
Effekt einer L-Carnitinzulage auf Leistungsparameter und ... · das energiereiches Phosphat mit...
179
Aus dem Physiologischen Institut und dem Institut für Tierernährung der Tierärztlichen Hochschule Hannover Effekt einer L-Carnitinzulage auf Leistungsparameter und den Muskelcarnitingehalt bei jungen Trabern im Laufe eines Trainings INAUGURAL-DISSERTATION zur Erlangung des Grades eines DOCTOR MEDICINAE VETERINARIAE durch die Tierärztliche Hochschule Hannover Vorgelegt von Cornelia Chrobok aus Mülheim an der Ruhr Hannover 2000
Transcript of Effekt einer L-Carnitinzulage auf Leistungsparameter und ... · das energiereiches Phosphat mit...

Aus dem Physiologischen Institut und dem Institut für Tierernährungder Tierärztlichen Hochschule Hannover
Effekt einer L-Carnitinzulageauf Leistungsparameter und den Muskelcarnitingehalt
bei jungen Trabern im Laufe eines Trainings
INAUGURAL-DISSERTATIONzur Erlangung des Grades eines
DOCTOR MEDICINAE VETERINARIAEdurch die Tierärztliche Hochschule Hannover
Vorgelegt vonCornelia Chrobok
aus Mülheim an der Ruhr
Hannover 2000

Wissenschaftliche Betreuung: Univ.-Prof. Dr. J. Harmeyer
Univ.-Prof. Dr. M. Coenen
1. Gutachter: Univ.-Prof. Dr. J. Harmeyer und Univ. -Prof. Dr. M. Coenen
2. Gutachter: Prof. Dr. E. Kallweit
Tag der mündlichen Prüfung: 21.11.2000

Meinen ElternMeinen Eltern


Inhaltsverzeichnis
Kapitel Seite
I. Einleitung..........................................................................................................9
II. Schrifttum .....................................................................................................10
1. Grundlagen des Energiestoffwechsels .............................................................................. 10
2. L-Carnitin: Chemische Struktur und Biosynthese ......................................................... 15
3. Funktionen von L-Carnitin ............................................................................................... 18
4. Carnitinstoffwechsel........................................................................................................... 22
4.1. Absorption und Elimination .......................................................................................... 22
4.2. Carnitingehalte im Körper............................................................................................. 23
4.3. Altersabhängigkeit ........................................................................................................ 25
4.4. Tagesschwankungen im Plasmacarnitin........................................................................ 26
5. Carnitingehalte in Futter- und Lebensmitteln ................................................................ 27
6. Trainings- und belastungsbedingte Einflüsse auf den Carnitinstatus........................... 27
7. Energiestoffwechsel des Pferdes während der Belastung und nach dem Training ..... 29
8. Einfluss von Carnitinzulagen ............................................................................................ 33
9. Abschließende Bemerkung................................................................................................ 41
III. Material und Methoden .............................................................................42
1. Versuchsziel und Fragestellung ........................................................................................ 42
2. Versuchsaufbau .................................................................................................................. 42
3. Versuchstiere ...................................................................................................................... 44
4. Fütterung............................................................................................................................. 45
5. Versuchsdurchführung ...................................................................................................... 47
5.1. Stufentest ....................................................................................................................... 47

5.2. Trainingsphase .............................................................................................................. 49
5.2.1. Kurzzeitbelastung................................................................................................... 50
5.2.2. Langzeitbelastung................................................................................................... 51
5.3. Ruheperiode .................................................................................................................. 52
5.4. Probennahmen............................................................................................................... 53
6. Messungen und Analyse biochemischer Parameter........................................................ 56
6.1. L-Carnitin im Plasma .................................................................................................... 56
6.2. L-Carnitin in der Muskulatur ........................................................................................ 58
6.3. Herzfrequenz ................................................................................................................. 60
6.4. Lactat im Vollblut ......................................................................................................... 60
6.5. Glucose im Plasma ........................................................................................................ 63
6.6. Glykogen in der Muskulatur ......................................................................................... 63
6.7. Freie Fettsäuren im Plasma ........................................................................................... 64
6.8. Harnstoff im Plasma...................................................................................................... 65
6.9. Hämatokrit..................................................................................................................... 66
6.10. Gesamtprotein im Plasma............................................................................................ 66
6.11. Körpergewicht ............................................................................................................. 66
7. Statistik................................................................................................................................ 67
IV. Ergebnisse ....................................................................................................69
1. L-Carnitin im Plasma ........................................................................................................ 69
2. L-Carnitin in der Muskulatur........................................................................................... 73
3. Herzfrequenz ...................................................................................................................... 78
4. Lactat im Vollblut .............................................................................................................. 83
5. Glucose im Plasma ............................................................................................................. 89
6. Glykogen in der Muskulatur............................................................................................. 92
7. Freie Fettsäuren im Plasma............................................................................................... 94
8. Harnstoff im Plasma .......................................................................................................... 96
9. Hämatokrit.......................................................................................................................... 97
10. Gesamtprotein im Plasma ............................................................................................... 97

V. Diskussion ......................................................................................................99
1. Kritik der Versuchseinstellung ......................................................................................... 99
1.1. Versuchsdurchführung .................................................................................................. 99
1.2. Tiergut ......................................................................................................................... 102
2. Diskussion der Ergebnisse ............................................................................................... 103
2.1. L-Carnitin im Plasma .................................................................................................. 103
2.2. L-Carnitin in der Muskulatur ...................................................................................... 105
2.3. Herzfrequenz ............................................................................................................... 108
2.4. Lactat ........................................................................................................................... 110
2.5. Glucose im Plasma ...................................................................................................... 111
2.6. Glykogen in der Muskulatur ....................................................................................... 117
2.7. Freie Fettsäuren im Plasma ......................................................................................... 119
3. Schlußfolgerungen............................................................................................................ 127
VI. Zusammenfassung.....................................................................................129
VII. Summary ..................................................................................................132
VIII. Literaturverzeichnis ..............................................................................134
IX. Tabellenanhang .........................................................................................160

Abkürzungsverzeichnis Carnitin L-Carnitin CoA Coenzym A DE digestible energy, verdauliche Energie FC Freies Carnitin FFA Free fatty acids FFS Freie Fettsäuren GC Gesamtcarnitin HF Herzfrequenz KGW Körpergewicht n. Bel. Nach Belastung PDH Pyruvat-Dehydrogenase x arithmetischer Mittelwert SD Standardabweichung SpM Schläge pro Minute TPP Gesamtprotein im Plasma TM Trockenmasse uS ursprüngliche Substanz v Geschwindigkeit v2,5 Geschwindigkeit, die rechnerisch eine Lactatkonzentration von 2,5
mmol/l Blut hervorruft
v4 Geschwindigkeit, die rechnerisch eine Lactatkonzentration von 4
mmol/l Blut hervorruft

I. Einleitung________________________________________________________________________
9
I. Einleitung
L-Carnitin ist ein lebenswichtiges körpereigenes Aminosäurenderivat, dessen Bedarf durch
Eigensynthese und durch alimentäre Zufuhr gedeckt wird. Pflanzliche Futtermittel sind arm
an Carnitin, während tierische Futter- und Nahrungsmittel meist viel Carnitin enthalten. Das
gilt besonders für Herz- und Skelettmuskulatur. Als Pflanzenfresser sind Pferde somit
hauptsächlich auf die körpereigene Synthese von Carnitin angewiesen.
Carnitin fungiert im Körper als Carrier für den Transport von langkettigen Fettsäuren aus dem
Cytosol durch die innere Mitochondrienmembran zum Ort der b-Oxidation. Ferner fungiert es
als Regulator des Acetyl-CoA/CoA Verhältnisses in der Mitochondrienmatrix und im
Cytosol. Carnitin spielt dadurch eine wichtige Rolle für die Fettsäurenoxidation und für die
Funktion des Citratcyclus. Es wurde daher vermutet, dass zusätzliche Gaben von Carnitin die
körperliche Leistung z. B. durch Stimulation der Fettsäurenoxidation bei Tieren und
Menschen steigern könnten. Bei Tauben, Ratten, Hunden und Menschen wurde über einen
solchen positiven Einfluss von Carnitin auf die Leistungsfähigkeit berichtet (BORGHIJS u.
DE WILDE 1992; DUBELAAR et al. 1994). Beim Pferd lassen dazu durchgeführte
Untersuchungen noch keinen eindeutigen Schluss zu. Hinweise auf mögliche positive Effekte
auch beim Pferd liegen aus Untersuchungen von IBEN et al. (1992) und FALASCHINI u.
TROMBETTA (1994) vor.
Wichtig für die Erzielung eines positiven Effektes von Carnitinzulagen auf motorische
Leistungen könnte sein, dass es durch solche Zulagen zu einer Erhöhung des Carnitingehaltes
in der Muskulatur kommt. In Untersuchungen bei Ratten konnte nach sechswöchigen oralen
Gaben von 500 mg/kg pro Tag eine Zunahme des Carnitingehaltes in der Muskulatur um
74 % nachgewiesen werden. Ob ein solcher Effekt auch beim Pferd zu erzielen ist, ist bisher
nicht bekannt. Der Transport von Carnitin aus dem Blutplasma über das Plasmalemma in die
Muskelzellen ist ein langsamer energieabhängiger Prozess.
In diesem Projekt wurde bei Sportpferden der Effekt einer mehrmonatigen oralen
Carnitinzulage in Verbindung mit einem fünfwöchigen Training auf den Carnitingehalt in der
Muskulatur und auf Leistungsparameter untersucht.

II. Schrifttum ___________________________________________________________________________
10
II. Schrifttum:
1. Grundlagen des Energiestoffwechsels
Muskuläre Tätigkeit erfordert die Umwandlung von chemischer Bindungsenergie in
mechanische Arbeit. Es handelt sich bei dieser Umwandlung in der Muskulatur um einen
komplexen Vorgang, der durch viele Faktoren beeinflusst wird. Die Akkumulation von
Stoffwechselendprodukten kann z. B. die Bereitstellung der Energie limitieren.
Die von der Muskulatur primär verwertbare Energiequelle ist Adenosintriphosphat (ATP), das
in Adenosindiphosphat (ADP) und freies Phosphat gespalten wird. Der Abbau der
verschiedenen Energie liefernden Substrate dient dieser ATP-Synthese.
Der ruhende Muskel hat nur geringe ATP-Mengen gespeichert (22-28 mmol/kg TM) (SNOW
et al. 1985; ESSEN-GUSTAVSSON et al. 1995), die selbst in Ruhe nur für drei Minuten
(HOLLMANN u. HETTINGER 1990), bei maximaler Belastung nur für eine Sekunde
ausreichen (ÅSTRAND u. ROHDAHL 1986).
Zusätzlich zu diesem ATP-Vorrat besitzt die Muskulatur im Gegensatz zu den meisten
anderen Organen einen zusätzlichen energiereichen Phosphatspeicher – das Kreatinphosphat,
das energiereiches Phosphat mit Hilfe der Kreatinkinase auf ADP übertragen kann. Der
Energiespeicher in Form von Kreatinphosphat ist drei bis vier mal so groß wie der in Form
von ATP. HARRIS et al. (1987) ermittelte beim Pferd eine durchschnittliche
Kreatinphosphat-Konzentration in der Muskulatur von 58,4 - 62,2 mmol/kg TM.
ATP ADP + Pi
Energie Arbeit
ATP ADP + Pi
Energie Arbeit

II. Schrifttum ___________________________________________________________________________
11
Kreatinkinase
Kreatinphosphat + ADP Kreatin +ATP
In Situationen eines akuten ATP-Mangels kann der Muskel darüber hinaus aus zwei
Molekülen ADP ein Molekül ATP zurückgewinnen. Dabei entsteht dann ein Molekül AMP,
das weiter abgebaut wird. Diese Reaktion wird durch die Myokinase katalysiert und läuft auf
niedriger Stufe bei jeder Belastung ab (Myokinasereaktion) (MC MIKEN 1983).
Myokinase
2 ADP ATP + AMP
Die verschiedenen energiereichen Phosphatverbindungen können nur kurzfristig die
Kontraktionsarbeit des Muskels unterhalten. Daher müssen andere Energie liefernde Substrate
eine Resynthese von ADP zu ATP langfristig ermöglichen. Hauptsubstrate, aus denen der
Muskel ATP gewinnt, sind Glucose und freie Fettsäuren einschließlich deren Speicherformen,
Glykogen und Triglyceride sowie einige unter bestimmten Bedingungen beim Abbau
entstehende Zwischenprodukte (Lactat, Ketonkörper) (KEUL u. BERG 1985). Auch
Aminosäuren können in geringerem Umfang der Energiebereitstellung dienen.
Von den Energie liefernden Substraten kann die Glucose unter zwei verschiedenen
Voraussetzungen abgebaut und zur Synthese energiereicher Phosphatverbindungen genutzt
werden, anaerob und aerob. Diese Wahlmöglichkeit bietet sich nur beim Abbau von Glucose.
Bei der Glykolyse wird Glucose bzw. deren Speicherform Glykogen ohne Verbrauch von
Sauerstoff zu Pyruvat abgebaut. Dabei wird ATP in der Phosphoglycerat- und in der
Pyruvatkinasereaktion gebildet. Dieser Abbauweg ist jedoch durch die Akkumulation von
Pyruvat, dem damit verbundenen Anstieg von Lactat und der dadurch ausgelösten pH-Wert
Senkung limitiert. Beim anaeroben Abbau von einem Mol Glucose werden zwei Mol ATP
und zwei Mol Lactat gebildet.

II. Schrifttum ___________________________________________________________________________
12
Eine weitere und wesentlich effektivere Möglichkeit zur Bereitstellung von ATP besitzt der
Muskel durch den oxidativen (aeroben) Abbau von Kohlenhydraten und auch von Fetten.
Dabei wird ADP in Anwesenheit von Sauerstoff in der Atmungskette in den Mitochondrien in
viel größerem Umfang zu ATP rephosphoryliert (HOLLMANN u. HETTINGER 1990). Es
entstehen bei der aeroben Umsetzung von einem Mol Glucose 38 Mol ATP. Dieser
Energiegewinn ist also, verglichen mit dem anaeroben Abbau der Glucose (zwei Mol ATP),
erheblich effektiver. Die vollständige Oxidation von einem Molekül Palmitat, einer C18-
kettigen FFS, liefert 129 Mol ATP.
Abgesehen vom Belastungsbeginn, an dem die aerobe ATP-Synthese noch nicht in Gang
gekommen ist, laufen bei muskulärer Arbeit aerobe und anaerobe Prozesse simultan ab. Die
aerobe Energiebereitstellung erreicht bei spontan einsetzender Maximalbelastung erst nach 10
bis 15 Sekunden einen nennenswerten Umfang und nimmt von da an ständig zu. Beim Pferd
haben TYLER et al. (1996) gezeigt, dass der aerobe Anteil an der ATP-Synthese in einem
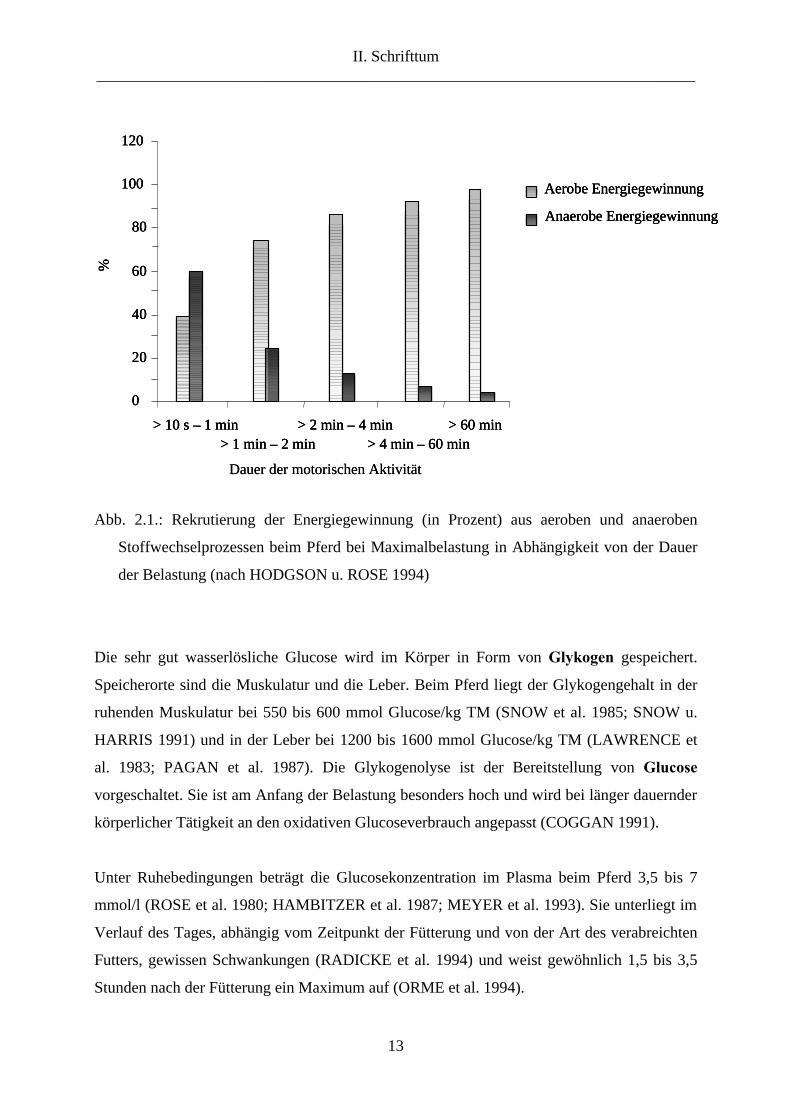
sich über ein bis zwei Minuten erstreckenden Rennen mehr als 70 % ausmacht. Abbildung
2.1. illustriert, wie sich die auf aerobe und anaerobe Prozesse stützende Energiegewinnung
beim Pferd unter Maximalbelastung verteilt in Abhängigkeit von der Dauer der Belastung.
II. Schrifttum ___________________________________________________________________________
13
Abb. 2.1.: Rekrutierung der Energiegewinnung (in Prozent) aus aeroben und anaeroben
Stoffwechselprozessen beim Pferd bei Maximalbelastung in Abhängigkeit von der Dauer
der Belastung (nach HODGSON u. ROSE 1994)
Die sehr gut wasserlösliche Glucose wird im Körper in Form von Glykogen gespeichert.
Speicherorte sind die Muskulatur und die Leber. Beim Pferd liegt der Glykogengehalt in der
ruhenden Muskulatur bei 550 bis 600 mmol Glucose/kg TM (SNOW et al. 1985; SNOW u.
HARRIS 1991) und in der Leber bei 1200 bis 1600 mmol Glucose/kg TM (LAWRENCE et
al. 1983; PAGAN et al. 1987). Die Glykogenolyse ist der Bereitstellung von Glucose
vorgeschaltet. Sie ist am Anfang der Belastung besonders hoch und wird bei länger dauernder
körperlicher Tätigkeit an den oxidativen Glucoseverbrauch angepasst (COGGAN 1991).
Unter Ruhebedingungen beträgt die Glucosekonzentration im Plasma beim Pferd 3,5 bis 7
mmol/l (ROSE et al. 1980; HAMBITZER et al. 1987; MEYER et al. 1993). Sie unterliegt im
Verlauf des Tages, abhängig vom Zeitpunkt der Fütterung und von der Art des verabreichten
Futters, gewissen Schwankungen (RADICKE et al. 1994) und weist gewöhnlich 1,5 bis 3,5
Stunden nach der Fütterung ein Maximum auf (ORME et al. 1994).
Aerobe Energiegewinnung
0
20
40
60
80
100
120
Anaerobe Energiegewinnung
%
> 10 s – 1 min > 2 min – 4 min > 60 min > 1 min – 2 min > 4 min – 60 min
Dauer der motorischen Aktivität
Aerobe Energiegewinnung
0
20
40
60
80
100
120
Anaerobe Energiegewinnung
%
> 10 s – 1 min > 2 min – 4 min > 60 min > 1 min – 2 min > 4 min – 60 min
Aerobe Energiegewinnung
0
20
40
60
80
100
120
Anaerobe Energiegewinnung
%
> 10 s – 1 min > 2 min – 4 min > 60 min > 1 min – 2 min > 4 min – 60 min
Dauer der motorischen Aktivität

II. Schrifttum ___________________________________________________________________________
14
Fettsäuren sind im Körper als Triglyceride gespeichert und im Fettgewebe sowie in der
Muskulatur deponiert. Die Trigycerid-Konzentration in der Muskulatur beträgt beim Pferd 27
bis 30 mmol/kg TM und hängt vom Verfettungszustand der Tiere ab (ESSEN-
GUSTAVSSON et al. 1989).
Durch Lipolyse werden freie Fettsäuren (FFS) aus Triglyceriden der Fettdepots in das
Plasma abgegeben oder direkt in der Muskulatur freigesetzt. Sie stellen ebenfalls eine
wichtige Energiequelle für die Muskelarbeit dar (JONES et al. 1980). Bei lang andauernden,
mäßig hohen Belastungen gewinnt die Muskulatur ihre Energie hauptsächlich durch die
Oxidation von FFS (JONES et al. 1980). Die Konzentration der FFS im Plasma schwankt
beim Pferd bereits unter Ruhebedingungen zwischen 20 und 500 µmol/l (LUCKE u. HALL
1980 a, b, c; ORME et al. 1994; HYYPPÄ et al. 1997). ORME et al. (1994) berichteten über
einen 4,5-fachen Anstieg der FFS-Konzentration im Plasma zwischen 4 und 10 Uhr morgens.
Aminosäuren werden in geringem Umfang zur Energiebereitstellung genutzt. Bei ihrem
Abbau entsteht zunächst Ammoniak, welches zum größten Teil durch Transaminierung auf
Pyruvat übertragen wird. Dabei entsteht Alanin (FEHLIG 1973; PÖSÖ et al. 1987). Die N-
freien C-Ketten der Aminosäuren werden an verschiedenen Stellen in den Citratcyclus
eingeschleust. Bei länger andauernden Belastungen etwa ab 60 Minuten werden Aminosäuren
in zunehmenden Maß zur Gewinnung von Energie herangezogen (HARALAMBIE u. BERG
1976).
Über einen weiten Bereich von Belastungsintensitäten wird in der Muskulatur Energie
parallel aus der Verbrennung von Glucose und Fettsäuren bereitgestellt (RANDEL et al.
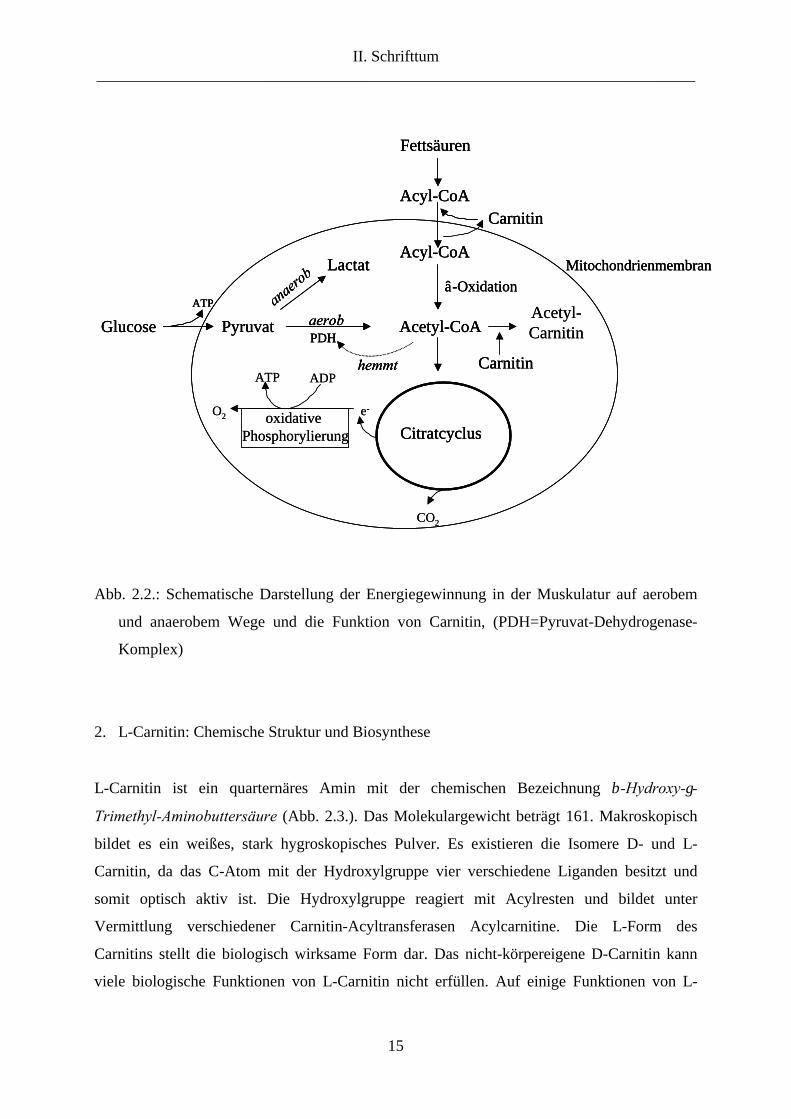
1963). Abbildung 2.2. veranschaulicht die daran beteiligten Stoffwechselwege.
II. Schrifttum ___________________________________________________________________________
15
Abb. 2.2.: Schematische Darstellung der Energiegewinnung in der Muskulatur auf aerobem
und anaerobem Wege und die Funktion von Carnitin, (PDH=Pyruvat-Dehydrogenase-
Komplex)
2. L-Carnitin: Chemische Struktur und Biosynthese
L-Carnitin ist ein quarternäres Amin mit der chemischen Bezeichnung β-Hydroxy-γ-
Trimethyl-Aminobuttersäure (Abb. 2.3.). Das Molekulargewicht beträgt 161. Makroskopisch
bildet es ein weißes, stark hygroskopisches Pulver. Es existieren die Isomere D- und L-
Carnitin, da das C-Atom mit der Hydroxylgruppe vier verschiedene Liganden besitzt und
somit optisch aktiv ist. Die Hydroxylgruppe reagiert mit Acylresten und bildet unter
Vermittlung verschiedener Carnitin-Acyltransferasen Acylcarnitine. Die L-Form des
Carnitins stellt die biologisch wirksame Form dar. Das nicht-körpereigene D-Carnitin kann
viele biologische Funktionen von L-Carnitin nicht erfüllen. Auf einige Funktionen von L-
Citratcyclus
Acetyl-CoAPyruvat aerob
Lactat
anaerob
PDH
â-OxidationMitochondrienmembran
hemmt
Acyl-CoA
Acyl-CoA
Fettsäuren
Carnitin
Carnitin
GlucoseATP
Acetyl-Carnitin
CO2
e-oxidative
Phosphorylierung
O2
ADPATP
Citratcyclus
Acetyl-CoAPyruvat aerob
Lactat
anaerob
PDH
â-OxidationMitochondrienmembran
hemmt
Acyl-CoA
Acyl-CoA
Fettsäuren
Carnitin
Carnitin
GlucoseATP
Citratcyclus
Acetyl-CoAPyruvat aerob
Lactat
anaerob
PDH
â-OxidationMitochondrienmembran
hemmt
Acyl-CoA
Acyl-CoA
Fettsäuren
Carnitin
Carnitin
GlucoseATP
Acetyl-Carnitin
CO2
e-oxidative
Phosphorylierung
O2
ADPATP

II. Schrifttum ___________________________________________________________________________
16
Carnitin wirkt es hemmend, z. B. auf die Carnitinacyltransferasen (MEIER 1987). D-Carnitin
muss daher als eine toxische Substanz angesehen werden.
CH3 OH
CH3 – N+ - CH2 - CH - CH2 - COO-
CH3
Abb. 2.3.: Strukturformel von L-Carnitin (β-Hydroxy-γ-Trimethyl-Aminobuttersäure)
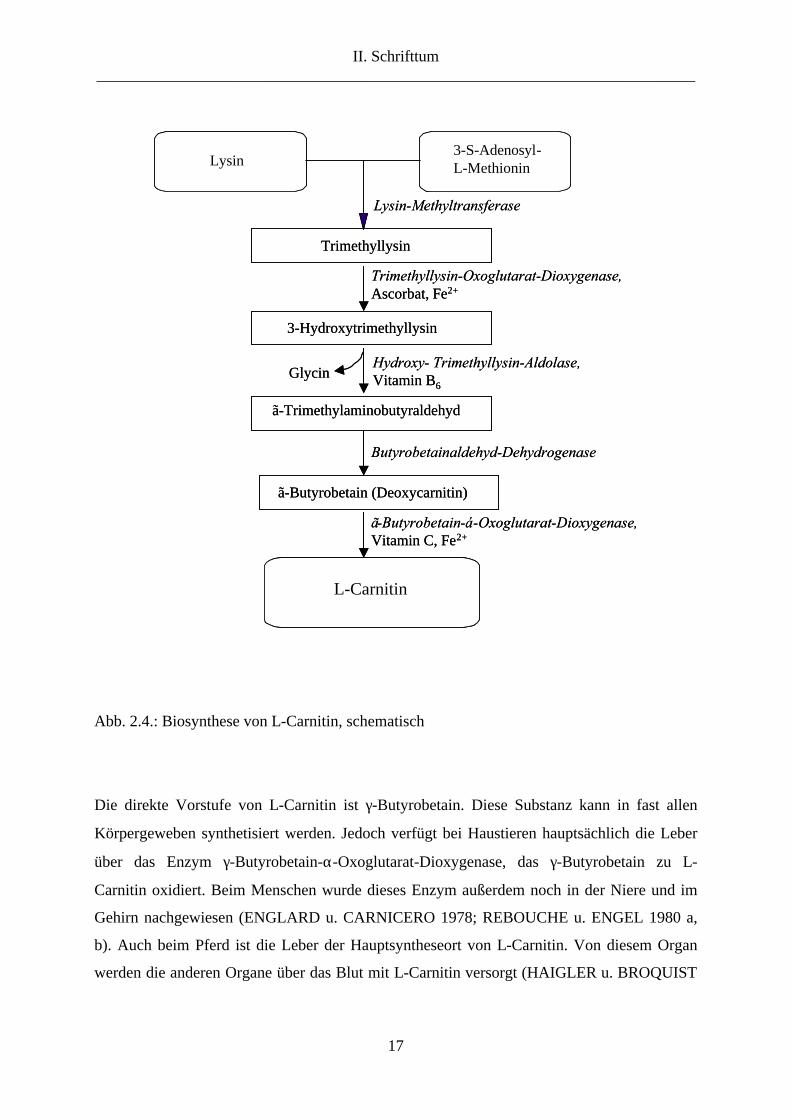
Carnitin wird aus Lysin und dem als Methylgruppendonator fungierenden Methionin über
mehrere Reaktionsschritte synthetisiert (BREMER 1961; LINDSTEDT u. LINDSTEDT
1961). Für seine Synthese ist ferner die Anwesenheit von Niacin, Vitamin B6, Ascorbinsäure
und Eisen notwendig (NEUMANN 1996) (Abb. 2.4.). Ein Mangel an diesen als Coenzym
bzw. Cofaktor wirkenden Stoffen kann daher die Carnitinbiosynthese im Körper
beeinträchtigen.
II. Schrifttum ___________________________________________________________________________
17
Abb. 2.4.: Biosynthese von L-Carnitin, schematisch
Die direkte Vorstufe von L-Carnitin ist γ-Butyrobetain. Diese Substanz kann in fast allen
Körpergeweben synthetisiert werden. Jedoch verfügt bei Haustieren hauptsächlich die Leber
über das Enzym γ-Butyrobetain-α-Oxoglutarat-Dioxygenase, das γ-Butyrobetain zu L-
Carnitin oxidiert. Beim Menschen wurde dieses Enzym außerdem noch in der Niere und im
Gehirn nachgewiesen (ENGLARD u. CARNICERO 1978; REBOUCHE u. ENGEL 1980 a,
b). Auch beim Pferd ist die Leber der Hauptsyntheseort von L-Carnitin. Von diesem Organ
werden die anderen Organe über das Blut mit L-Carnitin versorgt (HAIGLER u. BROQUIST
ã-Butyrobetain (Deoxycarnitin)
L-Carnitin
ã-Butyrobetain-á-Oxoglutarat-Dioxygenase,Vitamin C, Fe2+
Lysin-Methyltransferase
Trimethyllysin
3-Hydroxytrimethyllysin
ã-Trimethylaminobutyraldehyd
Trimethyllysin-Oxoglutarat-Dioxygenase,Ascorbat, Fe2+
Glycin
Butyrobetainaldehyd-Dehydrogenase
3-S-Adenosyl-L-MethioninLysin
Hydroxy- Trimethyllysin-Aldolase,Vitamin B6
ã-Butyrobetain (Deoxycarnitin)
L-CarnitinL-Carnitin
ã-Butyrobetain-á-Oxoglutarat-Dioxygenase,Vitamin C, Fe2+
Lysin-Methyltransferase
Trimethyllysin
3-Hydroxytrimethyllysin
ã-Trimethylaminobutyraldehyd
Trimethyllysin-Oxoglutarat-Dioxygenase,Ascorbat, Fe2+
Glycin
Butyrobetainaldehyd-Dehydrogenase
3-S-Adenosyl-L-MethioninLysin
Hydroxy- Trimethyllysin-Aldolase,Vitamin B6

II. Schrifttum ___________________________________________________________________________
18
1974; BROOKS u. MCINTOSH 1975; CEDERBLAD u. LINDSTEDT 1976; ZASPEL et al.
1980). Somit ist auch die Skelettmuskulatur auf die Synthese von L-Carnitin durch die Leber
angewiesen. Die Aufnahme aus dem Blut erfolgt mit Hilfe eines aktiven Natrium-abhängigen
Transportmechanismus entgegen einem Konzentrations-gradienten (BRASS 1992). Die
Synthesekapazität der Leber für Carnitin wird bei der Ratte von CEDERBLAD und
LINDSTEDT (1976) auf 2 µmol/100 g (3,2 g/kg) Körpergewicht pro Tag geschätzt. Der
Gehalt im Körper der Ratte beträgt 35 µmol/100 g (56,4 g/kg) Körpergewicht. Der tägliche
Umsatz soll dabei etwa 7 % (2,6 µmol/100 g, 4,2 g/kg Körpergewicht) betragen.
3. Funktionen von L-Carnitin
In Studien über den Wirkungsmechanismus von Carnitin berichteten FRIEDMANN und
FRAENKEL (1955) über eine reversible Transacetylierung zwischen L-Carnitin und
Coenzym A. Diese Reaktion steht im Zusammenhang mit dem Transport aktivierter
Fettsäuren aus dem Cytosol in die Mitochondrien. Zur gleichen Zeit zeigte FRITZ (1955),
dass Carnitin in Leberhomogenaten von Ratten die Fettsäurenoxidation stimuliert. Die
Autoren schrieben dem Carnitin daraufhin eine Funktion beim Transport von Fettsäuren aus
dem Cytosol in die Mitochondrien zu. Diese Rolle von Carnitin im Intermediärstoffwechsel
als Überträger von mittel- und langkettigen Fettsäuren aus dem Cytosol der Zelle in das
Mitochondrium, dem Ort der β-Oxidation, wurde von MARQUIS und FRITZ (1965) erstmals
nachgewiesen. Sie wird bis heute als eine der wichtigsten biologischen Funktionen von L-
Carnitin angesehen.
Für den Transport der aktivierten langkettigen Fettsäuren durch die innere
Mitochondrienmembran werden die an CoA gebundenen cytosolischen Fettsäuren mit Hilfe
einer Carnitin-Acyltransferase (I) vom CoA an Carnitin transferiert. Eine Carnitin-
Acylcarnitin-Translokase transportiert den Carnitin-Fettsäure-Ester im Austausch gegen freies
intramitochondriales Carnitin in die Mitochondrienmatrix (PANDE 1975). Dort wird durch
die Carnitin-Acyltransferase (II) der Acylrest wieder auf CoA übertragen. Das freie Carnitin
wird wieder in das Cytosol zurücktransportiert und steht dort für die Übernahme eines

II. Schrifttum ___________________________________________________________________________
19
weiteren aktivierten Acylrestes zur Verfügung (PANDE 1975; RAMSAY u. TUBBS 1975;
PAVIN u. PANDE 1979) (Abb. 2.5.).
Abb. 2.5.: Schematische Darstellung der Carrier-Funktion von L-Carnitin für den Transport
aktivierter Fettsäuren aus dem Cytosol in die Mitochondrien (Carnitin-Shuttle)
Unter bestimmten metabolischen Bedingungen wie Ketose, Diabetes, Hunger oder sportlichen
Hochleistungen im anaeroben Bereich akkumuliert Acetyl–CoA in den Mitochondrien
(CARTER et al. 1981), da die Kapazität des Citratcyclus für den Abbau der unter diesen
Bedingungen gebildeten Acetyl-CoA-Mengen nicht ausreicht. Carnitin übernimmt unter
diesen Bedingungen die Funktion eines Acetylpuffers. Ein Vorteil dieser Pufferfunktionen
besteht darin, dass CoA nach der Übertragung des Acetylrestes auf Carnitin wieder für
katabole Stoffwechselvorgänge zur Verfügung steht (Abb. 2.6.). Das sich intramitochondrial
anreichernde Acetyl-CoA rekrutiert sich aus dem Abbau von Pyruvat oder stammt aus dem
Fettabbau. Die Speicherung von aktivierten Acetylresten in Form von Acetylcarnitin beugt
einer Hemmung der Pyruvatdehydrogenase vor und vermindert den cytotoxischen Effekt, der
mit einer Oxidation des Pyruvats zu Lactat und der daraus resultierenden pH-Wert
Acyl-CoA Acetyl-CoAAcyl-CoAL-Carnitin
Coenzym AAcyl-Carnitin
Freie Fettsäuren
Citrat-zyklus
Coenzym A
Muskelzelle
Mitochondrium
Acyl-CoA Acetyl-CoAAcyl-CoAL-Carnitin
Coenzym AAcyl-Carnitin
Freie Fettsäuren
Citrat-zyklus
Coenzym A
Muskelzelle
Mitochondrium
II. Schrifttum ___________________________________________________________________________
20
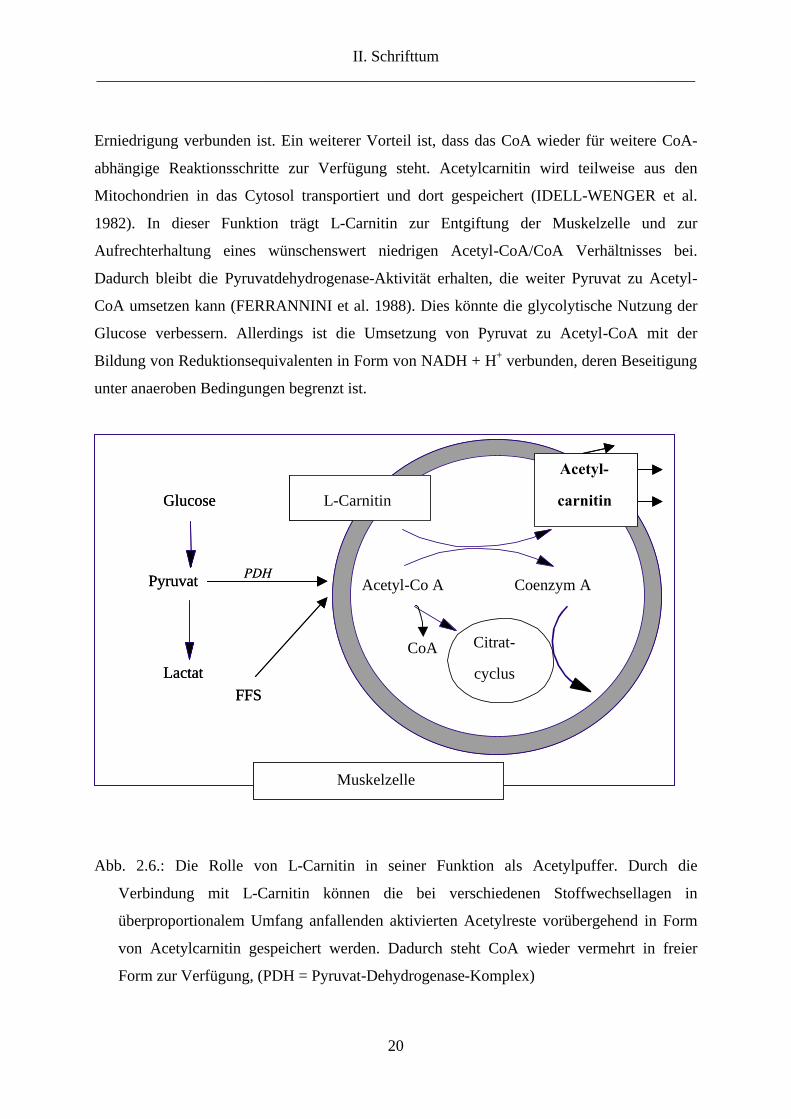
Erniedrigung verbunden ist. Ein weiterer Vorteil ist, dass das CoA wieder für weitere CoA-
abhängige Reaktionsschritte zur Verfügung steht. Acetylcarnitin wird teilweise aus den
Mitochondrien in das Cytosol transportiert und dort gespeichert (IDELL-WENGER et al.
1982). In dieser Funktion trägt L-Carnitin zur Entgiftung der Muskelzelle und zur
Aufrechterhaltung eines wünschenswert niedrigen Acetyl-CoA/CoA Verhältnisses bei.
Dadurch bleibt die Pyruvatdehydrogenase-Aktivität erhalten, die weiter Pyruvat zu Acetyl-
CoA umsetzen kann (FERRANNINI et al. 1988). Dies könnte die glycolytische Nutzung der
Glucose verbessern. Allerdings ist die Umsetzung von Pyruvat zu Acetyl-CoA mit der
Bildung von Reduktionsequivalenten in Form von NADH + H+ verbunden, deren Beseitigung
unter anaeroben Bedingungen begrenzt ist.
Abb. 2.6.: Die Rolle von L-Carnitin in seiner Funktion als Acetylpuffer. Durch die
Verbindung mit L-Carnitin können die bei verschiedenen Stoffwechsellagen in
überproportionalem Umfang anfallenden aktivierten Acetylreste vorübergehend in Form
von Acetylcarnitin gespeichert werden. Dadurch steht CoA wieder vermehrt in freier
Form zur Verfügung, (PDH = Pyruvat-Dehydrogenase-Komplex)
Glucose
Pyruvat Acetyl-Co A
Citrat-
cyclus
Coenzym A
Acetyl-
carnitinL-Carnitin
Lactat
Muskelzelle
FFS
CoA
PDH
Glucose
Pyruvat Acetyl-Co A
Citrat-
cyclus
Coenzym A
Acetyl-
carnitinL-Carnitin
Lactat
Muskelzelle
FFS
CoA
PDH

II. Schrifttum ___________________________________________________________________________
21
Es lassen sich die folgenden Funktionen von Carnitin charakterisieren:
a. katalytische Funktion: Transport von langkettigen aktivierten Fettsäuren in die
Mitochondrien für die β-Oxidation (L-Carnitin wird nicht verbraucht)
b. metabolische Funktion: Abpufferung überschüssiger aktivierter Acetylreste, dadurch
Stimulation der Pyruvat-Dehydrogenaseaktivität, (L-Carnitin wird
verbraucht)
Bei der katalytischen Funktion wird Carnitin nur in verhältnismäßig geringen Mengen
benötigt und nicht verbraucht, da es nach dem Transport von FFS durch die
Mitochiondrienmembran wieder in das Cytosol zurück befördert wird. Es steht dann für den
Transport von Fettsäuren wieder zur Verfügung. Bei der metabolischen Funktion wird freies
Carnitin zu Acetylcarnitin umgesetzt und dabei in größeren Mengen benötigt und gebunden.
Die Konzentration an freiem Carnitin in der ruhenden Muskulatur (4500 µmol/kg) ist 10 bis
100-fach größer als die vieler anderer am Energieumsatz beteiligter Intermediärprodukte, wie
z. B. die von Acetyl-CoA (20 µmol/kg), Citrat (250 µmol/kg) oder Pyruvat (45 µmol/kg).
Carnitin kommt in mmolaren Konzentrationen vor, während die meisten anderen an der
Energiegewinnung beteiligten Substrate im µmolaren Bereich liegen. Diese hohe
Konzentration an Carnitin ist erforderlich, um als Speicher zu fungieren.
Außer den bisher genannten Funktionen sind noch weitere biochemische Funktionen von
Carnitin beschrieben worden. CIPOLLA et al. (1999) wiesen in vitro an Endothelien und
glatten Muskelzellen menschlicher Arterien einen vasodilatierenden Effekt von Propionyl-L-
Carnitin nach. Nach Zugabe von Propionyl-L-Carnitin in einer Konzentration von 0,1 bis 100
µmol/l zu Inkubationslösungen mit Arterien, bei denen der Lumendurchmesser und der
Wanddruck registriert wurde, vergrößerte sich der Gefäßdurchmesser dosisabhängig um bis
zu 91 %. Vor der Messung befanden sich die Gefäße in einem Zustand der Vasokonstriktion.
BETTINI et al. (1985) machten die Beobachtung, dass Plasmacarnitin die Ausschüttung des
„Endothelial Derived Relaxing Factors“ (EDRF), einem körpereigenen Vasodilatator,
verstärkt. Dieser durchblutungsfördernde Effekt kommt besonders beim untrainierten Muskel
zum Tragen, wie die Studie von DUBELAAR et al. (1991) gezeigt hat. Diese Autoren haben
bei Hunden mittels Elektrostimulation den Musculus latissimus dorsi 12 Wochen lang

II. Schrifttum ___________________________________________________________________________
22
kontinuierlich mit einer sich alle zwei Wochen bis 60 Herz steigernden Frequenz gereizt
(„trainiert“). Zu Beginn und am Ende dieses Trainings wurde die Muskelkraft gemessen.
Dazu wurde den Tieren unter Narkose für acht Minuten gleichzeitig eine Lösung mit 0,15
mmol L-Carnitin pro Kilogramm oder eine physiologische Kochsalzlösung infundiert. Bei
den Tieren vor Elektrostimulation führte die Carnitininfusion im Vergleich zur Infusion von
physiologischer Kochsalzlösung zu einer Zunahme der Muskelkraft um 15 %, bei den
trainierten Tieren führte die Carnitininfusion im Vergleich zu physiologischer
Kochsalzlösung zu einer Zunahme um 3 %. Die Autoren schließen einen Effekt von Carnitin
in der Muskulatur aufgrund der kurzen Supplementationszeit (acht Minuten) aus und
begründen die oben aufgeführten Unterschiede mit einem vasodilatierenden Effekt.
4. Carnitinstoffwechsel
4.1. Absorption und Elimination
Außer durch endogene Synthese wird dem Körper noch mit der Nahrung Carnitin zugeführt.
Oral aufgenommenes Carnitin wird im Dünndarm absorbiert (HARRIS et al. 1995 a, b). Die
Absorption findet durch passive Diffusion und durch einen Natrium-abhängigen aktiven
Transport statt (SHAW et al. 1983; GUDJONSSON et al. 1985 a; LI et al. 1992). Beide
Vorgänge laufen im Duodenum und Jejunum ab. Im Ileum ist nur noch passive Diffusion
möglich (SHAW et al. 1983).
Bis jetzt wurden keine Untersuchungen über den Mechanismus der intestinalen Absorption
beim Pferd durchgeführt. FOSTER et al. (1988) gaben Pferden oral steigende Dosen L-
Carnitin (0 bis 60 g/d). Fraktionen von 3,5 bis 7,5 % der supplementierten Dosis wurden
innerhalb der ersten der ersten 24 Stunden wieder im Urin eliminiert. Daraus schlossen die
Autoren, dass die Absorptionsfähigkeit von oral verabreichtem Carnitin beim Pferd gering ist.
In weiteren Untersuchungen bei Pferden von HARRIS et al. (1995b) mit oral verabreichten
Carnitindosen von 10 g/d kam es nur zu einem geringen Anstieg der renalen Ausscheidung
von Carnitin während 24 Stunden. Bis zu dieser Dosis scheint die Nierenschwelle nicht
überschritten worden zu sein. Die Autoren folgerten daraus, dass oral verabreichtes Carnitin

II. Schrifttum ___________________________________________________________________________
23
auch im Gewebe gespeichert wird. Bei intravenöser Verabreichung von 10 g Carnitin wurden
etwa 80 – 90 % der Dosis mit dem Urin ausgeschieden. Die Nierenschwelle wurde in diesem
Falle überschritten. Ab welcher Plasmakonzentration es bei Pferden zu einer erhöhten renalen
Ausscheidung von Carnitin kommt, ist zur Zeit noch nicht bekannt. Die relative
Bioverfügbarkeit, die die Menge der am Wirkort verfügbaren Dosis beschreibt, wurde in
diesem zuletzt zitierten Versuch auf 7 % geschätzt.
Auf dem Blutweg gelangt das Carnitin aus dem Intestinum in die verschiedenen Gewebe und
Organe. Die Leber ist Synthese- und Speicherort für Carnitin. Aus der Leber kann es zu
einem sehr kleinen Teil in die Galle gelangen und mit ihr wieder in den Dünndarm, wo ein
Teil des Carnitins erneut absorbiert werden kann (enterohepatischer Kreislauf)
(GUDJONSON et al. 1985 b). Die Carnitinausscheidung über die Fäces ist beim Menschen
sehr gering (< 1 µmol/d) (SCHEK 1994).
Der Hauptausscheidungsweg des Carnitins ist die Niere (RUDMAN et al. 1977; MITCHEL
1978, BREMER 1983). L-Carnitn und Carnitinester sind in den Glomeruli frei filtrierbar.
Freies Carnitin wird beim Menschen mit normalem Carnitinstatus zu mehr als 98 % resorbiert
(ENGEL et al. 1981, LI et al 1992). Die tubuläre Resorption geht bei steigender
Plasmakonzentration von Carnitin prozentual zurück. (CARLIN et al 1986; BIEBER 1988).
Bei Carnitinzulagen steigt die renale Carnitinausscheidung deshalb relativ stark an (BAKER
et al. 1992).
4.2. Carnitingehalte im Körper
Carnitin ist in Grammengen im Körper gespeichert. Etwa 95 % dieser Menge befinden sich
beim Menschen im Muskelgewebe (Skelettmuskulatur und Herz) (ENGEL u. REBOUCHE
1984; SCHOLTE u. DE JONGE 1987). Mit Ausnahme des epididymalen Gewebes
(BROOKS 1980; CARTER et al. 1980) ist die Carnitinkonzentration in der Muskulatur am
höchsten. In den übrigen Geweben des Körpers (Niere, Leber u. a.) einschließlich der
extrazellulären Flüssigkeit befinden sich, wie die obigen Zahlen erkennen lassen, nur etwa ca.
5 % des Gesamtkörpercarnitins (Tab. 2.1.).

II. Schrifttum ___________________________________________________________________________
24
Tab. 2.1.: Prozentuale Verteilung von Carnitin im Körper und Konzentrationen von Carnitin
in einigen Geweben beim Hund, GC = Gesamtcarnitin
Verteilung im Körper % Literatur
Muskulatur 95 – 98
Sonstige Organe < 5
REBOUCHE u. ENGEL
(1983)
Extralzellulärflüssigkeit 0,3
Konzentration in Geweben
Plasma µmol/l
Gesamtcarnitin (GC) 24,9 ± 6,6 KEENE et al. (1991)
Freies Carnitin 21,9 ± 7,0
Skelettmuskel (GC) mmol/kg uS
M. quadriceps 1,62 – 7,38
M biceps 1,67 – 6,43
REBOUCHE u. ENGEL
(1983)
Leber (GC) 0,12 – 0,36
Niere (GC) 0,16 – 0,58
Die Konzentration des Gesamtcarnitins im Plasma bei Pferden schwankt zwischen 15 und 55
µmol/l (FOSTER et al. 1988, 1989 a, b; FOSTER u. HARRIS 1992). Es bestehen große
individuelle Unterschiede. Die Konzentration ist u. a. abhängig von der Zeit der
Probennahme. In einer Studie von LAMBERTZ (1999) betrug die Carnitinkonzentration im
Plasma im Tagesmittel bei 3 bis 5 jährigen Pferden 25 µmol/l. Ein Anteil von 70 bis 80 % des
Gesamtcarnitins im Plasma entfielen unter Ruhebedingungen auf freies Carnitin (FOSTER et
al. 1988; HARRIS et al. 1995 a).
Im Musculus glutaeus medius ist die Gesamtcarnitinkonzentration ca. 300 mal größer als im
Plasma (HARRIS et al. 1995 b). Beim ausgewachsenen Pferd erreicht die
Gesamtcarnitinkonzentration in der Muskulatur Werte von 29-33 mmol/kg TM (bei einem
TM-Gehalt des Muskels von 25 % entsprechend 7 – 8 mmol/kg uS) (FOSTER u. HARRIS
1987 b). Im Ruhezustand lagen 80 % des Gesamtcarnitins als freies Carnitin vor, 15 % als
kurzkettige Acylcarnitine (Acetylcarnitin) und 5 - 10 % als langkettige Acylcarnitine.

II. Schrifttum ___________________________________________________________________________
25
4.3. Altersabhängigkeit
In verschiedenen Untersuchungen wurde der Gehalt an Carnitin im Plasma in Abhängigkeit
vom Alter bei verschiedenen Spezies einschließlich des Menschen gemessen. Bei diesen
Untersuchungen zeigte sich, dass junge Tiere im Vergleich zu älteren niedrigere
Konzentrationen an Carnitin im Plasma aufweisen. Dieses wird dadurch erklärt, dass die
körpereigene Synthese bei jungen Tieren für Carnitin noch begrenzt ist (BATTISTELLA et
al. 1980; BORUM 1983). Daraus lässt sich jedoch nicht unbedingt auf einen Carnitinmangel
schließen, da die begrenzte körpereigene Synthesefähigkeit durch einen relativ hohen Gehalt
an Carnitin in der Milch ausgeglichen wird. Bei Kühen wiesen ROOS et al. (1992) zum
Zeitpunkt der Geburt in der Milch einen Carnitingehalt von 537 µmol/l nach, der bis 6
Wochen nach der Geburt auf 183 µmol/l absank. Die Milch enthält somit 10 bis 25 mal so
viel Carnitin wie Blutplasma. Ähnliches gilt auch für das Pferd. Die Carnitinkonzentration im
Plasma steigt beim Pferd in den ersten drei Lebensjahren relativ stark an. Anschließend bleibt
sie annähernd stabil. Bei Fohlen und Jährlingen ist die Carnitinkonzentration im Plasma nur
halb so groß wie beim ausgewachsenen Pferd (FOSTER et al. 1989 a).
Das Verhältnis von freiem zu Gesamtcarnitin im Plasma wie auch deren Konzentrationen sind
unabhängig vom Geschlecht, nicht aber vom Trainingszustand. Drei- bis sechsjährige
trainierte Pferde besaßen im Plasma 30 % mehr freies Carnitin als untrainierte (FOSTER et al.
1989 a). Die Autoren deuten diesen Anstieg als Ausdruck eines erhöhten Bedarfs und einer
verstärkten Eigensynthese. Bei jüngeren Pferden ist ein solcher Trainingseinfluss auf das freie
Plasmacarnitin nicht zu beobachten (FOSTER et al. 1989 a).
II. Schrifttum ___________________________________________________________________________
26
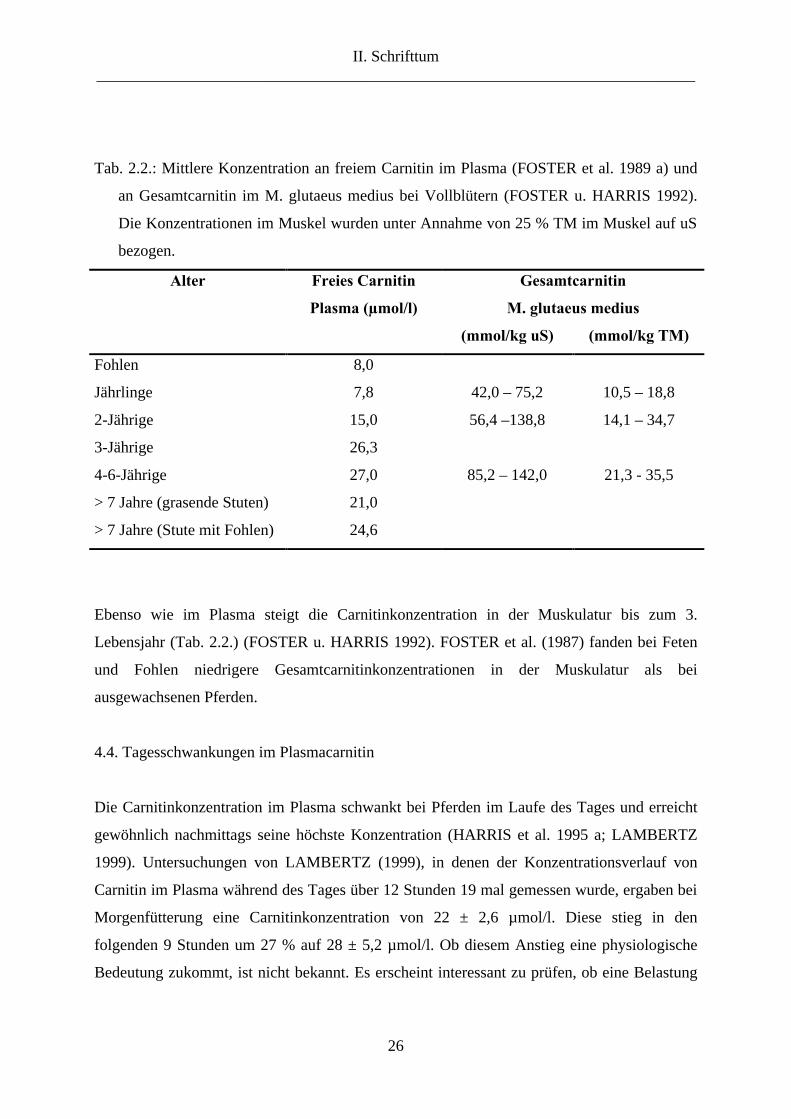
Tab. 2.2.: Mittlere Konzentration an freiem Carnitin im Plasma (FOSTER et al. 1989 a) und
an Gesamtcarnitin im M. glutaeus medius bei Vollblütern (FOSTER u. HARRIS 1992).
Die Konzentrationen im Muskel wurden unter Annahme von 25 % TM im Muskel auf uS
bezogen.
Alter Freies Carnitin
Plasma (µmol/l)
Gesamtcarnitin
M. glutaeus medius
(mmol/kg uS) (mmol/kg TM)
Fohlen 8,0
Jährlinge 7,8 42,0 – 75,2 10,5 – 18,8
2-Jährige 15,0 56,4 –138,8 14,1 – 34,7
3-Jährige 26,3
4-6-Jährige 27,0 85,2 – 142,0 21,3 - 35,5
> 7 Jahre (grasende Stuten) 21,0
> 7 Jahre (Stute mit Fohlen) 24,6
Ebenso wie im Plasma steigt die Carnitinkonzentration in der Muskulatur bis zum 3.
Lebensjahr (Tab. 2.2.) (FOSTER u. HARRIS 1992). FOSTER et al. (1987) fanden bei Feten
und Fohlen niedrigere Gesamtcarnitinkonzentrationen in der Muskulatur als bei
ausgewachsenen Pferden.
4.4. Tagesschwankungen im Plasmacarnitin
Die Carnitinkonzentration im Plasma schwankt bei Pferden im Laufe des Tages und erreicht
gewöhnlich nachmittags seine höchste Konzentration (HARRIS et al. 1995 a; LAMBERTZ
1999). Untersuchungen von LAMBERTZ (1999), in denen der Konzentrationsverlauf von
Carnitin im Plasma während des Tages über 12 Stunden 19 mal gemessen wurde, ergaben bei
Morgenfütterung eine Carnitinkonzentration von 22 ± 2,6 µmol/l. Diese stieg in den
folgenden 9 Stunden um 27 % auf 28 ± 5,2 µmol/l. Ob diesem Anstieg eine physiologische
Bedeutung zukommt, ist nicht bekannt. Es erscheint interessant zu prüfen, ob eine Belastung

II. Schrifttum ___________________________________________________________________________
27
der Tiere am Nachmittag, wenn das Carnitin im Plasma am höchsten ist, die
Leistungsfähigkeit verbessert.
5. Carnitingehalte in Futter- und Lebensmitteln
Pflanzliche Nahrungsmittel enthalten nur geringe Mengen Carnitin (< 10 mg/kg) (PANTER u.
MUDD 1969; MITCHELL 1978; BORUM et al. 1979). Tierische Produkte sind dagegen
meist reich an Carnitin (Tab. 2.3.).
Tab. 2.3.: Gehalte an L-Carnitin in pflanzlichen und tierischen Futter- und Lebensmitteln
(mg/kg uS) (HARMEYER et al. 1998)
Pflanzliche Produkte L-Carnitingehalt Tierische Produkte L-Carnitingehalt
Winterweizen 3,1 Milchaustauscher 223-236
Maismehl 4,1 Fischmehl 61
Wintergerste 2,8 Molkepulver 430-920
Sojaextraktionsschrot 9,7 Schweinefleisch (TM) 353-733
Weizenkleie 7,4 Pferdefleisch 270-1250
6. Trainings- und belastungsbedingte Einflüsse auf den Carnitinstatus
Im folgenden wird dargestellt, wie sich einzelne oder wiederholte Belastungen (Training) auf
den Carnitinstatus auswirken. Studien an Männern und Ratten (CIMAN et al. 1980;
LENNON et al. 1983; SANGUQ et al. 1984; NEGRAO et al. 1987) haben gezeigt, dass die
Konzentration an Carnitin im Plasma und in der Muskulatur durch Training ansteigt. Dies
betrifft sowohl die Menge an freiem als auch die an Acetylcarnitin. Nach Untersuchungen von
FOSTER et al. (1989 a) sind die Carnitinspiegel im Plasma bei trainierten drei- bis
sechsjährigen Vollblütern höher als bei gleichaltrigen untrainierten Vollblütern. Ob dieser
Anstieg auf einer vermehrten Synthese oder einer verminderten renalen Ausscheidung beruht,
ist nicht geklärt.

