Integrative Analysis of Head and Neck Cancer ... · typically using a combination of surgery,...
13
Biology of Human Tumors Integrative Analysis of Head and Neck Cancer Identifies Two Biologically Distinct HPV and Three Non-HPV Subtypes Michaela K. Keck 1,2 , Zhixiang Zuo 1 , Arun Khattri 1 , Thomas P. Stricker 3 , Christopher D. Brown 4 , Matin Imanguli 5 , Damian Rieke 1 , Katharina Endhardt 1 , Petra Fang 1 , Johannes Br€ agelmann 1 , Rebecca DeBoer 1 , Mohamed El-Dinali 1 , Serdal Aktolga 1 , Zhengdeng Lei 6 , Patrick Tan 6,7 , Steve G. Rozen 6 , Ravi Salgia 1,8 , Ralph R. Weichselbaum 1,8 , Mark W. Lingen 3,8 , Michael D. Story 9 , K. Kian Ang 10 , Ezra E.W. Cohen 11 , Kevin P. White 8,12 , Everett E. Vokes 1,8 , and Tanguy Y. Seiwert 1,8,12 Abstract Purpose: Current classification of head and neck squamous cell carcinomas (HNSCC) based on anatomic site and stage fails to capture biologic heterogeneity or adequately inform treatment. Experimental Design: Here, we use gene expression-based consensus clustering, copy number profiling, and human papil- lomavirus (HPV) status on a clinically homogenous cohort of 134 locoregionally advanced HNSCCs with 44% HPV þ tumors together with additional cohorts, which in total comprise 938 tumors, to identify HNSCC subtypes and discover several sub- type-specific, translationally relevant characteristics. Results: We identified five subtypes of HNSCC, including two biologically distinct HPV subtypes. One HPV þ and one HPV subtype show a prominent immune and mesenchymal pheno- type. Prominent tumor infiltration with CD8 þ lymphocytes char- acterizes this inflamed/mesenchymal subtype, independent of HPV status. Compared with other subtypes, the two HPV sub- types show low expression and no copy number events for EGFR/HER ligands. In contrast, the basal subtype is uniquely characterized by a prominent EGFR/HER signaling phenotype, negative HPV-status, as well as strong hypoxic differentiation not seen in other subtypes. Conclusion: Our five-subtype classification provides a com- prehensive overview of HPV þ as well as HPV HNSCC biology with significant translational implications for biomarker devel- opment and personalized care for patients with HNSCC. Clin Cancer Res; 21(4); 870–81. Ó2014 AACR. Introduction Head and neck squamous cell carcinoma (HNSCC) is the sixth most common nonskin cancer worldwide with an annual inci- dence of approximately 600,000 cases and a mortality rate of 40% to 50% (1, 2). The major known risk factors are environmental exposures to tobacco products, alcohol, and infection with high- risk human papillomaviruses (HPV). The incidence of HPV þ tumors is rising rapidly and HPV status is now the strongest prognostic marker (3). Although HNSCC is now widely viewed as composed of two distinct clinical entities, HPV þ and HPV , a therapeutically rel- evant molecular classification system remains elusive. Prior attempts at profiling this clinically heterogeneous disease have been hampered by lack of information about HPV status and/or absence of significant numbers of HPV þ cases, small sample size, and lack of annotation (4, 5). Currently, neither molecular classification nor validated biomarkers are used in clinical prac- tice. Instead all patients with HNSCC are treated independent of the underlying biology based on stage and anatomic location, typically using a combination of surgery, radiation, and chemo- therapy (6). Cetuximab, an anti-EGFR antibody, is the only approved targeted therapy for HNSCC, but predictive biomarkers remain to be identified and overall cetuximab has a response rate of 13% (7). In the current study, we investigate a fully clinically annotated, very homogenous patient cohort of locoregionally advanced HNSCC (including 44% HPV þ tumors), all treated uniformly with organ-preserving chemoradiotherapy together with all avail- able other HNC cohorts, including the cancer genome atlas head 1 Department of Medicine, The University of Chicago, Chicago, Illinois. 2 Institute of Biological Chemistry and Nutrition, The University of Hohenheim, Stuttgart, Germany. 3 Department of Pathology, The Uni- versity of Chicago, Chicago, Illinois. 4 Department of Human Genetics, The University of Chicago, Chicago, Illinois. 5 Medical Center, Depart- ment of Otolaryngology-Head and Neck Surgery, The University of Texas Southwestern, Dallas, Texas. 6 Duke-NUS Graduate Medical School, Singapore. 7 Genome Institute of Singapore, Singapore. 8 The University of Chicago Comprehensive Cancer Center, Chicago, Illinois. 9 Southwestern Medical Center, Department of Radiation Oncology, The University of Texas, Dallas,Texas. 10 MD Anderson Cancer Center, The University of Texas, Houston,Texas. 11 University of California San Diego, La Jolla, California. 12 Institute for Genomics and Systems Biol- ogy, The University of Chicago, Chicago, Illinois. Note: Supplementary data for this article are available at Clinical Cancer Research Online (http://clincancerres.aacrjournals.org/). M.K. Keck, Z. Zuo, and A. Khattri contributed equally to this article. Corresponding Author: Tanguy Seiwert, Knapp Center for Biomedical Discov- ery at the University of Chicago, 900 E. 57th Street, 7110, Chicago, IL 60637. Phone: 773-702-2452; Fax: 773-702-3002; E-mail: [email protected] doi: 10.1158/1078-0432.CCR-14-2481 Ó2014 American Association for Cancer Research. Clinical Cancer Research Clin Cancer Res; 21(4) February 15, 2015 870 on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481
Transcript of Integrative Analysis of Head and Neck Cancer ... · typically using a combination of surgery,...

Biology of Human Tumors
Integrative Analysis of Head and Neck CancerIdentifies TwoBiologically Distinct HPV and ThreeNon-HPV SubtypesMichaela K. Keck1,2, Zhixiang Zuo1, Arun Khattri1, Thomas P. Stricker3,Christopher D. Brown4, Matin Imanguli5, Damian Rieke1, Katharina Endhardt1,Petra Fang1, Johannes Br€agelmann1, Rebecca DeBoer1, Mohamed El-Dinali1,Serdal Aktolga1, Zhengdeng Lei6, Patrick Tan6,7, Steve G. Rozen6, Ravi Salgia1,8,Ralph R.Weichselbaum1,8, Mark W. Lingen3,8, Michael D. Story9, K. Kian Ang10,Ezra E.W. Cohen11, Kevin P.White8,12, Everett E. Vokes1,8, and Tanguy Y. Seiwert1,8,12
Abstract
Purpose:Current classification of head and neck squamous cellcarcinomas (HNSCC) based on anatomic site and stage fails tocapture biologic heterogeneity or adequately inform treatment.
Experimental Design: Here, we use gene expression-basedconsensus clustering, copy number profiling, and human papil-lomavirus (HPV) status on a clinically homogenous cohort of 134locoregionally advanced HNSCCs with 44% HPVþ tumorstogether with additional cohorts, which in total comprise 938tumors, to identify HNSCC subtypes and discover several sub-type-specific, translationally relevant characteristics.
Results: We identified five subtypes of HNSCC, including twobiologically distinct HPV subtypes. One HPVþ and one HPV�
subtype show a prominent immune and mesenchymal pheno-
type. Prominent tumor infiltration with CD8þ lymphocytes char-acterizes this inflamed/mesenchymal subtype, independent ofHPV status. Compared with other subtypes, the two HPV sub-types show low expression and no copy number events forEGFR/HER ligands. In contrast, the basal subtype is uniquelycharacterized by a prominent EGFR/HER signaling phenotype,negative HPV-status, as well as strong hypoxic differentiation notseen in other subtypes.
Conclusion: Our five-subtype classification provides a com-prehensive overview of HPVþ as well as HPV� HNSCC biologywith significant translational implications for biomarker devel-opment and personalized care for patients with HNSCC. ClinCancer Res; 21(4); 870–81. �2014 AACR.
IntroductionHead and neck squamous cell carcinoma (HNSCC) is the sixth
most common nonskin cancer worldwide with an annual inci-
dence of approximately 600,000 cases and amortality rate of 40%to 50% (1, 2). The major known risk factors are environmentalexposures to tobacco products, alcohol, and infection with high-risk human papillomaviruses (HPV). The incidence of HPVþ
tumors is rising rapidly and HPV status is now the strongestprognostic marker (3).
Although HNSCC is now widely viewed as composed of twodistinct clinical entities, HPVþ and HPV�, a therapeutically rel-evant molecular classification system remains elusive. Priorattempts at profiling this clinically heterogeneous disease havebeen hampered by lack of information about HPV status and/orabsence of significant numbers of HPVþ cases, small sample size,and lack of annotation (4, 5). Currently, neither molecularclassification nor validated biomarkers are used in clinical prac-tice. Instead all patients with HNSCC are treated independent ofthe underlying biology based on stage and anatomic location,typically using a combination of surgery, radiation, and chemo-therapy (6). Cetuximab, an anti-EGFR antibody, is the onlyapproved targeted therapy for HNSCC, but predictive biomarkersremain to be identified and overall cetuximab has a response rateof 13% (7).
