Aus der Klinik für Geflügel der Experimentelle Studie ä fileAus der Klinik für Geflügel der...
130
Aus der Klinik für Geflügel der Tierärztlichen Hochschule Hannover Experimentelle Studie über die Verträglichkeit und Wirksamkeit eines inaktivierten Mycoplasma gallisepticum - MG - Öl-Adjuvans-Impfstoffes bei Legehennen I N A U G U R A L - D I S S E R T A T I O N Zur Erlangung des Grades Eines Doktors der Veterinärmedizin (Dr. med. vet.) durch die Tierärztliche Hochschule Hannover Vorgelegt von Hanns-Dietrich Graack aus Itzehoe Hannover 2001
Transcript of Aus der Klinik für Geflügel der Experimentelle Studie ä fileAus der Klinik für Geflügel der...

Aus der Klinik für Geflügel der Tierärztlichen Hochschule Hannover
Experimentelle Studie über die Verträglichkeit und Wirksamkeit
eines inaktivierten Mycoplasma gallisepticum - MG -
Öl-Adjuvans-Impfstoffes bei Legehennen
I N A U G U R A L - D I S S E R T A T I O N
Zur Erlangung des Grades Eines Doktors der Veterinärmedizin
(Dr. med. vet.) durch die Tierärztliche Hochschule Hannover
Vorgelegt von Hanns-Dietrich Graack
aus Itzehoe
Hannover 2001

Wissenschaftliche Betreuung: Univ.-Prof. Dr. K.-H. Hinz 1. Gutachter Univ.-Prof. Dr. K.-H. Hinz
2. Gutachter Apl. Prof. Dr. med. vet. Dr. med. vet. habil. K. Pohlmeyer
Tag der mündlichen Prüfung: 19. November 2001

�����������

�
�
�
�
�
�
�
������ ����������������� ��
�������������������
������������������������������������������������
�������������������������������������������������
���� ����!���������������������������"����
�
��#�������$���%�&�'()�

Inhaltsverzeichnis Seite
1. Einleitung 11
2. Schrifttum 13
2.1. Historisches über die ersten Vakzinationsversuche gegen
Mycoplasma gallisepticum (MG) 13
2.2. Wirksamkeit von Mycoplasma gallisepticum - Totimpfstoffen 20
2.3. Mycoplasma gallisepticum - bedingte immunologische Reaktionen 42
2.4. Aerogene Inokulation mittels Aerosol 62
3. Material und Methoden 66
3.1. Allgemeiner Versuchsaufbau 66
3.2. Voruntersuchungen 66
3.3. Hauptuntersuchungen 67
3.3.1 Verträglichkeitsprüfung 67
3.3.2. Wirksamkeitsprüfung 68
3.4. Herkunft und Haltung der Versuchstiere 68
3.5. Mycoplasma gallisepticum - Stämme 68
3.5.1. Herkunft 69
3.5.2. Anzüchtung und Vermehrung 70
3.6. Vakzination 72
3.7. Experimentelle Belastungsinfizierung (EBI) 74
3.7.1. Infizierungskammer nach W. HOLLÄNDER (1995) 74
3.7.2. Pariboy - Ultraschallvernebler 75
3.7.3. Präparation und Applikation des Inokulums als Aerosol 75
3.8. Klinische Untersuchungen 77
3.8.1. Allgemeinbefinden 77
3.8.2. Lokale Impfstoffreaktion 78
3.8.3. Legeleistung 78
3.9. Serologisch nachweisbare Reaktionen 78

3.9.1. Serum-Schnellagglutinationstest (SSA) 79
3.9.2. Hämagglutinationshemmungstest (HAHT) 79
3.9.3. Enzymgebundener Immunadsorptionstest zum Nachweis von
Antikörpern ( ELISA) 81
3.10. Pathologisch-anatomische Untersuchungen 81
3.11. Kulturell-bakteriologische Untersuchungen 83
3.12. Statistische Auswertung 83
4. Ergebnisse 85
4.1. Voruntersuchungen 85
4.1.1. Zum Zeitpunkt der Einstallung 85
4.1.2. Nach der Vakzination 85
4.1.2.1. Versuch A 85
4.1.2.2. Versuche B und C 86
4.1.2.3. Versuch D 86
4.1.2.4. Versuch E 87
4.2. Hauptuntersuchungen 87
4.2.1. Verträglichkeitsprüfungen 87
4.2.2. Wirksamkeitsprüfungen 89
4.2.2.1. Legeleistung im Versuch1 89
4.2.2.2. Legeleistung im Versuch 2 90
4.2.2.3. Legeleistung im Versuch 3 91
4.3. Ergebnisse der serologischen Untersuchungen 93
4.3.1. Serumschnellagglutination (SSA) 93
4.3.2. Hämagglutinationshemmungstest (HAHT) 95
4.3.3. Enzymgebundener Immunadsorptionstest zum Nachweis von
Antikörpern (ELISA) 97
4.4. Ergebnisse der pathologisch-anatomischen Untersuchungen 98
4.5. Ergebnisse der kulturell-bakteriologischen Untersuchungen 102
5. Diskussion 104

6. Zusammenfassung 111
7. Summary 112
8. Literaturübersicht 113
9. Anhang 128
9.1. Verwendete Nährmedien und Lösungen 128

Abkürzungsverzeichnis µm Mikrometer Abb. Abbildung AE Aviäre Enzephalomyelitis AIB / AIBV Aviäre Infektiöse Bronchitis / Aviäres Infektiöses Bronchitis-Virus AID50 Dosis eines infektiösen Agens, welche bei 50 % der Versuchs-
tiere im Luftsack (airsac) eine pathologische Veränderungen herbeiführt
AK Antikörper bzw. beziehungsweise °C Grad Celsius cm2 Quadratzentimeter CCU Colour changing unit / Stoffwechseltiter CD Cluster of differentiation / charakteristische Oberflächenantigene
auf T-Lymphozyten CRD Chronic respiratory disease d. h. dass heißt DNA / DNS Desoxyribonukleinsäure EBI experimentelle Belastungsinfizierung E. coli Escherichia coli EID Dosis eines infektiösen Agens, welche ausreicht, um Eier
zu infizieren ELISA Enzyme-linked immunosorbant assay / enzymgebundener
Immunadsorptionstest zum Nachweis von Antikörpern g Gramm x g durch Zentrifugalbeschleunigung erzeugtes Vielfaches der Erd- beschleunigung (g= 9,81 m/sec) GMT Geometrischer Mittelwert H. Haemophilus HA Hämadsorption HAH / HAHT Hämagglutinationshemmungstest IBD Gumboro-Krankheit / Infectious Bursal Disease IB / IBV Infektiöse Bronchitis / Infektiöses Bronchitis-Virus ID50 Dosis einen infektiösen Agens, welche bei 50 % der Versuchs-
tiere zu einer Infektion führt I.E. Internationale Einheiten iCGN Iota carrageenan i.c. intrazoelomal IgA Immunglobulin der Klasse A IgG Immunglobulin der Klasse G IgM Immunglobulin der Klasse M IIPA Indirect immunoperoxidase assay i.n. intranasal i.t. intratracheal i.v. intravenös KBE Kolonie-bildende Einheit

kDA Kilodalton l Liter LAH Lohmann Animal Health (Pharmazeutisches Unternehmen) LMI Leukozytenmigrationshemmungstest LP Lipoprotein LSE Luftsackentzündung Lw. Lebenswoche M. Mycoplasma mm Millimeter Mab monoclonal antibody / monoklonaler Antikörper MID Minimale Infektiöse Dosis MG Mycoplasma gallisepticum MGn Mycoplasma gallinarum mg Milligramm ml Milliliter MS Mycoplasma synoviae m/s Meter pro Sekunde m2 Quadratmeter N. Nervus Nn. Nervi n Anzahl der Meßwerte ND / NDV Newcastle Disease / Newcastle Disease Virus p Irrtumswahrscheinlichkeit PAGE Polyacrylamid-Gel-Elektrophorese PCR Polymerase chain reaction pH negativer dekadischer Logarithmus der Wasserstoffionen-
konzentration p.i. post infectionem p.m. post mortem PPLO pleuropneumonia-like organisms eingetragenes Warenzeichen RDE receptor-destroying-enzyme RIA radio-immune-assay RVI reduced virulence isolate / Isolat mit reduzierter Virulenz s.c. subkutan SDS Sodium-dodecyl-sulfat / Natrium-dodecyl-sulfat s.o. siehe oben SPF specific pathogen free / frei von bestimmten pathogen Erregern STP Salmonella typhimurium-Zellwandprotein SSA Serum-Schnellagglutination TA tube agglutination / Röhrchenagglutination TCR t-cell-receptor / T-Zell-Rezeptor ts temperatursensibel Vlp variable lipoprotein / variables lipophiles Protein Vsp variable surface protein / variables Oberflächenprotein


11
1. Einleitung
Mycoplasma gallisepticum (MG) ist als Erreger der Mykoplasmose des Huhnes
weltweit verbreitet. Latente Infektionen mit MG können eine erhöhte Embryonal-
sterblichkeit, Entwicklungsstörungen der Jungtiere und Legeleistungsminderungen
bis zu 10 % zur Folge haben. Nach klinischen Erkrankungen können erhöhte
Abgangsraten und Legeleistungsminderungen bis zu 20 % auftreten. Hierdurch
entstehen der Geflügelwirtschaft bedeutende Verluste (JOHNSON 1983, MALLISON
1983 a, b, LEVISOHN und KLEVEN 2000), die allein in den USA mit jährlich 118
Millionen Dollar beziffert wurden (MALLISON und CARPENTER 1981).
Die effektivste Bekämpfung ist die Schaffung von MG-freien Herden. Zuchtherden
sind weitgehend MG-frei (STIPKOVITIS 1985, HINZ 1988), in Deutschland konnte
aber seit 1985 wieder eine Zunahme von MG-Neuinfektionen in Ablegebetrieben
festgestellt werden (HINZ und RYLL 1996). Die Eradikation des Erregers ist vor
allem in solchen Ablegebetrieben schwierig, in denen die Infektketten durch das
permanente Nebeneinander von Legehennen verschiedener Altersgruppen (multiple-
age) nicht unterbrochen wird; dies hat zur Folge, dass neu eingestallte MG-freie
Junghennen sich von den vorhandenen MG-infizierten Althennen anstecken. Auch
die regionale Zunahme von „single-age“-Betrieben und die nachlassende Sorgfalt im
Rahmen der Sicherung des Gesundheitszustandes der Herden (Biosecurity) bieten
dem Erreger die Möglichkeit (KALETA u. SIEGMANN 1993), in der Population zu
persistieren (HILL 1995, HINZ und RYLL 1996, KLEVEN 1994).
Eine Minderung wirtschaftlicher Verluste durch eine MG-assozierte
Legeleistungsminderung und schlechtere Futterverwertung ist durch Vakzination der
Junghennen vor der Umstallung in infizierte Ablegebetriebe möglich. Hierzu stehen
sowohl Lebend- als auch Totimpfstoffe zur Verfügung (KLEVEN 1994). Die Impfung
induziert eine spezifische Partialimmunität, die zwar eine klinisch manifeste
Mykoplasmose unterbindet, jedoch reicht der induzierte Impfschutz nicht aus, um bei
den vakzinierten Tieren eine Infektion mit einem MG-Feldstamm zu verhindern.
Vakzinierte Legehennen sind jedoch in signifikantem Maße besser vor einer MG-
bedingten Leistungsminderung geschützt als nicht vakzinierte Tiere.

12
In den USA ist die Impfung mit dem MG-Stamm F als Lebendimpfstoff möglich
(LEVISOHN und KLEVEN 1981, GLISSON 1987). Der Einsatz dieser Lebendvakzine
ist jedoch auf Grund der Pathogenität des Impfstammes für Puten, Broiler und auch
im geringeren Maße für Legehennen (CARPENTER et al. 1981) problematisch, und
daher ist sie in Deutschland nicht zugelassen (Hinz 1988). In der Bundesrepublik ist
jedoch die Vakzination mit inaktiviertem Impfstoff möglich. Zur Zeit sind in
Deutschland zwei Impfstoffe zugelassen.
Ziel dieser experimentellen Arbeit war es, die neuentwickelte inaktivierte Öl-
Adjuvans-Vakzine „Talovac 104 MG“ der Firma LAH (Lohmann Animal Health) auf
ihre Verträglichkeit und Wirksamkeit bei Legehennen zu prüfen.

13
2. Schrifttum
Schwerpunktmäßig soll hier auf die Entwicklung von Totimpfstoffen gegen
Mycoplasma gallisepticum eingegangen werden, da eine Literaturübersicht über die
Entwicklung von Lebendimpfstoffen bereits von ODENTHAL (1996) im Rahmen einer
Dissertation vorgelegt wurde.
2.1. Historisches zu ersten Vakzinationsversuchen gegen MG
J. B. NELSON untersuchte in den dreißiger Jahren dieses Jahrhunderts die
Ursachen von Atemwegserkrankungen bei Hühnern, Laborratten und Mäusen. Von
1933 bis 1938 veröffentlichte er ca. 10 Arbeiten, die sich mit der ursächlichen
Klärung respiratorischer Erkrankungen des Geflügels befassen (LANCASTER und
FABRICANT 1988). 1935 beschrieb NELSON erstmalig den Erreger von Schnupfen
beim Huhn, welcher sich durch seinen Krankheitsverlauf von dem durch H.
paragallinarum ausgelösten Geflügelschnupfen unterschied. Er konnte von diesen
Hühnern nasales Exsudat gewinnen und durch eine experimentelle Verabreichung
des erregerhaltigen Inokulums diese Erkrankung reproduzieren. Bei dieser
Schnupfenform traten erst nach einer bis vier Wochen bei einzelnen Versuchstieren
die ersten Krankheitssymptome auf. Diese hielten über einen Zeitraum von zwei
Monaten oder länger an. In dem von den Tieren ausgeschiedenen Exsudat konnte H.
paragallinarum nicht nachgewiesen werden. Das isolierte Exsudat der erkrankten
Hühner enthielt jedoch stets andere Bakterien, welche von NELSON (1935) jedoch
als sekundäre Erreger gewertet wurden. Es gelang ihm, mit einem nach Filtration
bakterienfreien Exsudat bei Hühnern den typisch protrahierten Krankheitsverlauf zu
reproduzieren. Mikroskopisch ließen sich in diesem Exsudat sehr kleine,
gramnegative, kokkoide Körperchen, welche gewöhnlich extrazellulär lagen,
nachweisen. Die Größe der Körperchen gab der Autor mit 0,1 – 0,5 µm an.
Im gleichen Jahr schlug KLIENEBERGER (zitiert nach: LANCASTER und
FABRICANT 1988) den Begriff „pleuropneumonia-like organisms“ (PPLO) für diese
Organismen vor. 1939 gelang es NELSON, das Wachstum der „coccobacilliform

14
bodies“ in „inaktiviertem“ Überstand von sedimentiertem Embryonalgewebe
nachzuweisen.
DELAPLANE und STUART (1943) beschrieben die Erkrankung einer Legeherde von
5000 Hennen, deren Legeleistung nach einer Infektion mit einem unbekannten
Erreger auf 30 % abfiel und die klinische, respiratorische Erscheinungen aufwiesen.
Sie verwandten erstmals den Begriff „chronic respiratory disease“ (CRD) für diese
ERkrankung, ohne dass sie einen Zusammenhang zu den PPLO herstellen konnten.
VAN HERICK und EATON gelang 1945 die Isolierung von PPLO aus embryonierten
Hühnereiern und die Anzüchtung in einer Bouillon mit Pferdeserum und Glukose. Sie
beobachteten, dass nach einer viertägigen Bebrütung (37 °C) in der Bouillon keine
lebenden Organismen mehr nachzuweisen waren. Dies führten sie auf die Metabo-
lisierung der Glukose und das damit verbundene Absinken des pH-Wertes zurück.
Mit der erregerhaltigen Bouillon konnten sie eine Agglutination von Hühner-
erythrozyten auslösen. Die Zugabe von Serum eines zuvor hyperimmunisierten
Kaninchens verhinderte diese Agglutinationsreaktion.
Erst 1952 konnten MARKHAM und WONG jedoch den Zusammenhang zwischen
diesem Erreger (PPLO) und der „chronic respiratory disease“ (CRD) der Hühner und
der „Infektiösen Sinusitis“ der Puten nachweisen, indem sie verschiedene Isolate von
erkrankten Tieren kulturell, morphologisch und in klinischen Versuchen verglichen.
DOMERMUTH (1957) beschrieb die PPLO als Ursache des chronischen
Geflügelschnupfens bei Hühnern. Er untersuchte, ob durch eine Impfung eine
Schutzwirkung gegen Luftsackentzündungen induziert werden kann. Nach einer
intramuskulären bzw. subkutanen Applikation von 0,1 bis 0,2 ml Suspension
lebender PPLO des Winchester-Stammes bei jungen Hühnern wurden diese in
einem Zeitabstand von vier bis neun Wochen nach dieser Impfung mit dem
homologen Stamm experimentell infiziert. Die Applikation des Inokulums erfolgte in
den Brustluftsack. Eine Woche nach dieser experimentellen Belastungsinfizierung
(EBI) wurden die Tiere getötet und pathologisch-anatomisch untersucht. Die
Luftsackveränderungen bewertete DOMERMUTH (1957) mit Hilfe einer vierstufigen
Skala und stellte fest, dass bei den geimpften Hühnern die Luftsackentzündungen
„weniger stark“ ausgeprägt waren.

15
ADLER (1958) impfte Puten mit einem inaktivierten bzw. attenuierten (90. Passage)
PPLO-Stamm S6. Vier Wochen nach der intramuskulären Injektion konnte er keine
spezifischen Titer agglutinierender Antikörper feststellen. Zu diesem Zeitpunkt
infizierte er die Puten durch eine intraperitoneale Injektion mit dem homologen
PPLO-Stamm. 21 Tage später wurden die Puten getötet und pathologisch-
anatomisch sowie serologisch untersucht. Er stellte bei allen Tieren ausgedehnte
Luftsackentzündungen fest. Diese Luftsackentzündungen erschienen ADLER (1958)
bei den Puten, die mit dem attenuierten Impfstoff intraperitoneal vakziniert worden
waren, „weniger stark“ ausgebildet. Bei serologischen Untersuchungen wurden nach
experimenteller Infizierung in allen untersuchten Blutproben spezifische Antikörper
gegen den PPLO-Stamm nachgewiesen.
EDWARD und KANAREK (1960) führten für die „pleuropneumonia-like organisms“
(PPLO), welche die „Infektiöse Sinusitis“ der Puten und die „chronic respiratory
disease“ (CRD) der Hühner auslösten, den Namen Mycoplasma gallisepticum (MG)
ein.
ADLER et al. (1960) suchten nach einer Möglichkeit, CRD-freie Hühner und Puten
durch Impfung mit Lebend- oder Totimpfstoffen vor einer Infektion durch
Mykoplasmen zu schützen. Zur Impfstoffherstellung und Infizierung verwandten sie
verschiedene Verdünnungen von Suspensionen des MG-Stammes S6. Die von
ihnen verwandten Lebendimpfstoffe enthielten 109 KBE/ml. Die Inaktivierung erfolgte
durch eine 30-minütige Ultraschallbehandlung. Die EBI der Versuchstiere erfolgte
durch Inokulation in den linken abdominalen Luftsack, die pathologisch-
anatomischen Veränderungen bewerteten sie mittels einer vierstufigen
Bewertungsskala auf Grundlage eines numerischen Punktesystems, dessen
gemittelte Werte sie als Luftsackentzündungs-Wert (LSE-Wert) angaben. ADLER et
al.(1960) stellten in ihren Experimenten fest, dass 106,5 KBE einer
Mykoplasmensuspension der 5. in vitro-Passage des S6-Stammes nötig sind, um 50
% der sieben Wochen alten Hühnerküken zu infizieren. Zehn Legehennen im Alter
von fünf Monaten infizierten ADLER et al. (1960) über die Eierstöcke mit dem MG-
Stamm S6. Daraufhin sank die Legeleistung dieser Hennen um 16 %. Vier Monate
später wurde eine EBI mit dem MG-Stamm F durchgeführt; nun fiel die Legeleistung

16
der Kontrollgruppe um 12 % ab, während die der bereits infizierten Gruppe konstant
blieb. Nach intravenöser Applikation von inaktiviertem Impfstoff konnten ADLER et al.
(1960) zwar bei diesen Tieren die höchsten Titer spezifischer Antikörper gegen MG
ermitteln, jedoch traten in dieser Versuchsgruppe nach experimenteller Infizierung
die ausgedehntesten Luftsackentzündungen auf, deren LSE-Wert sie für diese
Gruppe mit ≥ 2 angaben. Hühner, die i.m. geimpft worden waren, erkrankten in
einem geringeren Ausmaß (LSE-Wert: ≤ 1,5).
Mc MARTIN und ADLER (1961) arbeiteten in ihren Experimenten mit bei
Versuchsbeginn vier Wochen alten, SPF-freien Hühnerküken. Mit Hilfe von
logarithmisch verdünnten Mykoplasmensuspensionen, die zwischen 101 bis 107 KBE
enthielten, ermittelten sie die Dosis, die bei einer direkten Inokulation in den linken
abdominalen Luftsack, respektive einer intranasalen Applikation, bei 50 % der
Versuchstiere im Luftsack eine pathologische Veränderung herbeiführt (AID50). Zwei
Wochen nach Verabreichung des Inokulums wurden die Tiere getötet und
pathologisch-anatomisch untersucht. Sie ermittelten für die direkte Inokulation in den
Luftsack eine AID50 von 101,8 und für die intranasale Applikation eine AID50 von 102,9
Mykoplasmen. In weiteren Experimenten stellten sie fest, dass die intranasale
Applikation von 103 lebenden Mykoplasmen des S6-Stammes einen Schutz vor einer
Erkrankung nach einer erneuten experimentellen Infizierung mit dem homologen
Stamm bewirkte. In einem anderen Versuch impften sie Hühner durch die intranasale
Gabe von 108 Mykoplasmen und infizierten sie 21 Tage nach der Impfung durch
Inokulation des homologen Stammes in den linken abdominalen Luftsack. Bei den
Impflingen waren auch nach einer Infizierungsdosis von 300 x AID50 keine
Luftsackentzündungen festzustellen.
WARREN et al. (1968) vakzinierten mehrere Gruppen MG-freier Hühnerküken im
Alter von einem bis 14 Tagen mit einem inaktivierten MG-Impfstoff (MG-Stamm
293C-5261). Die Applikation des Impfstoffes erfolgte entweder subkutan oder
intranasal, einmalig oder zweimalig in einem Abstand von zwei Wochen. Die EBI mit
dem homologen Stamm erfolgte im Alter von 14 bis 28 Tagen mit 0,25 bzw. 0,5 ml
einer Suspension mit je 105,5 KBE/ml zwei Wochen post vaccinationem. Bei 89% der
nicht geimpften Kontrolltiere konnten post mortem Luftsackentzündungen festgestellt

17
werden. Der Schutz vor entzündlichen Veränderungen betrug in den geimpften
Gruppen zwischen 10 und 70%. Dieser Schutz vor einer Erkrankung zeigte sich nicht
nur in der Abnahme der Morbidität, sondern auch in einer Verminderung entzündlich
veränderter Lungen bei den Küken der Impfgruppe.
ADLER und LAMAS DA SILVA (1970) untersuchten die Wirksamkeit inaktivierter
MG-Impfstoffe in Abhängigkeit von der Applikationsroute. Für diese Versuche
verwendeten sie MG-freie Hühner. Sie stellten nach ein- bzw. zweimaliger
intravenöser Gabe des inaktivierten Impfstoffes einen Schutz vor
Luftsackentzündungen nach Belastungsinfizierung mit 109 Mykoplasmen des S6-
Stammes je Tier fest. Die durchschnittlichen LSE-Werte lagen mit 0,9 bzw. 1,0
deutlich unter denen der Kontrollgruppe (≥ 2). Die intranasale Gabe des inaktivierten
Impfstoffes erzeugte keinen Schutz. Bei Versuchen mit lebenden Mykoplasmen,
konnte eine Schutzwirkung, unabhängig von der Applikationsart, feststellt werden,
jedoch war die Belastbarkeit dieser Vakzination abhängig von der Anzahl der
verimpften Mykoplasmen. Aufgrund dieser Ergebnisse gelangten ADLER und
LAMAS DA SILVA (1970) zu der Auffassung, dass durch eine i.v. Impfung mit
inaktivierten Mykoplasmen ein Schutz gegen Luftsackentzündungen erzielt werden
kann, wenn die Belastungsinfizierung nicht später als 25 Tage nach der Impfung
erfolgt. Die Wirksamkeit der lokalen Applikation von inaktivierten MG-Adjuvans-
Impfstoffen untersuchten HAYATSU et al. (1974) bei MG-freien Mastküken. Sie
verwendeten für die Impfstoffherstellung und die EBI den virulenten MG-Stamm S6-
K. In verschiedenen Versuchsanordnungen untersuchten sie die Wirksamkeit von
Impfstoffen mit oder ohne Zusatz verschiedener Adjuvantien in Abhängigkeit von der
Applikationsweise, der Menge des verwendeten Impfstoffes bzw. der Menge der bei
der EBI verwendeten Mykoplasmen sowie die Dauer einer eventuell erzielten
Wirkung. Als Parameter für die Effizienz der Impfung wurden die post mortem
festgestellten Organveränderungen, die histologischen Befunde sowie die
Reisolationsrate von MG herangezogen. Hierzu wurden die Organveränderungen in
einer Skala (siehe ADLER 1960) bewertet. Die dreimalige, intramuskuläre Gabe
verschiedener Impfstoffarten erzeugte keinen Schutz vor einer Infektion der
Atemwege mit MG. Die intranasale und aerogene Applikation des Impfstoffes

18
hingegen führte zu einer Verminderung des Ausmaßes der Luftsackentzündungen
und/oder der Besiedlung des Tieres durch MG (HAYATSU et al. 1974). Nach der
Inokulation von 5,0 x 109 inaktivierten Mykoplasmen mittels Aerosol konnten sie
einen vollständigen Schutz vor einer Infektion und Luftsackentzündungen nach einer
EBI mit 7,4 x 103 KBE feststellen. Diesen Schutz beobachteten sie auch bei
geimpften Hühnern, die sie vier Wochen lang mit artifiziell infizierten Hühnern
zusammenhielten. In einem weiteren Experiment testeten sie die Dauer des
Impfschutzes in Abhängigkeit von ihrem verwendeten Impfkonzept, das eine
dreimalige Vakzination im Abstand von jeweils zehn Tagen mit 5,0 x 109 inaktivierten
Mykoplasmen vorsah. HAYATSU et al. (1974) konnten bei drei von neun Hühnern
auch sechs Monate nach der Impfung noch einen Schutz vor Luftsackentzündungen
nach einer EBI mit MG feststellen.
Diese Ergebnisse konnten HAYATSU et al. (1975) in einem kontrollierten
Feldversuch bestätigen. Sie impften via Aerosol 1760 Küken im Alter von drei, 14
und 25 Tagen mit 2,0 x 1010 inaktivierten Mykoplasmen des S6-K Stammes. 7170
Küken wurden als Kontrollgruppe unter identischen Bedingungen gehalten. Bei den
geimpften Küken war keine erhöhte Abgangsrate zu verzeichnen, die
Gewichtsentwicklung entsprach am 95. Tag der Zunahme der Kontrollgruppe. Bis
zum 74. Lebenstag konnten keine Mykoplasmen reisoliert werden. Dies war jedoch
in der Kontrollgruppe am 84. und 134. Lebenstag möglich. In den Luftsäcken konnten
bei der pathologisch-anatomischen Untersuchung am 84. und 134. Versuchstag
keine signifikanten Unterschiede im Ausmaß der Luftsackentzündung festgestellt
werden. Jedoch konnten HAYATSU et al. (1975) am 84. Versuchstag bei sechs von
zehn Hühnern der Kontrollgruppe Mykoplasmen reisolieren. Dies war bei keinem der
zehn untersuchten Küken der Impfgruppe der Fall. Nach einer aerogenen
Verabreichung von NDV am 45. Versuchstag zeigten sich in der Kontrollgruppe
deutlichere respiratorische Krankheitserscheinungen als in der geimpften Gruppe. Es
konnte nicht geklärt werden, ob diese Erkrankung Folge einer natürlichen Infektion
mit MG, der Applikation von NDV oder einer Kombination von beidem war. Im
Ergebnis wurde die Impfung mittels eines Aerosols als Möglichkeit bewertet, Hühner
vor einer natürlichen Infektion durch MG zu schützen.

19
Rimmler et al. (1978) untersuchten die Schutzwirkung von inaktivierten Vakzinen
gegen „Geflügelschnupfen“, welcher durch Haemophilus paragallinarum (HG)
und/oder MG hervorgerufen wurde. Es wurden in mehreren Versuchsanordnungen
verschiedene Kombinationen von MG- und HG-Impfstoffen untersucht. Zur
Herstellung der MG-Vakzine wurde der R-Stamm verwendet; die für die Versuche
verwendeten Hühner waren bei Versuchsbeginn 16 Wochen alt. Die Impfung erfolgte
im Alter von 16 bzw. 19 Wochen ein- bzw. zweimalig subkutan im Nacken. Maßstab
für die Schutzwirkung war die Häufigkeit von klinischen manifesten Erkrankungen im
Bereich der oberen Atemwege und der Luftsäcke. In ihren Versuchen konnten sie die
von NELSON (1933, 1938) als Typ I und Typ III klassifizierten Schnupfenformen bei
den Versuchshühnern reproduzieren. Eine zweimalige Impfung mit der kombinierten
MG/HG-Vakzine schützte die Tiere am besten vor einer Erkrankung nach der EBI mit
dem MG-Stamm R (3,1 x 108 KBE/ml) und dem HG-Stamm W. In der Impfgruppe
stellten sie nur bei drei von zehn Hühnern nicht genau definierte respiratorische
Erscheinungen fest, hingegen traten solche bei sieben von zehn Tieren der
Kontrollgruppe auf. Bei der pathologisch-anatomischen Untersuchung fanden
RIMMLER et al. (1978) lediglich bei zwei Hühnern der Impf- aber bei sieben Hühnern
der Kontrollgruppe eine Luftsackentzündung.

20
2.2. Wirksamkeit von MG-Totimpfstoffen
1979 untersuchte YODER, ob eine einmalige Impfung von MG-freien Legehennen im
Alter von 18 Wochen mit inaktivierten Vakzinen unterschiedlicher
Herstellungsverfahren einen Schutz vor einer klinischen Erkrankung oder einer
Legeleistungsminderung nach einer artifiziellen Infizierung bewirkte. Nach
intranasaler EBI in der 28. Lebenswoche mit 0,2 ml einer 24-Stunden-Bouillonkultur
des R-Stammes konnte er weder in der Impf- noch in der Kontrollgruppe Anzeichen
einer klinischen Erkrankung oder einer Minderung der Legeleistung feststellen. Den
Anstieg der Antikörpertiter nach der Infizierung mit MG wertete YODER (1979) nicht
als Zeichen für einen Schutz gegen eine Legeleistungsminderung durch Mycoplasma
gallisepticum.
PANIGRAHEY et al. (1981) testeten die Wirksamkeit von drei inaktivierten MG-
Impfstoffen unterschiedlicher Zusammensetzung an Hand klinischer Erkrankungen
nach der Belastungsinfizierung mit dem MG-Stamm R via „eye drop“. 150 MG-freie
Eintagsküken wurden in drei Impf- und eine Kontrollgruppe geteilt. Im Alter von fünf
Wochen erfolgte die erste Vakzination. Die Hälfte jeder Impfgruppe wurde sechs
Wochen später ein zweites Mal vakziniert. Zwei Wochen nach der Zweitimpfung
erfolgte die EBI. Bei den Hühnern, die zweimal mit einer Wasser in Öl-Vakzine
geimpft worden waren, wurden keine sichtbaren Anzeichen einer klinisch manifesten
Erkrankung festgestellt. Die signifikant höheren Titer spezifischer Antikörper in der
Gruppe, die zweimal mit einer Öl-Emulsions-Vakzine geimpft worden war, und der
fehlende weitere Titeranstieg nach der Infizierung der Küken bewerteten sie als
Impfschutz gegen MG.
Im Rahmen des AAAP Mycoplasmosis Symposiums berichtete YODER (1983) über
Untersuchungen, in denen die Schutzwirkung inaktivierter MG-Öl-Adjuvansimpfstoffe
vor Legeleistungsminderungen bei Hennen nach Impfung in der 18. Lebenswoche
(Lw) geprüft wurden. In der 28. bis 30. Lebenswoche wurden die Hennen zunächst
mit IBV und zwei bis drei Tage später mit MG infiziert. In zwei von vier
Versuchsdurchgängen war die Legeleistung der nicht geimpften Kontrollgruppen in
der zweiten Woche nach der experimentellen Infizierung „ein wenig geringer“.

21
YODER (1983) beschrieb, dass die Zusatzbelastung durch IBV notwendig war, um
„gute“ Auswertungen, d.h. statistisch verwertbare Unterschiede in der Legeleistung
zu erzielen. Lokale Impfreaktionen traten nach der subkutanen Injektion im
Nackenbereich nicht auf.
HILDEBRAND et al. (1983) untersuchten die Wirksamkeit einer inaktivierten Öl-
Adjuvans-Vakzine, „MG-Bac®“ (Salsbury Lab. Charles City, Iowa USA), hinsichtlich
ihres Schutzes gegen die Mykoplasmose bei Küken und vor
Legeleistungsminderungen bei Hennen. Sie verwendeten für die Laborstudien SPF-
Küken, welche im Alter von drei Wochen geimpft und vier Wochen später in den
rechten Infraorbitalsinus mit verschiedenen MG-Stämmen (R, S-6, PG-31 und 1150)
infiziert wurden. Sieben bis 14 Tage p. i. erfolgte die Untersuchung. Als Kriterium für
eine klinische Erkrankung wurde das Austreten mukösen Exsudates aus den
Nasenöffnungen gewertet. Bei den geimpften Küken stellte man einen 90 bis 95-
prozentigen Schutz vor einer klinischen Erkrankung fest. In den ungeimpften
Kontrollgruppen fanden sie bei 44% der Küken Luftsackentzündungen, in der
Impfgruppe hingegen nur bei 10 % der Versuchstiere. HILDEBRAND et al. (1983)
gelang die Reisolation von MG aus der Trachea bei allen Tieren der Kontrollgruppen.
Bei den Probanden der Impfgruppen war die Reisolation von MG auch nach einer
Subkultivierung nur bei 76 % der Tiere möglich. Für die Feldversuche wurden
110000 MG-freie Legehennen zweier Zuchtlinien verwendet. Den Tieren der
Impfgruppen wurde jeweils 0,5 ml der MG-Öl-Adjuvans-Vakzine subkutan im
Nackenbereich appliziert. 50 % der Hennen erhielten nur eine einmalige Impfung im
Alter von 14 Wochen, die andere Hälfte wurde im Alter von 18 Wochen erneut
vakziniert. Die Autoren empfahlen die Vakzination im unteren Drittel des Halses, da
sie Ödeme im Augenbereich infolge Impfstoffapplikation im oberen Halsabschnitt
beobachtet hatten. Eine Herde von ca. 165000 Hennen wurde in der zehnten
Lebenswoche mit lebenden Mykoplasmen des F-Stammes geimpft, um zwischen
Lebend- und Totimpfstoffen vergleichen zu können. Sie verglichen ihre Ergebnisse
mit der Legeleistung, die jährlich als Stall- bzw. Zuchtlinienstandards errechnet
wurde. 72 Hennen stallten sie als ungeimpfte Kontrollgruppe mit auf. Die einmal
geimpften Legehennen zeigten einen Schutz von 50 % bis 89 % vor einer klinisch

22
manifesten Mykoplasmose nach einer MG-Feldinfektion. Bei den Hühnern, die
zweimal geimpft worden waren, betrug der „Schutz“ zwischen 70 % und 100 %,
während 90 % bis 100 % der Kontrolltiere im Verlaufe des 64 Wochen währenden
Versuches erkrankten. Die Legeleistung der zweimalig mit MG-Bac® geimpften
Hennen lag mit 233,5 Eiern pro aufgestallter Henne über der Legeleistung der Herde,
die mit einer Dosis des F-Stammes (220,7 Eier/Henne) geimpft worden war. Den
Herdenstandard gaben sie mit 222 Eiern je aufgestallter Henne an. Auch die
Futterverwertung der mit MG-Bac® vakzinierten Herden lag über der mit dem F-
Stamm vakzinierten. HILDEBRAND et al. (1983) schätzten die untersuchte Vakzine
als sicher und wirksam ein.
Grundlage für die Effizienz einer Impfung mit einer inaktivierten Öl-Adjuvans-Vakzine
gegen MG stellte für YODER et al. (1984) der Schutz von Broilerküken vor
Luftsackentzündungen dar. Verschiedene Gruppen SPF-freie Küken wurden am
ersten, siebten oder 14. Lebenstag mit je 0,5 ml bzw. 0,1 ml der Öl-Adjuvans-
Vakzine subkutan im Nacken geimpft. Im Alter von sechs Wochen erfolgte die EBI
der Impf- und Kontrollgruppen. Zunächst wurden die Küken intratracheal mit IBV und
zwei Tage später mittels eines erregerhaltigen Aerosols mit dem MG-Stamm R
infiziert. Gleichzeitig wurden die Küken in eine kalte Umgebung gebracht. 21 Tage
nach dem Challenge wurden die Tiere getötet und pathologisch-anatomisch
untersucht. Die Schwere der Luftsackveränderungen wurden in einer Skala von 0
(keine Veränderungen) bis 4 (schwere Luftsackentzündungen mit viel Exsudat im
Bereich aller Luftsäcke) erfaßt und der LSE-Wert als arithmetischer Mittelwert
angegeben (siehe ADLER 1960). In dem ersten ihrer Experimente stellten YODER et
al. (1984) fest, dass Küken, die im Alter von einer bzw. zwei Wochen geimpft worden
waren, im Vergleich mit den Kontrollgruppen, einen signifikant höheren Schutz vor
Luftsackentzündungen aufwiesen. Ebenso erkrankten von 60 Küken, welche sie im
Alter von 15 Tagen geimpft hatten, nur 4 (6,6, %) Tiere, während in der
Kontrollgruppe bei 17 von 60 Küken (28,3 %) eine Luftsackentzündung festgestellt
wurde. Nach einer Impfung mit 0,1 ml der Vakzine erkrankten nur 8,3 %, in der
Kontrollgruppe hatten 35 % der Küken Luftsackentzündungen. Im selben Versuch

23
konnte in einer Gruppe, die mit 0,5 ml geimpft worden war, ein Schutz von 98,3 %
vor Luftsackentzündungen erzielt werden.
ZOLLI (1984) berichtete über Feldversuche mit einer inaktivierten Öl-Adjuvans-
Vakzine. Im Zusammenhang mit den Angaben zum verwendeten Impfstoff, der
Impfweise und dem Zeitpunkt der Infizierung verwies er auf die Publikation von
HILDEBRAND et al. (1983). Zwei Herden mit je 40000 Hühnern wurden im Alter von
16 Wochen einmalig geimpft. Es konnte ein Schutz vor einer
Legeleistungsminderung nach intrasinuidaler Belastungsinfizierung mit dem MG-
Stamm R festgestellt werden. Eine geimpfte Legehennenherde (n= 20000) wies im
Vergleich mit einer ungeimpften Kontrollherde (n= 20000) bis zum Alter von 47
Wochen eine um zwei Eier höhere Legeleistung auf. Dies stimmte mit dem Ergebnis
einer anderen Studie überein, in welcher zwei Herden mit je 75000 Hennen
verglichen wurden. Eine dieser Herden wurde einmal geimpft, in der ungeimpften
Herde kam es zu einem Ausbruch einer durch MG-bedingten CRD, so daß dieser
Herde von der 33. bis zur 60. Lebenswoche mit Tylosin medikamentiertes Futter
verabreicht wurde. Die geimpfte Herde erreichte einen um 4,6 % höheren
Legeleistungspeak und legte bis zur 60. Lebenswoche im Durchschnitt sieben Eier
mehr. Im Vergleich der Legeleistung von insgesamt 600000 Legehennen wurden
fünf Herden mit jeweils 60000 Hennen mit einer inaktivierten Öl-Adjuvans-Vakzine
und fünf Herden derselben Größe mit dem F-Stamm von MG geimpft. In vier von fünf
dieser Studien legten die Herden, die mit der inaktivierten Öl-Adjuvans-Vakzine
geimpft worden waren, mehr Eier als die jeweiligen Vergleichsgruppen.
GLISSON und KLEVEN (1984) untersuchten die Wirksamkeit unterschiedlicher MG-
Impfungen im Hinblick auf eine transovarielle Übertragung von Mykoplasmen und
eine Minderung der Legeleistung nach einer artifiziellen Infizierung mit dem MG-
Stamm R. 150 MG-freie Legehennen teilten sie in fünf Gruppen auf und verglichen
eine Kontrollgruppe mit vier Impfgruppen, die unterschiedlich geimpft wurden:
1. Vakzination im Alter von 16 Wochen mit dem F-Stamm (3x106 KBE/Tier) via
eye-drop.
2. kombinierte Vakzination mit dem F-Stamm (3x106 KBE/Tier) in der 16. Woche
und mit der inaktivierten Vakzine „MG-bac®“ im Alter von 20 Wochen.

