Cover 2 - anatomie.med.uni-muenchen.de · die Placenta eines Labornagers mikroskopiert, wird irri-...
36
NeadeA NEU 2010 MUNICH-ANATOMY-COMPETENCE.ORG Neues aus der Anstalt
Transcript of Cover 2 - anatomie.med.uni-muenchen.de · die Placenta eines Labornagers mikroskopiert, wird irri-...

NeadeA
NEU2010
MUNICH-ANATOMY-COMPETENCE.ORG
Neues aus der Anstalt

NeadeA – Neues aus der Anstalt – Vol. 1 Dezember 2010 Nr. 1
C. Schmitz, H.-G. Frank Neues aus der Anstalt (NeadeA) – Informationen aus der Anatomie und für die Anatomie 1-3 H.-G. Frank, C. Schmitz Der Mensch und „seine“ Placenta: Was bedeuten die beiden wirklich füreinander? 4-8 C. Schmitz „Design-based“ Stereologie – eine ganz kurze Einführung 9-13 A.K.E. Horn-Bochtler, K. Lienbacher, C. Zeeh, A. Messoudi, C. Glombik Nur bewegte Augen ermöglichen eine klare Sicht 14-18 C. Buhrmann, M. Shakibaei Stammzell Tissue Engineering und Elektronenmikroskopie: Die Rolle der Phytopharmaka auf die Differenzierung mesenchymaler Stammzellen zu Chondrozyten in einem Osteoarthritis Modell in vitro 19-23 A. Mayerhofer Neues von NGF und GDNF 24-25 M. Kiessling 7482 Nissl-Chloroform Upper East Side: Ein Labor-Erlebnisbericht 26-29 H.-G. Frank
Neues aus der Anstalt (NeadeA) – TEXnicalities 30-33

Neues aus der Anstalt (NeadeA)Informationen aus der Anatomie und für die Anatomie
C. Schmitz, H.-G. Frank1,∗
Anatomische Anstalt, Lehrstuhl II, Ludwig-Maximilians-Universität, Pettenkoferstrasse 11, 80336 München, Germany
Abstract
Die Anatomische Anstalt der Ludwig-Maximilians-Universität ist ein sehr traditionsreiches anatomisches Institut, in demviele bedeutende Anatomen ihre Wirkungsstätte hatten. Auch weiterhin wird hier nach Erkenntnisgewinn und Vermeh-rung des klinischen Nutzens in den Feldern der Medizin gestrebt werden, in denen mit anderen Methoden als denen derMorphologie kein substanzieller Erkenntnisgewinn möglich ist. Das schließt den fruchtbaren und konstruktiven Dialogzu den aktuellen Themenfeldern ein. Diesem Ziel verpflichtet, stellen wir hier ein informelles und im wesentlichen aufdie interne Kommunikation gerichtetes neues Medium vor, das den Titel „Neues aus der Anstalt” nicht ganz frei vonhumoristischen Bezügen tragen soll. Auch wenn der Namen augenzwinkernd vorgetragen wird, geht es um das ernste Ziel,die interne Kommunikation und das gegenseitige wissenschaftliche Verständnis zu pflegen. Beiträge von Doktorandenund Habilitanden zu ihren jeweiligen Projekten sowie generell interessante Themen, die von den Professoren vorgestelltwerden können, sollen den Strauss der wissenschaftlichen Themen innerhalb der Anatomischen Anstalt dokumentierenund auf breiterer Basis lebendig machen. Das Layout ist dem wissenschaftlicher Journale angepasst und standardisiert.Das Forum dient auch dazu, den jeweils neuen Mitarbeiterinnen und Mitarbeitern in den Laboren - hier vor allemden Doktoranden - die frühzeitige Gelegenheit und Verpflichtung zur schriftlichen Darstellung ihres Themas zu geben,direkt gebunden an typische wissenschaftsübliche Konventionen. In diesem Sinne wird NeadeA auch eine gute Platt-form zum Üben wissenschaftlichen Schreibens, die nicht unmittelbar dem Druck des externen „Peer Review” Systemsausgesetzt ist. Ein weiteres Anliegen von NeadeA ist es, wissenschaftliche Inhalte und Themen dem technischen undnicht-wissenschaftlichen Personal in verständlicher Sprache vorzustellen und auch umgekehrt diesen Mitarbeiterinnenund Mitarbeitern ein Forum zu bieten. NeadeA wurde vom Lehrstuhl II der Anatomischen Anstalt initiiert, wird aberhoffentlich eine breitere Verwendung im Haus finden. Es ist besonders zu wünschen, dass der Ernst der Wissenschaft denSpaß an der Wissenschaft und die nicht unwichtige Fähigkeit, auch über sich selbst lachen zu können, nicht verdrängt.
1. Welche Ziele sollen mit NeadeA erreicht wer-den?
• Alle Mitarbeiterinnen und Mitarbeiter sollen das Leis-tungsspektrum der Anatomischen Anstalt in ganzerBreite vorgestellt bekommen und erfassen können.
• Die Identifikation mit der Anatomischen Anstalt alsGanzes, ihrer Arbeit und ihren Zielen, soll gestärktwerden.
• Ein regelmässiger und strukturierter, sachorientierterund freier Informationsfluss über die Grenzen der wis-senschaftlichen Teildisziplinen der Anatomie hinwegsoll etabliert werden.
• Die Mitarbeiterinnen und Mitarbeiter, insbesondereder wissenschaftliche Nachwuchs sollen eine Plattformerhalten, die sich als erster Übungsplatz für wissen-schaftliches Publizieren eignet.
∗Email: [email protected]: Andrea Asikoglu
• Ehemalige Mitarbeiterinnen und Mitarbeiter sowieDoktoranden der Anatomischen Anstalt sollen dieMöglichkeit haben, mit den aktuellen Entwicklungenin der Anatomischen Anstalt in Verbindung zu blei-ben.
• Andere wissenschaftliche Institutionen - zuallererstnatürlich die anderen Einrichtungen der Medizini-schen Fakultät der Ludwig-Maximilians-Universität -sollen einen Einblick in die Arbeit und Leistung derAnatomischen Anstalt erhalten.
• Die Einzigartigkeit der Anatomischen Anstalt - es gibttatsächlich keine zweite Institution dieses Namens inDeutschland - soll ihren Ausdruck auch auf der in-haltlichen und kommunikativen Ebene finden.
2. Warum eine Zeitschrift, kein Kolloquium?
Große und häufig auch heterogene wissenschaftliche Ins-titute wie die Anatomische Anstalt haben es natürlicher-weise immer wieder schwer, die Kommunikation unter den
Internal Communication: Neues aus der Anstalt (NeadeA) 30. November 2010

4 WIE HOCH IST DER WISSENSCHAFTLICHE ANSPRUCH?
verschiedenen Wissenschaftlerinnen und Wissenschaftlern,technischen und nicht-technischen Mitarbeitern mit ihrenverschiedenen Hintergründen und Aufgaben sinnvoll zu ge-stalten. Je größer und heterogener eine Institution ange-legt ist, umso schwieriger wird dies. Auf Arbeitsgruppen-oder Lehrstuhlebene werden häufig Vorträge als Werkzeugzur Kommunikation genutzt. Diese haben den Vorteil dermündlichen und spontanen Präsentation und des persön-lichen Auftritts der Vortragenden sowie vor allem der un-mittelbaren Diskussion. Sie vergrößern aber immer wie-der auch die Heterogenität, weil zur inhaltlichen Hetero-genität die persönlichen Unterschiede zwischen Vortragen-den addiert werden müssen. Hinzu kommt, dass alle Hö-rer zum Zeitpunkt des Vortrages auch Zeit haben müssenund deswegen keine anderen Aufgaben wahrnehmen kön-nen. Dies schließt - wegen anderer Verpflichtungen - im-mer wieder Mitarbeiterinnen und Mitarbeiter aus und er-schwert die täglichen Arbeitsabläufe durch kollektive undlängere Unterbrechung. Für die technischen Mitarbeiterin-nen und Mitarbeiter sind allzu spezifische wissenschaftli-che Vorträge gelegentlich von ähnlichem Interesse wie län-gere Dia-abende im Familienkreis zum letzten Sommerur-laub. Vorträge aber, die ihre Adressaten - wegen divergen-ter Verständisvoraussetzungen und Interessen - gar nichterreichen können, sind weder für den Vortragenden nochfür die Zuhörer eine Bereicherung. Obwohl also Kolloqui-en - gezielt eingesetzt - sehr nützlich sein können, sind sieals universelles Kommunikationsmittel nicht immer opti-mal geeignet.Beiträge, die in einem schriftlichen Mitteilungsblatt zir-kuliert werden, haben einige dieser Nachteile nicht. Je-der Beitrag kann auf spezifische Adressatengruppen zu-geschnitten sein und durchaus auch von Mitarbeitern ausdiesen Gruppen (Wissenschaftlerinnen und Wissenschaft-ler, technische und nicht-technische Mitarbeiterinnen undMitarbeiter, Studentinnen und Studenten, Doktorandin-nen und Doktoranden, etc.) für die jeweiligen Gruppenerstellt werden. Diese Form schliesst niemanden aus undlädt alle ein. Mitarbeiterinnen und Mitarbeiter können dasLesen solcher Informationen frei in ihren Arbeitsablauf ein-passen und müssen Artikel, die Sie nicht interessieren, auchnicht lesen. Auch wenn eine Information schwierig zu er-fassen oder in einer Fremdsprache geschrieben ist, hat manbeim Lesen die nötige Zeit zur Verfügung, alles zu verste-hen. Beim Vortrag ist man schneller „abgehängt” und kannnicht einfach zurückblättern. Insgesamt also erscheint unsein schriftliches Mitteilungsblatt als eine entspanntere undeinfacher in die Arbeit zu integrierende Informationsmög-lichkeit als ein Vortrag, auch wenn dieser immer seinenbewährten Platz verteidigen wird, vor allem wenn es pri-mär um die spontane wissenschaftliche Diskussion geht.
3. Welche Themen kommen in Frage?
An einer universitären Einrichtung sollen Nachrichtenüber die Wissenschaft natürlich eine wichtige Rolle spie-len, aber dies stellt keine Beschränkung dar. Auch Be-
richte zu Änderungen z.B. der Rechtslage bei Ausbildung-Berufs- und Standesfragen, zu ethischen und auch finan-ziellen Fragen können eine Rolle spielen. Neue Lehr- undUnterrichtsveranstaltungen oder Unterrichtskonzepte kön-nen der gesamten Anatomischen Anstalt vorgestellt wer-den, und die technischen und sonstigen Mitarbeiterinnenund Mitarbeiter können Themen und Fortschritte aus ih-ren jeweiligen Arbeitsgebieten und Verantwortungsberei-chen vorstellen. Infrastrukturmassnahmen - zum Beispielbeim Umbau oder im Bereich der Datennetze - könnenerläutert werden.
Für die wissenschaftlichen Belange der AnatomischenAnstalt wird NeadeA eine besondere Funktion bei der In-formation über Forschungsthemen, Verwendungsmöglich-keiten von Grossgeräten, Information über Kooperationenund deren jeweiligen Fortschritt sowie aktuell bewilligteDrittmittelprojekte zugedacht. Am Lehrstuhl II wird eszur Gewohnheit werden, dass Doktorandinnen und Dokto-randen und neu hinzukommende Assistentinnen und As-sistenten - nach adäquater Einarbeitungszeit in ihr Thema- eine Übersichtsarbeit zu ihrem Themengebiet und ihrerjeweiligen spezifischen Zielstellung der gesamten Anato-mischen Anstalt in NeadeA vorstellen. Primär wird so diePerson und das Thema als Teil des Ganzen vorgestellt.Dies geschieht aber auch mit der Absicht, die im Bereichdes wissenschaftlichen Schreibens in der Regel relativ un-erfahrenen medizinischen Doktorandinnen und Doktoran-den frühzeitig an das Schreiben heranzuführen und dies ineinem niederschwelligen internen Zirkel zu tun, der trotz-dem natürlich schon eine gewisse Erwartungshaltung pfle-gen wird. Das Einstellen eines Artikels in NeadeA wird nurintern von den jeweiligen Betreuern begleitet, nicht einemexternen Peer Review System unterworfen und stellt dahereher eine ausbildungsbegleitende Betreuungsmassnahmefür Doktorandinnen und Doktoranden, vor allem aus demBereich der Medizin, dar. Das Zeitschriften-ähnliche Lay-out von NeadeA gibt darüber hinaus auch einen adäqua-ten Vorgeschmack darauf, was tatsächlich aus einer gutbetreuten und umgesetzten Promotionsarbeit in der Weltder Wissenschaft werden sollte. Für diejenigen - vor allemunter den Doktorandinnen und Doktoranden - , die sichdas auf der computertechnischen Ebene zutrauen, kann indem für NeadeA etablierten Satzsystem auch direkt imoffiziellen Template der LMU für Promotionen zur elek-tronischen Publikation gearbeitet werden, was eine Reihevon Vorteilen für die Einreichung von Promotionsschriftenhat.
4. Wie hoch ist der wissenschaftliche Anspruch?
Bei wissenschaftlichen Mitteilungen soll es darum ge-hen, auch die technischen und nicht-technischen Mitarbei-terinnen und Mitarbeiter in verständlicher Sprache überdie jeweiligen Themen zu informieren. Der wissenschaftli-che Anspruch schmälert sich dadurch aber nicht, weil dieMöglichkeit zur Flucht in eine entlegene Nomenklatur der
2

6 WIE OFT WIRD NEADEA ERSCHEINEN UND WIE SOLL ES ZIRKULIERT WERDEN?
jeweiligen fachlichen Nische - den dann nur noch die Fach-leute verstehen können - nicht bestehen soll. Damit müssenwissenschaftliche Aussagen klar, verständlich und einfachsein, was kein Zeichen von niedrigem wissenschaftlichenNiveau ist. Mehr als auf Details sollen also wissenschaftli-che Beiträge auf den „roten Faden”, auf die Geschichte, dieStrategie und die Zielrichtung eines Projektes oder The-mas konzentrieren und diese erläutern. Arbeiten, die be-stimmte Themen überblicksartig zusammenfassen, sollendas Destillat der wissenschaftlichen Literaturarbeit dar-stellen, was nur gelingen kann, wenn qualitativ hochwerti-ge Vorarbeit im stillen Kämmerlein geleistet wurde.
5. Welche Autoren kommen in Betracht?
Alle Mitarbeiterinnen und Mitarbeiter der Anatomi-schen Anstalt sind herzlich eingeladen, Beiträge für Nea-deA zu verfassen und zu erstellen. Gerne können zu spe-zifischen Themen - beispielsweise rechtlichen oder ethi-schen Themen - auch externe Autoren eingeladen werden.Um einen kontinuierlichen Fluss an Beiträgen zu ermög-lichen, werden Professoren und das übrige wissenschaftli-che Personal einen zahlenmässig sicher bedeutenden Anteilder Beiträge übernehmen. Aber neue Präparationsmetho-den und Vorgehensweisen, Kommentare zu wichtigen Er-eignissen (beispielsweise die ja irgendwann in hoffentlichnicht allzu ferner Zukunft anstehende Wiedereröffnung derSchausammlung), organisatorisches und allgemein für alleinteressante Aspekte können und sollten auch vom nicht-wissenschaftlichen Personal eingebracht werden.
6. Wie oft wird NeadeA erscheinen und wie soll eszirkuliert werden?
Es gibt zur Zeit keine Vorgaben für eine regelmässige Er-scheinungsweise, aber es ist angestrebt, zwei bis vier malim Jahr eine neue Ausgabe zusammenzustellen und zu zir-kulieren. Dies wird natürlich auch vom Eingang neuer Bei-träge aus dem Kreis der Mitarbeiterinnen und Mitarbeiterabhängen. Im Grundsatz soll eine neue Ausgabe von Nea-deA immer dann erscheinen, wenn mindestens 4 Beiträgevorliegen. Grundsätzlich soll ausser zu besonderen Anläs-sen kein Papier verwendet werden, sondern auf elektro-nischem Wege ein PDF-Dokument zirkuliert werden. FürArchiv-Zwecke und für auswärtige Interessenten, denen dieAnatomische Anstalt und ihre Arbeit ja auch mit diesemWerkzeug vorgestellt werden kann, können jederzeit dieggf. benötigten Print-Exemplare erstellt werden.
3