In the current study, we investigate a fully clinically annotated,very homogenous patient cohort of locoregionally advancedHNSCC (including 44% HPVþ tumors), all treated uniformlywith organ-preserving chemoradiotherapy together with all avail-able other HNC cohorts, including the cancer genome atlas head
1Department of Medicine, The University of Chicago, Chicago, Illinois.2Institute of Biological Chemistry and Nutrition, The University ofHohenheim, Stuttgart, Germany. 3Department of Pathology, The Uni-versity of Chicago, Chicago, Illinois. 4Department of Human Genetics,The University of Chicago, Chicago, Illinois. 5Medical Center, Depart-ment of Otolaryngology-Head and Neck Surgery, The University ofTexas Southwestern, Dallas, Texas. 6Duke-NUS Graduate MedicalSchool, Singapore. 7Genome Institute of Singapore, Singapore. 8TheUniversity of Chicago ComprehensiveCancer Center, Chicago, Illinois.9Southwestern Medical Center, Department of Radiation Oncology,The University of Texas, Dallas, Texas. 10MD Anderson Cancer Center,The University of Texas, Houston, Texas. 11University of California SanDiego, La Jolla, California. 12Institute for Genomics and Systems Biol-ogy, The University of Chicago, Chicago, Illinois.
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
M.K. Keck, Z. Zuo, and A. Khattri contributed equally to this article.
Corresponding Author: Tanguy Seiwert, Knapp Center for Biomedical Discov-ery at the University of Chicago, 900 E. 57th Street, 7110, Chicago, IL 60637.Phone: 773-702-2452; Fax: 773-702-3002; E-mail:[email protected]
doi: 10.1158/1078-0432.CCR-14-2481
�2014 American Association for Cancer Research.
ClinicalCancerResearch
Clin Cancer Res; 21(4) February 15, 2015870
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

and neck cancer cohort (TCGA), to identify HNSCC subtypes andtheir biologic and prognostic characteristics. We integrate geneexpression-based consensus clustering, cross-platform approachesfor validation, copy number profiling, and HPV status to discoversubtype-specific, translationally relevant biology.
The resulting new taxonomy of HNSCC closely correlateswith multiple HNC-relevant oncogenic processes and a biolo-gy-based taxonomy of HNSCC. In particular, we identify twosubtypes of HPVþ head and neck cancer and two subtypes (oneHPVþ and one HPV�) with a prominent immune phenotypecharacterized by CD8þ T-cell infiltration, as well as a subtypewith near exclusive HER ligand/EGFR copy number-drivensignaling. As such, our classification is broadly applicable tobiomarker and therapy development for head and neck cancerand will inform future clinical and preclinical studies, bio-marker development, and personalized HNC care.
Materials and MethodsPatient samples
OCT blocks of frozen tissue samples were obtained fromthe University of Chicago Head and Neck Cancer tissue bank(Chicago, IL; Institutional Review Board-approved protocolUCCCC#8980; Supplementary Experimental Procedures andSupplementary Fig. S1).
Nucleic acid extractionA total of 171 locoregionally advanced HNSCC specimens
obtained before treatment with concurrent chemoradiotherapy(all patients received organ-preserving chemoradiotherapywith curative intent) were selected for this study. Our aimwas to select at least 100 cases to be able to perform consensusclustering later. Samples were collected and banked between1997 and 2010, but our focus was on recent cases. A sectionwas cut from OCT-frozen blocks and stained with hematoxylinand eosin (H&E). An expert HNC pathologist reviewed slides
to determine and circle the area with the highest tumorcontent. Samples with microscopic tumor or tumor content<60% were excluded from further analysis. Guided by theH&E-stained slides, the region with the highest tumor contentwas cut from the OCT blocks, pulverized using CryoPrep(Covaris) and homogenized in lysis buffer from an All-PrepRNA/DNA/Protein Mini kit (Qiagen) or from an RNA/DNA/Protein Purification Kit (Norgen Biotek) using an Ultra-sonicator (Covaris). DNA, RNA, and protein were isolatedfrom each sample using the respective kit and following themanufacturer's protocol.
Gene expression and copy number analysisGene expression profiling was done using Agilent 4 � 44
Kv2 expression arrays on 134 samples (SupplementaryFig. S1 and Supplementary Experimental Procedures). To eva-luate copy number aberration (CNA) in HNSCC, we selecteda panel of 86 and subsequently 147 candidate genes knownto have frequent CNA in cancer and analyzed these using theNanostring nCounter assay (NanoString Technologies; ref. 8;Supplementary Experimental Procedures). Data and materialsavailability: Gene Expression Omnibus Accession number:GSE40774 (Embargoed until publication; http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?token¼jjsnbqwecyoyirg&acc¼GSE40774).
Discovery of subtypesTo discover gene expression-based subtypes, we used newly
generated gene expression data (Agilent, n ¼ 134), as well as adata based on Illumina BeadArrays (n ¼ 131; Yordy and collea-gues; in preparation), and publicly available HNSCC microarraydata from two studies based on Affymetrix HG133 arrays (n ¼106; refs. 9, 10; see Supplementary Experimental Procedures fordetails).
IHC and immunofluorescenceIHC and immunofluorescence were performed using routine
methods (Supplementary Experimental Procedures).
ResultsIdentification and validation of three HNSCC supergroups
Microarray gene expression profiles of 371 HNSCC sampleswere derived from several datasets (Supplementary Table S1),including our cohort (n ¼ 134, Agilent; Supplementary Fig.S1), an Illumina cohort (Yordy and colleagues; unpublishedresults), and two Affymetrix cohorts (9, 11). These cross-platform datasets were used as the discovery set for theclassification of HNSCC samples. All datasets were analyzedas described in the Supplementary Experimental Proceduresand outlined in Supplementary Fig. S2A. This resulted in 821reliable and informative genes (Supplementary Table S2),based on which five subtypes were derived separately in eachdataset (Supplementary Fig. S2B). These five subtypes slightlydiffer across the three datasets, but fall into three very stablecross-platform groups (termed supergroups) on the basis ofhierarchical clustering of their centroids (Fig. 1A, B, and C).All centroids in the three cross-platform supergroups show astrongly positive silhouette width (average ¼ 0.47), indicatingthat centroids fit their respective cluster (Fig. 1B; ref. 12). Thehigh correlation of our three supergroups and previously
Translational Relevance
Head and neck cancer (HNC) is composed of humanpapillomavirus HPVþ and HPV� tumors. However, thereremains significant heterogeneity in clinical behavior (e.g.,response to therapies, including anti-EGFR or anti–PD-1immunotherapy) and further biologic subclassification andelucidation of specific biologic characteristics are lacking. Onelimitation of available classifications is the absence of anylarger (and representative) cohort of HPVþ tumors or the lackof validation across multiple datasets/platforms. We identifyfive HNC subtypes, including two distinct HPV subtypes withdifferential biology across multiple HNC cohorts, and showvalidity of subtypes and associated biology across all currentlyavailable HNC datasets, including The Cancer Genome Atlas(TCGA) HNC cohort: In particular, we report (i) an immune(and mesenchymal) phenotype present in a group of HNCtumors independent of HPV status and (ii) a group with non–HPV-associated tumors showing a prominent HER-drivenphenotype as well as hypoxia, which are candidate biomarkersfor respective therapies (e.g., PD-1, EGFR/HER, or hypoxia-targeting agents).
