Fischzuchtlinien für standortgerechte Aquakultur! · 100 mg/l Benzokain bzw. 0,1‐0,5 ml/...
52
PILOTPROJEKT IM RAHMEN DES OPERATIONELLEN PROGRAMMS DES EUROPÄISCHEN FISCHEREIFONDS (EFF) Wissenschaftlicher Abschlussbericht zum Vorhaben VI 560/7308 Fischzuchtlinien für standortgerechte Aquakultur! Biotechnologische Prüfung auf Robustheit selektierter Regenbogenforellen (Stamm BORN) auf Eignung als Standortlinie und Tiermodell in differenten regionalen Aquakulturanlagen Projektlaufzeit: 01.04.2012 bis 30.11.2015 Antragsteller: Campus bioFISCH M‐V Projekt‐ und Campuskoordinator: PD Dr. habil. Tom Goldammer Leiter AG Fischgenetik FB Molekularbiologie Leibniz‐Institut für Nutztierbiologie (FBN) Wilhelm‐Stahl‐Allee 2 D‐18196 Dummerstorf Telefon: +49 (0)38 208 68 708 Telefax: +49 (0)38 208 68 702 E‐Mail: tomgoldammer@fbn‐dummerstorf.de
Transcript of Fischzuchtlinien für standortgerechte Aquakultur! · 100 mg/l Benzokain bzw. 0,1‐0,5 ml/...

PILOTPROJEKT IM RAHMEN DES
OPERATIONELLEN PROGRAMMS DES
EUROPÄISCHEN FISCHEREIFONDS (EFF)
Wissenschaftlicher Abschlussbericht zum Vorhaben VI 560/7308
Fischzuchtlinien für standortgerechte Aquakultur! Biotechnologische Prüfung auf Robustheit selektierter Regenbogenforellen (Stamm BORN) auf
Eignung als Standortlinie und Tiermodell in differenten regionalen Aquakulturanlagen
Projektlaufzeit: 01.04.2012 bis 30.11.2015 Antragsteller: Campus bioFISCH M‐V Projekt‐ und Campuskoordinator: PD Dr. habil. Tom Goldammer
Leiter AG Fischgenetik FB Molekularbiologie Leibniz‐Institut für Nutztierbiologie (FBN) Wilhelm‐Stahl‐Allee 2 D‐18196 Dummerstorf Telefon: +49 (0)38 208 68 708 Telefax: +49 (0)38 208 68 702 E‐Mail: tomgoldammer@fbn‐dummerstorf.de

2
Kooperationspartner im Campus bioFISCH M‐V:
1. Leibniz‐Institut für Nutztierbiologie, FBN, Dummerstorf
Dr. T. Goldammer, Dr. A. Rebl, Prof. K. Wimmers, Prof. M. Schwerin
2. BFI für Tiergesundheit, FLI, Riems, Institut für Infektionsmedizin
Dr. B. Köllner, Prof. T.C. Mettenleiter
Südufer 10, D‐17493 Greifswald, Insel Riems
Email: bernd.kö[email protected]
3. LFA Landwirtschaft und Fischerei M‐V, LFAMV, Institut für Fischerei
Dipl. Biol. C. Kühn, Prof. C. Gienapp, Dr. Ralf Bochert
Fischerweg 408, 18069 Rostock
Email: [email protected]
4. Praxispartner
BIMES Binnenfischerei GmbH, Teichanlage Frauenmark
Geschäftsführer Herr G. Thies
Forellenzucht Uhthoff GmbH, Neubrandenburg
Geschäftsführer Herr H. Uhthoff
Weitere Praxispartner außerhalb des Verbundes:
AQUA Fischzucht Wesenberg
Geschäftsführer Herr Thomas Splett
Fischerei Werner Loch, Hohen‐Sprenz
Geschäftsführer Herr Werner Loch
Forellenzucht Dobbin, Dobbin
Geschäftsführer Herr Siegfried Wuttge

3
Inhaltsverzeichnis
1 Einleitung 5
2 Ziele, Versuchsansatz, Arbeitspakete 5
3 Fische, Material, Methoden 6
3.1 Fische 6
3.2 Methodenspektrum 7
3.3 Phänotypanalysen und Umweltparameter 8
3.4 Geschlechtsbestimmung 10
3.5 Zellen 11
3.6 Gewebeproben 12
3.7 Bioinformatik 12
4 Ergebnisse und Diskussion 13
4.1 Forellenproduktion 13
4.1.1 Forellenproduktion Vergleich 1 13
4.1.2 Forellenproduktion Vergleich 2 16
4.1.3 Forellenproduktion Vergleich 3 21
4.1.4 Fazit der vergleichenden Forellenproduktion in Aquakulturanlagen in M‐V 25
4.1.5. Empfehlung: Die Bornforelle ist für die regionale Aquakultur geeignet 26
4.2 Biotechnologische Charakterisierung des Merkmals Robustheit 26
4.2.1 Publikationen – Ergebnisse und Diskussion 27
4.2.1.1 Publikation 1 – Rebl et al. (2012) CBP Part B Biochem Mol Biol 163(1):65‐73 27
4.2.1.2 Publikation 2 – Verleih et al. (2013) Gene 512(2):251‐258 28
4.2.1.3 Publikation 3 – Köbis et al. (2013) Mol Biol Rep 40(2):1955‐1966 29
4.2.1.4 Publikation 4 – Korytář T et al. (2013) Fish Shellfish Immunol 35(4):1192‐1199 31
4.2.1.5 Publikation 5 – Rebl et al. (2013) Mar Biotechnol (NY) 15(4):445‐460 33
4.2.1.6 Publikation 6 – Verleih et al. (2015) Mar Biotechnol (NY) 17(5):576‐592 33
4.2.1.7 Publikation 7 – Borchel A. et al. (2014) Springerplus 3:510 eCollection 36
4.2.1.8 Publikation 8 – Borchel A. (2015) Diss Universität Rostock: pp100 + Anhang 38
4.2.2 Weitere Projektergebnisse und Diskussion 39
4.2.2.1 Holistische Transkriptomanalysen 39
4.2.2.2 Transkriptomanalysen zur Identifizierung von DNA‐Polymorphismen 40

4
4.2.2.3 Besatzdichteexperimente 43
4.2.2.4 Zelluläre Analysen 45
4.2.3 Fazit der biotechnologischen Analysen zur Bornforelle 46
5 Die Bornforelle als Modell u. Standortlinie ‐ Entwicklung von Zuchtrichtlinien 46
6 Wissenstransfer 47

5
1. Einleitung
Verschiedene modellartig konzipierte Vorgängerprojekte gaben Hinweise dafür, dass die regional seit
1975 auf Überleben im Brackwasser gezüchtete Regenbogenforelle (Oncorhynchus mykiss) – Stamm
BORN – sich besser für die standortgerechte Aquakultur eignet als Importforellen. Als mögliche Ursache
wurde angenommen, dass sie optimal an regionale Umweltbedingungen adaptiert und dies genetisch
manifestiert ist. Verschiedentlich wurden Hinweise auf eine erhöhte Widerstandsfähigkeit und ein hohes
produktives Adaptationsvermögen der Bornforelle gefunden. Dies wurde unter dem Merkmal Robustheit
zusammengefasst und definiert. Unter anderem reagierte beispielsweise das Immunsystem schneller auf
Pathogene und die Mortalität war deutlich geringer. Wachstum und Schlachtgewicht waren unter
Stresseinfluss (Temperatur) signifikant erhöht. Dazu durchgeführte initiale molekulare und
immunologische Studien zeigten, dass zahlreiche kaum aufgeklärte genetische Faktoren und
Signalkaskaden in verschiedenen Organen und Geweben der Regenbogenforelle ursächlich an diesen
metabolischen Phänomenen beteiligt sein könnten. Eine Verifizierung der vorliegenden Studien im
Produktionsmaßstab gab es bisher nicht. Mit dem Ziel zu prüfen, ob die regionale Regenbogenforellen‐
Zuchtlinie Born sich für die Aufzucht in lokalen Aquakulturanlagen besser eignet als üblicherweise
genutzte Importforellen einer Wachstumsselektionslinie aus den USA, fanden deshalb im Rahmen des
Pilotprojektes die definierte parallele Produktion und eine biotechnologische Analyse wirtschaftlich
relevanter Fischmengen der Forellenzuchtlinien in technologisch unterschiedlich betriebenen regionalen
Aquakulturanlagen mit zusätzlich differentem Selektionsdruck statt. Grundsätzlich wurde so in dem über
den EFF und das Ministerium für Landwirtschaft, Umweltschutz und Verbraucherschutz M‐V geförderten
Pilotprojekt die technische Durchführbarkeit und Wirtschaftlichkeit einer innovativen Technik unter
realitätsnahen Bedingungen geprüft, um technische bzw. wirtschaftliche Kenntnisse über die oben
genannte Technik, zu gewinnen und zu verbreiten.
Das Projekt war ein Kooperationsprojekt, welches der Campus bioFISH M‐V organisiert und koordiniert
hat. An der Projektbearbeitung beteiligt waren als interdisziplinäre Forschungspartner das Leibniz‐
Institut für Nutztierbiologie (FBN), das Bundesforschungsinstitut für Tiergesundheit (FLI) und die
Landesforschungsanstalt für Landwirtschaft und Fischerei M‐V (LFA M‐V). Mehrere fischereiliche
Betriebe in M‐V waren als Praxispartner in das Forschungsvorhaben implementiert.
2. Ziele, Versuchsansatz, Arbeitspakete
Das wesentliche Ziel des vorliegenden Projektes bestand darin, langjährige wissenschaftliche Vorarbeiten
des Institutes für Fischerei der LFA M‐V zur Bornforelle unter Produktionsbedingungen zu verifizieren

6
und Aussagen zum möglichen Einsatz der Bornforelle in der Aquakultur Mecklenburg‐Vorpommerns und
darüber hinaus zu treffen. Dazu erfolgte erstmalig eine definierte parallele Produktion und Analyse
wirtschaftlich relevanter Fischmengen von Born‐ und Importforelle in unterschiedlich betriebenen
regionalen Aquakulturanlagen in Mecklenburg‐Vorpommern. Abbildung 1 zeigt schematisiert den dazu
gewählten Versuchsansatz. Die Aufzucht der Forellen wurde unter den jeweiligen technischen
Gegebenheiten der Fischzuchtanlagen durchgeführt, die lokalen natürlichen Umweltbedingungen und
der damit verbundene differente Selektionsdruck wurden berücksichtigt und standardisierte
Fütterungsbedingungen und Futtermittel wurden gewährleistet. Eine phänotypische Charakterisierung
der Fische in den jeweiligen Aquakulturanlagen wurde durchgeführt.
Abb. 1: Versuchsansatz Pilotprojekt Bornforelle.
Ein weiteres Ziel des Projektes bestand darin, mit biotechnologischen Ansätzen genetische,
immunologische und auch zelluläre Ursachen für die differente Robustheit der verglichenen Zuchtlinien
zu erhalten. Dazu erfolgten umfassende globale Transkriptomanalysen, die Auskunft über die Aktivität
von Genen unter definierten Haltungsbedingungen gaben. Davon abgeleitet, wurden Gennetzwerke
erstellt, die dazu beitragen, das Zusammenspiel der Gene zu interpretieren und Rückschlüsse auf eine
bestimmte Performance der Fische zu ziehen. Ausgewählte Parameter wurden molekularbiologisch,
immunologisch und in Zellmodellen weiter charakterisiert und so auch grundlegende Unterschiede
zwischen den Forellenlinien aufgeklärt.
Der Campus bioFISCH M‐V hatte im Hinblick auf die gestellten Ziele folgende Arbeitspakete (AP)
geschnürt und vollständig bearbeitet:
AP I Phänotypische Analysen unter Produktionsbedingungen;
AP II Vergleichende Produktion und Charakterisierung von Born‐ und Importforelle;
AP III Biotechnologische Charakterisierung des Merkmals Robustheit in vitro und in vivo;
AP IV Funktionelle Charakterisierung nach Stresseinfluss in vitro und in vivo;
AP V Dateninterpretation – Modell, Standortlinie, Zuchtrichtlinien.

7
3. Fische, Material, Methoden
3.1 Fische
Für die Untersuchungen verwendete Regenbogenforellen (Abb. 2) stammten im Falle der regionalen
Zuchtlinie Born aus dem Institut für Fischerei der LFA M‐V. Mit diesen Fischen erfolgt seit 1975 über
Generationen hinweg eine Selektion mit dem Zuchtziel: Überleben im Brackwasser (Anders, 1986). Als
weitere Zuchtlinie wurde eine wachstumsselektierte Regenbogenforellenlinie aus den USA
(www.troutlodge.com) verwendet. Diese stellt eine häufig eingesetzte Zuchtlinie in der deutschen und
weltweiten Aquakultur dar. Augenpunkteier dieser sterilen, rein weiblichen Linie wurden parallel mit
Eiern der Bornforelle im Institut für Fischerei der LFA (Born) erbrütet, vorgestreckt und wiederholt je
1000 Fingerlinge zu verschiedenen Aquakulturanlagen überführt und bis zu einem Schlachtgewicht von
ca. 350 g bis 400°g wachsen gelassen. Aufzucht und Vorstrecken erfolgten mit natürlichen
Schwankungen der Umweltparamater. Gefüttert wurde handelsübliches Forellenfutter.
Abb. 2: Darstellung unterschiedlicher Entwicklungsstadien der Regenbogenforelle. Von Links: Augenpunkteier; Dottersacklarven; Fingerling in unterschiedlichen Wasserqualitäten (oben Trinkwasser, unten Boddenwasser); Adulte Fische (oben eine dunkler gefärbte Forelle der Zuchtlinie Born und unten eine typisch silbrig glänzende Forelle der Wachstumslinie aus den USA). Fotos: Tom Goldammer
3.2 Methodenspektrum
Für die Projektbearbeitung kamen umfassende molekularbiologische, immunologische,
zellphysiologische und bioinformatische Methoden zur Anwendung. Aufgrund ihres Umfangs werden sie
an dieser Stelle nur stichpunktartig aufgelistet. Der Großteil der Methoden ist standardisiert und
orientiert sich an den Vorgaben der Hersteller für entsprechende Medien, Chemikalien, DNA‐, RNA‐Kits
etc.. Abwandlungen von den jeweiligen Methoden sind in detaillierter Form in den angehängten
Publikationen zum Projekt beschrieben oder es gibt Verweise auf entsprechende Literatur.

8
Methoden auf Phänotypebene:
u.a. Bestimmung Gewicht, Durchschnittsgewicht, Länge, Durchschnittslänge, Besatz in kg und
Anzahl Fische, Anzahl Verluste, Futtermenge in kg, Gesamt‐Stückzahl nach Erreichen des
Schlachtgewichts, Gesamtmasse (gewogen und kalkuliert), Zuwachs in kg, Fütterungsquotient
Methoden auf Genom‐ bzw. DNA‐Ebene:
u.a. DNA‐Isolation, Genome Walking, Polymerase‐Kettenreaktion (PCR), Schnelle
Amplifikation von DNA‐Enden (RACE‐Technologie), DNA‐Sequenzierung
Methoden auf Transkriptom‐ bzw. RNA‐Ebene:
u.a. RNA‐Isolation, cDNA‐Synthese, semiquantitiative PCR, Echtzeit‐PCR (qPCR) mit Lightcycler
96 (Roche), DNA‐Chip‐Analysen (Salmon 4x44K, 8x60K Agilent‐Array, Salmon 16k cDNA‐
Array), cDNA‐Sequenzierung, Transkriptomsequenzierung
Methoden auf Proteinebene:
u.a. GFP‐gekoppelte Proteinexpression, Western‐Blot
Methoden auf Zellebene:
u.a. FACS‐Fluoreszenz aktivierte Zellsortierung, verschiedene Fischzellkultivierungsmethoden,
Mikroskopie/ zelluläre Fluoreszenzmikroskopie mit Zeiss AxioObserver Z1 Mikroskop.
3.3 Phänotypanalysen und Umweltparameter
Die vergleichenden Untersuchungen wurden an verschiedenen Aquakulturstandorten in M‐V
durchgeführt in denen regelmäßig i.d.R. Importforellen im Produktionsmaßstab gehalten werden und
somit sämtliche technischen Gegebenheiten für die Forellenaufzucht unter den vorhandenen
Umweltbedingungen auf neuestem Kenntnisstand vorlagen. Beispiele für Anlagen mit differenten
Wasserqualitäten waren: eine Kaltwasserdurchflussanlage mit Rinnen (KWA) sowie temperierbare
Großmodule (GM) mit Filtersystem am Fischereiinstitut der LFA in Born, Netzkäfiganlagen in
Oberflächengewässern (NKA), Teichanlagen (TA) sowie Rinnenanlagen (RA1 und RA2) im Durchfluss von
Oberflächengewässern (Abb. 3). Die Anlage RA1 wurde am Projektbeginn zunächst als Standort für die
parallele Haltung verschiedener Zuchtlinien evaluiert.

9
A B
C
D
Abb. 3.: Standorte der parallelen Forellenhaltung. A) Durchflussanlage am Fischereiinstitut der LFA in Born; B) Netzkäfiganlage in einem Oberflächengewässer; C) Teichanlage; D) Rinnenanlage im Durchfluss eines Oberflächengewässers. Fotos: Ralf Bochert
Zum Versuchsbeginn wurden Satzfische mit Frischmassen von 2,5 g ‐6 g bzw. größer 100 g an die
entsprechenden Aquakulturstandorte verbracht. Der Besatz erfolgte mit praxisüblichen Dichten
entsprechend der gelieferten Fischmassen. Die parallele Mast der Forellen erfolgte unter den
praxisüblichen Bedingungen der jeweiligen Fischzüchter. Tägliche Mess‐ und Dokumentationswerte
waren die Wassertemperatur, der Sauerstoffgehalt, die Futtermenge und die Stückverluste. Wöchentlich
wurden Wasserproben entnommen und u.a. die Gehalte an Nitrat, Nitrit und Ammonium bestimmt.
Monatlich erfolgten an allen Standorten Messungen (Frischmasse, Totallänge) an jeweils ca. 10‐50
Tieren. Nach Erreichen des Zielgewichtes von > 350 g FM wurden die Gesamtmassen bestimmt und die
Frischmassen und Totallängen von 50 Tieren erfasst. Für eine Schlachtkörperanalyse wurden jeweils
zehn Tiere pro Gruppe zum Versuchsende entnommen und frisch bearbeitet.
Für die statistische Auswertung in R (Version 2.1.3.1.) wurden die Daten mit dem Shapiro‐Wilk‐Test auf
Normalverteilung überprüft. Bei Vorliegen von normalverteilten Stichproben wurden die Mittelwerte mit

