
Anleitung zum Biochemischen Praktikum für Mediziner · Praktikum zu entleeren und kurz mit Wasser...
75
Anleitung zum Biochemischen Praktikum für Mediziner Universität zu Köln 2
Transcript of Anleitung zum Biochemischen Praktikum für Mediziner · Praktikum zu entleeren und kurz mit Wasser...

Anleitung zum
Biochemischen Praktikum
für Mediziner
Universität zu Köln
2

3
Inhaltsverzeichnis
„Zehn Gebote“, Einführung, allgemeine Hinweise, Unfallverhütung, Pipetten, pH-Meter
Versuch 1: Aminosäuren und Proteine I
Lambert-Beer'sches Gesetz
Quantitative Biuret-Reaktion
Titrationskurve von Aminosäuren
Proteinstrukturen (Teil I)
Versuch 2: Aminosäuren und Proteine II
SDS-Polyacrylamidgelelektrophorese
Western-Blot
Proteinstrukturen (Teil II)
Versuch 3: Enzyme
Enzymkinetik der sauren Phosphatase
Acetylcholinesterase-Bestimmung im Serum
Hemmung des Enzyms mit (DIFP)
Reaktivierung mit PAM
Laktatdehydrogenase (opt. Test)
Versuch 4: Lipide
Nachweisreaktionen von Lipiden
DC-Trennung von Myelin-Lipiden
Gaschromatographie von FSM-Estern
DC von Steroidhormonen
4
Versuch 5: Immunologie
ELISA (enzyme linked immunosorbent assay), Enzymgekoppelter Immunnachweis
Western-Blot
Test zur Schwangerschaftsfrüherkennung aus Urin
Versuch 6: Blut
Gelfiltration, Spektroskopie und
die quantitative Bestimmung des Hämoglobins
Dialyse-Versuch
Enzymatische Bestimmung der Glucose
Polarimetrie
Versuch 7: Nukleinsäuren I
Löslichkeit der Harnsäure
Xanthinoxidase-Reaktion und ihre kompetitive Hemmung
Bestimmung der Harnsäure mit Hilfe der UV-Spektroskopie
Restriktionsendonukleasen
Polymerasekettenreaktion (PCR)
Versuch 8: Nukleinsäuren II
Regulation der Gen-Aktivität am Beispiel des lac Operons
DNA-Isolierung
Analyse von Erbkrankheiten am Beispiel der Cystischen Fibrose

5 Die ZEHN Gebote
1. Unfallverhütung: Beachten Sie bitte die Hinweise im Praktikumsbuch!
2. Es gibt keinen Fehltag! Bei Krankheit bringen Sie bitte ein Attest, und erfragen, wann der versäumte Versuch nachgeholt werden kann.
3. Sollten Sie keine ausreichende Vorbereitung nachweisen können, müssen Sie den Versuch an einem anderen Tag wiederholen.
4. Analysen: Mindestens 6 von 9 Analysen müssen richtig sein. Die Analysen müssen von jedem Studenten einzeln durchgeführt werden und die Ergebnisse in die ausliegenden Listen eingetragen werden. (Die Menge der ausgegebenen Analysenlösung ist begrenzt).
5. Sauberes Arbeiten: Bitte benutzen Sie nicht die Pipetten, um die Reagenzien aus der Vorratsflasche zu entnehmen. Die ungefähr benötigte Menge soll in ein Becherglas gegossen werden, und daraus pipettiert werden. Restlösungen bitte wegschütten und nicht in die Originalflaschen zurückgeben.
6. Arbeitsplatz: Bitte hinterlassen Sie Ihren Arbeitsplatz abends sauber und ordentlich. Jede Gruppe meldet sich beim Assistenten ab, damit er den Platz abnimmt.
7. Ende des Praktikums 17.00 Uhr
8. Protokollhefte (Meßdaten, Berechnungen, Diagramme) bitte knapp abfassen. Die Anleitung nicht abschreiben! Montagsgruppe: Do. einwerfen Mo. zurück Mittwochsgruppe: Fr. einwerfen Mi. zurück
9. Kleiderschränke in den Waschräumen können benutzt werden (eigenes Vorhängeschloß mitbringen!).
10. Die erfolreiche Teilnahme am Praktikum wird nur denjenigen bescheinigt, die alle Versuche absolviert haben und ein Referat gehalten haben. Andernfalls muss das Praktikum wiederholt werden
6 ALLGEMEINE HINWEISE
Die vorliegende Anleitung für das biochemische Praktikum an der Universität zu
Köln versucht, den Studenten mit einem breiten Spektrum biochemischer
Methoden von unterschiedlichem Schwierigkeitsgrad vertraut zu machen. Eine
Beschreibung der theoretischen Grundlagen ergänzt die ausführlich beschriebenen
Versuche.
Jeder Versuch muss vom Studenten theoretisch gründlich vorbereitet und die
Methodik vor Beginn genau durchdacht werden. Anders ist ein erfolgreiches
Experimentieren nicht denkbar.
Jedem Versuch ist eine Zusammenstellung der Wissensgebiete vorangestellt. Sie
werden in der Vorlesung in Anlehnung an die Approbationsordnung ausführlich
dargestellt. Diese Gebiete sollten die Studenten beherrschen.
Jeder Versuch mit allen Experimenten muss an einem Praktikumstag beendet
werden. Ausnahme ist der Versuch 2. Die verfügbare Zeit sollte daher
zweckmäßig eingeteilt werden. Versuche mit langer Dauer sollten als erste
begonnen werden; Wartezeiten bei länger laufenden Versuchen können für
kürzere Experimente genutzt werden. Die Praktikumszeit ist einzuhalten. Bei
zügiger Durchführung verbleibt in allen Versuchen Zeit zur sorgfältigen Aus-
wertung der Ergebnisse und Besprechung der theoretischen Grundlagen.
Ein Versuch gilt als abgeschlossen, wenn die experimentellen Ergebnisse vom
Assistenten überprüft und testiert, der Arbeitsplatz und die benutzten Geräte
aufgeräumt und zur Reinigung vorbereitet sind.
Über alle Versuche ist Protokoll zu führen. Das Protokollheft muss enthalten:
1) Datum und Bezeichnung des Versuchs.
2) Fragestellung, evtl. Reaktionsgleichungen
3) In knappem Stil, aber lückenlos, die experimentellen Ergebnisse (Chro-
matogramme, Beobachtungen, Meßzahlen) und die Auswertung dieser
Ergebnisse (Diagramme, Berechnungen). Es sollten alle zum Nachvollziehen
des Versuchs notwendigen Angaben, insbesondere Konzentrations- und
Mengenangaben gemacht werden.

7 4) Abschätzung der Fehler mit anschließender Diskussion, inklusive Vergleich
mit in der Klinik auftretenden Werten.
Es wird besonderer Wert auf eine übersichtliche und saubere Anordnung der
Werte im Protokollheft gelegt. Chromatogramme, Diagramme sollen eingeheftet
oder eingeklebt werden, "fliegende Blätter" werden nicht akzeptiert..
Die regelmäßige, erfolgreiche Teilnahme am biochemischen Praktikum wird
ferner nach einem zufriedenstellend gehaltenen Referat (s.u.) bescheinigt.
Lösungen werden in der benötigten Menge aus der Vorratsflasche in ein Rea-
genzglas oder in ein Becherglas gegossen; aus der Vorratsflasche sollen Lösungen
nie mit einer Pipette entnommen werden. Feste Substanzen werden mit sauberem
Spatel entnommen. Man arbeite grundsätzlich mit möglichst kleinen
Substanzmengen. Beim Arbeiten mit Eppendorfreaktionsgefäßen achten Sie bitte
darauf, dass sich die Lösung am Boden des Gefäßes befindet (Gegebenenfalls kurz
abzentrifugieren!)
Die erforderlichen Geräte müssen sorgsam benutzt werden. Zentrifugen bitte nur
betreiben, wenn sie vorher austariert worden sind. Die Glasgeräte sind nach dem
Praktikum zu entleeren und kurz mit Wasser vorzureinigen; die Sauberkeit der
Geräte ist Vorbedingung für zuverlässige Ergebnisse.
Unvorbereitete Studenten müssen den Versuch wiederholen.
Referate
Neben den acht experimentellen Versuchsnachmittagen werden zusätzlich an drei
Tagen Seminare abgehalten, in denen die Studenten jeweils vier Referate über
zum Praktikumsabschnitt passende Themen halten. Die nicht mehr im Praktikum
enthaltenen Versuchsthemen „Puffer“, „Bioenergetik“ und „Kohlenhydrate“ sind
auch Bestandteil dieser Seminare.
Die Referatthemen werden vorher von den Assistenten vergeben. Jedes Referat
dauert 20 Minuten und wird von zwei Studenten gehalten, die beide ihren Teil
8 eigenständig vortragen. Anschließend wird 10 Minuten diskutiert und die
Leistung der Vortragenden bewertet. Jeder Student hat im Verlauf des
Blockunterrichts ein Referat zu halten. Ohne Referat wird der Schein zur
erfolgreichen Teilnahme am Biochemischen Praktikum nicht ausgestellt.

9
HINWEISE ZUR UNFALLVERHÜTUNG
Im Labor ist das Tragen eines Baumwollkittels und einer Schutzbrille Pflicht!
Pipettieren
Die am Arbeitsplatz bereitliegenden automatischen Pipetten, Schliffpipetten,
Pipettierhilfen oder Spritzen sollen zur Dosierung entsprechender Lösungen
benutzt werden. Es darf auf keinen Fall mit dem Mund pipettiert werden!
Konzentrierte Säuren wie z.B. HNO3, H2SO4, CH3COOH, HCl oder Laugen wie
z.B. NaOH, KOH, NH4OH sowie giftige (Cyanid-Lösungen, DIFP) oder
infektiöse Lösungen wie Serum, E. coli-Suspensionen können ein erhebliches
Gefahrenpotenzial darstellen. Daher stehen automatische Pipetten s. u. und
Pipettierhilfen zur Verfügung: Blaue Pipettierhilfen für 1 ml Glaspipetten, rote
Pipettierhilfen für 5, 10 oder 20ml Glaspipetten. Die Pipetten werden vorsichtig
in den Gummiadaptor gesteckt und die Flüssigkeit wird mit Hilfe des Rändelrads
aufgezogen. Zum Ablassen der Flüssigkeit wird der seitlich angebrachte Schalter
gedrückt. Vermeiden Sie unter allen Umständen, dass Flüssigkeit in die
Pipettierhilfe gelangt.
Tipp: Ziehen Sie zuerst einen Überschuss an Flüssigkeit auf, und stellen Sie
dann das genaue Volumen mit Hilfe des Ablassschalters ein.
Jeder Student ist angehalten, in eigener Verantwortung mit den Chemikalien
umsichtig und fachgerecht so umzugehen, dass selbst verhältnismäßig unge-
fährliche Lösungen nicht auf den Körper oder in den Mund gelangen können.
Essen, Trinken, Rauchen und die Anwendung von Kosmetika sind strikt
untersagt. Der Umgang mit Chemikalien ist nicht narrensicher und verlangt
Vorsicht, Verantwortungsbewußtsein und entsprechende Sachkenntnisse.
10 Gebrauch der automatischen Pipetten
Um auch im Mikroliter-Bereich genau und sicher pipettieren zu können, stehen
Ihnen mechanische Pipettierhilfen (Abb. 1) der Firmen Gilson (PIPETMAN) oder
Biozym (PreCision) zur Verfügung. Je nach Modell können Sie damit zwischen 2
und 5000µl pipettieren. Das unsachgemäße Arbeiten mit diesen
Präzisonsgeräten führt zur Zerstörung der Pipettierhilfen, die hohe Kosten
verursachen. Beachten Sie bitte diese Arbeitshinweise und üben Sie vor dem
ersten Einsatz der Pipetten den Gebrauch mit entionisiertem Wasser.
Abb. 1 Automatische Pipettierhilfen
Die Pipetten haben ein digitales Mikrometer, das das Volumen angibt. Das
Volumen wird durch Drehen einer Einstellschraube (bei der PreCision identisch
mit dem Druckknopf, bei PIPETMAN, schwarze geriffelte Schraube) eingestellt.
Dabei dürfen die Einstellbereiche der einzelnen Pipetten auf keinen Fall über-
oder unterschritten werden:

11 Pipetman P20 2 - 20µl Pipetman P5000 1000 -5000µl
Pipetman P100 20 -100µl Precision rot 5 - 50µl
Pipetman P 200 50 - 200µl Precision blau 100 - 1000µl
Pipetman P 1000 200 - 1000µl
Die Digitalanzeigen der PreCision Pipetten werden von links nach rechts
abgelesen und zeigen den tatsächlichen Wert in µl an. Bei der Precision rot sind
0,5µl Schritte einstellbar. Die Digitalanzeige der Pipetman-Pipetten werden von
oben nach unten abgelesen und bestehen aus 3 Ziffern. Bei P20, P100 und P200
bedeuten die schwarzen Ziffern Mikroliter und die roten Ziffern zehntel
Mikroliter, bei P1000 und P5000 bedeuten die roten Ziffern Milliliter und die
schwarzen Ziffern Mikroliter.
Für Pipetman P20, P100, P200 und Precision rot werden gelbe, für Pipetman 1000
und Precision blau werden blaue und für Pipetman P5000 werden transparente
Einwegpipettenspitzen aus Polypropylen verwendet. Die passenden Spitzen
werden auf den Schaft der Pipette aufgesteckt und mit leichtem Drehen so fest
angedrückt, dass eine absolute Dichtheit gewährleistet ist. Dabei fassen Sie die
Spitzen immer nur am oberen Ende an, niemals an der Spitze. ACHTUNG:
NIEMALS FLÜSSIGKEITEN OHNE SPITZE AUFNEHMEN!
Füllen
Den Druckknopf bis zum ersten Druckpunkt eindrücken (Abb. 2A). Die Pipette
senkrecht halten, und die Spitze in die Probenflüssigkeit eintauchen. Den
Druckknopf dann langsam und gleichmäßig zurückgleiten lassen (Abb. 2B) Eine
Sekunde warten, und dann die Spitze aus der Flüssigkeit nehmen.
Entleeren
Das Ende der Spitze in einem Winkel von 10 - 40 Grad gegen die Innenwand des
Gefäßes halten. Den Druckknopf langsam bis zum ersten Druckpunkt
herunterdrücken (Abb. 2C). Eine Sekunde warten. Den Druckknopf bis zum
zweiten Druckpunkt herunterdrücken, um restliche Flüssigkeit auszustoßen. (Abb.
2D) Die Pipette mit ganz gedrücktem Druckknopf herausnehmen. Den
Druckknopf loslassen (Abb. 2E)
12 Die Spitze durch Drücken des Spitzenabwerfers (P5000 nur manuell) abwerfen.
Die Spitze muss nur gewechselt werden, wenn eine andere Flüssigkeit pipettiert
oder die Volumeneinstellung geändert wird.
Abb. 2 A-E
Gebrauch der pH-Meter
Die pH-Meter müssen vor Arbeitsbeginn geeicht werden (Versuch 1). Dazu stehen
zwei Eichlösungen mit pH7 und pH4 zur Verfügung. Zuerst wird die
Glaselektrode mit dest. Wasser gespült, dann (zerstörungsfrei) in die pH7-Lösung
eingetaucht und der pH-Wert ggf. mit dem Assymetriedrehknopf auf 7 einstellt.
Dann wird die Elektrode mit Wasser gespült und in die pH4-Lösung eingetaucht.
Erreicht der abgelesene Wert 3,9 -4,1, ist die Genauigkeit ausreichend für die in
diesem Praktikum durchgeführten Versuche. Nur wenn der pH-Wert darunter oder
darüber liegt, muss die Steilheit (2. Drehknopf) entsprechend erniedrigt oder
erhöht werden. Nach dem Anpassen der Steilheit wird die Eichung wie oben
beschrieben erneut durchgeführt, zuerst bei pH7 dann bei pH4.

1-1
Versuch 1: Aminosäuren und Proteine I
Versuche:
Lambert-Beer'sches Gesetz
Quantitative Biuret-Reaktion
Titrationskurve von Aminosäuren
Analysen: 1. Bestimmung von unbekannten Aminosäuren durch Titration (Ausnahme:
Gemeinsame Analyse)
2. Proteinbestimmung nach Biuret
Wissensgebiete
Lambert-Beer'sches Gesetz, Photometrie
Aminosäuren: Struktur, Kurzschreibweise, chemische Eigenschaften, essentielle
AS
Enzymatischer Mechanismus der AS-Transaminierung, Decarboxylierung zu
biogenen Aminen, oxidative Desaminierung
Interpretation einer Titrationskurve, isoelektrischer Punkt, pK-Wert, pH-Wert,
Dissoziation des Wassers
Harnstoff-Zyklus und Harnstoff-Biosynthese
Peptide und Proteine: Primär-, Sekundär-, Tertiär- und Quartärstruktur
Bindungsarten in Proteinen: Peptidbindung, Disulfidbrücken
Proteine des Blut-Serums, des Blut-Plasmas
Serum-Normalwerte: Gesamtproteinmenge, prozentuale Zusammensetzung der
Proteine
Allgemeine und spezifische Nachweisreaktionen der Aminosäuren und Proteine
(SDS)- Gelelektrophorese
Trennmethoden:
Ionenaustauscher- und Affinitäts-Chromatographie, Gelfiltration.
1-2 Aminosäuren und Proteine I
Einführung Photometrie
Der Farbcharakter eines Farbstoffs kommt dadurch zustande, dass der Farbstoff
die für ihn charakteristischen Wellenlängen des sichtbaren Lichts absorbiert. Je
konzentrierter die Farbstofflösung ist, desto größer ist die Absorption oder
Lichtschwächung.
Das Prinzip der Photometrie beruht auf der Schwächung eines monochroma-
tischen Lichtstrahls, der durch eine Lösung geschickt wird. Dabei ist die
Abschwächung des monochromatischen Lichtstrahls proportional zur
Konzentration des gelösten Stoffes und der Dicke der Schicht. Das einfallende
Licht wird beim Durchtritt durch eine differentielle Schicht dx um dI geschwächt.
Die Lichtschwächung gehorcht der Funktion
-dI = K⋅I⋅dx (K = Substanzkonstante) Das bestimmte Integral lautet dann:
⌡⌠
Io
I
1
I . dI = -K 1 dx .
x=0
d
∫ (d = Schichtdicke)
Daraus folgt: ln I - ln Io = -K⋅ d
und: ln (IIo
) = ln D = -K⋅d (D=Durchlässigkeit)
Daraus ergibt sich das Lambert'sche Gesetz:
IIo
= e-K⋅d = 10-m⋅d (m = K⋅ln10)
m ist ein für die absorbierende Materie und die Wellenlänge typischer Faktor, und
ist in Lösungen in der Regel der Konzentration c proportional:
m = ε⋅c. Die Proportionalitätskonstante ε ist der molare Extinktionskoeffizient. Den
reziproken Wert IoI bevorzugt man gegenüber der Durchlässigkeit I
Io ,weil sein
Wert anwächst, je größer die Absorption wird. Dessen dekadischer Logarithmus
wird als die Extinktion E bezeichnet:

Einführung 1-3
IoI = 10md = 10εcd
log IoI = ε • c • d = E Lambert-Beer'sches Gesetz
Der molare Extinktionskoeffizient
ε = Ε
c • d [l × mol-1 × cm-1] E = Extinktion
c = Konzentration (mol/l) d = Schichtdicke (cm)
Die Extinktion E, welche auch als Absorption oder optische Dichte bezeichnet
wird, ist der Schichtdicke d und der Konzentration c proportional. Die Extinktion ist die Messgröße bei allen photometrischen Untersuchungen. Die Proportionalität
zwischen Extinktion und Konzentration, die das Beer’sche Gesetz verlangt, gilt
nicht in allen Fällen, vor allem nicht mehr in höheren Konzentrationsbereichen.
Da das durchfallende Licht auch durch Reflexion an den Oberflächen der
Küvettenwände geschwächt wird, unter Umständen auch durch das Lösungsmittel,
muss zum Vergleich die Extinktion einer gleich großen, mit Lösungsmittel
beschickten Küvette als Nullwert gemessen werden. Standardlösungen bekannter
Zusammensetzung sind zur Anlage der Eichkurve bzw. zur Ermittlung von ε
notwendig.
Erstellen einer Eichkurve: Eine Reihe von Lösungen bekannter Konzentrationen werden gemessen und die
erhaltenen Extinktionen auf der Y-Achse gegen die Konzentrationen auf der X-
Achse aufgetragen. An dieser Eichkurve lässt sich für jeden Wert von E die
zugehörige Substanzmenge ablesen. Man kann auch aus jedem gemessenen Eichwert (Extinktion Ey und Y mg Substanz) mit Hilfe des Dreisatzes die zuge-
hörige Substanzmenge X einer Probe mit der Extinktion Ex im Reaktionsansatz
berechnen.
1-4 Aminosäuren und Proteine I
30002000100000,0
0,1
0,2
0,3
0,4
0,5
0,6
mg/l
E
Abbildung 1-1: Eichkurve für eine photometrische Analyse
Die Abbildung 1-2 zeigt den schematischen Aufbau eines Photometers. Von einer
Lichtquelle (hier eine Wolfram-Glühlampe für den sichtbaren und eine
Deuteriumlampe für den UV-Bereich) werden, nachdem ein dünner Strahl durch
eine Spaltblende ausgesondert wurde, bestimmte Wellenlängen durch einen
Monochromator (Filter, Gitter oder Prismen) selektiert. Für z.B. orangefarbene
Lösungen wird ein Filter der Komplementärfarbe blau verwendet. Diese
monochromatische (einfarbige) Strahlung wird durch eine Linse gebündelt und
gelangt durch einen Ausgangsspalt zur Küvette, einem quaderförmigen Gefäß für
die zu bestimmende Lösung in der das durch die Lösung gehende Licht je nach
Konzentration des Farbstoffes geschwächt wird. Je nach Wellenlänge werden
unterschiedliche Küvetten verwendet, z. B Quarzküvetten für den UV-Bereich,
Glas- bzw. Plastikküvetten für den sichtbaren Bereich (ca. 1-5 ml Inhalt, genau
angegebenen Innenabmessungen, meist 1.0 cm "Schichtdicke"),. Das Licht gelangt
schließlich zum Detektor (Photozelle oder Photodioden, die Licht in Strom
wandeln) und das Messergebnis kann auf verschiedene Weise angezeigt werden
(Amperemeter/ Display/ Computer/ Schreiber/ Drucker).
Das im Praktikum verwendete Spektralphotometer ULTROSPEC1000 ist ein
einfach zu benutzendes, mikroprozessorgesteuertes Gerät, das Sie zur Messung
der Absorption von Lösungen bei von Ihnen vorgewählten Wellenlängen ver-

Einführung 1-5
wenden. Die Geräte sind bei Beginn des Praktikums bereits eingeschaltetet und
betriebsbereit. Drücken Sie die λ-Taste auf der Tastatur, geben Sie die gewünschte
Wellenlänge ein und bestätigen Sie mit der F3-Taste (↵). Dann füllen Sie eine
Küvette mit dem „Leerwert“ (z. B. Wasser oder ein Reagenzienmix ohne zu
messende Substanz), stellen sie in den Strahlengang (Orientierung beachten!),
schließen den Deckel des Geräts und drücken Sie die -Taste. Der Leerwert
wird automatisch auf 0,000 eingestellt. Nehmen Sie zur Messung entweder
dieselbe Küvette oder bestimmen Sie den Küvettenfehler, indem Sie die
Messküvette ebenfalls mit dem Leerwert füllen und die Extinktion bestimmen.
Stellen Sie dann die Küvette mit ihrer Messlösung in den Strahlengang und
notieren Sie das Ergebnis.
Lampe Spalt Mono- Linse Spalt Küvette Detektor Anzeige
chromator Abbildung 1-2
1-6 Aminosäuren und Proteine I
Aminosäuren Die meisten Aminosäuren sind α-Aminocarbonsäuren, z. B. Glycin oder Valin:
H2N CH
CH3
COOH
H3C CH3
COOHCH
CH
H2N
20 verschiedene Aminosäuren werden in ein Protein während der Synthese am
Ribosom eingebaut. Sie werden in der Dreibuchstaben-Kurzschreibweise nach-
stehend angegeben:
Ala Arg Asn Asp Cys Gln Glu Gly His Ile
Leu Lys Met Phe Pro Ser Thr Tyr Trp Val
Neben den zwanzig kodierten Aminosäuren enthalten viele Proteine noch andere
Aminosäuren. Sie werden posttranslational enzymatisch verändert. Diese Proteine
haben spezifische Funktionen. Überlegen Sie sich, welche der Aminosäuren
verändert sind und welche Funktionen diese Proteine erfüllen. Die Aminosäuren
selbst unterliegen metabolischen Vorgängen im Körper. Überlegen Sie sich,
welche Reaktionen diese sind und welche Produkte (u. a. Neurotransmitter) dabei
entstehen.
Die unterschiedlichen Seitenketten der Aminosäuren prägen neben den Amino-
und Carboxylgruppen ihre Eigenschaften. Sie entscheiden über ihre chemischen
Eigenschaften, beispielsweise sauer, basisch oder neutral. Ferner klassifiziert man
noch die aromatischen, die polaren bzw. apolaren Aminosäuren. Diese
Unterschiede können zu ihrer Trennung genutzt werden.
Im Teilversuch I lernen Sie die Eigenschaften der Aminosäuren kennen. In
Proteinen treten dagegen die Summe der Eigenschaften der Seitenketten hervor.
Im Teilversuch II werden Sie Plasmaproteine untersuchen, die wegen dieser
unterschiedlichen Eigenschaften durch Ionenaustauscherchromatographie
aufgetrennt wurden.

Einführung 1-7
Säure-Base-Eigenschaften von Aminosäuren:
Da die Aminosäuren mindestens eine Carboxylgruppe und eine Aminogruppe
besitzen, liegt eine Monoamino-monocarbonsäure in wässriger Lösung in drei
Ionisationsstufen vor:
R C
H
NH3
COOH R C
H
COO
NH3
R C
H
NH2
COO
K2K1
AS-AS+-AS
+
- -
++
Das Mengenverhältnis der drei Ionisationsformen ist abhängig von den pK-
Werten der beiden funktionellen Gruppen und dem pH-Wert. Für die Dissoziation
der Carboxylgruppe gilt die Henderson-Hasselbalch'sche Gleichung:
pH = pK1 + log AS
AS
±
+ Gleichung 1
für die Aminogruppe:
pH = pK2 + log AS
AS
−
± Gleichung 2
Die pK-Werte der Aminosäuren versieht man mit Indices: Dabei beziehen sich: der pK1 auf die α-Carboxylgruppe, der pK2 auf die α-Aminogruppe und der pKR
bezieht sich stets auf die funktionelle Gruppe der Seitenkette.
Tabelle 1-1: pK-Werte der ionisierenden Gruppen von α-Aminosäuren bei 25 oC:
Aminosäure
pK1
-COOH pK2
-NH3+
pKR Seitenkette
Glycin 2,35 9,78
Asparaginsäure 1,99 9,90 3,90
Glutaminsäure 2,10 9,47 4,07
Arginin 1,82 8,99 12,48
Lysin 2,16 9,18 10,79
Tyrosin 2,20 9,17 10,13
Histidin 1,82 9,11 6,00
1-8 Aminosäuren und Proteine I
Isoelektrischer Punkt:
Der isoelektrische Punkt ist als derjenige pH-Wert definiert, bei dem die Summe
der positiven Ladungen einer Substanz gleich der der negativen ist.
Da das Zwitterion nach außen neutral ist, muss dann also gelten:
[ ] [ ]AS AS+ = −
Hieraus und aus den Gleichungen 1 und 2 folgt:
pI = pK + pK
2 1 2
Der pI von Monoaminodicarbonsäuren (Asp, Glu) ergibt sich aus:
pI = pK + pK
2 1 3
der pI von Diaminomonocarbonsäuren (Lys, Arg) aus:
pI = pK + pK
2 2 3
Das Endprodukt des Aminosäurestoffwechsels.
Harnstoff, das Endprodukt des Aminosäure-Stoffwechsels beim Menschen, wird
durch Urease (kommt in Sojabohnen und Bakterien vor) hydrolytisch in CO2 und
NH3 gespalten. Auf der Wirkung der Bakterien-Urease beruht die
"ammoniakalische Gärung" von Urin. Für die Bestimmung von Harnstoff in
biologischem Material (Blut, Harn) hat Urease große Bedeutung, da sie streng
spezifisch ist und das entstehende Ammoniak nach verschiedenen Methoden
genau bestimmt werden kann (keine Durchführung in diesem Praktikum).
pKR
pKR

Biuret 2-9
I. Quantitative Biuret-Reaktion
Proteine und Peptide bilden mit Cu2+-Ionen in alkalischer Lösung (Biuret-Rea-
genz) rot bis violett gefärbte Komplexverbindungen, freie Aminosäuren jedoch
nicht. Die quantitative Biuret-Probe ist eine der häufigsten Proteinbestim-
mungsmethode. Der Name der Reaktion stammt von der einfachsten Verbindung,
die mit dem Biuret-Reagenz (Cu2+ in alkalischer Lösung) die Komplexverbindung
liefert, dem Biuret. Es entsteht durch trockenes Erhitzen von Harnstoff:
O
NH2
C
NH2
CO
O C
NH
NH2
NH2
+ NH32
Cu2+-Ionen liegen gewöhnlich als Tetrahydrat vor. Diese vier Wassermoleküle
werden durch Aminogruppen verdrängt. Es sind also mindestens zwei
Peptidbindungen für den Komplex erforderlich:
CO
NH C
ONH2
wie im Biuretoder 2 Säureamidreste Lösungen: Analyse unbekannte Proteinlösung Protein-Standardlösung 10 g/l Biuret-Reagenz (in PVC-Flasche):
1,5 g CuSO4⋅5 H2O + 6 g Kalium-Natrium-Tartrat in 500 ml Wasser lösen; dazu 300 ml 10 %ige NaOH. Auffüllen auf 1 l.
Zur Herstellung einer Eichkurve werden sechs Proben (0,0 (Leerwert); 0,4; 0,8;
1,2; 1,6 und 2 ml) einer Standard-Proteinlösung in Reagenzgläser pipettiert, und
mit dest. Wasser auf 2 ml aufgefüllt. In zwei andere Reagenzgläser werden je 2 ml
der Analysenlösung pipettiert. Alle Proben werden mit 8 ml Biuret-Reagenz gut
gemischt und zur Ausbildung der Farbe 30 min stehengelassen. Gemessen wird in
1 cm-Küvetten bei 578 nm gegen den Leerwert (2 ml dest. Wasser und 8 ml
Biuret-Reagenz). Denken Sie immer daran Doppelbestimmungen durchzuführen:
Einzelmessungen haben oft Lotteriecharakter!
1-10 Aminosäuren und Proteine I
Auswertung:
Als Ordinate wird die Extinktion, als Abszisse die mg Protein/Liter Lösung
eingetragen.
a) Bestimmen Sie aus der Eichkurve den Proteingehalt der Analysenlösung,
indem Sie den zur gemessenen Extinktion gehörenden Wert in mg
Protein/Liter ablesen, oder
b) Berechnen Sie mit jedem Punkt, den Sie für die Erstellung der Eichkurve
verwendet haben, durch Dreisatz den Proteingehalt der Probe. Vergleichen Sie
den Mittelwert mit dem bei a) bestimmten Wert.
c) Geben Sie das Ergebnis in mg Protein pro Liter Analysenlösung an.
II. pH-Titrationskurve einer Aminosäure
Durch schrittweise Zugabe von HCl oder NaOH wird in einer wässrigen Lösung
die Konzentration von H+
-Ionen erhöht oder erniedrigt. Durch die chemischen
Eigenschaften der gelösten Aminosäure können diese Veränderungen zum Teil
abgefangen werden. Der pH-Wert bleibt streckenweise relativ konstant
(Pufferkapazität) und schlägt erst an charakteristischen Punkten um.
Bei der Titration einer Aminosäure findet man mindestens zwei Umschlagspunkte
(NH4+, COO-), je nach Seitenkette auch einen dritten. Die Titrationskurven von
Arginin, Glutaminsäure und Glycin sind hier dargestellt. Achtung Fehlerquelle:
Die Umschlagspunkte von Gly und Glu liegen sehr nahe beieinander! Markieren
Sie bitte die entsprechenden pK-Werte der drei Aminosäuren in der Kurve und
machen Sie sich bitte Gedanken darüber, warum die Kurven wie in der Abbildung
1-2 aussehen.