Integrative Analysis Identifies Five Distinct HNC Subtypes
www.aacrjournals.org Clin Cancer Res; 21(4) February 15, 2015 871
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481
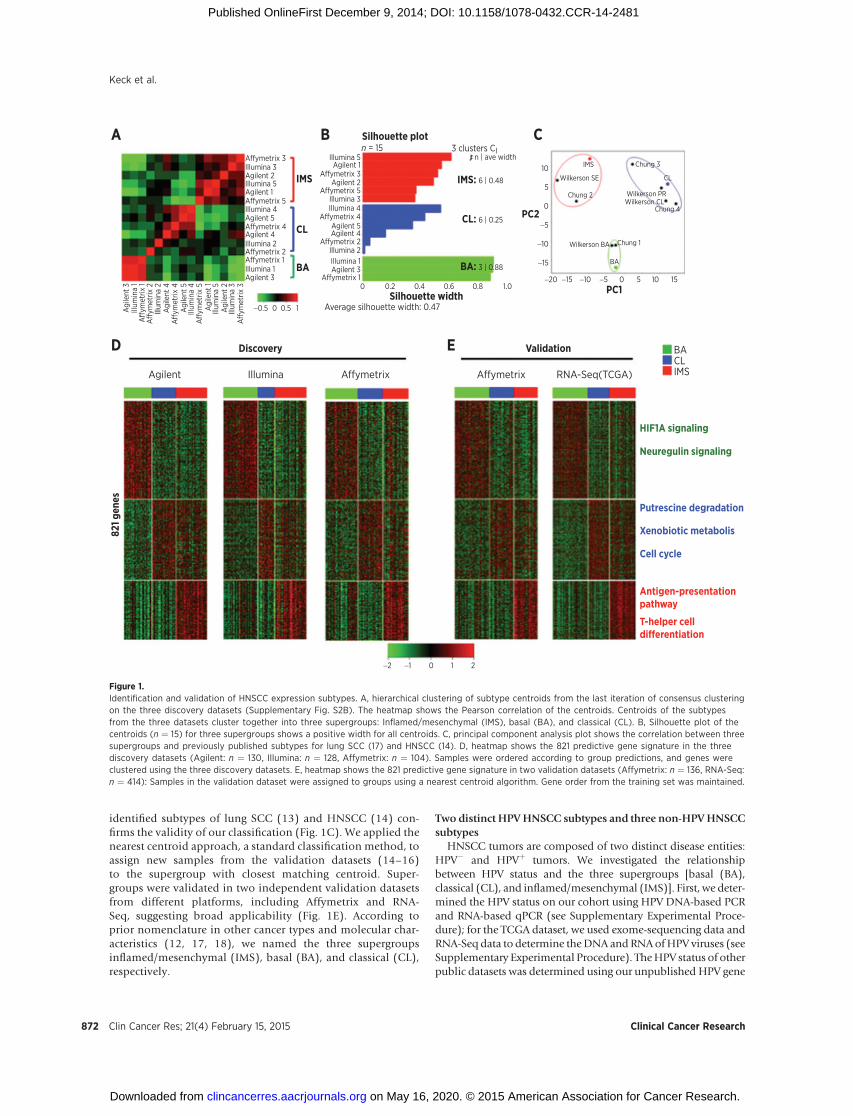
identified subtypes of lung SCC (13) and HNSCC (14) con-firms the validity of our classification (Fig. 1C). We applied thenearest centroid approach, a standard classification method, toassign new samples from the validation datasets (14–16)to the supergroup with closest matching centroid. Super-groups were validated in two independent validation datasetsfrom different platforms, including Affymetrix and RNA-Seq, suggesting broad applicability (Fig. 1E). According toprior nomenclature in other cancer types and molecular char-acteristics (12, 17, 18), we named the three supergroupsinflamed/mesenchymal (IMS), basal (BA), and classical (CL),respectively.
Two distinct HPVHNSCC subtypes and three non-HPVHNSCCsubtypes
HNSCC tumors are composed of two distinct disease entities:HPV� and HPVþ tumors. We investigated the relationshipbetween HPV status and the three supergroups [basal (BA),classical (CL), and inflamed/mesenchymal (IMS)]. First, we deter-mined the HPV status on our cohort using HPV DNA-based PCRand RNA-based qPCR (see Supplementary Experimental Proce-dure); for the TCGA dataset, we used exome-sequencing data andRNA-Seq data to determine theDNA and RNAofHPV viruses (seeSupplementary Experimental Procedure). TheHPV status of otherpublic datasets was determined using our unpublished HPV gene
Affymetrix 3
Affymetrix 3
Affymetrix 5
Affymetrix 4
Affymetrix 2
Affymetrix 1
Affymetrix 5
Affymetrix 4
Affymetrix 2Affymetrix 1
Illumina 3Illumina 5
10
5
0
−5
−10
−15
−15 −10 −5 0PC1
5 10 15
BACLIMS
−20
Illumina 3Illumina 4
Illumina 2Illumina 1
n = 15 3 clusters CI
IMS: 6 | 0.48
PC2
n | ave width
Wilkerson SE
Wilkerson PRWilkerson CL
Wilkerson BA
CL
Chung 2
Chung 3
Chung 4
Chung 1
BA
IMS
CL: 6 | 0.25
BA: 3 | 0.88
IMS
Silhouette plotA
D E
B C
CL
BA
Illumina 4
Illumina 2
Illumina 1
Illumina 5Agilent 2
Agilent 1
Agilent 2
Agilent 5Agilent 4
Agilent 3
Agilent 5
Agilent 4
Aff
ymet
rix 1
Aff
ymet
rix 2
Aff
ymet
rix 4
Aff
ymet
rix 5
Aff
ymet
rix 3
Illum
ina
1
Illum
ina
2
Illum
ina
4
Agi
lent
3
Affymetrix Affymetrix RNA-Seq(TCGA)IlluminaAgilent
Agi
lent
4
Agi
lent
5
Illum
ina
5A
gile
nt 1
Illum
ina
3A
gile
nt 2
Agilent 3
−0.5 0.5
0 0.2
Average silhouette width: 0.47
0.4Silhouette width
Discovery
821 g
enes
Validation
HIF1A signaling
Neuregulin signaling
Putrescine degradation
Xenobiotic metabolis
Cell cycle
Antigen-presentationpathway
T-helper celldifferentiation
0.6 0.8 1.0
10
−2 −1 0 1 2
Agilent 1
Figure 1.Identification and validation of HNSCC expression subtypes. A, hierarchical clustering of subtype centroids from the last iteration of consensus clusteringon the three discovery datasets (Supplementary Fig. S2B). The heatmap shows the Pearson correlation of the centroids. Centroids of the subtypesfrom the three datasets cluster together into three supergroups: Inflamed/mesenchymal (IMS), basal (BA), and classical (CL). B, Silhouette plot of thecentroids (n ¼ 15) for three supergroups shows a positive width for all centroids. C, principal component analysis plot shows the correlation between threesupergroups and previously published subtypes for lung SCC (17) and HNSCC (14). D, heatmap shows the 821 predictive gene signature in the threediscovery datasets (Agilent: n ¼ 130, Illumina: n ¼ 128, Affymetrix: n ¼ 104). Samples were ordered according to group predictions, and genes wereclustered using the three discovery datasets. E, heatmap shows the 821 predictive gene signature in two validation datasets (Affymetrix: n ¼ 136, RNA-Seq:n ¼ 414): Samples in the validation dataset were assigned to groups using a nearest centroid algorithm. Gene order from the training set was maintained.
Keck et al.
Clin Cancer Res; 21(4) February 15, 2015 Clinical Cancer Research872
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

expression signature. In all the datasets, HPVþ tumors were notgathered into one group, but fall into two distinct groups: (i) theinflamed/mesenchymal (IMS) supergroup and (ii) the classical(CL) supergroup. No HPVþ tumors fell in the basal supergroup(Fig. 2A). It is evident that HPVþ HNSCCs are composed of
two distinct gene expression subtypes, namely, IMS-HPV andCL-HPV. Interestingly, there is biologic overlap of these HPVþ
samples in a supergroup andHPV� tumors in the same supergroup(e.g., immune or mesenchymal differentiation). The non-HPVsubtypes were named as basal, CL-nonHPV, and IMS-nonHPV.
−1 1 2−2 0
neg
neg neg neg neg neg
pos neg pos neg pos
neg posneg posneg
0 10 0
SLC2A1SLC16A1H1F1ALAMC2COL17A1ITGB1AREGNRG1EGFRCDH3KRT16KRT17MCM2MCM10CDC7CDKN2AE2F2RPA2AKR1C1AKR1C3ALDH3A1ICOSCD8ALAG3HLA-DRAVIMMMP9
SLC2A1SLC16A1H1F1ALAMC2COL17A1ITGB1AREGNRG1EGFRCDH3KRT16KRT17MCM2MCM10CDC7CDKN2AE2F2RPA2AKR1C1AKR1C3ALDH3A1ICOSCD8ALAG3HLA-DRAVIMMMP9
50 0 10 0 10 20 30 40 0 20 4030 50100 15030
GroupBACLIMS
HPV Tobacconeverneg
pos lightheavyNA
50
pos
neg pos neg pos
neg posneg pos
AgilentA
B
C
Agilent
Agilent
Hypoxia-up(Winter et al.)Hypoxia-down(Winter et al.)Epithelial(Taube et al.)Mesenchymal(Taube et al)Proliferation(Whitfield et al.)
CL
BA
IMS
Sample number Sample number Sample number
Hypoxia
Cell adhesion
HER signaling
Epithelial
Cell cycle
Sample number Sample number
Affymetrix (D) Affymetrix (V)RNA-Seq(TCGA)
RNA-Seq(TCGA)
RNA-Seq(TCGA)
Illumina
Xenobiotic metabolism
Immune response
Mesenchymal
BA CL-HPV CL-nonHPV IMS-HPV IMS-nonHPV BA CL-HPV CL-nonHPV IMS-HPV IMS-nonHPV
Figure 2.Stratification of HNC tumors by supergroups and HPV status. A, the HNC tumors in both the discovery datasets (Agilent, Illumina, Affymetrix; Discovery) andvalidation datasets (RNA-Seq, Affymetrix; Validation) are stratified into five subtypes according to supergroups and HPV status. HPV tumors are subdividedinto classical (CL) and inflamed/mesenchymal (IMS), and completely absent in basal (BA). B, selection of HNC-relevant genes/pathways according toingenuity pathway annotation shows that supergroups/subtypes are characterized by distinct and differential biologic processes. Smoking history anddifferential expression of genes related to smoking are also displayed. C, significantly associated pathways with subtypes. The pathway activity of hypoxic(up and down), epithelial, mesenchymal, and proliferation for each patient was obtained by single sample gene set enrichment analysis (ssGSEA). Heatmaps areused to show the difference of pathway activities between subtypes in Agilent and TCGA cohorts.
