B1 10 Arsentransfer Abschlussbericht · LABO-Vorhaben B 1.10 31.5.2012 2 1 EINLEITUNG ahu AG Von...
101
Abschlussbericht Vorhaben B 1.10 „Arsentransfer aus belasteten Böden in Nahrungs- und Futterpflanzen“ des Länderfinanzierungsprogramms Wasser, Boden, Abfall, Teil Boden der Bund/Länder-Arbeitsgemeinschaft Bodenschutz (LABO) Datum: 31. Mai 2012 Auftraggeber: Ministerium für Landwirtschaft, Umwelt und Verbraucherschutz Mecklenburg-Vorpommern Auftrag vom: 20. August 2009 Ansprechpartner: Dr. Ingo Müller (Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie) Marie Heusler (Ministerium für Landwirtschaft, Umwelt und Verbraucherschutz Mecklenburg-Vorpommern) Auftragnehmer: ahu AG Wasser · Boden · Geomatik, Aachen Technische Universität Dresden Projektbearbeitung: Dipl.-Geogr. Carolin Kaufmann-Boll, Dr. Silke Höke Dr. Silvia Lazar (ahu AG) Dr. Carsten Brackhage, Prof. Dr. E. Gert Dudel (TU Dresden) Aktenzeichen: 10089 / LABOAS Ausfertigung Nr.: Abschlussbericht ahu AG Wasser · Boden · Geomatik, Vorstand: Dr. H.-Georg Meiners Kirberichshofer Weg 6, 52066 Aachen, Telefon: +49 241 900011-0, Fax: +49 241 900011-9 TU Dresden, Institut für Allgemeine Ökologie und Umweltschutz, Prof. Dr. E. Gert Dudel Pienner Straße 7, 01735 Tharandt, Telefon: +49 35203 3831390, Fax: +49 35203 3831399
Transcript of B1 10 Arsentransfer Abschlussbericht · LABO-Vorhaben B 1.10 31.5.2012 2 1 EINLEITUNG ahu AG Von...

Abschlussbericht
Vorhaben B 1.10 „Arsentransfer aus belasteten Böden in Nahrungs- und Futterpflanzen“ des Länderfinanzierungsprogramms Wasser, Boden, Abfall, Teil Boden der Bund/Länder-Arbeitsgemeinschaft Bodenschutz (LABO)
Datum: 31. Mai 2012
Auftraggeber: Ministerium für Landwirtschaft, Umwelt und Verbraucherschutz Mecklenburg-Vorpommern
Auftrag vom: 20. August 2009
Ansprechpartner: Dr. Ingo Müller (Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie) Marie Heusler (Ministerium für Landwirtschaft, Umwelt und Verbraucherschutz Mecklenburg-Vorpommern)
Auftragnehmer: ahu AG Wasser · Boden · Geomatik, Aachen Technische Universität Dresden
Projektbearbeitung: Dipl.-Geogr. Carolin Kaufmann-Boll, Dr. Silke Höke Dr. Silvia Lazar (ahu AG) Dr. Carsten Brackhage, Prof. Dr. E. Gert Dudel (TU Dresden)
Aktenzeichen: 10089 / LABOAS
Ausfertigung Nr.: Abschlussbericht
ahu AG Wasser · Boden · Geomatik, Vorstand: Dr. H.-Georg Meiners
Kirberichshofer Weg 6, 52066 Aachen, Telefon: +49 241 900011-0, Fax: +49 241 900011-9
TU Dresden, Institut für Allgemeine Ökologie und Umweltschutz, Prof. Dr. E. Gert Dudel
Pienner Straße 7, 01735 Tharandt, Telefon: +49 35203 3831390, Fax: +49 35203 3831399

LABO-Vorhaben B 1.10 31.5.2012
1
INHALT
1 Einleitung 2
2 Wissenschaftlicher Stand zum Arsentransfer Boden – Pflanze 3
2.1 Einfluss der P-Verfügbarkeit 4
2.2 Akkumulation und Speziierung von Arsen 4
2.3 Aufnahmemechanismen für Arsen 7
2.4 Toxizität und Toleranzmechanismen 10
2.5 Bedeutung von Mykorrhiza-Pilzen 12
2.6 Schlussfolgerungen 13
3 Erfahrungen der Vollzugspraxis 15
3.1 Bezugsrahmen und -literatur 15 3.2 Vorliegende Erfahrungen mit Arsen in Boden und Nutzpflanze 15
3.3 Defizite 25 3.4 Schlussfolgerungen 27
4 Aussagekraft und Aufwand von Arsenspezies-untersuchungen 28
5 Konzeption und Ergebnisse der Gefäß- und Feldversuche 33
5.1 Gefäßversuche – Material und Methoden 33 5.1.1 Versuchsdesign 34
5.1.2 Substrat – Beschaffung, Vorbereitung und Charakterisierung 36
5.1.3 Substrat – Messparameter und Aufbereitung 39
5.1.4 Pflanzen - Pflanzung und Kulturbedingungen 40
5.1.5 Pflanzen - Messparameter und Aufarbeitung 42
5.1.6 Statistische Auswertung 43
5.2 Gefäßversuche – Ergebnisse Grünlandnutzung 45 5.2.1 Veränderung pH-Wert und Elementgehalte im Substrat 46
5.2.2 Veränderung der Bodenlösung 48
5.2.3 Einfluss der Versuchsvarianten auf Biomasse und Sproß/Wurzel-Verhältnis 52
5.2.4 Einfluss der Versuchsvarianten auf den Elementgehalt der Pflanzen 54
5.2.5 Transfer und Verteilung in der Pflanze 60
5.3 Gefäßversuche – Ergebnisse Ackernutzung 62 5.3.1 Veränderungen pH-Wert und Elementgehalt im Substrat 62
5.3.2 Veränderungen der Bodenlösung 65
5.3.3 Einfluss der Versuchsvarianten auf Biomasse und Sproß/Wurzel-Verhältnis 72
5.3.4 Einfluss der Versuchsvarianten auf den Elementgehalt der Pflanzen 76
5.3.5 Transfer und Verteilung in der Pflanze 84
5.4 Feldversuche 88
5.5 Feldversuche – Ergebnisse Bodenanalyse 89 5.6 Feldversuche – Ergebnisse Pflanzenanalysen und Transfer 91
6 Fazit 93
7 Literatur 95
8 Abkürzungsverzeichnis 100

LABO-Vorhaben B 1.10 31.5.2012
2
1 EINLEITUNG
ahu AG
Von Juli 2010 bis Mai 2012 wurde das Vorhaben B 1.10 „Arsentransfer aus belasteten Böden in Nahrungs- und Futterpflanzen“ bearbeitet. Die Förderung erfolgte im Rahmen des Länderfinanzierungsprogramms Wasser, Boden, Abfall, Teil Boden der Bund/Länder-Arbeitsgemeinschaft Bodenschutz (LABO). Ausgangspunkt für die Bearbeitung waren die Ergebnisse des LABO-Vorhabens B 4.07 „Ermittlung und Beurteilung des Mobilisierungspotenzials von Arsen in Böden“.
Ziel des Vorhabens war es, weiterführende praxisgerechte Hinweise, z.B. Bewirtschaftungsempfehlungen, für die Beurteilung eines möglichen Übergangs von Arsen vom Boden in Nahrungs- und Futterpflanzen zu geben. Soweit hinreichend konkrete Gefährdungen identifiziert werden, sollten diese mit praktischen Lösungsansätzen minimiert werden. Das wesentliche Endprodukt des Vorhabens ist ein vollzugs- und anwendungsorientierter Leitfaden, der auf dem aktuellen wissenschaftlichen Kenntnisstand sowie auf vorhandenen Erfahrungen beim Umgang mit Bodenbelastungen durch Arsen in Bund und Ländern aufbaut. Darüber hinaus wurden vertiefende, Gefäß- und Feldversuche konzipiert und durchgeführt, um die vorhandenen Erkenntnisse weiter zu erhärten. Die Ergebnisse der Versuche werden im vorliegenden Bericht ausführlich beschrieben und sind in die Kenaussagen des Leitfadens eingeflossen.
Auf Anregung des unterstützenden und projektbegleitenden Arbeitskreises wurden in den Leitfaden Musterempfehlungen für Landwirte integriert, die von den Unteren Bodenschutzbehörden und Landwirtschaftsbehörden als Vorlage für eine lokale Empfehlungsbroschüre verwendet werden können.
Der vorliegende Abschlussbericht dient zur Ergebnisdokumentation und ergänzt den erarbeiteten Leitfaden Arsentransfer aus Böden in Nahrungs- und Futterpflanzen – Gefahrenbeurteilung und Maßnahmen. Er enthält die Ergebnisse der Literaturrecherche, die vorliegenden Erfahrungen aus der Vollzugspraxis der Länder, eine Gegenüberstellung des Aufwands und des Nutzens von Arsenspeziesuntersuchungen sowie das Konzept zur Durchführung der Gefäß- und Feldversuche und ihre Ergebnisse.

LABO-Vorhaben B 1.10 31.5.2012
3
2 WISSENSCHAFTLICHER STAND ZUM ARSENTRANSFER BODEN – PFLANZE
TU Dresden
Die wissenschaftlichen Untersuchungen widmen sich in der Regel den mechanistischen Erklärungen der As-Aufnahme und -Akkumulation in Pflanzen sowie den entsprechenden Toleranzmechanismen und der Toxizität. Die für die Einschätzung des Transfers im Nahrungsnetz so wichtige Quantifizierung steht fast immer im Hintergrund. Daher sind aus diesen Studien zwar die Einflussgrößen für einen Arsentransfer zu ermitteln, oft aber nicht die Größenordnung, mit der diese wirksam werden. Dies hängt selbstverständlich auch damit zusammen, dass das komplexe Zusammenwirken der vielen Faktoren in Abhängigkeit der spezifischen Standortverhältnisse keine generalisierenden Aussagen zulässt.
Die meisten wissenschaftlichen Untersuchungen zu Arsen in Pflanzen gibt es über Reis (Oryza sp.). Als großflächig angebaute Nahrungspflanze in Gebieten mit erhöhter Arsenbelastung steht er im Fokus der weltweiten Sorge um Gefahren durch erhöhten Arsentransfer im Nahrungsnetz. Dagegen sind für weitere Getreidearten und Futtergräser nur wenige Untersuchungen bekannt.
Arsen liegt in der pflanzlichen Umwelt (Boden, Atmosphäre/Staub, Wasser) in verschiedenen Spezies vor. Die häufigsten sind Arsenat(III) (auch Arsenit) und Arsenat(V). Entsprechend der Speziierung z. B. im Boden bzw. in der Bodenlösung wird Arsen durch die Pflanzen unterschiedlich aufgenommen. Dies wirkt sich auf das Akkumulationsverhalten aus, zeigt sich in unterschiedlichen Aufnahmemechanismen für unterschiedliche Arsenspezies und steht damit auch unter dem Einfluss unterschiedlicher Boden- bzw. Umgebungsparameter. Bodenparameter, die von den meisten Untersuchungen als wesentlich eingestuft werden, sind:
� Redoxpotenzial,
� pH-Wert,
� P-Gehalt (pflanzenverfügbarer P, damit P-Düngung),
� Verfügbare andere Nährstoffe (wichtig für die Stöchiometrie in den Pflanzen),
� Menge an organischem Kohlenstoff im Boden.
Weitere direkte oder indirekte Einflussgrößen resultieren daraus (u. a. Arsenspeziierung) bzw. aus der gegenseitigen Beeinflussung von Boden und Pflanze. Im Folgenden wird zusammenfassend auf die vielen empirischen Studien zum Einfluss der P-Verfügbarkeit auf das Aufnahme- und Akkumulationsverhalten von Arsen durch Pflanzen eingegangen, bevor weitere Aspekte des Arsentransfers vom Boden in die Pflanzen behandelt werden.

LABO-Vorhaben B 1.10 31.5.2012
4
Wissenschaftliche Erkenntnisse zur Freisetzbarkeit von As aus dem Boden unter verschiedenen Standortbedingungen – insbesondere des Redox-potenzials – wurden bereits im Vorgänger-Projekt der LABO B 4.07 „Ermitt-lung und Beurteilung des Mobilisierungspotenzials“ aufgearbeitet (siehe Overesch et al. 2008). Rinklebe & Du Laing 2011 zeigen ein Modell, wie unter Einbezug von biotischen und abiotischen Redoxvorgängen sowie von Eisen, die Mobilität, Dynamik, Bioverfügbarkeit, Toxizität, letztlich also das Vorkommen und der Verbleib von Arsen in der Umwelt gesteuert werden. Diese biogeochemischen Prozesse führen zur Bildung oder Zerstörung arsenhaltiger Trägerphasen und verändern die Spezies und Bindungsformen des Arsens. Praxisrelevante Aspekte hierzu wurden in den Leitfaden Arsentransfer aus Böden in Nahrungs- und Futterpflanzen – Gefahrenbeurteilung und Maßnahmen eingearbeitet.
2.1 Einfluss der P-Verfügbarkeit
Die Abhängigkeit der Arsenaufnahme und Translokation/Akkumulation von der P-Verfügbarkeit im Substrat/in der Nährlösung wird in vielen Studien beschrieben (z. B. Meharg et al. 1994a; Meharg & Macnair 1994; Meharg et al. 1994b; Geng et al. 2006; Zhao et al. 2010). In den meisten Fällen wurde die P-Verfügbarkeit nicht im Einzelnen nachgewiesen, sondern nur eine P-Düngung vollzogen. Das Ergebnis bleibt trotzdem in fast allen Fällen das Gleiche: Erhöhte P-Verfügbarkeit bzw. stärkere P-Düngung führt zu einer verminderten Aufnahme und Akkumulation von Arsen in der Pflanze (kompetitive Hemmung). Die mechanistischen Erklärungen dafür werden in den nächsten Kapiteln beschrieben. In welchem Umfang beispielsweise eine P-Düngung bei landwirtschaftlichen Nutzpflanzen zur Reduzierung der Arsenaufnahme führt, wurde von Serfling & Klose 2008 getestet. Sie konn-ten eine um 41 % geringere Arsenkonzentration in der Gesamtpflanze von Sommergerste bei Vergleich zwischen P-gedüngten und nicht gedüngten Varianten feststellen. Allerdings waren für die As-Konzentrationen im Korn keine Unterschiede feststellbar. Eine extrem weit über den Pflanzenbedarf hinaus gehende „Sanierungsdüngung“ mit Phosphat scheint hingegen einen gegenteiligen Effekt auszulösen: Marschner et al. (2010) berichten in diesem Zusammenhang über eine deutliche Steigerung der As-Mobilität im Boden und einer erhöhten Aufnahme in Gemüsepflanzen, insb. Kopfsalat nach Einbringen großer Mengen an Triplesuperphosphat.
2.2 Akkumulation und Speziierung von Arsen
Artspezifische Unterschiede
Verschiedene Pflanzenarten zeigen ein sehr unterschiedliches Aufnahme- und Akkumulationsverhalten bezüglich Arsen. Auf nicht kontaminierten Böden liegen die Pflanzengehalte in oberirdischen Kompartimenten zumeist unter 1 mg kg-1 Trockengewicht (TG) (Cullen & Reimer 1989; Kabata-Pendias & Pendias 1992). Untersucht man Boden und oberirdische

LABO-Vorhaben B 1.10 31.5.2012
5
Pflanzenkompartimente am gleichen Standort, werden normalerweise Transferfaktoren (TF) von < 0,1 gefunden. Viele Pflanzen einschließlich Nahrungs- und Futterpflanzen vermeiden daher offensichtlich die Aufnahme. Wichtiger aber noch ist die in der Regel geringe Translokation von den Wurzeln in oberirdische Kompartimente. Auf der anderen Seite gibt es als Arsen-Hyperakkumulatoren bezeichnete Pflanzen, die auf kontaminierten Standorten bis zu 2 % ihrer oberirdischen Biomasse an Arsen akkumulieren (Ma et al. 2001; Zhao et al. 2009). Dieses Phänomen ist bisher beschränkt auf bestimmte Farnarten (z. B. die Saumfarnart Pteris vittata). Daneben sind einige Pflanzenarten bekannt, die relativ tolerant gegenüber erhöhten Arsenkonzentrationen im Gewebe sind, wie Schachtelhalm-(Equisetum)-Arten (Meharg 2003), Douglasie (Pseudotsuga menziesie) (Haug et al. 2004) oder auch die Brassicaceae Isatis capadocica (Karimi et al. 2009). Diese Arten kommen auf kontaminierten Standorten vor und zeigen dann erhöhte As-Konzentrationen (Toleranz, s. Kapitel 2.4). Untersuchungen, inwieweit diese Arten als Zeigerpflanzen für As-belastete Flächen dienen können, fehlen derzeit. Alle bisher genannten Arten kommen auch auf unbelasteten Flächen vor. Hinweise für eine entsprechende Belastung erhält man möglicherweise nur über die Artenzusammensetzung an einem Standort. Dabei ist die Überlagerung mit Effekten durch andere Kontaminanten zu berücksichtigen und vielfach schwierig. Ergänzend ist zu erwähnen, dass auch bei Wasserpflanzen (aquatischen Makrophyten) Robinson et al. 2006 und Mkandawire & Dudel 2005 auf As-belasteten Standorten in Neuseeland und Deutschland erhöhte As-Konzentrationen feststellen konnten. Eingehende Untersuchungen inklusive Gefäßversuchen im Gewächshaus zeigten, dass dies allerdings ein Resultat physiko-chemischer Adsorptionsprozesse an der Oberfläche von untergetauchten Pflanzenteilen und Wurzeln ist, was durch die Bildung u. a. von Fe-Präzipitaten („Fe plaques“) eben dort ermöglicht wird.
Fazit: Es gibt große artspezifische Unterschiede, aber es sind nur wenige „Hyperakkumulatoren“ bekannt. Die meisten Pflanzenarten schränken zumindest den Transfer in die oberirdischen Organe anteilig ein.
Sortenspezifische Unterschiede
Es gibt immer wieder Hinweise in der Literatur auf sortenspezifische Unterschiede im Verhalten bezüglich Arsenaufnahme und -akkumulation, wobei sich bisher vorliegende Arbeiten zumeist auf Reis, Sommergerste und Winterweizen beschränken. Wesentliche Ergebnisse liefern Untersuchungen zu Reis (Zhang et al. 2008; Norton et al. 2009a; Norton et al. 2009b). Die Arsenkonzentrationen in Reiskörnern von 13 Sorten unterschieden sich auf unterschiedlichen Standorten in Bangladesh, Indien und China jeweils beim Vergleich an einem Standort signifikant bei Konzentrationen von 0,11 – 0,84 mg kg-1 (Norton et al. 2009a). In Deutschland wurden von Serfling & Klose 2008 Gefäßversuche mit 20 Winterweizensorten und 10 Sommergerstesorten durchgeführt. Die Arsenkonzentration in der Gesamtpflanze variierte zwischen den Sommergerstesorten bei gleichem Substrat nach 4 Wochen Wachstum

LABO-Vorhaben B 1.10 31.5.2012
6
zwischen 5,5 – 8,1 mg kg-1, wobei kein signifikanter Unterschied zwischen den Sorten festzustellen war. Zwischen den Winterweizensorten waren dagegen die As-Konzentrationen signifikant unterschiedlich. Sie lagen substratabhängig zwischen 3,9 und 7,0 bzw. 5,6 und 8,3 mg kg-1. Bei einem Sortenunterschied von bis zu 44 % kann so die Sortenwahl und damit die genetische Prädisposition einen nicht zu vernachlässigenden Einfluss auf den As-Gehalt in relevanten Nutzpflanzen haben. Nicht klar zum Ausdruck in diesem Bericht kommt, ob es sich bei Bezug auf „Gesamtpflanze“ um den gesamten oberirdischen Teil handelt, oder wirklich die gesamte Pflanze gemeint ist (einschließlich Wurzel, und wenn mit Wurzel, ob diese dann gewaschen wurde). Die As-Konzentrationen im Korn von Winterweizen sind ebenfalls sortenabhängig signifikant unterschiedlich mit Werten von 0,6 – 1,0 mg kg-1 TM. Dies entspricht auch in der Größenordnung den Ergebnissen bei Reiskörnern. In Gefäßversuchen sowohl als Hydrokultur-Ansatz als auch Substrat-Ansatz mit zwei Sorten Winterweizen fanden Geng et al. 2006 hoch signifikante Unterschiede der As-Konzentrationen zwischen den Sorten sowohl für die oberirdischen als auch die unterirdischen Pflanzenteile. Schließlich ist zu erwähnen, dass auch zwei Sorten Tomaten nach Exposition mit unterschiedlichen As-Spezies ein signifikant unterschiedliches Verhalten zeigten. Dies betraf sowohl das Pflanzenwachstum als auch das Akkumulationsverhalten von Arsen in den einzelnen Pflanzenteilen.
Fazit: Die Sortenwahl scheint eine Einflussgröße zur Verringerung der As-Konzentrationen in Getreide und anderen Nutzpflanzen zu sein. Das bisher untersuchte Sortenspektrum sollte allerdings ergänzt und auf weitere relevante Nutzpflanzen (z. B. Mais) ausgedehnt werden.
Arsenspeziierung in Pflanzen
Studien einer ganzen Reihe von Pflanzen sowohl im terrestrischen als auch im aquatischen Bereich (Wasserpflanzen) an Standorten mit erhöhten As-Konzentrationen in der Umwelt haben gezeigt, dass – quantitativ gesehen – im Wesentlichen anorganische Arsenspezies (Asi) in den Pflanzen zu finden sind (Koch et al. 2000; Kuehnelt et al. 2000; Geiszinger et al. 2002). Sowohl Arsenat(III) als auch Arsenat(V) werden dabei nachgewiesen und das Verhältnis dieser beiden anorganischen Spezies variiert von Pflanzenart zu Pflanzenart. Allerdings werden von Zhao et al. 2010 die Resultate z. T. angezweifelt, da durch entsprechende Probengewinnung (chemische Extraktionsverfahren) und Aufbereitung (Gefriertrocknung) Arsenat(III) leicht zu Arsenat(V) oxidiert wird. Andere Untersuchungen unter kontrollierten Bedingungen zeigen, dass vornehmlich Arsenat(III) als anorganische Spezies in Pflanzen vorliegt (Pickering et al. 2000; Dhankher et al. 2002; Raab et al. 2005). Die Untersuchungen an Acker-Schmalwand (Arabidopsis thaliana) als auch Ruten-Kohl (Indischer Senf) (Brassica juncea) zeigen darüber hinaus, dass Arsenat(III) in Komplexbindung mit den Sulfhydryl-Gruppen thiol-reicher Peptide wie Glutathion und entsprechenden Phytochelatinen (pflanzliche Chelatbildner) vorliegt (Pickering et al. 2000; Dhankher et al. 2002).

LABO-Vorhaben B 1.10 31.5.2012
7
Neben den anorganischen Spezies gibt es eine Reihe methylierter As-Spezies wie Monomethylarsonsäure (MMA), Dimethylarsinsäure (DMA) und Trimethylarsinoxid (TMAO). Die Konzentrationen der methylierten Arsenspezies sind in der Regel niedrig, können aber z. B. bei alleiniger Verfügbarkeit dieser Spezies in der Bodenlösung bis über 70 % des gesamten Arsens in der Pflanze ausmachen (Zhao et al. 2010). Schon in frühen Studien zur Arsenspeziierung in Pflanzen weisen Nissen & Benson 1982 darauf hin, dass methylierte Spezies in den pflanzlichen Zellen auch selbst entstehen können, speziell wenn Stress durch Nährstoffmangel (in diesem Fall durch P und N) entsteht.
Schließlich werden in höheren Pflanzen neben einigen unbekannten As-Spezies noch Arsenozucker, Arsenocholine und Arsenobetaine gefunden (Koch et al. 2000; Kuehnelt et al. 2000). Während in marinen Algen Arsen hauptsächlich als Arsenozucker vorliegt (Castlehouse et al. 2003), sind die Konzentrationen in höheren Pflanzen niedrig und es ist unklar, ob diese in den pflanzlichen Zellen gebildet oder aufgenommen werden (Zhao et al. 2010).
Fazit: Quantitativ spielen die anorganischen As-Spezies mit bis zu 90 % Anteil am gesamten As-Gehalt die wesentliche Rolle im pflanzlichen Gewebe, gefolgt von den methylierten Spezies MMA und DMA. Die Verteilung der analytisch nachweisbaren Spezies wird auch durch die Probenahme und -aufbereitung maßgeblich beeinflusst. Daher sind viele Untersuchungsergebnisse nicht direkt vergleichbar.
2.3 Aufnahmemechanismen für Arsen
Arsenspeziierung in der Rhizosphäre
Das Milieu der Rhizosphäre kann stark unterschiedlich zu den umgebenden Bodenverhältnissen sein. Dies ist besonders unter anaeroben Verhältnissen und damit für Feuchtgebietspflanzen gegeben, wie wir sie auch in als Grünland genutzten Auenbereichen vorfinden. Unter wassergesättigten Bedingungen liegt Arsen hauptsächlich als Arsenat(III) vor. In der Rhizosphäre kann Arsenat(III) dagegen mit Hilfe des über Aerenchymgewebe von Makrophyten transportierten und dann durch Rhizome und Wurzeln ausgeschiedenen Sauerstoffs zu Arsenat(V) oxidiert werden. Zusätzlich wird Eisen oxidiert und präzipitiert als sog. „Fe plaque“ (zumeist Fe(OH)3) auf der Wurzeloberfläche. Arsenat(V) zeigt eine hohe Affinität zum „Fe plaque“, was einen signifikanten Effekt auf die Absorptionskinetik von Arsen hat (Chen et al. 2005). Bei Reiswurzeln mit „Fe plaque“ wurde eine verminderte Aufnahme von Arsenat(V), aber eine verstärkte Aufnahme von Arsenat(III) festgestellt.
Unter aeroben Bedingungen ist Arsenat(V) die im Wesentlichen von den Pflanzen aufgenommene anorganische Arsenspezies. Allerdings gibt es Hinweise auf das Vorhandensein auch von Arsenat(III) unter diesen

LABO-Vorhaben B 1.10 31.5.2012
8
Bedingungen (Ultra et al. 2007; Vetterlein et al. 2007), was als Folge von Wurzelausscheidungen (u. a. organische Säuren, saure Phosphatasen) diskutiert wird (Xu et al. 2007; Logoteta et al. 2009). Ausgeschiedenes Arsenat(III) wird in der Rhizosphäre entweder zu Arsenat(V) oxidiert oder wieder resorbiert.
Fazit: Die Analyse der Bodenverhältnisse führt nicht in jedem Fall zu einer genauen Einschätzung der möglichen Aufnahme durch die Pflanze. Im Zweifelsfall ist eine Pflanzenanalyse anzuschließen.
Aufnahme und Transport in der Pflanze
Die im Wesentlichen von den Pflanzen über die Wurzeln aufgenommenen Arsenspezies sind Arsenat(V), Arsenat(III), Monomethylarsonsäure (MMA) und Dimethylarsinsäure (DMA).
Arsenat(V) wird über Phosphattransportersysteme (Systeme u. a. bestehend aus Membranproteinen zum Transport von Phosphat durch die Zellmembran) aufgenommen. Zu diesem Schluss kommt man zum einen durch physiologische Untersuchungen, die eine starke Reduzierung der Arsenaufnahme durch Anwesenheit von Phosphat belegen (Ullrich-Eberius et al. 1989; Abedin et al. 2002; Mkandawire et al. 2006). Zum anderen zeigen Mutanten des Acker-Schmalwands (Arabidopsis thaliana) mit defektem Phosphattransportsystem stärkere Toleranz gegenüber Arsenat(V) (Gonzalez et al. 2005; Catarecha et al. 2007). Verschiedene Phosphattransportersysteme zeigen unterschiedliche Affinitäten zu Arsenat(V). Beispielsweise wird für die Saumfarnart Pteris vittata (Arsen-Hyperakkumulator) eine höhere Affinität für Arsenat(V) gegenüber Pflanzen ohne Hyperakkumulator-Eigenschaften beschrieben (Wang et al. 2002; Poynton et al. 2004).
Im Gegensatz zu Arsenat(V) ist die arsenige Säure (H3AsO3) des Arsenat(III) bei pH-Werten < 8 zu mehr als 94 % nicht dissoziiert. Pflanzen nehmen daher Arsenat(III) vornehmlich als As(OH)3 auf. Die Eintrittsstellen sind, ähnlich wie bei Mikroorganismen und auch tierischen Zellen, intrinsische Proteine des Plasmalemmas (Zellmembran), eine Unterfamilie der Aquaporine. Für die im Zusammenhang mit Arsentransport als NIPs (nodulin 26-like intrinsic proteins) beschriebenen Aquaporine konnte in mehreren Studien die Durchlässigkeit für Arsenat(III) beschrieben werden (Bleeker et al. 2006; Isayenkov & Maathuis 2008; Ma et al. 2008; Kamiya et al. 2009). Dabei handelt es sich um eine ganze Reihe verschiedener NIPs, die diese Eigenschaft zeigen. Eine Übersicht dazu mit Referenzen findet sich bei Zhao et al. 2010. Wichtig für den Transport ist die Tatsache, dass spezielle NIPs an der proximalen Seite von Exodermis- und Endodermiszellen für die Abgabe von Arsenat(III) in Richtung Xylem (Gefäßteil der Leitbahnen) und damit den Ferntransport verantwortlich sind. Dies wurde zumindest so für Reis nachgewiesen (Ma et al. 2008). Interessant ist, dass diese NIPs eigentlich die Aufnahme von Si aktiv steuern und somit eine Beeinflussung der As-Aufnahme durch Si möglich

LABO-Vorhaben B 1.10 31.5.2012
9
ist, was mittlerweile durch einige Untersuchungen mit Si-Düngung belegt wurde (Bogdan & Schenk 2008; Li et al. 2009b).
Methylierte Arsenspezies wie MMA und DMA entstehen im Wesentlichen durch die Aktivität von Mikroorganismen im Substrat/Sediment. Die Aufnahme dieser Spezies, zumindest unter den kontrollierten Bedingungen einer Hydrokultur, wurde in mehreren Studien nachgewiesen (Marin et al. 1992; Marin et al. 1993; Carbonell-Barrachina et al. 1998; Carbonell-Barrachina et al. 1999; Abedin et al. 2002). Eine Aufnahme in die pflanzlichen Zellen durch Diffusion durch das Plasmalemma (Zellmembran) kommt allerdings kaum in Frage (Cullen & Nelson 1992). Li et al. 2009a konnten an Reis zeigen, dass undissoziierte methylierte Arsenspezies über das Aquaporin Lsi1 in die Wurzelzellen aufgenommen werden. Da MMA und DMA geringere Dissoziationskonstanten als Arsenat(III) haben, ist die Aufnahme stark vom äußeren pH-Wert abhängig. Je geringer der pH-Wert, desto höher die Aufnahme von MMA und DMA, was einhergeht mit einem höheren Anteil an undissoziierten Molekülen.
Im Xylemsaft und damit im Ferntransport der meisten untersuchten Pflanzen dominiert Arsenat(III). Dies führt zu der Vermutung, dass dies auch die Spezies ist, die hauptsächlich von den Wurzelzellen an den Xylemsaft abgegeben wird (Zhao et al. 2009). Auch bei alleiniger Gabe von Arsenat(V) wird hauptsächlich Arsenat(III) im Xylem transportiert, was mit der Tatsache zusammenpasst, dass Wurzeln über eine hohe Kapazität zur Reduktion von Arsenat(V) verfügen (Pickering et al. 2000; Xu et al. 2007). Wenig ist bekannt über den Transport im Phloem.
Fazit: Die Untersuchung von Arsenspezies in der Bodenlösung gibt zwar einen Hinweis, ob mehr methylierte oder anorganische Arsenspezies aufgenommen werden bzw. werden können, hat aber keine Bedeutung für eine quantitative Aussage zum Aufnahmeverhalten und den tatsächlich in der Pflanze vorhandenen mehr oder weniger toxischen Spezies.
Arsenmetabolismus in der Pflanze
Die wesentlichen in der Pflanze stattfindenden metabolischen Prozesse sind sicherlich die Oxidation/Reduktion anorganischer Arsenspezies und die hier schon erwähnte Reduzierung von Arsenat(V) zu Arsenat(III) sowie die Methylierung. Daneben spielen die im Zusammenhang mit der Toxizität bzw. Detoxifikation stehenden Bindungen/Komplexierungen z. B. mit Sulfhydryl-Gruppen von Proteinen eine Rolle, auf die aber im nächsten Kapitel eingegangen wird.
Die Methylierung von anorganischen Arsenspezies in der Pflanze wurde schon von Nissen & Benson 1982 wahrscheinlich gemacht (s. Kap. 2.2). Sie setzten radioaktives 74As (Arsenat(V)) in Gefäßversuchen als Düngelösung ein und fanden unter P- und N-limitierten Bedingungen 74As in der Pflanze auch in methylierten Spezies wieder. Bei ausreichender P- und N-Versorgung konnten kaum methylierte Spezies nachgewiesen

LABO-Vorhaben B 1.10 31.5.2012
10
werden. In einigen neueren Untersuchungen unter Hydrokultur-Bedingungen wurden jeweils anorganische Arsenspezies der Lösung zugesetzt und in pflanzlichem Gewebe als auch im Xylemsaft konnten anschließend methylierte As-Spezies in geringen Mengen nachgewiesen werden (Quaghebeur & Rengel 2003; Mihucz et al. 2005; Raab et al. 2005). Wie die Methylierung tatsächlich abläuft, kann von den aufgeklärten Reaktionsfolgen bei Pilzen abgeleitet werden (Bentley & Chasteen 2002). Mit Arsenat(III) als Ausgangssubstrat folgen bis zum volatilen Trimethylarsin drei vergleichbare Methylierungsschritte. Diese werden über die S-adenosyl-L-methyltransferase katalysiert, wobei jeweils S-adenosyl-L-methionin (SAM) als Methyl-Donor fungiert. Die Aktivität einer SAM-abhängigen As-Methyltransferase konnte auch in Blattextrakten von Rotem Straußgras (Agrostis capillaris, früher Agrostis tenuis) festgestellt werden (Wu et al. 2002), einem Süßgras, das auch in Deutschland auf eher trockenen Grünlandflächen weit verbreitet ist. Es entsteht im ersten Schritt MMA(V). In den Mikroorganismen (Pilzen) ist nach Reduzierung zu MMA(III) der nächste Methylierungsschritt zu DMA(V) nachgewiesen. Nach erneuter Reduktion (DMA(III)) wird noch einmal zu TMAO(V) methyliert, was wiederum zum volatilen TMA(III) reduziert wird (Qin et al. 2009). In Pflanzen ist bisher nur von Reiswurzeln bekannt, dass sie MMA(V) zu MMA(III) reduzieren können (Li et al. 2009a). Nicht bekannt ist, ob Pflanzen volatile Komponenten wie TMA(III) metabolisieren (Zhao et al. 2010) und so möglicherweise Arsen verlieren.
Inwieweit die auch in Pflanzen zu findenden Arsenozucker, Arsenocholine und Arsenobetaine in den Pflanzen selbst metabolisiert werden oder als solche aufgenommen werden, ist bisher nicht geklärt (Geiszinger et al. 2002).
Fazit: Offensichtlich ist die Methylierung in der Pflanze quantitativ von geringer Bedeutung. Damit ist eine Reduktion durch Volatilisierung (Ausgasung in die Atmosphäre) wahrscheinlich mengenmäßig vernachlässigbar. Die energieaufwendige Reduktion von Arsenat(V) zu Arsenat(III) führt möglicherweise zu reduziertem Wachstum.
2.4 Toxizität und Toleranzmechanismen
Die Toxizität von Arsen ist in starkem Maße abhängig von seiner Speziierung in der Pflanze. Arsenat(V) als Phosphat-Analogon hat einen starken Einfluss auf den Phosphatmetabolismus und damit z. B. auf die Phosphorylierung und die ATP Synthese (Zhao et al. 2010). Dies führt zur Störung des Energiehaushalts der Pflanzen und damit beispielsweise zu Wachstumsreduktionen. Für Arsenat(III) ist ein Effekt auf die Struktur und katalytische Funktion von Proteinen/Enzymen durch Bindung an Sulfhydryl-Gruppen beschrieben (Ullrich-Eberius et al. 1989). Dies wiederum hat Einfluss auf die biochemischen Prozesse innerhalb der einzelnen Zellen und kann ebenfalls zu Wachstumsreduktionen bis hin zum Absterben von Zellen und dann Geweben (im Extremfall zum Absterben der ganzen Pflanze) führen. Es wird vermutet, dass die meisten beschriebenen toxischen Effekte durch Arsenat(V) eigentlich auf die Wirkung von

LABO-Vorhaben B 1.10 31.5.2012
11
Arsenat(III) zurückgehen, da Arsenat(V) in der Pflanze relativ schnell zu Arsenat(III) reduziert wird (s. o.) und damit keine relevanten Arsenat(V)-Konzentrationen im Cytoplasma der Zellen erreicht werden (Bertolero et al. 1987).
Die Exposition von Pflanzen mit anorganischen Arsenspezies führt zur Bildung von Sauerstoffradikalen. Dies induziert oxidativen Stress und führt zu entsprechenden Reaktionen der pflanzlichen Zellen (Mylona et al. 1998; Requejo & Tena 2005). Beispielsweise kommt es durch erhöhte Arsenat(III)-Konzentrationen zur vermehrten Bildung von Enzymen des antioxidativen Systems (Superoxid-Dismutase, Katalase, Gluthathion-S-Transferase), aber auch nicht-enzymatischer Antioxidantien wie Ascorbat und Glutathion. Glutathion dient gleichzeitig als Vorläufer zur Bildung von Phytochelatinen, deren Bildung ebenfalls durch anorganische Arsenspezies induziert wird. Dies wiederum verringert die Menge an Glutathion, die als Antioxidans zur Verfügung steht. Schließlich ist beschrieben, dass die anorganischen Arsenspezies in Pflanzen weiter zu methylierten Spezies (z. B. MMA, DMA) metabolisiert werden können, was zu weiterem oxidativen Stress führt. Wie die Stressreaktionen artspezifisch ablaufen, ist im Einzelfall noch zu untersuchen.
Toxizitätsgrenzen für Arsen ohne Differenzierung in die vorliegenden Arsenspezies liegen für die meisten Pflanzen zwischen 1 und 100 mg Arsen kg-1 TG in den oberirdischen Kompartimenten (Kabata-Pendias & Pendias 1992). Davon ausgenommen sind Hyperakkumulatoren wie der Saumfarn Pteris vittata, die erst ab 5.000 – 10.000 mg kg-1 Arsen in den Wedeln Anzeichen von Toxizität zeigen (Zhao et al. 2002; Singh et al. 2006; Wei & Chen 2006).
Toleranzmechanismen beginnen mit der Vermeidung der Arsenaufnahme. Dies ist von einigen Pflanzen auf As-kontaminierten Standorten beschrieben, die die Aktivität des Phosphor-Transporter-Systems unterdrücken und so weniger Arsen (in diesem Fall Arsenat(V)) aufnehmen (Meharg & Hartley-Whitaker 2002). Wird Arsen in größeren Mengen aufge-nommen, stehen mehrere Möglichkeiten der Detoxifikation zur Verfügung:
Zum einen kann Arsen komplexiert werden. Arsen induziert sehr stark die Phytoochelatin-Synthese (Sneller et al. 1999; Schmöger et al. 2000) (s. o.). Wird die Phytochelatin-Synthese künstlich blockiert (z. B. durch L-buthionin-sulfoxim), erreicht man dadurch eine Hypersensitivität sowohl für Arsenat(V) als auch für Arsenat(III) (Schmöger et al. 2000; Schat et al. 2002; Bleeker et al. 2006). Ähnliches wird auch von einem Arabidopsis-Mutanten ohne Phytechelatin-Synthese beschrieben (Ha et al. 1999). Im Wesentlichen sind es die Sulfhydryl-Gruppen des Cysteins der Phytochelatine, die mit Arsenat(III) komplexieren. Die Detoxifizierung über Komplexbildung mit Phytochelatinen gilt im Wesentlichen für Nicht-Hyperakkumulatoren. Dagegen besitzen die hyperakkumulierenden Farne wie Pteris vittata andere Toleranzmechnismen (s. u.).
Die Phytochelatin-Arsenat(III)-Komplexe können dann als weiterer Schritt in der Detoxifikation in die Vakuole transportiert und dort gespeichert werden

LABO-Vorhaben B 1.10 31.5.2012
12
(Kompartimentierung). Für Hefe wurde ein entsprechendes Transportersystem identifiziert. Am Tonoplast von Wurzelzellen des wolligen Honiggrases (Holcus lanatus) wurde der Transport entsprechender Komplexe ebenfalls beobachtet (Bleeker et al. 2006). Wie genau der Transport abläuft und welche Proteine beteiligt sind, ist bisher nicht vollständig geklärt.
In den Zellen der Wedel der Saumfarnart Pteris vittata ist Arsen auch in den Vakuolen abgelagert, allerdings als anorganisches Arsenat(III) (Lombi et al. 2002; Pickering et al. 2006). Es wird ein stark energieabhängiger Transport vermutet, der bisher nicht aufgeklärt wurde, aber wahrscheinlich den Schlüssel für die Hyperakkumulation darstellt.
Als letzter möglicher Toleranzmechnismus kommt die aktive Ausscheidung von Arsen durch Wurzelzellen in Frage. Dies wurde als Möglichkeit in einer neueren Studie mit dem wolligen Honiggras (Holcus lanatus) für Arsenat(III) dargestellt (Logoteta et al. 2009).
Fazit: Die meisten höheren Pflanzen besitzen keine ausgeprägten Toleranzmechanismen gegenüber Arsen bzw. höheren Arsenkonzentrationen in der Umwelt. Entsprechend sind die Toxizitätsgrenzen niedrig und zudem abhängig von der Arsenspeziierung in der Pflanze.
2.5 Bedeutung von Mykorrhiza-Pilzen
In der Regel sind Pflanzen, die auf Standorten mit As-Kontaminierung wachsen, mykorrhiziert (Meharg et al. 1999; Sharples et al. 2000a; Sharples et al. 2000b). Von Meharg et al. 1994b konnte gezeigt werden, dass resistente Phänotypen des wolligen Honiggrases (Holcus lanatus) gegenüber nicht-resistenten Phänotypen signifikant stärker mykorrhiziert waren. In Untersuchungen von Sharples et al. 2000a wurde nachgewiesen, dass der Pilzsymbiont Hymenoscyphus ericae eine entscheidende Rolle bei der Kolonisation von arsenkontaminierten Bergbaustandorten (z.B. Halden, Aufbereitungsplätze) durch Besenheide (Calluna vulgaris) spielt. Der Mechanismus beruht dabei auf einer verstärkten Ausscheidung von Arsenat(III) durch den Pilzsymbionten, ein Mechanismus für Arsenresistenz, wie er auch für Bakterien und Hefe beschrieben wurde (Meharg & Hartley-Whitaker 2002).
Dagegen werden für die Saumfarnart Pteris vittata als Hyperakkumulator in mykorrhizierten Varianten noch einmal erhöhte Arsenaufnahmen gegenüber nicht mykorrhizierten Varianten beschreiben (Al Agely et al. 2005; Liu et al. 2005).
Schließlich stellen Serfling & Klose 2008 fest, dass die Mykorrhizierung von Sommergerste zwar zu einer leichten Erhöhung der Arsenkonzentration in

LABO-Vorhaben B 1.10 31.5.2012
13
der Gesamtpflanze geführt hat, aber keine Unterschiede für die Arsengehalte im Korn festzustellen sind.
Fazit: Die derzeit vorliegenden Erkenntnisse zeigen positive und negative Effekte der Mykorrhiza auf die Arsenaufnahme und reichen nicht aus, um Schlussfolgerungen für die Gefahrenabwehr abzuleiten. Zudem ist die Mykorrhizierung in der Praxis für die Grünlandnutzung nicht zu beeinflussen (vgl. Kap. 2.6).
2.6 Schlussfolgerungen
Die P-Düngung und hier speziell der Einfluss auf unterschiedlichen Standorten (mit entsprechend unterschiedlichen Verhältnissen) ist art- und sortenspezifisch z. T. widersprüchlich belegt und daher weiter zu untersuchen und deren Effekte weiter zu evaluieren.
Wenig bekannt ist bisher über die Wechselwirkung von verringertem Redoxpotenzial und P-Düngung. Hierzu sind weitere Untersuchungen dringend nötig. Speziell die Quantifizierung dieser beiden Komponenten unter unterschiedlichen Bedingungen (Pflanzenart/-sorte, Höhe und Qualität der Arsenbelastung) ist zu untersuchen. Die angestrebten Gefäßversuche sollen hierzu einen ersten Beitrag leisten.
Die Spezies-Analyse in den Pflanzen ist nur sinnvoll zur Unterscheidung der toxikologisch relevanteren anorganischen Arsenspezies und den weiteren Arsenverbindungen. Der Zusammenhang zwischen dem Vorkommen bestimmter Arsenspezies im Boden und in der Pflanze ist nicht eindeutig herzustellen, da es im Zuge der Aufnahme und Translokation in der Pflanze zu einer veränderten Speziierung kommen kann (speziell Reduktion von Arsenat(V) zu Arsenat(III). Außerdem ist die Methylierung von Arsen durch Pflanzen zwar nachgewiesen, hat aber möglicherweise quantitativ keine Bedeutung.
Interessant sind die wenigen Hinweise auf den Einfluss anderer Stressoren, wie weiterer Nährstoffmangel durch Makro- und Mikronährstoffe. Über die Wirkung komplexerer Belastungen mit Arsen als einem Stressor neben anderen, z. B. Schwermetallen, gibt es bisher keine eindeutigen Hinweise und dies ist deswegen in Zukunft ein weiteres weites Forschungsfeld.
Toleranzmechanismen gegenüber erhöhten Arsenkonzentrationen in der Umwelt, wie Vermeidung, Komplexierung oder Ausscheidung, besitzen die meisten höheren Pflanzen nicht. Die Toxizitätsgrenzen sind niedrig und abhängig von der Arsenspeziierung in der Pflanze.
Schließlich hat die Mykorrhizierung zwar einen Effekt, ist aber in der Praxis für die Grünlandnutzung nicht zu beeinflussen, da sich standortbedingt immer eine entsprechende Pilzflora einstellt. Bei z. B. Getreidenutzung

LABO-Vorhaben B 1.10 31.5.2012
14
(Korngewinnung) ist die Mykorrhizierung möglicherweise ohne Bedeutung, da es für die Arsengehalte im Erntegut keine oder nur geringe Unterschiede gibt.

LABO-Vorhaben B 1.10 31.5.2012
15
3 ERFAHRUNGEN DER VOLLZUGSPRAXIS
ahu AG
Für einen Überblick über die Optionen des behördlichen Handelns und das rechtskonforme Vorgehen, wurden die Erfahrungen der Vollzugsbehörden des Bodenschutzes und landwirtschaftlicher Institutionen ausgewertet.
3.1 Bezugsrahmen und -literatur
Um einen einheitlichen Bezugsrahmen für den Ablauf von Gefährdungsabschätzungen und den Untersuchungsablauf zu definieren, baut der Leitfaden Arsentransfer aus Böden in Nahrungs- und Futterpflanzen – Gefahrenbeurteilung und Maßnahmen in erster Linie auf die Vorgaben des BBodSchG 1998 und der BBodSchV 1999 auf. Allgemeine Arbeitshilfen zur Bewertung des Pfades Βoden – Nutzpflanze bei schädlichen Bodenveränderungen auf landwirtschaftlich genutzten Flächen wurden zusammengestellt, um im Leitfaden darauf zu verweisen. Enthaltene Hinweise für Arsen sind in den Leitfaden eingeflossen. Der Leitfaden für den Arsentransfer Boden – Nutzpflanze wird als Ergänzung zu diesen Arbeitshilfen angesehen und ersetzt diese nicht.
3.2 Vorliegende Erfahrungen mit Arsen in Boden und Nutzpflanze
Die Erfahrungen der Vollzugsbehörden bei der Bearbeitung von Fragen zum Arsentransfer Boden – Nutzpflanze wurden auf Grundlage der in Tabelle 3-1 aufgeführten Berichte und ergänzenden Recherchen bei ausgewählten Stellen der Länder1 ausgewertet.
In vier Bundesländern liegen umfangreiche, speziell auf Arsenbelastungen in Böden ausgerichtete Untersuchungen und Handlungskonzepte für die Bodenschutzbehörden vor (BY, BW, NW, SN). Handlungsempfehlungen für betroffene Landwirte zum Umgang mit erhöhten Arsengehalten in Böden stehen darüber hinaus in BY (Erdinger, Freisinger, Dachauer Moos), BW (Regierungsbezirke Karlsruhe, Freiburg), NW (Kreise Coesfeld Borken) und in SN (Landesamt für Umwelt, Landwirtschaft und Geologie, Sächsische Landesanstalt für Landwirtschaft) zur Verfügung (vgl. Tab. 3-1). Die
1 Regierungspräsidium Freiburg - Referate Gewässer/Boden und Futtermittelüberwachung (BW), Niedersächsisches Ministerium für Umwelt und Klimaschutz, Niedersächsisches Landesamt für Bergbau, Energie und Geologie, Landwirtschaftskammer Niedersachsen, Staatliche Betriebsgesellschaft für Umwelt und Landwirtschaft (SN), Landesdirektion Chemnitz (SN), Landesamt für Umwelt und Verbraucherschutz NRW, Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie, Bayerisches Landesamt für Umwelt.

