Institut für Mikrobiologie und Tierseuchen der ... · der Actinomycetales und der dazugehörigen...
153
Institut für Mikrobiologie und Tierseuchen der Tierärztlichen Hochschule Hannover Paratuberkulose-Diagnostik in Milch: Erreger- und Antikörpernachweis mittels PCR und ELISA INAUGURAL-DISSERTATION zur Erlangung des Grades eines DOCTOR MEDICINAE VETERINARIAE durch die Tierärztliche Hochschule Hannover Vorgelegt von Corinna Winterhoff aus Hagen/Westf. Hannover 2000
Transcript of Institut für Mikrobiologie und Tierseuchen der ... · der Actinomycetales und der dazugehörigen...

Institut für Mikrobiologie und Tierseuchen
der Tierärztlichen Hochschule Hannover
Paratuberkulose-Diagnostik in Milch:
Erreger- und Antikörpernachweis
mittels PCR und ELISA
INAUGURAL-DISSERTATION
zur Erlangung des Grades eines
DOCTOR MEDICINAE VETERINARIAE
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Corinna Winterhoff
aus Hagen/Westf.
Hannover 2000

Wissenschaftliche Betreuung: Univ.-Prof. Dr. G. F. Gerlach
1. Gutachter: Univ.-Prof. Dr. G. F. Gerlach
2. Gutachter: Univ.-Prof. Dr. Dr. h.c. Ch. Ring
Tag der mündlichen Prüfung: 29.11.2000
Diese Arbeit wurde durch die Niedersächsische Tierseuchenkasse und die Landesvereinigung
Milchwirtschaft Niedersachsen e.V. gefördert.

„Da es meine Absicht ist, etwas Brauchbares für den zu schreiben, der Interesse dafür
hat, schien es mir zweckmäßiger, dem wirklichen Wesen der Dinge nachzugehen als
deren Phantasiebild.“
(Niccolo Machiavelli)
Meiner Mutter
und
meinem Vater
gewidmet.


Teile der vorliegenden Dissertation wurden bereits auf folgenden Tagungen vorgestellt:
Homuth, M., C. Winterhoff, K. Strutzberg (2000):
„Ergebnisse eines Ringversuchs zur serologischen, mikroskopischen und kulturellen Untersu-
chung auf Paratuberkulose des Rindes in Niedersachsen“
Vortrag auf der 1. Arbeitstagung des veterinärmedizinischen Referenzlabors für Tuberkulose
28./29. März in Jena
Winterhoff, C., M. Homuth, K. Strutzberg, G.-F. Gerlach (2000):
„PCR zum diagnostischen Nachweis des Erregers der Paratuberkulose des Rindes aus Milch“
Poster, DVG-Tagung, Fachgruppe Bakteriologie und Mykologie
15.-17. Juni in Leipzig


Inhalt
1 Einleitung.....................................................................................................10
2 Schrifttum....................................................................................................112.1 Mycobacterium paratuberculosis ............................................................................ 11
2.1.1 Taxonomie und Morphologie........................................................................... 11
2.2 Paratuberkulose ........................................................................................................ 12
2.2.1 Pathogenese und Krankheitsbild...................................................................... 12
2.2.2 Immunologie .................................................................................................... 13
2.2.3 Pathologie und Histologie................................................................................ 13
2.3 Epidemiologie .......................................................................................................... 14
2.3.1 Tenazität........................................................................................................... 14
2.3.2 Wirtsspektrum.................................................................................................. 16
2.3.3 Geographische Verbreitung ............................................................................. 17
2.3.4 Empfänglichkeit der Wirtstiere........................................................................ 18
2.3.5 Erregerausscheidung ........................................................................................ 19
2.3.6 Übertragungswege ........................................................................................... 20
2.3.7 Wirtschaftliche Bedeutung............................................................................... 21
2.4 Bekämpfung und Sanierung..................................................................................... 22
2.4.1 Therapie ........................................................................................................... 22
2.4.2 Impfung............................................................................................................ 22
2.4.3 Sanierung ......................................................................................................... 23
2.5 Nachweis und Diagnose........................................................................................... 24
2.5.1 Direkter Erregernachweis ................................................................................ 24
2.5.2 Serologische Verfahren.................................................................................... 26
2.5.3 Polymerase Kettenreaktion (PCR) ................................................................... 29
3 Material und Methoden .............................................................................343.1 Material .................................................................................................................... 34
3.1.1 Bakterienstämme und Medien ......................................................................... 34
3.1.2 Probengewinnung............................................................................................. 35
3.2 Methoden ................................................................................................................. 36

3.2.1 Anzucht des Erregers ....................................................................................... 36
3.2.2 Anzucht anderer Mykobakterien...................................................................... 36
3.2.3 Bestimmung der Erregerdichte ........................................................................ 36
3.2.4 Mikroskopischer und kultureller Nachweis von M. paratuberculosis............. 37
3.2.5 Antigenherstellung ........................................................................................... 38
3.2.6 Präparation von Fab-Fragmenten ...................................................................... 38
3.2.7 DNA-Präparation von Mykobakterien............................................................. 39
3.2.8 Bestimmung der DNA-Konzentration ............................................................. 40
3.2.9 SDS-Polyacrylamidgelelektrophorese (SDS-PAGE) ...................................... 40
3.2.10 Western-Blotting.............................................................................................. 41
3.2.11 Agarosegelelektrophorese................................................................................ 42
3.2.12 Enzyme-Linked Immunosorbent Assay (ELISA)............................................ 43
3.2.13 Andere Paratuberkulose-ELISA-Testverfahren............................................... 45
3.2.14 Polymerasekettenreaktion (PCR)..................................................................... 46
3.2.15 Methoden der statistischen Auswertung .......................................................... 49
4 Ergebnisse....................................................................................................524.1 Adaption des SVANOVIR®-ELISA für Blutserum an Milchserum........................ 52
4.1.1 Auswahl des ELISA......................................................................................... 52
4.1.2 Darstellung der Bindungsstelle des Svanovir®-Konjugates............................. 54
4.1.3 Milchaufreinigungsmethode ............................................................................ 55
4.1.4 Reproduzierbarkeit........................................................................................... 56
4.1.5 Evaluierung des Milch-ELISA mittels ROC-Analyse ..................................... 57
4.1.6 Korrelation des Blut- und Milch-ELISA bei festgelegten „Cut off“-
Werten des Milch-ELISA ................................................................................ 65
4.2 Entwicklung einer Milch-PCR................................................................................. 68
4.2.1 Methode zur Milchaufbereitung....................................................................... 68
4.2.2 Etablierung der PCR ........................................................................................ 70
4.3 Validierung von Milchserologie und Milch-PCR in einer
Verlaufsuntersuchung .............................................................................................. 76
4.3.1 Untersuchunsergebnisse................................................................................... 77

5 Diskussion....................................................................................................915.1 Adaption des Svanovir®- ELISA für Blutserum an Milchserum............................. 91
5.2 Entwicklung einer Milch-PCR................................................................................. 96
5.3 Validierung von Milchserologie und Milch-PCR in einer
Verlaufsuntersuchung .............................................................................................. 99
6 Zusammenfassung ....................................................................................102
7 Summary....................................................................................................103
8 Literaturverzeichnis .................................................................................104
9 Anhang.......................................................................................................1299.1 Tabellenanhang ...................................................................................................... 129
9.2 Kulturmedien zur Anzucht von M. avium ssp.paratuberculosis ........................... 139
9.3 Agarosegelelektrophorese ...................................................................................... 141
9.4 SDS-Polyacrylamidgelelektrophorese (SDS-PAGE)............................................. 142
9.5 Coomassie®-Blau-Färbung..................................................................................... 143
9.6 Proteintransfer von SDS-Gelen im Tankblotting-Verfahren ................................. 144
9.7 Aufreinigung von IgG............................................................................................ 145
9.8 ELISA .................................................................................................................... 145
9.9 Primer..................................................................................................................... 146
9.10 Chemikalien ........................................................................................................... 147
9.11 Enzyme................................................................................................................... 148
9.12 Abbildungsverzeichnis........................................................................................... 149
9.13 Tabellenverzeichnis ............................................................................................... 151
9.14 Abkürzungsverzeichnis .......................................................................................... 152

Einleitung
10
1 Einleitung
Mycobacterium paratuberculosis verursacht eine chronische, granulomatöse und nicht
therapierbare Enteritis bei Wiederkäuern, die Johne´sche Krankheit. Weltweit entstehen be-
deutende wirtschaftliche Verluste infolge von Rückgang in der Milchleistung, Abmagerung
und Fruchtbarkeitsstörungen. Die Paratuberkulose verbreitet sich innerhalb betroffener Be-
stände nur relativ langsam. Die Ansteckung erfolgt über die Aufnahme von erregerhaltigem
Kolostrum sowie Kot. Die Tiere infizieren sich in der Regel in den ersten Lebensmonaten und
zeigen danach oft über Jahre keinerlei klinische Symptome, obwohl eine intermittierende oder
kontinuierliche Erregerausscheidung über Kot und Milch erfolgt. Gerade die Ausscheidung
über Milch wird in letzter Zeit kritisch betrachtet, da eine sichere Abtötung der Erreger durch
Pasteurisierung in Frage gestellt worden ist (MILLAR et al. 1996).
Die wachsende wirtschaftliche Bedeutung der Paratuberkulose sowie die hohe Tenazität des
Erregers, die auch eine vielmonatige Persistenz in der Außenwelt ermöglicht, lassen eine
kontinuierliche Überwachung von Milchviehbetrieben sinnvoll erscheinen. Von einer derarti-
gen Maßnahme wurde bisher Abstand genommen. Dies wird unter anderem durch die auf-
wendige Probenentnahme (Kot und Blut) und Diagnostik (u.a. mehrmonatige Kotkultur) be-
gründet.
Basierend auf diesen Überlegungen war das Ziel der Arbeit, auf der Basis des leicht zugängli-
chen Materials „Milch“ kostengünstige, diagnostische Methoden zum Erreger- und Antikör-
pernachweis zu entwickeln. Durch Etablierung einer Milch-PCR und eines Milch-ELISA
konnte dieses Ziel erreicht werden. Beide Verfahren wurden abschließend in einer Ver-
laufsuntersuchung mit zehn an Paratuberkulose erkrankten Milchkühen durch Blutserologie,
Kotuntersuchungen sowie durch die Untersuchung des Ileocaecallymphknotens validiert.

Schriftum
11
2 Schrifttum
2.1 Mycobacterium paratuberculosis
Im Jahre 1895 wurde der Erreger bei einer Kuh mit chronischer Enteritis erstmals be-
schrieben (JOHNE und FRONTHINGHAM 1895). Die Autoren nahmen an, daß es sich bei
dem beschriebenen Fall um eine besondere Form von Darmtuberkulose handelt. Die Eigen-
ständigkeit dieser Erkrankung wurde elf Jahre später erkannt (BANG 1906). Das Krankheits-
bild wurde als „Enteritis chronica bovis pseudotuberculosa“ bezeichnet.
2.1.1 TAXONOMIE UND MORPHOLOGIE
Heute wird Mycobacterium paratuberculosis (M. paratuberculosis) in die Ordnung
der Actinomycetales und der dazugehörigen Familie der Mycobacteriaceae eingeordnet. Auf-
grund von biochemischen Charakteristika wird der Erreger als eine weitere Subspezies (ssp.)
von Mycobacterium avium, neben M. avium ssp. silvaticum und M. avium ssp. avium, klassi-
fiziert (M. avium ssp. paratuberculosis) (THOREL et al. 1990). Diese Klassifizierung wird in
Europa bevorzugt, in den USA dagegen wird M. paratuberculosis eher als eigene Spezies
angesehen (SWEENEY 1996).
In molekularbiologischen Untersuchungen an genomischer DNA zeigte sich, daß M. paratu-
berculosis eine einzigartige Insertionssequenz (IS900) besitzt (GREEN et al. 1989). Bei M.
avium ssp. silvaticum und M. avium ssp. avium, nicht aber bei M. paratuberculosis konnten
verwandte Sequenzen (IS901/IS902) gefunden werden (KLAUSEN et al. 1997; KUNZE et al.
1992).
M. paratuberculosis ist ein säurefestes, unbewegliches, schwach grampositives Stäbchen von
0,3-2 µm Breite und 0,3-0,5 µm Länge (BISPING und AMTSBERG 1988). Die Zellwand
besteht aus einer äußeren Peptidglykanschicht, es folgt eine Schicht mit langkettigen Fettsäu-
ren, die mit Arabinogalaktan gekoppelt sind. Nach außen wird die Zellwand durch eine
Schicht von Peptidoglycolipiden abgeschlossen (CLARKE 1997). Das Lipoarabinomannan ist
ein Glycolipid, welches in der Zellmembran verankert ist und von dort aus in die Zellwand
reicht (BRENNAN und NIKAIDO 1995).

Schriftum
12
Charakteristisch für M. paratuberculosis ist das langsame, Mycobactin-abhängige Wachstum.
Mycobactin ist ein zellwandassoziiertes Siderophor, welches von allen Mykobakterien mit
Ausnahme von M. leprae und M. paratuberculosis in vitro synthetisiert wird (WHEELER und
RATLEDGE 1994).
Bei einer Generationszeit von 1,3 bis 4,4 Tagen werden sichtbare Kolonien auf festen Nähr-
böden erst nach 8-12 Wochen erwartet (LAMBRECHT et al. 1988). Als Standard-Nährboden
für die Anzucht von M. paratuberculosis wird das eidotterhaltige Medium nach Herrold (Her-
rold’s egg yolk medium) angesehen (MERKAL et al. 1968), auch in flüssigen Nährmedien,
wie z.B. Watson-Reid-Medium, kann M. paratuberculosis kultiviert werden.
Die Subspezies M. paratuberculosis ist also nur durch eine einzelne phänotypische Eigen-
schaft (Mycobactinabhängigkeit) und eine einzelne genotypische Eigenschaft (IS 900) cha-
rakterisiert.
2.2 Paratuberkulose
2.2.1 PATHOGENESE UND KRANKHEITSBILD
Die Erreger der Paratuberkulose gelangen über die orale Aufnahme in den Darm, wo
sie durch die M-Zellen im Domeepithel der Peyer´schen Platten die Darmschleimhaut über-
winden (CHIODINI et al. 1984; MOMOTANI et al. 1988). Die Mykobakterien werden dar-
aufhin von den subepithelialen und intraepithelialen Makrophagen aufgenommen, in denen
sie lange Zeit überleben und sich vermehren können. Bedingt durch das intrazelluläre
Wachstum von M. paratuberculosis kommt es zum Absterben des Makrophagen und zur er-
neuten Aufnahme in weitere Phagozyten. Es entsteht in diesem Bereich eine chronische ka-
tarrhalische Entzündung begleitet von Hyperplasie (MERKAL et al. 1970). Die Infektion
breitet sich im Dünndarm aus und befällt großflächigere Regionen sowie angrenzende Kör-
perbereiche wie Leber, Niere und Milz. Es kommt zu einer Enteropathie mit Proteinverlust
und schließlich zur Hypoproteinämie. Später werden die überfüllten epitheloiden Zellen ins
Darmlumen abgestreift, und die Mykobakterien werden mit dem Kot ausgeschieden. Die ge-
fundenen Zellzahlen liegen bei 108 Zellen/g Kot bzw. 1012 Zellen/ Tag (CHIODINI et al.
1984).

Schriftum
13
Die Paratuberkulose hat eine lange Inkubationszeit (ca. 2-5 Jahre) und verläuft meist subkli-
nisch, wobei der Erreger intermittierend ausgeschieden werden kann. Symptomatisch werden
drei unterschiedliche klinische Stadien unterschieden. Im ersten, subklinischen Stadium
scheiden die Tiere den Erreger nicht aus und zeigen keinerlei klinische Symptome. Im näch-
sten Stadium wird der Erreger intermittierend mit dem Kot und der Milch ausgeschieden
(SWEENEY et al. 1992b; TAYLOR et al. 1981). Im dritten Stadium sind die Tiere klinisch
erkrankt. In diesem letzten Stadium ist eine Verdachtsdiagnose aufgrund des zuerst intermit-
tierenden, später anhaltenden, therapieresistenten Durchfalls mit fortschreitender Abmagerung
bei jedoch kaum gestörtem Allgemeinbefinden und erhaltener Freßlust meist leicht zu stellen.
Wesentlich schwieriger gestaltet sich die Erkennung von subklinisch infizierten Tieren, die
über Jahre hinweg die Erreger ausscheiden können.
2.2.2 IMMUNOLOGIE
Wenn die Erreger der Paratuberkulose phagozytiert werden, kommt es zur zellulären
Immunantwort. Die Freisetzung der Bakterien aus den abgestorbenen Makrophagen führt im
nächsten Stadium zu einer humoralen Immunantwort. Das Gleichgewicht verschiebt sich von
der zellvermittelten zur humoralen Immunreaktion, die schließlich vorherrscht (CHIODINI et
al. 1984). Im Finalstadium der Erkrankung, jedoch auch während anderer Phasen der Infekti-
on, kann der Zustand der Anergie eintreten. In diesem Zustand fehlen sämtliche Reaktionen
der Immunabwehr auf die antigenen Eigenschaften des Erregers (MERKAL et al. 1970).
2.2.3 PATHOLOGIE UND HISTOLOGIE
Die pathologischen Veränderungen zeigen das Bild einer chronisch-hyperplastischen
Enteritis. Besonders im Bereich des Ileums, aber auch im mittleren Jejunum, Caecum und
Colon ist eine deutliche Verdickung der Schleimhaut erkennbar. Im klassischen Fall zeigt die
Schleimhaut einen hirnwindungsartigen Verlauf. Die mesenterialen Lymphgefäße sind hoch-
gradig gestaut, die Lymphknoten dieses Bereiches sind stark geschwollen und können bis auf
das Fünffache vergrößert sein. Im Anschnitt zeigen sie eine mit granulomatösen Entzün-
dungsherden durchsetzte Schnittfläche.
Histologisch sind in der Darmwand große Mengen an epitheloiden Zellen und Makrophagen,
sowie einzelne Riesenzellen von Langhanstyp und eine mäßige Lymphoplasmeninfiltration zu

Schriftum
14
beobachten. Die Zellinfiltrate können die Krypten verdrängen und sich in Mucosa und Sub-
mucosa ausdehnen (POHLENZ 1991).
Bei der gelegentlich auftretenden generalisierten Infektion finden sich in verschiedenen inne-
ren Organen wie Leber, Milz, Niere, Lunge und Uterus miliare Herde, die histologisch aus
epitheloiden Zellen und wenigen Langhansschen Riesenzellen aufgebaut sind (CHIODINI et
al. 1984; ROSENBERGER 1978).
2.3 Epidemiologie
2.3.1 TENAZITÄT
Der Erreger der Paratuberkulose ist ein sehr widerstandsfähiger Keim, der in der Au-
ßenwelt lange überleben kann. So beträgt die Überlebensdauer in Mist und anaerober Gülle
bis zu 9 Monate (GAY und SHERMAN 1992; JØRGENSEN 1977). Im Weidekot infizierter
Rinder kann der Erreger noch nach 11 Monaten nachgewiesen werden (ROSENBERGER
1978). Die Nachweisdauer noch lebender Mykobakterien beträgt im Boden 11 Monate und im
Wasser 17 Monate (GAY und SHERMAN 1992). Die Überlebensdauer im Boden kann durch
saure pH-Bedingungen günstig beeinflußt werden (RICHARDS 1988). Ein ursächlicher Zu-
sammenhang zwischen saurem Boden-pH und erhöhten Prävalenzen für Paratuberkulose ist
noch nicht abschließend geklärt (JOHNSON-IFEARULUNDU und KANEENE 1997). Auch
gegen Austrocknung ist M. paratuberculosis äußerst resistent. So kann der Erreger eine 47
Monate anhaltende Austrocknung überleben. Direkte Sonneneinstrahlung tötet den Erreger
allerdings bereits nach 100 Stunden (GAY und SHERMAN 1992).
Auch in der Kälberaufzucht stellt die Tenazität des Erregers ein Problem dar. In kontrolliert
infizierten (102 bis 104 KBE/ml) Kolostrumproben, konnte nach Pasteurisierung (63°C für 30
min) in 2 von 18 Proben M. paratuberculosis kulturell nachgewiesen werden (MEYLAN et
al. 1996). Die Autoren empfehlen aber trotz einer Reduktion des IgG-Gehaltes um 12,3 %
eine Thermobehandlung von Kolostrum in Problembetrieben, da die Zahl der Mykobakterien
deutlich reduziert werden konnte.
Die hohe Thermotoleranz von M. paratuberculosis zeigt ein besonderes Problem in der Qua-
litätssicherung des Lebensmittels Milch auf. So wurde schon 1993 gezeigt, daß bei einer Er-
hitzung von Milch auf 63°C für 30 Minuten oder 72°C für 15 Sekunden in ca. 5-9 %, bzw. 3-

Schriftum
15
5 % der Milchproben M. paratuberculosis noch kulturell nachgewiesen werden konnte.
(CHIODINI und HERMON-TAYLOR 1993). Hierbei wurden zur Inokulation der Milchpro-
ben ca. 1 x 104 KBE/ml sowohl boviner als auch humaner Isolate verwendet. Isolate humanen
Ursprungs waren generell Hitze-resistenter als bovine Isolate. Im Laufe der nächsten Jahre
wurden vermehrt Untersuchungen zur Überlebensrate von M. paratuberculosis Isolaten in
pasteurisierter Milch vorgenommen. Bei Pasteurisierungsbedingungen von 63,5°C für 30 Mi-
nuten und 71,7°C für 15 Sekunden konnten in Milch Überlebensraten <1 % dokumentiert
werden (GRANT et al. 1996). Die eingesetzten Inokulationsdosen waren mit 107 und 104
KBE/ml relativ hoch, und zudem entsprachen die experimentellen Bedingungen nicht ganz
den kommerziellen Pasteurisierungsbedingungen. Aber auch mit niedrigeren Keimzahlen
(103-102 KBE/ml Milch) konnte durch HTST-Bedingungen (high temperature, short time:
71,7°C für 15 Sekunden) M. paratuberculosis nicht vollständig inaktiviert werden (GRANT
et al. 1998). Dieselben Autoren konnten ein Jahr später bei Pasteurisierungsbedingungen mit
steigender Temperatur bei gleichbleibender Einwirkzeit (72°,75°,78°,80°,85°,90°C für 15
Sekunden) und gleichbleibender Temperatur bei längerer Einwirkzeit (72°C für 20 Sekunden
und 72°C für 25 Sekunden) dokumentieren, daß bei einer eingesetzten Inokulationsdosis von
106 KBE/ml Milch sich längere Einwirkzeiten effektiver bei der Inaktivierung von
M. paratuberculosis auswirken als höhere Temperaturen (GRANT et al. 1999). Auch bei Ino-
kulationsdosen von nur 101 KBE/ml Milch kann M. paratuberculosis die HTST-
Pasteurisierung überleben (SUNG und COLLINS 1998). In einer weiteren Untersuchung
wurde eine Anlage, die industrielle Pasteurisationsbedingungen exakt nachzeichnet, verwen-
det (STABEL et al. 1997). Bei Einsatz von tiefgefrorenen und Ultraschall-behandelten My-
kobakterien konnten die Autoren nach Pasteurisierung keine überlebensfähigen Erreger
nachweisen. Die Bedeutung dieser Daten wird kontrovers diskutiert (GRANT 1998). Trotz-
dem kann nicht ausgeschlossen werden, daß M. paratuberculosis die kommerzielle Milch-
Pasteurisierung überlebt. So konnte in England mittels PCR in pasteurisierter Milch, die im
Einzelhandel erhältlich war, M. paratuberculosis nachgewiesen werden (MILLAR et al.
1996). Den Autoren gelang der Nachweis in 7 % der 312 in Süd- und Westengland gekauften
Proben.
Die Zahl der wirksamen Desinfektionsmitteln ist begrenzt. Die meisten Desinfektionsmittel
sind unwirksam gegen M. paratuberculosis, insbesondere dann, wenn sich der Erreger im Kot
befindet. Als wirksam haben sich Formaldehyd-haltige Desinfektionsmittel erwiesen.

Schriftum
16
2.3.2 WIRTSSPEKTRUM
Die Paratuberkulose ist eine Darmerkrankung der Wiederkäuer. Neben den Hauswie-
derkäuern wie Rind, Schaf und Ziege ist die Infektion auch bei Wild und Zoowiederkäuern
nachgewiesen worden (CHIODINI und VAN KRUININGEN 1983; CLARKE 1997; WEBER
et al. 1991). Ebenso wurde die Infektion bei Lamas, Kamelen, Antilopen, Büffeln, Yaks,
Gnus und anderen Wiederkäuern beschrieben (STEHMAN 1996; WILLIAMS und SPRA-
KER 1979). Bei niedriger Bestandsdichte des Wildes ist zwar eine Weiterverbreitung des Er-
regers unwahrscheinlich, die Wahrscheinlichkeit wächst aber mit höheren Bestandsdichten
etwa durch Gatterhaltung, so daß Wild eine potentielle Infektionsquelle für Hauswiederkäuer
darstellen kann (CHIODINI und VAN KRUININGEN 1983).
In Schottland konnte sowohl M. paratuberculosis im Kot von Kaninchen nachgewiesen als
auch die typischen pathologischen Befunde im Darm der Tiere dokumentiert werden (GREIG
et al. 1997).
Es gelang nach oraler Verabreichung des Erregers in größeren Mengen sowohl Schweine als
auch Pferde zu infizieren (LARSEN et al. 1972). Beim Pferd (LIENAUX 1913), beim
Schwein (RUNNELS 1955), beim Hund (VOGEL 1970) und beim Esel (VAN ULSEN 1970)
konnten auch natürliche Infektionen nachgewiesen werden. Ebenso konnte die Empfänglich-
keit von Hühnern gegenüber dem Bakterium demonstriert werden (LARSEN und MOON
1972). Bei all diesen Tieren kommt es zwar zu einer Vermehrung des Erregers, die klinische
Symptomatik einer Darmerkrankung ist jedoch nicht festzustellen (CHIODINI et al. 1984).
Bei infizierten Primaten hingegen zeigte sich ein deutliches klinisches Bild. In einer Herde
von 38 Stummelschwanz-Makaken waren 29 Tiere mit Mykobakterien, die in der PCR (IS
900) positiv waren und ein mycobactinabhängiges Wachstum aufzeigten, infiziert. Die Aus-
scheidung der Keime mit dem Kot konnte nachgewiesen werden. Innerhalb von fünf Jahren
starben 13 Tiere mit klinischen und pathologischen Erscheinungen ähnlich der Paratuberkulo-
se-Infektion beim Rind (MCCLURE et al. 1987).
Weiterhin wird diskutiert, ob unterschiedliche Biotypen von M. paratuberculosis innerhalb
der Hauswiederkäuerarten existieren. Unterschiede in den erforderlichen Kultivierungsbedin-
gungen für spezifisch bovine und ovine M. paratuberculosis Isolate deuten darauf hin. Trotz
hoher genetischer Homologie gelang der Nachweis drei unterschiedlicher Typen von M. pa-
ratuberculosis mit Hilfe von Hybridisierungstechniken (COLLINS et al. 1990; DE LISLE et
al. 1992). Unklarheit besteht darüber, inwieweit sich unterschiedliche Tierarten Spezies über-

Schriftum
17
greifend infizieren können. Es bestehen Hinweise darauf, daß sich die Hauswiederkäuer un-
tereinander anstecken können (STEHMAN 1996), da experimentelle Infektionen vom Rind
und Hirsch auf Schafe gelangen, und Rinder mit ovinen Stämmen infiziert werden konnten
(ANGUS und GILMOUR 1971; JUSTE et al. 1994; KLUGE et al. 1968; TAYLOR 1953).
Die Beziehung zwischen der bovinen Paratuberkulose und der Crohn’schen Erkrankung des
Menschen ist noch nicht geklärt. Die Crohn’sche Erkrankung wurde 1932 als eine eigenstän-
dige Darmerkrankung, abgegrenzt zu der Darmtuberkulose beschrieben (CROHN et al. 1932).
Sie äußert sich als eine granulomatöse Ileocolitis, von der insbesondere jüngere Menschen
betroffen sind. Schon im Jahr 1913 wurden Vermutungen geäußert, basierend auf der Ähn-
lichkeit der Krankheitsbilder von Morbus Crohn und Paratuberkulose, daß M. paratuberculo-
sis eine entscheidende Rolle in der Pathogenese von Morbus Crohn zukommt (CHIODINI
1989; TAMBOLI 1996). Einige Autoren bezeichnen Paratuberkulose als eine potentielle Zo-
onose (CHIODINI und ROSSITER 1996; HERMON-TAYLOR et al. 1990).
In den vergangenen 15 Jahren haben sich eine Vielzahl von Untersuchungen mit dem labor-
diagnostischen Nachweis von M. paratuberculosis bei Morbus Crohn-Patienten beschäftigt.
Es wurden ELISA-Tests, Western-Blot-Verfahren, PCR und mikrobiologische Kultivie-
rungstechniken eingesetzt. Die Ergebnisse sind insgesamt sehr heterogen. Dies spiegelt sich
in zwei Untersuchungen dieses Jahres wieder. Es wird einerseits ein eindeutiger Zusammen-
hang von M. paratuberculosis und Morbus Crohn gesehen, da der Erreger bei Patienten nach-
gewiesen werden konnte. Diese Patienten zeigten nach einer Kombinations-Therapie mit ins-
besondere gegen Mykobakterien wirksamen Antibiotika eine Verbesserung und in einigen
Fällen eine scheinbare Heilung (HERMON-TAYLOR et al. 2000). Andererseits wird nicht
bestritten, daß M. paratuberculosis und Morbus Crohn gemeinsam auftreten können, es ist
aber bis heute ungeklärt, ob M. paratuberculosis vor oder nach dem Ausbrechen der Crohn‘
schen Erkrankung sich im Darm des Patienten manifestierte. Auch sind die Koch’schen Po-
stulate nicht erfüllt (COOK 2000).
2.3.3 GEOGRAPHISCHE VERBREITUNG
Die Paratuberkulose ist eine weltweit verbreitete Darmerkrankung der Wiederkäuer.
Das erste Auftreten wurde aufgrund von eingesandten Untersuchungsmaterialien eines Tier-
arztes Harms aus Jever beschrieben (JOHNE und FRONTHINGHAM 1895). In Deutschland

Schriftum
18
ist die Paratuberkulose seit 1970 eine meldepflichtige Erkrankung. In der Zeit zwischen 1971
und 1981 traten 90 % aller in Deutschland gemeldeten Paratuberkulosefälle in Norddeutsch-
land und hier vor allem in der Küstenregion auf (SCHLIESSER und SCHAAL 1984). Die
Paratuberkulose ist aber in ganz Europa verbreitet, wobei neben Norddeutschland, besonders
die Niederlande, Großbritannien (CETINKAYA et al. 1996) und Dänemark (JØRGENSEN
1977; NIELSEN et al. 2000) von der Paratuberkulose betroffen sind. Aufgrund der Merzung
aller Paratuberkulose-infizierten Rinderherden ist Schweden das einzige Paratuberkulose-freie
Land innerhalb der Europäischen Gemeinschaft (VISKE et al. 1996). Für Italien schwankten
die Angaben für Herdenprävalenzen von 2,8 % bis 10-15 % (BELLETTI et al. 1992; KOR-
MENDY et al. 1989). Die Seroprävalenz innerhalb der Milchkühe in der Plateau de Diesse-
Region in Frankreich betrug auf der Grundlage eines kommerziellen ELISA-Verfahrens
(Commonwealth Serum Laboratoires) 5,99 % (MEYLAN et al. 1995). Die individuellen Prä-
valenzen innerhalb verschiedener Herden schwankten dabei von 0 -17 %. In vielen der ande-
ren europäischen Ländern wurden bisher keine flächendeckenden Untersuchungen durchge-
führt
Außerhalb Europas wurden in den USA umfangreiche Untersuchungen zum Vorkommen der
Paratuberkulose durchgeführt. In den Jahren 1983 und 1984 wurden die Lymphknoten des
Ileocaecalbereichs von klinisch gesund erscheinenden Schlachtrindern untersucht (MERKAL
et al. 1987). Die kulturellen Ergebnisse ergaben eine Prävalenz von 1,6 % bei Rindern und 2,9
% bei Milchkühen in insgesamt 32 Staaten der USA.
Von dem „Office International des Epizooties“ (OIE) wird die Paratuberkulose der Wieder-
käuer als eine Liste B-Erkrankung geführt. Damit wird sie als eine Erkrankung mit sozioöko-
nomischer Bedeutung und/oder Bedeutung für die menschliche Gesundheit eingestuft.
2.3.4 EMPFÄNGLICHKEIT DER WIRTSTIERE
Die Empfänglichkeit der Wirtstiere bei der Paratuberkulose-Erkrankung ist altersab-
hängig. Kälber unter 30 Tagen zeigen die höchste Empfänglichkeit, jedoch treten klinische
Erkrankungen in der Regel erst im Alter von 2 bis 5 Jahren auf. Bei sehr hohen Infektionsdo-
sen können auch Kälber unter 12 Monaten bereits klinisch erkranken (CLARKE 1997;
HIETALA 1992). Die Resistenz gegen eine Infektion mit dem Erreger wächst mit zunehmen-
den Alter des Tieres (LARSEN et al. 1975). Dies wird in Zusammenhang mit der starken,

Schriftum
19
zahlenmäßige Verringerung der M-Zellen bei adulten Tiere gebracht. So gelten Rinder über 2
Jahre in der Regel als resistent gegenüber einer Infektion. Die Schwere der klinischen Er-
scheinungen ist mit zunehmenden Alter bei der Erstinfektion deutlich herabgesetzt
(CHIODINI et al. 1984). Auch Risikofaktoren wie Intensivhaltung, schlechte Fütterung, sau-
rer Boden und Stressfaktoren, wie Transport und Abkalbung, wirken sich begünstigend auf
eine Infektion aus.
2.3.5 ERREGERAUSSCHEIDUNG
Die Erregerausscheidung mit dem Kot steht bei der Paratuberkulose im Vordergrund.
Klinisch erkrankte Rinder scheiden ca. 108 KBE/g Kot aus (CHIODINI und HERMON-
TAYLOR 1993). Auch subklinisch infizierte Tiere scheiden den Erreger intermittierend mit
dem Kot aus (COLLINS 1996). Dabei kann aber die Ausscheidungsrate wesentlich geringer
sein als in der klinischen Phase der Erkrankung. Die Tiere scheiden ca. 101 bis 102 Erreger/g
Kot aus (WHITLOCK et al. 1994). Da die Anzahl der ausgeschiedenen Erreger schwankt,
scheiden nur wenige subklinisch infizierte Rinder M. paratuberculosis in ausreichender Men-
ge für eine kulturelle Untersuchung aus (GAY und SHERMAN 1992).
Die Ausscheidung von M. paratuberculosis über Kolostrum und Milch scheint ebenfalls eine
große Bedeutung für die Verbreitung der Erkrankung zu besitzen. Etwa ein Drittel klinisch
erkrankter Kühe scheidet M. paratuberculosis mit der Milch aus (TAYLOR et al. 1981). In
diesem Versuch konnte durch die parallel durchgeführte Untersuchung auf koliforme Keime
eine Kontamination der Milch durch Kot ausgeschlossen werden. Bei der Untersuchung von
Euterlymphknoten und Milchproben subklinisch infizierter Kühe konnte in 27 % der
Lymphknoten und in 11,6 % der Milchproben M. paratuberculosis kulturell nachgewiesen
werden (SWEENEY et al. 1992b). Die Untersuchungen weisen darauf hin, daß M. paratuber-
culosis über hämatogene und/oder über lymphogene Infektionswege in das Euter und die be-
nachbarten Lymphknoten gelangt. Die Häufigkeit des kulturellen Nachweises in Lymphkno-
ten oder Milch korrelierte mit der Stärke der Ausscheidung im Kot. Die Autoren empfehlen
auf Grund dieser Ergebnisse, in von Paratuberkulose betroffenenen Betrieben, Kälber generell
mit Milchaustauscher zu füttern, da die Gefahr einer oralen Übertragung durch Milch von
subklinisch infizierten Kühen auf Kälber besteht.

