MPI News jan 99 - mpibpc.mpg.de · Dinitrophenyl-Moleküle (DNP, grün) bin-den. Wie Abb. 4 (unten)...
4
Langreichweitige Kräfte schließlich, insbe- sondere Coulomb-Kräfte (rot, blau) zwi- schen den (meist) partiell geladenen Ato- men (d+, d-) tragen wesentlich zur Stabilität einer Proteinstruktur bei. All diese Kräfte (und noch einige weitere, die hier nicht beschrieben werden) bestimmen die Bewe- gung jedes einzelnen Atoms und werden in einer MD-Simulation entsprechend be- rücksichtigt. Da sich die Kräfte mit der Atombewegung rasch ändern, müssen alle Kräfte in kurzen Zeitschritten (typischer- weise 10 -15 sec.) wiederholt berechnet wer- den. Nach 10 6 Integrationsschritten erhält man so eine Beschreibung der Bewegung eines jeden Atoms innerhalb der kurzen Zeitspanne einer Nanosekunde. Dies ist gegenwärtig die Länge einer typischen MD- Simulation, begrenzt durch die jeweils ver- fügbare Computerleistung. Der größte Teil des Rechenaufwands entfällt dabei auf die Berechnung der lang- reichweitigen Coulombkräfte. So erfährt etwa das grün markierte Atom in Abb. 3(a) Kräfte von allen anderen Atomen (rote Lini- en). Aus Effizienzgründen wird daher in den meisten Fällen nur die unmittelbare Umgebung des Atoms berücksichtigt [Abb. 3(b)]. Max-Planck-Institut für biophysikalische Chemie MPIbpc News 5. Jahrgang Göttingen Ausgabe Nr. 1/99 Januar 1999 1. Einleitung P raktisch alle Stoffwechselvorgänge im Körper werden durch hochspezialisierte Proteine bewerkstelligt oder gesteuert – man kann sie mit gutem Recht als die bio- chemischen ‘Nano-Maschinen’ der Zelle bezeichnen. Dies nicht nur wegen ihrer komplexen und gleichzeitig hochgeord- neten räumlichen Struktur – eindrucksvoll belegt durch inzwischen über 8700 gelöste Proteinstrukturen –, sondern insbesondere wegen der oft mit großer Präzision koordi- nierten atomaren Bewegungen, welche diese Vorgänge antreiben. Schon bald nach der erstmaligen Auf- klärung der räumlichen Struktur eines Pro- teins – des Sauerstofftransporters Myoglo- bin – durch Perutz und Kendrew [1] erkannte man, daß korrelierte atomare Bewegungen oft unverzichtbar für die Pro- teinfunktion sind: Wäre das Myoglobin- Molekül nämlich so starr wie die Abbildun- gen seiner Struktur in Lehrbüchern sugge- rieren, so könnte kein Sauerstoffmolekül die im Inneren des Proteins gelegene Bin- dungsstelle erreichen. Wie auch Moleku- lardynamikrechnungen zeigten [2], schafft erst die thermische Bewegung der Proteina- tome genügend Flexibilität. Jüngste Fortschritte insbesondere im Bereich von Einzelmolekülexperimenten, Spektroskopie und zeitaufgelöster Struktur- aufklärung erlaubten es, einigen ‘Nano- Maschinen’ bei der Arbeit zuzusehen. Sie offenbaren erstaunliche Mechanismen, etwa fein abgestimmte ‘induced-fit’-Bewe- gungen bei spezifischer Ligandenbindung, die mechanische Energieübertragung in der F1-ATPase über eine elastische ‘Kurbelwel- le’ oder das ‘Krabbeln’ von Myosin oder Kinesinen. Gegenwärtig sind wir außerstande, sol- che Präzisionsfeinmechaniken de novo zu konstruieren – aber vielleicht können wir deren Mechanismen verstehen oder sogar Eigenschaften ihrer Funktion berechnen und vorhersagen. Diesem Ziel dient in unserer Arbeitsgruppe die Methode der Molekulardynamik-(MD-)Simulation. Im vorliegenden Text wollen wir, nach einer kurzen Skizze dieser Methode, beispielhaft unsere Untersuchung des Bindungsmecha- nismus eines Antikörpers beschreiben. 