Titration 2-11
12
10
8
6
4
2
pH
ml NaOH
Glu
Gly
Arg
Abbildung 1-2: Die Titrationskurven von Arg, Gly und Glu Geräte: pH-Meter mit Elektrode Magnetrührer mit Teflonrührer Becherglas: 100 ml Vollpipette: 20 ml Büretten: 2 x 50 ml
Lösungen: 3 unbekannte Aminosäure-Lösungen (saure Lösungen, 5 g/l) 0,1 M NaOH
1-12 Aminosäuren und Proteine I
Aufgabe: Titration von drei unbekannten Aminosäuren mit NaOH
Ausführung: 30 ml einer unbekannten Aminosäure-Lösung (5 g/l) werden in ein 100 ml
Becherglas gegeben. Diese Lösung wird langsam auf dem Magnetrührer gerührt.
Mit dem pH-Meter (Eichung: siehe Einleitung) wird durch Eintauchen der
Elektrode in die Lösung der pH-Wert bestimmt (Vorsicht: Tief genug, aber nicht
zu weit eintauchen, damit der Rührfisch nicht die Elektrode beschädigt).
Aus einer 50 ml-Bürette wird in 1 ml Schritten 0,1 m NaOH-Lösung zugegeben
bis pH 11 erreicht ist, mindestens aber 30 ml. Der pH-Wert wird nach jeder
Zugabe abgelesen und in einer Wertetabelle festgehalten.
Anschließend werden Becherglas und Rührfisch mit demineralisiertem Wasser
ausgespült und der Versuch mit der nächsten unbekannten Aminosäure-Lösung
wiederholt.
Auswertung: Die Messwerte werden auf Millimeterpapier grafisch dargestellt. Vergleichen Sie
Ihre Kurven mit denen in Abb. 1-2 und geben Sie an, welche Analysen welchen
Aminosäuren entsprechen (Ergebnisse in Liste eintragen). Anhand der Kurve
können pK-Werte und pI-Werte abgeschätzt werden, die ebenfalls Rückschlüsse
auf die eingesetzten Aminosäuren ermöglichen. Wo liegen sie? Welche
chemischen Eigenschaften haben diese Aminosäuren?

Aufgaben 2-13
Übungsaufgaben:
1. Nur isoelektrisches Lysin, Lys°, ist Substrat der Lysindecarboxylase a) Formulieren Sie die Ionenpaare bei pKa1 = 2,18; pKa2 = 8,95; pKa3 = 10,54 b) Wie groß ist [Lys°] einer 10-3 M Lösung bei pH = 7,6?
2. Formulieren Sie die Struktur des Peptids H2N-Tyr-Arg-Trp-Ile-COOH a) zu welchem Pol wandert das Peptid in der Elektrophorese bei pH 4? b) Welche Form der Arg-Seitenkette liegt bei diesem pH vor? (pKa1 = 2,17; pKa2 (α-NH2) = 9,04; pKa3 (Seitenkette) = 12,48
3. Histidin hat einen pKa3 = 6,1 a) Formulieren Sie die beiden Ionenspezies bei pH 7,4, und b) Bestimmen Sie das Verhältnis von basischer zu saurer Form
4. a) γ-Aminobuttersäure entsteht aus _________ durch_____________ b) Ihr isoelektrischer Punkt beträgt 7,3. Eine Lösung von γ-Amino- buttersäure enthält bei pH 3,25 10 % Zwitterion.Wie groß sind die pK-Werte pK1 und pK2?
5. Zeichnen Sie die Titrationskurve von 0,05 M Alanin-Hydrochlorid in wässriger Lösung bei Zugabe von 0,1 M NaOH. (Anfangs-pH = 0,5). pKa1 = 2,35, pKa2 = 9,78. Ordinaten quantitativ bezeichnen!
6. Zeichnen Sie die Titrationskurve von 1 l 0,02 M Glycin-Hydrochlorid in wässriger Lösung, deren Anfangs-pH = 0,5 ist, unter Zugabe von 0,1 M NaOH. pK1 = 2,25, pK2 = 9,77. a) Bezeichnen Sie die Ordinate (pH) und Abszisse (ml 0,1 N NaOH) für jeden Punkt exakt. b) Berechnen Sie mindestens zwei Messpunkte links und rechts vom be-rechneten isoelektrischen Punkt.
7. Geben Sie den Bereich des isoelektrischen Punktes eines sauren bzw. basi-schen Proteins an.
8. Wieviele Protonen kann Lysin ausgehend vom pH 1 maximal abgeben? Wieviele saure bzw. basische Funktionen enthält Lysin?
9. Geben Sie die relative Lage der folgenden Peptide an, die in einer Papier-elektrophorese bei pH 7 aufgetrennt werden. A: Phe-Met-Gly, B: Leu-Gly-Arg und C: Glu-Ile-Ala.
1-14 Aminosäuren und Proteine I
10. a) Wie groß ist der pH-Wert einer 0,1 N HCl; einer 0,1 N NaOH; einer 0,1 N Essigsäure? b) Der pH-Wert des Blutes sei 7,4. Wie groß ist die [H+]? c) Wieviele H+/ml hat eine Lösung mit einem pH-Wert von 1,2?
11. a) Wieviel ml einer 28 %-igen HCl (Gew. %, MG = 36,5 g/mol, Dichte 1,15 g/ml) benötigt man, um 2 l einer 0,4 M HCl herzustellen? b) Wieviel ml 0,15 M KOH sind zur Neutralisation 180 g H2SO4 erfor-derlich? MG H2SO4 = 98 g/mol; MG KOH = 56 g/mol c) Wieviel ml 0,025 M H2SO4 benötigt man, um 525 ml 0,06 M KOH zu neutralisieren?
12. Die Pufferkapazität ist definiert als die Anzahl mol H+, die den pH-Wert eines Liter Puffers um 1 verändert. Berechnen Sie die Pufferkapazität eines 0,3 M Acetatpuffers von pH = 5,0 in saurer Richtung. Ka = 1,9 x10-5 mol/l
13. Tris Puffer: H2N-C(CH2OH)3 → H3N+-C(CH2OH)3 Sie sollen 500 ml eines 0,1 M Tris-Puffers vom pH 7,4 aus fester Tris-Base (MG = 121 g/mol, pK = 9) und 33 %-iger HCl (Gew. %; Dichte 1,165 g/ml) herstellen. Wieviele g bzw. ml benötigen Sie von jeder Komponente?
14. Wieviel g Glycin und wieviel g NaOH benötigen Sie, um einen 5 Liter eines 0,05 M Glycin-Puffers mit einem pH-Wert von 9,6 herzustellen? (pK1 = 2,5; pK2 = 9,6)

2-1
Versuch 2: Aminosäuren und Proteine II
Versuche:
SDS-Polyacrylamidgelelektrophorese
Western-Blot
Proteinstrukturen (Teil I+ II)
Analyse:
Molekulargewichtsbestimmung eines Proteins
Wissensgebiete Funktion von Proteinen: Enzyme, Transportmoleküle, Strukturelemente,
Antikörper, Hormone
Proteasen: ihre Bildung, Vorkommen, pH-Optimum
Spezifitäten: Warum spaltet Pepsin nicht bei pH7?
Lipoproteine, Klassifizierung und Komponenten, Trennmethoden und Patho-
biochemie
Beispiele von Proteinen mit Kofaktoren (prosthetischen Gruppen)
Posttranslationale Modifikationen
Sequenzanalyse
2-2 Aminosäuren und Proteine II
Proteolytischer Verdau
Medizinische Bedeutung: Proteolytische Prozesse sind für eine Vielzahl physiologischer Funktionen es-
sentiell: z.B. wird die Blutgerinnung durch eine Kaskade von proteolytischen
Schritten reguliert, durch die Enzyme (Zymogene) aktiviert werden und letzt-
endlich aus Fibrinogen das unlösliche Fibrin entsteht. Hämophilie kann durch das
Fehlen oder die Mutation eines der beteiligten Proteine, z.B. Faktor IX genannt,
hervorgerufen werden. Ein weiteres Beispiel ist die Aktivierung von Prohormonen
wie Proinsulin oder Wachstumsfaktoren durch Abspaltung eines Peptides.
Proteasen besitzen mehr oder weniger breite Spezifitäten für gewisse Peptid-
bindungen. In diesem Versuch sollen Sie die entstehenden Spaltprodukte von
Albumin aus Ei (Ovalbumin) und aus Serum nach Verdau mit Chymotrypsin ana-
lysieren.
Ovalbumin (OV) und Serumalbumin (SA) wurden vom Assistenten mit
Chymotrypsin verdaut. Die Proteinfragmente werden am heutigen Versuchstag
mit Hilfe der SDS-Polyacrrylamidgelelektrophorese getrennt und die Gele mit
Coomassie angefärbt.
Ionenaustauscherchromatographie von Proteinen
Unter Ionenaustauscher-Chromatographie versteht man eine Methode, bei der ein
lösliches Ion gegen ein gleich geladenes Ion an einer unlöslichen Trägersubstanz
ausgetauscht wird. Die im Labor verwendeten Ionenaustauscher sind unter
anderen synthetische Träger, z. B. Polymere auf Polystyrolbasis (mit
Quervernetzung). Häufig eingesetzt werden auch modifizierte Zuckerpolymere.
Die Kationenaustauscher enthalten Sulfonsäure (–SO3H), Carboxyl- (–COOH)
oder phenolische Gruppen als funktionelle Gruppe. Nach der
Dissoziationskonstante (pK) der ionischen Gruppen unterscheidet man stark saure
(–SO3H) und schwach saure (–COOH) Austauscher.

Ionenaustauscherchromatographie 2-3
Die Anionenaustauscher enthalten basische Gruppen, z. B. –N+(CH3)3 (stark
basisch) oder –N+H(CH3)2 (schwach basisch).
Wirkungsweise von Ionenaustauschern:
Kationenaustauscher:
TrägerTräger
+Protein++
SO3- Protein
+SO3- Na
+
Na+
Das auf die mit Kationenaustauscher (Na+-Form) gefüllte Säule gegebene positive
Ion, in diesem Falle Protein+, wird gegen das Na+ ausgetauscht. Natriumionen
erscheinen im Eluat der Säule.
Anionenaustauscher:
CH2
CH3
H3C N CH3
CH2
CH3
H3C N CH3+ +
OH-
Träger Träger
Protein-+
Protein-
OH-+
Das OH- der quarternären Ammoniumbase des Austauschers wird z. B. durch
Protein- ausgetauscht.
2-4 Aminosäuren und Proteine II
Die Affinität eines Ionenaustauschers für ein Protein hängt von der Natur der
geladenen Gruppe auf dem Träger ab und von der Natur und Anzahl der geladenen
Gruppen auf der Proteinoberfläche. Je größer die Zahl der Ladungen eines Ions,
um so fester wird das Protein an den Austauscher gebunden.
Die Konzentration der Ionen, die Ionenstärke, beeinflusst ebenfalls die Bindung.
In der Praxis spielen u.U. auch die Proteingröße und die Quervernetzung (Größe
der Löcher im Träger) eine Rolle. Die Ionenstärke wird berechnet nach:
I = 1
2 ∑ c⋅z2 (c = Konzentration; z = Ladungszahl)
Ionenaustauscher werden für viele Zwecke in der Medizin, Biochemie und
Technik verwendet: Als Medikament zum Natriumentzug; zur Herstellung Ca2+-
armer Milch in der Diätetik; als Diagnostikum zur sondenlosen Funktionsprüfung
der Magensaftsekretion. In der Biochemie werden Ionenaustauscher zur
chromatographischen Trennung einer Vielzahl von Molekülen verwendet, z.B.
von Proteinen oder Aminosäuren. In großem Umfang werden Ionenaustauscher
zum Entsalzen von Lösungen, insbesondere zur Wasserenthärtung, verwendet.
Der pI von Peptiden und Proteinen ergibt sich aus dem Verhältnis aller disso-
ziierbaren Gruppen. Die pI-Werte von Proteinen mit einem überwiegenden Anteil
an sauren Aminosäuren liegen im sauren pH-Bereich; Proteine mit einem größeren
Anteil an basischen Aminosäuren haben ihre pI-Werte im alkalischen. Liegt der
pH-Wert einer Pufferlösung unterhalb des pI-Wertes eines Proteins ist die
Gesamtladung des Proteins positiv; liegt er oberhalb, ist die Summe der Ladungen
negativ.
Im folgenden Versuch werden humane Plasmaproteine (siehe Tabelle 2.2)
verwendet, die durch Anionenaustausch-Chromatographie auf DEAE-Sephacel
aufgetrennt wurden. (Versuch wurde bereits vom Assistenten durchgeführt).
DEAE-Sephacel ist ein schwach basischer Anionenaustauscher aus Diethyl-
aminoethyl (DEAE = -O-CH2CH2N+H(CH2CH2)2) substituierter Cellulose.
Cellulose dient hier als Trägermaterial; die Trennung der Proteine erfolgt über
ionische Wechselwirkungen mit den positiv geladenen DEAE-Gruppen in Ab-
hängigkeit vom pH-Wert und der Ionenstärke des Puffers.

SDS-PAGE 2-5
Unter den Startbedingungen der Chromatographie (pH 8,6) sind die Plasma-
proteine negativ geladen und werden über elektrostatische Wechselwirkungen an
den Ionenaustauscher gebunden. Die Elution der Immunglobuline (pI 8,3) erfolgt
bei pH 7,0. Unter diesen Bedingungen bleiben die übrigen Plasmaproteine
aufgrund ihrer niedrigeren pI-Werte (z.B. Albumin pH 5,6) an die Säule gebunden
und können anschließend bei pH 4,2 eluiert werden. Die Analyse der Eluate) in
der SDS-Polyacrylamidgel Elektrophorese (SDS-PAGE).
I. SDS-Polyacrylamidgelelektrophorese
Medizinische Bedeutung: Polyacrylamidgelelektrophorese (PAGE) gehört zu den wichtigsten bioche-
mischen Untersuchungsmethoden. PAGE eignet sich sowohl zum Nachweis der
Reinheit von Proteinen als auch zum Nachweis von Enzymmustern bzw. Pro-
teinmustern. Desweiteren wird sie für Analyse der Größe von DNA-Fragmenten
verwendet wie z.B. für Sequenzierung oder die Analyse von Mutationen in ver-
erbbaren Krankheiten. Die hier durchzuführende SDS-PAGE ist eine spezielle
Methode zur Analyse von Proteinen.
Grundlagen: SDS (Sodiumdodecylsulfat) ist ein anionisches Detergenz, das die Proteine in der
Probe denaturiert und an diese denaturierten Proteine bindet. Die Proteine werden
somit unabhängig von ihrem pI-Wert negativ geladen und wandern zur Anode.
Die Wanderung wird dann nur noch von der Größe der Proteine bestimmt, die
durch den Molekularsiebeffekt des Polyacrylamidgeles zu einer unterschiedlichen
Wanderungsstrecke führt.
Acrylamid ist das Material der Wahl, um Proteine nach Größe elektrophoretisch
zu trennen. Das Acrylamid bildet zusammen mit Bisacrylamid ein quervernetzes
Netzwerk, wenn ein Radikalstarter zugesetzt wird. Bei Verwendung von hohen
Konzentrationen an Acrylamid und Bisacrylamid erhöht man den Grad der
Quervernetzung und verringert die Porengröße, so dass kleine Proteine gut
aufgetrennt werden. Für große Proteine verwendet man niedrigprozentige Gele.
2-6 Aminosäuren und Proteine II
Um eine optimale Auflösung der Proteine zu erhalten, werden zweischichtige Gele
verwendet. Den unteren Teil nennt man Trenngel, den oberen Sammelgel.
Letzteres hat eine geringe Konzentration an Acrylamid und einen anderen pH-
Wert. Dies erlaubt es den Proteinen einer Probe sich in scharfen Banden zu
„sammeln“, bevor sie im Trenngel aufgetrennt werden.
Aussagen über intra- oder intermolekulare Disulfidbrücken in Proteinen kann man
erhalten, wenn man die Proben vor Auftrag auf das Polyacrylamidgel mit β-
Mercaptoethanol reduziert und mit nicht reduzierten Proben vergleicht.
Geräte: Mighty Small Elekrophorese Kammer
Materialien: SDS-Polyacrylamid-Gele, gebrauchsfertig, 10% Acrylamid Laufpuffer: 20 mM Tris, 192 mM Glycin, 0.1% SDS, pH 8.3 Probenpuffer nicht-reduzierend: (4fach): 8% SDS, 40% Glycerin, 0.1%Bromphenolblau,
250mM TRIS/HCl, pH 6.8, Probenpuffer reduzierend: (4fach): wie nicht-reduzierend, plus 10% β-Mercaptoethanol Coomassie-Färbelösung: 0.2%Coomassie Blue R250 in 50%MeOH, 5% HOAc Entfärbelösung: 40% MeOH, 7% HOAc Fixierlösung: 7% HOAc

SDS-PAGE 2-7
Aufgaben: 1. Die Fraktionen der Ionenaustauscherchromatographie (vorbereitet) sollen im
reduzierten und nichtreduzierten Zustand aufgetrennt und die Hauptbanden
anhand ihres Molekulargewichtes identifiziert werden.
2. Als Analyse soll das Molekulargewicht eines unbekannten Proteines bestimmt
werden.
3. Unbehandeltes und mit Chymotrypsin behandeltes Serumalbumin und
Ovalbumin (vorbereitet) sollen aufgetrennt und die Molekulargewichte der
auftretenden Banden bestimmt werden.
4. Milchproteine sollen aufgetrennt und identifiziert werden.
5. Plasma und zwei Fraktionen der Ionenaustauscherchromatographie der
Plasmas sollen aufgetrennt werden, um sie für den immunologischen Nachweis
von IgG in Versuch 5 auf Nitrocellulose zu blotten.
Durchführung: 1. Probenvorbereitung:
Jeweils 15 µl Probe werden mit 5 µl Probenpuffer (reduzierend bzw nicht
reduzierend) in einem Eppendorf-Reaktionsgefäß versetzt (gelbe Pipetten-
spitzen verwenden), 5 min bei 100°C inkubiert und kurz abzentrifugiert.
2. Gelaufbau:
Da nicht polymerisiertes Acrylamid toxisch ist, erhalten Sie gebrauchsfertige,
auspolymerisierte Gele. Diese 1 mm dicken Gele wurden zwischen zwei
Glasplatten auspolymerisiert und verbleiben während der Auftrennung auch
zwischen den Glasplatten. Im Sammelgel befindet sich ein Kamm. Wird dieser
aus dem Gel herausgezogen, ergeben sich Aussparungen, in die die
Proteinproben aufgetragen werden.
• Die Gele müssen Sie mittels zweier roter Klammern in die Laufkammern
einsetzen, durchsichtige Seite nach vorne. Beachten Sie, dass zwei verschie-
dene Gele verwendet werden. Gel 1 hat eine Trenngelkonzentration von
10% Polyacrylamid, Gel 2 von 15%.
• Die Gelkammern mit Laufpuffer befüllen.
• Die Markierungsschablone auf der Glasplatte am Kamm ausrichten und den
Kamm vorsichtig herausziehen.
2-8 Aminosäuren und Proteine II
• Die vorbereiteten Proben werden mit den speziellen ausgezogenen Spitzen
(farblos) langsam in die Taschen eingefüllt. Die Molekulargewichtsmarker
können direkt, ohne Zugabe von Probenpuffer, aufgetragen werden (20µl).
Beladung des Gels 1: (Fraktionen der Ionenaustauscherchromatographie)
Gel 1, 10%
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
F0
nr
F1
nr
F2
nr
F3
nr
F4
nr
F5
nr
leer leer F0
r
F1
r
F2
r
F3
r
F4
r
F5
r
M
W
Beladung des Gels 2: Molekulargewichtsmarker MW, Analysen zur Molekular-
gewichtsbestimmung, proteolytische Verdaus von Serumalbumin (SA1, SA2) und
Ovalbumin (OV1,OV2) und den Serum/Plasma Proben für den Westernblot.
Achtung: Bis auf Proben Nr. 10-12 alle Proben reduzieren!
Gel 2, 15%
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
M
W
Ana-
lyse
1
r
Ana-
lyse
2
r
S
A
1
r
S
A
2
r
O
V
1
r
O
V
2
r
Milch-
pulver
r
leer F0
nr
F5
nr
F3
nr
leer F3
r
M
W
• Gelkammerdeckel aufsetzen (auf die richtige Polung achten),
Elektrophorese starten. 160 V, 200 mA, Dauer ca. 1 h, bis Blaumarker am
unteren Ende des Gels angelangt ist.
• Spannung abschalten, Deckel abnehmen.
• Mit der grossen 50 ml Spritze Gelpuffer in der Kammer abziehen (Ausguss)
und die Gele herausnehmen.
• Seitliche Trennstücke herausziehen und Glasplatte vorsichtig vom Gel trennen.
Das Gel 2 mit den Analysen und den Proben für den Westernblot mit einem
Skalpell in Spur 9 durchschneiden. Die Verwendung des Gelstückes für den
Westernblot wird unter B) beschrieben. Das Gelstück mit den Analyseproben

SDS-PAGE 2-9
und das Gel 1 mit den Ionenaustauscherfraktionen wird in eine Plastikschale
zu ungefähr 75ml (halbvolle Schale) Coomassie-Färbelösung gegeben und 15
min auf dem Schüttler gefärbt.
• Dann das Gel ~1 h in Entfärbelösung entfärben. Dazu Coomassie-Lösung in
entsprechende Recyclingflasche zurückfüllen, Gel kurz mit Wasser abspülen
und Entfärbelösung zugeben.
• Entfärbelösung in entsprechende Recyclingflasche abgießen und Fixierlösung
zugeben.
• Zusammen mit dem Assistenten das Gel mit der Videokamera fotografieren.
Auswertung:
1. Ionenaustauscherchromatographie von Plasma:
Beschreiben Sie die Hauptbanden in den verschiedenen Fraktionen und wie
sich das Molekulargewicht dieser Fraktionen bei Reduktion verhält.
2. Messen Sie die Laufstrecken der Markerproteine und ihres Analysenproteins
exakt aus.
Die Markerproteine sind: Phosphorylase B 97400 Da
Serumalbumin 67000 Da
Albumin aus Ei 45000 Da
Carboanhydrase 29000 Da
Lactalbumin 14200 Da
Tragen Sie die Laufstrecke gegen der Logarithmus des Molekulargewichtes
auf und verbinden Sie die Punkte. Berechnen Sie das Molekulargewicht ihres
Analysenproteins. Wichtig ist, dass Sie alle Markerbanden auf dem Gel
erkennen und eindeutig zuordnen können. Zwei Probleme können auftreten: 1.
Das Lactalbumin kann aus dem Gel herausgelaufen sein. 2. Abbaubanden der
übrigen Markerproteine entstehen. Bei Unklarheiten bitte Assistenten
befragen. Tipp: Serumalbumin-Bande als Anhaltspunkt benutzen!
3. Verfahren Sie analog mit Ovalbumin und dem mit Chymotrypsin geschnit-
tenen Ovalbumin. Die Aminosäuresequenz von Ovalbumin lautet:
2-10 Aminosäuren und Proteine II 1 GSIGAASMEF CFDVFKELKV HHANENIFYC PIAIMSALAM VYLGAKDSTR
51 TQINKVVRFD KLPGFGDSIE AQCGTSVNVH SSLRDILNQI TKPNDVYSFS
101 LASRLYAEER YPILPEYLQC VKELYRGGLE PINFQTAADQ ARELINSWVE
151 SQTNGIIRNV LQPSSVDSQT AMVLVNAIVF KGLWEKAFKD EDTQAMPFRV
201 TEQESKPVQM MYQIGLFRVA SMASEKMKIL ELPFASGTMS MLVLLPDEVS
251 GLEQLESIIN FEKLTEWTSS NVMEERKIKV YLPRMKMEEK YNLTSVLMAM
301 GITDVFSSSA NLSGISSAES LKISQAVHAA HAEINEAGRE VVGSAEAGVD
351 AASVSEEFRA DHPFLFCIKH IATNAVLFFG RCVSP
Chymotrypsin schneidet normalerweise hinter aromatischen Aminosäuren.
Wieviele Fragmente würden sie ausgehend von der Sequenz erwarten? Wie
erklären Sie die im Gel aufgetretenen Banden?
4. Beschreiben Sie die Hauptproteinbanden der Milchproteine. Welche Funktion
haben die Proteine der Milch?
Tabelle 2-1: Ein-Buchstaben-Code der Aminosäuren
A Ala Q Gln L Leu S Ser
R Arg E Glu K Lys T Thr
N Asn G Gly M Met Y Tyr
D Asp H His F Phe W Trp
C Cys I Ile P Pro V Val

