
Zellbiologiefiles.schulbuchzentrum-online.de/pdf/978-3-507-10980-3-1-l.pdf · Eukaryotische Zelle...
40
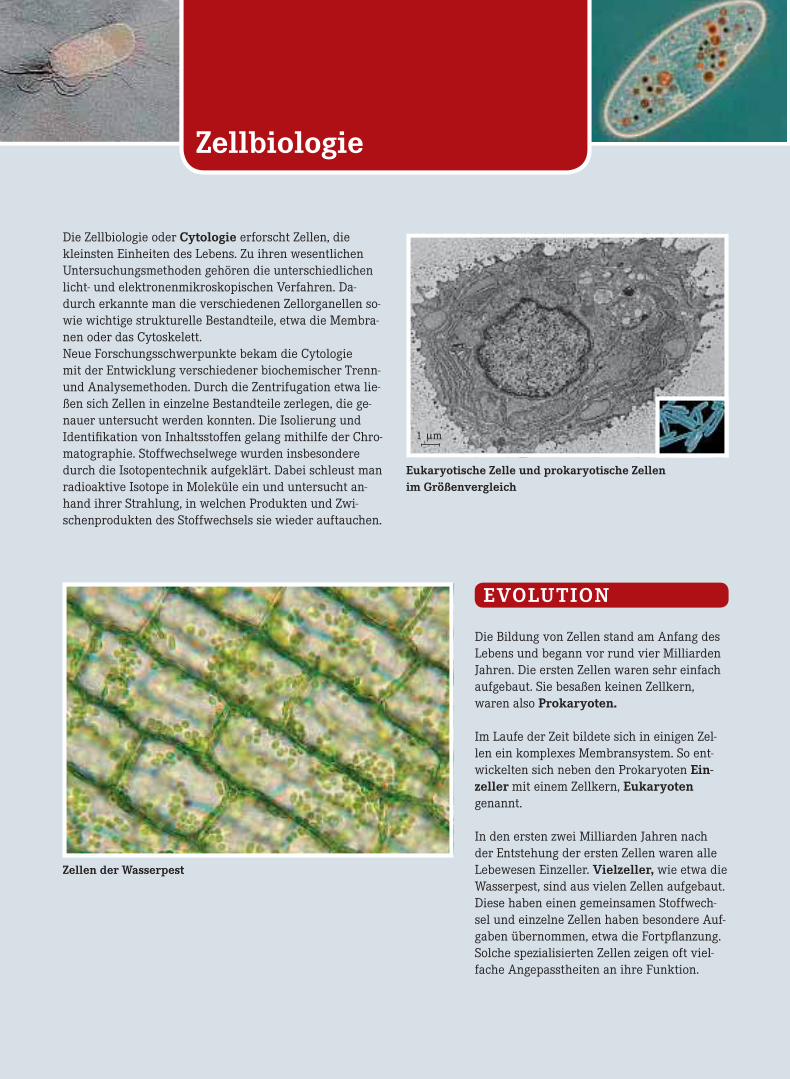
Zellbiologie Die Zellbiologie oder Cytologie erforscht Zellen, die kleinsten Einheiten des Lebens. Zu ihren wesentlichen Untersuchungsmethoden gehören die unterschiedlichen licht- und elektronenmikroskopischen Verfahren. Da- durch erkannte man die verschiedenen Zellorganellen so- wie wichtige strukturelle Bestandteile, etwa die Membra- nen oder das Cytoskelett. Neue Forschungsschwerpunkte bekam die Cytologie mit der Entwicklung verschiedener biochemischer Trenn- und Analysemethoden. Durch die Zentrifugation etwa lie- ßen sich Zellen in einzelne Bestandteile zerlegen, die ge- nauer untersucht werden konnten. Die Isolierung und Identifikation von Inhaltsstoffen gelang mithilfe der Chro- matographie. Stoffwechselwege wurden insbesondere durch die Isotopentechnik aufgeklärt. Dabei schleust man radioaktive Isotope in Moleküle ein und untersucht an- hand ihrer Strahlung, in welchen Produkten und Zwi- schenprodukten des Stoffwechsels sie wieder auftauchen. Eukaryotische Zelle und prokaryotische Zellen im Größenvergleich Zellen der Wasserpest EVOLUTION Die Bildung von Zellen stand am Anfang des Lebens und begann vor rund vier Milliarden Jahren. Die ersten Zellen waren sehr einfach aufgebaut. Sie besaßen keinen Zellkern, waren also Prokaryoten. Im Laufe der Zeit bildete sich in einigen Zel- len ein komplexes Membransystem. So ent- wickelten sich neben den Prokaryoten Ein- zeller mit einem Zellkern, Eukaryoten genannt. In den ersten zwei Milliarden Jahren nach der Entstehung der ersten Zellen waren alle Lebewesen Einzeller. Vielzeller, wie etwa die Wasserpest, sind aus vielen Zellen aufgebaut. Diese haben einen gemeinsamen Stoffwech- sel und einzelne Zellen haben besondere Auf- gaben übernommen, etwa die Fortpflanzung. Solche spezialisierten Zellen zeigen oft viel- fache Angepasstheiten an ihre Funktion. 1 μm
Transcript of Zellbiologiefiles.schulbuchzentrum-online.de/pdf/978-3-507-10980-3-1-l.pdf · Eukaryotische Zelle...
Zellbiologie
Die Zellbiologie oder Cytologie erforscht Zellen, diekleinsten Einheiten des Lebens. Zu ihren wesentlichenUntersuchungsmethoden gehören die unterschiedlichenlicht- und elektronenmikroskopischen Verfahren. Da-durch erkannte man die verschiedenen Zellorganellen so-wie wichtige strukturelle Bestandteile, etwa die Membra-nen oder das Cytoskelett.Neue Forschungsschwerpunkte bekam die Cytologiemit der Entwicklung verschiedener biochemischer Trenn-und Analysemethoden. Durch die Zentrifugation etwa lie-ßen sich Zellen in einzelne Bestandteile zerlegen, die ge-nauer untersucht werden konnten. Die Isolierung undIdentifikation von Inhaltsstoffen gelang mithilfe der Chro-matographie. Stoffwechselwege wurden insbesonderedurch die Isotopentechnik aufgeklärt. Dabei schleust manradioaktive Isotope in Moleküle ein und untersucht an-hand ihrer Strahlung, in welchen Produkten und Zwi-schenprodukten des Stoffwechsels sie wieder auftauchen.
Eukaryotische Zelle und prokaryotische Zellenim Größenvergleich
Zellen der Wasserpest
EVOLUTION
Die Bildung von Zellen stand am Anfang desLebens und begann vor rund vier MilliardenJahren. Die ersten Zellen waren sehr einfachaufgebaut. Sie besaßen keinen Zellkern,waren also Prokaryoten.
Im Laufe der Zeit bildete sich in einigen Zel-len ein komplexes Membransystem. So ent-wickelten sich neben den Prokaryoten Ein-zeller mit einem Zellkern, Eukaryotengenannt.
In den ersten zwei Milliarden Jahren nachder Entstehung der ersten Zellen waren alleLebewesen Einzeller. Vielzeller, wie etwa dieWasserpest, sind aus vielen Zellen aufgebaut.Diese haben einen gemeinsamen Stoffwech-sel und einzelne Zellen haben besondere Auf-gaben übernommen, etwa die Fortpflanzung.Solche spezialisierten Zellen zeigen oft viel-fache Angepasstheiten an ihre Funktion.
1 μm

Seit sich die Zellbiologie als Wissenschaft etabliert hat,wurde versucht, einzelne Zellen und Gewebe von Tierenoder Pflanzen in Nährlösungen am Leben zu erhalten,um sie zu untersuchen. Schon 1885 gelang es, Zellenaus Hühnerembryonen in einer Salzlösung für mehrereTage am Leben zu erhalten. Solche Zellen, die in einemNährmedium kultiviert werden, bezeichnet man alsZellkulturen. Sie sind ein wichtiges Arbeitsmaterialin der Zellbiologie.
Die ersten menschlichen Zellen, die man in Zellkulturenhalten konnte, waren die sogenannten HELA-Zellen.1951 hatte man sie als Probe aus dem Gebärmutterhals-tumor von Henrietta Larcks entnommen. Die Patientinstarb mit 31 Jahren an ihrer Krebserkrankung. IhreTumorzellen teilen sich in Nährlösungen noch heuteungehemmt weiter. Sie liefern Zellmaterial zuForschungszwecken und sind weltweit in zahlreichenLabors vorrätig. Viele Arzneimittel konnten mit HELA-Zellen entwickelt werden. Würde man heute alleHELA-Zellen zusammentragen, so wären es mehr Zellen,als Henrietta Larcks je hatte.
Anwendung finden die Erkenntnisse der Zell-biologie insbesondere in der Biotechnologie.Medikamente wie etwa Impfstoffe, Blutgerin-nungsfaktoren und Wachstumsfaktoren fürdie Blutbildung wie das Erythropoietin, auchEPO genannt, werden mittels Zellkulturen inBiofermentern in großen Mengen hergestellt.Zellkulturen sind auch die Grundlage fürdie Gewebezüchtung. Damit wird geschädig-tes Gewebe, etwa Knorpelgewebe in denGelenken, regeneriert oder zerstörtes Ge-webe, zum Beispiel verbrannte Haut, ersetzt.Auch die Züchtung von Herzklappen auspatienteneigenem Material ist bereits inersten Studien gelungen.Biofermenter
Hela-Zellen
ANWENDUNG
FORSCHUNG
Schwerpunkte der zellbiologischen Forschung liegenheutzutage auf der Zellteilung, der Differenzierung vonZellen und dem programmierten Zelltod, der Apoptose.Durch Apoptose werden Zellen, die der Körper nichtmehr benötigt, beseitigt. Dieser Prozess hat sich als einneues universelles Merkmal von eukaryotischen Zellenherausgestellt.

Zellbiologie16
1 Der Bau von Zellen
1.1 Das lichtmikroskopische Bildvon Zellen
Nachdem niederländische Brillenmacher ge-gen Ende des 16. Jahrhunderts die ersteneinfachen Mikroskope aus mehreren mitein-ander verbundenen Linsen gebaut hatten, er-öffnete sich Naturforschern eine ganz neueWelt: die Welt des Mikrokosmos. Der englischeUniversalgelehrte Robert Hooke veröffent-lichte 1665 eine Sammlung von Zeichnungenin seinem Buch „Micrographia“, darunter auchdie einer dünnen Korkscheibe, deren waben-artig angeordneten Kammern er als mikrosko-pisch kleine „Poren“ oder „Zellen“ bezeichnete.Hooke wurde durch seine Beobachtungenzu einem der Begründer der Cytologie oderZellbiologie.
Knapp 200 Jahre später, in den Jahren 1838und 1839, formulierten der Zoologe TheodorSchwann und der Botaniker MatthiasSchleiden die Zelltheorie, welche die Zelleals Grundbaustein von Tieren und Pflanzenbeschrieb. Sie wurde 1850 durch den Medi-ziner Rudolf Virchow erweitert. Berühmtwurde seine Formulierung „Omnis cellula ecellula.“: Jede Zelle geht aus einer anderenZelle hervor.
Die Zelltheorie wurde in der Folge so über-zeugend bestätigt, dass sie heute allgemeinanerkannt ist. Ihre wesentlichen Aussagenlauten:
(1) Alle Lebewesen bestehen aus Zellen.(2) Alle Zellen sind in ihrem Grundbauplan
und in ihrem Stoffwechsel im Wesent-lichen gleich.
(3) Alle Zellen entstehen aus Zellen durchZellteilung.
(4) Die Zelle ist die grundlegende Einheit fürdie Struktur und Funktion der Organis-men.
Unser Auge hat ein Auflösungsvermögenvon circa 100 μm (= 0,1 mm). Das heißt, eskann zwei nebeneinander liegende Punktenur dann getrennt wahrnehmen, wenn ihr Ab-stand mindestens 100 μm beträgt. Da die meis-ten Zellen kleiner als 100 μm sind, kann mansie ohne Hilfsmittel nicht sehen.
3 Verschiedene Zelltypen.A Chlamydomonas (pflanzlicher Einzeller);B Blattzellen der Wasserpest;C Nervenzelle (gefärbt);D Leberzellen (gefärbt)
1 ZweilinsigesMikroskop vonHooke
2 Zeichnung vonHooke. Zellstrukturvon Kork
A
B
C
Dμm = Mikrometernm = Nanometer
Zellbiologie 17
Begrenzt wird das Auflösungsvermögen nebenanatomischen Gegebenheiten unter anderemdurch den Sehwinkel. Je größer der Sehwin-kel, desto größer nimmt man das Objekt wahr.Die Lupe bewirkt dabei eine Vergrößerung desSehwinkels; das Objekt erscheint uns dadurchgrößer als es in Wirklichkeit ist (virtuellesBild).
Wenn man sehr kleine Strukturen erkennenwill, benötigt man die Hilfe eines Mikroskops.Die Vergrößerung des Objekts erfolgt hierdurch die Kombination von zwei Linsensyste-men, dem Objektiv und dem Okular. DasObjektiv erzeugt zunächst ein vergrößertesZwischenbild des Objekts. Dieses wird vomOkular, das wie eine Lupe wirkt, nochmalsvergrößert. Die Gesamtvergrößerung einesMikroskops ergibt sich aus dem Produkt vonObjektiv- und Okularvergrößerung.
Entscheidend für die Leistungsfähigkeit einesMikroskops ist aber weniger die Gesamtver-größerung, sondern sein Auflösungsvermögen.Dieses ist zum einen von der Qualität der Lin-sen, zum anderen von der Wellenlänge desverwendeten Lichtes abhängig. Dabei ent-spricht die maximale Auflösung aus physika-lischen Gründen der Hälfte der Wellenlängedes eingesetzten Lichtes. Bei blauem Lichtmit einer Wellenlänge von 400 nm beträgt dastheoretische Auflösungsvermögen des Licht-mikroskops (LM) circa 200 nm (= 0,2 μm).
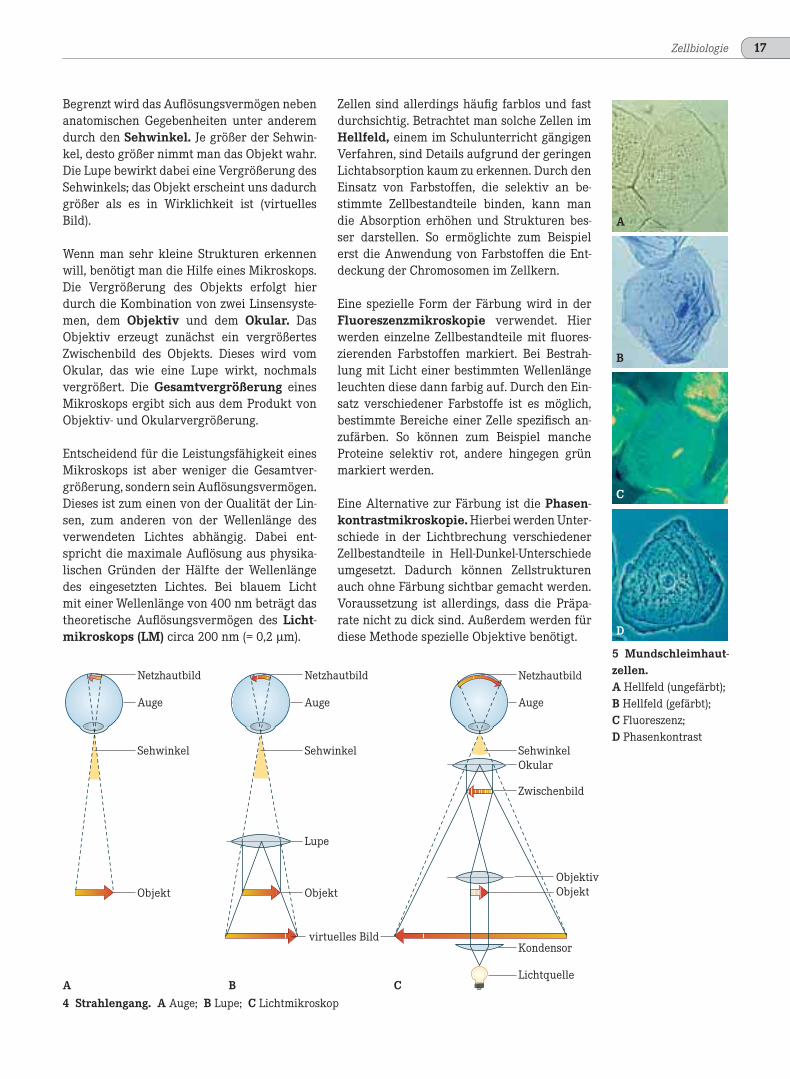
Zellen sind allerdings häufig farblos und fastdurchsichtig. Betrachtet man solche Zellen imHellfeld, einem im Schulunterricht gängigenVerfahren, sind Details aufgrund der geringenLichtabsorption kaum zu erkennen. Durch denEinsatz von Farbstoffen, die selektiv an be-stimmte Zellbestandteile binden, kann mandie Absorption erhöhen und Strukturen bes-ser darstellen. So ermöglichte zum Beispielerst die Anwendung von Farbstoffen die Ent-deckung der Chromosomen im Zellkern.
Eine spezielle Form der Färbung wird in derFluoreszenzmikroskopie verwendet. Hierwerden einzelne Zellbestandteile mit fluores-zierenden Farbstoffen markiert. Bei Bestrah-lung mit Licht einer bestimmten Wellenlängeleuchten diese dann farbig auf. Durch den Ein-satz verschiedener Farbstoffe ist es möglich,bestimmte Bereiche einer Zelle spezifisch an-zufärben. So können zum Beispiel mancheProteine selektiv rot, andere hingegen grünmarkiert werden.
Eine Alternative zur Färbung ist die Phasen-kontrastmikroskopie. Hierbei werden Unter-schiede in der Lichtbrechung verschiedenerZellbestandteile in Hell-Dunkel-Unterschiedeumgesetzt. Dadurch können Zellstrukturenauch ohne Färbung sichtbar gemacht werden.Voraussetzung ist allerdings, dass die Präpa-rate nicht zu dick sind. Außerdem werden fürdiese Methode spezielle Objektive benötigt.
4 Strahlengang. A Auge; B Lupe; C Lichtmikroskop
5 Mundschleimhaut-zellen.A Hellfeld (ungefärbt);B Hellfeld (gefärbt);C Fluoreszenz;D Phasenkontrast
A
B
C
D

Zellbiologie18
Da das Auflösungsvermögen guter Lichtmi-kroskope auf etwa 200 nm begrenzt ist, kön-nen zwar einzelne Zellorganellen wie der Zell-kern oder Mitochondrien erkannt werden,ihre Feinstruktur jedoch nicht. Erst die Ent-wicklung des Elektronenmikroskops (EM)durch Ernst Ruska im Jahr 1931 erlaubte dieDarstellung von Strukturen und Details, dieim Lichtmikroskop nicht sichtbar sind.
In der Elektronenmikroskopie benutzt manElektronenstrahlen, die eine deutlich kürze-re Wellenlänge als die des sichtbaren Lichtshaben. Das Auflösungsvermögen erhöht sichdadurch um das 2000-Fache gegenüber demLichtmikroskop; es können Strukturen bis zu0,1 nm aufgelöst werden.
Da Elektronen von Luftteilchen abgelenktwerden, muss im Elektronenmikroskop einHochvakuum erzeugt werden. Der von derElektronenquelle erzeugte Elektronenstrahlwird mithilfe elektromagnetischer Felder, diewie die Linsen im Lichtmikroskop wirken,auf das Objekt gelenkt. Das entstehendevergrößerte Zwischenbild wird durch ein elek-tromagnetisches Okular, Projektiv genannt,nachvergrößert. Das sichtbare Bild entsteht,wenn die Elektronen auf den Leuchtschirmtreffen.
1.2 Das elektronenmikroskopischeBild von Zellen
Um Objekte im Elektronenmikroskop unter-suchen zu können, müssen diese zunächstchemisch fixiert werden, damit die Zell-strukturen möglichst unverändert erhaltenbleiben. Anschließend werden die Präparateentwässert, in Kunstharz eingebettet und miteinem Ultramikrotom in 50 bis 100 nm dünneScheiben geschnitten. Ein menschliches Haarkann so in 1000 bis 2000 Längsschnitte zer-legt werden.
Bei der Transmissionselektronenmikro-skopie (TEM) durchdringt der Elektronen-strahl, ähnlich wie der Lichtstrahl im Licht-mikroskop, das Präparat. Vergleichbar mitder Färbung bei der Lichtmikroskopie mussim TEM der Kontrast der Präparate verstärktwerden. Hierzu werden die Schnitte mitSchwermetallsalzen behandelt. Durch dieKombination von zweidimensionalen Bildernvieler aufeinanderfolgender Dünnschnittelässt sich die räumliche Gestalt eines Orga-nells rekonstruieren.
Mit dem Rasterelektronenmikroskop (REM)können räumliche Bilder der Oberfläche ei-nes Objekts erzeugt werden. Hierzu wird zu-nächst die Oberfläche des Objekts mit einerdünnen Schwermetallschicht bedampft. An-schließend wird ein dünner Elektronenstrahl,die Primärelektronen, über das meistensmassive Objekt geführt. Dabei werden Elektro-nen zurückgestreut oder aus dem Objektherausgeschleudert, die Sekundärelektro-nen. Diese werden registriert und auf einemMonitor dargestellt. Durch zeilenweise Abtas-tung, das sogenannte Rastern, entsteht so einräumliches Bild von der Objektoberfläche.
Durch die Kombination von TEM- und REM-Aufnahmen kann schließlich der prinzipielleAufbau einer Zelle nachgebildet werden. Dasso entstandene dreidimensionale Modell kannallerdings die Dynamik, die in einer lebendenZelle herrscht, nicht verdeutlichen.
Erläutern Sie, warum im Elektronen-mikroskop keine Frischpräparate unter-sucht werden können.
2 Transmissionselektronenmikroskop.A im Labor; B Strahlengang im TEM
1 Sinneszellen ausdem Ohr einesOchsenfrosches.A LM;B TEM;C REM
A
B
C
A B