10
dem T‐Test analysiert. Im abweichenden Fall wurde der Mann‐Whitney U‐Test durchgeführt. Das
Signifikanzlevel wurde auf p < 0,05 festgelegt.
Von Fischen, die moderaten Stressoren ausgesetzt waren, erfolgte für nachfolgende biotechnologische
Analysen u.a. die Erfassung von Gewicht, Länge, Gesundheitszustand, Mortalität, Erfassung von
Futtermittel, ‐mengen, O2‐Gehalt, Wassertemperatur und Besatzdichte. Die Fischtötung wurde im
Wasserbad durchgeführt ‐ entweder mit Benzokain >250 mg/l (Altmann et al., 2015) oder
Phenoxyethanol >2,5 ml/l (Rebl et al., 2012): Die Betäubung von Fischen wurde entsprechend mit 25‐
100 mg/l Benzokain bzw. 0,1‐0,5 ml/ Phenoxylethanol durchgeführt.
Die folgende Übersicht spiegelt beispielhaft den schematischen Ablauf von Versuchsansätzen der
experimentellen Vergleiche zwischen den untersuchten Forellenlinien wieder, die den
biotechnologischen Folgeanalysen vorangestellt waren (Abb. 4).
Abb. 4: Experimentelle Ansätze im Projekt. Oben links – Schema für generelle Vergleiche gesunder Fische beider Zuchtlinien und nachfolgende Kandidatengenanalysen bzw. zelluläre Ansätze; Oben Rechts – Darstellung des zeitlichen Ablaufs der Infektionsexperimente. TCO = Importforelle, BORN = Bornforelle. Unten – Weitere Versuchsansätze mit Belastung der Fische durch Temperaturveränderung , Besatz oder kombiniert Besatz‐Temperatur.
3.4 Geschlechtsbestimmung
Die Importforellen sind im Gegensatz zur Bornforelle rein weiblich. Es bestand deshalb die
Notwendigkeit, bei den beprobten Tieren der Linie Born das Geschlecht zu bestimmen, um eventuell
auftretende geschlechtsspezifische Unterschiede bei den später durchgeführten Analysen
berücksichtigen zu können. Die Geschlechter der beprobten Forellen ließen sich mittels PCR‐Verfahren
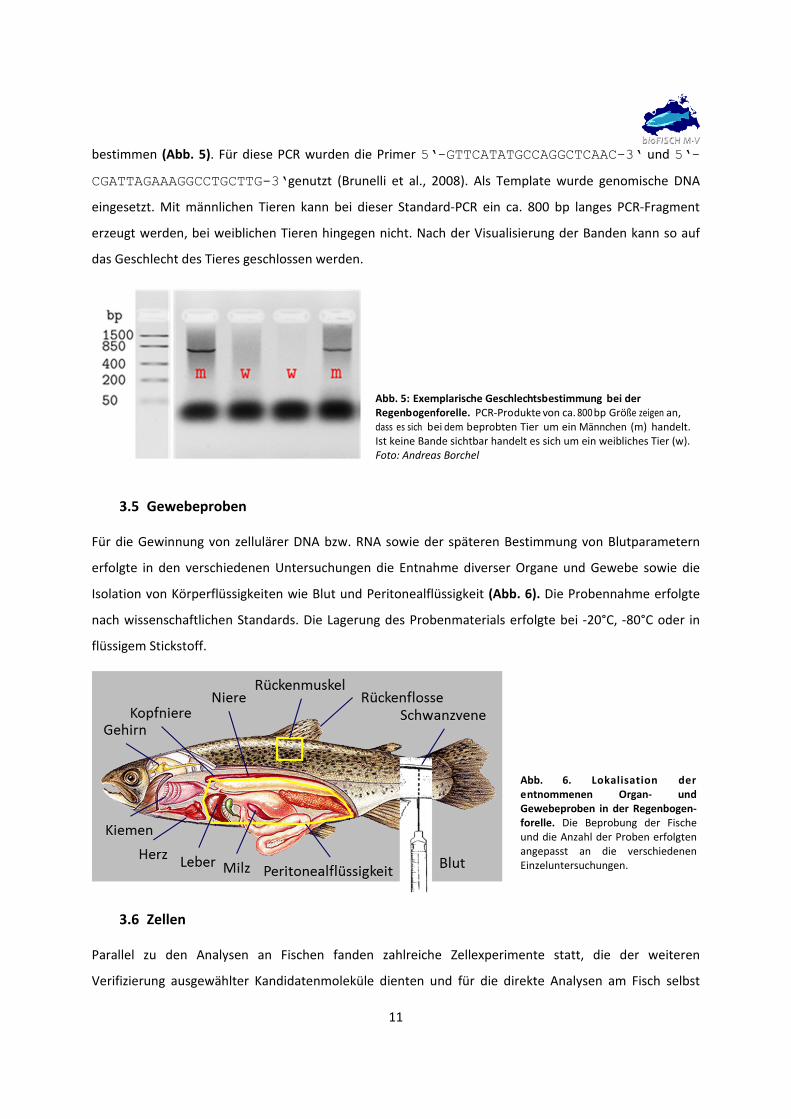
11
bestimmen (Abb. 5). Für diese PCR wurden die Primer 5‘-GTTCATATGCCAGGCTCAAC-3‘ und 5‘-
CGATTAGAAAGGCCTGCTTG-3‘genutzt (Brunelli et al., 2008). Als Template wurde genomische DNA
eingesetzt. Mit männlichen Tieren kann bei dieser Standard‐PCR ein ca. 800 bp langes PCR‐Fragment
erzeugt werden, bei weiblichen Tieren hingegen nicht. Nach der Visualisierung der Banden kann so auf
das Geschlecht des Tieres geschlossen werden.
Abb. 5: Exemplarische Geschlechtsbestimmung bei der Regenbogenforelle. PCR‐Produkte von ca. 800 bp Größe zeigen an, dass es sich bei dem beprobten Tier um ein Männchen (m) handelt. Ist keine Bande sichtbar handelt es sich um ein weibliches Tier (w). Foto: Andreas Borchel
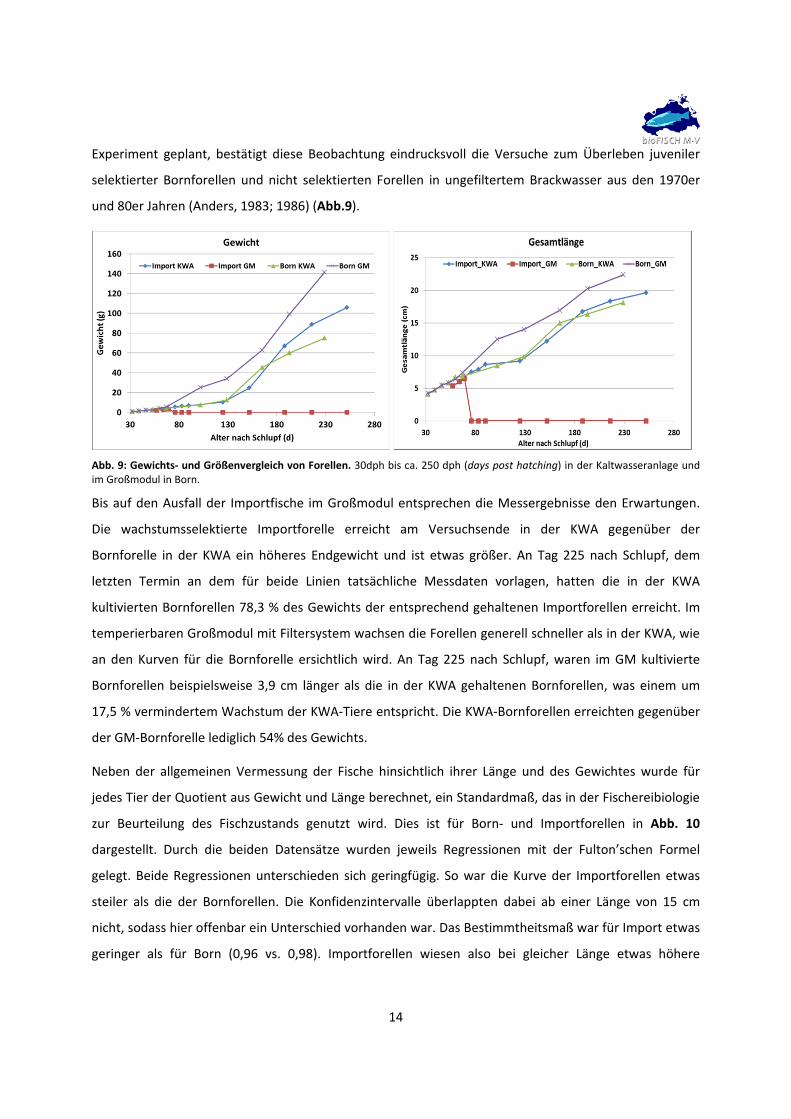
3.5 Gewebeproben
Für die Gewinnung von zellulärer DNA bzw. RNA sowie der späteren Bestimmung von Blutparametern
erfolgte in den verschiedenen Untersuchungen die Entnahme diverser Organe und Gewebe sowie die
Isolation von Körperflüssigkeiten wie Blut und Peritonealflüssigkeit (Abb. 6). Die Probennahme erfolgte
nach wissenschaftlichen Standards. Die Lagerung des Probenmaterials erfolgte bei ‐20°C, ‐80°C oder in
flüssigem Stickstoff.
Abb. 6. Lokalisation der entnommenen Organ‐ und Gewebeproben in der Regenbogen‐forelle. Die Beprobung der Fische und die Anzahl der Proben erfolgten angepasst an die verschiedenen Einzeluntersuchungen.
3.6 Zellen
Parallel zu den Analysen an Fischen fanden zahlreiche Zellexperimente statt, die der weiteren
Verifizierung ausgewählter Kandidatenmoleküle dienten und für die direkte Analysen am Fisch selbst

12
versuchstechnisch nicht zwingend notwendig bzw. im Rahmen des Projektes Untersuchungen am
lebenden Tier zu aufwendig waren. Für die Zellexperimente wurden im Wesentlichen die Zelllinien
RTgill‐W1 (Regenbogenforelle, Kiemenepithelzellen) und HEK‐293 (Humane embryonale
Nierenepithelzellen) eingesetzt (Abb. 7). Nach Infektionsexperimenten wurden für weitere Analysen
auch Leukozytenzellen aus der Bauchhöhle von Forellen mittels FACS isoliert und verschiedene Zelltypen
separiert – u.a. B‐,T‐Lymphozyten, Myeolidzellen, Trombozyten. Darüber hinaus wurden für
Kurzzeitversuche primäre Zelllinien von beiden Forellenlinien erstellt.
Abb.7: Verwendete Zelllinien. Exemplarische lichtmikroskopische Darstellung der Zelllinie HEK‐293 (links) und der Zelllinie RTgill‐W1 (rechts). Fotos: Alexander Rebl
3.7 Bioinformatik
Für die Auswertung der molekularen, immunologischen und zellulären Daten kamen u.a. folgende
bioinformatischen Methoden zur Anwendung: Sequenzanalyse mit UGene (Okonechnikov et al., 2012);
Exon‐Intron‐Grenzen mit Spidey (Wheelan et al., 2001) und MGAlign (Lee et al., 2003);
Stammbaumerstellung und multiples Sequenzalignment mit ClustalW (Goujon et al., 2010);
Clusteranalyse (Heatmaps) mit gplot (Warnes et al., 2014) mit Programmiersprache R; Clusterbildung mit
UPGMA‐Methode und amap (Lucas, 2014); IPA (www.ingenuity.com); weiterhin Excel, Sigmaplot, BLAST,
Primer 3 u.a.

13
4. Ergebnisse und Diskussion
4.1 Forellenproduktion
4.1.1 Forellenproduktion Vergleich 1
Im Zeitraum von April 2012 ‐2015 erfolgte die parallele Aufzucht von Born‐ und Importforellen in einer
Brackwasser gespeisten Kaltwasserdurchflussanlage (KWA) sowie in ebenfalls Brackwasser gespeisten
temperierbaren Großmodulen (GM) mit Vorfilterung. Gestartet wurde mit ca. 30000 bzw. 20000 Eiern
der Import‐ bzw. Bornforelle. Dabei ist zu berücksichtigen, dass die Importeier steril und rein weiblich
sowie befruchtet als Augenpunkteier eingesetzt wurden, während die Eier der Bornforelle vor Ort
befruchtet wurden und die Nachkommen gemischtgeschlechtlich waren. Die Augenpunkteier der
Importforellen waren größer als die der Bornforellen. Erbrütung und Anzucht der Fischlarven erfolgten
für ca. 12 Wochen in Süßwasser (Trinkwasser) in einem Temperaturbereich von 8‐16°C. Die Bornforellen
schlüpften nach 38 Tagen und waren nach 41 Tagen vollständig geschlüpft. Die Importforellen schlüpften
nach 40 Tagen und waren nach 43 Tagen vollständig geschlüpft. Unter Berücksichtigung der Tatsache,
dass die Importforellen bereits als befruchtete Augenpunkteier geliefert werden, lässt sich unter den
lokalen Gegebenheiten auf eine kürzere Schlupfzeit der Bornforelle schließen. Für alle Forellen begann
ca. 3 Wochen nach dem Schlupf die Fütterung und es erfolgte eine Umsetzung und Aufteilung der Fische
in verschiedene Produktionsanlagen. Von den befruchteten Eiern bis zum 100‐sten Tag nach dem Schlupf
wurden wöchentlich Proben für die DNA‐ und RNA‐Isolation genommen (jeweils n=10) (Abb. 8) sowie
Gewicht und Gesamtlänge der Fische erfasst. Danach wurde monatlich beprobt. Die Phänotypanalysen
sind in den Abb. 9 und 10 zusammengefasst. Nach der Umsetzung der Forellen ins GM trat für 48h ein
Filterproblem auf, weshalb 98% der Importfische starben und eine weitere Datenerfassung an dieser
Fischgruppe unmöglich wurde. Es ist an dieser Stelle zu erwähnen, dass die Bornforellen das
Filterproblem mit marginalen Verlusten von ca. 4% überstanden. Wenn auch nicht als wissenschaftliches
Abb. 8: RNA‐Isolation aus Regenbogenforellen. Das Beispiel zeigt gewonnene RNA aus Importforellen verschiedener Entwicklungsstadien in optimaler Qualität für Transkriptomanalysen. Die sichtbaren Banden entsprechen der 18S rRNA (unten) bzw. der 28S rRNA (oben). Die RNA‐Konzentration lag allgemein bei allen Proben beider Linien zwischen 1 und 4 µg/µl.
14
Experiment geplant, bestätigt diese Beobachtung eindrucksvoll die Versuche zum Überleben juveniler
selektierter Bornforellen und nicht selektierten Forellen in ungefiltertem Brackwasser aus den 1970er
und 80er Jahren (Anders, 1983; 1986) (Abb.9).
Abb. 9: Gewichts‐ und Größenvergleich von Forellen. 30dph bis ca. 250 dph (days post hatching) in der Kaltwasseranlage und im Großmodul in Born.
Bis auf den Ausfall der Importfische im Großmodul entsprechen die Messergebnisse den Erwartungen.
Die wachstumsselektierte Importforelle erreicht am Versuchsende in der KWA gegenüber der
Bornforelle in der KWA ein höheres Endgewicht und ist etwas größer. An Tag 225 nach Schlupf, dem
letzten Termin an dem für beide Linien tatsächliche Messdaten vorlagen, hatten die in der KWA
kultivierten Bornforellen 78,3 % des Gewichts der entsprechend gehaltenen Importforellen erreicht. Im
temperierbaren Großmodul mit Filtersystem wachsen die Forellen generell schneller als in der KWA, wie
an den Kurven für die Bornforelle ersichtlich wird. An Tag 225 nach Schlupf, waren im GM kultivierte
Bornforellen beispielsweise 3,9 cm länger als die in der KWA gehaltenen Bornforellen, was einem um
17,5 % vermindertem Wachstum der KWA‐Tiere entspricht. Die KWA‐Bornforellen erreichten gegenüber
der GM‐Bornforelle lediglich 54% des Gewichts.
Neben der allgemeinen Vermessung der Fische hinsichtlich ihrer Länge und des Gewichtes wurde für
jedes Tier der Quotient aus Gewicht und Länge berechnet, ein Standardmaß, das in der Fischereibiologie
zur Beurteilung des Fischzustands genutzt wird. Dies ist für Born‐ und Importforellen in Abb. 10
dargestellt. Durch die beiden Datensätze wurden jeweils Regressionen mit der Fulton’schen Formel
gelegt. Beide Regressionen unterschieden sich geringfügig. So war die Kurve der Importforellen etwas
steiler als die der Bornforellen. Die Konfidenzintervalle überlappten dabei ab einer Länge von 15 cm
nicht, sodass hier offenbar ein Unterschied vorhanden war. Das Bestimmtheitsmaß war für Import etwas
geringer als für Born (0,96 vs. 0,98). Importforellen wiesen also bei gleicher Länge etwas höhere
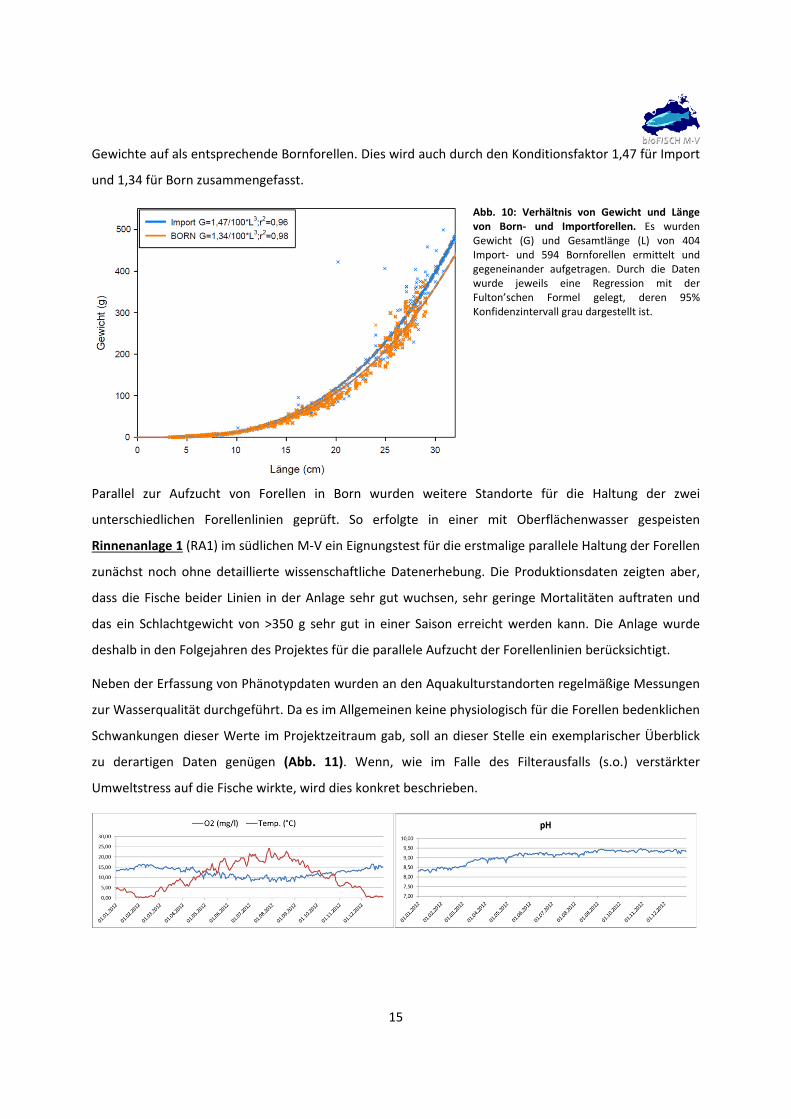
15
Gewichte auf als entsprechende Bornforellen. Dies wird auch durch den Konditionsfaktor 1,47 für Import
und 1,34 für Born zusammengefasst.
Abb. 10: Verhältnis von Gewicht und Länge von Born‐ und Importforellen. Es wurden Gewicht (G) und Gesamtlänge (L) von 404 Import‐ und 594 Bornforellen ermittelt und gegeneinander aufgetragen. Durch die Daten wurde jeweils eine Regression mit der Fulton’schen Formel gelegt, deren 95% Konfidenzintervall grau dargestellt ist.
Parallel zur Aufzucht von Forellen in Born wurden weitere Standorte für die Haltung der zwei
unterschiedlichen Forellenlinien geprüft. So erfolgte in einer mit Oberflächenwasser gespeisten
Rinnenanlage 1 (RA1) im südlichen M‐V ein Eignungstest für die erstmalige parallele Haltung der Forellen
zunächst noch ohne detaillierte wissenschaftliche Datenerhebung. Die Produktionsdaten zeigten aber,
dass die Fische beider Linien in der Anlage sehr gut wuchsen, sehr geringe Mortalitäten auftraten und
das ein Schlachtgewicht von >350 g sehr gut in einer Saison erreicht werden kann. Die Anlage wurde
deshalb in den Folgejahren des Projektes für die parallele Aufzucht der Forellenlinien berücksichtigt.
Neben der Erfassung von Phänotypdaten wurden an den Aquakulturstandorten regelmäßige Messungen
zur Wasserqualität durchgeführt. Da es im Allgemeinen keine physiologisch für die Forellen bedenklichen
Schwankungen dieser Werte im Projektzeitraum gab, soll an dieser Stelle ein exemplarischer Überblick
zu derartigen Daten genügen (Abb. 11). Wenn, wie im Falle des Filterausfalls (s.o.) verstärkter
Umweltstress auf die Fische wirkte, wird dies konkret beschrieben.
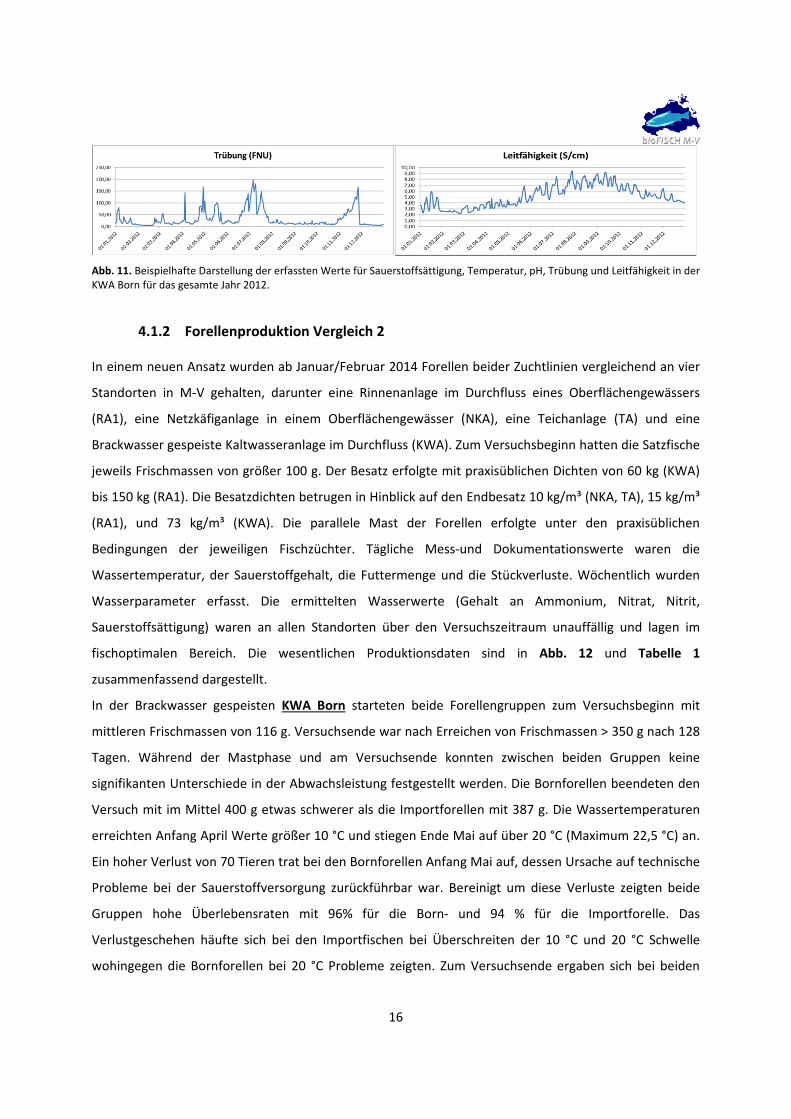
16
Abb. 11. Beispielhafte Darstellung der erfassten Werte für Sauerstoffsättigung, Temperatur, pH, Trübung und Leitfähigkeit in der KWA Born für das gesamte Jahr 2012.
4.1.2 Forellenproduktion Vergleich 2
In einem neuen Ansatz wurden ab Januar/Februar 2014 Forellen beider Zuchtlinien vergleichend an vier
Standorten in M‐V gehalten, darunter eine Rinnenanlage im Durchfluss eines Oberflächengewässers
(RA1), eine Netzkäfiganlage in einem Oberflächengewässer (NKA), eine Teichanlage (TA) und eine
Brackwasser gespeiste Kaltwasseranlage im Durchfluss (KWA). Zum Versuchsbeginn hatten die Satzfische
jeweils Frischmassen von größer 100 g. Der Besatz erfolgte mit praxisüblichen Dichten von 60 kg (KWA)
bis 150 kg (RA1). Die Besatzdichten betrugen in Hinblick auf den Endbesatz 10 kg/m³ (NKA, TA), 15 kg/m³
(RA1), und 73 kg/m³ (KWA). Die parallele Mast der Forellen erfolgte unter den praxisüblichen
Bedingungen der jeweiligen Fischzüchter. Tägliche Mess‐und Dokumentationswerte waren die
Wassertemperatur, der Sauerstoffgehalt, die Futtermenge und die Stückverluste. Wöchentlich wurden
Wasserparameter erfasst. Die ermittelten Wasserwerte (Gehalt an Ammonium, Nitrat, Nitrit,
Sauerstoffsättigung) waren an allen Standorten über den Versuchszeitraum unauffällig und lagen im
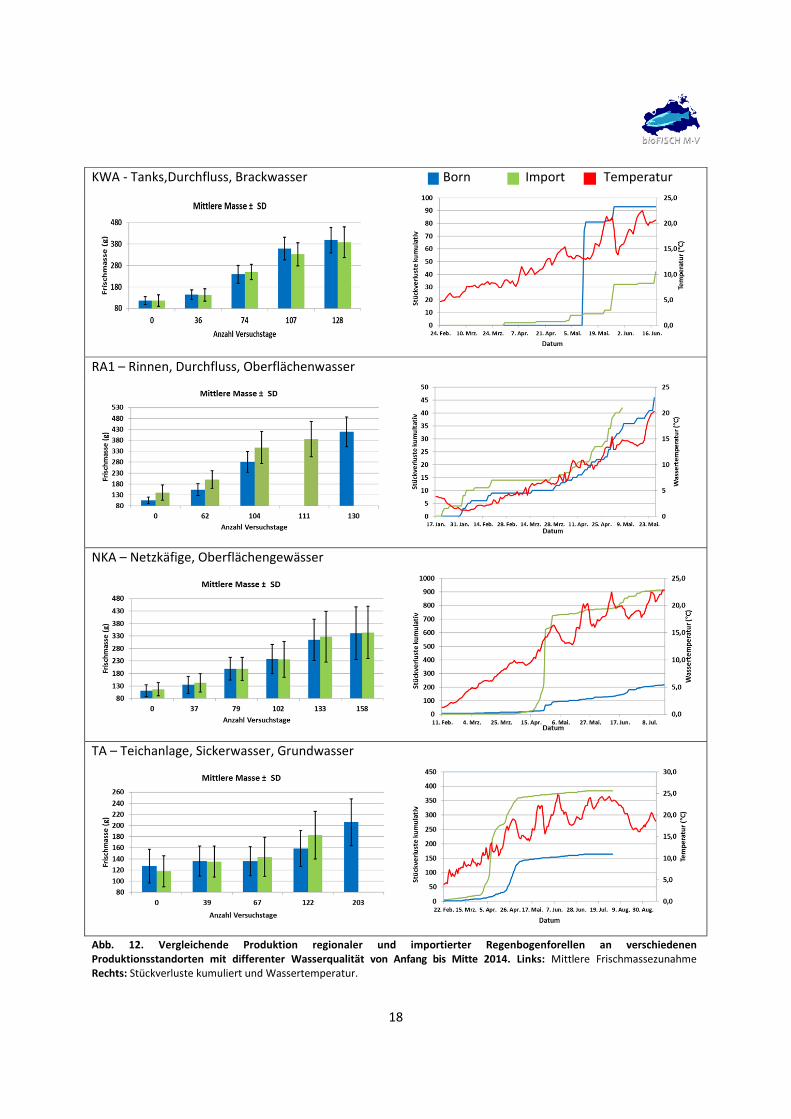
fischoptimalen Bereich. Die wesentlichen Produktionsdaten sind in Abb. 12 und Tabelle 1
zusammenfassend dargestellt.
In der Brackwasser gespeisten KWA Born starteten beide Forellengruppen zum Versuchsbeginn mit
mittleren Frischmassen von 116 g. Versuchsende war nach Erreichen von Frischmassen > 350 g nach 128
Tagen. Während der Mastphase und am Versuchsende konnten zwischen beiden Gruppen keine
signifikanten Unterschiede in der Abwachsleistung festgestellt werden. Die Bornforellen beendeten den
Versuch mit im Mittel 400 g etwas schwerer als die Importforellen mit 387 g. Die Wassertemperaturen
erreichten Anfang April Werte größer 10 °C und stiegen Ende Mai auf über 20 °C (Maximum 22,5 °C) an.
Ein hoher Verlust von 70 Tieren trat bei den Bornforellen Anfang Mai auf, dessen Ursache auf technische
Probleme bei der Sauerstoffversorgung zurückführbar war. Bereinigt um diese Verluste zeigten beide
Gruppen hohe Überlebensraten mit 96% für die Born‐ und 94 % für die Importforelle. Das
Verlustgeschehen häufte sich bei den Importfischen bei Überschreiten der 10 °C und 20 °C Schwelle
wohingegen die Bornforellen bei 20 °C Probleme zeigten. Zum Versuchsende ergaben sich bei beiden