SDS-PAGE 2-11
Tabelle 2-2: Wichtige Proteine des Blutplasmas (g/l)
Frauen Männer Kinder
Präalbumin) 0,15 - 0,30 0,20 - 0,30 0,04 - 0,12
Albumin 38,00 -51,00 40,00 - 52,00 28,00 - 41,00
α1-Antitrypsin 0,95 - 1,85 0,85 - 1,85 0,60 - 1,60
Orosomukoid 0,50 - 1,00 0,55 - 1,15 0,10 - 0,40
α2-Makroglobulin 2,05 - 4,30 1,80 - 3,40 2,35 - 5,30
Haptoglobin 0,50 - 2,70 0,55 - 3,20 < 0,20
Ceruloplasmin 0,25 - 0,55 0,20 - 0,45 0,05 - 0,25
Transferrin 1,65 - 2,90 1,54 - 2,75 0,90 - 1,90
Hemopexin 0,60 - 0,95 0,65 - 0,95 0,04 - 0,30
C3-Komplementfaktor 0,80 - 1,85 0,90 - 1,90 0,70 - 1,15
C4-Komplementfaktor 0,20 - 1,00 0,30 - 1,25
Fibrinogen 1,90 - 4,70 1,80 - 4,50
IgG 7,20 - 15,50 6,70 - 14,70 5,50 - 12,70
IgA 0,50 - 3,00 0,50 - 3,00 < 0,10
IgM 0,20 - 2,10 0,15 - 1,20 0,02 - 0,12
2-12 Aminosäuren und Proteine II
II. Blotting der Proteine auf Nitrocellulose (Vorbereitung für Versuch 5)
Grundlagen: Dieser Versuch ist die Vorbereitung für den immunologischen Versuch 5. Die
medizinische Bedeutung ist dort erläutert. Proteine, die mittels SDS-PAGE
getrennt wurden, werden in dem sogenannten Blotting-Verfahren auf Nitro-
cellulose übertragen, auf der dann immunologische Nachweise durchgeführt
werden können. Die Übertragung erfolgt wie bei der SDS-PAGE elektrophore-
tisch, d.h. die mit SDS beladenen Proteine werden von einem elektrischen Feld
aus dem Gel heraus auf die Nitrocellulose-Membran transferiert. Da die Proteine
negativ geladen sind, ist beim Aufbau der Transfereinheit darauf zu achten, dass
sich die Nitrocellulose auf den Anodenseite befindet.
Geräte: Nitrocellulose Transphor Blotting Kammer
Lösungen: Transferpuffer: 50 mM Borsäure pH 8.5, 5% Methanol Ponceau S TBS + Tween TBS + 5% Milchpulver
Durchführung:
1. Blotten:
Nach Beendigung der Gelelektrophorese wird das für den Western-Blot vorge-
sehene Gelstück (Spur 10-15)-in eine Schale gelegt, die Transfer-Puffer enthält.
Die Halterung für die Blotapparatur wird in eine Photoschale getaucht, die
ebenfalls Transfer-Puffer enthält. Dabei ist auf folgende Anordnung zu achten:
• Die Halterung wird mit der grauen Seite (zukünftige Anode) zuunterst in die
Schale gelegt (Abb. 2-1).
• Auf diese wird nacheinander ein Schwamm, ein Whatman-Papier, die
Nitrocellulose-Membran (mit Namen beschriften!), das Gel, wieder ein What-
man-Papier, ein Schwamm und zuletzt die schwarze Seite (zukünftige

Western-Blot 2-13
Kathode) der Halterung aufgelegt und eingerastet. Die einzelnen Schichten
müssen jeweils luftblasenfrei aufgelegt werden.
• Dieser Sandwichaufbau wird in die Blotkammer überführt (Anode zu Anode,
Kathode zu Kathode) und eine Stunde lang bei 1000 mA geblottet.
Abbildung 2-1: Schale mit schematischem Aufbau des Blot-Sandwiches
2. Ponceau S-Färbung (Zum Anfärben der Markerproteine):
• Durch Zugabe von 10 ml Ponceau S werden die Proteine auf der Membran
reversibel gefärbt, die Proteine des Markers werden mit Kugelschreiber vor-
sichtig markiert.
• Die Membran wird mit 25 ml TBS/Tween wieder vollständig entfärbt.
• Die Membranen werden eine Stunde lang in 5% Milchpulver geschüttelt, um
alle noch freien Proteinbindungsstellen zu blockieren.
• Das Milchpulver wird mit TBS abgewaschen und die beschrifteten Mem-
branen bis zum Versuch 5 eingefroren.
2-14 Aminosäuren und Proteine II
III. Dreidimensionale Strukturen von Proteinen
Dieser Teilversuch soll Ihnen einen Einblick in die dreidimensionale Struktur von
Proteinen vermitteln. Die 3D-Struktur von Proteinen kann mittels Röntgen-
kristallographie oder Kernresonanzspektroskopie (NMR) gelöst werden. Für die
Röntgenkristallographie muss man Proteinkristalle (ungefähr 1mm3) züchten, in
denen die Proteine wie in einem Kochsalzkristall in einer definierten Anordnung
sitzen. Diese Kristalle werden mit einem Röntgenstrahl bestrahlt und aus dem
Beugungsmuster des Strahles kann man die exakten Koordinaten aller Atome des
Proteins berechnen. Aufgrund der Wellenlänge des üblicherweise verwendeten
Röntgenlichtes können Wasserstoffatome nicht detektiert werden.
Die Kernresonanzspektroskopie erlaubt die Auflösung von Proteinstrukturen in
wässriger Lösung. Die Proteine werden in ein Magnetfeld gebracht und mit einem
Radiowellen bestrahlt. Die Strahlung regen die Atomkerne bei einer Re-
sonanzfrequenz an. Die Resonanzenergie, die von den Atomkernen nach der
Anregung wieder abgegeben wird, wird detektiert und zur Berechnung der Ko-
ordinaten der Atome verwendet. Zur Zeit ist die Anwendung der Kernreso-
nanzspektroskopie auf Proteine mit einem Molekulargewicht kleiner 40 kDa
limitiert. Auch Elektronenmikroskopie wird zur Strukturanalyse verwendet, die
Auflösung ist jedoch wegen der geringeren Energie der Elektronen viel schlechter
und nur die Umrisse von Proteinen können so sichtbar gemacht werden.
Medizinische Bedeutung: Einblicke in die Struktur von Proteinen tragen essentiell zum Verständnis mole-
kularer Vorgänge bei. Viele vererbbare Krankheiten werden durch einzelne
Punktmutationen in einem Gen hervorgerufen, die im Protein zum Austausch einer
Aminosäure führen. Warum diese mutierte Aminosäure einen Phänotyp
hervorrufen kann, kann oftmals nur anhand der dreidimensionalen Struktur
verstanden werden. So sitzt z.B. die eingetauschte Aminosäure Valin im Sichel-
zellhämoglobin auf der Oberfläche des Proteins und passt in eine hydrophobe
Tasche eines zweiten Hämoglobins. Dies führt zu einer Polymerisation des
Hämoglobins und der charakteristischen Sichelzellform der Erythrozyten.

Proteinstrukturen 2-15
Große Hoffnungen werden z.Z. auf das spezifische Design von Pharmaka anhand
der Struktur der Zielproteine gesetzt, dies zeigt sich z.B. daran, dass alle großen
pharmazeutischen Firmen Strukturlabors unterhalten. Man versucht z.B.
spezifische Inhibitoren für Proteasen aus HIV zu entwickeln, indem man gewisse
Aminosäuren eines bekannten, aber schwach bindenden Inhibitors am Computer
austauscht und untersucht, ob so zusätzliche Bindungen zwischen der Proteinase
und dem Inhibitor gebildet werden.
Grundlagen: Die grundlegende Bindung in Proteinen ist die Peptidbindung zwischen der
Aminogruppe und der Carbonsäuregruppe zweier Aminosäuren.
Die Bindung zwischem dem Kohlenstoff und dem Stickstoff hat aufgrund ihres
mesomeren Charakters Doppelbindungscharakter und deshalb sind alle Peptid-
bindungen planar (die vier Atome C, O, N und H liegen in einer Ebene).
O O-
¦ ←→ |
-C-N- -C=N+-
| |
H H
Das sogenannte Rückgrat der Proteine ergibt sich aus der Abfolge der planaren
Peptidbindungen, die jeweils durch das α-C-Atom, das die Seitenkette trägt,
getrennt sind. Die Bindungen zum α-C-Atom sind drehbar:
H O H O H O H O
| ¦ | ¦ | ¦ | ¦
-N——C——C——N——C——C——N——C——C——N——C——C-
| | | | | | | |
H R H R H R H R
AS1 AS2 AS3 AS4
2-16 Aminosäuren und Proteine II
Die Interaktion benachbarter Aminosäuren führt auf einer ersten Ebene zur Aus-
bildung von sogenannten Sekundärstrukturen, wie α-Helices, β-Faltblättern, β-
turns u.a.. Diese kommen durch Wasserstoffbrücken zwischen den Sauerstoff- und
den Wasserstoffatomen der Peptidbindung zustande. Welche Sekundärstruktur
dabei von einem Peptidstück angenommen wird, hängt von der Natur der
Seitenketten (Größe, Ladung) ab, die die relativen Stellungen der Peptidbindungen
zueinander bestimmt. Je nach Verdrehung der aufeinanderfolgenden planaren
Peptidbindungen ergeben sich so α-Helices, β-Faltblätter usw.
Die nächste Ebene, die Tertiärstruktur, beeinhaltet die räumliche Anordnung der
Sekundärstrukturen. Diese wird durch die Wechselwirkung der Sekundär-
strukturelemente bestimmt. An diesen Interaktionen sind sowohl das Peptid-
rückgrat als auch die Seitenketten beteiligt (ionische, van der Waals-, und Was-
serstoff-Brücken-Bindungen).
Im Falle von multimeren Proteinen, d.h. solchen die aus mehreren Untereinheiten
aufgebaut sind, gibt es eine weitere strukturelle Ebene, die Quartärstruktur. Mit ihr
wird die räumliche Anordnung der Tertiärstrukturen der Untereinheiten
beschrieben.
Denaturierung
Sekundär-, Tertiär- und Quartärstrukturen der Proteine sind nicht unter allen
Bedingungen stabil. Unter Denaturierung versteht man die Auflösung aller dieser
Strukturen, so dass das Protein als ungefaltete Polypeptidkette vorliegt. Dies kann
u.a. durch Temperaturerhöhung, pH-Wert-Änderung, Ionenstärkeänderung oder
sogenannte chaotrope Reagentien wie z.B. Harnstoff geschehen. So ist eine der
Hauptfunktionen der Salzsäure neben ihrer bakteriziden Wirkung die
Denaturierung von Proteinen im Magen.
Im ersten Teil dieses Versuchs werden Ihnen vom Assistenten verschiedene
Sekundärstrukturelemente und einige Proteine mit Hilfe des Programmes Rasmol
vorgestellt. Im zweiten Teil sollen Sie als Analyse eine Sekundär-, Tertiär- und -
wenn vorhanden - Quartärstruktur-Beschreibung eines kleinen Proteins
vornehmen.

Proteinstrukturen 2-17
Ausführung: Das Programm Rasmol ermöglicht es, 3-dimensionale Strukturen zu bewegen, zu
drehen und einzelne Elemente getrennt anzuschauen. Beim Öffnen des Pro-
grammes öffnet sich zum einen das Fenster (Rasmol Version 2.6 in oberster
Zeile), in dem die Struktur dargestellt wird und ein zweites Fenster (Rasmol
Command Line), in dem man vordefinierte Programme erstellen und abrufen
kann.
Übersicht über die wichtigsten Befehle im Rasmol Version 2.6- Fenster
(mit der Maus anzuwählen)
Display-Modus:
Rasmol erlaubt 7 Darstellungsformen, davon sind 3 interessant: 1. Sticks: alle Atome, Stabform
2. Spacefill: alle Atome in Kugelform: zeigt die Oberfläche eines Proteins
3. Cartoons schöne Darstellung der Sekundärstrukturelemente
Colours:
Hier sind drei Darstellungen wichtig:
1. CPK: Standardfärbung: O rot, N blau, C grau, S gelb, H weiß
2. Chain: färbt in multimeren Proteinen die einzelnen Untereinheiten ver-
schieden
3. Structure: färbt α-Helices rot, β-Stränge gelb, β -turns (Schleifen) blau,
random (ungeordnet) grau:
1. Öffnen einer Struktur:
• Vom Assistenten erhalten Sie einen Dateinamen, unter dem das Protein,
das Sie analysieren sollen, abgespeichert ist.
• Mit der Maus: File Open, dann entsprechendes File auswählen.
• Mit der Maus: Display Cartoon:
• Mit der Maus: Colour Structure:
• In dieser Darstellung sollen Sekundär-, Tertiär- und Quartärstruktur be-
schrieben werden.
2-18 Aminosäuren und Proteine II
• Vor dem Öffnen einer neuen Struktur, alte Struktur schließen mit: File
Close
2. Drehen einer Struktur:
Mit linker Maustaste Struktur anklicken und mit festgehaltener Taste ziehen
3. Verschieben einer Struktur:
Mit rechter Maustaste Struktur anklicken und mit festgehaltener Taste ziehen
4. Zoomen:
a) Vergrößern: Shift-Taste gedrückt halten und mit linker Maustaste auf die
Struktur klicken und nach unten ziehen.
b) Verkleinern: Shift-Taste gedrückt halten und mit linker Maustaste auf die
Struktur klicken und nach oben ziehen.

Proteinstrukturen 2-19
Aufgabe: Bestimmung der Struktur eines Proteins
Sie sollen die Sekundär, Tertiär- und -wenn vorhanden- die Quartärstruktur eines
Proteines beschreiben. Sie erhalten vom Assistenten die Nummer eines Proteins
und öffnen dies mit Hilfe des Programmes Rasmol.
Durchführung (s.o.). Die Beschreibung der Strukturelemente im Protokollheft
wird durch den Assistenten überprüft.
Beispielauswertung:
Protein xxx:
Sekundärstrukturelemente vom Amino- zum Carboxyterminus:
H3N+-Helix-Schleife-Helix-Schleife-Faltblatt-Schleife-Faltblatt-Schleife-Faltblatt-
COO-
Tertiärstruktur:
Monomeres globuläres Protein, das aus zwei Helices und einem antiparallelen -
Faltblatt gebildet wird. Das β-Faltblatt besteht aus drei Strängen.
2-20 Aminosäuren und Proteine II
Übungsaufgaben:
1. Wenn Sie ein Ei kochen, welche Art der Konformationsänderung der Proteine erfolgt dabei?
2. Geben Sie zwei Nachweismethoden für Proteine an.
3. Wie ist die Primär-, Sekundär-, Tertiär- und Quartärstruktur von Proteinen definiert?
4. Nennen Sie zwei Formen von Sekundärstrukturen von Proteinen.
5. Zwischen welchen chemischen Gruppen werden die Wasserstoffbrücken-
bindungen in der α-Helix von Proteinen gebildet?
6. Zeichnen sie zwei aufeinander folgende Peptidbindungen einer Peptidkette. Welche Bindungen sind beweglich?
7. Was versteht man unter einem hydrophoben Kern eines Proteins? Welche Aminosäuren haben aromatische Reste?
8. Mit welchen Methoden kann man das Molekulargewicht eines Proteins bestimmen?
9. Nennen Sie je ein Protein aus dem Blutplasma mit: a) Transportfunktion, b) Abwehrfunktion, c) Enzymatischer Funktion
10. Wie wird Trypsin aktiviert?
11. Welche Rolle spielt Pyridoxalphosphat bei der Transaminierungsreaktion?
12. Welche zellulären Kompartimente sind am Harnstoffzyklus beteiligt? Welche beiden Möglichkeiten hat eine Aminosäure die Aminogruppe für die Harnstoffsynthese zur Verfügung zu stellen?

Aufgaben 2-21
13. Sequenzanalyse eines Peptids:
↓ ↓ ↓ ↓ H2N-Ala-Met-Asp-Ile-Arg-Cys-Phe-Leu-COOH S ³ S H2N-His-Arg-Gly-Lys-Cys-Ser-Gly-COOH
↑ ↑
Schreiben Sie an den Pfeilen die zugehörigen Reagenzien.
Mercaptoäthanol SH-CH2-CH2-OH Phenylisothiocyanat Phenyl-N=C=S Sangerreagenz: Fluoro-dinitro-benzol Dansylchlorid Bromcyan BrCN Proteasen: Trypsin, Chymotrypsin, Pepsin, Thermolysin, Thrombin, Car-
boxypeptidase, Aminopeptidase
14. Ein Peptid reagiert mit Dinitrofluorbenzol nur in der Seitenkette des Lysins des Peptids. Durch Thrombinspaltung (Thrombin schneidet zwischen Arg-Gly) entsteht ein Peptid der Stöchiometrie:
Ala1 Tyr1 Gly1 Lys1 Ser1 Phe1 Arg1 Pro1
Mit dem Edman Reagenz wird aus diesem Gly PTH abgespalten. Schreiben Sie die Strukturformel des Derivats:
Wird das Thrombin-gespaltene Peptid mit Trypsin behandelt, so entstehen zwei Peptide der Stöchiometrie:
Ser1 Tyr1 Pro1 Arg1 und Phe1 Gly1 Ala1 Lys1
Chymotrypsin-Behandlung des Thrombin-gespaltenen Peptids liefert zwei Dipeptide mit der Zusammensetzung Gly1 Phe1 und Arg1 Ser1 und das übrige Tetrapeptid. Formulieren Sie die Sequenz des Peptids in der Drei-buchstaben-Schreibweise.
2-22 Aminosäuren und Proteine II
15. Die vollständige Hydrolyse eines Peptids ergibt:
1 Gly, 1 Ala, 2 Cys, 1 Lys, 1 Glu, 1 Ile, 1 Thr, 1 Phe, 1 Val
Reduktion mit Mercaptoäthanol ergibt zwei kleinere Peptide A und B. A enthält: Ala, Cys, Glu, Gly, Ile, Lys
Carboxypeptidase setzt Ile frei: Edman-Abbau mit Phenylisothiocyanat ergibt das PTH-Derivat von
Gly im 1-Cyclus, Ala im 2-Cyclus.
Trypsin setzt zwei Peptide frei: a) enthält Glu und Ile b) Ala, Cys, Gly, Lys
B enthält: Cys, Phe, Thr, Val Carboxypeptidase setzt Val frei; Chymotrypsin: Val und ein Tripeptid mit Cys, Phe, Thr; Edman-Abbau ergibt PTH-Thr.
Wie lautet die Sequenz des gesamten Peptids?
16. Ein Peptid hat die folgende stöchiometrische Zusammensetzung:
1 Ala, 1 Arg, 2 Asp, 2 Glu, 3 Gly, 1 Leu, 3 Val
Die folgenden Peptide wurden nach partieller Spaltung isoliert: 1. Asp-Glu-Val-Gly-Gly-Glu-Ala 2. Val-Asp-Val-Asp-Glu 3. Val-Asp-Val 4. Glu-Ala-Leu-Gly-Arg 5. Val-Gly-Gly-Glu-Ala-Leu-Gly-Arg 6. Leu-Gly-Arg Wie lautet die Aminosäuresequenz des Ausgangspeptids?

3-1
Versuch 3: Enzyme
Versuche:
Enzymkinetik der sauren Phosphatase
Acetylcholinesterase-Bestimmung im Serum
Hemmung des Enzyms mit Diisopropylfluorophosphat (DIFP)
Reaktivierung mit Pyridin-2-Aldoxim-Methyljodid (PAM)
Laktatdehydrogenase (opt. Test)
Wissensgebiete
Einteilung der Enzyme in Enzymklassen
Aktivierungsenergie
Enzymsubstrat-Komplex
Reaktionen 0. und 1. Ordnung, Reaktionsgeschwindigkeit, pH- und Temperatur-
Einfluss, Substrat- und Enzymkonzentrations-Diagramme, Lineweaver-
Burk-Diagramme, kompetitive und nicht kompetitive Hemmung, Beispiele Km-Bestimmung, Substratspezifität, Hemmsubstanzen, Mechanismus der
Hemmung, Wechselzahl
Quantitative Bestimmung der enzymatischen Aktivität
Einheit der Enzymaktivität: Katal (neu), Units (alt)
Isoenzyme
Multienzym-Komplexe und multifunktionelle Enzyme
Regulation der Enzymaktivität, Allosterie, negative Rückkopplung, Beispiele
Saure Phosphatase, Acetylcholinesterase, Neurotransmitter, Inhibitoren und
Enthemmer
Optischer Test (Formeln, optische Eigenschaften sowie Coenzymfunktionen von
NAD+, NADP+ und FAD)
Kombinierter optischer Test (Kofaktor-gekoppelte Reaktion)
3-2 Enzyme
Einführung
Bei reversiblen chemischen Reaktionen stellt sich ein Gleichgewicht ein. Im
Gleichgewichtszustand ist die Geschwindigkeit der Hin- gleich der der Rück-
reaktion. Katalysatoren erhöhen die Reaktionsgeschwindigkeit und beschleu-
nigen damit die Gleichgewichtseinstellung; sie beeinflussen aber nicht die Lage des Gleichgewichts. Enzyme sind Proteine, die als Katalysatoren fungieren.
Die International Union of Biochemistry hat 1961 und 1964 Regeln für die No-
menklatur der Enzyme aufgestellt. Die Enzyme werden in sechs Hauptklassen
eingeteilt, die jeweils gleiche chemische Reaktionstypen katalysieren.
1. Oxidoreduktasen übertragen Wasserstoff zwischen zwei Reaktionspartnern.
Zu ihnen gehören die Dehydrogenasen, Hydroxylasen und Oxigenasen.
2. Transferasen katalysieren die Übertragung von Atomgruppen oder ganzen
Molekülen zwischen zwei Reaktionspartnern. Untergruppen dieser Klasse sind
die "Kinasen": Carboxyl-Transferasen, Amino-Transferasen, Glycosyl-Trans-
ferasen und Acyl-Transferasen.
3. Hydrolasen sind Ester-, Peptid- und Glycosylbindungen spaltenden En-
zyme. Bekannte Enzyme wie Pepsin, Chymotrypsin, Trypsin, Renin und Papain
gehören dazu.
4. Lyasen katalysieren die nichthydrolytische Spaltung kovalenter Bindungen:
Decarboxylasen, Aldolasen, Dehydratasen.
5. Isomerasen epimerisieren ein optisch aktives C-Atom oder racemisieren
optisch aktive Verbindungen.
6. Ligasen führen zur Verknüpfung zweier Substratmoleküle unter gleich-
zeitiger Spaltung von ATP in AMP und Pyrophosphat. Wichtige Vertreter dieser
Gruppe sind die Synthetasen, z.B. Aminoacyl-tRNS-Synthetasen, Acyl-CoA-
Synthetasen oder auch die Carboxylasen.
Die Enzymaktivität bestimmt man, indem man Enzyme und Substrat bei bes-
timmter Temperatur, bestimmtem pH-Wert, definierter Substrat- und Enzym-
konzentration sowie in Gegenwart erforderlicher Kofaktoren inkubiert und die
Reaktion in Abhängigkeit von der Zeit verfolgt.

Enzymkinetik der sauren Phosphatase 3-3
Bei einer Reaktion 1. Ordnung hängt die Reaktionsgeschwindigkeit nur von der
Konzentration des Substrats ab.
Bestimmung der Geschwindigkeitskonstanten k einer Reaktion 1. Ordnung
v = [ ]
−d S
dt= k⋅[S]
[ ]−
d S
dS = k⋅dt
v = Reaktionsgeschwindigkeit [S] = Substratkonzentration k = Geschwindigkeitskonstante t = Zeit
Wenn x die Substratmenge ist, die in der Zeit t umgesetzt wurde, und a die
Substrat-Anfangskonzentration bei t = 0 ist, so gilt:
[ ]−
−
∫d S
a
a x
dS= k⋅ dt
t
0∫
a = Anfangskonzentration des Substrats
x = umgesetzte Substratmenge in der Zeit t
ln (a - x) - ln a= k⋅(t - 0)
k ⋅ t = ln a
a - x
Bei der Halbwertszeit t 1
2
ist x = a
2, daraus ergibt sich
t 1
2
= ln 2
k
3-4 Enzyme
Die enzymatischen Reaktionen sind Reaktionen 2. Ordnung: Die Reaktionsge-
schwindigkeit hängt von Konzentrationen zweier Komponenten ab. Man kann
die Behandlung dieses Systems vereinfachen, indem man: 1. Substrat im Überschuss einsetzt oder
2. die Enzymkonzentration konstant hält.
1. Die Reaktionsgeschwindigkeit ist proportional der Enzymkonzentration E bei konstanter Substratkonzentration S (Überschuss).
v = -d[S]
dt = k⋅[E]⋅[S] = k '⋅[E]
[E] = Enzymkonzentration (mg Protein)
Nimmt man pro Enzymmolekül eine Wirkgruppe an, so bezeichnet man die
Anzahl von Substratmolekülen, die pro Molekül Enzym in einer Minute umge-
setzt werden, als Wechselzahl oder "turn-over number" des Enzyms. (z.B.:
Fumarase 105, Acetylcholinesterase 2 x 107 mol Substrat/mol Enzym x Min).
2. Abhängigkeit der Reaktionsgeschwindigkeit von der Substratkonzentration
Hält man die Enzymkonzentration [E] konstant und steigert die Substratkon-
zentration [S], so erhält man folgendes Bild:
v0
[S]
I
II
III
Abbildung 3-1: Reaktionsgeschwindigkeit bei konstanter Enzym- und varia-bler Substratkonzentration

Enzymkinetik der sauren Phosphatase 3-5
Mit steigender Substratkonzentration wird ein Plateau erreicht, d.h. das Enzym
ist mit Substrat gesättigt. Die maximale Reaktionsgeschwindigkeit ist erreicht,
und eine weitere Zugabe von Substrat bei konstanter Enzymkonzentration stei-
gert die Umsatzgeschwindigkeit nicht mehr. Eine solche zweiphasige Kurve
kann man erklären, wenn man eine Komplexbildung zwischen Enzym und Sub-
strat (Enzym-Substrat-Komplex, ES) annimmt, bevor das Reaktionsprodukt
gebildet wird. Die abnehmende Reaktionsgeschwindigkeit (III) erklärt man
durch Substrathemmung: Zu viele Substratmoleküle vermindern die Effizienz
der Enzym-Substrat-Wechselwirkung.
In der Praxis sind jedoch die beiden Extremfälle selten anzutreffen. Die Be-
rechnung der Interaktion wird dadurch komplizierter. Unter Voraussetzung einer
konstanten Konzentration des Enzym-Substrat-Komplexes beschreibt eine
Gleichung von Michaelis und Menten die Wechselwirkung:
k1 k3
k4 k2 E + P ES E + S
P = Reaktionsprodukt
k1⋅[E]⋅[S] - k2⋅[ES] = k3⋅[ES] - k4⋅[E]⋅[P]
Nettogeschwindigkeit der = Nettogeschwindigkeit des
Bildung von [ES] Zerfalls von [ES]
Die Einführung der Konstanten Km (der Michaelis-Konstanten), der maximalen
Reaktionsgeschwindigkeit vmax und die Umformulierung nach vo ergibt:
1
vo
= K
vm
max
• 1
S +
1
v max
(y = m⋅x + b)
3-6 Enzyme
v 0 1
1 K m –
1 V max
1 [S]
Abbildung 3-2: Lineweaver-Burk-Plot
Trägt man 1
vo
als Ordinate gegen 1
Sals Abszisse auf, resultiert eine Gerade
(Lineweaver-Burk-Plot). Die Gerade schneidet die Ordinate bei 1
v max
und die
Abszisse bei - 1
K m
, ihre Steigung ist K
vm
max
. Experimentell erhält man die
Gerade durch Messungen der Anfangsgeschwindigkeit bei verschiedenen Sub-
stratkonzentrationen und konstanter Enzymmenge. Die reziproken Anfangs-
geschwindigkeiten werden gegen die reziproken Substratmengen aufgetragen.
Eine andere Darstellungsart der Gleichung erhält man, wenn man beide Seiten
der Gleichung mit [S] multipliziert. Zeichnet man vo gegen [S] auf, so erhält
man eine Hyperbel. Bei dieser Darstellungsart wird die Bedeutung von Km
deutlich, die bei der Ableitung der Gleichung eingeführt wurde.
Km ist demnach die Substratkonzentration, bei der vo = 1
2vmax ist.

Enzymkinetik der sauren Phosphatase 3-7
12
Vmax
v0
Km [S]
Abbildung 3-3: Abhängigkeit der Reaktionsgeschwindigkeit von der Sub-stratkonzentration
Geräte: Wasserbad 37 °C, mit Kontaktthermometer Einsatz für Reagenzgläser Filter-Photometer Filter 623 nm , 2 × 1 cm Küvetten Reagenzgläser 30, im Reagenzglasständer Vollpipetten 5 ml Messpipetten 1 ml, 5 ml
Lösungen Enzym: 1 mg saure Phosphatase in 100 ml Wasser Phenol-Standard-Lösung: 0,5 mM Dinatriumphenylphosphat (DPP): 0,005 M, mit 2 N HCl auf pH 5,3 eingestellt. Ethylendiamin-Zitratpuffer: 0,1 M, pH 5,9
(0,1 M Lösung von Ethylendiamin, pH mit fester Zitronensäure einstellen). Na2CO3-Lösung: 7 %ig Folin-Reagenz: 1:1 mit Wasser verdünnt
3-8 Enzyme
I. Der optische Test: Bestimmung von Pyruvat
Vorbemerkung: Die Pyridinnukleotide NAD+ und NADP+ können reversibel Wasserstoff auf-
nehmen. Diese Fähigkeit bedingt ihre Funktion als Coenzym vieler Dehydro-
genasen (Oxidoreduktasen). Bei der Hydrierung wird der aromatische Charakter
des Nikotinsäureamid-Ringsystems aufgehoben. Das entstandene Dihydro-
pyridin-Ringsystem besitzt eine spezifische Lichtabsorption mit einem Maxi-
mum von 340 nm, die bei NAD+ fehlt.
5004003002000,0
0,2
0,4
0,6
0,8
1,0
1,2
NAD+NADH
nm
E
Abbildung 3-9: Absorptionsspektren von NADH und NAD+
Man kann daher die Umwandlung von NAD+ in NADH und umgekehrt mit
dem Photometer messen (optischer Test).
Die Konzentration des gebildeten bzw. verbrauchten NADH oder die Konzen-
tration der entsprechenden Reaktionspartner der Enzyme werden mit Hilfe des
Lambert-Beerschen Gesetzes berechnet.