Zellbiologie 19
3 EM-Bilder und schematische Darstellung einer tierischen Zelle (Größenangaben: 1mm = 10–3m, 1μm = 10–6m, 1nm = 10–9m)
Zellbiologie20
1.3 Vergleich von Prokaryotenund Eukaryoten
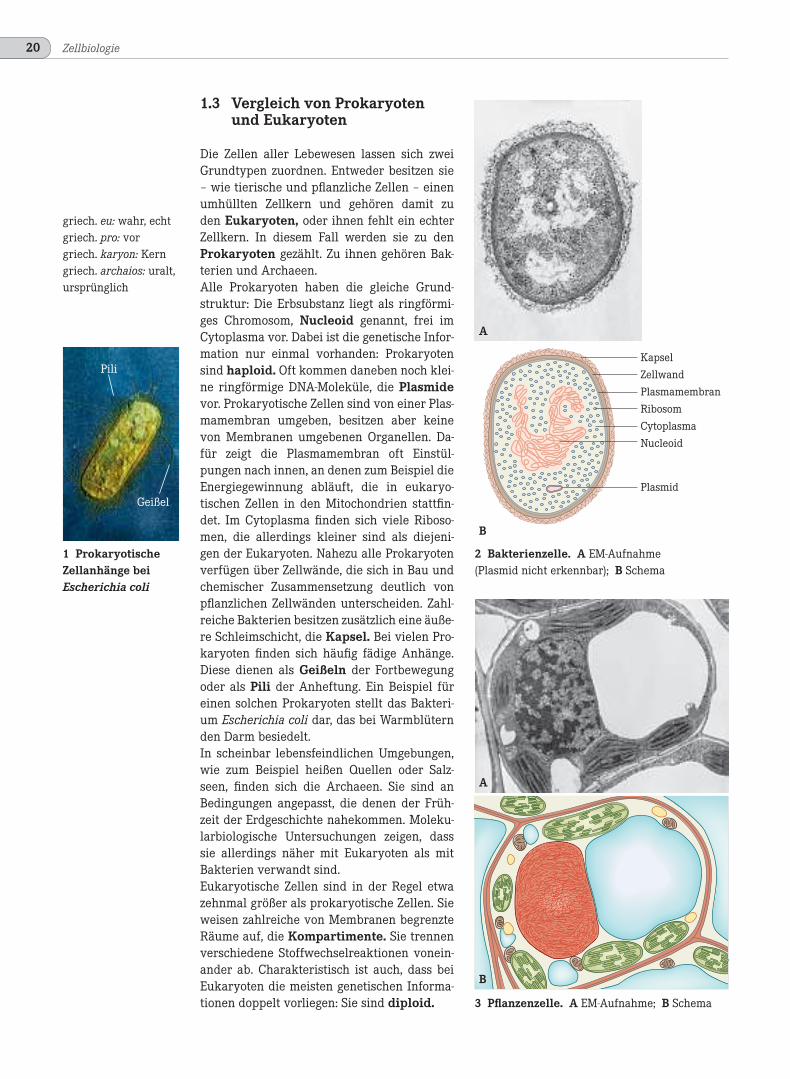
Die Zellen aller Lebewesen lassen sich zweiGrundtypen zuordnen. Entweder besitzen sie– wie tierische und pflanzliche Zellen – einenumhüllten Zellkern und gehören damit zuden Eukaryoten, oder ihnen fehlt ein echterZellkern. In diesem Fall werden sie zu denProkaryoten gezählt. Zu ihnen gehören Bak-terien und Archaeen.Alle Prokaryoten haben die gleiche Grund-struktur: Die Erbsubstanz liegt als ringförmi-ges Chromosom, Nucleoid genannt, frei imCytoplasma vor. Dabei ist die genetische Infor-mation nur einmal vorhanden: Prokaryotensind haploid. Oft kommen daneben noch klei-ne ringförmige DNA-Moleküle, die Plasmidevor. Prokaryotische Zellen sind von einer Plas-mamembran umgeben, besitzen aber keinevon Membranen umgebenen Organellen. Da-für zeigt die Plasmamembran oft Einstül-pungen nach innen, an denen zum Beispiel dieEnergiegewinnung abläuft, die in eukaryo-tischen Zellen in den Mitochondrien stattfin-det. Im Cytoplasma finden sich viele Riboso-men, die allerdings kleiner sind als diejeni-gen der Eukaryoten. Nahezu alle Prokaryotenverfügen über Zellwände, die sich in Bau undchemischer Zusammensetzung deutlich vonpflanzlichen Zellwänden unterscheiden. Zahl-reiche Bakterien besitzen zusätzlich eine äuße-re Schleimschicht, die Kapsel. Bei vielen Pro-karyoten finden sich häufig fädige Anhänge.Diese dienen als Geißeln der Fortbewegungoder als Pili der Anheftung. Ein Beispiel füreinen solchen Prokaryoten stellt das Bakteri-um Escherichia coli dar, das bei Warmblüternden Darm besiedelt.In scheinbar lebensfeindlichen Umgebungen,wie zum Beispiel heißen Quellen oder Salz-seen, finden sich die Archaeen. Sie sind anBedingungen angepasst, die denen der Früh-zeit der Erdgeschichte nahekommen. Moleku-larbiologische Untersuchungen zeigen, dasssie allerdings näher mit Eukaryoten als mitBakterien verwandt sind.Eukaryotische Zellen sind in der Regel etwazehnmal größer als prokaryotische Zellen. Sieweisen zahlreiche von Membranen begrenzteRäume auf, die Kompartimente. Sie trennenverschiedene Stoffwechselreaktionen vonein-ander ab. Charakteristisch ist auch, dass beiEukaryoten die meisten genetischen Informa-tionen doppelt vorliegen: Sie sind diploid.
griech. eu: wahr, echtgriech. pro: vorgriech. karyon: Kerngriech. archaios: uralt,ursprünglich
2 Bakterienzelle. A EM-Aufnahme(Plasmid nicht erkennbar); B Schema
3 Pflanzenzelle. A EM-Aufnahme; B Schema
Ribosom
Cytoplasma
Zellwand
Plasmamembran
Nucleoid
Plasmid
Kapsel
A
B
A
B
1 ProkaryotischeZellanhänge beiEscherichia coli
Pili
Geißel
Zellbiologie 21
Endocytose eines weiteren Bakteriums(Energiegewinnung aus der Fotosynthese)
Pilze Tierische Zellen Pflanzliche Zellen
Mitochondrien Chloro-plast
1.4 Die Endosymbionten-Theorie
Wie sich im Laufe der Evolution aus pro-karyotischen Zellen komplexe Zellen miteinem Zellkern und verschiedenen Zellorga-nellen entwickelt haben, ist auch heutzutagenoch eine fundamentale Frage der Biologie.
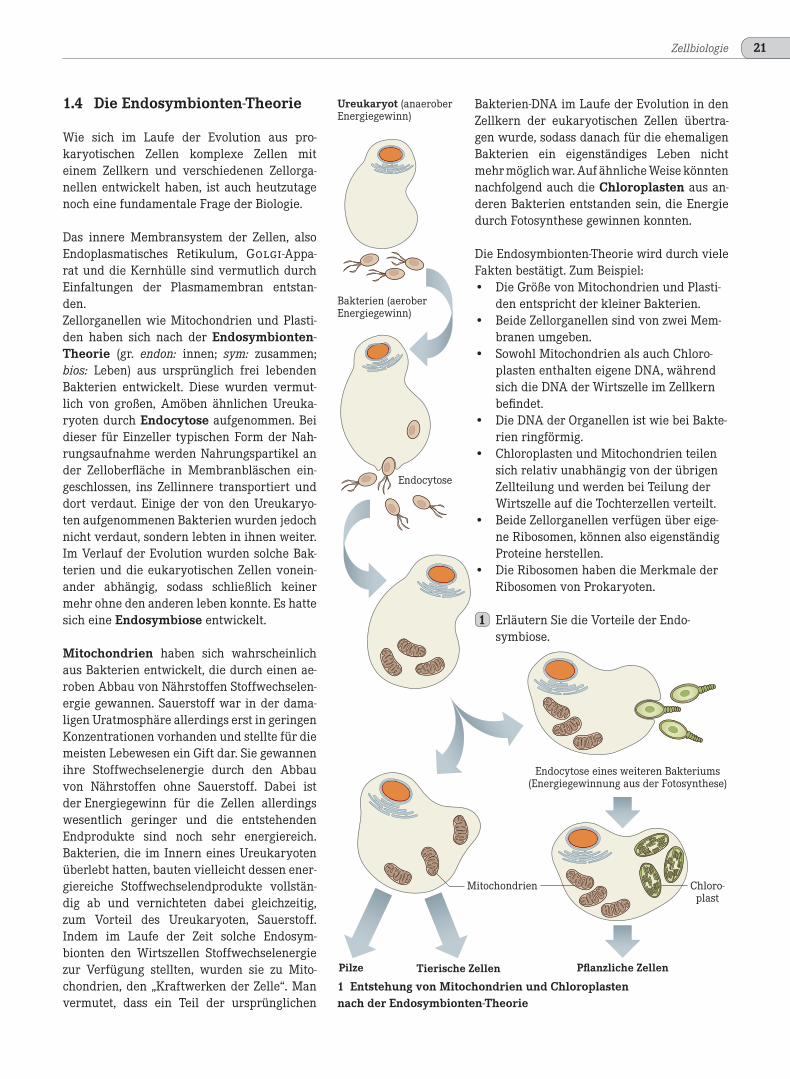
Das innere Membransystem der Zellen, alsoEndoplasmatisches Retikulum, Golgi-Appa-rat und die Kernhülle sind vermutlich durchEinfaltungen der Plasmamembran entstan-den.Zellorganellen wie Mitochondrien und Plasti-den haben sich nach der Endosymbionten-Theorie (gr. endon: innen; sym: zusammen;bios: Leben) aus ursprünglich frei lebendenBakterien entwickelt. Diese wurden vermut-lich von großen, Amöben ähnlichen Ureuka-ryoten durch Endocytose aufgenommen. Beidieser für Einzeller typischen Form der Nah-rungsaufnahme werden Nahrungspartikel ander Zelloberfläche in Membranbläschen ein-geschlossen, ins Zellinnere transportiert unddort verdaut. Einige der von den Ureukaryo-ten aufgenommenen Bakterien wurden jedochnicht verdaut, sondern lebten in ihnen weiter.Im Verlauf der Evolution wurden solche Bak-terien und die eukaryotischen Zellen vonein-ander abhängig, sodass schließlich keinermehr ohne den anderen leben konnte. Es hattesich eine Endosymbiose entwickelt.
Mitochondrien haben sich wahrscheinlichaus Bakterien entwickelt, die durch einen ae-roben Abbau von Nährstoffen Stoffwechselen-ergie gewannen. Sauerstoff war in der dama-ligen Uratmosphäre allerdings erst in geringenKonzentrationen vorhanden und stellte für diemeisten Lebewesen ein Gift dar. Sie gewannenihre Stoffwechselenergie durch den Abbauvon Nährstoffen ohne Sauerstoff. Dabei istder Energiegewinn für die Zellen allerdingswesentlich geringer und die entstehendenEndprodukte sind noch sehr energiereich.Bakterien, die im Innern eines Ureukaryotenüberlebt hatten, bauten vielleicht dessen ener-giereiche Stoffwechselendprodukte vollstän-dig ab und vernichteten dabei gleichzeitig,zum Vorteil des Ureukaryoten, Sauerstoff.Indem im Laufe der Zeit solche Endosym-bionten den Wirtszellen Stoffwechselenergiezur Verfügung stellten, wurden sie zu Mito-chondrien, den „Kraftwerken der Zelle“. Manvermutet, dass ein Teil der ursprünglichen
1 Entstehung von Mitochondrien und Chloroplastennach der Endosymbionten-Theorie
Bakterien-DNA im Laufe der Evolution in denZellkern der eukaryotischen Zellen übertra-gen wurde, sodass danach für die ehemaligenBakterien ein eigenständiges Leben nichtmehr möglich war. Auf ähnliche Weise könntennachfolgend auch die Chloroplasten aus an-deren Bakterien entstanden sein, die Energiedurch Fotosynthese gewinnen konnten.
Die Endosymbionten-Theorie wird durch vieleFakten bestätigt. Zum Beispiel:• Die Größe von Mitochondrien und Plasti-
den entspricht der kleiner Bakterien.• Beide Zellorganellen sind von zwei Mem-
branen umgeben.• Sowohl Mitochondrien als auch Chloro-
plasten enthalten eigene DNA, währendsich die DNA der Wirtszelle im Zellkernbefindet.
• Die DNA der Organellen ist wie bei Bakte-rien ringförmig.
• Chloroplasten und Mitochondrien teilensich relativ unabhängig von der übrigenZellteilung und werden bei Teilung derWirtszelle auf die Tochterzellen verteilt.
• Beide Zellorganellen verfügen über eige-ne Ribosomen, können also eigenständigProteine herstellen.
• Die Ribosomen haben die Merkmale derRibosomen von Prokaryoten.
Erläutern Sie die Vorteile der Endo-symbiose.
Ureukaryot (anaeroberEnergiegewinn)
Endocytose
Bakterien (aeroberEnergiegewinn)

Zellbiologie22
1 Gewebetypen beim Pflanzenblatt (C, D = LM-Aufnahmen).A Laubblatt (Schema); B Blattadern (Leitbündel); C Palisadenparenchym; D untere Epidermis mit Spaltöffnungen
1.5 Spezialisierung von Zellen
Rund drei Milliarden Jahre lang war die Erdeausschließlich von einzelligen Lebewesen be-wohnt. Gegen Ende des Präkambriums, voretwa 700 Millionen Jahren, tauchten erstmalsmehrzellige Organismen auf. Im weiteren Ver-lauf der Evolution entwickelten sich dann Or-ganismen, die durch eine Vielfalt unterschied-licher Zelltypen mit jeweils unterschiedlicherFunktion gekennzeichnet waren. Diese Spe-zialisierung ermöglichte eine effektivereArbeitsteilung.
In Lebewesen, wie zum Beispiel in höherenPflanzen und Tieren, bilden Zellen des glei-chen Typs mit ähnlicher Funktion ein Ge-webe. In Pflanzen unterscheidet man dreiverschiedene Gewebetypen: Das Abschluss-gewebe, die Epidermis, bedeckt den Korpusder Pflanze und dient ihrem Schutz. Sie be-steht aus dicht gepackten Zellen, die eineoder mehrere Schichten ausbilden können.Die Epidermis kann auch weitere spezialisier-
te Zellen enthalten. So bilden abgewandelteEpidermiszellen die sogenannten Schließzel-len in den Blättern. Durch die Veränderungihrer Form werden die Spaltöffnungen im Blattgeschlossen oder geöffnet. Über diese kanndie Pflanze ihren Gasaustausch mit der Um-gebungsluft sowie den Wasserverlust kontrol-lieren. Die Epidermis ist oftmals von einerWachsschicht, der Cuticula, überzogen. Dieseschränkt zusätzlich den Wasserverlust ein.Das Grundgewebe, auch Parenchym genannt,wird im Blatt von zwei Schichten fotosyn-thetisch aktivem Gewebe gebildet. Die obereSchicht besteht aus langgestreckten, zylindri-schen Zellen, die das Palisadenparenchymbilden. Es dient größtenteils der Fotosynthese.Die untere Schicht weist unregelmäßig ge-formte Zellen auf, die von großen Interzellular-räumen umgeben sind. Im Vergleich zum Pali-sadenparenchym enthält dieses sogenannteSchwammparenchym etwa drei- bis fünfmalweniger Chloroplasten. Seine Hauptaufgabeist die Durchlüftung des Parenchyms. Überseine Interzellularräume gelangt das für dieFotosynthese benötigte Kohlenstoffdioxid bisin das Palisadenparenchym.Die Leitbündel bilden das Leitgewebe einerPflanze. Über sie gelangen Wasser und daringelöste Mineralstoffe von den Wurzeln zu denBlattzellen. Im Gegenzug werden Fotosynthe-seprodukte aus den Blattzellen über die Leit-bündel in den restlichen Pflanzenkörper trans-portiert.Mehrere Gewebetypen ergänzen sich und wir-ken gemeinsam als Organ. Die Organe der Blü-tenpflanzen sind Wurzel, Spross und Blatt.Mehrere Organe, die gemeinsam ein funktio-nelles Ganzes bilden, wie zum Beispiel bei derBlüte einer Pflanze, nennt man Organsystem.Ein Organismus setzt sich aus verschiedenenOrganen und Organsystemen zusammen, diesich in ihren Aufgaben ergänzen.
A
B C D
Zellbiologie 23
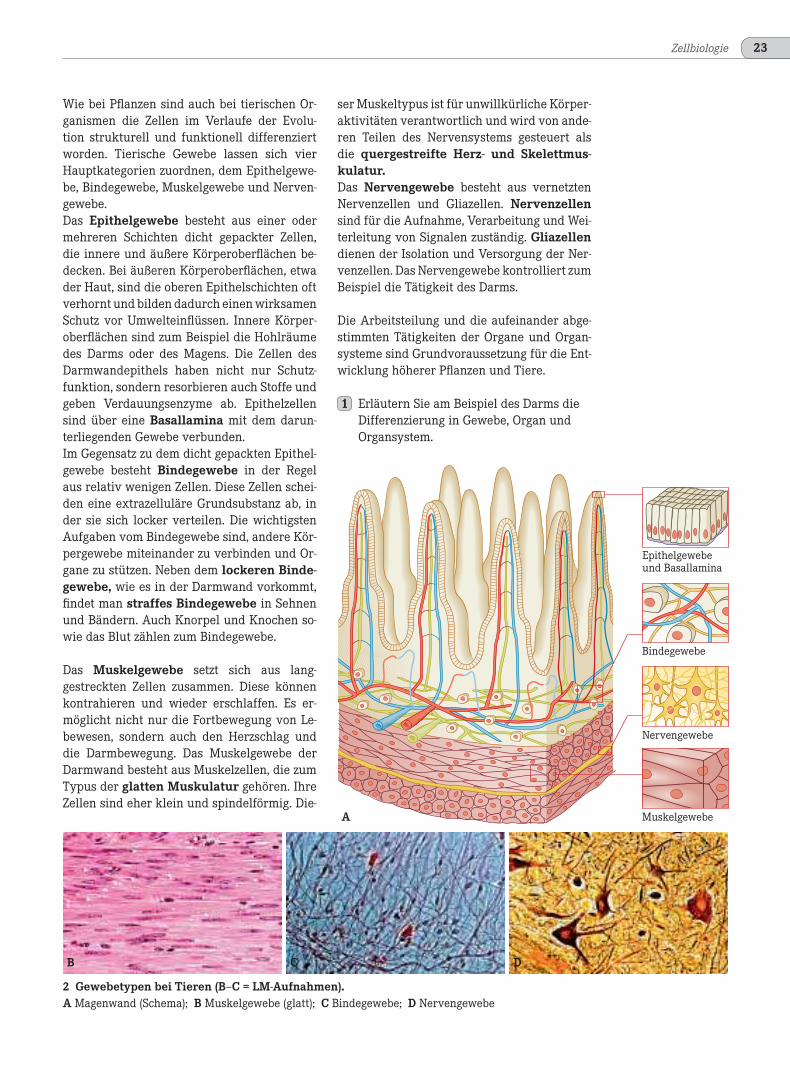
2 Gewebetypen bei Tieren (B–C = LM-Aufnahmen).A Magenwand (Schema); B Muskelgewebe (glatt); C Bindegewebe; D Nervengewebe
Wie bei Pflanzen sind auch bei tierischen Or-ganismen die Zellen im Verlaufe der Evolu-tion strukturell und funktionell differenziertworden. Tierische Gewebe lassen sich vierHauptkategorien zuordnen, dem Epithelgewe-be, Bindegewebe, Muskelgewebe und Nerven-gewebe.Das Epithelgewebe besteht aus einer odermehreren Schichten dicht gepackter Zellen,die innere und äußere Körperoberflächen be-decken. Bei äußeren Körperoberflächen, etwader Haut, sind die oberen Epithelschichten oftverhornt und bilden dadurch einen wirksamenSchutz vor Umwelteinflüssen. Innere Körper-oberflächen sind zum Beispiel die Hohlräumedes Darms oder des Magens. Die Zellen desDarmwandepithels haben nicht nur Schutz-funktion, sondern resorbieren auch Stoffe undgeben Verdauungsenzyme ab. Epithelzellensind über eine Basallamina mit dem darun-terliegenden Gewebe verbunden.Im Gegensatz zu dem dicht gepackten Epithel-gewebe besteht Bindegewebe in der Regelaus relativ wenigen Zellen. Diese Zellen schei-den eine extrazelluläre Grundsubstanz ab, inder sie sich locker verteilen. Die wichtigstenAufgaben vom Bindegewebe sind, andere Kör-pergewebe miteinander zu verbinden und Or-gane zu stützen. Neben dem lockeren Binde-gewebe, wie es in der Darmwand vorkommt,findet man straffes Bindegewebe in Sehnenund Bändern. Auch Knorpel und Knochen so-wie das Blut zählen zum Bindegewebe.
Das Muskelgewebe setzt sich aus lang-gestreckten Zellen zusammen. Diese könnenkontrahieren und wieder erschlaffen. Es er-möglicht nicht nur die Fortbewegung von Le-bewesen, sondern auch den Herzschlag unddie Darmbewegung. Das Muskelgewebe derDarmwand besteht aus Muskelzellen, die zumTypus der glatten Muskulatur gehören. IhreZellen sind eher klein und spindelförmig. Die-
ser Muskeltypus ist für unwillkürliche Körper-aktivitäten verantwortlich und wird von ande-ren Teilen des Nervensystems gesteuert alsdie quergestreifte Herz- und Skelettmus-kulatur.Das Nervengewebe besteht aus vernetztenNervenzellen und Gliazellen. Nervenzellensind für die Aufnahme, Verarbeitung und Wei-terleitung von Signalen zuständig. Gliazellendienen der Isolation und Versorgung der Ner-venzellen. Das Nervengewebe kontrolliert zumBeispiel die Tätigkeit des Darms.
Die Arbeitsteilung und die aufeinander abge-stimmten Tätigkeiten der Organe und Organ-systeme sind Grundvoraussetzung für die Ent-wicklung höherer Pflanzen und Tiere.
Erläutern Sie am Beispiel des Darms dieDifferenzierung in Gewebe, Organ undOrgansystem.
A
B C D

