
Neurobiologische Untersuchungen im Praetectum von ... · In den Vertebratenklassen Mammalia, Aves,...
152
Neurobiologische Untersuchungen im Praetectum von Knochenfischen Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften der Fakultät für Biologie der Ruhr- Universität Bochum angefertigt am Lehrstuhl für Allgemeine Zoologie und Neurobiologie vorgelegt von Matthias Klar aus Dortmund Bochum 2004
Transcript of Neurobiologische Untersuchungen im Praetectum von ... · In den Vertebratenklassen Mammalia, Aves,...

Neurobiologische Untersuchungen im Praetectum
von Knochenfischen
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie
der Ruhr- Universität Bochum
angefertigt am
Lehrstuhl für Allgemeine Zoologie und Neurobiologie
vorgelegt von
Matthias Klar
aus
Dortmund
Bochum 2004

Dissertation eingereicht am: 02. Februar 2004
Betreuer: Prof. Dr. K.-P. Hoffmann
(Lehrstuhl Allgemeine Zoologie und Neurobiologie, Fakultät Biologie)
Koreferent: Prof. Dr. W. H. Kirchner
(AG Verhaltensbiologie und Didaktik der Biologie, Fakultät Biologie)
Dekan: Prof. Dr. Dr. H. Hatt
(Lehrstuhl Zellphysiologie, Fakultät Biologie)

Inhaltsverzeichnis I. Elektrophysiologische Einzelzellableitungen im Praetectum
von Salmo gairdneri und Esox lucius
1. Einleitung .................................................................................................. 1
1.1 Das Praetectum als Ort richtungsselektiver Neurone ...................................................1
2. Material und Methoden ........................................................................... 6
2.1 Versuchstiere ................................................................................................................6
2.2 Fischhälterung ..............................................................................................................6
2.3 Anästhesie, Präparation und Fixierung .........................................................................7
2.4 Elektrophysiologischer Versuchsaufbau ......................................................................8
2.4.1 Der Ableitstand .......................................................................................................8
2.4.2 Visuelle Stimulation .............................................................................................10
2.4.3 Stimulussteuerung und elektrophysiologische Datenregistrierung ......................11
2.5 Erstellen des Penetrationsrasters ................................................................................11
2.6 Bestimmung der rezeptiven Felder .............................................................................13
2.7 Datenanalyse und Auswertung ...................................................................................13
2.7.1 Statistische Verfahren ...........................................................................................14
2.7.2 Histologie und Verifizierung des Ableitortes .......................................................14
3. Ergebnisse ................................................................................................16
3.1 Lokalisation richtungsselektiver Neurone im Praetectum von Salmo gairdneri
und Esox lucius ...........................................................................................................16
3.2 Praetectale richtungsselektive Neurone ......................................................................19
3.3 Untersuchungen zum Vorkommen weiterer richtungsselektiver Neurone .................24
3.4 Verteilung der Vorzugsrichtungsverteilung aller abgeleiteter Neurone .....................29
3.5 Entscheidet die Anzahl richtungsselektiver Neurone über eine Symmetrie bzw.
Asymmetrie des monokularen horizontalen Reflexes? ...............................................32
3.6 Populationsantwort .....................................................................................................33

3.7 Weitere Charakterisierungen der prätektalen Neuronen von Salmo gairdneri
und Esox lucius ...........................................................................................................34
3.7.1 Neuronale Aktivität in Abhängigkeit von der Reizgeschwindigkeit.....................34
3.7.2 Analyse der Latenzzeit .........................................................................................36
3.7.3 Größe und Verteilung der rezeptiven Felder richtungsselektiver Neurone...........37
3.8 Rekonstruktion des Ableitortes richtungsselektiver Neurone im Praetectum
von Salmo gairdneri und Esox lucius .........................................................................39
3.8.1 Histologische Lokalisation der praetectalen richtungsselektiven Neurone ..........39
3.8.2 Existiert eine topographische Ordnung? ...............................................................41
4. Diskussion ................................................................................................45 4.1 Auswahl der Versuchstiere und die Wahl der Methode..............................................45
4.2 Elektrophysiologische Ableitungen im Praetectum ...................................................46
4.2.1 Richtungsselektive Neurone .................................................................................47
4.2.2 Fanden die ersten Ableitungen tatsächlich im Praetectum statt? . ........................48
4.3 Richtungspräferenz der richtungsselektiven Neurone ................................................51
4.4 Praetectale richtungsselektive Neurone und optokinetisches Verhalten .....................54
4.5 Geschwindigkeitsverhalten der richtungsselektiven Neurone ....................................56
4.6 Lage der richtungsselektiven Neurone im Praetectum ...............................................57
4.7 Topographische Organisation der Vorzugsrichtungen ...............................................59
II. Verhaltensexperimente zum visuomotorischen System an
Salmo gairdneri 1. Einleitung .................................................................................................60
2. Material und Methoden ..........................................................................63 2.1 Versuchstiere ..............................................................................................................63
2.2 Verhaltensexperimente ...............................................................................................63
2.2.1 Kälte - Immobilisierung und Fixation ..................................................................63
2.2.2 Versuchsaufbau für die Verhaltensexperimente zum optokinetischen
und vestibulo-okulären Reflex .................................................................................64