Schriftum
20
Auch in Kolostrumproben von subklinisch infizierten Kühen konnte M. paratuberculosis
kulturell nachgewiesen werden (STREETER et al. 1995). Dabei konnte der Erreger in 36,4 %
der Kolostrumproben von Kühen, die M. paratuberculosis in starken Maße mit dem Kot aus-
scheiden, und in 16 % der Kolostrumproben von geringgradig ausscheidenen Kühen nachge-
wiesen werden. Da diese Werte wesentlich höher sind als die bei entsprechenden Untersu-
chungen in Milchproben, stellt das Kolostrum eine gefährliche Infektionsquelle für Kälber
auch bei subklinisch infizierten Kühen dar.
Weiterhin ist belegt, daß M. paratuberculosis aus Uterusspülflüssigkeit klinisch erkrankter
Kühe isoliert werden kann (ROHDE und SHULAW 1990). Ebenso konnte gezeigt werden,
daß klinisch erkrankte Bullen (LARSEN und KOPECKY 1970) und auch subklinisch infi-
zierte Bullen (LARSEN et al. 1981) M. paratuberculosis mit dem Samen ausscheiden.
2.3.6 ÜBERTRAGUNGSWEGE
Die Infektion der Tiere erfolgt vor allem postnatal durch orale Aufnahme des Erregers
im Kälberalter, über erregerhaltiges Kolostrum oder über kotverschmierte Zitzen. Es liegen
jedoch keine Angaben über die Höhe der infektiösen Dosis vor (SWEENEY 1996). Tiere, die
den Erreger mit dem Kot ausscheiden, können direkt Weiden, Futter oder auch Wasserstellen
kontaminieren. Ferner kann der Landwirt direkt oder indirekt, über kontaminierte Geräte, die
Umgebung des Kalbes mit M. paratuberculosis kontaminieren (SWEENEY 1996).
Auch die intrauterine Infektion scheint eine bedeutsame Rolle zu spielen. Für Feten von kli-
nisch erkrankten Kühen besteht ein Risiko von 26,4 %, daß sie sich intrauterin mit dem Erre-
ger der Paratuberkulose infizieren (SEITZ et al. 1989). Dieses Risiko wird für subklinisch
infizierte Tiere mit knapp 10 % angegeben (SWEENEY et al. 1992c). Der Nachweis gelang
allerdings nur bei Tieren, die den Erreger auch in großen Mengen mit dem Kot ausschieden.
Die intrauterin infizierten Kälber entwickeln keine meßbare Immunantwort (LARSEN et al.
1975). Aus der Untersuchung der BVD-Ätiologie ist bekannt, daß die im zweiten Drittel der
Trächtigkeit infizierten Kälber zu immuntoleranten Ausscheidern werden. Diese Möglichkeit
muß auch bei der intrauterinen Infektion mit M. paratuberculosis in Betracht gezogen wer-
den. Beim Embryotransfer ist eine Übertragung der Erkrankung vom infizierten Empfänger-

Schriftum
21
tier auf den Fetus möglich. Empfängertiere sollten daher aus nachweislich Paratuberkulose-
freien Betrieben stammen.
Im Samen von infizierten Bullen konnte M. paratuberculosis nachgewiesen werden. Jedoch
konnte experimentell eine Übertragung des Erregers durch den Deckakt oder durch künstliche
Besamung nicht bewiesen werden (SWEENEY 1996). Für die künstliche Besamung besteht
ein nur geringes Risiko, wenn Kotproben der Spenderbullen regelmäßig kulturell untersucht
werden (LARSEN et al. 1981). Allerdings wurde auf die möglichen Gefahren einer Übertra-
gung der Paratuberkulose durch die künstliche Besamung hingewiesen (PHILPOTT 1993).
2.3.7 WIRTSCHAFTLICHE BEDEUTUNG
Wirtschaftliche Schäden entstehen einmal durch den direkten Verlust oder den gerin-
gen Schlachterlös von Tieren, die klinisch an Paratuberkulose erkrankt sind. Aber insbesonde-
re subklinisch infizierte Tiere können durch eine geringere Milchleistung und verminderte
Fertilität wirtschaftliche Schäden verursachen (HUTCHINSON 1996). Der Abfall in der
Milchleistung bei Tieren, die klinisch an Paratuberkulose erkrankt sind, ist beachtlich. So
produzieren erkrankte Kühe im Durchschnitt über 15 % weniger Milch als nicht infizierte
Kühe. Subklinisch infizierte Rinder produzieren durchschnittlich ca. 5-15 % weniger Milch
als nicht infizierte Rinder (RIEMANN und ABBAS 1983; WILSON et al. 1993). Im Ver-
gleich zu vorangegangenen Laktationen konnte für ein klinisch erkranktes Tier ein Rückgang
von bis zu 19,5 % (BENEDICTUS et al. 1987) und für ein subklinisch infiziertes Tier ein
Rückgang von 6 % ermittelt werden (HUTCHINSON 1996). Beim Vergleich der Milchlei-
stung ELISA-positiv mit ELISA-negativ getesteter Kühe war der Rückgang in der ELISA-
positiven Versuchsgruppe signifikant (NORDLUND et al. 1996).
Die Erkrankung nimmt auch Einfluß auf die Milchinhaltsstoffe. So betrug der Rückgang des
Fett- und Proteingehaltes in der Milch 21 % für klinisch erkrankte und 17,5 % für subklinisch
erkrankte Rinder (BENEDICTUS et al. 1987).
Ebenso darf die Anzahl der Tiere, die aufgrund von Sekundärerkrankungen, wie Mastitis und
Sterilität, geschlachtet werden, nicht unterschätzt werden (MERKAL et al. 1975). Ein Zu-
sammenhang zwischen klinischer Mastitis und Paratuberkulose konnte jedoch nicht doku-
mentiert werden (WILSON et al. 1993). Allerdings konnten andere Autoren demonstrieren,

Schriftum
22
daß bei der serologischen Untersuchung von 304 Milchviehherden in Ontario ein serologisch
positives Ergebnis mit einem erhöhten Zellgehalt der Milch korreliert war (MCNAB et al.
1991b). Auch die Angaben über Zwischenkalbezeiten sind widersprüchlich. Die Zwischen-
kalbezeiten infizierter Kühe betragen nach einer Untersuchung 15,18 Monate bei serologisch
positiven Kühen im Gegensatz zu 13,45 Monaten bei negativ getesteten Kühen (RIEMANN
und ABBAS 1983). Im Gegensatz dazu wurde in einer weiteren Studie eine Zwischenkalbe-
zeit von 12,65 Monaten bei kulturell positiven Kühen und 13,08 Monaten bei kulturell nega-
tiven Tieren ermittelt (DINSMORE 1986).
2.4 Bekämpfung und Sanierung
2.4.1 THERAPIE
Die Paratuberkulose ist nicht heilbar. Daher ist eine Therapie, die eine vorübergehende
Besserung der klinischen Symptome bewirkt, nur bei genetisch wertvollen Tieren angezeigt.
Diese müssen während der folgenden Zeit getrennt gehalten werden, da sie weiterhin den Er-
reger mit dem Kot ausscheiden. Die Therapie ermöglicht es, von den Tieren hochwertige Em-
bryo- oder Samenproben zu entnehmen (ST JEAN 1996).
Die Zielzellen für M. paratuberculosis sind die intestinalen Makrophagen, deshalb müssen
wirksame Therapeutika zellwandgängig sein. Nach Isoniazid-Behandlung für 26 Tage waren
Kühen bis zu 6 Monate nach dem Ende der Therapie klinisch unauffällig (BALDWIN 1976).
Auch Clofazimin induziert eine klinische Remission, verhindert aber nicht die Ausscheidung
des Erregers. Im Gegensatz dazu zeigen mit Streptomycin oder Dapson behandelte Tiere be-
reits 3 bis 4 Wochen nach dem Ende der Therapie wieder klinische Symptome. Derzeit stellt
Isoniazid, evtl. in Kombination mit Rifampicin, das Mittel der Wahl dar (ST JEAN 1996). Die
Kosten dieser Behandlung sind untragbar, und die Rückstände in dem Lebensmittel Fleisch
sind als bedenklich anzusehen.
2.4.2 IMPFUNG
Erstmals im Jahr 1926 wurde eine Impfung gegen Paratuberkulose beschrieben
(VALLEE und RINJARD 1926). Zur Zeit wird in den USA ein Hitze-inaktivierter Impfstoff

Schriftum
23
(SWEENEY 1996), in Europa auch ein Lebendimpfstoff verwendet. Die bisher verwendeten
Impfstoffe bieten zwar Schutz vor klinischen Erscheinungen, verhindern aber nicht die Infek-
tion sowie die spätere Ausscheidung des Erregers (CHIODINI 1996). Allerdings konnte teil-
weise beobachtet werden, daß geimpfte Tiere den Erreger in geringeren Mengen ausscheiden
als nicht geimpfte Kühe. Als problematisch erwiesen sich jedoch die starke Kreuzreaktion mit
bovinem Tuberkulin und die großen Granulome, die sich an der Injektionsstelle bilden kön-
nen.
Anhand eines Impfversuchs in zwei Milchviehbetrieben in den Niederlanden konnte doku-
mentiert werden, daß die Zahl der aufgrund klinischer Paratuberkulose geschlachteten Rinder
als auch die Zahl der subklinischen Fälle gesenkt werden konnte (WENTINK et al. 1994).
Die Impfung soll durch die Verminderung des Infektionsdrucks und die Reduktion der sub-
klinischen und klinische Fälle einen wirtschaftlichen Vorteil bieten (VAN SCHAIK et al.
1996).
Bei einem Impfversuch in Ungarn konnte die Ausscheidungsrate von M. paratuberculosis im
Kot deutlich abgesenkt, aber der Erreger nicht eliminiert werden (KORMENDY 1994). Da-
hingegen stieg die Anzahl positiver und verdächtiger Reaktionen mit bovinem und aviärem
Tuberkulin im Tuberkulintest.
2.4.3 SANIERUNG
In Deutschland wurden von einigen Bundesländern Bestrebungen zur Sanierung Pa-
ratuberkulose-infizierter Rinderbestände eingeleitet.
In Nordrhein-Westfalen ist seit 1992 eine „Richtlinie für die Sanierung von mit Paratuberku-
lose infizierten Rinderbeständen“ in Kraft.
In Niedersachsen wird seit 1990 ein durch die Niedersächsische Tierseuchenkasse geförderter
Versuch zur Bestandssanierung Paratuberkulose-infizierter Milchviehherden durchgeführt
(REHM 1999). Die „Satzung über die Gewährung von Beihilfen bei der Paratuberkulose der
Rinder“ wurde resultierend aus den Ergebnissen im Jahr 1998 überarbeitet und genehmigt.

Schriftum
24
2.5 Nachweis und Diagnose
2.5.1 DIREKTER ERREGERNACHWEIS
2.5.1.1 Mikroskopischer Erregernachweis
Der mikroskopische Erregernachweis wird als schnell durchzuführende Methode bei
der Untersuchung von Kot- und Rektumschleimhautproben sowie postmortal bei der Untersu-
chung von Mesenteriallymphknoten angewendet. Zur Darstellung von Mykobakterien wird
die ZIEHL-NEELSEN Färbung (DREWS 1983) als Methode der Wahl angesehen. Charakte-
ristisch für M. paratuberculosis sind schlanke, deutlich rot gefärbte Stäbchen, die in Nestern
liegen. Diese typische Lagerung ist durch die Ausbildung interzellulärer Filamente bedingt
(MERKAL 1973). Einzeln liegende, säurefeste Stäbchen können bei der mikroskopischen
Untersuchung nicht gewertet werden, da es sich meist um atypische, saprophytäre Mykobak-
terien handelt, die auch im Kot gesunder Tiere vorkommen. Die Mikroskopie eignet sich zur
Bestätigung eines klinischen Befundes. Sie ist aber aufgrund mangelnder Sensitivität zur Er-
kennung subklinisch erkrankter Tiere ungeeignet (MERKAL et al. 1968). Andere Autoren
empfehlen das mikroskopische Verfahren nicht nur aufgrund der schnellen Durchführbarkeit,
sondern auch zur Absicherung der Diagnose bei klinisch erkrankten Tieren in Verbindung mit
serologischen Methoden (SCHLIESSER und SCHAAL 1984). Der direkte mikroskopische
Nachweis von M. paratuberculosis gelingt allerdings nur in ca. 25 bis 35 % der kulturell-
positiv getesteten Kotproben, bzw. 60 % der kulturell positiv getesteten Lymphknoten
(HIETALA 1992).
2.5.1.2 Kultureller Erregernachweis
Erschwerend für die Anzüchtung von M. paratuberculosis in vitro sind die hohen
Nährbodenansprüche des Erregers und sein extrem langsames, strikt an das Vorhandensein
von Mycobactin gebundenes Wachstum. Wegen der langen Bebrütungszeit müssen Begleit-
keime durch Zusatzstoffe im Nährboden gehemmt werden. Auch das Probenmaterial muß vor
der Beimpfung dekontaminiert werden.

Schriftum
25
Die kulturelle Untersuchung von Ileocaecallymphkoten gilt bis heute als der zuverlässigste
Test für den Nachweis von M. paratuberculosis und wird deshalb auch als „Goldstandard“
bezeichnet (MERKAL et al. 1987). Allerdings birgt diese Methode keine 100 % Sensitivität
(WHITLOCK et al. 1997). So gelang in 18 % der Kühe, die einen positiven Befund in der
kulturellen Kotuntersuchung aufwiesen, kein Nachweis aus den Ileocaecallymphknoten.
Bei der kulturellen Anzucht werden flüssige oder feste Nährmedien mit und ohne Zusatz von
Mycobactin mit Lymphknotenmaterial inokuliert und für mindestens 3 Monate bei 37°C be-
brütet. Die Diagnose, daß es sich um M. paratuberculosis handelt, kann gestellt werden, wenn
langsam wachsende, Mycobactin-abhängige, in Haufen liegende, säurefeste Stäbchen nach-
gewiesen werden.
Der kulturelle Nachweis des Erregers in Kotproben gelingt nur, wenn mindestens 10-100
vermehrungsfähige Keime pro Gramm Kot vorhanden sind (WHIPPLE et al. 1992). Der
Nachweis von M. paratuberculosis im Kot von erkrankten Tieren gilt als 100 % spezifisch für
das Vorliegen der Erkrankung (COLLINS 1996). In einer anderen Studie konnte demonstriert
werden, daß ein kultureller Nachweis der Darmpassanten aus dem Kot nach oraler Aufnahme
des Erregers in größeren Mengen möglich ist (SWEENEY et al. 1992a). Die Sensitivität des
kulturellen Kotnachweises wird auf ca. 50 % geschätzt, da ca. die Hälfte der subklinisch infi-
zierten Tiere den Erreger nicht oder nur intermittierend ausscheiden (COLLINS 1996; STA-
BEL 1997).
Die für den kulturellen Nachweis von M. paratuberculosis aus dem Kot beschriebenen Ver-
fahren unterscheiden sich in der Art der Dekontamination zur Unterdrückung der Begleitflora,
in der eventuell durchgeführten Konzentrierung des Erreger durch Sedimentation, Zentrifuga-
tion oder Filtration und der Art des verwendeten Anzuchtmediums (KIM et al. 1989; RIDGE
1993; STABEL 1997; WHIPPLE et al. 1992).
Zur Reduzierung der Begleitflora wird am häufigsten Hexadecylpyridiniumchlorid (HPC)
verwendet (COLLINS 1996). Aber auch Oxalsäure, NaOH und Natriumhypochlorid werden
eingesetzt. Die Zentrifugation der Proben erhöht die Kontaminationsgefahr mit Begleitkeimen
(WHIPPLE et al. 1992). Dahingegen kann ein Sedimentationsverfahren Verluste der Kulturen
durch Kontaminationen verringern.
Die Anzucht der Kulturen erfolgt auf Herrold’s egg yolk Medium ohne und mit Mycobactin-
zusatz (HEY und HEYM). Inzwischen sind Medien auch kommerziell erhältlich (3.1.1).

Schriftum
26
Eine Alternative zur klassischen Kultivierung stellt das BACTEC System1 dar. Das dort ein-
gesetzte Medium arbeitet auf der Basis eines Flüssigmediums, dem 14C-markiertes Palmitat
zugesetzt ist. Das Wachstum von M. paratuberculosis kann anhand der Freisetzung von 14CO2
gemessen werden. Diese Technik in Kombination mit einem Konzentrierungsverfahren er-
laubt einen schnelleren und sensitiveren Nachweis als die klassische Kultivierung (COLLINS
1996; SOCKETT et al. 1992a; WHITTINGTON et al. 1998). Ein Nachteil der Methode, die
Erzeugung von radioaktiven Abfall, wird durch ein neues BACTEC System ausgeschaltet2.
Dieser Vollautomat mißt die Abnahme der O2-Konzentration durch Veränderung von Fluo-
reszenzsignalen in einem modifizierten Middlebrook-Medium, die aufgrund des Wachtums
der Mykobakterien entsteht. Die kulturelle Diagnostik wird mittels dieses Systems bei klini-
schen Materialien bis auf 24 Tage verkürzt (KÜHN und HELBING 2000).
2.5.2 SEROLOGISCHE VERFAHREN
2.5.2.1 Komplementbindungsreaktion (KBR)
Die KBR ist der erste serologische Test, der zur Diagnostik der Paratuberkulose be-
schrieben wurde (BANG und ANDERSEN 1913; TWORT und INGRAM 1912). Bei der
KBR handelt es sich um eine der ältesten serologischen Diagnostikmethoden, und entspre-
chend zahlreich und unterschiedlich sind die Angaben über Spezifität und Sensitivität. Wäh-
rend in einer Studie von einer niedrigen Spezifität und Sensitivität ausgegangen wird
(COCITO et al. 1994), halten andere Autoren die KBR für durchaus vergleichbar mit den
anderen serologischen Verfahren (COLLINS 1996). Die mangelnde Sensitivität ist der
Schwachpunkt der KBR, so daß dieses Verfahren nur als Screening-Test angewendet werden
sollte (BELLETTI et al. 1992). In Vergleichsuntersuchungen konnte die KBR die Sensitivität
von neueren ELISA-Systemen nicht erreichen (MCNAB et al. 1991a).
Die KBR ist international anerkannt, und ein negatives Testergebnis ist in vielen Ländern
Voraussetzung für den Import von Rindern (COCITO et al. 1994; COLLINS und SOCKETT
1 BACTEC 460 TB Becton Dickinson Laboratories, Inc., Sparks, MD2 BACTEC MGIT 960 Becton Dickinson Laboratories, Inc., Sparks, MD

Schriftum
27
1993). Der Test ist nicht tierartspezifisch und durch seinen komplexen Aufbau nicht interna-
tional standardisiert.
2.5.2.2 Agargelimmunodiffusionstest (AGIDT)
Mit diesem einfachen und kostengünstigen Verfahren kann mit geringem Aufwand in-
nerhalb von 24 bis 48 Stunden eine Diagnose gestellt werden. Aber bei dem Vergleich von
vier serologischen Testverfahren konnte bei einer Spezifität des Tests von 100 % nur eine
Sensitivität von 26 % - die niedrigste Sensitivität aller untersuchten Verfahren – erreicht wer-
den (SOCKETT et al. 1992b). Bei einem weiteren Vergleich des AGIDT mit einem ELISA-
Verfahren wurde ebenfalls eine niedrigere Sensitivität des AGIDT ermittelt (CLARKE et al.
1996).
2.5.2.3 Enzyme-Linked Immunosorbent Assay (ELISA)
2.5.2.3.1 Nachweis von Antikörpern gegen M. paratuberculosis in Blut
Der diagnostische Wert des ELISA bei der Paratuberkulose ist hoch, da die Tiere lan-
ge vor dem Auftreten klinischer Symptome erkannt werden. Der Antikörperspiegel steigt
dann bis zum Auftreten klinischer Anzeichen an (COLLINS 1996).
Diese ELISA-Technik wurde erstmals im Jahr 1971 beschrieben (ENGWALL und PERL-
MANN 1971). Zur serologischen Diagnostik der Paratuberkulose wurde das Verfahren sieben
Jahre später eingeführt (JØRGENSEN und JENSEN 1978).
In den darauffolgenden Jahren wurden einige unterschiedliche ELISA-Verfahren entwickelt.
Bei der Erkennung subklinisch erkrankter Tiere weisen viele der Tests eine nur geringe Sen-
sitivität und Spezifität auf (BECH-NIELSEN et al. 1992; COLLINS et al. 1991). Die Spezi-
fität konnte durch Präabsorption der diagnostischen Seren mit einer Suspension von M. phlei
gesteigert werden. (YOKOMIZO et al. 1985). Andere Autoren demonstrierten, daß die Präab-
sorption mit M. phlei keine wesentlichen Vorteile erbringt (HILBINK et al. 1994).
Die beschriebenen ELISA-Methoden unterscheiden sich hinsichtlich der Präparation und der
Qualität des Antigens, welches zum Beschichten der Mikrotiterplatten benutzt wird. Antige-
ne, die einige Autoren (COLLINS und SOCKETT 1993; YOKOMIZO et al. 1988) in ihren

Schriftum
28
Arbeiten einsetzten, wurden aus einem M. paratuberculosis Stamm 18 isoliert, der sich später
aufgrund molekularbiologischer Differenzierung als M. avium Stamm 18 herausstellte
(CHIODINI 1993). Ein rekombinantes Polypeptid wurde als Antigen in einem anderen ELI-
SA-System eingesetzt (VANNUFFEL et al. 1994). Es handelt sich hierbei um ein immunoge-
nes 34-kDA-Protein. Die Kreuzreaktionen mit M. bovis und dem nahverwandten M. avium
gingen deutlich zurück (DE KESEL et al. 1993).
Auch ein Zellwandbestandteil, das Lipoarabinomannan (LAM), wird als Antigen bei einigen
ELISA-Systemen eingesetzt (JARK 1996; SUGDEN et al. 1986; SUGDEN et al. 1989;
SWEENEY et al. 1994).
Eine vergleichende Studie zeigte für den LAM-ELISA eine signifikant höhere Sensitivität als
für einen ELISA mit einem aus dem M. avium Stamm 18 isolierten Antigen (REICHEL et al.
1999).
Die drei zur Zeit in Deutschland kommerziell erhältlichen ELISA-Systeme sind die von Bun-
desinstitut für gesundheitlichen Verbraucherschutz und Veterinärmedizin (BgVV) zugelasse-
ne Testkits der Firmen Idexx, Wörrstadt (HerdChek™ Mycobacterium paratuberculosis),
Riemser Arzneimittel GmbH, Insel Riems (ELISA zur Serodiagnostik der bovinen Paratuber-
kulose) und Svanova, Schweden (Svanovir® Paratuberkulose ELISA).
Der „HerdChek™ Mycobacterium paratuberculosis“- ELISA hat, mit Bezug auf die kultu-
relle Kotuntersuchung, eine Sensitivität von 57 % und eine Spezifität von 99,7 %
(ROSSITER und BURHANS 1996). In diesem ELISA-System werden die Seren mit M.
phlei-Extrakt vorabsorbiert. Er gilt in den USA als das zur Zeit sensitivste serologische Test-
system (COLLINS 1996). Das im Test verwendete Festphasenantigen stammt aus dem M.
avium Stamm 18.
Der „ELISA zur Serodiagnostik der bovinen Paratuberkulose“ (Riemser Arzneimittel GmbH,
Insel Riems) beruht auf einem ELISA-System, das in einem Feldversuch mit 327 Kühen eine
Sensitivität von 57 % und eine Spezifität von 98 % aufwies (MILNER et al. 1990). Auch in
diesem ELISA-System werden die Seren mit M. phlei-Extrakt vorabsorbiert.
Der „Svanovir® Paratuberkulose ELISA“, ein auf LAM basierender ELISA, wurde als einzi-
ger der drei ELISA-Systeme mit Bezug auf die kulturelle Untersuchung der Ileocaecal-
lymphknoten evaluiert. Er hat in ersten Untersuchungen eine scheinbare Spezifität von 99 %
und eine scheinbare Sensitivität von 93,75 % (JARK 1996; JARK et al. 1997).

Schriftum
29
2.5.2.3.2 Nachweis von Antikörpern gegen M. paratuberculosis in Milch
Ein ELISA zur Detektion von Antikörpern gegen M. paratuberculosis in Milchproben
stellt aufgrund der leichten Probennahme eine kostengünstige Alternative zum Blut-ELISA
dar (RICHARDS 1990).
Auf der Basis eines LAM-ELISA konnte eine gute Korrelation zwischen Blut- und Milch-
ELISA ermittelt werden (SWEENEY et al. 1994). Milch- und Blutproben von 764 Kühen aus
13 Betrieben, von denen 2 Betriebe bisher keine Paratuberkulose-Erkrankungen zu verzeich-
nen hatten, wurden mittels LAM-ELISA getestet. Ebenso wurden Kotproben dieser Tiere
kulturell auf M. paratuberculosis untersucht. Der kulturelle Nachweis von M. paratuberculo-
sis wurde als „Goldstandard“ angesehen. Die Inter-Assay-Variation betrug für den Milch-
ELISA 0,19 und für den Blut-ELISA 0,15. Die Validierung des Milch-ELISA erfolgte anhand
der ROC-Kurve. Es ergaben sich bei einer Sensitivität von 50 % bzw. 60 % eine Spezifität
von 87 % bzw. 83 %. Der Korrelationskoeffizient betrug r = 0,71. Dies weist auf eine gute
Korrelation zwischen Blut- und Milch-ELISA hin.
Ein anderer Milch-ELISA wurde auf der Basis eines Protoplasma-Antigens (PPA-3) entwik-
kelt (HARDIN und THORNE 1996). Die Autoren konnten nachweisen, daß eine Vorabsorp-
tion der Milch mit M. phlei-Extrakt keine signifikanten Unterschiede der ELISA-Ergebnisse
ergab. Die Korrelation der Ergebnisse des Milch- und Blut-ELISA war sehr gering (r = 0,08).
Anhand von Untersuchungen an Tankmilchproben mittels eines kommerziell erhältlichen
ELISA-Systems wurde die Prävalenz in dänischen Milchviehherden bestimmt (NIELSEN et
al. 2000). Als Antigen wurden Bestandteile des M. avium Stammes 18 eingesetzt. Die Auto-
ren ermittelten eine Sensitivität von 97 % und eine Spezifität von 86 %. Der ELISA war nur
wenig zuverlässig, da geringe Schwankungen des „Cut off“ Wertes die Prävalenz extrem ver-
änderten.
2.5.3 POLYMERASE KETTENREAKTION (PCR)
Die PCR ist eine schnelle Methode zur „In-Vitro-Amplifikation“ von Nukleinsäuren.
Diese ermöglicht es, einen bestimmten Zielabschnitt (Target) einer vorliegenden Nukleinsäu-
re (Template-DNA), z. B. eines Krankheitserregers, selektiv zu vermehren. Das Orginalproto-
koll für die PCR wurde erstmals 1985 veröffentlicht (SAIKI et al. 1985).

Schriftum
30
2.5.3.1 Lysis und DNA-Präparation von Mykobakterien für die PCR
Die mechanische Lysis von Mykobakterien mit Glas- oder Zirkoniumbeads in einem
„Bead beater“ wird von mehreren Autoren beschrieben (BLEUMINK-PLUYM et al. 1994;
CHALLANS et al. 1994; COLLINS et al. 1993). Die Bakteriensuspension wird mit den
Beads gemischt und in dem „Bead beater“ für einige Minuten bei einer Frequenz von 100 Hz
kräftig geschüttelt. Die Zellwände werden so zerstört und die DNA freigesetzt.
Auch alleiniges Kochen der Suspension für 15 Minuten (MOSS et al. 1991) bzw. 20 Minuten
(HURLEY et al. 1993) setzte geeignete DNA zur Verwendung in der PCR frei.
Andere Autoren präparierten die DNA von Mykobakterien enzymatisch und setzten dazu
Subtilisin, Lysozym sowie SDS und Pronase ein (VISUVANATHAN et al. 1989). Ebenfalls
eine enzymatische Methode mit Lysozym und Proteinase K, bei der nach Cäsiumchloridbe-
handlung 30 Volumina destilliertes Wasser hinzugegeben wurde, um durch osmotischen
Schock die Zellwände zu sprengen, ergab eine Ausbeute von 1,2-2 mg DNA/g Zellfeuchtge-
wicht (BOSE et al. 1993).
2.5.3.2 DNA-Extraktion
Meist wird das Untersuchungsmaterial nach dem Zellaufschluß einer weiteren Be-
handlung zur Reinigung und gleichzeitigen Konzentrierung der Nukleinsäuren unterzogen.
Eine Chloroform-Phenolextraktion ist die klassische Methode (SAMBROOK et al. 1989). Die
hydrophilen DNA-Moleküle reichern sich in der wäßrigen Phase an und können aus dieser
zurückgewonnen werden. Die Präzipitation der gelösten DNA erfolgt mit Hilfe von Ethanol.
Zum Nachweis von M. tuberculosis in Gewebeproben wurde nach enzymatischen Aufschluß
der Proben die DNA durch Chloroform-Phenolextraktion mit anschließender Ethanolfällung
gewonnen (TAN et al. 1997).
Die Grundlage für die Aufreinigung von Nukleinsäuren über eine Silikamatrix ist die Beob-
achtung, daß DNA in Gegenwart chaotroper Agentien wie Natriumjodid und Natriumperchlo-
rat oder Guanidin-Thiocyanat bzw. –hydrochlorid an Silika (Siliziumdioxid)- bzw. Glasparti-
kel bindet (BOOM et al. 1990). Liegen hohe Konzentrationen chaotroper Stoffe vor, werden
Nukleinsäuren an die Silika-Partikel gebunden und lösen sich von diesen bei niedrigen Kon-
zentrationen der chaotropen Substanzen wieder ab. Durch diese selektive Bindung kann DNA
von Störstoffen befreit und danach durch Schwächung der Bindung wieder von der Matrix

Schriftum
31
gelöst werden. Trotz dieser DNA-Aufreinigungsmethode wurde in einer weiteren Untersu-
chung zum Nachweis von M. tuberculosis demonstriert, daß einige Proben noch PCR-
Inhibitoren enthielten (NOORDHOEK et al. 1995).
Zunehmend werden auch DNA-Extraktionskits auf der Basis von Chloroform-Extraktion oder
DNA-bindenden Matrices angeboten und eingesetzt. Die DNA-Extraktionskits sind oft zeit-
sparend und bieten aufgrund des standardisierten Verfahrens eine gute Reproduzierbarkeit.
DNA-bindende Matrices sind darüber hinaus mit einer geringeren anfallenden Menge an toxi-
schem Sondermüll verbunden. Es werden DNA-Extraktionskits für nahezu alle DNA-haltigen
Medien angeboten.
2.5.3.3 Aufreinigung von Milchproben für die PCR
Um aus dem komplexen, die PCR vollständig hemmenden Medium Milch (POWELL
et al. 1994) Erreger-DNA zu isolieren, werden mehrere Möglichkeiten genutzt.
Zum Nachweis von Coxiella burnettii aus Milch wurde nach Proteinase K-Abbau der Protein-
strukturen die DNA mittels Chloroform-Phenolextraktion gewonnen (WILLEMS et al. 1994).
Die Autoren erreichen mit dieser Methode eine Nachweisgrenze von einem Erreger pro ml
Milch. Ebenfalls zum Nachweis von Coxiella burnettii aus Milch wurde von einem anderen
Autor nach Beschallung, Entrahmung, Auslabung und Ultrazentrifugation der Molke zur
DNA-Extraktion ein kommerziell erhältliches DNA-Extraktionskit (Qiamp® Tissue Kit, Qia-
gen, Hamburg) eingesetzt (EDINGLOH et al. 1999; EDINGLOH 1999). Die Nachweisgrenze
lag bei 4 Erregern pro ml Milch.
Dasselbe DNA-Extraktionskit wurde in einer Arbeit zum Nachweis von Mycobacterium tu-
berculosis in Milch, Lymphknoten und Nasenabstrichen eingesetzt (VITALE et al. 1998). Die
Milchproben wurden enzymatisch vorverdaut und die DNA laut Anleitung extrahiert.
Erstmals im Jahr 1996 wurde der Nachweis von M. paratuberculosis in Milchproben be-
schrieben (MILLAR et al. 1996). In dieser Untersuchung wurden die Milchproben durch
Zentrifugation in Pellet, Fett und Milchserum fraktioniert. Die Mykobakterien wurden durch
20 minütiges Kochen der unterschiedlichen Milchfraktionen aufgeschlossen und der Über-
stand nach Zentrifugation direkt in der PCR eingesetzt. Andere Autoren beschrieben den
Nachweis von M. paratuberculosis in Milchproben, die enzymatisch vorbehandelt wurden
(HERMAN et al. 1999).

Schriftum
32
2.5.3.4 Ausschluß falscher Ergebnisse in der PCR
Häufig auftretende Probleme beim Einsatz der PCR sind falsch negative Ergebnisse
infolge einer Hemmung der Polymerase und falsch positive Ergebnisse durch Kontamination
der PCR-Ansätze mit der gesuchten DNA.
Eine zentrale Rolle bei der Hemmung der Polymerase spielt dabei die Verfügbarkeit des Co-
faktors Magnesium (ROLFS et al. 1992). Hemmstoffe binden häufig freies Magnesium. Zu
den Hemmstoffen, die aus der Probe bei der Aufbereitung nicht ausreichend eliminiert wur-
den, gehören EDTA, Proteine, Heparin u.v.a.. Auch können Reste der Lösungen, die zur
DNA-Isolierung eingesetzt wurden, wie SDS und Proteinase K, die PCR empfindlich stören.
Zwar kann die Konzentration der Hemmstoffe durch Verdünnung der DNA-Lösung gesenkt
werden, aber somit sinkt auch die Sensitivität der PCR. Empfehlenswert ist die erneute Auf-
reinigung der Probe. Als Maßnahmen zur Kontrolle der PCR-Ansätze sollte zu jedem Ansatz
ein Vergleichsansatz mit einem Gemisch aus Proben-DNA und zugesetzter amplifizierbarer
DNA erstellt werden.
Durch Kontaminationen sowie durch unspezifische Reaktionen kann es zu falsch positiven
Ergebnissen kommen. Die Kontaminationsgefahr ist besonders hoch in der Diagnostik, da es
zu einer Anreicherung von DNA im Labor kommt. Die Kontrolle der PCR-Ansätze kann
durch einen Ansatz, dem statt DNA Wasser zugesetzt wurde, gewährleistet werden.
Daraus ergibt sich, daß vor allem für den Erregernachweis aus klinischem Probenmaterial
eine genaue Kenntnis über Hemmstoffe und Kontaminationsquellen erforderlich ist (IEVEN
und GOOSSENS 1997). Es kommt bei einer möglichst einfach gestalteten Prozedur der
DNA-Extraktion auf die sichere Reproduzierbarkeit der Ergebnisse an.
2.5.3.5 PCR zum Nachweis des Erregers aus klinischem Untersuchungsmaterial
Ein spezifischer Nachweis von M. paratuberculosis in Kot mittels PCR stellt zwar ei-
ne schnelle Alternative für den Nachweis von mittel- bis hochgradig ausscheidenden Kühen
dar; er ist jedoch als Nachweis von nur mit wenigen Mykobakterien belastetem Kot ungeeig-
net, da mindestens 104 KBE/g Kot zum Nachweis benötigt werden (VARY et al. 1990;
WHIPPLE et al. 1992).
Ein kommerziell erhältliches Kit (DNA-Probe®-Test, Idexx, Wörrstadt) mit dem ein IS900
Fragment einer Länge von 229 bp aus aufbereiteten Kotproben vervielfältigt und dann mit

Schriftum
33
einer Gensonde nachgewiesen wird, wurde in einer vergleichenden Untersuchung als der
kulturellen Untersuchung unterlegen gewertet, da eine Sensitivität von 55 % gegenüber 89 %
und 74 % in zwei verschiedenen kulturellen Verfahren zu verzeichnen war (SOCKETT et al.
1992a). Andere Autoren konnten in Untersuchungen demonstrieren, daß mittels DNA-
Probe®-Test lediglich ein Drittel der kulturell positiven Kotproben identifiziert werden konn-
ten (WHIPPLE et al. 1992). Neuere Untersuchungen unterstützen diese Ergebnisse (ZIMMER
et al. 1999). In einer anderen Untersuchung konnte in der PCR bei allen Proben mit einem
Gehalt von > 1600 KBE/g Kot M. paratuberculosis-spezifische DNA nachgewiesen werden.
Bereits ab einem Gehalt von 480 KBE/g Kot sank die Sensitivität allerdings auf 60 %
(COLLINS et al. 1993).
Für einen weiteren PCR-gestützten Nachweis von M. paratuberculosis aus dem Kot ermittel-
ten die Autoren eine Sensitivität von 34 % und eine Spezifität von 100 % (SOCKETT et al.
1992a).
Der Nachweis des IS900 von M. paratuberculosis aus Milch wurde erstmals 1996 beschrie-
ben (MILLAR et al. 1996). Die Autoren untersuchten 312 pasteurisierte Milchproben in der
Zeit von September 1991 bis März 1993 und darüber hinaus noch Milchproben von sicher
nicht infizierten und infizierten Kühen. Die Nachweisgrenze lag bei 200 bis 300 Keimen pro
ml Milch. Bei den künstlich mit lebenden Erregern kontaminierten Milchproben konnte nach
Zentrifugation der Erreger mittels PCR im Pellet oder im Rahm nachgewiesen werden. Den
Autoren war es möglich in 7 % der pasteurisierten Milchproben M. paratuberculosis nachzu-
weisen.
Bei Einsatz einer nested-PCR kann die Nachweisgrenze auf 30 KBE/ml Milch gesenkt wer-
den (HERMAN et al. 1999). Auch hier konnte der Erreger im Milchfett nachgewiesen wer-
den.
Für einen PCR-gestützten Nachweis des IS900 von M. paratuberculosis aus Lymphknoten
wurde eine Methode zur Zelllyse auf der Basis der Zerstörung des Erregers mit Zirkonium-
beads in einem „Bead beater“ beschrieben (CHALLANS et al. 1994). Es folgt eine Chloro-
form-Phenolextraktion. Die Nachweisgrenze lag bei 35-45 Keimen/g Gewebe.

Material und Methoden
34
3 Material und Methoden
Rezepte für Lösungen, Puffer und Medien sind jeweils im Anhang aufgeführt.
3.1 Material
3.1.1 BAKTERIENSTÄMME UND MEDIEN
Die Untersuchungen wurden mit dem M. paratuberculosis-Stamm 6783 aus der
Stammsammlung des Instituts für Mikrobiologie und Tierseuchen durchgeführt. Es handelt
sich hierbei um einen Mykobakterienstamm, der in Norddeutschland isoliert wurde. Der
Stamm zeigt ein mycobactinabhängiges Wachstum und kann zusätzlich durch den Nachweis
des Insertionselementes IS900 mittels PCR als M. paratuberculosis identifiziert werden. Der
Bakterienstamm ist bei der Deutschen Gesellschaft für Mikroorganismen und Zellkulturen
(DSM, Braunschweig) unter der DSM-Nr. 10129 hinterlegt.
Tabelle 1: Herkunft der Bakterienstämme
Isolat Herkunft
M. avium ssp. paratuberculosis
Isolat 6783
klinisches Isolat
(DSM 10129)
M. avium ssp. avium
DSM 44158
Deutsche Gesellschaft für Mikroor-
ganismen und Zellkulturen (DSM,
Braunschweig)
M. phlei
NCTC 8151
National Collection of Type Cultu-
res (NCTC, Central Public Health
Laboratory, Colindale, UK)
Alle Isolate wurden zunächst aus Stammkulturen des Institutes auf Middlebrook Me-
dium (9.2) angezogen. M. paratuberculosis 6783 und M. phlei wurden dann in Watson-Reid-
Medium (9.2), M. avium 44158 in Long Medium (9.2) subkultiviert.

Material und Methoden
35
3.1.2 PROBENGEWINNUNG
3.1.2.1 Blutproben
Blutproben wurden durch Punktion der Vena coccygea mediana gewonnen und nach
etwa einer Stunde Lagerzeit bei Raumtemperatur bei mindestens 1500 x g3 für 10 Minuten
zentrifugiert. Das Serum wurde abpipettiert und bei –20°C4 längerfristig gelagert.
3.1.2.2 Milchproben
Es wurden zweimal 10 ml als Viertelgemelkprobe zu Beginn der Melkzeit steril ent-
nommen. Milchproben wurden bis zu einer Woche bei 4°C und längerfristig bei –20°C4gelagert.
3.1.2.3 Kotproben
Kotproben wurden aus dem Enddarm der Tiere entnommen. Die Proben wurden bei
–20°C4gelagert.
3.1.2.4 Organproben
Nach der Schlachtung wurden die Ileocaecallymphknoten entnommen. Die
Lymphknoten wurden bei –20°C4 gelagert.
3 C312, Jouan, Unterhaching4 Liebherr, Ochsenhausen

Material und Methoden
36
3.2 Methoden
3.2.1 ANZUCHT DES ERREGERS
Für die Kultivierung von M. paratuberculosis wurden 800 ml-Zellkulturflaschen
(NUNC, Wiesbaden) mit Watson-Reid-Medium verwendet. Von einer Reinkultur wurden
Bakterien mit einer Impföse entnommen und in ca. 2 ml n-Pentan vereinzelt. Die Mykobakte-
riensuspension wurde mit einer Glaspipette auf die Oberfläche des Mediums aufgetragen. Die
Bakterien blieben nach der Verdunstung des n-Pentans an der Oberfläche liegen. Das
Wachstum vollzog sich inselartig. Die Inkubation erfolgte unter aeroben Bedingungen bei
37°C5. Nach ca. zwei Monaten konnten die Bakterien geerntet werden.
3.2.2 ANZUCHT ANDERER MYKOBAKTERIEN
M. avium wurde in Flüssigmedium nach Long und M. phlei in Watson-Reid-Medium
exclusive Mycobactin bei 37°C aerob angezüchtet5.
3.2.3 BESTIMMUNG DER ERREGERDICHTE
(DREWS 1983)
Die Bakterienzelldichte wurde durch Messung der optischen Dichte von Flüssigkultu-
ren bei 660 nm (OD660) gegen unbeimpftes Medium ermittelt6. Nach Ernte der Mykobakterien
wurde die OD660 mit sterilem PBS (9.8) auf 0,1 eingestellt. Die Zählung erfolgte in der
THOMA-ZEISS-Zählkammer7. Die Zellzahl wurde gemäß der Formel: Zellzahl gesamt x
15,625 x Vorverdünnung x 1000 = Bakterien pro ml ermittelt.
5 Brutschrank, Memmert, Schwabach6 Spectralphotometer Ultrospec III, Pharmacia, Freiburg7 Mikroskop: Dialux 22 EB, Leitz, Wetzlar

Material und Methoden
37
3.2.4 MIKROSKOPISCHER UND KULTURELLER NACHWEIS VON M. PARATUBERCULOSIS
3.2.4.1 Mikroskopie
Die mikroskopische Untersuchung von Kotproben und Lymphknoten auf säurefeste
Stäbchen wurde mittels ZIEHL-NEELSEN Färbung durchgeführt. Die Kotproben wurden
dünn auf einen Glasobjektträger ausgestrichen, luftgetrocknet und hitzefixiert. Bei den
Lymphknotenproben wurde eine frische Schnittfläche des Lymphknotens auf einen Objekt-
träger ausgestrichen, ebenfalls luftgetrocknet und hitzefixiert. Die Ausstriche wurden nach
ZIEHL-NEELSEN (DREWS 1983) gefärbt und mikroskopiert7.
3.2.4.2 Kultur
Zur kulturellen Untersuchung von Kotproben wurden ca. 3 g Kot in 5 ml 0,75 % HPC
(Hexapyridiumchlorid)-Lösung suspendiert. Nach Inkubation über Nacht bei Raumtemperatur
wurden drei Schrägagarröhrchen (Enclitt, Oelzschau), eines mit Löwenstein-Jensen-Agar und
zwei mit Mycobactin-Zusatz zur Überprüfung der Mycobactinabhängigkeit, mit dem Boden-
satz beimpft.
Die Anzucht von M. paratuberculosis aus Lymphknoten erfolgte durch Ausstreichen frischer
Schnittflächen von abgeflammten und unter sterilen Bedingungen aufgeschnittenen
Lymphknoten auf die Nährbodenoberflächen. Sofern sich Kontaminationen zeigten, wurden
die Lymphknoten wie Kotproben mit 0,75 % HPC dekontaminiert.
Sichtbare Kolonien traten nach frühestens zehn Wochen auf und wurden mikroskopisch7
mittels ZIEHL-NEELSEN Färbung auf das Vorhandensein säurefester Stäbchen untersucht.