Zellbiologie24
�
��
��
�
���� ���
����������
�
�����������
1.5 Bau der Biomembranen
Jede lebende Zelle wird durch eine Zellmem-bran begrenzt. Im Cytoplasma der Zelle be-grenzen wiederum Membranen zahlreicheReaktionsräume, Kompartimente genannt.So sind etwa Vakuolen, Chloroplasten und Mi-tochondrien von Membranen umgeben. Insge-samt besteht 60 bis 90 Prozent der Trocken-masse einer Zelle aus Membranen.
Membranen, die Bestandteile von Zellen sind,bezeichnet man als Biomembranen. Sie be-stehen hauptsächlich aus Lipiden und Pro-teinen. Bei den Membranlipiden handelt essich um Phospholipide. Diese Moleküle sindamphipathisch, das heißt sie besitzen gleich-zeitig einen wasserliebenden, hydrophilenund einen wasserabstoßenden, hydrophobenBereich. Aufgrund dieses Aufbaus lagern sichPhospholipide in hochgeordneten Molekülver-bänden zusammen. Auf einer Wasserober-fläche zum Beispiel bilden sie eine Schicht,die nur aus einer einzigen Lage von Molekülenbesteht. In dieser sogenannten monomole-kularen Schicht tauchen die hydrophilen Be-reiche der Phospholipide ins Wasser ein unddie hydrophoben Bereiche ragen aus demWasser heraus.
In Biomembranen sind Phospholipide in einerDoppelschicht angeordnet. Darin sind ihre hy-drophilen Bereiche jeweils nach außen, zurwässrigen Umgebung hin ausgerichtet. Diehydrophoben Bereiche sind nach innen ge-lagert, sodass sie kaum mit Wasser in Kon-takt kommen.
Unter Berücksichtigung dieser und weitererErkenntnisse konnten Seymour Singer undGarth Nicolson 1972 ein Membran-Modellvorschlagen, das bis heute Arbeitsgrundlageder Cytologie ist. Ihr Modell beschreibt, dassdie Phospholipid-Doppelschicht mehr oderweniger flüssig ist, vergleichbar mit einem Öl-film auf der Oberfläche eines Sees. Die Mem-branproteine schwimmen darin unregelmäßigverteilt „wie Eisberge in der See“. Deshalbwird das Modell auch Fluid-Mosaik-Modellgenannt. Manche Proteine sind auf die Phos-pholipid-Doppelschicht aufgelagert. Sie wer-den als periphere Proteine bezeichnet. ImVergleich dazu tauchen integrale Proteinemehr oder weniger tief in die Lipid-Doppel-schicht ein. Einige integrale Proteine reichensogar von der einen bis zur anderen Membran-seite. Mehrere Proteine können auch als Pro-teinkomplexe zusammengelagert sein.
Membranproteine haben verschiedene Funk-tionen, dazu gehören der Stofftransport sowiedie Verknüpfung von Zellen miteinander durchsogenannte zellverbindende Proteine.
Auf der Außenseite von Zellmembranen findetman oft noch Kohlenhydrate. Sie sind entwe-der an Membranproteine oder an Phospho-lipide gebunden. Demzufolge unterscheidetman zwischen Glykoproteinen und Glyko-lipiden. Diese Kohlenhydratmoleküle habenals Erkennungsmerkmale der Zellen einegroße Bedeutung. Außerdem verleihen sietierischen Zellen ihre Festigkeit.
Skizzieren und erläutern Sie die An-ordnung von Phospholipiden auf einerÖloberfläche.Sticht man mit einer dünnen Nadel ineine Zelle, so bleibt an der Einstichstellenach dem Herausziehen der Nadel keinLoch. Erklären Sie diese Beobachtung.
3 Fluid-Mosaik-Modell einer Biomembran. A Phospholipid-Doppelschicht; B integrales Protein; C peripheres Protein;D Glykoprotein; E Glykolipid; F zellverbindendes Protein; G Proteinkomplex
2 Anordnung vonPhospholipiden aufeiner Wasserober-fläche
1 Phospholipid
��������� ���������� ����������� �������!��"���

Zellbiologie 25
METHODE Arbeiten mit Modellen
Modelle haben im naturwissenschaft-lichen Erkenntnisprozess eine wichti-ge Funktion. Wissenschaftler schlagenModelle unter anderem vor, um damitbisherige Erkenntnisse und Beobach-tungen zusammenzufassen, zu ver-einfachen und zu veranschaulichen.Ein Modell ist keine Kopie des Origi-nals. Es zeichnet sich durch Abstrak-tion aus: Bestimmte Merkmale desOriginals werden bewusst vernach-lässigt, andere hervorgehoben. ImSinne einer Theorie oder Hypothesewerden Modelle über das Originalkonstruiert. Neue Erkenntnissekönnen Modelle stützen, verfeinern,aber auch grundlegend verändern.Beispielhaft wird dies an der Erfor-schung der Biomembranen deut-lich.
Langmuir stellte 1917 erstmals ausamphipathischen Fettsäuren stabileSchichten auf einer Wasseroberflächeher. Aus dem Vergleich der Schicht-dicke und der Molekülgröße schlosser, dass die Schichten nur aus einereinzigen Lage von Molekülen beste-hen.
Mit dem gleichen Verfahren bildetenGorther und Grendel 1925 ausPhospholipiden, die sie aus den Zell-membranen einer bestimmten Anzahlvon Roten Blutkörperchen extrahierthatten, eine monomolekulare Schichtauf einer Wasseroberfläche. BeimAusmessen dieser Schicht stellten siefest, dass ihre Fläche genau doppeltso groß war wie die Oberflächen derverwendeten Roten Blutkörperchen.Daraus folgerten sie, dass die Phos-pholipide in Zellmembranen alsDoppelschicht angeordnet seinmüssen. Auf der Grundlage dieserErkenntnisse wurde 1935 ein erstesMembran-Modell, das Sandwich-Modell, vorgestellt. Es stellt die Bio-membran als Phospholipid-Doppel-schicht dar, auf der auf beidenSeiten jeweils eine Schicht aus kugel-förmigen Proteinen aufgelagert ist.
1 Sandwich-Modell
�����������
������ ����� ����������
�����������
3 Erweitertes Sandwich-Modell
������������
2 Biomembran (EM-Aufnahme)
Die ersten elektronenmikroskopischenUntersuchungen von Membranen ausden 50er Jahren schienen das Sand-wich-Modell zu bestätigen, da die Bil-der zunächst falsch gedeutet wurden.Man erkannte zwei dunkle Linien miteiner Dicke von jeweils 2 nm, zwi-schen denen sich eine etwa 3 nm di-cke hellere Linie befand. Die hellereLinie deutete man zunächst als dashydrophobe Innere der Membran, diebeiden äußeren dunklen Linien alsBereiche aus Proteinen und hydro-philen Bereichen der Phospholipide.Heute weiß man, dass die eigentlicheMembran nur der Bereich zwischenden dunklen Schichten ist. Die bei-den dunklen Linien entstehen durchdas Kontrastmittel, das der Membranbeidseitig aufgelagert ist.
teinen besitzen. Da man davon aus-ging, dass alle Biomembranen iden-tisch aufgebaut sind, sprach manauch von der Einheitsmembran.
Im weiteren Verlauf der Membran-forschung ergaben sich immer mehrWidersprüche zu diesen Modell-vorstellungen. Zum Beispiel ergabenchemische Analysen, dass das Ver-hältnis von Lipiden zu Proteinen beiMembranen verschiedener Herkunftauch sehr unterschiedlich sein kann.Vor allem wurde die Lage der Protei-ne bezweifelt. Membranen mit aufge-lagerten Proteinschichten könntennicht so flexibel sein, wie man es beiBiomembranen beobachtete.
Nachdem man mithilfe besondererMikroskopiertechniken festgestellthatte, dass Membranproteine unter-schiedlich tief in die Phospholipid-Doppelschicht eintauchen und sehrunregelmäßig in Membranen verteiltsein können, schlugen SeymourSinger und Garth Nicolson 1972das Fluid-Mosaik-Modell vor. Natür-lich wird auch dieses Membran-Modell ständig überprüft, ob es neugewonnenen Erkenntnissen und Be-obachtungen widerspricht. Es wirdweiterentwickelt und muss vielleichtauch eines Tages grundsätzlich um-gestaltet werden. Zurzeit ist es aberdie geeignetste Arbeitsgrundlage fürdie Erforschung von Membranen undZellen.
Um den Transport von hydrophilenStoffen durch Biomembranen erklä-ren zu können, wurde das Sandwich-Modell erweitert. Man vermutete,dass Biomembranen Kanäle aus Pro-

Zellbiologie26
1.7 Stofftransport durchBiomembranen
Zellen und Zellkompartimente müssen einer-seits vor dem Eindringen vieler Stoffe ge-schützt werden, andererseits sind sie auf einenständigen Stoffaustausch mit ihrer Umgebungangewiesen. Biomembranen ermöglichen denkontrollierten Stoffaustausch der Zellen undZellkompartimente. So sind sie für bestimmteMoleküle, wie etwa Wasser, gut durchlässig,während andere Moleküle und Ionen sie nichtüberwinden können. Man bezeichnet Biomem-branen deshalb als semipermeabel.
Trennt eine semipermeable Membran zweiKompartimente, wobei ein KompartimentWasser, das andere eine Zuckerlösung enthält,nimmt im Verlauf der Zeit das Volumen derZuckerlösung zu, während das Volumen desWassers abnimmt. Aus dieser Beobachtunglässt sich schließen, dass Wasser durch dieMembran in die Zuckerlösung dringt. Aberwarum bewegen sich Wassermoleküle in dasKompartiment mit der Zuckerlösung hinein?
2 Stofftransport ( = Zuckerlösung, = Wasser). A Diffusion; B Osmose
Gibt man einige Tropfen rote Tinte in ein Ge-fäß mit Wasser, ohne dass die Lösung umge-rührt oder bewegt wird, kann man nacheiniger Zeit feststellen, dass das Wasser über-all gleichmäßig rot gefärbt ist.
In Flüssigkeiten und Gasen befinden sich dieTeilchen ständig in regelloser Bewegung. ImLauf der Zeit verteilen sie sich dadurch gleich-mäßig in dem ihnen zur Verfügung stehendenRaum. Dieser Vorgang wird als Diffusion be-zeichnet. Die roten Tintenmoleküle verteilensich langsam aufgrund ihrer Eigenbewegunggleichmäßig in dem Wasser. Sie diffundierendabei von Bereichen, in denen sie in hohenKonzentrationen vorliegen, dahin, wo ihreKonzentration gering ist, nämlich ins Wasser.Auch die Wasserteilchen diffundieren in Be-reiche, in denen ihre Konzentration geringerist, also zwischen die Tintenteilchen. Das Er-gebnis ist ein Konzentrationsausgleich.
Befindet sich zwischen der Zuckerlösung unddem Wasser eine semipermeable Membran,können zwar die Wassermoleküle durch dieMembran diffundieren, die großen Zucker-moleküle aber nicht. Die Wassermoleküle ver-teilen sich gleichmäßig in dem ihnen zur Ver-fügung stehenden Raum, also in den beidenKompartimenten, während sich die Zucker-moleküle nur in dem Kompartiment bewegenkönnen, in welches sie eingefüllt wurden.Dies erklärt die Volumenzunahme der Zucker-lösung. Diese Diffusion durch eine semiper-meable Membran wird Osmose genannt. Dif-fundieren Wassermoleküle in Kompartimentehinein, kommt es dort neben einer Volumen-vergrößerung auch zu einer Druckerhöhung.
Für die Diffusion muss eine Zelle keine Stoff-wechselenergie aufwenden. Deshalb gehörtdie Diffusion zu den sogenannten passivenTransportprozessen.
Nur sehr kleine unpolare Moleküle wieSauerstoff und Kohlenstoffdioxid können diePhospholipid-Doppelschicht der Membranendurch einfache Diffusion überwinden. Dabeigilt, je fettlöslicher ein Molekül ist, destoschneller diffundiert es durch die Phospho-lipid-Doppelschicht. Polare Wassermolekülekönnen im Vergleich dazu nur sehr schlechtdurch die Phospholipid-Doppelschicht diffun-dieren. Viele Zellen sind aber auf einen schnel-len Wassertransport angewiesen. Er wird mit-
�������������� ���
�
��������
�
� ��� ������
lat. semi: halblat. permeo: etwasdurchwandern
1 Versuch zurDiffusion

Zellbiologie 27
hilfe integraler Membranproteine möglich, dievon einem zentralen Kanal durchzogen wer-den. Durch ihn kann Wasser ungehindert voneiner Membranseite zur anderen diffundieren.Solche Wasserkanalproteine, auch Aquapori-ne genannt, sind so gebaut, dass nur Wasser-moleküle ihren Kanal durchqueren können.Durch andere Kanalproteine können ver-schiedene gelöste Moleküle und Ionen voneiner zur anderen Membranseite diffundie-ren.
Für die Reizaufnahme und Erregungsweiter-leitung durch Sinnes- und Nervenzellen habengesteuerte Ionenkanäle eine große Bedeu-tung. Sie funktionieren ähnlich wie Tore, diedurch ein Signal geöffnet und geschlossenwerden. Ionenkanäle in den Membranen kön-nen durch Signalmoleküle wie Hormone oderdurch Ladungsveränderungen an der Mem-bran gesteuert werden. Ist ein Kanal erst ein-mal geöffnet, können in Abhängigkeit vomKonzentrationsgradienten viele Ionen gleich-zeitig hindurchströmen.
Andere Transportproteine in der Membranhaben charakteristische Bindungsstellen fürMoleküle, die durch die Membran transpor-tiert werden sollen. Ähnlich wie ein Schlüsselzum Schloss passen muss, können nur genaupassende Moleküle daran binden. Dadurchverändert sich die räumliche Struktur desTransportproteins so, dass dabei das gebun-dene Molekül durch die Membran geschleustwird. Solche spezifischen Transportproteine
werden als Carrier, der Transportvorgang alsCarriertransport bezeichnet.
Manche Carrier besitzen Bindungsstellen fürmehrere Substrate. Zum Beispiel binden diefür Glucose spezifischen Carrier neben einemGlucosemolekül gleichzeitig Natrium-Ionen.Nur wenn alle Bindungsstellen besetzt wer-den, erfolgt der Transport. Dieser sogenann-te Cotransport verläuft beim Symport ingleicher Richtung, beim Antiport in entgegen-gesetzter Richtung.
Carrier können ihre Moleküle nur durch dieMembran transportieren, wenn der Stoff aufeiner Seite in höherer Konzentration vor-kommt als auf der anderen Seite. Der Trans-port erfolgt also, wie auch bei der Diffusion,mit dem Konzentrationsgefälle. Deshalb mussdie Zelle dafür keine Stoffwechselenergieaufwenden. Sowohl die carrier- als auchdie kanalvermittelte Diffusion zählen zumpassiven Transport.
Viele Stoffe, wie etwa Glucose, können inZellen aufgenommen werden, obwohl in denZellen bereits eine höhere Glucosekonzentra-tion vorliegt als außerhalb. Für den Carrier-transport entgegen eines Konzentrationsge-fälles müssen Zellen Stoffwechselenergie be-reitstellen. Deshalb spricht man dann voneinem aktiven Transport. Die Energie wirdbenötigt, um den Stoff gegen den Konzentra-tionsgradienten zu befördern.
3 Transportprozesse durch Biomembranen
�������
��������������� ����
��������������������� ��������������� � ����
������� ��������� ������ ���������
� ������� �
�� ����� ����
�����������
�����
�
������������
4 Symport
5 Antiport

Zellbiologie28
1 Plasmolyse und Deplasmo-lyse bei Ligusterbeerenzellen
In lichtmikroskopischen Aufnahmenvon Zellen der Ligusterbeere erkenntman, dass sie fast vollständig voneiner zentralen Vakuole ausgefülltsind. Der Inhalt der Vakuole istdurch den Pflanzenfarbstoff Ligulinblauviolett gefärbt.
Befinden sich die Zellen in einer wäss-rigen Umgebung, ist das Zellplasmanur als schmaler Saum zwischenZellwand und Vakuole zu erkennen.Die Zellmembran liegt, wie im Sche-ma A dargestellt, eng an der Zell-wand an. Legt man die Zellen inkonzentrierte Zucker- oder Salz-Lösungen, beobachtet man, dass derProtoplast, so bezeichnet man dieZelle ohne Zellwand, schrumpft. Erlöst sich dabei, wie im Schema B dar-gestellt, von der Zellwand ab. Dabeiintensiviert sich die Farbe in der Va-kuole. Dieser Vorgang wird als Plas-molyse bezeichnet. Gibt man solcheplasmolysierten Zellen in Wasser, soerfolgt der umgekehrte Vorgang, dieDeplasmolyse. Der Protoplast ge-langt dabei wieder in seinen ur-sprünglichen Zustand zurück.
a) Benennen Sie die gekennzeichne-ten Zellstrukturen in den Schemata Aund B.b) Deuten Sie die Beobachtungen beider Plasmolyse und Deplasmolyse.c) Betrachten Sie die Animation zurPlasmolyse bei Zwiebelzellen undvergleichen Sie diese mit der Plasmo-lyse bei den Ligusterbeerenzellen.
2 Osmose-Modell
a) Erklären Sie mithilfe des abge-bildeten Osmose-Modells die Vor-gänge an der Zellmembran und amTonoplasten sowohl bei der Plasmo-lyse als auch bei der Deplasmolyse.b) Überprüfen Sie, welche Aspektedurch das Modell nur begrenzt ver-anschaulicht werden.
3 GrenzplasmolyseDen Zustand einer Zelle, bei dem sichdie Zellmembran nur minimal vonder Zellwand abgelöst hat, bezeichnetman als Grenzplasmolyse. Im Zell-verband spricht man von Grenz-plasmolyse, wenn 50 Prozent der Zel-len im plasmolysierten Zustandvorliegen. Anhand einer Grenzplas-molyse kann man die Konzentra-tionen von gelösten Stoffen im Zell-plasma und Zellsaft bestimmen.Planen Sie ein Experiment, mit demfestgestellt werden kann, wie hochdie Glucosekonzentration in den Zel-len der Ligusterbeeren ist.
4 Aufbau der BiomembranGibt man Zellen der Ligusterbeerein Aceton oder Alkohol, so kannman beobachten, dass der FarbstoffLigulin aus der Vakuole und aus derZelle in das umgebende Mediumausläuft.
Deuten Sie diese Beobachtung undskizzieren Sie in diesem Zusammen-hang den Aufbau einer Biomembran.
5 AquaporineQuecksilber-Ionen verstopfen dieAquaporine in Biomembranen.a) Erklären Sie die Funktion vonAquaporinen in Biomembranen.Vergleichen Sie in diesem Zusam-menhang die kanalvermittelte mitder carriervermittelten Diffusion.b) Beschreiben und erklären Sie,welches Ergebnis Sie bei einemPlasmolyse- und Deplasmolyse-versuch erwarten, wenn die Zellender Ligusterbeere kurz vor demVersuch in eine Quecksilberchlorid-Lösung getaucht werden.
6 Stofftransport durchBiomembranen
Die Abbildung zeigt modellhaft denStofftransport an der Biomembraneines Kompartiments. In Schema Bbefindet sich im Kompartiment einEnzym, welches den Stoff umwandelt.a) Vergleichen Sie den Stofftransportin Schema A und B und erklären Siedie Unterschiede.b) Betrachten Sie die Animation zumIonenfallenexperiment und schrei-ben Sie dazu ein Versuchsprotokoll.c) Vergleichen Sie das Ionenfallen-experiment mit dieser modellhaftenDarstellung des Stofftransports.
�������
� �
�� ��� �
AUFGABEN Plasmolyse und Deplasmolyse bei Ligusterbeerenzellen