LABO-Vorhaben B 1.10 31.5.2012
16
Handlungsempfehlungen beziehen häufig auch andere Wirkungspfade wie z. B. Boden – Mensch ein und erweitern damit den Kreis der Betroffenen (z. B. Haus- und Gartenbesitzer).
Im Vordergrund der Berichte stehen zumeist die Untersuchung des Bodens und der Mobilisierbarkeit des darin gebundenen Arsens, die Arsenquellen und der Umgang mit der Belastung durch angepasste Maßnahmen zur Gefahrenabwehr. Einzelne Untersuchungen von Pflanzenmaterial liegen sowohl in BY, BW und NW vor, während in SN systematische Feld- und Gefäßuntersuchungen zur Sortenabhängigkeit der Arsenaufnahme von Futter- und Getreidepflanzen durchgeführt wurden.
Zur Einschätzung der Aussagekraft der Ammoniumnitrat-Extraktion gemäß BBodSchV bei Arsen wurden von den Vertretern der Länder in der projektbegleitenden Arbeitsgruppe Überschreitungen des Prüfwertes für den Schadstoffübergang Boden - Nutzpflanze auf Ackerbauflächen im Hinblick auf Wachstumsbeeinträchtigungen recherchiert.
Die Erfahrungen aus der Vollzugspraxis werden nachfolgend zusammengefasst.
Tab. 3-1: Berichte und Empfehlungen insbesondere für Arsen in Boden und Pflanzen aus der Vollzugspraxis der Länder
Land Bericht Inhaltl. Bezug
Hinweise zum Umgang mit naturbedingt erhöhten Arsengehalten in Böden des Erdinger, Freisinger und Dachauer Mooses (für Landwirte, Verbraucher, Bauherren) erstellt von Regierung von Oberbayern (2008) in Zusammenarbeit mit den bayerischen Fachbehörden LfU, LfL, LGL.
Handlungsempfehlungen für Betroffene
Handlungsempfehlungen für kommunale und staatliche Behörden zum Umgang mit arsenbelasteten Böden im Erdinger, Freisinger und Dachauer Moos erstellt von Regierung von Oberbayern (2004) in Zusammenarbeit mit den bayerischen Fachbehörden LfU, LfL, LGL. Dieses Merkblatt ist im bayerischen behördeninternen Netz verfügbar.
Handlungsempfehlungen für Betroffene
Untersuchungen über die Arsenaufnahme von Grünlandpflanzen auf belasteten Böden im Erdinger Moos im Jahresbericht Jahresbericht 2004 und 2005 der Landesanstalt für Landwirtschaft Institut für Agrarökologie, ökologischen Landbau und Bodenschutz
Untersuchungen Pflanzenaufnahme
BY
Flughafen München - Planfeststellungsverfahren 3. Start- und Landebahn. Boden- und Rohstoffmanagement (2007)
Gefährdungsabschätzung Boden – Mensch und Boden-GW; nicht Boden –Nutzpflanze
Merkblatt für den Gemüseanbau auf schwermetallbelasteten Böden (Stand Januar 2011). Landratsamt Emmendingen.
Handlungsempfehlungen für Landwirtschaft
Merkblatt für den Anbau von Nahrungs- und Futterpflanzen auf schwermetallbelasteten Böden. Landratsamt Emmendingen (Stand 2011)
Pflichtenhinweise für Landwirte
BW
Arsen in Böden und Gesteinen im Regierungsbezirk Karlsruhe. Status, Bewertung, Konsequenzen herausgegeben von LUBW Landesanstalt für Umwelt, Messungen und Naturschutz Baden-Württemberg (2009)
Regionale Erfassung von Verdachtsflächen und Gefahrenbeurteilung
LABO-Vorhaben B 1.10 31.5.2012
17
Land Bericht Inhaltl. Bezug
Arsen in Böden – Hinweise zum Umgang mit naturbedingt erhöhten Arsengehalten im Oberboden des Kreises Borken (2010). Handlungsempfehlungen zur verschmutzungsarmen Nutzpflanzenernte. Herausgegeben vom Kreis Borken und der LWK Nordrhein-Westfalen, Kreisstelle Borken, Bezirksstelle für Agrarstruktur Münsterland.
Handlungsempfehlungen Gefahrenabwehr Landwirtschaft
Arsen – Hinweise zum Umgang mit naturbedingt erhöhten Arsengehalten in Böden (2006). Herausgegeben vom Kreis Coesfeld und der LWK Nordrhein-Westfalen, Kreisstelle Coesfeld, Bezirksstelle für Agrarstruktur Münsterland.
Handlungsempfehlungen für Betroffene
Bodenkundliche Untersuchung, Probenahme und Analytik von Flächen mit prognostizierter Arsenanreicherung (2010). Geologischer Dienst NRW. Dezember 2010.
Bodenchemische Untersuchung
Bodenschutz; Schädliche Bodenveränderungen durch geogene Arsenvorkommen (2008). Stellungnahme des LANUV NRW an den Kreis Recklinghausen bzgl. des Stadtgebietes von Datteln (unervöffentlicht).
Gefährdungsabschätzung, Amtsinterne Stellungnahme
Arsen in Böden des Kreises Borken – Untersuchung von Pflanzenproben (2009). Gutachten der IFUA Projekt-GmbH im Auftrag des Kreises Borken. November 2009.
Weitere Sachverhaltsermittlung, Amtsinternes Gutachten
Nachuntersuchungen zur digitalen Bodenbelastungskarte, Clörather Mühle im Kreis Viersen. Gutachten der ahu AG im Auftrag des Kreises Viersen, Amt für Wasser- und Abfallwirtschaft und Kreisstraßen. März 2007.
Gefährdungsabschätzung, Amtsinternes Gutachten
NW
Fachgespräch „Maßnahmen bei großflächigen schädlichen Bodenveränderungen – Wirkungspfad Boden – Pflanze und Direktpfad Boden – Mensch – am 30. September und 1. Oktober 2003 beim BEW Essen. Dokumentation herausgegeben vom MUNLV, AAV und LUA NRW (2003).
Ergebnisse eines Fachgesprächs mit besonderen Hinweisen zu Arsen
SN Arsentransfer Boden – Pflanze – Entwicklung von Maßnahmen zur Verhinderung des Arsentransfers im System Boden – Pflanze (2008). Schriftenreihe des Landesamtes für Umwelt, Landwirtschaft und Geologie Heft 32/2008.
Maßnahmen Boden –Pflanze
LABO Ermittlung und Beurteilung des Mobilisierungspotenzials von Arsen in Böden. Abschlussbericht zum Projekt B 4.07 im Länderfinanzierungsprogramm „Wasser, Boden und Abfall“ (Overesch et al. 2008)
Untersuchungen Freisetzung aus Böden/ Redox
Arsen-Gesamtgehalte im Boden
Geeignete Voraussetzungen für eine As-Anreicherung im Boden finden sich offensichtlich in:
- Niederungsböden mit As-Anreicherungen aus dem Grundwasser, - Böden der Flussauen (Auenböden) und - Böden aus Ausgangssubstraten mit erhöhten As-Gehalten.
Es treten verstärkt hohe As-Gehalte in den Talauen alter Bergbauregionen der Mittelgebirge auf. Die Belastungen sind unabhängig davon, ob der Bergbau in keltischer, römischer oder jüngerer Zeit stattfand. Solche

LABO-Vorhaben B 1.10 31.5.2012
18
Belastungen finden sich z.B. in den Regierungsbezirken Freiburg und Karlsruhe, aber auch in Sachsen-Anhalt.
In belasteten Auenböden wird der Maßnahmenwert der BBodSchV von 50 mg kg-1 im Boden bei Grünlandnutzung häufig überschritten (z.B. in SN, vgl. Kardel et al. 2011).
Die KW-löslichen As-Gehalte in Ackerböden liegen meist geringer als unter Grünland. Die Arsengehalte im Boden schwanken kleinräumig stark in Abhängigkeit der Standortbedingungen. Arsen ist im Boden häufig mit Eisen vergesellschaftet.
Ammoniumnitratextrahierbare Arsen-Gehalte im Boden
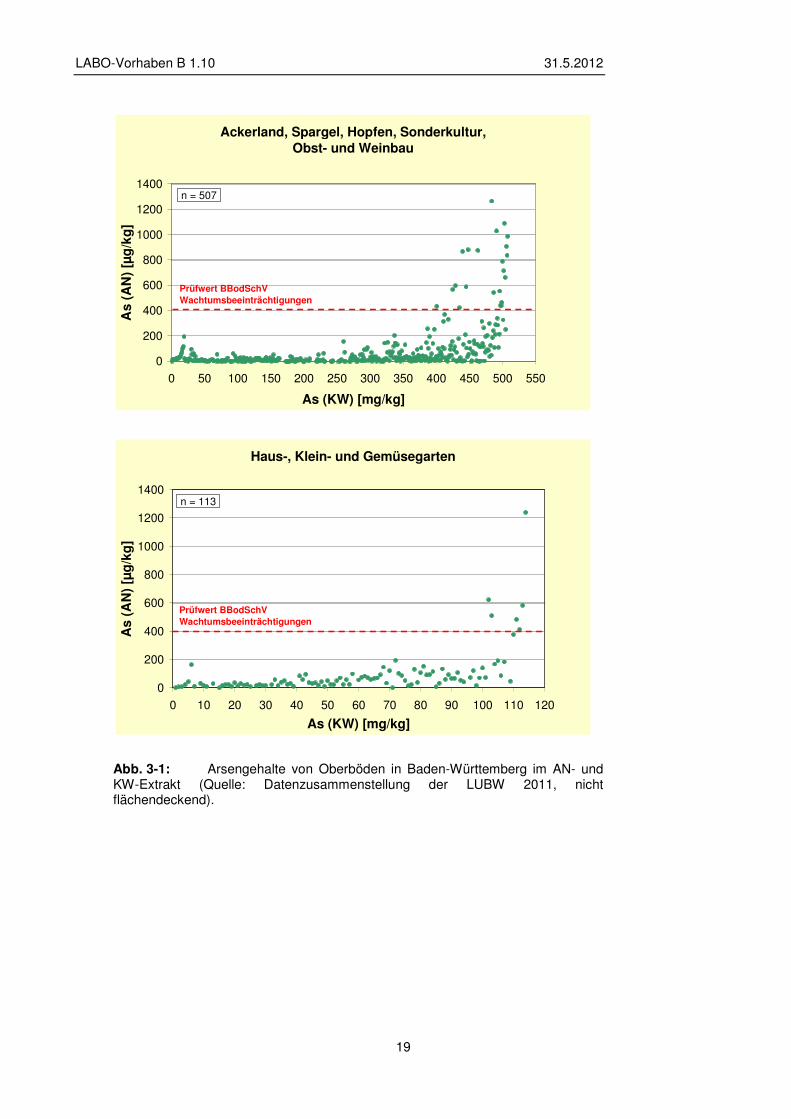
Überschreitungen des Prüfwertes im AN-Extrakt für den Schadstoffübergang Boden - Nutzpflanze auf Ackerbauflächen im Hinblick auf Wachtumsbeeinträchtigungen treten in Baden-Württemberg auf Ackerflächen und Sonderkulturen bei KW-extrahierbaren Gehalten über ca. 400 mg kg-1 auf. In Gärten sind bei Gesamtgehalten ab 100 mg kg-1
vereinzelt Überschreitungen feststellbar (vgl. Abb. 3-1).
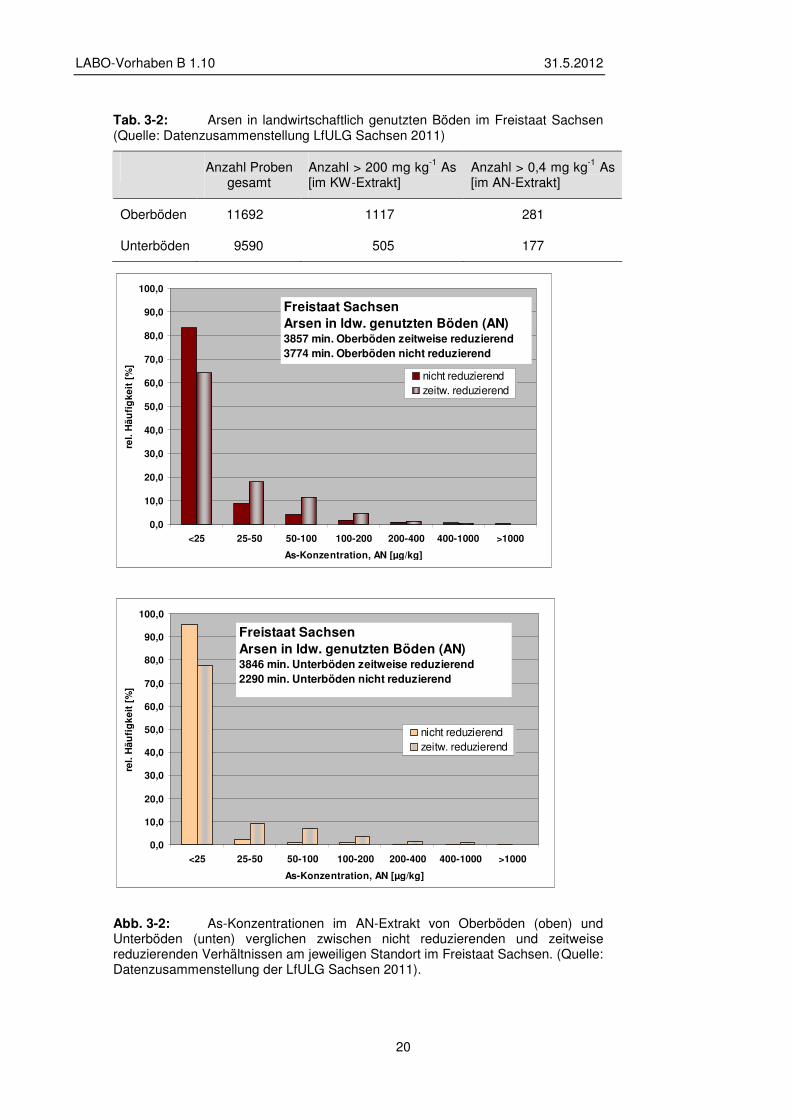
Im Freistaat Sachsen finden sich in landwirtschaftlich genutzten Böden in rund 10 % der bei der LfULG vorliegenden Oberbodenproben und in rund 5 % der Unterbodenproben mehr als 200 mg kg-1 As (Prüfwert Pflanzenqualität). Der Prüfwert für mobile Arsengehalte für Wachstumsbeeinträchtigungen von 0,4 mg kg-1 im Ammoniumnitrat-extrakt wird in den Oberböden in 2,4 % und in den Unterböden in 1,8 % der Gesamtproben überschritten (vgl. Tab. 3-2). Die Abb. 3-2 zeigt, dass eine Differenzierung mit Blick auf Bodentypen, bei denen zeitweise reduzierende Bedingungen erwartet werden können (Auenböden, Gleye, Anmoore, Moore, Reduktosole) für die mineralischen Horizonte möglich ist. Es wird deutlich, dass sich in diesen Böden häufiger höhere mobile Gehalte finden als in Böden, die kein reduktives Milieu erwarten lassen. Diese Aussage trifft auf Ober- wie Unterböden gleichermaßen zu. Ein Zusammenhang zwischen den KW- und AN-Gehalten lässt sich jedoch nicht herstellen (Mittlg. LfULG Sachsen 2011).
LABO-Vorhaben B 1.10 31.5.2012
19
Ackerland, Spargel, Hopfen, Sonderkultur, Obst- und Weinbau
0
200
400
600
800
1000
1200
1400
0 50 100 150 200 250 300 350 400 450 500 550
As (KW) [mg/kg]
As
(AN
) [µ
g/k
g]
Prüfwert BBodSchVWachtumsbeeinträchtigungen
n = 507
Haus-, Klein- und Gemüsegarten
0
200
400
600
800
1000
1200
1400
0 10 20 30 40 50 60 70 80 90 100 110 120
As (KW) [mg/kg]
As
(AN
) [µ
g/k
g]
Prüfwert BBodSchVWachtumsbeeinträchtigungen
n = 113
Abb. 3-1: Arsengehalte von Oberböden in Baden-Württemberg im AN- und KW-Extrakt (Quelle: Datenzusammenstellung der LUBW 2011, nicht flächendeckend).
LABO-Vorhaben B 1.10 31.5.2012
20
Tab. 3-2: Arsen in landwirtschaftlich genutzten Böden im Freistaat Sachsen (Quelle: Datenzusammenstellung LfULG Sachsen 2011)
Anzahl Proben gesamt
Anzahl > 200 mg kg-1 As [im KW-Extrakt]
Anzahl > 0,4 mg kg-1 As [im AN-Extrakt]
Oberböden 11692 1117 281
Unterböden 9590 505 177
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
90,0
100,0
<25 25-50 50-100 100-200 200-400 400-1000 >1000
As-Konzentration, AN [µg/kg]
rel.
Hä
ufi
gke
it [
%]
nicht reduzierendzeitw. reduzierend
Freistaat SachsenArsen in ldw. genutzten Böden (AN)3857 min. Oberböden zeitweise reduzierend3774 min. Oberböden nicht reduzierend
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
90,0
100,0
<25 25-50 50-100 100-200 200-400 400-1000 >1000
As-Konzentration, AN [µg/kg]
rel.
Hä
ufi
gk
eit
[%
]
nicht reduzierendzeitw. reduzierend
Freistaat SachsenArsen in ldw. genutzten Böden (AN)3846 min. Unterböden zeitweise reduzierend2290 min. Unterböden nicht reduzierend
Abb. 3-2: As-Konzentrationen im AN-Extrakt von Oberböden (oben) und Unterböden (unten) verglichen zwischen nicht reduzierenden und zeitweise reduzierenden Verhältnissen am jeweiligen Standort im Freistaat Sachsen. (Quelle: Datenzusammenstellung der LfULG Sachsen 2011).

LABO-Vorhaben B 1.10 31.5.2012
21
In landwirtschaftlich und forstlich genutzten Böden Bayerns treten auf landwirtschaftlich genutzten Flächen keine Überschreitungen des Prüfwertes für Wachstumsbeeinträchtigungen im Ackerbau von 0,4 mg kg-1 im AN auf. In der zugrunde gelegten, nicht flächendeckenden Datenzusammenstellung der LfU Bayern (2011) liegen für Ackerflächen jedoch nur vier Messwerte mit KW-Gehalten über 200 mg kg-1 und nur ein Wert über 400 mg kg-1 vor. Von insgesamt 1.530 Proben zeigen 10 Proben mehr als 0,4 mg kg-1 As im AN (max. 1,1 mg kg-1), wobei davon neun von Waldstandorten aus dem Oberboden stammen. Von diesen neun Waldstandorten zeigt keine Probe mehr als 50 mg kg-1 As im KW. Neben den Waldstandorten zeigt eine Torfprobe unter Ödland mehr als 0,4 mg kg-1 As im AN. Auch diese Torfprobe weist keine auffällig hohen Arsengehalte > 50 mg kg-1 im KW auf.
Bei Betrachtung aller verfügbare Datensätze von landwirtschaftlich genutzten Böden betragen die KW-Maxima unter Acker 940 mg kg-1 und unter Grünland 1610 mg kg-1. Es konnten keine Zusammenhänge zwischen den AN- und KW-Gehalten für alle Datensätze (n=1.530) sowie für die Nutzungen Acker (n=195) und Grünland (n=381) gefunden werden. Die AN-Maxima betragen unter Acker 0,28 mg kg-1 und unter Grünland 0,26 mg kg-1. Die hohen KW-Gehalte im Grünland stammen aus dem Erdinger und Freisinger Moos. Dort führt ein arsenhaltiger Grundwasserstrom zu den Anreicherungen im Boden. Da die Grundwässer häufig auch eisenhaltig sind und zu einer stabilen Festlegung des Arsens führen, könnte dies die geringen AN-Werte unter Grünland erklären. Möglicherweise nimmt auch ein hoher pH-Wert im Grundwasserstrom darauf Einfluss.
Es konnten weiterhin keine Zusammenhänge zwischen KW- und den AN-Gehalten in Oberböden (n=499), Unterböden (n=466), Torfen (n=127) sowie oxidierten (n=47) und reduzierten Gley-Horizonten (n=67) gefunden werden. In den Oberbodenhorizonten wurden mit Ausnahme einiger ebenfalls oberflächennaher Torfhorizonte die höchsten AN-Gehalte gemessen, die jedoch stets unterhalb des Prüfwertes von 0,4 mg kg-1 für Wachtumsbeeinträchtigungen liegen (ggf. Sonderfall Erdinger/Freisinger Moos aufgrund der stabilen Anreicherung von As aus dem Grundwasser im Oberboden). In den Unterbodenhorizonten2 wurden relativ geringe KW- (max. ca. 117 mg kg-1) und AN-Konzentrationen (max. 0,1 mg kg-1) beobachtet. In den Torfhorizonten liegen die KW-Werte mit zwei Ausnahmen unter 300 mg kg-1 und die AN-Werte unter 0,3 mg kg-1. In den Go-Horizonten liegen mit ≤ 0,1 mg kg-1 geringe AN-Gehalte vor. Auch die As-KW-Werte sind mit nur einem Datensatz über 200 mg/kg moderat und insgesamt liegen nur vier Proben über dem Maßnahmenwert von 50 mg kg-1 für Grünlandnutzung bzw. dem Prüfwert auf Ackerflächen und in Nutzgärten bei zeitweise reduzierenden Bedingungen. Ähnlich verhält es sich mit den Gr-Horizonten. Hier wird der Maßnahmenwert von 50 mg kg-1 nur in einer Probe überschritten und die AN-Gehalte betragen max. 0,1 mg kg-1.
2 In den Unterbodenhorizonten wurden alle A-, G-, H- und C-Horizonte sowie reliktische und fossile Horizonte ausgeschlossen.

LABO-Vorhaben B 1.10 31.5.2012
22
Für 1.278 Datensätze wurden zudem die Zusammenhänge zwischen AN- und KW-Gehalten von Arsen zu Fe-Dithionit, Fe-Oxalat, Feo/Fed-Quotient, pH-Wert und Corg betrachtet. Auch hier sind keine Zusammenhänge nachweisbar. Es deutet sich an, dass eine Zunahme der KW-Gehalte mit zunehmenden Fed-Gehalten erfolgt und dass sich erhöhte AN-Aktivitäten zeigen, wenn die Fe-Konzentrationen (Oxalat und Dithionit) sehr gering sind; gesicherte monokausale Beziehungen sind hier jedoch nicht ableitbar.
Auch nach den Erfahrungen der LfL Bayern werden bei Maßnahmenwertüberschreitung auf Grünland im AN-Extrakt sehr geringe Arsengehalte gemessen (< 0,5 mg kg-1) (LfL Bayern 2005).
In Nordrhein-Westfalen liegen im Fachinformationssystem Stoffliche Bodenbelastung (FIS StoBo) 39 As-Analysen (AN) von Ackerböden vor. Sie stammen überwiegend aus dem Raum Bochum und dem Einzugsgebiet der Ruhr. Alle Werte liegen im Bereich zwischen 0,01 und 0,1 mg kg-1, d.h. deutlich unter dem Prüfwert Boden-Pflanze für Wachstumsbeeinträchtigungen von 0,4 mg kg-1. Der maximale KW-Gehalt liegt bei 120 mg kg-1 (Mittlg. LANUV NRW 2011).
Arsentransfer Boden-Pflanze
Die Verschmutzung mit anhaftendem Boden stellt bei belüfteten Böden den maßgeblichen Aufnahmepfad für Arsen dar und es besteht ein geringes Risiko der Überschreitung des FutM-Höchstgehaltes über den systemischen Pfad.
Bei gering verschmutztem Grünlandaufwuchs besteht kein oder nur ein schwacher Zusammenhang zwischen den Arsengehalten (KW) im Boden und den Arsengehalten im Aufwuchs und auch kein Zusammenhang der Arsengehalte (AN) mit den Arsengehalten im Aufwuchs.
Eine Überschreitung des Höchstgehaltes der Futtermittelverordnung (FutMV) für Einzelfuttermittel von 2 mg kg-1 bei 88 % TM kann auf Standorten mit Arsenanreicherungen und einer Freisetzungsgefährdung aufgrund von zeitweise reduzierenden Bedingungen nicht ausgeschlossen werden. Allerdings können auch auf hoch belasteten Böden Lebens- und Futtermittelpflanzen angebaut und geerntet werden, die die bestehenden Grenzwerte einhalten. Überschreitungen des Arsen-Höchstgehaltes werden im Rahmen von Futtermitteluntersuchungen selten festgestellt (Erfahrungen aus SN: Kardel et al. 2011, mündl. Mittlg Landesdirektion Chemnitz; Erfahrungen aus BW: mündl. Mittlg. Regierungspräsidium Freiburg). Im Kreis Borken wurden in fünf untersuchten Grasproben messbare Arsenkonzentrationen festgestellt. In zwei Fällen wurde dabei der Höchstgehalt gemäß Futtermittelverordnung von 2 mg/kg deutlich überschritten. Demgegenüber sind im Kreis Coesfeld keine Überschreitungen im Futtermittel festgestellt worden (Erfahrungen aus NW: IFUA 2008).

LABO-Vorhaben B 1.10 31.5.2012
23
Pflanzen nehmen Arsen i. d. R. aus dem Boden nicht oder nur in sehr geringem Umfang über die Wurzel auf; eine Aufnahme über die Wurzel ist jedoch in schlecht durchlüfteten Böden möglich. Anhaltspunkte für eine Gefährdung der Futter- und Lebensmittelsicherheit werden nur kleinflächig festgestellt. In Lebensmitteln bzw. verzehrbaren Pflanzenteilen wurden messbare Arsengehalte vereinzelt z.B. bei Weizen geringfügig über der Nachweisgrenze (Erdinger, Freisinger und Dachauer Moos, BY), bei Rapskorn trotz Bodenbelastung < 0,03 mg/kg und Weizenkorn max. 0,06 mg/kg (88 % TM) (Ortenaukreis, BW) festgestellt. Zum Teil sind jedoch in Lebensmitteln keine Arsengehalte messbar (Kreis Coesfeld, NW).
Es deutet sich an, dass die Pflanzenart und -sorte einen starken Einfluss auf die Arsenaufnahme über die Wurzel hat (vgl. Kap. 2.2 und Serfling & Klose 2008).
Das aufgenommene Arsen verteilt sich in den verschiedenen Pflanzen-teilen unterschiedlich (Wurzel > Blätter oder Stängel > Früchte oder Körner).
Wegen der vielfältigen Sorptionsmöglichkeiten, der pH- und Redox-Abhängigkeit der Freisetzung, der heterogenen Verteilung im Boden und der ständig wechselnden Arsenspezies ist es schwierig, die Wirkung von Maßnahmen zur Verringerung des Arsentransfers vom Boden in Nutzpflanzen zu untersuchen.
Umgang mit Arsenbelastungen im Vollzug und Maßnahmen
Aufgrund der Vielzahl unterschiedlicher Einflussgrößen ist es nicht möglich, von den Schadstoffgehalten im Boden hinreichend sicher auf die Gehalte im Ernteprodukt zu schließen (Kardel et al. 2011, LfL Sachsen 2008).
Durch belastete Talfüllungen in alten Bergbauregionen liegt ein Erfordernis zur besonderen Zusammenarbeit zwischen Bodenschutz- und Altlastenbehörden mit den Stellen der Gesundheitsverwaltung, der Abfallrechtsbehörde, der Futtermittelrechtsbehörde, Landwirtschafts-, Lebensmittelrechts- sowie Baurechtsbehörde vor. Die Einbindung und Mitwirkung der Gemeinden sowie die Information ihrer Bürger ist von besonderer Bedeutung.
Zur Vereinfachung des Umgangs mit großflächigen As-Belastungen wird in Baden-Württemberg an der Entwicklung gebietsbezogener Regelungen gearbeitet, durch die unter vertretbarem Aufwand die gebotene Sicherheit für die Bodenbewirtschafter, -nutzer, Bürger, Kommunen und Verwaltung gewährleistet sowie der Vorsorge vor dem Entstehen schädlicher Bodenveränderungen Rechnung getragen wird (Regierungspräsidium Freiburg). Mit vergleichbaren Aufgaben sieht man sich in den Bergbaugebieten des Harzes und Sachsens konfrontiert.

LABO-Vorhaben B 1.10 31.5.2012
24
Auf gefährdeten Standorten sind regelmäßige und repräsentative Pflanzenuntersuchungen erforderlich und werden den betroffenen Flächennutzern zur Anwendung empfohlen (LfL Sachsen 2006a,b, LUA NRW 2006, Mittlg. Landratsamt Emmendingen). Die Vor-Ernte-Untersuchung stellt in belasteten Gebieten eine effektive Maßnahme dar, unerwünschte Belastungen z.B. von Getreide frühzeitig zu erkennen und zu minimieren (in Sachsen seit 2004 empfohlen, vgl. Kardel et al. 2011).
Das Regierungspräsidium Freiburg hat auf schwermetallbelastetem Grünland in zwei Tälern Kalkungsversuche durchgeführt und jeweils zu vier Terminen die Gesamtgehalte (KW) und mobilen Arsengehalte (AN) im Boden und die Gehalte im Aufwuchs untersuchen lassen. Die Bodenprobeahmetermine lagen überwiegend im Sommer und Herbst. Bei Gesamtgehalten von 27 bis 71 mg kg-1 As im KW treten zum letzten Beprobungstermin Anfang September in beiden Tälern mobile Arsengehalte von über 0,4 mg kg-1 auf. An den drei vorherigen Terminen lagen in beiden Untersuchungsgebieten die mobilen Arsengehalte unter 0,025 mg kg-1. Die Ursache für die höheren Gehalte zum letzten Beprobungstermin ist unklar. Eine Abnahme mobiler Arsengehalte oder der Arsengehalte im Aufwuchs durch die Kalkung deutet sich (aus den Werten für As) nur bedingt an. Bei Cd, Zn und Pb zeigt sich nach der Kalkung eindeutig eine Reduktion der AN-Gehalte. Ein Zusammenhang zwischen mobilem Bodengehalt und Arsengehalt im Aufwuchs ist nicht erkennbar. Dieser potenzielle Zusammenhang wird vermutlich überdeckt durch standortspezifische Einflüsse, Probenahmetermin, Verschmutzungsgrad u.a. Auch der deutliche Anstieg der Arsengehalte im AN schlägt sich in keiner Weise in den Pflanzengehalten nieder. Zweifel an der Verhältnismäßigkeit von kategorischen Nutzungsverboten aufgrund mobiler Bodengehalte wurden damit erhärtet. Die Qualität von Futteraufwüchsen lässt sich nicht ausschließlich anhand von Bodengehalten beurteilen. Die Verwaltung benötigt für sachgerechte Entscheidungen, die dem Einzelfall gerecht werden, Ermessensspielräume. Insgesamt untermauern die Versuche, dass bei Grünlandnutzung die Heranziehung von Bodengesamtgehalten nach BBodSchV für die Bewertung des Pfades Boden-Pflanze im Blick auf die futtermittelrechtlichen Grenzwerte sinnvoll ist. Auch auf Böden, die die Maßnahmenwerte nach der BBodSchV überschreiten, ist bei Anlage von futtermittelrechtlichen Maßstäben häufig die Verwertung des Aufwuchses möglich. Eine Prognose allein anhand von Bodenparametern, ob eine Futternutzung möglich ist, erscheint jedoch vage (Mittlg. Regierungspräsidium Freiburg 2011).
Im Fall hoher Arsenbelastungen im Boden wird z.B. im Bodenplanungsgebiet „Raum Freiberg“ in Sachsen und im Regierungsbezirk Freiburg die Verantwortung des Landwirts hinsichtlich seiner Pflichten als Lebensmittel- und Futtermittelunternehmer eindeutig klargestellt. Dazu dient ein Merkblatt bzw. die Verordnung zur Festlegung des Bodenplanungsgebietes. Liegt ein Grund zur Annahme vor, dass ein abgegebenes Lebensmittel nicht den gesetzlichen Anforderungen genügt, besteht im Regierungsbezirk Freiburg eine aktive Mitteilungspflicht an die Behörden. Es wird deutlich hervorgehoben, dass bereits ein fahrlässiger Verstoß gegen die Bestimmungen des Lebensmittelrechts eine Straftat darstellt. Hinsichtlich des Futtermittelrechts wird darauf hingewiesen, dass

LABO-Vorhaben B 1.10 31.5.2012
25
wer fahrlässig oder vorsätzlich gegen die Vorschriften verstößt, ordnungswidrig handelt (Mittlg. Regierungspräsidium Freiburg 2011, Landesdirektion Chemnitz 2011).
Zur Verminderung der Verschmutzung von Ernteprodukten sind jeweils angepasste Maßnahmen für unterschiedliche Bewirtschaftungssysteme geeignet; sie sind zusammengestellt in Elsässer et al. 2004 (LABO-Vorhaben B 4.03). Daneben können Anbau-, Dünge- und Nutzungsänderungen zur Verminderung einer systemischen Arsenaufnahme in Ernteprodukte beitragen (am umfassendsten aufgeführt in Serfling & Klose/LfL Sachsen 2008):
- Auswahl von Sorten mit möglichst geringer As-Aufnahme;
- P- Düngung bis 25 kg ha-1 unabhängig vom P-Gehalt (nach CAL Methode);
- Erhöhung des pH-Wertes im Boden bis in einen neutralen Bereich (≈ pH 6 -7);
- Vermeidung von Staunässe;
- Beseitigung von Verdichtungen beispielsweise durch tiefwurzelnde Zwischenfrüchte;
- gute Durchlüftung des Oberbodens, wenn sich dort die höchsten As-Konzentrationen befinden;
- Meiden älterer bodennaher Pflanzenteile bei der Verwendung als Futter.
3.3 Defizite
Wenige Vollzugserfahrungen liegen vor für:
- die Einflussfaktoren, die eine systemische Aufnahme steuern wie z. B. Nährstoffzustand und Sortenabhängigkeit (Ausnahme: Untersuchungen der LfL Sachsen zu Sortenabhängigkeit, Mykorrhizierung und Phosphor-Düngung, siehe Serfling & Klose / LfL Sachsen 2008),
- die Definition von Kriterien für die Gefahrenabschätzung (z. B. wie lange herrschen im Boden reduzierende Bedingungen und wie wird dies bewertet),
- die Anwendung alternativer Extraktionsverfahren zum Ammoniumnitrat zur Ermittlung mobiler (leicht freisetzbarer) Arsenanteile im Boden wie z.B. Ammoniumsulfat (Wenzel et al. 2001), Ammoniumacetat oder Ammoniumoxalat (Mattusch & Wennrich 1998),

LABO-Vorhaben B 1.10 31.5.2012
26
- den Einsatz von weiterführenden, i. d. R. aufwändigen Untersuchungs-ansätzen wie z. B. sequentieller Extraktion zur Ermittlung unterschiedlich mobiler Arsenanteile im Boden (von schwach adsorbiert bis residual in Mineralen nach Wenzel et al. 2001) oder Sickerwasserbeprobung,
- die Reproduzierbarkeit von Arsenuntersuchungen in Pflanzen je nach Probenahmeverfahren (z. B. auch zum Einfluss von Waschverfahren),
- die Anwendung einer angepassten Probenentnahme aus reduzierten Horizonten (die Ergebnisse des LABO-Projekts B 4.07 deuten darauf hin, dass die Probenvorbehandlung unter Schutzgas sich nicht maßgeblich auf die Ergebnisse für mobile Arsengehalte im Boden auswirkt) und
- den Einfluss des Witterungsverlaufs (z. B. stellt sich die Frage nach einer höheren Arsenanreicherung in Pflanzen in sehr nassen Jahren).
Keine Erfahrungen aus der Vollzugspraxis sind bekannt für
- die Untersuchung und Bewertung von verschiedenen Arsenspezies,
- die praktische Umsetzung von Maßnahmen mit veränderten Anbaustra-tegien mit Pflanzen geringen Anreicherungspotenzials (Anbauempfehlungen für Getreide- und Gemüsesorten leiten sich derzeit insbesondere aus den lebensmittelrechtlichen Höchstwerten für Blei und Cadmium ab, z.B. Landratsamt Emmendingen 2001) sowie
- die Nutzung von Indikatoren wie Zeigerpflanzen und Pflanzenschäden zur Erfassung von Verdachtsflächen und zur Gefahrenbeurteilung.
Unterschiedliche Erfahrungen sind festzustellen im Hinblick auf
- angewendete Methoden zur Erfassung von Verdachtsflächen und zur räumlichen Differenzierung, Stichprobenhafte Analysen von Ober- und Unterböden im Bereich naturbedingt erhöhter Arsengehalte im Bereich arsenführender Gesteine (BW, Reg.bez. Karlsruhe); Bodenanalysen und flächenhafte Darstellung von Arsengehalten der Oberböden in Digitalen Bodenbelastungskarten (NW); Ausweisung von Bodenplanungsgebieten (Harz/Goslar - Neufassung 2011), Raum Freiberg / Erzgebirge (2011)
- die angewendeten Methoden zur Ermittlung des Verschmutzungsgrads in Gefährdungsabschätzungen, Unterschiedliche Konventionen: Während in BW und NW wie in Elsässer et al. (2004) 3 % als unvermeidbar gilt (LUBW 2011, Kreis Borken 2009, Kreis Coesfeld 2009), wird in BB unter günstigen Bedingungen ein Anteil von 1 % angenommen (LUA Brandenburg 2010).
- die angewendeten Methoden zur Ermittlung / Abschätzung eines Transfers Boden – Pflanze, Messung mobiler Anteile oder sequentielle Extraktion � Abschätzung mit Regressionsfunktionen; untersuchte Pflanzenteile

LABO-Vorhaben B 1.10 31.5.2012
27
- die Handlungsempfehlungen unter Berücksichtigung der pH-Abhängigkeit der Arsenfreisetzung. z. B. Empfehlung von Ziel-pH-Werten in NW-Kreisen Coesfeld und Borken; Empfehlung der Demobilisierung der Schadstoffe durch Kalkung auf pH 6,5 bis 6,7 auf Grünland im Oberharz (LWK NI); kein Nutzen einer Aufkalkung für Arsen in aid 2009 und unveröffentlichten Versuchsergebnissen des RP Freiburg; Empfehlung der Erhöhung des pH-Wertes im Boden bis in einen neutralen Bereich in Serfling & Klose/LfL Sachsen 2008; Hinweis, dass eine Aufkalkung sogar eine Erhöhung der Arsenmobilität in Böden bewirken kann in MUNLV, AAV & LUA NRW 2003 sowie Kiesewalter & Röhricht 2008.
3.4 Schlussfolgerungen
Aus den Vollzugserfahrungen bei der Erfassung von Verdachtsflächen, der Gefahrenbeurteilung (Orientierende Untersuchung, Detailuntersuchung) sowie der Gefahrenabwehr wurden – unter Berücksichtigung der Erkenntnisse der Literaturrecherche und den Ergebnissen der Gefäß- und Feldversuche – Empfehlungen für den Leitfaden Arsentransfer aus Böden in Nahrungs- und Futterpflanzen abgeleitet.
Als Schlussfolgerung der Diskussion der Werteregelungen der BBodSchV in der projektbegleitenden Arbeitsgruppe wurde festgestellt, dass der Maßnahmenwert für Grünland von 50 mg kg-1 (KW) und der Prüfwert für Ackerbau von 200 bzw. abgesenkt von 50 mg kg-1 (KW) sachgerecht sind. Der Prüfwert für Acker im Hinblick auf mögliche Wachstums-beeinträchtigungen in Höhe von 0,4 mg/kg (AN) wird nach derzeitigen Erfahrungen als bedingt sachgerecht angesehen (i.d.R. zu geringe Extraktionsausbeute und geringer Zusammenhang mit As-Konzentrationen in Pflanzenkompartimenten). Der Wert ist jedoch bei der derzeit bestehenden Rechtslage im Vollzug anzuwenden.
LABO-Vorhaben B 1.10 31.5.2012
28
4 AUSSAGEKRAFT UND AUFWAND VON ARSENSPEZIES-UNTERSUCHUNGEN
ahu AG, TU Dresden
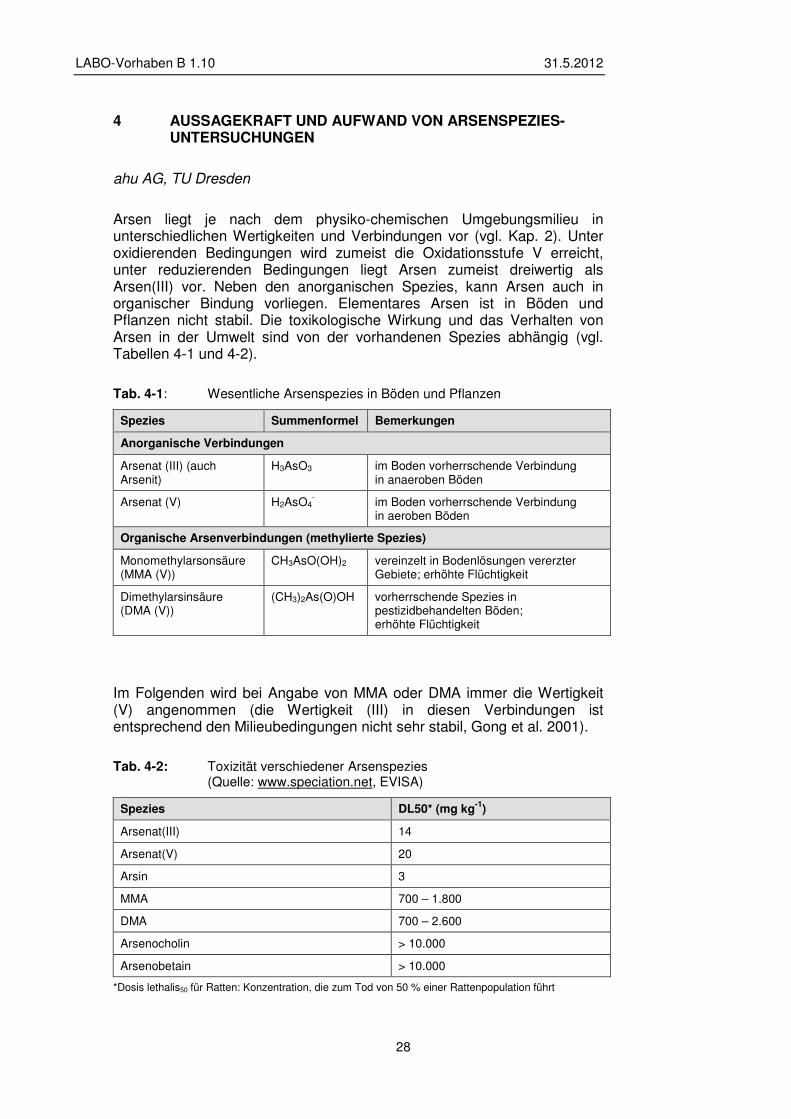
Arsen liegt je nach dem physiko-chemischen Umgebungsmilieu in unterschiedlichen Wertigkeiten und Verbindungen vor (vgl. Kap. 2). Unter oxidierenden Bedingungen wird zumeist die Oxidationsstufe V erreicht, unter reduzierenden Bedingungen liegt Arsen zumeist dreiwertig als Arsen(III) vor. Neben den anorganischen Spezies, kann Arsen auch in organischer Bindung vorliegen. Elementares Arsen ist in Böden und Pflanzen nicht stabil. Die toxikologische Wirkung und das Verhalten von Arsen in der Umwelt sind von der vorhandenen Spezies abhängig (vgl. Tabellen 4-1 und 4-2).
Tab. 4-1: Wesentliche Arsenspezies in Böden und Pflanzen
Spezies Summenformel Bemerkungen
Anorganische Verbindungen
Arsenat (III) (auch Arsenit)
H3AsO3 im Boden vorherrschende Verbindung in anaeroben Böden
Arsenat (V) H2AsO4- im Boden vorherrschende Verbindung
in aeroben Böden
Organische Arsenverbindungen (methylierte Spezies)
Monomethylarsonsäure (MMA (V))
CH3AsO(OH)2 vereinzelt in Bodenlösungen vererzter Gebiete; erhöhte Flüchtigkeit
Dimethylarsinsäure (DMA (V))
(CH3)2As(O)OH vorherrschende Spezies in pestizidbehandelten Böden; erhöhte Flüchtigkeit
Im Folgenden wird bei Angabe von MMA oder DMA immer die Wertigkeit (V) angenommen (die Wertigkeit (III) in diesen Verbindungen ist entsprechend den Milieubedingungen nicht sehr stabil, Gong et al. 2001).
Tab. 4-2: Toxizität verschiedener Arsenspezies (Quelle: www.speciation.net, EVISA)
Spezies DL50* (mg kg-1)
Arsenat(III) 14
Arsenat(V) 20
Arsin 3
MMA 700 – 1.800
DMA 700 – 2.600
Arsenocholin > 10.000
Arsenobetain > 10.000
*Dosis lethalis50 für Ratten: Konzentration, die zum Tod von 50 % einer Rattenpopulation führt

LABO-Vorhaben B 1.10 31.5.2012
29
Im Folgenden wird die Aussagekraft von Speziesanalysen für Arsen dem dafür erforderlichen Aufwand gegenübergestellt. Die Praktikabilität im Rahmen von bodenschutzrechtlichen Amtsermittlungen (§ 9 Abs. 1 BBodSchG) bzw. im Rahmen von Detailuntersuchungen (§ 9 Abs. 2 BBodSchG) wird geprüft und bewertet.
Anwendungsbereiche
Die Notwendigkeit der Unterscheidung von Arsenspezies bei der Bewertung des toxikologischen oder ökotoxikologischen Potenzials ist im medizinischen Bereich allgemein anerkannt. Eine häufige Anwendung ist die As-Speziierung in Urinproben. Zur Praxis der Lebens-mitteluntersuchung gehört die Bestimmung von anorganischem Arsen in Algen und des als Arsenobetain vorliegenden Arsens in Fischereierzeugnissen und in Krusten-, Schalen- und Weichtieren (vgl. Amtliche Sammlung von Untersuchungsverfahren nach § 64 Lebensmittel- und Futtermittelgesetzbuch LFGB). Hinsichtlich der Arsenspezies-untersuchung in Boden und Pflanzen sind die Erkenntnisse jedoch derzeit noch lückenhaft.
Im Ergebnis der Untersuchung eines breiten Spektrums von Grünlandpflanzenarten (insgesamt 29) bezüglich ihres Gehaltes an organischen und anorganischen Arsenspezies sind bis zu 90 % des extrahierbaren As-Gehaltes aus den toxischen Spezies As(III) und As(V) zusammengesetzt und nur ca. 10 % aus mindertoxischen organischen Verbindungen (MMA und DMA, unbekannte organische Spezies). Dabei konnte keine Abhängigkeit des Vorkommens organischer Spezies vom Standort und von der Pflanzenart festgestellt werden. Die an der Pflanze anhaftenden Bodenpartikel erhöhte nicht den Gehalt organischer Spezies in der Pflanze (LfL Sachsen 2006b).
Indirekte Rückschlüsse von Bodeneigenschaften auf die im Boden oder der Pflanze vorliegenden Arsenspezies sind schwierig. So können mittels Pt-Elektroden im ungestörten Boden gemessene Redoxpotenziale nicht für quantitative Aussagen über auftretende Arsenspezies herangezogen werden. Dies liegt zum einen daran, dass generell durch Redox-Elektroden Redoxspezies nur dann erfasst werden, wenn sie in der Messlösung ausreichend hoch konzentriert sind und wenn das beteiligte Redoxpaar elektrodenwirksam ist, d. h. in ausreichender Geschwindigkeit Elektronen auf die Elektrode überträgt. Zum anderen weichen besonders unter reduzierenden Bedingungen die Redoxpotenziale, welche in der Bodenlösung gemessen wurden, deutlich (bis zu 400 mV) von den im ungestörten Boden gemessenen Werten ab (Overesch et al. 2008). Zudem ist der Zusammenhang zwischen dem Vorkommen bestimmter Arsenspezies im Boden und in der Pflanze nicht eindeutig, da es im Zuge der Aufnahme und Translokation in der Pflanze zu einer veränderten Speziierung kommen kann (vgl. Kap. 2).

LABO-Vorhaben B 1.10 31.5.2012
30
Verfahren zur Arsen-Speziesbestimmung
Die Bestimmung von As-Spezies ist prinzipiell in der wässrigen Phase (z. B. Bodenlösung, Grundwasser, Eluat) möglich. Analysen an Pflanzen oder der Bodenfestphase erfordern eine vorangehende Elution, bei der die As-Spezies erhalten bleiben. Oxidativ aufgeschlossene Proben kommen für die As-Speziierung nicht in Frage. Die Bestimmung einzelner organischer As-Spezies ist derzeit äußerst aufwändig und in der Laborpraxis nicht standardisiert. In der Regel wird As(III) und As(V) bestimmt und der Rest als organisch angesprochen. As-Speziierung in Boden und Pflanze ist Gegenstand aktueller Forschung und kommt derzeit vor allem auf Rüstungsaltlasten zum Einsatz (arsenorganische Kampfstoffe). Nur wenige Laboratorien führen derzeit Analysen zur As-Speziierung durch.
Ein generelles genormtes Verfahren zur Arsen-Speziesbestimmung existiert bislang nicht. Aktuell in der Praxis eingesetzte Verfahren sind:
� Arsenanalyse mit Hydridtechnik nach EN ISO 11969, wobei der in der Norm vorgesehene spezieshomogenisierende Aufschluss durch eine speziesselektive Hydridgenerierung nach Anderson et al. (1986) ersetzt ist. Definierte Millieubedingungen ermöglichen die selektive Erfassung von As (III). Anwendungsbeispiel: Overesch et al (2008). As(V) dort als Differenz zum Gesamt-Arsen.
� Ionenchromatographische Trennung mit plasma-massenspektrometrischer Detektion. Anwendungsbeispiel: LfL Sachsen / Haßler und Klose 2006b. Dort wurde die Methode nach Kohlmeyer et al. 2002 eingesetzt. Diese funktioniert mit einem sauren Eluenten unter Zuhilfenahme eines Ionenpaar-Reagenzes und ermöglicht die Erfassung von As(III), As(V) und mehrerer organischer Spezies. Basis-Arbeiten zu dieser Technik: Mattusch & Wennrich 1998 und Francesconi et al. 2002. (Die Methode ist an der TU Dresden verfügbar.)
� Photometrische Arsenanalyse mit Silberdiethylthiocarbamat nach DIN EN 26595, wobei der in der Norm vorgesehene spezieshomogenisierende Aufschluss durch eine speziesselektive Hydridgenerierung ersetzt ist (z. B. Labor Wessling, Hannover).
As-Speziierung ist zum Teil mit einem sehr viel höheren logistischen Aufwand für Probenvorbereitung, Analyse, Technik und Sicherheit im Vergleich zur Gesamtarsenanalyse verbunden. Die Kosten für Arsenspeziesuntersuchungen in Boden- und Wasserproben bzw. Pflanzen schwanken erheblich, da schon bei der Extraktion/Aufbereitung je nach Probe verschiedene analytische Probleme zu umgehen sind (z. B. Analyse unter Schutzgasatmosphäre, da Arsenspezies nicht redoxstabil sind). Zudem hängt der jeweilige Preis stark vom untersuchten Kompartiment ab (Wasser am preiswertesten, Boden je nach Eluierbarkeit, Pflanzen am aufwändigsten). Zudem preisbestimmend sind die zu bestimmenden Bindungsformen (organisch >> anorganisch), die Extraktion und

LABO-Vorhaben B 1.10 31.5.2012
31
Eluierbarkeit, die Anzahl analysierter Proben, die eingesetzte Technik (z. B. ICP-MS, HPLC) und der Einsatzbereich (Forschung � Gutachterliche Praxis).
Über Erfahrungen mit der Analyse von As-Spezies verfügen3:
� Universitäten und Forschungseinrichtungen:
o TU Dresden (Prof. Dudel)
o TU Freiberg (Entwicklung Routine-Methode Pflanze)
o Prof. Küper, München (Entwicklung Routine-Methode Pflanze)
o Universität Bayreuth, Bayreuther Zentrum für Ökologie und Umweltforschung, Prof. Matzner, G. Ilgen
o Universität Bayreuth, Umweltgeochemie, Prof. Planer-Friedrich
o BOKU Wien, Prof. Gunda Koellensperger (Übergang Boden – Pflanze, Ribosomübergang)
o UFZ Leipzig (FB Ökotoxikologie, Umwelt Analytik, Dr. Mattusch, Dr. Rainer Wennrich; Dr. Daus)
o Universität Köln (Geographisches Institut, Prof. Tim Mansfeldt)
o TU Bergakademie Freiberg (Geologie, Prof. Broder Merkel, Dipl.-Chem. Peter Volke)
o TU Berlin (Gesamtarsen über wellenlängen-dispersive Röntgenfluoreszenzspektroskopie (WD-RFA), Brandt 2003)
� Privatwirtschaftliche und behördliche Labore: o Bundesanstalt für Materialforschung und -prüfung BAM (Prof. Irene
Nehls, As-Proteinwechselwirkung)
o Bundesanstalt für Geowissenschaften und Rohstoffe BGR (Dr. Jens Stummeyer, Harazim Bodo; Bergbaualtlasten, Methodenentwicklung)
o IFE Analytik Leipzig (Dr. Michael Hanrieder)
o Labor Wessling, Hannover (Hr. Habersaat, Hr. Symura in Bochum)
o Toxlab Zentrallabors für Toxikologische Analysen Universität Duisburg-Essen (http://www.toxlab.de/analytik/)
o Staatliche Betriebsgesellschaft für Umwelt und Landwirtschaft BfUL (GB 6, Dr. Ralf Klose)
Bewertung von Aussagekraft und Anwendung
Die Unterschiede im chemischen Verhalten und in der Toxizität verschiedener Arsenverbindungen (vgl. Tab. 4-1) führen zu der Annahme, dass es für eine Gefahrenbeurteilung nützlich ist, die Konzentrationen der einzelnen Spezies zu bestimmen statt nur den Gesamtgehalt an Arsen. Der potenzielle Anwendungsbereich von Speziesanalysen liegt in der Detailuntersuchung, wenn z.B. in einer abschließenden Gefährdungs-abschätzung die Belastungsursache(n), die Möglichkeiten der
3 Die Zusammenstellung basiert auf einer Literaturrecherche und telefonischen Anfragen. Sie erhebt keinen Anspruch auf Vollständigkeit.

