
BBBBBBBBB - europarl.europa.eu · B Landwirtschaft und ländliche Entwicklung Kultur und Bildung...
76
B Landwirtschaft und ländliche Entwicklung Kultur und Bildung Regionale Entwicklung Verkehr und Fremdenverkehr GENERALDIREKTION INTERNE POLITIKBEREICHE FACHABTEILUNG STRUKTUR- UND KOHäSIONSPOLITIK Fischerei
Transcript of BBBBBBBBB - europarl.europa.eu · B Landwirtschaft und ländliche Entwicklung Kultur und Bildung...

B
Landwirtschaft und ländliche Entwicklung
Kultur und Bildung
Fischerei
Regionale Entwicklung
Verkehr und Fremdenverkehr
B Rolle
Die Fachabteilungen sind Forschungsreferate, die die Ausschüsse, interparlamentarischen Delegationen und andere parlamentarische Einrichtungen beraten.
PolitikbereicheLandwirtschaft und ländliche EntwicklungKultur und BildungFischereiRegionale EntwicklungVerkehr und Fremdenverkehr
DokumenteSiehe Website des Europäischen Parlaments: http://www.europarl.europa.eu/studies
FachabteilungStruktur- und kohäSionSpolitik
Bildnachweise: istock international inc., Photodisk, Phovoir
Generaldirektion interne Politikbereiche
Generaldirektion interne Politikbereiche
FachabteilungStruktur- und kohäSionSpolitik
Verkehr und Fremdenverkehr
Regionale Entwicklung
Fischerei
Kultur und Bildung
Landwirtschaft und ländliche Entwicklung
DRAFT ! DRAFT ! DRAFT ! DRAFT ! DRAFT ! DRAFT PRE-RELEASE! PRE-RELEASE! PRE-RELEASE!


GENERALDIREKTION INTERNE POLITIKBEREICHE DER UNION
FACHABTEILUNG B: STRUKTUR- UND KOHÄSIONSPOLITIK
FISCHEREI
FÖRDERT DIE ÜBERFISCHUNG DAS AUFTRETEN
VON ALGENBLÜTEN?
THEMENPAPIER

Dieses Dokument wurde vom Fischereiausschuss des Europäischen Parlaments angefordert. VERFASSER Britas Klemens ERIKSSON Department of Marine Benthic Ecology and Evolution, Centre for Ecological and Evolutionary Studies, University of Groningen Niederlande VERANTWORTLICHE VERWALTUNGSBEAMTIN Irina POPESCU Fachabteilung Struktur- und Kohäsionspolitik Europäisches Parlament E-Mail: [email protected] REDAKTIONSASSISTENZ Virginija KELMELYTE SPRACHFASSUNGEN Original: EN Übersetzungen: DE, ES, FR, IT, PT ÜBER DEN HERAUSGEBER Kontakt zur Fachabteilung oder Bestellung des monatlichen Newsletters: [email protected] Redaktionsschluss: November 2011. Brüssel, © Europäisches Parlament, 2011. Dieses Dokument ist im Internet abrufbar unter: http://www.europarl.europa.eu/studies HAFTUNGSAUSSCHLUSS Die hier vertretenen Auffassungen geben die Meinung des Verfassers wieder und entsprechen nicht unbedingt dem offiziellen Standpunkt des Europäischen Parlaments. Nachdruck und Übersetzung – außer zu kommerziellen Zwecken – mit Quellenangabe gestattet, sofern der Herausgeber vorab unterrichtet und ihm ein Exemplar übermittelt wird.

GENERALDIREKTION INTERNE POLITIKBEREICHE DER UNION
FACHABTEILUNG B: STRUKTUR- UND KOHÄSIONSPOLITIK
FISCHEREI
FÖRDERT DIE ÜBERFISCHUNG DAS AUFTRETEN
VON ALGENBLÜTEN?
THEMENPAPIER
Inhalt In diesem Themenpapier werden wissenschaftliche Hinweise auf einen Zusammenhang zwischen Überfischung und Algenblüte präsentiert. Außerdem werden mehrere europäische Fallstudien vorgestellt, die diese Hypothese untermauern. Die Überfischung trägt zum wachsenden Problem der Algenblüte in Europa bei. Die Übernutzung von Beständen vor der Küste hat zur Veränderung der Struktur vieler Meeresökosysteme geführt, was wiederum die Bildung von Algenbiomasse fördert. Inzwischen greifen die Auswirkungen der Überfischung von den küstennahen Nahrungsnetzen auf die Küstenökosysteme über und verursachen eine Beeinträchtigung der Wasserqualität sowie den Verlust von Lebensräumen im küstennahen Bereich.
IP/B/PECH/IC/2011-105 November 2011 PE 474.461 DE


Fördert die Überfischung das Auftreten von Algenblüten?
INHALTSVERZEICHNIS
LISTE DER ABKÜRZUNGEN 5
VERZEICHNIS DER ABBILDUNGEN 7
TABELLENVERZEICHNIS 9
ZUSAMMENFASSUNG 11
1. EINLEITUNG 15
2. ARTEN UND AUSWIRKUNGEN DER ALGENBLÜTE 19
2.1. Was bedeutet Algenblüte? 19 2.2. Umweltauswirkungen der Algenblüte 19 2.3. Schädliche Algengruppen 20 2.4. Allgemeine gesellschaftliche Probleme und Kosten 22 2.5. Konkrete Auswirkungen auf die Fischereien in Europa 23
3. DIE ÖKOLOGIE DER ALGENBLÜTEN 27
3.1. Ein natürlicher Prozess, der aufgrund der Eutrophierung zunimmt 27 3.2. Der Einfluss von Nährstoffen und Algenfraß auf die Algenblüte 30 3.3. Rückgang der Spitzenprädatoren bei gleichzeitiger Algenzunahme 32 3.4. Abhängigkeit trophischer Kaskaden von Ressourcen und
Prädatorenvielfalt 34
4. TRÄGT DIE ÜBERFISCHUNG ZUM AUFTRETEN VON ALGENBLÜTEN IN EUROPA BEI? 37
4.1. Darstellung der Auswirkungen von Überfischung und Eutrophierung im Modell 37
4.2. Fallstudie: Nordsee 40 4.3. Fallstudie: Schwarzes Meer 43 4.4. Fallstudie: Kattegat 46 4.5. Fallstudie: Ostsee im küstennahen Bereich 50 4.6. Fallstudie: Ostsee im Küstenbereich 54
5. SCHLUSSFOLGERUNGEN UND EMPFEHLUNG 59
LITERATURVERZEICHNIS 63
3

Fachabteilung B: Struktur- und Kohäsionspolitik
4

Fördert die Überfischung das Auftreten von Algenblüten?
LISTE DER ABKÜRZUNGEN
ASP Amnesic Shellfish Poisoning (amnestische Schalentiervergiftung)
CFP Ciguatera Fish Poisoning (Ciguatera-Fischvergiftung)
CPUE Catch Per Unit Effort (Fang je Aufwandseinheit)
DSP Diarrhetic Shellfish Poisoning (diarrhöische Schalentiervergiftung)
EMB Ecosystem-Based Management (ökosystembasierte Bewirtschaftung)
FK Fish Killing algal blooms (Fischsterben durch Algenblüten)
GLM General Linear Model (allgemeines Linerarmodell)
HAB Harmful Algal Bloom (schädliche Algenblüte)
HBT High Biomass Toxic algal blooms (Bildung großer Biomassemengen in Form giftiger Algen)
HBNT High Biomass Non-Toxic algal blooms (Bildung großer Biomassemengen in Form ungiftiger Algen)
NAO Nordatlantikoszillation
PSP Paralytic Shellfish Poisoning (paralytische Schalentiervergiftung)
NSP Neurotoxic Shellfish Poisoning (neurotoxische Schalentiervergiftung)
ST Seafood Toxic algal blooms (für Meeresfrüchte toxische Algenblüten)
5

Fachabteilung B: Struktur- und Kohäsionspolitik
6

Fördert die Überfischung das Auftreten von Algenblüten?
VERZEICHNIS DER ABBILDUNGEN Abbildung 1: Potenzielle Auswirkungen der Überfischung auf die Struktur mariner Nahrungsnetze, die einen Zusammenhang zwischen Fischerei und Algenblüten belegen 16
Abbildung 2: Gruppen von Organismen, die schädliche Algenblüten auslösen, und verschiedene Arten von schädlichen Algenblüten 19
Abbildung 3: Anstieg des Sommerpeaks bei Algenbiomasse im zeitlichen Verlauf im Schwarzen Meer 27
Abbildung 4: Dauer der Algenblütensaison im Wattenmeer 29
Abbildung 5: Räumliche Ausdehnung zum Höhepunkt der durch Cyanobakterien ausgelösten Algenblüte in der Ostsee im Zeitraum 1997 bis 2009. 30
Abbildung 6: Der relative Einfluss von Grasern auf die Algenbiomasse in benthischen und pelagischen Systemen 32
Abbildung 7: Fischereibedingte Rückgänge von Spitzenprädatoren fallen mit sprunghaften Steigerungen des ständigen Algenbestands im Nordatlantik, in der Ostsee und im Schwarzen Meer zusammen. 33
Abbildung 8: Die durch Nährstoffanreicherung bedingte erhöhte Biomasseproduktion findet je nach Anzahl der Trophieebenen im Ökosystem auf verschiedenen trophischen Ebenen statt. 36
Abbildung 9: Generisches Nahrungsnetz eines küstennahen pelagischen Ökosystems in einem Gebiet mit gemäßigtem Klima 38
Abbildung 10: Generisches Nahrungsnetz eines küstennahen pelagischen Ökosystems in einem Gebiet mit gemäßigtem Klima 39
Abbildung 11: Trends bei den Anlandungen großer und kleiner Fische aus der Nordsee 41
Abbildung 12: Trends bei den Anlandungen großer pelagischer Arten aus der Nordsee 41
Abbildung 13: Trends bei den Anlandungen großer demersaler Arten aus der Nordsee 42
Abbildung 14: Trends bei der Algendichte in der Nordsee 42
Abbildung 15: Trends bei den Anlandungen aus der mittleren trophischen Ebene der Nordsee 42
Abbildung 16: Trends bei den Anlandungen von Thun- und Fächerfischen in russischen, rumänischen und türkischen Schwarzmeerfischerei 45
Abbildung 17: Trends bei den Anlandungen kleiner und mittelgroßer pelagisches Arten aus dem Schwarzen Meer 45
7

Fachabteilung B: Struktur- und Kohäsionspolitik
Abbildung 18: Trends bei der mittleren trophischen Ebene der Anlandungen aus dem Schwarzen Meer 45
Abbildung 19: Trends bei der relativen Biomasse von Zooplankton und Phytoplankton im Schwarzen Meer 46
Abbildung 20: Trends bei Anlandungen von Kabeljau und anderen großen Raubfischen im Kattegat 48
Abbildung 21: Trends bei der Gesamtbiomasse von Kabeljau im Kattegat 48
Abbildung 22: Trends bei der Dichte von Strandkrabben und mittelgroßen Fischen, die im Rahmen der Küstenüberwachung in Ringhals am Kattegat ermittelt wurde 48
Abbildung 23: Verhältnis zwischen küstenfernen Kabeljaupopulationen und Küstenpopulationen mittelgroßer Räuber = Gesamtdichte von Strandkrabben, Lippfischen und Schwarzgrundeln 49
Abbildung 24: Verhältnis zwischen Frühjahrstemperaturen und Küstenpopulationen mittelgroßer Räuber = Gesamtdichte von Strandkrabben, Lippfischen und Schwarzgrandeln 49
Abbildung 25: Trends bei dominierenden Fischpopulationen in der Ostsee 51
Abbildung 26: Zoo- und Phytoplanktonbiomasse in der Gotlandsee, südliche Ostsee 51
Abbildung 27: Intensität der Frühjahrsblüte in der Gotlandsee, südliche Ostsee 51
Abbildung 28: Relative Bedeutung verschiedener biologischer Faktoren und Umgebungsfatoren für die Bestimmung der Sprottendichte in der Ostsee. GLM-Modellierung auf der Grundlage einer absteigenden Selektion mit Mallows Cp-Statistik als Auswahlkriterien (Näheres siehe Originalpublikation) 52
Abbildung 29: Relative Bedeutung verschiedener biologischer Faktoren und Umgebungsfatoren für die Bestimmung der Dichte von Zooplankton und Phytoplankton in der südlichen Ostsee. GLM-Modellierung auf der Grundlage einer absteigenden Selektion mit Mallows Cp-Statistik als Auswahlkriterien (Näheres siehe ursprüngliche Publikation) 53
Abbildung 30: Rückgang beim Barsch an der schwedischen Küste in der südlichen Ostsee 55
Abbildung 31: Rückgang beim Hecht an der schwedischen Küste in der südlichen Ostsee 55
Abbildung 32: Trends bei der Dichte größerer Küstenräuber im Kalmarsund in der südlichen Ostsee 56
Abbildung 33: Trends bei der Dichte von Kabeljau, Sprotten und Stichlingen in küstenfernen Gebieten 56
Abbildung 34: Stichlingsdichte je Fang mit Schleppnetz von Hand und prozentualer Anteil der mit filamentösen Algen zugewucherten Fläche in Abhängigkeit von der Dichte der Spitzenprädatoren 57
Abbildung 35: Entfernung von Barschen und Hechten bei gleichzeitiger Eutrophierung führt zur vermehrten Bildung von blühenden filamentösen Algen 57
8

Fördert die Überfischung das Auftreten von Algenblüten?
TABELLENVERZEICHNIS Tabelle 1: Berichte über Algenblüten im Mittelmeer und im Schwarzen Meer 28
Tabelle 2: Verhältnis zwischen größeren pelagischen Räubern (z. B. Biomasse von Thun- und Fächerfischen) und niedrigeren trophischen Ebenen im Schwarzen Meer im Zeitverlauf 46
Tabelle 3. Verhältnis zwischen benachbarten trophischen Ebenen in der Ostsee im Zeitraum 1974-2006. Die Angaben zur Biomasse von Kabeljau und Sprotten wurden den Gesamtschätzungen für die Ostsee entnommen, die Zahlen für die Zoo- und Phytoplanktonbiomasse beruhen auf Überwachungsdaten aus der südlichen Ostsee (Gotlandsee) 52
9

Fachabteilung B: Struktur- und Kohäsionspolitik
10

Fördert die Überfischung das Auftreten von Algenblüten?
ZUSAMMENFASSUNG
Hintergrund Algenblüte ist ein Sammelbegriff für das schnelle und übermäßige Wachstum von Algen und photosynthetisierenden Bakterien. Die für Algenblüten verantwortlichen spezifischen Organismen sind an sich nur entfernt miteinander verwandt und gehören zu einer höchst vielschichtigen Gruppe von photosynthetisch aktiven Wasserorganismen. Algenblüten können aufgrund ihrer negativen Auswirkungen auf wertvolle Teile von Ökosystemen ein erhebliches Problem für den Menschen darstellen – in solchen Fällen wird von einer „schädlichen Algenblüte“ (HAB) gesprochen. Die vorliegenden Daten für europäische Meeresgewässer zeigen, dass die sozioökonomischen Folgen schädlicher Algenblüten insgesamt auf mindestens 850 Mio. EUR pro Jahr zu beziffern sind. Diese Zahl schließt jedoch nicht alle Länder Europas ein, und nicht erfasste Fälle bleiben ebenfalls unberücksichtigt. Die Häufigkeit von Algenblüten hat mit der globalen Eutrophierung zugenommen, und in Europa kommen die negativen Effekte schädlicher Algenblüten seit den 1950er Jahren immer stärker zum Tragen. Die nachteiligen Auswirkungen von Algenblüten können grob in zwei Gruppen unterteilt werden.
Dichte Algenblüten: Große sichtbare Ansammlungen von Algen oder Bakterien, die eine Verfärbung des Wassers bewirken und Störungen aufgrund ihres bloßen Wucherns verursachen. Große Biomasseansammlungen in Form von Algen können sich in unterschiedlicher Weise nachteilig auf das Ökosystem auswirken, so beispielsweise durch die Entstehung treibender Algenteppiche und eine übermäßige Belastung durch Algenschleim, der das Wasser trübt und zusammen mit Schaum an den Stränden abgelagert wird, oder auch durch eine anschließende Anoxie infolge der Zersetzung großer Mengen organischer Materie.
Toxinproduzenten: Hochpotente Toxine werden von einer Gruppe einzelliger Algen produziert, wobei die Gruppe der Dinoflagellaten die bedeutendste Rolle spielt. Diese Toxine sind für eine hohe Mortalität bei Fischen und Schalentieren verantwortlich und können sich bei Akkumulation im Nahrungsnetz massiv auf die menschliche Gesundheit auswirken, insbesondere in Form von Schalentiervergiftungen. Die für derartige Vergiftungen verantwortlichen Algenblüten sind oft nicht mit bloßem Auge erkennbar.
Andere Arten haben beträchtliche direkte Auswirkungen auf Tier und Mensch, wobei hier eine Kombination aus dichter Algenblüte und geringer Toxizität zum Tragen kommt.
In den vergangenen zehn Jahren hat sich zunehmend die Erkenntnis durchgesetzt, dass die Überfischung mitverantwortlich für das Problem der Algenblüte sein könnte. In etlichen marinen Systemen gingen die Entstehung größerer Biomassemengen und die Häufung von Algenblüten mit dramatischen Rückgängen der kommerziell befischten Bestände an größeren Raubfischen einher. Eine Gemeinsamkeit der betroffenen Systeme besteht darin, dass nach den Verlusten an größeren Raubfischen 1) eine signifikante Zunahme bei deren Beutepopulationen, d. h. kleineren Fischen, und 2) eine Abnahme bei Grasern, der Beute kleinerer Fische, die selbst auch Algen fressen, zu verzeichnen war. Daher besteht ein potenzieller Zusammenhang zwischen den rückläufigen Beständen der kommerziell
11

Fachabteilung B: Struktur- und Kohäsionspolitik
befischten Arten und der Zunahme von Algenblüten, die sich aus Veränderungen des Verhältnisses zwischen Raub- und Beutefischen ergibt, wodurch letztlich der Fressdruck auf Algen nachlässt.
Ziel Das allgemeine Ziel dieses Themenpapiers besteht darin, kurz den Hintergrund darzulegen, der für das Verständnis des Zusammenhangs zwischen Algenblüte und Fischerei erforderlich ist, und anhand wissenschaftlicher Erkenntnisse aufzuzeigen, dass die Überfischung zum größer werdenden Problem der Algenblüten beitragen könnte. In dem Themenpapier werden europäische Fallstudien vorgestellt, die Anhaltspunkte für einen Zusammenhang zwischen Algenblüte und Überfischung bieten, und es wird erörtert, inwieweit und unter welchen Bedingungen die Erschöpfung von Fischbeständen Algenblüten verursachen oder fördern kann. In diesem Themenpapier werden vier konkrete Fragen behandelt:
Welche Konsequenzen haben Algenblüten für die Fischerei?
Bestehen Synergieeffekte zwischen Überfischung und Eutrophierung?
Bei welchen Fischarten fördert die Überfischung die Algenblüte?
Welche Arten der Algenblüte stehen mit der Überfischung im Zusammenhang?
Wichtigste Erkenntnisse Schädliche Algenblüten haben negative Auswirkungen auf die Fischerei in Europa. Schätzungen zufolge entstehen der europäischen Fischerei durch schädliche Algenblüten Gesamtkosten von mindestens 177 Mio. EUR pro Jahr. Darin eingeschlossen sind die nachteiligen Auswirkungen auf die kommerzielle Fischerei, soziale Verluste und die Kosten der Überwachung von Algenblüten zwecks Feststellung einer Toxinbelastung von Meeresfrüchten Toxische Algenblüten verursachen hohe Kosten in der Aquakultur. Die direkten Verluste an marktfähigen Erzeugnissen im Muschelsektor belaufen sich auf 15 bis 62 Mio. EUR pro Jahr. Algenblüten bereiten Probleme für jede Art von Küstenfischerei. Treibende Algenteppiche und schleimbildende Algen geraten in die Fanggeräte und verringern deren Wirksamkeit. Die nach einer Algenblüte auftretende Anoxie verursacht eine erhebliche Mortalität unter Aquakultur- und Wildfischbeständen. Zudem beeinträchtigt sie die Reproduktion entlang der Küsten, darunter die wichtigsten Rekrutierungsgebiete zahlreicher Fischpopulationen im küstennahen Bereich. Die Überfischung von größeren, an der Spitze der Nahrungskette stehenden Raubfischen trägt dazu bei, dass Algenblüten häufiger und mit größerer Intensität auftreten. Algenblüten sind Naturphänomene, die durch eine Reihe von Umweltfaktoren beeinflusst werden. Die Überfischung trägt zum Problem der Algenblüte bei, da sie in einen wichtigen biologischen Mechanismus zur Kontrolle übermäßigen Wachstums eingreift. Das Verschwinden von Spitzenprädatoren aufgrund von Überfischung führt zu Veränderungen in der Struktur von Nahrungsnetzen dahingehend, dass die Verbreitung kleinerer Fische generell gefördert wird und die Dichte von Algengrasern wie Zooplankton oder wirbellosen Herbivoren abnimmt. Neben abiotischen Faktoren halten Algengraser den Algenwuchs unter Kontrolle, so dass sich Algenblüten leichter ausbreiten können, wenn bei den Grasern ein Rückgang zu verzeichnen ist. Inzwischen greifen die Auswirkungen der Überfischung von den küstennahen Nahrungsnetzen auf die Küstenökosysteme über und verursachen eine
12

Fördert die Überfischung das Auftreten von Algenblüten?
Beeinträchtigung der Wasserqualität sowie den Verlust von Lebensräumen im küstennahen Bereich. In der Ostsee und im Schwarzen Meer besteht eine sehr hohe Wahrscheinlichkeit, dass die Überfischung zum größer werdenden Problem schädlicher Algenblüten beigetragen hat. Ungeachtet der Komplexität der Ökologie und Regulierung von Algenblüten ergeben sich aus der Analyse von Zeitreihen und zeitgleich auftretenden Ereignissen starke Anhaltspunkte dafür, dass die Übernutzung zur Zunahme von Algenblüten in der Ostsee und im Schwarzen Meer und zum Anstieg der wirtschaftlichen Kosten beigetragen hat, die infolge der umfassenden Veränderungen in der Struktur dieser marinen Ökosysteme entstehen. In der Nordsee hat die Überfischung wahrscheinlich zum größer werdenden Problem schädlicher Algenblüten beigetragen. Die Modellierung und Auswertung verfügbarer Daten zur Nordsee liefert Hinweise, dass die zunehmende Algenbelastung mit fischereibedingten Veränderungen in der Struktur der Fischgemeinschaft zusammenhängen könnte. Für das Kattegat fehlen Daten zu Tendenzen bei der Algenblüte, doch deuten Versuche darauf hin, dass der dokumentierte langfristige Rückgang beim Kabeljau nachteilige Auswirkungen auf Seegrasgemeinschaften hatte, da es zu einer zunehmenden Ausbreitung von filamentösen Algen kam. Allgemein bestehen Probleme, was die Verfügbarkeit von Daten zu europäischen Meeren anbelangt; aufgrund dessen ist eine Einschätzung der Auswirkungen der Fischerei nur bedingt möglich. Es fehlt generell an Trendanalysen für mehr als eine Gruppe von Organismen aus mehreren Meeresbecken in Europa, aus denen Probleme mit der Überfischung und auch mit Algenblüten gemeldet werden. Daraus lässt sich schließen, dass wichtige Informationen fehlen, weil viele europäische Organisationen und Institutionen wertvolle Daten, die für Vergleiche herangezogen werden könnten, nicht veröffentlichen. Überfischung und Eutrophierung erzeugen Synergieeffekte. Veränderungen in Gemeinschaften von Spitzenprädatoren und Algenblüten sind eine Begleiterscheinung der Eutrophierung. Die europaweite Eutrophierung hat die Algenblüte zweifellos begünstigt, doch Veränderungen in der Zusammensetzung der Fischgemeinschaft haben das Problem schädlicher Algenblüten weiter verschärft. Angesichts des heutigen hohen Nährstoffgehalts in der Ostsee lassen sich Trends in der Entwicklung der Algenbiomasse eher auf Trends in der Entwicklung kommerziell befischter Bestände zurückführen als auf die Nährstoffsituation, wobei sich abzeichnet, dass die Überfischung inzwischen in erheblichem Maße zur Algenblüte beiträgt. Das Bestehen von Synergieeffekten zwischen Überfischung und Nährstoffeinträgen wird durch Feldversuche bestätigt, die eindeutig belegen, dass das Wachstum filamentöser Algen sowohl durch die Zufuhr von Nährstoffen als auch durch das Verschwinden von Spitzenprädatoren begünstigt wird und beide Komponenten - Nährstoffeintrag und Raubfischfang – gemeinsam stärker wirken als jede für sich genommen. Die Überfischung dürfte Algenblüten hauptsächlich dann fördern, wenn eine einzige Art befischt wird, die zahlen- und funktionsmäßig in der Fischgemeinschaft eine Vorrangstellung einnimmt. Die Auswirkungen der Entnahme von Raubfischen auf die Beutearten kommen im Allgemeinen wesentlich stärker zum Tragen, wenn die Gemeinschaft der Räuber von einer einzigen Art dominiert wird, wie das bei den demersalen Kabeljau-Populationen in der Ostsee und im Kattegat oder bei pelagischen Anchovis-Gemeinschaften im Schwarzen Meer der Fall ist.
13

Fachabteilung B: Struktur- und Kohäsionspolitik
Der Zusammenhang zwischen dichten Algenblüten und Überfischung ist am deutlichsten ausgeprägt. Die verfügbaren Daten deuten darauf hin, dass es sich bei der nichttoxischen dichten Algenblüte um die Art von Algenblüte handelt, die am stärksten auf eine Überfischung anspricht. Daher dürfte die Überfischung hauptsächlich dichte Algenblüten fördern und Probleme wie 1) treibende Algenteppiche, 2) Schleim- und Schaumbildung und 3) Anoxie nach der Algenblüte hervorrufen. Allerdings zeigen generische Analysen mariner Nahrungsnetze in gemäßigten Zonen, dass eine intensivierte Fischerei, die sich auch auf kleinere Grundfischarten richtet, auch toxische Algenblüten und das vermehrte Auftreten von Quallen fördern kann. Dies spiegelt sich im Nahrungsnetz des Schwarzen Meeres wider, wo eine dramatische Zunahme der Quallenpopulation zu verzeichnen war, nachdem die Intensivbefischung von Anchovis in den 1990er Jahren zusammengebrochen war.
Empfehlung Im vorliegenden Themenpapier wird aufgezeigt, dass die traditionelle Bewirtschaftung von Meeresressourcen gravierende Schwachstellen hat, werden doch oftmals die Interaktionen sowohl innerhalb von Nahrungsnetzen als auch zwischen Nahrungsnetzen vor und an der Küste außer Acht gelassen. Kaskadeneffekte der Überfischung verändern die Beziehungen zwischen Raub- und Beutearten und bewirken damit, dass schädliche Algenblüten häufiger und mit größerer Intensität auftreten; dies wiederum schmälert die Rentabilität der Fischereiindustrie und kann negative langfristige Auswirkungen auf wichtige kommerziell befischte Bestände haben. Allerdings zahlt nicht allein der Fischereisektor den Preis dafür; betroffen sind auch die Küstengemeinden, in denen wachsende Probleme aufgrund von Algenblüten zu einem nennenswerten Rückgang der Tourismuseinnahmen führen und Kosten verursachen, die durch die negativen Auswirkungen auf die menschliche Gesundheit bedingt sind. Es hat sich erwiesen, dass ein ökosystembasiertes Bewirtschaftungskonzept (EBM) benötigt wird, das sowohl 1) den Auswirkungen der Fischerei auf das übrige Ökosystem als auch 2) den Wechselwirkungen zwischen den Lebensräumen vor und an der Küste Rechnung trägt.
14

Fördert die Überfischung das Auftreten von Algenblüten?
1. EINLEITUNG Algenblüte ist ein Sammelbegriff für das schnelle und übermäßige Wachstum von Algen und photosynthetisierenden Bakterien. Algenblüten können aufgrund ihrer negativen Auswirkungen auf wertvolle Teile von Ökosystemen ein erhebliches Problem für den Menschen darstellen – in solchen Fällen wird von einer „schädlichen Algenblüte“ (HAB) gesprochen (Hoagland und Scatasta, 2006). Zu solchen negativen Effekten gehören die Ablagerung von Algen an Stränden und in Fanggeräten, das Entstehen anoxischer Bedingungen, unter denen pelagische Invertrebraten und Grundfische verenden, und die Toxizität von Meerwasser und Nahrung aus dem Meer. Es besteht kein Zweifel daran, dass anthropogene Nährstoffeinträge überall in der Welt mit einem gehäuften Auftreten schädlicher Algenblüten einhergehen (Hallegraeff, 1993; Sellner et al., 2003; Glibert und Burkholder, 2006; Heisler et al., 2008; Smith und Schindler, 2009). In den vergangenen zehn Jahren hat sich jedoch zunehmend die Erkenntnis durchgesetzt, dass die Erschöpfung von Fischbeständen auf den höheren trophischen Ebenen mitverantwortlich für das Problem der Algenblüte sein könnte (Frank et al., 2005; Scheffer et al., 2005; Daskalov et al., 2007; Casini et al., 2008; Moksnes et al., 2008; Eriksson et al., 2009; Eriksson et al., 2011). In etlichen marinen Systemen traten die Entstehung größerer Biomassemengen und die Häufung von Algenblüten zeitgleich mit dramatischen Rückgängen bei den kommerziell befischten Beständen an größeren Raubfischen auf. Eine Gemeinsamkeit der betroffenen Systeme besteht darin, dass nach den Verlusten bei größeren Raubfischen 1) eine signifikante Zunahme bei deren Beutepopulationen, d. h. kleineren Fischen, und 2) eine Abnahme bei Grasern, der Beute kleinerer Fische, die selbst auch Algen fressen, zu verzeichnen war. Daher besteht ein potenzieller Zusammenhang zwischen den rückläufigen Beständen der kommerziell befischten Arten und der Zunahme von Algenblüten, die sich aus Veränderungen des Verhältnisses zwischen Raub- und Beutefischen ergibt, wodurch letztlich der Fressdruck auf Algen nachlässt (Abbildung 1). In diesem Themenpapier wird kurz der wissenschaftliche Hintergrund dargelegt, der für das Verständnis des Zusammenhangs zwischen Algenblüten und Fischerei erforderlich ist, und es werden wissenschaftliche Erkenntnisse vorgestellt, die darauf hindeuten, dass die Überfischung Algenblüten fördert. Algen sind Primärproduzenten an der Basis der meisten marinen Nahrungsnetze und treten durch Energieübertragung in Interaktion mit höheren trophischen Ebenen. Das in Algen gespeicherte Sonnenlicht wird im Nahrungsnetz nach oben weitergeleitet und steuert letztlich die Produktion wertvoller Fischbestände. Diese Beschreibung einer einfachen trophischen Pyramide erscheint möglicherweise trivial, doch handelt es sich bei Pyramiden dieser Art nicht um eine einfache lineare Kette von Abläufen, in der die Menge verfügbarer abiotischer Ressourcen in ein vorbestimmtes organisches Produkt auf höheren Trophieebenen umgewandelt wird. Vielmehr bringen komplexe Interaktionen zwischen verschiedenen biologischen Organismen neue Eigenschaften hervor, die für den Menschen wünschenswert oder nicht wünschenswert sein können. Aus Sicht der Bewirtschaftung gilt es zwei sehr wichtige Merkmale mariner Nahrungsnetze zu erörtern, die die Nachhaltigkeit eines Ökosystems gewährleisten, das wertvolle Bestände kommerziell interessanter Fische hervorbringt: 1) Sämtliche Konsumenten sind auf die Menge und Qualität der Nahrung angewiesen. Daher ergibt sich die Struktur des Systems, d. h. die Produktion und Zusammensetzung der Fischgemeinschaft, nicht nur aus der Dichte, sondern auch aus der Zusammensetzung der Algengemeinschaft. 2) Die oberen Trophieebenen wirken auf die unteren Trophieebenen ein, indem sie sie konsumieren, d. h. die Raubfische dezimieren die Zahl ihrer Beutefische, die wiederum durch das Fressen von Pflanzen und Algen eine Abnahme der primären Biomasse bewirken. Diese in beide Richtungen funktionierenden Interaktionen im Nahrungsnetz haben eine schon lange währende wissenschaftliche
15