2.2.3 Reizmodalitäten ....................................................................................................66
2.2.3.1 Vertikaler optokinetischer Reflex ...................................................................66
2.2.3.2 Kompensatorische Augenbewegungen des vestibulo-okulären Reflexes ......66
2.2.3.3 Monokularer, horizontaler optokinetischer Reflex im Läsionsexperiment ....66
2.2.4 Messdatenerfassung, Analyse und Auswertung ...................................................67
2.2.4.1 Vertikaler und horizontaler optokinetischer Reflex .......................................68
2.2.4.2 Vestibulo-okulärer Reflex ..............................................................................68
2.2.5 Statistische Verfahren zur Signifikanzbestimmung .............................................69
2.2.5.1 Vestibulo-okulärer Reflex ..............................................................................69
2.2.5.2 Läsionsexperiment ..........................................................................................70
2.3 Elektrophysiologisches Läsionsexperiment ...............................................................70
2.3.1 Fischanästhesie und Präparation ...........................................................................70
2.3.2 Versuchsaufbau und Versuchsdurchführung ........................................................71
2.3.3 Bestimmung der neuronalen Richtungscharakteristik ..........................................71
2.3.4 Histologische Lokalisation des Läsionsortes ........................................................71
3. Ergebnisse ................................................................................................72
3.1 Untersuchungen zum vertikalen optokinetischen Reflex an Salmo gairdneri ...........72
3.2 Visueller Einfluss auf die kompensatorischen Augenbewegungen beim
vestibulo- okulären Reflex .........................................................................................76
3.2.1 Der vertikale vestibulo- okuläre Reflex ...............................................................76
3.2.2 Der horizontale vestibulo- okuläre Reflex ............................................................84
3.2.3 Der vertikale und der horizontale vestibulo- okuläre Reflex im Dunklen ...........86
3.3 Optokinetische Augenbewegungen vor und nach Läsion des dorsalen
Praetectums .................................................................................................................89
3.3.1 Elektrophysiologische Läsionsexperiment ...........................................................89
3.3.2 Verhaltensbeobachtung nach Läsion dorsalen Praetectums .................................91
3.3.3 Der monokulare horizontale optokinetische Reflex nach der Läsion ...................91
3.3.4 Analyse der Augenfolgebewegungsereignisse während der
Stimulationsphasen ...............................................................................................94
3.3.4.1 Anzahl der Augenfolgebewegungen ..............................................................94
3.3.4.2 Amplitude der Augenfolgebewegungen .........................................................96

3.3.4.3. Geschwindigkeit der Augenfolgebewegung ..................................................98
3.3.5 Spontane Augenbewegungen .............................................................................100
3.3.7 Histologische Lokalisation des Läsionsortes ......................................................101
4. Diskussion ..............................................................................................104
4.1 Der vertikale optokinetische Reflex bei Salmo gairdneri ........................................104
4.1.2 Gründe für das Ausbleiben des vertikalen optokinetischen Reflexes ................105
4.1.2.1 Anordnung der extraokulären Muskeln ........................................................105
4.1.2.2 Die Wahl des adäquaten visuellen Reizes ....................................................106
4.1.2.3 Anpassung an das Habitat .............................................................................107
4.2 Der visuelle Einfluss auf das vestibulo- okuläre System .........................................108
4.2.1 Der vertikale vestibulo- okuläre Reflex .............................................................108
4.2.2 Der horizontale vestibulo-okuläre Reflex ...........................................................112
4.3.3 Die Kopplung des optokinetischen und vestibulo-okulären Systems ................113
4.3 Auswirkungen einer unilateralen praetectalen Läsion auf den
optokinetischen Reflex .............................................................................................115
4.3.1 Die Voruntersuchung zum mhOKR und das Läsionsexperiment ......................115
4.3.2 Verhaltensbeobachtung nach der praetctalen Läsion .........................................116
4.3.3 Der monokulare horizontale optokinetische Nystagmus nach
der Inaktivierung ................................................................................................117
4.3.4 Welche Anteile des horizontalen optokinetischen Reflexes sind
während der visuellen Stimulation betroffen ......................................................118
4.3.5 Das Model der optokinetischen Projektion für das echte Konochenfische ........119
III. Zusammenfassung ........................................................................121
IV. Literaturverzeichnis ..................................................................... 125
V. Abkürzungen ..................................................................................... 143