LABO-Vorhaben B 1.10 31.5.2012
32
Schadstoffausbreitung und die Exposition von Schutzgütern zu beurteilen sowie geeignete Gefahrenabwehrmaßnahmen festzulegen sind.
Sinnvoll wäre eine Speziesuntersuchung jedoch nur dann, wenn zu erwarten ist, dass Arsenspezies unterschiedlicher Toxizität in Pflanzen oder Arsenspezies unterschiedlicher Aufnahmeeigenschaften im Bodenwasser auftreten. Theoretisch könnte z. B. ein Gefahrenverdacht ausgeräumt werden, wenn das Arsen im Ernteprodukt ausschließlich in Spezies mit geringer Toxizität vorliegt (z. B. als organisch gebundene Spezies). Gleichwohl könnte theoretisch ein Gefahrenverdacht durch einen Nachweis von maßgeblichen Anteilen hoch toxischer Arsenspezies (z. B. As(III) und As(V)) im Ernteprodukt belegt werden. Wie jedoch in Kapitel 2 beschrieben gibt die Untersuchung von Arsenspezies in der Bodenlösung zwar einen Hinweis, ob mehr methylierte oder anorganische Arsenspezies aufgenommen werden bzw. werden können, sie lässt aber keine quantitativen Aussagen zum Aufnahmeverhalten in die Pflanze zu.
In Landpflanzen kommen nach aktuellem Kenntnisstand im Wesentlichen die beiden toxischen anorganischen Spezies As(III) und As(V) vor. Die weniger toxischen organisch gebundenen Spezies (insbes. MMA und DMA) treten nur in geringen Anteilen auf (vgl. Kap. 2.2). Für die in gemäßigtem Klima angebauten Getreide- und Grünlandpflanzen beinhaltet die Auftrennung von anorganisch und organisch gebundenen As-Spezies in Pflanzen nach derzeitigem Stand somit nur eine geringe Aussagekraft. Der Kenntnisstand sollte allerdings durch weitere Forschungsaktivitäten erweitert werden. Im Hinblick auf die hohen Kosten und die bislang fehlende Standardisierung der Probenvorbereitungs- und Analyseverfahren wird eine As-Speziierung in der Bodenlösung und in Ernteprodukten derzeit als nicht verhältnismäßig angesehen. Auch die Identifizierung von As(III) und As(V) erbringt keinen bewertbaren Mehrwert für die Gefahrenbeurteilung, denn die Konzentrationen in Pflanzen, ab deren Höhe einzelne As-Spezies toxisch wirken, sind bislang nicht bekannt.
Im rechtlichen Umfeld von Futtermittel- und Lebensmittelproduktion sowie im Bodenschutz fehlen derzeit Beurteilungs- und Grenzwerte für verschiedene Arsenspezies sowohl in der Bodenlösung als auch in Pflanzen. Eine toxikologisch begründete und reproduzierbare Beurteilung der Ergebnisse einer As-Speziesanalytik in Bodenlösung und Ernteprodukten in der Praxis ist somit derzeit nicht erreichbar. Es ist jedoch möglich, dass sich eine Auftrennung von anorganisch und organisch gebundenen Arsenspezies künftig in den rechtlichen Regelungen zur Futtermittelsicherheit niederschlagen wird4. Erst wenn sich dies konkret abzeichnet, eine Beurteilungsmethodik und entsprechende standardisierte Untersuchungsverfahren vorliegen, ist eine Anwendung dieser Verfahren in der Praxis zu empfehlen.
4 siehe z. B. Aktivitäten der CEN/Technical Committee 327 „Animal feedingstuffs – Methods of sampling ans analysis“ zur Methodenentwicklung zur Bestimmung von anorganischem Arsen (As(III) + As(V))

LABO-Vorhaben B 1.10 31.5.2012
33
5 KONZEPTION UND ERGEBNISSE DER GEFÄß- UND FELDVERSUCHE
TU Dresden
Die Konzeption der Gefäß- und Feldversuche wurde in enger Abstimmung mit dem Projektbetreuer und dem projektbegleitenden Arbeitskreis erstellt. Den Schwerpunkt der Arbeiten bildeten die Untersuchungen mittels Gefäßversuchen, die durch angepasste Felduntersuchungen ergänzt wurden.
Ziel der Untersuchungen war eine exemplarische Ergänzung von Erkenntnissen zum Einfluss des Redoxpotenzials im Boden / in der Boden-lösung auf die Aufnahme von Arsen durch die Pflanzen. Hier bestehen derzeit noch deutliche Wissenslücken (s. Kap. 2). Die Zielstellung ist mit folgenden Kernfragen unterlegt:
1. Führt ein verringertes Redoxpotenzial im Boden zu einer verstärkten Aufnahme von Arsen bei Getreide als auch bei Grünlandpflanzen?
2. Wird der Effekt durch ein verändertes Redoxpotenzial im Boden beeinflusst durch die Verfügbarkeit von Phosphat (Düngung)?
3. Sind die Effekte durch Redoxpotenzial und P-Verfügbarkeit überlagert von anderen Effekten (z. B. N-Düngung, anderen Bodeneigenschaften)?
5.1 Gefäßversuche – Material und Methoden
Um den Zielstellungen und gleichzeitig dem Aspekt unterschiedlicher Arsenbelastungen zu entsprechen, wurde beschlossen, für die Gefäßversuche Substrate unterschiedlicher Standorte aus verschiedenen Bundesländern einzubeziehen. Vorzugsweise sollten Standorte ausgewählt werden, zu denen Voruntersuchungen vorliegen und die gleichzeitig momentan über eine entsprechende Messausstattung verfügen. Schließlich war die derzeitige Nutzung als Grünland oder Ackerfläche ein weiteres Auswahlkriterium.

LABO-Vorhaben B 1.10 31.5.2012
34
Drei Standorte für die Entnahme von Bodenmaterial für die Gefäßversuche wurden ausgewählt:
1. Colditz in der Nähe einer Bodendauerbeobachtungsfläche (SN, BDF 33, Grünland; Kontakt über Fr. Barth, LfULG SN);
2. Niederschopfheim im Ortenaukreis (BW, Acker; Kontakt über Hr. Nöltner, LUBW);
3. Heubachniederung (NW, Grünland; Kontakt über Hr. Neite, LANUV NRW).
5.1.1 Versuchsdesign
Es wurde vereinbart, Gefäßversuche mit Bezug zur Grünlandnutzung von jenen mit Bezug zu Ackernutzung im Versuchsdesign zu trennen.
Der Gefäßversuch mit Bezug zur Grünlandnutzung beschränkt sich auf den Einsatz von Deutschem Weidelgras (Lolium perenne, Sorte „KERATOS“). Als Variablen (Faktoren) mit wenigstens zwei Faktorstufen wurden Bodenfeuchte und Substrat der drei unterschiedlichen Standorte festgelegt. Das Versuchsdesign ist in Tab. 5-1 dargestellt.
Tab. 5-1: Versuchsdesign Gefäßversuch mit Bezug Grünlandnutzung. Ausgewählte Pflanzenart: Lolium perenne (Sorte KERATOS). Es ist die im Weiteren verwendete Bezeichnung (Bez.) und die Anzahl der Wiederholungen (n) angegeben.
Bodenfeuchte Bez. Standort (Substrat) Bez. n
mäßig feucht 70 Colditz (SN) Colditz 5 (60 – 80 % Feldkapazität) Heubachniederung (NRW) Heubach 5
Niederschopfheim (BW) Schopfheim 5
staunass 100 Colditz (SN) Colditz 5 (periodischer Überstau) Heubachniederung (NRW) Heubach 5
Niederschopfheim (BW) Schopfheim 5
Der Gefäßversuch mit Bezug zur Ackernutzung beschränkt sich auf den Einsatz von Winterweizen (Triticum aestivum; Sorte „AKTEUR“). Die Sortenwahl fand auf der Grundlage von Ergebnissen von Serfling & Klose (2008) statt. In dieser Untersuchung wird das Aufnahme- und Akkumulationsverhalten von Arsen durch verschiedene Winterweizen- und Sommergerstesorten beschrieben. Es zeigte sich, dass für die Sorte „AKTEUR“ im Vergleich der verschiedenen Sorten hohe Arsenkonzentrationen in oberirdischen Pflanzenkompartimenten nachgewiesen werden konnten. Gleichzeitig ist diese Sorte im Anbau sehr weit verbreitet. Beide Fakten zusammengenommen führten zur Auswahlentscheidung.
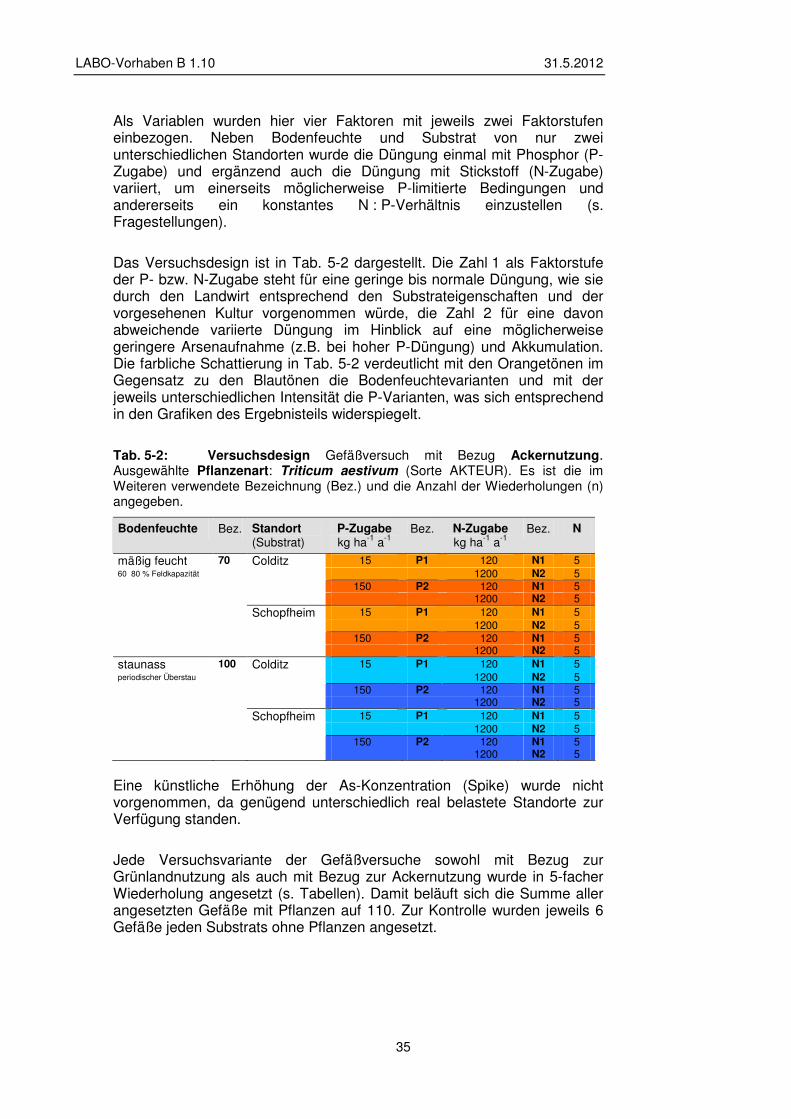
LABO-Vorhaben B 1.10 31.5.2012
35
Als Variablen wurden hier vier Faktoren mit jeweils zwei Faktorstufen einbezogen. Neben Bodenfeuchte und Substrat von nur zwei unterschiedlichen Standorten wurde die Düngung einmal mit Phosphor (P-Zugabe) und ergänzend auch die Düngung mit Stickstoff (N-Zugabe) variiert, um einerseits möglicherweise P-limitierte Bedingungen und andererseits ein konstantes N : P-Verhältnis einzustellen (s. Fragestellungen).
Das Versuchsdesign ist in Tab. 5-2 dargestellt. Die Zahl 1 als Faktorstufe der P- bzw. N-Zugabe steht für eine geringe bis normale Düngung, wie sie durch den Landwirt entsprechend den Substrateigenschaften und der vorgesehenen Kultur vorgenommen würde, die Zahl 2 für eine davon abweichende variierte Düngung im Hinblick auf eine möglicherweise geringere Arsenaufnahme (z.B. bei hoher P-Düngung) und Akkumulation. Die farbliche Schattierung in Tab. 5-2 verdeutlicht mit den Orangetönen im Gegensatz zu den Blautönen die Bodenfeuchtevarianten und mit der jeweils unterschiedlichen Intensität die P-Varianten, was sich entsprechend in den Grafiken des Ergebnisteils widerspiegelt.
Tab. 5-2: Versuchsdesign Gefäßversuch mit Bezug Ackernutzung. Ausgewählte Pflanzenart: Triticum aestivum (Sorte AKTEUR). Es ist die im Weiteren verwendete Bezeichnung (Bez.) und die Anzahl der Wiederholungen (n) angegeben.
Bodenfeuchte Bez. Standort (Substrat)
P-Zugabe kg ha-1 a-1
Bez. N-Zugabe kg ha-1 a-1
Bez. N
mäßig feucht 70 Colditz 15 P1 120 N1 5 60 80 % Feldkapazität 1200 N2 5 150 P2 120 N1 5 1200 N2 5 Schopfheim 15 P1 120 N1 5 1200 N2 5 150 P2 120 N1 5 1200 N2 5 staunass 100 Colditz 15 P1 120 N1 5 periodischer Überstau 1200 N2 5 150 P2 120 N1 5 1200 N2 5 Schopfheim 15 P1 120 N1 5 1200 N2 5 150 P2 120 N1 5 1200 N2 5
Eine künstliche Erhöhung der As-Konzentration (Spike) wurde nicht vorgenommen, da genügend unterschiedlich real belastete Standorte zur Verfügung standen.
Jede Versuchsvariante der Gefäßversuche sowohl mit Bezug zur Grünlandnutzung als auch mit Bezug zur Ackernutzung wurde in 5-facher Wiederholung angesetzt (s. Tabellen). Damit beläuft sich die Summe aller angesetzten Gefäße mit Pflanzen auf 110. Zur Kontrolle wurden jeweils 6 Gefäße jeden Substrats ohne Pflanzen angesetzt.

LABO-Vorhaben B 1.10 31.5.2012
36
Für die Etablierung der Gefäßversuche wurden die notwendigen orga-nisatorischen und logistischen Vorarbeiten im August und September 2010 durchgeführt.
5.1.2 Substrat – Beschaffung, Vorbereitung und Charakterisierung
Die Gewinnung der Substrate von den vereinbarten Standorten erwies sich als organisatorische, logistische und zeitliche Herausforderung. Eine schnelle Substratbeschaffung war von eminenter Wichtigkeit, um die Gefäßversuche noch im Oktober 2010 etablieren und damit den Winterweizen als junge Schösslinge überwintern lassen zu können (Vernalisierung). Tatsächlich wurden die Substrate am 22.09.10 (Colditz), 03.10.10 (Heubachniederung) und 18.10.10 (Niederschopfheim) geerntet und nach Tharandt auf das Ökologische Prüffeld verbracht.
Vom Standort Colditz wurde das Substrat aus einer Tiefe von 10–60 cm entnommen. Dies entspricht den am Aufschluss der Bodendauerbeoachtungsfläche 33 beschriebenen Horizonte aAh – IV aM. Allerdings wurde am aktuellen Probenahmestandort für diesen Versuch (ca 100 m entfernt) eine stärkere Mächtigkeit des II aM Horizonts ohne die beschriebenen linsenförmigen Holzkohleeinlagerungen festgestellt. Am Standort Heubachniederung wurde das Substrat aus einer Tiefe von 15 - 60 cm gewonnen während am Probenahmestandort Niederschopfheim das Substrat aus 0-30 cm Tiefe zwischen den Pflanzreihen eines Maisackers geerntet wurde. Der Standort hier wurde so gewählt, dass er in unmittelbarer Nähe (ca. 10 m Entfernung) zu einem vorher angelegten Aufschluss mit ausführlicher Bodencharakterisierung lag (Vergleichswerte).
Colditz Heubachniederung Niederschopfheim
Abb. 5-1: Gewinnung der Substrate von den festgelegten Standorten in den Bundesländern Sachsen (Colditz), Nordrhein-Westfalen (Heubachniederung) und Baden-Württemberg (Niederschopfheim).
Zur Vorbereitung des Versuchsansatzes wurden die Substrate auf 1 cm gesiebt. Dazu mussten die schluffig-tonigen Substrate (vgl. Tab. 5-3) von den Standorten Heubachniederung und Niederschopfheim durch Ausbreiten auf gesäuberten PP-Platten zunächst in ihrem Feuchtegehalt
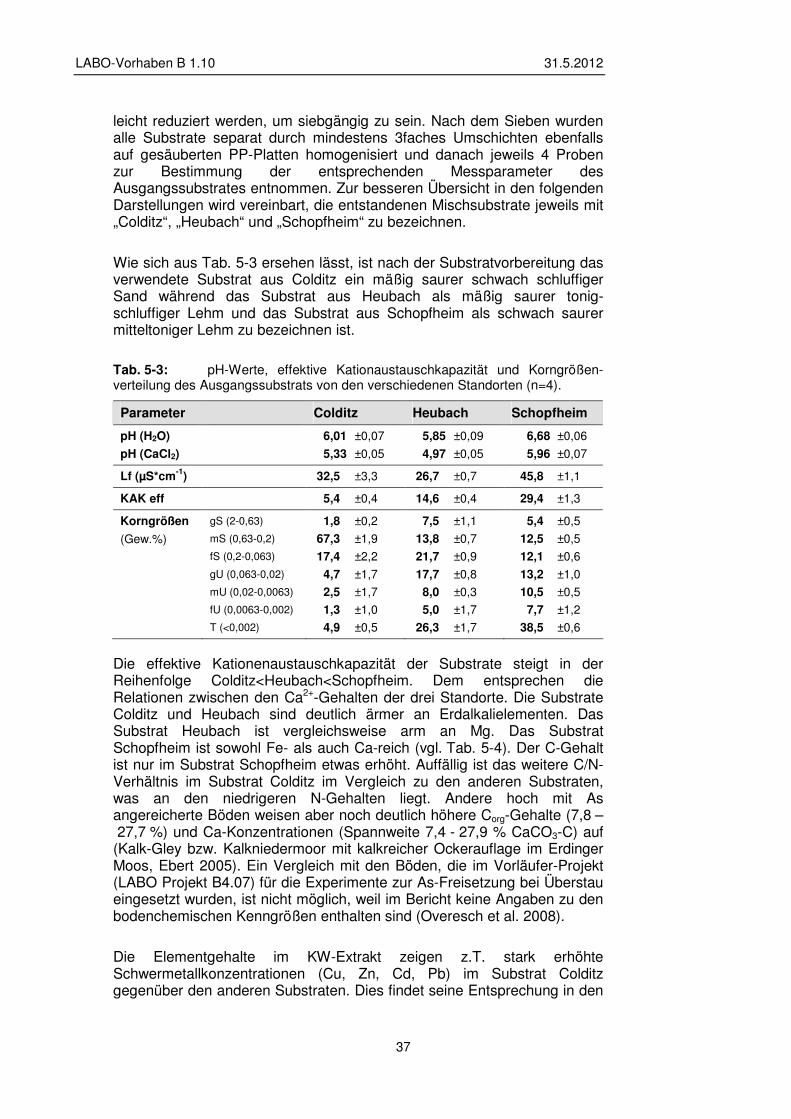
LABO-Vorhaben B 1.10 31.5.2012
37
leicht reduziert werden, um siebgängig zu sein. Nach dem Sieben wurden alle Substrate separat durch mindestens 3faches Umschichten ebenfalls auf gesäuberten PP-Platten homogenisiert und danach jeweils 4 Proben zur Bestimmung der entsprechenden Messparameter des Ausgangssubstrates entnommen. Zur besseren Übersicht in den folgenden Darstellungen wird vereinbart, die entstandenen Mischsubstrate jeweils mit „Colditz“, „Heubach“ und „Schopfheim“ zu bezeichnen.
Wie sich aus Tab. 5-3 ersehen lässt, ist nach der Substratvorbereitung das verwendete Substrat aus Colditz ein mäßig saurer schwach schluffiger Sand während das Substrat aus Heubach als mäßig saurer tonig-schluffiger Lehm und das Substrat aus Schopfheim als schwach saurer mitteltoniger Lehm zu bezeichnen ist.
Tab. 5-3: pH-Werte, effektive Kationaustauschkapazität und Korngrößen-verteilung des Ausgangssubstrats von den verschiedenen Standorten (n=4).
Parameter Colditz Heubach Schopfheim
pH (H2O) 6,01 ±0,07 5,85 ±0,09 6,68 ±0,06
pH (CaCl2) 5,33 ±0,05 4,97 ±0,05 5,96 ±0,07
Lf (µS*cm-1) 32,5 ±3,3 26,7 ±0,7 45,8 ±1,1
KAK eff 5,4 ±0,4 14,6 ±0,4 29,4 ±1,3
Korngrößen gS (2-0,63) 1,8 ±0,2 7,5 ±1,1 5,4 ±0,5
(Gew.%) mS (0,63-0,2) 67,3 ±1,9 13,8 ±0,7 12,5 ±0,5
fS (0,2-0,063) 17,4 ±2,2 21,7 ±0,9 12,1 ±0,6
gU (0,063-0,02) 4,7 ±1,7 17,7 ±0,8 13,2 ±1,0
mU (0,02-0,0063) 2,5 ±1,7 8,0 ±0,3 10,5 ±0,5
fU (0,0063-0,002) 1,3 ±1,0 5,0 ±1,7 7,7 ±1,2
T (<0,002) 4,9 ±0,5 26,3 ±1,7 38,5 ±0,6
Die effektive Kationenaustauschkapazität der Substrate steigt in der Reihenfolge Colditz<Heubach<Schopfheim. Dem entsprechen die Relationen zwischen den Ca2+-Gehalten der drei Standorte. Die Substrate Colditz und Heubach sind deutlich ärmer an Erdalkalielementen. Das Substrat Heubach ist vergleichsweise arm an Mg. Das Substrat Schopfheim ist sowohl Fe- als auch Ca-reich (vgl. Tab. 5-4). Der C-Gehalt ist nur im Substrat Schopfheim etwas erhöht. Auffällig ist das weitere C/N-Verhältnis im Substrat Colditz im Vergleich zu den anderen Substraten, was an den niedrigeren N-Gehalten liegt. Andere hoch mit As angereicherte Böden weisen aber noch deutlich höhere Corg-Gehalte (7,8 – 27,7 %) und Ca-Konzentrationen (Spannweite 7,4 - 27,9 % CaCO3-C) auf (Kalk-Gley bzw. Kalkniedermoor mit kalkreicher Ockerauflage im Erdinger Moos, Ebert 2005). Ein Vergleich mit den Böden, die im Vorläufer-Projekt (LABO Projekt B4.07) für die Experimente zur As-Freisetzung bei Überstau eingesetzt wurden, ist nicht möglich, weil im Bericht keine Angaben zu den bodenchemischen Kenngrößen enthalten sind (Overesch et al. 2008).
Die Elementgehalte im KW-Extrakt zeigen z.T. stark erhöhte Schwermetallkonzentrationen (Cu, Zn, Cd, Pb) im Substrat Colditz gegenüber den anderen Substraten. Dies findet seine Entsprechung in den
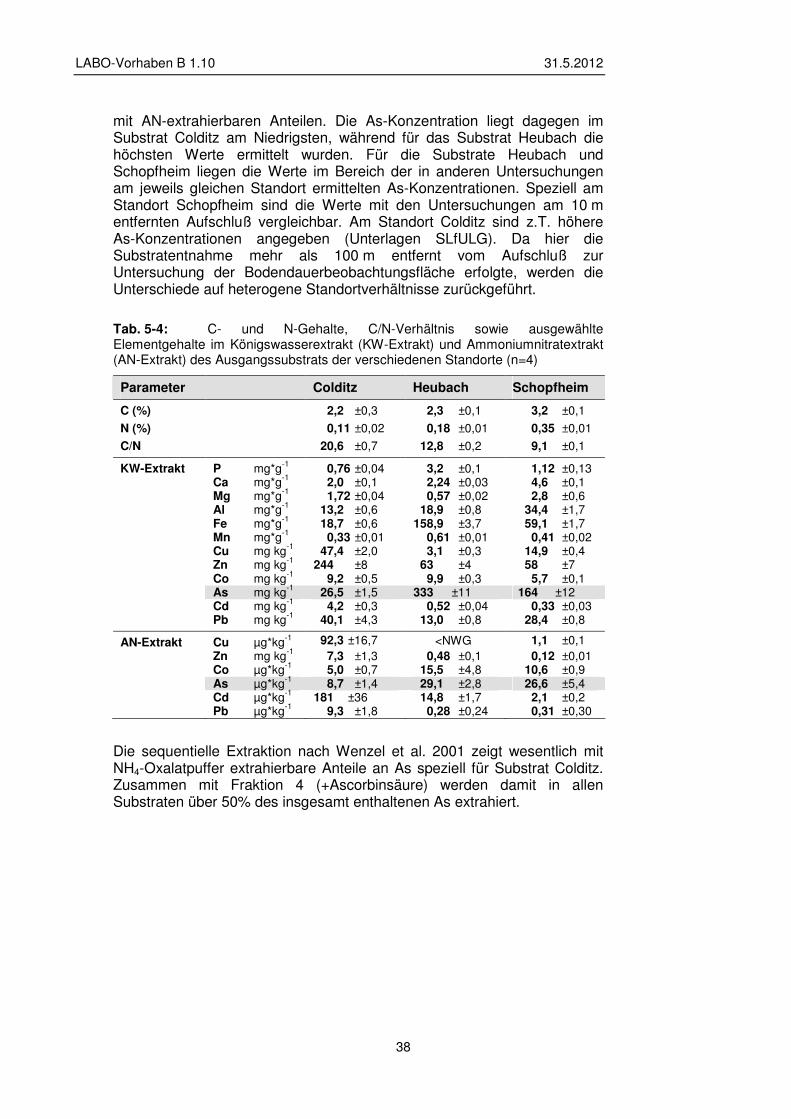
LABO-Vorhaben B 1.10 31.5.2012
38
mit AN-extrahierbaren Anteilen. Die As-Konzentration liegt dagegen im Substrat Colditz am Niedrigsten, während für das Substrat Heubach die höchsten Werte ermittelt wurden. Für die Substrate Heubach und Schopfheim liegen die Werte im Bereich der in anderen Untersuchungen am jeweils gleichen Standort ermittelten As-Konzentrationen. Speziell am Standort Schopfheim sind die Werte mit den Untersuchungen am 10 m entfernten Aufschluß vergleichbar. Am Standort Colditz sind z.T. höhere As-Konzentrationen angegeben (Unterlagen SLfULG). Da hier die Substratentnahme mehr als 100 m entfernt vom Aufschluß zur Untersuchung der Bodendauerbeobachtungsfläche erfolgte, werden die Unterschiede auf heterogene Standortverhältnisse zurückgeführt.
Tab. 5-4: C- und N-Gehalte, C/N-Verhältnis sowie ausgewählte Elementgehalte im Königswasserextrakt (KW-Extrakt) und Ammoniumnitratextrakt (AN-Extrakt) des Ausgangssubstrats der verschiedenen Standorte (n=4)
Parameter Colditz Heubach Schopfheim
C (%) 2,2 ±0,3 2,3 ±0,1 3,2 ±0,1
N (%) 0,11 ±0,02 0,18 ±0,01 0,35 ±0,01
C/N 20,6 ±0,7 12,8 ±0,2 9,1 ±0,1
KW-Extrakt P mg*g-1 0,76 ±0,04 3,2 ±0,1 1,12 ±0,13 Ca mg*g-1 2,0 ±0,1 2,24 ±0,03 4,6 ±0,1 Mg mg*g-1 1,72 ±0,04 0,57 ±0,02 2,8 ±0,6 Al mg*g-1 13,2 ±0,6 18,9 ±0,8 34,4 ±1,7 Fe mg*g-1 18,7 ±0,6 158,9 ±3,7 59,1 ±1,7 Mn mg*g-1 0,33 ±0,01 0,61 ±0,01 0,41 ±0,02 Cu mg kg-1 47,4 ±2,0 3,1 ±0,3 14,9 ±0,4 Zn mg kg-1 244 ±8 63 ±4 58 ±7 Co mg kg-1 9,2 ±0,5 9,9 ±0,3 5,7 ±0,1 As mg kg-1 26,5 ±1,5 333 ±11 164 ±12 Cd mg kg-1 4,2 ±0,3 0,52 ±0,04 0,33 ±0,03 Pb mg kg-1 40,1 ±4,3 13,0 ±0,8 28,4 ±0,8
AN-Extrakt Cu µg*kg-1 92,3 ±16,7 <NWG 1,1 ±0,1 Zn mg kg-1 7,3 ±1,3 0,48 ±0,1 0,12 ±0,01 Co µg*kg-1 5,0 ±0,7 15,5 ±4,8 10,6 ±0,9 As µg*kg-1 8,7 ±1,4 29,1 ±2,8 26,6 ±5,4 Cd µg*kg-1 181 ±36 14,8 ±1,7 2,1 ±0,2 Pb µg*kg-1 9,3 ±1,8 0,28 ±0,24 0,31 ±0,30
Die sequentielle Extraktion nach Wenzel et al. 2001 zeigt wesentlich mit NH4-Oxalatpuffer extrahierbare Anteile an As speziell für Substrat Colditz. Zusammen mit Fraktion 4 (+Ascorbinsäure) werden damit in allen Substraten über 50% des insgesamt enthaltenen As extrahiert.

LABO-Vorhaben B 1.10 31.5.2012
39
0
50
100
150
200
250
300
350A
s[m
g*kg
TG
]-1
Colditz Heubach Schopfheim
0
20
40
60
80
100
%
Colditz Heubach Schopfheim
HNO /H O3 2 2
NH -Oxalatpuffer + Ascorbinsäure4
NH -Oxalatpuffer4
(NH )H PO4 2 4
(NH ) SO4 2 4
Abb. 5-2: Ergebnisse für die As-Konzentrationen nach sequentieller Extraktion (Wenzel et al. 2001) für die einzelnen Ausgangssubstrate.
Nach Vorbereitung und Probenahme wurden die Substrate in weiße PP-Pflanzgefäße mit einem Volumen von 13 L verfüllt und in einem Gewächshaus zur Pflanzung bereitgestellt.
5.1.3 Substrat – Messparameter und Aufbereitung
Die Messparameter im Substrat werden im Einzelnen in Tab. 5-5 aufgelistet. Ausgewählte Parameter wurden einmalig am Ausgangssubstrat ermittelt. Zum Zeitpunkt der Ernte wurde die Elementbestimmung im KW-Extrakt und im AN-Extrakt sowie die pH-Wert-Bestimmung wiederholt, um Veränderungen zu dokumentieren. Während des Versuchs wurden nur die Parameter Feuchte, Temperatur und Redoxpotenzial kontinuierlich bestimmt. Die Feuchte und Temperatur wurde an ausgewählten Gefäßen mit TDR-Sonden und Datenaufnahme über ein Loggersystem ermittelt. Alle anderen Gefäße wurden täglich mit FDR-Sonden (ML-2x Theta-Sonden mit HH2, Fa. Delta-T) kontrolliert. Ebenfalls an ausgewählten Gefäßen wurden zur kontinuierlichen Messung des Redoxpotenzials Redox-Elektroden mit einem Platinstab (Ø 1 mm, Länge 5 mm) nach Mansfeldt 2003 verwendet (Bezugselektrode: Ag/AgCl-Referenzelektrode), die an einen Datenlogger angeschlossen wurden.
LABO-Vorhaben B 1.10 31.5.2012
40
Tab. 5-5: Messparameter Substrat mit Angabe der Methode sowohl zur Durchführung wie auch zur Analyse. t0: Messung vor Versuchsbeginn; tn: Messung am Versuchsende.
Allgemeine Parameter t0 t1 t2 … tn Methode/Analyse
pH-Wert X X BZE HFA* Leitfähigkeit (Lf) X X BZE HFA* KAKeff X BZE HFA* Korngrößenverteilung X BZE HFA* Feuchte kont. Kont. kont. kont. kont. Temperatur kont. Kont. kont. kont. kont. Redoxpotenzial kont. Kont. kont. kont. kont. C_total X DIN ISO 10694 N_total X DIN ISO 13878
KW-Extrakt (nach EN 13657)
As_gesamt X X DIN EN ISO 17294-2 P, Ca, Mg X X Fe, Mn, Cu, Zn, Co, Cd, Pb X X DIN EN ISO 11885
AN-Extrakt nach Anhang 1 BBodSchV
As_gesamt X X DIN EN ISO 17294-2 Cu, Zn, Co, Cd, Pb X X DIN EN ISO 11885
Sequentielle Extraktion
As_gesamt X Wenzel 2001
*BZE HFA: Bodenzustandserhebung – Handbuch Forstliche Analytik 2009
Bodenlösungsproben wurden mit Hilfe von 10 cm langen Mikro-Saugkerzen gewonnen (Porengröße 0,1 µm, Fa. Eijkelkamp). In jeder Versuchsvariante wurden damit 3 Gefäße bestückt (Budgetlimitation). Aus den Varianten mit Überstau konnten in der Regel durchgängig Bodenlösungsproben gewonnen werden. In den Varianten unter mäßig feuchten Bedingungen wurden nur zu Beginn des Versuchs Mengen > 2 ml erhalten. Danach waren speziell von den Substraten Heubach und Schopfheim keine analysierbaren Probenmengen mehr zu gewinnen.
Folgende Messparameter wurden in der Bodenlösung bestimmt:
Parameter Methode Sulfat, Nitrat, Phosphat, Chlorid DIN 38405 As(III)*, As (V)* IC-ICPMS-Koppl. As_gesamt, Ca, Mg, Fe, Mn, Cu, Zn, Co, Cd, Pb DIN EN ISO 11885
*nur ausgewählte Proben
5.1.4 Pflanzen - Pflanzung und Kulturbedingungen
Das Deutsche Weidelgras (Lolium perenne „KERATOS“) als auch der Winterweizen (Triticum aestivum „AKTEUR“) wurden als Versuchssaatgut von den entsprechenden Anbietern bezogen.
In jedes Gefäß wurden anfänglich 20 Samen Anfang November gesetzt. Der Winterweizen zeigte eine gute bis sehr gute Keimfähigkeit (>95%) mit erstem Spitzen nach 14 Tagen. In diesem Zustand konnten die Gefäße eingewintert werden. Dagegen war die Keimfähigkeit des Weidelgrases deutlich geringer (<70%, einige Gefäße mit Totalausfall). Daher musste hier
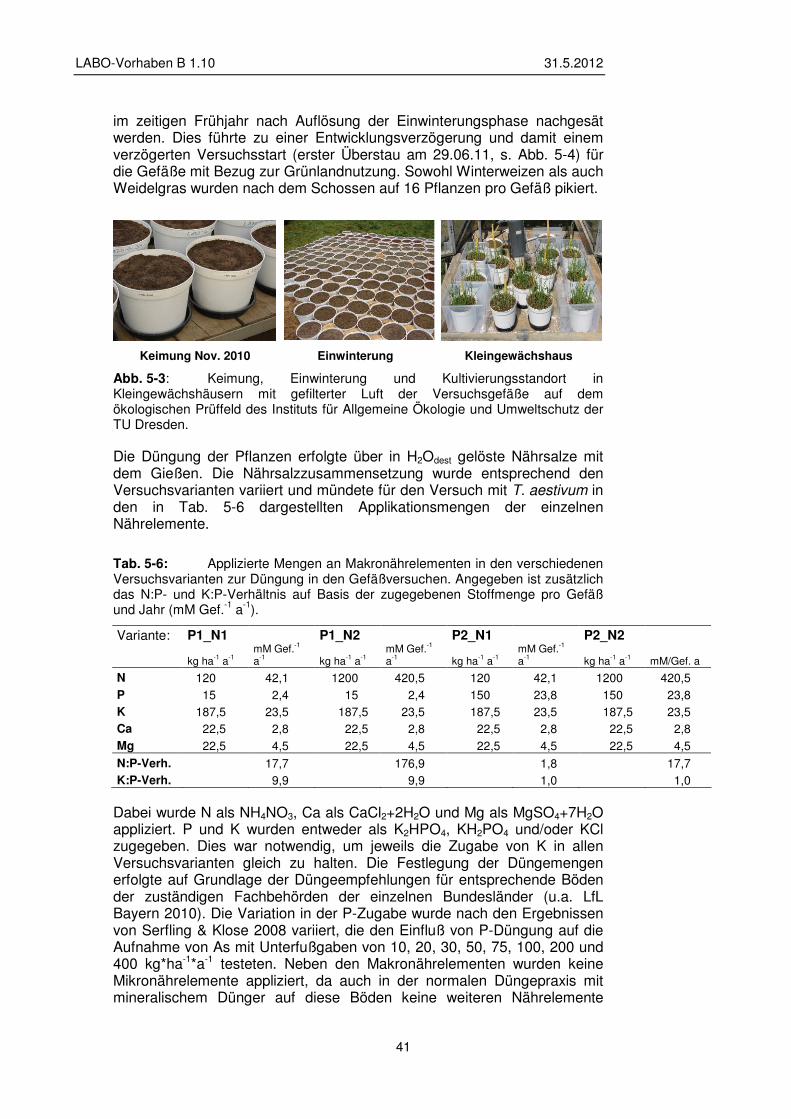
LABO-Vorhaben B 1.10 31.5.2012
41
im zeitigen Frühjahr nach Auflösung der Einwinterungsphase nachgesät werden. Dies führte zu einer Entwicklungsverzögerung und damit einem verzögerten Versuchsstart (erster Überstau am 29.06.11, s. Abb. 5-4) für die Gefäße mit Bezug zur Grünlandnutzung. Sowohl Winterweizen als auch Weidelgras wurden nach dem Schossen auf 16 Pflanzen pro Gefäß pikiert.
Keimung Nov. 2010 Einwinterung Kleingewächshaus
Abb. 5-3: Keimung, Einwinterung und Kultivierungsstandort in Kleingewächshäusern mit gefilterter Luft der Versuchsgefäße auf dem ökologischen Prüffeld des Instituts für Allgemeine Ökologie und Umweltschutz der TU Dresden.
Die Düngung der Pflanzen erfolgte über in H2Odest gelöste Nährsalze mit dem Gießen. Die Nährsalzzusammensetzung wurde entsprechend den Versuchsvarianten variiert und mündete für den Versuch mit T. aestivum in den in Tab. 5-6 dargestellten Applikationsmengen der einzelnen Nährelemente.
Tab. 5-6: Applizierte Mengen an Makronährelementen in den verschiedenen Versuchsvarianten zur Düngung in den Gefäßversuchen. Angegeben ist zusätzlich das N:P- und K:P-Verhältnis auf Basis der zugegebenen Stoffmenge pro Gefäß und Jahr (mM Gef.-1 a-1).
Variante: P1_N1 P1_N2 P2_N1 P2_N2
kg ha-1 a-1 mM Gef.-1 a-1 kg ha-1 a-1
mM Gef.-1 a-1 kg ha-1 a-1
mM Gef.-1 a-1 kg ha-1 a-1 mM/Gef. a
N 120 42,1 1200 420,5 120 42,1 1200 420,5 P 15 2,4 15 2,4 150 23,8 150 23,8 K 187,5 23,5 187,5 23,5 187,5 23,5 187,5 23,5 Ca 22,5 2,8 22,5 2,8 22,5 2,8 22,5 2,8 Mg 22,5 4,5 22,5 4,5 22,5 4,5 22,5 4,5
N:P-Verh. 17,7 176,9 1,8 17,7 K:P-Verh. 9,9 9,9 1,0 1,0
Dabei wurde N als NH4NO3, Ca als CaCl2+2H2O und Mg als MgSO4+7H2O appliziert. P und K wurden entweder als K2HPO4, KH2PO4 und/oder KCl zugegeben. Dies war notwendig, um jeweils die Zugabe von K in allen Versuchsvarianten gleich zu halten. Die Festlegung der Düngemengen erfolgte auf Grundlage der Düngeempfehlungen für entsprechende Böden der zuständigen Fachbehörden der einzelnen Bundesländer (u.a. LfL Bayern 2010). Die Variation in der P-Zugabe wurde nach den Ergebnissen von Serfling & Klose 2008 variiert, die den Einfluß von P-Düngung auf die Aufnahme von As mit Unterfußgaben von 10, 20, 30, 50, 75, 100, 200 und 400 kg*ha-1*a-1 testeten. Neben den Makronährelementen wurden keine Mikronährelemente appliziert, da auch in der normalen Düngepraxis mit mineralischem Dünger auf diese Böden keine weiteren Nährelemente

LABO-Vorhaben B 1.10 31.5.2012
42
aufgebracht würden. Außerdem deuten die Werte im KW-Extrakt bzw. AN-Extrakt für die ermittelten Mikronährelemente nicht auf eine Unterversorgung hin (vgl. Tab. 5-4).
Die Applikation von 2/3 der Nährelementmengen erfolgte im Versuch mit T. aestivum am 21.04.2011. Das restliche Drittel wurde am 23.05.2011 zugegeben. Zur Düngung des Versuchs mit L. perenne wurden die Nährelementmengen der Variante P1_N1 einmalig am 01.06.2011 appliziert.
In den folgenden Abbildungen sind die Versuchsabläufe einschließlich Düngetermine für die Versuche zur Grünlandnutzung mit L. perenne sowie zur Ackernutzung mit T. aestivum zusammenfassend dargestellt.
29.3. 15.4. 29.4. 15.5. 29.5. 15.6. 29.6. 15.7. 29.7. 15.8. 29.8.
2011
Aus
win
teru
ng
1.M
ahd
Dün
gung
Neu
saat
3.M
ahd
(Ern
te)
24h
Übe
rsta
u
2.M
ahd
24h
Übe
rsta
u
24h
Übe
rsta
u
24h
Übe
rsta
u
24h
Übe
rsta
u
24h
Übe
rsta
u
24h
Übe
rsta
u
24h
Übe
rsta
u
Abb. 5-4: Versuchsablauf im Versuch zur Grünlandnutzung mit L. perenne.
Aus
win
teru
ng
3hÜ
bers
tau
2.D
üngu
ng
1.D
üngu
ng
Ern
te
48h
Übe
rsta
u
24h
Übe
rsta
u
3hÜ
bers
tau
24h
Übe
rsta
u
29.3. 15.4. 29.4. 15.5. 29.5. 15.6. 29.6.
2011 Abb. 5-5: Versuchsablauf im Versuch zur Ackernutzung mit T. aestivum.
5.1.5 Pflanzen - Messparameter und Aufarbeitung
Zu den verschiedenen Mahdterminen des Grünlandversuchs wurden die Pflanzen in 3-5 cm Höhe über dem Substrat abgeschnitten und als Proben in Papiertüten verpackt (Sproßproben). Zur 3. Mahd erfolgte gleichzeitig die Ernte der restlichen beiden Kompartimente Stoppeln (Sproß von
LABO-Vorhaben B 1.10 31.5.2012
43
Schnittstelle bis zum Wurzelansatz) und Wurzeln. Das Mahdgut (Sproß ohne Stoppeln) der 3. Mahd wurde in 2 Fraktionen geteilt und eine Fraktion gründlich mit H2Odest. gewaschen, die andere Fraktion ungewaschen belassen. Die Wurzeln wurden ebenfalls vor der weiteren Aufbereitung gründlich mit H2Odest. gewaschen.
Die Ernte der Pflanzen von T. aestivum erfolgte jeweils zur Totreife. Oberirdisch erfolgte die Trennung zunächst in Ähren, Blätter und Stängel (3 cm über dem Substrat abgeschnitten). Im Labor wurden dann die Körner von den Spelzen getrennt. Für Stoppeln und Wurzeln wurde genauso wie bei L. perenne verfahren. Die Probenvorbereitung der Pflanzenproben zur weiteren Analyse erfolgte entsprechend den Methoden des Handbuchs Forstliche Analytik 2005 (HFA 2005) in der Ergänzungsfassung 2009. Ergänzend erwähnt sei nur, dass die Proben generell gefriergetrocknet wurden, in Ausnahmefällen bei 70°C bis zur Gewichtskonstanz. Der Aufschluß der Proben erfolgte mittels mikrowellen-gestütztem Druckaufschluß (MARS Express, Fa. CEM). In Vorbereitung zur Messung der As-Spezies in den oberirdischen Kompartimenten Körner und Blätter bzw. Sproß zum Zeitpunkt der Ernte wurden die Proben nach Zhu et al. 2008 extrahiert und innerhalb der nächsten 48h vermessen.
Folgende Messparameter wurden an den Pflanzenproben bestimmt:
L. perenne T. aestivum Parameter Sproß Stoppeln Wurzeln Körner Spelzen Blätter Stängel Wurzeln
Frischgewicht X X X X X X X X Trockengewicht X X X X X X X X As_gesamt X X X X X X As(III)*, As(V)* X X X C, N X X P, Ca, Mg, Fe, Mn X X X X X X Cu, Zn, Co X X X X X X Cd, Pb X X X X X X
* nur ausgewählte Proben
Die Elementgehalte wurden entsprechend DIN EN ISO 11885 (P, Ca, Mg, Fe, Mn, Cu, Zn, Co, Cd, Pb), DIN EN ISO 17294-2 (As_gesamt), DIN ISO 10694 (C), DIN ISO 13878 (N) bestimmt. Die As-Speziesanalytik folgt den Angaben in Kap. 4 (IC-ICPMS-Kopplung).
Zur Qualitätssicherung der Probenaufbereitung und der Analyseverfahren wurden Standardreferenzmaterialien eingesetzt (für Pflanzenproben: GBW7604 (Pappelblätter), NIST 1575 (Kiefernnadeln), BCR101 (Weidelgras); für Bodenproben: GBW 7404).
5.1.6 Statistische Auswertung
Die Berechnung der statistischen Tests erfolgte mit dem Programmpaket SPSS für Windows (Version 11.5.1). Für die Darstellung der Signifikanzen wird folgende Vereinbarung getroffen:

LABO-Vorhaben B 1.10 31.5.2012
44
Tab. 5-7: Verwendete Signifikanzniveaus, deren Bezeichnung und Symbole
Signifikanzniveau ((1-p)•100) Bezeichnung Symbol p > 0,05 < 95 % nicht signifikant n.s. p ≤ 0,05 ≥ 95 % signifikant * p ≤ 0,01 ≥ 99 % sehr signifikant ** p ≤ 0,001 ≥ 99,9 % hoch signifikant ***
Zur Prüfung der Daten von den Gefäß- und Freilandversuchen auf Normalverteilung wurde der zweiseitige Kolmogorov-Smirnov-Test für eine Stichprobe verwendet (Siegel 1985). Dieser Test prüft den Grad der Übereinstimmung zwischen einer kumulativen Häufigkeitsverteilung entsprechend einer theoretischen Verteilung und einer beobachteten kumulierten Häufigkeitsverteilung. Konnte keine Normalverteilung festgestellt werden, wurden nichtparametrische Tests verwendet.
Regressionsanalyse und Diagramme
Regressionsgeraden sowie sämtliche Diagramme wurden mit dem Programm XACT (Fa. SCILAB, Hamburg) erstellt. Das Programm gibt den Korrelationskoeffizienten, die Varianz und die Standardabweichung der Residuen sowie die Koeffizienten der Regressionsgeraden a und b aus. Die neben Linien- und Balkendiagrammen verwendeten Boxplot-Diagramme geben den Median (Querstrich), den Quartilbereich (grauer Balken) sowie die Extremwerte (Endpunkte der Längsstriche) wider. Dieser Diagrammtyp wurde gewählt, wenn unterschiedliche Stichprobengrößen mit teilweise nicht absicherbarer Normalverteilung zusammen dargestellt werden.
Nichtparametrische Tests
Wurde keine Normalverteilung gefunden und konnte auch durch Lineartransformation (Logarithmierung) für die Werte der Variablen keine Normalverteilung herbeigeführt werden, wurden nichtparametrische Tests verwendet. In den hier vorgestellten Freilanduntersuchungen sowie in den Gefäßversuchen konnten in den allermeisten Fällen Normalverteilungen wahrscheinlich gemacht werden. Daher werden auch in der deskriptiven Statistik in den meisten Fällen arithmetische Mittelwerte mit Standardabweichungen angegeben. Allerdings wird von Bortz 1993; Bortz 2005 und Siegel 1985 generell empfohlen, bei sehr kleinen Stichprobenumfängen auf nichtparametrische statistische Tests zurückzugreifen. Im Falle von k unabhängigen Stichproben wurde mit der Rangvarianzanalyse nach Kruskal-Wallis (H-Test oder auch Kruskal-Wallis-Test) die Nullhypothese untersucht, ob die Stichproben aus der gleichen Population stammen. Wird die Nullhypothese abgelehnt, wurden paarweise Vergleiche zur Differenzierung der Einzelunterschiede angeschlossen. Für den Paar-Vergleich zweier unabhängiger Stichproben wurde der U-Test nach Mann-Whitney verwendet. Mehrere abhängige Stichproben wurden mit dem Friedman-Test untersucht. Dieser rangbasierte Test stellt das nichtparametrische Äquivalent zu einer einfaktoriellen Varianzanalyse mit Messwiederholungen für den Sonderfall nur einer Faktorstufe dar. Einzelvergleiche bei abhängigen Stichproben wurden mit dem Wilcoxon-Vorzeichenrang-Test (Wilcoxon-Test) untersucht. Als nichtparametrisches