Material und Methoden
38
3.2.5 ANTIGENHERSTELLUNG
(JARK 1996; RINGENA 1995)
M. paratuberculosis wurde hierzu in Flüssigmedium nach Watson-Reid angezüchtet.
Nach ca. 3 Monaten wurden die Bakterien durch Zentrifugation bei 3000 x g8 geerntet. Das
Pellet wurde gewogen und mit 1 ml A. dest pro 0,6 g Bakterienfeuchtmasse resuspendiert und
bei 100°C im Wasserbad für 10 Minuten inaktiviert. Die Mykobakteriensuspension wurde
mittels French Press9 bei einem Druck von 15.000 psi aufgeschlossen. Die groben Zellbe-
standteile wurden durch Zentrifugation (6500 x g) pelletiert. Der erhaltene Überstand wurde
mit dem identischen Volumen Proteinase K-Puffer (9.7) versetzt. Die Proteinase K wurde 15
Minuten bei 37°C10 inkubiert und danach in einer Konzentration von 100 mg/g Bakterienma-
sse eingesetzt. Der Verdau wurde für eine Stunde im Hitzeblock bei 37°C durchgeführt und
durch Erhitzen auf 100°C für 5 Minuten beendet. Die Suspension wurde erneut bei 6500 x g
zentrifugiert. Es folgte eine Filtration (Porengröße 0,45µm, Millex®HA, Millipore) des Über-
standes. Das Filtrat wurde mittels Ultrafiltration weiter aufgereinigt (Centriplus®30, AMI-
CON®). Das Retentat wurde bei -80°C gelagert.
3.2.6 PRÄPARATION VON FAB-FRAGMENTEN
(HARLOW und LANE 1988)
Die Immunglobuline der IgG Klasse des positiven Kontrollserums aus dem Svanovir®
Paratuberkulose ELISA-Testkit (Svanova, Schweden) wurden mittels Papain gespalten und
elektrophoretisch aufgetrennt, um im Western Blot zu prüfen, an welches Fragment der
monoklonale Antikörper des Testkits (Svanova, Schweden) bindet. Hierzu wurden 2 ml des
Serums mittels dreimaliger Zentrifugation8 in einem Zentrifugations Filter (Centriplus®30,
AMICON®) in den Puffer für die Papain-Spaltung (9.7) umgepuffert und das Volumen auf
200 µl aufgefüllt. 100 µl immobilisiertes Papain wurden zweimal mit dem Puffer gewaschen
und 100 µl IgG-Lösung hinzugegeben. Es folgte eine Inkubation unter Schütteln für 5 Stun-
8 Sorvall RC, Du Pont Ind., Bad Homburg9 French Pressure Cell, American Instrument Company, Inc., Silver Spring, Maryland

Material und Methoden
39
den bei 37°C. Danach wurden 600 µl 10 mM Tris-HCl pH 7,5 zugegeben, der Verdau ge-
mischt und zentrifugiert (9000 x g, 5 Minuten). Die ABICAP ProteinA-Mikroaffinitätssäule
wurde mit 10 mM Tris-HCl pH 7,5 äquilibriert und dann der Überstand auf die Säule appli-
ziert. Die nicht gebundenen Fab-Fragmente wurden mit 10 mM Tris-HCl pH 7,5 von der Säule
gewaschen. Gebundene Fc-Fragmente und nicht verdaute IgG-Moleküle wurden mit 450 µl
Elutionspuffer (9.7) eluiert und durch Vorlage von 50 µl Neutralisierungspuffer (9.7) neutrali-
siert.
Die Fab-Fragmente in der Waschlösung konnten durch einen Zentrifugations Filter (Centri-
plus®30, AMICON®) aufkonzentriert werden. Die Ausgangs-IgG-Lösung, die Fab-Fragmente
und die Fc-Fragmente wurden unter nicht reduzierenden Bedingungen aufbereitet und in zwei
SDS-Gelelektrophoresen (3.2.9) aufgetrennt. In einem Gel wurden die aufgetrennten Proteine
mittels Coomassie-Blau®-Färbung (3.2.9.2) dargestellt; von einem weiteren wurde ein We-
stern Blot (3.2.10) erstellt und mit dem monoklonalen Antikörper entwickelt.
3.2.7 DNA-PRÄPARATION VON MYKOBAKTERIEN
(BOSE et al. 1993)
Etwa 100 mg Bakterienpellet wurden zweimalig mit TE-Puffer (9.3) gewaschen. Dar-
aufhin wurde das gleiche Volumen gesättigte CsCl-Lösung mit 1 % Triton-X zugegeben.
Nach gründlichen Mischen (10 Minuten) wurde die Suspension mit dem 30fachen Volumen
A. dest. aufgefüllt. Nach Zentrifugation8 (4000 x g, 15 Minuten) wurde der Überstand mit
Lysozym (Endkonzentration 1 mg/ml) für eine Stunde auf Eis verdaut. Nach Zugabe von SDS
(Endkonzentration 0,5 %) und Proteinase K (Endkonzentration 100 µg/ml) folgte eine Inku-
bation für 4 Stunden im Wasserbad bei 55°C. Der Zellsuspension wurde 1 ml Guanidium-
thiocyanat (0,5 g/ml) zugegeben und bei 65°C für 30 Minuten unter Schütteln inkubiert. Dann
wurden die Zelltrümmer von der Lösung durch Zentrifugation (4000 x g, 15 Minuten) ent-
fernt. Aus dem Überstand wurde die DNA mittels Chloroform-Phenol-Extraktion gewonnen
(SAMBROOK et al. 1989).
10 Blockthermostat BT 100, Landgraf Hannover

Material und Methoden
40
3.2.8 BESTIMMUNG DER DNA-KONZENTRATION
(SAMBROOK et al. 1989)
Die DNA-Konzentration wurde spektralphotometrisch bestimmt6. Die DNA-Lösungen
wurden bei 260 und 280 nm gemessen. Die Meßwerte bei 260 nm erlaubten die Kalkulation
der DNA-Konzentration in der Probe. Mit Hilfe des Quotienten aus den bei 260 und 280 nm
gemessenen optischen Dichten wurde die Reinheit der DNA-Lösung abgeschätzt. Reine Prä-
parationen haben Werte zwischen 1,8-2,0.
3.2.9 SDS-POLYACRYLAMIDGELELEKTROPHORESE (SDS-PAGE)
(modif. nach LÄMMLI 1970)
Die elektrophoretische Auftrennung von Proteinen erfolgte mit einem Elektrophorese-
System11 unter Verwendung von 10 % Polyacrylamidgelen und Kämmen für 10 Proben (Bio-
Rad, München). Die entstandenen Proteinprofile wurden mittels Coomassie®-Blau angefärbt
und im Western Blot immunologisch charakterisiert. Die vorbereiteten Proben wurden in ei-
nem Volumen von 20 µl mit einer Mikroliterspritze12 in die Geltaschen appliziert. Die Elek-
trophorese erfolgte mit SDS-Laufpuffer (9.4) bei 150 V für 60 Minuten13.
3.2.9.1 Vorbereitung der Proben für die SDS-Page
Die Proben wurden 1:2 mit nicht reduzierenden Probenpuffer (2 x) versetzt (9.4), für
30 Minuten bei 37°C inkubiert und in die Geltaschen appliziert.
Als Molekulargewichtsstandard wurde ein vorgefärbter Protein-Standard mit den Molekular-
massen 19, 28, 34, 47, 81, 104 kDA (BioRad Prestained SDS-Page Standard®, Low Range)
nach Herstellerangaben präpariert und auf jedes Gel mit aufgetragen.
11 Mini-Protean II Elektrophorese-System®, BioRad, München12 Hamilton®, Sigma, München13 Powerpac 300 Power Supply, BioRad, München

Material und Methoden
41
3.2.9.2 Coomassie-Blau®-Färbung
Die elektrophoretisch in der SDS-Page aufgetrennten Proteine können mit einer
Nachweisgrenze von 100 ng/Bande mit dem Farbstoff Coomassie-Blau® sichtbar gemacht-
werden.
Rezepte für Färbe- und Entfärbelösung sind im Anhang aufgeführt (9.5). Die Gele wurden für
15 Minuten in der Färbelösung geschwenkt. Zur Entfärbung wurde die Färbelösung abgespült
und die Entfärbelösung mehrmals gewechselt. Die gefärbten Gele wurden konserviert14.
3.2.10 WESTERN-BLOTTING
Die mittels SDS-Page hergestellten Proteinprofile wurden elektrophoretisch auf eine
Membran transferiert und immunologisch nachgewiesen.
3.2.10.1 Transfer der Proteine im Tankblotting-Verfahren
(modif. nach TOWBIN et al. 1979)
Der Transfer der im SDS-Gel aufgetrennten Proteine, wurde mittels Transfer-Zelle15
auf eine Nitrozellulosemembran (Protan BA85 0,45 µm, Schleicher und Schuell, Dassel)
durchgeführt. Die Blotkassette wurde gemäß der Angaben des Herstellers im Submarine-
Verfahren zusammengebaut. Der elektrophoretische Transfer der Proteine erfolgte bei einer
angelegten Spannung von 50 V mit einer Eiskühlungseinheit innerhalb von 50 Minuten13.
3.2.10.2 Nachweis immunogener Proteine
(SAMBROOK et al. 1989)
Nach dem Transfer wurden die Proteine auf der Nitrozellulosemembran mit dem
monoklonalen Antikörper des Svanovir®-ELISA detektiert.
14 GelAir Drying System, BioRad, München15 Transfer-Zelle Mini Trans Blot, BioRad, München

Material und Methoden
42
Zur Blockierung unspezifischer Bindungsstellen wurde die Nitrozellulosemembran über
Nacht bei 4°C in Blockpuffer (9.6) inkubiert. Nach 3 x 5 Minuten waschen in Waschpuffer
(9.6) schloß sich die Inkubation für 1 Stunde bei Raumtemperatur mit dem monoklonalen,
HRP-konjugierten Antikörper (Verdünnung 1:500) an. Nach Inkubation wurde der Blot 2 x 5
Minuten mit Waschpuffer und 2 x 10 Minuten mit Substratpuffer (9.6) gewaschen. Die Ent-
wicklung nach Zugabe von Substrat wurde unter Sichtkontrolle durchgeführt und mit Lei-
tungswasser abgestoppt. Alle Wasch- und Inkubationsschritte wurden bei Raumtemperatur
auf einem Orbital-Schüttler16 vorgenommen.
Der Blot wurde zwischen Filterpapier (Gel-Blotting-Papier, Schleicher und Schuell, Dassel)
getrocknet.
3.2.11 AGAROSEGELELEKTROPHORESE
(SAMBROOK et al. 1989)
Für die Elektrophorese wurden 1,5 % Agarose-Gele hergestellt. Dem Gel wurden vor
dem Gießen 1 µl Ethidiumbromid [10 mg/ml] je 50 ml zugegeben. Als Laufpuffer diente 0,5
x TBE-Puffer (9.3). Die DNA-Proben wurden mit einem 1/10 Volumen 6 x Probenpuffer
(9.3) gemischt und 20 µl in die Geltaschen geladen. Als Größenstandard wurde ein 100 bp
Ladder (Gene Ruler™ 100 bp DNA Ladder, ready-to-use, MBI Fermentas, St. Leon Roth)
eingesetzt. Die Elektrophorese wurde bei 180 V für 90 Minuten in einer horizontalen Gel-
kammer17 durchgeführt13.
Das Gel wurde unter UV-Licht (254 nm) beurteilt und dokumentiert18.
16 3011, GFL, Burgwedel17 Gator™, Gelsystem Modell A2, peqlab, Erlangen18 MultiAnalyst-System, BioRad, München

Material und Methoden
43
3.2.12 ENZYME-LINKED IMMUNOSORBENT ASSAY (ELISA)
(JARK 1996)
Zur quantitativen Messung von bovinen Immunglobulinen gegen M. paratuberculosis
im Blutserum wurde der von der Firma Svanova, Schweden hergestellte und vom BgVV amt-
lich zugelassene Test (Svanovir® Paratuberkulose ELISA) verwendet.
3.2.12.1 Durchführung des Svanovir® Paratuberkulose ELISA (Svanova, Schweden) für
Blutserum
Das Testkit beinhaltet mit Lipoarabinomannan (LAM) beschichteten Mikrotiterplatten,
positives und negatives Kontrollserum und PBS-Tween(PBST)-Tabletten. Weiterhin befindet
sich im Kit Anti-Rind-IgG-HRP-Konjugat in lyophilisierter Form, als Substratlösung dient
Tetramethylbenzidin und als Stoplösung 2 M Schwefelsäure. Der Test wurde nach der beilie-
genden Gebrauchsanleitung durchgeführt19 20 21.
3.2.12.2 Auswertung des Svanovir® Paratuberkulose ELISA
(modif. nach WRIGHT et al. 1993)
Mit Hilfe eines PC-gestützten ELISA-Programmes22 wurden aus den gemessenen Ex-
tinktionen ELISA-Einheiten (EU) nach der Referenz-Standard-Methode berechnet.
Das positive Kontrollserum stellt das Referenzserum dar, welches eine fiktive ELISA-
Aktivität von 100 EU erhält. Die Extinktionen der zu untersuchenden Seren werden mit der
Extinktion des Referenzserums verglichen und damit eine zum Referenzserum relative ELI-
SA-Aktivität berechnet. Von den Extinktionswerten der positiven Kontrolle wurden die des
Leerwertes subtrahiert. Die korrigierten Extinktionswerte und die dazugehörigen Titerstufen
wurden dekadisch logarithmiert und die Regressionsgerade berechnet. Aus dieser Regressi-
19 Plattenschüttler: MTS 4, Omnilab, Gehrden20 ELISA-Washer: 96 PW, Tecan, Salzburg, Austria21 ELISA-Photometer: Spectr III, SLT, Crailsheim22 Tecan Easy Win®, Salzburg, Austria

Material und Methoden
44
onsgeraden wird die Steigung, der y-Achsenabschnitt und der Variationskoeffizient berech-
net. Voraussetzungen für die Auswertung einer Platte waren, daß mindestens drei Verdün-
nungsstufen des Referenzstandards meßbar waren, die berechnete Regressionsgerade eine
Steigung von 0,7-1,1 aufwies und der Variationskoeffizient nicht größer als 20 war. Nun wur-
den die Mittelwerte der Proben ebenfalls logarithmiert und die EU nach folgendem Algorith-
mus berechnet:
����
� −−•=−
dardtanS
dardtanSobePrSteigung
hnittAchsenabscy)ODlog(obePr
10
100Verdünnung)EU(EinheitenELISA
3.2.12.3 Bewertungsschema des Svanovir® Paratuberkulose ELISA
ELISA-Aktivitäten: ≤ 20 EU negativ
21-40 EU zweifelhaft
> 40 EU positiv
3.2.12.4 Vorbehandlung der Milchproben
(MANZ et al. 1986)
Die folgenden vier Methoden wurden miteinander verglichen:
1. Milch (10 ml) wurde für 36 Stunden bei 37°C inkubiert. Das Milchserum wurde durch
Zentrifugation (4000 x g)3 gewonnen.
2. Milch (10 ml) wurde für 48 Stunden bei 20°C inkubiert. Das Milchserum wurde durch
Zentrifugation (4000 x g)3 gewonnen.
3. Zur Milchserumgewinnung wurde 10 ml Milch, die auf 37°C erwärmt worden war, mit
Rennin (Sigma, München) in einer Konzentration von 1 IU versetzt. Die Inkubation er-
folgte bei 37°C für 30 Minuten. Das Milchserum wurde durch Zentrifugation (4000 x g)3
gewonnen.
4. 10 ml Milch wurden durch Zentrifugation (4000 x g)3 entrahmt (MACMILLAN et al.
1990).

Material und Methoden
45
3.2.12.5 Durchführung des Svanovir® Paratuberkulose ELISA für Milch
Die Durchführung des Milch-ELISA unterschied sich nur in der eingesetzten Proben-
verdünnung von der des Blut-ELISA. Milchproben wurden in einer Verdünnung von 1:10
eingesetzt. Die weiteren Arbeitsschritte erfolgten wie in der Anleitung beschrieben19 20 21.
3.2.13 ANDERE PARATUBERKULOSE-ELISA-TESTVERFAHREN
3.2.13.1 Durchführung des HerdChek™ Mycobacterium paratuberculosis
(Idexx, Wörrstadt)
Das Testkit beinhaltet mit M. paratuberculosis-Antigenen beschichtete Mikrotiter-
platten, positives und negatives Kontrollserum, Waschkonzentrat, Anti-Rind-IgG-HRP-
Konjugat und M. phlei-Absorbens, mit dem die Proben und Kontrollen vorinkubiert werden.
Als Substratlösung dient Tetramethylbenzidin und als Stoplösung 1 M Salzsäure. Der Test
wurde nach der beiliegenden Gebrauchsanleitung durchgeführt21.
3.2.13.2 Durchführung des ELISA zur Serodiagnostik der bovinen Paratuberkulose®,
(Riemser Arzneimittel GmbH, Insel Riems)
Das Testkit beeinhaltet Mikrotiterplatten, die alternierend mit Paratuberkulose-
Antigen und Kontrollantigen beschichtet sind, positives und negatives Kontrollserum und
Waschkonzentrat. Weiterhin befindet sich im Kit Anti-Rind-IgG-HRP-Konjugat, sowie M.
phlei-Verdünnungspuffer, mit dem die Proben und Kontrollen vorinkubiert werden. Als Sub-
stratlösung dient Tetramethylbenzidin und als Stoplösung 0,5 M Schwefelsäure. Der Test
wurde nach der beiliegenden Gebrauchsanleitung durchgeführt19 21.

Material und Methoden
46
3.2.14 POLYMERASEKETTENREAKTION (PCR)
3.2.14.1 Milchprobenaufbereitung für die PCR
1 ml einer Milchprobe wurde für 10 Minuten bei 100°C im Heizblock10 erhitzt. Nach
Abkühlung der Milch im Eisbad und Zugabe von Lysozym (Endkonzentration: 1 mg/ml)
wurde diese für 1 Stunde im Eisbad inkubiert. Proteinase K (Endkonzentration: 1 mg/ml)
wurde der auf 55°C erhitzten Milch zusammen mit SDS (Endkonzentration: 1%) und EDTA
(Endkonzentration: 0,1 molar) zugegeben. Die Inkubation erfolgte über Nacht bei 55°C. Die
Phasentrennung der Milch gelang durch Zentrifugation23 (7000 rpm, 10 Minuten). Zur DNA-
Extraktion wurden 200 µl der wäßrigen Phase eingesetzt.
3.2.14.2 DNA-Extraktion mit dem R.Z.N.A Cell and Tissue Kit (peqlab, Erlangen)
Die Durchführungsvorschrift des Extraktionskits wurde der Milchaufreinigung ange-
paßt. Da der Proteinase K-Verdau an der gesamten Milchprobe durchgeführt wurde, entfiel
die Zugabe des im Kit enthaltenen TL-Puffers. An dieser Stelle wurden 200 µl der wäßrigen
Phase der vorbehandelten Milch (3.2.12.4) mit 220 µl BL-Puffer zur Zelllyse gründlich ge-
mischt und für 30 Minuten bei 70°C10 inkubiert. Bei Zugabe von 220 µl absoluten Ethanols
(-20°C) trat weißer Niederschlag auf, der mit dem gesamten Ansatz auf die Zentrifugensäule
gegeben wurde. Der Ansatz wurde ultrafiltriert23 (10.000 rpm, 1 Minute). Das Filtrat wurde
verworfen, der Filter zweimal mit 750 µl Waschpuffer gewaschen (12.000 rpm, 1 Minute)
und in einem abschließenden Zentrifugationsschritt getrocknet (14.000 rpm, 2 Minute). Die
Zentrifugensäule wurde in einem sterilen Reaktionsgefäß im Hitzeblock auf 70°C erwärmt,
mit 50 µl auf 70°C erwärmten Elutionspuffer versetzt und für 3 Minuten bei 70°C inkubiert.
Das Eluat wurde durch Zentrifugation (14.000 rpm, 1 Minute) gewonnen und der Elutions-
schritt wiederholt. 5 µl des Eluats wurden in der PCR eingesetzt.
23 Centrifuge 5415 C, Eppendorf, Hamburg

Material und Methoden
47
3.2.14.3 DNA-Extraktion mit dem GeneClean II®Kit, Dianova, Hamburg
Diese Aufreinigung wurde angeschlossen, wenn die interne Funktionskontrolle ein ne-
gatives Ergebnis zeigte. Das Eluat aus den R.Z.N.A Cell and Tissue Kit (peqlab, Erlangen)
wurde mit dem dreifachen Volumen NaJ aufgefüllt. 5 µl der gut gemischten Glasmilch zuge-
geben und für 1 Stunde auf Eis inkubiert, die Reaktion wurde regelmäßig geschwenkt. Die
Glasmilch mit der gebundenen DNA wurde bei 14.000 rpm für 30 Sekunden pelletiert23 und
der Überstand verworfen. Das Pellet wurde dreimal mit 500 µl eiskaltem New Wash® gewa-
schen und bei 14.000 rpm für 30 Sekunden zentrifugiert. Nach dem letzten Waschschritt wur-
de der Überstand sehr sorgfältig abgenommen, erneut für 3 Minuten zentrifugiert und wieder-
um der Überstand abpipettiert. Das Pellet wurde in 20 µl A. bidest gelöst und für 3 Minuten
bei 55°C inkubiert10. Nach Zentrifugieren für 30 Sekunden bei 14.000 rpm wurde der Über-
stand, der jetzt die gelöste DNA enthielt, in ein neues Reaktionsgefäß überführt.
3.2.14.4 PCR-Kontrollen
Als positive Kontroll-Lösung diente aufgereinigte M. paratuberculosis-DNA (BOSE
et al. 1993). Als Negativkontrolle wurde einem Ansatz statt der Matrizen-DNA 5 µl A. bidest
zugesetzt. Mit Hilfe der Negativkontrolle konnte so ein falsch-positives PCR-Ergebnis auf
Grund von Kontamination des Mastermix ausgeschlossen werden.
3.2.14.5 PCR-Ansatz und Profil
Die PCR erfolgte in 50 µl Ansätzen, als Polymerase wurde die Taq-DNA-
Polymerase® von Gibco, Eggenstein in einer Konzentration von 2,5 U pro Ansatz eingesetzt.
Die verwendeten Primer sind im Anhang aufgelistet. Pro PCR-Ansatz wurden 5 µM jedes
Primers, 1,5 mM MgCl2, 0,2 mM jedes Desoxynukleotids (peqlab, Erlangen) und der mit dem
Enzym mitgelieferte Reaktionspuffer in einfacher Konzentration eingesetzt. A. bidest, Puffer,
MgCl2 und Nukleotide wurden für alle gemeinsam zu bearbeitenden Ansätze auf Eis zusam-
mengemischt und die Primer hinzugegeben. Nach Zugabe der Matrizen-DNA wurde der An-
satz halbiert und als interne Funktionskontrolle 1µl aufgereinigte M. paratuberculosis-DNA
zu einer Hälfte hinzugegeben.

Material und Methoden
48
Bei dem mittels Thermocycler24 erstellten PCR-Profil handelt es sich um ein touchdown Pro-
fil, das mit einem Denaturierungsschritt bei 94°C für 3 Minuten beginnt.
Tabelle 2: PCR-Profil
Anzahl der Zyklen Denaturierung Annealing Extension
5 30‘‘ 94°C 30‘‘ 65°C 30‘‘ 72°C
5 30‘‘ 94°C 30‘‘ 64°C 30‘‘ 72°C
5 30‘‘ 94°C 30‘‘ 63°C 30‘‘ 72°C
20 30‘‘ 94°C 30‘‘ 62°C 30‘‘ 72°C
Nach dem letzten Zyklus schloß sich eine zehnminütige Extension an. Anschließend wurde
auf 4°C gekühlt. Die Ansätze wurde bis zur Elektrophorese bei 4°C oder bei –20°C aufbe-
wahrt.
3.2.14.6 DNA-Aufreinigung aus PCR-Ansätzen mit dem NucleoSpin Extract (Macherey-
Nagel, Düren)
Der PCR-Ansatz wurde mit TE-Puffer auf ein Volumen von 100 µl aufgefüllt. Nach
Zugabe von 400 µl des NT 2 Puffers wurde der Ansatz auf einen Filter, der sich in einer Zen-
trifugensäule befindet, gegeben. Der Ansatz wurde durch den Filter zentrifugiert (10.000 rpm,
1 Minute)23. Das Filtrat wurde verworfen, der Filter zweimal mit 600 µl Waschpuffer gewa-
schen (12.000 rpm, 1 Minute) und in einem abschließenden Zentrifugationsschritt getrocknet
(14.000 rpm, 2 Minute). Die an den Filter gebundene DNA wurde mit A. bidest durch Zentri-
fugation (14.000 rpm, 1 Minute) eluiert.
3.2.14.7 Sequenzierung
Alle Sequenzierreaktionen wurden von der Firma SequLab, Göttingen durchgeführt.
24 Primus 25/96, MWG, Ebersberg

Material und Methoden
49
3.2.14.8 Computeranalyse der Sequenzen
Die Sequenzdatenanalyse wurde am UNIX-Rechner „Genius“ im Krebsforschungs-
zentrum Heidelberg via Telnet mit dem Programmpaket „HUSAR 4.0“ durchgeführt.
3.2.15 METHODEN DER STATISTISCHEN AUSWERTUNG
(HARMS 1998)
3.2.15.1 Korrelationskoeffizienten
Der Korrelationskoeffizient macht eine Aussage darüber, wie eng der lineare statisti-
sche Zusammenhang zwischen zwei Größen ist. Der Korrelationskoeffizient kann einen Wert
zwischen +1 und -1 annehmen. Ein positives Vorzeichen bedeutet, daß bei Zunahme der x-
Werte auch die y-Werte steigen. Wenn der Korrelationskoeffizient den Wert +1 oder –1 hat,
läßt sich y aus x errechnen, ohne daß die zufallsbedingten Abweichungen eine Rolle spielen.
3.2.15.1.1 Produktmoment-Korrelationskoeffizient nach Pearson:
Voraussetzung für die Berechnung ist, daß die Variablen aus normalverteilten Grundgesamt-
heiten stammen und in einer linearen Abhängigkeit zueinander stehen.
yxPearson SAQSAQ
SAPr•
=
SAP: Summe der Abweichungsprodukte
SAQ: Summe der Abweichungsquadrate
3.2.15.1.2 Spearmanscher Rangkorrelationskoeffizient:
nn
D61r
3
2i
Spearman−
•−=
n: Anzahl der Wertpaare
Di: Rangdifferenz des einzelnen Wertpaares

Material und Methoden
50
3.2.15.2 ROC (relative operating characteristic)-Analyse
(JENSEN und POULSEN 1992)
In der ROC-Kurve werden Diagnostik-Sensitivität (y-Achse) und 100-Diagnostik-
Spezifität (x-Achse) bei verschiedenen Schwellenwerten („Cut off“) gegeneinander aufgetra-
gen. Der Abstand der Kurve zur y = x-Achse wird über die Formel:
richtig Positive – falsch Positive————————————— berechnet.
2
3.2.15.3 Parameter zur Bewertung diagnostischer Verfahren
Die Diagnostik-Sensitivität (D-SN), Diagnostik-Spezifität (D-SP) und die Vorhersa-
gewerte (prädiktive Werte) werden zur Bewertung diagnostischer Verfahren herangezogen.
Mit der Einrichtung eines „Goldstandards“ wird eine Basis zur Bewertung der diagnostischen
Verfahren geschaffen. Bei der Bewertung des Milch-ELISA wurde der Blut-ELISA als
„Goldstandard“ zugrundegelegt. Eine Vierfeldertafel (Tabelle 7) beschreibt die möglichen 4
unterschiedlichen Merkmalsausprägungen. So können bezogen auf den „Gold Standard“,
richtig positive (a), falsch positive (b), richtig negative (d) und falsch negative (c) Ergebnisse
vorkommen. Die Parameter D-SN, D-SP und die prädiktiven Werte beschreiben die Verhält-
nisse dieser unterschiedlichen Merkmalsausprägungen.

Material und Methoden
51
Abbildung 1: Vierfeldertafel zur Schätzung von D-SN/D-SP und der Vorhersagewerte
„Gold Standard“
Diagnose positiv negativ
positiv a b positiver Vorhersagewert:a /(a + b)
negativ c d negativer Vorhersagewert:d /(d + c)
D-Sensitivität:a/(a + c)
D-Spezifität:d/(d + b)
D-SN: Die D-SN beschreibt die Empfindlichkeit eines Meßverfahrens, erkrankte Indi-
viduen als erkrankt zu erkennen. Sie drückt den Anteil der testpositiven Tiere
unter allen infizierten Tieren aus.
D-SP: Die D-SP beschreibt die Wahrscheinlichkeit, mit der ein Gesunder als gesund
erkannt wird. Sie drückt den Anteil der testnegativen Tieren unter allen nicht
infizierten Tieren aus.
Prädiktive Werte:
Die prädiktiven Werte geben jeweils die Wahrscheinlichkeit an, mit der eine
aus einem Test-Ergebnis gezogene Schlußfolgerung richtig ist.
Der positive prädiktive Wert sagt aus, mit welcher Wahrscheinlichkeit die be-
treffende Krankheit vorliegt, wenn der dazugehörige Test positiv ausfällt.
Der negative prädiktive Wert sagt aus, mit welcher Wahrscheinlichkeit die be-
treffende Krankheit nicht vorliegt, wenn der dazugehörige Test negativ ausfällt.

Ergebnisse
52
4 Ergebnisse
Das Ziel der vorliegenden Arbeit war die Erarbeitung zweier Verfahren zur routine-
mäßigen Paratuberkulose-Diagnostik in Milch. Diese sollen zu einer Verbesserung der Tier-
gesundheit und zur Qualitätssicherung des Lebensmittel Milch beitragen. Zum einen wurde
der von JARK (1996) entwickelte und inzwischen von der Firma SVANOVA, Schweden her-
gestellte ELISA (SVANOVIR®-ELISA) für Blutserum an Milch adaptiert. Desweiteren wur-
de eine Methode zum diagnostischen direkten Nachweis von M. paratuberculosis spezifischer
DNA in Kuhmilch auf der Basis einer PCR entwickelt (4.2). Beide Methoden wurden an Pro-
ben einer Gruppe von 10 Milchkühen in einer Verlaufsuntersuchung überprüft (4.3).
4.1 Adaption des SVANOVIR®-ELISA für Blutserum an Milchserum
4.1.1 AUSWAHL DES ELISA
Zur Zeit sind drei ELISA-Testkits zur Untersuchung von Blutserum auf Antikörper
gegen M. paratuberculosis im Handel erhältlich.
Zwei der ELISA-Testverfahren führen eine Vorinkubation mit M. phlei-Antigen durch
(HerdChek™ M.pt Ab, Idexx, Wörrstadt und ELISA zur Serodiagnostik der bovinen
Paratuberkulose, Riemser Arzneimittel GmbH, Insel Riems). Beide Testverfahren nutzen
ein M. paratuberculosis Ganzzellantigen. Im Svanovir®- ELISA (Svanova, Schweden) wird
das zellwandständige Lipoarabinomannan als Antigen verwendet.
Zur Auswahl des ELISA, der als Grundlage des Milch-ELISA dienen sollte, wurden die Er-
gebnisse eines Ringversuchs, der im Auftrag der Niedersächsischen Tierseuchenkasse durch-
geführt wurde, herangezogen.
Der Ringversuch wurde in Form einer Verlaufsuntersuchung über einen Zeitraum von 6 Wo-
chen an 10 Milchkühen (1-10), denen bei der späteren Schlachtung der Ileocaecallymphkno-
ten entnommen wurde, durchgeführt. Die Kühe wurden ausgesucht, da sie aufgrund ihrer kli-
nischen Symptome als Paratuberkulose-verdächtig einzustufen waren. Es kamen die Paratu-
berkulose-ELISA-Testkits der Firmen Idexx, Riemser Arzneimittel GmbH und Svanova zum
Einsatz. Beteiligt an diesem Ringversuch war die Gesellschaft für Innovative Veterinärdia-

Ergebnisse
53
gnostik (IVD) und sechs Untersuchungsstellen in Niedersachsen (A-F). Die Ergebnisse sind
in Tabellen im Anhang (9.1) dargestellt.
In der kulturellen Untersuchung der Ileocaecallymphknoten konnte bei sechs der zehn kli-
nisch verdächtigen Tiere M. paratuberculosis nachgewiesen werden (209, 238, 239, 37M,
80M, 76M). Aufgrund der klinischen Erscheinungen der Paratuberkulose mußte das Tier mit
der Nummer 239 in der 5. Woche des Ringversuchs und das Tier mit der Nummer 80 im Zeit-
raum der Verlaufsuntersuchung (4.3.1) geschlachtet werden. Bei einem der verbleibenden vier
Tiere konnte der Erreger in der Verlaufsuntersuchung mittels Milch-PCR nachgewiesen wer-
den (235).
Der HerdChek™ M.pt Ab-ELISA (Idexx, Wörrstadt) erkennt in der 1.-3. Woche fünf (209,
238, 239, 37M, 80M) der sechs Lymphknoten-Kultur positiven Tiere als positiv. In den fol-
genden Wochen wird das Tier mit der Nummer 37M zunächst von drei Untersuchungsstellen
(A, E, F in der 4. Woche), vier Untersuchungsstellen (IVD, A, E, F in der 5. Woche) und
schließlich sechs Untersuchungsstellen (IVD, A, B, C, E, F in der 6.Woche) als negativ gete-
stet. Die übrigen klinisch verdächtigen Tiere sowie das Milch-PCR positive Tier (235) sind
im HerdChek™ M.pt Ab-ELISA negativ. Beim Vergleich der Ergebnisse der Untersuchungs-
stellen zeigte der ELISA eine gute Reproduzierbarkeit.
Der ELISA zur Serodiagnostik der bovinen Paratuberkulose (Riemser Arzneimittel
GmbH, Insel Riems) erkennt in der 1.-5. Woche vier (209, 238, 239, 80M) der sechs
Lymphknoten-Kultur positiven Tiere als positiv. In der 6. Woche wird die Kuh mit der Num-
mer 238 von drei Untersuchungsstellen (IVD, A, B) als serologisch zweifelhaft und von einer
Untersuchungsstelle (D) als negativ ermittelt. Die übrigen klinisch verdächtigen Tiere sowie
das Milch-PCR positive Tier (235) werden nicht erkannt. In die Bewertung des Ringversuchs
wurden die stark abweichenden Ergebnisse der Untersuchungsstelle C nicht einbezogen, da
davon ausgegangen werden muß, daß dort ein Durchführungs- oder Auswertungsfehler vor-
lag. Beim Vergleich der Ergebnisse der übrigen Untersuchungsstellen zeigt der ELISA eine
gute Reproduzierbarkeit.
Der Svanovir®- ELISA (Svanova, Schweden) erkennt schon in der 1. Woche alle sechs
Lymphknoten-Kultur positiven Tiere (209, 238, 239, 37M, 80M, 76M) und weiterhin das
Milch-PCR positive Tier (235). Bei der Kuh mit der Nummer 238 zeichnet sich im Verlauf
der 2.-6. Woche ein Abfall der Antikörper gegen M. paratuberculosis im Blut ab. In der 2.
Woche ermitteln drei Untersuchungsstellen (IVD, A, F), in der 3. Woche fünf Untersuchungs-

Ergebnisse
54
stellen (IVD, A, B, C, F), in der 4. und 5. Woche sechs Untersuchungsstellen (IVD, A, B, C,
D, F) einen zweifelhaften Befund. Diese sechs Untersuchungsstellen beurteilten das Tier in
der 6. Woche als negativ und die Untersuchungsstelle E als zweifelhaft. Von den übrigen kli-
nisch verdächtigen Tieren wird die Kuh mit der Nummer 282 im gesamten Untersuchungs-
zeitraum von allen teilnehmenden Untersuchungsstellen als negativ beurteilt. Zwei weitere
klinisch verdächtige Tiere (37, 284) werden überwiegend von allen Untersuchungsstellen als
zweifelhaft ermittelt. Beim Vergleich der Ergebnisse der Untersuchungsstellen zeigt der Sva-
novir®- ELISA eine gute Reproduzierbarkeit.
Der Svanovir®-ELISA wurde als Grundlage zur Entwicklung des Milch-ELISA ausgewählt,
da er als einziger der eingesetzten ELISA-Systeme alle Lymphknoten-Kultur oder/und Milch-
PCR positiven Tiere erkennt und zwar schon in der ersten Woche des Ringversuchs. Er zeigt
eine gute Reproduzierbarkeit. Außerdem zeichnet der Svanovir®-ELISA sich durch seine ein-
fache und wenig zeitintensive Durchführung aus. Im Gegensatz zu den beiden anderen einge-
setzten ELISA-Systemen mußten bei diesem Test die Seren nicht vorinkubiert werden. Wei-
terhin ist der Svanovir®-ELISA das kostengünstigste Produkt.
4.1.2 DARSTELLUNG DER BINDUNGSSTELLE DES SVANOVIR ®-KONJUGATES
Zur Überprüfung, ob der sich im ELISA-Testkit der Firma Svanova, Schweden, be-
findliche monoklonale Sekundärantikörper auch zur Detektion der Milchantikörper eignet,
wurde das IgG des positiven Kontrollserums aufgereinigt, mit Papain gespalten und schließ-
lich getestet, an welches Fragment des Immunglobulins der monoklonale Antikörper bindet.
Das mit Coomassie®-Blau gefärbte SDS-Gel zeigt die Auftrennung der Spaltprodukte des IgG
(Abbildung 2: A Spur 1) im Vergleich zum intakten IgG-Molekül (Spur 3) und dem Retentat
nach der Zentrifugation (Centriplus® 30, Amicon®) (Spur 2). Es ist eine Bande bei ca. 50 kDa
zu erkennen. Es handelt sich dabei um die Fab–Fragmente (HARLOW und LANE 1988), die
aufgrund des gleichen Molekulargewichts nicht zu unterscheiden sind. Eine weitere Bande
befindet sich ca. in Höhe der 24 kDa Bande des Molekularmassenstandards. Dabei handelte
es sich um das Fc–Fragment mit einem Molekulargewicht von ca. 25 kDa.
Der mit dem monoklonalen Sekundärantikörper entwickelte entsprechende Western-Blot
(Abbildung 2 B Spur 1) zeigt nur im Bereich des Fc-Fragments eine deutlich sichtbar gefärbte

Ergebnisse
55
Bande. Daraus konnte gefolgert werden, daß der Antikörper an das Fc-Fragment des Im-
munglobulins bindet.
Abbildung 2: Abb. A: Coomassie-Blau® gefärbtes 10 % Acrylamidgel
Abb. B: Western-Blot mit monoklonalem Sekundärantikörper entwickelt
M: Proteinstandard (BioRad Prestained SDS-Page Standard®, Low Range)
1: Papain gespaltenes IgG des positiven Kontrollserums
2: Retentat nach Zentrifugation
3: Gesamt IgG
4.1.3 MILCHAUFREINIGUNGSMETHODE
In Vorversuchen zeigte sich, daß die Milchaufreinigungsmethode mit Rennin zwei an-
deren Methoden (3.2.12.4 unter 1. und 2.) vorzuziehen war, da das so aufgereinigte Milchse-
rum klar war und sich gut zum Einsatz im ELISA eignete. Die Methode war allerdings sehr
zeit- und arbeitsaufwendig. Auch die Trennung der Milchphasen nach Zentrifugation gestal-
tete sich schwierig.
In Anlehnung an die Durchführungsvorschrift eines anderen Milch-ELISA (Brucellen-
ELISA, Svanova, Schweden) wurde die dort beschriebene Aufreinigung durch Zentrifugation
(5000 rpm,15 Minuten) mit der Rennin-Methode verglichen. Hierbei wurden vierzig Milch-
proben mit beiden Methoden aufgereinigt und im ELISA getestet. Der Svanovir® –ELISA
wurde laut Anleitung durchgeführt (3.2.12.1).