Fachabteilung B: Struktur- und Kohäsionspolitik
Debatte darüber ausgelöst, ob Bottom-up-Effekte (Energieübertragung von den Ressourcen an der Basis) oder Top-down-Effekte (Konsumtion auf höheren Trophieebenen) die Fülle an Pflanzen und Algen in verschiedenen Ökosystemen beeinflussen. Heute wissen wir, dass zwischen Konsumenten, Beute und Ressourcen oftmals eine gegenseitige Abhängigkeit besteht, d. h. dass die Akkumulation von Algen durch Erhöhung der Ressourcen von der Intensität des Verbrauchsdrucks abhängt, der von Grasern ausgeht (Worm et al., 2002). Abbildung 1: Potenzielle Auswirkungen der Überfischung auf die Struktur
mariner Nahrungsnetze, die einen Zusammenhang zwischen Fischerei und Algenblüten belegen
phytoplankton
zooplankton
planktivorous fish
piscivorous fish
sustainable fishing
phytoplankton
zooplankton
planktivorous fish
piscivorous fish
overfishingIncreased fishing
pressure
decreases in large predatory fish
increases in smaller predatory fish
decreases in organisms that eat algae ‐ grazers
increases in primary producing algae
Quelle: Entlehnt aus Scheffer et al. (2005), frei nach Frank et al. (2005).
Die dokumentierte Interaktion zwischen aquatischen Ressourcen und Konsumenten hat Anlass zu der Vermutung gegeben, dass durch Überfischung bedingte Veränderungen in marinen Nahrungsnetzen Algenblüten auslösen könnten. Wichtig ist hier der Hinweis, dass das Verschwinden von Konsumenten nicht unbedingt eine Steigerung der Produktivität von Primärproduzenten bedeutet. Die Primärerzeugung wird durch Primärressourcen wie Licht und Nährstoffe geregelt und ist von Umgebungsbedingungen wie der Temperatur abhängig. Die Entnahme von Fischen kurbelt also den Algenwuchs im Allgemeinen nicht an. Darum geht es bei dieser Diskussion gar nicht. Vielmehr hat die Wissenschaft den Nachweis dafür erbracht, dass Konsumenten die Struktur von Nahrungsnetzen innerhalb der Grenzen beeinflussen können, die durch bestimmte Grund- und Umgebungsbedingungen gesetzt werden. Angesichts der Erkenntnis, dass Interaktionen im Nahrungsnetz an der Gestaltung von Ökosystemen beteiligt sind und Auswirkungen auf deren Funktion und Fähigkeit zur Bereitstellung von Dienstleistungen und Gütern haben, gab es einen allgemeinen Ruf nach einer ökosystembasierten Bewirtschaftung (EBM) von Meeresressourcen. Unter EBM ist ein adaptives Bewirtschaftungskonzept zu verstehen, bei dem die Komplexität der Interaktionen in und zwischen ökologischen und sozialen Systemen im Mittelpunkt steht,
16

Fördert die Überfischung das Auftreten von Algenblüten?
wobei der Tatsache Rechnung getragen wird, dass die Vielfalt der Arten und ihrer Merkmale eine wichtige Grundlage für die Leistungsfähigkeit und Stabilität von Ökosystemen ist (Christensen et al., 1996). Bislang liegen nur begrenzte wissenschaftliche Belege für den Einfluss der Fischerei auf die Algenentwicklung vor. Dies ist der Tatsache zuzuschreiben, dass allgemein nur ein geringes Interesse vorhanden ist, die Verbindungen und Beziehungen zwischen den trophischen Ebenen im Rahmen von Meeresüberwachungsprogrammen zu erkunden. Eine Überwachung von Algenblüten und deren Konsumenten (Zooplankton und wirbellose Graser) findet nirgendwo besonders häufig statt, wodurch die Länge verfügbarer Zeitreihen bedenklich beschränkt ist. Auch den kleineren Fischen als wichtigem Bindeglied zwischen größeren, kommerziell interessanten Fischen und Algen wurde seit jeher nur geringe Aufmerksamkeit gewidmet. Zudem sind nur wenige veröffentlichte Ergebnisse oder frei zugängliche Datenbanken vorhanden, die Aufschluss über die Überwachung von Fischgemeinschaften mit Algen und den entsprechenden Herbivoren geben und eindeutige Trendvergleiche zwischen diesen Gruppen zulassen würden. Dies ist auf die Trennlinien zwischen den einzelnen Bewirtschaftungssektoren (z. B. Fischerei/Wasserqualität) und die Abgrenzung zwischen den verschiedenen akademischen Fachbereichen (z. B. Zoologie/Botanik) zurückzuführen und weist darauf hin, dass möglicherweise die Organisationsstruktur der Bewirtschaftung verändert werden muss, um eine angemessene Bewertung der Auswirkungen der Fischerei auf Ebene des Ökosystems zu ermöglichen (Olsson et al., 2008; Osterblom et al., 2010). Viel problematischer ist jedoch, dass Organisationen, die entsprechende Informationen erheben, diese nicht an externe Forscher weitergeben. Organisationsstrukturen sowie private Interessen verhindern also oftmals eine übergreifende Betrachtung von Auswirkungen auf Ökosysteme, bei der mehrere Gruppen von Organismen und Trophieebenen erfasst werden. Gleichwohl deuten immer mehr wissenschaftliche Indizienbeweise darauf hin, dass die Übernutzung kommerziell befischter Bestände unter günstigen Umweltbedingungen die Entwicklung von Algenblüten fördern kann, da ein Bestandteil des Nahrungsnetzes wegfällt, der die Akkumulation von Algenbiomasse eindämmt. In diesem Themenpapier wird eine kurze Erläuterung zur Ökologie von Algenblüten gegeben, soweit dies für das Verständnis der von ihnen ausgehenden Gefahr und ihrer Funktionsweise erforderlich ist. Anschließend werden einschlägige Fallstudien und die jüngsten Forschungen über den Zusammenhang zwischen Algenblüten und Fischerei vorgestellt. Im zweiten Kapitel werden Algenblüten und ihre negativen Folgen für die Gesellschaft behandelt, darunter insbesondere die Auswirkungen von Algenblüten auf die Fischerei. Das dritte Kapitel behandelt die Ökologie von Algenblüten; es werden Tendenzen und Zusammenhänge zwischen Algenblüten und Fischerei aufgezeigt und ökologische Faktoren genannt, die einen solchen Zusammenhang begünstigen können. Kapitel 4 enthält Ausführungen zu möglichen Synergieeffekten von Überfischung, übermäßigen Nährstoffeinleitungen und Algenblüten in Europa und liefert anhand von Fallbeispielen Belege dafür, dass die Überfischung Auslöser von Algenblüten sein kann.
17

Fachabteilung B: Struktur- und Kohäsionspolitik
18

Fördert die Überfischung das Auftreten von Algenblüten?
2. ARTEN UND AUSWIRKUNGEN DER ALGENBLÜTE
2.1. Was bedeutet Algenblüte? Algenblüten werden durch viele verschiedene Algenarten ausgelöst, die nur entfernt miteinander verwandt sind. Bei den Lebensformen kann es sich um Einzeller, koloniebildende oder filamentöse Algen, aber auch um Planktonalgen oder benthische Algen handeln. Zu den weitverbreiteten blühenden Algengruppen zählen Dinoflagellaten, sonstige Flagellaten, Diatomeen, Cyanobakterien sowie filamentöse Grün-, Braun- und Rotalgen (Graneli und Turner, 2006). Aufgrund der breit gefächerten Taxonomie erweist sich eine genaue Definition des Begriffs „Algenblüte“ als schwierig. Es bedarf also einer kontextbezogenen Begriffsbestimmung. Daher können sich bei den Warnstufen für eine Algenblüte in Überwachungsprogrammen Werte ergeben, die von 100-300 Zellen/l im Fall von Dinoflagellaten bis 150-200 000 Zellen/l im Fall von Diatomeen und Cyanobakterien reichen (Weltgesundheitsorganisation, 2002; Hinder et al., 2011). Für die Zwecke dieses Themenpapiers werden Algenblüten als große Algenansammlungen (einschließlich Cyanobakterien) mit schädlichen Auswirkungen auf das umgebende Ökosystem definiert.
2.2. Umweltauswirkungen der Algenblüte Aufgrund der breit gefächerten taxonomischen Zusammensetzung haben Algenblüten auch sehr unterschiedliche schädliche Wirkungen. Allerdings können die negativen Effekte in folgende zwei Hauptarten untergliedert werden: Auswirkungen, die mit der hohen Dichte an sich zusammenhängen, und Toxinproduktion/chemische Effekte (Abbildung 2) (Smayda, 1997; Nastasi, 2010). Abbildung 2: Gruppen von Organismen, die schädliche Algenblüten auslösen, und
verschiedene Arten von schädlichen Algenblüten
Harmful algal blooms
diatoms
dino‐flagellates
flagellatescyano‐bacteria
filamen‐tous algae
Producing toxins/
”red tides”
shellfish poisioning
direct lethal toxicity
ambush predation
Producing high biomass/
”green tides”
mechanical disturbance
shading
anoxia
clogging the water & overgrowth
mucus & foam formation
Quelle: Graneli et al. (1996)
19

Fachabteilung B: Struktur- und Kohäsionspolitik
Dichte Algenblüten, die eine Verfärbung des Wassers bewirken und aufgrund ihres bloßen Wucherns lästig sind, gehen Berichten zufolge auf viele verschiedene Arten von blühenden Algen zurück, doch charakteristische Auslöser sind Cyanobakterien, einige Flagellaten (Phaeocystis, Crysocromulina), Diatomeen und filamentöse Algen (grüne Tide). Große Biomasseansammlungen in Form von Algen können sich in unterschiedlicher Weise nachteilig auf das Ökosystem auswirken. Mechanische Schäden wie durch Partikel ausgelöste Reizungen können Infektionen an den Kiemen von Fischen und Krustentieren hervorrufen, und die Verletzung der Kiemen durch Diatomeen kann sogar zu Atemversagen führen (Smayda, 1997). Wieder andere Organismen können durch extrazelluläre Polymere Schaden nehmen, wenn sich beispielsweise die Kiemen zusetzen und eine übermäßige Belastung durch Algenschleim auftritt, der die Gewässer trübt und zusammen mit Schaum an Stränden abgelagert wird (Smayda, 2006). Bei der Zersetzung der großen Mengen angehäufter organischer Materie nach der Blüte wird möglicherweise der gesamte verfügbare Sauerstoff verbraucht, wodurch eine lokale Anoxie auftritt, die tödlich für alle am Meeresboden lebenden Organismen ist und bewirkt, dass Vertreter der mobilen Fauna wie Fische und Krustentiere flüchten oder verenden (Hallegraeff, 1993; Vahteri et al., 2000; Cloern, 2001; Breitburg, 2002). Zu den Toxinproduzenten gehört eine Gruppe von 60-80 Mikroalgenarten (einzellige Algen), von denen drei Viertel Dinoflagellaten sind (Smayda, 1997; Van Dolah, 2000; Hinder et al., 2011). Algentoxine sind äußerst potent und verursachen eine hohe Mortalität bei Fischen und Schalentieren. Bei Akkumulation im Nahrungsnetz können sie schwere gesundheitliche Schäden hervorrufen, insbesondere in Form von Schalentiervergiftungen. Bei diesen Toxinproduzenten reichen geringe Zelldichten aus, um gefährliche Toxinspiegel zu erreichen, und aus diesem Grund sind toxische Algenblüten oft nicht mit bloßem Auge erkennbar. Cyanobakterien und sonstige Flagellaten (z. B. Chrysocromulina, Prymnesium) lösen hingegen üblicherweise sichtbare Blüten aus und produzieren ebenfalls Toxine mit erheblichen direkten negativen Auswirkungen auf Mensch und Tier (Weltgesundheitsorganisation, 2002; Edvardsen und Imai, 2006; Peaerl und Fulton III, 2006).
2.3. Schädliche Algengruppen Es gibt fünf Hauptkategorien blühender Algen, die aus der Überwachung von Algenblüten und/oder der wissenschaftlichen Literatur zu diesem Thema bekannt sind. Im folgenden Abschnitt werden sie kurz vorgestellt. Detailliertere und umfassendere Informationen sind den Studien von Smayda (2006) sowie Graneli und Turner (2006) zu entnehmen, die sich speziell mit Algen befassen.
2.3.1. Dinoflagellaten Dinoflagellaten sind einzellige Organismen, die aus einer Geißelzelle mit begrenzter, aber dennoch beachtlicher Beweglichkeit bestehen (Burkholder et al., 2006). Sie sind für vier der fünf am häufigsten festgestellten Gruppen von Schalentiervergiftungen verantwortlich: die paralytische Schalentiervergiftung (PSP), die neurotoxische Schalentiervergiftung (NPS), die Ciguatera-Fischvergiftung (CFP) und die diarrhöische Schalentiervergiftung (DSP) (Smayda, 1997; Van Dolah, 2000). In Europa werden Dinoflagellaten-Blüten, die zur Vergiftung von Meeresfrüchten führen, vor allem von den Atlantikküsten gemeldet (Smayda, 2006). Gelegentlich finden sich Berichte über einen Zusammenhang zwischen Dinoflagellaten und durch dichte Algenblüten hervorgerufenen Störungen, wobei als Beispiele die anoxische Blüte von Ceratium tripos in New York, die zum Fischsterben in einem Gebiet von mehr als einer Million Hektar führte, sowie das Blühen anderer
20

Fördert die Überfischung das Auftreten von Algenblüten?
schleimbildender Algen (vor allem Gonyaulax fragilis) im Mittelmeer und im Marmarameer genannt werden können (Tufekci et al., 2010). Karenia mikimotoi ist ein weiterer sehr weit verbreiteter Dinoflagellat, der als große Gefahr für die Aquakultur gilt. Zu den bekannten schädigenden Folgen gehören die hohe Mortalität von Schalentieren an der französischen Küste sowie von Schalentieren und Zuchtlachs in Norwegen (Smayda, 2006). Aus dem Mittelmeerraum werden regelmäßig Probleme mit dichten Blüten der Ostreopsis-Arten gemeldet.
2.3.2. Sonstige Flagellaten Zu den sonstigen Flagellaten, die schädliche Blüten mit erheblichen negativen Auswirkungen auf den Menschen auslösen, zählen hauptsächlich die Klassen Prymnesiophyceae und Raphidophyceae (Edvardsen und Imai, 2006). Prymnesiophyten sind eine Klasse von Flagellaten mit weltweiter Verbreitung, die gelegentlich schädliche Blüten verursachen; in Europa werden schädliche Blüten vor allem aus der Nordsee gemeldet. Prymnesium parvus kann Dichten von bis zu 800 Mio. Zellen/l erreichen (Smayda, 2006) und löst regelmäßig örtlich begrenzte Fischsterben entlang der britischen Küste aus, tritt aber auch in hohen Dichten bis hin in die südliche Ostsee auf. Chrysochromulina polylepis ist ein giftiger Vertreter der Prymnesiophyceae, der ebenfalls zu Fischsterben führt. 1988 richtete die Blüte von C. polylepis an den Atlantikküsten in Schweden und Norwegen in einem 75 000 km2 großen Gebiet schwere Schäden unter wildlebenden Organismen und in der Fischaquakultur an (Maestrini und Graneli, 1991; Granéli et al., 1996; Dahl et al., 2005). Die Algenart Phaeocystis gehört zu den Prymnesiophyceae und blüht immer wieder im Nordatlantik, wobei die Blüte mit der Bildung großer Schleimmengen einhergeht, die als auf der Wasseroberfläche schwimmende und an die Strände gespülte Schaumklumpen sichtbar sind (Lancelot et al., 1998; Smayda, 2006). 2001 verendeten 10 Mio. kg Muscheln infolge einer durch die Blüte von Phaeocystis ausgelösten Anoxie in der Oosterschelde, einem Meeresarm an der niederländischen Atlantikküste (Peperzak und Poelman, 2008). Die Raphidophyten umfassen drei Taxa, die als Auslöser für Fischsterben bekannt sind: Chattonella-Arten, Heterosigma akashiwo und Fibrocapsa japonica. Meldungen über Fischsterben aufgrund der Blüte von Raphidophyten liegen sowohl von der Adriaküste als auch von den französischen, niederländischen und britischen Küsten vor (de Boer, 2006). Mehrere Massensterben von Fischen in Nordeuropa wurden zunächst mit Chattonella verruculosa in Verbindung gebracht; später stellte sich jedoch heraus, dass der Silicoflagellat Pseudochattonella verruculosa dafür verantwortlich war. Die erste Blüte dieser Art, die zum Fischsterben führte, wurde in April/Mai 1998 im Skagerrak in der Nordsee beobachtet. Seither wurden mehrere Fischsterben in diesem Gebiet auf Pseudochattonella zurückgeführt.
2.3.3. Diatomeen Diatomeen-Blüten sind im Allgemeinen nicht toxisch, doch werden häufig schädliche Auswirkungen auf Zuchtfische in Europa gemeldet, wenn dichte Blüten der ca. 14 Kieselalgenarten auftreten (Smayda, 2006). Diatomeen verfügen über Frusteln – äußere Skelette aus hartem Siliziumdioxid – und weisen oftmals einen Dorn auf, der während der Blüte mechanische Kiemenverletzungen verursachen kann (Smayda, 1997). Häufig wird auch über Folgewirkungen nach der Blüte berichtet, die auf Anoxie und Schleimbildung zurückzuführen sind (Smayda, 2006). Zudem wird die amnestische Schalentiervergiftung (ASP) durch eine Kieselalge (Pseudo-nitzschia multiseries) hervorgerufen. Regelmäßig
21

Fachabteilung B: Struktur- und Kohäsionspolitik
finden sich Berichte über das Auftreten von Pseudo-nitzschia multiseries-Blüten an der europäischen Nordseeküste, die zu vorübergehenden Schließungen der Kamm- und Miesmuschelfischerei führen (Smayda, 2006).
2.3.4. Cyanobakterien Cyanobakterien werden häufig als Blaualgen bezeichnet, doch handelt es sich hierbei um Photosynthese betreibende Bakterien, die Kolonien bilden können. Cyanobakterien-Blüten treten am häufigsten in Brackwasserhabitaten und interkontinentalen Meeren wie der Ostsee, im Marmarameer und in Meeresarmen auf (Peaerl und Fulton III, 2006). Üblicherweise handelt es sich um dichte Blüten mit mehr als 200 000 Einheiten/l, die aber auch Toxine produzieren. Es gibt drei weit verbreitete Gruppen von Cyanotoxinen: Hepatotoxine, die Leberschäden verursachen, Neurotoxine, die das Nervensystem schädigen, und Dermatoxine, die bei Kontakt Reizungen und allergische Reaktionen hervorrufen (Weltgesundheitsorganisation, 2002).
2.3.5. Filamentöse Algen Filamentöse Algen sind für die sogenannten grünen Tiden im küstennahen Bereich verantwortlich und weisen eine Grün-, Rot- oder Braunfärbung auf; sie schließen Arten der Gattungen Cladophora, Ectocarpus, Pilayella, Ulothrix, Ulva und Urospora ebenso ein wie koloniebildende Diatomeen der Gattung Melosira (Eriksson et al., 2009; Eriksson et al., 2011). Sie sind nicht giftig, bilden aber dicht wuchernde Biomasse und riesige treibende Algenteppiche, die benthische Makrophyten verdrängen und in Buchten eine räumlich begrenzte Anoxie auslösen. Zudem setzen sich Blüten filamentöser Algen in Fanggeräten, Booten und Anlagen an Stränden fest und stellen eine Gefahr für den lokalen Tourismus dar. In Europa werden Schäden durch treibende Algenteppiche und Blüten filamentöser Algen vor allem aus der Ostsee, dem Kattegat und dem Skagerrak gemeldet (Bonsdorff et al., 1997; Pihl et al., 1999; Vahteri et al., 2000; Baden et al., 2003; Eriksson et al., 2009; Eriksson et al., 2011), es finden sich aber auch Berichte über Schäden durch Strandablagerungen an der Atlantikküste (Weltgesundheitsorganisation, 2002).
2.4. Allgemeine gesellschaftliche Probleme und Kosten Es gibt eine Reihe erwiesener negativer gesellschaftlicher Auswirkungen von Algenblüten, die wesentliche wirtschaftliche Folgen für Küstengemeinden haben (Hoagland et al., 2002; Weltgesundheitsorganisation, 2002; ECOHARM, 2003; Hoagland und Scatasta, 2006). Toxische Algenblüten lösen beim Menschen bei Berühren, Verschlucken und Einatmen von Sprühtröpfchen gesundheitliche Probleme aus (Weltgesundheitsorganisation, 2002). Dies hat nachteilige wirtschaftliche Folgen für den Fremdenverkehr und die Meeresfrüchteindustrie (Hoagland et al., 2002; ECOHARM, 2003; Hoagland und Scatasta, 2006). Infolge der Zersetzung großer Ansammlungen von Algenbiomasse kommt es zur Anoxie mit erheblichen negativen Auswirkungen auf Fischbestände und Schalentiere (Hallegraeff, 1993; Breitburg, 2002). Fische in frühen Entwicklungsstadien wie Larven und Eier können von einer Anoxie besonders betroffen sein, weil sie nur bedingt bewegungsfähig sind (Breitburg, 2002). Dichte Algenblüten, die mit der Entstehung großer Biomassemengen einhergehen, können die Wirksamkeit von Fanggeräten dadurch beeinträchtigen, dass die Netze schwer und durch die Verstopfung beschädigt werden, wozu die Algenwucherung ebenso beiträgt wie die Schleimbildung (Weltgesundheitsorganisation, 2002). Große Ansammlungen von Algenbiomasse führen auch dazu, dass Küstengewässer und Strände für Menschen unattraktiv werden, weil das
22

Fördert die Überfischung das Auftreten von Algenblüten?
Wasser deutlich getrübt ist und weil durch die Zersetzung großer Mengen an organischem Material ein fauliger Geruch entsteht (Weltgesundheitsorganisation, 2002). Die Europäische Kommission stellte Fördermittel für das Programm ECOHARM (2003) bereit, das der Ermittlung der sozioökonomischen Verluste gewidmet war, die durch schädliche Algenblüten in Europa entstehen. Für die Zwecke von ECOHARM wurden bei schädlichen Algenblüten folgende Kategorien unterschieden: für Meeresfrüchte toxische Blüten (ST), Fischsterben durch Algenblüten (FK), Bildung großer Biomassemengen in Form giftiger Algen (HBT) und Bildung großer Biomassemengen in Form ungiftiger Algen (HBNT). Wie die Ergebnisse zeigen, sind für Meeresfrüchte toxische Blüten die am häufigste gemeldete HAB-Störung in Europa, gefolgt von der Bildung großer Biomassemengen in Form ungiftiger Algen. Im Rahmen von ECOHARM wurde errechnet, dass sich die durch HAB in Europa pro Jahr verursachten wirtschaftlichen Verluste auf durchschnittlich 813 Mio. USD (USD von 2005) belaufen. Dabei erfolgte eine Aufschlüsselung nach Auswirkungen auf die öffentliche Gesundheit (11 Mio. USD), kommerzielle Fischereien (147 Mio. USD), Erholung und Fremdenverkehr (637 Mio. USD) und Kosten für die Überwachung und Bewertung toxischer Blüten (18 Mio. USD) (siehe auch Hoagland und Scatasta, 2006). Die Auswirkungen waren ungleichmäßig auf Europa verteilt, wobei im Rahmen von ECOHARM je nach Art der Blüte, zu der die meisten Meldungen vorlagen, zwischen HBNT- und ST-Ländern unterschieden wurde. Zu den HBNT-Ländern gehörten Deutschland, Estland, Finnland, Frankreich, Lettland, die Niederlande, Polen und Schweden. Dies deutet darauf hin, dass Probleme mit nichttoxischen Algenblüten unter Bildung großer Biomassemengen das größte Problem in den Ostseeländern, einschließlich des Wattenmeers und der Meeresarme an der niederländischen Küste, darstellen. In Frankreich hing das HBNT-Problem hauptsächlich mit Ansammlungen filamentöser Algen an Stränden zusammen, es wurde aber auch eine Vielzahl von für Meeresfrüchte toxischen Blüten gemeldet (fünfthöchste Rate in Europa). Zu den ST-Ländern zählten Dänemark, Irland, Italien, Norwegen, Portugal, Spanien und das Vereinigte Königreich. Dies deutet darauf hin, dass die Vergiftung von Meeresfrüchten nicht nur in Frankreich, sondern überhaupt im Atlantikgebiet und im Mittelmeerraum das Hauptproblem darstellt. Es ist darauf hinzuweisen, dass das Schwarze Meer und das Marmarameer nicht in die Bewertung einbezogen waren.
2.5. Konkrete Auswirkungen auf die Fischereien in Europa Bei den Gesamtkosten, die den Fischereien in Europa durch schädliche Algenblüten entstehen, sind die direkten negativen Auswirkungen auf kommerzielle Fischereien, soziale Verluste und die Kosten berücksichtigt, die für die Überwachung von Algenblüten anfallen, um Meeresfrüchtevergiftungen zu verhindern. Laut ECOHARM (2003) sind die jährlichen Gesamtkosten für die europäischen Fischereien mit geschätzten 177 Mio. EUR zu veranschlagen. Es ist schwierig, bestimmte Arten als Verursacher schädlicher Algenblüten auszumachen, die von den Fischereien mit mehr oder minder großem Aufwand zu bewältigen sind, weil die Kosten für sämtliche schädigenden Ereignisse, die durch die vorstehend genannten Algengruppen verursacht werden, zusammen erfasst werden. Allerdings ist eine Aufschlüsselung bei bestimmten Beispielen, für die eine ausreichende wissenschaftlich Dokumentation vorliegt, nach Art der Auswirkungen und damit eine allgemeine Lageeinschätzung möglich, sofern bedacht wird, dass weitere Gefahren und viele lokale Ausnahmen bestehen.
23

Fachabteilung B: Struktur- und Kohäsionspolitik
2.5.1. Für Meeresfrüchte toxische Algenblüten Algenblüten zählen neben krankheits- und wetterbedingten Schäden zu den drei wichtigsten Ursachen für Verluste in der Aquakultur (Rutter, 2010), und 99 % aller Berichte über für Meeresfrüchte toxische Blüten in Europa betreffen Schalentiere (ECOHARM, 2003). Die direkten Einnahmeverluste des Muschelaquakultursektors belaufen sich in Europa – ohne Kosten für die Überwachung und Sozialausgaben – auf 15 bis 62 Mio. EUR pro Jahr. Für Meeresfrüchte toxische Blüten treten vor allem an den europäischen Atlantik- und Mittelmeerküsten auf, wobei natürlich die Länder mit nennenswerter Aquakultur am stärksten betroffen sind (ECOHARM, 2003). Die größten Probleme bereiten Blüten verschiedener Dinoflagellaten, wie zum Beispiel Dinophysis-Arten, Alexandrium-Arten und Karenia mikimotoi, die Meeresfrüchtevergiftungen auslösen und zum Sterben von wildlebenden Beständen und Zuchtbeständen führen können (Smayda, 2006). Große Besorgnis löst auch die toxinproduzierende Diatomee Pseudo-nitzschia multiseries aus, die regelmäßig an der europäischen Nordseeküste blüht (Smayda, 2006). Alle diese Arten führen dazu, dass die Schalentierfischereien an den europäischen Küsten in regelmäßigen Abständen geschlossen werden. Auch die von den Raphidophyten Heterosigma akashiwo und Fibrocapsa japonica ausgehenden Bedrohungen für Meeresfrüchte rücken zunehmend ins Bewusstsein (de Boer, 2006).
2.5.2. Algenblüten mit Entstehung großer Biomassemengen Die größten Probleme mit Algenblüten, die mit einem hohem Biomasseanfall einhergehen, bestehen offenbar in interkontinentalen Binnenmeeren und Meeresarmen (ECOHARM, 2003). Eine wichtige Ausnahme dürften die Ansammlungen von Pheocystis und filamentösen Algen an den Nordseeküsten Frankreichs, Belgiens und der Niederlande sein (Weltgesundheitsorganisation, 2002). Algenblüten mit Entstehung großer Biomassemengen können Auswirkungen verschiedener Art auf Fischereien haben, darunter mechanische Störungen, Verhungern und Ersticken (siehe oben, Smayda, 1997; Van Dolah, 2000). Allerdings werden in europäischen Fallstudien das Wuchern treibender Algenteppiche und eine übermäßige Schleimbildung sowie Anoxie nach der Blüte am häufigsten als Ursache für erhebliche wirtschaftliche Probleme im Fischereisektor genannt.
Treibende Algenteppiche
Berichte über dokumentierte nachteilige Effekte treibender Algenteppiche auf die Fischerei stammen hauptsächlich aus der Ostseeregion. So meldeten beispielsweise 2005 in Finnland 65 % der Berufsfischer, die mit stationären Fanggeräten (Fischfallen oder Kiemennetze) arbeiten, und 35 % der Fischer, die Schleppnetze einsetzen, dass sie wirtschaftlichen Schaden aufgrund treibender Cyanobakterien-Teppiche erlitten hätten (Viitasalo et al., 2005). Während der Blüte hätte die Reinigung ihrer Ausrüstung viel Zeit in Anspruch genommen, und die Fangmengen seien zurückgegangen, weil die Fische nicht in die mit Algen verstopften Fanggeräte gingen. Treibende Fadenalgenteppiche können ebenfalls zu einem weiträumigen Rückgang der Qualität von Rekrutierungsgebieten für Fische führen, die flache Buchten mit flacher Vegetation als Aufwuchsgebiete nutzen (Pihl et al., 2006; Ljunggren et al., 2010). Die Qualität des Lebensraums hat großen Einfluss auf die Rekrutierung (Alheit und Niquen, 2004; Leggett und Frank, 2008). Im Kattegat und im Skagerrak haben starke Rückgänge der Seegrasbeete (Zostera marina), die durch die zunehmende Ausbreitung filamentöser Algen bedingt sind, Schäden in wichtigen Rekrutierungsgebieten von Populationen des Kabeljau (Gadus morhua) vor der Küste und im küstennahen Bereich angerichtet (Pihl et al., 1999; Baden et al., 2003; Pihl et al., 2006). In den flachen Buchten der Ostsee
24