2. Molekulardynamiksimulation - Prinzip und Tricks Wie in Abb. 2 veranschaulicht, wirken in einem Makromolekül interatomare Kräf- te unterschiedlichen Typs. Chemische Bin- dungskräfte, hier dargestellt durch Federn, zwingen etwa gebundene Atome in ihren Gleichgewichtsabstand (grüne Pfeile) oder Gleichgewichtswinkel (magentafarbene Pfei- le). Die Pauli-Abstoßung (graue Pfeile) ver- hindert, daß sich Atome durchdringen. Inhalt Theoretische Molekulare Biophysik 1-4 Neueinstellungen 5 Abgänge 6 Gäste 6 Sicherheitsdatenblätter 6 Impressum 6 Promotionen 7 GWDG informiert 7 Publikationen 7-8 Suzanne Jovin 8 Fortsetzung auf S. 2 Berichte aus dem Institut Berichte aus dem Institut Proteindynamik von Ligand/Rezeptor- Bindungen Helmut Grubmüller und Berthold Heymann Arbeitsgruppe Theoretische Molekulare Biophysik (071)
Transcript of MPI News jan 99 - mpibpc.mpg.de · Dinitrophenyl-Moleküle (DNP, grün) bin-den. Wie Abb. 4 (unten)...

Langreichweitige Kräfte schließlich, insbe-sondere Coulomb-Kräfte (rot, blau) zwi-schen den (meist) partiell geladenen Ato-men (d+, d-) tragen wesentlich zur Stabilitäteiner Proteinstruktur bei. All diese Kräfte(und noch einige weitere, die hier nichtbeschrieben werden) bestimmen die Bewe-gung jedes einzelnen Atoms und werdenin einer MD-Simulation entsprechend be-rücksichtigt. Da sich die Kräfte mit derAtombewegung rasch ändern, müssen alleKräfte in kurzen Zeitschritten (typischer-weise 10-15 sec.) wiederholt berechnet wer-den. Nach 106 Integrationsschritten erhältman so eine Beschreibung der Bewegungeines jeden Atoms innerhalb der kurzenZeitspanne einer Nanosekunde. Dies istgegenwärtig die Länge einer typischen MD-Simulation, begrenzt durch die jeweils ver-fügbare Computerleistung.
Der größte Teil des Rechenaufwandsentfällt dabei auf die Berechnung der lang-reichweitigen Coulombkräfte. So erfährtetwa das grün markierte Atom in Abb. 3(a)Kräfte von allen anderen Atomen (rote Lini-en). Aus Effizienzgründen wird daher inden meisten Fällen nur die unmittelbareUmgebung des Atoms berücksichtigt[Abb. 3(b)].
MMaaxx--PPllaanncckk--IInnssttiittuutt ffüürr bbiioopphhyyssiikkaalliisscchhee CChheemmiiee
MPIbpc News5. Jahrgang Göttingen Ausgabe Nr. 1/99 Januar 1999
1. Einleitung
Praktisch alle Stoffwechselvorgänge imKörper werden durch hochspezialisierte
Proteine bewerkstelligt oder gesteuert –man kann sie mit gutem Recht als die bio-chemischen ‘Nano-Maschinen’ der Zellebezeichnen. Dies nicht nur wegen ihrerkomplexen und gleichzeitig hochgeord-neten räumlichen Struktur – eindrucksvollbelegt durch inzwischen über 8700 gelösteProteinstrukturen –, sondern insbesonderewegen der oft mit großer Präzision koordi-nierten atomaren Bewegungen, welchediese Vorgänge antreiben.