I. Elektrophysiologie 1. Einleitung
I. Elektrophysiologische Einzelzellableitungen im
Praetectum von Salmo gairdneri und Esox lucius
1. Einleitung Mit der Entwicklung von Augen, die ein scharfes Abbild der Umwelt auf einer Schicht von
Sehzellen entwerfen, besitzen Tiere ein wichtiges Fernsinnesorgan, das es ihnen erlaubt,
Objekte in ihrer Umgebung zu erkennen. Da die meisten dieser Tier auch die Fähigkeit
entwickelt haben, sich in ihrem Lebensraum oder ihre Augen zu bewegen, entsteht ein
Problem. Das Abbild der Umwelt bewegt sich dabei auf der Sinneszellschicht, es
„verwackelt“. Daher haben diese Tiere Reflexe für die Ausführung kompensatorischer
Augenbewegungen entwickelt, die das retinale Bild stabilisieren. Man nimmt sogar an,
dass bei Vertebraten der Evolutionsdruck für die Ausbildung eines effektiven Augen-
bewegungsapparates in der Notwendigkeit der Blickstabilisierung bestand. Alle
Vertebraten mit Augen haben sechs Augenmuskel, die durch komplexe neuronale
Schaltkreise angesteuert werden. Dieses okulomotorische System wird durch die
Sinnesorgane Auge und Gleichgewichtssystem über Eigenbewegungen des Tieres
informiert und erzeugt die kompensatorische Aktion der Augenmuskeln damit das Bild auf
der Retina stabil bleibt. Der über die visuelle Information gesteuerte Reflex wird
optokinetischer Reflex (OKR), der über vestibuläre Information gesteuerte Reflex wird
vetibulo-okulärer Reflex (VOR) genannt.
1.1 Das Praetectum als Ort richtungsselektiver Neurone In den Vertebratenklassen Mammalia, Aves, Reptilia und Amphibia wurden weitreichende,
neuroanatomische, cytoarchitektonische und elektrophysiologische Untersuchungen zur
genauen Lokalisation und Charakterisierung praetectaler richtungsselektiver Neurone
durchgeführt, die maßgeblich an der langsamen Augenfolgephase des horizontalen
optokinetischen Reflexes (hOKR) beteiligt sind.
Solche Neurone liegen bei Säugern im Nukleus des optischen Traktes (Nucleus tractus
optici, NOT) und bei Aves, Reptilia und Amphibia im Nucleus lentiformis mesencephali
(nLM) (Amphibia: Katte & Hoffmann, 1980; Fite, 1985; Montgomery et al., 1985;
1