24
3. zweimalige Vakzination mit „MG-bac®“ in der 16. und 20. Lebenswoche
4. einmalige Impfung mit „MG-bac®“ in der 20. Lebenswoche.
Die Applikation des inaktivierten Impfstoffes erfolgte jeweils subkutan. Zum Zeitpunkt
des Produktionsmaximums in der 28. Lebenswoche wurden alle Tiere durch ein
erregerhaltiges Aerosol und durch Gabe von 1,5 x 107 Mykoplasmen je Henne in den
linken Infraorbitalsinus infiziert. In allen vier Impfgruppen konnten GLISSON und
KLEVEN (1984) feststellen, dass im Vergleich mit der Kontrollgruppe die
Übertragung von MG erst zu einem späteren Zeitpunkt nach der EBI begann. In der
Gruppe, die zweimal mit dem inaktivierten Impfstoff geimpft worden war, konnten sie
diese Übertragung erst nach 82 Tagen nachweisen. Warum gerade diese
Impfgruppe gegen Ende des Versuches in der 15. bis 25. Woche p.i. den höchsten
prozentualen Anteil infizierter Eier aufwies (3,7 % - 12,6 %), konnten GLISSON und
KLEVEN (1984) nicht erklären. Die Legeleistung der vakzinierten Gruppen lag im
Versuchszeitraum von der 19. - 54. Lebenswoche 5 bis 15 Eier über der der
Kontrollgruppe. Gründe, warum die Impfgruppe, welche sie mit der Kombination F-
Stamm und MG-bac® vakziniert hatten, über den gesamten Versuchszeitraum in der
Legeleistung zurückblieb, konnten nicht gefunden werden. Alle Impfgruppen waren
jedoch vor einer Legeleistungsminderung nach experimenteller Infizierung mit dem
MG-Stamm R geschützt. Allein in der Kontrollgruppe kam es bis zur 30.
Lebenswoche zu einen Abfall der Legeleistung von 90 % auf 20 %.
GLISSON und KLEVEN versuchten 1985, diese Ergebnisse in entsprechend
gestalteten Studien zu bestätigen. Sie verglichen eine ungeimpfte Kontrollgruppe mit
drei unterschiedlich vakzinierten Gruppen von jeweils 30 MG-freien Legehennen: Die
erste wurde zweimalig in der 17. und 21. Lebenswoche mit MG-Bac® (Salsbury Lab.
Charles City, Iowa, USA) geimpft, in der zweiten Impfgruppe wurde diese Impfung in
der 30., 41. und 52. Lebenswoche wiederholt. Die dritte Gruppe wurde erst zwei
Wochen nach der Belastungsinfektion geimpft. Diese Belastungsinfektion via Aerosol
und Injektion von 2,3 x 107 KBE/Henne in den rechten Infraorbitalsinus erfolgte in der
28. Lebenswoche zum Zeitpunkt des Legeleistungsmaximums. Die zweimalige
Impfung mit MG-Bac® (Salsbury Lab. Charles City, Iowa, USA) schützte fast
vollständig gegen die transovarielle Übertragung nach der Infizierung mit MG. Nur in

25
einem von 4725 Eiern konnten GLISSON und KLEVEN (1985) den MG-Stamm R
nachweisen. Die Boosterung der Impfung verhinderte diese Übertragung von MG
vollständig. Im Vergleich mit der Kontrollgruppe war dieser Schutz signifikant. Im
Unterschied zu den Ergebnissen ihrer Studien aus dem Jahr 1984 erfolgte auch im
weiteren Verlauf ihres Versuches bis zur 60. Lebenswoche keine transovarielle
Übertragung von MG. Nach der Belastungsinfektion fiel die Legeleistung in den
beiden zu diesem Zeitpunkt ungeimpften Gruppen von 85 % auf 30 % ab. Diese
Legeleistungsminderung betrug in den beiden, zum Zeitpunkt der Infizierung bereits
geimpften Gruppen nur 15 % bzw. 30 %. Eine einmalige Impfung von zuvor natürlich
infizierten Hennen mit MG-Bac® (Salsbury Lab. Charles City, Iowa, USA) verminderte
die transovarielle Übertragung von Mycoplasma gallisepticum, verhinderte sie aber
nicht vollständig. GLISSON und KLEVEN (1985) fanden in diesem Versuch in 4,1 %
der Eier der Kontrollgruppe Mykoplasmen. Durch die Impfung p.i. verringerte sich der
Anteil der infizierten Eier auf 1,2 %.
YODER und HOPKINS (1985) prüften einen inaktivierten Öl-Emulsions-Impfstoff in
Bezug auf seine Schutzwirkung vor einer MG-bedingten Minderung der Legeleistung
bei Legehennen, die sie aus kommerziell gehaltenden Herden und einem
Versuchslabor erhielten. Die Ergebnisse ihrer Versuche zeigten ein unterschiedliches
Bild. In ihrem ersten Experiment hatten sie 32 Hühner in der elften und 19.
Lebenswoche mit 0,5 ml des Impfstoffes subkutan im Nacken geimpft. Impf- und
Kontrollgruppe wurden in der 29. Lebenswoche mit 0,2 ml einer MG-Bouillon (Stamm
R) infiziert. YODER und HOPKINS (1985) konnten im Laufe dieses Versuches keine
Unterschiede in der Legeleistung feststellen. Dies war auch bei einem Vergleich von
in der 20. Lebenswoche geimpften 48 Hennen mit ihren unter identischen
Bedingungen gehaltenden Kontrolltieren der Fall. Beide Gruppen wurden in der 28.
Lebenswoche zunächst mit IBV infiziert. Zwei Tage danach wurde den Hühnern 0,4
ml einer MG-Kultur des R-Stammes verabreicht. Die Infizierung mit MG erfolgt
intrakonjunktival, intranasal und intratracheal. Vor dieser Infizierung wiesen beide
Gruppen eine Legeleistung von ca. 92 % auf, diese fiel zunächst auf 85 % ab, um
dann in der Legeleistung wieder auf 87 % anzusteigen. Die 48 Legehennen, die
YODER und HOPKINS (1985) im Anschluß an ein Experiment mit IBV in der 33.

26
Lebenswoche gegen MG geimpft hatten, zeigten nach einer kombinierten Infizierung
mit IBV und MG in der 38. Lebenswoche keinen Abfall in der Legeleistung. In der
entsprechenden Kontrollgruppe verminderte sich die Legeleistung von ca. 90 % auf
76 %. Nach einer Impfung gegen MG in der 20. Lebenswoche kam es bei Hennen
nach einer kombinierten Infizierung mit IBV und MG zum Zeitpunkt des
Legeleistungsmaximums in der 29. Lebenswoche nur zu einer Verringerung der
Legeleistung von 93 % auf 91 %. In einem weiteren Versuch wurden Hennen durch
ein erregerhaltiges Aerosol infiziert. Die Legeleistung der vakzinierten Gruppe betrug
vor der experimentellen Infizierung 85 %, die der Kontrollgruppe 82 %. Eine dritte
Gruppe, welche weder geimpft noch infiziert worden war, zeigte im Verlaufe des
Versuches eine Legeleistung von 78 % bis 80 %. Im Zeitraum von zwei Wochen
nach der EBI mit MG fiel die Legeleistung der Kontrollgruppe auf 66 % ab. Die
vakzinierte Gruppe zeigte nur einen leichten Legeleistungsrückgang auf 80 %,
erholte sich aber binnen drei Wochen auf 83 % bis 85 %. Die Legeleistung der
Kontrollgruppe stieg lediglich wieder auf 76 % bis 79 % an. Anzeichen einer
klinischen Erkrankung zeigten sich nur bei den Hennen der ungeimpften
Kontrollgruppe, ebenso konnten in dieser Gruppe eine Zunahme von Abweichungen
in der Größe, Form und Schalenqualität der Eier festgestellt werden. YODER und
HOPKINS (1985) konnten keine Unterschiede bei der Reisolation von MG feststellen.
KLEVEN (1985) untersuchte, ob nach einer Impfung mit der kommerziell erhältlichen
Vakzine MG-Bac® (Salsbury Lab. Charles City, Iowa, USA) sich die Anzahl von
Mykoplasmen in der Trachealschleimhaut nach einer experimentellen
Belastungsinfizierung mit MG vermindert. Hierzu bildete er drei Gruppen mit je 75
SPF-Küken, die aus normalen Feldherden stammten. Die erste Gruppe wurde in der
15. und 19. Lebenswoche, die zweite Gruppe nur einmalig in der 19. Lebenswoche
geimpft. Die Impfdosis betrug jeweils 0,5 ml und wurde subkutan verabreicht. Eine
Gruppe von 75 Hühnern blieb als Kontrollgruppe ungeimpft. In der 23. Lebenswoche
wurden alle Hühner mit je 1,5 x 105 KBE/Tier des MG-Stammes R intratracheal
infiziert. Von jeweils 20 Tieren jeder Gruppe wurden in der 15., 19. und 23. Woche
Blut entnommen und mittels SSA und HAH auf MG-Antikörper untersucht. Zufällig
ausgewählte Tiere wurden im Verlauf des Versuches seziert. Bei den vakzinierten

27
Hühnern konnte bis zur achten Woche p.i. eine quantitativ geringere Kolonisation der
Trachea mit Mykoplasmen festgestellt werden, jedoch waren die Unterschiede im
Verhältnis zur Kontrollgruppe nicht signifikant. Ab der achten Woche p.i. bis zum
Versuchsende 28 Wochen p.i. konnte KLEVEN (1985) keine Unterschiede mehr
feststellen.
In Fortführung seiner Arbeiten aus dem Jahre 1983 überprüfte HILDEBRAND (1985)
den Schutz vor einer Mykoplasmose bei Küken und Legehennen nach einer Impfung
mit der inaktivierten Öl-Adjuvans-Vakzine MG-Bac® (Salsbury Lab. Charles City,
Iowa, USA). Für seine Laborstudien verwendete er sowohl SPF-Küken als auch
kommerziell erhältliche Broilerküken. Diese wurden in den Versuchen
unterschiedlichen Vakzinations- und Infektionsprozeduren unterzogen. Die EBI
erfolgte in den rechten Infraorbitalsinus mit dem MG-Stamm R, die Infektionsdosis
lag zwischen 2 x 104 und 1 x 105 KBE/Tier. Jeweils sieben Tage nach der Inokulation
des Agens wurden die Hühner auf klinische Erkrankung, d. h. auf die
Nachweisbarkeit eines nasalen Exsudates untersucht. In diesen Versuchen zeigte
sich, daß die SPF-Küken, welche in der ersten Lebenswoche geimpft worden waren,
vor einer MG-bedingten Erkrankung geschützt waren. Dieser Schutz war bei Gabe
der vollen Impfstoffdosis von 0,5 ml mit 95 % Schutz vier Wochen nach der
Infizierung besser ausgeprägt als bei der Gabe von nur 0,2 ml (80 %). Die Impfung
von zuvor mit MG-Feldstamm infizierten Hühnern bewirkte einen vollständigen
Schutz vor einer vier Wochen später stattfindenden EBI mit 1 x 105 Mykoplasmen.
Die Dokumentation der Feldversuche erscheinen identisch mit denen, die ZOLLI
(1984) veröffentlichte. Bei dem Vergleich der mit MG-Bac® (Salsbury Lab. Charles
City, Iowa, USA) geimpften Herden mit den Herden, die mit der Lebendvakzine
(Conn. F-Stamm) geimpft wurden, unterließ HILDEBRAND (1985) allerdings die
Darstellung des Feldversuches, in dem die mit dem MG-Stamm F geimpfte Herde in
dem Zeitraum von 60 Wochen eine um sieben Eier höhere Legeleistung aufwies.
Die vertikale Übertragung von Mykoplasmen stellte für SASIPREEYAJAN et al.
(1985) das Arbeitsfeld dar. Zur Herstellung des inaktivierten Öl-Adjuvans-Impfstoffes
verwendete er den MG-Stamm R. In einem vierwöchigem Abstand erfolgte eine
zweimalige Vakzination von seropositiven Elterntiere im Alter von 12 bis 20 Wochen.

28
Die Dosis betrug 0,5 ml und wurde subkutan verabreicht. Diese Vakzination erfolgte
in zwei aufeinander folgenden Generationen. Die Hühner der dritten Generation
wurden nicht vakziniert. Von den Legehennen der zweiten Generation wurden in der
33. Lebenswoche 36 Eier kulturell auf MG untersucht. In keinem der untersuchten
Eier fanden SASIPREEYAJAN et al. (1985) Mykoplasmen. Von den Hühnern der
dritten Generation wurden am ersten Lebenstag, in der sechsten, elften, und 31.
Lebenswoche stichprobenartig Blutproben für die serologische Untersuchung
entnommen. In der 16. Lebenswoche entnahmen SASIPREEYAJAN et al. (1985)
von allen 700 Hühnern dieser Studie Blutproben. In keinem der untersuchten Seren
konnten sie spezifische Antikörper gegen MG nachweisen. SASIPREEYAJAN et al.
(1985) hielten es für möglich, durch dieses Impfverfahren MG aus Kleinbeständen
und Ziervogelzuchten zu eliminieren.
Der Schutz vor einer Kolonisation der Trachea mit Mykoplasmen des R-Stammes
beschäftigte TALKINGTON und KLEVEN (1985). Hierzu untersuchten sie 164
kommerziell erhältliche MG-freie Legehennen. Für ihre Experimente teilten sie die
Hennen in drei Gruppen auf. Die erste Gruppe wurde im Alter von 16 und 20 Wochen
mit 0,5 ml MG-Bac® (Salsbury Lab. Charles City, Iowa, USA) intramuskulär geimpft.
Die zweite Gruppe erhielt lediglich eine Impfung in der 20. Lebenswoche, die dritte
Gruppe diente als ungeimpfte Kontrolle. In der 24. Lebenswoche wurden die Hühner
mit 0,1 ml verschiedener Verdünnungen einer Kultur des R-Stammes von MG
intratracheal infiziert. Die Anzahl der Mykoplasmen betrug in der 10-4 Verdünnung
15000 KBE/Tier und in der 10-9 Verdünnung 0,15 KBE/Tier. Drei Tage nach der
erfolgten Infizierung wurden die Tiere getötet und kulturell untersucht. Im Vergleich
der einzelnen Gruppen konnte festgestellt werden, dass bereits 15 KBE als
„Minimale Infektiöse Dosis“ (MID) ausreichten, um die Hühner der Kontrollgruppe mit
MG zu infizieren. Die MID lag für die einmalig geimpften Hühner bei 150 KBE bzw.
bei 1500 KBE nach der zweifachen Vakzination. Ebenso betrug die ID50 für die
einzelnen Gruppen 794, 2512 und 5012 KBE.
Im Gegensatz zu HILDEBRAND (1985) konnten KHAN et al. (1986) nicht feststellen,
daß eine einmalige Impfung mit dem Impfstoff MG-Bac® (Salsbury Lab., Charles City,
Iowa, USA) bei den geimpften Hennen zu einem signifikanten Schutz vor einer

29
Legeleistungsminderung durch eine Infektion mit Mycoplasma gallisepticum führte. In
einen Betrieb, in welchem Legehennen in fünf verschiedenen, latent mit MG
infizierten Altersgruppen gehalten wurden, wurden einen Monat nach der Ausstallung
der ältesten Gruppe 7700 neue Legehennen eingestallt. Die benachbarten Gruppen
waren vor der Einstallung der neuen Legehennen mit MG infiziert. 3400 Hennen
wurden zum Zeitpunkt der Aufstallung in der 19. Lebenswoche mit Impfstoff MG-
Bac® (Salsbury Lab., Charles City, Iowa, USA) vakziniert. 4300 Hühner dienten als
ungeimpfte Kontrollgruppe. Durch den Nachweis spezifischer Antikörper gegen MG
im Serum der ungeimpften Kontrolltiere in der 26. Lebenswoche stellen KHAN et al.
(1986) fest, dass die Legehennen sich auf natürlichem Weg infiziert hatten. Der
kulturelle Nachweis von MG gelang ihnen jedoch erst in der 48. Lebenswoche in
beiden Gruppen. Zeitgleich stellten sie in beiden Gruppen Anzeichen einer
respiratorischen Erkrankung fest, ohne diese genauer zu beschreiben. Signifikante
Unterschiede in der Legeleistung waren im Verlaufe des Versuches nicht zu
erkennen.
KARACA und LAM (1986) verglichen die Auswirkung einer Vakzination mit dem
temperatursensitiven MG-Stamm TS 100 in seiner lebenden Form mit der
inaktivierten Variante des Impfstoffes in Bezug auf einen Schutz vor
Luftsackentzündungen. Für diese Versuche verwendeten sie kommerziell erhältliche,
MG-freie Küken. Im Alter von einer Woche wurden auf unterschiedliche Weise
geimpft: intranasal mit 0,1 ml TS 100 lebend (3x107 KBE/0,1ml), subkutan mit 0,1 ml
TS 100 lebend (3x107 KBE/0,1ml), intranasal mit 0,2 ml TS 100 inaktiviert (2,25x109
KBE/0,1 ml) und subkutan mit 0,5 ml TS 100 inaktiviert (2,25x109 KBE/0,1 ml). Zwei
Gruppen blieben zur Kontrolle der Versuchsergebnisse ungeimpft. Drei Wochen
nach der Impfung wurden die Küken durch die Applikation von 1x104 KBE des MG-
Stammes S6 in die Luftsäcke infiziert. Eine Woche später wurden die Probanden
getötet und pathologisch-anatomisch untersucht. KARACA und LAM (1986) fanden
deutliche Unterschiede in der Schutzwirkung der einzelnen Impfverfahren. So wies
nur eines von 54 Küken, welches intranasal mit der Lebendvariante des Impfstoffes
TS 100 geimpft worden war, Luftsackentzündungen auf. Durch die subkutane
Applikation der Lebendvariante erzielten sie eine Schutzwirkung von 81 %,

30
respektive 85 % bei Verwendung des inaktivierten Impfstoffes. Die intranasale
Verabreichung der inaktivierten Impfstoffvariante schützte dagegen nur
unzureichend (44 %) gegen Luftsackentzündungen. In den ungeimpften
Kontrollgruppen wiesen 93 % bzw. 70 % der Küken eine Woche nach der EBI eine
Luftsackentzündung auf. Die Bemühungen von KARACA und LAM (1986), aus
unveränderten Luftsäcken von Küken, die intranasal mit der Lebendvariante von TS
100 geimpft worden waren, Mykoplasmen des S6-Stammes zu isolieren, schlugen
fehl.
Mit demselben Versuchsaufbau testeten KARACA und LAM (1987) in zwei
Versuchsdurchgängen auch die Schutzwirkung des Impfstoffes MG-Bac® (Salsbury
Lab. Charles City, Iowa, USA). Sie verabreichten eine Impfstoffmenge von 0,5 ml
subkutan im Nacken. Drei Wochen nach der Impfung wurden die Küken mit 1x104
Mykoplasmen des S6-Stammes experimentell infiziert. Eine Woche später wurden
die Küken getötet und pathologisch-anatomisch untersucht. In den Impfgruppen
stellten KARACA und LAM (1987) lediglich bei 10 % bzw. 9 % der Küken
Luftsackentzündungen fest. Für die Kontrollgruppe gaben sie den Anteil der Küken
mit Luftsackentzündungen mit 63 % und 83 % an, diese Unterschiede sind in beiden
Versuchen statistisch hochsignifikant.
YAGIHASHI et al. (1987) untersuchten die Schutzwirkung einer inaktivierten
Aluminiumhydroxid-Vakzine. Für den Versuch wurden 96 SPF-Hühner der Rasse
„White Leghorn“ aus der eigenen Versuchsaufzucht verwendet, welche bei
Versuchsbeginn fünf Wochen alt waren. Die Impfgruppen wurden zweimalig geimpft,
eine Gruppe zweimal im Abstand von 14 Tagen intramuskulär, die andere
intramuskulär und intratracheal. Die Kontrollgruppe wurde nicht vakziniert. Zur
Herstellung der Vakzine und zur Infizierung wurde der SAS-Stamm verwendet, die
Konzentration der Mykoplasmen vor der Inaktivierung mit 0,1 % Formalin betrug
annähernd 109 KBE/ml. Eine Woche nach der letzten Impfung wurden alle Tiere mit
0,5 ml Mykoplasmensuspension (107 KBE/Tier) intratracheal infiziert. Vier, sieben
und 14 Tage nach der Infizierung wurden Hühner aus den Versuchsgruppen getötet,
die Tracheen in Formalin fixiert und mikroskopisch untersucht. Die Veränderungen in
der Trachealschleimhaut wurden in einer Skala von 0 bis 3 bewertet. Bei beiden

31
Impfgruppen konnten deutlich weniger entzündliche Läsionen in der Trachea durch
Mycoplasma gallisepticum festgestellt werden. Sieben Tage p.i. lag der
durchschnittliche Wert für die festgestellten Trachealläsionen für die zweimal
intramuskulär geimpfte Gruppe bei 1,0 und für die intramuskulär und intratracheal
geimpfte Gruppe bei 0,3. Im Vergleich mit der Kontrollgruppe (2,0) ergab sich für die
kombiniert geimpfte Gruppe ein signifikanter Schutz gegen entzündliche Läsionen in
der Trachealschleimhaut. Gleichzeitig wurden in den Trachealspülproben auch
signifikant weniger Mykoplasmen reisoliert.
Die Schutzwirkung von liposomalen Vakzinen vor einer Infektion der Trachea nach
einer experimentellen Infizierung mit Mycoplasma gallisepticum untersuchten
BARBOUR et al. (1987). In dem Versuch wurden Hühnergruppen, die mit sechs
unterschiedlichen liposomalen Impfstoffpräparationen und zwei Öl-Emulsions-
Vakzinen geimpft worden waren, mit den Hühnern einer ungeimpften Kontrollgruppe
verglichen. Jeweils eine von zwei identischen Impfstoffpräparationen wurde mit
Ultraschall behandelt. Bei diesem Versuch wurden 56 MG-freie Küken aus
kommerzieller Aufzucht verwendet. Zur Herstellung der Vakzine wurde der R-Stamm
benutzt. Die Impfung der Hühner erfolgte im Alter von 13 und 17 Wochen mit jeweils
einer Impfstoffmenge von 0,5 ml/Tier. Die Belastungsinfizierung erfolgte mit dem
homologen Stamm, intratracheal mit 17800 KBE/Tier. 72 Stunden nach der EBI
wurden die Hühner getötet und seziert. BARBOUR et al. (1987) fertigten
histologische Schnitte der Trachea zwischen dem ersten und fünften Trachealring
an, machten aber keine Angaben zur Beurteilung dieser histo-pathologischen
Untersuchungen. Die Schutzwirkung der einzelnen Impfstoffpräparationen war sehr
unterschiedlich, drei Gruppen zeigten einen guten Schutz gegen die Infektion der
Trachealschleimhaut. Hierzu gehörten zwei Gruppen (Gruppen 2 und 3), die mit
liposomalen Impfstoffen geimpft worden waren. 14 Tage nach der Infizierung
konnten nur bei jeweils einem von sechs Hühnern (16,67 %) MG aus der Trachea
isoliert werden. Auch in der Gruppe 8, die mit der nicht mit Ultraschall behandelten
Öl-Emulsions-Vakzine geimpft worden war, fand sich nur bei drei von acht Hühnern
(37,5 %) MG. Die Gruppe 3 war die einzige, in welcher bei keinem Huhn
Veränderungen in der Trachealschleimhaut festgestellt werden konnte. Ebenso wies

32
diese Gruppe die im Verhältnis zur Kontrollgruppe signifikant höchsten
Antikörpertiter im Respirationstrakt gegen MG auf.
SASIPREEYAJAN et al. (1987) prüften, ob durch eine inaktivierte MG-Vakzine ein
Schutz vor Legeleistungsminderung und transovarieller Übertragung nach einer EBI
mit MG erzielt werden kann. Für diese Untersuchungen verwendeten sie 49 MS- und
MG-freie Legehennen unbekannter Herkunft. Eine Impfgruppe wurde in der 19. und
23., die andere nur in der 23. Lebenswoche im Bereich der Schädelbasis subkutan
mit 0,5 ml geimpft. Zur Herstellung der Vakzine und für die Infizierung wurde der MG-
Stamm R verwendet. In der 27. Lebenswoche wurden beide Impfgruppen und die
Kontrollgruppe infiziert. Es wurden jeweils 0,1 ml Inokulum in den linken kaudalen
thorakalen Luftsack und 0,1 ml in den linken Infraorbitalsinus verabreicht. Die
Konzentration der Mykoplasmen in der Suspension betrug 1,3 x 107 KBE/ml. Zum
Zeitpunkt der Infizierung lag die Legeleistung bei allen drei Gruppen bei 90 %.
Binnen zweier Wochen fiel die Legeleistung der Kontrollgruppe auf 40 % ab und
stieg bis zum Versuchsende, sieben Wochen nach der EBI, nur auf 69 % an. In
beiden Impfgruppen war nur eine Legeleistungsminderung um ca. 5 % festzustellen.
Die Impfgruppen zeigten nach der experimentellen Infizierung eine signifikant höhere
Legeleistung. Im Zeitraum von sieben Wochen p.i. lag die Legeleistung der einmal
vakzinierten Gruppe mit 84,63 % über der jeweiligen der nur zweimal geimpften
Gruppe (79,97 %) und der Kontrollgruppe (60,33 %). In der Kontrollgruppe stellten
vier Hennen ihre Legetätigkeit ein, in den Impfgruppen war dies nur bei einem Tier
der Fall. Auch bei der Übertragung von Mykoplasmen auf die Eier zeigten sich
signifikante Unterschiede zwischen den einzelnen Gruppen. So konnten
SASIPREEYAJAN et al. (1987) bei 45,9 % der Eier der Kontrollgruppe MG von der
Dottermembran isolieren. Dieser kulturelle Nachweis war nur bei 38,55 % der Eier
der einmal vakzinierten Gruppe bzw. bei 17,65 % der zweimal vakzinierten Gruppe
möglich. Sieben Wochen nach der Infizierung fanden SASIPREEYAJAN et al. (1987)
nur in einem von zehn untersuchten Eileitern der zweimal geimpften Hennen MG. In
der einmal geimpften Gruppe war dies bei zwei, in der Kontrollgruppe bei acht von
zehn Eileitern der Fall. Auch BARBOUR und NEWMAN (1990) untersuchten die
Wirksamkeit verschiedener Präparationen von MG-Impfstoffen anhand der

33
Legeleistung und Eiübertragung von MG nach einer Belastungsinfizierung mit dem
homologen Stamm. Sie verglichen die Wirkung von zwei Öl-Emulsions-Vakzinen und
zwei liposomalen, positiv geladenen Vakzinen. Jeweils eine dieser Präparationen
erhielt einen Zusatz von Salmonella typhimurium - Zellwandprotein (STP). Die
Vakzination der 32 MG-freien Legehennen erfolgte in der 13. und 17. Lebenswoche.
Zwei Kontrollgruppen wurden nicht geimpft und wiederum eine von diesen auch nicht
infiziert. Die EBI mit dem R-Stamm wurde in der 21. Lebenswoche durch die Injektion
von jeweils 1,3 x 105 KBE/Tier in den rechten und linken Brustluftsack durchgeführt.
Die Legeleistung der einzelnen Gruppen wurde für den Zeitraum von zwei Monaten
p.i. dokumentiert. Die Vittelinische Membran der Eier, die jeweils an den beiden
letzten Tagen einer Woche gelegt wurden, wurde kulturell auf MG untersucht.
Angaben zur Legeleistung der Hennen vor der EBI machten sie nicht. Im ersten
Monat nach der Infizierung lag die Legeleistung der mit der liposomalen Vakzine
ohne STP geimpften Hennen bei 87,1 % und die der mit der Öl-Emulsions-Vakzine
ohne STP bei 82,1 %. Dieser Abfall war nicht signifikant gegenüber der Legeleistung
der ungeimpften und nicht infizierten Kontrollgruppe (88,0 %). Im Vergleich mit einer
infizierten Kontrollgruppe war dies ein signifikanter besserer Schutz vor einer
Legeleistungsminderung. Die Legeleistung der beiden Gruppen, bei denen das
Salmonella typhimurium - Zellwandprotein zum Impfstoff hinzugefügt worden war, fiel
im ersten Monat nach der Infektion auf ca. 65 % ab. Diese Unterschiede
verringerten sich im zweiten Monat p.i.. Die transovarielle Übertragung von MG war
in allen geimpften Gruppen im zweiten Monat signifikant geringer als in der
Kontrollgruppe. BARBOUR und NEWMAN (1990) werteten dies als Anzeichen dafür,
dass durch Impfung der Hennen eine Reduktion der transovariellen Übertragung von
MG erzielt worden ist.
REFAI et al. (1990) untersuchten die Schutzwirkung einer inaktivierten MG-
Adjuvans-Vakzine (Aluminium-Kalium-Sulfat) in Abhängigkeit von der
Applikationsroute. Als Maßstab für die Bewertung gaben sie klinische Symptome und
post mortem festgestellte Veränderungen an, die sie jedoch nicht genauer
beschrieben. Für die Experimente verwendeten sie 540 kommerziell erhältliche
Mycoplasma spp.-freie Hühner. Sie verglichen die Schutzwirkung des Impfstoffes bei

34
Tieren, die zum Zeitpunkt der Impfung unterschiedlich alt waren: einen Tag (n=160),
eine Woche (n=120), zwei Wochen (n=100), vier Wochen (n=80) und sechs Wochen
(n=42). Jede dieser Altersstufen wurde wiederum in Subgruppen unterteilt, die sich
durch die Applikationsart (i.m.,s.c.,i.n.) unterschieden. Ein Teil der Hühner blieb als
Kontrollgruppe ungeimpft. Die Hühner wurden mit 0,2 ml des Impfstoffes geimpft und
zwei Wochen danach mit dem homologen MG-Stamm S6 infiziert. Angaben darüber,
welches Volumen der Suspension mit 2x108 KBE/ml für die Infizierung verwandt
wurde, machten REFAI et al. (1990) nicht. Sie stellten fest, daß der „beste Schutz“
durch die intramuskuläre Applikation des Impfstoffes bei Hühnern im Alter von einer
Woche zu erzielen war. Bei 93,3 % der Hühner fanden sie p.m. keine Läsionen.
REDMANN und GÖBEL (1991) berichteten von unterschiedlichen Erfolgen bei der
Impfung mit einer inaktivierten Vakzine, um Legeleistungsminderungen durch MG zu
reduzieren. In vier von ihnen ausgewählten Betrieben (Betriebe A, B, C, D), in denen
Hennen unterschiedlichen Alters gehalten wurden, lag die Legeleistung deutlich
unter der Norm, die von den Zuchtgesellschaften angegeben wurden. MG-
Infektionen waren in diesen Herden mittels SSA mehrfach nachgewiesen worden. In
Betrieb „D“ stellten sie überdies in der 34. und 35. Lebenswoche einer Legegruppe
eine klinisch manifeste Mykoplasmose fest. Die Verluste durch die latente Infektion
mit MG wurden für den Betrieb „A“ mit ca. DM 22500.- im Zeitraum der letzten fünf
Jahre angegeben. Der Geflügelgesundheitsdienst Gießen empfahl den
Betriebsleitern, nur noch Junghennen in ihre Betriebe einzustallen, die drei bis vier
Wochen vor der Ankunft in ihrem Betrieb einmal mit einer inaktivierten MG-Vakzine
geimpft worden waren. Maßstab für den Erfolg der Impfung war die Legeleistung. Die
Ergebnisse wurden nur anhand der Legeleistungsstandards der Zuchtgesellschaften
bewertet. Im Betrieb „A“ verlief die Legeleistungskurve der geimpften Herde trotz
eines geringen Einbruchs (ca. 4 %) in der 28. Lebenswoche entsprechend der
vorgegebenen Standardkurve. Insgesamt lag die Legeleistung der geimpften Herden
im Zeitraum von 52 Wochen 14 % über den Legeleistungen der zuvor in diesem
Betrieb gehaltenen nicht vakzinierten Gruppen. Auch im Betrieb „B“ entsprach die
Legeleistung zweier neu eingestallter geimpfter Gruppen den erwarteten
Normwerten. Durch die Einstallung geimpfter Junghennen in den Betrieb „C“

35
verbesserte sich die Situation, jedoch ohne dass diese Herden die Normwerte
erreichten. Ein Grund hierfür war möglicherweise der Ausbruch der tumorösen Form
der Marek´schen Krankheit, die erhebliche Verluste verursachte. Im Betrieb „D“
hingegen hatte die Impfung nicht den gewünschten Effekt. Auch bei den beiden neu
eingestallten geimpften Herden kam es in der 30. Lebenswoche zu einem Rückgang
der Legeleistung um 5 bis 10 %. REDMANN und GÖBEL (1991) vermuteten, dass
die unterschiedliche Virulenz der MG-Stämme für den Mißerfolg in Betrieb „D“
verantwortlich gewesen ist.
BEHR et al. (1991) berichteten über Feldversuche, in denen Legehennen durch die
Impfung mit der inaktivierten Öl-Adjuvans-Vakzine „Talovac 104 MG“ (TAD
Pharmazeutisches Werk GmbH, Cuxhaven) vor Legeleistungsminderungen infolge
einer natürlichen Infizierung durch MG geschützt werden sollten. Die Versuche
wurden in Betrieben durchgeführt, in denen eine MG- und MS-Infektion serologisch
zuvor nachgewiesen worden waren. In den drei Ablegebetrieben (H, B, L) wurden
jeweils zeitgleich braune und weiße Hennen in zwei (L) respektive drei (H, B)
Altersgruppen gehalten. Die Junghennen stammten aus MG-freien
Aufzuchtbetrieben. In den Betrieb H wurden für die Versuche 340, in den Bestand B
1404 und in den Bestand L 400 Hennen eingestallt. Die Hälfte von ihnen wurde in
der 16. und 20. Lebenswoche (Betrieb H) bzw. 12./13. und 17./18. Lebenswoche
(Betriebe B, L) mit 0,5 ml des Impfstoffes subkutan im Nacken geimpft. Die andere
Hälfte der einzustallenden Hennen verblieb jeweils als Kontrollgruppe ungeimpft. Die
Legeleistung sowie die Tierverluste wurden in allen drei Beständen täglich
protokolliert. Zusätzlich wurden in den Beständen zunächst in drei-, später in
sechswöchigem Abstand Blutproben entnommen, um den Zeitpunkt einer MG-
Feldinfektion bei den ungeimpften Kontrolltieren feststellen zu können. Im Betrieb H
verzögerte sich der Legebeginn bei den geimpften Hennen. Die Legeleistungskurven
der Gruppen glichen sich aber an, so daß zwischen der 26. und 46. Lebenswoche
keine Unterschiede mehr im Verlauf der Legeleistungskurven zu erkennen waren.
Eine Infizierung mit MG konnten BEHR et al. (1991) bei den braunen Legehennen ab
der 26. und bei den weißen Legehennen ab der 32. Lebenswoche serologisch
nachweisen (HINZ et al. 1991). In der 47. Woche beginnend kam es - bei den

36
weißen Hennen deutlicher als bei den braunen - zu einer Legeleistungsminderung in
den ungeimpften Kontrollgruppen. BEHR et al. (1991) vermuteten, dass die bei der
Ausstallung einer älteren Hennengruppe verbundene Belastung der Tiere
disponierend eine Rolle gespielt hat. Bis zur Ausstallung der Versuchsgruppe in
Betrieb H in der 68. Lebenswoche legten die geimpften weißen Hennen mit 252
Eiern 22 Eier mehr als die Kontrollgruppe. Der Unterschied bei den braunen Hennen
betrug 254 zu 256 Eier. Auch im Betrieb B zeigten sich Unterschiede in der
Legeleistung nur zwischen der Impf- und der Kontrollgruppe bei den weißen Hennen.
Auch in diesem Betrieb kam es zu einem Einbruch der Legeleistung der ungeimpften
weißen Hennen ab der 43. Lebenswoche. In den 65 Legewochen legten die
geimpften weißen Hennen 337 Eier und damit 22 mehr als die Hennen der
ungeimpften Kontrollgruppe. Auch in diesem Betrieb gab es bei den braunen
Legehennen kaum Unterschiede in der Legeleistung zwischen geimpften (301) und
ungeimpften Hennen (297). Im Betrieb L kam es dagegen zu einer langanhaltenden
Legeleistungsminderung der ungeimpften braunen Hennen. Ab der 35. Lebenswoche
konnten in allen entnommenen Blutproben spezifische Antikörper gegen MG
nachgewiesen werden. Nach der Einstallung neuer Junghennen in den Betrieb kam
es in allen Versuchsgruppen zu einem Rückgang der Legeleistung, von welchem
sich die braunen ungeimpften Hennen nicht wieder erholten. So legten die geimpften
braunen Hennen in 66 Legewochen 27 Eier mehr (355 zu 328) als die ungeimpften
Kontrolltiere. Bei den weißen Hennen war in diesem Betrieb kein Unterschied
zwischen geimpften (366) und ungeimpften (365) Tieren zu erkennen. Diese
Ergebnisse sprechen nach Ansicht von BEHR et al. (1991) dafür, dass die Impfung
vor einer Legeleistungsminderung durch MG geschützt hat. Die Unterschiede
zwischen den Legeleistungskurven der braunen und weißen Legehennen ließen sich
durch die bekannte, unterschiedliche Sensibilität von braunen und weißen
Legehennen gegenüber definierten Stressoren infektiöser (NDV, AIBV, E. coli) und
nicht infektiöser Art (Temperatur, Staub, Schadgase) erklären.
YAGIHASHI et al. (1992) verglichen Hühner zweier verschiedener Zuchtlinien, die
„high responder line“ GSP mit der „low responder line“ BM-C, deren schwächere
Antikörperbildung auf eine Infizierung mit MG bekannt ist. Die MG-freien Küken

37
bezogen sie im Alter von sechs Wochen aus einer speziellen Laboratoriumsaufzucht.
Zunächst wurden die Hühner mit inaktivierter Adjuvans-Vakzine intramuskulär
geimpft. 14 Tage später erfolgte eine Boosterung durch eine intratracheale Gabe des
Impfstoffes ohne Adjuvans. Wiederum eine Woche nach der zweiten Impfung wurden
die geimpften Hühner und ungeimpfte Kontrolltiere gleichermaßen durch die
intratracheale Gabe von 0,5 ml Suspension mit 1x106 Mykoplasmen des SAS-
Stammes infiziert. Die histo-pathologischen Veränderungen in den Tracheen und die
Veränderungen der Luftsäcke wurden post mortem in einer vierstufigen
Bewertungsskala auf Grundlage eines numerischen Punktesystems klassifiziert. Zum
Vergleich der Impf- und Kontrollgruppen wurde das arithmetische Mittel der
einzelnen Werte gebildet. Die von YAGIHASHI et al. (1992) vorgenommene Art der
Vakzination schützte die geimpften Hühner beider Linien weitgehend vor einer
Infektion der Luftsäcke durch MG. So konnten bei allen Hühnern der Kontrollgruppe
vier Tage nach der Inokulation in allen untersuchten Luftsäcken Mykoplasmen
reisoliert werden. Dies gelang YAGIHASHI et al. (1992) jedoch nur bei zwei Hühnern
der GSP- und bei vier Hühnern der BM-C-Linie (n=6). Auch Luftsackentzündungen
stellten sie sieben Tage nach der Infizierung bei den geimpften Hühnern seltener
fest. Von sechs untersuchten Hühnern der GSP-Impfgruppe wies nur eines
Anzeichen einer Luftsackentzündung auf. In der Kontrollgruppe traten bei allen sechs
Hühnern Luftsackentzündungen auf. Allgemein war der Impferfolg in der GSP-Linie
„deutlicher“ ausgebildet als in der BM-C-Linie. Dies deckte sich auch mit den
Ergebnissen der serologischen Untersuchungen. YAGIHASHI et al. (1992) vertraten
die Auffassung, dass lokale Antikörper eine wichtige Rolle bei der Immunabwehr vor
einer respiratorischen Erkrankung infolge einer EBI mit MG spielen.
ELFAKI et al. (1992) untersuchten die Wirksamkeit einer inaktivierten MG-Vakzine
mit und ohne Iota carrageenan (iCGN) als Adjuvans in verschiedenen
Konzentrationen im Vergleich mit der kommerziell erhältlichen Vakzine MG-bac®
(Salsbury Lab., Charles City, Iowa, USA) und einer Kontrollgruppe. Darüber hinaus
prüften sie den Einfluß der Applikationsart auf den Impferfolg. Für die Versuche
verwendeten sie Mycoplasma spp.-freie männliche Eintagsküken, die sie aus einer
örtlichen Zucht erhielten. Die Impfungen erfolgten in der dritten und fünften

38
Lebenswoche. Die MG-Vakzine mit iCGN wurde bei der ersten Impfung intrazölomal
(i.c.) und bei der zweiten Vakzination intrabursal (i.b.) verabreicht. In allen drei
Versuchen wurden die Küken in der siebten Lebenswoche via Aerosol mit dem MG-
Stamm R belastet. Für die serologischen Untersuchungen wurden jeweils zwei
Wochen nach den Impfungen bzw. zwei Wochen nach der Belastungsinfizierung
Blutproben und Trachealspülproben entnommen. Für die kulturelle Untersuchung auf
MG nach der Infizierung wurde ein Trachealabstrich von den Hühnern gemacht. 14
Tage nach der EBI wurden die Küken getötet und die Luftsackveränderungen im
Rahmen einer pathologisch-anatomischen Untersuchung protokolliert sowie in einer
Skala von 0 bis 4 bewertet. Für jede Gruppe wurde ein Mittelwert gebildet und dieser
durch den theoretisch zu erzielenden LSE-Maximalwert dividiert. Dabei entspricht der
LSE-Wert von 1,0 den maximal möglichen pathologisch-anatomischen
Veränderungen. Die intrazölomale Impfung mit dem inaktivierten MG-Impfstoff mit
0,2 % iCGN und der anschließenden intrabursalen Boosterung schützte die Küken
am besten vor der Belastungsinfizierung. Der LSE-Wert betrug für die geimpfte
Gruppe nur 0,15. Bei 75 % dieser Küken konnten ELFAKI et al. (1992) keine
Luftsackentzündung feststellen. Dieser Wert war im Vergleich mit der Kontrollgruppe
signifikant besser. Die Impfung mit MG-bac® (Salsbury Lab., Charles City, Iowa,
USA) schützte 50 % der Küken vor Luftsackentzündungen; den LSE dieser Gruppen
gaben sie mit 0,40 an.
Die Auswirkung von Vakzination und EBI mit Mycoplasma gallisepticum bei MG-
freien Legehennen untersuchten EVANS et al. (1992 a). Sie verglichen in drei
Experimenten mit verschiedenen inaktivierten Vakzinen oder mit Lebendvakzinen
geimpfte Gruppen mit einer Kontrollgruppe. Hierzu wurden die Impfgruppen einmalig
in der zehnten Lebenswoche bzw. zum Teil zweimalig in der 10. und 16.
Lebenswoche vakziniert. Die Tiere wurden in der 29. bzw. 30. Lebenswoche mit dem
MG-Stamm R in form eines erregerhaltigen Aerosols infiziert; die hierzu verwendete
Mykoplasmensuspension enthielt 107 KBE je Huhn. Die Hühner aller Gruppen
zeigten nach dieser Belastungsinfizierung einen Einbruch in der Legeleistung binnen
drei bis sieben Tagen. Die Legeleistung der drei Kontrollgruppen betrug im
Durchschnitt vor der Infizierung 88,7 % und fiel infolge der EBI mit MG auf 26 % ab.