Ergebnisse
56
Bei beiden Aufreinigungsmethoden war kein wesentlicher Unterschied bei den optischen
Dichten der Milchproben zu beobachten. Im Folgenden wurde die weniger arbeitsintensive
Zentrifugationsmethode angewendet.
4.1.4 REPRODUZIERBARKEIT
4.1.4.1 Inter Assay-Variation
Die Reproduzierbarkeit von Tag zu Tag wurde anhand von 20 Milchproben an 8 ver-
schiedenen Tagen überprüft. Dabei handelte es sich sowohl um Proben von Tieren, die blutse-
rologisch im ELISA positiv reagierten, als auch um Proben von Tieren, die negativ reagierten.
Die Tabelle 3 stellt das Ergebnis dieses Versuchs dar. Die für jede Milchprobe erhaltenen
Untersuchungsergebnisse (Extinktionen) sind im Anhang aufgeführt. Der Mittelwert, die
Standardabweichung und der Variationskoeffizient in der nachfolgenden Tabelle wurden aus
den Extinktionen (9.1) berechnet.
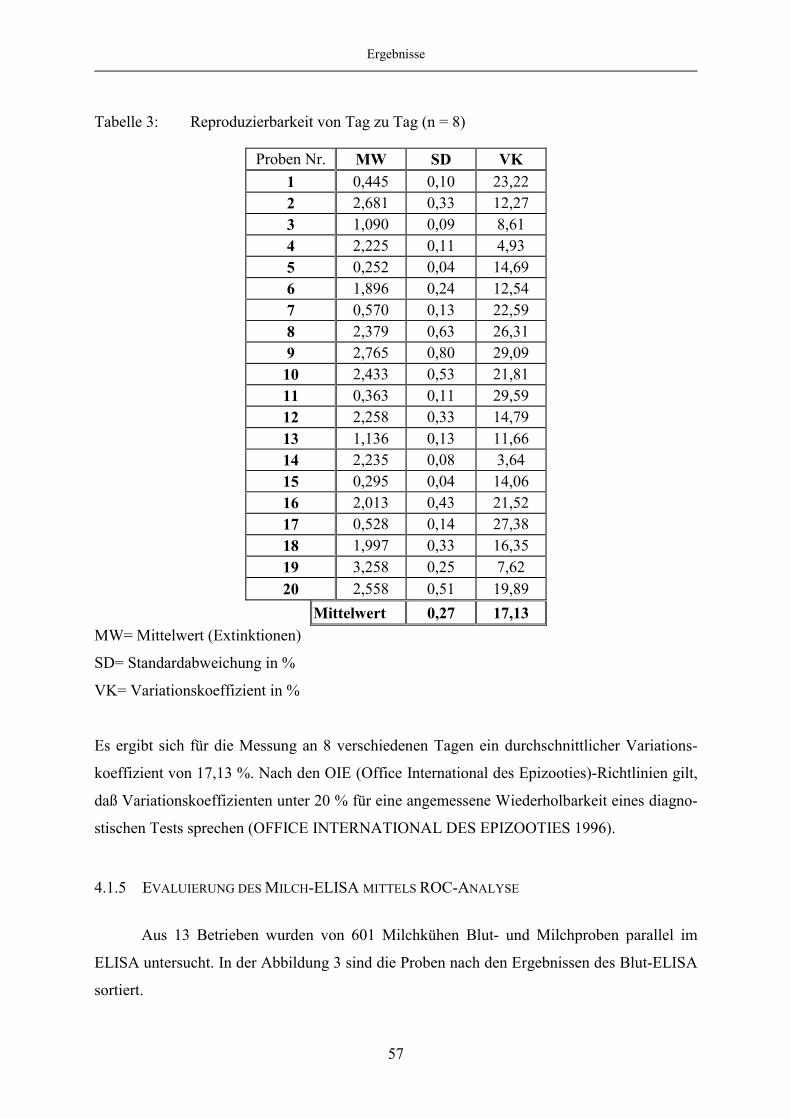
Ergebnisse
57
Tabelle 3: Reproduzierbarkeit von Tag zu Tag (n = 8)
Proben Nr. MW SD VK1 0,445 0,10 23,222 2,681 0,33 12,273 1,090 0,09 8,614 2,225 0,11 4,935 0,252 0,04 14,696 1,896 0,24 12,547 0,570 0,13 22,598 2,379 0,63 26,319 2,765 0,80 29,0910 2,433 0,53 21,8111 0,363 0,11 29,5912 2,258 0,33 14,7913 1,136 0,13 11,6614 2,235 0,08 3,6415 0,295 0,04 14,0616 2,013 0,43 21,5217 0,528 0,14 27,3818 1,997 0,33 16,3519 3,258 0,25 7,6220 2,558 0,51 19,89
Mittelwert 0,27 17,13MW= Mittelwert (Extinktionen)
SD= Standardabweichung in %
VK= Variationskoeffizient in %
Es ergibt sich für die Messung an 8 verschiedenen Tagen ein durchschnittlicher Variations-
koeffizient von 17,13 %. Nach den OIE (Office International des Epizooties)-Richtlinien gilt,
daß Variationskoeffizienten unter 20 % für eine angemessene Wiederholbarkeit eines diagno-
stischen Tests sprechen (OFFICE INTERNATIONAL DES EPIZOOTIES 1996).
4.1.5 EVALUIERUNG DES MILCH-ELISA MITTELS ROC-ANALYSE
Aus 13 Betrieben wurden von 601 Milchkühen Blut- und Milchproben parallel im
ELISA untersucht. In der Abbildung 3 sind die Proben nach den Ergebnissen des Blut-ELISA
sortiert.
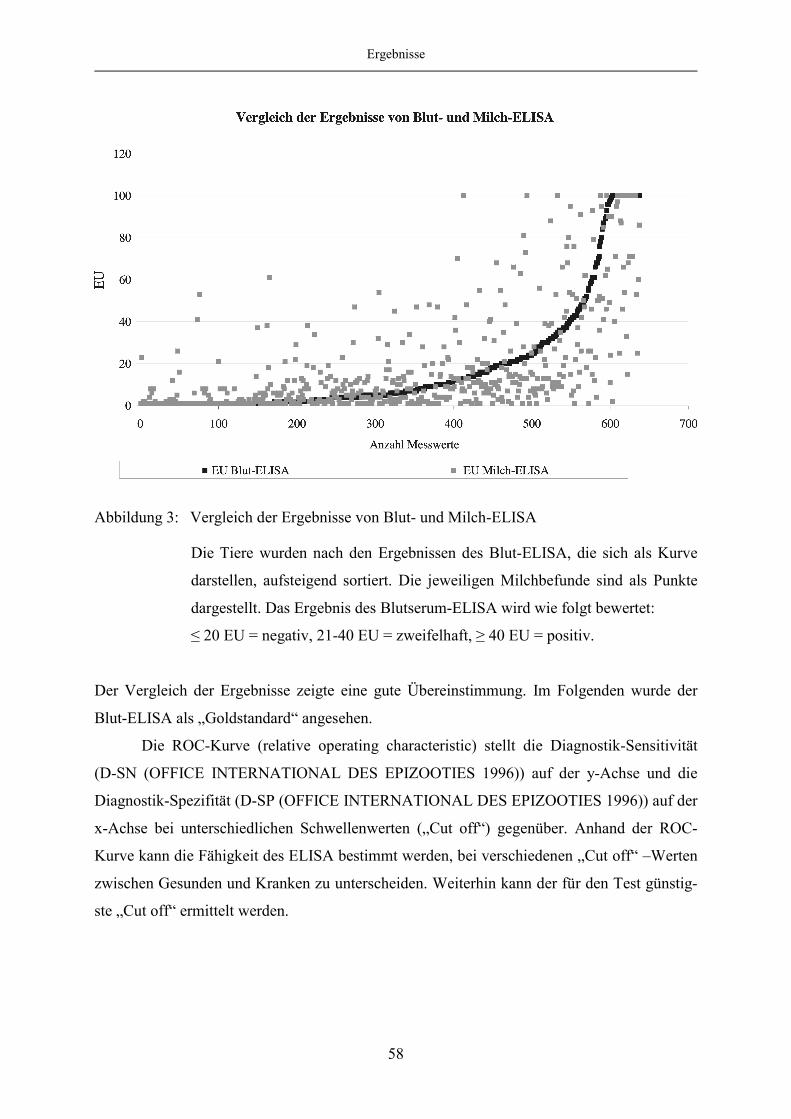
Ergebnisse
58
Abbildung 3: Vergleich der Ergebnisse von Blut- und Milch-ELISA
Die Tiere wurden nach den Ergebnissen des Blut-ELISA, die sich als Kurve
darstellen, aufsteigend sortiert. Die jeweiligen Milchbefunde sind als Punkte
dargestellt. Das Ergebnis des Blutserum-ELISA wird wie folgt bewertet:
≤ 20 EU = negativ, 21-40 EU = zweifelhaft, ≥ 40 EU = positiv.
Der Vergleich der Ergebnisse zeigte eine gute Übereinstimmung. Im Folgenden wurde der
Blut-ELISA als „Goldstandard“ angesehen.
Die ROC-Kurve (relative operating characteristic) stellt die Diagnostik-Sensitivität
(D-SN (OFFICE INTERNATIONAL DES EPIZOOTIES 1996)) auf der y-Achse und die
Diagnostik-Spezifität (D-SP (OFFICE INTERNATIONAL DES EPIZOOTIES 1996)) auf der
x-Achse bei unterschiedlichen Schwellenwerten („Cut off“) gegenüber. Anhand der ROC-
Kurve kann die Fähigkeit des ELISA bestimmt werden, bei verschiedenen „Cut off“ –Werten
zwischen Gesunden und Kranken zu unterscheiden. Weiterhin kann der für den Test günstig-
ste „Cut off“ ermittelt werden.
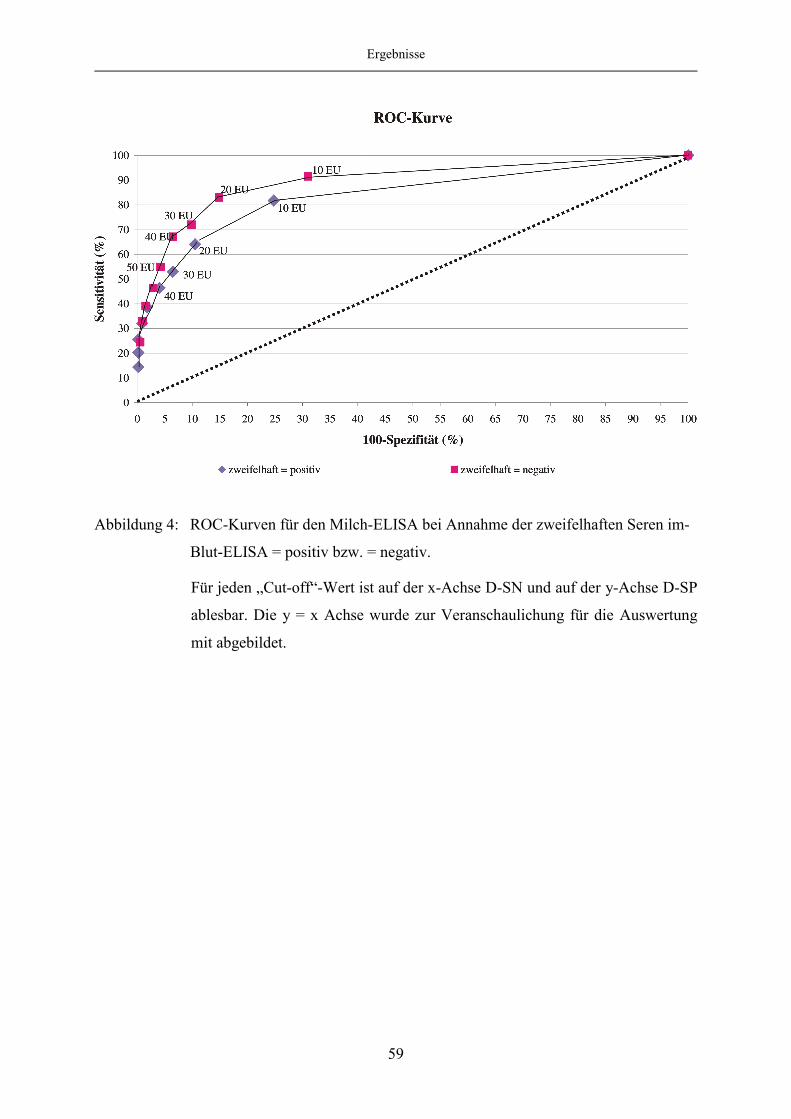
Ergebnisse
59
Abbildung 4: ROC-Kurven für den Milch-ELISA bei Annahme der zweifelhaften Seren im-
Blut-ELISA = positiv bzw. = negativ.
Für jeden „Cut-off“-Wert ist auf der x-Achse D-SN und auf der y-Achse D-SP
ablesbar. Die y = x Achse wurde zur Veranschaulichung für die Auswertung
mit abgebildet.
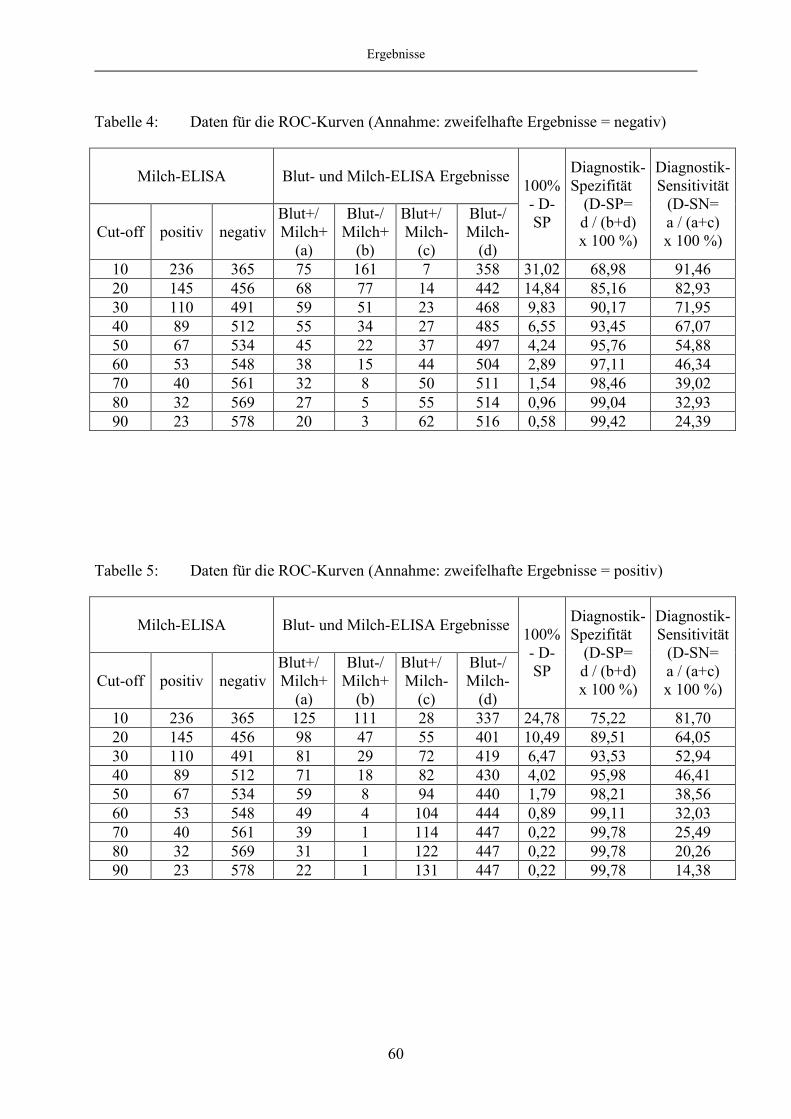
Ergebnisse
60
Tabelle 4: Daten für die ROC-Kurven (Annahme: zweifelhafte Ergebnisse = negativ)
Milch-ELISA Blut- und Milch-ELISA Ergebnisse
Cut-off positiv negativBlut+/Milch+
(a)
Blut-/Milch+
(b)
Blut+/Milch-
(c)
Blut-/Milch-
(d)
100%- D-SP
Diagnostik-Spezifität
(D-SP=d / (b+d)x 100 %)
Diagnostik-Sensitivität
(D-SN=a / (a+c)x 100 %)
10 236 365 75 161 7 358 31,02 68,98 91,4620 145 456 68 77 14 442 14,84 85,16 82,9330 110 491 59 51 23 468 9,83 90,17 71,9540 89 512 55 34 27 485 6,55 93,45 67,0750 67 534 45 22 37 497 4,24 95,76 54,8860 53 548 38 15 44 504 2,89 97,11 46,3470 40 561 32 8 50 511 1,54 98,46 39,0280 32 569 27 5 55 514 0,96 99,04 32,9390 23 578 20 3 62 516 0,58 99,42 24,39
Tabelle 5: Daten für die ROC-Kurven (Annahme: zweifelhafte Ergebnisse = positiv)
Milch-ELISA Blut- und Milch-ELISA Ergebnisse
Cut-off positiv negativBlut+/Milch+
(a)
Blut-/Milch+
(b)
Blut+/Milch-
(c)
Blut-/Milch-
(d)
100%- D-SP
Diagnostik-Spezifität
(D-SP=d / (b+d)x 100 %)
Diagnostik-Sensitivität
(D-SN=a / (a+c)x 100 %)
10 236 365 125 111 28 337 24,78 75,22 81,7020 145 456 98 47 55 401 10,49 89,51 64,0530 110 491 81 29 72 419 6,47 93,53 52,9440 89 512 71 18 82 430 4,02 95,98 46,4150 67 534 59 8 94 440 1,79 98,21 38,5660 53 548 49 4 104 444 0,89 99,11 32,0370 40 561 39 1 114 447 0,22 99,78 25,4980 32 569 31 1 122 447 0,22 99,78 20,2690 23 578 22 1 131 447 0,22 99,78 14,38
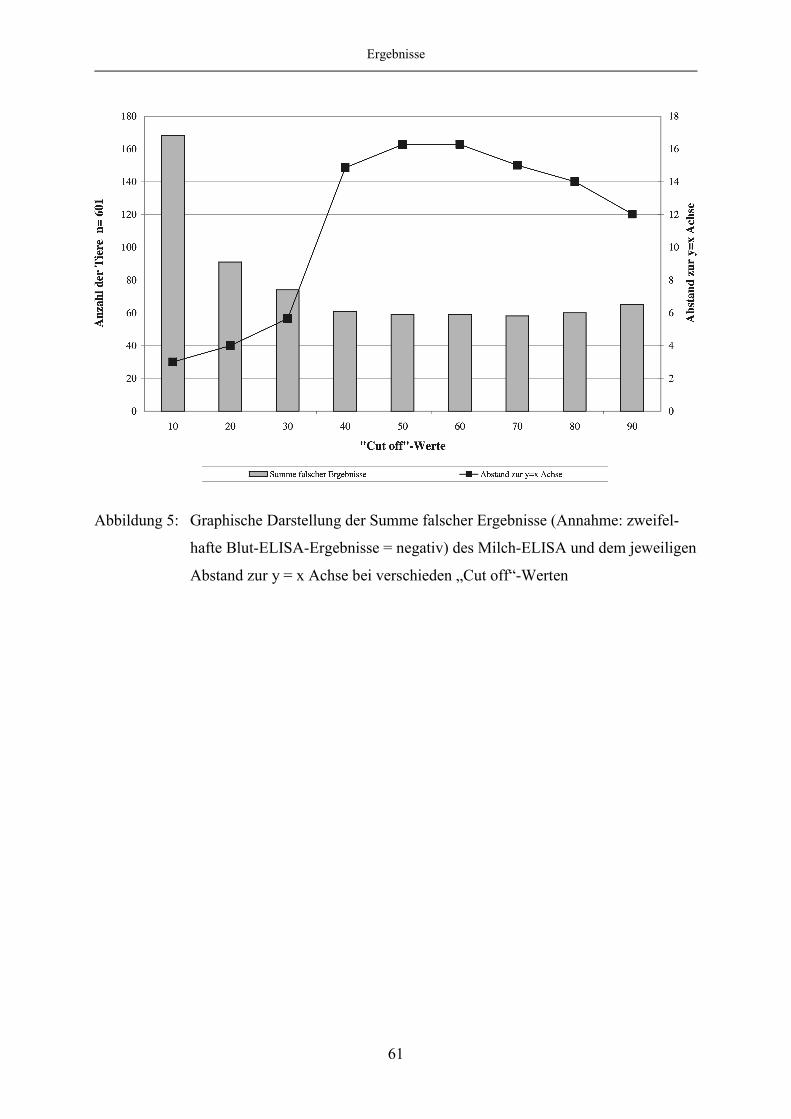
Ergebnisse
61
Abbildung 5: Graphische Darstellung der Summe falscher Ergebnisse (Annahme: zweifel-
hafte Blut-ELISA-Ergebnisse = negativ) des Milch-ELISA und dem jeweiligen
Abstand zur y = x Achse bei verschieden „Cut off“-Werten
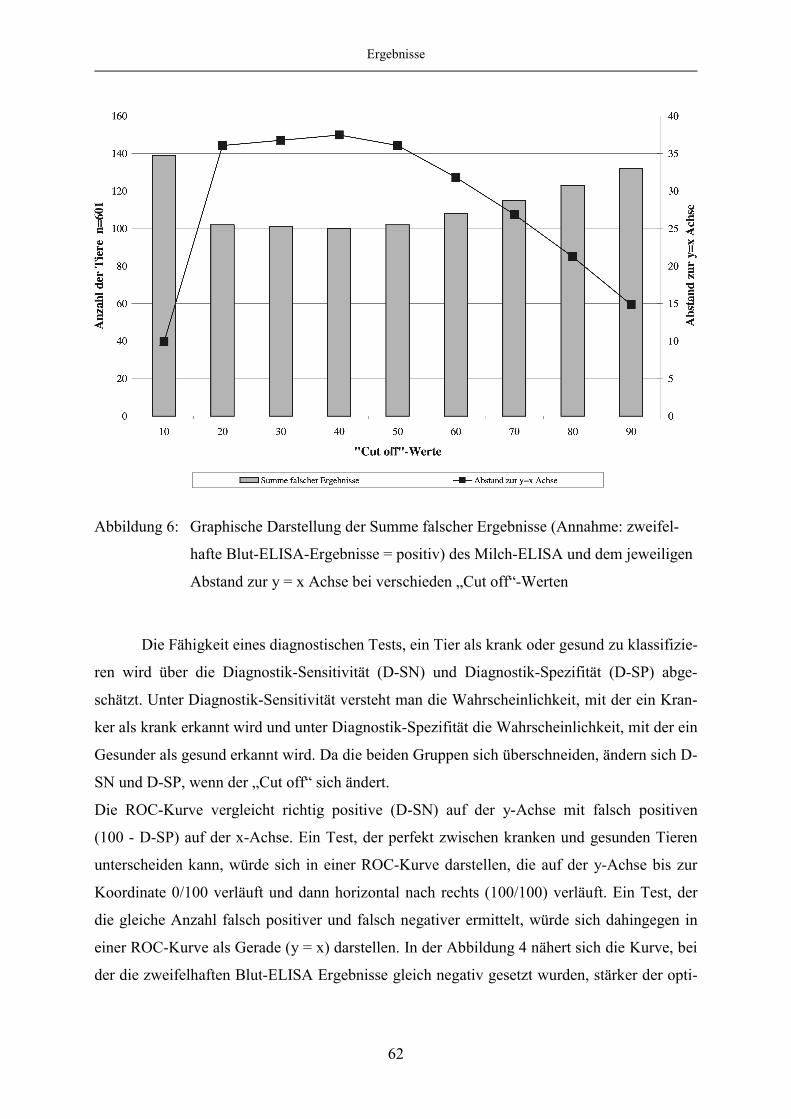
Ergebnisse
62
Abbildung 6: Graphische Darstellung der Summe falscher Ergebnisse (Annahme: zweifel-
hafte Blut-ELISA-Ergebnisse = positiv) des Milch-ELISA und dem jeweiligen
Abstand zur y = x Achse bei verschieden „Cut off“-Werten
Die Fähigkeit eines diagnostischen Tests, ein Tier als krank oder gesund zu klassifizie-
ren wird über die Diagnostik-Sensitivität (D-SN) und Diagnostik-Spezifität (D-SP) abge-
schätzt. Unter Diagnostik-Sensitivität versteht man die Wahrscheinlichkeit, mit der ein Kran-
ker als krank erkannt wird und unter Diagnostik-Spezifität die Wahrscheinlichkeit, mit der ein
Gesunder als gesund erkannt wird. Da die beiden Gruppen sich überschneiden, ändern sich D-
SN und D-SP, wenn der „Cut off“ sich ändert.
Die ROC-Kurve vergleicht richtig positive (D-SN) auf der y-Achse mit falsch positiven
(100 - D-SP) auf der x-Achse. Ein Test, der perfekt zwischen kranken und gesunden Tieren
unterscheiden kann, würde sich in einer ROC-Kurve darstellen, die auf der y-Achse bis zur
Koordinate 0/100 verläuft und dann horizontal nach rechts (100/100) verläuft. Ein Test, der
die gleiche Anzahl falsch positiver und falsch negativer ermittelt, würde sich dahingegen in
einer ROC-Kurve als Gerade (y = x) darstellen. In der Abbildung 4 nähert sich die Kurve, bei
der die zweifelhaften Blut-ELISA Ergebnisse gleich negativ gesetzt wurden, stärker der opti-

Ergebnisse
63
malen ROC-Kurve, als die Kurve, bei der die zweifelhaften Blut-ELISA Ergebnisse gleich
positiv gesetzt wurden, und damit dem Koordinatenpunkt 0/100, der die höchste D-SN und D-
SP widerspiegelt.
4.1.5.1 Bestimmung des „Cut off“
An einer ROC-Kurve kann der optimale „Cut off“-Wert nicht unmittelbar abgelesen
werden, da bei der Bestimmung des optimalen „Cut off“-Wertes noch andere Faktoren mit
einbezogen werden sollten.
Es ist aber möglich, den „Cut off“-Wert mit der höchsten D-SN und D-SP anhand der Kurve
zu bestimmen. Dieser Wert hat den kürzesten Abstand (3.2.15.2) zum Koordinatenpunkt
0/100 oder den größten Abstand zur y = x-Achse. Ein weiterer Hinweis für einen geeigneten
„Cut off“-Wert gibt die Summe der falschen Ergebnisse, also der falsch negativen und der
falsch positven. Bei dem ausgewählten „Cut off“-Wert sollte diese Summe möglichst gering
sein.
Die Abbildung 5 und Abbildung 6 stellen die Summe der falschen Ergebnisse und den Ab-
stand zur y = x-Achse für beide Kurven gegenüber.
Die Kurve, bei der die zweifelhaften Blut-ELISA Ergebnisse gleich negativ gesetzt wurden,
zeigte den größten Abstand zur y = x-Achse bei den „Cut off“-Werten 50 und 60. Bei diesen
„Cut off“-Werten wies auch die Summe der falschen Ergebnisse den geringsten Wert mit 59
auf. Bei dem „Cut off“-Wert 40 betrug die Summe der falschen Ergebnisse 61. Bei dem Ver-
gleich von D-SN und D-SP für die „Cut off“-Werte 40, 50 und 60 in Tabelle 4 zeigte sich,
daß bei einem „Cut off“-Wert von 60 die D-SN nur 46,34 % und die D-SP 97,11 % betrug,
während bis zum „Cut off“-Wert 40 die D-SN stark auf 67,07 % anstieg und die D-SP im
Verhältnis dazu nur gering abfiel (93,45 %). Bei einem „Cut off“-Wert von 50 betrug die D-
SN 54,88 % und die D-SP 95,76 %.
Die Kurve, bei der die zweifelhaften Blut-ELISA Ergebnisse gleich positiv gesetzt wurden,
zeigte den größten Abstand zur y = x-Achse bei „Cut off“-Wert 40. Bei „Cut off“-Wert 40
wies auch die Summe der falschen Ergebnisse den geringsten Wert mit 100 auf. Die D-SN für
den „Cut off“-Wert 40 beträgt 95,98 % und D-SP 46,41 %.
Bei der Auswahl des „Cut off“-Wertes wurde die Kurve, bei der die zweifelhaften Blut-
ELISA Ergebnisse gleich negativ gesetzt wurden, stärker berücksichtigt, da sie den besseren

Ergebnisse
64
Kurvenverlauf zeigte. Demnach sollte der ausgewählte „Cut off“ zwischen den „Cut off“-
Werten 40 und 50 liegen.
Mit der Methode zur „Cut off“-Wert-Berechnung nach (TIJSSEN 1985) ließen sich diese Er-
gebnisse bestätigen. Bei dieser Berechnung wird für die positiven und negativen Ergebnisse
eine Normalverteilung angenommen. Der „Cut off“-Wert berechnet sich aus dem für eine
negative Population erhaltenen arithmetischen Mittelwert zu dem eine einfache, zweifache
oder dreifache Standardabweichung addiert wird.
Aus den berechneten ELISA-Aktivitäten der negativen Population (448 Tiere) wurden der
arithmetische Mittelwert (MW = 8,4) sowie die Standardabweichung (SD = 12,4) errechnet.
Der Schwellenwert für ein negatives Ergebnis (EU 21) errechnet sich aus der Summe des
arithmetischen Mittelwertes (MW = 8,4) zuzüglich der einfachen Standardabweichung (SD =
12,4). Der „Cut off“-Wert für ein positives Ergebnis (EU 46) errechnet sich aus dem arithme-
tischen Mittelwert (MW = 8,4) plus der dreifachen Standardabweichung (3*SD = 37,2). Die
erhaltenen Werte wurden jeweils aufgerundet. Die ELISA-Aktivitäten, die zwischen diesen
beiden „Cut off“-Werten liegen, wurden als zweifelhaft beurteilt.
Tabelle 6: „Cut off“-Werte des Milch-ELISA (TIJSSEN 1985)
ELISA-Aktivitäten
≤ 21 = negativ
22-45 = zweifelhaft
≥ 46 = positiv
Auf der Grundlage dieser „Cut-off“-Werte wurden Proben mit 1-21 ELISA-Units als negativ,
mit 22-45 ELISA-Units als zweifelhaft und ab 46 ELISA-Units als positiv bewertet.

Ergebnisse
65
Tabelle 7: Vierfeldertafel zum Vergleich des Milch-ELISA mit dem Blut-ELISA auf der
Grundlage der berechneten „Cut-off“-Werte
Milch-ELISA
Blut-ELISA negativ positiv Summe
negativ 405 14 419
positiv 14 53 67
Summe 419 67 486
Aus der Tabelle 7 ergibt sich für den Milch-ELISA bezogen auf den Blut-ELISA eine Spezi-
fität von 96,6 % und eine Sensitivität von 79,1 %. Die zweifelhaften Ergebnisse wurden in die
Berechnung nicht mit einbezogen. Desweiteren ergibt sich bei einer Prävalenz von 13,7 % ein
positiver prädiktiver Wert von 96,6 % und ein negativer prädiktiver Wert von 79,1 %.
4.1.6 KORRELATION DES BLUT- UND MILCH-ELISA BEI FESTGELEGTEN „CUT OFF“-WERTEN
des Milch-ELISA
Die Ergebnisse der Paralleluntersuchungen wurden in die Stufen negativ, zweifelhaft
und positiv eingeteilt und im Folgenden miteinander verglichen. Die dabei eingesetzten
Schwellenwerte („Cut off“) beruhen auf der Berechnung in Kapitel 4.1.5.1.
In der Tabelle 8 sind die serologischen Untersuchungsergebnisse von Blut und Milch zusam-
mengefaßt. Die Spalten stellen eine Einteilung in die negativen, zweifelhaften oder positiven
Bereiche des ELISA dar.
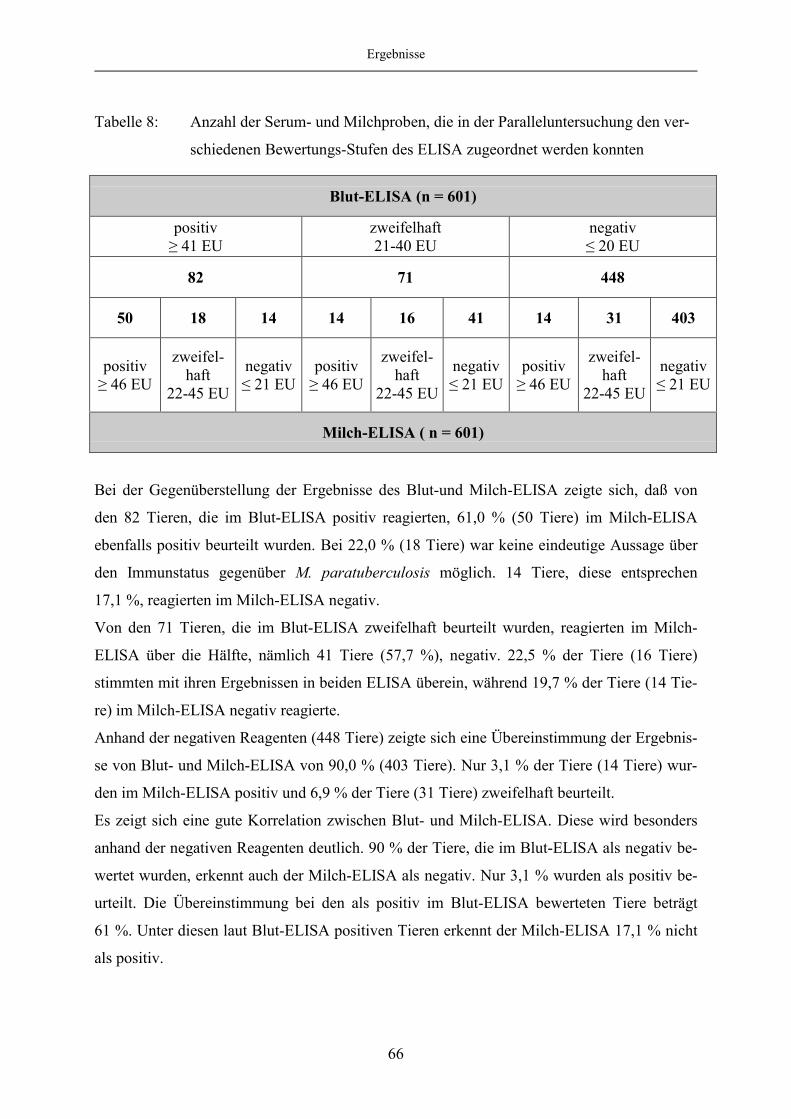
Ergebnisse
66
Tabelle 8: Anzahl der Serum- und Milchproben, die in der Paralleluntersuchung den ver-
schiedenen Bewertungs-Stufen des ELISA zugeordnet werden konnten
Blut-ELISA (n = 601)
positiv≥ 41 EU
zweifelhaft21-40 EU
negativ≤ 20 EU
82 71 448
50 18 14 14 16 41 14 31 403
positiv≥ 46 EU
zweifel-haft
22-45 EU
negativ≤ 21 EU
positiv≥ 46 EU
zweifel-haft
22-45 EU
negativ≤ 21 EU
positiv≥ 46 EU
zweifel-haft
22-45 EU
negativ≤ 21 EU
Milch-ELISA ( n = 601)
Bei der Gegenüberstellung der Ergebnisse des Blut-und Milch-ELISA zeigte sich, daß von
den 82 Tieren, die im Blut-ELISA positiv reagierten, 61,0 % (50 Tiere) im Milch-ELISA
ebenfalls positiv beurteilt wurden. Bei 22,0 % (18 Tiere) war keine eindeutige Aussage über
den Immunstatus gegenüber M. paratuberculosis möglich. 14 Tiere, diese entsprechen
17,1 %, reagierten im Milch-ELISA negativ.
Von den 71 Tieren, die im Blut-ELISA zweifelhaft beurteilt wurden, reagierten im Milch-
ELISA über die Hälfte, nämlich 41 Tiere (57,7 %), negativ. 22,5 % der Tiere (16 Tiere)
stimmten mit ihren Ergebnissen in beiden ELISA überein, während 19,7 % der Tiere (14 Tie-
re) im Milch-ELISA negativ reagierte.
Anhand der negativen Reagenten (448 Tiere) zeigte sich eine Übereinstimmung der Ergebnis-
se von Blut- und Milch-ELISA von 90,0 % (403 Tiere). Nur 3,1 % der Tiere (14 Tiere) wur-
den im Milch-ELISA positiv und 6,9 % der Tiere (31 Tiere) zweifelhaft beurteilt.
Es zeigt sich eine gute Korrelation zwischen Blut- und Milch-ELISA. Diese wird besonders
anhand der negativen Reagenten deutlich. 90 % der Tiere, die im Blut-ELISA als negativ be-
wertet wurden, erkennt auch der Milch-ELISA als negativ. Nur 3,1 % wurden als positiv be-
urteilt. Die Übereinstimmung bei den als positiv im Blut-ELISA bewerteten Tiere beträgt
61 %. Unter diesen laut Blut-ELISA positiven Tieren erkennt der Milch-ELISA 17,1 % nicht
als positiv.
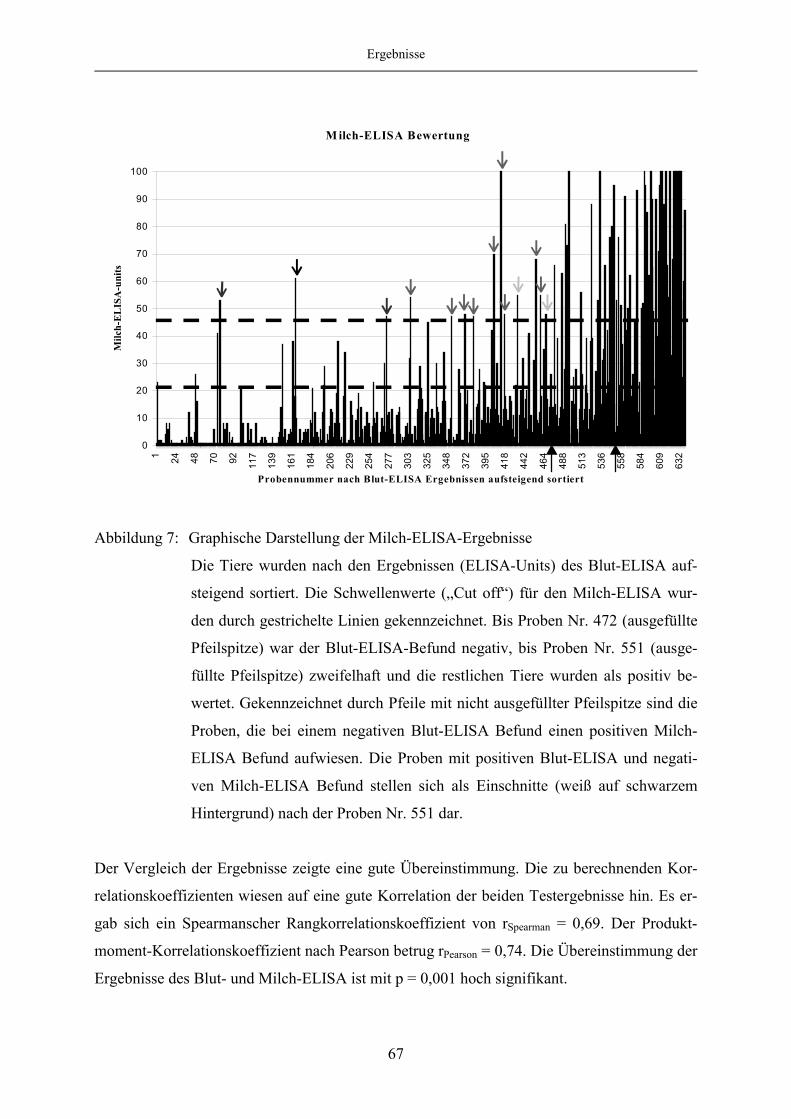
Ergebnisse
67
M ilch-ELISA Bewertung
0
10
20
30
40
50
60
70
80
90
100
1 24 48 70 92 117
139
161
184
206
229
254
277
303
325
348
372
395
418
442
464
488
513
536
558
584
609
632
Probennummer nach Blut-ELISA Ergebnissen aufsteigend sortiert
Milc
h-E
LIS
A-u
nits
Abbildung 7: Graphische Darstellung der Milch-ELISA-Ergebnisse
Die Tiere wurden nach den Ergebnissen (ELISA-Units) des Blut-ELISA auf-
steigend sortiert. Die Schwellenwerte („Cut off“) für den Milch-ELISA wur-
den durch gestrichelte Linien gekennzeichnet. Bis Proben Nr. 472 (ausgefüllte
Pfeilspitze) war der Blut-ELISA-Befund negativ, bis Proben Nr. 551 (ausge-
füllte Pfeilspitze) zweifelhaft und die restlichen Tiere wurden als positiv be-
wertet. Gekennzeichnet durch Pfeile mit nicht ausgefüllter Pfeilspitze sind die
Proben, die bei einem negativen Blut-ELISA Befund einen positiven Milch-
ELISA Befund aufwiesen. Die Proben mit positiven Blut-ELISA und negati-
ven Milch-ELISA Befund stellen sich als Einschnitte (weiß auf schwarzem
Hintergrund) nach der Proben Nr. 551 dar.
Der Vergleich der Ergebnisse zeigte eine gute Übereinstimmung. Die zu berechnenden Kor-
relationskoeffizienten wiesen auf eine gute Korrelation der beiden Testergebnisse hin. Es er-
gab sich ein Spearmanscher Rangkorrelationskoeffizient von rSpearman = 0,69. Der Produkt-
moment-Korrelationskoeffizient nach Pearson betrug rPearson = 0,74. Die Übereinstimmung der
Ergebnisse des Blut- und Milch-ELISA ist mit p = 0,001 hoch signifikant.

Ergebnisse
68
4.2 Entwicklung einer Milch-PCR
4.2.1 METHODE ZUR MILCHAUFBEREITUNG
Auf der Grundlage des Verfahren zur DNA-Aufreinigung aus Mykobakterien (BOSE
et al. 1993) wurde ein Verfahren zur Milchprobenaufbereitung entwickelt. Die Probenaufbe-
reitung erfolgte durch einen enzymatischen Verdau und anschließende Entrahmung der
Milch. Da die PCR für den diagnostischen Einsatz entwickelt wurde, sollte eine standardi-
sierte Methode zur DNA-Extraktion eingesetzt werden (R.Z.N.A Cell and Tissue Kit, peqlab,
Erlangen).
Im Folgenden werden zunächst zwei Versuche geschildert, die aus unterschiedlichen Gründen
nicht einer optimalen Milchaufreinigung für den Einsatz in der PCR entsprachen.
In dem ersten Versuch wurde 1 ml Milch mit Rennin versetzt (3.12.2.4) und das Milchserum
durch Zentrifugation gewonnen. 200 µl des Milchserums wurden enzymatisch mit Lysozym
(Endkonzentration: 1 mg/ml) für 1 Stunde bei 4°C und mit Proteinase K (Endkonzentration: 1
mg/ml) über Nacht bei 55°C behandelt. Danach wurde der Ansatz mittels R.Z.N.A Cell and
Tissue Kit (3.2.14.2) aufgereinigt. In der angeschlossenen PCR konnte mit der infizierten
Milch kein Amplifikat erlangt werden. Es mußte daher davon ausgegangen werden, daß das
Milchserum Erreger nicht oder in zu geringer Anzahl enthielt.
Im zweiten Versuch wurde 1 ml Milch zentrifugiert (14000 rpm, 30 Minuten). Das Pellet ließ
sich in 200 µl des TL- Puffer des R.Z.N.A Cell and Tissue Kit nur schwer resuspendieren.
Der anschließende Verdau mit Lysozym und Proteinase K wurde wie oben beschrieben
durchgeführt. Bei Zentrifugation (10000 rpm, 1 Minute) der Milch durch die im R.Z.N.A Cell
and Tissue Kit enthaltenen Mini-Säulen kam es immer wieder zu Verstopfungen der Säulen,
da auch nach Enzymverdau eine vollständige Homogenisierung des Milchpellets nicht mög-
lich war. Die DNA-Elution von den verstopften Mini-Säulen gelang nicht.
Die optimale Milchaufbereitung schließlich wurde mit Milch einer an Paratuberkulose er-
krankten Kuh entwickelt. 1 ml Milch wurde für 10 Minuten bei 100°C aufgekocht. Vorversu-
che hatten gezeigt, daß der hohe Proteingehalt in Kolostralmilch dazu führte, daß die Milch
nach dem Kochschritt nicht weiterverarbeitet werden konnte. Deshalb wurde Kolostralmilch
nicht erhitzt. Nach Abkühlung der Milch im Eisbad und Zugabe von Lysozym (Endkonzen-
tration: 1 mg/ml) wurde diese für 1 Stunde im Eisbad inkubiert. Es folgte ein weiterer enzy-

Ergebnisse
69
matischer Verdau. Proteinase K (Endkonzentration: 1 mg/ml) wurde der auf 55°C erhitzten
Milch zusammen mit SDS (Endkonzentration: 1%) und EDTA (Endkonzentration: 0,1 Molar)
zugegeben. Die Inkubation erfolgte über Nacht bei 55°C. Die Phasentrennung der Milch ge-
lang durch Zentrifugation (7000 rpm, 10 Minuten). Zur DNA-Extraktion wurden 200 µl der
wäßrigen Phase eingesetzt.
4.2.1.1 Entwicklung einer standardisierten Methode zur DNA-Extraktion
Für die DNA-Extraktion aus Milch konnte der R.Z.N.A Cell and Tissue Kit der Firma
peqlab, Erlangen erfolgreich angepasst werden.
1. Die Inkubationszeit mit dem zur Zellyse eingesetzten BL-Puffer wurde von 10 Minu-
ten auf 30 Minuten verlängert.
2. Die Elution der DNA von der Säulenmatrix wurde zweimalig mit jeweils 50 µl Eluti-
onspuffer durchgeführt.
Im Übrigen wurde bei der DNA-Extraktion nach der Herstellervorschrift verfahren.
4.2.1.2 Aufbereitung der Milchproben mit negativer interner Funktionskontrolle
Im Falle einer negativen internen Funktionskontrolle wurde eine weitere Aufreinigung
mit dem Gene clean II Kit (DIANOVA, Hamburg) der DNA-Extraktion angeschlossen. Im
Rahmen der Verlaufsuntersuchung (4.3.1) konnte bei 6 Milchproben mit negativer interner
Funktionskontrolle nach der zweiten Aufreinigung ein auswertbares Ergebnis in der PCR er-
mittelt werden.
4.2.1.3 Lagerung der Milchproben
Rohmilch konnte noch nach einer Lagerzeit von fünf bis sieben Tagen bei 4°C aufge-
arbeitet werden. Auch bei –20°C gelagerte Milch konnte ohne Einschränkung nach der be-
schriebenen Methode aufbereitet werden.