Enzymkinetik der sauren Phosphatase 3-9
1510500,1
0,2
0,3
0,4
Ext
inkt
ion
Zeit [min] A
Abbildung 3-10: Pyruvat-Bestimmung mit dem optischen Test
Prinzip: Pyruvat (Pyr) wird von NADH in Gegenwart von Laktatdehydrogenase (LDH)
nach folgender Gleichung zu L+-Laktat (Lakt) reduziert.
CH3-CO-COO- + NADH + H+ → CH3-CH(OH)-COO- + NAD+
Die Gleichgewichtskonstante dieser Reaktion ist bei 25 °C:
K = [Lakt] • [NAD ]
[Pyr] • [NADH] • [H ]
+
+= 3,6 • 1011 l/mol
Pyruvat wird praktisch vollständig zu Laktat reduziert.
Lösungen:
Citrat-Puffer pH 6,0
Laktat-Dehydrogenase: 0,025 ml Konzentrat (10 mg/2 ml) ad 10 ml H2O
NADH-Lösung 1 mM
Pyruvat Analysenlösung
Ausführung: Das Photometer wird mit Wasser als Leerwert bei 340 nm auf 0,000 eingestellt.
In eine 1 cm-Küvette werden 1,5 ml Citrat-Puffer pH 6,0; 0,2 ml Natrium-
pyruvatlösung und 0,2 ml 1 mM NADH pipettiert. Nach Mischen mit dem
Plastikspatel wird die Extinktion viermal in Abständen von 1 min gemessen und
3-10 Enzyme
notiert. Dann werden 0,1 ml LDH zur Probenküvette gegeben und sofort in
Abständen von 1 min die Extinktionen bei 340 nm über eine Dauer von 10 min
gemmesen und notiert.
Berechnung: Bei photometrischen Bestimmungen wird die Menge der unbekannten Probe mit
Hilfe eines in seiner Menge bekannten Standardwertes oder mit Hilfe einer
Eichkurve ermittelt. Hier steht als Messergebnis nur die Extinktionsdifferenz der
unbekannten Lösung zur Verfügung. Man braucht deshalb die Angabe des
molaren Extinktionskoeffizienten und die Volumina des Reaktionsansatzes und
der darin enthaltenen Probemenge. Die Berechnung geht vom Lambert-
Beer'schen Gesetz aus. Berücksichtigen Sie bei der Berechnung der
Pyruvatkonzentration c (Einheit: mol/l) den Verdünnungsfaktor.
∆E=ε·d·c (∆Ε=ΕΑnfang-EEnde; ε = 6,22⋅103 mol-1⋅cm-1⋅l; d=1cm)
Berechnen Sie die Pyruvatkonzentration in g/l (MW Pyruvat = 88 g/mol).
Tragen Sie in einer Graphik die Pyruvatkonzentration gegen die Zeit (min) auf.

Enzymkinetik der sauren Phosphatase 3-11
II. Enzymkinetik der sauren Phosphatase
Die saure Phosphatase spaltet organische Phospatester bei einem pH-Optimum
zwischen pH 4,5 bis 6,0. Sie ist ein lysosomales Enzym und kommt in allen
Organen vor.
Die im menschlichen Blutplasma enthaltenen sauren Phosphatasen stammen aus
den verschiedensten Organen oder Zellen. Sie unterscheiden sich durch ihre
Hemmbarkeit durch bestimmte Substanzen. So wird z.B. die aus der Prostata
stammende saure Phosphatase selektiv durch L-Tartrat gehemmt, während die
aus Erythrozyten, Knochen oder Milz stammende saure Phosphatase nicht durch
L-Tartrat gehemmt wird. Aus der Differenz zwischen der Gesamtphos-
phataseaktivität des Serums und der tartratgehemmten Phosphatase lässt sich der
Anteil der Prostata-Phosphatase im Serum berechnen.
Medizinische Bedeutung: Die Gesamtphosphatase ist erhöht bei akuter myeloischer Leukämie, bei Leber-
parenchymschäden, bei Lipidspeicherkrankheiten, bei denen die Niere betroffen
ist (Lipideinlagerung). Die mit L-Tartrat hemmbare spezifische Prostata-
Phosphatase ist erhöht bei Erkrankungen der Prostata (Karzinom, Entzündung).
Im folgenden Versuch wird die Michaelis-Menten-Konstante der sauren Phos-
phatase aus Kartoffeln bestimmt. Als Substrat dient Dinatriumphenylphosphat
(DPP). Das freigesetzte Phenol wird mit dem Folin-Ciocalteu-Reagenz quanti-
tativ bestimmt.
O P
O
O
O OH-
Phosphatase
saure2 Na+
+ HPO42- + 2 Na +
2--
3-12 Enzyme
1) Standardkurve mit Phenol
Ausführung: In sechs Reagenzgläser pipettiert man jeweils 0; 0,1; 0,2; 0,3; 0,4 und 0,5 ml der
0,5 mM Phenol-Standardlösung und ergänzt die Proben mit Wasser auf ein
Volumen von 1 ml. Die Probe, die nur Wasser enthält, dient als Leerwert. Dann
gibt man zu jeder Probe 5 ml einer 10 %igen Na2CO3-Lösung. Die Proben
werden gut durchgeschüttelt, anschließend mit 1 ml Folin-Reagenz versetzt
und nach dem Durchmischen 15 min. bei Raumtemperatur stehengelassen. Die
Extinktion wird bei 623 nm im Photometer abgelesen.
Ausführung: Die Werte werden in Tabelle 3-1 eingetragen. Berechnen Sie die Phenolkonzen-
tration (Einheit: mM) unter Einbeziehung des jeweiligen Verdünnungsfaktors
und tragen Sie die Werte gegen die Extinktionen auf.
Tabelle 3-1: Messergebnisse zur Anfertigung der Abb. 4-4 und 4-5
Glas-Nr
µmol DPP/
ml
E(gemessen) nach Min. E Eich-kurv
e
% Hydrolyse nach Inku-bation
1 5 ∆5-1 10 15 1 5 10 15
1 0,0 2 0,5 3 1,0 4 1,5 5 2,0 6 2,5
2) Bestimmung der Michaelis-Konstanten
Ausführung: Vorbereitend beschriftet man 24 Reagenzgläser (Angaben im Seminar
beachten!) und pipettiert in jedes 5 ml einer 10 %igen Na2CO3-Lösung.
Für die Enzymreaktion gibt man in sechs weitere Reagenzgläser jeweils 0; 0,1;
0,2; 0,3; 0,4 und 0,5 ml der 5 mM DPP-Lösung. Die Proben ergänzt man mit

Enzymkinetik der sauren Phosphatase 3-13
% Hydro- lyse
5 10 15 20 25 30
0,25µmol/min 0,15µmol/min
0,05µmol/min
min
Wasser auf ein Volumen von 1 ml. Dazu pipettiert man 7 ml Citrat-Puffer. Die
sechs Inkubationsansätze werden in ein Wasserbad von 37 °C gestellt. Nach
5 min pipettiert man nacheinander in jedes der 6 Gläser 2 ml der sauren
Phosphatase-Lösung und mischt gut. Achtung: Ab jetzt läuft die Zeit!
Je 1 ml wird nach genau 1, 5, 10 und 15 min aus den sechs Reaktionsansätzen
entnommen und sofort in die entsprechend vorbereiteten Reagenzgläser mit 5 ml
10 %iger Na2CO3-Lösung pipettiert. Dadurch wird die Reaktion gestoppt. Zum
Schluss wird je 1 ml Folin-Reagenz in alle Reagenzgläser gegeben, gut
durchgeschüttelt und nach 15 min die Extinktion bei 623 nm gemessen und in
Tabelle 3-1 eingetragen.
Berechnungen: a) Abhängigkeit der Hydrolyse von der Inkubationszeit
Die Extinktionen der Phenol-Eichkurve entsprechen einer 100 %-igen Hydro-
lyse des Substrates. Der Hydrolysegrad in Prozent wird nach der Formel:
Egemessen
E Eichkurve . 100 = % Hydrolyse
berechnet, in die Tabelle 3-1 eingetragen und graphisch gegen die Inkubations-
zeit aufgetragen (Abbildung 3-4).
Abbildung 3-4:
Prozentuale Abhängigkeit der Hydrolyse von der Inkubationszeit
3-14 Enzyme
b) Abhängigkeit der Anfangsgeschwindigkeit von der Substratkonzentration
Die Anfangsgeschwindigkeit (Hydrolysegrad/Min.) kann man aus dem linearen
Teil der Kurven in Abb. 3-4 ermitteln und auf µmol/Min. umrechnen. Einfacher
ist es, die Extinktionsdifferenzen der ersten Messungen (zwischen ein und fünf
Minuten), die den Anfangsgeschwindigkeiten vo entsprechen, zu verwenden.
Diese Differenzen werden in der Tabelle 3-1 eingetragen und gegen die
Substratkonzentration aufgetragen (vgl. Abb. 3-5). Bestimmen Sie aus Ihrem
Graphen die Substratsättigungskonzentration.
V0
µMol DPP/ml
0,04
0,03
0,02
0,01
0,05 0,15 0,25
∆ E 5-1
Abbildung 3-5: Abhängigkeit der Anfangsgeschwindigkeit von der Substrat-konzentration
c) Zeichnen des Lineweaver-Burk-Diagramms
Die Michaelis-Menten-Konstante KM wird graphisch ermittelt. Tragen Sie dazu
1/v0 gegen 1/[S] auf und bestimmen Sie anhand des Graphen Vmax und KM.

Enzymkinetik der sauren Phosphatase 3-15
Tabelle 3-2: Wertetabelle für Lineweaver-Burk-Diagramm
[S] 1/[S] (∆E5-∆E1) vo 1/vo
0,5 2,0
1,0 1,0
1,5 0,67
2,0 0,5
2,5 0,4
3) Einfluss steigender Enzymkonzentration:
Vorbereitend pipettiert man in fünf Reagenzgläser je 5 ml einer 10%igen
Na2CO3-Lösung. Dann werden nach Tabelle 3-3 die Reaktionsansätze in fünf
neue Reagenzgläser pipettiert.
Tabelle 3-3: Volumina der Reagenzien in ml
Nach der Inkubationszeit wird jeweils 1 ml aus den fünf Reagenzgläser
entnommen und sofort in die vorbereiteten Reagenzgläser mit 5 ml 10%iger
Na2CO3 pipettiert. Nach Zugabe von 1 ml Folin-Reagenz und gutem
Durchmischen wartet man 15 min und misst die Extinktionen bei 623 nm. Dies
entspricht direkt dem Hydrolysegrad in Prozent oder den Molen gebildeten
Phenols. Tragen Sie bitte auf Millimeterpapier die Extinktionen gegen die
Enzymmengen auf.
Reagenzglas-Nr.
1
(Nullwert)
2
3
4
5
Citratpuffer pH 6,0 [ml] 3 3 3 3 3
H2Odest [ml] 3 2,5 2 1 0
saure Phosphatase [ml] 0 0,5 1 2 3
mischen und 5 min Inkubation im Wasserbad (37°C)
0,25 mM DPP-Lösung [ml] 0,5 0,5 0,5 0,5 0,5
mischen und 10 min Inkubation im Wasserbad (37°C)
Enzyme 3-16
III. Best. der Serum-Acetylcholinesterase-Aktivität
Axon
Synaptische Vesikel
Synaptischer Spalt (50 nm)
Präsynaptische Membran
Richtung des Nervenimpulses
Postsynaptische Membran
Abbildung 3-6: Schema einer Synapse
Vorbemerkungen: Breitet sich die Erregung in Form eines Aktionspotenzials über ein Axon - kon-
tinuierlich bei myelinfreien und saltatorisch über myelinisierte Neuriten - bis hin
zu den terminalen Synapsen aus, so wird dort eine chemische Transmitter-
Substanz, z. B. Acetylcholin, aus synaptischen Vesikeln in den Synapsenspalt
freigesetzt.
+
+ + +
+ +
+ +
+
polarisierte synaptische Membran,
∆ E ca. -75 mV
hohe [K ], niedrige [Na ] +
+
Acetylcholin
K +
Na +
depolarisierte postsynaptische Membran, ∆ E ca. 0 mV
Abbildung 3-7: Depolarisation der postsynaptischen Membran durch Ace-tylcholin

Optischer Test 3-17
Die am längsten bekannte und am besten untersuchte Transmittersubstanz ist
das Acetylcholin, andere sind die γ-Aminobuttersäure und die Glutaminsäure.
Der berühmte Versuch von Otto Loewi (1921) bewies am perfundierten Frosch-
herzen, dass bei Vagusreizung Acetylcholin an den Endigungen postgang-
lionärer präsynaptischer Nervenfasern entsteht. Dale und Feldberg (1930) zeig-
ten, dass Acetylcholin an vielen Synapsen des peripheren Nervensystems der
Säuger entsteht. Auch an den motorischen Endplatten des Skelettmuskels wird
Acetylcholin freigesetzt.
Acetylcholin wird präsynaptisch in cholinergen Neuronen durch Cholin-Acetat-
Transferase nach folgender Gleichung synthetisiert:
CH3-CO~SCoA + HO-CH2-CH2-N+(CH3)3 →
CH3-COO-CH2-CH2-N+(CH3)3 + CoASH
Das Acetylcholin wird dann in Vesikeln gespeichert und bei der Depolarisation
der präsynaptischen Membran in "Quanten" in den Synapsenspalt ausgeschüttet.
Nach Bindung an den Acetylcholinrezeptor der postsynaptischen Membran mit
Öffnung der Natriumkanäle (Depolarisation) wird es durch die Cholinesterase
hydrolysiert. Das Ruhepotenzial wird wieder aufgebaut.
CH3-COO-CH2-CH2-N+(CH3)3 + H2O →
CH3-COOH + HO-CH2-CH2-N+(CH3)3
Das Enzym hat im aktiven Zentrum zwei charakteristische Wirkungszentren:
1) die anionische Bindungsstelle, an die der quaternäre Stickstoff des Cholins
bindet,
2) die Esterase-Gruppe, die Elektronen auf den Acetatrest übertragen kann,
wodurch ein acetyliertes Enzym entsteht, das anschließend hydrolysiert
wird.
3-18 Enzyme
OH
Ser
(CH3)3-N-CH2-CH2-O-C=O
CH3
-+
E +AcCh EAc + Ch
EAc + H2O E + AcOH
Acetylcholinesterase gehört, wie die Phosphatasen, zur Klasse der Hydrolasen.
Man unterscheidet zwei Typen von Cholinesterasen. Wie aus der Tabelle her-
vorgeht, enthält vor allem Nervengewebe einen hohen Gehalt an echter Acetyl-
cholinesterase. Die Rolle der Pseudocholinesterase dürfte im intermediären
Stoffwechsel des Cholins und verwandter Verbindungen sowie in Entgiftungs-
vorgängen (Procain-Esterase, Aspirin-Esterase) zu suchen sein.
Tabelle 3-4: Eigenschaften der Cholinesterase-Typen
Kriterium Acetylcholinesterase
(E C 3.1.1.7.) (E C 3.1.1.8)
Synonym echte Cholinesterase Pseudo-Cholinesterase
Vorkommen Gehirn, Nerven, Erythrozyten, Leber, Pankreas, Blutserum
Schlangengifte
pH-Optimum 7,8 8,5
Spezifität spaltet nur Ester der Essigsäure weiter Spezifitätsbereich
Die Aktivität beider Cholinesterase-Typen kann durch spezifische Inhibitoren
teilweise oder völlig gehemmt werden. Das Enzym mit Aktivitäten um 2000 U/l
im Serum wird in der Leber synthetisiert. Daher sind bei schweren Leberer-
krankungen die Werte im Serum erniedrigt. Pharmakologische und physio-
logische Studien haben zur Entwicklung vieler Enzym-Inhibitoren geführt, von
denen einige sehr toxisch sind. Cholinesterasen werden von zahlreichen Insek-
tiziden (z.B. E 605, p-Nitrophenyl-di-ethylester der Thiophosphorsäure) und
von Physostigmin stark gehemmt. Praktische Bedeutung haben vor allem fol-
gende Gruppen von Cholinesterase-Hemmern:

Optischer Test 3-19
Enzym
CH2OH +
O
H3CCH
CH3
PF O
O
CHH3C
O
H3CCH
CH3
CH3
PO O + HF
O
CHH3C CH3
H2C
Enzym
a) Medikamente, die auf Grund einer gewissen Strukturanalogie die Acetyl-
cholinspaltung kompetitiv hemmen. Dazu gehören die als Parasympathiko-
mimetika verwendeten Pharmaka wie Prostigmin, Biotin, Eserin.
b) Organische Phosphorsäure- oder Thiophosphorsäure-Ester, welche als In-
sektizide in der Landwirtschaft verbreitet Anwendung finden, z. B. Parathion.
Ähnlich in Bau und Wirkung sind die in der experimentellen Forschung ver-
wendeten Cholinesterase-Blocker DFP oder DIFP (= Diisopropylfluorophos-
phat). Es handelt sich dabei um äußerst gefährliche Nervengifte.
c) Nervengifte vom Typ der Trilone, deren Verwendung als chemische Kampf-
stoffe (Sarin, Soman, Tabun) nach Genfer-Konvention verboten ist.
CH
O
P
O
CH
CH3
H3C
OO
H3C
H2C
CH3
Enzym
N
CH3
CH+ HON
Enzym
CH2OH
CH
O
P
O
CH
CH3
H3C
O+ O
H3C
CH3
N
CH3
CN
H
+
+
3-20 Enzyme
P
OH5C2 O
H5C2 O
NO2O
P
OH5C2 S
H5C2 O
NO2O
HC O P O CH
O
F
H3C
CH3H3C
CH3
N
N
H3C
CH3
CH3
C
O
NH OH3C
(H3C)3N
H2C
H2C
O
C
H2C CH2
C
O
CH2
CH2
N(CH3)3
O O
O
C
NCH3
NH3C CH3
H3C
CH3
O
N
CH3
CH NOH
+
Deblocker
Pyridin-2-aldoxim- methojodid (PAM)
Prostigmin+
Succinyldicholin
++
p-Nitrophenyl-diethylester der Phosphorsäure
p-Nitrophenyl-diethylester der Thio-phosphorsäure: E 605 = Parathion
Diisopropylfluorophosphat
Physostigmin-Eserin
Während die Wirkung von Stoffen der Gruppe a) auf einer reversiblen Verän-
derung beruht, kommt es bei denen der Gruppe b) zu einer festen Bindung des
Blockers an das Serin des Enzymproteins. Die völlige Hemmung der Acetyl-
cholinesterase im zentralen und peripheren Nervensystem durch solche "Bloc-
ker" führt zu einem stetigen Ansteigen der Acetylcholinkonzentration im Be-
reich der cholinergischen Synapsen und Nervenendigungen. Die Folge ist eine
starke Parasympathikusreizung und eine Störung der neuromuskulären Reiz-

Optischer Test 3-21 übertragung. Der Tod infolge Atemlähmung kann innerhalb einer Stunde ein-
treten.
Die Formel einiger Cholinesterase-Substrate und Inhibitoren sowie des De-
blockers PAM werden auf der Vorseite aufgeführt. Da Vergiftungsfälle durch
Inhibitoren relativ häufig sind (Verwechslungen, Unachtsamkeit, Suizidver-
suche), sei kurz auf den biochemischen Aspekt von Diagnose und Therapie
dieser Vergiftung eingegangen.
Die Diagnose hat in erster Linie auf Grund des allgemeinen klinischen Bildes
und des Augenbefundes (kleine, stecknadelkopfgroße, reaktionslose Pupillen!)
zu erfolgen. Als Ergänzung vermag auch die Bestimmung der Cholinesterase-
Aktivität des Serums im Schnelltest (Testpapierstreifen) wertvolle Hinweise zu
geben. Bei der Behandlung derartiger Vergiftungsfälle sollen neben Atropin (in
hoher Dosierung!) spezifisch auf den Cholinesterase-Inhibitor-Komplex ein-
wirkende Substanzen wie Picolinhydroxansäure oder PAM (Pyridin-2-Aldoxim-
Methojodid) verwendet werden. Der therapeutische Effekt von PAM beruht auf
einer Befreiung des Enzyms vom anhaftenden Inhibitor.
Auch zur Erkennung der nicht allzu seltenen Fälle von Dyscholinesterasämie
leistet die Bestimmung der Cholinesterase-Aktivität im Serum wertvolle Dien-
ste. Bei dieser genetisch determinierten Anomalie wird eine atypische Variante
von Cholinesterase mit einem unterschiedlichen Spezifitätsbereich synthetisiert.
Die strukturanaloge Verbindung Succinyldicholin wird beispielsweise infolge
einer etwa 100fach geringeren Affinität von dem atypischen Enzym kaum um-
gesetzt. Die Träger dieser Anomalie sind nicht in der Lage, Succinyldicholin zu
spalten bzw. zu inaktivieren. Wird diese Substanz als Muskelrelaxans bei der
Operationsvorbereitung injiziert, kommt es zu einem lebensbedrohenden
Zustand (Apnoe). Die Untersuchung der Serum-Cholinesterase auf atypisches
Verhalten gehört daher zu einer gewissenhaften Operationsvorbereitung, sofern
die Verabreichung von Succinyldicholin beabsichtigt ist.
Prinzip:
Als Substrat dient Acetylthiocholinjodid und als Indikator für freigesetztes
Thiocholin wird 5,5'-Dithio-bis-2-nitrobenzoesäure verwendet, die zur gelb ge-
färbten 5-Mercapto-2-nitrobenzoesäure reduziert wird. Zur Bestimmung der
3-22 Enzyme
Pseudocholinesteraseaktivität wird der Thiocholinester der Buttersäure verwen-
det.
SHOOC
O2N
S
NO2
CH2
SHCOOH
+ H 2C CH2 N(CH3)3
SHOOC
O2N
S CH2 N
+ HS
(CH3)3
NO2
COOH
5-Mercapto-2-nitrobenzoesäure
5,5'-Dithio-bis-2-nitro-benzoesäure
+
+
In alkalischem Medium vertieft sich die gelbe Farbe der 5-Mercapto-2-nitro-
benzoesäure unter Bildung der chinoiden Verbindung:
S
N
O
O
COOH
Na+
Geräte: Eppendorf-Pipetten, Hamilton-Spritzen Laborwecker Photometer, Filter 405 nm, 2 × 1 cm-Plastikküvetten
Lösungen: Acetylthiocholiniodid 5 mM (CH3–CO–S–CH2–CH2–N(CH3)3+ I– 5,5'-Dithio-bis-(2-nitrobenzoesäure) in 50 mM Phosphatpuffer 0,25 mM, pH 7,2 Diisopropylfluorophosphat 0,1 mM PAM 10 mM Serum

Optischer Test 3-23
Im Folgenden wird die Aktivität, Hemmung und Reaktivierung der Acetylcholinesterase
bestimmt. Der Inhibitor DIFP ist eine hoch toxische Substanz. Achten Sie bitte
darauf, dass die Haut nicht mit der Flüssigkeit in Berührung kommt.
Ausführung: Nach gutem Mischen wird nach einer Minute und dann in einem Zeitraum von
30 min alle fünf Minuten die Extinktion der drei Reaktionsküvetten (2-4, siehe
Tabelle 3-5) bei 405 nm gemessen (Achtung die Zeit läuft nach Zugabe des
Substrats Acetylcholiniodid!). Vor jeder Messung muss der Leerwert mit
Küvette-Nr. 1 erneut auf Null eingestellt werden, da das Serum mit dem
Reagenz reagiert. Tragen Sie die Werte in Tabelle 3-6 ein.
Tabelle 3-5: Durchführung der Aktivität, Hemmung und Reaktivierung der AChE
Küvetten-Nr. 1 Leerwert
2 Aktivität
3 Hemmung
4 Reaktivierung
0,25 mM 5,5‘-Dithio-bis-(2-nitro-benzoesäure) in 50 mM Phosphatpuffer
3 ml 3 ml 3 ml 3 ml
Serum (enthält Acetylcholinesterase) 20 µl 20 µl 20 µl 20 µl
DIFP (Hemmstoff) 50 µl - 50 µl 50 µl
jeweils gut mischen!
5 min nach mischen: PAM (Enthemmer) 50 µl - - 50 µl
15 min nach mischen: 5mMAcetylthiocholinjodid (Substrat)
- 100 µl 100 µl 100 µl
Tragen Sie bitte die abgelesenen Extinktionen gegen die Zeit auf (vgl. Abb. 3-
8). Aus dem Graphen wird die ∆E/min. abgelesen (Steigungsdreieck).
Berechnen Sie die Acetylcholinesterase-Aktivität c/min (Einheit: Mol/min) mit
Hilfe des Lambert-Beer`schen Gesetzes.
ε = 13,3 x 103 [l x mol-1 x cm-1]
3-24 Enzyme
Geben Sie die Acetylcholinesterase-Aktivität in U/l an (1 U = 1 µMol/min).
Tabelle 3-6: Wertetabelle für die Zeit-Umsatzkurve
Zeit nach Substratzugabe
Werte für Aktivität
Werte für Hemmung
Werte für Reaktivierung
1 min [E(405nm)1] 3 min [E(405nm)3] 5 min [E(405nm)5] 10 min [E(405nm)10] 15 min [E(405nm)15] 20 min [E(405nm)20] 25 min [E(405nm)25] 30 min [E(405nm)30]
30 20 10 0 0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4 E
1
2
3
Zeit [min]
Abbildung 3-8: 1) Aktivität der Acetylcholinesterase
2) Hemmung mit DIFP
3) Reaktivierung mit PAM

Aufgaben 3-25
Übungsaufgaben:
1. Eine reine Enzymlösung (100 µg Enzym/ml) hat eine Aktivität von 50
U/ml. MG des Enzyms 80.000. Wie groß ist die Wechselzahl?
2 Hexokinase hat im Gehirn ein Km von 10-5 mol/l und in der Leber ein Km von 10-3 mol/l.
a) In welchem der beiden Organe läuft bei ständig abnehmender Glc-
Konzentration die Reaktion länger mit maximaler Geschwindigkeit ab?
b) Wie groß muss die Glucose-Konzentration in mg/ml intrazellulärer
Flüssigkeit in beiden Organen sein, damit die Reaktion mit halbmaximaler
Geschwindigkeit abläuft ? (MG der Glucose = 180)
3. Formulieren Sie die Reaktionsgleichung der
a) Alkoholdehydrogenase (ADH) mit Coenzym.
b) Die Reaktion zeigte mit verschiedenen Konzentrationen Äthanol [S]
folgende Geschwindigkeiten:
[Äthanol] mM Vo (∆E340/5 min)
0,75 0,094
1,00 0,115
1,50 0,148
3,00 0,206
12,00 0,293
Ermitteln Sie graphisch Km für Äthanol und Vmax der ADH.
c) Einen Umsatz von 0,356⋅105 µmol NADH/l⋅min⋅mg Protein errechnet
man aus Vmax. 1 % der für die Messung verwendete Proteinfraktion war
ADH. MG = 80.000. Berechnen Sie die Wechselzahl der ADH
4. Eine Reaktion 1. Ordnung beginnt mit einer Anfangskonzentration des
Substrates von 10-5 mol/l getestet. Sie ist eine Reaktion 1. Ordnung. Nach
6 Minuten ist die Hälfte des Substrates umgesetzt.
a) Wie groß ist k;
b) Wie groß ist die Konzentration nach 10 Minuten?
3-26 Enzyme
5. In einer Isomerase Reaktion werden 2,5 mg Enzym vom Molekularge-
wicht 125.000 eingesetzt. Km wurde zu 3⋅10-3 mol/l bestimmt und Vmax
zu 275 µmol/Min.
a) Wie groß ist die Wechselzahl?
b) Berechnen Sie die Anfangsgeschwindigkeit bei einer Substratkonzen-
tration von 7,5 mmol/l
6. a) Für welche Reaktion wird Vitamin K als Cofaktor benötigt?
b) Für welchen physiologischen Vorgang ist diese Reaktion von Bedeu-
tung?
7. a) Wie beinflusst ein erhöhter ADP-Gehalt die Umlaufgeschwindigkeit im
Citratzyklus?
b) Auf welche Enzyme des Citratzyklus wirkt ADP?
8. a) Welches Organell der Zelle enthält vor allem Hydrolasen?
b) Nennen Sie drei Stoffklassen, die dort hydrolysiert werden.
9. Stellen Sie den Effekt, den ein Enzym auf die Aktivierungenergie einer
Reaktion hat, in einer einfachen Graphik dar.
10. Wie beinflusst ein kompetitiver Inhibitor Vmax und KM einer enzymati-
schen Reaktion?
11. a) Nennen Sie einen Inhibitor der Acetylcholinesterase, der eine kovalente
Bindung mit dem Enzym eingeht.
b) Nennen Sie die betroffene Aminosäure.