Schon bald nach der erstmaligen Auf-klärung der räumlichen Struktur eines Pro-teins – des Sauerstofftransporters Myoglo-bin – durch Perutz und Kendrew [1]erkannte man, daß korrelierte atomareBewegungen oft unverzichtbar für die Pro-teinfunktion sind: Wäre das Myoglobin-Molekül nämlich so starr wie die Abbildun-gen seiner Struktur in Lehrbüchern sugge-rieren, so könnte kein Sauerstoffmoleküldie im Inneren des Proteins gelegene Bin-dungsstelle erreichen. Wie auch Moleku-lardynamikrechnungen zeigten [2], schaffterst die thermische Bewegung der Proteina-tome genügend Flexibilität.
Jüngste Fortschritte insbesondere imBereich von Einzelmolekülexperimenten,Spektroskopie und zeitaufgelöster Struktur-aufklärung erlaubten es, einigen ‘Nano-Maschinen’ bei der Arbeit zuzusehen. Sie
offenbaren erstaunliche Mechanismen,etwa fein abgestimmte ‘induced-fit’-Bewe-gungen bei spezifischer Ligandenbindung,die mechanische Energieübertragung in derF1-ATPase über eine elastische ‘Kurbelwel-le’ oder das ‘Krabbeln’ von Myosin oderKinesinen.
Gegenwärtig sind wir außerstande, sol-che Präzisionsfeinmechaniken de novo zukonstruieren – aber vielleicht können wirderen Mechanismen verstehen oder sogarEigenschaften ihrer Funktion berechnenund vorhersagen. Diesem Ziel dient inunserer Arbeitsgruppe die Methode derMolekulardynamik-(MD-)Simulation. Imvorliegenden Text wollen wir, nach einerkurzen Skizze dieser Methode, beispielhaftunsere Untersuchung des Bindungsmecha-nismus eines Antikörpers beschreiben.
2. Molekulardynamiksimulation - Prinzipund Tricks
Wie in Abb. 2 veranschaulicht, wirkenin einem Makromolekül interatomare Kräf-te unterschiedlichen Typs. Chemische Bin-dungskräfte, hier dargestellt durch Federn,zwingen etwa gebundene Atome in ihrenGleichgewichtsabstand (grüne Pfeile) oderGleichgewichtswinkel (magentafarbene Pfei-le). Die Pauli-Abstoßung (graue Pfeile) ver-hindert, daß sich Atome durchdringen.
Inhalt
Theoretische Molekulare Biophysik 1-4
Neueinstellungen 5
Abgänge 6
Gäste 6
Sicherheitsdatenblätter 6
Impressum 6
Promotionen 7
GWDG informiert 7
Publikationen 7-8
Suzanne Jovin 8
Fortsetzung auf S. 2
Berichte aus dem Institut
Berichte aus dem Institut
Proteindynamik von Ligand/Rezeptor-BindungenHelmut Grubmüller und Berthold HeymannArbeitsgruppe Theoretische Molekulare Biophysik (071)

Seite 2
Fortsetzung von S.1
Es zeigte sich jedoch, daß dadurchgerade die uns interessierende – weil funk-tionsrelevante – Dynamik konformativerUmlagerungen in Proteinen häufig dra-stisch verfälscht wird [3,4], weshalb wireinen anderen Weg eingeschlagen haben[5]. In unseren Simulationen werden dieProteinatome hierarchisch gruppiert[Abb. 3(c)] und die zugehörigen Partialla-dungen zu Multipolen [farbige Flächen inAbb. 3(d)] zusammengefaßt. Die Wechsel-wirkung dieser Multipole mit dem heraus-gegriffenen (grünen) Atom (farbige Linien)kann nun sehr viel effizienter berechnetwerden [5]. Diese und weitere effizienz-steigernde Methoden [6,7] haben wir fürParallelrechner implementiert [8], so daßwir nun die Leistungsfähigkeit solcherRechner für ausgedehnte Simulationennutzen können.