39
Der Unterschied in der Legeleistungsminderung war im ersten Versuch zwischen
Kontroll- und Impfgruppe nicht signifikant. Im Vergleich der Wirksamkeit zwischen
einem inaktivierten Impfstoff, der aus einem „Isolat reduzierter Virulenz (RVI)“
hergestellt worden war, der kommerziell erhältlichen Vakzine MG-bac® (Salsbury
Lab., Charles City, Iowa, USA) und einer Kontrollgruppe fiel die Legeleistung der
Kontrollgruppe am stärksten (von 85,14 % auf 39,8 %) ab. In der Gruppe, die mit
dem inaktivierten RVI-Impfstoff vakziniert worden war, verzeichneten EVANS et al.
(1992 a) eine Legeleistungsminderung von 81,5 % auf 50,6 %. Die dritte Gruppe
(MG-bac®) wies vor der Infizierung die höchste Legeleistung (90,07 %) auf. Sie fiel in
der Legeleistung auf 61,9 % ab. Diese Gruppe erholte sich am schnellsten von der
Legeleistungsminderung nach der EBI und erreichte wieder einen Wert von 84,23 %.
Dieser Schutz vor Legeleistungsminderung nach EBI durch MG war im Vergleich mit
der Kontrollgruppe signifikant.
ELFAKI et al. (1993) verglichen die Wirksamkeit von Mykoplasmenimpfstoffen in
Abhängigkeit von der Art der Applikation. Sie verwendeten MG-freie männliche
Küken aus kommerzieller Aufzucht. In einzelnen Gruppen wurden Kombinationen
von intrazölomaler (i.c.), intratrachealer (i.t.), intranasaler (i.n.), intravenöser (i.v.),
subkutaner (s.c.) und oraler Vakzination mit MG-Impfstoffen mit und ohne Iota
carrageenan (iCGN) als Adjuvans im Vergleich mit einer kommerziell erhältlichen
Vakzine und einer ungeimpften Kontrollgruppe geprüft. Die Erstimpfung der Küken
erfolgte in der dritten, die zweite Vakzination in der fünften Lebenswoche. Die
Versuchstiere wurden in der siebten Lebenswoche aerogen infiziert. In der zur
Infizierung verwendeten Mykoplasmensuspension (MG-Stamm R) befanden sich
mindestens 2,9x108 KBE/ml-1. Die Menge, die je Küken zur Infizierung verwandt
wurde, gaben ELFAKI et al. (1993) nicht an. Zwei Wochen nach der
Belastungsinfektion wurden die Küken getötet und pathologisch-anatomisch
untersucht. Die Veränderungen der Luftsäcke wurden auf einer Skala von 0 bis 4
bewertet und der LSE-Wert für jede Gruppe berechnet (ELFAKI et al. 1992). Im
Verlaufe des Versuches wurden von den Küken Blut- und Trachealspülproben
gewonnen und Abstriche aus der Trachea entnommen. Die Vakzination der Tiere
bewirkte eine deutlichen Schutz vor Luftsackentzündungen nach einer EBI mit dem

40
MG-Stamm R. Der LSE-Wert der Kontrollgruppe betrug 1,00 und erreichte damit den
maximal möglichen Wert. Für die Gruppen, die s.c. mit der kommerziell erhältliche
Vakzine geimpft worden waren, gaben ELFAKI et al. (1993) den LSE-Wert mit 0,05
an. Die Impferfolge mit dem Impfstoff, der iCGN als Adjuvans enthielt, waren
abhängig von der Applikationsart: So betrug der LSE nach i.c./i.n. Impfung 0,00,
nach i.c. Impfung 0,15 und nach i.c./i.t. Impfung 0,14. Der Impfschutz dieser vier
Gruppen war im Vergleich zur Kontrollgruppe signifikant höher. Die Anzahl der
reisolierten Mykoplasmen war in der Kontrollgruppe signifikant höher als in den
geimpften Gruppen. Dieser Unterschied fiel zwischen der Kontrollgruppe (7,41 log10
KBE/ml-1) und der mit der kommerziell erhältlichen Vakzine geimpften Gruppe (5,71
log10 KBE/ml-1) am deutlichsten aus.
ABD-EL-MOTELIB und KLEVEN (1993) untersuchten den induzierbaren Schutz
durch verschiedene Impfstoffe vor Luftsackentzündungen nach einer EBI mit dem
MG-Stamm R. Als Versuchstiere nutzten sie insgesamt 349 MG-freie Hühner der
Rasse „White Leghorn“. Sie verglichen die Effizienz dreier Lebendvakzinen der MG-
Stämme F, ts 11 und 6/85 und eines inaktivierten Impfstoffs (Intervet America) im
Vergleich mit einer ungeimpften Kontrollgruppe. Es wurden in dieser Studie - nicht
näher beschriebene - respiratorische Erscheinungen nach der Vakzination, die
serologische Immunantwort, die Persistenz der Impfstämme und das Ausmaß von
Luftsackentzündungen nach einer EBI bewertet. Die Küken wurden im Alter von 10
Tagen geimpft. Dies geschah bei den drei Lebendvakzinen via eye-drop und bei der
inaktivierten Vakzine durch eine subkutane Injektion. 30, 60 und 90 Tage nach der
Impfung wurden Hühner aus jeder Gruppe via Aerosol infiziert. Die für diese EBI
genutzte Suspension des R-Stammes enthielt 3,62x108 CCU/ml. 10 Tage p.i. wurden
die Hühner getötet und seziert. Die festgestellten pathologisch-anatomischen
Veränderungen der Luftsäcke wurden mit Hilfe einer Skala von 0 (keine Läsionen)
bis 4 (schwere Läsionen) klassifiziert. Nach der Vakzination mit dem F-Stamm kam
es bei den Hühnern dieser Gruppe zu „leichten respiratorischen Erscheinungen“.
Ähnliche Symptome stellten ABD-EL-MOTELIB und KLEVEN (1993) auch in der ts
11-, 6/85- und Kontrollgruppe nach der EBI fest. Die Reisolierung von Mykoplasmen
des F-Stammes gelang auch 90 Tage nach der Impfung aus den Tracheen aller

41
untersuchten Hühner. Mykoplasmen des ts-11-Stammes waren zu diesem Zeitpunkt
nur noch bei zwei von fünf Tieren und bei Mykoplasmen des 6/85-Stammes bei
keinem Tier nachzuweisen. Die Vakzination mit dem F-Stamm schützte die Hühner
„am besten“ vor Luftsackentzündungen. Die LSE-Werte dieser Gruppe waren in allen
drei Fällen mit 0,2, 0,2 und 0,1 am niedrigsten. Die Werte für die mit der inaktivierten
Vakzine geimpften Hühner lagen mit 1,2, 1,7 und 3,0 zum Teil sogar über denen der
Kontrollgruppe (1,5, 1,7 und 2,3). Die LSE-Werte der ts-11- bzw. 6/85-Gruppe lagen
bei 0,6, 1,9 und 1,3 bzw. 0,9, 1,2 und 2,0. In der Addition aller drei
Versuchsabschnitte ergab sich ein signifikant besserer Schutz vor
Luftsackentzündungen durch die Vakzination mit dem F-Stamm. ABD-EL-MOTELIB
und KLEVEN (1993) vertraten die Ansicht, dass die höhere Virulenz des
verwendeten F-Stammes mit der besseren Schutzwirkung vor Luftsackentzündungen
korreliert. Einen Schutz vor einer Infektion konnten sie nicht nachweisen. Die durch
die Impfung mit der inaktivierten Vakzine induzierten hohen Antikörpertiter im Serum
korrelierten nicht mit dem Schutz vor Luftsackentzündungen.
NAKAMURA et al. (1995) untersuchten den Einfluß der Applikationsart des
Impfstoffes auf die serologische Reaktion und den Schutz vor Trachealläsionen nach
einer Infizierung mit dem homologen MG-Stamm. Hierzu wurden SPF-Küken im Alter
von vier und sechs Wochen mit einer inaktivierten Vakzine, hergestellt mit dem SAS-
Stamm, intramuskulär, intratracheal, per os (in drei verschiedenen Konzentrationen)
und in den Kropf geimpft. Nach zwei Wochen wurden die Küken mit 1,0 ml
Phosphatpufferlösung, in welcher 106,6 KBE des MG-Stammes SAS suspendiert
waren, intratracheal infiziert und weitere zwei Wochen später getötet. Die
Schleimhautveränderungen der Tracheen wurden patho-histologisch untersucht, die
festgestellten Veränderungen wurden in einer Skala von 0 bis 3 klassifiziert und
Ergebnisse > 2 als positiv bewertet. Die intratracheale und intramuskuläre Impfung
der Küken ergab den besten Schutz (100%). Auch die orale Impfung mit 1011
inaktivierten Mykoplasmen schützte die Küken zu 100%, doch konnten NAKAMURA
et al. (1995) nicht ausschließen, dass ein Teil des verimpften Volumens (5 ml) bei
der Atmung in den Respirationstrakt gelangt war und dieses Impfverfahren somit
einer intratrachealen Vakzination entsprach. Diese Vermutung entsprach auch dem

42
Ergebnis, welches sie durch die direkte Applikation der Vakzine in den Kropf erzielt
hatten. Bei vier von sechs Küken, die sie auf diese Weise geimpft hatten, bildeten
sich Trachealläsionen aus. Dies entsprach den Werten, die bei der ungeimpften
Kontrollgruppe festgestellt worden waren.
2.3. MG-bedingte immunologische Reaktionen
Im folgenden Kapitel werden die Reaktionen des Immunsystems von Hühnern auf
eine Infektion mit Mycoplasma gallisepticum sowie die Befähigung von MG, sich
diesen Reaktionen zu entziehen, dargestellt.
ROBERTS et al. (1967) verglichen verschiedene Methoden (SSA, HAH und
Röhrchenagglutination (TA)) zum Nachweis spezifischer Antikörper gegen MG.
Hierzu infizierten sie MG- und MS-freie Hühner mit dem MG-Stamm Hy. Sie konnten
in der SSA, im HAH und der TA bis zur 28. Woche p.i. spezifische Antikörper gegen
MG nachweisen. Gleichzeitig stellen sie in den Untersuchungen der Seren der nicht
infizierten Kontrolltiere ihnen unerklärliche positive Reaktionen in der SSA und TA
fest. Ähnliche unspezifische Reaktionen im HAH ließen sich durch die Behandlung
der Blutserumprobe mit einem „receptor-destroying enzyme“ (RDE) von Vibrio
cholerae verhindern. Als Ursache für die von ihnen gefundenen unspezifischen
Reaktionen vermuteten ROBERTS et al. (1967) Kontaminationen des Serums oder
Kreuzreaktionen mit anderen Bakterien.
Bis zu vier Wochen nach einer Impfung von zehn Wochen alten, MG-freien Küken
gegen Erysipelothrix rhusiopathiae konnten auch VARDAMAN und YODER (1971)
unspezifische Agglutinationsreaktionen in der SSA gegen MG feststellen. Bei der
Überprüfung dieser Ergebnisse im HAHT wies jedoch keines der Seren spezifische
Antikörper gegen MG auf.
CULLEN und TIMMS (1972) berichteten von ähnlichen falsch positiven Ergebnissen.
Sie hatten MG-freie Hühner aus einer Laboraufzucht im Alter von drei, neun und 16
Wochen intramuskulär mit verschiedenen Vakzinen (ND, ND+IB, ND+IB+AE, AE)
geimpft oder ihnen reines Adjuvans bzw. eine Kombination von Adjuvans und Protein

43
injiziert. In der elften Lebenswoche stellten sie beispielsweise in der Gruppe, die mit
ND-Impfstoff vakziniert worden war, in neun von zwölf untersuchten
Blutserumproben eine positive Reaktion mit dem MG-Antigen fest. Auch in den
Serumproben von Hühnern, denen lediglich reines Adjuvans bzw. ein Gemisch von
Adjuvans und Protein injiziert worden war, fanden sie im Alter von 18 Wochen bei
jeweils drei Hühnern agglutinierende Antikörper, die in der SSA mit dem MG-Antigen
reagierten. In keinem der zur Kontrolle dieser Ergebnisse im HAH untersuchten
Seren konnten CULLEN und TIMMS (1972) spezifische Antikörper gegen MG
nachweisen.
Auch YODER (1989) untersuchte das Auftreten von unspezifischen Agglutinations-
reaktionen in der SSA für MG und MS nach Impfungen mit acht verschiedenen
Vakzinen. Für seine Versuche verwendete er acht Wochen alte SPF-freie Hühner
(Mycoplasma spp., NDV, IBV, AE und Marek). Die Agglutinationsreaktionen in der
SSA wurden in einer Skala von 0 bis 4 bewertet. Nach einer Vakzination mit
verschiedenen Kombinationsimpfstoffen (NDV+IBV+IBD+Reo-Virus, IBD+NDV) und
monovalenten Vakzinen gegen IBD, beispielsweise Coryza-vac® (Salsbury Lab.
Charles City, Iowa, USA), kam es bei der Untersuchung der Seren in der SSA mit
MG-Antigen zu „starken“ unspezifischen Reaktionen. Versuche zur Eliminination
dieser Reaktivität schlugen fehl, jedoch war das Auftreten nur vorübergehend
nachzuweisen. Sechs Wochen nach der Impfung mit Coryza-vac® (Salsbury Lab.
Charles City, Iowa, USA) war der Titer der unspezifischen Reaktionen wieder von 3,2
auf 0,2 abgefallen. Zu keinem Zeitpunkt seiner Versuche konnte YODER (1989) im
HAH spezifische Antikörpertiter gegen MG feststellen.
Bei weiteren Arbeiten über die serologischen Reaktionen nach einer MG-Infektion
verglich ROBERTS (1969) die Antikörperreaktion der MG-Stämme S 6, F und WS7
in der SSA und im HAH. Hierzu verwendete er sechs Wochen alte, MG- und MS-freie
Hühner. In drei Gruppen mit jeweils 14 Tieren aufgeteilt, wurden sie mit 0,5 x 107
Mykoplasmen der drei genannten Stämme intranasal infiziert. Im wöchentlichem
Abstand wurden je Gruppe zwei Hühner getötet und das Blut serologisch untersucht.
In den beiden ersten Wochen p.i. stellte ROBERTS (1969) überwiegend positive
Reaktionen in der SSA fest. Diese Antigen-Antikörperreaktion war abhängig von den

44
hochmolekularen Immunglobulinen des Typs IgM. Er fand in der SSA Reaktionen
sowohl mit den homologen als auch mit den heterologen Antigenen. Der HAH-Test
reagierte dagegen stammspezifischer. Diese Reaktion ist abhängig von den
niedermolekularen Immunglobulinen des Typs IgG. So reagierte das Serum der mit
dem F-Stamm infizierten Hühner zu keinem Zeitpunkt des Versuches mit dem
Antigen des WS7-Stammes.
Ähnliche Kreuzreaktionen bei hochvirulenten Stämmen beobachten LIN und
KLEVEN (1982 b) im HAHT. Nach einer Impfung bzw. einer aerogenen Infizierung
mit den MG-Stämmen R, F und S6 stellten sie fest, daß Agglutinationsreaktionen
auch bei heterologen Antigenpräparationen mit den MG-Stämmen S6, A 5969, 730
und 503 auftraten. Dagegen reagierten die Seren von Hühnern, die mit den MG-
Stämmen 730 und 503 geimpft worden waren, nur mit dem entsprechenden
homologen Antigen.
KLEVEN (1975) und HINZ (1977, 1978) empfahlen den Einsatz der SSA als
Suchtest für Infektionen in Legehennenherden mit MG. Zur Verifizierung positiver
Ergebnisse sollten solche Proben mit dem HAH überprüft werden. HINZ (1977)
beschrieb, dass positive HA-Hemmtiter immer mit einer positiven Reaktion in der
SSA verbunden sind, dies aber im umgekehrten Fall nicht gilt. In der SSA waren bei
der Verwendung von heterologen Antigenen verzögerte oder schwach positive
Reaktionen festzustellen. In keinem Fall konnte HINZ (1977) im HAH bei Seren, die
er mit RDE behandelt hatte, Reaktionen mit heterologen Antigenen erkennen.
Bereits eine Woche nach einer EBI via eye-drop mit dem R-Stamm von MG konnten
AVAKIAN und LEY (1993) bei allen Versuchshühnern agglutinierende Antikörper in
der SSA nachweisen. Nach einer intrakonjunktivalen Applikation der inaktivierten
MG-Vakzine (MG-bac®, Salsbury Lab.) konnten ABD-EL-MOTELIB und KLEVEN
(1993) zwei Wochen nach der Impfung spezifische Antikörpertiter (IgM, IgG) im SSA,
HAH und ELISA nachweisen. Im Vergleich zu drei Lebendvakzinen (F, ts-11, 6/85)
erfolgte der Nachweis der spezifischen Antikörper zwei Wochen früher und war
länger möglich, in ihrem Versuch bis zur zwölften Woche. Einen Zusammenhang
zwischen den Antikörpertitern und einem Schutz gegen die Adhäsion der
Mykoplasmen im Respirationstrakt nach einer EBI mit dem R-Stamm konnten sie

45
jedoch nicht feststellen. KHAN et al. (1986) fanden in einer von ihnen geimpften
Hennengruppe (n = 3400) drei Wochen nach der Impfung mit der Vakzine MG-bac®,
Salsbury Lab. die ersten Antikörper in der SSA. Im Verlaufe ihrer Untersuchungen
(50 Wochen) wies die geimpfte Gruppe im Vergleich zur Kontrollgruppe signifikant
höhere Antikörpertiter im SSA-Verdünnungstest auf.
Mit der Frage, wie lange nach einer Impfung mit einer inaktivierten MG-Vakzine
humorale Antikörper gegen MG nachzuweisen sind, beschäftigten sich HINZ et al.
(1991). Vier Wochen nach der ersten Impfung wiesen sie das erste Auftreten von
agglutinierenden Antikörpern in der SSA nach. In einer Gruppe reagierten zu diesem
Zeitpunkt 77 % und in zwei anderen Gruppen 100 % der untersuchten Seren positiv.
Erste negative Reaktionen stellten sie ein Jahr nach der Zweitvakzination fest, wobei
über 90 % der untersuchten Seren auch 78 respektive 85 Wochen nach der zweiten
Impfung in der SSA positiv reagierten. In diesen Seren konnten sie auch mit Hilfe des
ELISA spezifische Antikörper gegen MG nachweisen. Der Nachweis spezifisch HA-
hemmender Antikörper schwankte vier Wochen nach der Erstimpfung zwischen 0%
und 66 %. Zwar konnten sie vier Wochen nach der Zweitvakzination in allen
Versuchsgruppen positive Reaktionen nachweisen, doch lag die Nachweishäufigkeit
zwischen 20 % und 100%. Im Gegensatz zu den agglutinierenden Antikörpern
konnten HINZ et al. (1991) HA-hemmende Antikörper nur bis maximal 27 Wochen
nach der Zweitimpfung nachweisen.
Diesen Zeitraum für den Nachweis HA-hemmender Antikörper gegen MG gaben
auch GLISSON und KLEVEN (1984) an. Sie konnten nach Impfungen in der 16. und
20. Lebenswoche sowie einer experimentellen Infektion in der 28. Lebenswoche bis
zur 55. Lebenswoche spezifische Antikörpertiter im HAH nachweisen.
BENCINA und DORRER (1983) überprüften die Sicherheit serologischer
Nachweismethoden bei lang zurückliegenden Infektionen mit MG. Sie untersuchten
Hühner, die aus einer latent infizierten Herde stammten. Diese Tiere hatten von der
fünften bis zur 13. Lebenswoche deutliche respiratorische Erscheinungen und
Nasenausfluß gezeigt. Zum Zeitpunkt der Erkrankung konnte MG aus der Trachea
isoliert werden. Im Alter von 57 Wochen wurden bei den Hühnern Abstriche aus der
Trachea entnommen. Bei 12 Hühnern konnten mittels indirektem

46
Fluoreszenzantiglobulintest MG Serotyp A nachgewiesen werden. Bei den
serologischen Kontrolluntersuchungen stellten sie fest, daß z. B. in der SSA in
Abhängigkeit von dem verwendeten Antigen bei neun von zwölf getesteten Seren
kein Nachweis für die latente MG-Infektion erbracht werden konnte. Im HAHT stellten
sie nur Antikörpertiter zwischen 1:32 und 1:128 fest, wobei das arithmetische Mittel
bei 1:72 lag. Eine MG-Infektion war bei ihrem Versuchsaufbau jedoch erst ab einem
Titer von ≥ 1:80 sicher zu erkennen.
Auch die Untersuchungen von KLEVEN et al. (1988) zeigten, daß in der Diagnostik
von MG Lücken auftreten können. Blutserumproben von Hühnern, die mit 16
verschiedenen MG-Stämmen infiziert worden waren, wurden von ihnen im HAH mit
insgesamt 19 unterschiedlichen Antigenpräparationen untersucht. Prinzipiell stellten
sie fest, dass im HAHT mit den homologen Antigenen die höchsten Titer zu erreichen
waren. Sie waren überrascht, dass gerade der in vielen Labors zur
Antigenherstellung verwandte Stamm A5969 derjenige war, bei welchem sie die
geringsten Antikörpertiter fanden. Bei anderen verwendeten Antigenen waren alle
untersuchten Seren negativ, so dass diese Proben bei der Untersuchung eines MG-
Ausbruches als negativ gewertet worden wären. Mit keinem der von ihnen
verwendeten Antigene konnten alle MG-Stämme nachgewiesen werden.
Die Untersuchungen von BENICINA et al. (1991) zeigten diagnostische Lücken bei
latenten Infektionen mit mehreren Mycoplasma spp. auf. Bei einer kulturellen
Untersuchung von acht einjährigen Hühnern konnten sie MS, MG und MGn
nachweisen. Die Blutserumproben der Hühner zeigten in der SSA für M. synoviae
deutliche Agglutinationsreaktionen. Die spezifischen Titer für MS im HAHT
schwankten zwischen 1:40 und 1:160. Auch mit dem IIPA konnten sie spezifische
Antikörpertiter (IgG, IgM) für MS bis zu einer Höhe von 1:2560 nachweisen. Mit dem
MGn-Antigen konnten sie jedoch nur in einem der getesteten Seren eine schwache
Agglutinationsreaktion erkennen. Ein Nachweis spezifischer Antikörper im Serum
gegen MG gelang ihnen weder in der SSA noch im HAH oder der IIPA. Aus allen
acht Milzen und drei Harder´schen Drüsen war jedoch mittels IIPA ein Nachweis
spezifischer Antikörper möglich. Die Autoren fanden, dass eine vorangehende

47
Infektion mit MS die Schwere einer nachfolgenden klinischen Erkrankung durch MG
und die Höhe der Antikörpertiter mindert.
RODRIGUEZ und KLEVEN (1980 a) verglichen die Höhe spezifischer Antikörpertiter
im HAH nach einer EBI mittels eines erregerhaltigen Aerosols mit unterschiedlich
pathogenen MG-Stämmen. Die im Alter von 30 Tagen mit dem R-Stamm infizierten
Küken wiesen signifikant höhere Antikörpertiter im HAH auf als die Vergleichsgruppe,
die mit dem F-Stamm infiziert worden war. Auch nach einer weiteren EBI, die 14
Tage später erfolgte, lagen die Titer HA-hemmender Antikörper in der mit dem R-
Stamm infizierten Gruppe höher (GMT 212,5) als in der Gruppe (GMT 127), die
erneut mit dem F-Stamm von MG infiziert worden war (RODRIGUEZ und KLEVEN
1980 b).
SOERIPTO et al. (1989 b) untersuchten die Wirksamkeit einer Impfung von sieben
Wochen alten SPF-Küken mit dem MG-Stamm 80083, bzw. der 50. (80083M) und
der 100. (80083H) Passage dieses Stammes. Die Hühner, die über Trinkwasser(tw)
oder eye drop(ed) mit dem MG-Stamm 80083 vakziniert worden waren, zeigten eine
serologische Reaktion im HAHT (GMT 1:15(tw,ed) und im ELISA (GMT 105(tw) bzw.
139(ed)), wobei die Titer deutlich über denen der beiden anderen Gruppen lagen.
Keines der mit dem Ausgangsstamm geimpften Hühner erkrankte an einer
Luftsackentzündung nach einer EBI mit dem virulenten Stamm Ap3AS. Serologisch
konnten SOERIPTO et al. (1989 b) nach der EBI einen deutlichen Anstieg der
Antikörpertiter feststellen. Die Hühner, die mit 80083M vakziniert wurden, wiesen vor
der EBI nur niedrige serologische Titer auf, waren aber vor Luftsackentzündungen
geschützt. Bei den Tieren, welche mit dem Stamm 80083H geimpft worden waren,
konnten sie vor der EBI keine Antikörper nachweisen. Diese Hühner erkrankten nach
der Belastungsinfizierung an einer Mykoplasmose. Aus denen im Rahmen der
pathologisch-anatomischen Untersuchungen festgestellten, auf einer Skala von 0 bis
4 bewerteten Luftsackveränderungen, ergaben sich Mittelwerte von ≥ 3,5.
SOERIPTO et al. (1989 b) vermuteten einen Zusammenhang zwischen der Höhe der
Antikörpertiter und dem Schutz vor Luftsackentzündungen, wobei die Ausbildung der
Antikörpertiter von der Virulenz des Impfstammes abhängig sein soll. So konnten sie
bei 37 Wochen alten Hühnern 13 Wochen nach der Impfung hohe Antikörpertiter

48
gegen den Impfstamm MG-Stamm 80083 nachweisen. Ein Schutz vor
Luftsackentzündungen durch eine EBI war zu diesem Zeitpunkt nur gegen den
homologen Stamm vollständig ausgebildet.
Einen Schutz vor pathologisch-anatomischen Veränderungen der Tracheal-
schleimhaut stellten NAKAMURA et al. (1995) grundsätzlich nur bei den Hühnern
fest, bei denen sie in der serologischen Untersuchung auch agglutinierende
Antikörper in der SSA fanden. In den Blutserumproben von neun Hühnern, die sie im
Alter von vier und sechs Wochen intratracheal mit einem inaktiviertem Impfstoff (MG-
Stamm SAS) vakziniert hatten, stellten sie vier Wochen nach der zweiten Gabe
agglutinierende Antikörper fest. Zu diesem Zeitpunkt wurden die Hühner mit 106,6
Mykoplasmen des homologen Stammes infiziert. Bei keinem dieser Hühner konnten
sie bei der pathologisch-anatomischen Untersuchung zwei Wochen p.i.
Trachealläsionen diagnostizieren.
Der von YODER et al. (1984) festgestellte signifikante Schutz gegen
Luftsackentzündungen bei Broilern deckte sich mit den Ergebnissen ihrer
serologischen Untersuchungen. So ermittelten sie in einem Versuch bei geimpften
Hühnern vor der EBI spezifische Antikörper gegen MG. Der geometrische Mittelwert
der im HAH untersuchten Seren lag bei 1:25, während in der Kontrollgruppe nur ein
unspezifischer Titer von ≤ 1:10 gemessen wurde. Auch in einem zweiten Versuch
stellten sie bei geimpften Tieren vor der Infizierung spezifische HAH-Titer (GMT 1:40)
fest. Nach der EBI lagen die Titer der Tiere von Impf- und Kontrollgruppe bei 1:320,
jedoch wiesen die Hühner der Kontrollgruppe signifikant häufiger
Luftsackentzündungen auf.
Auch TALKINGTON und KLEVEN (1985) fanden bei zwölf Wochen alten,
kommerziell erworbenen Hühnern nach einer einmaligen Impfung mit inaktiviertem
MG-Impfstoff humorale Antikörpertiter im HAHT (GMT 1:68). Nach einer Boosterung
stieg der GMT auf 1:112 an. Alle zweimal geimpften Hühner wiesen mindestens
einen spezifischen Antikörpertiter von ≥ 1:40 auf. Alle untersuchten Seren zeigten in
der SSA Agglutinationsreaktionen. Für die erfolgreiche Infizierung der geimpften
Hühner waren im Durchschnitt eine größere Anzahl von lebenden Mykoplasmen
notwendig. So mußten Tiere, die zweimal vakziniert worden waren, mit 1500

49
KBE/Tier infiziert werden, damit drei Tage p.i. MG aus der Trachea zu reisolieren
war. Bei den einmal geimpften Tieren bzw. der Kontrollgruppe waren für eine
Infektion nur 150 respektive 15 KBE/Tier notwendig. TALKINGTON und KLEVEN
(1985) konnten jedoch bei der Reisolation keinen Zusammenhang zwischen der
Höhe der serologischen Titer und der Anzahl der gefundenen Mykoplasmen
feststellen. Die Anzahl der Mykoplasmen stieg nach ihren Angaben auch bei
geimpften Hühnern nach einer Infektion zunächst stark an, um dann langsam
abzufallen.
Auch BARBOUR et al. (1987) stellten einen Zusammenhang zwischen einer zuvor
erfolgten Immunisierung (s. o.), lokal sezernierten Antikörpern und einem Schutz vor
einer Infektion der Trachea nach experimenteller Infizierung mit dem R-Stamm von
MG fest. Beim Vergleich verschiedener Impfstoffpräparationen wies allein die Gruppe
mit dem signifikant höchsten lokalen Antikörpertiter keine mikroskopisch
nachweisbaren Läsionen der Trachealschleimhaut auf. Bei der Untersuchung der
Überempfindlichkeitsreaktion vom Typ IV zeigten drei zuvor mit liposomaler
Adjuvans-Vakzine grundimmunisierte Gruppen 24 Stunden nach einer intradermalen
Applikation von 0,9 mg MG-Äquivalent einen im Vergleich mit der Kontrollgruppe
signifikant stärker verdickten Kehllappen auf.
YODER und HOPKINS (1985) beschrieben bei geimpften Legehennen, die entweder
aus einer Versuchstierhaltung oder aus kommerziellen Beständen stammten, einen
Schutz vor einer Legeleistungsminderung (s.o.). Die geimpften Hennen (Experiment
6) wiesen nach der EBI mit dem MG-Stamm R im Durchschnitt einen deutlich
höheren Titer (GMT: 557) im HAH auf als die der Kontrollgruppe (GMT: 320). Im
Vergleich der subkutanen*1 Impfung mit der intratrachealen*2 stellten sie jeweils vier
Wochen nach der ersten bzw. zweiten Impfung bei subkutaner Verabreichung
höhere Antikörpertiter im HAH fest (GMT*1: 1:155 und 1:150; GMT*2: 1:32 und 1:45).
In diesem Versuch lagen sechs Wochen nach der Infizierung die HAH-Titer beider
Impf- und der Kontrollgruppe bei etwa 1:300.
Eine Korrelation zwischen der Anzahl der nach experimenteller Infizierung
(105 KBE/Tier) reisolierten Mykoplasmen (SAS-Stamm) und der Schwere der
Trachealläsionen stellten YAGIHASHI und TAJIMA (1986) fest. Gleichzeitig wiesen

50
sie nach der Infektion einen Anstieg lokaler (IgG, IgA, IgM) Antikörper nach, die bei
IgA (log102,6 ±0,4) und IgM (log102,0 ±0) zwei Wochen p.i. und bei IgG (log103,1 ±0,3)
drei Wochen p.i. ihre höchsten Werte erreichten. Die Titer der im Blutserum
nachgewiesenen Antikörper erreichten ebenfalls nach ca. drei Wochen ihre höchsten
Werte (IgG log103,8, IgA log103,7), die dann bis zur zwölften Woche auf hohem
Niveau konstant blieben. Drei bzw. 14 Tage nach einer zweiten EBI konnten sie aus
den Tracheen der Tiere signifikant weniger Mykoplasmen reisolieren als nach der
ersten. Diese Hühner zeigten geringere entzündliche Veränderungen der Trachea
und eine schnellere Elimination der Mykoplasmen. YAGIHASHI und TAJIMA (1986)
bewerteten dies als Indiz dafür, dass lokale Antikörper eine Rolle bei der
Infektionsabwehr gegen MG spielen.
Dagegen beobachtete KLEVEN (1985) direkt nach Belastungsinfektion geimpfter
Hühner einen starken Abfall der Serumantikörpertiter im HAHT. So fiel der im
geometrischen Mittel errechnete HAH-Titer bei den Tieren, die zweimal geimpft
worden waren, von 1:149,3 zum Zeitpunkt der Infizierung mit MG auf 1:22,0 zwei
Wochen p.i. ab. Zurückgeführt wurde dies auf die Neutralisation der bereits
vorhandenen Antikörper durch die in den Organismus eindringenden Mykoplasmen.
Die Frage, welche Bedeutung maternale Antikörper gegen MG und MS für Küken
haben, beschäftigte SNELL und CULLEN (1977). Sie konnten in Blutserumproben
der Küken bis zum 18. Tag maternale Antikörper in der SSA und dem HAHT
nachweisen. Jedoch nahm die Anzahl der Küken, die über maternale Antikörper
verfügten, sehr schnell ab. So konnten sie am vierten Tag nach dem Schlupf bei 35%
der Küken keine maternalen Antikörper in der SSA mehr finden. LIN und KLEVEN
(1984) verglichen Küken MG-positiver und MG-freier Abstammung in Bezug auf die
Bedeutung maternaler Antikörper. Nach experimenteller Infizierung mit dem MG-
Stamm R am dritten und 14. Lebenstag konnten sie keine signifikanten Unterschiede
im Ausmaß der Luftsackentzündungen feststellen. Von den Probanden, die im Alter
von sieben Tagen infiziert worden waren, wiesen die Küken der MG-positiven
Hennen signifikant niedrigere LSE-Werte auf als die der MG-freien Hennen. Das
Vorhandensein maternaler Antikörper beeinflußte die Entwicklung der Antikörpertiter
nach einer Impfung mit dem MG-Stamm F am ersten Lebenstag nicht.

51
Die Art und den Umfang von zellulärer, humoraler und lokaler Immunantwort nach
Infizierung mit MG (S 6) untersuchten CHHABRA und GOEL (1980) bei SPF-
Hühnern, die bei Versuchsbeginn sieben Wochen alt waren. Bereits eine Woche
nach einer EBI konnten sie eine zelluläre Immunantwort mit dem
Leukozytenmigrationshemmungstest (LMI) nachweisen, wobei sie die höchsten
Werte (36,4%) der Migrationshemmung sieben Wochen p. i. feststellten. Ebenfalls
als Anzeichen einer zellulären Immunantwort werteten sie eine lokale
Hypersensibilitätsreaktion in der 17. Lebenswoche. Hierzu injizierten sie sieben
Hühnern der bereits MG-positiven Gruppe und fünf Hühner der Kontrollgruppe durch
eine intradermale Injektion von 0,1 ml Antigen (MG-Stamm S6) in den rechten
Kehllappen. Die Dickenzunahme der Haut binnen 48 Stunden war in der zuvor
infizierten Gruppe signifikant größer als in der Kontrollgruppe. Die humorale
Antikörperreaktion untersuchten sie per HAHT. Sie stellten schon eine Woche nach
der Infizierung Antikörper fest. Der Titer erreichte seinen Maximalwert ebenfalls in
der siebten Woche p. i.. CHHABRA und GOEL (1980) stellten eine hochsignifikante
Korrelation zwischen LMI und HAHT fest. Die Antikörperreaktion im Serum war
geprägt durch einen signifikant höheren Titer von Immunglobulinen des Typs IgG im
Vergleich zu den Kontrolltieren. Mit Trachealspülproben ermittelten sie die Titer lokal
sezernierter Antikörper vom Typ IgG und IgA. Hierbei stellten sie einen Anstieg der
Titer fest, wobei das Maximum in der sechsten Woche p.i. lag. Der Beginn der
Genesung der Hühner von der experimentell erzeugten Mykoplasmose erfolgte
zeitgleich mit der Abnahme der Anzahl der aus der Trachea zu reisolierenden
Mykoplasmen, der Abnahme der Trachealläsionen und den höchsten Antikörpertitern
in den serologischen Untersuchungen.
BENCINA und DORRER (1989) untersuchten Seren und Schleim aus dem oberen
Respirationstrakt von natürlich und experimentell infizierten Hühnern auf Antikörper
(IgG) gegen MG und MS mittels indirektem Immunoperoxidase-Test (IIPA). In 90 %
der Fälle stimmten die Ergebnisse des Antikörpernachweises im IIPA für Serum und
Schleim überein. Jedoch fanden BENICINA und DORRER (1989) bei drei Hühnern
lediglich Antikörper im Schleim aus dem oberen Respirationstrakt. Auch die generelle
Übereinstimmung des Nachweises von Immunglobulinen (IgG) in der Harder´schen

52
Drüse und dem Schleim aus der Choanenspalte ließen sie eine partielle lokale
Antikörperproduktion vermuten. Den Ursprung der von ihnen in der Bursa cloacalis
gefundenen MG-Antikörper vermochten sie nicht zu klären.
Anzeichen für die Bedeutung der Bursa cloacalis bei der Immunantwort von Hühnern
auf Mykoplasmen belegen die Untersuchungsergebnisse von VARDAMAN et al.
(1973). Hühner, denen die Bursa cloacalis chirurgisch entfernt worden war, wiesen
nach einer Infektion mit M. synoviae signifikant niedrigere Antikörpertiter im HAH auf.
Zudem stellten sie bei diesen Tieren nach einer aerogenen Belastungsinfektion bei
21 von 32 Hühnern Luftsackentzündungen mit fibrinösem Exsudat fest, während
entsprechende Veränderungen nur bei drei Tieren der Kontrollgruppe gefunden
wurden.
Auch LAM und LIN (1984) entfernten bei Eintagsküken die Bursa cloacalis bzw. den
Thymus. Am zweiten Lebenstag wurden die Küken mit 1x1010 Mykoplasmen (TS
100) je Tier geimpft. Nach drei Wochen erfolgte eine EBI mit dem MG-Stamm S6 in
die Luftsäcke. Nach einer Woche wurden die Küken getötet, das Blut zur
serologischen Untersuchung gewonnen und die Tiere pathologisch-anatomisch
untersucht. Im Vergleich zu einer nicht bursektomierten und geimpften
Kontrollgruppe (15%) wiesen ungeimpfte (47 %) und geimpfte aber bursektomierte
Küken (50%) signifikant häufiger Luftsackentzündungen auf. Durch die Entfernung
des Thymus ergaben sich keine signifikanten Unterschiede in der Häufigkeit von
Luftsackentzündungen. Beim Antikörpernachweis per serologischer Untersuchung in
der SSA konnten LAM und LIN (1984) keine signifikanten Unterschiede zwischen
burs- bzw. thymektomierten Tieren und ihren jeweiligen Kontrollgruppen feststellen.
Sie vertraten die Ansicht, dass die Ergebnisse ihrer Untersuchungen auf einen B-
Zell-vermittelten Schutzmechanismus vor Luftsackentzündungen hinwiesen.
LAM et al. (1986) stellten bei SPF-Küken nach einer EBI mit dem MG-Stamm TS-100
eine „milde“ zelluläre Reaktion im oberen Atemtrakt fest. Küken, die sie mit dem MG-
Stamm S6 infiziert hatten, zeigten „deutlichere“ Reaktionen. Serologisch konnten sie
keine Unterschiede der Titer in der SSA, im HAHT und Radioimmunassay feststellen.
Nur bei 50 % der Küken, die sie zuvor bursektomiert und experimentell infiziert
hatten, fanden sie in Trachealspülproben im Radioimmunassay (RIA) spezifische

53
Antikörpertiter (IgG) gegen MG. Dagegen konnten sie bei allen nicht
bursektomierten, aber infizierten Küken der Kontrollgruppe spezifische Antikörper
(IgG) im RIA nachweisen. Die Tatsache, daß bei den bursektomierten Tieren zwar
humorale Antikörper nachzuweisen waren, sie jedoch lokal in der Trachea keine
spezifischen Antikörper fanden, deuteten LAM et al. (1986) als einen Hinweis auf
eine B-Zell-vermittelte lokale Antikörperproduktion in der Trachealschleimhaut.
ELFAKI et al. (1992, 1993) beschrieben die Immunreaktionen von kommerziell
erworbenen Küken, die sie im Alter von drei und sieben Wochen mit einer
inaktivierten Vakzine, die mit iota carrageenan (iCGN) als Adjuvans hergestellt
worden war, impften. Sie verglichen verschiedene Applikationsrouten bei der
Impfstoffgabe (s. o.). Küken, die nach der Impfung einen signifikanten Schutz vor
Luftsackentzündungen aufwiesen, hatten zeitgleich auch signifikant höhere Titer
spezifischer Immunglobuline (IgG, IgA) im Serum. Nach der EBI stiegen die
Antikörpertiter (IgG, IgA) in den Trachealspülproben deutlich von <1,0 log10 auf 3,03
± 0,058 log10 respektive < 1,0 log10 auf 3,03 ±0,130 log10 an. Dieser Anstieg der
Antikörpertiter war signifikant höher als der Anstieg der Antikörpertiter
Kontrollgruppe. Zwei Wochen nach der Infizierung konnten aus den Tracheen der
geimpften Hühner signifikant weniger Mykoplasmen reisoliert werden.
Nach einer EBI von 15 MG-freien Hühnern im Alter von 35 Tagen mit dem MG-
Stamm R konnten AVAKIAN und LEY (1993) in Spülproben der oberen und unteren
Atemwege Immunglobuline der Klassen IgG, IgM, und IgA nachweisen. In einer von
ihnen angelegten Trachealringkultur konnten sie eine signifikant höhere
Beweglichkeit der Zilien feststellen, wenn sie diese Kultur mit der Spülflüssigkeit aus
den oberen Atemwegen zuvor infizierter Hühner behandelt hatten. Desweiteren
hemmten die Spülflüssigkeiten aus den oberen und unteren Atemwegen die
Anheftung von Mykoplasmen in vitro, in Abhängigkeit von der in der Spülflüssigkeit
mittels ELISA nachgewiesen Antikörpermenge. Dies könnte auf einen
Abwehrmechanismus hinweisen, der in vivo die Adhäsion der Mykoplasmen im
Respirationstrakt beeinflußt. Den Ursprung der von ihnen nachgewiesenen
Antikörper konnten die Autoren nicht belegen.

