
Vertiefendes Seminar zur Vorlesung Biochemie I · Aufgabe 1 Glyoxalat-Zyklus • Ermöglicht die...
14
Vertiefendes Seminar zur Vorlesung Biochemie I 22.01.2016 Bearbeitung Übungsblatt 10 Gerhild van Echten-Deckert Fon. +49-228-732703 Homepage: http://www.limes-institut-bonn.de/forschung/arbeitsgruppen/unit-3/
Transcript of Vertiefendes Seminar zur Vorlesung Biochemie I · Aufgabe 1 Glyoxalat-Zyklus • Ermöglicht die...

Vertiefendes Seminar zur Vorlesung Biochemie I
22.01.2016
Bearbeitung Übungsblatt 10g g
Gerhild van Echten-DeckertFon. +49-228-732703
Homepage:
http://www.limes-institut-bonn.de/forschung/arbeitsgruppen/unit-3/

Glyoxalat-ZyklusAufgabe 1
• Ermöglicht die Synthese von Glukose aus Acetyl-CoA
• in Pflanzen, Pilzen, Mikroorganismen
• Erzeugung von Zucker aus gespeichertem Fett zur Keimun
i O ll Gl i dt P i• eigenes Organell: Glyoxisom, verwandt zum Peroxisom
• verwendet auch Enzyme des Citrat-Cyclus• verwendet auch Enzyme des Citrat-Cyclus
• benötigt zwei zusätzliche Enzymeg y• Isocitrat-Lyase
• Malat-SynthaseA h it d Gl C i Sä ti i t t itt
2• Anwesenheit des Glyox.-C. in Säugetieren ist umstritten
Christoph Thiele / Biochemie 1 / WS2015/16

Glyoxalat-ZyklusGluconeog. Cytopl.
Glyoxalat-C. GlyoxisomGlyoxisom
ICL S i d Gl l• ICL erzeugt Succinat und Glyoxalat• Succinat wird in Mito. transportiert• Succinat Oxaloacetat• Oxaloacetat GlukoneogeneseO a oacetat G u o eoge ese• Glyoxalat + AcCoA erzeugen Malat
(Malat-Synthase)• Isocitrat wird aus Malat regeneriert
• Die Decarboxylierungen des Citrat.-C werden umgangen
Citrat-C. Mito.
3

Glyoxalat-Zyklus: Organisation
Glyoxisomey
4
Eastmond & Graham, TRENDS in Plant Science, Vol.6 No.2 February 2001
Christoph Thiele / Biochemie 1 / WS2015/16

Glyoxalat-Zyklus
Bilanz
4 x Ac-CoA + 2 FAD + 4 NAD+ 2 PEP + 2 CO2 + 4 NADH + 2 FADH2 + 2 CoA2 PEP + 2 NADH + 2 ATP Glukose + 2 NAD+ + 2 ADP2 PEP + 2 NADH + 2 ATP Glukose + 2 NAD+ + 2 ADP
4 Ac-CoA + 2 ATP + 2 NAD+ + 2 FAD 1 Glukose + 2 NADH + 2 FADH2 + 2 ADP + 2 CoA + 2 CO2
5

Glyoxalat-Zyklus: gesamte C-Bilanz
4 Ac-CoA + 2 ATP + 2 NAD+ + 2 FAD Glukose + 2 NADH + 2 FADH2 + 2 ADP + 2 CoA + 2 CO2
nur C Bilanz und x 4 ergibt: 16 Ac CoA 4 Glukose + 8 COnur C-Bilanz, und x 4 ergibt: 16 Ac-CoA 4 Glukose + 8 CO2
Fettsäure beta-Oxidation: 2 Palmitat 16 Ac-CoA
F tt ä bi th (k l R h i h BC 2 lid 119)Fettsäurebiosynthese (komplexe Rechnung, siehe BC 2 slide 119)
9 Glukose 2 Palmitat + 22 CO2
k l tt Z kl Gl k ( Ph t th ) F tt ä (i S ) A t l C A Gl kkompletter Zyklus: Glukose (aus Photosynthese) Fettsäure (im Samen) Acetyl-CoA Glukose (während der Keimung für Zellwandsynthese etc.)
9 Glukose 4 Glukose + 30 CO2 (+ Reduktionsequivalente / ATP)
6Christoph Thiele / Biochemie 1 / WS2015/16