3. Beispiel Antiköper/Antigen-Bindung
Damit waren wir in der Lage, die nicht-kovalente Bindung eines Antiköper/Anti-gen-Komplexes zu studieren; einige Ergeb-nisse sollen hier beispielhaft geschildertwerden.
Der Komplex (Abb. 4, oben) besteht ausdem monoklonalen Antikörper AN02(rot/blau), dessen hypervariable Regionen
Abbildung 2: Interatomare Kräfte (Pfeile, ‘Federn’) ineinem Makromolekül; Beschreibung siehe Text.
Abbildung 3: (a) Coulombkräfte (rot) wirken auf jedes geladene Atom eines Proteins (z.B. dasgrün markierte). Hierarchische Multipolmethoden vermeiden das üblicherweise notwendigeAbschneiden (b) der langreichweitigen Wechselwirkungen. Dazu werden alle Atome des Pro-teins (schwarz) hierarchisch gruppiert (c) und deren Ladungen zu Multipolen [(d), farbigeFlächen] zusammengefaßt, deren Wechselwirkungen sich sehr effizient berechnen lassen.
Abbildung 1: Untersuchung nichtkovalenter Ligand/Rezeptor-Bindungendurch Molekulardynamiksimulationen der erzwungenen Bindungstren-nung. Zu Beginn der Simulation (oberes Bild) befindet sich ein Biotin-molekül (gelb) in der Bindungstasche eines Streptavidinmoleküls (rot).Ein Atom des Biotins wird an ein harmonisches Potential gekoppelt (imBild symbolisch durch eine Feder dargestellt), das gleichförmig vomStreptavidin wegbewegt wird; der Schwerpunkt des Streptavidins wirddabei fixiert. Übersteigt die auf das Biotin so ausgeübte Zugkraft die Bin-dungskraft, die das Molekül zunächst in der Bindungstasche zurückhält(Bild mitte), löst sich das Biotin vom Streptavidin. Die Abreißkraft kannaus der Ausdehnug der ‘Feder’ abgelesen werden. Ist der Komplexgetrennt (unteres Bild), kann das Biotin dem Zug folgen, und die Zugkraftnimmt wieder ab. Entsprechend ist die Abreißkraft das Maximum derwährend des Trennungsprozesses aufgewendeten Zugkraft.
Fortsetzung auf Seite 3
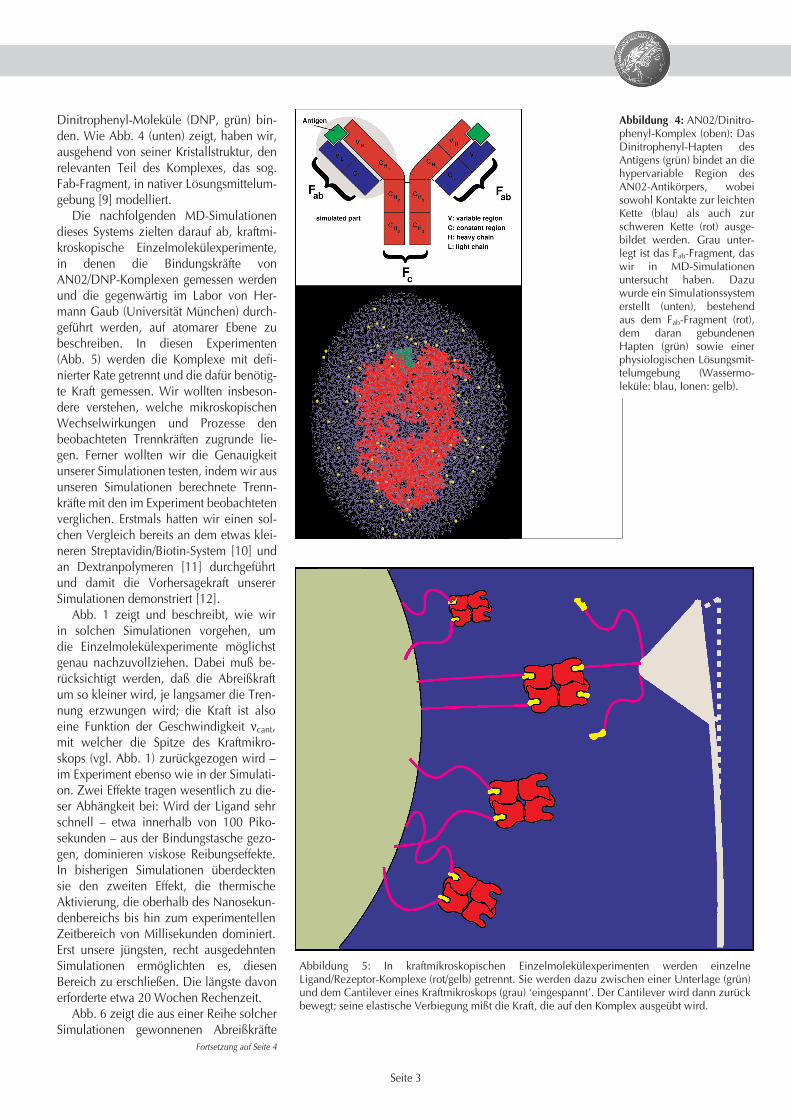
Dinitrophenyl-Moleküle (DNP, grün) bin-den. Wie Abb. 4 (unten) zeigt, haben wir,ausgehend von seiner Kristallstruktur, denrelevanten Teil des Komplexes, das sog.Fab-Fragment, in nativer Lösungsmittelum-gebung [9] modelliert.