54
YAGIHASHI et al. (1987) untersuchten die Wirksamkeit einer inaktivierten Öl-
Adjuvansvakzine bei SPF-Hühnern. Hierbei testeten sie unterschiedliche
Applikationswege. Eine Gruppe wurde zweimal intramuskulär geimpft, bei der
zweiten Gruppe folgte auf die intramuskuläre eine intratracheale Impfung. Beide
Gruppen wiesen zum Zeitpunkt der Infektion mit dem MG-Stamm SAS ähnliche Titer
(ELISA (IgG, IgA)) in den Trachealspülproben und im Serum auf, jedoch war die
Schutzwirkung bei den intratracheal nachgeimpften Tieren höher. Sieben Tage nach
der Infizierung konnten sie bei den i.m. und i.t. geimpften Tieren signifikant weniger
Mykoplasmen reisolieren und stellten signifikant weniger Läsionen in der Trachea
fest. Der bessere Schutz der intratracheal geimpften Tiere deutete ihrer Meinung
nach darauf hin, dass bei den Hühnern auch eine lokale zellvermittelte Abwehr eine
Rolle beim Schutz gegen eine MG-Infektion spielt. Sie vermuteten hierbei eine lokale
Antigenproduktion. Letztendlich konnten sie in dieser Studie keinen Nachweis über
die Herkunft der Antikörper führen, nahmen aber an, dass es sich sowohl um lokal
sezernierte Antikörper als auch um transsudierte Immunglobuline handeln könnte.
Die Abhängigkeit des Impfschutzes von der Höhe bestehender Antikörpertiter und
der immunologischen Fähigkeit der Hühner, auf eine Impfung zu reagieren,
untersuchten YAGIHASHI et al. (1992). Zwei verschiedene Hühnerlinien wurden als
Versuchstiere ausgewählt. Hennen der GSP-Linie sind als „high responder“ bekannt.
Als weitere Versuchstiere wurden Hennen einer Linie gewählt, die als „low-responder
line“ (BM-C) bekannt ist. Die Hühner beider Linien wurden mit inaktivierter
Aluminium-Hydroxid-Vakzine zunächst intramuskulär, dann intratracheal geimpft. Die
EBI erfolgte mit dem MG-Stamm SAS. Die GSP-Hühner zeigten deutlich höhere
Antikörpertiter (IgG und IgA) im ELISA. So zeigten diese Tiere 14 Tage nach der
ersten Impfung einen Wert von 2,7 ± 0,3 log10 Antikörper des Typs IgG im Serum.
Die Hühner der BM-C Linie hatten zur gleichen Zeit einen signifikant niedrigeren Titer
von 0,3 ± 0,6 log10. Vier Tage nach der EBI wurden in den Trachealspülproben der
GSP-Hühner signifikant höhere Antikörpertiter des Typs IgG gefunden. Bei diesen
Probanden konnten sie auch in Tränenflüssigkeit bzw. im Speichel signifikant
häufiger Immunglobuline vom Typ IgG und IgA feststellen. Beide Linien zeigten nach
der EBI einen Schutz vor einer Infektion der Trachea mit MG. Der „Grad der

55
Schutzwirkung“ ist jedoch bei der GSP-Linie ausgeprägter. Die Ergebnisse dieser
Untersuchungen deuten darauf hin, dass die Qualität der Immunabwehr mit der
Menge der lokal vorhandenen Antikörper korreliert ist. BRANTON et al. (1997)
berichteten über die Veränderungen des weißen Blutbildes nach einer EBI von MG-
freien Hühnern im Alter von sechs Wochen mit 0,4 ml einer Mykoplasmensuspension
(F-Stamm). In der akuten Phase innerhalb der ersten vier Wochen nach der
Inokulation der Mykoplasmen via eye-drop, stellten sie bei der infizierten Gruppe
Heterophilie, von ihnen als Verschiebung des Verhältnisses zwischen
Gesamtleukozytenzahl und Lymphozytenzahl definiert, sowie Lymphopenie,
Monozytose, Eosinopenie, und Basopenie fest. Im Vergleich mit einer nicht infizierten
Kontrollgruppe waren diese Unterschiede signifikant. Histopathologisch konnten sie
bei den infizierten Hühnern eine Hyperplasie der Lymphzentren in Milz und
Knochenmark feststellen.
GAUNSON et al. (2000) untersuchten erstmals die Anzahl und Verteilung von T-
Lymphozytzen in der Trachea von Hühnern nach einer EBI mit dem virulenten MG-
Stamm R. Signifikante Unterschiede in der Häufigkeit von zytotoxischen T-
Lymphozyten (CD 8) im oberen, mittleren und unteren Bereich der Trachea konnten
sie nicht feststellen. Während die T-Helferzellen (CD 4) gleichmäßig in der
Trachealschleimhaut verteilt waren, lagen die zytotoxischen T-Lymphozyten in
„follikelähnlichen“ Haufen zusammen. Im gesamten Verlauf des Experiments konnten
sie hohe Konzentrationen von T- Helferzellen und zytotoxischen T-Lymphozyten in
der Trachealschleimhaut feststellen, die sich jedoch in der Häufigkeit ihrer
spezifischen T-Zell-��������������� +�������+������������������������
Anzahl der Mykoplasmen in der Trachealschleimhaut nahm in der zweiten Woche p.i.
ab, währenddessen die Zahl der T-Lymphozyten gleichzeitig noch anstieg.
GAUNSON et al. (2000) bewerteten die spezifische Aktivierung der zytotoxischen T-
Lymphozyten als wichtigen Abwehrmechanismus des Wirtes gegen eine Infektion
durch MG.
Die Fähigkeit von MG in nicht phagozytierenden Zellen einzudringen und sich so der
Immunabwehr des Wirtes zu entziehen, stellt eine weitere Überlebensstrategie von
MG dar. WINNER et al. (2000) konnten diese Fähigkeit von MG in vitro bei

56
menschlichen Epithelzellen und Fibroblasten aus Hühnerembryonen nachweisen.
Binnen 20 Minuten gelangten Mykoplasmen aus dem interzellularen Bereich in die
Zellen hinein und konnten hier auch durch eine antibiotische Behandlung mit
Gentamycin nicht abgetötet werden. Da sie gleichzeitig ihre Fähigkeit, diese Zellen
wieder zu verlassen, nicht einbüßten, sahen WINNER et al. (2000) dies als eine der
Ursachen chronischer und generalisierter Infektionen an.
KHAN et al. (1986) stellten bei der Untersuchung der Proteinzusammensetzung
verschiedener MG-Stämme (S6, F, A5969) in der Natrium-dodecyl-sulfat - Polyacryl-
amid-Gel-Elektrophorese (SDS-PAGE) fest, dass sich die einzelnen MG-Stämme in
ihrem Bandenmuster unterschieden. Anhand geringer Abweichungen in den
Bandenmustern konnten sie auch Subpopulationen eines Stammes unterschiedlicher
Herkunft von einander abgrenzen. KHAN et al. (1986) hoben die Bedeutung der
SDS-PAGE für epidemiologische Studien hervor.
Auch YOGEV et al. (1988) konnten mit Hilfe von RNA-Gensonden unterschiedliche
Genotypen innerhalb eines MG-Stammes nachweisen. Ihre Untersuchungen
deuteten darauf hin, daß einige Genomabschnitte bei allen Stämmen von MG
identisch sind und andere stammspezifisch und somit verantwortlich für die Spezifität
des „genetischen Fingerabdrucks“ eines Stammes zeichnen. Aber auch bei
Subpopulationen eines Stammes waren unterschiedliche Bandenmuster zu
erkennen, so dass beispielsweise eine Abgrenzung von Feldisolaten des F-Stammes
gegenüber dem Impfstamm möglich war.
BARBOUR und NEWMAN (1990) untersuchten die Bildung spezifischer Antikörper
nach Impfung von Mycoplasma-spp.-freien Hühnern mit MG-„sub-unit-Vakzinen“.
Nach Impfung mit einer „sub-unit-Vakzine“ im Alter von 13 und 17 Wochen, die das
Adhäsinprotein von MG enthielt, konnten sie die Bildung spezifischer Antikörper
nachweisen. Diese Antikörper konnten sie im Western-Blot als Proteinbande im
Molekulargewichtsbereich von 75 kDA darstellen. Gleichzeitig fanden sie in ihren
Untersuchungen auch Kreuzreaktionen drei Wochen nach der ersten bzw. zweiten
Impfung im Bereich von 85 kDA respektive 56 kDA. Auch nach Impfung mit MG-
Vakzinen, die aus dem ganzen Erreger hergestellt waren, konnten Antikörper mit
einem Molekulargewicht von 75 kDA nachgewiesen werden. Sie werteten dies als

57
Hinweis auf die immunologische Bedeutung des Adhäsins für die Antikörper-
entwicklung und die damit verbundene Wirksamkeit einer Vakzine gegen die
Anheftung und Infektion mit MG.
Bereits 1988 berichteten WATSON et al. über die Heterogenität des Antigen-
Komplexes V-1 bei Mycoplasma pulmonis. Mit monoklonalen und monospezifischen
Antikörpern untersuchten sie 18 verschiedene Stämme von M. pulmonis. Alle
unterschieden sich in ihren Bandenmustern voneinander. Bei der Untersuchung von
subklonierten Populationen von vier Mycoplasma pulmonis-Stämmen konnten sie im
Immunblotting Unterschiede in der Struktur und im antigenetischen Charakter dieser
Subpopulationen nachweisen. Die Häufigkeit der Phasenvariation des Oberflächen-
antigen-Komplexes V-1 wurde mit 2 x 10-3 pro Zelle und Generation angegeben.
Diese Variationsmöglichkeit der Majorantigene der Zellmembran bewerteten die
Autoren als bedeutende Möglichkeit von M. pulmonis, sich der Wirtsabwehr zu
entziehen.
ROSENGARTEN und WISE (1990) stellten bei Mycoplasma hyorhinis, einer für das
Schwein pathogenen Mykoplasmenart, im Verlaufe von über 100 Kulturpassagen
Veränderungen im Erscheinungsbild der Kolonien fest. Hierbei konnten sie drei
unterschiedliche Kolonieformen identifizieren. Bei der Untersuchung von Sub-
populationen stellten sie fest, dass der Wechsel von einer in die andere Form
möglich ist. Sie konnten feststellen, dass die phänotypische Instabilität durch einen
hochfrequenten Wechsel von membranständigen Oberflächenproteinen bedingt ist.
Diesen Drift der Oberflächenstruktur konnte in einer Studie über das als „Variable
lipoprotein (Vlp) systems“ bezeichneten Komplexes bei M. hyorhinis bestätigt
werden. Die antigenetischen Variationsmöglichkeiten von drei membranständigen
Lipoproteinen (VlpA, VlpB, VlpC), von denen jedes unabhängig von den anderen
innerhalb einer isogenen Population hochfrequente Phasenvariationen aufwies,
wurden beschrieben.
ROSENGAREN und WISE (1991) fanden 12 bis 30 spontane Größenvariationen je
Vlp, die unabhängig voneinander auftraten. So konnten sie 104 verschiedene
strukturelle Unterschiede der Vlp´s dokumentieren. Den Nachweis von Antikörpern
gegen diese Vlp`s im Serum von Schweinen, welche sie zuvor experimentell mit M.

58
hyorhinis infiziert hatten, werteten sie als Hinweis darauf, daß das System der Vlp´s
auch in vivo auftritt.
Dagegen konnten EVANS et al. (1992 b) bei der Untersuchung der genetischen
Stabilität der MG-Stämme S6 und 6/85 (MGI; Intervet America Inc., Millsboro, Del.)
kaum Unterschiede im Verlauf von zehn in vivo Passagen feststellen. Sie verglichen
die mittels Restriktionsendonukleasen gelösten DNS-Banden in der Darstellung mit
der SDS-PAGE vor bzw. nach drei, sechs und zehn Passagen in Hühnern und
Puten. Bis auf eine deutlichere Ausprägung einer Proteinbande im Bereich von 86
kDa konnten sie keine Veränderungen der DNA-Bandenmuster im Bereich von 46,5
kDA, 50-54 kDA, 58-64 kDA und 105-140 kDA feststellen und werteten dies als
Anzeichen für eine genetische Stabilität der MG-Stämme S6 und 6/85.
PANANGALA et al. (1992) entwickelten monoklonale Antikörper (MAb) gegen den
Mycoplasma gallisepticum-Stamm PG 31. Mit diesen untersuchten sie die
antigenetischen Profile von fünf bekannten (PG 31, R, S6, F, A5969) Stämmen und
sechs Feldisolaten. Ein monoklonaler Antikörper (G12) reagierte mit allen Stämmen
und Isolaten und zeigte dabei in der SDS-PAGE ein einheitliches Bandenmuster. Mit
dem monoklonalen Antikörper G9 reagierten ebenfalls alle Stämme, jedoch
schwankte die Molekularmasse der nachgewiesenen Antigene zwischen 90 und 98
kDA. Die MAb´s G10 und G11 reagierten dagegen selektiv nur mit einigen MG-
Stämmen bzw. Isolaten. PANANGALA et al. (1992) konnten mit goldkonjugierten
Antigenen die Lokalisation der Antigene in der Zellmembran von MG nachweisen.
Der MAb G9 reagierte in ihrem Versuchsaufbau mit Oberflächenantigenen und war
an der Agglutinationsreaktion von MG beteiligt. Der Grad der Agglutinationsreaktion
war bei den einzelnen MG-Stämmen unterschiedlich stark. Es wurde vermutet, dass
die unterschiedliche Expression der Epitope für die antigenetischen Unterschiede der
einzelnen MG-Stämme verantwortlich ist.
Die Bedeutung des Hämagglutinins, einem lipophilen Oberflächenprotein von MG mit
einem Molekulargewicht von 64 kDA, für die Immunantwort untersuchten FORSYTH
et al. (1992). Durch Immunoelektronenmikroskopie konnten sie die Bindung dieses
Proteins an die Basis des terminalen Endes des Fab-Fragmentes feststellen. Obwohl
dieses Protein (LP 64) nur 1,7 % der Gesamtproteinmasse darstellt, trägt es zu 7,4

59
% zur Antigenbildung (IgG) vier Wochen p.i. bei. Sie konnten mittels Immunoblotting
das Hämagglutinin auf der Membran der MG-Stämme R, F, PG 31 und A 5969
nachweisen; dies gelang ihnen jedoch nicht bei M. synoviae. Anti-LP 64-Serum zuvor
infizierter Kaninchen verhinderte die Hämagglutination von Hühnererythrozyten bis
zu einer Verdünnung von 1:256. Durch eine 14C-Markierung von MG konnten
FORSYTH et al. (1992) nachweisen, daß Anti-LP 64-Serum von Kaninchen die
Anheftung von MG an die Epithelzellen der Trachea um 62 % vermindert. Sie werten
dies als Hinweis auf die Bedeutung des LP 64 als Adhäsionsfaktor.
MARKHAM et al. (1993) isolierten beim Mycoplasma gallisepticum-Stamm S6 mit
Hilfe eines monoklonalen Antikörpers (MAb) in der Affinitätschromatographie ein
Protein („pMGA“) mit einem Molekulargewicht von 67 kDA. Dieses Oberflächen-
protein wird mit den hämagglutinierenden Eigenschaften von MG in Verbindung
gebracht. Sie versuchten mit Hilfe der DNS-Restriktionsanalyse das Gen, welches
das pMGA kodiert, zu isolieren. Sie fanden jedoch nur zwei in ihrer Sequenz sehr
ähnliche Gene, die das von ihnen isolierte „pMGA“ jedoch nicht kodierten.
MARKHAM et al. (1992) vermuteten eine Genfamilie, die für die Exprimierung sehr
ähnlicher Oberflächenstrukturen verantwortlich ist.
ROSENGARTEN et al. (1994) untersuchten die Protein- und Antigenprofile von elf
M. bovis-Isolaten. In der SDS-PAGE stellten sie deutliche Unterschiede der
Bandenmuster zwischen den einzelnen Stämmen fest. Diese antigenetischen
Unterschiede traten unabhängig von der geographischen Herkunft, dem klinischen
Krankheitsbild oder der Art der Probenentnahme auf. ROSENGARTEN et al. (1994)
beobachteten diesen hochfrequenten Wechsel in der Ausbildung von Epitopen auch
bei von ihnen klonierten Subpopulationen des M. bovis-Stammes PG 45.
Auch BEHRENS et al. (1994) untersuchten subklonierte Nachkommen des M. bovis-
Stammes PG45 mit Hilfe von monoklonalen Antikörpern (MAb´s). Hierbei
identifizierten sie eine „Familie“ von MAb´s und strukturell ähnlichen, variablen
Oberflächenproteinen (variable surface proteins; Vsps). Drei Mitglieder dieser
„Familie“ konnten sie identifizieren, sie bezeichneten sie als VspA, VspB und VspC.
Diese Oberflächenproteine veränderten unabhängig voneinander ihre Größe;
BEHRENS et al. (1994) vertreten die Ansicht, dass diese Fähigkeit zur

60
Oberflächenvariatiion eine Möglichkeit für M. bovis darstellt, sich der Wirtsabwehr zu
entziehen.
YOGEV et al. (1994) verwandten in ihrem Experiment denselben monoklonalen
Antikörper (1E5), den BEHRENS et al. (1994) zur Darstellung von variablen Epitopen
auf Oberflächenproteinen bei M. bovis genutzt hatten. Sie untersuchten 15
verschiedene MG-Stämme, u .a. F, R, S6, PG31, 6/85 und A5969. Bei diesen
Stämmen konnten sie auf dem Oberflächenprotein (PvpA) dieses Epitop erneut
nachweisen. Sie stellten fest, dass das PvpA ein integraler Bestandteil der
Zellmembran ist und das Epitop aus dieser Oberfläche herausragt. YOGEV et al.
(1994) konnten eine hochfrequente phänotypische Veränderung des Proteins
erkennen, für die sie eine reversible Veränderung des Genotyps in Form eines „Ein-
Aus-Ein-Mechanismus“ verantwortlich machten. Im Gegensatz zu den Versuchen
von BEHRENS et al. (1994), die mit dem MAb 1E5 lipophile Oberflächenproteine
nachgewiesen hatten, wies das Antigen bei den MG-Stämmen einen hydrophilen
Charakter auf.
GLEW et al. (2000) untersuchten die molekulare Struktur des Oberflächenproteins
(pMGA) in vivo. Hierzu infizierten sie Hühner mit dem MG-Stamm S 6. Zwei Tage p.i.
hatte sich bei 40 % der Mykoplasmen die phänotypische Struktur dieses Ober-
flächenproteins verändert. Sechs Tage p.i. konnten GLEW et al. (2000) diese
reversiblen Strukturveränderungen bei über 50 % der untersuchten Oberflächen-
proteine feststellen. Dieser Wechsel der Struktur der Oberflächenproteine fand auch
ohne das Vorhandensein spezifischer Antikörper statt und schien der spezifischen
Immunabwehr des Wirtes vorauszugehen. GLEW et al. (2000) sahen diese Fähigkeit
von MG als großen Vorteil für die Besiedlung von Wirtsorganismen an.
Auch BENICINA et al. (1994) verwendeten monoklonale Antikörper (n = 12) gegen
Epitope von MG-Stämmen (R, F, S6 (208) und PET2) zur Untersuchung der
Oberflächenantigene von insgesamt 22 MG-Stämmen. Zwei MAB´s reagierten mit
Epitopen einer Molekularmasse von 56 kDA bei allen untersuchten MG-Stämmen,
sechs MAB´s reagierten zwar mit fast allen Stämmen, jedoch nicht mit den MG-
Stämmen K 503 und K 703. Drei MAB´s erkannten drei unterschiedliche
Oberflächenantigene auf Proteinen von etwa 67 kDA, und ein MAb band an variable

61
Epitope auf Proteinen mit dem Molekulargewicht von 33 bzw. 80 kDA. Mittels
Darstellung durch zweidimensionale Immunoblots konnten sie Unterschiede in der
Ladung der Proteine nachweisen, die bei verschiedenen MG-Stämmen diese
variablen Oberflächen-antigene trugen. Einige subklonierte Kolonien wechseln die
mit den 67 kDA und 80 kDA assoziierten Epitope. Dieser phänotypische, reversible
Wechsel der Antigenstruktur trat bei einem bis fünf Prozent der Population der
nächsten Generation auf und führte zur Bildung von Subpopulationen; es wurde
vermutet, dass dieser Wechsel der Epitope wahrscheinlich auch in vivo vorkommt
und es MG so möglich wird, der Immunantwort des Wirts zu entgehen.
Die Ausprägung verschiedener Oberflächenepitope untersuchten GARCIA et al.
(1994) bei den Mycoplasma gallisepticum-Stämmen A5969, R, S6, F, 6/85, ts-11,
K503, K703 und K730 mit sieben verschiedenen monoklonalen Antikörpern (MAb).
Keiner der MAb´s reagierte in der IIPA mit den drei schwachvirulenten Stämmen
K503, K703 und K730. Im Western-Blot banden zwei MAb´s mit einem Protein (44
kDA) und ein MAb mit einem Protein (119 kDA) bei allen MG-Stämmen. Drei MAb´s,
die ihren Ursprung im S6-Stamm hatten, reagierten mit einem Protein, das ein
Molekulargewicht von 69 kDA aufwies. GARCIA et al. (1994) konnten an Hand
subklonierter Populationen in vitro einen Wechsel in der Ausprägung der Epitope
nachweisen.
LEVISOHN et al. (1995) untersuchten im Western-Blot die unterschiedlichen
antigenetischen Profile von MG-Isolaten (R-Stamm). Nach einer artifiziellen
aerogenen Infizierung reisolierten sie die Mykoplasmen mittels Abstrich drei und acht
Wochen p.i. aus den Tracheen der Versuchshühner. Die Bandenmuster der durch
die MAb´s identifizierten Oberflächenproteine veränderten sich im Verlaufe des
Versuches von der Inokulation beginnend bis acht Wochen p.i. deutlich. Die Autoren
sehen dies als Nachweis dafür, dass MG auch in vivo die Fähigkeit besitzt, seine
Oberflächenstruktur verändern und sich so der Wirtsabwehr entziehen zu können.
ELLAKANY et al. (1998) konnten in zwei Experimenten die Sensitivität eines
Immunoblottingverfahrens zum Nachweis von Antikörpern gegen MG in Tracheal-
spülproben und aus dem Blutserum belegen. Der Nachweis erfolgte mit Hilfe von
monoklonalen Antikörpern gegen IgG, IgM und IgA von Hühnern. Sie infizierten

62
Hühner im Alter von zehn Tagen mit dem MG-Stamm 1226. Zwei Wochen nach
dieser Belastungsinfektion untersuchten sie die Trachealspülproben und stellten fest,
daß die monoklonalen Antikörper gegen IgA mit acht Polypeptiden von MG
reagierten. Die Antikörper gegen IgG und IgM reagierten mit jeweils 3 Polypeptiden.
Im Serum konnten sie zu diesem Zeitpunkt keine IgA-Antikörper nachweisen, doch
reagierten die IgM-Antikörper mit acht Polypeptiden und die IgG-Antikörper mit 16
Polypeptiden von MG. In weiteren Untersuchungen stellten sie fest, daß nach einer
Infektion zunächst die Immunglobuline des Typs IgA in den Trachealspülproben
dominant waren und daß im Serum der Typ IgM vorherrschte. Ab der zweiten Woche
p.i. traten jedoch sowohl im Serum als auch in den Trachealspülproben die
Immunoglobuline des Typs IgG in den Vordergrund.
Die Möglichkeit einen Impfstoff gegen MG, auf Basis des Hühnerpockenvirus zu
entwickeln beschäftigte YOSHIDA et al. (2000). Hierzu isolierten sie den MAb 35A6
gegen den MG-Stamm R aus Mäusen. Dieser MAb bindet an ein Membranprotein
mit einen Molekulargewicht von 120 kDa und ist in der Lage sowohl das Wachstum
als auch den Stoffwechsel von MG in vitro zu unterbinden. Nach einer
Gensequenzierung und Genisolierung war es ihnen möglich, dass für das
Membranprotein entsprechende Gen (MGC 3) in das Genom von Hühnerpockenvirus
zu transferieren. Ein Hühnerpockenvirus war in der Lage das von dem MGC 3-Gen
terminierte Protein auf der Zelloberfläche zu exprimieren. YOSHIDA et al. (2000)
hielten es für möglich auf diese Weise einen Impfstoff gegen MG zu entwickeln.
2.4. Aerogene Infektion mittels Aerosol
Im Allgemeinen erfolgt eine Infizierung von Hühnern mit MG durch Inspiration die
anschließender Anheftung der Mykoplasmen an die Schleimhaut, vor allem der
oberen Atemwege (STIPKOVITS 1985). Bei der Beurteilung der Effizienz von
Impfstoffen im Experiment ist es notwendig, das natürliche Infektionsgeschehen
nachzuahmen (WHITHEAR 1996). JARNYCH (1976) faßte die Grundlagen der
Anwendung von Aerosolen in der Veterinärmedizin zusammen. Soll die
Verabreichung eines Aerosols zur Infektion eines Tieres über die Atemwege führen,

63
so ist die Schwebfähigkeit des Erregers und damit seine Fähigkeit, tief in die
Atmungsorgane einzudringen, entscheidend. Dies ist abhängig von der Größe der
Tröpfchen und der Konzentration des Aerosols. Bei einer zu hohen Anzahl von
Tröpfchen je Raumeinheit kommt es infolge der Brown´schen Bewegung durch
Kollision der Tröpfchen zu einer Größenzunahme der einzelnen Tropfen durch
„Koagulation“ (JARNYCH 1976). Ein konzentriertes Aerosol enthält daher,
unabhängig von seiner Ausganskonzentration, schon nach einigen Minuten nicht
mehr als 107 Tropfen / cm3.
Je kleiner die Tropfen sind, desto größer ist ihre Schwebfähigkeit. Aerosole, deren
Teilchen einen Durchmesser von 100 µm aufweisen, sind instabil und können daher
nicht in der Veterinärmedizin eingesetzt werden. Bei Teilchen mit einem
Durchmesser von weniger als 10-5 cm übertrifft die Brown´sche Molekularbewegung
die Schwerkraft. Diese Aerosole sedimentieren daher nur wenig. Sie werden als
hochdisperse Aerosole bezeichnet. Grobdisperse Aerosole besitzen einen
Teilchendurchmesser von 10-4 cm und weisen eine deutliche Sedimentationstendenz
auf. Aerosole mit der Teilchengröße von 10-4 bis 10-5 cm nehmen eine
Zwischenstellung ein. Besonders Teilchen mit einem Durchmesser von 0,2 µm
werden in Atmungsorganen und Filtern besonders schlecht zurückgehalten und
können daher auch tief in die Atmungsorgane eindringen.
Gleichzeitig weisen Stoffe, die als Aerosol vorliegen, eine stark vergrößerte
Oberfläche auf. Auf Grund dieser stark vergrößerten Oberfläche steigt die
Oberflächenaktivität an. Ein Beispiel hierfür ist die exponentielle Wirkungssteigerung
bei fein verteilten Desinfektionslösungen.
Die Anforderungen an Bauart und Betrieb einer Aerosol-Infektions-Box beschrieben
BEARD und EASTERDAY (1965). Es handelte sich hierbei um einen
Luftdruckvernebler mit zwei Kammern. Eine Kammer diente zur Vernebelung des
Agens, die andere zur zusätzlichen Versorgung der Tiere mit Luft. Die
Gesamtkapazität der Anlage betrug 8 l Luft/Minute. In das System war ein Filter
integriert, welcher einen Porendurchmesser von 0,8 +/- 0,05 µm aufwies. In der
Aerosolkammer herrschte negativer Luftdruck, um das Entweichen des Agens in die
Raumluft zu verhindern. Eine Vernebelungsdauer von 20 Minuten war bei diesem

64
Versuchsaufbau ausreichend, um eine Sättigung der Luft mit dem Agens zu
erreichen. Die Partikelgröße war nach ihrer Auffassung entscheidend für die
Eindringtiefe des Aerosols in den Respirationstrakt. Zur Infektion in den unteren
Abschnitten des Respirationstraktes verwendeten sie eine Partikelgröße von 4 - 5
µm. BEARD und EASTERDAY (1965) vermochten mit diesem Versuchsaufbau
erfolgreich Hühner mit Newcastle Disease Virus zu infizieren.
KLEVEN et al. (1972) beschrieben die erfolgreiche Infizierung von Eintagsküken
mittels Aerosol durch Mycoplasma synoviae, ohne genauer auf die technischen
Daten der Geräte einzugehen. Im ersten Versuch wurde ein „Sprayer“ für Impfungen
und in zwei weiteren Versuchen der „Vaponefrin standard nebulizer“ (Vaponefrin
Company, Edison, N.J.), verwendet. HAYATSU et al. (1974) nutzten eine 150 l große
Kammer zur aerogenen Impfung von Hühnern mit inaktiviertem MG-Impfstoff. Der
Vernebler (Toyoshima Co.) hatte eine Kapazität von 0,25 ml/min und arbeitete mit
einem Druck von 0,7 kg/cm². Sie gaben für diesen Versuch eine Impfdosis von 5 x
109 Zellen an. Für die Infizierung der Hühner verwendeten sie
Mykoplasmenkonzentrationen zwischen 7,4 x 103 bis 7,4 x 105 KBE/ml. Für die
Impfung von 1760 Broilern in einem Feldversuch verwendeten HAYATSU et al.
(1975) den „Sprayer“ MS-10 (Matsushita Electric Industrial Company, Ltd, Kadoma,
Japan). Der Sprayer hatte eine Durchflußrate von 10 ml/min, dies entsprach einer
Impfstoffmenge von 0,22 ml/Huhn in einem Zeitraum von 40 Minuten. Die
Konzentration des Impfstoffes gaben sie in diesem Versuch mit 2 x 1010 Zellen an.
Auf den Versuchsaufbau von KLEVEN (1972) bezogen sich LEVISOHN et al. (1986),
ABD-EL-MOTELIB und KLEVEN (1993) und ORTIZ et al. (1995). Zur Infektion der
Tiere verwendeten sie jeweils den MG-Stamm R, bzw. LEVISOHN et al. (1986) auch
die Stämme F, S-6, 227 und A 5969 in einem vergleichenden Experiment. Die
Konzentration der Mykoplasmen in der Suspension gaben ABD-EL-MOTELIB und
KLEVEN (1993) mit 5,0 x 108 KBE / ml und ORTIZ et al. (1995) mit 3,8 x 108 KBE /
ml an. RODRIGUEZ und KLEVEN (1980 a, b) verwendeten einen „Devilbiss No. 42
standard nebulizer“, um 14 Tage alte Küken mit dem F-Stamm (3 x 106 KBE / ml)
bzw. dem R-Stamm (4 x 108 oder 1,5 x 106 KBE / ml) zu infizieren.

65
CARPETER et al. (1981) benutzten in ihren Versuchen zur Vakzination von Hühnern
mit dem F-Stamm den Vernebler „HCS1-2AA“ der Firma Micro-Gen equipment, San
Antonio, Texas. Die Teilchengröße betrug 5 µm, für 1000 Hühner wurden 100 ml
einer Mykoplasmensuspension mit 109 KBE / ml versprüht.
LIN und KLEVEN (1982 a, b, 1984 a, b,) bezogen sich in ihrem Versuchsaufbau
wiederum auf RODRIGUEZ und KLEVEN (1980 a, b). Es gelang, mit dem Vernebler
„Devilbiss No. 42“ eine Infektion mit dem R-Stamm zu erzeugen und die
anschließende transovarielle Übertragung von MG auf Eier nachzuweisen. Auch
ELFAKI et al. (1992 und 1993) verwendeten diesen Vernebler, um Hühner mit dem
R-Stamm zu infizieren.
WHITHEAR et al. (1996) beschrieben ein Gerät zur aerogenen Infizierung mit MG,
um die Effizienz von MG-Vakzinen testen zu können. Grundlage für ihr
Infektionsmodell war die Arbeit von BEARD und EASTERDAY (1965). Die Kammer
faßte ein Volumen von 80 l bei einer Luftflußrate von 40 l/min, so dass in zwei
Minuten jeweils der Austausch der Luft erfolgte. Der Vernebler war eine Anfertigung
der Chemischen Werkstätten der Universität Melboure, Parkville und arbeitete mit
einer Luftrate von 15 l/min. Zur Infektion wurde die Luftrate der Frischluft auf 25 l/min
gesenkt. Die Teilchengröße betrug 1 bis 10 µm, wobei der Anteil der Teilchen < 5 µm
über 50 % lag. Für die Infektionsversuche verwendeten WHITHEAR et al. (1996) den
virulenten australischen Feldstamm Ap3AS. Hühner, die für 10 Minuten mit Aerosol
(40 l/min) infiziert wurden, welches 102 - 103 CCU/l enthielt, zeigten deutliche
Luftsackentzündungen und serologische Reaktionen. In der Trachea waren jedoch
keine pathologisch-anatomisch oder histologisch erkennbaren Läsionen
nachzuweisen.

66
3. Material und Methoden
3.1. Allgemeiner Versuchsaufbau
Zunächst wurde in Voruntersuchungen geprüft, ob die Hennen für die
Verträglichkeits- und Wirksamkeitsprüfung geeignet sind. Danach wurde die Hälfte
der aufgestallten Hennen in der 14. und 18. Lebenswoche mit der zu prüfenden MG-
Vakzine geimpft. Nachdem sowohl die Impfgruppe als auch die ungeimpfte
Kontrollgruppe eine Legeleistung von mindestens 70 % erreicht hatten, wurden beide
Gruppen mit dem Mycoplasma gallisepticum-Stamm R in Form eines erregerhaltigen
Aerosols inokuliert. Die Entwicklung der Legeleistung vor und nach der artifiziellen
Infizierung wurde dokumentiert und es wurden Blutproben zur serologischen
Untersuchung gewonnen. Nach einer Beobachtungszeit von mindestens vier
Wochen p.i. wurden die Tiere getötet. Es erfolgte eine pathologisch-anatomische
sowie eine kulturell-bakteriologische Untersuchung veränderter Organe. Zur
kulturellen Reisolation von MG wurden Proben aus Trachea, Lunge und Luftsäcken
gewonnen. Die zeitlichen Abläufe der einzelnen Versuchsdurchgänge sind in der
Tabelle 1 dargestellt.
Tabelle 1: Zeitlicher Ablauf der Versuchsdurchgänge 1, 2 und 3
Versuch 1 Versuch 2 Versuch 3 1. Impfung*1 14. Lebenswoche 14. Lebenswoche 14. Lebenswoche 2. Impfung*2 18. Lebenswoche 18. Lebenswoche 18. Lebenswoche Infizierung*3 24. Lebenswoche 26. Lebenswoche 25. Lebenswoche
Versuchsende*4 28. Lebenswoche 30. Lebenswoche 29. Lebenswoche *1 Zeitpunkt der Aufstallung in die Klinik, 1. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme, Tötung und Sektion aller Versuchstiere
3.2. Voruntersuchungen
Zur Durchführung der Versuche wurden je Versuchsdurchgang 80 Legehennen im
Alter von 14 Wochen per Zufallsprinzip aus einer Herde kommerzieller Junghennen
entnommen und in die Klinik für Geflügel nach Hannover verbracht. 40 Hennen

67
bildeten die Impfgruppe, die anderen 40 die Kontrollgruppe. Die Aufteilung in diese
beiden Gruppen erfolgte nach dem Zufallsprinzip. Die Hennen wurden bei
Versuchsbeginn mit farbigen numerierten Flügelmarken gekennzeichnet. Hierbei
wurden die Hennen einer kurzen Allgemeinuntersuchung unterzogen. Je
Versuchsgruppe wurde bei jeweils 20 zufällig ausgewählten Hennen zum Zeitpunkt
der Aufstallung 10 ml je Tier Blut entnommen, um den MG- und MS-Status der Tiere
mittels Serumschnellagglutination feststellen zu können. In der 15. Lebenswoche
wurde je Gruppe eine gepoolte Kotprobe parasitologisch untersucht. Ziel der
Voruntersuchungen war es festzustellen, ob die Legehennen für den Hauptversuch
geeignet sind. Als Kriterien zur Beurteilung des Gesundheitszustandes wurden das
Allgemeinbefinden (Sensibilität [Aufmerksamkeit], Verhalten, Futter- und
Wasseraufnahme, Befiederung), die Ausbildung und Färbung des Kammes und die
Gewichtsentwicklung herangezogen. Zeigten Hennen eine Störung des
Allgemeinbefindens, erfolgte eine weitergehende Untersuchung des Tieres. Bei
einem Verdacht auf schwerwiegende Erkrankungen wurden diese Tiere getötet und
pathologisch-anatomisch untersucht. Die Voruntersuchung endete mit Beginn der
Legetätigkeit.
3.3. Hauptuntersuchungen
3.3.1. Verträglichkeitsprüfung
Als Kriterien für die Verträglichkeit des Impfstoffes wurde das Allgemeinbefinden
(Sensibilität, [Aufmerksamkeit], Verhalten, Futter- und Wasseraufnahme,
Befiederung, Gewichtsentwicklung) und die Legeleistung (nur bei adulten Hennen)
sowie die lokalen Reaktionen am Ort der Impfstoffapplikationen herangezogen. Die
Hennen wurden durch ein Kontrollfenster beobachtet sowie einige bei der täglichen
Stallkontrolle nach dem Zufallsprinzip einer kurzen Allgemeinuntersuchung
unterzogen und die Legeleistung dokumentiert. Da durch äußere Störungen, wie z.
B. klinische Untersuchungen, Gewichtsermittlung, Stallarbeiten oder Umstallungen
die Legeleistung von Hennen in erheblichem Maße negativ beeinflusst wird, wurde
nach Beginn der Legetätigkeit die klinische Untersuchung nur noch bei auffällig
gestörtem Allgemeinbefinden durchgeführt.

68
3.3.2. Wirksamkeitsprüfung
Die Überprüfung der Wirksamkeit wurde an der für den Impfstoff vorgesehenen
Zielart, Legehennen, vorgenommen. Daher war der Vergleich der Legeleistung
zwischen den geimpften Hennen und der ungeimpften Kontrollgruppe einziger
entscheidender Parameter. Ab der 18. Lebenswoche wurde die Legeleistung der
Hennen täglich dokumentiert. Bei Erreichen von mindestens 70 % Legeleistung
erfolgte die EBI beider Gruppen mit dem MG-Stamm R via Aerosol . Die Entwicklung
der Legeleistung wurde weiterhin dokumentiert und der Versuch nach einem
Beobachtungszeitraum von mindestens 28 Tagen p.i. beendet.
3.4. Herkunft und Haltung der Versuchstiere
Für die Voruntersuchungen und die Durchführung der Verträglichkeits- und
Legeleistungsprüfung wurden kommerziell erhältliche „Lohmann brown“-Legehennen
des Bodenhaltungstyps verwendet. Sie wurden von der Firma „Geflügelzuchtbetriebe
Gudendorf-Ankum GmbH & Co.“ als MG- und MS-freie Junghennen bezogen. Zum
Zeitpunkt der Aufstallung befanden sich die Hennen in der 14. Lebenswoche. Sie
entstammten den Legehennenaufzuchtbetrieben N. (Voruntersuchungsgruppe A),
G/S. (Voruntersuchungsgruppen B und C) W.-L. (Voruntersuchungsgruppe D) und N.
(Voruntersuchungsgruppe E). Die Tiere wurden in zwei identischen Versuchsställen
der Klinik für Geflügel in Bodenhaltung mit Sägespänen als Einstreu aufgestallt. Die
Grundfläche des Stalls betrug 9 m2. Weiterhin befand sich in dem Raum eine
Sitzstange von 2,5 Metern Länge und 10 cm Durchmesser. In der 18. Lebenswoche
wurden übereinander 2 x 5 Legenester Marke „Eierglück“ der Firma Siepmann,
Herdecke, aufgestellt.
Da die Ställe über keine Klimatisierung verfügten, war es in den Sommermonaten
nicht möglich, die Temperatur in den Ställen bei Außentemperaturen von über 18° C
auf diesem Niveau zu halten, so dass es im ersten Versuchsdurchgang zu
Temperaturschwankungen kam. Durch die Möglichkeit, die Zuluft zu erwärmen, war
es jedoch möglich, bei niedrigen Außentemperaturen die Raumtemperatur in den
Versuchsställen bei konstant 18° C zu halten.