Integrative Analysis Identifies Five Distinct HNC Subtypes
www.aacrjournals.org Clin Cancer Res; 21(4) February 15, 2015 873
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

Molecular and genetic characteristics of HNSCC subtypesBasal subtype—BA. One distinctive feature of the basal subtypeis the significant enrichment for hypoxia signaling, representedby hypoxia-responsive genes such as HIF1A, CA9, and VEGF(Fig. 2B and Supplementary Fig. S3A). In addition, the geneexpression profile of the basal subtype closely matches a pub-lished hypoxia gene signature (Fig. 2C) which validates thehypoxic phenotype (19). The second key characteristic of thebasal subtype is strong enrichment for neuregulin signaling,including EGFR, AREG (amphiregulin), and NRG1 (neuregulin1/heregulin; Fig. 2B and Supplementary Fig. S3A). Overexpres-sion of epithelial markers such as P-cadherin (CDH3) and cyto-keratins (KRT1, KRT9) is another distinctive characteristic ofthe basal subtype (Fig. 2B and Supplementary Fig. S3A) similarto basal breast cancer (20). Moreover, a published EMT signa-ture also demonstrates the elevated epithelial pathway activitiesin the basal subtype (Fig. 2C; ref. 21). Consistent with the highexpression of cytokeratins in the basal subtype, the morphologydata also demonstrated that basal tumors are highly keratinizingand well differentiated (Supplementary Fig. S4).
Classical subtypes—CL-HPV and CL-nonHPV. The most distinc-tive feature of the classical supergroup is the significantenrichment for putrescine (polyamine) degradation pathway(Supplementary Fig. S3B), which is relevant for detoxification,
for example, related to tobacco use. Increased polyaminelevels are associated with increased cell proliferation (22).Consistently, a published proliferation signature also indicatesthat the classical supergroup has a higher proliferation ratecompared with the other groups (Supplementary Fig. S4;ref. 23).
Although the CL-HPV and CL-nonHPV subtypes share sim-ilarities, they are still two distinct disease entities, reflected inmany biologic pathways. Cell-cycle genes, such as mini-chro-mosome maintenance proteins (MCM2 and MCM10), celldivision cycle protein kinase (CDC7), and -related genes(CDKN2A, E2F2, and RPA2) are overexpressed in the CL-HPVsubtype. The two subtypes also show significant difference intobacco use with 74% heavy smokers in CL-nonHPV comparedwith 42% heavy smokers in CL-HPV (Fig. 2B and Table 1, P ¼0.01, Fisher exact test). Consistent with smoking status, xeno-biotic metabolism pathway genes AKR1C1, AKR1C3, andALDH3A1, which are known to be associated with smoking,are enriched in CL-nonHPV (Fig. 2B; refs. 24, 25).
Inflamed/mesenchymal subtypes—IMS-HPV and IMS-nonHPV.The distinguishing features of the IMS group is expression ofimmune response genes like CD8, ICOS, LAG3, and HLA-DRA(Fig. 2B) related to the infiltration of CD8þ T lymphocytes intumors.
Table 1. Patient and sample overview
Total Basal CL-HPV CL-nonHPV IMS-HPV IMS-nonHPV
Patients, n 130 43 19 19 36 13Age, y (median) 57 56 56 60 59 57GenderFemale 23 10 2 7 3 1Male 107 33 17 12 33 12
% TumorMedian 70 70 70 70 70 70Interquartile range 60–77.5 60–72.5 60–70 60–80 60–80 60–70
HPV statusPositive 55 0 19 0 36 0Negative 75 43 0 19 0 13
Anatomic siteLarynx 29 10 0 11 0 8Oral cavity 25 18 1 3 1 2Oropharynx 75 14 18 5 35 3Othera 1 1 0 0 0 0
Tobacco useNever 22 (17%) 5 (11%) 4 (21%) 1 (5%) 11 (31%) 1 (7.5%)Light 36 (28%) 11 (26%) 7 (37%) 4 (21%) 13 (36%) 1 (7.5%)Heavy 72 (55%) 27 (63%) 8 (42%) 14 (74%) 12 (33%) 11 (85%)
Alcohol useNever 16 (12%) 4 (9%) 3 (16%) 0 (0%) 8 (22%) 1 (7.5%)Light 35 (27%) 16 (37%) 4 (21%) 3 (16%) 11 (31%) 1 (7.5%)Heavy 79 (61%) 23 (54%) 12 (63%) 16 (84%) 17 (47%) 11 (85%)
T stageT0–T2 43 9 6 3 20 5T3–T4 87 34 13 16 16 8
N stageN0 12 10 1 1 0 0N1 9 4 1 1 2 1N2 90 22 12 15 31 10N3 19 7 5 2 3 2
Tumor stageI–II 2 2 0 0 0 0III 3 1 0 0 2 0IV 124 40 18 19 34 13
NOTE: 4 out of 134 samples (3%) were undefined and did not met criteria to allow classification into only one subtype.aParanasal sinus.
Keck et al.
Clin Cancer Res; 21(4) February 15, 2015 Clinical Cancer Research874
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481
Mesenchymal genes such as vimentin (VIM), matrix metallo-proteinases (MMP9), and S100A4 also show increased expressionin the IMS group (Fig. 2B), which typically associates withincreased metastatic risk (8, 26). Epithelial markers such as P-cadherin (CDH3) and cytokeratins (KRT1, KRT9) are downregu-lated, suggesting epithelial-to-mesenchymal transition (EMT; Fig.2B; ref. 21). Consistently, a published EMT signature (21) isupregulated in the IMS group and there is downregulation ofepithelial differentiation markers (Fig. 2C).
The two subtypes in the IMS group show a significantdifference in cell-cycle pathways and smoking-associated path-ways (Fig. 2B). Similar to the CL-HPV subtype, the IMS-HPVsubtype has significantly higher cell-cycle pathway activities,in which HPV is known to play a critical role. The IMS-HPV
subtype shows a higher proliferation rate indicated by a pub-lished proliferation signature (Fig. 2C; ref. 23) and IMS-HPVsubtype tumors are nonkeratinizing and poorly differentiatedaccording to the morphology review by light microscopy (Sup-plementary Fig. S3).