I. Elektrophysiologie 1. Einleitung
Reptilia: Schröder, 1981; Aves: Wylie & Forst, 1990; Mammalia: Collewijn, 1975;
Hoffmann & Schoppmann, 1975). Klassenübergreifend zeigen Neurone innerhalb dieses
Kerngebietes typische Antworteigenschaften.
Die visuelle Information erreicht diese Kerne, abhängig vom Dekussationsgrad der
Sehnerven vorwiegend von der kontralateralen Retina. Die Stärke des ipsilateralen
Eingangs variiert mit der Lage der Augen im Kopf und der retinalen Foveation (Tauber &
Atkin, 1968; Hoffmann, 1985).
Generell wird die Information einer Bildverschiebung im Gesichtsfeld nach links von den
Ganglienzellen der rechten Retina registriert, monosynaptisch zu den Neuronen des linken
NOT bzw. nLM übertragen und dort weiter verarbeitet. Bildverschiebungsneurone des
NOT und nLM sind charakteristischerweise richtungsselektiv.
Große rezeptive Felder werden benötigt, um langsame Augenfolgebewegungen ohne
Abbruch durchzuführen. Bei Detektion kleiner Objekte während der langsamen
Augenfolgephase, die sich in einer von der Hintergrundbewegung entgegengesetzten
Richtung bewegen, könnten zu einem Abbruch des OKRs führen. Damit würde das System
fehlerhaft arbeiten, weil unter natürlichen Bedingungen sicherlich keine konstanten
visuellen Bedingungen vorherrschen.
Ein weiteres, wichtiges Charakteristikum ist die Abhängigkeit der neuronalen Antwort in
diesen Kernen von der Geschwindigkeit des präsentierten visuellen Reizes. Um einer
retinalen Bildverschiebung entgegenzuwirken, muss nicht nur die Richtung der
Reizbewegung detektiert werden, sondern auch deren Geschwindigkeit. Dies wird von den
richtungsselektiven Neuronen über eine Erniedrigung oder Erhöhung der Aktivität erreicht,
die letztendlich zur Ausführung der Augenbewegung mit der richtigen Geschwindigkeit
führt.
Der optokinetische Reflex kann nicht nur in horizontaler Bewegungsrichtung ausgeführt
werden. Visuelle Richtungsinformationen über vertikale Bildverschiebungen werden
primär von Kernen des akzessorischen optischen Systems (AOS) im Mesencephalon
verarbeitet. Bei Säugern sind dies der laterale terminale Nukleus (LTN) und der mediale
terminale Nukleus (MTN).
Bei Aves, Reptilia und Amphibia wurde der „Nucleus of the basal optic root“ (nBOR) im
Mesencephalon als dem LTN und MTN der Mammalia vergleichbares Areal identifiziert.
2

I. Elektrophysiologie 1. Einleitung
Im nBOR wurden richtungsselektive Neurone gefunden, die neben vertikalen
Bildverschiebungen auch eine horizontale kontraversive Stimulusrichtung kodieren. Der
nBOR enthält also drei Neuronenpopulationen mit unterschiedlichen Vorzugsrichtungen.
Die rezeptiven Feldeigenschaften wie Richtungs- und Geschwindigkeitsselektivität dieser
Neuronenpopulationen gleichen denen des NOT und nLM.
Die Bildverschiebungsneurone des NOT und nLM sind das visuomotorische Bindeglied im
Schaltkreis für den horizontalen OKR (hOKR) und VOR. Anatomisch konnten zwei
Projektionswege im optokinetischen System identifiziert werden.
Die für die langsame Folgephase des hOKR verantwortliche Projektion zieht ipsilateral in
den Hirnstamm zum Nucleus praepositus hypoglossi (NPH) und / oder zum Nucleus
reticularis tegmenti pontis (NRTP) (Maekawa et al., 1981; Cazin et al., 1980, 1982, 1984;
Magnin et al., 1983, 1989; Fuchs et al., 1992; Watanabe et al., 1993; Mustari et al., 1994).
Diese Kerne projizieren ihrerseits zu den kontralateralen Vestibulariskernen. Von den
Vestibulariskernen verläuft der Projektionsweg zu den Augenmuskelkernen, die
schliesslich die entsprechenden äußeren Augenmuskeln innervieren.
Außerdem projizieren Neurone des NOT zur ipsilateralen Oliva inferior (IO); von dort
ziehen Kletterfasern zum Kleinhirn und den cerebellären Kernen. Diese Projektion dient
der Rekalibirierung des vestibulo - okulären Reflexes (Ito et al., 1974; Ito, 1982, 1993).
In ersten Verhaltensuntersuchungen fand Easter (1972) heraus, dass der OKR des
Goldfisches (Carassius auratus) mit dem anderer Vertebraten vergleichbar ist. Mit
anterograden Markierungstechniken identifizierten Finger und Karten (1978) am Goldfisch
zwei praetectale Bereiche. Die Autoren gaben ihnen die Bezeichnung P1 und P2 und sahen
diese als homologe Struktur des AOS der Mammalia, Aves, Reptilia und Amphibia an. In
späteren Arbeiten wurden diese verwirrenden anatomischen Bezeichnungen praetectaler
Areale revidiert. Den Autoren Vanegas und Ito (1983) und Fite (1985) gelang es, sechs
primäre visuelle Kerne im Praetectum von Knochenfischen anatomisch zu identifizieren.
Die Abbildung I. 1.1 zeigt diese praetectalen Kerngebiete an einem schematisierten
Frontalschnitt durch das Gehirn eines Knochenfisches.
3