17
Versuchsgruppen ähnliche Leistungsparameter. Die täglichen Wachstumsraten betrugen 0,94 (Import)
und 0,97 %/Tag (Bornforelle). Für den Futterquotienten ergaben sich mäßige Werte die bei den
Bornforellen mit 1,5 schlechter waren als der Fütterungsquotient von 1,4 für die Importfische. Die
Analyse der Schlachtkörper ergab kaum Unterschiede. Die Schlachtkörperanteile betrugen 88 – 89 % der
Frischmassen. Der Filetanteil war bei den Importforellen mit 44 % signifikant höher als in der
Vergleichsgruppe, wohingegen die Bornforellen mit 3,5 % etwas mehr Eingeweidefett besaßen. Gonaden
waren nur unmerklich ausgebildet. Die Konditionsfaktoren lagen bei 1,3‐1,4.
Für die Rinnenanlage 1 standen zu Versuchsbeginn Mitte Januar keine einheitlichen Satzfische zur
Verfügung. Die mittleren Frischmassen der Bornforelle lagen mit 105 g deutlich unter den Werten der
Importfische mit 141 g. Dies führte zu unterschiedlichen Versuchsenden Anfang Mai nach 111 Tagen für
die Importforellen, die zu diesem Zeitpunkt ein mittleres Gewicht von 386 g aufwiesen und nach 130
Tagen für die Bornforellen mit dann im Mittel 418 g. Während der Mastphase blieben die
Wachstumsvorteile der Importforellen weiterhin sichtbar. Die Wassertemperaturen sanken bis Ende
Januar noch einmal ab, erreichten Anfang April Werte größer 10°C und stiegen erst zum Versuchsende
im Mai auf über 20 °C an. In beiden Gruppen traten schleichende Verluste auf. Die Überlebensraten
betrugen bei beiden Gruppen 96 %. Das Verlustgeschehen zeigte im Verlauf keine Spitzen. Für die
Futterquotienten ergaben sich zum Versuchsende sehr gute Werte von 1,0. Die tägliche Wachstumsrate
lag für die Bornforelle mit 1,1 %/Tag etwas höher als für die Importfische mit 0,9 %/Tag. Die
Schlachtkörperanalysen ergaben kaum Unterschiede. Die Schlachtkörperanteile betrugen 85 – 86 % der
Frischmassen. Der Filetanteil (ohne Haut) war bei den Importforellen mit 50 % signifikant höher als in der
Vergleichsgruppe mit 46%. Die Tiere waren mit Eingeweidefettwerten von 1,4‐2 % fettarm. Gonaden
waren bei den Bornforellen nur unmerklich ausgebildet. Die Konditionsfaktoren lagen bei 1,3‐1,5 für
Born‐ bzw. Importforellen.
Der Besatz der Versuchsgruppen in die Netzkäfiganlage erfolgte Anfang Februar. Die mittleren
Frischmassen der Bornforellen lagen mit 111 g nur unwesentlich höher als in der Vergleichsgruppe mit
117 g. Mitte Juli wurde der Versuch nach 158 Tagen beendet. Beide Vergleichsgruppen hatten im Mittel
fast 350 g Stückgewicht erreicht. Während der Mastphase zeigten sich keine Unterschiede beim
Wachstum, jedoch traten bei den Importforellen massive Verluste auf. Mit dem Anstieg der
Wassertemperatur über 10°C setzte eine kurzzeitig hohe Sterblichkeit von mehr als der Hälfte der Tiere
in dieser Versuchsgruppe ein. In beiden Versuchsgruppen stellten sich in den Folgewochen erhöhte
schleichende Verluste ein. Die Verluste stiegen erneut Mitte Juni nach einer Periode mit
Wassertemperaturen über 20 °C. Diese Verluste waren bei den Bornforellen geringer als bei den
Importfischen. Insgesamt ergaben sich Überlebensraten von 80 % für die Bornforelle bzw. nur 19 % für
18
KWA ‐ Tanks,Durchfluss, Brackwasser Born Import Temperatur
RA1 – Rinnen, Durchfluss, Oberflächenwasser
NKA – Netzkäfige, Oberflächengewässer
TA – Teichanlage, Sickerwasser, Grundwasser
Abb. 12. Vergleichende Produktion regionaler und importierter Regenbogenforellen an verschiedenen Produktionsstandorten mit differenter Wasserqualität von Anfang bis Mitte 2014. Links: Mittlere Frischmassezunahme Rechts: Stückverluste kumuliert und Wassertemperatur.
19
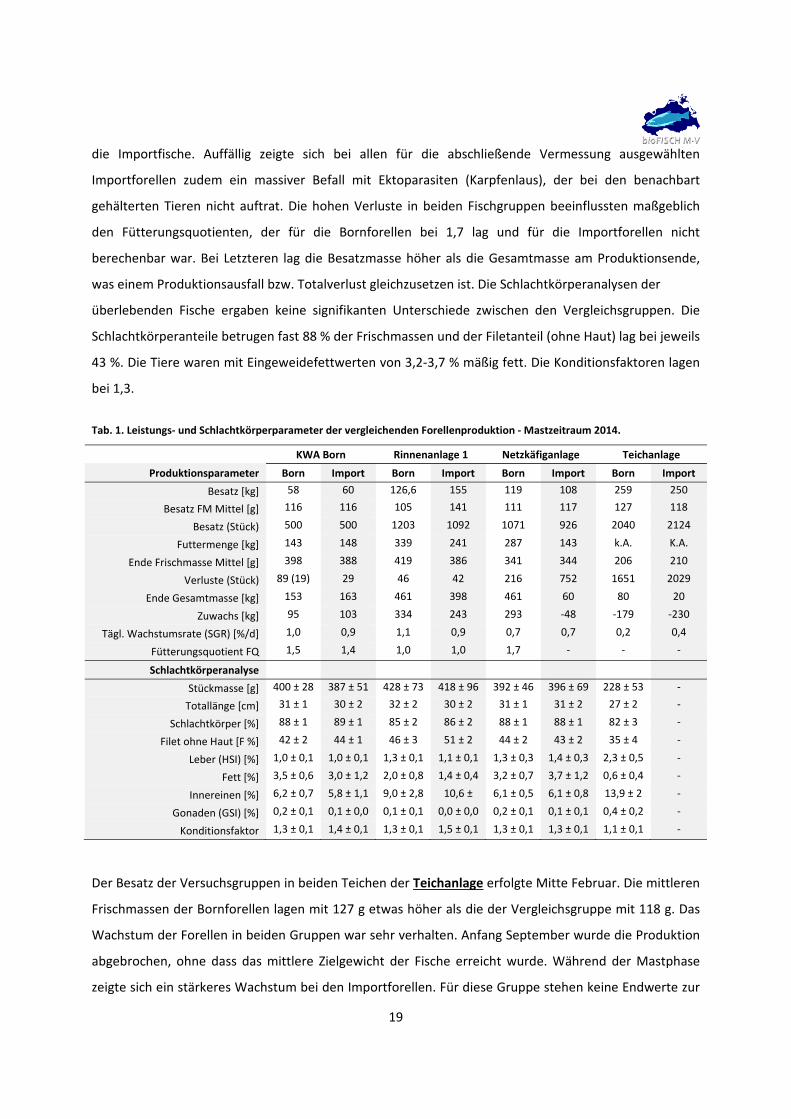
die Importfische. Auffällig zeigte sich bei allen für die abschließende Vermessung ausgewählten
Importforellen zudem ein massiver Befall mit Ektoparasiten (Karpfenlaus), der bei den benachbart
gehälterten Tieren nicht auftrat. Die hohen Verluste in beiden Fischgruppen beeinflussten maßgeblich
den Fütterungsquotienten, der für die Bornforellen bei 1,7 lag und für die Importforellen nicht
berechenbar war. Bei Letzteren lag die Besatzmasse höher als die Gesamtmasse am Produktionsende,
was einem Produktionsausfall bzw. Totalverlust gleichzusetzen ist. Die Schlachtkörperanalysen der
überlebenden Fische ergaben keine signifikanten Unterschiede zwischen den Vergleichsgruppen. Die
Schlachtkörperanteile betrugen fast 88 % der Frischmassen und der Filetanteil (ohne Haut) lag bei jeweils
43 %. Die Tiere waren mit Eingeweidefettwerten von 3,2‐3,7 % mäßig fett. Die Konditionsfaktoren lagen
bei 1,3.
Tab. 1. Leistungs‐ und Schlachtkörperparameter der vergleichenden Forellenproduktion ‐ Mastzeitraum 2014.
KWA Born Rinnenanlage 1 Netzkäfiganlage Teichanlage
Produktionsparameter Born Import Born Import Born Import Born Import
Besatz [kg] 58 60 126,6 155 119 108 259 250
Besatz FM Mittel [g] 116 116 105 141 111 117 127 118
Besatz (Stück) 500 500 1203 1092 1071 926 2040 2124
Futtermenge [kg] 143 148 339 241 287 143 k.A. K.A.
Ende Frischmasse Mittel [g] 398 388 419 386 341 344 206 210
Verluste (Stück) 89 (19) 29 46 42 216 752 1651 2029
Ende Gesamtmasse [kg] 153 163 461 398 461 60 80 20
Zuwachs [kg] 95 103 334 243 293 ‐48 ‐179 ‐230
Tägl. Wachstumsrate (SGR) [%/d] 1,0 0,9 1,1 0,9 0,7 0,7 0,2 0,4
Fütterungsquotient FQ 1,5 1,4 1,0 1,0 1,7 ‐ ‐ ‐
Schlachtkörperanalyse
Stückmasse [g] 400 ± 28 387 ± 51 428 ± 73 418 ± 96 392 ± 46 396 ± 69 228 ± 53 ‐
Totallänge [cm] 31 ± 1 30 ± 2 32 ± 2 30 ± 2 31 ± 1 31 ± 2 27 ± 2 ‐
Schlachtkörper [%] 88 ± 1 89 ± 1 85 ± 2 86 ± 2 88 ± 1 88 ± 1 82 ± 3 ‐
Filet ohne Haut [F %] 42 ± 2 44 ± 1 46 ± 3 51 ± 2 44 ± 2 43 ± 2 35 ± 4 ‐
Leber (HSI) [%] 1,0 ± 0,1 1,0 ± 0,1 1,3 ± 0,1 1,1 ± 0,1 1,3 ± 0,3 1,4 ± 0,3 2,3 ± 0,5 ‐
Fett [%] 3,5 ± 0,6 3,0 ± 1,2 2,0 ± 0,8 1,4 ± 0,4 3,2 ± 0,7 3,7 ± 1,2 0,6 ± 0,4 ‐
Innereinen [%] 6,2 ± 0,7 5,8 ± 1,1 9,0 ± 2,8 10,6 ± 6,1 ± 0,5 6,1 ± 0,8 13,9 ± 2 ‐
Gonaden (GSI) [%] 0,2 ± 0,1 0,1 ± 0,0 0,1 ± 0,1 0,0 ± 0,0 0,2 ± 0,1 0,1 ± 0,1 0,4 ± 0,2 ‐
Konditionsfaktor 1,3 ± 0,1 1,4 ± 0,1 1,3 ± 0,1 1,5 ± 0,1 1,3 ± 0,1 1,3 ± 0,1 1,1 ± 0,1 ‐
Der Besatz der Versuchsgruppen in beiden Teichen der Teichanlage erfolgte Mitte Februar. Die mittleren
Frischmassen der Bornforellen lagen mit 127 g etwas höher als die der Vergleichsgruppe mit 118 g. Das
Wachstum der Forellen in beiden Gruppen war sehr verhalten. Anfang September wurde die Produktion
abgebrochen, ohne dass das mittlere Zielgewicht der Fische erreicht wurde. Während der Mastphase
zeigte sich ein stärkeres Wachstum bei den Importforellen. Für diese Gruppe stehen keine Endwerte zur