Aufgabe 2

Penicilline greifen im Bereich der Zellteilung in denPenicilline greifen im Bereich der Zellteilung in den Stoffwechsel der Bakterien ein und blockieren die Synthese der bakteriellen Zellwand: Der Beta-Lactam-Ring der meisten gPenicillin-Antibiotika öffnet sich im Zytosol des exponierten Bakteriums und bindet in der geöffneten Form an das bakterielle Enzym D-Alanin-Transpeptidase. Auf diese Weise kann dieses Enzym nicht mehr die Alaninreste der Zellwand verknüpfen das befallene Bakterium stirbt dadurch schließlichverknüpfen, das befallene Bakterium stirbt dadurch schließlich ab.Durch die Wirkung im Bereich der bakteriellen Teilungsphase g g pist eine gleichzeitige Gabe von teilungshemmenden sogenannten Bakteriostatika kontraindiziert.

Tetracyclin verhindert die Anlagerung von Aminoacyl-tRNA an dierRNA in der 30-S-yUntereinheit des Bakterien-Ribosoms. Dadurch wird die Translation und letztlich die Proteinbiosynthese gestoppt.DieToxizität könnte auf eine Inaktivierung der 30-S Ribosomen der in den eukaryotischen WirtszellenS-Ribosomen der in den eukaryotischen Wirtszellen vorhandenen Mitochondrien zurückzuführen sein.

Ciprofloxacin ist ein synthetisches Antibiotikum mit breitem Wirkspektrum aus der Gruppe der Fluorchinolone. Die Substanz wurde 1981 von der Firma Bayer entwickelt und 1983 patentiert. [7][8] Antibiotika dieser Gr ppe hemmen die G rase on Bakterien nddieser Gruppe hemmen die Gyrase von Bakterien und damit ihre DNA-Replikation und ihre Zellteilung. Sie wirken dadurch bakterizid, und zwar vor allem gegenwirken dadurch bakterizid, und zwar vor allem gegen gramnegative Keime.

Folgende Mechanismen werden zur Erklärung der preferential exclusion herangezogen [56-59]:
Aufgabe 3
o ge de ec a s e we de u ä u g de p e e e t a e c us o e a ge oge [56 59]:1. Das Protein verhält sich inert. Die Interaktion zwischen Protein und Hilfsstoff werden einzig durch die Beschaffenheit der Hilfsstofflösung beeinflusst.�2 Infolge unterschiedlicher Größe von Hilfsstoffmolekül und Wasser können die kleinen Wassermoleküle i di ht Ob flä h b d k P t i i h l di öß Hilf t ff l kül E li t ieine dichtere Oberflächenbedeckung am Protein erreichen als die größeren Hilfsstoffmoleküle. Es liegt ein
sterischer Ausschluss der Hilfsstoffmoleküle vor.�2 Durch den Hilfsstoffzusatz wird die Oberflächenspannung der Lösung verändert. An der Grenzfläche zum hydrierten Protein werden die Hilfsstoffmoleküle bevorzugt ausgeschlossen, und es kommt zu einer y g g ,Ungleichverteilung in der Lösung. Die native Form des Proteins wird durch Hilfsstoffzusatz stabilisiert, welche die Grenzflächenspannung von Wasser erhöhen.2. Die chemische Eigenschaft der Proteinoberfläche bestimmt die Anziehung oder Abstoßung zwischen P t i d Hilf t fflöProtein und Hilfsstofflösung.�2 Durch Abstoßung der Hilfsstoffmoleküle von der Proteinoberfläche aufgrund gleicher Ladungen steht an der Proteinoberfläche mehr Platz für Wassermoleküle zur Verfügung. Das Protein wird bevorzugt hydratisiert und die native Struktur durch preferential exclusion stabilisiert.p�2 Hilfsstoffe wie Glycerol werden wegen eines solvophoben Effektes von der Proteinober- fläche ausgeschlossen. Der Kontakt zwischen den unpolaren Regionen des Proteins und der Glycerol-Wasser-Mischung ist entropisch noch ungünstiger als der Kontakt mit Wasser. Aus diesem Grund entfernen sich Gl l l kül d P t i b flä h d hi t l i it W i h tGlycerolmoleküle von der Proteinoberfläche und hinterlassen eine mit Wasser angereicherte Proteinoberfläche. Glycerol zeigt dieses Verhalten, obwohl es eine Affinität zu polaren Regionen der Proteine aufweist.
S.N.Timasheff, Stabilization of protein structure by solvent additives, Pharm.Biotechnol., Vol.3, Stab. Prot. Pharm., Part B, 1992, p.265-285.