II. Schrifttum ___________________________________________________________________________
28
FOSTER und HARRIS (1992) untersuchten die Carnitinkonzentration im M. glutaeus medius
bei trainierten und untrainierten zweijährigen Vollblütern. Sie fanden in dieser Altersgruppe
keinen Anstieg an Gesamtcarnitin im Laufe eines niedrigen bis mäßigen sechsmonatigen
Trainings. Allerdings wies das Gesamtcarnitin bei trainierten eine geringere Variationsbreite
auf als bei untrainierten (Tab. 2.4.). Die Autoren sehen den Grund in einer zunehmenden
Mitochondriendichte.
Tab. 2.4.: Gesamtcarnitinkonzentration des M. glutaeus medius bei zweijährigen Vollblütern
in unterschiedlichem Trainingszustand (FOSTER u. HARRIS 1992)
Trainingsstand Gesamtcarnitin
M. glutaeus medius
(mmol/kg TM)
Untrainiert 18,5 – 34,7
Trainiert (niedrige Belastung) 14,1 – 24,2
Trainiert (mäßige Belastung) 22,9 – 26,9
Im Gegensatz zu einer trainingsbedingten Anpassung hat eine akute Belastung keinen
Einfluss auf den Gehalt an Gesamtcarnitin des Muskels (FOSTER u. HARRIS 1987 a, b;
FOSTER et al. 1988; HARRIS u. FOSTER 1990; FOSTER u. HARRIS 1992). Allerdings
kommt es, wie schon erwähnt, im Laufe einer Belastung zu einem Abfall an freiem Carnitin
und einem parallelen Anstieg an Acetylcarnitin (FOSTER u. HARRIS 1987 a, b; CARLIN et
al. 1990; HARRIS u. FOSTER 1990). Der Anstieg an Acetylcarnitin ist Ausdruck der
metabolischen (Puffer)Funktion von L-Carnitin.
Im Abstand von mindestens drei Tagen absolvierten sechs Vollblüter sieben
Laufbandbelastungen. Jede Belastung bestand aus einer 15-minütigen Aufwärmphase, einer
zweiminütigen Belastung bei 6 m/s Laufbandgeschwindigkeit und einer zweiminütigen
Belastung bei entweder 6, 7, 8, 9, 10, 11 oder 12 m/s und 5 %-iger Steigung des Laufbandes.
Die Acetylcarnitinkonzentration in der Glutaeusmuskulatur erhöhte sich schon bei moderater
Belastung (6 m/s für vier Minuten) auf das Doppelte des Ruhewertes. Das freie Carnitin sank

II. Schrifttum ___________________________________________________________________________
29
entsprechend (HARRIS u. FOSTER 1990). Ferner unterzogen FOSTER und HARRIS
(1987a) zwei Vollblüter einer Laufbandbelastung von vier Minuten bei einer
Laufbandgeschwindigkeit von 3,2 m/s und vier Minuten bei 8 oder 10 m/s mit einer Steigung
von 5 % und einer sich anschließenden Erholungsphase von 12 Minuten im Schritt bei 1,6 m/s
ohne Steigung. Die Ruhekonzentration an Gesamtcarnitin in der Muskulatur betrug 29,5
mmol/kg TM mit 88 % freiem, 7 % Acetyl- und 5 % Acylcarnitin. Die Belastung änderte die
Konzentration an Gesamtcarnitin nicht. Das freie Carnitin sank um 19,7 mmol/kg TM, das
Acetylcarnitin stieg um 21,5 mmol/kg TM. Während der Erholung kehrte sich der Trend
wieder um. Nach 12 Minuten Erholung waren 72 % der Ruhekonzentration an freiem Carnitin
wieder erreicht.
Die höchste Konzentration an Acetylcarnitin in der Glutaenmuskulatur war kurz vor dem
Anstieg des Plasmalactats erreicht (HARRIS und FOSTER 1990). Dieses zeitliche
Nacheinander im Anstieg von Acetylcarnitin und Lactat könnte bedeuten, dass der
Acetylüberschuß zunächst durch Carnitin abgepuffert und danach als Pyruvat zu Lactat
umgesetzt wird.
7. Der Energiestoffwechsel des Pferdes während der Belastung und nach dem Training
- belastungsabhängige Veränderungen
Die Anpassung des Energiestoffwechsels an eine Belastung hängt von der Dauer und der
Höhe der Belastung ab. Am Anfang der Belastung wird die Energie auf anaerobem Wege
gewonnen. Zuerst werden die ATP-Vorräte und dann das Kreatinphosphat genutzt. Parallel
dazu steigt die Glykolyserate.
Die Ruhelactatkonzentration im Vollblut von 0,5 bis 1,5 mmol/l steigt abhängig von der
Belastung zuerst langsam (linear) und dann exponentiell an. Dieser Übergang wird als „aerob-
anaerobe Schwelle“ angesehen (SNOW u. VALBERG 1994). Von da an übersteigt die
Lactatneubildung erheblich die Kapazität der Eliminierung (HARRIS et al. 1987). Bei
kurzzeitigen Belastungen, z. B. Gallopprennen über eine Distanz von 1200 – 2400 m, wird die

II. Schrifttum ___________________________________________________________________________
30
aerob-anaerobe Schwelle regelmäßig überschritten. Es kommt dabei zu Lactatanstiegen bis 30
mmol/l (SNOW et al. 1983). Auch bei Trabrennen wird diese Schwelle regelmäßig
überschritten und es treten Lactatkonzentrationen von 23 mmol/l auf (KRZYWANEK 1974).
Anders ist es bei Distanzritten (bis 210 km), bei denen das Lactat im Blut etwa zwischen zwei
und sieben mmol/l liegt (GROSSKOPF u. VAN RENSBURG 1983).
Voraussetzung für die hohe Lactogenese ist eine ebenso starke Glykogenolyserate. Sie geht
der Lactogenese zeitlich voraus und ist zu Beginn jeder Belastung am größten (COGGAN
1991). Dementsprechend sinkt die Glykogenkonzentration im Muskel bei 800 bis 2000 m
Galopprennen um 20-35 % (HARRIS et al. 1987; SNOW u. HARRIS 1991). Aber auch bei
Distanzritten über 40 bis 160 km nimmt der Muskelglykogengehalt um bis zu 75 % ab
(SNOW u. BAXTER 1981; SNOW et al. 1982). Grosse Glykogenspeicher der Muskulatur
und hohe Konzentrationen an FFS im Plasma können die Glucoseentnahme aus dem Blut
verringern (COGGAN 1991). Dennoch steigt die Glucoseaufnahme aus dem Plasma in die
Muskulatur exponentiell mit der Belastung (COGGAN et al. 1995 a). Der Plasmaglucosepool
wird von der Leber gespeist, die die Glucose durch Glykogenolyse und auch durch
Resynthese von Glucose aus Lactat und Alanin bereitstellt. Bei kurzen sehr hohen
Belastungen (Rennen von 1000 bis 2400 m) steigt die Plasmaglucose von vier bis fünf mmol/l
auf 10 bis 12 mmol/l an (JUDSON et al. 1983; SNOW et al. 1983; HARRIS et al. 1987;
FREESTONE et al. 1991). SNOW et al. (1983) führen den Plasmaglucoseanstieg bei
intensiven Belastungen auf einen überproportionalen Glucoseeinstrom im Vergleich zum
Glucoseverbrauch zurück. Im Gegensatz zu kurzen intensiven Belastungen fällt bei
Dauerbelastungen (Distanzritten) mit niedrigerer Intensität die Glucosekonzentration im
Plasma ab (LUCKE u. HALL 1980 a, b; HAMBITZER u. BENT 1988; JAHN et al. 1996).
GROSSKOPF u. VAN RENSBURG (1983) und SLOET VAN OLDRUITENBORGH-
OOSTERBAAN et al. (1991) berichten bei einem Distanzritt über 80 bis 100 km von einem
Anstieg der Plasmagucosekonzentration während der ersten 30-60 km und danach von einem
Abfall. Sie führen den Abfall auf eine Entleerung der Leberglykogenspeicher zurück (SNOW
et al. 1982; ROSE et al. 1983 a).
Mit zunehmender Belastungsdauer erfolgt die Energiebereitstellung mehr und mehr aerob.
Dies ist nicht mit einem pH-Abfall der Muskelzelle verbunden und die Energieausbeute ist

II. Schrifttum ___________________________________________________________________________
31
bezogen auf das verbrauchte Substrat wesentlich höher. Darüber hinaus ist unter diesen
Bedingungen die Energiereserve in Form von FFS sehr viel höher. Der prozentuale Anteil der
FFS, die zur Energiegewinnung genutzt werden, steigt mit zunehmender Belastungsdauer.
Bei Pferden stiegt die Plasmakonzentration der FFS nach Distanzritten (60-100 km) um 300
bis 1000 % auf Werte bis zu 2000 µmol/l (LUCKE u. HALL 1980 b, c; HAMBITZER u.
BENT 1988; SLOET VAN OLDRUITENBORGH-OOSTERBAAN et al. 1991). Nach
intensiven kurzen Belastungen (Galopprennen bis 2000 m) steigen die FFS im Plasma
dagegen nur wenig an (SNOW et al. 1979; PÖSÖ et al. 1989; ZIMMERMANN et al. 1992).
Geht die Dauerbelastung jedoch bis zur Erschöpfung, kommt es schließlich zu einem Abfall
der FFS-Konzentration im Plasma (JONES et al. 1980).
- trainingsabhängige Veränderungen
Neben den kurzfristigen Anpassungsvorgängen des Muskels während der Belastung treten im
Laufe eines Trainings langfristige Änderungen im Muskel ein. Jedes Training verbessert die
aerobe und die anaerobe Energiebereitstellung der Muskulatur.
Zur Einschätzung eines Trainingserfolges eignet sich besonders die Messung der
Blutlactatkonzentration bei definierter Belastung (HODGSON u. ROSE 1994; LINDNER
1997). Die Verbesserung der Leistungsfähigkeit kommt in einer niedrigeren Blut- und
Muskellactatkonzentration bei gleicher submaximaler Belastung zum Ausdruck. Ein
wichtiges Merkmal trainingsbedingter Anpassungen der Muskulatur ist die Zunahme der
Energiegewinnung aus Fetten zu Lasten von Glucose (GREEN et al. 1992).
BROOKS u. DONOVAN (1983) konnten bei Ratten nachweisen, dass Trainierte schneller
FFS aus Körperfett bereitstellen als Untrainierte. Die Einsparung des Muskelglykogens und
die niedrigere Muskellactatkonzentration zeigen, dass außer der schnelleren Bereitstellung
auch vermehrt FFS verwertet werden und somit die Glykolyse vermindert wird, also die
Fähigkeit der FFS-Verwertung im Trainingsverlauf ansteigt (BROOKS u. DONOVAN 1983;
HOLLOSZY u. COYLE 1984; GREEN et al. 1991). In Studien von JANSSON und KAIJSER
(1987) und HURLEY et al. (1986) wurde bei Menschen gezeigt, dass nicht vermehrt FFS aus
dem Plasma oxidiert werden, sondern intramuskuläre Triglyceride zur Energiegewinnung

II. Schrifttum ___________________________________________________________________________
32
herangezogen wurden. Die Aufnahme von FFS aus dem Plasma in die Muskulatur war bei
Untrainierten genauso hoch wie bei Trainierten. Auch in diesen Studien wurde bei den
Trainierten weniger Muskelglykogen und Blutglucose verbraucht. In Studien von KOIVISTO
et al. (1982) bei Menschen und ZENDZIAN-PIOTROWSKA und GORSKI (1993) bei Ratten
war der Anstieg der FFS im Plasma bei Trainierten niedriger als bei Untrainierten, was die
Autoren auf einen erhöhten Verbrauch an FFS aus dem Plasma zurückführen. Im Gegensatz
zu den vorher genannten Studien sind hierbei keine Kalkulationen der Umsatzraten
durchgeführt worden, und die Autoren stellen nur Vermutungen an, wieso der FFS-Spiegel im
Plasma bei Trainierten niedriger war als bei Untrainierten. Die FFS-Aufnahme durch den
Muskel sinkt bei Sprinttrainierten während der Belastung trotz höhere FFS-Konzentrationen
im Plasma, was bei Ausdauersportlern nicht der Fall ist (TURCOTTE et al. 1992; KIENS
1997). KIENS (1997) begründet dieses unterschiedliche Verhalten durch eine Verbesserung
der carrierabhängigen Transportfähigkeit durch Ausdauertraining, nicht aber durch anderes
Training.
Im Zusammenhang damit ist der Plasmaglucoseverbrauch beim Trainierten niedriger als beim
Untrainierten (COGGAN 1991). Die Glucoseaufnahme durch die Muskulatur, die
Glykogenolyse und Entnahme der Glucose aus dem Plasma sinken nach Training beim
Menschen, wodurch Trainierte bei gleicher Belastung im Vergleich mit Untrainierten einen
höheren Plasmaglucosespiegel haben (JANSON u. KAIJSER 1987; RICHTER et al. 1998;
BERGMAN et al. 1999). Die verminderte Glucoseaufnahme durch die Muskulatur kommt
durch eine verminderte Translokation der Glucosetransportproteine (GLUT-4) aus der Zelle
ins Sarkolemma zustande. Interessanterweise steigt die Anzahl der GLUT-4 im Muskel und
die Hexokinaseaktivität, welche intrazellulär die Glucose-6-Phosphat Konzentration bei
Belastung senkt, im Trainingsverlauf (TURCOTTE et al. 1992; RICHTER et al. 1998).
Vermutlich verbessert der Anstieg der GLUT-4 bei Trainierten die Transportfähigkeit der
Glucose aus dem Plasma in die Muskulatur in der Erholungsphase.
Der Glykogengehalt der Muskulatur kann durch Training beeinflusst werden. Nach einer
sechswöchigen Trainingsperiode waren bei sechs Pferden die Glykogengehalte in der
Muskulatur um 12 % angestiegen (GANSEN et al. 1997). SNOW und HARRIS (1991)
wiesen tendentiell einen Anstieg der Ruheglykogenkonzentration nach einem 14-tägigen

II. Schrifttum ___________________________________________________________________________
33
Training nach, wohingegen ESSEN-GUSTAVSSON et al. (1989) ein Absinken im Verlauf
eines fünfwöchigen Trainings beobachteten. In anderen Untersuchungen konnte der
Glykogengehalt nicht durch Training beeinflusst werden (GUY u. SNOW 1977; WILSON et
al. 1987; FOREMAN et al. 1990 a). Der Glykogenverbrauch bei Belastung wird durch
Training gesenkt. COGGAN et al. (1993) hat einen um 36 % niedrigeren Verbrauch an
Muskelglykogen bei Männern nach einem 12-wöchigen Training bei einer Belastung von 75
% VO2max über 40 Minuten Laufen und von 30 – 90 % VO2max über ca. 45 Minuten Radfahren
jeweils drei Tage pro Woche gemessen. Auch andere Studien bestätigen diese Ergebnisse
(HERMANSSEN et al. 1967; HURLEY et al. 1986; JANSSON u. KAIJSER 1987).
Die Herzfrequenz wird durch Training gesenkt. GOTTLIEB-VEDI et al. (1995) stellten eine
signifikante Senkung der Herzfrequenz bei Pferden bei derselben Belastung nach einem
vierwöchigen Laufbandtraining drei Tage pro Woche bei v4 fest. Echographisch wurde in
einer Studie mit sieben Vollblütern nach einem 18-wöchigen Renntraining eine Vergrößerung
des Durchmessers des linken Ventrikels und eine Herzwandverdickung gemessen (YOUNG
1999). Das Körpergewicht der Tiere änderte sich während dieser Studie nicht. Die
Herzhypertrophie als Folge des Trainings führte zu einem größeren Schlagvolumen. Die
Herzfrequenz sinkt bei gleicher Belastung nach Training.
8. Einfluss von Carnitinzulagen
Gestützt auf die metabolische und katalytische Funktion von Carnitin könnte man fragen, ob
eine Supplementation mit Carnitin die körperliche Leistungsfähigkeit verbessern kann. Bei
Tieren und Menschen wurden daher eine Reihe von Untersuchungen durchgeführt, um den
Einfluss einer Carnitinsupplementierung auf die Leistungsfähigkeit zu prüfen. Beim
Menschen wird L-Carnitin schon länger als Leistungsförderer angesehen und in diesem Sinne
verwendet. Die wissenschaftlichen Ergebnisse, die bisher zu dieser Frage vorliegen, lassen
jedoch noch keine eindeutigen Schlüsse zu.
II. Schrifttum ___________________________________________________________________________
34
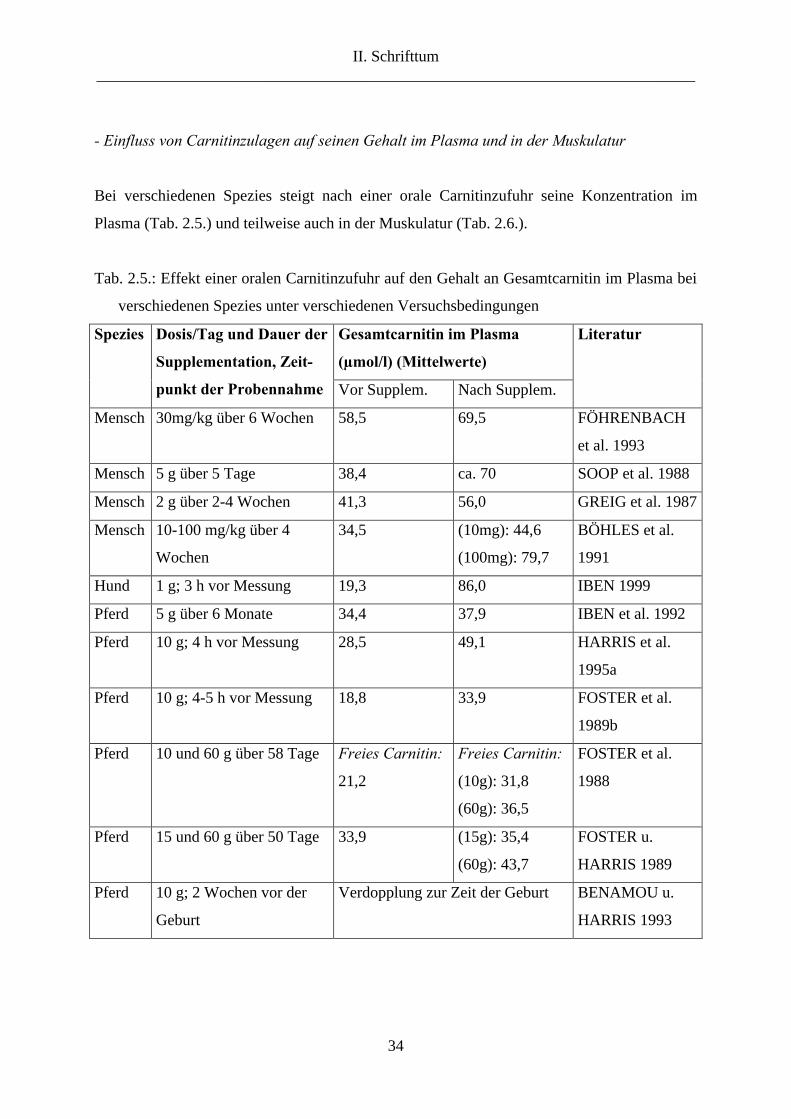
- Einfluss von Carnitinzulagen auf seinen Gehalt im Plasma und in der Muskulatur
Bei verschiedenen Spezies steigt nach einer orale Carnitinzufuhr seine Konzentration im
Plasma (Tab. 2.5.) und teilweise auch in der Muskulatur (Tab. 2.6.).
Tab. 2.5.: Effekt einer oralen Carnitinzufuhr auf den Gehalt an Gesamtcarnitin im Plasma bei
verschiedenen Spezies unter verschiedenen Versuchsbedingungen
Gesamtcarnitin im Plasma
(µmol/l) (Mittelwerte)
Spezies Dosis/Tag und Dauer der
Supplementation, Zeit-
punkt der Probennahme Vor Supplem. Nach Supplem.
Literatur
Mensch 30mg/kg über 6 Wochen 58,5 69,5 FÖHRENBACH
et al. 1993
Mensch 5 g über 5 Tage 38,4 ca. 70 SOOP et al. 1988
Mensch 2 g über 2-4 Wochen 41,3 56,0 GREIG et al. 1987
Mensch 10-100 mg/kg über 4
Wochen
34,5 (10mg): 44,6
(100mg): 79,7
BÖHLES et al.
1991
Hund 1 g; 3 h vor Messung 19,3 86,0 IBEN 1999
Pferd 5 g über 6 Monate 34,4 37,9 IBEN et al. 1992
Pferd 10 g; 4 h vor Messung 28,5 49,1 HARRIS et al.
1995a
Pferd 10 g; 4-5 h vor Messung 18,8 33,9 FOSTER et al.
1989b
Pferd 10 und 60 g über 58 Tage
Freies Carnitin:
21,2
Freies Carnitin:
(10g): 31,8
(60g): 36,5
FOSTER et al.
1988
Pferd 15 und 60 g über 50 Tage 33,9 (15g): 35,4
(60g): 43,7
FOSTER u.
HARRIS 1989
Pferd 10 g; 2 Wochen vor der
Geburt
Verdopplung zur Zeit der Geburt BENAMOU u.
HARRIS 1993

II. Schrifttum ___________________________________________________________________________
35
FOSTER et al. (1988) supplementierten Vollblüter oral 58 Tage lang mit 1x10 bis 3x20 g
Carnitin täglich. Zwei bis vier Stunden nach der Gabe war ein Peak in der
Plasmakonzentration an Gesamtcarnitin messbar. Bei einmaliger Gabe von 10 g Carnitin stieg
der Plasmacarnitinspiegel sieben Stunden nach Gabe auf 31,8 µmol/l, bei 2x30 g stieg er nur
wenig höher auf 36,5 µmol/l. Der Anstieg im Plasmacarnitin ist nicht proportional zur
Zulagehöhe. Es liegen starke transiente Schwankungen bei hohen Carnitingaben vor und die
Bioverfügbarkeit nimmt mit zunehmender Zulagehöhe ab. Es stellt sich die Frage, welche
Zulagehöhe bei Supplementation empfehlenswert ist. FOSTER und HARRIS (1989)
supplementierten fünf Pferde über einen Zeitraum von 50 Tagen mit 15, 30 und 60 g Carnitin
pro Tag auf drei Dosen verteilt. Die Autoren empfahlen eine Dosis von 3x5 g pro Tag, da eine
höhere Zulage den Plasmaspiegel kaum weiter ansteigen ließ. Die Autoren vermuten, dass der
Absorptionsmechanismus des Darms bei 3x5 g gesättigt war und empfahlen eine niedrige
jedoch häufige Verabreichung von Carnitin.
Verglichen mit den Konzentrationsanstiegen im Plasma sind die Konzentrationszunahmen in
der Muskulatur geringer und teilweise nicht nachweisbar. FOSTER et al. (1988) konnten
keinen signifikanten Anstieg der Gesamtcarnitinkonzentration in der Muskulatur beim Pferd
bei einer Supplementation von 10 - 60 g pro Tag über 58 Tage feststellen. Die
Carnitinkonzentration in der Muskulatur konnte selbst bei i. v. Injektionen von 10 g Carnitin
über 26 Tage nicht signifikant erhöht werden (HARRIS et al. 1995 a). Anders als bei der
Ratte scheint die Muskulatur auf Carnitinzulagen nach bisher vorliegenden Messungen beim
Menschen und beim Pferd wenig anzusprechen. Tabelle 2.6. zeigt beispielhaft die
Gesamtcarnitinkonzentration in der Muskulatur nach Zulagen bei verschiedenen Spezies.
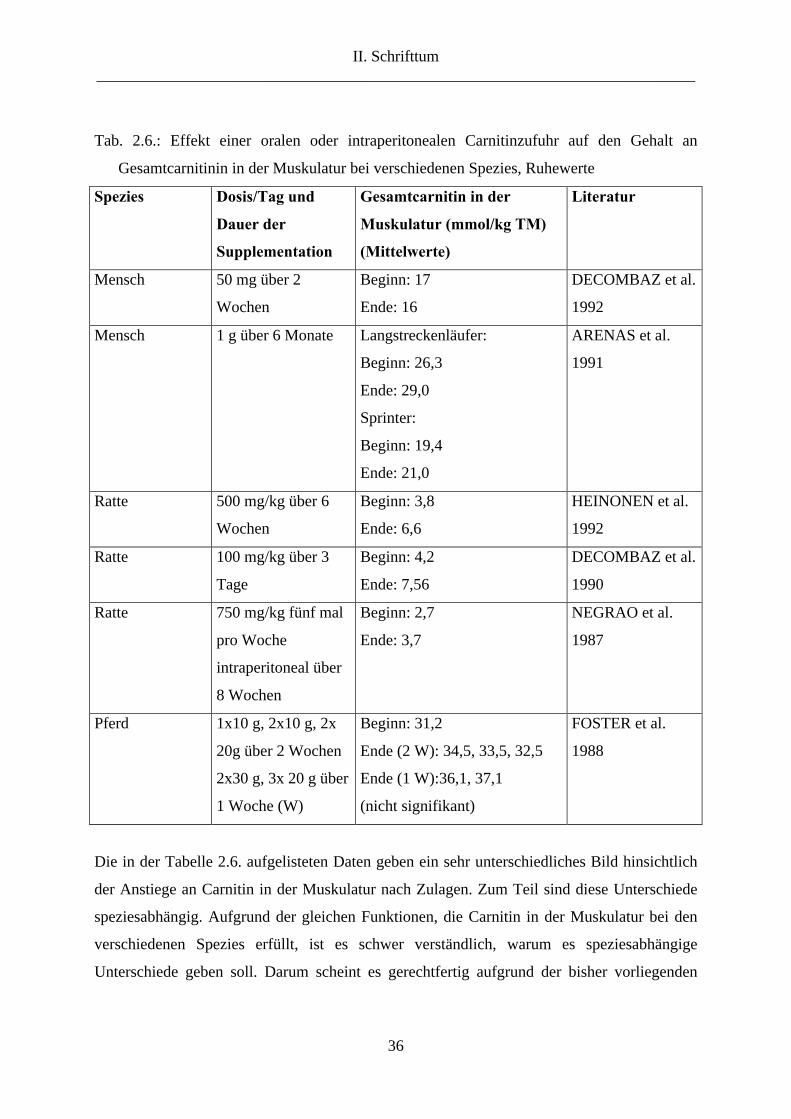
II. Schrifttum ___________________________________________________________________________
36
Tab. 2.6.: Effekt einer oralen oder intraperitonealen Carnitinzufuhr auf den Gehalt an
Gesamtcarnitinin in der Muskulatur bei verschiedenen Spezies, Ruhewerte
Spezies Dosis/Tag und
Dauer der
Supplementation
Gesamtcarnitin in der
Muskulatur (mmol/kg TM)
(Mittelwerte)
Literatur
Mensch 50 mg über 2
Wochen
Beginn: 17
Ende: 16
DECOMBAZ et al.
1992
Mensch 1 g über 6 Monate Langstreckenläufer:
Beginn: 26,3
Ende: 29,0
Sprinter:
Beginn: 19,4
Ende: 21,0
ARENAS et al.
1991
Ratte 500 mg/kg über 6
Wochen
Beginn: 3,8
Ende: 6,6
HEINONEN et al.
1992
Ratte 100 mg/kg über 3
Tage
Beginn: 4,2
Ende: 7,56
DECOMBAZ et al.
1990
Ratte 750 mg/kg fünf mal
pro Woche
intraperitoneal über
8 Wochen
Beginn: 2,7
Ende: 3,7
NEGRAO et al.
1987
Pferd 1x10 g, 2x10 g, 2x
20g über 2 Wochen
2x30 g, 3x 20 g über
1 Woche (W)
Beginn: 31,2
Ende (2 W): 34,5, 33,5, 32,5
Ende (1 W):36,1, 37,1
(nicht signifikant)
FOSTER et al.
1988
Die in der Tabelle 2.6. aufgelisteten Daten geben ein sehr unterschiedliches Bild hinsichtlich
der Anstiege an Carnitin in der Muskulatur nach Zulagen. Zum Teil sind diese Unterschiede
speziesabhängig. Aufgrund der gleichen Funktionen, die Carnitin in der Muskulatur bei den
verschiedenen Spezies erfüllt, ist es schwer verständlich, warum es speziesabhängige
Unterschiede geben soll. Darum scheint es gerechtfertig aufgrund der bisher vorliegenden

II. Schrifttum ___________________________________________________________________________
37
Daten, auch bei den Tieren, bei denen bisher kein deutlicher Effekt gemessen wurde,
Versuche unter standardisierten Bedingungen zu wiederholen.
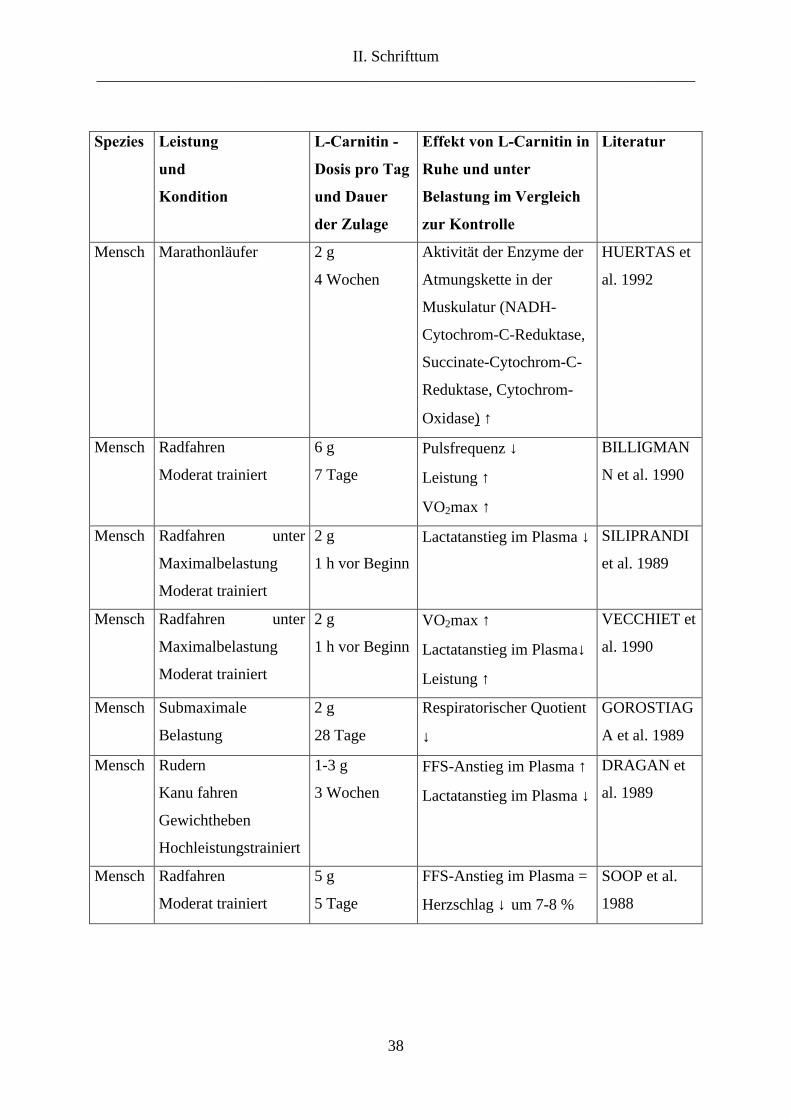
- Einfluss von Carnitingaben auf Leistungs- und biochemische Parameter
Schon unter Ruhebedingungen beeinflusst Carnitin einige Leistungsparameter. SUZUKI et al.
(1981) wiesen bei mit Carnitin supplementierten Hunden (8-minütige Infusion mit 80
mg/kg/min) eine um 17 % niedrigere Herzfrequenz und eine um 60 % höhere
Coronararteriendurchblutung nach als bei den Kontrolltieren. Diese Daten lassen vermuten,
dass Carnitin einen direkt vasodilatierenden und einen positiv inotropen Effekt hat.
In der nachfolgenden Tabelle 2.7. sind die Effekte von Carnitingaben auf verschiedene
Leistungsparameter und biochemische Merkmale zusammengestellt. Die Ergebnisse sind zum
Teil uneinheitlich. Sie hängen offenbar von der Dauer und der Höhe der Supplementation und
der Höhe der Belastung ab.
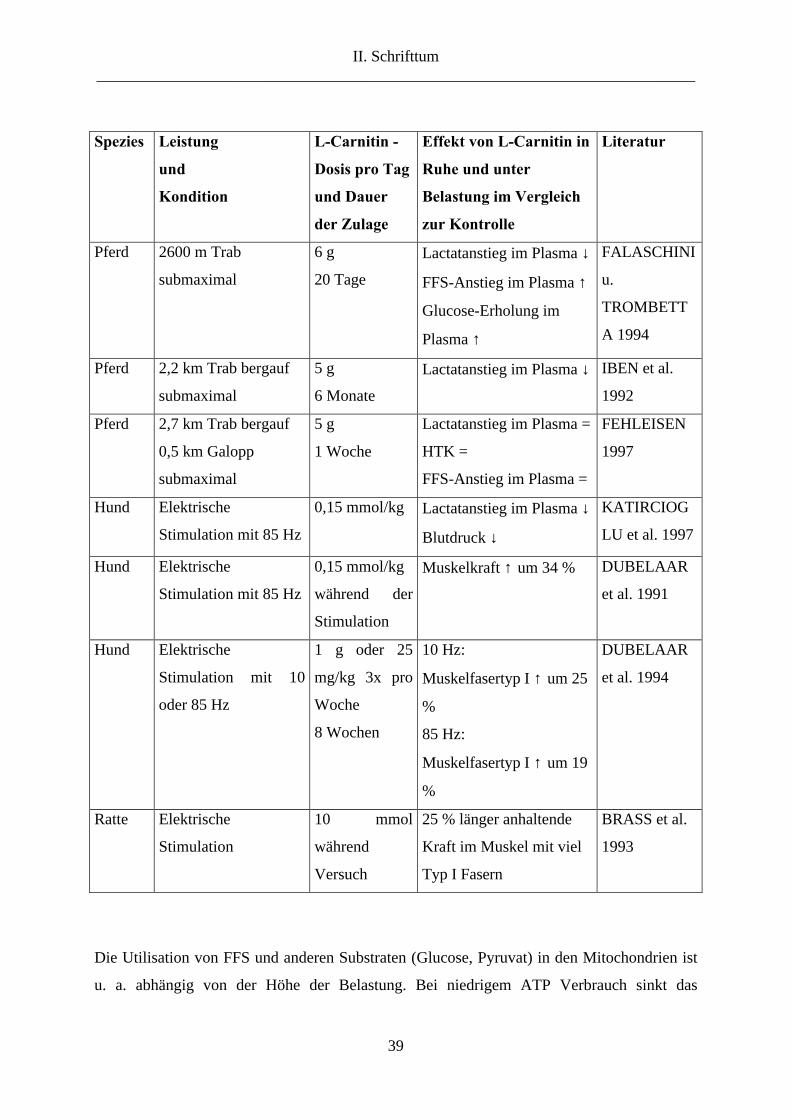
Tab. 2.7.: Effekte von Carnitingaben auf leistungsabhängige Funktionsmerkmale in Ruhe und
bei Belastung im Vergleich zu Kontrollgruppen. ↓ = niedriger, ↑ = höher, = = kein
Unterschied zur Kontrolle
Spezies Leistung
und
Kondition
L-Carnitin -
Dosis pro Tag
und Dauer
der Zulage
Effekt von L-Carnitin in
Ruhe und unter
Belastung im Vergleich
zur Kontrolle
Literatur
Taube trainiert 45-90 mg
7 Tage
In Ruhe
FFS im Plasma ↓
Bei Belastung
FFS-Anstieg im Plasma ↓
(dosisabhängig)
Lactatanstieg im Plasma ↓
bei 90 mg
BORGHIJS u.
DE WILDE
1992
II. Schrifttum ___________________________________________________________________________
38
Spezies Leistung
und
Kondition
L-Carnitin -
Dosis pro Tag
und Dauer
der Zulage
Effekt von L-Carnitin in
Ruhe und unter
Belastung im Vergleich
zur Kontrolle
Literatur
Mensch Marathonläufer 2 g
4 Wochen
Aktivität der Enzyme der
Atmungskette in der
Muskulatur (NADH-
Cytochrom-C-Reduktase,
Succinate-Cytochrom-C-
Reduktase, Cytochrom-
Oxidase) ↑
HUERTAS et
al. 1992
Mensch Radfahren
Moderat trainiert
6 g
7 Tage
Pulsfrequenz ↓
Leistung ↑
VO2max ↑
BILLIGMAN
N et al. 1990
Mensch Radfahren unter
Maximalbelastung
Moderat trainiert
2 g
1 h vor Beginn
Lactatanstieg im Plasma ↓ SILIPRANDI
et al. 1989
Mensch Radfahren unter
Maximalbelastung
Moderat trainiert
2 g
1 h vor Beginn
VO2max ↑
Lactatanstieg im Plasma↓
Leistung ↑
VECCHIET et
al. 1990
Mensch Submaximale
Belastung
2 g
28 Tage
Respiratorischer Quotient
↓
GOROSTIAG
A et al. 1989
Mensch Rudern
Kanu fahren
Gewichtheben
Hochleistungstrainiert
1-3 g
3 Wochen
FFS-Anstieg im Plasma ↑
Lactatanstieg im Plasma ↓
DRAGAN et
al. 1989
Mensch Radfahren
Moderat trainiert
5 g
5 Tage
FFS-Anstieg im Plasma =
Herzschlag ↓ um 7-8 %
SOOP et al.
1988
II. Schrifttum ___________________________________________________________________________
39
Spezies Leistung
und
Kondition
L-Carnitin -
Dosis pro Tag
und Dauer
der Zulage
Effekt von L-Carnitin in
Ruhe und unter
Belastung im Vergleich
zur Kontrolle
Literatur
Pferd 2600 m Trab
submaximal
6 g
20 Tage
Lactatanstieg im Plasma ↓
FFS-Anstieg im Plasma ↑
Glucose-Erholung im
Plasma ↑
FALASCHINI
u.
TROMBETT
A 1994
Pferd 2,2 km Trab bergauf
submaximal
5 g
6 Monate
Lactatanstieg im Plasma ↓ IBEN et al.
1992
Pferd 2,7 km Trab bergauf
0,5 km Galopp
submaximal
5 g
1 Woche
Lactatanstieg im Plasma =
HTK =
FFS-Anstieg im Plasma =
FEHLEISEN
1997
Hund Elektrische
Stimulation mit 85 Hz
0,15 mmol/kg Lactatanstieg im Plasma ↓
Blutdruck ↓
KATIRCIOG
LU et al. 1997
Hund Elektrische
Stimulation mit 85 Hz
0,15 mmol/kg
während der
Stimulation
Muskelkraft ↑ um 34 % DUBELAAR
et al. 1991
Hund Elektrische
Stimulation mit 10
oder 85 Hz
1 g oder 25
mg/kg 3x pro
Woche
8 Wochen
10 Hz:
Muskelfasertyp I ↑ um 25
%
85 Hz:
Muskelfasertyp I ↑ um 19
%
DUBELAAR
et al. 1994
Ratte Elektrische
Stimulation
10 mmol
während
Versuch
25 % länger anhaltende
Kraft im Muskel mit viel
Typ I Fasern
BRASS et al.
1993
Die Utilisation von FFS und anderen Substraten (Glucose, Pyruvat) in den Mitochondrien ist
u. a. abhängig von der Höhe der Belastung. Bei niedrigem ATP Verbrauch sinkt das

II. Schrifttum ___________________________________________________________________________
40
Redoxpotential in den Mitochondrien und verlangsamt die Substratoxidation
(Atmungskettenkontrolle). Die oxidative Decarboxylierung von Pyruvat zu Acetyl-CoA und
seine weitere Verbrennung im Citratcyclus werden gehemmt, wenn das NADH/NAD
Verhältnis in den Mitochondrien steigt (BREMER 1969; LUMENG et al. 1976). Ein hohes
NADH/NAD Verhältnis und viel Acetyl-CoA hemmt auch die β-Oxidation der FS,
insbesondere die Aktivität der 3-Hydroxyacyl-CoA-Dehydrogenase und der Thiolase
(BREMER et al. 1984). Im humanen Skelettmuskel stimuliert ein niedriges Acetyl-CoA/CoA
Verhältnis die Pyruvat- und die FS-Oxidation (UZIEL et al. 1988).
BORGHIJS und DE WILDE (1992) supplementierten 12 Brieftauben oral mit 0, 45 und 90
mg Carnitin jeweils über sieben Tage. Bei einer Belastung in Form einer elektrischen
Stimulation der Pectoralismuskulatur über sechs Stunden fiel der Anstieg der FFS und des
Lactats im Plasma gegenüber unsupplementierten Kontrollen niedriger aus. Die Autoren
erklären diesen Effekt mit einer vermehrten Aufnahme und verstärkten Oxidation von FFS in
der Muskulatur und einer verminderten Glucoseverbrennung im Vergleich zu
unsupplementierten Kontrollen.
In einer Studie mit 10 moderat trainierten Männern mit einer Carnitinsupplementation von 2 g
1 Stunde vor einem Fahrradbelastungstest wiesen VECCHIET et al. (1990) einen
verminderten belastungsinduzierten Lactatanstieg und einen höheren Sauerstoffverbrauch
während der Belastung gegenüber denselben Männern ohne Supplementation nach. Sie zogen
den Schluss, dass durch die hohe O2-Aufnahme nach Carnitingabe die aerobe
Energiebereitstellung im Muskel begünstigt wird.
SILIPRANDI et al. (1990) beobachteten einen niedrigeren Lactat- und Pyruvatspiegel und
einen höheren Acetylcarnitin Spiegel im Plasma bei mit 2 g Carnitin eine Stunde vor
Belastung supplementierten Männern nach einem Fahrradbelastungstest bis zur Erschöpfung
im Vergleich zu denselben unsupplementierten Testpersonen. Die Autoren vermuten eine
Stimulation der Pyruvatdehydrogenase (PDH) Aktivität durch Carnitin. Diese Stimulation
könnte durch ein durch Carnitin vermitteltes niedrigeres Acetyl-CoA/CoA-Verhältnis
hervorgerufen werden. Dadurch wurde weniger Pyruvat zu Lactat umgesetzt.

II. Schrifttum ___________________________________________________________________________
41
Neben diesen positiven Effekten einer Carnitinuzulage wird auch über Versuche berichtet, in
denen L-Carnitin keinen Einfluss auf die Leistung ausübt (DECOMBAZ et al. 1992; IBEN
1998). IBEN (1999) konnte bei Beaglen keinen Einfluss einer Carnitinsupplementation von 1
g drei Stunden vor einer 20 minütigen Laufbandbelastung bei 6 km/h auf die Herzfrequenz
und die Lactatkonzentration im Blut feststellen. In einer Studie von FEHLEISEN (1997)
absolvierten vier Pferde eine Teststrecke von 2,7 km im Trab über 280 Höhenmeter und
danach 0,5 km im Galopp in neun Minuten. Es wurde kein Einfluss einer Carnitinzulage von
5 g/Tier und Tag über eine Woche auf das Plasmalactat und die Pulsfrequenz festgestellt.
Allerdings wurde bei diesen Pferden auch kein Anstieg der Carnitinspiegel im Plasma
festgestellt.
9. Abschließende Bemerkung
Aufgrund seiner spezifischen biochemischen Funktionen für die Bereitstellung von Energie
genießt Carnitin im humanen Leistungssport den Ruf eines Leistungsförderers. Im
Pferdeleistungssport sind die Ergebnisse von dazu publizierten Untersuchungen bisher
uneinheitlich. Die unterschiedlichen Befunde lassen keinen eindeutigen Schluss über einen
Einfluss einer L-Carnitinsupplementierung auf Leistungsmerkmale bei Pferden zu. Auch die
Frage, ob sich beim Pferd der Carnitingehalt in der Muskulatur durch eine längere
Carnitinzulage erhöht, ist bisher nicht eindeutig geklärt. Im Hinblick auf mögliche
leistungsabhängige Effekte von Carnitin kommt der Dauer und der Höhe einer solche Zulage
vielleicht eine Bedeutung zu. Aufgrund der biochemischen Funktionen von Carnitin und im
Hinblick auf viele positive Befunde beim Menschen und anderen Spezies erscheint ein
positiver Effekt auch beim Pferd unter bestimmten Bedingungen möglich. Voraussetzung
dafür ist jedoch wohl, dass sein Gehalt im Muskel durch eine längere Carnitingabe zunimmt.
Es war ein Ziel dieser Untersuchungen bei etwa 2-jährigen Pferden, die in Verbindung mit
einem Trainingsprogramm täglich 10 g Carnitin erhielten, zu untersuchen, ob diese Zulage die
Carnitinkonzentration in der Muskulatur erhöht und ob sich dadurch Effekte auf die
Leistungsfähigkeit ergeben.

III. Material und Methoden ________________________________________________________________________
42
III. Material und Methoden
1. Versuchsziel und Fragestellung
Ziel der Untersuchung war festzustellen, ob bei Pferden eine orale Zulage von L-Carnitin in
Verbindung mit einem körperlichen Training einen Effekt auf Leistungsparameter wie
Herzfrequenz, Blutlactat und andere Parameter des Energiestoffwechsels wie Glucose, freie
Fettsäuren und Harnstoff ausübt. Ferner war von Interesse zu prüfen, ob eine sich über
mehrere Monate erstreckende orale Supplementierung mit L-Carnitin den Carnitingehalt der
Muskulatur beeinflusst.
Es handelt sich um ein gemeinsames Forschungsvorhaben des Physiologischen Instituts und
des Instituts für Tierernährung, an dessen Durchführung außerdem noch Frau U. Wedemeyer
beteiligt war.
2. Versuchsaufbau
Der Versuch bestand aus einer Trainingsperiode von sechseinhalb Wochen Dauer und einer
anschließenden Ruheperiode (Detrainingsperiode) von 10 Wochen. Zu Beginn der
Trainingsperiode wurde zur Einstufung des individuellen Leistungsvermögens der Tiere ein
Stufentest durchgeführt, der am Ende des Trainingsprogramms wiederholt wurde.
Anschließend an den zweiten Stufentest wurden die Tiere zwei weiteren Trainingsbelastungen
unterzogen. Das Trainingsprogramm bestand aus alternierenden Kurz- und
Langzeitbelastungen mit dazwischenliegenden eintägigen Pausen. Zur Durchführung der
Versuche stand ein Hochgeschwindigkeitslaufband (Mustang, Fa. Kagra, Schweiz) zur
Verfügung, das sich in einem halboffenen, unklimatisierten Raum befand.
Beim Stufentest wurden die Tiere stufenweise einer ansteigenden körperlichen Belastung
unterzogen, indem sie auf dem Laufband nach einem festgelegten
Geschwindigkeitsprogramm für eine Gesamtdauer von 15 Minuten immer schneller laufen

III. Material und Methoden ________________________________________________________________________
43
mussten. Der während der einzelnen Belastungsstufen und am Ende der Belastung
vorliegende Blutlactatspiegel wurde sofort gemessen. Die Beziehung zwischen der
Laufbandgeschwindigkeit und dem sich unter Gleichgewichtsbedingungen einstellenden
Lactatspiegel im Blut bildete die Grundlage zur Abschätzung des körperlichen
Leistungsvermögens der einzelnen Tiere.
Am Ende der 39-tägigen Trainingsperiode nach 16 einzelnen Trainingsbelastungen (Abb.
3.6.) wurde der Leistungszustand durch einen zweiten gleich angelegten Stufentest erneut
ermittelt und der Trainingsfortschritt anhand der Lactat-Geschwindigkeitskurve abgeschätzt.
Vier Pferde erhielten am Abend nach dem ersten Stufentest über eine Dauer von vier Monaten
bis zum Ende der zehnwöchigen Ruhephase (Abb. 3.6.) eine orale Carnitinzulage. Vier Pferde
dienten als unsupplementierte Kontrollen.
Während des Trainings wurden die Tiere jeden zweiten Tag, alternierend in Form einer Kurz-
und einer Langzeitbelastung, trainiert.
Die Kurzzeitbelastung entsprach in ihrem Grundkonzept einem Stufentest. Auch bei dieser
Belastung wurde die Laufbandgeschwindigkeit im Rahmen einer Belastungsdauer von 15
Minuten stufenweise erhöht. Die Anfangsgeschwindigkeit des Laufbands wurde jedoch so
gewählt, dass sich, basierend auf den Daten des zuvor absolvierten Stufentests, ein
Blutlactatspiegel von 4 mmol/l einstellen sollte (v4). Die Laufbandgeschwindigkeit wurde von
dieser Anfangsgeschwindigkeit an in einminütigen Abständen um 0,3 m/s erhöht. Die Dauer
der Belastung wurde so gewählt, dass alle Tiere am Ende der Belastung einen
Blutlactatspiegel von mindestens 12 mmol/l aufwiesen, also auf jeden Fall anaerob Energie
gewinnen müssen. Die für diese Belastung notwendige Energie wurde für jedes Pferd
entsprechend seiner Laufgeschwindigkeit nach den Vorgaben der Gesellschaft für
Ernährungsphysiologie der Haustiere (1994) berechnet (Tab. 3.3.).
Während der Langzeitbelastung war die Laufgeschwindigkeit der Tiere konstant. Hier wurde
die Laufgeschwindigkeit so gewählt, dass sich, basierend auf den Daten des Stufentests, ein
Blutlactatspiegel von 2,5 mmol/l einstellen sollte (v2,5). Die Belastung wurde so lange
aufrecht erhalten, bis der Energieverbrauch dem der Kurzzeitbelastung entsprach (Tab. 3.3.).

III. Material und Methoden ________________________________________________________________________
44
Übergreifendes Ziel des Trainingsprogramms war es, die Muskulatur der Tiere in etwa
gleichgroßem Umfang einer Belastung mit vorwiegend anaerober (Kurzzeitbelastung) oder
aerober (Langzeitbelastung) Energiebereitstellung zu unterwerfen.
Die Abbildung 3.1. gibt einen groben Überblick über den zeitlichen Ablauf des
Versuchsprogramms.
Abb. 3.1.: Zeitlicher Ablauf des Versuchsprogramms
3. Versuchstiere
Für den Versuch standen acht Traber (4 Stuten und 4 Wallache) im Alter von zwei Jahren zur
Verfügung. Alle Tiere stammten von einem Hengst ab und wurden bis zum Versuchsbeginn
gleich aufgezogen. Vor Beginn des Trainings wurden die Tiere zwei Monate an das Laufband
und an ihre Umgebung gewöhnt. Die Tiere wurden in Boxen auf Stroh gehalten und erhielten
täglich Zugang zu einem Sandauslauf. Sie wurden nach dem Zufallsprinzip in zwei Gruppen
aufgeteilt. Einer Gruppe wurden täglich 10 g L-Carnitin, entsprechend 20 g Carniking (Fa.
Lonzagroup, Basel), mit dem Futter verabreicht (Carnitingruppe). Die andere Gruppe erhielt
kein Carnitin und diente als Kontrollgruppe. Die Supplementierung begann am Abend nach
dem ersten Stufentest. Zu Beginn der Versuche waren alle Tiere klinisch gesund.
Ruheperiode(10 Wochen)
Trainingsprogramm(8 Kurz- u.
8 Langzeitbelastungen)berechnet nach dem
1. Stufentest
1 Kurz- u. 1 Langzeit-belastung,berechnetnach dem
2. Stufentest
1. Stufen-test
2. Stufen-
test
16,5 Wochen
Ruheperiode(10 Wochen)
Trainingsprogramm(8 Kurz- u.
8 Langzeitbelastungen)berechnet nach dem
1. Stufentest
1 Kurz- u. 1 Langzeit-belastung,berechnetnach dem
2. Stufentest
1. Stufen-test
2. Stufen-
test
16,5 Wochen
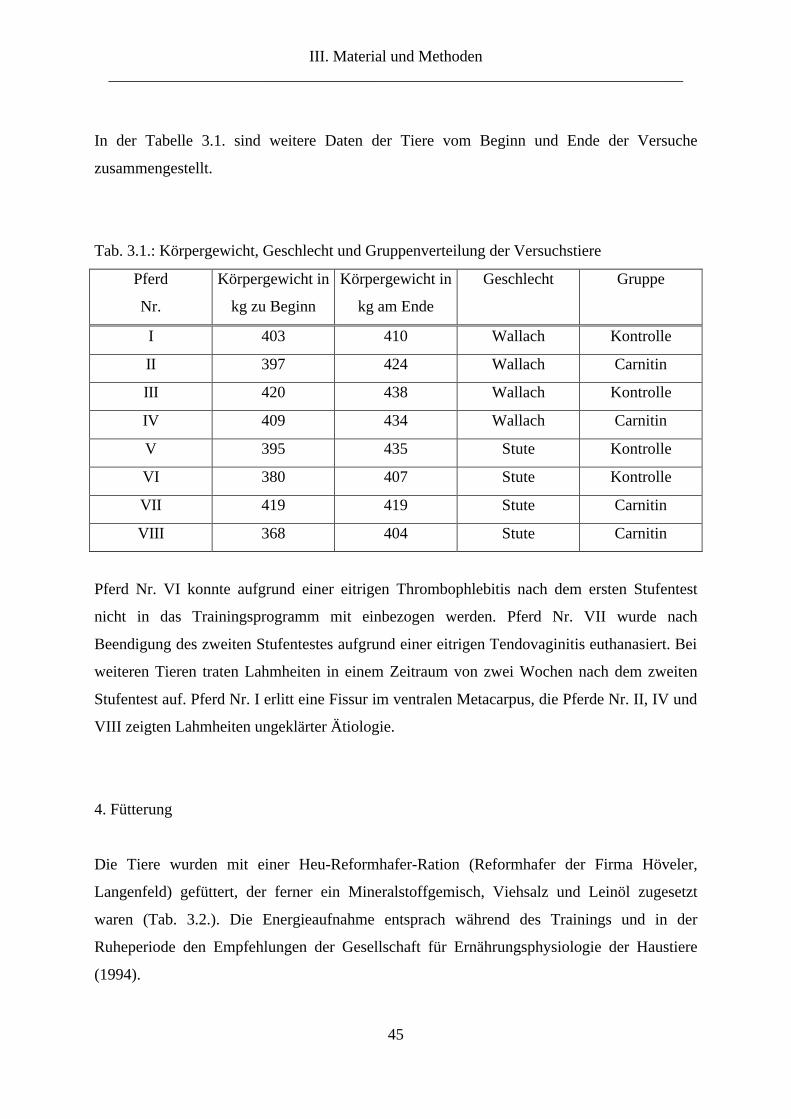
III. Material und Methoden ________________________________________________________________________
45
In der Tabelle 3.1. sind weitere Daten der Tiere vom Beginn und Ende der Versuche
zusammengestellt.
Tab. 3.1.: Körpergewicht, Geschlecht und Gruppenverteilung der Versuchstiere
Pferd
Nr.
Körpergewicht in
kg zu Beginn
Körpergewicht in
kg am Ende
Geschlecht Gruppe
I 403 410 Wallach Kontrolle
II 397 424 Wallach Carnitin
III 420 438 Wallach Kontrolle
IV 409 434 Wallach Carnitin
V 395 435 Stute Kontrolle
VI 380 407 Stute Kontrolle
VII 419 419 Stute Carnitin
VIII 368 404 Stute Carnitin
Pferd Nr. VI konnte aufgrund einer eitrigen Thrombophlebitis nach dem ersten Stufentest
nicht in das Trainingsprogramm mit einbezogen werden. Pferd Nr. VII wurde nach
Beendigung des zweiten Stufentestes aufgrund einer eitrigen Tendovaginitis euthanasiert. Bei
weiteren Tieren traten Lahmheiten in einem Zeitraum von zwei Wochen nach dem zweiten
Stufentest auf. Pferd Nr. I erlitt eine Fissur im ventralen Metacarpus, die Pferde Nr. II, IV und
VIII zeigten Lahmheiten ungeklärter Ätiologie.
4. Fütterung
Die Tiere wurden mit einer Heu-Reformhafer-Ration (Reformhafer der Firma Höveler,
Langenfeld) gefüttert, der ferner ein Mineralstoffgemisch, Viehsalz und Leinöl zugesetzt
waren (Tab. 3.2.). Die Energieaufnahme entsprach während des Trainings und in der
Ruheperiode den Empfehlungen der Gesellschaft für Ernährungsphysiologie der Haustiere
(1994).
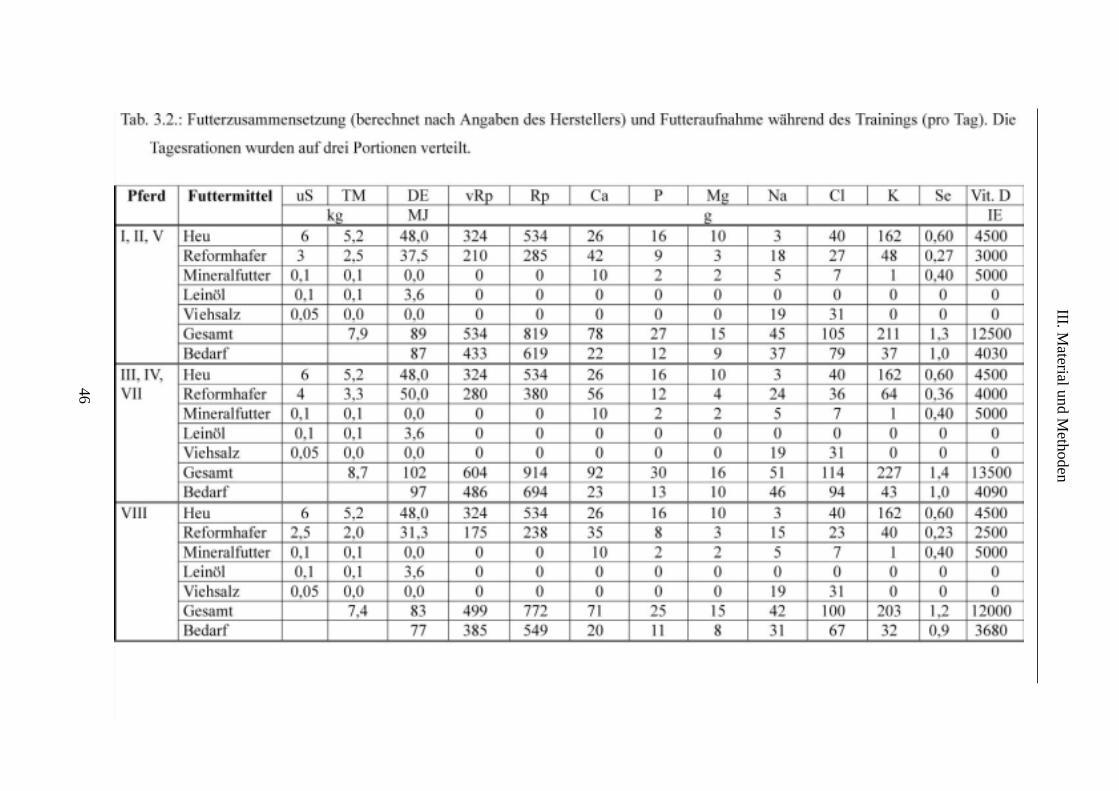
III. Material und M
ethoden ________________________________________________________________________
46