Der Mensch und „seine” Placenta: Was bedeuten die beiden wirklich füreinander?
H.-G. Frank1,∗, C. SchmitzAnatomische Anstalt, Lehrstuhl II, Ludwig-Maximilians-Universität, Pettenkoferstrasse 11, 80336 München, Germany
Abstract
Die Placenta des Menschen als großes parenchymatöses Organ ist in der forschenden Medizin lange Zeit eines der amwenigsten beachteten und bearbeiteten Themen gewesen. Neben dem geburtshilflich-neonatologischen Interesse sowieInteresse von wenigen Grundlagenforschern schien die Placenta für die anderen medizinischen Disziplinen lange Zeitohne Relevanz. Die Frage, was die neunmonatige Abhängigkeit menschlichen Lebens von diesem Organ und seinerFunktion wirklich für das ganze Leben bedeutet, wurde in letzter Zeit neu gestellt, nachdem gezeigt werden konnte,dass pränatale Mangelzustände lebenslang wirkende Spuren im Leben der Nachkommen hinterlassen können. Dabeiwurde dieses Phänomen zunächst am Beispiel der metabolischer Verarbeitungsmuster entdeckt, obwohl dem Befundan sich generelle biologische Bedeutung zukommen dürfte. Hier gilt unser Interesse vor allem der Gehirnentwicklung,die in der zweiten Hälfte der Schwangerschaft mit der Myelinisierung weiter Gehirnabschnitte wesentliche funktionelleReifungsschritte durchläuft, während die Placenta immer höhere Leistungen bei laufendem Umbau des Organs erbringenmuss. Um die möglichen Auswirkungen dieses Lebensabschnittes auf das spätere Leben und die Wechselwirkung zwischenplacentarer Reifung und Gehirnentwicklung so zu verstehen, dass klinische Relevanz entstehen kann, sind neue undmoderne quantitative Verfahren der Histologie sowohl für die Analyse placentaren Gewebes wie auch für die Analyse desGehirnmaterials von Bedeutung und teilweise noch zu adaptieren und zu entwickeln.
1. Was ist so faszinierend an der menschlichen Pla-centa?
1.1. Zum Einstieg eine Anekdote
Als ich mich nach meiner Habilitation zu einem Themader Placentaforschung bei den Professoren der Medizini-schen Fakultät der RWTH Aachen vorstellen musste, wareines der Vorstellungsgespräche beim damaligen Dekan derFakultät und Lehrstuhlinhaber für Psychiatrie zu führen.Seine - durchaus provokant gemeinte - Frage war, warumdenn überhaupt jemand an diesem Organ forschen sollte,das unmittelbar nach der Geburt seinen Dienst getan hatund bei allen Ereignissen des späteren Lebens keine primärwahrnehmbare Rolle spielt. Die Antwort, die ich auf dieseFrage gab, war politisch vielleicht nicht die geschickteste:Ich wies darauf hin, dass die Placenta das einzige großeparenchymatöse Organ des menschlichen Körpers ist, dasgänzlich ohne Innervation und ohne eigene Nervenzellenauskommt, sich entwickelt und seine Funktionen erfüllt.Sie wäre darum das beste Beispiel dafür, dass menschli-ches Leben überhaupt und „an sich” auch ganz ohne Ner-venzellen auskommen könne. Trotz des politischen Missge-schickes macht die Antwort aber doch unmittelbar deut-lich, dass dieses explosiv wachsende „Organ für neun Mo-nate” Besonderheiten aufweist, die es mindestens für die
∗Korrespondierender Autor; Email: [email protected]
1Redaktion: Andrea Asikoglu
Forschung, aber eben auch für die klinische Medizin - undich hoffe, das unten näher auslegen zu können - bedeutsammachen.
1.2. Unsere Placenta unterscheidet uns vom (Modell-)Tier
Neben der Tatsache, dass dieses Organ mit einem vol-len Organgewicht von ca. 500 g und seine Entwicklungvöllig frei sind von lokoregionalen neuronalen Einflüssen,ist die Placenta auch das Organ, das für den Menschenim Speziesvergleich am kennzeichnendsten ist, in mancherHinsicht kennzeichnender als selbst der Stolz der mensch-lichen Rasse, das Gehirn. Während das Gehirn des Men-schen und die Gehirne z.B. der häufig als Modellorganis-men benutzten Labornager viele offensichtliche Analogienaufweisen und die Parallelen durch das ganze Reich derSäugetiere hindurch in diesem wichtigen Organ - bei al-len Unterschieden im Detail - auffällig groß sind, ist dasbei der Placenta gänzlich anders [1, 2, 3, 4]. Wer sich andie Zotten der menschlichen Placenta (s. auch Abb. 1a)und ihre sich wandelnde Erscheinungsform im Laufe derSchwangerschaft gewöhnt hat und dann zum ersten Maldie Placenta eines Labornagers mikroskopiert, wird irri-tiert fragen, ob er denn überhaupt das richtige Präparatunter dem Mikroskop hat, so anders und nicht wiederzuer-kennen ist die Histologie dieser Placenten im Vergleich zurmenschlichen Placenta. Die Placenten der Labornager [5]sind labyrinthäre Placenten (s. auch Abb. 1b), in denen inhoch organisierter und hocheffizienter Form Trophoblast-kanäle (durchströmt von mütterlichem Blut) und Blutge-
Internal Communication: Neues aus der Anstalt (NeadeA) 30. November 2010
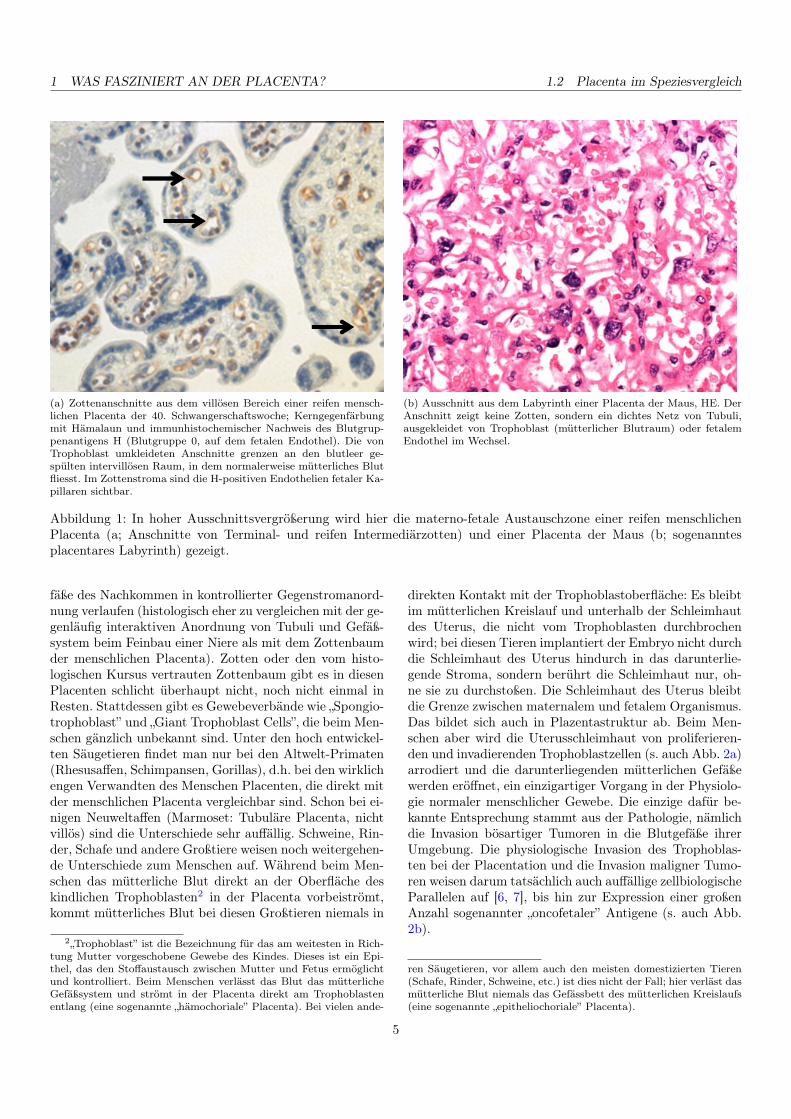
1 WAS FASZINIERT AN DER PLACENTA? 1.2 Placenta im Speziesvergleich
(a) Zottenanschnitte aus dem villösen Bereich einer reifen mensch-lichen Placenta der 40. Schwangerschaftswoche; Kerngegenfärbungmit Hämalaun und immunhistochemischer Nachweis des Blutgrup-penantigens H (Blutgruppe 0, auf dem fetalen Endothel). Die vonTrophoblast umkleideten Anschnitte grenzen an den blutleer ge-spülten intervillösen Raum, in dem normalerweise mütterliches Blutfliesst. Im Zottenstroma sind die H-positiven Endothelien fetaler Ka-pillaren sichtbar.
(b) Ausschnitt aus dem Labyrinth einer Placenta der Maus, HE. DerAnschnitt zeigt keine Zotten, sondern ein dichtes Netz von Tubuli,ausgekleidet von Trophoblast (mütterlicher Blutraum) oder fetalemEndothel im Wechsel.
Abbildung 1: In hoher Ausschnittsvergrößerung wird hier die materno-fetale Austauschzone einer reifen menschlichenPlacenta (a; Anschnitte von Terminal- und reifen Intermediärzotten) und einer Placenta der Maus (b; sogenanntesplacentares Labyrinth) gezeigt.
fäße des Nachkommen in kontrollierter Gegenstromanord-nung verlaufen (histologisch eher zu vergleichen mit der ge-genläufig interaktiven Anordnung von Tubuli und Gefäß-system beim Feinbau einer Niere als mit dem Zottenbaumder menschlichen Placenta). Zotten oder den vom histo-logischen Kursus vertrauten Zottenbaum gibt es in diesenPlacenten schlicht überhaupt nicht, noch nicht einmal inResten. Stattdessen gibt es Gewebeverbände wie „Spongio-trophoblast” und „Giant Trophoblast Cells”, die beim Men-schen gänzlich unbekannt sind. Unter den hoch entwickel-ten Säugetieren findet man nur bei den Altwelt-Primaten(Rhesusaffen, Schimpansen, Gorillas), d.h. bei den wirklichengen Verwandten des Menschen Placenten, die direkt mitder menschlichen Placenta vergleichbar sind. Schon bei ei-nigen Neuweltaffen (Marmoset: Tubuläre Placenta, nichtvillös) sind die Unterschiede sehr auffällig. Schweine, Rin-der, Schafe und andere Großtiere weisen noch weitergehen-de Unterschiede zum Menschen auf. Während beim Men-schen das mütterliche Blut direkt an der Oberfläche deskindlichen Trophoblasten2 in der Placenta vorbeiströmt,kommt mütterliches Blut bei diesen Großtieren niemals in
2„Trophoblast” ist die Bezeichnung für das am weitesten in Rich-tung Mutter vorgeschobene Gewebe des Kindes. Dieses ist ein Epi-thel, das den Stoffaustausch zwischen Mutter und Fetus ermöglichtund kontrolliert. Beim Menschen verlässt das Blut das mütterlicheGefäßsystem und strömt in der Placenta direkt am Trophoblastenentlang (eine sogenannte „hämochoriale” Placenta). Bei vielen ande-
direkten Kontakt mit der Trophoblastoberfläche: Es bleibtim mütterlichen Kreislauf und unterhalb der Schleimhautdes Uterus, die nicht vom Trophoblasten durchbrochenwird; bei diesen Tieren implantiert der Embryo nicht durchdie Schleimhaut des Uterus hindurch in das darunterlie-gende Stroma, sondern berührt die Schleimhaut nur, oh-ne sie zu durchstoßen. Die Schleimhaut des Uterus bleibtdie Grenze zwischen maternalem und fetalem Organismus.Das bildet sich auch in Plazentastruktur ab. Beim Men-schen aber wird die Uterusschleimhaut von proliferieren-den und invadierenden Trophoblastzellen (s. auch Abb. 2a)arrodiert und die darunterliegenden mütterlichen Gefäßewerden eröffnet, ein einzigartiger Vorgang in der Physiolo-gie normaler menschlicher Gewebe. Die einzige dafür be-kannte Entsprechung stammt aus der Pathologie, nämlichdie Invasion bösartiger Tumoren in die Blutgefäße ihrerUmgebung. Die physiologische Invasion des Trophoblas-ten bei der Placentation und die Invasion maligner Tumo-ren weisen darum tatsächlich auch auffällige zellbiologischeParallelen auf [6, 7], bis hin zur Expression einer großenAnzahl sogenannter „oncofetaler” Antigene (s. auch Abb.2b).
ren Säugetieren, vor allem auch den meisten domestizierten Tieren(Schafe, Rinder, Schweine, etc.) ist dies nicht der Fall; hier verläst dasmütterliche Blut niemals das Gefässbett des mütterlichen Kreislaufs(eine sogenannte „epitheliochoriale” Placenta).
5

2 WAS BEDEUTET DIE PLACENTA FÜR DEN MENSCHEN?
1.3. Die Placenta des Menschen verändert sich währendder Schwangerschaft
Während bei vielen Labornagern die Grundstruktur derPlacenta während der zweiten Hälfte der Schwangerschafterhalten bleibt und im wesentlichen eine Veränderung derGröße der Placenta (sprich: der Länge der gegenläufigenTrophoblastkanäle und Kapillaren im Labyrinth) als Ad-aptation an die wachsenden Anforderungen des Nachkom-men zu beobachten ist, verändert sich beim Menschen ca.ab Mitte der Schwangerschaft darüber hinaus der Baudes Zottenbaumes qualitativ. Bestimmte Zottentypen, diefür die ersten Monate dominierend waren („unreife In-termediärzotten”) verschwinden zunehmend und Zottenty-pen, die bisher nicht vorkamen, werden zur dominierendenStruktur („reife Intermediärzotten” und „Terminalzotten”;[8, 9, 10]. Jedem Mitarbeiter der Anatomischen Anstalt istspontan klar, was es bedeutet, wenn ein Umbau währenddes laufenden Betriebes zu geschehen hat. Es bedeutet einhöheres Risiko für unerwartete Fehlfunktion. Genau einsolcher „Umbau im laufenden Betrieb” aber findet bei je-der menschlichen Schwangerschaft in dem Organ statt, vondem unser intrauterines Wohl und Wehe abhängt. Auch istklar, dass die großen Speziesunterschiede zwischen Labor-nagern und dem Menschen den Wert tierexpertimentellerAnsätze ganz wesentlich limitieren. Wirklich relevante undin der Medizin anwendbare Ergebnisse können hier - allerVoraussicht nach - nur an menschlichem Material gewon-nen werden, selbst wenn tierexperimentelle Modellsystememöglicherweise trotzdem eingesetzt werden müssen.
1.4. Macht die Placenta den Menschen?Biologisch ist dieser große Speziesunterschied im Bereich
eines wichtigen Reproduktionsorgans nicht völlig unerklär-lich, denn die Trennung von Entwicklungslinien in der Evo-lution in verschiedene Arten erfolgt ganz wesentlich da-durch, dass Reproduktionsmechanismen (und bei Säuge-tieren ist einer dieser Mechanismen die Placentation) nichtmehr zur Deckung gebracht werden können. Sobald dieFortpflanzung aufgrund solcher Differenzen unmöglich ge-worden ist, ist der Prozess der Bildung zweier neuer, ver-wandter Spezies unumkehrbar geworden und damit einewesentliche Voraussetzung für die Definition einer neuenSpezies erfüllt. Darum ist es durchaus verständlich, dassdie Placenta des Menschen so auffällig geringe strukturel-le Überlappungen mit den Placenten anderer Säugetierezeigt. Sie ist eben im wahrsten Sinn des Begriffes Teil des-sen, was „spezifisch” menschlich ist.
2. Was bedeutet die Placenta für den Menschen?
Oben wurde ausgeführt, dass die menschliche Plazentaetwas besonderes unter den Organen des Menschen ist, unddies vor allem im Vergleich zu verschiedenen Tierarten.Eine Frage ist, ob sich aus diesen Besonderheiten, die fürden Menschen und wenige Primatenspezies recht typischsind, auch weitergehende Konsequenzen ergeben.
(a) Nachweis von Mib-1 (Proli-ferationsmarker →) in den Zell-kernen von Trophoblastzellen derZellsäule. Die Proliferationsrateist ausserordentlich hoch. Hierlösen sich die Trophoblastzellenvon den Zotten (dann als ex-travillöse Trophoblastzellen be-zeichnet) und invadieren in Rich-tung mütterliche Gefäße des Pla-centabettes.
(b) Nachweis eines oncofetalenAntigens (oncofetales Fibronek-tin; Ak BC-1), das als Produktder invadierenden Trophoblastzel-len unpolar zwischen den Zellenaufgehäuft wird.
Abbildung 2: Folgeschnitte einer Verankerungszone (soge-nannte trophoblastische Zellsäule) im Bereich einer Ba-salplatte einer menschlichen Plazenta der 10. Schwanger-schaftswoche.
Zum einen ist zu fragen, ob die „spezifischen” Eigenar-ten der Placentation beim Menschen etwas zu tun habenmit den Eigenheiten, auf die die Gattung Homo Sapiensbesonders stolz ist, vorweg natürlich mit der „Sapientia”,der Weisheit, die zu besitzen sie sich schon vom Namen herrühmt. Ganz auszuschliessen ist jedenfalls nicht, dass wäh-rend des intrauterinen Lebens wesentliche Grundlagen derGehirnentwicklung in Abhängigkeit von einer optimalenplacentaren Funktion stehen. Die relativ lange intrauterinePhase der Gehirnentwicklung beim Menschen im Vergleichzu anderen Spezies könnte ein mit-konstituierender Faktorfür die wesentlichen evolutiven Vorteile in der Leistungs-fähigkeit des Gehirns sein. Im Unterschied zu z.B. denLabornagern finden wichtige Teile der Gehirnentwicklungbeim Menschen noch intrauterin (sozusagen „im Nest”)statt, während sie bei Labornagern erst nach der Geburtstattfinden (Review siehe in [11]). Damit ergeben sich ganzandere Möglichkeiten der Interaktion zwischen einer opti-malen Placentafunktion und der Entwicklung des Gehirnswährend der pränatalen Phase, vor allem in der zweitenHälfte der Schwangerschaft. Wie oben ausgeführt, unter-liegt die Placenta während dieser Periode einem struktu-rellen Wandel, der sicher einer besonderen funktionellenBelastung und damit einer auch medizinisch riskanterenPhase entspricht. Wie in der Medizin eigentlich immerim Rahmen komplexer Prozesse ist auch hier zu erwar-ten, dass es zu placentaren Dysfunktionen verschiedenen
6

3 WIE IST DIE BEDEUTUNG DER PLACENTA ZU ERMESSEN?
PI Hazard Ratio (95% CI) Fälle / Männer≤25 1,66 (1,11-2,48) 104 / 109327 1,44 (0,97-2,13) 135 / 164329 1,18 (0,78-1,78) 84 / 1260>29 1,00 (Referenz) 31 / 578
P for trend 0,0006
Tabelle 1: Relative Erkrankungsrisiken (Hazard Ratios)für koronare Herzerkrankung bei Männern in Abhängig-keit vom Geburtsgewicht. In der Tabelle werden die Zahlender sogenannten Helsinki-Kohorte benutzt. Der Ponderal-Index PI berechnet wich aus (PI = m/l3), wobei m dasKörpergewicht und l die Körperlänge ist. Dies dient alsstandardisiertes Maß des Körpergewichtes bei Neugebore-nen. Die Tabelle wurde auszugsweise aus [17] übernom-men.
Schweregrades kommen kann und damit möglicherweise zuWechselwirkungen mit der parallel ablaufenden Gehirn-entwicklung. Diese werden nicht zwangsläufig primär Vi-talfunktionen betreffen, sondern möglicherweise sogar be-vorzugt diejenigen Gehirnregionen, die humanspezifischeFunktionen höherer Ordnung übernehmen sollen und imwesentlichen während der zweiten Hälfte der Schwanger-schaft in Reifungsvorgänge verwickelt sind. Analog zu denaus der Teratologie bestens bekannten Fakten wäre auchhier zu eruieren, ob es einen Zusammenhang zwischen Orteiner Funktionsstörung im Gehirn und dem Zeitpunkt derpränatal aufgetretenen Dysfunktion geben könnte. Unterdem Schlagwort des „Prenatal Programming” ist in denletzten Jahrzehnten die Wahrnehmung dafür gewachsen,dass pränatale Ereignisse möglicherweise langfristige ge-sundheitliche Konsequenzen für das Kind haben können[12, 13, 14, 15]. Am weitestgehenden konnte bisher erhär-tet werden, dass intrauterine Mangelsituationen eine sta-bile „Spar-Stoffwechsellage” festzurren, die lebenslang alsendogener metabolischer „Trend” weiterbesteht und dannin der heute in der ersten Welt regelhaft bestehenden le-benslangen Überernährungssituation eine ungünstige Vor-aussetzung, nämlich eine potenzielle Gesundheitsgefähr-dung, darstellt. Der Kern dieser Gesundheitsgefährdungsind langsam entstehende und chronische Fehlentwicklun-gen der metabolischen Lage, die zum Entstehen biogra-phisch relativ spät auftretender Gesundheitsprobleme imkardiovaskulären Bereich beitragen können [16, 14]. Datenaus einer solchen epidemiologischen Studienkohorte sindauszugsweise in Tabelle 1 dargestellt.
Die Datenlage bezüglich Auswirkungen intrauterinerProblemsituationen auf die Gehirnentwicklung und ihrermöglichen Konsequenzen [18, 19, 20, 21]ist - noch - dürfti-ger, obwohl die Gehirnentwicklung als Gegenstand der zel-lulären und klinischen Neurowissenschaften ein in besonde-rer Weise sensitiver und auch prädestinierter Bereich fürdie Entwicklung möglicher Langzeitfolgen ist. Allerdingsist es auch sehr schwer, in diesem Bereich die pränatal-postnatale Transmission sicher zu erfassen. Die meisten
Forschungsvorhaben aus dem Bereich des Prenatal Pro-gramming kämpfen mit dem Problem der Ungenauigkeitpränataler Daten. Bisher beruhen die hier zugrundeliegen-den Studien in der Regel auf der Erhebung von Datenaus Geburtsregistern, die u.a. Kindsgewicht, Kopfumfangund Abdominalumfang des Kindes, Placentagewicht, Pla-centainsertionsfläche („placental surface”) etc. beinhalten.Aus diesen epidemiologischen Daten lassen sich nur Rück-schlüsse auf sehr grobe intrauterine Mangelsituationen zie-hen; es sind wegen der breiten Überlagerung mit Norm-verteilungen große Fallzahlen notwendig, um Risikowahr-scheinlichkeiten sicher und mit ausreichender statistischerAussagekraft berechnen zu können. Trotz dieser Limita-tionen bestehender Studien ist klar, dass die Epidemiolo-gie hier einen wesentlichen Beitrag zur Weiterentwicklungwissenschaftlicher Konzepte und Fragestellungen geleistethat.
3. Wie kann man die Bedeutung der Placenta fürden Menschen ermessen?
Was bisher ganz wesentlich in der Diskussion um diemögliche Bedeutung pränataler Veränderungen für daspostnatale Leben fehlt, sind geeignete Methoden, die prä-natalen Veränderungen viel genauer als bisher zu erfas-sen. Dabei geht es nicht um diejenigen Veränderungen, dieso schwerwiegend sind, dass sie das Leben des Kindes di-rekt bedrohen. Für diese Situationen ist die moderne Me-dizin durchaus gerüstet. Es geht vielmehr um diejenigenSituationen, in denen eine diskrete, nicht direkt lebens-bedrohliche und möglicherweise nur zeitweilig vorhandeneStörung der Placentafunktion zu Veränderungen mit lang-fristigen, sich häufig erst spät postnatal manifestierendenNormabweichungen führt. Es geht ausserdem nicht nur umdie Veränderungen in der Placenta, sondern auch um diekorrespondierenden Veränderungen in den potenziell einenspäteren Schaden mediierenden Gehirnstrukturen.
Es ist aber mit den heute zur Verfügung stehendenwissenschaftlichen Methoden praktisch unmöglich, diesefunktionellen Veränderungen in situ direkt zu erfassen.Vielmehr erscheint es sinnvoll, nach diskreten strukturel-len Veränderungen zu suchen, die funktionellen Verän-derungen zugrunde liegen und diese somit indirekt an-zeigen könnten. Dieses Konzept basiert auf dem zentra-len Struktur-Funktions-Paradigma der modernen Medizin,das sicherlich für diese Fragestellung ebenfalls valide an-wenbar ist. Die Tatsache, dass Tiermodelle im Falle der Be-sonderheiten der späten intrauterinen Lebensphase nichtoptimal zur Beantwortung der anstehenden Fragen geeig-net - teilweise wegen der enormen Speziesdifferenzen sogarungeeignet - sind, macht die Bedeutung der (human-) ana-tomischen Strukturforschung in diesem Gebiet nicht gerin-ger. Langfristig wird es nötig sein, prospektive Studien zuinitiieren, die imstande sind, früh erhobene placentare undneuroanatomische Befunde mit späteren Entwicklungen zukorrelieren. Dies kann nur in Zusammenarbeit mit den kli-nischen Disziplinen erfolgen. Eine weitere Möglichkeit be-
7

3 WIE IST DIE BEDEUTUNG DER PLACENTA ZU ERMESSEN?
steht darin, die Archive placentarer Biopsien und Gewe-beproben, die - beispiels aus juristischen Gründen in denUSA - als Banken von Paraffinmaterial angelegt wurden,retrospektiv zu analysieren und dies mit dem Gesundheits-zustand der inzwischen in ihrem Lebensalter fortgeschrit-tenen Nachkommen zu korrelieren. Für alle diese Analysenmüssen auf der Seite der Analyse placentaren Materialsund auch auf der Seite der Analyse von Gehirnmaterialnoch wesentlich methodische Arbeiten bei der Verfeine-rung existierender Methoden und ihrer Adaptation auf diepränatale Situation geleistet werden. Diese Arbeitsrich-tung stellt einen langfristig ausgerichteten Schwerpunktder Arbeit am Lehrstuhl II der Anatomischen Anstalt darund soll nach dem Aufbau der erforderlichen Infrastrukturauch rasch drittmittelfähig und drittmittelgestützt weiter-entwickelt werden.
[1] O. Grosser, Vergleichende Anatomie und entwicklungsgeschich-te der Eihäute und der placenta, Wilhelm Braumüller„ Vienna,1909.
[2] O. Grosser, Frühentwicklung, Eihautbildung und placentationdes Menschen und der Säugetiere„ J.F. Bergmann, München,1927.
[3] E. Ramsey, The placenta : human and animal, Praeger, NewYork N.Y., 1982.
[4] H. Mossman, Vertebrate fetal membranes : comparative onto-geny and morphology, evolution, phylogenetic significance, ba-sic functions, research opportunities, Rutgers University Press,New Brunswick N.J., 1987.
[5] E. Ramsey, The placenta of laboratory animals and man, HoltRinehart and Winston, [New York, 1975.
[6] H. G. Frank, F. Malekzadeh, S. Kertschanska, C. Crescimanno,M. Castellucci, I. Lang, G. Desoye, P. Kaufmann, Immunohi-stochemistry of two different types of placental fibrinoid., ActaAnat (Basel) 150 (1) (1994) 55–68.
[7] H. G. Frank, B. Huppertz, S. Kertschanska, D. Blanchard,D. Roelcke, P. Kaufmann, Anti-adhesive glycosylation offibronectin-like molecules in human placental matrix-type fibri-noid., Histochem Cell Biol 104 (4) (1995) 317–329.
[8] K. Benirschke, Pathology of the human placenta, 5th Edition,Springer, New York, 2006.
[9] D. K. Sen, P. Kaufmann, G. Schweikhart, Classification of hu-man placental villi. ii. morphometry., Cell Tissue Res 200 (3)(1979) 425–434.
[10] P. Kaufmann, D. K. Sen, G. Schweikhart, Classification of hu-man placental villi. i. histology., Cell Tissue Res 200 (3) (1979)409–423.
[11] B. Clancy, R. B. Darlington, B. L. Finlay, Translating develop-mental time across mammalian species., Neuroscience 105 (1)(2001) 7–17.
[12] D. J. Barker, P. D. Winter, C. Osmond, B. Margetts, S. J. Sim-monds, Weight in infancy and death from ischaemic heart di-sease., Lancet 2 (8663) (1989) 577–580.
[13] D. J. Barker, C. Osmond, C. M. Law, The intrauterine and earlypostnatal origins of cardiovascular disease and chronic bronchi-tis., J Epidemiol Community Health 43 (3) (1989) 237–240.
[14] D. J. P. Barker, J. Gelow, K. Thornburg, C. Osmond, E. Ka-jantie, J. G. Eriksson, The early origins of chronic heart failure:impaired placental growth and initiation of insulin resistance inchildhood., Eur J Heart Fail 12 (8) (2010) 819–825.URL http://dx.doi.org/10.1093/eurjhf/hfq069
[15] D. J. P. Barker, D. T. Lackland, Prenatal influences on strokemortality in england and wales., Stroke 34 (7) (2003) 1598–1602.URL http://dx.doi.org/10.1161/01.STR.0000077257.27430.7E
[16] K. L. Thornburg, P. F. O’Tierney, S. Louey, Review: The pla-centa is a programming agent for cardiovascular disease, Pla-centa 31 Suppl (2010) S54–59, PMID: 20149453.
URL http://www.ncbi.nlm.nih.gov/pubmed/20149453[17] D. J. Barker, Fetal Origins of Adult Disease, 3rd Edition, Vol. 1,
Saunders, 2004.[18] H. Kunugi, S. Nanko, R. M. Murray, Obstetric complications
and schizophrenia: prenatal underdevelopment and subsequentneurodevelopmental impairment., Br J Psychiatry Suppl 40(2001) s25–s29.
[19] H. Kunugi, N. Takei, S. Nanko, K. Saito, H. Kazamatsuri,Head circumference at birth and schizophrenia., Br J Psych-iatry 165 (4) (1994) 551–552.
[20] R. J. V. Lieshout, L. P. Voruganti, Diabetes mellitus duringpregnancy and increased risk of schizophrenia in offspring: areview of the evidence and putative mechanisms., J PsychiatryNeurosci 33 (5) (2008) 395–404.
[21] R. M. Murray, P. Jones, E. O’Callaghan, Fetal brain develop-ment and later schizophrenia., Ciba Found Symp 156 (1991)155–63; discussion 163–70.
8