20
Verfügung. Die Bornforellen hatten zum Versuchsende nach mehr als 200 Tagen nur eine mittlere Masse
von 205 g und waren zudem massiv mit Ektoparasiten (Karpfenlaus) befallen. In der Teichanlage müssen
beide Versuchsgruppen als Totalverlust eingestuft werden. Von den anfänglich jeweils besetzten ca. 250
kg je Gruppe konnten im September insgesamt nur jeweils 60 bzw. 80 kg abgefischt werden. Die
ermittelten täglichen Wachstumsraten waren extrem gering und betrugen für die Bornforellen lediglich
0,2 %/Tag und für die Importforellen (bis Tag 122) 0,4 %/Tag. Für die finalen Schlachtkörperanalysen
standen keine Importforellen mehr zur Verfügung. Die Schlachtkörperanteile der Bornforellen betrugen
82 % der Frischmassen und der Filetanteil (ohne Haut) lag bei nur 35 %. Die Tiere hatten kaum
Eingeweidefett (0,6 %), einen hohen Leberindex HSI=2,3 % sowie einen schwachen Konditionsfaktor von
1,1.
Im Ergebnis dieser vergleichenden Forellenproduktion mit regionalen Bornforellen und Importforellen
lässt sich feststellen, dass die ermittelten Abwachsleistungen und Leistungsparameter der Fische zum
Versuchsende in den Aquakulturanlagen KWA, RA1 und NKA praxisübliche Werte ergaben. Lediglich in
der Teichanlage wurden schon im Versuchsverlauf stark abweichende Messwerte festgehalten. Die
Forellen zeigten kaum Wachstum und benötigten mehr als 200 Tage, um auf Stückmassen von im Mittel
mehr als 200 g abzuwachsen. Diese Stückmassen wurden an allen anderen Standorten bereits nach 2‐3
Monaten erreicht. In Verbindung mit dem beobachteten extrem hohen Verlustgeschehen, den extrem
niedrigen täglichen Wachstumsraten und den mäßigen Konditionsfaktoren der Fische ist von einer
weniger optimalen Hälterung auszugehen und somit sind die Ergebnisse aus der Teichanlage für die
weitere vergleichende Betrachtung nicht reproduzierbar. An den übrigen Standorten konnte nach 111‐
158 Tagen die Mast der Speiseforellen erfolgreich abgeschlossen werden. Trotzdem muss auch dort das
Verlustgeschehen für zukünftige Produktionsansätze diskutiert und berücksichtigt werden.
Für das jeweilige Verlustgeschehen, als ein wichtiger Marker der Robustheit gegenüber den
Standortfaktoren, ergaben sich im Ergebnis unterschiedliche Bilder. Lediglich an den Standorten KWA
und RA1 zeigten sich niedrige Verlustraten von 3‐5 % wobei zwischen den Vergleichsgruppen kaum
Unterschiede auftraten. Höhere Verluste bei der Bornforelle in der KWA Born resultierten aus einem
technischen Defekt. Im Vergleich sind jedoch auch die Verlustraten von 3‐5% als relativ hoch
einzustufen, denn Jansen et al. (2004, 2007) gaben bei der Speiseforellenmast lediglich Mortalitäten von
1,3 % bzw. 0,2 % für Bornforellen an. Die Verluste in der Netzkäfiganlage waren bei der Bornforelle
wesentlich höher, jedoch in der Vergleichsgruppe mit > 80 % Verlusten gravierend. An diesem Standort
zeigte die Bornforelle deutlich bessere Überlebensraten. Im Gegensatz zu den kontinuierlichen Verlusten
in der Rinnenanlage RA1, konzentrierten sich die Verluste in der KWA Born und in der Netzkäfiganlage

21
NKA zu den Zeitpunkten des Überschreitens der Wassertemperatur > 10 °C und > 20 °C. Zu diesen
Zeitpunkten stiegen vor allem in der Gruppe der Importforellen die Mortalitäten deutlich an. Für die
Rinnenanlage konnte diese Beobachtung nicht festgehalten werden, da die Wassertemperatur erst nach
Versuchsende über 20 °C anstieg. Für beide Versuchsgruppen in der Teichanlage ergaben sich
Totalverluste.
Aus den vorliegenden Daten ließ sich schlussfolgern, dass KWA Born und RA1 sich generell sehr gut für
die Forellenproduktion eignen. Sowohl die Zuchtlinie Born als auch die Importlinie erscheinen für die
Produktion an diesen Standorten geeignet. Die Ergebnisse in der NKA deuten hingegen auf weniger
geeignete Haltungsbedingungen zumindest für die Importforellen hin. Die Bornforelle ist an diesem
Standort deutlich im Vorteil. Dennoch sind die Verluste auch bei diesen Fischen zu groß. Die
Forellenhaltung in Teichanlagen ohne Manipulationsmöglichkeiten der Umweltbedingungen (z.B.
Belüftung, künstlicher Wasseraustausch) in Extremsituationen erscheint hingegen als nicht ratsam.
4.1.3 Forellenproduktion Vergleich 3
In einem weiteren Ansatz wurde basierend auf den Ergebnissen der vorherigen Produktionsansätze ein
paralleler Mastansatz für die Regenbogenforellenlinien in der Durchflussanlage KWA in Born, sowie in
zwei weiteren regionalen Aquakulturanlagen mit Rinnensystem durchgeführt. Während die Anlage in
Born wiederum mit Brackwasser gespeist wurde, nutzten die Rinnenanlagen (RA1 und RA2)
Oberflächenwasser für die Aufzucht. Der Besatz erfolgte im Juni 2014. Zum Versuchsbeginn Mitte Juni
2014 hatten die Satzfische mittlere Frischmassen (FM) von 2,7 g Bornforelle bzw. 5,1 g Importforelle. Der
Besatz erfolgte in Mengen von je 500 Stück (KWA) und 1000 Stück (RA1, RA2). Die parallele Mast der
Forellen und die Datenerfassung erfolgten unter den praxisüblichen Bedingungen der jeweiligen
Fischzüchter. Gefüttert wurde handelsübliches Forellenfutter. An allen Standorten erfolgte eine
Zwischenmessung (Frischmasse, Totallänge) an jeweils 50 Tieren. Nach Erreichen des Zielgewichtes von >
350‐400 g wurden die Gesamtmassen bestimmt und die Frischmassen und Totallängen von 50 Tieren
erfasst. Für eine Schlachtkörperanalyse wurden jeweils zehn Tiere pro Gruppe zum Versuchsende
entnommen und bearbeitet. Die Ergebnisse der Forellenproduktion and den drei Standorten sind in
Abbildung 13 und Tabelle 2 zusammengefasst. Abb. 13 gibt neben Daten zur Wachstumsleistung
Auskunft über die Mortalitäten und die Wassertemperatur während der Produktion.
In der KWA Born wurde die Forellenproduktion im Mai 2015 nach 321 Tagen (Importforelle) bzw. 345
Tagen (Bornforelle) beendet. Im Abwachszeitraum traten keine Umweltmesswerte auf, die außerhalb
des physiologischen Toleranzbereiches von Regenbogenforellen lag. Dementsprechend wuchsen die

22
Forellen der wachstumsselektierten Importforellen etwas besser und wiesen zum Messzeitpunkt
innerhalb der Mastphase höhere Gewichte auf als die Bornforellen und erreichten am Versuchsende das
Schlachtgewicht von > 350 g vor der Vergleichsgruppe. Zu berücksichtigen ist dabei, dass die Satzfische
der Bornforelle im Mittel 1,4 g weniger wogen als die Importfische, was zu Beginn der Aufzucht einer
Differenz von ca. 14 Tagen entspricht. Die Bornforellen (465 g) waren am Ende der Mast wiederum
schwerer als die Importfische (392 g), weil sie in der absoluten Differenz ca. 10 Tage länger wachsen
konnten (Abb. 13). Und dies in einem optimalen Mastzeitraum mit Temperaturen von 15°C. Es ist
anzunehmen, dass die Schlachtgewichte der Fische beider Linien bei einem exakt gleichen Erbrütungs‐
und Aufzuchtzeitraum in der KWA Born sehr ähnlich gewesen wären. Dies untermauern auch die sehr
ähnlichen Leistungsparameter, die sich zum Versuchsende bei beiden Versuchsgruppen ergaben. Die
täglichen Wachstumsraten betrugen 1,5 %/d (Born) und 1,5 %/d (Import). Zum Beginn der Mast (Juli‐
August 2014) traten bei erhöhten Wassertemperaturen von bis zu 25°C in beiden Fischgruppen
schleichende Verlust auf mit höheren Verlusten bei der Bornforelle. Hier setzten sich die Verluste über
den Sommer fort und stiegen im Oktober 2014 noch etwas an. Ab Dezember traten kaum noch Verluste
auf. Im Gegensatz dazu setzte Mitte August für einige Wochen ein massives Sterben bei den
Importforellen ein. Ab Oktober traten kaum noch Verluste auf. Die Überlebensraten waren bei der
Bornforelle mit 79 % signifikant höher als bei den Importforellen (42 %). Aufgrund der hohen Mortalität
bei den Importforellen weisen die Bornforellen mit einem finalen Gesamtgewicht von 179 kg gegenüber
den Importforellen mit 87 kg ein Produktionsergebnis auf, das um das 2‐fache höher liegt, als das der
Importforellen. Diese Zahlen spiegeln sich ebenso im Fütterungsquotienten wieder, der für die
Bornforelle bei sehr guten 0,9 für die Importforelle jedoch nur bei 1,9 lag (Tabelle 2). Bei der Analyse
der Schlachtkörper ergaben sich kaum Unterschiede. Die unterschiedlichen mittleren Stückmassen
resultieren aus unterschiedlichen Probetagen. Die Schlachtkörperanteile betrugen ca. 84 % der
Frischmassen. Die Leber war bei den RFBo etwas größer ausgebildet. Der Filetanteil war bei den
Importfischen mit 45 % signifikant höher als in der Vergleichsgruppe. Beide Gruppen zeigten wenig
Eingeweidefett (1,9‐2,0 %). Die Konditionsfaktoren lagen bei 1,3‐1,4.

23
KWA ‐ Tanks,Durchfluss, Brackwasser
RA1 – Rinnen, Durchfluss, Oberflächenwasser
RA2 – Rinnen, Durchfluss, Oberflächenwasser
Abb. 13. Vergleichende Produktion regionaler und importierter Regenbogenforellen an verschiedenen Produktionsstandorten mit differenter Wasserqualität im Zeitraum 2014‐2015. (Links) Mittlere Frischmassezunahme von Regenbogenforellen. (Rechts) Stückverluste kumuliert und Wassertemperatur. Für RA2 fehlt die Dokumentation der Verluste. SD‐ Standardabweichung.
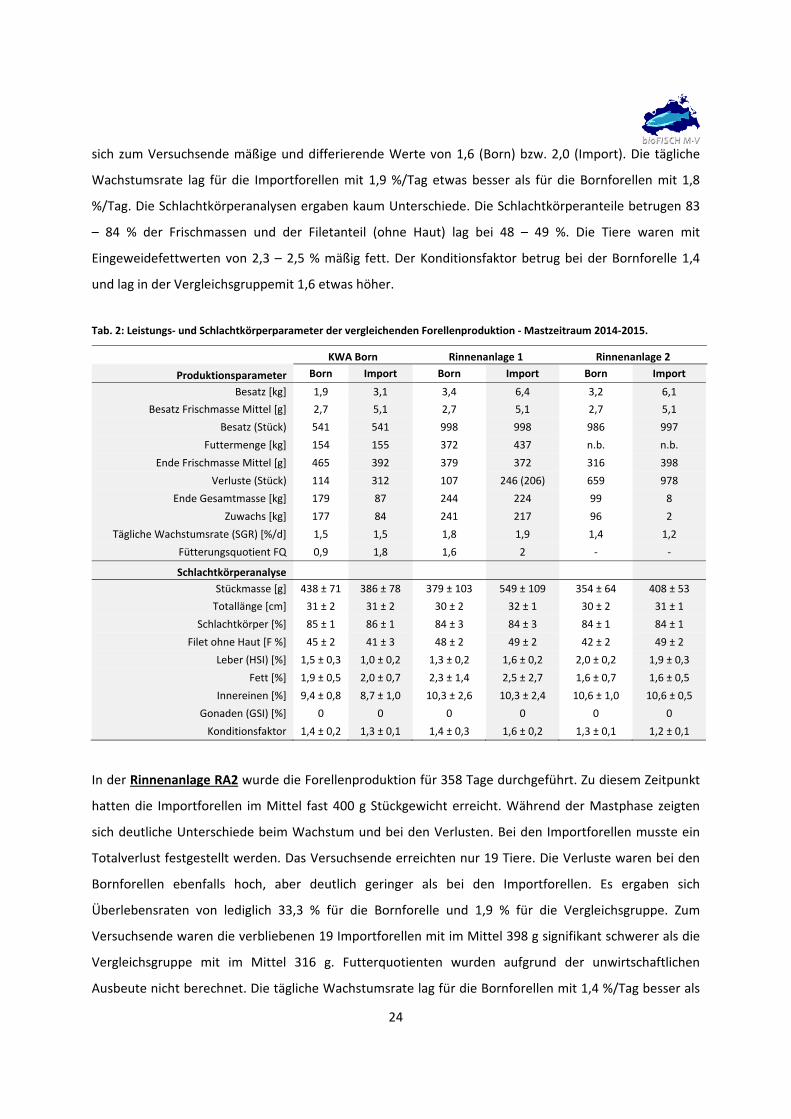
In der Rinnenanlage RA1 erreichten die Importforellen bereits nach 227 Tagen ein Gewicht von beinahe
380 g und übertrafen damit das avisierte Zielgewicht von >350 g. Ein ähnliches mittleres Stückgewicht
erreichten die Bornforellen erst nach 282 Tagen. Während der Mastphase zeigten sich Unterschiede
beim Wachstum. Ende Januar waren die Importforellen im Mittel bereits 64 g schwerer als die
Vergleichsgruppe. Bis Mitte August 2014, mit ansteigenden und hohen Wassertemperaturen, stellten
sich in beiden Versuchsgruppen stetige Verluste ein. Diese waren bei den Importforellen deutlicher
ausgeprägt. Ende Dezember wurde versehentlich eine größere Anzahl Importforellen (n = 40) aus dem
Versuch entfernt. In beiden Gruppen traten weiter über den Versuchszeitraum verteilt schleichende
Verluste auf. Die Überlebensraten waren unterschiedlich und betrugen 89 % für die Bornforellen und 79
% (bereinigt um die entnommenen Forellen) für die Importforellen. Für die Futterquotienten ergaben
24
sich zum Versuchsende mäßige und differierende Werte von 1,6 (Born) bzw. 2,0 (Import). Die tägliche
Wachstumsrate lag für die Importforellen mit 1,9 %/Tag etwas besser als für die Bornforellen mit 1,8
%/Tag. Die Schlachtkörperanalysen ergaben kaum Unterschiede. Die Schlachtkörperanteile betrugen 83
– 84 % der Frischmassen und der Filetanteil (ohne Haut) lag bei 48 – 49 %. Die Tiere waren mit
Eingeweidefettwerten von 2,3 – 2,5 % mäßig fett. Der Konditionsfaktor betrug bei der Bornforelle 1,4
und lag in der Vergleichsgruppemit 1,6 etwas höher.
Tab. 2: Leistungs‐ und Schlachtkörperparameter der vergleichenden Forellenproduktion ‐ Mastzeitraum 2014‐2015.
KWA Born Rinnenanlage 1 Rinnenanlage 2
Produktionsparameter Born Import Born Import Born Import
Besatz [kg] 1,9 3,1 3,4 6,4 3,2 6,1
Besatz Frischmasse Mittel [g] 2,7 5,1 2,7 5,1 2,7 5,1
Besatz (Stück) 541 541 998 998 986 997
Futtermenge [kg] 154 155 372 437 n.b. n.b.
Ende Frischmasse Mittel [g] 465 392 379 372 316 398
Verluste (Stück) 114 312 107 246 (206) 659 978
Ende Gesamtmasse [kg] 179 87 244 224 99 8
Zuwachs [kg] 177 84 241 217 96 2
Tägliche Wachstumsrate (SGR) [%/d] 1,5 1,5 1,8 1,9 1,4 1,2
Fütterungsquotient FQ 0,9 1,8 1,6 2 ‐ ‐
Schlachtkörperanalyse
Stückmasse [g] 438 ± 71 386 ± 78 379 ± 103 549 ± 109 354 ± 64 408 ± 53
Totallänge [cm] 31 ± 2 31 ± 2 30 ± 2 32 ± 1 30 ± 2 31 ± 1
Schlachtkörper [%] 85 ± 1 86 ± 1 84 ± 3 84 ± 3 84 ± 1 84 ± 1
Filet ohne Haut [F %] 45 ± 2 41 ± 3 48 ± 2 49 ± 2 42 ± 2 49 ± 2
Leber (HSI) [%] 1,5 ± 0,3 1,0 ± 0,2 1,3 ± 0,2 1,6 ± 0,2 2,0 ± 0,2 1,9 ± 0,3
Fett [%] 1,9 ± 0,5 2,0 ± 0,7 2,3 ± 1,4 2,5 ± 2,7 1,6 ± 0,7 1,6 ± 0,5
Innereinen [%] 9,4 ± 0,8 8,7 ± 1,0 10,3 ± 2,6 10,3 ± 2,4 10,6 ± 1,0 10,6 ± 0,5
Gonaden (GSI) [%] 0 0 0 0 0 0
Konditionsfaktor 1,4 ± 0,2 1,3 ± 0,1 1,4 ± 0,3 1,6 ± 0,2 1,3 ± 0,1 1,2 ± 0,1
In der Rinnenanlage RA2 wurde die Forellenproduktion für 358 Tage durchgeführt. Zu diesem Zeitpunkt
hatten die Importforellen im Mittel fast 400 g Stückgewicht erreicht. Während der Mastphase zeigten
sich deutliche Unterschiede beim Wachstum und bei den Verlusten. Bei den Importforellen musste ein
Totalverlust festgestellt werden. Das Versuchsende erreichten nur 19 Tiere. Die Verluste waren bei den
Bornforellen ebenfalls hoch, aber deutlich geringer als bei den Importforellen. Es ergaben sich
Überlebensraten von lediglich 33,3 % für die Bornforelle und 1,9 % für die Vergleichsgruppe. Zum
Versuchsende waren die verbliebenen 19 Importforellen mit im Mittel 398 g signifikant schwerer als die
Vergleichsgruppe mit im Mittel 316 g. Futterquotienten wurden aufgrund der unwirtschaftlichen
Ausbeute nicht berechnet. Die tägliche Wachstumsrate lag für die Bornforellen mit 1,4 %/Tag besser als