Zellbiologie 29
1 Plasmolyse und Deplasmo-lyse bei EpidermiszellenMaterial: rote Küchenzwiebel;Pinzette; Rasierklinge; Objektträger;Deckgläschen; in Streifen geschnitte-nes Filtrierpapier; Tropfpipette;1-molare Kaliumnitrat-Lösung;Mikroskop; WasserDurchführung: Schneiden Sie indie untere Epidermis einer Zwiebel-schuppe mit einer Rasierklingemehrere kleineRechtecke.
Fassen Sie eines der Rechtecke mitder Pinzette an einer Ecke an undziehen Sie es vorsichtig von derZwiebelschuppe ab. Übertragen Siees in einen Wassertropfen auf einemObjektträger, sodass die Oberseiteoben liegt, und decken Sie es miteinem Deckgläschen ab. Saugen Siedas überschüssige Wasser an derSeite des Deckgläschens mit Filtrier-papier ab. Mikroskopieren Sie dasPräparat mindestens bei 300-facherVergrößerung. Geben Sie dann einenTropfen Kaliumnitrat-Lösung nebendas Deckgläschen und saugen Sie esvon der gegenüberliegenden Seitemit Filtrierpapier unter das Deck-gläschen.
Mikroskopieren Sie nach etwa zehnMinuten das Präparat bei mindestens300-facher Vergrößerung.Aufgaben:a) Zeichnen Sie eine Zelle aus demZellverband, die sich in Wasser be-
findet, und eine Zelle, die sich in Kali-umnitrat-Lösung befindet. BeschriftenSie die erkennbaren Strukturen.b) Erklären Sie auf der Grundlageder Osmose, wie die Unterschiedezwischen den Zellen in Wasser und inder Kaliumnitrat-Lösung entstehen.c) Informieren Sie sich anhand derAnimation zur Plasmolyse undDeplasmolyse über diese Vorgängeund erklären Sie, welche Vorgängeim Endstadium einer Plasmolyse amTonoplasten ablaufen.d) Planen Sie ein Experiment füreine Deplasmolyse bei IhremZwiebelzellenpräparat. Führen Siedas Experiment danach durch.
2 Turgordruck im Modell
Strömt Wasser in den Protoplasteneiner Pflanzenzelle, so nimmt er anVolumen zu. Dabei drückt er auf dieZellwand. Dieser Druck, Turgorgenannt, nimmt solange zu, bis derGegendruck durch die Zellwandverhindert, dass sich der Protoplastweiter ausdehnen kann. Der Turgorspielt eine entscheidende Rolle fürdie Stabilität und Steifheit vonPflanzen und pflanzlichen Geweben.
Material: Luftballon; dünnwandigePlastikflasche (In Boden und Wändewurden Löcher gestochen.)Durchführung: Basteln Sie aus denangegebenen Materialien ein Modelleiner Pflanzenzelle.Aufgaben:a) Vergleichen Sie eine Pflanzen-zelle mit dem Modell. Geben Sie an,welche Bestandteile des Modellsund einer Pflanzenzelle sichentsprechen.b) Erklären Sie anhand des Modells,was man in den Naturwissenschaftenunter einem Modell versteht.c) Erklären Sie mit dem Modell denTurgordruck, die Plasmolyse und dieDeplasmolyse.d) Beschreiben Sie, welche Trans-portvorgänge am Plasmalemma ab-laufen, wenn der Turgordruck undder Gegendruck durch die Zellwandgleich groß sind.
3 TurgorverlustMaterial: frisch abgeschnitteneSprosse mit Laubblättern einerkrautigen Pflanze, z.B. FleißigesLieschen; zwei Bechergläser (100 ml);gesättigte Kochsalz-Lösung; WasserDurchführung: Stellen Sie einenSpross in Wasser, den anderen indie gesättigte Kochsalz-Lösung.Aufgabe:Beschreiben und erklären Sie dasVersuchsergebnis, das nach etwa24 Stunden zu beobachten ist.
1 Epidermiszellen der roten Küchenzwiebel (LM-Aufnahme). Die Zellen sindfast vollständig von der Vakuole ausgefüllt. Der Vakuoleninhalt ist durch Pflanzen-farbstoffe, sogenannte Anthocyane, rot gefärbt.
�������
������
��� �� �������
������������
������������
PRAKTIKUM Plasmolyse bei Pflanzenzellen

Zellbiologie30
1.8 Zellbestandteile
Der Zellkern oder Nucleus (Größe: 3–15 μm)ist abgesehen von der Zentralvakuole in man-chen Pflanzenzellen das größte Zellorganelleiner eukaryotischen Zelle. Er enthält das Erb-material, das für die Zellteilung und für dieSteuerung der Stoffwechselvorgänge verant-wortlich ist, und gilt somit als Steuerzentraleder Zelle.
Der Kern ist vom Cytoplasma durch die Kern-hülle abgegrenzt. Diese besteht aus einer in-neren und einer äußeren Membran und stehtmit dem Endoplasmatischen Retikulum (ER)der Zelle in enger Verbindung. Über die Kern-poren in der Kernhülle ist ein kontrollierterStoffaustausch zwischen dem Kerninnerenund dem umgebenden Zellplasma möglich. ImInnern des Zellkerns befindet sich, eingebettetin eine wässrige Grundsubstanz, ein Gewirraus fädigen Strukturen, das als Chromatinbezeichnet wird. Es besteht aus Proteinen undNucleinsäuren und enthält die genetische In-formation der Zelle. Das Chromatin verdichtetsich vor jeder Zellteilung, sodass klar vonein-ander getrennte Chromosomen sichtbar wer-den. Weiter kommen im Zellkern ein odermehrere Zellkernkörperchen oder Nucleolivor. Sie sind beteiligt an der Bildung der Ribo-somen, den Orten der Proteinbiosynthese.
Ribosomen (Größe: 15–30 nm) sind sehr klei-ne rundliche Zellorganellen, die frei im Cyto-plasma vorkommen oder an die röhrenför-migen Strukturen des EndoplasmatischenRetikulums gebunden sind. Ribosomen syn-thetisieren mithilfe von Bauanleitungen ausdem Zellkern die Zellproteine. Besonders vieleRibosomen kommen in Zellen vor, die ständigviele Proteine synthetisieren, wie etwa Drü-senzellen. So kann eine menschliche Leberzel-le mehrere Millionen Ribosomen besitzen.Freie Ribosomen produzieren Proteine für denEigenbedarf der Zelle. Am ER gebundene Ribo-somen synthetisieren hauptsächlich Proteine,die in die Zellmembranen eingebaut oder vonder Zelle abgegeben werden.
Mitochondrien (Größe: 1–10 μm) sind stäb-chenförmige Zellorganellen. Die Zahl der Mito-chondrien pro Zelle ist abhängig von der Stoff-wechselaktivität der Zelle. Eine menschlicheLeberzelle enthält in der Regel mehr als 1000Mitochondrien. Mitochondrien vermehrensich durch Zweiteilung. Sie werden von zweiMembranen begrenzt, einer inneren und äu-ßeren. Die innere Membran ist zur Ober-flächenvergrößerung in viele Falten gelegt, diesich ins Innere des Mitochondriums erstre-cken. Die Grundsubstanz, die ein Mitochon-drium ausfüllt wird Matrix genannt. Sie ent-hält DNA, Ribosomen und zahlreiche Enzyme.
1 Zellkern. A EM-Aufnahme; B Schema; C REM-Aufnahme; D Kernporen (EM-Aufnahme)
μm = Mikrometer1 μm = 10–6 m;entspricht demMillionstel einesMeters
nm = Nanometer1 nm = 10–9 m;entspricht demMilliardstel einesMeters
1 μmA B
Nucleoli
Kernhülle
Chromatin
ERRibosom
C D
Kernpore
1 μm 100 nm
Zellbiologie 31
Mitochondrien werden auch als „Kraftwerkeder Zellen“ bezeichnet, denn in ihnen findenStoff- und Energieumwandlungen statt. Ener-giereiche Stoffe werden in den Mitochondrienmithilfe von Sauerstoff enzymatisch abgebaut.Diesen Prozess bezeichnet man als Zell-atmung. Die dabei frei werdende Energie wirdin Form von Adenosintriphosphat, abge-kürzt ATP, gespeichert. Die Bildung von ATPfindet an der inneren Mitochondrienmembranstatt. ATP-Moleküle werden ins Cytoplasmabefördert und können dort überall bei ener-gieverbrauchenden Reaktionen Energie be-reitstellen. Zum Beispiel liefert ATP die Ener-gie für den aktiven Carriertransport.
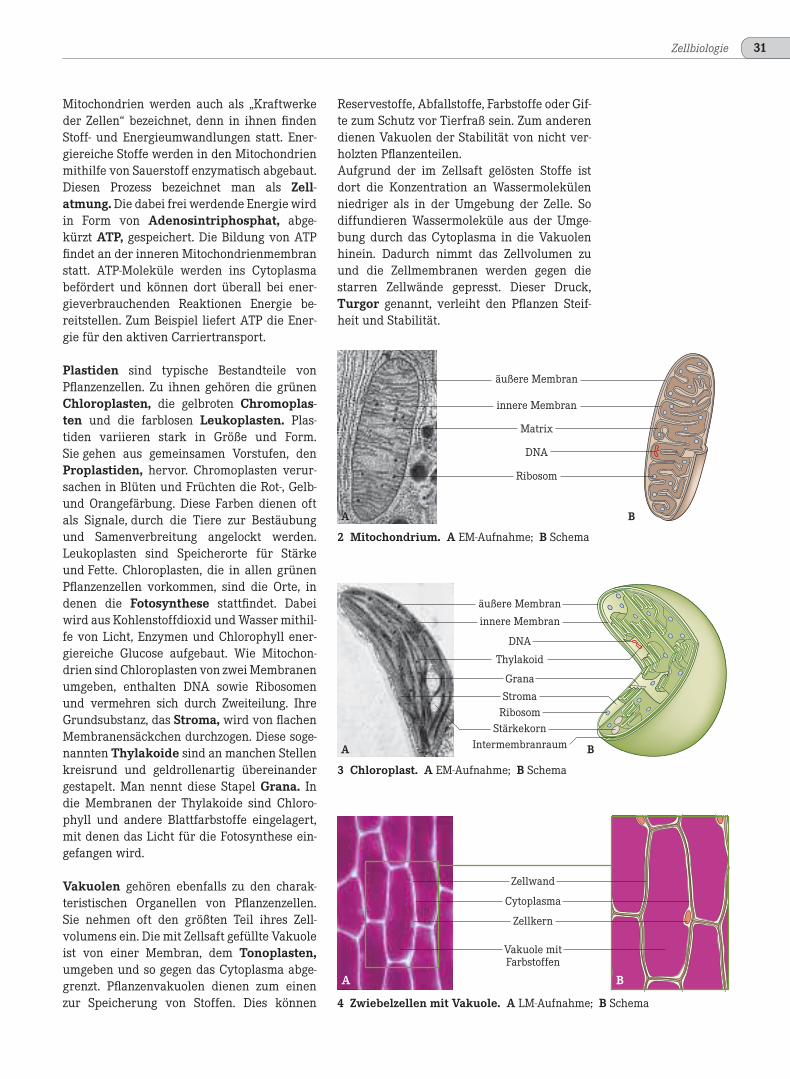
Plastiden sind typische Bestandteile vonPflanzenzellen. Zu ihnen gehören die grünenChloroplasten, die gelbroten Chromoplas-ten und die farblosen Leukoplasten. Plas-tiden variieren stark in Größe und Form.Sie gehen aus gemeinsamen Vorstufen, denProplastiden, hervor. Chromoplasten verur-sachen in Blüten und Früchten die Rot-, Gelb-und Orangefärbung. Diese Farben dienen oftals Signale, durch die Tiere zur Bestäubungund Samenverbreitung angelockt werden.Leukoplasten sind Speicherorte für Stärkeund Fette. Chloroplasten, die in allen grünenPflanzenzellen vorkommen, sind die Orte, indenen die Fotosynthese stattfindet. Dabeiwird aus Kohlenstoffdioxid und Wasser mithil-fe von Licht, Enzymen und Chlorophyll ener-giereiche Glucose aufgebaut. Wie Mitochon-drien sind Chloroplasten von zwei Membranenumgeben, enthalten DNA sowie Ribosomenund vermehren sich durch Zweiteilung. IhreGrundsubstanz, das Stroma, wird von flachenMembranensäckchen durchzogen. Diese soge-nannten Thylakoide sind an manchen Stellenkreisrund und geldrollenartig übereinandergestapelt. Man nennt diese Stapel Grana. Indie Membranen der Thylakoide sind Chloro-phyll und andere Blattfarbstoffe eingelagert,mit denen das Licht für die Fotosynthese ein-gefangen wird.
Vakuolen gehören ebenfalls zu den charak-teristischen Organellen von Pflanzenzellen.Sie nehmen oft den größten Teil ihres Zell-volumens ein. Die mit Zellsaft gefüllte Vakuoleist von einer Membran, dem Tonoplasten,umgeben und so gegen das Cytoplasma abge-grenzt. Pflanzenvakuolen dienen zum einenzur Speicherung von Stoffen. Dies können
Reservestoffe, Abfallstoffe, Farbstoffe oder Gif-te zum Schutz vor Tierfraß sein. Zum anderendienen Vakuolen der Stabilität von nicht ver-holzten Pflanzenteilen.Aufgrund der im Zellsaft gelösten Stoffe istdort die Konzentration an Wassermolekülenniedriger als in der Umgebung der Zelle. Sodiffundieren Wassermoleküle aus der Umge-bung durch das Cytoplasma in die Vakuolenhinein. Dadurch nimmt das Zellvolumen zuund die Zellmembranen werden gegen diestarren Zellwände gepresst. Dieser Druck,Turgor genannt, verleiht den Pflanzen Steif-heit und Stabilität.
2 Mitochondrium. A EM-Aufnahme; B Schema
3 Chloroplast. A EM-Aufnahme; B Schema
4 Zwiebelzellen mit Vakuole. A LM-Aufnahme; B Schema
BA
äußere Membran
innere Membran
Matrix
DNA
Ribosom
BA
äußere Membran
innere Membran
Stroma
Thylakoid
DNA
RibosomStärkekorn
Intermembranraum
Grana
Zellwand
Cytoplasma
Zellkern
Vakuole mitFarbstoffen
BA

Zellbiologie32
Bei eukaryotischen Zellen wird ein großer Teildes Zellvolumens von einem ausgedehnten in-neren Membransystem ausgefüllt. Es bestehtim Wesentlichen aus dem Golgi-Apparatund dem Endoplasmatischen Retikulum(ER). Zwischen diesen bewegen sich kleine,von Membranen umgebene Bläschen, dieVesikel, hin und her.
Das Wort „Endoplasmatisches Retikulum“heißt übersetzt „innerplasmatisches Netz-werk“. Es besteht aus miteinander verbun-denen, membranbegrenzten Röhren und ab-geflachten Säckchen und durchzieht dasCytoplasma der gesamten Zelle. In EM-Aufnah-men erkennt man, dass es sich bis in die äuße-
re Membran der Kernhülle fortsetzt. In vielenBereichen sind auf seiner Oberfläche Riboso-men angeheftet. Man kann daher zwischendem rauen, ribosomentragenden ER und demglatten ER, das nicht mit Ribosomen besetztist, differenzieren. Raues und glattes ER unter-scheiden sich in ihren Aufgaben im Zellstoff-wechsel.Das glatte ER ist insbesondere am Abbau vonGiften oder auch Medikamenten beteiligt.Außerdem hat es in bestimmten Zelltypen dieAufgabe, Lipide und Hormone zu synthetisie-ren.Das raue ER arbeitet eng mit den auf seinerOberfläche angelagerten Ribosomen zusam-men. Diese synthetisieren Polypeptidkettenund geben sie in den Innenraum des ERs ab.Dort werden sie in mehreren Schritten verän-dert und gefaltet, sodass daraus Proteine mitihrer typischen Raumstruktur entstehen.
Der Transport der Proteine zu den Bestim-mungsorten in der Zelle erfolgt dann zu einemTeil im Innenraum des Röhrensystems desrauen ERs, zu einem anderen Teil durch Vesi-kel. Für den Vesikeltransport werden die Pro-teine an der Oberfläche des ERs in kleineMembranaussackungen gegeben. Diese ver-größern sich und werden dann als Vesikel ab-geschnürt. Solche ER-Vesikel können mithilfevon Mikrotubuli – fädigen Proteinen, die dasCytoplasma durchziehen – zur Zelloberflächebewegt werden und dort mit der Zellmembranverschmelzen. Auf diese Weise gelangen dieProteine aus dem Vesikelinneren ins Außen-medium der Zelle. Andere ER-Vesikel beför-dern Proteine zum Golgi-Apparat.
Der Golgi-Apparat besteht aus abgeflachtenMembransäckchen, den sogenannten Zister-nen, sowie aus Golgi-Vesikeln. Die Zisternensind oft dicht gestapelt. Solche Membran-stapel werden Dictyosomen genannt. Die Ge-samtheit aller Dictyosomen einer Zelle ist derGolgi-Apparat. Er ist eine Schaltstelle desStofftransports. Insbesondere ist er am Exportvon Stoffen aus der Zelle beteiligt. Der Golgi-Apparat übernimmt Proteine vom ER. Dazuverschmelzen Vesikel, die vom ER abgeschnürtwerden, mit Zisternen des Golgi-Apparats.Dabei gelangen die Proteine aus den Golgi-Vesikeln in den Innenraum der Zisternen.Manche Proteine werden dort noch weiter ver-ändert. Vor allem aber werden Proteine dortsortiert und konzentriert. Danach werden sie
5 Endoplasmatisches Retikulum.A EM-Aufnahme (Querschnitt); B Schema
6 Dictyosom. A EM-Aufnahme (Querschnitt); B Schema
B
B
A
A
entstehendesGolgi-Vesikel
Golgi-Vesikel ER-Vesikel
Zisterne
Zellbiologie 33
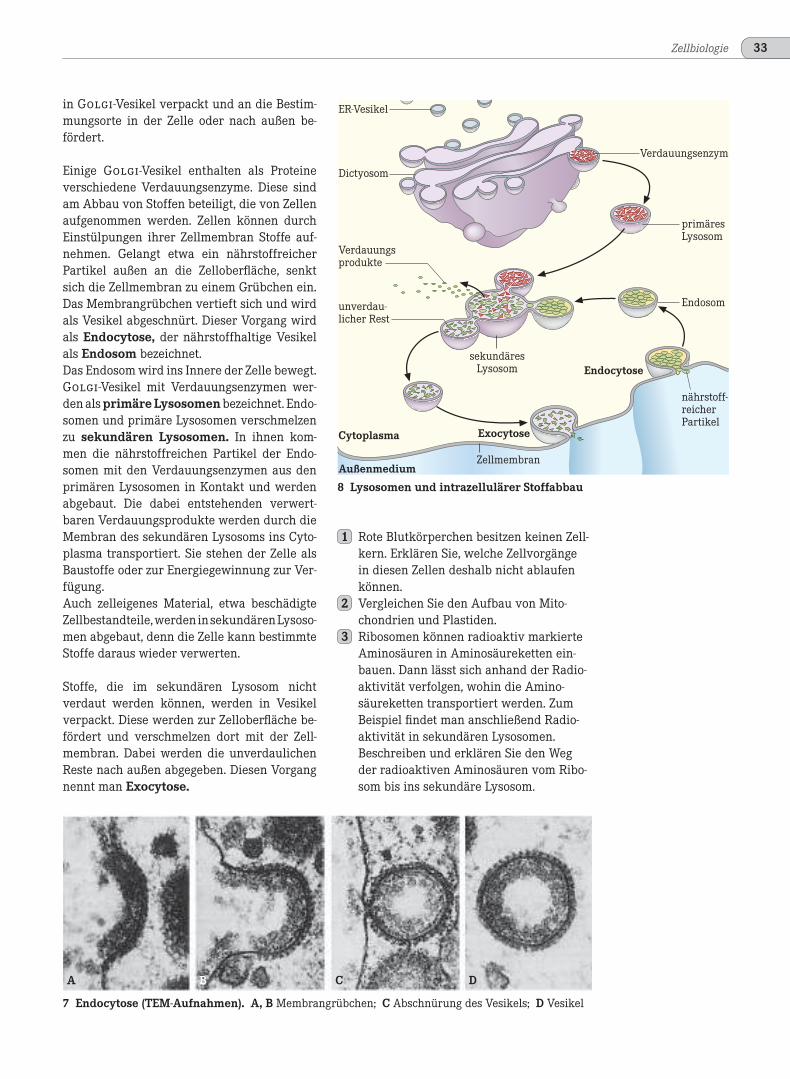
in Golgi-Vesikel verpackt und an die Bestim-mungsorte in der Zelle oder nach außen be-fördert.
Einige Golgi-Vesikel enthalten als Proteineverschiedene Verdauungsenzyme. Diese sindam Abbau von Stoffen beteiligt, die von Zellenaufgenommen werden. Zellen können durchEinstülpungen ihrer Zellmembran Stoffe auf-nehmen. Gelangt etwa ein nährstoffreicherPartikel außen an die Zelloberfläche, senktsich die Zellmembran zu einem Grübchen ein.Das Membrangrübchen vertieft sich und wirdals Vesikel abgeschnürt. Dieser Vorgang wirdals Endocytose, der nährstoffhaltige Vesikelals Endosom bezeichnet.Das Endosom wird ins Innere der Zelle bewegt.Golgi-Vesikel mit Verdauungsenzymen wer-den als primäre Lysosomen bezeichnet. Endo-somen und primäre Lysosomen verschmelzenzu sekundären Lysosomen. In ihnen kom-men die nährstoffreichen Partikel der Endo-somen mit den Verdauungsenzymen aus denprimären Lysosomen in Kontakt und werdenabgebaut. Die dabei entstehenden verwert-baren Verdauungsprodukte werden durch dieMembran des sekundären Lysosoms ins Cyto-plasma transportiert. Sie stehen der Zelle alsBaustoffe oder zur Energiegewinnung zur Ver-fügung.Auch zelleigenes Material, etwa beschädigteZellbestandteile,werdeninsekundärenLysoso-men abgebaut, denn die Zelle kann bestimmteStoffe daraus wieder verwerten.
Stoffe, die im sekundären Lysosom nichtverdaut werden können, werden in Vesikelverpackt. Diese werden zur Zelloberfläche be-fördert und verschmelzen dort mit der Zell-membran. Dabei werden die unverdaulichenReste nach außen abgegeben. Diesen Vorgangnennt man Exocytose.
8 Lysosomen und intrazellulärer Stoffabbau
Rote Blutkörperchen besitzen keinen Zell-kern. Erklären Sie, welche Zellvorgängein diesen Zellen deshalb nicht ablaufenkönnen.Vergleichen Sie den Aufbau von Mito-chondrien und Plastiden.Ribosomen können radioaktiv markierteAminosäuren in Aminosäureketten ein-bauen. Dann lässt sich anhand der Radio-aktivität verfolgen, wohin die Amino-säureketten transportiert werden. ZumBeispiel findet man anschließend Radio-aktivität in sekundären Lysosomen.Beschreiben und erklären Sie den Wegder radioaktiven Aminosäuren vom Ribo-som bis ins sekundäre Lysosom.
primäresLysosom
sekundäresLysosom
Endosom
nährstoff-reicherPartikel
Verdauungsenzym
ER-Vesikel
Dictyosom
Außenmedium
Cytoplasma
Zellmembran
Exocytose
Endocytose
unverdau-licher Rest
Verdauungsprodukte
7 Endocytose (TEM-Aufnahmen). A, B Membrangrübchen; C Abschnürung des Vesikels; D Vesikel
A B C D