„Design-based“ Stereologie – eine ganz kurze Einführung
C. Schmitz∗
Anatomische Anstalt, Lehrstuhl II, Ludwig-Maximilians-Universität München, Pettenkoferstrasse 11, 80336 München
Abstract
„Design-based“ Stereologie ist eine Sammlung von Methoden zur Bestimmung von Anzahl, Größe, Raumausrichtungund anderer Variabler sogenannter biologischer „events“ (z.B. Zellkerne, Zellen, Gefässe, etc.) in einem „Referenzraum“(z.B. einer Gehirnregion). Die konsequente Anwendung design-based stereologischer Methoden kann eine entscheidendeRolle bei der Überprüfung experimenteller Hypothesen in der modernen biomedizinischen Forschung spielen, und hatim Bereich der quantitativen Histologie in den letzten 20 Jahren zu einer erheblichen Verbesserung der Aussagekraftentsprechender Studien in der Literatur geführt. Viele angesehene Fachzeitschriften verlangen heute bei der Publikationquantitativ histologischer Daten die Anwendung design-based stereologischer Methoden. Am Lehrstuhl II der Anatomi-schen Anstalt wird die design-based Stereologie eine zentrale Rolle im Methodenspektrum der Arbeitsgruppe einnehmen.
Die Anwendung design-based stereologischer Methodenbei modernen biomedizinischen Studien erlaubt Forscherndie effektive und effiziente Erfassung von quantitativ his-tologischen Daten. Der Name „design-based“ verweist aufdie annahme-freie, mathematische und statistische Naturder entsprechenden Methoden, im Gegensatz zu „model-based“ Methoden, bei denen Annahmen z.B. über dieGröße, Form etc. der untersuchten biologischen eventsgemacht werden.
Das Prinzip der design-based Stereologie und ihreVorteile seien an dem folgenden Beispiel erläutert:
Beispiel: Bestimmung der Anzahl von Purkinjezellen imKleinhirn der Ratte
Lassen Sie uns annehmen, dass ein Forscherdie Hypothese testen möchte, dass eine bes-timmte experimentelle Behandlung von Ratten(z.B. in der Toxikologie) zum Verlust von Purkin-jezellen im Kleinhirn führt. Um diese Hypothesezu testen, stellt der Forscher im Labor 5 µm dickeParaffinschnitte der Kleinhirne von sechs behan-delten und sechs unbehandelten Ratten her undfärbt sie mit Cresylviolett (einer Nissl-Färbung).Dann sucht er unter einem Standardmikroskopmit einem 40er Objektiv einige repräsentativemikroskopische Gesichtsfelder pro Tier. Er wähltjeweils 10 Gesichtsfelder pro Tier, die die besteFärbung der Purkinjezellen zeigen, und zähltalle Purkinjezellen, die er in den Gesichtsfeldernidentifizieren kann. Zu diesem Zweck verwen-det er ein Okular, das über dem Gesichtsfeldein Rechteck zeigt. Anschliessend teilt er die
∗E-Mail: [email protected]
Anzahl der gezählten Purkinjezellen pro Klein-hirn durch die Größe des Areals, das untersuchtwurde (die Größe dieses Areals ergibt sich ausdem Produkt der Größe des Rechtecks im Okularund der Anzahl der untersuchten Gesichtsfelder– in unserem Beispiel also 10). Anhand dieserDaten berechnet der Forscher die durchschnitt-liche Dichte von Purkinjezellen im Kleinhirn vonbehandelten und unbehandelten Tieren, und ver-gleicht den Unterschied zwischen den Gruppenstatistisch mit Hilfe des Student-t Tests. DieAnalyse zeigt eine leichte, statistisch aber nichtsignifikante (d.h. p > 0,05) Reduktion der mitt-leren Dichte von Purkinjezellen im Kleinhirn derRatte als Reaktion auf die experimentelle Be-handlung. Basierend auf dieser Analyse kommtder Forscher zu dem Schluss, dass die experi-mentelle Behandlung keinen Verlust von Purkin-jezellen im Kleinhirn der Ratte auslöst.
Unzählige Studien dieser Art können in der Literatur ge-funden werden. Dies wirft die Frage auf, wie man über-haupt auf die Idee kommen könnte, dass mit dieser Ana-lyse etwas falsch sein sollte, und wie man diese Studie an-ders ausführen sollte. Es ist in der Tat zunächst nichtsfalsch am Zählen in diesem Beispiel, abgesehen vom Stand-punkt der design-based Stereologie aus. Die Diskus-sion sollte stattdessen darauf fokussiert werden, ob diebeschriebene Daten-Analysen-Prozedur tatsächlich einenstatistisch signifikanten Unterschied zwischen den Grup-pen zeigen würde, falls er denn existiert. Oder, anders aus-gedrückt, die Methodik sollte so optimiert werden, dass dieNicht-Ablehnung der Nullhypothese (Nullhypothese: keinUnterschied zwischen den Gruppen) tatsächlich auch dasbiologische Ergebnis ist (biologisches Ergebnis: kein Ver-lust von Purkinjezellen im Kleinhirn).
Internal Communication: Neues aus der Anstalt (NeadeA) November 30, 2010

Große Unterschiede zwischen den Gruppen mit ver-gleichsweise geringer interindividueller Variabilität könnenwahrscheinlich mit der im obigen Beispiel beschriebenenZählmethode beobachtet werden, aber eher diskrete – bio-logisch aber wichtige – Veränderungen nach der experi-mentellen Behandlung werden so wahrscheinlich nicht ge-funden. Um dies nachvollziehen zu können, muss manberücksichtigen, dass die beschriebene Untersuchung anhistologischen Schnitten von Gehirngewebe durchgeführtund deshalb von den folgenden Faktoren beeinflusst wurde:(i) systematische Fehler bei der Suche „repräsentativer“Gesichtsfelder, (ii) Zuverlässigkeit und (iii) Gültigkeit vonZellzählungen und Zelldichtemessungen.
Wir werden nun jeden dieser Faktoren kritisch disku-tieren und sehen, wie sie mit design-based Stereologie op-timiert werden können:
• Die Klassifikation eines bestimmten Gewebeschnittsaus einem Referenzraum (hier: Kleinhirn der Ratte)als „repräsentativ“ impliziert, dass die Ergebnisse,die an diesem Schnitt erzielt werden, mehr oderweniger identisch mit dem Durchschnitt der Ergeb-nisse sind, die erzielt würden, wenn alle Schnitte,die den Referenzraum umfassen, in der gleichenWeise analysiert würden. Dies wurde in dem obengenannten Beispiel jedoch nicht getestet, sondernvielmehr einfach angenommen bzw. vorausgesetzt.In der Literatur wurde jedoch gezeigt, dass Ergeb-nisse von Zellzählungen, die an einzelnen „repräsen-tativen“ Schnitten aus dem Kleinhirn der Rattedurchgeführt wurden, sich erheblich von den Ergeb-nissen solcher Zellzählungen unterscheiden können,die auf der Analyse einer „systematisch-zufällig“ aus-gewählten Schnittserie basieren („systematic and ran-dom sampling“; „SRS“; der Begriff SRS wird weiterunten erläutert) [1]. Von daher wird in der design-based Stereologie die Analyse einer SRS-Serie vonSchnitten durch den Referenzraum als Goldstandardangesehen [2]. Das Prinzip der Erstellung einer SRS-Schnittserie durch das Kleinhirn einer Ratte ist in Ab-bildung 1A gezeigt.
• Ebenso problematisch ist die Auswahl bestimmtermikroskopischer Gesichtsfelder, die auf lokaler Fär-bequalität basieren (oder jedes andere subjektiveKriterium). Ein bestimmtes „Set“ mikroskopischerGesichtsfelder als „repräsentativ“ zu klassifizieren im-pliziert, dass die Ergebnisse, die an diesem Set vonGesichtsfeldern gewonnen werden, mehr oder wenigeridentisch sind mit dem Durchschnitt der Ergeb-nisse, die gewonnen würden, wenn alle Regionen desSchnitts in der gleichen Weise analysiert würden.Auch dies wurde im oben beschriebenen Beispiel nichtgetestet, sondern einfach angenommen bzw. voraus-gesetzt. Um dies zu vermeiden, untersucht man in derdesign-based Stereologie grundsätzlich eine SRS-Serievon mikroskopischen Gesichtsfeldern, die tatächlichrepräsentativ für den gesamten untersuchten Schnitt
ist. Die entsprechende Vorgehensweise ist in Abbil-dung 1C gezeigt.
• In Paraffinschnitten haben die Purkinjezellen imKleinhirn der Ratte einen mittleren Perikaryen-Durchmesser von ca. 20 µm. In dem obenbeschriebenen Beispiel hat der Forscher daher tat-sächlich gar keine Purkinjezellen gezählt, sondernPurkinjezellanschnitte (auch Zellprofile genannt), dadie Schnitte (5 µm dick) ja viel dünner waren alsder mittlere Durchmesser der Perikaryen. Dabeimuss aber beachtet werden, dass die Wahrschein-lichkeit, ein Zellprofil einer Zelle im Schnitt zu finden,mit zunehmender Größe der Zelle steigt (vgl. [1]).Auch dies mag im oben beschriebenen Beispiel keinenEinfluss auf die Ergebnisse der quantitativen Unter-suchung gehabt haben – vorausgesetzt, die mittlereGröße der Perikaryen der Purkinjezellen hatte sichdurch die experimentelle Behandlung nicht verändert.Aber auch hier - dies wurde nicht getestet, sonderneinfach angenommen bzw. vorausgesetzt. In derdesign-based Stereologie umgeht man dieses Prob-lem dadurch, dass man keine Zellen bzw. Zellprofilezählt, sondern sogenannte „repräsentative Punkte“ in-nerhalb der Zellen, wobei jede Zelle genau einen dieser„repräsentativen Punkte“ aufweisen muss, keinesfallsaber mehr als einen dieser Punkte. Um dies zu er-reichen, werden in der design-based Stereologie meis-tens auch dicke Schnitte (erheblich dicker als 5 µm)analysiert, und zwar mit sogenannten „unbiased vir-tual counting spaces“, die virtuell in den dickenSchnitten angelegt werden. Wie dies realisiert wird,ist in Abbildung 1D prinzipiell gezeigt.
• Schliesslich muss erwähnt werden, dass die Be-stimmung von Zelldichten in Studien, bei denendie Hypothese einer Änderung der mittleren Zel-lzahl überprüft werden soll, äusserst problematischsein kann. Eines der eindrucksvollsten Beispiele indiesem Zusammenhang ist wahrscheinlich die Re-duzierung der mittleren Gesamtzahl von Purkin-jezellen im Kleinhirn der Ratte um ca. 50% alsFolge einer bestimmten pränatalen Bestrahlung, ohnedass dies zu einer Änderung der mittleren Dichte derPurkinjezellen geführt hätte [1]. Hintergrund diesesPhänomens ist, dass sich bei der untersuchten präna-talen Bestrahlung auch das Volumen des Kleinhirnsder Ratte (d.h. des Referenzraums) um ca. 50%reduziert hatte. Nochmals, dieses Phänomen mussauf die Ergebnisse des oben beschriebenen Beispielskeinerlei Einfluss gehabt haben, solange sich die mitt-lere Größe des Kleinhirns durch die experimentelleBehandlung nicht geändert hatte. Aber – wir wis-sen es halt nicht; es wurde einfach vorausgesetzt bzw.angenommen.
Zusammenfassend können wir festhalten, dass die Ergeb-nisse im oben beschriebenen Beispiel auf vier nicht über-
10

prüften Annahmen basieren:
• Einzelne Schnitte aus dem Kleinhirn sind „repräsen-tativ“ für das gesamte Organ.
• Einzelne Gesichtsfelder aus diesen Schnitten sind „re-präsentativ“ für den gesamten Schnitt.
• Die mittlere Größe der Purkinjezell-Perikaryen hatsich durch die experimentelle Behandlung nicht ver-ändert.
• Die mittlere Größe des Kleinhirns hat sich durch dieexperimentelle Behandlung nicht verändert.
Dies ist jedoch ein äusserst unbefriedigender Zustand – je-de dieser Annahmen könnte richtig oder falsch sein, unddemnach das Ergebnis der quantitativ histologischen Un-tersuchung beeinflusst haben. Tatsächlich kann also ebennicht gesagt werden, in wie weit die Ergebnisse der quan-titativ histologischen Untersuchung tatsächlich die biolo-gische Realität wiedergeben. Unter wissenschaftlichen Ge-sichtspunkten ist eine solche Situation jedoch abzulehnen.Wenn man es sich genau überlegt, sind die Ergebnisse imoben beschriebenen Beispiel wissenschaftlich wertlos
Der Schlüssel zum Erfolg in der design-based Stereolo-gie liegt darin, jedem biologischen event (d.h., jeder Purk-injezelle) die gleiche Wahrscheinlichkeit einzuräumen, imRahmen einer Stichprobenuntersuchung einer Anzahl vonGesichtsfeldern gezählt zu werden (Infratest und andereMeinungsforschungsinstitute arbeiten übrigens nach dengleichen statistischen Kriterien). Dies geschieht durch dieKombination eines mehrstufigen SRS-Auswahlprozesses(Schnitte, Gesichtfelder), die Analyse dicker Schnitte mitvirtuellen „counting spaces“, und der (mathematischen)Reduzierung jedes biologischen events auf einen Punkt(der dann z.B. keine Größe, keine Form und auch keineRaumrichtung mehr besitzt).
Tatsächlich kann man die Gesamtzahl von biologischenevents in einem Referenzraum auf zwei verschiedene Artenbestimmen. Bei der ersten Methode (der sogenannten„Optical Fractionator“ Methode) bestimmt man die An-zahl von events in einer SRS-Stichprobe des Referen-zraums, und errechnet die Gesamtzahl von events ausder gezählten Anzahl der events und der Wahrschein-lichkeit, einen bestimmten Teilbereich des Referenzraumsuntersucht zu haben. Bei der zweiten Methode (der so-genannten „VRef × NV “ Methode) bestimmt man dieGesamtzahl von events als Produkt der Dichte der eventsin einem bestimmten Teilvolumen des Referenzraums undder absoluten Größe des Referenzraums (vgl. [3]). Insge-samt ist die „Optical Fractionator“ Methode in ihrer An-wendung etwas einfacher als die „VRef × NV “ Methode,und liefert dabei auch noch etwas präzisere Ergebnisse([3]).
Alle bisher diskutierten Aspekte beziehen sich auf dieValidität von Resultaten quantitativ histologischer Stu-dien. Aber wie ist es um die Reliabilität der Ergebnissesolcher Studien bestellt, d.h., der Wiederholbarkeit der
erzielten Ergebnisse? Was wäre dem Forscher in unseremobigen Beispiel passiert, wenn er seine Analyse an an-deren Schnitten wiederholt hätte, oder andere Gesichts-felder ausgezählt hätte? Wäre er zu derselben Schlussfol-gerung gekommen, oder eventuell zu einer anderen? Auchdiese Frage kann nicht beantwortet werden, was der im obi-gen Beispiel beschriebenen Methodik eigentlich den „kissof death“ gibt. Im Gegensatz dazu stehen in der design-based Stereologie eine Reihe von mächtigen, statistischenMethoden zur Verfügung, um genau dieser Fragestellungnachzugehen. Computersimulationen (z.B. [4, 5]) habendie Validität dieser Methoden aufgezeigt. Diese Metho-den sind auch Grundlage dafür, dass Wissenschaftler dieFrage beantworten können, was gewesen wäre, wenn sieeine andere SRS Serie von Schnitten bzw. eine andereSRS-Serie von Gesichtsfeldern analysiert hätten. Es sindgenau diese Zusatzwerkzeuge, die der design-based Stere-ologie ihren festen Platz in der modernen quantitativenHistologie gesichert haben – denn die Auswahl von Schnit-ten und Gesichtsfeldern nach SRS-Kriterien bringt es mitsich, dass die finalen Schlussfolgerungen unabhängig davonsind, ob man nun eine bestimmte oder eine andere SRS-Serie von Schnitten bzw. Gesichtsfeldern analysiert hätte.
Zusammengefasst kann man sagen, dass design-basedStereologie ein gewaltiges Angebot an modernsten Meth-oden in der quantitativen Histologie bietet, das nurein einziges Ziel vor Augen hat, nämlich die Gültigkeitvon quantitativ histologischen Forschungsergebnissensicherzustellen. In unserem obigen Beispiel hätte einemoderne design-based stereologische Analyse demnach wiefolgt ausgesehen:
Ein besserer Weg zur Bestimmung der Gesamtzahl vonPurkinjezellen im Kleinhirn der Ratte
Lassen Sie uns nochmals davon ausgehen,dass ein Forscher die Hypothese testen möchte,dass eine bestimmte experimentelle Behandlungvon Ratten (z.B. in der Toxikologie) zum Ver-lust von Purkinjezellen im Kleinhirn führt. Umdiese Hypothese zu testen, erstellt er von jeweilssechs behandelten und unbehandelten Rattenvollständige Serien 50 µm dicker Frontalschnittedurch das Kleinhirn (vorzugsweise Gefrier-schnitte), und wählt für jede Ratte eine SRS-Serie jedes fünften Schnitts aus (zufällig ist dabeidie Wahl des ersten Schnitts der SRS-Serie ausden ersten fünf Schnitten des Kleinhirns, z.B.Schnitt Nr. 3; systematisch ist dann die Auswahlder SRS-Serie jeden fünften Schnitts, d.h., dieSchnitte Nr. 3, 8, 13, 18, usw.). Der Forscherfärbt die Schnitte mit einer modifizierten Nissl-Färbung, die für 50 µm dicke Schnitte opti-miert wurde. Dann analysiert der Forscher jedenSchnitt der SRS-Serien mit einem stereologischenArbeitsplatz, der (u.a.) aus einem Mikroskop,einer Digitalkamera, einem motorisierten Objekt-trägertisch, einer motorisierten Fokussiereinheit
11

sowie einem Computer mit Stereologie-Softwarebesteht (Abbildung 2). Der Forscher umfährtdie Konturen des Kleinhirns bei niedrigem Ab-bildungsmaßstab mit der Maus auf dem Monitor,und die Software berechnet automatisch die Posi-tion einer SRS-Serie von Gesichtsfeldern (in Ab-hängigkeit von der gewünschten Gesamtzahl vonzu untersuchenden Gesichtsfeldern und der An-zahl der Schnitte pro Kleinhirn). Anschliessendfährt die Software bei hohem Abbildungsmaßstabjedes einzelne Gesichtsfeld der SRS-Serie vonGesichtsfeldern an, und überlagert es mit einemsogenannten „unbiased counting frame“ (Abbil-dung 1D), dessen Dimensionen von dem Forscherfrei gewählt werden können und in einer Pilot-studie so bestimmt wurden, dass für jedes Klein-hirn eine bestimmte Anzahl von Zellen gezähltwird (z.B. im Mittel 750 Zellen pro Kleinhirn).Beim Durchfokussieren durch den dicken Schnittwarnt die Software den Forscher optisch (undevtl. auch akustisch), wenn die Fokusebeneausserhalb des „unbiased virtual counting spaces“(d.h., höher oder tiefer) liegt. Der Forscher klicktnun jede Purkinjezelle an, die alle gewähltenZählkriterien erfüllt. Nach Abschluss der Ana-lyse aller Schnitte eines Kleinhirns berechnetdie Software automatisch die „estimated“ (dasdeutsche Wort „geschätzt“ ist in diesem Zusam-menhang unzutreffend) Gesamtzahl von Purk-injezellen im untersuchten Kleinhirn, zusammenmit einer „predicted“ (das deutsche Wort „voraus-gesagt“ ist in diesem Zusammenhang ebenfallsunzutreffend) Präzision der Messung. Nach Ab-schluss der Analyse aller Kleinhirne berechnetder Forscher die durchschnittliche Gesamtzahlvon Purkinjezellen im Kleinhirn von behandel-ten und unbehandelten Tieren, und vergleichtden Unterschied zwischen den Gruppen statis-tisch mit Hilfe des Student-t Tests. Die Ana-lyse zeigt eine statistisch signifikante (d.h. p< 0,05) Reduktion der mittleren Anzahl vonPurkinjezellen im Kleinhirn um etwa 20% derRatte als Reaktion auf die experimentelle Be-handlung. Basierend auf dieser Analyse kommtder Forscher zu dem Schluss, dass die experi-mentelle Behandlung einen Verlust von Purkin-jezellen im Kleinhirn der Ratte auslöst. In derentsprechenden Publikation der Ergebnisse kannder Forscher darlegen, dass die Ergebnisse un-abhängig von den untersuchten Schnitten bzw.den untersuchten Gesichtsfeldern sind, und auchnicht auf möglichen Größenänderungen der Purk-injezellen bzw. des Kleinhirns als Ergebnis derexperimentellen Behandlung basieren. Vielmehrhandelt es sich bei dem gefundenen Verlust vonca. 20% der Purkinjezellen im Kleinhirn derRatte um eine biologische Reaktion auf die exper-
imentelle Behandlung. Diese Ergebnisse habeneinen erheblichen Einfluss auf die Bewertungder Ergebnisse der zugrundeliegenden toxikolo-gischen Untersuchung.[1] C. Schmitz, M. Born, P. Dolezel, B. P. F. Rutten,
L. de Saint-Georges, P. R. Hof, H. Korr, Prenatalprotracted irradiation at very low dose rate inducessevere neuronal loss in rat hippocampus and cerebel-lum., Neuroscience 130 (4) (2005) 935–948.URL http://dx.doi.org/10.1016/j.neuroscience.2004.08.034
[2] J. Glaser, G. Green, S. Hendricks, Stereology for Bio-logical Research with a Focus on Neuroscience., MBFPress, Williston, 2007.
[3] C. Schmitz, P. R. Hof, Design-based stereology in neu-roscience., Neuroscience 130 (4) (2005) 813–831.URL http://dx.doi.org/10.1016/j.neuroscience.2004.08.050
[4] C. Schmitz, Variation of fractionator estimates andits prediction., Anat Embryol (Berl) 198 (5) (1998)371–397.
[5] E. M. Glaser, P. D. Wilson, The coefficient of errorof optical fractionator population size estimates: acomputer simulation comparing three estimators., JMicrosc 192 (Pt 2) (1998) 163–171.
12