LABO-Vorhaben B 1.10 31.5.2012
45
Korrelationsmaß wurde der Rang-Korrelationskoeffizient rS nach Spearman verwendet.
Zwei- und Dreifaktorielle Varianzanalyse (ANOVA)
Für die Gefäßversuche mit in den meisten Fällen n=5 konnten in der Regel normalverteilte Werte festgestellt werden. Da in fast allen Fällen Varianzhomogenität (Levene-Test) gegeben war, wurde für den Versuch mit Bezug zur Grünlandnutzung für die Untersuchung der Effekte der Feuchtevariante (Feuchte) und des Substrats (Substrat) als unabhängige Variablen (Faktoren) auf die jeweilige abhängige Variable eine zweifaktorielle Varianzanalyse gerechnet. Im Versuch mit Bezug zur Ackernutzung wurde zur Untersuchung der Effekte der Feuchtevariante (Feuchte), der P-Zugabe und der N-Zugabe als unabhängige Variablen (Faktoren) auf die jeweilige abhängige Variable eine dreifaktorielle Varianzanalyse angewendet. Als Ergebnis angegeben werden die aus den Quadratsummen sich ergebende Varianzaufklärung bezüglich der untersuchten abhängigen Variablen durch die unabhängigen Variablen (Haupteffekte) und deren Interaktion (Interaktionseffekt) in Prozent (erklärte Streuung – „Erkl. Streuung“) sowie der zur Signifikanzprüfung notwendige F-Wert einschließlich Signifikanz (Signif.). Die Varianzanalyse selbst gibt nur an, ob insgesamt Unterschiede bezogen auf die einzelnen Faktoren bestehen. Einzelvergleiche der jeweiligen Faktorstufen zur Überprüfung von a priori Hypothesen wurden a posteriori mit dem konservativen Scheffé-Test durchgeführt. Dieser Post-Hoc-Test (wie auch andere Tests dieser Art) berücksichtigt die Tatsache, dass das α-Fehlerniveau der Varianzanalyse nicht überschritten werden darf und eliminiert so die Gefahr, dass es zu einer α-Fehlerkumulation kommt.
In Ausnahmefällen wurden die Mittelwerte zweier Stichproben mit dem t-Test verglichen. Dieser überprüft die Nullhypothese, dass zwei unabhängige Stichproben aus Populationen stammen, deren Erwartungswerte gleich sind. Neben der Unabhängigkeit der Stichproben wird für den t-Test speziell bei kleinen Stichproben Normalverteilung und Varianzhomogenität vorausgesetzt, was in den Fällen der Anwendung des t-Tests gegeben war. Zur Überprüfung von Zusammenhangshypothesen wurde für den Gefäßversuch die Produkt-Moment-Korrelation verwendet (s.a. Bortz 2005). Soweit nicht anders angegeben, wurden alle Tests richtungsneutral durchgeführt, d.h. zweiseitig getestet.
5.2 Gefäßversuche – Ergebnisse Grünlandnutzung
Im Ergebnisteil zum Gefäßversuch mit Bezug zur Grünlandnutzung wird zunächst auf die Substrat- und Bodenlösungsparameter eingegangen, bevor die biometrischen Parameter der Pflanzen sowie deren Elementgehalte behandelt werden.
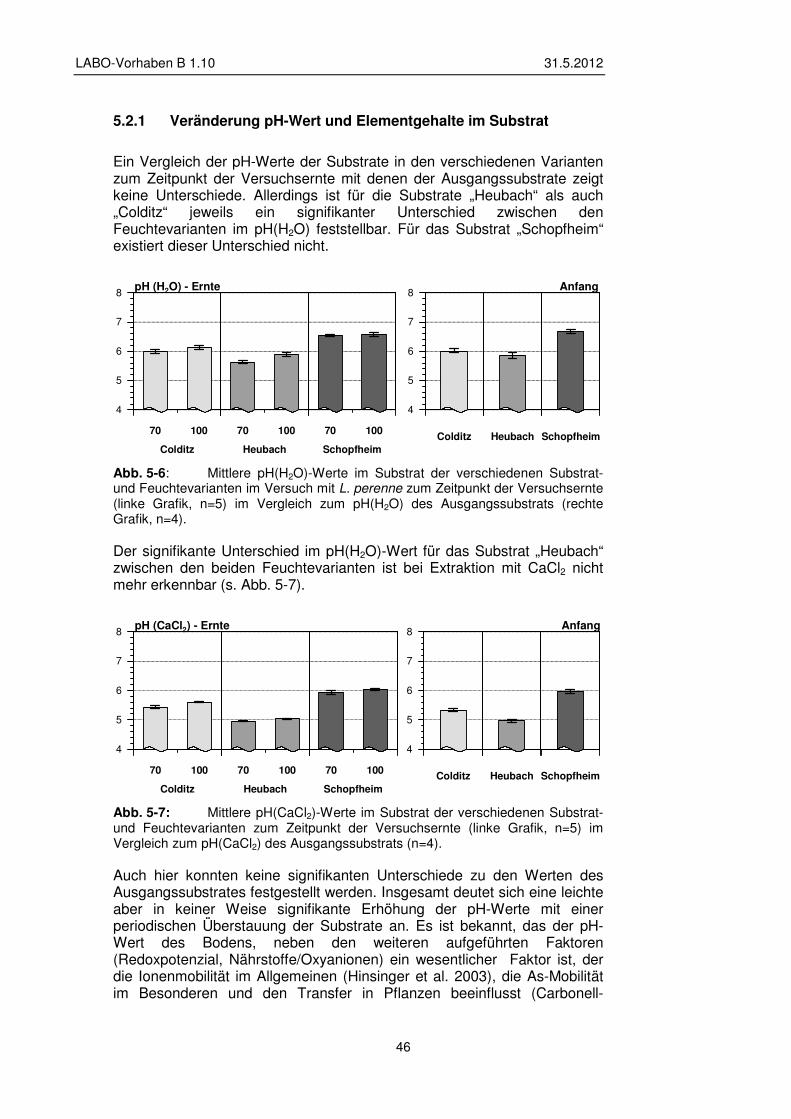
LABO-Vorhaben B 1.10 31.5.2012
46
5.2.1 Veränderung pH-Wert und Elementgehalte im Substrat
Ein Vergleich der pH-Werte der Substrate in den verschiedenen Varianten zum Zeitpunkt der Versuchsernte mit denen der Ausgangssubstrate zeigt keine Unterschiede. Allerdings ist für die Substrate „Heubach“ als auch „Colditz“ jeweils ein signifikanter Unterschied zwischen den Feuchtevarianten im pH(H2O) feststellbar. Für das Substrat „Schopfheim“ existiert dieser Unterschied nicht.
4
5
6
7
8
70 100
Colditz
70 100
Heubach
70 100
Schopfheim
pH (H2O) - Ernte
4
5
6
7
8
Colditz Heubach Schopfheim
Anfang
Abb. 5-6: Mittlere pH(H2O)-Werte im Substrat der verschiedenen Substrat- und Feuchtevarianten im Versuch mit L. perenne zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zum pH(H2O) des Ausgangssubstrats (rechte Grafik, n=4).
Der signifikante Unterschied im pH(H2O)-Wert für das Substrat „Heubach“ zwischen den beiden Feuchtevarianten ist bei Extraktion mit CaCl2 nicht mehr erkennbar (s. Abb. 5-7).
4
5
6
7
8
70 100
Colditz
70 100
Heubach
70 100
Schopfheim
4
5
6
7
8
Colditz Heubach Schopfheim
pH (CaCl2) - Ernte Anfang
Abb. 5-7: Mittlere pH(CaCl2)-Werte im Substrat der verschiedenen Substrat- und Feuchtevarianten zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zum pH(CaCl2) des Ausgangssubstrats (n=4).
Auch hier konnten keine signifikanten Unterschiede zu den Werten des Ausgangssubstrates festgestellt werden. Insgesamt deutet sich eine leichte aber in keiner Weise signifikante Erhöhung der pH-Werte mit einer periodischen Überstauung der Substrate an. Es ist bekannt, das der pH-Wert des Bodens, neben den weiteren aufgeführten Faktoren (Redoxpotenzial, Nährstoffe/Oxyanionen) ein wesentlicher Faktor ist, der die Ionenmobilität im Allgemeinen (Hinsinger et al. 2003), die As-Mobilität im Besonderen und den Transfer in Pflanzen beeinflusst (Carbonell-

LABO-Vorhaben B 1.10 31.5.2012
47
Barrachina et al. 2000). Eine regulär signifikante Erhöhung der pH-Werte im Boden in den Varianten mit periodischer Wassersättigung wurde nur in den etwas kohlenstoffärmeren Böden mit weiterem C/N-Verhältnis (Colditz, Heubach) nachgewiesen. pH-Wert-Veränderungen um mehr als eine Einheit wurden sowohl in gut durchlüfteten als auch anoxischen Böden beschrieben. Sie werden durch induzierten Nährstoffmangel (Phosphat oder Fe, Hinsinger 2001) und durch die Form der N-Ernährung (Ammonium vs. Nitrat: Marschner 2002) verursacht. Auch eine oxidative Versauerung infolge der Sauerstoffausscheidung der Wurzeln und/oder Anreicherung organischer Säuren u.a. infolge fermentativer Energiegewinnung in Wurzeln und der Rhizosphäre bei längerem Überstau ist möglich (Blossfeld et al. 2011). Auf den radialen Sauerstofftransfer von den Wurzeln in das Milieu dürften sowohl die Porosität der Wurzeln als auch die Durchlässigkeit des Bodens einen Einfluss haben, wovon wiederum der Arsentransfer in die Pflanzen abhängig ist (Li et al. 2011). Bezüglich dieser Parameter unterscheidet sich insbesondere das Substrat Colditz relevant vom Substrattyp Schopfheim. In letzterem wurde in keinem Fall eine signifikante pH-Veränderung beobachtet. Es ist anzunehmen, das nicht nur (i) die Nitrifikation in diesem Bodentyp verstärkt wird und infolgedessen die Pflanzen eher Nitrat als Ammonium aufnehmen (müssen), was zur pH-Anhebung führt, und (ii) durch die bessere Sauerstoffversorgung auch die Respiration im Wurzelraum und die dadurch verursachte Alkalinisierung verstärkt wird, sondern dass auch durch die reduzierenden Bedingungen (iii) verstärkt DOC und Fe(II) freigesetzt wird (s. Kap.5.2.2). Diese Freisetzung ist offensichtlich eine Reaktion auf reduzierende Prozesse bei „circum-neutralen“ (leichtsauren bis leicht basischen) Milieuverhältnissen, wo die Deprotonierung der Hydroxylgruppen an den Mineraloberflächen zur Verminderung der Oberflächenladung führt. Es kann angenommen werden, das demzufolge die Fe(OH)n-Reduktion nicht der einzige Prozess ist der zur Erhöhung der Spurenelementmobilität führt (Grybos et al. 2007).
Im KW-Extrakt sind für die Substrate Colditz und Schopfheim keine bzw. leicht unterschiedliche(n) As-Konzentrationen zwischen den Werten im Ausgangssubstrat und den Werten zum Zeitpunkt der Ernte festzustellen (Abb. 5-8).
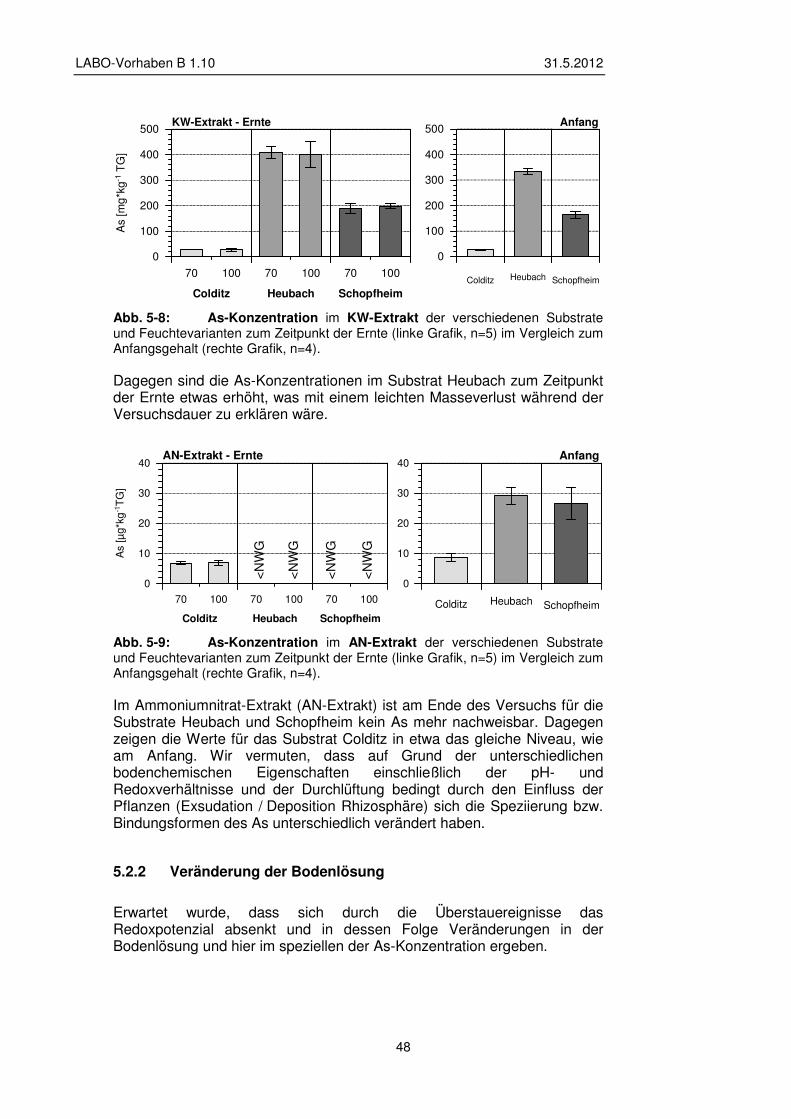
LABO-Vorhaben B 1.10 31.5.2012
48
0
100
200
300
400
500A
s[m
g*kg
TG
]-1
70 100
Colditz
70 100
Heubach
70 100
Schopfheim
0
100
200
300
400
500
Colditz Heubach Schopfheim
AnfangKW-Extrakt - Ernte
Abb. 5-8: As-Konzentration im KW-Extrakt der verschiedenen Substrate und Feuchtevarianten zum Zeitpunkt der Ernte (linke Grafik, n=5) im Vergleich zum Anfangsgehalt (rechte Grafik, n=4).
Dagegen sind die As-Konzentrationen im Substrat Heubach zum Zeitpunkt der Ernte etwas erhöht, was mit einem leichten Masseverlust während der Versuchsdauer zu erklären wäre.
0
10
20
30
40
As
[µg*
kgT
G]
-1
70 100
Colditz
70 100
Heubach
70 100
Schopfheim
<N
WG
<N
WG
<N
WG
<N
WG
0
10
20
30
40
Colditz Heubach Schopfheim
AN-Extrakt - Ernte Anfang
Abb. 5-9: As-Konzentration im AN-Extrakt der verschiedenen Substrate und Feuchtevarianten zum Zeitpunkt der Ernte (linke Grafik, n=5) im Vergleich zum Anfangsgehalt (rechte Grafik, n=4).
Im Ammoniumnitrat-Extrakt (AN-Extrakt) ist am Ende des Versuchs für die Substrate Heubach und Schopfheim kein As mehr nachweisbar. Dagegen zeigen die Werte für das Substrat Colditz in etwa das gleiche Niveau, wie am Anfang. Wir vermuten, dass auf Grund der unterschiedlichen bodenchemischen Eigenschaften einschließlich der pH- und Redoxverhältnisse und der Durchlüftung bedingt durch den Einfluss der Pflanzen (Exsudation / Deposition Rhizosphäre) sich die Speziierung bzw. Bindungsformen des As unterschiedlich verändert haben.
5.2.2 Veränderung der Bodenlösung
Erwartet wurde, dass sich durch die Überstauereignisse das Redoxpotenzial absenkt und in dessen Folge Veränderungen in der Bodenlösung und hier im speziellen der As-Konzentration ergeben.
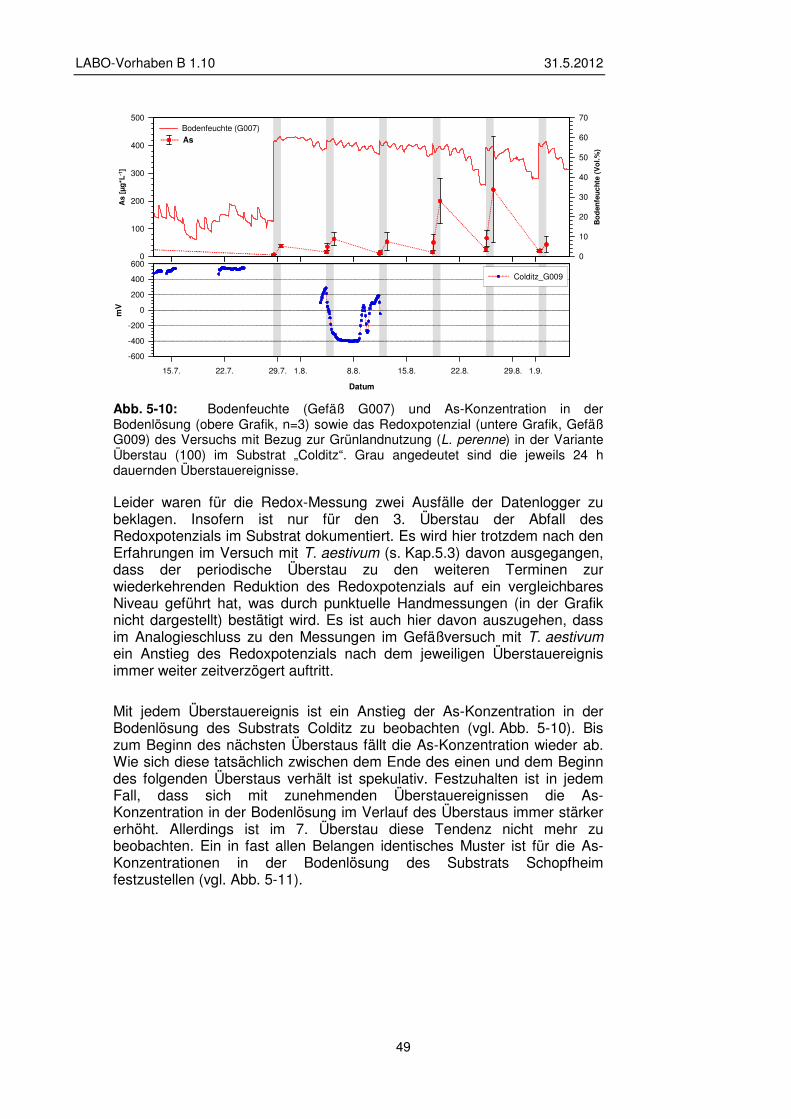
LABO-Vorhaben B 1.10 31.5.2012
49
0
100
200
300
400
500A
s[µ
g*L
]-1
As
0
10
20
30
40
50
60
70
Bo
den
feu
chte
(Vo
l.%)
Bodenfeuchte (G007)
-600
-400
-200
0
200
400
600
mV
15.7. 22.7. 29.7. 1.8. 8.8. 15.8. 22.8. 29.8. 1.9.
Datum
Colditz_G009
Abb. 5-10: Bodenfeuchte (Gefäß G007) und As-Konzentration in der Bodenlösung (obere Grafik, n=3) sowie das Redoxpotenzial (untere Grafik, Gefäß G009) des Versuchs mit Bezug zur Grünlandnutzung (L. perenne) in der Variante Überstau (100) im Substrat „Colditz“. Grau angedeutet sind die jeweils 24 h dauernden Überstauereignisse.
Leider waren für die Redox-Messung zwei Ausfälle der Datenlogger zu beklagen. Insofern ist nur für den 3. Überstau der Abfall des Redoxpotenzials im Substrat dokumentiert. Es wird hier trotzdem nach den Erfahrungen im Versuch mit T. aestivum (s. Kap.5.3) davon ausgegangen, dass der periodische Überstau zu den weiteren Terminen zur wiederkehrenden Reduktion des Redoxpotenzials auf ein vergleichbares Niveau geführt hat, was durch punktuelle Handmessungen (in der Grafik nicht dargestellt) bestätigt wird. Es ist auch hier davon auszugehen, dass im Analogieschluss zu den Messungen im Gefäßversuch mit T. aestivum ein Anstieg des Redoxpotenzials nach dem jeweiligen Überstauereignis immer weiter zeitverzögert auftritt.
Mit jedem Überstauereignis ist ein Anstieg der As-Konzentration in der Bodenlösung des Substrats Colditz zu beobachten (vgl. Abb. 5-10). Bis zum Beginn des nächsten Überstaus fällt die As-Konzentration wieder ab. Wie sich diese tatsächlich zwischen dem Ende des einen und dem Beginn des folgenden Überstaus verhält ist spekulativ. Festzuhalten ist in jedem Fall, dass sich mit zunehmenden Überstauereignissen die As-Konzentration in der Bodenlösung im Verlauf des Überstaus immer stärker erhöht. Allerdings ist im 7. Überstau diese Tendenz nicht mehr zu beobachten. Ein in fast allen Belangen identisches Muster ist für die As-Konzentrationen in der Bodenlösung des Substrats Schopfheim festzustellen (vgl. Abb. 5-11).
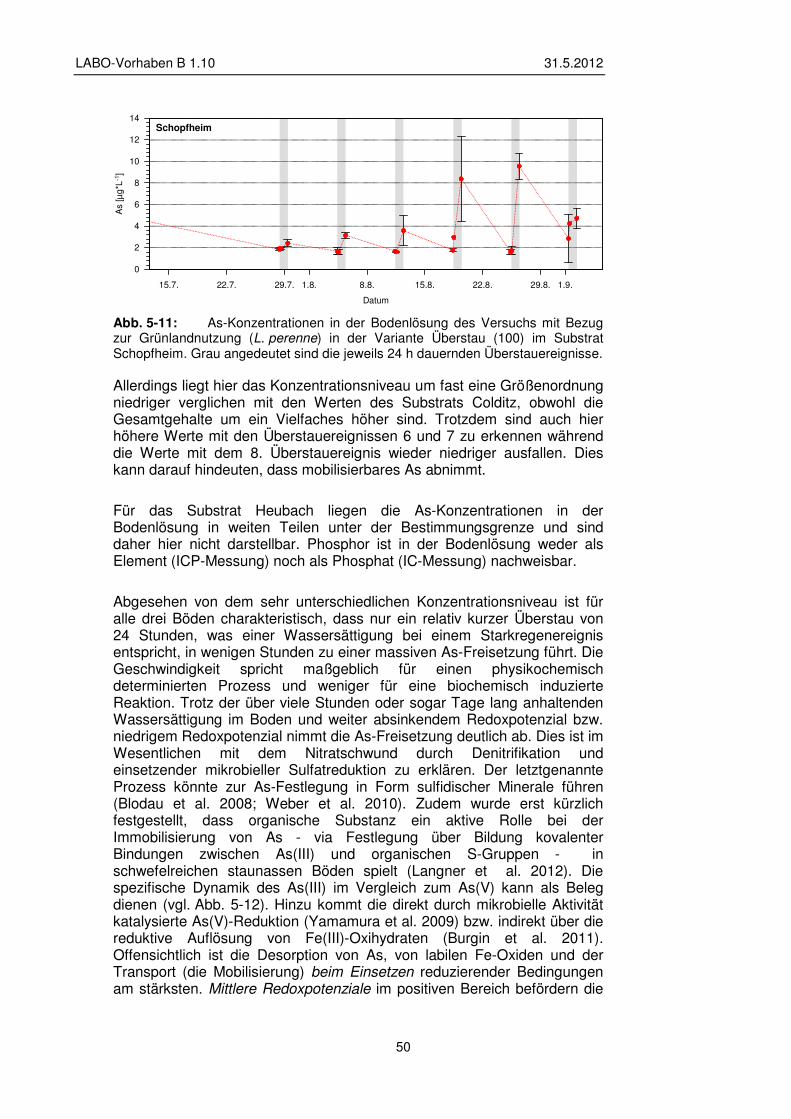
LABO-Vorhaben B 1.10 31.5.2012
50
0
2
4
6
8
10
12
14A
s[µ
g*L
]-1
15.7. 22.7. 29.7. 1.8. 8.8. 15.8. 22.8. 29.8. 1.9.
Datum
Schopfheim
Abb. 5-11: As-Konzentrationen in der Bodenlösung des Versuchs mit Bezug zur Grünlandnutzung (L. perenne) in der Variante Überstau (100) im Substrat Schopfheim. Grau angedeutet sind die jeweils 24 h dauernden Überstauereignisse.
Allerdings liegt hier das Konzentrationsniveau um fast eine Größenordnung niedriger verglichen mit den Werten des Substrats Colditz, obwohl die Gesamtgehalte um ein Vielfaches höher sind. Trotzdem sind auch hier höhere Werte mit den Überstauereignissen 6 und 7 zu erkennen während die Werte mit dem 8. Überstauereignis wieder niedriger ausfallen. Dies kann darauf hindeuten, dass mobilisierbares As abnimmt.
Für das Substrat Heubach liegen die As-Konzentrationen in der Bodenlösung in weiten Teilen unter der Bestimmungsgrenze und sind daher hier nicht darstellbar. Phosphor ist in der Bodenlösung weder als Element (ICP-Messung) noch als Phosphat (IC-Messung) nachweisbar.
Abgesehen von dem sehr unterschiedlichen Konzentrationsniveau ist für alle drei Böden charakteristisch, dass nur ein relativ kurzer Überstau von 24 Stunden, was einer Wassersättigung bei einem Starkregenereignis entspricht, in wenigen Stunden zu einer massiven As-Freisetzung führt. Die Geschwindigkeit spricht maßgeblich für einen physikochemisch determinierten Prozess und weniger für eine biochemisch induzierte Reaktion. Trotz der über viele Stunden oder sogar Tage lang anhaltenden Wassersättigung im Boden und weiter absinkendem Redoxpotenzial bzw. niedrigem Redoxpotenzial nimmt die As-Freisetzung deutlich ab. Dies ist im Wesentlichen mit dem Nitratschwund durch Denitrifikation und einsetzender mikrobieller Sulfatreduktion zu erklären. Der letztgenannte Prozess könnte zur As-Festlegung in Form sulfidischer Minerale führen (Blodau et al. 2008; Weber et al. 2010). Zudem wurde erst kürzlich festgestellt, dass organische Substanz ein aktive Rolle bei der Immobilisierung von As - via Festlegung über Bildung kovalenter Bindungen zwischen As(III) und organischen S-Gruppen - in schwefelreichen staunassen Böden spielt (Langner et al. 2012). Die spezifische Dynamik des As(III) im Vergleich zum As(V) kann als Beleg dienen (vgl. Abb. 5-12). Hinzu kommt die direkt durch mikrobielle Aktivität katalysierte As(V)-Reduktion (Yamamura et al. 2009) bzw. indirekt über die reduktive Auflösung von Fe(III)-Oxihydraten (Burgin et al. 2011). Offensichtlich ist die Desorption von As, von labilen Fe-Oxiden und der Transport (die Mobilisierung) beim Einsetzen reduzierender Bedingungen am stärksten. Mittlere Redoxpotenziale im positiven Bereich befördern die
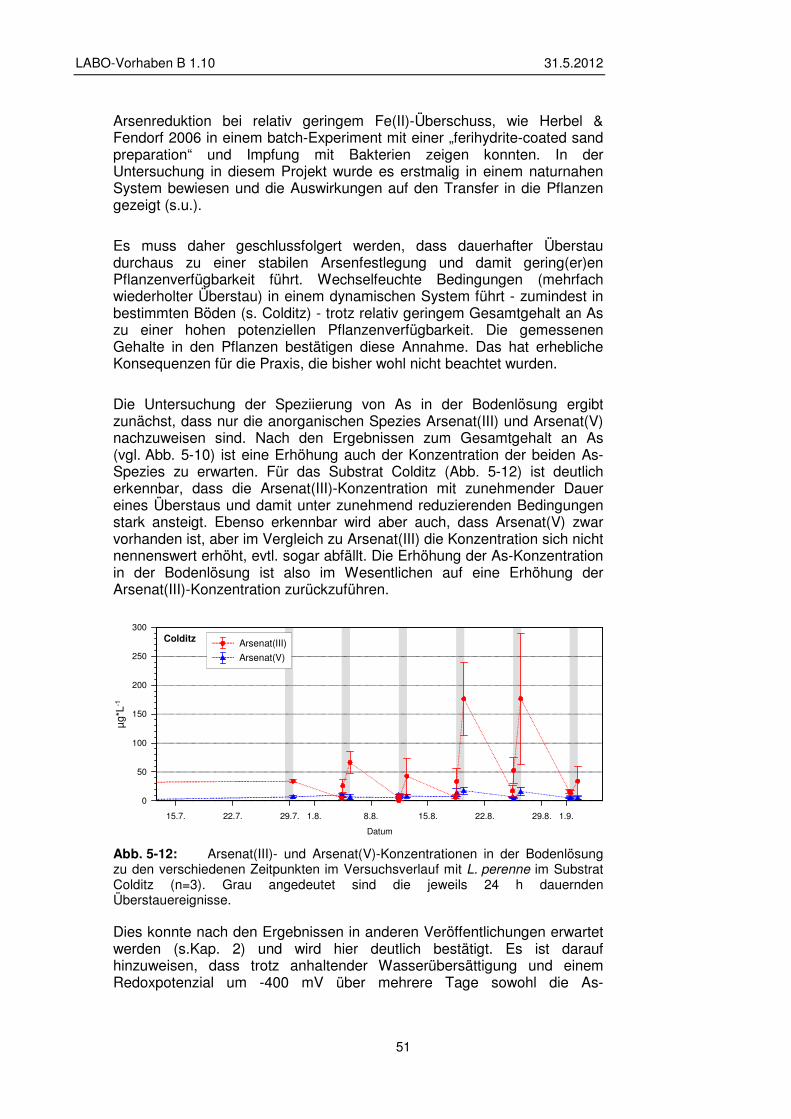
LABO-Vorhaben B 1.10 31.5.2012
51
Arsenreduktion bei relativ geringem Fe(II)-Überschuss, wie Herbel & Fendorf 2006 in einem batch-Experiment mit einer „ferihydrite-coated sand preparation“ und Impfung mit Bakterien zeigen konnten. In der Untersuchung in diesem Projekt wurde es erstmalig in einem naturnahen System bewiesen und die Auswirkungen auf den Transfer in die Pflanzen gezeigt (s.u.).
Es muss daher geschlussfolgert werden, dass dauerhafter Überstau durchaus zu einer stabilen Arsenfestlegung und damit gering(er)en Pflanzenverfügbarkeit führt. Wechselfeuchte Bedingungen (mehrfach wiederholter Überstau) in einem dynamischen System führt - zumindest in bestimmten Böden (s. Colditz) - trotz relativ geringem Gesamtgehalt an As zu einer hohen potenziellen Pflanzenverfügbarkeit. Die gemessenen Gehalte in den Pflanzen bestätigen diese Annahme. Das hat erhebliche Konsequenzen für die Praxis, die bisher wohl nicht beachtet wurden.
Die Untersuchung der Speziierung von As in der Bodenlösung ergibt zunächst, dass nur die anorganischen Spezies Arsenat(III) und Arsenat(V) nachzuweisen sind. Nach den Ergebnissen zum Gesamtgehalt an As (vgl. Abb. 5-10) ist eine Erhöhung auch der Konzentration der beiden As-Spezies zu erwarten. Für das Substrat Colditz (Abb. 5-12) ist deutlich erkennbar, dass die Arsenat(III)-Konzentration mit zunehmender Dauer eines Überstaus und damit unter zunehmend reduzierenden Bedingungen stark ansteigt. Ebenso erkennbar wird aber auch, dass Arsenat(V) zwar vorhanden ist, aber im Vergleich zu Arsenat(III) die Konzentration sich nicht nennenswert erhöht, evtl. sogar abfällt. Die Erhöhung der As-Konzentration in der Bodenlösung ist also im Wesentlichen auf eine Erhöhung der Arsenat(III)-Konzentration zurückzuführen.
0
50
100
150
200
250
300
µg*
L-1
15.7. 22.7. 29.7. 1.8. 8.8. 15.8. 22.8. 29.8. 1.9.
Datum
Arsenat(III)
Arsenat(V)
Colditz
Abb. 5-12: Arsenat(III)- und Arsenat(V)-Konzentrationen in der Bodenlösung zu den verschiedenen Zeitpunkten im Versuchsverlauf mit L. perenne im Substrat Colditz (n=3). Grau angedeutet sind die jeweils 24 h dauernden Überstauereignisse.
Dies konnte nach den Ergebnissen in anderen Veröffentlichungen erwartet werden (s.Kap. 2) und wird hier deutlich bestätigt. Es ist darauf hinzuweisen, dass trotz anhaltender Wasserübersättigung und einem Redoxpotenzial um -400 mV über mehrere Tage sowohl die As-
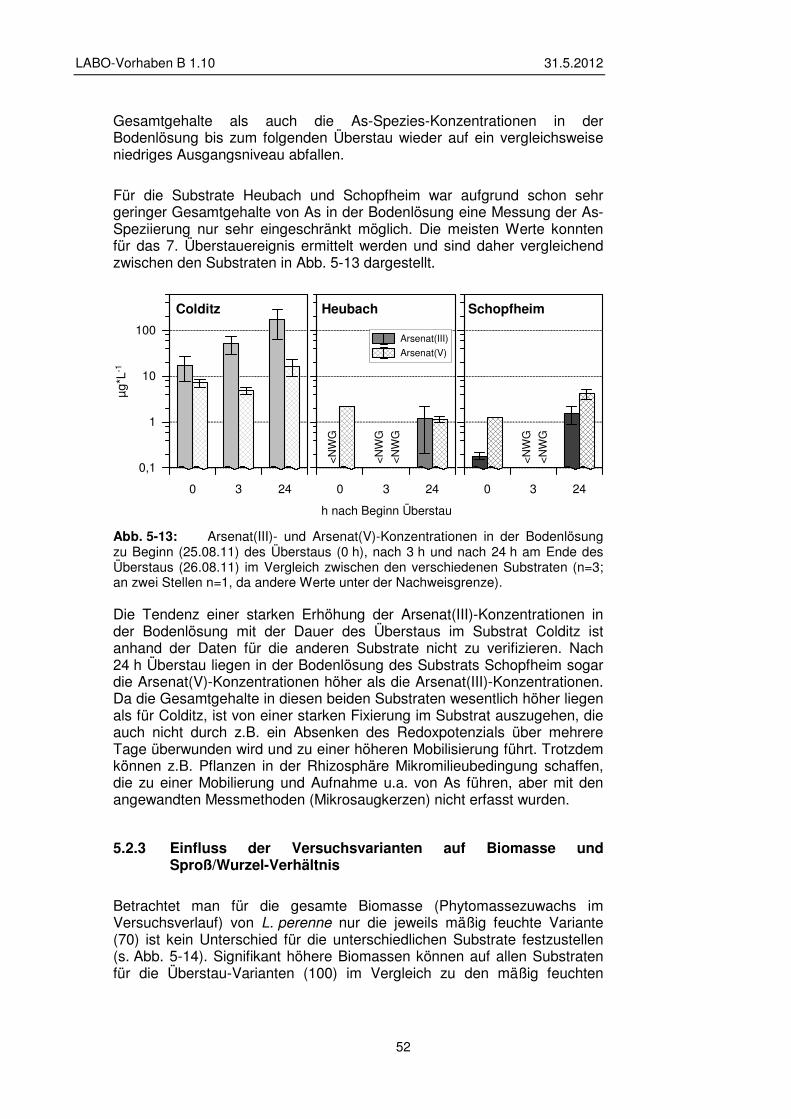
LABO-Vorhaben B 1.10 31.5.2012
52
Gesamtgehalte als auch die As-Spezies-Konzentrationen in der Bodenlösung bis zum folgenden Überstau wieder auf ein vergleichsweise niedriges Ausgangsniveau abfallen.
Für die Substrate Heubach und Schopfheim war aufgrund schon sehr geringer Gesamtgehalte von As in der Bodenlösung eine Messung der As-Speziierung nur sehr eingeschränkt möglich. Die meisten Werte konnten für das 7. Überstauereignis ermittelt werden und sind daher vergleichend zwischen den Substraten in Abb. 5-13 dargestellt.
0,1
1
10
100
µg*
L-1
0 3 24 0 3 24
Arsenat(III)
Arsenat(V)
0 3 24
Colditz SchopfheimHeubach
h nach Beginn Überstau
<N
WG
<N
WG
<N
WG
<N
WG
<N
WG
Abb. 5-13: Arsenat(III)- und Arsenat(V)-Konzentrationen in der Bodenlösung zu Beginn (25.08.11) des Überstaus (0 h), nach 3 h und nach 24 h am Ende des Überstaus (26.08.11) im Vergleich zwischen den verschiedenen Substraten (n=3; an zwei Stellen n=1, da andere Werte unter der Nachweisgrenze).
Die Tendenz einer starken Erhöhung der Arsenat(III)-Konzentrationen in der Bodenlösung mit der Dauer des Überstaus im Substrat Colditz ist anhand der Daten für die anderen Substrate nicht zu verifizieren. Nach 24 h Überstau liegen in der Bodenlösung des Substrats Schopfheim sogar die Arsenat(V)-Konzentrationen höher als die Arsenat(III)-Konzentrationen. Da die Gesamtgehalte in diesen beiden Substraten wesentlich höher liegen als für Colditz, ist von einer starken Fixierung im Substrat auszugehen, die auch nicht durch z.B. ein Absenken des Redoxpotenzials über mehrere Tage überwunden wird und zu einer höheren Mobilisierung führt. Trotzdem können z.B. Pflanzen in der Rhizosphäre Mikromilieubedingung schaffen, die zu einer Mobilierung und Aufnahme u.a. von As führen, aber mit den angewandten Messmethoden (Mikrosaugkerzen) nicht erfasst wurden.
5.2.3 Einfluss der Versuchsvarianten auf Biomasse und Sproß/Wurzel-Verhältnis
Betrachtet man für die gesamte Biomasse (Phytomassezuwachs im Versuchsverlauf) von L. perenne nur die jeweils mäßig feuchte Variante (70) ist kein Unterschied für die unterschiedlichen Substrate festzustellen (s. Abb. 5-14). Signifikant höhere Biomassen können auf allen Substraten für die Überstau-Varianten (100) im Vergleich zu den mäßig feuchten
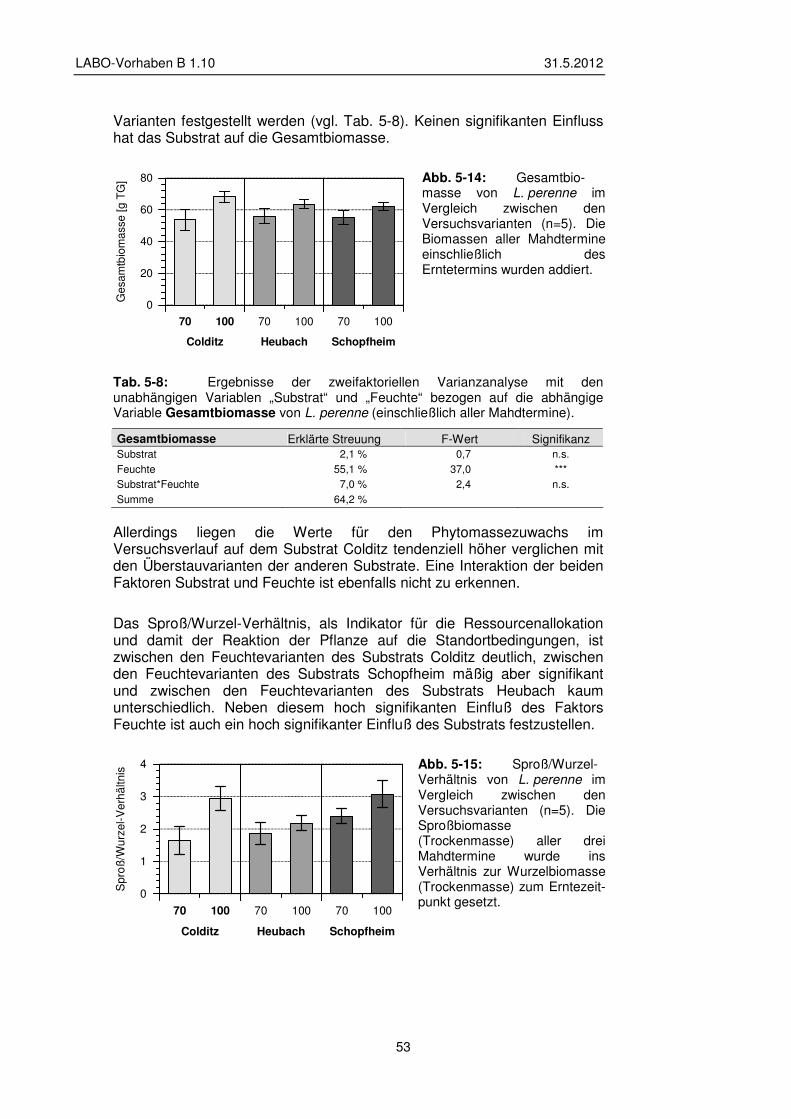
LABO-Vorhaben B 1.10 31.5.2012
53
Varianten festgestellt werden (vgl. Tab. 5-8). Keinen signifikanten Einfluss hat das Substrat auf die Gesamtbiomasse.
0
20
40
60
80
Ges
amtb
iom
asse
[gT
G]
70 100
Colditz
70 100
Heubach
70 100
Schopfheim
Abb. 5-14: Gesamtbio-masse von L. perenne im Vergleich zwischen den Versuchsvarianten (n=5). Die Biomassen aller Mahdtermine einschließlich des Erntetermins wurden addiert.
Tab. 5-8: Ergebnisse der zweifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Substrat“ und „Feuchte“ bezogen auf die abhängige Variable Gesamtbiomasse von L. perenne (einschließlich aller Mahdtermine).
Gesamtbiomasse Erklärte Streuung F-Wert Signifikanz Substrat 2,1 % 0,7 n.s.
Feuchte 55,1 % 37,0 *** Substrat*Feuchte 7,0 % 2,4 n.s.
Summe 64,2 %
Allerdings liegen die Werte für den Phytomassezuwachs im Versuchsverlauf auf dem Substrat Colditz tendenziell höher verglichen mit den Überstauvarianten der anderen Substrate. Eine Interaktion der beiden Faktoren Substrat und Feuchte ist ebenfalls nicht zu erkennen.
Das Sproß/Wurzel-Verhältnis, als Indikator für die Ressourcenallokation und damit der Reaktion der Pflanze auf die Standortbedingungen, ist zwischen den Feuchtevarianten des Substrats Colditz deutlich, zwischen den Feuchtevarianten des Substrats Schopfheim mäßig aber signifikant und zwischen den Feuchtevarianten des Substrats Heubach kaum unterschiedlich. Neben diesem hoch signifikanten Einfluß des Faktors Feuchte ist auch ein hoch signifikanter Einfluß des Substrats festzustellen.
0
1
2
3
4
Spr
oß/W
urze
l-Ver
hältn
is
70 100
Colditz
70 100
Heubach
70 100
Schopfheim
Abb. 5-15: Sproß/Wurzel-Verhältnis von L. perenne im Vergleich zwischen den Versuchsvarianten (n=5). Die Sproßbiomasse (Trockenmasse) aller drei Mahdtermine wurde ins Verhältnis zur Wurzelbiomasse (Trockenmasse) zum Erntezeit-punkt gesetzt.
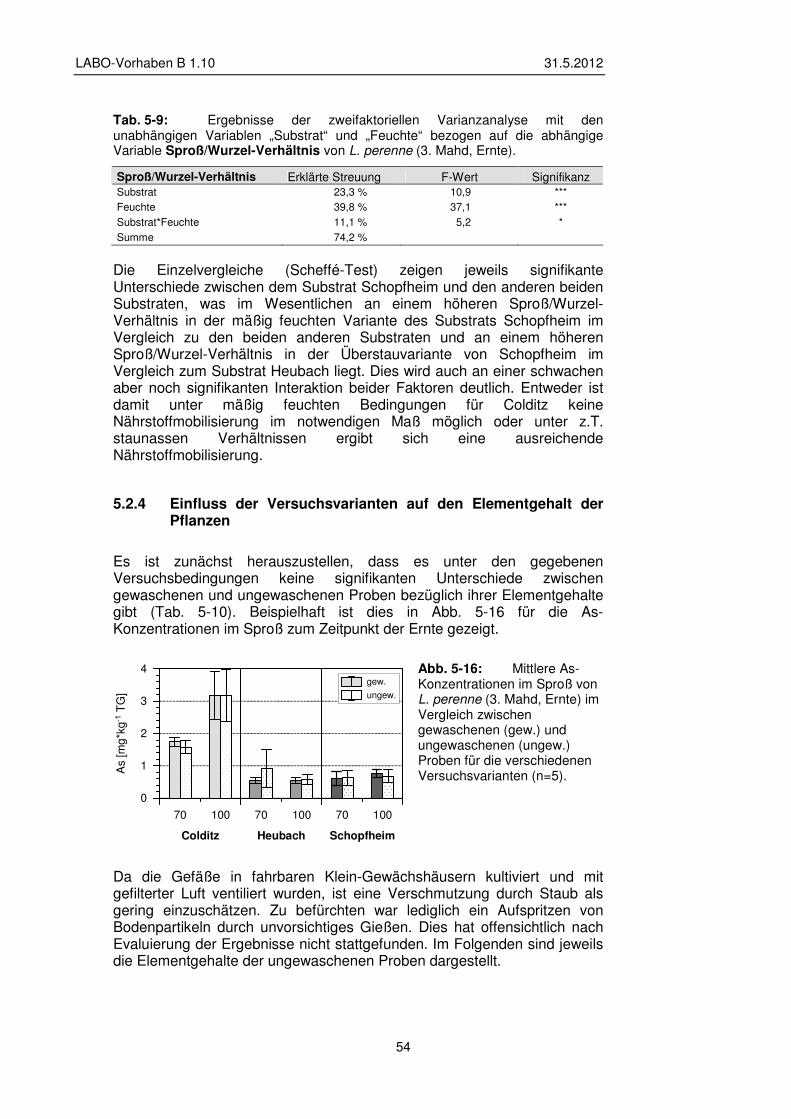
LABO-Vorhaben B 1.10 31.5.2012
54
Tab. 5-9: Ergebnisse der zweifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Substrat“ und „Feuchte“ bezogen auf die abhängige Variable Sproß/Wurzel-Verhältnis von L. perenne (3. Mahd, Ernte).
Sproß/Wurzel-Verhältnis Erklärte Streuung F-Wert Signifikanz Substrat 23,3 % 10,9 *** Feuchte 39,8 % 37,1 ***
Substrat*Feuchte 11,1 % 5,2 *
Summe 74,2 %
Die Einzelvergleiche (Scheffé-Test) zeigen jeweils signifikante Unterschiede zwischen dem Substrat Schopfheim und den anderen beiden Substraten, was im Wesentlichen an einem höheren Sproß/Wurzel-Verhältnis in der mäßig feuchten Variante des Substrats Schopfheim im Vergleich zu den beiden anderen Substraten und an einem höheren Sproß/Wurzel-Verhältnis in der Überstauvariante von Schopfheim im Vergleich zum Substrat Heubach liegt. Dies wird auch an einer schwachen aber noch signifikanten Interaktion beider Faktoren deutlich. Entweder ist damit unter mäßig feuchten Bedingungen für Colditz keine Nährstoffmobilisierung im notwendigen Maß möglich oder unter z.T. staunassen Verhältnissen ergibt sich eine ausreichende Nährstoffmobilisierung.
5.2.4 Einfluss der Versuchsvarianten auf den Elementgehalt der Pflanzen
Es ist zunächst herauszustellen, dass es unter den gegebenen Versuchsbedingungen keine signifikanten Unterschiede zwischen gewaschenen und ungewaschenen Proben bezüglich ihrer Elementgehalte gibt (Tab. 5-10). Beispielhaft ist dies in Abb. 5-16 für die As-Konzentrationen im Sproß zum Zeitpunkt der Ernte gezeigt.
70 100
Schopfheim
0
1
2
3
4
As
[mg*
kgT
G]
-1
70 100
Colditz
gew.
ungew.
70 100
Heubach
Abb. 5-16: Mittlere As-Konzentrationen im Sproß von L. perenne (3. Mahd, Ernte) im Vergleich zwischen gewaschenen (gew.) und ungewaschenen (ungew.) Proben für die verschiedenen Versuchsvarianten (n=5).
Da die Gefäße in fahrbaren Klein-Gewächshäusern kultiviert und mit gefilterter Luft ventiliert wurden, ist eine Verschmutzung durch Staub als gering einzuschätzen. Zu befürchten war lediglich ein Aufspritzen von Bodenpartikeln durch unvorsichtiges Gießen. Dies hat offensichtlich nach Evaluierung der Ergebnisse nicht stattgefunden. Im Folgenden sind jeweils die Elementgehalte der ungewaschenen Proben dargestellt.
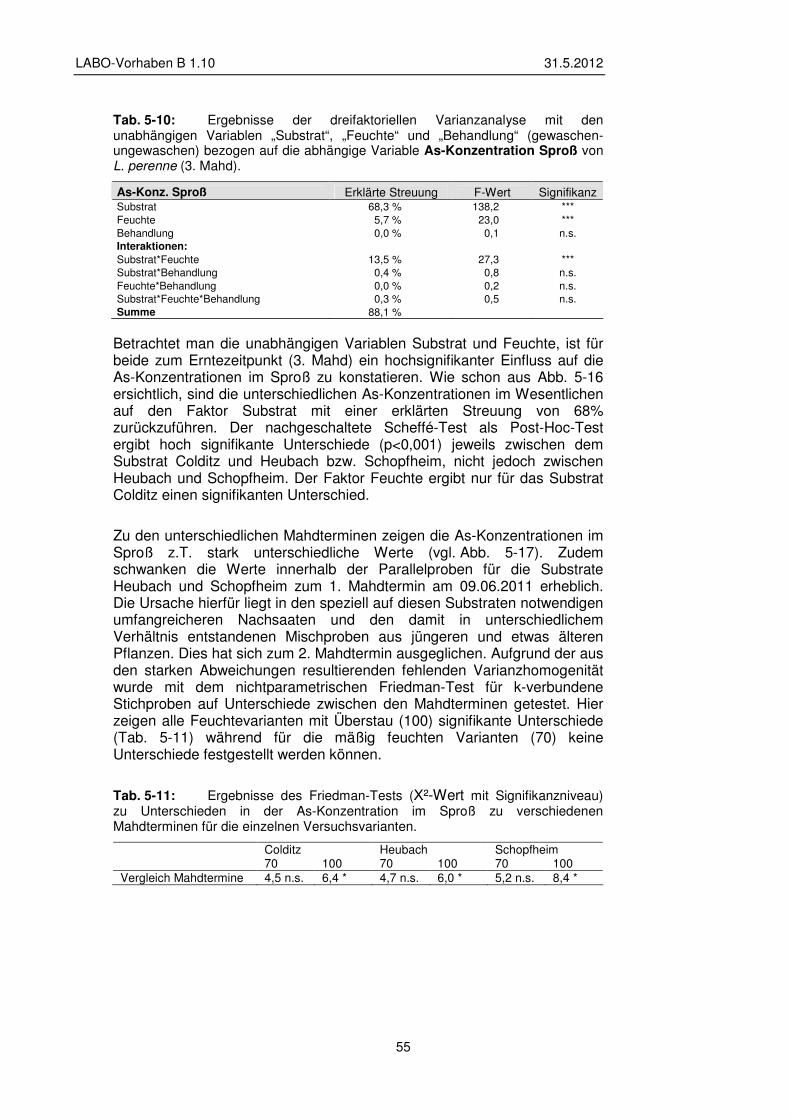
LABO-Vorhaben B 1.10 31.5.2012
55
Tab. 5-10: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Substrat“, „Feuchte“ und „Behandlung“ (gewaschen-ungewaschen) bezogen auf die abhängige Variable As-Konzentration Sproß von L. perenne (3. Mahd).
As-Konz. Sproß Erklärte Streuung F-Wert Signifikanz Substrat 68,3 % 138,2 *** Feuchte 5,7 % 23,0 *** Behandlung 0,0 % 0,1 n.s. Interaktionen: Substrat*Feuchte 13,5 % 27,3 *** Substrat*Behandlung 0,4 % 0,8 n.s. Feuchte*Behandlung 0,0 % 0,2 n.s. Substrat*Feuchte*Behandlung 0,3 % 0,5 n.s. Summe 88,1 %
Betrachtet man die unabhängigen Variablen Substrat und Feuchte, ist für beide zum Erntezeitpunkt (3. Mahd) ein hochsignifikanter Einfluss auf die As-Konzentrationen im Sproß zu konstatieren. Wie schon aus Abb. 5-16 ersichtlich, sind die unterschiedlichen As-Konzentrationen im Wesentlichen auf den Faktor Substrat mit einer erklärten Streuung von 68% zurückzuführen. Der nachgeschaltete Scheffé-Test als Post-Hoc-Test ergibt hoch signifikante Unterschiede (p<0,001) jeweils zwischen dem Substrat Colditz und Heubach bzw. Schopfheim, nicht jedoch zwischen Heubach und Schopfheim. Der Faktor Feuchte ergibt nur für das Substrat Colditz einen signifikanten Unterschied.
Zu den unterschiedlichen Mahdterminen zeigen die As-Konzentrationen im Sproß z.T. stark unterschiedliche Werte (vgl. Abb. 5-17). Zudem schwanken die Werte innerhalb der Parallelproben für die Substrate Heubach und Schopfheim zum 1. Mahdtermin am 09.06.2011 erheblich. Die Ursache hierfür liegt in den speziell auf diesen Substraten notwendigen umfangreicheren Nachsaaten und den damit in unterschiedlichem Verhältnis entstandenen Mischproben aus jüngeren und etwas älteren Pflanzen. Dies hat sich zum 2. Mahdtermin ausgeglichen. Aufgrund der aus den starken Abweichungen resultierenden fehlenden Varianzhomogenität wurde mit dem nichtparametrischen Friedman-Test für k-verbundene Stichproben auf Unterschiede zwischen den Mahdterminen getestet. Hier zeigen alle Feuchtevarianten mit Überstau (100) signifikante Unterschiede (Tab. 5-11) während für die mäßig feuchten Varianten (70) keine Unterschiede festgestellt werden können.
Tab. 5-11: Ergebnisse des Friedman-Tests (Χ²-Wert mit Signifikanzniveau) zu Unterschieden in der As-Konzentration im Sproß zu verschiedenen Mahdterminen für die einzelnen Versuchsvarianten.
Colditz Heubach Schopfheim 70 100 70 100 70 100 Vergleich Mahdtermine 4,5 n.s. 6,4 * 4,7 n.s. 6,0 * 5,2 n.s. 8,4 *
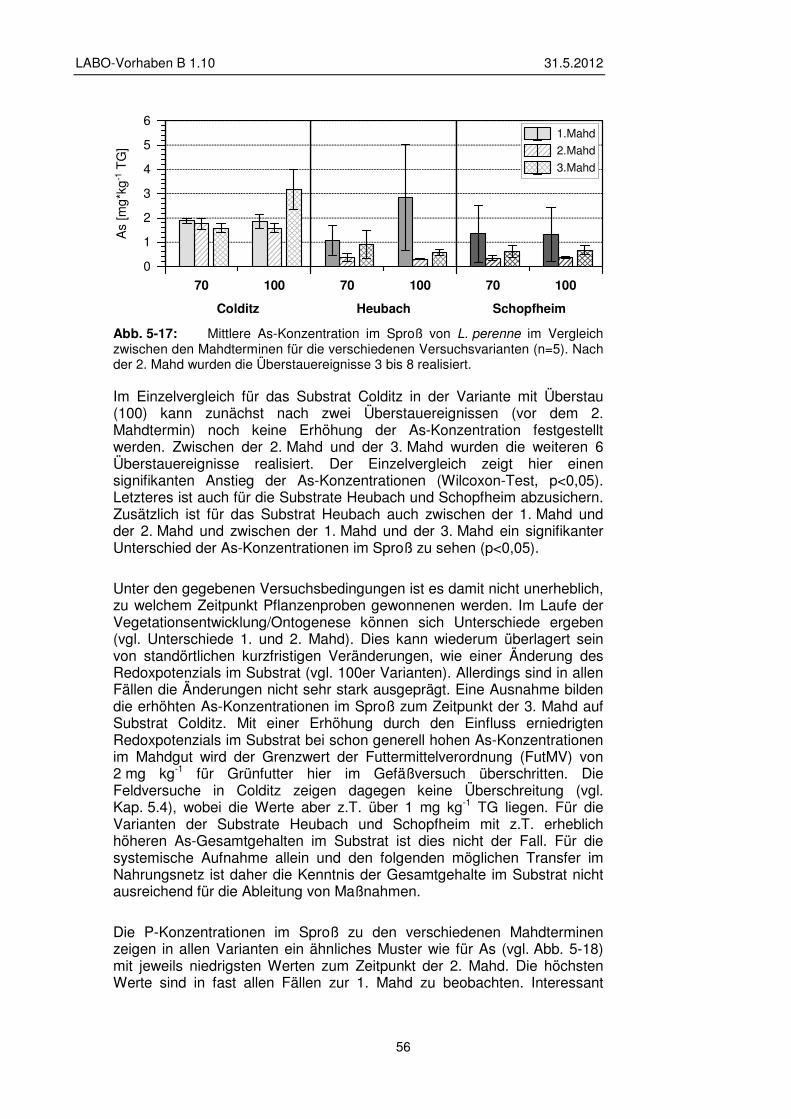
LABO-Vorhaben B 1.10 31.5.2012
56
70 100
Heubach
70 100
Schopfheim
0
1
2
3
4
5
6A
s[m
g*kg
TG
]-1
70 100
Colditz
1.Mahd
2.Mahd
3.Mahd
Abb. 5-17: Mittlere As-Konzentration im Sproß von L. perenne im Vergleich zwischen den Mahdterminen für die verschiedenen Versuchsvarianten (n=5). Nach der 2. Mahd wurden die Überstauereignisse 3 bis 8 realisiert.
Im Einzelvergleich für das Substrat Colditz in der Variante mit Überstau (100) kann zunächst nach zwei Überstauereignissen (vor dem 2. Mahdtermin) noch keine Erhöhung der As-Konzentration festgestellt werden. Zwischen der 2. Mahd und der 3. Mahd wurden die weiteren 6 Überstauereignisse realisiert. Der Einzelvergleich zeigt hier einen signifikanten Anstieg der As-Konzentrationen (Wilcoxon-Test, p<0,05). Letzteres ist auch für die Substrate Heubach und Schopfheim abzusichern. Zusätzlich ist für das Substrat Heubach auch zwischen der 1. Mahd und der 2. Mahd und zwischen der 1. Mahd und der 3. Mahd ein signifikanter Unterschied der As-Konzentrationen im Sproß zu sehen (p<0,05).
Unter den gegebenen Versuchsbedingungen ist es damit nicht unerheblich, zu welchem Zeitpunkt Pflanzenproben gewonnenen werden. Im Laufe der Vegetationsentwicklung/Ontogenese können sich Unterschiede ergeben (vgl. Unterschiede 1. und 2. Mahd). Dies kann wiederum überlagert sein von standörtlichen kurzfristigen Veränderungen, wie einer Änderung des Redoxpotenzials im Substrat (vgl. 100er Varianten). Allerdings sind in allen Fällen die Änderungen nicht sehr stark ausgeprägt. Eine Ausnahme bilden die erhöhten As-Konzentrationen im Sproß zum Zeitpunkt der 3. Mahd auf Substrat Colditz. Mit einer Erhöhung durch den Einfluss erniedrigten Redoxpotenzials im Substrat bei schon generell hohen As-Konzentrationen im Mahdgut wird der Grenzwert der Futtermittelverordnung (FutMV) von 2 mg kg-1 für Grünfutter hier im Gefäßversuch überschritten. Die Feldversuche in Colditz zeigen dagegen keine Überschreitung (vgl. Kap. 5.4), wobei die Werte aber z.T. über 1 mg kg-1 TG liegen. Für die Varianten der Substrate Heubach und Schopfheim mit z.T. erheblich höheren As-Gesamtgehalten im Substrat ist dies nicht der Fall. Für die systemische Aufnahme allein und den folgenden möglichen Transfer im Nahrungsnetz ist daher die Kenntnis der Gesamtgehalte im Substrat nicht ausreichend für die Ableitung von Maßnahmen.
Die P-Konzentrationen im Sproß zu den verschiedenen Mahdterminen zeigen in allen Varianten ein ähnliches Muster wie für As (vgl. Abb. 5-18) mit jeweils niedrigsten Werten zum Zeitpunkt der 2. Mahd. Die höchsten Werte sind in fast allen Fällen zur 1. Mahd zu beobachten. Interessant
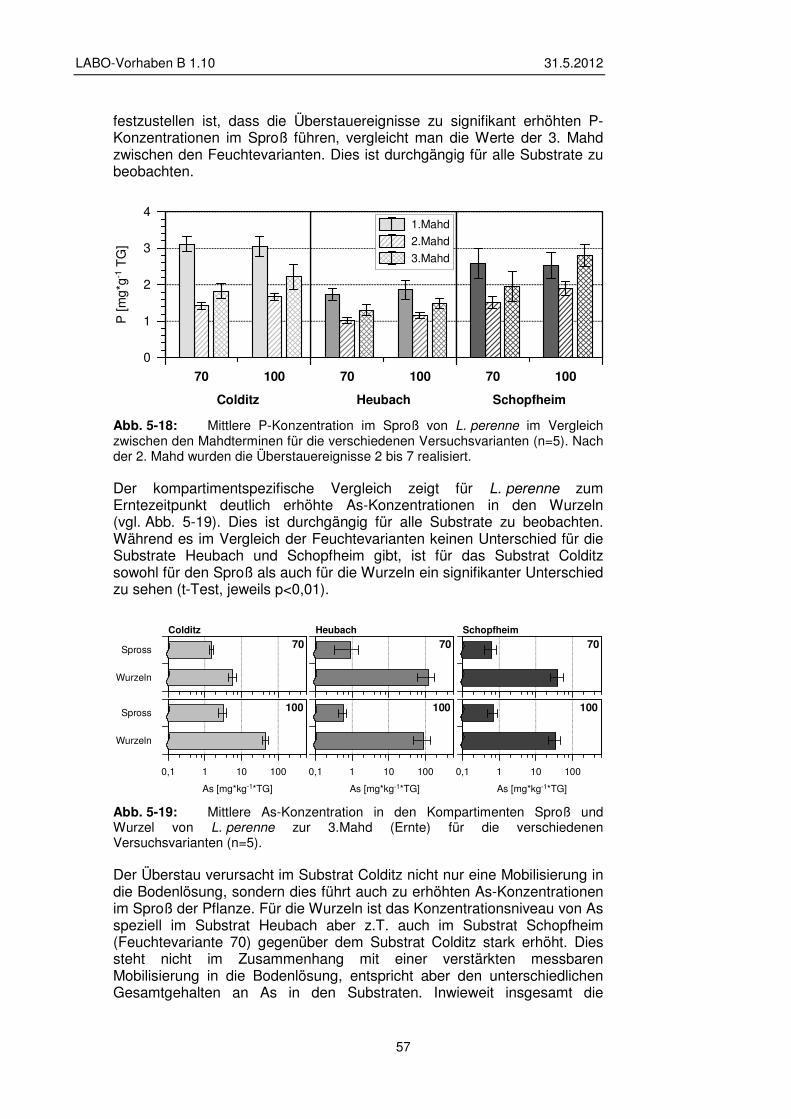
LABO-Vorhaben B 1.10 31.5.2012
57
festzustellen ist, dass die Überstauereignisse zu signifikant erhöhten P-Konzentrationen im Sproß führen, vergleicht man die Werte der 3. Mahd zwischen den Feuchtevarianten. Dies ist durchgängig für alle Substrate zu beobachten.
70 100
Heubach
0
1
2
3
4
P[m
g*g
TG
]-1
70 100
Colditz
1.Mahd
2.Mahd
3.Mahd
70 100
Schopfheim Abb. 5-18: Mittlere P-Konzentration im Sproß von L. perenne im Vergleich zwischen den Mahdterminen für die verschiedenen Versuchsvarianten (n=5). Nach der 2. Mahd wurden die Überstauereignisse 2 bis 7 realisiert.
Der kompartimentspezifische Vergleich zeigt für L. perenne zum Erntezeitpunkt deutlich erhöhte As-Konzentrationen in den Wurzeln (vgl. Abb. 5-19). Dies ist durchgängig für alle Substrate zu beobachten. Während es im Vergleich der Feuchtevarianten keinen Unterschied für die Substrate Heubach und Schopfheim gibt, ist für das Substrat Colditz sowohl für den Sproß als auch für die Wurzeln ein signifikanter Unterschied zu sehen (t-Test, jeweils p<0,01).
Spross
Wurzeln
Colditz
70
0,1 1 10 100
As [mg*kg *TG]-1
Spross
Wurzeln
100
Heubach
70
0,1 1 10 100
As [mg*kg *TG]-1
100
Schopfheim
70
0,1 1 10 100
As [mg*kg *TG]-1
100
Abb. 5-19: Mittlere As-Konzentration in den Kompartimenten Sproß und Wurzel von L. perenne zur 3.Mahd (Ernte) für die verschiedenen Versuchsvarianten (n=5).
Der Überstau verursacht im Substrat Colditz nicht nur eine Mobilisierung in die Bodenlösung, sondern dies führt auch zu erhöhten As-Konzentrationen im Sproß der Pflanze. Für die Wurzeln ist das Konzentrationsniveau von As speziell im Substrat Heubach aber z.T. auch im Substrat Schopfheim (Feuchtevariante 70) gegenüber dem Substrat Colditz stark erhöht. Dies steht nicht im Zusammenhang mit einer verstärkten messbaren Mobilisierung in die Bodenlösung, entspricht aber den unterschiedlichen Gesamtgehalten an As in den Substraten. Inwieweit insgesamt die