Zellbiologie 35
1.9 Angewandte Biologie:Zellkulturen in der Medizin
Die meisten Pflanzen- und Tierzellen könnenunter geeigneten Bedingungen in einem Kul-turmedium leben und sich vermehren. Sol-che Zellkulturen kommen immer mehr inder Medizin zur Anwendung. Primäre Zell-kulturen enthalten Einzelzellen, die einemOrgan direkt entnommen und für die Kulturaufbereitet werden. Tierische Zellen teilensich in Kultur aber nur etwa 50- bis 100-mal.Durch Eingriffe, wie etwa das Einschleusenvon Tumorgenen, kann man erreichen, dasssich auch tierische Zellen in Zellkultur perma-nent weiterteilen. Sekundäre Zellkulturenenthalten solche manipulierten Zellen.
Zellkulturen ersetzen in zunehmendem MaßeTierversuche, in denen zum Beispiel die Ver-träglichkeit von Pharmaka, Chemikalien undBiomaterial, wie etwa Kontaktlinsen und Pro-thesen, getestet werden. Kulturen aus mensch-lichen Leberzellen nutzt man insbesondere,um die Wirksamkeit von potentiellen Medika-menten zu untersuchen und zu prüfen, ob beiihrem Abbau im Zellstoffwechsel schädlicheStoffe gebildet werden. Die Ergebnisse aus sol-chen Testverfahren lassen sich viel besser aufMenschen übertragen, als Ergebnisse aus Tier-versuchen.
Große Bedeutung haben Zellkulturen für dieVermehrung von Viren, die man für die Her-stellung von Impfstoffen benötigt. Da Virensich nur in lebenden Zellen vermehren kön-nen, mussten sie bislang entweder in Tieren
oder in befruchteten Hühnereiern vermehrtwerden. Ihre Vermehrung in Zellkulturen be-schleunigt und vereinfacht die Impfstoffher-stellung wesentlich.
Jedes Jahr muss gegen Grippeviren ein neuerImpfstoff entwickelt werden, da sich die Ober-flächenproteine des Virus schnell verändernund eine vorherige Impfung nicht vor einerInfektion durch eine neue Virusvarianteschützt. Seit über 50 Jahren nutzt man fürdie Vermehrung der neuen Virusvariantenbebrütete Hühnereier aus einer sterilen An-zucht. In Deutschland wurde 2007 der ersteGrippeimpfstoff aus einer Zellkultur zugelas-sen. Mit diesem Verfahren kann schneller undflexibler als bisher auf einen erhöhten Bedarfan Impfstoff reagiert werden. So müssen etwabei einer drohenden Epidemie die speziellenHühnereier nicht erst besorgt werden, son-dern aus wenigen tiefgefrorenen Zellen lassensich sehr schnell große Zellkulturen gewin-nen. Dadurch hat sich die Impfstoffproduktionvon neun auf vier Monate verkürzt.
Auch verschiedene andere Impfstoffe werdenbereits mithilfe von Zellkulturen hergestellt,meist jedoch noch mit primären Zellkulturen.Dabei werden ständig wieder Lebewesen, zumBeispiel Hühner oder Affen, gebraucht, um tei-lungsfähige Zellen zu gewinnen. Im Gegensatzdazu handelt es sich bei den Zellen, in denenGrippeviren gezüchtet werden, um sich per-manent weiter teilende Zellen. Solche sekun-dären Zellkulturen werden auch schon für dieProduktion von Polio- und Tollwut-Impfstoffeneingesetzt.
2 Grippeviren.A EM-Aufnahme(gefärbt);B Schema einesGrippevirus
1 Produktion des Grippeimpfstoffes. A Die Zellkultur wird aufgetaut und die Zellen werden zur Vermehrung angeregt.B Nach Zugabe und Vermehrung der Grippeviren in den Zellen werden diese zerstört und die Viren gelangen in die umgebendeLösung. C Die Viren werden aus der Lösung isoliert. D Nur Bruchstücke aus der Virusoberfläche werden verwendet.E Nach den Qualitätskontrollen wird der Impfstoff abgefüllt.
������������
������������ ������
������������ ����
������������ ����
��������� ���������������������
������������ �������
� � � � �
�����������
A
B
Zellbiologie36
AUFGABEN (vernetzt) Erythrocyten
1 ZellorganelleDie Zellorganellen von Erythrocytenwerden im Verlauf ihrer Reifung ab-gebaut. Aber ihr Zellinneres ist voneinem ausgeprägten Cytoskelettdurchzogen. Dieses ist unmittelbarunter der Zellmembran besondersstark ausgebildet und an vielen Stel-len mit ihren integralen Proteinenverknüpft. Es stabilisiert ihre Linsen-form und erlaubt gleichzeitig ihreelastische Verformbarkeit. Im Cyto-plasma liegt eine hohe Konzentrationan Hämoglobin vor, ein Protein, dasdie Atemgase bindet.a) Vergleichen Sie Erythrocyten mitanderen tierischen Zellen hinsicht-lich der Zellbestandteile und dendamit verbundenen Funktionen.b) Erklären Sie am Beispiel der Ery-throcyten das Basiskonzept Strukturund Funktion.
2 Erythrocytenmembran
Auf der Erythrocytenmembran be-finden sich verschiedene Glykolipideund Glykoproteine, welche die Blut-gruppenspezifitäten, wie etwa dasAB0-System, bestimmen. Gegen dieseAntigene befinden sich im Blut jederanderen Blutgruppe Antikörper.Sie bewirken, dass Erythrocytenmit den entsprechenden Antigenenverklumpen. Nur bei Blutgruppe 0liegen keine Antigene auf derErythrocytenmembran vor.a) Skizzieren und beschriften Sieden Bau einer Erythrocytenmembranunter Berücksichtigung ihrer Ver-knüpfung mit dem Cytoskelett.
b) Erklären Sie das Schema zurVerträglichkeit von Spender- undEmpfängerblut.
3 ErythrocytenresistenzIn einem Versuch gibt man Erythro-cyten in 20 Reagenzgläser mit an-steigenden Salzkonzentrationen von0 Prozent bis 5 Prozent. Von den Ver-suchsergebnissen sind der Übersicht-lichkeit wegen nur drei dargestellt:
Zusätzliche mikroskopische Unter-suchungen zeigen zum Teil veränder-te Zellformen der Erythrocyten inden verschiedenen Salz-Lösungen.Die Bildfolge D bis F zeigt jeweilseinen normalen und zwei Erythro-cyten mit abnormer Zellform:
a) Formulieren Sie mögliche Fragen,die mit dieser Versuchsreihe geklärtwerden sollten.
b) Deuten Sie die Beobachtungenin den drei Reagenzgläsern unterBerücksichtigung der LM-Aufnah-men. Ordnen Sie dazu die LM-Aufnah-men D bis F den Reagenzgläsern mitden entsprechenden Salzkonzentrati-onen A bis C zu.
4 TransportprozesseIm Plasma des Erythrocyten liegt einesehr hohe Hämoglobinkonzentrationvor. Zum Ausgleich wird die Ionen-konzentration durch Natrium-Kalium-Pumpen niedrig gehalten.
a) Beschreiben Sie anhand der Ab-bildung die Funktion der aktivenNatrium-Kalium-Pumpe.b) Erläutern Sie, warum eine Zer-störung der Erythrocyten erfolgt,wenn man ihre Natrium-Kalium-Pumpen hemmt.
5 Glucoseaufnahme
Die Abbildung zeigt die Aufnahmevon Glucose in Erythrocyten inAbhängigkeit von der Glucose-konzentration im Aufnahmemedium.a) Erstellen Sie eine Anleitung fürdie Durchführung des Versuches.b) Deuten Sie das Versuchsergebnis.
���
��
����
�
��� �
� � �� �
�
��
�
�
������
���
��
���������������� ������������� ���������
������ ����
����������
�����
��� �����
�!����"�� #$$
%$$
&$$
$$ %' '$
D
E
F
A B C

Stoffwechselbiologie54
1 Enzyme bewirken Stoffwechsel
1.1 Enzyme sind Biokatalysatoren
In jeder Zelle finden während des Stoffwech-sels unzählige chemische Reaktionen statt.Diese Reaktionen laufen stets unter Energie-umsetzung ab, da das Lösen und Knüpfenchemischer Bindungen Energieumwandlun-gen erfordert. Für viele chemische Reaktionengilt dabei der Grundsatz, dass sie nur dannspontan ablaufen, wenn ihre Ausgangsstoffemehr Energie enthalten als die entstehendenProdukte.
Spontane Reaktionen können aber so langsamverlaufen, dass sie kaum wahrnehmbar sind.Eine wässrige Saccharose-Lösung kann manbeispielsweise jahrelang bei Raumtemperaturstehen lassen, ohne dass eine nennenswerteReaktion zu den energieärmeren ProduktenGlucose und Fructose erfolgt. Der Grundhierfür liegt in einer Energiebarriere, die zwi-schen Ausgangsstoffen und Endproduktenliegt. Zunächst muss durch Zufuhr einesEnergiebetrages die Energiebarriere überwun-den werden, bevor die Reaktion ohne weitereEnergiezufuhr selbstständig ablaufen kann.Die anfängliche Energiezufuhr zum Start derReaktion nennt man Aktivierungsenergie.Sie kann durch Zufuhr von Wärmeenergie be-reitgestellt werden. In Zellen würden dadurchaber alle Reaktionen gleichzeitig beschleunigtwerden. Außerdem würden beispielsweiseProteine denaturiert werden. Ein geregelterStoffwechsel wäre unmöglich. Unter Zellbe-dingungen werden daher Stoffwechselreak-tionen in Gang gesetzt beziehungsweise be-
2 Substratspezifität. A Schlüssel-Schloss-Modell; B Modell der induzierten Passform
3 Energieprofil chemischer Reaktionen
schleunigt, indem Wirkstoffe die erforderlicheAktivierungsenergie herabsetzen. Der Wirk-stoff selbst geht unverändert aus der Reaktionhervor. Solche Biokatalysatoren ermöglichenes, dass bestimmte Stoffwechselreaktionen beiKörpertemperatur ablaufen können. Die in le-benden Zellen wirkenden Biokatalysatorenheißen Enzyme.
Im Vergleich zu den aus der Technik bekann-ten Katalysatoren wie Platin oder Nickel be-schleunigen Enzyme Reaktionen um ein Viel-faches. Zudem reagieren sie häufig nur miteinem Ausgangsstoff, dem Substrat. Beispiels-weise spaltet das Enzym Saccharase nur dasDisaccharid Saccharose, nicht aber andereDisaccharide wie Maltose. Enzyme sind sub-stratspezifisch.Die meisten Enzyme sind Proteine, die voneiner oder mehreren Polypeptidketten gebil-det werden. Sie verleihen dem Enzym eine in-dividuelle dreidimensionale Gestalt. Das Sub-strat bindet nur an einer bestimmten Regiondes Enzyms, dem aktiven Zentrum. Die Sub-stratspezifität beruht darauf, dass zwischenSubstrat-Molekül und dem aktiven ZentrumWechselwirkungen möglich sind, sodass dasSubstrat von dem aktiven Zentrum angezogenund gebunden werden kann. Modellhaft kannman sich vorstellen, dass ein Substrat in dasaktive Zentrum des Enzyms passt, wie einSchlüssel in das dazugehörige Schloss. Manspricht vom Schlüssel-Schloss-Modell.
aktivesZentrum
Substrat Substrat
Substrat
+ +
+
–
– –
Enzym
+– –
–
–
–
–
–
–
–
–
Enzym Enzym
Enzym
A B
Substrat
+ +
+
+
+
+
+
+
+
+
–
1 Schlüssel-Schloss-Modell
Ener
gie
Reaktionsverlauf
Aktivierungs-energie fürnicht-katalysierteReaktion
Aktivierungs-energie fürkatalysierteReaktion
EnergieniveauAusgangs-stoffe (z.B.Saccharose)
Energie-gewinn
Energieniveau Endprodukte(z.B. Glucose und Fructose)

Stoffwechselbiologie 55
Diese Modellvorstellung berücksichtigt abernicht, dass sich sowohl die Raumstruktur desSubstrat-Moleküls als auch die des Enzym-Moleküls durch Wechselwirkungen zwischenSubstrat und aktivem Zentrum verändern.Man hat daher das starre Schlüssel-Schloss-Modell um das Modell der induziertenPassform (engl. induced-fit) erweitert. Diewechselseitige Anpassung von Substrat undEnzym kann man sich modellhaft veranschau-lichen: Beim Ankleiden passt sich die Formeiner Hose der Körperform an, gleichzeitigverleiht sie dem Träger eine bestimmte Form.
Durch die Bindung des Substrat-Molekülsan das aktive Zentrum des Enzym-Molekülsentsteht ein Enzym-Substrat-Komplex. DasSubstrat-Molekül befindet sich dabei in eineminstabilen Zustand, in dem bestimmte Bin-dungen gespannt oder gelockert sind. So kön-nen diese leichter gelöst und andere neu ge-bildet werden. Das Substrat wird dadurchchemisch verändert.
Im Substrat-Molekül können nur ganz be-stimmte Bindungen gelöst oder geknüpft wer-den. Das Enzym hat also eine ganz bestimmteWirkung auf das Substrat-Molekül. Andersausgedrückt kann das Enzym nur eine be-stimmte Reaktion katalysieren. Man sagt, En-zyme sind wirkungs- oder reaktionsspezi-fisch. Zum Beispiel spaltet das EnzymSaccharase das Disaccharid Saccharose stetsunter Wasseranlagerung in Glucose und Fruc-tose. Andere denkbare Reaktionen werdennicht katalysiert.
Aus dem Enzym-Substrat-Komplex werden dasentstandene Produkt und das Enzym freige-setzt. Das Enzym geht dabei unverändert ausder Reaktion hervor. Bereits kleinste Enzym-mengen genügen daher, um große Substrat-mengen umzusetzen. Beispielsweise kann eineinziges Saccharase-Molekül innerhalb einerSekunde mehrere Tausend Saccharose-Mole-küle umsetzen.
Enzyme steuern und regeln chemische Reak-tionen in lebenden Zellen. Sie katalysierensowohl anabole Stoffwechselschritte, bei de-nen kleine Moleküle zu großen Molekülen auf-gebaut werden, als auch katabole Stoffwech-selschritte, bei denen große Moleküle abgebautwerden. Enzyme sind damit die universellenWerkzeuge des Stoffwechsels. 4 Katalysierter Zyklus
5 Modell derinduzierten Passform
Begründen Sie anhand von Abbildung 3,warum die freigesetzte Energiemenge un-abhängig davon ist, ob die Reaktion durchWärmeenergie oder Enzymkatalyse er-möglicht wurde.Methanolvergiftungen wurden früherbehandelt, indem man dem Patientengroße Mengen Ethanol (Trinkalkohol)zuführte. Beide Alkohole werden imKörper zunächst durch die Alkohol-Dehydrogenase oxidiert. Aus Ethanolentsteht dabei Ethanal, Methanol reagiertzum giftigen Methanal. Im weiterenVerlauf werden beide Substanzen zu Koh-lenstoffdioxid und Wasser abgebaut.a) Beschreiben Sie die Substrat- undWirkungsspezifität der Alkohol-Dehydro-genase.b) Begründen Sie die Therapie der Metha-nolvergiftung mit Ethanol.Erläutern Sie anhand der Abbildung 2das Modell der induzierten Passform undvergleichen Sie es mit dem Schlüssel-Schloss-Modell.Erklären Sie, warum der Austausch einerAminosäure im Enzym-Molekül zum voll-ständigen Funktionsverlust des Enzymsführen kann, aber auch ohne Auswirkungbleiben kann.
Substrat(Saccharose)
Enzym(Saccharase)
Enzym-Substrat-Komplex
aktivesZentrum
Glucose
Fructose
H2O
OH
+
OH