Figure 1: Essentielle Schritte bei der design-based stereologischenAnalyse einer Gehirnregion (hier: die rechte Kleinhirnhälfte einerneun Monate alten Ratte). Nähere Erläuterungen zu den Details desVorgehens finden sich unten.
Erläuterungen zu Abbildung 1:Die Ratte wurde in tiefer Narkose durch transkardiale
Perfusion mit 4% Paraformaldehyd in phosphat-gepufferterKochsalzlösung (pH 7.4) getötet (alle Versuche waren von derTierversuchskommission der Universität Maastricht [Nieder-lande], wo die entsprechenden Tierversuche durchgeführt wur-den, genehmigt). Das Gehirn wurde aus der Schädelkalotte prä-pariert und in der gleichen Fixierlösung für zwei Wochen bei 4°C nachfixiert. A: SRS-Serie von 50 µm dicken Frontalschnit-ten (Gefrierschnitte) durch die rechte Kleinhirnhälfte (Nissl-Färbung). Die gesamte Kleinhirnhälfte ist erfasst; der Ab-stand der (Oberkanten der) Schnitte zueinander beträgt 250µm (d.h., jeder fünfte Schnitt wurde gefärbt). B: Schnitt Nr.7 der SRS-Schnittserie bei höherer VerGrößerung. Über den
Schnitt wurde ein rechtwinkliges Gitter mit gleichförmigem Ab-stand der Gitterlinien in den Richtungen X und Y projiziert.Die Position des Gitters relativ zum Schnitt wurde zufälliggewählt. Im Ergebnis wurde die Position der Kreuzungspunktedes Gitters relativ zu den Strukturen des Kleinhirns system-atisch (gleichförmiger Abstand der Gitterlinien) und zugle-ich zufällig (zufällige Position des Gitters auf dem Schnitt)gewählt. Aus (i) der Anzahl der Kreuzungspunkte des Git-ters, die auf der Granularzellschicht des Kleinhirns (schwarzin B und C) zu liegen kommen und (ii) des Abstands derGitterlinien in den Richtungen X und Y zueinander kann die„estimated“ Fläche der Granularzellschicht in Schnitt Nr. 7berechnet werden. Das Produkt der Summe aller „estimated“Flächen der Granularzellschicht aller Schnitte in der SRS-Schnittserie und dem gleichförmigen Abstand der (Oberkantender) Schnitte zueinander (250 µm) ergibt das „estimated“ Vol-umen der Granularzellschicht des untersuchten Kleinhirns. C:ebenfalls Schnitt Nr. 7 der SRS-Schnittserie bei höherer Ver-Größerung, jetzt aber mit einem anderen rechtwinkligen Git-ter mit gleichförmigem Abstand der Gitterlinien überlagert,das die Position einer SRS-Serie von Gesichtsfeldern vorgibt(graue Rechtecke). D: eines dieser Gesichtsfelder bei höhererVerGrößerung. Gezeigt ist die Molekularzellschicht (ML), diePurkinjezellschicht (PL) und die Granularzellschicht (GL) desKleinhirns. Fünf Anschnitte von Purkinjezellen wurden indiesem Gesichtsfeld gefunden (Pfeile). Um die Purkinjezellen„unbiased“ zu zählen (d.h., unabhängig von ihrer Größe, Form,Lage und räumlichen Anordnung), ist das Gesichtsfeld miteinem „unbiased counting frame“ überlagert, der die Grund-fläche eines „unbiased virtual counting spaces“ bildet (Detailsbei [3] sowie bei [2]). Der Balken entspricht 8 mm in A, 2,5 mmin B und C, sowie 20 µm in D. Bildquelle: [2] (mit freundlicherGenehmigung des Verlags)
Figure 2: Ein typischer stereologischer Arbeitsplatz (MBF Bios-cience, Williston, VT, USA). A: Forschungsmikroskop. B: Digital-kamera. C: Motorisierter Objektträgertisch. D: Kontrolleinheit. E:Dreiachsiger Joystick. F: Computer mit Stereologiesoftware. Die mo-torisierte Fokussiereinheit ist nicht gezeigt. Bildquelle: MBF Bios-cience.
13

Nur bewegte Augen ermöglichen eine klare Sicht
A. K. E. Horn-Bochtler, K. Lienbacher, C. Zeeh, A. Messoudi, C. Glombik∗
Anatomische Anstalt, Lehrstuhl I, Ludwig-Maximilians-Universität, Pettenkoferstr. 11, 80336 München
Abstract
Augenbewegungen sind essentiell, um bewegte Blickziele auf der Fovea centralis unserer Netzhaut zu fixieren, womit dieWahrnehmung eines scharfen Bildes gewährleistet ist. Die besonderen Erfordernisse an einerseits sehr schnelle Bewegun-gen der Augen und andererseits an Haltefunktion bei Fixation schlagen sich bereits in der sehr komplexen Anatomie deräußeren Augenmuskeln nieder. Fünf verschiedene Augenbewegungstypen werden über anatomisch voneinander getrennteBahnen generiert, deren selektive Schädigung zum Ausfall eines Augenbewegungstyps, z.B. vertikale Blickbewegungen,führen kann, während die anderen Augenbewegungen erhalten bleiben. Das Wissen um die neuronalen Netzwerke für dieverschiedenen Augenbewegungen ermöglicht eine einfache Diagnose des Arztes zur Lokalisation der Schädigung bei Pati-enten mit Augenbewegungsstörungen. Die Identifikation der für Augenbewegungen relevanten Hirnstammareale erfolgenmit Physiologen im Tierexperiment an Rhesusaffen, die neuronalen Verbindungen dieser Areale werden mit Trakt-Tracer-Methoden untersucht. In einem nächsten Schritt werden die funktionellen Hirnstammareale beim Affen histochemischcharaktierisiert, was mit den gleichen Markern die Identifizierung der homologen Zellgruppen beim Menschen erlaubt.Korrelativ klinisch-pathologisch-anatomische Untersuchungen an post-mortem Hirnstämmen von Patienten mit doku-mentierten Augenbewegungsstörungen bei degenerativen Erkrankungen ermöglichen neben dem Verständnis für dasStörungsbild der Erkrankung Erkenntnisse über die Progressionslinie der Degeneration.
1. Fünf Typen von Augenbewegungen sorgen fürein scharfes Bild
Der Sehsinn ist eines der wichtigsten Sinnesfunktionenunseres Körpers. Ein klares Bild erhalten wir jedoch nur,wenn visuelle Objekte auf der Fovea centralis, dem Ort derhöchsten Zapfendichte, auf unserer Netzhaut festgehaltenwerden. Hinzu kommt, daß die visuellen Objekte gleicher-maßen mit der Fovea centralis beider Augen erfasst wer-den müssen, um binokulares Sehen zu ermöglichen – dieGrundlage für räumliches Sehen. Augenbewegungen spie-len nun eine entscheidende Rolle für die Wahrnehmungscharfer Bilder der Umwelt, weil sie dafür sorgen, die Blick-ziele sowohl bei Eigenbewegungen des Körpers, wie auchBewegung der visuellen Umwelt, auf der Fovea centralisfest zu halten [1]. Zum Beispiel kompensieren kleine Au-genbewegungen fortwährend die Auf- und Abwärtsbewe-gungen unseres Kopfes, während wir gehen, womit auchim Laufen ein scharfes Bild der Umwelt wahrgenommenwird. Ohne diese kleinen kompensatorischen Augenbewe-gungen, die wir meist gar nicht realisieren, würden wir dieUmwelt verwackelt und verschwommen wie durch eine Vi-deokamera beim Laufen sehen.
Beim Säuger lassen sich fünf Typen von Augenbewegun-gen voneinander unterscheiden [2, 3] (Abb. 1):
1. Der entwicklungsgeschichtlich sehr alte vestibulo-okuläre Reflex wird über Reizung der Bogengänge
∗Korrespondenz an: [email protected]
ausgelöst. Wenn wir unseren Kopf z.B. nach rechtsdrehen, dann bewegen sich unsere Augen automatischin die Gegenrichtung, bis sie zum „Anschlag“ gelan-gen, und durch eine schnelle Rückstellbewegung wie-der zurück bewegt werden. Die periodische Abfolgedieser langsamen kompensatorischen Augenbewegun-gen und schnellen Rückstellsakkaden bezeichnet manals Nystagmus.
2. Der vestibulo-okuläre Reflex wird im täglichen Le-ben durch den optokinetischen Reflex ergänzt. In die-sem Fall werden kompensatorische Augenbewegungendurch große bewegte visuelle Reizmuster ausgelöst.Jeder von uns hat schon einmal im Zug periodischauftretende Augenbewegungen bei seinem Gegenüberbeobachtet (als es noch keine Großraumwagen gab),während dieser aus dem Fenster auf die vorbei ziehen-de Landschaft starrt [4].
3. Während Sie diese Zeilen lesen, „springen“ Ihre Au-gen von einem Wort zum nächsten und führen dabeisogenannte Sakkaden (Blicksprünge) aus. Wenn wirunsere visuelle Umgebung erkunden, so dienen Sakka-den dazu, unsere Augen immer von einem Fixierpunktmit einer schnellen Bewegung zum nächsten zu brin-gen. Während dieser Sakkade sorgt das Gehirn dafür,dass wir blind sind, um nicht durch das bewegte Ab-bild auf der Netzhaut wiederum eine Augenbewegung(optokinetischen Reflex) auszulösen. Willentlich aus-geführte Sakkaden erfordert das Vorhandensein einerFovea centralis, wie es bei uns der Fall ist, nicht aber
Internal Communication: Neues aus der Anstalt (NeadeA) 30. November 2010

2 AUGENMUSKELN ZEIGEN EINEN SEHR KOMPLEXEN AUFBAU
z.B. bei der Ratte oder dem Kaninchen, auf der dasBlickziel fixiert werden soll. Im Rahmen des vestibulo-okulären und optokinetischen Reflexes treten schnelleRückstellsakkaden bei allen Wirbeltieren auf [1].
4. Langsame Augenfolgebewegungen („smooth pursuit“)machen ebenfalls eine Fovea centralis erforderlich underlauben die Fixation von kleinen bewegten Blickzie-len, z.B. ein Vogel, der am Himmel fliegt.
5. Im Unterschied zu allen vorgenannten Augenbewe-gungstypen sind Vergenzbewegungen als einzige Formdiskonjugiert, d.h. die Augen bewegen sich aufeinan-der zu, während sie sonst immer parallel bewegt wer-den. Wir setzen sie immer ein, wenn wir etwas in derNähe fixieren, z.B. beim Lesen.
Die Augenbewegungen werden in anatomisch voneinandergetrennten Arealen und Bahnen generiert, die erst auf Ebe-ne der Motoneurone als gemeinsamer Endstrecke konver-gieren (Abb. 1).
2. Augenmuskeln zeigen einen sehr komplexenAufbau
Vier gerade und zwei schräge äußere Augenmuskeln sor-gen dafür, dass wir unsere Augen innerhalb der Augen-höhle bewegen können, dabei steht sicher die horizonta-le Bewegung im Vordergrund. Für konjugierte (parallele)Augenbewegungen müssen immer die jeweiligen Muskelnbeider Augen zusammenarbeiten, während gleichzeitig dieGegenmuskeln beider Augen gehemmt werden. KleinsteAbweichungen der Parallelität der Augenstellung und Au-genbewegungen beider Augen führen zur Wahrnehmungstörender Doppelbilder (s. Abb 2). Wir können das an ei-nem einfachen Eigenversuch nachvollziehen, indem wir beiAnsicht eines Objektes mit dem Finger seitlich auf einenAugapfel drücken und damit seine Sehachse verstellen.
Für diese Fähigkeit, einerseits das Auge sehr rasch zubewegen, andererseits aber eine Feinjustierung bei Fixier-bewegungen durchführen zu können und diese zu halten,weist der Augenmuskel einen sehr komplexen Aufbau auf,der sich von der quergestreiften Muskulatur unseres Ske-letts stark unterscheidet. Der äußere Augenmuskel weistzwei Schichten auf, eine äußere, der Augenhöhle zugewand-te, orbitale Schicht und eine innere, dem Augapfel anlie-gende, globale Schicht [5, 6]. Nur die Muskelfasern derglobalen Schicht inserieren über eine Sehne am Augap-fel, während die orbitale Schicht über „Pulleys“ mit demorbitalen Bindegewebe verbunden ist [7] (s. auch Abb. 3).
Außerdem enthalten die äußeren Augenmuskeln mindes-tens 6 verschiedene Muskelfasertypen, die sich an Hand ih-rer Innervierung in zwei Hauptkategorien einteilen lassen:einfach innervierte Muskelfasern mit „twitch-„ Eigenschaf-ten, die den Skelettmuskelfasern entsprechen, und multipelinnervierte Muskelfasern mit „non-twitch“-Eigenschaften,die beim Säuger sehr ungewöhnlich sind, und nur nochim Musculus vocalis des Kehlkopfes und Musculus tensor
Abbildung 2: A: Seitenansicht auf das rechte Auge nach Entfernungder seitlichen knöchernen Orbita und des Fettgewebes und Drüsen.B: Schematische Darstellung zur Lage der orbitalen und globalenSchicht der äußeren Augenmuskeln. IO: M obliquus inferior; IR: M.rectus inferior; LP: M. levator palpebrae; LR: M. rectus lateralis; SR:M. rectus superior (aus Sobotta, Atlas der Anatomie, 21. Auflage)
Abbildung 3: A, B: Benachbarte Querschnitte (Sterne zur Orientie-rung) eines äußeren Augenmuskels (M. obliquus superior) in einerVan Giesson-Färbung (B) und Immunfärbung auf eine Isoform vonMyosin-Schwerketten (slow), mit der nur die tonischen Muskelfasernangefärbt werden (dunkelbraun). Die gestrichelte Linie markiert dieGrenze zwischen orbitaler und globaler Schicht.
tympani des Mittelohrs vorkommen, und sonst eher als ty-pisch für Amphibien beschrieben werden (Übersicht: [6], s.auch Abb. 4).
Im Unterschied zum Skelettmuskel bestehen immernoch Unklarheiten bezüglich der Informationsverarbei-tung sensorischer Signale vom Augenmuskel. Klassischepropriozeptive Organe, wie Muskelspindeln oder Golgi-Sehnenorgane sind höchst variabel nur in einigen Tierspe-zies vorhanden, oft nur rudimentär ausgebildet, meist ganzfehlend [8, 9]. Unsere Arbeitsgruppe beschäftigt sich indiesem Zusammenhang mit einem besonderen Nervenen-digungskomplex, den sogenannten Palisadenendigungen,die bisher nur im Augenmuskel nachgewiesen wurden[8, 10, 11].
Abbildung 4: Detailansichten von Augenmuskellängsschnitten einesMenschen mit immuncytochemischer Darstellung der Innervationüber SNAP-25. A: große „en-plaque“-Endigung an „twitch-Faser“ B:multiple „en-grappe“-Endigungen an dünner „non-twitch“-Faser.
15