LABO-Vorhaben B 1.10 31.5.2012
58
erhöhten Konzentrationen in den Wurzeln auf tatsächlich in die Wurzelzellen inkorporiertes As zurückzuführen ist oder ein erheblicher Teil auch an Zellwandbestandteilen anhaftet oder sogar nur an Wurzeloberflächen präzipitiert (Fe-Plaque-Bildung) ist, kann hier nicht unterschieden werden.
Spross
Wurzeln
Colditz
70
0,01 0,1 1 10
Fe [mg*g *TG]-1
Spross
Wurzeln
100
Heubach
70
0,01 0,1 1 10
Fe [mg*g *TG]-1
100
Schopfheim
70
0,01 0,1 1 10
Fe [mg*g *TG]-1
100
Abb. 5-20: Mittlere Fe-Konzentration in den Kompartimenten Sproß und Wurzel von L. perenne zur 3. Mahd (Ernte) für die verschiedenen Versuchsvarianten (n=5).
Ein Hinweis ist allerdings über die Fe-Konzentrationen in den Kompartimenten der Pflanzen zu erhalten. Diese zeigen das gleiche Muster wie die As-Konzentrationen mit stark erhöhten Gehalten in/an den Wurzeln (z.T. >10 mg g-1 TG !) für alle Substrate. Das unterschiedliche Niveau der Fe-Gehalte entspricht wiederum den unterschiedlichen Gesamtgehalten an Fe in den Substraten (vgl. Tab. 5-4). Auch für Fe ist kein Unterschied in den Feuchtevarianten für die Substrate Heubach und Schopfheim erkennbar. Auf Substrat Colditz zeigen sich dagegen für Fe wie schon für As signifikante Unterschiede zwischen den Feuchtevarianten. Gleichzeitig lässt sich hier ein hoch signifikanter Zusammenhang zwischen beiden Elementkonzentrationen in den Wurzeln herstellen (s. Abb. 5-21). Dies ist ein deutlicher Hinweis darauf, dass durch eine beschriebene Mobilisierung von Fe unter teilweise reduzierenden Bedingungen mit anschließender Fällung in/an Wurzeln unter oxischen Bedingungen in der Rhizosphäre gleichzeitig mobilisiertes As gebunden und nicht in oberirdische Kompartimente transportiert wird (s. Kap. 2). Da die Wurzeln gründlich gewaschen wurden, kann man vermuten, dass wesentliche Prozesse der Fe-As-Wechselwirkung im Apoplasten der Wurzelrinde stattfinden. Dies ist speziell für die Substrate Heubach und Schopfheim mit sowohl erhöhten Fe-Gehalten als auch erhöhten As-Gehalten wahrscheinlich. Fe wird von den Pflanzen einerseits stark reguliert, da es intrazellulär an der Hydroxyl-Radikal-Bildung beteiligt ist. Andererseits kann es auf vielen Böden ein Mangelfaktor sein (geringe Pflanzeverfügbarkeit), was beispielsweise von Gräsern mit der Ausscheidung von Phytosiderophoren (Chelator) und Wiederaufnahme von Fe-Phytosiderophoren kompensiert wird (Kim & Guerinot 2007). Unter bestimmten Umständen bilden sich im Apoplasten der Wurzelrinde Fe-Pools (durch Fe-Fällungen) aus (Bienfait et al. 1985), was hier der Fall sein dürfte. Diese treten dann in Wechselwirkung mit As und führen zu den ermittelten Zusammenhängen.

LABO-Vorhaben B 1.10 31.5.2012
59
0,0
0,2
0,4
0,6
0,8A
s[m
mol
*kg-
1T
G]
0 50 100 150
Fe [mmol*kg-1 TG]
R²=0,945***
0,0
0,2
0,4
0,6
0,8
25 50 75 100
Fe [mmol*kg TG]-1
R²=0,945**
R²=0,977**
Wurzel
Abb. 5-21: Zusammenhang zwischen der As- und der Fe-Konzentration in Wurzeln von L. perenne auf Substrat Colditz zur Ernte (3.Mahd). Linke Grafik: alle Werte; rechte Grafik: ausgefüllte Marker = Überstauvariante (100), hohle Marker = mäßig feuchte Variante (70).
Stellt man den Zusammenhang zwischen As- und Fe-Konzentrationen für das Substrat Colditz getrennt für beide Feuchtevarianten her, ist ein unterschiedlicher Anstieg der Regressionsgeraden zu erkennen (rechte Grafik in Abb. 5-21).
Andere Untersuchungen legen ebenfalls die Vermutung nahe, dass As tatsächlich unter staunassen/wechselfeuchten Verhältnissen in erheblichem Maße von außen anhaftet und nicht aufgenommen ist (Zhao et al. 2009). Dies wird ebenfalls mit Fe-Plaque-Bildung begründet, zu dem speziell As(V) eine hohe Affinität zeigt.
Die As-Speziierung in der Bodenlösung spiegelt sich nicht in der As-Speziierung im Sproß wider. Im Sproß sind höhere As(V)-Konzentrationen zu finden, während in der Bodenlösung speziell während der Überstauereignisse die As(III)-Konzentration ansteigt.
70 100
Heubach
0,0
0,2
0,4
mg*
kgT
G-1
1
2
70 100
Colditz
As(III)As(V)
70 100
Schopfheim
Abb. 5-22: Mittlere As(III)- und As(V)-Konzentrationen im Sproß (3.Mahd) von L. perenne im Vergleich zwischen den Versuchsvarianten (n=5).
Die Befunde hier stehen nicht im Einklang mit den Hinweisen aus der Literatur. Dort wird vermutet, dass hauptsächlich As(III) in der Pflanze vorkommt (neben möglicherweise organischen As-Verbindungen). Inwieweit dies auf die Probenvorbereitung und das Extraktionsverfahren zurückzuführen ist, muss diskutiert werden.

LABO-Vorhaben B 1.10 31.5.2012
60
5.2.5 Transfer und Verteilung in der Pflanze
Sproß
Wurzeln
Colditz
70Heubach
70Schopfheim
70
1 10 100
TF
Sproß
Wurzeln
100
1 10 100
TF
100
1 10 100
TF
70
Abb. 5-23: Mittlere Transferfaktoren (TF) Boden – Pflanze für As bezogen auf die As-Konzentrationen nach KW-Extrakt und die As-Konzentrationen in Sproß und Wurzeln für alle Versuchsvarianten.
Ein Zusammenhang zwischen den As-Konzentrationen im AN-Extrakt und den As-Konzentrationen in Sproß und Wurzel der einzelnen Versuchsvarianten zum Zeitpunkt der Ernte konnte nur für das Substrat Colditz gerechnet werden. Für die anderen Substrate lagen die As-Konzentrationen im AN-Extrakt unter der Nachweisgrenze (s. Abb. 5-9). Für das Substrat Colditz ist weder für den Sproß noch für die Wurzel ein signifikanter Zusammenhang herzustellen (Sproß: r=-0,234 p>0,05; Wurzel: r=0,104 p>0,05).
Die Verteilung innerhalb der Pflanzenkompartimente ist sowohl mit den kompartimentspezifischen As-Konzentrationen als auch mit den berechneten Transferfaktoren für die einzelnen Kompartimente angedeutet. Allerdings wurde bislang der Biomassebezug nicht hergestellt. Nach Kalkulation der As-Stoffmenge auf der Grundlage von As-Konzentrationen und Biomasse lässt sich der prozentuale Anteile von As in den einzelnen Kompartimenten und damit die genaue interne Verteilung berechnen.
0
20
40
60
80
100
%
70 100
Colditz
70 100
Heubach
70 100
Schopfheim
WurzelSproß
Anteil an As in
Abb. 5-24: Mittlerer prozentualer Anteil der As-Stoffmenge in Sproß (Summe aller Mahdtermine) und Wurzeln an der gesamten As-Stoffmenge in den L. perenne-Pflanzen für die unterschiedlichen Versuchsvarianten.
Für die Substrate Heubach und Schopfheim sind mehr als 90% der gesamten As-Stoffmenge der Pflanzen in den Wurzeln zu finden. Die höheren As-Konzentrationen in den Wurzeln werden im Stoffmengenvergleich nicht durch die höhere Sproßbiomassen ausgeglichen (vgl. Abb. 5-15). Dies gilt ebenfalls für die Varianten mit