Stoffwechselbiologie56
Änderungen der Temperatur und des pH-Wertes beeinflussen die Enzymaktivität. Siehaben Auswirkungen auf die Raumstrukturder Enzyme und bestimmen damit wesentlichderen Aktivität.
Der Zusammenhang zwischen der Aktivitäteines Enzyms und der Temperatur wird durcheine Optimumkurve verdeutlicht. Bei stei-genden Temperaturen erhöht sich die Reak-tionsgeschwindigkeit zunächst stark. Die Be-obachtung, dass die Geschwindigkeit einerReaktion mit steigender Temperatur zunimmt,ist bei vielen chemischen Reaktionen zu ma-chen. So führt eine Temperaturerhöhung um10°C im Allgemeinen zu einer Verdoppelungder Reaktionsgeschwindigkeit. Man sprichtvon der Reaktions-Geschwindigkeits-Tem-peratur-Regel, kurz RGT-Regel.Der beobachtete Zusammenhang lässt sichinsbesondere mit der wachsenden Teilchen-bewegung erklären. Substrat- und Enzym-Moleküle treffen infolgedessen mit größererWahrscheinlichkeit aufeinander, sodass esschneller zur Bildung von Enzym-Substrat-Komplexen und somit zu einem höheren Stoff-umsatz pro Zeiteinheit kommt. Die höchsteAktivität erreicht ein Enzym beim Tempe-raturoptimum.Bei höheren Temperaturen verringert sich dieEnzymaktivität wieder. Enzym- und Substrat-Moleküle bewegen sich jetzt zu schnell, sodassSubstrat-Moleküle nicht im aktiven Zentrumgebunden werden können. Außerdem nimmtim Enzym-Molekül die Bewegung der Poly-peptidkette zu. Dadurch werden Wechselwir-
2 Enzymaktivität in Abhängigkeit von derTemperatur
Enzy
mak
tivi
tät[
rela
tive
Einh
eite
n]
Temperatur [°C]2010 30 40 50 60
Reaktions-geschwindigkeit Inaktivierung
1.2 Beeinflussung der Enzymaktivität
1 pH-Optima verschiedener Verdauungsenzyme des Menschen
Enzy
mak
tivi
tät[
rela
tive
Einh
eite
n]
Pepsin
Amylase
Trypsin
pH-Wert1 2 3 4 5 6 7 8 9 10 11 12 13 14
kungen zwischen den Aminosäureresten derPolypeptidkette zerstört. Dies hat eine Verän-derung der Raumstruktur des Enzym-Mole-küls zur Folge. Das Enzym denaturiert undkann kein Substrat mehr im aktiven Zentrumbinden und umsetzen.
Untersucht man die Abhängigkeit der Enzym-aktivität vom pH-Wert, so erhält man auchhier eine Optimumkurve. Jedes Enzym zeigtein charakteristisches pH-Optimum. Eine Ab-weichung von diesem pH-Wert führt zu einerVerringerung der Enzymaktivität. Erniedrigtman beispielsweise den pH-Wert, so lagernsich vermehrt H+-Ionen an negativ geladeneAminosäurereste des Proteins. Diese werdendadurch ungeladen und können nicht mehrmit anderen geladenen Bereichen der Poly-peptidkette in Wechselwirkung treten. Dietypische räumliche Struktur des Proteins än-dert sich. Das Enzym denaturiert und kannseine Funktion nicht mehr erfüllen.
Grünes Gemüse wird beim Lagern all-mählich braun, da der grüne FarbstoffChlorophyll durch ein zelleigenes Enzymabgebaut wird. Begründen Sie, warumman zum Beispiel Erbsen oder Brokkolivor dem Einfrieren kurz in kochendesWasser tauchen sollte.Ordnen Sie die Enzyme aus Abbildung 1den verschiedenen Abschnitten desmenschlichen Verdauungstraktes zu undbegründen Sie Ihre Entscheidung.
Stoffwechselbiologie 57
1.3 Cofaktoren
Nur wenige Enzyme bestehen ausschließlichaus Proteinen. Die Mehrzahl benötigt nebendem eigentlichen Substrat weitere Bestandtei-le für die Katalyse. Diese im Vergleich zu Enzy-men kleinen Moleküle oder Ionen nennt manCofaktoren.
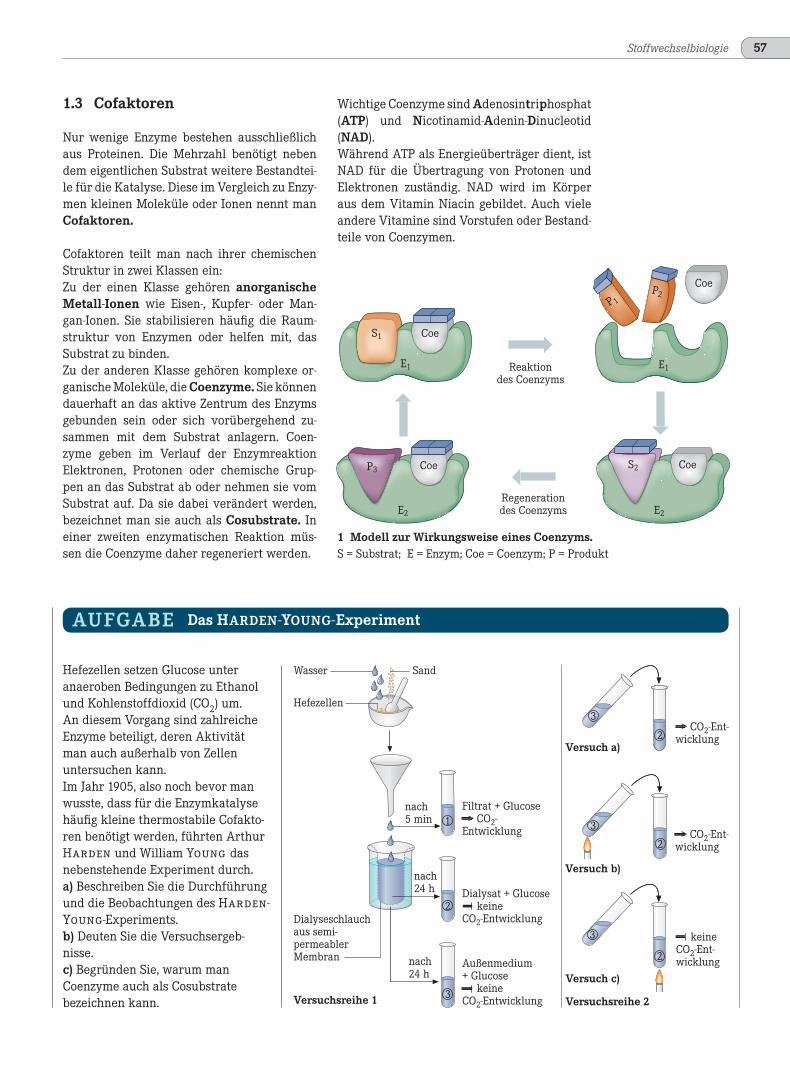
Cofaktoren teilt man nach ihrer chemischenStruktur in zwei Klassen ein:Zu der einen Klasse gehören anorganischeMetall-Ionen wie Eisen-, Kupfer- oder Man-gan-Ionen. Sie stabilisieren häufig die Raum-struktur von Enzymen oder helfen mit, dasSubstrat zu binden.Zu der anderen Klasse gehören komplexe or-ganische Moleküle, die Coenzyme. Sie könnendauerhaft an das aktive Zentrum des Enzymsgebunden sein oder sich vorübergehend zu-sammen mit dem Substrat anlagern. Coen-zyme geben im Verlauf der EnzymreaktionElektronen, Protonen oder chemische Grup-pen an das Substrat ab oder nehmen sie vomSubstrat auf. Da sie dabei verändert werden,bezeichnet man sie auch als Cosubstrate. Ineiner zweiten enzymatischen Reaktion müs-sen die Coenzyme daher regeneriert werden.
Wichtige Coenzyme sind Adenosintriphosphat(ATP) und Nicotinamid-Adenin-Dinucleotid(NAD).Während ATP als Energieüberträger dient, istNAD für die Übertragung von Protonen undElektronen zuständig. NAD wird im Körperaus dem Vitamin Niacin gebildet. Auch vieleandere Vitamine sind Vorstufen oder Bestand-teile von Coenzymen.
1 Modell zur Wirkungsweise eines Coenzyms.S = Substrat; E = Enzym; Coe = Coenzym; P = Produkt
Reaktiondes Coenzyms
Regenerationdes Coenzyms
CoeS1
S2
E1 E1
E2 E2
P3
P1P2
Coe
Coe
Coe
Hefezellen setzen Glucose unteranaeroben Bedingungen zu Ethanolund Kohlenstoffdioxid (CO2) um.An diesem Vorgang sind zahlreicheEnzyme beteiligt, deren Aktivitätman auch außerhalb von Zellenuntersuchen kann.Im Jahr 1905, also noch bevor manwusste, dass für die Enzymkatalysehäufig kleine thermostabile Cofakto-ren benötigt werden, führten ArthurHarden und William Young dasnebenstehende Experiment durch.a) Beschreiben Sie die Durchführungund die Beobachtungen des Harden-Young-Experiments.b) Deuten Sie die Versuchsergeb-nisse.c) Begründen Sie, warum manCoenzyme auch als Cosubstratebezeichnen kann.
AUFGABE Das HARDEN-YOUNG-Experiment
Stoffwechselbiologie58
Substratkonzentration [S]
Rea
ktio
nsge
schw
indi
gkei
t[v]
A BSubstratkonzentration [S]
Rea
ktio
nsge
schw
indi
gkei
t[v]
ungehemmteReaktion
kompetitivgehemmte Reaktion
ungehemmteReaktion
nichtkompetitivgehemmte Reaktion
S
I
I
Enzym Enzym
S = SubstratI = Inhibitor
S = SubstratI = Inhibitor
S
I
I
Enzym Enzym
���������� ������� ���
� �
����
� ��
����
���
���
��
����
�
�
�
�
�
�
�������������
1.4 Reaktionsgeschwindigkeit undHemmung der Enzymaktivität
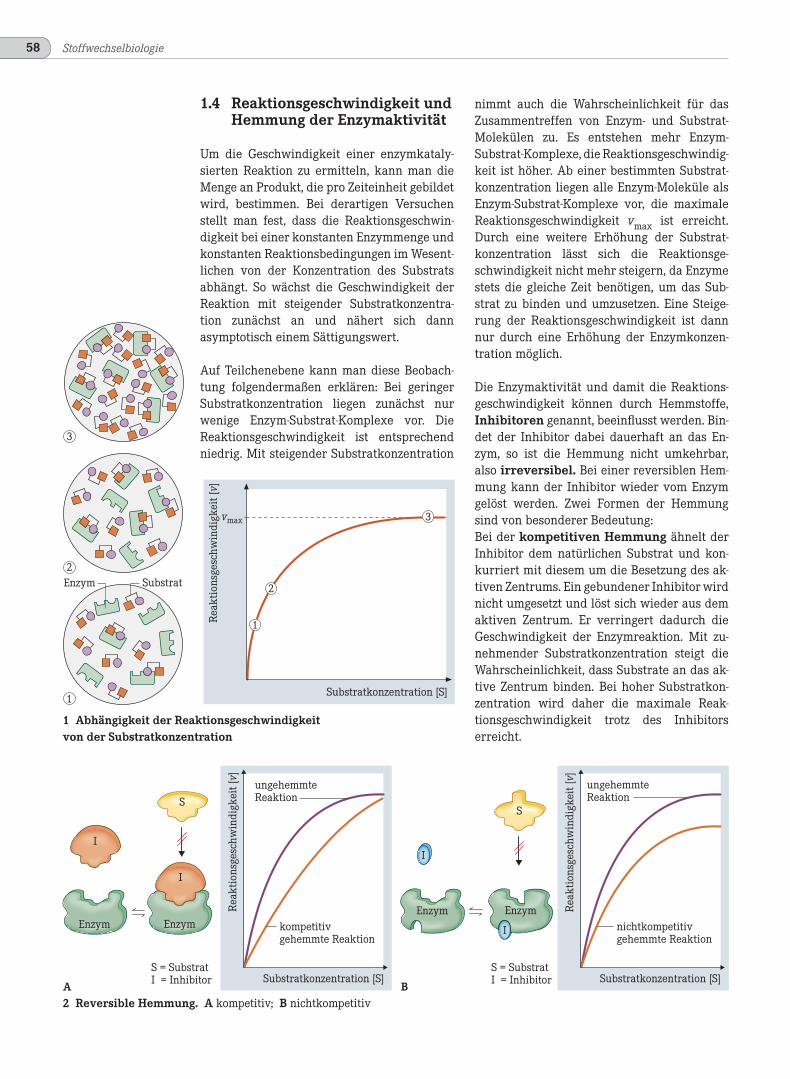
Um die Geschwindigkeit einer enzymkataly-sierten Reaktion zu ermitteln, kann man dieMenge an Produkt, die pro Zeiteinheit gebildetwird, bestimmen. Bei derartigen Versuchenstellt man fest, dass die Reaktionsgeschwin-digkeit bei einer konstanten Enzymmenge undkonstanten Reaktionsbedingungen im Wesent-lichen von der Konzentration des Substratsabhängt. So wächst die Geschwindigkeit derReaktion mit steigender Substratkonzentra-tion zunächst an und nähert sich dannasymptotisch einem Sättigungswert.
Auf Teilchenebene kann man diese Beobach-tung folgendermaßen erklären: Bei geringerSubstratkonzentration liegen zunächst nurwenige Enzym-Substrat-Komplexe vor. DieReaktionsgeschwindigkeit ist entsprechendniedrig. Mit steigender Substratkonzentration
1 Abhängigkeit der Reaktionsgeschwindigkeitvon der Substratkonzentration
2 Reversible Hemmung. A kompetitiv; B nichtkompetitiv
nimmt auch die Wahrscheinlichkeit für dasZusammentreffen von Enzym- und Substrat-Molekülen zu. Es entstehen mehr Enzym-Substrat-Komplexe, die Reaktionsgeschwindig-keit ist höher. Ab einer bestimmten Substrat-konzentration liegen alle Enzym-Moleküle alsEnzym-Substrat-Komplexe vor, die maximaleReaktionsgeschwindigkeit vmax ist erreicht.Durch eine weitere Erhöhung der Substrat-konzentration lässt sich die Reaktionsge-schwindigkeit nicht mehr steigern, da Enzymestets die gleiche Zeit benötigen, um das Sub-strat zu binden und umzusetzen. Eine Steige-rung der Reaktionsgeschwindigkeit ist dannnur durch eine Erhöhung der Enzymkonzen-tration möglich.
Die Enzymaktivität und damit die Reaktions-geschwindigkeit können durch Hemmstoffe,Inhibitoren genannt, beeinflusst werden. Bin-det der Inhibitor dabei dauerhaft an das En-zym, so ist die Hemmung nicht umkehrbar,also irreversibel. Bei einer reversiblen Hem-mung kann der Inhibitor wieder vom Enzymgelöst werden. Zwei Formen der Hemmungsind von besonderer Bedeutung:Bei der kompetitiven Hemmung ähnelt derInhibitor dem natürlichen Substrat und kon-kurriert mit diesem um die Besetzung des ak-tiven Zentrums. Ein gebundener Inhibitor wirdnicht umgesetzt und löst sich wieder aus demaktiven Zentrum. Er verringert dadurch dieGeschwindigkeit der Enzymreaktion. Mit zu-nehmender Substratkonzentration steigt dieWahrscheinlichkeit, dass Substrate an das ak-tive Zentrum binden. Bei hoher Substratkon-zentration wird daher die maximale Reak-tionsgeschwindigkeit trotz des Inhibitorserreicht.

Stoffwechselbiologie
Gicht ist eine Stoffwechselkrankheit,bei der es infolge hoher Harnsäure-konzentrationen im Blut zu Ablage-rungen von Harnsäurekristallen inGelenken und Geweben kommt. Diebetroffenen Gelenke sind gerötet,angeschwollen und schmerzen starkbei Berührung.
und -ausscheidung besteht, ist beiden meisten Gichtkranken die Aus-scheidung über die Nieren gestört.Eine erhöhte Harnsäurekonzentra-tion im Blut ist die Folge. Ein bewähr-tes Medikament zur Gichtbehand-lung ist Allopurinol.
a) Stellen Sie die Versuchsergebnissegrafisch dar und erläutern Sie diese.b) Erklären Sie die Wirkungsweisevon Allopurinol unter Berücksichti-gung der Strukturformeln.
2 Allopurinol
Substrat-konzen-
tration [S]in μmol/l
Reaktions-geschwindigkeit vin μmol/(l · min)
ohneAllopurinol
mitAllopurinol
0,5 1,8 0,4
1,0 2,8 0,8
2,0 4,0 1,3
3,0 4,8 1,9
5,0 5,7 3,0
10,0 6,5 4,7
15,0 6,7 6,2
20,0 6,8 6,8
3 Versuchsergebnisse
1 Abbauweg der Purine
��
�
�
�
�
�
��
�
�
�
�
���
��
�
�
�
�
���
�
�
������� ����� ���
���
��
��� �
��� ����� ��� ������� ����������� ���
������� ���
��� ��� ���
59
Bei der nichtkompetitiven Hemmung be-steht zwischen dem Inhibitor und dem Substratkeine Ähnlichkeit. Der Inhibitor bindet nichtam aktiven Zentrum, sondern an einer ande-ren Stelle. Er verursacht dadurch eine Ände-rung der Raumstruktur des Enzyms. Das ak-tive Zentrum wird dabei so verändert, dass eskein Substrat mehr binden kann. Durch nicht-kompetitive Inhibitoren wird damit immer einTeil der Enzym-Moleküle blockiert. Auch wennsich der Inhibitor ablöst, bindet er sofort aneinem anderen Enzym-Molekül. Da hierdurchdie Zahl aktiver Enzyme verringert wird, kanndie maximale Reaktionsgeschwindigkeit einerungehemmten Reaktion nicht erreicht wer-den. Auch kann die hemmende Wirkung nichtdurch eine erhöhte Substratkonzentration auf-gehoben werden, denn Inhibitor und Substrat
konkurrieren bei dieser Form der Hemmungnicht um das aktive Zentrum.
Kompetitive und nichtkompetitive Inhibitorenkönnen auch dauerhaft an Enzyme binden. Indiesen Fällen liegt eine irreversible Hem-mung vor. So wirkt beispielsweise das Anti-biotikum Penicillin, indem es eine kovalenteBindung mit einem Aminosäurerest im ak-tiven Zentrum des Enzyms Transpeptidase ein-geht. Dadurch wird die Synthese bakteriellerZellwände verhindert, sodass die Bakteriengetötet werden.Schwermetall-Ionen wie Cyanid-, Quecksilber-oder Blei-Ionen binden außerhalb des aktivenZentrums und verändern dabei die Raum-struktur des Enzyms. Sie wirken damit alsEnzymgifte.
AUFGABEN Enzymhemmung in der Medizin
Harnsäure entsteht beim Abbau vonPurinen. Purine sind Bestandteilevon Nucleinsäuren, die vom Körpergebildet werden. Der Mensch nimmtauch große Mengen an Purinen mitder Nahrung auf. Diese werden inmehreren Schritten zunächst in was-serlösliche Verbindungen überführtund anschließend durch das EnzymXanthinoxidase in die schwerlöslicheHarnsäure, die mit dem Urin ausge-schieden wird.
Während bei Gesunden ein Gleich-gewicht zwischen Harnsäurebildung

Stoffwechselbiologie 61
Stärkebrei aus Mais,Kartoffeln oder Weizen
Maltodextrinsirup
Maltosesirup Glucosesirup
Glucose-isomerase
GlucoamylasePullulanase
seisomeraseras
Fructosesirup
1. Stärke-verflüssigung
2. Verzuckerung
3. Isomerisierung
1 Enzymatische Stärkeverzuckerung(Schema)
1.6 Angewandte Biologie: Enzymefür Industrie und Haushalt
Enzyme spielen nicht nur für den Stoffwechselvon Lebewesen eine herausragende Rolle. Ihrebesonderen Eigenschaften nutzt man auch beivielen biochemischen Prozessen, zum Beispielin der Lebensmittel- oder Waschmittelindus-trie. Die benötigten Enzyme gewinnt man ingroßen Mengen und hoher Reinheit zumeistaus gentechnisch veränderten Bakterien.
Im Lebensmittelbereich besitzt die enzymati-sche Stärkeverzuckerung große wirtschaft-liche Bedeutung. In einem mehrstufigen Pro-zess wird pflanzliche Stärke – etwa aus Mais,Kartoffeln oder Weizen – in verschiedeneKohlenhydrate umgewandelt. Man erhält soSüßungsmittel aus preiswerter Stärke undist nicht auf die Saccharosegewinnung ausZuckerrohr oder Zuckerrüben angewiesen.
Im ersten Schritt dieses Verfahrens, der Stär-keverflüssigung, wird Stärke durch eineα-Amylase in Bruchstücke zerlegt. Es entstehtMaltodextrin-Sirup, ein Gemisch aus Maltoseund längeren Ketten aus Glucosemolekülen.Bei der Verzuckerung wird dieser Sirupdurch verschiedene Enzyme weiter zerlegt. Jenach Wahl des Enzyms erhält man beispiels-weise Maltosesirup oder Glucosesirup, einGemisch aus Glucose und anderen Zuckern.Glucosesirup hat mittlerweile Saccharose ganzoder teilweise in vielen Süß- und Backwarenersetzt. Da Fructose eine deutlich höhere Süß-kraft als Glucose besitzt, wird bei der Isomeri-sierung ein Teil der Glucose-Moleküle durchdas Enzym Glucoseisomerase in Fructose-Moleküle umgewandelt. Der entstandene Siruphat einen Fructose-Anteil von bis zu 55 Pro-zent und besitzt annähernd die gleiche Süß-kraft wie Saccharose. In den USA hat dieseraus Maisstärke gewonnene HFC-Sirup (engl.high fructose corn) Saccharose als Süßungs-mittel weitgehend verdrängt.
Ein weiteres, wirtschaftlich bedeutsames An-wendungsgebiet für Enzyme sind Waschmittel.Beim Waschvorgang werden Fettverschmut-zungen durch Tenside vom Gewebe gelöst.Tenside versagen aber weitgehend bei eiweiß-haltigen Verschmutzungen wie zum BeispielEigelb, Milch oder Blut. Mit Proteasen, vondenen etwa 200 mg pro kg Waschpulver zuge-setzt werden, kann man dieses Problem lösen.
Proteasen spalten an bestimmten Stellen Pep-tidbindungen, sodass die eiweißhaltigen Ver-schmutzungen aus dem Gewebe gelöst werdenkönnen.
Neben Proteasen setzt man Waschmittelnhäufig auch Amylasen zu, um stärkehaltigeFlecken zu entfernen. Der Zusatz von Lipasendient zum Fettabbau bei niedrigen Waschtem-peraturen. Cellulasen bauen von der Baum-wollfaser abstehende Mikrofasern ab, sodasssich die Wäsche anschließend weicher an-fühlt.
Was
chak
tivi
tät[
rel.
Einh
eit]
Waschmittelmit Enzymen
ohne Enzyme
Waschtemperatur [°C]0 20 40 60 80 100
AUFGABE Waschmittelenzyme
Leiten Sie aus dernebenstehendenAbbildungEmpfehlungen fürdie VerwendungenzymhaltigerWaschmittel ab.
Stoffwechselbiologie62
AUFGABEN Enzyme
1 Konzentrations-Zeit-Abhängigkeit einer Enzym-reaktion
Die Abbildung gibt die Konzentrati-onsänderungen von freiem Enzym(E), freiem Substrat (S) und die desEnzym-Substrat-Komplexes (ES) imzeitlichen Verlauf einer Enzymreak-tion wieder.a) Erklären Sie den Verlauf derKurven und geben Sie an, wie sichdie Konzentration des gebildetenProdukts entwickeln wird. Gehen Siedavon aus, dass aus einem Substrat-Molekül ein Produkt-Molekül ent-steht.b) Erläutern Sie, wie sich die Kon-zentrationen von Enzym, Enzym-Substrat-Komplex und Substrat biszum Ende der Reaktion weiterentwickeln werden.
2 Bestimmung derReaktionsgeschwindigkeitDas Enzym Katalase kommt in denZellen nahezu aller Lebewesen vorund bewirkt die Spaltung des Zell-giftes Wasserstoffperoxid (H2O2) zuWasser (H2O) und Sauerstoff (O2).In einem Experiment wurde derEinfluss der Substratkonzentrationauf die Geschwindigkeit derKatalasereaktion bestimmt.Zur Verfügung standen eine Katalase-und eine konzentrierte Wasserstoff-peroxid-Lösung.
Kon
zent
rati
on[r
el.E
inhe
iten
]
Zeit [rel. Einheiten]
(S)
(ES)
(E)
Die Messungen wurden mit folgenderApparatur durchgeführt, die im Fol-genden skizziert ist:
Bei der Durchführung des Experi-ments erhielt man folgende Mess-ergebnisse:
Substrat-konzentration
[%]
Reaktions-geschwindigkeit
[ml O2/5 min]
0,50,75
1,01,52,03,06,012
19,2223,1827,4529,9431.9834,6040,6733,23
a) Definieren Sie den Begriff Reakti-onsgeschwindigkeit und erklären Sie,wie man diese mit den gegebenenMaterialien experimentell bestimmt.b) Stellen Sie die Versuchsergebnissegrafisch dar und erklären Sie denKurvenverlauf.c) Erläutern Sie, wie sich die Ge-schwindigkeit der Reaktion erhöhenließe.d) Katalase kann durch Hydroxyl-amin gehemmt werden. ErläuternSie, wie man experimentell die Fragebeantworten kann, ob der Hemmstoffkompetitiv oder nichtkompetitivwirkt.
3 Regulation des Stoffwechsels
In einem Versuch wurde die Frageuntersucht, wie die Synthese derAminosäuren Threonin und Lysinreguliert wird. Dazu wurde die proZeiteinheit gebildete Menge an Phos-phoasparaginsäure in Abhängigkeitvon der Lysinkonzentrationohne Threonin und mit Threoninbestimmt.
a) Werten Sie den Versuch aus undprüfen Sie, wie das Enzym Aspartat-kinase reguliert wird.b) Skizzieren Sie ein Modell derAspartatkinase mit allen Bindungs-stellen, dem zugehörigen Substratund den Effektor-Molekülen.c) Erklären Sie den Vorteil dieserEnzymregulation durch Lysin undThreonin.
Kon
zent
rati
onan
Phos
phoa
spar
agin
-sä
ure
pro
Zeite
inhe
it[r
el.E
inhe
iten]
Konzentration an Lysin[rel. Einheiten]
ohne Threonin
mit Threonin
0
��������� ����� ����
��������
��������������
�������������������
�������������������
�������
��������
�������
�����

