
Charakterisierung und Lokalisation der Toxoplasma gondii ... · Katalase, Peroxidasen und...
70
Aus dem Institut für Hygiene und Mikrobiologie der Universität Würzburg Vorstand: Prof. Dr. med. M. Frosch Charakterisierung und Lokalisation der Toxoplasma gondii Katalase: Peroxisomen in Apicomplexa? Inaugural – Dissertation zur Erlangung der Doktorwürde der Medizinischen Fakultät der Bayerischen Julius-Maximilians-Universität zu Würzburg vorgelegt von Achim J. Kaasch aus Rottweil Würzburg, Juli 2002
-
Upload
duongkhanh -
Category
Documents
-
view
214 -
download
0
Transcript of Charakterisierung und Lokalisation der Toxoplasma gondii ... · Katalase, Peroxidasen und...

Aus dem Institut für Hygiene und Mikrobiologie
der Universität Würzburg
Vorstand: Prof. Dr. med. M. Frosch
Charakterisierung und Lokalisation der Toxoplasma gondiiKatalase: Peroxisomen in Apicomplexa?
Inaugural – Dissertation
zur Erlangung der Doktorwürde der
Medizinischen Fakultät
der
Bayerischen Julius-Maximilians-Universität zu Würzburg
vorgelegt von
Achim J. Kaasch
aus Rottweil
Würzburg, Juli 2002

Referent: Prof. Dr. med M. Frosch
Koreferent: Prof. Dr. rer. nat. H. Moll
Dekan: Prof. Dr. med V. ter Meulen
Tag der mündlichen Prüfung: 27. Juni 2003
Der Promovend ist Arzt

Inhaltsverzeichnis
1 Einleitung 1
1.1 Die Gattung Toxoplasma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
1.2 Medizinische Bedeutung von Toxoplasma gondii . . . . . . . . . . . . . . . . 4
1.3 Antioxidative Schutzmechanismen von T. gondii . . . . . . . . . . . . . . . 5
1.4 Das Enzym Katalase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
1.5 Peroxisomen – Morphologie und enzymatische Ausstattung . . . . . . . . 9
1.6 Peroxisomen – Biogenese und Protein Targeting . . . . . . . . . . . . . . . 10
1.7 Spezialisierte Peroxisomen: Glyoxysomen und Glykosomen . . . . . . . . 12
2 Ergebnisse 15
2.1 Klonierung und Sequenzierung der Toxoplasma gondii Katalase . . . . . . . 15
2.2 Analyse der Sequenz . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
2.3 Western Blot mit Antiserum gegen T. gondii Katalase . . . . . . . . . . . . . 25
2.4 Lokalisation in der Immunfluoreszenz . . . . . . . . . . . . . . . . . . . . . 26
2.5 Lokalisation mittels Histochemie . . . . . . . . . . . . . . . . . . . . . . . . 28
2.6 Expression von Katalase in CHO-Zellen . . . . . . . . . . . . . . . . . . . . 28
2.7 Expression eines Reportergens in CHO-Zellen . . . . . . . . . . . . . . . . 29
3 Materialien und Methoden 31
3.1 Bakterienstämme . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.2 Parasitenstämme . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.3 Lambda-Phagen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.4 Zellinien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.5 Plasmide . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
3.6 Primäre Antikörper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
3.7 Sekundäre Antikörper . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
3.8 Kulturbedingungen für Zellinien . . . . . . . . . . . . . . . . . . . . . . . . 32
3.9 Passagieren der Zellinien . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
3.10 Passage der Parasiten in der Zellkultur . . . . . . . . . . . . . . . . . . . . . 33
3.11 In vivo Exzision . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
3.12 Praktisches Vorgehen bei der in vivo Excision . . . . . . . . . . . . . . . . . 35
3.13 „Rapid Amplification of cDNA Ends“ (5’ RACE) . . . . . . . . . . . . . . . 35
3.14 Sequenzierung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
3.15 Herstellung des Antiserums gegen T. gondii Katalase . . . . . . . . . . . . . 36
3.16 Antikörper Aufreinigung . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38

3.17 Western Blot . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
3.18 Immunfluoreszenz . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.19 DAB Präzipitation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.20 Klonierung der Expressions-Konstrukte . . . . . . . . . . . . . . . . . . . . 40
3.21 Transfektion der CHO-Zellen . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.22 Allgemeine molekularbiologische Techniken . . . . . . . . . . . . . . . . . 41
3.23 Oligonukleotide und Chemikalien . . . . . . . . . . . . . . . . . . . . . . . 41
3.24 Protein-Modellierung mit Swissmodel . . . . . . . . . . . . . . . . . . . . . 41
3.25 Verwendete Software . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
3.26 Puffer und Lösungen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
3.27 Benutzte Geräte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
4 Diskussion 46
5 Zusammenfassung 50
6 Abkürzungsverzeichnis 52

1 Einleitung
1.1 Die Gattung Toxoplasma
Der eukaryontische Erreger Toxoplasma gondii gehört zu den Apicomplexa, einem Phylum
der Protozoa. Dieser Gruppe gehören ca. 5000 verschiedene Spezies an, die weitreichen-
de Bedeutung in der Medizin und Biologie besitzen [51]. Weitere wichtige Vertreter der
Apicomplexa sind Plasmodium, Sarcocystis, Babesia, Eimeria, Perkinsus und Theileria.
Insgesamt zählt Toxoplasma gondii zu den am weitesten verbreiteten Krankheitserre-
gern mit einer regional unterschiedlichen Durchseuchungsrate von 40-70 %. Plasmodium
hingegen, der Erreger der Malaria, stellt mit 300-500 Millionen Fällen die weltweit häu-
figste Infektionskrankheit dar. Die Infektion von Tieren mit Eimeria, Babesia, Theileria und
Perkinsus führt zu großen Verlusten in der Viehhaltung [19, 27, 89].
Das gemeinsame taxonomisches Merkmal der Apicomplexa ist der „apikale Komplex“,
eine aus mehreren Zellorganellen bestehende Struktur am vorderen Ende der Parasiten,
die bei der Invasion der Wirtszelle eine entscheidende Rolle spielt. Der apikale Komplex
besteht aus dem Konoid, Rhoptrien und Mikronemen (Abbildung 1). Weitere elektro-
nenmikroskopisch nachweisbare Zellorganellen von T. gondii sind der Apikoplast, ein
sekundär erworbener Chloroplast [87], ein einzelnes Mitochondrium, der Golgi-Apparat
und elektronendichte, sekretorische Granula, die sogenannten „dense granules“ [50]. Der
obligat intrazelluläre Einzeller ist ca. 5-6µm lang, 2-3 µm breit und leicht gekrümmt, was
in der Namensgebung berücksichtigt wurde (toxon = Bogen).
Der Lebenszyklus von Toxoplasma (Abb. 2) wechselt fakultativ zwischen geschlecht-
licher Vermehrung (Gamogonie) im Darmepithel des Endwirts (Katzen) und der unge-
schlechtlichen Vermehrung (Schizogonie) in einer Reihe von verschiedenen Zwischen-
wirten. Im Darmepithel einer infizierten Katze entstehen Oozysten, die mit dem Kot aus-
geschieden werden (Abb. 2, 1). In einer Oozyste reifen zwei Sporozysten mit jeweils vier
Sporozoiten heran (Abb. 2, 2). Nach oraler Aufnahme durch einen Zwischenwirt, wer-
den die Sporozoiten im Darm freigesetzt und dringen in extraintestinalen Zellen ein.
Innerhalb dieser Zellen bildet sich eine Pseudozyste (Vakuole), in der sich die Parasiten
ungeschlechtlich vermehren (Abb. 2, 4). Die Teilung findet in einem 6-8 h lang dauern-
dem Zyklus statt und führt zu Tachyzoiten, die Vakuole rasch anfüllen. Nach einigen
Teilungsschritten bricht die Wirtszelle auf und die Tachyzoiten werden freigesetzt.
Über Blut und Lymphe gelangen Tachyzoiten in Hirn- und Muskelgewebe, wo sie
sich zu Bradyzoiten, der langsam wachsenden Form, weiterentwickeln können. Brady-
zoiten bilden Gewebszysten, die bis zu mehreren hundert Parasiten enthalten können
(Abb. 2, 6). Durch Genuß von rohem Fleisch werden weitere Zwischenwirte infiziert, was
1

Abbildung 1: ElektronenmikroskopischeAbbildung eines einzelnen Parasiten ineiner Wirtszelle. Parasitäre Vakuole (PV),perivakuoläre Membran (PVM), Rhoptrien(R), Mikronemen (M), Dense Granules (DG),Endoplasmatisches Retikulum (ER), Golgi(G), Nukleus (N), Mitochondrium (Mt). Zubeachten sind auch die membranassoziiertenMitochondrien der Wirtszelle (hMt). Nicht zuerkennen ist der Apikoplast.
wiederum zur Entstehung von Tachyzoiten führt (Abb. 2, 8). Eine Infektion des Endwirts
kann entweder über die Aufnahme von Oozysten, Pseudozysten oder Gewebszysten er-
folgen.
Die Tachyzoiten von T. gondii dringen aktiv in die Wirtszellen ein [28]. Dieser Vorgang
dauert ca. 20-30 Sekunden. Zuerst dockt der Parasit mit seinem vorderen Ende an der
Oberfläche der Wirtszelle an. Dann entsteht die „moving junction“, eine elektronendichte
Verbindung zwischen der Parasiten und Wirtszellmembran, die für Membranproteine
undurchlässig ist [60]. Durch aktives Eindringen des Parasiten in die Zelle entsteht eine
Vakuole, in der T. gondii sich vermehren kann.
Bei der Invasion spielt das sekretorische System von T. gondii eine entscheidende Rol-
le [30,44]. Die Produkte von Mikronemen, Rhoptrien und Granula densa werden sequen-
tiell sezerniert und tragen zur Bildung und Erhaltung der Vakuole bei [29].
Die Exozytose der Mikronemen wird durch Erhöhung des intraparasitären Kalzium-
Spiegels ausgelöst [13]. Die sezernierten Produkte der Mikronemen dienen der Fortbe-
wegung des Parasiten und Erkennung von Oberflächen, indem sie eine Verbindung zwi-
schen dem Parasiten und extrazellulären Oberflächen schaffen [94]. Mit der Exozytose
der Rhoptrien beginnt die Einstülpung der Wirtszellmembran und die Bildung der Va-
kuole. Die Rhoptrien haben einen hohen Cholesterol- und Phospholipid-Gehalt und tra-
gen vermutlich zur Bildung der vakuolären Membran bei. Die aus den Rhoptrien se-
2

Abbildung 2: Lebenszyklus vonToxoplasma gondii. Einzelheitenfinden sich im Text. Abkürzun-gen: EN Teilung durch Endodyo-genie, HC Wirtszelle, N Nukleus,NH Wirtszellnukleus, OC Oozyste,PC primäre Zystenwand, PV para-sitäre Vakuole, RB Residualkörper,SP Sporozoit, SPC Sporozoitenzy-ste. Übersicht nach Mehlhorn [57].
zernierten Proteine werden entweder in die vakuoläre Membran eingebaut oder liegen
in löslicher Form innerhalb der Vakuole vor. Eines dieser Proteine, Rop2, vermittelt die
Assoziation von Mitochondrien der Wirtszelle mit der vakuolären Membran über einen
neuartigen Mechanismus [98, 99]. Die Sekretion aus den Granula densa trägt zur Aus-
bildung des tubulovesikulären Netzwerks bei [96]. Dieses Netz besteht aus Membranen,
die sich innerhalb der Vakuolenmembran fingerförmig ausstülpen. Eine Funktion dieses
Netzwerks ist bisher nicht bekannt.
Innerhalb der Vakuole vermehren sich die Tachyzoiten durch Endodyogenie. Endo-
dyogenie ist eine spezialisierte Form der Zellteilung, bei der zwei Tochterzellen innerhalb
der Mutterzelle entstehen. Während der Bildung der Tochterzellen wird die Mutterzelle
zerstört [26]. Unter entsprechenden Umweltbedingungen entwickeln sich Bradyzoiten.
Bradyzoiten, die langsam wachsende Form von T. gondii, liegen in Gewebszysten vor,
die viele hundert Parasiten enthalten können. Trigger für die Bildung von Gewebszy-
sten in vitro sind Stressoren wie Erhöhung des pH-Wertes oder der Temperatur [100]. Der
Induktor für die Zystenbildung in vivo ist jedoch nicht bekannt.
Andere Organismen, wie Legionella pneumophila oder Mycobacterium ssp., haben ver-
3

schiedene Strategien entwickelt, um dem lysosomalen System des Wirts zu entgehen. Sie
beeinflussen entweder die Fusion der Phagosomen mit den Lysosomen, entkommen aus
der Vakuole ins Zytoplasma oder verhindern die Ansäuerung des Phagosoms [97]. T.
gondii umgeht durch die aktive Invasion das endosomale System der Wirtszelle [42]. Im
weiteren Verlauf kommt es zu keiner Fusion des lysosomalen Kompartimentes mit der
parasitären Vakuole – die Vakuole bleibt für die Wirtszelle unsichtbar.
Die Nährstoff-Aufnahme von T. gondii ist nicht vollständig bekannt. Lösliche Mole-
küle bis zu einer Größe von 1300 Da, wie z. B. Aminosäuren oder Nukleotide, können
über Poren in der Membran in die Vakuole übertreten [90]. Die Aufnahme von Lipiden
und Cholesterol, letzteres erhält der Parasit vollständig von der Wirtszelle, ist nicht ge-
klärt [17].
1.2 Medizinische Bedeutung von Toxoplasma gondii
In den 1980er Jahren war die Toxoplasma-Enzephalitis häufigste Todesursache bei HIV-
Erkrankten. Dies hat sich erst nach Einführung der „highly active anti-retroviral therapy“
(HAART) und der Entwicklung neuer Antibiotika wie Pyrimethamin verändert.
Eine Übertragung auf den Menschen geschieht über Genuß von rohem Fleisch infi-
zierter Tiere, über oozystenhaltigen Katzenkot (z. B. ungewaschener Salat) oder diapla-
zentar bei einer Erstinfektion während der Schwangerschaft. In der Regel verläuft ei-
ne Infektion subklinisch. Gelegentlich treten Lymphknotenschwellungen, Fieber, sowie
Kopf- und Muskelschmerzen auf. Unter Immunsuppression, z. B. bei Zytostatika Be-
handlung oder im Verlauf einer HIV Erkrankung, manifestiert sich die Toxoplasmose
als fulminante Enzephalitis mit eventueller septischer Streuung in Herz, Leber und Milz.
Im Rahmen einer Erstinfektion in der Schwangerschaft führt die diaplazentare Über-
tragung von T. gondii Tachyzoiten zur Fetopathia toxoplasmotica. Hierbei kommt es zu
Früh- oder Totgeburt, Hepatosplenomegalie, Ikterus, Myokarditis oder einer interstitiel-
len Pneumonie. Leichtere Fälle gehen mit der typischen enzephalitischen Trias bestehend
aus Hydrozephalus, Chorioretinitis und periventrikulären Verkalkungen einher.
Der Nachweis einer frischen Toxoplasma Infektion gelingt serologisch über den An-
stieg des IgM-Titers. Dieser kann jedoch bei immunsupprimierten Patienten ausbleiben.
Hier kann ein direkter Erregernachweis aus Blut oder Liquor hilfreich sein. Schwangere
Frauen sollten prophylaktisch auf das Vorliegen von IgG Antikörpern untersucht wer-
den, und bei Serokonversion einer Therapie zugeführt werden. Therapie der ersten Wahl
ist Pyrimethamin kombiniert mit Sulfadiazin oder Clindamycin. Vor der 20. Schwanger-
schaftswoche wird mit Spiramycin behandelt [38].
4

1.3 Antioxidative Schutzmechanismen von T. gondii
Die Fähigkeit verschiedener intrazellulärer Parasiten, in Makrophagen zu überleben be-
ruht auf einer Reihe von unterschiedlichen Mechanismen [11, 58]. Generell kann man
mindestens vier Strategien abgrenzen: Rückzug des Parasiten in ein sicheres intrazellu-
läres Kompartiment, Unterdrückung der Produktion antimikrobieller Substanzen und
die Modulation der Wirtszelle auf der Ebene der Apoptose [71], der Zytokin-Produktion
[6, 12] und der Antigenpräsentation [53]. Auf die Modulation der Immunantwort durch
T. gondii soll an dieser Stelle nicht eingegangen werden.
Die Vakuole, in der T. gondii sich aufhält, ist ein idealer Schutzort, da sie einerseits
innerhalb der Zelle liegt und so vor extrazellulär freigesetzten Substanzen geschützt
ist, sowie andererseits keinen Kontakt zum lysosomalen System der Wirtszelle besitzt
[14, 97]. Letzteres wird dadurch erklärt, daß T. gondii nicht über Phagozytose aufge-
nommen wird [42, 61] und bei der Invasion alle Wirtszellproteine von der Vakuolen-
membran ausgeschlossen werden [60]. Da somit auch die Rezeptoren für die Vesikel-
fusion (SNARE-Proteine) fehlen, ist die Vakuole für die Wirtszelle als Kompartiment im
Membran-Verkehr unsichtbar.
Die wichtigsten antimikrobiellen Substanzen bei der Immunantwort auf eine T. gon-
dii Infektion sind freie Radikale [3, 66]. Dazu gehören Wasserstoffperoxid (H2O2 ), das
Superoxid-Radikal (O−2 ) und das Hydroxyl-Radikal (OH ·). Diese Sauerstoffradikale wer-
den durch Neutrophile und Makrophagen während des „respiratory burst“ freigesetzt
und schädigen DNA, Lipide und Proteine [54].
Beim „respiratory burst“ ist der Sauerstoff- und Glukoseverbrauch der Makrophagen
exzessiv gesteigert. Der Sauerstoff wird über die NADPH-Oxidase zu O−2 umgesetzt,
während die Glukose über den Hexose-Monophosphat-Shunt das verbrauchte NADPH
regeneriert.
O2 +NADPH −→ O−2 +NADP+ +H+
Der überwiegende Teil des entstehenden O−2 reagiert weiter zu H2O2 :
2O−2 + 2H+ −→ H2O2 +O2
Über die metallkatalysierte Haber-Weiss Reaktion entsteht das Hydroxyl-Radikal (OH ·):
O−2 +H2O2
Fe2+−→ OH ·+OH− +O2
Das Hydroxyl-Radikal kann nun Lipide, Proteine, Kohlenhydrate und sogar DNA
angreifen und irreversibel schädigen. Besonders Membranlipide können an ihren mehr-
fach ungesättigten Fettsäuren angegriffen werden, was eine Transformation von cis nach
5

trans und damit eine Formänderung bewirkt. Häufig wird eine Kettenreaktion ausgelöst,
die zur Polymerisierung der Fettsäuren führt.
Schutzmechanismen gegen freie Radikale umfassen Antioxidantien (Vitamin E, β-
Karotin, Ascorbinsäure und Glutathion) und Enzymsysteme wie Superoxid-Dismutase,
Katalase, Peroxidasen und Peroxidoxin. Antioxidantien reagieren mit freien Sauerstoffra-
dikalen und neutralisieren sie. Superoxid-Dismutase (SOD) eliminert das Superoxid-Ra-
dikal (O−2 ). Katalase und Peroxidasen setzen H2O2 um.
O−2 +O2 + 2H −→ H2O2 +O2 (SOD)
H2O2 +H2O2 −→ 2H2O +O2 (Katalase)
H2O2 +R−H2 −→ 2H2O+ R (Peroxidase)
Peroxidoxine (oder Peroxiredoxine) sind eine relative neu entdeckte Familie antioxi-
dativer Enzyme [56]. Sie können unterschiedliche Peroxide, so auch Wasserstoffperoxid,
spalten. Ihre Besonderheit besteht darin, daß sie, im Gegensatz zu allen anderen bis-
her bekannten antioxidativen Enzymen, keine Metalle im aktiven Zentrum enthalten.
Ihr Reaktions-Mechanismus ist noch nicht eindeutig geklärt, jedoch spielen redox-aktive
Cysteine eine entscheidende Rolle.
Die Toleranz von T. gondii gegenüber H2O2 ist ungewöhnlich hoch [40, 65–68]. Diese
Beobachtung führte schon vor 15 Jahren zum enzymatischen und elektrophoretischen
Nachweis von Superoxid-Dismutase und Katalase [69, 95]. Die entsprechenden Gene
wurden jedoch erst neuerdings kloniert [23, 45, 74].
Auf die Existenz des Glutathion-Systems läßt sich anhand einer EST-Sequenz für
Glutathion-Reduktase schließen (Genbank Acc. Nr. W00127), die eine Ähnlichkeit von
58 % mit der Glutathion-Reduktase von Plasmodium falciparum aufweist. Das T. gondii Per-
oxidoxin wurde ebenfalls beim EST-Projekt identifiziert und nachfolgend kloniert [102].
Ihr Beitrag zum Überleben des Parasiten ist noch nicht geklärt.
1.4 Das Enzym Katalase
Das Enzym Katalase kommt in fast allen aeroben Lebewesen vor und katalysiert die Zer-
setzung von Wasserstoffperoxid zu Wasser und Sauerstoff. Diese Reaktion verhindert
die Entstehung von Sauerstoffradikalen und schützt die Zelle vor deren toxischen Wir-
kung. Ein Beispiel hierfür ist das seltene Krankheitsbild der Akatalasämie (Morbus Kata-
hara). Dieser autosomal-dominant vererbten Krankheit liegt eine verminderte Katalase-
Funktion in den Erythrozyten zugrunde. Sie führt infolge fehlendem Abbaus des von
vergrünenden Streptokokken gebildeten Wasserstoffperoxids zu einer Stomatitis ulcero-
sa [75].
6