I. Elektrophysiologie 1. Einleitung
Abb. I. 1.1:
TeO = Tectum
Diese Kern
Nucleus co
lateralis (NG
Spezies aus
Kerne NPT
und Ausprä
nachgewies
Experiment
praetectale
die Area
okulomotor
Kerngebiete
Diese anato
und diente
Homologisi
echten Kn
charakterisi
Funktionell
das neurona
Lokalisation der sechs visuellen praetectalen Kerngebiete eines generalisierten Teleosteers.
opticum, V3 = dritter Vertrikel. Quelle: Fite, 1985.
Retina
TeO
NDL
NPT
NCP
NC
NGL
APT
V3
gebiete sind der Nucleus praetectalis (NPT), Nucleus dorsolateralis (NDL),
mmissurae posterioris (NCP), Nucleus corticalis (NC), Nucleus geniculatus
L) und die Area praetectalis (APT). In Untersuchungen an unterschiedlichen
den Familien Cyprinidea und Percidea konnte das konstante Vorkommen der
, NDL, NCP, APT gezeigt, aber auch eine große Variabilität in Vorkommen
gung der praetectalen Kerngebiete NC und NP zwischen den Fischfamilien
en werden (Vanegas & Ito, 1983; Ito & Murakami, 1984). In weiterführenden
en wurden die Projektionen der sechs praetectalen Kerne untersucht. Zwei
Kerngebiete, der NPT und NCP, projizieren zum Cerebellum. Ein Kerngebiet,
praetectalis (APT) projiziert ebenfalls zum Cerebellum und zu den
ischen Kernen. Aufgrund dieser Projektionswege werden diese drei
(NPT, NCP, APT) anatomisch zum AOS der Teleostei gezählt.
mischen Untersuchungen wurden an verschiedenen Fischspezies durchgeführt
n neben der Lokalisation visueller Areale auch dem Versuch einer
erung zu den anderen Vertebratenklassen. Dies gestaltete sich schwierig, da die
ochenfische (Teleostei) durch sehr hohe Spezialisierung und Divergenz
ert sind.
konnte mit elektrophysiologischen Methoden in der Überordnung Teleostei
le Korrelat zur Generierung der langsamen Folgephase des hOKRs bisher nicht
4

I. Elektrophysiologie 1. Einleitung
nachgewiesen werden. Aus diesem Grund werden in diesem ersten Kapitel folgende
Fragestellungen untersucht:
- Existieren richtungsselektive Neurone im Praetectum von Forelle (Salmo gairdneri)
und Hecht (Esox lucius)?
- Wenn ja, welche weiteren Antworteigenschaften besitzen diese richtungsselektiven
Neurone?
- Können Vergleiche zu Befunden aus den anderen Vertebratenklassen gezogen
werden?
- Wie kann eine funktionelle Beteiligung solcher richtungsselektiver Neurone am
OKR gezeigt werden?
5

I. Elektrophysiologie 2. Material & Methoden
2. Material und Methoden
2.1 Versuchstiere Die elektrophysiologischen Experimente und Verhaltensuntersuchungen in dieser Arbeit
wurden an weiblichen und männlichen Regenbogenforellen (Salmo gairdneri) und Hechten
(Esox lucius) durchgeführt. Die Versuchstiere hatten ein Alter von einem bis zweieinhalb
Jahren und wogen 140 g bis 380 g. Die Regenbogenforellen stammten aus der Fischzucht
Gebr. Viedt, Am Masling 54, 58456 Witten, die Hechte aus der Fischzucht Peschkes,
Beckrather Str. 115, 41119 Mönchengladbach.
Alle Fische waren ohne Krankheitsbefall, agil und zeigten keine auffälligen Defizite in
ihrem Verhalten. Die Versuche wurden von den lokalen Behörden und der Ethikkommission
genehmigt. Ihre Durchführung erfolgte gemäß dem deutschen Tierschutzgesetz und den
europäischen Richtlinien (S6 609 EEC) sowie den NIH Richtlinien zur Durchführung von
Tierversuchen.
Die elektrophysiologischen Untersuchungen wurden an 24 Salmo gairdneri und 8 Esox
lucius durchgeführt.
2.2 Fischhälterung Die Fische wurden in Aquarien mit der Größe 180 cm x 60 cm x 40 cm gehältert. Pro
Hälterungsbecken wurden maximal vier Tiere eingesetzt, um innerartlichen Stress
aufgrund eines Überbesatzes zu vermeiden.
In den Aquarien von Salmo gairdneri wurden größere Steine eingebracht, eine ständige
Wasserströmung erzeugt und für eine zusätzliche Luftzufuhr gesorgt.
Wegen des bekannten Kannibalismus in der Familie Esocidae wurden die Hechte einzeln
gehältert. Jedem Tier stand ein Areal von 45 cm x 60 cm x 40 cm zur Verfügung.
Die Wassertemperatur betrug stets 12°C (+/- 1°C). Die Fische waren auf einen Tag/Nacht
Rhythmus von 12 Stunden eingestellt.
6