5 KLINISCHE ASPEKTE
Abbildung 5: Detailansichten vom Nucleus oculomotorius des Rhe-susaffen nach Doppelimmunfluoreszenz auf Cholinacetyltransfera-se zur Darstellung der Motoneurone (rot) und Chondroitinsulfat-Proteoglykan (grün) zur Darstellung der perineuronalen Netze. Be-achte, dass die peripheren Motoneurone der multipel innerviertenMuskelfasern (vergleiche mit B) keine perineuronalen Netze tragen.B,C: Selektive Markierung der peripheren Motoneurone im Nucleusoculomotorius nach Tracerinjektion in den Muskelsehnenübergangdes M. rectus medialis.
3. Motoneurone
Die äußeren Augenmuskeln werden von Motoneuro-nen innerviert, die in drei Hirnstammkernen liegen, demNucleus oculomotorius und Nucleus trochlearis im Mit-telhirn und dem Nucleus abducens im pontomedullärenÜbergang (Abb. 6A). Mit Hilfe von unterschiedlich plat-zierten Trakt-Tracern in die äußeren Augenmuskeln vonRhesusaffen und Ratten konnte unsere Arbeitsgruppe zei-gen, dass die Motoneurone der multipel-innervierten non-twitch Muskelfasern in eigenen Gruppen außerhalb derklassischen Augenmuskelkerne liegen, dass sie sich histo-chemisch von den Motoneuronen der twitch-Muskelfasernunterscheiden, und dass sie auch andere Eingänge erhalten([12, 13, 14, 15, 16], s. auch Abb. 5).
Die Motoneurone vertikal ziehender Augenmuskeln wer-den von anderen Transmittereingängen kontrolliert als dieder horizontal ziehenden Augenmuskeln [17]. Interessan-terweise bestehen auch Unterschiede in der Histochemieder Eingänge auf die Motoneurone der Augenheber ver-sus Augensenker [18], deren Quellen zum Teil identifi-ziert wurden [19]. Auf Grundlage dieser Befunde am Affenund entsprechenden Immunfärbungen an Humanschnittenkonnte die anatomische Karte des Nucleus oculomotoriusund Nucleus abducens beim Menschen aktualisiert wer-den [20, 21]. Dabei konnte z.B. auch gezeigt werden, dassder klassische Edinger-Westphal-Kern als assoziierter Kerndes Nucleus oculomotorius, beim Menschen nicht Sitz derpräganglionären Neurone für den Pupillenreflex ist [22].
4. Die Auslösung verschiedenener Augenbewe-gungstypen erfolgt über anatomisch voneinan-der getrennte prämotorische Bahnen
Wie eingangs beschrieben werden verschiedene Augen-bewegungen in anatomisch voneinander getrennten Area-len und Bahnen generiert (Abb. 1). Das hat zur Folge,
dass bei Schädigung einer dieser prämotorischen Arealeoder Bahnen ein bestimmter Augenbewegungstyp ausfällt,sich in einer Parese äußert, aber der gleiche, scheinbar ge-lähmte Muskel über ein anderes Bewegungsprogramm an-gesteuert werden kann. Die genaue Kenntnis der prämo-torischen Bahnen des Augenbewegungssystems ist daherfür den Kliniker ein wertvolles Hilfsmittel, um ohne grö-ßeren Geräteeinsatz eine rasche Diagnose über die Ursa-che, ob z.B. zentrale oder periphere Läsion, von Augenbe-wegungsstörungen zu stellen. Als Beispiel sei ein Patientgenannt, der darüber klagt, dass er beim Treppensteigenzunehmend Schwierigkeiten hat, da er seinen Blick nichtmehr nach unten richten kann, d.h. eine vertikale Blick-parese nach unten hat. Auf die Bitte des Arztes, seinenKopf doch einmal nach hinten zu legen, führt der Pati-ent korrekterweise vertikale Augenbewegungen nach untenaus. Das zeigt, dass weder die Augenmuskeln noch derenMotoneurone geschädigt sind, sondern die den Motoneuro-nen vorgeschalteten Verbindungen für vertikale Sakkaden.Bei Nutzung anderer prämotorischer Strukturen und Bah-nen, hier die vestibulo-okulären Bahnen zur Auslösung desvestibulo-okulären Reflexes, können die Motoneurone undderen Muskeln korrekt aktiviert werden. Dem Arzt signa-lisiert das in diesem Fall eine zentrale Schädigung, vermut-lich im Bereich des Mittelhirns, dem er jetzt mit weiterenTests oder bildgebenden Verfahren auf die Spur kommenkann.
5. Klinische Aspekte
In Fortsetzung der Pionierarbeiten von Jean Büttner-Ennever, die bis 2008 an der Anatomischen Anstalt tätigwar, und in langjähriger Kollaboration mit Physiologenam Affen viele für Augenbewegungen wichtige funktionel-le Hirnstammareale anatomisch identifiziert hat, wie z.B.das Hirnstammzentrum zur Auslösung von Horizontalsak-kaden (PPRF) [23] und Vertikalsakkaden (RIMLF) [24],ist ein Schwerpunkt unserer Forschung die histochemi-sche Charakterisierung der funktionellen Neuronengrup-pen beim Affen. Mit Hilfe der gefundenen Marker kön-nen dann auch die funktionellen Neuronengruppen desAugen- und Lidbewegungssystems in Hirnstammschnit-ten beim Menschen auf zellulärer Ebene identifiziert wer-den [25, 26, 27, 28]. Das wiederum bietet die Möglich-keit, post-mortem Untersuchungen an Gehirnen von Pa-tienten, die unter bestimmten Augenbewegungsstörungenlitten, gezielt auf degenerative Veränderungen an identifi-zierten funktionellen Neuronen durchzuführen ([29, 30, 31],s. auch Abb. 6).
Im Rahmen des am Klinikum Großhadern neu eingerich-teten vom BMFB geförderten „Integrierten Forschungs-und Behandlungszentrums für Schwindel und Okulomoto-rikstörungen“ werden zur Zeit in einem ersten Projekt inZusammenarbeit mit der Neurologischen Klinik und demInstitut für Neuropathologie post-mortem Untersuchungenvon Fällen mit progressiver supranukleärer Parese (PSP)
16
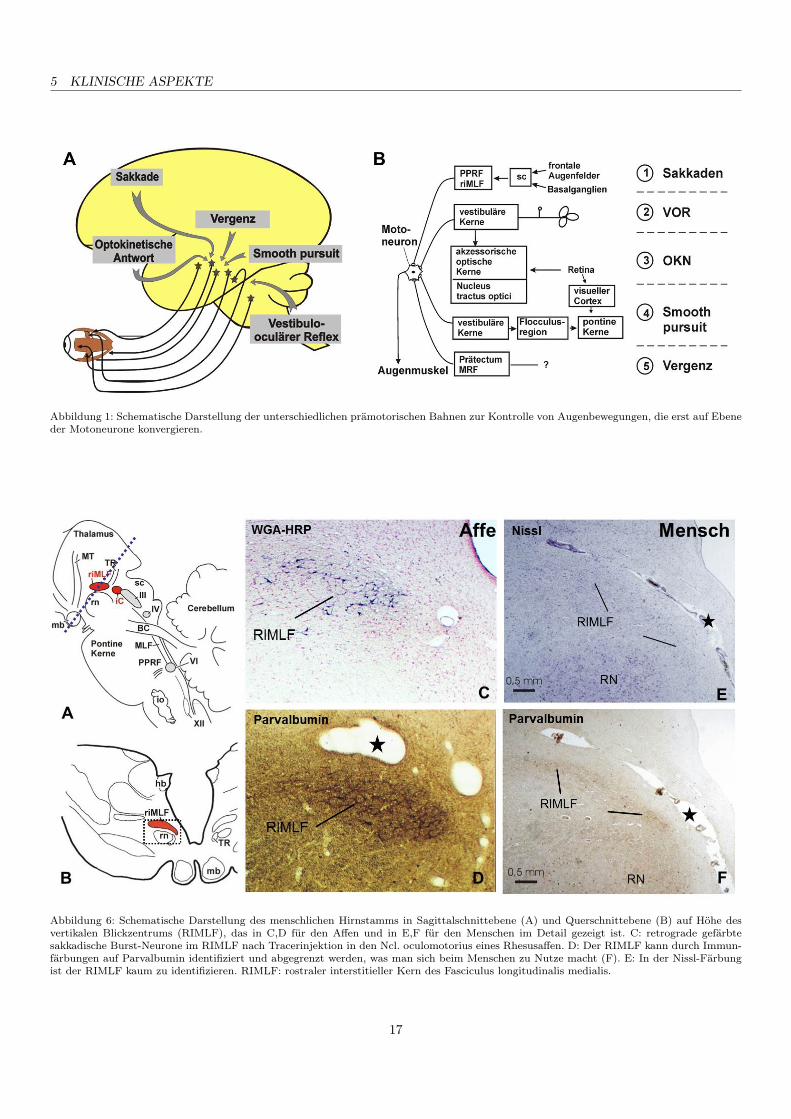
5 KLINISCHE ASPEKTE
Abbildung 1: Schematische Darstellung der unterschiedlichen prämotorischen Bahnen zur Kontrolle von Augenbewegungen, die erst auf Ebeneder Motoneurone konvergieren.
Abbildung 6: Schematische Darstellung des menschlichen Hirnstamms in Sagittalschnittebene (A) und Querschnittebene (B) auf Höhe desvertikalen Blickzentrums (RIMLF), das in C,D für den Affen und in E,F für den Menschen im Detail gezeigt ist. C: retrograde gefärbtesakkadische Burst-Neurone im RIMLF nach Tracerinjektion in den Ncl. oculomotorius eines Rhesusaffen. D: Der RIMLF kann durch Immun-färbungen auf Parvalbumin identifiziert und abgegrenzt werden, was man sich beim Menschen zu Nutze macht (F). E: In der Nissl-Färbungist der RIMLF kaum zu identifizieren. RIMLF: rostraler interstitieller Kern des Fasciculus longitudinalis medialis.
17

5 KLINISCHE ASPEKTE
untersucht, deren Leitsymptom eine früh auftretende ver-tikale Blicklähmung war. Die pathologisch-anatomischeAnalyse von Fällen in unterschiedlichen Krankheitsstadi-en kann Aufschluss über die Wege des Voranschreitens derErkankung liefern. Langfristig sollen die Untersuchungendann auch auf andere degenerative Erkrankungen erwei-tert werden, wie amyotrophe Lateralsklerose (ALS), einerfortschreitenden Degeneration motorischer Bahnen, wobeidie Augenbewegungen am längsten erhalten bleiben, odercortico-basaler Degeneration.
[1] R. J. Leigh, D. Zee, The Neurology of Eye-Movements, OxfordUniversity Press, New York, 2006.
[2] J. Büttner-Ennever, A. Horn, Reticular formation: eye move-ments, gaze and blinks, The Human Nervous System, ElsevierAcademic Press, Amsterdam, 2004, Ch. 16, pp. 479–510.
[3] U. Büttner, J. A. Büttner-Ennever, Present concepts of oculo-motor organization., Prog Brain Res 151 (2006) 1–42.URL http://dx.doi.org/10.1016/S0079-6123(05)51001-X
[4] J. A. Büttner-Ennever, Patterns of connectivity in the vestibu-lar nuclei., Ann N Y Acad Sci 656 (1992) 363–378.
[5] S. Y. Oh, V. Poukens, J. L. Demer, Quantitative analysis ofrectus extraocular muscle layers in monkey and humans., InvestOphthalmol Vis Sci 42 (1) (2001) 10–16.
[6] R. F. Spencer, J. D. Porter, Biological organization of the ex-traocular muscles., Prog Brain Res 151 (2006) 43–80.URL http://dx.doi.org/10.1016/S0079-6123(05)51002-1
[7] J. L. Demer, The orbital pulley system: a revolution in conceptsof orbital anatomy., Ann N Y Acad Sci 956 (2002) 17–32.
[8] I. M. Donaldson, The functions of the proprioceptors of the eyemuscles., Philos Trans R Soc Lond B Biol Sci 355 (1404) (2000)1685–1754.URL http://dx.doi.org/10.1098/rstb.2000.0732
[9] J. A. Büttner-Ennever, K. Z. Konakci, R. Blumer, Sensory con-trol of extraocular muscles., Prog Brain Res 151 (2006) 81–93.URL http://dx.doi.org/10.1016/S0079-6123(05)51003-3
[10] A. C. Eberhorn, A. K. E. Horn, N. Eberhorn, P. Fischer, K.-P.Boergen, J. A. Büttner-Ennever, Palisade endings in extraocu-lar eye muscles revealed by snap-25 immunoreactivity., J Anat206 (3) (2005) 307–315.URL http://dx.doi.org/10.1111/j.1469-7580.2005.00378.x
[11] K. Lienbacher, M. Mustari, H. Ying, J. Büttner-Ennever,A. Horn, Do the palisade endings in extraocular muscles arisefrom neurons in the motonuclei?, Invest Ophthalmol Vis Sci(InPress).
[12] G. Ugolini, F. Klam, M. D. Dans, D. Dubayle, A.-M. Brandi,J. Büttner-Ennever, W. Graf, Horizontal eye movement net-works in primates as revealed by retrograde transneuronal trans-fer of rabies virus: differences in monosynaptic input to "slow"and "fast" abducens motoneurons., J Comp Neurol 498 (6)(2006) 762–785.URL http://dx.doi.org/10.1002/cne.21092
[13] J. A. Büttner-Ennever, A. K. E. Horn, The neuroanatomicalbasis of oculomotor disorders: the dual motor control of ex-traocular muscles and its possible role in proprioception., CurrOpin Neurol 15 (1) (2002) 35–43.
[14] J. A. Büttner-Ennever, A. K. Horn, H. Scherberger,P. D’Ascanio, Motoneurons of twitch and nontwitch extraoc-ular muscle fibers in the abducens, trochlear, and oculomotornuclei of monkeys., J Comp Neurol 438 (3) (2001) 318–335.
[15] A. C. Eberhorn, P. Ardeleanu, J. A. Büttner-Ennever, A. K. E.Horn, Histochemical differences between motoneurons supply-ing multiply and singly innervated extraocular muscle fibers., JComp Neurol 491 (4) (2005) 352–366.URL http://dx.doi.org/10.1002/cne.20715
[16] A. C. Eberhorn, J. A. Büttner-Ennever, A. K. E. Horn, Identifi-cation of motoneurons supplying multiply- or singly-innervatedextraocular muscle fibers in the rat., Neuroscience 137 (3)(2006) 891–903.
URL http://dx.doi.org/10.1016/j.neuroscience.2005.10.038
[17] R. F. Spencer, S. F. Wang, R. Baker, The pathways and func-tions of gaba in the oculomotor system., Prog Brain Res 90(1992) 307–331.
[18] C. Schulze, M. Mustari, G. Holstein, A. Horn, Transmitterinputs to different motoneuron subgroups in the oculomotorand trochlear nucleus in monkey, Soc Neurosci Abstr 35 (2009)356.9.
[19] J. Ahlfeld, M. Mustari, G. Holstein, A. Horn, Sources of calre-tinin input to the upgaze motoneurons of the oculomotor nu-cleus in monkey, Soc Neurosci Abstr 36 (2010) 894.2.
[20] A. Horn, E. C. Ngwa, J. Büttner-Ennever, Histochemistry andlocation of motoneurons usbgroups in the oculomotor nucleusof human, Soc Neursci Abst 35 (2009) 356.10.
[21] A. Eberhorn, A. Horng, W. Härtig, J. Büttner-Ennever,A. Horn, Distribution of functional cell groups in the abducensnucleus of monkey and man, Soc Neurosci Abstr 30 (2004) 881.6.
[22] A. K. Horn, A. Eberhorn, W. Härtig, P. Ardeleanu, A. Mes-soudi, J. A. Büttner-Ennever, Perioculomotor cell groups inmonkey and man defined by their histochemical and func-tional properties: reappraisal of the edinger-westphal nucleus.,J Comp Neurol 507 (3) (2008) 1317–1335.URL http://dx.doi.org/10.1002/cne.21598
[23] J. A. Büttner-Ennever, V. Henn, An autoradiographic study ofthe pathways from the pontine reticular formation involved inhorizontal eye movements., Brain Res 108 (1) (1976) 155–164.
[24] J. A. Büttner-Ennever, U. Büttner, A cell group associated withvertical eye movements in the rostral mesencephalic reticularformation of the monkey., Brain Res 151 (1) (1978) 31–47.
[25] A. Horn, J. Büttner-Ennever, U. Büttner, Saccadic premotorneurons in the brainstem: functional neuroanatomy and clinicalimplications, Neuro-Ophthalm 16 (1996) 229–240.
[26] A. K. Horn, J. A. Büttner-Ennever, Y. Suzuki, V. Henn, Histo-logical identification of premotor neurons for horizontal saccadesin monkey and man by parvalbumin immunostaining., J CompNeurol 359 (2) (1995) 350–363.URL http://dx.doi.org/10.1002/cne.903590212
[27] A. K. Horn, J. A. Büttner-Ennever, M. Gayde, A. Messoudi,Neuroanatomical identification of mesencephalic premotor neu-rons coordinating eyelid with upgaze in the monkey and man.,J Comp Neurol 420 (1) (2000) 19–34.
[28] A. K. E. Horn, The reticular formation., Prog Brain Res 151(2006) 127–155.URL http://dx.doi.org/10.1016/S0079-6123(05)51005-7
[29] L. Averbuch-Heller, C. Helmchen, A. K. Horn, R. J. Leigh, J. A.Büttner-Ennerver, Slow vertical saccades in motor neuron dis-ease: correlation of structure and function., Ann Neurol 44 (4)(1998) 641–648.URL http://dx.doi.org/10.1002/ana.410440410
[30] J. A. Büttner-Ennever, C. Jenkins, H. Armin-Parsa, A. K. Horn,J. S. Elston, A neuroanatomical analysis of lid-eye coordinationin cases of ptosis and downgaze paralysis., Clin Neuropathol15 (6) (1996) 313–318.
[31] S. Geiner, A. K. E. Horn, N. H. Wadia, H. Sakai, J. A.Büttner-Ennever, The neuroanatomical basis of slow saccadesin spinocerebellar ataxia type 2 (wadia-subtype)., Prog BrainRes 171 (2008) 575–581.URL http://dx.doi.org/10.1016/S0079-6123(08)00683-3
18

Stammzell Tissue Engineering und Elektronenmikroskopie:Die Rolle der Phytopharmaka auf die Differenzierung mesenchymaler Stammzellen zu
Chondrozyten in einem Osteoarthritis Modell in vitro
C. Buhrmann und M. Shakibaei1,∗
Institut für Anatomie, Lehrstuhl I, Ludwig-Maximilians Universität München,Arbeitsgruppe: Muskuloskeletales System, Pettenkoferstrasse 11, 80336 München, Germany
Abstract
Verletzungen am Gelenkknorpel führen zu degenerativen Gelenkveränderungen (Osteoarthrose/Arthritis) mit darausresultierenden Leistungsminderungen und stellen die Orthopädie vor eine große Herausforderung [1]. Der Hauptgrundhierfür ist die geringe Regenerationsfähigkeit des Knorpels, da das Knorpelgewebe avaskulär, alymphatisch und aneu-ral ist. Während der Embryonalentwicklung entsteht Knorpel durch Kondensation und Differenzierung mesenchymalerStammzellen (MSC) zu Chondrozyten. Neue Studien haben ergeben, dass auch im adulten Gelenkknorpel MSC vorkom-men und ihre Anzahl in arthrotischem Knorpel sogar noch erhöht ist [2]. Dass es trotzdem im arthrotischen Knorpel nichtzu einer Knorpeldefektheilung kommt, liegt möglicherweise an dem hohen Gehalt kataboler und pro-inflammatorischerCytokine (IL-1β und TNF-α), die zu einer fehlerhaften chondrogenen Differenzierung der MSC und damit zu Verkalkungenund Faserknorpelbildung führen. In der Arbeitsgruppe wird seit Jahren die Hypothese geprüft, dass der Transkriptions-faktor NF-κB als zentrales downstream-Target für pro-inflammatorische Zytokine therapeutisch für die Arthrosetherapienutzbar ist. Experimente mit Phytopharmaka (z.B. Curcumin/Resveratrol), untoxischen Wirkstoffen (Polyphenolen)und effektiven Hemmstoffen von NF-κB, haben gezeigt, dass diese die Zytokin-induzierten katabolen Effekte in Chon-drozyten abschwächen können und hierbei NF-κB als Haupttarget für die Phytopharmaka fungiert. Die Wirkung vonCurcumin/Resveratrol auf Chondrozyten wurde bisher insbesondere im Hinblick auf einen möglichen Einsatz in derArthrosetherapie noch nicht genauer untersucht. Die hierbei gewonnenen Erkenntnisse über die Feinmodulation derChondrogenese, können neue Impulse für die prophylaktische Therapie der Osteoarthritis und zur in vitro Herstellungvon belastbarem funktionellem Knorpelgewebe zur Transplantation geben.
1. Osteoarthritis:
Osteoarthritis ist eine fortschreitende degenerativeGelenkerkrankung, die zu volkswirtschaftlichen Proble-men weltweit führt. Verletzungen am Gelenkknor-pel führen zu degenerativen Gelenkveränderungen (Os-teoarthrose/Arthritis) mit daraus resultierenden Leis-tungsminderungen [1]. In der Pathogenese der Os-teoarthrose/Arthritis spielt besonders die Produktion undFreisetzung kataboler Zytokine, wie Interleukin-1β (IL-1β)und Tumor Necrosis Factor-alpha (TNF-α), eine zentraleRolle [3, 4]. IL-1β und TNF-α begünstigen unter an-derem durch Aktivierung des ubiquitären Transkriptions-faktors Nuclear Faktor-kappaB (NF-κB) die ProduktionKnorpelmatrix degradierender Enzyme, wie der Matrix-Metalloproteinasen (MMPs) und die Aktivierung weitererpro-inflammatorischer Enzyme, wie der Cyclooxygenase-2 (Cox-2), die zu weiterem Abbau der Knorpelmatrix
∗E-Mail: [email protected]: Der besondere Dank der Autoren gilt der technis-
chen Unterstützung von Frau K. Sperling.
und Schmerzen bei den Patienten führen [5]. Desweit-eren führen die katabolen Zytokine zu einer Aktivierungder Apoptosemechansimen über eine Aktivierung des pro-apoptotischen Enzyms Caspase-3 und Spaltung des DNAReparaturenzyms Poly(ADP-ribose)polymerase (PARP)[6, 7].
2. Mesenchymale Stammzellen (MSC)
Mesenchymale Stammzellen (MSC) sind undifferen-zierte Zellen, die man im Embryo, Fetus und dem adul-ten Individuum findet. Im Gegensatz zu embryonalenStammzellen weisen Stammzellen, die aus dem adultenIndividuum gewonnen werden, keine Omnipotenz mehrauf, sondern sind pluripotent und damit eingeschränkt dif-ferenzierbar. Stammzellen wurden mittlerweile aus vie-len Geweben unter anderem aus dem Knochenmark, demBlut, der Leber, dem Fett, dem Pankreas und dem ZNSisoliert [9, 10]. Aus dem Knochenmark lassen sich Stamm-zellen isolieren, welche sich in Gewebezellen differenzierenlassen [11, 12]. Diese mesenchymalen Stammzellen machenca. 1-2% der mononukleären Zellen des Knochenmarks
Internal Communication: Neues aus der Anstalt (NeadeA) November 30, 2010

3 PHYTOPHARMAKA
Figure 1: Curcumin alleine induziert keine Chondrogenese in MSCs 14 Tage high-density Kultur. Apoptose und/oder Nekrose wurde inunbehandelten MSCs Kulturen (a), in 4 Stunden mit Curcumin vorbehandelten Kulturen (c) oder in 4 Stunden mit Curcumin vorbehandeltenKulturen, die dann 14 Tage weiter mit Curcumin behandelt wurden (e) beobachtet. Im Gegensatz dazu, führte ein gleichzeitige chondrogeneStimulation mit Wachstumsfaktoren sowohl in MSC Kulturen (b), in 4 Stunden mit Curcumin vorbehandelten Kulturen (d) oder in 4 Stundenmit Curcumin vorbehandelten Kulturen die dann 14 Tage weiter mit Curcumin behandelt wurden (f) zur Chondrogenese. Vergrösserung:6000x; Balken = 1 µm; C=Chondrozyten; F=Fibroblastenähnliche Zellen; M=EZM [8]
aus und können über mehrere Passagen expandiert wer-den, ohne dass sie ihr proliferatives Potential verlieren[13, 14]. Ihre Morphologie ist sehr heterogen und geht vonlanggestreckten, fibroblasten-ähnlichen Zellen über flachausgebreitete Zellen hin zu kurzen polygonalen Zellen.Ihre Population ist bezüglich ihrer Oberfächenmarkersehr inhomogen, es lassen sich eine Reihe von Markern,die auf mesenchymalem, endothelialem und epithelialemGewebe gefunden werden, nachweisen [15]. Mesenchy-male Stammzellen sollten aber immer CD105+, CD73+und CD90+ positiv und CD45-, CD34- und für anderehemato-poetische Stammzellmarker negativ sein [14, 16].Ihr wichtigstes Definitionsmerkmal ist die Fähigkeit sichin vitro in verschiedene mesenchymale Gewebe zu differen-zieren [15, 17, 18]. Zum Nachweis des osteogenen Poten-tials der gewonnenen MSC, werden Osteocalcin, KollagenTyp I und Calcium Ablagerungen mittels von Kossa Fär-bung, Transmissions- und Immuno-elektronenmikroskopiein der neugebildeten Matrix nachgewiesen [13]. Dasadipogene Potential der MSC wird durch eine fürFettzellen charakteristische Menge an eingelagerten Fett-tröpfchen mittels Oil Red O Färbung und Transmissions-elektronenmikroskopie in den Zellen nachgewiesen. Das
chondrogene Potential der MSC zeigt sich durch En-twicklung typischer Knorpelzellknötchen mit großen, run-den typischen Knorpelzellen und spezifischen Knorpelma-trixablagerungen (Knorpel spezifische Proteoglykane undKollagen Typ II). Diese werden mittels Alcian Blau Fär-bung, Transmissions- und Immuno-elektronenmikroskopienachgewiesen [13]. Desweiteren werden die isolierten MSCsehr erfolgreich in dem dreidimensionalen Organoidkul-tursystem kultiviert. Der besondere Vorteil von MSCfür die Regenerative Medizin und das Tissue Engineer-ing liegt somit in ihrer relativ einfachen Gewinnung ausdem Knochenmark oder Fettgewebe, ihrem hohen prolif-erativen Potential und ihrer mesenchymalen Pluripotenz[14].
3. Phytopharmaka
Potente Wirkstoffe, die für die prophylaktische Therapieder Osteoarthritis und Arthrose in Betracht kommen, sindPhytopharmaka wie z. B. Curcumin und Resveratrol .Diese Phytopharmaka haben eine lange Geschichte alsArzneimittel, so wird Curcumin in der indischen Ayurveda
20