Biochemisch unterscheidet man zwischen zwei verschiedenen Typen der Katalase:
Mangan-Katalasen und Häm-Katalasen. Die Mn-Katalasen sind Hexamere, enthalten ein
zentrales Mangan-Atom im aktiven Zentrum und kommen nur in Prokaryoten vor [15].
Die Häm-Katalasen hingegen sind ubiquitär verbreitet und haben ein Molekulargewicht
von 200-330 kDa. Jede Untereinheit des tetrameren Enzyms enthält ein Häm-Molekül.
Dieses besteht aus einem zentralen Eisenatom und Protoporphyrin IX.
Die klassische katalytische Aktivität der Katalase ist die Spaltung von zwei Molekü-
len Wasserstoffperoxid:
2H2O2 −→ H2O +O2
Zusätzlich zeigt Katalase unter bestimmten Bedingungen eine Peroxidase-Aktivität.
Die postulierte Reaktion von Katalase (E ), einem kleinen Peroxid als Substrat (S ) und
einem Wasserstoff-Donator (AH2) findet folgendermaßen statt:
E + S −→ ES (Katalase-Peroxid Komplex)
ES + AH2 −→ E + SH2 +A
Die Reaktionskinetik der Katalase ist ungewöhnlich, da bereits geringe Konzentratio-
nen des Substrates H2O2 die Aktivität des Enzyms hemmen. Betrachtet man den Reak-
tionsmechanismus der Zersetzung von H2O2 genauer, fallen zwei Teilschritte auf [33].
Zuerst erfolgt eine Oxidation von zwei Elektronen:
E − FeIII +H2O2 −→ E − FeV=O +H2O
Der oxidierte Zustand (FeV =O ) wird „Compound I“ genannt und entspricht einer
oxoferryl-Gruppe mit einem π-Porphyrin Radikal. Der zweite Schritt der Reaktion ist
eine zweimalige Reduktion des „Compound I“ durch ein weiteres Molekül H2O2 :
E − FeV +H2O2 −→ E − FeIII=O +H2O +O2
Bei kontinuierlichem Flux von H2O2 wird der „Compound I“ langsam zum „Com-
pound II“ reduziert. Dieser „Compound II“ ist gegenüber H2O2 enzymatisch inaktiv.
Wird der Zufluß von H2O2 jedoch gestoppt, kehrt das Enzym spontan zum Ausgangs-
zustand zurück.
Viele Katalasen enthalten NADPH als Kofaktor, der in der Peripherie des Enzyms fest
gebunden ist. Die Oxidation von NADPH zu NADP+ verhindert die Inaktivierung des
Enzyms durch H2O2 und beschleunigt die Reaktion [39, 46]. Ein Modell für den Reakti-
onsmechanismus [33], der diesen Eigenschaften Rechenschaft trägt ist in Abb. 3 darge-
stellt.
7

FerricCatalase(E-FeIII)
Compound I(E-FeV)
Compound II(E-FeIV=O)
NADP+
NADPH
(2 e-)
Reaktion 1
H2O2 H2O
Reaktion 2
Reaktion 3
NADP+
NADPH
A*
AH
(1 e-)
(2 e-)
(1 e-) AH
A*
H2O2
(2 e-)
(2 e-)
O2 + H2O
Abbildung 3: Möglicher Reaktionsmechanismus der Katalase bei konstantem Substratflux. Re-aktion 1: Compound I oxidiert das gebundene NADPH zu NADP+. Reaktion 2: Nach Oxidationdes NADPH wird Compound I durch einen endogenen oder hinzukommenden Elektronendona-tor (AH) zu Compound II reduziert. Reaktion 3: Der inaktive Compound II wird durch spontaneReduktion oder durch Zugabe von NADPH in den Ausgangszustand (Ferric Catalase) zurück-versetzt.
8

Die Katalase war eines der ersten Enzyme, das aufgereinigt und kristallisiert werden
konnte [104]. Die Kristallstruktur der bovinen Katalase wurde zuerst 1981 mit einer Auf-
lösung von 2.5 Å erstellt [70,85]. Zur Zeit sind 24 Kristallstrukturen von 7 verschiedenen
Organismen in der Entrez-Structure Datenbank (National Institute of Health, Bethesda)
verzeichnet. Da die Aminosäuresequenz des Proteins über die Arten hinweg hinreichend
konserviert ist, können modellbildende Algorithmen (z. B. Swissmodel) zu einer relativ
präzisen Strukturvorhersage benutzt werden.
1.5 Peroxisomen – Morphologie und enzymatische Ausstattung
Die Peroxisomen sind subzelluläre Organellen, die erstmals im Jahre 1954 von Rhodin
elektronenmikroskopisch im proximalen Tubulus der Mäuseniere als „Microbodies“ be-
schrieben wurden [86]. In der nachfolgenden Dekade zeigte de Duve durch biochemische
und morphologische Studien, daß Katalase, Urat-Oxidase und D-Aminosäuren-Oxidase
in einem eigenständigen subzellulären Kompartiment vorliegen, das den Microbodies
entspricht [5, 18]. Dieses Kompartiment wurde Peroxisom genannt, um auf das bioche-
mische Zusammenspiel zwischen H2O2 -bildenden Oxidasen und Katalase hinzuweisen
(Abbildung 4). In den darauf folgenden Jahren wurden Peroxisomen in fast allen euka-
ryontischen Zellen nachgewiesen (mit Ausnahme der Trichomonaden).
Morphologisch sind Peroxisomen durch eine fein granulierte Matrix und eine sie um-
gebende einfache Membran gekennzeichnet. In einigen Spezies enthält die Matrix kristal-
line Strukturen. Die Membran enthält ca. 1,2 nm große Poren, die für Moleküle bis 800 Da
durchlässig sind. Häufig sind Peroxisomen rund oder oval, in manchen Zelltypen können
sie allerdings auch miteinander verbunden sein oder in anderen komplexen Formationen
vorliegen [117]. Der Durchmesser runder Peroxisomen liegt abhängig vom Zelltyp zwi-
schen 100-500 nm. In den Leberzellen der Ratte sind sie mit einem Volumenanteil von
1,5 % vertreten [114]. Innerhalb einer Zelle sind die Peroxisomen in der Regel gleichmä-
ßig verteilt, es wird jedoch eine Assoziation mit dem Endoplasmatischen Retikulum (ER)
H2O
KATALASE
H2O2 + H2O2
reduziertesSubstrat
oxidiertesProdukt
O2
OXIDASE
Abbildung 4: Prinzip der peroxiso-malen Oxidation. Substrate derperoxisomalen Oxidasen werdenunter Bildung von H2O2 oxidiert.H2O2 wird anschließend durchKatalase enzymatisch in O2 undH2O gespalten. Das Ineinander-greifen dieser Reaktionen verhin-dert die Entstehung von toxischenSauerstoff-Radikalen.
9

Abbildung 5: Peroxisomen verschiedener Vo-gelarten. Peroxisomen sind sehr polymorpheZellorganellen. Häufig finden sich kristallineEinschlüsse (64, 66, 67), die bei Vögeln zum Teilals tubuläre Strukturen auftreten (62). Eine As-soziation mit dem ER (67) und Marginalplatten(66, rechts oben) treten auch auf. (Abbildungaus [10]).
gelegentlich beschrieben (Abb. 5).
Die enzymatische Ausstattung der Peroxisomen variiert sehr stark zwischen verschie-
denen Zelltypen. Insgesamt sind etwa 50 verschiedene Reaktionen beschrieben worden.
Interessanterweise haben die meisten peroxisomalen Enzyme Gegenstücke in anderen
Zellkompartimenten, die gleiche oder ähnliche Reaktionen katalysieren, jedoch ohne das
toxische Nebenprodukt H2O2 zu bilden. Eine Auswahl von biochemischen Reaktionen,
die in Peroxisomen zu finden sind, sind in Tabelle 1 dargestellt [84].
1.6 Peroxisomen – Biogenese und Protein Targeting
Die Entstehung von Peroxisomen wird noch kontrovers diskutiert [103]. Nach dem von
Lazarow und Fujiki postulierten Modell entstehen neue Peroxisomen nicht de novo, son-
dern nur durch Teilung oder Abspaltung von bestehenden Peroxisomen [49]. Dieses Mo-
dell begründet sich aus der Beobachtung, daß Matrix- und Membranproteine der Pero-
xisomen im Zytosol synthetisiert werden und dann posttranslational in die Organellen
transportiert werden. Daraus leitet sich eine Interaktion zwischen Import-Signalen auf
dem importierten Protein und Rezeptoren auf der peroxisomalen Oberfläche, sowie die
10

Oxidation Synthese Metabolismus
kurz- und langkettige Fett-säurenProstaglandineXenobiotikaSpermidin und SperminHarnsäureD-AminosäurenEthanol, MethanolNitritIsozitrat
CholesterolDolicholGlycerolipidelangkettige FettsäurenGallensäuren
Wasserstoffperoxidfreie RadikaleEpoxideAlaninGlyoxylatL-AminosäurenPurinePolyamineEicosanoide
Tabelle 1: Biochemische Stoffwechselvorgänge in den Peroxisomen.
Existenz einer Import-Maschinerie ab. Zwei dieser „peroxisomal targeting signals“ (PTS1
und PTS2) konnten bisher für Matrix-Proteine definiert und die dazu korrespondieren-
den Rezeptoren (Pex5p und Pex7p) isoliert werden [34, 77, 105].
Das PTS1 wurde zuerst am Luciferase-Genprodukt der Glühwürmchen entdeckt. Es
ist ein aus drei Aminosäuren bestehendes Signal am C-Terminus des Proteins mit fol-
gender Konsensus-Sequenz: (S/T/A/G/C/N) - (R/K/H) - (L/I/V/M/A/F/I) 1. Nicht
alle dieser Kombinationen sind jedoch Import-Signale und auch die Region unmittelbar
vor dem Signal hat einen noch nicht vollkommen geklärten Einfluß auf die Effizienz des
Imports [63, 64, 82].
Das PTS2-Signal liegt am N-terminus und hat die Konsensus-Sequenz (X)2-(R/K)-
(L/V/I)-(Q)-(X)5-(H/Q)-(L/A). Ein Beispiel ist das Importmotiv der peroxisomale Thio-
lase von Saccharomyces cerevisiae mit der Sequenz XXRLQX5HL [77, 105]. Die bisher be-
kannten Interaktionen von PTS1 und PTS2 mit den Import-Proteinen ist in Abb. 6 darge-
stellt.
Targeting-Signale, die die Membran-Proteine in die peroxisomale Membranen ein-
bauen (mPTS1), konnten bisher noch nicht exakt definiert werden. Unter anderem wur-
de im PEX3-Genprodukt von Pichia pastoris eine 40 Aminosäuren lange, N-terminale Se-
quenz [116] und ein 20 Aminosäuren langer Abschnitt im PMP47-Genprodukt von Can-
dida boidini identifiziert [31]. Beide Sequenzen enthalten fünf basische Aminosäuren hin-
tereinander, die von teilweise konservierten Regionen flankiert werden. Ein Konsensus
Motiv läßt sich aus der bisherigen Datenlage allerdings nicht ableiten.
In den letzten Jahren jedoch gab es Hinweise, daß das ER eine wichtige Rolle in der
Biogenese der Peroxisomen spielt [48, 111]. Nach dem Modell von Titorenko und Rachu-
binski gelangen einige Membran-Proteine (Pex2p und Pex16p) vom ER, wo sie glyko-
1PROSITE, Swiss Institute of Bioinformatics
11

Pex5p
Pex13p
PTS2
Pex7p
PTS1 - Katalase
PTS1
PTS2 - Thiolase
Docking & Import
Pex14p
Abbildung 6: Importsignalefür Matrixproteine. Der post-translationale Proteinimportin die Peroxisomen wird überzwei verschiedene peroxiso-male Targeting-Signale (PTS)vermittelt. Das C-terminalePTS1 (z. B. Katalase) interagiertmit seinem Rezeptor Pex5p,das N-terminale PTS2 (z. B.Thiolase) mit Pex7p. Es bildetsich ein Protein-Komplex mitPex13p und Pex14p, der in dasOrganell transportiert wird.
syliert werden, mittels kleiner sekretorischer Vesikel [109]zu den Peroxisomen (Abb. 7)
[110]. Hierbei entstehen zwei verschiedene transiente Kompartimente (PV1 und PV2),
die Katalase bzw. Thiolase enthalten und somit die beiden Importwege (PTS1 und PTS2)
widerspiegeln. Durch die Verschmelzung von PV1 und PV2 mit wachsenden Peroxiso-
men werden die Proteine in der peroxisomalen Membran integriert. Der Vesikeltransport
zwischen ER und Peroxisomen basiert auf Interaktionen der beteiligten Membranen mit
den Coatomeren COPI und COPII und wird von ARF reguliert [79].
1.7 Spezialisierte Peroxisomen: Glyoxysomen und Glykosomen
Die unterschiedlichen Anforderungen an die metabolischen Leistungen hat spezialisierte
Peroxisomen in verschiedenen Tier- und Pflanzenzellen hervorgebracht. Prominenteste
Beispiele sind die Glyoxysomen der Pflanzenzellen und die Glykosomen der Kinetopla-
stidae.
Die Glyoxysomen sind der Ort des Glyoxylat-Zyklus. Bei diesem bisher nur in Pflan-
zen nachgewiesenen Zyklus wird Acetat zu Succinat umgewandelt, was im weitesten
Sinne einem Umbau von Fettsäuren zu Zuckern entspricht. Die beiden Hauptenzyme
des Glyoxylat-Zyklus, Isozitrat-Lyase und Malat-Synthase, konnten bisher nicht sicher
in Toxoplasma gondii nachgewiesen werden. Glyoxysomen unterliegen einer hohen Wan-
delbarkeit. Sie können während ihres Lebenszyklus unterschiedliche Enzyme enthalten
und sich von Glyoxysomen zu Blatt-Peroxisomen und wieder zurück verwandeln [37].
Eine weitere wichtige Gruppe der spezialisierten Peroxisomen sind die Glykosomen
der parasitären Protozoen der Ordnung Kinetoplastida, zu der unter anderem Trypanosoma
12

PV1
PV2
ReifePeroxisomen
COP I & ARF vermitteltes Retrieval
CatalasePTS1
ThiolasePTS2
Pex16p
Pex2p
Budding Fusion
COP IIER
Abbildung 7: Biogenese von Peroxisomen. Das Modell von Titorenko und Rachubinski postuliertdie Entstehung des peroxisomalen Kompartimentes aus dem endoplasmatischen Retikulum (ER)und verknüpft die zwei Importwege für Matrixproteine (PTS1 und PTS2) mit zwei verschiede-nen vesikulären Kompartimenten (PV1 und PV2), die unreifen peroxisomalen Kompartimentenentsprechen. Vesikel, die Pex2p und Pex16p als Membranproteine enthalten, spalten sich vom ERab und werden mit peroxisomalen Matrixproteinen beladen. Die Kompartimente PV1 und PV2verschmelzen mit schon bestehenden peroxisomalen Vesikeln und bilden reife Peroxisomen.
und Leishmania gehören. In den Glykosomen sind Enzyme der Glykolyse kompartimen-
talisiert [108]. Es wird spekuliert daß dadurch die biochemischen Reaktionen effizienter
ablaufen [76].
Die Glykolyse ist für Kinetoplastida von Bedeutung, wenn sie sich im Blut des Wirts-
organismus aufhalten, da dort hohe Glukosespiegel einen Abbau zu Pyruvat fördern. Im
Vergleich zur oxidativen Phosphorylierung ist die Energiegewinnung bei der Glykolyse
geringer, was nach dieser Hypothese durch eine Kompartimentalisierung ausgeglichen
wird. Befindet sich der Parasit wieder im Insektenvektor, werden glykosomale Enzyme
vermindert exprimiert und die ATP-Erzeugung auf oxidative Phosphorylierung in den
Mitochondrien umgestellt.
Neuere Arbeiten haben allerdings die Sichtweise verschoben [59]. Bakker et al. konn-
ten anhand einer Computermodellierung zeigen, daß die Kompartimentalisierung kei-
neswegs den Substratflux erhöht [4]. Durch virtuelle Entfernung der glykosomalen Mem-
bran in ihrem Modell entstand eine Akkumulation von Hexose-6-Phosphaten im Zytosol,
die vermutlich zu einem osmotischen Schaden führt. Blattner et al. demonstrierten, daß
Expression von Phosphoglycerat-Kinase im Zytosol zum Tod der amastigoten Parasiten
führt [8].
Die Biogenese der Glykosomen ähnelt, soweit untersucht, der Biogenese der Pero-
xisomen [78, 101]. Bei einigen glykosomalen Enzymen konnte gezeigt werden, daß auch
13

hier PTS1-Signale als Import-Signale wirken. Der zugehörige Rezeptor wurde kloniert
[20]. Jedoch sind die Sequenzen, die PTS1-abhängigen Import bewirken, nicht so strin-
gent, wie in den bisher untersuchten Systemen. So gibt es PTS1-Sequenzen, die Import
in Glykosomen bewirken, nicht jedoch in Peroxisomen der Säugetiere [9]. Die Analyse
der Kristallstruktur der Trypanosoma brucei Aldolase läßt vermuten, daß hier ein PTS2
vermittelter Import in die Glykosomen vorliegt [16].
14

2 Ergebnisse
2.1 Klonierung und Sequenzierung der Toxoplasma gondii Katalase
Die Existenz von Katalase in Toxoplasma gondii wurde vor knapp fünfzehn Jahren enzy-
matisch und elektrophoretisch nachgewiesen [95]. Weitere Schritte zur Charakterisierung
dieses Enzyms sind jedoch nie unternommen worden. Dies veranlaßte uns, gezielt nach
Katalase Sequenzen in der „expressed sequence tag“ (EST) Datenbank für das Toxoplasma
gondii Genom 2 zu suchen [1, 113]. Dort sind zwei Sequenzen verzeichnet (W63499 und
W99737), die Ähnlichkeit mit Katalasen anderer Species aufweisen.
Aus den beiden λ-Phagen Klonen der EST Datenbank wurde das Insert, das die T.
gondii spezifische Sequenz trägt, mit einem Helferphagen (VCSM13) in vivo exzidiert
und sequenziert. Ausgehend von dieser Sequenz wurde mittels „Rapid Amplification
of cDNA Ends“ (5‘RACE) weitere 300 Basenpaare gewonnen. Da jedoch keine größeren
Fragmente kloniert werden konnten, wurde mittels „nested“ reverser Transkriptase PCR
(RT-PCR) mit einem degenerierten Primer (Fdeg) und drei spezifischen Primern (R3, R4,
R5) weitere 700 Basenpaare kloniert und sequenziert. Durch eine weitere 5‘RACE konn-
ten weitere Basenpaare über das Startcodon hinaus bestimmt werden. Die Basenabfolge
der Katalase wurde mittels Sequenzierung vier unabhängiger Klone eines per RT-PCR
(F4, R0) gewonnen Fragmentes von 2480 bp bestätigt. Die zur Klonierung verwendeten
Methoden sind in Abb. 8 dargestellt. Die verwendeten Oligonukleotide sind in einer Ta-
belle in Materialien und Methoden auf S. 42 zusammengefaßt.
R7, R6
Fdeg
Phagen Sequenz
5’RACE
RT-PCR mit degenerierten Primern
5’RACE
R0R2, R1R5, R4 R3
F1160 1685 2480
R9
Abbildung 8: Methoden zur Klonierung der T. gondii Katalase. T. gondii Katalase wurde mitHilfe von verschiedenen Techniken kloniert. Die Balken zeigen an, welche Methode für welchenAbschnitt der Sequenz verwendet wurde. Die 5‘ und 3‘ UTRs (untranslated regions) sind weiß,die kodierende Region schwarz dargestellt. Die Pfeile geben die Position der Primer an.
2http://www.cbil.upenn.edu/ParaDBs/Toxoplasma
15