Fördert die Überfischung das Auftreten von Algenblüten?
beeinträchtigt das Wuchern treibender Fadenalgenteppiche an einigen Stellen die Rekrutierung des Barsches, der an der Küste der vorherrschende Raubfisch ist (Perca fluviatilis) und in Gewässern mit flacher Vegetation laicht (Ljunggren et al., 2010).
Blüte schleimbildender Algen
Negative Auswirkungen der Schleimbildung auf Fanggeräte werden hauptsächlich aus dem Adriatischen Meer und dem Marmarameer gemeldet (Zengin et al., 2009; Turk et al., 2010; Zengın et al., 2011). Im Mittelmeer und im Marmarameer findet sich in Algenblüten, die in Form einer gallertartigen cremigen Masse auftreten, eine Fülle verschiedener Arten, einschließlich Cyanobakterien, Diatomeen und Dinoflagellaten (Tufekci et al., 2010; Turk et al., 2010). Vor allem die Diatomeen Skeletonema costatum und Cylindrotheca closterium und der Dinoflagellat Gonyaulax fragilis zählen zu den gängigsten schleimbildenden Arten (Tufekci et al., 2010). Zwar wird seit Jahrhunderten über Blüten schleimbildender Algen im Adriatischen Meer berichtet, doch seit einigen Jahrzehnten häufen sich die Berichte (Turk et al., 2010). Im Marmarameer gingen die Anlandungen der Fischerei während der heftigen Blüten schleimbildender Algen in den Jahren 2007 und 2008 gegenüber 2006 um fast 50 % von 75 000 Tonnen auf 44 450 bzw. 38 400 Tonnen zurück (Zengin et al., 2009; Zengın et al., 2011). Die Fanggeräte waren aufgrund der mit Quallen vermischten gallertartigen Algenmasse, die sich darin verfangen hatte, so schwer, dass die Netze rissen und nicht eingeholt werden konnten. Davon waren Fischereien jeglicher Art im Marmarameer betroffen, wobei Schätzungen zufolge 91 % des jährlichen Gewinns verloren gingen (Zengin et al., 2009; Zengın et al., 2011).
Anoxie nach der Algenblüte
Anoxie tritt vor allem in interkontinentalen Binnenmeeren und Meeresarmen auf. Eine Abnahme des Sauerstoffangebots aufgrund der Zersetzung großer Algenbiomassemengen tritt schneller ein, wenn sich im Sommer eine Dichtestratifizierung zeigt, die den Wasseraustausch am Meeresboden behindert (Cloern, 2001; Breitburg, 2002). Die Anoxie stellt eine ernste Bedrohung für sämtliche Aquakulturen und Gemeinschaften benthischer Invertrebraten dar. So verendeten beispielsweise 2001 10 Mio. kg Zuchtmuscheln (Mytilus edulis) infolge einer durch die Blüte von Phaeocystis ausgelösten Anoxie in der Oosterschelde, einem Meeresarm an der niederländischen Atlantikküste; dies verursachte Schätzungen zufolge Verluste in Höhe von 15 bis 20 Mio. EUR (Peperzak und Poelman, 2008). Die Anoxie gefährdet aber auch wildlebende Fischbestände. In der Regel weichen erwachsene Fische an die Gewässeroberfläche aus, wenn Anoxie am Grund auftritt (Breitburg, 2002). Gelegentlich sind jedoch auch flache Gewässer von der Sauerstofferschöpfung betroffen. 1997 griff eine durch die Blüte des Dinoflagellaten Prorocentrum minimum ausgelöste Anoxie auf einer Länge von 25 km rasch auf die gesamte Wassersäule im Mariager-Fjord in Norddänemark über und führte zum Tod von Fischen und Invertrebraten (Fallesen et al., 2000). Anoxie stellt eine potenziell große Gefahr für die Rekrutierung wildlebender Fischbestände dar. Treibende Algenteppiche können in flachen Buchten zu einer lokalen Anoxie mit anschließendem Sterben der Eier und Larven von Küstenfischen führen (Pihl et al., 1996; Ljunggren et al., 2010). Außerdem können die Eier pelagischer Arten in sauerstofflose Schichten des Gewässers absinken (Breitburg, 2002). Vor allem aufgrund der Stratifizierung der Ostsee können Kabeljaueier nur dann überleben, wenn der Salzgehalt des Wassers so hoch ist, dass die Eier schwimmfähig sind (und nicht auf den Grund
25

Fachabteilung B: Struktur- und Kohäsionspolitik
sinken), und wenn der Sauerstoffgehalt ausreichend hoch ist. Dies wird als Reproduktionsvolumen eines Gewässers bezeichnet (Österblom et al., 2007). Während der massiven Rückgänge des Kabeljaubestands Ende der 1980er Jahre wiesen die Bodenwasserschichten einen sehr niedrigen Salzgehalt auf und 95 % der Kabeljaueier fanden sich in Schichten mit einem Sauerstoffgehalt, der für ein Überleben nicht ausreichte (Breitburg, 2002; Österblom et al., 2007). Der Salzgehalt und der Sauerstoffgehalt des Ostseewassers hängen vom Zustrom sauerstoffreichen Wassers mit hohem Salzgehalt aus dem Atlantik ab, aber die Zunahme des Anteils an sauerstofflosem Wasser am Meeresgrund wird auch durch die umfassende Zersetzung von Plankton gefördert (HELCOM, 2009). Algenblüten tragen also möglicherweis direkt zu der rückläufigen Rekrutierung von Kabeljau bei, doch wurde dieser Zusammenhang bislang kaum erforscht.
26
Fördert die Überfischung das Auftreten von Algenblüten?
3. DIE ÖKOLOGIE DER ALGENBLÜTEN
3.1. Ein natürlicher Prozess, der aufgrund der Eutrophierung zunimmt
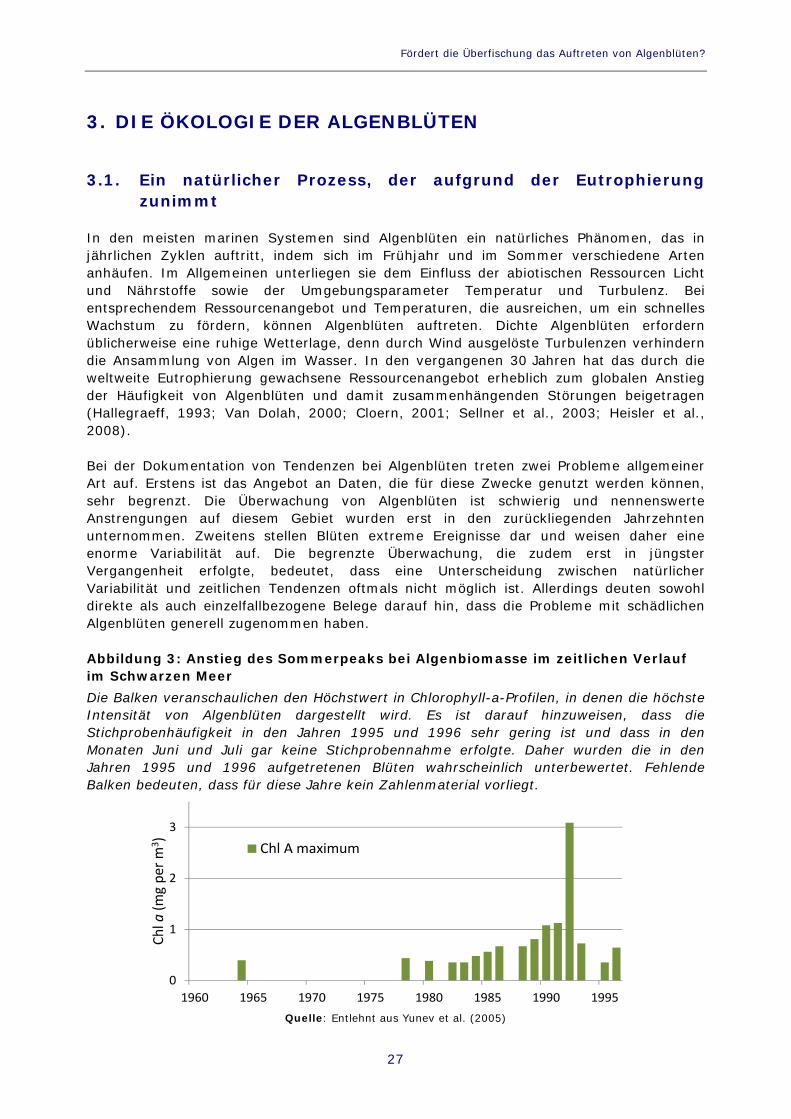
In den meisten marinen Systemen sind Algenblüten ein natürliches Phänomen, das in jährlichen Zyklen auftritt, indem sich im Frühjahr und im Sommer verschiedene Arten anhäufen. Im Allgemeinen unterliegen sie dem Einfluss der abiotischen Ressourcen Licht und Nährstoffe sowie der Umgebungsparameter Temperatur und Turbulenz. Bei entsprechendem Ressourcenangebot und Temperaturen, die ausreichen, um ein schnelles Wachstum zu fördern, können Algenblüten auftreten. Dichte Algenblüten erfordern üblicherweise eine ruhige Wetterlage, denn durch Wind ausgelöste Turbulenzen verhindern die Ansammlung von Algen im Wasser. In den vergangenen 30 Jahren hat das durch die weltweite Eutrophierung gewachsene Ressourcenangebot erheblich zum globalen Anstieg der Häufigkeit von Algenblüten und damit zusammenhängenden Störungen beigetragen (Hallegraeff, 1993; Van Dolah, 2000; Cloern, 2001; Sellner et al., 2003; Heisler et al., 2008). Bei der Dokumentation von Tendenzen bei Algenblüten treten zwei Probleme allgemeiner Art auf. Erstens ist das Angebot an Daten, die für diese Zwecke genutzt werden können, sehr begrenzt. Die Überwachung von Algenblüten ist schwierig und nennenswerte Anstrengungen auf diesem Gebiet wurden erst in den zurückliegenden Jahrzehnten unternommen. Zweitens stellen Blüten extreme Ereignisse dar und weisen daher eine enorme Variabilität auf. Die begrenzte Überwachung, die zudem erst in jüngster Vergangenheit erfolgte, bedeutet, dass eine Unterscheidung zwischen natürlicher Variabilität und zeitlichen Tendenzen oftmals nicht möglich ist. Allerdings deuten sowohl direkte als auch einzelfallbezogene Belege darauf hin, dass die Probleme mit schädlichen Algenblüten generell zugenommen haben. Abbildung 3: Anstieg des Sommerpeaks bei Algenbiomasse im zeitlichen Verlauf im Schwarzen Meer Die Balken veranschaulichen den Höchstwert in Chlorophyll-a-Profilen, in denen die höchste Intensität von Algenblüten dargestellt wird. Es ist darauf hinzuweisen, dass die Stichprobenhäufigkeit in den Jahren 1995 und 1996 sehr gering ist und dass in den Monaten Juni und Juli gar keine Stichprobennahme erfolgte. Daher wurden die in den Jahren 1995 und 1996 aufgetretenen Blüten wahrscheinlich unterbewertet. Fehlende Balken bedeuten, dass für diese Jahre kein Zahlenmaterial vorliegt.
0
1
2
3
1960 1965 1970 1975 1980 1985 1990 1995
Chl a
(mg per m
3 ) Chl A maximum
Quelle: Entlehnt aus Yunev et al. (2005)
27
Fachabteilung B: Struktur- und Kohäsionspolitik
Im Schwarzen Meer fiel die Zunahme von Blüten mit hohen Algenbiomassen mit dem Beginn der heutigen Eutrophierung zusammen (Zaitsev, 1992). Die durchschnittliche Phytoplanktonbiomasse verdoppelte sich in den 1980er Jahren und die Häufigkeit und Intensität von Phytoplanktonblüten haben seit den 1970er Jahren dramatisch zugenommen (Zaitsev, 1992; Kideys, 1994; Daskalov, 2002; Yunev et al., 2002; Yunev et al., 2005) (Abbildung 3). Der relative Anteil von Dinoflagellaten stieg von 10 % in den 1960er und 1970er Jahren auf 30 % in den 80er Jahren; und auch die Zahl der roten Tiden sowie die Dichte einiger schädlicher Dinoflagellaten-Arten (vor allem Prorocentrum cordatum und Noctiluca miliaris) nahmen erheblich zu (Kideys, 1994). Tabelle 1: Berichte über Algenblüten im Mittelmeer und im Schwarzen Meer
Jahr Algenarten, die laut Bericht für die Blüte verantwortlich
waren
Ende der 1960er Jahre
Nördliches Adriatisches Meer: Prorocentrum
1970er Jahre Nördliches Adriatisches Meer: Noctiluca, Gonyaulax, Prorocentrum, Gyrodinium, Glenodinium
1980er Jahre
Nördliches Adriatisches Meer: Katodinium, Noctiluca, Glenodinium, Prorocentrum, Gyrodinium, Gonyaulax, Scrippsiella, Massarthia Spanien 1984: Gonyaulax Schwarzes Meer 1986: Prorocentrum Mittelmeer 1989: Gymnodinium
1990er Jahre
Schwarzes Meer: Pseudo-nitzschia, Nitzschia, Cheatoceros, Ditylum, Cylindrotheca, Rhizosolenia, Heterocapsa, Protoperidinium, Scripsiella, Emiliania, Gonyaulax, Prorocentrum Schwarzes Meer 1993: Dinoflagellaten Tunesien 1994: Karenia Spanien 1994 und 1996-1999: Alexandrium Griechenland 1994 und 1997: Prorocentrum, Noctiluca, Erythropsidinium Frankreich 1998: Alexandrium Griechenland 1998: Chattonella Frankreich 1999: Prorocentrum Tyrrhenisches Meer 1998, 2000-2001: Ostreopsis
Ab 2000
Schwarzes Meer: Skeletonema, Cerataulina, Prorocentrum, Gymnodinium Spanien 2003: Alexandrium, Gymnodinium Griechenland 2000-2004: Prorocentrum, Noctiluca, Gymnodinium, Alexandrium, Dinophysis, Pseudo-nitzschia Ligurisches Meer 2005-2006: Ostreopsis Tunesien 2006: Coolia Nördliches Adriatisches Meer 2007: Ostreopsis Italien 2010: Ostreopsis Puglia 2010 Sizilien 2010
Quelle: Nastasi (2010)
28
Fördert die Überfischung das Auftreten von Algenblüten?
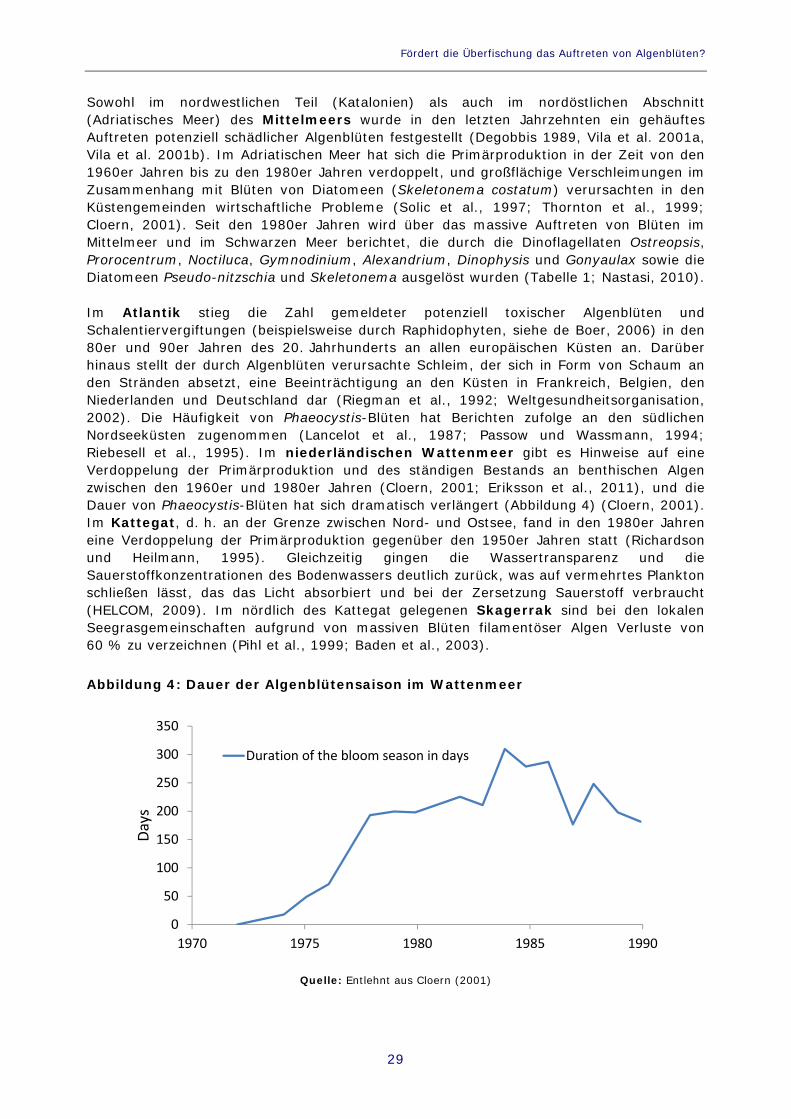
Sowohl im nordwestlichen Teil (Katalonien) als auch im nordöstlichen Abschnitt (Adriatisches Meer) des Mittelmeers wurde in den letzten Jahrzehnten ein gehäuftes Auftreten potenziell schädlicher Algenblüten festgestellt (Degobbis 1989, Vila et al. 2001a, Vila et al. 2001b). Im Adriatischen Meer hat sich die Primärproduktion in der Zeit von den 1960er Jahren bis zu den 1980er Jahren verdoppelt, und großflächige Verschleimungen im Zusammenhang mit Blüten von Diatomeen (Skeletonema costatum) verursachten in den Küstengemeinden wirtschaftliche Probleme (Solic et al., 1997; Thornton et al., 1999; Cloern, 2001). Seit den 1980er Jahren wird über das massive Auftreten von Blüten im Mittelmeer und im Schwarzen Meer berichtet, die durch die Dinoflagellaten Ostreopsis, Prorocentrum, Noctiluca, Gymnodinium, Alexandrium, Dinophysis und Gonyaulax sowie die Diatomeen Pseudo-nitzschia und Skeletonema ausgelöst wurden (Tabelle 1; Nastasi, 2010). Im Atlantik stieg die Zahl gemeldeter potenziell toxischer Algenblüten und Schalentiervergiftungen (beispielsweise durch Raphidophyten, siehe de Boer, 2006) in den 80er und 90er Jahren des 20. Jahrhunderts an allen europäischen Küsten an. Darüber hinaus stellt der durch Algenblüten verursachte Schleim, der sich in Form von Schaum an den Stränden absetzt, eine Beeinträchtigung an den Küsten in Frankreich, Belgien, den Niederlanden und Deutschland dar (Riegman et al., 1992; Weltgesundheitsorganisation, 2002). Die Häufigkeit von Phaeocystis-Blüten hat Berichten zufolge an den südlichen Nordseeküsten zugenommen (Lancelot et al., 1987; Passow und Wassmann, 1994; Riebesell et al., 1995). Im niederländischen Wattenmeer gibt es Hinweise auf eine Verdoppelung der Primärproduktion und des ständigen Bestands an benthischen Algen zwischen den 1960er und 1980er Jahren (Cloern, 2001; Eriksson et al., 2011), und die Dauer von Phaeocystis-Blüten hat sich dramatisch verlängert (Abbildung 4) (Cloern, 2001). Im Kattegat, d. h. an der Grenze zwischen Nord- und Ostsee, fand in den 1980er Jahren eine Verdoppelung der Primärproduktion gegenüber den 1950er Jahren statt (Richardson und Heilmann, 1995). Gleichzeitig gingen die Wassertransparenz und die Sauerstoffkonzentrationen des Bodenwassers deutlich zurück, was auf vermehrtes Plankton schließen lässt, das das Licht absorbiert und bei der Zersetzung Sauerstoff verbraucht (HELCOM, 2009). Im nördlich des Kattegat gelegenen Skagerrak sind bei den lokalen Seegrasgemeinschaften aufgrund von massiven Blüten filamentöser Algen Verluste von 60 % zu verzeichnen (Pihl et al., 1999; Baden et al., 2003). Abbildung 4: Dauer der Algenblütensaison im Wattenmeer
0
50
100
150
200
250
300
350
1970 1975 1980 1985 1990
Days
Duration of the bloom season in days
Quelle: Entlehnt aus Cloern (2001)
29
Fachabteilung B: Struktur- und Kohäsionspolitik
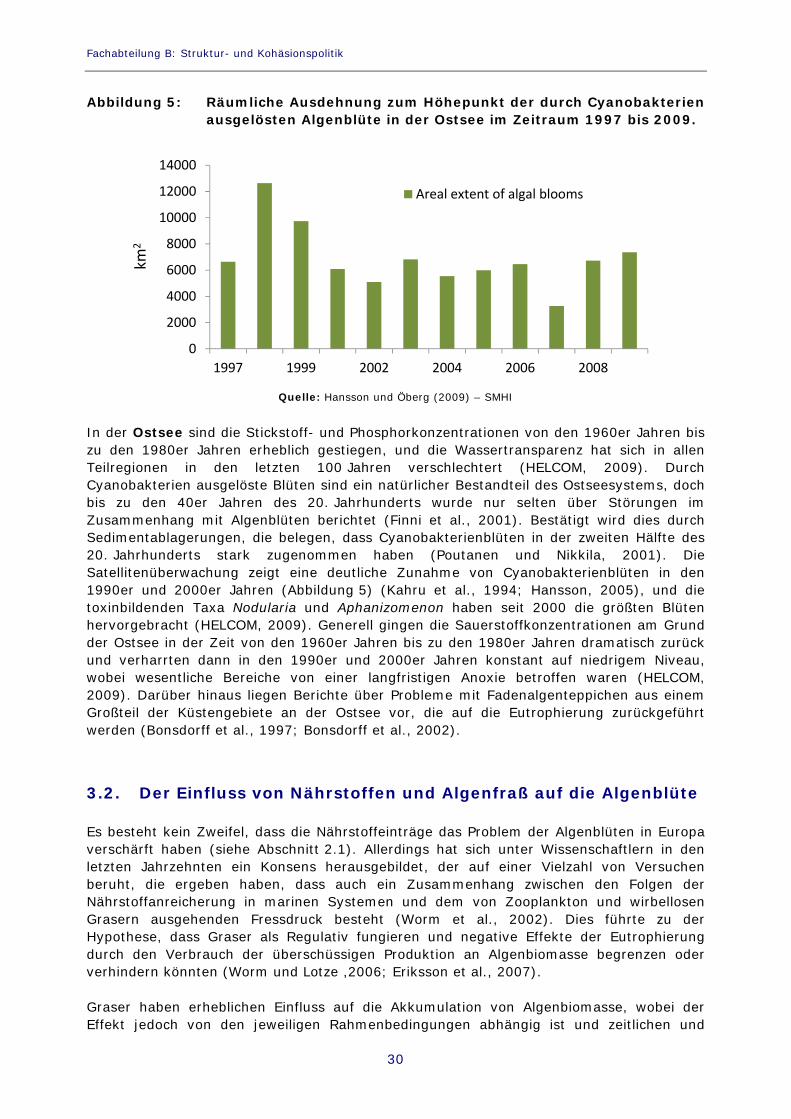
Abbildung 5: Räumliche Ausdehnung zum Höhepunkt der durch Cyanobakterien ausgelösten Algenblüte in der Ostsee im Zeitraum 1997 bis 2009.
0
2000
4000
6000
8000
10000
12000
14000
1997 1999 2002 2004 2006 2008
km2
Areal extent of algal blooms
Quelle: Hansson und Öberg (2009) – SMHI
In der Ostsee sind die Stickstoff- und Phosphorkonzentrationen von den 1960er Jahren bis zu den 1980er Jahren erheblich gestiegen, und die Wassertransparenz hat sich in allen Teilregionen in den letzten 100 Jahren verschlechtert (HELCOM, 2009). Durch Cyanobakterien ausgelöste Blüten sind ein natürlicher Bestandteil des Ostseesystems, doch bis zu den 40er Jahren des 20. Jahrhunderts wurde nur selten über Störungen im Zusammenhang mit Algenblüten berichtet (Finni et al., 2001). Bestätigt wird dies durch Sedimentablagerungen, die belegen, dass Cyanobakterienblüten in der zweiten Hälfte des 20. Jahrhunderts stark zugenommen haben (Poutanen und Nikkila, 2001). Die Satellitenüberwachung zeigt eine deutliche Zunahme von Cyanobakterienblüten in den 1990er und 2000er Jahren (Abbildung 5) (Kahru et al., 1994; Hansson, 2005), und die toxinbildenden Taxa Nodularia und Aphanizomenon haben seit 2000 die größten Blüten hervorgebracht (HELCOM, 2009). Generell gingen die Sauerstoffkonzentrationen am Grund der Ostsee in der Zeit von den 1960er Jahren bis zu den 1980er Jahren dramatisch zurück und verharrten dann in den 1990er und 2000er Jahren konstant auf niedrigem Niveau, wobei wesentliche Bereiche von einer langfristigen Anoxie betroffen waren (HELCOM, 2009). Darüber hinaus liegen Berichte über Probleme mit Fadenalgenteppichen aus einem Großteil der Küstengebiete an der Ostsee vor, die auf die Eutrophierung zurückgeführt werden (Bonsdorff et al., 1997; Bonsdorff et al., 2002).
3.2. Der Einfluss von Nährstoffen und Algenfraß auf die Algenblüte Es besteht kein Zweifel, dass die Nährstoffeinträge das Problem der Algenblüten in Europa verschärft haben (siehe Abschnitt 2.1). Allerdings hat sich unter Wissenschaftlern in den letzten Jahrzehnten ein Konsens herausgebildet, der auf einer Vielzahl von Versuchen beruht, die ergeben haben, dass auch ein Zusammenhang zwischen den Folgen der Nährstoffanreicherung in marinen Systemen und dem von Zooplankton und wirbellosen Grasern ausgehenden Fressdruck besteht (Worm et al., 2002). Dies führte zu der Hypothese, dass Graser als Regulativ fungieren und negative Effekte der Eutrophierung durch den Verbrauch der überschüssigen Produktion an Algenbiomasse begrenzen oder verhindern könnten (Worm und Lotze ,2006; Eriksson et al., 2007). Graser haben erheblichen Einfluss auf die Akkumulation von Algenbiomasse, wobei der Effekt jedoch von den jeweiligen Rahmenbedingungen abhängig ist und zeitlichen und
30