4 DIE BEDEUTUNG DER PHYTOPHARMAKA FÜR DIE PROPHYLAXE UND BEHANDLUNG DEROSTEOARTHRITIS
Figure 2: Curcumin hemmt die IL-1b Aktivität, und ermöglicht dadurch Wachstumsfaktor induzierte Chondrogenese in den MSCs 14 Tagehigh-density Kultur. In unbehandelten MSC-Kulturen (a), mit IL-1b und dem chondrogenem Medium behandelten Kulturen (b) oder in 4Stunden mit Curcumin vorstimulierten und dann mit IL-1b stimulierten Kulturen (c) kam es zur Apoptose. Im Gegensatz dazu kam es inden Kulturen die 4 Stunden mit Curcumin vorstimuliert wurden und dann entweder sowohl mit IL-1b und chondrogenem Medium behandeltwurden (d) oder sowohl mit IL-1b, chondrogenem Medium und Curcumin behnadelt wurden (e) zu Chondrogenese. Vergrösserung: 6000x;Balken = 1 µm; C=Chondrozyten; F=Fibroblastenähnliche Zellen; M=EZM[8].
Medizin seit langem bei Osteoarthrose/Arthritis einge-setzt und erste wissenschaftlich fundierte Hinweise aufeine Wirkung bei Osteoarthrose/Arthritis wurden 1980berichtet [19, 20]. Die Behandlung des osteoarthritischenKnorpels mit Wirkstoffen, welche die Wirkung katabolerZytokine hemmen und gleichzeitig die adäquate chondro-gene Differenzierung der MSC begünstigen, könnte neue,erfolgreiche Therapiemöglichkeiten in der Behandlung derOsteoarthrose/Arthritis schaffen. Für Resveratrol wurdebereits im Tiermodell eine positive Wirkung auf die Os-teoarthrose/Arthritis beschrieben [21, 22, 23, 24].
3.1. Curcumin (Diferuloylmethane)
Curcumin, ein Inhaltsstoff der Pflanze Curcuma longa,führt zu einer Hemmung der Aktivierung von NF-κB.Es verhindert wahrscheinlich die Phosphorylierung unddamit den Abbau von IκBα durch Hemmung der IκB-Kinase [25, 26]. Curcumin zeigt durch seine Wirkung aufNF-κB anti-inflammatorische Eigenschaften und hemmtdie Expression von Enzymen wie Lipoxygenase, COX-2,iNOS, uPA sowie von MMPs [27, 28, 29]. In verschiede-nen Zelltypen wurden auch andere durch Curcumin her-vorgerufene Effekte beobachtet wie z.B. Hemmung an-derer Transkriptions-faktoren, wie AP-1 und Egr-1, Hem-mung der JNK (c-Jun N-terminal kinase), Protein Tyrosinsowie Serine/Threonin Kinasen und Förderung der Zelld-ifferenzierung [30, 31, 32]. Außerdem zeigt Curcumin anti-oxidative Eigenschaften [33]. Zur Zeit findet Curcumin inder Tumortherapie Einsatz. In Versuchen am Menschenzeigte es keine die Dosis limitierende Toxizität. Somitstellt sich die Frage, ob dieser weitgehend untoxische, imCurry-Gewürz enthaltene Hemmstoff für den Einsatz inder Arthrosetherapie geeignet sein könnte.
3.2. Resveratrol (trans-3,5, 4´-trihydroxystilbene)
Resveratrol ist eine Polyphenol-Schutzverbindung, diedurch rote Trauben (und einige andere Pflanzen) pro-duziert wird, um sich gegen Parasiten zu schützen [34, 35].Es ist wissenschaftlich wegen seiner vielfältigen positiven
pharmakologischen Wirkungen und nicht zuletzt wegendes „französischen Paradoxes“ bekannt: Trotz hohem Kon-sum von gesättigten Fettsäuren und Nikotin weisen Fran-zosen durch den regelmäßigen Genuss von Rotwein, derResveratrol und weitere interessante Polyphenole enthält,eine wesentlich geringere Rate (<40%) an Herz-Kreislauf-Erkrankungen auf. Im Auftrag der WHO ergaben sich ineiner Zehnjahresstudie (MONICA-Studie), durchgeführtin 41 Städten auf der ganzen Welt, extreme Abweichungenin den Ländern mit geringem oder sehr hohem Weinkon-sum. So war in den USA sowie Kanada und England dasInfarktrisiko im Schnitt 3mal (Männer) bis sogar 12mal(Frauen) höher als im Weinland Frankreich. Damit istResveratrol wirksamer als Vitamin E. Resveratrol wirktgegen mehrere Arten von freien Radikalen, um die Ox-idation von Lipoproteinen mit geringer Dichte (LDL) zuverhindern, hemmt die Aggregation von Blutplättchen undblockiert die Wirkung des Thrombins und anderen Gerin-nungsfaktoren. Desweiteren begünstigt es die Produktionvon Stickoxid (NO), das die Arterien entspannt und er-weitert.
4. Die Bedeutung der Phytopharmaka für die Pro-phylaxe und Behandlung der Osteoarthritis
In unserer Arbeitsgruppe beschäftigen wir uns seitlangem auch mit Phytopharmaka in unserem Arthritis-Modell. In verschiedenen Studien mit Chondrozytenkonnten wir in unserem dreidimensionalen Kultur-Modell(High Density Kultur) bereits den anti-inflammatorischenEffekt verschiedener Phytopharmaka, wie Curcuminund Resveratrol, und ihre Wirkungsmechanismen aufmit IL-1β- stimulierte Chondrozyten nachweisen [6,22, 36, 37]. Insbesondere konnten wir nachweisen,dass sowohl Curcumin als auch Resveratrol die Ak-tivierung des ubiquitären Transkriptionsfaktors NF-κBund somit die Produktion und Aktivierung der Matrix-metalloproteinasen (MMPs) und die Aktivierung weitererpro-inflammatorischer Enzyme, wie der Cyclooxygenase-2 (Cox-2) in IL-1β stimulierten Chondrozyten hemmen
21
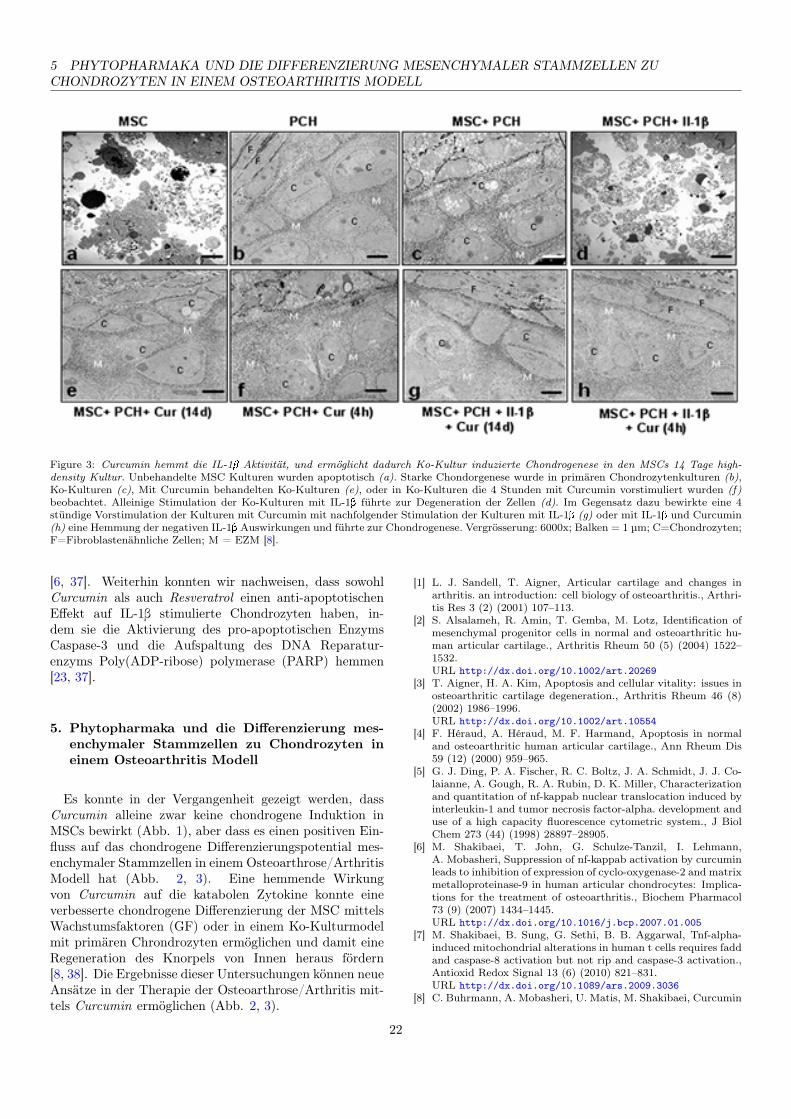
5 PHYTOPHARMAKA UND DIE DIFFERENZIERUNG MESENCHYMALER STAMMZELLEN ZUCHONDROZYTEN IN EINEM OSTEOARTHRITIS MODELL
Figure 3: Curcumin hemmt die IL-1b Aktivität, und ermöglicht dadurch Ko-Kultur induzierte Chondrogenese in den MSCs 14 Tage high-density Kultur. Unbehandelte MSC Kulturen wurden apoptotisch (a). Starke Chondorgenese wurde in primären Chondrozytenkulturen (b),Ko-Kulturen (c), Mit Curcumin behandelten Ko-Kulturen (e), oder in Ko-Kulturen die 4 Stunden mit Curcumin vorstimuliert wurden (f)beobachtet. Alleinige Stimulation der Ko-Kulturen mit IL-1b führte zur Degeneration der Zellen (d). Im Gegensatz dazu bewirkte eine 4stündige Vorstimulation der Kulturen mit Curcumin mit nachfolgender Stimulation der Kulturen mit IL-1b (g) oder mit IL-1b und Curcumin(h) eine Hemmung der negativen IL-1b Auswirkungen und führte zur Chondrogenese. Vergrösserung: 6000x; Balken = 1 µm; C=Chondrozyten;F=Fibroblastenähnliche Zellen; M = EZM [8].
[6, 37]. Weiterhin konnten wir nachweisen, dass sowohlCurcumin als auch Resveratrol einen anti-apoptotischenEffekt auf IL-1β stimulierte Chondrozyten haben, in-dem sie die Aktivierung des pro-apoptotischen EnzymsCaspase-3 und die Aufspaltung des DNA Reparatur-enzyms Poly(ADP-ribose) polymerase (PARP) hemmen[23, 37].
5. Phytopharmaka und die Differenzierung mes-enchymaler Stammzellen zu Chondrozyten ineinem Osteoarthritis Modell
Es konnte in der Vergangenheit gezeigt werden, dassCurcumin alleine zwar keine chondrogene Induktion inMSCs bewirkt (Abb. 1), aber dass es einen positiven Ein-fluss auf das chondrogene Differenzierungspotential mes-enchymaler Stammzellen in einem Osteoarthrose/ArthritisModell hat (Abb. 2, 3). Eine hemmende Wirkungvon Curcumin auf die katabolen Zytokine konnte eineverbesserte chondrogene Differenzierung der MSC mittelsWachstumsfaktoren (GF) oder in einem Ko-Kulturmodelmit primären Chrondrozyten ermöglichen und damit eineRegeneration des Knorpels von Innen heraus fördern[8, 38]. Die Ergebnisse dieser Untersuchungen können neueAnsätze in der Therapie der Osteoarthrose/Arthritis mit-tels Curcumin ermöglichen (Abb. 2, 3).
[1] L. J. Sandell, T. Aigner, Articular cartilage and changes inarthritis. an introduction: cell biology of osteoarthritis., Arthri-tis Res 3 (2) (2001) 107–113.
[2] S. Alsalameh, R. Amin, T. Gemba, M. Lotz, Identification ofmesenchymal progenitor cells in normal and osteoarthritic hu-man articular cartilage., Arthritis Rheum 50 (5) (2004) 1522–1532.URL http://dx.doi.org/10.1002/art.20269
[3] T. Aigner, H. A. Kim, Apoptosis and cellular vitality: issues inosteoarthritic cartilage degeneration., Arthritis Rheum 46 (8)(2002) 1986–1996.URL http://dx.doi.org/10.1002/art.10554
[4] F. Héraud, A. Héraud, M. F. Harmand, Apoptosis in normaland osteoarthritic human articular cartilage., Ann Rheum Dis59 (12) (2000) 959–965.
[5] G. J. Ding, P. A. Fischer, R. C. Boltz, J. A. Schmidt, J. J. Co-laianne, A. Gough, R. A. Rubin, D. K. Miller, Characterizationand quantitation of nf-kappab nuclear translocation induced byinterleukin-1 and tumor necrosis factor-alpha. development anduse of a high capacity fluorescence cytometric system., J BiolChem 273 (44) (1998) 28897–28905.
[6] M. Shakibaei, T. John, G. Schulze-Tanzil, I. Lehmann,A. Mobasheri, Suppression of nf-kappab activation by curcuminleads to inhibition of expression of cyclo-oxygenase-2 and matrixmetalloproteinase-9 in human articular chondrocytes: Implica-tions for the treatment of osteoarthritis., Biochem Pharmacol73 (9) (2007) 1434–1445.URL http://dx.doi.org/10.1016/j.bcp.2007.01.005
[7] M. Shakibaei, B. Sung, G. Sethi, B. B. Aggarwal, Tnf-alpha-induced mitochondrial alterations in human t cells requires faddand caspase-8 activation but not rip and caspase-3 activation.,Antioxid Redox Signal 13 (6) (2010) 821–831.URL http://dx.doi.org/10.1089/ars.2009.3036
[8] C. Buhrmann, A. Mobasheri, U. Matis, M. Shakibaei, Curcumin
22

5 PHYTOPHARMAKA UND DIE DIFFERENZIERUNG MESENCHYMALER STAMMZELLEN ZUCHONDROZYTEN IN EINEM OSTEOARTHRITIS MODELL
mediated suppression of nuclear factor-?b promotes chondro-genic differentiation of mesenchymal stem cells in a high-densityco-culture microenvironment., Arthritis Res Ther 12 (4) (2010)R127.URL http://dx.doi.org/10.1186/ar3065
[9] R. Cancedda, B. Dozin, P. Giannoni, R. Quarto, Tissue en-gineering and cell therapy of cartilage and bone., Matrix Biol22 (1) (2003) 81–91.
[10] E. Fuchs, T. Tumbar, G. Guasch, Socializing with the neighbors:stem cells and their niche., Cell 116 (6) (2004) 769–778.
[11] A. I. Caplan, Mesenchymal stem cells., J Orthop Res 9 (5)(1991) 641–650.URL http://dx.doi.org/10.1002/jor.1100090504
[12] B. Short, N. Brouard, T. Occhiodoro-Scott, A. Ramakrishnan,P. J. Simmons, Mesenchymal stem cells., Arch Med Res 34 (6)(2003) 565–571.
[13] C. Csaki, U. Matis, A. Mobasheri, H. Ye, M. Shakibaei, Chon-drogenesis, osteogenesis and adipogenesis of canine mesenchy-mal stem cells: a biochemical, morphological and ultrastruc-tural study., Histochem Cell Biol 128 (6) (2007) 507–520.URL http://dx.doi.org/10.1007/s00418-007-0337-z
[14] M. F. Pittenger, A. M. Mackay, S. C. Beck, R. K. Jaiswal,R. Douglas, J. D. Mosca, M. A. Moorman, D. W. Simonetti,S. Craig, D. R. Marshak, Multilineage potential of adult humanmesenchymal stem cells., Science 284 (5411) (1999) 143–147.
[15] P. A. Conget, J. J. Minguell, Phenotypical and functional prop-erties of human bone marrow mesenchymal progenitor cells., JCell Physiol 181 (1) (1999) 67–73.URL http://dx.doi.org/3.0.CO;2-C
[16] M. Dominici, K. L. Blanc, I. Mueller, I. Slaper-Cortenbach,F. Marini, D. Krause, R. Deans, A. Keating, D. Prockop,E. Horwitz, Minimal criteria for defining multipotent mesenchy-mal stromal cells. the international society for cellular therapyposition statement., Cytotherapy 8 (4) (2006) 315–317.URL http://dx.doi.org/10.1080/14653240600855905
[17] L. A. Fortier, P. J. Brofman, A. J. Nixon, H. O. Mohammed,Disparate chondrocyte metabolism in three-dimensional fibrincultures derived from autogenous or commercially manufac-tured fibrinogen., Am J Vet Res 59 (4) (1998) 514–520.
[18] W. R. Otto, J. Rao, Tomorrow’s skeleton staff: mesenchymalstem cells and the repair of bone and cartilage., Cell Prolif 37 (1)(2004) 97–110.
[19] S. D. Deodhar, R. Sethi, R. C. Srimal, Preliminary study on an-tirheumatic activity of curcumin (diferuloyl methane)., IndianJ Med Res 71 (1980) 632–634.
[20] D. Khanna, G. Sethi, K. S. Ahn, M. K. Pandey, A. B. Kun-numakkara, B. Sung, A. Aggarwal, B. B. Aggarwal, Naturalproducts as a gold mine for arthritis treatment., Curr OpinPharmacol 7 (3) (2007) 344–351.URL http://dx.doi.org/10.1016/j.coph.2007.03.002
[21] N. Elmali, I. Esenkaya, A. Harma, K. Ertem, Y. Turkoz,B. Mizrak, Effect of resveratrol in experimental osteoarthritisin rabbits., Inflamm Res 54 (4) (2005) 158–162.URL http://dx.doi.org/10.1007/s00011-004-1341-6
[22] C. Csaki, N. Keshishzadeh, K. Fischer, M. Shakibaei, Regula-tion of inflammation signalling by resveratrol in human chon-drocytes in vitro., Biochem Pharmacol 75 (3) (2008) 677–687.URL http://dx.doi.org/10.1016/j.bcp.2007.09.014
[23] M. Shakibaei, T. John, C. Seifarth, A. Mobasheri, Resveratrolinhibits il-1 beta-induced stimulation of caspase-3 and cleavageof parp in human articular chondrocytes in vitro., Ann N YAcad Sci 1095 (2007) 554–563.URL http://dx.doi.org/10.1196/annals.1397.060
[24] N. Elmali, O. Baysal, A. Harma, I. Esenkaya, B. Mizrak, Effectsof resveratrol in inflammatory arthritis., Inflammation 30 (1-2)(2007) 1–6.URL http://dx.doi.org/10.1007/s10753-006-9012-0
[25] A. C. Bharti, B. B. Aggarwal, Nuclear factor-kappa b and can-cer: its role in prevention and therapy., Biochem Pharmacol64 (5-6) (2002) 883–888.
[26] Y.-J. Surh, Cancer chemoprevention with dietary phytochemi-
cals., Nat Rev Cancer 3 (10) (2003) 768–780.URL http://dx.doi.org/10.1038/nrc1189
[27] A. Liacini, J. Sylvester, W. Q. Li, W. Huang,F. Dehnade, M. Ahmad, M. Zafarullah, Induction of ma-trix metalloproteinase-13 gene expression by tnf-alpha ismediated by map kinases, ap-1, and nf-kappab transcriptionfactors in articular chondrocytes., Exp Cell Res 288 (1) (2003)208–217.
[28] A. Liacini, J. Sylvester, W. Q. Li, M. Zafarullah, Inhibition ofinterleukin-1-stimulated map kinases, activating protein-1 (ap-1) and nuclear factor kappa b (nf-kappa b) transcription factorsdown-regulates matrix metalloproteinase gene expression in ar-ticular chondrocytes., Matrix Biol 21 (3) (2002) 251–262.
[29] J. K. Lin, M. H. Pan, S. Y. Lin-Shiau, Recent studies on the bio-functions and biotransformations of curcumin., Biofactors 13 (1-4) (2000) 153–158.
[30] Y. R. Chen, T. H. Tan, Inhibition of the c-jun n-terminal kinase(jnk) signaling pathway by curcumin., Oncogene 17 (2) (1998)173–178.URL http://dx.doi.org/10.1038/sj.onc.1201941
[31] U. R. Pendurthi, L. V. Rao, Suppression of transcription factoregr-1 by curcumin., Thromb Res 97 (4) (2000) 179–189.
[32] D. Thaloor, K. J. Miller, J. Gephart, P. O. Mitchell, G. K.Pavlath, Systemic administration of the nf-kappab inhibitorcurcumin stimulates muscle regeneration after traumatic in-jury., Am J Physiol 277 (2 Pt 1) (1999) C320–C329.
[33] B. B. Aggarwal, A. Kumar, A. C. Bharti, Anticancer potentialof curcumin: preclinical and clinical studies., Anticancer Res23 (1A) (2003) 363–398.
[34] J. Leiro, J. A. Arranz, A. Paramá, M. F. Alvarez, M. L.Sanmartín, In vitro effects of the polyphenols resveratrol,mangiferin and (-)-epigallocatechin-3-gallate on the scuticocili-ate fish pathogen philasterides dicentrarchi., Dis Aquat Organ59 (2) (2004) 171–174.
[35] J. Leiro, E. Cano, F. M. Ubeira, F. Orallo, M. L. Sanmartín, Invitro effects of resveratrol on the viability and infectivity of themicrosporidian encephalitozoon cuniculi., Antimicrob AgentsChemother 48 (7) (2004) 2497–2501.URL http://dx.doi.org/10.1128/AAC.48.7.2497-2501.2004
[36] G. Schulze-Tanzil, A. Mobasheri, J. Sendzik, T. John, M. Shak-ibaei, Effects of curcumin (diferuloylmethane) on nuclear factorkappab signaling in interleukin-1beta-stimulated chondrocytes.,Ann N Y Acad Sci 1030 (2004) 578–586.URL http://dx.doi.org/10.1196/annals.1329.067
[37] M. Shakibaei, G. Schulze-Tanzil, T. John, A. Mobasheri, Cur-cumin protects human chondrocytes from il-l1beta-induced in-hibition of collagen type ii and beta1-integrin expression andactivation of caspase-3: an immunomorphological study., AnnAnat 187 (5-6) (2005) 487–497.
[38] C. Csaki, A. Mobasheri, M. Shakibaei, Synergistic chondropro-tective effects of curcumin and resveratrol in human artic-ular chondrocytes: inhibition of il-1beta-induced nf-kappab-mediated inflammation and apoptosis., Arthritis Res Ther11 (6) (2009) R165.URL http://dx.doi.org/10.1186/ar2850
23