Patient characteristics and survivalOur cohort of 134 HNSCC samples with 44% HPVþ is repre-
sentative for the patient population presenting to a large referralcenter for advanced HNC. There is an increasing proportion ofHPVþ cases (Table 1). All patients in this new cohort had locor-egionally advanced disease and were treated with concurrentchemoradiotherapy. Consistent with previous reports, patientswith oropharyngeal tumors are mainly in the two HPV subtypes:
AgilentA
B
0
1.0
0.8
0.6
0.4
0.2
0.0
0.2 0.45-year survival
0 500 1,000 1,500 2,000
0.6 0.8 0 0.2 0.45-year survival
0.6 0 0.2 0.45-year survival
0.60.8
IMS-HPV
IMS-nonHPV
CL-HPV
CL-nonHPV
CL-nonHPV (19)
IMS-nonHPV (13)IMS-HPV (36)
CL-HPV (19)
BA
TCGA
Agilent overall survival
Illumina overall survival Combined overall survival
Ove
rall
surv
ival
1.0
0.8
0.6
0.4
0.2
0.0
Ove
rall
surv
ival
1.0
0.8
0.6
0.4
0.2
0.0
Ove
rall
surv
ival
Survival time in days0
P = 0.036
BA (42)
P = 0.589 P = 8e-06
P = 0.003
500 1,000 1,500 2,000Survival time in days
0 500 1,000 1,500 2,000Survival time in days
0 500 1,000 1,500 2,000Survival time in days
1.0
0.8
0.6
0.4
0.2
0.0
Ove
rall
surv
ival
TCGA overall survival
Illumina
CL-nonHPV (18)
IMS-nonHPV (22)IMS-HPV (9)
CL-HPV (6)
BA (44) CL-nonHPV (107)
IMS-nonHPV (123)IMS-HPV (76)
CL-HPV (47)
BA (235)
CL-nonHPV (69)
IMS-nonHPV (88)IMS-HPV (25)
CL-HPV (21)
BA (147)
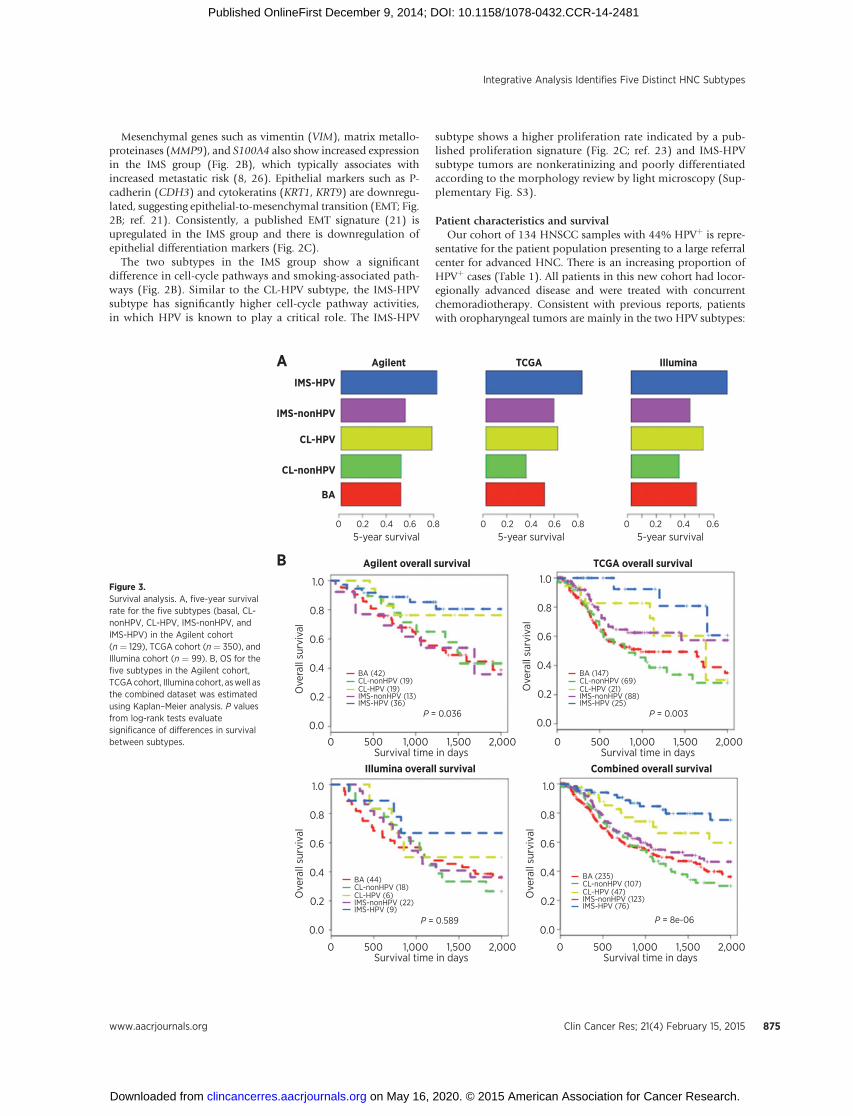
Figure 3.Survival analysis. A, five-year survivalrate for the five subtypes (basal, CL-nonHPV, CL-HPV, IMS-nonHPV, andIMS-HPV) in the Agilent cohort(n ¼ 129), TCGA cohort (n ¼ 350), andIllumina cohort (n ¼ 99). B, OS for thefive subtypes in the Agilent cohort,TCGAcohort, Illumina cohort, aswell asthe combined dataset was estimatedusing Kaplan–Meier analysis. P valuesfrom log-rank tests evaluatesignificance of differences in survivalbetween subtypes.
Integrative Analysis Identifies Five Distinct HNC Subtypes
www.aacrjournals.org Clin Cancer Res; 21(4) February 15, 2015 875
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

Loss Gscore Gain GscoreA
C
B
CL-HPV IMS-HPV CL-HPV IMS-HPV CL-HPV IMS-HPV
Copy
num
ber
Discovery set (PIK3CA) Validation set (TP63) Validation set (SOX2)
Chr3q26 (gain): PIK3CA, TP63, SOX2
BA
Tobacco
Subgroup
NeverLightHeavy
CL-HPVCL-nonHPVIMS-HPVIMS-nonHPV
−1.0
8 –
6 –
4 –
2 –
8 –
6 –
4 –
2 –
10.0 –
7.5 –
5.0 –
2.5 –
−0.5 0.0
−2
−1
0
1
2
0.5 1.0
chr1
chr2
chr3
chr4
chr5
chr6
chr7
chr8
chr9
chr10
chr11
chr12
chr13
chr14
chr15
chr17
chr18chr19
chr20chr22
Keck et al.
Clin Cancer Res; 21(4) February 15, 2015 Clinical Cancer Research876
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

IMS-HPV and CL-HPV (71%, P ¼ 4.19 � 10�11). In contrast,oral cavity tumors are overrepresented in the basal group (72%,P¼ 1.04� 10�5). Seventy-four percent of the CL-nonHPV tumorsand 85% of IMS-nonHPV are heavy smokers, which is signifi-cantly higher than other subtypes.
Five-year survival rate was assessed for our Agilent, TCGA,and Illumina cohorts, respectively. In all the cohorts, the HPVsubtypes have a significantly higher 5-year survival than non-HPV subtypes. Furthermore, between the two HPV subtypes,the IMS-HPV subtype shows a trend toward higher 5-yearsurvival than CL-HPV (Fig. 3A) consistent across all cohorts.We used Kaplan–Meier estimator to measure the overallsurvival (OS for the subtypes in the above three cohorts,separately. The log-rank test was used to evaluate theKaplan–Meier curve difference among the subtypes. We foundthat OS differs significantly among the five subtypes in ourAgilent cohort (P ¼ 0.037) and the TCGA cohort (P ¼ 0.003)but not in the Illumina cohort (P ¼ 0.589; Fig. 3B). Further-more, the Kaplan–Meier analysis on the combined dataset ofthe three cohorts showed a clearer difference in OS of the fivesubtypes (P ¼ 8e-06), which was more significant than anycohort independently (Fig. 3B). Importantly, the IMS-HPVsubtype is different from the CL-HPV subtype by better sur-vival (Fig 3A and B) suggesting a potential positive impact ofimmune response.
Copy number analysis exhibits distinct alterations per subtypeCopy number analysis was executed in all samples. Initially,
101 samples were analyzed targeting 75 genes previouslydescribed to have CNA (27–29). A GISTIC-derived strategy wasused to identify significant CNA. Significant copy number gainswere detected in 12 genes, including PIK3CA (3q26.3), VEGFA(6p21), EGFR (7p11),MYC (8q24), and CCND1 (11q13; Fig. 4Aand B and Supplementary Table S3). Significant copy numberlosses were detected in 16 genes, including CDKN2A (9p21) andRB1 (13q14; Fig. 4B and Supplementary Tables S3). Subsequent-ly, we aimed to further investigate these aberrant regions byscanning 72 additional genes within copy number altered regionsin 55 of the samples. We identified that TP63, SOX2 (3q26-28),and PIK3CA (3q26.3; Supplementary Table S4) are coamplified inmost tumors.
Furthermore, CNAs for a lot of oncogenes and tumor sup-pressors are found to have strong associations with subtypes(Supplementary Table S5). Amplification of EGFR (7p12),CCND1 (11q13), and FADD (11q13), as well as deletion ofFHIT (3p14) and CDKN2A (9p2) with few exceptions are onlyobserved in the nonHPV subtypes, particularly in the basalsubtype (Supplementary Table S5). Amplification of MYC(8q24) and ITGB4 (17q25) is significantly enriched in thebasal subtype (Supplementary Table S5). ITGB4 is also over-expressed in the basal subtype, and it is reported that ITGB4mediates cell adhesion and plays a crucial role on the initiation,progression, and metastasis of solid tumors (16, 30).
The copy number gains of genes (PRKCI, PIK3CA, andDCUN1D1) in the 3q26-28 region can be seen in all subtypes,but are significantly higher in the classical subtypes (Supplemen-tary Table S5). Although the frequency of E2F3 (6p22) amplifi-cation in our cohort is only 5%, most of the amplification eventsoccur in the classical subtypes, including both CL-HPV andCL-nonHPV subtypes (Supplementary Table S5). Consistently,E2F3 is overexpressed in classical subtypes. E2F3 encodes atranscription factor important for cell-cycle regulation and DNAreplication, and its amplification and overexpression are knownto associate with invasive tumor growth and rapid tumor cellproliferation in urinary bladder cancer (31). Importantly, the twoHPV subtypes differ significantly in these regions, suggestingdifferent biology of two HPV subtypes. The amplification of3q26-28 was validated by different markers such as TP63 andSOX2 (Fig. 4C).
Using IHC, we validated the ability of key markers (SOX2,Cyclin D1/CCND1, p16/CDKN2A, and TP63) to differentiate sub-types (Supplementary Fig. S5).