25
für die Importfische mit 1,2 %/Tag. Die Konditionsfaktoren waren vergleichbar (Born 1,3 und Import 1,2).
Die Schlachtkörperanalysen ergaben nur in einem Merkmal signifikante Unterschiede zwischen den
Vergleichsgruppen. Die Schlachtkörperanteile betrugen 84 % der Frischmassen, der Filetanteil (ohne
Haut) betrug bei der Bornforelle im Mittel 42 % und lag bei der Vergleichsgruppe mit 49 % deutlich
höher. Die Tiere waren mit Eingeweidefettwerten von 1,6 % sehr mager.
Die vergleichende Produktion von Forellen in KWA bzw. RA1 und RA2 liefert bezogen auf das
Fischwachstum und damit im Zusammenhang stehende Leistungsparameter und Schlachtkörperwerte
praxisübliche Ergebnisse. An allen drei Standorten waren die wachstumsselektierten Importfische zum
Versuchsende im Mittel deutlich schwerer bzw. erreichten das avisierte Zielgewicht von >350 g deutlich
eher als die robuste Bornforelle. Zwischen den Standorten gab es jedoch deutliche Unterschiede im
Fischwachstum. In der RA1 erreichten die Forellen Ihr Schlachtgewicht 65‐100 Tage eher als in KWA und
RA2. Ebenso waren in der RA1 die Verluste in beiden Linien verhältnismäßig gering. Damit scheint die
RA1 wie schon im Validierungsansatz und dem vorherigen Produktionsansatz als Produktionsanlage
sowohl für Born‐ als auch Importforelle sehr gut geeignet. Werden alle hier betrachteten
Produktionsparameter einbezogen, zeigte die seit beinahe 40 Jahren im Brackwasser gezüchtete
Bornforelle in der KWA final die wirtschaftlichste Performance mit einem Futterumsatzquotienten von
0,9. Zu berücksichtigen ist aber bei einer Wirtschaftlichkeitsstudie ihr etwas langsameres Wachstum,
weshalb sie die Produktionsanlagen länger blockiert. Bezogen auf die hohen Mortalitäten der
Importforelle gegenüber der Bornforelle erscheint es im Sinne des Tierwohls dennoch sinnvoll, an den
Standorten KWA und RA1 bevorzugt die robustere Bornforelle einzusetzen. In der RA2 ist entsprechend
der vorliegenden extrem schlechten Ergebnislage (Totalverlust) keine Forellenaufzucht zu empfehlen.
4.1.4 Fazit der vergleichenden Forellenproduktion in Aquakulturanlagen in M‐V
Die Standorte KWA Born und RA1, sowie mit Einschränkungen NKA erscheinen generell für die
Forellenproduktion geeignet. Für die weiteren geprüften Anlagen TA und RA2 kann dies auf Basis der
vorliegenden Ergebnisse nicht empfohlen werden. Die wachstumsselektierten Importfische zeigen
gegenüber der robusten Zuchtlinie Born unter den geprüften lokalen Bedingungen Vorteile bei der
Abwachsleistung solange die Bedingungen optimal sind. Tritt in einer Produktionsanlage Umweltstress in
Form von Temperaturanstieg oder auch Parasiten auf, bestätigen sich frühere Versuche im
Modellmaßstab (DIREFO I und II; Goldammer et al., 2009‐2015), die zeigen, dass die Bornforelle
letztendlich im Mittelwert bessere Produktionsleistungen zeigt. Dementsprechend empfiehlt sich

26
basierend auf den reinen Produktionsleistungen die fertile Bornforelle als lokale Zuchtlinie eher für die
regionale Aquakulturproduktion in M‐V als die sterilen, nicht anpassbaren Importforellen.
4.1.5 Empfehlung: Die Bornforelle ist für die regionale Aquakultur geeignet
Mit Blick auf potentielle Zuchtmerkmale, die die Bornforelle für die regionale Aquakultur gegenüber
Vergleichslinien begünstigen, lassen sich die vorliegenden Phänotypdaten so zusammenfassen, dass die
Zucht auf das Merkmal Robustheit mit geringerer Mortalität der Fische korreliert ist, und dass die
Bornforellen schneller vollständig schlüpfen. Von Nachteil ist ihr messbar langsameres Wachstum.
Optimale Temperaturbedingungen können diesen Nachteil ausgleichen. In der Summe ist die Nutzung
der regionalen Zuchtlinie zu empfehlen. Die regelmäßige natürliche Selektion und Reproduktion der
Forellenlinie am Standort Born ist die Voraussetzung für die Erhaltung der noch unaufgeklärten
merkmalsbeeinflussenden genetischen Faktoren. Gleichzeitig ist es notwendig, neben der Robustheit
weitere relevante Zuchtziele, wie ein schnelleres Wachstum, zur Verbesserung der Wirtschaftlichkeit der
Bornforellenproduktion zu berücksichtigen.
4.2 Biotechnologische Charakterisierung des Merkmals Robustheit
Das Projekt sah vor, verschiedenste genetische, molekulare und immunologische Daten im Vergleich der
unterschiedlichen Forellenlinien zu erheben und diese Untersuchungen für ausgewählte Kandidatengene
bzw. –moleküle auch auf die zelluläre Ebene zu erweitern. Damit sollte ein wesentlicher Beitrag zur
Aufklärung der Stoffwechselvorgänge erfolgen, die für die Robustheit und damit in erster Linie für das
Stressadaptationspotential der Bornforelle verantwortlich sind.
Zu diesem Zweck wurden den Arbeitspaketen entsprechend in vivo und in vitro Belastungsexperimente
mit den different resistenten Forellenlinien Born und Import und auf Zellebene durchgeführt.
Zu Versuchszwecken wurden Fische sowohl in 300l Frischwasseraquarien im FLI (Infektionsexperimente)
und in 300l Brackwasseraquarien des Inst. f. Fischerei (Born) gehältert und dabei verschiedenen
natürlichen Stressoren ausgesetzt (Abb. 14).

27
Abb. 14. Parallele Haltungsmöglichkeit für die untersuchten Forellenlinien Born und Import. Beispiel: Versuchsbecken des Inst. für Fischerei in Born. Foto: Tom Goldammer
4.2.1 Publikationen – Ergebnisse und Diskussion
Von einer Reihe der durchgeführten Experimente sind die Ergebnisse bereits in peer reviewed Journalen
detailreich publiziert bzw. befinden sich im Stadium der Begutachtung oder sind eingereicht. Im
Folgenden werden diese Ergebnisse deshalb nur in ihrer Kernaussage vorgestellt und darüber hinaus auf
die Originalmanuskripte verwiesen.
4.2.1.1 Publikation 1
Rebl A, Verleih M, Köllner B, Korytář T, Goldammer T. Duplicated NELL2 genes show different expression
patterns in two rainbow trout strains after temperature and pathogen challenge. Comp Biochem Physiol
B Biochem Mol Biol. 2012 Sep;163(1):65‐73. doi: 10.1016/j.cbpb.2012.05.001. Epub 2012 May 8.
Globale Transkriptomanalysen, die im Anschluss an Temperaturstress bzw. eine Infektion mit dem
Modell‐Fischpathogen Aeromonas salmonicida durchgeführt wurden, identifizierten das Gen NELL2 als
Kandidatengen und Marker für eine differente Stressreaktion der Born‐ und der Importforelle. NELL2
kodiert den neural epidermis growth factor like ‐ like 2. Das Gen war in der Forelle nicht bekannt und
wurde deshalb hinsichtlich seiner DNA‐Struktur (Accession‐Nummer FN556022.1) zunächst aufgeklärt.
Die im Rahmen der eigenen Analysen identifizierte Genessequenz dient heute in der Genomdatenbank
unter der Nummer NM_001199152 als Referenzsequenz für das Gen NELL2 in der Regenbogenforelle

28
(Gene ID: 100526797). Die RACE‐PCR identifizierte eine weitere Genvariante in der Forelle, so dass die
Genvarianten NELL2A und NELL2B beschrieben sind. Genduplikationen sind aufgrund einer im Gegensatz
zu anderen Vertebraten zusätzlichen Ploidisierung des Genoms bei Salmoniden häufig. Die quantitativen
Analysen identifizierten eine entwicklungs‐ und gewebespezifische Genexpression der Genvarianten.
Ebenso wurde eine linienspezifische Expression von NELL2A nach Temperaturstress gefunden. So zeigte
z.B. der weiße Muskel der Bornforelle gegenüber den Importforellen bei 8°C eine signifikant erhöhte
Expression an NELL2A sowie generell in beiden Linien eine Erhöhung beim Anstieg der Temperatur von
8° auf 23°C. Sieben Tage nach einer Infektion gab es eine signifikante Expressionszunahme in der
immunrelevanten Kopfniere bei der Bornforelle, die in der Importforelle nicht auftrat sowie eine
Zunahme der Expression in beiden Linien nach 14 Tagen im Muskel. Die Daten ließen die Vermutung zu,
dass die erhöhte Expression von NELL2A unter beiden geprüften Stressbedingungen für das Überleben
der Forellen generell bedeutsam sein könnte und somit die linienspezifisch höhere Expression in der
Bornforelle einen Hinweis auf die verbesserte Robustheit gibt.
4.2.1.2 Publikation 2
Verleih M, Rebl A, Köllner B, Korytář T, Köbis JM, Kühn C, Wimmers K, Goldammer T. Iron‐sulfur cluster
scaffold (ISCU) gene is duplicated in salmonid fish and tissue and temperature dependent expressed in
rainbow trout. Gene. 2013 Jan 10;512(2):251‐8. doi: 10.1016/j.gene.2012.10.037. Epub 2012 Nov 5.
In dieser Studie geht es um die molekulargenetische Charakterisierung des Gens ISCU (iron‐sulfur cluster
scaffold). Aufgrund von Literaturinformationen sowie aufgrund der differenten Genexpression von ISCU,
die in früheren Untersuchungen an gesunden und nicht gestressten Fischen der Forellenlinien Born und
Import nachgewiesen wurde, war das Gen in die Liste der Kandidatengene für das Merkmal Robustheit
aufgenommen worden. Ungleich regulierte Gene der Born‐ und Importforelle könnten relevant für die
Ausprägung der erhöhten Robustheit der lokalen Zuchtlinie sein. Das Protein ISCU reguliert wichtige
Stoffwechsel‐ und Ionentransportprozesse in der Zelle über die Bindung und Weitergabe von [Fe‐S]‐
Clustern. In der Studie wurde die Duplikation des Gens ISCU in Regenbogenforelle und Ostseeschnäpel
nachgewiesen und die entsprechenden Gensequenzen charakterisiert und publiziert (Reference
Sequence: NM_001171861). Zusätzlich wurde ein spezifisches Expressionsprofil der Genvarianten in der
Forelle erstellt. Insgesamt konnte trotz partiell differenter Genexpression in Born‐ und Importforellen
zwei Wochen nach einer peritoneal injizierten Dosis von Aeromonas salmonicida Bakterien der
vermutete Einfluss des Gens auf die differente Robustheit der Born‐ und Importforellen nach einer
Infektion nicht bestätigt werden.

29
4.2.1.3 Publikation 3
Köbis JM, Rebl A, Kühn C, Goldammer T. Comparison of splenic transcriptome activity of two rainbow
trout strains differing in robustness under regional aquaculture conditions. Mol Biol Rep. 2013
Feb;40(2):1955‐66. doi: 10.1007/s11033‐012‐2252‐1. Epub 2012 Oct 20.
Die Milz als Organ ist im Fisch wesentlich an der Immunantwort nach Infektionen beteiligt. Ziel der
Studie war es, zu zeigen, ob es in diesem Organ bereits zwischen gesunden Fischen der untersuchten
Forellenlinien Unterschiede in der Expression von Genen bzw. der Aktivität von Signalkaskaden gibt, die
sich generalisieren lassen und die im Hinblick auf das Immunsystem der Fische auf eine linienspezifische
Aktivität hindeuten. Dazu wurden holistische Transkriptomanalysen durchgeführt (Abb. 15).
Abb. 15. Beispielschema für den Ablauf holistischen Transkriptomanalysen. Im Beispiel werden die Transkriptome gesunder Forellen der Zuchtlinien Born und Import verglichen und anschließend erfolgt eine Verifizierung der Genexpression ausgewählter Gene mit dem Lightcycler 96 (Roche, Germany).
Die Transkriptomanalyse zwischen den Forellenlinien identifizierte 807 different exprimierte Features
(bekannte und kodierende unbekannte Gensequenzen) in der Milz mit einem sogenannten fold change >
1,5 (FC > 1,5; p ≤ 0.001). 237 der identifizierten Gene sind in viele grundlegende Prozesse im Organismus
involviert (Abb. 16). Neun Prozent dieser Gene sind an immunrelevanten Reaktionen des Organismus
beteiligt und wurden genauer betrachtet und ausgewählte Gene im Rahmen umfangreicher
Strukturanalysen charakterisiert. Ebenso erfolgte über die Erstellung so genannter IPA networks
(Ingenuity pathway analysis tool) eine funktionale Charakterisierung, die das mögliche Zusammenspiel
der Gene im Organismus prognostiziert. Diese Daten helfen, unbekannte funktionale Zusammenhänge
von Genen aufzuklären und dienen darüber hinaus auch der Hypothesenfindung für neue
Forschungsansätze (Abbildung siehe Originalmanuskript).
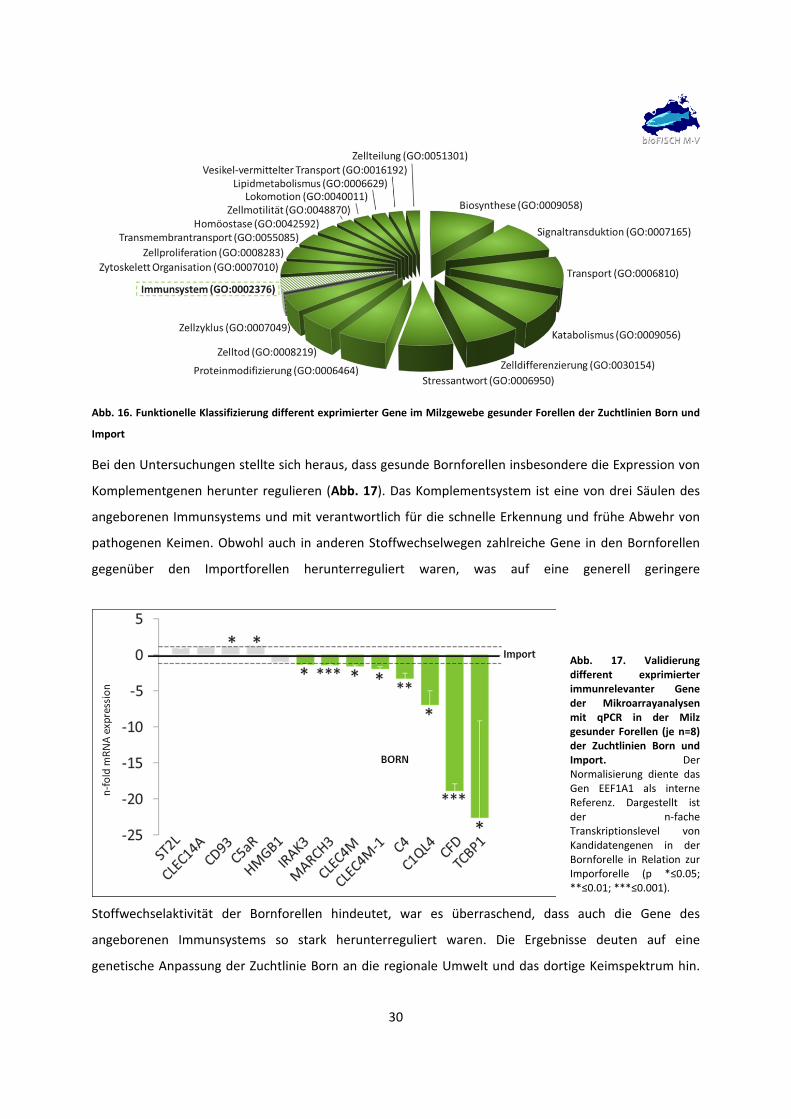
30
Abb. 16. Funktionelle Klassifizierung different exprimierter Gene im Milzgewebe gesunder Forellen der Zuchtlinien Born und
Import
Bei den Untersuchungen stellte sich heraus, dass gesunde Bornforellen insbesondere die Expression von
Komplementgenen herunter regulieren (Abb. 17). Das Komplementsystem ist eine von drei Säulen des
angeborenen Immunsystems und mit verantwortlich für die schnelle Erkennung und frühe Abwehr von
pathogenen Keimen. Obwohl auch in anderen Stoffwechselwegen zahlreiche Gene in den Bornforellen
gegenüber den Importforellen herunterreguliert waren, was auf eine generell geringere
Stoffwechselaktivität der Bornforellen hindeutet, war es überraschend, dass auch die Gene des
angeborenen Immunsystems so stark herunterreguliert waren. Die Ergebnisse deuten auf eine
genetische Anpassung der Zuchtlinie Born an die regionale Umwelt und das dortige Keimspektrum hin.
Abb. 17. Validierung different exprimierter immunrelevanter Gene der Mikroarrayanalysen mit qPCR in der Milz gesunder Forellen (je n=8) der Zuchtlinien Born und Import. Der Normalisierung diente das Gen EEF1A1 als interne Referenz. Dargestellt ist der n‐fache Transkriptionslevel von Kandidatengenen in der Bornforelle in Relation zur Imporforelle (p *≤0.05; **≤0.01; ***≤0.001).

31
Umgekehrt antworten möglicherweise die gesunden Importforellen in ihnen „unbekannten“ Gewässern
auf dort vorhandene „unbekannte“ Pathogene mit einer erhöhten Aktivität des Komplementsystems.
Weiterführende Infektionsstudien mit dem Erreger Aeromonas salmonicida zu dieser Thematik sowie in
anderen immunrelevanten Geweben sind im Rahmen einer Promotionsarbeit zur „Charakterisierung von
Genen des angeborenen Immunsystems in der Regenbogenforelle nach holistischer Transkriptomanalyse
in der Milz“ (Köbis, Dissertation Universität Rostock 2013: pp121 + Anhang) zusammengefasst. Diese
untermauern z.T. die obige Hypothese. Da die Daten inhaltlich teilweise auch anderen Projekten
zugeordnet sind, wird an dieser Stelle auf ihre erweiterte Darstellung verzichtet.
4.2.1.4 Publikation 4
Korytář T, Jaros J, Verleih M, Rebl A, Kotterba G, Kühn C, Goldammer T, Köllner B. Novel insights into the
peritoneal inflammation of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2013
Oct;35(4):1192‐9. doi: 10.1016/j.fsi.2013.07.032. Epub 2013 Jul 31.
Im Rahmen dieser immunologischen Studie wurde erstmalig analysiert, wie sich Leukozyten nach einer
Stimulierung von Regenbogenforellen mit dem Modell‐Pathogen Aeromonas salmonicida verhalten. Für
den initialen Stimulationsversuch wurden ca. 10 Monate alte Forellen der Importzuchtlinie infiziert und
jeweils 4 Fische an 5 Zeitpunkten (6, 12, 24, 48, 72 h) beprobt. Die Studie sollte insbesondere erste
Informationen zur Zellkinetik und –dynamik geben. Für die Zellanalysen kam die Fluoreszenzaktivierte
Durchflusszytometrie (FACS) zur Anwendung. Die Analysen nach einer Stimulierung zeigten eine
gewebespezifische Zellkinetik für das Peritoneum im Vergleich zu den immunrelevanten Geweben
Thymus, Milz und Kopfniere. Die Betrachtung der Zellkinetik nach Immunisierung (tote Bakterien) bzw.
Infektion im zeitlichen Verlauf von 72 Stunden ergab einen massiven Wechsel in der Population der
Myeolidzellen und Lymphoidzellen (Abb. 18). In den ersten 24 h steigt zunächst die Anzahl der
Myeolidzellen stark an, deren Verantwortung u.a. in der Pathogenerkennung und sofortigen Vernichtung
z.B. durch Phagozytose liegt. Damit kann eine Infektion gleich an ihrem Beginn unterdrückt werden.
Dieses Ergebnis zeigt auch, dass am Anfang der Stimulation vor allem die Myeolidzellen und nicht wie
zuvor vermutet von Beginn an auch die Lymphoidzellen die Erregerinvasion stoppen. Im weiteren
Verlauf, nach etwa 48‐72 h, nehmen dann die lymphoiden Leukozyten zu, deren Aufgabe u.a. darin
besteht, dass erworbene Immunsystem zu aktivieren, um eine Infektion nachhaltig zu bekämpfen.
Bemerkenswert ist auch, dass die Myeolidzellen nach der Infektion gegenüber der Immunisierung um
das Dreifache zunehmen. Detaillierte Informationen sind im Manuskript dargelegt.
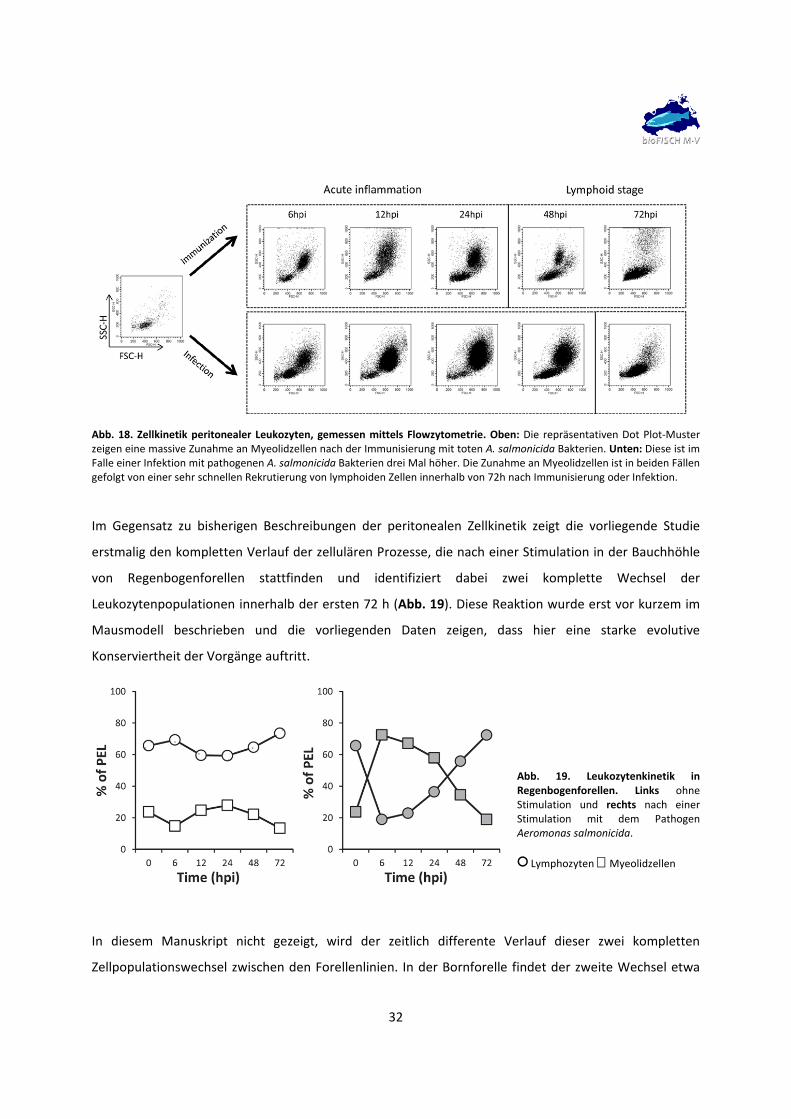
32
Abb. 18. Zellkinetik peritonealer Leukozyten, gemessen mittels Flowzytometrie. Oben: Die repräsentativen Dot Plot‐Muster zeigen eine massive Zunahme an Myeolidzellen nach der Immunisierung mit toten A. salmonicida Bakterien. Unten: Diese ist im Falle einer Infektion mit pathogenen A. salmonicida Bakterien drei Mal höher. Die Zunahme an Myeolidzellen ist in beiden Fällen gefolgt von einer sehr schnellen Rekrutierung von lymphoiden Zellen innerhalb von 72h nach Immunisierung oder Infektion.
Im Gegensatz zu bisherigen Beschreibungen der peritonealen Zellkinetik zeigt die vorliegende Studie
erstmalig den kompletten Verlauf der zellulären Prozesse, die nach einer Stimulation in der Bauchhöhle
von Regenbogenforellen stattfinden und identifiziert dabei zwei komplette Wechsel der
Leukozytenpopulationen innerhalb der ersten 72 h (Abb. 19). Diese Reaktion wurde erst vor kurzem im
Mausmodell beschrieben und die vorliegenden Daten zeigen, dass hier eine starke evolutive
Konserviertheit der Vorgänge auftritt.
Abb. 19. Leukozytenkinetik in Regenbogenforellen. Links ohne Stimulation und rechts nach einer Stimulation mit dem Pathogen Aeromonas salmonicida.
○ Lymphozyten Myeolidzellen
In diesem Manuskript nicht gezeigt, wird der zeitlich differente Verlauf dieser zwei kompletten
Zellpopulationswechsel zwischen den Forellenlinien. In der Bornforelle findet der zweite Wechsel etwa