4-1
Versuch 4: Lipide
Versuche:
Nachweisreaktionen von Lipiden
DC-Trennung von Myelin-Lipiden
Gaschromatographie von Fettsäuremethylestern
Dünnschichtchromatographie von Steroidhormonen
2 Analysen:
1. Aceton in Wasser
2. Hormongemisch
Wissensgebiete
Chemische Strukturen der gesättigten und ungesättigten Fettsäuren
Allgemeine Struktur der Glyceride, des Cholesterins, der Cholesterinester,
der Phospolipide, der Sphingolipide
Membranbausteine, Aufbau von Membranen
Lipasen und Phospholipasen
Biosynthese der Fettsäuren und Lipide, β-Oxidation, Energiebilanz
Biosynthese und Umwandlung des Cholesterins zu Gallensäuren und Steroid-
hormonen, Regulation der Cholesterin-Biosynthese
Galle, Zusammensetzung und Funktion
Prostaglandinfamilie: Prostaglandine, Leukotriene, Thromboxane
Steroidhormone: allgemeine Struktur und Funktion
Seifen und Verseifung
Prinzip der Gaschromatographie, Dünnschichtchromatographie
4-2 Lipide
I. Nachweisreaktionen von Lipiden
1) Fettsäuren
Doppelbindungen in ungesättigten Fettsäuren können durch die Addition von
Brom nachgewiesen werden: Zwei Tropfen Olivenöl werden in etwa 3 ml
Chloroform gelöst und tropfenweise eine 2%-ige Bromlösung in Chloroform
zugegeben. Die braune Farbe der Bromlösung entfärbt sich spontan. Überlegen
Sie sich, in welchen Naturprodukten Doppelbindungen vorkommen und warum
einige Fettsäuren essentiell sind.
2) Ketonkörper
Im Urin eines Diabetikers erscheinen nicht nur Glukose, sondern durch die
verstärkte Lipolyse auch Ketonkörper, u. a. Aceton, das auch über die Atemluft
abgegeben werden kann (charakteristischer Geruch im Krankenzimmer!).
Aceton-Nachweise:
a) Natriumnitroprussidprobe
Natriumnitroprussidlösung bildet mit Aceton folgenden rot gefärbten Komplex:
CH3-CO-CH3 + [Fe2+(CN)5NO+] 2- + 2 OH- →
[Fe2+(CN)5NO+CH-CO-CH3] 4- + 2 H2O
Beim Ansäuern mit Eisessig wird ein Proton an den Komplex angelagert und die
Farbe vertieft sich:
[Fe2+(CN)5NO+ CH2-CO-CH3]3-
Ausführung: Zu 3 ml der Analysenlösung gibt man fünf Tropfen frische Nariumnitroprussid-
lösung und 1 ml NaOH. Anschließend wird mit wenigen Tropfen Eisessig
schwach angesäuert.

Lipidnachweisreaktionen 4-3
Der Nachweis von Aceton mit Hilfe von Natriumnitroprussidlösung kann aber
durch andere Metaboliten gestört werden. Überlegen Sie sich, welche diese sein
könnten.
b) Jodoformprobe
CH3-CO-CH3 + 3 I2 + 3 NaOH → CH3-CO-CI3 + 3 NaI + 3 H2O
CH3-CO-CI3 + NaOH → CHI3 + CH3COONa
Ausführung: Zu 3 ml Acetonlösung und 1 ml Jod-Jodkalium-Lösung gibt man tropfenweise
2 N NaOH, bis Entfärbung eintritt. Bei positivem Ausfall der Reaktion tritt ein
gelber Niederschlag und typischer Jodoform-Geruch auf.
Geräte: Reagenzglasgestell mit 10 Reagenzgläsern Spatel
Chemikalien: Aceton 1%ig Brom 2% in Chloroform Eisessig Jod-Jodkalium-Lösung Natriumnitroprussidlösung 5%ig NaOH 2 N Olivenöl
4-4 Lipide
II. Trennung von Myelin-Lipiden durch Dünnschicht-Chromatographie
Geräte: 1 Dünnschichtplatte 10 × 10 cm, beschichtet mit Kieselgel H Entwicklungstrog
Lösungen: Fließmittel: Chloroform/Methanol/Wasser = 65:25:4 Lipid-Extrakt aus Myelin Testlösungen (2 mg/ml) : Cholesterin, Lecithin, Kephalin, Cerebrosid, Sulfatid
Normalwerte im Serum (pro 100 ml) Gesamtlipide 0,6-1 g Triglyceride 70-180 mg Gesamtcholesterin 160-230 mg Verestertes Cholesterin 120-180 mg Phospholipide 160-230 mg freie Fettsäuren 10-35 mg β-Lipoproteine bis 550 mg Bilirubin 0,5-1 mg Gesamtprotein 6-8 g Harnsäure 2-6,8 mg Glucose 60-100 mg Natrium 140 mVal pro Liter Kalium 5 Calcium 5 Magnesium 2 HCO3
– 27 Cl– 103 Protein– 17
Lipidzusammensetzung des Myelins des menschlichen Gehirns.
PC, Phosphatidylcholin 10,0 % PE, Phosphatidylethanolamin 20,0 % PI, Phosphatidylinositol 1,0 % PS, Phosphatidylserin 8,5 % SPM, Sphingomyelin 9,0 % Cerebrosid 20,8 % Sulfatid 5,2 % Cholesterin 26,0 % Lysophosphatidylcholin Spuren

Myelinlipid DC 4-5
Auf einer mit Kieselgel beschichteten Glasplatte werden nach dem Prinzip der
Verteilungschromatographie verschiedene Lipide aufgetrennt und anschließend
mit spezifischen Färbereagenzien nachgewiesen. Die Verteilung findet zwischen
der stationären, polaren Hydrathülle des Kieselgels und dem mobilen, unpolaren
Laufmittel statt. Daher laufen die unpolaren Lipide nahe der Laufmittelfront und
je stärker der polare Charakter des Lipids ist, um so stärker ist ihre Wech-
selwirkung mit der polaren, stationären Phase und umso kürzer ist die Laufstrecke
dieser Lipide. Die Phosphatgruppe der Phospholipide kann mit Hilfe von
Molybdänblau (Zinzadse) nachgewiesen werden, Ninhydrin erfasst die Aminogruppen von Phosphatidylethanolamin und Phosphatidylserin, H2SO4-
Eisessig (Salkowsky) färbt Cholesterin und das Anthron-Reagenz Kohlenhydrate.
Als Laufmittel dient Chloroform-Methanol-Wasser 65:25:4.
1 2 3 4
PE Sulfatid
Cerebrosid
Cholesterin
PC PS SPM Lyso-PC
Salkowsky
Anthron
Phosphor
Ninhydrin
Phosphor
Phosphor
Ninhydrin
Anthron
Auftrags- stelle
Abbildung 4-1: Dünnschichtchromatogramm der Myelinlipide
Die Proben werden punktförmig mit der Spitze einer Pasteurpipette aufgetragen.
Dabei sollte die Spitze 3 mal hintereinander mit jeweils 3-5 mm Probe gefüllt und
so aufgetragen werden, dass der Durchmesser der Flecken nicht größer als 3-4 mm
4-6 Lipide
ist. Dabei ist darauf zu achten, dass die Auftragspunkte so weit vom unteren Rand
der Platte entfernt sind, dass sie nicht im Laufmittel „ertrinken“.
Aufgetragen werden: 1. Phosphatidylcholin (Lecithin) 2. Cerebrosid und Sulfatid (Probenlösung handwarm machen!) 3. Lipidextrakt aus Myelin 4. Cholesterin
Pasteurpipette zwischendurch mit Aceton reinigen. Der Chromatographietrog wird
verschlossen und die Chromatographie so lange durchgeführt, bis die
Laufmittelfront bis auf ca. 1 cm Abstand den oberen Rand der Platte erreicht hat.
Nachdem die Platten im Abzug getrocknet sind, werden sie nacheinander:
1. mit Ninhydrin-Lösung besprüht und 15-20 Min. bei Raumtemperatur liegen
gelassen.
2. mit Zinzadse-Reagenz eingesprüht
3. mit konz. H2SO4-Eisessig 1:1 besprüht und bei 120°C im Trockenschrank
über 15 min. erhitzt
4. mit Anthron-Reagenz besprüht und erneut 10-15 Min bei 90 °C im Trocken-
schrank erhitzt
Notieren Sie jeweils nach jeder Färbung die Lage der Flecken.

GC 4-7 III. Gaschromatographie der Fettsäuremethylester
Triglyceride sind Energiespeicher des Körpers und kommen z. B. im Fettgewebe
oder in Lipoproteinen vor. Den mit tierischer und pflanzlicher Nahrung aufge-
nommenen Triglyceriden werden unterschiedliche Qualitäten zugeschrieben.
Objektiv unterscheiden sie sich nur in der Zusammensetzung ihrer Fettsäuren. Die
Zusammensetzung der Fettsäuren lässt sich mit Hilfe der Gaschromatographie
quantitativ ermitteln. Überlegen Sie sich, warum man zunächst die Fettsäuren in
Fettsäureester umwandeln muss.
Geräte: Gaschromatograph, Erlenmeyerkolben 100 ml , 20 ml Wasserbad zur Verseifung und Veresterung Wasserbad zum Abdampfen von Lösungsmitteln, unter dem Abzug
Chemikalien: Bortrifluorid/Methanol 1:5 Petrolether Na2SO4 wasserfrei NaOH 2 N Olivenöl Methanolische KOH 20% HCL Chloroform
1) Verseifung von Triglyceriden:
0,1 ml Olivenöl oder ca. 0,1 g Speisefett aus dem Haushalt (Margarine, Butter,
Salatöl, Speck usw., bitte von zu Hause mitbringen!) und 5 ml 15%-ige
methanolische KOH (Methanol/Wasser = 9:1) werden in einem 100 ml-
Erlenmeyerkolben mit Deckel 30 min in einem elektrisch beheizten, siedenden
Wasserbad erhitzt. Der kugelförmige Deckel wirkt wie ein primitiver
Rückflusskühler und verhindert das Verdampfen des Methanols. Anschließend
kühlt man ab und säuert mit 10 ml ca. 20%-iger HCl an. Die freigesetzten
Fettsäuren werden in einem Schütteltrichter mit 20 ml Petrolether ausgeschüttelt.
Die untere Phase wird verworfen, dabei muss der Glasstopfen abgenommem
werden. Die obere Phase (Petrolether) wird dann im Schütteltrichter 3 mal mit je
10 ml Wasser gewaschen und in einem 20 ml-Erlenmeyerkolben mit 2-3
Spatelspitzen wasserfreiem Na2SO4 versetzt („getrocknet“), um die Wasserreste
4-8 Lipide
zu binden. Nach kurzer Zeit und wiederholtem Umschwenken wird die Lösung in
einen 20 ml Erlenmeyerkolben filtriert und im Abzug auf einem ebenfalls
elektrisch beheizten Wasserbad zur fast vollständigen Trockne eingedampft.
2) Veresterung der freien Fettsäuren mit 2% Bortrifluorid-Etherat in
Methanol:
• 1 ml der BF3-Methanollösung wird in den Erlenmeyerkolben gegeben, die
eingedampften Fettsäuren durch Schwenken gelöst und sofort in ein Sovirelglas
(=Reagenzglas mit Schraubverschluss) überführt.
• Sovirelglas fest verschließen! 15 min. im siedenden Wasserbad erhitzen.
• 2 ml H2O zugeben, anschließend mit 2 ml Petrolether durch Schütteln extra-
hieren.
• Abhebern der Petroletherphase mit Pasteurpipette, Umfüllen in ein Reagenzglas
und gegebenenfalls mit Na2SO4 trocknen.
• In ein Reagenzglas filtrieren und im Wasserbad im Abzug zur Trockne ein-
dampfen.
• Rückstand in 0,5 ml CHCl3 lösen, ca. 0,1 µl aufspritzen. Die genau
aufzuspritzende Menge muß jeweils ausgetestet werden. (Bei Verlusten während
der Veresterung, CHCl3-Volumen verringern)

GC 4-9
Abbildung 4-2: Schematische Darstellung eines Gaschromatographen
Die Gaschromatographie ist eine Verteilungschromatographie. Verdampfbare
Substanzen verteilen sich zwischen einer flüssigen und einer gasförmigen Phase.
Als flüssige stationäre Phase werden für Fettsäuremethylester Polyester zwischen
Dicarbonsäuren (z.B. Bernsteinsäure) und Glykolen oder Siliconöle und höhere
Kohlenwasserstoffe verwendet. Die Trennphase wird auf ein inertes, feinkörniges
Trägermaterial (z. B. Kieselgur 0,1-0,2 mm Ø) aufgetragen. Trägermaterial mit
aufgezogener Trennphase ist das Füllmaterial der Gaschromatographiesäule. Sie
wird von der gasförmigen Phase, dem Trägergas, durchströmt, wodurch die
Substanzen der zu analysierenden Probe getrennt in den Detektor transportiert
werden. Dort werden elektrische Signale erzeugt, die über einen Verstärker vom
Schreiber aufgezeichnet werden.
Die GC ist in der Lage, die Trennung komplizierter Gemische in µg-Mengen exakt
qualitativ und quantitativ durchzuführen. Jede Verbindung hat eine
charakteristische Retentionszeit (Verweilzeit in der Säule). Der Vergleich mit
Testsubstanzen erlaubt die Identifizierung der Komponenten eines Gemisches, in
unserem Fall der einzelnen Fettsäuremethylester. Das Gaschromatogramm wird
quantitativ nach der Dreiecksmethode ausgewertet:
F = h•b
4-10 Lipide
Den Flächeninhalt berechnet man durch Multiplikation der Peakhöhe (h) und der
Peakbreite (b) auf halber Höhe.
Abbildung 4-3: Gaschromatogramm eines Fettsäuremethylestergemisches

Steroidhormon DC 4-11 IV. Dünnschichtchromatographie von Steroid-
hormonen.
Die Steroidhormone Progesteron, Cortison, Testosteron, Östradiol und Cortexon
sollen getrennt und parallel dazu die Komponenten eines Gemisches bestimmt
werden.
O
O OHC
CH2OH
O
O
C
CH2OH
O
O
C
CH3
O
HO
OH
O
OH
Testosteron C 19Östradiol C 18
ProgesteronCortexonCortison
Abbildung 4-4: Strukturformeln der untersuchten Steroidhormone
Geräte: Chromatographietrog Dünnschichtplatten 10 × 10 cm mit Kieselgel H Sprüher zum Anfärben Teflonschläuche Trockenschrank 120 °C UV-Lampe
Lösungen: Steroidhormonlösungen einzeln Laufmittel: Chloroform/Aceton 9:1 Sprühreagenzien: 1) 50% H2SO4 in Methanol
2) 0,1%ige 2,4-Dinitrophenylhydrazin in Ethanol + 1 ml konz. HCl
4-12 Lipide
Ausführung: Mit Hilfe einer Pasteurpipette werden die Testsubstanzen und die Analysen in
Abständen von 1 cm aufgetragen (siehe 4 II und Abb. 4-5). Die Spitze der
Pasteurpipette sollte zwischendurch mit entionisiertem Wasser gespült werden.
Als Laufmittel dient Chloroform/Aceton 9:1. Nach Lufttrocknung (Abzug) wird
die Platte zuerst mit 50% H2SO4 in Methanol und dann mit 0,1% 2,4-Dinitro-
phenylhydrazin in Ethanol + HCl eingesprüht und zur Farbentwicklung 10 Min. in
einem Trockenschrank von 120°C gelegt. Notieren Sie die Farben der Flecken und
beobachten Sie ihre Fluoreszenz unter der UV-Lampe. Bestimmen Sie die beiden
Steroidhormone in Ihrer Analyse. Berechnen Sie die Rf-Werte.
Front
1 2 3 4 5
Auftrags-stelle
8 cm
A1 A2
1,5 cm
Abbildung 4-5: Dünnschichtchromatogramm von Steroidhormonen
1 Östradiol 2 Cortexon A1 Analyse 1 3 Cortison A2 Analyse 2 4 Testosteron 5 Progesteron

Aufgaben 4-13
Übungsaufgaben:
1. Wie kann der Abbau von Triglyceriden zur Glukoneogenese beitragen?
2. Wie beeinflusst der Prozentsatz der ungesättigten Fettsäuren der Membran-lipide die Fluidität einer Membran?
3. Warum sind Plasmamembranen für geladene Moleküle undurchlässig?
4. Nennen Sie drei Substanzen, die aus Membranbausteinen entstehen und an der Übertragung von Signalen mitwirken.
5. N-Acetylneuraminsäure (Sialinsäure) ist Bestandteil von: a) ...........und b) ..........
6. a) Nennen sie das Schlüsselenzym der Biosynthese von Fettsäuren b) Formulieren Sie die von ihm katalysierte Reaktion (in Worten). c) Nennen Sie je ein positives und ein negatives Effektormolekül.
7. Nennen Sie Namen der drei Ketonkörper:
8. Wo sind in der Zelle die Rezeptoren für a) Insulin, b) Cortisol und c) die Schilddrüsenhormone T3/T4 lokalisiert?

5-1
Versuch 5: Immunologie
Versuche:
ELISA (enzyme linked immunosorbent assay), Enzymgekoppelter Im-
munnachweis
Western-Blot
Test zur Schwangerschaftsfrüherkennung aus Urin
Analyse:
IgG-Test mittels ELISA, 2 Proben (A und B)
Wissensgebiete
Allgemeine Struktur und grundsätzlicher Aufbau von Antikörpern
Klassen der Immunglobuline
Polyklonale, monoklonale Antikörper
Prinzip der humoralen und zellulären Immunantwort
Wechselwirkung zwischen Antigen und Antikörper
Komplementsystem
Polyklonale und monoklonale Antikörper
Prinzip der Herstellung monoklonaler Antikörper
Prinzip von ELISA, RIA, Westernblot
Blut, Serum, Plasma
Blutgruppendeterminanten
5-2 Immunologie
Medizinische Grundlage:
Antikörper sind Proteine, die vom Immunsystem des Körpers produziert werden,
um eine hochspezifische extrazelluläre Abwehr gegen Infektionen aufzubauen.
Außerhalb des Körpers, im Labor, sind Antikörper unentbehrliche Werkzeuge
geworden, die in einer Vielzahl von Methoden die Bindungseigenschaften der
Antikörper zur qualitativen und quantitativen Analyse von Antigenen nutzen. Im
Praktikum werden Sie anhand von drei Beispielen einen ersten Einblick
bekommen.
I. ELISA
Die Markierung von Antikörpern mit Enzymen ist eine weit verbreitete Methode
zur Detektion der Antikörper und damit der daran gebundenen Antigene und hat
die radioaktiven Markierungsmethoden mit dem Nachweis im RIA (radio immuno
assay) in vielen Bereichen verdrängt.
Der Aufbau eines ELISA kann auf verschiedene Art und Weise erfolgen und
hängt von den Anforderungen ab, die an ihn gestellt werden. Generell kann man
zwei verschiedene Verfahren unterscheiden:
A: Direkt-ELISA,
B: Inhibitions-ELISA.
Im Praktikum werden Sie aus Zeitgründen die einfachste Form eines Direkt-
ELISA durchführen. Sie sollen humanes IgG mit Hilfe eines affinitätsgereinigten,
mit Meerettich-Peroxidase gekoppelten Antikörpers aus Kaninchen nachweisen.
Normalerweise wird zuerst ein nicht gekoppelter Antikörper eingesetzt, der dann
mit Hilfe eines Enzym-gekoppelten Zweitantikörpers nachgewiesen wird.
Alle Immunglobuline besitzen die gleiche Grundstruktur aus vier Untereinheiten,
zwei leichten Ketten (je 23 kDa) und zwei schweren Ketten (je 53 -75 kDa). Die
Untereinheiten lagern sich über Disulfidbrücken und hydrophobe Wech-
selwirkungen zusammen und bilden eine Y-förmige Struktur. Die sezernierten
Immunglobuline des Menschen lassen sich in 5 Klassen einteilen, IgA, IgD, IgE,
IgG und IgM, die sich im Typ ihrer schweren Ketten unterscheiden. IgD, IgE und
IgG bestehen nur aus Monomeren, IgA besteht aus Monomeren, Dimeren und

ELISA 5-3
Trimeren, während IgM aus Pentameren der Y-förmigen Struktur besteht. Die
Multimere sind dabei sowohl untereinander als auch mit einer sogenannten J-Kette
(J=joining) über Disulfidbrücken verbunden.
In einem direkten ELISA sollen Sie IgG im Plasma nachweisen, als Leerwert dient
TBS. In der Analyse sollen Sie ermitteln, ob in den beiden Proben, die Sie
erhalten, humanes IgG vorhanden ist oder nicht. Dazu werden in einem ersten
Schritt die Plasmaproteine irreversibel an eine ELISA-Mikrotestplatte gebunden.
Nach dem Blockieren unspezifischer Bindungsstellen mit Milchproteinen wird ein
in Kaninchen hergestellter Peroxidase-gekoppelter Antikörper zugegeben, der
gegen humanes IgG gerichtet ist. Der Nachweis der gebundenen Antikörper
erfolgt dann durch die enzymatische Reaktion. Als Substrat dient Tetra-
methylbenzidin, dessen Umsatz photometrisch bei 450 nm detektiert werden kann.
Materialien und Geräte: ELISA-Reader 96er Microtiter-Platte Pipetten: Precision rot/blau 500 ml Spritzflasche 15 ml Plastikgefäß mit Schraubdeckel Eppendorf-Reaktionsgefäße
Lösungen: Plasma (für die Verdünnungsreihe) Protein-Analysen-Lösungen A und B TBS TBS/Tween 20, 0,02 %(Detergenz) 5% Milchpulver in TBS Kaninchen-Anti-Human-IgG Antikörper (Peroxidase gekoppelt; 1:40000) Citrat pH 6.0 10 mg/ml Tetramethylbenzidin-Lösung in DMSO (carcinogen) 3% ige H2O2-Lösung 20%ige H2SO4
5-4 Immunologie
Durchführung:
1. Beschichten
Die Löcher einer 96-Loch (12x8) Microtiter-Platte werden mit jeweils 100 µl
Antigenlösung beschichtet. Hierzu wird eine Plasma-Verdünnungsreihe in
TBS-Puffer (siehe Tabelle) hergestellt, zusammen mit den unverdünnten
Analysen A und B (ebenfalls je 100µl) wie folgt aufgetragen (alles in
Doppelbestimmung) und eine Stunde bei Raumtemperatur (RT) inkubiert.
Bitte benutzen Sie immer nur 3 nebeneinanderliegende Spalten und vermeiden
Sie Proteinkontaminationen der freien Plätze. Die Platten werden in der
nächsten Woche von Ihren Kommilitonen weiterverwendet. Zur Sicherheit
lassen Sie jede 4. Spalte frei. Bitte markieren Sie die benutzten Spalten mit
Filzstift.
Also zu benutzen: Spalten 1,2,3 oder 5,6,7 oder 9,10,11.
Verdünnung Plasma (Doppelbestimmung)
Analysen
Leerwert AX1,
1:104 AX2 Student X
1:5x104 BX1
1:105 BX2
1:5x105 AY1
1: 106 AY2 Student Y
1:5x106 BY1
1: 107 BY2
2. Waschen
Die Lösungen werden über dem Waschbecken ausgegossen und dann dreimal
mit TBS/T-Puffer gewaschen. Man füllt die Löcher mit dem Waschpuffer aus
einer Polyethylen-Spritzflasche, und schüttet sie ebenfalls über dem
Waschbecken aus. Anhaftende Tropfen können durch Ausklopfen entfernt
werden.

ELISA 5-5
3. Blockieren
Um die restlichen Proteinbindungsstellen am Plastik zu blockieren, inkubiert
man 30 Minuten lang bei RT mit 200 µl Blockierungsreagenz (5% Milch-
pulver in TBS).
4. Waschen
Blockierungslösung entfernen und dreimal (s. o.) mit TBS/T waschen.
5. Peroxidase-gekoppelter Antikörper
je 100 µl Antikörper (Anti-human IgG, 1:40.000 in TBS) pro Loch auftragen
und 45 Minuten lang bei RT inkubieren.
6. Waschen
Antikörperlösung entfernen, dreimal (s. o.) mit TBS/T und anschließend
zweimal mit H2O (entsalzt) waschen.
7. Detektion
Ansetzen von Substratlösung in 15 ml Plastikgefäßen:
4,5 ml aqua dest., 0,5 ml 1M Na-Actat Puffer (pH 6), 31 µl Tetramethylbenzi-
din-Lösung (10 mg/ml DMSO), 5 µl 3 %ige H2O2-Lösung.
jeweils 100 µl Substratlösung werden in jedes Loch pipettiert. Danach min-
destens 5-10 Minuten lang inkubieren, bis eine Blaufärbung zu sehen ist.
8. Stoppen der Farbentwicklung
je 100 µl 20%ige H2SO4 pro Loch hinzugeben.
9. Messen
im ELISA-Reader bei 450 nm (mit Hilfe des Assistenten)
10. Auswerten
Tragen Sie die gemessenen Absorptionswerte gegen den Logarithmus der
Antigenverdünnung auf. Diskutieren Sie den Kurvenverlauf. Geben Sie das
Ergebnis der Analyse an (positiv oder negativ), im Vergleich mit dem Standard
aus der Plasmaverdünnungsreihe
Immunologie 5-6
II. Western Blot
Prinzip:
Die Trennung von Proteinen über SDS-Gelelektrophorese ist eine Standard-
methode zur Analyse von Proteinen. Um eine spezifische Antigenbindung der
getrennten Proteine nachzuweisen, überträgt man die Proteine durch Anlegung
eines elektrischen Felds auf eine Nitrocellulose-Membran. Diese kann dann mit
einem spezifischen Antikörper gegen ein bestimmtes Protein inkubiert werden und
durch die gekoppelte enzymatische Aktivität eines ersten oder zweiten
Antikörpers nachgewiesen werden. Damit wird nicht nur wie beim ELISA eine
Antigen-Antikörper-Bindung nachgewiesen, sondern zusätzlich auch noch eine
Information über die Größe des Proteins erhalten. Der Western Blot dient z. B. als
Bestätigung des Nachweises einer HIV-Infektion, wenn in einem ELISA
Antikörper gegen HIV nachgewiesen wurden. Im Western Blot werden virale
Proteine im SDS-Gel der Größe nach aufgetrennt und auf dem Blot die Antikörper
der Patienten gegen die verschiedenen Proteine nachgewiesen. Beim Immuno-Blotting kann die Trennung eines Proteingemisches mittels SDS-
PAGE mit der Spezifität vom immunologischen Nachweis gekoppelt werden.
Dabei unterscheidet man folgende sechs Schritte:
1. Proteingemisch in Probenpuffer 2. Trenunng der Proteine durch Gele-
lektophorese 3. Transfer auf Nitrocellulose
4. Blocken unspezifischer Bindungen 5. Antikörperaddition 6. Detektion

Western-Blot 5-7 Dieses Experiment schließt sich an Versuch 2 (Aminosäuren und Proteine II) an.
Die Blotprozedur wird dort beschrieben. Materialen und Geräte: geblottete Nitrocellulose (Versuch 2) Plastikschale Schüttler
Lösungen: TBS-Puffer: 0.15 M NaCl, 50 mM Tris pH 7.4 TBS/T (0.01 % Tween 20 in TBS) TBS/T + 5% Milchpulver Kaninchen-Anti-Human-IgG Antikörper (Peroxidase gekoppelt; 1:15.000) 30%ige H2O2-Lösung 0.3 % 4-Chloronaphtol (in Methanol) (carcinogen!)
Durchführung: − die Membran wird eine Stunde lang mit 10 ml Kaninchen-Anti-Human-IgG
(Peroxidase gekoppelt) (1:15.000) inkubiert und dann
− dreimal für 5-10 min mit jeweils 10 ml TBS/T gewaschen.
− zweimal kurz mit Wasser gewaschen
− Farbreaktion: Der Antikörper ist mit Peroxidase gekoppelt, die ein farbloses
Substrat (4-Chloronaphtol) zu einem Farbstoff umsetzen kann. Hierzu stellt
man kurz vor Gebrauch folgende Lösung her:
0,3 ml 0.3% 4-Chloronaphtol
4,7 ml TBS
5 µl 30%ige H2O2-Lösung
Nach Zugabe dieser Lösung erscheint auf der Membran eine farbige Bande, die
selektiv die Lage und damit die Existenz des Antigens anzeigt.
Die Färbereaktion wird anschließend mit TBS-Puffer abgestoppt.
Auswertung:
Die Beobachtungen nach der Detektion mit Chlornaphthol werden festgehalten
und im Vergleich zur Poinceau S-Färbung kommentiert. Hierzu werden, vom
unteren Rand der Nitrocellulose ausgehend, sowohl die Markerbanden ausge-
messen, als auch die Lage der prominentesten Proteinbanden.
Immunologie 5-8 III. Test zur Schwangerschaftsfrüherkennung aus Urin
Medizinische Grundlage:
HCG, humanes Choriongonadotropin, ist ein Glykoproteinhormon, das bereits
kurz nach der Befruchtung von der Eizelle sezerniert wird. HCG regt den Gelb-
körper (corpus luteum) dazu an, sich nicht zurückzubilden, sondern weiterhin
Progesteron zu produzieren, so dass keine Menstruation stattfindet. Bei normalem
Schwangerschaftsverlauf kann HCG im Serum bereits 7 Tage nach der
Befruchtung nachgewiesen werden. HCG ist daher ein hervorragender Indikator
zur Früherkennung einer Schwangerschaft, es kann auch im Urin nachgewiesen
werden. Die Konzentration steigt während der Frühphase der Schwangerschaft
rasch an. Ein positives Ergebnis kann außer durch eine Schwangerschaft nur durch
ein sehr selten vorkommendes Chorionkarzinom entstehen, bei dem ebenfalls
große Mengen HCG sezerniert werden.
HCG besteht aus zwei Untereinheiten, α und β. Die α Untereinheit hat eine große
Homologie zu den α-Ketten der Hypophysenhormone LH, FSH und TSH,
außerdem sind 94 der 115 Aminosäuren der β-Untereinheit mit denen des β-LH
identisch. Daher ist es unmöglich einen immunologischen Nachweis mittels eines
polyklonalen Antikörpers durchzuführen, da Kreuzreaktivitäten der Antikörper
eine sichere Bestimmung der Hormone unmöglich machen würde. Nur ein
monoklonaler Antikörper, der gegen das für HCG spezifische Epitop in der β-
Kette gerichtet ist, kann zur Diagnostik eingesetzt werden.
Prinzip
Der Versuch wird mit einem hochempfindlichen kommerziell erhältlichen Test
(„STORCH-TEST“ oder „SCHWANGER JA:NEIN“) durchgeführt. Es handelt
sich hierbei um einen Sandwich-Immun-Assay, der aus einer Kombination von
monoklonalen und polyklonalen Antikörpern besteht. Durch einen
chromatographischen Schritt werden die beiden Antikörper mit dem Antigen
zusammengebracht und die Aggregation durch die Goldmarkierung sichtbar
gemacht. Die Kontrolle für eine korrekte Testdurchführung ist durch die Reaktion
des markierten Antikörpers mit einem weiteren Antikörper gewährleistet. Die
Nachweisgrenze des Tests liegt bei 10 IU HCG/l Urin.