Die nachfolgenden MD-Simulationendieses Systems zielten darauf ab, kraftmi-kroskopische Einzelmolekülexperimente,in denen die Bindungskräfte vonAN02/DNP-Komplexen gemessen werdenund die gegenwärtig im Labor von Her-mann Gaub (Universität München) durch-geführt werden, auf atomarer Ebene zubeschreiben. In diesen Experimenten(Abb. 5) werden die Komplexe mit defi-nierter Rate getrennt und die dafür benötig-te Kraft gemessen. Wir wollten insbeson-dere verstehen, welche mikroskopischenWechselwirkungen und Prozesse denbeobachteten Trennkräften zugrunde lie-gen. Ferner wollten wir die Genauigkeitunserer Simulationen testen, indem wir ausunseren Simulationen berechnete Trenn-kräfte mit den im Experiment beobachtetenverglichen. Erstmals hatten wir einen sol-chen Vergleich bereits an dem etwas klei-neren Streptavidin/Biotin-System [10] undan Dextranpolymeren [11] durchgeführtund damit die Vorhersagekraft unsererSimulationen demonstriert [12].
Abb. 1 zeigt und beschreibt, wie wirin solchen Simulationen vorgehen, umdie Einzelmolekülexperimente möglichstgenau nachzuvollziehen. Dabei muß be-rücksichtigt werden, daß die Abreißkraftum so kleiner wird, je langsamer die Tren-nung erzwungen wird; die Kraft ist alsoeine Funktion der Geschwindigkeit νcant,mit welcher die Spitze des Kraftmikro-skops (vgl. Abb. 1) zurückgezogen wird –im Experiment ebenso wie in der Simulati-on. Zwei Effekte tragen wesentlich zu die-ser Abhängkeit bei: Wird der Ligand sehrschnell – etwa innerhalb von 100 Piko-sekunden – aus der Bindungstasche gezo-gen, dominieren viskose Reibungseffekte.In bisherigen Simulationen überdecktensie den zweiten Effekt, die thermischeAktivierung, die oberhalb des Nanosekun-denbereichs bis hin zum experimentellenZeitbereich von Millisekunden dominiert.Erst unsere jüngsten, recht ausgedehntenSimulationen ermöglichten es, diesenBereich zu erschließen. Die längste davonerforderte etwa 20 Wochen Rechenzeit.