III. Material und Methoden ________________________________________________________________________
47
In der Ruhephase wurde die Hafermenge auf 1,5 kg und das Viehsalz auf 20 g reduziert.
Die Tiere wurden jeweils um 8, 14 und 19 Uhr gefüttert. An den Trainingstagen wurden die
Tiere jeweils drei Stunden vor der zu verschiedenen Uhrzeiten stattfindenden Belastung
gefüttert. Wasser stand aus Selbsttränken ad libitum zur Verfügung. Das L-Carnitin wurde in
zwei Dosen morgens und abends verabreicht.
5. Versuchsdurchführung
Alle Pferde wurden im gleichen Zeitrahmen trainiert.
5.1. Stufentest
Vor Beginn der stufenweise zunehmenden Belastung wurden die Tiere 5 Minuten bei 2 m/s
und 10 Minuten bei 3,5 m/s auf dem Laufband bewegt (Aufwärmphase). Danach wurde die
erste Blutprobe aus der Vena jugularis externa entnommen. Der Stufentest bestand aus 6
Stufen à 5 Minuten mit einer jeweils einminütigen Pause zur Blutentnahme. Während der
ersten Stufe betrug die Laufgeschwindigkeit 5 m/s. Sie wurde jeweils nach 5 Minuten
Laufarbeit und einer Minute Pause um 1 m/s erhöht, so dass die Pferde eine
Endgeschwindigkeit von 10 m/s erreichten. Vier und 24 Stunden nach der Belastung wurde je
eine weitere Blutproben entnommen. Vor dem Aufwärmen und nach der letzten
Belastungsstufe wurde Muskelgewebe aus dem Musculus glutaeus medius entnommen. Es
wurden zwei Stufentests im Abstand von fünf Wochen durchgeführt. Abbildung 3.2.
verdeutlicht den zeitlichen Ablauf des Stufentests.
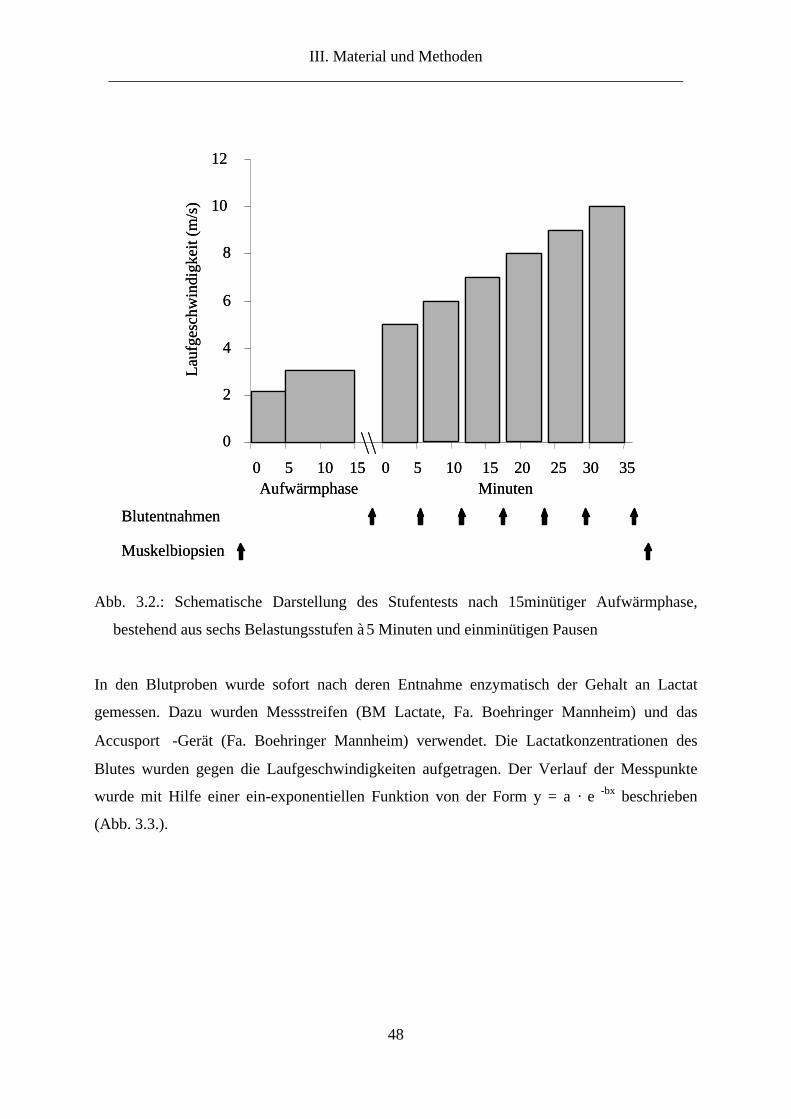
III. Material und Methoden ________________________________________________________________________
48
Abb. 3.2.: Schematische Darstellung des Stufentests nach 15minütiger Aufwärmphase,
bestehend aus sechs Belastungsstufen à 5 Minuten und einminütigen Pausen
In den Blutproben wurde sofort nach deren Entnahme enzymatisch der Gehalt an Lactat
gemessen. Dazu wurden Messstreifen (BM Lactate, Fa. Boehringer Mannheim) und das
Accusport-Gerät (Fa. Boehringer Mannheim) verwendet. Die Lactatkonzentrationen des
Blutes wurden gegen die Laufgeschwindigkeiten aufgetragen. Der Verlauf der Messpunkte
wurde mit Hilfe einer ein-exponentiellen Funktion von der Form y = a · e -bx beschrieben
(Abb. 3.3.).
0
2
4
6
8
10
12
Blutentnahmen
Muskelbiopsien
Lauf
gesc
hwin
digk
eit (
m/s
)
0 5 10 15 0 5 10 15 20 25 30 35MinutenAufwärmphase
0
2
4
6
8
10
12
Blutentnahmen
Muskelbiopsien
Lauf
gesc
hwin
digk
eit (
m/s
)
0 5 10 15 0 5 10 15 20 25 30 35MinutenAufwärmphase
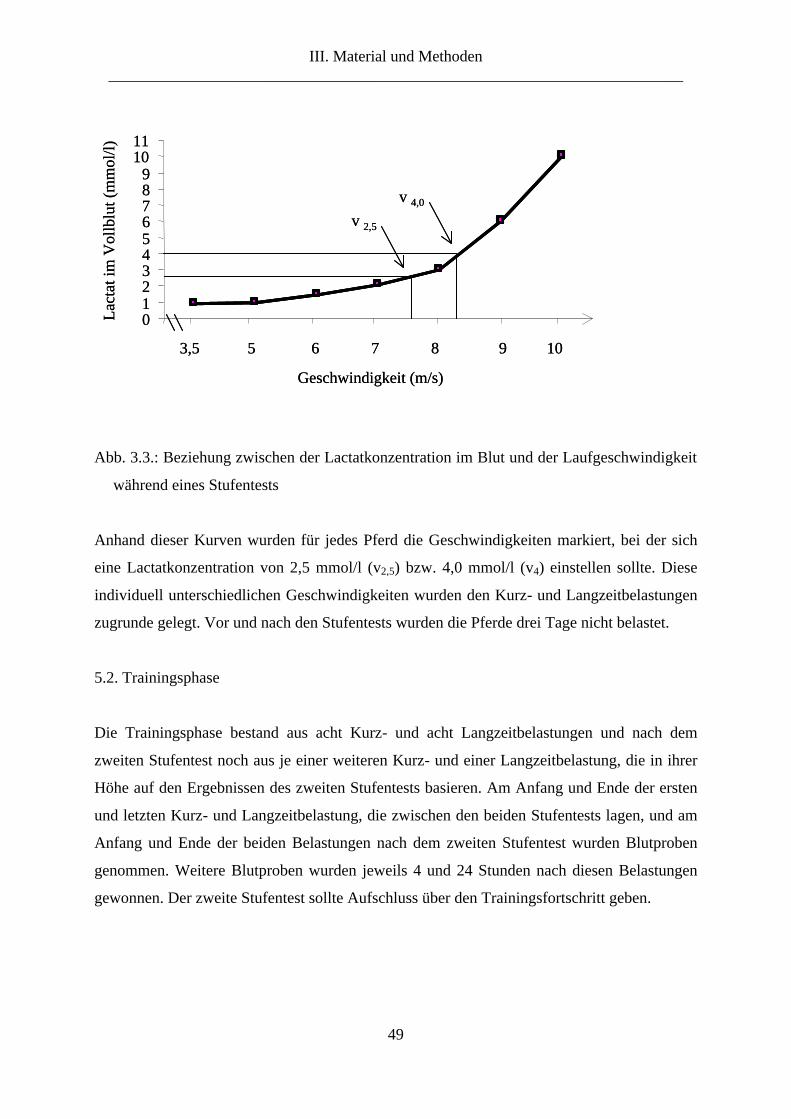
III. Material und Methoden ________________________________________________________________________
49
Abb. 3.3.: Beziehung zwischen der Lactatkonzentration im Blut und der Laufgeschwindigkeit
während eines Stufentests
Anhand dieser Kurven wurden für jedes Pferd die Geschwindigkeiten markiert, bei der sich
eine Lactatkonzentration von 2,5 mmol/l (v2,5) bzw. 4,0 mmol/l (v4) einstellen sollte. Diese
individuell unterschiedlichen Geschwindigkeiten wurden den Kurz- und Langzeitbelastungen
zugrunde gelegt. Vor und nach den Stufentests wurden die Pferde drei Tage nicht belastet.
5.2. Trainingsphase
Die Trainingsphase bestand aus acht Kurz- und acht Langzeitbelastungen und nach dem
zweiten Stufentest noch aus je einer weiteren Kurz- und einer Langzeitbelastung, die in ihrer
Höhe auf den Ergebnissen des zweiten Stufentests basieren. Am Anfang und Ende der ersten
und letzten Kurz- und Langzeitbelastung, die zwischen den beiden Stufentests lagen, und am
Anfang und Ende der beiden Belastungen nach dem zweiten Stufentest wurden Blutproben
genommen. Weitere Blutproben wurden jeweils 4 und 24 Stunden nach diesen Belastungen
gewonnen. Der zweite Stufentest sollte Aufschluss über den Trainingsfortschritt geben.
0123456789
1011
3,5 5 6 7 8 9 10
Geschwindigkeit (m/s)
Lact
atim
Vol
lblu
t (m
mol
/l)
v 2,5
v 4,0
0123456789
1011
3,5 5 6 7 8 9 10
Geschwindigkeit (m/s)
Lact
atim
Vol
lblu
t (m
mol
/l)
v 2,5
v 4,0
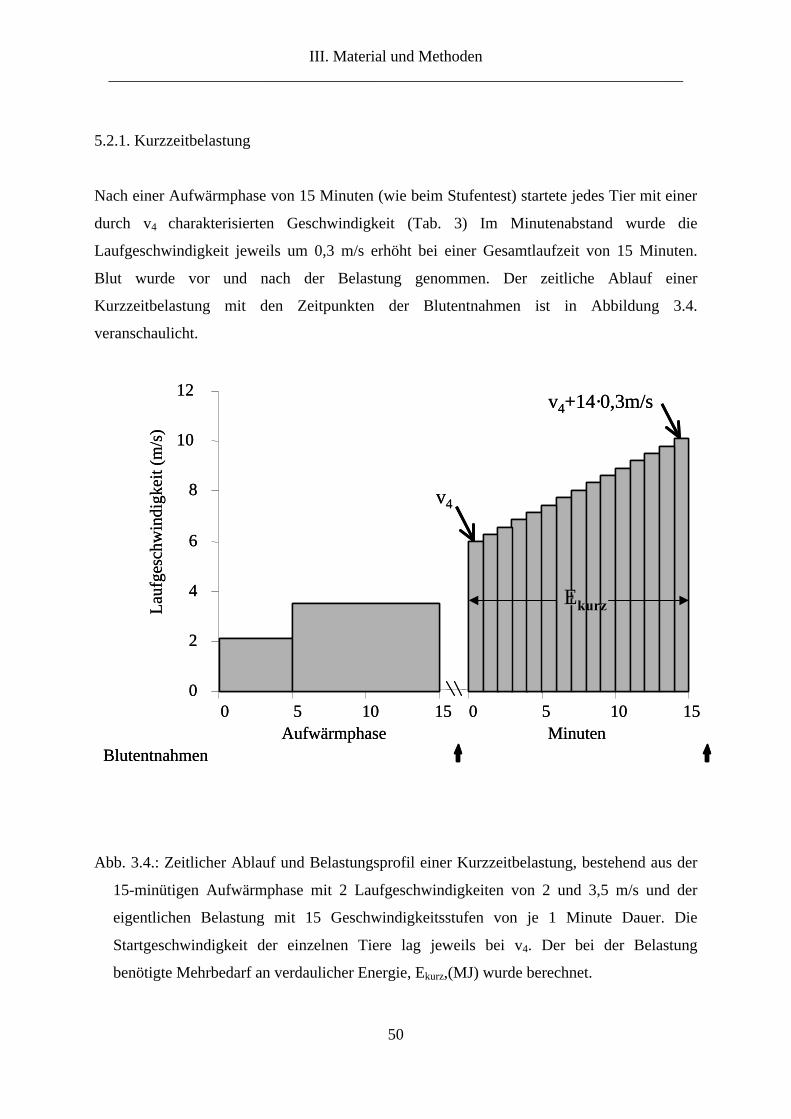
III. Material und Methoden ________________________________________________________________________
50
5.2.1. Kurzzeitbelastung
Nach einer Aufwärmphase von 15 Minuten (wie beim Stufentest) startete jedes Tier mit einer
durch v4 charakterisierten Geschwindigkeit (Tab. 3) Im Minutenabstand wurde die
Laufgeschwindigkeit jeweils um 0,3 m/s erhöht bei einer Gesamtlaufzeit von 15 Minuten.
Blut wurde vor und nach der Belastung genommen. Der zeitliche Ablauf einer
Kurzzeitbelastung mit den Zeitpunkten der Blutentnahmen ist in Abbildung 3.4.
veranschaulicht.
Abb. 3.4.: Zeitlicher Ablauf und Belastungsprofil einer Kurzzeitbelastung, bestehend aus der
15-minütigen Aufwärmphase mit 2 Laufgeschwindigkeiten von 2 und 3,5 m/s und der
eigentlichen Belastung mit 15 Geschwindigkeitsstufen von je 1 Minute Dauer. Die
Startgeschwindigkeit der einzelnen Tiere lag jeweils bei v4. Der bei der Belastung
benötigte Mehrbedarf an verdaulicher Energie, Ekurz,(MJ) wurde berechnet.
v4+14·0,3m/s
0
2
4
6
8
10
12
MinutenBlutentnahmen
Lauf
gesc
hwin
digk
eit (
m/s
)
v4
0 5 10 15 0 5 10 15Aufwärmphase
EEkurzkurz
v4+14·0,3m/s
0
2
4
6
8
10
12
MinutenBlutentnahmen
Lauf
gesc
hwin
digk
eit (
m/s
)
v4
0 5 10 15 0 5 10 15Aufwärmphase
EEkurzkurz
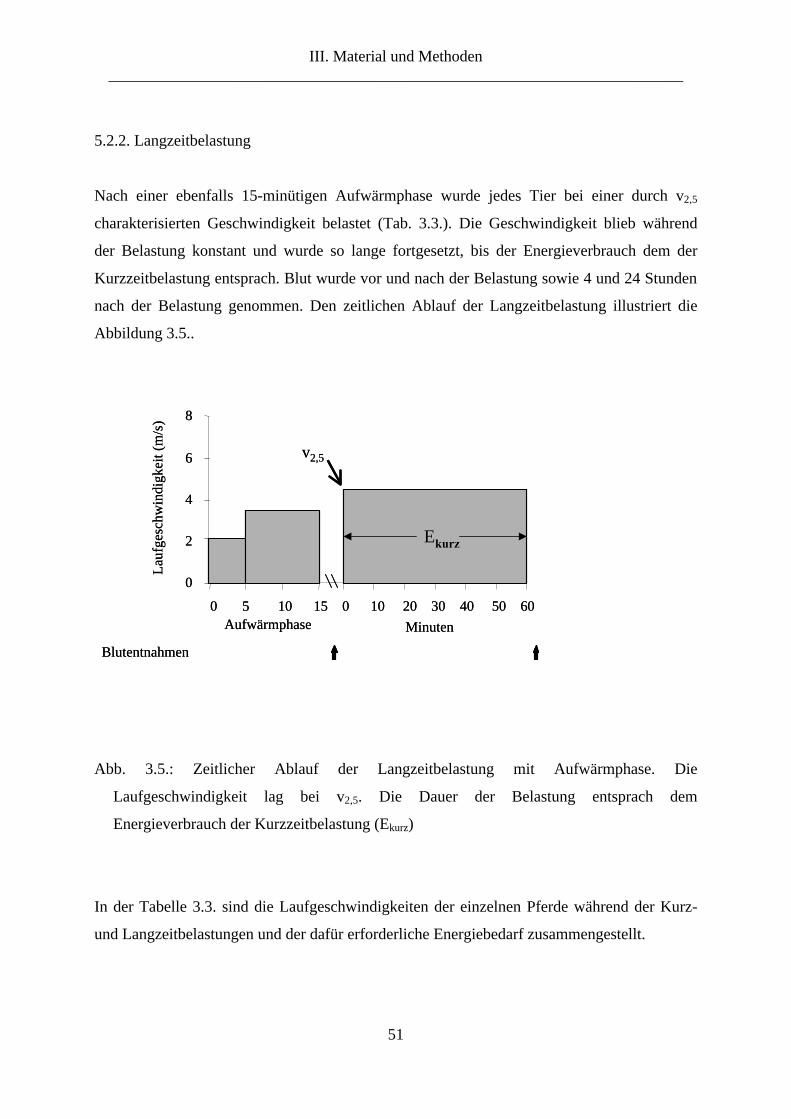
III. Material und Methoden ________________________________________________________________________
51
5.2.2. Langzeitbelastung
Nach einer ebenfalls 15-minütigen Aufwärmphase wurde jedes Tier bei einer durch v2,5
charakterisierten Geschwindigkeit belastet (Tab. 3.3.). Die Geschwindigkeit blieb während
der Belastung konstant und wurde so lange fortgesetzt, bis der Energieverbrauch dem der
Kurzzeitbelastung entsprach. Blut wurde vor und nach der Belastung sowie 4 und 24 Stunden
nach der Belastung genommen. Den zeitlichen Ablauf der Langzeitbelastung illustriert die
Abbildung 3.5..
Abb. 3.5.: Zeitlicher Ablauf der Langzeitbelastung mit Aufwärmphase. Die
Laufgeschwindigkeit lag bei v2,5. Die Dauer der Belastung entsprach dem
Energieverbrauch der Kurzzeitbelastung (Ekurz)
In der Tabelle 3.3. sind die Laufgeschwindigkeiten der einzelnen Pferde während der Kurz-
und Langzeitbelastungen und der dafür erforderliche Energiebedarf zusammengestellt.
0
2
4
6
8
Blutentnahmen
Lauf
gesc
hwin
digk
eit (
m/s
)
v2,5
0 5 10 15 0 10 20 30 40 50 60
EEkurzkurz
Aufwärmphase Minuten
0
2
4
6
8
Blutentnahmen
Lauf
gesc
hwin
digk
eit (
m/s
)
v2,5
0 5 10 15 0 10 20 30 40 50 60
EEkurzkurz
Aufwärmphase Minuten
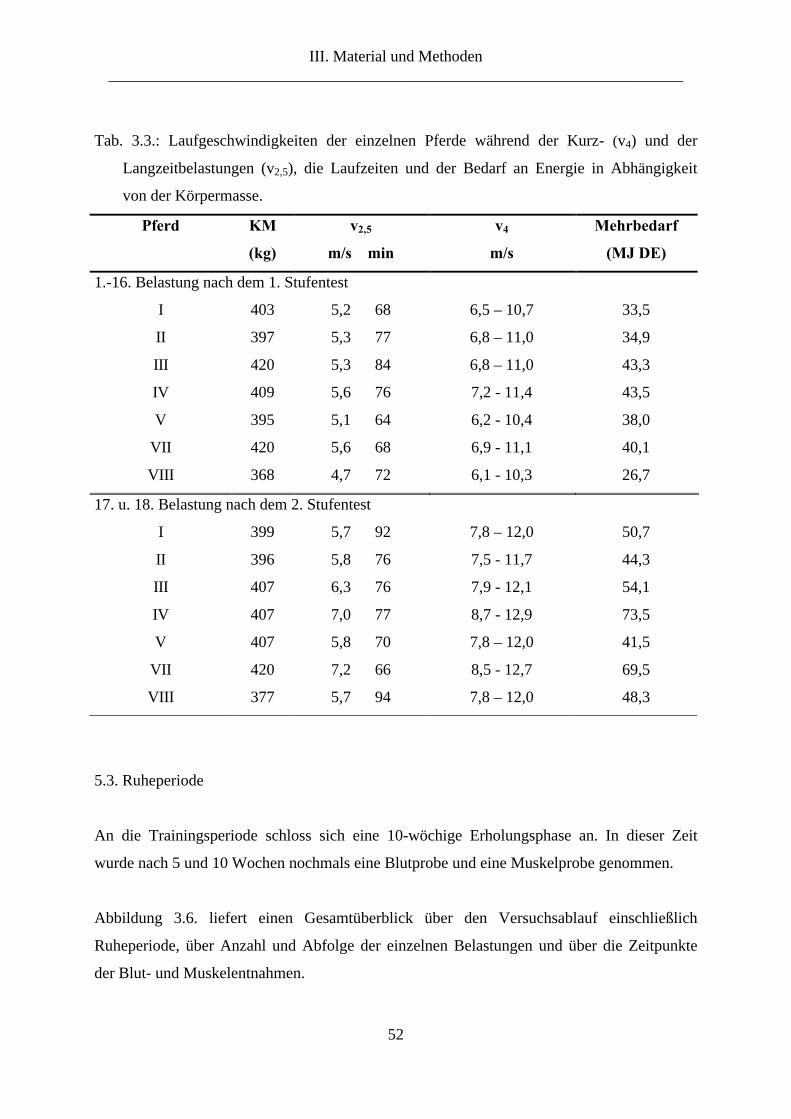
III. Material und Methoden ________________________________________________________________________
52
Tab. 3.3.: Laufgeschwindigkeiten der einzelnen Pferde während der Kurz- (v4) und der
Langzeitbelastungen (v2,5), die Laufzeiten und der Bedarf an Energie in Abhängigkeit
von der Körpermasse.
Pferd KM
(kg)
v2,5
m/s min
v4
m/s
Mehrbedarf
(MJ DE)
1.-16. Belastung nach dem 1. Stufentest
I 403 5,2 68 6,5 – 10,7 33,5
II 397 5,3 77 6,8 – 11,0 34,9
III 420 5,3 84 6,8 – 11,0 43,3
IV 409 5,6 76 7,2 - 11,4 43,5
V 395 5,1 64 6,2 - 10,4 38,0
VII 420 5,6 68 6,9 - 11,1 40,1
VIII 368 4,7 72 6,1 - 10,3 26,7
17. u. 18. Belastung nach dem 2. Stufentest
I 399 5,7 92 7,8 – 12,0 50,7
II 396 5,8 76 7,5 - 11,7 44,3
III 407 6,3 76 7,9 - 12,1 54,1
IV 407 7,0 77 8,7 - 12,9 73,5
V 407 5,8 70 7,8 – 12,0 41,5
VII 420 7,2 66 8,5 - 12,7 69,5
VIII 377 5,7 94 7,8 – 12,0 48,3
5.3. Ruheperiode
An die Trainingsperiode schloss sich eine 10-wöchige Erholungsphase an. In dieser Zeit
wurde nach 5 und 10 Wochen nochmals eine Blutprobe und eine Muskelprobe genommen.
Abbildung 3.6. liefert einen Gesamtüberblick über den Versuchsablauf einschließlich
Ruheperiode, über Anzahl und Abfolge der einzelnen Belastungen und über die Zeitpunkte
der Blut- und Muskelentnahmen.

III. Material und Methoden ________________________________________________________________________
53
Abb.3.6.: Übersicht über den zeitlichen Ablauf des Trainingsprogramms, beginnend mit dem
1. Stufentest, der anschließenden Trainingsperiode, dem 2. Stufentest, den beiden letzten
Kurz- und Langzeitbelastungen und der anschließenden Ruheperiode. Die Zeitpunkte der
Blut- und Muskelentnahmen sind oberhalb des Balkens durch Pfeile markiert
5.4. Probennahmen
Tabelle 3.4. gibt eine Übersicht über die Zeitpunkte der Probenentnahmen und die
untersuchten Parameter während der unterschiedlichen Belastungen und in der Ruheperiode.
1. 5. 9. 13. 17. 21. 25. 29. 33. 39. 43. 45. 1. 5. 10.
S K L K L K L K L K L K L K L K L S K L
S = Stufentest; K = Kurzzeitbelastung; L = Langzeitbelastung; MB = Muskelbiopsie; BE = Blutentnahme
RuheperiodeTrainingsperiode
MB+BE BE BE BE BE MB+BE BE BE MB+BE MB+BE
Tage Wochen
1. 5. 9. 13. 17. 21. 25. 29. 33. 39. 43. 45. 1. 5. 10.
S K L K L K L K L K L K L K L K L S K L
S = Stufentest; K = Kurzzeitbelastung; L = Langzeitbelastung; MB = Muskelbiopsie; BE = Blutentnahme
RuheperiodeTrainingsperiode
MB+BE BE BE BE BE MB+BE BE BE MB+BE MB+BE
Tage Wochen
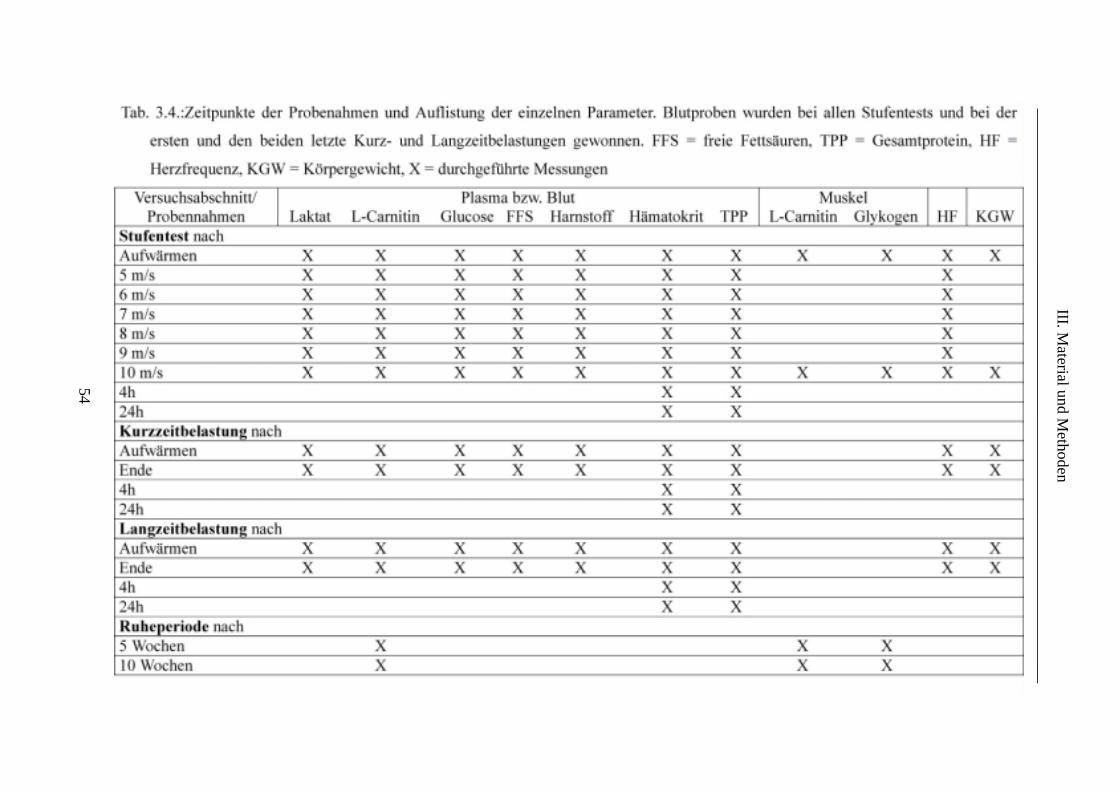
III. Material und M
ethoden ________________________________________________________________________
54

III. Material und Methoden ________________________________________________________________________
55
Blut
Bei jedem Stufentest, der ersten Kurz- und Langzeitbelastung, die dem Stufentest folgte,
sowie den letzten Kurz- und Langzeitbelastungen, die einem Stufentest vorausgingen, wurden
Blutproben entnommen. Ferner wurden 4 und 24 Stunden nach diesen Belastungen
Blutproben gewonnen. Schließlich wurden nochmals in der Ruhepause nach 5 und 10
Wochen zwei Blutproben genommen.
Für die Blutentnahme wurde ein Verweilkatheter (Fa. Braun, 14 G, 45 cm Länge) nach Rasur
und Desinfektion der Haut in die Vena jugularis externa rechts oder links geschoben und mit
einem Einzelknopfheft aus Catgut an der Haut fixiert. Der Katheter war mit einem
Verlängerungsstück (Fa. Vygon, Aachen) und einem Absperrhahn (Luer-Lock, Fa. Vygon,
Aachen) versehen. Für die Entnahmen wurden Monovetten (Fa. Sarstedt, Nümbrecht)
benutzt. Der Katheter wurde nach jeder Entnahme mit 10 ml isotonischer Kochsalzlösung
gespült. Nach der letzten Blutentnahme wurde der Katheter entfernt. Die Blutprobe, die 24
Stunden nach der Belastung entnommen wurde, wurde mir einem Vacutainer (Fa. Becton
Dickinson) gezogen.
Für die Bestimmung der Blutparameter Glucose, Carnitin, Harnstoff, Gesamtprotein und
Lactat wurden 9 ml Monovetten mit Lithium-Heparin (15 I.E. Heparin/ml Blut, Fa.
Sarstedt, Nümbrecht) verwendet. Für die Bestimmung der freien Fettsäuren und des
Hämatokrits wurden 9 ml Monovetten mit EDTA (1,2-2 mg EDTA/ml Blut, Fa. Sarstedt,
Nümbrecht) verwendet. Die Proben wurden bei 3000 g für 10 Minuten zentrifugiert, Das
Plasma wurde abpipettiert und in Eppendorfgefäßen in 1 ml Portionen bei –20°C bis zur
weiteren Untersuchung eingefroren.
Muskel
Zu Beginn und am Ende jedes Stufentests und in der 5. und 10. Woche der Ruhephase wurden
der linke und rechte Musculus glutaeus medius nach der Methode von LINDHOLM und

III. Material und Methoden ________________________________________________________________________
56
PIEHL (1974) bioptiert. In den gewonnenen Gewebeproben wurde der Glykogen- und der
Carnitingehalt bestimmt. Vor den Stufentests wurde die linke und nach den Stufentests die
rechte Seite des Tieres für die Biopsie ausgewählt. Die Entnahmestelle für das Bioptat befand
sich am Ende des cranialen Drittels der Verbindungslinie zwischen Hüfthöcker und
Schweifansatz. Nach Rasur, Reinigung und Desinfektion der Haut wurden 2 ml Lidocain
2%ig (Fa. Albrecht, Aulendorf) ohne Sperrkörper intracutan an der geplanten Incisionsstelle
injiziert und mit einem Skalpell ein 1 cm langer Hautschnitt durchgeführt ebenso wie die
Durchstoßung der darunterliegenden Faszie. An der Schnittstelle wurde mit einer Nadel
(Außendurchmesser 0,6 cm) nach BERGSTRÖM, die mit einem Abstandhalter von 6 cm
versehen war, etwa 100 bis 300 mg Muskelgewebe entnommen. Die Biopsien wurden sofort
in ein Kryogefäß verbracht, in flüssigem Stickstoff bei –70°C eingefroren und bis zur
weiteren Verarbeitung gelagert.
6. Messungen und Analyse biochemischer Parameter
6.1. L-Carnitin im Plasma
Die Konzentration von freiem und Gesamtcarnitin im Plasma wurde mit einem modifizierten
radiometrischen Enzymtest bestimmt (MCGARRY und FOSTER 1976). Dieses
Messverfahren basiert auf der enzymatischen Acetylierung von freiem Carnitin mit Tritium
markiertem Acetyl-CoA zu [3H]Acetylcarnitin. Das freie Carnitin im Plasma wurde in
Doppelbestimmungen nach Zusatz des mit Tritium markierten Acetyl-CoA mit Hilfe der
Carnitinacetyltransferase (CAT) (Fa. Roche Diagnistics GmbH, Mannheim) zu Tritium
markiertem Acetylcarnitin umgesetzt:
CAT
↓
[3H]–Acetyl–CoA + Carnitin [3H]–Acetylcarnitin + CoA–SH

III. Material und Methoden ________________________________________________________________________
57
Bei der oben beschriebenen Reaktion wird [3H]–Acetyl–CoA im Überschuss zugegeben. Um
das Reaktionsgleichgewicht so zu beeinflussen, dass möglichst die gesamte Carnitinmenge
der Probe zu Acetylcarnitin umgesetzt wird, wurde das entstandene CoA mittels eines
Thiolreagenzes (N-ethylmaleimid, NEM) gebunden. Mit Hilfe eines Anionenaustauschers
(DOWEX 1X8-400; 6MBH in der H-Form, Fa. Sigma Aldrich Chemie GmbH, Deisenhofen)
wurde das nach Abschluss der Reaktion noch vorhandene [3H]–Acetyl–CoA aus dem
Gemisch entfernt. Die Aktivität des im Überstand vorliegenden [3H]–Acetylcarnitins diente
als Maß für den Carnitingehalt der Probe und wurde mit einem Flüssigkeitsszintillationszähler
(Tri-Carb 2500, Fa. Canberra-Packard, Dreieich) gemessen. Die Carnitinmenge wurde aus
der Radioaktivität mit Hilfe einer Eichkurve berechnet.
Das Gesamtcarnitin im Plasma besteht aus dem freien Carnitin und den Carnitinestern. Durch
alkalische Hydrolyse wurde das an Ac(et)yl gebundene Carnitin gespalten und anschließend
mit der oben beschriebenen Methode gemessen. Zur Messung des Gesamtcarnitins wurden
300 µl Plasma mit 600 µl 0,2 molarer Kaliumhydroxidlösung vermischt und eine Stunde bei
37 °C inkubiert. Anschließend wurde die Probe mit 600 µl 0,2 molarer Salzsäure neutralisiert.
Der Anteil der Carnitinester ergab sich aus der Differenz des Gehaltes an Gesamtcarnitin und
an freiem Carnitin.
Bei jedem Assay wurde eine Standardplasmaprobe vom Pferd mitgemessen. Die
Variationsbreite dieser Proben zwischen allen Assays lag beim freien Carnitin bei 6,9 %
(n=27), beim Gesamtcarnitin bei 8,5 % (n = 27). Zu dieser Berechnung wurden die
Mittelwerte der Doppelbestimmungen jedes Assays gemittelt und die prozentuale
Abweichung berechnet. Die mittlere prozentuale Abweichung aller Doppelbestimmungen des
Plasmas lag bei 3,8 ± 2,9 %. Zur Berechnung dieser Werte wurden die Standardabweichungen
der Doppelbestimmungen in Prozent der Mittelwerte gebildet. Daraus wurde die mittlere
prozentuale Standardabweichung berechnet (Variationskoeffizient).

III. Material und Methoden ________________________________________________________________________
58
6.2. L-Carnitin in der Muskulatur
Die Konzentration von freiem und Gesamtcarnitin in der Muskulatur wurde anhand eines
modifizierten radiometrischen Enzymtests bestimmt (siehe 6.1.).
Da in der Literatur Angaben über die Aufbereitung des Muskelgewebes für diesen Test
fehlten, wurden im Vorfeld Versuche zur Aufbereitung von in Milligramm Mengen
vorliegendem Muskelgewebe durchgeführt. Aus den Bioptaten standen Gewebeproben von
150 bis 300 mg zur Verfügung. Da in diesem Gewebe außer Carnitin noch Glykogen
bestimmt werden sollte, war es das Ziel, die für die Carnitinmessung einzusetzende
Bioptatmenge auf ein Minimum zu begrenzen. In den Vorversuchen wurden aus
Pferdemuskeln, die von Autopsien zur Verfügung standen, Stückchen von 300 mg gewonnen.
Diese wurden bei –20 °C eingefroren. Als erstes wurde versucht, die gefrorenen
Muskelstückchen in einem Mörser zu zerkleinern. Dieses gelang nicht, da das Gewebe zu
rasch auftaute und am Boden des Mörsers und am Pistill festklebte. Außerdem blieb die
Zerkleinerung unvollständig, da auch nach längerer Bearbeitung noch immer unversehrte
Muskelfaserfragmente zu erkennen waren. In einem zweiten Vorversuch wurde das gefrorene
Gewebe im Mörser mit flüssigem Stickstoff übergossen und anschließend gemörsert. Bei
diesem Zerkleinerungsversuch sprangen Teile des spröden Gewebematerials aus dem Mörser
und eine kontrollierte, vollständige Zerkleinerung der Probe erwies sich als nicht
durchführbar. Nach diesen Erfahrungen wurden die kleinen Muskelstückchen zunächst
gefriergetrocknet und anschließend bei Zimmertemperatur im Mörser fein zerrieben. Dabei
blieb jedoch eine nicht unbeträchtliche Menge des zerkleinerten Materials relativ fest in der
Mörserschale und an der Oberfläche des Pistills haften, das sich mechanisch nicht vollständig
genug zurückgewinnen ließ. Eine befriedigende Rückgewinnung dieser Gewebeanteile gelang
nur durch Abspülen mit 10 ml deionisiertem Wasser. Das wiederum war mit einer nicht zu
tolerierenden Verdünnung der Probe verbunden. Dadurch erschien dieser Versuch der
Probenaufbereitung auch als nicht brauchbar. Als gangbarer Weg zur Messung von Carnitin
in repräsentativen Gewebeproben von 10 mg TS, die aus einer größeren Stichprobe von etwa
150 bis 300 mg uS (ca. 37,5 bis 75 mg TS) entnommen wurden, erwies sich schließlich das
Zerkleinern der gefriergetrockneten Stichproben im Mörser unter flüssigem Stickstoff. Das
Gewebe ließ sich zu staubförmig feiner Korngröße vermahlen, blieb nicht an der Oberfläche

III. Material und Methoden ________________________________________________________________________
59
haften und konnte mit Hilfe des flüssigen Stickstoffs nahezu quantitativ in entsprechende
Aufbewahrungsbehälter überführt und bei – 20 °C gelagert werden.
Als nächstes wurde geprüft, ob Aliquots von 10 mg aus auf diese Weise hergestelltem
Gewebspulver als repräsentativ für die gesamte Probe zur Messung von Carnitin angesehen
werden konnten. Dazu wurde pulverisiertes Gewebe von fünf verschiedenen Zerkleinerungen
zu einer Probe vereinigt und gut vermischt. Aus diesem Gemisch wurden jeweils drei
Aliquots von 10, 20, 50 und 100 mg abgewogen und auf ihren Gehalt an freiem und
Gesamtcarnitin untersucht. Der Variationskoeffizient der bei diesem Test gewonnenen 12
Messergebnissen betrug 3,6 %. Ein Aliquot von 10 mg, das aus einem größeren Bioptat nach
der hier beschriebenen Aufbereitung entnommen wird, kann daher als repräsentativ für die
Bestimmung von freiem und Gesamtcarnitin in dieser Probe angesehen werden.
Die in diesem Versuchsvorhaben gewonnenen, gefrorenen Bioptate wurden zunächst von
makroskopisch sichtbarem Fett, Bindewebe und Blut befreit. Danach wurden die
Gewebsfragmente gefriergetrocknet (α2-4 Christ LDC-IM), unter flüssigem Stickstoff wie
oben beschrieben homogenisiert und in einem 10 ml Glasgefäß aufbewahrt. Ein Aliquot von
10 mg wurden daraus abgewogen, mit 2 ml deionisiertem Wasser vermischt und 15 Minuten
mit Ultraschall behandelt. Die Proben wurden 5 Minuten bei 5000 g zentrifugiert. Außer dem
Gehalt an freien Carnitin und an Gesamtcarnitin wurden bei den Carnitinestern die Fraktionen
der kurzkettigen und langkettigen Ester getrennt erfasst. Dabei wurde der Umstand genutzt,
daß die kurzkettigen Carnitinester wasserlöslich sind und in den wässrigen Extraktüberstand
übergehen, die langkettigen Carnitinester dagegen in den zerkleinerten Gewebestücken der
Muskulatur zurückbleiben. Der wässrige Extraktüberstand (ohne Sediment) und das in
Wasser suspendierte Muskelhomogenat wurden getrennt hydrolysiert. Der Gehalt an
Gesamtcarnitin im wässrigen Extraktüberstand umfasste das freie Carnitin und die
kurzkettigen Carnitinester. Der Gehalt an Gesamtcarnitin im wässrigen Gewebehomogenat
umfasste das freie Carnitin und alle kurz- und langkettigen Carnitinester. Ein Aliquot von 400
µl des Überstandes wurde mit 800 µl deionisiertem Wasser vermischt und zur Bestimmung
des freien Carnitins (FC) verwendet. Ein zweites 600 µl Aliquot des Überstandes wurde zur
Bestimmung des freien Carnitins und der wasserlöslichen Carnitinester (GC1) mit 1,2 ml
KOH vermischt, eine Stunde bei 37 °C hydrolisiert und anschließend mit 1,2 HCl

III. Material und Methoden ________________________________________________________________________
60
neutralisiert. Der Rest der Probe, bestehend aus 1 ml Überstand und Muskelpulvereinwaage,
wurde ebenfalls hydrolisiert. Der Gehalt an Carnitin in diesem Hydrolysat repräsentierte den
Gehalt an Gesamtcarnitin (freies Carnitin und kurz- und lankettige Carnitinester, GC2). Es
wurde daraus unter Berücksichtigung der zuvor durchgeführten Entnahmen berechnet. In den
radioenzymatischen Assay wurde zur Messung von freiem Carnitin 200 µl und zur Messung
von GC1 und GC2 jeweils 300 µl eingesetzt.
Die Berechnung der Konzentration an Gesamtcarnitin (GC) ergab sich aus:
(GC2-GC1)/2+GC1 = GC
Die Konzentration der kurzkettigen Carnitinester (KKCE) errechnete sich mit der Gleichung:
GC1-FC=KKCE
Die Konzentration der langkettigen Carnitinester (LKCE) war gegeben durch die Gleichung:
GC-GC1=LKCE
Die mittlere prozentuale Abweichung aller Doppelbestimmungen von Carnitin in der
Muskulatur lag bei 1,8 ± 0,4 % (n = 50) (zur Variationbreite des Standards und zur
Berechunung dieser Werte siehe Plasma).
6.3. Herzfrequenz
Die Herzfrequenz wurde telemetrisch (Polar Horsetrainer Herzfrequenz-Messgerät, Fa.
Baumann & Haldi SA, Schweiz) gemessen. Die Messeinrichtung besteht aus einem Sender
(Elektroden) und einem Empfänger (Polar Vantage XL). Zur Aufnahme der
elektromagnetischen Signale wurden zwei Elektroden am linken Widerist und am ventralen
Brustkorb des Pferdes befestigt. Nach Beendigung der Messung wurden die im Empfänger
gespeicherten Daten mit Hilfe eines „Polar-Interface“ auf einem Computer übertragen.
6.4. Lactat im Vollblut
Lactat wurde im heparinisierten Vollblut mit dem Accusport-Gerät (Fa. Boehringer
Mannheim) gemessen. Das Gerät wurde für Lactatmessungen im Humanblut entwickelt. Nach
Untersuchungen von LINDNER (1996) und WILLIAMSON et al. (1996) ist es aber auch für
Messungen im Pferdeblut einsetzbar. Laut Angaben des Herstellers ist das Gerät für
Lactatkonzentrationen zwischen 0,8 und 26 mmol/l ausgelegt. Bei den Untersuchungen im

III. Material und Methoden ________________________________________________________________________
61
Rahmen dieses Projektes lagen die Konzentrationen innerhalb dieses Bereiches. Bei einem
Vergleich der Messergebnisse des Accusport-Gerätes mit denen einer nass-chemischen
Methode (EPOS 5060, Eppendorf Instruments, Hamburg), ergab sich eine Korrelation von bei
r = 0,95. Auch die mit beiden Methoden gemessenen absoluten Konzentrationen
unterschieden sich nicht signifikant voneinander. Der Variationskoeffizient bei Messung von
16 Blutproben mit fünf verschiedenen Accusport-Geräten lag bei 2,48 – 14,37 %. Bei der
Messung zweier von Boehringer Mannheim mitgelieferter Standards betrug der
Variationskoffizient laut Herstellerangaben 4,42 – 7,0 % (n= unbekannt). In vergleichenden
Messungen mit diesem Gerät ergab sich bei Untersuchungen von LINDNER (1996) ein
Intraassay-Variationskoeffizient von 5,56 bis 7,23 %. Die sich aus diesen Messungen
ergebenden Messgenauigkeiten erschienen ausreichend, um das Gerät im Rahmen dieses
Projektes zu verwenden. Zusätzlich wurde in 7 Plasmaproben die Lactatkonzentration mit
einem KODAK EKTACHEM DT 60 Analyser mit Hilfe von Lactat-Plättchen (Fa. Ortho-
Clinical Diagnostics, Hamburg) im heparinisierten Plasma gemessen. Die Ergebnisse dieser
Untersuchung wurden mit denen im Vollblut verglichen. Es ergab sich eine mittlere
prozentuale Abweichung von 8 % (Abb. 3.7.).
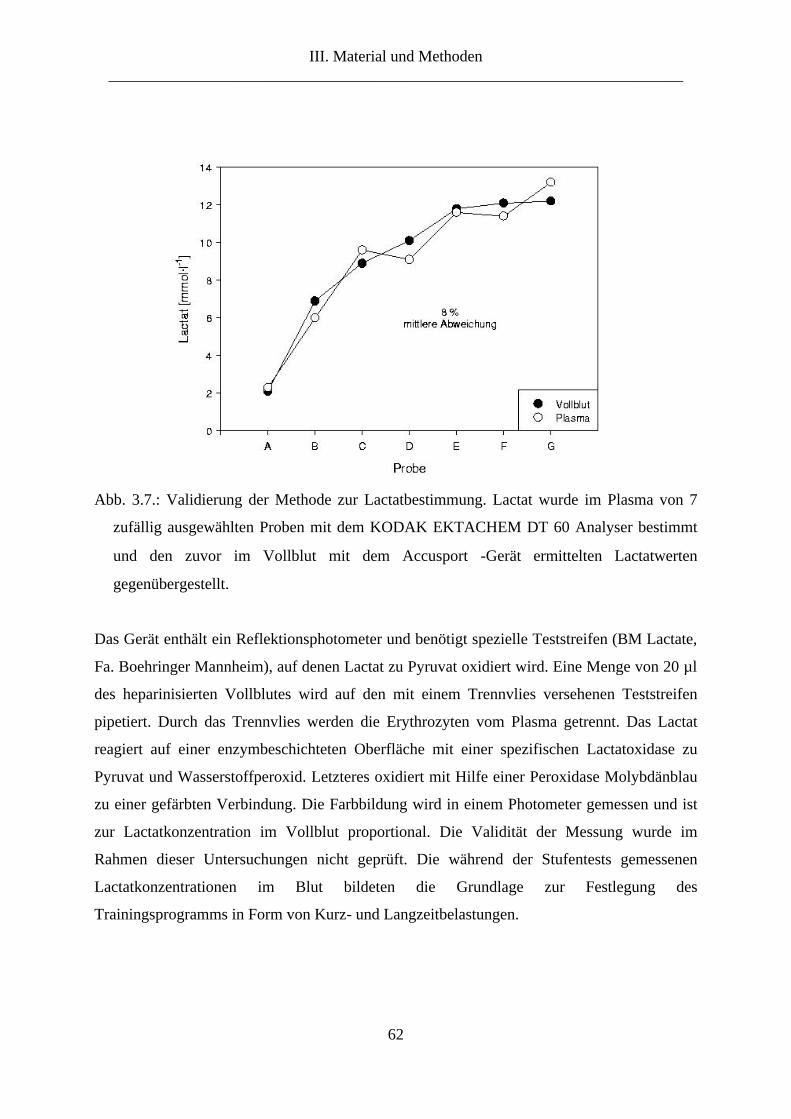
III. Material und Methoden ________________________________________________________________________
62
Abb. 3.7.: Validierung der Methode zur Lactatbestimmung. Lactat wurde im Plasma von 7
zufällig ausgewählten Proben mit dem KODAK EKTACHEM DT 60 Analyser bestimmt
und den zuvor im Vollblut mit dem Accusport-Gerät ermittelten Lactatwerten
gegenübergestellt.
Das Gerät enthält ein Reflektionsphotometer und benötigt spezielle Teststreifen (BM Lactate,
Fa. Boehringer Mannheim), auf denen Lactat zu Pyruvat oxidiert wird. Eine Menge von 20 µl
des heparinisierten Vollblutes wird auf den mit einem Trennvlies versehenen Teststreifen
pipetiert. Durch das Trennvlies werden die Erythrozyten vom Plasma getrennt. Das Lactat
reagiert auf einer enzymbeschichteten Oberfläche mit einer spezifischen Lactatoxidase zu
Pyruvat und Wasserstoffperoxid. Letzteres oxidiert mit Hilfe einer Peroxidase Molybdänblau
zu einer gefärbten Verbindung. Die Farbbildung wird in einem Photometer gemessen und ist
zur Lactatkonzentration im Vollblut proportional. Die Validität der Messung wurde im
Rahmen dieser Untersuchungen nicht geprüft. Die während der Stufentests gemessenen
Lactatkonzentrationen im Blut bildeten die Grundlage zur Festlegung des
Trainingsprogramms in Form von Kurz- und Langzeitbelastungen.

III. Material und Methoden ________________________________________________________________________
63
6.5. Glucose im Plasma
Die Glucose wurde mit einem KODAK EKTACHEM DT 60 Analyser mit Hilfe von
Glucose-Plättchen (Fa. Ortho-Clinical Diagnostics, Hamburg) im heparinisierten Plasma
gemessen. Entsprechende Plättchen, die anhand eines Strichcodes von dem Gerät identifiziert
wurden, wurden mit 10 µl Plasma betropft. Das Plättchen besteht aus mehreren Schichten, die
auf einem Träger aufgebracht sind und alle für den Test benötigten Reagenzien enthalten. Der
Nachweis basiert auf einer durch Glucoseoxidase katalysierten Oxidation der Glucose und der
damit verbundenen Bildung von Wasserstoffperoxid. Dieser wird durch Bildung eines
Farbstoffs für die Bestimmung genutzt. Die Glucose im Plasma wird durch die
Glucoseoxidase zu D-Gluconsäure oxidiert. Das dabei entstehende H2O2 bildet unter
Vermittlung von Peroxidase aus 1,7-Dihydroxynaphtalin und 4-Aminoantipyrin einen
Farbstoff, der bei 555 nm Wellenlänge gemessen wird. Nach einer Inkubationszeit von 5
Minuten bei 37°C ist die Farbreflexion proportional zur Glucosekonzentration. Als
Bezugsgröße dienten Wieß-Schwarz-Bezugswerte, ein Kalibrationsmodell und gespeicherte
Kalibrationsparameter. Das Gerät berechnet den Wert und druckt das Ergebnis aus.
Das Gerät wurde einmal pro Woche mit einer Kalibrationslösung (Fa. Ortho-Clinical
Diagnostics, Hamburg) geeicht. Eine Plasmaprobe (Probenstandard) wurde zur Kontrolle der
Genauigkeit des Gerätes täglich zu Beginn gemessen. Die Glucosekonzentration des
Probenstandards betrug 4,5 ± 0,08 mmol/l (n = 3). Weitere Validitätsparameter der Methode
wurden nicht bestimmt.
6.6. Glykogen in der Muskulatur
Die Glykogenbestimmung in der Muskulatur erfolgte nach der Methode von LINDHOLM
(1979). Das Messprinzip beruht auf der Hydrolyse des Glykogens zu Glucose und deren
anschließender enzymatischen Bestimmung.
Glykogen in ca. 10 mg des Muskelpulver (siehe Carnitinbestimmung) wurde mit 1 ml 1 M
Salzsäure 2 Stunden bei 100°C hydrolysiert. Eine Menge von 50 µl des Hydrolysates wurde
zur Enteiweißung mit 950 µl 0,36 M Perchlorsäure vermischt und 2 Minuten bei 10.000 g

III. Material und Methoden ________________________________________________________________________
64
zentrifugiert. Die Bestimmung der Glucose erfolgte nach der GOD-Perid-Methode (Test –
Kombination Glucose, Fa. Boehringer Mannheim) entsprechend den Angaben von WERNER
et al. (1970). Bei dieser Methode wird Glucose durch Glucoseoxidase (GOD) zu
Glukonolakton oxidiert, das in wässriger Lösung in Glukonsäure übergeht. Dabei entsteht
H2O2, das in Gegenwart von Peroxidase 2,2-Azino-di [3-ethyl-benzthiazolinsulfonsäure-(6)]-
diammoniumsalz (ABTS) zu einer blaugrün gefärbten Verbindung oxidiert. Die Intensität
des Farbstoffs ist der Glucosekonzentration proportional. Sie wird bei einer Wellenlänge von
578 nm gegen einen Leerwert im Spektralphotometer (Spectrophotometer, Pharmacia LKB)
gemessen. Mit Hilfe eines Standards wird die Glucosekonzentration berechnet. Die Validität
der Methode wurde nicht getestet.
6.7. Freie Fettsäuren im Plasma
Die freien Fettsäuren (FFS) wurden im EDTA-Plasma direkt bestimmt. Für diese Messung
darf kein Heparin zugesetzt werden, da Heparin die Lipoproteinlipase stimuliert und aus den
Lipoproteinen zusätzlich freie Fettsäuren freisetzt. Für die Messungen wurde ein
enzymatischer Farbtest der Fa. Wako Chemicals GmbH (Neuss) verwendet. Das Testprinzip
beruht auf einer Aktivierung der FFS zu Acyl-CoA-Estern in Anwesenheit von ATP und
Acyl-CoA-Synthase. Die Acyl-CoA-Thioester werden unter Vermittlung von Acyl-CoA-
Oxidase in Anwesenheit von Sauerstoff zu 2,3-trans-Enoyl-CoA reduziert. Dabei entsteht
Wasserstoffperoxid. In Anwesenheit von Peroxidase kondensiert H2O2 einen Farbkomplex
oxidativ, der aus einem Anilinderivat (3-Methyl-N-Ethyl-N-(β-Hydroxyethyl)-Anilin) und 4-
Aminoantipyrin besteht. Der rotgefärbte Farbstoff absorbiert Licht mit einer Wellenlänge von
550 nm. Ascorbinsäure, die mit H2O2 interferiert, wurde durch Zugabe von Ascorbat-Oxidase
vorher aus dem Reaktionsgemisch entfernt. Nach Abzug des Leerwertes wird die
Konzentration an FFS aus der Extinktion im Verhältnis zu einer Standardlösung berechnet.
Zur Überprüfung der Genauigkeit der Messungen wurde täglich vor der Messung der
Probenstandard gemessen. Die Abweichung dieses Standards betrug 2,4 % (n = 267).
Neben dieser Methode wurden 20 Proben der FFS im Plasma noch kolorimetrisch mit der
Methode von ITAYA (1977) bestimmt, da einige Werte zweifelhaft erschienen. Diese

III. Material und Methoden ________________________________________________________________________
65
Methode beruht auf dem Prinzip, dass freie Fettsäuren mit Kupfer-Ionen unter der Zugabe
von Diphenylcarbazid und Diphenylcarbazon einen blau-violetten Farbkomplex bilden,
dessen Farbintensität der Konzentration der freien Fettsäuren im Plasma proportional ist. Zur
Einstellung des pH-Wertes und zur Extration der freien Fettsäuren wurden 100 µl
Probenmaterial in 0,5 ml Phosphatpuffer (0,5 M, pH 7,0), 0,4 ml NaCl-Lösung (0,15 M) und
in 4 ml Chloroform gegeben. Die Proben wurden 30 Minuten im Rüttler belassen und
anschließend 10 Minuten bei 3000 g zentrifugiert. Die obere Schicht wurde abgesaugt. Auf
die untere Chloroformschicht mit den darin enthaltenen FFS wurden 2 ml einer Kupfer-
Triethanollösung (Cu-TEA) gegeben. Diese Lösung bestand aus 1 M Triethanolamin, 1 M
Essigsäure und aus einer 6,45 %igen Kupfernitratlösung. Die Lösungen verblieben 30
Minuten im Rüttler und wurden anschließend bei 3000 g zentrifugiert. Die obere Schicht
wurde abgesaugt. Ein Aliquot von zwei ml der unteren Chloroformschicht mit den FFS wurde
in ein neues Glasröhrchen verbracht und 0,75 ml Farbreagenz hinzugefügt. Die Farbreagenz
setzte sich aus 0,5 % Diphenylcarbazon und 0,5 % Diphenylcarbazid in Methanol zusammen.
Die stabile Farblösung wurde bei 546 nm mit einem Spektralphotometer (Spectrophotometer
Pharmacia LBK) gegen einen Reagenzienleerwert gemessen.
Die mit dieser Methode erhaltenen Messwerte waren durchschnittlich um 10 % niedriger als
die Ergebnisse der zuerst beschriebenen Methode. Sie waren aber nicht signifikant
voneinander verschieden. In Relation zueinander waren sie gleich. Da die Messung der FFS
im Plasma hauptsächlich einem Vergleich zwischen den Gruppen dienen sollte, wurden die
Messwerte der ersten Methode verwendet, da die routinemäßige Validitätskontrolle dieser
Methode zuverlässige Messdaten erbrachte.
6.8. Harnstoff im Plasma
Harnstoff wurde mit einem KODAK EKTACHEM DT 60 Analyser mit Hilfe von Harnstoff-
Plättchen (Fa. Ortho-Clinical Diagnostics, Hamburg) im heparinisierten Plasma bestimmt.
Entsprechende Plättchen, die anhand eines Strichcodes von dem Gerät identifiziert werden,
wurden mit 10 µl Plasma betropft. Das Plättchen besteht aus mehreren Schichten, die auf
einem Träger aufgebracht sind und alle für den Test benötigten Reagenzien enthalten. Der
Nachweis basiert auf einer durch Urease katalysierten Hydrolyse von Harnstoff zu Ammoniak

III. Material und Methoden ________________________________________________________________________
66
und Kohlendioxid. Das Ammoniak wird mit einem vom Hersteller nicht näher bezeichneten
„Indikator“ zu einem Farbstoff umgesetzt, der reflektometrisch bei 660 nm gemessen wird.
Das Gerät wurde einmal pro Woche mit einer vom Hersteller zur Verfügung gestellten
Kalibrationslösung geeicht. Eine Plasmaprobe (Probenstandard) wurde zur Kontrolle der
Genauigkeit des Gerätes täglich zu Beginn gemessen. Die Harnstoffkonzentration des
Probenstandards betrug 4,9 ± 0,08 mmol/l (n = 3). Nach Angaben des Herstellers beträgt die
Intra- und Interassayvarianz entsprechend 2,4 und 2,6 %. Weitere Validitätsparameter der
Methode wurden nicht erfasst.
6.9. Hämatokrit
Der Hämatokrit wurde mit Hilfe einer Mikrohämatokritzentrifuge in EDTA-Vollblut
gemessen. Die Kapillarröhrchen wurden zu ¾ gefüllt und mit Spezialkitt verschlossen und
sechs Minuten zentrifugiert. Das Ergebnis wurde in einem Auswertegerät abgelesen und in %
angegeben. Da es sich bei dieser Methode um eine wenig störanfällige Standardmethode
handelt, wurden keine Messungen zur Überprüfung der Präzision vorgenommen.
6.10. Gesamtprotein im Plasma
Mit Hilfe einer refraktiometrischen Messung wurde das Gesamtprotein im Plasma bestimmt.
Das Messprinzip besteht in der Brechung vom sichtbaren Licht beim Übergang in ein optisch
unterschiedlich dichtes Medium. Die optische Dichte der Probe (Plasma) ist zum
Proteingehalt proportional. Das Verhältnis zwischen dem Sinus des Einfallwinkels (i) des
Lichtes (sin(i)) im Glas und dem Sinus des Ausfallwinkels (e) (sin(e)) bildet den
Refraktionsindex. Er dient der Quantifizierung des Proteingehaltes im Plasma. Das Verfahren
wurde als Standardlabormethode im Rahmen dieser Arbeit eingesetzt. Die Validität der
Methode wurde nicht überprüft.
6.11. Körpergewicht
Die Pferde wurden zu Anfang und nach den Stufentests und zu Anfang und zu Ende der
beprobten Belastungen sowie vor der Probennahme in der Ruheperiode gewogen. Die

III. Material und Methoden ________________________________________________________________________
67
Wägung erfolgte auf einer festinstallierten Viehwaage und einem Gewichtsbereich zwischen
25 kg und 600 kg und einer Genauigkeit von 0,2 kg (Fa. Bitzer GmbH, Hildesheim).
7. Statistik
Die statistische Auswertung der Versuchsdaten erfolgte mit Hilfe der Statistiksoftware Sigma
Stat (Versin 1.0, SPSS Science Software, GmbH, Erkenrat). Der Probenumfang „N“
bezeichnet die Anzahl der Versuchstiere, die zur Berechnung der Mittelwerte herangezogen
wurden. Als signifikant wurden Irrtumswahrscheinlichkeiten kleiner 5 % (p < 0,05) und als
hochsignifikant Irrtumswahrscheinlichkeiten kleiner 1 % (p < 0,01) bezeichnet.
Der Gehalt an Gesamtcarnitin und an freiem Carnitin im Plasma am Anfang und am Ende der
Belastung und die basale und belastungsabhängige Konzentration der FFS im Plasma am
Anfang bzw. am Ende des Trainings wurden mit Hilfe einer 2-faktoriellen Varianzanalyse auf
Signifikanz geprüft. Der Carnitinstatus bildete den ersten und die Belastung jeweils den
zweiten Faktor.
Die Veränderungen der Messwerte (Herzfrequenz, Lactat, Glucose, L-Carnitin im Plasma und
in der Muskulatur, FFS, Harnstoff, Hämatokrit, Gesamtprotein, Muskelglykogen) vor und
nach dem Training (Trainingseffekt) wurden auf die folgende Weise auf Signifikanz geprüft.
Es wurden die Differenzen der Parameter bei jeweils gleicher Belastung vor und nach dem
Training zwischen Gruppen (mit und ohne Carnitin) auf Signifikanz getestet. In gleicher
Weise wurden die Differenzen der einzelnen Parameter, ausgedrückt in Prozent (Werte am
Trainingsanfang gleich 100 %), zwischen den Gruppen auf Signifikanz geprüft.
Vor der Anwendung der statistischen Verfahren wurden die Werte aller Parameter zunächst
auf Normalveteilung und gleiche Varianz geprüft. Die Prüfung ergab, dass alle Messwerte
nicht, die Differenzen zwischen den entsprechenden Messpunkten aber, normal verteilt waren.
Die oben definierten, normal verteilten Differenzen mit gleicher Varianz wurden mit Hilfe
einer 2-faktoriellen Varianzanalyse auf Signifikanz geprüft. Bei den während des Stufentests

III. Material und Methoden ________________________________________________________________________
68
erhobenen Daten war der Carnitinstatus (mit bzw. ohne Zulage) der erste Faktor und die
Laufgeschwindigkeit der Tiere der zweite Faktor. Bei den vor und nach den Kurz- und
Langzeitbelastungen erhobenen Daten bildete der Carnitinstatus ebenfalls den ersten Faktor
und Anfang und Ende der Belastung den zweiten Faktor. Die Signifikanzprüfung erfolgte für
alle Werte mit Hilfe des Student-Newman-Keuls-Tests.
Die Ergebnisse der statistischen Tests wurden nur, soweit sie statistisch gesicherte Effekte
ergaben, in die Ergebnisse mit aufgenommen.

IV. Ergebnisse___________________________________________________________________________
69
IV. Ergebnisse
1. L-Carnitin im Plasma
Neben der Frage, ob eine orale Carnitinzulage bei Pferden durch Training Leistungsparameter
beeinflußt, war es ein Hauptanliegen dieser Untersuchung, den Einfluß einer Carnitinzulage auf
den Carnitinstatus der Tiere zu untersuchen. Dazu wurde die Carnitinkonzentration im Plasma
und in Bioptaten des Musculus glutaeus medius bei sieben Pferden gemessen. In Abbildung 4.1.
ist der zeitlich Verlauf der Gesamtcarnitinkonzentration im Plasma bei den vier mit Carnitin
supplementierten und den drei Kontrollpferden während des vier Monate währenden
Versuchsprogramms dargestellt. Ferner zeigt die Abbildung 4.1. die Konzentrationen an
Plasmacarnitin während kürzerer Zeiträume im Laufe der Stufentests sowie vor und nach
einzelnen Kurz- und Langzeitbelastungen.
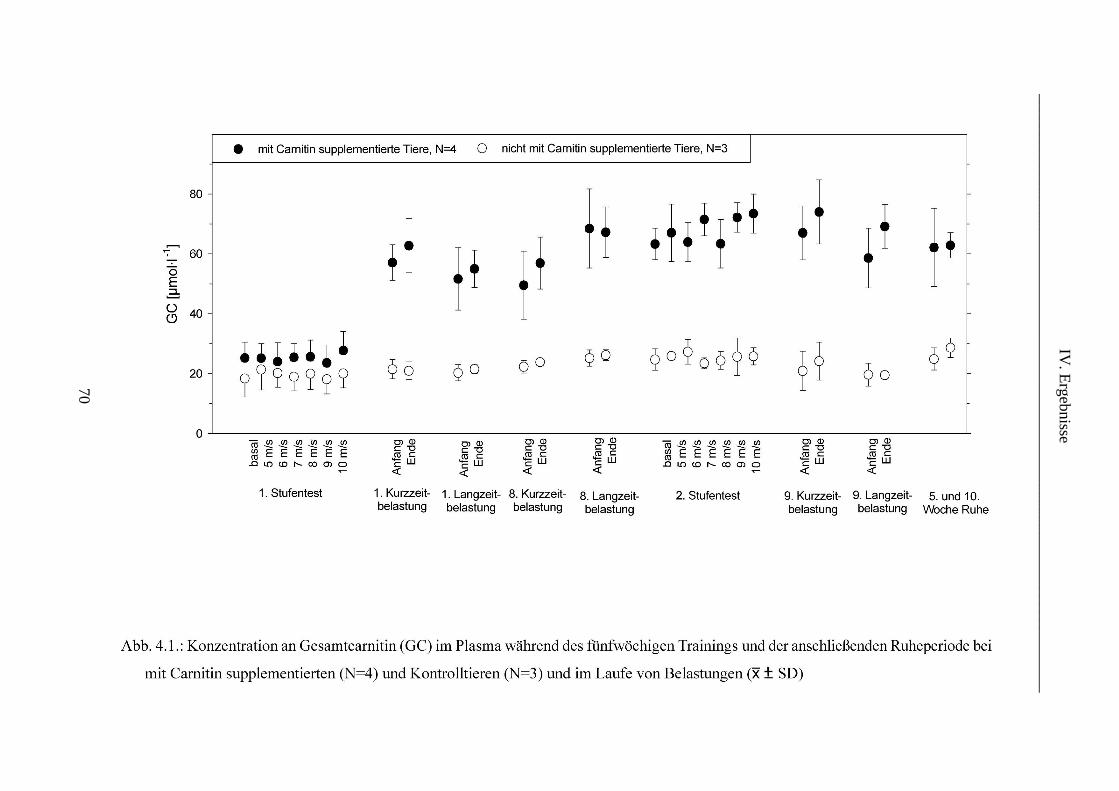
IV. Ergebnisse
___________________________________________________________________________
70

IV. Ergebnisse___________________________________________________________________________
71
Die Konzentration von Gesamtcarnitin im Plasma betrug bei den Kontrollpferden am Anfang
des Versuchszeitraumes 19,6 ± 4,7 :mol/l. Dieser Wert wird durch die sieben während des
Stufentests gemessenen Werte repräsentiert. Er blieb über den viermonatigen Versuchszeitraum
etwa auf diesem Niveau (22,7 ± 4,4 :mol/l). Beim ersten Stufentest war noch keines der Pferde
mit Carnitin supplementiert. Zu diesem Zeitpunkt betrug die mittlere Konzentration an
Gesamtcarnitin bei den Pferden, für die nach diesem Test die Supplementationsperiode begann,
25,3 ± 5,3 :mol/l und lag signifikant um 16 % höher als bei den Kontrolltieren. Bereits vier
Tage nach Beginn der Supplementation, also während der ersten Kurzzeitbelastung, wies diese
Gruppe einen deutlich um 137 % erhöhten Plasmaspiegel an Gesamtcarnitin auf (59,9 ± 7,7
:mol/l). Bei den supplementierten Pferden blieb der bereits bei der ersten Kurzzeitbelastung
nachweisbare erhöhte Spiegel mit einigen Schwankungen ebenfalls während des gesamten
Versuchszeitraums bestehen. Er betrug im Mittel 63,7 ± 10,2 :mol/l.
Während der drei Kurzzeitbelastungen, während zweier Langzeitbelastungen und während des
zweiten Stufentests stieg die Konzentration an Gesamtcarnitin im Plasma bei den mit Carnitin
supplementierten Tieren um durchschnittlich 13 %. Während der achten Langzeitbelastung blieb
sie gleich. Bei den Kontrolltieren blieb die Konzentration an Gesamtcarnitin im Plasma im Laufe
aller Belastungen etwa gleich. Ferner fällt auf, dass die Standardabweichung der
Carnitinkonzentration bei den supplementierten Pferden (± 7,6 :mol/l) und bei den
Kontrollpferden (± 3,7 :mol/l) mit entsprechend 15,4 und 17,0 % des Mittelwertes etwa gleich
war.
Da körperliche Belastungen, sofern sie die aerobe Schwelle überschreiten, mit einer deutlichen
Zunahme der Carnitinester auf Kosten des freien Carnitins in der Muskulatur verbunden sind,
war von Interesse, die Konzentration an freiem Carnitin im Plasma bei diesen Tiere,
insbesondere in Abhängigkeit von den Belastungen, zu messen. Die Ergebnisse dieser
Messungen sind in Abbildung 4.2. nach dem gleichen Schema wie in Abbildung 4.1. dargestellt.
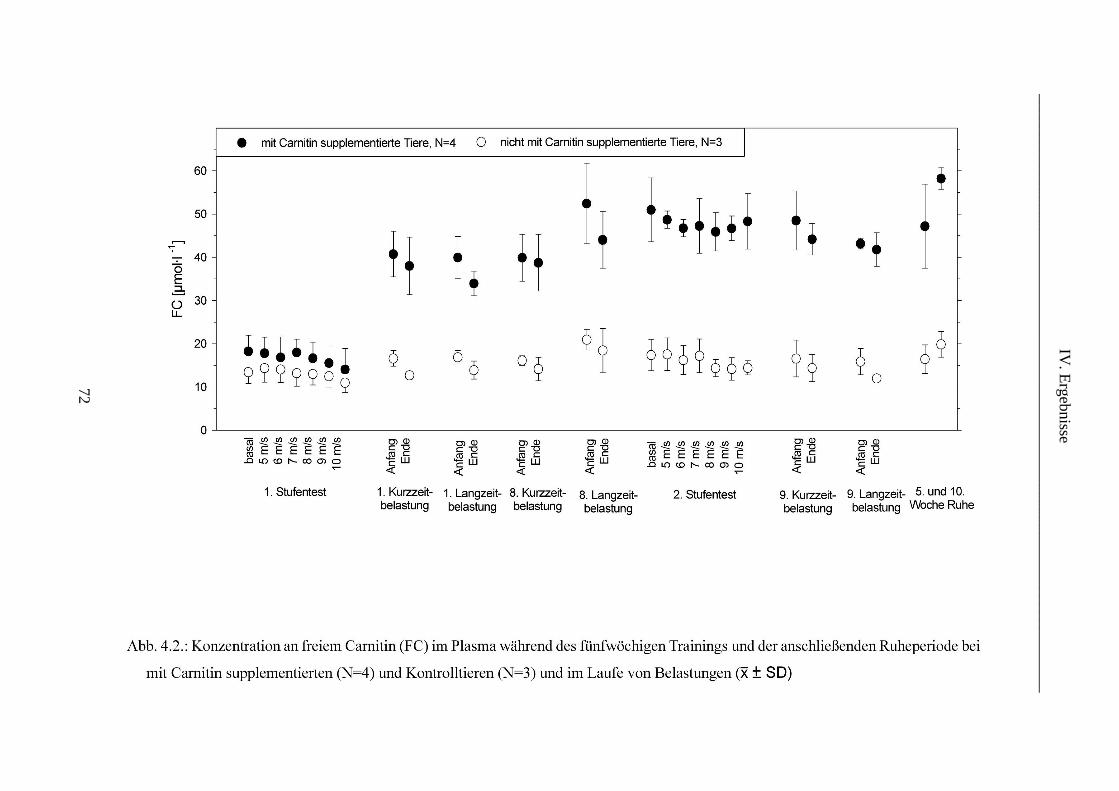
IV. Ergebnisse
___________________________________________________________________________
72

IV. Ergebnisse___________________________________________________________________________
73
In seinem Grundmuster entsprach das freie Carnitin im Plasma dem des Gesamtcarnitins. Bei
den Kontrolltieren blieb es während der viermonatigen Meßperiode auf einem etwa gleichen
Niveau. Bei den mit Carnitin supplementierten Tieren nahm das freie Carnitin ebenfalls bereits
nach vier Tagen um 122 % zu und blieb von da an über den ganzen Versuchszeitraum auf dieser
Höhe. Bei den supplementierten Pferden bestand nach Beginn der Supplementation bei Ruhe
73 % des Gesamtcarnitins aus freiem Carnitin. Bei den Kontolltieren waren es 75 %.
Auffällig war jedoch, dass, im Gegensatz zum Gesamtcarnitin, das freie Carnitin im Plasma bei
allen Tieren am Ende der drei Kurz- und der drei Langzeitbelastungen immer geringfügig
niedriger lag als am Anfang der jeweiligen Belastung. Am Anfang der sechs Belastungen betrug
die Konzentration bei den mit Carnitin supplementieren Tieren im Mittel 44,2 ± 7,4 :mol/l und
am Ende 40,0 ± 6,0 :mol/l. Setzt man den Wert am Anfang der Belastung gleich Hundert, dann
lag die Konzentration am Ende im Mittel bei 90 %. Der Abfall war mit p=0,04 signifikant. Auch
bei den Kontrolltieren war die Konzentration an freiem Carnitin am Ende der Kurz- und
Langzeitbelastungen im Vergleich zum Belastungsbeginn in allen sechs Fällen auf 83 %
abgefallen (17,2 ± 2,8 vs. 14,3 ± 3,2 :mol/l) (Konzentration zu Beginn der Belastung = 100 %).
Auch dieser Abfall war mit p=0,006 signifikant. Der Konzentrationsabfall an freiem Carnitin im
Plasma am Ende aller Belastungen (einschließlich beider Stufentests) betrug bei den mit Carnitin
supplementierten Tieren durchschnittlich 14 ± 3 % und bei den Kontrolltieren 17 ± 4 % der
jeweiligen Anfangskonzentration.
2. L-Carnitin in der Muskulatur
Es war ein Hauptanliegen dieser Arbeit, der Frage nachzugehen, ob eine orale Carnitingabe auch
den Carnitingehalt in der Muskulatur beeinflußt. Dazu erhielten vier Tiere über den
Versuchszeitraum von vier Monaten täglich 10 g L-Carnitin, beginnend nach dem ersten
Stufentest. Insgesamt wurden pro Pferd acht Bioptate genommen, von denen sechs unter
Ruhebedingungen und zwei nach Belastung gewonnen wurden. Die Abbildung 4.3. zeigt die
Konzentration an Gesamtcarnitin im Musculus glutaeus medius bei den mit Carnitin
supplementierten und den Kontrolltieren während des Versuchszeitraums.
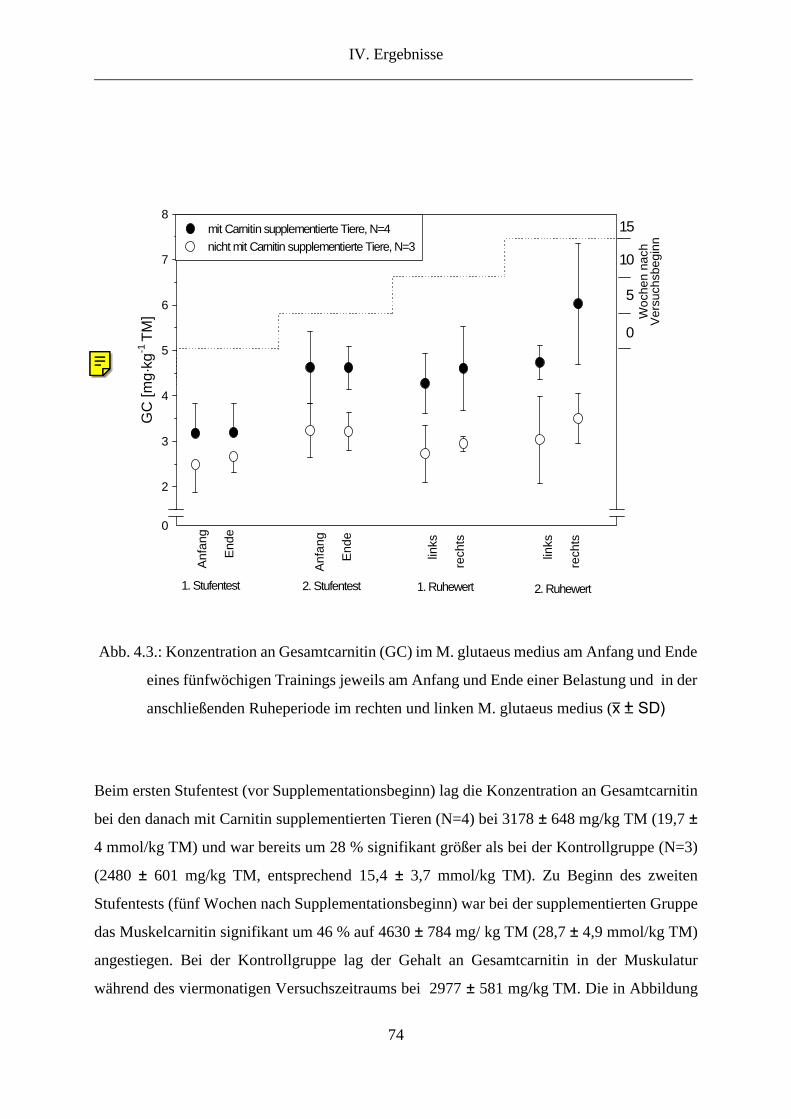
IV. Ergebnisse___________________________________________________________________________
74
GC
[mg·
kg-1
TM]
0
2
3
4
5
6
7
8
1. Stufentest 2. Stufentest
Anf
ang
End
e
Anf
ang
End
e
links
rech
ts
links
rech
ts
mit Carnitin supplementierte Tiere, N=4nicht mit Carnitin supplementierte Tiere, N=3
2. Ruhewert1. Ruhewert
15
0
5
10
Woc
hen
nach
Ver
such
sbeg
inn
Abb. 4.3.: Konzentration an Gesamtcarnitin (GC) im M. glutaeus medius am Anfang und Ende
eines fünfwöchigen Trainings jeweils am Anfang und Ende einer Belastung und in der
anschließenden Ruheperiode im rechten und linken M. glutaeus medius (x # SD)
Beim ersten Stufentest (vor Supplementationsbeginn) lag die Konzentration an Gesamtcarnitin
bei den danach mit Carnitin supplementierten Tieren (N=4) bei 3178 ± 648 mg/kg TM (19,7 ±
4 mmol/kg TM) und war bereits um 28 % signifikant größer als bei der Kontrollgruppe (N=3)
(2480 ± 601 mg/kg TM, entsprechend 15,4 ± 3,7 mmol/kg TM). Zu Beginn des zweiten
Stufentests (fünf Wochen nach Supplementationsbeginn) war bei der supplementierten Gruppe
das Muskelcarnitin signifikant um 46 % auf 4630 ± 784 mg/ kg TM (28,7 ± 4,9 mmol/kg TM)
angestiegen. Bei der Kontrollgruppe lag der Gehalt an Gesamtcarnitin in der Muskulatur
während des viermonatigen Versuchszeitraums bei 2977 ± 581 mg/kg TM. Die in Abbildung
bibliothek
Erratum In der linke Beschriftung der Abb. 4.3 muß "mg" durch "g" ersetzt werden.