Ergebnisse
70
4.2.2 ETABLIERUNG DER PCR
Die PCR wurde als touchdown-PCR mit den Primern nach DORAN et al. 1994 eta-
bliert. Der Ansatz hatte ein Volumen von 50 µl und setzte sich wie folgt zusammen: 20 mM
Tris-HCl, 50 mM KCl, 1,5 mM MgCl2, 0,2 mM dNTPs, Primerpaar je 5µM, 2,5 U Taq-DNA-
Polymerase (Gibco, Eggenstein).
4.2.2.1 Bestimmung der Nachweisgrenze
Zur Ermittlung der Nachweisgrenzen wurde eine Suspension mit definierter Erreger-
dichte benötigt. Dafür wurde von einer Erregersuspension in PBS eine Verdünnungsreihe
angefertigt, die von einer Suspension mit einer optischen Dichte von 0,1 bei 660 nm ausging.
Diese Verdünnungsstufe enthielt bei mehrmaliger Zählung in der THOMA-Kammer eine Er-
regerdichte von 107 M. paratuberculosis pro ml. Durch Zentrifugieren (14000 rpm, 30 Minu-
ten) wurden die Keime pelletiert und die Milch nachweislich nicht infizierter Kühe experi-
mentell damit kontaminiert.
Im Folgenden wurde die Nachweisgrenze wiederholt an Milch von Kühen verschiedener
Laktations- und Eutergesundheitsstadien ermittelt. Zur Absicherung der erhaltenen Nach-
weisgrenzen wurden die Bestimmungen mehrmals durchgeführt. Die Tabelle 8 zeigt den Ver-
gleich der Empfindlichkeit der PCR. In Milchproben der Hochlaktation war bei 14 Wieder-
holungen eine Nachweisgrenze von mindestens 103 Erregern pro ml Milch verifizierbar. 103
Erreger pro ml Milch konnten auch in einer bakteriell stark belasteten Mastitis-Milch repro-
duzierbar nachgewiesen werden. Für Kolostrum wurde eine Nachweisgrenze von 104 Erre-
gern pro ml Milch ermittelt.
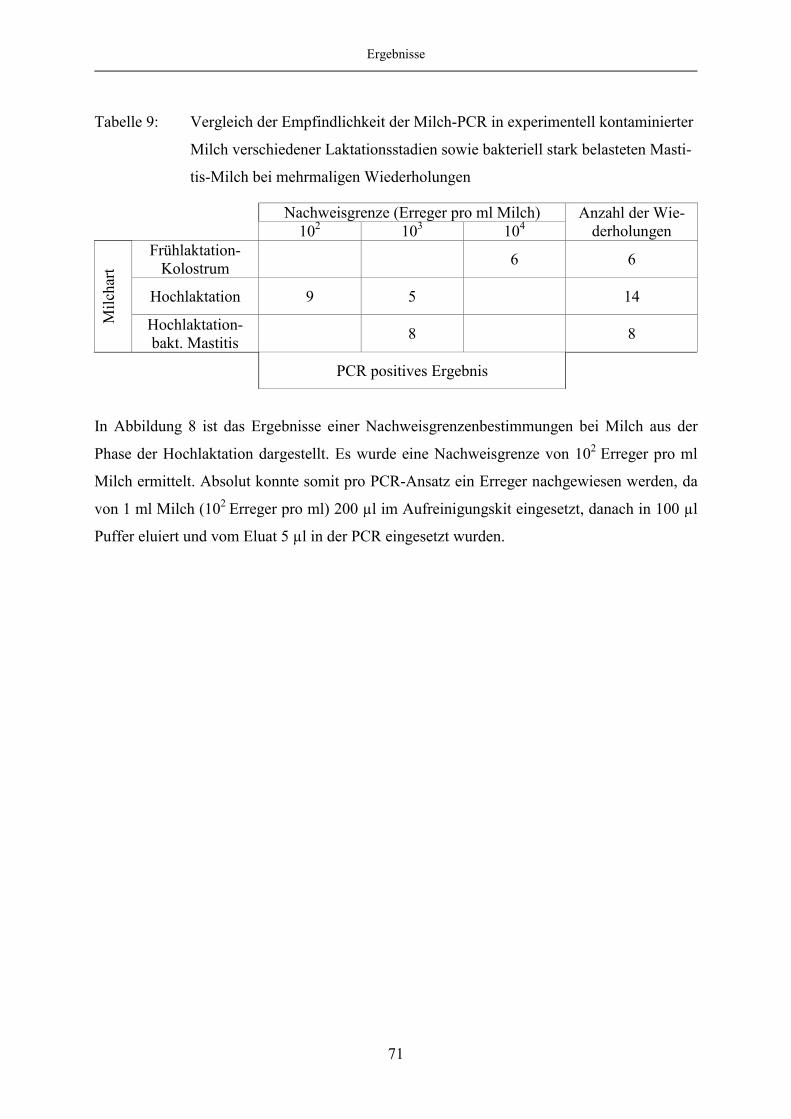
Ergebnisse
71
Tabelle 9: Vergleich der Empfindlichkeit der Milch-PCR in experimentell kontaminierter
Milch verschiedener Laktationsstadien sowie bakteriell stark belasteten Masti-
tis-Milch bei mehrmaligen Wiederholungen
Nachweisgrenze (Erreger pro ml Milch)102 103 104
Anzahl der Wie-derholungen
Frühlaktation-Kolostrum 6 6
Hochlaktation 9 5 14
Milc
hart
Hochlaktation-bakt. Mastitis 8 8
PCR positives Ergebnis
In Abbildung 8 ist das Ergebnisse einer Nachweisgrenzenbestimmungen bei Milch aus der
Phase der Hochlaktation dargestellt. Es wurde eine Nachweisgrenze von 102 Erreger pro ml
Milch ermittelt. Absolut konnte somit pro PCR-Ansatz ein Erreger nachgewiesen werden, da
von 1 ml Milch (102 Erreger pro ml) 200 µl im Aufreinigungskit eingesetzt, danach in 100 µl
Puffer eluiert und vom Eluat 5 µl in der PCR eingesetzt wurden.

Ergebnisse
72
Abbildung 8: Ermittlung der Nachweisgrenze experimentell kontaminierter Milch (Hoch-
laktation)
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2: Erregerdichte 10 / ml MilchSpur 3: Erregerdichte 102 / ml MilchSpur 4: Erregerdichte 103 / ml MilchSpur 5: Erregerdichte 104 / ml MilchSpur 6: Erregerdichte 105 / ml MilchSpur 7: Erregerdichte 106 / ml MilchSpur 8: Erregerdichte 107 / ml Milch
4.2.2.2 Interne Funktionskontrolle
Für den diagnostischen Einsatz von PCR-Tests wird neben einer standardisierbaren
Probenaufbereitung eine interne Funktionskontrolle gefordert (IEVEN und GOOSSENS
1997).
Interne Funktionskontrollen können im Rahmen einer Multiplex-PCR (EDINGLOH et al.
1999) eine Aussage über die Funktionalität jedes einzelnen PCR-Ansatzes ermöglichen.

Ergebnisse
73
Positiver Milch wurde Mycobacterium phlei und in weiteren Vorversuchen Mycobacterium
avium vor der Aufreinigung zugesetzt, um in der anschließenden PCR mit einem weiteren
Primerpaar ein zweites Amplifikat zu erhalten. Die Erregerdichten wurden, wie unter 3.2.3,
beschrieben bestimmt. Bei M. phlei wurde eine Region nachgewiesen, die nur einmal im Ge-
nom vorkommt (Primer 264/283) (SCHEIBL 1996). Die Erregerdichte von M. phlei in der
Milch mußte 109 Keime/ml Milch betragen, um in der PCR ein Amplifikat zu erhalten. In
dieser Keimkonzentration war es nicht mehr möglich die M. paratuberculosis spezifische
DNA zu amplifizieren.
Bei M. avium mußte die Erregerdichte 105 Keime/ml Milch betragen. Es handelte sich bei
dem Amplifikat um einen Ausschnitt eines Insertionselements (IS 901), das mehrmals im
Genom vorhanden ist. Doch auch hier war es nicht möglich gleichzeitig M. paratuberculosis
spezifische DNA zu amplifizieren. In den beschriebenen Vorversuchen gelang es nicht, in
einem einzigen PCR-Ansatz zwei Amplifikate zu erhalten.
Schließlich wurde als interne Funktionskontrolle M. paratuberculosis-DNA zusätzlich zum
Template in der PCR eingesetzt, indem ein vollständiger PCR-Ansatz geteilt und eine Hälfte
mit M. paratuberculosis-DNA versetzt wurde.
Mittels Titration der Template-DNA in der PCR konnte die geeignete Konzentration für die
interne Funktionskontrolle ermittelt werden. Nach spektralphotometrischer Bestimmung er-
gab sich eine DNA-Konzentration der Ausgangslösung von 0,75 µg/µl. Die geeignete Kon-
zentration für die interne Funktionskontrolle betrug bei einer Verdünnung der Ausgangslö-
sung von 1:105 7,5 pg/µl. Von der verdünnten Lösung wurde 1 µl pro PCR-Ansatz eingesetzt.

Ergebnisse
74
Abbildung 9: Bestimmung der optimalen Konzentration an M. paratuberculosis-DNA für
die interne Funktionskontrolle
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2: Verdünnung 1: 7,5x104
Spur 3: Verdünnung 1: 105
Spur 4: Verdünnung 1: 5x105
Spur 5: Verdünnung 1: 106
Abbildung 10 zeigt die Ergebnisse einer PCR-Analyse dreier verschiedener Milchproben. In
der links aufgetragenen Probe (Spur 2 und Spur 3) konnte M. paratuberculosis spezifische
DNA nachgewiesen werden. Die interne Funktionskontrolle ist positiv. In der mittleren Probe
(Spur 4 und Spur 5) konnte keine M. paratuberculosis spezifische DNA nachgewiesen wer-
den. Auch hier ist die interne Funktionskontrolle positiv. In der rechten Probe (Spur 6 und
Spur 7) wurde die interne Funktionskontrolle nicht amplifiziert. Diese Probe wurde erneut
aufgereinigt.

Ergebnisse
75
Abbildung 10: PCR-Analyse dreier ausgewählter Milchproben zur Interpretation der internen
Funktionskontrolle
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2: Probe der Kuh 239 von 10.07.1999 mit interner Funktionskontrolle
(Spur 3)Spur 4: Probe der Kuh 282 von 10.07.1999 mit interner Funktionskontrolle
(Spur 5)Spur 6: Probe der Kuh 235 von 11.09.1999 mit interner Funktionskontrolle
(Spur 7)
Als negativ gewertet wurde eine Milchprobe, wenn nur der Ansatz mit der internen Funk-
tionskontrolle ein Amplifikat in der Größe von 548 bp aufwies. Wenn sowohl in dem Ansatz,
indem sich nur die Probe befand, als auch in dem Ansatz mit Probe und interner Funktions-
kontrolle das PCR-Produkt amplifiziert wurde, wurde die Probe als positiv bewertet. Die Pro-
ben, in denen keine interne Funktionskontrolle amplifiziert werden konnte, wurden einer er-
neuten Aufreinigung unterzogen.
4.2.2.3 Sequenzierung zufällig ausgewählter PCR-Produkte
Die zur Bestätigung der Amplifikate durchgeführte Sequenzierung von zwei zufällig
ausgewählten PCR-Produkten bestätigte die angegebene Länge des Amplifikates von 548 bp
zum Nachweis von M. paratuberculosis (DORAN et al. 1994). Die Sequenzen der ausge-
wählten Amplifikate stimmten mit den Datenbankeinträgen überein.

Ergebnisse
76
4.3 Validierung von Milchserologie und Milch-PCR in einer Verlaufsuntersuchung
Im folgenden Versuchsabschnitt wurden die beiden im Rahmen dieser Arbeit ent-
wickelten Nachweismethoden an Milchproben von zehn klinisch verdächtigen Tieren aus
zwei verschiedenen Beständen angewendet. Die Tiere wurden anhand einer Eingangsuntersu-
chung (Blutserologie und Kotkultur) zwei Monate vor Versuchsbeginn ausgewählt. Die Ver-
laufsuntersuchung begann im Juli 1999 und endete mit der Entnahme der Ileocaecal-
lymphknoten bei der Schlachtung der Tiere im November 1999. In diesem Untersuchungs-
zeitraum wurden wöchentlich Blutproben, Kotproben und Viertelgemelkproben genommen.
Die Viertelgemelkproben für den Nachweis von M. paratuberculosis spezifischer DNA mit-
tels PCR wurden zu gleichen Anteilen gemischt (Einzelgemelkproben). Im Fall eines negati-
ven Ergebnisses in der Einzelgemelkprobe wurden die Viertelgemelkproben in der Reihenfol-
ge vorne rechts, vorne links, hinten rechts und hinten links nachgetestet. Bei negativen inter-
nen Funktionskontrollen wurde die PCR wiederholt und gegebenenfalls eine zweite Aufreini-
gung durchgeführt.
Im Folgenden werden die Ergebnisse der Einzeltiere von jeweils fünf Terminen exemplarisch
gezeigt, gefolgt von einer Übersichtstabelle, die die Ergebnisse aller Tiere im Verlauf der
Untersuchung zusammenfaßt. Aus Gründen der Übersichtlichkeit wurden die Daten der Tiere
in Steckbriefform dargestellt.

Ergebnisse
77
4.3.1 UNTERSUCHUNSERGEBNISSE
Kuh Nr. 209Geburtsdatum: 07.12.1992 Abkalbetermin: 02.11.1998 / 31.10.99 (während der Verlaufsuntersuchung)
Schlachttermin: 17.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war positiv.M. paratuberculosis konnte kulturell im Kot nicht nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA wurde regelmäßig in der Milch sowie im Kolostrum (31.10.99)mittels PCR nachgewiesen. Die milch- und blutserologische Untersuchung war positiv. M. paratuber-culosis konnte kulturell im Kot nicht nachgewiesen werden. Der kulturelle Nachweis des Erregers imIleocaecallymphknoten war positiv.
Entnahme-datum 10.7.99 27.7.99 31.10.99 5.11.99 1) 11.11.991)
Milch-ELISA[[[[EU]]]] 93 100 93 72 60
Blut-ELISA[[[[EU]]]] 84 73 65 55 81
Kotkultur - - - - -
Kulturbefund Ileocaecallymphknoten: positiv
Abbildung 11: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 209
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 209Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41
1) Diese Proben wurden wegen negativer interner Funktionskontrolle nachuntersucht.

Ergebnisse
78
Kuh Nr. 235Geburtsdatum: 19.09.1999 Abkalbetermin: 23.12.1998 Schlachttermin: 17.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war positiv.M. paratuberculosis konnte kulturell im Kot nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:In der zweiten Hälfte des Untersuchungszeitraumes (ab 04.09.99) war es mittels Milch-PCR möglich M.paratuberculosis spezifische DNA nachzuweisen. Zu diesem Zeitpunkt stieg der Antikörperspiegel inder Milch in den fraglichen (04.09.99/23.09.99) bzw. positiven (11.09.99) Bereich. Die blutserologischeUntersuchung war positiv. M. paratuberculosis konnte kulturell im Kot nachgewiesen werden. Derkulturelle Nachweis des Erregers im Ileocaecallymphknoten war negativ.
Entnahme-datum 10.7.99 02.8.99 04.9.99 11.9.991) 23.9.99
Milch-ELISA[[[[EU]]]] 13 15 42 51 48
Blut-ELISA[[[[EU]]]] 61 75 68 70 69
Kotkultur - + + + +
Kulturbefund Ileocaecallymphknoten: negativ
Abbildung 12: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 235
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 235Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41
1) Diese Proben wurden wegen negativer interner Funktionskontrolle nachuntersucht.
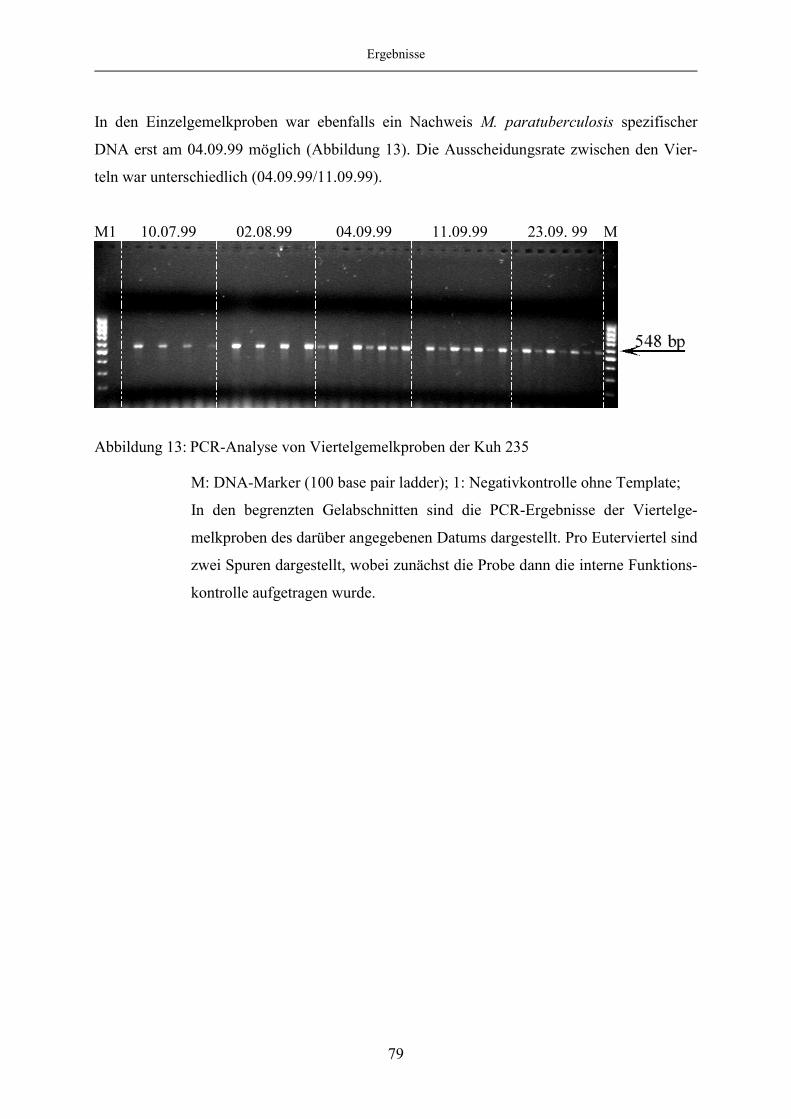
Ergebnisse
79
In den Einzelgemelkproben war ebenfalls ein Nachweis M. paratuberculosis spezifischer
DNA erst am 04.09.99 möglich (Abbildung 13). Die Ausscheidungsrate zwischen den Vier-
teln war unterschiedlich (04.09.99/11.09.99).
M1 10.07.99 02.08.99 04.09.99 11.09.99 23.09. 99 M
Abbildung 13: PCR-Analyse von Viertelgemelkproben der Kuh 235
M: DNA-Marker (100 base pair ladder); 1: Negativkontrolle ohne Template;
In den begrenzten Gelabschnitten sind die PCR-Ergebnisse der Viertelge-
melkproben des darüber angegebenen Datums dargestellt. Pro Euterviertel sind
zwei Spuren dargestellt, wobei zunächst die Probe dann die interne Funktions-
kontrolle aufgetragen wurde.

Ergebnisse
80
Kuh Nr. 238Geburtsdatum: 05.04.1994 Abkalbetermin: 21.05.1998 (Abort: 12.08.1999) Schlachttermin: 18.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war zweifelhaft.M. paratuberculosis konnte kulturell im Kot nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR nicht nachgewiesen werden. Diemilchserologische Untersuchung war negativ, die Blutserologie zweifelhaft. Der kulturelle Nachweisdes Erregers im Kot und Ileocaecallymphknoten war negativ
Entnahme-datum 03.9.991) 11.9.991) 17.9.99 23.9.99 20.11.99
Milch-ELISA[[[[EU]]]] 36 44 39 37 28
Blut-ELISA[[[[EU]]]] 56 68 75 85 57
Kotkultur - - - - -
Kulturbefund Ileocaecallymphknoten: positiv
Abbildung 14: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 238
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 238Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41
1) Diese Proben wurden wegen negativer interner Funktionskontrolle nachuntersucht.
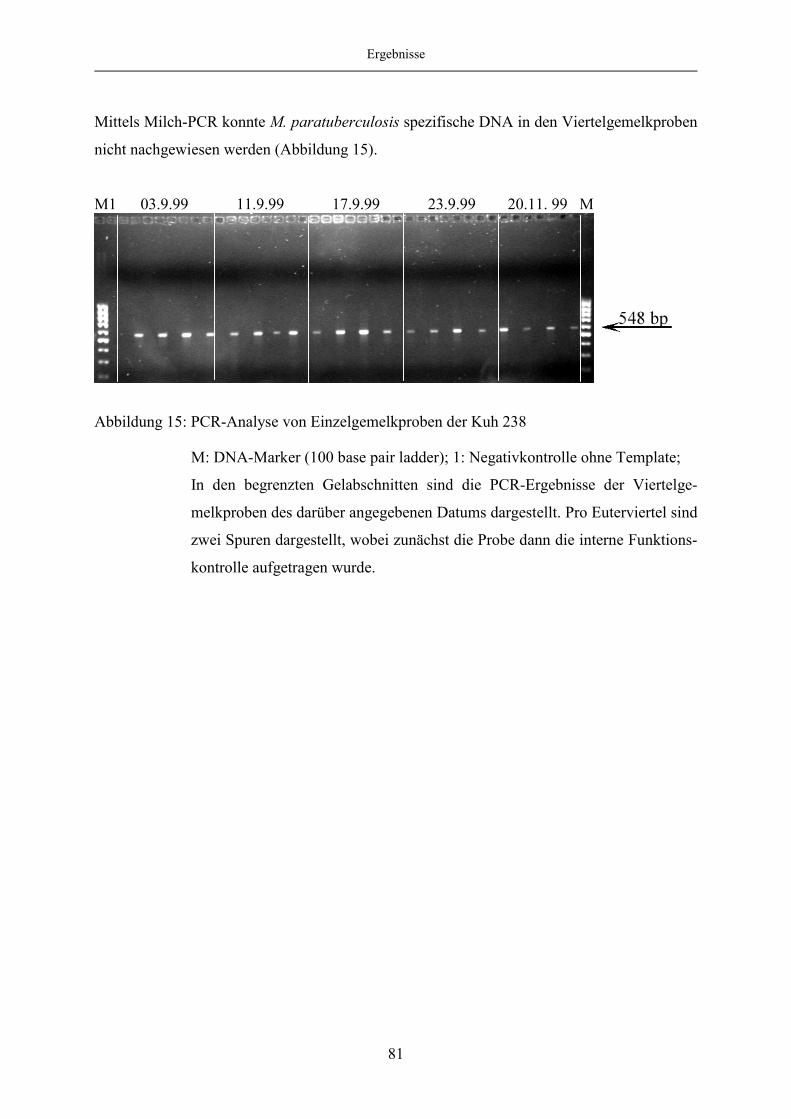
Ergebnisse
81
Mittels Milch-PCR konnte M. paratuberculosis spezifische DNA in den Viertelgemelkproben
nicht nachgewiesen werden (Abbildung 15).
M1 03.9.99 11.9.99 17.9.99 23.9.99 20.11. 99 M
Abbildung 15: PCR-Analyse von Einzelgemelkproben der Kuh 238
M: DNA-Marker (100 base pair ladder); 1: Negativkontrolle ohne Template;
In den begrenzten Gelabschnitten sind die PCR-Ergebnisse der Viertelge-
melkproben des darüber angegebenen Datums dargestellt. Pro Euterviertel sind
zwei Spuren dargestellt, wobei zunächst die Probe dann die interne Funktions-
kontrolle aufgetragen wurde.

Ergebnisse
82
Kuh Nr. 239Geburtsdatum: 30.10.1999 Abkalbetermin: 21.04.1999
Schlachttermin: 11.08.1999 aufgrund klinischer Paratuberkulose
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war positiv.M. paratuberculosis konnte kulturell im Kot nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR regelmäßig nachgewiesen werden. Diemilch- und blutserologische Untersuchung war positiv. Der kulturelle Nachweis des Erregers im Kotund Ileocaecallymphknoten war positiv.
Entnahme-datum 10.7.99 27.7.99 07.8.99 09.8.99 11.8.99
Milch-ELISA[[[[EU]]]] 51 62 62 68 72
Blut-ELISA[[[[EU]]]] 48 50 51 46 55
Kotkultur + + + + +
Kulturbefund Ileocaecallymphknoten: positiv
Abbildung 16: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 239
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 239Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41

Ergebnisse
83
Kuh Nr. 282Geburtsdatum: 08.11.1996 Abkalbetermin: 23.03.1999 Schlachttermin: 16.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war negativ.M. paratuberculosis konnte kulturell im Kot nicht nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR nicht nachgewiesen werden. Diemilch- und blutserologische Untersuchung war negativ. Der kulturelle Nachweis des Erregers im Kotund Ileocaecallymphknoten war negativ.Mittels Milch-PCR war M. paratuberculosis spezifische DNA in den Viertelgemelken nicht nachweis-bar.
Entnahme-datum 10.7.99 27.7.99 05.8.99 12.8.99 23.9.99
Milch-ELISA[[[[EU]]]] 1 1 1 1 1
Blut-ELISA[[[[EU]]]] 9 10 7 8 10
Kotkultur - - - - -
Kulturbefund Ileocaecallymphknoten: negativ
Abbildung 17: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 282
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 282Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41

Ergebnisse
84
Kuh Nr. 284Geburtsdatum: 25.14.1996 Abkalbetermin: 01.05.1999 Schlachttermin: 18.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war negativ.M. paratuberculosis konnte kulturell im Kot nicht nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR nicht nachgewiesen werden. Diemilch- und blutserologische Untersuchung war negativ. Der kulturelle Nachweis des Erregers im Kotund Ileocaecallymphknoten war negativ. Mittels Milch-PCR war M. paratuberculosis spezifische DNAin den Viertelgemelken nicht nachweisbar.
Entnahme-datum 10.7.99 17.7.99 27.7.99 07.8.99 11.9.99
Milch-ELISA[[[[EU]]]] 2 2 2 2 2
Blut-ELISA[[[[EU]]]] 20 16 15 16 19
Kotkultur - - - - -
Kulturbefund Ileocaecallymphknoten: negativ
Abbildung 18: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 284
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 284Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41

Ergebnisse
85
Kuh Nr. 76Geburtsdatum: 15.10.1993 Abkalbetermin: 13.09.1998 Schlachttermin: 16.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war positiv.M. paratuberculosis konnte kulturell im Kot nicht nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR regelmäßig nachgewiesen werden. Diemilch- und blutserologische Untersuchung war positiv. Der kulturelle Nachweis des Erregers im Kot(27.7.99/07.8.99/12.8.99) und Ileocaecallymphknoten war positiv.
Entnahme-datum 10.7.99 17.7.99 27.7.99 07.8.99 12.8.99
Milch-ELISA[[[[EU]]]] 52 59 50 67 50
Blut-ELISA[[[[EU]]]] 77 72 76 78 76
Kotkultur - - + + +
Kulturbefund Ileocaecallymphknoten: positiv
Abbildung 19: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 76
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 76Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41

Ergebnisse
86
Kuh Nr. 37Geburtsdatum: 30.01.1991 Abkalbetermin: 17.04.1999 Schlachttermin: 16.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war negativ.M. paratuberculosis konnte kulturell im Kot nicht nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR nicht nachgewiesen werden. Diemilch- und blutserologische Untersuchung war negativ. Der kulturelle Nachweis des Erregers im Kotund Ileocaecallymphknoten war negativ.Mittels Milch-PCR war M. paratuberculosis spezifische DNA in den Viertelgemelken nicht nachweis-bar.
Entnahme-datum 10.7.99 17.7.99 27.7.99 07.8.99 11.9.99
Milch-ELISA[[[[EU]]]] 15 15 17 16 10
Blut-ELISA[[[[EU]]]] 16 14 19 10 10
Kotkultur - - - - -
Kulturbefund Ileocaecallymphknoten: negativ
Abbildung 20: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 37
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 37Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41

Ergebnisse
87
Kuh Nr. 37 MGeburtsdatum: 08.01.1992 Abkalbetermin: 09.08.1998 Schlachttermin: 19.11.1999
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war positiv.M. paratuberculosis konnte kulturell im Kot nicht nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR nicht nachgewiesen werden. Die se-rologische Untersuchung der Milch ergab ein zweifelhaftes, die des Blutserums ein positives Ergebnis.An zwei Terminen (07.08.99/12.08.99) konnte der Erreger kulturell im Kot nachgewiesen werden. Derkulturelle Nachweis im Lymphknoten war positiv.Mittels Milch-PCR war M. paratuberculosis spezifische DNA in den Viertelgemelken nicht nachweis-bar.
Entnahme-datum 10.7.99 17.7.99 27.7.99 07.8.991) 12.8.99
Milch-ELISA[[[[EU]]]] 29 40 39 39 28
Blut-ELISA[[[[EU]]]] 85 80 85 67 66
Kotkultur - - - + +
Kulturbefund Ileocaecallymphknoten: positiv
Abbildung 21: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 37 M
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 37 MSpur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41
1) Diese Probe wurde wegen negativer interner Funktionskontrolle nachuntersucht.

Ergebnisse
88
Kuh Nr. 80Geburtsdatum: 20.12.1993 Abkalbetermin: 20.08.1999
Schlachttermin: 16.09.1999 aufgrund klinischer Paratuberkulose
Ergebnisse der Eingangsuntersuchung:Die blutserologische Untersuchung war positiv.M. paratuberculosis konnte kulturell im Kot nicht nachgewiesen werden.
Ergebnisse der Verlaufsuntersuchung:M. paratuberculosis spezifische DNA konnte mittels Milch-PCR regelmäßig nachgewiesen werden. Diemilch- und blutserologische Untersuchung war positiv. Der kulturelle Nachweis des Erregers im Kotund Ileocaecallymphknoten war positiv.
Entnahme-datum 20.8.99 25.8.99 01.8.99 11.9.99 16.9.99
Milch-ELISA[[[[EU]]]] 100 100 79 90 98
Blut-ELISA[[[[EU]]]] 67 95 100 100 100
Kotkultur + + + + +
Kulturbefund Ileocaecallymphknoten: positiv
Abbildung 22: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-PCR, ELISA), im Blutse-
rum, Kot und Lymphknoten der Kuh Nr. 80
M: DNA-Marker (100 base pair ladder)Spur 1: Negativkontrolle ohne TemplateSpur 2,4,6,8,10: Einzelgemelkproben der Kuh Nr. 80Spur 3,5,7,9,11: interne FunktionskontrolleMilch-ELISA/ Blut-ELISA: negativ EU ≤ 21 / EU ≤ 20
zweifelhaft EU 22-45 / EU 21-40positiv EU ≥ 46 / EU ≥ 41
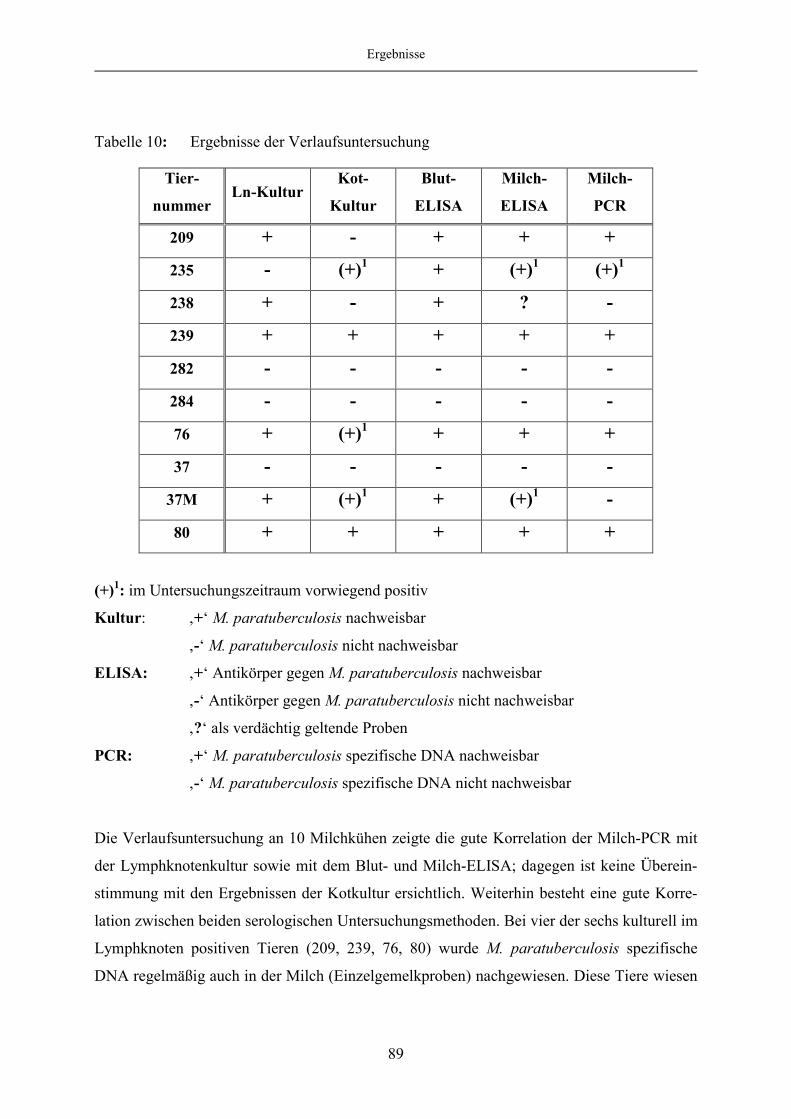
Ergebnisse
89
Tabelle 10: Ergebnisse der Verlaufsuntersuchung
Tier-
nummerLn-Kultur
Kot-
Kultur
Blut-
ELISA
Milch-
ELISA
Milch-
PCR
209 + - + + +
235 - (+)1 + (+)1 (+)1
238 + - + ? -
239 + + + + +
282 - - - - -
284 - - - - -
76 + (+)1 + + +
37 - - - - -
37M + (+)1 + (+)1 -
80 + + + + +
(+)1: im Untersuchungszeitraum vorwiegend positiv
Kultur: ‚+‘ M. paratuberculosis nachweisbar
‚-‘ M. paratuberculosis nicht nachweisbar
ELISA: ‚+‘ Antikörper gegen M. paratuberculosis nachweisbar
‚-‘ Antikörper gegen M. paratuberculosis nicht nachweisbar
‚?‘ als verdächtig geltende Proben
PCR: ‚+‘ M. paratuberculosis spezifische DNA nachweisbar
‚-‘ M. paratuberculosis spezifische DNA nicht nachweisbar
Die Verlaufsuntersuchung an 10 Milchkühen zeigte die gute Korrelation der Milch-PCR mit
der Lymphknotenkultur sowie mit dem Blut- und Milch-ELISA; dagegen ist keine Überein-
stimmung mit den Ergebnissen der Kotkultur ersichtlich. Weiterhin besteht eine gute Korre-
lation zwischen beiden serologischen Untersuchungsmethoden. Bei vier der sechs kulturell im
Lymphknoten positiven Tieren (209, 239, 76, 80) wurde M. paratuberculosis spezifische
DNA regelmäßig auch in der Milch (Einzelgemelkproben) nachgewiesen. Diese Tiere wiesen

Ergebnisse
90
im gesamten Untersuchungszeitraum im Blut- und Milch-ELISA einen als positiv zu bewer-
tenden Antikörperspiegel auf. Bei zwei in der Lymphknotenkultur positiven Tieren (238, 37
M) konnte keine M. paratuberculosis spezifische DNA in der Milch nachgewiesen werden;
die blutserologische Untersuchung war positiv, die milchserologische Untersuchung überwie-
gend positiv. Bei einem weiteren Tier (235), das serologisch positiv, aber im Ileocaecal-
lymphknoten kulturell negativ war, war es in Einzelgemelkproben in der zweiten Hälfte des
Untersuchungszeitraumes möglich mittels Milch-PCR M. paratuberculosis spezifische DNA
nachzuweisen. Die Nachtestung der Viertelgemelkproben im Fall negativer Einzelgemelkpro-
ben zeigte unterschiedliche Erreger-Ausscheidungsraten innerhalb der Euterviertel.