LABO-Vorhaben B 1.10 31.5.2012
61
Substrat Colditz. Allerdings erreicht hier die Stoffmenge As im Sproß einen Anteil von 20-30%. Dies ist auch das einzige Substrat, für das Unterschiede zwischen den Feuchtevarianten existieren. Unter mäßig feuchten Bedingungen wird prozentual mehr As in den Sproß verlagert.
In der Berechnung des Transfers im Nahrungsnetz spielt der Anteil der Verschmutzung durch Staubablagerungen und aufgespritzten Boden eine wesentliche Rolle. Der Maßnahmewert der BBodSchV für As im Boden mit Grünlandnutzung von 50 mg kg-1 wurde unter der Maßgabe einer 3%igen Verschmutzung des Grünschnitts mit anhaftendem Bodenmaterial bei gleichzeitig angenommenem geringen Transfer über die Wurzelaufnahme festgelegt. Legt man diese Annahme für den Versuch mit L. perenne zugrunde und berechnet eine theoretische Verschmutzung des Mahdguts (Sproß) mit den entsprechenden Substraten, ist die in Abb. 5-25 dargestellte Verteilung zwischen dem Anteil von As durch systemische Aufnahme und dem Anteil durch Verschmutzung zu sehen.
0
20
40
60
80
100
%
70 100
Colditz
70 100
Heubach
Substrat (Verschmutzung)Sproß (systemisch)
70 100
Schopfheim
70 100
Colditz
70 100
Colditz
1.Mahd 3.Mahd2.Mahd
70 100
Heubach
70 100
Heubach
1.Mahd 3.Mahd2.Mahd
70 100
Schopfheim
70 100
Schopfheim
1.Mahd 3.Mahd2.Mahd
Abb. 5-25: Prozentualer Anteil von As im Sproß (systemische Aufnahme) und anhaftendem Substrat (Verschmutzung) im Mahdgut bei angenommener Verschmutzung von 3% durch das Substrat für die unterschiedlichen Versuchsvarianten getrennt nach Mahden.
Entsprechend den As-Konzentrationen im Substrat würden hier also auf Substrat Colditz nur zwischen 20 und 30% des As durch Verschmutzung im Nahrungsnetz weitergegeben. Mehr als 70% würde durch eine systemische Aufnahme und Akkumulation in oberirdischen Pflanzenkompartimenten das Nahrungsnetz erreichen. Dieser Anteil erhöht sich auf fast 80% unter teilweise reduzierenden Bedingungen im Substrat (Variante 100 zur 3.Mahd), was für erhöhte As-Konzentrationen im Mahdgut sorgt.
Ganz anders sieht es auf den Substraten Heubach und Schopfheim aus. Hier kommt der wesentliche Beitrag (80 - <90%) durch die Verschmutzung mit kontaminiertem Substrat zustande, was eindeutig auf die höheren As-Konzentrationen im Substrat zurückzuführen ist.
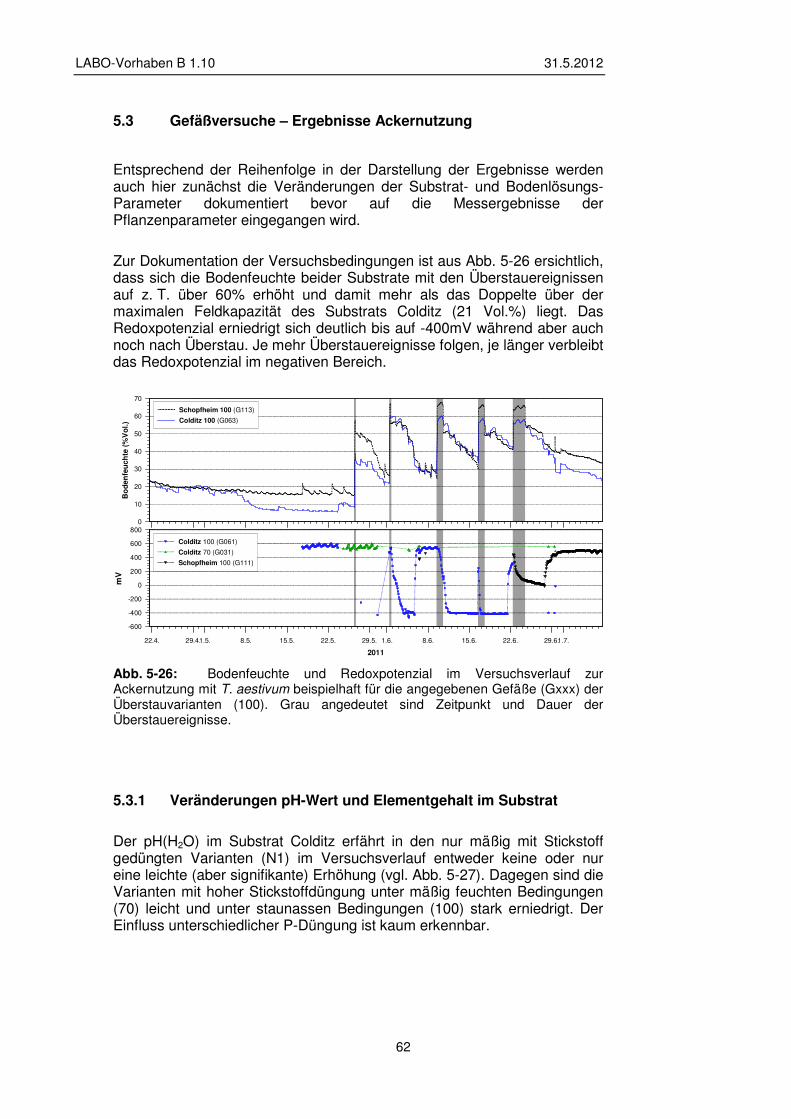
LABO-Vorhaben B 1.10 31.5.2012
62
5.3 Gefäßversuche – Ergebnisse Ackernutzung
Entsprechend der Reihenfolge in der Darstellung der Ergebnisse werden auch hier zunächst die Veränderungen der Substrat- und Bodenlösungs-Parameter dokumentiert bevor auf die Messergebnisse der Pflanzenparameter eingegangen wird.
Zur Dokumentation der Versuchsbedingungen ist aus Abb. 5-26 ersichtlich, dass sich die Bodenfeuchte beider Substrate mit den Überstauereignissen auf z. T. über 60% erhöht und damit mehr als das Doppelte über der maximalen Feldkapazität des Substrats Colditz (21 Vol.%) liegt. Das Redoxpotenzial erniedrigt sich deutlich bis auf -400mV während aber auch noch nach Überstau. Je mehr Überstauereignisse folgen, je länger verbleibt das Redoxpotenzial im negativen Bereich.
0
10
20
30
40
50
60
70
Bo
den
feu
chte
(%V
ol.)
Schopfheim 100 (G113)
Colditz 100 (G063)
-600
-400
-200
0
200
400
600
800
mV
22.4. 29.4.1.5. 8.5. 15.5. 22.5. 29.5. 1.6. 8.6. 15.6. 22.6. 29.6.1.7.
2011
Colditz 100 (G061)
Colditz 70 (G031)
Schopfheim 100 (G111)
Abb. 5-26: Bodenfeuchte und Redoxpotenzial im Versuchsverlauf zur Ackernutzung mit T. aestivum beispielhaft für die angegebenen Gefäße (Gxxx) der Überstauvarianten (100). Grau angedeutet sind Zeitpunkt und Dauer der Überstauereignisse.
5.3.1 Veränderungen pH-Wert und Elementgehalt im Substrat
Der pH(H2O) im Substrat Colditz erfährt in den nur mäßig mit Stickstoff gedüngten Varianten (N1) im Versuchsverlauf entweder keine oder nur eine leichte (aber signifikante) Erhöhung (vgl. Abb. 5-27). Dagegen sind die Varianten mit hoher Stickstoffdüngung unter mäßig feuchten Bedingungen (70) leicht und unter staunassen Bedingungen (100) stark erniedrigt. Der Einfluss unterschiedlicher P-Düngung ist kaum erkennbar.
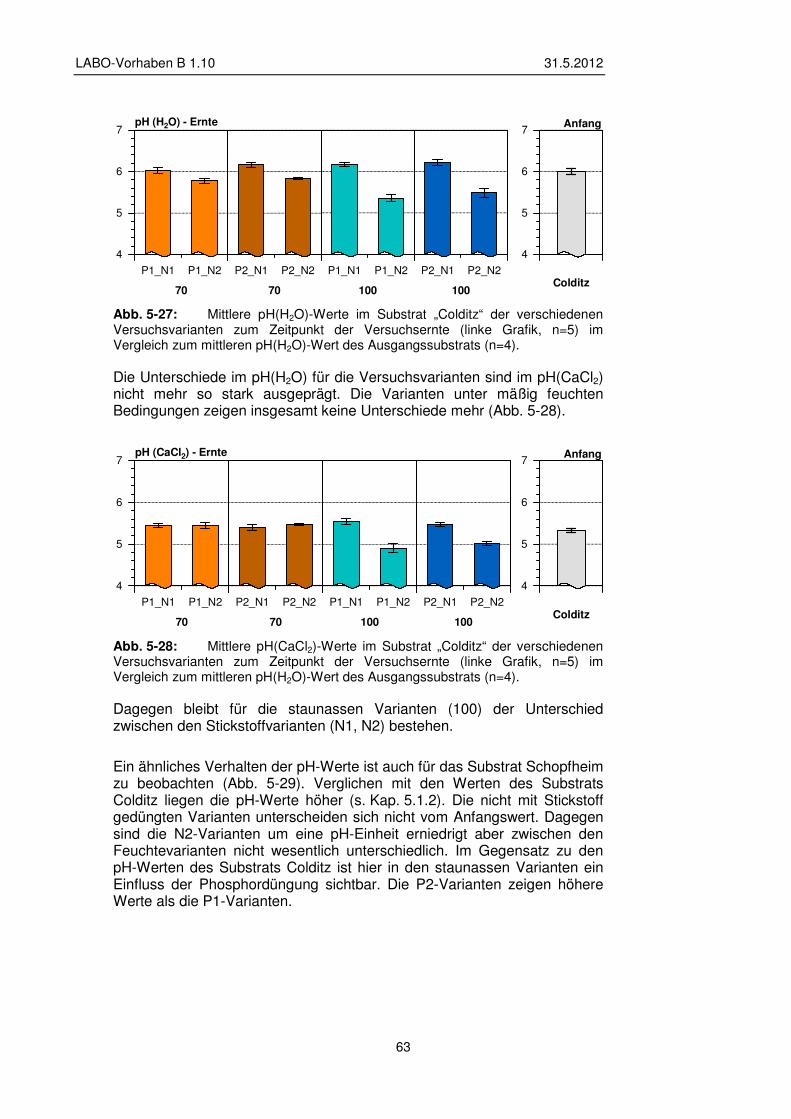
LABO-Vorhaben B 1.10 31.5.2012
63
4
5
6
7
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
4
5
6
7
Colditz
pH (H2O) - Ernte Anfang
Abb. 5-27: Mittlere pH(H2O)-Werte im Substrat „Colditz“ der verschiedenen Versuchsvarianten zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zum mittleren pH(H2O)-Wert des Ausgangssubstrats (n=4).
Die Unterschiede im pH(H2O) für die Versuchsvarianten sind im pH(CaCl2) nicht mehr so stark ausgeprägt. Die Varianten unter mäßig feuchten Bedingungen zeigen insgesamt keine Unterschiede mehr (Abb. 5-28).
4
5
6
7
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
pH (CaCl2) - Ernte Anfang
4
5
6
7
Colditz
Abb. 5-28: Mittlere pH(CaCl2)-Werte im Substrat „Colditz“ der verschiedenen Versuchsvarianten zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zum mittleren pH(H2O)-Wert des Ausgangssubstrats (n=4).
Dagegen bleibt für die staunassen Varianten (100) der Unterschied zwischen den Stickstoffvarianten (N1, N2) bestehen.
Ein ähnliches Verhalten der pH-Werte ist auch für das Substrat Schopfheim zu beobachten (Abb. 5-29). Verglichen mit den Werten des Substrats Colditz liegen die pH-Werte höher (s. Kap. 5.1.2). Die nicht mit Stickstoff gedüngten Varianten unterscheiden sich nicht vom Anfangswert. Dagegen sind die N2-Varianten um eine pH-Einheit erniedrigt aber zwischen den Feuchtevarianten nicht wesentlich unterschiedlich. Im Gegensatz zu den pH-Werten des Substrats Colditz ist hier in den staunassen Varianten ein Einfluss der Phosphordüngung sichtbar. Die P2-Varianten zeigen höhere Werte als die P1-Varianten.
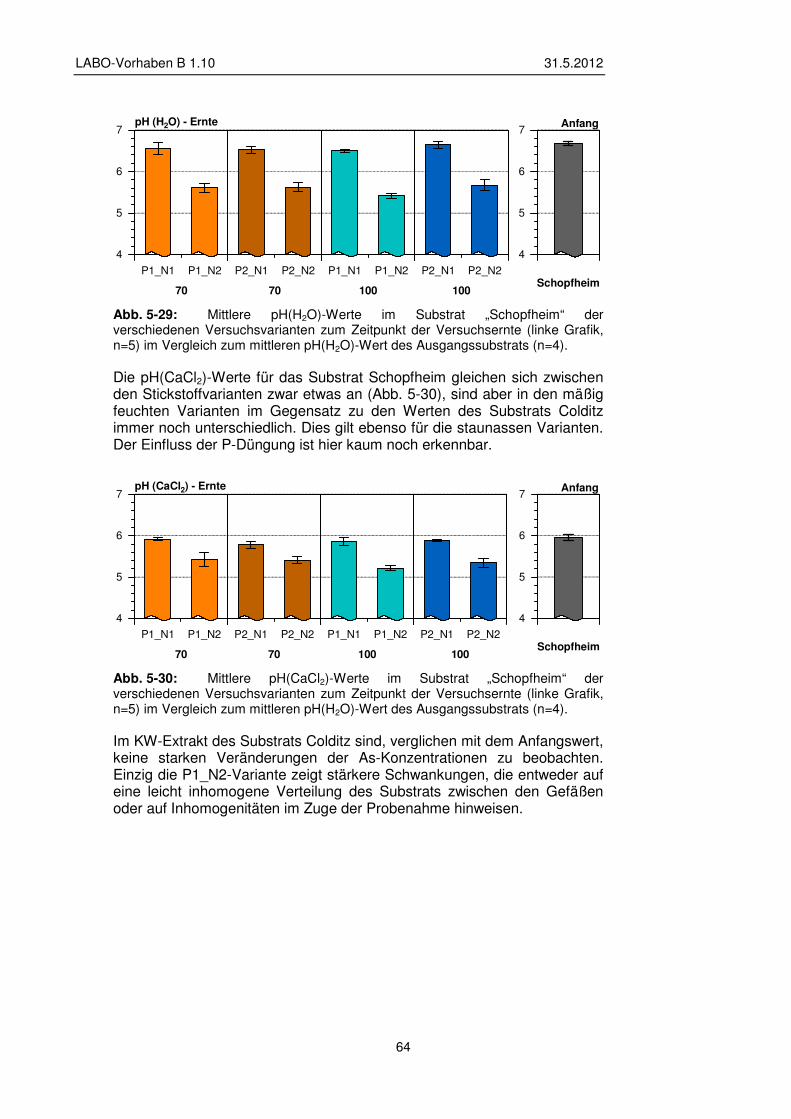
LABO-Vorhaben B 1.10 31.5.2012
64
4
5
6
7
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
4
5
6
7
Schopfheim
pH (H2O) - Ernte Anfang
Abb. 5-29: Mittlere pH(H2O)-Werte im Substrat „Schopfheim“ der verschiedenen Versuchsvarianten zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zum mittleren pH(H2O)-Wert des Ausgangssubstrats (n=4).
Die pH(CaCl2)-Werte für das Substrat Schopfheim gleichen sich zwischen den Stickstoffvarianten zwar etwas an (Abb. 5-30), sind aber in den mäßig feuchten Varianten im Gegensatz zu den Werten des Substrats Colditz immer noch unterschiedlich. Dies gilt ebenso für die staunassen Varianten. Der Einfluss der P-Düngung ist hier kaum noch erkennbar.
4
5
6
7
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
4
5
6
7
Schopfheim
pH (CaCl2) - Ernte Anfang
Abb. 5-30: Mittlere pH(CaCl2)-Werte im Substrat „Schopfheim“ der verschiedenen Versuchsvarianten zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zum mittleren pH(H2O)-Wert des Ausgangssubstrats (n=4).
Im KW-Extrakt des Substrats Colditz sind, verglichen mit dem Anfangswert, keine starken Veränderungen der As-Konzentrationen zu beobachten. Einzig die P1_N2-Variante zeigt stärkere Schwankungen, die entweder auf eine leicht inhomogene Verteilung des Substrats zwischen den Gefäßen oder auf Inhomogenitäten im Zuge der Probenahme hinweisen.
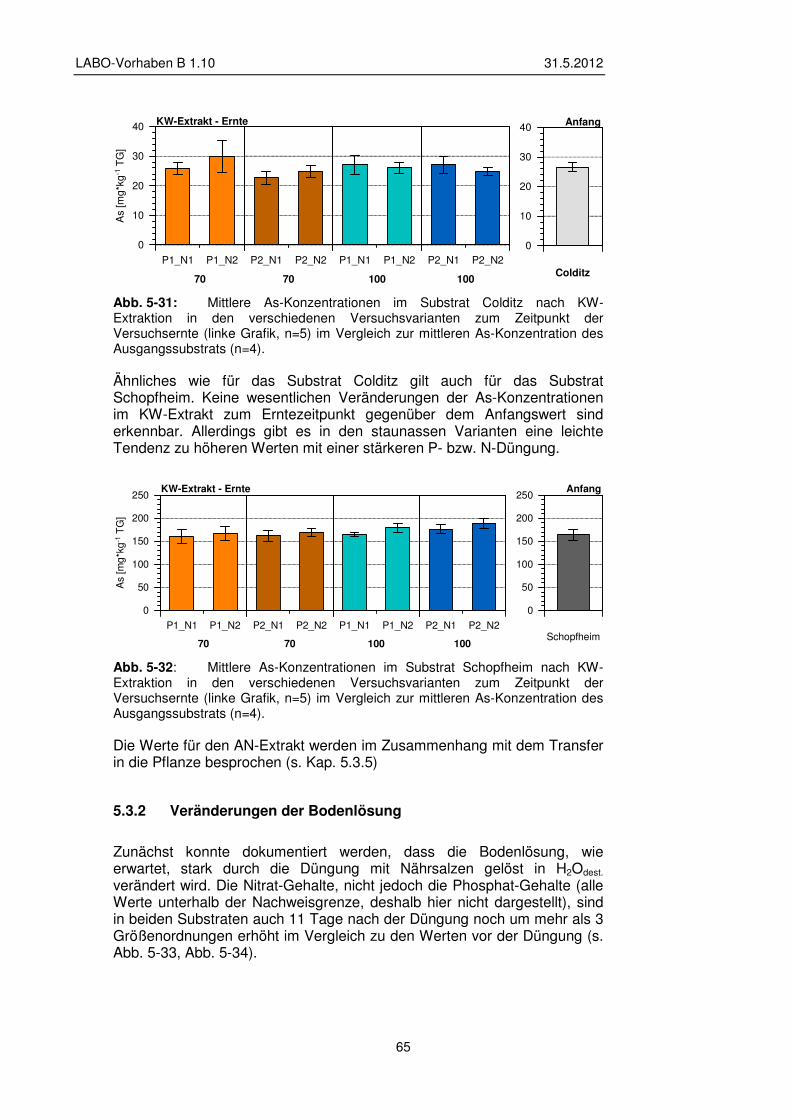
LABO-Vorhaben B 1.10 31.5.2012
65
0
10
20
30
40A
s[m
g*kg
TG
]-1
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
KW-Extrakt - Ernte
0
10
20
30
40
Colditz
Anfang
Abb. 5-31: Mittlere As-Konzentrationen im Substrat Colditz nach KW-Extraktion in den verschiedenen Versuchsvarianten zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zur mittleren As-Konzentration des Ausgangssubstrats (n=4).
Ähnliches wie für das Substrat Colditz gilt auch für das Substrat Schopfheim. Keine wesentlichen Veränderungen der As-Konzentrationen im KW-Extrakt zum Erntezeitpunkt gegenüber dem Anfangswert sind erkennbar. Allerdings gibt es in den staunassen Varianten eine leichte Tendenz zu höheren Werten mit einer stärkeren P- bzw. N-Düngung.
0
50
100
150
200
250
As
[mg*
kgT
G]
-1
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
KW-Extrakt - Ernte
0
50
100
150
200
250
Schopfheim
Anfang
Abb. 5-32: Mittlere As-Konzentrationen im Substrat Schopfheim nach KW-Extraktion in den verschiedenen Versuchsvarianten zum Zeitpunkt der Versuchsernte (linke Grafik, n=5) im Vergleich zur mittleren As-Konzentration des Ausgangssubstrats (n=4).
Die Werte für den AN-Extrakt werden im Zusammenhang mit dem Transfer in die Pflanze besprochen (s. Kap. 5.3.5)
5.3.2 Veränderungen der Bodenlösung
Zunächst konnte dokumentiert werden, dass die Bodenlösung, wie erwartet, stark durch die Düngung mit Nährsalzen gelöst in H2Odest. verändert wird. Die Nitrat-Gehalte, nicht jedoch die Phosphat-Gehalte (alle Werte unterhalb der Nachweisgrenze, deshalb hier nicht dargestellt), sind in beiden Substraten auch 11 Tage nach der Düngung noch um mehr als 3 Größenordnungen erhöht im Vergleich zu den Werten vor der Düngung (s. Abb. 5-33, Abb. 5-34).
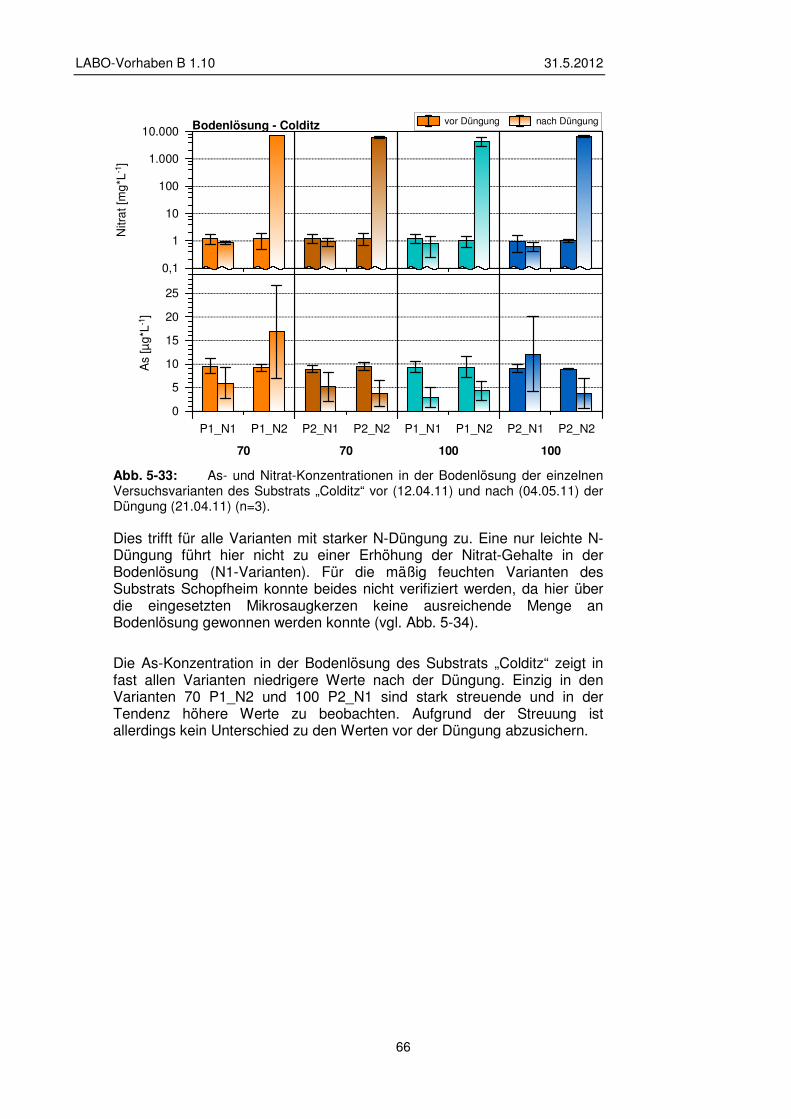
LABO-Vorhaben B 1.10 31.5.2012
66
P2_N1 P2_N2
70
0,1
1
10
100
1.000
10.000N
itra
t[m
g*L
]-1
P1_N1 P1_N2
70
vor Düngung nach Düngung
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Bodenlösung - Colditz
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
0
5
10
15
20
25
As
[µg*
L]
-1
P1_N1 P1_N2
70 Abb. 5-33: As- und Nitrat-Konzentrationen in der Bodenlösung der einzelnen Versuchsvarianten des Substrats „Colditz“ vor (12.04.11) und nach (04.05.11) der Düngung (21.04.11) (n=3).
Dies trifft für alle Varianten mit starker N-Düngung zu. Eine nur leichte N-Düngung führt hier nicht zu einer Erhöhung der Nitrat-Gehalte in der Bodenlösung (N1-Varianten). Für die mäßig feuchten Varianten des Substrats Schopfheim konnte beides nicht verifiziert werden, da hier über die eingesetzten Mikrosaugkerzen keine ausreichende Menge an Bodenlösung gewonnen werden konnte (vgl. Abb. 5-34).
Die As-Konzentration in der Bodenlösung des Substrats „Colditz“ zeigt in fast allen Varianten niedrigere Werte nach der Düngung. Einzig in den Varianten 70 P1_N2 und 100 P2_N1 sind stark streuende und in der Tendenz höhere Werte zu beobachten. Aufgrund der Streuung ist allerdings kein Unterschied zu den Werten vor der Düngung abzusichern.
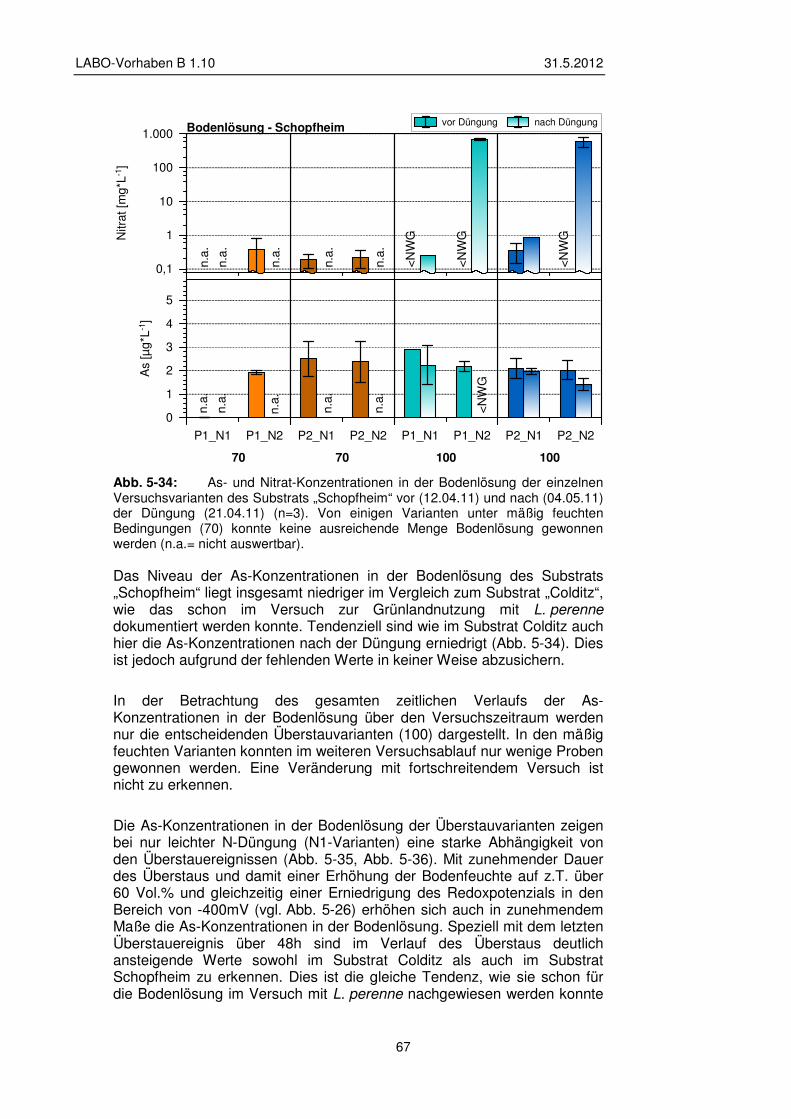
LABO-Vorhaben B 1.10 31.5.2012
67
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
0,1
1
10
100
1.000N
itrat
[mg*
L]
-1
P1_N1 P1_N2
70
Bodenlösung - Schopfheim
<N
WG
n.a.
n.a.
n.a.
n.a.
n.a.
<N
WG
<N
WG
P2_N1 P2_N2
100
P1_N1 P1_N2
100
vor Düngung nach Düngung
0
1
2
3
4
5
As
[µg*
L]
-1
P1_N1 P1_N2
70
P2_N1 P2_N2
70
<N
WG
n.a.
n.a.
n.a.
n.a.
n.a.
Abb. 5-34: As- und Nitrat-Konzentrationen in der Bodenlösung der einzelnen Versuchsvarianten des Substrats „Schopfheim“ vor (12.04.11) und nach (04.05.11) der Düngung (21.04.11) (n=3). Von einigen Varianten unter mäßig feuchten Bedingungen (70) konnte keine ausreichende Menge Bodenlösung gewonnen werden (n.a.= nicht auswertbar).
Das Niveau der As-Konzentrationen in der Bodenlösung des Substrats „Schopfheim“ liegt insgesamt niedriger im Vergleich zum Substrat „Colditz“, wie das schon im Versuch zur Grünlandnutzung mit L. perenne dokumentiert werden konnte. Tendenziell sind wie im Substrat Colditz auch hier die As-Konzentrationen nach der Düngung erniedrigt (Abb. 5-34). Dies ist jedoch aufgrund der fehlenden Werte in keiner Weise abzusichern.
In der Betrachtung des gesamten zeitlichen Verlaufs der As-Konzentrationen in der Bodenlösung über den Versuchszeitraum werden nur die entscheidenden Überstauvarianten (100) dargestellt. In den mäßig feuchten Varianten konnten im weiteren Versuchsablauf nur wenige Proben gewonnen werden. Eine Veränderung mit fortschreitendem Versuch ist nicht zu erkennen.
Die As-Konzentrationen in der Bodenlösung der Überstauvarianten zeigen bei nur leichter N-Düngung (N1-Varianten) eine starke Abhängigkeit von den Überstauereignissen (Abb. 5-35, Abb. 5-36). Mit zunehmender Dauer des Überstaus und damit einer Erhöhung der Bodenfeuchte auf z.T. über 60 Vol.% und gleichzeitig einer Erniedrigung des Redoxpotenzials in den Bereich von -400mV (vgl. Abb. 5-26) erhöhen sich auch in zunehmendem Maße die As-Konzentrationen in der Bodenlösung. Speziell mit dem letzten Überstauereignis über 48h sind im Verlauf des Überstaus deutlich ansteigende Werte sowohl im Substrat Colditz als auch im Substrat Schopfheim zu erkennen. Dies ist die gleiche Tendenz, wie sie schon für die Bodenlösung im Versuch mit L. perenne nachgewiesen werden konnte
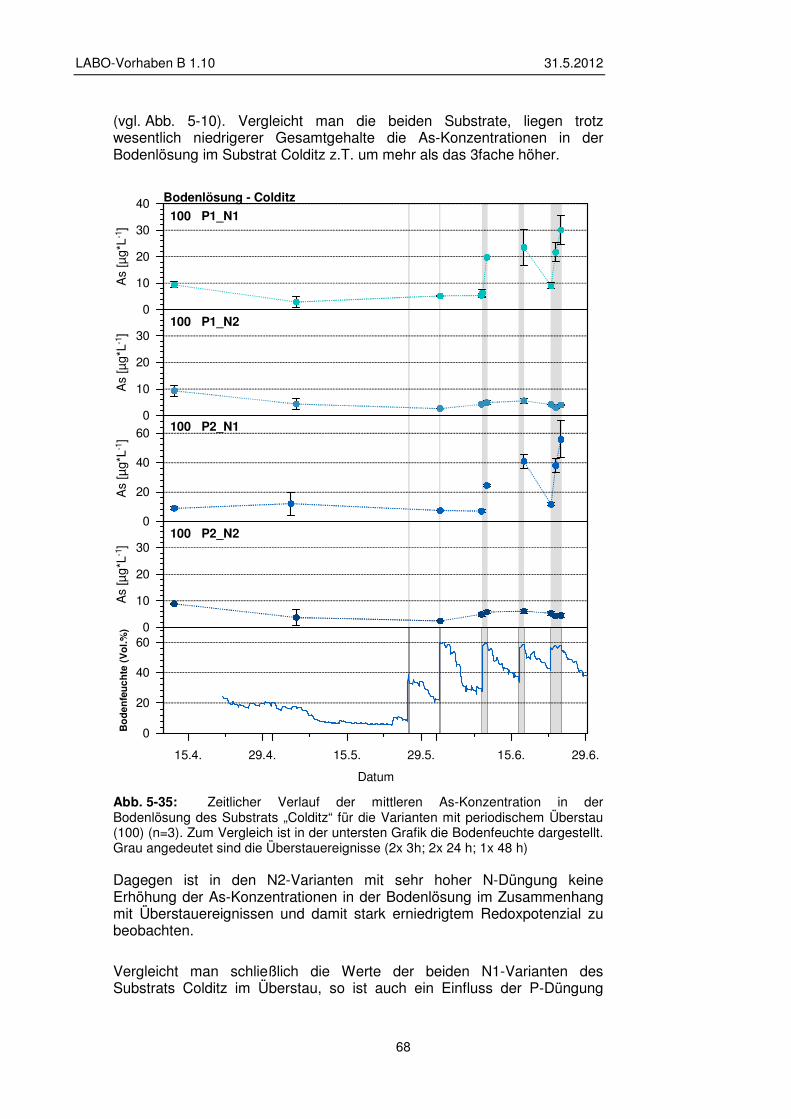
LABO-Vorhaben B 1.10 31.5.2012
68
(vgl. Abb. 5-10). Vergleicht man die beiden Substrate, liegen trotz wesentlich niedrigerer Gesamtgehalte die As-Konzentrationen in der Bodenlösung im Substrat Colditz z.T. um mehr als das 3fache höher.
0
10
20
30
40
As
[µg*
L]
-1
100 P1_N1
0
10
20
30
As
[µg*
L]
-1
0
20
40
60
As
[µg*
L]
-1
0
10
20
30
As
[µg*
L]
-1
Bodenlösung - Colditz
100 P2_N1
100 P2_N2
100 P1_N2
0
20
40
60
Bo
den
feu
ch
te(V
ol.
%)
15.4. 29.4. 15.5. 29.5. 15.6. 29.6.
Datum Abb. 5-35: Zeitlicher Verlauf der mittleren As-Konzentration in der Bodenlösung des Substrats „Colditz“ für die Varianten mit periodischem Überstau (100) (n=3). Zum Vergleich ist in der untersten Grafik die Bodenfeuchte dargestellt. Grau angedeutet sind die Überstauereignisse (2x 3h; 2x 24 h; 1x 48 h)
Dagegen ist in den N2-Varianten mit sehr hoher N-Düngung keine Erhöhung der As-Konzentrationen in der Bodenlösung im Zusammenhang mit Überstauereignissen und damit stark erniedrigtem Redoxpotenzial zu beobachten.
Vergleicht man schließlich die Werte der beiden N1-Varianten des Substrats Colditz im Überstau, so ist auch ein Einfluss der P-Düngung
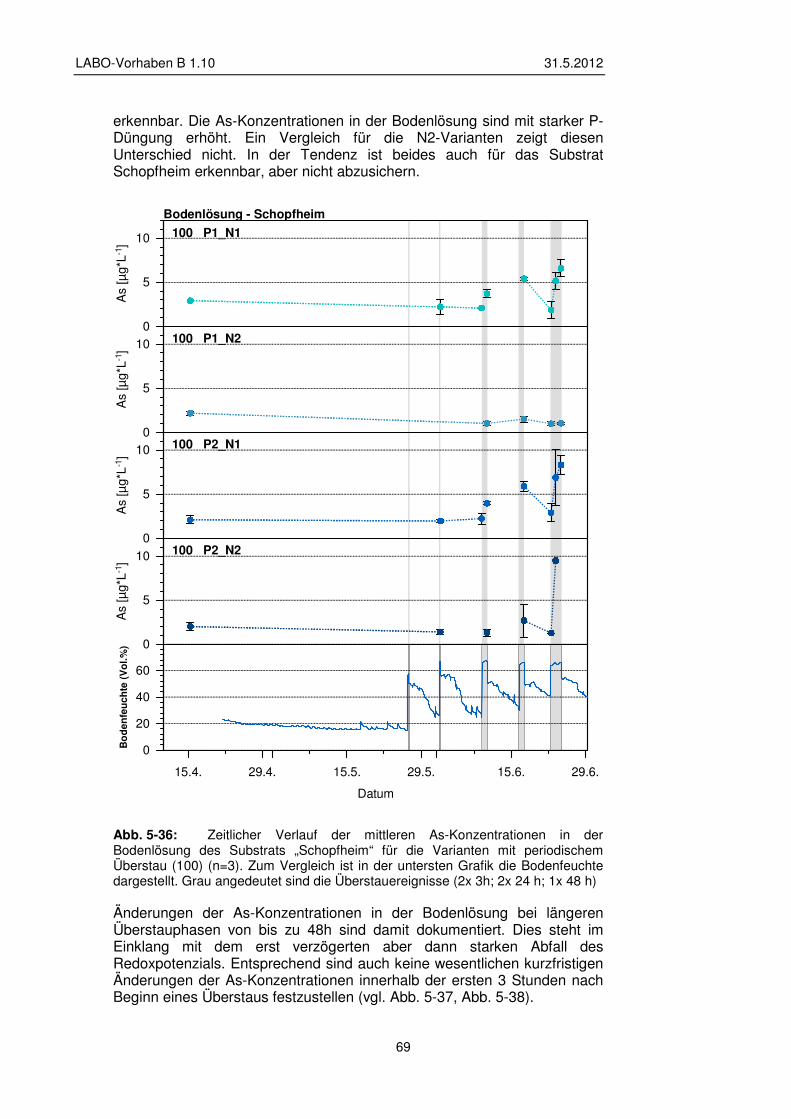
LABO-Vorhaben B 1.10 31.5.2012
69
erkennbar. Die As-Konzentrationen in der Bodenlösung sind mit starker P-Düngung erhöht. Ein Vergleich für die N2-Varianten zeigt diesen Unterschied nicht. In der Tendenz ist beides auch für das Substrat Schopfheim erkennbar, aber nicht abzusichern.
0
5
10
As
[µg*
L]
-1
0
5
10
As
[µg*
L]
-1
0
5
10
As
[µg*
L]
-1
0
5
10
As
[µg*
L]
-1
Bodenlösung - Schopfheim
100 P1_N1
100 P2_N2
100 P2_N1
100 P1_N2
0
20
40
60
Bo
den
feu
chte
(Vo
l.%
)
15.4. 29.4. 15.5. 29.5. 15.6. 29.6.
Datum
Abb. 5-36: Zeitlicher Verlauf der mittleren As-Konzentrationen in der Bodenlösung des Substrats „Schopfheim“ für die Varianten mit periodischem Überstau (100) (n=3). Zum Vergleich ist in der untersten Grafik die Bodenfeuchte dargestellt. Grau angedeutet sind die Überstauereignisse (2x 3h; 2x 24 h; 1x 48 h)
Änderungen der As-Konzentrationen in der Bodenlösung bei längeren Überstauphasen von bis zu 48h sind damit dokumentiert. Dies steht im Einklang mit dem erst verzögerten aber dann starken Abfall des Redoxpotenzials. Entsprechend sind auch keine wesentlichen kurzfristigen Änderungen der As-Konzentrationen innerhalb der ersten 3 Stunden nach Beginn eines Überstaus festzustellen (vgl. Abb. 5-37, Abb. 5-38).
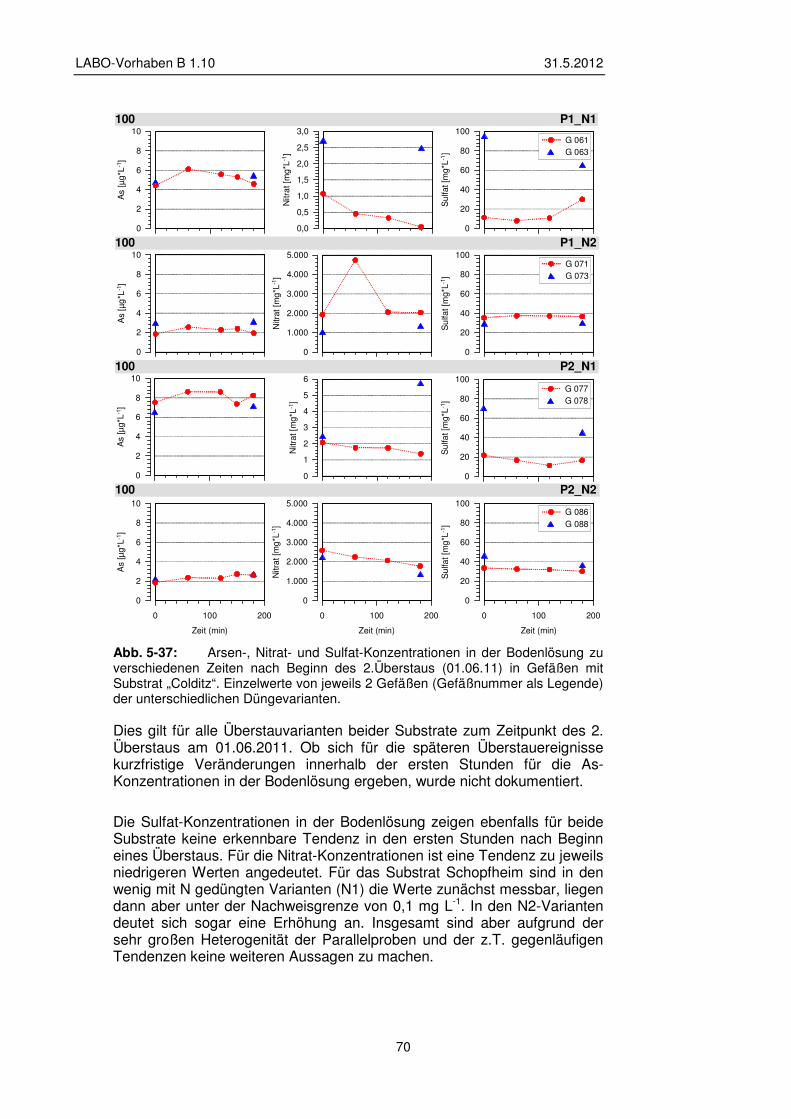
LABO-Vorhaben B 1.10 31.5.2012
70
100 P1_N1
100 P1_N2
100 P2_N1
100 P2_N2
0,0
0,5
1,0
1,5
2,0
2,5
3,0
Nitr
at[m
g*L
]-1
0
1.000
2.000
3.000
4.000
5.000
Nitr
at[m
g*L
]-1
0
1
2
3
4
5
6
Nitr
at[m
g*L
]-1
0
20
40
60
80
100
Sul
fat[
mg*
L]
-1
G 061G 063
0
20
40
60
80
100
Su
lfat[
mg*
L]
-1
G 071G 073
0
20
40
60
80
100
Sul
fat[
mg*
L]
-1
G 077G 078
0
20
40
60
80
100
Sul
fat[
mg*
L]
-1
0 100 200
Zeit (min)
G 086
G 088
0
2
4
6
8
10A
s[µ
g*L
]-1
0
2
4
6
8
10
As
[µg
*L]
-1
0
2
4
6
8
10
As
[µg*
L]
-1
0 100 200
Zeit (min)
0
1.000
2.000
3.000
4.000
5.000
Nitr
at[m
g*L
]-1
0 100 200
Zeit (min)
0
2
4
6
8
10
As
[µg*
L]
-1
Abb. 5-37: Arsen-, Nitrat- und Sulfat-Konzentrationen in der Bodenlösung zu verschiedenen Zeiten nach Beginn des 2.Überstaus (01.06.11) in Gefäßen mit Substrat „Colditz“. Einzelwerte von jeweils 2 Gefäßen (Gefäßnummer als Legende) der unterschiedlichen Düngevarianten.
Dies gilt für alle Überstauvarianten beider Substrate zum Zeitpunkt des 2. Überstaus am 01.06.2011. Ob sich für die späteren Überstauereignisse kurzfristige Veränderungen innerhalb der ersten Stunden für die As-Konzentrationen in der Bodenlösung ergeben, wurde nicht dokumentiert.
Die Sulfat-Konzentrationen in der Bodenlösung zeigen ebenfalls für beide Substrate keine erkennbare Tendenz in den ersten Stunden nach Beginn eines Überstaus. Für die Nitrat-Konzentrationen ist eine Tendenz zu jeweils niedrigeren Werten angedeutet. Für das Substrat Schopfheim sind in den wenig mit N gedüngten Varianten (N1) die Werte zunächst messbar, liegen dann aber unter der Nachweisgrenze von 0,1 mg L-1. In den N2-Varianten deutet sich sogar eine Erhöhung an. Insgesamt sind aber aufgrund der sehr großen Heterogenität der Parallelproben und der z.T. gegenläufigen Tendenzen keine weiteren Aussagen zu machen.
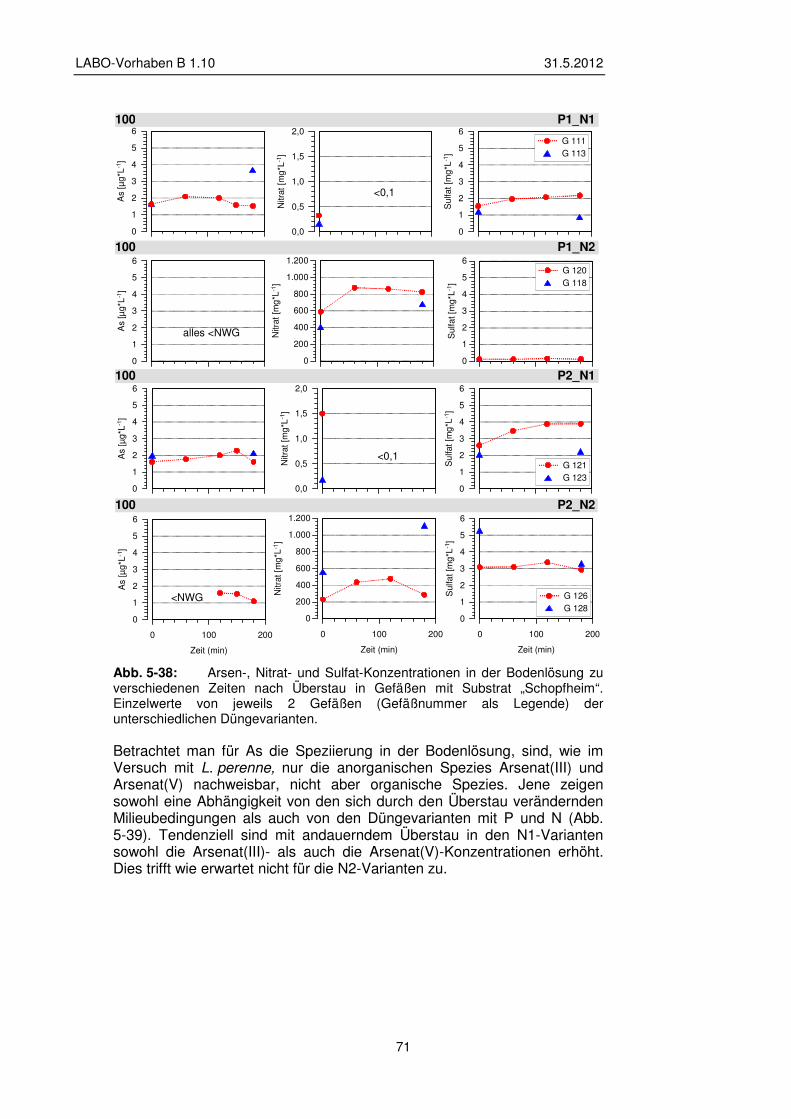
LABO-Vorhaben B 1.10 31.5.2012
71
100 P1_N1
100 P1_N2
100 P2_N1
100 P2_N2
0
1
2
3
4
5
6
Sul
fat[
mg*
L]
-1
G 111G 113
0,0
0,5
1,0
1,5
2,0
Nitr
at[m
g*L
]-1
<0,1
0
1
2
3
4
5
6A
s[µ
g*L
]-1
0
1
2
3
4
5
6
As
[µg*
L]
-1
alles <NWG
0
200
400
600
800
1.000
1.200
Nitr
at[m
g*L
]-1
0
1
2
3
4
5
6
Sul
fat[
mg*
L]
-1
G 120
G 118
0
200
400
600
800
1.000
1.200
Nitr
at[m
g*L
]-1
0 100 200
Zeit (min)
0
1
2
3
4
5
6
As
[µg*
L]
-1
0 100 200
Zeit (min)
0
1
2
3
4
5
6
Sul
fat[
mg*
L]
-1
0 100 200
Zeit (min)
G 126G 128
<NWG
0,0
0,5
1,0
1,5
2,0
Nitr
at[m
g*L
]-1
<0,1
0
1
2
3
4
5
6
Sul
fat[
mg*
L]
-1
G 121G 123
0
1
2
3
4
5
6
As
[µg*
L]
-1
Abb. 5-38: Arsen-, Nitrat- und Sulfat-Konzentrationen in der Bodenlösung zu verschiedenen Zeiten nach Überstau in Gefäßen mit Substrat „Schopfheim“. Einzelwerte von jeweils 2 Gefäßen (Gefäßnummer als Legende) der unterschiedlichen Düngevarianten.
Betrachtet man für As die Speziierung in der Bodenlösung, sind, wie im Versuch mit L. perenne, nur die anorganischen Spezies Arsenat(III) und Arsenat(V) nachweisbar, nicht aber organische Spezies. Jene zeigen sowohl eine Abhängigkeit von den sich durch den Überstau verändernden Milieubedingungen als auch von den Düngevarianten mit P und N (Abb. 5-39). Tendenziell sind mit andauerndem Überstau in den N1-Varianten sowohl die Arsenat(III)- als auch die Arsenat(V)-Konzentrationen erhöht. Dies trifft wie erwartet nicht für die N2-Varianten zu.
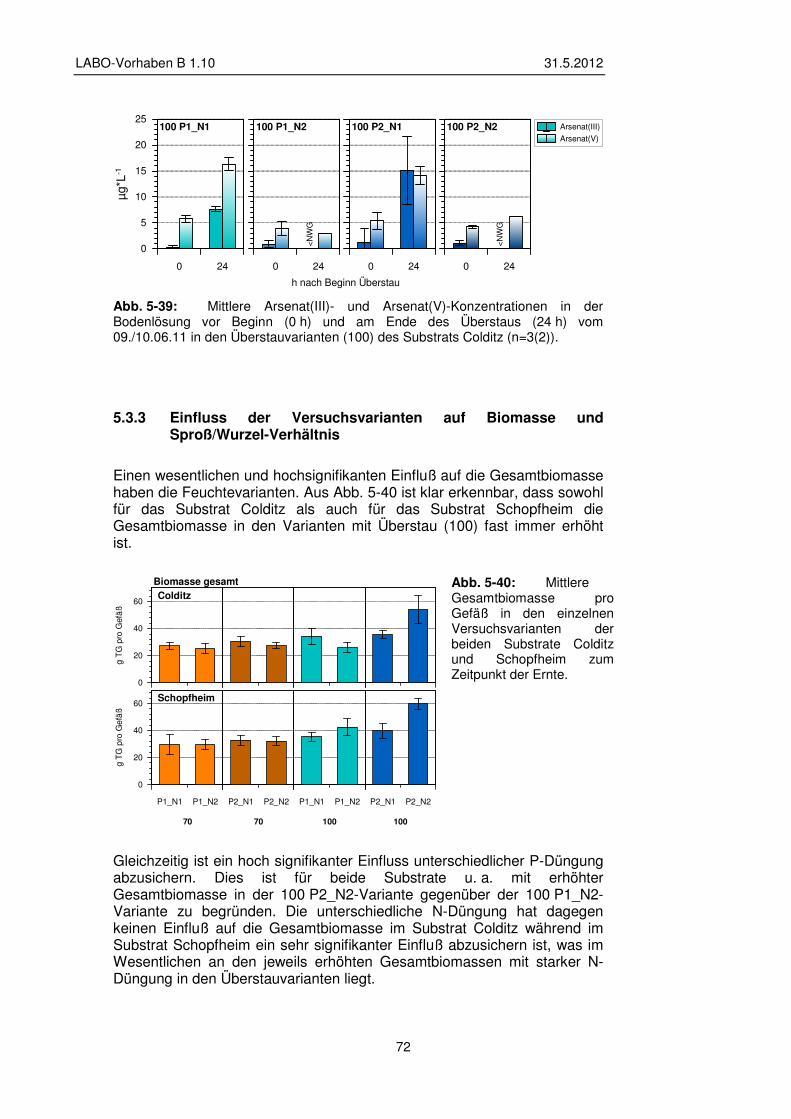
LABO-Vorhaben B 1.10 31.5.2012
72
0 240 240 24
0
5
10
15
20
25µg
*L-1
0 24
Arsenat(III)
Arsenat(V)100 P1_N2100 P1_N1
<N
WG
100 P2_N1 100 P2_N2
<N
WG
h nach Beginn Überstau Abb. 5-39: Mittlere Arsenat(III)- und Arsenat(V)-Konzentrationen in der Bodenlösung vor Beginn (0 h) und am Ende des Überstaus (24 h) vom 09./10.06.11 in den Überstauvarianten (100) des Substrats Colditz (n=3(2)).
5.3.3 Einfluss der Versuchsvarianten auf Biomasse und Sproß/Wurzel-Verhältnis
Einen wesentlichen und hochsignifikanten Einfluß auf die Gesamtbiomasse haben die Feuchtevarianten. Aus Abb. 5-40 ist klar erkennbar, dass sowohl für das Substrat Colditz als auch für das Substrat Schopfheim die Gesamtbiomasse in den Varianten mit Überstau (100) fast immer erhöht ist.
0
20
40
60
gT
Gpr
oG
efäß
Biomasse gesamt
0
20
40
60
gT
Gp
roG
efäß
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Schopfheim
Colditz
Abb. 5-40: Mittlere Gesamtbiomasse pro Gefäß in den einzelnen Versuchsvarianten der beiden Substrate Colditz und Schopfheim zum Zeitpunkt der Ernte.
Gleichzeitig ist ein hoch signifikanter Einfluss unterschiedlicher P-Düngung abzusichern. Dies ist für beide Substrate u. a. mit erhöhter Gesamtbiomasse in der 100 P2_N2-Variante gegenüber der 100 P1_N2-Variante zu begründen. Die unterschiedliche N-Düngung hat dagegen keinen Einfluß auf die Gesamtbiomasse im Substrat Colditz während im Substrat Schopfheim ein sehr signifikanter Einfluß abzusichern ist, was im Wesentlichen an den jeweils erhöhten Gesamtbiomassen mit starker N-Düngung in den Überstauvarianten liegt.
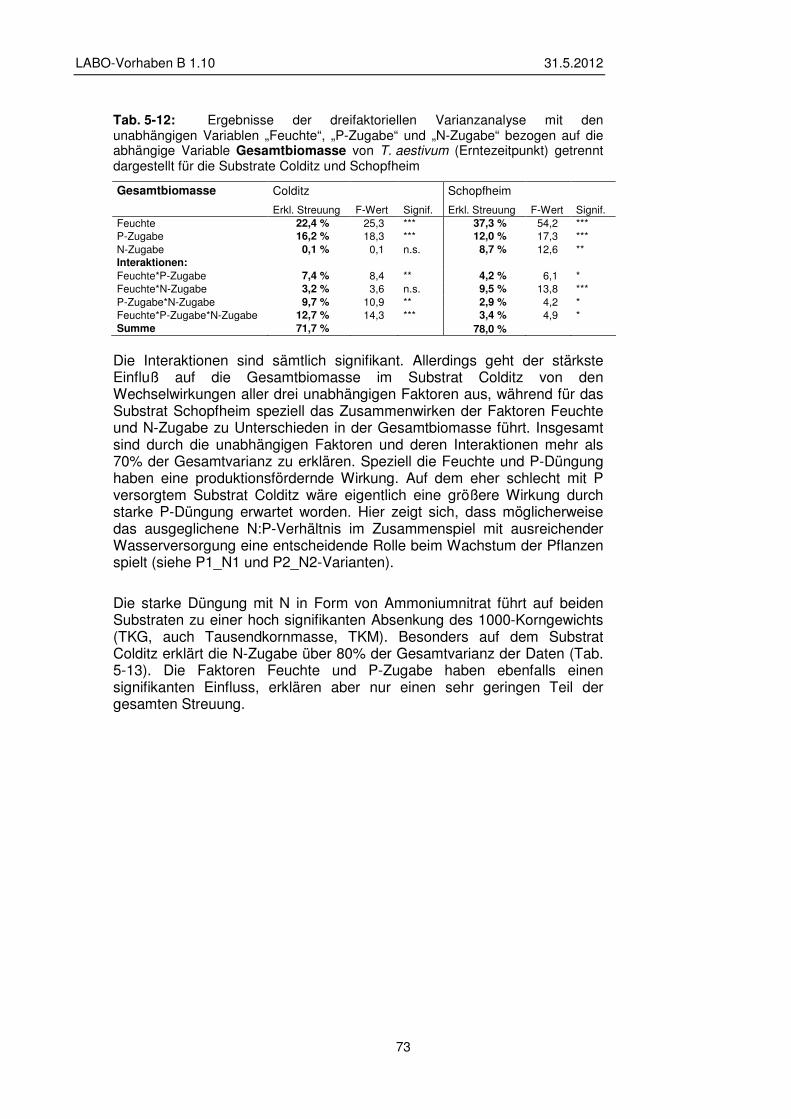
LABO-Vorhaben B 1.10 31.5.2012
73
Tab. 5-12: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Feuchte“, „P-Zugabe“ und „N-Zugabe“ bezogen auf die abhängige Variable Gesamtbiomasse von T. aestivum (Erntezeitpunkt) getrennt dargestellt für die Substrate Colditz und Schopfheim
Gesamtbiomasse Colditz Schopfheim
Erkl. Streuung F-Wert Signif. Erkl. Streuung F-Wert Signif. Feuchte 22,4 % 25,3 *** 37,3 % 54,2 *** P-Zugabe 16,2 % 18,3 *** 12,0 % 17,3 *** N-Zugabe 0,1 % 0,1 n.s. 8,7 % 12,6 ** Interaktionen: Feuchte*P-Zugabe 7,4 % 8,4 ** 4,2 % 6,1 * Feuchte*N-Zugabe 3,2 % 3,6 n.s. 9,5 % 13,8 *** P-Zugabe*N-Zugabe 9,7 % 10,9 ** 2,9 % 4,2 * Feuchte*P-Zugabe*N-Zugabe 12,7 % 14,3 *** 3,4 % 4,9 * Summe 71,7 % 78,0 %
Die Interaktionen sind sämtlich signifikant. Allerdings geht der stärkste Einfluß auf die Gesamtbiomasse im Substrat Colditz von den Wechselwirkungen aller drei unabhängigen Faktoren aus, während für das Substrat Schopfheim speziell das Zusammenwirken der Faktoren Feuchte und N-Zugabe zu Unterschieden in der Gesamtbiomasse führt. Insgesamt sind durch die unabhängigen Faktoren und deren Interaktionen mehr als 70% der Gesamtvarianz zu erklären. Speziell die Feuchte und P-Düngung haben eine produktionsfördernde Wirkung. Auf dem eher schlecht mit P versorgtem Substrat Colditz wäre eigentlich eine größere Wirkung durch starke P-Düngung erwartet worden. Hier zeigt sich, dass möglicherweise das ausgeglichene N:P-Verhältnis im Zusammenspiel mit ausreichender Wasserversorgung eine entscheidende Rolle beim Wachstum der Pflanzen spielt (siehe P1_N1 und P2_N2-Varianten).
Die starke Düngung mit N in Form von Ammoniumnitrat führt auf beiden Substraten zu einer hoch signifikanten Absenkung des 1000-Korngewichts (TKG, auch Tausendkornmasse, TKM). Besonders auf dem Substrat Colditz erklärt die N-Zugabe über 80% der Gesamtvarianz der Daten (Tab. 5-13). Die Faktoren Feuchte und P-Zugabe haben ebenfalls einen signifikanten Einfluss, erklären aber nur einen sehr geringen Teil der gesamten Streuung.
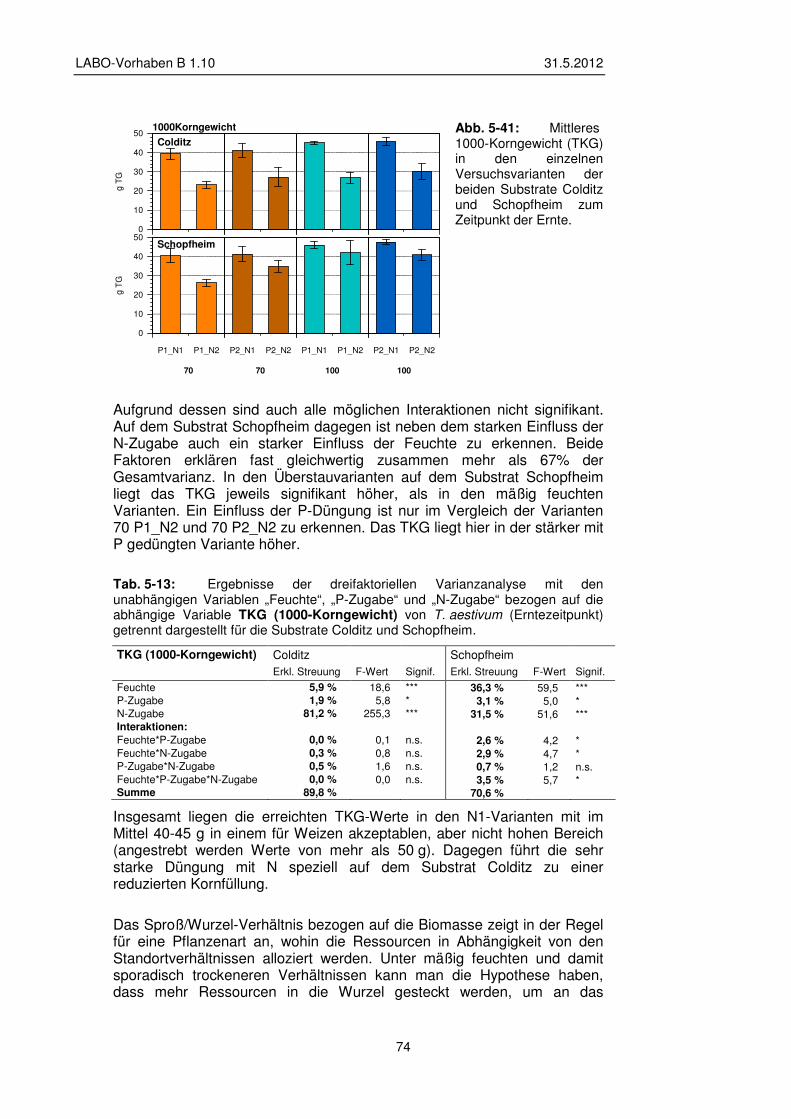
LABO-Vorhaben B 1.10 31.5.2012
74
0
10
20
30
40
50
gT
G
Colditz
1000Korngewicht
0
10
20
30
40
50
gT
G
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Schopfheim
Abb. 5-41: Mittleres 1000-Korngewicht (TKG) in den einzelnen Versuchsvarianten der beiden Substrate Colditz und Schopfheim zum Zeitpunkt der Ernte.
Aufgrund dessen sind auch alle möglichen Interaktionen nicht signifikant. Auf dem Substrat Schopfheim dagegen ist neben dem starken Einfluss der N-Zugabe auch ein starker Einfluss der Feuchte zu erkennen. Beide Faktoren erklären fast gleichwertig zusammen mehr als 67% der Gesamtvarianz. In den Überstauvarianten auf dem Substrat Schopfheim liegt das TKG jeweils signifikant höher, als in den mäßig feuchten Varianten. Ein Einfluss der P-Düngung ist nur im Vergleich der Varianten 70 P1_N2 und 70 P2_N2 zu erkennen. Das TKG liegt hier in der stärker mit P gedüngten Variante höher.
Tab. 5-13: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Feuchte“, „P-Zugabe“ und „N-Zugabe“ bezogen auf die abhängige Variable TKG (1000-Korngewicht) von T. aestivum (Erntezeitpunkt) getrennt dargestellt für die Substrate Colditz und Schopfheim.
TKG (1000-Korngewicht) Colditz Schopfheim Erkl. Streuung F-Wert Signif. Erkl. Streuung F-Wert Signif.
Feuchte 5,9 % 18,6 *** 36,3 % 59,5 *** P-Zugabe 1,9 % 5,8 * 3,1 % 5,0 * N-Zugabe 81,2 % 255,3 *** 31,5 % 51,6 *** Interaktionen: Feuchte*P-Zugabe 0,0 % 0,1 n.s. 2,6 % 4,2 * Feuchte*N-Zugabe 0,3 % 0,8 n.s. 2,9 % 4,7 * P-Zugabe*N-Zugabe 0,5 % 1,6 n.s. 0,7 % 1,2 n.s. Feuchte*P-Zugabe*N-Zugabe 0,0 % 0,0 n.s. 3,5 % 5,7 * Summe 89,8 % 70,6 %
Insgesamt liegen die erreichten TKG-Werte in den N1-Varianten mit im Mittel 40-45 g in einem für Weizen akzeptablen, aber nicht hohen Bereich (angestrebt werden Werte von mehr als 50 g). Dagegen führt die sehr starke Düngung mit N speziell auf dem Substrat Colditz zu einer reduzierten Kornfüllung.
Das Sproß/Wurzel-Verhältnis bezogen auf die Biomasse zeigt in der Regel für eine Pflanzenart an, wohin die Ressourcen in Abhängigkeit von den Standortverhältnissen alloziert werden. Unter mäßig feuchten und damit sporadisch trockeneren Verhältnissen kann man die Hypothese haben, dass mehr Ressourcen in die Wurzel gesteckt werden, um an das
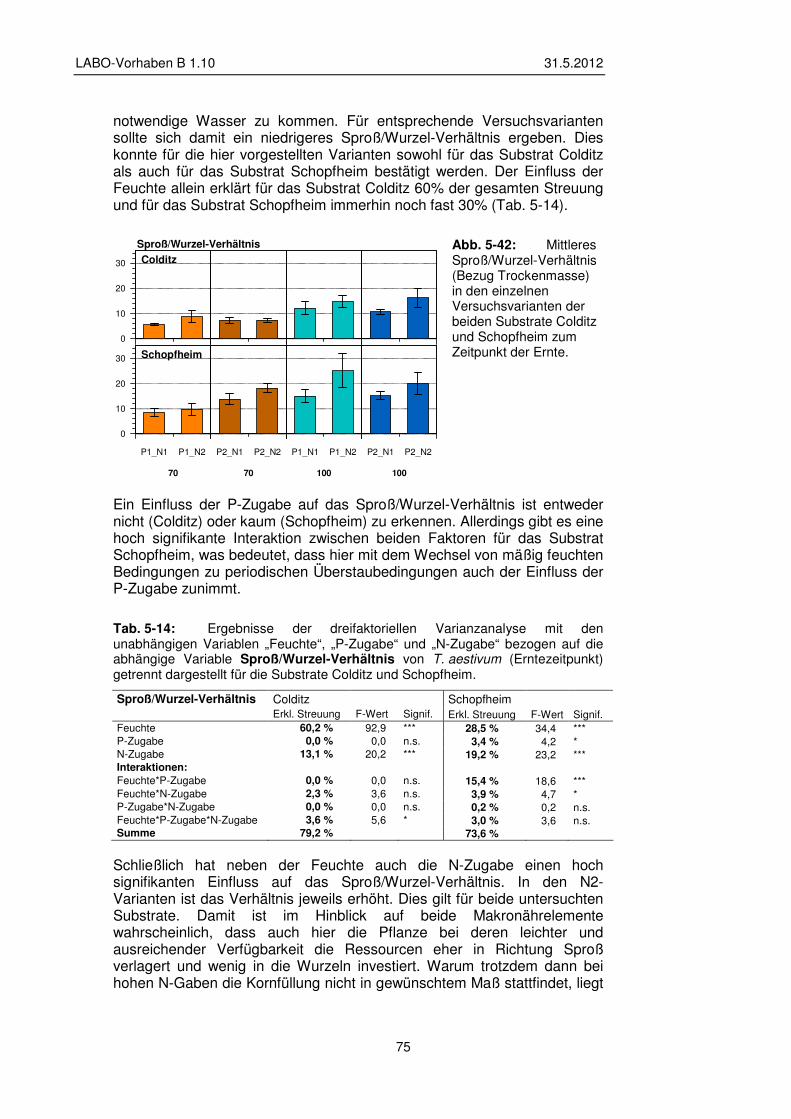
LABO-Vorhaben B 1.10 31.5.2012
75
notwendige Wasser zu kommen. Für entsprechende Versuchsvarianten sollte sich damit ein niedrigeres Sproß/Wurzel-Verhältnis ergeben. Dies konnte für die hier vorgestellten Varianten sowohl für das Substrat Colditz als auch für das Substrat Schopfheim bestätigt werden. Der Einfluss der Feuchte allein erklärt für das Substrat Colditz 60% der gesamten Streuung und für das Substrat Schopfheim immerhin noch fast 30% (Tab. 5-14).
0
10
20
30
0
10
20
30
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Sproß/Wurzel-Verhältnis
Colditz
Schopfheim
Abb. 5-42: Mittleres Sproß/Wurzel-Verhältnis (Bezug Trockenmasse) in den einzelnen Versuchsvarianten der beiden Substrate Colditz und Schopfheim zum Zeitpunkt der Ernte.
Ein Einfluss der P-Zugabe auf das Sproß/Wurzel-Verhältnis ist entweder nicht (Colditz) oder kaum (Schopfheim) zu erkennen. Allerdings gibt es eine hoch signifikante Interaktion zwischen beiden Faktoren für das Substrat Schopfheim, was bedeutet, dass hier mit dem Wechsel von mäßig feuchten Bedingungen zu periodischen Überstaubedingungen auch der Einfluss der P-Zugabe zunimmt.
Tab. 5-14: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Feuchte“, „P-Zugabe“ und „N-Zugabe“ bezogen auf die abhängige Variable Sproß/Wurzel-Verhältnis von T. aestivum (Erntezeitpunkt) getrennt dargestellt für die Substrate Colditz und Schopfheim.
Sproß/Wurzel-Verhältnis Colditz Schopfheim Erkl. Streuung F-Wert Signif. Erkl. Streuung F-Wert Signif. Feuchte 60,2 % 92,9 *** 28,5 % 34,4 *** P-Zugabe 0,0 % 0,0 n.s. 3,4 % 4,2 * N-Zugabe 13,1 % 20,2 *** 19,2 % 23,2 *** Interaktionen: Feuchte*P-Zugabe 0,0 % 0,0 n.s. 15,4 % 18,6 *** Feuchte*N-Zugabe 2,3 % 3,6 n.s. 3,9 % 4,7 * P-Zugabe*N-Zugabe 0,0 % 0,0 n.s. 0,2 % 0,2 n.s. Feuchte*P-Zugabe*N-Zugabe 3,6 % 5,6 * 3,0 % 3,6 n.s. Summe 79,2 % 73,6 %
Schließlich hat neben der Feuchte auch die N-Zugabe einen hoch signifikanten Einfluss auf das Sproß/Wurzel-Verhältnis. In den N2-Varianten ist das Verhältnis jeweils erhöht. Dies gilt für beide untersuchten Substrate. Damit ist im Hinblick auf beide Makronährelemente wahrscheinlich, dass auch hier die Pflanze bei deren leichter und ausreichender Verfügbarkeit die Ressourcen eher in Richtung Sproß verlagert und wenig in die Wurzeln investiert. Warum trotzdem dann bei hohen N-Gaben die Kornfüllung nicht in gewünschtem Maß stattfindet, liegt
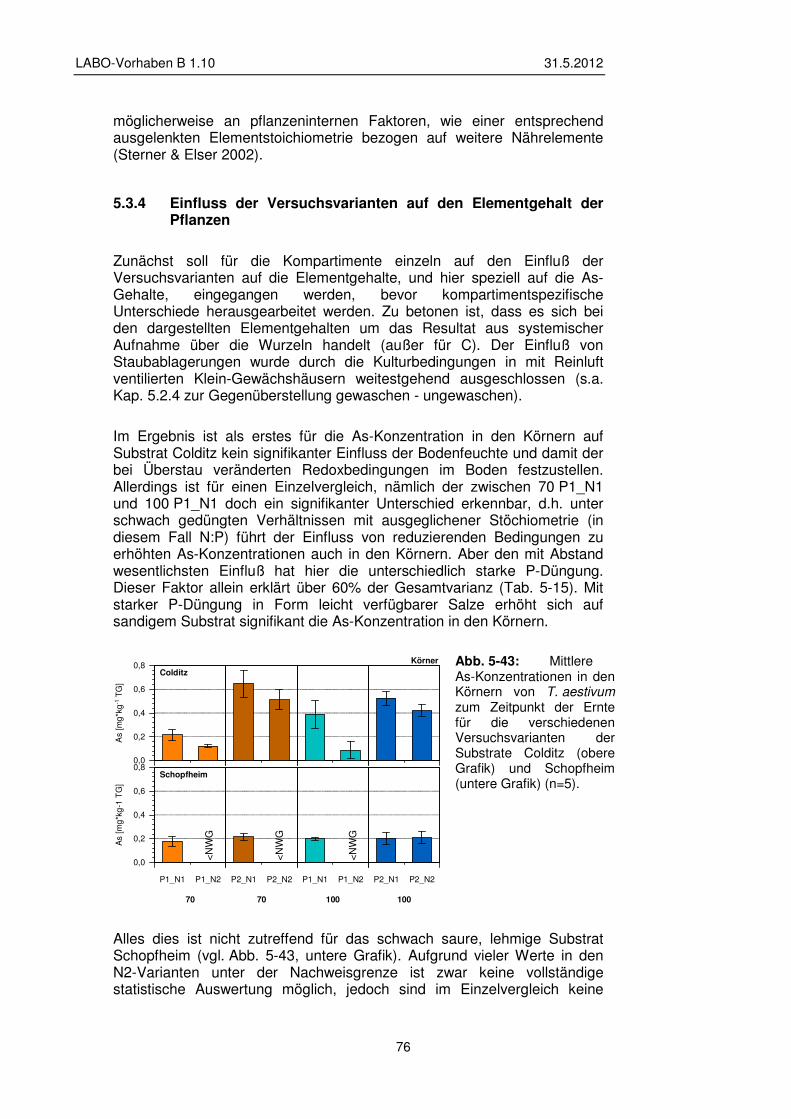
LABO-Vorhaben B 1.10 31.5.2012
76
möglicherweise an pflanzeninternen Faktoren, wie einer entsprechend ausgelenkten Elementstoichiometrie bezogen auf weitere Nährelemente (Sterner & Elser 2002).
5.3.4 Einfluss der Versuchsvarianten auf den Elementgehalt der Pflanzen
Zunächst soll für die Kompartimente einzeln auf den Einfluß der Versuchsvarianten auf die Elementgehalte, und hier speziell auf die As-Gehalte, eingegangen werden, bevor kompartimentspezifische Unterschiede herausgearbeitet werden. Zu betonen ist, dass es sich bei den dargestellten Elementgehalten um das Resultat aus systemischer Aufnahme über die Wurzeln handelt (außer für C). Der Einfluß von Staubablagerungen wurde durch die Kulturbedingungen in mit Reinluft ventilierten Klein-Gewächshäusern weitestgehend ausgeschlossen (s.a. Kap. 5.2.4 zur Gegenüberstellung gewaschen - ungewaschen).
Im Ergebnis ist als erstes für die As-Konzentration in den Körnern auf Substrat Colditz kein signifikanter Einfluss der Bodenfeuchte und damit der bei Überstau veränderten Redoxbedingungen im Boden festzustellen. Allerdings ist für einen Einzelvergleich, nämlich der zwischen 70 P1_N1 und 100 P1_N1 doch ein signifikanter Unterschied erkennbar, d.h. unter schwach gedüngten Verhältnissen mit ausgeglichener Stöchiometrie (in diesem Fall N:P) führt der Einfluss von reduzierenden Bedingungen zu erhöhten As-Konzentrationen auch in den Körnern. Aber den mit Abstand wesentlichsten Einfluß hat hier die unterschiedlich starke P-Düngung. Dieser Faktor allein erklärt über 60% der Gesamtvarianz (Tab. 5-15). Mit starker P-Düngung in Form leicht verfügbarer Salze erhöht sich auf sandigem Substrat signifikant die As-Konzentration in den Körnern.
0,0
0,2
0,4
0,6
0,8
As
[mg*
kgT
G]
-1
0,0
0,2
0,4
0,6
0,8
As
[mg*
kg-1
TG
]
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
<N
WG
<N
WG
<N
WG
Schopfheim
ColditzKörner
Abb. 5-43: Mittlere As-Konzentrationen in den Körnern von T. aestivum zum Zeitpunkt der Ernte für die verschiedenen Versuchsvarianten der Substrate Colditz (obere Grafik) und Schopfheim (untere Grafik) (n=5).
Alles dies ist nicht zutreffend für das schwach saure, lehmige Substrat Schopfheim (vgl. Abb. 5-43, untere Grafik). Aufgrund vieler Werte in den N2-Varianten unter der Nachweisgrenze ist zwar keine vollständige statistische Auswertung möglich, jedoch sind im Einzelvergleich keine
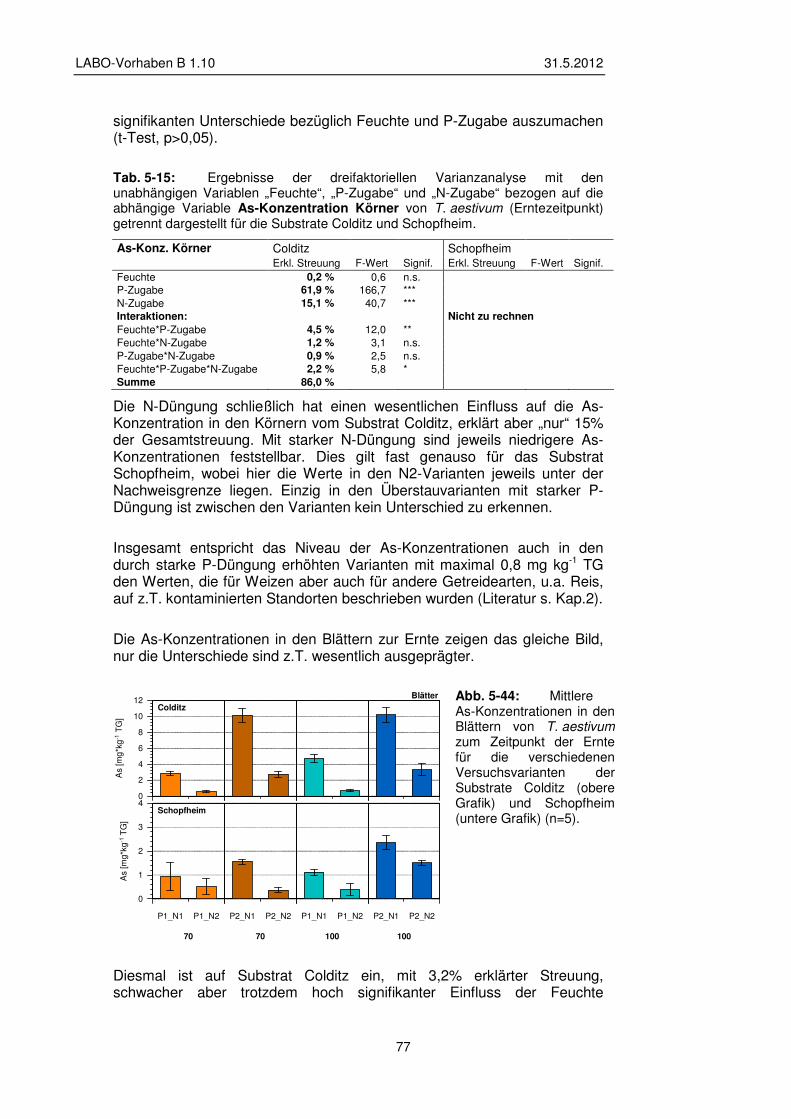
LABO-Vorhaben B 1.10 31.5.2012
77
signifikanten Unterschiede bezüglich Feuchte und P-Zugabe auszumachen (t-Test, p>0,05).
Tab. 5-15: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Feuchte“, „P-Zugabe“ und „N-Zugabe“ bezogen auf die abhängige Variable As-Konzentration Körner von T. aestivum (Erntezeitpunkt) getrennt dargestellt für die Substrate Colditz und Schopfheim.
As-Konz. Körner Colditz Schopfheim Erkl. Streuung F-Wert Signif. Erkl. Streuung F-Wert Signif. Feuchte 0,2 % 0,6 n.s. P-Zugabe 61,9 % 166,7 *** N-Zugabe 15,1 % 40,7 *** Interaktionen: Nicht zu rechnen Feuchte*P-Zugabe 4,5 % 12,0 ** Feuchte*N-Zugabe 1,2 % 3,1 n.s. P-Zugabe*N-Zugabe 0,9 % 2,5 n.s. Feuchte*P-Zugabe*N-Zugabe 2,2 % 5,8 * Summe 86,0 %
Die N-Düngung schließlich hat einen wesentlichen Einfluss auf die As-Konzentration in den Körnern vom Substrat Colditz, erklärt aber „nur“ 15% der Gesamtstreuung. Mit starker N-Düngung sind jeweils niedrigere As-Konzentrationen feststellbar. Dies gilt fast genauso für das Substrat Schopfheim, wobei hier die Werte in den N2-Varianten jeweils unter der Nachweisgrenze liegen. Einzig in den Überstauvarianten mit starker P-Düngung ist zwischen den Varianten kein Unterschied zu erkennen.
Insgesamt entspricht das Niveau der As-Konzentrationen auch in den durch starke P-Düngung erhöhten Varianten mit maximal 0,8 mg kg-1 TG den Werten, die für Weizen aber auch für andere Getreidearten, u.a. Reis, auf z.T. kontaminierten Standorten beschrieben wurden (Literatur s. Kap.2).
Die As-Konzentrationen in den Blättern zur Ernte zeigen das gleiche Bild, nur die Unterschiede sind z.T. wesentlich ausgeprägter.
0
2
4
6
8
10
12
As
[mg*
kgT
G]
-1
Colditz
0
1
2
3
4
As
[mg*
kgT
G]
-1
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Schopfheim
Blätter
Abb. 5-44: Mittlere As-Konzentrationen in den Blättern von T. aestivum zum Zeitpunkt der Ernte für die verschiedenen Versuchsvarianten der Substrate Colditz (obere Grafik) und Schopfheim (untere Grafik) (n=5).
Diesmal ist auf Substrat Colditz ein, mit 3,2% erklärter Streuung, schwacher aber trotzdem hoch signifikanter Einfluss der Feuchte
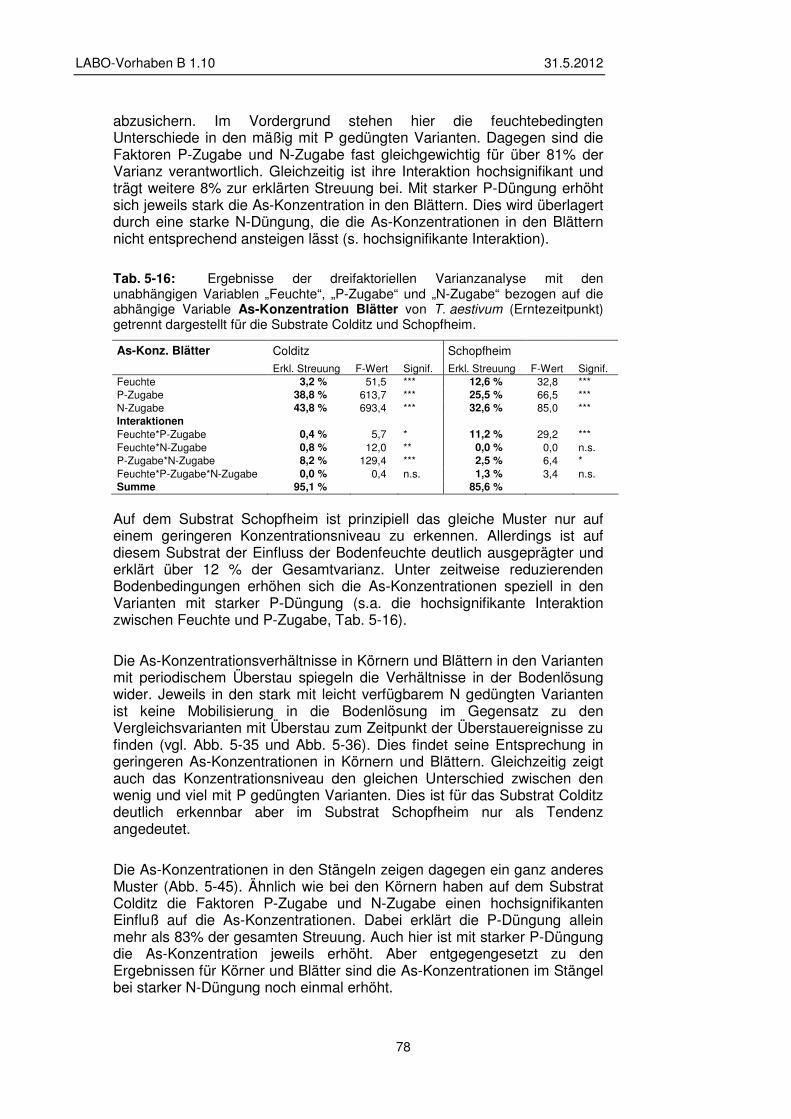
LABO-Vorhaben B 1.10 31.5.2012
78
abzusichern. Im Vordergrund stehen hier die feuchtebedingten Unterschiede in den mäßig mit P gedüngten Varianten. Dagegen sind die Faktoren P-Zugabe und N-Zugabe fast gleichgewichtig für über 81% der Varianz verantwortlich. Gleichzeitig ist ihre Interaktion hochsignifikant und trägt weitere 8% zur erklärten Streuung bei. Mit starker P-Düngung erhöht sich jeweils stark die As-Konzentration in den Blättern. Dies wird überlagert durch eine starke N-Düngung, die die As-Konzentrationen in den Blättern nicht entsprechend ansteigen lässt (s. hochsignifikante Interaktion).
Tab. 5-16: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Feuchte“, „P-Zugabe“ und „N-Zugabe“ bezogen auf die abhängige Variable As-Konzentration Blätter von T. aestivum (Erntezeitpunkt) getrennt dargestellt für die Substrate Colditz und Schopfheim.
As-Konz. Blätter Colditz Schopfheim
Erkl. Streuung F-Wert Signif. Erkl. Streuung F-Wert Signif. Feuchte 3,2 % 51,5 *** 12,6 % 32,8 *** P-Zugabe 38,8 % 613,7 *** 25,5 % 66,5 *** N-Zugabe 43,8 % 693,4 *** 32,6 % 85,0 *** Interaktionen Feuchte*P-Zugabe 0,4 % 5,7 * 11,2 % 29,2 *** Feuchte*N-Zugabe 0,8 % 12,0 ** 0,0 % 0,0 n.s. P-Zugabe*N-Zugabe 8,2 % 129,4 *** 2,5 % 6,4 * Feuchte*P-Zugabe*N-Zugabe 0,0 % 0,4 n.s. 1,3 % 3,4 n.s. Summe 95,1 % 85,6 %
Auf dem Substrat Schopfheim ist prinzipiell das gleiche Muster nur auf einem geringeren Konzentrationsniveau zu erkennen. Allerdings ist auf diesem Substrat der Einfluss der Bodenfeuchte deutlich ausgeprägter und erklärt über 12 % der Gesamtvarianz. Unter zeitweise reduzierenden Bodenbedingungen erhöhen sich die As-Konzentrationen speziell in den Varianten mit starker P-Düngung (s.a. die hochsignifikante Interaktion zwischen Feuchte und P-Zugabe, Tab. 5-16).
Die As-Konzentrationsverhältnisse in Körnern und Blättern in den Varianten mit periodischem Überstau spiegeln die Verhältnisse in der Bodenlösung wider. Jeweils in den stark mit leicht verfügbarem N gedüngten Varianten ist keine Mobilisierung in die Bodenlösung im Gegensatz zu den Vergleichsvarianten mit Überstau zum Zeitpunkt der Überstauereignisse zu finden (vgl. Abb. 5-35 und Abb. 5-36). Dies findet seine Entsprechung in geringeren As-Konzentrationen in Körnern und Blättern. Gleichzeitig zeigt auch das Konzentrationsniveau den gleichen Unterschied zwischen den wenig und viel mit P gedüngten Varianten. Dies ist für das Substrat Colditz deutlich erkennbar aber im Substrat Schopfheim nur als Tendenz angedeutet.
Die As-Konzentrationen in den Stängeln zeigen dagegen ein ganz anderes Muster (Abb. 5-45). Ähnlich wie bei den Körnern haben auf dem Substrat Colditz die Faktoren P-Zugabe und N-Zugabe einen hochsignifikanten Einfluß auf die As-Konzentrationen. Dabei erklärt die P-Düngung allein mehr als 83% der gesamten Streuung. Auch hier ist mit starker P-Düngung die As-Konzentration jeweils erhöht. Aber entgegengesetzt zu den Ergebnissen für Körner und Blätter sind die As-Konzentrationen im Stängel bei starker N-Düngung noch einmal erhöht.
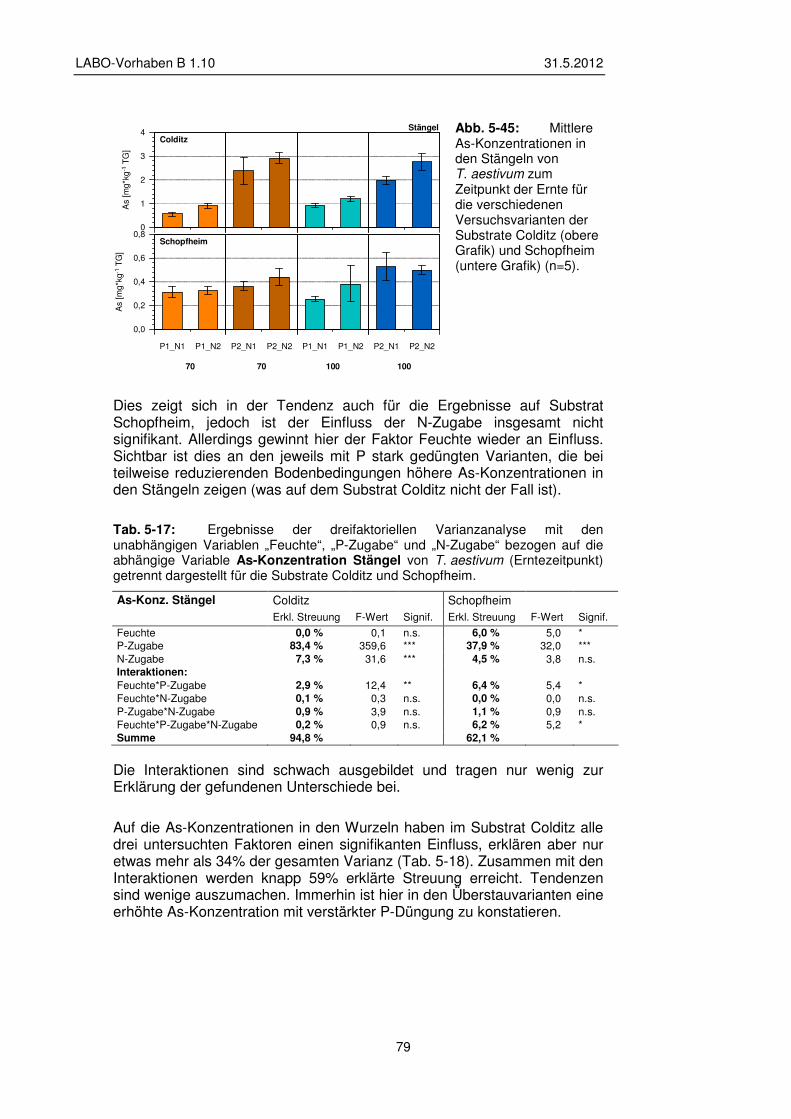
LABO-Vorhaben B 1.10 31.5.2012
79
0
1
2
3
4A
s[m
g*kg
TG
]-1
Colditz
0,0
0,2
0,4
0,6
0,8
As
[mg*
kgT
G]
-1
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Schopfheim
Stängel
Abb. 5-45: Mittlere As-Konzentrationen in den Stängeln von T. aestivum zum Zeitpunkt der Ernte für die verschiedenen Versuchsvarianten der Substrate Colditz (obere Grafik) und Schopfheim (untere Grafik) (n=5).
Dies zeigt sich in der Tendenz auch für die Ergebnisse auf Substrat Schopfheim, jedoch ist der Einfluss der N-Zugabe insgesamt nicht signifikant. Allerdings gewinnt hier der Faktor Feuchte wieder an Einfluss. Sichtbar ist dies an den jeweils mit P stark gedüngten Varianten, die bei teilweise reduzierenden Bodenbedingungen höhere As-Konzentrationen in den Stängeln zeigen (was auf dem Substrat Colditz nicht der Fall ist).
Tab. 5-17: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Feuchte“, „P-Zugabe“ und „N-Zugabe“ bezogen auf die abhängige Variable As-Konzentration Stängel von T. aestivum (Erntezeitpunkt) getrennt dargestellt für die Substrate Colditz und Schopfheim.
As-Konz. Stängel Colditz Schopfheim Erkl. Streuung F-Wert Signif. Erkl. Streuung F-Wert Signif.
Feuchte 0,0 % 0,1 n.s. 6,0 % 5,0 * P-Zugabe 83,4 % 359,6 *** 37,9 % 32,0 *** N-Zugabe 7,3 % 31,6 *** 4,5 % 3,8 n.s. Interaktionen: Feuchte*P-Zugabe 2,9 % 12,4 ** 6,4 % 5,4 * Feuchte*N-Zugabe 0,1 % 0,3 n.s. 0,0 % 0,0 n.s. P-Zugabe*N-Zugabe 0,9 % 3,9 n.s. 1,1 % 0,9 n.s. Feuchte*P-Zugabe*N-Zugabe 0,2 % 0,9 n.s. 6,2 % 5,2 * Summe 94,8 % 62,1 %
Die Interaktionen sind schwach ausgebildet und tragen nur wenig zur Erklärung der gefundenen Unterschiede bei.
Auf die As-Konzentrationen in den Wurzeln haben im Substrat Colditz alle drei untersuchten Faktoren einen signifikanten Einfluss, erklären aber nur etwas mehr als 34% der gesamten Varianz (Tab. 5-18). Zusammen mit den Interaktionen werden knapp 59% erklärte Streuung erreicht. Tendenzen sind wenige auszumachen. Immerhin ist hier in den Überstauvarianten eine erhöhte As-Konzentration mit verstärkter P-Düngung zu konstatieren.
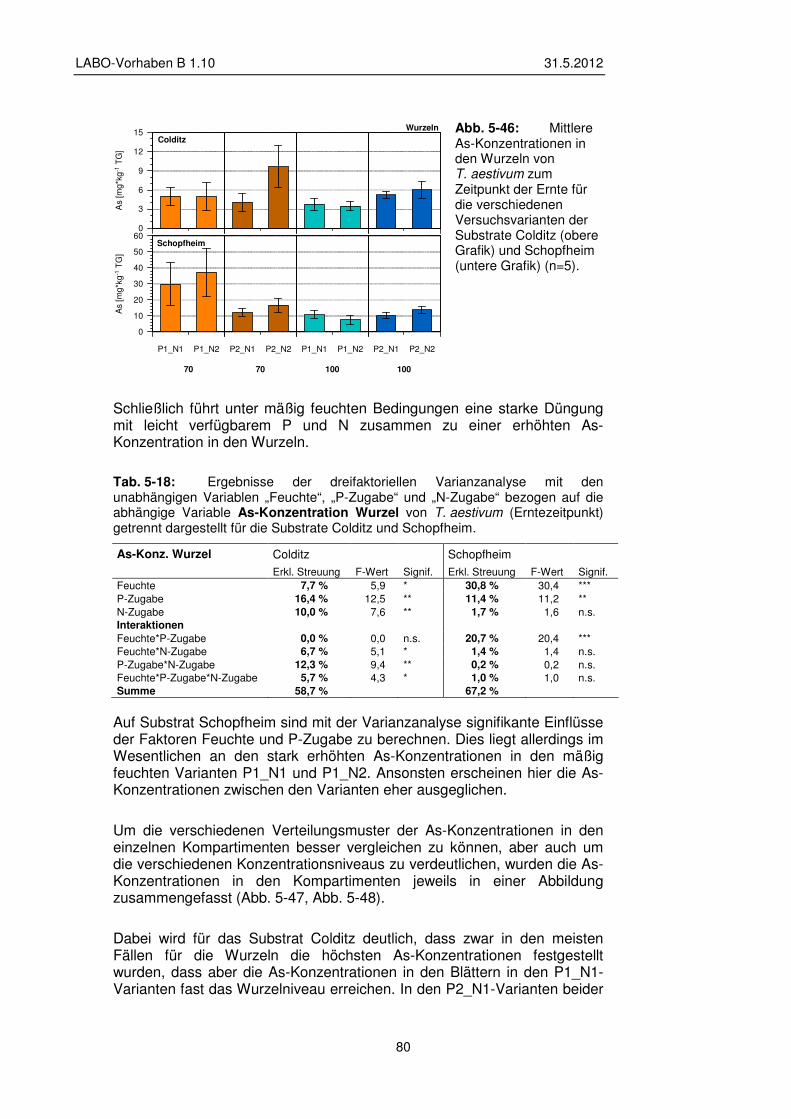
LABO-Vorhaben B 1.10 31.5.2012
80
0
3
6
9
12
15
As
[mg*
kgT
G]
-1
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
ColditzWurzeln
0
10
20
30
40
50
60
As
[mg*
kgT
G]
-1
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Schopfheim
Abb. 5-46: Mittlere As-Konzentrationen in den Wurzeln von T. aestivum zum Zeitpunkt der Ernte für die verschiedenen Versuchsvarianten der Substrate Colditz (obere Grafik) und Schopfheim (untere Grafik) (n=5).
Schließlich führt unter mäßig feuchten Bedingungen eine starke Düngung mit leicht verfügbarem P und N zusammen zu einer erhöhten As-Konzentration in den Wurzeln.
Tab. 5-18: Ergebnisse der dreifaktoriellen Varianzanalyse mit den unabhängigen Variablen „Feuchte“, „P-Zugabe“ und „N-Zugabe“ bezogen auf die abhängige Variable As-Konzentration Wurzel von T. aestivum (Erntezeitpunkt) getrennt dargestellt für die Substrate Colditz und Schopfheim.
As-Konz. Wurzel Colditz Schopfheim
Erkl. Streuung F-Wert Signif. Erkl. Streuung F-Wert Signif. Feuchte 7,7 % 5,9 * 30,8 % 30,4 *** P-Zugabe 16,4 % 12,5 ** 11,4 % 11,2 ** N-Zugabe 10,0 % 7,6 ** 1,7 % 1,6 n.s. Interaktionen Feuchte*P-Zugabe 0,0 % 0,0 n.s. 20,7 % 20,4 *** Feuchte*N-Zugabe 6,7 % 5,1 * 1,4 % 1,4 n.s. P-Zugabe*N-Zugabe 12,3 % 9,4 ** 0,2 % 0,2 n.s. Feuchte*P-Zugabe*N-Zugabe 5,7 % 4,3 * 1,0 % 1,0 n.s. Summe 58,7 % 67,2 %
Auf Substrat Schopfheim sind mit der Varianzanalyse signifikante Einflüsse der Faktoren Feuchte und P-Zugabe zu berechnen. Dies liegt allerdings im Wesentlichen an den stark erhöhten As-Konzentrationen in den mäßig feuchten Varianten P1_N1 und P1_N2. Ansonsten erscheinen hier die As-Konzentrationen zwischen den Varianten eher ausgeglichen.
Um die verschiedenen Verteilungsmuster der As-Konzentrationen in den einzelnen Kompartimenten besser vergleichen zu können, aber auch um die verschiedenen Konzentrationsniveaus zu verdeutlichen, wurden die As-Konzentrationen in den Kompartimenten jeweils in einer Abbildung zusammengefasst (Abb. 5-47, Abb. 5-48).
Dabei wird für das Substrat Colditz deutlich, dass zwar in den meisten Fällen für die Wurzeln die höchsten As-Konzentrationen festgestellt wurden, dass aber die As-Konzentrationen in den Blättern in den P1_N1-Varianten fast das Wurzelniveau erreichen. In den P2_N1-Varianten beider