Fördert die Überfischung das Auftreten von Algenblüten?
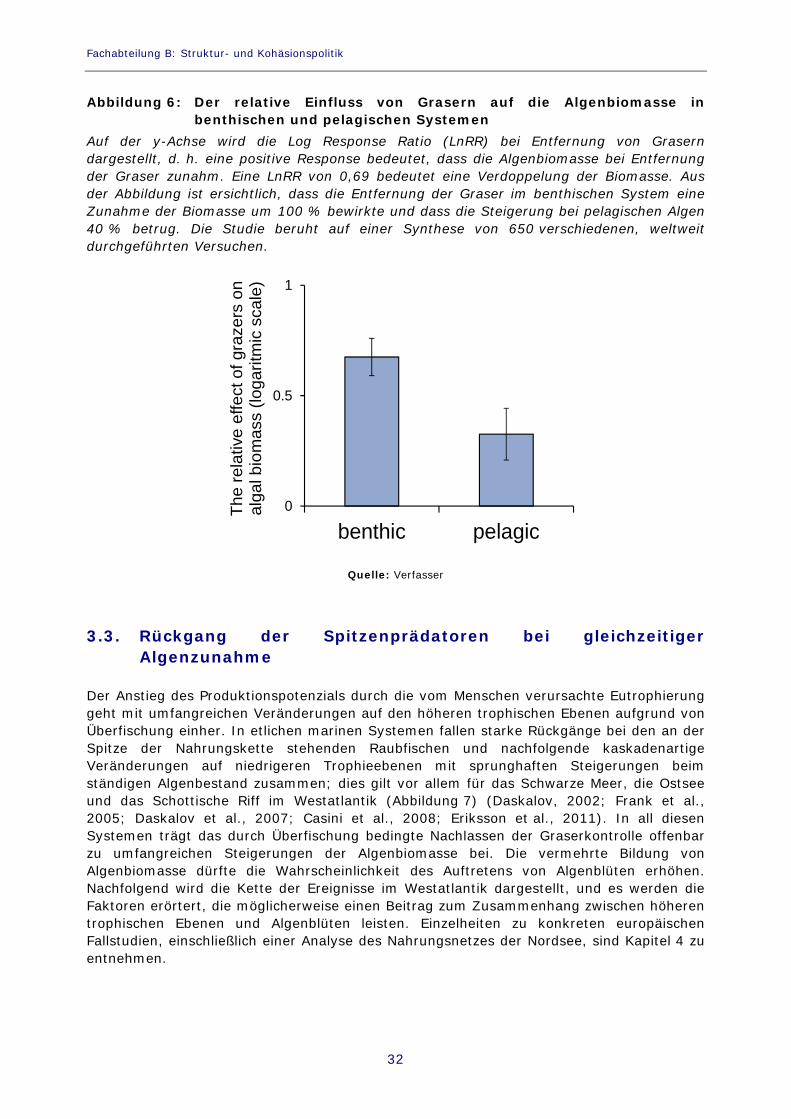
räumlichen Schwankungen unterliegt. Ökologische Synthesen, bei denen die durchschnittlichen Auswirkungen anhand zahlreicher verschiedener Versuche aus verschiedenen Gebieten berechnet werden, haben gezeigt, dass das Verschwinden von Grasern oder zusätzliche Nährstoffeinträge zu einer Erhöhung – häufig sogar zu einer Verdoppelung –- der Algenbiomassemenge in marinen Systemen jeglicher Art führen (Worm et al., 2002; Gruner et al., 2008). In einer Synthesestudie von Gruner et al. (2008) bewirkte die Entfernung von Grasern eine Zunahme der relativen Auswirkung von Nährstoffen an Felsenstränden (signifikante Interaktionswirkung). Dies bedeutet zum Beispiel, dass Nährstoffe bei Anwesenheit von Grasern eine Zunahme der Algenbiomasse um 40 % bewirken, während die Steigerung bei Abwesenheit von Grasern 80 % beträgt. Auf weichen Gründen führten Nährstoffe unabhängig von der An- oder Abwesenheit von Grasern im Durchschnitt zu einer Verdoppelung der Biomasse, aber dennoch reduzierten die Graser die gesamte Biomasse um die Hälfte. Worm und Lotze (2006) führten in eutrophierten Buchten an der Ostsee und in Nova Scotia einen koordinierten Versuch mit Nährstoffen und Grasern durch und stellten fest, dass Graser in allen untersuchten Gebieten eine Abnahme der durch Fadenalgenblüten gebildeten Biomasse um ca. 80 % bewirken, die Nährstoffe aber trotz hohen Fressdrucks dennoch die Biomasseakkumulation begünstigen. Die negativen Auswirkungen der Eutrophierung können also durch das Grasen verhindert werden, doch fällt die Nährstoffanreicherung mitunter stärker ins Gewicht als die Kontrolle durch Graser, und die Algenbiomasse beginnt zu wachsen. Gleichwohl führt das Grasen auf dem üblichen natürlichen Niveau im Allgemeinen dazu, dass das Algenaufkommen deutlich geringer ist als ohne die Anwesenheit von Grasern. Ausgehend von der Gesamtheit der weltweit durchgeführten Versuche mit Grasern ist eine allgemeine Bewertung der Kontrolle durch Graser in benthischen und pelagischen Ökosystemen möglich. Insgesamt 650 Versuche in 507 benthischen und 143 pelagischen Systemen ergeben, dass die Anwesenheit von Grasern eine Begrenzung der benthischen Algenbiomasse auf ca. 50 % der ständigen Biomasse bedeutet, die sich bei Entfernung von Grasern bildet, und dass es bei der pelagischen Algenbiomasse ca. 70 % sind (Abbildung 6). Die Wissenschaft ist sich demnach weitgehend einig, dass Graser dazu beitragen, die Küstengemeinden gegen die negativen Folgen der Eutrophierung zu wappnen.
31
Fachabteilung B: Struktur- und Kohäsionspolitik
Abbildung 6: Der relative Einfluss von Grasern auf die Algenbiomasse in benthischen und pelagischen Systemen
Auf der y-Achse wird die Log Response Ratio (LnRR) bei Entfernung von Grasern dargestellt, d. h. eine positive Response bedeutet, dass die Algenbiomasse bei Entfernung der Graser zunahm. Eine LnRR von 0,69 bedeutet eine Verdoppelung der Biomasse. Aus der Abbildung ist ersichtlich, dass die Entfernung der Graser im benthischen System eine Zunahme der Biomasse um 100 % bewirkte und dass die Steigerung bei pelagischen Algen 40 % betrug. Die Studie beruht auf einer Synthese von 650 verschiedenen, weltweit durchgeführten Versuchen.
0
0.5
1
benthic pelagic
The r
ela
tive e
ffect
of gra
zers
on
alg
al b
iom
ass
(lo
ga
ritm
ic s
cale
)
Quelle: Verfasser
3.3. Rückgang der Spitzenprädatoren bei gleichzeitiger Algenzunahme
Der Anstieg des Produktionspotenzials durch die vom Menschen verursachte Eutrophierung geht mit umfangreichen Veränderungen auf den höheren trophischen Ebenen aufgrund von Überfischung einher. In etlichen marinen Systemen fallen starke Rückgänge bei den an der Spitze der Nahrungskette stehenden Raubfischen und nachfolgende kaskadenartige Veränderungen auf niedrigeren Trophieebenen mit sprunghaften Steigerungen beim ständigen Algenbestand zusammen; dies gilt vor allem für das Schwarze Meer, die Ostsee und das Schottische Riff im Westatlantik (Abbildung 7) (Daskalov, 2002; Frank et al., 2005; Daskalov et al., 2007; Casini et al., 2008; Eriksson et al., 2011). In all diesen Systemen trägt das durch Überfischung bedingte Nachlassen der Graserkontrolle offenbar zu umfangreichen Steigerungen der Algenbiomasse bei. Die vermehrte Bildung von Algenbiomasse dürfte die Wahrscheinlichkeit des Auftretens von Algenblüten erhöhen. Nachfolgend wird die Kette der Ereignisse im Westatlantik dargestellt, und es werden die Faktoren erörtert, die möglicherweise einen Beitrag zum Zusammenhang zwischen höheren trophischen Ebenen und Algenblüten leisten. Einzelheiten zu konkreten europäischen Fallstudien, einschließlich einer Analyse des Nahrungsnetzes der Nordsee, sind Kapitel 4 zu entnehmen.
32

Fördert die Überfischung das Auftreten von Algenblüten?
Abbildung 7: Fischereibedingte Rückgänge von Spitzenprädatoren fallen mit sprunghaften Steigerungen des ständigen Algenbestands im Nordatlantik, in der Ostsee und im Schwarzen Meer zusammen.
phytoplankton
larger zooplankton
planktivorous fish
demersal top predators
(mainly cod)
overfishingIncreased fishing
pressure
decreases in large predatory fish
increases in smaller predatory fish
decreases in organisms that eat algae ‐ grazers
increases in primary producing algae
phytoplankton/ dinoflagellates
larger zooplankton
planktivorous fish (mainly anchovy)
pelagic top predators
overfishing
Scotian shelf Black Sea
phytoplankton
larger zooplankton
planktivorous fish (sprat)
cod
overfishing
Baltic Sea
Quelle: Frank et al. (2005), Daskalov (2007), Casini et al. (2008) Auf dem Schottischen Riff im Nordwestatlantik (Nova Scotia, Kanada) kam es im Zeitraum 1985 – 1995 zu einem Zusammenbruch der massiv befischten benthischen Fischgemeinschaft und parallel dazu zu einer allgemeinen Veränderung des Phytoplanktonaufkommens von gering (1960er und 1970er Jahre) nach hoch (1990er und 200er Jahre) (Frank et al., 2005). In der benthischen Fischgemeinschaft nahm der Kabeljau (Gadus morhua) eine Spitzenposition ein, doch auch bei anderen größeren demersalen Fischarten wie dem Schellfisch (Melanogrammus aeglefinus), Seehechten (Urophycis tenuis, Merluccius bilinearis) und dem Seelachs (Pollachius virens) waren Rückgänge zu verzeichnen. Nach dem weitgehenden Wegfall der wichtigsten größeren Räuber wuchsen die Bestände einer Vielzahl ihrer Beutearten wie auch die Gesamtbiomasse kleiner pelagischer Fische, Krabben und Garnelen (Frank et al., 2005). Im Zuge dieser Verschiebung zugunsten kleinerer planktivorer Fische vollzog sich auch in der Zooplanktongemeinschaft ein Wechsel von hohen zu geringen Dichten großer Arten (Mountain und Kane, 2010; Pierik et al., 2011). Größeres Zooplankton ist zudem die bevorzugte Nahrungsquelle der kleineren pelagischen Fische, und Anfang der 1980er Jahre war die Dichte von größerem Zooplankton noch um 45 % höher als in den 1990er Jahren, während die Dichten von kleinerem Zooplankton konstant blieben. Korrelationsanalysen ergaben, dass die jährlichen Veränderungen bei benthischen Fischen seit den 1970er Jahren negativ bei kleinen pelagischen Fischen und Phytoplankton und positiv bei größerem Zooplankton zu Buche schlugen: Es bestand eine signifikante negative Korrelation zwischen der Biomasse benthischer Fische und kleiner pelagischer Fische (r=-0,61, n=33), eine signifikante positive Korrelation zwischen der Biomasse benthischer Fische und größerem Zooplankton (r=0,45, n=23) und eine signifikante negative Korrelation zwischen der Biomasse benthischer Fische und Phytoplankton (r=-0,72, n=24) (Frank et al., 2005). Diese reziproken Veränderungen in der Abundanz der Trophieebenen deuten darauf hin, dass der Rückgang der größeren demersalen Fische eine Zunahme der Dichte von
33

Fachabteilung B: Struktur- und Kohäsionspolitik
Phytoplankton gefördert haben könnte, indem indirekt die Dichte größerer Zooplanktonarten verringert wurde und folglich der allgemeine Fressdruck durch Graser nachließ. Veränderungen in der Struktur des Nahrungsnetzes, bei denen ein Rückgang der Spitzenprädatoren zu Veränderungen auf den verschiedenen trophischen Ebenen führt – z. B. Zunahme kleinerer Räuber, Abnahme von Herbivoren und Zunahme von Pflanzen oder Algenbiomasse -, werden trophische Kaskaden genannt (Pace et al., 1999). Es gibt nur wenige Experimente in einer für das jeweilige Ökosystem relevanten Größenordnung, die einen kausalen Zusammenhang zwischen Räubern und Algen in marinen Systemen herstellen. Dennoch nimmt die Zahl der Indizienbeweise für systemweite marine trophische Kaskaden zu (Estes et al., 1998; Jackson et al., 2001; Daskalov, 2002; Frank et al., 2005; Casini et al., 2008; Eriksson et al., 2011), und eine Reihe kleinerer Versuche untermauert die Bedeutung von Fischräubern für die Menge der Algenbiomasse (Shurin et al., 2002; Borer et al., 2005; Borer et al., 2006; Eriksson et al., 2009; Sieben et al., 2011a, 2011b). Trophische Kaskaden sind aus der Bewirtschaftung von Seen allgemein bekannt; dort haben Versuche mit ganzen Seen gezeigt, dass die Freisetzung von Spitzenprädatoren in Seen, die zuvor durch Phytoplankton getrübt waren, eine deutliche Reduzierung der Algenbiomasse und eine Klärung des Wassers bewirken kann (Carpenter et al., 2001). Daher hat sich die biologische Manipulation höherer Trophieebenen als nützliches Instrument zur Eindämmung der Entstehung von Algenblüten in eutrophierten Seen erwiesen (Sondergaard et al., 2007).
3.4. Abhängigkeit trophischer Kaskaden von Ressourcen und Prädatorenvielfalt
Die Auswirkungen des Rückgangs von Raubfischen auf das Ökosystem sind in den letzten Jahrzehnten im Rahmen von Modellversuchen und Feldstudien gründlich erforscht worden. Heute weiß man, dass kaskadenartige Veränderungen in der Struktur von Nahrungsnetzen nach Rückgängen von Raubfischen auf einer höheren trophischen Ebene in einer Vielzahl verschiedener Ökosysteme üblich sind, darunter in Seen (Carpenter et al., 1985), Flüssen (Power, 1990), Zuckertangwäldern (Estes et al., 1998), Flussmündungen (Jackson., 2001), felsigen Wattystemen (Menge, 2000), Festlandsockeln (Frank et al., 2007) und auch im offenen Ozean (Ward und Myers, 2005). Allmählich setzt sich aber auch die Erkenntnis durch, dass trophische Kaskaden außerdem auf komplexe Wechselwirkungen zwischen Umweltbedingungen, Ressourcenangebot, Merkmalen einzelner Arten und der Komplexität des Nahrungsnetzes zurückgehen (Strong, 1992; Polis und Strong, 1996; Menge, 2000; Oksanen und Oksanen, 2000; Shurin et al., 2002; Borer et al., 2005; Borer et al., 2006; Hopcraft et al., 2010). Daher sind die Einflüsse von Räubern auf die Zusammensetzung eines Nahrungsnetzes sehr stark kontextabhängig, und die Eigenschaften des Ökosystems bestimmen etwaige Reaktionen auf die Überfischung von Spitzenprädatoren. Es lassen sich jedoch einige generelle Feststellungen treffen, die darauf hindeuten könnten, welche Systeme anfällig für trophische Kaskaden sind und an welcher Stelle eine Überfischung zur Entstehung von Algenblüten beitragen könnte. Trophische Kaskaden werden durch die Produktivität verstärkt (Pace et al., 1999), was bedeutet, dass eine Wechselwirkung zwischen Rückgängen von Spitzenprädatoren und der Eutrophierung der Meere bestehen könnte. Bei trophischen Kaskaden wird nämlich unterstellt, dass auf jeder zweiten Trophieebene eine Ressourcenkontrolle stattfindet und Konsumenten als Regulativ fungieren (Oksanen und Oksanen, 2000); dies bedeutet ein positives Verhältnis zwischen der Primärproduktion und der jeweils zweiten Trophieebene. Daher dürften in einem marinen System mit vier funktionierenden trophischen Ebenen -
34

Fördert die Überfischung das Auftreten von Algenblüten?
(1) Algen, 2) Herbivoren, 3) kleinere Räuber, die sich von Pflanzenfressern ernähren und 4) Räuber an der Spitze, die kleinere Räuber fressen - Herbivoren und Spitzenprädatoren zahlenmäßig zunehmen, wenn sich die Nährstoffanreicherung erhöht. Bei einer Reduzierung auf drei funktionierende trophische Ebenen aufgrund der Überfischung von Spitzenprädatoren wiederum dürften Algen und kleinere Räuber zunehmen, wenn sich die Nährstoffanreicherung erhöht (Oksanen und Oksanen, 2000; Eriksson, 2011) (Abbildung 8). Dies zeigen auch dokumentierte Beispiele für Kaskadeneffekte im Schwarzen Meer und in der Ostsee, bei denen ein Rückgang der Spitzenprädatoren mit einer Zunahme der Algenbiomasse einherging (siehe Fallstudien; Daskalov, 2002; Casini et al., 2008), wenn Überfischung und Eutrophierung gleichzeitig auftraten. Terrestrische und marine Feldstudien untermauern das Bestehen eines positiven Zusammenhangs zwischen hoher Produktivität und trophischen Kaskaden im großen Maßstab (Sinclair et al., 2000; Elmhagen und Rushton, 2007; Vasas et al., 2007; Hopcraft et al., 2010), und kleinmaßstäbliche Versuche im marinen Benthos stützen speziell die These, dass die Nährstoffanreicherung das Entstehen trophischer Kaskaden von den Spitzenprädatoren bis hinunter zu den Algen fördert (Moksnes et al., 2008; Eriksson et al., 2009; Sieben et al., 2011b). Beispielsweise entwickelten Vasas et al. (2007) das Modell eines Nahrungsnetzes auf der Grundlage veröffentlichter Nordseestudien und wiesen nach, dass Überfischung und Eutrophierung Synergieeffekte haben, die schädliche Algenblüten fördern (siehe Abschnitt 4.2, Fallstudie: Nordsee). Sieben et al. (2011b) zeigten, dass die Entnahme von benthischen Spitzenprädatoren mittels Käfigen eine trophische Kaskade in Gang setzte, durch die die Biomasse benthischer Algen drastisch anstieg, was jedoch nur in Kombination mit einer Nährstoffanreicherung der Fall war (siehe Abschnitt 4.6, Fallstudie: Ostsee). Wie sich die Entnahme von Spitzenprädatoren auf die Entwicklung von Algenblüten auswirkt, hängt also von der Ressourcenverfügbarkeit ab. Trophische Kaskaden wirken im marinen Benthos stärker als im marinen Plankton (Strong, 1992; Shurin et al., 2002; Borer et al., 2005). Im Allgemeinen kommen positive Auswirkungen auf das Wachstum der Pflanzen- oder Algenbiomasse, die auf die Entfernung von Herbivoren durch Prädation zurückzuführen sind, im Wasser stärker zum Tragen als an Land (Strong, 1992). Dies ist jedoch hauptsächlich die Ursache sehr starker Kaskadeneffekte auf Algen im marinen Benthos (Shurin et al., 2002). Bei einer Betrachtung aller seinerzeit veröffentlichten Studien zu trophischen Kaskaden im Meer führten Borer et al. (2005) den Nachweis, dass mittelgroße Räuber, die Herbivoren fressen, signifikante positive Auswirkungen auf das marine Phytoplankton und benthische Algen hatten; doch während die Planktonbiomasse infolge der Prädation um ca. 30 % zunahm, stieg die Biomasse benthischer Algen um ein Mehrfaches an. Daher ist zu erwarten, dass die Entfernung von Spitzenprädatoren deutlich stärkere Auswirkungen auf Blüten filamentöser Algen als auf Phytoplanktonblüten hat. Des Weiteren ist mit trophischen Kaskaden mit starken wechselseitigen Räuber-Beute-Effekten hauptsächlich in Systemen zu rechnen, in denen die Stärke der Interaktion von einigen wenigen funktional dominierenden Arten abhängt (Strong, 1992; Pace et al., 1999). Ein Überblick über 108 Versuche, bei denen Prädatoren entnommen wurden, zeigt, dass die Räuber im Durchschnitt 57 % der Beuteindividuen oder der Biomasse entfernten (persönliche Daten). Hier besteht ein hohes Maß an Übereinstimmung mit der durchschnittlichen Entfernung von Produzenten-Biomasse durch Herbivoren, die in einer Synthese von 191 Versuchen analysiert wurde (Gruner et al., 2008). Allerdings war der Effekt der Prädation auf die Biomasse der auf einer niedrigeren trophischen Ebene angesiedelten Beute stets nur in Systemen negativ, in denen eine Räuberart die Räubergemeinschaft dominierte. In Gemeinschaften mit einem dominierenden Räuber hatte die Prädation im Durchschnitt das Verschwinden von zwei Dritteln der Beutebiomasse zur
35

Fachabteilung B: Struktur- und Kohäsionspolitik
Folge, während bei größerer Räubervielfalt keine signifikanten negativen Effekte der Prädation festgestellt wurden. Dies deutet darauf hin, dass zwar Räuber offenkundig Biomasse entnehmen, der von ihnen ausgehende Prädationsdruck aber häufig nicht ausreicht, um die Beutebiomasse unter Kontrolle zu halten, wenn die Räubervielfalt groß ist; dies ist vermutlich negativen Interaktionen zwischen den Räubern zuzuschreiben. Aus diesem Grund sind stärkere Auswirkungen der Überfischung in marinen Systemen zu erwarten, in denen die Fischgemeinschaft von einigen wenigen, zahlenmäßig extrem großen und funktional wichtigen Arten dominiert wird. Hier besteht ein hohes Maß an Übereinstimmung mit dem System im Schwarzen Meer, wo der Anchovis eine Vorrangstellung in der pelagischen Fischgemeinschaft einnimmt, und mit dem Ostsee-System, wo der Kabeljau die Gemeinschaft der Spitzenprädatoren dominiert und Clupeidae (Sprotte und Hering) in der plantivoren Fischgemeinschaft eine Vorrangstellung einnehmen (siehe Fallstudien in den Abschnitten 4.3 und 4.5). Abbildung 8: Die durch Nährstoffanreicherung bedingte erhöhte
Biomasseproduktion findet je nach Anzahl der Trophieebenen im Ökosystem auf verschiedenen trophischen Ebenen statt.
Smaller predators
Algae
nutrients
Herbivores
Herbivores
Smaller predators
Top‐predators
Algae
4 trophic levels
3 trophic levels
Quelle: Frank et al., (2005); Daskalov (2007); Casini et al. (2008)
36

Fördert die Überfischung das Auftreten von Algenblüten?
4. TRÄGT DIE ÜBERFISCHUNG ZUM AUFTRETEN VON ALGENBLÜTEN IN EUROPA BEI?
Aus methodischer Sicht ist es nahezu unmöglich, schlüssige Beweise für die Auswirkungen derart großräumiger Prozesse wie der Überfischung zu finden. Überlagert werden die Effekte der Fischerei von einer Reihe wichtiger ökologischer Variablen, und es finden Interaktionen zwischen diesen Effekten und Variablen statt. Da Systeme von der Größe eines Ozeanbeckens nicht manipuliert werden können, müssen Trendanalysen und eine auf Felddaten beruhende Modellierung in Verbindung mit kleinmaßstäblichen Versuchen genutzt werden, um die wahrscheinlichsten Szenarien zu untersuchen und die wahrscheinlichsten Auswirkungen zu bewerten. Belege für den Einfluss der Fischerei auf Algenblüten zeigen, dass die Überfischung höchstwahrscheinlich zu Algenblüten in einigen europäischen Meeressystemen beiträgt, auch wenn es sich vor allem um Indizienbeweise handelt. Daher muss der Schluss gezogen werden, dass die Überfischung zum Auftreten von Algenblüten beiträgt, der Umfang und die Schwere der Blüte im Verlaufe der Saison aber zeitlich, räumlich, zwischen den marinen Systemen und innerhalb dieser Systeme variiert.
4.1. Darstellung der Auswirkungen von Überfischung und Eutrophierung im Modell
Um gezielt die kombinierten Auswirkungen von Überfischung und Eutrophierung der Meere zu überprüfen, schufen Vasas et al. (2007) ein Netzmodell eines generischen küstennahen pelagischen Ökosystems in einem Gebiet mit gemäßigtem Klima. Als Modell fungierte ein topologisches Nahrungsnetz, d. h. eine vereinfachte Darstellung dessen, wie die Beziehungen zwischen verschiedenen wichtigen Artengruppen aussehen und wer wen frisst. Dieses Modell bildet die Grundstruktur des Ökosystems ab und kann genutzt werden, um das allgemeine Verhalten des Systems beim Auftreten klar umrissener Störungen zu untersuchen, ist aber nicht geeignet, um spezifische Dichten oder ein etwaiges dynamisches Verhalten vorherzusagen. Die Ermittlung der aus funktioneller Sicht wichtigsten Nährstoffpools und Artengruppen sowie der Beziehungen zwischen Artengruppen erfolgte anhand einer allgemeinen Auswertung von 25 Studien zum Thema trophischer Transfer, d. h. Verbrauch eines Nährstoffpools bzw. einer Artengruppe oder Beitrag zu einem Nährstoffpool durch Nährstoffeintrag oder Sedimentation, sowie auf der Grundlage von Felddaten aus der Nordsee (Rousseau et al., 2000). Daher sind die Ergebnisse dieser Analyse für alle Küstenökosysteme in Europa, besonders aber für die Nordsee von Relevanz. Im Wege der Literaturrecherche ermittelten Vasas et al. (2007) 19 Artengruppen und 7 Nährstoffpools, die Einfluss auf das Funktionieren pelagischer Gemeinschaften im küstennahen Bereich von Gebieten mit gemäßigtem Klima haben. Dazu gehörten vier blühende Algengruppen: 1) große Diatomeen, 2) große nichttoxische Dinoflagellaten, 3) ungenießbare Algen einschließlich schädlicher Algen und Bakterien, die toxische Algenblüten oder Blüten ungenießbarer Algen hervorrufen; in die Analyse waren zum Beispiel schädliche Algenarten wie Cyanobakterien (Nodularia spumigena), Dinoflagellaten (Noctiluca, Alexandrium-Arten, Karenia brevis), Prymnesiophyten (Prymnesium parvus, Chrysochromulina polylepis, Phaeocystis-Arten) und Raphidophyten (Heterosigma akashiwo) einbezogen. Wichtig für diese Analyse war, dass das Netz auch großes Zooplankton, das größere genießbare Diatomeen und Dinoflagellaten frisst (Gruppen 1 und 2), planktivore Fische (kleinere pelagische Fische, die sich von Plankton ernähren), piscivore Fische (größere pelagische Fische, die Fische fressen) und Quallen, d. h.
37
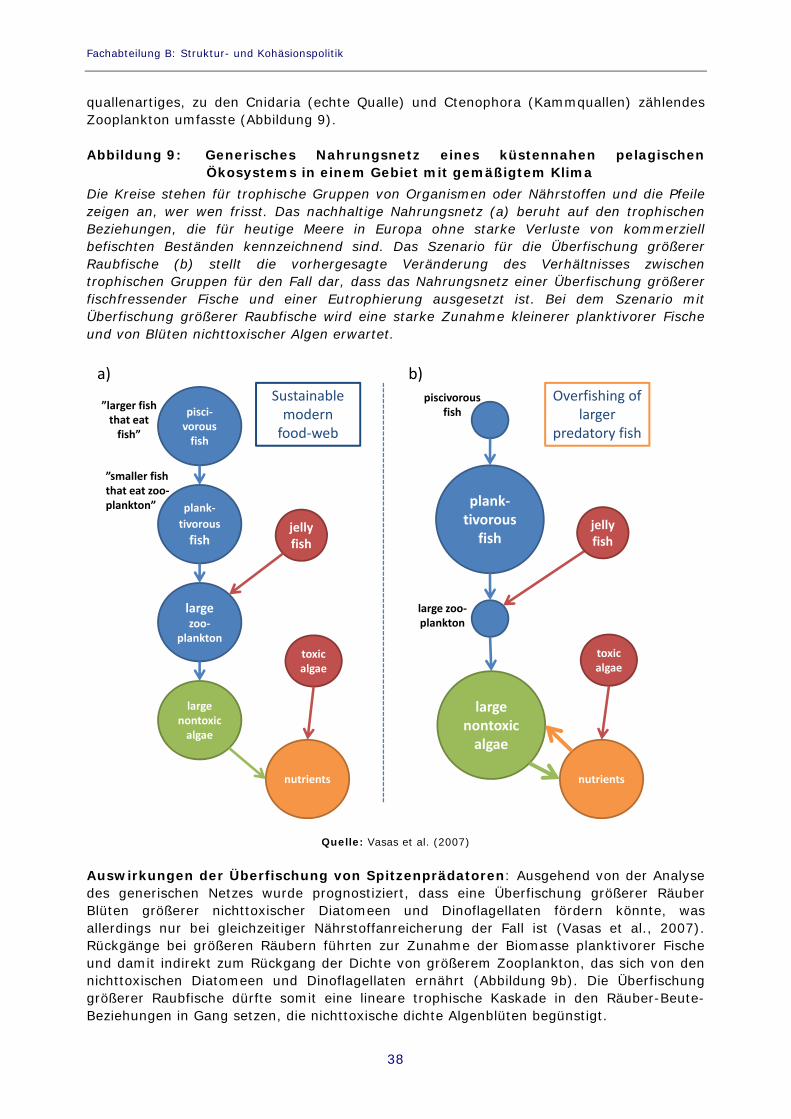
Fachabteilung B: Struktur- und Kohäsionspolitik
quallenartiges, zu den Cnidaria (echte Qualle) und Ctenophora (Kammquallen) zählendes Zooplankton umfasste (Abbildung 9). Abbildung 9: Generisches Nahrungsnetz eines küstennahen pelagischen
Ökosystems in einem Gebiet mit gemäßigtem Klima Die Kreise stehen für trophische Gruppen von Organismen oder Nährstoffen und die Pfeile zeigen an, wer wen frisst. Das nachhaltige Nahrungsnetz (a) beruht auf den trophischen Beziehungen, die für heutige Meere in Europa ohne starke Verluste von kommerziell befischten Beständen kennzeichnend sind. Das Szenario für die Überfischung größerer Raubfische (b) stellt die vorhergesagte Veränderung des Verhältnisses zwischen trophischen Gruppen für den Fall dar, dass das Nahrungsnetz einer Überfischung größerer fischfressender Fische und einer Eutrophierung ausgesetzt ist. Bei dem Szenario mit Überfischung größerer Raubfische wird eine starke Zunahme kleinerer planktivorer Fische und von Blüten nichttoxischer Algen erwartet.
pisci‐vorous fish
plank‐
tivorous
fish
large zoo‐
plankton
large nontoxic algae
toxic algae
jelly fish
nutrients
plank‐tivorous fish
large nontoxic algae
nutrients
jelly fish
toxic algae
large zoo‐plankton
piscivorous fish
Overfishing of larger
predatory fish
Sustainablemodern food‐web
”larger fish that eat fish”
”smaller fish that eat zoo‐plankton”
a) b)
Quelle: Vasas et al. (2007) Auswirkungen der Überfischung von Spitzenprädatoren: Ausgehend von der Analyse des generischen Netzes wurde prognostiziert, dass eine Überfischung größerer Räuber Blüten größerer nichttoxischer Diatomeen und Dinoflagellaten fördern könnte, was allerdings nur bei gleichzeitiger Nährstoffanreicherung der Fall ist (Vasas et al., 2007). Rückgänge bei größeren Räubern führten zur Zunahme der Biomasse planktivorer Fische und damit indirekt zum Rückgang der Dichte von größerem Zooplankton, das sich von den nichttoxischen Diatomeen und Dinoflagellaten ernährt (Abbildung 9b). Die Überfischung größerer Raubfische dürfte somit eine lineare trophische Kaskade in den Räuber-Beute-Beziehungen in Gang setzen, die nichttoxische dichte Algenblüten begünstigt.
38
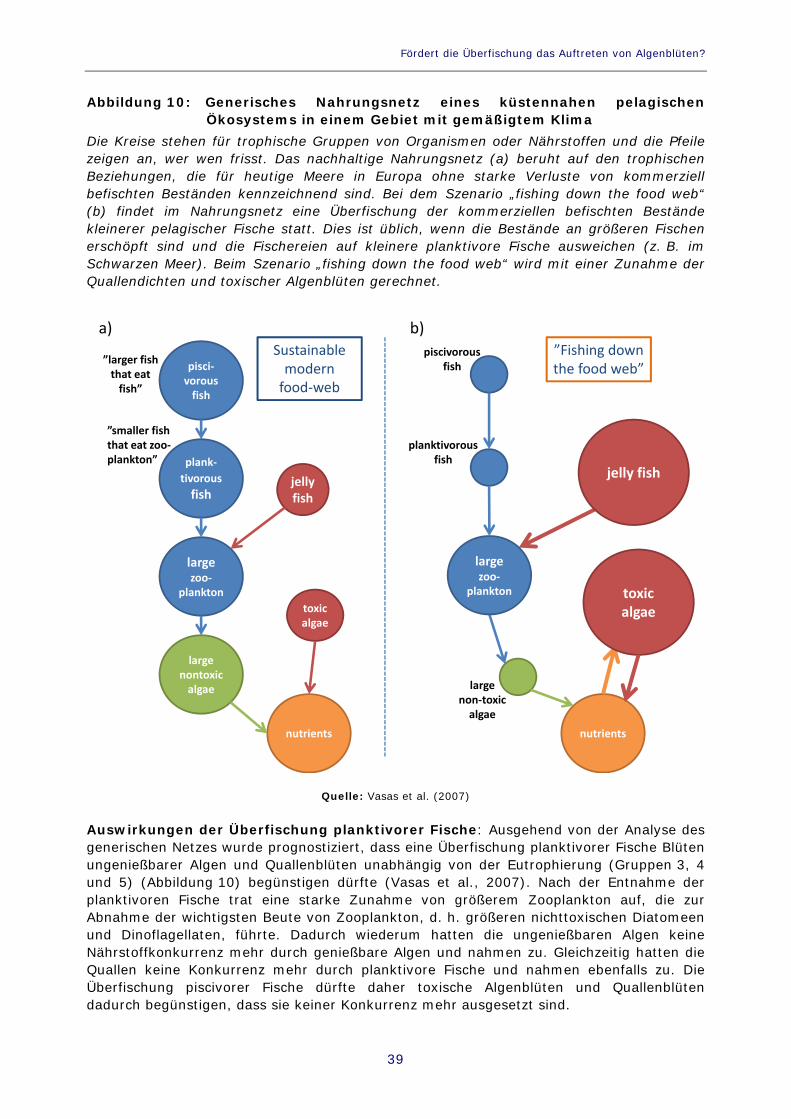
Fördert die Überfischung das Auftreten von Algenblüten?
Abbildung 10: Generisches Nahrungsnetz eines küstennahen pelagischen Ökosystems in einem Gebiet mit gemäßigtem Klima
Die Kreise stehen für trophische Gruppen von Organismen oder Nährstoffen und die Pfeile zeigen an, wer wen frisst. Das nachhaltige Nahrungsnetz (a) beruht auf den trophischen Beziehungen, die für heutige Meere in Europa ohne starke Verluste von kommerziell befischten Beständen kennzeichnend sind. Bei dem Szenario „fishing down the food web“ (b) findet im Nahrungsnetz eine Überfischung der kommerziellen befischten Bestände kleinerer pelagischer Fische statt. Dies ist üblich, wenn die Bestände an größeren Fischen erschöpft sind und die Fischereien auf kleinere planktivore Fische ausweichen (z. B. im Schwarzen Meer). Beim Szenario „fishing down the food web“ wird mit einer Zunahme der Quallendichten und toxischer Algenblüten gerechnet.
nutrients
jelly fish
toxic algae
large zoo‐
plankton
largenon‐toxicalgae
piscivorous fish
planktivorous fish
”Fishing down the food web”pisci‐
vorous fish
plank‐
tivorous
fish
large zoo‐
plankton
large nontoxic algae
toxic algae
jelly fish
nutrients
Sustainablemodern food‐web
”larger fish that eat fish”
”smaller fish that eat zoo‐plankton”
a) b)
Quelle: Vasas et al. (2007) Auswirkungen der Überfischung planktivorer Fische: Ausgehend von der Analyse des generischen Netzes wurde prognostiziert, dass eine Überfischung planktivorer Fische Blüten ungenießbarer Algen und Quallenblüten unabhängig von der Eutrophierung (Gruppen 3, 4 und 5) (Abbildung 10) begünstigen dürfte (Vasas et al., 2007). Nach der Entnahme der planktivoren Fische trat eine starke Zunahme von größerem Zooplankton auf, die zur Abnahme der wichtigsten Beute von Zooplankton, d. h. größeren nichttoxischen Diatomeen und Dinoflagellaten, führte. Dadurch wiederum hatten die ungenießbaren Algen keine Nährstoffkonkurrenz mehr durch genießbare Algen und nahmen zu. Gleichzeitig hatten die Quallen keine Konkurrenz mehr durch planktivore Fische und nahmen ebenfalls zu. Die Überfischung piscivorer Fische dürfte daher toxische Algenblüten und Quallenblüten dadurch begünstigen, dass sie keiner Konkurrenz mehr ausgesetzt sind.
39