IV. Ergebnisse___________________________________________________________________________
75
4.3. erkennbaren Schwankungen erscheinen zufallsbedingt. Bei den mit Carnitin
supplementierten Tieren waren die Ruhewerte auch nach fünf Wochen etwa auf dem gleichen
Niveau wie beim zweiten Stufentest. Der hohe Mittelwert in der rechten Muskulatur nach zehn
Wochen Ruhe wird durch einen Messwert beim Pferd Nr. IV (6071 mg/kg TM) hervorgerufen,
der als Ausreisserwert interpretiert werden kann. Während der Ruheperiode liegen die Werte der
rechten Seite bei beiden Gruppen tendenziell höher als die der linken. Im Mittel lag die
Carnitinkonzentration in der Muskulatur während der viermonatigen Supplementationsperiode
bei den mit Carnitin supplementierten Tieren bei 4676 ± 865 mg/kg TM. Dies stellte einen
Anstieg von 57 % gegenüber der Kontrollgruppe dar.
Die Konzentration an freiem Carnitin und an Gesamtcarnitin in der Muskulatur verdeutlicht
Abbildung 4.4..
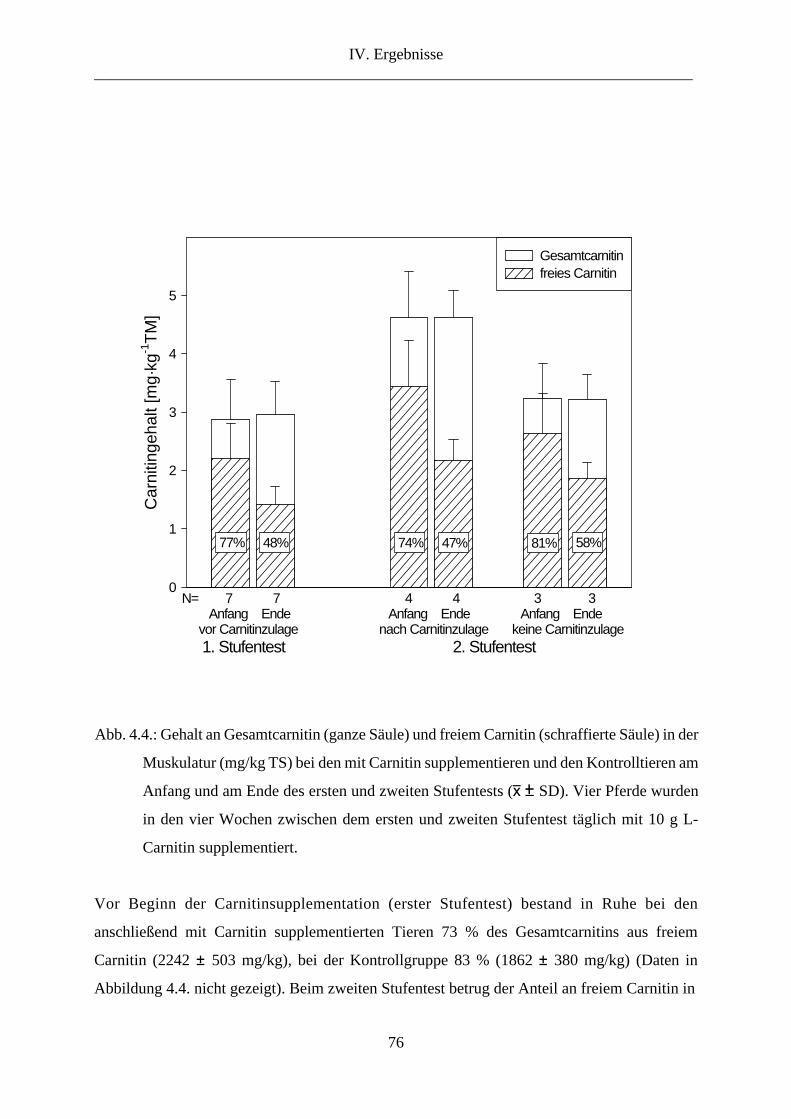
IV. Ergebnisse___________________________________________________________________________
76
Car
nitin
geha
lt [m
g·kg
-1TM
]
0
1
2
3
4
5
Gesamtcarnitinfreies Carnitin
N= 7 7 4 4 3 3 Anfang Ende Anfang Ende Anfang Ende vor Carnitinzulage nach Carnitinzulage keine Carnitinzulage 1. Stufentest 2. Stufentest
77% 48% 74% 47% 81% 58%
Abb. 4.4.: Gehalt an Gesamtcarnitin (ganze Säule) und freiem Carnitin (schraffierte Säule) in der
Muskulatur (mg/kg TS) bei den mit Carnitin supplementieren und den Kontrolltieren am
Anfang und am Ende des ersten und zweiten Stufentests (x # SD). Vier Pferde wurden
in den vier Wochen zwischen dem ersten und zweiten Stufentest täglich mit 10 g L-
Carnitin supplementiert.
Vor Beginn der Carnitinsupplementation (erster Stufentest) bestand in Ruhe bei den
anschließend mit Carnitin supplementierten Tieren 73 % des Gesamtcarnitins aus freiem
Carnitin (2242 ± 503 mg/kg), bei der Kontrollgruppe 83 % (1862 ± 380 mg/kg) (Daten in
Abbildung 4.4. nicht gezeigt). Beim zweiten Stufentest betrug der Anteil an freiem Carnitin in
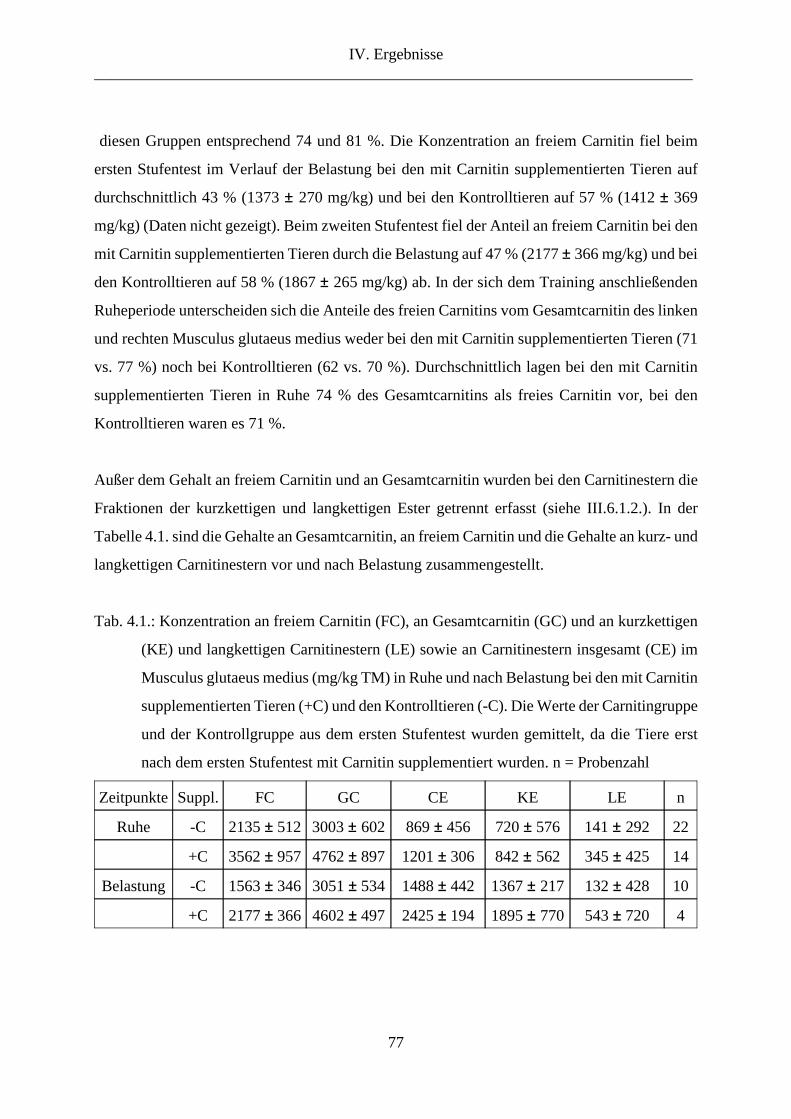
IV. Ergebnisse___________________________________________________________________________
77
diesen Gruppen entsprechend 74 und 81 %. Die Konzentration an freiem Carnitin fiel beim
ersten Stufentest im Verlauf der Belastung bei den mit Carnitin supplementierten Tieren auf
durchschnittlich 43 % (1373 ± 270 mg/kg) und bei den Kontrolltieren auf 57 % (1412 ± 369
mg/kg) (Daten nicht gezeigt). Beim zweiten Stufentest fiel der Anteil an freiem Carnitin bei den
mit Carnitin supplementierten Tieren durch die Belastung auf 47 % (2177 ± 366 mg/kg) und bei
den Kontrolltieren auf 58 % (1867 ± 265 mg/kg) ab. In der sich dem Training anschließenden
Ruheperiode unterscheiden sich die Anteile des freien Carnitins vom Gesamtcarnitin des linken
und rechten Musculus glutaeus medius weder bei den mit Carnitin supplementierten Tieren (71
vs. 77 %) noch bei Kontrolltieren (62 vs. 70 %). Durchschnittlich lagen bei den mit Carnitin
supplementierten Tieren in Ruhe 74 % des Gesamtcarnitins als freies Carnitin vor, bei den
Kontrolltieren waren es 71 %.
Außer dem Gehalt an freiem Carnitin und an Gesamtcarnitin wurden bei den Carnitinestern die
Fraktionen der kurzkettigen und langkettigen Ester getrennt erfasst (siehe III.6.1.2.). In der
Tabelle 4.1. sind die Gehalte an Gesamtcarnitin, an freiem Carnitin und die Gehalte an kurz- und
langkettigen Carnitinestern vor und nach Belastung zusammengestellt.
Tab. 4.1.: Konzentration an freiem Carnitin (FC), an Gesamtcarnitin (GC) und an kurzkettigen
(KE) und langkettigen Carnitinestern (LE) sowie an Carnitinestern insgesamt (CE) im
Musculus glutaeus medius (mg/kg TM) in Ruhe und nach Belastung bei den mit Carnitin
supplementierten Tieren (+C) und den Kontrolltieren (-C). Die Werte der Carnitingruppe
und der Kontrollgruppe aus dem ersten Stufentest wurden gemittelt, da die Tiere erst
nach dem ersten Stufentest mit Carnitin supplementiert wurden. n = Probenzahl
Zeitpunkte Suppl. FC GC CE KE LE n
Ruhe -C 2135 ± 512 3003 ± 602 869 ± 456 720 ± 576 141 ± 292 22
+C 3562 ± 957 4762 ± 897 1201 ± 306 842 ± 562 345 ± 425 14
Belastung -C 1563 ± 346 3051 ± 534 1488 ± 442 1367 ± 217 132 ± 428 10
+C 2177 ± 366 4602 ± 497 2425 ± 194 1895 ± 770 543 ± 720 4

IV. Ergebnisse___________________________________________________________________________
78
In Ruhe bestanden ca. 75 % und nach Belastung ca. 83 % der Carnitinester aus kurzkettigen
Estern. Bei den mit Carnitin supplementierten Tieren stieg der Anteil der kurzkettigen
Carnitinester durch Belastung von 70 auf 78 % und bei den Kontrolltieren von 83 auf 92 %
(Carnitinester insgesamt = 100 %). Diese Ergebnisse sind naturgemäß mit einer nicht
unerheblichen Streuung behaftet, da nur 5 % des Gesamtcarnitins als langkettige Carnitinester
vorliegen. Diese Menge entsprach in etwa der Größe des methodischen Fehlers.
3. Herzfrequenz
Ein wichtiges Merkmal zur Beurteilung trainingsbedingter Anpassungsvorgänge und möglicher
Effekte einer Carnitinzulage bildet die Herzfrequenz unter verschiedenen standardisierten
Bedingungen einer Belastung. Die Abbildung 4.5. zeigt die Zunahme der Herzfrequenz während
der beiden in fünfwöchigem Abstand durchgeführten Stufentests bei den vier mit Carnitin
supplementierten (links) und den drei Kontrolltieren (rechts).
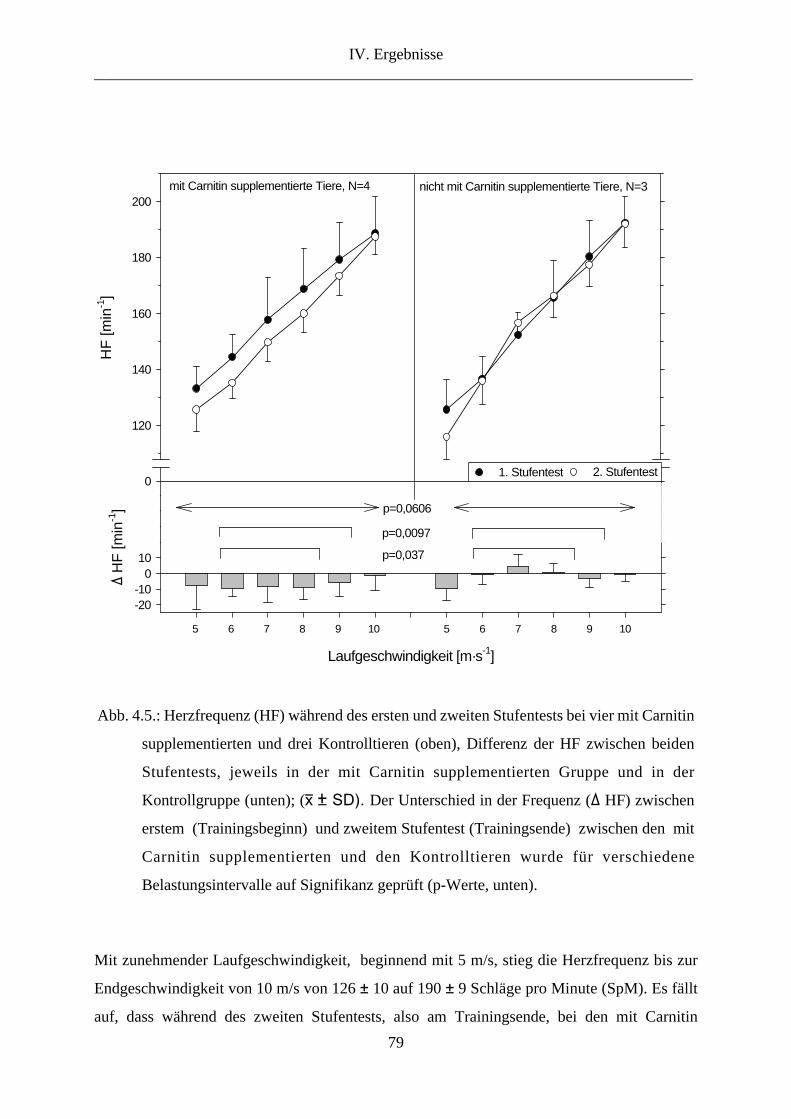
IV. Ergebnisse___________________________________________________________________________
79
HF
[min
-1]
0
120
140
160
180
200
Laufgeschwindigkeit [m·s-1]
5 6 7 8 9 10 5 6 7 8 9 10
d H
F [m
in-1
]
-20-10
01020304050
mit Carnitin supplementierte Tiere, N=4 nicht mit Carnitin supplementierte Tiere, N=3
1. Stufentest 2. Stufentest
p=0,0606
p=0,0097
p=0,037
Abb. 4.5.: Herzfrequenz (HF) während des ersten und zweiten Stufentests bei vier mit Carnitin
supplementierten und drei Kontrolltieren (oben), Differenz der HF zwischen beiden
Stufentests, jeweils in der mit Carnitin supplementierten Gruppe und in der
Kontrollgruppe (unten); (x # SD). Der Unterschied in der Frequenz (d HF) zwischen
erstem (Trainingsbeginn) und zweitem Stufentest (Trainingsende) zwischen den mit
Carnitin supplementierten und den Kontrolltieren wurde für verschiedene
Belastungsintervalle auf Signifikanz geprüft (p-Werte, unten).
Mit zunehmender Laufgeschwindigkeit, beginnend mit 5 m/s, stieg die Herzfrequenz bis zur
Endgeschwindigkeit von 10 m/s von 126 ± 10 auf 190 ± 9 Schläge pro Minute (SpM). Es fällt
auf, dass während des zweiten Stufentests, also am Trainingsende, bei den mit Carnitin

IV. Ergebnisse___________________________________________________________________________
80
supplementierten Tieren die Herzfrequenz im Mittel um 7 ± 3 SpM niedriger lag als während
des ersten Stufentests (Trainingsanfang). In dem zwischen beiden Stufentests liegenden
Zeitraum von fünf Wochen hatten die Tiere täglich 10 g L-Carnitin mit dem Futter
aufgenommen. Der frequenzsenkende Effekt war bis zu einer Laufgeschwindigkeit von 8 m/s
mit 8 ± 1 SpM am größten (Abb. 4.5. unten). Mit der weiteren Erhöhung der
Laufgeschwindigkeit auf 9 m/s und 10 m/s verringerte er sich und war bei 10 m/s nicht mehr
nachweisbar.
Bei den Kontrolltieren lag die Herzfrequenz, mit Ausnahme der ersten Belastungsstufe (5 m/s),
bei beiden Stufentests etwa auf der gleichen Höhe. Die trainingsinduzierte Reduktion der
Herzfrequenz war bei den mit Carnitin supplementierten Tieren mit 7 SpM über den gesamten
Belastungsbereich von 5 m/s bis 10 m/s ausgeprägter als bei den Kontrollpferden (2 SpM,
p=0,06). Für enger gefasste Belastungsintervalle innerhalb des Tests war der Unterschied mit
p<0,05 signifikant (Abb. 4.5. unten).
Während der Langzeitbelastungen zeigte sich bei der Herzfrequenz ein ähnlicher Trend wie bei
den Stufentests. In der mit Carnitin supplementierten Gruppe war die sich auf ein steady state
einstellende Herzfrequenz am Ende des fünfwöchigen Trainings (achte Langzeitbelastung) im
Mittel um 8 SpM niedriger als zu Beginn des Trainings (erste Langzeitbelastung) (130 vs. 122
SpM) (Tab. 4.2.). Bei den Kontrollpferden betrug die Abnahme der Herzfrequenz im steady state
nur 4 SpM (128 vs. 124/min).
Ein in die gleiche Richtung weisender Effekt der Carnitinzulage auf die Herzfrequenz war auch
während der Kurzzeitbelastungen zu beobachten. Unter diesen Bedingungen war bei den mit
Carnitin supplementierten Tieren am Ende des Trainings (achte Kurzzeitbelastung) ein
Rückgang der Maximalfrequenz gegenüber Trainingsbeginn (erste Kurzzeitbelastung) um 13
SpM eingetreten (209 vs. 196 SpM). Bei den Kontrolltieren betrug der trainingsbedingte
Rückgang unter diesen Bedingungen nur 4 SpM ( 202 vs. 198 SpM) (Tab. 4.2.).
Für die neunte Kurz- und Langzeitbelastung waren die Pferde aufgrund der
Lactatkonzentrationen während des zweiten Stufentests individuell auf ein höheres
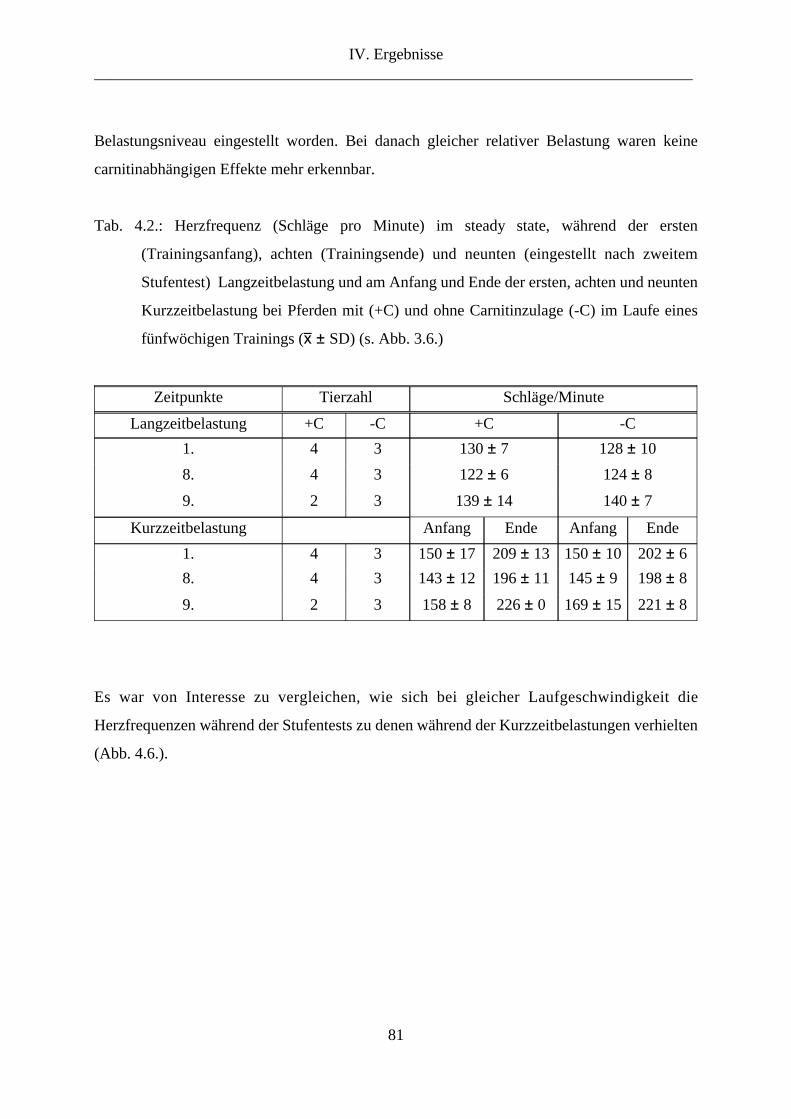
IV. Ergebnisse___________________________________________________________________________
81
Belastungsniveau eingestellt worden. Bei danach gleicher relativer Belastung waren keine
carnitinabhängigen Effekte mehr erkennbar.
Tab. 4.2.: Herzfrequenz (Schläge pro Minute) im steady state, während der ersten
(Trainingsanfang), achten (Trainingsende) und neunten (eingestellt nach zweitem
Stufentest) Langzeitbelastung und am Anfang und Ende der ersten, achten und neunten
Kurzzeitbelastung bei Pferden mit (+C) und ohne Carnitinzulage (-C) im Laufe eines
fünfwöchigen Trainings (x ± SD) (s. Abb. 3.6.)
Zeitpunkte Tierzahl Schläge/Minute
Langzeitbelastung +C -C +C -C
1. 4 3 130 ± 7 128 ± 10
8. 4 3 122 ± 6 124 ± 8
9. 2 3 139 ± 14 140 ± 7
Kurzzeitbelastung Anfang Ende Anfang Ende
1. 4 3 150 ± 17 209 ± 13 150 ± 10 202 ± 68. 4 3 143 ± 12 196 ± 11 145 ± 9 198 ± 8
9. 2 3 158 ± 8 226 ± 0 169 ± 15 221 ± 8
Es war von Interesse zu vergleichen, wie sich bei gleicher Laufgeschwindigkeit die
Herzfrequenzen während der Stufentests zu denen während der Kurzzeitbelastungen verhielten
(Abb. 4.6.).
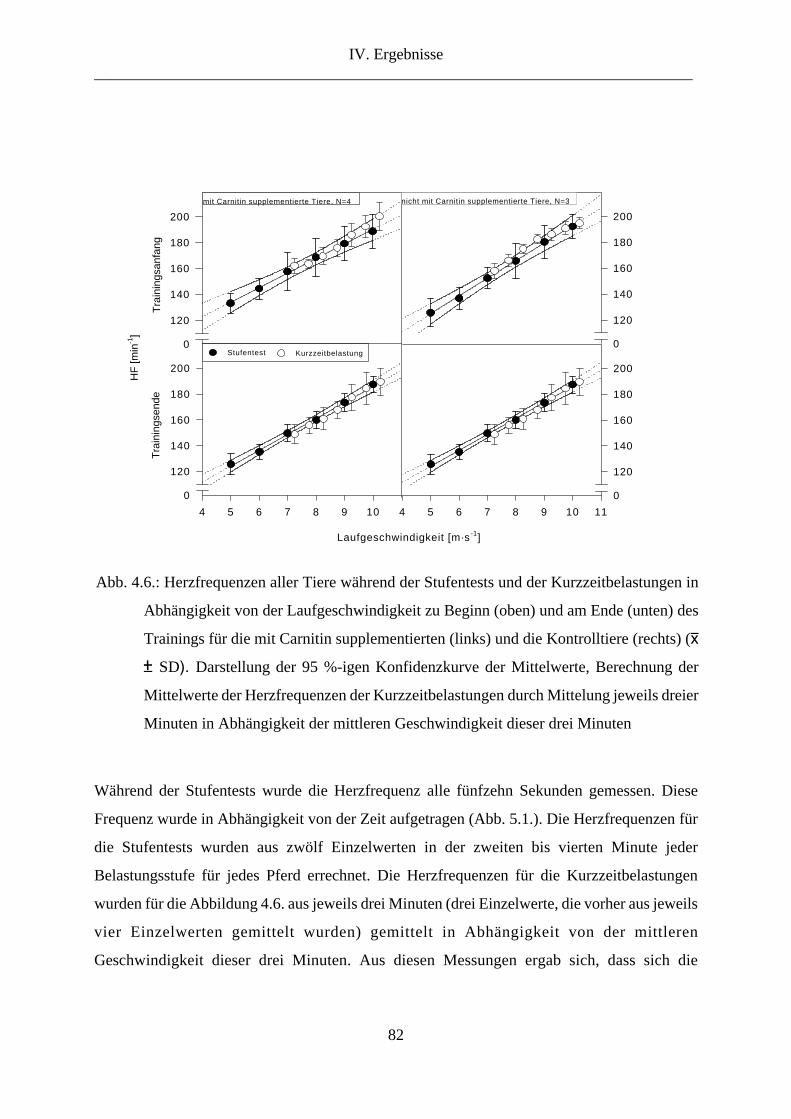
IV. Ergebnisse___________________________________________________________________________
82
HF
[min
-1]
0
120
140
160
180
200
0
120
140
160
180
200
4 5 6 7 8 9 10
0
120
140
160
180
200
Laufgeschwindigkeit [m·s -1]
4 5 6 7 8 9 10 110
120
140
160
180
200
Trai
ning
sanf
ang
Trai
ning
send
e
mit Carnitin supplementierte Tiere, N=4 nicht mit Carnitin supplementierte Tiere, N=3
Stufentest Kurzzeitbelastung
Abb. 4.6.: Herzfrequenzen aller Tiere während der Stufentests und der Kurzzeitbelastungen in
Abhängigkeit von der Laufgeschwindigkeit zu Beginn (oben) und am Ende (unten) des
Trainings für die mit Carnitin supplementierten (links) und die Kontrolltiere (rechts) (x
# SD). Darstellung der 95 %-igen Konfidenzkurve der Mittelwerte, Berechnung der
Mittelwerte der Herzfrequenzen der Kurzzeitbelastungen durch Mittelung jeweils dreier
Minuten in Abhängigkeit der mittleren Geschwindigkeit dieser drei Minuten
Während der Stufentests wurde die Herzfrequenz alle fünfzehn Sekunden gemessen. Diese
Frequenz wurde in Abhängigkeit von der Zeit aufgetragen (Abb. 5.1.). Die Herzfrequenzen für
die Stufentests wurden aus zwölf Einzelwerten in der zweiten bis vierten Minute jeder
Belastungsstufe für jedes Pferd errechnet. Die Herzfrequenzen für die Kurzzeitbelastungen
wurden für die Abbildung 4.6. aus jeweils drei Minuten (drei Einzelwerte, die vorher aus jeweils
vier Einzelwerten gemittelt wurden) gemittelt in Abhängigkeit von der mittleren
Geschwindigkeit dieser drei Minuten. Aus diesen Messungen ergab sich, dass sich die

IV. Ergebnisse___________________________________________________________________________
83
Herzfrequenz bei jeder Belastungsstufe in der Regel nach weniger als einer Minute auf ein der
jeweiligen Laufgeschwindigkeit entsprechendes steady state einstellte.
Es war von Interesse zu prüfen, ob die Herzfrequenzen, die sich in den einzelnen
Belastungsstufen während der Kurzzeitbelastungen einstellten, denen entsprachen, die die Tiere
während des Stufentests aufwiesen. Entsprechend wurden die Herzfrequenzen der ersten
Kurzzeitbelastung mit den Herzfrequenzen des ersten Stufentests und die Herzfrequenzen der
achten Kurzzeitbelastung mit denen des zweiten Stufentests bei entsprechender
Laufgeschwindigkeit verglichen (Abb. 4.6.). Die mittleren Herzfrequenzen aus beiden
Belastungsarten, aufgetragen in Abhängigkeit von der dazugehörigen Laufgeschwindigkeit der
Tiere, sind weitgehend deckungsgleich.
4. Lactat im Vollblut
Mit zunehmender Laufgeschwindigkeit nahm die Lactatkonzentration im Vollblut während des
Stufentests, beginnend mit 2,2 ± 0,3 mmol/l, zunächst kaum messbar zu. Erst ab einer
Geschwindigkeit von 7 m/s wurde der Anstieg steiler und beschleunigte sich von da an stetig mit
der linearen Zunahme der Laufgeschwindigkeit (Abb. 4.7.).
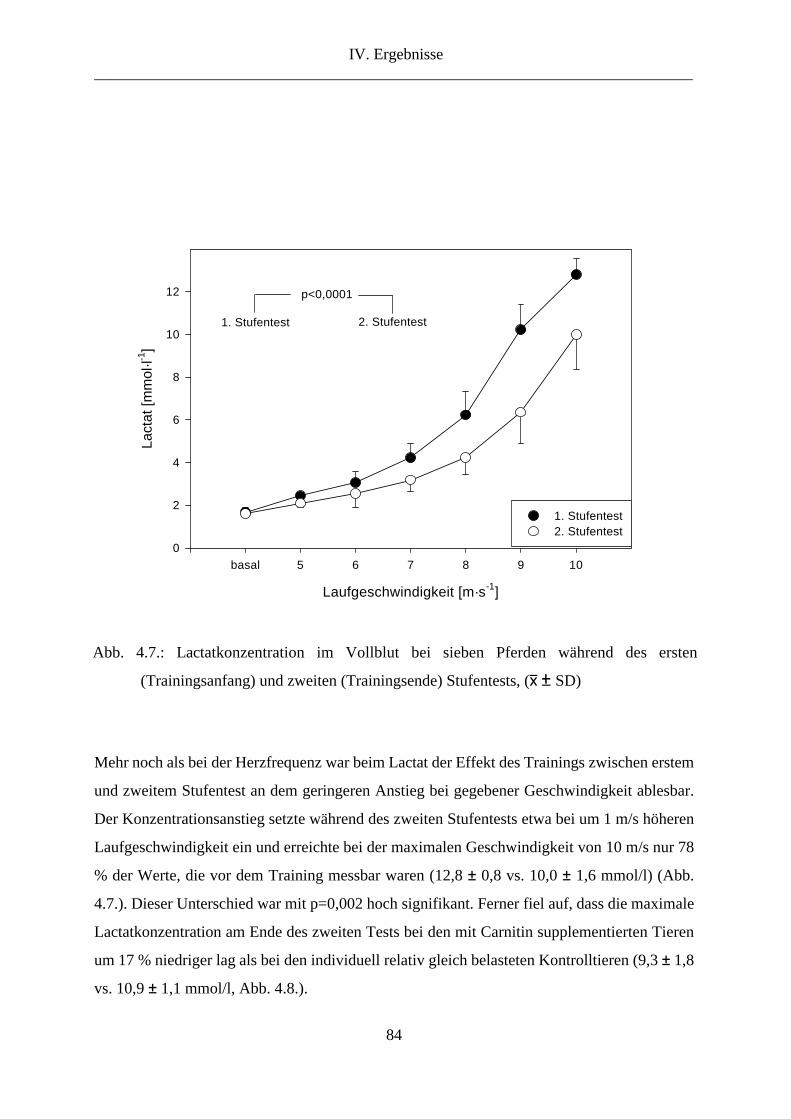
IV. Ergebnisse___________________________________________________________________________
84
Laufgeschwindigkeit [m·s-1]
basal 5 6 7 8 9 10
Lact
at [m
mol
·l-1]
0
2
4
6
8
10
12
1. Stufentest2. Stufentest
1. Stufentest 2. Stufentest
p<0,0001
Abb. 4.7.: Lactatkonzentration im Vollblut bei sieben Pferden während des ersten
(Trainingsanfang) und zweiten (Trainingsende) Stufentests, (x # SD)
Mehr noch als bei der Herzfrequenz war beim Lactat der Effekt des Trainings zwischen erstem
und zweitem Stufentest an dem geringeren Anstieg bei gegebener Geschwindigkeit ablesbar.
Der Konzentrationsanstieg setzte während des zweiten Stufentests etwa bei um 1 m/s höheren
Laufgeschwindigkeit ein und erreichte bei der maximalen Geschwindigkeit von 10 m/s nur 78
% der Werte, die vor dem Training messbar waren (12,8 ± 0,8 vs. 10,0 ± 1,6 mmol/l) (Abb.
4.7.). Dieser Unterschied war mit p=0,002 hoch signifikant. Ferner fiel auf, dass die maximale
Lactatkonzentration am Ende des zweiten Tests bei den mit Carnitin supplementierten Tieren
um 17 % niedriger lag als bei den individuell relativ gleich belasteten Kontrolltieren (9,3 ± 1,8
vs. 10,9 ± 1,1 mmol/l, Abb. 4.8.).
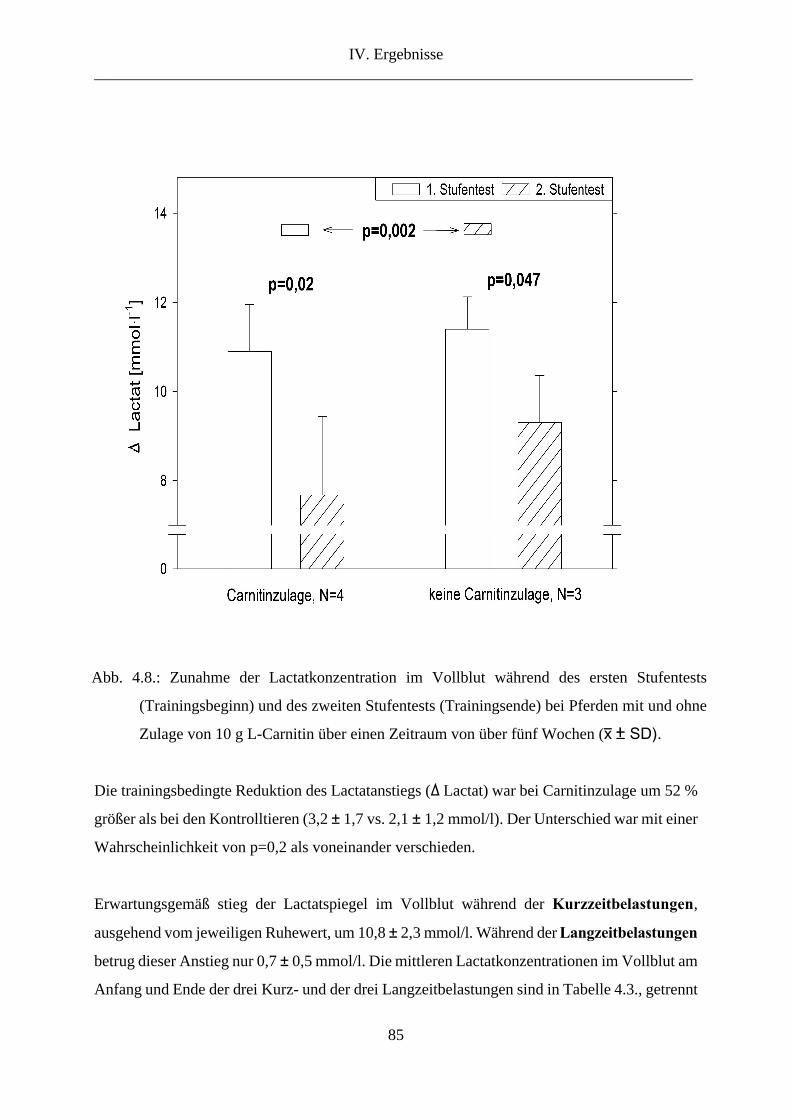
IV. Ergebnisse___________________________________________________________________________
85
Abb. 4.8.: Zunahme der Lactatkonzentration im Vollblut während des ersten Stufentests
(Trainingsbeginn) und des zweiten Stufentests (Trainingsende) bei Pferden mit und ohne
Zulage von 10 g L-Carnitin über einen Zeitraum von über fünf Wochen (x # SD).
Die trainingsbedingte Reduktion des Lactatanstiegs (d Lactat) war bei Carnitinzulage um 52 %
größer als bei den Kontrolltieren (3,2 ± 1,7 vs. 2,1 ± 1,2 mmol/l). Der Unterschied war mit einer
Wahrscheinlichkeit von p=0,2 als voneinander verschieden.
Erwartungsgemäß stieg der Lactatspiegel im Vollblut während der Kurzzeitbelastungen,
ausgehend vom jeweiligen Ruhewert, um 10,8 ± 2,3 mmol/l. Während der Langzeitbelastungen
betrug dieser Anstieg nur 0,7 ± 0,5 mmol/l. Die mittleren Lactatkonzentrationen im Vollblut am
Anfang und Ende der drei Kurz- und der drei Langzeitbelastungen sind in Tabelle 4.3., getrennt
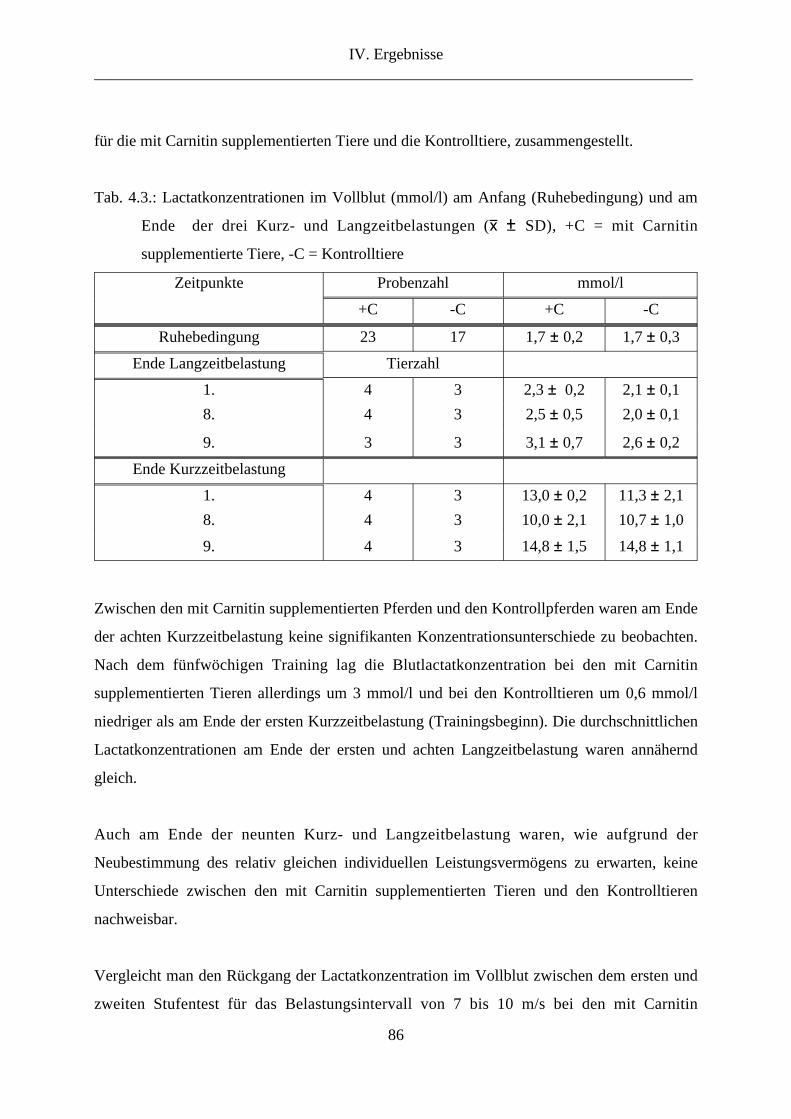
IV. Ergebnisse___________________________________________________________________________
86
für die mit Carnitin supplementierten Tiere und die Kontrolltiere, zusammengestellt.
Tab. 4.3.: Lactatkonzentrationen im Vollblut (mmol/l) am Anfang (Ruhebedingung) und am
Ende der drei Kurz- und Langzeitbelastungen (x # SD), +C = mit Carnitin
supplementierte Tiere, -C = Kontrolltiere
Zeitpunkte Probenzahl mmol/l
+C -C +C -C
Ruhebedingung 23 17 1,7 ± 0,2 1,7 ± 0,3
Ende Langzeitbelastung Tierzahl
1. 4 3 2,3 ± 0,2 2,1 ± 0,18. 4 3 2,5 ± 0,5 2,0 ± 0,1
9. 3 3 3,1 ± 0,7 2,6 ± 0,2
Ende Kurzzeitbelastung
1. 4 3 13,0 ± 0,2 11,3 ± 2,18. 4 3 10,0 ± 2,1 10,7 ± 1,0
9. 4 3 14,8 ± 1,5 14,8 ± 1,1
Zwischen den mit Carnitin supplementierten Pferden und den Kontrollpferden waren am Ende
der achten Kurzzeitbelastung keine signifikanten Konzentrationsunterschiede zu beobachten.
Nach dem fünfwöchigen Training lag die Blutlactatkonzentration bei den mit Carnitin
supplementierten Tieren allerdings um 3 mmol/l und bei den Kontrolltieren um 0,6 mmol/l
niedriger als am Ende der ersten Kurzzeitbelastung (Trainingsbeginn). Die durchschnittlichen
Lactatkonzentrationen am Ende der ersten und achten Langzeitbelastung waren annähernd
gleich.
Auch am Ende der neunten Kurz- und Langzeitbelastung waren, wie aufgrund der
Neubestimmung des relativ gleichen individuellen Leistungsvermögens zu erwarten, keine
Unterschiede zwischen den mit Carnitin supplementierten Tieren und den Kontrolltieren
nachweisbar.
Vergleicht man den Rückgang der Lactatkonzentration im Vollblut zwischen dem ersten und
zweiten Stufentest für das Belastungsintervall von 7 bis 10 m/s bei den mit Carnitin

IV. Ergebnisse___________________________________________________________________________
87
supplementierten Tieren mit den Kontrolltieren, ist festzustellen, dass dieser bei den mit Carnitin
supplementierten Tieren prozentual signifikant (p<0,05) größer ist als bei den Kontrolltieren
(Abb. 4.9. unten). Auch am Ende der Kurzzeitbelastung war der trainingsbedingte Rückgang der
Lactatkonzentration im Vollblut bei den mit Carnitin supplementierten Tieren zwischen der
ersten und achten Kurzzeitbelastung mit p=0,03 signifikant. Bei den Kontrolltieren war unter
diesen Bedingungen kein Unterschied feststellbar (Abb. 4.6. oben).
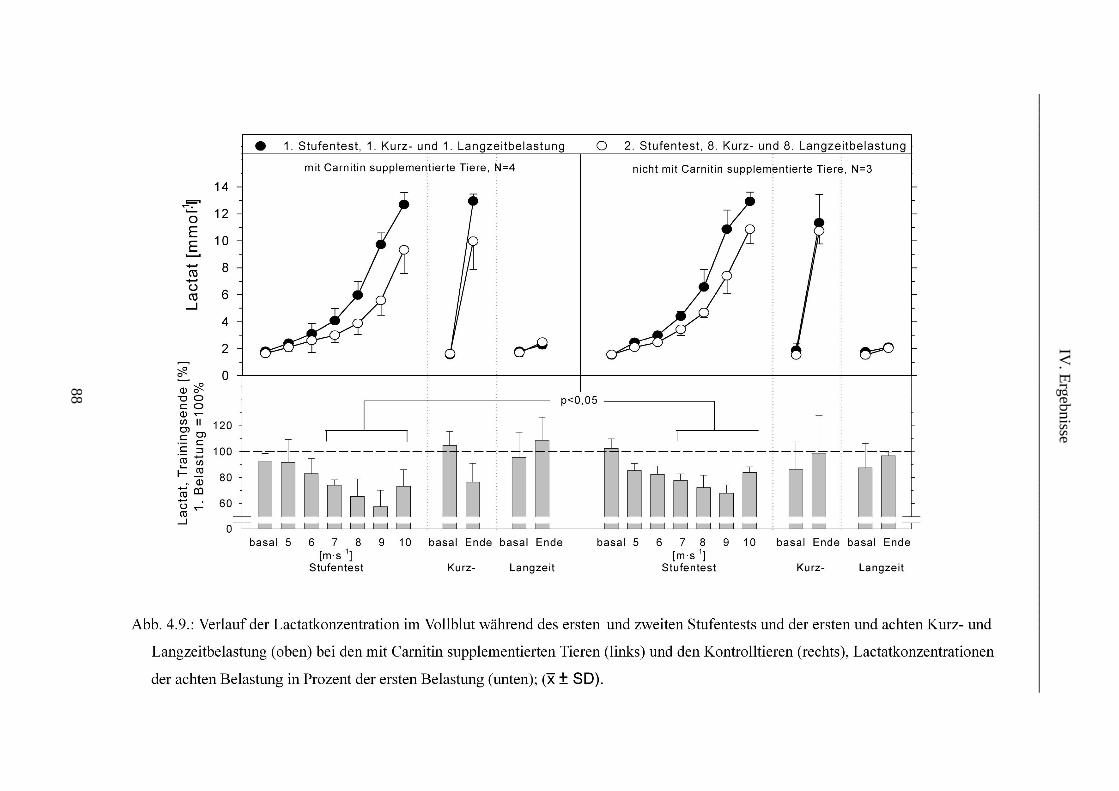
IV. Ergebnisse
___________________________________________________________________________
88

IV. Ergebnisse___________________________________________________________________________
89
Wie bei der Herzfrequenz war es auch bei der Lactatkonzentration von Interesse zu sehen,
welche Lactatkonzentration sich am Ende der ersten und achten Kurzzeitbelastung im Vergleich
zum ersten und zweiten Stufentest bei entsprechend gleicher Laufgeschwindigkeit einstellte.
Trotz unterschiedlicher Belastungsprofile während der Kurzzeitbelastung einerseits und der
Stufentests andererseits (siehe Kapitel III.5) war die Lactatkonzentration im Vollblut bei fünf
Pferden am Ende der ersten und achten Kurzzeitbelastung etwa auf dem gleichen Niveau wie
während der etwa zeitgleich durchgeführten Stufentests bei gleicher Laufgeschwindigkeit.
Bei zwei Pferden lag die Lactatkonzentration im Vollblut am Ende der Kurzzeitbelastung jedoch
deutlich niedriger als während des Stufentests bei entsprechend gleicher Laufgeschwindigkeit.
Ein solcher Unterschied wurde einmal zu Beginn des Trainings bei dem Kontrolltier Nr. V
beobachtet. Dieses Tier wies während des ersten Stufentests bei einer Laufgeschwindigkeit von
10 m/s eine Lactatkonzentration im Vollblut von 13,7 mmol/l auf und am Ende der ersten
Kurzzeitbelastung bei gleicher Laufgeschwindigkeit eine Konzentration von 8,9 mmol/l. Die
zweite Ausnahme betraf das mit Carnitin supplementierte Tier Nr. VIII am Ende des Trainings.
Bei diesem Tier lag die Lactatkonzentration im Vollblut am Ende des zweiten Stufentests bei
10,1 mmol/l und am Ende der achten Kurzzeitbelastung bei einer Laufgeschwindigkeit von 10
m/s bei 6,9 mmol/l.
5. Glucose im Plasma
Da die Glucose zu Beginn jeder körperlichen Belastung und darüber hinaus in sehr großem
Umfang bei der anaeroben Energiegewinnung als Energielieferant dient, war es von Interesse,
die Veränderungen der Plasmaglucosekonzentration im Rahmen dieses Projektes in Verbindung
mit den einzelnen Belastungen zu messen. Unter Ruhebedingungen wurde bei allen Tieren eine
mittlere Glucosekonzentration im Plasma von 4,4 ± 0,5 mmol/l gemessen. Dieser Ruhewert
veränderte sich über die Dauer der Versuche nicht. Es gab keine Unterschiede zwischen den mit
Carnitin supplementierten und den Kontrolltieren. Abbildung 4.10. stellt den Verlauf der
Glucosekonzentration im Plasma während beider Stufentests dar.
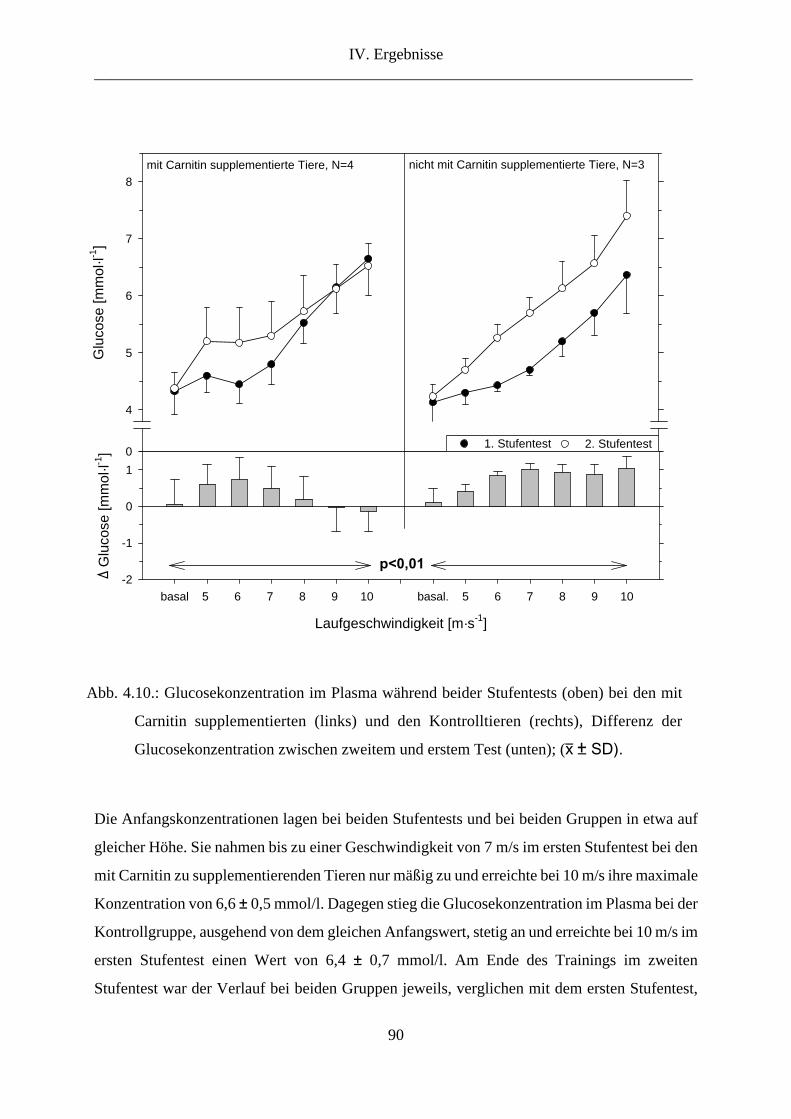
IV. Ergebnisse___________________________________________________________________________
90
Glu
cose
[mm
ol·l-1
]
0
4
5
6
7
8
Laufgeschwindigkeit [m·s-1]
basal 5 6 7 8 9 10 basal. 5 6 7 8 9 10
d G
luco
se [m
mol
·l-1]
-2
-1
0
1
mit Carnitin supplementierte Tiere, N=4 nicht mit Carnitin supplementierte Tiere, N=3
p<0,01
1. Stufentest 2. Stufentest
Abb. 4.10.: Glucosekonzentration im Plasma während beider Stufentests (oben) bei den mit
Carnitin supplementierten (links) und den Kontrolltieren (rechts), Differenz der
Glucosekonzentration zwischen zweitem und erstem Test (unten); (x # SD).
Die Anfangskonzentrationen lagen bei beiden Stufentests und bei beiden Gruppen in etwa auf
gleicher Höhe. Sie nahmen bis zu einer Geschwindigkeit von 7 m/s im ersten Stufentest bei den
mit Carnitin zu supplementierenden Tieren nur mäßig zu und erreichte bei 10 m/s ihre maximale
Konzentration von 6,6 ± 0,5 mmol/l. Dagegen stieg die Glucosekonzentration im Plasma bei der
Kontrollgruppe, ausgehend von dem gleichen Anfangswert, stetig an und erreichte bei 10 m/s im
ersten Stufentest einen Wert von 6,4 ± 0,7 mmol/l. Am Ende des Trainings im zweiten
Stufentest war der Verlauf bei beiden Gruppen jeweils, verglichen mit dem ersten Stufentest,

IV. Ergebnisse___________________________________________________________________________
91
ähnlich. Die Glucosekonzentration nahm während der ersten Belastungsstufen bis etwa 7 m/s bei
den mit Carnitin supplementierten Tieren, abgesehen von der ersten Belastungsstufe, nicht zu
und erreichte danach durch einen relativ flachen Anstieg bis 10 m/s denselben Endwert wie
während des ersten Stufentests. Es bestand kein Unterschied in der Endkonzentration zwischen
beiden Stufentests bei den mit Carnitin supplementierten Tieren zu Anfang und Ende des
Trainings. Bei den Kontrolltieren blieb die Glucosekonzentration am Ende des Trainings über
die gesamte Belastungsspanne deutlich oberhalb der sich am Anfang des Trainings einstellenden
Konzentrationen und erreichte im zweiten Stufentest eine Endkonzentration von 7,4 ± 0,6
mmol/l. Die Konzentrationszunahmen am Ende des Trainings sind über den gesamten
Belastungsbereich während des zweiten Stufentests bei den Kontrolltieren hochsignifikant
(p<0,01) größer als bei den mit Carnitin supplementierten Tieren (Abb. 4.10. unten).
Die Glucosekonzentration im Plasma stieg auch bei den anderen Belastungen, abhängig von der
Belastungsart, an. Bei den Langzeitbelastungen war bei allen Tieren ein Anstieg von 1,5 ± 1,1
mmol/l und bei den Kurzzeitbelastungen ein Anstieg von 2,4 ± 1,3 mmol/l zu verzeichnen.
Die mittleren Glucosekonzentrationen im Plasma am Anfang und Ende der drei Kurz- und der
drei Langzeitbelastungen sind in Tabelle 4.4., getrennt für die mit Carnitin supplementierten
Tiere und die Kontrolltiere, zusammengestellt.
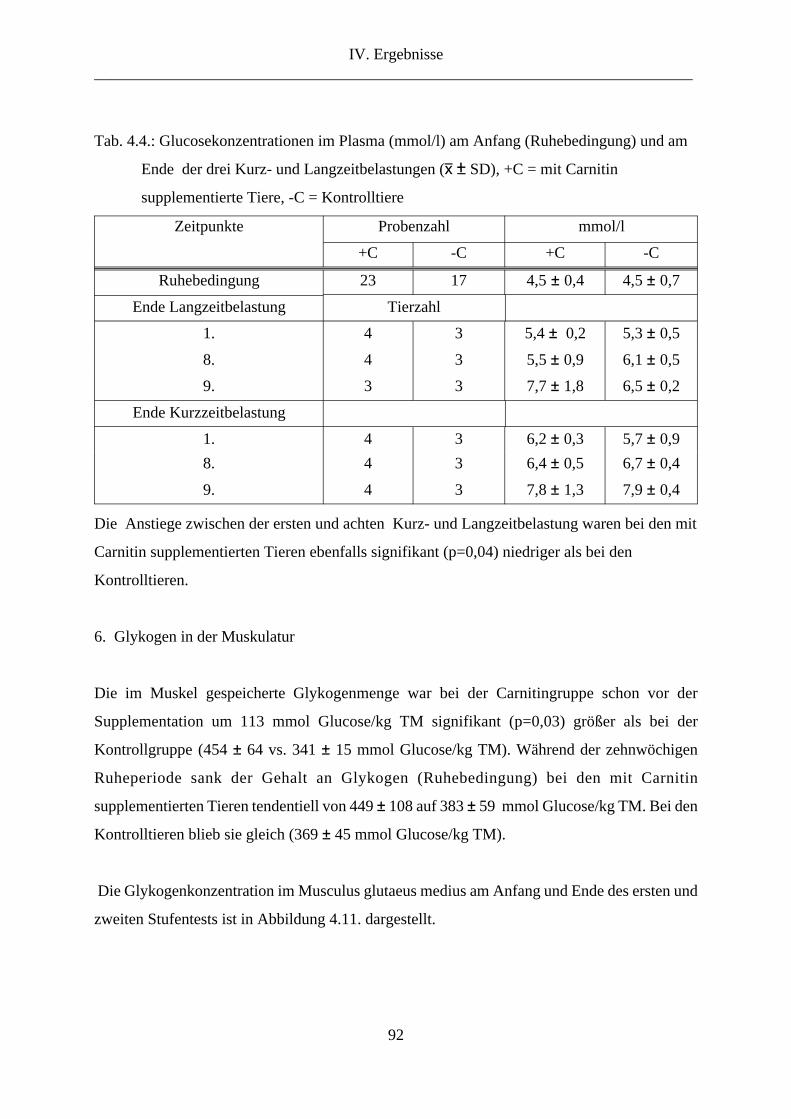
IV. Ergebnisse___________________________________________________________________________
92
Tab. 4.4.: Glucosekonzentrationen im Plasma (mmol/l) am Anfang (Ruhebedingung) und am
Ende der drei Kurz- und Langzeitbelastungen (x # SD), +C = mit Carnitin
supplementierte Tiere, -C = Kontrolltiere
Zeitpunkte Probenzahl mmol/l
+C -C +C -C
Ruhebedingung 23 17 4,5 ± 0,4 4,5 ± 0,7
Ende Langzeitbelastung Tierzahl
1. 4 3 5,4 ± 0,2 5,3 ± 0,5
8. 4 3 5,5 ± 0,9 6,1 ± 0,5
9. 3 3 7,7 ± 1,8 6,5 ± 0,2
Ende Kurzzeitbelastung
1. 4 3 6,2 ± 0,3 5,7 ± 0,98. 4 3 6,4 ± 0,5 6,7 ± 0,4
9. 4 3 7,8 ± 1,3 7,9 ± 0,4
Die Anstiege zwischen der ersten und achten Kurz- und Langzeitbelastung waren bei den mit
Carnitin supplementierten Tieren ebenfalls signifikant (p=0,04) niedriger als bei den
Kontrolltieren.
6. Glykogen in der Muskulatur
Die im Muskel gespeicherte Glykogenmenge war bei der Carnitingruppe schon vor der
Supplementation um 113 mmol Glucose/kg TM signifikant (p=0,03) größer als bei der
Kontrollgruppe (454 ± 64 vs. 341 ± 15 mmol Glucose/kg TM). Während der zehnwöchigen
Ruheperiode sank der Gehalt an Glykogen (Ruhebedingung) bei den mit Carnitin
supplementierten Tieren tendentiell von 449 ± 108 auf 383 ± 59 mmol Glucose/kg TM. Bei den
Kontrolltieren blieb sie gleich (369 ± 45 mmol Glucose/kg TM).
Die Glykogenkonzentration im Musculus glutaeus medius am Anfang und Ende des ersten und
zweiten Stufentests ist in Abbildung 4.11. dargestellt.
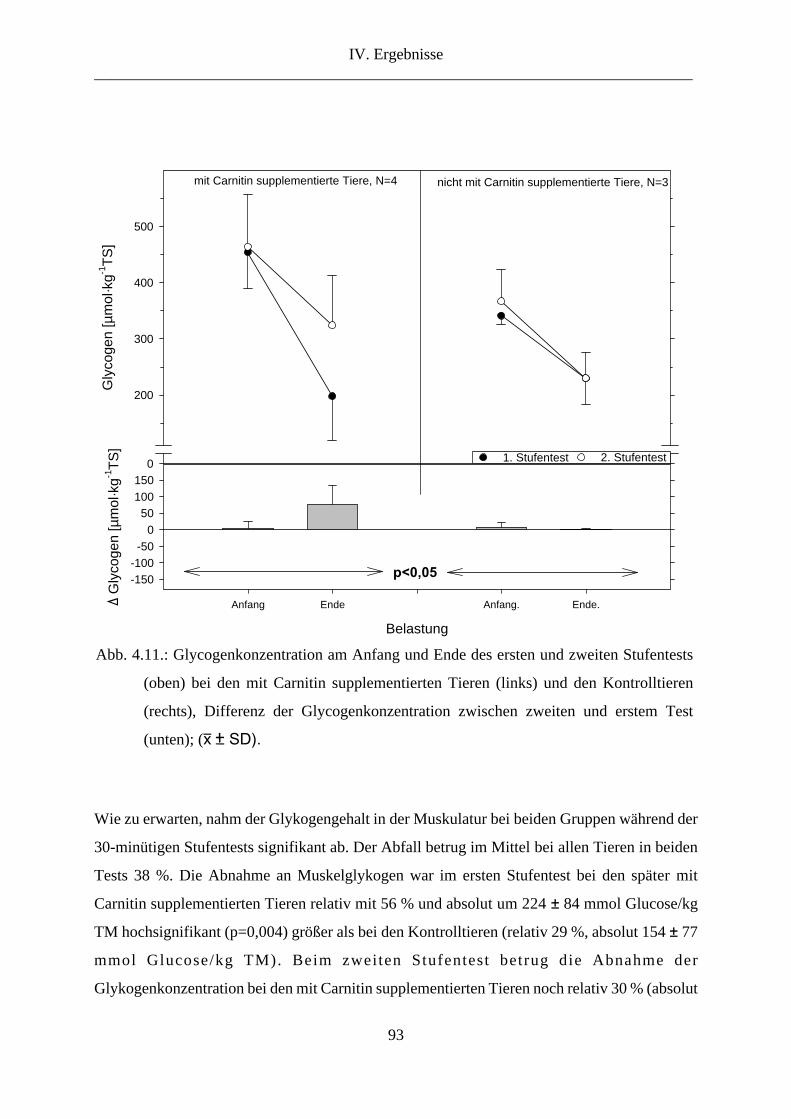
IV. Ergebnisse___________________________________________________________________________
93
Belastung
Anfang Ende Anfang. Ende.d G
lyco
gen
[µm
ol·k
g-1TS
]
-150-100-50
050
100150
Gly
coge
n [µ
mol
·kg-1
TS]
0
200
300
400
500
mit Carnitin supplementierte Tiere, N=4 nicht mit Carnitin supplementierte Tiere, N=3
p<0,05
1. Stufentest 2. Stufentest
Abb. 4.11.: Glycogenkonzentration am Anfang und Ende des ersten und zweiten Stufentests
(oben) bei den mit Carnitin supplementierten Tieren (links) und den Kontrolltieren
(rechts), Differenz der Glycogenkonzentration zwischen zweiten und erstem Test
(unten); (x # SD).
Wie zu erwarten, nahm der Glykogengehalt in der Muskulatur bei beiden Gruppen während der
30-minütigen Stufentests signifikant ab. Der Abfall betrug im Mittel bei allen Tieren in beiden
Tests 38 %. Die Abnahme an Muskelglykogen war im ersten Stufentest bei den später mit
Carnitin supplementierten Tieren relativ mit 56 % und absolut um 224 ± 84 mmol Glucose/kg
TM hochsignifikant (p=0,004) größer als bei den Kontrolltieren (relativ 29 %, absolut 154 ± 77
mmol Glucose/kg TM). Beim zweiten Stufentest betrug die Abnahme der
Glykogenkonzentration bei den mit Carnitin supplementierten Tieren noch relativ 30 % (absolut