Schwangerschaftstest 5-9 Aufbau
Auf einer hydrophilen Membran sind drei verschiedene Bereiche zu unter-
scheiden:
1. der Kontrollbereich (C), beschichtet mit polyklonalem Ziegen-Antimaus-IgG
Antikörper (IgG),
2. der Testbereich (T), beschichtet mit polyklonalem Ziegen-Anti-HCG-Anti-
körper (IgG) und
3. der Probenauftragsbereich (P). Dieser Bereich enthält den getrockneten, mit
kolloidalem Gold konjugierten monoklonalen Maus-Anti-HCG-Antikörper auf
Papierstreifen, der mit der Membran verbunden ist.
Kontrolle (C) Testergebnis (T) Probenauftrag (P)
Durchführung:
Stellen Sie das Testkit mit den Sichtfenstern nach oben auf den Labortisch. Füllen
Sie die Pipette mit dem Testurin. Geben Sie exakt fünf Tropfen Testurin (ohne
Luftbläschen) in das Probenauftragsloch. Warten Sie eine Minute und lesen Sie
das Testergebnis im Sichtfenster ab. Der Test ist nicht verwertbar, wenn sich keine
bzw. später als nach 5 Minuten eine Linie im Sichtfenster zeigt.
Aufgaben:
Protokollieren Sie das Testergebnis
1. Erklären Sie genau, wie Ihr Testergebnis zustande gekommen ist. Warum ist
der T-Bereich positiv bzw. negativ? Warum ist der C-Bereich positiv?
2. Erklären Sie, warum bei einer nur mit LH versetzten Probe keine Reaktion im
T-Bereich festgestellt werden kann, obwohl die dort vorhandenen polyklonalen
Antikörper auch mit LH reagieren.

6-1
Versuch 6: Blut
Versuche:
Gelfiltration, Spektroskopie und
die quantitative Bestimmung des Hämoglobins
Dialyse-Versuch
Enzymatische Bestimmung der Glucose
Polarimetrie
Analysen:
1. Hämoglobin-Bestimmung
2. Bestimmung von Glukose, Fruktose, Saccharose oder Xylose mittels
Polarimetrie
Wissensgebiete
Puffersysteme des Blutes, Azidose, Alkalose
Hämoglobin als Puffer, 2,3-Bisphosphoglycerat
Sauerstoff-Transport, Kohlendioxidtransport
Myoglobin-, Hämoglobin-Struktur, Biosynthese, Porphyrin-Strukturen
Katabolismus des Hämoglobins, Bilirubin-Chemie
Transport, Glucuronidierung, Ikterusformen
Kolloid-osmotischer Druck, Dialyse Enzymatische Grundlage des GOD-Testes
(Coenzyme von GOD und POD, Funktion und Coenzym von Katalase)
Optische Aktivität, Polarimetrie
6-2 Blut
I. Gelfiltration, quantitative Bestimmung und Spek-troskopie von verschiedenen Formen des Hämo-globins
Die Funktion des Hämoglobins besteht im Transport von O2. Hämoglobin enthält
als prosthetische Gruppe Häm, das im Zentrum eines Porphyrinrings ein Fe2+-Ion
in Chelatform gebunden hält. Dieses Fe2+-Ion ist in der Koordination mit sechs
Liganden am stabilsten. Vier davon werden von den Pyrrol-Stickstoffatomen des
Porphyrinrings und ein weiterer durch einen Histidin-Stickstoff aus der
Polypeptidkette der Globine geliefert. Die sechste Position kann durch
verschiedene Liganden besetzt werden und bestimmt das Absorptionsspektrum
des Hämoglobins. Im Oxihämoglobin z. B. ist die sechste Position mit
molekularem Sauerstoff besetzt. Neben den für die unterschiedlichen Liganden
1. Oxihämoglobin ____
2. Desoxihämoglobin ........
3. CO-Hämoglobin ------
Abbildung 6-1: Absorptionscharakteristika von Hämoglobin mit unterschied-
lichen Liganden.

Spektralphotometrie 6-3
charakteristischen Absorptionsbanden zwischen 500 und 600 nm besitzen alle
Hämoglobine eine gemeinsame Soret-Bande bei 415 - 430 nm.
Charakteristika der einzelnen Hämoglobinarten:
1. Cyan-Methämoglobin ([CN-Hb(III)]): Das Fe2+ wird mit Hexacyanoferrat(III)
zum Fe3+ oxidiert. Diese Form transportiert keinen Sauerstoff und besitzt eine
braune Farbe eine Absorptionsbande bei 541 nm und ist eine stabile
Verbindung
2. Oxihämoglobin ([O2-Hb(II)]): scharlachrot, zwei Absorptionsbanden im
gelbgrünen bei 576 - 540 nm
3. Desoxihämoglobin oder "reduziertes" Hb ([Hb(II)]): purpurrot, eine Ab-
sorptionsbande bei 555 nm
4. Kohlenmonoxid-Hämoglobin ([CO-HB(II)]): kirschrot, Absorptionsbanden bei
570 nm und 541 nm
Geräte: Glas-Chromatographiesäule, gefüllt mit Sephadex G25, grob Gummibällchen, Pasteurpipette Messpipetten 5 ml, 1 ml Reagenzglasständer mit Reagenzgläsern Spektralphotometer, Glasküvetten
Lösungen: Transformationslösung (=Drabkin'sche Lösung) : 1 g NaHCO3, 50 mg KCN und 200 g K3[FeCN6]/l Heparin-Gesamtblut wird als Hämolysat zur Verfügung gestellt. Kaliumhexacyanoferrat(III) 5% in Puffer Natriumdithionit 1% in Puffer Phosphat-Puffer 20 mM, pH 7,0
6-4 Blut
1) Gelfiltration
Prinzip: Gelfiltration trennt Substanzen nach Molekulargewicht. Für die Gelfiltration
werden quervernetzte Polymere als Gelfiltrations-Medien verwendet. Wegen eines
Ausschlußeffekts eluieren große Moleküle vor kleineren. In unserem Experiment
wird das Hämolysat mit Dithionit reduziert und das aus dem Methämoglobin
entstandene Desoxihämoglobin wird bei der Passage über die Säule durch die
Größenfraktionierung vom überschüssigen Dithionit abgetrennt. Das
Desoxihämoglobin wird durch Luftsauerstoff oxigeniert und als Oxihämoglobin
eluiert.
Ausführung: Eine vorbereitete 25 ml-Säule wurde an ihrem Ausfluss mit Glaswolle abgedichtet
und mit einer Suspension von gequollenem Sephadex G25 bis zu einer Höhe von
8 cm gefüllt. Überpüfen Sie, ob die gepackte Säule frei von Luftblasen ist!
Die Säule wird zuerst mit 10 ml Phosphat-Puffer gewaschen. Die Oberfläche der
Säulenfüllung sollte eben sein. Wenn der Pufferspiegel gerade bis auf die Höhe
des Säulenmaterials abgesunken ist, wird eine frisch hergestellte Mischung aus 0,2
ml Natriumdithionit-Lösung und 0,5 ml Hämolysat auf die Säule aufgetragen,
ohne Säulenmaterial aufzuwirbeln. Man lässt die Lösung in das Säulenmaterial
einsinken. Dann wäscht man zunächst mit 1 ml Puffer nach und füllt anschließend
die Säule mit Puffer (ebenfalls möglichst verwirbelungsfrei). Die rotgefärbte
Oxihämoglobin-Fraktion wird in einem Minimum Puffervolumen, ausreichend
für eine Küvettenfüllung, aufgefangen.
2) Spektralphotometrie des Hämoglobins Ausführung: Das Oxihämoglobin, Methämoglobin und das Desoxihämoglobin werden für
spektroskopische Messungen verwendet.
a) Für das Oxihämoglobin wird das Eluat der Gelfiltration verwendet
b) Methämoglobin: ca. 0,5 ml Hämolysat mit 1,5 ml Hexacyanoferrat
c) Desoxihämoglobin: 0,5 ml Hämolysat mit 1,5 ml Dithionit-Lösung
Die Farbe des Hexacyanoferrats und die der drei Formen des Hämoglobins sollten
beobachtet werden. Die drei Lösungen werden unter Anleitung in einem mit einem
Drucker verbundenen ULTROSPEC 1000 zwischen 600 bis 450 nm gegen Wasser

Spektralphotometrie 6-5
gemessen und die Ergebnisse in 5 nm Schritten ausgedruckt. Zeichnen Sie die
Spektren auf Millimeterpapier.
3) Bestimmung von Carboxihämoglobin
Ausführung: Zu 0,5 ml Hämolysat werden 1 ml Phosphat-Puffer und 1 ml Dithionitlösung
zugefügt. Dann mischen und innerhalb von 30 Sekunden in einer Küvette die
Extinktionen im Spektralphotometer bei 555 und 480 nm messen. 0,2 ml CO-
gesättigter Phosphatpuffer pH 7 werden zu der Probe gegeben. Sofort die Ex-
tinktionen bei 555 und 480 nm messen! Trotz der 200-fach höheren Affinität des
Kohlenmonoxids zum Hb sollte eine Äquilibrierung mit Luftsauerstoff vermieden
werden.
Charakteristisch für den CO-Gehalt im Hb ist das Extinktionsverhältnis E555/E480.
100 % Desoxi-Hb: E555/E480 = 3,15 ± 0,05
100 % CO-Hb: E555/E480 = 1,94 ± 0,05
Nichtraucher haben 0,21-2 %, Raucher 0,7 %-6,5 % CO-Hb.
4) Quantitative Bestimmung des Hämoglobins als Hämoglobin-Cyanid
Ausführung: In zwei Reagenzgläser legt man je 5 ml dest. Wasser vor. Mit einer 50 µl Spritze
werden 0,05 ml Blut dazugegeben und sorgfältig vermischt. Die Spritze sofort mit
dest. Wasser spülen. Danach gibt man 5 ml Transformations-Lösung dazu (gut
umschütteln). Die Extinktion der Doppelbestimmung wird nach genau 30 min
gegen den Leerwert (5 ml Wasser + 5 ml Transformationslösung) bei 546 nm
gemessen.
Auswertung: Berechnen Sie die Konzentration des Hämoglobins in g/100 ml Blut!
MG = 68.000 Da; Extinktionskoeffizient: ε = 44⋅103 l⋅mol-1⋅cm-1
6-6 Blut
II. Dialyse-Versuch
Prinzip: Niedermolekulare Stoffe lassen sich durch Diffusion durch eine semipermeable
Membran in Richtung des Konzentrationsgefälles von kolloidalen Stoffen trennen.
Die Kolloide können wegen ihrer Größe die Membranporen nicht passieren. Die
Dialysegeschwindigkeit ist abhängig von:
1) dem Verhältnis Membranoberfläche zum Flüssigkeitsvolumen
2) dem Konzentrationsgefälle der niedermolekularen Stoffe auf beiden Seiten der
Membran.
Die Dialyse findet nicht nur im biochemischen Laboratorium zur Entsalzung von
Proteinen Anwendung. Ihre klinische Bedeutung liegt in der Anwendung der
Dialyse in der künstlichen Niere.
Geräte: Rührmotor, 250 ml Becherglas, Magnetrührfisch Vollpipette 25 ml Pasteurpipette Bürette
Lösungen: Hg(NO3)2 (171,3 mg/l) 0,001 N Jod-Jodkaliumlösung 0,1 N Stärke-NaCl-Lösung:
200 ml Stärkelösung (0,5 %ig) + 50 ml 1 M NaCl (58,44 g NaCl/l)
Ausführung: 5 ml Stärke-NaCl-Lösung werden in einen zuvor in Wasser eingeweichten Dia-
lyseschlauch pipettiert. Die Enden des Schlauchs werden dabei mit Plastik-
klammern verschlossen. Danach wird der Schlauch insgesamt eine Stunde dialy-
siert. Als Dialysebad wird ein mit 250 ml dest. Wasser gefülltes Becherglas
verwendet. Mit einem Magnetrührer wird das Dialysat gerührt.
Nach 5, 10, 15, 20, 30, 45 und 60 Min. werden je 5 ml Dialysat entnommen und
die Cl–-Menge im Dialysat nach Zugabe von sechs Tropfen Diphenylcarbazon-

Glucosebestimmung 6-7
lösung durch Titration mit 0,001 N Hg(NO3)2-Lösung bestimmt. Die lachsrote
Farbe des Indikators schlägt in tiefviolett um.
Die mmol-Menge des Rest-Chlorids im Schlauch wird daraus berechnet. Nach
Beendigung der Entnahmen werden Schlauchinhalt bzw. 5 ml des Dialysats mit
zwei Tropfen Jod-Jodkaliumlösung versetzt. Beobachtung?
Zeichnen Sie ein Diagramm:
Abszisse: Zeit (Minuten), Ordinate: Restmenge Cl– im Schlauch in mmol.
III. Enzymatische Bestimmung der wahren Glucose
Eine schnelle, sichere und spezifische Bestimmungsmethode der Blutglucose im
Serum beruht auf folgender Reaktion: Die Glucoseoxidase (GOD), ein FAD-
Enzym aus Penicillium notatum, katalysiert die Oxidation von D-Glucose zum
Gluconolacton, das mit Wasser zu Gluconsäure hydrolysiert:
GOD Glucose + H2O + O2 → Gluconsäure + H2O2
Das dabei entstehende H2O2 wird in einer gekoppelten Indikatorreaktion weiter
umgesetzt, indem es durch Peroxidase (POD) katalytisch einen Wasserstoff-
donator wie z. B. das 2,2'-Azino-di-[3-ethyl-benzthiazolinsulfonsäure-6]-
diammoniumsalz oxidiert.
N
C2H5
N
S=N–N=
S3S4+NH O
–4
+NHSO 3–
C2H5
Ammoniumsalz der ABTS: 2,2'-Azino-di-[3-ethyl-benzthiazolinsulfonsäure-6]
6-8 Blut
Lösungen
Glucose-Standard: 100 mg Glucose in 100 ml H2O, bereitgestellt wird die Mischung von
1 ml Glucose Standard + 10 ml Perchlorsäure
Glucose-Reagenz: 2 mg Peroxidase, 18 mg Glucose-Oxidase und 100 mg 2,2'-Azino-di-
[3-ethyl-benzthiazolinsulfonsäure-6]-diammoniumsalz (H2-Donator)
in 100 ml 0,1 M Phosphatpuffer (pH 7).
Perchlorsäure: 0,33 M, 47 g 70 %ige HClO4 in 1000 ml H2O
Durchführung:
1. Die Enteiweißung wurde vom Assistenten wie folgt vorbereitet:
1 ml Blut wurde mit 0,1 ml Citratlösung versetzt, um die Gerinnung zu
verhindern. Danach wurden 10 ml 0,33 M Perchlorsäure zugesetzt, die Lösung in
ein Zentrifugenglas umgefüllt und 20 Minuten zentrifugiert. Der Überstand wurde
vorsichtig in ein Reagenzglas überführt.
Das Reagenzglas mit dem Überstand steht dann für Sie am Arbeitsplatz bereit!
2. Bereiten Sie folgende Ansätze in fünf Reagenzgläsern vor:
1 Reagenzien-Leerwert 0,1 ml Wasser
2 Serumproben je 0,1 ml Zentrifugenüberstand
2 Glucose-Standardwerte je 0,1 ml Glucose-Standardlösung
je 5 ml Glucose-Reagenz dazugeben, sofort schütteln! Die Proben werden
35 Minuten bei Raumtemperatur stehengelassen und dann die Extinktionen im
Photometer bei 436 nm gegen den Reagenzien-Leerwert gemessen, d.h. Sie stellen
zuerst die Extinktion des Leerwerts auf 0,000 und messen dann Ihre Proben.
Wenden Sie zur Ergebnisberechnung den Dreisatz an und berücksichtigen Sie
dabei, dass der Glucosestandard (100 mg/100 ml), wie das Blut, mit dem gleichen
Volumen Perchlorsäure versetzt wurde. Geben Sie das Ergebnis in mg
Glucose/100 ml Blut und mmol Glucose/l Blut an.

Polarimetrie 6-9
Berücksichtigen Sie dabei, dass das Blut vor der Enteiweißung durch Zugabe von
Citratlösung ungerinnbar gemacht wurde (Verdünnungsfaktor!). Vergleichen Sie
das Ergebnis mit den Normalwerten.
IV. Polarimetrie
Prinzip: Von vielen biochemisch wichtigen Verbindungen (z.B. Aminosäuren, Zucker) gibt
es theoretisch zwei räumlich nicht identische, stereoisomere Formen, die sich wie
Bild und Spiegelbild verhalten (vgl. rechter und linker Schuh). Grund hierfür ist in
der Regel ein im Molekül vorhandenes Kohlenstoffatom mit vier
unterschiedlichen Substituenten, das dem Molekül eine Asymmetrie verleiht. In
der Natur kommt aber nie das Gemisch beider Formen, das sogenannte Racemat
vor, sondern immer nur eine der beiden Formen (Enantiomere), z. B. die L-
Aminosäuren oder D-Glukose. Die Bezeichnungen D-, L- (nach Emil Fischer),
beziehen sich nur auf die sterische Konfiguration der Moleküle und keinesfalls auf
optische Eigenschaften; denn eine solche asymmetrische Verbindung kann die
Ebene des polarisierten Lichtes nach rechts oder links drehen. Eine Substanz, die
das planpolarisierte Licht im Uhrzeigersinn dreht, nennt man rechtsdrehend (+).
Linksdrehende Substanzen (-) drehen die Ebene nach links.
Die Größe und Richtung der Drehung sind spezifisch für jede Verbindung. Der
spezifische Drehwinkel einer Substanz [α]D20 errechnet man aus derm
gemessenen Drehwinkel α, der Schichtdicke l (in Dezimetern!) und der
Konzentration c (in g/100 ml Lösung!):
[α]D20 =
α 100
l c
••
grad 100
dm g / 100ml
••
6-10 Blut
planpolarisiertes Licht
optisch aktive Substanz
Ebene des planpolarisiertes Lichts ist gedreht
Abbildung 6-2: Schematische Veranschaulichung der Drehung der Ebene des
planpolarisierten Lichtes durch eine optisch aktive Substanz.
Der spezifische Drehwinkel ist von der Wellenlänge, der Temperatur und dem
Lösemittel abhängig. Diese Parameter müssen immer mit angegeben werden. Als
Lichtquelle wird die D-Linie einer Natriumspektrallampe (589 nm) benutzt. Der
Drehwinkel wird mit Hilfe des Polarimeters bestimmt. Das Nicolsche Prisma
besteht aus zwei in einer bestimmten Ebene geschnittenen und durch
Kanadabalsam verkitteten Flussspatkristallen, die nur den ausserordentlichen
Strahl als planpolarisiertes Licht durchlassen, während der ordentliche Strahl
durch Totalreflexion an der Kittschicht ausgelöscht wird. Nach Durchgang des auf
dieser Weise am Polarisator-Prisma planpolarisierten Lichts durch die optisch
aktive Substanz in der Küvette trifft es auf das Analysator-Prisma, das nun gedreht
werden muss, um im Einstellfeld des Okulars wieder gleichen Lichtdurchgang wie
bei der Nulleinstellung und Eichung des Polarimeters gegen das reine
Lösungsmittel zu bekommen, also die gleiche Helligkeit in den beiden äußeren
und dem mittleren Feld. Den Drehwinkel und die Richtung liest man an der
graduierten Drehscheibe ab, die mit dem Analysator gekoppelt ist.

Polarimetrie 6-11
1 2 3 4 5 6
Abbildung 6-3: Schematische Zeichnung eines Polarimeters
1) Lichtquelle mit Farbfilter (Natriumdampflampe/D-Linie)
2) Polarisator (Nicolsches Prisma)
3) Küvette mit Lösung der optisch aktiven Substanz
4) Analysator, versehen mit graduierter Drehscheibenskala
5) Okular
6) Beobachterauge
Ausführung: Die spezifischen Drehwinkel der beiden Testlösungen (Glukose und Fruktose) und
der Analysenlösung sollen ermittelt werden. Eine Substanz oder Lösung ist
rechtsdrehend, wenn die beiden Außenfelder des dreigeteilten Einstellfeldes
dunkler als das mittlere Feld sind. In diesem Fall dreht man die Skala so, dass sie
bei der Einstellung auf eine gleiche Helligkeitsstufe nach links wandert. Bei
Linksdrehung erscheint das mittlere Feld dunkler. Hier erfolgt beim Abgleich der
Helligkeit eine Verschiebung der Skala nach rechts.
a) Die Nulleinstellung (gleiche Lichtstärke der drei Felder) wird am Polarimeter
an der Analysator-Skala geprüft.
b) Das Polarimeterrohr wird mit der zu bestimmenden Lösung luftblasenfrei
gefüllt. Ein luftblasenfreies Rohr bekommt man am einfachsten dann, wenn
sich der Flüssigkeitsspiegel der Lösung etwas über dem plangeschliffenen
Ende des Rohres wölbt. Nun wird das Deckglas aufgezogen und das Rohr
verschlossen. Es ist darauf zu achten, dass das Polarimeterrohr sauber und
trocken in das Gerät eingelegt wird!
c) Durch Drehen des Analysators in der oben angegebenen Richtung wird das
Hell-Dunkel-Feld auf die gleiche Helligkeitsstufe gebracht und an der Skala
6-12 Blut
mit Hilfe des Nonius der Drehwinkel auf 0,05 Grad genau abgelesen. Aus dem
gemessenen Wert wird der spezifische Drehwinkel berechnet.
rechts links
Abbildung 6-4: Im Okular sichtbare Einstell- und Ablesefelder
Analyse: Berechnen Sie mit Hilfe der Formel auf Seite 6-9 den spezifischen Drehwinkel
und geben Sie im Vergleich mit den in der Tabelle 6-1 angegeben Werten an, um
welchen Zucker es sich handelt.
Die Konzentrationen der Analysenlösungen betragen 9 g/100 ml.
Tabelle 6-1
Xylose Glucose Fructose Saccharose
Spez. Drehwinkel
[α]D20
+18,7 +52,8 -92 +66,5

Aufgaben 6-13
Übungsaufgaben: 1. Wie beeinflusst 2,3-Diphosphoglycerat die Sauerstoff-Affinität des Hämo-
globins?
2. Welche Funktion erfüllt Glutathion im Erythrocyten?
3. Zeichnen Sie die O2-Sättigungskurve von Hämoglobin in Abhängigkeit von
pO2 und ihre Veränderung durch 2,3-Bisphosphoglycerat.
4. a)Welches Endprodukt des Porphyrinabbaus wird in der Leber gebildet? b)Wie
verändert sich der Serumgehalt dieser Verbindung bei:
A) einem Gallengangsverschluss
B) bei massiver Hämolyse?
5. Warum entstehen Ödeme bei längerem Hungern?
6. Welche Porengrößen einer Membran wären für die Dialyse eines Nieren-
kranken geeignet: 500 Da, 50.000 Da oder 500.000 Da ?
7. In welcher Reihenfolge eluieren Carboanhydrase (29000 Da), Lactalbumin
(14200 Da), Phosphorylase B (97400 Da), Albumin aus Ei (45000 Da) und
Serumalbumin (67000 Da) von einer Gelfiltrationssäule?
8. Die Glukosekonzentration im Serum eines Patienten beträgt 6 mmol/l.
Berechnen Sie die Konzentration in mg/100 ml.

7-1
Versuch 7: Nukleinsäuren I
Versuche:
Löslichkeit der Harnsäure
Xanthinoxidase-Reaktion und ihre kompetitive Hemmung
Bestimmung der Harnsäure mit Hilfe der UV-Spektroskopie
Restriktionsendonukleasen
Polymerasekettenreaktion (PCR)
Analyse:
Hypoxanthin in 50 mM Phosphatpuffer pH 7,5
Wissensgebiete
RNA- und DNA-Struktur
Biosynthese der Pyrimidin- und Purinbasen, Salvage Pathway
Purin-Abbau, Harnsäure, Xanthinoxidase, Uricase, kompetitive Hemmung, Gicht
Stickstoffbilanz
Restriktionsendonukleasen, rekombinante DNA
PCR
7-2 Nucleinsäuren I
Grundbestandteile der Nukleinsäuren sind die Nukleotide, die wiederum aus
folgenden Bestandteilen aufgebaut sind:
a) Pentose b) Phosphat
c) N-haltige heterozyklische Basen:
Purinreihe: Adenin, Guanin
Pyrimidinreihe: Cytosin, Uracil (RNA), Thymin (DNA).
N
NHO
Ribose
O
P O
OH
-O
OH
N
N N
N
NH2
Ribose
O
P
OH
O-O
N
NHO
Ribose
O
P O
OH
-O
NH2
N
N N
N
OH
Ribose
O
P
OH
O-O
H2N
GMPCMPAMPUMP
Die Nukleotide sind in DNA und RNA durch Phosphorsäurediesterbindungen
verknüpft. Die geringe Löslichkeit von Nukleinsäuren in Wasser-Alkohol-Ge-
mischen bzw. ihre Löslichkeit in konzentrierter Kochsalzlösung nutzt man zu ihrer
Isolierung.
Während die Endprodukte des Pyrimidinstoffwechsels, β-Alanin und β-Ami-
noisobutyrat, Aminosäuren sind und verstoffwechselt werden können, ist Harn-
säure beim Menschen und den anderen Primaten das Endprodukt des Purin-
stoffwechsels (also auch des mit der Nahrung als Nukleinsäuren aufgenommenen
Purins). Harnsäure entsteht durch schrittweise Oxidation der Purinbasen
Hypoxanthin und Xanthin mit molekularem Sauerstoff unter der Wirkung von
Xanthinoxidase.
Harnsäure und Mononatriumurat sind schwer löslich. Bei Stoffwechselstörungen
kann diese geringe Löslichkeit zu Ablagerungen (Gicht, Nierensteine) führen.