2.2 Analyse der Sequenz
Die vollständige DNA Sequenz ist in Abb. 9 dargestellt. Die cDNA kodiert für ein 502
Aminosäuren langes Protein mit einem molekularen Gewicht von 57,2 kDa. Ein Vergleich
der Aminosäuresequenz mit anderen eukaryotischen Katalasen zeigt starke Homologien
(Tabelle 2), während die DNA Sequenz eine vergleichsweise geringe Ähnlichkeit mit an-
deren Katalasen aufweist. Ein Alignment von Katalasen verschiedener Species mit der T.
gondii Katalase ist in Abb. 10 gezeigt.
Eine weitergehende Analyse der Aminosäuresequenz bestätigt, daß es sich in der Tat
um Katalase handelt. Beide Katalase Konsensus-Sequenzen 3 sind vorhanden: Die Signa-
tur des aktiven Zentrums (As 53-69) und die proximale Häm-Bindungsstelle (As 343-
351).
Konsensus Sequenz T. gondii SequenzProximale HämBindungsstelle
R-[LIVMFSTAN]-F-[GASTNP]-Y-x-D-[AST]-[QEH]
RMFSYIDTH
Signatur desaktiven Zentrums
[IF]-x-[RH]-x(4)-[EQ]-R-x(2)-H-x(2)-[GAS]-[GASTF]-[GAST]
FDRERIPERVVHAKGGG
Dreidimensionale Modelle eines Proteins können aus der Aminosäuresequenz durch
Vergleich mit bekannten Strukturen verwandter Proteine errechnet werden. Diese Struk-
turvorhersagen sind erstaunlich präzise [36,80,81]. Mit Hilfe geeigneter Software erfolgte
eine dreidimensionale Modellierung der T. gondii Katalase. Als Modellvorlage dienten
bekannte Strukturen der bovinen Katalase. Die folgenden Stereobilder können durch
leichtes Einwärtsschielen betrachtet werden. (Näheres zur Modellierung in Materialien
und Methoden).
Die Homologie zwischen der bovinen Katalase und der T. gondii Katalase beträgt
63 %, was für eine 3D-Modellierung gut geeignet ist. Die geringe Abweichung der Koh-
3PROSITE, Swiss Institute of Bioinformatics
Organismus GenBank Nr. Identische As. Positive As. Wahrscheinlichkeit
B. taurus S49452 63 % 75 % 10 −170
H. sapiens P04040 61 % 74 % 10 −168
C. elegans 690941A 57 % 72 % 10 −153
P. mirabilis P42321 53 % 69 % 10 −141
S. cerevisiae 115709 49 % 65 % 10 −124
Tabelle 2: Ergebnisse eines Alignments mittels PSI-Blast [2] von Katalasen fünf ausgewählter Or-ganismen.
16

GTG CGG CTC CTC GAA GGA CTG AGT GAA ACG CTG AGG TGC GCG TCT CGT GAA GGC CAC TGT 60TGC TTG TCC TCA CGG TGA AGG TCG GCC AGC CAG CTT TTG ACG GTA CGG AGA TTC TTC TGT 120TTT TGA CCC CGA AGG GAA ACG TGC GCG CCG TCA GCG AAA ATG ACT CAG GTT CCG CCC GTA 180
M T Q V P P VACA TTC CAG CAG TAT GGA CCG GTT ATC ACG ACC TCT GCT GGG AAC CCA GTC GAT GAC AAC 240T F Q Q Y G P V I T T S A G N P V D D NCAA AAC TCG GTA ACT GCT GGA CCC TAC GGC CCC GCG ATC CTG AGT AAC TTC CAT CTC ATC 300Q N S V T A G P Y G P A I L S N F H L IGAC AAG CTG GCC CAC TTC GAT CGA GAA CGC ATT CCC GAG CGC GTT GTG CAT GCC AAG GGC 360D K L A H F D R E R I P E R V V H A K GGGT GGC GCC TTT GGC TAT TTC GAG GTC ACT CAT GAC ATC ACC AGA TTC TGC AAA GCG AAG 420G G A F G Y F E V T H D I T R F C K A KCTG TTC GAG AAA ATC GGA AAG CGG ACG CCT GTG TTT GCA CGT TTT TCG ACG GTG GCT GGC 480L F E K I G K R T P V F A R F S T V A GGAG TCG GGG AGT GCG GAC ACC AGG CGC GAC CCG CGC GGC TTC GCA CTC AAG TTC TAC ACG 540E S G S A D T R R D P R G F A L K F Y TGAG GAG GGG AAC TGG GAC ATG GTT GGA AAC AAC ACT CCT ATC TTT TTC GTT CGA GAC GCC 600E E G N W D M V G N N T P I F F V R D AATA AAG TTC CCC GAC TTC ATT CAT ACG CAG AAG CGG CAT CCG CAA ACG CAC CTC CAC GAC 660I K F P D F I H T Q K R H P Q T H L H DCCG AAC ATG GTG TGG GAT TTC TTC TCG CTC GTT CCG GAA AGT GTG CAT CAA GTG ACC TTC 720P N M V W D F F S L V P E S V H Q V T FCTG TAC ACG GAC CGT GGC ACA CCG GAC GGC TTC CGC CAT ATG AAT GGC TAC GGG AGT CAC 780L Y T D R G T P D G F R H M N G Y G S HACG TTC AAA TTC ATC AAC AAA GAC AAC GAA GCC TTT TAT GTC AAG TGG CAC TTC AAG ACA 840T F K F I N K D N E A F Y V K W H F K TAAC CAA GGA ATC AAG AAC CTG AAC AGA CAG CGC GCA AAA GAA CTC GAG AGC GAA GAT CCT 900N Q G I K N L N R Q R A K E L E S E D PGAC TAC GCG GTT CGT GAC CTG TTC AAC GCC ATT GCC AAG CGA GAG TTT CCC TCT TGG ACC 960D Y A V R D L F N A I A K R E F P S W TTTC TGT ATT CAG GTA ATG CCC TTG AAA GAT GCA GAG ACG TAC AAA TGG AAT GTC TTT GAC 1020F C I Q V M P L K D A E T Y K W N V F DGTG ACC AAA GTG TGG CCG CAC GGA GAC TAC CCA CTC ATC CCG GTC GGA AAA CTC GTG CTG 1080V T K V W P H G D Y P L I P V G K L V LGAT CGC AAC CCA GAA AAC TAT TTC CAA GAC GTG GAA CAA GCA GCT TTT GCG CCA GCT CAT 1140D R N P E N Y F Q D V E Q A A F A P A HATG GTG CCA GGG ATT GAA CCG AGC GAA GAT CGC ATG CTG CAG GGA CGC ATG TTC TCC TAC 1200M V P G I E P S E D R M L Q G R M F S YATC GAC ACT CAT CGG CAC AGA CTT GGC GCC AAT TAC CAT CAG ATT CCT GTG AAC CGC CCG 1260I D T H R H R L G A N Y H Q I P V N R PTGG AAC GCC CGT GGG GGC GAC TAC TCC GTC CGC GAT GGT CCA ATG TGC GTC GAC GGG AAC 1320W N A R G G D Y S V R D G P M C V D G NAAA GGG TCT CAG CTG AAT TAC GAA CCG AAC AGT GTG GAC GGC TTT CCA AAG GAA GAC AGA 1380K G S Q L N Y E P N S V D G F P K E D RAAT GCT GCT GTG TCG GGA ACG ACC ACA GTG AGT GGC ACG GTC GCG TGC CAT CCT CAA GAG 1440N A A V S G T T T V S G T V A C H P Q ECAC CCC AAC AGT GAC TTT GAG CAG CCA GGG AAC TTT TAT AGG ACG GTC TTG AGT GAG CCG 1500H P N S D F E Q P G N F Y R T V L S E PGAG AGG GAA GCG CTC ATT GGC AAC ATC GCC GAG CAT CTC AGG CAG GCG AGA CGA GAT ATT 1560E R E A L I G N I A E H L R Q A R R D ICAA GAG CGA CAG GTG AAG ATT TTT TAC AAG TGC GAT CCA GAG TAT GGC GAG CGC GTC GCT 1620Q E R Q V K I F Y K C D P E Y G E R V ACGA GCC ATC GGT TTG CCG ACT GCG GCG TGC TAC CCA GCC AAG ATG TAG GCG TGC GTT GTC 1680R A I G L P T A A C Y P A K M *CCG CTT GAG CCA GAA GAC CAA CAA TAG AGG GAC TAA ATA TGC GCA AGG AAG GAA GAC AGG 1760ACC CTT TCG AAG ACG GAA AAC ACG CAG TCT GAA TTC TTG GCG CTG AGG GGA CCA ACG GGT 1820TCG TCG CCT TGT GTG ACA ACG ACG AGT AAA AAG ACA AAG CGA ACA GCG ATG ACC GTA GCG 1880TTC GTC CAC CTG CGT AAA TGT GGC TTA TCT CTG TCA CGA TGC GCA CAA GCG TTT GGT CAT 1940CGC GTA ATT ACA GAT AGT TGT GTA GTG ACT TGC TGC TGT TGC GTC TTC CAA CAC TTG TAG 2000TTA GAG AAA TGC ATG TGT ATG GGT AAG CTT GCA TGA ACT GCT ACA TAT CCA TGG TAA ACC 2060AAT TTG ACT GCA GGT GTC GGC GTT TAC GAC GTC AGT TTC ACA TTC TTG TTC TTG ATT TCC 2120TTC TTG CAA TCA CTT GCA CTC TGG CAA TTT TTG GAA GGG AAA GGA AAT GTG AGG GTG GGA 2180GGG CTC AGC ATC CTC TTC CAG TGG TCC TGC CGT GTC CAG TTT GCG CCT AAA GGT AGA GAT 2240CGC GGG GGG GGT CTC AGG AAT CTA TCC GCG TTT TTT CTC AAA GGA TTT GCA TTT ACT GTT 2300TCT GAA AGG TGA AAG ACC GGG TGT ATG TGC ACC CAC CTG AGC CAA CCC AGC ATT TCG TAA 2360TTC CAT CTG CAG CGT GTT TCC CCC TTT TCA AAG CGG ACC TTT TGT GAG AAA ACG TTC CCG 2420CGG GAA CAG CAC TTT CTG AAC GAG AAA AAA AAA AAA AAA AAA AAA AAA AAA AAA AAA AAA 2480
Abbildung 9: cDNA Sequenz der T. gondii Katalase. Die komplette cDNA Sequenz der T. gondiiKatalase ist in der „Genbank“- Datenbank unter der Zugriffsnummer AF136344 gespeichert. Dieunterstrichenen Basenpaare entsprechen der Position der zur Klonierung verwendeten Primer.
17

1 10 20 30 40T.gondii MTQVPPVTFQQYGPVITTSAGNPVDDNQNSVTAGPYGPAILSNFHLIDKH.sapiens MADSRDPASDQMQHWKEQRAA.KAD.L..G.....G.KL.VI.V..R..LLVQDVVFT.EC.elegans MPNDPSDNQLKTYKET--YPKPQ.....N.A.IYSKTAVL...RR..MLMQDVVYM.ES.cerevisae MSKLGQEKNEVNYSDVREDR.V.N.T.N.INEPFVTQRI.EH..LL.QDYN...S
50 60 70 80 90 100T.gondii LAHFDRERIPERVVHAKGGGAFGYFEVTHDITRFCKAKLFEKIGKRTPVFARFSTVAGESH.sapiens M.................A.............KYS...V..H...K..IAV.........C.elegans M.................A..H.........SKY...DI.N.V..Q..LLI.....G...S.cerevisae ....N..N..Q.NP..H.S.........D...DI.GSAM.S......KCLT.....G.DK
# ># : # : / ++- --- /
110 120 130 140 150 160T.gondii GSADTRRDPRGFALKFYTEEGNWDMVGNNTPIFFVRDAIKFPDFIHTQKRHPQTHLHDPNH.sapiens .....V.......V.....D....L.........I..P.L..S...S...N.....K..DC.elegans .....A.......I..........L.........I..P.H..N.......N.....K...S.cerevisae .....V.......T........L.W.Y....V..I..PS...H.......N...N.R.AD
++ # / \\ ++ + ++ ++> >
170 180 190 200 210 220T.gondii MVWDFFSLVPE---SVHQVTFLYTDRGTPDGFRHMNGYGSHTFKFINKDNEAFYVKWHFKH.sapiens .....W..R..---.L...S..FS...I...H............LV.ANG..V.C.F.Y.C.elegans .IF..WLHR..---AL...M..FS...L...Y............MV...GK.I...F.F.S.cerevisae .F...LTT-..NQVAI...MI.FS.....ANY.S.H..SG..Y.WS..NGDWH..QV.I.
o + \+ o o o \ < o \230 240 250 260 270 280
T.gondii TNQGIKNLNRQRAKELESEDPDYAVRDLFNAIAKREFPSWTFCIQVMPLKDAETYKWNVFH.sapiens .D......SVED.AR.SQ.....GI........TGKY....FY....TFNQ...FPF.P.C.elegans PT..V...TVEK.GQ.A.S....SI.......E.GD..V.KMF....TFEQ..KWEF.P.S.cerevisae .D......TIEE.TKIAGSN...CQQ...E..QNGNY....VY..T.TER..KKLPFS..
\290 300 310 320 330 340
T.gondii DVTKVWPHGDYPLIPVGKLVLDRNPENYFQDVEQAAFAPAHMVPGIEPSEDRMLQGRMFSH.sapiens .L......K............N...V...AE...I..D.SN.P....A.P.K.....L.AC.elegans ..............E...M..N...R...AE...S..C...I.....F.P.K.....I..S.cerevisae .L.....Q.QF..RR...I..NE..L.F.AQ........STT..YQ.A.A.PV..A.L..
\o\o # < #
350 360 370 380 390 400T.gondii YIDTHRHRLGANYHQIPVNRPWNARGGDYSVRDGPMCVDGNKGSQLNYEPNSVDGFPKEDH.sapiens .P........P..LH....C.YR..VAN.Q-......MQD.Q.GA...Y...FG--AP.QC.elegans .T...F....P..I.L...C.YRS.AHNTQ-...A.AY-D.QQHA..FF...FN--YGKTS.cerevisae .A.A..Y...P.F......C.YASKFFNPAI.....N.N..F..E.T.LA.DKSYTYIQQ
# - :/
410 420 430 440 450 460T.gondii RNAAVSGTTTVSGTVACHPQEHP--NSDFEQPGNFYRTVLSEP-EREALIGNIAEHLRQAH.sapiens QPS.LEHSIQY..E.RRFNTAND--D-NVT.VRA..VN..N.E-Q.KR.CE...G..KD.C.elegans .PDVKDT.FPAT.D.DRYESGDD--.-NYD..RQ.WEK..DTG-A..RMCQ.F.GP.GECS.cerevisae DRPIQQHQEVWN.PAIPYHWATSPGDV..V.AR.L..VLGKQ.GQQKN.AY..GI.VEG.
o \ o\ \470 480 490 500
T.gondii RRDIQERQVKIFYKCDPEYGERVARAIGLPTAACYPAKMH.sapiens QIF..KKA..N.TEVH.D..SHIQALLDKYNAEKPKNAIHTFVQSGSHLAAREKANLC.elegans HDF.IKGMIDH.S.VH.DF.A..KALIQKQARSHIS.cerevisae CPQ..Q.VYDM.ARV.KGLS.AIKKVAEAKHASELSSNSKF
Abbildung 10: Sequenzvergleich verschiedener Katalasen. Katalasen der folgenden Spezies sindin einem Alignment dargestellt: T. gondii (GenBank Zugriffsnummer AF136344 ), H. sapiens (NCBZugriffsnummer 179950), C. elegans (NCB Zugriffsnummer 1078837) und S. cerevisae (NCB Zu-griffsnummer 115709). Punkte zeigen an, daß die As. mit denen der T. gondii Katalase identischsind, wohingegen ein Bindestrich eine Lücke anzeigt. Die Symbole unter dem Alignment wei-sen auf mögliche Funktionen der As. hin: Distale Häm Bindungsstelle ”>”, Proximale Häm Bin-dungsstelle ”<”; Bindung der Propionat-Seitenketten des Häm ”:”; As. in der Nähe des aktivenZentrums ”# ”; Bindungspartner des NADPH ”o”; As., die am katalytischen Mechanismus teil-nehmen ”/”; Säurereste in der Umgebung des NADPH ”\”; As. des Substrat-Kanals ”+”; As., diemit den anderen Homomeren interagieren ”-”. PTS-Signale sind fett gedruckt. Die Signatur desAktiven Zentrums (aa 53-69) und die Proximale Häm-Bindungs-Signatur (aa 343-351) sind unter-strichen. Das Alignment wurde mittels ClustalW1.73 durchgeführt [107].
18

Funktion Aminosäuren
Aktives Zentrum distale Hämbindungspartner: F142, F150, F53
proximale Hämbindungspartner: H207, R343
Interaktion mit Propionatgruppe: H207, R343
Umgebung des aktiven Zentrums Y347, D54, H64, V135, F323, S346A und L50M
Katalytischer Mechanismus H64, N137, S103, Y347
NADPH Bindungsstelle R192, T190, N202, H294, H183, Q435, W292, H224, V443
Umgebung des NADPH E448, F187, T139, P140, Y204, K226, Q271, P293, V291,
F439, L444, T190S und E448Q
Substratkanal V105, A106, D117, P118, F142, F143, F150, F153, I154,
V185, L188, Q157, und K158
Tetramerinteraktion L50, D54, F53, D349 und R55
PTS1 A500, K501, M502
Tabelle 3: Funktionell relevante Aminosäurereste der T. gondii Katalase.
lenstoffgerüste in der Übereinanderprojektion dieser Katalasen in Abb. 11 unterstreicht
dies.
Die Anordnung der Sekundärstrukturen weist einige Besonderheiten auf (Abb. 13).
Acht antiparallele β-Faltblätter schirmen die Hämgruppe auf einer Seite ab. Die ande-
re Seite des Häms ist frei zugänglich und bildet den Substratkanal (s. u.). Die NADPH -
und die Hämgruppe befinden sich in relativer Nähe, was ein Zusammenwirken beim ka-
talytischen Mechanismus wahrscheinlich macht. Dabei soll NADPH die Rolle des Elek-
tronendonators übernehmen und die Bildung des inaktiven „Compound II“ rückgängig
machen [7]. Der lange „wrapping loop“, der keiner eindeutigen Sekundärstruktur zu-
geordnet werden kann, ist auf der rechten Seite zu sehen. Er dient der Interaktion mit
den benachbarten Monomeren [43]. Der C -Terminus ist in dieser Abbildung oben links
als α-Helix zu sehen. Er bildet das peroxisomale Targeting-Signal. Da Katalase erst nach
der Proteinfaltung und Zusammenbau des Tetramers in die Peroxisomen transportiert
wird [112], ist seine exponierte Lage nachvollziehbar.
Die Darstellung der Oberfläche eines Katalase Monomers in Abb. 12 zeigt, wie kom-
pakt die Katalase in ihrem Kernbereich gefaltet ist. In der oberen Mitte ist ein Teil des
NADPH -Moleküls zu sehen, das von der Oberfläche zugänglich ist.
Ein Vergleich der T. gondii Katalase Sequenz mit anderen Katalasen zeigt, daß nicht
nur die Sekundärstrukturen ähnlich sind. Aminosäuren mit entscheidenden Funktionen
sind hoch konserviert. Eine Übersicht über diese Aminosäuren bietet Tabelle 3.
Aminosäurereste der T. gondii Katalase, die das aktive Zentrum bilden, sind identisch
mit den entsprechenden Aminosäuren der humanen Katalase. Alle weiteren Aminosäu-
19