Zusatz von Glycerol. Bei vielen Proteinen führt der Zusatz von Gl l (bi 50 % / ) lä H ltb k it b i ti fGlycerol (bis zu 50 % w/v) zu längerer Haltbarkeit bei tiefen Temperaturen, vor allem, weil Glycerol/Proteingemische auch bei tiefen Temperaturen nicht einfrieren Glucose Saccharosetiefen Temperaturen nicht einfrieren. Glucose, Saccharose, Fructose und Sorbitol scheinen einen ähnlichen Effekt auf die Stabilität eines Proteins in Lösung zu haben
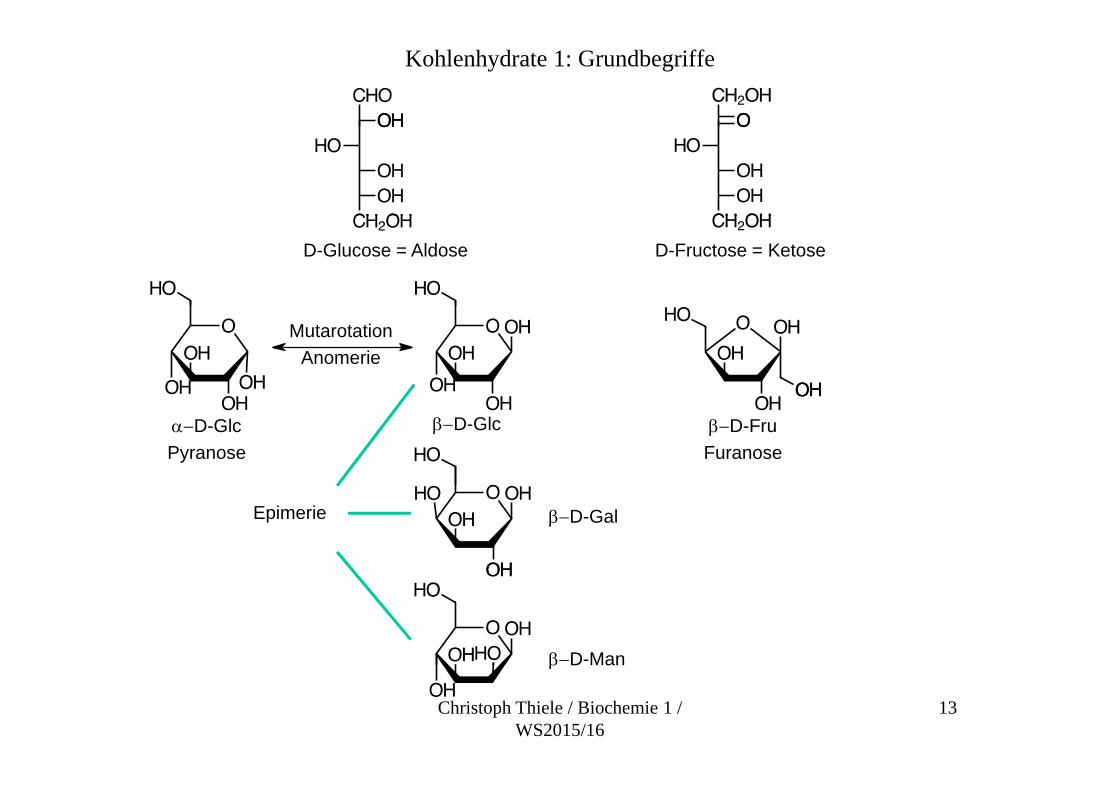
Kohlenhydrate 1: Grundbegriffe
D-Glucose = Aldose D-Fructose = Ketose
MutarotationAnomerie
D-GlcPyranose
D-Glc D-FruFuranose
Epimerie D-Gal
D-Man
Christoph Thiele / Biochemie 1 / WS2015/16
13
D Man
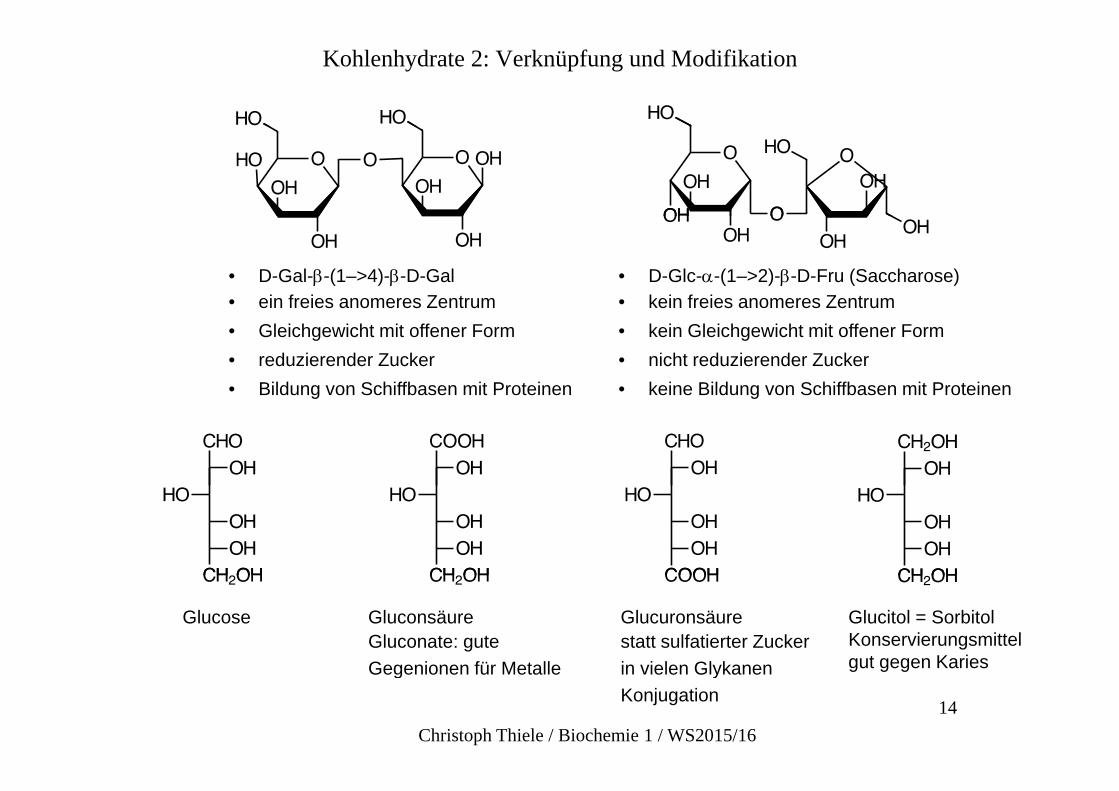
Kohlenhydrate 2: Verknüpfung und Modifikation
• D-Gal--(1–>4)--D-Gal• ein freies anomeres Zentrum
• D-Glc--(1–>2)--D-Fru (Saccharose)• kein freies anomeres Zentrumein freies anomeres Zentrum
• Gleichgewicht mit offener Form• reduzierender Zucker• Bildung von Schiffbasen mit Proteinen
kein freies anomeres Zentrum• kein Gleichgewicht mit offener Form• nicht reduzierender Zucker• keine Bildung von Schiffbasen mit Proteineng g
Glucose GluconsäureGluconate: gute Gegenionen für Metalle
Glucuronsäurestatt sulfatierter Zucker in vielen Glykanen
Glucitol = SorbitolKonservierungsmittelgut gegen Karies
14
Gegenionen für Metalle in vielen GlykanenKonjugation
gut gegen Karies
Christoph Thiele / Biochemie 1 / WS2015/16


