Fachabteilung B: Struktur- und Kohäsionspolitik
Abschließend kann festgestellt werden, dass das von Vasas et al. (2007) entwickelte Modell eines generischen Nahrungsnetzes eines küstennahen pelagischen Ökosystems in einem Gebiet mit gemäßigtem Klima folgende Vorhersagen ermöglicht: 1) Die Überfischung größerer Raubfische in Verbindung mit einer Eutrophierung dürfte die Entstehung nichttoxischer Algenblüten mit hohem Biomasseaufkommen erleichtern, und 2) die Überfischung kleinerer planktivorer Fische – „fishing down the food web“ – dürfte die Entstehung toxischer Algenblüten und eine Zunahme der Dichten von gallertartigem Zooplankton begünstigen. Diese einfache Analyse dient als Schlüssel zum Verständnis des Beitrags der Überfischung zu den zeitlichen Trends in den nachfolgenden Fallstudien.
4.2. Fallstudie: Nordsee In der Nordsee sind durch Überfischung bedingte Rückgänge bei größeren Raubfischen seit den 1970er Jahren dokumentiert (Pauly et al., 1998; Christensen et al., 2003). 1980 setzte jedoch auch eine allgemeine Veränderung des Ökosystems Nordsee ein, die mit Änderungen in der Zusammensetzung der Fischgemeinschaft und Zunahmen der Dichte von Phytoplankton einherging (Reid et al., 2001; Weijerman et al., 2005; McQuatters-Gollop et al., 2007). Die Fänge von Raubfischen mit einer Länge von mindestens 90 cm waren seit 1970 von einem Spitzenwert von 1 000 000 Tonnen ständig weiter gesunken und erreichten im Jahr 2000 nur noch 200 000 Tonnen (Abbildung 11). Der Thun- und Fächerfischfang sowie die Befischung größerer pelagischer Räuber kam bereits Anfang der 1960er Jahre zum Erliegen, während die Hai-und die Rochenfischerei in dieser Zeit ihren Höhepunkt erreichten, gefolgt von einem kontinuierlichen Rückgang ab 1970 (Abbildung 12). Heute zählen zu den kommerziell wichtigsten größeren Räubern die demersalen Arten Kabeljau und Seelachs (Pollachius virens). Bei den Kabeljau- und Seelachsfängen wurden in den 1970er Jahren Spitzenwerte verzeichnet; seinerzeit wurden 2,5 Mal größere Mengen als heute gefangen. Seit etwa 1980 sind die Fangmengen bei Kabeljau und Seelachs stark rückläufig (Abbildung 13). Mitte der 1970er Jahre veränderte sich die Zusammensetzung der Fänge deutlich, weil der Rückgang bei größeren Fischen mit einer starken Zunahme der Fänge von Fischen mit einer Länge unter 30 cm einherging (Abbildung 11). Neben der allgemeinen Abnahme der Bestände an größeren Räubern trägt auch die starke Zunahme der industriellen Sandaalfischerei zu dieser Schwerpunktverlagerung auf kleinere Fische bei (seaaroundus.org). Um das Jahr 1980 herum veränderte sich die Struktur des Ökosystems Nordsee vor allem aufgrund der rapiden Zunahme der Algenbiomasse (Abbildung 14) (McQuatters-Gollop et al., 2007). Ein Vergleich der Zeiträume 1947-1979 und 1980-2003 ergibt eine im Durchschnitt 13 % gestiegene Konzentration von Phytoplanktonpigmenten im Wasser der offenen Nordsee und eine um 21 % gestiegene Konzentration in den Küstenregionen der Nordsee (McQuatters-Gollop et al., 2007). Analysen großskaliger Klimavariablen haben Anlass zu der Vermutung gegeben, dass etliche potenzielle Umgebungsvariablen, die Auswirkungen auf die gesamte Region Nordeuropa haben, diese Veränderung in der biologischen Struktur ausgelöst haben könnten; zu nennen wären hier vor allem der Salzgehalt, die Wetterbedingungen und großräumige Veränderungen der Wasserklarheit (Reid et al., 2001; Weijerman et al., 2005; McQuatters-Gollop et al., 2007). Dies hat zu der Annahme geführt, dass Veränderungen bei großskaligen Klimabedingungen einen großräumigen Regimewechsel in der Nordsee verursacht haben, von dem sämtliche trophischen Ebenen – vom Plankton über Seefische bis hin zu benthischen Invertebraten und Küstenfischen – betroffen waren (Beaugrand, 2004). Allerdings fällt die Verschiebung auch mit Einbußen auf der mittleren Trophieebene des Fangs zusammen, die dem Einfluss der kombinierten Effekte einer stärkeren Befischung kleinerer Fische und des Rückgangs größerer Räuber unterliegt (Abbildung 15). So sind parallele Veränderungen in der Fisch- und Phytoplanktongemeinschaft erkennbar, indem eine intensivere Fischerei und
40
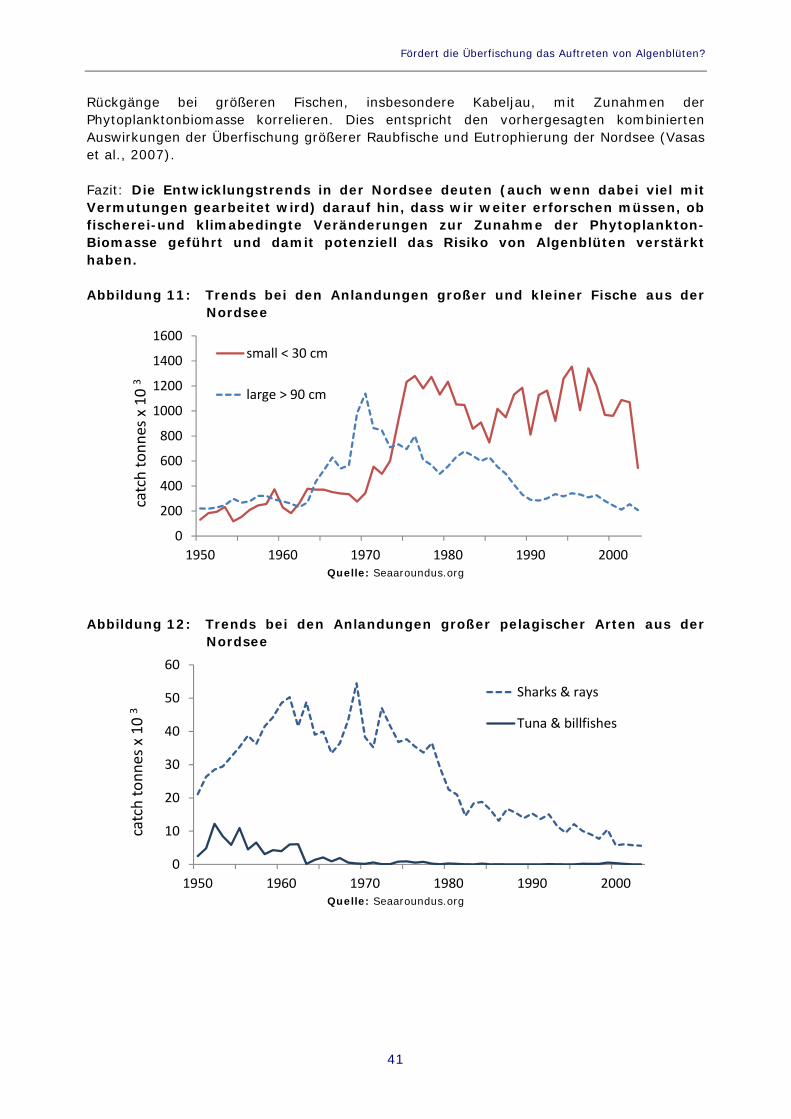
Fördert die Überfischung das Auftreten von Algenblüten?
Rückgänge bei größeren Fischen, insbesondere Kabeljau, mit Zunahmen der Phytoplanktonbiomasse korrelieren. Dies entspricht den vorhergesagten kombinierten Auswirkungen der Überfischung größerer Raubfische und Eutrophierung der Nordsee (Vasas et al., 2007). Fazit: Die Entwicklungstrends in der Nordsee deuten (auch wenn dabei viel mit Vermutungen gearbeitet wird) darauf hin, dass wir weiter erforschen müssen, ob fischerei-und klimabedingte Veränderungen zur Zunahme der Phytoplankton-Biomasse geführt und damit potenziell das Risiko von Algenblüten verstärkt haben. Abbildung 11: Trends bei den Anlandungen großer und kleiner Fische aus der
Nordsee
0
200
400
600
800
1000
1200
1400
1600
1950 1960 1970 1980 1990 2000
catch tonnes x 10 3
small < 30 cm
large > 90 cm
Quelle: Seaaroundus.org
Abbildung 12: Trends bei den Anlandungen großer pelagischer Arten aus der
Nordsee
0
10
20
30
40
50
60
1950 1960 1970 1980 1990 2000
catch tonnes x 10
3
Sharks & rays
Tuna & billfishes
Quelle: Seaaroundus.org
41
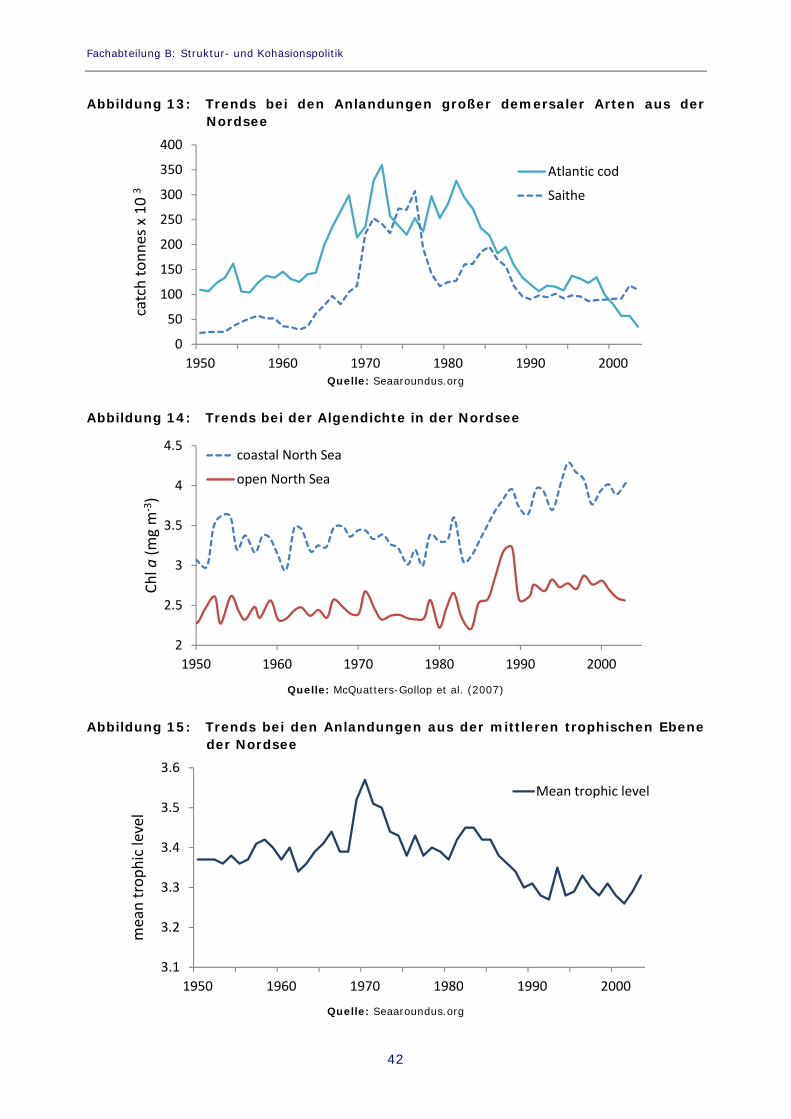
Fachabteilung B: Struktur- und Kohäsionspolitik
Abbildung 13: Trends bei den Anlandungen großer demersaler Arten aus der Nordsee
0
50
100
150
200
250
300
350
400
1950 1960 1970 1980 1990 2000
catch tonnes x 10 3
Atlantic cod
Saithe
Quelle: Seaaroundus.org
Abbildung 14: Trends bei der Algendichte in der Nordsee
2
2.5
3
3.5
4
4.5
1950 1960 1970 1980 1990 2000
Chl a
(mg m
‐3)
coastal North Sea
open North Sea
Quelle: McQuatters-Gollop et al. (2007) Abbildung 15: Trends bei den Anlandungen aus der mittleren trophischen Ebene
der Nordsee
3.1
3.2
3.3
3.4
3.5
3.6
1950 1960 1970 1980 1990 2000
mean trophic level
Mean trophic level
Quelle: Seaaroundus.org
42

Fördert die Überfischung das Auftreten von Algenblüten?
4.3. Fallstudie: Schwarzes Meer Im Schwarzen Meer folgte auf einen starken Rückgang bei größeren Räubern, dessen Höhepunkt Anfang der 1970er Jahre verzeichnet wurde, ein starker Anstieg der Phytoplanktonbiomasse, der Anfang der 1980er mit einer relativen Steigerung von 100 % einen Spitzenwert erreichte (Daskalov, 2002; Daskalov et al., 2007). Seit der Veränderung in der Zusammensetzung der Fischgemeinschaft in den 1970er Jahren hat die Häufigkeit und Intensität von Phytoplanktonblüten dramatisch zugenommen (Zaitsev, 1992; Kideys, 1994; Daskalov, 2002; Yunev et al., 2002; Yunev et al., 2005). Während der gesamten 1960er Jahre wurden die wichtigsten Spitzenprädatoren, darunter Delfine und große wandernde Arten wie der Bonito (Sarda sarda), Thunfisch und Schwertfisch, massiv befischt (Daskalov, 2002). Allerdings brach die Fischerei größerer Arten Ende der 60er Jahre zusammen, und die betreffenden Bestände waren weitestgehend erschöpft oder verschwanden. Heute ist den Fangstatistiken zu entnehmen, dass sich die Thun- und Fächerfischbestände langsam wieder erholen, was vor allem daran liegt, dass die türkischen und die russischen Fischereien vermehrt den atlantischen Gelbflossenthun befischen (Abbildung 16). Nach dem Rückgang der großen pelagischen Arten waren bei deren wichtigsten Beutearten, d. h. kleinen und mittelgroßen pelagischen Arten wie dem Anchovis (Engraulis encrasiccolus), Sprotten (Sprattus sprattus) und den Mittelmeerstöckern (Trachurus mediterraneus), Anfang der 1970er Jahre starke Zuwächse zu verzeichnen; dadurch gerieten sie ins Blickfeld der kommerziellen Fischereien (Abbildung 17) (Kideys, 1994; Daskalov, 2002). Daraufhin sank die durchschnittliche trophische Ebene der Fänge im Schwarzen Meer in der zweiten Hälfte des 20. Jahrhunderts deutlich ab, wodurch in der Zusammensetzung der Fischgemeinschaft ein Wechsel von großen zu mittelgroßen und kleineren pelagischen Arten gefördert wurde (Abbildung 18). Nach dem Wechsel in der Fischgemeinschaft von größeren Räubern zu kleineren planktivoren Fischen ging die Zooplanktonbiomasse stark zurück und beim Phytoplankton war eine entsprechende Steigerung zu verzeichnen (Abbildung 19) (Daskalov, 2002; Daskalov et al., 2007). Planktivore Fische ernähren sich hauptsächlich von größerem Zooplankton, und Veränderungen in der Zusammensetzung von Zooplanktongemeinschaften deuten darauf hin, dass der Biomasserückgang teilweise dadurch bedingt war, dass größere Arten durch kleinere Arten von Krustentierplankton verdrängt wurden (Zaitsev, 1992; Kideys, 1994). Auch der Charakter der Phytoplanktongemeinschaft veränderte sich, da die relative Dichte von größerem Phytoplankton, bei dem Dinoflagellaten eine Vorrangstellung innehatten, stärker zunahm als die von kleinerem Phytoplankton, bei dem Diatomeen dominierten (Kideys, 1994; Daskalov, 2002). Vor allem im Schwarzen Meer ist ein zweites Kaskadenereignis erkennbar, durch das die Dichte der Phytoplanktonbiomasse und die Häufigkeit von Algenblüten in den 1990er Jahren sogar noch zugenommen haben. Nachdem die kommerziellen Fischereien in den 1960er Jahren den Wechsel von größeren Spitzenprädatoren zu kleineren pelagischen Arten vollzogen hatten, wuchs die Anchovisfischerei deutlich, und der Anchovis avancierte zum kommerziell wichtigsten planktivoren Fisch in der türkischen Fischerei. Im Zeitraum 1988-1991 gingen die Anchovisfänge auf 25 % der Fangmengen der 1980er Jahre zurück. Der Einbruch bei den Anchovisfängen wird durch einen drastischen Abfall der Gesamtbiomasse kleinerer planktivorer Fische im Schwarzen Meer bestätigt (Daskalov et al., 2007). Allerdings hat sich die Zooplanktongemeinschaft nicht erholt. Stattdessen war bei zwei Arten von gallertartigem Zooplankton, die Konkurrenten der planktivoren Fische im Hinblick auf deren wichtigste Zooplanktonbeute sind, eine starke Zunahme zu verzeichnen. Dies deutet darauf hin, dass die Fischerei Algenblüten im Schwarzen Meer im Zuge zweier
43

Fachabteilung B: Struktur- und Kohäsionspolitik
Überfischungsereignisse gefördert haben könnte, die beide dazu führten, dass der von Zooplankton ausgehende Fressdruck auf das Phytoplankton nachließ. Es liegen deutliche Indizien dafür vor, dass Überfischung und Eutrophierung zu einer trophischen Kaskade beigetragen haben, durch die eine Zunahme der Phytoplanktonmasse im Schwarzen Meer ausgelöst wurde (Daskalov, 2002; Daskalov et al., 2007). Erstens ergibt sich eine inverse zeitliche Korrelation zwischen benachbarten trophischen Ebenen (1950-1960 bis zum Jahr 2003) und eine positive Korrelation zwischen nicht benachbarten Trophieebenen (Tabelle 2) (Daskalov, 2007): Vor allem besteht ein signifikantes negatives Verhältnis zwischen der Dichte größerer pelagischer Raubfische und der Phytoplanktonbiomasse (n=43, r=-0,39). Zweitens entwickelte Daskalov (2002) ein Modell eines Nahrungsnetzes (ein dynamisches Massenbilanzmodell) des pelagischen Systems in den 1960er Jahren, indem er die mittlere Biomasse, die Produktion, die Verbrauchsraten und die Nahrung von 15 wichtigen Gruppen im Nahrungsnetz kartierte. Dazu gehörten Delfine, größere Räuber, planktivore Fische, Fischlarven, gallertartiges Zooplankton verschiedener Art sowie großes und kleines Zoo- und Phytoplankton. Daskalov (2002) führte dann Modellmodelsimulationen der Fischerei und der Eutrophierung durch. Ein Szenario, bei dem nur die Fischerei untersucht und von der Annahme ausgegangen wurde, dass sich der fischereiliche Druck innerhalb eines Zeitraums von 30 Jahren verdreifacht hat, spiegelte die jeweiligen Veränderungen auf den trophischen Ebenen, die in den 1970er und 1980er Jahren im Nahrungsnetz des Schwarzen Meeres beobachtet wurden, korrekt wider, einschließlich 1) der Zunahmen bei planktivoren Fischen, 2) der Abnahmen bei der Zooplanktonbiomasse, die durch die sinkende relative Dichte größerer Zooplanktonarten bedingt waren, und 3) des Anstiegs der Phytoplanktonbiomasse, der auf die Steigerung der relativen Dichte größerer Phytoplanktonarten zurückzuführen war. Hinzuweisen ist darauf, dass der unterstellte fischereiliche Druck eine Mindestschätzung darstellt; so soll eine Überzeichnung der Auswirkungen vermieden werden. Allerdings wurde mit dem Modell, bei dem nur die Fischerei betrachtet wurde, ein Anstieg der Phytoplanktonbiomasse von ca. 30 % ermittelt, der deutlich hinter der festgestellten Zunahme von 100 % liegt. Wenn man jedoch eine dreifache Zunahme der Fischerei und eine vermehrte Primärproduktion des Systems aufgrund von Eutrophierung unterstellt, wird richtig ein Anstieg der Phytoplanktonbiomasse von 95 bis 100 % ermittelt. Fazit: Angesichts 1) der Häufung von Algenblüten in den 1970er Jahren (Zaitsev, 1992; Kideys, 1994; Daskalov, 2002; Yunev et al., 2002; Yunev et al., 2005), 2) der parallelen Veränderungen in der Zusammensetzung der Fischgemeinschaft und in den Beziehungen zwischen Räubern und Beute (Abbildungen 16 bis 19) (Daskalov, 2002; Daskalov et al., 2007) und 3) der Ergebnisse des von Daskalov (2002) entwickelten Nahrungsnetzmodells, die darauf hindeuten, dass die nachlassende Kontrolle durch größere Raubfische in Kombination mit der Eutrophierung zu einer verstärkten Bildung von Phytoplanktonbiomasse führt, ist es durchaus wahrscheinlich, dass sowohl die Überfischung als auch die Eutrophierung zur Entstehung von Algenblüten im Schwarzen Meer beitragen. Bei den Algen, für die eine Zunahme aufgrund von Überfischung im Schwarzen Meer vorhergesagt wurde, handelte es sich hauptsächlich um großes Phytoplankton wie die größeren, rote Tiden verursachenden Dinoflagellaten Prorocentrum cordatum und Noctiluca miliaris, die in den 1970er Jahren, als die Fischerei auf größere pelagische Fische zusammenbrach, ebenfalls stark zunahmen (Kideys, 1994; Daskalov, 2002).
44
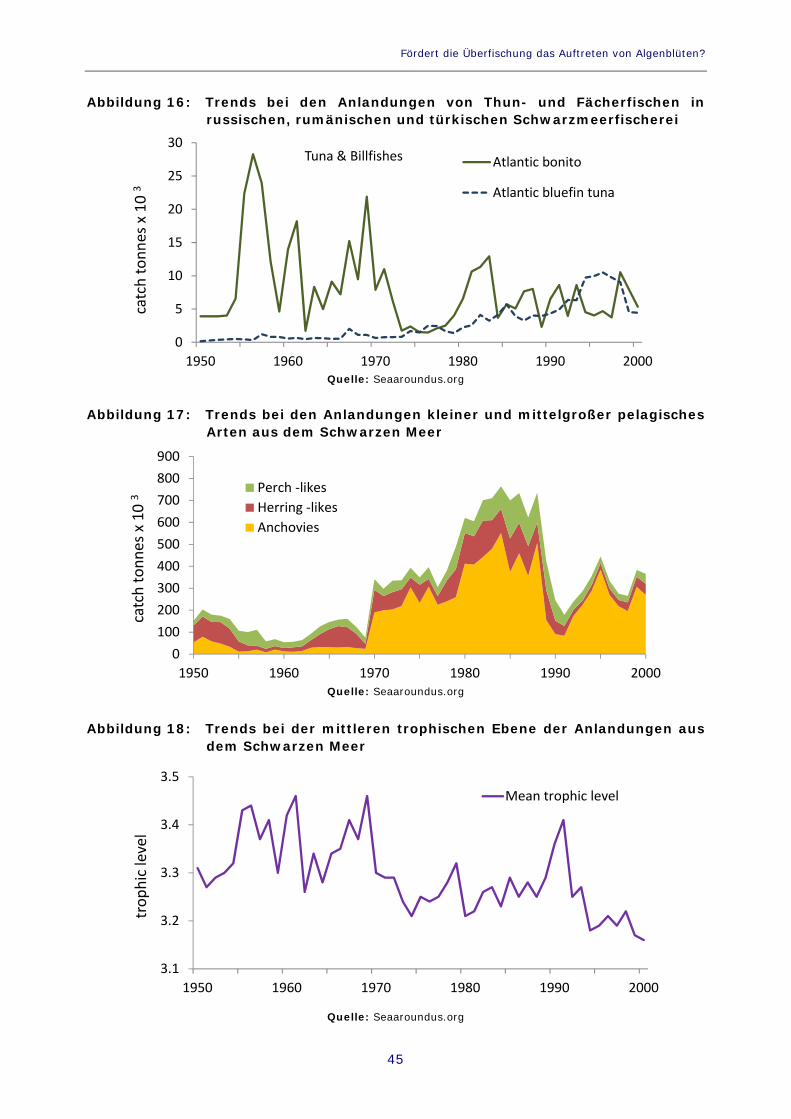
Fördert die Überfischung das Auftreten von Algenblüten?
Abbildung 16: Trends bei den Anlandungen von Thun- und Fächerfischen in russischen, rumänischen und türkischen Schwarzmeerfischerei
0
5
10
15
20
25
30
1950 1960 1970 1980 1990 2000
catch tonnes x 10 3
Tuna & Billfishes Atlantic bonito
Atlantic bluefin tuna
Quelle: Seaaroundus.org
Abbildung 17: Trends bei den Anlandungen kleiner und mittelgroßer pelagisches
Arten aus dem Schwarzen Meer
0
100
200
300
400
500
600
700
800
900
1950 1960 1970 1980 1990 2000
catch tonnes x 10
3
Perch ‐likes
Herring ‐likes
Anchovies
Quelle: Seaaroundus.org
Abbildung 18: Trends bei der mittleren trophischen Ebene der Anlandungen aus
dem Schwarzen Meer
3.1
3.2
3.3
3.4
3.5
1950 1960 1970 1980 1990 2000
trophic level
Mean trophic level
Quelle: Seaaroundus.org
45
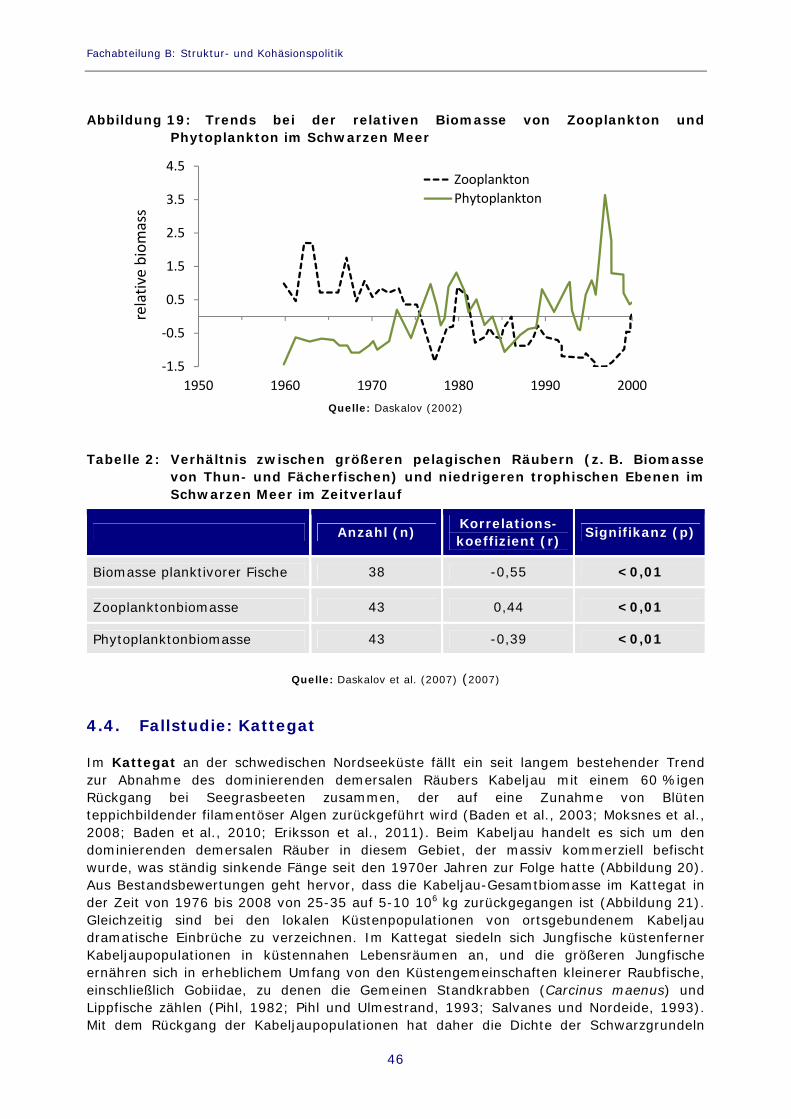
Fachabteilung B: Struktur- und Kohäsionspolitik
Abbildung 19: Trends bei der relativen Biomasse von Zooplankton und
Phytoplankton im Schwarzen Meer
‐1.5
‐0.5
0.5
1.5
2.5
3.5
4.5
1950 1960 1970 1980 1990 2000
relative biomass
Zooplankton
Phytoplankton
Quelle: Daskalov (2002)
Tabelle 2: Verhältnis zwischen größeren pelagischen Räubern (z. B. Biomasse
von Thun- und Fächerfischen) und niedrigeren trophischen Ebenen im Schwarzen Meer im Zeitverlauf
Anzahl (n) Korrelations-koeffizient (r)
Signifikanz (p)
Biomasse planktivorer Fische 38 -0,55 <0,01
Zooplanktonbiomasse 43 0,44 <0,01
Phytoplanktonbiomasse 43 -0,39 <0,01
Quelle: Daskalov et al. (2007) (2007)
4.4. Fallstudie: Kattegat Im Kattegat an der schwedischen Nordseeküste fällt ein seit langem bestehender Trend zur Abnahme des dominierenden demersalen Räubers Kabeljau mit einem 60 %igen Rückgang bei Seegrasbeeten zusammen, der auf eine Zunahme von Blüten teppichbildender filamentöser Algen zurückgeführt wird (Baden et al., 2003; Moksnes et al., 2008; Baden et al., 2010; Eriksson et al., 2011). Beim Kabeljau handelt es sich um den dominierenden demersalen Räuber in diesem Gebiet, der massiv kommerziell befischt wurde, was ständig sinkende Fänge seit den 1970er Jahren zur Folge hatte (Abbildung 20). Aus Bestandsbewertungen geht hervor, dass die Kabeljau-Gesamtbiomasse im Kattegat in der Zeit von 1976 bis 2008 von 25-35 auf 5-10 106 kg zurückgegangen ist (Abbildung 21). Gleichzeitig sind bei den lokalen Küstenpopulationen von ortsgebundenem Kabeljau dramatische Einbrüche zu verzeichnen. Im Kattegat siedeln sich Jungfische küstenferner Kabeljaupopulationen in küstennahen Lebensräumen an, und die größeren Jungfische ernähren sich in erheblichem Umfang von den Küstengemeinschaften kleinerer Raubfische, einschließlich Gobiidae, zu denen die Gemeinen Standkrabben (Carcinus maenus) und Lippfische zählen (Pihl, 1982; Pihl und Ulmestrand, 1993; Salvanes und Nordeide, 1993). Mit dem Rückgang der Kabeljaupopulationen hat daher die Dichte der Schwarzgrundeln
46