IV. Ergebnisse___________________________________________________________________________
94
169 ± 33 mmol Glucose/kg TM) und bei den Kontrolltieren relativ 37 % (absolut 98 ± 72 mmol
Glucose/kg TM). Der Abfall des Muskelglykogens war bei den mit Carnitin supplementierten
Tieren im zweiten Stufentest hochsignifikant (p=0,009) geringer als im ersten.
7. Freie Fettsäuren im Plasma
Freie Fettsäuren stellen ebenso wie Glucose bei körperlicher Belastung eine wichtige
Energiequelle dar. Sie können jedoch nur unter aeroben Bedingungen energetisch genutzt
werden. Aufgrund der katalytischen Funktion von Carnitin wäre es denkbar, dass durch eine
Carnitinzulage die energetische Nutzung von FFS durch den arbeitenden Muskel beeinflusst
wird. Aus diesem Grunde wurde die Konzentration der FFS im Plasma während der Stufentests
gemessen. Die Ergebnisse sind in Abbildung 4.12. dargestellt. Die Anfangskonzentration an FFS
betrug bei dem später mit Carnitin supplementierten Pferd Nr. IV beim ersten Stufentest 1317
µmol/l. Bei den übrigen dieser Tiere lag sie im Mittel bei 419 ± 314 µmol/l. Der Wert von Pferd
Nr. IV wurde als Ausreißer angesprochen und nicht mit in die Berechnung einbezogen.
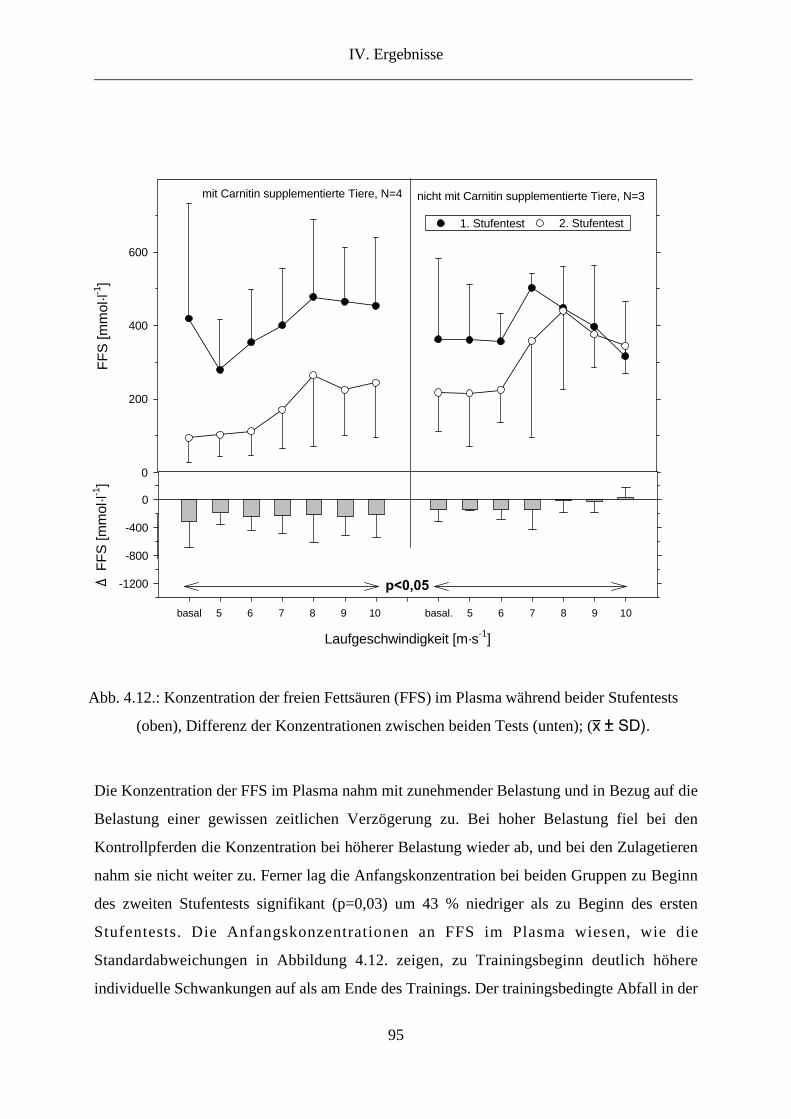
IV. Ergebnisse___________________________________________________________________________
95
FFS
[mm
ol·l-1
]
0
200
400
600
Plot 1 Plot 2
Laufgeschwindigkeit [m·s-1]
basal 5 6 7 8 9 10 basal. 5 6 7 8 9 10
d F
FS [m
mol
·l-1]
-1200
-800
-400
0
mit Carnitin supplementierte Tiere, N=4 nicht mit Carnitin supplementierte Tiere, N=3
p<0,05
1. Stufentest 2. Stufentest
Abb. 4.12.: Konzentration der freien Fettsäuren (FFS) im Plasma während beider Stufentests
(oben), Differenz der Konzentrationen zwischen beiden Tests (unten); (x # SD).
Die Konzentration der FFS im Plasma nahm mit zunehmender Belastung und in Bezug auf die
Belastung einer gewissen zeitlichen Verzögerung zu. Bei hoher Belastung fiel bei den
Kontrollpferden die Konzentration bei höherer Belastung wieder ab, und bei den Zulagetieren
nahm sie nicht weiter zu. Ferner lag die Anfangskonzentration bei beiden Gruppen zu Beginn
des zweiten Stufentests signifikant (p=0,03) um 43 % niedriger als zu Beginn des ersten
Stufentests. Die Anfangskonzentrationen an FFS im Plasma wiesen, wie die
Standardabweichungen in Abbildung 4.12. zeigen, zu Trainingsbeginn deutlich höhere
individuelle Schwankungen auf als am Ende des Trainings. Der trainingsbedingte Abfall in der
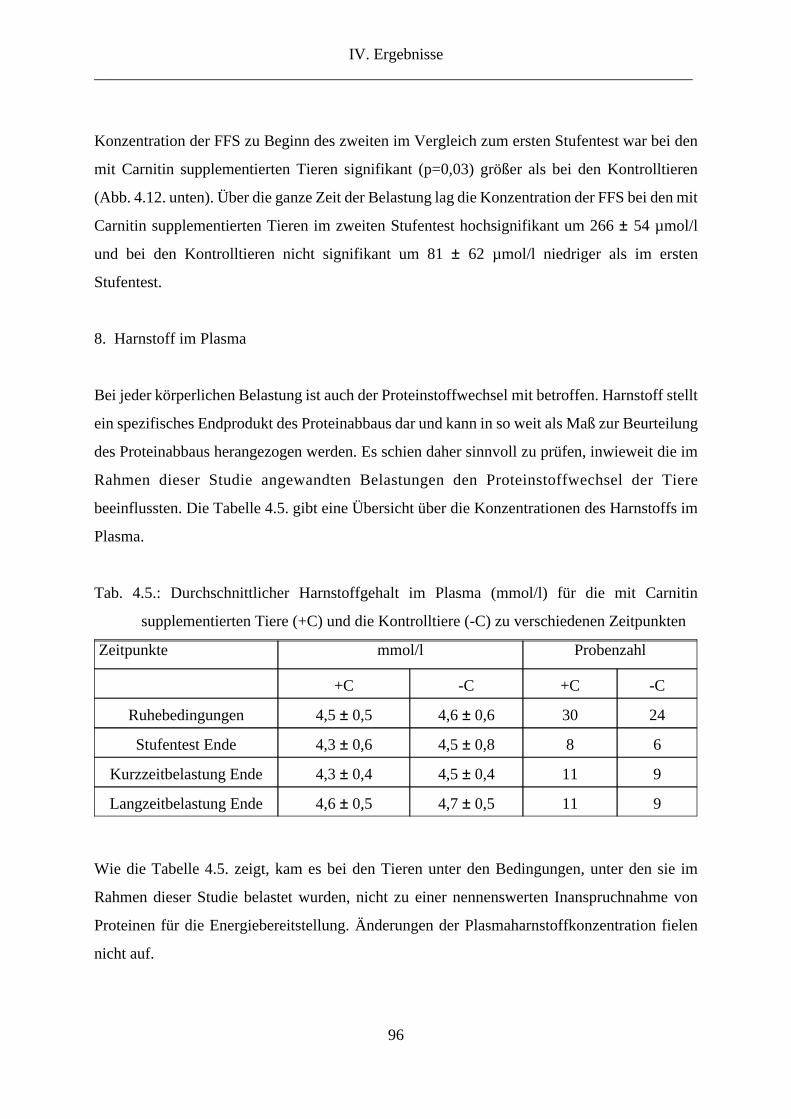
IV. Ergebnisse___________________________________________________________________________
96
Konzentration der FFS zu Beginn des zweiten im Vergleich zum ersten Stufentest war bei den
mit Carnitin supplementierten Tieren signifikant (p=0,03) größer als bei den Kontrolltieren
(Abb. 4.12. unten). Über die ganze Zeit der Belastung lag die Konzentration der FFS bei den mit
Carnitin supplementierten Tieren im zweiten Stufentest hochsignifikant um 266 ± 54 µmol/l
und bei den Kontrolltieren nicht signifikant um 81 ± 62 µmol/l niedriger als im ersten
Stufentest.
8. Harnstoff im Plasma
Bei jeder körperlichen Belastung ist auch der Proteinstoffwechsel mit betroffen. Harnstoff stellt
ein spezifisches Endprodukt des Proteinabbaus dar und kann in so weit als Maß zur Beurteilung
des Proteinabbaus herangezogen werden. Es schien daher sinnvoll zu prüfen, inwieweit die im
Rahmen dieser Studie angewandten Belastungen den Proteinstoffwechsel der Tiere
beeinflussten. Die Tabelle 4.5. gibt eine Übersicht über die Konzentrationen des Harnstoffs im
Plasma.
Tab. 4.5.: Durchschnittlicher Harnstoffgehalt im Plasma (mmol/l) für die mit Carnitin
supplementierten Tiere (+C) und die Kontrolltiere (-C) zu verschiedenen Zeitpunkten
Zeitpunkte mmol/l Probenzahl
+C -C +C -C
Ruhebedingungen 4,5 ± 0,5 4,6 ± 0,6 30 24
Stufentest Ende 4,3 ± 0,6 4,5 ± 0,8 8 6
Kurzzeitbelastung Ende 4,3 ± 0,4 4,5 ± 0,4 11 9
Langzeitbelastung Ende 4,6 ± 0,5 4,7 ± 0,5 11 9
Wie die Tabelle 4.5. zeigt, kam es bei den Tieren unter den Bedingungen, unter den sie im
Rahmen dieser Studie belastet wurden, nicht zu einer nennenswerten Inanspruchnahme von
Proteinen für die Energiebereitstellung. Änderungen der Plasmaharnstoffkonzentration fielen
nicht auf.

IV. Ergebnisse___________________________________________________________________________
97
9. Hämatokrit
Durch Entspeicherung der Milz und Wasserverluste durch Schwitzen steigt der Hämatokrit. Bei
Änderungen des Hämatokrits infolge der Entspeicherungspolyglobulie und von Wasserverlusten
um mehr als 5 % ist es ratsam zu prüfen, ob im Plasma gemessene Parameter entsprechend
korrigiert werden müssen. Der mittlere Hämatokritgehalt unter Ruhebedingungen lag bei allen
Tieren bei 34 ± 2 %. Er stieg signifikant auf 41 ± 4 % bei den Kurz- und Langzeitbelastungen
und auf 42 ± 2 % beim Stufentest. Vier Stunden nach den Belastungen waren die Ruhewerte
wieder erreicht. Bei gleichen Belastungen unterschied sich der Hämatokritgehalt zwischen den
mit Carnitin supplementierten Pferden und den Kontrollpferden nicht. Tabelle 4.6. gibt eine
Übersicht über den Hämatokrit bei den mit Carnitin supplementierten und den Kontrolltieren
unter verschiedenen Belastungsbedingungen.
Tab. 4.6.: Hämatokritgehalt (%) bei Pferden in Ruhe und nach Belastung, mit Carnitin
supplementierte Tiere (+C), Kontrolltiere (-C); (x # SD)
Zeitpunkte % Anzahl der Messungen
+C -C +C -C
Ruhebedingungen 35 ± 2 34 ± 2 31 24
Stufentest Ende 42 ± 2 43 ± 2 8 6
Kurzzeitbelastung Ende 43 ± 2 41 ± 3 12 9
Langzeitbelastung Ende 40 ± 3 37 ± 4 11 9
10. Gesamtprotein im Plasma
Die Messung des Gehaltes an Gesamtprotein gibt Hinweise auf mögliche Wasserverschiebungen
und Wasserverluste aus dem intravasalen Raum. Diese Messungen ermöglichen es, bei
Änderungen des Hämatokritwertes im Laufe einer Belastung zwischen dem Einstrom von
Blutzellen und Nettowasserverschiebungen zu unterscheiden. Die Tabelle 4.7. gibt eine
Übersicht über die Konzentrationen des Gesamtproteins im Plasma am Anfang und am Ende
verschiedener Belastungen.
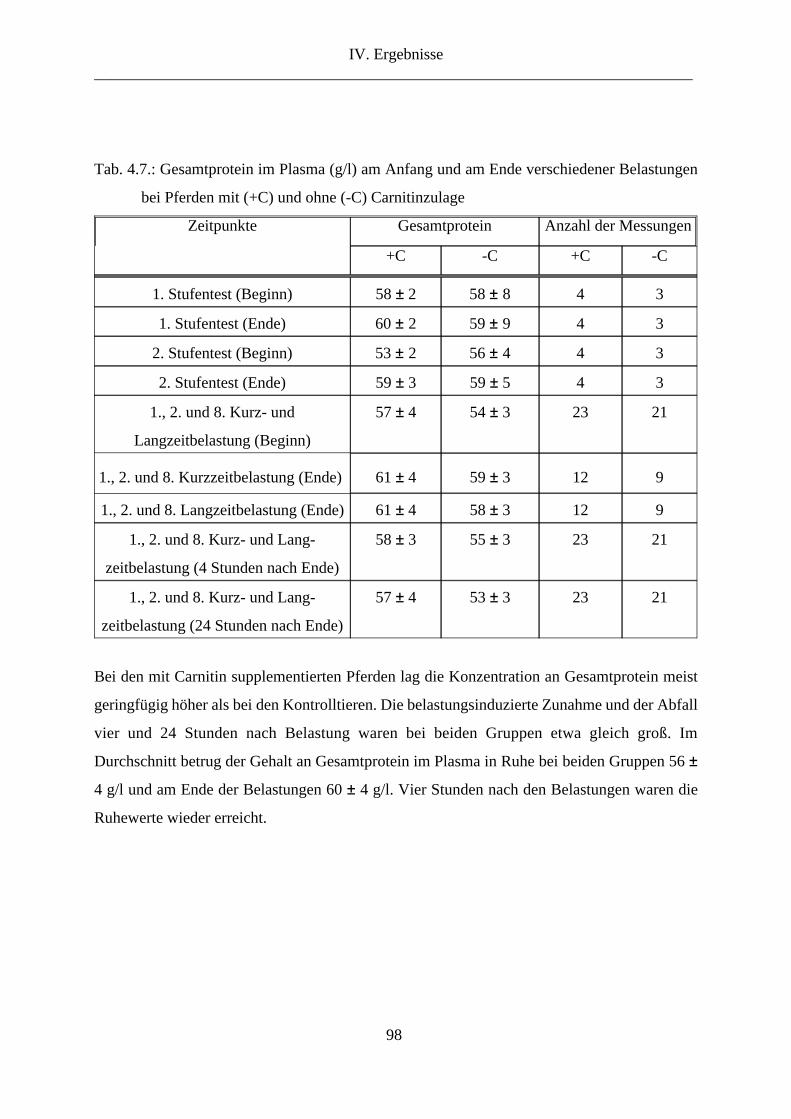
IV. Ergebnisse___________________________________________________________________________
98
Tab. 4.7.: Gesamtprotein im Plasma (g/l) am Anfang und am Ende verschiedener Belastungen
bei Pferden mit (+C) und ohne (-C) Carnitinzulage
Zeitpunkte Gesamtprotein Anzahl der Messungen
+C -C +C -C
1. Stufentest (Beginn) 58 ± 2 58 ± 8 4 3
1. Stufentest (Ende) 60 ± 2 59 ± 9 4 3
2. Stufentest (Beginn) 53 ± 2 56 ± 4 4 3
2. Stufentest (Ende) 59 ± 3 59 ± 5 4 3
1., 2. und 8. Kurz- und
Langzeitbelastung (Beginn)
57 ± 4 54 ± 3 23 21
1., 2. und 8. Kurzzeitbelastung (Ende) 61 ± 4 59 ± 3 12 9
1., 2. und 8. Langzeitbelastung (Ende) 61 ± 4 58 ± 3 12 9
1., 2. und 8. Kurz- und Lang-
zeitbelastung (4 Stunden nach Ende)
58 ± 3 55 ± 3 23 21
1., 2. und 8. Kurz- und Lang-
zeitbelastung (24 Stunden nach Ende)
57 ± 4 53 ± 3 23 21
Bei den mit Carnitin supplementierten Pferden lag die Konzentration an Gesamtprotein meist
geringfügig höher als bei den Kontrolltieren. Die belastungsinduzierte Zunahme und der Abfall
vier und 24 Stunden nach Belastung waren bei beiden Gruppen etwa gleich groß. Im
Durchschnitt betrug der Gehalt an Gesamtprotein im Plasma in Ruhe bei beiden Gruppen 56 ±
4 g/l und am Ende der Belastungen 60 ± 4 g/l. Vier Stunden nach den Belastungen waren die
Ruhewerte wieder erreicht.

V. Diskussion ___________________________________________________________________________
99
V. Diskussion
In der vorliegenden Studie sollte geprüft werden, inwieweit eine orale Carnitinzulage bei
Pferden die körperliche Leistungsfähigkeit in Hinblick auf die Herzfrequenz und den
Energiestoffwechsel beeinflusst. Es war außerdem von Interesse zu prüfen, ob orale
Carnitinzulagen die Carnitinkonzentration in der Muskulatur erhöhen. Aufgrund der
biochemischen Funktionen von Carnitin im Energiestoffwechsel erschien es denkbar, dass
Carnitinzulagen die körperliche Leistung steigern. Von Tauben liegen Ergebnisse über eine
Verbesserung der körperlichen Leistungsfähigkeit durch Verschiebung der muskulären
Energiegewinnung von der anaeroben auf die aerobe Ebene vor (BORGHIJS u. DE WILDE
1992). Auch beim Hund konnte eine Muskelkraftsteigerung durch Carnitinzulagen erzielt
werden (DUBELAAR et al. 1991). Bei Menschen wurde ein positiver Effekt von Carnitin auf
die Leistungsfähigkeit in Form einer Herzfrequenzsenkung und eines niedrigeren
Lactatanstiegs bei gleicher Belastung nachgewiesen (SOOP et al. 1988; DRAGAN et al.
1989; SILIPRANDI et al. 1989; BILLIGMANN et al. 1990; VECCHIET et al. 1990).
Carnitin wird beim Menschen schon als Leistungsförderer eingesetzt. Vom Pferd liegen dazu
nur wenige Untersuchungen vor. Um einen Carnitineffekt zu beurteilen, sind die
Belastungsintensität und die Dauer der Supplementation vermutlich von Bedeutung. Bisher ist
nicht bekannt, ob und bei welcher Belastung sich eine Carnitinzulage positiv auf die
körperliche Leistung bei Pferden auswirkt.
1. Kritik der Versuchseinstellung
1.1. Versuchsdurchführung
In dieser Studie wurden die Tiere durch einen standardisierten Stufentest individuell belastet.
Anhand der während dieses Tests geschwindigkeitsabhängig vorliegenden
Lactatkonzentration im Blut wurde die Höhe der Belastung für die Kurz- und
Langzeitbelastungen berechnet. Außer der Lactatkonzentration stellte die Herzfrequenz eine
weitere Möglichkeit zur Beurteilung der Leistungsfähigkeit der Tiere dar (ERICKSON et al.

V. Diskussion ___________________________________________________________________________
100
1990; EVANS et al. 1993). Allerdings ist die Herzfrequenz außer vom Grad der Belastung
von verschiedenen anderen Faktoren, z. B. von der Psyche der Tiere, abhängig und daher
nicht immer geeignet, die Belastungsintensität objektiv zu beurteilen. Die maximale
Sauerstoffaufnahmekapazität, welche häufig beim Menschen zur Beurteilung der
Leistungsfähigkeit eingesetzt wird, findet auch zunehmend beim Pferd Anwendung (ART u.
LEKEUX 1993; TYLER et al. 1996). Nachteil dieser Methode ist der relativ hohe apparative
Aufwand, der damit verbunden ist. Deshalb schien es bei dieser Untersuchung ratsam, die
Lactatkonzentration zur Beurteilung der Belastungsintensität zu nutzen.
Vor Beginn des Stufentests wurden die Tiere aufgewärmt, da häufig ohne Aufwärmphase zu
Beginn der Belastung ein höherer Lactatspiegel beobachtet wird als im weiteren Verlauf
(MARTIN et al. 1975; TYLER et al. 1996). Dieses Phänomen kommt wahrscheinlich dadurch
zustande, dass der aerobe Stoffwechsel eine gewisse Zeit zur belastungsabhängigen
Energiebereitstellung benötigt (WEBER et al. 1987; LINDNER 1997). Wie der
Konzentrationsverlauf des Lactats während des Stufentests zeigt, reichte die von uns gewählte
Aufwärmphase aus, um einen aussagekräftigen Anstieg des Lactats während der Belastung zu
erzielen.
Gewählt wurde in diesem Versuch eine Dauer der Belastungsstufen von fünf Minuten. Diese
erwies sich als ausreichend, um die Herzfrequenz in jeder Belastungsstufe auf einen
gleichbleibenden Wert einzustellen. Die Herzfrequenz stieg schnell (nach ungefähr 30
Sekunden), entsprechend der Belastung, an (KRYWANEK et al. 1970; LINDHOLM u.
SALTIN 1974).
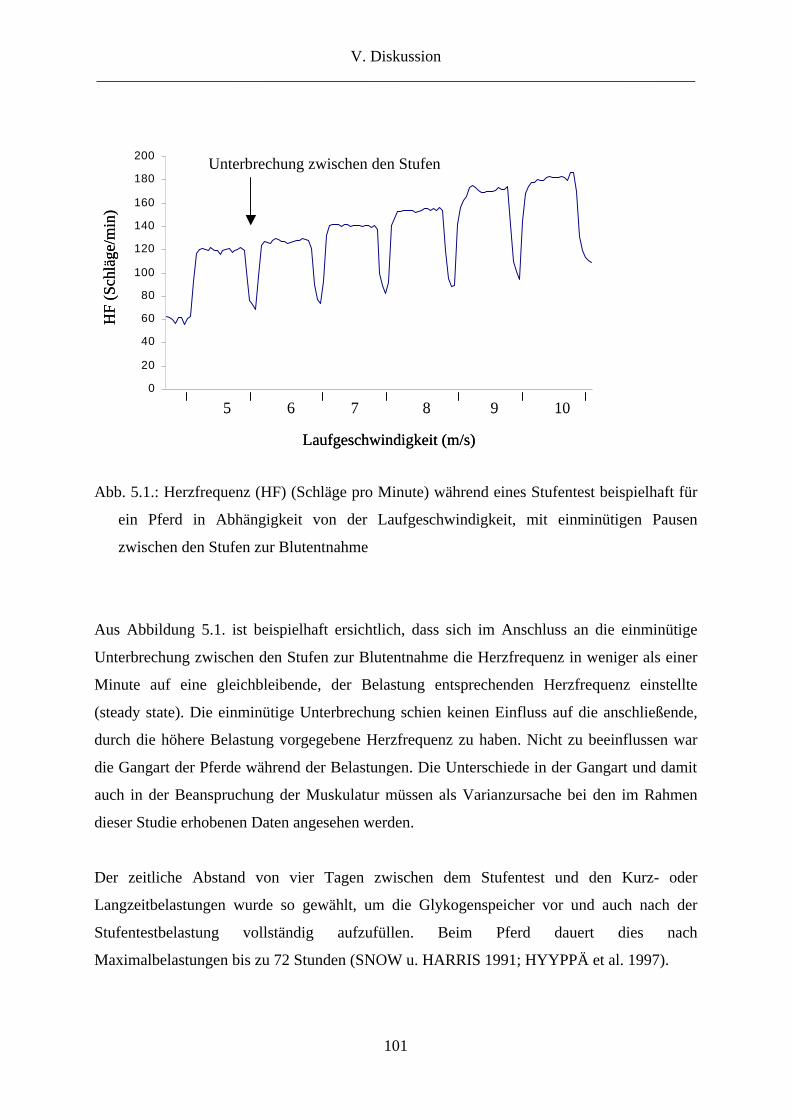
V. Diskussion ___________________________________________________________________________
101
Abb. 5.1.: Herzfrequenz (HF) (Schläge pro Minute) während eines Stufentest beispielhaft für
ein Pferd in Abhängigkeit von der Laufgeschwindigkeit, mit einminütigen Pausen
zwischen den Stufen zur Blutentnahme
Aus Abbildung 5.1. ist beispielhaft ersichtlich, dass sich im Anschluss an die einminütige
Unterbrechung zwischen den Stufen zur Blutentnahme die Herzfrequenz in weniger als einer
Minute auf eine gleichbleibende, der Belastung entsprechenden Herzfrequenz einstellte
(steady state). Die einminütige Unterbrechung schien keinen Einfluss auf die anschließende,
durch die höhere Belastung vorgegebene Herzfrequenz zu haben. Nicht zu beeinflussen war
die Gangart der Pferde während der Belastungen. Die Unterschiede in der Gangart und damit
auch in der Beanspruchung der Muskulatur müssen als Varianzursache bei den im Rahmen
dieser Studie erhobenen Daten angesehen werden.
Der zeitliche Abstand von vier Tagen zwischen dem Stufentest und den Kurz- oder
Langzeitbelastungen wurde so gewählt, um die Glykogenspeicher vor und auch nach der
Stufentestbelastung vollständig aufzufüllen. Beim Pferd dauert dies nach
Maximalbelastungen bis zu 72 Stunden (SNOW u. HARRIS 1991; HYYPPÄ et al. 1997).
0
20
40
60
80
100
120
140
160
180
200
Laufgeschwindigkeit (m/s)
5 6 7 8 9 10
HF
(Sch
läge
/min
)
Unterbrechung zwischen den Stufen
0
20
40
60
80
100
120
140
160
180
200
Laufgeschwindigkeit (m/s)
5 6 7 8 9 10
HF
(Sch
läge
/min
)
Unterbrechung zwischen den Stufen

V. Diskussion ___________________________________________________________________________
102
1.2. Tiergut
Für die Durchführung der Versuche standen acht zweijährige, vom selben Hengst
abstammende und unter gleichen Aufzuchtbedingungen gehaltene Traber zur Verfügung. Da
die Carnitinkonzentration im Plasma altersabhängig ist (FOSTER et al. 1988), ist die
Gleichaltrigkeit der Tiere vorteilhaft. Ein etwaiger Einfluss der Fütterung vor Versuchsbeginn
mit unterschiedlich carnitinhaltigem Futter konnte in diesem Falle ebenfalls ausgeschlossen
werden. Während des Versuchs waren die Fütterungs- und Haltungsbedingungen über den
gesamten Versuchszeitraum für alle Tiere gleich. Da der Abstand zwischen der letzten
Fütterung und der Belastung immer drei Stunden betrug, wurde der Verlauf der Glucose und
der FFS im Plasma dadurch nicht in unkontrollierter Weise beeinflusst. Der hohe
Verwandtschaftsgrad der Tiere ist auch als ein varianzbegrenzender Faktor bei der
Gewinnung der Messdaten anzusehen.
Auffällig und letztlich auch unerklärlich war, dass sich die beiden Pferdegruppen zu Beginn
der Studie, also vor Beginn der Carnitinsupplementation, in ihrem Carnitingehalt im Plasma
und in der Muskulatur bereits geringfügig aber signifikant unterschieden. Der Gehalt an
Carnitin im Plasma lag bei den Tieren, die anschließend die Supplementationsgruppe
bildeten, zu Anfang im Plasma um 16 % und in der Muskulatur um 28 % höher als bei den
Kontrollpferden (Abb. 4.1. und 4.3.). Dieser Unterschied war jedoch, gemessen am
Supplementationseffekt, so klein, dass er keinen Einfluss auf die Beurteilung des
Zulageeffektes hatte. Wahrscheinlich wäre der Effekt der Zulage absolut noch größer
ausgefallen, wenn die Anfangskonzentrationen an Carnitin in der Zulagegruppe auf der Höhe
der Kontrollpferde gelegen hätten. Außer in dem Carnitingehalt unterschieden sich die beiden
Gruppen noch zufallsbedingt in ihrem Gehalt an Muskelglykogen sowie in der Höhe der
Depletion des Glykogens während des ersten Stufentests, was eine Interpretation dieses
Wertes im Trainingsverlauf und nach Carnitingabe erschwert.

V. Diskussion ___________________________________________________________________________
103
2. Diskussion der Ergebnisse
2.1. L-Carnitin im Plasma
-basale Carnitinkonzentrationen
Die während des ersten Stufentests im Plasma gemessenen Konzentrationen an Carnitin
zeigen, dass die Konzentration an Gesamtcarnitin im Laufe der Belastung gering anstieg.
Beim freien Carnitin war bei der Supplementationsgruppe bei hoher Belastung ein
geringfügiger Trend nach unten zu erkennen. Dies deutet darauf hin, dass die Konzentration
der Carnitinester unter diesen Bedingungen geringfügig angestiegen war. Die zu
Trainingsbeginn in beiden Gruppen gemessenen Gehalte an Gesamtcarnitin und auch an
freiem Carnitin lagen in einem Bereich, der auch von anderen Autoren angegeben wird.
FOSTER et al. (1989 a) ermittelten bei Jährlingen einen Gesamtcarnitingehalt im Plasma von
18,3 µmol/l. IBEN et al. (1992) berichteten bei siebenjährigen Pferden von einer
Konzentration an Gesamtcarnitin von 23 µmol/l. In einer Studie von LAMBERTZ (1999)
betrug die Gesamtcarnitinkonzentration im Plasma im Tagesmittel bei drei- bis fünfjährigen
Pferden 25 µmol/l mit Schwankungen zwischen 18 und 40 µmol/l. Diese Daten sprechen
dafür, dass der Konzentrationsunterschied zwischen beiden Gruppen zu Anfang dieses
Versuchs wohl zufallsbedingt ist.
-Effekt der Carnitinzulage
Aus der Literatur (siehe Kapitel II.8) ist bekannt, dass orale Carnitingaben beim Pferd den
Plasmacarnitinspiegel anheben. In der hier vorliegenden Untersuchung erhöhte sich die
Konzentration an Gesamtcarnitin bereits nach vier Tagen Supplemetation mit 10 g Carnitin
täglich auf das 2,4-fache (von 25 auf 59 µmol/l). Dieser hohe Gehalt blieb dann über den
weiteren Versuchszeitraum bestehen. In einer Studie von FOSTER et al. (1989 b) wurde von
einem 1,8-fachen Anstieg von 19 auf 34 µmol/l bei gleich hoher oraler Zulage berichtet. Die
Pferde erhielten auch 10 g Carnitin, jeweils vier bis fünf Stunden vor der Messung.

V. Diskussion ___________________________________________________________________________
104
Allein durch Training (Kontrolltiere) stieg die Konzentration an Gesamtcarnitin im Plasma
tendentiell an. ARENAS et al. (1991) und BORDIN et al. (1992) fanden demgegenüber bei
trainierten und untrainierten Menschen im Plasma keinen Unterschied in der Konzentration an
Gesamtcarnitin. Anders war es in Untersuchungen mit Pferden bei FOSTER et al. (1989 a).
Diese Autoren berichten über eine um ca. 30 % höhere Konzentration an freiem Carnitin im
Plasma bei trainierten drei bis sechs Jahre alten Pferden im Vergleich zu gleichaltrigen
untrainierten Pferden. Ob dieser Unterschied auf Speziesunterschieden beruht, kann zur Zeit
nicht beurteilt werden.
-Einfluss der Belastung auf den Carnitingehalt
In den eigenen Untersuchungen stieg der Gehalt an Gesamtcarnitin im Plasma im Laufe der
Belastungen bei den mit Carnitin supplementierten Tieren in sechs von sieben Fällen um
durchschnittlich 13 % (hochsignifikant) an. Auch bei den Kontrolltieren war wie schon beim
ersten Stufentest eine solche Tendenz erkennbar (Anstieg um 8 %). Möglicherweise beruht
dieser belastungsbedingte Anstieg des Carnitins auf einer vermehrten Durchlässigkeit der
Muskulatur für Carnitin während der Belastung. Auch einige Muskelenzyme gelangen bei
Belastung vermehrt in das Blut (SICILIANO et al. 1997). Der große
Konzentrationsunterschied beim Carnitin zwischen Muskulatur und Plasma (100 bis 500 zu 1)
und die sehr unterschiedlichen Poolgrößen (Muskulatur : Plasma > 99 : 1) führt dazu, dass
schon minimale Mengen Carnitin, wenn sie aus der Muskulatur austreten, zu messbaren
Konzentrationszunahmen im Plasma führen. IBEN et al. (1992) beobachteten auch einen
Anstieg der Konzentration an Gesamtcarnitin im Plasma im Laufe einer siebenminütigen
Trabbelastung bergauf sowohl bei den Pferden, die mit 5 g pro Tag supplementiert wurden
(von 34,4 auf 38,0 µmol/l) als auch bei den Kontrolltieren (von 23,0 auf 24,2 µmol/l).
Ebenso wie das Gesamtcarnitin wurde auch das freie Carnitin im Plasma durch die orale
Zulage angehoben. Das Verhältnis von freiem zu Gesamtcarnitin blieb jedoch unter
Ruhebedingungen über den Versuchszeitraum bei beiden Gruppen annähernd gleich (77 ±
6 % freies Carnitin vom Gesamtcarnitin). In Untersuchungen von FOSTER et al. (1988) und
HARRIS et al. (1995 a) entfielen ein Anteil von 70 bis 80 % des Gesamtcarnitins unter
Ruhebedingungen auf freies Carnitin. Das freie Carnitin im Plasma war im Gegensatz zum

V. Diskussion ___________________________________________________________________________
105
Gesamtcarnitin am Ende der Belastung immer erniedrigt. Dadurch sank bei den
Kontrollpferden und bei den supplementierten Pferden das Verhältnis zwischen freiem und
Gesamtcarnitin im Mittel von 80 % (vor Belastung) auf 60 % (nach Belastung). Auch in einer
Studie von FOSTER und HARRIS (1987 b) fiel das freie Carnitin im Plasma, allerdings erst
während der Erholungsphase nach 10 Minuten. Dies war nach einer nur zweiminütigen
Belastung mit einer Laufgeschwindigkeit von 12 m/s der Fall. Der Konzentrationsabfall an
freiem Carnitin im Plasma am Ende einer Belastung spricht jedoch gegen die Annahme, dass
es durch Belastung infolge einer Schädigung des Sarkolemms zu einem Austritt von Carnitin
aus der Muskulatur in die extrazelluläre Flüssigkeit kommt. Wäre das der Fall, müsste die
Konzentration an freiem Carnitin im Plasma nach Belastung auch zunehmen. Der Befund ist
mit der Annahme vereinbar, dass der arbeitende Muskel verstärkt freies Carnitin aus dem
Plasma aufnimmt und dafür Carnitinester an das Plasma abgibt. Unberücksichtigt bleibt bei
dieser Überlegung die Rolle der Leber, deren Einfluss auf die Carnitinkonzentration im
Plasma während und nach Belastung noch nicht abgeschätzt werden kann.
2.2. L-Carnitin in der Muskulatur
-basale Carnitinkonzentrationen
Die in der eigenen Studie gemessene Gesamtcarnitinkonzentration im Musculus glutaeus
medius in Ruhe vor dem Training und vor Carnitinsupplementation lag bei 14,7 ± 4,3
mmol/kg TM. Dieser Wert liegt an der unteren Grenze des von FOSTER und HARRIS (1992)
für zweijährige Vollblüter angegebenen Bereichs (14,1 – 34,7 mmol/kg TM). In der eigenen
Studie stieg bei den Kontrollpferden das Gesamtcarnitin in der Muskulatur im
Trainingsverlauf vom ersten bis zum zweiten Stufentest um 12 % (nicht signifikant). Für
einen solchen Befund gibt es in der Literatur bisher keine Bestätigung. FOSTER und
HARRIS (1992) fanden nur eine geringere Variationsbreite in der Konzentration von
Gesamtcarnitin bei trainierten zweijährigen Vollblütern im Vergleich zu untrainierten. Es
erscheint aber nicht abwegig zu vermuten, dass der Carnitinbedarf des Muskels durch
Training ansteigt und dass der hier beobachtete Effekt tatsächlich eingetreten ist. Im
Interspeziesvergleich gibt es Beispiele, die zeigen, dass der Gehalt an Carnitin im Muskel
stark abhängig ist von der motorischen Aktivität, der die Tiere ausgesetzt sind (Tab. 5.1.).
V. Diskussion ___________________________________________________________________________
106
Tab. 5.1.: Gehalte an Gesamtcarnitin in der Muskulatur bei verschiedenen Spezies,
vergleichbare Tierarten nebeneinander (GUSTAVSEN 2000)
Bewegungsaktive
Spezies
Gesamtcarnitin
(mg/kg uS)
Bewegungsarme
Spezies
Gesamtcarnitin
(mg/kg uS)
Hase 1200 Kaninchen 244
Wildschwein 460 Hausschwein 240
Hirsch 1930 Hausrind 1430
Wildtaube 211 Huhn 80
Diese Gehalte an Carnitin lassen es plausibel erscheinen, dass auch Training den Gehalt an
Carnitin in der Muskulatur anheben kann.
-Effekt der Carnitinzulage
Die Carnitinsupplementation führte in dieser Studie in fünf Wochen zu einem signifikanten
Anstieg des Muskelcarnitins um 46 %. Bei untrainierten Pferden haben FOSTER et al. (1988)
in einem ähnlichen Versuch mit oralen Carnitinzulagen von 10 bis 60 g pro Tag über acht
Wochen nur einen Anstieg von 3 bis 5 % erzielt. Auch nach einer täglichen intravenösen
Injektion von 10 g Carnitin über 26 Tage war der Muskelcarnitingehalt nicht angestiegen
(HARRIS et al 1995 a). In dieser Studie wurden die Tiere ebenfalls nicht belastet.
Andererseits wurde bei Langstreckenläufern und Sprintern nach einer Supplementation von
1 g Carnitin täglich über sechs Monate bei Fortführung des Trainings ein
Konzentrationsanstieg von 10 % (ARENAS et al. 1991) und bei Ratten (HEINONEN et al.
1992) nach täglicher Supplementation von 500 mg/kg über sechs Wochen und
Schwimmtraining ein Anstieg um 74 % gemessen. Die eigenen Untersuchungen zeigen
eindeutig und zum ersten Mal, dass auch beim Pferd eine tägliche Zulage von 10 g Carnitin
über fünf Wochen in Verbindung mit einem Training den Carnitingehalt in der Muskulatur
deutlich nicht signifikant erhöht. Welche Bedeutung dem Training in diesem Kontext genau
zukommt, kann aus den bisher vorliegenden Daten nicht beurteilt werden.

V. Diskussion ___________________________________________________________________________
107
-Einfluss der Belastung auf den Carnitingehalt
In Ruhe bestand das Muskelcarnitin zu 73 % aus freiem Carnitin und zu 21 % aus
kurzkettigen Carnitinestern. Der Rest waren langkettige Carnitinester (Tab. 4.1.). Ähnliche
Werte wurden auch in einer Studie von FOSTER und HARRIS (1987 b) gemessen. Durch die
Belastung kam es zu sehr deutlichen Verschiebungen im Konzentrationsverhältnis zwischen
freiem Carnitin und den Carnitinestern. Der Anteil an freiem Carnitin sank von 73 % auf
50 % und der der kurzkettigen Carnitinester stieg von 21 auf 43 %. Die Konzentration der
langkettigen Carnitinester unterlag keinen größeren belastungsinduzierten Änderungen. Die
belastungsbedingte Abnahme an freiem Carnitin und die entsprechende Zunahme an
kurzkettigen Carnitinestern war prozentual bei den mit Carnitin supplementierten Pferden und
den Kontrollpferden gleich. Das entsprach absolut jedoch bei den supplementierten Pferden
einer Abnahme von 1262 mg/kg TM und bei den Kontrollpferden von 753 mg/kg TM.
Offenbar war das mehr an Carnitin, das bei den supplementierten Pferden in der Muskulatur
vorhanden war, funktionell voll in den belastungsinduzierten Muskelstoffwechsel
eingebunden.
Diese Untersuchung gibt keine Auskunft darüber, wie sich jeweils in Abhängigkeit von der
jeweiligen Höhe der Belastung das Verhältnis zwischen freiem Carnitin und Carnitinestern
veränderte. HARRIS und FOSTER (1990) berichten, dass schon ein vierminütiger Galopp mit
einer Geschwindigkeit von 6 m/s zu einer Verdopplung der Acetylcarnitinkonzentration in der
Muskulatur führte. Bei höheren Belastungen als sie im Rahmen dieser Studie angewendet
wurden nahm das freie Carnitin im Muskel weiter ab und die Carnitinester stiegen weiter an.
FOSTER und HARRIS (1987 a) stellten bei einer 12-minütigen Belastung von 3,2 bis 10 m/s
mit 5 % iger Steigung fest, dass das freie Carnitin von 88 % auf 20 % des Gesamtcarnitins
abgefallen und dass das Acetylcarnitin in gleichem Maße angestiegen war. CARLIN et al.
(1990) haben berechnet, dass bei Pferden das Konzentrationsverhältnis von Acetylcarnitin zu
Acetyl-CoA in der Muskulatur nach extremer Belastung 350 : 1 beträgt. Die Umwandlung
von freiem Carnitin in Acetylcarnitin in der arbeitenden Muskulatur steht im Einklang mit der
Pufferfunktion von Carnitin, die einer Akkumulation von Acetyl-CoA entgegenwirkt und das
intramitochondriale Verhältnis von Acetly-CoA zu CoA auf einem funktionell
wünschenswert niedrigen Wert hält.

V. Diskussion ___________________________________________________________________________
108
Die Ergebnisse der Tabelle 4.1. zeigten, dass die Konzentration an langkettigen Carnitinestern
bei den mit Carnitin supplementierten Tieren unter Ruhebedingungen gegenüber den
Kontrolltieren zugenommen hatten (141 vs. 345 mg/kg TM) und dass diese bei Belastung
weiter anstiegen (auf 543 mg/kg TM). Bei den Kontrolltieren änderte sich diese
Konzentration dagegen nicht messbar. Eine Belastung stellt für den Muskel immer einen
metabolischen Stress dar, bei dem auch längerkettige verzweigte organische Säuren in
gewissem Umfang anfallen. Es ist daher als günstig zu beurteilen, wenn die Muskulatur
Vorraussetzungen bietet, die eine vorrübergehende Speicherung und Entgiftung dieser
Substanzen ermöglicht und den Muskelstoffwechsel dadurch entlasten (ALLEN et al. 1982;
SCHOLTE u. DE JONGE 1987).
2.3. Herzfrequenz
Bei den Kontrollpferden war in der Herzfrequenz kein Trainingseffekt erkennbar. Weder bei
den Stufentests noch bei den Kurz- und Langzeitbelastungen wies die Schlagfrequenz bei
gleicher absoluter Belastung zwischen Anfang und Ende des fünfwöchigen Trainings einen
Unterschied auf (Abb. 4.5., Tab. 4.2.). Eine Ausnahme bildete die Herzfrequenz am Anfang
der Belastung beim Stufentest, also bei geringer Laufgeschwindigkeit (5 m/s), die tendentiell
nach dem Training etwas geringer war. Aber schon bei der nächsten und allen weiteren
Belastungsstufen war die Herzfrequenz genauso hoch wie zu Beginn des Trainings. Diese
Beobachtung stimmt mit Ergebnissen anderer Untersucher überein. Sie zeigen
übereinstimmend, dass beim Pferd, anders als beim Menschen, die Herzfrequenz kaum mit
einer Reduktion auf Trainingseinflüsse reagiert (SKARDA et al. 1976; ROSE et al. 1983 b;
EVANS u. ROSE 1988; FOREMAN et al. 1990 b). Einige Untersucher haben allerdings eine
geringgradige trainingsbedingte Frequenzabnahme gefunden (GOTTLIEB-VEDI et al. 1995;
YOUNG 1999). Sie war jedoch sehr viel niedriger als beim Menschen.
Bei den mit Carnitin supplementierten Pferden war die Herzfrequenz während des Stufentests
am Ende des Trainings bei fast allen Belastungsstufen und auch bei den Kurz- und
Langzeitbelastungen bei gleicher absoluter Belastung deutlich niedriger als am Anfang des
Trainings. Mit zunehmender Belastung während des Stufentests wurde dieser Effekt immer
kleiner. Über den Belastungsbereich von 5 bis 9 m/s war die Frequenz bei den mit Carnitin

V. Diskussion ___________________________________________________________________________
109
supplementierten Tieren am Ende des Trainings signifikant um 5 % niedriger als vor dem
Training. Auch in Studien mit Menschen wird von einer frequenzsenkenden Wirkung von
Carnitin berichtet (SOOP et al. 1988; BILLIGMANN et al. 1990). Diese könnte mit der seit
langem bekannten vasodilatatorischen Wirkung von Carnitin in einem ursächlichen
Zusammenhang stehen (CIPOLLA et al. 1999) und wurde in Form einer verbesserten
Durchblutung des Herzmuskels direkt nachgewiesen (SUZUKI et al. 1981). Besonders auf
das ischämische Herz hat Carnitin einen positiv inotropen Effekt (CAPONNETTO u.
BRUNELLI 1995). Dieser Effekt ist sowohl in vivo als auch in vitro nachweisbar (SUZUKI
et al. 1981; PAULSON et al. 1984, 1986). Ein entsprechender cardialer Effekt bei Belastung
könnte beim Pferd, zumindest im unteren und mittleren Belastungsbereich, die
Sauerstoffzufuhr begünstigen und die Kontraktilität des Herzmuskels fördern. Folge einer
verstärkten Muskelkraft wäre ein größeres Schlagvolumen mit einer entsprechenden
Reduktion der Frequenz bei gegebener Belastung. Beim Pferd kam dieser Effekt von Carnitin
mit zunehmender Belastung immer weniger zum Tragen und war bei 10 m/s
Laufgeschwindigkeit nicht mehr vorhanden. Der im unteren bis mittleren Belastungsbereich
besonders deutliche Effekt von Carnitin wird weiter dadurch belegt, dass die
Frequenzunterschiede bei den Zulagepferden am Anfang der Kurzzeitbelastung (Tab. 4.2.)
auch deutlich größer waren als am Ende der Belastung. Die Tatsache, dass der Effekt bei
höherer Belastung verschwand, deutet darauf hin, dass die durch Carnitin vermittelten
günstigen Einflüsse auf das Herz bei höheren Belastungen und mit zunehmendem
Sauerstoffmangel nicht mehr wirken und die Schlagfrequenz des Herzens dann offenbar durch
andere regulative Einflüsse dominiert werden. Mit zunehmender Belastung übernehmen durch
den Sympatikus gesteuerte Anpassungsvorgänge mehr und mehr die Steuerung der
Herzfunktion (HARMEYER 2000). Angesichts der relativ geringen Zahl an Versuchstieren,
die für diese Untersuchung zur Verfügung standen, erscheint es jedoch wünschenswert, den
signifikanten Effekt von Carnitin durch weitere Messungen zu ergänzen.
Die Frequenzzunahme des Herzens zwischen Anfang und Ende der Stufentestbelastung
unterscheidet sich zwischen beiden Gruppen nicht. Dieser Umstand stützt zusätzlich die
Vermutung, dass es sich bei der Frequenzabnahme innerhalb derselben Gruppe, d. h. bei
denselben Tieren vor und nach dem Training, um einen Carnitineffekt handelte. Bei der
niedrigeren Frequenz in der Kontrollgruppe am Anfang des zweiten Stufentests bei 5 m/s

V. Diskussion ___________________________________________________________________________
110
Laufgeschwindigkeit im Vergleich zum ersten Stufentest ist nicht klar, ob dies als ein
Zufallsbefund oder als ein Trainingseffekt, der nur bei niedriger Laufgeschwindigkeit auftritt,
angesehen werden muss.
Die gute Übereinstimmung der Herzfrequenzen in Abhängigkeit von der Laufgeschwindigkeit
unabhängig von der Belastungsart (Stufentest, Kurz-, Langzeitbelastung) am Trainingsanfang
und Trainingsende (Abb. 4.6.) verdeutlicht die gute Erfassbarkeit und Aussagekraft dieses
Merkmals bei einer gegebenen Belastung. Bei den hier beschriebenen Versuchen wurden die
Tiere während der körperlichen Arbeit psychisch wenig belastet.
2.4. Lactat
Die niedrigere Blutlactatkonzentration bei gleicher absoluter Belastung bestätigt die Eignung
dieses Parameters beim Pferd, um trainingsbedingte Leistungsfortschritte zu quantifizieren
(BOURGELA et al. 1991; CASINI u. GREPPI 1996; LINDNER 1997; EATON et al. 1999).
Bei den Langzeitbelastungen zeigte das Blutlactat den Trainingsfortschritt nicht, da bei dieser
Belastungsintensität das Blutlactat noch nicht ansprach. Nach Untersuchungen von
LINDNER (1997) eignet sich das Blutlactat erst etwa ab einer Konzentration von 4 mmol/l,
um Trainingseffekte zu erfassen. Der belastungsabhängige Verlauf des Blutlactats vor und
nach dem Training in der hier vorliegenden Untersuchung (Abb. 4.7.) zeigt jedoch, dass der
Trainingseffekt nicht erst ab einer Lactatkonzentration von 4 mmol/l in Erscheinung tritt.
Auch schon unterhalb 4 mmol/l ist ein trainingsabhängiger Unterschied deutlich sichtbar, der
sich ab einer Lactatkonzentration von > 4 mmol/l noch zunehmend weiter vergrößert (bis
etwa 8 mmol/l). Bis zu einer Geschwindigkeit von 7 m/s stieg das Blutlactat annähernd linear.
Danach folgte ein exponentieller Anstieg. Dieser Übergang lag hier, wie in vielen anderen
Untersuchungen (BOURGELA et al. 1991; KRZYWANEK et al. 1996; EATON et al. 1999)
zwischen 2 und 4 mmol/l. Er wird als „anaerobe Schwelle“ bezeichnet (HOLLMANN u.
HETTINGER 1990; SNOW u. VALBERG 1994) und dokumentiert den von dieser Belastung
an exponentiell zunehmenden Anteil an Energie, der durch Glycolyse bereitgestellt wird.
Durch das Training wurde diese Schwelle um eine Belastungsstufe von etwa 1 m/s
Laufgeschwindigkeit angehoben.

V. Diskussion ___________________________________________________________________________
111
Bei den mit Carnitin supplementierten Tieren war die Konzentrationsdifferenz bei gleicher
absoluter Belastung zwischen dem ersten und zweiten Stufentest größer als bei den
Kontrolltieren. Wenn die mit Carnitin supplementierten Pferde nach dem Training bei
gleicher absoluter Belastung genauso viel Glucose umsetzten wie die Kontrolltiere, spräche
dieser Befund dafür, dass bei den supplementierten Tieren ein größerer Teil des Pyruvats
weiter zu Acetyl-CoA umgesetzt und anschließend als Acetylcarnitin gebunden wurde als bei
den Kontrolltieren. Die niedrigeren Plasmaglucosespiegel der supplementierten Tiere (Abb.
4.10.) nach dem Training sprechen dafür, dass diese Pferde nach dem Training bei gleicher
absoluter Belastung mehr Glucose nutzen als die Kontrolltiere (s. u.).
Für die niedrigere Lactatkonzentration des Pferdes Nr. VIII am Ende der achten
Kurzzeitbelastung und des Pferdes Nr. V am Ende der ersten Kurzzeitbelastung im Vergleich
zu einer entsprechend hohen Belastung im Verlauf des Stufentests konnten keine
versuchstechnischen Ursachen erkannt werden. Eine zweite Messung des Lactats in der
gleichen Probe drei Monate später ergab die gleiche Konzentration wie die erste Messung.
Die bei den beiden Tieren zum fraglichen Zeitpunkt gemessenen Herzfrequenzen am Ende
der Kurzzeitbelastungen lagen auf der für diese Laufgeschwindigkeit typischen Höhe, so dass
Zweifel an der Höhe der Belastung zum Messzeitpunkt nicht gegeben sind. Eine sinnvolle
Deutung dieses Phänomens ist daher aus der vorliegenden Datenlage nicht möglich. Bei der
Messung des Blutlactats in Verbindung mit körperlicher Belastung kommen solche
„Ausreißer“ offenbar vor. Ferner nimmt der Blutlactatspiegel im Laufe eines Trainings bei
gleicher absoluter Belastung ab, die Herzfrequenz jedoch nicht bzw. nicht wesentlich. Wenn
es im Rahmen eines Trainingsprogramms z. B. unter Feldbedingungen darum geht, die Höhe
einer bestimmten Belastung genauer festzustellen, ist das bei psychisch unbeeinflussten
Pferden durch Messung der Herzfrequenz wesentlich genauer möglich als durch Messung des
Blutlactats.
2.5. Glucose im Plasma
Während des Stufentests stieg mit zunehmender Laufgeschwindigkeit die
Glucosekonzentration im Plasma an (Abb. 4.10.). Ein solcher belastungsinduzierter
Konzentrationsanstieg ist aus anderen Versuchen mit Pferden bereits bekannt (FREESTONE

V. Diskussion ___________________________________________________________________________
112
et al. 1991; HODGSON u. ROSE 1994; GEOR et al. 2000 a, b). Zu Beginn des Trainings
(erster Stufentest) betrug der Konzentrationsanstieg zwischen Beginn und Ende der Belastung
gut 2 mmol/l. Der belastungsabhängige Konzentrationsanstieg war nicht linear. Bis zu einer
Laufgeschwindigkeit von etwa 7 m/s war der Konzentrationsanstieg mit etwa 0,5 mmol/l sehr
moderat. Danach verstärkte er sich mit zunehmender Belastung stetig. Zwischen den beiden
Tiergruppen, die nach diesem ersten Test unterschiedlich mit Carnitin supplementiert wurden,
bestand im Konzentrationsverlauf zu diesem Zeitpunkt kein signifikanter Unterschied.
Nach Abschluss des vierwöchigen Trainings wies der Verlauf der Plasmaglucose bei absolut
gleicher Laufbelastung wie beim ersten Stufentest jedoch Unterschiede zum ersten Stufentest
auf. Bei den drei nicht mit Carnitin supplementierten Tieren lag die Glucosekonzentration
nach dem Training durchweg während aller Belastungsstufen etwa 0,5 mmol/l höher als
während des ersten Stufentests. Bei den vier mit Carnitin supplementierten Pferden war dieses
trainingsbedingte höhere Konzentrationsniveau nur bis zu einer Laufgeschwindigkeit von 7
m/s zu beobachten. Danach stieg bei diesen Tieren die Glucosekonzentration mit
zunehmender Belastung deutlich weniger stark an als vor dem Training und fiel bereits bei
einer Laufgeschwindigkeit von 7 – 8 m/s wieder auf das niedrigere Konzentrationsniveau des
ersten Stufentests zurück. Der Richtungswechsel im Konzentrationsverlauf der Plasmaglucose
trat bei den mit Carnitin supplementierten Tieren bei einer Belastung von 7 m/s
Laufgeschwindigkeit ein. Bei dieser Belastung zeigte die belastungsinduzierte Zunahme der
Lactatkonzentration im Vollblut deutlich nach oben (Abb. 4.7.).
Über den Effekt eines Trainings auf den glucoseabhängigen Energiestoffwechsel des Muskels
und auf die Glucosekonzentration im Plasma wurden in der Vergangenheit verschiedene
Untersuchungen angestellt. Die meisten dieser Untersuchungen ergaben, dass die
Glucosekonzentration im Plasma nach einem Training stärker anstieg bzw. weniger stark
abfiel als vor dem Training, wenn der Organismus der gleichen absoluten Belastung
ausgesetzt wurde. Solche Befunde liegen von Pferden (SNOW u. MACKENZIE 1977;
HODGSON u. ROSE 1994; GEOR et al. 2000 a) und in größerem Umfang von Ratten
(GALBAO et al. 1977; DOHM et al. 1983; ZENDZIAN-PIOTROWSKA u. GORSKI 1993)
und Menschen (JANSSON u. KAIJSER 1987; RICHTER et al. 1998; BERGMAN et al. 1999

V. Diskussion ___________________________________________________________________________
113
a) vor. Der durch Belastung induzierte Anstieg der Glucosekonzentration des Plasmas fällt im
Interspeziesvergleich beim Pferd jedoch meist größer aus als z. B. bei Mensch und Ratte.
Der Trainingseffekt auf den belastungsinduzierten Glucoseanstieg ist Folge eines
zunehmenden Ungleichgewichts zwischen dem Glucoseeinstrom (glucose appearance rate,
Ra) und dem Glucoseabstrom (rate of glucose disposal, Rd) in den bzw. aus dem
extrazellulären Raum. Ein solches Ungleichgewicht in der Glucoregulation wurde, wenn auch
nicht so ausgeprägt wie beim Pferd, schon vor über 10 Jahren auch beim Menschen
beobachtet und beschrieben (KJAER et al. 1986; COGGAN et al. 1990). Der Hauptgrund für
dieses Ungleichgewicht wird darin gesehen, dass die Muskulatur durch ein mehrwöchiges bis
mehrmonatiges Training bei gleichbleibender Ra mehr und mehr befähigt ist, besonders
während einer die anaerobe Schwelle nicht überschreitenden Belastung, Energie aus der
Oxidation von Fettsäuren zu gewinnen und dabei Glucose einzusparen (HURLEY et al. 1986;
KJAER et al. 1986; BERGMAN et al. 1999 a).
Dieser „Stoffwechselshift“ wird von einer Reihe von Anpassungsvorgängen in der
Muskulatur und in anderen Körperorganen begleitet. Durch Training nimmt bei Ratten
(GALBO et al. 1977; ZENDZIAN-PIOTROWSKA u. GORSKI 1993) und Menschen
(MCCONELL et al. 1994; BERGMAN et al. 1999 b) der Ruheglykogengehalt in der Leber
und in der Muskulatur deutlich zu. Außerdem ist bei Trainierten der Glykogengehalt in der
Muskulatur (GREEN et al. 1991; PHILLIPS et al. 1995) nach einer bestimmten Belastung
noch deutlich höher als bei Untrainierten. Die Menge Glucose, die aus dem Muskelglykogen
bei gleicher Belastung bereitgestellt wird, nimmt durch Training bei Menschen und Ratten ab
(HERMANSSEN et al. 1967; HURLEY et al. 1986; JANSSON u. KAIJSER 1987;
COGGAN et al. 1990, 1993). Auch die Leber setzt beim Menschen nach einer
Trainingsperiode bei gleicher absoluter Belastung wie vor dem Training durch Glykogenolyse
und durch Gluconeogenese weniger Glucose frei als vor dem Training (COGGAN et al. 1995
b; PHILLIPS et al. 1995; BERGMAN et al. 1999 b). Dies ist der Fall, obwohl der hepatische
Blutfluss durch Training um 20 % ansteigt (ROWELL et al. 1986). Der Glucoseumsatz des
Plasmas insgesamt geht beim Menschen durch Training bei gleichbleibender Belastung
zurück (BERGMAN et al. 1999 a).