N
C
WL
Abbildung 11: Projektion des Kohlenstoffrückgrats verschiedener Katalasen. Das Kohlen-stoffrückgrat der T. gondii Katalase (grün) wurde auf die Modellvorlagen der verschiedenen Un-tereinheiten der bovinen Katalase projiziert. Beachtenswert ist die geringe Abweichung im zen-tralen Bereich des Proteins. Der N- und C-Terminus sind entsprechend bezeichnet. WL: „wrap-ping loop“ rot: Häm und NADPH.
ren aus der Umgebung des aktiven Zentrums sind ebenfalls konserviert, mit Ausnahme
geringer Abweichungen (S346A und L50M) [33]. Weiterhin sind Aminosäuren konser-
viert, die im katalytischen Mechanismus eine Rolle spielen [52]. Drei der Aminosäuren
in der Nähe des aktiven Zentrums (L50, F53, D54) gehören zum benachbarten Monomer.
Die sterischen Verhältnisse im aktiven Zentrum sind in Abb. 14 dargestellt.
Einige Katalasen, wie z. B. humane und bovine Katalase, binden NADPH als Cofak-
tor. NADPH soll dabei unter anderem die Bildung des „Compound II“, ein inaktiver
Zustand im katalytischen Zyklus, verhindern [39, 46]. Konservierte Aminosäuren lassen
darauf schließen, daß T. gondii Katalase auch NADPH binden kann (Abb. 15). Die Säu-
rereste der möglichen NADPH -Bindungsstelle und deren Umgebung sind identisch mit
jenen der humanen Katalase mit Ausnahme von T190S und E448Q [33].
Das Substrat H2O2 diffundiert zum aktiven Zentrum der Katalase durch einen circa
30 Å langen Kanal (Abb. 16) [33]. Aminosäuren, die vermutlich die Wände dieses Kanals
bilden, sind identisch mit der humanen Sequenz. Dies weist darauf hin, daß die Band-
20
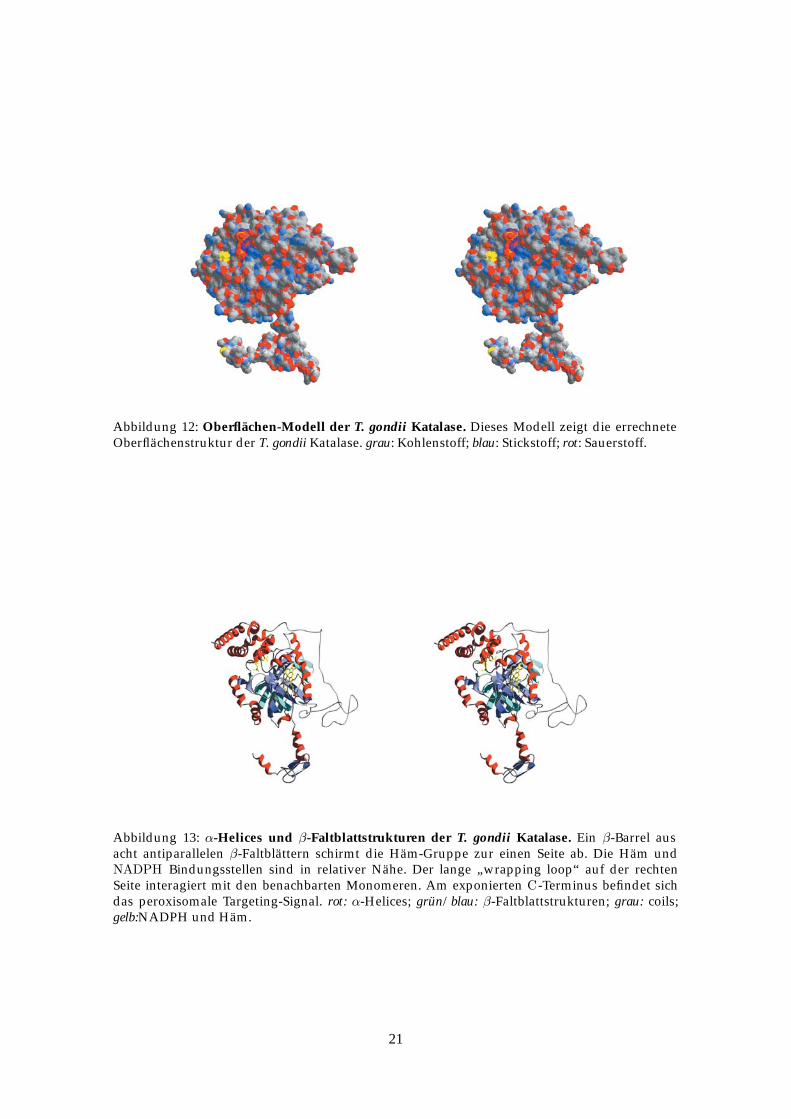
Abbildung 12: Oberflächen-Modell der T. gondii Katalase. Dieses Modell zeigt die errechneteOberflächenstruktur der T. gondii Katalase. grau: Kohlenstoff; blau: Stickstoff; rot: Sauerstoff.
Abbildung 13: α-Helices und β-Faltblattstrukturen der T. gondii Katalase. Ein β-Barrel ausacht antiparallelen β-Faltblättern schirmt die Häm-Gruppe zur einen Seite ab. Die Häm undNADPH Bindungsstellen sind in relativer Nähe. Der lange „wrapping loop“ auf der rechtenSeite interagiert mit den benachbarten Monomeren. Am exponierten C-Terminus befindet sichdas peroxisomale Targeting-Signal. rot: α-Helices; grün/blau: β-Faltblattstrukturen; grau: coils;gelb:NADPH und Häm.
21

Abbildung 14: Aktives Zentrum der T. gondii Katalase. rot: Häm; pink: Aminosäuren des be-nachbarten Monomers; blau: Aminosäuren, die am katalytischen Mechanismus teilnehmen; grün:Bindungspartner des Häm; grau: weitere Aminosäuren aus der Umgebung des Häm.
Abbildung 15: Aminosäuren aus der Umgebung der NADPH-Bindungsstelle der T. gondii Ka-talase. rot: NADPH ; grün: NADPH -Bindungspartner; grau: Nähere Umgebung des NADPH .
22

Abbildung 16: Substrat-Kanal der T. gondii Katalase. Aminosäuren, die den Substratkanal bil-den, der von der Oberfläche des Proteins zum Häm (gelb) führt sind hier im Kugelmodell darge-stellt (blau). α-Helices und β-Faltblätter sind analog zu Abb. 13 dargestellt.
breite möglicher Substrate ähnlich ist [118].
Die meisten Katalasen sind Homotetramere, mit Ausnahme der prokaryontischen
Mangan-Katalasen. Aminosäuren, die in Proteus mirabilis Katalase Interaktionen zwi-
schen den einzelnen Monomeren vermitteln, sind identisch mit den entsprechenden Säu-
reresten der T. gondii Katalase (L50, D54, F53, D349 und R55). Humane und bovine Kata-
lase haben zusätzlich einen längeren C-terminus, der die gegenseitige Bindung der ein-
zelnen Monomere stabilisiert [33]. In Abb. 17 sind die vier Untereinheiten unterschiedlich
eingefärbt, um die Lage der einzelnen Monomere zueinander deutlich zu machen.
Weiterhin besitzt T. gondii Katalase ein peroxisomales Targeting Signal (PTS1) am C-
Terminus. Die letzten drei Aminosäuren (-AKM) stimmen mit dem Konsensusmotiv für
peroxisomale Targeting Signale (PTS) der Klasse 1 überein. Experimentell wurde bereits
in Affennierenzellen (CV1) gezeigt, daß –AKM als PTS1 wirken kann, wenn es an Chlor-
amphenicol-Acetyltransferase gekoppelt ist [106]. Die exponierte Lage des PTS1 in der
3D-Struktur ist Vorbedingung für den peroxisomalen Transport.
23

Abbildung 17: T. gondii Katalase als Homotetramer. Unten: Die vier Monomere sind in den Far-ben grün, blau, pink und orange dargestellt, die Häm-Gruppe in gelb, das NADPH in rot. Oben: DieLage der einzelnen Monomere zueinander wird durch Betrachtung der Sekundärstrukturelemen-te deutlich.
24

RH Vero
102
81
47
32
30
27
GST-Kat
alase
rKat
in C
HO
Abbildung 18: Western Blot mit Antiserum gegen ein T. gondii-Katalase-GST Fusionsprotein.Das Antiserum gegen die T. gondii Katalase reagiert mit den Parasitenlysat (Spur 1), das aus einerVero-Zellkultur stammt. Die negative Kontrolle mit einer nicht-infizierten Vero-Zellkultur (Spur2) zeigt keine Reaktion. Als positive Kontrolle fungiert das GST-Katalase Fusionsprotein (Spur 3)und die in CHO-Zellen exprimierte rekombinante T. gondii-Katalase.
2.3 Western Blot mit Antiserum gegen T. gondii Katalase
Für die Produktion eines Antiserums wurde ein Fusionsprotein aus der Glutathion-S-
Transferase (GST) von Schistosoma mansoni mit den Aminosäuren 335-502 der T. gondii
Katalase hergestellt (Molekulargewicht: 47 kDa). Diese Region der Katalase wurde als
Antigen gewählt, da sie die größten Unterschiede gegenüber der humanen Katalase Se-
quenz aufweist. Die Herstellung und Testung des Antiserums ist im Kapitel Materialien
und Methoden genauer beschrieben.
Wie erwartet, reagiert das Antiserum mit dem rekombinantem GST-Katalase Fusions-
protein, das bei 47 kDa läuft (Abb. 18, Spur 3). Eine einzelne Bande bei 63 kDa erkennt
das Antiserum im Lysat ganzer Parasiten (Abb. 18, Spur 1). Die Größe dieses Proteins
ist im Einklang mit der vorhergesagten Größe von T. gondii Katalase (57,2 kDa). Zudem
läuft es in derselben Höhe wie rekombinante in CHO-Zellen exprimierte T. gondii Ka-
talase (Abb. 18, Spur 4). Kein Signal gibt es im Lysat der unveränderten Wirtszellen
(Abb. 18, Spur 2) und bei Inkubation mit prä-Immun-Serum (nicht dargestellt). Somit
reagiert das Antiserum mit einem 63 kDa T. gondii Protein, das wahrscheinlich der Kata-
lase entspricht.
25

Abbildung 19: Immunfluoreszenz der T. gondii Katalase. Dargestellt ist ein Immunfluoreszenz-Assay mit einem aufgereinigten Antikörper gegen die T. gondii Katalase an intra- und extrazellu-lären Parasiten. a: Intrazelluläre Parasiten replizieren sich in der Wirtszelle und bilden dabei eineRosette in der Vakuole. Das anteriore Ende der Parasiten zeigt nach außen. b: Eine Schemazeich-nung der Parasiten in der Vakuole verdeutlicht deren Position in der Phasenkontrastdarstellung.In einem Parasiten wurde zusätzlich der Nukleus (N) und die Katalase enthaltenden Strukturendargestellt. c und d: In extrazellulären Parasiten erscheint das Signal kompakter als in intrazellu-lären Parasiten, was möglicherweise jedoch ein Fixationsartefakt ist. Skalierung 2µm.
2.4 Lokalisation in der Immunfluoreszenz
In der Immunfluoreszenz markiert das anti-GST-Katalase Antiserum vesikuläre Struk-
turen im vorderen Ende des Parasiten (Abb. 19). Zusätzlich besteht ein diffuses Signal
im Inneren des Parasiten. Dieses Signal ist spezifisch für Katalase, da es verschwindet,
wenn das Antiserum zuerst mit rekombinanten Protein inkubiert wird. Vermutlich ent-
spricht diese diffuse Verteilung einem Pool von Katalase im Zytosol des Parasiten. Der
Immunfluoreszenz-Assay mit einem kommerziell erhältlichen monoklonalen Antikörper
gegen die Glutathion-S-Transferase ergibt kein Signal in T. gondii.
Werden zwei verschiedene Antikörper bei der Immunfluoreszenz verwendet, kann
eine Kolokalisation von Katalase mit bisher bekannten Zellorganellen ausgeschlossen
werden. In Abb. 20 überlappt das Signal für Katalase (grün) mit keiner der Zellorga-
nellen (Rhoptrien, Mikronemen, Granula densa und den Apikoplasten). Daraus läßt sich
schließen, daß Katalase in einem eigenständigen Kompartiment innerhalb von Toxoplas-
ma gondii vorkommt.
26

Abbildung 20: Kolokalisation der T. gondii Katalase mit verschiedenen subzellulären Orga-nellen. Katalase (in grün) kolokalisiert nicht mit den bekannten subzellulären Kompartimentenvon T. gondii. Mikronemen (a), Rhoptrien (c) und Dense granules (e) (hier rot dargestellt) wur-den mit monoklonalen Antikörpern markiert. Die entsprechenden Phasenkontrastbilder sind in(b) und (d) gezeigt. Die Anfärbung der Apikoplast DNA (f, Pfeile) wurde mit 4’,6-Diamidino-2-Phenylindol erreicht. Die Pfeilspitzen in (f ) zeigen auf die Nuklei der Parasiten. Skalierung 2µm.
27

Abbildung 21: Katalase Nachweis mitder DAB Präzipitationstechnik. Elek-tronenmikroskopische Aufnahmen in-trazellulärer Parasiten (a und b). MMitochondrium; N, Nukleus; p, Pero-xisom (Skalierung 0,5µm). Das Inset in azeigt eine vergrößerte Darstellung einesPeroxisoms (Skalierung 0,5µm). In derLichtmikroskopie erscheint das DAB-Präzipitat als dunkle Spots im Phasen-kontrastbild (c, Pfeile). In der Nomarski-Optik (d) stellen sich Parasiten besserdar.
2.5 Lokalisation mittels Histochemie
Katalase kann durch die DAB-Präzipitationstechnik licht- und elektronenmikroskopisch
nachgewiesen werden [22]. Diaminobenzidin (DAB) bildet mittelsH2O2 und Katalase ein
instabiles Zwischenprodukt, das spontan zu einem braunen Niederschlag oxidiert, wenn
folgende Bedingungen vorliegen: eine milde Fixierung, hohe Substrat (H2O2 ) Konzentra-
tion und eine alkalische Umgebung. Spezifisch hemmbar ist dieser Prozess mit Katalase
Inhibitoren, wie z. B. 20 mM 3-Amino-1,2,4-Triazol (AT) [21].
In der Lichtmikroskopie kann das Reaktionsprodukt im Inneren des Parasiten gefun-
den werden (Abb. 21 b, c). Dieses Präzipitat ist spezifisch, da es bei gleichzeitiger Inku-
bation mit AT nicht auftritt. Elektronenmikroskopisch finden sich 100 bis 300 nm große,
spezifische Präzipitate in juxtanukleärer Position, die meist eine sehr geordnete, kristall-
ähnliche Struktur aufweisen (Abb. 21, a, b und Inset). Zudem tritt das Reaktionsprodukt
in den Cristae der Mitochondrien auf. Wird das Substrat H2O2 in der Präzipitationsreak-
tion weggelassen oder der Inhibitor AT zugegeben, verschwinden die spezifischen Prä-
zipitate, das Signal in den Cristae der Mitochondrien jedoch bleibt. Folglich ist das Signal
in den Mitochondrien ein unspezifisches Reaktionsprodukt und kann wahrscheinlich auf
dort ansässige Peroxidasen [22] zurückgeführt werden.
2.6 Expression von Katalase in CHO-Zellen
Die funktionelle Bedeutung des C -terminalen peroxisomalen Targeting Signals der T.
gondii Katalase kann durch Expression rekombinanter Proteine in Säugetierzellen und
28

Feststellung der Lokalisation mittels Immunfluoreszenz, getestet werden. Dafür eigenen
sich CHO-Zellen im besonderen, da sie ein gut definiertes peroxisomales Kompartiment
aufweisen [119]. Die Lokalisation des peroxisomalen Kompartiments in der Immunfluo-
reszenz kann mit einem anti-SKL Antikörper gezeigt werden (Abb. 22e). Dieser Antikör-
per, der das C -terminale Tripeptid S-K-L erkennt, reagiert mit Peroxisomen vieler ver-
schiedener Organismen [35].
Transiente Expression der kompletten T. gondii Katalase führt zu punktförmigen Si-
gnalen im Zellkörper, die mit einer peroxisomalen Lokalisation im Einklang stehen (Abb.
22c). Werden jedoch die letzten drei Aminosäuren der Katalase entfernt, verteilt sich das
Signal diffus über die ganze Zelle, was charakteristisch für die zu erwartende zytosoli-
sche Lokalisation ist (Abb. 22d). Somit ist das PTS1 der T. gondii Katalase, zumindest in
einem heterologem System, funktionell von Bedeutung.
2.7 Expression eines Reportergens in CHO-Zellen
Die Frage, ob ein PTS1-Motiv ausreichend ist, um Katalase in Peroxisomen zu importie-
ren, kann durch einen Suffizienztest beantwortet werden. Hierbei wird das vermutliche
PTS1-Motiv an ein Reportergen angehängt und dieses in CHO-Zellen exprimiert.
Expression von Chloramphenicol-Acetyltransferase (CAT) in CHO-Zellen führt zu
einer zytosolischen Lokalisation (Abb. 22b) [106]. CAT-SKL hingegen, das CAT-Gen mit
dem PTS1-Motiv –SKL, ergibt ein peroxisomales Muster (nicht dargestellt), da das Prote-
in in die Peroxisomen importiert wird [106].
Fusioniert man die drei letzten Aminosäuren der T. gondii Katalase (-AKM) an das
CAT-Gen, ergibt sich ebenso eine peroxisomale Lokalisation (Abb. 22a). Diese Experi-
mente zeigen, daß das PTS1 der T. gondii Katalase ausreicht, um den Import eines zyto-
solisches Reporterproteins in die Peroxisomen zu veranlassen.
29

CAT AKM CAT
T. gondii Katalase AKM T. gondii Katalase ∆500-502Abbildung 22: Import der T. gondii Ka-talase und CAT in CHO-Zellen. Dieperoxisomale Lokalisation des CAT-AKM (a) und die diffuse zytosolischeVerteilung der nativen CAT ohne PTS(b) in der Immunfluoreszenz. In CHO-Zellen exprimierte T. gondii Katalasezeigt eine gepunktete Verteilung (c).Deletion der letzten drei Aminosäurenführt zu einer zytosolischen Lokalisati-on (d). In (e) ist das typische peroxiso-male Signal des anti-SKL Antikörpersgezeigt. Skalierung 5µm.
30

3 Materialien und Methoden
3.1 Bakterienstämme
DH5α (Gibco BRL): F−−φ80d lacZ ∆M15∆(lacZYA-argF) U169 endA1 recA1 hsdR17(r−Km+K)
deoR thi-1 supE44 λ− gyrA96 relA1
XL1 Blue (Stratagene, La Jolla, CA): recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac
[F’ proAB lacIqZ ∆M15 Tn10 (Tetr)]
3.2 Parasitenstämme
In der vorliegenden Arbeit wurde ausschließlich der RH-Stamm verwendet. Diese Para-
sitenlinie wird seit über 50 Jahren (Sabin und Feldman 1946) im Labor passagiert und hat
die Fähigkeit zur Zystenbildung in vivo weitgehend verloren [88]. In vitro jedoch ist eine
Induktion der Zystenbildung z. B. durch Hitzeschock noch möglich [100].
3.3 Lambda-Phagen
Folgende Lambda-Phagen-Klone aus der T. gondii EST Datenbank wurden verwendet:
dbEST ID gi Nummer Accession EST ID Library
571279 1370813 W63499 TgESTzy67b01.r1 ME49 tachyzoite
607975 1435613 W99737 TgESTzz06g03.r1 ME49 tachyzoite
3.4 Zellinien
Folgende Zellinien wurden in der vorliegenden Arbeit verwendet:
Vero-Zellen African green monkey kidney cells (ATCC-Nr. 4: CCL81)
CHO-Zellen Chinese hamster ovary cells K1 (ATCC-Nr.: CCL 61)
HFF-Zellen Human foreskin fibroblasts. Die Primärkultur dieser Fibroblasten
stammt aus der Yale University Dermatology Core Facility. In dieser
Arbeit verwendete Zellen waren in der 10. Passsage.
4ATCC: American Type Culture Collection
31

3.5 Plasmide
Folgende Plasmide wurden in der vorliegenden Arbeit verwendet:
Klonierung pGEM-T (Promega, Madison, WI)
pBluescript II SK+ (Stratagene, La Jolla, CA)
Expression in E. coli pGEX-4T-1 (Amersham Pharmacia Biotech, Piscataway,
NJ)
Expression in CHO-Zellen DNA3.1/Zeo(+) (Invitrogen Carlsbad, CA)
3.6 Primäre Antikörper
Folgende monoklonale Antikörper wurden freundlicherweise von J. F. Dubremetz, Lille,
Frankreich, zur Verfügung gestellt:
T3 4A7 monoklonaler Maus-Antikörper gegen das Rop2,3,4-Antigen
T6 2H11 monoklonaler Maus-Antikörper gegen das Gra3-Antigen
D2 R3 monoklonaler Maus-Antikörper gegen das Mic2-Antigen
Folgende polyklonale Antiseren wurden in dieser Arbeit verwendet:
Rabbit anti-CAT (5prime 3prime Inc., Boulder, CO)
Rabbit anti-SKL (Zymed Laboratories Inc., San Francisco, CA)
3.7 Sekundäre Antikörper
Folgende sekundäre Antikörper wurden in dieser Arbeit verwendet:
FITC linked goat anti-rabbit-IgG (Calbiochem-Novachem Corp. La Jolla, CA)
Rhodamine-linked goat anti-mouse-IgG (Roche Molecular Biochemicals)
HRP-conjugated goat anti-rabbit-IgG (Calbiochem-Novachem Corp. La Jolla, CA)
3.8 Kulturbedingungen für Zellinien
Alle Inkubationen fanden bei 37˚ C in 5 % CO2 Atmosphäre statt. Alle Zellkulturmedien
und Zusätze wurden von der Media Core Facility des Department of Cell Biology (Yale
University, New Haven, CT) hergestellt.
Vero-Zellen Minimum Eagle Medium (MEM) + 7 % FBS + Penicillin/Streptomycin
HFF-Zellen αMEM + Glutamin + 7 % FBS + Penicillin/Streptomycin
CHO-Zellen αMEM + Glutamin + 3.5 % FBS + Penicillin/Streptomycin
32