I. Elektrophysiologie 2. Material & Methoden
2.3 Anästhesie, Präparation und Fixierung Die initiale Anästhesie aller Fische, die in den elektrophysiologischen Experimenten
eingesetzt wurden, erfolgte mit MS-222 (3-Aminobenzoid-Acid-Ethyl-Ester-
Methanesulfonate, Sigma). In einem 15 l fassenden Styroporbehälter wurde eine 0,5 %- ige
MS-222 Lösung mit Eiswasser (4°C) angesetzt. Die Versuchstiere befanden sich ca. 15
Minuten lang in diesem Narkosebad, bis eine vollständige Anästhesie eintrat. Als
Kontrolle diente das Ausbleiben der kompensatorischen Augenbewegungen bei Drehung
des Fisches um seine Körperlängsachse, die Reduktion der Kiemenschlagfrequenz und die
Relaxation der gesamten Rumpfmuskulatur.
Im Anschluss wurden die anästhesierten Fische mit Ausnahme von Kopf und
Kiemenregion in feuchtes Zellulosepapier gehüllt, um einer vermehrten Schleimbildung
durch physiologischen Stress während des elektrophysiologischen Experimentes
entgegenzuwirken. Die Fische wurden dann in eine Fixationshalterung eingesetzt. Diese
bestand aus einer Plexiglasgrundplatte (2 cm x 30 cm x 1 cm) mit zwei verschiebbaren
Seitenhalterungen (5 cm x 25 cm), die für jede untersuchte Fischart individuell an die
Rumpfform angepasst war. Sie reichten beidseitig eng anliegend von der Schwanzflosse
bis kurz hinter die Kiemenregion. An der Grundplatte war eine Maulhalterung befestigt, in
der ein Tubus integriert war. Dieser wurde den Fischen in das Maul eingeführt und war an
die Form der Gaumenplatte der untersuchten Fischart angepasst.
Die fixierten Fische wurden dann in ein Präparationsbecken (20 cm x 30 cm x 20 cm)
überführt. Umgehend wurde das eisgekühlte Narkosewasser bis zum dorsalen Rand der
Augen in das Becken eingefüllt, so dass die Kiemen im Wasser waren und der dorsale
Kopfbereich aus dem MS-222 Narkosewasser ragte.
Durch den Tubus wurde den Kiemen sauerstoffangereichertes Narkosewasser zugeführt.
Zur Vermeidung unwillkürlicher Körper- und Augenbewegungen während der Präparation
und des elektrophysiologischen Experiments wurde den Versuchstieren 0,6 ml
Alcuroniumchlorid (Alloferin®, 1 mg/ml, ICN) i. m. appliziert. Die Injektionsorte waren
beidseitig der Körpermittellinie in der rostralen, medialen und caudalen Rumpfregion. Mit
dieser Menge konnte eine anhaltende Immobilisierung über 15 bis 20 Stunden erreicht
werden. Je nach Versuchsdauer (bis zu 40 Stunden) wurde 0,3 ml Alloferin® nachinjiziert.
Vor der Kraniotomie und alle 2 – 3 Stunden während des elektrophysiologischen
7