Abb. 6 zeigt die aus einer Reihe solcherSimulationen gewonnenen Abreißkräfte
Seite 3
Fortsetzung auf Seite 4
Abbildung 4: AN02/Dinitro-phenyl-Komplex (oben): DasDinitrophenyl-Hapten desAntigens (grün) bindet an diehypervariable Region desAN02-Antikörpers, wobeisowohl Kontakte zur leichtenKette (blau) als auch zurschweren Kette (rot) ausge-bildet werden. Grau unter-legt ist das Fab-Fragment, daswir in MD-Simulationenuntersucht haben. Dazuwurde ein Simulationssystemerstellt (unten), bestehendaus dem Fab-Fragment (rot),dem daran gebundenenHapten (grün) sowie einerphysiologischen Lösungsmit-telumgebung (Wassermo-leküle: blau, Ionen: gelb).
Abbildung 5: In kraftmikroskopischen Einzelmolekülexperimenten werden einzelneLigand/Rezeptor-Komplexe (rot/gelb) getrennt. Sie werden dazu zwischen einer Unterlage (grün)und dem Cantilever eines Kraftmikroskops (grau) ‘eingespannt’. Der Cantilever wird dann zurückbewegt; seine elastische Verbiegung mißt die Kraft, die auf den Komplex ausgeübt wird.

Literatur
[1] J. C. Kendrew, R. E. Dickerson, B. E.Strandberg, R. G. Hart, D. R. Davies, D. C. Phil-lips, and V. C. Shore. Structure of myoglobin: Athree-dimensional fourier synthesis at 2Å resolu-tion. Nature (London) 185, 422-427, 1960.[2] R. M. Levy, R. P. Sheridan, J. W. Keepers,G. S. Dubey, S. Swaminathan, and M. Karplus.Molecular dynamics of myoglobin at 298°K.Biophys. J. 48, 509-518, 1985.[3] Helmut Grubmüller and Paul Tavan.Molecular dynamics of conformational substatesfor a simplified protein model. J. Chem. Phys.101, 5047-5057, 1994.[4] Helmut Grubmüller and Paul Tavan. Mul-tiple time step algorithms for molecular dyna-mics simulations of proteins: How good arethey? J. Comp. Chem.19, 1534-1552, 1998.[5] M. Eichinger, H. Grubmüller, H. Heller,and P. Tavan. FAMUSAMM: A new algorithm forrapid evaluation of electrostatic interactions inmolecular dynamics simulations. J. Comp Chem.18, 1729-1749, 1997.[6] Helmut Grubmüller, Helmut Heller, And-reas Windemuth, and Klaus Schulten. Generali-zed Verlet algorithm for efficient molecular
Seite 4
strukturelle Vielfalt möglicher Bindungsre-aktionswege (besser: Trennreaktionswege),die von unseren Simulationen nahegelegtwird. Dies um so mehr, als eine vergleich-bare Heterogenität in früheren Abreiß-Simulationen, etwa des Streptavidin/Bio-tin-Systems, nicht aufgetreten war. Ent-sprechend untersuchen wir gegenwärtigdie funktionelle Bedeutung dieser Hetero-genität, etwa im Licht des dadurch beding-ten entropischen Beitrags zur freien Ener-gie entlang des Reaktionswegs.
(Punkte) als Funktion der Ziehgeschwin-digkeit. Im Vergleich zwischen der loga-rithmischen und der linearen (Inset) Auftra-gung sind die beiden Regimes deutlich zuerkennen. Ebenfalls dargestellt ist eineAbschätzung der experimentell zu erwar-tenden Abreißkraft (weiße Raute), die aufeiner Interpolation zwischen der sponta-nen Trennrate (Kraft null) und den aus denSimulationen gewonnenen Kräften beruht.Vorläufige Ergebnisse der kraftmikroskopi-schen Messungen in München deuten dar-auf hin, daß unsere Vorhersage zutrifft.