33
48 h nach einer Infektion statt, in der Importforelle erst 72h danach. Der Erreger Aeromonas salmonicida
ist in verschiedensten Subspezies weltweit vertreten. Die hier verwendete Subspezies des Erregers war
für die Immunsysteme beider Forellenlinien unbekannt. Der zeitliche Versatz kann deshalb auf ein
generell intakteres, schnelleres, adaptiveres Immunsystem bei der Bornforelle hindeuten. Ergebnisse zu
dieser Thematik werden für die Publikation vorbereitet.
4.2.1.5 Publikation 5
Rebl A, Verleih M, Köbis JM, Kühn C, Wimmers K, Köllner B, Goldammer T. Transcriptome profiling of gill
tissue in regionally bred and globally farmed rainbow trout strains reveals different strategies for coping
with thermal stress. Mar Biotechnol (NY). 2013 Aug;15(4):445‐60. doi: 10.1007/s10126‐013‐9501‐8. Epub
2013 Apr 3.
4.2.1.6 Publikation 6
Verleih M, Borchel A, Krasnov A, Rebl A, Korytář T, Kühn C, Goldammer T. Impact of Thermal Stress on
Kidney‐Specific Gene Expression in Farmed Regional and Imported Rainbow Trout. Mar Biotechnol (NY).
2015 Oct;17(5):576‐92. doi: 10.1007/s10126‐015‐9640‐1. Epub 2015 May 28.
Die Studien 5 und 6 entstanden im Ergebnis wiederholter holistischer Transkriptomanalysen mit 4x44K
Salmoniden‐Mikroarrays (Abb. 20) nach moderatem Temperaturstress von Regenbogenforellen der
Linien Born und Import. Ausgangspunkt der Untersuchungen war, dass in Abhängigkeit von der Lokalität
und Wasserversorgung bzw. Wasserqualität in einer Aquakulturanlage saisonal differente
Wassertemperaturschwankungen auftreten, deren Amplituden aufgrund globaler Klimaveränderungen
in Zukunft noch zunehmen. In beiden Studien wurden in parallelen Ansätzen gesunde ca. 10 Monate alte
Forellen an Temperaturen von 8°C, 15°C bzw. 23°C adaptiert und beprobt. Das Ziel der Analysen bestand
in der Identifizierung von Genen, die generell temperaturabhängig exprimiert werden und solchen, die
zwischen den Zuchtlinien unterschiedlich reguliert sind.

34
Abb. 20. Heat maps der Top‐10 auf‐ bzw. abregulierten Gene nach Temperaturstress. Die Pfeile markieren Kandidatengene, die näher charakterisiert worden sind.
Bei diesen Analysen wurden u.a. klassische Stressgene, wie etwa verschiedene Hitzeschockproteingene
exprimiert, die der positiven Validierung des jeweiligen Versuchsansatzes dienten. Weiterhin wurden
zahlreiche Gene gefunden, die bisher nicht im Zusammenhang mit Temperaturstress diskutiert wurden.
Viele dieser Gene sind sogenannte Akute‐Phase‐Gene, die üblicherweise in der angeborenen
Immunabwehr fungieren, also wiederum der aktiven Erstbeseitigung eindringender Erreger dienen. Hier
waren diese Gene aber beachtenswerter weise nach Temperaturstress ebenso hoch reguliert. Andere
Gene werden aktiv im Zusammenhang mit dem Zelltod (Abb. 21).
Abb. 21. Potentielle Interaktion temperaturregulierter Gene (fett, rot), die in die Apoptose involviert sind. Links Zuchtlinie Born; Rechts Zuchtlinie Import. IPA (Ingenuity® Systems, www.ingenuity.com) generierte Netzwerke. Durchgezogene und
35
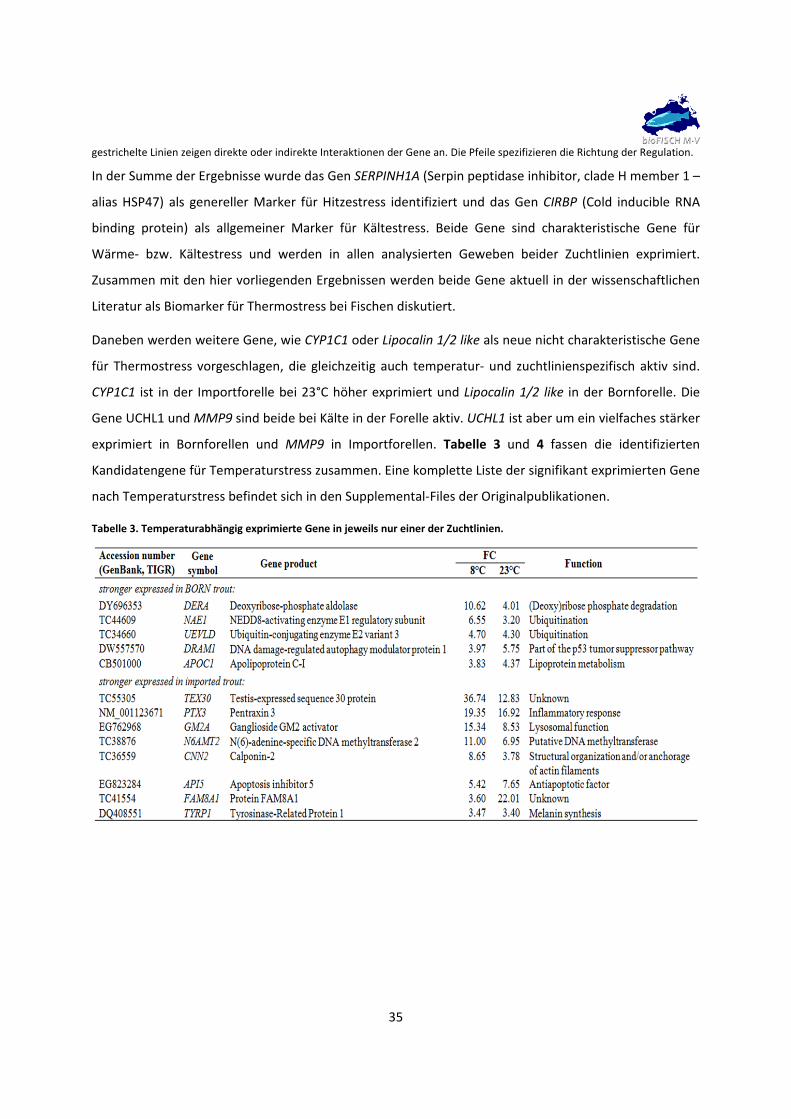
In der Summe der Ergebnisse wurde das Gen SERPINH1A (Serpin peptidase inhibitor, clade H member 1 –
alias HSP47) als genereller Marker für Hitzestress identifiziert und das Gen CIRBP (Cold inducible RNA
binding protein) als allgemeiner Marker für Kältestress. Beide Gene sind charakteristische Gene für
Wärme‐ bzw. Kältestress und werden in allen analysierten Geweben beider Zuchtlinien exprimiert.
Zusammen mit den hier vorliegenden Ergebnissen werden beide Gene aktuell in der wissenschaftlichen
Literatur als Biomarker für Thermostress bei Fischen diskutiert.
Daneben werden weitere Gene, wie CYP1C1 oder Lipocalin 1/2 like als neue nicht charakteristische Gene
für Thermostress vorgeschlagen, die gleichzeitig auch temperatur‐ und zuchtlinienspezifisch aktiv sind.
CYP1C1 ist in der Importforelle bei 23°C höher exprimiert und Lipocalin 1/2 like in der Bornforelle. Die
Gene UCHL1 und MMP9 sind beide bei Kälte in der Forelle aktiv. UCHL1 ist aber um ein vielfaches stärker
exprimiert in Bornforellen und MMP9 in Importforellen. Tabelle 3 und 4 fassen die identifizierten
Kandidatengene für Temperaturstress zusammen. Eine komplette Liste der signifikant exprimierten Gene
nach Temperaturstress befindet sich in den Supplemental‐Files der Originalpublikationen.
Tabelle 3. Temperaturabhängig exprimierte Gene in jeweils nur einer der Zuchtlinien.
gestrichelte Linien zeigen direkte oder indirekte Interaktionen der Gene an. Die Pfeile spezifizieren die Richtung der Regulation.

36
Tabelle 4. Temperaturabhängig exprimierte Gene in beiden Zuchtlinien bei 8°C und 23°C.
4.2.1.7 Publikation 7
Borchel A, Verleih M, Rebl A, Kühn C, Goldammer T. Creatine metabolism differs between mammals and
rainbow trout (Oncorhynchus mykiss). Springerplus. 2014 Sep 9;3:510. doi: 10.1186/2193‐1801‐3‐510.
eCollection 2014
Die Idee dieser Studie bestand darin ausgewählte Moleküle des Energiestoffwechsels biotechnologisch
näher zu charakterisieren, um zu prüfen, ob es u.a. zuchtlinienspezifische Differenzen bei der
Aufrechterhaltung der Homöostase der Regenbogenforellen gibt. Die Studie widmet sich der Analyse von
Molekülen im Kreatinkreislauf. Dieser stellt die Energie bereit, die in den ersten ca. 5 Sekunden von
Flucht oder Beutefang benötigt wird. Kreatin und Kreatinphosphat agieren mit Hilfe von Kreatinkinasen
(CKs) als räumlicher und zeitlicher Energiepuffer und ermöglichen den Energietransport zwischen Zellen

37
und Organen. Die Kreatinaufnahme erfolgt über die Nahrung oder seine Synthese über einen Prozess,
der durch GATM und GAMT katalysiert wird. Gefunden wird Kreatin in Geweben mit hohem
Energiebedarf, wie Muskel oder im Gehirn. Gene, die in dieser Signalkaskade eine wichtige Rolle spielen,
sind CKM, CKB, GATM und GAMT. Funktionale Untersuchungen zu dieser Thematik in Fischen gibt es
kaum. Im Vergleich von etwa 10 Monate alten gesunden Forellen der Zuchtlinien Born und Import
wurden hinsichtlich der Expression obiger Gene keine signifikanten Unterschiede gefunden. Bei
Einbeziehung von moderatem Temperaturstress (8°C bzw. 23°C) traten zwischen den Gruppen teilweise
signifikante, aber dennoch nur geringe Unterschiede auf. Ebenso traten zwischen den verschiedenen
Temperaturen in einigen Geweben für einige der Gene signifikante Differenzen auf. Insgesamt
betrachtet, ist das Kreatinsystem der Zuchtlinien sehr ähnlich, die nachgewiesenen Differenzen könnten
dennoch auch linienspezifische Ursachen haben. Gefunden wurden aber hochsignifikante
gewebespezifische Unterschiede in der Genexpression. Bemerkenswert ist dabei, dass die Ergebnisse
teilweise konträr zu bisherigen Studien an Säugetieren (Abb. 22) , aber auch bei z.B. Zebrafischen sind.
Im Säuger wird die Kreatinproduktion über Leber und Niere und nur teilweise über den Muskel
organisiert und das fertige Kreatin‐Protein dann in den Muskel transportiert. Bei der Regenbogenforelle
hingegen findet nicht nur die Verwendung sondern auch ein Großteil der Kreatinsynthese direkt im
Zielorgan, dem weißen Muskel, statt. Diese Ergebnisse werfen ein neues Licht auf die evolutive
Entwicklung des Kreatinsystems bei Vertebraten. In der Zwischenzeit wurden die Ergebnisse durch
eigene Studien an weiteren Fischen, Säugern, Vögeln und Reptilien etc. untermauert (Daten noch
unveröffentlicht).
Abb. 22. Schematisch vereinfachte Darstellung der Kreatinsynthese und Genexpression in Maus und Regenbogenforelle.

38
4.2.1.8 Publikation 8
Borchel A. Charakterisierung von Transkriptom und Phänotyp different robuster Zuchtlinien der
Regenbogenforelle mit Fokus auf das Kreatin‐System der Fische. Dissertation Universität Rostock 2015:
pp100 + Anhang, 2015.
An dieser Stelle wird nur die Zusammenfassung der Promotionsarbeit wiedergegeben, die vollständig im
Rahmen des Projektes angefertigt wurde. Die in der Bibliothek der Universität Rostock hinterlegte Arbeit
kann bei Bedarf auch als pdf‐Datei zur Verfügung gestellt werden.
Zusammenfassung
In der vorliegenden Arbeit wurden die beiden Forellenzuchtlinien Born und Import hinsichtlich
molekularer und phänotypischer Unterschiede untersucht. Bei der Bornforelle handelt es sich um eine
lokale Zuchtlinie, die als robuster als die Importforelle und besonders gut angepasst an die lokalen
Umweltbedingungen gilt. Die vorliegende Arbeit konnte neue Erkenntnisse zu den Unterschieden von
Born‐ und Importforelle erbringen. Ausgehend von einer holistischen Transkriptomanalyse wurden vier
Gene identifiziert, welche in vier Geweben unter verschiedensten Besatzdichte‐ und
Temperaturbedingungen different zwischen beiden Linien exprimiert waren. CEPT1A sowie GIMAP7
waren stärker in der Bornforelle exprimiert, während RARRES3 und TPM1A stärker in der Importforelle
exprimiert wurden. Damit wurden erstmals Gene identifiziert, welche unabhängig von den konkreten
Umweltbedingungen, in denen die Tiere lebten, in vier Geweben gleichzeitig different exprimiert waren.
Dies deutet auf genetische Unterschiede zwischen den beiden Forellenlinien hin.
40 Jahre Zucht mit dem Selektionsziel Überleben in Brackwasser haben offenbar die Herausbildung
dieser genetischen Unterschiede bewirkt. Die identifizierten Gene könnten relevant für die Robustheit
der Bornforelle sein, auch wenn die Kausalität in dieser Arbeit nicht nachgewiesen werden konnte.
Dennoch können diese Gene als Biomarker zur Unterscheidung der beiden Forellenlinien dienen.
Weiterhin konnte das Kreatinsystem der Fische grundlegend charakterisiert werden. In insgesamt fünf
verschiedenen Fischspezies konnten Gene des Kreatinsystems ansequenziert werden. Für die
Regenbogenforelle wurden die gesamten codierenden Bereiche der Gene GATM, GAMT und CKM
identifiziert. Weiterhin wurde in der Forelle erstmalig für einen Salmoniden die Sequenz des
Kreatintransporters SLC6A8 bestimmt. Auf Basis der Sequenzinformation konnte dann die Genexpression
untersucht werden. Fische sind offenbar dazu in der Lage, Kreatin, einen zellulären Energiespeicher, im
Muskel zu synthetisieren. Die Genexpressionsprofile waren für alle Fischarten vergleichbar und deuteten
im Vergleich zu Niere und Leber auf eine große Relevanz des Muskels hin. In Säugetieren hingegen