Stoffwechselbiologie 63
PRAKTIKUM Enzyme
1 Enzymeigenschaften
Das Enzym Urease spaltet Harnstoff in Ammoniak.
Eine dem Harnstoffchemisch ähnlicheSubstanz ist N-Methyl-harnstoff.Material: 2%ige Harnstoff-Lösung; 50%ige Harnstoff-Lösung; 2%ige N-Methylharnstoff-Lösung (Xn); Urease;0,1%ige Urease-Suspension; Phenolphthalein-Lösung (F);Tropfpipetten; Spatel; Brenner; 6 Reagenzgläser;ReagenzglasständerDurchführung: Füllen Sie in Reagenzglas 1 und 2 je-weils 2 ml der verdünnten Harnstoff-Lösung, fügen Siezwei Tropfen Phenolphthalein-Lösung zu beiden Ansät-zen hinzu und schütteln Sie um.Geben Sie dann 1 ml der Urease-Suspension in Reagenz-glas 2.Geben Sie in Reagenzglas 3 drei Spatelspitzen Harnstoff,erhitzen Sie die Probe über dem Brenner und führen Sieanschließend durch Zufächeln eine Geruchsprobedurch.Geben Sie 2 ml verdünnter Harnstoff-Lösung in Reagenz-glas 4 und 2 ml gesättigter Harnstoff-Lösung in Reagenz-glas 5.Füllen Sie 2 ml der N-Methylharnstoff-Lösung in Reagenz-glas 6.Geben Sie zwei Tropfen Phenolphthalein-Lösung und an-schließend zeitgleich jeweils 1 ml der Urease-Suspensionzu den Reagenzgläsern 4 bis 6.
Reagenzglas 1 2 3 4 5 6
2%ige Harnstoff-Lösung (ml) 2 2 – 2 – –
50%ige Harnstoff-Lösung (ml) – – – – 2 –
Harnstoff (Spatelspitzen) – – 3 – – –
N-Methylharnstoff-Lösung (Tr.) – – – – – 2
Phenolphthalein-Lösung (Tr.) 2 2 – 2 2 2
Urease-Suspension (ml) – 1 – 1 1 1
Aufgabe: Notieren Sie Ihre Versuchsbeobachtungenund erklären Sie diese.
2 Einfluss von Hitze auf die EnzymwirkungUrsache für die Verfärbung von angeschnittenem Obstoder Kartoffeln ist die Reaktion pflanzlicher Phenole mitSauerstoff. Die Reaktion wird durch das pflanzeneigeneEnzym Phenoloxidase katalysiert.Material: Banane; Becherglas; Brenner; StativklammerDurchführung: Tauchen Sieeine Banane etwa zehn Se-kunden lang bis zur Hälftein kochendes Wasser. HaltenSie anschließend eine zweiteBanane für etwa 5 bis 10 Se-kunden in der gezeigten Weisein die nicht leuchtende Brenner-flamme.Die Schale darf dabei nicht verkohlen.Aufgabe: Beschreiben und erklären Sie die Versuchs-beobachtung.
3 Untersuchung der Enzymhemmung
Acarbose hemmt den enzymatischen Abbau von Stärke zuGlucose. Unter dem Handelsnamen Glucobay® wird es inder Diabetes-Therapie eingesetzt.Material: 1 Tablette Glucobay®; Iod-Kaliumiodid-Lösung;1%ige Amylose-Lösung; 1%ige Amylase-Lösung; Mörsermit Pistill; 6 Reagenzgläser; Reagenzglasständer; 3 Becher-gläser; Mikropipetten; WasserDurchführung:Zerreiben Sie eine Tablette Glucobay® im Mörser undschlämmen Sie diese in 20 ml Wasser auf. Befüllen Siesechs Reagenzgläser mit jeweils 5 ml der 1%igen Amylose-Lösung. Geben Sie in Reagenzglas 3 bis 6 steigende Men-gen (25, 50, 100, 150 μl) Glucobay®-Suspension. Geben Siezu Reagenzglas 2 bis 6 rasch jeweils 0,5 ml Amylase-Lösung. Geben Sie nach fünf Minuten jeweils zwei Trop-fen Iod-Kaliumiodid-Lösung in alle Reagenzgläser.
Reagenzglas 1 2 3 4 5 6
Amylose-Lösung (ml) 5 5 5 5 5 5
Glucobay-Suspension (μl) – – 25 50 100 150
Amylase-Lösung (ml) – 0,5 0,5 0,5 0,5 0,5
Iod-Kaliumiodid-Lösung (Tr.) 2 2 2 2 2 2
Aufgabe: Beschreiben und deuten Sie die Versuchs-ergebnisse.

Stoffwechselbiologie 95
AUFGABEN (forschen und erkennen) HILL-Reaktion
Frage Aus welchen Ausgangsstoffen der Fotosynthesestammt der freigesetzte Sauerstoff?
6 CO2 + 6 H2O LichtenergieChlorophyll 6 O2 + C6H12O6
Hypothesen Der bei der Fotosynthese freigesetzte Sauerstoff stammt:• aus Kohlenstoffdioxid.• aus dem Wasser.• zum Teil aus dem Wasser und zum anderen Teil
aus dem Kohlenstoffdioxid.
Methode Für den Versuch werden isolierte Chloroplasten inkohlenstoffdioxidfreiem Wasser aufgeschwemmt.Als Elektronenakzeptor wird DCPIP verwendet.DCPIP ist im oxidierten Zustand blau, im reduziertenZustand (DCPIPH2) farblos. Folgende Versuche werdenangesetzt:
An-satz
isolierte Chloroplastenim CO2-freiem Wasser
DCPIP Licht
1 7 ml 0,2 ml abgedunkelt*
2 7 ml 0,2 ml belichtet
3 7 ml — belichtet
* (Glas mit Alufolie umwickelt)
Ergebnis Beschreibung:
Deutung:
Erkenntnis Das Versuchsergebnis zeigt, dass Sauerstoff auch inAbwesenheit von Kohlenstoffdioxid freigesetzt werdenkann und in isolierten Chloroplasten Teilschritte derFotosynthese ablaufen können.Damit ist die Hypothese, „der bei der Fotosynthese frei-gesetzte Sauerstoff stammt aus Kohlenstoffdioxid“,widerlegt und die Hypothese, „der freigesetzte Sauerstoffstammt aus dem Wasser“, bestätigt.
�
������ ��� �� ��� �����
�
�
������ ��� �� ��� �����
�
�
�
Der dargestellte Versuch wurde 1937von Robert Hill durchgeführt undwar der erste experimentelle Beweisdafür, dass der bei der Fotosynthesegrüner Pflanzen freigesetzte Sauer-stoff aus der Spaltung des Wassersstammt. Der Versuch zeigt, dass inChloroplasten bei Belichtung die fol-gende Reaktion erfolgt.
H2O + A LichtenergieChloroplasten AH2 + 1_
2 O2
(A = Elektronenakzeptor)
Hill verwendete als Elektronen-akzeptoren Eisen-Ionen. In dem hierdargestellten Versuch nutzt mandafür den Farbstoff DCPIP, bei demman die reduzierte und oxidierteForm eindeutig an der Farbe unter-scheiden kann. Auf Grund der Ver-suchserkenntnis, dass der bei derFotosynthese freigesetzte Sauerstoffaus dem Wasser stammt, lautet dasReaktionsschema der Fotosynthese:
6 CO2 + 12 H2OC6H12O6 + 6 O2 + 6 H2O
a) Beschreiben und deuten Sie dasVersuchsergebnis der dargestelltenHill-Reaktion.b) Erklären Sie, warum durch denVersuch die Hypothese, „der bei derFotosynthese freigesetzte Sauerstoffstammt zum Teil aus dem Wasserund zum anderen Teil aus demKohlenstoffdioxid“, widerlegt wird.c) Prüfen Sie, ob man in dem Ver-such auch eine Entfärbung desDCPIPs beobachten könnte, wennman anstelle der Chloroplasten eineChlorophyll-Lösung verwendenwürde.d) Skizzieren Sie, ähnlich der Ab-bildung 1 Seite 93, den Elektronen-transport von Wasser nach DCPIP.e) Das Herbizid DCMU hemmt denElektronentransport zwischen demFotosystem II und I. Welches Ver-suchsergebnis erwarten Sie bei derHill-Reaktion, wenn man in die dreiVersuchsansätze DCMU geben wür-de? Begründen Sie Ihre Vermutung.
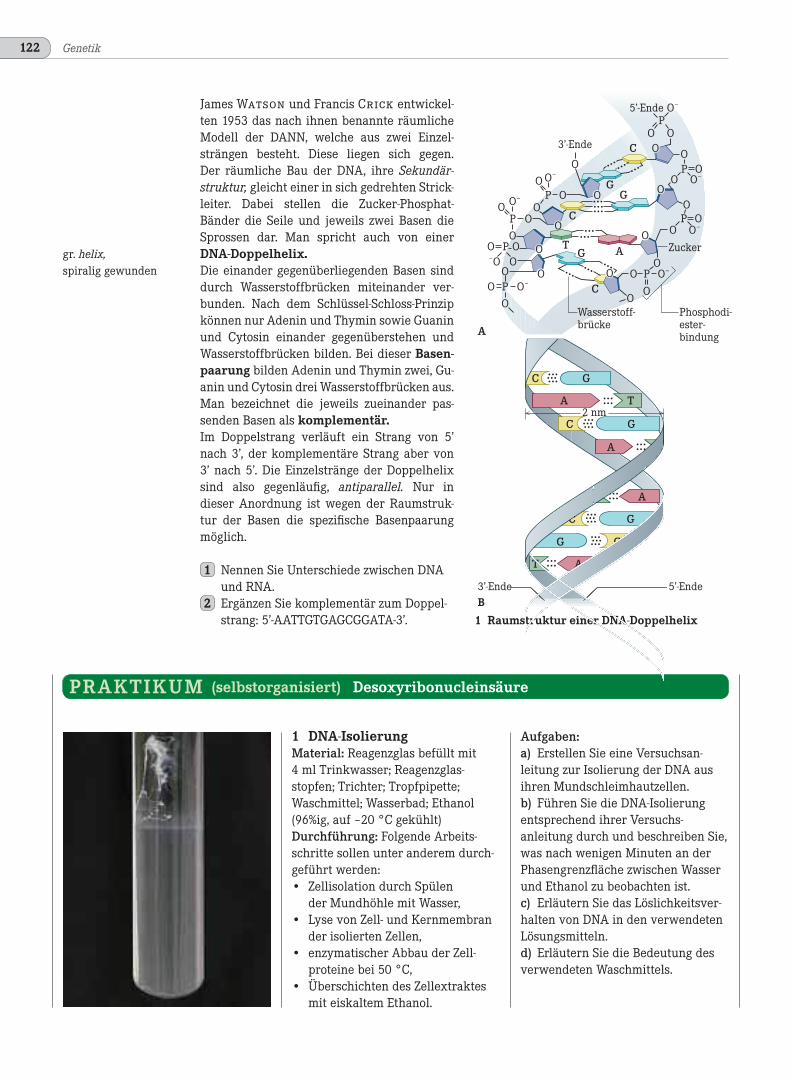
Genetik122
James Watson und Francis Crick entwickel-ten 1953 das nach ihnen benannte räumlicheModell der DANN, welche aus zwei Einzel-strängen besteht. Diese liegen sich gegen.Der räumliche Bau der DNA, ihre Sekundär-struktur, gleicht einer in sich gedrehten Strick-leiter. Dabei stellen die Zucker-Phosphat-Bänder die Seile und jeweils zwei Basen dieSprossen dar. Man spricht auch von einerDNA-Doppelhelix.Die einander gegenüberliegenden Basen sinddurch Wasserstoffbrücken miteinander ver-bunden. Nach dem Schlüssel-Schloss-Prinzipkönnen nur Adenin und Thymin sowie Guaninund Cytosin einander gegenüberstehen undWasserstoffbrücken bilden. Bei dieser Basen-paarung bilden Adenin und Thymin zwei, Gu-anin und Cytosin drei Wasserstoffbrücken aus.Man bezeichnet die jeweils zueinander pas-senden Basen als komplementär.Im Doppelstrang verläuft ein Strang von 5’nach 3’, der komplementäre Strang aber von3’ nach 5’. Die Einzelstränge der Doppelhelixsind also gegenläufig, antiparallel. Nur indieser Anordnung ist wegen der Raumstruk-tur der Basen die spezifische Basenpaarungmöglich.
Nennen Sie Unterschiede zwischen DNAund RNA.Ergänzen Sie komplementär zum Doppel-strang: 5’-AATTGTGAGCGGATA-3’.
PRAKTIKUM (selbstorganisiert) Desoxyribonucleinsäure
1 DNA-IsolierungMaterial: Reagenzglas befüllt mit4 ml Trinkwasser; Reagenzglas-stopfen; Trichter; Tropfpipette;Waschmittel; Wasserbad; Ethanol(96%ig, auf –20 °C gekühlt)Durchführung: Folgende Arbeits-schritte sollen unter anderem durch-geführt werden:• Zellisolation durch Spülen
der Mundhöhle mit Wasser,• Lyse von Zell- und Kernmembran
der isolierten Zellen,• enzymatischer Abbau der Zell-
proteine bei 50 °C,• Überschichten des Zellextraktes
mit eiskaltem Ethanol.
Aufgaben:a) Erstellen Sie eine Versuchsan-leitung zur Isolierung der DNA ausihren Mundschleimhautzellen.b) Führen Sie die DNA-Isolierungentsprechend ihrer Versuchs-anleitung durch und beschreiben Sie,was nach wenigen Minuten an derPhasengrenzfläche zwischen Wasserund Ethanol zu beobachten ist.c) Erläutern Sie das Löslichkeitsver-halten von DNA in den verwendetenLösungsmitteln.d) Erläutern Sie die Bedeutung desverwendeten Waschmittels.
1 Raumstruktur einer DNA-Doppelhelix
gr. helix,spiralig gewunden
�������
��������
�
�
�
� �����
�
�
��
������������������
�����������������������
��
�
�
�
�
�
�
������� ��������
� �
� �
� �
�
�
� �
��
��
� ��

Genetik126
METHODE PCR
Mit der Polymerasekettenreaktion(engl. polymerase chain reaction,PCR) beschrieb Kary Mullis 1983erstmals ein Verfahren zur gezieltenVervielfältigung, der Amplifikationvon DNA-Abschnitten in vitro. Dabeiist die Basis der Vervielfältigung dieDNA-Neusynthese. Benötigt werdendaher für eine PCR alle Komponen-ten, die auch bei der Replikation inZellen benötigt werden.
Eine Ausgangs-DNA wird in ein Reak-tionsgefäß zusammen mit einer hitze-stabilen DNA-Polymerase, einemGemisch aus den vier Desoxyribo-nucleosid-Triphosphaten und zweiPrimer-Molekülen gegeben.Primer sind einzelsträngige DNA-Moleküle von 15 bis 25 NucleotidenLänge. Sie sind in ihrer Basense-quenz komplementär zum Anfangund Ende des DNA-Abschnittes, dervervielfältigt werden soll. Da die Pri-mer chemisch synthetisiert werden,kann die PCR nur zur Vervielfälti-gung von DNA-Abschnitten eingesetztwerden, deren Anfangs- und End-sequenzen bekannt sind. Das PCR-
Reaktionsgefäß wird dann in einprogrammierbares Heiz-/Kühlgerät,einen Thermocycler gestellt, dessenTemperatur sehr rasch verändertwerden kann.
Ein PCR-Reaktionszyklus besteht ausdrei Temperaturschritten. Im erstenSchritt, der Denaturierung, wirdder Reaktionsansatz auf etwa 95°Cerhitzt, um doppelsträngige DNA-Moleküle in Einzelstränge zu tren-nen. Danach kühlt der Thermocyclerauf eine Temperatur zwischen 40 und60°C ab. Bei dieser Temperaturlagern sich die Primer komplementäran die Ausgangs-DNA oder die be-reits entstandenen PCR-Produktean. Dieser Schritt wird Annealinggenannt. Im dritten Schritt wird dieTemperatur auf 72°C, dem Tempe-raturoptimum der Taq-Polymeraseerhöht. Dieses Enzym wird aus demBakterium Thermus aquaticus, dasin heißen Quellen lebt, gewonnen.Es ist hitzestabil und übersteht diehohen Temperaturen während derPCR. Bei 72°C verlängert es die bei-den Primer an ihren 3’-Enden und
synthetisiert so jeweils einen neuenkomplementären Strang. Dieser Vor-gang wird Polymerisation genannt.Am Ende eines solchen dreischritti-gen Zyklus hat sich die Zahl der DNA-Vorlagen für den folgenden Zyklus imReaktionsansatz verdoppelt.
Im ersten PCR-Zyklus beginnt dieDNA-Polymerase am 3’-Ende derPrimer komplementär zur Ausgangs-DNA mit der Synthese. Die Polymeri-sation endet erst, wenn durch dieTemperaturerhöhung auf etwa 95°C,zu Beginn des zweiten Zyklus, dieDNA-Denaturierung erfolgt. Da diedabei synthetisierten Stränge nur aneinem Ende durch den Primer klarbegrenzt werden, sind sie länger alsder zu vervielfältigende DNA-Ab-schnitt. Auch diese längeren Strängefungieren neben der Ausgangs-DNAab dem zweiten Zyklus als Vorlageder Polymerase. Dienen diese PCR-Produkte als Vorlage, sind die neu-synthetisierten Stränge an der einenSeite durch den Primer und an deranderen Seite durch das Ende derVorlage begrenzt. Somit entstehen
��������������
����� �����
�������
����� �
�� ��
�������
��������
�� ��������� ��������������
����� �
�� ��������� ��������������
����� �
�� ���������
�����
������ ������� �������� �� �������� ��
� ����������� ��� ����� ������
���� � �!" �#
� ����������� ��� ����� ������
���� � �!" �#
� ����������� ���� ����� ������
���� � �!" �#
$% &%
&% $%
���'
$% &%
&% $%
�
�
1 Polymerasekettenreaktion. A PCR-Zyklus; B Prinzip der PCR