V. Diskussion ___________________________________________________________________________
114
Die Aufnahme von Glucose aus dem Blut durch den arbeitenden Muskel sinkt beim
Menschen durch Training bei gleicher absoluter Belastung wie vorher ebenfalls ab
(JANSSON u. KAIJSER 1987; RICHTER et al. 1998; BERGMAN et al. 1999 a). Der
belastungsinduzierte Anstieg der Glucosekonzentration im Muskel ist bei Untrainierten höher
als bei Trainierten (RICHTER et al. 1998; BERGMAN et al. 1999 b). Trotz einer
trainingsbedingten Zunahme in der Konzentration der GLUT-4-Transportproteine im
trainierten Muskel (TURCOTTE et al. 1992; MCCONELL et al. 1994; RICHTER et al. 1998)
kommt es zu einem solchen Rückgang bei der Glucoseaufnahme. Nach den oben zitierten
Untersuchungen sind nach einem Training somit beide, der Glucoseeinstrom (Ra) und der
Glucoseabstrom (Rd), vermindert, jeweils bei gleich hoher Belastung wie vor dem Training.
Trotzdem stieg nach einem Training in den oben zitierten Versuchen die
Plasmaglucosekonzentration nach dem Training stärker an als vor dem Training. Daraus
ergibt sich, dass trainingsbedingt der Glucoseabstrom, Rd, weniger stark reduziert wurde als
der Glucoseeinstrom, Ra. Diese Schlussfolgerung stützt sich insbesondere auf Messungen bei
Menschen und Ratten. Pferde zeigen jedoch trainingsbedingt den gleichen Effekt auf die
Plasmaglucose wie Menschen und Ratten (Abb. 4.10.). Insoweit erscheint es gerechtfertigt,
diese Befunde qualitativ auch auf Pferde zu übertragen.
In dem hier beschriebenen Versuch stieg bei den vier mit Carnitin supplementierten Pferden
die Plasmaglucose nach fünfwöchigem Training im oberen Belastungsbereich deutlich
weniger stark an als bei den drei Kontrollpferden (Abb. 4.10.). Diese Beobachtung spricht
zunächst dafür, dass die Carnitinzulage einen Einfluss auf den Kohlenhydratstoffwechsel der
Tiere ausgeübt hat. Der Effekt könnte grundsätzlich auf einer verstärkten Aufnahme von
Glucose in die Muskulatur (oder in andere Gewebe) oder auf einer verminderten
hepatischen Glucosefreisetzung im Vergleich zu den Kontrolltieren beruhen.
Gegen eine erhöhte Glucoseaufnahme in die arbeitende Muskulatur spräche zunächst der bei
diesen Tieren tendentiell niedrigere Anstieg der Lactatkonzentration im Blut im Vergleich zu
den Kontrolltieren (Abb. 4.8.). Andererseits könnte die Muskulatur der Zulagetiere dennoch
mehr Glucose aufgenommen haben als die Kontrolltiere. Dann nämlich, wenn das durch
Glycolyse entstehende Pyruvat nicht in gleichem Umfang zu Lactat reduziert wurde, sondern
unter Vermittlung des PDH-Komplexes oxidativ zu Acetyl-CoA decarboxyliert wurde. Da bei

V. Diskussion ___________________________________________________________________________
115
den mit Carnitin supplementierten Tieren 64 % mehr Carnitin in der Muskulatur zu
Verfügung stand (Abb. 4.3.), war die Pufferkapazität für aktivierte Acetylgruppen bei diesen
Tieren sicherlich erheblich größer als bei den Kontrolltieren.
Die Carnitinzulage könnte somit auch den Glucoseabbau und im Zuge damit die
Glucoseaufnahme in die Muskulatur stimulieren. Durch seine Funktion als Acetylpuffer sinkt
bei einer Carnitinanreicherung in der Muskulatur bei Belastung das intramitochondriale
Acetyl-CoA zu CoA Verhältnis im Vergleich zu den Kontrolltieren. Die relativ höhere
Konzentration an freiem Coenzym A hemmt die PDH-Kinase und stimuliert dadurch den
PDH-Komplex (SUGDEN u. HOLNESS 1994). Dies produziert aus Pyruvat mehr Acetyl-
CoA, dessen Acetyl-Rest durch Carnitin-Acyl-Transferasen an Carnitin gebunden und
abgepuffert werden könnte. Das auf diese Weise intramitochondrial vermehrt anfallende
Acetylcarnitin könnte über bidirektional fungierende Carnitin-Acylcarnitin-Translokasen in
das Cytosol (ABDEL-ALEEM et al. 1996) und von dort in kleinen Mengen durch das
Sarkolemma über bisher nicht bekannte Transportmechanismen auch in den extrazellulären
Raum abgegeben werden. Für diese Vermutung spricht z. B., dass die Konzentration an
Gesamtcarnitin im Plasma am Ende der Belastungen (Abb. 4.1.) signifikant zu- und die
Konzentration an freiem Carnitin (Abb. 4.2.) signifikant abgenommen hatte. Zudem war aber
die Konzentration der Carnitinestern im Plasma am Ende der Belastung bei den
supplementierten Pferden erheblich mehr angestiegen als bei den Kontrolltieren. Es gibt
ferner Hinweise, dass das aus dem Pyruvatabbau stammende Acetyl-CoA leichter für die
Carnitin-Acetyl-Transferase verfügbar ist als das aus der β-Oxidation stammende Acetyl-
CoA (LYSIAK et al. 1986; RUSSELL u. TAEGTMEYER 1992; ABDEL-ALEEM et al
1996).
In isolierten Langendorff- Herzen von Ratten hemmte eine Zugabe von freiem CoA zur
Perfusionslösung die Oxidation von Palmitat (LOPASCHUK et al. 1986; SADDIK et al
1993). Ferner stimulierte eine Zugabe von Carnitin zur Perfusionslösung im Herzen unter den
gleichen Bedingungen die Glucoseoxidation (LOPASCHUK et al. 1994). In anderen
Versuchen mit perfundierten Rattenherzen reduzierte eine Zugabe von Palmitat die
Glykolyserate des Herzens um ein Drittel. Wurde der Perfusionslösung jedoch zusätzlich zum

V. Diskussion ___________________________________________________________________________
116
Palmitat noch L-Carnitin zugesetzt, blieb die Hemmung der Glykolyserate aus (BRODERIK
et al. 1992).
Ein Blick auf die Redoxsituation der Zelle bei hoher (anaerober) Belastung spricht ebenfalls
dafür, dass das Muskelcarnitin eher dafür genutzt wird, das aus dem Kohlenhydratabbau
stammende Acetyl-CoA, als das aus der β-Oxidation der Fettsäuren anfallende Acetyl-CoA
zu transacetylieren. Ein β-Oxidationscyclus belastet die Zelle mit vier
Reduktionsäquivalenten (1 FADH2 und 1 NADH + H+). Dies bringt, wenn die reduzierten
Coenzyme aufgrund des bestehenden Sauerstoffmangels nicht mehr oxidiert werden können,
keinen Gewinn an ATP. Die Bildung eines Moleküls Acetyl-CoA aus Pyruvat belastet die
Zelle nur mit zwei Reduktionsäquivalenten (NADH + H+). Die dadurch erreichte Absenkung
der Pyruvatkonzentration ermöglicht aber den Fortgang der Glykolyse mit einem daraus
resultierenden Gewinn an ATP. Diese Überlegung steht im Einklang mit einer seit langem
bekannten Beobachtung am ischämischen Herzen. Dort bewirkt die Zufuhr von Carnitin ohne
Änderung anderer Bedingungen eine signifikante Verbesserung der Kontraktionskraft
(LOPASCHUK et al. 1994; DE JONG u. FERRARI 1995).
Bezogen auf die Befunde dieser Studie bei Pferden, ist diese Betrachtung zunächst reine
Spekulation. Die am Herzmuskel erhobenen Befunde lassen sich nicht ohne weiteres auf den
Skelettmuskel übertragen. Die Energieversorgung mit verschiedenen Substraten und die der
Energiebereitstellung zugrunde liegenden, regulativen Mechanismen sind zwischen Herz- und
Skelettmuskel sehr verschieden. Der geringere Anstieg der Plasmaglucose bei den
supplementierten Pferden im Vergleich zu den Kontrollpferden ließe sich jedoch auf der
Grundlage dieser experimentell belegten Beobachtungen sinnvoll deuten.
Die zweite (theoretische) Möglichkeit, die als Erklärung für den der verminderten
Glucoseanstieg im Plasma bei den mit Carnitin supplementierten Pferden nach dem Training
bei hoher Belastung dienen könnte, ist die Annahme eines verminderten hepatischen
Glucoseeinstroms, Ra. Über einen solchen Effekt von Carnitin auf die Leber ist bisher jedoch
nichts bekannt. Die wenigen Untersuchungen zu diesem Thema sprechen eher dafür, dass eine
Carnitinzulage die hepatische Gluconeogeneserate nicht negativ beeinflusst. In isolierten

V. Diskussion ___________________________________________________________________________
117
Rattenhepatozyten wurde die Glucoseproduktion durch Zugabe von Propionat gehemmt.
Durch gleichzeitige Zugabe von L-Carnitin zur Inkubationslösung wurde dieser hemmende
Effekt von Propionat auf die hepatische Glucoseproduktion wieder aufgehoben (BRASS u.
BEYERINCK 1987). Bekannt ist, dass Carnitingaben in vitro (MCGARRY u. FOSTER
1979) und vermutlich auch in vivo (BREMER u. OSMONDSEN 1984) in der Leber die
Oxidation FFS stimulieren. Der Glucosestoffwechsel insgesamt könnte durch diese Effekte
nur insoweit betroffen werden, als durch die erhöhte Fettsäurenoxidation in der Leber weniger
Energie aus Glucose für den Leberstoffwechsel bereitgestellt werden muss. Die bisherigen
Untersuchungen über den Effekt von Carnitin auf den Glucosestoffwechsel im arbeitenden
Muskel und in der Leber sprechen jedoch dafür, dass der weniger steile belastungsinduzierte
Anstieg der Glucosekonzentration im Plasma bei den mit Carnitin supplementierten Pferden
auf einem stärkeren Glucoseabstrom, Rd, in die Muskulatur und nicht auf einer verminderte
Glucosefreisetzung aus der Leber beruht.
2.6. Glykogen in der Muskulatur
Eine zentrale Rolle im Hinblick auf die Energiebereitstellung während der Belastung besitzen
die intramuskulären Glykogenspeicher. Der durchschnittliche Glykogengehalt in der
ruhenden Muskulatur wird beim Pferd mit 550-600 mmol Glucose/kg TM angegeben (SNOW
et al. 1985; SNOW u. HARRIS 1991). Bei der eigenen Untersuchung wurde vor Beginn der
Belastungen (erster und zweiter Stufentest) durchschnittlich eine tendentiell niedrigere
Muskelglykogenkonzentration von 414 mmol Glucose/kg TM gemessen. Die Zulagetiere
hatten im Vergleich zu den Kontrolltieren schon vor der Supplementation eine signifikant
höhere Konzentration an Glykogen in der Muskulatur. Dieser Unterschied zwischen den
Gruppen erschwert die Erfassung eines eventuellen Carnitineffektes.
In der eigenen Untersuchung änderte sich die Ruheglykogenkonzentration in der Muskulatur
im Trainingsverlauf weder bei den Kontrolltieren noch bei den supplementierten Tieren.
SNOW und HARRIS (1991) beobachteten während einer 14-tägigen Trainingsperiode bei
Galoppern, dass nach intensiven Kurzzeitbelastungen von 1000 bzw. 1600 m Sprints
innerhalb von drei Tagen die Glykogenspeicher auf immer höhere Ruhewerte eingestellt
wurden. Im Gegensatz dazu beobachteten ESSEN-GUSTAVSSON et al. (1989) ein Absinken

V. Diskussion ___________________________________________________________________________
118
um 10 bis 15 % nach ein bis zwei Wochen intensivem Training täglich auf einem
Hochgeschwindigkeitlaufband. Bei dem überwiegenden Teil der zu dieser Frage
durchgeführten Untersuchungen konnte der Glykogengehalt durch Training nicht beeinflusst
werden (GUY u. SNOW 1977; WILSON et al. 1987; FOREMAN et al. 1990 a). Diese
scheinbar widersprüchlichen Ergebnisse könnten auf der unterschiedlichen Art des Trainings
beruhen. Denkbar ist hingegen auch, dass, je nach Art und Dauer eines Trainings,
unterschiedliche Anteile der Muskulatur beansprucht und stimuliert werden (KISSENBECK
1999). Ferner besteht die Möglichkeit, dass bei der Biopsie nicht immer die gleichen Anteile
des Muskels zugänglich sind. Diese Varianzursachen sind in die Betrachtung der
divergierenden Befunde über den trainingsabhängigen Ruheglykogengehalt mit
einzubeziehen. Für die richtige Erfassung des Ruheglykogengehaltes muss natürlich beachtet
werden, dass nach einer Belastung bis zu 72 Stunden verstreichen, bevor das Muskelglykogen
seinen Ruhewert wieder erreicht hat.
Während der Stufentests (intensive kurze Belastungen) bei der eigenen Untersuchung sank
die Glykogenkonzentration in der Muskulatur im Mittel um 38 %. NIMMO und SNOW
(1983) und HARRIS et al. (1987) ermittelten nach intensiven Kurzzeitbelastungen einen
Abfall der Glykogenkonzentration von 20 – 35 %. Allerdings fiel der Glykogengehalt bei den
Zulagetieren im ersten Stufentest ausgehend von dem höheren Niveau in Ruhe um 56 % ab,
bei den Kontrolltieren nur um 29 %. Beide Gruppen erreichten nach der Belastung einen
vergleichbaren Depletionsgrad. Naheliegend wäre, dass bei hohen Glykogenausgangswerten
mehr Glykogen mobilisiert würde als bei niedrigeren Ausgangswerten. Die eigenen Befunde
schließen einen solchen Zusammenhang nicht unbedingt aus (Abb. 5.2.).
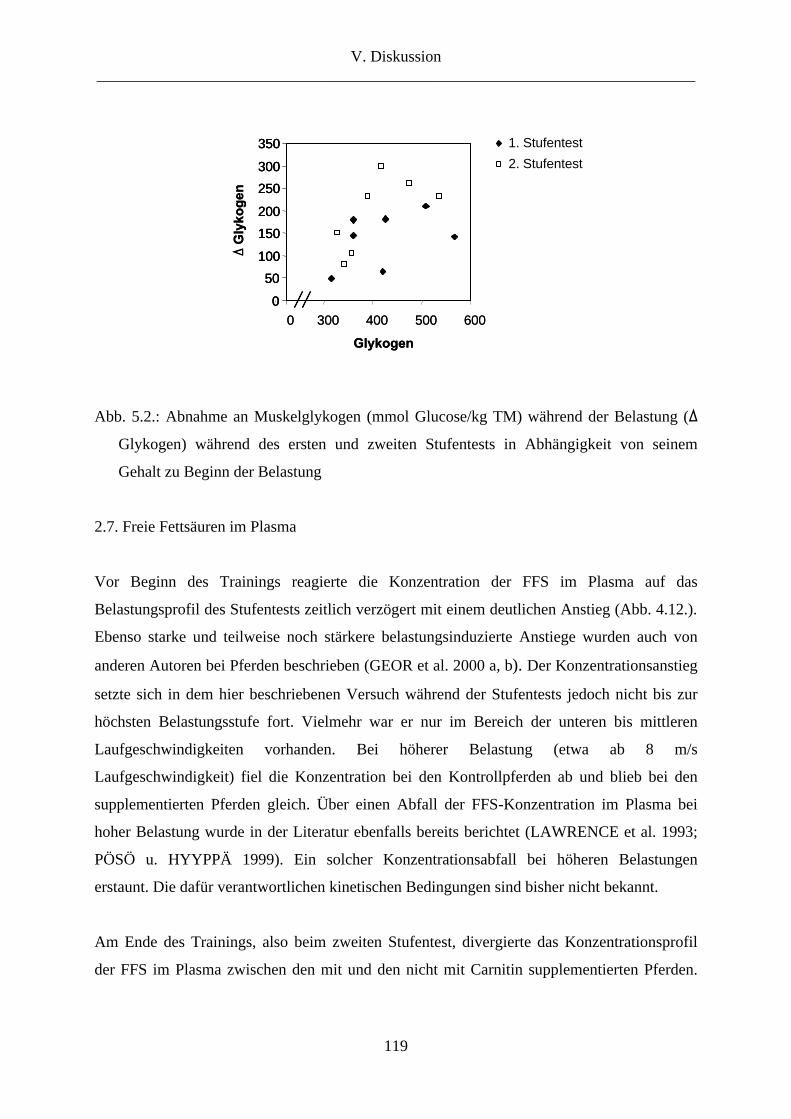
V. Diskussion ___________________________________________________________________________
119
Abb. 5.2.: Abnahme an Muskelglykogen (mmol Glucose/kg TM) während der Belastung (d
Glykogen) während des ersten und zweiten Stufentests in Abhängigkeit von seinem
Gehalt zu Beginn der Belastung
2.7. Freie Fettsäuren im Plasma
Vor Beginn des Trainings reagierte die Konzentration der FFS im Plasma auf das
Belastungsprofil des Stufentests zeitlich verzögert mit einem deutlichen Anstieg (Abb. 4.12.).
Ebenso starke und teilweise noch stärkere belastungsinduzierte Anstiege wurden auch von
anderen Autoren bei Pferden beschrieben (GEOR et al. 2000 a, b). Der Konzentrationsanstieg
setzte sich in dem hier beschriebenen Versuch während der Stufentests jedoch nicht bis zur
höchsten Belastungsstufe fort. Vielmehr war er nur im Bereich der unteren bis mittleren
Laufgeschwindigkeiten vorhanden. Bei höherer Belastung (etwa ab 8 m/s
Laufgeschwindigkeit) fiel die Konzentration bei den Kontrollpferden ab und blieb bei den
supplementierten Pferden gleich. Über einen Abfall der FFS-Konzentration im Plasma bei
hoher Belastung wurde in der Literatur ebenfalls bereits berichtet (LAWRENCE et al. 1993;
PÖSÖ u. HYYPPÄ 1999). Ein solcher Konzentrationsabfall bei höheren Belastungen
erstaunt. Die dafür verantwortlichen kinetischen Bedingungen sind bisher nicht bekannt.
Am Ende des Trainings, also beim zweiten Stufentest, divergierte das Konzentrationsprofil
der FFS im Plasma zwischen den mit und den nicht mit Carnitin supplementierten Pferden.
0
50
100
150
200
250
300
350
0 300 400 500 600
Glykogen
dG
lyko
gen
1. Stufentest
2. Stufentest
0
50
100
150
200
250
300
350
0
50
100
150
200
250
300
350
0 300 400 500 600
Glykogen
dG
lyko
gen
1. Stufentest
2. Stufentest

V. Diskussion ___________________________________________________________________________
120
Bei den drei Kontrollpferden lag die Konzentration nur im unteren Belastungsbereich (bis
etwa 7 m/s Laufgeschwindigkeit) deutlich unter der Konzentration zum Trainingsbeginn. Aus
Versuchen von Menschen ist bekannt, dass der Anstieg der Konzentration der FFS im Plasma
nach einem Training bei gleicher absoluter Belastung wie vor dem Training signifikant um 20
bis 40 % geringer ausfällt (PHILLIPS et al. 1995). Training begünstigt die Verbrennung von
Fettsäuren und führt zur Einsparung bei der Glucose. Dieser Effekt ist bei den
Kontrollpferden am Ende des Trainings aber nur bei niedriger Laufgeschwindigkeit zu
beobachten. Bei mittlerer Laufgeschwindigkeit (von 6 – 8 m/s) stieg bei diesen Tieren die
FFS-Konzentration nach dem Training sogar stärker an als vor dem Training (Abb. 4.12.).
Bei den mit Carnitin supplementierten Pferden war der belastungsabhängige Verlauf der FFS-
Konzentration im Plasma am Anfang und am Ende des Trainings seiner Form nach etwa
gleich. Am Ende des Trainings lag die Konzentration während des Stufentests auf einem
signifikant niedrigeren Niveau als bei den Kontrolltieren. Mit zunehmender Belastung stieg
die Konzentration zwar an, der Anstieg war aber deutlich geringer als bei den
Kontrollpferden. So blieb die Konzentration während des gesamten Tests bis zum Ende
signifikant niedriger als am Anfang des Trainings. Das Konzentrationsniveau der FFS nach
Supplementation mit Carnitin unterschied sich somit signifikant von dem der Kontrollpferde.
Dieses Ergebnis überrascht zunächst. Hätte man doch bei diesen Tieren – angesichts des
geringeren Anstiegs der Plasmaglucose am Ende des Trainings (Abb. 4.10.) und des
vermuteten höheren Glucoseverbrauchs - erwartet, dass die Konzentration der FFS im Plasma
höher liegen würde als bei den Kontrollpferden. Ein höherer Glucoseverbrauch sollte die
Aufnahme FFS entsprechend vermindern (KATZ et al. 1991) und - eine gleiche Lipolyserate
wie vor dem Training vorausgesetzt - den belastungsinduzierten Konzentrationsanstieg
entsprechend vergrößern. Diese Vermutung bestätigte sich jedoch nicht.
Versuche mit Menschen (HURLEY et al. 1986; PHILLIPS et al. 1995) und Ratten (RENNIE
et al. 1976; GÓRSKI et al. 1988) haben ergeben, dass die Konzentration der FFS im Plasma
mit zunehmender Belastung ansteigt. Dieser Befund wird durch die hier vorgelegten Daten
mit Pferden grundsätzlich bestätigt. Der niedrigere basale Spiegel der FFS nach dem Training
im Vergleich zum Trainingsbeginn bleibt bei gleicher absoluter Belastung zunächst in beiden

V. Diskussion ___________________________________________________________________________
121
Pferdegruppen niedrig. Dieser Befund steht im Einklang mit der Vorstellung einer höheren
Fettsäurenoxidationsrate im trainierten Muskel im Vergleich zum untrainierten. Er ist auch
aus Untersuchungen vom Menschen bekannt (KOIVISTO et al. 1982; HURLEY et al. 1986).
In einer neueren Untersuchung mit 14 bis 27-jährigen ausdauertrainierten jungen Männern lag
die Konzentration an FFS im Plasma dagegen bei einer relativ hohen Belastung von 75 –
80 % VO2max nach dem Training signifikant höher als vor dem Training (COGGAN et al.
2000). Aufgrund dieser Beobachtung ist es daher nicht überraschend, dass der
trainingsinduzierte niedrigere FFS-Spiegel mit zunehmend höherer Belastung verschwindet.
Bei einer Deutung dieser trainings- und belastungsabhängigen Konzentrationsschwankungen
müssen verschiedene Prozesse berücksichtigt werden. So lassen
Konzentrationsschwankungen der FFS im Plasma nur in sehr beschränktem Umfang
Rückschlüsse auf die von der arbeitenden Muskulatur genutzten Mengen an FFS zu. Nur ein
relativ kleiner Teil der zur Energieversorgung im Muskel vorgesehenen FFS wird in Form
FFS direkt aus dem Plasma aufgenommen. Der größere Teil stammt aus der
Lipoproteinfraktion des Plasmas. Die Fettsäuren der Lipoproteine werden durch die Aktivität
der Lipoprotein-Lipase erst im Muskel frei und an Ort und Stelle direkt aus dem Plasma
aufgenommen. Dies führt nicht zu einer Reduktion der FFS-Konzentration im Plasma. Die
Aufnahme der Fettsäuren durch die Muskulatur ist eng gekoppelt an die Lipoprotein-
Lipaseaktivität in den Muskelkapillaren, die durch Training zunimmt (HUTTUNEN 1982;
ROMIJN et al. 1993). Durch diesen trainings- und belastungsabhängigen Einstrom von
Fettsäuren aus den Lipoproteinen in die Muskulatur wird die Konzentration von FFS im
Plasma nicht betroffen. Sie wird unabhängig davon durch deren Zu- und Abstrom geregelt. In
diesem Zusammenhang ist es von Interesse, dass der Einstrom von FFS in das Plasma beim
Menschen bei gleicher absoluter Belastung durch Training zurückgeht (MARTIN et al. 1994).
Bei den mit Carnitin supplementierten Pferden lag die Konzentration der FFS am Ende des
Trainings basal und während der unteren Belastungsstufen tendentiell noch niedriger als bei
den Kontrolltieren. Andererseits überrascht nicht, dass eine alimentäre Zufuhr von Carnitin
die Konzentration der FFS und der Triglyceride im Plasma senkt (FROHLICH et al. 1978;
AHMAD et al. 1989; BENEVENGA et al. 1989). Der Effekt beruht zum Teil auf einer
Stimulation der basalen Fettsäurenoxidation, vor allem in Leber, Herzmuskel und vermutlich

V. Diskussion ___________________________________________________________________________
122
in der Skelettmuskulatur (FROHLICH et al. 1978; REGITZ et al. 1982). Aufgrund dieses
Effektes weisen Carnitinzulagen bei hyperlipidämischen Erkrankungen einen Plasmalipid und
Fettsäuren senkenden Effekt auf (POLA et al. 1980; GLÖGGLER et al. 1989). Bei den mit
Carnitin supplementierten Pferden war die Konzentration der FFS im Plasma während des
zweiten Stufentests, also am Ende des Trainings, bei allen Belastungsstufen signifikant
niedriger als bei während des ersten Stufentests. Zur Beurteilung dieses Unterschieds muss
auch die sehr viel größere Abnahme an Muskelglykogen bei diesen Tieren während des ersten
Stufentests im Vergleich zum zweiten Stufentest in Betracht gezogen werden. Falls der
höhere Verbrauch an Muskelglykogen während des ersten Stufentests eine geringere
Entnahme von FFS aus dem Plasma nach sich zieht, könnte das den etwas stärkeren Anstieg
der FFS im ersten Stufentest am Ende der Belastung im Vergleich zum zweiten Stufentest
ebenfalls erklären.
Mit zunehmender körperlicher Belastung während des zweiten Stufentests war in dieser
Studie der Konzentrationsanstieg an FFS im Plasma bei den mit Carnitin supplementierten
Pferden jedoch deutlich moderater als bei den Kontrollpferden, so dass der
Konzentrationsabstand zwischen Trainingsanfang und –ende während des gesamten Tests
erhalten blieb. Bei den Kontrolltieren wurde diese Schere dagegen langsam geschlossen (Abb.
4.12.).
Theoretisch kommen für den geringeren Anstieg der FFS bei den Zulagetieren, wie schon bei
der Plasmaglucose dargelegt, zwei Möglichkeiten in Betracht. Entweder wurde durch
Training, jeweils im Vergleich mit den Kontrolltieren, die Lipolyserate und dadurch der
Einstrom FFS reduziert oder der Abstrom aus dem Plasma wurde stimuliert.
1. Möglichkeit
Zur Zeit sind keine Untersuchungen bekannt, die auf eine durch Carnitin vermittelte
Hemmung der Lipolyserate und auf eine Reduktion eines Einstroms FFS in das Plasma
schließen ließen.

V. Diskussion ___________________________________________________________________________
123
2. Möglichkeit
Für die Vermutung, dass durch eine Carnitinzulage nach einem Training der Abstrom FFS aus
dem Plasma stimuliert werden kann, sprechen drei Beobachtungen.
1) Seit längerem ist bekannt, dass Carnitin die basale Fettsäurenoxidation in vielen
Körpergeweben stimuliert (FROHLICH et al. 1978; REGITZ et al. 1982). Somit ist denkbar,
dass die niedrigere Fettsäurenoxidation nach dem Training im arbeitenden Muskel bei den
Zulagetieren durch eine höhere Fettsäurenoxidationsrate in anderen Geweben einschließlich
der Leber kompensiert wurde. Beim Menschen entfallen bei hoher körperlicher Belastung
etwa 95 % des Fettsäurenumsatzes des Plasmas auf die arbeitende Muskulatur. Bei
Untrainierten sind es aber nur 65 % (JANSSON u. KAIJSER 1987). Im Rahmen dieses
Regulationsspielraums könnte eine Carnitinzulage die Relation des Fettsäurenabstroms aus
dem Plasma in die arbeitende Muskulatur und in andere Gewebe beeinflussen (Abb. 5.2.a).
Eine Reihe von Arbeiten bei verschiedenen Spezies belegen ferner, dass die Aufnahmerate
von FFS in die arbeitende Muskulatur eng und positiv mit deren Konzentration im Plasma
korreliert (HAGENFELDT 1975; HURLEY et al. 1986). Dies würde dafür sprechen, dass die
Arbeitsmuskulatur der mit Carnitin supplementierten Tiere weniger FFS aus dem Plasma
aufnimmt als die der Kontrolltiere.
2) Als zweites muss berücksichtigt werden, dass von den im arbeitenden Muskel oxidierten
Fettsäuren nur ein kleiner Teil aus dem FFS-Pool des Plasmas entnommen wird. Der weitaus
größere Teil rekrutiert sich aus den Lipoproteinen des Plasmas, insbesondere aus den
Lipoproteinen der HDL-Fraktion (LOPASCHUK et al. 1994). Die Konzentration der HDL-
Fraktion und die der Triglyceride im Plasma nimmt beim Menschen bei Belastung und durch
Training sowohl in Ruhe als auch bei Belastung um bis zu 30 % zu (HUTTUNEN 1982;
THOMPSON et al. 1991; ROMIJN et al. 1993; PODL et al. 1994). Die in den HDL
enthaltenen Fettsäuren werden erst durch die endotheliale Lipoprotein-Lipase frei (ROMIJN
et al. 1993) und gelangen durch das Kapillarendothel bzw. durch die Zwischenzellräume aber
auf direktem Wege in die Muskelzelle (EISENBERG 1984). Dieser Fettsäurenflux beeinflusst
die Konzentration der FFS im Plasma somit nicht. Es ist denkbar, dass die bei den
Zulagetieren vermutete, geringere Fettsäurenoxidationsrate im arbeitenden Muskel sich in

V. Diskussion ___________________________________________________________________________
124
erster Linie in einer Reduktion von FFS aus der Lipoprotein-Fraktion des Plasmas
niederschlägt und dass umgekehrt FFS aus dem Plasma sogar mehr entnommen wurden
(ROSSI u. SILIPRANDI 1982) (Abb. 5.2.b).
3) Schließlich kann noch ein dritter Aspekt in Betracht gezogen werden. Verschiedene
Untersuchungen haben ferner gezeigt, dass der Anteil der aus dem Plasma in die Muskulatur
(Herz- und Skelettmuskel) und in andere Gewebe (Leber- und Fettgewebe) aufgenommenen
FFS nur zu einem geringen Prozentsatz (ca. 30 %) direkt für die oxidative Energiegewinnung
genutzt wird (WOLFE et al 1990). Der größere Anteil wird im Gewebe wieder zu
Triglyceriden aufgebaut (Triglycerid-Fettsäuren-Cyclus) (KLEIN et al. 1989; WOLFE et al
1990). Eine solche Lipogenese findet auch im arbeitenden Muskel statt. Die Konzentration
der FFS im Plasma wird indirekt auch von der Rate dieses Fettsäuren-Recyclings beeinflusst.
Untersuchungen beim Menschen zu dieser Frage haben gezeigt, dass der Plasmaspiegel der
FFS sinkt, wenn die Reesterifizierungsrate zunimmt (ROMIJN et al. 1993). Bei Ratten
verdoppelt sich z. B. bei hoher körperlicher Belastung der Fettgehalt der Leber (GÓRSKI et al
1988). Auch dieser Mechanismus könnte durch eine Zulage von Carnitin modifiziert werden
und dadurch Einfluss auf die Konzentration der FFS im Plasma nehmen (Abb. 5.2.c). Der
Anteil, der über Triglyceride rezirkulierenden Fettsäuren kann hormonell kurzfristig, z. B. bei
Belastungen, stark geändert werden (WOLFE et al 1990). Auf diese Weise kann der
arbeitende Muskel die für eine Belastung notwendige Menge an Fettsäuren rasch verfügbar
machen (WOLFE et al 1990; ROMIJN et al. 1993). Die Reesterifizierungsrate der FFS des
Plasmas nimmt beim Menschen durch Ausdauertraining bereits unter Ruhebedingungen um
das zwei- bis vierfache gegenüber untrainierten zu (ROMIJN et al. 1993). Vieles spricht
dafür, dass die niedrigere basale Konzentration an FFS im Plasma nach dem Training auch
bei den Pferden mit auf einem ähnlichen Effekt beruht.
Aus dieser Erörterung wird klar, dass der größere Teil an Energie, der bei Trainierten im
Vergleich zu Untrainierten aus der Fettsäurenoxidation stammt, überwiegend durch eine
verstärkte Lipolyserate von Muskelfett und durch einen erhöhten Einstrom von Fettsäuren aus
Lipoproteinen bereitgestellt wird (HURLEY et al. 1986) und somit nicht direkt aus dem FFS-
Pool des Plasmas stammt. Es ist daher naheliegend zu vermuten, dass der unmittelbar aus den
FFS des Plasmas stammende Anteil an Energie, der für die Muskelarbeit aufgebracht wurde,

V. Diskussion ___________________________________________________________________________
125
bei den mit Carnitin supplementierten Tieren höher lag, der aus Muskelfett und Lipoproteinen
stammende Anteil aber niedriger lag als bei den Kontrolltieren. Langkettige aktivierte
Fettsäuren, wie Palmitoyl-CoA, hemmen im Herzmuskel die Lipoproteinlipase
(MCDONOUGH u. NEELY 1988). Ferner hemmen hohe Konzentration an Acyl-Carnitin im
Herzmuskel von Ratten die Lipolyseaktivität (HULSMANN et al. 1982). Falls solche Effekte
von Carnitin auch in der Skelettmuskulatur auftreten, würde diese die Aufnahme von
Fettsäuren in die Zellen nicht vermindern, die β-Oxidation der Fettsäuren aber nicht
stimulieren. GÓRSKI (1990) weist in einer Übersichtsarbeit über den Triglyceridstoffwechsel
bei Belastung darauf hin, dass während einer Belastung neben ausgeprägten tierartlichen
Unterschieden keine Korrelation besteht zwischen der Rate des Triglyceridabbaus im Muskel
und der Konzentration der FFS im Plasma. Dieser Befund lässt Raum für den Einfluss von
Carnitin auf den Energiestoffwechsel des Muskels und auf die Rekrutierung der Energie aus
verschiedenen Kohlenhydrat- und Fettquellen. Es erschient somit nicht von vornherein
widersprüchlich, dass bei den Zulagetieren nach dem Training sowohl mehr Glucose als auch
mehr FFS aus dem Plasma aufgenommen wurden. Bei diesen Tieren könnten mehr FFS in
nicht aktive Gewebe geflossen sein. Es könnten aber auch mehr FFS von der arbeitenden
Muskulatur aufgenommen worden sein, wenn bei diesen Tieren die Reesterifizierungsrate
höher und der Lipoprotein-Lipase vermittelte Einstrom niedriger war (Abb. 5.2.). Diese
Effekte würden in gewisser Weise eine Parallele darstellen zu der Zunahme der
Glykogenspeicher in der Muskulatur nach einem Training sowohl vor Belastung wie auch
nach einer Belastung (PHILLIPS et al. 1995; BERGMAN et al. 1999 b). Ein solcher Effekt
stünde auch nicht im Widerspruch zu der Beobachtung, dass der Gehalt an Gesamtkörperfett
durch Training generell reduziert wird (PELS et al 1985). Der Fettgehalt im Musculus
glutaeus medius wurde in dieser Studie nicht bestimmt. Es erscheint rückblickend auch nicht
gesichert, ob der hier diskutierte Effekt von Carnitin durch eine Messung des
Muskelfettgehaltes ausreichend genau hätte erfasst werden können.
Die drei möglichen Mechanismen, die nach Supplementation von Carnitin zu einer höheren
Entnahme von FFS aus dem Plasma bei gleichzeitiger Reduktion der Fettsäurenoxidationsrate
führen können, sind in Abbildung 5.3. schematisch zusammengefasst.
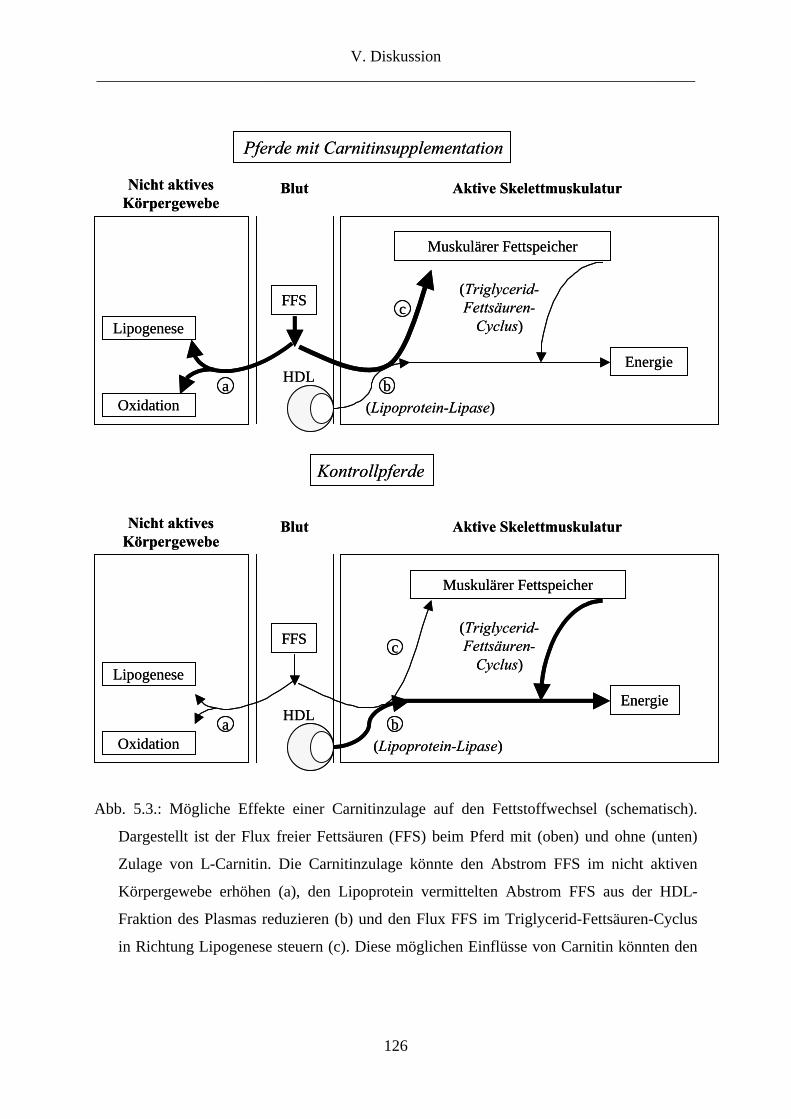
V. Diskussion ___________________________________________________________________________
126
Abb. 5.3.: Mögliche Effekte einer Carnitinzulage auf den Fettstoffwechsel (schematisch).
Dargestellt ist der Flux freier Fettsäuren (FFS) beim Pferd mit (oben) und ohne (unten)
Zulage von L-Carnitin. Die Carnitinzulage könnte den Abstrom FFS im nicht aktiven
Körpergewebe erhöhen (a), den Lipoprotein vermittelten Abstrom FFS aus der HDL-
Fraktion des Plasmas reduzieren (b) und den Flux FFS im Triglycerid-Fettsäuren-Cyclus
in Richtung Lipogenese steuern (c). Diese möglichen Einflüsse von Carnitin könnten den
Lipogenese
Oxidationa
c
b
FFS
HDL
(Lipoprotein-Lipase)
Muskulärer Fettspeicher
Energie
(Triglycerid-Fettsäuren-
Cyclus)
Nicht aktivesKörpergewebe
Blut Aktive Skelettmuskulatur
Lipogenese
Oxidationa
c
b(Lipoprotein-Lipase)
(Triglycerid-Fettsäuren-
Cyclus)
Nicht aktivesKörpergewebe
Blut Aktive Skelettmuskulatur
Kontrollpferde
Pferde mit Carnitinsupplementation
HDL
FFS
Muskulärer Fettspeicher
Energie
Lipogenese
Oxidationa
c
b
FFS
HDL
(Lipoprotein-Lipase)
Muskulärer Fettspeicher
Energie
(Triglycerid-Fettsäuren-
Cyclus)
Nicht aktivesKörpergewebe
Blut Aktive Skelettmuskulatur
Lipogenese
Oxidationa
c
b(Lipoprotein-Lipase)
(Triglycerid-Fettsäuren-
Cyclus)
Nicht aktivesKörpergewebe
Blut Aktive Skelettmuskulatur
Kontrollpferde
Pferde mit Carnitinsupplementation
HDL
FFS
Muskulärer Fettspeicher
Energie

V. Diskussion ___________________________________________________________________________
127
Nettoabstrom von FFS aus dem Plasma erhöhen (dicke Linien) und gleichzeitig die
Oxidationsrate der FFS in der aktiven Skelettmuskulatur reduzieren (dünne Linien).
Das niedrigere belastungsabhängige Konzentrationsprofil der FFS im Plasma bei den
trainierten mit Carnitin supplementierten Pferden im Vergleich zu den Kontrollpferden könnte
somit:
a) auf einer höheren Fettsäurenoxidationsrate in nicht aktiven Geweben z. B. in der
Leber
b) auf einer verminderten Aufnahme von Fettsäuren aus der Lipoprotein-Fraktion des
Plasma und
c) auf einer höheren intrazellulären Lipogeneserate im Rahmen des Triglycerid-
Fettsäuren-Cyclus
beruhen.
3. Schlussfolgerungen
1) Eine Supplementation von 10 g L-Carnitin pro Tag über vier Monate erhöhte die
Carnitinkonzentration im Plasma bereits nach vier Tagen um das 2,4-fache (von 25 auf 59
µmol/l). Der Gehalt an Gesamtcarnitin in der Muskulatur am Versuchsanfang von 19,7
mmol/kg TS erhöhte sich in fünf Wochen in Verbindung mit einem Training um 46 %. Wie
bei anderen Tierarten und beim Menschen nahm somit auch beim Pferd der Carnitingehalt der
Muskulatur bei einer entsprechenden Zulage und gleichzeitigem Training beträchtlich zu.
2) Die Carnitinzulage reduzierte in Verbindung mit einem fünfwöchigen Training über einen
weiten Belastungsbereich signifikant die Herzfrequenz. Dieser Effekt steht im Einklang mit
experimentell gut belegten inotropen Effekten von Carnitin am ischämischen Herzen.
3) Die Carnitinzulage verminderte den belastungsinduzierten Lactatanstieg im Plasma und
reduzierte somit die Lactacidose. Gleichzeitig erhöhte die Carnitinzulage die
belastungsinduzierte Synthese von Acetyl-Carnitin in der Arbeitsmuskulatur. Es wird
vermutet, dass der Rückgang der Milchsäurebildung mit der verstärkten
Transacetylierungsrate des Carnitins in einem funktionellen Zusammenhang steht.

V. Diskussion ___________________________________________________________________________
128
4) Die Carnitinzulage verminderte den belastungsinduzierten Anstieg der Plasmaglucose. Für
das Zustandekommen dieses Effektes wird vermutet, dass die Glucoseutilisation durch die
Carnitinzulage stimuliert und dass das durch Glycolyse vermehrt anfallende Pyruvat teilweise
nach Decarboxylierung an Carnitin gebunden wurde.
5) Die Carnitinzulage senkte die Konzentration der FFS im Plasma und reduzierte den
belastungsinduzierten Anstieg der FFS im Plasma. Es wird vermutet, dass, neben anderen
Wirkungen auf den Fettstoffwechsel, durch die Carnitinzulage der Abstrom der FFS aus dem
Plasma stimuliert wurde.
Die durch die Carnitinzulage in Verbindung mit einem fünfwöchigen Training beobachteten
Effekt wie die Reduktion der Herzfrequenz, der geringere belastungsinduzierte Lactatanstieg,
der geringere belastungsinduzierte Anstieg der Plasmaglucose und der FFS im Plasma, sind
insgesamt im Sinne einer Verbesserung der körperlichen Fitness als günstig zu beurteilen. Ob
diese Effekte von Carnitin das körperliche Leistungsvermögen der Tiere unter bestimmten
Wettkampfbedingungen verbessern, erscheint möglich, wurde im Rahmen dieser Studie aber
nicht untersucht.

VI. Zusammenfassung ___________________________________________________________________________
129
VI. Zusammenfassung
Im Bereich des Sports ist die Nachfrage nach Substanzen, die die körperliche
Leistungsfähigkeit steigern, ungebrochen. L-Carnitin, einer körpereigenen Substanz, wird
aufgrund ihrer besonderen biochemischen Wirkungen eine solche leistungssteigernde
Funktion zugesprochen. Untersuchungen bei Menschen, Pferden, Hunden, Tauben stützen nur
teilweise die an diese Substanz geknüpften Erwartungen.
Zweifel an der Wirksamkeit von L-Carnitin werden unter anderem durch die Auffassung
begründet, dass es nicht möglich sei, durch orale Zufuhr den Carnitingehalt der Muskulatur
über das normale Maß hinaus zu erhöhen. Befürworter von Carnitin halten dieses Argument
für experimentell nicht bewiesen und weisen darauf hin, dass Carnitin sich vermutlich nur in
der Muskulatur anreichere, wenn es in ausreichend hoher Dosis und über einen genügend
langen Zeitraum (am besten in Verbindung mit einem Training) verabreicht werde.
Ziel der hier vorliegenden Arbeit war es, bei Sportpferden (Trabern) zu untersuchen, ob der
Carnitingehalt im Plasma und in der Muskulatur im Laufe einer viermonatigen Zulageperiode
ansteigt. Während der ersten sechseinhalb Wochen wurden die Pferde außerdem trainiert. Ein
weiteres Ziel der Untersuchung war festzustellen, ob die Carnitinzulage während des
Trainings die Herzfrequenz und leistungsabhängige Parameter des Blutes beeinflusst.
Für die Versuche standen Traber im Alter von zwei Jahren zur Verfügung. Sie stammten alle
vom gleichen Hengst. Vier Tiere erhielten täglich in zwei Portionen 10 g L-Carnitin mit dem
Futter (+C). Drei Tiere dienten als Kontrollen (-C). Das Trainingsprogramm bestand aus
individuell abgestimmten, alternierenden Laufbandbelastungen auf aerober (v2,5) und
anaerober (v4) Stufe mit je einem Tag Pause zwischen den Belastungen (Abb. 3.6.). Das
Leistungsvermögen der Tiere wurde zur Beurteilung des Trainingseffektes zu Anfang und am
Ende des Trainings mit einem Stufentest mit Hilfe der belastungsabhängigen
Blutlactatkonzentration erfasst.

VI. Zusammenfassung ___________________________________________________________________________
130
Die Studie erbrachte die folgenden Ergebnisse:
1. Durch die Carnitinzulage war die Konzentration an Gesamtcarnitin im Plasma bereits
nach vier Tagen etwa auf das 2,5-fache angestiegen (25 vs. 60 µmol/l) und blieb über
gesamte Zulage auf dem gleichen Niveau (Abb. 4.1.).
2. In der Muskulatur (M. glutaeus medius) hatte der Gehalt an Gesamtcarnitin nach
fünfwöchiger Zulage um etwa 50 % zugenommen (4630 vs. 3180 mmol/kg TS). Er
blieb auch während der Ruheperiode auf diesem Niveau (Abb. 4.3.).
3. Bei den +C-Pferden war die Herzfrequenz am Ende des Trainings bei gleicher
Laufgeschwindigkeit wie vor dem Training (5 bis 9 m/s) signifikant um ca. 7 Schläge
pro Minute niedriger. Bei den –C-Pferden war die Herzfrequenz bei gleicher
Laufgeschwindigkeit vor und nach dem Training gleich (Abb. 4.5.).
4. Bei gleicher Laufgeschwindigkeit (10 m/s) war die Lactatkonzentration im Blut nach
vierwöchigem Training bei allen Tieren um 22 % niedriger als vor dem Training (10,0
vs. 12,8 mmol/l). Diese trainingsbedingte Reduktion der Lactatkonzentration war bei
den +C-Pferden ausgeprägter als bei den –C-Tieren (Abb. 4.8.).
5. Der belastungsinduzierte Anstieg der Plasmaglucose war am Ende des Trainings bei
den +C-Pferden signifikant geringer als bei den –C-Pferden (Abb. 4.10.).
6. Die Konzentration der FFS im Plasma war bei den +C-Pferden nach dem Training im
Gegensatz zu den –C-Pferden bei allen Belastungsstufen signifikant niedriger als vor
dem Training (Abb. 4.12.).
7. Der Ruheglykogengehalt des Muskels wies keine carnitin- und trainingsabhängigen
Unterschiede auf. Der belastungsinduzierte Abfall des Muskelglykogens schien durch
die Carnitinzulage nicht beeinflusst zu werden.

VI. Zusammenfassung ___________________________________________________________________________
131
Die Ergebnisse dieser Studie zeigen, dass die Muskulatur des Pferdes in Verbindung mit
einem Training bei einer Dosis von 10 g pro Tag in nennenswerten Mengen aus dem Plasma
aufnimmt und deponiert. Die Carnitinzulage erhöhte signifikant die belastungsinduzierte
Kontraktionskraft des Herzens. Die Effekte der Carnitinzulage auf die Konzentration der
Glucose und der FFS im Plasma während der Belastung sind mit der Annahme vereinbart,
dass L-Carnitin sowohl die Elimination von Glucose als auch von FFS aus dem Plasma
erhöht. Diese Effekte könnten mit Einsparungen von Muskelglykogen und –fett kombiniert
sein. Sie sind insgesamt im Hinblick auf eine Steigerung der körperlichen Fitness als günstig
zu beurteilen.
Ob – und, wenn ja – bei welchen Belastungsarten und in welchem Umfang eine
Carnitinzulage auch unter Wettkampfbedingungen Leistungssteigerungen erbringt, müssen
weitere Versuche klären.

VII. Summary ___________________________________________________________________________
132
Cornelia Chrobok: Combined effects of physical training and carnitine supplementation on
carnitine content in muscle and on certain plasma parameters during exercise in young trotters
VII. Summary
In sports, there is continuing interest to increase physical fitness by use of certain food
constituents or additives. Due to its biochemical properties, L-carnitine, a normal constituent
of the body, is said to improve physical performance. Such expectations have, so far, only
partially been met by studies carried out on humans, horses, dogs or pigeons.
Doubts in general have been expressed in the potential effectiveness of L-carnitine of
improving physical performance. It has been argued that it would probably be impossible to
increase the content of carnitine in muscle simply by oral supplementation of carnitine. Others
believe, that this might be well possible, provided the oral dose of carnitine is high enough
and is offered for a sufficient length of time. They also argue that the best results are achieved
when carnitine is given in combination with training.
It was the aim of the present study to measure the content of total and free carnitine in plasma
and skeletal muscle of horses over the course of a four month supplementation period. During
the first five weeks of carnitine supplementation the horses were also trained. Examination of
the effect of carnitine and training on heart rate and on some plasma parameters was
attempted.
Seven two-year-old trotters, all descendants from one stallion, were randomly allotted to a
supplemented group (+C) (four animals) and a control group (-C) (three animals). The
animals in the supplemented group were fed with daily amounts of 10 g of L-carnitine offered
in two portions with the morning and the afternoon feed. The initial five week training
program consisted of alternating treadmill exercises at an aerobic level (v2,5) and at an
anaerobic level (starting with v4). There was one day of rest between each set of exercise. The
physical fitness of the horses was estimated at the beginning and end of the training period
from the lactate concentration in blood obtained during a graded exercise test.
The following results were obtained from the study.

VII. Summary ___________________________________________________________________________
133
1. The concentration of carnitine in blood plasma had increased 2,5-fold (from 25 to 60
µmol/l) four days after the start of carnitine supplementation and remained at that high
level during the whole four month period of carnitine supplementation.
2. The content of total carnitine in the skeletal muscle (M. glutaeus medius) had
increased by about 50 % (from 3180 to 4630 mmol/kg dry matter) after four weeks of
carnitine supplementation combined with training. The concentration in muscle
remained constant at this high level during the remaining three months of detraining.
3. Comparing the +C horses to the –C horses at the end of training and with the same
absolute power output as before training:
a) the heart rate during exercise was significantly lower by seven beats per
second in the supplemented animals and was unchanged in control animals
b) the lactate concentration in blood during exercise was 22 % lower
c) the exercise-induced increase in plasma glucose was significantly lower and
d) the concentration of FFA before and during exercise was significantly lower.
4. The resting glycogen stores in M. glutaeus medius was not affected by carnitine and
training. The exercise induced decrease of muscular glycogen content appeared also
not to be affected by L-carnitine supplementation.
The study has shown that, concomitantly with training, the skeletal muscle of horses is
capable of considerably increasing its carnitine content when the animals are supplemented
with 10 g of carnitine per day. Carnitine supplementation improved cardiac power output over
a certain range of exercise intensities. The effects of carnitine on exercise-related
concentrations of glucose and FFA in plasma could be explained by assuming an increased
rate of glucose disposal and an increased rate of uptake of FFA from plasma, both, probably
indirectly mediated by carnitine. The cardiac and haematological effects of carnitine are
regarded as being positive with respect of improving the physical fitness of the animals.
Whether, and if so, to what extent, such effects of carnitine can improve competitive physical
performance of horses is still unknown.

VIII. Literaturverzeichnis ___________________________________________________________________________
134
VIII. Literaturverzeichnis ABDEL-ALEEM, S., M. A. NADA, M. SAYED-AHMED, S. C. HENDRICKSON, J. ST LOUIS, H. P. WALTHALL u. J. E. LOWE (1996): Regulation of fatty acid oxidation by acetyl-CoA generated from glucose utilization in isolated myocytes. J. Mol. Cell. Cardiol. 28(5), 825-833 AHMAD, S., A. DASGUPTA u. M. A. KENNY (1989): Fatty acid abnormalities in hemodialysis patients: Effect of L-carnitine administration. Kidney International 36(27), 243-246 ALLEN, R. J., D. B. HANSCH u. H. L. C. WU (1982): Hypocarnitinemia in disorders of organic acid metabolism. Lancet 2(8296), 500-501 ÅSTRAND, P. O., u. K. ROHDAHL (1986): Textbook of Work Physiology. 3. Aufl., McGraw-Hill Comp., New York ARENAS, J., J. R. RICOY, A. R. ENCINAS, P. POLA, S. D´IDDIO, M. ZEVIANI, S. DIDONATO u. M. CORSI (1991): Carnitine in muscle, serum and urine of nonprofessional athletes effects of physical exercise, training and L-carnitine administration. Muscle and Nerve 14, 598-604 ART, T., u. P. LEKEUX (1993): Training-induced modifications in cardiorespiratory and ventilatory measurements in Thoroughbred horses. Equine vet. J. 25(6), 532-536 BAKER, H., O. FRANK, B. DEANGELIS u. E. R. BAKER (1992): Absorption and excretion of L-carnitine during single or multiple dosings in humans. Int. J. Vitam. Nutr. Res. 63, 22-26 BATTISTELLA, P. A., L. VERGANI, F. DONZELLI, F. F. RUBALTELLI u. C. ANGELINI (1980): Plasma and urine carnitine levels during development. Pediatr. Res. 14, 1379-1381 BENAMOU, A. E., u. R. C. HARRIS (1993): Effect of carnitine supplement to the dam on plasma carnitine concentration in the sucking foal. Equine vet. J. 25, 49-52

VIII. Literaturverzeichnis ___________________________________________________________________________
135
BENEVENGA, N. J., J. K. STEINMAN-GOLDSWORTHY, T. D. CRENSHAW u. J. OLDE (1989): Utilitation of medium-chain triglycerides by neonatal piglets: I. Effects on milk consumption and body fuel utilization J. Anim. Sci. 67, 3331-3339 BERGMAN, B. C., G. E. BUTTERFIELD, E. E. WOLFEL, G. D. LOPASCHUK, G. A. CASAZZA, M. A. HORNING u. G. A. BROOKS (1999 a): Muscle net glucose uptake and glucose kinetics after endurance training in men. Am. J. Physiol. 277(40), E81-E92 BERGMAN, B. C., G. E. BUTTERFIELD, E. E. WOLFEL, G. A. CASAZZA, G. D. LOPASCHUK, u. G. A. BROOKS (1999 b): Evaluation of exercise and training on muscle lipid metabolism. Am. J. Physiol. 276(39), E106-E117 BETTINI, V., C. CATOZZO, R. MARTINO, F. MAYELLARO, L. MUNARI, V. TEGAZZIN u. P. TON (1985): Changes in acetylcholine contractions induced by carnitine in coronary vessels isolated ''in vitro''. Acta Vitaminol. Enzymol. 7(1-2), 61-69 BIEBER, L. L. (1988): Carnitine. Ann. Rev. Biochem. 57, 261-283 BILLIGMANN, P.-W., U. KÜNZEL u. S. BERTSCH (1990): Wie wirkt sich L-Carnitin auf die physische Maximalbelastung aus? Therapiewoche 40, 1866-1872 BÖHLES, H., D. MICHALK u. E. VON WENDT-GÖKNUR (1991): Der Einfluss einer L-Carnitin-Supplementierung auf den Lipidstoffwechsel niereninsuffizienter, dialysepflichtiger Kinder und Jugendlicher. Infusionstherapie 18, 224-226 BORDIN, D., D. BOTTECCHIA, V. BETTINI, R. ARAGNO u. L. SARTORELLI (1992): Effects of middle-intensity exercise on carnitine and β-hydroxybutyrate plasmatic concentration in men and women. J. Sports Med. Phys. Fitness 32, 394-399 BORGHIJS, H. K., u. R. O. DE WILDE (1992): The influence of two different dosages of L-carnitine on some blood parameters. J. Vet. Nutr. 1, 31-35

VIII. Literaturverzeichnis ___________________________________________________________________________
136
BORUM, P. R. (1983): Carnitine. Ann. Review Nutr. 3, 233-259 BORUM, P. R., C. M. YORK u. H. P. BROQUIST (1979): Carnitine content of liquid formulas and special diets. Am. J. Clin. Nutr. 32, 2272-2276 BOURGELA, M., D. BLAIS u. M. MARCOUX (1991): Reproducibility and validity of VLA4 in Standardbred pacer horses on track. In: S. G. B. PERSSON, A. LINDHOLM u. L. B. JEFFCOTT (Edit.): Equine Exercise Physiology 3. ICEEP Publications, Davis, California, S. 196-201 BLOOM, S. R., R. H. JOHNSON, D. M. PARK, M. J. RENNIE u. S. R. SULAIMAN (1976): Differences in the metabolic and hormonal response to exercise between racing cyclists and untrained individuals. J. Physiol. 258, 1-18 BRASS, E. (1992): L-carnitine and its role in medicine: From function to therapy. In: R. FERRARI, G. DIMAURO u. G. SHERWOOD (Edit.): Carnitine Transport. Academic Press, London, New York, S. 21-36 BRASS, E. P., u. R. A. BEYERINCK (1987): Interactions of propionate and carnitine metabolism in isolated rat hepatocytes. Metabolism 36, 781-787 BRASS, E. P., A. M. SCARROW, L. J. RUFF, K. A. MASTERSON u. E. VAN LUNTEREN (1993): Carnitine delays rat skeletal muscle fatigue in vitro. J. Appl. Physiol. 75(4), 1595-1600 BREMER, J. (1961): Biosynthesis of carnitine in vivo. Biochim. Biophys. Acta 48, 622-624 BREMER, J. (1969): Pyruvate dehydrogenase, substrate specificity and product inhibition. Eur. J. Biochem. 8, 535-540 BREMER, J. (1983): Carnitine-metabolism and functions. Physiol. Rev. 63, 1420-1480

VIII. Literaturverzeichnis ___________________________________________________________________________
137
BREMER, J., E. J. DAVIS u. B. BORREBAEK (1984): Factors influencing the carnitine-dependent oxidation of fatty acids in the heart. In: R. FERRARI, A. M. KATZ, A. SHUG u. O. VISIOLI (Edit.): Myocardial Ischemia and Lipid Metabolism. Plenum Press, New York, S. 15-26 BREMER, J., u. H. OSMONDSEN (1984): Fatty acid oxidation and its regulation. In: S. NUMA (Edit.): Fatty Acid Metabolism and Its Regulation. Elsevier Science Publishers, Amsterdam, S. 113-154 BRODERICK, T. L., H. A. QUINNEY u. G. D. LOPASCHUK (1992): Carnitine stimulation of glucose oxidation in the fatty acid perfused isolated working rat heart. J. Biol. Chem. 267(6), 3758-3763 BROOKS, D. E. (1980): Carnitine in the male reproductive tract and its relation to the metabolism of the epididymidis and spermatozoa. In: R. A. FRENKEL u. J. D. MCGARRY (Edit.): Carnitine Biosynthesis, Metabolism and Functions. Academic Press, New York, S. 219-235 BROOKS, D. E., u. C. M. DONOVAN (1983): Effect of endurance training on glucose kinetics during exercise. Am. J. Physiol. 244, E505-E512 BROOKS, D. E., u. J. E. A. MCINTOSH (1975): Turnover of carnitine by rat tissue. Biochem. J. 148, 439-445 CAPONNETTO, S., u. C. BRUNELLI (1995): Hemodynamic and metabolic effect of propionyl-L-canitine in patients with heart failure. In: J. W. DE JONG u. R. FERRARI (Edit.): The Carnitine System. Kluwer Academic Publishers, Dordrecht, Niederlande, S. 337-351 CARLIN, J. I., R. C. HARRIS, G. CEDERBLAD, D. C. TEODOSIU, D. H. SNOW u. E. HULTMAN (1990): Association between muscle acetyl-CoA and acetylcarnitine levels in the exercising horse. J. Appl. Physiol. 69, 42-45 CARLIN, J. I., W. REDDAR u. M. SANIAK (1986): Carnitine metabolism during prolonged exercise and recovery in humans. J. Appl. Physiol. 55, 489-495

VIII. Literaturverzeichnis ___________________________________________________________________________
138
CARTER, A. L., D. L. F. LENNON u. F. W. STRATMAN (1981): Increased acetyl-carnitine in rat skeletal muscle as a result of high-intensity short-duration exercise. FEBS Lett. 126, 21-24 CARTER, A. L., F. W. STRATMAN, S. M. HUTSON u. H. A. LARDY (1980): The role of carnitine and its esters in sperm metabolism. In: R. A. FRAENKEL u. J. D. MCGARRY (Edit.): Carnitine and Metabolism and Functions. Academic Press, New York, S. 251-263 CASINI, L., u. G. F. GREPPI (1996): Correlation of racing performance with fitness parameters after exercise tests on treadmill and on track in Standardbred racehorses. Pferdeheilkunde 12, 466-469 CEDERBLAD, G., u. S. LINDSTEDT (1976): Metabolism of labelled carnitine in the rat. Arch. Biochem. Biophys. 175, 173-180 CIMAN, M., V. RIZZOLI u. N. SILIPRANDI (1980): Effect of physical training on carnitine concentration in liver, heart and gastrocnemius muscle of rat. Int. J. Vit. Nutr. Res. 50, 40-43 CIPOLLA, M. J., A. NICOLOFF, T. REBELLO, A. AMATO u. J. M. PORTER (1999): Propionyl-L-carnitine dilates human subcutaneous arteries through an endothelium-dependent mechanism. J. Vasc. Surg. 29(6), 1097-1103 COGGAN, A. R. (1991): Plasma glucose metabolism during exercise in humans. Sports Medicine 11, 102-124 COGGAN, A. R., W. M. KOHRT, R. J. SPINA, D. M. BIER u. J. O. HOLLOSZY (1990): Endurance training decreases plasma glucose turnover and oxidation during moderate-intesity exercise in men. J. Appl. Physiol. 68(3), 990-996 COGGAN, A. R., R. J. SPINA, W. M. KOHRT u. J. O. HOLLOSZY (1993): Effect of prolonged exercise on muscle citrate concentration before and after endurance training in men. Am. J. Physiol. 264(27), E215-E220

VIII. Literaturverzeichnis ___________________________________________________________________________
139
COGGAN, A. R., C. A. RAGUSO, B. D. WILLIAMS, L. S. SIDOSI u. A. GASTALDELLI (1995 a): Glucose kinetics during high-intensity exercise in endurance-trained and untrained humans. J. Appl. Physiol. 78, 1203-1207 COGGAN, A. R., S. C. SWANSON, L. A. MENDENHALL, D. L. HABASH u. C. L. KIEN (1995 b): Effect of endurance training on hepatic glycogenolysis and gluconeogenesis during prolonged exercise in men. Endocrinol. Metab. 31, E375-E383 COGGAN, A. R., C. A. RAGUSO, A. GASTALDELLI, L. S. SIDOSSIS u. C. W. YECKEL (2000): Fat metabolism during high-intensity exercise in endurance-trained and untrained men. Metabolism 49(1), 122-128 DECOMBAZ, J. E., B. REFFET u. Y. BLOEMHARD (1990): Effect of L-carnitine and stimulated lipolysis on muscle substrates in the exercising rat. Experienta 46 (5), 457-458 DECOMBAZ, J., B. GMUENDER, G. SIERRO u. P. CERRETELLI (1992): Muscle carnitine after strenuous endurance exercise. J. Appl. Physiol. 72(2), 423-427 DE JONG, J. W., u. R. FERRARI (1995): Therapeutic efficacy of L-carnitine at the myocardial level. In: The Carnitine System. Kluwer Academic Publishers, Dordrecht, Niederlande, S. 169-249 DOHM, G. L., E. B. TAPSCOTT, H. A. BARAKAT u. G. J. KASPEREK (1983): Influence of fasting on glycogen depletion in rats during exercise. J. Appl. Physiol. 55(3), 830-833 DRAGAN, I. G., A. VASILIU, E. GEORGESCU u. N. EREMIA (1989): Studies concerning chronic and acute effects of L-carnitina in elite athletes. Rev. roum. Morphol. Embryol. Physiol. 26(2), 111-129 DUBELAAR, M.-L., C. M. H. B. LUCAS u. W. C. HÜLSMANN (1991): Acute effect of L-carnitine on skeletal muscle force tests in dogs. Am. J. Physiol. 260, E189-E193 DUBELAAR, M.-L., J. F. C. GLATZ, Y. F. DE JONG, F. H. VAN DER VEEN u. W. C. HÜLSMANN (1994): L-carnitine combined with minimal electrical stimulation promotes type transformation of canine latissimus dorsi. J. Appl. Physiol. 76, 1636-1642

VIII. Literaturverzeichnis ___________________________________________________________________________
140
EATON, M. D., D. R. HODGSON, D. L. EVANS u. R. J. ROSE (1999): Effects of low- and moderate-intensity training on metabolic response to exercise in Thoroughbreds. Equine vet. J. Suppl. 30, 521-527 EISENBERG, S. (1984): High density lipoprotein metabolism. J. Lipid Res. 25, 1017-1058 ENGEL, A. G. u. C. J. REBOUCHE (1984): Carnitine metabolism and inborn errors. J. Inherit. Metab. Dis. 7 (Suppl 1), 38-43 ENGEL, A. G., C. J. REBOUCHE, D. M. WILSON, A. M. GLASGOW, C. A. ROMSHE u. R. P. CRUSE (1981): Primary systemic carnitine deficiency. II. Renal handling of carnitine. Neurology 31, 819-825 ENGLARD, S., u. H. H. CARNICERO (1978): Gamma-Butyrobetaine hydroxylation to carnitine in mammlian kidney. Arch. Biochem. Biophys. 190, 357-364 ERICKSON, H. H., B. K. ERICKSON, C. S. LUNDIN, J. R. GILLESPIE u. J. R. COFFMAN (1990): Performance indices for the evalution of the equine athlete. Proc. Am. Ass. Equine Prac. 36, 457-469 ESSEN-GUSTAVSSON, B., D. MC MIKEN, K. KARLSTRÖM, A. LINDHOLM u. S. G. B. PERSSON (1989): Muscular adaptations of horses during intensive training and detraining. Equine vet. J. 21, 27-33 ESSEN-GUSTAVSSON, B., G. NYMAN u. P. D. WAGNER (1995): Muscle and blood metabolic response to intense exercise during acute hypoxia and hyperoxia. Equine vet. J. 18, 181-187 EVANS, D. L., u. R. J. ROSE (1988): Cardiovascular and respiratory response to submaximal exercise training in the Thoroughbred horse. Pflügers Arch. 411(3), 316-321 EVANS, D. L., R. C. HARRIS u. D. H. SNOW (1993): Correlation of racing performance with blood lactate and heart rate after exercise in Thoroughbred horses. Equine vet. J. 25, 441-445

VIII. Literaturverzeichnis ___________________________________________________________________________
141
FALASCHINI, A., u. M. F. TROMBETTA (1994): Use of diets containing soya oil and L-carnitine in Trotting-horses. Zoot. Nutr. Anim. 20, 253-262 FEHLEISEN, B. (1997): Einsatz von Carnitin im Training von Sportpferden. Wien, Veterinärm. Univers., Diss. FEHLIG, P. (1973): The glucose-alanine cycle. Metabolism 2, 179-199 FERRANNINI, E., G. BUZZIGOLI, S. BEVILACQUA, C. BONI, D. DEL CHIARO, M. OLEGGINI, L. BRANDI u. F. MACCARI (1988): Interactions of carnitine with insulin stimulated glucose metabolism in humans. Am. J. Physiol. 255, 946-952 FÖHRENBACH, R., W. MÄRZ, H. LOHREER, R. SIEKMEIER, A. EVANGELIOU u. H. J. BÖHLES (1993): Der Einfluss von L-Carnitin auf den Lipidstoffwechsel von Hochleistungssportlern. Deutsche Zeitschrift für Sportmedizin 8, 349-356 FOREMAN, J. H., W. M. BAYLY, J. R. ALLEN, H. MATOBA, B. D. GRANT u. P. D. GOLLNICK (1990 a): Muscle responses of Thoroughbreds to conventional race training and detraining. Am. J. Vet. Res. 51(6), 909-913 FOREMAN, J. H., W. M. BAYLY, B. D. GRANT u. P. D. GOLLNICK (1990 b): Standardized exercise test and daily heart rate response of Thoroughbreds undergoing conventional race training and detraining. Am. J. Vet. Res. 51(6), 914- 920 FOSTER, C. V. L., u. R. C. HARRIS (1987 a): Formation of acetylcarnitine in muscle of horses during high intensity exercise. Eur. J. Appl. Physiol. 56, 639-642 FOSTER, C. V. L., u. R. C. HARRIS (1987 b): Changes in the free and bound carnitine in muscle with maximal sprint exercise in the Thoroughbred horse. In: J. R. GILLESPIE u. N. E. ROBINSON (Edit.): Equine Exercise Physiology 2. ICEEP Publications, Davis, California, S. 332-340 FOSTER, C. V. L., u. R. C. HARRIS (1989): Plasma carnitine concentrations in the horse following oral supplementation using a triple dose regime. Equine vet. J. 24, 52-57