Fördert die Überfischung das Auftreten von Algenblüten?
(Gobius niger), der Strandkrabben und der dominierenden Lippfische Goldmaid (Symphodus melops) und Klippenbarsch (Ctenolabrus rupestris) an der Küste ständig zugenommen (Abbildung 22) (Eriksson et al., 2011). Schwarzgrundeln, Strandkrabben und Lippfische verfügen alle über das Potenzial, die Dichte von Krustentieren und Gastropoden („Mesograsern“) zu regulieren, die sich von den Algen ernähren, die Fadenalgenblüten bilden (Norderhaug et al., 2005; Newcombe und Taylor, 2010). Vergleiche der in Seegrasbeeten lebenden Tiergemeinschaften, die in den 1980er Jahren begannen und bis in das erste Jahrzehnt des 21. Jahrhunderts hinein fortgesetzt wurden, haben übereinstimmend ergeben, dass die Gemeinschaft der Krustentier-Mesograser in den 1980er Jahren eine große Dichte in Seegrasbeeten aufwies, ab 2000 aber vollständig daraus verschwand (Baden et al., 2011). Gleichzeitig wurde beim Kabeljau eine vierfache Verringerung der Populationen und bei kleineren Räubern wie den Schwarzgrundeln sogar eine sechsfache Abnahme festgestellt. Die Überfischung des Kabeljaus hat daher möglicherweise Kaskadeneffekte ausgelöst, die Blüten filamentöser Algen fördern, indem indirekt der Fressdruck vermindert wird, der von den wirbellosen Grasern ausgeht (Moksnes et al., 2008; Baden et al., 2010; Eriksson et al., 2011). Sowohl Indizien als auch Versuchsergebnisse stützen die Hypothese, dass die Veränderungen in der Struktur des Nahrungsnetzes im Kattegat, die auf die fischereibedingten Rückgänge des Kabeljau zurückzuführen sind, Anteil an der zunehmenden Dichte von Algenblüten und den großflächigen Rückgängen von Seegras haben. Erstens besteht eine signifikante Korrelation zwischen dem Rückgang bei Kabeljau in küstenfernen Gewässern und der Zunahme bei kleineren Küstenfischen (Abbildung 23). Des Weiteren ist eine deutliche Korrelation zwischen hohen Temperaturen in Küstengewässern im Frühjahr und größeren Dichten kleinerer Küstenfische (Abbildung 24) erkennbar. Es gibt jedoch keine Tendenz zum Temperaturanstieg in den Küstengewässern, in denen der Fisch überwacht wird, und die Korrelation zum Rückgang beim Kabeljau ist statistisch stärker – zu 47 % wird die Veränderung bei den Gesamtdichten kleinerer Raubfische dem Kabeljau zugeschrieben und nur zu 28 % der Temperatur (vgl. R-Werte in den Abbildungen 23 und 24). Demnach spielt zwar die Temperatur eine wichtige Rolle für das Überleben von Küstenfischen, doch größer ist die Wahrscheinlichkeit, dass der Rückgang beim Kabeljau den Anstieg bei kleineren Raubfischen verursacht hat (Eriksson et al., 2011). Zudem haben Versuche deutliche Anhaltspunkte dafür geliefert, dass kleinere Raubfische die Bildung größerer Biomassemengen filamentöser Algen dadurch fördern, dass sie sich von grasenden Krustentieren im System ernähren (Moksnes et al., 2008; Baden et al., 2010). Die Entnahme von Grasern und Räubern mithilfe von Käfigen verschiedener Art vor Ort ergab, dass die natürliche Dichte kleinerer Räuber in Seegrasbeeten zu einer Abnahme von Krustentier-Mesograsern um 98 % führt und das Seegraswachstum dadurch negativ beeinflusst, dass die Menge filamentöser Algen steigt, die in den Küstengebieten des Kattegat störende Teppiche bilden.
47
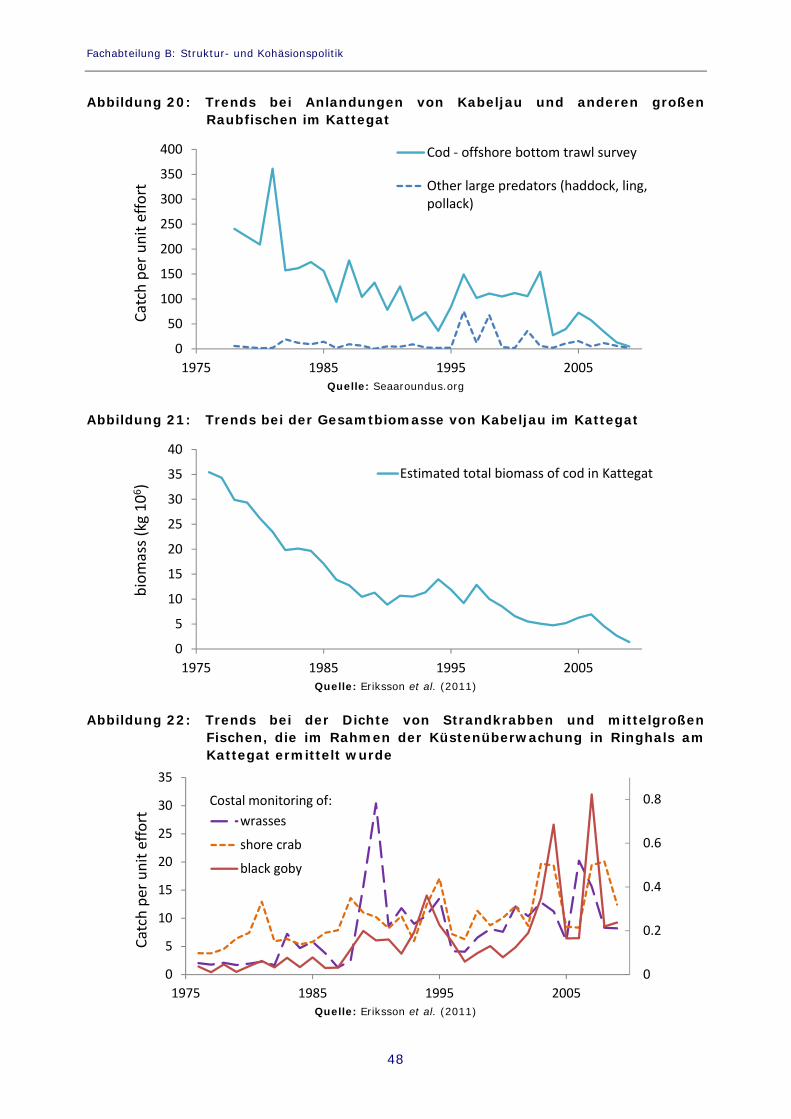
Fachabteilung B: Struktur- und Kohäsionspolitik
Abbildung 20: Trends bei Anlandungen von Kabeljau und anderen großen Raubfischen im Kattegat
0
50
100
150
200
250
300
350
400
1975 1985 1995 2005
Catch per unit effort
Cod ‐ offshore bottom trawl survey
Other large predators (haddock, ling,pollack)
Quelle: Seaaroundus.org
Abbildung 21: Trends bei der Gesamtbiomasse von Kabeljau im Kattegat
0
5
10
15
20
25
30
35
40
1975 1985 1995 2005
biomass (kg 106)
Estimated total biomass of cod in Kattegat
Quelle: Eriksson et al. (2011)
Abbildung 22: Trends bei der Dichte von Strandkrabben und mittelgroßen
Fischen, die im Rahmen der Küstenüberwachung in Ringhals am Kattegat ermittelt wurde
0
0.2
0.4
0.6
0.8
0
5
10
15
20
25
30
35
1975 1985 1995 2005
Catch per unit effort
Costal monitoring of:
wrasses
shore crab
black goby
Quelle: Eriksson et al. (2011)
48
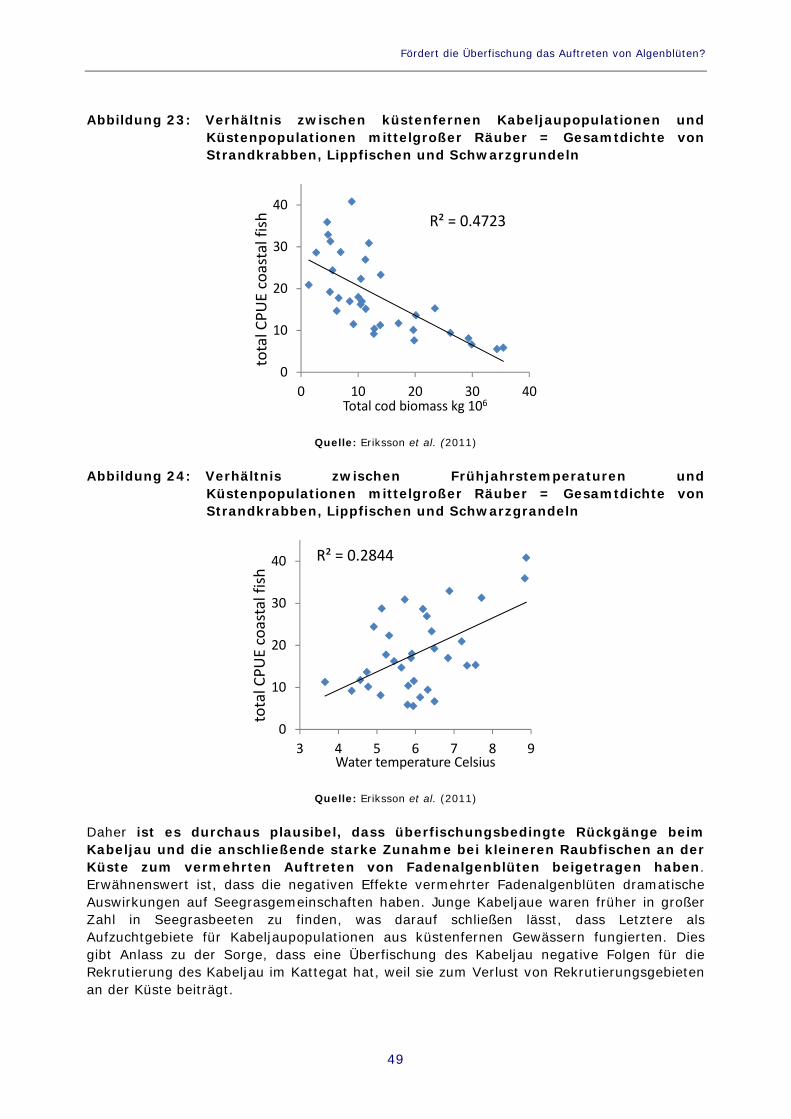
Fördert die Überfischung das Auftreten von Algenblüten?
Abbildung 23: Verhältnis zwischen küstenfernen Kabeljaupopulationen und
Küstenpopulationen mittelgroßer Räuber = Gesamtdichte von Strandkrabben, Lippfischen und Schwarzgrundeln
R² = 0.4723
0
10
20
30
40
0 10 20 30 4
total CPUE coastal fish
Total cod biomass kg 1060
Quelle: Eriksson et al. (2011) Abbildung 24: Verhältnis zwischen Frühjahrstemperaturen und
Küstenpopulationen mittelgroßer Räuber = Gesamtdichte von Strandkrabben, Lippfischen und Schwarzgrandeln
R² = 0.2844
0
10
20
30
40
3 4 5 6 7 8 9
total CPUE coastal fish
Water temperature Celsius
Quelle: Eriksson et al. (2011) Daher ist es durchaus plausibel, dass überfischungsbedingte Rückgänge beim Kabeljau und die anschließende starke Zunahme bei kleineren Raubfischen an der Küste zum vermehrten Auftreten von Fadenalgenblüten beigetragen haben. Erwähnenswert ist, dass die negativen Effekte vermehrter Fadenalgenblüten dramatische Auswirkungen auf Seegrasgemeinschaften haben. Junge Kabeljaue waren früher in großer Zahl in Seegrasbeeten zu finden, was darauf schließen lässt, dass Letztere als Aufzuchtgebiete für Kabeljaupopulationen aus küstenfernen Gewässern fungierten. Dies gibt Anlass zu der Sorge, dass eine Überfischung des Kabeljau negative Folgen für die Rekrutierung des Kabeljau im Kattegat hat, weil sie zum Verlust von Rekrutierungsgebieten an der Küste beiträgt.
49

Fachabteilung B: Struktur- und Kohäsionspolitik
4.5. Fallstudie: Ostsee im küstennahen Bereich In der Ostsee hat sich das küstenferne Nahrungsnetz seit den 1980er Jahren dramatisch verändert: Der Bestand an Kabeljau als dominierender demersaler Fisch ist stark rückläufig, und bei der Phytoplanktonbiomasse sind ebenso Steigerungen zu verzeichnen wie bei der Häufigkeit von Algenblüten (Kahru et al., 1994; Poutanen und Nikkila, 2001; Alheit et al., 2005; Hansson, 2005; Casini et al., 2008; Möllmann et al., 2008; HELCOM, 2009). Zunächst, d. h. nach umfangreicher Befischung des Kabeljaus in den 1970er Jahren, gingen die Kabeljaupopulationen in den küstenfernen Gewässern der Ostsee in den 80er Jahren um 75 % zurück (Abbildung 25) (ICES, 2010). Auf diesen Rückgang folgte eine Zunahme bei der Sprotte (Sprattus sprattus), der den wichtigsten Beutefisch für Kabeljau darstellt und zum dominierenden planktivoren Tiefseefisch avancierte (Österblom et al., 2007; Casini et al., 2008). Zum Zooplankton liegen nur wenige Erkenntnisse vor, doch zeigen Daten aus der Gotlandsee in der südlichen Ostsee, dass die Sommerbiomasse der wichtigsten Zooplanktonbeutearten der Sprotte (etliche große Copepoden und Cladocera (Crustacea)), in den 90er Jahren verglichen mit den 1980er Jahren um 50 % zurückging (Casini et al., 2008). Gleichzeitig verdoppelte sich die anhand der Chlorophyllkonzentrationen im Wasser geschätzte Phytoplanktonbiomasse (Abbildung 26). Des Weiteren war in den 1990er Jahren in der Gotlandsee im Vergleich zu den 80er Jahren eine deutliche Zunahme der Phytoplanktonbiomasse in Oberflächengewässern während der Frühjahrsblüte von März bis Mai zu verzeichnen (Abbildung 27) (Wasmund et al., 1998). Es gibt starke Indizien dafür, dass auch tiefere trophische Ebenen in der Ostsee von Veränderungen in der Zusammensetzung der Fischgemeinschaft betroffen waren (Österblom et al., 2007, Casini et al., 2008). Vor allem bestand zwischen Kabeljau- und Sprottenbiomasse, Sprotten- und Zooplanktonbiomasse, Zooplankton- und Phytoplanktonbiomasse eine deutlich ausgeprägte gegenseitige inverse Abhängigkeit (Zahlenmaterial von Casini et al. (2008), unter Verwendung der Daten für die Zoo- und Phytoplanktonbiomasse aus der Gotlandsee in der südlichen Ostsee; Tabelle 3). Des Weiteren wurde, wie für eine trophische Kaskade vorhergesagt, ein positiver Zusammenhang zwischen nicht benachbarten trophischen Ebenen ermittelt: die geringe Kabeljaubiomasse ging mit einer geringen Zooplanktonbiomasse einher, und die hohe Sprottenbiomasse fiel mit einer hohen Phytoplanktonbiomasse zusammen. Zweitens verglichen Casini et al. (2008) den Einfluss jeder höheren trophischen Ebene auf die niedrigere trophische Ebene mit dem Effekt verschiedener Umgebungsvariablen, einschließlich Temperatur, Salzgehalt und Klimaschwankungen insgesamt (NAO-Winterindex). Der Vergleich ergab durchgängig, dass die Biomasse oder Dichte der höheren trophischen Ebene der ausschlaggebende Faktor für die Erklärung der Abweichung auf der niedrigeren Trophieebene war: Die Kabeljaudichte lieferte die Erklärung für 90 % der jährlichen Abweichung bei Sprotten; aus der Sprottendichte erklärte sich zu 83 % die jährliche Abweichung bei der Zooplanktonbiomasse; und die Zooplanktonbiomasse lieferte die Erklärung für 91 % der jährlichen Variation bei Phytoplankton (Abbildungen 28 und 29). Abschließend kann festgestellt werden, dass Rückgänge beim Kabeljau aufgrund des nachlassenden Prädatordrucks zu einer massiven Zunahme bei Sprotten in der offenen Ostsee geführt haben und dass dies wahrscheinlich ab Beginn der 1990er Jahre eine trophische Kaskade mit generell geringeren Zooplanktonbiomassen und höheren Phytoplanktonbiomassen in Gang gesetzt hat. Die Zunahme der Phytoplanktonbiomasse fällt mit einer deutlichen Zunahme der Intensität der Frühjahrsblüte zusammen (Abbildung 27), und in den 1990er und 200er Jahren wurde auch eine Häufung und Zunahme der Intensität von Cyanobakterienblüten in der Ostsee festgestellt (Abbildung 5) (Kahru et al., 1994; Poutanen und Nikkila, 2001; Hansson, 2005; HELCOM, 2009). Daher
50
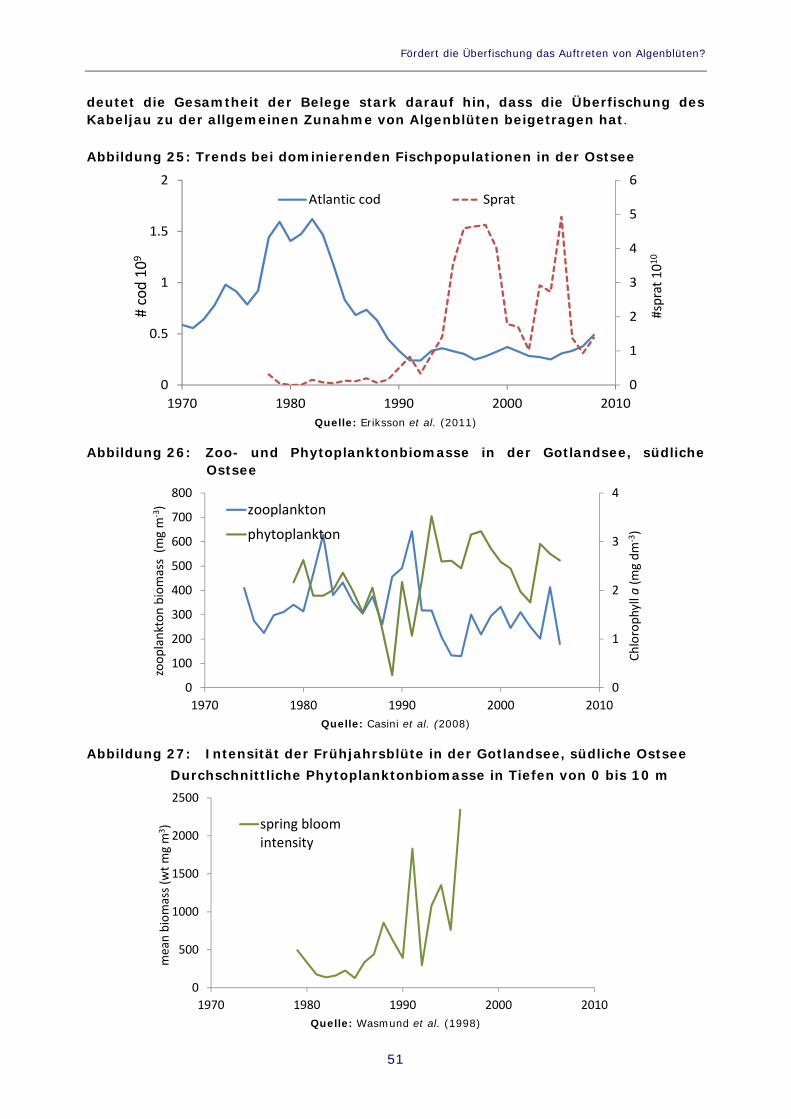
Fördert die Überfischung das Auftreten von Algenblüten?
deutet die Gesamtheit der Belege stark darauf hin, dass die Überfischung des Kabeljau zu der allgemeinen Zunahme von Algenblüten beigetragen hat. Abbildung 25: Trends bei dominierenden Fischpopulationen in der Ostsee
0
1
2
3
4
5
6
0
0.5
1
1.5
2
1970 1980 1990 2000 2010
#sprat 10
10
# cod 109
Atlantic cod Sprat
Quelle: Eriksson et al. (2011)
Abbildung 26: Zoo- und Phytoplanktonbiomasse in der Gotlandsee, südliche
Ostsee
0
1
2
3
4
0
100
200
300
400
500
600
700
800
1970 1980 1990 2000 2010
Chlorophyll a
(mg dm
‐3)
zooplankton biomass (mg m
‐3) zooplankton
phytoplankton
Quelle: Casini et al. (2008)
Abbildung 27: Intensität der Frühjahrsblüte in der Gotlandsee, südliche Ostsee
Durchschnittliche Phytoplanktonbiomasse in Tiefen von 0 bis 10 m
0
500
1000
1500
2000
2500
1970 1980 1990 2000 2010
mean biomass (w
t mg m
3) spring bloom
intensity
Quelle: Wasmund et al. (1998)
51
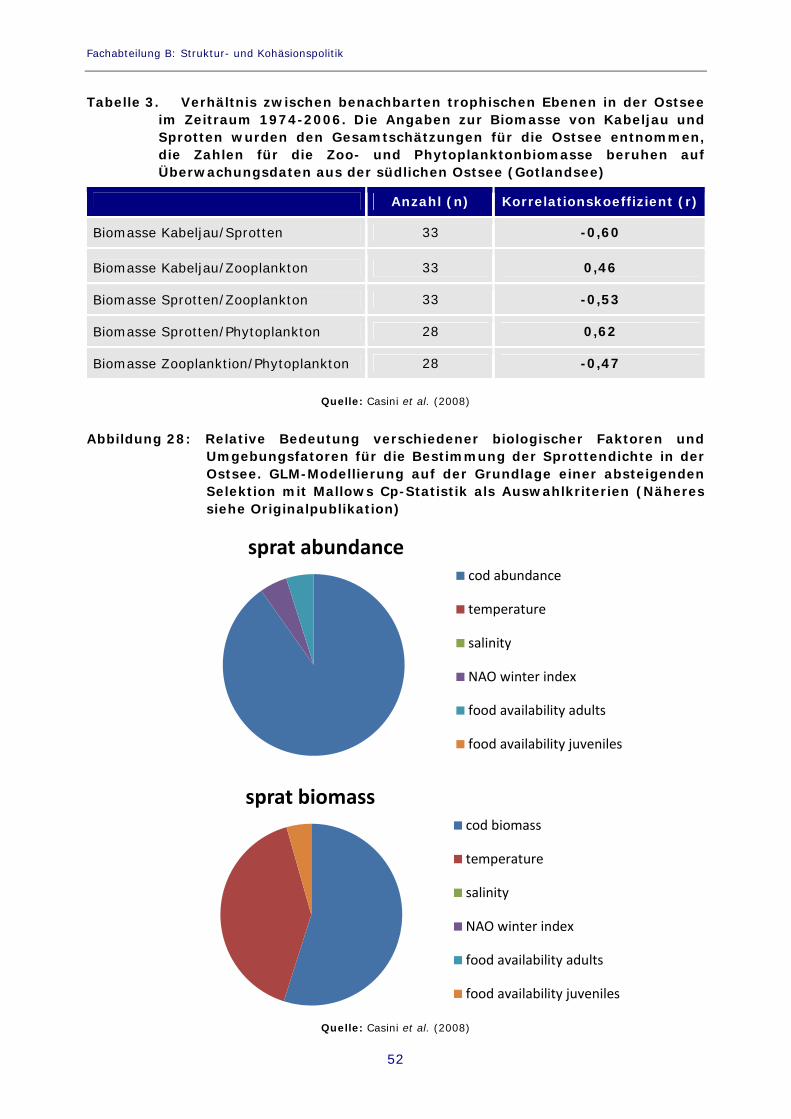
Fachabteilung B: Struktur- und Kohäsionspolitik
Tabelle 3. Verhältnis zwischen benachbarten trophischen Ebenen in der Ostsee im Zeitraum 1974-2006. Die Angaben zur Biomasse von Kabeljau und Sprotten wurden den Gesamtschätzungen für die Ostsee entnommen, die Zahlen für die Zoo- und Phytoplanktonbiomasse beruhen auf Überwachungsdaten aus der südlichen Ostsee (Gotlandsee)
Anzahl (n) Korrelationskoeffizient (r)
Biomasse Kabeljau/Sprotten 33 -0,60
Biomasse Kabeljau/Zooplankton 33 0,46
Biomasse Sprotten/Zooplankton 33 -0,53
Biomasse Sprotten/Phytoplankton 28 0,62
Biomasse Zooplanktion/Phytoplankton 28 -0,47
Quelle: Casini et al. (2008)
Abbildung 28: Relative Bedeutung verschiedener biologischer Faktoren und
Umgebungsfatoren für die Bestimmung der Sprottendichte in der Ostsee. GLM-Modellierung auf der Grundlage einer absteigenden Selektion mit Mallows Cp-Statistik als Auswahlkriterien (Näheres siehe Originalpublikation)
sprat abundancecod abundance
temperature
salinity
NAO winter index
food availability adults
food availability juveniles
sprat biomasscod biomass
temperature
salinity
NAO winter index
food availability adults
food availability juveniles
Quelle: Casini et al. (2008)
52
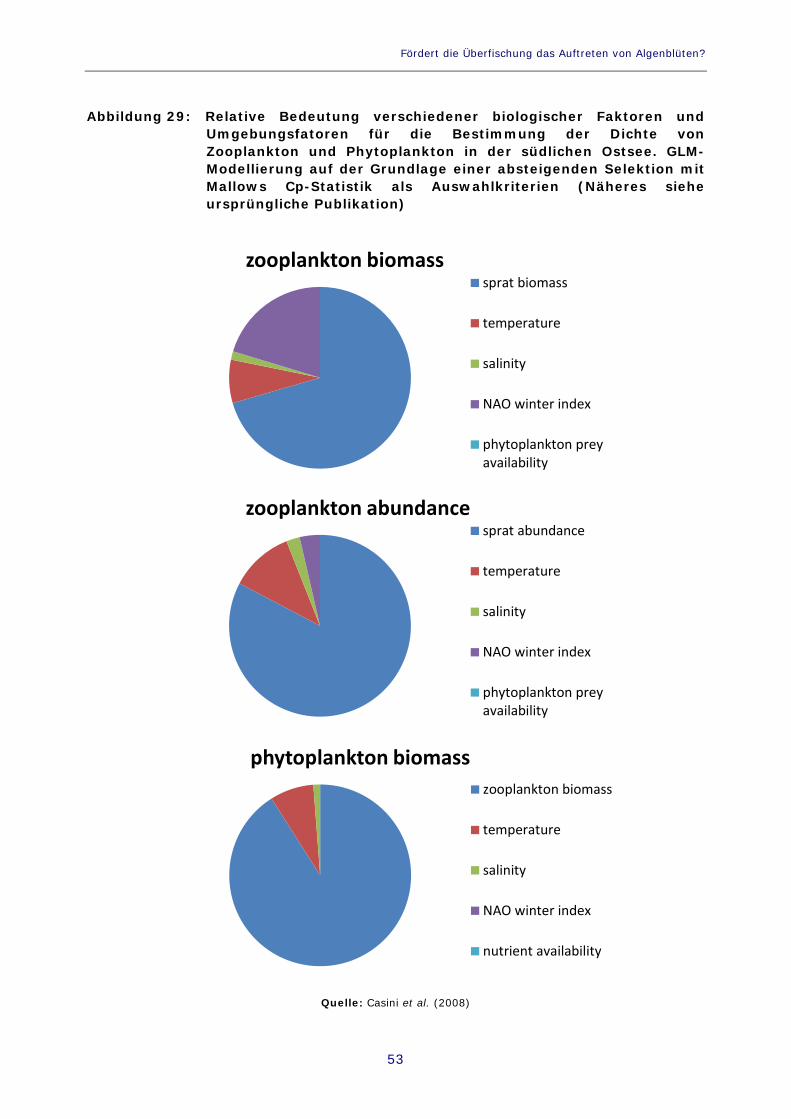
Fördert die Überfischung das Auftreten von Algenblüten?
Abbildung 29: Relative Bedeutung verschiedener biologischer Faktoren und
Umgebungsfatoren für die Bestimmung der Dichte von Zooplankton und Phytoplankton in der südlichen Ostsee. GLM-Modellierung auf der Grundlage einer absteigenden Selektion mit Mallows Cp-Statistik als Auswahlkriterien (Näheres siehe ursprüngliche Publikation)
zooplankton biomasssprat biomass
temperature
salinity
NAO winter index
phytoplankton preyavailability
zooplankton abundancesprat abundance
temperature
salinity
NAO winter index
phytoplankton preyavailability
phytoplankton biomass
zooplankton biomass
temperature
salinity
NAO winter index
nutrient availability
Quelle: Casini et al. (2008)
53