3.9 Passagieren der Zellinien
Um eine konfluente Zellkultur auf weitere Kulturschalen (Falcon, Ø 10 cm) aufzuteilen,
wurden die anheftenden Zellen mit 5 ml sterilem PBS-Puffer mehrmals gewaschen. Nach
kurzer Inkubation (5 min) wurde der PBS-Puffer entfernt und durch 1 ml Trypsin-Lösung
(Gibco-BRL) ersetzt. Nach kurzer Inkubation bei 37˚ C für 2 min wurden die Zellen in
12 ml angewärmten Kulturmedium resuspendiert und im Verhältnis 1 : 4 bis 1 : 6 auf neue
Kulturschalen verteilt. Alternativ wurden 2 ml dieser Suspension auf einer 6-well Platte
gleichmäßig verteilt. Die durchschnittliche Zeit bis zur erneuten Konfluenz der Zellen
betrug für CHO- und Vero-Zellen etwa 2-3 Tage, für HFF-Zellen eine Woche.
3.10 Passage der Parasiten in der Zellkultur
Etwa 5x106 Parasiten wurden zu einer 80 % konfluenten Vero-Zellkultur in einer 10 cm
Kulturschale hinzugegeben und für 2-3 Tage bei 37˚ C in 5 % CO2 Atmosphäre inkubiert.
Während dieser Zeit vermehren sich die Parasiten in den Wirtszellen. Das Kulturmedium
wurde abgesaugt, die Zellen mit PBS-Puffer gewaschen, mit einem Teflon-beschichteten
Schaber abgekratzt und in 2 ml PBS-Puffer resuspendiert. Die Zellsuspension wurde mit
Hilfe einer Spritze (3 ml Luer Lock) durch eine 19 Gauge Nadel getrieben. Dadurch wer-
den die Wirtszellen zerstört, während die Parasiten intakt bleiben. Das Lysat wurde ab-
zentrifugiert (5 min bei 1500 rpm in einem Beckmann Swingout-Rotor), das Sediment in
1 ml PBS-Puffer resuspendiert und die Parasitenzahl in einer Zählkammer bestimmt.
3.11 In vivo Exzision
Die Klone aus der EST Datenbank (W63499 und W99737) lagen in einem Lamda ZAP II
Vektor (Stratagene, La Jolla, CA) vor. Durch Koinfektion mit einem Helferphagen kann
das pBluescript Phagemid, das die EST-Klone enthält, aus dem Lambda ZAP II Vektor
(Abb. 23) ausgeschnitten werden und als Plasmid in E. coli propagiert werden [93].
Diese „In vivo Exzision“ ist abhängig von DNA Sequenzen im Lambda ZAP II Vektor,
sowie von Hilfsproteinen eines M13 Phagen oder F1 Phagen. Die Proteine des F1 Pha-
gen erkennen eine Region der DNA, die normalerweise der „origin of replication“ für
den positiven DNA-Strang in einem F1 Phagen ist. Dieser Bereich kann jedoch in eine
Initiations- und eine Terminations-Region aufgeteilt werden [25]. Zwischen diese beiden
Regionen ist im Lambda Zap II Vektor der pBluescript Vektor als Träger der ESTs kloniert.
Um nun die λ-Phagen DNA den Proteinen des F1 Phagen zugänglich zu machen,
wird ein E. coli Stamm mit beiden Phagen gleichzeitig infiziert. In E. coli erkennen nun
33

A - J c1857pBluescript + EST-DNAT I
Exzision mit F1-Phagen
pBluescript SK+
EST Klon
lacZ
Amp
EST-DNA
coscos
Initiator
Terminatorf1 ori
Abbildung 23: Prinzipder in vivo Exzision desLambda ZAP II Vektors.Mittels des λ-Phagen kanneine DNA Bibliothek er-stellt werden. Ko–Infektioneines Wirtsbakterium miteinem F1 Phagen schnei-det den Bereich zwischender Initiator- (I) undTerminator-Region (T) her-aus und ligiert ihn, so daßsich ein funktionsfähigespBluescript Plasmid bildet.
die F1 „Hilfs“-Proteine die Initiator-Region im λ-Phagen. Sie setzen einen „nick“ in der
Initiator DNA und replizieren einen Strang bis zur Terminatorregion. Das Gen II Produkt
des M13 Phagen bildet nun zirkuläre DNA, die nur noch das pBluescript Plasmid, die
EST Sequenz, einen F1-origin, die Terminator- und Initiatorregion enthält, während alle
anderen Sequenzen des λ-Phagen nicht mehr enthalten sind.
Die F1 Terminatorregion ermöglicht es nun, daß das Phagemid verpackt und von E.
coli sezerniert wird. Die E. coli Bakterien im Überstand werden nun durch Erhitzung auf
70˚ C abgetötet, während das Phagemid diese Temperatur übersteht. Um doppelsträngi-
ge DNA zu erzeugen, werden frische E. coli Zellen mit dem Phagemid (das sich ja selbst
nicht replizieren kann) inkubiert und auf Ampicillin ausgestrichen. Nur die Bakterien,
die das Phagemid als Plasmid tragen, sind Ampicillin resistent und können zur DNA
Isolation verwendet werden.
Als E. coli Stamm wurde XL1-Blue verwendet, da dieser Stamm das F‘-Episome trägt,
welches hier zwei Funktionen hat: Einerseits trägt es ein lacZ Gen mit der ∆M15 Mu-
tation, das eine blau/weiss-Selektion mit dem Lambda Zap II Vektor gestattet (weiße
Kolonien haben DNA in der „Multiple Cloning Site“ von pBluescript), andererseits ent-
hält das F‘-Plasmid die Gene für die Expression der F‘ Pili, die für eine Infektion mit
einem filamentösen Phagen (F1 oder M13) benötigt werden. Zudem trägt es das TN10
Tetrazyklin-Resistenz Gen, so daß mit Tetrazyklin für das F‘-Episome selektiert werden
kann.
34

RNA cDNA tailed cDNA
5’ 3’ 3’ 5’3’CCCC 5’
CCCC
CCCC
CCCC
PCR
3’CCCC 5’5’GGGG 3’
spezifisches Produkt
TdTrT
Pol
Abbildung 24: Prinzip der 5’ RACE.Bei der 5’ RACE wird RNA durch re-verser Transkriptase (rT) umgeschrie-ben und die entstandene cDNA en-zymatisch mittels Terminaler desoxy-nukleotid Transferase (TdT) um ca. 20Cytosin-Nukleotiden verlängert. Mit-tels Polymerasekettenreaktion (PCR)kann dann das spezifische Produktamplifiziert werden.
3.12 Praktisches Vorgehen bei der in vivo Excision
Eine 5 ml Schüttel-Kultur XL1-Blue wurde mit VCSM13 (kanr) infiziert, der gegenüber
dem klassischem F1-Phagen den Vorteil besitzt, eine Kanamycin Resistenz zu tragen. Mit
Phagen infizierte Bakterien wurden mit Kanamycin selektiert (In 5 ml LB: 1 Kolonie XL1-
Blue, 20 λ einer 20 % Maltose-Lösung, 50 λ von 1 M MgSO4, nach 3 h wird Kanamycin
hinzugegeben). Die Zellen wurden mit 1000 x g abzentrifugiert und in 2 ml SM Puffer re-
suspendiert. Alle Bakterien tragen nun den Helferphagen in sich. Es wurden 200 λ einer
Lambda-Phagen-Suspension hinzugegeben und 20 min bei 37˚ C inkubiert, damit sich die
Phagen an die Bakterien anheften können. Nach Zugabe von 5 ml 2xYT Medium wur-
de nochmals für 3 h bei 37˚ C inkubiert. In dieser Zeit integriert sich der Lambda Phage
im Chromosom und der Bluescript Vektor wird vom Helferphagen ausgeschnitten. Die
Bakterien wurden abzentrifugiert (4000 x g für 5 min) und 20 min auf 70˚ C erhitzt. Nach
Abkühlung wurden 200 λ des Überstands mit 200 λ XL1-Blue Zellen für 15 Min. bei 37˚ C
inkubiert. Während dieser Inkubation infiziert das Phagemid E. coli und repliziert sich als
Plasmid. Danach werden 10 λ und 100 λ auf Ampicillin Platten ausgestrichen. Kolonien
wurden isoliert, eine Plasmid-Präparation durchgeführt und die Plasmid-DNA sequen-
ziert.
3.13 „Rapid Amplification of cDNA Ends“ (5’ RACE)
Mittels 5’ RACE kann die Sequenz einer mRNA upstream von einem bekannten Teilstück
bestimmt werden. Hierbei werden enzymatisch mehrere Cytosin-Nukleotide an das 3’-
Ende der cDNA angehängt und dieses mit einem komplementären Primer in mehreren
nested PCRs amplifiziert, was die nachfolgende Klonierung und das Screening verein-
facht (Abb. 24). Die einzelnen Schritte sind im folgenden kurz dargestellt.
35

Zur RNA-Isolierung wurde TRIzol Reagenz nach den Angaben des Herstellers (Li-
fe Technologies, Rockville, MD) verwendet. Die mRNA wurde mit Superscript II rever-
se Transkriptase (Life Technology, Rockville, MD) in cDNA umgeschrieben, dann die
mRNA mit RNAseH degradiert und die cDNA über Qiaquick-Säulen (Qiagen, Valen-
cia, CA) aufgereinigt. Bei der sogenannten „Tailing“-Reaktion wird durch die Terminale-
deoxynukleotidyl-Transferase (TdT) eine Kette von entsprechend zugegebenen Nukleo-
tiden an das 3’-Ende der cDNA angehängt. Die Länge dieses poly-Nukleotid-Stückes
kann über die Reaktionsdauer gesteuert werden. Hier wurde mit Cytosin und einer Re-
aktionsdauer von 12 Minuten gearbeitet, was erfahrungsgemäß einen poly-C Stretch von
durchschnittlich 15-20 Basen erzeugt. Durch nested PCR wird die cDNA amplifiziert,
wobei spezifische Primer auf der einen und ein poly-dG Primer auf der anderen Seite
benutzt wird. Das entstehende Reaktionsprodukt wird auf einem Agarose-Gel aufgetra-
gen, ausgeschnitten, aufgereinigt und in den pGEM-T Vektor ligiert. Durch Screening der
Kolonien mittels PCR werden positive Klone identifiziert, das Plasmid aufgereinigt und
die Sequenz analysiert.
3.14 Sequenzierung
Die Basenabfolge der Katalase wurde mittels Sequenzierung vier unabhängiger Klone ei-
nes per RT-PCR (F4, R0) gewonnen Fragmentes von 2480 bp bestätigt. Es wurden jeweils
beide Stränge eines DNA Stücks mittels Dideoxy-Sequenzierung analysiert. Die Sequen-
zierung der DNA wurde vom W.M. Keck Sequencing center, Yale University School of
Medicine, durchgeführt.
3.15 Herstellung des Antiserums gegen T. gondii Katalase
Mittels Polymerase Kettereaktion (PCR) wurde das entsprechende Stück der T. gondii
Katalase amplifiziert (Primer: ExF und ExR) und in die BamHI und XmaI Schnittstelle
des Vektors pGEX-4T-1 kloniert. Das entstandene GST-Katalase-Fusionsprotein wurde
in Escherichia coli exprimiert, die Synthese mit Isopropyl-1-thio-β-D-galactopyranosoid
(IPTG) induziert und das Fusionsprotein über Glutathion-Sepharose Perlen aufgereinigt
(Abb. 25). Im folgenden wird dies näher beschrieben.
Eine 6 ml Übernachtkultur wurde mit LB Medium und Ampicillin auf 300 ml ver-
dünnt und bis zum Erreichen einer optischen Dichte von 0,6 bei 37˚ C geschüttelt. Nach
Zugabe von 0.5 mM IPTG zur Induktion der Fusionsprotein-Synthese wurden weitere 4 h
geschüttelt. Die Bakterien wurden abzentrifugiert, und mit 20 ml eiskaltem STE gewa-
schen. Die eine Hälfte des Ansatzes wurde für spätere Verwendung eingefroren, mit der
36
+ IPTG
- IPTG
GST-fusio
n
47 kD
102 kD
81 kD
47 kD
32 kD
+ IPTG
Eluat
1
Eluat
2
Centri
prep-3
Centri
con-3
0
Standar
d
a b
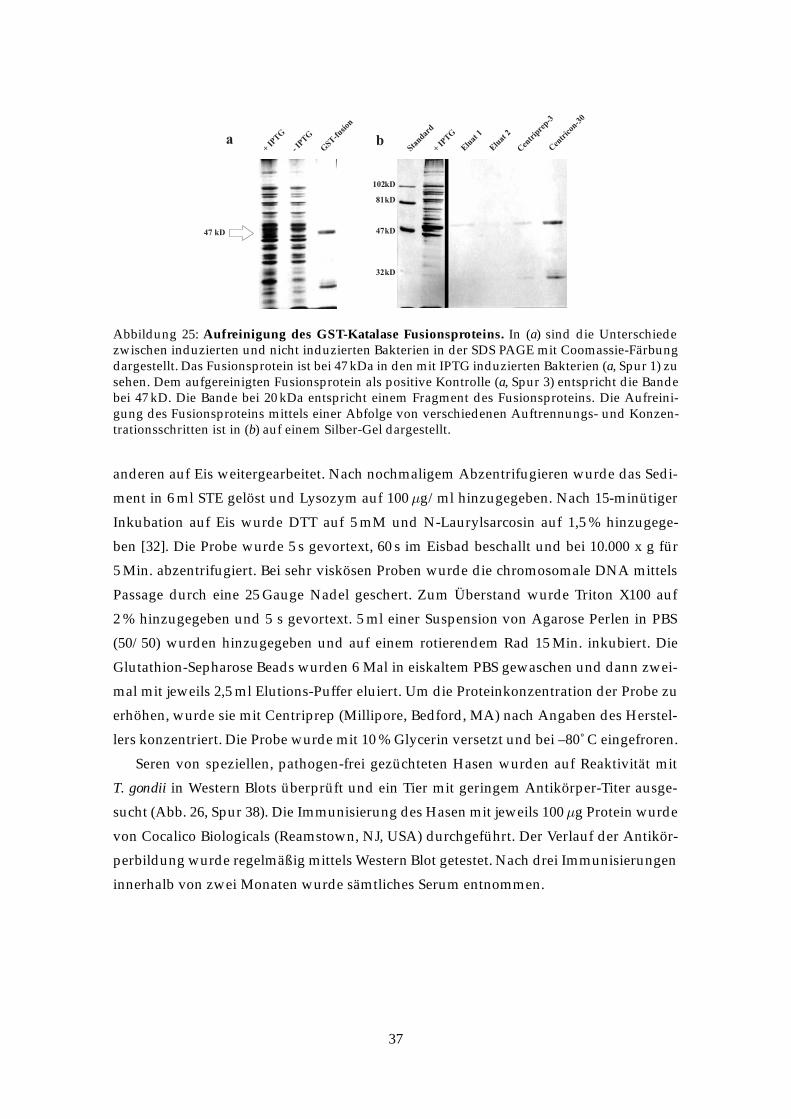
Abbildung 25: Aufreinigung des GST-Katalase Fusionsproteins. In (a) sind die Unterschiedezwischen induzierten und nicht induzierten Bakterien in der SDS PAGE mit Coomassie-Färbungdargestellt. Das Fusionsprotein ist bei 47 kDa in den mit IPTG induzierten Bakterien (a, Spur 1) zusehen. Dem aufgereinigten Fusionsprotein als positive Kontrolle (a, Spur 3) entspricht die Bandebei 47 kD. Die Bande bei 20 kDa entspricht einem Fragment des Fusionsproteins. Die Aufreini-gung des Fusionsproteins mittels einer Abfolge von verschiedenen Auftrennungs- und Konzen-trationsschritten ist in (b) auf einem Silber-Gel dargestellt.
anderen auf Eis weitergearbeitet. Nach nochmaligem Abzentrifugieren wurde das Sedi-
ment in 6 ml STE gelöst und Lysozym auf 100 µg/ml hinzugegeben. Nach 15-minütiger
Inkubation auf Eis wurde DTT auf 5 mM und N-Laurylsarcosin auf 1,5 % hinzugege-
ben [32]. Die Probe wurde 5 s gevortext, 60 s im Eisbad beschallt und bei 10.000 x g für
5 Min. abzentrifugiert. Bei sehr viskösen Proben wurde die chromosomale DNA mittels
Passage durch eine 25 Gauge Nadel geschert. Zum Überstand wurde Triton X100 auf
2 % hinzugegeben und 5 s gevortext. 5 ml einer Suspension von Agarose Perlen in PBS
(50/50) wurden hinzugegeben und auf einem rotierendem Rad 15 Min. inkubiert. Die
Glutathion-Sepharose Beads wurden 6 Mal in eiskaltem PBS gewaschen und dann zwei-
mal mit jeweils 2,5 ml Elutions-Puffer eluiert. Um die Proteinkonzentration der Probe zu
erhöhen, wurde sie mit Centriprep (Millipore, Bedford, MA) nach Angaben des Herstel-
lers konzentriert. Die Probe wurde mit 10 % Glycerin versetzt und bei –80˚ C eingefroren.
Seren von speziellen, pathogen-frei gezüchteten Hasen wurden auf Reaktivität mit
T. gondii in Western Blots überprüft und ein Tier mit geringem Antikörper-Titer ausge-
sucht (Abb. 26, Spur 38). Die Immunisierung des Hasen mit jeweils 100µg Protein wurde
von Cocalico Biologicals (Reamstown, NJ, USA) durchgeführt. Der Verlauf der Antikör-
perbildung wurde regelmäßig mittels Western Blot getestet. Nach drei Immunisierungen
innerhalb von zwei Monaten wurde sämtliches Serum entnommen.
37
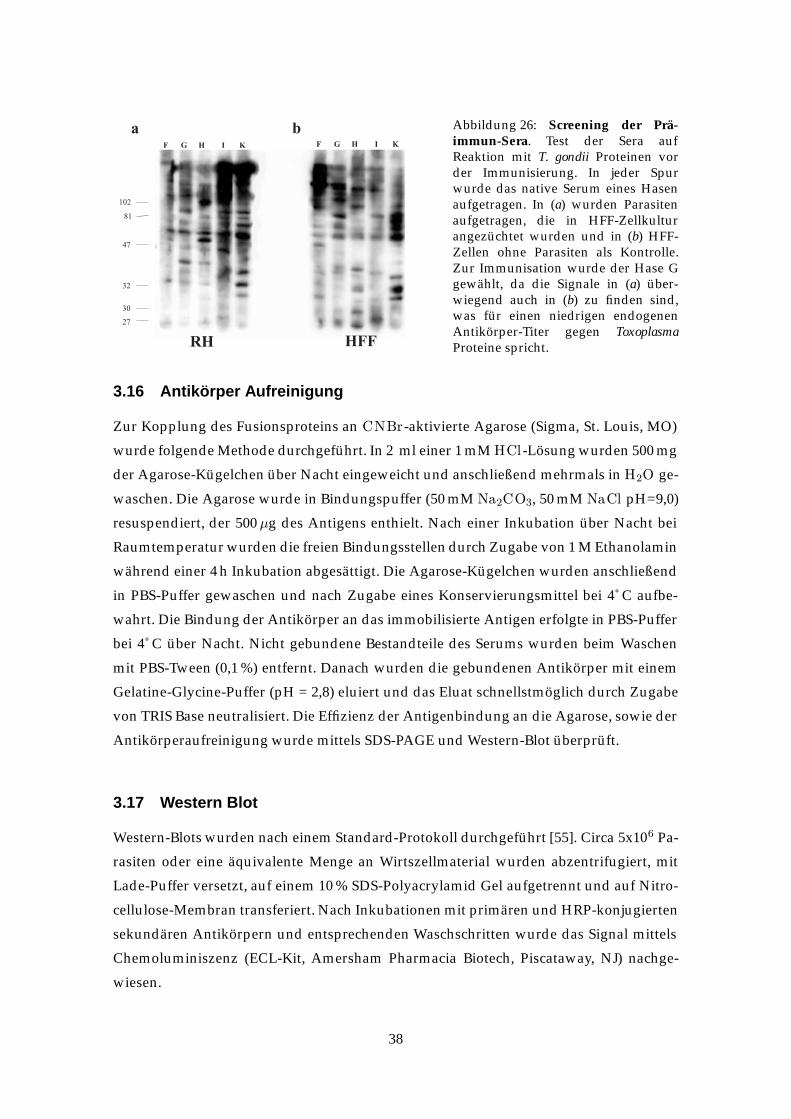
102
81
47
32
30
27
RH HFF
F G H I K F G H I K
a b Abbildung 26: Screening der Prä-immun-Sera. Test der Sera aufReaktion mit T. gondii Proteinen vorder Immunisierung. In jeder Spurwurde das native Serum eines Hasenaufgetragen. In (a) wurden Parasitenaufgetragen, die in HFF-Zellkulturangezüchtet wurden und in (b) HFF-Zellen ohne Parasiten als Kontrolle.Zur Immunisation wurde der Hase Ggewählt, da die Signale in (a) über-wiegend auch in (b) zu finden sind,was für einen niedrigen endogenenAntikörper-Titer gegen ToxoplasmaProteine spricht.
3.16 Antikörper Aufreinigung
Zur Kopplung des Fusionsproteins an CNBr -aktivierte Agarose (Sigma, St. Louis, MO)
wurde folgende Methode durchgeführt. In 2 ml einer 1 mM HCl -Lösung wurden 500 mg
der Agarose-Kügelchen über Nacht eingeweicht und anschließend mehrmals in H2O ge-
waschen. Die Agarose wurde in Bindungspuffer (50 mM Na2CO3, 50 mM NaCl pH=9,0)
resuspendiert, der 500µg des Antigens enthielt. Nach einer Inkubation über Nacht bei
Raumtemperatur wurden die freien Bindungsstellen durch Zugabe von 1 M Ethanolamin
während einer 4 h Inkubation abgesättigt. Die Agarose-Kügelchen wurden anschließend
in PBS-Puffer gewaschen und nach Zugabe eines Konservierungsmittel bei 4˚ C aufbe-
wahrt. Die Bindung der Antikörper an das immobilisierte Antigen erfolgte in PBS-Puffer
bei 4˚ C über Nacht. Nicht gebundene Bestandteile des Serums wurden beim Waschen
mit PBS-Tween (0,1 %) entfernt. Danach wurden die gebundenen Antikörper mit einem
Gelatine-Glycine-Puffer (pH = 2,8) eluiert und das Eluat schnellstmöglich durch Zugabe
von TRIS Base neutralisiert. Die Effizienz der Antigenbindung an die Agarose, sowie der
Antikörperaufreinigung wurde mittels SDS-PAGE und Western-Blot überprüft.
3.17 Western Blot
Western-Blots wurden nach einem Standard-Protokoll durchgeführt [55]. Circa 5x106 Pa-
rasiten oder eine äquivalente Menge an Wirtszellmaterial wurden abzentrifugiert, mit
Lade-Puffer versetzt, auf einem 10 % SDS-Polyacrylamid Gel aufgetrennt und auf Nitro-
cellulose-Membran transferiert. Nach Inkubationen mit primären und HRP-konjugierten
sekundären Antikörpern und entsprechenden Waschschritten wurde das Signal mittels
Chemoluminiszenz (ECL-Kit, Amersham Pharmacia Biotech, Piscataway, NJ) nachge-
wiesen.
38