VIII. Literaturverzeichnis ___________________________________________________________________________
142
FOSTER, C. V. L., u. R. C. HARRIS (1992): Total carnitine content of the middle gluteal muscle of Thoroughbred horses: normal values, variability and effect of acute exercise. Equine vet. J. 24, 52-57 FOSTER, C. V. L., R. C. HARRIS u. P. D. ROSSDALE (1987): Tissue concentration of carnitine in fetuses, neonates and adult horses. J. Reprod. Fert. 35, 739-740 FOSTER, C. V. L., R. C. HARRIS u. D. H. SNOW (1988): The effect of oral L-carnitine supplementation on the muscle and plasma concentrations in the Thoroughbred horse. Comp. Biochem. Physiol. 91A, 827-835 FOSTER, C. V. L., R. C. HARRIS u. E. J.-M. POURET (1989 a): Survey of plasma free carnitine levels in 74 Thoroughbred horses at stud and in training. Equine vet. J. 21, 139-141 FOSTER, C. V. L., R. C. HARRIS u. E. J.-M. POURET (1989 b): Effect of oral L-carnitine on its concentration in the plasma of yearling Thoroughbred horses. Vet. Rec. 125, 125-128 FREESTONE, J. F., K. J. WOLFSHEIMER, S. G. KAMERLING, C. CHURCH, J. HAMARA u. C. BAGWELL (1991): Exercise induced hormonal and metabolic changes in Thoroughbred horses: effects of conditioning and acepromazine. Equine vet. J. 23, 219-223 FRIEDMAN, S., u. G. FRAENKEL (1955): Reversible enzymatic acetylation of carnitine. Arch. Biochem. Biophys. 59, 491-501 FRITZ, I. B. (1955): The effect of muscle extracts on the oxidation of palmatic acid by liver slices and homogenates. Acta Physiol. Scand. 34, 367-385 FROHLICH, J., D. W. SECCOMBE, P. HAHN, P. DODEK u. I. HYNIE (1978): Effect of fasting on free and esterified carnitine levels in human serum and urin: correlation with serum levels of free fatty acids and β-hydroxybutyrate. Metabolism 27(5), 555-561 GALBO, H., E. A. RICHTER, J. J. HOLST u. N. J. CHRISTENSEN (1977): Diminished hormonal response to exercise in trained rats. J. Appl. Physiol. 43(6), 953-958

VIII. Literaturverzeichnis ___________________________________________________________________________
143
GEH: GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE DER HAUSTIERE (1994): Empfehlungen zur Energie- und Nährstoffversorgung des Pferdes. DLG-Verlag, Frankfurt/Main GEOR, R. J., K. W. HINCHCLIFF, L. J. MCCUTCHEON u. R. A. SAMS (2000 a): Epinephrine inhibits exogenous glucose utilization in exercising horses. J. Appl. Physiol. 88, 1777-1790 GEOR, R. J., K. W. HINCHCLIFF u. R. A. SAMS (2000 b): Glucose infusion attenuates endogenous glucose production and enhances glucose use of horses during exercise. J. Appl. Physiol. 88, 1765-1776 GLÖGGLER, A., M. BULLA u. P. FÜRST (1989): Effect of low dose supplementation of L-carnitine on lipid metabolism in hemodialysed children. Kidney Intern. 36(27), 256-258 GOROSTIAGA, E. M., C. A. MAURER u. J. P. ECLACHE (1989): Decrease in respiratory quotient during exercise following L-carnitine supplementation. Int. J. Sports Med. 10, 169-174 GÓRSKI, J. (1990): Muscle triglyceride metabolism during exercise. Can. J. Physiol. Pharmacol. 70, 123-131 GÓRSKI, J., M. NOWACKA, Z. NAMIOT u. U. PUCH (1988): Effect of prolonged exercise on the level of triglycerides in the rat liver. Eur. J. Appl. Physiol. 57, 554-557 GOTTLIEB-VEDI, M., S. PERSSON, H. ERICKSON u. E. KORBUTIAK (1995): Cardiovascular, respiratory and metabolic effects of interval training at VLA4. Zentralbl. Veterinärmed. A. 42 (3), 165-175 GREEN, H. J., S. JONES, M. E. BALL-BOURNET, D. SMITH u. B. W. FARRANCE (1991): Early muscular and metabolic adaptations to prolonged exercise training in humans. J. Appl. Physiol. 70, 2032-2038 GREEN, H. J., R. HELYA, M. E. BALL-BOURNET, N. KOWALCHUK, S. SYMON u. B. FARRANCE (1992): Metabolic adaptations to training precede changes in muscular mitochondrial capacity. J. Appl. Physiol. 72, 484-491

VIII. Literaturverzeichnis ___________________________________________________________________________
144
GREIG, C., K. M,. FINCH, A. D. JONES, M. COOPER, A. J. SARGEANT u. C. A. FORTE (1987): The effect of oral supplementation with L-carnitine on maximum and submaximum exercise capacity. Eur. J. Appl. Physiol. 56, 457-460 GROSSKOPF, J. F. W., u. J. J. VAN RENSBURG (1983): Some observations on the haematology and blood chemistry of horses competing in 80 km endurance rides. In: D. H. SNOW, S. G. B. PERSSON u. R. J. ROSE (Edit.): Equine Exercise Physiology 1. Granta Editions, Cambridge, S. 425-431 GUDJONSSON, H., B. U. LI, A. L. SHUG u. W. A. OLSEN (1985 a): In vivo studies of intestinal carnitine absorption in rats. Gastroenterology 88, 1880-1887 GUDJONSSON, H., B. U. K. LI, A. L. SHUG u. W. A. OLSEN (1985 b): Studies of carnitine metabolism in relation of intestinal absorption. Am. J. Physiol. 248, 306-319 GUSTAVSEN, H. S. M. (2000): Bestimmung des L-Carnitingehaltes in rohen und behandelten pflanzlichen und tierischen Lebensmitteln. Hannover, Tierärztl. Hochsch., Diss. in Vorbereitung GUY, P. S., u. D. H. SNOW (1977): The effect of training and detraining on muscle composition in the horse. J. Physiol. 269, 33-51 HAGENFELDT, L. (1975): Turnover of individual free fatty acids in man. Federation Proc. 34, 2236-2240 HAIGLER, H. T., u. H. P. BROQUIST (1974): Carnitine synthesis in rat tissue slices. Biochem. Biophys. Res. Commun. 56, 676-681 HAMBITZER, R., E. BENT, C. FAISST u. H. SOMMER (1987): Belastungsinduzierte Veränderungen im Blutprofil von Araberpferden nach einem Distanzritt. Prakt. Tierarzt 8, 9-13 HAMBITZER, R., u. E. BENT (1988): Effect of long distance exercise on free fatty acids in blood plasma of Arab horses. J. vet. Med. A. 35, 622-625

VIII. Literaturverzeichnis ___________________________________________________________________________
145
HARALAMBIE, G., u. A. BERG (1976): Serum urea and amino nitrogen changes with exercise duration. Eur. J. Appl. Physiol. 36, 39 HARMEYER, J. (2000): Regulation der Herztätigkeit. In: W. VON ENGELHARDT u. G. BREVES (Edit): Physiologie der Haustiere. Enkeverlag, Stuttgart, S. 150-155 HARMEYER, J., C. SCHLUMBOHM u. M. BAUMGARTNER (1998): Der Gehalt an L-Carnitin in Futter- und Lebensmitteln pflanzlicher und tierischer Herkunft und der Einfluß von Herstellungsverfahren auf dessen Gehalt. In: Einfluß von Erzeugung und Verarbeitung auf die Qualität landwirtschaftlicher Produkte. VDLUFA Verlag, Darmstadt, S. 489-492 HARRIS, R. C., u. C. V. L. FOSTER (1990): Changes in muscle free carnitine and acetylcarnitine with increasing work intesity. Eur. J Appl. Physiol. 60, 81-85 HARRIS, R. C., C. V. L. FOSTER und D. H. SNOW (1995 a): Plasma carnitine concentration and uptake into muscle following oral and intravenous administration. In: N. E. Robinson (Edit.): Equine Exercise Physiology 4. ICEEP Publications, Kooralbyn, Australia, S. 382-387 HARRIS, R. C., C. V. L. FOSTER and D. H. SNOW (1995 b): Muscle carnitine content and plasma concentration in Thoroughbred horses following oral and intravenous administration. Equine vet. J. Suppl. 18, 42 HARRIS, R. C., D. J. MARLIN u. D. H. SNOW (1987): Metabolic response to maximal exercise of 800 and 2000 m in the Thoroughbred horse. J. Appl. Physiol. 63, 12-19 HEINONEN, O. J., J. TAKALA u. M. H. KVIST (1992): Effect of carnitine loading on long-chain fatty acid oxidation, maximal exercise capacity, and nitrogen balance. Eur. J. Appl. Physiol. 65(1), 13-17 HERMANSSEN, L., E. HULTMAN u. B. SALTIN (1967): Muscle glycogen during prolonged serve exercise. Acta Physiol. Scand. 71, 129-139 HODGSON, D. R., u. R. J. ROSE (1994): The Athletic Horse - Principles and Practice of Equine Sports Medicine. W. B. Saunders Comp., Philadelphia

VIII. Literaturverzeichnis ___________________________________________________________________________
146
HOLLMANN, W., u. H. HETTINGER (1990): Sportmedizin: Arbeits- und Trainingsgrundlagen. 3. Aufl., Schattauer, Stuttgart, New York HOLLOSZY, O. J., u. E. F. COYLE (1984): Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J. Appl. Phys. 56, 831-838 HUERTAS, R., Y. CAMPOS, E. DIAZ, J. ESTEBAN, L. VECHIETTI, G. MONTANARI u. S. D`IDDIO (1992): Respiratory chain enzymes in muscle of endurance athletes: effect of L-carnitine. Biochem. Biophys. R. Comm. 1, 102-107 HULSMANN, W. C., H. STAM u. F. MACCARI (1982): The effect of excess (acyl) carnitine on lipid metabolism in rat heart. Biochim. Biophys. Acta 713(1), 39-45 HURLEY, B. F., P. M. NEMETH, W. H. MARTIN III, J. M. HAGBERG, G. P. DALSKY u. J. O. HOLLOSZY (1986): Muscle triglyceride utilisation during exercise: effect of training. J. Appl. Physiol. 60, 562-567 HUTTUNEN, J. K. (1982): Physical activity and plasma lipids and lipoproteins. Ann. Clin. Res. 14, Suppl. 34, 124.129 HYYPPÄ, S., L. A. RÄSÄNEN u. A. R. PÖSÖ (1997): Resynthesis of glycogen in skeletal muscle of Standardbred Trotters after repeated bouts of exercise. Am. J. Vet. Res. 2, 162-166 IBEN, C. (1998): Einsatz von Carnitin beim Schlittenhund. Wien. Tierärztl. Mschr. 85, 334-339 IBEN, C. (1999): Effects of L-carnitine administration on treadmill test performance of untrained dogs. J. Anim. Physiol. a. Anim. Nutr. 82, 66-79 IBEN, C., G. BERGMEISTER, E. SADILA u. J. LEIBETSEDER (1992): Einfluß von oralen Carnitingaben auf einige Blut- und Leistungsparameter beim Pferd. Pferdeheilkunde (Sonderausgabe), 150-152

VIII. Literaturverzeichnis ___________________________________________________________________________
147
IDELL-WENGER, J. A., L. W. GROTYOHANN u. J. R. NEELY (1982): Regulation of fatty acid utilisation in heart. Role of the carnitine-acetyl-CoA transferase and carnitine-acetyl carnitine translocase system. J. Mol. Cell. Card. 14, 413-417 ITAYA, K. (1977): A more sensitive and stable colorimetric determination of free fatty acids in blood. J. Lipid Res. 18, 663-665 JAHN, P., H. HÄRTLOVA, M. MAL, R. KABER u. J. HANAK (1996): PCV and plasma biochemistry in relation to fitness of horses competing in endurance rides. Pferdeheilkunde 12, 506-509 JANSSON, E., u. L. KAIJSER (1987): Substrate utilisation and enzymes in skeletal muscle of extremely endurance-trained men. J. Appl. Physiol. 662, 999-1005 JONES, N. L., G. J. F. HEIGENHAUSER, A. KUKSIS, C. G. MATSOS, J. R. SUTTON u. C. J. TOEWS (1980): Fat metabolism in heavy exercise. Clin. Sci. 59, 469-478 JUDSON, G.J., H. C. FRAUENFELDER u. G. J. MOONES (1983): Biochemical changes in Thoroughbred racehorses following submaximal and maximal exercise. In: D. H. SNOW, S. G. B. PERSSON u. R. J. ROSE (Edit.): Equine Exercise Physiology 1. Granta Editions, Cambridge, S. 408-415 KATIRCIOGLU, P. A. GRANDJEAN, S. KUCUKER, Z. SARITAS, S. YAVAS, O. TASDEMIR u. K. BAYAZIT (1997): Effects of carnitine on preconditioned latissimus dorsi muscle at different burst frequencies. J. Card. Surg. 12(2), 120-125 KATZ, A., K. SAHLIN u. S. BROBERG (1991): Regulation of glucose utilization in human skeletal muscle during moderate dynamic exercise. Am. J. Physiol. 260(23), E411-E415 KEENE, B. W., D. P. PANCIERA, C. E. ATKINS, V. REGITZ, M. J. SCHMIDT u. A. L. SHUG (1991): Myocardial L-carnitine deficiency in a family of dogs with dilated cardiomyopathy. J. Am. Vet. Med. Assoc. 198, 647-650 KEUL, J., u. A. BERG (1985): Energiestoffwechsel und körperliche Leistung. In: W. HOLLMANN (Edit.): Zentrale Themen der Sportmedizin. Springer Verlag, Berlin, S. 197-243

VIII. Literaturverzeichnis ___________________________________________________________________________
148
KIENS, B. (1997): Effects of endurance training on fatty acid metabolism: local adaptations. Med. Sci. Sports Exerc. 29, 640-645 KISSENBECK, S. (1999): Einfluß eines Trainings auf den Glykogengehalt und den Glykogenverbrauch im M. glutaeus medius von Pferden. Bonn, Hohe Landwirtsch. Fak. der Univ., Diss. KJAER, M., P. A. FARRELL, N. J. CHRISTENSEN u. H. GALBO (1986): Increased epinephrine response and inaccurate glucoregulation in exercising athlets. J. Appl. Physiol. 61(5), 1693-1700 KLEIN, S. E. J. PETERS, O. B. HOLLAND u. R. R. WOLFE (1989): Effect of short- and long-term β-adrenergic blockade on lipolysis during fasting in humans. Am. J. Physiol. 257(20), E-65-E73 KOIVISTO, V., R. HENDLER, E. NADEL u. P. FELIG (1982): Influence of physical training on the fuel-hormone response to prolonged low intensity exercise. Metabolism 31 (2), 192-197 KRZYWANEK, H. (1974): Lactic acid concentrations and pH values in Trotters after racing. J. S. Afr. Vet. Ass. 45, 355-360 KRZYWANEK, H., E. MOHR, J. MILL u. M. SCHARPENACK (1996): Veränderung von Serumenzymen, Lactat-, und Hämoglobinkonzentrationen im Blut junger Trabrennpferde durch Trainingsbelastung. J. Vet. Med. 43, 345-352 KRZYWANEK, H., G. WITTKE, A. BAYER u. P. PORMAN (1970): The heart rate of Thoroughbred horses during a race. Equine vet. J. 2, 115-117 LAMBERTZ, A. (1999): Untersuchungen zum L-Carnitinstoffwechsel beim Pferd. Hannover, Tierärztl. Hochsch., Diss. LAWRENCE, L. M., L. V. SODERHOLM, A. ROBERTS, J. M WILLIAMS u. H. HINTZ (1993): Feeding status effects glucose metabolism in exercising horse. J. Nutr. 123, 2152-2157

VIII. Literaturverzeichnis ___________________________________________________________________________
149
LENNON, D. L. F., F. W. STRATMAN, E. SHRAGO, F. J. NAGLE, M. MADDEN, P. HANSON u. A. L. CARTER (1983): Effects of acute moderate intensity exercise on carnitine metabolism in men and women. J. Appl. Physiol. 55, 489-495 LI, B., M. L. LLOYD, H. GUDJONSSON, A. L. SHUG u. W. A. OLSEN (1992): The effect of enteral carnitine administrations in humans. Am. J. Clin. Nutr. 55, 838-845 LINDHOLM, A. (1979): Substrate utilisation and muscle fiber types in Standardbred Trotters during exercise. Proc. Am. Assoc. Equine Pract. 25, 329-336 LINDHOLM, A., u. K. PIEHL (1974): Fibre composition, enzyme activity and concentrations of metabolites and electrolytes in muscle of Standardbred horses. Acta Vet. Scand. 15, 287-309 LINDHOLM, A., u. B. SALTIN (1974): The physiological and biochemical response of Standardbred horses to exercise of varying speed and duration. Acta Vet. Scand. 15, 310-324 LINDNER, A. (1996): Measurement of plasma lactate concentration with Accusport. Equine vet. J. 28, 403-405 LINDNER, A. (1997): Grundlagen des Laktatstoffwechsels. In: Laktat und Leistung. Reihe Basis- und Praxiswissen über Pferde, S. 86-89 LINDSTEDT, G., u. S. LINDSTEDT (1961): On the biosynthesis and degradation of carnitine. Biochem. Biophys. Res. Comm. 6, 319-323 LOPASCHUK, G. D., D. D. BELKE, J. GAMBLE, T. ITOI u. B. O. SCHÖNEKESS (1994): Regulation of fatty acid oxidation in the mammalian heart in health and disease. Biochim. Biophys. Acta 1213, 263-276 LOPASCHUK, G. D., C. A. HANSEN u. J. R. NEELY (1986): Fatty acid metabolism in hearts containing elevated levels of CoA. Am J. Phsiol. 250, H351-H359

VIII. Literaturverzeichnis ___________________________________________________________________________
150
LUCKE, J. N., u. G. M. HALL (1980 a): A biochemical study of the Arab Horse Society´s marathon race. Vet. Rec. 107, 523-525 LUCKE, J. N., u. G. M. HALL (1980 b): Further studies on the metabolic effects of long distance riding: Golden Horseshoe Ride 1979. Equine vet. J. 12, 189-192 LUCKE, J. N., u. G. M. HALL (1980 c): Long distance exercise in the horse: Golden Horseshoe Ride 1978. Vet. Rec. 106, 405-407 LUMENG, L., J. BREMER u. E. J. DAVIS (1976): Suppression of the mitochondrial oxidation of (-)-palmityl-carnitine by the malate-aspartate and α-glycerophosphate shuttles. J. Biol. Chem. 251, 277-284 LYSIAK, W., P. P. TOTH, C. H. SUELTER u. L. L. BIEBER (1986): Quantitation of the efflux of acylcarnitines from rat heart, brain, and liver mitochondria. J. Biol. Chem. 261(29), 13698-16703 MARQUIS, N. R., u. I. B. FRITZ (1965): Effects of carnitine, acetylcarnitne and carnitineacetyltranferase in tissues of the reproductive system of the male rat. J. Biol. Chem. 240, 2197-2202 MARTIN, W. H. I., G. P. DALSKY, B. F. HURLEY, D. E. MATHEWS, D. M. BIER, J. M. HAGBERG, M. A. ROGERS, D. S. KING u. J. O. HOLLOSZY (1994): Effect of endurance training on free fatty acid turnover and oxidation during exercise. Am. J. Physiol. 265(28), E708-E714 MARTIN, B. J., S. ROBINSON, D. L. WEIGMAN u. L. H. AULICK (1975): Effect of warm-up on metabolic responses to strenuous exercise. Med. Sci. Sports Exerc. 7, 146-149 MCCONELL, G., M. MCCOY, J. PROIETTO u. M. HARGREAVES (1994): Skeletal muscle GLUT-4 and glucose uptake during exercise in humans. J. Appl. Physiol. 77(3), 1565-1568 MCDONOUGH, K. H., u. J. R. NEELY (1988): Inhibition of myocardial lipase by palmityl CoA. J. Mol. Cell Cardiol. Suppl. 2, 31-39

VIII. Literaturverzeichnis ___________________________________________________________________________
151
MCGARRY, J. D., u. D. W. FOSTER (1976): An improved and simplified radioisotopic assay for the determination of free and esterified carnitine. J. Lipid Res. 17, 277-281 MCGARRY, J. D., u. D. W. FOSTER (1979): In support of the roles of malonyl CoA and carnitine acyltransferase 1 in the regulation of hepatic fatty acid oxidation and ketogenesis. J. Biol. Chem. 254, 8163-8167 MC MIKEN, D. F. (1983): An energetic basis of equine performance. Equine vet. J. 15, 123-133 MEIER, P. J. (1987): Carnitin in der Medizin. In: R. GITZELMANN, K. BAERLOCHER u. B. STEINMANN (Edit.): Carnitin in der Medizin. Schattauer Verlag, Stuttgart, S. 101-104 MEYER, H., B. STADERMANN, S. RADICKE, E. KIENZLE u. ANYARI (1993): Untersuchung zum Einfluß der Futterart auf Füllung und Zusammensetzung des Verdauungskanals sowie postprandiale Blut- und Harnparameter. Pferdeheilkunde 9, 15-25 MITCHELL, M. E. (1978): Carnitine metabolism in human subjects. Am. J. Clin. Nutr. 31, 293-306 NEGRAO, C. E., L. L. JI, J. E. SCHAUER, F. J. NAGLE u. H. A. LARDY (1987): Carnitine supplemention and depletion: tissue carnitine and enzymes in fatty acid oxidation. J. Appl. Physiol. 63, 315-321 NEUMANN, G. (1996): Wirkung von L-Carnitin auf die sportliche Leistungsfähigkeit. Leipziger Carnitin-Symposium 27-28 März NIMMO, M. A., u. D. H. SNOW (1983): Changes in muscle glycogen, lactate and pyruvate concentrations in the Thoroughbred horse following maximal exercise. In: D. H. SNOW, S. G. B. PERSSON u. R. J. ROSE (Edit.): Equine Exercise Physiology 1. Granta Editions, Cambridge, S. 237-244 ORME, C. E., M. DUNNETT u. R. C. HARRIS (1994): Variation in the concentration of long chain free fatty acids in equine plasma over 24 hours. Br. Vet. J. 150, 339-342

VIII. Literaturverzeichnis ___________________________________________________________________________
152
PAGAN, J. D., B. ESSEN-GUSTAVSSON, A. LINDHOLM u. J. THORNTON (1987): The effect of dietary energy source on exercise performande on Standardbred horses. In: J. R. GILLESPIE u. N. E. ROBINSON (Edit.): Equine Exercise Physiology 2. ICEEP Publications, Davis, California, S. 686-456 PANDE, S. V. (1975): A mitochondrial carnitine acylcarnitine translocase system. Proc. Nat. Acad. Sci. 72, 883-887 PANTER, R. A., u. J. B. MUDD (1969): Carnitine levels in some higher plants. FEBS Lett. 5, 169-170 PARVIN, R., u. S. V. PANDE (1979): Enhancement of mitochondrial carnitine and carnitine acylcarnitine translocase-mediated transport of fatty acids into liver mitochondria under ketogenetic conditions. J. Biol. Chem. 254, 5423-5429 PAULSEN, J. D., M. J. SCHMIDT, J. S. TRAXLER, M. T. RAMACCI u. A. L. SHUG (1984): Improvement of myocardium function in diabetic rats after treatment with L-carnitine. Metab. Clin. Exp. 4, 358-363 PAULSEN, J. D., J. S. TRAXLER u. M. J. SCHMIDT (1986): Protection of the ischaemic myocardium by L-propionyl-carnitine: effects on the recovery of the cardiac output after ischaemia and reperfusion, carnitine transport, and fatty acid oxidation. Cardiovasc. Res. 20, 536-541 PELS, A. E., T. P. WHITE u. W. D. BLOCK (1985): Effects of exercise training on plasma lipids and lipoproteins in rats. J. Appl. Physiol 58(2), 612-618 PHILLIPS, S. M., H. J. GREEN, M. A. TARNOPOLSKY u. S. M. GRANT (1995): Decreased glucose turnover after short-term training is unaccompanied by changes in muscle oxidative potential. Am. J. Physiol. 269(32), E222-E230 PODL, T. R., J. M. ZMUDA, S. M. YURGALEVITCH, M. C. FAHRENBACH, L. L. BAUSSERMAN, R. B. TERRY u. P. D. THOMPSON (1994): Lipoprotein lipase activity and plasma triglyceride clearance are elevated in endurance-trained women. Metabolism 43(7), 808-813

VIII. Literaturverzeichnis ___________________________________________________________________________
153
POLA, P., L. SAVI, M. GRILLI, R. FLORE u. M. SERRICCHIO (1980): Carnitine in the therapy of dyslipidemic patients. Cur. Ther. Res. 27, 208-215 PÖSÖ, A. R., u. S. HYYPPÄ (1999): Metabolic and hormonal changes after exercise in relation to muscle glycogen concentrations. Equine vet. J., Suppl. 30, 332-336 PÖSÖ, A. R., T. SOVERI, M. ALAVIUHKOLA, L. LINDQVIST u. L. ALAKUIJALA, P. H. MÄENPÄA u. H. E. OKSANEN (1987): Metabolic responses to exercise in the racehorse: changes in plasma alanine concentrations. J. Appl. Physiol. 63, 2195-2200 PÖSÖ, A. R., E. VILIJANEN-TARIFA, T. SOVERI u. H. E. OKSANEN (1989): Exercise-induced transient hyperlipidemia in the racehorse. J. vet. Med. A. 36, 603-611 RADICKE, S., H. MEYER u. E. KIENZLE (1994): Über den Einfluß von Futterart und Fütterungszeitpunkt auf den Blutglucosespiegel bei Pferden. Pferdeheilkunde 10, 187-190 RAMSAY, R. R., u. P. K. TUBBS (1975): The mechanism of fatty acid uptake by heart mitochondria: an acylcarnitine-carnitine exchange. FEBS Lett. 54, 21-25 RANDEL, P. J., P. B. GARLAND, C. N. HALES u. E. A. NEWSHOLME (1963): The glucose fatty acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1, 785-789 REBOUCHE, C. J., u. A. G. ENGEL (1980 a): Tissue distribution of carnitine biosynthetic enzymes in man. Biochim. Biophys. Acta. 630, 22-29 REBOUCHE, C. J., u. A. G. ENGEL (1980 b): Significance of renal gamma-butyrobetaine hydroxylase for carnitine biosynthesis in man. J. Biol. Chem. 255, 8700-8705 REBOUCHE, C. J., u. A. G. ENGEL (1983): Kinetic compartment analysis of carnitine metabolism in the dog. Arch. Biochem. Biophys. 220, 60-70

VIII. Literaturverzeichnis ___________________________________________________________________________
154
REGITZ, V., R. J. HODACH u. A. L. SHUG (1982): Carnitin-Mangel: Eine behandelbare Ursache kindlicher Kardiomyopathien. Klin. Wochenschr. 60(8), 393-400 RENNIE, M. J., W. W. WINDER u. J. O. HOLLOSZY (1976): A sparing effect of increased plasma fatty acids on muscle and liver glycogen content in exercising rat. Biochem. J. 156, 647-655 RICHTER, E. A., P. JENSEN, B. KIENS u. S. KRISTIANSEN (1998): Sarcolemmal glucose transport and GLUT-4 translocation during exercise are diminished by endurance training. Am. J. Physiol. 274(37), E555-E95 ROMIJN, J. A., S. KLEIN, E. F. COYLE, L. S. SIDOSSIS u. R. P. WOLFE (1993): Strenuous endurance training increases lipolysis and triglyceride-fatty acid cycle at rest. J. Appl. Physiol. 75 (1), 108-113 ROOS, N., M. DEVRESE, H. SCHULTE-COERNE u. C. A. BARTH (1992): L-Carnitin in Milch von monozygoten Zwillingskühen. Kieler Milchwirtsch. Forschungsber. 44, 363-370 ROSE, R. J., J. R. ALLEN, D. R. HODGSON, J. H. STEWART u. W. CHAN (1983 b): Response to submaximal treadmill exercise and training in the horse: changes in haematology, arterial blood gas and acid base measurements, plasma biochemical values and heart rate. Vet. Rec. 113(26-27), 608-612 ROSE, R. J., D. R. HODGSON, D. SAMPSON, D. u. W. CHAN (1983 a): Changes in plasma biochemistry in horses competing in a 160 km endurance ride. Austr. Vet. J. 67, 101-105 ROSE, R. J., J. ILKIW, K. S. ARNOLD, J. W. BACKHOUSE u. D. SAMPSON (1980): Plasma biochemistry in the horse during 3-day event competition. Equine vet. J. 12, 132-136 ROSSI, C. S., u. N. SILIPRANDI (1982): Effect of carnitine on serum HDL-cholesterol: report on 2 cases. John Hopkins Med. J. 150, 51-54 ROWELL, L. B., B. SALTIN, B. KIENS u. N. J. CHRISTENSEN (1986): Is peak quadriceps blood flow in humans even higher during exercise with hypoxemia. Am. J. Physiol. 251, H1038-H1044 RUDMAN, D. C. W., J. D. SEWELL u. J. D. ANSLEY (1977): Deficiency of carnitine in cachectic cirrhotic patients. J. Clin. Invest. 60, 716-723

VIII. Literaturverzeichnis ___________________________________________________________________________
155
RUSSEL, R. R., u. H. TAEGTMEYER (1992): Coenzym A sequestration in rat hearts oxidizing ketone bodies. J. Clin. Invest. 89, 968-973 SADDIK, M., J. GAMBLE, L. A. WITTERS u. G. D. LOPASCHUK (1993): Acetyl-CoA carboxylase regulation of fatty acid oxidation in the heart. J. Biol. Chem. 268, 25836-25845 SANGUQ, M., D. J. PAULSON, R. FREGOSI, W. REDDAN u. A. L. SHUG (1984): Effects of training on maximum oxygen consumption, mitochondrial respiration, cardiac performance and carnitine tissue levels. Fed. Proc. 43, 293 SCHEK, A. (1994): L-Carnitin: Sinn und Unsinn der Substitution einer körpereigenen Substanz. Ernährungsumschau 41, 9-15 SCHOLTE, H. R. u. P. C. DE JONGE (1987): Metabolism, function and transport of carnitine in health and disease. In: R. GITZELMANN, K. BAERLOCHER u. B. STEINMANN (Edit.): Carnitin in der Medizin. Verlag Schattauer, Stuttgart, New York, S. 21-59 SHAW, R.D., B. U. K. LI, A. L. HAMILTON, A. L. SHUG u. W. A. OLSEN (1983): Carnitine transport in rat small intestine. Am. J. Physiol. 245, G376-G381 SICILIANO, P. D., A. L. PARKER u. L. M. LAWRENCE (1997): Effect of dietary vitamin E supplementation on the integrity of skeletal muscle in exercises horses. J. Anim. Sci. 75(6),1553-1560 SILIPRANDI, N., F. DI LISA, G. PIERALISI, P. RIPARI, F.Ö MACCARI, R. MENABO, M. A. GIAMBERARDINO u. L. VECCHIET (1990): Metabolic changes induced by maximal exercise in human subjects following L-carnitine administration. Biochim. Biophys. Acta 1034, 17-21 SILIPRANDI, N., L. VECCHIET u. F. DI LISA (1989): Carnitine administration in physical exercise. In: G. BENZI u. J. LIBBEY (Edit.): Advances in myochemistry. 2. Aufl., Eurotext Ltd., S. 273-278

VIII. Literaturverzeichnis ___________________________________________________________________________
156
SKARDA, R. T., W. W. MUIR, D. W. MILNE u. A. A. GABEL (1976): Effects of training on resting and postexercise ECG in Standardbred horses, using a standardized exercise test. Am. J. Vet. Res. 37(12), 1485-1488 SLOET VAN OLDRUITENBORGH-OOSTERBAAN, M. M., T. WENSING, A. BARNEVELD u. J. BREUKING (1991): Heart rate, blood biochemistry and performance of horses competing in a 100 km endurance ride. Vet. Rec. 128, 175-179 SNOW, D. H., u. P. BAXTER (1981): Muscle fiber composition and glycogen depletion in horses competing in an endurance ride. Vet. Rec. 108, 374-378 SNOW, D. H., u. R. C. HARRIS (1991): Effects of daily exercise on muscle glycogen in the Thoroughbred Racehorse. In: S. G. B. PERSSON, A. LINDHOLM u. L. B. JEFFCOTT (Edit.): Equine Exercise Physiology 3. ICEEP Publications, Davis, California, S. 299-304 SNOW, D. H., R. C. HARRIS u. S. GASH (1985): Metabolic response of equine muscle to intermittent maximal exercise. J. Appl. Phys. 58, 1689-1697 SNOW, D. H., M. G. KERR, M. A. NIMMO u. E. M. ABBOT (1982): Alterations in blood, sweat, urine and muscle composition during prolonged exercise in the horse. Vet. Rec. 110, 377-384 SNOW, D. H., u. G. MACKENZIE (1977): Some metabolic effects of maximal exercise in the horse and adaptations with training. Equine vet. J. 9(3), 134-140 SNOW, D. H., D. K. MASON, S. W. RICKETTS u. T. A. DOUGLAS (1983): Post-race blood biochemistry in Thoroughbreds. In: D. H. SNOW, S. G. B. PERSSON u. R. J. ROSE (Edit.): Equine Exercise Physiology 1. Granta Editions, Cambridge, S. 389-407 SNOW, D. H., R. J. SUMMERS u. P. S. GUY (1979): The actions of the β-adrenoceptor blocking agents propranolol and metoprolol in the maximal exercised horse. Res. Vet. Sci. 27, 22-29

VIII. Literaturverzeichnis ___________________________________________________________________________
157
SNOW, D. H., u. VALBERG (1994): Muscle anatomy, physiology and adaptations to exercise and training. In: D. R. HODGSON u. R. J. ROSE (Edit.): The Athletic Horse – Principales and Practice of Equine Sports Medicine. W. B. Saunders Comp., Philadelphia, S. 145-179 SOOP, M., O. BJORKMAN, G. CEDERBLAD, L. HAGENFELDT u. J. WAHREN (1988): Influence of carnitine supplementation on muscle substrate and carnitine metabolism during exercise. J. Appl. Physiol. 64, 2394-2399 SUGDEN, M. C., u. M. J. HOLNESS (1994): Interactive regulation of the pyruvat dehydrogenase complex and the carnitine palmitoyltransferase system. Faseb. J. 8(1), 54-61 SUZUKI, Y., T. KAMIKAWA u. N. YAMAZAKI (1981): Effect of L-carnitine on cardiac hemodynamics. Jpn. Heart. J. 22(2), 219-225 THOMPSON, P. D., E. M. CULLINANE, S. P. SADY, M. M. FLYNN, C. B. CHENEVERT u. P. N. HERBERT (1991): High density lipoprotein metabolism in endurance athletes and sedentary men. Circulation 84, 140-152 TURCOTTE, L. P., E. A. RITCHER u. B. KIENS (1992): Increased plasa FFA uptake and oxidation during prolonged exercise in trained vs. untrained humans. Am. J. Physiol. 262, E791-E799 TYLER, C. M., D. R. HODGSON u. R. J. ROSE (1996): Effect of a warm-up on energy supply during high intensity exercise in horses. Equine vet. J. 28, 117-120 VECCHIET, L., F. DI LISA, G. PIERALISI, P. RIPARI, R. MENABO, M. A. GIAMBERARDINO u. N. SILIPRANDI (1990): Influence of L-carnitine administration on maximal physical exercise. Eur. J. Appl. Physiol. 61, 486-490 UZIEL, G., B. BARAVAGALIA u. S. DIDONATO (1988): Carnitine stimulation of pyruvate dehydrogenase complex (PDHC) in isolated human skeletal muscle mitochondria. Muscle Nerve 11, 720-724

VIII. Literaturverzeichnis ___________________________________________________________________________
158
WEBER, J. M., W. S. PARKHOUSE, G. P. DOBSON, J. C. HARMAN, D. H. SNOW u. P. W. HOCHACHKA (1987): Onset of submaximal exercise in Thoroughbred horses. Ca. J. Zoo. 65, 2513-2518 WEICKER, H. (1994): Interaktion zwischen aerober und anaerober Energieproduktion, Laktatproduktion, Release und Elimination. In: D. CLASIN, H. WEICKER u. D. BÖNING: Stellenwert der Laktatbestimmung in der Leistungsdiagnostik. Gustav Fischer Verlag, Stuttgart, Jena, New York, S. 11-25 WERNER, W., H. G. REY u. Z. WIELINGER (1970): Über die Eigenschaften eines neuen Chromogens für die Blutzuckerbestimmung nach der GOD/POD-Methode. Z. Analyt. Chem. 252, 224 WILLIAMSON, C. C., E. A. JAMES, M. P. JAMES, C. D. MAY u. P. J. CASEY (1996): Horse plasma lactate determination: comparison of wet and dry chemistry methods and the effect of storage. Equine vet. J. 28, 406-408 WILSON, R. G., J. R. THRONTON, S. INGLIS u. J. AINSCOW (1987): Skeletal muscle adaptation in racehorses following high intensity interval training. In: J. R. GILLESPIE u. N. E. ROBINSON (Edit.): Equine Exercise Physiology 2. ICEEP Publications, Davis, California, S. 367-375 WOLFE, R. R., S. KLEIN, F. CARRARO u. J. M. WEBER (1990): Role of triglyceride-fatty acid cycling in controlling fat metabolism in humans during and after exercise. Am. J. Physiol. 258(11), E382-E389 YOUNG, L. E. (1999): Cardiac responses to training in 2-year-old Thoroughbreds: an echocardiographic study. Equine vet. J. Suppl. 30, 195-198 ZASPEL, B. J., K. J. SHERIDAN u. L. M. HENDERSON (1980): Transport and metabolism of carnitine precursors in various organs of the rat. Biochim. Biophys. Acta. 631, 192-202 ZENDZIAN-PIOTROWSKA, M., u. J. GORSKI (1993): Metabolic adaptation to daily exercise of moderate intensity to exhaustion in the rat. Eur. J. Appl. Physiol. 67 (1), 77-82