69
Die Junghennen erhielten bis zur 18. Lebenswoche pelletiertes Junghennenaufzucht-
futter in einem Hennenfuttertrog mit einer Länge von einem Meter. Die Futtermenge
steigerte sich von 72 g je Henne in der 14. Lebenswoche auf 84 g in der 18.
Lebenswoche. Ab der 18. Lebenswoche erhielten die Hennen pelletiertes
Legehennenfutter, bis zur 22. Lebenswoche 100 g je Henne pro Tag, danach wurde
ad libitum gefüttert. Bei der täglichen Stallkontrolle wurde verbliebenes Futter aus
den Trögen entfernt. Wasser wurde durch eine automatische Standard-Hängetränke
stets ad libitum angeboten. Die Tränken und Tröge wurden täglich gereinigt.
Alleinfuttermittel für Legehennen: Inhaltsstoffe:
Rohprotein 16,50 % Methionin 0,36 % Rohfett 5,00 % Rohfaser 4,50 % Rohasche 12,50 % Calzium 3,40 % Phosphor 0,55 % Natrium 0,15 % MJ ME 11,00
Zusatzstoffe je kg Mischfutter:
Vitamin A 13500 I. E. Vitamin E 16 mg Vitamin D3 2700 I.E. Kupfer 28 mg Zinkbacitracin 50 mg Antioxydans (Buthylhydroxytoluol) Canthaxanthin
Die Beleuchtungsintensität in den Ställen betrug bis zur 16. Lebenswoche der
Hennen 5 Lux, in der 17. Woche 10 Lux und ab der 18. Lebenswoche bis zum
Versuchsende 15 Lux. Die Beleuchtungsdauer richtete sich nach dem
Beleuchtungsprogramm der Lohmann Tierzucht GmbH. Sie steigerte sich von acht
Stunden pro Tag in der 14. Lebenswoche auf schließlich 15 Stunden täglich ab der
23. Lebenswoche.
3.5. Mycoplasma gallisepticum - Stämme
3.5.1. Herkunft
R (K 781) 1990 S.H. KLEVEN, Poultry Research Center, Department of Avian
Medicine, University of Georgia, Athens, Georgia 30605, USA

70
A 5969 1990 S.H. KLEVEN, Poultry Research Center, Department of Avian
Medicine, University of Georgia, Athens, Georgia 30605, USA
3.5.2. Anzüchtung und Vermehrung
Herstellung der Mycoplasma-Nährmedien
Für die Anzüchtung von MG wurde das Flüssigmedium nach FREY et al. (1968)
verwendet. Die genaue Zusammensetzung des Mediums ist im Anhang mitgeteilt.
Für die Zubereitung von 500 ml Flüssigmedium wurden die einzelnen Substanzen
abgewogen und in 450 ml Ampuwa (Fresenius AG) mittels eines Magnetrührers
gelöst und mit 1N NaOH-Lösung der pH-Wert auf 7,8 eingestellt. Anschließend
wurde die Lösung mit Hilfe autoklavierter (121 °C, 15 min) Seitz-Filter (0,2 µm) der
Firma Schleicher & Schuell sterilfiltriert. Danach wurden 50 ml inaktiviertes (56 °C, 35
min) und sterilfiltriertes Schweineserum zugegeben und die Bouillon durchmischt.
Die Aufbewahrung der fertigen Nährmedien erfolgte in sterilen 10 ml
Polystyrolröhrchen mit Schraubverschluß (Firma Greiner), die mit je 5 ml Bouillon
beschickt wurden, oder in autoklavierten Erlenmeyerkolben (100 ml) mit
Schraubverschluß für maximal eine Woche bei 7 °C. Die Herstellung größerer MG-
Antigenmengen als Inokulum erfolgte in je 500 ml Mykoplasmenbouillon, abgefüllt in
sterilen Erlenmeyerkolben (1000 ml), die mit sterilen Zellulosestopfen verschlossen
wurden. Zur Herstellung des Festmediums nach FREY et al. (1968) wurde zunächst
15 g Noble-Agar (Difco) in 450 ml Ampuwa gelöst und eine Stunde stehengelassen,
bevor diese Suspension bei 121 °C für 15 Minuten autoklaviert wurde. Anschließend
wurde 450 ml flüssiger Agar mit 450 ml Flüssigmedium ohne Phenolrot (s. o.) bei 50°
C vermischt und sterilfiltriertes, inaktiviertes Schweineserum (100 ml) hinzugefügt. Je
10 ml wurden mit autoklavierten Cornwall-Spritzen in Petrischalen gefüllt, wobei die
Luftbläschen an der Oberfläche mit dem Bunsenbrenner entfernt wurden. Die
Mycoplasma-Agarplatten wurden in Plastiktüten eingeschweißt und maximal eine
Woche bei 7 °C gelagert. Zur Sterilitätskontrolle der Bouillon wurde je ein Tropfen mit
einer sterilen Pipette auf MG-Agarböden und Blutagar verbracht, und anschließend
bei 37° C aerob und mikroaerob bebrütet. Zur Kontrolle der MG-Agarplatten wurden
diese stichprobenartig wie oben beschrieben bebrütet.

71
Anzüchtung der MG-Stämme
Die verwendeten Stämme waren als Lyophilisat verfügbar. Der Inhalt der
lyophilisierten Röhrchen wurde mit 0,5 ml physiologischer NaCl-Lösung
resuspendiert. Die Anzüchtung erfolgte im Overlay-Medium in Polystyrolröhrchen
(Firma Greiner), die mit 1 ml MG-Agar beschickt und mit 5 ml MG-Flüssigmedium
überschichtet worden waren, bei 37°C ± 1 °C. Nach 24 Stunden erfolgte die erste
Kontrolle auf den Umschlag des Farbindikators. Bei einer Vermehrung der
Mykoplasmen kommt es durch die Metabolisierung von Glukose und der damit
verbundenen Freisetzung von H+-Ionen zu einem Absinken des pH-Wertes. Diese
Änderung des pH-Wertes bewirkt einen Farbumschlag des Indikators von rot nach
orange bis gelb. Die optische Kontrolle erfolgte unter standardisierten
Lichtverhältnissen vor demselben Hintergrund. Nach der kulturellen Anzüchtung
wurde die Einsaatkultur jeweils im Mengenverhältnis 1 : 10 verimpft. Subkulturen
wurden jeweils mit 0,05 ml der Flüssigkultur angelegt. Die Bebrütung erfolgte unter
mikroaeroben Bedingungen in einen Exsikkator bei 37 °C +/- 1 °C bis zu 72 Stunden.
Alle Arbeiten mit Mycoplasmen erfolgten in einem seperatem Sterillabor.
Herstellung des Inokulums
Zur Herstellung des zur Infizierung benötigten Inokulums wurde der Mycoplasma
gallisepticum-Stamm R verwendet und zunächst Wachstumsstudien mit
verschiedenen Beimpfungsmengen durchgeführt. So war es möglich, die
Wachstumsdynamik des Stammes zu verfolgen, um die für die artifizielle Infizierung
benötigte Menge des Inokulums zum gewünschten Zeitpunkt zur Verfügung zu
haben. Bei der Herstellung des Inokulums wurde pro Versuchsdurchgang schließlich
mit 6 x 50 ml Einsaatkultur 6 x 500 ml auf 37 °C vorgewärmtes MG-Flüssigmedium
beimpft und bei 37 °C bebrütet. Bei einem pH-Wert von 7,2 wurde die Kultur aus dem
Brutschrank genommen und in der Zentrifuge „Sorvall Superspeed RC-2-B“ mit dem
Rotor „GS-3“ bei 4 °C für 30 Minuten mit 10000 x g zentrifugiert. Der Überstand
wurde verworfen und die sedimentierten Mykoplasmen mit insgesamt 30 ml
Phosphatpufferlösung resuspendiert. Jeweils ein Milliliter wurde zur Bestimmung der
Anzahl der KBE/ml verwendet.

72
Herstellung des MG-HA-Antigens
Die Herstellung des Antigens für den HAH-Test erfolgte wie oben beschrieben unter
Verwendung des MG-Stammes A 5969. Nach sieben Passagen wurde 50 ml
Einsaatkultur auf 500 ml Flüssigmedium überimpft und bis zu einem pH-Wert von 7,0
bebrütet. Hierauf wurde die Kultur bei 4° C für 30 Minuten bei 10000 x g in der
Zentrifuge „Sorvall Superspeed RC-2-B“ mit dem Rotor „GS-3“ zentrifugiert. Der
Überstand wurde dekantiert und das Mykoplasmenpellet in 5 ml
Phosphatpufferlösung resuspendiert.
Bestimmung des vermehrungsfähigen Mykoplasmengehalts
Zur Bestimmung der Anzahl der koloniebildenden Einheiten (KBE) je Milliliter wurde
jeweils von der MG-Suspension, die zur experimentellen Infizierung der Hennen
vorgesehen war, ein Milliliter abgenommen und eine Verdünnungsreihe hergestellt.
Jeweils 0,05 ml mykoplasmenhaltige Suspension wurden in Zehnerstufen (log10)
jeweils im Verhältnis 1:10 in MG-Bouillon verdünnt und mit einem Schüttelautomaten
vermischt. Mit einer Eppendorf-Pipette wurden nun wiederum 0,05 ml Suspension
jeder Verdünnungsstufe in die Mitte der Agarplatten verbracht und ausgestrichen.
Die Agarplatten wurden dann unter mikroaeroben und humiden Klima im Exsikkator
bei 37° C +/- 1° C für zwei Tage bebrütet. Die Agarplatten, bei denen die Kolonien
weitgehend einzeln lagen ( 20 bis 100 Kolonien), wurden zur Auszählung
herangezogen. Die Auszählung erfolgte unter einem Stereomikroskop bei 20-facher
Vergrößerung.
3.6. Vakzination
In den drei Versuchsdurchgängen 1, 2 und 3 wurden die Chargen 5, 6 und 7 des
Impfstoffes „TALOVAC 104 MG“ der Firma Lohmann Animal Health GmbH & Co. KG
geprüft.
Nach drei Tagen Lagerung des Impfstoffes bei 6° C im Kühlschrank zeigte sich bei
den drei Chargen eine unterschiedliche Entmischung des Impfstoffes. In allen drei
Flaschen der Charge 5 bildete sich die von LAH beschriebene hellgelbliche, 2-3-mm
dünne, Schicht heraus, welche nach leichtem Schütteln wieder aufzulösen war. Der

73
Inhalt der Flaschen der Chargen 6 und 7 wies eine andere Schichtung auf. Zuoberst
befand sich ein hellweißlicher, 6-8 mm dicker Streifen einer Flüssigkeit, die im
Tröpfchentest im 4°C kalten Wasser den Charakter einer öligen Substanz aufwies.
Darunter folgte eine Emulsion, die adspektorisch ohne Anzeichen einer Entmischung
war. Am Boden der Flaschen befand sich ein ca. 2 mm dünner Film fast klarer
Flüssigkeit. Durch leichtes Schütteln ließen sich auch diese Bestandteile wieder
vermischen. Ursachen für die unterschiedliche Entmischung konnten von der Firma
LAH nicht angegeben werden.
Abweichend von dem von LAH empfohlenen Impfschema, 1. Impfung 6. - 8.
Lebenswoche und 2. Impfung 14. - 16. Lebenswoche, erfolgten die Vakzinationen in
dieser Versuchsanordnung in der 14. und 18. Lebenswoche.
Die Impfstoffmenge betrug 0,5 ml je Henne, die Impfung erfolgte mit einer
Revolverspritze mit Luer-Konus. Zur Impfung wurden Einmalkanülen 1,1 x 25 mm
verwendet, die nach jeder zehnten Henne erneuert wurden. Die Impfstoffapplikation
erfolgte subkutan, kaudal im Bereich der Halsbasis.
Nach den Vakzinationen wurden lokale Veränderungen an der Injektionsstelle nach
dem folgendem Beurteilungsschlüssel von Dr. Opitz, (1982) University of Maine,
Cooperative Extension, Orono Maine, USA, bewertet:
Grad 0 = keine Veränderungen
Grad 1 = Reste von Öl, keine Entzündungsreaktion
Grad 2 = einzelne Granulome, weniger als 2 cm Durchmesser
Grad 3 = einzelne oder multiple Granulome - im Durchmesser von 2 cm oder
mehr
Grad 4 = große Granulome, diffuse Schwellungen mit Nekrosebildungen; 4 bis
6 cm im Durchmesser, die das Allgemeinbefinden des Impflings durch
Druck auf Organe wie Nerven, Blutgefäße oder Schlund stören.
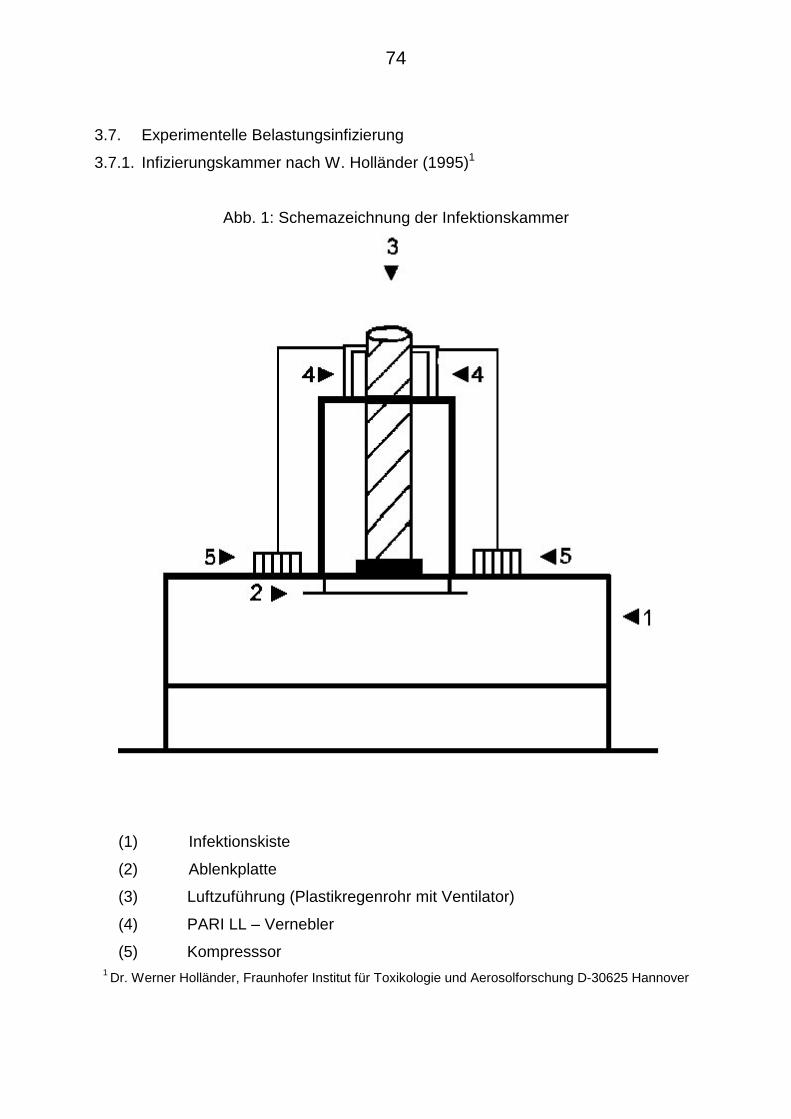
74
3.7. Experimentelle Belastungsinfizierung
3.7.1. Infizierungskammer nach W. Holländer (1995)1
Abb. 1: Schemazeichnung der Infektionskammer
(1) Infektionskiste
(2) Ablenkplatte
(3) Luftzuführung (Plastikregenrohr mit Ventilator)
(4) PARI LL – Vernebler
(5) Kompresssor 1 Dr. Werner Holländer, Fraunhofer Institut für Toxikologie und Aerosolforschung D-30625 Hannover

75
Für die Inokulation der Hühner mit dem erregerhaltigen Aerosol wurde aus 8 mm
starken Spanholzplatten eine Kiste (1) mit einer Grundfläche von 2 m x 1 m und
einer Höhe von 0,5 m gebaut. Der Kistendeckel war im Verhältnis ¼ zu ¾ geteilt. Um
die Hühner in die Kiste setzen zu können, war der größere Teil des Deckels
aufklappbar. In der Mitte befand sich ein Loch mit einem Durchmesser von 10 cm
und unterhalb davon, auf der Deckelunterseite, war parallel ein Brett (2) angebracht,
welches zur Luftablenkung und damit zur gleichmäßigen Verteilung des
erregerhaltigen Aerosols diente. Die Luftzufuhr (3) erfolgte über ein 1 m langes
Plastikregenrohr (Hagebaumarkt) mit einem Durchmesser von 10 cm, an dessen
unteren Ende sich ein 12-Volt Ventilator (Conrad Electronics) befand. Dieser wurde
mit einem Eisenbahntrafo (Märklin) auf minimale Drehzahl reduziert, so dass die
Luftaustrittsgeschwindigkeit am Ausgang des Rohres 2,2 m/s betrug. Am oberen
Rand des Rohres waren zwei Öffnungen gebohrt, welche die Mundstücke der beiden
„PARI LL - Vernebler“ (4) aufnahmen; über Schläuche wurde aus den Kompressoren
(5) die benötigte Druckluft zugeführt.
3.7.2 Pariboy-Ultraschallvernebler
Für die aerogene Inokulation mit den Mycoplasmen des R-Stammes wurden zwei
PARI LL - Vernebler , PARI GmbH, Moosstraße 9, D-82319 Starnberg, mit dem Pari
Boy als Kompressor gewählt. Der Pari Boy wird im Bereich der Humanmedizin zur
Behandlung von Atemwegserkrankungen eingesetzt.
Bei dem PARI LL - Vernebler handelt es sich um einen Ultraschallvernebler, der mit
Druckluft betrieben wird. Der mediane Massendurchmesser (MMD) beträgt 4,8 µm,
und die Auswurfleistung beträgt 0,42 g / min (0,9 % NaCl-Lösung). Der Pari Boy
Kompressor liefert einen Betriebsdruck von 0,75 bar.
3.7.3. Präparation und Applikation des Inokulums als Aerosol
Die Mykoplasmen (Durchmesser von 0,25-0,5 µm) liegen in Form einer Suspension
in einer Phosphatpufferlösung vor. Der Pari Boy vernebelt diese Suspension, wobei
52 % der Tröpfchen einen Durchmesser < 5 µm aufweisen. Dies hat zur Folge, dass
die einzelnen Mykoplasmen in einem Wassertropfen eingehüllt vorliegen. Die Größe
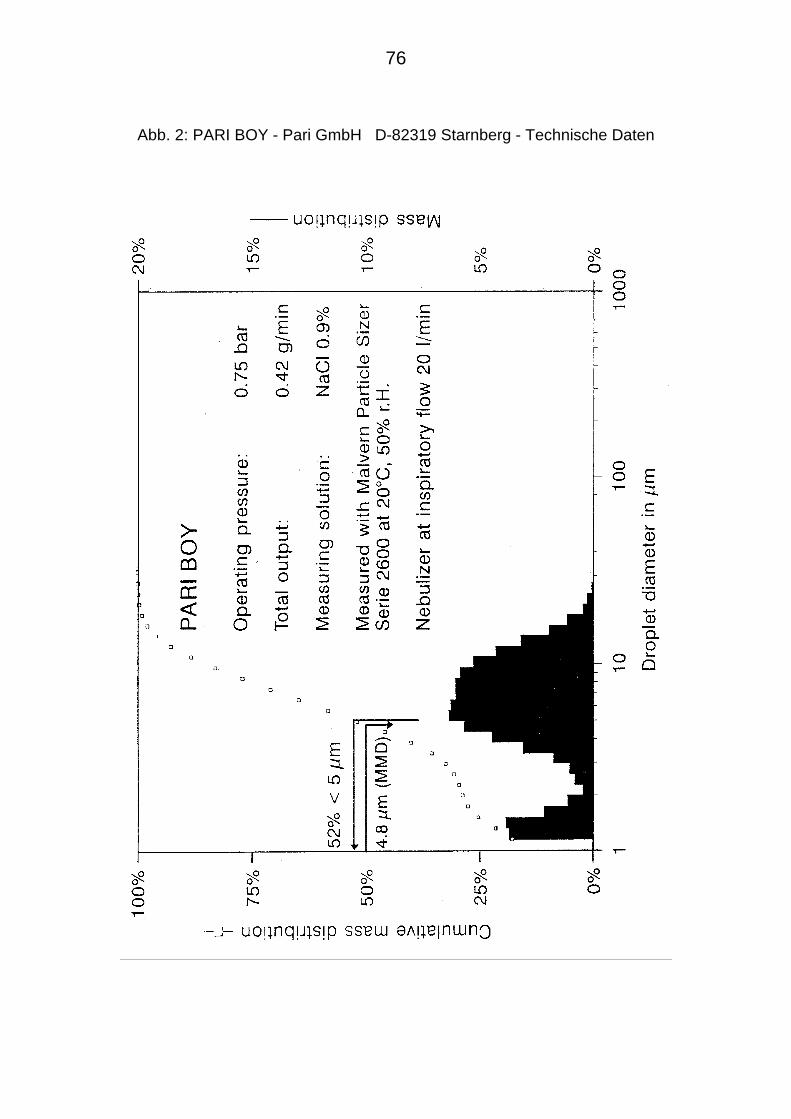
76
Abb. 2: PARI BOY - Pari GmbH D-82319 Starnberg - Technische Daten

77
dieser Teilchen reduziert die Schwebfähigkeit und die Eindringtiefe der Mykoplasmen
in die Bronchien, Lungen und Luftsäcke (siehe 2.4.). Durch den Luftsog, der durch
den Ventilator erzeugt wird, werden die Mykoplasmen, die aus den Pari-Boy
Ultraschallverneblern in das Rohr eintreten, in die Infektionskammer transportiert.
Während dieses Vorganges verdunstet die Wasserhülle der meisten Mykoplasmen.
Hierdurch erhöht sich wiederum die Schwebfähigkeit und Eindringtiefe der
Mykoplasmen. Eine Schädigung der Lebensfähigkeit der Mykoplasmen durch die
Ultraschallvernebelung ist durch die kurze Zeit der direkten Einwirkung nicht zu
befürchten.
Die Präparation des Inokulums erfolgte wie oben beschrieben. Die Hennen jeder
Gruppe wurden jeweils gemeinsam aus ihrem Stall in die Infektionskammer
verbracht und dort mit je 15 ml Mykoplasmensuspension inokuliert. Jeder der beiden
PARI - LL - Vernebler wurde mit 7,5 ml gefüllt. Nachdem diese Menge vernebelt war,
wurden beide PARI - LL - Vernebler noch einmal mit je 7,5 ml Phosphatpufferlösung
befüllt. Nach Abschluß der Vernebelung verblieben die Hühner für weitere 15
Minuten in der Infektionskammer, bevor sie wieder in die zwischenzeitlich gereinigten
Ställe verbracht wurden.
Für die zur Belastungsinfektion der Hennen gebrauchten Suspensionen wurden
durch Verdünnungsreihen folgende Titer ermittelt:
Tabelle 2: Infizierungsdosen für die experimentellen Belastunginfizierung
Versuchsdurchgang 1 - Charge 5 2 x 15 ml mit je 3,2 x 107 KBE / ml Versuchsdurchgang 2 - Charge 6 2 x 15 ml mit je 4,5 x 107 KBE / ml Versuchsdurchgang 3 - Charge 7 2 x 15 ml mit je 4,8 x 107 KBE / ml
3.8. Klinische Untersuchungen
3.8.1. Allgemeinbefinden
Die Legetätigkeit der Hennen ist ein sehr empfindlicher Indikator für Störungen der
Hömöostase. Daher wurde das Allgemeinbefinden (Sensibilität [Aufmerksamkeit],
Verhalten, Futter- und Wasseraufnahme, Befiederung) täglich nach einer mindestens
zehnminütige Beobachtung durch ein verdecktes Stallfenster beurteilt. Eine

78
weitergehende Untersuchung der einzelnen Tiere beschränkte sich auf die Hennen,
die bei den täglichen Stallarbeiten (Eier sammeln, Reinigung von Trog und Tränke
sowie Befüllung des Troges mit Futter) in die Hand genommen werden mussten. Bei
diesen Tieren wurden Augen, Nasenöffnungen und der Ernährungszustand
untersucht.
3.8.2. Lokale Impfstoffreaktion
Im Rahmen der Untersuchung des Allgemeinbefindens (3.7.1.) wurde bei den
Hennen der Impfgruppen, die bei der täglichen Stallkontrolle in die Hand genommen
wurden, das untere Halsdrittel im Bereich der Impfstoffapplikation durchgetastet.
3.8.3. Legeleistung
Die Eier wurden während der täglichen Stallkontrolle eingesammelt und die
Legeleistung einer jeden Gruppe notiert.
3.9. Serologisch nachweisbare Reaktionen
Zur serologischen Untersuchung wurde von den Hennen dreimal Blut gewonnen. Die
erste Blutentnahme erfolgte zum Zeitpunkt der zweiten Vakzination, die zweite zum
Zeitpunkt der Infizierung und die dritte zum Zeitpunkt der Tötung bei Versuchsende
(siehe Tabelle 1). Während des Versuchszeitraumes erfolgte die Blutentnahme aus
der Vena basilaris, bei Versuchsende aus der Vena jugularis. Der Blutkuchen wurde
anschließend mit einem Glasstab von der Wand der Auffanggefäße gelöst, bei 37 °C
für eine Stunde im Brutschrank inkubiert und dann für zehn Minuten zentrifugiert
(Megafuge 1,0; Heraeus 3000 U/min. entsprechend 3120 x g).
Unmittelbar nach der Serumgewinnung erfolgte die Untersuchung in der
Serumschnellagglutination. Anschließend wurden die Serumröhrchen mit sterilen
Zellulosestopfen verschlossen und für 24 Stunden bei - 18 °C eingefroren. Bis zur
Durchführung weiterer serologischer Untersuchungen (ELISA, HAH), die nach dem
Ende des dritten Versuchsdurchganges erfolgten, wurden die Seren bei - 70 °C
gelagert.

79
3.9.1. Serum-Schnellagglutinations-Test
Der Test wurde entsprechend der Gebrauchsinformation des Herstellers und nur mit
frisch gewonnenen Seren durchgeführt. Zunächst wurden Serum, Antigen sowie
Tüpfelplatte, Glasstäbe und Pipetten auf Raumtemperatur erwärmt. Ein Tropfen des
Serums wurde jeweils mit einer sterilen Glaspipette auf eine Tüpfelplatte verbracht,
mit Hilfe eines Glasstabes mit je einem Tropfen MG- bzw. MS-Antigen (Mycoplasma-
Gallisepticum- und Mycoplasma-Synnoviae-Antigen Vemie, Vemie Veterinär Chemie
GmbH, Kempen) vermischt. Die Tüpfelplatte wurde für 30 Sekunden stehengelassen
und dann für 90 Sekunden vorsichtig geschwenkt. Erfolgte innerhalb von 120
Sekunden eine Agglutinationsreaktion, so wurde dies als positives Ergebnis, d.h. als
Nachweis von spezifischen agglutinierenden Antikörpern gegen MG bzw. MS
gewertet. Zeitlich spätere oder schwache Agglutinationsreaktionen wurden als
zweifelhaftes Ergebnis angesehen. Je nach Menge der im Serum vorhandenen
Antikörper kam es zu einer unterschiedlichen Größe der Flocken, die aber nicht zur
quantitativen Bewertung herangezogen wurden.
Als Maßstab für die Bewertung der Agglutinationsreaktionen wurde folgende
Einteilung zu Grunde gelegt:
Keine Antigen-Antikörper (Flockung) Reaktion -
Schwache und/oder verzögerte Reaktion (Flockung nach mehr als 2 min.) (+)
Positive Reaktion (Flockung innerhalb von 2 min.) +
Bei jeder Untersuchung wurden Negativkontrollen durchgeführt. Reines Antigen
wurde auf die Tüpfelplatte getropft und auf Flockenbildung kontrolliert.
3.9.2. Hämagglutinationshemmungstest (HAHT)
Der Hämagglutinationshemmungstest wurde entsprechend der Beschreibung von
HINZ (1977) im Mikrotestverfahren durchgeführt. Zunächst wurde eine 0,75 %ige
Erythrozytensuspension hergestellt. Hierzu wurden 3 ml Alsever´sche Lösung und 7
ml Hühnerblut in einem Reagenzröhrchen gemischt. Anschließend wurde diese
Suspension bei 3000 U/min für zehn Minuten zentrifugiert und der Überstand
verworfen. Zweimal wurde der Bodensatz mit Phosphatpuffer aufgeschwemmt und

80
erneut zentrifugiert, so daß schließlich ein Erythrozytenkonzentrat vorlag. 0,75 ml
wurden mit 99,25 ml Phosphatpufferlösung zur Erythrozytensuspension gemischt.
Da bei der Verwendung von frisch hergestelltem HA-Antigen keine unspezifischen
Reaktionen auftreten, kann auch ohne eine Behandlung mit RDE (receptor
destroying enzyme) (ROBERTS u. OLESIUK 1967) bereits ein Hemmtiter von 1:5 als
spezifisch anzusehen ist (Hinz 1977).
Zunächst wurde durch eine Titrationsreihe der HA-Antigenendtiter bestimmt. Hierzu
wurde eine geometrische Verdünnungsreihe, beginnend mit 1:2, 1:4 usw. erstellt. Zu
jeder Stufe der Verdünnungsreihe (0,025 ml) wurden ein Tropfen physiologischer
NaCl-Lösung und zwei Tropfen (0,05 ml) einer 0,5 % Hühnererythrozytenlösung
gegeben und alles gut durchmischt. Nach 45-minütigem Stehenlassen bei
Zimmertemperatur wurde die Bewertung durchgeführt. Als HA-Antigenendtiter wurde
die Verdünnungsstufe gewertet, in der eine vollständige Hämagglutination bei
gleichzeitiger einwandfreier Erythrozytenkontrolle vorlag. Für die Antigensuspension
wurde ein HA-Antigenendtiter von 1:64 ermittelt. Im HAHT fand eine
Antigengebrauchsverdünnung von 1:16 Anwendung, entsprechend 4 HA-Einheiten in
0,025 ml.
Die Seren wurden nach der β-Methode beginnend mit 1:5, 1:10 usw. in
physiologischer Kochsalzlösung verdünnt. Nach Zugabe von Antigen und
Erythrozytensuspension wurde die Mikrotiterplatte (Nunc-Mikrotiterplatten aus
Polystyrol mit U-förmigen Vertiefungen) für eine Minute geschüttelt und für eine
Stunde bei Zimmertemperatur stehengelassen. Danach erfolgte die Ablesung der
Ergebnisse. Als Hemmtiter wurde die Serumverdünnung gewertet, bei der eine
vollständige Hemmung des HA vorlag und der knopfförmige Erythrozytenbodensatz
bei Schräghaltung der Titerplatten auslief.
Da bei der Berechnung des geometrischen Mittelwertes aufgrund der Einteilung nach
der β-Methode große Schwankungen entstehen, wurden die einzelnen
Verdünnungen mit Titerstufen gleichgesetzt. Hierbei entsprach die Verdünnung von
1:5 der Titerstufe 1, die Verdünnung von 1:10 der Stufe 2 usw.. Aus den Werten der
einzelnen Proben wurde dann der geometrische Mittelwert (GMT) der Gruppe
berechnet.

81
3.9.3. Enzymgebundener Immunadsorptionstest zum Nachweis von Antikörpern
(ELISA)
Für die Durchführung des ELISA wurde der ProFlok® „Mycoplasma gallisepticum
antibody test kit“ der Firma Kirkegaard and Perry Laboratories, Gaithersburg
Maryland, USA, entsprechend der Gebrauchsanweisung verwendet. Die
Extinktionsbestimmung wurde mit einem Photometer bei einer Wellenlänge von 405
nm vorgenommen und mittels KPL-ELISA Programm am Computer ausgewertet.
Proben mit einem Titer von ≥ 744 sind als spezifisch positiv zu bewerten.
Bei fraglichen Ergebnissen in der SSA (MG/MS - Kreuzreaktionen) wurde zur
Klärung zusätzlich der entsprechende MS-ELISA durchgeführt.
3.10. Pathologisch-anatomische Untersuchungen
Traten im Verlauf der Vor- bzw. Hauptuntersuchungen gravierende Störungen des
Allgemeinbefindens einzelner Tiere oder Anzeichen einer Erkrankung mehrerer
Hennen einer Gruppe auf, wurden diese Tiere während des Versuchsdurchganges
getötet und seziert. Am Ende der Versuche wurden dann alle verbliebenen Hennen
getötet und seziert. Die Sektion erfolgte entsprechend des Sektionsganges nach
HINZ et al. (1993). Veränderungen an den Organen wurden protokolliert und nach
ihrem Schweregrad und dem Grad ihrer Ausbreitung mit Hilfe einer vierstufigen
Bewertungsskala numerisch kodiert. Zum Vergleich der Gruppen wurde jeweils das
arithmetische Mittel gebildet. Die Bewertung der entzündlichen Veränderungen der
Luftsäcke (LSE-Score-Werte) in den Kontroll- und Impfgruppen wurde in Anlehnung
an das System von YODER et al. (1984) vorgenommen.
Die folgende Bewertung der Organveränderungen wurden durchgeführt:
Herzbeutel:
geringgradig: grau-gelbliche Trübung und einfache Dickenzunahme des Peri-
kards, vor allem im Bereich der Verbindungsstelle zu den
thorakalen kranialen Luftsäcken.

82
Lunge:
geringgradig: Stauungslunge
mittelgradig: herdförmige Verdichtungsherde
Luftsäcke:
geringgradig: diffuse, grau-gelbe Trübung einer Luftsackwand, teilweise
verbunden mit herdförmigen Auflagerungen eines dünnen
Fibrinfilms.
mittelgradig: grau-gelbe Trübung und bis zweifache Verdickung der Luftsack-
wände mit großflächigen Auflagerungen von fibrino-purulentem
bis fibrinösem Exsudat bei einem oder zwei Luftsäcken.
hochgradig: entzündlich verdickte grau-gelbe Luftsäcke mit Ansammlungen
von fibrinösem Exsudat im Luftsacklumen bei zwei oder mehr
Luftsäcken
Leber, Milz:
geringgradig: Hyperämie
mittelgradig: Vergrößerung um 1/3 des Gewichts und Abflachung der
Leberränder.
hochgradig: Vergrößerung und Verdoppelung des Gewichtes bei Vorliegen
einer Stauungshyperämie.
Eierstock:
geringgradig: ein bis zwei Fibrinflocken zwischen den Follikeln eines aktiven
Eierstocks
mittelgradig: drei bis vier Fibrinflocken zwischen den Follikeln eines aktiven
Eierstocks
hochgradig: Mesosalpinx und die dem Eierstock benachbarten Luftsäcke
zeigen ausgeprägte fibrinöse entzündiche Veränderungen und
zahlreiche Fibrinflocken zwischen den Tertiärfollikeln des aktiven
Eierstocks; oder Vorhandensein von einem inaktiven,
degenerativ veränderten Eierstock.

83
3.11. Kulturell-bakteriologische Untersuchungen
Kulturell auf MG wurden ein ca. 5 cm langes Teilstück der Trachea aus dem Bereich
des kranialen Brusteinganges, ein Lungenflügel und ein Teil der thorakalen und
abdominalen Luftsäcke untersucht. Bei den Versuchsdurchgängen 1 und 3 wurde die
Schleimhaut der Tracheen mit einer jeweils zuvor ausgeglühten Platinöse
abgeschabt und anschließend auf MG-Agar ausgestrichen. Die Tracheaproben der
Hennen des zweiten Versuchsdurchganges wurden mit sterilen Wattetupfern
genommen. Bei diesen Proben erfolgte zusätzlich eine Untersuchung mittels PCR-
DNS-Sondentechnik auf MG-DNS. (SALISCH et al. 1998). Zum Nachweis von MG
aus Lunge und Luftsack wurden Proben von frisch entnommenen Organen auf MG-
Agar übertragen und ausgestrichen.
Im Rahmen der pathologisch-anatomischen Untersuchungen wurden Proben von
Herz, Leberlappen, Milz, Teilen der Lunge und Luftsäcke sowie veränderte und
unveränderte Ovarien mit sterilem Besteck entnommen und auf Blut - und
Drigalskiagar verimpft. Beim Herz, der Leber, der Milz und der Lunge wurden frisch
angelegte Schnittflächen auf den Nährböden ausgestrichen. Von den Luftsäcken
wurde mit einer ausgeglühten Platinöse eine Probe entnommen und verimpft. Die
Blutagarplatten wurden bei 37° C für 24 Stunden unter mikroaeroben Verhältnissen
angezüchtet. Die Bebrütung der Drigalskinährböden erfolgte ebenfalls bei 37° C
unter aeroben Bedingungen. Die weitere Differenzierung und Beurteilung erfolgte wie
bei BLOME (1994) beschrieben.
3.12. Statistische Auswertung
Die biostatistisch vergleichende Auswertung der Legeleistungen der Impf- mit der
Kontrollgruppe und die Untersuchungen auf die Signifikanz wurden mit Hilfe des
SAS® (Statistical Analysis Systems, Version 6.04 1989) Computerprogramms im Chi-
Quadrat-Test mit freundlicher Unterstützung des Institutes für Biometrie der
Tierärztlichen Hochschule Hannover, durchgeführt. Die biostatistische Auswertung
der ELISA-, HAH- und der LSE-Score-Werte erfolgte mit dem SPSS Programm
Version 9.0 (1999). Die Signifikanzprüfung der errechneten ELISA-Werte wurde im

84
T-Test, die der HAH- und der LSE-Score-Werte im Mann-Whitney-U-Test
durchgeführt.

85
4. Ergebnisse
4.1. Voruntersuchungen
4.1.1. Zum Zeitpunkt der Einstallung
Zum Zeitpunkt der Aufstallung in der Klinik konnte bei keiner der Hennen eine
Störung des Allgemeinbefindens festgestellt werden.
In den Kotproben, deren Untersuchung in der 15. Lebenswoche der Hennen erfolgte,
konnten mikroskopisch keine Parasiten bzw. deren Eier nachgewiesen werden.
Die Seren aller Hennen, die bei der Ankunft in der Klinik mittels SSA untersucht
worden waren, wiesen keine spezifischen agglutinierenden Antikörper gegen das
Mycoplasma gallisepticum-Antigen auf. Im Versuch A konnten jedoch bei 14 von 20
untersuchten Blutserumproben in der SSA bei der Verwendung von Mycoplasma
synoviae-Antigen eine positive Reaktion festgestellt werden. Auch die Junghennen
der Versuche B, C und D zeigten in der SSA für MS positive Reaktionen. Bei der
Voruntersuchungsgruppe D wurden bei zwölf von 20 Blutserumproben Antikörper
gegen Mycoplasma synoviae in der SSA festgestellt. Nur die
Voruntersuchungsgruppe E wies in der SSA keine agglutinierenden Antikörper gegen
MS-Antigen auf. Die Spezifität der positiven Reaktionen in der SSA konnten in allen
Fällen im enzymgebundenen Immunadsorptionstest zum Nachweis von Antikörpern
(ELISA) mit dem Pro-Flok® „Mycoplasma synoviae antibody test kit“ der Firma
Kirkegaard and Perry Laboratories verifiziert werden.
4.1.2. Nach der Vakzination
4.1.2.1. Versuch A
Die Junghennen der Impf- und Kontrollgruppe zeigten im Verlaufe der
Voruntersuchungen ein ungestörtes Allgemeinbefinden, keines der Tiere ließ eine
Entwicklungsstörung erkennen. Der Beginn der Legetätigkeit erfolgte in beiden
Gruppen zeitgleich.
Die Hennen des Versuchs A wurden daher im Rahmen der Hauptuntersuchungen
(Versuch 1) zur Prüfung der Verträglichkeit und Wirksamkeit der Impfstoffcharge 5
verwendet.

86
4.1.2.2. Versuche B und C
Die Junghennen beider Versuchsdurchgänge wurden zeitgleich aufgestallt. Hierbei
zeigten einige Hennen bereits kurz nach der Aufstallung in der Klinik für Geflügel
eine deutliche Störung des Allgemeinbefindens. In der Impfgruppe des Versuches B
verendeten fünf Hennen und in der Impfgruppe des Versuches C zwei Hennen. Bei
fünf von diesen Tieren wurden pathologisch-anatomische Veränderungen wie bei der
Marek´schen Krankheit gefunden. Histologisch konnte diese Befunde nicht bestätigt
werden. Da diese Hennen für die Verträglichkeits- und Wirksamkeitsprüfungen in den
Hauptuntersuchungen nicht zu verwenden waren, wurden die verbliebenen Hennen
der Voruntersuchungsgruppen B und C getötet. Eine Henne zeigte das klinische Bild
der Marek`schen Krankheit, bei insgesamt 20 Tieren der Impfgruppen (n = 80)
wurden im Rahmen der pathologisch-anatomischen Untersuchungen an den Nn.
ischiadici eine Verdickung und der Verlust der Querstreifung festgestellt wie sie für
die Marek´sche Lähme typisch ist. Hennen aus den Kontrollgruppen der Versuche B
und C waren von diesen Veränderungen nicht betroffen.
4.1.2.3. Versuch D
Bis zur zweiten Vakzination zeigten sowohl die Impf- als auch die Kontrollgruppe ein
ungestörtes Allgemeinbefinden. Die Entwicklung der Tiere in Bezug auf die
Gewichtszunahme und die Ausbildung der Kämme war gleichmäßig. Nach der
Wiederholungsimpfung fiel bei den Hennen der Impfgruppe eine leichte
Beeinträchtigung des Allgemeinbefindens auf. Sie bewegten sich weniger als die
Hennen der Kontrollgruppe, reagierten mit geringerer Intensität auf optische und
akustische Reize, wie z. B. Handbewegungen oder Fingerschnipsen und zeigten
einen geringgradigen Federverlust. Vier Hennen, die am deutlichsten in der
Entwicklung zurückblieben, wurden getötet. In der pathologisch-anatomischen
Untersuchung konnten Veränderungen, ins-besondere solche wie sie für die
Marek´sche Krankheit typisch sind, nicht festgestellt werden. Drei der sezierten
Hennen wiesen einen aktiven Legedarm auf. Der Beginn der Legetätigkeit verzögerte
sich in der Impfgruppe um zehn Tage.