Diskussion
91
5 Diskussion
In Niedersachsen wird seit 1990 ein durch die Niedersächsische Tierseuchenkasse ge-
förderter Versuch zur Bestandssanierung Paratuberkulose-infizierter Milchviehherden durch-
geführt. Über einen Zeitraum von 5 Jahren werden in genau definierten Zeitabständen einer-
seits die Blutproben aller über zwei Jahre alten Milchkühe serologisch mittels ELISA und
andererseits die Kotproben kulturell untersucht. Die Probenentnahme wird von Tierärzten
durchgeführt und gestaltet sich besonders in den Sommermonaten schwierig.
In den am Sanierungsprogramm beteiligten Herden hat die Paratuberkuloseinzidenz deutlich
abgenommen und wird am Ende des Programms vermutlich zwischen 2 % und 4 % liegen. In
diesem Prävalenzbereich treten keine klinisch kranken Tiere mehr auf, und Leistungseinbußen
sind für den Landwirt nicht spürbar. Daher stößt in diesen Betrieben eine Fortsetzung der re-
lativ zeit- und kostenintensiven Blut- und Kotuntersuchung auf geringe Akzeptanz.
Um langfristig eine flächendeckende Tilgung der Paratuberkulose zu erreichen, ist zusätzlich
zur Sanierung besonders stark betroffener Betriebe auch die Ermittlung eines Paratuberkulo-
sestatus bisher nicht untersuchter Betriebe erforderlich. Da die Prävalenz in solchen Betrieben
zwischen 0 % und 25 % liegt, ist durch stichprobenartige Untersuchungen von Tieren in ei-
nem Betrieb keine sichere Aussage über den Herdenstatus zu machen. Andererseits ist die
flächendeckende, kombinierte Blut- und Kotuntersuchung aller Tiere nicht finanzierbar. Eine
mögliche Lösung – sowohl zur Aufrechterhaltung des Gesundheitsstatus nach 5-jähriger Teil-
nahme am Sanierungsprogramm der Niedersächsischen Tierseuchenkasse, wie auch zur Erhe-
bung des Paratuberkulosestatus in bisher nicht untersuchten Herden – wäre die Einführung
eines ELISA auf der Basis von Milchserum eventuell kombiniert mit einem schnellen Erre-
gernachweis aus Milch mittels PCR.
5.1 Adaption des Svanovir®- ELISA für Blutserum an Milchserum
Im Rahmen dieser Arbeit wurde, basierend auf einem Blutserum-ELISA-System
(JARK 1996), ein Milch-ELISA entwickelt. Für die Entwicklung war entscheidend, daß eine
Korrelation der IgG Konzentration in Milch und Blutserum besteht, da der größte Anteil der

Diskussion
92
in der Milch enthaltenen Immunglobuline mit dem Blut in die Milchdrüse transportiert wird
(CAFFIN et al. 1983; CAFFIN und POUTREL 1987). Diese Korrelation zwischen Blut- und
Milch-Antikörperspiegeln konnte auch für M. paratuberculosis spezifische Antikörper auf der
Basis eines LAM-ELISA bereits ermittelt werden (SWEENEY et al. 1994). Bei der Validie-
rung dieses ELISA wurde der kulturelle Nachweis von M. paratuberculosis in Kot als „Gold-
standard“ angesehen.
Um das für die Etablierung des Milch-ELISA am besten geeignete Testsystem zu identifizie-
ren wurde zunächst ein Ringversuch, an dem sieben Untersuchungsstellen in Niedersachsen
beteiligt waren, durchgeführt und ausgewertet. In diesem Versuch wurden die drei zur Zeit in
Deutschland erhältlichen Paratuberkulose-ELISA-Testkits anhand von 60 Seren, die von 10
klinisch verdächtigen Tieren zu verschiedenen Zeitpunkten genommen worden waren, mit-
einander verglichen. Mittels Kultur des Ileocaecallymphknotens konnte bei sechs Tieren M.
paratuberculosis nachgewiesen werden. Bei einem weiteren Tier wurde in der Milchuntersu-
chung die Ausscheidung des Erregers mit der Milch mittels PCR bewiesen. Bei drei der kli-
nisch verdächtigen Tiere war ein kultureller Nachweis des Erregers im Ileocaecallymphkno-
ten nicht möglich. Bei der Bewertung dieses Ergebnisses ist zu beachten, daß sogar die kultu-
relle Untersuchung des Ileocaecallymphknotens keinen absolut sicheren „Goldstandard“ dar-
stellt, sondern nur die sicherste unter Schlachthofbedingungen noch praktikable Möglichkeit.
Als „Goldstandard“ wird dabei das Testverfahren bezeichnet, das die Basis für die Beurtei-
lung eines diagnostischen Tests darstellt. Der „Goldstandard“ sollte also eine sichere Aussage
darüber erlauben, ob eine Krankheit vorliegt oder ein Tier gesund ist. Es konnte jedoch in
anderen Untersuchungen bewiesen werden, daß bei 18 % von an Paratuberkulose erkrankten
Tieren kein Nachweis des Erregers im Ileocaecallymphknoten gelang (WHITLOCK und BU-
ERGELT 1996). In einer weiteren Untersuchung konnte demonstriert werden, daß nur bei 65
% der Kotkultur positiven Tiere der Erreger kulturell im Ileocaecallymphknoten nachweisbar
war (PAVLIK et al. 1999).
In dem genannten Ringversuch zeigte der ELISA zur Serodiagnostik der bovinen Paratuber-
kulose (Riemser Arzneimittel GmbH, Insel Riems) insgesamt eine gute Reproduzierbarkeit
bei den Ergebnissen der Untersuchungsstellen. Eine Untersuchungsstelle zeigte allerdings
stark abweichende Ergebnisse; es ist jedoch nicht auszuschließen, daß es sich bei diesen Er-
gebnissen um einen Durchführungs- oder Auswertungsfehler handelt. Der ELISA erkennt nur

Diskussion
93
vier der sechs Lymphknoten-Kultur positiven Tiere. Die übrigen klinisch verdächtigen Tiere
sowie das Milch-PCR positive Tier werden vom ELISA nicht erkannt. Die Abweichung von
der angegebenen Sensitivität von 57 % ist dadurch zu erklären, daß es sich bei den unter-
suchten Tieren nicht um eine zufällige Stichprobe handelt und daß die Validierung dieses
ELISA auf der Basis der Kotkultur als „Goldstandard“ vorgenommen wurde (MILNER et al.
1990). Der kulturelle Nachweis des Erregers im Kot ist jedoch als „Goldstandard“ für die Pa-
ratuberkulosediagnostik absolut ungeeignet, da die Tiere im subklinischen Stadium den Erre-
ger nicht und später auch nur intermittierend ausscheiden (COLLINS 1996). Die mangelnde
Eignung der kulturellen Kotuntersuchung als Referenzgröße wird auch durch andere Arbeiten
belegt (MEYER ZU VILSENDORF 1995). Dieser Autor stellt in seinen Untersuchungen fest,
daß die Kotkultur gegenüber der kulturellen Untersuchung des Ileocaecallymphknotens nur
eine Sensitivität von 33 % besitzt.
Ebenso wurde die Validierung des HerdChek™ M.pt Ab-ELISA (Idexx, Wörrstadt) anhand
der Kotkultur vorgenommen (ROSSITER und BURHANS 1996). Auch dieser Test erkennt
bei Beginn des Ringversuchs ein „Goldstandard“ (Ileocaecallymphknoten-Kultur) positives
Tier nicht. Gegen Ende des Ringversuchs wird noch ein weiteres „Goldstandard“ (Ileocae-
callymphknoten-Kultur) positives Tier von dem ELISA nicht als positiv erkannt. Die übrigen
klinisch verdächtigen Tiere sowie das Milch-PCR positive Tier werden vom ELISA nicht
erkannt. Die Reproduzierbarkeit der Ergebnisse in den sieben Untersuchunsstellen ist unein-
geschränkt gut.
Als einziger der eingesetzten ELISA-Tests erkennt der Svanovir®- ELISA (Svanova, Schwe-
den) jedes der Lymphknoten-Kultur positiven Tiere sowie das nur in der Milch-PCR positive
Tier (235) und dies schon in der ersten Woche der Untersuchung. Von den übrigen klinisch
verdächtigen Tieren wird eine Kuh im gesamten Untersuchungszeitraum von allen teilneh-
menden Untersuchungsstellen als negativ beurteilt. Zwei weitere klinisch verdächtige Tiere
werden überwiegend von allen Untersuchungsstellen als zweifelhaft ermittelt.
Die Reproduzierbarkeit der Ergebnisse der Untersuchungsstellen ist beim Svanovir®- ELISA
uneingeschränkt gut. Die positive Beurteilung aller „Goldstandard“ (Ileocaecallymphknoten-
Kultur) positiven Tiere und eine frühe Erkennung auch schon subklinisch erkrankter Tiere
stimmt mit den früheren Ergebnissen zur Validierung des ELISA mittels Ileocaecal-
lymphknoten-Kultur überein (JARK 1996; REHM 1999). Auch konnten beide Autoren de-
monstrieren, daß der Svanovir®- ELISA insbesondere Tiere im relativ frühen Stadium der

Diskussion
94
Infektion, bevor die Tiere den Erreger kontinuierlich im Kot ausscheiden, erkennt. In einer
Dissertation wurde an Proben von Kühen in Nordrhein-Westfalen der HerdChek™ M.pt Ab-
ELISA mit dem Svanovir®- ELISA verglichen (VOM SCHLOß 2000); dabei wurde ermittelt,
daß der Svanovir®- ELISA infizierte Kühe signifikant eher erkennt als der HerdChek™ M.pt
Ab-ELISA.
Für die Sicherung eines Sanierungserfolges ist gerade die frühe Erkennung subklinisch infi-
zierter Tiere von großer Bedeutung. Ebenso ist es für die Qualitätssicherung des Lebensmit-
tels Milch wichtig, die Kühe vor der Ausscheidung des Erregers zu erkennen. Diese Kriterien
waren nur mit dem Svanovir®- ELISA zu erfüllen. Zusätzlich zu den fachlichen Kriterien hat
der Svanovir®- ELISA einen Vorteil im „handling“ (keine Vorinkubation) und er stellt das
kostengünstigste Produkt auf dem Markt dar. Aus diesen Gründen wurde der Svanovir®-
ELISA als Basis zur Entwicklung eines Milch-ELISA ausgewählt.
Im ELISA-Kit des Svanovir®- ELISA ist ein konjugierter, monoklonaler anti-Rind-IgG-
Antikörper enthalten, dessen Spezifität für IgG Subtypen und deren Bindungsdomäne nicht
bekannt sind. Da Kuhmilch, anders als die Milch anderer Tierarten, überwiegend Immunglo-
bulin IgG1 enthält, erschien es wahrscheinlich, daß der monoklonale Antikörper auch für den
Milch-ELISA einsetzbar war. Allerdings mußte dazu sicher gestellt werden, daß der mono-
klonale Antikörper unabhängig von den IgG-Subklassen am Fc-Fragmente bindet. Darum
wurde nach enzymatischen Verdau mit Papain des IgG, welches aus dem positiven Kon-
trollserum des Svanovir®- ELISA-Testkits aufgereinigt wurde, gezeigt, daß der monoklonale
Antikörper an das Fc-Fragment bindet und somit ohne Einschränkung für den Milch-ELISA
verwendet werden kann.
Weiterhin war es für die Etablierung eines Milch-ELISA erforderlich, eine schnelle und re-
produzierbare Methode zur Milchaufreinigung zu etablieren. Die beste Trennung des Milchse-
rums wurde durch die Behandlung der Milch mit dem Labferment Rennin erreicht (MANZ et
al. 1986). Diese extrem zeitintensive und aufwendige Methode wurde mit der in anderen
Milch-ELISA - z.B. zur Brucellen Diagnostik - angewandten Methode (MACMILLAN et al.
1990) verglichen. Die untersuchten Milchproben wiesen keine Unterschiede in den Extinktio-
nen im Svanovir®- ELISA auf, so daß mit der Zentrifugationsmethode eine einfache und
schnelle Milchaufreinigung für den Milch-ELISA etabliert werden konnte.

Diskussion
95
Bei der Etablierung eines serologischen Testverfahrens ist die Zuverlässigkeit, gemessen an-
hand der Reproduzierbarkeit, ein entscheidendes Kriterium. Die Reproduzierbarkeit von Tag
zu Tag wurde an 20 Milchproben an 8 verschiedenen Tagen überprüft. Es ergab sich ein
durchschnittlicher Variationskoeffizient von 17,13 %. Damit entspricht der Test den Forde-
rungen der OIE (Office International des Epizooties) (OFFICE INTERNATIONAL DES
EPIZOOTIES 1996).
Wenn die Reproduzierbarkeit eines serologischen Testverfahrens sichergestellt ist, muß als
nächster Schritt bei der Etablierung eine Validierung mithilfe eines anerkannten Referenzver-
fahrens erfolgen. Für den Milch-ELISA wurde dies anhand der blutserologischen Untersu-
chung von 601 Tieren mittels Svanovir®- ELISA durchgeführt, da dieser ELISA vom BgVV
zugelassen und auf der Basis der Lymphknotenkultur validiert worden war. Die kulturelle
Untersuchung von Kotproben, die zur Beurteilung anderer ELISA-Systeme diente, wurde aus
den schon genannten Gründen, wie die fehlende Ausscheidung bei subklinisch infizierten
Tieren und die nur intermittierende Ausscheidung bei erkrankten Tieren, nicht in die Validie-
rung des Milch-ELISA einbezogen.
Die Bestimmung des optimalen „Cut off“ wurde mit Hilfe einer ROC-Analyse durchgeführt
(JENSEN und POULSEN 1992). Hierbei werden Sensitivität und Spezifität bei verschiedenen
Schwellenwerten in einer Kurve dargestellt. Die Schwellenwerte mit der höchsten Sensitivität
und Spezifität konnten durch den größten Abstand zur y = x-Achse bestimmt werden. Weiter-
hin wurden die „falsch positiven“ und „falsch negativen“ Ergebnisse für die jeweiligen „Cut
off“-Werte in die Bestimmung mit einbezogen. Die so bestimmten „Cut off“-Werte waren ≤
21 (negativ) und ≥ 46 (positiv) mit einer Diagnostik-Spezifität (D-SP) von 96,6 % und einer
Diagnostik-Sensitivität (D-SN) von 79,1 %. Bei diesen „Cut off“-Werten waren 10,3 % der
untersuchten Proben als zweifelhaft zu beurteilen. Dieser zweifelhafte Bereich wurde in die
Bewertung des Milch-ELISA mit übernommen, da bei Betrieben mit hoher Prävalenz erwartet
wird, daß auch Tiere ohne persistierende Infektion etwas höhere Antikörperspiegel (gemessen
in ELISA-Einheiten) aufweisen. Die Tiere mit zweifelhaften Befund könnten somit entspre-
chend der Prävalenzen der Betriebe bewertet werden. Beim Vergleich der D-SP und D-SN
verschiedener Milch-ELISA zur Diagnostik der Paratuberkulose sind die D-SP und D-SN der
vorliegenden Arbeit als positiv zu bewerten. So konnte bei einem anderen ELISA zum Nach-

Diskussion
96
weis von M. paratuberculosis in der Milch nur eine D-SP von 83 % und eine D-SN von 60 %
erreicht werden (SWEENEY et al. 1994); ein weiterer Milch-ELISA erreicht eine D-SP von
89,7 % und eine D-SN von 71,4 % (JAKOBSEN et al. 2000).
Damit wurde im Rahmen dieser Arbeit ein Milch-ELISA etabliert, der sich für die Untersu-
chung von Einzeltieren eignet. Der Test zeigt eine gegenüber dem Blutserum-ELISA geringe-
re Sensitivität; daher ist sein Einsatz für laufende Sanierungsverfahren vorsichtig zu beurtei-
len. Er scheint jedoch unter Berücksichtigung der Kosten-Nutzen Relation als „Nachsorge-
untersuchung“ in Betrieben nach Abschluß des Sanierungsprogramms empfehlenswert. Wei-
terhin erscheint der Test zur Erhebung eines Herdenstatus geeignet. Zusätzlich haben erste
Untersuchungen gezeigt, daß die Entwicklung eines Tankmilch-ELISA auf der Basis dieses
Milch-ELISA möglich scheint. Jedoch muß systematisch überprüft werden, inwieweit das
Ergebnis der Tankmilchuntersuchung eines Betriebes mit dem Ergebnis einer Einzeltierunter-
suchung übereinstimmt. Gegebenenfalls könnten auf der Basis eines solchen Test zuminde-
stens Herden mit hoher Prävalenz sicher und kostengünstig identifiziert werden. Eine derarti-
ge Untersuchung wäre wichtig beispielweise vor Exportverkäufen, bei denen der beamtete
Tierarzt die „Paratuberkulosefreiheit“ bescheinigen muß.
5.2 Entwicklung einer Milch-PCR
Für die vorliegende Untersuchung wurde zunächst eine Methode zur Aufbereitung von
Milchproben entwickelt. Die einzige bisher beschriebene Methode wurde nicht übernommen,
da sie nur zur Aufreinigung pasteurisierter Milchproben eingesetzt wurde (MILLAR et al.
1996) und in den Vorversuchen bei Verwendung nicht pasteurisierter Milch einen sehr hohen
Anteil an falsch negativen Ergebnissen ergab.
Stattdessen wurde ein enzymatischer Aufschluß von Mykobakterien, mit dem eine hohe Aus-
beute an DNA pro Gramm Bakterienfeuchtmasse erreicht werden konnte (BOSE et al. 1993)
als Grundlage gewählt. In Anlehnung an diese Methode wurde dann ein Verfahren zur DNA-
Aufreinigung aus M. paratuberculosis entwickelt. Dabei zeigte sich, daß bei der Auftrennung
von künstlich kontaminierter Milch durch Zentrifugation in Pellet, Milchserum und Rahm
keine Erreger im Milchserum und im Rahm nachweisbar waren. Die alleinige Weiterverar-
beitung des Pellets führte aber zu Verstopfungen der Zentrifugationssäule im Extraktionskit.
Im Folgenden wurde daher jeweils 1 ml Milch nach dem Aufkochen ohne Phasentrennung

Diskussion
97
enzymatisch mit Lysozym und Proteinase K verdaut. Diese Probenaufbereitung wurde mit
dem R.Z.N.A Cell and Tissue Kit (peqlab, Erlangen) kombiniert. Die Vorteile dieses Verfah-
rens liegen in der Möglichkeit einer standardisierten Anwendung, eines sicheren Extrakti-
onserfolges sowie der schnellen und einfachen Durchführung. Die zeitaufwendige und an den
Einsatz von Phenol und Chloroform gebundene klassische DNA-Extraktion (SAMBROOK et
al. 1989) wurde mit diesem System umgangen.
Die Bestimmung der Nachweisgrenze wurde mit künstlich kontaminierten Milchproben von
nachweislich nicht infizierten Kühen durchgeführt. Die erreichte Nachweisgrenze differierte
in Abhängigkeit von der Laktationsperiode. Die höchste Nachweisgrenze (102) konnte bei
Milch aus der Hochlaktation, die nicht durch andere Keime belastet war, erreicht werden. Bei
diesen Proben wird die Nachweisgrenze weder durch hohen Proteingehalt noch durch Fremd-
DNA oder eine Kontamination mit Bakterien oder Pilzen negativ beeinflußt. Die Untersu-
chung von Kolostrum gestaltete sich dagegen aufwendiger. Der normalerweise am Beginn der
Aufreinigung durchgeführte Kochschritt mußte hierbei entfallen, da das Kolostrum nach dem
Aufkochen durch den hohen Proteingehalt „schnittfest“ war und sich zur weiteren Aufreini-
gung nicht eignete. Bei der alleinigen Aufreinigung mit dem R.Z.N.A Cell and Tissue Kit
(peqlab, Erlangen) war es nicht zuverlässig möglich, die Proteine und andere PCR-
Hemmstoffe in ausreichendem Maße aus der Probe zu entfernen. Dies zeigte sich durch die
negative interne Funktionskontrolle in der PCR. Mittels weiterer Aufreinigung mit dem Ge-
neClean II®Kit, Dianova, Hamburg, die mit dem Eluat aus der ersten Aufreinigung durchge-
führt wurde, konnten diese PCR-Hemmstoffe zuverlässig aus der Probe entfernt werden. Al-
lerdings war die DNA-Ausbeute nach diesen beiden Aufreinigungsschritten geringer. Daher
konnte bei Kolostrum nur eine Nachweisgrenze von 104 Keimen/ml Kolostrum erreicht wer-
den.
Im Vergleich zu veröffentlichten Berichten sind diese erzielten Nachweisgrenzen positiv zu
beurteilen. So konnte in der Untersuchung von künstlich mit M. paratuberculosis kontami-
nierten Proben nur bei Verwendung pasteurisierter Milch eine Nachweisgrenze von 200-300
Erregern/ml Milch angegeben werden (MILLAR et al. 1996). Die sehr niedrige Nachweis-
grenze von 4 Erregern pro ml Milch, die in einer Coxiellen-PCR erreicht wurde (EDINGLOH
et al. 1999) wird dadurch erlangt, daß die Methodik eine Aufkonzentration des Erregers um
den Faktor 100 mittels Ultrazentrifugation vorsieht. Unter Berücksichtigung dieser in der

Diskussion
98
Routinediagnostik nicht praktikablen Methode liegt die hier vorgestellt Milch-PCR mit ihrer
Nachweisgrenze im akzeptablen Bereich.
Ohne interne Funktionskontrolle kann bei einem negativen PCR-Ergebnis eine Hemmung der
Polymerasefunktion als Ursache nicht ausgeschlossen werden. Derartige falsch negative Er-
gebnisse sind jedoch für die diagnostische Arbeit nicht akzeptabel. Diese Problematik läßt
sich prinzipiell am einfachsten mit Hilfe einer Multiplex-PCR ausschließen (EDINGLOH
1999). Hierbei ist grundsätzlich nur der Einsatz eines weiteren Primerpaars und entsprechen-
der Kontroll-DNA notwendig. Vorversuche zeigten jedoch, daß die Zugabe anderer Myko-
bakterien als interner Kontrolle zur Milch sowohl die Aufreinigung als auch die Amplifikati-
on der M. paratuberculosis spezifischen DNA stark störten oder sogar völlig blockierten. Es
ist bekannt, daß die Polymerase durch verschiedene Inhaltsstoffe gehemmt wird (ROLFS et
al. 1992). Hierzu zählt auch eine hohe Konzentration an Fremd-DNA, die bei der Zugabe an-
derer Mykobakterien auftrat.
Daher wurde anstelle einer Multiplex-PCR eine Teilung der zu untersuchenden Probe mit
Zugabe von M. paratuberculosis-DNA zu einem Ansatz vorgenommen. Mit diesem Ansatz
konnte die Hemmung der PCR durch Inhaltsstoffe in der Probe sicher ausgeschlossen werden,
ohne daß eine Beeinträchtigung der Sensitivität erfolgte.
Im Rahmen des Sanierungsprogramms der Niedersächsischen Tierseuchenkasse wird der Er-
regernachweis kulturell aus Kot geführt. Dieser Nachweis nimmt aufgrund der langen Gene-
rationszeit des Erregers Wochen in Anspruch. Der Nachweis von M. paratuberculosis in Kot
mittels PCR ist zur Zeit keine Alternative, da er sich nur für den Nachweis von mittel- bis
hochgradig ausscheidenden Kühen eignet (mindestens 104 KBE/g Kot) (VARY et al. 1990;
WHIPPLE et al. 1992).
Die Nachweisgrenze der hier vorgestellten Milch-PCR liegt jedoch deutlich unter der der Kot-
PCR und ist vergleichbar mit der Nachweisgrenze der routinemäßig durchgeführten kulturel-
len Kotuntersuchung (103 KBE/g Kot) (COLLINS 1996). Die in der Literatur angegebene
Nachweisgrenze für die kulturelle Kotuntersuchung von 10-100 KBE/g Kot (WHIPPLE et al.
1992) wird jedoch dadurch erreicht, daß die Kotproben parallel auf mehrere Anzuchtmedien
(5-10) aufgebracht wurden. Damit könnte die Milch-PCR eine mögliche Alternative zur kul-
turellen Kotuntersuchung im Rahmen des Sanierungprogrammes darstellen.

Diskussion
99
5.3 Validierung von Milchserologie und Milch-PCR in einer Verlaufsuntersuchung
Die entwickelten Methoden wurden abschließend an mehreren Hundert Milchproben, die im
Rahmen einer fünf Monate dauernden Verlaufsuntersuchung mit zehn klinisch verdächtigen
Milchkühen gewonnen worden waren, validiert. Diesen Tieren wurden in zweiwöchentlichem
Abstand Blut-, Milch- und Kotproben zur vergleichenden serologischen bzw. kulturellen Un-
tersuchung entnommen. Bei der Schlachtung wurde zusätzlich der Ileocaecallymphknoten
entnommen und kulturell auf M. paratuberculosis untersucht. Von jeder Kuh lagen somit von
ca. 20 Terminen Viertelgemelkproben vor, die sowohl einzeln als auch zu gleichen Anteilen
gemischt als Einzelgemelkprobe im Milch-ELISA untersucht wurden. Die Ergebnisse der
Viertel- und Einzelgemelkproben stimmen wie erwartet überein. Die Einzelgemelkprobe kann
somit als geeignetes Untersuchungsmaterial für den Milch-ELISA angesehen werden. In der
Milch-PCR zeigten sich bei der Untersuchung der Viertel- und Einzelgemelkproben Unter-
schiede in der Ausscheidungsrate von M. paratuberculosis. Diese Unterschiede zwischen den
Eutervierteln sind in dem Aufbau der Milchdrüse begründet und von anderen Erkrankungen
her bekannt. Jedes Viertel stellt aufgrund der Entwicklung und damit der Ausbildung des
Hohlraumsystems eine selbständige Einheit dar (WENDT et al. 1994). Bei der Mastitis
phlegmonosa, die Folge eines primär im Körper vorhandenen Infektionsherdes ist, gelangen
die Erreger, wie auch der Erreger der Paratuberkulose, über den hämatogenen oder lympho-
genen Infektionsweg ins Euter (GRUNERT 1995). Häufig ist bei dieser Form der Mastitis nur
ein Euterviertel betroffen. Auch bei Salmonella dublin-Infektionen konnte beobachtet werden,
daß sich die Erregerausscheidung auf ein Euterviertel beschränkte (SMITH et al. 1989). Die
alleinige Untersuchung der Einzelgemelkproben mit der Milch-PCR reicht somit nicht aus. So
konnte auch im Rahmen der vorliegenden Arbeit gezeigt werden, daß bei nachweisbarer Erre-
gerausscheidung in nur einem Viertel in der Einzelgemelkprobe aufgrund der Verdünnung
keine M. paratuberculosis spezifische DNA mehr nachweisbar war. Damit ist die Untersu-
chung der Einzelgemelkproben zwar für Screening-Zwecke empfehlenswert; bei negativen
PCR-Befund und klinischen Verdacht sind jedoch die Viertelgemelkproben einzeln zu unter-
suchen.
In der Verlaufsuntersuchung wurde die gute Korrelation zwischen Blut- und Milch-ELISA
bestätigt. Diese Korrelation konnte mit den Korrelationskoeffizienten nach Spearman (r Spear-
man = 0,69) und Pearson (r Pearson = 0,74) rechnerisch ausgedrückt werden (HARMS 1998).
Diese Korrelation ist vergleichbar zu einem anderen Milch-ELISA zur Diagnostik der Paratu-

Diskussion
100
berkulose (SWEENEY et al. 1994) für den eine Korrelation von r = 0,71 bestimmt wurde.
Auch für andere Milch-ELISA-Systeme (Salmonella dublin) wurden vergleichbare Korrela-
tionen (r = 0,63-0,71 für verschiedene Versuchsgruppen) beim Vergleich der serologischen
Blut- und Milch-Ergebnisse angegeben (SMITH et al. 1989).
Mit der Milch-PCR konnte bei vier der sechs „Goldstandard“ (Ileocaecallymphknoten-Kultur)
positiven Tieren die Erregerausscheidung mit der Milch bewiesen werden. Der fehlende
Nachweis des Erregers in der Milch bei zwei „Goldstandard“ positiven Tieren könnte durch
eine intermittierende Ausscheidung des Erregers mit der Milch bedingt sein; eine intermittie-
rende Ausscheidung wurde bei Salmonella dublin-Infektionen angenommen (SMITH et al.
1989). Insgesamt wird die gute Sensitivität der hier vorgestellten Milch-PCR durch den Ver-
gleich mit früheren Untersuchungen untermauert, in denen gezeigt wurde, daß auch mittels
kultureller Milchuntersuchung der Erreger bei nur 35 % der klinisch erkrankten (TAYLOR et
al. 1981) und 11,6 % der subklinisch infizierten Tiere nachweisbar war (SWEENEY et al.
1992b).
Der Vergleich der Milch-PCR und der kulturellen Kotuntersuchung zeigte, daß die Erre-
gerausscheidung mit der Milch und die Ausscheidung über den Kot nicht in einem direkten
Zusammenhang stehen. So war bei einem Tier der Erreger im Kot kulturell nachweisbar, aber
mittels PCR an diesem Termin in der Milch nicht nachzuweisen. Bei einem anderen Tier
wurde der Erreger mittels Milch-PCR nachgewiesen während zum selben Zeitpunkt ein kultu-
reller Nachweis im Kot nicht möglich war. Dieser Befund ist mit den unterschiedlichen We-
gen, auf denen der Erreger in die Milch und in den Kot gelangt, zu erklären. So tritt der Erre-
ger in die Milch auf lymphogenem und/oder hämatogenem Weg ein (SWEENEY et al.
1992b); in den Kot hingegen gelangen die Mykobakterien bei Abstreifung der befallenen
epitheloiden Zellen ins Darmlumen (CHIODINI et al. 1984).
Zusammenfassend ist zu sagen, daß die Ergebnisse des Milch-ELISA und der Milch-PCR
durch blutserologische Befunde sowie durch die kulturelle Untersuchung von Kotproben und
Ileocaecallymphknoten bestätigt werden konnten. Damit ist gezeigt worden, daß die Milch-
PCR und der Milch-ELISA im Rahmen der Paratuberkulose-Diagnostik wertvolle neue dia-
gnostische Werkzeuge zur frühzeitigen, sicheren und kostengünstigen Diagnose dieser Er-

Diskussion
101
krankung darstellen und insbesondere im Rahmen der Nachsorge sanierter Betriebe eingesetzt
werden können. In zukünftigen Untersuchungen bleibt zu prüfen, ob die in der Beihilfesat-
zung der Tierseuchenkasse vorgeschriebene, kulturelle Kotuntersuchung durch die Milch-
PCR ersetzt werden kann, oder ob eventuell sogar eine Sanierung anhand einer Kombination
von Milch-ELISA und Milch-PCR möglich ist. Weiterhin ist die Sensitivität des Milch-
ELISA bei Tankmilchuntersuchungen zu ermitteln, um festzustellen, inwieweit sich ein sol-
ches Testverfahren für flächendeckende Statuserhebungen im Rahmen der regelmäßig durch-
geführten Routineuntersuchungen auf Brucellose und Leukose eignet.

Zusammenfassung
102
6 Zusammenfassung
Die Paratuberkulose des Rindes ist eine weltweit verbreitete Erkrankung. Die Tiere in-
fizieren sich in der Regel in den ersten Lebensmonaten, zeigen aber danach oft über Jahre
keinerlei klinische Symptome, obwohl sie den Erreger mit dem Kot und der Milch ausschei-
den.
In der vorliegenden Arbeit wurden zwei Verfahren zur Diagnose der Paratuberkulose aus
Milch etabliert. Das eine Verfahren erlaubt einen Antikörpernachweis mittels ELISA und das
andere einen Erregernachweis mittels PCR.
Für den Antikörpernachweis wurde aufgrund der Ergebnisse eines Ringversuchs, in dem drei
Testsysteme verglichen wurden, der Svanovir® Paratuberkulose-ELISA (Firma Svanova,
Schweden) ausgewählt. Dieser ELISA konnte für die Untersuchung von Milch auf Antikörper
gegen M. paratuberculosis erfolgreich adaptiert werden. Die Inter-Assay-Variation betrug für
den Milch-ELISA 17 %. Die parallele, serologische Untersuchung der Blut- und Milchproben
von insgesamt 601 Milchkühen im Svanovir® ELISA zeigte eine signifikante Übereinstim-
mung der Ergebnisse beider Methoden. Der optimale „Cut off“ mit einer Spezifität von 96%
und eine Sensitivität von 79% wurde anhand der ROC-Analyse bestimmt.
Der Erregernachweis wurde mittels touchdown-PCR mit Funktionskontrolle etabliert. Zur
Steigerung der Sensitivität wurde eine DNA-Präparation mit einem Präparationskit (R.Z.N.A
Cell and Tissue Kit, Firma peqlab, Erlangen) entwickelt, der ein Enzymverdau der Milch mit
Lysozym und Proteinase K vorangeht. Die reproduzierbare Nachweisgrenze in künstlich
kontaminierten Milchproben beträgt 103 Keime pro ml Milch und 104 Erreger pro ml Ko-
lostrum. Die interne Funktionskontrolle erlaubt den für den diagnostischen Einsatz von PCR-
Tests erforderlichen Ausschluß falsch negativer Ergebnisse.
In einer von der Niedersächsischen Tierseuchenkasse unterstützten Verlaufsuntersuchung mit
zehn Milchkühen, von denen sechs Tiere nachweislich mit M. paratuberculosis infiziert wa-
ren, konnten die Ergebnisse des Milch-ELISA und der Milch-PCR durch Blutserologie, kultu-
relle Kotuntersuchungen sowie durch die kulturelle Untersuchung des Ileocaecallymphkno-
tens bestätigt werden.

Summery
103
7 Summary
Corinna Winterhoff: Diagnosis of paratuberculosis in milk: pathogen and antibody detection
by PCR and ELISA
Bovine paratuberculosis is a disease which occurs world-wide. Cattle are usually in-
fected during the first few months of life. Although they shed the pathogen with milk and fae-
ces, they may not show any clinical signs for years after the primary infection.
The thesis presented here introduces two techniques to diagnose paratuberculosis from milk.
The first method allows antibody detection by means of ELISA, the other allows detection of
the pathogen by means of PCR. For antibody detection the Svanovir® Paratuberculosis-
ELISA (by Svanova, Sweden) was chosen, based on the results of a ring test comparing three
test systems. This ELISA could be successfully adapted for testing milk for antibody against
M. paratuberculosis. Inter Assay Variation of the milk ELISA was 17 %. A parallel serologi-
cal milk and blood antibody analysis of 601 dairy cows by means of Svanovir® ELISA indi-
cated that the results obtained with both methods were clearly correlated. The optimal "cut
off" for the milk ELISA with a specificity of 96 % and a sensitivity of 79 % was determined
by ROC analysis. For detection of M. paratuberculosis a touchdown PCR with function con-
trol was established. To increase sensitivity, a DNA preparation kit (R.Z.N.A. Cell and Tissue
Kit by peqlab, Erlangen) was used, it was preceded by an enzyme digestion of milk with lyso-
zyme and proteinase K. The reproducible limit of detectability in artificially contaminated
milk samples is 10³ bacteria per ml milk and 104 bacteria per ml colostrum. The internal con-
trol, which is required for the diagnostic use of PCR tests, allows the exclusion of false-
negative results. Finally the reproducibility and diagnostic value of both test was confirmed in
a 4 month experiment taken weekly milk, blood and faecal samples from 10 dairy cows. The
true M. paratuberculosis status („Goldstandard“) of these animals was assessed by
lymphnode culture at the end of the experiment.

Literaturverzeichnis
104
8 Literaturverzeichnis
ANGUS, K. W. und N. J. GILMOUR (1971):
Effect of the rimino phenazine B663 (G30320) on Mycobacterium johnei infection and rein-
fection in sheep. II. Pathology
J. Comp Pathol. 81, 227-34
BALDWIN, E. W. (1976):
Isoniazid therapy in two cases of Johne's disease
Vet. Med. Small Anim Clin. 71, 1359-62
BANG, B. (1906):
Chronische pseudotuberkulöse Darmentzündung beim Rinde
Tieraerztl. Wochenschrift 42, 759-63
BANG, O. und C. W. ANDERSEN (1913):
Einige Untersuchungen über komplementbindende Antistoffe bei experimenteller und sponta-
ner Tuberkulose sowie paratuberkulöser Darmentzündung des Rindes
Zbl. Bakt. Hyg. I Abt. Orig. 69, 517-38
BECH-NIELSEN, S., J. B. JØRGENSEN, P. AHRENS, und N. C. FELD (1992):
Diagnostic accuracy of a Mycobacterium phlei-absorbed serum ELISA for diagnosis of bo-
vine paratberculosis in dairy cows
J. Clin. Microbiol. 30, 613-8
BELLETTI, G., N. ARRIGONI, und G. SALI (1992):
Johne's disease in cattle: clinical and epidemiological investigations and experiences of a
control program
Tierärztliche Umschau 47, 328-30
BENEDICTUS, G., A. A. DIJKHUIZEN, und J. STELWAGEN (1987):
Economic losses due to paratuberculosis in dairy cattle
Vet. Rec. 121, 142-6

Literaturverzeichnis
105
BISPING, W. und G. AMTSBERG (1988):
Farbatlas zur Diagnose bakterieller Infektionserreger der Tiere
Verlag Parey, Berlin, Hamburg, 119-20
BLEUMINK-PLUYM, N. M., D. J. HOUWERS, und S. D. PEETERS-VAN GENT (1994):
Sample preparation for M paratuberculosis polymerase chain reaction in faeces and tissues
[letter]
Vet. Rec. 135, 120
BOOM, R., C. J. SOL, M. M. SALIMANS, C. L. JANSEN, P. M. WERTHEIM-VAN DIL-
LEN, und N. J. VAN DER (1990):
Rapid and simple method for purification of nucleic acids
J. Clin. Microbiol. 28, 495-503
BOSE, M., A. CHANDER, und R. H. DAS (1993):
A rapid and gentle method for the isolation of genomic DNA from mycobacteria
Nucleic Acids Res. 21, 2529-30
BRENNAN, P. J. und H. NIKAIDO (1995):
The envelope of mycobacteria
Annu. Rev. Biochem. 64, 29-63
CAFFIN, J. P. und B. POUTREL (1987):
Physiological and pathological factors influencing bovine immunoglobulin G2 concentration
in milk
J. Dairy Sci 71, 2035-43
CAFFIN, J. P., B. POUTREL, und P. RAINARD (1983):
Physiological and pathological factors influencing bovine immunoglobulin G1 concentration
in milk
J. Dairy Sci 66, 2161-6

Literaturverzeichnis
106
CETINKAYA, B., K. EGAN, D. A. HARBOUR, und K. L. MORGAN (1996):
An abattoir-based study of the prevalence of subclinical Johne's disease in adult cattle in south
west England
Epidemiol. Infect. 116, 373-9
CHALLANS, J. A., K. STEVENSON, J. D. REID, und J. M. SHARP (1994):
A rapid method for the extraction and detection of Mycobacterium avium subspecies paratu-
berculosis from clinical specimens
Vet. Rec. 134, 95-6
CHIODINI, R. J. (1989):
Crohn's disease and the mycobacterioses: a review and comparison of two disease entities
Clin. Microbiol. Rev. 2, 90-117
CHIODINI, R. J. (1993):
Abolish Mycobacterium paratuberculosis strain 18 [letter; comment]
J. Clin. Microbiol. 31, 1956-8
CHIODINI, R. J. (1996):
Immunology: resistance to paratuberculosis
Vet. Clin. North Am. Food Anim Pract. 12, 313-43
CHIODINI, R. J. und J. HERMON-TAYLOR (1993):
The thermal resistance of Mycobacterium paratuberculosis in raw milk under conditions si-
mulating pasteurization
J. Vet. Diagn. Invest 5, 629-31
CHIODINI, R. J. und C. A. ROSSITER (1996):
Paratuberculosis: a potential zoonosis?
Vet. Clin. North Am. Food Anim Pract. 12, 457-67
CHIODINI, R. J. und H. J. VAN KRUININGEN (1983):
Eastern white-tailed deer as a reservoir of ruminant paratuberculosis
J. Am. Vet. Med. Assoc. 182, 168-9

Literaturverzeichnis
107
CHIODINI, R. J., H. J. VAN KRUININGEN, und R. S. MERKAL (1984):
Ruminant paratuberculosis (Johne's disease): the current status and future prospects
Cornell Vet. 74, 218-62
CLARKE, C. J. (1997):
The pathology and pathogenesis of paratuberculosis in ruminants and other species
J. Comp Pathol. 116, 217-61
CLARKE, C. J., I. A. PATTERSON, K. E. ARMSTRONG, und J. C. LOW (1996):
Comparison of the absorbed ELISA and agar gel immunodiffusion test with clinicopathologi-
cal findings in ovine clinical paratuberculosis
Vet. Rec. 139, 618-21
COCITO, C., P. GILOT, M. COENE, M. DE KESEL, P. POUPART, und P. VANNUFFEL
(1994):
Paratuberculosis
Clin. Microbiol. Rev. 7, 328-45
COLLINS, D. M., D. M. GABRIC, und G. W. DE LISLE (1990):
Identification of two groups of Mycobacterium paratuberculosis strains by restriction endo-
nuclease analysis and DNA hybridization
J. Clin. Microbiol. 28, 1591-6
COLLINS, D. M., F. HILBINK, D. M. WEST, B. D. HOSIE, M. M. COOKE, und G. W. DE
LISLE (1993):
Investigation of Mycobacterium paratuberculosis in sheep by faecal culture, DNA characteri-
sation and the polymerase chain reaction
Vet. Rec. 133, 599-600
COLLINS, M. T. (1996):
Diagnosis of paratuberculosis
Vet. Clin. North Am. Food Anim Pract. 12, 357-71

Literaturverzeichnis
108
COLLINS, M. T. und D. C. SOCKETT (1993):
Accuracy and economics of the USDA-licensed enzyme-linked immunosorbent assay for bo-
vine paratuberculosis
J. Am. Vet. Med. Assoc. 203, 1456-63
COLLINS, M. T., D. C. SOCKETT, S. RIDGE, und J. C. COX (1991):
Evaluation of a commercial enzyme-linked immunosorbent assay for Johne's disease
J. Clin. Microbiol. 29, 272-6
COOK, L. S. (2000):
A causal role for Mycobacterium avium subspecies paratuberculosis in Crohn's disease?
Can. J. Gastroenterol. 14, 479-80
CROHN, B., L. GINZBURG, und G. OPPENHEIMER (1932):
Regional ileitis, a pathological and clinical entity
J. Am. Med. Assoc. 99, 1323-9
DE KESEL, M., P. GILOT, M.-C. MISONNE, M. COENE, und C. COCITO (1993):
Cloning and expression of portions of the 34-kilodalton-protein gene of Mycobacterium pa-
ratuberculosis: Its application to serological analysis of Johne's disease
J. Clin. Microbiol. 31, 947-54
DE LISLE, G. W., D. M. COLLINS, und H. F. HUCHZERMEYER (1992):
Characterization of ovine strains of Mycobacterium paratuberculosis by restriction endo-
nuclease analysis and DNA hybridization
Onderstepoort J. Vet. Res. 59, 163-5
DINSMORE, R. P. (1986):
Effects of subclinical paratuberculosis in a research dairy herd
Proceedings of the 3rd Northeast Region Meeting on Paratuberculosis, Ithaka, NY,20,
DORAN, T. J., J. K. DAVIES, A. J. RADFORD, und A. L. HODGSON (1994):
Putative functional domain within ORF2 on the Mycobacterium insertion sequences IS900
and IS902
Immunol. Cell Biol. 72, 427-34