Antikörper-Konzentrationen:
T. gondii Katalase Antiserum 1 : 500
HRP-konjugierter goat anti-rabbit IgG Antikörper 1 : 2000
3.18 Immunfluoreszenz
Für den Immunfluoreszenz-Assay wurden Zellen auf runden Deckgläschen mit 15 mM
Durchmesser über Nacht herangezogen. Nach dreimaligem Waschen in PBS++ (1mM
CaCl2 und 1 mM MgCl2 als Zusatz zu PBS-Puffer) innerhalb von 5 min mit ca. 2 ml/well
wurden die Zellen in 3 % Paraformaldehyd bei Raumtemperatur fixiert. Dann folgte wie-
derholtes dreimaliges Waschen und anschließend eine Permeabilisierung der Membran
mit 0.1 % Triton X-100 in PBS++ für 5 Minuten. Nach anschließendem zweimaligem Wa-
schen in PBS++ wurden durch Inkubation für 30 Minuten mit 50 % Ziegen-Serum freie
Bindungsstellen geblockt. Die Antikörper wurden in 3 % BSA in PBS++ verdünnt, die
Deckgläschen umgekehrt auf einen Tropfen dieser Lösung gegeben und 30 Minuten bei
Raumtemeperatur inkubiert. Nach dreimaligen Waschen in PBS++ wurde mit dem zwei-
ten Antikörper auf die gleiche Weise inkubiert. Nach 30 Minuten wurden drei weitere
Waschschritte mit PBS++ vorgenommen und die Deckgläser mittels Mowiol 40-88 (Sig-
ma St. Louis, MO) auf einem Objektträger upside-down montiert. Nach 4 h Aushärtung
können die Präparate betrachtet werden.
Alternativ kann die Fixation für 15 min in eiskaltem Aceton erfolgen. Dabei entfällt
der anschließende Permeabilisierungsschritt mit TritonX-100. Eine Färbung des Apiko-
plast läßt sich durch Zugabe von wenigen Kristallen 4’,6-Diamidino-2-Phenylindol in
die Einbettungsflüssigkeit erreichen.
Antikörperkonzentrationen für den Immunfluoreszenz Assay
T. gondii Katalase Antiserum 1 : 250
aufgereinigtes T. gondii Katalase Antiserum 1 : 5
Rabbit anti-CAT 1 : 500
Rabbit anti-SKL 1 : 500
T3 4A7 1 : 250
T6 2H11 1 : 250
D2 R3 1 : 250
FITC linked goat anti-rabbit IgG 1 : 500
Rhodamine-linked goat anti-mouse-IgG 1 : 500
39

3.19 DAB Präzipitation
Die Parasiten wurden in der Kulturschale für 15 min mit 1,5 % Glutaraldehyd in 0,1 M PI-
PES Puffer (pH 7,4) fixiert, zweimal in PBS und einmal in Teorell-Stenhagen (T/S) Puffer
(pH 10,5) gewaschen. Nach einer zweistündigen Inkubation mit 2 mM DAB in T/S wur-
de H2O2 in einer End-Konzentration von 1,5 % für 1-3 h hinzugegeben, danach jeweils
einmal mit T/S und 100 mM Natrium-Cacodylat (pH 7,4) gewaschen und die Probe über
Nacht bei 4˚ C in 2,5 % Glutaraldehyd und 100 mM Natrium-Cacodylat fixiert. Die Pro-
be wurde mittels eines Teflon Schabers in ein Eppendorf-Cap überführt und in EPON
eingebettet. Im Microtom wurden 60-80 nm dicke Schnitte angefertigt und auf Formvar
beschichteten Nickel Trägern montiert. Die Schnitte wurden nicht mit Bleizitrat gegen-
gefärbt, damit das DAB Präzipitat leichter erkannt werden kann. Zur Kontrolle wurden
Inkubationen ohne H2O2 oder in Anwesenheit eines Katalase Inhibitors (z. B. 20 mM 3-
Amino-1,2,4-Triazol) durchgeführt.
Sämtliche Materialien für die Elektronenmikroskopie wurden von er Firma Electron
Microscopy Sciences, Fort Washington, PA bezogen. Für die Lichtmikroskopie wurde obi-
ge Prozedur auf Objektträgern durchgeführt, die nach der Inkubation mit H2O2 unter-
sucht werden können.
3.20 Klonierung der Expressions-Konstrukte
Mit Hilfe der Polymerase-Kettenreaktion (PCR) wurde die DNA-Sequenz der Chlor-
amphenicol-Acetyltransferase um die Aminosäuren -AKM am C -terminus erweitert, so-
wie an den Enden eine HindIII und KpnI Restriktionsenzym-Schnittstelle eingefügt. Das
entstandene PCR-Produkt wurde in den pcDNA3.1/Zeo(+)-Vektor eingefügt, der für
die Expression des CAT-AKM Genprodukts in Säugerzellen einen Zytomegalie-Virus-
Promoter enthält.
Auf die gleiche Weise wurde die T. gondii Katalase in die NheI und BamHI Restrikti-
onsenzym-Schnittstellen des pcDNA3.1/Zeo(+)-Vektors eingefügt. Die unterschiedliche
Länge der T. gondii Katalase wurde durch Verwendung verschiedener Primer erreicht:
NheCat und AKMBgl für die volle Länge, NheCat und CYPBgl für die drei Aminosäuren
kürzere Version.
3.21 Transfektion der CHO-Zellen
Zur Transfektion der Säugerzellen wurde Superfect (Qiagen, Valencia, CA) nach den An-
gaben des Herstellers verwendet. Die Transfektion wurde in Kulturschalen durchgeführt,
40

die Zellen nach 24-48 h auf Deckgläser umgesetzt und 24-48 h später mittels Immunfluo-
reszenz angefärbt.
3.22 Allgemeine molekularbiologische Techniken
Die Plasmid-Präparationen wurden nach dem alkalische Lyse-Verfahren durchgeführt
[55], bzw. Mini-, Midi- und Maxiprep-Kits (Qiagen, Valencia, CA) benutzt. Als Enzyme
für die Polymerase-Kettenreaktion (PCR) wurde Taq-Polymerase und PfU-Polymerase
(New England Biolabs, MA) nach Angaben des Herstellers verwendet.
Alle Restriktionsendonukleasen, die T4-DNA-Ligase und die Calf-Intestinal-Phos-
phatase wurden von New England Biolabs, MA, bezogen und nach Angabe des Her-
stellers verwendet.
Die Auftrennung von DNA erfolgte auf Gelen aus Seakem LE Agarose oder Sea-
plaque Agarose (Sigma, St. Louis, MO). Die DNA wurde mit Ethidiumbromid unter
UV-Licht dargestellt. DNA wurde aus Seaplaque Agarose-Gelen mit Phenol extrahiert
und auf einer Qiaquick-Säule (Qiagen, Valencia, CA) aufgereinigt. Proteine wurden auf
Acrylamid-Gelen mittels SDS-PAGE nach einem Standardverfahren aufgetrennt [55].
Transformations-kompetente E. coli wurden mit der CaCl-Methode hergestellt und
die Transformation mit Hitzeschock eingeleitet [55].
3.23 Oligonukleotide und Chemikalien
Sämtliche Oligonukleotide (siehe Tabelle 4) wurden von der W.M. Keck Foundation Bio-
technology Resource Labaratroy (Yale University, New Haven, CT) bezogen. Alle wei-
teren Chemikalien, deren Hersteller nicht aufgeführt ist, wurden von Sigma (St. Louis,
MO) bezogen.
3.24 Protein-Modellierung mit Swissmodel
Die Modellierung der T. gondii Katalase wurde mit Hilfe des Swiss-Model-Servers 5 vor-
genommen. Dieser Server vergleicht die eingegebene Aminosäuresequenz mit den Se-
quenzen experimentell erfaßter 3D-Strukturen mittels BlastP2 und sucht geeignete Vor-
lagen für ein 3D-Modell aus. Anschließend wird mittels ProModII die eingegebene Se-
quenz anhand der dreidimensionalen Vorlagen modelliert und eine Energieminimierung
5http://www.expasy.ch/swissmod/SWISS-MODEL.html
41

Name Sequenz
R0 TTCTCGTTCAGAAAGTGCTG
R1 CATTTCTGTCTTCCTTTGGAAAGC
R2 TCGCCTGCCTGAGATGC
R3 GGGAAACTCTCGCTTGGC
R4 CATTACCTGAATACAGAAGG
R5 GGCGTTGAACAGGTCACG
R6 GTCGCGCCTGGTGTCC
R7 CACTCCCCGACTCGCC
R9 GGAAGTTACTCAGGATCGC
F1 GCTGCAGGGACGCATGTTCTC
F4 GTGCGGCTCCTCGAAGG
Fdeg CCnGArmGnGTnGTnCAyGC
ExF CGGGATCCCCGAGCGAAGATCGCATGC
ExR TCCCCCGGGACAACGCACGCCTAC
NheCat CTTCGTCAGCTAGCATGACTCAGGTTCCGCCCG
AKMNot CGTTTAGCGGCCGCAGATCTACATCTTGGCTGGGTAG
CYPNot CGTTTAGCGGCCGCAGATCTATGGGTAGCACGCCGCAG
CATFHind CCCAAGCTTAATGCATGAGAAAAAAATCACTGG
CATAKMKpn GGGGTACCTTACATCTTTGCTCCCGCCCCGCCCTGCC
Tabelle 4: In dieser Arbeit verwendete Primer. Schnittstellen für Restriktionsenzyme sind fett ge-druckt. n = A,C,G,T; r = A,G; m = A,C; y = C,T.
mit Gromos96 durchgeführt [36, 80, 81]. Die Ausgabe erfolgt als PDB-Datei, die alle mo-
dellierten Atomkoordinaten enthält. Diese Datei kann nun mit Hilfe eines PDB-Viewers
dargestellt werden.
Die Aussagekraft der 3D-Modelle hängt von mehreren Einflußgrößen ab. Man unter-
scheidet zwischen der correctness, die von der Qualität des Alignments abhängt, und der
accuracy, die Abweichung des errechneten Modells von der experimentell gewonnenen
Struktur beschreibt. Bei mangelnder correctness treten Fehler beim Alignment auf und
die entsprechende Region wird räumlich falsch modelliert. Diese Fehler können durch
ein sorgfältiges Alignment vermieden werden. Zum Teil fallen diese durch stereoche-
misch unmögliche Konformationen auf. Die Abweichung eines errechneten Modells von
einer experimentell bestimmten Struktur, accuracy, kann zwei Ursachen haben. Einerseits
Fehler im Modelling-Verfahren, andererseits die Variabilität in den Verfahren der experi-
mentellen Strukturbestimmung.
Um eine genauere Vorstellung von der accuracy zu bekommen wurden im 3DCrunch-
Projekt 1200 Modell/Struktur-Paare verglichen [91]. Es zeigte sich, daß eine Überein-
42

stimmung der Aminosäuresequenz zwischen Modell und Vorlage von über 70 % zu äu-
ßerst präzisen Strukturvorhersagen führt. Diese Modelle sind aufgrund ihrer Präzision
u. a. für Compound-Optimierung im Drug Design geeignet. Modelle mit korrekten Ali-
gnment trotz geringerer Sequenzübereinstimmung sind noch immer nützliche Werkzeu-
ge bei „rational mutagenesis“ Experimenten. Modelle mit inkorrekten Alignment haben
ihre Aussagekraft in den Regionen mit falschen Alignment verloren, können aber für die
anderen Regionen durchaus korrekt modelliert sein.
Die Modellstruktur der Aminosäuren 10 bis 490 der T. gondii Katalase wurde mit-
tels dem Swiss-Model-Server anhand folgender Strukturen der bovinen Katalase aus
der ExNRL-3D Datenbank errechnet: 4BLCA, 8CATA, 7CATA, 4BLCD, 4BLCC. Die hohe
Identität der Aminosäuren zwischen T. gondii und boviner Katalase (63 %) läßt auf eine
hohe Qualität des Modells schließen.
Die fehlenden N- und C -terminalen Aminosäuren wurden mit dem Swiss PDB View-
er von Hand hinzugefügt. Da keinerlei Daten über die dreidimensionale Anordnung
dieser Aminosäuren vorhanden ist, sind Fehler in diesem Bereich des Modells unver-
meidlich. Das Rendering der Stereo-Abbildungen wurde mittels Raytracing (Pov-Ray)
durchgeführt. Ein 3D-Effekt läßt sich bei den Stereobildern durch leichtes Einwärtsschie-
len erreichen.
3.25 Verwendete Software
Die vorliegende Arbeit wurde mit dem Textsatzprogramm LATEX 2ε und BIBTEX 0. 99 c aus
der Distribution MiKTEX 2. 0 6 gesetzt. Abbildungen wurden mit Adobe Photoshop 5. 5
und Adobe Illustrator 8. 0 bearbeitet. Die 3D-Modellierung der T. gondii Katalase wurde
mit Swiss-PdbViewer v 3. 7 7 durchgeführt und mit Pov-Ray 3. 1 8 nachbearbeitet.
6http://www.miktex.de7http://www.expasy.ch/spdbv/mainpage.html8http://www.pov-ray.org
43

3.26 Puffer und Lösungen
10 x PBS 800 ml dH2O , 80 g NaCl , 2 g KCl , 14.4 g Na2HPO4, 2.4 g KH2PO4,
auf 1 Liter auffüllen, pH auf 7,2 bringen, 1 x Lösung autoklavieren
und bei Raumtemperatur aufbewahren.
PBS++ 1 l 1 x PBS, 1 ml 1 M MgCl2 und 1 ml 1 M CaCl2 hinzugeben, bei
Raumtemperatur lagern.
TE 10 mM Tris, 1 mM EDTA: 0.06 g Tris, 0.19 g Na2EDTA in 50 ml dH2O ,
pH auf 7,6 einstellen.
20 x TBE 108 g Tris, 55 g Borsäure in 900 ml dH2O , 80 ml 0,25 M EDTA pH 8,0
hinzugeben, pH auf 8,4 bringen und auf 1 l mit dH2O auffüllen.
EDTA 9.3 g NaEDTA in 100 ml dH2O , 10 N NaOH zugeben bis zu einem
pH von 8,4.
STE 10 mM TRIS, 150 mM NaCl , 1 mM EDTA.
Elutions-Puffer 75 mM Hepes (pH 7,4), 150 mM NaCl , 10 mM reduziertes GSH,
5 mM DTT, 2 % N-Octylguanid.
44

3.27 Benutzte Geräte
Die folgende Tabelle führt einige bisher nicht näher bezeichnete Geräte an. Kleinere La-
borgeräte und Massenartikel, wie z. B. Pipettenspitzen, wurden ausgelassen.
Gerät Bezeichnung Hersteller/Vertrieb
Elektronenmikroskop Philips 101 Philips Inc., Peawick, NY
Fluoreszenz-Mikroskop Microphot-FxA Nikon, JP
Ölimmersionsobjektiv PlanApo100/1.40 Nikon, JP
Quecksilberdampflampe HB10101 AF Nikon, JP
Kamera Sensys II Photometrics, NJ
Spektrometer Genesys 5 Spectronic Instruments, Roche-
ster, NY
SpeedVac DNA110 Savant Instruments Inc., Far-
mingdale, NY
Transilluminator FBTIV88 Fisher Scientific
PCR-Gerät Thermal-Cycler III Hybaid, NJ
Gene Pulser Biorad 2010 Bio-Rad Inc. Hercules, CA
Sonicator Modell G112SP1G Laboratory Supplies Co. Inc.,
Hicksville, NY
Tabletop-Zentrifuge Eppendorf 5415c Fisher Scientific
Swingout Rotor Modell GPR Beckman Inc, Wakefield, MA
45

4 Diskussion
Die Katalase ist ein ubiquitär vorkommendes Enzym das in vielen Organismen eine
wichtige Bedeutung für den Stoffwechsel hat. Sie katalysiert die Umwandlung von Was-
serstoffperoxid in Wasser und Sauerstoff und verringert somit die Bildung von freien
Radikalen. Dadurch schützt sie vor deren toxischen Wirkung auf DNA, Proteine und Li-
pide [54]. Da Wasserstoffperoxid leicht über biologische Membranen diffundieren kann,
ist es auch ein wesentlicher Bestandteil der Immunabwehr und tötet z. B. Bakterien in den
Phagosomen ab [41]. Reaktive Sauerstoffradikale sind ein wesentlicher Abwehrmecha-
nismus zur Bekämpfung einer Toxoplasma gondii Infektion [40,65–68]. Untersuchungen an
Ucp2-Knockout Mäusen, die resistent gegen Infektionen mit Toxoplasma sind, untermau-
ern dies [3]. Die Makrophagen dieser Knockout-Mäusen produzieren größere Mengen
freier Radikale als Wildtyp-Makrophagen und können Toxoplasma gondii fünffach besser
abtöten.
Ein Vergleich der Protein-Expression zwischen einem Wildtyp und einem attenuier-
ten T. gondii Stamm, ersterer wurde in Mäusen passagiert, letzterer ausschließlich in Zell-
kultur, ergab eine verminderte Expression der Katalase in dem attenuierten Stamm [73].
Dies ist ein weiterer Hinweis darauf, daß Katalase ein Pathogenitätsfaktor ist.
Gegen die Sauerstoffradikale der Makrophagen schützt sich T. gondii mit einer Reihe
von Enzymen. Die schon vor langer Zeit beobachtete hohe Resistenz gegen H2O2 [66]
erklärt sich aus dem Zusammenspiel von Katalase [45, 95] Superoxid-Dismutase [74, 95],
Peroxidasen und Peroxiredoxin [102].
In der vorliegenden Arbeit haben wir die Katalase von Toxoplasma gondii kloniert und
genauer untersucht. Die Ergebnisse unserer Sequenzierung bestätigten Ding et al. in einer
zeitgleichen, unabhängigen Arbeit [23]. Außerdem konnte diese Gruppe mit Northern-
und Southern-Blot-Analyse die Transkription der Katalase DNA nachweisen.
Bei unseren Untersuchungen zeigte sich im Western-Blot mit einem polyklonalen An-
tiserum gegen ein Katalase-GST-Fusionsprotein ein Protein in der Größe von ca. 63 kDa.
Dies steht in guter Übereinstimmung mit der berechneten Größe. Somit gehen wir davon
aus, daß das Antiserum in der Tat mit Katalase reagiert.
Die starke Konservierung von Katalase verschiedener Organismen über die Artgren-
zen hinweg begünstigt eine strukturelle Analyse. So können Voraussagen über struk-
turelle Zusammenhänge durch Analogieschluss mit hoher Wahrscheinlichkeit getroffen
werden. Die absolute Konservierung der Aminosäuren, die für die Bindung des Häm-
und NADPH -Moleküls verantwortlich sind und das aktive Zentrum bilden, läßt auf eine
Ähnlichkeit des katalytischen Reaktionsmechanismus schließen. Aminosäurereste, die
46