I. Elektrophysiologie 2. Material & Methoden
Experiments wurde 0,1 ml Prilocainhydrochlorid–Lösung (Xylonest®, 5 mg/ml, ASTRA)
im Wundbereich verabreicht, um eine zusätzliche Lokalanästhesie zu erreichen.
Die Kraniotomie begann mit der Markierung eines 1 cm x 1 cm Hautbereichs im dorsalen
Kopfbereich oberhalb der linken Gehirnhemisphäre. In diesem Bereich wurde die Haut und
das darrunterliegende Muskelgewebe entfernt und mit Hilfe eines Zahnarztbohrers
(Chiyoda, Etelna) der Schädelknochen im Bereich des darunter befindlichen Tectum
opticum (TeO) abgetragen. Anschließend konnte die Dura mater und das Fettgewebe,
welches das Gehirn umgibt, im dorsalen Bereich entfernt werden, so dass der rostrale
Bereich des linken und Teilbereiche des rechten Tectum opticum frei zugänglich vorlag.
Mit einem Plexiglasstab (∅ = 0,5 cm, Länge 3 cm), der von der linken Seite horizontal auf
dem Nasale mit Sekundenkleber befestigt wurde, wurde der Kopf des Versuchstieres
fixiert. Anschließend wurde das präparierte Versuchstier in der Fixationshalterung in den
Ableitstand in der Plexiglashalbkugel überführt.
2.4 Elektrophysiologischer Versuchsaufbau Die Beschreibung des elektrophysiologischen experimentellen Versuchsaufbaus wird in
drei Bereiche gegliedert:
- Ableitstand
- visuelle Stimulation
- Stimulussteuerung und Datenregistrierung
2.4.1 Der Ableitstand Die in der Fischhalterung fixierten Versuchstiere wurden über einen Halter, der mit dem
Rahmengestell der Plexiglashalbkugel verbunden war, mit dem rechten Auge zentriert in
die Plexiglashalbkugel eingesetzt. Dieser Verbund, das Versuchstier in der
Fixationshalterung und die Ableithalterung, an der sich in drei Raumachsen (X, Y, Z)
angeordnete Mikromanipulatoren (Mikochima) und ein Mikrofeinvortrieb (Burleigh, PZ
550) befanden, wurde genau um 45° in der Körperlängsachse des Fisches nach rechts
8

I. Elektrophysiologie 2. Material & Methoden
geneigt. Dadurch befand sich die rechte Kopfhälfte im und die linke Kopfhälfte außerhalb
des Wassers. Der Abstand des rechten Auges zur opaken (undurchsichtigen) Oberfläche
der Plexiglashalbkugel betrug allseits 20 cm. Mit dieser Anordnung war es möglich, die
elektrophysiologischen Ableitungen in der linken Gehirnhemisphäre durchzuführen und
einen möglichst großen Bereich des rechten Gesichtsfelds homogen mit einem
Zufallspunktemuster, das mit einem Planetarium auf die Innenseite der Plexiglashalbkugel
projiziert wurde, visuell zu stimulieren. (Abb. I. 2.1).
10 cm
20 cm
10 cmcwccw
0°
90°
270°315°
135° 45°
225°
180° nasal
nasal-unten
nasal-obenoben
unten
temporal
temporal-unten
temporal-oben
CCW CWPause Pause
2 s 3 s 2 s 3 s
Verlauf der visuellen Stimulation
A B
Wasserlinie
Grundplatte der Fixationshalterung
Beatmungstubus
Nasenhalterung zur Kopffixierung
Ableitelektrode
Abb. I. 2.1: In der Teilabbildung A ist eine schematische Darstellung des elektrophysiologischen
Ableitstandes mit der Position des Fisches in der Plexiglashalbkugel, der Position des Planetariums und
dessen Drehrichtung (cw im Uhrzeigersinn, ccw gegen den Uhrzeigersinn) sowie die Rotationsachse, aus der
sich die acht Stimulationsrichtungen ergeben, gezeigt. Die Teilabbildung B zeigt eine Frontalansicht des
Versuchtieres in der Fixationshalterung im Plexiglasbecken.
Die Wasserstandregulierung in der Plexiglashalbkugel erfolgte durch den Wasserabfluss
aus der Plexiglashalbkugel und Weiterleitung zu einem Kryostaten. Das Gerät regelte die
Wassertemperatur auf konstante 12°C. Das Wasser wurde mittels Druckluft mit Sauerstoff
angereichert. Die abgeflossene Wassermenge wurde wieder aufbereitet und den
Versuchstieren über den im Maul befindlichen Tubus erneut zugeleitet (Salmo gairdneri =
300 ml/min, Esox lucius = 185 ml/min).
Die mit glasisolierten Wolfrahmdrahtelektroden (Impedanz 2,5 bis 3,5 MΩ, Eigenbau des
Instituts für Allg. Zoologie und Neurobiologie, Herr H. Korbmacher) registrierten
neuronalen Signale wurden über einen Vorverstärker (200 bis 400 - fache Verstärkung)
digitalisiert mit einem Glasfaserkabel zu einem Ableitendverstärker (2000 - fache Ver-
stärkung) weitergeleitet. Vor der Endverstärkung wurde das digitale Signal in ein analoges
9