Fachabteilung B: Struktur- und Kohäsionspolitik
4.6. Fallstudie: Ostsee im Küstenbereich Ebenso wie im Fall des Kattegat gibt es deutliche Anhaltspunkte von der Küste der zentralen Ostsee, dass Fadenalgenblüten als direkte Folge der Kabeljaufischerei in küstenfernen Gewässern zugenommen haben, doch geht dies in diesem Fall mit lokalen Rückgängen bei den Küstenpopulationen von Barsch und Hecht einher (Eriksson et al., 2009; Ljunggren et al., 2010; Eriksson et al., 2011; Sieben et al., 2011a, 2011b). Seit 1990 sind die Fänge bei den dominierenden größeren Räubern im küstennahen Bereich - Barsch (Perca fluviatilis) und Hecht (Esox lucius) - an der schwedischen Ostseeküste kontinuierlich gesunken (Abbildungen 30 und 31). Die Rückgänge bei Barsch und Hecht sind örtlich begrenzt, da sich die Populationen in einigen Gebieten offenbar in einem guten Zustand befinden, in anderen Gebieten aber vollständig verschwunden sind. Ein Indiz dafür sind auch die Ausfälle bei der Rekrutierung, die sich in den offenen Inselgebieten konzentrieren, während die Rekrutierung und starke Populationen von Barsch und Hecht in den geschützten Gebieten an der Küste offenbar noch begünstigt werden. Dementsprechend ergibt sich aus der langfristigen Überwachung des Kalmarsund, eines Meeresarms vor der schwedischen Ostseeküste, eine allgemeine Abnahme größerer Räuber, einschließlich ehemals großer Küstenpopulationen des Kabeljau (Abbildung 32) (Eriksson et al., 2011). Somit sind von den Veränderungen in der Struktur der Fischgemeinschaft in den offenen Gewässern der Ostsee auch küstennahe Gebiete betroffen. In den 70er und 80er Jahren des 20. Jahrhunderts wurden üblicherweise hohe Kabeljaudichten im ufernahen Bereich gemeldet, doch seit Anfang der 1990er Jahre ist der Kabeljau verschwunden. Gegenwärtig beherrscht der dreistachlige Stichling (Gasterosteus aculeatus) – ein kleinerer Raubfisch, der sich von Krustentieren und Gastropoden, die Mesograser sind, ernährt – vollständig das Bild in geschützten Küstengemeinschaften (Eriksson et al., 2009; Ljunggren et al., 2010). Stichlinge spielen in Küstenüberwachungsprogrammen kaum eine Rolle, weil die Mehrheit nach der Rekrutierungszeit im Sommer in küstenferne Gebiete abwandert; an den Küsten werden Stichlinge hauptsächlich im Mai und Juni gesichtet (unveröffentlichte Daten). Mithilfe der Echolotung in küstenfernen Gebieten wurde jedoch festgestellt, dass die Stichlingspopulationen in den Jahren 2006-2010 sprunghaft angestiegen sind (Abbildung 33) (Ljunggren et al., 2010). Daher könnte eine starke Zunahme bei den Stichlingen sowohl eine Folge des nachlassenden Prädatordrucks geschwächter Küstenpopulationen von Kabeljau, Barsch und Hecht als auch eine Folge des Rückgangs von Kabeljaupopulationen in küstenfernen Gebieten sein. Einige Anhaltspunkte sprechen dafür, dass die hohen Stichlingsdichten auf die geschrumpften Populationen von Barsch und Hecht zurückzuführen sind und dass diese Verschiebung in der Zusammensetzung der Fischgemeinschaft indirekt das Auftreten von Fadenalgenblüten fördert. Weder findet eine Überwachung wirbelloser Graser statt noch liegen genügend Zeitreihen für filamentöse Algen vor, um zeitliche Trends zu analysieren. Allerdings treten an der schwedischen Ostseeküste vermehrt filamentöse Algen auf und nimmt die Dichte von Stichlingen zu, wenn die Dichte von Barsch und Hecht abnimmt (Eriksson et al., 2009). In Gebieten mit geringer Dichte von Barsch und Hecht nehmen Stichlinge in Fischgemeinschaften eine dominierende Stellung ein und nahezu 50 % der flachen Buchten sind mit filamentösen Algen zugewuchert (Abbildung 34). In Gebieten mit hoher Barsch- und Hechtdichte kommen Stichlinge selten vor, und lediglich 10 % der Buchten sind zugewuchert. In der Ostsee wurde im Rahmen von Versuchen auch ein kausaler Zusammenhang zwischen Küstenräubern und Algen nachgewiesen. In einer Bucht mit vielen Stichlingen, in der häufig filamentöse Algen wuchern, wurde durch eine groß angelegte Aktion, bei der
54
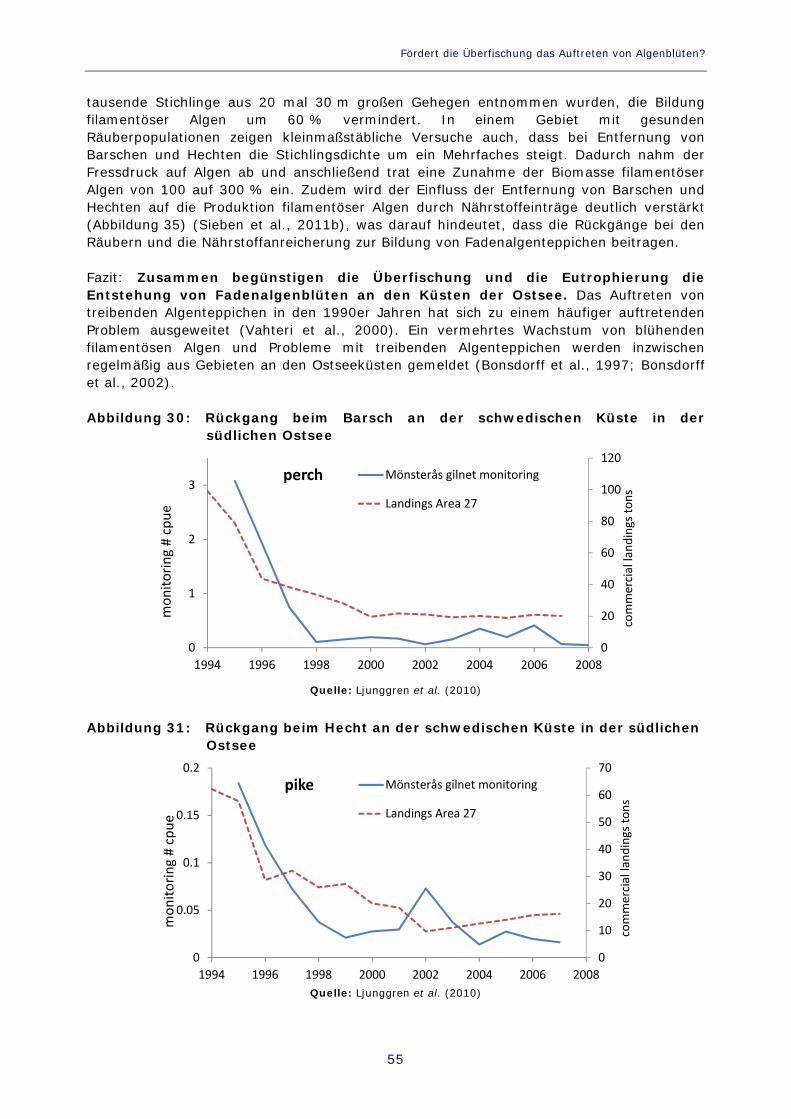
Fördert die Überfischung das Auftreten von Algenblüten?
tausende Stichlinge aus 20 mal 30 m großen Gehegen entnommen wurden, die Bildung filamentöser Algen um 60 % vermindert. In einem Gebiet mit gesunden Räuberpopulationen zeigen kleinmaßstäbliche Versuche auch, dass bei Entfernung von Barschen und Hechten die Stichlingsdichte um ein Mehrfaches steigt. Dadurch nahm der Fressdruck auf Algen ab und anschließend trat eine Zunahme der Biomasse filamentöser Algen von 100 auf 300 % ein. Zudem wird der Einfluss der Entfernung von Barschen und Hechten auf die Produktion filamentöser Algen durch Nährstoffeinträge deutlich verstärkt (Abbildung 35) (Sieben et al., 2011b), was darauf hindeutet, dass die Rückgänge bei den Räubern und die Nährstoffanreicherung zur Bildung von Fadenalgenteppichen beitragen. Fazit: Zusammen begünstigen die Überfischung und die Eutrophierung die Entstehung von Fadenalgenblüten an den Küsten der Ostsee. Das Auftreten von treibenden Algenteppichen in den 1990er Jahren hat sich zu einem häufiger auftretenden Problem ausgeweitet (Vahteri et al., 2000). Ein vermehrtes Wachstum von blühenden filamentösen Algen und Probleme mit treibenden Algenteppichen werden inzwischen regelmäßig aus Gebieten an den Ostseeküsten gemeldet (Bonsdorff et al., 1997; Bonsdorff et al., 2002). Abbildung 30: Rückgang beim Barsch an der schwedischen Küste in der
südlichen Ostsee
0
20
40
60
80
100
120
0
1
2
3
1994 1996 1998 2000 2002 2004 2006 2008
commercial landings tons
monitoring # cpue
perch Mönsterås gilnet monitoring
Landings Area 27
Quelle: Ljunggren et al. (2010) Abbildung 31: Rückgang beim Hecht an der schwedischen Küste in der südlichen
Ostsee
0
10
20
30
40
50
60
70
0
0.05
0.1
0.15
0.2
1994 1996 1998 2000 2002 2004 2006 2008
commercial landings tons
monitoring # cpue
pike Mönsterås gilnet monitoring
Landings Area 27
Quelle: Ljunggren et al. (2010)
55
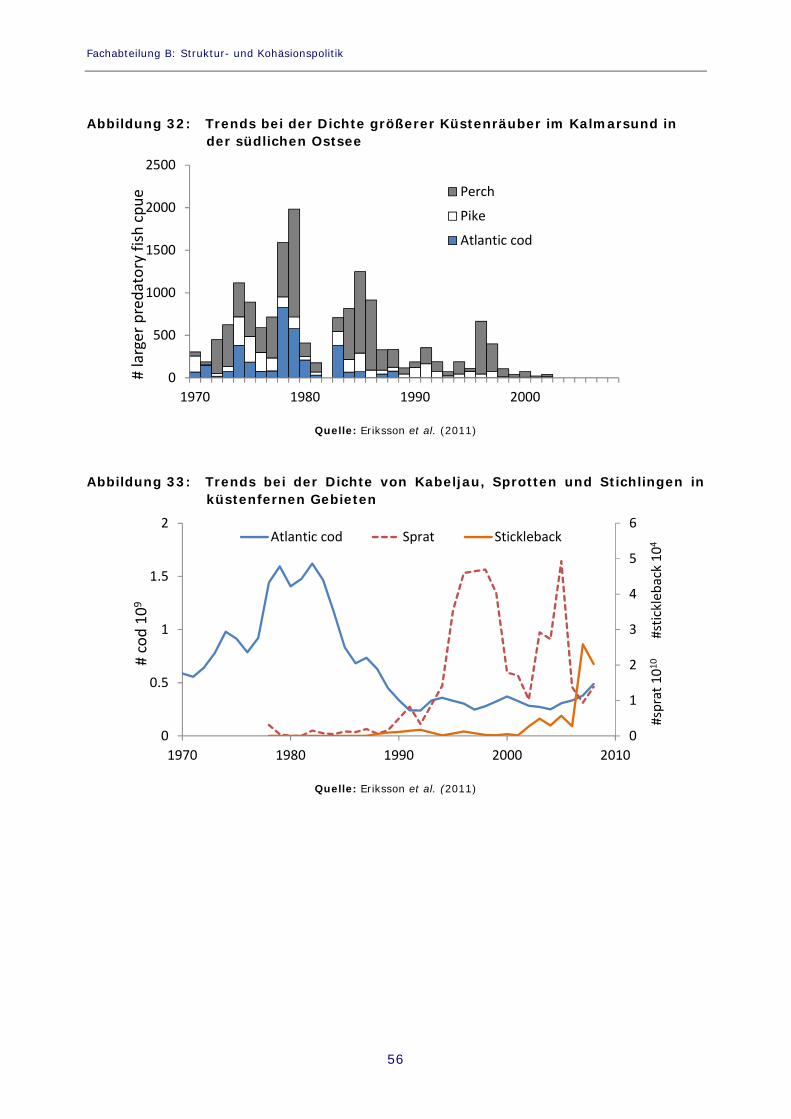
Fachabteilung B: Struktur- und Kohäsionspolitik
Abbildung 32: Trends bei der Dichte größerer Küstenräuber im Kalmarsund in
der südlichen Ostsee
0
500
1000
1500
2000
2500
1970 1980 1990 2000
# larger predatory fish cpue Perch
Pike
Atlantic cod
Quelle: Eriksson et al. (2011) Abbildung 33: Trends bei der Dichte von Kabeljau, Sprotten und Stichlingen in
küstenfernen Gebieten
0
1
2
3
4
5
6
0
0.5
1
1.5
2
1970 1980 1990 2000 2010
#sprat 1010 #stickleback 104
# cod 109
Atlantic cod Sprat Stickleback
Quelle: Eriksson et al. (2011)
56
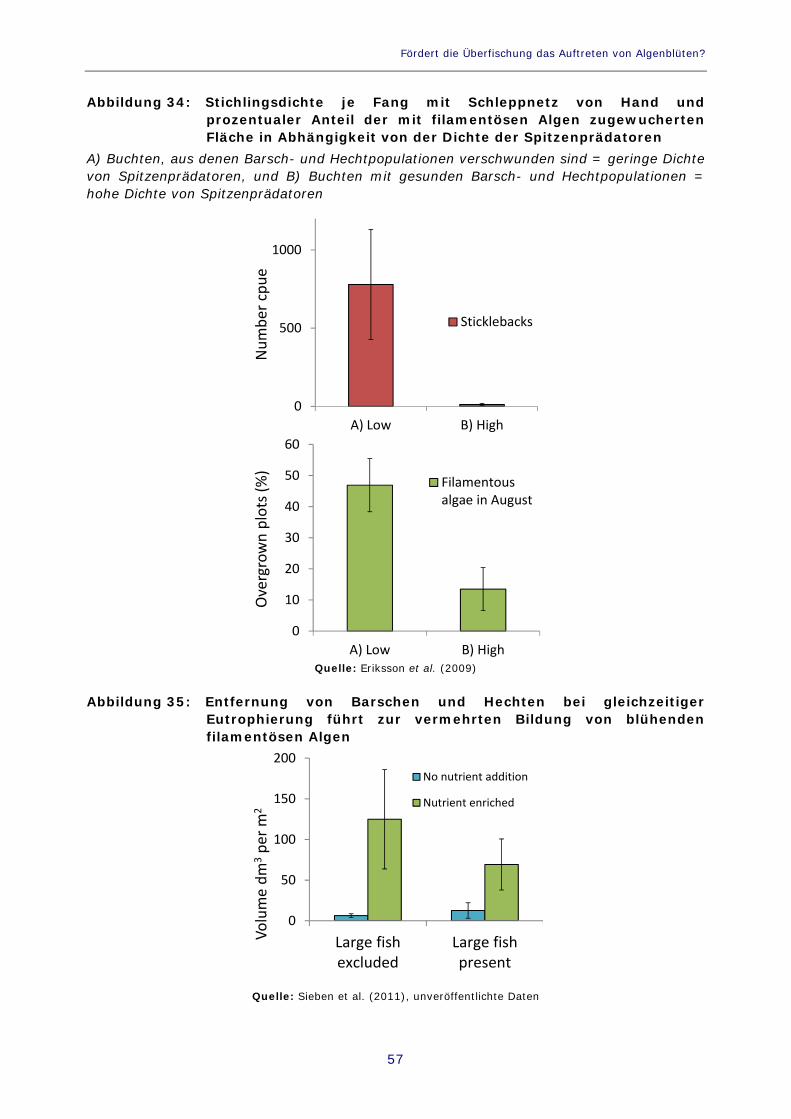
Fördert die Überfischung das Auftreten von Algenblüten?
Abbildung 34: Stichlingsdichte je Fang mit Schleppnetz von Hand und prozentualer Anteil der mit filamentösen Algen zugewucherten Fläche in Abhängigkeit von der Dichte der Spitzenprädatoren
A) Buchten, aus denen Barsch- und Hechtpopulationen verschwunden sind = geringe Dichte von Spitzenprädatoren, und B) Buchten mit gesunden Barsch- und Hechtpopulationen = hohe Dichte von Spitzenprädatoren
0
500
1000
A) Low B) High
Number cpue
Sticklebacks
0
10
20
30
40
50
60
A) Low B) High
Overgrown plots (%)
Filamentousalgae in August
Quelle: Eriksson et al. (2009)
Abbildung 35: Entfernung von Barschen und Hechten bei gleichzeitiger
Eutrophierung führt zur vermehrten Bildung von blühenden filamentösen Algen
0
50
100
150
200
Large fishexcluded
Large fishpresent
Volume dm
3per m
2
No nutrient addition
Nutrient enriched
Quelle: Sieben et al. (2011), unveröffentlichte Daten
57

Fachabteilung B: Struktur- und Kohäsionspolitik
58

Fördert die Überfischung das Auftreten von Algenblüten?
5. SCHLUSSFOLGERUNGEN UND EMPFEHLUNG Diese Untersuchung liefert deutliche Anhaltspunkte dafür, dass die Überfischung von größeren, an der Spitze der Nahrungskette stehenden Raubfischen dazu beiträgt, dass Algenblüten in Europa häufiger und mit größerer Intensität auftreten: Algenblüten sind Naturphänomene, die durch eine Reihe von Umweltfaktoren ausgelöst werden. Die Überfischung trägt zum Problem der Algenblüten bei, indem sie in einen wichtigen biologischen Mechanismus zur Kontrolle übermäßigen Wachstums eingreift. Das Verschwinden von Spitzenprädatoren infolge der Überfischung führt zu Veränderungen in der Struktur von Nahrungsnetzen, die im Allgemeinen die Ausbreitung kleinerer Fische fördern und Verluste bei der Dichte von Algengrasern wie Zooplankton oder wirbellosen Herbivoren zur Folge haben. Neben abiotischen Faktoren halten Graser den Algenbiomassewuchs unter Kontrolle, und wenn bei den Grasern ein Rückgang zu verzeichnen ist, können sich Algenblüten leichter ausbreiten. Daher variieren die Auswirkungen der Fischerei auf Algenblüten je nach System und Jahreszeit und in den einzelnen Jahren. Dies ist ein wichtiger Hinweis, wird doch dadurch die Ermittlung und Bewältigung von Auswirkungen der Fischerei auf Algenblüten zusätzlich erschwert. Die Reaktionen von Algen stehen nicht in einem linearen Zusammenhang mit dem Aufkommen an Spitzenprädatoren, sondern hängen auch von anderen Umständen ab, die etwaige Effekte einer Überfischung mehr oder minder verstärken können. Daher ist der Zugriff auf abgleichbare Langzeitdaten, die alle trophischen Ebenen im marinen Nahrungsnetz erfassen, wichtig für die Klärung der Frage, inwieweit die Fischerei zu Algenblüten beiträgt. In der Ostsee und im Schwarzen Meer hat die Überfischung Veränderungen in der Struktur der Ökosysteme verursacht und höchstwahrscheinlich zum größer werdenden Problem schädlicher Algenblüten beigetragen: Ungeachtet der Komplexität der Ökologie und Regulierung von Algenblüten gibt es starke Indizien dafür, dass die Überfischung zu den größer werdenden Problemen mit Algenblüten bei gleichzeitigen Rückgängen der kommerziellen Bestände an größeren Raubfischen und erheblichen Zunahmen der Algenbiomasse und schädlicher Algenblüten in etlichen europäischen Meeren beigetragen hat. Was die Ostsee und das Schwarze Meer anbelangt, untermauern Analysen von Zeitreihen und zeitgleich auftretenden Ereignissen die Annahme, dass die Überfischung zum größer werdenden Problem der Algenblüten und zum Anstieg der dadurch verursachten wirtschaftlichen Kosten beigetragen hat. Vor allem deuten Tendenzen in der Ostsee darauf hin, dass von den nachteiligen Auswirkungen der Überfischung küstenferner Bestände inzwischen auch Küstenökosysteme betroffen sind, wodurch die Wasserqualität beeinträchtigt wird und Lebensräume im küstennahen Bereich verloren gehen. Für das Kattegat fehlen Daten zu Tendenzen bei der Algenblüte, doch deuten Versuche darauf hin, dass der dokumentierte langfristige Rückgang der Kabeljaupopulationen in küstenfernen Gebieten und Küstengebieten nachteilige Auswirkungen auf Seegrasgemeinschaften in Form der zunehmenden Ausbreitung von filamentösen Algen hatte. Für die Nordsee werden anhand einer Modellierung und der Analyse verfügbarer Daten zunehmende Algenbelastungen vorhergesagt; in diesem Zusammenhang wird auf dokumentierte, durch die Fischerei verursachte Veränderungen in der Struktur der Fischgemeinschaft verwiesen. Es fehlt generell an Trendanalysen für mehr als nur eine Gruppe von Organismen aus mehreren Meeresbecken in Europa, aus denen Probleme gemeldet werden, die sowohl die
59

Fachabteilung B: Struktur- und Kohäsionspolitik
Überfischung als auch Algenblüten betreffen. Daraus lässt sich schließen, dass wichtige Informationen fehlen, weil Daten für Vergleichszwecke nicht öffentlich verfügbar sind. Die Analysen zu den Ökosystemen der Ostsee, des Schwarzen Meeres und der Nordsee veranschaulichen, dass die Verbindungen zwischen den trophischen Ebenen untersucht werden müssen, um die Auswirkungen der Fischereien verstehen zu können. Überfischung und Eutrophierung erzeugen Synergieeffekte: Das bedeutet doppelte Probleme! Veränderungen in den Gemeinschaften der Spitzenprädatoren und Algenblüten treten zeitgleich mit der Eutrophierung auf. Es besteht kein Zweifel daran, dass die europaweite Eutrophierung Algenblüten gefördert hat, doch später haben die Folgen der veränderten Zusammensetzung von Fischgemeinschaften das Problem schädlicher Algenblüten weiter verschärft. So traten beispielsweise ab dem Einsetzen der Eutrophierung in den 1960er/1970er Jahren in der Ostsee vermehrt Probleme mit anoxischen Meeresgründen auf, die durch eine stärkere Algenbildung verursacht wurden. Was die letzten Jahrzehnte anbelangt, lassen sich die jährlichen Schwankungen bei der Phytoplanktonbiomasse wesentlich besser anhand des Grasens erklären als anhand der Nährstoffsituation (geringe Zooplanktonbiomasse = hohe Phytoplanktonbiomasse; Abbildung 29). In der nährstoffbelasteten Ostsee hat die Überfischung beispielsweise zum Anstieg der Algenbiomasse beigetragen. Zudem belegen Feldversuche eindeutig, dass das Wachstum filamentöser Algen sowohl durch die Zufuhr von Nährstoffen als auch durch das Verschwinden von Spitzenprädatoren begünstigt wird und beide Komponenten - Nährstoffeintrag und Raubfischfang – gemeinsam oft wesentlich stärker wirken als jede für sich genommen. Der Zusammenhang zwischen dichten Algenblüten an den Küsten und Überfischung ist am deutlichsten ausgeprägt Es gibt keinen eindeutigen Beweis dafür, dass bestimmten Arten oder Algenblüten empfindlicher oder unempfindlicher auf Veränderungen in der Zusammensetzung von Fischgemeinschaften reagieren. Aus den verfügbaren Daten lassen sich jedoch einige allgemeine Tendenzen ablesen. Bei den schädlichen Algenblüten weisen nichttoxische dichte Algenblüten die größte Empfindlichkeit gegenüber einer Überfischung auf, da toxische oder ungenießbare Blüten weniger stark vom Grasen betroffen sein dürften. Dementsprechend kommen die schlüssigsten Beweise für Überfischungseffekte aus Küstenbuchten, in denen Rückgänge bei größeren Raubfischen nichttoxische Blüten teppichbildender filamentöser Algen begünstigen. Modellergebnisse, die auf verfügbarer Erkenntnissen über Küstenmeere in gemäßigten Zonen und Daten aus der Nordsee beruhen, belegen auch, dass größere nichttoxische Diatomeen und Dinoflagellaten die wichtigsten der Gruppen darstellen, bei denen aufgrund der Überfischung größerer Raubfische eine Zunahme eintreten würde. Daher dürfte die Überfischung hauptsächlich dichte Algenblüten fördern und Probleme wie 1) treibende Algenteppiche, 2) Schleim- und Schaumbildung und 3) Anoxie nach der Algenblüte hervorrufen. Allerdings ergab die von Vasas et al. (2007) durchgeführte Analyse des Nahrungsnetzes der Nordsee auch, dass die Intensivfischerei unerwartete Wirkungen hervorbringen kann, wenn in die Konkurrenzhierarchien zwischen Algen und Planktivoren eingegriffen wird. Die Ergebnisse deuten darauf hin, dass vermehrt toxische Algenblüten und Quallen auftreten, wenn man zulässt, dass kommerzielle Fischereien rückläufige Fänge bei größeren Raubfischen durch die intensivere Befischung kleinerer planktivorer Fische („fishing down the food web“) ausgleichen. Dies zeigt sich auch am Nahrungsnetz des Schwarzen Meeres,
60

Fördert die Überfischung das Auftreten von Algenblüten?
wo nach dem Zusammenbruch der Anchovis-Intensivfischerei in den 1990er Jahren die Quallenpopulation dramatisch zunahm. Die Überfischung dürfte Algenblüten hauptsächlich dann fördern, wenn eine einzige Art befischt wird, die zahlen- und funktionsmäßig in der Fischgemeinschaft eine Vorrangstellung einnimmt Die Auswirkungen der Entnahme von Raubfischen auf die Beutearten kommen im Allgemeinen wesentlich stärker zum Tragen, wenn die Gemeinschaft der Räuber von einer einzigen Art dominiert wird. Das belegen die am besten dokumentierten europäischen Fallstudien. In der Ostsee und im Kattegat besteht ein Zusammenhang zwischen den Auswirkungen und Rückgängen bei einem einzigen Räuber (dem Kabeljau), der in den Gemeinschaften demersaler Räuber an der Spitze der Nahrungskette stand. Im Schwarzen Meer waren weitere Spitzenprädatoren an den anfänglichen Veränderungen des Ökosystems in den 1960er und 1970er Jahren beteiligt. Die stärksten Auswirkungen auf das Ökosystem, einschließlich eines vermehrten Algenwuchses, traten auf, als die pelagische Gemeinschaft kleinerer Fische um das Jahr 1990 herum zusammenbrach. In dieser Gemeinschaft dominierte eine einzige, massiv befischte Anchovisart. Dies war ein Beleg für die Notwendigkeit der Nutzung der Biodiversität als Bewirtschaftungsstrategie, als Instrument zum Erhalt einer diversifizierten Fischerei und artenreicher Fischgemeinschaften mit als Puffer fungierenden marinen Systemen zur Abwehr schädlicher Kaskadeneffekte, die Algenblüten fördern. Schädliche Algenblüten haben negative Auswirkungen auf die Fischerei in Europa Schätzungen zufolge entstehen der europäischen Fischerei durch schädliche Algenblüten Gesamtkosten von mindestens 177 Mio. EUR pro Jahr. Darin eingeschlossen sind die nachteiligen Auswirkungen auf die kommerzielle Fischerei, soziale Verluste und die Kosten der Überwachung von Algenblüten zwecks Feststellung einer Toxinbelastung von Meeresfrüchten. Toxische Algenblüten verursachen auch hohe Kosten im Aquakultursektor. Die direkten Verluste an marktfähigen Erzeugnissen im Muschelsektor belaufen sich auf 15 bis 62 Mio. EUR pro Jahr. Dichte Algenblüten bereiten Probleme für jede Art von Küstenfischerei. Treibende Algenteppiche und schleimbildende Algen geraten in die Fanggeräte und verringern deren Wirksamkeit. Die nach einer Algenblüte auftretende Anoxie verursacht eine erhebliche Mortalität bei Aquakultur- und Wildfischbeständen. Zudem beeinträchtigt sie die Reproduktion entlang der Küste, wovon auch die wichtigsten Rekrutierungsgebiete zahlreicher Fischpopulationen in küstenfernen Gebieten betroffen sind. In der Ostsee kann die erhöhte Anoxie am Meeresgrund sogar die Rekrutierung des Kabeljaubestands in küstenfernen Gebieten beeinträchtigen. Empfehlung: ökosystembasierte Bewirtschaftung Im vorliegenden Themenpapier wird aufgezeigt, dass die traditionelle Bewirtschaftung von Meeresressourcen gravierende Schwachstellen hat, werden doch oftmals die Interaktionen sowohl innerhalb von Nahrungsnetzen als auch zwischen Nahrungsnetzen vor und an der Küste außer Acht gelassen. Kaskadeneffekte der Überfischung verändern die Beziehungen zwischen Raub- und Beutearten und bewirken damit, dass schädliche Algenblüten häufiger und mit größerer Intensität auftreten; dies wiederum schmälert die Rentabilität der Fischereiindustrie und kann negative langfristige Auswirkungen auf wichtige kommerziell befischte Bestände haben. Negative Rückwirkungen können somit die langfristige Nachhaltigkeit mariner Ressourcen beeinträchtigen. Allerdings zahlt nicht allein der Fischereisektor den Preis dafür; betroffen sind auch die Küstengemeinden, in denen wachsende Probleme aufgrund von Algenblüten zu einem nennenswerten Rückgang der
61