87
Die Hennen des Versuchs D wurden im Rahmen der Hauptuntersuchungen
(Versuch 2) zur Prüfung der Verträglichkeit und Wirksamkeit der Impfstoffcharge 6
verwendet.
4.1.2.4. Versuch E
Die Hennen beider Gruppen zeigten ein ungestörtes Allgemeinbefinden und eine
gleichmäßige Entwicklung aller Tiere, jedoch trat bei Hennen der Impfgruppe
vereinzelt nach der zweiten Vakzination ein geringgradiger Federverlust auf.
Die Hennen des Versuchs E wurden im Rahmen der Hauptuntersuchungen (Versuch
3) zur Prüfung der Verträglichkeit und Wirksamkeit der Impfstoffcharge 7 verwendet.
4.2. Hauptuntersuchung
4.2.1. Verträglichkeitsprüfung
Nach der Impfstoffapplikation wurde bei den Hennen des Versuchs 1 keine Störung
des Allgemeinbefindens festgestellt. Die Hennen waren aufmerksam und zeigten ein
ungestörte Motilität. Eine Beeinträchtigung der Futter- bzw. Wasseraufnahme konnte
nicht beobachtet werden.
Bei den Hennen der Impfgruppe des Versuchs 2 kam es nach der
Wiederholungsimpfung bei einigen Hennen zu einem geringgradigem Federverlust.
Die Hennen dieser Gruppe zeigten, im Vergleich zu den Hennen der Kontrollgruppe,
eine verminderte Bewegungsaktivität. In dieser Gruppe verzögerte sich der
Legebeginn um zehn Tage. Die Hennen der Impfgruppe im Versuch 3 zeigten ein
ungestörtes Allgemeinbefinden. Nur bei einigen Tieren war ein leicht vermehrter
Federverlust zu beobachten. Die Gruppe war aufmerksam und bewegungsfreudig.
Bei der pathologisch-anatomischen Untersuchung am Versuchsende waren an den
Injektionsstellen der Legehennen in den drei Versuchsdurchgängen im
unterschiedlichen Ausmaß lokale Gewebsreaktionen feststellbar. In dem Versuch 1
waren die lokalen Reaktionen im Bereich der Injektionsstelle des Impfstoffes im
Vergleich mit den beiden anderen Versuchsdurchgängen, am geringsten ausgeprägt.
In den meisten Fällen (n = 32) waren nur noch Reste des Ölemulsions-Impfstoffes
festzustellen oder es konnten vereinzelte kleine Granulome gefunden werden. Bei
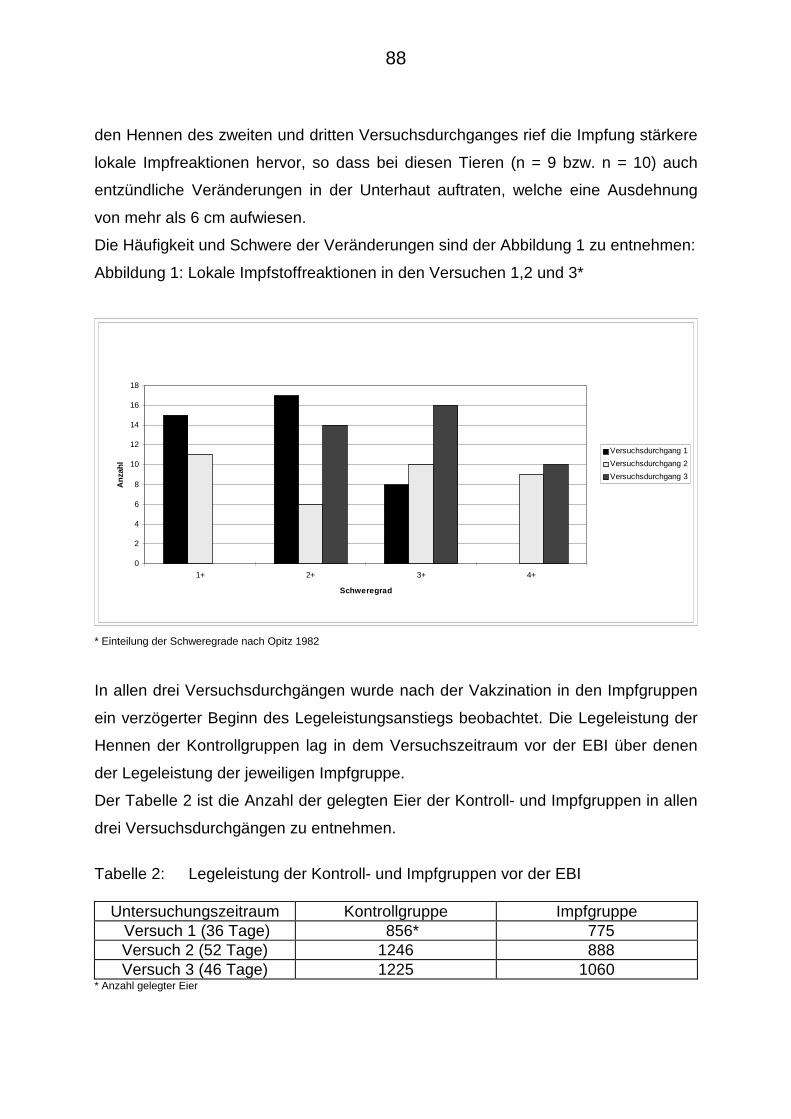
88
den Hennen des zweiten und dritten Versuchsdurchganges rief die Impfung stärkere
lokale Impfreaktionen hervor, so dass bei diesen Tieren (n = 9 bzw. n = 10) auch
entzündliche Veränderungen in der Unterhaut auftraten, welche eine Ausdehnung
von mehr als 6 cm aufwiesen.
Die Häufigkeit und Schwere der Veränderungen sind der Abbildung 1 zu entnehmen:
Abbildung 1: Lokale Impfstoffreaktionen in den Versuchen 1,2 und 3*
0
2
4
6
8
10
12
14
16
18
1+ 2+ 3+ 4+
Schweregrad
An
zah
l
Versuchsdurchgang 1
Versuchsdurchgang 2
Versuchsdurchgang 3
* Einteilung der Schweregrade nach Opitz 1982
In allen drei Versuchsdurchgängen wurde nach der Vakzination in den Impfgruppen
ein verzögerter Beginn des Legeleistungsanstiegs beobachtet. Die Legeleistung der
Hennen der Kontrollgruppen lag in dem Versuchszeitraum vor der EBI über denen
der Legeleistung der jeweiligen Impfgruppe.
Der Tabelle 2 ist die Anzahl der gelegten Eier der Kontroll- und Impfgruppen in allen
drei Versuchsdurchgängen zu entnehmen.
Tabelle 2: Legeleistung der Kontroll- und Impfgruppen vor der EBI
Untersuchungszeitraum Kontrollgruppe Impfgruppe Versuch 1 (36 Tage) 856* 775 Versuch 2 (52 Tage) 1246 888 Versuch 3 (46 Tage) 1225 1060
* Anzahl gelegter Eier

89
4.2.2. Wirksamkeitsprüfung
Die Wirksamkeit des Impfstoffes ließ sich biostatischtisch nur beurteilen, wenn die
vor der MG-Belastungsinfizierung vorhandenen Legeleistungsdifferenzen zwischen
Kontroll- und Impfgruppe mitberücksichtigt wurden. Dabei blieben die Zeiträume des
anfänglich starken Legeleistungsanstiegs bei der Berechnung der prozentualen
Legeleistung unberücksichtigt, da es als Folge der Belastung durch die Impfung in
den vakzinierten Gruppen zu einer Verzögerung des Legebeginns kam.
Zum Zeitpunkt der experimentellen Belastungsinfizierung mit dem MG-Stamm R
betrug die Legeleistung von Kontroll- und Impfgruppen jedoch mindestens 70 %.
4.2.2.1. Legeleistung im Versuch 1
Die Legeleistung beider Gruppen des ersten Versuchsdurchganges stiegen
zeitgleich von der 19. bis zur 21. Lebenswoche bis auf ca. 70 % der Legeleistung an.
Bis zur 24. Lebenswoche schwankte die Legeleistung beider Gruppen zwischen 70
und 90 %. Hierbei lag die Legeleistung der Kontrollgruppe im Durchschnitt um 7,61
% über der Leistung der Impfgruppe. Nach der EBI fiel die Legeleistung beider
Gruppen in einem Zeitraum von 14 Tagen zunächst auf ca. 60 % ab, um von diesem
Wert aus wieder auf eine Niveau von ca. 80 % anzusteigen. Die Legeleistung der
Impfgruppe übertraf nach der EBI die Legeleistung der Kontrollgruppe um 1,76 %.
In der Tabelle 3 ist die Entwicklung der Legeleistung vor und nach der EBI
dargestellt.
Tabelle 3: Prozentualer Legeleistungvergleich der Impf- und Kontrollgruppe
Versuch 1
Zeitraum Kontrollgruppe Impfgruppe Differenz vor der EBI (23 Tage) 80,45 % 72,84 % - 7,61 %
nach der EBI (29 Tage) 70,14 % 71,90 % + 1,76 % Differenz - 10,31 % - 0,94 % + 9,37 %
Die Entwicklung der Legeleistung im Vergleich vor und nach der
Belastungsinfizierung ist in der Abbildung 2 dargestellt.
Der Tabelle 3 und der Abbildung 2 ist zu entnehmen, dass die Legeleistung der
Kontrollgruppe nach der Belastungsinfektion um rund 10 % abfiel, während die

90
Legeleistung der Impfgruppe nahezu stabil blieb. Die biostatistische Auswertung
ergab für die Impfgruppe nach der EBI eine schwachsignifikant bessere Entwicklung
der Legeleistung (p ≤ 0.066).
Abb. 2: Legeleistungsveränderung im Versuch 1
Versuchsdurchgang 1
Prozentuale Legeleistungsveränderung im Vergleich der Zeiträume 23 Tage vor und 29 Tage nach der EBI
- 10,31%
- 0,94%
-12,00%
-10,00%
-8,00%
-6,00%
-4,00%
-2,00%
0,00% 1
Leg
elei
stun
gsv
erän
der
ung
Kontrollgruppe Impfgruppe
4.2.2.2. Legeleistung im Versuch 2
Beim Vergleich der Gruppenleistungen in diesem Versuch fällt der um zehn Tage
verzögerte Eintritt des Legebeginns in der Impfgruppe auf. Auch wenn der verzögerte
Legeleistungsbeginn nicht berücksichtigt wird, bleibt die Legeleistung der Impfgruppe
in den 29 Tagen vor der EBI um ca. 12 % unter der der Kontrollgruppe. In der
Tabelle 4 ist die Entwicklung der Legeleistung vor und nach der EBI dargestellt.
Mit der Belastungsinfizierung der Gruppen ändert sich dieses Bild. Während die
Legeleistung der Kontrollgruppe nach der EBI um gut 4 % abfällt, steigt die
Legeleistung der Impfgruppe weiter an.
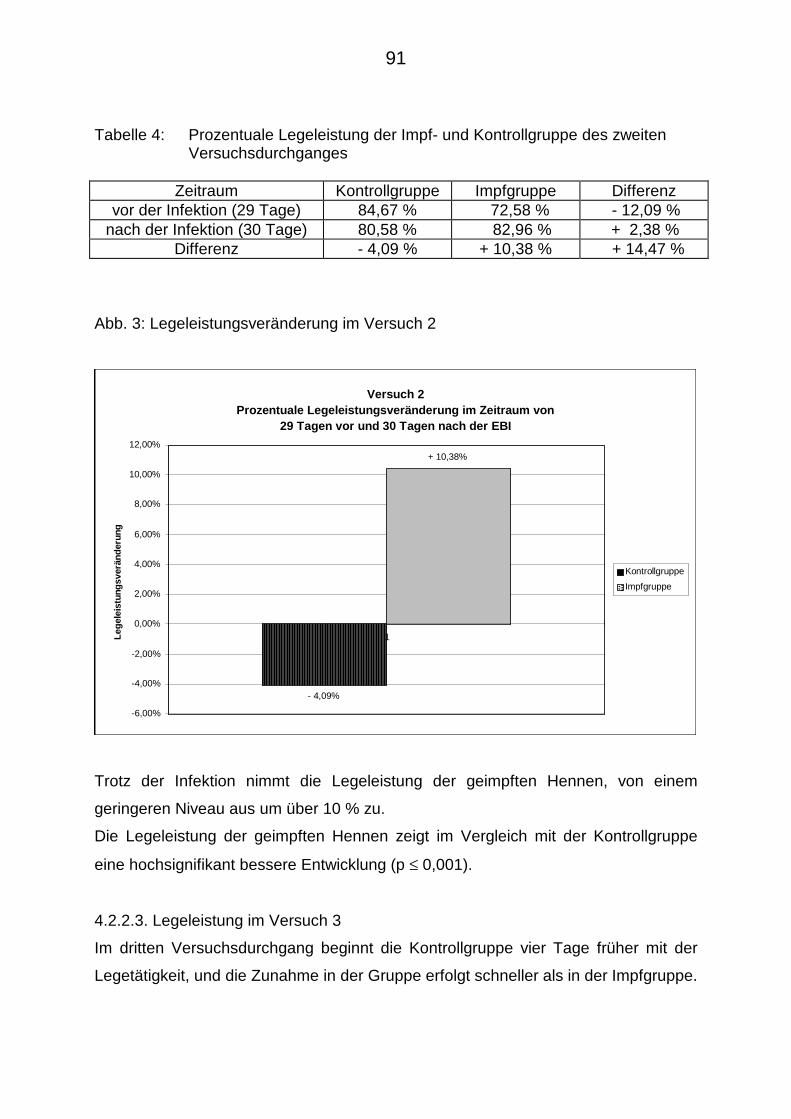
91
Tabelle 4: Prozentuale Legeleistung der Impf- und Kontrollgruppe des zweiten Versuchsdurchganges
Zeitraum Kontrollgruppe Impfgruppe Differenz vor der Infektion (29 Tage) 84,67 % 72,58 % - 12,09 %
nach der Infektion (30 Tage) 80,58 % 82,96 % + 2,38 % Differenz - 4,09 % + 10,38 % + 14,47 %
Abb. 3: Legeleistungsveränderung im Versuch 2
Versuch 2
Prozentuale Legeleistungsveränderung im Zeitraum von 29 Tagen vor und 30 Tagen nach der EBI
- 4,09%
+ 10,38%
-6,00%
-4,00%
-2,00%
0,00%
2,00%
4,00%
6,00%
8,00%
10,00%
12,00%
1 Leg
elei
stun
gsv
erän
der
ung
Kontrollgruppe Impfgruppe
Trotz der Infektion nimmt die Legeleistung der geimpften Hennen, von einem
geringeren Niveau aus um über 10 % zu.
Die Legeleistung der geimpften Hennen zeigt im Vergleich mit der Kontrollgruppe
eine hochsignifikant bessere Entwicklung (p ≤ 0,001).
4.2.2.3. Legeleistung im Versuch 3
Im dritten Versuchsdurchgang beginnt die Kontrollgruppe vier Tage früher mit der
Legetätigkeit, und die Zunahme in der Gruppe erfolgt schneller als in der Impfgruppe.
92
In der 22. Lw. liegt die Legeleistung beider Gruppen über 80 %, wobei die
Legeleistung der Impfgruppe geringfügig über der der Kontrollgruppe liegt. Zum
Zeitpunkt der EBI beträgt die Legeleistung in beiden Gruppen 95 %, um nach der EBI
auf 80 % abzufallen. Die Legeleistung der Impfgruppe steigt binnen 14 Tagen wieder
auf ca. 90 % an und liegt damit im Durchschnitt um 4,03 % über der Legeleistung vor
der EBI. Die durchschnittliche Legeleistung der Kontrollgruppe fällt dagegen nach der
Infizierung um 7,47 % ab und erreicht 14 Tage nach der experimentellen Infizierung
eine Legeleistung von ca. 80 %. In der Tabelle 5 ist die Entwicklung der Legeleistung
vor und nach der EBI dargestellt.
Tabelle 5: Prozentuale Legeleistung der Impf- und Kontrollgruppe des dritten Versuchsdurchganges
Zeitraum Kontrollgruppe Impfgruppe Differenz vor der EBI (28 Tage) 86,55 % 83,88 % - 2,67 %
nach der EBI (30 Tage) 79,08 % 87,91 % + 8,83 % Differenz - 7,47 % + 4,03 % + 11,5 %
In der Abbildung 4 ist die Entwicklung der Legeleistung vor und nach der EBI grafisch
dargestellt.
Die Hennen der Kontrollgruppe zeigen eine größere und länger anhaltende
Depression der Legeleistung. Die Gruppe bleibt deutlich unter dem Niveau, welches
sie vor der Belastungsinfektion erreicht hatte.
Dieser Unterschied in der Entwicklung der Legeleistung der Kontroll- und Impfgruppe
ist hochsignifikant (p ≤ 0,001).
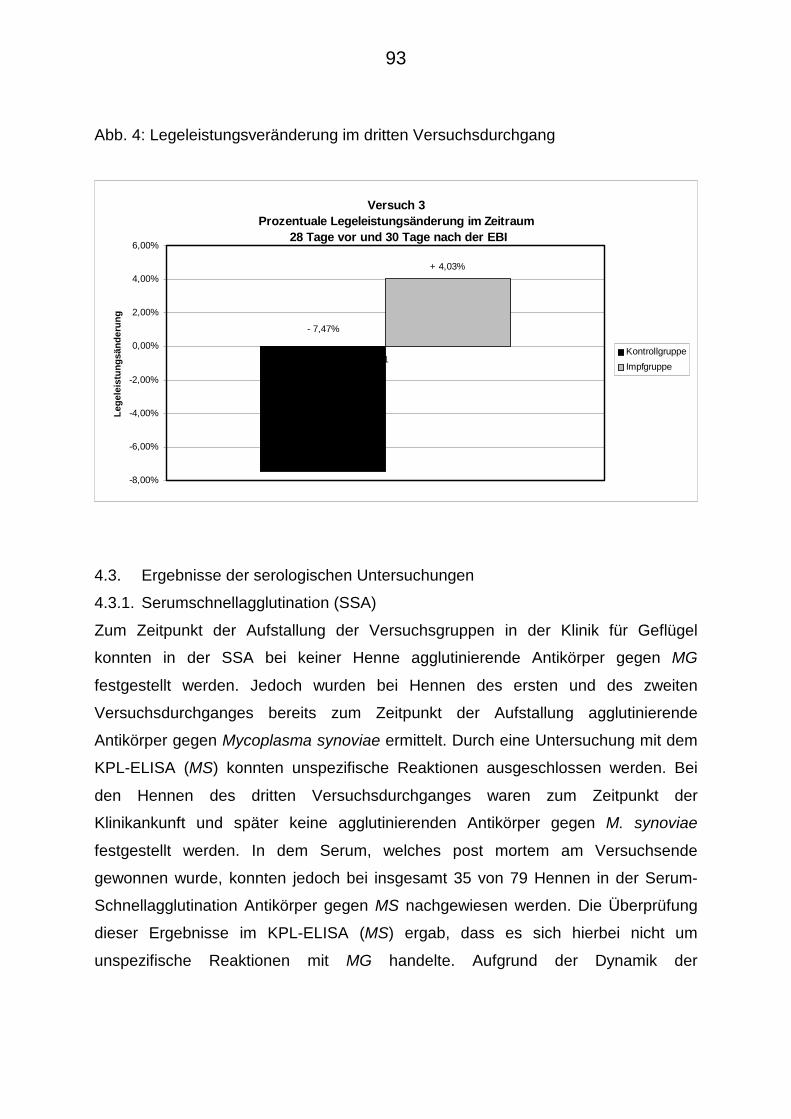
93
Abb. 4: Legeleistungsveränderung im dritten Versuchsdurchgang
Versuch 3Prozentuale Legeleistungsänderung im Zeitraum
28 Tage vor und 30 Tage nach der EBI
- 7,47%
+ 4,03%
-8,00%
-6,00%
-4,00%
-2,00%
0,00%
2,00%
4,00%
6,00%
1
Leg
elei
stu
ng
sän
der
un
g
Kontrollgruppe
Impfgruppe
4.3. Ergebnisse der serologischen Untersuchungen
4.3.1. Serumschnellagglutination (SSA)
Zum Zeitpunkt der Aufstallung der Versuchsgruppen in der Klinik für Geflügel
konnten in der SSA bei keiner Henne agglutinierende Antikörper gegen MG
festgestellt werden. Jedoch wurden bei Hennen des ersten und des zweiten
Versuchsdurchganges bereits zum Zeitpunkt der Aufstallung agglutinierende
Antikörper gegen Mycoplasma synoviae ermittelt. Durch eine Untersuchung mit dem
KPL-ELISA (MS) konnten unspezifische Reaktionen ausgeschlossen werden. Bei
den Hennen des dritten Versuchsdurchganges waren zum Zeitpunkt der
Klinikankunft und später keine agglutinierenden Antikörper gegen M. synoviae
festgestellt werden. In dem Serum, welches post mortem am Versuchsende
gewonnen wurde, konnten jedoch bei insgesamt 35 von 79 Hennen in der Serum-
Schnellagglutination Antikörper gegen MS nachgewiesen werden. Die Überprüfung
dieser Ergebnisse im KPL-ELISA (MS) ergab, dass es sich hierbei nicht um
unspezifische Reaktionen mit MG handelte. Aufgrund der Dynamik der
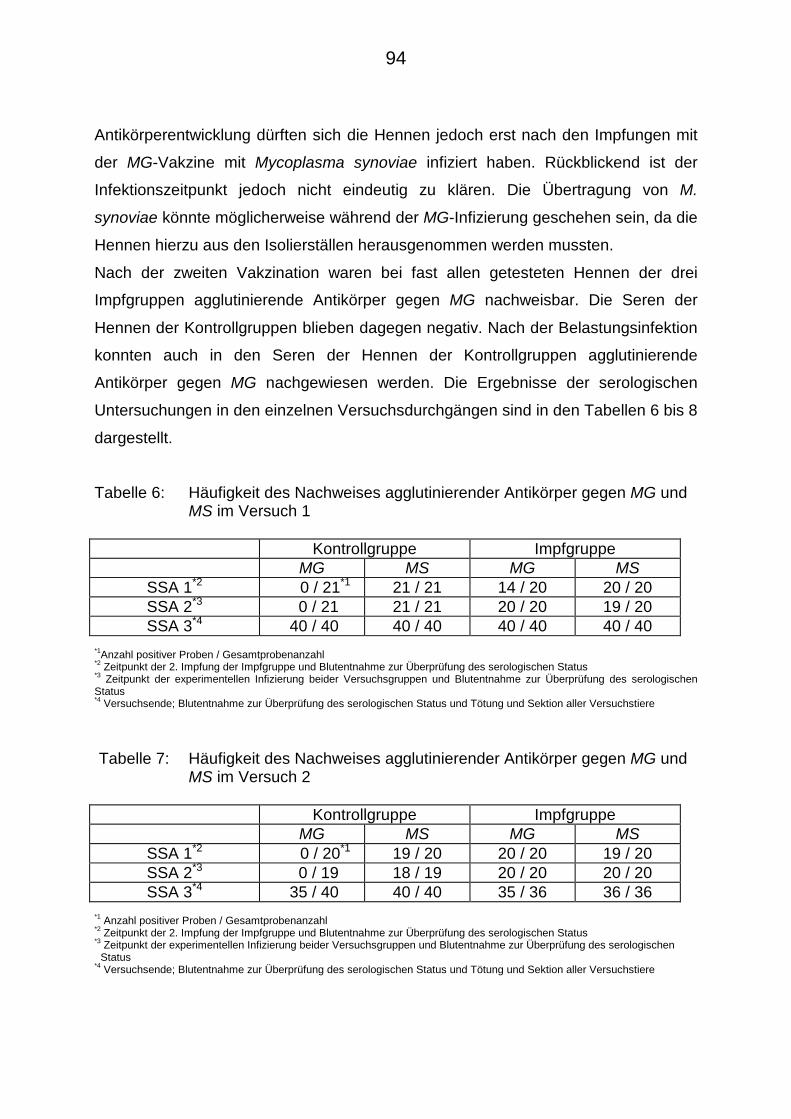
94
Antikörperentwicklung dürften sich die Hennen jedoch erst nach den Impfungen mit
der MG-Vakzine mit Mycoplasma synoviae infiziert haben. Rückblickend ist der
Infektionszeitpunkt jedoch nicht eindeutig zu klären. Die Übertragung von M.
synoviae könnte möglicherweise während der MG-Infizierung geschehen sein, da die
Hennen hierzu aus den Isolierställen herausgenommen werden mussten.
Nach der zweiten Vakzination waren bei fast allen getesteten Hennen der drei
Impfgruppen agglutinierende Antikörper gegen MG nachweisbar. Die Seren der
Hennen der Kontrollgruppen blieben dagegen negativ. Nach der Belastungsinfektion
konnten auch in den Seren der Hennen der Kontrollgruppen agglutinierende
Antikörper gegen MG nachgewiesen werden. Die Ergebnisse der serologischen
Untersuchungen in den einzelnen Versuchsdurchgängen sind in den Tabellen 6 bis 8
dargestellt.
Tabelle 6: Häufigkeit des Nachweises agglutinierender Antikörper gegen MG und MS im Versuch 1
Kontrollgruppe Impfgruppe MG MS MG MS
SSA 1*2 0 / 21*1 21 / 21 14 / 20 20 / 20 SSA 2*3 0 / 21 21 / 21 20 / 20 19 / 20 SSA 3*4 40 / 40 40 / 40 40 / 40 40 / 40
*1Anzahl positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
Tabelle 7: Häufigkeit des Nachweises agglutinierender Antikörper gegen MG und MS im Versuch 2
Kontrollgruppe Impfgruppe MG MS MG MS
SSA 1*2 0 / 20*1 19 / 20 20 / 20 19 / 20 SSA 2*3 0 / 19 18 / 19 20 / 20 20 / 20 SSA 3*4 35 / 40 40 / 40 35 / 36 36 / 36
*1 Anzahl positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
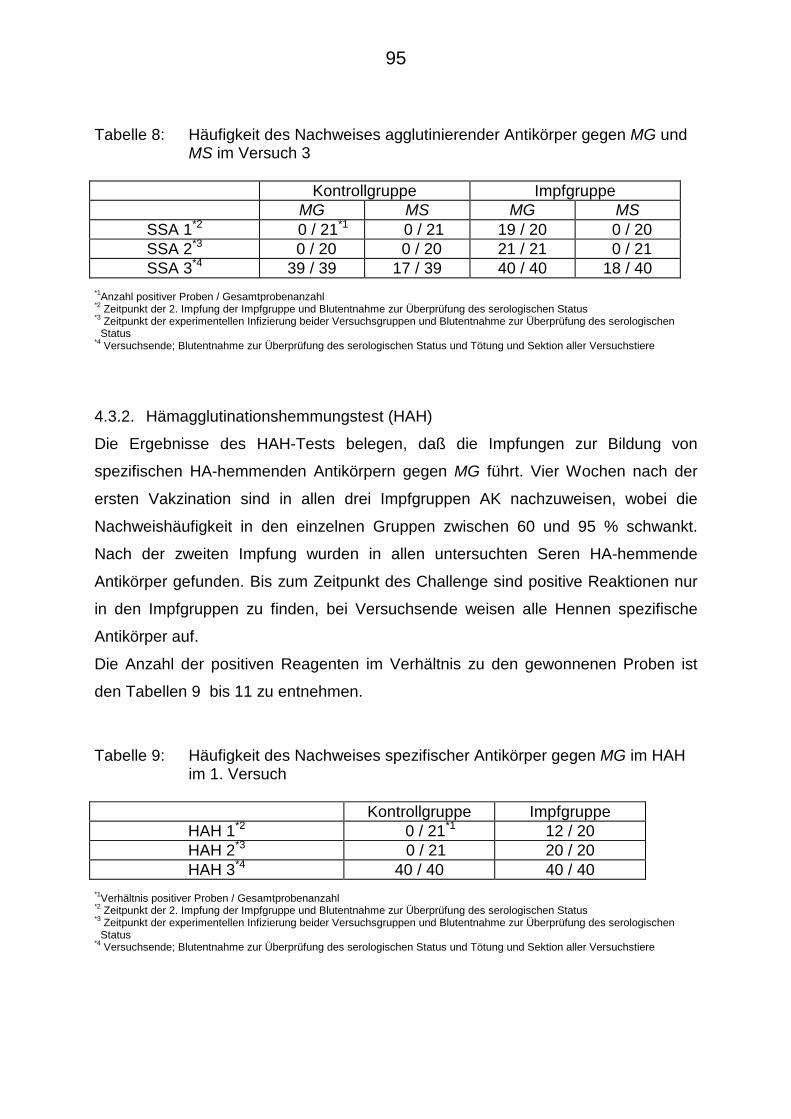
95
Tabelle 8: Häufigkeit des Nachweises agglutinierender Antikörper gegen MG und MS im Versuch 3
Kontrollgruppe Impfgruppe MG MS MG MS
SSA 1*2 0 / 21*1 0 / 21 19 / 20 0 / 20 SSA 2*3 0 / 20 0 / 20 21 / 21 0 / 21 SSA 3*4 39 / 39 17 / 39 40 / 40 18 / 40
*1Anzahl positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
4.3.2. Hämagglutinationshemmungstest (HAH)
Die Ergebnisse des HAH-Tests belegen, daß die Impfungen zur Bildung von
spezifischen HA-hemmenden Antikörpern gegen MG führt. Vier Wochen nach der
ersten Vakzination sind in allen drei Impfgruppen AK nachzuweisen, wobei die
Nachweishäufigkeit in den einzelnen Gruppen zwischen 60 und 95 % schwankt.
Nach der zweiten Impfung wurden in allen untersuchten Seren HA-hemmende
Antikörper gefunden. Bis zum Zeitpunkt des Challenge sind positive Reaktionen nur
in den Impfgruppen zu finden, bei Versuchsende weisen alle Hennen spezifische
Antikörper auf.
Die Anzahl der positiven Reagenten im Verhältnis zu den gewonnenen Proben ist
den Tabellen 9 bis 11 zu entnehmen.
Tabelle 9: Häufigkeit des Nachweises spezifischer Antikörper gegen MG im HAH
im 1. Versuch
Kontrollgruppe Impfgruppe HAH 1*2 0 / 21*1 12 / 20 HAH 2*3 0 / 21 20 / 20 HAH 3*4 40 / 40 40 / 40
*1Verhältnis positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
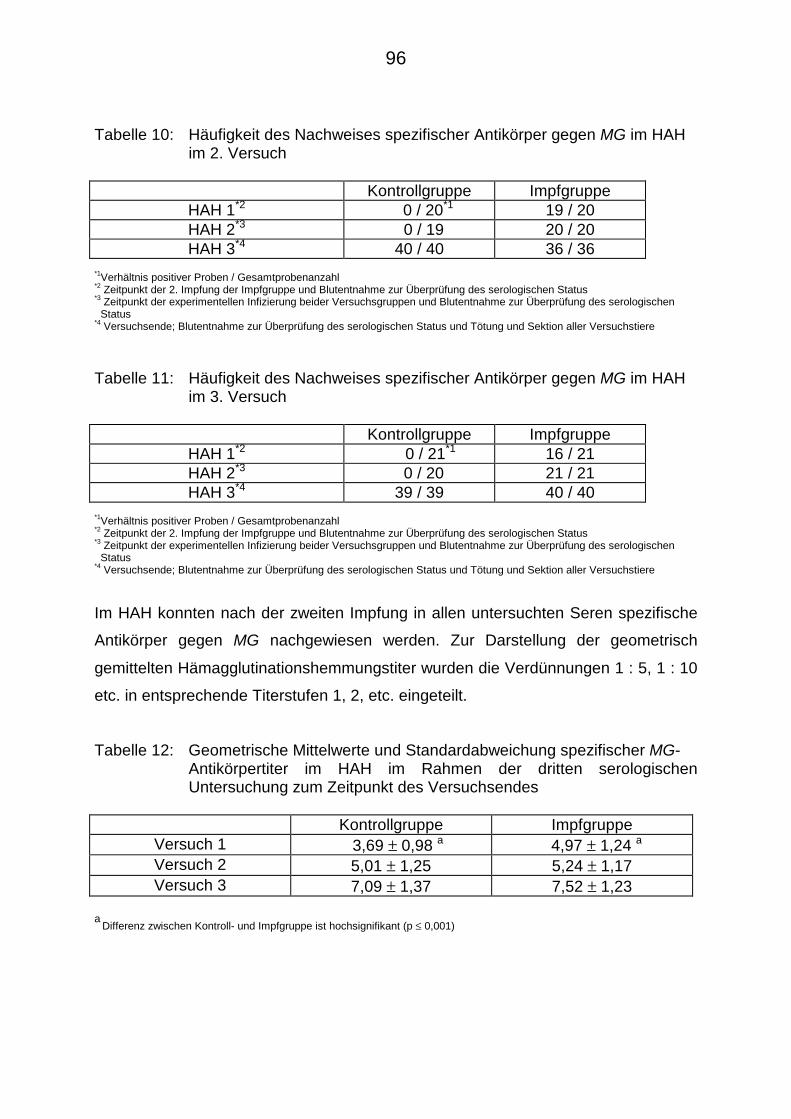
96
Tabelle 10: Häufigkeit des Nachweises spezifischer Antikörper gegen MG im HAH im 2. Versuch
Kontrollgruppe Impfgruppe HAH 1*2 0 / 20*1 19 / 20 HAH 2*3 0 / 19 20 / 20 HAH 3*4 40 / 40 36 / 36
*1Verhältnis positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
Tabelle 11: Häufigkeit des Nachweises spezifischer Antikörper gegen MG im HAH
im 3. Versuch
Kontrollgruppe Impfgruppe HAH 1*2 0 / 21*1 16 / 21 HAH 2*3 0 / 20 21 / 21 HAH 3*4 39 / 39 40 / 40
*1Verhältnis positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
Im HAH konnten nach der zweiten Impfung in allen untersuchten Seren spezifische
Antikörper gegen MG nachgewiesen werden. Zur Darstellung der geometrisch
gemittelten Hämagglutinationshemmungstiter wurden die Verdünnungen 1 : 5, 1 : 10
etc. in entsprechende Titerstufen 1, 2, etc. eingeteilt.
Tabelle 12: Geometrische Mittelwerte und Standardabweichung spezifischer MG- Antikörpertiter im HAH im Rahmen der dritten serologischen Untersuchung zum Zeitpunkt des Versuchsendes Kontrollgruppe Impfgruppe
Versuch 1 3,69 ± 0,98 a 4,97 ± 1,24 a Versuch 2 5,01 ± 1,25 5,24 ± 1,17 Versuch 3 7,09 ± 1,37 7,52 ± 1,23
a Differenz zwischen Kontroll- und Impfgruppe ist hochsignifikant (p ≤ 0,001)
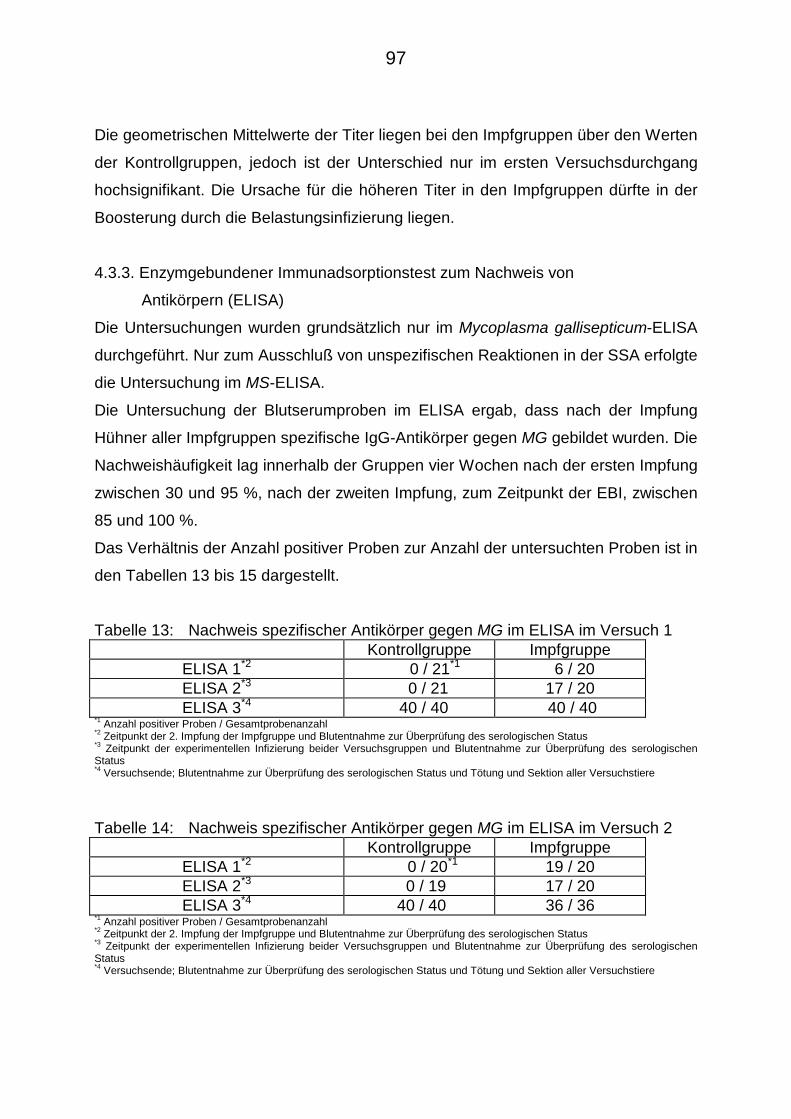
97
Die geometrischen Mittelwerte der Titer liegen bei den Impfgruppen über den Werten
der Kontrollgruppen, jedoch ist der Unterschied nur im ersten Versuchsdurchgang
hochsignifikant. Die Ursache für die höheren Titer in den Impfgruppen dürfte in der
Boosterung durch die Belastungsinfizierung liegen.
4.3.3. Enzymgebundener Immunadsorptionstest zum Nachweis von
Antikörpern (ELISA)
Die Untersuchungen wurden grundsätzlich nur im Mycoplasma gallisepticum-ELISA
durchgeführt. Nur zum Ausschluß von unspezifischen Reaktionen in der SSA erfolgte
die Untersuchung im MS-ELISA.
Die Untersuchung der Blutserumproben im ELISA ergab, dass nach der Impfung
Hühner aller Impfgruppen spezifische IgG-Antikörper gegen MG gebildet wurden. Die
Nachweishäufigkeit lag innerhalb der Gruppen vier Wochen nach der ersten Impfung
zwischen 30 und 95 %, nach der zweiten Impfung, zum Zeitpunkt der EBI, zwischen
85 und 100 %.
Das Verhältnis der Anzahl positiver Proben zur Anzahl der untersuchten Proben ist in
den Tabellen 13 bis 15 dargestellt.
Tabelle 13: Nachweis spezifischer Antikörper gegen MG im ELISA im Versuch 1 Kontrollgruppe Impfgruppe
ELISA 1*2 0 / 21*1 6 / 20 ELISA 2*3 0 / 21 17 / 20 ELISA 3*4 40 / 40 40 / 40
*1 Anzahl positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
Tabelle 14: Nachweis spezifischer Antikörper gegen MG im ELISA im Versuch 2 Kontrollgruppe Impfgruppe
ELISA 1*2 0 / 20*1 19 / 20 ELISA 2*3 0 / 19 17 / 20 ELISA 3*4 40 / 40 36 / 36
*1 Anzahl positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
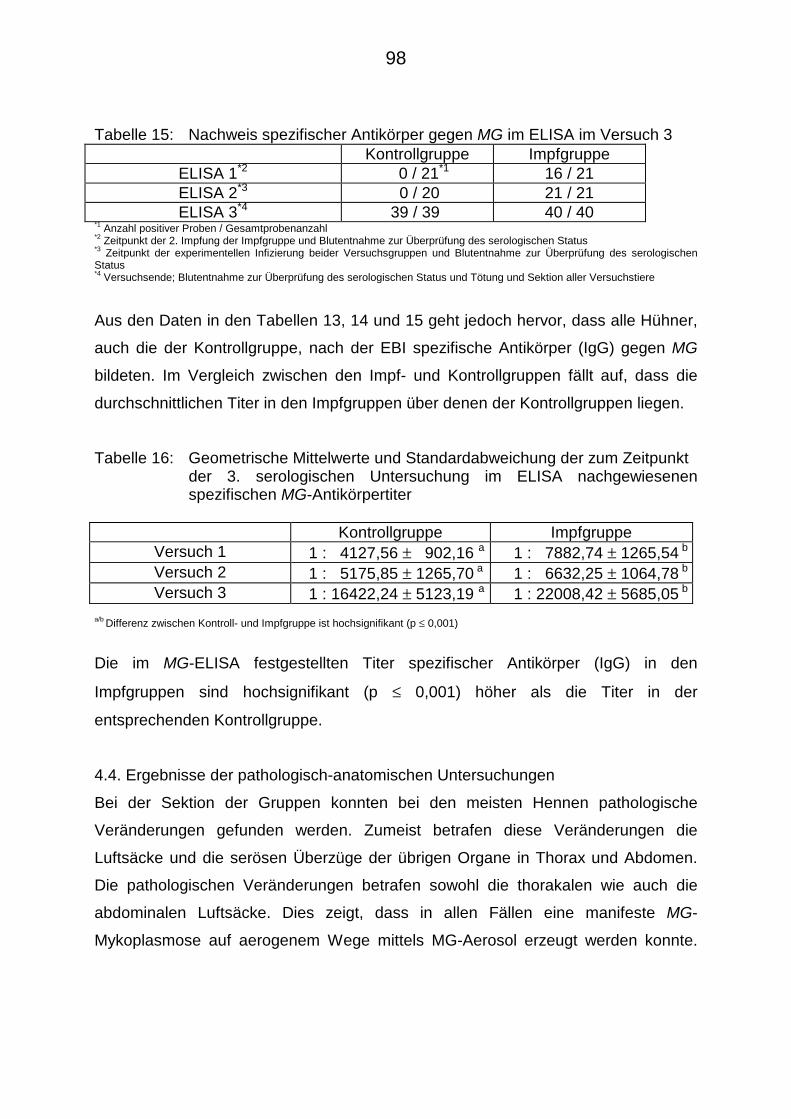
98
Tabelle 15: Nachweis spezifischer Antikörper gegen MG im ELISA im Versuch 3 Kontrollgruppe Impfgruppe
ELISA 1*2 0 / 21*1 16 / 21 ELISA 2*3 0 / 20 21 / 21 ELISA 3*4 39 / 39 40 / 40
*1 Anzahl positiver Proben / Gesamtprobenanzahl *2 Zeitpunkt der 2. Impfung der Impfgruppe und Blutentnahme zur Überprüfung des serologischen Status *3 Zeitpunkt der experimentellen Infizierung beider Versuchsgruppen und Blutentnahme zur Überprüfung des serologischen Status *4 Versuchsende; Blutentnahme zur Überprüfung des serologischen Status und Tötung und Sektion aller Versuchstiere
Aus den Daten in den Tabellen 13, 14 und 15 geht jedoch hervor, dass alle Hühner,
auch die der Kontrollgruppe, nach der EBI spezifische Antikörper (IgG) gegen MG
bildeten. Im Vergleich zwischen den Impf- und Kontrollgruppen fällt auf, dass die
durchschnittlichen Titer in den Impfgruppen über denen der Kontrollgruppen liegen.
Tabelle 16: Geometrische Mittelwerte und Standardabweichung der zum Zeitpunkt der 3. serologischen Untersuchung im ELISA nachgewiesenen spezifischen MG-Antikörpertiter Kontrollgruppe Impfgruppe
Versuch 1 1 : 4127,56 ± 902,16 a 1 : 7882,74 ± 1265,54 b Versuch 2 1 : 5175,85 ± 1265,70 a 1 : 6632,25 ± 1064,78 b Versuch 3 1 : 16422,24 ± 5123,19 a 1 : 22008,42 ± 5685,05 b
a/b Differenz zwischen Kontroll- und Impfgruppe ist hochsignifikant (p ≤ 0,001)
Die im MG-ELISA festgestellten Titer spezifischer Antikörper (IgG) in den
Impfgruppen sind hochsignifikant (p ≤ 0,001) höher als die Titer in der
entsprechenden Kontrollgruppe.
4.4. Ergebnisse der pathologisch-anatomischen Untersuchungen
Bei der Sektion der Gruppen konnten bei den meisten Hennen pathologische
Veränderungen gefunden werden. Zumeist betrafen diese Veränderungen die
Luftsäcke und die serösen Überzüge der übrigen Organe in Thorax und Abdomen.
Die pathologischen Veränderungen betrafen sowohl die thorakalen wie auch die
abdominalen Luftsäcke. Dies zeigt, dass in allen Fällen eine manifeste MG-
Mykoplasmose auf aerogenem Wege mittels MG-Aerosol erzeugt werden konnte.
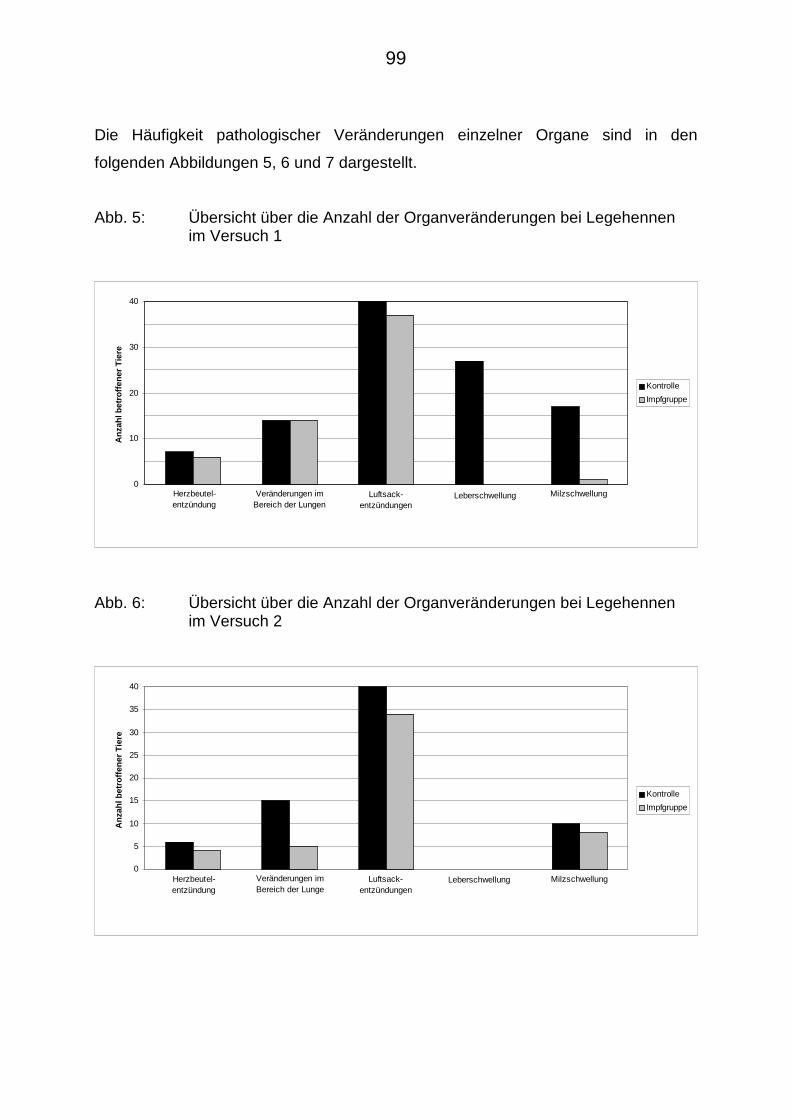
99
Die Häufigkeit pathologischer Veränderungen einzelner Organe sind in den
folgenden Abbildungen 5, 6 und 7 dargestellt.
Abb. 5: Übersicht über die Anzahl der Organveränderungen bei Legehennen im Versuch 1
Herzbeutel-entzündung
Veränderungen im Bereich der Lungen
MilzschwellungLuftsack-entzündungen
Leberschwellung0
10
20
30
40
An
zah
l bet
roff
ener
Tie
re
Kontrolle
Impfgruppe
Abb. 6: Übersicht über die Anzahl der Organveränderungen bei Legehennen
im Versuch 2
MilzschwellungLeberschwellungVeränderungen im Bereich der Lunge
Herzbeutel-entzündung
Luftsack-entzündungen
0
5
10
15
20
25
30
35
40
An
zah
l bet
roff
ener
Tie
re
Kontrolle
Impfgruppe
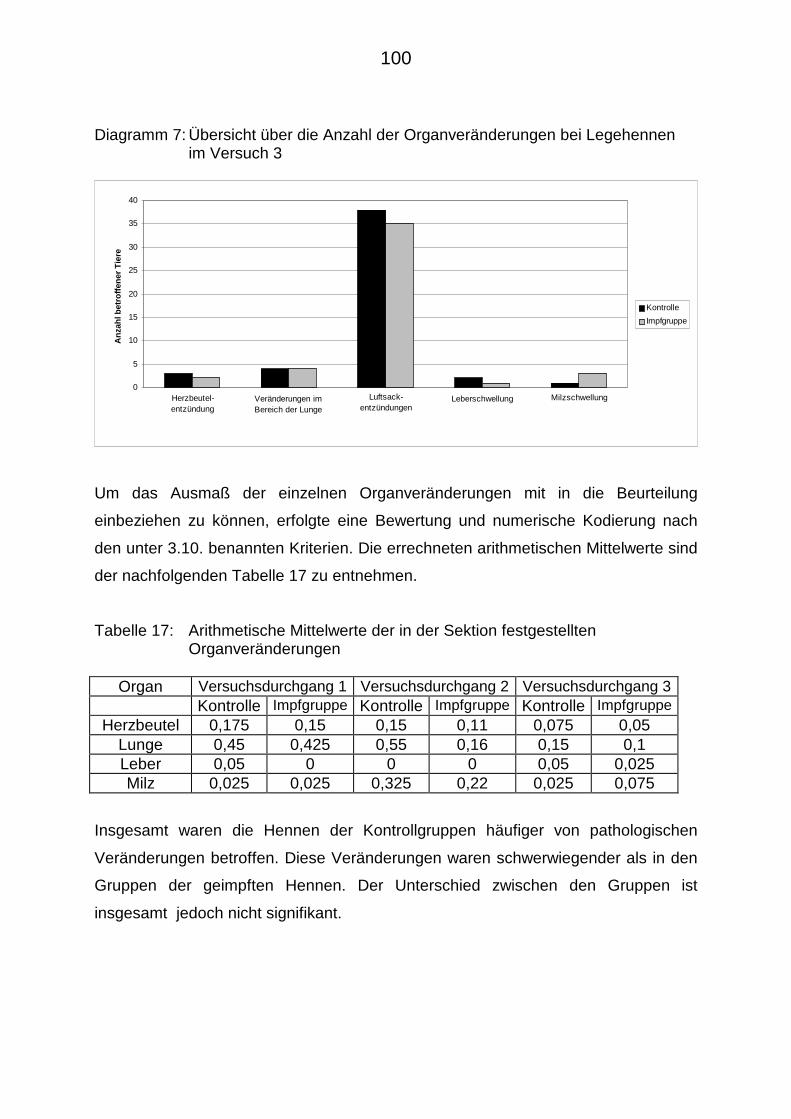
100
Diagramm 7: Übersicht über die Anzahl der Organveränderungen bei Legehennen im Versuch 3
Veränderungen imBereich der Lunge
Luftsack-entzündungen
Leberschwellung MilzschwellungHerzbeutel-entzündung
0
5
10
15
20
25
30
35
40
An
zah
l bet
roff
ener
Tie
re
Kontrolle
Impfgruppe
Um das Ausmaß der einzelnen Organveränderungen mit in die Beurteilung
einbeziehen zu können, erfolgte eine Bewertung und numerische Kodierung nach
den unter 3.10. benannten Kriterien. Die errechneten arithmetischen Mittelwerte sind
der nachfolgenden Tabelle 17 zu entnehmen.
Tabelle 17: Arithmetische Mittelwerte der in der Sektion festgestellten Organveränderungen
Organ Versuchsdurchgang 1 Versuchsdurchgang 2 Versuchsdurchgang 3 Kontrolle Impfgruppe Kontrolle Impfgruppe Kontrolle Impfgruppe
Herzbeutel 0,175 0,15 0,15 0,11 0,075 0,05 Lunge 0,45 0,425 0,55 0,16 0,15 0,1 Leber 0,05 0 0 0 0,05 0,025 Milz 0,025 0,025 0,325 0,22 0,025 0,075
Insgesamt waren die Hennen der Kontrollgruppen häufiger von pathologischen
Veränderungen betroffen. Diese Veränderungen waren schwerwiegender als in den
Gruppen der geimpften Hennen. Der Unterschied zwischen den Gruppen ist
insgesamt jedoch nicht signifikant.
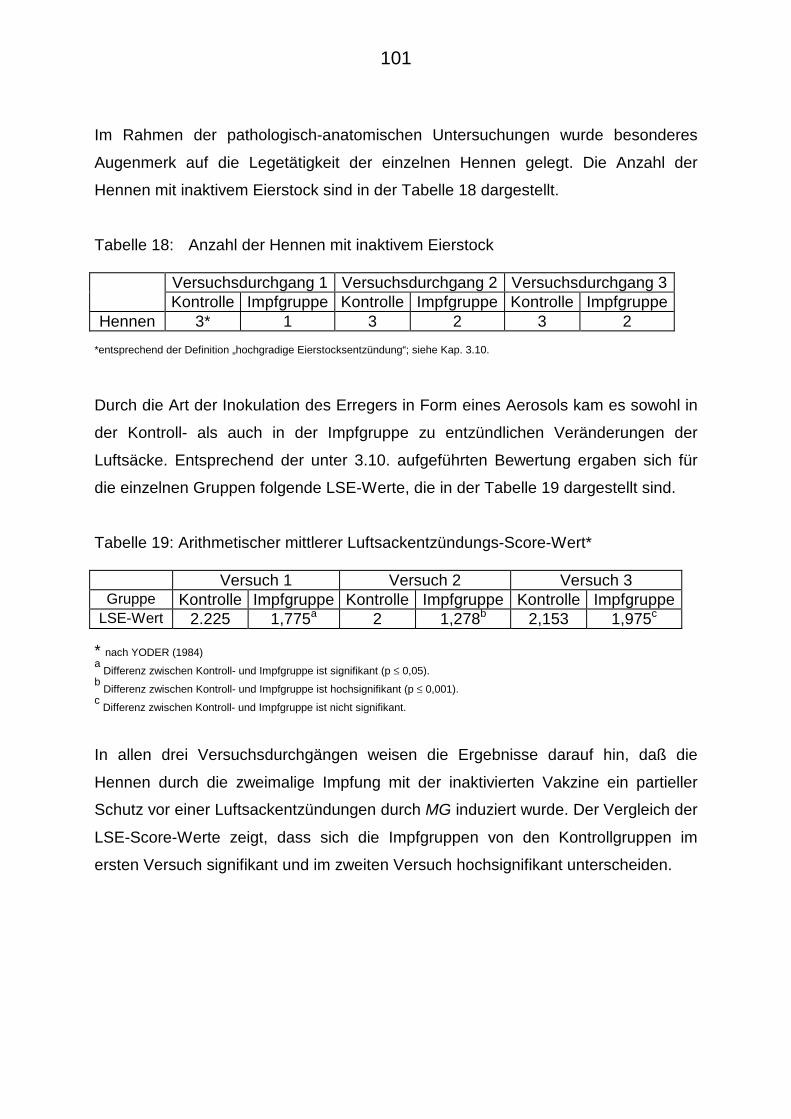
101
Im Rahmen der pathologisch-anatomischen Untersuchungen wurde besonderes
Augenmerk auf die Legetätigkeit der einzelnen Hennen gelegt. Die Anzahl der
Hennen mit inaktivem Eierstock sind in der Tabelle 18 dargestellt.
Tabelle 18: Anzahl der Hennen mit inaktivem Eierstock
Versuchsdurchgang 1 Versuchsdurchgang 2 Versuchsdurchgang 3 Kontrolle Impfgruppe Kontrolle Impfgruppe Kontrolle Impfgruppe
Hennen 3* 1 3 2 3 2 *entsprechend der Definition „hochgradige Eierstocksentzündung“; siehe Kap. 3.10.
Durch die Art der Inokulation des Erregers in Form eines Aerosols kam es sowohl in
der Kontroll- als auch in der Impfgruppe zu entzündlichen Veränderungen der
Luftsäcke. Entsprechend der unter 3.10. aufgeführten Bewertung ergaben sich für
die einzelnen Gruppen folgende LSE-Werte, die in der Tabelle 19 dargestellt sind.
Tabelle 19: Arithmetischer mittlerer Luftsackentzündungs-Score-Wert*
Versuch 1 Versuch 2 Versuch 3 Gruppe Kontrolle Impfgruppe Kontrolle Impfgruppe Kontrolle Impfgruppe
LSE-Wert 2.225 1,775a 2 1,278b 2,153 1,975c
* nach YODER (1984) a Differenz zwischen Kontroll- und Impfgruppe ist signifikant (p ≤ 0,05).
b Differenz zwischen Kontroll- und Impfgruppe ist hochsignifikant (p ≤ 0,001).
c Differenz zwischen Kontroll- und Impfgruppe ist nicht signifikant.
In allen drei Versuchsdurchgängen weisen die Ergebnisse darauf hin, daß die
Hennen durch die zweimalige Impfung mit der inaktivierten Vakzine ein partieller
Schutz vor einer Luftsackentzündungen durch MG induziert wurde. Der Vergleich der
LSE-Score-Werte zeigt, dass sich die Impfgruppen von den Kontrollgruppen im
ersten Versuch signifikant und im zweiten Versuch hochsignifikant unterscheiden.
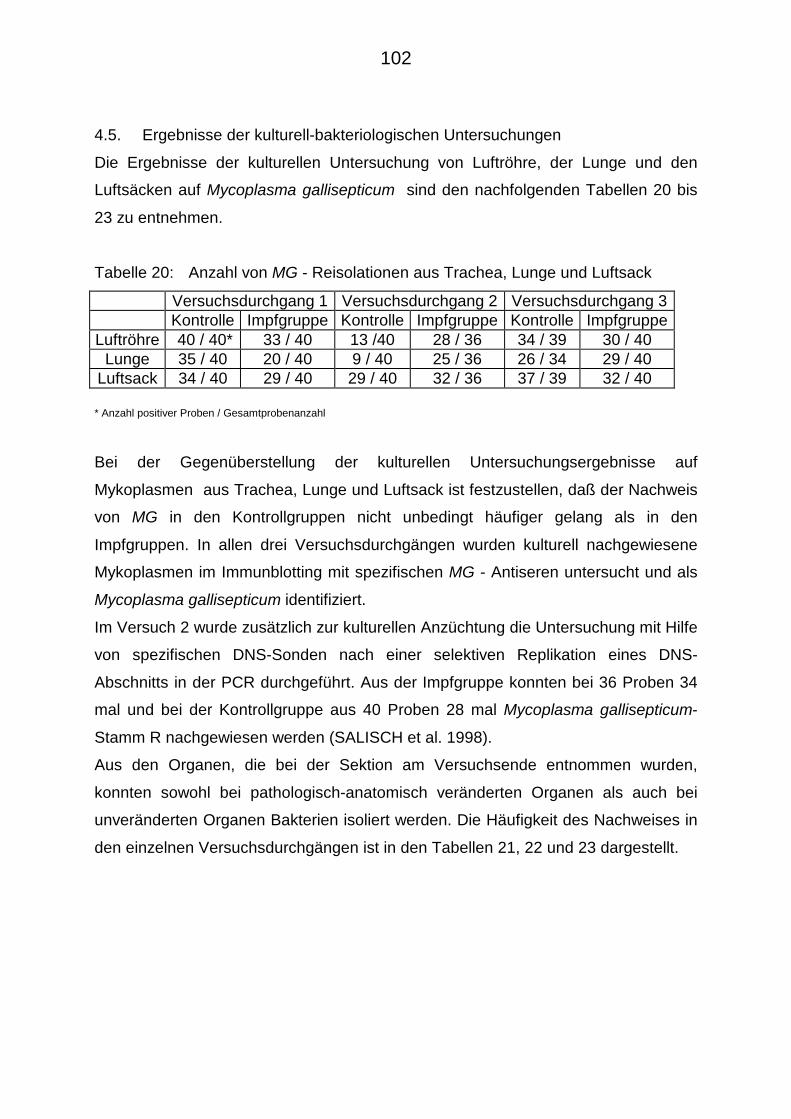
102
4.5. Ergebnisse der kulturell-bakteriologischen Untersuchungen
Die Ergebnisse der kulturellen Untersuchung von Luftröhre, der Lunge und den
Luftsäcken auf Mycoplasma gallisepticum sind den nachfolgenden Tabellen 20 bis
23 zu entnehmen.
Tabelle 20: Anzahl von MG - Reisolationen aus Trachea, Lunge und Luftsack
Versuchsdurchgang 1 Versuchsdurchgang 2 Versuchsdurchgang 3 Kontrolle Impfgruppe Kontrolle Impfgruppe Kontrolle Impfgruppe
Luftröhre 40 / 40* 33 / 40 13 /40 28 / 36 34 / 39 30 / 40 Lunge 35 / 40 20 / 40 9 / 40 25 / 36 26 / 34 29 / 40
Luftsack 34 / 40 29 / 40 29 / 40 32 / 36 37 / 39 32 / 40
* Anzahl positiver Proben / Gesamtprobenanzahl
Bei der Gegenüberstellung der kulturellen Untersuchungsergebnisse auf
Mykoplasmen aus Trachea, Lunge und Luftsack ist festzustellen, daß der Nachweis
von MG in den Kontrollgruppen nicht unbedingt häufiger gelang als in den
Impfgruppen. In allen drei Versuchsdurchgängen wurden kulturell nachgewiesene
Mykoplasmen im Immunblotting mit spezifischen MG - Antiseren untersucht und als
Mycoplasma gallisepticum identifiziert.
Im Versuch 2 wurde zusätzlich zur kulturellen Anzüchtung die Untersuchung mit Hilfe
von spezifischen DNS-Sonden nach einer selektiven Replikation eines DNS-
Abschnitts in der PCR durchgeführt. Aus der Impfgruppe konnten bei 36 Proben 34
mal und bei der Kontrollgruppe aus 40 Proben 28 mal Mycoplasma gallisepticum-
Stamm R nachgewiesen werden (SALISCH et al. 1998).
Aus den Organen, die bei der Sektion am Versuchsende entnommen wurden,
konnten sowohl bei pathologisch-anatomisch veränderten Organen als auch bei
unveränderten Organen Bakterien isoliert werden. Die Häufigkeit des Nachweises in
den einzelnen Versuchsdurchgängen ist in den Tabellen 21, 22 und 23 dargestellt.
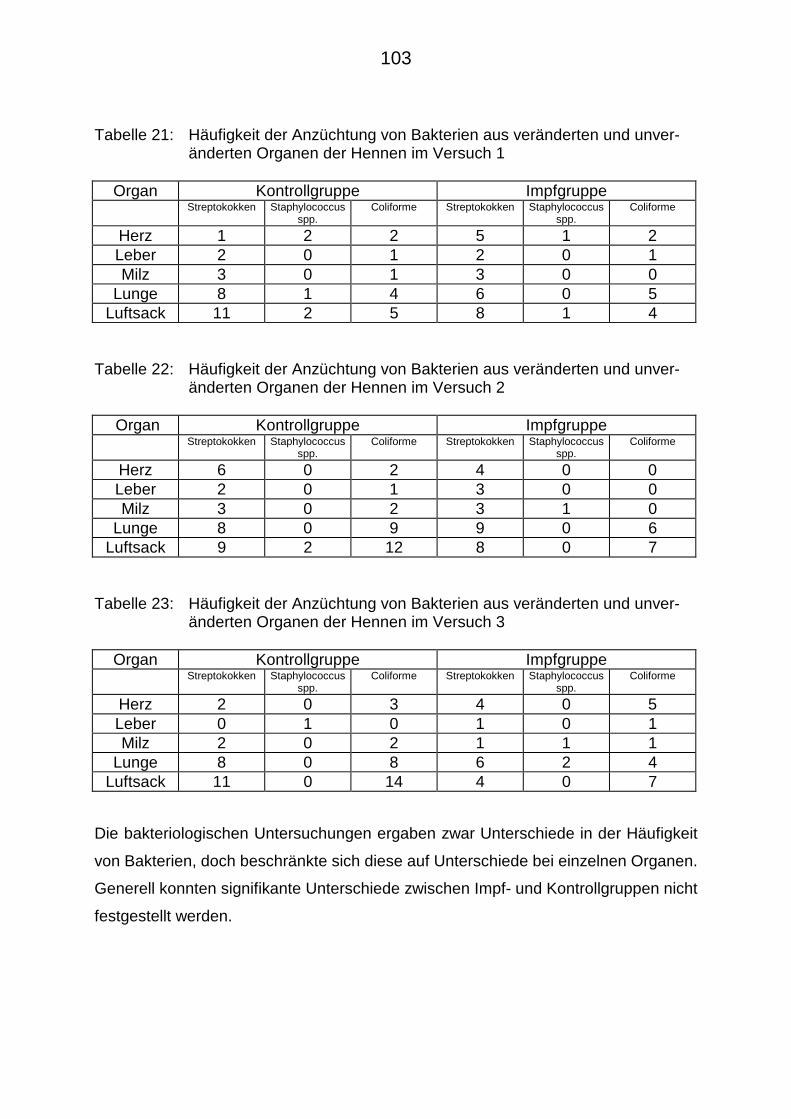
103
Tabelle 21: Häufigkeit der Anzüchtung von Bakterien aus veränderten und unver- änderten Organen der Hennen im Versuch 1
Organ Kontrollgruppe Impfgruppe Streptokokken Staphylococcus
spp. Coliforme Streptokokken Staphylococcus
spp. Coliforme
Herz 1 2 2 5 1 2 Leber 2 0 1 2 0 1 Milz 3 0 1 3 0 0
Lunge 8 1 4 6 0 5 Luftsack 11 2 5 8 1 4
Tabelle 22: Häufigkeit der Anzüchtung von Bakterien aus veränderten und unver-
änderten Organen der Hennen im Versuch 2
Organ Kontrollgruppe Impfgruppe Streptokokken Staphylococcus
spp. Coliforme Streptokokken Staphylococcus
spp. Coliforme
Herz 6 0 2 4 0 0 Leber 2 0 1 3 0 0 Milz 3 0 2 3 1 0
Lunge 8 0 9 9 0 6 Luftsack 9 2 12 8 0 7
Tabelle 23: Häufigkeit der Anzüchtung von Bakterien aus veränderten und unver-
änderten Organen der Hennen im Versuch 3
Organ Kontrollgruppe Impfgruppe Streptokokken Staphylococcus
spp. Coliforme Streptokokken Staphylococcus
spp. Coliforme
Herz 2 0 3 4 0 5 Leber 0 1 0 1 0 1 Milz 2 0 2 1 1 1
Lunge 8 0 8 6 2 4 Luftsack 11 0 14 4 0 7
Die bakteriologischen Untersuchungen ergaben zwar Unterschiede in der Häufigkeit
von Bakterien, doch beschränkte sich diese auf Unterschiede bei einzelnen Organen.
Generell konnten signifikante Unterschiede zwischen Impf- und Kontrollgruppen nicht
festgestellt werden.