39
zeigten sich Niere und Leber wichtiger für diesen Prozess. Evolutionär scheint sich diese unterschiedliche
Kreatinsynthese schon sehr früh etabliert zu haben. Während alle Fische sehr ähnliche
Genexpressionsprofile der Kreatingene zeigten, unterschieden sich diejenigen von Paarhufern und Maus,
was auf eine relativ neue Anpassung im Kreatinsystem der verschiedenen Säuger‐Gruppen hindeutet.
Die zwei bisherigen Untersuchungen zum Kreatinsystem von Fischen hatten den Skelettmuskel nicht in
die Untersuchung mit einbezogen. Daher war eine muskuläre Kreatinsynthese im Fisch bisher noch nicht
postuliert worden. Beim Vergleich der Kreatinsysteme der beiden Forellenlinien gab es ebenfalls
Hinweise auf Unterschiede. Zumindest bei warmen Temperaturen schien die muskuläre Kreatinsynthese
auf Basis der Genexpression der dafür benötigten Enzyme in Bornforellen unterhalb derjenigen von
Importforellen zu liegen. Weiterhin gab es Hinweise auf eine unterschiedliche Zuckersynthese in den
beiden Linien, was ebenfalls auf Unterschiede im Energiehaushalt der Forellenlinien hindeutet.
Auch im Phänotyp, der grundsätzlich neben der Umwelt auch ein Produkt des Genotyps ist, zeigten sich
Unterschiede. Bezüglich des Verhaltens weisen beide Forellenlinien eine unterschiedliche Präferenz
hinsichtlich der Schwimmtiefe auf. Es muss weiter untersucht werden, wodurch diese hervorgerufen
werden. Zudem wurde die Wachstumsleistung der Tiere verglichen. Bornforellen wuchsen etwas
langsamer als Importforellen, welche bei gleicher Länge auch etwas schwerer waren. Ein Befund, der die
Attraktivität der Bornforelle für die kommerzielle Aquakultur verringern dürfte, wobei nachfolgende
Experimente jedoch noch den Geschlechtsfaktor stärker berücksichtigen müssten.
Insgesamt konnte somit in dieser Arbeit gezeigt werden, dass sich die Regenbogenforellen‐Zuchtlinien
Born und Import sowohl hinsichtlich ihres Transkriptoms als auch ihres Phänotyps voneinander
unterscheiden.
4.2.2 Weitere Projektergebnisse und Diskussion
Im Projektzeitraum wurden weitere Ergebnisse produziert, die hier kurz zusammengefasst werden. Die
Daten befinden sich in der Auswertung und sind derzeit noch unveröffentlicht.
4.2.2.1 Holistische Transkriptomanalysen
Im Rahmen des Projektes wurden zahlreiche weitere globale Genexpressionsstudien nach Infektion,
Besatzdichtestress oder Temperaturstress durchgeführt, die grundsätzlich ausgewertet wurden, die aber
aufgrund ihrer Parallelität und der Art der Durchführung viele weitere Vergleichsmöglichkeiten der
Daten erlauben. Bisher lag eine gewebespezifische Betrachtung der Daten unter ganz konkreten
Umweltbedingungen im Fokus der Analysen. Abb. 23 zeigt jedoch deutlich auf, dass die Expression
40
gleicher Gene in den verschiedenen Geweben sowohl große Unterschiede als auch Ähnlichkeiten
aufweist im Vergleich der Gewebe miteinander aufweist. Die funktionale globale Analyse derartiger
Quervernetzungen ist Bestandteil zukünftiger Betrachtungen.
Abb. 23. Beispiel einer Heat map für die 100 Top auf‐ und abregulierten Gene eines kompletten experimentellen Datensatzes von Born‐ und Importforellenproben nach
Temperaturstress.
4.2.2.2 Transkriptomanalysen zur Identifizierung von DNA‐Polymorphismen
Für die gezielte genetische Selektion und Entwicklung genetisch definierter Zuchtlinien bildet der
Nachweis von DNA‐Polymorphismen (single nucleotide polymorphism – SNP), die z.B. mit dem Merkmal
Robustheit gekoppelt und im Idealfall für das Merkmal kausal sind, eine wichtige Grundlage. Um diesem
Ziel bei der Bornforelle näher zu kommen, wurden wiederholt vergleichende de novo
Transkriptomsequenzierungen auf dem Genome Analyzer GA IIX (Illumina) zwischen gesunden
ungestressten Born‐ und Importforellen durchgeführt (Abb. 24). Mit dieser sogenannten paired end
Sequenzierung, mit der gleichzeitig die gewebespezifische Geneexpression und Sequenzpolymorphismen
identifizierbar sind, gelang es, die zum Probezeitpunkt exprimierte und isolierte RNA von verschiedenen
Geweben (Muskel, Kieme, Kopfniere, Leber, Herz und Milz) vielfach zu sequenzieren. Im Ergebnis
wurden für die Forellen Hunderte Millionen kurzer DNA‐Fragmente (reads von jeweils 125bp Länge)
erfasst. Die Assemblierung der reads mit Hilfe der Bioinformatik ist noch nicht abgeschlossen.

41
Haupursache dafür ist eine fehlende hochwertige Genomsequenz für die Regenbogenforelle in den
öffentlichen Datenbanken wodurch die Assemblierung – insbesondere die Contig‐Bildung
(Zusammenlagerung größerer Fragmente) – erschwert wird. Eine weitere Ursache bestand in der etwas
zu geringen Anzahl an reads je Tier und Gewebe, wofür Ungenauigkeiten bei der Probenvorbereitung
(library preparation) verantwortlich sein könnten. Für Fische müssen die entsprechenden Protokolle
aufgrund fehlender Grunddaten im Gegensatz zu Säugern zum Teil empirisch angepasst werden. Zum
Projektende wurden deshalb einige Transkriptomsequenzierungsexperimente wiederholt.
Abb. 24: Beispielschema für die vergleichende Transkriptomanalyse vn Born‐ und Importforellen. Die Hauptkomponentenanalyse beweist eine eindeutige linienspezifische Genexpression (Rot: Bornforellen n=8; Blau: Importforellen n=8; Gewebe: Kopfniere). Das Bild rechts daneben zeigt die Genexpressionswolke aller gemessenen DNA‐Sequenzen (reads) basierend auf der Anzahl zueinander identischer bzw. überlappender reads. Rechts außen sind Cluster solcher reads gezeigt. Herausgezoomt werden SNP in der Sequenz deutlich.
Wie nützlich diese Analysemethode ist, zeigen u.a. die Ergebnisse zur Identifizierung allgemeiner
Biomarker für Stress in Regenbogenforellen. Hier wurde, wie in Abschnitt 4.2.1.8 beschrieben, u.a. das
Gen GIMAP7 (GTPase, immunity‐associated protein Family Member 7) als so ein potentieller Biomarker
identifiziert. Schaut man sich den Vergleich der DNA‐Sequenz dieses Gens zwischen den beiden
Zuchtlinien an, wird das Potential der Methode deutlich. Das Gen GIMAP7 wird in beiden Zuchtlinien bei
jeglicher Art von Stress (Infektion, Temperatur, Besatz) hochreguliert (Abb. 25). Mit Hilfe der qPCR in
einem Fluidigm‐Quantifizierungsgerät unter Nutzung von Mikofluidchips (192x24 Proben, Fluidigm)
konnte die Illumina‐Transkriptomsequenzierung verifiziert werden und es wurde gezeigt, dass die
GIMAP‐Genexpression der Bornforellen unter beinahe allen geprüften Stressbedingungen signifikant
stärker ist als bei den Importforellen.
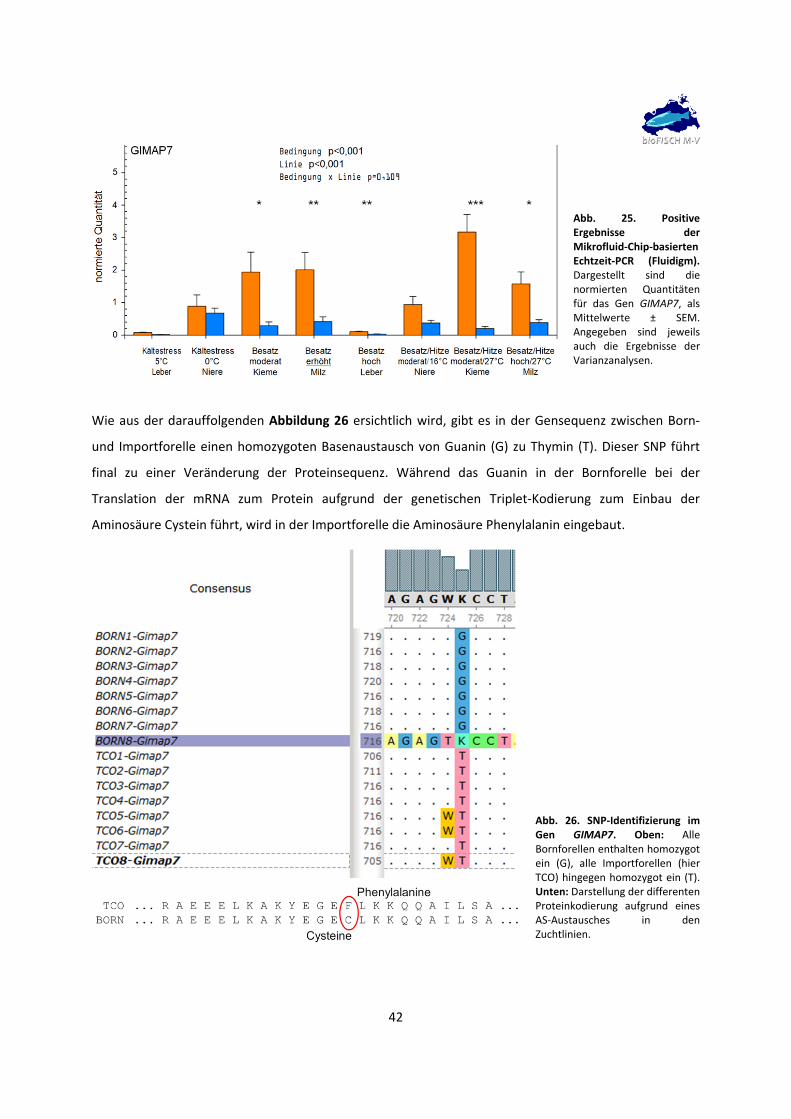
42
Abb. 25. Positive Ergebnisse der Mikrofluid‐Chip‐basierten Echtzeit‐PCR (Fluidigm). Dargestellt sind die normierten Quantitäten für das Gen GIMAP7, als Mittelwerte ± SEM. Angegeben sind jeweils auch die Ergebnisse der Varianzanalysen.
Wie aus der darauffolgenden Abbildung 26 ersichtlich wird, gibt es in der Gensequenz zwischen Born‐
und Importforelle einen homozygoten Basenaustausch von Guanin (G) zu Thymin (T). Dieser SNP führt
final zu einer Veränderung der Proteinsequenz. Während das Guanin in der Bornforelle bei der
Translation der mRNA zum Protein aufgrund der genetischen Triplet‐Kodierung zum Einbau der
Aminosäure Cystein führt, wird in der Importforelle die Aminosäure Phenylalanin eingebaut.
Abb. 26. SNP‐Identifizierung im Gen GIMAP7. Oben: Alle Bornforellen enthalten homozygot ein (G), alle Importforellen (hier TCO) hingegen homozygot ein (T). Unten: Darstellung der differenten Proteinkodierung aufgrund eines AS‐Austausches in den Zuchtlinien.
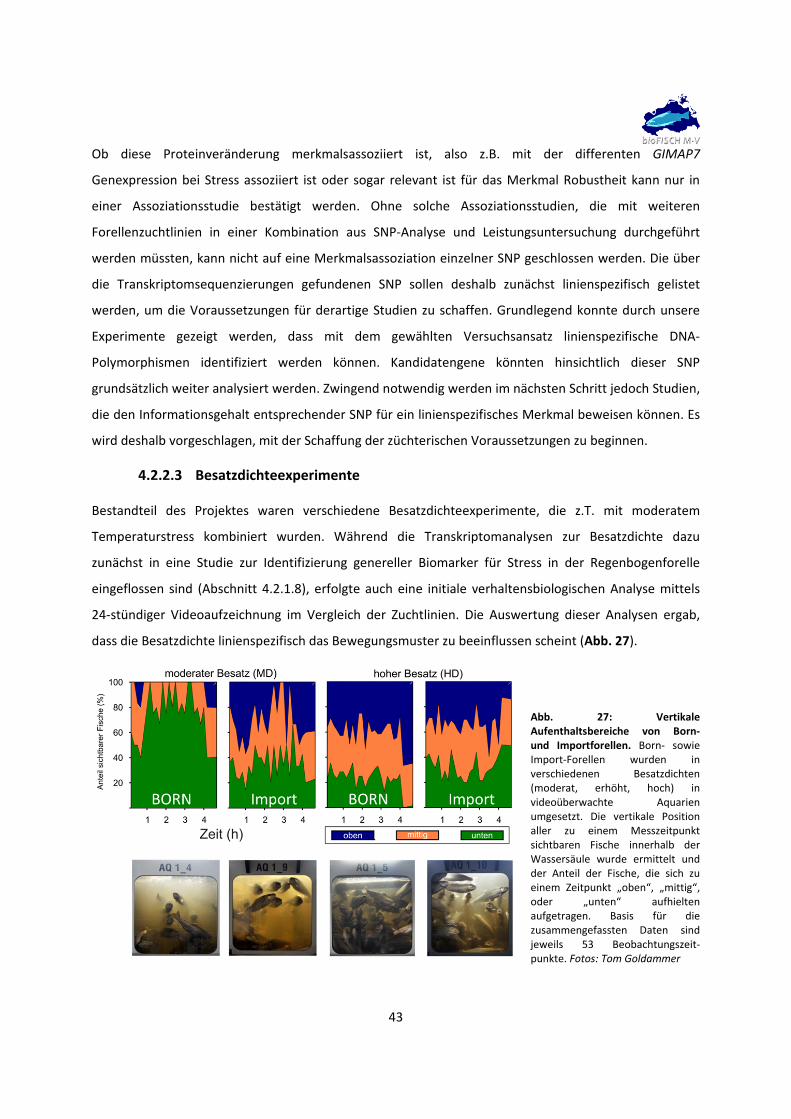
43
Ob diese Proteinveränderung merkmalsassoziiert ist, also z.B. mit der differenten GIMAP7
Genexpression bei Stress assoziiert ist oder sogar relevant ist für das Merkmal Robustheit kann nur in
einer Assoziationsstudie bestätigt werden. Ohne solche Assoziationsstudien, die mit weiteren
Forellenzuchtlinien in einer Kombination aus SNP‐Analyse und Leistungsuntersuchung durchgeführt
werden müssten, kann nicht auf eine Merkmalsassoziation einzelner SNP geschlossen werden. Die über
die Transkriptomsequenzierungen gefundenen SNP sollen deshalb zunächst linienspezifisch gelistet
werden, um die Voraussetzungen für derartige Studien zu schaffen. Grundlegend konnte durch unsere
Experimente gezeigt werden, dass mit dem gewählten Versuchsansatz linienspezifische DNA‐
Polymorphismen identifiziert werden können. Kandidatengene könnten hinsichtlich dieser SNP
grundsätzlich weiter analysiert werden. Zwingend notwendig werden im nächsten Schritt jedoch Studien,
die den Informationsgehalt entsprechender SNP für ein linienspezifisches Merkmal beweisen können. Es
wird deshalb vorgeschlagen, mit der Schaffung der züchterischen Voraussetzungen zu beginnen.
4.2.2.3 Besatzdichteexperimente
Bestandteil des Projektes waren verschiedene Besatzdichteexperimente, die z.T. mit moderatem
Temperaturstress kombiniert wurden. Während die Transkriptomanalysen zur Besatzdichte dazu
zunächst in eine Studie zur Identifizierung genereller Biomarker für Stress in der Regenbogenforelle
eingeflossen sind (Abschnitt 4.2.1.8), erfolgte auch eine initiale verhaltensbiologischen Analyse mittels
24‐stündiger Videoaufzeichnung im Vergleich der Zuchtlinien. Die Auswertung dieser Analysen ergab,
dass die Besatzdichte linienspezifisch das Bewegungsmuster zu beeinflussen scheint (Abb. 27).
Abb. 27: Vertikale Aufenthaltsbereiche von Born‐ und Importforellen. Born‐ sowie Import‐Forellen wurden in verschiedenen Besatzdichten (moderat, erhöht, hoch) in videoüberwachte Aquarien umgesetzt. Die vertikale Position aller zu einem Messzeitpunkt sichtbaren Fische innerhalb der Wassersäule wurde ermittelt und der Anteil der Fische, die sich zu einem Zeitpunkt „oben“, „mittig“, oder „unten“ aufhielten aufgetragen. Basis für die zusammengefassten Daten sind jeweils 53 Beobachtungszeit‐punkte. Fotos: Tom Goldammer

44
In Hinblick auf das Verhalten der beiden Regenbogenforellenlinien wurde im Rahmen des
Besatzdichteversuchs untersucht, inwieweit sich die beiden Linien hinsichtlich ihres vertikalen
Aufenthaltsorts in der Wassersäule unterscheiden. Bei moderatem und erhöhtem Besatz waren die
beobachteten Bornforellen ungleichmäßig in der Wassersäule verteilt. 83 % bzw. 66 % der beobachteten
Fische schwammen im untersten Drittel des Beckens, im obersten hingegen nur 2 % bzw. 4 %. Bei
moderatem Besatz war der Unterschied zwischen Born und Import signifikant (p<0,001 für „oben“ und
„unten“, p=0,023 für „mittig“; Holm‐ Sidak‐Test, nach zweifaktorieller Varianzanalyse mit signifikanten
Interaktionen zwischen Linie und Besatzdichte). Bei erhöhtem Besatz lag der Anteil der oben
schwimmenden Fische der Bornlinie signifikant unter dem der Importlinie, im Gegenzug war der Anteil
der unten schwimmenden Fische signifikant erhöht (jeweils p<0,001). Bei hohem Besatz hingegen gab es
keine Unterschiede zwischen den Linien und die Anteile lagen jeweils in etwa bei 33 %. Jedoch muss
hierzu angemerkt werden, dass bei hohem Besatz die Dichte so hoch war, dass stets der komplette
Sichtbereich der Kamera mit Fischen gefüllt war.
Zur Beurteilung der Verhaltensmuster der Tiere erwies sich der gewählte Versuchsaufbau als
unzureichend. Eine Auswertung der Aktivitäten der Fische konnte daher nicht vorgenommen werden.
Die Schwimmtiefe von Salmoniden ist Gegenstand vieler Untersuchungen. Für Lachse konnte gezeigt
werden, dass diese in Netzkäfigen im Meer ihre Körpertemperatur regulieren, indem sie ihre
Schwimmtiefe anpassen (Johansson et al., 2009). Für die Forellen in diesem Versuch kommt diese
Erklärung kaum in Frage, da die Temperatur in den relativ flachen Aquarien bei guter Zirkulation keine
allzu starken Temperaturgradienten ausgebildet haben dürfte. Ein weiterer Zusammenhang wurde
zwischen Schwimmtiefen von Fischen und der Verfügbarkeit von gelöstem Sauerstoff in
unterschiedlichen Wassertiefen hergestellt (Kramer, 1987), wobei auch dies für das Experiment eher
unwahrscheinlich erscheint. Wahrscheinlicher erscheint der Faktor der Lichtvermeidung. Lachse meiden
hohe Lichtintensitäten. Bei starkem Licht schwimmen sie in größere Tiefen (Huse und Holm, 1993). Als
Ursache für die Lichtvermeidung wird die gesteigerte Gefahr genannt, in das Blickfeld von Prädatoren zu
geraten (Fernö et al., 1995). Insgesamt erscheint die vertikale Fischposition dabei als Trade‐off zwischen
Lichtvermeidung und Nahrungsaufnahme, zumal in höheren Wasserschichten mehr Nahrung vorhanden
ist. Möglicherweise könnten sich unterschiedliche Lichtpräferenzen auch durch die unterschiedlichen
Lebensumstände der beiden Zuchtlinien in mehr oder minder trübem Wasser erklären lassen, die sich
genetisch manifestieren haben könnten. Um zu überprüfen, inwieweit sich die Lichtvermeidung von
Import‐ und Bornforelle tatsächlich unterscheiden, und ob sie die beobachteten Verhaltensunterschiede
erklären können, müssten weitere Versuche durchgeführt werden. Denkbar wären die Beobachtung der

45
Fischposition im Dunkeln sowie die Beobachtung bei verschiedenen Lichtintensitäten. Dies könnte
weitere Hinweise über Ursachen für das unterschiedliche Verhalten beider Forellenlinien liefern.
4.2.2.4 Zelluläre Analysen
Für einzelne Kandidatengene war es hilfreich und notwendig zelluläre Analysen für eine verbesserte
Funktionsaufklärung durchzuführen. Für derartige Analysen kam das im Rahmen des Projektes
angeschaffte Fluoreszenzmikroskopsystem für Lebendzellanalysen mehrfach zur Anwendung. Unter
anderem wurde im Rahmen der Untersuchungen zum Kreatinsystem ein in der Forelle noch
unbeschriebener Kreatintransporter SLC6A8 identifiziert. Neben der Aufklärung seiner Gen‐ und
potentiellen Proteinstruktur wurde im Zellexperiment die Membranständigkeit des SLC6A8‐Proteins
überprüft. Um die vorhergesagte Lokalisation in der Membran zu überprüfen, wurde dazu ein SLC6A8‐
GFP‐Fusionsgen hergestellt und zur Transfektion von HEK‐293‐Zellen verwendet. Die Proteinlokalisation
ist exemplarisch in Abb. 28 dargestellt. Deutlich zu erkennen ist, dass nur ein geringer Anteil der Zellen
eine Fluoreszenz zeigte. Diese grüne Fluoreszenz war aber deutlich auf die Zellmembran fokussiert,
innerhalb der Zellen gab es nur einige kleinere Stellen, an denen Fluoreszenz vorhanden war. Zu
Sicherstellung des Ergebnisses erfolgte ein so genannter Western Blot (nicht dargestellt) (Borchel, 2015).
Abb. 28. Zelluläre Lokalisation des SLC6A8‐GFP‐Fusionsproteins. Oben links: Zellkernanfärbung mit DAPI; oben rechts: Lichtmikroskopische Aufnahme; unten links: GFP‐Fluoreszenz; unten rechts: fusioniertes Bild. Fotos: Andreas Borchel