Literaturverzeichnis
109
DREWS, G. (1983):
Mikrobiologisches Praktikum
4. neubearbeitete Auflage, Springer Verlag, Berlin, Heidelberg,
EDINGLOH, M. (1999):
Untersuchung zur Anwendbarkeit der Polymerase Kettenreaktion in der Routinediagnostik
des Q-Fiebers beim Rind unter besonderer Berücksichtigung des Erregernachweises aus
Milch
Berlin, Fachbereich Veterinärmedizin der Freien Universität, Diss.
EDINGLOH, M., C. C. MERCK, und E. MANZ (1999):
[Multiplex PCR for the diagnostic detection of Coxiella burnetii in cow's milk]
Berl Munch. Tierarztl. Wochenschr. 112, 5-9
ENGWALL, E. und P. PERLMANN (1971):
Enzyme-Linked Immunosorbent Assay (ELISA). Quantitative assay of Immunglobulin G
Immunochemistry 8, 871-4
GAY, J. M. und D. M. SHERMAN (1992):
Factors in the epidemiologie and control of ruminant paratuberculosis
Vet. Med. 87, 1133-9
GRANT, I. R. (1998):
Does Mycobacterium paratuberculosis survive current pasteurization conditions? [letter;
comment]
Appl. Environ. Microbiol. 64, 2760-1
GRANT, I. R., H. J. BALL, S. D. NEILL, und M. T. ROWE (1996):
Inactivation of Mycobacterium paratuberculosis in cows' milk at pasteurization temperatures
Appl. Environ. Microbiol. 62, 631-6
GRANT, I. R., H. J. BALL, und M. T. ROWE (1998):
Effect of high-temperature, short-time (HTST) pasteurization on milk containing low num-
bers of Mycobacterium paratuberculosis
Lett. Appl. Microbiol. 26, 166-70

Literaturverzeichnis
110
GRANT, I. R., H. J. BALL, und M. T. ROWE (1999):
Effect of higher pasteurization temperatures, and longer holding times at 72 degrees C, on the
inactivation of Mycobacterium paratuberculosis in milk
Lett. Appl. Microbiol. 28, 461-5
GREEN, E. P., M. L. TIZARD, M. T. MOSS, J. THOMPSON, D. J. WINTERBOURNE, J. J.
MCFADDEN, und J. HERMON-TAYLOR (1989):
Sequence and characteristics of IS900, an insertion element identified in a human Crohn's
disease isolate of Mycobacterium paratuberculosis
Nucleic Acids Res. 17, 9063-73
GREIG, A., K. STEVENSON, V. PEREZ, A. A. PIRIE, J. M. GRANT, und J. M. SHARP
(1997):
Paratuberculosis in wild rabbits (Oryctolagus cuniculus) [see comments]
Vet. Rec. 140, 141-3
GRUNERT, E. (1995):
Buiatrik
Verlag M. & H. Schaper Alfeld, Hannover Band 1, 54-5
HARDIN, L. E. und J. G. THORNE (1996):
Comparison of milk with serum ELISA for the detection of paratuberculosis in dairy cows
J. Am. Vet. Med. Assoc. 209, 120-2
HARLOW, E. und D. LANE (1988):
Antibodies: A Laboratory Manual
Cold Spring Harbor, New York,
HARMS, V. (1998):
Biomathematik, Statistik und Dokumentation
Verlag Harms, Kiel,
HERMAN, L., W. REYBROECK, E. D'HAESE, und H. J. NELIS (1999):
Detection and Enumeration of Mycobacterium paratuberculosis in Milk
Proceedings of 6th Int. Coll. Paratubercul., Melbourne, Australia, 75

Literaturverzeichnis
111
HERMON-TAYLOR, J., T. J. BULL, J. M. SHERIDAN, J. CHENG, M. L. STELLAKIS,
und N. SUMAR (2000):
Causation of Crohn's disease by Mycobacterium avium subspecies paratuberculosis
Can. J. Gastroenterol. 14, 521-39
HERMON-TAYLOR, J., M. MOSS, M. TIZARD, Z. MALIK, und J. SANDERSON (1990):
Molecular biology of Crohn's disease mycobacteria
Baillieres Clin. Gastroenterol. 4, 23-42
HIETALA, S. K. (1992):
The options in diagnosing ruminant paratuberculosis
Vet. Med. 87, 1122-32
HILBINK, F., D. M. WEST, G. W. DE LISLE, R. KITTELBERGER, B. D. HOSIE, J. HUT-
TON, M. M. COOKE, und M. PENROSE (1994):
Comparison of a complement fixation test, a gel diffusion test and two absorbed and unabsor-
bed ELISAs for the diagnosis of paratuberculosis in sheep
Vet. Microbiol. 41, 107-16
HURLEY, S. S., G. A. SPLITTER, und R. A. WELCH (1993):
Rapid lysis technique for mycobacterial species
J. Clin. Microbiol. 25, 3216-22
HUTCHINSON, L. J. (1996):
Economic impact of paratuberculosis
Vet. Clin. North Am. Food Anim Pract. 12, 373-81
IEVEN, M. und H. GOOSSENS (1997):
Relevance of nucleic acid amplification techniques for diagnosis of respiratory tract infections
in the clinical laboratory
Clin. Microbiol. Rev. 10, 242-56
JAKOBSEN, M. B., L. ALBAN, und S. S. NIELSEN (2000):
A cross-sectional study of paratuberculosis in 1155 danish dairy cows
Prev. Vet. Med. 46, 15-27

Literaturverzeichnis
112
JARK, U. (1996):
Etablierung eines ELISA zur Erkennung subklinischer Paratuberkulose-Infektionen beim
Rind
Hannover, Tierärztl. Hochsch., Diss.
JARK, U., I. RINGENA, B. FRANZ, G. F. GERLACH, M. BEYERBACH, und B. FRANZ
(1997):
Development of an ELISA technique for serodiagnosis of bovine paratuberculosis
Vet. Microbiol. 57, 189-98
JENSEN, A. L. und J. S. POULSEN (1992):
Evaluation of diagnostic tests using relative operating characteristic (ROC) curves and the
differential positive rate. An example using the total serum bile acid concentration and the
alanine aminotransferase activity in the diagnosis of canine hepatobiliary diseases
Zentralbl. Veterinarmed. [A] 39, 656-68
JOHNE, H. A. und L. FRONTHINGHAM (1895):
Ein eigenthümlicher Fall von Tuberculose beim Rind
Dtsch. Z. Thiermed. Vergl. Path. 21, 438-54
JOHNSON-IFEARULUNDU, Y. J. und J. B. KANEENE (1997):
Relationship between soil type and Mycobacterium paratuberculosis
J. Am. Vet. Med. Assoc. 210, 1735-40
JØRGENSEN, J. B. (1977):
Survival of Mycobacterium paratuberculosis in slurry
Nord. Vet. Med. 29, 267-70
JØRGENSEN, J. B. und A. L. JENSEN (1978):
Enzyme-Linked Immunosorbent Assay (ELISA) for detection of antibodies to Mycobacteri-
um paratuberculosis in cattle
Acta Vet. Scand. 19, 310-2

Literaturverzeichnis
113
JUSTE, R. A., J. F. GARCIA MARIN, B. PERIS, C. S. SAEZ DE OCARIZ, und J. J. BA-
DIOLA (1994):
Experimental infection of vaccinated and non-vaccinated lambs with Mycobacterium paratu-
berculosis
J. Comp Pathol. 110, 185-94
KIM, Y. G., S. BECH-NIELSEN, J. C. GORDON, R. D. SLEMONS, und E. SPANGLER
(1989):
Comparison of two methods for isolation of Mycobacterium paratuberculosis from bovine
fecal samples
Am. J. Vet. Res. 50, 1110-3
KLAUSEN, J., V. PEREZ, S. B. GIESE, J. F. GARCIA MARIN, und P. AHRENS (1997):
Immunological detection of sheep experimentally infected with strains of Mycobacterium
avium subspecies containing insertion sequence IS901/IS902 and a 40 kDa protein
Vet. Microbiol. 57, 181-7
KLUGE, J. P., R. S. MERKAL, W. S. MONLUX, A. B. LARSEN, K. E. KOPECKY, F. K.
RAMSEY, und R. P. LEHMANN (1968):
Experimental paratuberculosis in sheep after oral, intratracheal, or intravenous inoculation
lesions and demonstration of etiologic agent
Am. J. Vet. Res. 29, 953-62
KORMENDY, B. (1994):
The effect of vaccination on the prevalence of paratuberculosis in large dairy herds
Vet. Microbiol. 41, 117-25
KORMENDY, B., T. KOPAL, T. BALINT, M. SZILAGYI, und L. BEKI (1989):
Economic losses caused by paratuberculosis in a dairy herd: case report
Acta Vet. Hung. 37, 45-53
KUNZE, Z. M., F. PORTAELS, und J. J. MCFADDEN (1992):
Biologically distinct subtypes of Mycobacterium avium differ in possession of insertion se-
quence IS901 [see comments]
J. Clin. Microbiol. 30, 2366-72

Literaturverzeichnis
114
KÜHN, T. und H. HELBING (2000):
Neue methodische Ansätze in der Kultivierung von Mycobacterium paratuberculosis und
erste Erfahrungen mit dem System MGIT in der veterinärmedizinischen Diagnostik
1. Arbeitstagung des veterinärmedizinischen Referenzlabors für Tuberkulose, Jena,
LAMBRECHT, R. S., J. F. CARRIERE, und M. T. COLLINS (1988):
A model for analyzing growth kinetics of a slowly growing Mycobacterium sp
Appl. Environ. Microbiol. 54, 910-6
LARSEN, A. B. und K. E. KOPECKY (1970):
Mycobacterium paratuberculosis in reproductive organs and semen of bulls
Am. J. Vet. Res. 31, 255-8
LARSEN, A. B., R. S. MERKAL, und R. C. CUTLIP (1975):
Age of cattle as related to resistance to infection with Mycobacterium paratuberculosis
Am. J. Vet. Res. 36, 255-7
LARSEN, A. B. und H. W. MOON (1972):
Experimental Mycobacterium paratuberculosis infection in chickens
Am. J. Vet. Res. 33, 1231-5
LARSEN, A. B., H. W. MOON, und R. S. MERKAL (1972):
Susceptibility of horses to Mycobacterium paratuberculosis
Am. J. Vet. Res. 33, 2185-9
LARSEN, A. B., O. H. STALHEIM, D. E. HUGHES, L. H. APPELL, W. D. RICHARDS,
und E. M. HIMES (1981):
Mycobacterium paratuberculosis in the semen and genital organs of a semen-donor bull
J. Am. Vet. Med. Assoc. 179, 169-71
LÄMMLI, U. K. (1970):
Cleavage of structural proteins during the assembly of the head of bacteriophage T4
Nature 227, 680-5

Literaturverzeichnis
115
LIENAUX, E. (1913):
Un cas d'enterite hypertrophiante chez le cheval
Ann. Med. vet 62, 197-200
MACMILLAN, A. P., I. GREISER-WILKE, V. MOENING, und L. A. MATHIAS (1990):
A competition enzyme immunoassay for brucellosis diagnosis
Dtsch. tierärztl. Wschr. 97, 83-5
MANZ, D., D. WIEGAND, und F. BEHRENS (1986):
Untersuchung zur Konzentration spezifischer Antikörper gegen das Virus der enzootischen
bovinen Leukose in Einzel-und Sammelgemelken
Dtsch. tierärztl. Wschr. 93, 107-10
MCCLURE, H. M., R. J. CHIODINI, D. C. ANDERSON, R. B. SWENSON, W. R. THAY-
ER, und J. A. COUTU (1987):
Mycobacterium paratuberculosis infection in a colony of stumptail macaques (Macaca arctoi-
des)
J. Infect. Dis. 155, 1011-9
MCNAB, W. B., A. H. MEEK, J. R. DUNCAN, B. W. BROOKS, A. A. VAN DREUMEL,
S. W. MARTIN, K. H. NIELSEN, E. A. SUGDEN, und C. TURCOTTE (1991a):
An evaluation of selected screening tests for bovine paratuberculosis
Can. J. Vet. Res. 55, 252-9
MCNAB, W. B., A. H. MEEK, S. W. MARTIN, und J. R. DUNCAN (1991b):
Associations between dairy production indices and lipoarabinomannan enzyme-immunoassay
results for paratuberculosis
Can. J. Vet. Res. 55, 356-61
MERKAL, R. S. (1973):
Laboratory diagnosis of bovine paratuberculosis
J. Am. Vet. Med. Assoc. 163, 1100-2

Literaturverzeichnis
116
MERKAL, R. S., K. E. KOPECKY, und A. B. LARSEN (1970):
Immunologic mechanisms in bovine paratuberculosis
Am. J. Vet. Res. 31, 475-85
MERKAL, R. S., A. B. LARSEN, und G. D. BOOTH (1975):
Analysis of the effect of inapparent bovine paratuberculosis
Am. J. Vet. Res. 36, 837-8
MERKAL, R. S., A. B. LARSEN, und K. E. KOPECKY (1968):
Comparison of examination and test methods for early detection of paratuberculous cattle
Am. J. Vet. Res. 29, 1533-8
MERKAL, R. S., D. L. WHIPPLE, J. M. SACKS, und G. R. SNYDER (1987):
Prevalence of Mycobacterium paratuberculosis in ileocecal lymph nodes of cattle culled in
the United States
J. Am. Vet. Med. Assoc. 190, 676-80
MEYER ZU VILSENDORF, U. (1995):
Untersuchungen zur Bekämpfung der Paratuberkulose mittels Merzung serologisch oder kul-
turell positiver Rinder
Hannover, Tierärztl. Hochsch., Diss.
MEYLAN, M., J. NICOLET, A. BUSATO, A. BURNENS, und J. MARTIG (1995):
[The prevalence of paratuberculosis in the Plateau de Diesse region]
Schweiz. Arch. Tierheilkd. 137, 22-5
MEYLAN, M., D. M. RINGS, W. P. SHULAW, J. J. KOWALSKI, S. BECH-NIELSEN, und
G. F. HOFFSIS (1996):
Survival of Mycobacterium paratuberculosis and preservation of immunoglobulin G in bo-
vine colostrum under experimental conditions simulating pasteurization
Am. J. Vet. Res. 57, 1580-5

Literaturverzeichnis
117
MILLAR, D., J. FORD, J. SANDERSON, S. WITHEY, M. TIZARD, T. DORAN, und J.
HERMON-TAYLOR (1996):
IS900 PCR to detect Mycobacterium paratuberculosis in retail supplies of whole pasteurized
cows' milk in England and Wales
Appl. Environ. Microbiol. 62, 3446-52
MILNER, A. R., W. N. MACK, K. J. COATES, J. HILL, I. GILL, und P. SHELDRICK
(1990):
The sensitivity and specificity of a modified ELISA for the diagnosis of Johne's disease from
a field trial in cattle
Vet. Microbiol. 25, 193-8
MOMOTANI, E., D. L. WHIPPLE, A. B. THIERMANN, und N. F. CHEVILLE (1988):
Role of M cells and macrophages in the entrance of Mycobacterium paratuberculosis into
domes of ileal Peyer's patches in calves
Vet. Pathol. 25, 131-7
MOSS, M., E. GREEN, M. TIZARD, Z. MALIK, und J. HERMON-TAYLOR (1991):
Specific detection of Mycobacterium paratuberculosis bei DNA hybridization with a frag-
ment of the insertion element IS900
Gut 32, 395-8
NIELSEN, S. S., S. M. THAMSBORG, H. HOUE, und V. BITSCH (2000):
Bulk-tank milk ELISA antibodies for estimating the prevalence of paratuberculosis in Danish
dairy herds
Prev. Vet. Med. 44, 1-7
NOORDHOEK, G. T., J. A. KAAN, S. MULDER, H. WILKE, und A. H. KOLK (1995):
Routine application of the polymerase chain reaction for detection of Mycobacterium tuber-
culosis in clinical samples
J. Clin. Pathol. 48, 810-4

Literaturverzeichnis
118
NORDLUND, K. V., W. J. GOODGER, J. PELLETIER, und M. T. COLLINS (1996):
Associations between subclinical paratuberculosis and milk production, milk components,
and somatic cell counts in dairy herds
J. Am. Vet. Med. Assoc. 208, 1872-6
OFFICE INTERNATIONAL DES EPIZOOTIES (1996):
Manual of standards for diagnostic tests and vaccines
, 8-15
PAVLIK, I., L. MATLOVA, J. BARTL, P. SVASTOVA, L. DVORSKA, und R. H. WHIT-
LOCK (1999):
Parallel faecal und organ Mycobacterium avium subsp. paratuberculosis culture showing or-
ganism distribution in different breeds of cattle
Proceedings of 6th Int. Coll. Paratubercul., Melbourne, Australia, 642-56
PHILPOTT, M. (1993):
The dangers of disease transmission by artificial insemination and embryo transfer [published
erratum appears in Br Vet J 1994 Mar- Apr;150(2):209]
Br. Vet. J. 149, 339-69
POHLENZ, J. (1991):
Krankheiten der Verdauungsorgane
SCHULZ, L. C. Pathologie der Haustiere Band 1, 1. Auflage
Verlag Gustav Fischer, Jena, 312-3
POWELL, H. A., C. M. GOODING, S. D. GARRETT, M. B. LUND, und R. A. MCKEE
(1994):
Proteinase inhibition of the detection of Listeria monocytogenes in milk using the polymerase
chain reaction
Lett. Appl. Microbiol. 18, 59-61
REHM, T. (1999):
Etablierung und Standardisierung eines Sanierungsprogrammes zur Bekämpfung der Paratu-
berkulose in infizierten Rinderbeständen
Hannover, Tierärztl. Hochsch., Diss.

Literaturverzeichnis
119
REICHEL, M. P., R. KITTELBERGER, M. E. PENROSE, R. M. MEYNELL, D. COUSINS,
T. ELLIS, L. M. MUTHARIA, E. A. SUGDEN, A. H. JOHNS, und G. W. DE LISLE (1999):
Comparison of serological tests and faecal culture for the detection of Mycobacterium avium
subsp. paratuberculosis infection in cattle and analysis of the antigens involved
Vet. Microbiol. 66, 135-50
RICHARDS, W. D. (1988):
The apparent effect of environmental acidity on the incidence of paratuberculosis
MERKAL, R. S. u. M. F. THOREL Second International Colloquium on Paratuberculosis,
Laboratoire Central de Recherches Veterinaires, Maison-Alfort,France
, 342-9
RICHARDS, W. D. (1990):
Milk ELISA: a new inexpensive test for detecting paratuberculosis in dairy herds
Paratuberculosis Newsletter 2, 7-8
RIDGE, S. E. (1993):
Cultivation of Mycobacterium paratuberculosis from bovine fecal samples by using elements
of the Roche MB Check system
J. Clin. Microbiol. 31, 400-5
RIEMANN, H. P. und B. ABBAS (1983):
Diagnosis and control of bovine paratuberculosis (Johne's disease)
Adv. Vet. Sci. Comp Med. 27, 481-506
RINGENA, I. (1995):
Untersuchungen zur Diagnostik der Paratuberkulose des Rindes unter besonderer Berück-
sichtigung der Sanierung von Problembeständen
Hannover, Tierärztl. Hochsch., Diss.
ROHDE, R. F. und W. P. SHULAW (1990):
Isolation of Mycobacterium paratuberculosis from the uterine flush fluids of cows with clini-
cal paratuberculosis
J. Am. Vet. Med. Assoc. 197, 1482-3

Literaturverzeichnis
120
ROLFS, A., I. SCHULLER, U. FINKH, und I. WEBER-ROLFS (1992):
PCR: Clinical diagnostics and research
Verlag Springer, Berlin/Heidelberg,
ROSENBERGER, G. (1978):
Infektionskrankheiten des Verdauungsapparates
ROSENBERGER, G. Krankheiten des Rindes, 2. Auflage
Verlag Parey, Berlin, Hamburg, 756-60
ROSSITER, C. A. und W. S. BURHANS (1996):
Farm-specific approach to paratuberculosis (Johne's disease) control
Vet. Clin. North Am. Food Anim Pract. 12, 383-415
RUNNELS, R. A. (1955):
Case 17-paratuberculosis in a pig
J. Am. Vet. Med. Assoc. 127, 523-4
SAIKI, R. K., S. SCHARF, F. FALOONA, K. B. MULLIS, G. T. HORN, A. H. EHRLICH,
und N. ARNHEIM (1985):
Enzymatic amplification of beta-globulin genomic sequences and restriction site analysis for
diagnosis of sickle cell anemia
Science 230, 1350-4
SAMBROOK, J., F. FRITSCH, und T. MANIATIS (1989):
Molecular Cloning: A Laboratory Manual, 2nd ed.
Cold Spring Harbor, New York,
SCHEIBL, P. (1996):
Differenzierung klinischer Mycobacterium paratuberculosis-Isolate mittels 16S-23-S rDNS-
Spacer-Analyse und Random Amplified Polymorphic DNA-PCR (RAPD-PCR)
Hannover, Tierärztl. Hochsch., Diss.

Literaturverzeichnis
121
SCHLIESSER, T. und E. SCHAAL (1984):
Prevalence and diagnosis of paratuberculosis in the Federal Republic of Germany
Commission of the European Communities, Report EUR 9000, EN, 103-14
SEITZ, S. E., L. E. HEIDER, W. D. HEUSTON, S. BECH-NIELSEN, D. M. RINGS, und L.
SPANGLER (1989):
Bovine fetal infection with Mycobacterium paratuberculosis
J. Am. Vet. Med. Assoc. 194, 1423-6
SMITH, B. P., D. G. OLIVER, P. SINGH, G. DILLING, und P. A. MARVIN (1989):
Detection of Salmonella dublin mammary gland infection in carrier cows, using an enzyme-
linked immunosorbent assay for antibody in milk or serum
Am. J. Vet. Res. 50, 1352-60
SOCKETT, D. C., D. J. CARR, und M. T. COLLINS (1992a):
Evaluation of conventional and radiometric fecal culture and a commercial DNA probe for
diagnosis of Mycobacterium paratuberculosis infections in cattle
Can. J. Vet. Res. 56, 148-53
SOCKETT, D. C., D. J. CARR, W. D. RICHARDS, und M. T. COLLINS (1992b):
A repository of specimens for comparison of diagnostic testing procedures for bovine paratu-
berculosis
J. Vet. Diagn. Invest 4, 188-91
ST JEAN, G. (1996):
Treatment of clinical paratuberculosis in cattle
Vet. Clin. North Am. Food Anim Pract. 12, 417-30
STABEL, J. R. (1997):
An improved method for cultivation of Mycobacterium paratuberculosis from bovine fecal
samples and comparison to three other methods
J. Vet. Diagn. Invest 9, 375-80

Literaturverzeichnis
122
STABEL, J. R., E. M. STEADHAM, und C. A. BOLIN (1997):
Heat inactivation of Mycobacterium paratuberculosis in raw milk: are current pasteurization
conditions effective? [see comments]
Appl. Environ. Microbiol. 63, 4975-7
STEHMAN, S. M. (1996):
Paratuberculosis in small ruminants, deer, and South American camelids
Vet. Clin. North Am. Food Anim Pract. 12, 441-55
STREETER, R. N., G. F. HOFFSIS, S. BECH-NIELSEN, W. P. SHULAW, und D. M.
RINGS (1995):
Isolation of Mycobacterium paratuberculosis from colostrum and milk of subclinically infec-
ted cows
Am. J. Vet. Res. 56, 1322-4
SUGDEN, E. A., A. H. CORNER, B. S. SAMAGH, B. W. BROOKS, C. TURCOTTE, K. H.
NIELSEN, R. W. STEWART, und J. R. DUNCAN (1989):
Serodiagnosis of ovine paratuberculosis, using lipoarabinomannan in an enzyme-linked im-
munosorbent assay
Am. J. Vet. Res. 6, 850-4
SUGDEN, E. A., B. S. SAMAGH, D. R. BUNDLE, und J. R. DUNCAN (1986):
Lipoarabinomannan and lipid-free arabinomannan antigens of Mycobacterium paratuberculo-
sis
Infect. Immun. 55, 762-70
SUNG, N. und M. T. COLLINS (1998):
Thermal tolerance of Mycobacterium paratuberculosis
Appl. Environ. Microbiol. 64, 999-1005
SWEENEY, R. W. (1996):
Transmission of paratuberculosis
Vet. Clin. North Am. Food Anim Pract. 12, 305-12

Literaturverzeichnis
123
SWEENEY, R. W., R. H. WHITLOCK, C. L. BUCKLEY, P. SPENCER, A. E. ROSEN-
BERGER, und L. J. HUTCHINSON (1994):
Diagnosis of paratuberculosis in dairy cattle, using enzyme-linked immunosorbent assay for
detection of antibodies against Mycobacterium paratuberculosis in milk
Am. J. Vet. Res. 55, 905-9
SWEENEY, R. W., R. H. WHITLOCK, A. N. HAMIR, A. E. ROSENBERGER, und S. A.
HERR (1992a):
Isolation of Mycobacterium paratuberculosis after oral inoculation in uninfected cattle
Am. J. Vet. Res. 53, 1312-4
SWEENEY, R. W., R. H. WHITLOCK, und A. E. ROSENBERGER (1992b):
Mycobacterium paratuberculosis cultured from milk and supramammary lymph nodes of in-
fected asymptomatic cows
J. Clin. Microbiol. 30, 166-71
SWEENEY, R. W., R. H. WHITLOCK, und A. E. ROSENBERGER (1992c):
Mycobacterium paratuberculosis isolated from fetuses of infected cows not manifesting signs
of the disease
Am. J. Vet. Res. 53, 477-80
TAMBOLI, C. P. (1996):
A hypothesis for explaining the geographical distribution of Crohn's disease
Can. J. Gastroenterol. 10, 173-7
TAN, M. F., W. C. NG, S. H. CHAN, und W. C. TAN (1997):
Comparative usefulness of PCR in the detection of Mycobacterium tuberculosis in different
clinical specimens
J. Med. Microbiol. 46, 164-9
TAYLOR, A. W. (1953):
The experimental infection of cattle with varieties of Mycobacterium johnei isolated from
sheep
J. Comp Pathol. 63, 368-73

Literaturverzeichnis
124
TAYLOR, T. K., C. R. WILKS, und D. S. MCQUEEN (1981):
Isolation of Mycobacterium paratuberculosis from the milk of a cow with Johne's disease
Vet. Rec. 109, 532-3
THOREL, M. F., M. KRICHEVSKY, und V. V. LEVY-FREBAULT (1990):
Numerical taxonomy of mycobactin-dependent mycobacteria, emended description of My-
cobacterium avium, and description of Mycobacterium avium subsp. avium subsp. nov., My-
cobacterium avium subsp. paratuberculosis subsp. nov., and Mycobacterium avium subsp.
silvaticum subsp. nov
Int. J. Syst. Bacteriol. 40, 254-60
TIJSSEN, P. (1985):
Laboratory techniques: Practice and Theory of Enzyme Immunoassays
Elsevier, Amsterdam, Niederlande, 393-7
TOWBIN, H., T. STAEHELIN, und J. GORDON (1979):
Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: proce-
dure and some applications
Proc. Natl. Acad. Sci. U. S. A 76, 4350-4
TWORT, F. W. und G. L. Y. INGRAM (1912):
A method for isolating and cultivating the Mycobacterium enteritidis chronicae pseudotuber-
culosae bovis johne and some experiments on the preparation of a diagnostic vaccine for
pseudotuberculous enterititis of bovines
Proc. Roy. Soc. London 84, 517-43
VALLEE, H. und P. RINJARD (1926):
Etudes sur l'enterite paratuberculeuse des bovides
Revue Generale Med. Vet. 1-9
VAN SCHAIK, G., C. H. KALIS, G. BENEDICTUS, A. A. DIJKHUIZEN, und R. B.
HUIRNE (1996):
Cost-benefit analysis of vaccination against paratuberculosis in dairy cattle
Vet. Rec. 139, 624-7

Literaturverzeichnis
125
VAN ULSEN, F. W. (1970):
Paratuberculosis in a pygmy ass, Equus asinus
Tijdschr. Diergeneesk. 95, 446-8
VANNUFFEL, P., P. GILOT, B. LIMBOURG, B. NAERHUYZEN, C. DIETERICH, M.
COENE, L. MACHTELINCKX, und C. COCITO (1994):
Development of species-specific enzyme-linked immunosorbent assay for diagnosis of Joh-
ne's disease in cattle
J. Clin. Microbiol. 32, 1211-6
VARY, P. H., P. R. ANDERSEN, E. GREEN, J. HERMON-TAYLOR, und J. J. MCFAD-
DEN (1990):
Use of highly specific DNA probes and the polymerase chain reaction to detect Mycobacteri-
um paratuberculosis in Johne's disease
J. Clin. Microbiol. 28, 933-7
VISKE, D., B. LARSSON, A. ENGVALL, und G. BÖLSKE (1996):
Paratuberculosis in Sweden
The Paratuberculosis Newsletter 8-2, 29
VISUVANATHAN, S., M. MOSS, J. L. STANFORD, J. HERMON-TAYLOR, und J. J.
MCFADDEN (1989):
Simple enzymic method for isolation of DNA from diverse bacteria
J. Microbiol. Methods 10, 59-64
VITALE, F., G. CAPRA, L. MAXIA, S. REALE, G. VESCO, und S. CARACAPPA (1998):
Detection of Mycobacterium tuberculosis complex in cattle by PCR using milk, lymph node
aspirates, and nasal swabs
J. Clin. Microbiol. 36, 1050-5
VOGEL, O. (1970):
Paratuberkulose bei einem Hund
Berl Munch. Tierärztl. Wochenschr. 90, 419-21

Literaturverzeichnis
126
VOM SCHLOSS, A. (2000):
Auswertung eines Sanierungsverfahrens zur Bekämpfung der Paratuberkulose in Rinderbe-
ständen in Nordrhein-Westfalen
Hannover, Tierärztl. Hochsch., Diss., eingereicht,
WEBER, A., M. ROTH, und R. GÜRKE-WERNER (1991):
Paratuberkulose beim Damwild
Vet. 9, 21-3
WENDT, K., H. BOSTEDT, H. MIELKE, und H. W. FUCHS (1994):
Euter- und Gesäugekrankheiten
Verlag Gustav Fischer, Jena, 20
WENTINK, G. H., J. H. BONGERS, A. A. ZEEUWEN, und F. H. JAARTSVELD (1994):
Incidence of paratuberculosis after vaccination against M. paratuberculosis in two infected
dairy herds
Zentralbl. Veterinarmed. [B] 41, 517-22
WHEELER, P. R. und C. RATLEDGE (1994):
Metabolism of Mycobacterium paratuberculosis
BLOOM, R. B. Tuberculosis: Pathogenesis, protection and control
ASM Press, Washington, DC, 360-3
WHIPPLE, D. L., P. A. KAPKE, und P. R. ANDERSEN (1992):
Comparison of a commercial DNA probe test and three cultivation procedures for detection of
Mycobacterium paratuberculosis in bovine feces
J. Vet. Diagn. Invest 4, 23-7
WHITLOCK, R. H. und C. BUERGELT (1996):
Preclinical and clinical manifestations of paratuberculosis (including pathology)
Vet. Clin. North Am. Food Anim Pract. 12, 345-56

Literaturverzeichnis
127
WHITLOCK, R. H., L. J. HUTCHINSON, R. W. SWEENEY, P. SPENCER, A. E. ROSEN-
BERGER, und M. A. VAN BUSKIRK (1994):
Pattern of detection of M. paratuberculosis infected cattle in ten dairy herds cultured every
six month for four years
Proceedings Int. Coll. Paratubercul., Cambridge, 47-53
WHITLOCK, R. H., A. E. ROSENBERGER, R. W. SWEENEY, und P. SPENCER (1997):
Distribution of M. paratuberculosis in tissues of cattle from herds infected with Johne's disea-
se
Proceedings of 5th Int. Coll. Paratubercul., Wisconsin, 168-74
WHITTINGTON, R. J., I. MARSH, M. J. TURNER, S. MCALLISTER, E. CHOY, G. J.
EAMENS, D. J. MARSHALL, und S. OTTAWAY (1998):
Rapid detection of Mycobacterium paratuberculosis in clinical samples from ruminants and
in spiked environmental samples by modified BACTEC 12B radiometric culture and direct
confirmation by IS900 PCR
J. Clin. Microbiol. 36, 701-7
WILLEMS, H., D. THIELE, R. FRÖLICH-RITTER, und H. KRAUSS (1994):
Detection of Coxiella burnetii in cow's milk using the polymerase chain reaction
J. Vet. Med. 41, 580-7
WILLIAMS, E. S. und T. R. SPRAKER (1979):
Paratuberculosis in free-ranging bighorn sheep and a Rocky Mountain goat with a brief re-
view of the disease in wild species
Proceedings Annu. meet. Am. Assoc. Zoo vet. 122-4
WILSON, D. J., C. ROSSITER, H. R. HAN, und P. M. SEARS (1993):
Association of Mycobacterium paratuberculosis infection with reduced mastitis, but with
decreased milk production and increased cull rate in clinically normal dairy cows
Am. J. Vet. Res. 54, 1851-7

Literaturverzeichnis
128
WRIGHT, P. F., E. NILSSON, E. M. VAN ROOIJ, M. LELENTA, und M. H. JEGGO
(1993):
Standardisation and validation of enzyme-linked immunosorbent assay techniques for the
detection of antibody in infectious disease diagnosis
Rev. Sci. Tech. 12, 435-50
YOKOMIZO, Y., I. NISHIMORI, M. KISHIMA, und H. YUGI (1988):
Evaluation of an enzyme-linked immunosorbent assay (ELISA) fo the diagnosis of bovine
paratuberculosis. A proposal of replacing teh complement fixation test with the ELISA as the
official diagnostic test for paratuberculosis in Japan
Proceedings of 2nd Int. Coll. Paratubercul., Maisons-Alfort, France, 206-14
YOKOMIZO, Y., H. YUGI, und R. S. MERKAL (1985):
A method for avoiding false-positive reactions in an enzyme-linked immunosorbent assay
(ELISA) for the diagnosis of bovine paratuberculosis
Nippon Juigaku. Zasshi 47, 111-9
ZIMMER, K., K. G. DRAGER, W. KLAWONN, und R. G. HESS (1999):
Contribution to the diagnosis of Johne's disease in cattle. Comparative studies on the validity
of Ziehl-Neelsen staining, faecal culture and a commercially available DNA-Probe test in
detecting Mycobacterium paratuberculosis in faeces from cattle
Zentralbl. Veterinarmed. [B] 46, 137-40

Anhang
129
9 Anhang
9.1 Tabellenanhang
Tabelle 11: Serumergebnisse des HerdChek™ Mycobacterium paratuberculosis (Idexx,
Wörrstadt) aus dem Ringversuch
Zehn Tiere mit bekanntem Lymphknotenbefund (Ln kult.) wurden von 7 Untersuchungsstel-
len (IVD GmbH und Institute A-F) über einen Zeitraum von 6 Wochen mittels Blut-ELISA
(Idexx, Wörrstadt) untersucht. Das Tier 239 wurde in der 5. Woche aufgrund klinischer Pa-
ratuberkulose geschlachtet. Farblich hervorgehoben sind die von dem kulturellen Lymphkno-
tenergebnis („Goldstandard“) abweichenden serologischen Befunde. Die Ergebnisse der
zweiten Messung wurden nur bei Abweichungen angegeben.
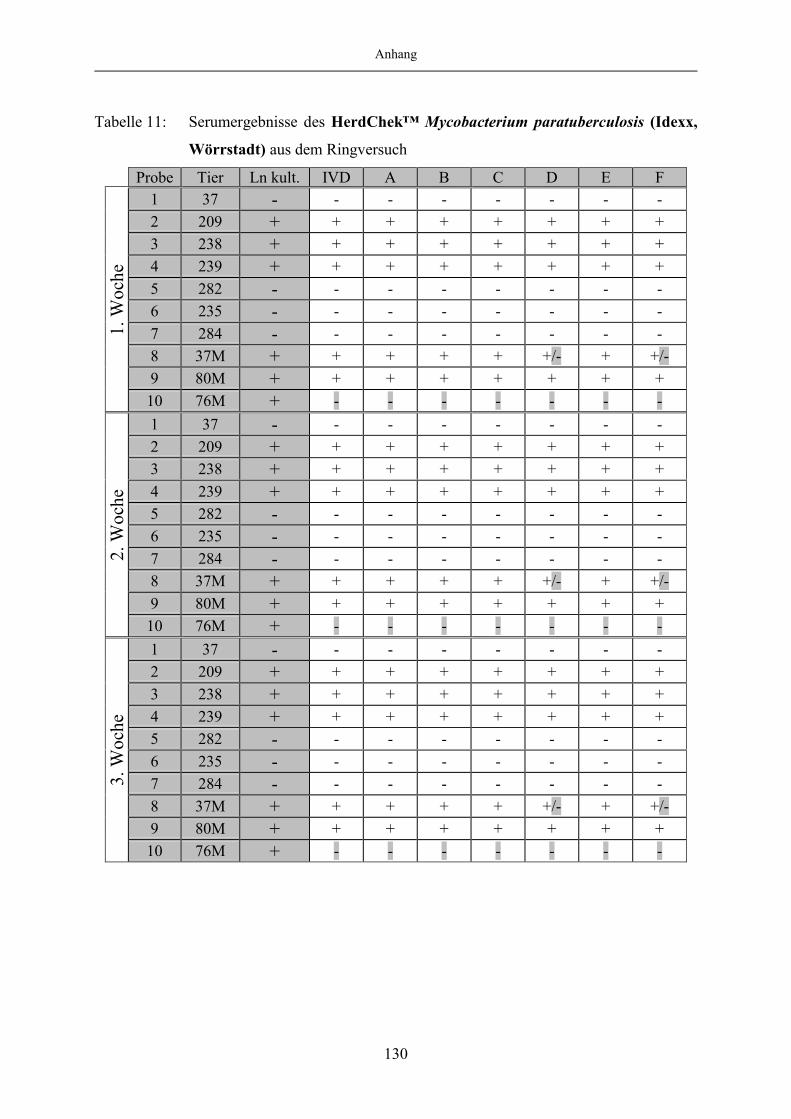
Anhang
130
Tabelle 11: Serumergebnisse des HerdChek™ Mycobacterium paratuberculosis (Idexx,
Wörrstadt) aus dem Ringversuch
Probe Tier Ln kult. IVD A B C D E F1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - - - - -7 284 - - - - - - - -8 37M + + + + + +/- + +/-9 80M + + + + + + + +
1. W
oche
10 76M + - - - - - - -1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - - - - -7 284 - - - - - - - -8 37M + + + + + +/- + +/-9 80M + + + + + + + +
2. W
oche
10 76M + - - - - - - -1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - - - - -7 284 - - - - - - - -8 37M + + + + + +/- + +/-9 80M + + + + + + + +
3. W
oche
10 76M + - - - - - - -
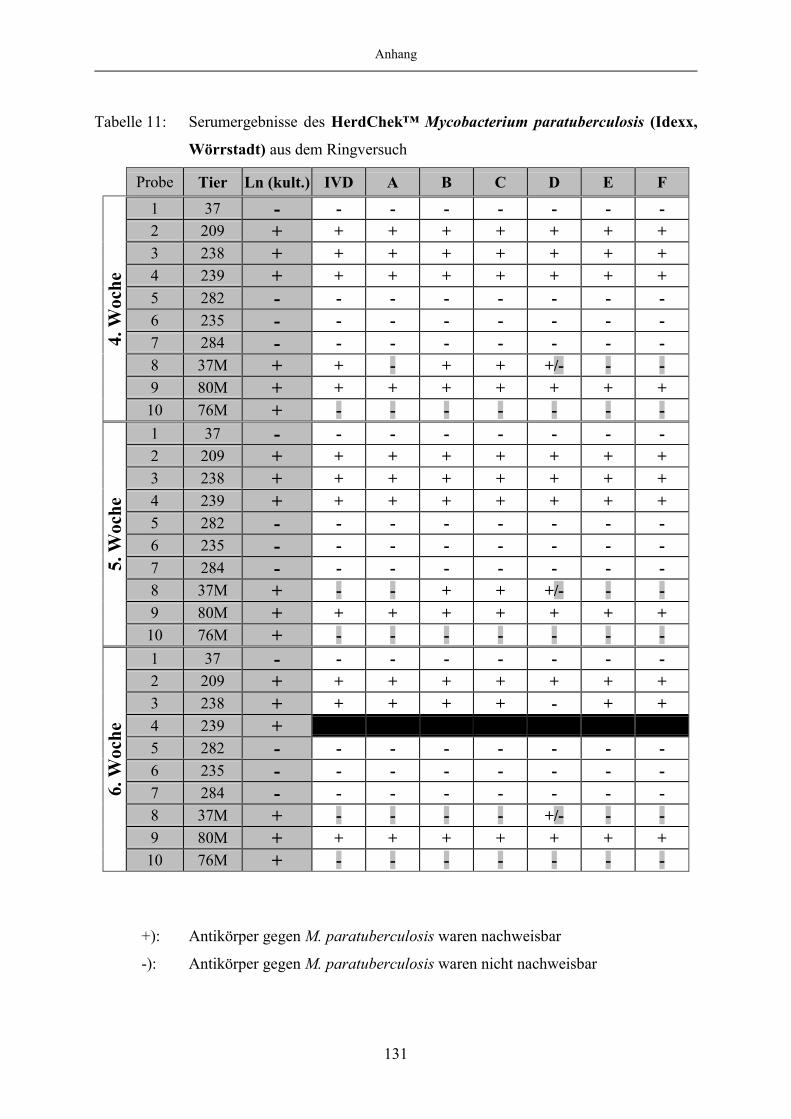
Anhang
131
Tabelle 11: Serumergebnisse des HerdChek™ Mycobacterium paratuberculosis (Idexx,
Wörrstadt) aus dem Ringversuch
Probe Tier Ln (kult.) IVD A B C D E F1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - - - - -7 284 - - - - - - - -8 37M + + - + + +/- - -9 80M + + + + + + + +
4. W
oche
10 76M + - - - - - - -1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - - - - -7 284 - - - - - - - -8 37M + - - + + +/- - -9 80M + + + + + + + +
5. W
oche
10 76M + - - - - - - -1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + - + +4 239 +5 282 - - - - - - - -6 235 - - - - - - - -7 284 - - - - - - - -8 37M + - - - - +/- - -9 80M + + + + + + + +
6. W
oche
10 76M + - - - - - - -
+): Antikörper gegen M. paratuberculosis waren nachweisbar
-): Antikörper gegen M. paratuberculosis waren nicht nachweisbar