Cytotoxic T-cell infiltration and subtypesOne of the key findings based on gene expression is the
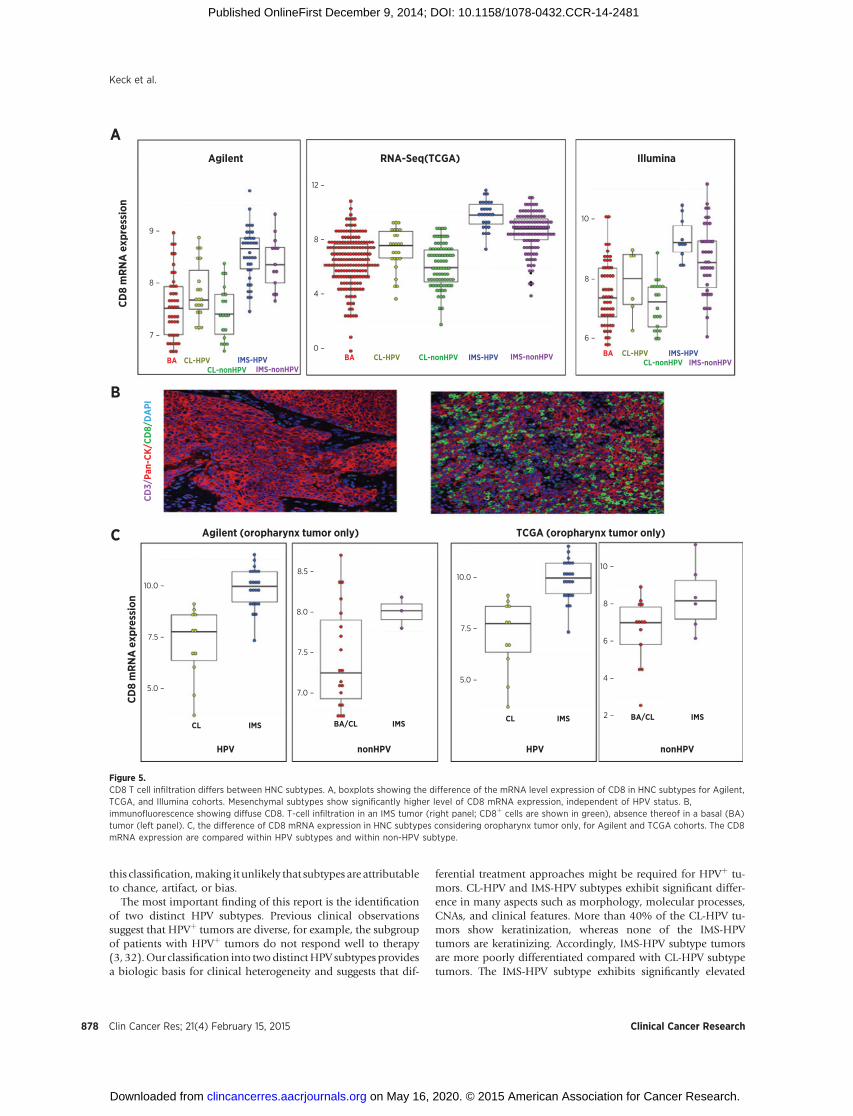
discovery of immune-related marker expression (CD8A/B) inIMS tumors (Fig. 5A). We performed multicolor immunoflu-orescence to identify tumor-infiltrating CD8þ lymphocytes inthese tumors (Fig. 5B). The enrichment of cytotoxic T-cellinfiltration in mesenchymal tumors is present even whenconsidering oropharynx tumors only, suggesting independenceof anatomic location and lack of contamination from normallymphoid tissue (Fig. 5C).
DiscussionAlthough previous attempts at expression profiling of HNSCC
identified distinct tumor subtypes (11, 14, 19), a therapeuticallyrelevant molecular classification remains elusive due to lack ofinformation about HPV status and/or absence of significantnumbers of HPVþ cases, small sample size, and lack of predictivebiomarkers. The HNSCC subtypes proposed in our study areidentified in an unsupervised, cross-cohort and cross-platformway, which is supported by four discovery cohorts and fourindependent validation cohorts, which together total 938patients. We showed that our subtypes are remarkably similarto those found in LUSC (17) and also have a strong correlationwith the previous identified HNSCC subtypes (14). Moreover,our expression profiling includes a large and representativenumber of HPVþ tumors, allowing us to identify two biologi-cally distinct HPVþ subtypes.
Consequently, we now propose that HNSCC can be classifiedinto five distinct subtypes—two HPV and three non-HPV sub-types. The close correlation of the five HNSCC subtypes withmorphologic characteristics, molecular processes, survival, andcopy number changes supports a biologic and clinical basis for
Figure 4.Copy number (CN) analysis in the five subtypes. A, genome-wide plot of copy number G-scores across the samples is shown for each subtype. The subtypes arerepresented by different colors. B, copy number for 75 genes in 101 samples (discovery set). Heatmap shows samples in columns, genes in rows ordered bychromosomal region. The copy number is normalized to�2 to 2, where�2 means high-level deletion,�1 means low-level deletion, 1 means low-level amplification,and 2 means high-level amplification. C, box plots show validation of chr3q26 amplification using different marks in this region. PIK3CA is used as markers in adiscovery dataset with 101 samples. TP63 and SOX2 are used as the markers in a validation dataset with 55 samples.
Integrative Analysis Identifies Five Distinct HNC Subtypes
www.aacrjournals.org Clin Cancer Res; 21(4) February 15, 2015 877
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481
this classification,making it unlikely that subtypes are attributableto chance, artifact, or bias.
The most important finding of this report is the identificationof two distinct HPV subtypes. Previous clinical observationssuggest that HPVþ tumors are diverse, for example, the subgroupof patients with HPVþ tumors do not respond well to therapy(3, 32).Our classification into twodistinctHPV subtypes providesa biologic basis for clinical heterogeneity and suggests that dif-
ferential treatment approaches might be required for HPVþ tu-mors. CL-HPV and IMS-HPV subtypes exhibit significant differ-ence in many aspects such as morphology, molecular processes,CNAs, and clinical features. More than 40% of the CL-HPV tu-mors show keratinization, whereas none of the IMS-HPVtumors are keratinizing. Accordingly, IMS-HPV subtype tumorsare more poorly differentiated compared with CL-HPV subtypetumors. The IMS-HPV subtype exhibits significantly elevated
CL IMS
HPV nonHPV HPV nonHPV
CL IMSBA/CL IMS
BA/CL IMS
Agilent
A
B
C Agilent (oropharynx tumor only) TCGA (oropharynx tumor only)
9 –
8 –
7 –
10.0 –
7.5 –
5.0 –
10.0 –
7.5 –
5.0 –
10 –
8 –
6 –
4 –
2 –
8.5 –
8.0 –
7.5 –
7.0 –
12 –
10 –
8 –
6 –
8 –
4 –
0 –
CD8
mRN
A e
xpre
ssio
nCD
8 m
RNA
exp
ress
ion
RNA-Seq(TCGA) Illumina
CL-nonHPVCL-nonHPVBA CL-HPV
CD3/
Pan-
CK/C
D8/
DA
PI
BA CL-HPV BA CL-HPV CL-nonHPVIMS-HPV
IMS-HPVIMS-HPVIMS-nonHPV
IMS-nonHPVIMS-nonHPV
Figure 5.CD8 T cell infiltration differs between HNC subtypes. A, boxplots showing the difference of the mRNA level expression of CD8 in HNC subtypes for Agilent,TCGA, and Illumina cohorts. Mesenchymal subtypes show significantly higher level of CD8 mRNA expression, independent of HPV status. B,immunofluorescence showing diffuse CD8. T-cell infiltration in an IMS tumor (right panel; CD8þ cells are shown in green), absence thereof in a basal (BA)tumor (left panel). C, the difference of CD8 mRNA expression in HNC subtypes considering oropharynx tumor only, for Agilent and TCGA cohorts. The CD8mRNA expression are compared within HPV subtypes and within non-HPV subtype.
Keck et al.
Clin Cancer Res; 21(4) February 15, 2015 Clinical Cancer Research878
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

expression of mesenchymal markers, whereas the CL-HPV sub-type shows higher proliferation. Compared with the IMS-HPVsubtype, the CL-HPV subtype presents significantly more canon-ical genomic aberrations associated with squamous cell carcino-ma, such as amplification of 3q26-27. For example, E2F3 ampli-fication, which is associated with cell proliferation, is only foundin the CL-HPV subtype but not in the IMS-HPV subtype, whichcorrelates with the proliferation signature seen by expression inthe CL-HPV subtype. The differences in the biologic patternsfound between the two HPV subtypes are supported by a trendtoward better survival in the IMS-HPV subtype comparedwith theCL-HPV subtype, and this observation holds true across allcohorts, although it does not correlate as strongly with prognosisas, for example, tobacco use.