Genetik 127
erst ab dem zweiten PCR-ZyklusDNA-Abschnitte mit der gewünsch-ten, durch die Lage der Primerdefinierten Länge. In den folgendenZyklen erhöht sich die Anzahl derDNA-Abschnitte mit der gewünschtenLänge exponenziell und wird damitnachweisbar, während die Anzahlder „längeren“ PCR-Produkte nurlinear ansteigt.
In der Praxis werden 20 bis 30 PCR-Zyklen durchlaufen. Ein einzelnerZyklus dauert nur etwa fünf Minu-ten. Dies bedeutet, dass innerhalbweniger Stunden viele MillionenKopien eines bestimmten DNA-Ab-schnittes hergestellt werden können.
Die PCR ist eine Methode, die aufvielen Gebieten angewendet wird,
zum Beispiel bei der Suche nachGendefekten, der Ursache etwa vonErbkrankheiten und Krebs sowie beider kriminalistischen Analytik.
Die PCR ist eine extrem empfindlicheMethode: Sie kann eine einzige DNA-Sequenz in einer Probe aufspürenund sie so stark vervielfältigen, dasssie nachweisbar wird. Dazu müssendie bei der PCR gewonnenen ver-schieden langen DNA-Abschnittedurch Gelelektrophorese getrenntwerden. Bei der Gelelektrophoreseberuht die Trennung auf der unter-schiedlich schnellen Wanderungelektrisch geladener Moleküle zwi-schen den Elektroden eines Gleich-spannungsfeldes. Nucleinsäuren sindbei physiologischem pH-Wert durchihre Phosphatgruppen negativ gela-
den. Sie wandern im elektrischen Feldzum Pluspol, also in Richtung Anode.Verwendet man ein poröses Agaro-segel, so werden Nucleinsäuren striktnach Größe getrennt: Das Gel wirktwie ein Sieb, durch dessen Porenkleine DNA-Fragmente schneller alsgroße Fragmente wandern. Gleich-artige Stücke konzentrieren sich da-bei zu Banden. Diese DNA-Bandensind erst einmal unsichtbar. Manmacht sie durch eine Färbung etwamit Ethidiumbromid sichtbar, dassich zwischen die Basenpaare derDNA-Doppelhelix schiebt und unterUV-Strahlung orange fluoresziert.Um die Größe der vervielfältigtenDNA-Fragmente abschätzen zukönnen, lässt man DNA-Fragmentemit bekannter Größe im Gel mit-laufen.
1 Chorea HuntingtonChorea Huntington ist eine mono-gene Erbkrankheit, die mit einerErkrankung des Gehirns einhergeht.Sie beginnt mit dem Verlust der Be-wegungskontrolle und steigert sichbis hin zur Demenz. Die Diagnose er-folgt durch eine genetische Analyseeiner Blutprobe. Dabei wird die PCRmit anschließender gelelektrophore-tischer Auftrennung der erhaltenenDNA-Produkte eingesetzt.Der Gentest beruht darauf, dassChorea Huntington mit einer be-stimmten Anzahl von CAG-Triplettsim entsprechenden Gen korreliert,die mehrfach hintereinander vor-kommen. Abbildung 1 zeigt einenFamilienstammbaum über dreiGenerationen (I, II, III) und das Er-gebnis der Gelelektrophorese, dasElektropherogramm, der mittelsPCR erhaltenen DNA-Produkte deruntersuchten Familienmitglieder.
a) Erstellen Sie ein typisches Tempe-ratur-/Zeit-Diagramm bis zum Endedes zweiten Zyklus einer PCR und be-gründen Sie die gewählten Tempera-turstufen und deren Dauer.b) In der Regel werden die theore-tisch möglichen Ausbeuten an PCR-Produkten nicht erreicht. Nennen Siedafür zwei mögliche Gründe.c) Berechnen Sie die hypothetischeAnzahl der vorhandenen DNA-Kopienausgehend von einem DNA-Molekülnach 20 PCR-Zyklen.
�
������� �
� ��
� �
�� �������� ��� ���� ���� ���� ����
��
���
�
� � �
�
1 Testergebnisse Chorea Huntington. A Stammbaum; B Elektropherogramm
AUFGABEN PCR
d) Leiten Sie aus der Darstellung desElektropherogramms in der folgendenAbbildung B die Laufrichtung derPCR-Produkte und die Polung deselektrischen Feldes ab.e) Stellen Sie einen Zusammenhangzwischen der Anzahl der CAG-Wieder-holungseinheiten im Gen und demAuftreten der Krankheit her und lei-ten Sie einen möglichen Vererbungs-modus ab. Geben Sie die theoretischmöglichen Genotypen aller unter-suchten Familienmitglieder an.
A B
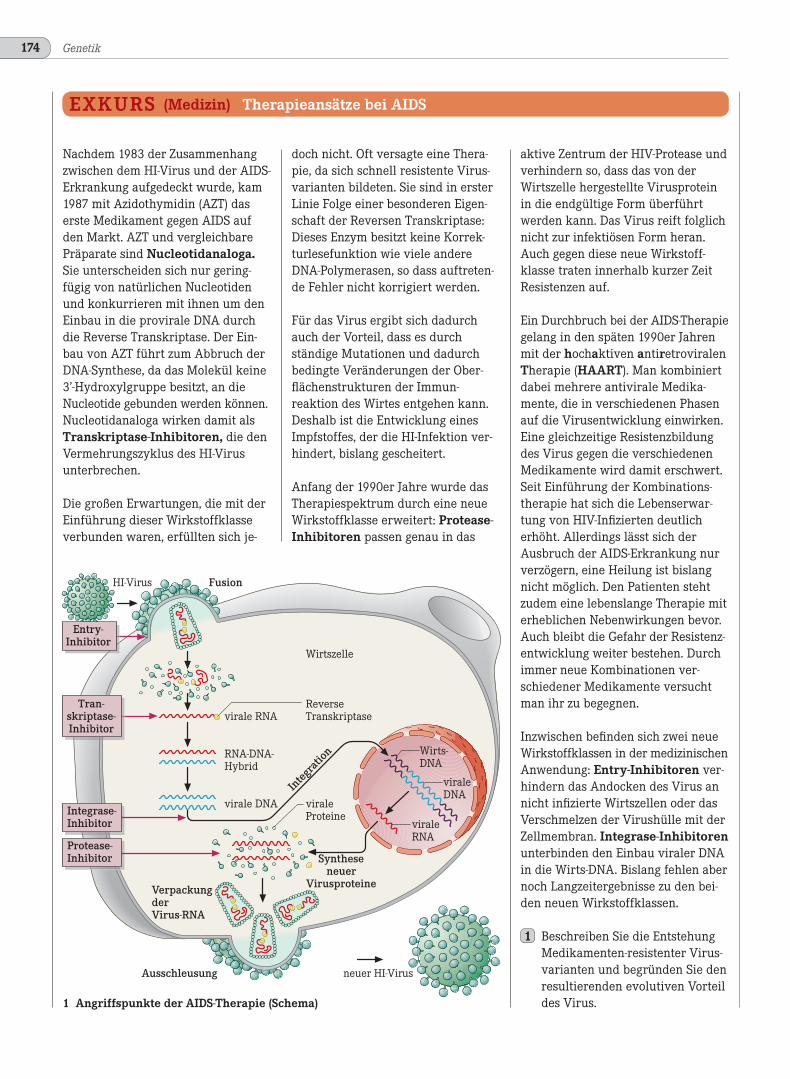
Genetik174
EXKURS (Medizin) Therapieansätze bei AIDS
�����������
����
������ �
���� �����
������ �
�������
�������
�����������
������� ����
����� �������
��������������������
����������
��������������������
��������������
�������
�������
����������� �
�������������������� �
�������������� �
�� ������������ �
�������
Nachdem 1983 der Zusammenhangzwischen dem HI-Virus und der AIDS-Erkrankung aufgedeckt wurde, kam1987 mit Azidothymidin (AZT) daserste Medikament gegen AIDS aufden Markt. AZT und vergleichbarePräparate sind Nucleotidanaloga.Sie unterscheiden sich nur gering-fügig von natürlichen Nucleotidenund konkurrieren mit ihnen um denEinbau in die provirale DNA durchdie Reverse Transkriptase. Der Ein-bau von AZT führt zum Abbruch derDNA-Synthese, da das Molekül keine3’-Hydroxylgruppe besitzt, an dieNucleotide gebunden werden können.Nucleotidanaloga wirken damit alsTranskriptase-Inhibitoren, die denVermehrungszyklus des HI-Virusunterbrechen.
Die großen Erwartungen, die mit derEinführung dieser Wirkstoffklasseverbunden waren, erfüllten sich je-
doch nicht. Oft versagte eine Thera-pie, da sich schnell resistente Virus-varianten bildeten. Sie sind in ersterLinie Folge einer besonderen Eigen-schaft der Reversen Transkriptase:Dieses Enzym besitzt keine Korrek-turlesefunktion wie viele andereDNA-Polymerasen, so dass auftreten-de Fehler nicht korrigiert werden.
Für das Virus ergibt sich dadurchauch der Vorteil, dass es durchständige Mutationen und dadurchbedingte Veränderungen der Ober-flächenstrukturen der Immun-reaktion des Wirtes entgehen kann.Deshalb ist die Entwicklung einesImpfstoffes, der die HI-Infektion ver-hindert, bislang gescheitert.
Anfang der 1990er Jahre wurde dasTherapiespektrum durch eine neueWirkstoffklasse erweitert: Protease-Inhibitoren passen genau in das
aktive Zentrum der HIV-Protease undverhindern so, dass das von derWirtszelle hergestellte Virusproteinin die endgültige Form überführtwerden kann. Das Virus reift folglichnicht zur infektiösen Form heran.Auch gegen diese neue Wirkstoff-klasse traten innerhalb kurzer ZeitResistenzen auf.
Ein Durchbruch bei der AIDS-Therapiegelang in den späten 1990er Jahrenmit der hochaktiven antiretroviralenTherapie (HAART). Man kombiniertdabei mehrere antivirale Medika-mente, die in verschiedenen Phasenauf die Virusentwicklung einwirken.Eine gleichzeitige Resistenzbildungdes Virus gegen die verschiedenenMedikamente wird damit erschwert.Seit Einführung der Kombinations-therapie hat sich die Lebenserwar-tung von HIV-Infizierten deutlicherhöht. Allerdings lässt sich derAusbruch der AIDS-Erkrankung nurverzögern, eine Heilung ist bislangnicht möglich. Den Patienten stehtzudem eine lebenslange Therapie miterheblichen Nebenwirkungen bevor.Auch bleibt die Gefahr der Resistenz-entwicklung weiter bestehen. Durchimmer neue Kombinationen ver-schiedener Medikamente versuchtman ihr zu begegnen.
Inzwischen befinden sich zwei neueWirkstoffklassen in der medizinischenAnwendung: Entry-Inhibitoren ver-hindern das Andocken des Virus annicht infizierte Wirtszellen oder dasVerschmelzen der Virushülle mit derZellmembran. Integrase-Inhibitorenunterbinden den Einbau viraler DNAin die Wirts-DNA. Bislang fehlen abernoch Langzeitergebnisse zu den bei-den neuen Wirkstoffklassen.
Beschreiben Sie die EntstehungMedikamenten-resistenter Virus-varianten und begründen Sie denresultierenden evolutiven Vorteildes Virus.1 Angriffspunkte der AIDS-Therapie (Schema)
Immunbiologie
Der 14. Mai 1796 gilt als die Geburtsstundeder Immunbiologie. An diesem Tag impfte derenglische Landarzt Edward Jenner einengesunden achtjährigen Jungen mit Gewebs-flüssigkeit, die er einer Pustel von einer mitKuhpocken infizierten Milchmagd entnom-men hatte.
Nachdem der Junge die harmlose Kuhpocken-erkrankung überstanden hatte, infizierte ihnJenner sechs Wochen später mit echtenPocken. Auch diese Infektion überstand derJunge ohne schwerwiegende Symptome.
Jenner ging dieses Wagnis ein, weil er be-obachtet hatte, dass Melkerinnen, die sichmit den für Menschen harmlos verlaufendenKuhpocken infiziert hatten, von den damalshäufig auftretenden und oftmals tödlichverlaufenden Pockeninfektionen verschontblieben oder nur leichte Krankheitsverläufezeigten.
Wie man heute weiß, hatten die den Pocken-viren ähnlichen aber harmlosen Erreger der
Kuhpocken eine Immunreaktion ausgelöst.Dabei wurden Antikörper und Gedächtnis-zellen gebildet, die für eine lebenslangeImmunität auch gegen die Erreger der gefähr-lichen Pocken sorgten. Edward Jenner giltheute als Begründer der aktiven Immunisie-rung und der Immunologie.
Durchführung einer Impfung
Phagocytose von Bakterien durch Fresszellen
EVOLUTION
Im Verlauf der Evolution haben Lebewesen unterschied-liche Abwehrsysteme entwickelt, um sich effizient vorInfektionen zu schützen.
Bakterien besitzen zum Schutz vor einer PhageninfektionRestriktionsenzyme, mit deren Hilfe sie das Erbmaterial desPhagen zerschneiden und dadurch unschädlich machen.
Wirbellose verfügen über eine unspezifische Abwehr.Eindringlinge, wie etwa Bakterien, vernichten sie vor-wiegend durch Phagocytose mittels Fresszellen.
Wirbeltiere verfügen zusätzlich zum unspezifischen Abwehr-system noch über ein spezifisches Immunsystem. Dabeihandelt es sich um ein komplexes System aus Molekülenund Zellen, durch das körperfremde Strukturen, wie etwaBakterien und Viren, erkannt und zerstört werden.

FORSCHUNG
Zentraler Forschungsgegenstand der Immunbiologie ist das Immun-system. Manchmal entgleist es aus noch ungeklärten Gründen undwendet sich gegen den eigenen Körper. Autoimmunkrankheiten,wie etwa Jugenddiabetes, können die Folge sein.
Patienten mit dem erworbenen Immunschwäche-Syndrom AIDSbesitzen eine hohe Anfälligkeit für Infektionserkrankungen, die mitfortschreitender Immunschwäche auch zum Tod führen. An derEntwicklung von Medikamenten zur Behandlung von HIV-infiziertenMenschen und einem HIV-Impfstoff wird intensiv geforscht.
Auch bei Krebserkrankungen spielt das Immunsystem eine wichtigeRolle. Patienten, die etwa nach einer Organtransplantation Medika-mente einnehmen müssen, die das Immunsystem hemmen, so dassdas Organ nicht abgestoßen wird, zeigen eine deutlich erhöhteHäufigkeit bestimmter Krebserkrankungen. Daraus gewann mandie Erkenntnis, dass ein intaktes Immunsystem auch Krebszellenkontrolliert und sie inaktivieren kann, so dass kein Tumor entsteht.
ANWENDUNG
Seit September 2006 gibt es eine Impfung gegen HumanePapillomaviren, kurz HPV. Die Viren werden durchsexuellen Kontakt übertragen und können bei FrauenGebärmutterhalskrebs auslösen.
Ohne die Impfung infizieren sich sieben von zehn Frauenmindestens einmal im Leben mit HPV. Es gibt etwa 100verschiedene Typen von HPV. Mädchen im Alter von12 bis 17 Jahren wird empfohlen, sich gegen HPV impfenzu lassen. Die Impfung schützt vor allem gegen die amhäufigsten vorkommenden Typen 16 und 18.
Angesichts der Vielzahl der potenziellen krebserregendenHPV-Typen gibt es aber keinen sicheren Schutz vor Gebär-mutterhalskrebs. Wer sich impfen lässt, darf sich nicht inSicherheit wiegen. Regelmäßige Vorsorgeuntersuchungensind weiterhin erforderlich.Humanes Papillomavirus (Rekonstruktion)
Abtöten einer Krebszelle durch eineKillerzelle

BASISWISSEN MIT BASISKONZEPTEN Steuerung und Regelung
Alle Organismen müssen bestimmteZustände aufrecht erhalten oder aufVeränderungen reagieren. Um dieszu gewährleisten, müssen lebendeSysteme auf verschiedenen System-ebenen zur Regulation fähig sein.
Bei der Regulation unterscheidet manRegelung und Steuerung. Bei derRegelung wird die Regelgröße, wiezum Beispiel die Körpertemperaturbei endothermen Tieren, weitgehendunabhängig von der Umgebungstem-peratur konstant gehalten. Dabeispielt die negative Rückkopplung,eine zentrale Rolle. Jede Abweichungder Regelgröße vom Sollwert löstautomatisch Vorgänge aus, die derAbweichung entgegenwirken. Es liegtein geschlossener Regelkreis vor.
Die konstant zu haltende Regelgrößewird durch einen Fühler fortlaufendgemessen. Dieser meldet den Istwertan den Regler. Dort wird er mit demSollwert verglichen, der von einerFührungsgröße vorgegeben wird. Beieiner Abweichung des Istwertes vomSollwert durch Störgrößen erteilt derRegler Befehle an die Stellglieder, diedaraufhin den Abweichungen ent-gegenwirken.
Bei der Steuerung wird ein Zustandoder Vorgang nicht konstant gehal-ten, sondern durch Signale in einebestimmte Richtung beeinflusst. ZumBeispiel steuert Licht das Längen-wachstum von Pflanzen.
Steuerung und Regelung finden aufden verschiedenen Systemebenenstatt. Auf zellulärer Ebene werdenzum Beispiel die Aktivitäten vonGenen und Enzymen reguliert.Bei vielzelligen Organismen findetman Regelkreise auf der Ebene derGewebe und Organe, etwa bei derRegulation der Pupillenweite desAuges oder auf der Ebene des gesam-ten Organismus, bei der Regulationder Körpertemperatur. Selbst aufder Ebene der Biozönose findet Regu-lation statt. So wird etwa die Dichteder beteiligten Populationen oder dasVerhältnis von Räuber- und Beute-tieren reguliert.
Zelluläre RegulationDie Regulation der Genaktivität kannauf verschiedenen Ebenen erfolgen.Sie ist bei Eukaryoten erheblich kom-plizierter als bei Prokaryoten. So gibtes bei Eukaryoten innerhalb einesGens Exons und Introns. Durch alter-natives Spleißen können aus einerprä-mRNA mit mehreren Exons undIntrons unterschiedlich reife mRNA-Moleküle entstehen, die in verschie-dene Peptide mit jeweils unterschied-licher Funktion übersetzt werden.
Bei den Prokaryoten ist das lac-Operon von Escherichia coli das ambesten untersuchte Beispiel für dieRegulation der Transkription. Wirdvon dem Bakterium Lactose auf-genommen, lagert sich ein Lactose-Molekül an die Bindungsstelle imLac-Repressor-Protein an. Dadurchändert sich dessen Raumgestaltreversibel, so dass der Repressornicht mehr an den Operator bindenund so die RNA-Polymerase blockie-ren kann. Demzufolge kann die
RNA-Polymerase nun an den Promo-tor binden und die Strukturgenetranskribieren. Lactose induziert alsodie Synthese für die Lactose abbau-enden Enzyme.
Durch diese Regulation wird gewähr-leistet, dass die Bakterien nur dannEnergie für die Synthese der Lactoseabbauenden Enzyme verbrauchen,wenn energiereiche Lactose zum Ab-bau zur Verfügung steht.
Enzyme können in ihrer Aktivitätdurch andere Stoffe gehemmt odergefördert werden. Dadurch ist eineRegulation von Stoffwechselprozes-sen möglich. In der Glykolyse führtdas Enzym Phosphofructokinasedie Umwandlung von Fructose-6-Phosphat in Fructose-1,6-bisphosphatdurch. Der weitere Glucoseabbauführt dann über Pyruvat zum Acetyl-CoA, welches im Citratzyklus abge-baut wird.
�����������
��������
������
������������� ��������
����������
������������� ������
�������������� ���������
���
���
�
���������� �����
���������� ���
� �� ��
��
�� ���!"����
#�� �$����%�&�$ �'����(�)��*+
� ������)��� ����
" ��
&�$��� ���������,�-!"�
&�$ �'����(���)��*+
���������� ��
��
����)��� ����
�� ���!"����
&�$���
��
Neurobiologie286
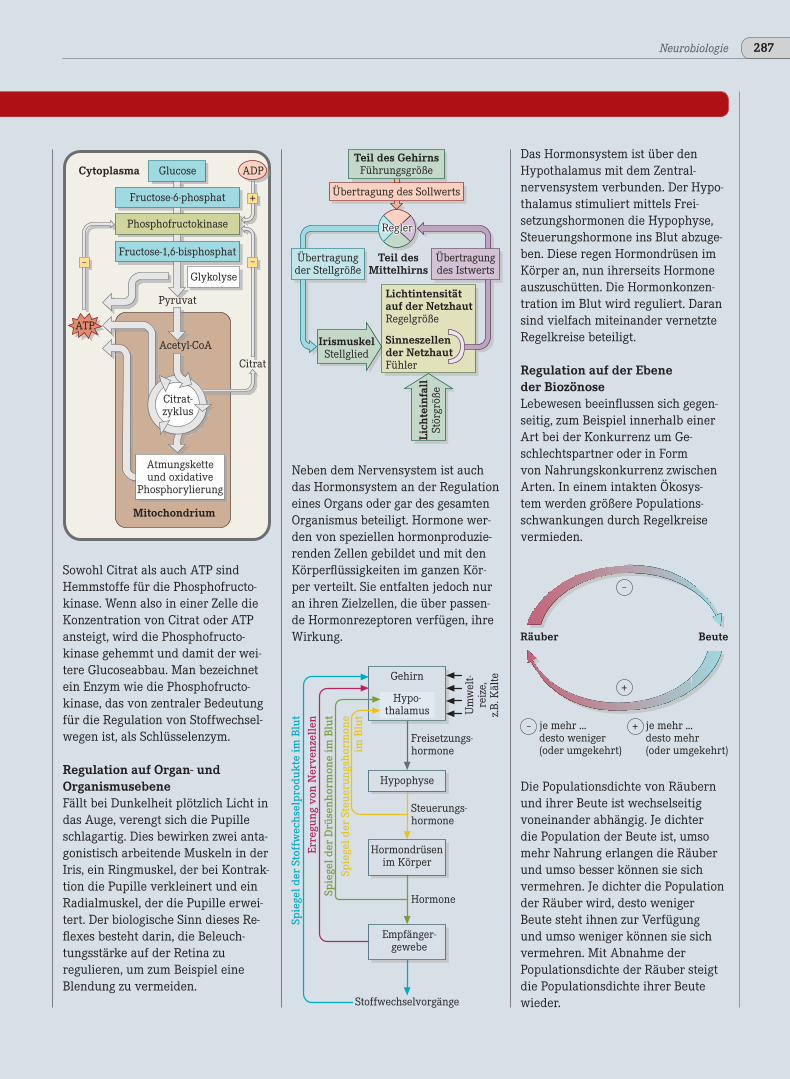
Sowohl Citrat als auch ATP sindHemmstoffe für die Phosphofructo-kinase. Wenn also in einer Zelle dieKonzentration von Citrat oder ATPansteigt, wird die Phosphofructo-kinase gehemmt und damit der wei-tere Glucoseabbau. Man bezeichnetein Enzym wie die Phosphofructo-kinase, das von zentraler Bedeutungfür die Regulation von Stoffwechsel-wegen ist, als Schlüsselenzym.
Regulation auf Organ- undOrganismusebeneFällt bei Dunkelheit plötzlich Licht indas Auge, verengt sich die Pupilleschlagartig. Dies bewirken zwei anta-gonistisch arbeitende Muskeln in derIris, ein Ringmuskel, der bei Kontrak-tion die Pupille verkleinert und einRadialmuskel, der die Pupille erwei-tert. Der biologische Sinn dieses Re-flexes besteht darin, die Beleuch-tungsstärke auf der Retina zuregulieren, um zum Beispiel eineBlendung zu vermeiden.
Neben dem Nervensystem ist auchdas Hormonsystem an der Regulationeines Organs oder gar des gesamtenOrganismus beteiligt. Hormone wer-den von speziellen hormonproduzie-renden Zellen gebildet und mit denKörperflüssigkeiten im ganzen Kör-per verteilt. Sie entfalten jedoch nuran ihren Zielzellen, die über passen-de Hormonrezeptoren verfügen, ihreWirkung.
Das Hormonsystem ist über denHypothalamus mit dem Zentral-nervensystem verbunden. Der Hypo-thalamus stimuliert mittels Frei-setzungshormonen die Hypophyse,Steuerungshormone ins Blut abzuge-ben. Diese regen Hormondrüsen imKörper an, nun ihrerseits Hormoneauszuschütten. Die Hormonkonzen-tration im Blut wird reguliert. Daransind vielfach miteinander vernetzteRegelkreise beteiligt.
Regulation auf der Ebeneder BiozönoseLebewesen beeinflussen sich gegen-seitig, zum Beispiel innerhalb einerArt bei der Konkurrenz um Ge-schlechtspartner oder in Formvon Nahrungskonkurrenz zwischenArten. In einem intakten Ökosys-tem werden größere Populations-schwankungen durch Regelkreisevermieden.
Die Populationsdichte von Räubernund ihrer Beute ist wechselseitigvoneinander abhängig. Je dichterdie Population der Beute ist, umsomehr Nahrung erlangen die Räuberund umso besser können sie sichvermehren. Je dichter die Populationder Räuber wird, desto wenigerBeute steht ihnen zur Verfügungund umso weniger können sie sichvermehren. Mit Abnahme derPopulationsdichte der Räuber steigtdie Populationsdichte ihrer Beutewieder.
��������
���������
���� �������
�� ��������
��������������� ��� ���
���������������
����������������� �
��������������� ��
���������� ��!������� �
�������
����� �
�"�
�#�
$
%
�� �
���������
$
&!���� ���� ��� '���(����
���� �� �������)��������*+�
���� ������������
&!���� ������� ,��(����
&!���� ������� '������*+�
�� �������������� ��� ��������-������*+�
�������������� ���������)����
�������'��������
�� ���������
'�*�
��*+
�
--����������-�����
�����
.������ � ���
.��������
.�������)���� /*����
'����(����������0���
1�
(��
����
��
�232/
0���
����������� �� �������� ������!"���
�����# ���#�$����
���������%�&��� �! ��!"���
������������������ �! �
�!"���
�������������������
'�����������������
.������
4���0�������(�!�
������ ����
$
%
5� ���� 222����� (�����6���� ���������7
5� ���� 222����� ����6���� ���������7
%$
Neurobiologie 287


