Neues von NGF und GDNF
A. Mayerhofer∗
Zellbiologie, Ludwig-Maximilians-Universität, Biedersteiner Straße 29, 80802 München
Abstract
Über die zellulären Grundlagen der humanen Reproduktion ist wenig bekannt. Untersuchungen an isolierten humanenZellen des Ovars (Granulosazellen) und des Hodens (testikuläre peritubuläre Zellen) sind ein Ansatz, den wir verfolgen,um neue Einblicke in das normale (Fertilität) und gestörte Geschehen (Infertilität) bei Frau und Mann zu erhalten.Dabei sind wir in den letzten Jahren auf neuronale Moleküle und Mechanismen im Ovar und im Hoden gestoßen, dievon Ionenkanälen über Neurotransmitter bis zu Neurotrophinen reichen. Der Nervenwachstumsfaktor, NGF ("Nervegrowth factor"), wird beispielsweise von Granulosazellen des Ovars gebildet. Er ist zudem in der Follikelflüssigkeit vonPatientinnen mit dem Polyzystischen Ovar-Syndrom (PCOS) erhöht. Dass ein Zuviel an NGF ein PCOS erzeugenkann, wurde durch einen systemischen Ansatz nachgewiesen. Eine transgene Maus, die NGF im Ovar verstärkt bildet,entwickelt einen PCOS Phänotyp. GDNF ("Glial cell line derived neurotrophic factor") ist wichtig für die Regulation derStammzellproliferation des Hodens (Stammzelle der Spermatogenese = spermatogoniale Stammzelle). Wir fanden, dassdie Stammzellen des Hodens beim Mann in unmittelbarer Nachbarschaft zu den peritubulären, myoiden Zellen liegen.An isolierten peritubulären Zellen und ex-vivo konnten wir zeigen, dass diese bisher wenig untersuchten Zellen einenBeitrag zur Stammzellnische des menschlichen Hodens leisten, indem sie konstitutiv GDNF produzieren.
1. Einleitung
Die zellulären Grundlagen der humanen Reproduktionsind nur rudimentär bekannt. Da viele grundlegende Un-terschiede zwischen der Spezies Mensch und Tieren bei derReproduktion bestehen, kann man nicht einfach aus demTierexperiment auf den Menschen schließen. Der Teufelsteckt hier - wie immer - im Detail. Untersuchungen amMenschen sind daher notwendig, um Einblicke in das nor-male Geschehen bei Fertilität und um Verständnis für dasgestörte Geschehen bei Infertilität von Frau und Mann zuerhalten. Wie lässt sich das aber umsetzten?
Wir greifen in unserer Arbeit auf zwei zelluläre Model-le zurück, primäre humane ovarielle Granulosazellen ausdem sprungreifen Follikel von Patientinnen, und humanetestikuläre peritubuläre Zellen, also myoide Zellen aus derWand der Hodenkanälchen. Primärkulturen beider Zellar-ten, die nur durch Zusammenarbeit mit klinischen Part-nern möglich sind, erlauben einen experimentellen Ansatz,um zumindest Aspekte des menschlichen Ovars und Ho-dens kennen zu lernen. Die so gewonnenen Ergebnisse müs-sen natürlich immer wieder am wirklichen Geschehen imOvar und Hoden überprüft werden und im Idealfall zusätz-lich mit einem systemischen Ansatz experimentell getestetwerden.
Bei unseren Untersuchungen sind wir in den letzten Jah-ren auf neuronale Moleküle und Mechanismen im Ovar und
∗Für Rückfragen, Kooperationsideen oder Informationen zu un-seren Projekten, E-Mail: [email protected]
im Hoden gestoßen, die von Ionenkanälen (z.B. [1, 2, 3, 4,5, 6]) über Neurotransmitter (z.B. [7, 8, 9, 10, 11, 12, 13])bis zu Neurotrophinen [14, 15, 16] reichen.
Zwei Beispiele zu Neutrotrophinen aus letzter Zeit sollendies verdeutlichen.
2. Beispielhafte Ergebnisse
2.1. Nerven-Wachstumsfakor NGF ("nerve growth fac-tor")
Der Nerven-Wachstumsfakor NGF ("nerve growth fac-tor") wird von humanen Granulosazellen bereits im Folli-kel und in kultivierten Zellen in hoher Konzentration ge-bildet. NGF-Rezeptoren, die Granulosazellen steuern kön-nen, kommen zahlreich im Ovar und im Follikel vor, undwir versuchen derzeit die Konsequenzen der Aktivierungaufzuklären.
Granulosazellen von Frauen mit dem häufigen "Polyzys-tischen Ovar-Syndrom (PCOS)", das zu Unfruchtbarkeitführt, bilden, wie wir fanden, deutlich mehr NGF. Könntees daher vielleicht eine Rolle bei dieser Erkrankung spie-len? Diese Frage, also ob ein Zuviel an NGF PCOS erzeu-gen kann, wurde in Zusammenarbeit mit einer Gruppe inden USA durch einen systemischen Ansatz getestet: Einetransgene Maus, die NGF im Ovar verstärkt bildet, ent-wickelt tatsächlich einen typischen PCOS Phänotyp [15].NGF ist also vermutlich von fundamentaler Bedeutung imOvar.
Internal Communication: Neues aus der Anstalt (NeadeA) 30. November 2010

4 DANKSAGUNG
2.2. GDNF ("Glial cell line derived neurotrophic factor")
NGF wird auch von humanen testikulären peritubulä-ren Zellen (HTPCs) gebildet [14] aber seine Bedeutungist noch unbekannt. HTPCs bilden daneben noch weite-re neurotrophe Faktoren. GDNF ("Glial cell line derivedneurotrophic factor") ist so ein Faktor, der von Sertolizel-len produziert wird und die Stammzelle der Spermatoge-nese (spermatogoniale Stammzelle) steuert. Dies belegenUntersuchungen an Nagern, bei denen die Wand der Sa-menkanälchen jedoch aus nur einer Lage von myoiden Zel-len besteht. Dagegen finden sich beim Mann drei bis fünfSchichten von Zellen, was auf eine möglicherweise andereBedeutung dieser Zellen hinweisen mag.
Wir fanden nun kürzlich, dass die Zellen dieser Wandeinen Beitrag zur Stammzellnische des menschlichen Ho-dens leisten können [16]. Sie produzieren, wie die Serto-lizelle, GDNF und unmittelbar benachbarte spermatogo-niale Stammzellen besitzen, wie wir fanden, einen Rezep-tor für GDNF (GFRα1). Die Freisetzung von GDNF er-folgt konstitutiv und so könnten die peritubulären Zellen,als Nachbarn der Stammzellen, diese auf parakrinem Wegsteuern. Ob sie auch über andere Stoffe einen Beitrag fürdie Stammzellnische leisten können, wollen wir im Rahmeneines beantragten Projektes weiter untersuchen.
3. Ausblick
Die Namen der Neurotrophine spiegeln die Geschichteihrer Entdeckung wider. Es wird klar, dass sie nicht nurim Nervensystem wichtig sind, sondern eine Bedeutung inOrgankompartimenten haben, die völlig ohne Innervati-on auskommen, also im ovariellen Follikel und im Samen-kanälchen. Dass dort zudem andere aus dem Nervensytembekannte Mechanismen greifen (Neurotransmitter, derenRezeptoren und Ionenkanäle), ist eine Überraschung. Wel-chen Stellenwert sie bei der normalen und gestörten Funk-tion der Gonaden haben, gilt es herauszuarbeiten.
4. Danksagung
Unsere Forschung wird von der DFG in Einzelprojek-ten (z.B. MA 1080/16-3; MA1080/17-3), vom DAAD undder Friedrich-Baur Stiftung gefördert. Wir sind Mitgliedim Reprozentrum der LMU (Reprozentrum) und arbeiteneng zusammen mit klinischen Partnern in München undGruppen in Finnland, Chile, Argentinien und den USA.[1] A. Bulling, F. D. Berg, U. Berg, D. M. Duffy, R. L. Stouffer,
S. R. Ojeda, M. Gratzl, A. Mayerhofer, Identification of an ova-rian voltage-activated Na+-channel type: hints to involvementin luteolysis., Mol Endocrinol 14 (7) (2000) 1064–1074.
[2] L. Kunz, A. Thalhammer, F. D. Berg, U. Berg, D. M. Duffy,R. L. Stouffer, G. A. Dissen, S. R. Ojeda, A. Mayerhofer, Ca2+-activated, large conductance K+ channel in the ovary: identi-fication, characterization, and functional involvement in steroi-dogenesis., J Clin Endocrinol Metab 87 (12) (2002) 5566–5574.
[3] L. Kunz, R. Rämsch, A. Krieger, K. A. Young, G. A. Dissen,R. L. Stouffer, S. R. Ojeda, A. Mayerhofer, Voltage-dependentK+ channel acts as sex steroid sensor in endocrine cells of the
human ovary., J Cell Physiol 206 (1) (2006) 167–174.URL http://dx.doi.org/10.1002/jcp.20453
[4] L. Kunz, J. S. Richter, A. Mayerhofer, The adenosine 5’-triphosphate-sensitive potassium channel in endocrine cells ofthe human ovary: role in membrane potential generation andsteroidogenesis., J Clin Endocrinol Metab 91 (5) (2006) 1950–1955.URL http://dx.doi.org/10.1210/jc.2005-2269
[5] L. Kunz, C. Roggors, A. Mayerhofer, Ovarian acetylcholine andovarian KCNQ channels: insights into cellular regulatory sys-tems of steroidogenic granulosa cells., Life Sci 80 (24-25) (2007)2195–2198.URL http://dx.doi.org/10.1016/j.lfs.2007.01.022
[6] M. H. Traut, D. Berg, U. Berg, A. Mayerhofer, L. Kunz, Iden-tification and characterization of Ca2+-activated K+ channelsin granulosa cells of the human ovary., Reprod Biol Endocrinol7 (2009) 28.URL http://dx.doi.org/10.1186/1477-7827-7-28
[7] A. Mayerhofer, G. A. Dissen, M. E. Costa, S. R. Ojeda, A rolefor neurotransmitters in early follicular development: inducti-on of functional follicle-stimulating hormone receptors in newlyformed follicles of the rat ovary., Endocrinology 138 (8) (1997)3320–3329.
[8] A. Mayerhofer, G. D. Smith, M. Danilchik, J. E. Levine, D. P.Wolf, G. A. Dissen, S. R. Ojeda, Oocytes are a source of catecho-lamines in the primate ovary: evidence for a cell-cell regulatoryloop., Proc Natl Acad Sci U S A 95 (18) (1998) 10990–10995.
[9] A. Mayerhofer, S. Fritz, S. Mani, T. R. Kumar, A. Thalhammer,P. Ingrassia, A. A. Fienberg, P. Greengard, Ovarian functionand morphology after deletion of the DARPP-32 gene in mice.,Exp Clin Endocrinol Diabetes 112 (8) (2004) 451–457.URL http://dx.doi.org/10.1055/s-2004-821205
[10] A. Mayerhofer, L. Kunz, A. Krieger, B. Proskocil, E. Spindel,A. Amsterdam, G. A. Dissen, S. R. Ojeda, I. Wessler, FSH re-gulates acetycholine production by ovarian granulosa cells., Re-prod Biol Endocrinol 4 (2006) 37.URL http://dx.doi.org/10.1186/1477-7827-4-37
[11] V. Rey-Ares, N. Lazarov, D. Berg, U. Berg, L. Kunz, A. Mayer-hofer, Dopamine receptor repertoire of human granulosa cells.,Reprod Biol Endocrinol 5 (2007) 40.URL http://dx.doi.org/10.1186/1477-7827-5-40
[12] M. Greiner, A. Paredes, V. Rey-Ares, S. Saller, A. Mayerhofer,H. E. Lara, Catecholamine uptake, storage, and regulated re-lease by ovarian granulosa cells., Endocrinology 149 (10) (2008)4988–4996.URL http://dx.doi.org/10.1210/en.2007-1661
[13] S. Saller, L. Kunz, G. A. Dissen, R. Stouffer, S. R. Ojeda,D. Berg, U. Berg, A. Mayerhofer, Oxytocin receptors in theprimate ovary: molecular identity and link to apoptosis in hu-man granulosa cells., Hum Reprod 25 (4) (2010) 969–976.URL http://dx.doi.org/10.1093/humrep/dep467
[14] C. Schell, M. Albrecht, C. Mayer, J. U. Schwarzer, M. B. Frun-gieri, A. Mayerhofer, Exploring human testicular peritubularcells: identification of secretory products and regulation by tu-mor necrosis factor-alpha., Endocrinology 149 (4) (2008) 1678–1686.URL http://dx.doi.org/10.1210/en.2007-1064
[15] G. A. Dissen, C. Garcia-Rudaz, A. Paredes, C. Mayer, A. Mayer-hofer, S. R. Ojeda, Excessive ovarian production of nerve growthfactor facilitates development of cystic ovarian morphology inmice and is a feature of polycystic ovarian syndrome in humans.,Endocrinology 150 (6) (2009) 2906–2914.URL http://dx.doi.org/10.1210/en.2008-1575
[16] K. Spinnler, F. M. Köhn, U. Schwarzer, A. Mayerhofer, Glialcell line-derived neurotrophic factor is constitutively producedby human testicular peritubular cells and may contribute tothe spermatogonial stem cell niche in man., Hum Reprod 25 (9)(2010) 2181–2187.URL http://dx.doi.org/10.1093/humrep/deq170
25

7482 Nissl-Chloroform Upper East Side: Ein Labor-Erlebnisbericht
M. Kiessling1,∗
Anatomische Anstalt, Lehrstuhl II, Ludwig-Maximilians-Universität, Pettenkoferstr. 11, 80336 München
Abstract
Im Hinblick auf eine zeitnah umzusetzende Promotionsarbeit auf Grundlage der modernen design-basierten Stereologiewurde ich nach New York geschickt, um mir dort in vier Wochen Crash-Kurs praktische Laborerfahrung mit dieserMethode anzueignen und darin so sicher zu werden, dass ich nahtlos in München mit der Umsetzung an meinem eigenenThema, dem humanen Kleinhirn, beginnen konnte. Nach einer intensiven und arbeitsreichen Zeit jenseits des Atlantikhabe ich in München unmittelbar mit meiner Rückkehr aus New York begonnen, die dort erlernten Kenntnisse undFähigkeiten produktiv in unserem Arbeitsalltag zu integrieren. Tatsächlich helfen mir die Arbeitsschritte, die ich mir inNew York zu eigen machen konnte, in der täglichen Arbeit hier immens weiter und ich bin dankbar für die Erfahrung,die ich fern der Heimat sammeln durfte.
1. Vorgeschichte
Schon im Vorfeld der Übernahme des Lehrstuhls II deranatomischen Anstalt durch Herrn Professor Schmitz hat-te sich herausgestellt, dass mit der Neubesetzung des Lehr-stuhls auch neue Methoden – schwerpunktmäßig die quan-titative Morphologie auf Basis der Stereologie – Einzughalten würden.
Die Bereitstellung der dafür benötigten Infrastrukturbegann unmittelbar mit dem Wechsel der Lehrstuhllei-tung, die endgültige Ausstattung konnte aber unmöglichvon Anfang an zur Verfügung stehen. Da die Stereologie,deren Etablierung in München bereits begonnen hat, imLabor von Prof. Patrick Hof am Department of Neuros-cience der Mount Sinai School of Medicine in New York(USA) zur täglichen Arbeitsroutine gehört, und Herr Prof.Hof und Herr Prof. Schmitz zusammen mehr als 30 Ar-beiten publiziert haben, sowie zur raschen Einarbeitungin die Methodik frühzeitig während der Aufbauphase inMünchen, wurde ich, als Mitarbeiterin des Lehrstuhl II, indieses Kooperationslabor in New York entsandt, um dortErfahrungen mit der praktischen Anwendung stereologi-scher Methoden zu erwerben. Ich wurde dort direkt in einlaufendes Forschungsprojekt am Gehirn des Seehundes ein-bezogen und habe dabei die Grundlagen der Verarbeitungdes Gewebes Gehirn sowie die erforderlichen und adäqua-ten stereologischen Techniken vier Wochen ausgiebig ge-übt. Da sich das Gehirn von Seehunden zwar ein wenig
∗Email: [email protected]: Ich möchte an dieser Stelle Herrn Professor Patrick
Hof und seinen Kollegen für die freundliche Aufnahme in seinem La-bor, die Geduld seiner Mitarbeiter - besonders Camilla Butti fürdie tolle Betreuung - und die schöne Zeit dort danken. Und natür-lich auch Herrn Professor Christoph Schmitz und dem Lehrstuhl, diediese Reise erst möglich gemacht haben.
im Aufbau, grundsätzlich aber nicht in der Art der An-wendung der stereologischen Techniken vom menschlichenGehirn unterscheidet, konnte ich hier in München sofortmit der Arbeit beginnen und wende nun die in New Yorkerworbenen Kenntnisse bereits erfolgreich an.
2. Die neue Laborwelt – faszinierend und inter-essant
Am ersten Tag – die Zeiteinheit richtet sich noch nachder mir geläufigen europäischen Norm von Minuten undStunden.
Upper East Side, New York. Auf der kurzen Streckevon meinem Domizil an der Emergency-Anfahrtszone desMount-Sinai-Hospital zu meinem Arbeitsplatz bewundereich die Schlichtheit des vor mir liegenden Laborgebäudes.Eine halbe Stunde früher als die per e-Mail mit den Mit-arbeitern vereinbarte Zeit betrete ich das Gebäude durcheine Drehtüre und durchquere voll Tatendrang die Lobby.Wie selbstverständlich gehe ich am sich dort befindlichenDoorman vorbei – denke ich zumindest. Schon hier wirdjedermanns Zugangsberechtigung überprüft, die man an-hand einer ID-Card belegen muss.
Diese ominöse Karte, von deren Existenz ich bis zu die-sem Zeitpunkt nicht einmal etwas ahnte, kann ich natürlichnicht vorweisen. Also muss ich mich in eine Besucherlisteeintragen. Der Aufzug, der einen ohne Zwischenstop vomdort so bezeichneten 1. Stock, der unserem Erdgeschoßentspricht, in den 9. Stock katapultiert, spuckt mich vordem Labor aus. Da die Kollegin, mit der ich mich verab-redet hatte, noch nicht im Hause ist, parkt man mich ineinem Aufenthaltsraum. Hier betrachte ich ausgiebig denAusblick aus dem grossen Fenster und geniesse die Sichtauf die Stadt aus einer anderen Perspektive.
Internal Communication: Neues aus der Anstalt (NeadeA) 30. November 2010