Anhang
132
Tabelle 12: Serumergebnisse des ELISA zur Serodiagnostik der bovinen Paratuber-
kulose, (Riemser Arzneimittel GmbH, Insel Riems) aus dem Ringversuch
Zehn Tiere mit bekanntem Lymphknotenbefund (Ln kult.) wurden von 7 Untersuchungsstel-
len (IVD GmbH und Institute A-F) über einen Zeitraum von 6 Wochen mittels Blut-ELISA
(Riemser Arzneimittel GmbH, Insel Riems) untersucht. Das Tier 239 wurde in der 5. Woche
aufgrund klinischer Paratuberkulose geschlachtet. Farblich hervorgehoben sind die von dem
kulturellen Lymphknotenergebnis („Goldstandard“) abweichenden serologischen Befunde.
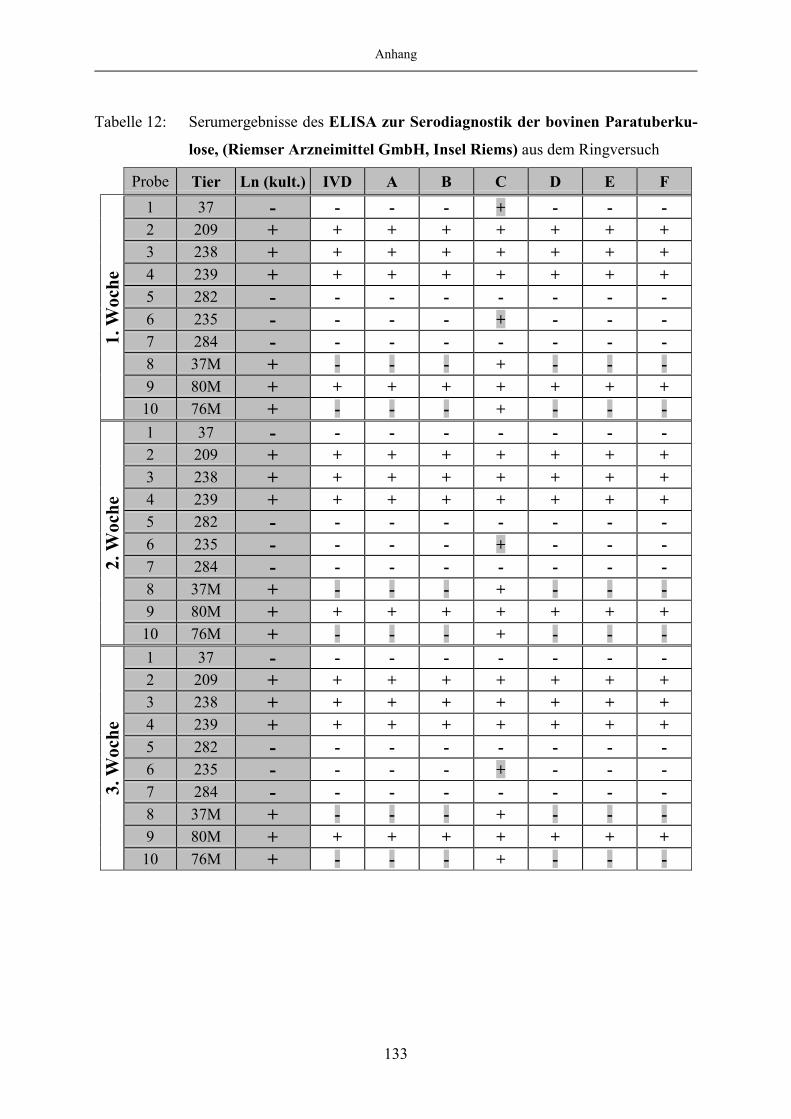
Anhang
133
Tabelle 12: Serumergebnisse des ELISA zur Serodiagnostik der bovinen Paratuberku-
lose, (Riemser Arzneimittel GmbH, Insel Riems) aus dem Ringversuch
Probe Tier Ln (kult.) IVD A B C D E F1 37 - - - - + - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - + - - -7 284 - - - - - - - -8 37M + - - - + - - -9 80M + + + + + + + +
1. W
oche
10 76M + - - - + - - -1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - + - - -7 284 - - - - - - - -8 37M + - - - + - - -9 80M + + + + + + + +
2. W
oche
10 76M + - - - + - - -1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - + - - -7 284 - - - - - - - -8 37M + - - - + - - -9 80M + + + + + + + +
3. W
oche
10 76M + - - - + - - -
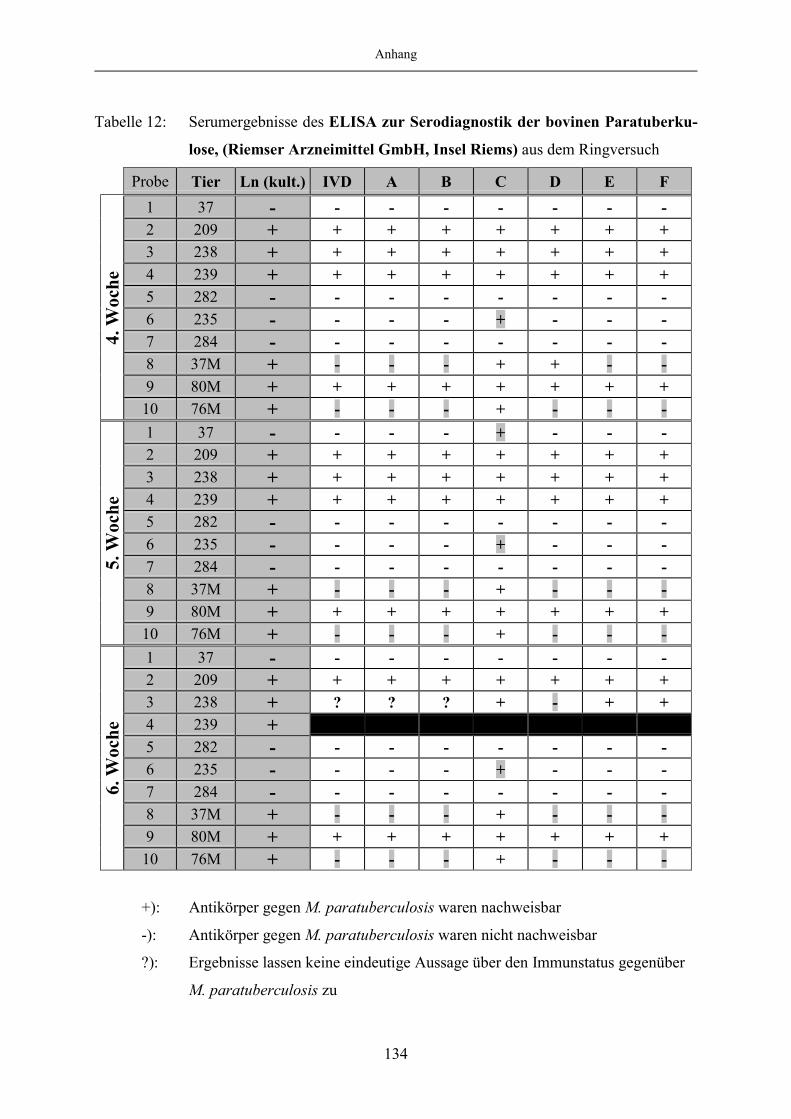
Anhang
134
Tabelle 12: Serumergebnisse des ELISA zur Serodiagnostik der bovinen Paratuberku-
lose, (Riemser Arzneimittel GmbH, Insel Riems) aus dem Ringversuch
Probe Tier Ln (kult.) IVD A B C D E F1 37 - - - - - - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - + - - -7 284 - - - - - - - -8 37M + - - - + + - -9 80M + + + + + + + +
4. W
oche
10 76M + - - - + - - -1 37 - - - - + - - -2 209 + + + + + + + +3 238 + + + + + + + +4 239 + + + + + + + +5 282 - - - - - - - -6 235 - - - - + - - -7 284 - - - - - - - -8 37M + - - - + - - -9 80M + + + + + + + +
5. W
oche
10 76M + - - - + - - -1 37 - - - - - - - -2 209 + + + + + + + +3 238 + ? ? ? + - + +4 239 +5 282 - - - - - - - -6 235 - - - - + - - -7 284 - - - - - - - -8 37M + - - - + - - -9 80M + + + + + + + +
6. W
oche
10 76M + - - - + - - -
+): Antikörper gegen M. paratuberculosis waren nachweisbar
-): Antikörper gegen M. paratuberculosis waren nicht nachweisbar
?): Ergebnisse lassen keine eindeutige Aussage über den Immunstatus gegenüber
M. paratuberculosis zu

Anhang
135
Tabelle 13: Serumergebnisse des Svanovir® Paratuberkulose ELISA, (Svanova, Schwe-
den) aus dem Ringversuch
Zehn Tiere mit bekanntem Lymphknotenbefund (Ln kult.) wurden von 7 Untersuchungsstel-
len (IVD GmbH und Institute A-F) über einen Zeitraum von 6 Wochen mittels Blut-ELISA
(Svanova, Schweden) untersucht. Das Tier 239 wurde in der 5. Woche aufgrund klinischer
Paratuberkulose geschlachtet. Farblich hervorgehoben sind die von dem kulturellen
Lymphknotenergebnis (Gold-Standard) abweichenden serologischen Befunde. Die Ergebnisse
der zweiten Messung wurden nur bei Abweichungen angegeben.
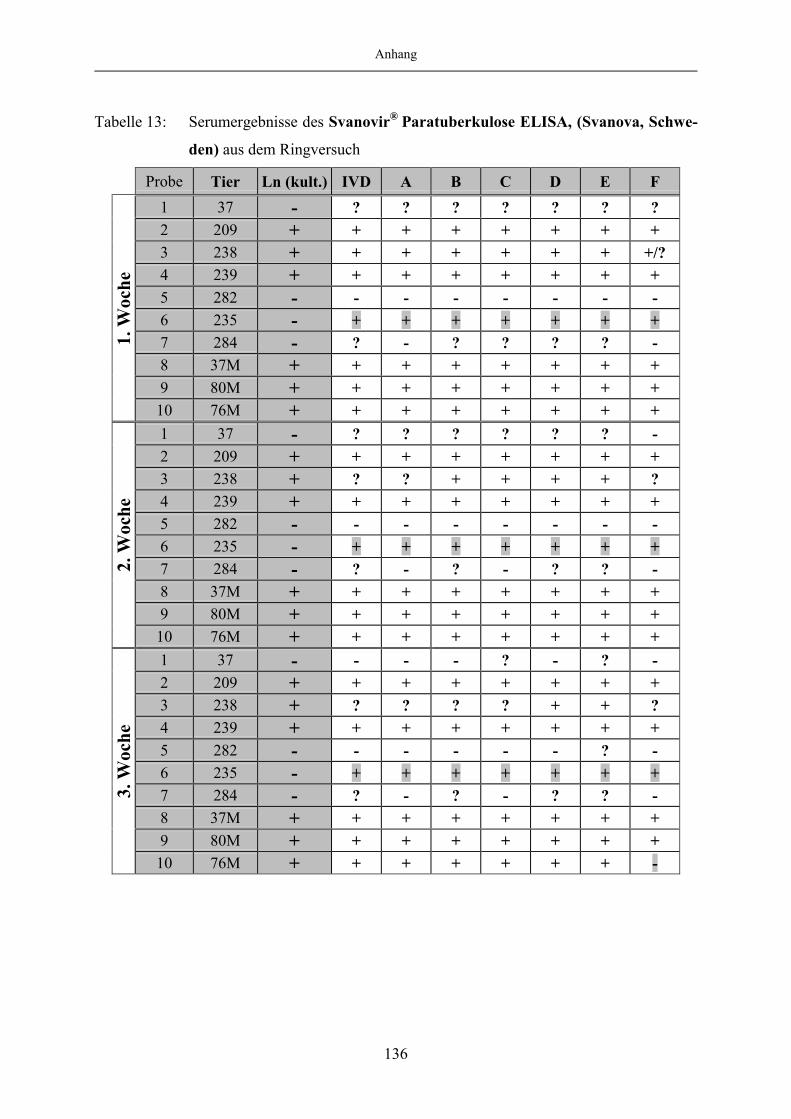
Anhang
136
Tabelle 13: Serumergebnisse des Svanovir® Paratuberkulose ELISA, (Svanova, Schwe-
den) aus dem Ringversuch
Probe Tier Ln (kult.) IVD A B C D E F1 37 - ? ? ? ? ? ? ?2 209 + + + + + + + +3 238 + + + + + + + +/?4 239 + + + + + + + +5 282 - - - - - - - -6 235 - + + + + + + +7 284 - ? - ? ? ? ? -8 37M + + + + + + + +9 80M + + + + + + + +
1. W
oche
10 76M + + + + + + + +1 37 - ? ? ? ? ? ? -2 209 + + + + + + + +3 238 + ? ? + + + + ?4 239 + + + + + + + +5 282 - - - - - - - -6 235 - + + + + + + +7 284 - ? - ? - ? ? -8 37M + + + + + + + +9 80M + + + + + + + +
2. W
oche
10 76M + + + + + + + +1 37 - - - - ? - ? -2 209 + + + + + + + +3 238 + ? ? ? ? + + ?4 239 + + + + + + + +5 282 - - - - - - ? -6 235 - + + + + + + +7 284 - ? - ? - ? ? -8 37M + + + + + + + +9 80M + + + + + + + +
3. W
oche
10 76M + + + + + + + -
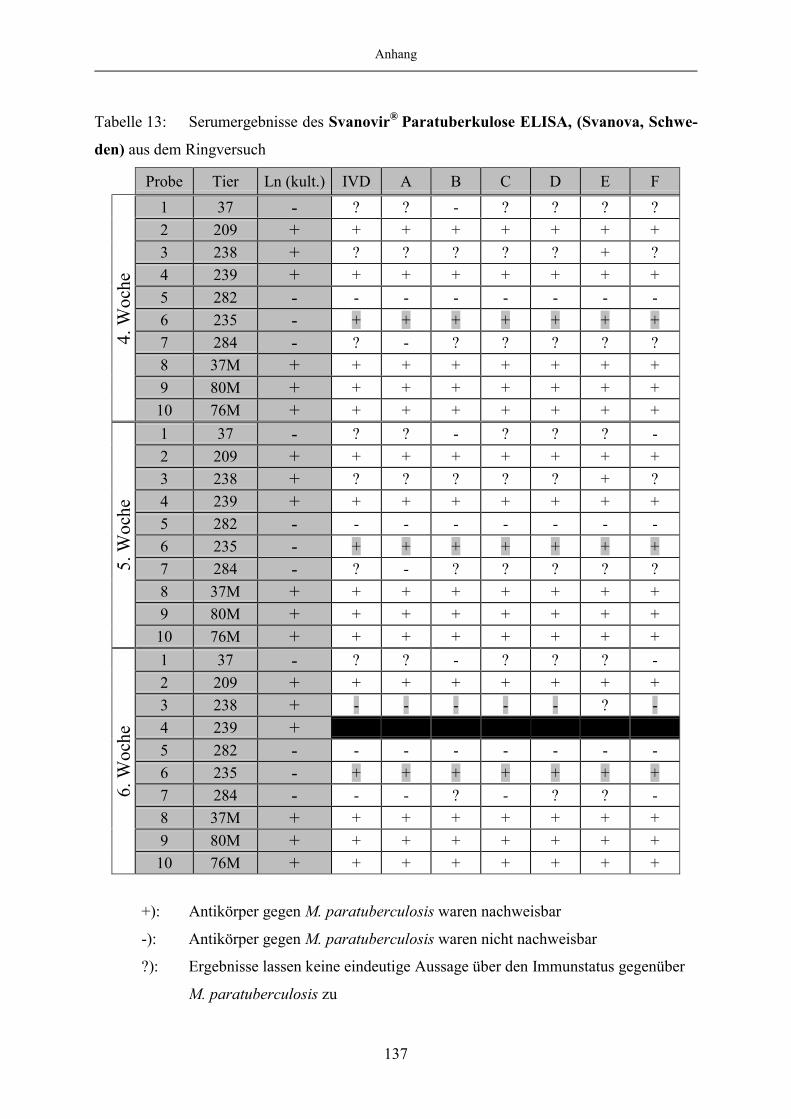
Anhang
137
Tabelle 13: Serumergebnisse des Svanovir® Paratuberkulose ELISA, (Svanova, Schwe-
den) aus dem Ringversuch
Probe Tier Ln (kult.) IVD A B C D E F1 37 - ? ? - ? ? ? ?2 209 + + + + + + + +3 238 + ? ? ? ? ? + ?4 239 + + + + + + + +5 282 - - - - - - - -6 235 - + + + + + + +7 284 - ? - ? ? ? ? ?8 37M + + + + + + + +9 80M + + + + + + + +
4. W
oche
10 76M + + + + + + + +1 37 - ? ? - ? ? ? -2 209 + + + + + + + +3 238 + ? ? ? ? ? + ?4 239 + + + + + + + +5 282 - - - - - - - -6 235 - + + + + + + +7 284 - ? - ? ? ? ? ?8 37M + + + + + + + +9 80M + + + + + + + +
5. W
oche
10 76M + + + + + + + +1 37 - ? ? - ? ? ? -2 209 + + + + + + + +3 238 + - - - - - ? -4 239 +5 282 - - - - - - - -6 235 - + + + + + + +7 284 - - - ? - ? ? -8 37M + + + + + + + +9 80M + + + + + + + +
6. W
oche
10 76M + + + + + + + +
+): Antikörper gegen M. paratuberculosis waren nachweisbar
-): Antikörper gegen M. paratuberculosis waren nicht nachweisbar
?): Ergebnisse lassen keine eindeutige Aussage über den Immunstatus gegenüber
M. paratuberculosis zu
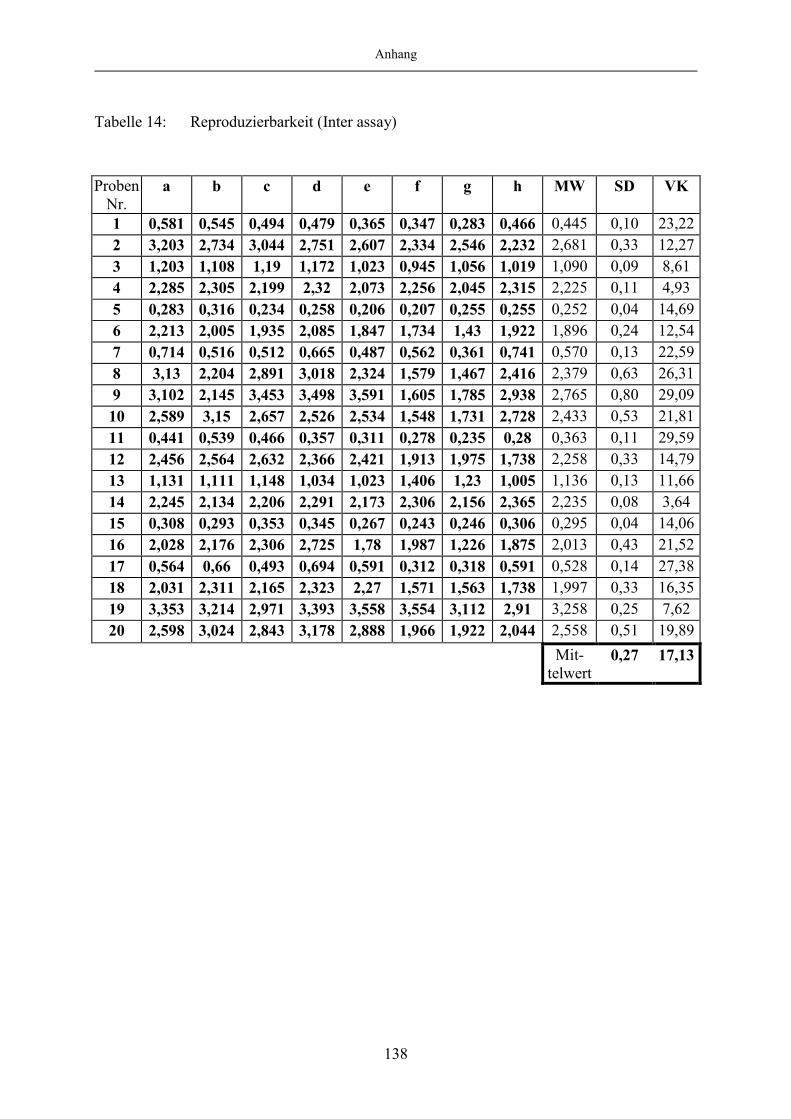
Anhang
138
Tabelle 14: Reproduzierbarkeit (Inter assay)
ProbenNr.
a b c d e f g h MW SD VK
1 0,581 0,545 0,494 0,479 0,365 0,347 0,283 0,466 0,445 0,10 23,222 3,203 2,734 3,044 2,751 2,607 2,334 2,546 2,232 2,681 0,33 12,273 1,203 1,108 1,19 1,172 1,023 0,945 1,056 1,019 1,090 0,09 8,614 2,285 2,305 2,199 2,32 2,073 2,256 2,045 2,315 2,225 0,11 4,935 0,283 0,316 0,234 0,258 0,206 0,207 0,255 0,255 0,252 0,04 14,696 2,213 2,005 1,935 2,085 1,847 1,734 1,43 1,922 1,896 0,24 12,547 0,714 0,516 0,512 0,665 0,487 0,562 0,361 0,741 0,570 0,13 22,598 3,13 2,204 2,891 3,018 2,324 1,579 1,467 2,416 2,379 0,63 26,319 3,102 2,145 3,453 3,498 3,591 1,605 1,785 2,938 2,765 0,80 29,0910 2,589 3,15 2,657 2,526 2,534 1,548 1,731 2,728 2,433 0,53 21,8111 0,441 0,539 0,466 0,357 0,311 0,278 0,235 0,28 0,363 0,11 29,5912 2,456 2,564 2,632 2,366 2,421 1,913 1,975 1,738 2,258 0,33 14,7913 1,131 1,111 1,148 1,034 1,023 1,406 1,23 1,005 1,136 0,13 11,6614 2,245 2,134 2,206 2,291 2,173 2,306 2,156 2,365 2,235 0,08 3,6415 0,308 0,293 0,353 0,345 0,267 0,243 0,246 0,306 0,295 0,04 14,0616 2,028 2,176 2,306 2,725 1,78 1,987 1,226 1,875 2,013 0,43 21,5217 0,564 0,66 0,493 0,694 0,591 0,312 0,318 0,591 0,528 0,14 27,3818 2,031 2,311 2,165 2,323 2,27 1,571 1,563 1,738 1,997 0,33 16,3519 3,353 3,214 2,971 3,393 3,558 3,554 3,112 2,91 3,258 0,25 7,6220 2,598 3,024 2,843 3,178 2,888 1,966 1,922 2,044 2,558 0,51 19,89
Mit-telwert
0,27 17,13

Anhang
139
9.2 Kulturmedien zur Anzucht von M. avium ssp.paratuberculosis
Herrold’s egg yolk-Medium mit Mycobactin (Heym)
Pepton 9,0 g
NaCl 4,5 g
Agar-Agar 15,3 g
Fleischextrakt 2,7 g
Glycerin 25 ml
A. dest. 870 ml
• pH-Wert 7,3 mit 1 M NaOH eingestellen
• 15 min bei 121°C autoklaviert
• auf 56°C abkühlen und folgende Substanzen zugegeben:
Natriumpyruvat 4,0 g
Cyclohexemid 0,75 g
Chloramphenicol 50 mg
Penicillin 200.000 I.E.
Mycobactin 2 mg
Malachitgrün-Lösung 2 % 5,1 ml
Dotter von 7 Eiern
Herrold’s egg yolk Medium ohne Mycobactin (Hey)
Dieses Medium entspricht dem oben beschriebenen Rezept, wobei der Zusatz von Mycobac-tin unterbleibt.

Anhang
140
Nährmedium nach Long
L-Asparagin 5,0 g
KH2PO4 3,0 g
NH4-dihydrogencitrat 5 g
MgSO4 x 7 H2O 1,0 g
NaCl 2,0 g
Na2CO3 0,9 g
FeCl3 50 mg
Glycerin 40 ml
A. dest . ad 1000 ml
• pH Wert 5,6-5,8 einstellen
• 15 min bei 121°C autoklaviert.
Watson-Reid-Medium
L-Asparagin 5 g
D-Glukose 10 g
Diammoniumzitrat 2 g
Eisenammoniumzitrat 75 mg
NaCl 2 g
KH2PO4 2 g
MgSO4 x 7 H2O 2 g
ZnSO4 x 7 H2O 2 mg
CaCl2 x 2 H2O 10 mg
Glycerin 63 ml
A. dest . ad 1000 ml

Anhang
141
• pH-Wert 5,6 mit NaOH einstellen
• autoklavieren
• nach Abkühlung auf 56°C Zugabe von:
Natriumpyruvat 4 g
Mycobactin 2 mg
9.3 Agarosegelelektrophorese
TE-Puffer
Tris pH 8 10 mM
EDTA pH 8 1 mM
TAE-Puffer (Tris-Acetat-Puffer), 50-fache Stammlösung
Tris 242,0 g
Essigsäure, 99%ig [w/v] 57,1 ml
0,5 M EDTA (pH 8.0) 100,0 ml
A. dest. ad 1000 ml
TBE-Puffer, 10-fache Stammlösung
Tris 108,0 g
Borsäure 55,0 g
EDTA 6,72 g
A. dest. ad 1000 ml
DNA-Ladungspuffer, 6-fache Stammlösung
Bromphenolblau 0,25% [w/v]
Xylenecyanol 0,25%
Glycerin 30% [v/v]

Anhang
142
9.4 SDS-Polyacrylamidgelelektrophorese (SDS-PAGE)
Bisacrylamid-Acrylamid-Lösung, 30%ig [w/v]
Acrylamid (4x) 29,20 g
Bisacrylamid (2x) 0,80 g
A. dest. 100 ml
• im Dunkeln über Nacht rühren
• sterilfiltrieren
• bei 4°C im Kühlschrank aufbewahren
Ammoniumpersulfat (APS)
Ammoniumpersulfat (APS) 10,0 g
A. dest. ad 100 ml
Trenngelpuffer, 1,5 M TRIS-Puffer
Tris (-base) 18,18 g
A. dest. ad 100 ml
• pH 8,8 mit Essigsäure einstellen
Sammelgelpuffer, 0,5 M TRIS-Puffer (SDS-PAGE, 1D)
Tris (-base) 6,06 g
A. dest. ad 100 ml
• pH 6,8 mit Essigsäure einstellen
Spaltpuffer, nicht reduzierend (2x)
0,5 M Tris, pH 6.8 1,5 ml
10% [w/v] SDS 6,0 ml
50´% [v/v] Glycerin 3,0 ml
1% [w/v] Bromphenolblau 100 µl

Anhang
143
Laufpuffer II für SDS-Gele (10x) (nach LAEMMLI)
Tris 30,0 g
Glycin 144,0 g
SDS 10,0 g
A. dest. ad 1000 ml
9.5 Coomassie®-Blau-Färbung
Färbelösung
Ethanol, 96%ig [v/v] 500,0 ml
Eisessig, 96%ig [v/v] 160,0 ml
Coomassie Brilliant Blue R250 10,0 g
A. dest. ad 2000 ml
Entfärbelösung
Methanol, 70%ig [v/v] 300,0 ml
Eisessig, 96%ig [v/v] 50,0 ml
A. dest. ad 600 ml
Geltrockungslösung
Ethanol 25%ig [v/v]
Eisessig 5%ig [v/v]
Glycerin 10% [v/v]

Anhang
144
9.6 Proteintransfer von SDS-Gelen im Tankblotting-Verfahren
Transferpuffer ohne SDS
Tris 3,0 g
Glycin 8,5 g
Methanol 200 ml
A. dest. ad 1000 ml
Blockpuffer (AP)
Tris 1,2 g
NaCl 8,8 g
A. dest. ad 800 ml
• nach Einstellung des pH-Wertes mit HCl auf 8,0 Zusatz von
Tween-20 0,5 ml
Gelatine 5,0 g
A. dest. ad 1000 ml
Waschpuffer, 10-fach (AP)
Tris 12,0 g
NaCl 88,0 g
A. dest. ad 800 ml
• nach Einstellung des pH-Wertes mit HCl auf 8,0 Zusatz von
Tween-20 5,0 ml

Anhang
145
9.7 Aufreinigung von IgG
Waschpuffer (Aufreinigung von IgG mit Abicap)
Kaliumphosphatpuffer, pH 9,0 50 mM
NaCl 150 mM
Tween 20 0,005%
Elutionspuffer (Aufreinigung von IgG mit Abicap)
Natriumcitratpuffer, pH 3,5 2 M
Proteinase K-Reaktionspuffer
10 mM Tris, pH 7.8 5 µl der 1 M Stammlösung
5 mM EDTA 5 µl der 0,5 M Stammlösung
0,5 % SDS 5 µl der 10% [w/v] Stammlösung
50 µg/ml Proteinase K 1,25 µl der Stammlösung
A. dest. ad 500 µl
Puffer für Papainverdau
Na2HPO4 20 mM
L-Cystein-HCl 20 mM
EDTA 10 mM
• pH 6,2
9.8 ELISA
PBS
NaCl 8,0 g
KH2PO4 0,2 g
Na2HPO4 x 12H2O 2,9 g
KCl 0,2 g
A. dest. ad 1000 ml

Anhang
146
PBS-Tween, pH 7.2
NaCl 8,0 g
KH2PO4 0,2 g
Na2HPO4 x 12H2O 2,9 g
KCl 0,2 g
Tween 20 0,5 ml
A. dest. ad 1000 ml
9.9 Primer
Primer 264 und 283 (SCHEIBL 1996)
264: TGC ACA CAG GCC ACA AGG GA
283: GAG TTT GAT CCT GGC TCA GGA
Die Annealingtemperatur beträgt 64°C.
Primer MK 5 und MK 6 (DORAN 1994)
MK5: TTC TTG AAG GGT GTT CGG GGC C
MK6: GCG ATG ATC GCA GCG TCT TTG G
Die Annealingtemperatur beträgt 62°C.

Anhang
147
9.10 Chemikalien
Acrylamid Fa. Serva, Heidelberg
Agar Nr. 1 Fa. Oxoid, Wessel
Agarose Fa. Gibco, Eggenstein
Ammoniumchlorid Fa. Sigma, Deisenhofen
Ammoniumdihydrogencitrat Fa. Sigma, Deisenhofen
Ammoniumperoxidsulfat (APS) Fa. Roth, Karlsruhe
Bisacrylamid Fa. Serva, Heidelberg
Borsäure Fa. Sigma, Deisenhofen
Bromphenolblau Fa. Sigma, Deisenhofen
Chloramphenicol Fa. Sigma, Deisenhofen
Coomassie® Brilliant Blue Fa. Sigma, Deisenhofen
Cyclohexemid Fa. Sigma, Deisenhofen
D-Glukose Fa. Sigma, Deisenhofen
Diammoniumcitrat Fa. Sigma, Deisenhofen
EDTA Fa. Sigma, Deisenhofen
Eisenammoniumcitrat Fa. Sigma, Deisenhofen
Eisenchlorid Fa. Sigma, Deisenhofen
Essigsäure Fa. Roth, Karlsruhe
Ethanol, vergällt Fa. Roth, Karlsruhe
Fleischextrakt Fa. Oxoid, Wessel
Gelatine Fa. Sigma, Deisenhofen
Glycin Fa. Sigma, Deisenhofen
Glyzerin Fa. Roth, Karlsruhe
Kaliumchlorid Fa. Sigma, Deisenhofen
Kaliumhydrogenphosphat Fa. Sigma, Deisenhofen
Kaliumphosphat Fa. Sigma, Deisenhofen
Kalziumchlorid Fa. Sigma, Deisenhofen
L-Asparagin Fa. Sigma, Deisenhofen

Anhang
148
Laurylsulfat (SDS) Fa. Sigma, Deisenhofen
Magnesiumsulfat Fa. Gibco, Eggenstein
Malachitgrün Fa. Sigma, Deisenhofen
Methanol Fa. Roth, Karlsruhe
Mycobactin Fa. Rhone Merieux
Natriumcarbonat Fa. Sigma, Deisenhofen
Natriumchlorid Fa. Sigma, Deisenhofen
Natriumcitrat Fa. Sigma, Deisenhofen
Natriumhydrogenphosphat Fa. Sigma, Deisenhofen
Natriumpyruvat Fa. Sigma, Deisenhofen
Natronlauge Fa. Sigma, Deisenhofen
Penicillin Fa. Sigma, Deisenhofen
Salzsäure Fa. Roth, Karlsruhe
Tris-(hydroxymethyl)-aminomethan Fa. Sigma, Deisenhofen
Tween 20 ® Fa. Sigma, Deisenhofen
Xylencyanol Fa. Sigma, Deisenhofen
Zinksulfat Fa. Sigma, Deisenhofen
9.11 Enzyme
Lysozym Fa. Roth, Karlsruhe
Proteinase K Fa. Roth, Karlsruhe
Taq-DNA Polymerase Fa. Gibco, Eggenstein

Abbildungsverzeichnis
149
9.12 Abbildungsverzeichnis
Abbildung 1: Vierfeldertafel zur Schätzung von D-SN/D-SP und der Vorhersagewerte ...... 51
Abbildung 2: Abb. A: Coomassie®-Blau gefärbtes 10 % Acrylamidgel
Abb. B: Western-Blot mit monoklonalem Sekundärantikörper entwickelt ..... 55
Abbildung 3: Vergleich der Ergebnisse von Blut- und Milch-ELISA ................................... 58
Abbildung 4: ROC-Kurven für den Milch-ELISA bei Annahme der zweifelhaften
Seren im Blut-ELISA = positiv bzw. = negativ. .............................................. 59
Abbildung 5: Graphische Darstellung der Summe falscher Ergebnisse (Annahme:
zweifelhafte Blut-ELISA-Ergebnisse = negativ) des Milch-ELISA und
dem jeweiligen Abstand zur y = x Achse bei verschieden „Cut off“-
Werten .............................................................................................................. 61
Abbildung 6: Graphische Darstellung der Summe falscher Ergebnisse (Annahme:
zweifelhafte Blut-ELISA-Ergebnisse = positiv) des Milch-ELISA und
dem jeweiligen Abstand zur y = x Achse bei verschieden „Cut off“-
Werten .............................................................................................................. 62
Abbildung 7: Graphische Darstellung der Milch-ELISA-Ergebnisse .................................... 67
Abbildung 8: Ermittlung der Nachweisgrenze experimentell kontaminierter Milch
(Hochlaktation)................................................................................................. 72
Abbildung 9: Bestimmung der optimalen Konzentration an M. paratuberculosis-DNA
für die interne Funktionskontrolle .................................................................... 74
Abbildung 10: PCR-Analyse dreier ausgewählter Milchproben zur Interpretation der
internen Funktionskontrolle ............................................................................. 75
Abbildung 11: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 209 .......... 77
Abbildung 12: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 235 .......... 78
Abbildung 13: PCR-Analyse von Viertelgemelkproben der Kuh 235 ..................................... 79
Abbildung 14: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 238 .......... 80
Abbildung 15: PCR-Analyse von Einzelgemelkproben der Kuh 238...................................... 81

Abbildungsverzeichnis
150
Abbildung 16: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 239 .......... 82
Abbildung 17: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 282 .......... 83
Abbildung 18: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 284 .......... 84
Abbildung 19: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 76 ............ 85
Abbildung 20: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 37 ............ 86
Abbildung 21: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 37 M........ 87
Abbildung 22: Ergebnisse der Verlaufsuntersuchung in Einzelgemelkproben (Milch-
PCR, ELISA), im Blutserum, Kot und Lymphknoten der Kuh Nr. 80 ............ 88

Tabellenverzeichnis
151
9.13 Tabellenverzeichnis
Tabelle 1: Herkunft der Bakterienstämme ........................................................................ 34
Tabelle 2: PCR-Profil ........................................................................................................ 48
Tabelle 3: Reproduzierbarkeit von Tag zu Tag (n = 8) ..................................................... 57
Tabelle 4: Daten für die ROC-Kurven (Annahme: zweifelhafte Ergebnisse =
negativ) ............................................................................................................. 60
Tabelle 5: Daten für die ROC-Kurven (Annahme: zweifelhafte Ergebnisse =
positiv).............................................................................................................. 60
Tabelle 6: „Cut off“-Werte des Milch-ELISA (TIJSSEN 1985)....................................... 64
Tabelle 7: Vierfeldertafel zum Vergleich des Milch-ELISA mit dem Blut-ELISA
auf der Grundlage der berechneten „Cut-off“-Werte ....................................... 65
Tabelle 8: Anzahl der Serum- und Milchproben, die in der Paralleluntersuchung
den verschiedenen Bewertungs-Stufen des ELISA zugeordnet werden
konnten ............................................................................................................. 66
Tabelle 9: Vergleich der Empfindlichkeit der Milch-PCR in experimentell
kontaminierter Milch verschiedener Laktationsstadien sowie bakteriell
stark belasteten Mastitis-Milch bei mehrmaligen Wiederholungen................. 71
Tabelle 10: Ergebnisse der Verlaufsuntersuchung .............................................................. 89
Tabelle 11: Serumergebnisse des HerdChek™ Mycobacterium paratuberculosis
(Idexx, Wörrstadt) aus dem Ringversuch .................................................... 129
Tabelle 12: Serumergebnisse des ELISA zur Serodiagnostik der bovinen
Paratuberkulose, (Riemser Arzneimittel GmbH, Insel Riems) aus
dem Ringversuch............................................................................................ 132
Tabelle 13: Serumergebnisse des Svanovir® Paratuberkulose ELISA, (Svanova,
Schweden) aus dem Ringversuch .................................................................. 135
Tabelle 14: Reproduzierbarkeit (Inter assay) .................................................................... 138

Abkürzungsverzeichnis
152
9.14 Abkürzungsverzeichnis
µ mikro (x10-6)A. dest. Aqua destillata (destilliertes Wasser)BgVV Bundesinstitut für gesundheitlichen Verbraucherschutz und Veterinärmedizinbzw. beziehungsweiseca. zirkaELISA Enzyme linked immuno sorbent assayEU engl. ELISA units, ELISA-Einheiteng Grammg Erdbeschleunigungskonstanteh Stunde (lateinisch: hora)I.E. Internationale EinheitenKBR KomplementbindungsreaktionkDa Kilodalton (103 Dalton)LAM Lipoarabinomannanl Literm milli (x10-3)M mol / lM. Mycobacteriummin Minute(n)n bei Berechnung des Mittelwertes die Anzahl der EinzelbeobachtungenNr. NummerPCR Polymerase-Ketten-Reaktion (engl.: polymerase chain reaction)RT Raumtemperaturrpm engl. rotation per minute (Umdrehungen pro Minute)s. siehesek Sekunde(n)ssp. lateinisch subspecies (Subspezies)Tris Trishydroxymethylaminomethanü.N. über Nachtz.B. zum Beispiel

Danksagungen
Danksagung
Herrn Prof. Dr. G.-F. Gerlach danke ich sehr herzlich für die Überlassung des interessanten,praxisnahen Themas, seinen fachlichen Rat und seine jederzeit gewährte freundliche Unter-stützung.
Mein ganz besonderer Dank gilt Frau Dr. Katrin Strutzberg und Herrn Dr. Matthias Homuth,Geschäftsführer der IVD GmbH, für die motivierende und freundschaftliche Betreuung injeder Phase dieser Arbeit, zu deren Gelingen ihre fachlichen Anregungen und ihr Engagemententscheidend beigetragen haben.
Frau Dr. Flebbe, Leiterin der Niedersächsischen Tierseuchenkasse, danke ich herzlich für diegute Kooperation und die freundliche Unterstützung.
Bei Herrn Dr. Beyerbach möchte ich mich für die sehr gute fachliche Unterstützung bei derstatistischen Auswertung bedanken.
Herrn Dr. Eicken danke ich für die gute Zusammenarbeit und die Unterstützung bei der Pro-bennahme.
Bedanken möchte ich mich auch bei Herrn Dr. Gehrisch, Tiergesundheitsdienst Thüringen,und bei Herrn Dr. Brentrup, Landwirtschftskammer Westfalen-Lippe, für die Bereitstellungvon Blut- und Milchproben.
Jörg Matusch danke ich herzlich für die Hilfe in allen PC-Angelegenheiten und für seine Ge-duld.
Weiterhin danke ich dem gesamten IVD Team Frau Dr. Katrin Strutzberg, Herrn Dr. MatthiasHomuth, Frau Dr. Astrid Tschentscher, Herrn Dr. Jan Böhmer und Yener Selcuk für dieHilfsbereitschaft und die freundliche Atmosphäre.
Mein Dank gilt ebenso den Institutsangehörigen und meinen Mitstreitern, insbesondere Birgit,Thomas, Sabine, Nina, Walaiporn, Anja, Janin und Mark für die moralische Unterstützungund manche fröhliche Stunde.