Another key finding of this report is the recognition of thedifferential expression of immune markers in the subtypes. Can-cer immune surveillance is considered to be a factor in the body'sability to prevent cancer (33). Evading immune surveillance hasbeen considered as an emerging hallmark of cancer (34). Impor-tantly, activity of checkpoint blockade was recently reported inHNC using anti–PD-1 therapies, and PD-L1 expression correlateswith an inflamed phenotype that is consistent with and validatesour inflamed/mesenchymal (IMS) subtypes (35, 36).
Furthermore, accumulating evidence indicates that cancerimmune suppression can be driven by hypoxia (37, 38). Wedetected that the basal subtype is highly enriched for HIF1Asignaling. Immunosuppressive factors such as VEGF (39) areupregulated in the basal subtype, and a lack of immune-relatedmarkers is evident. Alternative to hypoxia-induced activation,HIF1A can be activated under normoxic conditions by EGFRsignaling (40, 41),which is also found to be significantly activatedin the basal subtype tumors. EGFR amplification and MYCamplification are found in the basal subtypes. Taken together,the immunosuppression of the basal subtype could be driven byeither hypoxia or potentially also EGFR signaling, and furthervalidation will be important as immune-targeted therapies arebeing developed for HNC.
Currently, no clinically relevant EGFR biomarkers exist andanti-EGFR therapy is administered indiscriminately despite alow single-agent response rate of 13% (7, 42). Our results sug-gest that the basal subtype could serve as a candidate predictivebiomarker in evaluating the benefit of an anti-EGFR therapy, aswell as a hypoxia-targeting therapy.
In contrast with the absence of immune markers of the basalsubtype, the IMS subtypes, including both the IMS-HPV and theIMS-nonHPV subtypes, exhibit a strongly activated immunephenotype, validated by CD8þ T-cell infiltration on IHC. Infil-tration of tumors with CD8þ cytotoxic T lymphocytes has beenassociated with a favorable prognosis in several tumor types andmay be a predictive biomarker for cancer immunotherapy (43).Furthermore, novel immunomodulatory therapies (e.g., PD-1checkpoint targeting drugs) have shown activity in squamouscell carcinomas (44) and evidence of activity in HNSCC wasrecently reported. We demonstrate that both IMS subtypes inde-pendent of HPV status are characterized by CD8þ T-cell infiltra-tion (Fig. 5A and B) irrespective of anatomic location (Fig. 5C).Immune response against HPVþ is of particular interest andevidence of immune escape was reported recently (45, 46). Thisis the first report that HPVþ tumors have two subtypes that exhibita different immune phenotype. CD8þ T-cell infiltration is there-fore one possible explanation for the trend toward a more
favorable prognosis of IMS-HPV subtype tumors, and consistentwith findings in other cancer types. Although further investiga-tions are needed to explain the difference in immune responsebetween the two HPV subtypes, one can hypothesize that differ-ences in the host, tumor, and viral genetic background, as well astheir interplay, are etiologic.
Given the widespread use of IHC for diagnostic purposes inroutine pathology practice (e.g., p16, p63), we evaluated theability of protein expression to differentiate subtypes (Supple-mentary Fig. S5). Although additional optimization and valida-tion are necessary, development of either an IHCor a qPCR-basedassay could potentially also allow classification ofHNSCC similarto what is used for breast cancer (47).
In conclusion, we propose a new taxonomy of HNSCC basedon five subtypes with a comprehensive overview of HPVþ as wellas HPV� HNSCC biology validated across 938 HNSCC patienttumors. There are significant translational implications withrespect to immunotherapy, presence of two biologically distinctHPV subtypes, and HER-targeted therapies that will inform bio-marker development andpersonalized care efforts that are alreadybeing pursued.
Disclosure of Potential Conflicts of InterestT.Y. Seiwert is a consultant/advisory board member for Merck/MSD and
Bayer/Onyx. No potential conflicts of interest were disclosed by the otherauthors.
Authors' ContributionsConception and design:M.K. Keck, Z. Zuo, T.P. Stricker, M. El Dinali, R. Salgia,E.E.W. Cohen, K.P. White, E.E. Vokes, T.Y. SeiwertDevelopment of methodology: M.K. Keck, Z. Zuo, A. Khattri, T.P. Stricker,M. El Dinali, S. Aktolga, Z. Lei, S.G. Rozen, T.Y. SeiwertAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): M.K. Keck, T.P. Stricker, M. Imanguli, P. Fang,R.R. Weichselbaum, M.W. Lingen, M.D. Story, K.K. Ang, E.E.W. Cohen,K.P. White, E.E. Vokes, T.Y. SeiwertAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis):M.K. Keck, Z. Zuo, A. Khattri, T.P. Stricker, C. Brown,D. Rieke, J. Bragelmann, R. DeBoer, M. El Dinali, S. Aktolga, S.G. Rozen,R. Salgia, R.R. Weichselbaum, M.W. Lingen, E.E.W. Cohen, T.Y. SeiwertWriting, review, and/or revision of the manuscript: M.K. Keck, Z. Zuo,D. Rieke, J. Bragelmann, R. DeBoer, S. Aktolga, P. Tan, R. Salgia, R.R. Weich-selbaum, M.W. Lingen, M.D. Story, K.K. Ang, E.E.W. Cohen, K.P. White,E.E. Vokes, T.Y. SeiwertAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): M.K. Keck, D. Rieke, K. Endhardt, R. DeBoer,M. El Dinali, S. Aktolga, T.Y. SeiwertStudy supervision: E.E. Vokes, T.Y. SeiwertOther (funding): T.Y. Seiwert
AcknowledgmentsThe authors thank Drs. Janet Rowley and Yusuke Nakamura for their critical
feedback, suggestions, and guidance, the Gleason Family, and the AchatzResearch Fund for their support of HNC research (to T.Y. Seiwert).
Grant SupportThis work was supported by an ASCO Translational Professorship (to E.E.
Vokes).Dr. T.Y. Seiwertwas supported by The Flight AttendantMedical ResearchInstitute (FAMRI) Young Investigator Award.
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received October 14, 2014; accepted November 7, 2014; publishedOnlineFirst December 9, 2014.
www.aacrjournals.org Clin Cancer Res; 21(4) February 15, 2015 879
Integrative Analysis Identifies Five Distinct HNC Subtypes
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

References1. Siegel R, Ward E, Brawley O, Jemal A. Cancer statistics, 2011. CA Cancer J
Clin 2011;61:212–36.2. Vokes EE, Weichselbaum RR, Lippman SM, Hong WK. Head and neck
cancer. N Engl J Med 1993;328:184–94.3. Ang KK, Harris J, Wheeler R, Weber R, Rosenthal DI, Nguyen-Tan PF, et al.
Human papillomavirus and survival of patients with oropharyngeal can-cer. N Engl J Med 2010;363:24–35.
4. Leemans CR, Braakhuis BJM, Brakenhoff RH. The molecular biology ofhead and neck cancer. Nat Rev Cancer 2010;11:9–22.
5. Slebos RJC. Gene expression differences associated with human papillo-mavirus status in head and neck squamous cell carcinoma. Clin Cancer Res2006;12:701–9.
6. Posner MR, Hershock DM, Blajman CR, Mickiewicz E, Winquist E,Gorbounova V, et al. Cisplatin and fluorouracil alone or withdocetaxel in head and neck cancer. N Engl J Med 2007;357:1705–15.
7. Vermorken JB, Trigo J, Hitt R, Koralewski P, Diaz-Rubio E, Rolland F, et al.Open-label, uncontrolled, multicenter phase II study to evaluate theefficacy and toxicity of cetuximab as a single agent in patientswith recurrentand/or metastatic squamous cell carcinoma of the head and neck whofailed to respond to platinum-based therapy. J Clin Oncol 2007;25:2171–77.
8. Northcott PA, Shih DJH, Peacock J, Garzia L, Sorana Morrissy A, Zichner T,et al. Subgroup-specific structural variation across 1,000medulloblastomagenomes. Sci Rep 2012;488:49–56.
9. Cohen EEW, ZhuH, LingenMW,Martin LE, KuoWL, Choi EA, et al. A feed-forward loop involving protein kinase C and MicroRNAs regulates tumorcell cycle. Cancer Res 2009;69:65–74.
10. Pavon M, Parreno M, Tellez-Gabriel M, Sancho F, Lopez M, Cespedes M,et al. Gene expression signatures and molecular markers associated withclinical outcome in locally advanced head and neck carcinoma. Carcino-genesis 2012;33:1707–16.
11. Walter V, Yin X, Wilkerson MD, Cabanski CR, Zhao N, Du Y, et al.Molecular subtypes in head and neck cancer exhibit distinct patterns ofchromosomal gain and loss of canonical cancer genes. PLoS ONE 2013;8:e56823.
12. Verhaak RGW, Hoadley KA, Purdom E,Wang V, Qi Y, WilkersonMD, et al.Integrated genomic analysis identifies clinically relevant subtypes of glio-blastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, andNF1. Cancer Cell 2010;17:98–110.
13. Wilkerson MD, Hayes DN. ConsensusClusterPlus: a class discovery toolwith confidence assessments and item tracking. Bioinformatics 2010;26:1572–3.