3 EIN TYPISCHER LABORTAG ODER: EINE EINFÜHRUNG IN DIE NISSL-ZEITMESSUNG
Anders als erwartet, werde ich zur Begrüßung von denneuen Kollegen nicht etwa mit den örtlichen Gegebenhei-ten vertraut gemacht, sondern überraschenderweise mit ei-ner Liste versehen, umgehend wieder weggeschickt. Dieseenthält eine Abfolge von Orten, die ich nacheinander auf-zusuchen und deren Besuch ich mit Unterschriften zu bele-gen habe, bevor ich auch nur irgendein Glas, eine Flüssig-keit oder sonst etwas im Labor in die Hand nehmen darf –bevor ich das Labor sogar überhaupt betreten darf. Allemvoran habe ich eine Sicherheitseinweisung, die die unbe-dingte Voraussetzung für die ersehnte ID-Karte darstellt,zu absolvieren. Diese Belehrung beinhaltet ein ca. zehn-minütiges Video über das adäquate Verhalten im Brand-fall, dem ein über eine halbe Stunde dauernder Film überden verantwortungsvollen Umgang mit vertraulichen Pa-tientendaten folgt. Danach habe ich noch ein Meeting mitder Dame, die die Visa-Austauschunterlagen überwacht,um dann mit einer unterschriebenen Bestätigung, dass ichnun alle Unterlagen zusammen hätte, wieder zurück in dasGebäude zu eilen, wo besagte Filmchen laufen. Fototermin– und endlich bin ich mittags stolze Besitzerin meiner ID-Karte.
3. Ein typischer Labortag oder: eine Einführung indie Nissl-Zeitmessung
Ein Tag an meinem vorübergehenden Arbeitsplatz inUpper East Side, Manhattan könnte typischerweise wiefolgt ausgesehen haben.
An dem Wachhund in der Lobby bin ich mit meiner ID-Card ohne Probleme vorbei, der Aufzug hat mich in denneunten Stock befördert und nun befinde ich mich vor derLabortüre. Ebenso wie ein Tisch, der für die Ablage vonSpeisen und Getränken vorgesehen ist, die im Labor kei-nen Zutritt haben. Also nochmal einen kräftigen Schluckaus dem Pappbecher genommen, der den auf den Wach-heitsgrad direkt Einfluß nehmenden Kaffee enthält. Undrein geht es ins Labor, in „mein“ Büro. Leider stehe ichhier vor verschlossener Türe, da ich mal wieder die Zah-lenfolge vergessen habe, die den Zugang sichert. Hier sindalle Räume, ausser dem Labor selbst, codegesichert, na-türlich mit verschiedenen Nummern (Codes sind wirklichlästig, besonders dann, wenn man es gewöhnt ist, alle fürdie Arbeit wichtigen Türen mit einem einzigen Transpon-der zu öffnen). Dazu kommen dann noch die Passwörteran den verschiedenen Rechnern. Da kann man schon maldurcheinander kommen, zumal ich mir weder die Zahlen-folgen, noch die Passwörter selber ausgedacht habe. Also,ein erneuter Versuch, die richtige Codenummer einzuge-ben. Und noch einer - und endlich bin ich in dem Büro,das ich mir zwar mit einer Kollegin teile, das ich aber diemeiste Zeit alleine nutze. Wir haben in dem Raum zusam-men gut Platz, der Tisch läßt viel Freiheit für meine Un-terlagen und der Rechner ist mit allen von mir benötigtenProgrammen ausgestattet. Ich lasse meine dicke Jacke erstmal an. Das Zimmer gleicht, wie immer wenn ich morgenskomme, einer Kühltruhe. Also klebe ich ein Eispack auf
den Sensor, der die Temperatur erfasst, damit diesem dasZimmer kälter erscheint und die Kühlung früher stoppt.
Auf dem Weg zum meinem Laborarbeitsplatz, ohne Ja-cke, hier ist es wohlig warm, sammle ich noch mein sorgfäl-tig geführtes Laborbuch ein, das alle Daten meiner Versu-che und Mikroskopierarbeiten enthält. Auch hier genießtdas Laborbuch einen sehr hohen Stellenwert als Dokument.Es wurde mir am ersten Tag mit dem Hinweis überge-ben wurde, alles hineinzuschreiben, was mit dem Laborzusammenhängt – und zwar sofort und immer mit Kugel-schreiber, um Änderungen durch andere vorzubeugen. DasLaborbuch dient einem selbst als Informationsquelle überdie eigenen durchgeführten Arbeitsschritte und Versuche,gewinnt aber vor allem in Zeiten zunehmender manipu-lierter wissenschaftlicher Arbeiten als dokumentarischerNachweis der Vorgehensweisen und angegebenen Ergeb-nisse immer mehr an Bedeutung.
Ein Blick auf den Mikroskopbelegungsplan, der an derTür zum Mikroskopierraum angebracht ist, verrät mir,dass ich erst nachmittags einen Zeitslot an dieser vielverwendeten Vergrösserungsvorrichtung habe. Dieser Planenthält als Kalenderblatt die Tage einer Woche und manschreibt sich nach Absprache mit den Kollegen einfach inden Stunden ein, die man gerne am Mikroskop verbringenwürde. Die Auswahl der Uhrzeiten hängt meist von denpersönlichen Vorlieben ab. Die eine will nicht über Mit-tag, die andere will nicht vor zehn anfangen und nach fünfUhr nachmittags ist das Mikroskop eigentlich zuverlässigfrei. Ich versuche immer, den letzten Termin zu bekommen,da man beim Zellenzählen leicht die Zeit vergisst und zwi-schendrin aufzuhören kontaproduktiv ist. Aber auch nachder Festlegung der Zeiten wird der Plan in Absprache sehrflexibel gehandhabt. Da ich nun noch viel Zeit habe, be-schließe ich, noch ein paar Schnitte zu färben.
Im Kühlschrank finde ich sauber beschriftete Plastik-kästen, die viele fortlaufend nummerierte Eppendorfhüt-chen enthalten und alle mit je einem Gehirnschnitt be-stückt sind. Ich nehme mir die für mich relevanten Num-mern heraus und richte mir gemütlich meinen grosszügigbemessenen Arbeitsplatz ein, den ich für die Zeit meinesAufenthalts von einer Kollegin übernommen habe. DasLabor ist ein großer, eher länglicher Raum, der von ei-nem Gang durchzogen ist. Von diesem gehen parallel dieArbeitsplätze ab, die aus zwei großen Tischen für die La-bortätigkeiten und einem Schreibplatz bestehen.
Es fehlt Puffer. Also begebe ich mich um die Ecke zu dendort stehenden großen Kanistern, wo ich, je nach Puffer,anteilig zwei Substanzen zusammenmische. Nun beginntdie feinmotorikintensive Beschäftigung mit den im Puf-fer schwimmenden Gehirnschnitten und der Aufgabe, die-se möglichst als Ganzes mittig auf einem Objektträger zuplatzieren. Eine wahre Herausforderung. Die ersten Schnit-te sind eine Katastrophe und ich brauche ewig. Mittlerwei-le funktioniert es aber schon ganz gut und die Arbeit gehtzügig voran. Dann werden die Objektträger noch flach zumTrocknen ausgelegt und in den folgenden Stunden ist einesorgfältige Überwachung der Schnitte erforderlich. Diese
27

4 SCHLUSSBEMERKUNG UND FAZIT
dürfen nicht zu trocken werden, damit sie nicht kaputt ge-hen, und um alle gleichzeitig färben zu können, werden die,die schneller trocken immer wieder befeuchtet - mit demPinsel in der Hand male ich die entsprechenden Stellen mitPuffer phantasievoll an. Die Wartezeit überbrücke ich mitLiteraturrecherche in „meinem“ Büro, wo die Temperaturlangsam ein erträgliches Maß erreicht.
Die Schnitte sind bereit zum Färben. Gewappnet mitHandschuhen und Schutzbrille fülle ich die letzten für dieFärbung benötigten Gefäße. Und nun beginnt mein fremd-bestimmtes Leben nach der Uhr. Die vor mir liegende Fär-bereihe besteht aus Gefäßen mit verschiedenen Flüssigkei-ten in Abstufungen von durchsichtig bis tieflila. Jedes derGefäße ist mit dem jeweiligen Inhalt beschriftet und diePfeile geben die Richtung an, in der die Reihe fortzuset-zen ist. So schafft man es sogar unter erheblichem Koffe-inmangel, die richtige Reihenfolge einzuhalten. Wenn mannun auch noch der intellektuellen Leistung gewachsen ist,das richtige Färbeprotokoll zu verwenden, das mit ande-ren gut sichtbar über der Färbereihe angebracht ist, sollteschön lila gefärbten Schnitten nichts mehr imWege stehen.Nach einigen Tagen färben gebe ich Zeitspannen jetzt nurnoch in Nissl-Einheiten an. „Ich brauche noch kurz, 5 NisslEthanol“ was zehn Minuten entspricht. Oder „drei Nissl-Chloroform“, also eine viertel Stunde.
Nachmittag, Mikroskopierzeit. Nachdem ich die Schnit-te zusammengesucht habe, deren histologischem Erschei-nungsbild heute mein besonderes Interesse gilt – die Infor-mation, welche das sind, habe ich natürlich meinem Labor-buch entnommen - mache ich mich mit den entsprechen-den Objektträgern und dem unvermeidlichen Laborbuchbewaffnet auf den Weg zum Mikroskop – und stehe malwieder vor einer verschlossenen Türe. Der vorbeieilendeKollege hilft mir mit dem Code aus, was mir die Müheerspart, in mein Büro zurückzulaufen – wo ich mich auchvor einer verschlossenen Türe wiederfinden würde - und inmeinen Notizen nachzusehen. Ich taste mich in dem weni-gen Licht, das vom Gang hereinfällt an den Arbeitsplatz,finde den Power-Schalter des Computers und der anderenGeräte. In dem nun herrschenden Dämmerlicht platziereich unter einigen Mühen den Objektträger in der ledig-lich vom Lichtstrahl des Mikroskops erhellten und dahernur zu erahnenden Haltevorrichtung und klemme ihn fest.Köhlern, öffnen des Programms und Übertragung des Bil-des auf den Monitor. Den Bereich, der für meine Zählungrelevant ist, habe ich schon in einem vorhergehenden Ar-beitsschritt mittels Mouse am Monitor umrandet und nunmuss ich den von mir erstellten Rahmen mittels Joystickwieder richtig positioniert mit dem Bild, das mir der Mo-nitor von dem eingespannten Schnitt liefert, in Einklangbringen. Was nun folgt ist stundenlanges Markieren vonZellen, was Konzentration und Durchhaltevermögen erfor-dert.
Über dem Kennzeichnen der Zellen per Mouseklick amMonitor vergesse ich vollkommen die Zeit. Wie viele derKollegen hier im Labor, habe auch ich mir angewöhnt,immer Musik bei mir zu haben und mit dem richtigen
Beat geht das Zählen leicht von der Hand. Eine besondereAbwechslung verschafft die Software, die diverse Überra-schungen für mich bereithält. Die einheitliche Bemerkungder Kollegen „Das Problem hatte ich noch nie“ ist in kei-nem dieser Fälle zielführend. Stunden später krieche ichaus dem Zimmer, der sonnige Tag hat sich in dunkle Nachtverwandelt und die Kollegen sind verschwunden – bis aufeine. Ich spreche sie an, aber wie so oft, muss ich mich an-derweitig bemerkbar machen, da auch hier die unvermeid-lichen Kopfhörer die Schallwellenleitung des von mir Arti-kulierten ins Innenohr erheblich einschränken. Wir haltennoch einen kurzen fachlichen Austausch ab, dann packe ichmeine Sachen zusammen, in meinem Büro, das mich auchdiesmal wieder vor die Frage des richtigen Zugangscodesstellt, und trete den Heimweg an. Ich falle acht Stockwerkein die Tiefe und wünsche dem Doorman eine gute Nacht,als ich an ihm vorbeigehe und die sichtbar an der Hosenta-sche angebrachte ID-Karte von dort entferne und in meinerUmhängetasche verstaue.
4. Schlussbemerkung und Fazit
„In Manhattan finden sich genug kleine Welten für einekleine Weltreise“ ([1], Seite 68). New York, das als NewAmsterdam ([2], Seite 21) seinen Beginn gefunden hat,ist eine Großstadt wie jede andere, könnte man auf denersten Blick meinen. In dieser Stadt hatten verschiedeneVölkergruppen eigene Viertel, was sich schon in den Na-men wie Chinatown oder Little Italy widerspiegelt. Hierwird Umweltschutz zwar groß geschrieben, das Geschirrist aber nahezu ausschliesslich Einweg. In dieser Stadt, inder man, um in einen anderen Staat zu gelangen, nur denFluss überqueren muss und im Laborgebäude sogar die Re-strooms codegesichert sind, habe ich mit all seinen über-raschenden Facetten vier Wochen lang einen Lebens- undArbeitsplatz gefunden. Hier habe ich die Möglichkeit be-kommen, einen Einblick in das Leben und Forschen in eineranderen Stadt, einem anderen Land, sogar einem anderenKontinent zu erhalten.
Wie forscht man nun in New York? Der Vergleich der La-bors in New York und München läßt mich zu dem Schlussgelangen, dass wohl im Grunde viele Labors vergleichbarerGrößenordnung mit ähnlichen Forschungsschwerpunktengleichartig arbeiten. Zwar weichen in dem besuchten La-bor die verwendeten Protokolle und die Aufteilung der Ar-beitsbereiche geringfügig von unseren ab, die Sicherheits-bestimmungen gelten aber entsprechend und die Arbeits-weisen gleichen unseren. Gravierende Unterschiede konnteich nicht feststellen. Auch in New York kämpfen die Wis-senschaftler mit Deadlines und auch dort lieben die fleis-sigen Labormäuse Schokolade. Für mich war es eine lehr-und erlebnisreiche Zeit die mir eine hervorragende Mög-lichkeit geboten hat, die Aufbauphase in München effektivzu nützen. Hoch motiviert bin ich aus New York zurück-gekehrt und wurde nach meiner Ankunft ohne Übergangsofort mit einem Projekt am menschlichen Kleinhirn be-traut. Nun habe ich die Gelegenheit, das, was mich in New
28

4 SCHLUSSBEMERKUNG UND FAZIT
York vier Wochen lang beschäftigt hat, umzusetzen. Esist spannend zu beobachten, wie die Infrastruktur hier inMünchen rasend schnell entsteht und so auch die für dieForschung benötigten Voraussetzungen geschaffen werden.Das Projekt ist in vollem Gange und ich freue mich auf dieErgebnisse, die uns das Kleinhirn mittels der Forschungauf Grundlage der Stereologie liefern wird.[1] G. Otis, Lonely Planet New York, 2nd Edition, Mairdumont,
Ostfildern, 2009.[2] D. von Kruse-Etzbach, New York, 6th Edition, Reisebuchverlag
Iwanowski, 2010.
29

Neues aus der Anstalt (NeadeA)TEXnicalitiesI,II
H.-G. Frank1,∗
Anatomische Anstalt, Lehrstuhl II, Ludwig-Maximilians-Universität, Pettenkoferstrasse 11, 80336 München, Germany
Abstract
The layout system of NeadeA is modified from existing layout templates of elsevier journals. In principal, the generalguidelines of elsevier apply and it will be no problem to adapt any manuscript being written according to elsevierguidelines into the layout system used here. The basic layout system (TEXand LATEX) is used under the editor Lyx, allsoftware being used under Open Source licensing conditions. Compatibility with future developments and high-qualityPDF output is possible and guaranteed. Rich hyperlinking from the text to other text sections, figures, tables andreferences is implemented. Authors can use any word processing software to prepare manuscripts. The most importantrules for doing this are summarized below. Authors are encouraged to write their manuscripts directly in the layoutsystem under Lyx in order to avoid and minimize additional redactional work due to transfer of text and other elementsof a manuscript from a word processing file to Lyx. For staff of the Anatomische Anstalt the software can be installedlocally by the IT-Support crew.
1. Introductory Remarks
1.1. Basic Layout System (LATEX) and Editor (Lyx)
The layout as given in this first volume is not preparedin an expensive softwaretool, but is built with the soft-warepackage LATEX under the editor Lyx. Both packagesare open source software, which can be used under the re-spective licenses without fees. As template for this layout,a LATEXbased layout template of the publishing house el-sevier was used and modified accordingly. This producesa final high-quality layout, which is very close to a finalprinted professional version. However, the layout of else-vier journals is not matched completely due to the fact theproprietary elsevier fonts are not used for typesetting themanuscripts of NeadeA, but the look-a-like is very close toa professional print-layout.
2. Instructions to Authors
2.1. Use of wordprocessing software
It is important to know that the file can be saved inthe native format of the wordprocessor used or as RichText File (RTF). The text should be in single-columnformat. Do not try to format it according to the printlayout of NeadeA. This print layout is achieved with the
IEssentially, this contribution is equivalent to „instructions to au-thors” of a scientific journal.
IIA german version of TEXnicalities is in preparation∗Email: [email protected]: Andrea Asikoglu
LATEXsystem. However, it is required that all text sectionsare present and provided in such a way that the texts canbe transferred to the editor Lyx by a copy and paste pro-cedure. Thus, avoid fancy formatting. Text of title footnotes and author footnotes should be a single paragrapheach named e.g. title footnote 1, or author footnote, etcand the position in title or author name should be indi-cated by a superscript symbol. In particular, do not usethe wordprocessor’s options to justify text or to hyphen-ate words. However, do use bold face, italics, subscripts,superscripts etc. Do not embed "graphically designed (i.e.drawn in a drawing program or e.g. Powerpoint" equationsor tables, but prepare these using the wordprocessor’s fa-cility. When preparing tables, if you are using a table grid,use only one grid for each individual table and not a gridfor each row. If no grid is used, use tabs, not spaces, toalign columns. The electronic text should be prepared in away very similar to that of conventional manuscripts (seealso e.g.: http://www.elsevier.com/guidepublication). Donot import the figures into the text file but, instead, indi-cate their approximate locations directly in the electronictext and on the manuscript. See also the section on Elec-tronic illustrations. To avoid unnecessary errors you arestrongly advised to use the "spell-check" and "grammar-check" functions of your wordprocessor.
2.2. Use of Lyx as a replacement for Word Processors
The programs Lyx and LATEX as well as standard BibTexeditors for reference management are freely available andcan be installed at every work place of the AnatomischeAnstalt. Templates for article setup can be obtained by
Internal Communication: Neues aus der Anstalt (NeadeA) November 30, 2010

2 INSTRUCTIONS TO AUTHORS 2.3 Article structure
Figure 1: This is an example of a figure in one column-width
Hans-Georg Frank or Andrea Asikoglu. Both can also givea short introduction in the use of these options.
2.3. Article structure
For every contribution, the authors are free in chosingthe respective Divisions and Subdivisions of their article.In the following paragraphs, the sections, which ususallywould be included in a scientific article are mentioned asan example. A contribution dealing e.g. with a legal issuewill certainly have completely different divisions and sub-divisions. Contributions with a more essay-like ductus willhave another type of division and subdivision, if at all. Itis impossible for this instruction to cover all these aspectsin advance and completely So, the well-known example ofa scientific article is given here as an illustration as whatwould be expected from authors in general in analogy tothis example. Moreover, some options of the layout sys-tem with regard to frequently used text tools are shown insample texts to illustrate the options.
2.3.1. SubdivisionNumbered sections. Divide your article into clearly definedand numbered sections. Subsections should be numbered1.1 (then 1.1.1, 1.1.2, ...), 1.2, etc. (the abstract is notincluded in section numbering). Use this numbering alsofor internal cross-referencing: do not just refer to "thetext" or „see above/below”. Any subsection may be givena brief heading. Each heading should appear on its ownseparate line.
Introduction State the objectives of the work andprovide an adequate background, avoiding a detailed lit-erature survey or a summary of the results.
Material and methods Provide sufficient detail toallow the work to be reproduced. Methods already pub-lished should be indicated by a reference: only relevantmodifications should be described.
Theory/calculation A Theory section should ex-tend, not repeat, the background to the article alreadydealt with in the Introduction and lay the foundation forfurther work. In contrast, a Calculation section representsa practical development from a theoretical basis.
Results Results should be clear and concise.
Discussion This should explore the significance ofthe results of the work, not repeat them. A combinedResults and Discussion section is often appropriate. Avoidextensive citations and discussion of published literature.
Conclusions The main conclusions of the study maybe presented in a short Conclusions section, which maystand alone or form a subsection of a Discussion or Resultsand Discussion section.
Appendices If there is more than one appendix, theyshould be identified as A, B, etc. Formulae and equationsin appendices should be given separate numbering: Eq.(A.1), Eq. (A.2), etc.; in a subsequent appendix, Eq. (B.1)and so on. Similarly for tables and figures: Table A.1; Fig.A.1, etc.
Lists (Numbered or Not Numbered) It is noproblem to use lists with either numbers or symbols. Upto 4 hierarchical levels can be handled by the layout sys-tem automatically. Examples are given in this article. Anexample of lists is given below:
List (not numbered, example 1)
• List Item 1
• List Item 2
• List Item 3
. Lower hierarchy item 1
. Lower hierarchy item 2
List (numbered, example 2)
1. List Item 12. List Item 23. List Item 3
(a) Lower hierarchy item 1(b) Lower hierarchy item 2
31

2 INSTRUCTIONS TO AUTHORS 2.6 Copyright Aspects
2.3.2. Essential title page informationPlease provide in separate paragraphs the full tile, se-
quence of authors, author names and complete affiliationsand title footnotes as well as author footnotes. One au-thor should be declared to be the corresponding authorand his/her email adress should be provided. Please in-dicate, in which position or with which author name therespective footnotes will have to appear.
2.4. Figures
Single Figures can be set either on one-column width(see Figure 1) or on two-column width (not shown). Se-veral connected figures can be arranged such that theyform a panel, with two subfigures one row being set two-column width. Depending on the vertical space needed toset the respective figures, panels of 2xn figures are possible.An example of this with two variants of the sign of theLudwig-Maximilians-University in one rowd is shown inFigures 2a and 2b. All figures should be referenced in therunning text as is shown in this paragraph for the twoexample figures. In word processor based manuscripts forNeadeA, the place, where a reference has to be placed inthe final layout has to be indicated clearly.
Figures must be of professional quality. When possi-ble, please use or supply first-generation artwork. Fig-ures are encouraged in electronic format with a printedproof on glossy or highquality photographic printer paperand should be at least 5 inches wide. Figures may beblack and white line art, graphs, halftones (photographs),or color. TIFF, EPS, or PDF formats are recommendedfor electronic artwork. Line art should have a minimumresolution of 1000 dpi, halftone art a minimum of 300 dpi,and combination art (line/halftone) a minimum of 500 dpi.MS Office files (Word, PowerPoint, and Excel) are also ac-cepted if they meet certain standards. Color figures shouldbe submitted in actual size.
When possible all lettering should be within the frame-work of the illustration; likewise the key to symbols shouldbe on the face of the chart. The following standard sym-bols should be used as they are easily available to theprinter: + ο ˙ [] ˘. Actual magnification of all photomi-crographs should be provided. Dimension scale should beindicated.
For help with preparing electronic artwork for bothon-screen review and print the Elsevier Author Gateway(http://authors.elsevier.com) is a good source to visit.
2.5. Tables
Tables can be set in a similar format as figures. Theycan be written completely in any word-processing format,but the transfer of tables into the layout system used hereis more difficult than the transfer of figures. Thus, authorsshould read proofs with tables very carefully. Like figures,tables should be referenced carefully in the text. The ta-bles should be self-explanatory and it should be possible to
have them set on a different print page than the text refer-ring to them. Table structure should be kept as simple aspossible and fancy layout options should be avoided, wherepossible. Special requirements for table formatting shouldalways be discussed with the redaction team of NeadeA inadvance. Like figures, tables will be numbered consecu-tively, each table should be provided on a separate page,if prepared in a word processor manuscript. Tables shouldbe numbered consecutively in arabic numbers. Subtables(like subfigues) should be numbered with letters.
In principal, tables can be one-coumn-width, two-column-width or cover a whole page, which would causethe page to be turned 90° to become a landscape formatpage.
An example of a table in one-column-width (german lan-guage) is shown in table 1.
Tabelle 1: Beispieltabelle
System Medipix 1 Medipix 2
Detektordicke [µm] 300 300 700
Kantenwinkel [°] 3.55 2.71 7.99
räumliche Auflösung [µm] 4.26 10.17 10.56
MTF bei fmax 0.53 0.37 0.39
LSF räumliche Auflösungin µm 129.7 52.75 50.78
in % der Pixelgröße 76.3 95.9 92.3
2.6. Copyright AspectsAll copyrights of materials being used for internal com-
munication with NeadeA reside with the authors. NeadeAdoes not request transfer of copyrights. All copyright-related committments also stay with the authors of a con-tribution.
32

2 INSTRUCTIONS TO AUTHORS 2.6 Copyright Aspects
(a) This shows a subfigure of a 2x1 panel. Panels can cover highernumbers of subfigures, but should be in the format 2xX subfigures.
(b) This is an example of a subfigure in a panel 2x1
Figure 2: This is an example of one figure built from two subfigures being set in two-column-width
33

NeadeAAnatomische Anstaltder Ludwig-Maximilians-Universität MünchenPettenkoferstraße 11D-80336 MünchenTel: 089-5160-4810 oder 4820Fax: 089-5160-4897E-Mail: [email protected]: http:// www.munich-anatomy-competence.org
©be
i den
Aut
oren
, V.i.S.d.P.
: Han
s-G
eorg
Fra
nk