LABO-Vorhaben B 1.10 31.5.2012
81
Feuchtestufen sind die As-Konzentrationen deutlich gegenüber allen anderen Kompartimenten erhöht.
0 5 10 15
As [mg*kg TG]-1
100 P2_N2
0 5 10 15
As [mg*kg TG]-1
100 P2_N1
0 5 10 15
As [mg*kg TG]-1
100 P1_N2
0 5 10 15
As [mg*kg TG]-1
Korn
Blätter
Stängel
Wurzel
100 P1_N1
70 P2_N270 P2_N170 P1_N2Korn
Blätter
Stängel
Wurzel
70 P1_N1Colditz
Abb. 5-47: Mittlere As-Konzentration in den einzelnen Kompartimenten von T. aestivum für die verschiedenen Versuchsvarianten mit Substrat Colditz (n=5).
Dies bedeutet, dass mit einer starken P-Verfügbarkeit im Substrat das aufgenommene As zu hohen Konzentrationen in den Blättern führt. Zwar sind die As-Konzentrationen in den anderen Kompartimenten ebenfalls gegenüber den entsprechenden Vergleichsvarianten erhöht, aber weitaus weniger, als für die Blätter.
Für die Versuchvarianten mit Substrat Schopfheim sind dagegen generell die As-Konzentrationen in den Wurzeln am höchsten. Trotzdem sind auch hier speziell in den Blättern höhere As-Konzentrationen im Vergleich zu den restlichen Kompartimenten zu beobachten. Dies ist ebenfalls verstärkt in den Varianten mit höherer P-Düngung festzustellen.
Korn
Blätter
Stängel
Wurzel
70 P1_N1 70 P1_N2 70 P2_N1 70 P2_N2
0,1 1 10 100
As [mg*kg TG]-1
Korn
Blätter
Stängel
Wurzel
100 P1_N1
0,1 1 10 100
As [mg*kg TG]-1
100 P1_N2
0,1 1 10 100
As [mg*kg TG]-1
100 P2_N1
0,1 1 10 100
As [mg*kg TG]-1
100 P2_N2
Schopfheim
<NWG
<NWG
<NWG
Abb. 5-48: Mittlere As-Konzentration in den einzelnen Kompartimenten von T. aestivum für die verschiedenen Versuchsvarianten mit Substrat Schopfheim (n=5).
Für die Varianten mit Substrat Schopfheim ließe sich insgesamt ein mehr oder weniger einheitlicher Trend zur Abstufung der As-Konzentrationen zwischen den Kompartimenten formulieren:
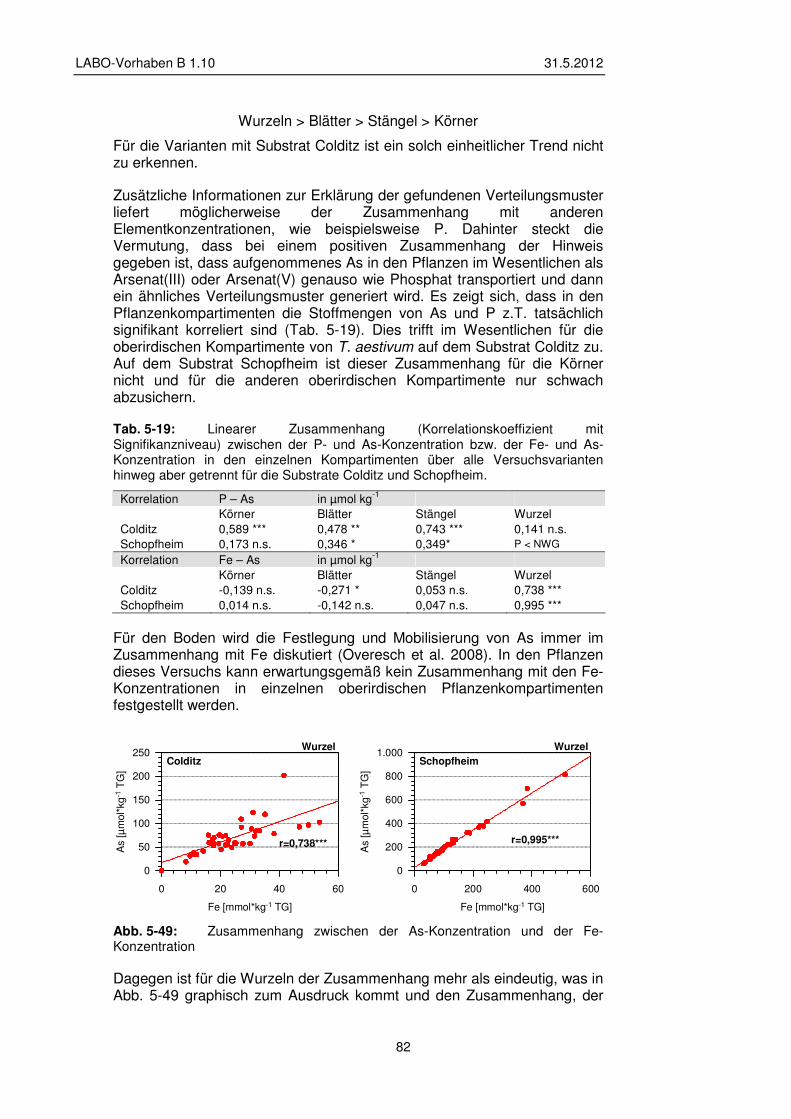
LABO-Vorhaben B 1.10 31.5.2012
82
Wurzeln > Blätter > Stängel > Körner
Für die Varianten mit Substrat Colditz ist ein solch einheitlicher Trend nicht zu erkennen.
Zusätzliche Informationen zur Erklärung der gefundenen Verteilungsmuster liefert möglicherweise der Zusammenhang mit anderen Elementkonzentrationen, wie beispielsweise P. Dahinter steckt die Vermutung, dass bei einem positiven Zusammenhang der Hinweis gegeben ist, dass aufgenommenes As in den Pflanzen im Wesentlichen als Arsenat(III) oder Arsenat(V) genauso wie Phosphat transportiert und dann ein ähnliches Verteilungsmuster generiert wird. Es zeigt sich, dass in den Pflanzenkompartimenten die Stoffmengen von As und P z.T. tatsächlich signifikant korreliert sind (Tab. 5-19). Dies trifft im Wesentlichen für die oberirdischen Kompartimente von T. aestivum auf dem Substrat Colditz zu. Auf dem Substrat Schopfheim ist dieser Zusammenhang für die Körner nicht und für die anderen oberirdischen Kompartimente nur schwach abzusichern.
Tab. 5-19: Linearer Zusammenhang (Korrelationskoeffizient mit Signifikanzniveau) zwischen der P- und As-Konzentration bzw. der Fe- und As-Konzentration in den einzelnen Kompartimenten über alle Versuchsvarianten hinweg aber getrennt für die Substrate Colditz und Schopfheim.
Korrelation P – As in µmol kg-1 Körner Blätter Stängel Wurzel Colditz 0,589 *** 0,478 ** 0,743 *** 0,141 n.s. Schopfheim 0,173 n.s. 0,346 * 0,349* P < NWG
Korrelation Fe – As in µmol kg-1 Körner Blätter Stängel Wurzel Colditz -0,139 n.s. -0,271 * 0,053 n.s. 0,738 *** Schopfheim 0,014 n.s. -0,142 n.s. 0,047 n.s. 0,995 ***
Für den Boden wird die Festlegung und Mobilisierung von As immer im Zusammenhang mit Fe diskutiert (Overesch et al. 2008). In den Pflanzen dieses Versuchs kann erwartungsgemäß kein Zusammenhang mit den Fe-Konzentrationen in einzelnen oberirdischen Pflanzenkompartimenten festgestellt werden.
0
200
400
600
800
1.000
As
[µm
ol*k
gT
G]
-1
0 200 400 600
Fe [mmol*kg TG]-1
r=0,995***
SchopfheimWurzel
0
50
100
150
200
250
As
[µm
ol*k
gT
G]
-1
0 20 40 60
Fe [mmol*kg TG]-1
r=0,738***
ColditzWurzel
Abb. 5-49: Zusammenhang zwischen der As-Konzentration und der Fe-Konzentration
Dagegen ist für die Wurzeln der Zusammenhang mehr als eindeutig, was in Abb. 5-49 graphisch zum Ausdruck kommt und den Zusammenhang, der
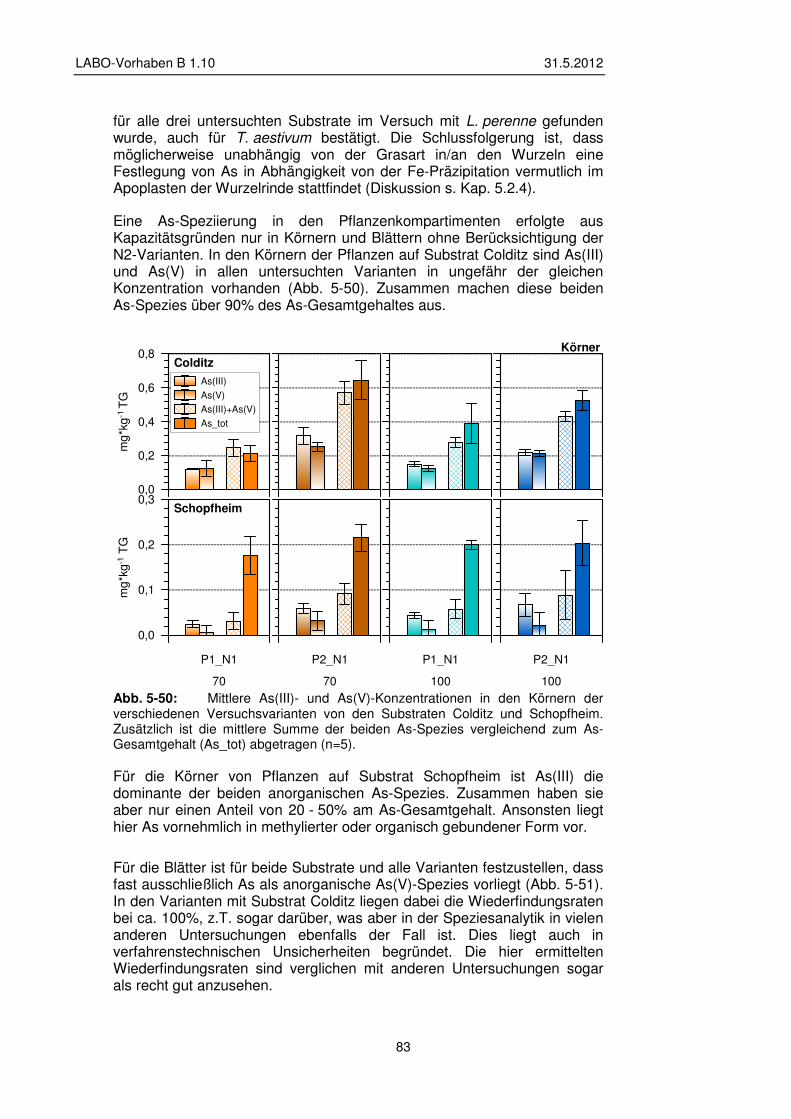
LABO-Vorhaben B 1.10 31.5.2012
83
für alle drei untersuchten Substrate im Versuch mit L. perenne gefunden wurde, auch für T. aestivum bestätigt. Die Schlussfolgerung ist, dass möglicherweise unabhängig von der Grasart in/an den Wurzeln eine Festlegung von As in Abhängigkeit von der Fe-Präzipitation vermutlich im Apoplasten der Wurzelrinde stattfindet (Diskussion s. Kap. 5.2.4).
Eine As-Speziierung in den Pflanzenkompartimenten erfolgte aus Kapazitätsgründen nur in Körnern und Blättern ohne Berücksichtigung der N2-Varianten. In den Körnern der Pflanzen auf Substrat Colditz sind As(III) und As(V) in allen untersuchten Varianten in ungefähr der gleichen Konzentration vorhanden (Abb. 5-50). Zusammen machen diese beiden As-Spezies über 90% des As-Gesamtgehaltes aus.
0,0
0,2
0,4
0,6
0,8
mg*
kgT
G-1
As(III)
As(V)
As(III)+As(V)
As_tot
ColditzKörner
0,0
0,1
0,2
0,3
mg
*kg
TG
-1
P1_N1
70
P2_N1
70
P1_N1
100
P2_N1
100
Schopfheim
Abb. 5-50: Mittlere As(III)- und As(V)-Konzentrationen in den Körnern der verschiedenen Versuchsvarianten von den Substraten Colditz und Schopfheim. Zusätzlich ist die mittlere Summe der beiden As-Spezies vergleichend zum As-Gesamtgehalt (As_tot) abgetragen (n=5).
Für die Körner von Pflanzen auf Substrat Schopfheim ist As(III) die dominante der beiden anorganischen As-Spezies. Zusammen haben sie aber nur einen Anteil von 20 - 50% am As-Gesamtgehalt. Ansonsten liegt hier As vornehmlich in methylierter oder organisch gebundener Form vor.
Für die Blätter ist für beide Substrate und alle Varianten festzustellen, dass fast ausschließlich As als anorganische As(V)-Spezies vorliegt (Abb. 5-51). In den Varianten mit Substrat Colditz liegen dabei die Wiederfindungsraten bei ca. 100%, z.T. sogar darüber, was aber in der Speziesanalytik in vielen anderen Untersuchungen ebenfalls der Fall ist. Dies liegt auch in verfahrenstechnischen Unsicherheiten begründet. Die hier ermittelten Wiederfindungsraten sind verglichen mit anderen Untersuchungen sogar als recht gut anzusehen.
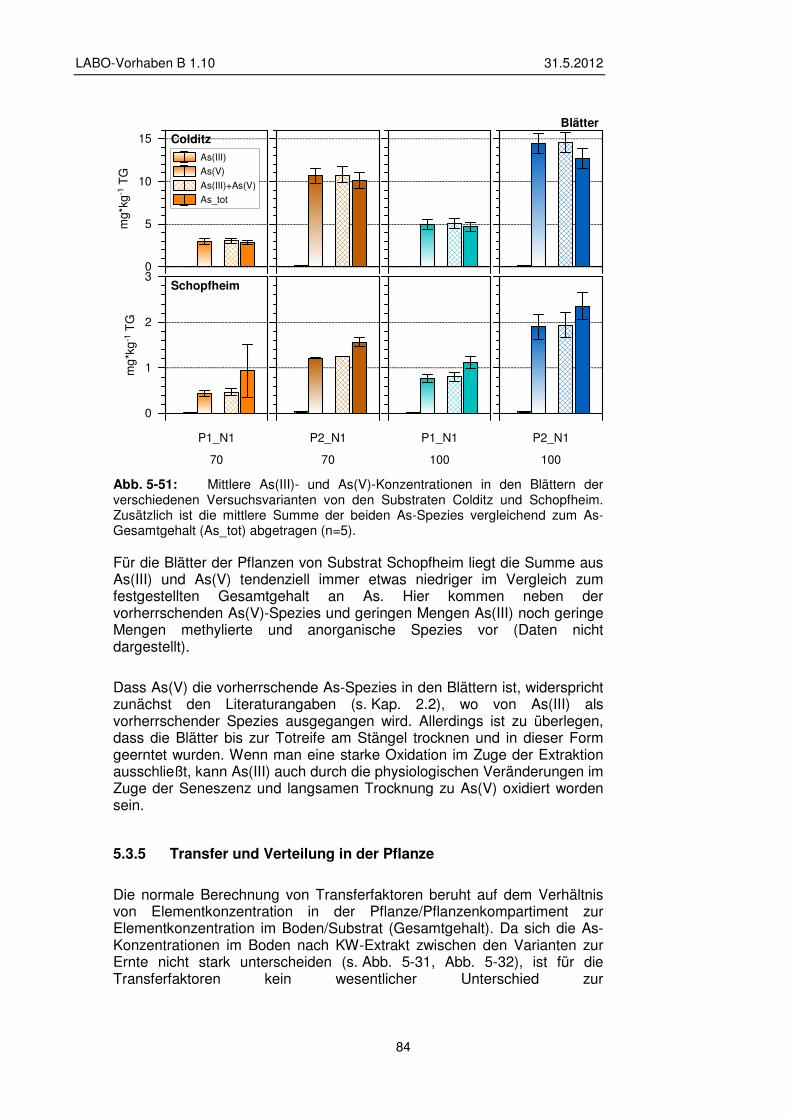
LABO-Vorhaben B 1.10 31.5.2012
84
0
5
10
15m
g*kg
TG
-1
As(III)
As(V)
As(III)+As(V)
As_tot
ColditzBlätter
P1_N1
100
0
1
2
3
mg*
kgT
G-1
P1_N1
70
P2_N1
70
P2_N1
100
Schopfheim
Abb. 5-51: Mittlere As(III)- und As(V)-Konzentrationen in den Blättern der verschiedenen Versuchsvarianten von den Substraten Colditz und Schopfheim. Zusätzlich ist die mittlere Summe der beiden As-Spezies vergleichend zum As-Gesamtgehalt (As_tot) abgetragen (n=5).
Für die Blätter der Pflanzen von Substrat Schopfheim liegt die Summe aus As(III) und As(V) tendenziell immer etwas niedriger im Vergleich zum festgestellten Gesamtgehalt an As. Hier kommen neben der vorherrschenden As(V)-Spezies und geringen Mengen As(III) noch geringe Mengen methylierte und anorganische Spezies vor (Daten nicht dargestellt).
Dass As(V) die vorherrschende As-Spezies in den Blättern ist, widerspricht zunächst den Literaturangaben (s. Kap. 2.2), wo von As(III) als vorherrschender Spezies ausgegangen wird. Allerdings ist zu überlegen, dass die Blätter bis zur Totreife am Stängel trocknen und in dieser Form geerntet wurden. Wenn man eine starke Oxidation im Zuge der Extraktion ausschließt, kann As(III) auch durch die physiologischen Veränderungen im Zuge der Seneszenz und langsamen Trocknung zu As(V) oxidiert worden sein.
5.3.5 Transfer und Verteilung in der Pflanze
Die normale Berechnung von Transferfaktoren beruht auf dem Verhältnis von Elementkonzentration in der Pflanze/Pflanzenkompartiment zur Elementkonzentration im Boden/Substrat (Gesamtgehalt). Da sich die As-Konzentrationen im Boden nach KW-Extrakt zwischen den Varianten zur Ernte nicht stark unterscheiden (s. Abb. 5-31, Abb. 5-32), ist für die Transferfaktoren kein wesentlicher Unterschied zur
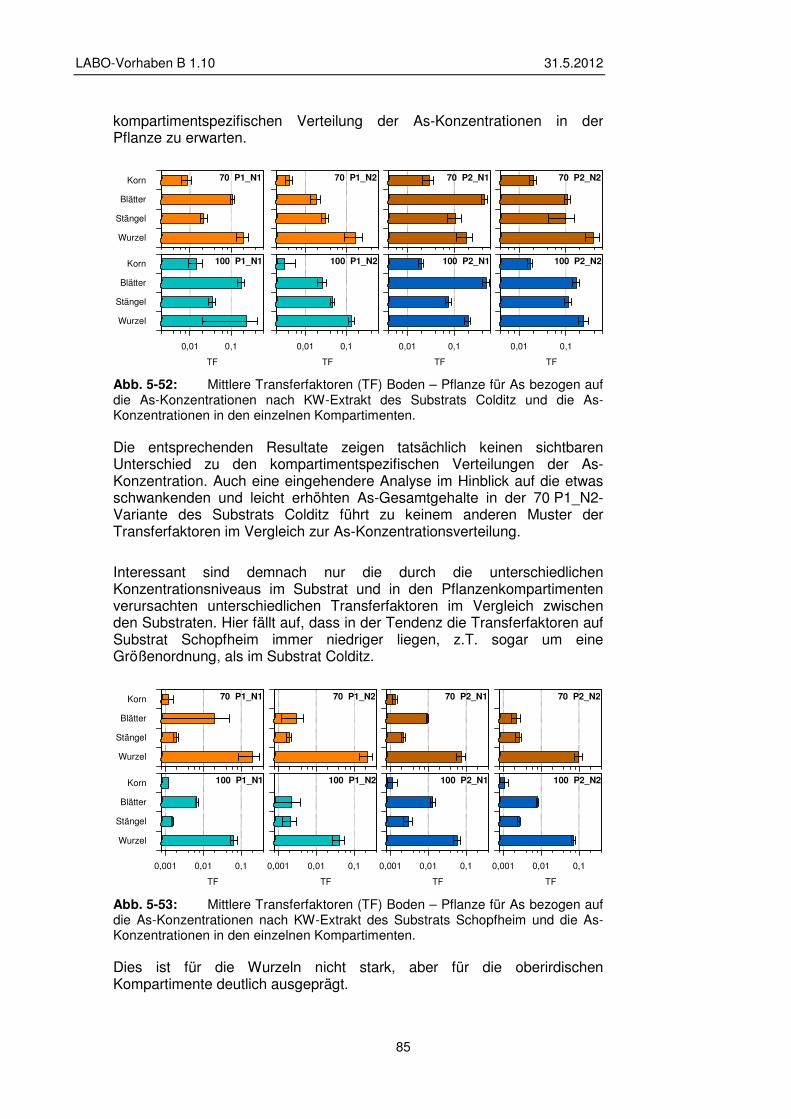
LABO-Vorhaben B 1.10 31.5.2012
85
kompartimentspezifischen Verteilung der As-Konzentrationen in der Pflanze zu erwarten.
Korn
Blätter
Stängel
Wurzel
70 P1_N1 70 P1_N2 70 P2_N1 70 P2_N2
0,01 0,1
TF
Korn
Blätter
Stängel
Wurzel
100 P1_N1
0,01 0,1
TF
100 P1_N2
0,01 0,1
TF
100 P2_N1
0,01 0,1
TF
100 P2_N2
Abb. 5-52: Mittlere Transferfaktoren (TF) Boden – Pflanze für As bezogen auf die As-Konzentrationen nach KW-Extrakt des Substrats Colditz und die As-Konzentrationen in den einzelnen Kompartimenten.
Die entsprechenden Resultate zeigen tatsächlich keinen sichtbaren Unterschied zu den kompartimentspezifischen Verteilungen der As-Konzentration. Auch eine eingehendere Analyse im Hinblick auf die etwas schwankenden und leicht erhöhten As-Gesamtgehalte in der 70 P1_N2-Variante des Substrats Colditz führt zu keinem anderen Muster der Transferfaktoren im Vergleich zur As-Konzentrationsverteilung.
Interessant sind demnach nur die durch die unterschiedlichen Konzentrationsniveaus im Substrat und in den Pflanzenkompartimenten verursachten unterschiedlichen Transferfaktoren im Vergleich zwischen den Substraten. Hier fällt auf, dass in der Tendenz die Transferfaktoren auf Substrat Schopfheim immer niedriger liegen, z.T. sogar um eine Größenordnung, als im Substrat Colditz.
Korn
Blätter
Stängel
Wurzel
70 P1_N1 70 P1_N2 70 P2_N1 70 P2_N2
0,001 0,01 0,1
TF
Korn
Blätter
Stängel
Wurzel
100 P1_N1
0,001 0,01 0,1
TF
100 P1_N2
0,001 0,01 0,1
TF
100 P2_N1
0,001 0,01 0,1
TF
100 P2_N2
Abb. 5-53: Mittlere Transferfaktoren (TF) Boden – Pflanze für As bezogen auf die As-Konzentrationen nach KW-Extrakt des Substrats Schopfheim und die As-Konzentrationen in den einzelnen Kompartimenten.
Dies ist für die Wurzeln nicht stark, aber für die oberirdischen Kompartimente deutlich ausgeprägt.