VIII. Literaturverzeichnis ___________________________________________________________________________
159
ZIMMERMAN, N. I., S. J. WICKLER, A. V. RODIEK u. M. A. HOWER (1992): Free fatty acids in exercising Arabian horses fed two common diets. J. Nutr. 12, 145-150
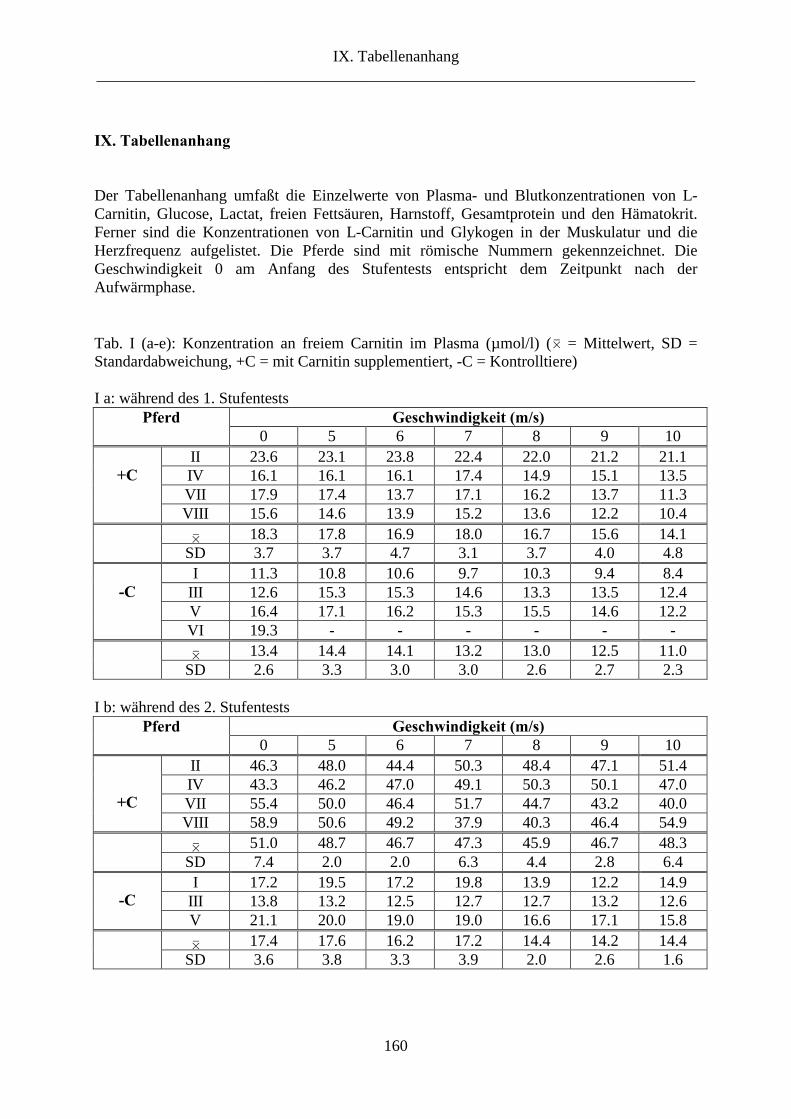
IX. Tabellenanhang ___________________________________________________________________________
160
IX. Tabellenanhang Der Tabellenanhang umfaßt die Einzelwerte von Plasma- und Blutkonzentrationen von L-Carnitin, Glucose, Lactat, freien Fettsäuren, Harnstoff, Gesamtprotein und den Hämatokrit. Ferner sind die Konzentrationen von L-Carnitin und Glykogen in der Muskulatur und die Herzfrequenz aufgelistet. Die Pferde sind mit römische Nummern gekennzeichnet. Die Geschwindigkeit 0 am Anfang des Stufentests entspricht dem Zeitpunkt nach der Aufwärmphase. Tab. I (a-e): Konzentration an freiem Carnitin im Plasma (µmol/l) (0 = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) I a: während des 1. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 23.6 23.1 23.8 22.4 22.0 21.2 21.1 IV 16.1 16.1 16.1 17.4 14.9 15.1 13.5 VII 17.9 17.4 13.7 17.1 16.2 13.7 11.3
+C VIII 15.6 14.6 13.9 15.2 13.6 12.2 10.4
0 18.3 17.8 16.9 18.0 16.7 15.6 14.1 SD 3.7 3.7 4.7 3.1 3.7 4.0 4.8
I 11.3 10.8 10.6 9.7 10.3 9.4 8.4 III 12.6 15.3 15.3 14.6 13.3 13.5 12.4 V 16.4 17.1 16.2 15.3 15.5 14.6 12.2
-C VI 19.3 - - - - - -
0 13.4 14.4 14.1 13.2 13.0 12.5 11.0 SD 2.6 3.3 3.0 3.0 2.6 2.7 2.3
I b: während des 2. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 46.3 48.0 44.4 50.3 48.4 47.1 51.4 IV 43.3 46.2 47.0 49.1 50.3 50.1 47.0 VII 55.4 50.0 46.4 51.7 44.7 43.2 40.0 +C
VIII 58.9 50.6 49.2 37.9 40.3 46.4 54.9 0 51.0 48.7 46.7 47.3 45.9 46.7 48.3
SD 7.4 2.0 2.0 6.3 4.4 2.8 6.4 I 17.2 19.5 17.2 19.8 13.9 12.2 14.9
III 13.8 13.2 12.5 12.7 12.7 13.2 12.6 -C V 21.1 20.0 19.0 19.0 16.6 17.1 15.8
0 17.4 17.6 16.2 17.2 14.4 14.2 14.4 SD 3.6 3.8 3.3 3.9 2.0 2.6 1.6
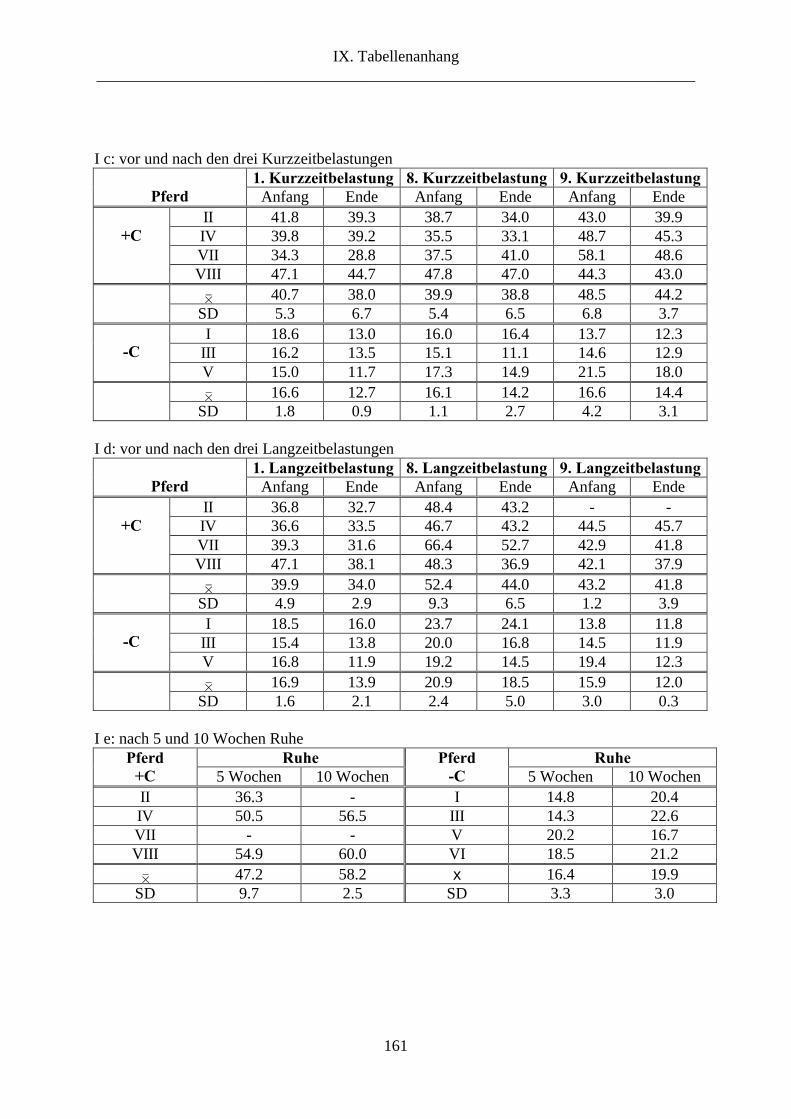
IX. Tabellenanhang ___________________________________________________________________________
161
I c: vor und nach den drei Kurzzeitbelastungen
1. Kurzzeitbelastung 8. Kurzzeitbelastung 9. Kurzzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 41.8 39.3 38.7 34.0 43.0 39.9 IV 39.8 39.2 35.5 33.1 48.7 45.3 VII 34.3 28.8 37.5 41.0 58.1 48.6
+C VIII 47.1 44.7 47.8 47.0 44.3 43.0
0 40.7 38.0 39.9 38.8 48.5 44.2 SD 5.3 6.7 5.4 6.5 6.8 3.7
I 18.6 13.0 16.0 16.4 13.7 12.3 III 16.2 13.5 15.1 11.1 14.6 12.9 -C
V 15.0 11.7 17.3 14.9 21.5 18.0 0 16.6 12.7 16.1 14.2 16.6 14.4
SD 1.8 0.9 1.1 2.7 4.2 3.1 I d: vor und nach den drei Langzeitbelastungen
1. Langzeitbelastung 8. Langzeitbelastung 9. Langzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 36.8 32.7 48.4 43.2 - - IV 36.6 33.5 46.7 43.2 44.5 45.7 VII 39.3 31.6 66.4 52.7 42.9 41.8
+C VIII 47.1 38.1 48.3 36.9 42.1 37.9
0 39.9 34.0 52.4 44.0 43.2 41.8 SD 4.9 2.9 9.3 6.5 1.2 3.9
I 18.5 16.0 23.7 24.1 13.8 11.8 III 15.4 13.8 20.0 16.8 14.5 11.9 -C
V 16.8 11.9 19.2 14.5 19.4 12.3 0 16.9 13.9 20.9 18.5 15.9 12.0
SD 1.6 2.1 2.4 5.0 3.0 0.3 I e: nach 5 und 10 Wochen Ruhe
Ruhe Ruhe Pferd +C 5 Wochen 10 Wochen
Pferd -C 5 Wochen 10 Wochen
II 36.3 - I 14.8 20.4 IV 50.5 56.5 III 14.3 22.6 VII - - V 20.2 16.7 VIII 54.9 60.0 VI 18.5 21.2 0 47.2 58.2 x 16.4 19.9
SD 9.7 2.5 SD 3.3 3.0
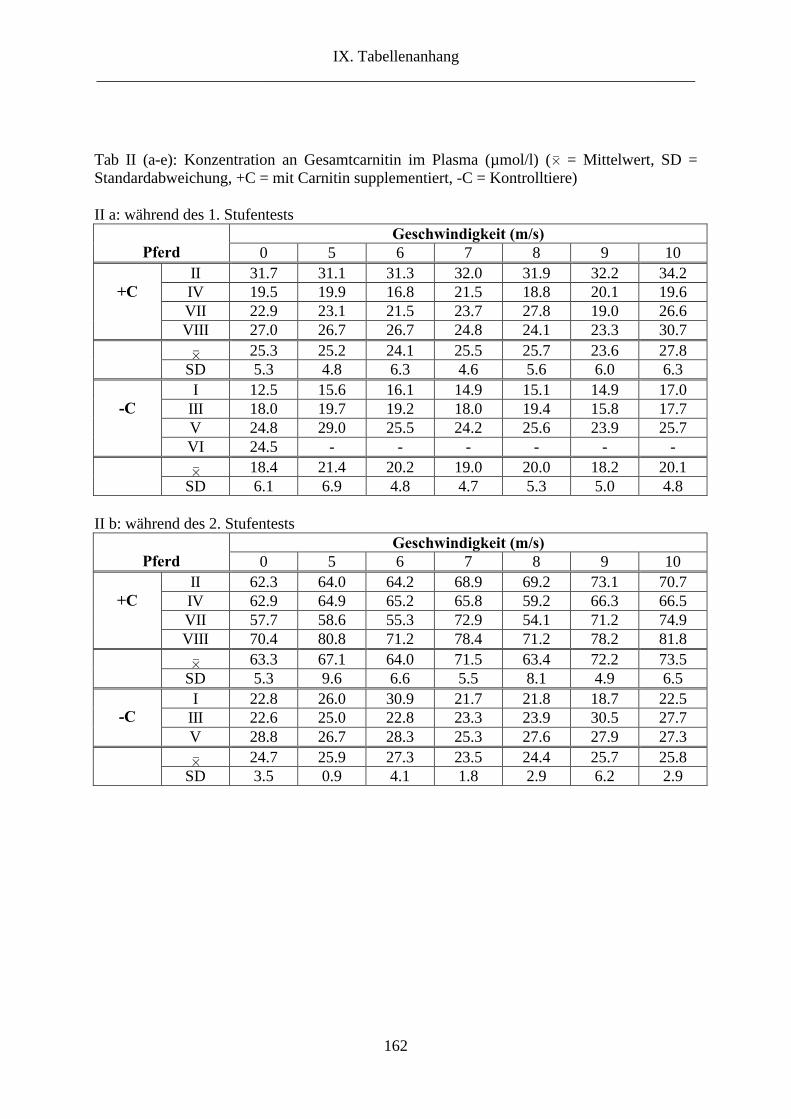
IX. Tabellenanhang ___________________________________________________________________________
162
Tab II (a-e): Konzentration an Gesamtcarnitin im Plasma (µmol/l) (0 = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) II a: während des 1. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 31.7 31.1 31.3 32.0 31.9 32.2 34.2 IV 19.5 19.9 16.8 21.5 18.8 20.1 19.6 VII 22.9 23.1 21.5 23.7 27.8 19.0 26.6
+C VIII 27.0 26.7 26.7 24.8 24.1 23.3 30.7
0 25.3 25.2 24.1 25.5 25.7 23.6 27.8 SD 5.3 4.8 6.3 4.6 5.6 6.0 6.3
I 12.5 15.6 16.1 14.9 15.1 14.9 17.0 III 18.0 19.7 19.2 18.0 19.4 15.8 17.7 V 24.8 29.0 25.5 24.2 25.6 23.9 25.7
-C VI 24.5 - - - - - -
0 18.4 21.4 20.2 19.0 20.0 18.2 20.1 SD 6.1 6.9 4.8 4.7 5.3 5.0 4.8
II b: während des 2. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 62.3 64.0 64.2 68.9 69.2 73.1 70.7 IV 62.9 64.9 65.2 65.8 59.2 66.3 66.5 VII 57.7 58.6 55.3 72.9 54.1 71.2 74.9
+C VIII 70.4 80.8 71.2 78.4 71.2 78.2 81.8
0 63.3 67.1 64.0 71.5 63.4 72.2 73.5 SD 5.3 9.6 6.6 5.5 8.1 4.9 6.5
I 22.8 26.0 30.9 21.7 21.8 18.7 22.5 III 22.6 25.0 22.8 23.3 23.9 30.5 27.7 -C
V 28.8 26.7 28.3 25.3 27.6 27.9 27.3 0 24.7 25.9 27.3 23.5 24.4 25.7 25.8
SD 3.5 0.9 4.1 1.8 2.9 6.2 2.9
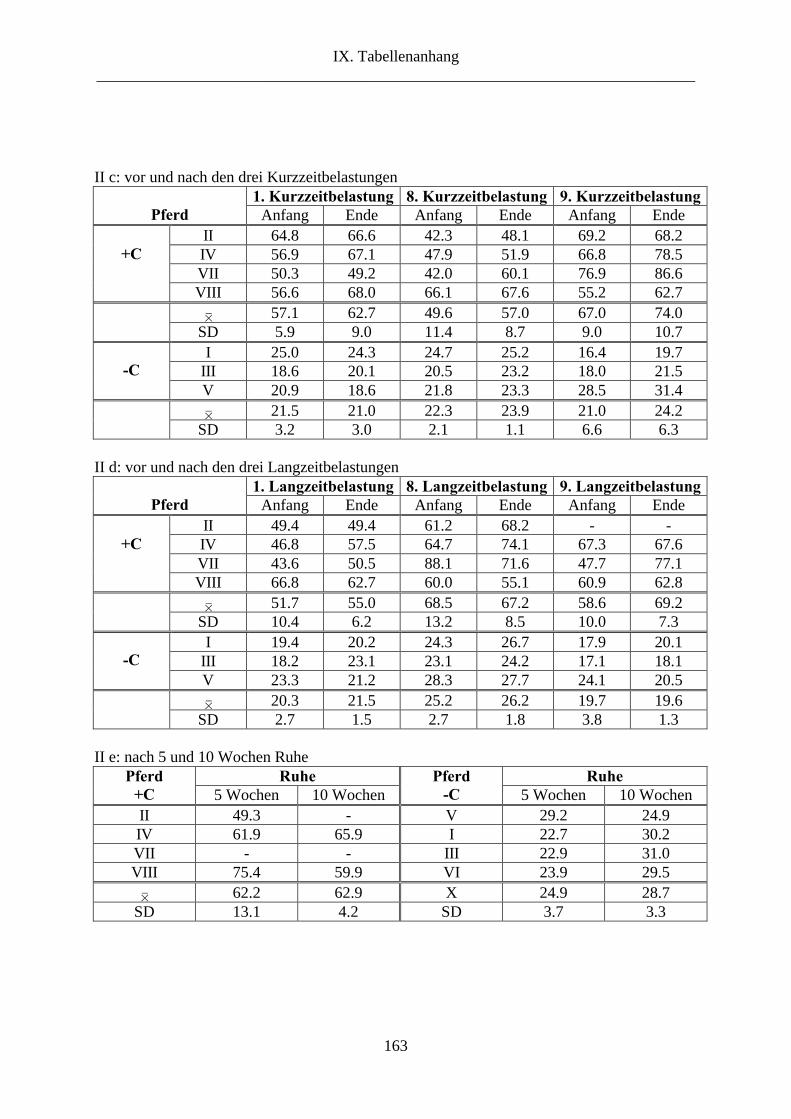
IX. Tabellenanhang ___________________________________________________________________________
163
II c: vor und nach den drei Kurzzeitbelastungen
1. Kurzzeitbelastung 8. Kurzzeitbelastung 9. Kurzzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 64.8 66.6 42.3 48.1 69.2 68.2 IV 56.9 67.1 47.9 51.9 66.8 78.5 VII 50.3 49.2 42.0 60.1 76.9 86.6
+C VIII 56.6 68.0 66.1 67.6 55.2 62.7
0 57.1 62.7 49.6 57.0 67.0 74.0 SD 5.9 9.0 11.4 8.7 9.0 10.7
I 25.0 24.3 24.7 25.2 16.4 19.7 III 18.6 20.1 20.5 23.2 18.0 21.5 -C
V 20.9 18.6 21.8 23.3 28.5 31.4 0 21.5 21.0 22.3 23.9 21.0 24.2
SD 3.2 3.0 2.1 1.1 6.6 6.3 II d: vor und nach den drei Langzeitbelastungen
1. Langzeitbelastung 8. Langzeitbelastung 9. Langzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 49.4 49.4 61.2 68.2 - - IV 46.8 57.5 64.7 74.1 67.3 67.6 VII 43.6 50.5 88.1 71.6 47.7 77.1
+C VIII 66.8 62.7 60.0 55.1 60.9 62.8
0 51.7 55.0 68.5 67.2 58.6 69.2 SD 10.4 6.2 13.2 8.5 10.0 7.3
I 19.4 20.2 24.3 26.7 17.9 20.1 III 18.2 23.1 23.1 24.2 17.1 18.1 -C
V 23.3 21.2 28.3 27.7 24.1 20.5 0 20.3 21.5 25.2 26.2 19.7 19.6
SD 2.7 1.5 2.7 1.8 3.8 1.3 II e: nach 5 und 10 Wochen Ruhe
Ruhe Ruhe Pferd +C 5 Wochen 10 Wochen
Pferd -C 5 Wochen 10 Wochen
II 49.3 - V 29.2 24.9 IV 61.9 65.9 I 22.7 30.2 VII - - III 22.9 31.0 VIII 75.4 59.9 VI 23.9 29.5 0 62.2 62.9 X 24.9 28.7
SD 13.1 4.2 SD 3.7 3.3
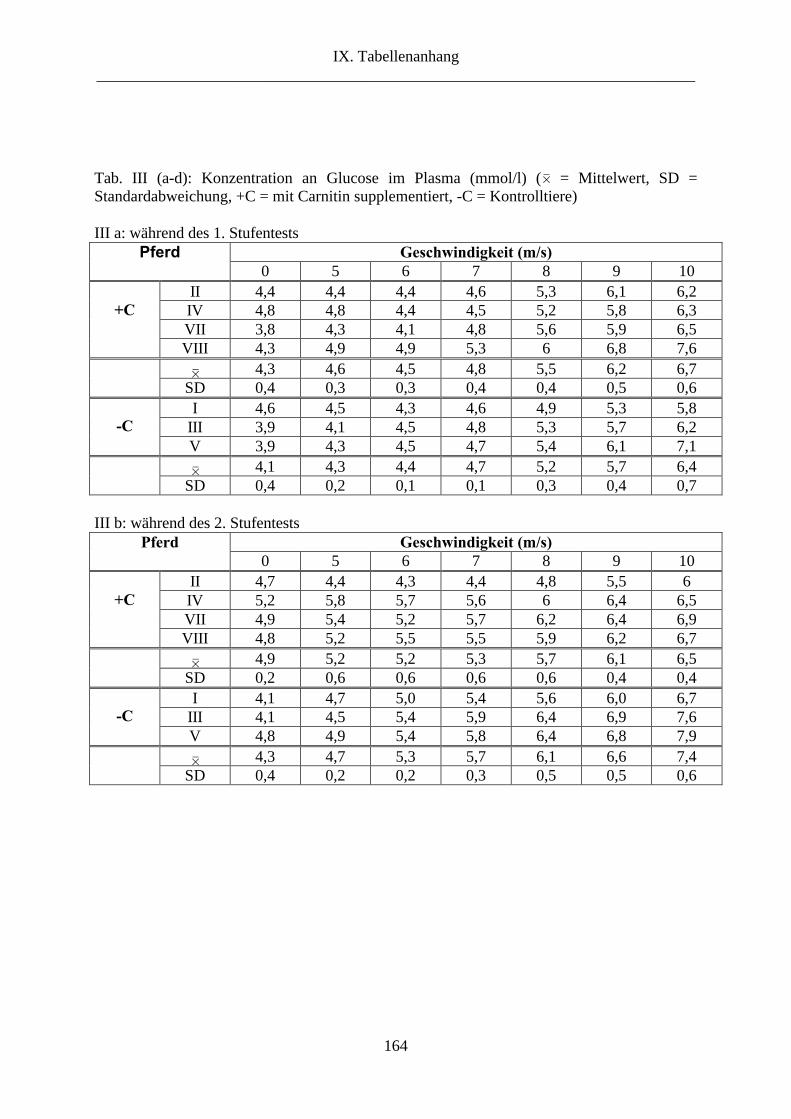
IX. Tabellenanhang ___________________________________________________________________________
164
Tab. III (a-d): Konzentration an Glucose im Plasma (mmol/l) (0 = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) III a: während des 1. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 4,4 4,4 4,4 4,6 5,3 6,1 6,2 IV 4,8 4,8 4,4 4,5 5,2 5,8 6,3 VII 3,8 4,3 4,1 4,8 5,6 5,9 6,5
+C VIII 4,3 4,9 4,9 5,3 6 6,8 7,6
0 4,3 4,6 4,5 4,8 5,5 6,2 6,7 SD 0,4 0,3 0,3 0,4 0,4 0,5 0,6
I 4,6 4,5 4,3 4,6 4,9 5,3 5,8 III 3,9 4,1 4,5 4,8 5,3 5,7 6,2 -C
V 3,9 4,3 4,5 4,7 5,4 6,1 7,1 0 4,1 4,3 4,4 4,7 5,2 5,7 6,4
SD 0,4 0,2 0,1 0,1 0,3 0,4 0,7 III b: während des 2. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 4,7 4,4 4,3 4,4 4,8 5,5 6 IV 5,2 5,8 5,7 5,6 6 6,4 6,5 VII 4,9 5,4 5,2 5,7 6,2 6,4 6,9
+C VIII 4,8 5,2 5,5 5,5 5,9 6,2 6,7
0 4,9 5,2 5,2 5,3 5,7 6,1 6,5 SD 0,2 0,6 0,6 0,6 0,6 0,4 0,4
I 4,1 4,7 5,0 5,4 5,6 6,0 6,7 III 4,1 4,5 5,4 5,9 6,4 6,9 7,6 -C
V 4,8 4,9 5,4 5,8 6,4 6,8 7,9 0 4,3 4,7 5,3 5,7 6,1 6,6 7,4
SD 0,4 0,2 0,2 0,3 0,5 0,5 0,6
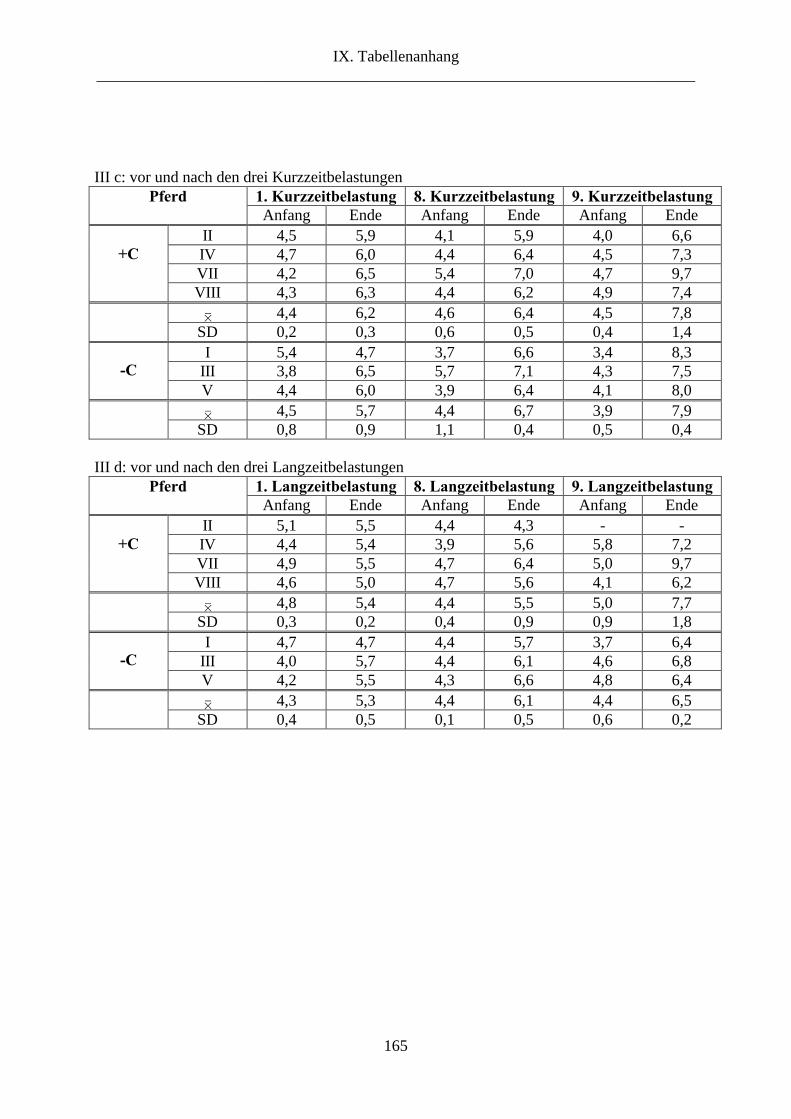
IX. Tabellenanhang ___________________________________________________________________________
165
III c: vor und nach den drei Kurzzeitbelastungen
1. Kurzzeitbelastung 8. Kurzzeitbelastung 9. Kurzzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 4,5 5,9 4,1 5,9 4,0 6,6 IV 4,7 6,0 4,4 6,4 4,5 7,3 VII 4,2 6,5 5,4 7,0 4,7 9,7
+C VIII 4,3 6,3 4,4 6,2 4,9 7,4
0 4,4 6,2 4,6 6,4 4,5 7,8 SD 0,2 0,3 0,6 0,5 0,4 1,4
I 5,4 4,7 3,7 6,6 3,4 8,3 III 3,8 6,5 5,7 7,1 4,3 7,5 -C
V 4,4 6,0 3,9 6,4 4,1 8,0 0 4,5 5,7 4,4 6,7 3,9 7,9
SD 0,8 0,9 1,1 0,4 0,5 0,4 III d: vor und nach den drei Langzeitbelastungen
1. Langzeitbelastung 8. Langzeitbelastung 9. Langzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 5,1 5,5 4,4 4,3 - - IV 4,4 5,4 3,9 5,6 5,8 7,2 VII 4,9 5,5 4,7 6,4 5,0 9,7
+C VIII 4,6 5,0 4,7 5,6 4,1 6,2
0 4,8 5,4 4,4 5,5 5,0 7,7 SD 0,3 0,2 0,4 0,9 0,9 1,8
I 4,7 4,7 4,4 5,7 3,7 6,4 III 4,0 5,7 4,4 6,1 4,6 6,8 -C
V 4,2 5,5 4,3 6,6 4,8 6,4 0 4,3 5,3 4,4 6,1 4,4 6,5
SD 0,4 0,5 0,1 0,5 0,6 0,2
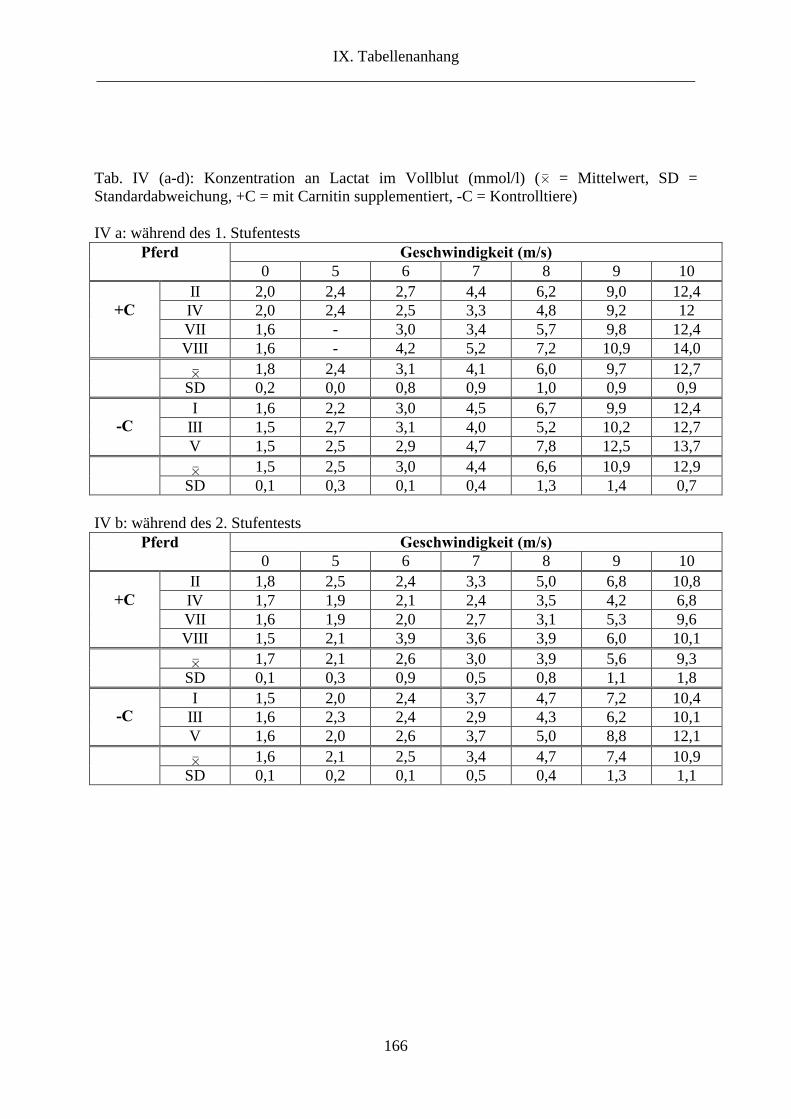
IX. Tabellenanhang ___________________________________________________________________________
166
Tab. IV (a-d): Konzentration an Lactat im Vollblut (mmol/l) (0 = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) IV a: während des 1. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 2,0 2,4 2,7 4,4 6,2 9,0 12,4 IV 2,0 2,4 2,5 3,3 4,8 9,2 12 VII 1,6 - 3,0 3,4 5,7 9,8 12,4
+C VIII 1,6 - 4,2 5,2 7,2 10,9 14,0
0 1,8 2,4 3,1 4,1 6,0 9,7 12,7 SD 0,2 0,0 0,8 0,9 1,0 0,9 0,9
I 1,6 2,2 3,0 4,5 6,7 9,9 12,4 III 1,5 2,7 3,1 4,0 5,2 10,2 12,7 -C
V 1,5 2,5 2,9 4,7 7,8 12,5 13,7 0 1,5 2,5 3,0 4,4 6,6 10,9 12,9
SD 0,1 0,3 0,1 0,4 1,3 1,4 0,7 IV b: während des 2. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 1,8 2,5 2,4 3,3 5,0 6,8 10,8 IV 1,7 1,9 2,1 2,4 3,5 4,2 6,8 VII 1,6 1,9 2,0 2,7 3,1 5,3 9,6
+C VIII 1,5 2,1 3,9 3,6 3,9 6,0 10,1
0 1,7 2,1 2,6 3,0 3,9 5,6 9,3 SD 0,1 0,3 0,9 0,5 0,8 1,1 1,8
I 1,5 2,0 2,4 3,7 4,7 7,2 10,4 III 1,6 2,3 2,4 2,9 4,3 6,2 10,1 -C
V 1,6 2,0 2,6 3,7 5,0 8,8 12,1 0 1,6 2,1 2,5 3,4 4,7 7,4 10,9
SD 0,1 0,2 0,1 0,5 0,4 1,3 1,1
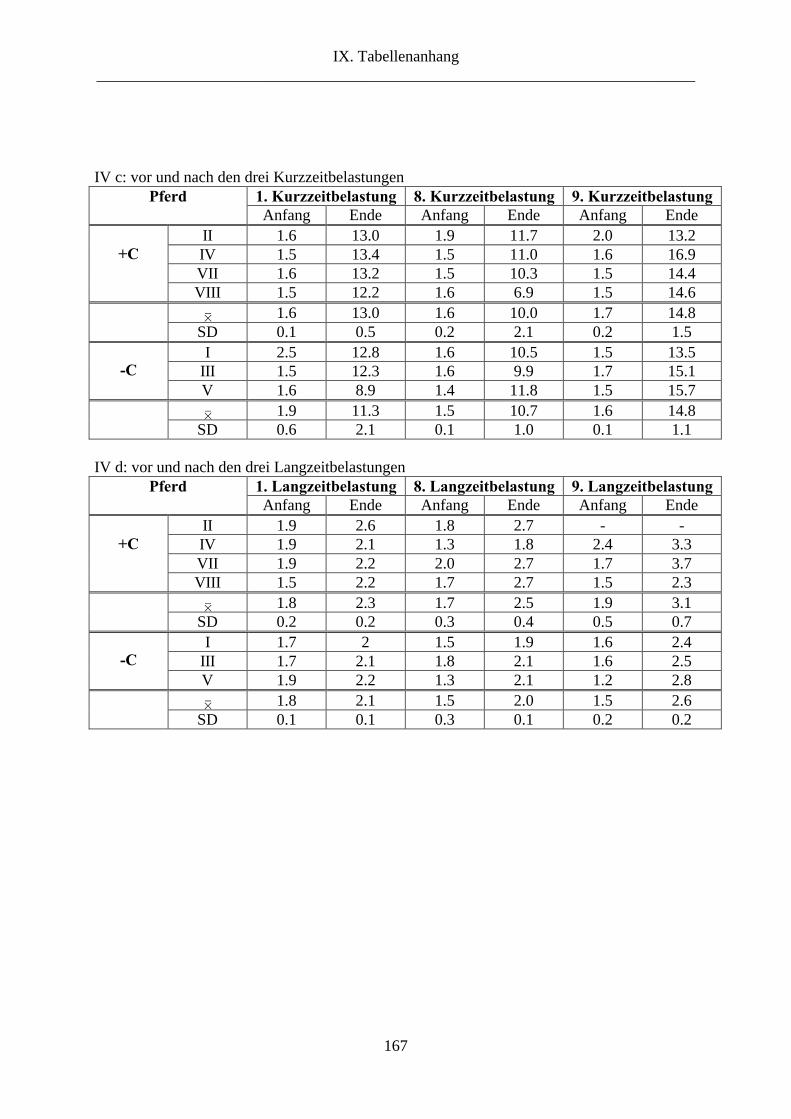
IX. Tabellenanhang ___________________________________________________________________________
167
IV c: vor und nach den drei Kurzzeitbelastungen
1. Kurzzeitbelastung 8. Kurzzeitbelastung 9. Kurzzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 1.6 13.0 1.9 11.7 2.0 13.2 IV 1.5 13.4 1.5 11.0 1.6 16.9 VII 1.6 13.2 1.5 10.3 1.5 14.4
+C VIII 1.5 12.2 1.6 6.9 1.5 14.6
0 1.6 13.0 1.6 10.0 1.7 14.8 SD 0.1 0.5 0.2 2.1 0.2 1.5
I 2.5 12.8 1.6 10.5 1.5 13.5 III 1.5 12.3 1.6 9.9 1.7 15.1 -C
V 1.6 8.9 1.4 11.8 1.5 15.7 0 1.9 11.3 1.5 10.7 1.6 14.8
SD 0.6 2.1 0.1 1.0 0.1 1.1 IV d: vor und nach den drei Langzeitbelastungen
1. Langzeitbelastung 8. Langzeitbelastung 9. Langzeitbelastung Pferd Anfang Ende Anfang Ende Anfang Ende
II 1.9 2.6 1.8 2.7 - - IV 1.9 2.1 1.3 1.8 2.4 3.3 VII 1.9 2.2 2.0 2.7 1.7 3.7
+C VIII 1.5 2.2 1.7 2.7 1.5 2.3
0 1.8 2.3 1.7 2.5 1.9 3.1 SD 0.2 0.2 0.3 0.4 0.5 0.7
I 1.7 2 1.5 1.9 1.6 2.4 III 1.7 2.1 1.8 2.1 1.6 2.5 -C
V 1.9 2.2 1.3 2.1 1.2 2.8 0 1.8 2.1 1.5 2.0 1.5 2.6
SD 0.1 0.1 0.3 0.1 0.2 0.2
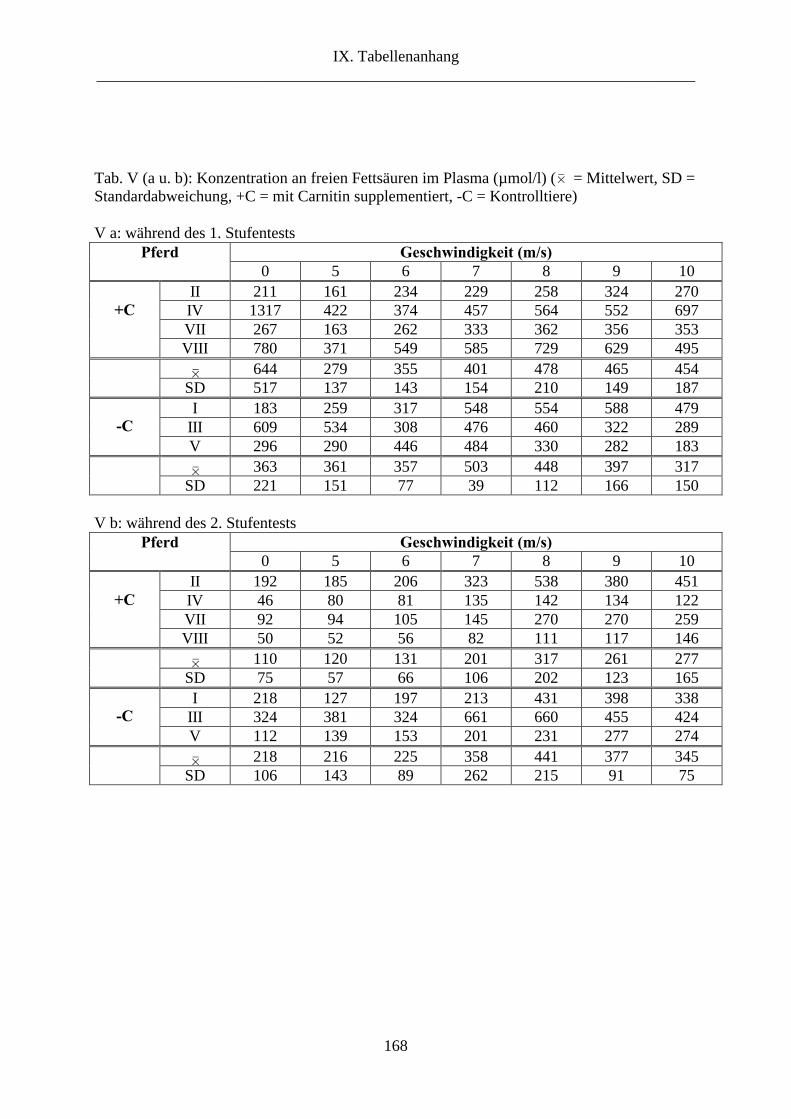
IX. Tabellenanhang ___________________________________________________________________________
168
Tab. V (a u. b): Konzentration an freien Fettsäuren im Plasma (µmol/l) (0 = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) V a: während des 1. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 211 161 234 229 258 324 270 IV 1317 422 374 457 564 552 697 VII 267 163 262 333 362 356 353
+C VIII 780 371 549 585 729 629 495
0 644 279 355 401 478 465 454 SD 517 137 143 154 210 149 187
I 183 259 317 548 554 588 479 III 609 534 308 476 460 322 289 -C
V 296 290 446 484 330 282 183 0 363 361 357 503 448 397 317
SD 221 151 77 39 112 166 150 V b: während des 2. Stufentests
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 192 185 206 323 538 380 451 IV 46 80 81 135 142 134 122 VII 92 94 105 145 270 270 259
+C VIII 50 52 56 82 111 117 146
0 110 120 131 201 317 261 277 SD 75 57 66 106 202 123 165
I 218 127 197 213 431 398 338 III 324 381 324 661 660 455 424 -C
V 112 139 153 201 231 277 274 0 218 216 225 358 441 377 345
SD 106 143 89 262 215 91 75
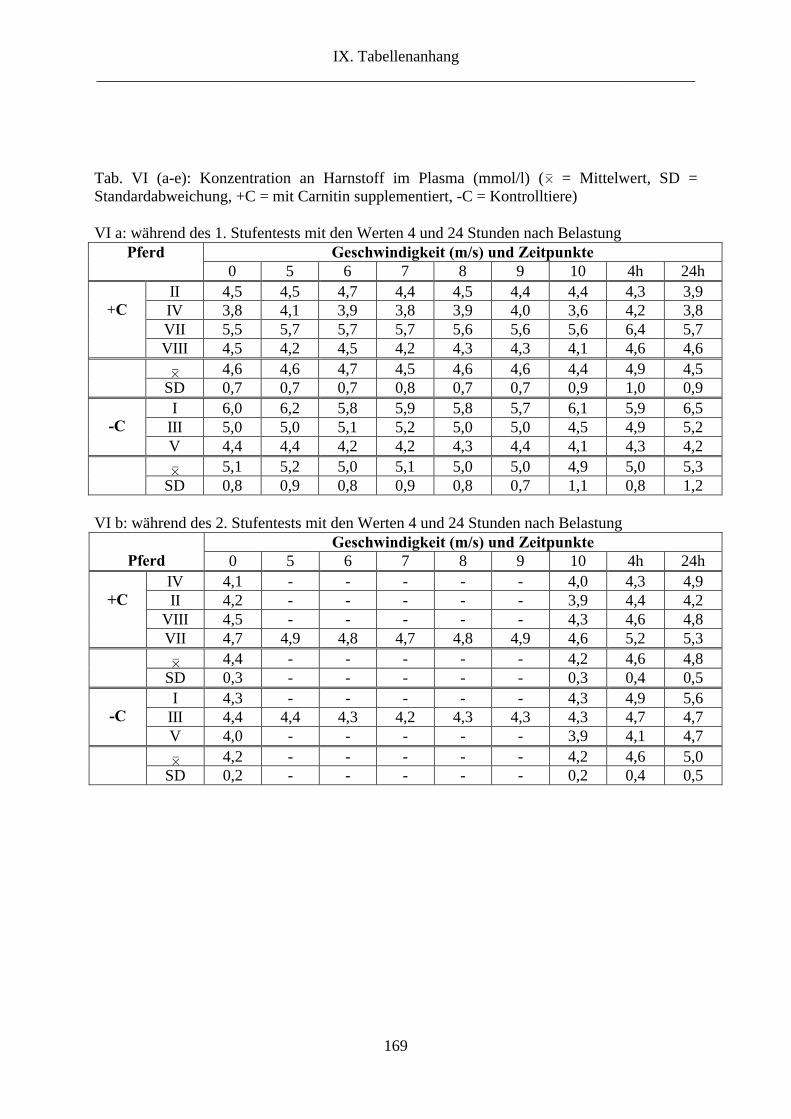
IX. Tabellenanhang ___________________________________________________________________________
169
Tab. VI (a-e): Konzentration an Harnstoff im Plasma (mmol/l) (0 = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) VI a: während des 1. Stufentests mit den Werten 4 und 24 Stunden nach Belastung
Geschwindigkeit (m/s) und Zeitpunkte Pferd 0 5 6 7 8 9 10 4h 24h
II 4,5 4,5 4,7 4,4 4,5 4,4 4,4 4,3 3,9 IV 3,8 4,1 3,9 3,8 3,9 4,0 3,6 4,2 3,8 VII 5,5 5,7 5,7 5,7 5,6 5,6 5,6 6,4 5,7
+C VIII 4,5 4,2 4,5 4,2 4,3 4,3 4,1 4,6 4,6
0 4,6 4,6 4,7 4,5 4,6 4,6 4,4 4,9 4,5 SD 0,7 0,7 0,7 0,8 0,7 0,7 0,9 1,0 0,9
I 6,0 6,2 5,8 5,9 5,8 5,7 6,1 5,9 6,5 III 5,0 5,0 5,1 5,2 5,0 5,0 4,5 4,9 5,2 -C
V 4,4 4,4 4,2 4,2 4,3 4,4 4,1 4,3 4,2 0 5,1 5,2 5,0 5,1 5,0 5,0 4,9 5,0 5,3
SD 0,8 0,9 0,8 0,9 0,8 0,7 1,1 0,8 1,2 VI b: während des 2. Stufentests mit den Werten 4 und 24 Stunden nach Belastung
Geschwindigkeit (m/s) und Zeitpunkte Pferd 0 5 6 7 8 9 10 4h 24h
IV 4,1 - - - - - 4,0 4,3 4,9 II 4,2 - - - - - 3,9 4,4 4,2
VIII 4,5 - - - - - 4,3 4,6 4,8 +C
VII 4,7 4,9 4,8 4,7 4,8 4,9 4,6 5,2 5,3
0 4,4 - - - - - 4,2 4,6 4,8 SD 0,3 - - - - - 0,3 0,4 0,5
I 4,3 - - - - - 4,3 4,9 5,6 III 4,4 4,4 4,3 4,2 4,3 4,3 4,3 4,7 4,7 -C
V 4,0 - - - - - 3,9 4,1 4,7 0 4,2 - - - - - 4,2 4,6 5,0
SD 0,2 - - - - - 0,2 0,4 0,5
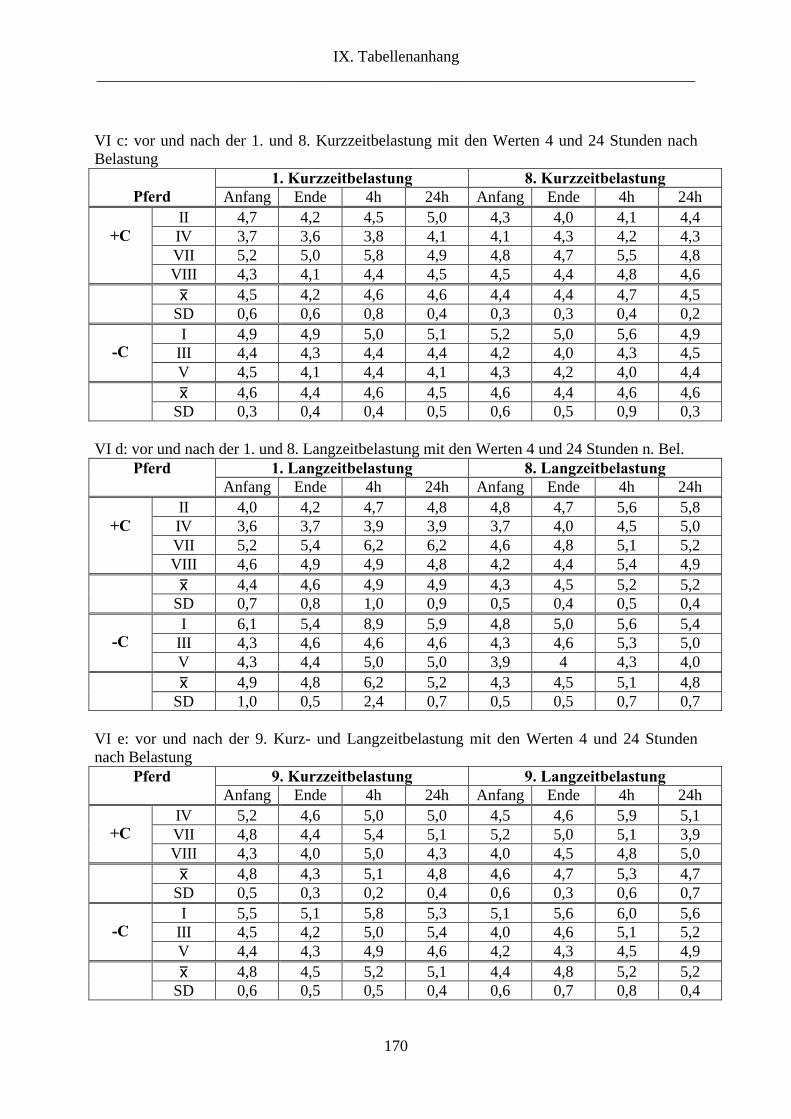
IX. Tabellenanhang ___________________________________________________________________________
170
VI c: vor und nach der 1. und 8. Kurzzeitbelastung mit den Werten 4 und 24 Stunden nach Belastung
1. Kurzzeitbelastung 8. Kurzzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 4,7 4,2 4,5 5,0 4,3 4,0 4,1 4,4 IV 3,7 3,6 3,8 4,1 4,1 4,3 4,2 4,3 VII 5,2 5,0 5,8 4,9 4,8 4,7 5,5 4,8
+C VIII 4,3 4,1 4,4 4,5 4,5 4,4 4,8 4,6
x 4,5 4,2 4,6 4,6 4,4 4,4 4,7 4,5 SD 0,6 0,6 0,8 0,4 0,3 0,3 0,4 0,2
I 4,9 4,9 5,0 5,1 5,2 5,0 5,6 4,9 III 4,4 4,3 4,4 4,4 4,2 4,0 4,3 4,5 -C
V 4,5 4,1 4,4 4,1 4,3 4,2 4,0 4,4 x 4,6 4,4 4,6 4,5 4,6 4,4 4,6 4,6
SD 0,3 0,4 0,4 0,5 0,6 0,5 0,9 0,3 VI d: vor und nach der 1. und 8. Langzeitbelastung mit den Werten 4 und 24 Stunden n. Bel.
1. Langzeitbelastung 8. Langzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 4,0 4,2 4,7 4,8 4,8 4,7 5,6 5,8 IV 3,6 3,7 3,9 3,9 3,7 4,0 4,5 5,0 VII 5,2 5,4 6,2 6,2 4,6 4,8 5,1 5,2
+C VIII 4,6 4,9 4,9 4,8 4,2 4,4 5,4 4,9
x 4,4 4,6 4,9 4,9 4,3 4,5 5,2 5,2 SD 0,7 0,8 1,0 0,9 0,5 0,4 0,5 0,4
I 6,1 5,4 8,9 5,9 4,8 5,0 5,6 5,4 III 4,3 4,6 4,6 4,6 4,3 4,6 5,3 5,0 -C
V 4,3 4,4 5,0 5,0 3,9 4 4,3 4,0 x 4,9 4,8 6,2 5,2 4,3 4,5 5,1 4,8
SD 1,0 0,5 2,4 0,7 0,5 0,5 0,7 0,7 VI e: vor und nach der 9. Kurz- und Langzeitbelastung mit den Werten 4 und 24 Stunden nach Belastung
9. Kurzzeitbelastung 9. Langzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
IV 5,2 4,6 5,0 5,0 4,5 4,6 5,9 5,1 VII 4,8 4,4 5,4 5,1 5,2 5,0 5,1 3,9 +C
VIII 4,3 4,0 5,0 4,3 4,0 4,5 4,8 5,0 x 4,8 4,3 5,1 4,8 4,6 4,7 5,3 4,7
SD 0,5 0,3 0,2 0,4 0,6 0,3 0,6 0,7 I 5,5 5,1 5,8 5,3 5,1 5,6 6,0 5,6
III 4,5 4,2 5,0 5,4 4,0 4,6 5,1 5,2 -C V 4,4 4,3 4,9 4,6 4,2 4,3 4,5 4,9
x 4,8 4,5 5,2 5,1 4,4 4,8 5,2 5,2 SD 0,6 0,5 0,5 0,4 0,6 0,7 0,8 0,4
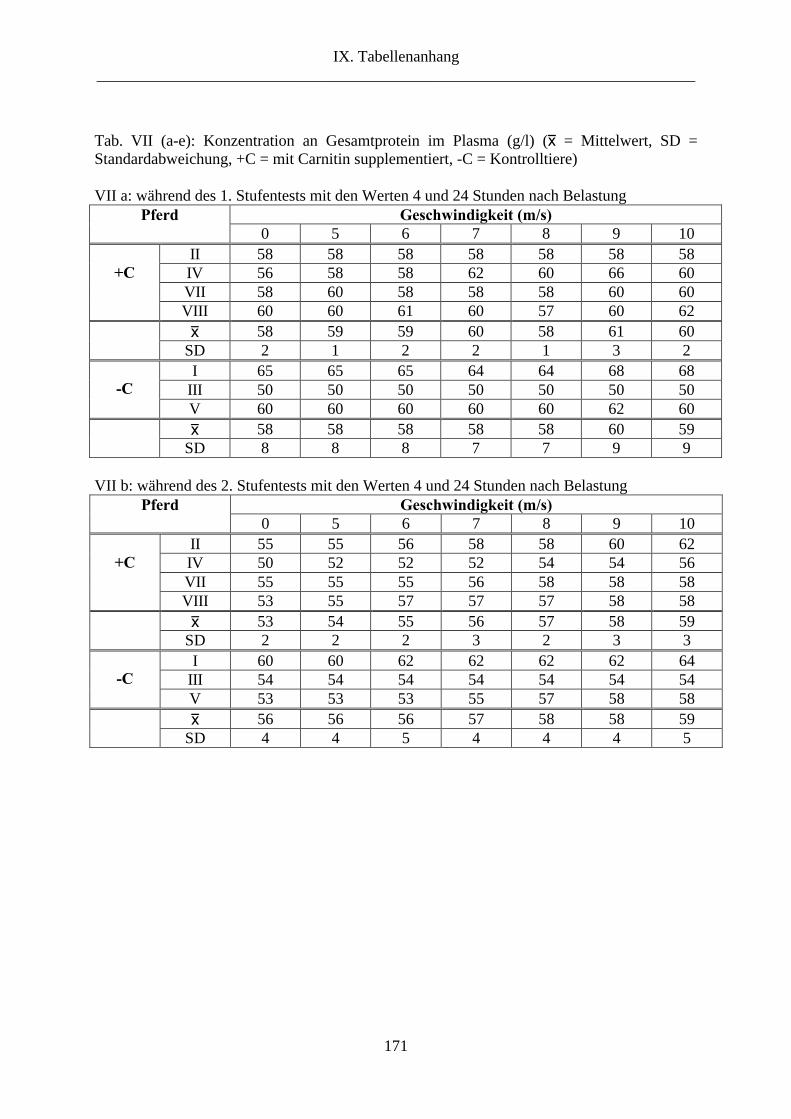
IX. Tabellenanhang ___________________________________________________________________________
171
Tab. VII (a-e): Konzentration an Gesamtprotein im Plasma (g/l) (x = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) VII a: während des 1. Stufentests mit den Werten 4 und 24 Stunden nach Belastung
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 58 58 58 58 58 58 58 IV 56 58 58 62 60 66 60 VII 58 60 58 58 58 60 60
+C VIII 60 60 61 60 57 60 62
x 58 59 59 60 58 61 60 SD 2 1 2 2 1 3 2
I 65 65 65 64 64 68 68 III 50 50 50 50 50 50 50 -C
V 60 60 60 60 60 62 60 x 58 58 58 58 58 60 59
SD 8 8 8 7 7 9 9 VII b: während des 2. Stufentests mit den Werten 4 und 24 Stunden nach Belastung
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 55 55 56 58 58 60 62 IV 50 52 52 52 54 54 56 VII 55 55 55 56 58 58 58
+C VIII 53 55 57 57 57 58 58
x 53 54 55 56 57 58 59 SD 2 2 2 3 2 3 3
I 60 60 62 62 62 62 64 III 54 54 54 54 54 54 54 -C
V 53 53 53 55 57 58 58 x 56 56 56 57 58 58 59
SD 4 4 5 4 4 4 5
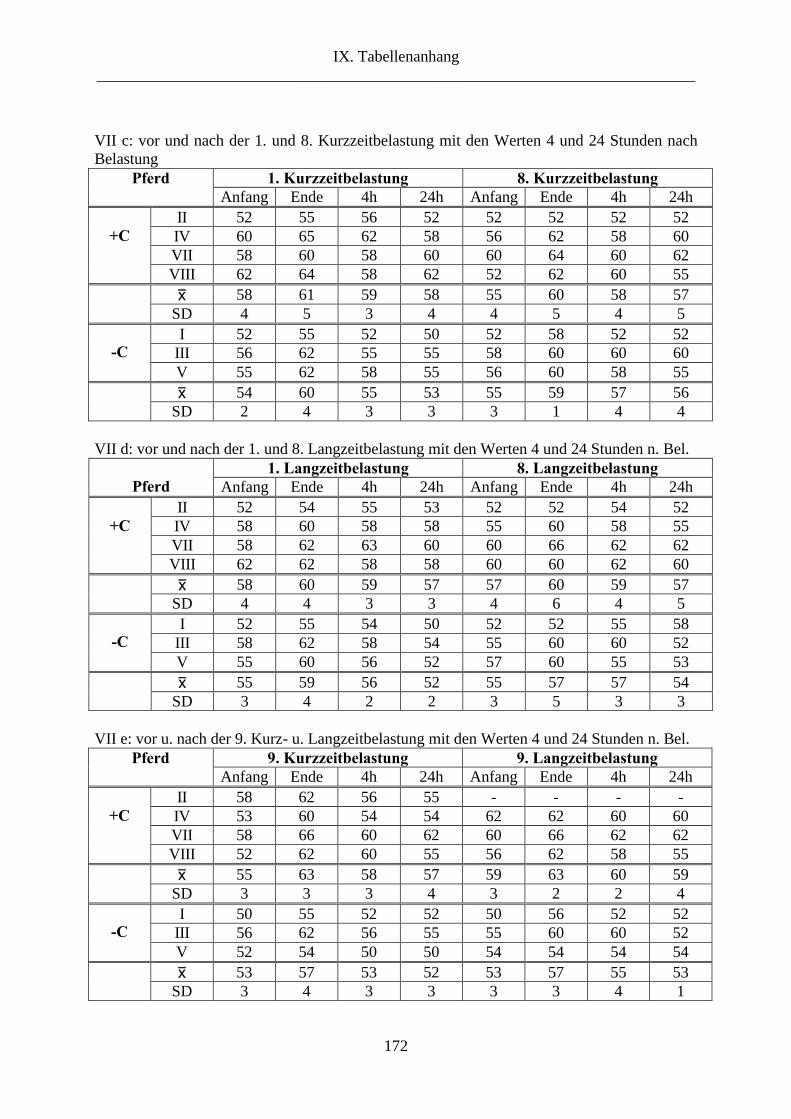
IX. Tabellenanhang ___________________________________________________________________________
172
VII c: vor und nach der 1. und 8. Kurzzeitbelastung mit den Werten 4 und 24 Stunden nach Belastung
1. Kurzzeitbelastung 8. Kurzzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 52 55 56 52 52 52 52 52 IV 60 65 62 58 56 62 58 60 VII 58 60 58 60 60 64 60 62
+C VIII 62 64 58 62 52 62 60 55
x 58 61 59 58 55 60 58 57 SD 4 5 3 4 4 5 4 5
I 52 55 52 50 52 58 52 52 III 56 62 55 55 58 60 60 60 -C
V 55 62 58 55 56 60 58 55 x 54 60 55 53 55 59 57 56
SD 2 4 3 3 3 1 4 4 VII d: vor und nach der 1. und 8. Langzeitbelastung mit den Werten 4 und 24 Stunden n. Bel.
1. Langzeitbelastung 8. Langzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 52 54 55 53 52 52 54 52 IV 58 60 58 58 55 60 58 55 VII 58 62 63 60 60 66 62 62
+C VIII 62 62 58 58 60 60 62 60
x 58 60 59 57 57 60 59 57 SD 4 4 3 3 4 6 4 5
I 52 55 54 50 52 52 55 58 III 58 62 58 54 55 60 60 52 -C
V 55 60 56 52 57 60 55 53 x 55 59 56 52 55 57 57 54
SD 3 4 2 2 3 5 3 3 VII e: vor u. nach der 9. Kurz- u. Langzeitbelastung mit den Werten 4 und 24 Stunden n. Bel.
9. Kurzzeitbelastung 9. Langzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 58 62 56 55 - - - - IV 53 60 54 54 62 62 60 60 VII 58 66 60 62 60 66 62 62
+C VIII 52 62 60 55 56 62 58 55
x 55 63 58 57 59 63 60 59 SD 3 3 3 4 3 2 2 4
I 50 55 52 52 50 56 52 52 III 56 62 56 55 55 60 60 52 -C
V 52 54 50 50 54 54 54 54 x 53 57 53 52 53 57 55 53
SD 3 4 3 3 3 3 4 1
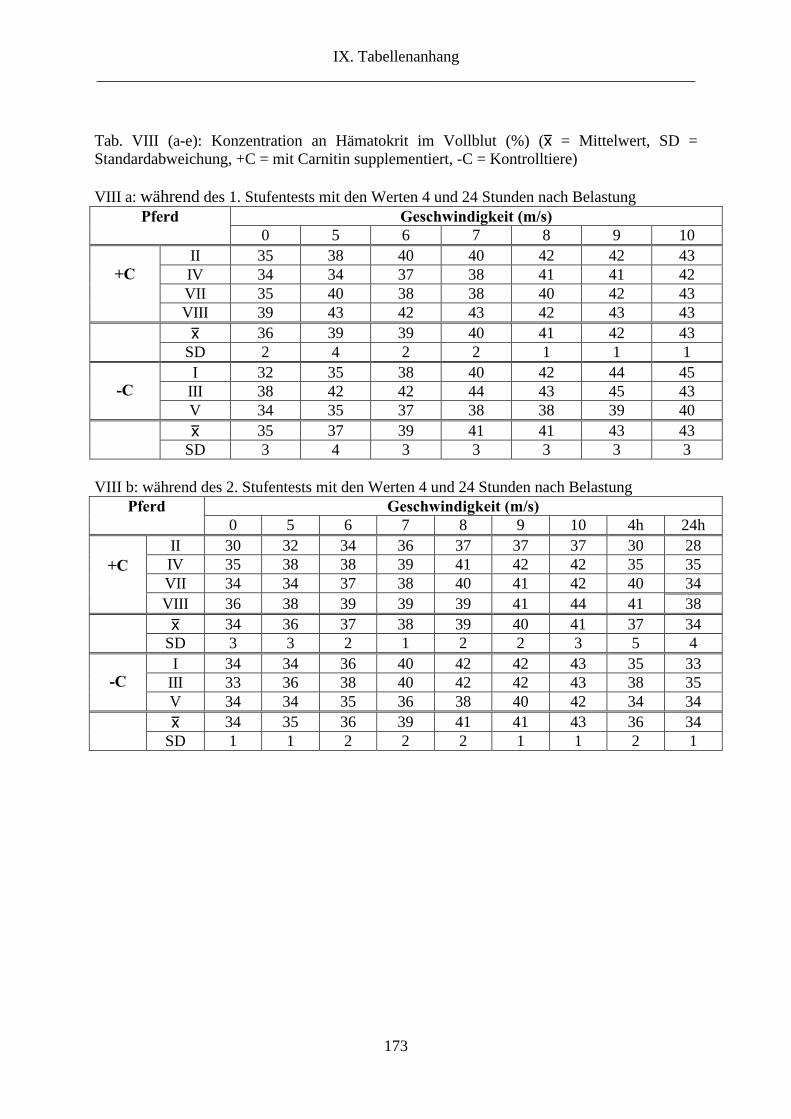
IX. Tabellenanhang ___________________________________________________________________________
173
Tab. VIII (a-e): Konzentration an Hämatokrit im Vollblut (%) (x = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) VIII a: während des 1. Stufentests mit den Werten 4 und 24 Stunden nach Belastung
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10
II 35 38 40 40 42 42 43 IV 34 34 37 38 41 41 42 VII 35 40 38 38 40 42 43
+C VIII 39 43 42 43 42 43 43
x 36 39 39 40 41 42 43 SD 2 4 2 2 1 1 1
I 32 35 38 40 42 44 45 III 38 42 42 44 43 45 43 -C
V 34 35 37 38 38 39 40 x 35 37 39 41 41 43 43
SD 3 4 3 3 3 3 3 VIII b: während des 2. Stufentests mit den Werten 4 und 24 Stunden nach Belastung
Geschwindigkeit (m/s) Pferd 0 5 6 7 8 9 10 4h 24h
II 30 32 34 36 37 37 37 30 28 IV 35 38 38 39 41 42 42 35 35 VII 34 34 37 38 40 41 42 40 34
+C VIII 36 38 39 39 39 41 44 41 38
x 34 36 37 38 39 40 41 37 34 SD 3 3 2 1 2 2 3 5 4
I 34 34 36 40 42 42 43 35 33 III 33 36 38 40 42 42 43 38 35 -C
V 34 34 35 36 38 40 42 34 34 x 34 35 36 39 41 41 43 36 34
SD 1 1 2 2 2 1 1 2 1
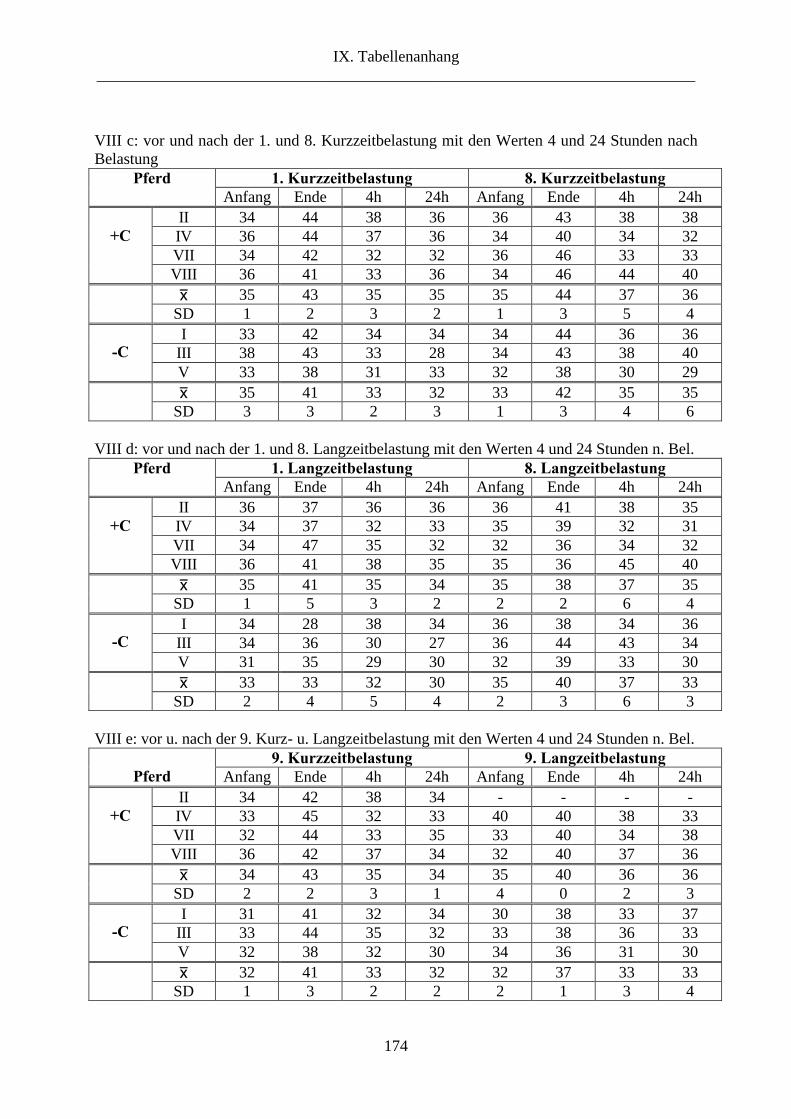
IX. Tabellenanhang ___________________________________________________________________________
174
VIII c: vor und nach der 1. und 8. Kurzzeitbelastung mit den Werten 4 und 24 Stunden nach Belastung
1. Kurzzeitbelastung 8. Kurzzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 34 44 38 36 36 43 38 38 IV 36 44 37 36 34 40 34 32 VII 34 42 32 32 36 46 33 33
+C VIII 36 41 33 36 34 46 44 40
x 35 43 35 35 35 44 37 36 SD 1 2 3 2 1 3 5 4
I 33 42 34 34 34 44 36 36 III 38 43 33 28 34 43 38 40 -C
V 33 38 31 33 32 38 30 29 x 35 41 33 32 33 42 35 35
SD 3 3 2 3 1 3 4 6 VIII d: vor und nach der 1. und 8. Langzeitbelastung mit den Werten 4 und 24 Stunden n. Bel.
1. Langzeitbelastung 8. Langzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 36 37 36 36 36 41 38 35 IV 34 37 32 33 35 39 32 31 VII 34 47 35 32 32 36 34 32
+C VIII 36 41 38 35 35 36 45 40
x 35 41 35 34 35 38 37 35 SD 1 5 3 2 2 2 6 4
I 34 28 38 34 36 38 34 36 III 34 36 30 27 36 44 43 34 -C
V 31 35 29 30 32 39 33 30 x 33 33 32 30 35 40 37 33
SD 2 4 5 4 2 3 6 3 VIII e: vor u. nach der 9. Kurz- u. Langzeitbelastung mit den Werten 4 und 24 Stunden n. Bel.
9. Kurzzeitbelastung 9. Langzeitbelastung Pferd Anfang Ende 4h 24h Anfang Ende 4h 24h
II 34 42 38 34 - - - - IV 33 45 32 33 40 40 38 33 VII 32 44 33 35 33 40 34 38
+C VIII 36 42 37 34 32 40 37 36
x 34 43 35 34 35 40 36 36 SD 2 2 3 1 4 0 2 3
I 31 41 32 34 30 38 33 37 III 33 44 35 32 33 38 36 33 -C
V 32 38 32 30 34 36 31 30 x 32 41 33 32 32 37 33 33
SD 1 3 2 2 2 1 3 4
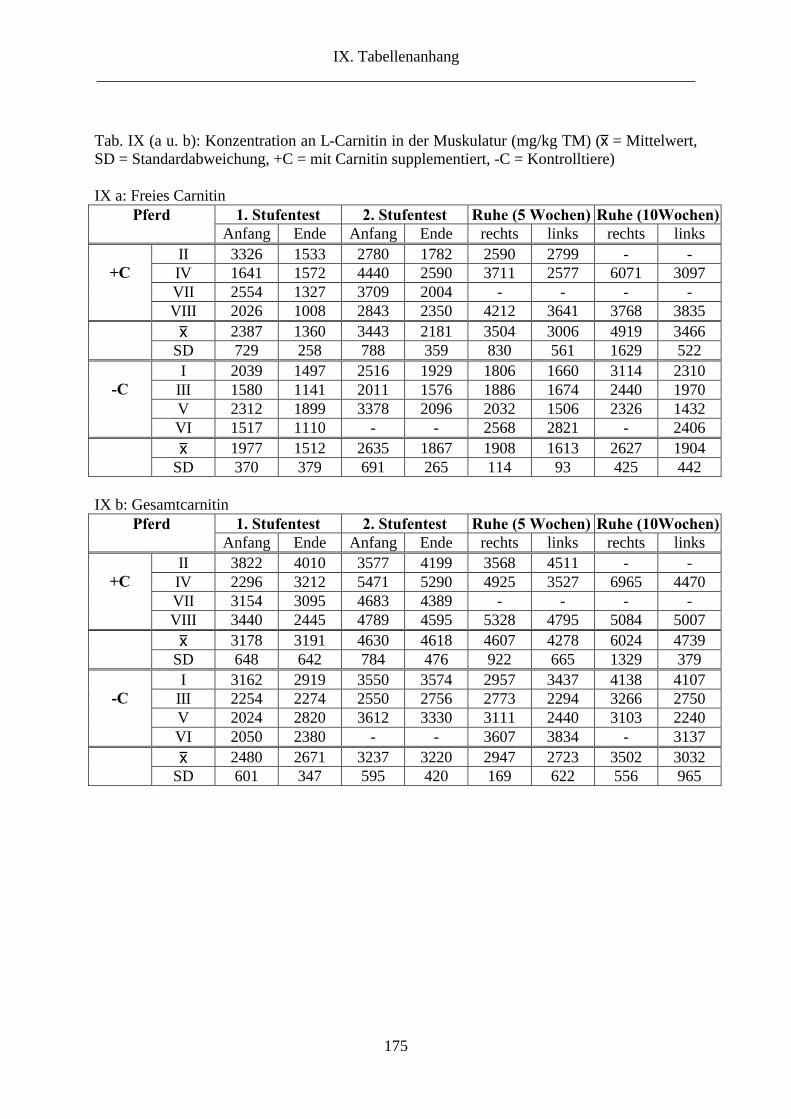
IX. Tabellenanhang ___________________________________________________________________________
175
Tab. IX (a u. b): Konzentration an L-Carnitin in der Muskulatur (mg/kg TM) (x = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) IX a: Freies Carnitin
1. Stufentest 2. Stufentest Ruhe (5 Wochen) Ruhe (10Wochen) Pferd Anfang Ende Anfang Ende rechts links rechts links
II 3326 1533 2780 1782 2590 2799 - - IV 1641 1572 4440 2590 3711 2577 6071 3097 VII 2554 1327 3709 2004 - - - -
+C VIII 2026 1008 2843 2350 4212 3641 3768 3835
x 2387 1360 3443 2181 3504 3006 4919 3466 SD 729 258 788 359 830 561 1629 522
I 2039 1497 2516 1929 1806 1660 3114 2310 III 1580 1141 2011 1576 1886 1674 2440 1970 V 2312 1899 3378 2096 2032 1506 2326 1432
-C VI 1517 1110 - - 2568 2821 - 2406
x 1977 1512 2635 1867 1908 1613 2627 1904 SD 370 379 691 265 114 93 425 442
IX b: Gesamtcarnitin
1. Stufentest 2. Stufentest Ruhe (5 Wochen) Ruhe (10Wochen) Pferd Anfang Ende Anfang Ende rechts links rechts links
II 3822 4010 3577 4199 3568 4511 - - IV 2296 3212 5471 5290 4925 3527 6965 4470 VII 3154 3095 4683 4389 - - - -
+C VIII 3440 2445 4789 4595 5328 4795 5084 5007
x 3178 3191 4630 4618 4607 4278 6024 4739 SD 648 642 784 476 922 665 1329 379
I 3162 2919 3550 3574 2957 3437 4138 4107 III 2254 2274 2550 2756 2773 2294 3266 2750 V 2024 2820 3612 3330 3111 2440 3103 2240
-C VI 2050 2380 - - 3607 3834 - 3137
x 2480 2671 3237 3220 2947 2723 3502 3032 SD 601 347 595 420 169 622 556 965
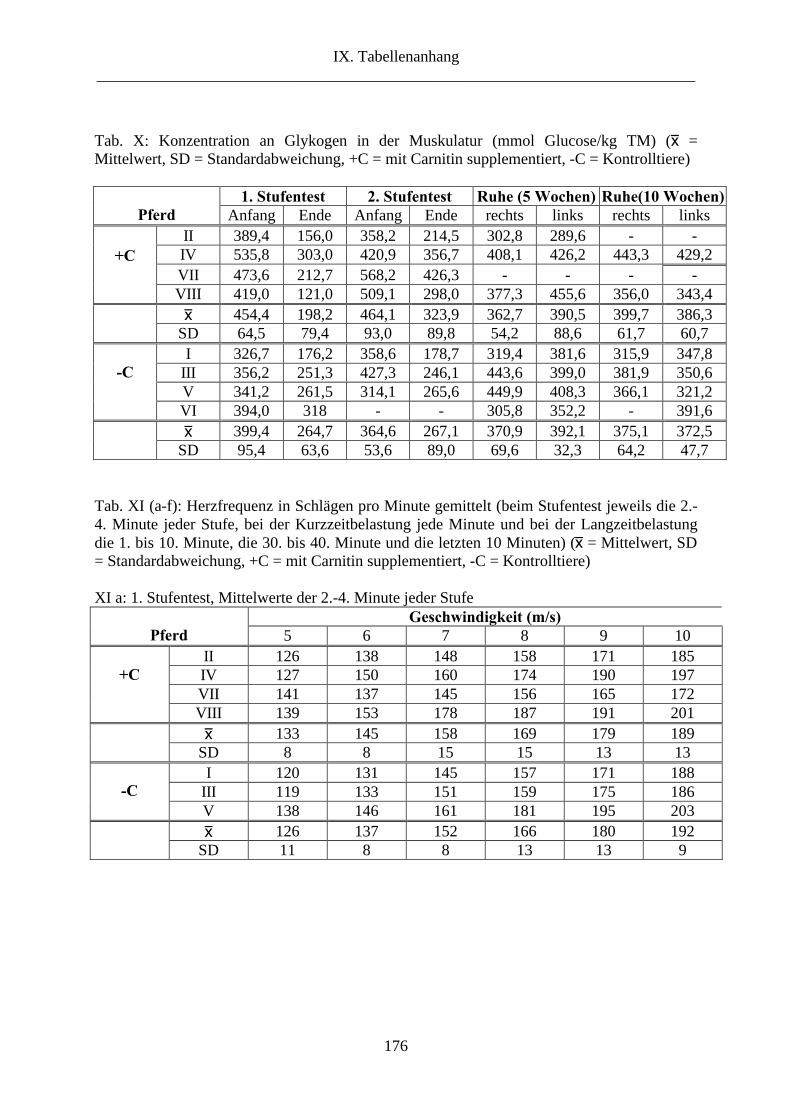
IX. Tabellenanhang ___________________________________________________________________________
176
Tab. X: Konzentration an Glykogen in der Muskulatur (mmol Glucose/kg TM) (x = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere)
1. Stufentest 2. Stufentest Ruhe (5 Wochen) Ruhe(10 Wochen) Pferd Anfang Ende Anfang Ende rechts links rechts links
II 389,4 156,0 358,2 214,5 302,8 289,6 - - IV 535,8 303,0 420,9 356,7 408,1 426,2 443,3 429,2 VII 473,6 212,7 568,2 426,3 - - - -
+C VIII 419,0 121,0 509,1 298,0 377,3 455,6 356,0 343,4
x 454,4 198,2 464,1 323,9 362,7 390,5 399,7 386,3 SD 64,5 79,4 93,0 89,8 54,2 88,6 61,7 60,7
I 326,7 176,2 358,6 178,7 319,4 381,6 315,9 347,8 III 356,2 251,3 427,3 246,1 443,6 399,0 381,9 350,6 V 341,2 261,5 314,1 265,6 449,9 408,3 366,1 321,2
-C VI 394,0 318 - - 305,8 352,2 - 391,6
x 399,4 264,7 364,6 267,1 370,9 392,1 375,1 372,5 SD 95,4 63,6 53,6 89,0 69,6 32,3 64,2 47,7
Tab. XI (a-f): Herzfrequenz in Schlägen pro Minute gemittelt (beim Stufentest jeweils die 2.-4. Minute jeder Stufe, bei der Kurzzeitbelastung jede Minute und bei der Langzeitbelastung die 1. bis 10. Minute, die 30. bis 40. Minute und die letzten 10 Minuten) (x = Mittelwert, SD = Standardabweichung, +C = mit Carnitin supplementiert, -C = Kontrolltiere) XI a: 1. Stufentest, Mittelwerte der 2.-4. Minute jeder Stufe
Geschwindigkeit (m/s) Pferd 5 6 7 8 9 10
II 126 138 148 158 171 185 IV 127 150 160 174 190 197 VII 141 137 145 156 165 172
+C VIII 139 153 178 187 191 201
x 133 145 158 169 179 189 SD 8 8 15 15 13 13
I 120 131 145 157 171 188 III 119 133 151 159 175 186 -C
V 138 146 161 181 195 203 x 126 137 152 166 180 192
SD 11 8 8 13 13 9
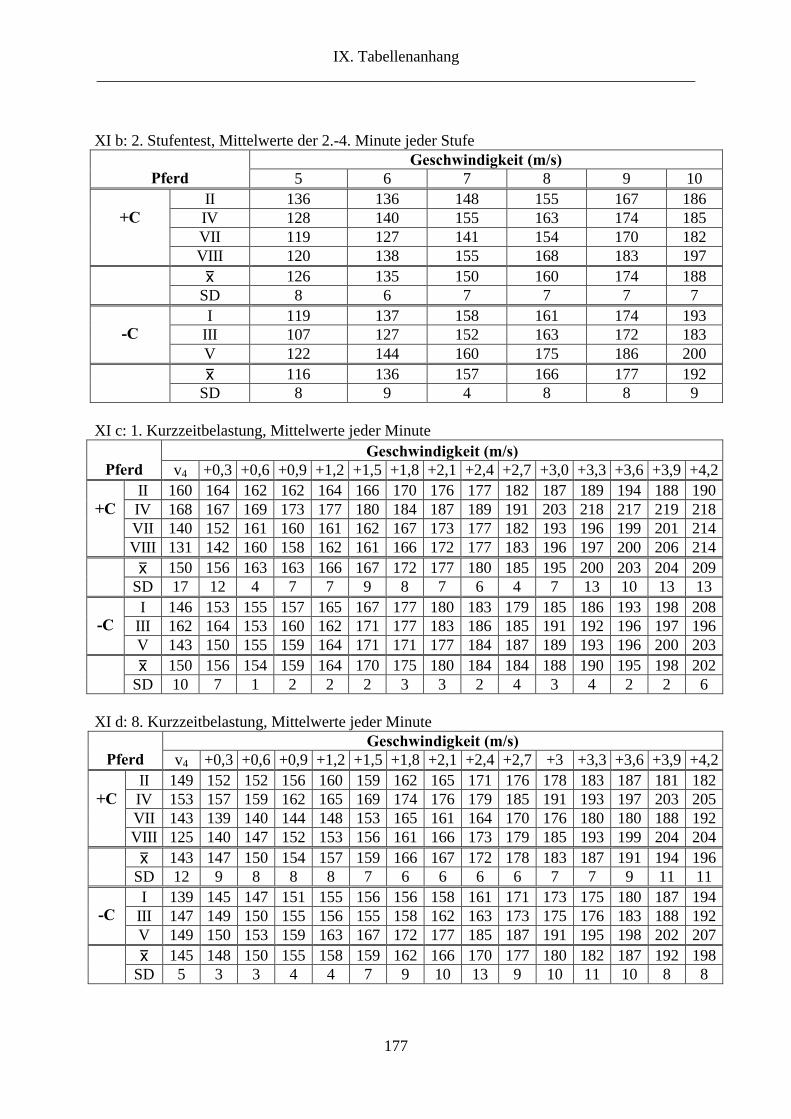
IX. Tabellenanhang ___________________________________________________________________________
177
XI b: 2. Stufentest, Mittelwerte der 2.-4. Minute jeder Stufe Geschwindigkeit (m/s)
Pferd 5 6 7 8 9 10 II 136 136 148 155 167 186 IV 128 140 155 163 174 185 VII 119 127 141 154 170 182
+C VIII 120 138 155 168 183 197
x 126 135 150 160 174 188 SD 8 6 7 7 7 7
I 119 137 158 161 174 193 III 107 127 152 163 172 183 -C
V 122 144 160 175 186 200 x 116 136 157 166 177 192
SD 8 9 4 8 8 9 XI c: 1. Kurzzeitbelastung, Mittelwerte jeder Minute
Geschwindigkeit (m/s) Pferd v4 +0,3 +0,6 +0,9 +1,2 +1,5 +1,8 +2,1 +2,4 +2,7 +3,0 +3,3 +3,6 +3,9 +4,2
II 160 164 162 162 164 166 170 176 177 182 187 189 194 188 190 IV 168 167 169 173 177 180 184 187 189 191 203 218 217 219 218 VII 140 152 161 160 161 162 167 173 177 182 193 196 199 201 214
+C VIII 131 142 160 158 162 161 166 172 177 183 196 197 200 206 214
x 150 156 163 163 166 167 172 177 180 185 195 200 203 204 209 SD 17 12 4 7 7 9 8 7 6 4 7 13 10 13 13
I 146 153 155 157 165 167 177 180 183 179 185 186 193 198 208 III 162 164 153 160 162 171 177 183 186 185 191 192 196 197 196 -C
V 143 150 155 159 164 171 171 177 184 187 189 193 196 200 203 x 150 156 154 159 164 170 175 180 184 184 188 190 195 198 202
SD 10 7 1 2 2 2 3 3 2 4 3 4 2 2 6 XI d: 8. Kurzzeitbelastung, Mittelwerte jeder Minute
Geschwindigkeit (m/s) Pferd v4 +0,3 +0,6 +0,9 +1,2 +1,5 +1,8 +2,1 +2,4 +2,7 +3 +3,3 +3,6 +3,9 +4,2
II 149 152 152 156 160 159 162 165 171 176 178 183 187 181 182 IV 153 157 159 162 165 169 174 176 179 185 191 193 197 203 205 VII 143 139 140 144 148 153 165 161 164 170 176 180 180 188 192
+C VIII 125 140 147 152 153 156 161 166 173 179 185 193 199 204 204
x 143 147 150 154 157 159 166 167 172 178 183 187 191 194 196 SD 12 9 8 8 8 7 6 6 6 6 7 7 9 11 11
I 139 145 147 151 155 156 156 158 161 171 173 175 180 187 194 III 147 149 150 155 156 155 158 162 163 173 175 176 183 188 192 -C
V 149 150 153 159 163 167 172 177 185 187 191 195 198 202 207 x 145 148 150 155 158 159 162 166 170 177 180 182 187 192 198
SD 5 3 3 4 4 7 9 10 13 9 10 11 10 8 8
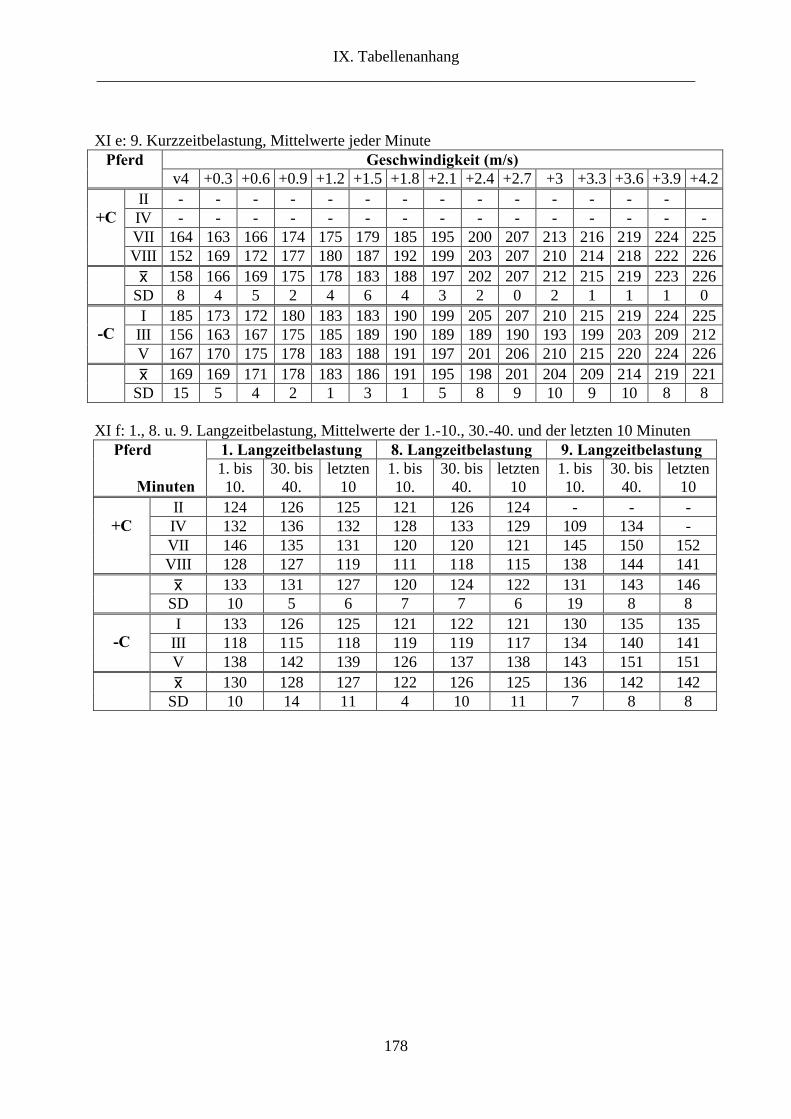
IX. Tabellenanhang ___________________________________________________________________________
178
XI e: 9. Kurzzeitbelastung, Mittelwerte jeder Minute Geschwindigkeit (m/s) Pferd
v4 +0.3 +0.6 +0.9 +1.2 +1.5 +1.8 +2.1 +2.4 +2.7 +3 +3.3 +3.6 +3.9 +4.2 II - - - - - - - - - - - - - - IV - - - - - - - - - - - - - - - VII 164 163 166 174 175 179 185 195 200 207 213 216 219 224 225
+C VIII 152 169 172 177 180 187 192 199 203 207 210 214 218 222 226
x 158 166 169 175 178 183 188 197 202 207 212 215 219 223 226 SD 8 4 5 2 4 6 4 3 2 0 2 1 1 1 0
I 185 173 172 180 183 183 190 199 205 207 210 215 219 224 225 III 156 163 167 175 185 189 190 189 189 190 193 199 203 209 212 -C
V 167 170 175 178 183 188 191 197 201 206 210 215 220 224 226 x 169 169 171 178 183 186 191 195 198 201 204 209 214 219 221
SD 15 5 4 2 1 3 1 5 8 9 10 9 10 8 8 XI f: 1., 8. u. 9. Langzeitbelastung, Mittelwerte der 1.-10., 30.-40. und der letzten 10 Minuten
1. Langzeitbelastung 8. Langzeitbelastung 9. Langzeitbelastung Pferd
Minuten 1. bis 10.
30. bis 40.
letzten 10
1. bis 10.
30. bis 40.
letzten 10
1. bis 10.
30. bis 40.
letzten 10
II 124 126 125 121 126 124 - - - IV 132 136 132 128 133 129 109 134 - VII 146 135 131 120 120 121 145 150 152
+C VIII 128 127 119 111 118 115 138 144 141
x 133 131 127 120 124 122 131 143 146 SD 10 5 6 7 7 6 19 8 8
I 133 126 125 121 122 121 130 135 135 III 118 115 118 119 119 117 134 140 141 -C
V 138 142 139 126 137 138 143 151 151 x 130 128 127 122 126 125 136 142 142
SD 10 14 11 4 10 11 7 8 8

Danksagung
Herrn Prof. Dr. J. Harmeyer danke ich für die Überlassung des Themas und der jederzeit
freundlichen und hilfsbereiten Unterstützung bei der Anfertigung dieser Arbeit.
Herrn Prof. Dr. M. Coenen möchte ich für die wissenschaftliche Betreuung und die
wertvollen Anregungen danken.
Besonderer Dank richtet sich auch an U. Wedemeyer, mit der ich bei der Durchführung der
praktischen Versuche sehr konstruktiv und harmonisch zusammengearbeitet habe.
Ganz besonders seien Dr. Vervuert und Dr. Sporleder gedankt, die in allen Lebenslagen mir
tatkräftig zur Seite standen und stets hilfreich mit ihren Anregungen den Werdegang der
Arbeit ebneten.
Den Mitarbeiterinnen und Mitarbeitern unserer Arbeitsgruppe danke ich für die gute
Arbeitsatmosphäre und die wertvollen Ratschläge ebenso wie für die Beratung bei den
Analysen.
Für die freundliche Unterstützung danke ich auch den Mitarbeiterinnen und Mitarbeitern der
Tierernährung, ganz besonders den Tierpflegern, durch deren wachsames Auge ein gutes
Gelingen der Arbeit möglich war.
Herzlich möchte ich mich auch bei all meinen Freunden bedanken, die mich bei der
Anfertigung dieser Arbeit unterstützt haben, sowohl fachkompetent als auch psychisch.
Ebenso sei meinen Eltern gedankt, durch deren finanzielle Unterstützung der schnelle
Werdegang dieser Arbeit erst möglich wurde.