Harnsäure 7-3
Interessant ist unter diesem Gesichtspunkt die pH-Abhängigkeit der
Harnsäurelöslichkeit (Abb. 7-1 Schema nach E. Aebi). Bei Vermeidung von
Fleischkost und Verwendung von Pflanzen-, sog. Basenkost, wird weniger
Harnsäure gebildet, und außerdem verschiebt sich der pH-Wert des Urins nach
rechts, es kann mehr Harnsäure ausgeschieden werden. Zur Therapie der Gicht
wird u.a. Allopurinol (1H-Pyrazolo-(3,4-d)-pyrimidin-4-ol), ein kompetitiver
Hemmstoff der Xanthinoxidase, angewandt.
Abbildung 7-1: Abhängigkeit der Harnsäurelöslichkeit vom pH-Wert
Der Gehalt des Serums an Harnsäure ist für die Diagnose und Überwachung der
Hyperurikämie von Bedeutung. Bei den meisten Säugetieren (nicht beim Men-
schen!) wird die Harnsäure durch das Enzym Uricase unter gleichzeitiger Bildung
von Wasserstoffperoxid weiter zu Allantoin abgebaut: Harnsäure-Normalwerte beim Erwachsenen: Urin: 500 - 580 mg/Tag 3000 - 3500 µmol/d Serum: 3,3 - 7,6 mg/100 ml (Mann) 200 - 450 µmol/l 1,7 - 6,2 mg/100 ml (Frau) 100 - 360 µmol/l
7-4 Nucleinsäuren I
I. Harnsäure Geräte: Messpipetten 1 ml, 5 ml Plastikküvetten 2 × 1 cm Spektralphotometer
Lösungen: Natronlauge, HCl 2N Na2CO3-Lösung 2N Natrium-Allopurinol -Lsg. 1,5 mM Boratpuffer 0,1 M; pH 9,5 Serum Substratpuffer-Lösung: 50 mM Phosphatpuffer pH 7,5; 100 µM EDTA Hypoxanthin (ist 10 min. mit Sauerstoff durchperlt) Uricaselösung: 1,8 kU/l Xanthinoxidaselösung: 1 g/l 1) Löslichkeit von Harnsäure
Die Harnsäure ist in Wasser fast unlöslich, in Na2CO3-Lösung gering löslich als
Mononatriumsalz, und in NaOH-Lösung gut löslich bei Erwärmen unter Bildung
des Dinatriumsalzes.
Ausführung: Je ca. 3 ml dest. Wasser, Na2CO3-Lösung und Natronlauge werden mit einer
Spatelspitze Harnsäure versetzt. Während sich Harnsäure in Lauge in der Hitze
(Wasserbad, Schutzbrille) unter Bildung von Dinatriumurat löst, ist dies in dest.
HN
N NH
N
O
HN
NH NH
N
O
O
HN
NH NH
NH
O
O
HN
N NN
O
H
O
1 und 2: Xanthinoxidase
21
Allopurinol
Hypoxanthin HarnsäureXanthin

Harnsäure 7-5
Wasser und Na2CO3-Lösung nicht der Fall. Die Dinatrium-Urat-Lösung wird anschließend mit wenigen Tropfen HCl versetzt. Die Harnsäure fällt als weißer, flockiger Niederschlag aus. 2) Xanthinoxidase-Reaktion und ihre kompetitive Hemmung
Prinzip: Die Oxidation von Hypoxanthin zu Xanthin und weiter zu Harnsäure durch die
Xanthinoxidase in Gegenwart von Sauerstoff wird spektrophotometrisch bei
297 nm verfolgt. Urate haben in alkalischer Lösung einen spezifischen Extink-
tionskoeffizienten von ε297 = 11,8 l/mmol.cm. Aus der Differenz der gemessenen
Endextinktion zur Anfangsextinktion lässt sich die Hypoxanthinmenge berechnen.
4030201000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Extin
ktio
n
1
2
Zeit [min] Abbildung 7-2: 1) Bildung von Harnsäure durch Oxidation von Hypo-xanthin
mit Xanthinoxidase 2) Hemmung der Xanthinoxidase mit Allopurinol
Ausführung: 3 ml einer sauerstoffgesättigten Hypoxanthinlösung werden in eine 1 cm-Plas-
tikküvette pipettiert. Die Extinktion wird zweimal gegen dest. Wasser bei 297 nm
gemessen. Durch Zugabe von 100 µl Xanthinoxidase startet man die Reaktion. In
7-6 Nucleinsäuren I
den ersten 5 Minuten wird die Extinktion wird alle 30 s gemessen und
anschließend noch nach 10, 15 und 20 Minuten.
Der Versuch wird wiederholt, jedoch werden vor dem Start mit Xanthinoxidase
100 µl Allopurinol-Natrium-Lösung in die Küvette pipettiert und gut gemischt
(Abb. 7-2, Kurve 2).
Auswertung: Die gemessenen Extinktionen werden in einem Diagramm gegen die Zeit aufge-
tragen. Aus den Zeitumsatzkurven für Xanthinoxidase werden die µmol Hypo-
xanthin/l Lösung, die Enzymkonzentration (U/l) und der Hemmungsgrad (%)
berechnet.
3) Bestimmung der Harnsäure mit Hilfe des UV-Tests
Prinzip: Die Umwandlung von Harnsäure zu Allantoin durch Uricase wird spektral-
photometrisch verfolgt.
HN
NH NH
NH
O
O
ONH2
NH
O
NH
NH
HO
O
AllantoinHarnsäure
Uricase
Ausführung: 200 µl Serum und 3 ml 0,1 M Boratpuffer pH 9,5 in einer Plastikküvette werden
bei 297 nm gegen dest. Wasser gemessen. Danach werden 200 µl Uricaselösung
eingerührt und der Extinktionsabfall verfolgt. Messwerte werden in Abständen
von 30 Sekunden bis zur Konstanz genommen. Aus der Differenz der
Anfangsextinktion und des konstanten Endwertes wird die Harnsäurekon-
zentration in µmol/l Serum berechnet.

Restriktionsendonukleasen 7-7
II. Restriktionsendonukleasen: Charakterisierung eines Plasmids
Restriktionsendonukleasen (RE) kommen in fast jedem Mikroorganismus vor und
spalten doppelsträngige DNA unter Bildung von Restriktionsfragmenten. Ein
DNA-Strang wird von einem RE spezifisch gespalten und liefert reproduzierbare
DNA-Fragmente. Ein Genom kann so in kleine DNA-Stücke, von denen jedes ein
ganzes Gen oder Teile davon enthält, zerlegt werden. Sie können isoliert, kloniert
und in ihrer Nukleotidsequenz analysiert werden.
Die Bakterien-DNA wird durch Methylierung von Basen vor dem Angriff durch
die eigene RE geschützt. Fremd-DNA, die durch Infektion, Konjugation oder
Transformation in die Zelle eindringt, wird abgebaut, falls sie nicht durch
Methylierung (katalysiert durch Methylasen mit S-Adenosylmethionin als Me-
thyldonator) modifiziert wird. Typ I-Endonukleasen, die an der Wirts-Restriktion
und –Modifikation beteiligt sind, sind große multimere Enzymkomplexe, die DNA
unter ATP-Verbrauch spalten und methylieren. Vom Typ II, kleinen monomeren
Proteinen, die Mg2+ für die enzymatische Aktivität benötigen, sind mehr als
250 RE bekannt. Sie erkennen spezifische Nukleotidsequenzen von 4 bis
8 Basenpaaren, die palindromartig angeordnet sind. Die Nomenklatur der
Enzymbezeichnungen sind aus dem Anfangsbuchstaben des Genus und den ersten
zwei oder drei Buchstaben der Spezies zusammengesetzt, aus der sie isoliert
wurden. Die römische Zahl deutet die Reihenfolge der Entdeckung des Enzyms in
diesem Stamm an:
EcoRI: Escherichia coli Stamm R: Palindrom aus Hexanukleotid
5' GAATTC 3' Dyaden-Symmetrie 3' CTTAAG 5' (zweifache Rotationssymmetrie)
Es entstehen 3'-überstehende Enden an der Schnittstelle, (sticky ends).
HinfI: Haemophilus influenzae Stamm f: Pentanukleotid
5' GANTC 3' CTNAG (sticky ends)
HaeIII: Haemophilus aegypticus: Tetranukleotid
5' GGCC liefert glatte Enden (blunt ends) 3' CCGG
7-8 Nucleinsäuren I
Statistisch gesehen treten diese Palindrome mit einer Häufigkeit von 1:44, 1:45,
1:46, d. h. in einem von 256, 1024 bzw. 4096 Fällen.
1:4N N = Länge der Restriktionssequenz
Medizinische Bedeutung: Jedes Gen kann in charakteristische Restriktions-
fragmente gespalten werden, wobei ein spezifisches Restriktionsmuster entsteht.
Mutationen des Gens sind am Verschwinden oder der Längenveränderung eines
oder mehrerer Fragmente, dem Restriktionsfragment-Längenpolymorphismus
(RFLP) erkennbar. Die Analyse der genomischen DNA von Patienten oder auch
des Föten in der pränatalen Diagnostik ist eine wichtige Methode bei der
Diagnostik von Erbkrankheiten.
Man geht dabei folgendermaßen vor: Aus Blut oder beliebigen anderen Kör-
perzellen wird DNA isoliert. Aliquots werden mit verschiedenen Restriktions-
enzymen geschnitten und die entstandenen Fragmente gelelektrophoretisch
aufgetrennt. Als Vergleich dient eine analog behandelte Probe von DNA eines
gesunden Individuums. Die Fragmente werden aus dem Gel auf eine Nylon-
membran übertragen (Southern blotting). Sie wird dabei denaturiert, so dass sie
einzelsträngig vorliegt. Die Nylonmembran wird mit einem ebenfalls einzel-
strängigen, radioaktiv markierten DNA-Fragment inkubiert, das zu einer Region
des Gens komplementär ist und daher mit diesem Bereich paart (hybridisiert), und
so die zu dem untersuchten Gen gehörigen Fragmente radioaktiv markiert. Auf
einem Röntgenfilms werden sie durch die Autoradiographie sichtbar gemacht.
DNA von verschiedenen Patienten werden auf strukturelle Änderungen des Gens
hin verglichen.
Im folgenden Versuch werden Restriktionsenzyme zur Analyse der DNA eines
Plasmids eingesetzt, das als Insert die cDNA des menschlichen Apolipoproteins
AI trägt. Apo AI (243 AS) aktiviert die Serum-Lecithin-Cholesterin-
Acyltransferase (LCAT) und ist ein Hauptstrukturprotein des Serum-HDLs, das
für den reversen Cholesterin-Transports aus den peripheren Geweben hin zur
Leber verantwortlich ist.

Restriktionsendonukleasen 7-9
Tabelle 7-1: Restriktionsenzyme und Fragmentlängen in Basenpaaren (bp)
Enzym Fragmentlänge (bp)
HindIII 905 5016 EcoRI 246 892 2130 2650
Abbildung 7-3 : pSV2 cat AI - Plasmid
Der menschliche Apo AI-cDNA-Klon wurde aus einer menschlichen Leber-
cDNA-Bank isoliert. Der Klon kodiert 18 N-terminale Aminosäurereste der
Signal- (Prä-) Sequenz, gefolgt von sechs Resten der Pro-Sequenz und 243
Aminosäureresten des reifen Apo AI. Die an beiden Enden mit der EcoRI-Re-
striktionssequenz versehene cDNA wurde über Adaptoren an beiden Seiten in die
HindIII-Restriktionsstelle des Plasmis pSV2 cat kloniert, wie in der Abb. 7-3
schematisch angedeutet.
7-10 Nucleinsäuren I
Nach der Isolierung des pSV2 cat-AI-Plasmids kann das Plasmid durch die
kombinierte Anwendung der Restriktionsenzyme EcoRI und HindIII charakte-
risiert werden. Es sollten die DNA-Fragmente entstehen, die in der Tabelle 7-1
aufgelistet sind. Sie werden in einem 1-%igen Agarosegel getrennt und durch
Ethidiumbromid (Hautkontakt vermeiden!), das mit DNA fluoreszierende
Komplexe bildet, in UV-Licht sichtbar gemacht. Die Fragmentgrößen werden durch Eichung mit DNA-Längenmarkern ermittelt. Die Rf-Werte der Marker-
banden werden gegen den dekadischen Logarithmus der Basenpaarzahl aufge-
tragen und die Größe der aus dem Klon entstandenen Fragmente mit Hilfe der
Eichgeraden bestimmt.
M H E
---
---
-
-
543
21,6
1
0,5
0,2
kb
Abbildung 7-4 : Agarosegelelektophorese des mit Hilfe von HindIII oder EcoRI verdautem pSV2 cat AI - Plasmids. M: Längenmarker, H: HindIII-Fragmente, E: EcoRI-Fragmente

Restriktionsendonukleasen 7-11
Geräte: Agarosegel, 1 % mit 0,5 µg Ethidiumbromid/ml Elektrophorese-Puffer: Tris-Borat pH 8,5 Flachbettelektrophoresekammer mit Spannungsgerät Saranfolie UV-Lampe
Lösungen: 10 × Restriktionspuffer: Indikator-Probenpuffer: 0,25 % Bromphenolblau, 20 % Ficoll 400 Restriktionsenzyme: EcoRI und HindIII Marker-DNA
Ausführung (Demonstration vom Assistenten): Die Enzyme sind stets im Eisbad aufzubewahren. Lösungen oder Geräte, die mit
DNA in Kontakt kommen, sollten nicht mit den Fingern berührt werden. In ein 1,5
ml Eppendorf-Röhrchen werden folgende Komponenten pipettiert:
2 µl 10 x Restriktionspuffer 14 µl H2O 2 µl DNA pSV2 cat AI-Lösung 2 µl EcoRI bzw. HindIII enthaltenden Enzymlösung,
gut gemischt, und eine Stunde lang bei 37 °C inkubiert. 5 µl Indikator-Proben-
puffer werden zugefügt und der Ansatz in die Probentasche des Agarose-Gels
vorsichtig mit einer Eppendorf-Pipette eingefüllt. Ein Gemisch von 10 µl H2O,
5 µl Indikator-Probenpuffer und 5 µl Längenmarker wird in die Nachbartasche
aufgetragen. Der Puffer in der Kammer schließt gerade mit der Oberkante des
Agarosegels ab. Die Elektrophorese wird bei 90 V durchgeführt, bis die
Bromphenolblaubande etwa 1 cm vom kathodischen Ende des Gels entfernt
gewandert ist. Das Spannungsgerät wird ausgeschaltet und das Gel, eingewickelt
in Saranfolie, unter dem UV-Licht betrachtet.
Mit Hilfe eines Videoaufzeichnungsgerät (siehe Versuch 2) wird das Gel do-
kumentiert. Die Abstände der fluoreszierenden Banden zum Start werden ge-
messen und die Größe der DNA Fragmente durch eine halblogarithmische
Auftragung bestimmt.
7-12 Nucleinsäuren I
III. PCR Kary B. Mullis, Nobelpreisvortrag 1993:
„And again, EUREKA!!!! I could do it over and over again. Every time I did it I
would double the signal. For those of you who got lost, we're back! I stopped the
car at mile marker 46,7 on Highway 128. In the glove compartment I found some
paper and a pen. I confirmed that two to the tenth power was about a thousand and
that two to the twentieth power was about a million, and that two to the thirtieth
power was around a billion, close to the number of base pairs in the human
genome. Once I had cycled this reaction thirty times I would be able to the
sequence of a sample with an immense signal and almost no background“
Die Polymerasekettenreaktion („polymerase chain reaction“) ist eine einfache Me-
thode zur Vervielfältigung von DNA in vitro und ist heute im molekularbiolo-
gischen Labor unverzichtbar geworden. Mit Hilfe einer vorgeschalteten Reversen-
Transkriptase-Reaktion ist auch die Vervielfältigung von RNA möglich (RT-
PCR). Die Anwendung der PCR in der Medizin ist inzwischen Routine bei der
Diagnostik von Infektionskrankheiten (Bakterien, Viren, Einzeller, Pilze), von
genetisch bedingten Erkrankungen und in der Forensik.
Die PCR ist eine DNA-Replikation im Reagenzglas, bei der eine DNA-abhängige
DNA-Polymerase eingesetzt wird. Als Matrize dient dabei jeweils einzelsträngige
DNA, die durch thermische Denaturierung doppelsträngiger DNA gewonnen wird.
Das Besondere dieser Methode ist der Einsatz einer thermostabilen Polymerase,
die einen raschen zyklischen Reaktionsablauf ermöglicht, durch den sich der
DNA-Gehalt pro Zyklus verdoppelt. Es ist also theoretisch möglich, aus einem
einzigen DNA-Fragment in 35 Zyklen 235
= 3,4x1010
Moleküle (34 Milliarden!)
zu generieren. Damit ist die Möglichkeit gegeben, einzelne Moleküle
nachzuweisen.
Welches Stück DNA amplifiziert werden soll, hängt von der Auswahl der Primer
ab, die die 5’- und 3’-Enden der amplifizierten DNA bilden. Zumindest die in den
Primer enthaltene Sequenz der zu amplifizierenden DNA (RNA) muss daher
bekannt sein. In späteren Zyklen entstehen nur noch Fragmente mit der genau
gewünschten Länge, die von den 5’-Enden der Primer festgelegt wird. Als Primer
werden chemisch synthetisierte, einzelstängige Oligonukleotide eingesetzt, die
eine Länge von ca. 20 Basen haben.

PCR 7-13
5'3'
5'
5'
5'
3'
3'
3'
3'
5'
5'
3'
3'5'
5'
5'
5'3'
3'
3'5'
5'
5'
5'
3'
3'
3'
3'5'
5'3'
3'
5'
5'3'
3'
1. Denaturierung2. Anlagerung der Primer
3. Kettenverlängerung
Wiederholung von 1. und 2.
Wiederholung von 3.
Wiederholung von 1. bis 3.
vielfacheWiederholung von 1. bis 3.
Bereich der Zielsequenz
PCR-Produkt, 2N Moleküle3'5'
1. Zyklus 21 = 2
2. Zyklus 22 = 4
3. Zyklus 23 = 8
N. Zyklus 2N=X
Abb. 7-5: PCR-Schema
7-14 Nucleinsäuren I
Im Einzelnen läuft eine PCR wie folgt ab (Abb.7-5): Die Matrizen-DNA
(template) wird zusammen mit den spezifischen Primern, dem Substrat, den
Desoxinukleotiden (dNTPs), der thermostabilen Polymerase (z.B. aus Thermus
aquaticus) und einem Mg2+
-Ionen enthaltenden Puffer in ein Eppendorfgefäß
gegeben. Der PCR-Ansatz wird dann in einem sogenannten Thermocycler einem
Programm mit einer Wiederholung von spezifischen Temperaturen für
Denaturierung, Anlagerung der Primer und Kettenverlängerung unterworfen. Die
Denaturierung findet bei 95°C statt, einer Temperatur, bei der die Polymerase eine
für die Versuchsdauer ausreichende Stabilität aufweist. Eine Dauer von 30-60 s ist
normalerweise ausreichend; nur die erstmalige Denaturierung zu Beginn der
Reaktion sollte 5 min betragen.
Zur Anlagerung der Primer (Annealing) wird die Temperatur soweit abgesenkt,
dass sich nur spezifische Primer mit der komplementären Sequenz anlagern
können. Die Temperatur sollte 5°C unter der sogenannten Schmelztemperatur
liegen, die man näherungsweise mit Hilfe der Formel
Tm=(ΣA+ΣT)x2°C+(ΣG+ΣC)x4°C berechnen kann. Auch hier sind 30-60 s
ausreichend.
Die Kettenverlängerung findet dann im Temperaturoptimum der Polymerase bei
72°C statt. Die Dauer dieses Schrittes hängt von der Länge des zu amplifi-
zierenden DNA Fragments ab. Als Faustregel gilt: 1 min pro 1000 Basenpaaren
(bp). Der finale Kettenverlängerungsschritt wird üblicherweise auf 5-10 min
ausgedehnt, um zu gewährleisten, dass auch alle DNA-Moleküle, die dann in
maximaler Menge vorliegen, vollständig verlängert werden.
Die Anzahl der Zyklen richtet sich nach der vorhandenen Menge der zu ampli-
fizierenden DNA und beträgt normalerweise 25-40 Zyklen. Die Länge der
Fragmente ist limitiert. Generell gilt: Je kürzer die Fragmente, umso unkompli-
zierter die PCR Reaktion. Bis zu einer Länge von ca. 3000 bp ist in den meisten
Fällen die oben beschriebene Standardmethode erfolgreich, Fragmente bis zu
10.000 bp können durch die Anwendung spezieller Polymerasen und die Opti-
mierung der spezifischen Reaktionsbedingungen hergestellt werden. Auch die
Herstellung von Fragmenten >10.000 bp ist beschrieben worden.
DNA, die in der PCR eingesetzt wird, kann aus den unterschiedlichsten Quellen
stammen. Es werden nur geringste Mengen an Gewebe, Blut, Speichel oder
Sperma benötigt, um genomische DNA zu amplifizieren. DNA von Erregern kann

Aufgaben 7-15 auch noch in einem großen Hintergrund von Fremd-DNA nachgewiesen werden.
Im Extremfall ist dadurch ein einziger Erreger unter 106 eukaryotischen Zellen
nachweisbar. Der Nachweis der DNA erfolgt normalerweise durch Elektrophorese
in Ethidiumbromid-gefärbten Agarosegelen.
Im vorliegenden Versuch sollen Sie mit Hilfe der PCR Ihr Geschlecht bestimmen.
Dazu soll ein repetitives DNA-Fragment amplifiziert werden, das auf dem Y-
Chromosom liegt und daher nur in „männlicher“ DNA und nicht in „weiblicher“
DNA nachgewiesen werden kann. Da ein repetitives DNA-Fragment (= in
mehreren Kopien vorliegend) amplifiziert wird, reichen 25 Zyklen aus, um
genügend DNA für eine Analyse mittels Agarosegel zu erhalten.
Geräte : Thermocycler BIOZYM PTC 200 Agarosegel-Flachbettkammer, Spannungsgerät Lösungen: PBS 0,1 MNaOH PCR-Mix Endkonzentration: 50 mM KCl; 10mM Tris, pH 8,3; 1,5 mM MgCl2; 0,2 mM dNTP’s; 0,2 µM Primer Y1/Y2; 1 U Taq-Polymerase pro Ansatz. Primer Y1: 5’ TCCACTTTATTCCAGGCCTGTCC 3’ Primer Y2: 5’ TTGAATGGAATGGGAACGAATGG 3’
1) Schnellpräparation von DNA aus Zellen der Mundschleimhaut
• Spülen Sie Ihren Mund mit Leitungswasser aus und verwerfen das Wasser.
• Spülen Sie anschließend den Mund sehr sorgfältig für 10s mit 10ml phos-phatgepufferter physiologischer Kochsalzlösung (PBS) und sammeln die Probe in einem 15ml Zentrifugenröhrchen.
• Zentrifugieren Sie die Zellen 10min. bei 2000 U/min. ab. Ein Zellpellet sollte eindeutig erkennbar sein.
• Gießen Sie den Überstand ab und resuspendieren Sie das Zellpellet mit ca. 10ml PBS.
• Zentrifugieren Sie erneut 10min bei 2000 U/min. Gießen Sie den Überstand ab und ziehen Sie den verbleibenden Überstand mit der Pipette ab.
• Resuspendieren Sie die Zellen mit 100µl 0,1M NaOH und überführen die Suspension in ein 1,5ml Eppendorfgefäß. Verschließen Sie das Gefäß und inkubieren Sie 5min bei 100°C im Heizblock. Lassen Sie das Gefäß kurz
7-16 Nucleinsäuren I
abkühlen und geben Sie 400 µl dest. Wasser dazu. (Vorsicht beim Öffnen, Schutzbrille tragen!)
• Zentrifugieren Sie für 2min in einer Mikrozentrifuge bei voller Geschwin-digkeit.
• Verwenden Sie den Überstand für die PCR.
2) PCR
Pipettieren Sie 20µl des PCR-Mix auf den Boden eines 250µl-PCR-Eppendorf-
gefäßes. (Vorsicht: Das Gefäß ist sehr dünnwandig und leicht zu beschädigen!).
Geben Sie dann 5µl der DNA-Lösung so zu den 20µl PCR-Mix, dass eine ein-
heitliche Lösung entsteht und verschließen Sie vorsichtig das Gefäß (evtl. kurz
zentrifugieren).
Stellen Sie das Gefäß in den Thermocycler. Das Programm wird dann vom
Assistenten gestartet.
Programm (Dauer ca. 70min.):
Start 5min 95 °C
30s 95 °C
25 Zyklen 30s 58 °C
30s 72 °C
Auffüllen 2min 72 °C
3) Agarose-Gelelektrophorese
Die Ansätze werden nach Beendigung der PCR mit 5µl Beladungspuffer ver-
mischt und mit Unterstützung des Assistenten auf ein mit Ethidiumbromid ver-
setztes, 2%iges Agarosegel aufgetragen. Zum Vergleich wird eine positiver und
ein negativer PCR-Ansatz mit aufgetragen. Die Elektrophorese wird bei 90 V
durchgeführt. Nachdem der Farbmarker ca. 1-2 cm ins Gel eingelaufen ist, wird
das Gel zur Dokumentation im UV-Durchlicht fotografiert.
Auswertung
Beschreiben Sie das Ergebnis und diskutieren Sie mögliche Fehler!

Aufgaben 7-17 Übungsaufgaben:
1. Erklären Sie kurz den biochemischen Hintergrund für den Einsatz von Al-
lopurinol bei Gicht.
2. Warum ist der Schmelzpunkt einer GC-reichen DNA höher als der einer AT-
reichen DNA?
3. Berechnen Sie, wieviele Kopien aus einem Molekül doppelsträngiger DNA
nach 25 Zyklen PCR entstanden sind.
4. Berechnen Sie die Schmelztemperatur der im Versuch eingesetzten Primer.
5. Wie viele DNA-Fragmente entstehen bei einem Restriktionsverdau, wenn 3
Erkennungssequenzen vorhanden sind:
a) bei einer zirkulären DNA (Plasmid)
b) bei einem linearen DNA-Fragment?
6. Welche RNA zeichnet sich durch einen hohen Gehalt an modifizierten Basen
aus?
7. In der DNA-Replikation wird die folgende Sequenz repliziert:
5’ A T T G C CT T G A G 3’.
Geben Sie die komplementäre Sequenz (mit 5’ und 3’ Orientierung) an.
8. Wie ist ein Nukleotid aufgebaut (Bausteine und Anordnung)?
Durch welche Art von chemischer Bindung sind die Nukleotide in Nuklein-
säuren kovalent miteinander verbunden?
Welche C-Atome der Ribose können in Nukleotiden mit einem Phosphatrest
verbunden sein?
9. Nennen Sie die beiden Hauptaufgaben des Pentosephosphat-Shunts.

8-1
Versuch 8: Nukleinsäuren II
Versuche:
Regulation der Gen-Aktivität am Beispiel des lac Operons
DNA-Isolierung
Analyse von Erbkrankheiten am Beispiel der Cystischen Fibrose
Wissensgebiete
DNA-Struktur, Replikation, Gen-Aufbau
RNA-Struktur, Transkription, reverse Transkription
tRNA-Struktur, Ribosomenstruktur
Proteinbiosynthese, Translation
Genetischer Code
Eukaryontische und prokaryontische Transkriptionskontrolle
Mutationen (Wildtyp, konstitutive Mutanten)
Enzym-Induktion und -Repression, Repressormolekül, Induktoren
Bakterienwand-Struktur und Biosynthese
Wirkung von Antibiotika auf Transkription (Rifamycin und Actinomycin) und
Translation (Streptomycin, Chloramphenicol, Tetracycline, Puromycin,
Erythromycin)
8-2 Nukleinsäuren II
I. Regulation der Genaktivität am Beispiel des lac operons
Einführung: Eine fein abgestimmte, sehr komplexe Kontrolle der Transkription bei Eu-
karyonten ermöglicht es, bestimmte Gene sehr selektiv zu exprimieren oder
abzuschalten. Die Mechanismen der Expressionskontrolle bei Prokaryonten sind
sehr viel einfacher, so dass auch reprimierte Gene immer noch eine basale
Expression zeigen. Die Promotoren, die durch die eukaryontische RNA-Poly-
merase II erkannt werden, sind erheblich länger und vielfältiger, als die Pro-
motoren prokaryontischer Gene, ebenso beeinflussen Enhancer und Silencer, die
weit entfernt vom Promotor liegen können, die Expression bei Eukaryonten.
Die Grundmechanismen der Expressionssteuerung ähneln sich aber, da sowohl bei
Eukaryonten, wie auch bei Prokaryonten Proteine selektiv an regulatorische Gen-
Sequenzen binden und so die Geschwindigkeit verändern, mit der die
Transkription initiiert wird.
Das vorliegende Experiment soll an einem einfachen Beispiel demonstrieren,
welche Mechanismen in Bakterienzellen wirksam sind. Sie garantieren, dass die
für den Anabolismus und Katabolismus erforderlichen Enzyme, sowie die zell-
eigenen Bausteine koordiniert und kontrolliert synthetisiert werden.
Die Regulation der Enzymaktivität bei der Synthese von kleinen Molekülen kann
z.B. über den "feed back"-Mechanismus kontrolliert werden. Hier reguliert das
Endprodukt die Aktivität des in einer Folge von Reaktionen am Anfang des
Synthesewegs stehenden allosterischen Enzyms.
↓ Enzym1 Enzym2 Enzym3
A → B → C → D