LABO-Vorhaben B 1.10 31.5.2012
86
Anders als zum Versuch mit L. perenne ausgeführt, konnte hier aufgrund von weniger Werten unter der Nachweisgrenze besser untersucht werden, ob von der As-Konzentration nach AN-Extrakt auf die As-Konzentration im Sproß bzw. in den einzelnen oberirdischen Pflanzenkompartimenten geschlossen werden kann.
Tab. 5-20: Zusammenhang (Korrelationskoeffizient mit Signifikanzniveau) zwischen den As-Konzentrationen in den einzelnen Kompartimenten und der As-Konzentration nach AN-Extrakt über alle Versuchsvarianten hinweg aber getrennt für die Substrate Colditz und Schopfheim.
Korrelation As-Konz. (AN-Extrakt) – As-Konz. (Kompartiment) Körner Blätter Stängel Wurzel Colditz 0,497 *** 0,232 n.s. 0,396 ** 0,346 * Schopfheim -0,169 n.s -0,217 n.s. 0,199 n.s. -0,029 n.s.
Tatsächlich gibt es signifikante Zusammenhänge für die Kompartimente Korn, Stängel und Wurzel auf Substrat Colditz. Allerdings sind diese Zusammenhänge für Stängel und Wurzel schwach bis sehr schwach ausgeprägt. Einzig für die Körner zeigt sich ein stärkerer Zusammenhang. Für Substrat Schopfheim ist in keiner Weise ein Zusammenhang herzustellen. Insgesamt lässt sich konstatieren, dass der AN-Extrakt nach diesen Ergebnissen keine zuverlässigen Rückschlüsse auf den Transfer von As in die Pflanze zulässt.
Die Verteilung innerhalb der Pflanzenkompartimente ist sowohl mit den kompartimentspezifischen As-Konzentrationen als auch mit den berechneten Transferfaktoren für die einzelnen Kompartimente angedeutet. Allerdings wurde bislang der Biomassebezug nicht hergestellt. Nach Kalkulation der As-Stoffmenge auf der Grundlage von As-Konzentrationen und Biomasse lässt sich der prozentuale Anteile von As in den einzelnen Kompartimenten und damit die genaue interne Verteilung berechnen. Hier zeigt sich, dass insgesamt für das Substrat Colditz das meiste As in fast allen Varianten im Sproß zu finden ist. Dagegen sind für das Substrat Schopfheim generell mehr als 50% der As-Stoffmenge in den Pflanzen in den Wurzeln akkumuliert. Die Translokation in den Sproß findet hier z.T. nur in geringem Maße statt.
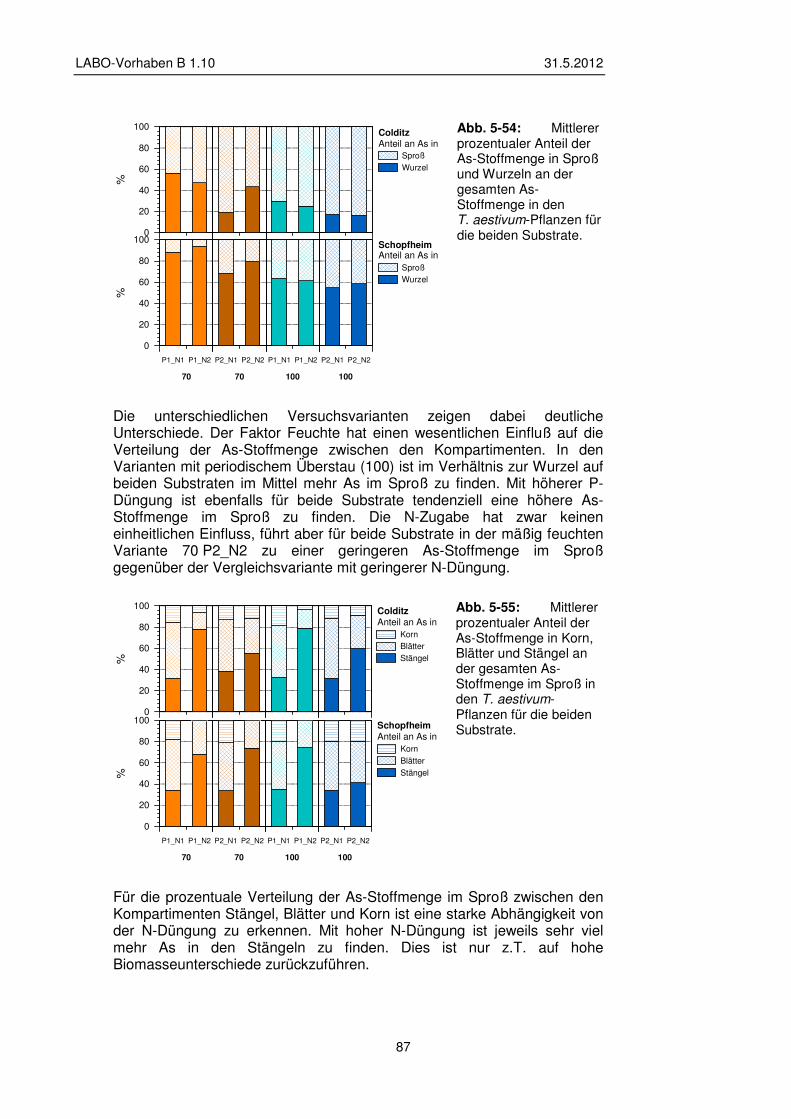
LABO-Vorhaben B 1.10 31.5.2012
87
0
20
40
60
80
100%
Wurzel
SproßAnteil an As in
0
20
40
60
80
100
%
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
WurzelSproß
Colditz
SchopfheimAnteil an As in
Abb. 5-54: Mittlerer prozentualer Anteil der As-Stoffmenge in Sproß und Wurzeln an der gesamten As-Stoffmenge in den T. aestivum-Pflanzen für die beiden Substrate.
Die unterschiedlichen Versuchsvarianten zeigen dabei deutliche Unterschiede. Der Faktor Feuchte hat einen wesentlichen Einfluß auf die Verteilung der As-Stoffmenge zwischen den Kompartimenten. In den Varianten mit periodischem Überstau (100) ist im Verhältnis zur Wurzel auf beiden Substraten im Mittel mehr As im Sproß zu finden. Mit höherer P-Düngung ist ebenfalls für beide Substrate tendenziell eine höhere As-Stoffmenge im Sproß zu finden. Die N-Zugabe hat zwar keinen einheitlichen Einfluss, führt aber für beide Substrate in der mäßig feuchten Variante 70 P2_N2 zu einer geringeren As-Stoffmenge im Sproß gegenüber der Vergleichsvariante mit geringerer N-Düngung.
0
20
40
60
80
100
% Stängel
BlätterKorn
Anteil an As inColditz
0
20
40
60
80
100
%
P1_N1 P1_N2
70
P2_N1 P2_N2
70
P1_N1 P1_N2
100
P2_N1 P2_N2
100
Stängel
Blätter
Korn
Anteil an As inSchopfheim
Abb. 5-55: Mittlerer prozentualer Anteil der As-Stoffmenge in Korn, Blätter und Stängel an der gesamten As-Stoffmenge im Sproß in den T. aestivum-Pflanzen für die beiden Substrate.
Für die prozentuale Verteilung der As-Stoffmenge im Sproß zwischen den Kompartimenten Stängel, Blätter und Korn ist eine starke Abhängigkeit von der N-Düngung zu erkennen. Mit hoher N-Düngung ist jeweils sehr viel mehr As in den Stängeln zu finden. Dies ist nur z.T. auf hohe Biomasseunterschiede zurückzuführen.

LABO-Vorhaben B 1.10 31.5.2012
88
5.4 Feldversuche
Die Felduntersuchungen beschränken sich auf den Standort Colditz (in der Nähe der Bodendauerbeobachtungsfläche 33) in Sachsen. Der Standort liegt in der Muldeaue und wird im Abstand von ca. 200 m von der Mulde als Grünland genutzt. Als solches wurde es vom Pächter im Untersuchungszeitraum 2011 dreimal zur Verwertung des Grünschnitts gemäht. In westlicher Richtung grenzt an das Grünlang nach einer Flurerhöhung Ackernutzung an. Hier wurde 2011 Mais angebaut.
Zur eingehenden Untersuchung des Transfers Boden – Pflanze wurde südlich der Bodendauerbeobachtungsfläche ein Transekt (Transekt I) von der Mulde bis in den Bereich der Ackernutzung etabliert. Die Etablierung eines 2. Transekts nördlich der Bodendauerbeobachtungsfläche konnte nicht realisiert werden. Die Transektführung folgt einem Überstaugradienten. Im Bereich vor einer dammartigen Erhöhung ist im Mittel einmal im Jahr mit einem Überstau durch ein Muldehochwasser zu rechnen. Hinter diesem Dammbereich ist nur zu außergewöhnlichen Hochwasserereignissen mit einem Überstau zu rechnen (s. Hochwasser 2002).
Entlang des Transekts I wurden Vegetationsaufnahmen zur Charakterisierung der Vegetation durchgeführt. Dies diente in erster Linie zur Evaluierung der Stetigkeit der Arten, was wiederum wichtig für Pflanzenauswahl zur Beprobung ist. Nur über das Transekt stetige Arten können zu Vergleichszwecken für eine Probenahme herangezogen werden. Zum zweiten sollte aber auch die Abundanz auf jeder einzelnen Versuchsfläche dokumentiert werden, um eine flächenhafte Abschätzung des As-Transfers zu ermöglichen. Entsprechend den Ergebnissen aus der Vegetationskartierung wurden dann die Probenahmepunkte festgelegt (s. Abb. 5-56, Tab. 5-21). Es wurden aufgrund der Untersuchungsergebnisse für die ersten Beprobungstermine 2 Pflanzenarten (Alopecurus pratensis Wiesenfuchsschwanz; Rumex acetosa Wiesen-Sauerampfer), für den letzten Beprobungstermin 3 Pflanzenarten (Alopecurus pratensis Wiesenfuchsschwanz; Rumex acetosa Wiesen-Sauerampfer, Glyceria fluitans Flutender Schwaden) ausgewählt.

LABO-Vorhaben B 1.10 31.5.2012
89
BDF 33
Transsekt I
I 1I 2
I 3I 4I 5I 6I 7
Abb. 5-56: Lage des Transsekts I, der Probenahmepunkte sowie der Bodendauerbeobachtungsfläche. Schraffiert ist der Bereich einer dammartigen Erhöhung eingezeichnet.
Tab. 5-21: Koordinaten für die Lage der Probepunkte
Probepunkte Hochwert Rechtswert I-1 5664460 4557117 I-2 5664467 4557109 I-3 5664477 4557103 I-4 5664481 4557091 I-5 5664488 4557077 I-6 5664496 4557073 I-7 5664499 4557060
Zu 3 verschiedenen Zeitpunkten (10.05.11, 30.06.11, 12.10.11) wurden Pflanzenproben, zu einem Zeitpunkt (12.10.11) Bodenproben als Mischproben von wenigstens 3 Stellen innerhalb einer Probefläche genommen. Eine erste Probenahme im Herbst 2010 scheiterte, da kurz vor dem anvisierten Termin überraschenderweise eine Mahd durch den Pächter stattfand.
Die Pflanzenproben wurden in 2 Fraktionen getrennt und die eine Fraktion noch am Tag der Probenahme mit H2Odest gründlich gewaschen, die andere Fraktion ungewaschen belassen. Die Proben wurden entsprechend den für die Gefäßversuche angegebenen Methoden aufgearbeitet und auf ihre Elementgehalte analysiert.
An den Bodenproben wurden die pH-Werte sowie die Elementgehalte nach AN-Extraktion und nach KW-Extraktion bestimmt (Methoden s. Kap. 5.1.3)
5.5 Feldversuche – Ergebnisse Bodenanalyse
Im Ergebnis zeigen die pH-Werte einen sowohl für pH(H2O) als auch für pH(CaCl2) einen Abfall mit Entfernung zur Mulde. Dieser Abfall ist eindeutig und reicht von Werten über 6,5 bis zu Werten von 5,7 im pH(H2O).
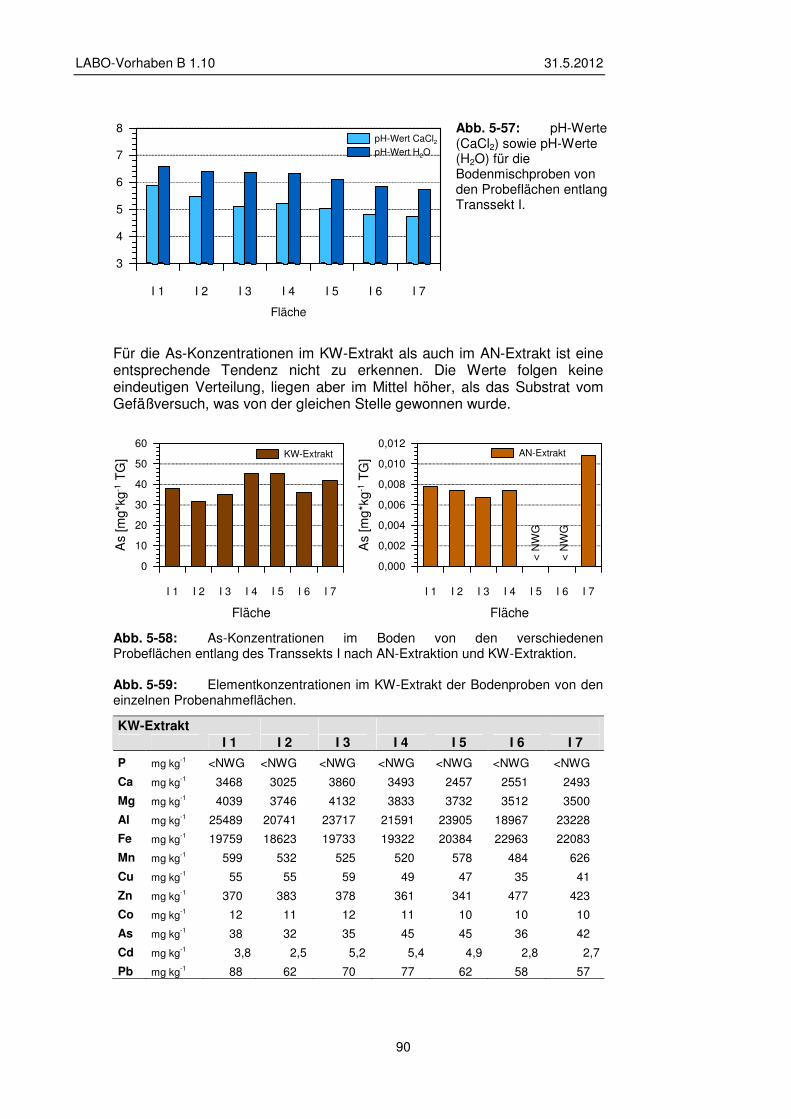
LABO-Vorhaben B 1.10 31.5.2012
90
3
4
5
6
7
8
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
pH-Wert CaCl2pH-Wert H O2
Abb. 5-57: pH-Werte (CaCl2) sowie pH-Werte (H2O) für die Bodenmischproben von den Probeflächen entlang Transsekt I.
Für die As-Konzentrationen im KW-Extrakt als auch im AN-Extrakt ist eine entsprechende Tendenz nicht zu erkennen. Die Werte folgen keine eindeutigen Verteilung, liegen aber im Mittel höher, als das Substrat vom Gefäßversuch, was von der gleichen Stelle gewonnen wurde.
0
10
20
30
40
50
60
As
[mg*
kgT
G]
-1
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
KW-Extrakt
0,000
0,002
0,004
0,006
0,008
0,010
0,012
As
[mg*
kgT
G]
-1
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
AN-Extrakt
<N
WG
<N
WG
Abb. 5-58: As-Konzentrationen im Boden von den verschiedenen Probeflächen entlang des Transsekts I nach AN-Extraktion und KW-Extraktion.
Abb. 5-59: Elementkonzentrationen im KW-Extrakt der Bodenproben von den einzelnen Probenahmeflächen.
KW-Extrakt I 1 I 2 I 3 I 4 I 5 I 6 I 7
P mg kg-1 <NWG <NWG <NWG <NWG <NWG <NWG <NWG
Ca mg kg-1 3468 3025 3860 3493 2457 2551 2493
Mg mg kg-1 4039 3746 4132 3833 3732 3512 3500
Al mg kg-1 25489 20741 23717 21591 23905 18967 23228
Fe mg kg-1 19759 18623 19733 19322 20384 22963 22083
Mn mg kg-1 599 532 525 520 578 484 626
Cu mg kg-1 55 55 59 49 47 35 41
Zn mg kg-1 370 383 378 361 341 477 423
Co mg kg-1 12 11 12 11 10 10 10
As mg kg-1 38 32 35 45 45 36 42
Cd mg kg-1 3,8 2,5 5,2 5,4 4,9 2,8 2,7
Pb mg kg-1 88 62 70 77 62 58 57
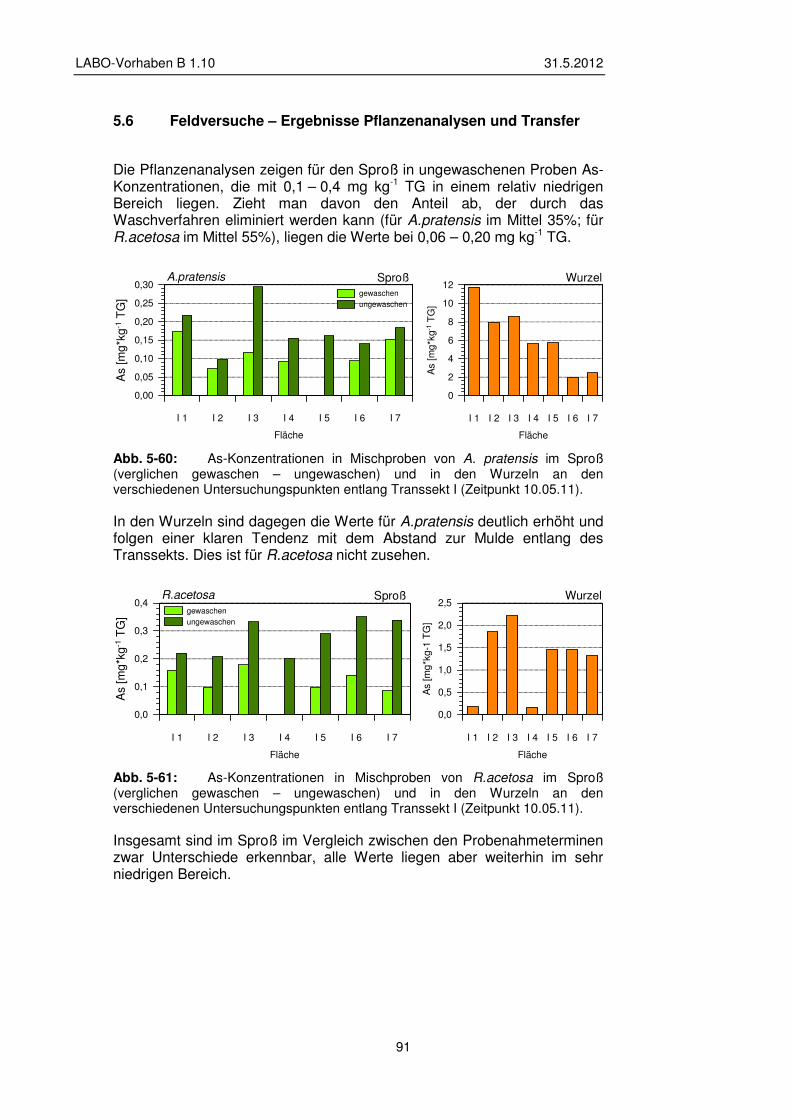
LABO-Vorhaben B 1.10 31.5.2012
91
5.6 Feldversuche – Ergebnisse Pflanzenanalysen und Transfer
Die Pflanzenanalysen zeigen für den Sproß in ungewaschenen Proben As-Konzentrationen, die mit 0,1 – 0,4 mg kg-1 TG in einem relativ niedrigen Bereich liegen. Zieht man davon den Anteil ab, der durch das Waschverfahren eliminiert werden kann (für A.pratensis im Mittel 35%; für R.acetosa im Mittel 55%), liegen die Werte bei 0,06 – 0,20 mg kg-1 TG.
0,00
0,05
0,10
0,15
0,20
0,25
0,30
As
[mg*
kgT
G]
-1
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
gewaschenungewaschen
A.pratensis Sproß
0
2
4
6
8
10
12
As
[mg*
kgT
G]
-1I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
Wurzel
Abb. 5-60: As-Konzentrationen in Mischproben von A. pratensis im Sproß (verglichen gewaschen – ungewaschen) und in den Wurzeln an den verschiedenen Untersuchungspunkten entlang Transsekt I (Zeitpunkt 10.05.11).
In den Wurzeln sind dagegen die Werte für A.pratensis deutlich erhöht und folgen einer klaren Tendenz mit dem Abstand zur Mulde entlang des Transsekts. Dies ist für R.acetosa nicht zusehen.
0,0
0,1
0,2
0,3
0,4
As
[mg*
kgT
G]
-1
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
gewaschenungewaschen
R.acetosa Sproß
0,0
0,5
1,0
1,5
2,0
2,5
As
[mg*
kg-1
TG
]
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
Wurzel
Abb. 5-61: As-Konzentrationen in Mischproben von R.acetosa im Sproß (verglichen gewaschen – ungewaschen) und in den Wurzeln an den verschiedenen Untersuchungspunkten entlang Transsekt I (Zeitpunkt 10.05.11).
Insgesamt sind im Sproß im Vergleich zwischen den Probenahmeterminen zwar Unterschiede erkennbar, alle Werte liegen aber weiterhin im sehr niedrigen Bereich.
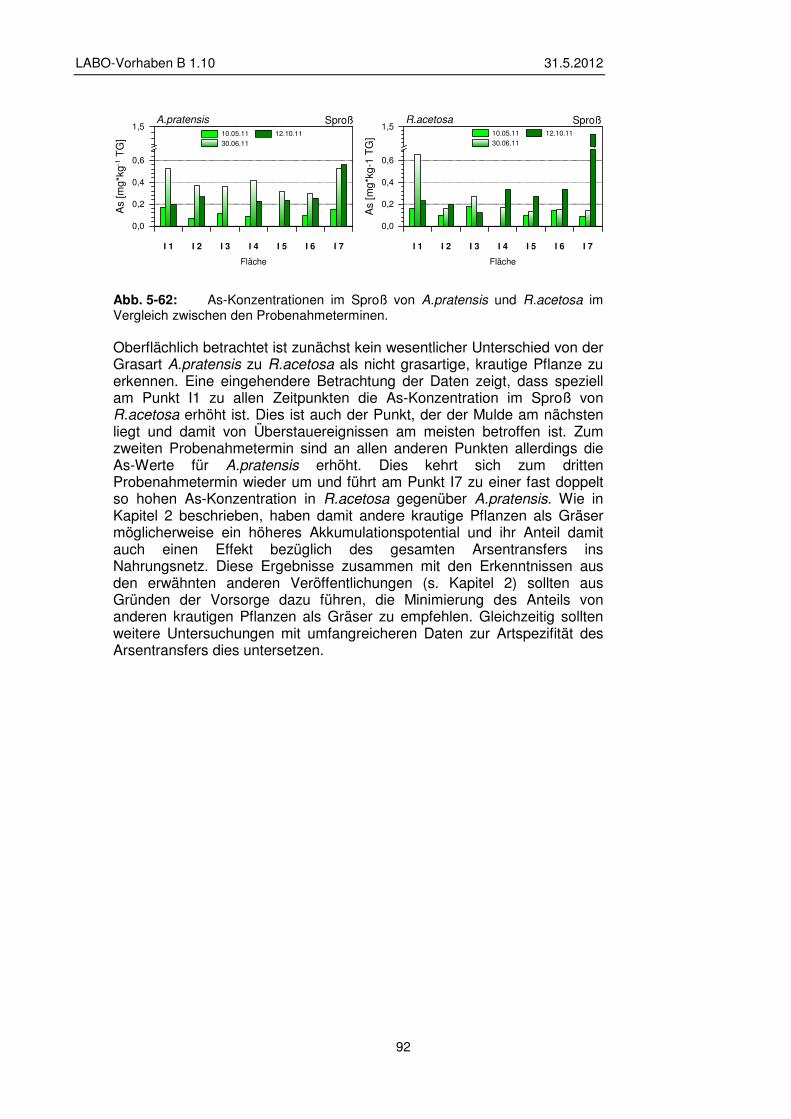
LABO-Vorhaben B 1.10 31.5.2012
92
0,0
0,2
0,4
0,6
As
[mg*
kgT
G]
-1
1,5
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
10.05.1130.06.11
12.10.11
A.pratensis Sproß
0,0
0,2
0,4
0,6
As
[mg*
kg-1
TG
]
1,5
I 1 I 2 I 3 I 4 I 5 I 6 I 7
Fläche
10.05.1130.06.11
12.10.11
R.acetosa Sproß
Abb. 5-62: As-Konzentrationen im Sproß von A.pratensis und R.acetosa im Vergleich zwischen den Probenahmeterminen.
Oberflächlich betrachtet ist zunächst kein wesentlicher Unterschied von der Grasart A.pratensis zu R.acetosa als nicht grasartige, krautige Pflanze zu erkennen. Eine eingehendere Betrachtung der Daten zeigt, dass speziell am Punkt I1 zu allen Zeitpunkten die As-Konzentration im Sproß von R.acetosa erhöht ist. Dies ist auch der Punkt, der der Mulde am nächsten liegt und damit von Überstauereignissen am meisten betroffen ist. Zum zweiten Probenahmetermin sind an allen anderen Punkten allerdings die As-Werte für A.pratensis erhöht. Dies kehrt sich zum dritten Probenahmetermin wieder um und führt am Punkt I7 zu einer fast doppelt so hohen As-Konzentration in R.acetosa gegenüber A.pratensis. Wie in Kapitel 2 beschrieben, haben damit andere krautige Pflanzen als Gräser möglicherweise ein höheres Akkumulationspotential und ihr Anteil damit auch einen Effekt bezüglich des gesamten Arsentransfers ins Nahrungsnetz. Diese Ergebnisse zusammen mit den Erkenntnissen aus den erwähnten anderen Veröffentlichungen (s. Kapitel 2) sollten aus Gründen der Vorsorge dazu führen, die Minimierung des Anteils von anderen krautigen Pflanzen als Gräser zu empfehlen. Gleichzeitig sollten weitere Untersuchungen mit umfangreicheren Daten zur Artspezifität des Arsentransfers dies untersetzen.

LABO-Vorhaben B 1.10 31.5.2012
93
6 FAZIT
ahu AG, TU Dresden
Die Ergebnisse des Vorhabens B 1.10 sind ein praxisorientierter Leitfaden Arsentransfer aus Böden in Nahrungs- und Futterpflanzen – Gefahrenbeurteilung und Maßnahmen und die vorliegende Ergebnisdokumentation. Damit wird die Erkenntnislage zu Arsen in den Bereichen der Erfassung von Verdachtsflächen, der orientierenden Untersuchung und Detailuntersuchung sowie für die Gefahrenabwehr verbessert. Die Gefäß- und Feldversuche liefern neue Erkenntnisse zur Aufnahme von Arsen in Pflanzen unter verschiedenen Standort-, Feuchte- und Düngebedingungen. Die zusammengefassten, vollzugsrelevanten Schlussfolgerungen aus diesem Abschlussbericht finden sich im Leitfaden.
Während das bodenchemische Verhalten von Arsen im Wesentlichen bekannt ist und mit dem Abschluss des Vorhabens exemplarische Versuchsergebnisse vorliegen, fehlen über einige Details von Prozessen und über die Aufnahme in Pflanzen immer noch Erkenntnisse. Daraus resultieren zum Teil Unsicherheiten bei der Gefahrenbeurteilung.
Klärungsbedarf besteht nach derzeitigem Kenntnisstand in Bezug auf folgende praxisrelevante Aspekte:
- Konkrete Anbauempfehlungen für Nahrungs- und Futterpflanzen in Abhängigkeit des Anreicherungvermögens; es besteht der Bedarf an Sortenempfehlungen zur Vermeidung der Aufnahme von Arsen und deren regelmäßiger Aktualisierung (z.B. jährlich),
- Temperaturabhängigkeit der Redoxtransformationen des Arsens,
- Methodische Unsicherheiten aufgrund fehlender Standarduntersuchungsverfahren (z.B. Eisen-/Mangan(hydr-)oxide, Arsenspezies, Probenentnahme in reduzierten Horizonten),
- Etablierung eines standardisierten Waschverfahrens zur näherungsweisen Quantifizierung der Arsenbelastung von Futtermitteln über systemische Aufnahme und über Verschmutzung,
- Wirkung von Bodenzusätzen wie Eisen, Kalk, organische Substanz
sowie in Bezug auf weitergehende Forschung hinsichtlich
- der Bedingungen für mikrobielle Methylierungsreaktionen und Umfang der Mobilisierung von Arsenverbindungen durch die Umwandlung in gasförmige oder gelöste Spezies,
- methodischen Schwierigkeiten bei der Identifizierung und Quantifizierung der verschiedenen Arsenspezies in Böden und den darin enthaltenen Lösungen und Gasen,

LABO-Vorhaben B 1.10 31.5.2012
94
- der Interaktionen mit Makronährstoffen und veränderter Nährstoffstöchiometrie (Imbalancen) � bisher nur N:P in begrenztem Umfang untersucht,
- der art- und sortenspezifische Kompensation des Stresses durch As,
- Auswirkungen der chemischen Erscheinungsformen von Nährstoffen (z.B. Ammonium- vs. Nitraternährung),
- Auswirkungen von Wechselfeuchte (Amplitude und Frequenz) im Vergleich zu Dauerfeuchte,
- der spezifischen Kompensation der Auswirkungen des Überstaus durch Arten und funktionelle Gruppen von Pflanzen (z.B. mit der Kapazität, den Wurzelraum selbst mit Sauerstoff zu versorgen oder alternativ Überstauphasen mit Gärung/Fermentation zu überbrücken),
- Einflüssen von multiplem oxidativen Stress, der nicht durch Metalle und Metalloide verursacht wird (u.a. UV, NOx, Bodenozon, Pathogene) und
- der Rolle pro- und symbiotischer Mikroben bezüglich Vermeidung, Toleranz und Akkumulation von Arsen.

LABO-Vorhaben B 1.10 31.5.2012
95
7 LITERATUR
Abedin, M. J., J. Feldmann and A. A. Meharg (2002): "Uptake kinetics of arsenic species in rice plants." Plant Physiology 128(3): 1120-1128.
Al Agely, Y., D. M. Sylvia and L. Q. Ma (2005): "Mycorhizae increase arsenic uptake by the hyperaccumulator Chinese brake fern (Pteris vittata L.)." Journal of Environmental Quality 34(6): 2181-2186.
Bentley, R. and T. G. Chasteen (2002): "Microbial methylation of metalloids: Arsenic, antimony, and bismuth." Microbiology and Molecular Biology Reviews 66(2): 250-+.
Bertolero, F., G. Pozzi, E. Sabbioni and U. Saffiotti (1987): "Cellular uptake and metabolic reduction of pentavalent to trivalent arsenic as determinants of cytotoxicity and morphological transformation." Carcinogenesis 8(6): 803-808.
Bienfait, H. F., W. Vandenbriel and N. T. Meslandmul (1985): "FREE SPACE IRON POOLS IN ROOTS - GENERATION AND MOBILIZATION." Plant Physiology 78(3): 596-600.
Bleeker, P. M., H. W. J. Hakvoort, M. Bliek, E. Souer and H. Schat (2006): "Enhanced arsenate reduction by a CDC25-like tyrosine phosphatase explains increased phytochelatin accumulation in arsenate-tolerant Holcus lanatus." Plant Journal 45(6): 917-929.
Blodau, C., B. Fulda, M. Bauer and K. H. Knorr (2008): "Arsenic speciation and turnover in intact organic soil mesocosms during experimental drought and rewetting." Geochimica Et Cosmochimica Acta 72(16): 3991-4007.
Bogdan, K. and M. K. Schenk (2008): "Arsenic in Rice (Oryza sativa L.) Related to Dynamics of Arsenic and Silicic Acid in Paddy Soils." Environmental Science & Technology 42(21): 7885-7890.
Bortz, J. (1993): Statistik für Sozialwissenschaftler. Berlin, Springer Verlag.
Bortz, J. (2005): Statistik für Human- und Sozialwissenschaftler. Heidelberg, Springer Medizin Verlag.
Carbonell-Barrachina, A. A., M. A. Aarabi, R. D. DeLaune, R. P. Gambrell and W. H. Patrick (1998): "The influence of arsenic chemical form and concentration on Spartina patens and Spartina alterniflora growth and tissue arsenic concentration." Plant and Soil 198(1): 33-43.
Carbonell-Barrachina, A. A., F. Burlo, E. Lopez and F. Martinez-Sanchez (1999): "Arsenic toxicity and accumulation in radish as affected by arsenic chemical speciation." Journal of Environmental Science and Health Part B-Pesticides Food Contaminants and Agricultural Wastes 34(4): 661-679.
Carbonell-Barrachina, A. A., A. Jugsujinda, F. Burlo, R. D. Delaune and W. H. Patrick (2000): "Arsenic chemistry in municipal sewage sludge as affected by redox potential and pH." Water Research 34(1): 216-224.
Castlehouse, H., C. Smith, A. Raab, C. Deacon, A. A. Meharg and J. Feldmann (2003): "Biotransformation and accumulation of arsenic in soil amended with seaweed." Environmental Science & Technology 37(5): 951-957.
Catarecha, P., M. D. Segura, J. M. Franco-Zorrilla, B. Garcia-Ponce, M. Lanza, R. Solano, J. Paz-Ares and A. Leyva (2007): "A mutant of the Arabidopsis phosphate transporter PHT1;1 displays enhanced arsenic accumulation." Plant Cell 19(3): 1123-1133.
Chen, Z., Y. G. Zhu, W. J. Liu and A. A. Meharg (2005): "Direct evidence showing the effect of root surface iron plaque on arsenite and arsenate uptake into rice (Oryza sativa) roots." New Phytologist 165(1): 91-97.
Cullen, W. R. and J. Nelson (1992): "The use of liposomes in predicting the biological mobility of arsenic compounds." Applied Organometallic Chemistry 6(2): 179-183.
Cullen, W. R. and K. J. Reimer (1989): "Arsenic speciation in the environment." Chemical Reviews 89(4): 713-764.

LABO-Vorhaben B 1.10 31.5.2012
96
Dhankher, O. P., Y. J. Li, B. P. Rosen, J. Shi, D. Salt, J. F. Senecoff, N. A. Sashti and R. B. Meagher (2002): "Engineering tolerance and hyperaccumulation of arsenic in plants by combining arsenate reductase and gamma-glutamylcysteine synthetase expression." Nature Biotechnology 20(11): 1140-1145.
Ebert, T. (2005). Untersuchungen über die Arsenaufnahme von Grünlandpflanzen auf belasteten Böden im Erdinger Moos. Fachbereich 3: Naturwissenschaften. Koblenz, Universität Koblenz-Landau: 122. Thesis
Francesconi, K., P. Visoottiviseth, W. Sridokchan and W. Goessler (2002): "Arsenic species in an arsenic hyperaccumulating fern, Pityrogramma calomelanos: a potential phytoremediator of arsenic-contaminated soils." Science of the Total Environment 284: 27-35.
Geiszinger, A., W. Goessler and W. Kosmus (2002): "Organoarsenic compounds in plants and soil on top of an ore vein." Applied Organometallic Chemistry 16(5): 245-249.
Geng, C. N., Y. G. Zhu, Y. P. Tong, S. E. Smith and F. A. Smith (2006): "Arsenate (As) uptake by and distribution in two cultivars of winter wheat (Triticum aestivum L.)." Chemosphere 62(4): 608-615.
Gong, Z., X. Lu, W. R. Cullen and X. Chris Le (2001): "Unstable trivalent arsenic metabolites, monomethylarsonous acid and dimethylarsinous acid." Journal of Analytical Atomic Spectrometry 16(12): 1409-1413.
Gonzalez, E., R. Solano, V. Rubio, A. Leyva and J. Paz-Ares (2005): "PHOSPHATE TRANSPORTER TRAFFIC FACILITATOR1 is a plant-specific SEC12-related protein that enables the endoplasmic reticulum exit of a high-affinity phosphate transporter in Arabidopsis." Plant Cell 17(12): 3500-3512.
Grybos, M., M. Davranche, G. Gruau and P. Petitjean (2007): "Is trace metal release in wetland soils controlled by organic matter mobility or Fe-oxyhydroxides reduction?" Journal of Colloid and Interface Science 314(2): 490-501.
Ha, S. B., A. P. Smith, R. Howden, W. M. Dietrich, S. Bugg, M. J. O'Connell, P. B. Goldsbrough and C. S. Cobbett (1999): "Phytochelatin synthase genes from Arabidopsis and the yeast Schizosaccharomyces pombe." Plant Cell 11(6): 1153-1163.
Haug, C. M., K. J. Reimer and W. R. Cullen (2004): "Arsenic uptake by the Douglas-fir (Pseudotsuga menziesie)." Applied Organometallic Chemistry 18(12): 626-630.
Herbel, M. and S. Fendorf (2006): "Biogeochemical processes controlling the speciation and transport of arsenic within iron coated sands." Chemical Geology 228(1-3): 16-32.
Hinsinger, P. (2001): "Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review." Plant and Soil 237(2): 173-195.
Hinsinger, P., C. Plassard, C. X. Tang and B. Jaillard (2003): "Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: A review." Plant and Soil 248(1-2): 43-59.
Isayenkov, S. V. and F. J. M. Maathuis (2008): "The Arabidopsis thaliana aquaglyceroporin AtNIP7;1 is a pathway for arsenite uptake." Febs Letters 582(11): 1625-1628.
Kabata-Pendias, A. and H. Pendias (1992): Trace elements in soils and plants. Boca Raton, CRC Press.
Kamiya, T., M. Tanaka, N. Mitani, J. F. Ma, M. Maeshima and T. Fujiwara (2009): "NIP1;1, an aquaporin homolog, determines the arsenite sensitivity of Arabidopsis thaliana." Journal of Biological Chemistry 284(4): 2114-2120.
Karimi, N., S. M. Ghaderian, A. Raab, J. Feldmann and A. A. Meharg (2009): "An arsenic-accumulating, hypertolerant brassica, Isatis capadocica." New Phytologist 184(1): 41-47.
Kim, S. A. and M. L. Guerinot (2007): "Mining iron: Iron uptake and transport in plants." Febs Letters 581(12): 2273-2280.
Koch, I., L. X. Wang, C. A. Ollson, W. R. Cullen and K. J. Reimer (2000): "The predominance of inorganic arsenic species in plants from Yellowknife, Northwest Territories, Canada." Environmental Science & Technology 34(1): 22-26.

LABO-Vorhaben B 1.10 31.5.2012
97
Kuehnelt, D., J. Lintschinger and W. Goessler (2000): "Arsenic compounds in terrestrial organisms. IV. Green plants and lichens from an old arsenic smelter site in Austria." Applied Organometallic Chemistry 14(8): 411-420.
Li, H., Z. H. Ye, Z. J. Wei and M. H. Wong (2011): "Root porosity and radial oxygen loss related to arsenic tolerance and uptake in wetland plants." Environmental Pollution 159(1): 30-37.
Li, R. Y., Y. Ago, W. J. Liu, N. Mitani, J. Feldmann, S. P. McGrath, J. F. Ma and F. J. Zhao (2009a): "The Rice Aquaporin Lsi1 Mediates Uptake of Methylated Arsenic Species." Plant Physiology 150(4): 2071-2080.
Li, R. Y., J. L. Stroud, J. F. Ma, S. P. McGrath and F. J. Zhao (2009b): "Mitigation of Arsenic Accumulation in Rice with Water Management and Silicon Fertilization." Environmental Science & Technology 43(10): 3778-3783.
Liu, Y., Y. G. Zhu, B. D. Chen, P. Christie and X. L. Li (2005): "Influence of the arbuscular mycorrhizal fungus Glomus mosseae on uptake of arsenate by the As hyperaccumulator fern Pteris vittata L." Mycorrhiza 15(3): 187-192.
Logoteta, B., X. Y. Xu, M. R. Macnair, S. P. McGrath and F. J. Zhao (2009): "Arsenite efflux is not enhanced in the arsenate-tolerant phenotype of Holcus lanatus." New Phytologist 183(2): 340-348.
Lombi, E., F. J. Zhao, M. Fuhrmann, L. Q. Ma and S. P. McGrath (2002): "Arsenic distribution and speciation in the fronds of the hyperaccumulator Pteris vittata." New Phytologist 156(2): 195-203.
Ma, J. F., N. Yamaji, N. Mitani, X. Y. Xu, Y. H. Su, S. P. McGrath and F. J. Zhao (2008): "Transporters of arsenite in rice and their role in arsenic accumulation in rice grain." Proceedings of the National Academy of Sciences of the United States of America 105(29): 9931-9935.
Ma, L. Q., K. M. Komar, C. Tu, W. H. Zhang, Y. Cai and E. D. Kennelley (2001): "A fern that hyperaccumulates arsenic " Nature 411(6836): 438-U433.
Marin, A. R., P. H. Masscheleyn and W. H. Patrick (1992): "The influence of chemical form and concentration of arsenic on rice growth and tissue arsenic concentration." Plant and Soil 139(2): 175-183.
Marin, A. R., P. H. Masscheleyn and W. H. Patrick (1993): "Soil redox-pH stability of arsenic species and its influence on arsenic uptake by rice." Plant and Soil 152(2): 245-253.
Marschner, H. (2002): Mineral Nutrition of Higher Plants. Amsterdam Boston Heidelberg, Academic Press.
Mattusch, J. and R. Wennrich (1998): "Determination of anionic, neutral, and cationic species of arsenic by ion chromatography with ICPMS detection in environmental samples." Analytical Chemistry 70(17): 3649-3655.
Meharg, A. A. (2003): "Variation in arsenic accumulation - hyperaccumulation in ferns and their allies." New Phytologist 157(1): 25-31.
Meharg, A. A., J. Bailey, K. Breadmore and M. R. Nacnair (1994a): "Biomass allocation, phosphorus-nutrition and vesicular-arbuscular mycorrhizal infection clones of Yorkshire fog, Holcus lanatus L. (Poaceae) that differ in their phosphate-uptake kinetics and tolerance to arsenate." Plant and Soil 160(1): 11-20.
Meharg, A. A., J. W. G. Cairney, A. H. Fitter and D. G. Raffaelli (1999): "Co-evolution of mycorrhizal symbionts and their hosts to metal-contaminated environments." Advances in Ecological Research 30: 69-112.
Meharg, A. A. and J. Hartley-Whitaker (2002): "Arsenic uptake and metabolism in arsenic resistant and nonresistant plant species." New Phytologist 154(1): 29-43.
Meharg, A. A. and M. R. Macnair (1994): "Relationship between plant phosphorus status and the kinetics of arsenate influx in clones of Deschampsia cespitosa (L.) Beauv. that differ in their tolerance to arsenate." Plant and Soil 162(1): 99-106.

LABO-Vorhaben B 1.10 31.5.2012
98
Meharg, A. A., J. Naylor and M. R. Macnair (1994b): "Phosphorus-nutrition of arsenate-tolerant and nontolerant phenotypes of Velvetgrass." Journal of Environmental Quality 23(2): 234-238.
Mihucz, V. G., E. Tatar, I. Virag, E. Cseh, F. Fodor and G. Zaray (2005): "Arsenic speciation in xylem sap of cucumber (Cucumis sativus L.)." Analytical and Bioanalytical Chemistry 383(3): 461-466.
Mkandawire, M. and E. G. Dudel (2005): "Accumulation of arsenic in Lemna gibba L. (duckweed) in tailing waters of two abandoned uranium mining sites in Saxony, Germany." Science of the Total Environment 336(1-3): 81-89.
Mkandawire, M., B. Taubert and E. G. Dudel (2006): "Limitations of growth-parameters in Lemna gibba bioassays for arsenic and uranium under variable phosphate availability." Ecotoxicology and Environmental Safety 65(1): 118-128.
Mylona, P. V., A. N. Polidoros and J. G. Scandalios (1998): "Modulation of antioxidant responses by arsenic in maize." Free Radical Biology and Medicine 25(4-5): 576-585.
Nissen, P. and A. A. Benson (1982): "Arsenic metabolism in freshwater and terrestrial plants." Physiologia Plantarum 54: 446-450.
Norton, G. J., G. L. Duan, T. Dasgupta, M. R. Islam, M. Lei, Y. G. Zhu, C. M. Deacon, A. C. Moran, S. Islam, F. J. Zhao, J. L. Stroud, S. P. McGrath, J. Feldmann, A. H. Price and A. A. Meharg (2009a): "Environmental and Genetic Control of Arsenic Accumulation and Speciation in Rice Grain: Comparing a Range of Common Cultivars Grown in Contaminated Sites Across Bangladesh, China, and India." Environmental Science & Technology 43(21): 8381-8386.
Norton, G. J., M. R. Islam, C. M. Deacon, F. J. Zhao, J. L. Stroud, S. P. McGrath, S. Islam, M. Jahiruddin, J. Feldmann, A. H. Price and A. A. Meharg (2009b): "Identification of low inorganic and total grain arsenic rice cultivars from Bangladesh." Environmental Science & Technology 43(15): 6070-6075.
Overesch, M., L. Düster, K. Greef, J. Rinklebe and T. Mansfeldt (2008). Ermittlung und Beurteilung des Mobilisierungspotenzials von Arsen in Böden. Länderfinanzierungsprogramm "Wasser, Boden und Abfall": 85. Report
Pickering, I. J., L. Gumaelius, H. H. Harris, R. C. Prince, G. Hirsch, J. A. Banks, D. E. Salt and G. N. George (2006): "Localizing the biochemical transformations of arsenate in a hyperaccumulating fern." Environmental Science & Technology 40(16): 5010-5014.
Pickering, I. J., R. C. Prince, M. J. George, R. D. Smith, G. N. George and D. E. Salt (2000): "Reduction and coordination of arsenic in Indian mustard." Plant Physiology 122(4): 1171-1177.
Poynton, C. Y., J. W. W. Huang, M. J. Blaylock, L. V. Kochian and M. P. Elless (2004): "Mechanisms of arsenic hyperaccumulation in Pteris species: root As influx and translocation." Planta 219(6): 1080-1088.
Qin, J., C. R. Lehr, C. G. Yuan, X. C. Le, T. R. McDermott and B. P. Rosen (2009): "Biotransformation of arsenic by a Yellowstone thermoacidophilic eukaryotic alga." Proceedings of the National Academy of Sciences of the United States of America 106(13): 5213-5217.
Quaghebeur, M. and Z. Rengel (2003): "The distribution of arsenate and arsenite in shoots and roots of Holcus lanatus is influenced by arsenic tolerance and arsenate and phosphate supply." Plant Physiology 132(3): 1600-1609.
Raab, A., H. Schat, A. A. Meharg and J. Feldmann (2005): "Uptake, translocation and transformation of arsenate and arsenite in sunflower (Helianthus annuus): formation of arsenic-phytochelatin complexes during exposure to high arsenic concentrations." New Phytologist 168(3): 551-558.
Requejo, R. and M. Tena (2005): "Proteome analysis of maize roots reveals that oxidative stress is a main contributing factor to plant arsenic toxicity." Phytochemistry 66(13): 1519-1528.

LABO-Vorhaben B 1.10 31.5.2012
99
Robinson, B., N. Kim, M. Marchetti, C. Moni, L. Schroeter, C. van den Dijssel, G. Milne and B. Clothier (2006): "Arsenic hyperaccumulation by aquatic macrophytes in the Taupo Volcanic Zone, New Zealand." Environmental and Experimental Botany 58(1-3): 206-215.
Schat, H., M. Llugany, R. Vooijs, J. Hartley-Whitaker and P. M. Bleeker (2002): "The role of phytochelatins in constitutive and adaptive heavy metal tolerances in hyperaccumulator and non-hyperaccumulator metallophytes." Journal of Experimental Botany 53(379): 2381-2392.
Schmöger, M. E. V., M. Oven and E. Grill (2000): "Detoxification of arsenic by phytochelatins in plants." Plant Physiology 122(3): 793-801.
Serfling, A. and R. Klose (2008): "Arsentransfer Boden-Pflanze - Entwicklung von Maßnahmen zur Verhinderung des Arsentransfers im System Boden-Pflanze." Schriftenreihe des Landesamtes für Umwelt, Landwirtschaft und Geologie 32: 1-37.
Sharples, J. M., A. A. Meharg, S. M. Chambers and J. W. G. Cairney (2000a): "Evolution - Symbiotic solution to arsenic contamination." Nature 404(6781): 951-952.
Sharples, J. M., A. A. Meharg, S. M. Chambers and J. W. G. Cairney (2000b): "Mechanism of arsenate resistance in the ericoid mycorrhizal fungus Hymenoscyphus ericae." Plant Physiology 124(3): 1327-1334.
Siegel, S. (1985): Nichtparametrische statistische Methoden. Eschborn, Fachbuchhandlung für Psychologie GmbH - Verlagsabteilung.
Singh, N., L. Q. Ma, M. Srivastava and B. Rathinasabapathi (2006): "Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L and Pteris ensiformis L." Plant Science 170(2): 274-282.
Sneller, F. E. C., L. M. Van Heerwaarden, F. J. L. Kraaijeveld-Smit, W. M. Ten Bookum, P. L. M. Koevoets, H. Schat and J. A. C. Verkleij (1999): "Toxicity of arsenate in Silene vulgaris, accumulation and degradation of arsenate-induced phytochelatins." New Phytologist 144(2): 223-232.
Sterner, R. W. and J. J. Elser (2002): Ecological Stoichiometry - The Biology of Elements form Molecules to the Biosphere. Princeton, Princeton University Press.
Ullrich-Eberius, C. I., A. Sanz and A. J. Novacky (1989): "Evaluation of arsenate-associated and vanadate-associated changes of electrical membrane-potential and phosphate-transport in Lemna gibba G1." Journal of Experimental Botany 40(210): 119-128.
Ultra, V. U., S. Tanaka, K. Sakurai and K. Iwasaki (2007): "Effects of arbuscular mycorrhiza and phosphorus application on arsenic toxicity in sunflower (Helianthus annuus L.) and on the transformation of arsenic in the rhizosphere." Plant and Soil 290(1-2): 29-41.
Vetterlein, D., K. Szegedi, J. Ackermann, J. Mattusch, H. U. Neue, H. Tanneberg and R. Jahn (2007): "Competitive mobilization of phosphate and arsenate associated with goethite by root activity." Journal of Environmental Quality 36(6): 1811-1820.
Wang, J. R., F. J. Zhao, A. A. Meharg, A. Raab, J. Feldmann and S. P. McGrath (2002): "Mechanisms of arsenic hyperaccumulation in Pteris vittata. Uptake kinetics, interactions with phosphate, and arsenic speciation." Plant Physiology 130(3): 1552-1561.
Weber, F. A., A. F. Hofacker, A. Voegelin and R. Kretzschmar (2010): "Temperature Dependence and Coupling of Iron and Arsenic Reduction and Release during Flooding of a Contaminated Soil." Environmental Science & Technology 44(1): 116-122.
Wei, C. Y. and T. B. Chen (2006): "Arsenic accumulation by two brake ferns growing on an arsenic mine and their potential in phytoremediation." Chemosphere 63(6): 1048-1053.
Wenzel, W. W., N. Kirchbaumer, T. Prohaska, G. Stingeder, E. Lombi and D. C. Adriano (2001): "Arsenic fractionation in soils using an improved sequential extraction procedure." Analytica Chimica Acta 436(2): 309-323.

LABO-Vorhaben B 1.10 31.5.2012
100
Wu, J. H., R. Zhang and R. M. Lilley (2002): "Methylation of arsenic in vitro by cell extracts from bentgrass (Agrostis tenuis): effect of acute exposure of plants to arsenate." Functional Plant Biology 29(1): 73-80.
Xu, X. Y., S. P. McGrath and F. J. Zhao (2007): "Rapid reduction of arsenate in the medium mediated by plant roots." New Phytologist 176: 590-599.
Yamamura, S., M. Watanabe, N. Yamamoto, K. Sei and M. Ike (2009): "Potential for microbially mediated redox transformations and mobilization of arsenic in uncontaminated soils." Chemosphere 77(2): 169-174.
Zhang, J., Y. G. Zhu, D. L. Zeng, W. D. Cheng, Q. Qian and G. L. Duan (2008): "Mapping quantitative trait loci associated with arsenic accumulation in rice (Oryza sativa)." New Phytologist 177(2): 350-355.
Zhao, F. J., S. J. Dunham and S. P. McGrath (2002): "Arsenic hyperaccumulation by different fern species." New Phytologist 156(1): 27-31.
Zhao, F. J., J. F. Ma, A. A. Meharg and S. P. McGrath (2009): "Arsenic uptake and metabolism in plants." New Phytologist 181(4): 777-794.
Zhao, F. J., S. P. McGrath and A. A. Meharg (2010): "Arsenic as a Food Chain Contaminant: Mechanisms of Plant Uptake and Metabolism and Mitigation Strategies." Annual Review of Plant Biology 61: 535-559.
Zhu, Y. G., G. X. Sun, M. Lei, M. Teng, Y. X. Liu, N. C. Chen, L. H. Wang, A. M. Carey, C. Deacon, A. Raab, A. A. Meharg and P. N. Williams (2008): "High percentage inorganic arsenic content of mining impacted and nonimpacted Chinese rice." Environmental Science & Technology 42(13): 5008-5013.
8 ABKÜRZUNGSVERZEICHNIS
AN Ammoniumnitrat
BY Bayern
BW Baden-Württemberg
BB Brandenburg
HH Hamburg
HE Hessen
KW Königswasser
MV Mecklenburg-Vorpommern
NI Niedersachsen
NW Nordrhein-Westfalen
RP Rheinland-Pfalz
SL Saarland
SN Sachsen
ST Sachsen-Anhalt
SH Schleswig-Holstein
TH Thüringen
TM / TG Trockenmasse / Trockengewicht