in anderen Katalasen mit den anderen Untereinheiten interagieren, sind auch in der T.
gondii Katalase vorhanden. Dies spricht für eine Organisation der Katalase als Homo-
tetramer, wie sie bei allen bisher bekannten eukaryontischen Katalasen vorkommt. Das
C -terminale PTS1 der T. gondii Katalase legt eine Lokalisation in einem peroxisomalen
Kompartiment nahe.
Die starke Homologie der T. gondii Katalase zu anderen Katalasen begünstigt auch
Aussagen über die Struktur des Enzyms im dreidimensionalen Raum. Durch Vergleich
mit bekannten Röntgendiffraktions- oder NMR-Strukturen kann eine Modellierung des
Proteins erfolgen. Hierfür sind zahlreiche Computeralgorithmen entwickelt worden. Ei-
ne Nachbearbeitung zur Verbesserung und Erweiterung eines computergenerierte Mo-
dells wird von Hand vorgenommen. Entscheidend für die Qualität der Modellierung ist
einerseits die Kenntnis der räumliche Auflösung der Vorlage und andererseits die Ähn-
lichkeit in der Aminosäuresequenz zwischen Modell und Vorlage. Im Falle der T. gondii
Katalase sind beide Einflußfaktoren günstig verteilt: Mit einer Homologie von 63 % zur
bovinen Katalase und einer Auflösung deren Kristallstruktur von 2.5 Å finden sich sehr
gute Voraussetzungen. Da sich die meisten Katalasen relativ einfach kristallisieren lassen,
stehen mehrere dreidimensionale Strukturen als Vorlage zur Verfügung [24, 33, 47, 85].
Durch die dreidimensionale Modellierung von Proteinen können die sterischen Ver-
hältnisse wesentlich anschaulicher dargestellt und besser interpretiert werden. Bei der
Sekundärstruktur der Katalase fällt ein β-Barrel aus acht antiparallelen β-Faltblattstruk-
turen auf, das das Häm-Molekül auf einer Seite abschirmt, während die andere Seite re-
lativ frei zugänglich ist. Ein Kanal führt direkt zum Eisenatom des Häms und dient dem
Anfluten des Substrats H2O2 . Die Modellierung des aktiven Zentrums und der NADPH -
Bindungsstelle illustriert die räumliche Anordnung der katalytischen Aminosäurereste.
Auch die Interaktionen zwischen den Monomeren sind durch eine 3D-Ansicht wesent-
lich anschaulicher dargestellt.
In den meisten eukaryontischen Zellen liegt Katalase in den Peroxisomen (Microbo-
dies) vor. Aus diesem Grunde ist sie zum Leitenzym dieses Kompartiments geworden. In
einigen Fällen jedoch, wie z. B. in Saccharomyces cerevisiae, finden sich zwei Isoformen, von
denen die eine in Peroxisomen und die andere im Zytosol vorliegt. In Toxoplasma haben
wir, sowie Ding et al., keine weitere Isoform der Katalase gefunden. Um die Lokalisation
der Katalase in T. gondii zu klären, haben wir die Verteilung licht- und elektronenmikro-
skopisch untersucht.
In der Immunfluoreszenz-Mikroskopie mit einem Antiserum gegen T. gondii Katalase
zeigte sich ein punktförmiges Signal im vorderen Ende des Parasiten, das an vesikuläre
Strukturen erinnert. Zudem lag ein diffuses, aber spezifisches Signal innerhalb des Para-
47

siten vor, das zytosolisch anmutet. Gleichzeitiges Anfärben verschiedener Zellorganellen
(Rhoptrien, Granula densa, Mikronemen und der Apikoplast) zeigte keine Signalüberla-
gerung. Daher muß davon ausgegangen werden, daß das vesikuläre Signal einem nicht
näher definierten Kompartiment entspricht.
Allerdings sind in Toxoplasma nicht alle Kompartimente definiert, die in Säugetierzel-
len bekannt sind. So gibt es kein überzeugendes Äquivalent für die Lysosomen und En-
dosomen. Obwohl es einige Hinweise für eine Rezeptor vermittelte Endozytose an der
„micropore“ (einem spezialisierten, eingestülpten Bereich der Zellmembran) gibt [72],
entzieht sich die Endozytose des Parasiten bisher der experimentellen Untersuchung
[92]. Auch der Nachweis von Lysosomen ist bisher nicht gelungen.
Die elektronenmikroskopische Darstellung der Katalase-Aktivität durch DAB-Präzi-
pitation zeigte 100-300 nm große, runde Ablagerungen. Diese Ablagerungen traten zu-
meist in juxtanukleärer Position auf und enthielten kristallähnliche Einschlüsse. Durch
gleichzeitige Inkubation mit einem Katalase-Inhibitor oder bei Auslassung des Substrats
(H2O2 ) verschwinden diese Ablagerungen, während das unspezifische Signal in den Mit-
ochondrien bestehen bleibt. Dies spricht für ein Katalase-spezifisches Signal in den run-
den Ablagerungen. Aus der geringen Häufigkeit der runden Präzipitate (1 auf 20-30 Pa-
rasitenanschnitte) kann man schließen, daß nur wenige Strukturen pro Parasit vorliegen.
Dies steht im Einklang mit den Immunfluoreszenz-Bildern auf denen jeweils 2-5 punkt-
förmige Signale pro Parasit zu sehen sind.
Unsere Ergebnisse legen nahe, daß wir ein bisher noch nicht näher definiertes, subzel-
luläres Kompartiment entdeckt haben. Es befindet sich im vorderen Ende der Parasiten,
in der Nähe des Golgi-Apparats. In diesem Bereich liegt vermutlich auch das Zentromer,
das Organisationszentrum der Mikrotubuli. Aus Experimenten an CHO-Zellen ist be-
kannt, daß Peroxisomen sich entlang Mikrotubuli bewegen können [83, 115]. Allerdings
findet man in vielen anderen eukaryontischen Zellen eine Vielzahl von Vesikeln in der
Nähe des Zentromers.
Das in der Fluoreszenz-Mikroskopie sichtbare, diffuse Signal innerhalb des Parasiten
spricht für die Existenz eines Katalase-Pools im Zytosol. Dies könnte durch eine Sätti-
gung der Importmaschinerie oder eine andere Isoform hervorgerufen sein. Ding et al.
haben allerdings keine weiteren Katalasen im T. gondii Genom entdeckt [23]. Der zyto-
solische Pool der Katalase könnte ein Schutzmechanismus gegen freie Radikale sein, der
das Überleben des Parasiten bei einer Immunreaktion des Wirtes sichert.
Entscheidend für die Definition eines peroxisomalen Kompartiments ist eine umge-
bende Membran. Die hier verwendeten elektronenmikroskopischen Methoden können
diese leider nicht überzeugend dargestellen, da keine Gegenfärbung mit Bleizitrat durch-
48

geführt wurde. Andererseits wird durch Auslassung der Gegenfärbung die Möglichkeit
falsch positive Resultate zu erreichen, stark verringert. Der Versuch, durch Immunogold-
labeling mit dem polyklonalen Antiserum gegen Katalase elektronenmikroskopisch eine
Klärung herbeizuführen, ist uns und anderen Arbeitsgruppen bisher nicht gelungen.
Ein weiteres Argument für eine peroxisomale Lokalisation der T. gondii Katalase ist
die Tatsache, daß sie in einem heterologen System (CHO-Zellen) in die Peroxisomen
transportiert wird. Die Funktionalität des PTS1 der T. gondii Katalase in CHO-Zellen er-
höht die Wahrscheinlichkeit, daß ein solches Signal in Toxoplasma auch funktionell in T.
gondii von Bedeutung ist. Eine endgültige Klärung der Bedeutung des PTS1 kann jedoch
nur durch Expression der rekombinanten Katalase in einem T. gondii Katalase knock-out
Stamm erzielt werden. Die dafür nötige Methodik ist seit einiger Zeit entwickelt [88].
Die Funktion eines peroxisomalen Kompartimentes in T. gondii ist zur Zeit nicht ab-
zuschätzen. Viele Stoffwechselwege bieten sich an. Von besonderem Interesse ist der Bei-
trag peroxisomaler Enzyme in der Lipid-Synthese und Metabolismus. Der Parasit scheint
unfähig zu sein, Cholesterol und Phospholipide de novo zu synthetisieren und bekommt
diese von der Wirtszelle [17]. Andererseits werden Enzyme der Fettsäurebiosynthese in
den Apikoplast importiert, ein Überrest eines Chloroplasten in vielen, wenn nicht allen
Apikomplexa [87]. Das Vorkommen von Enzymen des Glyoxylat-Zyklus wie Isozitrat-
Lyase und Malat-Synthetase [62], die in den Glyoxysomen der Pflanzen enthalten sind,
konnte von uns nicht eindeutig bestätigt werden, jedoch sind diese Enzyme potentiell in
Peroxisomen anzutreffen.
Mit der vorliegenden Arbeit konnte die Existenz eines peroxisomalen Kompartimen-
tes in T. gondii nicht zweifelsfrei geklärt werden, jedoch eröffnet sie die Möglichkeit in
neuer Weise über Stoffwechselvorgänge dieses Parasiten nachzudenken. Eine genauere
Untersuchung der Peroxisomen von T. gondii führt letztlich zu einem genaueren Ver-
ständnis des Metabolismus des Parasiten und potentiell zu neuen therapeutischen An-
sätzen.
49

5 Zusammenfassung
Toxoplasma gondii ist ein obligat intrazellulärer, einzelliger Parasit aus dem Phylum der
Apicomplexa. Infektionen des Menschen mit T. gondii verlaufen meist subklinisch. Nach
einer Infektion persistiert der Erreger für viele Jahre in Hirn- und Muskelgewebe. Durch
Reaktivierung des Erregers, z. B. durch eine Immunschwächekrankheit oder unter Im-
munsuppression, kann eine Enzephalitis mit septischer Streuung entstehen. Eine diapla-
zentare Infektion führt zur Fetopathia toxoplasmotica mit Früh- und Totgeburten oder zu
der typischen enzephalitischen Trias aus Chorioretinitis, Hydrozephalus und zerebralen
Verkalkungen.
Ein Mechanismus, der es T. gondii ermöglicht im Wirtsorganismus zu überleben, ist
die ungewöhnlich hohe Widerstandsfähigkeit gegenüber freien Radikalen. Die wichtig-
ste Quelle für freie Radikale bei der Abwehrreaktion des Wirtsorganismus ist Wasser-
stoffperoxid (H2O2 ). Es wird beim sogenannten „respiratory burst“ von Makrophagen
freigesetzt, diffundiert dann durch biologische Membranen und schädigt DNA, Lipide
und Proteine durch Zerfall in Sauerstoffradikale. Außerdem entsteht (H2O2 ) auch bei
normalen Stoffwechselvorgängen in den Persoxisomen der Zelle.
Das Enzym Katalase (EC 1.11.1.6) wandelt zwei Wasserstoffperoxidmoleküle in Was-
ser und Sauerstoff um und eliminiert somit toxisches Wasserstoffperoxid. Katalase liegt
zumeist in spezialisierten Zellorganellen, den Peroxisomen oder Microbodies, vor. Dort
dient es zum Abbau von bei metabolischen Prozessen entstehendem Wasserstoffperoxid.
Im Rahmen der vorliegenden Arbeit wurde die Katalase von Toxoplasma gondii klo-
niert und charakterisiert. Die Klonierung von T. gondii Katalase cDNA ergab ein Prote-
in mit 502 Aminosäuren und einem errechneten Gewicht von 57.2 kDa mit starker Ho-
mologie zu anderen eukaryontischen Katalasen. Ein polyklonales Antiserum gegen ein
GST-Fusionsprotein zeigte im Western-blot eine Bande bei ungefähr 63 kDa. Die Immun-
fluoreszenz zeigte ein vesikuläres Kompartiment im vorderen Ende des Parasiten. Die-
ses kann von anderen Zellorganellen (Mikronemen, Rhoptrien, Granula densa und dem
Apikoplast) durch doppelte Immunfluoreszenzmarkierung unterschieden werden. Zyto-
chemisch können Katalasen durch die DAB-Präzipitationstechnik nachgewiesen werden.
Hier zeigten sich vesikuläre Strukturen vor dem Nukleus in der Lichtmikroskopie und
runde, spezifische Präzipitate mit einem Durchmesser von 100 bis 300 nm in der Elektro-
nenmikroskopie.
Am C-terminus der T. gondii Katalase findet sich ein „peroxisomales Targeting Signal“
(PTS1) in den letzten 3 Aminosäuren (-AKM). Die Expression der vollständigen Katala-
se in CHO-Zellen resultiert in einer peroxisomalen Lokalisation, während ein Konstrukt
50

ohne die letzten 3 Aminosäuren im Zytosol verbleibt. Wird das PTS1 mit einem Repor-
terprotein (Chloramphenicol-Acetyltransferase) fusioniert, wechselt dessen Lokalisation
vom Zytosol zu den Peroxisomen. Damit wurde gezeigt, daß das PTS1 der T. gondii Kata-
lase in einem heterologen System sowohl im Kontext der Katalase als auch eines Repor-
terproteins den Import in Peroxisomen vermitteln kann.
Diese Ergebnisse sind die ersten Hinweise auf Peroxisomen in einem Parasiten der
Apikomplexa. Zugleich ist T. gondii, evolutionsbiologisch gesehen, der bisher niedrigste
Eukaryont in dem bisher Peroxisomen nachgewiesen wurden.
51

6 Abkürzungsverzeichnis
5’ RACE Rapid Amplification of cDNA Ends
CAT Chloramphenicol-Acetyltransferase
DAB Diamino-Benzidin
DAPI 4’6-Diamidino-2-phenylindol
DTT Dithiothreitol
EST Expressed Sequence Tag
FBS Fetales Kälberserum
GST Glutathion S-Transferase
IFA Immunfluoreszenz Assay
IPTG Isopropyl-1-thio-β-D-galactopyranosoid
PBS Phosphate buffered saline
PCR Polymerase Kettenreaktion
PTS Peroxisomales Targeting Signal
RT-PCR Reverse Transkriptase PCR
T/S Teorell-Stenhagen Puffer
52

Literatur
[1] Ajioka JW. Toxoplasma gondii: ESTs and gene discovery. Int J Parasitol, 28(7): 1025–31,
1998.
[2] Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, and Lipman
DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search
programs. Nucleic Acids Res, 25(17): 3389–402, 1997.
[3] Arsenijevic D, Onuma H, Pecqueur C, Raimbault S, Manning BS, Miroux B, Cou-
plan E, Alves-Guerra MC, Goubern M, Surwit R, Bouillaud F, Richard D, Collins S,
and Ricquier D. Disruption of the uncoupling protein-2 gene in mice reveals a role
in immunity and reactive oxygen species production. Nat Genet, 26(4): 435–9, 2000.
[4] Bakker BM, Mensonides FI, Teusink B, van Hoek P, Michels PA, and Westerhoff
HV. Compartmentation protects trypanosomes from the dangerous design of gly-
colysis. Proc Natl Acad Sci U S A, 97(5): 2087–92, 2000.
[5] Baudhuin P, Beaufay H, Rahman-Li Y, Sellinger OZ, Wattiaux R, Jacques P, and
De Duve C. Tissue fractionation studies. 17. Intracellular distribution of monoami-
ne oxidase, aspartate aminotransferase, alanine aminotransferase, D-amino acid
oxidase and catalase in rat-liver tissue. Biochem J, 92(1): 179–84, 1964.
[6] Bermudez LE RJ Covaro G. Infection of murine macrophages with Toxoplasma gon-
dii is associated with release of transforming growth factor beta and downregulati-
on of expression of tumor necrosis factor receptors. Infect Immun, 61(10): 4126–30,
1993.
[7] Bicout DJ, Field MJ, Gouet P, and Jouve HM. Simulations of electron transfer in the
NADPH-bound catalase from Proteus mirabilis PR. Biochim Biophys Acta, 1252(1):
172–6, 1995.
[8] Blattner J, Helfert S, Michels P, and Clayton C. Compartmentation of phosphogly-
cerate kinase in Trypanosoma brucei plays a critical role in parasite energy metabo-
lism. Proc Natl Acad Sci U S A, 95(20): 11596–600, 1998.
[9] Blattner J, Swinkels B, Dorsam H, Prospero T, Subramani S, and Clayton C. Gly-
cosome assembly in trypanosomes: variations in the acceptable degeneracy of a
COOH-terminal microbody targeting signal. J Cell Biol, 119(5): 1129–36, 1992.
[10] Bock P, Kramar R, and Pavelka M. Peroxisomes and related particles in animal tissues.
Springer-Verlag, Wien, New York, 1980.
53

[11] Bogdan C and Rollinghoff M. How do protozoan parasites survive inside macro-
phages? Parasitol Today, 15(1): 22–8, 1999.
[12] Butcher BA, Kim L, Johnson PF, and Denkers E. Toxoplasma gondii Tachyzoites In-
hibit Proinflammatory Cytokine Induction in Infected Macrophages by Preventing
Nuclear Translocation of the Transcription Factor NF-kappaB. J Immunol, 167(4):
2193–2201, 2001.
[13] Carruthers VB and Sibley LD. Mobilization of intracellular calcium stimulates mi-
croneme discharge in Toxoplasma gondii. Mol Microbiol, 31(2): 421–8, 1999.
[14] Chang HR and Pechere JC. Macrophage oxidative metabolism and intracellular
Toxoplasma gondii. Microb Pathog, 7(1): 37–44, 1989.
[15] Christianson DW. Structural chemistry and biology of manganese metalloenzymes.
Prog Biophys Mol Biol, 67(2-3): 217–52, 1997.
[16] Chudzik DM, Michels PA, de Walque S, and Hol WG. Structures of type 2 peroxiso-
mal targeting signals in two trypanosomatid aldolases. J Mol Biol, 300(4): 697–707,
2000.
[17] Coppens I, Sinai AP, and Joiner KA. Toxoplasma gondii exploits host low-density
lipoprotein receptor- mediated endocytosis for cholesterol acquisition. J Cell Biol,
149(1): 167–80, 2000.
[18] de Duve C and Baudhuin P. Peroxisomes (microbodies and related particles). Phy-
siological Reviews, 46: 323–357, 1966.
[19] de Graaf DC, Vanopdenbosch E, Ortega-Mora LM, Abbassi H, and Peeters JE. A
review of the importance of cryptosporidiosis in farm animals. Int J Parasitol, 29(8):
1269–87, 1999.
[20] de Walque S, Kiel JA, Veenhuis M, Opperdoes FR, and Michels PA. Cloning and
analysis of the PTS-1 receptor in Trypanosoma brucei. Mol Biochem Parasitol, 104(1):
106–19, 1999.
[21] Deimann W, Angermüller S, Stoward PJ, and Fahimi HD. Cytochemical methods
for Peroxidases. In PJ Stoward and AG Everson Pearse, Herausgeber, Histoche-
mistry, Band 3, Seiten 573–575. Churchill Livingstone, Edinburgh, 1991.
[22] Deimann W, Angermüller S, Stoward PJ, and Fahimi HD. Peroxidases. In PJ Sto-
ward and AG Everson Pearse, Herausgeber, Histochemistry, Band 3, Seiten 135–159.
Churchill Livingstone, Edinburgh, 1991.
54

[23] Ding M, Clayton C, and Soldati D. Toxoplasma gondii catalase: are there peroxisomes
in toxoplasma? J Cell Sci, 113(Pt 13): 2409–19, 2000.
[24] Dorset DL and Gilmore CJ. Direct methods in protein electron crystallography–
beef liver catalase in its fully hydrated form at room temperature. Acta Crystallogr
A, 55(Pt 3): 448–56, 1999.
[25] Dotto GP, Horiuchi K, and Zinder ND. The functional origin of bacteriophage f1
DNA replication. Its signals and domains. J Mol Biol, 172(4): 507–21, 1984.
[26] Dubey J and Kreier JP. Toxoplasma, Hammondia, Besnoita, Sarcocystis, and Other Tissue
Cyst-Forming Coccidia of Man and Animals, Band 3. Academic Press, NY, 1977.
[27] Dubey JP. Neosporosis in cattle: biology and economic impact. J Am Vet Med Assoc,
214(8): 1160–3, 1999.
[28] Dubremetz JF. Host cell invasion by Toxoplasma gondii. Trends Microbiol, 6(1): 27–30,
1998.
[29] Dubremetz JF, Achbarou A, Bermudes D, and Joiner KA. Kinetics and pattern of
organelle exocytosis during Toxoplasma gondii/host-cell interaction. Parasitol Res,
79(5): 402–8, 1993.
[30] Dubremetz JF and Schwartzman JD. Subcellular organelles of Toxoplasma gondii
and host cell invasion. Res Immunol, 144(1): 31–3, 1993.
[31] Dyer JM, McNew JA, and Goodman JM. The sorting sequence of the peroxisomal
integral membrane protein PMP47 is contained within a short hydrophilic loop. J
Cell Biol, 133(2): 269–80, 1996.
[32] Frangioni JV and Neel BG. Solubilization and purification of enzymatically active
glutathione S- transferase (pGEX) fusion proteins. Anal Biochem, 210(1): 179–87,
1993.
[33] Gouet P, Jouve HM, and Dideberg O. Crystal structure of Proteus mirabilis PR cata-
lase with and without bound NADPH. J Mol Biol, 249(5): 933–54, 1995.
[34] Gould SJ, Keller GA, Hosken N, Wilkinson J, and Subramani S. A conserved tri-
peptide sorts proteins to peroxisomes. J Cell Biol, 108(5): 1657–64, 1989.
[35] Gould SJ, Krisans S, Keller GA, and Subramani S. Antibodies directed against the
peroxisomal targeting signal of firefly luciferase recognize multiple mammalian
peroxisomal proteins. J Cell Biol, 110(1): 27–34, 1990.
55