14. Chung CH, Parker JS, Karaca G, Wu J, Funkhouser WK, Moore D,et al. Molecular classification of head and neck squamous cellcarcinomas using patterns of gene expression. Cancer Cell 2004;5:489–500.
15. Rickman DS, Millon R, De Reynies A, Thomas E, Wasylyk C, MullerD, et al. Prediction of future metastasis and molecular characteriza-tion of head and neck squamous-cell carcinoma based on transcrip-tome and genome analysis by microarrays. Oncogene 2008;27:6607–22.
16. Ye H, Yu T, Temam S, Ziober BL, Wang J, Schwartz JL, et al. Transcrip-tomic dissection of tongue squamous cell carcinoma. BMC Genomics2008;9:69.
17. WilkersonMD, Yin X, Hoadley KA, Liu Y, HaywardMC, Cabanski CR, et al.Lung squamous cell carcinoma mRNA expression subtypes are reproduc-ible, clinically important, and correspond to normal cell types. Clin CancerRes 2010;16:4864–75.
18. Sorlie T, Tibshirani R, Parker J, Hastie T,Marron JS, Nobel A, et al. Repeatedobservation of breast tumor subtypes in independent gene expression datasets. Proc Natl Acad Sci 2003;100:8418–23.
19. Winter SC, Buffa FM, Silva P, Miller C, Valentine HR, Turley H,et al. Relation of a hypoxia metagene derived from head andneck cancer to prognosis of multiple cancers. Cancer Res 2007;67:3441–9.
20. Ribeiro AS, Albergaria A, Sousa B, Correia AL, Bracke M, Seruca R, et al.Extracellular cleavage and shedding of P-cadherin: a mechanism underly-ing the invasive behaviour of breast cancer cells. Oncogene 2009;29:392–402.
21. Taube JH, Herschkowitz JI, Komurov K, Zhou AY, Gupta S, Yang J, et al.Core epithelial-to-mesenchymal transition interactome gene-expressionsignature is associated with claudin-low and metaplastic breast cancersubtypes. Proc Natl Acad Sci 2010;107:15449–54.
22. Gerner EW, Meyskens FL. Polyamines and cancer: old molecules, newunderstanding. Nat Rev Cancer 2004;4:781–92.
23. Whitfield ML, George LK, Grant GD, Perou CM. Common markers ofproliferation. Nat Rev Cancer 2006;6:99–106.
24. Jorgensen E, Stinson A, Shan L, Yang J, Gietl D, Albino AP. Cigarettesmoke induces endoplasmic reticulum stress and the unfolded proteinresponse in normal and malignant human lung cells. BMC Cancer2008;8:229.
25. Spira A, Beane J, Shah V, Liu G, Schembri F, Yang X, et al. Effects of cigarettesmoke on the human airway epithelial cell transcriptome. Proc Natl AcadSci 2004;101:10143–8.
26. Michael Zeisberg EGN. Biomarkers for epithelial-mesenchymal transi-tions. J Clin Invest 2009;119:1429.
27. Agrawal N, Frederick MJ, Pickering CR, Bettegowda C, Chang K, Li RJ,et al. Exome sequencing of head and neck squamous cell carcinomareveals inactivating mutations in NOTCH1. Science 2011;333:1154–7.
28. Beroukhim R, Getz G, Nghiemphu L, Barretina J, Hsueh T, Linhart D, et al.Assessing the significance of chromosomal aberrations in cancer: Meth-odology and application to glioma. Proc Natl Acad Sci 2007;104:20007–12.
29. Stransky N, Egloff AM, Tward AD, Kostic AD, Cibulskis K, Sivachenko A,et al. Themutational landscape of head andneck squamous cell carcinoma.Science 2011;333:1157–60.
30. Desgrosellier JS, Cheresh DA. Integrins in cancer: biological impli-cations and therapeutic opportunities. Nat Rev Cancer 2010;10:9–22.
31. Oeggerli M, Tomovska S, Schraml P, Calvano-Forte D, Schafroth S, SimonR, et al. E2F3 amplification and overexpression is associated with invasivetumor growth and rapid tumor cell proliferation in urinary bladder cancer.Oncogene 2004;23:5616–23.
32. Chaturvedi AK, Engels EA, Pfeiffer RM,Hernandez BY, XiaoW, Kim E, et al.Human papillomavirus and rising oropharyngeal cancer incidence in theunited states. J Clin Oncol 2011;29:4294–301.
33. Kim R, Emi M, Tanabe K. Cancer immunoediting from immune surveil-lance to immune escape. Immunology 2007;121:1–14.
34. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell2011;144:646–74.
35. Seiwert TY, Burtness B,Weiss J, Gluck I, Eder JP, Pai SI, et al. A phase Ib studyofMK-3475 in patients with human papillomavirus (HPV)-associated andnon-HPV–associated head and neck (H/N) cancer. J Clin Oncol 32:5s,2014 (suppl; abstr 6011).
36. Saloura V, Zuo Z, KoeppenH, KeckMK, Khattri A, BoeM, et al. Correlationof T-cell inflamed phenotype with mesenchymal subtype, expression ofPD-L1, and other immune checkpoints in head and neck cancer. J ClinOncol 32:5s, 2014 (suppl; abstr 6011).
37. Lee C-T, Mace T, Repasky EA. Hypoxia-driven immunosuppression: a newreason to use thermal therapy in the treatment of cancer? Int J Hyperther-mia 2010;26:232–46.
38. Duechler M, Peczek L, Zuk K, Zalesna I, Jeziorski A, Czyz M. The hetero-geneous immune microenvironment in breast cancer is affected by hyp-oxia-related genes. Lancet Oncol 2014;219:158–65.
39. Gavalas NG, Tsiatas M, Tsitsilonis O, Politi E, Ioannou K, Ziogas AC,et al. VEGF directly suppresses activation of T cells from ascites sec-ondary to ovarian cancer via VEGF receptor type 2. Br. J Cancer2012;107:1869–75.
40. Uniacke J, Holterman CE, Lachance G, Franovic A, Jacob MD, Fabian MR,et al. An oxygen-regulated switch in the protein synthesis machinery. Sci.Rep 2012;486:126–9.
41. Hoogsteen IJ, Marres HAM, van den Hoogen FJA, Rijken PFJW, Lok J,Bussink J, et al. Expression of EGFR under tumor hypoxia: identifi-cation of a subpopulation of tumor cells responsible for aggressivenessand treatment resistance. Int J Radiat Oncol Biol Phys 2012;84:807–14.
42. Stoehlmacher-Williams J, Villanueva C, Foa P, Rottey S, Winquist E,Licitra LF, et al. Safety and efficacy of panitumumab (pmab) in
Clin Cancer Res; 21(4) February 15, 2015 Clinical Cancer Research880
Keck et al.
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

HPV-positive (þ) and HPV-negative (�) recurrent/metastatic squa-mous cell carcinoma of the head and neck (R/M SCCHN): Analysisof the global phase III SPECTRUM trial. J Clin Oncol 30, 2012(suppl; abstr 5504^).
43. FridmanWH, Pag�es F, Saut�es-Fridman C, Galon J. The immune contexturein human tumours: impact on clinical outcome. Nat Rev Cancer2012;12:298–306.
44. Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermottDF, et al. Safety, activity, and immune correlates of anti–PD-1 antibodyin cancer. N Engl J Med 2012;366:2443–54.
45. Albers A. Antitumor activity of humanpapillomavirus type 16 E7-specific Tcells against virally infected squamous cell carcinoma of the head andneck.Cancer Res 2005;65:11146–55.
46. Lyford-Pike S, Peng S, Young GD, Taube JM, Westra WH, Akpeng B, et al.Evidence for a role of the PD-1:PD-L1 pathway in immune resistance ofHPV-associated head and neck squamous cell carcinoma. Cancer Res2013;73:1733–41.
47. Parker JS, Mullins M, Cheang MCU, Leung S, Voduc D, Vickery T, et al.Supervised risk predictor of breast cancer based on intrinsic subtypes. J ClinOncol 2009;27:1160–7.
www.aacrjournals.org Clin Cancer Res; 21(4) February 15, 2015 881
Integrative Analysis Identifies Five Distinct HNC Subtypes
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481

2015;21:870-881. Published OnlineFirst December 9, 2014.Clin Cancer Res Michaela K. Keck, Zhixiang Zuo, Arun Khattri, et al. Biologically Distinct HPV and Three Non-HPV SubtypesIntegrative Analysis of Head and Neck Cancer Identifies Two
Updated version
10.1158/1078-0432.CCR-14-2481doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2015/01/14/1078-0432.CCR-14-2481.DC2
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/21/4/870.full#ref-list-1
This article cites 44 articles, 15 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/21/4/870.full#related-urls
This article has been cited by 30 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/21/4/870To request permission to re-use all or part of this article, use this link
on May 16, 2020. © 2015 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1078-0432.CCR-14-2481