104
5. Diskussion
Ziel dieser Arbeit war es, die Verträglichkeit und Wirksamkeit der Öl-Adjuvans-
Vakzine „TALOVAC 104 MG“ der Firma LAH (Lohmann Animal Health) unter
experimentellen Bedingungen zu prüfen. Der Impfstoff wurde entwickelt, um
Legehennen vor einer MG-assoziierten Minderung der Legeleistung schützen.
Im Rahmen dieser experimentellen Prüfung wurden Junghennen in der 14. und 18.
Lebenswoche mit jeweils 0,5 ml des Impfstoffes subkutan, dorsal im unteren Drittel
des Halses, geimpft. Dieses Impfschema, bot sich, abweichend von der
Impfempfehlung der Herstellerfirma, an, weil in praxi der zweite Impftermin mit dem
Zeitpunkt der Umstallung der Junghennen vom Aufzucht- in den Ablegebetrieb
zusammenfällt und somit die Kosten der aufwendigen Einzeltiervakzination reduziert
werden können (JORDAN u. KLEVEN 1995). Auch von anderen Autoren wurde die
Impfung gegen MG zeitgleich in ähnlicher Weise durchgeführt (HILDEBRAND et al.
1983; GLISSON u. KLEVEN 1984, 1985; TAKLINGTON u. KLEVEN 1985;
SASIPREEYAJAN et al. 1987), ohne dass über eine Störung der Legetätigkeit zu
berichtet worden ist. Lediglich BARBOUR und NEWMAN (1990) und EVANS et al.
(1992) impften ihre Versuchshennen zu einem früheren Zeitpunkt, nämlich in der
zehnten und sechzehnten Lebenswoche.
Die pathologisch-anatomischen Untersuchungen der Hennen am Versuchsende
ergaben am Applikationsort des Impfstoffes entzündlich-granulomatöse Reaktionen,
die bei über 40 % der Tiere über das für diesen Impfstoff akzeptable Maß
hinausgingen. Der Einsatz von Adjuvantien, in diesem Fall Mineralöl, verzögert die
Freisetzung des Antigens und steigert die Makrophagentätigkeit, was wiederum zu
einer vermehrten Antigenpräsentation und Freisetzung von Lymphokinen führt
(BARBOUR et al. 1987; FINGER 1966; JANEWAY und TRAVERS 1997). Gerade bei
nur schwach immunogenen Antigenen ist so die Ausbildung eines langanhaltenden
Impfschutzes möglich (JANEWAY und TRAVERS 1997). STONE et al. (1978) und
YODER (1979) stellten nach Impfung mit einer MG-Öl-Adjuvansvakzine in den
Blutserumproben einen zwei- bis vierfach höheren Antikörpertiter, als in den Seren
der Hühner, die nur mit dem reinen Antigen geimpft worden waren, fest. HAYATSU

105
et al. (1974) konnten durch die Zugabe verschiedener Adjuvantien dagegen keinen
signifikanten Anstieg spezifischer Antikörpertiter gegen MG nachweisen.
Über lokale oder systemische Nebenwirkungen der Impfung, wie sie in den eigenen
Versuchen auftraten, haben nur wenige Autoren Angaben gemacht. HILDEBRAND
et al. (1983) berichteten, daß es nach Applikation eines inaktivierten MG-Impfstoffes
in der Halsmitte bei einigen Hennen zur Ausbildung periorbitaler Ödeme kam. Nach
Impfung in den M. pectoralis fanden DROUAL et al. (1990) undurchsichtige Zysten in
den Faszien, welche teilweise von lymphoiden Zellaggregationen umgeben waren.
Nach Impfungen in die Beinmuskulatur kam es zu lokalen Schwellungen, die sich
auch auf benachbarte Gelenke oder Sehnenscheiden ausdehnten (DROUAL et al.
1993). SASIPREEYAJAN et al. (1987) dagegen beschrieben solche Veränderungen
nicht, obwohl bei ihren Versuchen die Impfstoffapplikation direkt an der Schädelbasis
erfolgte.
In den Versuchen 2 und 3 kam es zusätzlich zu einer Verzögerung des Legebeginns
der geimpften Hennen. Dies wird durch die Anzahl der vor dem Zeitpunkt der
Infizierung gelegten Eier belegt. Lediglich BEHR et al. (1991) beobachteten bei
Einsatz eines inaktivierten Impfstoffes in einer Feldstudie eine Verzögerung des
Legebeginns bei geimpften Hennen. Durch eine Vakzination der Legehennen zu
einem früheren Zeitpunkt, wie ihn EVANS et al. (1992) und JORDAN und KLEVEN
(1995) REDMANN und GÖBEL (1991) ohne Angabe von Gründen empfahlen, hätte
die Verzögerung des Legebeginns wie im 2. und 3. Versuch eventuell vermieden
werden können. Ob die Impfung zur Beeinträchtigung des Allgemeinbefindens und
zur Beeinträchtigung der immunologischen Reaktionsfähigkeit der Hennen der
Impfgruppen in den Versuchen B und C und damit zur Manifestation der
Marek´schen Krankheit geführt hat, war nicht zu klären. Jedoch berichteten auch
KATZEN (1969) sowie REDMANN und GÖBEL (1991) über das Auftreten der
Marek´schen Krankheit nach Impfungen gegen Mycoplasma gallisepticum. Diese
Gesundheitsbeeinträchtigungen betrafen Hennen, die mit den Impfstoffchargen 6
und 7 geimpft worden waren. Diese Impfstoffchargen zeigten bei sachgerechter
Lagerung eine von den Herstellerangaben abweichende Entmischungscharakteristik.
Die ungenügende Verträglichkeit dieser Impfstoffchargen kann somit möglicherweise

106
auch durch eine produktionstechnische Komponente bedingt bzw. mitbeeinflusst
worden sein.
Kernanliegen der vorliegenden experimentellen Studie war die Prüfung der
Wirksamkeit der Vakzination mit dem Impfstoff „Talovac 104 MG“ anhand der
Legeleistung, die bei Legehennen ein hochsensibler Indikator für Störungen des
Gesund-heitszustandes darstellt. Mehr als vier Wochen nach der zweiten Impfung
und bei einem Legeleistungsniveau von mindestens 70 % erfolgte die aerogene
Infizierung mit dem MG-Stamm R. Ziel dieser Infizierung war nicht den Status einer
latenten MG-Infektion zu erzeugen, sondern den einer klinisch manifesten Störung
des Gesund-heitszustandes und die damit assoziierte Minderung der Legeleistung.
Bei alleiniger Infizierung mit MG ist dies nur durch Inokulation einer großen
Mykoplasmenanzahl möglich, wie sie unter natürlichen Bedingungen niemals
vorkommt. Die in diesen Versuchen vermittelte Exposition mit MG stellte somit eine
Maximalbelastung dar. Da Mycoplasma gallisepticum in der Außenwelt nur über eine
geringe Tenazität verfügt (HINZ 1993) und somit existenziell vom seinem Wirt
abhängig ist, hat eine Schädigung des Wirtes, die eventuell zum Tod desselben
führt, evolutionsbiologisch für MG keinen Vorteil. Die Persistenz im Wirt und die
vertikale und horizontale Übertragbarkeit garantieren MG das Überleben; daher ist
nach einer Infizierung mit MG die latente Infektion der Normalzustand (HINZ und
RYLL 1996). Stressoren infektiöser (NDV, AIBV, E. coli) und nicht infektiöser Art
(Temperatur, Staub, Schadgase) können das labile Gleichgewicht zwischen
Infektabwehr und MG stören (GIAMBRONE 1989) und zu einer Manifestation einer
Mykoplasmose führen (BEHR et al. 1991). So konnten YODER (1979) und YODER
und HOPKINS (1985), die ihre Versuchshühner nur mit 0,2 ml einer
Mykoplasmenkultur infizierten, bei dem Vergleich der Legeleistung keine
Unterschiede zwischen Impf- und Kontrollgruppe feststellen. Es ist anzunehmen,
dass sie eine Mykoplasmenmenge inokulierten, die lediglich zu einer Infektion, nicht
jedoch zu einer manifesten Erkrankung durch MG führte.
YODER (1983), YODER et al. (1984) und SOERIPTO (1989 a) belasteten die
Hühner zusätzlich mit IBV bzw. sie verbrachten die Hühner p.i. in eine kalte
Umgebung, um eine manifeste Mykoplasmose zu erzeugen. GLISSON und KLEVEN

107
(1984, 1985), EVANS (1992) und ABD-EL-MOTELIB und KLEVEN (1993)
vermochten hingegen durch die alleinige aerogenen Infizierung über ein Aerosol mit
dem MG-Stamm R (1,5x106 bis 3x108 Mykoplasmen je Henne)
Luftsackentzündungen und Legeleistungsminderungen hervorrufen. Eine Infizierung
mit einer derartigen MG-Menge entspricht der Maximalbelastung, wie sie auch in der
vorliegenden Studie durchgeführt wurde. Das Design der in den eigenen
Untersuchungen vorgenommenen experimentellen Belastungsinfizierung (1995/96)
entspricht dem Verfahren, das WHITHEAR et al. (1996) zur Infizierung von Hühnern
beschrieben haben.
Nach der experimentellen Belastungsinfizierung entwickelte sich die Legeleistung in
allen drei Impfgruppen besser als in der jeweils korrespondierenden Kontrollgruppe.
Der Unterschied zwischen der Impf- und der Kontrollgruppe war im 1. Versuch nur
schwachsignifikant, im 2. und 3. Versuch jedoch hochsignifikant. Mit einer sehr
ähnlichen Vakzine „TALOVAC 104 MG“ (andere Zusammensetzung des Adjuvans)
konnten BEHR et al. (1991, 1993) unter Feldbedingungen ebenfalls einen Schutz vor
einer Legeleistungsminderung induzieren. Während KHAN (1986) keine
Schutzwirkung nach einer einmaligen Impfung mit einer inaktivierten Öl-Adjuvans-
Vakzine feststellen konnte, bestätigen die Ergebnisse der vorliegenden Studie die
von HILDEBRAND et al. (1983), GLISSON und KLEVEN (1984, 1985), EVANS et al.
(1992) sowie SASIPREEJAJAN et al. (1987). Prozentual war die Entwicklung der
Legeleistung der geimpften Hennen nach der EBI um 9 % bis 14 % besser als die
der Kontrollgruppen. Diese Zahlen decken sich mit den Angaben von CARPENTER
et al. (1981), GLISSON (1987), MALLISON (1983 a) und MOHAMMED et al. (1987)
über MG-bedingte Legeleistungsminderungen. Ein antiinfektiöser Schutz durch die
Vakzination konnte nicht festgestellt werden. Bei der kulturellen Untersuchung von
Luftröhre, Lunge und Luftsäcken auf MG konnten in der Anzahl der reisolierten
Mykoplasmen keine signifikanten Unterschiede zwischen Impf- und Kontrollgruppen
festgestellt werden. KLEVEN (1985); TALKINGTON und KLEVEN (1985) und
YAGIHASHI (1987, 1992) berichten zwar, dass nach einer Impfung es zu einer
Verminderung der aus der Trachea zu reisolierenden Mykoplasmen kommt bzw. eine

108
größere Anzahl für eine Infektion notwendig ist; ein Schutz vor einer Infektion besteht
jedoch nicht (KLEVEN 1994).
Vor der ersten Vakzination konnten in keiner der untersuchten Blutserumproben
spezifische agglutinierende Antikörper gegen MG nachgewiesen werden. Da auch
die untersuchten Seren der Hennen der Kontrollgruppen bis zum Zeitpunkt der
experimentellen Belastungsinfizierung negativ waren, können die in diesen
Versuchen verwandten Hühner als MG-frei angesehen werden (HINZ 1977 und
1978; KLEVEN 1975). Nach Impfung mit einer MG-Vakzine kommt es bei MG-freien,
immunkompetenten Hühnern binnen drei bis vier Wochen zur Bildung spezifischer
agglutinierender Antikörper, die mittels Serum-Schnellagglutination nachgewiesen
werden können (AVAKIAN und LEY 1993; HINZ 1988; HINZ et al. 1991). In der
vorliegenden Studie konnten nach der zweiten Vakzination in allen in der SSA und im
HAH untersuchten Blutserumproben spezifische Antikörper gegen MG nachgewiesen
werden. Auch im ELISA wurden in allen Impfgruppen, jedoch nicht in allen
untersuchten Seren, spezifische Immunglobuline des Typs IgG gefunden. Auch dies
spricht für die Ausbildung einer protektiven Immunität und somit für einen Erfolg der
Impfung (GLISSON und KLEVEN 1984, 1985; KHAN et al. 1986; LIN und KLEVEN
1982 b; RODRIGUEZ und KLEVEN 1980 a, b). Nach der experimentellen Infizierung
stiegen die Serumtiter der geimpften Tiere deutlich an und lagen bei Versuchsende
über den in den Kontrollgruppen festgestellten Werten. Im 1. Versuch war die
Differenz zwischen der Impf- und Kontrollgruppe sowohl bei denen im HAH als auch
im ELISA ermittelten Werten hochsignifikant; in den Versuchen 2 und 3 zeigten sich
lediglich im ELISA hochsignifikante Unterschiede der Antikörpertiter. Einen ähnlichen
Anstieg der Antikörpertiter nach einer Infektion stellten SOERIPTO et al. (1989 b)
und YODER und HOPKINS (1985) fest. Bei ihnen lag der ermittelte Serumtiter der
Impfgruppe über dem der Kontrollgruppe. Dagegen wiesen die Blutserumproben der
geimpften Hühner in den Versuchen von SASIPREEYAJAN et al. (1987) und
YODER et al. (1984) nach einer EBI keine höheren Antikörpertiter als die der
Kontrolltiere auf.
NAKAMURA et al. (1995) konnten in ihren Untersuchungen grundsätzlich einen
Schutz vor MG nur dann feststellen, wenn sie auch spezifische Antikörper

109
nachweisen konnten. ELFAKI et al. (1992), SOERIPTO et al. (1989 b), YAGIHASHI
et al. (1992) und YODER et al. (1984) fanden eine positive Korrelation zwischen der
Höhe spezifischer Antikörpertiter gegen MG und dem Schutz vor
Luftsackentzündungen nach einer experimentellen Belastungsinfizierung. Dagegen
war eine solche Korrelation in den Versuchen von KARAKA und LAM (1986 a) nicht
festzustellen. ABD-EL-MOTELIB und KLEVEN (1993) sowie WHITHEAR et al.
(1990) konnten bei mit MG-Vakzine geimpften Legehennen ebenfalls keine
Korrelation zwischen der Höhe der ermittelten Antikörpertiter und der Häufigkeit und
Schwere von Luftsackveränderungen feststellen. Auch in der vorliegenden Studie
wiesen die geimpften Hennen höhere Antikörpertiter auf, jedoch waren bei der
pathologisch-anatomischen Untersuchung keine signifikanten Unterschiede im
Ausmaß und Schwere der Luftsackentzündungen zwischen Impf- und
Kontrollgruppen festzustellen. Trotz der maximalen Erregerbelastung war es möglich
einen Impfschutz nachzuweisen, obwohl in den Luftsäcken pathologisch-
anatomische Veränderungen festzustellen waren (HINZ 1988).
Die serologischen Untersuchungen ergaben einen deutliche Unterschied zwischen
der Höhe der spezifischen Antikörpertiter in den Versuche 1 und 2 im Vergleich zum
Versuch 3. Trotz der Aufbewahrung der Seren bei – 70° C könnte ein Grund hierfür
die durch die Dauer der Lagerung bedingte Abnahme der nachweisbaren
Antikörpertiter sein, da die Untersuchungen im HAH und ELISA erst nach Abschluß
des 3. Versuchs durchgeführt wurden (TIMMS 1971). Andererseits ist die Höhe der
Antikörpertiter auch abhängig von dem verwendeten Impfstamm (RODRIGUEZ und
KLEVEN 1980 b). BRADBURY (1975) und BENICINA und DORRER (1989) konnten
nachweisen, dass Hühner, bei denen eine latente Infektion mit M. synoviae vorlag,
nach einer EBI mit MG geringere spezifische Antikörpertiter entwickelten.
Die in den Versuchen 1 und 2 zum Zeitpunkt der Aufstallung in die Klinik bestehende
Infektion mit MS, stellte eine zusätzliche Belastung der Hennen dar. Zwar konnten
nach der Impfung und nach der Infizierung bei allen Hennen spezifische
Antikörpertiter gegen MG nachgewiesen werden, jedoch lagen diese Titer deutlich
unter den Werten, die bei den Hühnern in Versuch 3 ermittelt wurden; diese waren
zum Zeitpunkt der Aufstallung MS-frei, infizierten sich wahrscheinlich während sie

110
zur EBI aus den Isolierställen gebracht wurden mit MS. Bei einer kulturell
nachgewiesenen, latenten Infektion von Hühnern mit MG und MS konnten BENICINA
et al. (1991) zwar hohe spezifische Antikörpertiter gegen MS nachweisen, während
sie in der SSA und im HAH keine spezifischen Antikörper für MG fanden.
Ob die Remontierung einer Legehennenherde durch den Zukauf geimpfter aber MG-
freier Junghennen für den Besitzer einer latent MG infizierten Legehennenherde
(multiple-age) sinnvoll ist, entscheidet im wesentlichen die Wirtschaftlichkeit dieser
Maßnahme. Die Kosten für die Impfung trägt zunächst der Junghennenaufzüchter,
sie bewirken jedoch letztendlich einen höheren Preis des einzelnen Tieres für den
Legehennenhalter. Ausgehend von einer Verbesserung der durchschnittlichen
Legeleistung um 5% innerhalb der Legeperiode - entsprechend einer Anzahl von 12
bis 15 Eiern je Henne - und einem zu erzielenden Verkaufspreis je Ei von ca. 10 bis
15 Pfennig (bei einem hohen Anteil der Eigenvermarktung), ist ein Mehrerlös von bis
zu DM 2,25 je Henne möglich. Hierbei sind stärkere Legeleistungsminderungen
durch Cofaktoren infektiöser und nicht infektiöser Art, die bessere Futterverwertung
sowie die Verminderung der Eischalenqualität, wie sie bei latent MG-infizierten
Hennen zum Ende der Legeperiode auftreten, noch nicht berücksichtigt. Diesem
Mehrerlös entgegen stehen die Kosten des Junghennenaufzüchters, die sich für die
zweimalige Vakzination der Junghennen auf ca. 50 bis 60 Pfennig je Henne
belaufen. Diese Kosten setzen sich aus dem Preis für den Impfstoff sowie für die
Applikation zusammen und sind abhängig von der Anzahl der zu impfenden Tiere
und dem Management des Junghennenaufzüchters (Pöppel 2001).
Die Untersuchungsergebnisse belegen die Wirksamkeit des Impfstoffes „Talovac 104
MG“ gegen eine MG-bedingte Legeleistungsminderungen. Auch aus wirtschaftlichen
Gesichtspunkten ist die Impfung MG-freier Junghennen vor Einstallung in einen
latent mit MG-infizierten Legehennenbetrieb sinnvoll.