[36] Guex N and Peitsch MC. SWISS-MODEL and the Swiss-PdbViewer: an environ-
ment for comparative protein modeling. Electrophoresis, 18(15): 2714–23, 1997.
[37] Hayashi M, Toriyama K, Kondo M, Kato A, Mano S, De Bellis L, Hayashi-Ishimaru
Y, Yamaguchi K, Hayashi H, and M N. Functional transformation of plant pero-
xisomes. Cell Biochem Biophys, 32: 295–304, 2000.
[38] Herold G. Innere Medizin. Herold, 2001.
[39] Hillar A, Nicholls P, Switala J, and Loewen PC. NADPH binding and control of
catalase compound II formation: comparison of bovine, yeast, and Escherichia coli
enzymes. Biochem J, 300(Pt 2): 531–9, 1994.
[40] Hughes HP, Boik RJ, Gerhardt SA, and Speer CA. Susceptibility of Eimeria bovis
and Toxoplasma gondii to oxygen intermediates and a new mathematical model for
parasite killing. J Parasitol, 75(4): 489–97, 1989.
[41] Janeway C and Travers P. Immunobiology. Current Biology Ltd., London, 2. Auflage,
1996.
[42] Joiner KA, Fuhrman SA, Miettinen HM, Kasper LH, and Mellman I. Toxoplasma
gondii: fusion competence of parasitophorous vacuoles in Fc receptor-transfected
fibroblasts. Science, 249(4969): 641–6, 1990.
[43] Jouve HM, Gouet P, Boudjada N, Buisson G, Kahn R, and Duee E. Crystallization
and crystal packing of Proteus mirabilis PR catalase. J Mol Biol, 221(4): 1075–7, 1991.
[44] Kaasch AJ and Joiner KA. Protein-targeting determinants in the secretory pathway
of apicomplexan parasites. Curr Opin Microbiol, 3(4): 422–8, 2000.
[45] Kaasch AJ and Joiner KA. Targeting and subcellular localization of Toxoplasma gon-
dii catalase. Identification of peroxisomes in an apicomplexan parasite. J Biol Chem,
275(2): 1112–8, 2000.
[46] Kirkman HN, Rolfo M, Ferraris AM, and Gaetani GF. Mechanisms of protection
of catalase by NADPH. Kinetics and stoichiometry. J Biol Chem, 274(20): 13908–14,
1999.
[47] Ko TP, Safo MK, Musayev FN, Di Salvo ML, Wang C, Wu SH, and Abraham DJ.
Structure of human erythrocyte catalase. Acta Crystallogr D Biol Crystallogr, 56(Pt
2): 241–5, 2000.
56

[48] Kunau WH and Erdmann R. Peroxisome biogenesis: back to the endoplasmic reti-
culum? Curr Biol, 8(9): R299–302, 1998.
[49] Lazarow PB and Fujiki Y. Biogenesis of peroxisomes. Annu Rev Cell Biol, 1: 489–530,
1985.
[50] Leriche MA and Dubremetz JF. Exocytosis of Toxoplasma gondii dense granules into
the parasitophorous vacuole after host cell invasion. Parasitol Res, 76(7): 559–62,
1990.
[51] Levine ND. Progress in taxonomy of the Apicomplexan protozoa. J Protozool, 35(4):
518–20, 1988.
[52] Loewen P. Probing the structure of catalase HPII of Escherichia coli–a review. Gene,
179(1): 39–44, 1996.
[53] Luder C, Walter W, Beuerle B, Maeurer M, and Gross U. Toxoplasma gondii down-
regulates MHC class II gene expression and antigen presentation by murine macro-
phages via interference with nuclear translocation of STAT1alpha. Eur J Immunol,
31(5): 1475–84, 2001.
[54] Majno G and Joirs I. Cells, Tissues, and Disease. Blackwell Science, Cambridge MA,
1996.
[55] Maniatis T, Sambrook J, and Fritsch E. Molecular Cloning : A Laboratory Manual,
Band 1-3. Cold Spring Harbor Laboratory Press, 2nd edition Auflage, 1989.
[56] McGonigle S, Dalton JP, and James ER. Peroxidoxins: A New Antioxidant Family.
Parasitology Today, 14(4): 139–145, 1998.
[57] Mehlhorn HE. Parasitology in Focus. Springer Verlag, Berlin, 1988.
[58] Mehlotra RK. Antioxidant defense mechanisms in parasitic protozoa. Crit Rev
Microbiol, 22(4): 295–314, 1996.
[59] Michels PA, Hannaert V, and Bringaud F. Metabolic aspects of glycosomes in trypa-
nosomatidae - new data and views. Parasitol Today, 16(11): 482–9, 2000.
[60] Mordue DG, Desai N, Dustin M, and Sibley LD. Invasion by Toxoplasma gondii
establishes a moving junction that selectively excludes host cell plasma membrane
proteins on the basis of their membrane anchoring. J Exp Med, 190(12): 1783–92,
1999.
57

[61] Mordue DG, Hakansson S, Niesman I, and Sibley LD. Toxoplasma gondii resides in
a vacuole that avoids fusion with host cell endocytic and exocytic vesicular traf-
ficking pathways. Exp Parasitol, 92(2): 87–99, 1999.
[62] Mui E, Samuel B, Mack D, Roberts C, Pope C, Roberts F, Trelease D, Milhous W,
Kyle D, Tzipori S, and McLeod R. Isocitrate lyase and malate synthetase in T. gondii.
In 5th Toxoplasma Conference. Marshall, CA, 1999.
[63] Mullen RT, Lee MS, Flynn CR, and Trelease RN. Diverse amino acid residues func-
tion within the type 1 peroxisomal targeting signal. Implications for the role of ac-
cessory residues upstream of the type 1 peroxisomal targeting signal. Plant Physiol,
115(3): 881–9, 1997.
[64] Mullen RT, Lee MS, and Trelease RN. Identification of the peroxisomal targeting
signal for cottonseed catalase. Plant J, 12(2): 313–22, 1997.
[65] Murray HW. Cellular resistance to protozoal infection. Annu Rev Med, 37: 61–9,
1986.
[66] Murray HW and Cohn ZA. Macrophage oxygen-dependent antimicrobial activity.
I. Susceptibility of Toxoplasma gondii to oxygen intermediates. J Exp Med, 150(4):
938–49, 1979.
[67] Murray HW and Cohn ZA. Macrophage oxygen-dependent antimicrobial activity.
III. Enhanced oxidative metabolism as an expression of macrophage activation. J
Exp Med, 152(6): 1596–609, 1980.
[68] Murray HW, Juangbhanich CW, Nathan CF, and Cohn ZA. Macrophage oxygen-
dependent antimicrobial activity. II. The role of oxygen intermediates. J Exp Med,
150(4): 950–64, 1979.
[69] Murray HW, Nathan CF, and Cohn ZA. Macrophage oxygen-dependent antimi-
crobial activity. IV. Role of endogenous scavengers of oxygen intermediates. J Exp
Med, 152(6): 1610–24, 1980.
[70] Murthy MR, Reid TJd, Sicignano A, Tanaka N, and Rossmann MG. Structure of
beef liver catalase. J Mol Biol, 152(2): 465–99, 1981.
[71] Nash P, Purner M, Leon R, Clarke P, Duke R, and Curiel TJ. Toxoplasma gondii-
infected cells are resistant to multiple inducers of apoptosis. J Immunol, 160(4):
1824–30, 1998.
58

[72] Nichols BA, Chiappino ML, and Pavesio CE. Endocytosis at the micropore of Toxo-
plasma gondii. Parasitol Res, 80(2): 91–8, 1994.
[73] Nischnik N, Schade B, Dytnerska K, Dlugonska H, Reichmann G, and Fischer H.
Attenuation of mouse-virulent Toxoplasma gondii parasites is associated with a de-
crease in interleukin-12-inducing tachyzoite activity and reduced expression of ac-
tin, catalase and excretory proteins. Microbes and Infection, 3: 689–699, 2001.
[74] Odberg-Ferragut C, Renault JP, Viscogliosi E, Toursel C, Briche I, Engels A, Lepage
G, Morgenstern-Badarau I, Camus D, Tomavo S, and Dive D. Molecular cloning,
expression analysis and iron metal cofactor characterisation of a superoxide dis-
mutase from Toxoplasma gondii. Mol Biochem Parasitol, 106(1): 121–9, 2000.
[75] Ogata M. Acatalasemia. Hum Genet, 86(4): 331–40, 1991.
[76] Opperdoes FR, Baudhuin P, Coppens I, De Roe C, Edwards SW, Weijers PJ, and
Misset O. Purification, morphometric analysis, and characterization of the glyco-
somes (microbodies) of the protozoan hemoflagellate Trypanosoma brucei. J Cell Biol,
98(4): 1178–84, 1984.
[77] Osumi T, Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa
S, and Hashimoto T. Amino-terminal presequence of the precursor of peroxisomal
3-ketoacyl-CoA thiolase is a cleavable signal peptide for peroxisomal targeting. Bio-
chem Biophys Res Commun, 181(3): 947–54, 1991.
[78] Parsons M, Furuya T, Pal S, and Kessler P. Biogenesis and function of peroxisomes
and glycosomes. Mol Biochem Parasitol, 115(1): 19–28, 2001.
[79] Passreiter M, Anton M, Lay D, Frank R, Harter C, Wieland FT, Gorgas K, and Just
WW. Peroxisome biogenesis: involvement of ARF and coatomer. J Cell Biol, 141(2):
373–83, 1998.
[80] Peitsch MC. Protein modeling by E-mail. Bio/Technology, 13: 658–660, 1995.
[81] Peitsch MC. ProMod and Swiss-Model: Internet-based tools for automated compa-
rative protein modelling. Biochem Soc Trans, 24(1): 274–9, 1996.
[82] Purdue PE and Lazarow PB. Targeting of human catalase to peroxisomes is de-
pendent upon a novel COOH-terminal peroxisomal targeting sequence. J Cell Biol,
134(4): 849–62, 1996.
[83] Rapp S, Saffrich R, Anton M, Jakle U, Ansorge W, Gorgas K, and Just WW.
Microtubule-based peroxisome movement. J Cell Sci, 109(Pt 4): 837–49, 1996.
59

[84] Reddy and Janardan K. Peroxisomes. New York Acad. of Sciences, New York, 1996.
[85] Reid TJd, Murthy MR, Sicignano A, Tanaka N, Musick WD, and Rossmann MG.
Structure and heme environment of beef liver catalase at 2.5 A resolution. Proc Natl
Acad Sci U S A, 78(8): 4767–71, 1981.
[86] Rhodin J. Correlation of Ultrastructural Organization and Function in Normal and Expe-
rimentally changed Proximal Convoluted Tubule Cells of the Mouse Kidney. Aktiebologet
Godvil, Stockholm, 1954.
[87] Roos DS, Crawford MJ, Donald RG, Kissinger JC, Klimczak LJ, and Striepen B.
Origin, targeting, and function of the apicomplexan plastid. Curr Opin Microbiol,
2(4): 426–32, 1999.
[88] Roos DS, Donald RG, Morrissette NS, and Moulton AL. Molecular tools for genetic
dissection of the protozoan parasite Toxoplasma gondii. Methods Cell Biol, 45: 27–63,
1994.
[89] Ruff MD. Important parasites in poultry production systems. Vet Parasitol, 84(3-4):
337–47, 1999.
[90] Schwab JC, Beckers CJ, and Joiner KA. The parasitophorous vacuole membrane
surrounding intracellular Toxoplasma gondii functions as a molecular sieve. Proc
Natl Acad Sci U S A, 91(2): 509–13, 1994.
[91] Schwede T, Diemand A, Guex N, and Peitsch MC. Protein structure computing in
the genomic era. Res Microbiol, 151(2): 107–12, 2000.
[92] Shaw MK, Roos DS, and Tilney LG. Acidic compartments and rhoptry formation
in Toxoplasma gondii. Parasitology, 117(Pt 5): 435–43, 1998.
[93] Short JM, Fernandez JM, Sorge JA, and Huse WD. Lambda ZAP: a bacteriophage
lambda expression vector with in vivo excision properties. Nucleic Acids Res, 16(15):
7583–600, 1988.
[94] Sibley L, Hakansson S, and Carruthers V. Gliding Motility: An efficient mechanism
for cell penetration. Current Biology, 8(1): R12–R14, 1998.
[95] Sibley LD, Lawson R, and Weidner E. Superoxide dismutase and catalase in Toxo-
plasma gondii. Mol Biochem Parasitol, 19(1): 83–7, 1986.
60

[96] Sibley LD, Niesman IR, Parmley SF, and Cesbron-Delauw MF. Regulated secretion
of multi-lamellar vesicles leads to formation of a tubulo-vesicular network in host-
cell vacuoles occupied by Toxoplasma gondii. J Cell Sci, 108(Pt 4): 1669–77, 1995.
[97] Sinai AP and Joiner KA. Safe haven: the cell biology of nonfusogenic pathogen
vacuoles. Annu Rev Microbiol, 51: 415–62, 1997.
[98] Sinai AP and Joiner KA. The Toxoplasma gondii protein ROP2 mediates host orga-
nelle association with the parasitophorous vacuole membrane. J Cell Biol, 154(1):
95–108, 2001.
[99] Sinai AP, Webster P, and Joiner KA. Association of host cell endoplasmic reticulum
and mitochondria with the Toxoplasma gondii parasitophorous vacuole membrane:
a high affinity interaction. J Cell Sci, 110(Pt 17): 2117–28, 1997.
[100] Soete M, Camus D, and Dubremetz JF. Experimental induction of bradyzoite-
specific antigen expression and cyst formation by the RH strain of Toxoplasma gondii
in vitro. Exp Parasitol, 78(4): 361–70, 1994.
[101] Sommer JM and Wang CC. Targeting proteins to the glycosomes of African trypa-
nosomes. Annu Rev Microbiol, 48: 105–38, 1994.
[102] Son E, Song KJ, Shin JC, and Nam HW. Molecular cloning and characterization of
peroxiredoxin from Toxoplasma gondii. Korean J Parasitol, 39(2): 133–41, 2001.
[103] Subramani S. Components involved in peroxisome import, biogenesis, proliferati-
on, turnover, and movement. Physiol Rev, 78(1): 171–88, 1998.
[104] Sumner JB and Dounce AL. Crystalline Catalase. Science, 124: 417–424, 1937.
[105] Swinkels BW, Gould SJ, Bodnar AG, Rachubinski RA, and Subramani S. A novel,
cleavable peroxisomal targeting signal at the amino-terminus of the rat 3-ketoacyl-
CoA thiolase. Embo J, 10(11): 3255–62, 1991.
[106] Swinkels BW, Gould SJ, and Subramani S. Targeting efficiencies of various permu-
tations of the consensus C-terminal tripeptide peroxisomal targeting signal. FEBS
Lett, 305(2): 133–6, 1992.
[107] Thompson JD, Higgins DG, and Gibson TJ. CLUSTAL W: improving the sensitivity
of progressive multiple sequence alignment through sequence weighting, position-
specific gap penalties and weight matrix choice. Nucleic Acids Res, 22(22): 4673–80,
1994.
61

[108] Tielens AGM and Van Hellemond JJ. Differences in Energy metabolism Between
Trypanosomatidae. Parasitology Today, 14(7): 265–271, 1998.
[109] Titorenko VI, Chan H, and Rachubinski RA. Fusion of small peroxisomal vesicles
in vitro reconstructs an early step in the in vivo multistep peroxisome assembly
pathway of Yarrowia lipolytica. J Cell Biol, 148(1): 29–44, 2000.
[110] Titorenko VI and Rachubinski RA. The endoplasmic reticulum plays an essential
role in peroxisome biogenesis. Trends Biochem Sci, 23(7): 231–3, 1998.
[111] Titorenko VI and Rachubinski RA. Mutants of the yeast Yarrowia lipolytica defective
in protein exit from the endoplasmic reticulum are also defective in peroxisome
biogenesis. Mol Cell Biol, 18(5): 2789–803, 1998.
[112] Walton PA, Hill PE, and Subramani S. Import of stably folded proteins into pero-
xisomes. Mol Biol Cell, 6(6): 675–83, 1995.
[113] Wan KL, Blackwell JM, and Ajioka JW. Toxoplasma gondii expressed sequence tags:
insight into tachyzoite gene expression. Mol Biochem Parasitol, 75(2): 179–86, 1996.
[114] Weibel ER, Staubli W, Gnagi HR, and Hess FA. Correlated morphometric and bio-
chemical studies on the liver cell. I. Morphometric model, stereologic methods, and
normal morphometric data for rat liver. J Cell Biol, 42(1): 68–91, 1969.
[115] Wiemer EA, Wenzel T, Deerinck TJ, Ellisman MH, and Subramani S. Visualization
of the peroxisomal compartment in living mammalian cells: dynamic behavior and
association with microtubules. J Cell Biol, 136(1): 71–80, 1997.
[116] Wiemer EAC, Luers GH, Faber KN, Wenzel T, Veenhuis M, and Subramani S. Iso-
lation and characterization of Pas2p, a peroxisomal membrane protein essential
for peroxisome biogenesis in the methylotrophic yeast Pichia pastoris. J Biol Chem,
271(31): 18973–80, 1996.
[117] Yamamoto K and Fahimi HD. Three-dimensional reconstruction of a peroxisomal
reticulum in regenerating rat liver: evidence of interconnections between heteroge-
neous segments. J Cell Biol, 105(2): 713–22, 1987.
[118] Zamocky M, Herzog C, Nykyri LM, and Koller F. Site-directed mutagenesis of
the lower parts of the major substrate channel of yeast catalase A leads to highly
increased peroxidatic activity. FEBS Lett, 367(3): 241–5, 1995.
62

[119] Zoeller RA and Raetz CR. Isolation of animal cell mutants deficient in plasmalogen
biosynthesis and peroxisome assembly. Proc Natl Acad Sci U S A, 83(14): 5170–4,
1986.
63

Danksagung
An dieser Stelle möchte ich meinen Eltern danken, die mich immer unterstützt haben.
Ganz besonderer Dank gebührt Prof. Dr. Keith Joiner von der Yale University Medical
School, in dessen Labor der experimentelle Teil dieser Arbeit durchgeführt wurde, sowie
allen Mitgliedern des Labors, insbesondere Dr. Heinrich Hoppe, Dr. Anthony Sinai, Dr.
Verena Karsten, Dr. Timothy Stedman und Dr. Isabelle Coppens.
Herrn Prof. Dr. M. Frosch möchte ich für die kritische Durchsicht des Manuskripts und
die konstruktive Kritik danken.
Rottweil im Juli 2002, Achim Kaasch

Lebenslauf von Achim Kaasch
Geboren am 17.09.1973 in Fulda, Deutschland.
Ausbildung
15.04.1993 Abitur am Leibniz Gymnasium Rottweil.
01.08.1993-
30.10.1994
Zivildienst im stationären Pflegedienst des KKH Hiltrup.
01.11.1994 Beginn des Medizinstudiums an der Julius-Maximilians-Universität
Würzburg.
01.08.1996 Ärztliche Vorprüfung mit der Note „gut“.
20.08.1997 Erstes Staatsexamen mit der Note „gut“.
01.09.1997-
31.03.1999
Predoctoral Associate im Labor von Prof. Dr. K. Joiner, Infectious Di-
seases Section, Internal Medicine, Yale University School of Medicine,
New Haven (USA).
01.05.1999-
30.03.2000
Hilfswissenschaftler im Institut für Mikrobiologie und Hygiene der
Universität Würzburg.
29.03.2001 Zweites Staatsexamen mit der Note „gut“.
04.06.2002 Drittes Staatsexamen mit der Note „sehr gut“.
Klinische Ausbildung
23.04.2001-
12.08.2001
PJ-Tertial in der Neurologischen Uniklinik Würzburg.
01.09.2001-
31.12.2001
Chirurgisches PJ-Tertial im Kantonsspital Herisau (Schweiz).

01.01.2002-
28.02.2002
Medizinisches PJ-Tertial im St. Lukes Hospital, Gwardamangia (Malta).
04.03.2002-
28.04.2002
Medizinisches PJ-Tertial am Clinical Center der National Institutes of
Health, Bethesda (USA).
Studentische Aktivitäten
WS 1995 Studentischer Beirat im Uni-med Verlag.
WS 1996 Organisation des Kurses „Anatomie am Lebenden“.
WS 1996 Organisation einer biochemischen Seminarreihe für Medizinstudenten.
1999, 2000, 2001 Konzeption und Organisation des Promomed-Kongresses an der Uni-
versität Würzburg.
Stipendien und Preise
08/1997 FEBS Stipendium zur Teilnahme am Sommer-Kurs „Protein Targeting
and Secretion“ in Cargèse, Korsika (Frankreich).
01.02.1998-
30.03.1999
Stipendium der Yale University Medical School zur Durchführung der
Doktorarbeit.
seit 08/2000 Internet-Stipendium von „E-Fellows.net“.
11/2000 Preisträger beim Wettbewerb „CEO of the future“ von McKinsey & Co.
und dem Manager Magazin.
Hiermit bestätige ich, dass obige Angaben richtig sind.
Rottweil, Juni 2002












![Eimeria tenella sowie deren Nutzung in der Wirkstoffforschung · gondii und Cryptosporidium parvum) [MEHLHORN, 2008]. INLEITUNG E 3 Sporozoa (Apicomplexa) Perkinsea Sporozoea Gregarinia](https://static.fdokument.com/doc/165x107/5be5645d09d3f288458b923f/eimeria-tenella-sowie-deren-nutzung-in-der-wirkstoffforschung-gondii-und-cryptosporidium.jpg)