Wie erhofft, konnten wir aus unserenSimulationen ein mikroskopisches Modell(Abb. 7) des Abreißvorgangs gewinnen,welches im Detail das Trennen und auchein transientes Bilden von Wasserstoff-brücken (gestrichelte Linien) und van derWaals-Kontakten (nicht dargestellt) zwi-schen dem Liganden und dem Rezeptorzeigt. Dieses Modell, das hier nicht genau-er diskutiert werden kann, gestattet esetwa, Punktmutanten vorzuschlagen, wel-che die Spezifität und die Bindungskräfteder Antigenbindung erniedrigen oder auchsogar erhöhen sollten.
Bemerkenswert und in ihrem Umfangüberraschend erscheint uns die breite
Zwei der unzähligen möglichen Reakti-onswege sind in Abb. 7 dargestellt; sieunterscheiden sich sowohl in der Art undAbfolge intermediärer Bindungen zur Anti-körper-Bindungstasche als auch in derjeweiligen Abreißkraft. Letzterer Befundlegt nahe, daß die genaue Analyse der inEinzelmolekülexperimenten gemessenenAbreißkraftverteilungen einen kraftmikro-skopischen Zugang zur strukturellen Hete-rogenität erlauben sollte.
dynamics simulations with long-range interac-tions. Molec. Sim. 6, 121-142, 1991.[7] Helmut Grubmüller. Predicting slow struc-tural transitions in macromolecular systems:Conformational Flooding. Phys. Rev. E 52, 2893,1995.[8] Markus Eichinger, Helmut Grubmüler,and Helmut Heller. User Manual for EGO_VIII,Release 2.0 (electronic access: http://www.mpib-pc.gwdg.de/abteilungen/071/ego.html).[9] Helmut Grubmüller. Solvate: A program tocreate atomic solvent models, 1996. (electronicpublication: http://www.mpibpc.gwdg.de/abtei-lungen/071/solvate.html).[10] Helmut Grubmüller, Berthold Heymann,and Paul Tavan. Ligand binding: Molecularmechanics calculation of the streptavidin-biotinrupture force. Science 271 (5251), 997-999,1996.[11] M. Rief, F. Oesterhelt, B. Heymann, andH. E. Gaub. Single molecule force spectroscopyreveals conformational change in polysacchari-des. Science 275, 1295-1297, 1997.[12] H. J. C. Berendsen. Bio-molecular dyna-mics comes of age. Science 271, 954-955, 1996.
❏
Fortsetzung von S.3
Abb. 7: Strukturelle Heterogenitätder Trennung des AN02/DNP-Komplexes: Ausgehend vomgebundenen Zustand (oben, Mit-te), kann das Dinitrophenyl-Hap-ten (grau) ein ganzes Spektrumunterschiedlicher Trennungsreak-tionswege aus der Antikörper-Bindungstasche durchlaufen, wo-von die Abbildung je zweiMomentaufnahmen zweier mög-licher Reaktionswege (‘Heavychain route’, links; ‘Light chainroute’, rechts) zeigt. Die einzel-nen Reaktionswege unterschei-den sich in der Bewegung desHaptens, in der transienten Bil-dung kurzreichweitiger Wechsel-wirkungen zwischen Hapten undBindungstasche (etwa Wasser-stoffbrücken, gestrichelt; van derWaals-Kontakte, nicht gezeigt)sowie in den Konformationsbe-wegungen der Bindungstasche.
Abbildung 6: Berechnete Abreißkräfte (Punk-te) für unterschiedliche Ziehgeschwindig-keiten νcant (in logarithmischer Darstellung),entsprechend unterschiedlichen Zeitinterval-len tunbind, innerhalb derer die Bindungstren-nung erzwungen wurde. Die durchgezogeneLinie zeigt eine Anpassung unter Berücksich-tigung von Reibung und von thermisch akti-vierten Prozessen; die gestrichelte Linieberücksichtigt lediglich Reibungseffekte. EineAbschätzung der im Experiment zu erwarten-den Abreißkraft ist als Raute dargestellt. DerInset zeigt dieselben Daten in linearer Auftra-gung.