46
4.2.3 Fazit der biotechnologischen Analysen zur Bornforelle
Sämtliche Ergebnisse der molekularen und immunologischen Untersuchungsansätze zeigen deutliche
Unterschiede zwischen Born‐ und Importforellen auf. Wenngleich diese Unterschiede nur hypothetisch
mit der Robustheit der Bornforellen in Beziehung gebracht werden dürfen, da es keine
Assoziationsstudien mit den Zuchtlinien gibt, die die Kausalität der Unterschiede unterstützen, fügen sich
einige der Ergebnisse nahtlos in die Resultate der Phänotypanalysen ein, die in den Produktionsanlagen
gewonnen wurden. Die Robustheit der Bornforellen drückt sich vor allem durch höhere Überlebensraten
in Stresssituationen aus. Auf molekularer Ebene und mit Untersuchungen der Immunzellkinetik konnte
gezeigt werden, dass insbesondere das angeborene Immunsystem der Bornforellen in der regionalen
Umwelt z.T. völlig anders als das der Importforellen auf Stressoren reagiert. Schon im normalen nicht
gestressten Zustand zeigten sich teilweise extreme Unterschiede, wie die Herabregulation von
Komplementgenen. Auch wurde gezeigt, dass der Wechsel der Leukozytenzellpopulationen, die für die
Erstabwehr nach Pathogeneinwirkung verantwortlich sind, in Bornforellen schneller abläuft als in
Importforellen. Dies unterstützt zusammen mit weiteren Daten die Annahme, dass die Bornforellen
genetisch an die lokale Umgebung mit den dortigen Stressoren adaptiert sind. Ihr Immunsystem kann in
einem Ruhezustand verharren, während die Importforellen versuchen, unbekannte Stressoren aktiv zu
bekämpfen, was am Ende öfter erfolglos bleibt und in entsprechenden Mortalitäten mündet.
Die biotechnologischen Analysen unterstützen somit die phänotypischen Daten aus den
Produktionsanlagen und eine Nutzung der Bornforellen anstelle von Importforellen ist entsprechend der
Datenlage an schwierigen Standorten in jedem Fall vorzuziehen.
5 Die Bornforelle als Modell und Standortlinie ‐ Entwicklung von
Zuchtrichtlinien
Die vorliegenden und publizierten Daten belegen, dass die Zuchtlinie Born ein ausgezeichnetes
Nutzfischmodell für die Aquakulturforschung darstellt. Die Weiterführung der Zuchtlinie Born als Modell
für die Aquakultur von Regenbogenforellen wird empfohlen.
Als Standortlinie ist die Zuchtlinie Born in jedem Fall erhaltenswert, da sie in der Summe der Ergebnisse
unter den regionalen, oft nicht optimalen Gewässerqualitäten für eine Forellenaufzucht gegenüber
anderen Zuchtlinien die besseren Produktionsleistungen zeigt. Ihre Zucht muss weiter optimiert werden
im Hinblick auf die Charakterisierung der Robustheit, aber auch mit Blick auf ein besseres Wachstum.

47
Die Ergebnisse können zur Weiterentwicklung und Anpassung bestehender Zuchtrichtlinien genutzt
werden. Die vorliegenden Daten empfehlen sehr klar, die Entwicklung und Nutzung der regionalen
Zuchtlinien Born weiterzuführen, aber besser noch an das jeweilige Gewässer angepasste Zuchtlinien zu
entwickeln sofern offene oder halboffene Aquakultursysteme zum Einsatz kommen. Die Nutzung
importierter nicht regionaler Zuchtlinien empfiehlt sich für geschlossene Aquakulturanlagen. Zumindest
aber sollte für den Einsatz nicht regionaler Zuchtlinien die Gewässerqualität während der Aufzucht der
des Ursprungsgewässers der Zuchtlinie weitestgehend entsprechen. Dies kann natürlich nur für
abiotische Wasserparameter gelten, da u.a. regionale pathogene Keime, Parasiten oder Algentoxine
immer ein erhöhtes Risiko für genetisch nicht angepasste Zuchtlinien darstellen werden. Da auch in der
Aquakultur von Fischen das Tierwohl eine zunehmend größere Rolle spielt, und gerade im Hinblick auf
selbiges zahlreiche Hinweise auf Produktionsprobleme und Anpassungsschwierigkeiten der Forellen im
Projekt sichtbar wurden, ist es aus wissenschaftlicher Sicht zwingend, züchterisch tätig zu werden und
verschiedene regionale Zuchtlinien für unterschiedliche regionale Gewässerqualitäten zu entwickeln.
6 Wissenstransfer
Im Rahmen des Projektes kam es zu einer weitreichenden Weitergabe und Verbreitung der gewonnenen
Informationen.
Manuskripte (peer reviewed)
Rebl A, Verleih M, Köllner B, Korytář T, Goldammer T. Duplicated NELL2 genes show different
expression patterns in two rainbow trout strains after temperature and pathogen challenge.
Comp Biochem Physiol B Biochem Mol Biol. 2012 Sep;163(1):65‐73. doi:
10.1016/j.cbpb.2012.05.001. Epub 2012 May 8.
Verleih M, Rebl A, Köllner B, Korytář T, Köbis JM, Kühn C, Wimmers K, Goldammer T. Iron‐sulfur
cluster scaffold (ISCU) gene is duplicated in salmonid fish and tissue and temperature dependent
expressed in rainbow trout. Gene. 2013 Jan 10;512(2):251‐8. doi: 10.1016/j.gene.2012.10.037.
Epub 2012 Nov 5.
Köbis JM, Rebl A, Kühn C, Goldammer T. Comparison of splenic transcriptome activity of two
rainbow trout strains differing in robustness under regional aquaculture conditions. Mol Biol
Rep. 2013 Feb;40(2):1955‐66. doi: 10.1007/s11033‐012‐2252‐1. Epub 2012 Oct 20.
Rebl A, Verleih M, Köbis JM, Kühn C, Wimmers K, Köllner B, Goldammer T. Transcriptome
profiling of gill tissue in regionally bred and globally farmed rainbow trout strains reveals

48
different strategies for coping with thermal stress. Mar Biotechnol (NY). 2013 Aug;15(4):445‐60.
doi: 10.1007/s10126‐013‐9501‐8. Epub 2013 Apr 3.
Borchel A, Verleih M, Rebl A, Kühn C, Goldammer T. Creatine metabolism differs between
mammals and rainbow trout (Oncorhynchus mykiss). Springerplus. 2014 Sep 9;3:510. doi:
10.1186/2193‐1801‐3‐510. eCollection 2014
Korytář T, Jaros J, Verleih M, Rebl A, Kotterba G, Kühn C, Goldammer T, Köllner B. Novel insights
into the peritoneal inflammation of rainbow trout (Oncorhynchus mykiss). Fish Shellfish
Immunol. 2013 Oct;35(4):1192‐9. doi: 10.1016/j.fsi.2013.07.032. Epub 2013 Jul 31.
Verleih M, Borchel A, Krasnov A, Rebl A, Korytář T, Kühn C, Goldammer T. Impact of Thermal
Stress on Kidney‐Specific Gene Expression in Farmed Regional and Imported Rainbow Trout. Mar
Biotechnol (NY). 2015 Oct;17(5):576‐92. doi: 10.1007/s10126‐015‐9640‐1. Epub 2015 May 28.
Borchel A. Charakterisierung von Transkriptom und Phänotyp different robuster Zuchtlinien der
Regenbogenforelle mit Fokus auf das Kreatin‐System der Fische. Dissertation Universität Rostock
2015: pp100 + Anhang, 2015.
Weitere Schriften
Tom Goldammer, Alexander Rebl, Marieke Verleih, Bernd Köllner, Tomas Korytář, Carsten Kühn.
Standortgerechte Fischproduktion – Robuste Forellen für die Teichwirtschaft gezüchtet. For‐
schungsReport Spezial Ökologischer Landbau 2:10‐11, 2013.
Alexander Rebl, Tom Goldammer. Burnoutprophylaxe für Forellen – FBN‐Forscher erkunden die
geringe Stressempfindlichkeit der BORN‐Forelle, einer Zucht aus Mecklenburg‐Vorpommern.
Leibniz Nordost 18: 10‐11, 2014.
Andreas Borchel, Tom Goldammer. Widerstand gegen Aeromona – Warum ist die Bornforelle so
robust gegen Kranheiten? Eine Erkundung ihrer Gesundheitsgene. Leibniz Nordost 21: 8‐9, 2015.
Wesentliche Einträge in Genomdatenbanken
* You may view your GSE74332 study at:
http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE74332
* You may view your GSE75563 study at:
http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE75563
Hinter den Links sind projektbezogene Transkriptomdaten als verpackte *.txt.gz‐Dateien
abrufbar. Die darin befindlichen Daten sind im txt‐Format kodiert und können über FTP oder
HTTP heruntergeladen werden und dann in MS Excel für weitere Studien, Analysen und
Auswertungen, das Finden von Sequenzmotiven etc. einfach sichtbar gemacht werden.

49
Vorträge
A. Borchel, M. Verleih, A.Rebl, C. Kühn, T. Goldammer. Unterschiedliche Regulation des
Kreatinstoffwechsels bei differenten Temperaturen in zwei Regenbogenforellenlinien
(Oncorhynchus mykiss). Vortragstagung der DGfZ und GfT am 12.‐13. September 2012, Hal‐
le/Saale, Deutschland.
Andreas Borchel. Different regulation of creatine metabolism at different temperatures in two
strains of rainbow trout (Oncorhynchus mykiss). Tag des Doktoranden im FBN. 18.Oktober 2012,
Dummerstorf, Deutschland.
Tom Goldammer. Die Regenbogenforelle Stamm BORN ‐ Beispiel für die Entwicklung robuster
Nutzfischlinien für eine nachhaltige regionale Aquakultur. Seminar Senatsarbeitsgruppe
Ökologischer Landbau, FBN Dummerstorf, 24.‐25.Januar 2013, Dummerstorf, Deutschland.
Tom Goldammer. Animal health research with special reference to fish. Annual meeting of
European Forum of Farm Animal Breeders (EFFAB), FBN Dummerstorf, 24.‐25.April 2013,
Dummerstorf, Deutschland.
Andreas Borchel. Einblicke in das Kreatinsystem der Regenbogenforelle (Oncorhynchus mykiss):
Die besondere Rolle des Skelettmuskels. Vortragstagung der DGfZ und GfT am 04.‐05. September
2013, Göttingen, Deutschland.
Tom Goldammer. Robust aquaculture fish breeds ensure progressive gains, 5th edition of the
Ren‐dez‐Vous de Concarneau: where Industry meets Science in marine Biotechnology. 3.‐4. Oc‐
tober 2013, Concarneau, France.
Marieke Verleih. Die apoptotische Caspase‐Kaskade in der Regenbogenforelle ‐
Charakterisierung von Genen und Genvarianten. Kollquiumsreihe zur Nutztierforschung am FBN.
31.März 2014, Dummerstorf, Deutschland.
Tom Goldammer. Angewandte Genetik in der Aquakultur – Beispiel Regenbogenforelle Stamm
BORN. Gemeinsames Seminar der Nutztierwissenschaften der Justus‐Liebig‐Universität Giessen.
5.Mai 2014, Giessen, Deutschland.
Andreas Borchel. Untersuchungen zum Kreatinsystem der Regenbogenforelle (Oncorhynchus
my‐kiss). Aktuelle Forschungsarbeiten der Biochemie, Botanik, Mikrobiologie, Pflanzengenetik,
Pflanzenphysiologie an der Universität Rostock. 9.Mai 2014, Rostock, Deutschland.
Andreas Borchel, Marieke Verleih, Alexander Rebl, Tom Goldammer. Cold shock induced changes
in gene expression in rainbow trout (Oncorhynchus mykiss). Tag des Doktoranden im FBN.
13.Mai 2014, Dummerstorf, Deutschland.

50
Marieke Verleih, Alexander Rebl, Ronald M. Brunner, Judith Köbis, Andreas Borchel, Tomáš
Korytář, Bernd Köllner, Aleksei Krasnov, Carsten Kühn, Tom Goldammer. Global biotechnological
strategies for characterization of the trait robustness in rainbow trout. 11th Internatinal
Congress on the Biology of Fish, 3.‐7. August 2014, Heriot‐Watt University, Edinburgh, Scotland.
Andreas Borchel, Marieke Verleih, Alexander Rebl, Tom Goldammer. Vergleichende
Transkriptom‐analysen zur Kälteschockantwort der Regenbogenforelle (Oncorhynchus mykiss).
Vortragsta‐gung der DGfZ und GfT am 17.‐18.September 2014 im FBN. Dummerstorf,
Deutschland.
Tom Goldammer. Zuchtauswahl mittels genetischer Analyse: Die Born‐Forelle.
Fischereiforschung und Praxis, Informationsveranstaltung der LFA‐MV, 8.‐9.10.2014, Rostock,
Deutschland.
Tom Goldammer. Comparative molecular and immunological analyses of different robust
rainbow trout strains in aquaculture. Workshop Animal Immunogenetic and Molecular
Immunology, 30.‐31. October 2014, Kayseri, Turkey.
Andreas Borchel. Die BORN‐Forelle – Gute Gene gegen Ostsee‐Stress. Wettbewerb Wissenschaft
und Kommunikation 2015 – Rostock’s Eleven, Rostock denkt 365°. 10.‐11.6.2015, Universität
Rostock, Rostock, Deutschland
Andreas Borchel. Charakterisierung von Transkriptom und Phänotyp different robuster
Zuchtlinien der Regenbogenforelle mit Fokus auf das Kreatin‐System der Fische.
Promotionsverteidigung an der Mathematisch‐Naturwissenschaftlichen Fakultät der Universität
Rostock. 26.10.2015, Rostock, Deutschland.
Tom Goldammer, Alexander Rebl, Ronald M. Brunner, Judith Köbis, Marieke Verleih, Andreas
Borchel, Aleksei Krasnov, Tomáš Korytář, Carsten Kühn. Global transcriptome analyes in two
different selected rainbow trout strains for development of molecular biomarkers determining
fish welfare. 15th Aquaculture Europe Conference, 20.‐23.10.2015, Rotterdam, Netherlands.
Poster
Simone Altmann, Alexander Rebl, Marieke Verleih, Judith Köbis, Tomas Korytář, Carsten Kühn,
Bernd Köllner, Tom Goldammer. The rainbow trout strain BORN – Regional breeding resource
and fish model for comparative molecular genetic analyses of robustness. 1st International
Conference on Integrative Salmonid Biology (ICISB), 17.‐20.June 2012, Oslo, Norway.
Alexander Rebl, Marieke Verleih, Judith M. Köbis, Tomáš Korytář, Carsten Kühn, Bernd Köllner,
Klaus Wimmers, Tom Goldammer. Different strategies of coping with thermal stress in two

51
rainbow trout strains identified by transcriptome profiling in gill tissue 33rd Conference of the
International Society for Animal Genetics 15.‐20.July 2012, Cairns, Australia.
Marieke Verleih, Alexander Rebl, Bernd Köllner, Tomáš Korytář, Carsten Kühn, Klaus Wimmers,
Tom Goldammer. Duplicated rainbow trout genes ISCU, NELL2 and MARCH5 show gene variant
specific expression pattern. 33rd Conference of the International Society for Animal Genetics
15.‐20.July 2012, Cairns, Australia.
Marieke Verleih, Julia Brennmöhl, Andreas Borchel, Alexander Rebl, Ronald M. Brunner, Bernd
Köllner, Carsten Kühn, Tom Goldammer. Characterization of the Apoptotic Caspase Cascade in
Rainbow Trout. 11th International Congress on the Biology of Fish, 3.‐7. August 2014, Heriot‐
Watt University, Edinburgh, Scotland.
Andreas Borchel, Marieke Verleih, Alexander Rebl, Carsten Kühn, Tom Goldammer. Strong
expression of creatinerelated genes in the skeletal muscle of rainbow trout indicates differences
to mammals. 11th International Congress on the Biology of Fish, 3.‐7. August 2014, Heriot‐Watt
University, Edinburgh, Scotland.
Alexander Rebl, Marieke Verleih, Andreas Borchel, Mareen Nipkow, Tomáš Korytář, Henrike
Rebl, Aleksei Krasnov, Carsten Kühn, Ronald M. Brunner, Hans‐Martin Seyfert, Tom Goldammer.
Charakterisierung der frühen Immunantwort auf A. salmonicida in Lachsfischen6. Büsumer
Fischtag, 11.6.2015, mariCUBE Büsum, Deutschland.
Andreas Borchel, Marieke Verleih, Alexander Rebl, Carsten Kühn, Tom Goldammer. Expression
von Genen der Kreatinsynthese in Fischen im Vergleich zum Säuger. Deutscher Fischereitag, 25.‐
26.8.2015, Rostock, Deutschland.
Alexander Rebl, Marieke Verleih, Andreas Borchel, Mareen Nipkow, Tomáš Korytář, Henrike
Rebl, Aleksei Krasnov, Carsten Kühn, Ronald M. Brunner, Hans‐Martin Seyfert, Tom Goldammer.
Genombiologische Ansätze zur Analyse der differenten Robustheit von Salmoniden in der
regionalen Aquakultur. Deutscher Fischereitag, 25.‐26.8.2015, Rostock, Deutschland.
Marieke Verleih, Andreas Borchel, Aleksei Krasnov, Alexander Rebl, Tomáš Korytář, Carsten
Kühn, Tom Goldammer. Transcriptome data suggest SERPINH1 and CIRBP as molecular markers
for thermalstress characterization in rainbow trout (Oncorhynchus mykiss). 15th Aquaculture
Europe Conference, 20.‐23.10.2015, Rotterdam, Netherlands.
Ronald M. Brunner, Marieke Verleih, Judith Köbis, Andreas Borchel, Carsten Kühn, Frieder
Hadlich, Alexander Rebl, Tom Goldammer. Comparative transcriptome sequencing of different
robust rainbow trout lines (Oncorhynchus mykiss) for the identification of trait related candidate
genes. 15th Aquaculture Europe Conference, 20.‐23.10.2015, Rotterdam, Netherlands.

52
Vorlesungen ‐ Praktika
Im Rahmen des Wissenstransfers an die Universität Rostock erfolgte in den Sommersemestern
SS2012, SS2013, SS2014 und SS2015 die direkte Weitergabe von Erkenntnissen aus dem Projekt
an die Studenten des Masterstudienganges Aquakultur. Dies geschah im Rahmen der Vorlesung
sowie eines genetischen Praktikums über das Modul Technologie der Fischaquakultur – Teil
Fischgenetik.
Projektinformationen Online und Presse.
Informationen zum Projekt können über die Homepage WWW.Aquakultur‐MV.de abgerufen werden.
Über das Projekt erfolgten während der Projektlaufzeit verschiedene Pressemitteilungen auf
verschiedenen medialen Plattformen.
Die Ergebnisse des Projektes werden im Rahmen eines öffentlichen Kolloquiums in zusammengefasster
Form Anfang 2016 vorgestellt und parallel dazu in der Zeitschrift Fischerei und Fischmarkt publiziert.
Die Projektmitarbeiter bedanken sich beim Europäischen Fischereifond (EFF) und Ministerium für
Landwirtschaft, Umwelt und Verbraucherschutz des Landes Mecklenburg‐Vorpommern für die
Förderung des Pilotprojektes.
Die Projektkoordination bedankt sich bei allen Kooperationspartnern und den fischereilichen Betrieben
für die aktive Mitarbeit und Kooperation.
Dr. Tom Goldammer
Projektkkoordinator
Dummerstorf, 4.Dezember 2015