111
6. Zusammenfassung
In einer experimentellen Studie wurde die Verträglichkeit und Wirksamkeit der
inaktivierten Öl-Adjuvans-Vakzine „TALOVAC 104 MG“* der Firma Lohmann Animal
Health, 27472 Cuxhaven untersucht. Als Kriterium für die Wirksamkeit des
Impfstoffes wurde die Legeleistung vor und nach einer experimentellen
Belastungsinfizierung mit Mycoplasma gallisepticum herangezogen. In drei
Versuchsdurchgängen wurden drei Chargen des Impfstoffes getestet. Jeder
Versuchsdurchgang umfaßte je eine Impf- und Kontrollgruppe bestehend aus je 40
Hennen. Den Hennen der Impfgruppen wurde zweimal, nämlich in der 14. und 18.
Lebenswoche, 0,5 ml des Impfstoffes subkutan, dorsal im Bereich der Halsbasis,
verabreicht. Bei einer Legeleistung von mindestens 70 % erfolgte die
Belastungsinfizierung mit dem virulenten Mycoplasma gallisepticum-Stamm R über
ein erregerhaltiges Aerosol. Die Legeleistung beider Gruppen wurde vor und nach
der Infizierung dokumentiert. Zur Kontrolle der serologischen Immunreaktionen
wurden Blutproben genommen.
Bei allen geimpften Hennen entstanden lokale entzündlich-granulomatöse
Reaktionen, die bei über 40 % der Hennen über das für Impfstoff dieser Art
tolerierbare Maß hinausgingen. Bei einigen Hennen trat darüber hinaus eine leichte
Störung des Allgemeinbefindens auf, die zu einer Verzögerung des Legebeginns
führte. Die Legeleistung der geimpften Hennen lag nach der Belastungsinfektion
signifikant über der Legeleistung der Hennen der Kontrollgruppen. In zwei
Versuchsdurchgängen war dieser Unterschied hochsignifikant (p≤0,001).
Nach der zweiten Impfung konnten in allen untersuchten Blutserumproben der
geimpften Hennen spezifische agglutinierende und HA-hemmende Antikörper
nachgewiesen werden. Vier Wochen nach der Belastungsinfizierung lagen die im
ELISA ermittelten Serumtiter (GMT) der Impfgruppen hochsignifikant über denen der
Kontrollgruppen (p≤0,001).
*Die Adjuvanszusammensetzung des MG-Impfstoffes ist inzwischen geändert
worden.

112
7. Summary Hanns-Dietrich Graack 2001:
Experimental study on the safety and effectiveness of an inactivated Mycoplasma
gallisepticum (MG) oil-adjuvans-vaccine in layers.
This experimental study was thought to examine safety and effectiveness of the
inactivated oil-adjuvans-vaccine “Talovac 104 MG”* manufactured by Lohmann
Animal Health, Cuxhaven, Germany.
The criterion for the effectiveness was the egg yield before and after an experimental
infection of the population with Mycoplasma gallisepticum. Three lots of the vaccine
were tested in three experiments. Each experiment included one vaccination and one
control group of 40 hens, respectively. Hens of the vaccination group received 0,5 ml
of the vaccine subcutaneously in the caudal aspect of the neck at the 14th and 18th
week of age. When an egg yield of at least 70 % was reached both groups were
exposed to an airborne infection with the virulent R-strain of Mycoplasma
gallisepticum. The egg yields of both groups were recorded before and after the
infection. To control the serological immune response blood samples were taken.
In all vaccinated hen local inflammatory granulomatomatous reactions develop. In
40 % of these hens they were more pronounced tha is tolerated with this kind of
vaccine. Additionally some hens showed a slightly poorer general health, which
resulted in a delayed begin of egg production. After the experimental infection the
egg yield of the vaccinated hens was significantly higher than that of the control
groups. In two lots the difference was highly significant (p<0,001).
After the second vaccination in all examined blood samples of the vaccinated hens
specific agglutinating and haemagglutination-inhibiting antibodies were found. With a
high significance four weeks after the experimental infection the mean serum titers
(GMT) in ELISA of the vaccination groups were higher than in the control groups.
* The composition of the oil-adjuvans has meanwhile be changed.

113
8. Literaturübersicht ABD-EL-MOTELIB, T. Y., u. S. H. KLEVEN (1993): A comparative study of Mycoplasma gallisepticum vaccines in young chickens. Avian Dis. 37, 981 - 987 ADLER, H.E. (1958): A PPLO slide agglutination test for the detection of infectious sinusitis of turkeys. Poult. Sci. 37, 1116 - 1123 ADLER, H. E. u. J. M. L. DA SILVA (1970): Immunization against Mycoplasma gallisepticum. Avian Dis. 14, 763 - 769 ADLER, H. E., D. Mc MARTIN u. M. SHIFRINE (1960): Immunization against mycoplasma infections in poultry. Am. J. Vet. Res. 21, 482 - 485 AVAKIAN, A. P. u. D. H. LEY (1993): Protected immune response to Mycoplasma gallisepticum demonstrated in respiratory-tract washings from M. gallisepticum - infected chickens. Avian Dis. 37, 697 - 705 BARBOUR, E. K., u. J. A. NEWMAN (1990): Preliminary data of efficacy of Mycoplasma gallisepticum vaccines containing different adjuvants in laying hens. Vet. Immunol. Immunopathol. 26, 115 - 123 BARBOUR, E. K., J. A. NEWMAN, V. SIVANANDAN, D. A. HALVORSON u. J. SASIPREEYAJAN (1987): Protection and immunity in commercial chicken layers adminstered Mycoplasma gallisepticum liposomal bacterins. Avian Dis. 31, 723 - 729 BEARD, C. W., u. B. C. EASTERDAY (1965): An aerosol apparatus for the exposure of large and small animals: description and operating characteristics. Am. J. Vet. Res. 26, 174 - 182 BEHR, K., K.-H. HINZ, U. JOCHIMS, M. RYLL, H. LÜDERS u. W. LEHMACHER (1993): Über die Wirksamkeit einer inaktivierten Mycoplasma gallisepticum - Ölemulsionsvakzine bei Legehühnern unter Feldbedingungen. Arch. Geflügelk. 57, (1): 35 - 41

114
BEHR, K., K.-H. HINZ, H. LÜDERS u. M. RYLL (1991): Zur Wirksamkeit einer inaktivierten Mycoplasma-gallisepticum-Vakzine bei Legehennen unter Feldbedingungen. 41. DVG-Fachgespräch über Geflügelkrankheiten, Hannover Verlag DVG e. V., Gießen, DVG-Tagungsbericht 14 - 26 BEHRENS, A., M. HELLER, H. KIRCHHOFF, D. YOGEV u. R. ROSENGARTEN (1994): A family of phase- and size-variant membrane surface lipoprotein antigens (Vsps) of Mycoplasma bovis. Infect. Immun. 62, 5075 - 5084 BENCINA. D. und D. DORRER (1983): Demonstration of Mycoplasma gallisepticum in tracheas of health carrier chickens by flourescent-antibody procedure and the significance of certain serologic tests in estimating antibody response. Avian Dis. 28, 574 - 578 BENCINA. D. und D. DORRER (1989): Local antibody response against Mycoplasma gallisepticum and Mycoplasma synoviae in upper respiratory tract and in hoch joint of naturelly infected chicken. Praxis Veterinaria 37, 181 - 187 BENCINA, D., S. H. KLEVEN, M. G. ELFAKI, A. SNOJ, P. DOVC, D. DORRER und I. RUSS (1994): Variable expression of epitopes on the surface of Mycoplasma gallisepticum demonstrated with monoclonal antibodies. Avian Pathol. 23, 19 - 36 BENCINA, D., A. SYVETLIN, D. DORRER und T. TADINA-JAKSIC (1991): Humoral und local antibodies in chickens with mixed infection with three Mycoplasma species. Avian Pathol. 20, 325 – 334 BRADBURY, J. M. (1975): Concurrent infection of M. synoviae and M. gallisepticum in chickens. Proc. Of the 3rd conference on taxonomy and physiology of animal mycoplasmas Brno 1.-4. Juli 1974 S. 106 - 118 BRANTON, S. L., J. D. MAY, B. D. LOTT und W. R. MASLIN (1997): Various blood parameters in commercial hens acutely and chronically infected with Mycoplasma gallisepticum and Mycoplasma synoviae. Avian Dis. 41, 540 - 547

115
CARPENTER, T. E., E. T. MALLISON, K. F. MILLER, R. F. GENTRY u. L. D. SCHWARTZ (1981): Vaccination with F-strain Mycoplasma gallisepticum to reduce produktion losses in layer chickens. Avian Dis. 25, 404 - 409 CHHABRA, P. C. u. M. C. GOEL (1980): Immunological response of chickens to Mycoplasma gallisepticum infection. Avian Dis. 25, 279 - 293 CULLEN, G. A. and L. M. TIMMS (1972): Diagnosis of mycoplasma infection in poultry previously vaccinated with killed adjuvans vaccines. Br. Vet. J. 128, 94 - 100 DOMERMUTH, C. H. (1957): The response to avian PPLO vaccination I. Laboratory trials. Avian Dis. 1, 346 DROUAL, R., A. A. BICKFORD, B. R. CHARLTON u. D. R. KUNEY (1990): Investigation of problems associated with intramuscular breast injection of oil-adjuvanted killed vaccines in chickens. Avian Dis. 34, 473 - 478 DROUAL, R., A. A. BICKFORD u. G. J. CUTLER (1993): Local reaction and serological response in commercial layer chickens injected intramuscularly in the leg with oil adjuvanted Mycoplasma gallisepticum bacterin. Avian Dis. 37, 1001 - 1008 EDWARD, D. G. f. u. A. D. KANAREK (1960): Organisms of the pleuropneumonia group of avian origin: their classification into species. Ann. N. Y. Acad. Sci. 79, 696 - 702 ELFAKI, M. G., S. H. KLEVEN, L. H. Lin, u. W. L. RAGLAND (1992): Sequential intracoelomic and intrabursal immunization of chickens with inaktivated Mycoplasma gallisepticum bacterin and iota carrageenan adjuvant. Vaccine 10, 655 - 661 ELFAKI, M. G., S. H. KLEVEN, L. H. Lin, u. W. L. RAGLAND (1993): Protection against airsacculitis with sequential systemic and local immunization of chickens using killed mycoplasma bacterin with iota carrageenan adjuvant. Vaccine 11, 311 - 317

116
ELLAKANY, H., KATALIN FABIAN, I. NEMETH u. L. STIPKOVITS (1998): Antibody response detected by immunoblot in respiratory tract washings of chickens after infection with Mycoplasma gallisepticum. Avian Path. 27, 547 - 554 EVANS, R. D., Y .S . HAFEZ u. F. W. ORTHEL (1992 a): Mycoplasma gallisepticum vaccination-challenge: an egg production model. Avian Dis. 36, 956 - 963 EVANS, R. D., Y. S. HAFEZ und C. S. SCHREURS (1992 b): Demonstration of the genetic stability of a Mycoplasma gallisepticum stain following in vivo passage. Avian Dis. 36, 554 - 560 FINGER, Horst (1966): Die Adjuvanswirkung von Mykobakterien. Klin. Wochenschr. 19, 1105 - 1111 FORSYTH, M. H., M. E. TOURTELLOTTE u. S. J. GEARY (1992): Localisation of an immunodominant 64 kDa lipoprotein (LP 64) in the membrane of Mycoplasma gallisepticum and its role in cytadherence. Molekular Microbiology 6, 2099 - 2106 FREY, M. L., R. P. HANSON u. D. P. ANDERSON (1968): A medium for the isolation of avian mycoplasmas. Am. J. Vet. Res. 29, 2163 - 2171 GARCIA, M., M. G. ELFAKI u. S. H. KLEVEN (1994): Analysis of the variability in expression of Mycoplasma gallisepticum surface antigens. Vet. Microbiol. 42, 147 - 158 GAUNSON, J.E., C. J. PHILIP, K. G. WITHHEAR u. G. F. BROWNING (2000): Lymphocytic infiltration in the chicken trachea in response to Mycoplasma gallisepticum infection. Microbiology 146, 1223 – 1229 GIAMBRONE, J. J. (1989): Immunosuppression in poultry Monentum Vol. 1, MSD AG VET GLEW, M. D., G. F. BROWNING, P. F. MARKHAM und I. D. WALKER (2000): PMGA phenotypic variation in Mycoplasma gallisepticum occurs in vivo and is mediated by trinucleotide repeat length variation. Infect. Immun. 68, 6027 - 6033

117
GLISSON, J. R. (1987): Mycoplasmosis control, prevention in laying hens. Proc. 16th Annual Convention, Midwest Poultry Federation, February, 1987. Poult. Dig. 46, 288 - 289 GLISSON, J. R., u. S. H. KLEVEN (1984): Mycoplasma gallisepticum vaccination: effect on transmission and egg produktion. Avian Dis. 28, 406 - 415 GLISSON, J. R., u. S. H. KLEVEN (1985): Mycoplasma gallisepticum vaccination: further studies on egg transmission and egg produktion. Avian Dis. 29, 408 - 415 HAYATSU, E., H. SUGIYAMA, Y. KAWAKUBO, M.KIMURA u. M. YOSHIOKA (1974): Lokal immunizazion in chicken respiratory tract with killed Mycoplasma gallisepticum vaccine. Jap. J. Vet. Sci. 36, 311 - 319 HAYATSU, E., H. SUGIYAMA, K. KUME, Y. KAWAKUBO, M. KIMURA, M. YOSHIOKA, S. KANEKO, K. KOBAYASHI, T. YAMASAKI, u. Y. NISHIYAMA (1975): A field trial, using killed - Mycoplasma gallisepticum vaccine to protect against chicken respiratory mycoplasmosis. Am. J. Vet. Res. 36, (2) 217- 221 HILDEBRAND, D. (1985): Immunology and prophylaxis associated with the use of Mycoplasma gallisepticum bacterin in chickens. Clinica-Veterinaria 108, 89 - 94 HILDEBRAND, D. G., D. E. PAGE, and J. R. BERG (1983): Mycoplasma gallisepticum (MG)-laboratory and field studies evaluating the safety and efficacy of an inaktivated MG bacterin. Avian Dis. 27, 792 - 802 HILL, D. (1995): Mycoplasma field issues, diagnostics. Poultry Digest 32 - 33 HINZ, K. H. (1977): Untersuchungen zur Spezifität des HAH-Tests an experimentell mit Mycoplasma gallisepticum oder Mycoplasma synoviae infizierten Hühnerküken. Dtsch. tierärztl. Wschr. 84, 475 - 479

118
HINZ, K.-H. (1978): Zur serologischen Diagnose der Mycoplasma gallisepticum - und Mycoplasma synoviae -Infektion. Tierärztl. Umsch. 33, 336 – 339 HINZ, K.-H. (1988): Zur aktiven Immunisierung gegen Mycoplasma gallisepticum: Nutzen und Risiko eines Impfstoffeinsatzes. Tagungsbericht der DVG Tagung der Fachgruppe „Geflügelkrankheiten“ in Verbindung mit der WVPA und der Abt. Geflügelkrankheiten der I. Medizinischen Klinik, Veterinär-medizinische Universität Wien HINZ, K.-H. (1993): Kapitel 3. 3 Mykoplasmen. in: O. SIEGMANN (Hrsg.): Kompendium der Geflügelkrankheiten. Verlag Paul Parey, Berlin, Hamburg, 5. Aufl., 233 - 242 HINZ, K.-H., K.-P. BEHR, H. LÜDERS u. M. RYLL (1991): Dynamik der MG-Antikörperentwicklung bei Legehennen nach Impfung mit inaktivierter Mycoplasma gallisepticum (MG)-Vakzine unter Feldbedingungen. 41. DVG-Fachgespräch über Geflügelkrankheiten, Hannover Verlag DVG e. V., Gießen, DVG-Tagungsbericht 27 - 35 HINZ, K.-H., E. F. KALETA, J. M. KÖSTERS, H. LÜDERS, G. MONREAL und O. SIEGMANN (1993): Kapitel 2 Diagnose in: O. SIEGMANN (Hrsg.): Kompendium der Geflügelkrankheiten. Verlag Paul Parey, Berlin, Hamburg, 5. Aufl., 83 - 98 HINZ, K.-H. u. M. RYLL (1996): Mykoplasmen des Wirtschaftsgeflügels in der Diagnostik: Eine bleibende Heraus-forderung. 50. DVG-Fachgespräch über Geflügelkrankheiten, Hannover Verlag DVG e. V., Gießen, DVG-Tagungsbericht 71 - 78 HOLLÄNDER, W. (1995): Persönliche Mitteilungen Fraunhofer Institut für Toxikologie und Aerosolforschung Nikolai-Fuchs-Sraße 1, D-30625 Hannover JANEWAY, C. A. u. P. TRAVERS (1997): Immunologie Spektrum Akademischer Verlag

119
JARNYCH, V. S. (1976): Aerosole. Theorie und Praxis der Aerosolanwendung in der Veterinärnedizin und Tierproduktion. Hrsg. W. KURZWEG, Institut für angewandte Tierhygiene Eberswalde VEB Deutscher Landwirtschaftsverlag Berlin, GDR JOHNSON, D .C. (1983): Role of management and sanitation in controling mycoplasma outbreaks AAAP Mycoplasmosis symposium Avian Dis. 27, 342 - 343 JORDAN, F. u. S. H. KLEVEN (1995): Mycoplasmosis (Mycoplasma gallisepticum) OIE Manual: Amendment 1, April 1995 KALETA, E. F. u. O. SIEGMANN (1993): Kapitel 1.3. Spezielle Seuchenvorbeuge. in O. Siegmann (Hrsg.): Kompendium der Geflügelkrankheiten 5. Aufl. Paul Parey Verlag 1993: 75 - 82 KARAKA, K. u. K. M. LAM (1986 ): Effect of temperature-sensitive Mycoplasma gallisepticum vaccine preparations and routes of inoculation on resistence of white leghorns to challenge. Avian Dis. 30, 772 - 775 KARACA, K. u. K. M. LAM (1987): Efficacy on commercial Mycoplasma gallisepticum bacterin (MG-Bac®)in preventing air-sac lesions in chickens. Avian Dis. 31, 202 - 203 KATZEN, S., K. MATSUDA u. B. L. REID (1969): Amelioration of Marek´s disease by Mycoplasma gallisepticum Poultry Sci. 48, 1502 - 1506 KHAN, M. I., D. A. McMARTIN, R. YAMAMOTO u. H. B. ORTMAYER (1986): Observation on commercial layers vaccinated with Mycoplasma gallisepticum (MG) bacterin on a multiple-age site endemically infected with MG. Avian Dis. 30, 309 - 312 KLEVEN, S. H. (1975): Antibody response to avian mycoplasma. Am. J. vet. Res. 36, 563 - 565 KLEVEN, S. H. (1985): Tracheal populations of Mycoplasma gallisepticum after challenge of bacterin - vaccinated chickens. Avian Dis. 29, 1012 - 1017

120
KLEVEN, S. H. (1994): Summary of discussions of Avian mycoplasma team, international research program on comparative mycoplasmology, international organization of mycoplasmology, University of Ljubljana, Domzale, Slovenia September 29 - Oktober 1, 1993 Avian Pathol. 23, 587 - 594 KLEVEN, S. H., C. S. EDISON, D. P. ANDERSON u. O. J. FLETCHER (1972): Decrease of antibody response to Mycoplasma synoviae infected with Marek´s disease herpes-virus. Am. J. Vet. Res. 33, 2037 KLEVEN, S. H., C. J. MORROW und K. G. WHITHEAR (1988): Comparsion of Mycoplasma gallisepticum strains by hemagglutination-inhibition and restriction endonuclease analysis. Avian Dis. 32, 731 - 741 LAM, K. M., K. KARACA u. A. A. BICKFORD (1986): Response of chickens to inoculation with a temperature-sensitive mutant of Mycoplasma gallisepticum. Avian Dis. 30, 382 - 388 LAM, K.M. u. W. LIN (1984): Resistance of chickens immunized against Mycoplasma gallisepticum is mediated by bursal dependent lymphoid cells. Vet. Microbiol. 9, 509 - 514 LANCASTER, J. E. u. J. FABRICANT (1988): The history of avian medicine in the United States. IX. Events in the History of avian mycoplasmosis 1905-70. Avian Dis. 32, 607 - 623 LEVISOHN, S., M. J. DYKSTRA, M. Y. LIN u. S. H. KLEVEN (1986): Comparsion of in vivo and in vitro methods for pathogenicity evaluation for Mycoplasma gallisepticum in respiratory infection. Avian Pathol. 15, 233 – 246 LEVISOHN, S. u. S. H. KLEVEN (1981): Vaccination of chickens with nonpathogenic Mycoplasma gallisepticum as a means for displacement of pathogenic strains. Israel J. Med. Sci. 17, 669 – 673 LEVISOHN, S. u. S. H. KLEVEN (2000): Avian mycoplasmosis (Mycoplasma gallisepticum). Rev. sci. tech. Off. int. Epiz. 19, 425 - 442

121
LEVISOHN, S., R. ROSENGARTEN und D. YOGEV (1995): In vivo variation of Mycoplasma gallisepticum antigen expression in experimentally infected chickens. Vet. Mikrobiol. 45, 219 - 231 LIN, M. Y. u. S. H. KLEVEN (1982 a): Egg Transmission of two strains of Mycoplasma gallisepticum in chickens. Avian Dis. 26, 487- 495 LIN, M. Y. u. S. H. KLEVEN (1982 b): Cross-immunity and antigenetic relationships among five strains of Mycoplasma gallisepticum in young leghorn chickens. Avian Dis. 26, 496 - 507 LIN, M. Y., u. S. H. KLEVEN (1984 a): Transferred humural immunity in chickens to Mycoplasma gallisepticum. Avian Dis. 28, 79 - 87 LIN, M.Y. u. S. H. KLEVEN (1984 b): Evaluation of attenuated strains of Mycoplasma gallisepticum as vaccines in young chickens. Avian Dis. 28, 88 - 99 MALLISON, E. T. (1983 a): Live vaccine and eradication of Mycoplasma gallisepticum from multiple-age laying flocks. AAAP Mycoplasmosis Symposium Avian Dis. 27, 335 - 336 MALLISON, E. T. (1983 b): Planning and testing for mycoplasma eradication. Proc. 32th. Western Poultry Dis. Conf. and 17th. Poult. Hlth. Symp.: 29 - 31 MALLISON, E. T. u. T. E. CARPENTER (1981): The economics of MG control in laying flocks. Poult. Dig. 40, 365 - 368 MARKHAM, F. S. u. S. C. WONG (1952): Pleuropneumonia-like organisms in the etiologie of Turkey sinusitis and chronic respiratory disease of chickens. Poult. Sci. 31, 902 – 904 MARKHAM, P. F. , M. D. GLEW, K. G. WHITHEAR u. I. D. WALKER (1993): Molecular clonig of a member of the gene family that encodes pMGA, a hemagglutinin of Mycoplasma gallisepticum. Infect. Immun. 61, 903 - 909

122
McMARTIN, D. A. u. H. E. ADLER (1961): An immunologial phenomenon in chickens following infection with Mycoplasma gallisepticum. J. Comp. Path. 71, 311 - 324 MOHAMMED, H. O., T. E. CARPENTER, u. R. YAMAMOTO (1987): Economic impact of Mycoplasma gallisepticum and M. synoviae in commercial layer flocks Avian Dis. 31, 477 - 482 NAKAMURA, T., S. HOSHI, NAGASAWA u. S. UEDA (1995): The effect of route of inoculation on protection by killed vaccines in chickens. Avian Dis. 39, 507 - 513 NELSON, J. B. (1933): Studies on an uncomplicatedcoryza of domestic fowl. II. The relation of the „bacillary“ coryza to that produced by exsudate. J. Exptl. Med. 58, 297 - 304 NELSON, J. B. (1935): Cocco-bacilliform bodies associated with an infectious fowl coryza. Science 82, 43 - 44 NELSON, J. B. (1938): Studies on an uncomplicatedcoryza of domestic fowl. IX. The cooperative action of Hemophilus gallinarum and the coccobacilliform bodies in the coryza of rapid onset and long duration. J. Exptl. Med. 64, 847 - 855 NELSON, J. B. (1939): Growth of the fowl coryza bodies in tissue culture and blood agar. J. Expt. Med. 69, 199 – 209 ODENTHAL, L. (1996): Zur Eignung der Mycoplasma gallisepticum-Mutante TS 100 als Lebendimpfstoff bei Hühnern. Hannover, Tierärztl. Hochsch.; Diss. ORTIZ, A., R. FROYMAN u. S. H. KLEVEN (1995): Evaluation of enrofloxacin against egg transmission of Mycoplasma gallisepticum. Avian Dis. 39, 830 - 836 PANIGRAHY, B., L. C. GRUMBLES u. C. F. Hall (1981): Immunogenic potency of oil-emulsified Mycoplasma gallisepticum bacterins. Avian Dis. 35, 821 - 826

123
PANANGALA, V. S. M. A. MORSY, M. M. GRESHAM u. M. TOIVIO-KINNUCAN (1993): Antigen variation of Mycoplasma gallisepticum, as detected by use of monoclonal antibodies. Am. J. vet. Res. 53, 1139 – 1144 PÖPPEL, M. (2001): Persönliche Mitteilungen Drubbelstr. 2 33129 Delbrück REDMANN, T. u. T. GÖBEL (1991): Einfluß von Impfungen gegen Mycoplasma gallisepticum auf die Legeleistung von Legehennen. 41. DVG-Fachgespräch über Geflügelkrankheiten, Hannover Verlag DVG e. V., Gießen, DVG-Tagungsbericht 3 -13 REFAI, M., A. El-EBEEDY, A. G. HEGAZI, G. O. EDRIS, AIDA, M. ABDEL AZIZ u. NAWAL ALLAM (1990): Evaluation of the efficiency of inactivated Mycoplasma gallisepticum (formalin killed adjuvant) vaccine. Veterinary-Medical-Journal-Giza 38, 1, 19 - 27 RIMMLER, R. B., R. B. DAVIS, R. K. PAGE u. S. H. KLEVEN (1978): Infectious coryza preventing complicated coryza with Haemophilus gallinarum and Mycoplasma gallisepticum bacterin. Avian Dis. 22, 140 - 150 ROBERTS, D. H. (1969): Serological response produced in chickens by three strain of Mycoplasma gallisepticum. J. appl. Bact. 32, 395 - 401 ROBERTS, D. H., O. M: OLESIUK u. H. VAN ROECKEL (1967): Immunologic response of fowl to Mycoplasma gallisepticum and its relationship to latent infection. Am. J. Vet. Res. 28, 1135 - 1152 RODRIGUEZ, R., u. S. H. KLEVEN (1980 a): Pathogenicity of two strains of Mycoplasma gallisepticum in broilers Avian Dis. 24, 800 - 807 RODRIGUEZ, R., u. S. H. KLEVEN (1980 b) : Evaluation of a vaccine against Mycoplasma gallisepticum. Avian Dis. 24, 879 - 889

124
ROSENGARTEN, R., A. BEHRENS, A. STETEFELD, M. HELLER, M. AHRENS, K. SACHSE, D. YOGEV u. H. KIRCHHOFF (1994): Antigenetic heterogeneity among isolatesof Mycoplasma bovis is generated by high-frequent variation of diverse membrane surface proteins. Infect. Immun. 62, 5066 – 5074 ROSENGARTEN, R. u. K. S. WISE (1990): Phenotypic switching in mycoplasmas: Phase variation of diverse lipoproteins. Science 247, 315 - 318 ROSENGARTEN, R. u. K. M. WISE (1991): The Vlp system of Mycopasma hyorhinis: Combinatorial expresion of distinct size variant lipoproteins generating high-frequency surface antigenetic variation. J. bacteriol. 173, 4782 - 4793 SALISCH, H., K.-H. HINZ, H.-D. GRAACK und M. RYLL (1998): A comparsion of a commercial PCR-based test to culture methods for detection of Mycoplasma gallisepticum and Mycoplasma synoviae in concurrently infected chickens. Avian Pathol. 27, 142 - 147 SASIPREEYAJAN, J., D. A. HALVORSON u. J. A. NEWMAN (1985): Bacterin to control the vertical transmission of Mycoplasma gallisepticum in chickens. Avian Dis. 29, 1256 - 1259 SASIPREEYAJAN, J., D. A. HALVORSON u. J. A. NEWMAN (1987): Effect of Mycoplasma gallisepticum bacterin on egg-transmission and egg produktion. Avian Dis. 31, 776 - 781 SNELL, G. C., u. G. A. CULLEN (1977): The detection of maternal antibodies to Mycoplasma gallisepticum in chickens by rapid serum agglutination and hemagglutination inhibition test. Avian Pathol. 6, 181 - 185 SOERIPTO, K. G. WHITHEAR, G. S. COTTEW u. K. E. HARRIGAN (1989 a): Virulence and transmissibility of Mycoplasma gallisepticum. Austr. vet. J. 66, 3, 65 - 72 SOERIPTO, K. G. WHITHEAR, G. S. COTTEW u. K. E. HARRIGAN (1989 b): Immunogenicity of Mycoplasma gallisepticum. Austr. vet. J. 66, 3, 73-77 STIPKOVITS, L. (1985): Mycoplasmosis in poultry. Monatsh. Veterinärmed. 40, 165 - 170

125
STONE, H. D., M. BRUGH, S. R. HOPKINS, H. W. YODER u. C. W. BEARD (1978): Preparation of inactivated oil-emulsion vaccines with an avian viral or Mycoplasma antigens. Avian Dis. 22, 666 - 674 TAKLINGTON, F. D. u. S. H. KLEVEN (1985): Evaluation of protection against colonization of the chicken following administration of Mycoplasma gallisepticum bacterin. Avian Dis. 29, 998 - 1003 TIMMS, L. M. (1971): Effect of storage on the diagnosis of avian mycoplasmosis. Vet. Rec. 88, 698 VAN HERRICK, W. u. M. D. EATON (1945): An unidentified pleuropneumonia-like organism isolated during passages in chick embryos. J. bacteriol. 50, 47 - 55 VARDAMAN, T. H., K. LANDRETH, S. WHATELY, L. J. DREESEN und B. GLICK (1973): Resistance to Mycoplasma synoviae is bursal dependant. Infect. Immun. 8, 674 - 676 VARDAMAN, T. H. u. H. W. YODER (1970): Mycoplasma synoviae und Mycoplasma gallisepticum infections: differentiation by the hemagglutination-inhibition test. Poultr. Sci. 49, 157 - 161 VARDAMAN, T. H. u. H. W. YODER (1971): Determination of nonspecific serological reactions to avian mycoplasma antigens. Poultr. Sci. 50, 183 - 186 WARREN, J., B. SENTERFIT u. F. SIEIRO (1968): Inactivated culture vaccine against Mycoplasma gallisepticum infections in chickens. Am. J. vet. Res. 29, 1659 - 1664 WATSON, H. L., L. S. Mc Daniel, D. K. BLALOCK, M. T. FALLON und G. H. CASSELL (1988): Heterogenity among strains and a high rate of variation within strains of a major surface antigen of Mycoplasma pulmonis. Infect. Immun. 56, 1358 - 1363 WHITHEAR, K. G., K. E. HARRIGAN und S. H. KLEVEN (1996): Standardized method of aerosol challenge for testing the efficacy of Mycoplasma gallisepticum vaccines. Avian Dis. Vol. 40, 654 - 660

126
WHITHEAR, K. G., SOERIPTO, K. E. HARRIGAN u. E. CHIOCAS (1990): Safety of temperature sensitive mutant Mycoplasma gallisepticum vaccine. Aust. Vet. J. 67, 159 – 165 WINNER, F., R. ROSENGARTEN u. C. CITTI (2000): In vitro cell invasion of Mycoplasma gallisepticum. Infect. Immun. 68, 4238 - 4244 YAGIHASHI, T., T. NUNOYA, S. SANNAI und M. TAJIMA (1992): Comparison of imunity induced with a Mycoplasma gallisepticum bacterin between high and low-responder lines of chickens Avian Dis. Vol. 36,125-133 YAGIHASHI , T., T. NUNOYA und M. TAJIMA (1987): Immunity induced with an aluminium hydroxide-adsorbed Mycoplasma gallisepticum bacterin in chickens Avian Dis. Vol. 31, 149-155 YAGIHASHI, T. und M. TAJIMA (1986): Antibody responses in sera and respiratory secretons from chickens infected with Mycoplasma gallisepticum. Avian Dis. 30, 543 - 550 YODER, H. W., Jr. (1979): Serologic response of chickens vaccinated with inactivated preparations of Mycoplasma gallisepticum. Avian Dis. 23, 493 - 503 YODER, H. W., Jr. (1983): Laboratory studies with inactivated oil-emulsion Mycoplasma gallisepticum vaccines. AAAP Mycoplasmosis Symposium Avian Dis. 27, 339 - 340 YODER, H. W. (1989): Nonspecific reactions to mycoplasma serum plate antigens induced by inactivated poultry disease vaccines. Avian Dis. 33, 60 - 68 YODER, H. W. u. S. HOPKINS (1985): Efficacy of experimental inactivated Mycoplasma gallisepticum oil-emulsion bacterin in egg-layer-chickens. Avian Dis. 29, 322 - 334 YODER, H. W., S. R. HOPKINS u. B. W. MITCHELL (1984): Evaluation of inactivated Mycoplasma gallisepticum oil-emulsion bacterins for protection against airsacculitis in broilers. Avian Dis. 28, 224 – 234

127
YOGEV, D., D. MENAKER, K. STRUNZBERG, S. LEVISOHN, H. KIRCHHOFF, K.-H. HINZ u. R. ROSENGARTEN (1994): A surface epitope undergoing high-fequency phase variation is shared by Mycoplasma gallisepticum and Mycoplasma bovis. Infect. Immun. 62, 4962 - 4968 YOGEV, D., S. LEVISOHN, S. H. KLEVEN, D. HALACHIMI und S. RAZIN (1988): Ribosomal RNA gene probes to detect intraspecies heterogenity in Mycoplasma gallisepticum & M. synoviae. Avian Dis. 32, 220 – 231 YOSHIDA, S., A. FUJISAWA, Y. TSUZAKI u. S. SAITOH (2000) : Identification and expression of a Mycoplasma gallisepticum surface antigen recognized by a monoclonal antibody capble of inhibiting both browth and metabolism. Infect. Immun. 68, 3186 - 3192 ZOLLI, Z. (1984): Mycoplama gallisepticum - field evaluation on the use and efficacy of a MG bacterin. Proc. 33th. Western Poultry Conf. and 18th Poult. Hlth. Symp.: 49 - 51

128
9. Anhang 9.1. verwendete Nährmedien und Lösungen Flüssigmedium nach FREY et. al. (1968) Mycoplasma Broth Base 22, 3 g Vitamins 100 x for Basal Medium Eagle (Modified) Flow 0,25 ml Glukose (Merck) 3,0 g Penicillin-G Natriumsalz (Sigma) 1.000.000 I. E. Thallium (I)-acetat (RiedelDe-Haen) 200 mg Phenolrot 1 % Lsg. (Merck) 2,7 ml Schweineserum 100 ml Ampuwa ad 1000 ml für das Festmedium zusätzlich: Noble-Agar (Difco) 15 g es entfällt der Zusatz von Phenolrot Mycoplasma Broth Base : Pepton 140 (Gibco) 7,5 g Pepton 110 (Gibco) 2,5 g Yeast Extrakt (Gibco) 5,0 g NaCl (Merck) 5,0 g KCl (Merck) 0,4 g Magnesiumsulfat MgSO4 x 7 H2O (Merck) 0,2 g Dinatriumhydrogenphosphat Na2HPO4 (Riedel-De-Haen) 1,6 g Kaliumdihydrogenphosphat KH2PO4 (Merck) 0,1 g Alsever´s Lösung; ph-Wert: 6,1 Glukose (Merck) 20,5 g Natriumzitrat (Merck) 8,0 g Zitronensäure (Merck) 0,55 g Natriumchlorid (Merck) 4,2 g Aqua bidest. ad 1000 ml

129
0,01 M Phosphat-0,15 M NaCl-Puffer-Lösung Ausgangslösung 1: NaH2PO4 x H2O 1,38 g NaCl 8,77 g Aqua bidest. ad 1000 ml Ausgangslösung 2: Na2HPO4 1,42 g NaCl 8,77 g Aqua bidest. ad 1000 ml 0,01 M Phosphat-0,15 M NaCl-Puffer Ausgangslösung 1: 296 ml Ausgangslösung 2: 704 ml

Danksagungen
Die Erstellung dieser Arbeit und die Durchführung der Versuche wäre ohne die
Unterstützung und Hilfe anderer schwer möglich gewesen. Ich bedanke mich bei
allen, die mir geholfen haben.
Ich danke ganz besonders Herrn Prof. Dr. K.-H. Hinz für die freundliche Überlassung
des Themas und seine jederzeit gewährte geduldige Unterstützung und menschliche
und fachliche Hilfestellung bei der Anfertigung dieser Arbeit.
Ich danke der Firma Lohmann Annimal Health, Cuxhaven, für die finanzielle
Unterstützung des Projektes.
Ich danke allen Mitarbeitern der Klinik für Geflügel der Tierärztlichen Hochschule
Hannover, die mir bei der Durchführung des praktischen Teils der Arbeit
unterstützten, besonders möchte ich mich bei Frau Rita Leise und bei Frau Sonja
Bertram bedanken.
Ich danke Herrn Dr. Werner Holländer, Fraunhofer Institut für Toxikologie und
Aerosolforschung, Hannover, für die Informationen über die Konstruktion und Betrieb
der Infektionskammer.
Ich danke Herrn Dr. Heimberg und Herrn Dr. Rohn für die Hilfe bei der statistischen
Auswertung der Ergebnisse.
Bei meiner Familie, bei Frau Dr. Annette Gabriel und Frau Dr. Christine Lendl möchte
ich mich für die jederzeit gewährte Hilfe ganz besonders bedanken.