Induktion des lac Operons 8-3
Die feed-back-Hemmung stellt die Feinregulierung, die Enzymsynthesehemmung
eine Grobregulierung dar. Die Synthese wird über die Mechanismen der Re-
pression und Induktion der Proteinsynthese reguliert.
Die Synthese eines Enzyms kann so gesteuert werden, dass das Endprodukt einer
multienzymatischen Synthese die Synthese des ersten an einer Reaktionssequenz
beteiligten Enzyms und damit aller am Syntheseweg beteiligten Enzyme
unterdrückt. Man nennt dies Repression oder koordinierte Enzym-Repression.
Man findet sie vorwiegend bei der Synthese von Enzymen anabolischer
Reaktionen.
Als Enzym-Induktion bezeichnet man den Mechanismus, der eine beträchtliche
Steigerung der Enzymsynthese nur in Gegenwart des Substrats bewirkt. Hierbei
handelt es sich meist um Enzyme katabolischer Reaktionen. Enzyme, die weder
reprimierbar noch induzierbar sind, sondern ständig synthetisiert werden, nennt
man konstitutive Enzyme.
Die Induktion der Synthese von Enzymen katabolischer Reaktionen soll am
Beispiel des Laktose-Operons von E. coli erläutert werden. Die E. coli-Zelle kann
ihren Energiebedarf durch die Aufnahme vieler C-Quellen aus dem Nährmedium
decken, u.a. durch Laktose und Glukose. Für die Verwertung von Laktose werden
die erforderlichen Enzyme erst synthetisiert, wenn der Zelle β-Galaktoside
angeboten werden. Die β-Galaktosidase, die die Laktose zu Galaktose und
Glukose hydrolysiert, wird damit zu einem wesentlichen Enzym. Wächst E. coli
auf anderen C-Quellen, enthält die Bakterienzelle nur wenige β-Galaktosidase-
Moleküle, auf Laktose wachsend synthetisiert sie dagegen Tausende von
Enzymmolekülen.
β-Galaktosidase (z) ist also ein induzierbares Enzym. Gleichzeitig werden noch
zwei weitere Enzyme synthetisiert: die β-Galaktosid-Permease (y), die für den
Transport durch die Zellmembran erforderlich ist, und die β-Galaktosid-Trans-
acetylase (a), deren physiologische Rolle unbekannt ist (in vitro überträgt sie den
Acetylrest von Acetyl-CoA auf C6 des Thiogalaktosids). Der physiologische
Induktor ist die Allolaktose, die durch Transglycosylierung von Laktose durch die
wenigen vorhandenen β-Galaktosidase-Moleküle synthetisiert wird. Einige
Induktoren sind Induktoren, ohne selbst Substrat der Galaktosidase-Reaktion zu
sein. Ein solcher ist das Isopropylthiogalaktosid (IPTG).
8-4 Nukleinsäuren II
Die Information (Erbanlage) für die Primärstruktur der drei Enzyme der Laktose-
Verwertung (Laktose Operon) ist linear als "cluster" angelegt.
An dieser Stelle soll darauf hingewiesen werden, dass bei Eukaryonten die Gene
in der Regel einzeln angelegt sind und häufig Introns enthalten.
Laktose Operon Regulatorgen Steuerelemente Strukturgene
i p o z y a
Repressor Promotor Operator z: ß-Galaktosidase y: ß-Galaktosid-Permease a: ß-Galaktosid-Transacetylase
Die Information der DNA wird in die mRNA transkribiert. Die mRNA-Synthese
wird durch die RNA-Polymerase katalysiert, die an die Promotor (p)-Region
bindet und die RNA-Synthese startet, wenn der Operator, (o)-Region, frei ist.
Durch ein "Schlüsselprotein", den Repressor, kann der Operator blockiert werden.
Der Repressor ist das Produkt des Regulatorgens (i), der auf einem völlig anderen
DNA-Abschnitt lokalisiert sein kann.
i p o z y a
Bindungsstelle für Induktor
Bindungsstelle für Operator
Repressorprotein (R)
Ribosomen
m-RNS
mRNA

Induktion des lac Operons 8-5
Strukturelemente und Strukturgene stellen hier die funktionelle Einheit, das
Operon, dar. Die am Operon synthetisierte mRNA ist ein polycistronischer (die
Information mehrerer Gene enthaltender) Messenger.
Der Repressor hat zwei spezifische Bindungsstellen, eine für den Operator und
eine für den Induktor.
i p o z y a
Induktormolekül
Wenn Induktormoleküle anwesend sind, binden sie an die Induktorbindungsstelle,
wodurch die Konformation des Repressors verändert wird (allosterisches Protein).
Der Repressor wird frei, die Strukturgene sind nun frei für die Transkription.
Galaktosidase wird gebildet:
i p o z y a
Induktor-Repressor-Komplex
m-RNA
Ribosomen
Enzyme
8-6 Nukleinsäuren II
Die Repression der Synthese von Enzymen anabolischer Reaktionen durch ein
biosynthetisches Endprodukt lässt sich analog erklären. Freie Repressormoleküle
besitzen eine Konformation, die keine Bindung an den Operator erlaubt. Die
Transkription kann stattfinden:
i p o z y a
Repressor (inaktive Form)
Häuft sich das biosynthetische Endprodukt im Medium an, wirkt es als Korepres-
sor, indem es mit dem Repressor komplexiert:
i p o z y a
Korepressor-Repressor-Komplex
Dieser Komplex hat die geeignete Struktur, um an das Operatorgen zu binden. Die
Transkription wird blockiert, die Enzymsynthese reprimiert:
i p o z y a
aktiver Korepressor-Repressor-Komplex

Induktion des lac Operons 8-7
Es gibt also zwei Klassen von Repressor-Molekülen: Die eine ist an der Kontrolle
der Induktion, die andere an der Kontrolle der Repression durch Endprodukte
beteiligt.
Der Beginn der Transkription selbst wird durch ein anderes Protein, das soge-
nannte CAP-Protein (Catabolic Activating Protein) um das zwanzigfache be-
schleunigt. In Form eines cAMP-CAP-Komplexes bindet dieses Protein an die
DNA direkt vor der Anheftungsstelle der RNA-Polymerase. Das Polymerase-
Enzym allein kann offenbar die Startstelle nicht schnell genug binden.
Beim Wildstamm E. coli K12 ist das Enzym induzierbar. Es gibt Mutanten, die
das Enzym β-Galaktosidase auch ohne Induktor ständig bilden. Es handelt sich
hier um eine konstitutive Mutanten (E. coli lac i-o+z+a+) Die Mutation liegt
entweder im Regulatorgen (i-), so dass kein aktiver Repressor gebildet werden
kann, oder im Operatorgen (o) (z.B. E. coli lac i+ocz+a+; oc = constitutiv), so dass
der Operator nicht vom aktiven Repressor blockiert werden kann.
Im folgenden Experiment wird gezeigt, dass die Enzymsynthese der β-Galak-
tosidase im Wildstamm E. coli K12 (W), der normalerweise kaum mehr als ein
Molekül β-Galaktosidase pro Zelle enthält, durch das Galaktosid Isopro-
pylthiogalaktosid (IPTG) induziert wird. Diese Enzymsynthese wird mit der
Synthese in einer konstitutiven Mutante E. coli lac i-o+z+ verglichen.
CH SH3C
H3CC
C
C
C
C
CH2OH
OHH
HHO
HHO
O
H
Isopropylthiogalaktosid (IPTG)
Die Induktion wird mit IPTG als Substrat durchgeführt. Der Nachweis der ge-
steigerten Synthese des Enzyms β-Galaktosidase wird mit einem einfachen,
empfindlichen photometrischen Test mit dem Substrat o-Nitrophenyl-β-Ga-
laktosid (ONPG) geführt, das vom Enzym zu einem farbigen Produkt, o-Ni-
8-8 Nukleinsäuren II
trophenol (ONP), hydrolysiert wird. Das ONP vertieft seine gelbe Farbe (436 nm)
bei alkalischem pH.
O C
C
C
C
C
CH2OH
OHH
HHO
HHO
O
H
NO2 NO2
OH
+ Galactose
ONPgelb
o-Nitrophenyl-ß-galactosid (ONPG)
Geräte: Bakterienwachstumsröhrchen mit Belüftungsvorrichtung Laborwecker Spektralphotometer Glasküvetten Pipetten 10 ml, 5 ml, 2 ml, 1 ml Reagenzgläser 36 Schüttelwasserbad 37 °C Whirlmix
Lösungen: 200 ml von wachsenden Bakterienkulturen:
W: (Wildstamm) E. coli K12
C: (Constitutive Mutante) E. coli (lac i–o+z+)
Beide Kulturen wachsen in synthetischem M9-Medium mit 4⋅10-2 M Glycerin als Kohlenstoffquelle. Titer ca. 3⋅108 Bakterien/ml bei einer OD von 1,0 (623 nm). Medium: Bactotryptone 20g Yeastextrakt 5g NaCl 0,5 g aqua dest. ad 1 l Vor Gebrauch 20 ml 1 M MgSO4 zusetzen BME-Puffer (450 ml H2O, 50 ml 0,25 M Natriumphosphatpuffer pH 7 und 1 ml Mer-
captoethanol) Isopropylthio-β-Galaktosid (IPTG) 0,01 M Na2CO3 1 M
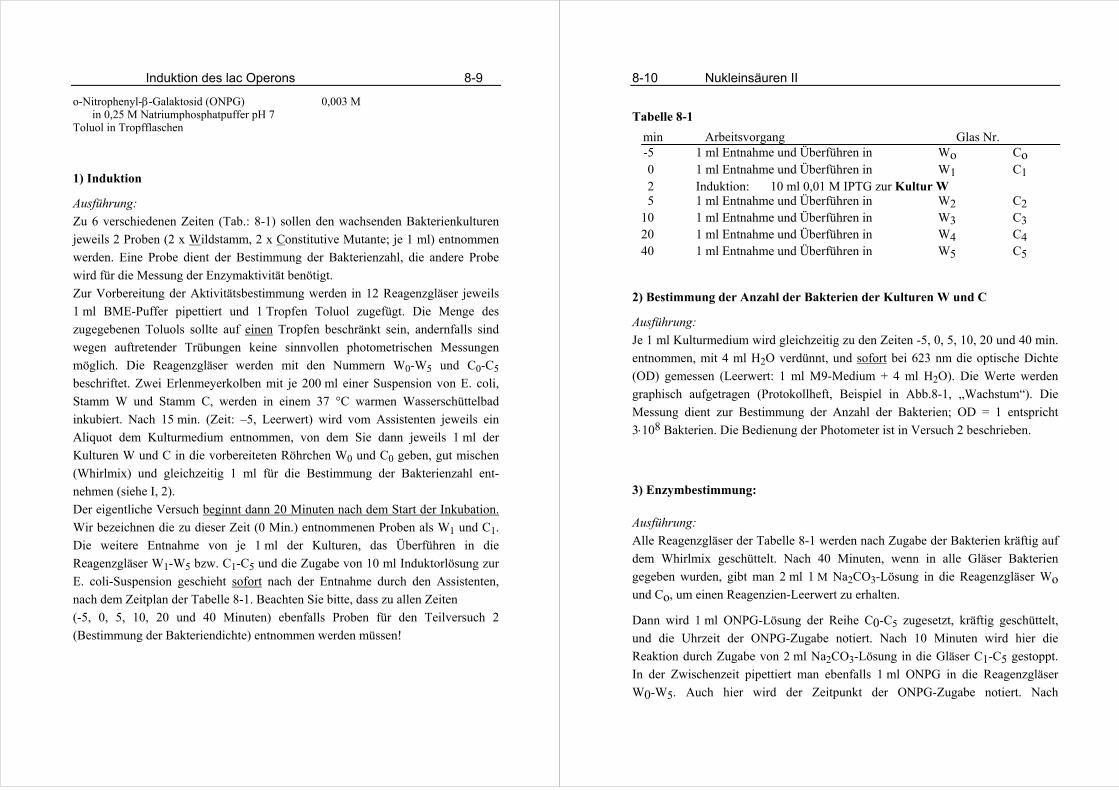
Induktion des lac Operons 8-9
o-Nitrophenyl-β-Galaktosid (ONPG) 0,003 M in 0,25 M Natriumphosphatpuffer pH 7
Toluol in Tropfflaschen
1) Induktion
Ausführung: Zu 6 verschiedenen Zeiten (Tab.: 8-1) sollen den wachsenden Bakterienkulturen
jeweils 2 Proben (2 x Wildstamm, 2 x Constitutive Mutante; je 1 ml) entnommen
werden. Eine Probe dient der Bestimmung der Bakterienzahl, die andere Probe
wird für die Messung der Enzymaktivität benötigt.
Zur Vorbereitung der Aktivitätsbestimmung werden in 12 Reagenzgläser jeweils
1 ml BME-Puffer pipettiert und 1 Tropfen Toluol zugefügt. Die Menge des
zugegebenen Toluols sollte auf einen Tropfen beschränkt sein, andernfalls sind
wegen auftretender Trübungen keine sinnvollen photometrischen Messungen
möglich. Die Reagenzgläser werden mit den Nummern W0-W5 und C0-C5
beschriftet. Zwei Erlenmeyerkolben mit je 200 ml einer Suspension von E. coli,
Stamm W und Stamm C, werden in einem 37 °C warmen Wasserschüttelbad
inkubiert. Nach 15 min. (Zeit: –5, Leerwert) wird vom Assistenten jeweils ein
Aliquot dem Kulturmedium entnommen, von dem Sie dann jeweils 1 ml der
Kulturen W und C in die vorbereiteten Röhrchen W0 und C0 geben, gut mischen
(Whirlmix) und gleichzeitig 1 ml für die Bestimmung der Bakterienzahl ent-
nehmen (siehe I, 2).
Der eigentliche Versuch beginnt dann 20 Minuten nach dem Start der Inkubation.
Wir bezeichnen die zu dieser Zeit (0 Min.) entnommenen Proben als W1 und C1.
Die weitere Entnahme von je 1 ml der Kulturen, das Überführen in die
Reagenzgläser W1-W5 bzw. C1-C5 und die Zugabe von 10 ml Induktorlösung zur
E. coli-Suspension geschieht sofort nach der Entnahme durch den Assistenten,
nach dem Zeitplan der Tabelle 8-1. Beachten Sie bitte, dass zu allen Zeiten
(-5, 0, 5, 10, 20 und 40 Minuten) ebenfalls Proben für den Teilversuch 2
(Bestimmung der Bakteriendichte) entnommen werden müssen!
8-10 Nukleinsäuren II
Tabelle 8-1
min Arbeitsvorgang Glas Nr. -5 1 ml Entnahme und Überführen in Wo Co 0 1 ml Entnahme und Überführen in W1 C1 2 Induktion: 10 ml 0,01 M IPTG zur Kultur W 5 1 ml Entnahme und Überführen in W2 C2 10 1 ml Entnahme und Überführen in W3 C3 20 1 ml Entnahme und Überführen in W4 C4 40 1 ml Entnahme und Überführen in W5 C5
2) Bestimmung der Anzahl der Bakterien der Kulturen W und C
Ausführung: Je 1 ml Kulturmedium wird gleichzeitig zu den Zeiten -5, 0, 5, 10, 20 und 40 min.
entnommen, mit 4 ml H2O verdünnt, und sofort bei 623 nm die optische Dichte
(OD) gemessen (Leerwert: 1 ml M9-Medium + 4 ml H2O). Die Werte werden
graphisch aufgetragen (Protokollheft, Beispiel in Abb.8-1, „Wachstum“). Die
Messung dient zur Bestimmung der Anzahl der Bakterien; OD = 1 entspricht
3⋅108 Bakterien. Die Bedienung der Photometer ist in Versuch 2 beschrieben.
3) Enzymbestimmung:
Ausführung: Alle Reagenzgläser der Tabelle 8-1 werden nach Zugabe der Bakterien kräftig auf
dem Whirlmix geschüttelt. Nach 40 Minuten, wenn in alle Gläser Bakterien
gegeben wurden, gibt man 2 ml 1 M Na2CO3-Lösung in die Reagenzgläser Wo
und Co, um einen Reagenzien-Leerwert zu erhalten.
Dann wird 1 ml ONPG-Lösung der Reihe C0-C5 zugesetzt, kräftig geschüttelt,
und die Uhrzeit der ONPG-Zugabe notiert. Nach 10 Minuten wird hier die
Reaktion durch Zugabe von 2 ml Na2CO3-Lösung in die Gläser C1-C5 gestoppt.
In der Zwischenzeit pipettiert man ebenfalls 1 ml ONPG in die Reagenzgläser
W0-W5. Auch hier wird der Zeitpunkt der ONPG-Zugabe notiert. Nach

Induktion des lac Operons 8-11
60 Minuten wird die Reaktion in den Proben durch Zufügen von 2 ml Na2CO3-
Lösung in die Gläser W1-W5 gestoppt.
Tabelle 8-2
Zeit
Glas- Nr. OD 436 nm U/min
(min) Verd. 1:5
Wildtyp Const. Mut. W. C. W. C. -5 Wo Co
0 W1 C1 5 W2 C2 10 W3 C3 20 W4 C4 40 W5 C5
40 30 20 10 0 -10 0
10
20
U
Zeit (Minuten)
Constitutive Mutante
Wachstum
Wildtyp
Abbildung 8-1: Bestimmung der β-Galaktosidase in den E. coli-Stäm men W und C. Induktion des Enzyms im Wildstamm W durch Zugabe eines β–Galaktosids (IPTG).
8-12 Nukleinsäuren II
Auswertung: Aus allen Reagenzgläsern (W1-W5 und C1-C5) entnimmt man 1 ml und verdünnt,
wenn nötig, mit 4 ml H2O. Die Extinktionen werden bei 436 nm gegen Wo bzw.
Co als Leerwert gemessen. Bitte keine Plastikküvetten benutzen. Das eingesetzte
Toluol greift den Kunststoff an. Eine Extinktion von 0,0075 entspricht der
Hydrolyse von 1 nmol ONPG. (Bedienung des Photometers wie in Versuch 6)
Die Einheit des Enzyms definieren wir hier als diejenige Menge, die 1 nmol
ONPG in 1 min. bei Zimmertemperatur und pH 7 hydrolysiert. Die Einheiten
(U/min) des Enzyms (Abb. 8–1) werden gegen die Zeit aufgetragen.

DNA-Isolierung 8-13
II. DNA-Isolierung
Das gesamte Genom eines Organismus ist in der DNA jeder kernhaltigen Kör-
perzelle als Kopie enthalten. Zur Analyse eines beliebigen Gens kann also DNA
aus leicht zugänglichen Zellen, wie Fibroblasten, Lymphozyten oder
Epithelzellen, herangezogen werden. (Schnellpräparation, mit Proteinen verun-
reinigt, siehe Versuch 7, PCR)
Um eine größere Menge relativ sauberer DNA zu gewinnen, sind einige zusätz-
liche Schritte nötig. Wir wollen hier der Einfachheit halber bakterielle chromo-
somale DNA isolieren. Bakterien enthalten häufig neben der genomischen DNA
noch zusätzlich Plasmide, ringförmige DNA-Moleküle, die z. B. Gene für
Antibiotika-Resistenzen tragen. Da Plasmide leicht künstlich in Bakterien einge-
schleust werden können, werden sie häufig in der rekombinanten DNA-
Technologie eingesetzt (siehe Versuch 7).
Die Zellwände der Bakterien werden zuerst mit Lysozym angedaut und an-
schließend werden die Zellen durch Detergenzien aufgeschlossen. Proteine werden
durch Chloroform denaturiert und durch Zentrifugation abgetrennt.
Material: Glasstab Schliffmesspipette 10 ml UV-Spektrometer Quarz-Küvette Vollpipette 2 ml Wasserbad 37 °C, 60 °C Zentrifugen 2 Zentrifugengläser ein 50 ml-Glas und zwei 10 ml-Gläser
Lösungen: Bakterienkultur 50 ml Chloroform-Isoamylalkohol 24:1 Isopropanol NaCl 0,15 M, enthält 0,1 M Na2EDTA, pH 8,0 Lysozym-Lösung 10 mg/ml NaClO4 3 M Natriumdodecylsulfat-Lösung (SDS) 12,5 %ig
8-14 Nukleinsäuren II
1) Isolierung
Ausführung: Bakterien einer 50 ml-Kultur (stehen bereit) werden in der Zentrifuge 10 min. bei
höchster Umdrehung abzentrifugiert. Der Überstand wird verworfen und die sedi-
mentierten Bakterien in 5 ml 0,15 M NaCl, 0,1 M Na2EDTA suspendiert. 0,5 ml
Lysozymlösung wird hinzugefügt, gut gemischt und 30 min. bei 37 °C inkubiert.
Durch Umschwenken wird die Suspension zwischenzeitlich gemischt. Zur
Vervollständigung der Lysis wird 1 ml 12,5% SDS-Lösung zugefügt und 10 min.
bei 60 °C inkubiert. Schließlich werden 2 ml 3M NaClO4 und 8 ml
Chloroform/Isoamylalkohol (24:1) zugefügt. Es wird gut geschüttelt, die Emulsion
auf 2 Zentrifugenröhrchen verteilt und die Phasen durch 10-minütige Zen-
trifugation getrennt. Anschließend wird die klare obere Phase abgenommen und
unter gelegentlichem Umrühren mit einem Glasstab in 50 ml Isopropanol ge-
gossen. Die DNA fällt aus und kann mit dem Glasstab herausgefischt werden.
Überschüssiges Isopropanol wird durch kurzes Halten in der Luft getrocknet.
2) Konzentrationsbestimmung der DNA
Ausführung: Die DNA wird in 1 ml Wasser gelöst. 0,1 ml davon werden mit 5 ml Wasser
verdünnt und die Absorption in einer Quarzküvette im Spektralphotometer bei
260 nm gemessen. Falls die Extinktion größer als 1 ist, muss die Probe verdünnt
werden. Zur Reinheitsbestimmung wird die Extinktion bei 280 nm gemessen.
Reine DNA hat ein Verhältnis der Extinktionen von E260/E280 von 2. Ein
kleineres Verhältnis zeigt die Verunreinigung mit Proteinen an, die bei 280 nm
absorbieren.
Berechnung der Ausbeute: OD260 = 1 entspricht 50 mg DNA/l

Molekulare Medizin 8-15
III. Analyse von Erbkrankheiten am Beispiel der cysti-schen Fibrose
Die cystische Fibrose ist eine autosomal-rezessive Erbkrankheit der exokrinen
Drüsen und der Schweißdrüsen. Die Krankheit beginnt bereits im Kindesalter.
Primär werden Verdauungs- und Atmungstrakte befallen. Die Hauptsymptome
sind chronische Infektionen des Atmungstrakts, Pankreasinsuffizienz, ausge-
sprochen viskose Schleimsekretionen und eine Empfindlichkeit für Hitzebela-
stung. Jeder 25.-30. ist heterozygoter Träger der Erbanlage, jedes 3000. Neu-
geborene ist erkrankt.
Das bei der cystische Fibrose betroffene Gen ist seit 1989 bekannt und seit dieser
Zeit sind viele Mutationen entdeckt worden, die die Krankheit verursachen. Das
Genprodukt wird als "Cystic Fibrosis Transmembrane-Conductance Regulator"
(CFTR) bezeichnet. Das membranständige Protein (170 kDa) ist an der Regulation
eines Chlorid-Ionenkanals beteiligt. Die Schwere der Erkrankung ist abhängig von
der Lokalisation der Mutation in bestimmten Domänen des Proteins. Neben vielen
transmembranalen Bereichen wurden durch Sequenzvergleiche auch zwei
nukleotidbindende Bereiche postuliert. 70% der Mutationen betreffen eine drei
Basenpaare umfassende Deletion, die einen Verlust des Phenylalanins an Position
508 bewirken (∆F508). Die restlichen 30 % der Mutationen sind über das Gen
verteilt. Für die exakte Bestimmung dieser Mutationen ist es notwendig die
kodierenden Bereiche des Gens, die der mRNA entsprechen, zu sequenzieren. Der
anschließende Vergleich der daraus abgeleiteten Primärstruktur des Proteins mit
der Primärstruktur des Wildtyps ergibt die genaue Lokalisation der Mutation.
Computer und Datenbanken sind wichtige Werkzeuge in der molekularen Medizin
geworden. Am Beispiel der cystischen Fibrose sollen Sie einen Einblick in deren
Einsatz bekommen. Im Zentrum für Angewandte Informatik der Universität zu
Köln (ZAIK) steht das „Wisconsin Sequence Analysis Program“, ein
Sequenzanalyseprogramm, allen Angehörigen der Universität zur Verfügung. Von
allen am UKLAN (Local Area Network der Universität zu Köln) angeschlossenen
Rechnern kann man über das Netz seine Arbeit am gendb-Server durchführen.
Alternativ kann die Analyse auch über das Internet erfolgen. Hier stehen z.B. die
Server am NCBI (National Center for Biotechnology Information, USA oder EBI,
8-16 Nukleinsäuren II
European Bioinformatics Institute, zur Verfügung.
(http://www.ncbi.nlm.nih.gov/blast/blast.cgi oder http://srs.ebi.ac.uk/)
Aufgabe: Die CFTR cDNA-Sequenz eines CF-Patienten (unter der Gruppennummer) wird
Ihnen zur Verfügung gestellt. Diese sollen Sie
1. in die Proteinsequenz translatieren, 2. mit der Sequenz des Wildtyps vergleichen,
die Sie in einer Proteinsequenzdatenbank abgerufen haben und 3. die Art der Mutation angeben.
Durchführung: Nach dem Einloggen in das WindowsNT-System (login liegt aus) des Computers
erscheint ein Auswahlmenü. Mit Hilfe der Maus wählen Sie
Start - Praktikum Biochemie - gendb.
Automatisch wird die Verbindung zum gendb-Server hergestellt. Es wird nach
dem USERNAME (akc37) gefragt. Das PASSWORT wird Ihnen vom Assistenten
mitgeteilt. Nach der Eingabe erscheint die Begrüßungsseite des gendb-Servers
und Sie befinden sich in Ihrem Arbeitsbereich im gendb. Der Rechner ist bereit,
Ihre Anweisungen entgegenzunehmen, wenn in der letzten Zeile folgendes
erscheint:
akc37@campfire%
Alle im folgenden fettgedruckten Befehle werden durch Drücken der RETURN-
Taste abgeschlossen und ausgeführt. Mit dem Befehl:
akc37@campfire% cd 37xx (xx ist Ihre Gruppennummer, 01, 02,
03, usw.)
gelagen Sie in Ihr Arbeitsverzeichnis. Mit:
akc37@campfire% dir
können Sie die vorhandenen Dateien Ihres Arbeitsbereichs auflisten.
Unter dem Namen cftr_gruppex.dna ist in einer Datei die cDNA Sequenz des
CF-Patienten abgespeichert. Die Datei können Sie sich mit den Befehlen:
akc37@campfire% pg cftr_gruppex.dna

Molekulare Medizin 8-17
am Bildschirm anschauen. Bitte notieren Sie sich den Beginn und das Ende der
DNA-Sequenz, die die Proteinsequenz oder einen Teil davon kodieren. Diese wer-
den in einer Zeile:
FT CDS nnn..nnnn
angegeben. Die Zeile ist eine „Feature Table“-Zeile und enthält den Beginn und
das Ende des Protein-kodierenden Bereichs. Diese beiden Zahlen werden Sie noch
für die Translation benötigen. Mit dem Befehl:
akc37@campfire% names swiss:cftr*
suchen Sie dann in einer Proteindatenbank nach dem Wildtyp des CFTR-Gens.
Das Sternchen bedeutet, dass alle Einträge, die in der Swiss-Protein-Datenbank im
Namen die Buchstabeanfolge „cftr“ enthalten, auf dem Bildschirm aufgelistet
werden sollen. Bestätigen Sie die Frage nach dem:
output file (*TERM*)
durch Drücken der RETURN-Taste. Unter dem Eintrag cftr_human ist die
menschliche Proteinsequenz in der SWISS-Proteindatenbank gespeichert. Das
Programm „names“ gibt alle zur Zeit bekannten CFTR-Gene an, auch die anderer
Organismen. Mit:
akc37@campfire% fetch swiss:cftr_human
kopieren Sie dann die Sequenz aus der Datenbank in Ihren Arbeitsbereich. Sie
können sich jetzt den Eintrag mit dem Befehl:
akc37@campfire% pg cftr_human.swnew
anschauen.
Zuerst erscheint eine Literaturliste, dann eine kurze Beschreibung der Krankheit.
Außerdem ist eine Internetadresse angegeben, bei der weitere Informationen
abgerufen werden können. In den Zeilen:
FT VARIANT 13 13 S -> F. usw.
werden einige der fast 900 gefundenen Mutationen angegeben, hier die Serin-
Phenylalanin Substitution (Austausch) in Position 13.
Nun translatieren Sie den kodierenden Bereich der CFTR-cDNA mit:
8-18 Nukleinsäuren II
akc37@campfire% translate cftr_gruppex.dna
die cDNA des CF-Patienten in die entsprechende Proteinsequenz. Die abgefragten
Einstellungen werden ohne Veränderungen durch die RETURN-Taste bestätigt.
Den Anfang und das Ende der Translation geben Sie entsprechend Ihrer Notizen
(s.o.) an. Als cftr_gruppex.pep wird die Proteinsequenz des Patienten
abgespeichert.
Mit dem Befehl:
akc37@campfire% pg cftr_gruppex.pep
kann die CFTR-Sequenz des Patienten angesehen werden.
Diese Sequenz wird mit Hilfe des Programms bestfit mit der Sequenz des
Wildtyps verglichen.
akc37@campfire%
bestfit cftr_human.swnew cftr_gruppex.pep
Die abgefragten Einstellungen werden wieder mit der RETURN-Taste bestätigt. Das
Vergleichsergebnis wird als cftr_human.pair-Datei abgelegt. Schauen Sie sich
die Datei an:
akc37@campfire% pg cftr_human.pair
Mit dem Befehl:
akc37@campfire% lpr –P cipm1ps cftr_human.pair
kann die Datei gedruckt und im Protokollheft abgeheftet werden. Die Sitzung
beenden Sie mit:
akc37@campfire% exit
Aufgabe:
Bitte markieren Sie die Mutationsstelle und geben die Art der Mutation an. Be-
nutzen Sie den genetischen Code und schlagen mögliche Basenaustausche vor.

Aufgaben 8-19
Übungsaufgaben:
1. Durch welchen Mechanismus kann ein einziges Gen die Expression von Proteinen unterschiedlicher Größe bewirken?
2. Woran binden Transkriptionsfaktoren? Was bewirken sie?
3. Durch welche strukturelle Komponente eines Proteins wird festgelegt, ob es sezerniert wird?
4. Geben Sie Mechanismen an, die der Entstehung der Antikörpervariabilität zugrundeliegen.
5. Zeichnen Sie auf DNA-Ebene drei prinzipiell unterschiedliche Beispiele für eine Mutation.
6. Formulieren Sie eine aus 6 Nukleotidpaaren bestehende Palindromstruktur.
7. Geben Sie Beispiele für folgende Mutationen an: Substitution, Deletion und Insertion.
8. Wie beeinflusst Penicillin den Metabolismus von Escherichia coli?


