Fachabteilung B: Struktur- und Kohäsionspolitik
Tourismuseinnahmen führen und Kosten verursachen, die durch die negativen Auswirkungen auf die menschliche Gesundheit bedingt sind. Es hat sich erwiesen, dass ein ökosystembasiertes Bewirtschaftungskonzept (EBM) benötigt wird, das eine geeignetere Grundlage für ein integriertes Fischerei- und Küstenmanagement ist. Mit dem künftigen EBM sollten folgende zwei Hauptziele verfolgt werden: 1) Berücksichtigung der Auswirkungen der Nutzung höherer trophischer Ebenen auf das Ökosystem und 2) räumliche Überlegungen, die auch den fließenden Übergängen zwischen Lebensräumen vor und an der Küste Rechnung tragen. Die hat zwei unmittelbare Konsequenzen: Erstens wird die Berücksichtigung der Auswirkungen von Fischereien auf das Ökosystem als erste Maßnahme eine Änderung der Bewirtschaftungsziele erfordern. Der Wiederaufbau von Räuberpopulationen dient nicht nur dazu, die Produktion der Zielarten zu maximieren, sondern sie auch auf ein Niveau anzuheben, auf dem ihre ökologische Funktion in küstenfernen Nahrungsnetzen und Küstennahrungsnetzen wiederhergestellt wird. Zu diesem Zweck muss die Dichte von Raubfischen, die an der Spitze der Nahrungskette stehen, so weit erhöht werden, bis sie wieder die Kontrolle über niedrigere Trophieebenen haben; dies schließt die stärkere Kontrolle von blühenden Algen durch Graser ein. Zweitens werden die Zusammenhänge zwischen den verschiedenen Bewirtschaftungszielen traditionell separater Bewirtschaftungssektoren sichtbar, wenn den wichtigen fließenden Übergängen zwischen küstenfernen Lebensräumen und Küstenhabitaten Rechnung getragen wird; dies betrifft solche Fragen wie die Bedeutung der Wiederherstellung der Funktion von Fischbeständen in küstenfernen Gewässern als Voraussetzung für die Verbesserung der Wasserqualität durch Verhinderung von Algenblüten und die Bedeutung eines stärkeren Schutzes wertvoller Küstenhabitate für die Rekrutierung zahlreicher küstenferner Bestände kommerziell wertvoller Fische.
62

Fördert die Überfischung das Auftreten von Algenblüten?
LITERATURVERZEICHNIS
Alheit J., Niquen, M., 2004. Regime shifts in the Humboldt Current ecosystem. Progress in Oceanography 60, 201-222.
Alheit J., Möllmann C., Dutz J., Kornilovs G., Loewe P., Mohrholz V., Wasmund N., 2005. Synchronous ecological regime shifts in the central Baltic and the North Sea in the late 1980s. ICES Journal of Marine Science 62, 1205-1215.
Baden S., Gullstrom M., Lunden B., Pihl L., Rosenberg R., 2003. Vanishing seagrass (Zostera marina, L.) in Swedish coastal waters. Ambio 32, 374-377.
Baden S., Bostrom C., Tobiasson S., Arponen H., Moksnes P.O., 2010. Relative importance of trophic interactions and nutrient enrichment in seagrass ecosystems: A broad-scale field experiment in the Baltic-Skagerrak area. Limnology and Oceanography 55, 1435-1448.
Baden S., Emanuelsson A., Pihl L., Svensson C.J., Åberg P., 2011. Shift in seagrass food web structure over decades linked to overfishing. ASLO Aquatic Sciences Meeting, 13-18 February 2011, San Juan, Puerto Rico, USA.
Beaugrand G., 2004. The North Sea regime shift: evidence, causes, mechanisms and consequences. Progress in Oceanography 60, 245-262.
Bonsdorff E., Blomqvist E. M., Mattila J., Norkko A., 1997. Coastal eutrophication: causes, consequences and perspectives in the archipelago areas of the northern Baltic Sea. Estuarine, Coastal and Shelf Science 44, 63-72.
Bonsdorff E., Ronnberg C., Aarnio, K., 2002. Some ecological properties in relation to eutrophication in the Baltic Sea. Hydrobiologia 475, 371-377.
Borer E.T., Seabloom E.W., Shurin J.B., Anderson K.E., Blanchette C.A., Broitman B., Cooper S.D., Halpern B.S., 2005. What determines the strength of a trophic cascade? Ecology 86, 528-537.
Borer E.T., Halpern B.S., Seabloom E.W., 2006. Asymmetry in community regulation: Effects of predators and productivity. Ecology 87, 2813-2820.
Breitburg D., 2002. Effects of hypoxia, and the balance between hypoxia and enrichment, on coastal fishes and fisheries. Estuaries 25, 767-781.
Burkholder J.M., Azanza R.V., Sako Y., 2006. The ecology of harmful dinoflagellates. In: Graneli E., Turner J.T. (editors). Ecology of harmful algal blooms. Springer.
Carpenter S.R., Kitchell J.F., Hodgson J.R., 1985. Cascading Trophic Interactions and Lake Productivity. Bioscience 35, 634-639.
Carpenter S.R., Cole J.J., Hodgson J.R., Kitchell J.F., Pace M.L., Bade D., Cottingham K.L., Essington T.E., Houser J.N., Schindler D.E., 2001. Trophic cascades, nutrients, and lake productivity: whole-lake experiments. Ecological Monographs 71, 163-186.
Casini M., Lovgren J., Hjelm J., Cardinale M., Molinero J.C., Kornilovs G., 2008. Multi-level trophic cascades in a heavily exploited open marine ecosystem. Proceedings of the Royal Society B-Biological Sciences 275, 1793-1801.
Christensen N.L., Bartuska A.M., Brown J.H., Carpenter S., Dantonio C., Francis R., Franklin J.F., MacMahon J.A., Noss R.F., Parsons D.J., Peterson C.H., Turner M.G., Woodmansee R.G., 1996. The report of the ecological society of America committee on the scientific basis for ecosystem management. Ecological Applications 6, 665-691.
63

Fachabteilung B: Struktur- und Kohäsionspolitik
Christensen V., Guenette S., Heymans J.J., Walters C.J., Watson R., Zeller D., Pauly D., 2003. Hundred-year decline of North Atlantic predatory fishes. Fish and Fisheries 4, 1-24.
Cloern J.E., 2001. Our evolving conceptual model of the coastal eutrophication problem. Marine Ecology Progress Series 210, 223-253.
Dahl E., Bagoien E., Edvardsen B., Stenseth N.C., 2005. The dynamics of Chrysochromulina species in the Skagerrak in relation to environmental conditions. Journal of Sea Research 54, 15-24.
Daskalov G.M., 2002. Overfishing drives atrophic cascade in the Black Sea. Marine Ecology Progress Series 225, 53-63.
Daskalov G.M., Grishin A.N., Rodionov S., Mihneva V., 2007. Trophic cascades triggered by overfishing reveal possible mechanisms of ecosystem regime shifts. Proceedings of the National Academy of Sciences of the United States of America 104, 10518-10523.
de Boer M.K., 2006. Fibrocapsa japonica (Raphidophyceae) in Dutch coastal waters. University of Groningen, Groningen.
Degobbis D., 1989. Increased eutrophication of the northern Adriatic Sea - 2nd act. Marine Pollution Bulletin 20, 452-457.
ECOHARM, 2003. The socio-economic impact of harmful algal blooms in European marine waters. http://www.bom.hik.se/ecoharm/index.html.
Edvardsen B., Imai I., 2006. The ecology of harmful flagellates. In: Graneli E., Turner J.T. (editors). Ecology of harmful algal blooms. Springer.
Elmhagen B., Rushton S.P., 2007. Trophic control of mesopredators in terrestrial ecosystems: top-down or bottom-up? Ecology Letters 10, 197-206.
Eriksson B.K., Rubach A., Hillebrand H., 2007. Dominance by a canopy forming seaweed modifies resource and consumer control of bloom-forming macroalgae. Oikos 116, 1211-1219.
Eriksson B.K., Ljunggren L., Sandstrom A., Johansson G., Mattila J., Rubach A., Raberg S., Snickars M., 2009. Declines in predatory fish promote bloom-forming macroalgae. Ecological Applications 19, 1975-1988.
Eriksson B.K., Sieben K., Eklöf J., Ljunggren L., Olsson J., Casini M., Bergström U., 2011. Effects of altered offshore food webs on coastal ecosystems emphasizes the need for cross-ecosystem management. Ambio 40, 786-797.
Estes J.A., Tinker M.T., Williams T.M., Doak D.F., 1998. Killer whale predation on sea otters linking oceanic and nearshore ecosystems. Science 282, 473-476.
Fallesen G., Andersen F., Larsen B., 2000. Life, death and revival of the hypertrophic Mariager Fjord, Denmark. Journal of Marine Systems 25, 313-321.
Finni T., Kononen K., Olsonen R., Wallstrom K., 2001. The history of cyanobacterial blooms in the Baltic Sea. Ambio 30, 172-178.
Frank K.T., Petrie B., Choi J.S., Leggett W.C., 2005. Trophic cascades in a formerly cod-dominated ecosystem. Science 308, 1621-1623.
Frank K.T., Petrie B., Shackell N.L., 2007. The ups and downs of trophic control in continental shelf ecosystems. Trends in Ecology and Evolution 22, 236-242.
Glibert P.M., Burkholder J.M., 2006. The complex relation between increases in fertilization of the earth, coastal eutrophication and profiliation of harmful algal blooms. In: Graneli E., Turner J.T. (editors). Ecology of harmful algal blooms. Springer.
64

Fördert die Überfischung das Auftreten von Algenblüten?
Granéli E., Paasche E., Maestrini S.Y., 1996. Three years after the Chrysochromulina polylepis bloom in Scandinavian waters in 1988: Some conclusions of recent research and monitoring. In: Smayda T.J., Shimizu Y. (editors). Toxic Phytoplankton Blooms in the Sea. Elsevier.
Graneli E., Turner J.T., 2006. Ecology of harmful algal blooms. Springer.
Gruner D. S., Smith J.E., Seabloom E.W., Sandin S.A., Ngai J.T., Hillebrand H., Harpole W.S., Elser J.J., Cleland E.E., Bracken M.E.S., Borer E.T., Bolker B.M., 2008. A cross-system synthesis of consumer and nutrient resource control on producer biomass. Ecology Letters 11, 740-755.
Hallegraeff G.M. 1993. A review of harmful algal blooms and their apparent global increase. Phycologia 32, 79-99.
Hansson M., 2005. Cyanobakterieblomningar i Östersjön, resultat från satellitövervakning 1997-2005. 0283-7714 ICES, Sveriges Meteorologiska och Hydrologiska Institut, Oceanografiska enheten, Göteborg.
Hansson M., Öberg J., 2009. Cyanobacterial blooms in the Baltic Sea. Swedish Meteorological and Hydrological Institute.
Heisler J., Glibert P.M., Burkholder J.M., Anderson D.M., Cochlan W., Dennison W.C., Dortch Q., Gobler C.J., Heil C.A., Humphries E., Lewitus A., Magnien R., Marshall H.G., Sellner K., Stockwell D.A., Stoecker D.K., Suddleson M., 2008. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 8, 3-13.
HELCOM, 2009. Eutrophication in the Baltic Sea - An integrated thematic assessment of the effects of nutrient enrichment ansd eutrophication in the Baltic Sea region. Baltic Sea Environmental Proceedings 115B.
Hinder S.L., Hays G.C., Brooks C.J., Davies C.J., Edwards M., Walne A.W., Gravenor M. B., 2011. Toxic marine microalgae and shellfish poisoning in the British Isles: history, review of epidemiology, and future implications. Environmental Health 10.
Hoagland P., Scatasta S., 2006. The economic effects of harmful algal blooms. In: Graneli E., Turner J.T. (editors). Ecology of harmful algal blooms. Springer.
Hoagland P., Anderson D.M., Kaoru Y., White A.W., 2002. The economic effects of harmful algal blooms in the United States: Estimates, assessment issues, and information needs. Estuaries 25, 819-837.
Hopcraft J.G. C., Olff H., Sinclair A.R.E., 2010. Herbivores, resources and risks: alternating regulation along primary environmental gradients in savannas. Trends in Ecology and Evolution 25, 119-128.
ICES, 2010. Report of the Baltic Fisheries Assessment Working Group (WGBFAS). ICES CM 2010/ACOM, 10.
Jackson J.B.C., Kirby M.X., Berger W.H., Bjorndal K.A., Botsford L.W., Bourque B.J., Bradbury R.H., Cooke R., Erlandson J., Estes J.A., Hughes T.P., Kidwell S., Lange C.B., Lenihan H.S., Pandolfi J.M., Peterson C.H., Steneck R.S., Tegner M.J., Warner R.R., 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science 293, 629-638.
Kahru M., Horstmann U., Rud O., 1994. Satellite detection of increased cyanobacteria blooms in the Baltic Sea - natural fluctuation or ecosystem change? Ambio 23, 469-472.
Kideys A.E., 1994. Recent dramatic changes in the Black Sea ecosystem - the reason for the sharp decline in Turkis anchovy fisheries. Journal of Marine Systems 5, 171-181.
65

Fachabteilung B: Struktur- und Kohäsionspolitik
Lancelot C., Billen G., Sournia A., Weisse T., Colijn F., Veldhuis M.J.W., Davies A., Wassman P., 1987. Phaeocystis blooms and nutrient enrichment in the continental coastal zones of the North Sea. Ambio 16, 38-46.
Lancelot C., Keller M.D., Rousseau V., Smith Jr. W.O., Mathot S., 1998. Autecology of the marine Haptophyte Phaeocystis sp. In: D.M. Anderson, A.D. Cembella, G.M. Hallegraeff (editors). Physiological Ecology of Harmful Algal Blooms. NATO ASI Series, Springer-Verlag, Berlin, 209-224.
Leggett W.C., Frank K.T., 2008. Paradigms in fisheries oceanography. Oceanography and Marine Biology: an Annual Review, 46.
Ljunggren L., Sandstrom A., Bergstrom U., Mattila J., Lappalainen A., Johansson G., Sundblad G., Casini M., Kaljuste O., Eriksson B.K., 2010. Recruitment failure of coastal predatory fish in the Baltic Sea coincident with an offshore ecosystem regime shift. ICES Journal of Marine Science 67, 1587-1595.
Maestrini S.Y., Graneli E., 1991. Environmental conditions and ecophysiological mechanisms which led to the 1988 chrysochromulina-polylepis bloom - an hypothesis. Oceanologica Acta 14, 397-413.
McQuatters-Gollop A., Raitsos D.E., Edwards M., Pradhan Y., Mee L.D., Lavender S.J., Attrill M.J., 2007. A long-term chlorophyll data set reveals regime shift in North Sea phytoplankton biomass unconnected to nutrient trends. Limnology and Oceanography 52, 635-648.
Menge B.A., 2000. Top-down and bottom-up community regulation in marine rocky intertidal habitats. Journal of Experimental Marine Biology and Ecology 250, 257-289.
Moksnes P.-O., Gullstrom M., Tryman K., Baden S., 2008. Trophic cascades in a temperate seagrass community. Oikos 117, 763-777.
Möllmann C., Muller-Karulis B., Kornilovs G., St John M.A., 2008. Effects of climate and overfishing on zooplankton dynamics and ecosystem structure: regime shifts, trophic cascade, and feedback coops in a simple ecosystem. ICES Journal of Marine Science 65, 302-310.
Mountain D.G., Kane J., 2010. Major changes in the Georges Bank ecosystem, 1980s to the 1990s. Marine Ecology Progress Series 398, 81-91.
Nastasi A., 2010. Algal and Jellyfish Blooms in the Mediterranean and Black Sea: a brief review. In: GFCM Workshop on Algal and Jellyfish Blooms in the Mediterranean and Black Sea: a brief review, Istanbul, Turkey.
Newcombe E.M., Taylor R.B., 2010. Trophic cascade in a seaweed-epifauna-fish food chain. Marine Ecology Progress Series 408, 161-167.
Norderhaug K.N., Christie H., Fossa J.H., Fredriksen S., 2005. Fish-macrofauna interactions in a kelp (Laminaria hyperborea) forest. Journal of the Marine Biological Association of the United Kingdom 85, 1279-1286.
Oksanen L., Oksanen, T., 2000. The logic and realism of the hypothesis of exploitation ecosystems. American Naturalist 155, 703-723.
Olsson P., Folke C., Hughes T.P., 2008. Navigating the transition to ecosystem-based management of the Great Barrier Reef, Australia. Proceedings of the National Academy of Sciences of the United States of America 105, 9489-9494.
Osterblom H., Gardmark A., Bergstrom L., Muller-Karulis B., Folke C., Lindegren M., Casini M., Olsson P., Diekmann R., Blenckner T., Humborg C., Mollmann C., 2010. Making the ecosystem approach operational-Can regime shifts in ecological- and governance systems facilitate the transition? Marine Policy 34, 1290-1299.
66

Fördert die Überfischung das Auftreten von Algenblüten?
Österblom H., Hansson S., Hjerne O., Larsson U., Wulff F., Elmgren R., Folke C., 2007. Human-induced trophic cascades and ecological regime shifts in the Baltic Sea. Ecosystems 10, 887-889.
Pace M.L., Cole J.J., Carpenter S.R., Kitchell J.F., 1999. Trophic cascades revealed in diverse ecosystems. Trends in Ecology and Evolution 14, 483-488.
Passow U., Wassmann P., 1994. On the trophic fate of Phaeocystis pouchetii (Hariot): IV. The formation of marine snow by pouchetii. Marine Ecology Progress Series 104, 153-161.
Pauly D., Christensen V., Dalsgaard J., Froese R., Torres F., 1998. Fishing down marine food webs. Science 279, 860-863.
Peaerl H.W., Fulton III R. S., 2006. Ecology of harmful cyanobacteria. In: Graneli E., Turner J.T. (editors). Ecology of harmful algal blooms. Springer.
Peperzak L., Poelman M., 2008. Mass mussel mortality in The Netherlands after a bloom of Phaeocystis globosa (prymnesiophyceae). Journal of Sea Research 60, 220-222.
Pierik M., van Ruijven J., Bezemer T.M., Geerts R., Berendse F., 2011. Recovery of plant species richness during long-term fertilization of a species-rich grassland. Ecology 92, 1393-1398.
Pihl L., 1982. Food-intake of young cod and flounder in a shallow bay on the Swedish west-coast. Netherlands Journal of Sea Research 15, 419-432.
Pihl L., Ulmestrand M., 1993. Migration pattern of juvenile cod (Gadus morhua) on the Swedish west coast. ICES J. Mar. Sci. 50, 63-70.
Pihl L., Magnusson G., Isaksson I., Wallentinus I., 1996. Distribution and growth dynamics of ephemeral macroalgae in shallow bays on the Swedish west coast. Journal of Sea Research 35, 169-180.
Pihl L., Svenson A.M., Moksnes P-O., Wennhage H., 1999. Distribution of green algal mats throughout shallow soft bottoms of the Swedish Skagerrak archipelago in relation to nutrient sources and wave exposure. Journal of Sea Research 41, 281-294.
Pihl L., Baden S., Kautsky N., Ronnback P., Soderqvist T., Troell M., Wennhage H., 2006. Shift in fish assemblage structure due to loss of seagrass Zostera marina habitats in Sweden. Estuarine Coastal and Shelf Science 67, 123-132.
Polis G.A., Strong D.R., 1996. Food web complexity and community dynamics. American Naturalist 147, 813-846.
Poutanen E.L., Nikkila K., 2001. Carotenoid pigments as tracers of cyanobacterial blooms in recent and post-glacial sediments of the Baltic Sea. Ambio 30, 179-183.
Power M.E., 1990. Effects of Fish in River Food Webs. Science 250, 811-814.
Reid, P.C., Borges M.D., Svendsen E., 2001. A regime shift in the North Sea circa 1988 linked to changes in the North Sea horse mackerel fishery. Fisheries Research 50, 163-171.
Richardson K., Heilmann J.P., 1995. Primary production in the Kattegat: Past and present. Ophelia 41, 317-328.
Riebesell U., Reigstad M., Wassmann P., Noji T., Passow U., 1995. On the trophic fate of Phaeocystis pouchetii (hariot): VI. Significance of Phaeocystis-derived mucus for vertical flux. Netherlands Journal of Sea Research 33, 193-203.
Riegman R., Noordeloos A.A.M., Cadee G.C., 1992. Phaeocystis blooms and eutrophication of the continental coastal zones of the North Sea. Marine Biology 112, 479-484.
67

Fachabteilung B: Struktur- und Kohäsionspolitik
Rousseau V., Becquevort S., Parent J.Y., Gasparini S., Daro M.H., Tackx M., Lancelot C., 2000. Trophic efficiency of the planktonic food web in a coastal ecosystem dominated by Phaeocystis colonies. Journal of Sea Research 43, 357-372.
Rutter T., 2010. Managing Risk and Uncertainty. In: Advancing the Aquaculture Agenda, Paris.
Salvanes A.G.V., Nordeide, J.T., 1993. Dominating sublittoral fish species in a west Norwegian fjord and their trophic links to cod (Gadus morhua L.). Sarsia 78, 221-234.
Scheffer M., Carpenter S., de Young B., 2005. Cascading effects of overfishing marine systems. Trends in Ecology and Evolution 20, 579-581.
Sellner K.G., Doucette G.J., Kirkpatrick G.J., 2003. Harmful algal blooms: causes, impacts and detection. Journal of Industrial Microbiology and Biotechnology 30, 383-406.
Shurin J.B., Borer E.T., Seabloom E.W., Anderson K., Blanchette C.A., Broitman B., Cooper S.D., Halpern, B.S., 2002. A cross-ecosystem comparison of the strength of trophic cascades. Ecology Letters 5, 785-791.
Sieben K., Ljunggren L., Bergstrom U., Eriksson B.K., 2011a. A meso-predator release of stickleback promotes recruitment of macroalgae in the Baltic Sea. Journal of Experimental Marine Biology and Ecology 397, 79-84.
Sieben K., Rippen A.D., Eriksson B.K., 2011b. Cascading effects from predator removal depend on resource availability in a benthic food web. Marine Biology 158, 391-400.
Sinclair A.R.E., Krebs C.J., Fryxell J.M., Turkington R., Boutin S., Boonstra R., Seccombe-Hett P., Lundberg P., Oksanen, L., 2000. Testing hypotheses of trophic level interactions: a boreal forest ecosystem. Oikos 89, 313-328.
Smayda T.J., 1997. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnology and Oceanography 42, 1137-1153.
Smayda T.J., 2006. Harmful Algal Bloom Communities in Scottish Coastal Waters: Relationship to Fish Farming and Regional Comparisons – A Review. Scottish Executive Environment Group Paper 2006/3 2005.
Smith V. H., Schindler D.W., 2009. Eutrophication science: where do we go from here? Trends in Ecology and Evolution 24, 201-207.
Solic M., Krstulovic N., Marasovic I., Baranovic A., Pucher Petkovic T., Vucetic T., 1997. Analysis of time series of planktonic communities in the Adriatic Sea: Distinguishing between natural and man-induced changes. Oceanologica Acta 20, 131-143.
Sondergaard M., Jeppesen E., Lauridsen T.L., Skov C., Van Nes E.H., Roijackers R., Lammens E., Portielje R., 2007. Lake restoration: successes, failures and long-term effects. Journal of Applied Ecology 44, 1095-1105.
Strong D.R., 1992. Are trophic cascades all wet? Differentiation and donor-control in speciose ecosystems. Ecology 73, 747-754.
Thornton D.C.O., Santillo D., Thake B., 1999. Prediction of Sporadic Mucilaginous Algal Blooms in the Northern Adriatic Sea. Marine Pollution Bulletin 38, 891-898.
Tufekci V., Balkis N., Beken C.P., Ediger D., Mantikci M., 2010. Phytoplankton composition and environmental conditions of a mucilage event in the Sea of Marmara. Turkish Journal of Biology 34, 199-210.
Turk V., Hagstrom A., Kovac N., Faganeli J., 2010. Composition and function of mucilage macroaggregates in the northern Adriatic. Aquatic Microbial Ecology 61, 279-289.
68

Fördert die Überfischung das Auftreten von Algenblüten?
Vahteri P., Makinen A., Salovius S., Vuorinen I., 2000. Are drifting algal mats conquering the bottom of the Archipelago Sea, SW Finland? Ambio 29, 338-343.
Van Dolah F.M., 2000. Marine algal toxins: Origins, health effects, and their increased occurrence. Environmental Health Perspectives 108, 133-141.
Vasas V., Lancelot C., Rousseau V., Jordán F., 2007. Eutrophication and overfishing in temperate nearshore pelagic food webs: a network perspective. Marine Ecology Progress Series 336, 1-14.
Viitasalo M., Peltonen H., Pääkkönen J.-P., 2005. Blue-green algal blooms interfere with the growth of young fish and cause fishermen financial losses. The Baltic Sea Portal http://www.itameriportaali.fi/en/ajankohtaista/mtl_uutisarkisto/2005/en_GB/210.
Vila M., Camp J., Garces E., Maso M., Delgado M., 2001a. High resolution spatio-temporal detection of potentially harmful dinoflagellates in confined waters of the NW Mediterranean. Journal of Plankton Research 23, 497-514.
Vila M., Garces E., Maso M., Camp J., 2001b. Is the distribution of the toxic dinoflagellate Alexandrium catenella expanding along the NW Mediterranean coast? Marine Ecology Progress Series 222, 73-83.
Ward P., Myers R.A., 2005. Shifts in open-ocean fish communities coinciding with the commencement of commercial fishing. Ecology 86, 835-847.
Wasmund N., Nausch G., Matthaus W., 1998. Phytoplankton spring blooms in the southern Baltic Sea - spatio-temporal development and long-term trends. Journal of Plankton Research 20, 1099-1117.
Weijerman M., Lindeboom H., Zuur A.F., 2005. Regime shifts in marine ecosystems of the North Sea and Wadden Sea. Marine Ecology Progress Series 298, 21-39.
World Health Organization, 2002. Eutrophication and Health. European Communities Report http://europa.eu.int.
Worm B., Lotze H.K., 2006. Effects of eutrophication, grazing, and algal blooms on rocky shores. Limnology and Oceanography 51, 569-579.
Worm B., Lotze H.K., Hillebrand H., Sommer U., 2002. Consumer versus resource control of species diversity and ecosystem functioning. Nature 417, 848-851.
Yunev O.A., Moncheva S., Carstensen J., 2005. Long-term variability of vertical chlorophyll a and nitrate profiles in the open Black Sea: eutrophication and climate change. Marine Ecology Progress Series 294, 95-107.
Yunev O.A., Vedernikov V.I., Basturk O., Yilmaz A., Kideys A.E., Moncheva S., Konovalov S.K., 2002. Long-term variations of surface chlorophyll a and primary production in the open Black Sea. Marine Ecology Progress Series 230, 11-28.
Zaitsev Y.P., 1992. Recent changes in the trophic structure of the Black Sea. Fisheries Oceanography 1:180-189.
Zengin M., Gungör H., Demirkol C., Yuksek A., 2009. Effects of Algal Bloom on the Fisheries of the Sea of Marmara through the 2007/2008 Fishing Period. In: FAO-GFCM SAC/SCMEE Working Group on Workshop on algal and jelly fish blooms, Istanbul.
Zengın M., Güngör H., Demırkol C., Yüksek A., 2011. Effects of mucilage bloom on the fisheries of the Sea of Marmara through the 2007/2008 fishing period. In: Report of the workshop on algal and jellyfish blooms in the Mediterranean and the Black Sea, Istanbul. Marseille, France.
69

Fachabteilung B: Struktur- und Kohäsionspolitik
70


B
Landwirtschaft und ländliche Entwicklung
Kultur und Bildung
Fischerei
Regionale Entwicklung
Verkehr und Fremdenverkehr
B Rolle
Die Fachabteilungen sind Forschungsreferate, die die Ausschüsse, interparlamentarischen Delegationen und andere parlamentarische Einrichtungen beraten.
PolitikbereicheLandwirtschaft und ländliche EntwicklungKultur und BildungFischereiRegionale EntwicklungVerkehr und Fremdenverkehr
DokumenteSiehe Website des Europäischen Parlaments: http://www.europarl.europa.eu/studies
FachabteilungStruktur- und kohäSionSpolitik
Bildnachweise: istock international inc., Photodisk, Phovoir
Generaldirektion interne Politikbereiche
Generaldirektion interne Politikbereiche
FachabteilungStruktur- und kohäSionSpolitik
Verkehr und Fremdenverkehr
Regionale Entwicklung
Fischerei
Kultur und Bildung
Landwirtschaft und ländliche Entwicklung
DRAFT ! DRAFT ! DRAFT ! DRAFT ! DRAFT ! DRAFT PRE-RELEASE! PRE-RELEASE! PRE-RELEASE!


