I. Elektrophysiologie 2. Material & Methoden
Signal zurückgewandelt (beide Geräte sind Eigenbau des Institut für Allg. Zoologie und
Neurobiologie). Das Ausgangssignal des Verstärkers wurde zu einem Oszillographen mit
integriertem Differenzverstärker und Filtereinheit (Tektronix, 5103N) und zu einem
Window - Diskriminator (W-P Instruments Inc., Model 120) weitergleitet. Der
Oszillograph stellte die abgeleitete neuronale Aktivität und die Schwellwert-Einstellung
des Window - Diskriminators zur Detektion der Aktionspotentiale (Spikes) der
Einzelzellableitungen dar. Das diskriminierte TTL – Signal wurde zu einem Flip/Flop (1
ms Auflösungsvermögen) Interface geleitet, das mit dem Computer zur Datenaufnahme
verbunden war. Zusätzlich wurden die gefilterten Ableitsignale der Elektrode und die
detektierten Spikes der Einzelzellableitung getrennt über Aktivlautsprecher (BOEKER,
25W) hörbar gemacht.
2.4.2 Visuelle Stimulation Die visuelle Großfeldstimulation erfolgte mit einem Planetarium. Mit einer Lichtquelle
innerhalb einer hohlen, zufällig durchlöcherten Metallkugel konnte ein
Zufallspunktemuster durch das Wasser auf die Innenseite der opaken Plexiglashalbkugel
projiziert werden. Der Abstand zwischen dem Kopf der Tiere und dem Planetarium betrug
10 cm. Die projizierten Lichtpunkte hatten auf der Oberfläche der Plexiglashalbkugel einen
Durchmesser von 1 cm. Über eine Ansteuerungseinheit konnte das Planetarium im und
gegen den Uhrzeigersinn gedreht werden. Zusätzlich konnte es vier unterschiedliche
Rotationspositionen einnehmen. Daraus ergaben sich acht visuelle Stimulusrichtungen, in
denen das rechte Auge des Versuchstiers zur Bestimmung der Richtungsempfindlichkeit
(Tuning-Kurven) stimuliert wurde: temporal ↔ nasal (180°↔0°), oben ↔ unten
(90°↔270°), nasal - oben ↔ temporal - unten (45°↔225°) und temporal - oben ↔ nasal -
unten (135°↔315°).
Eine einzelne Stimulationsphase (10 Sekunden) bestand aus: 2 Sekunden stationäres
Zufallspunktemuster, 3 Sekunden Bewegungsrichtung im Uhrzeigersinn, 2 Sekunden
stationäres Zufallspunktemuster und 3 Sekunden Bewegungsrichtung gegen den
Uhrzeigersinn (siehe Abb. I. 2.1). Dieses wurde zehn Mal pro Rotationsposition
(Stimulusrichtung) wiederholt. Die Stimulusgeschwindigkeit betrug konstant 10°/s.
10

I. Elektrophysiologie 2. Material & Methoden
Zur Untersuchung des geschwindigkeitsabhängigen Verhaltens der richtungsselektiven
Neurone wurde die Reizgeschwindigkeit von 0°/s bis 80°/s in jeweils 10°/s Schritten
variiert. Die Stimulationsdauer betrug acht Sekunden. Die Stimulusrichtung entsprach der
Vorzugsrichtung des jeweiligen Neurons.
2.4.3 Stimulussteuerung und elektrophysiologische Datenregistrierung Die zentrale Kontrolle der Ansteuerungseinheit des Planetariums zur visuellen Stimulation
und die Datenregistrierung der neuronalen Aktivität wurden von einem Computer (MW-
SOFT, Celeron 330 MHz) mit einer Multi I/O Karte (Computerboard, PIO 24-DAS 16,
Plug - In) geleistet. Für die elektrophysiologischen Untersuchungen wurde eine neue
Aufnahme- Software CORTEX (NIH, Version 5.3) etabliert.
Über programmierte Steuerungsfiles in CORTEX wurde zufallsgeneriert nacheinander eine
der vier Rotationspositionen des Planetariums mit Hilfe von Infrarot-Lichtschranken
angefahren. Zu jeder Rotationsposition erfolgte die visuelle Stimulation mit gleichzeitiger
Registrierung der vom Flip/Flop-Interface bereitgestellten TTL-Spikeaktivität in CORTEX
- Datenfiles. Gleichzeitig wurden Triggerdaten zu Beginn und Ende der Registrierung und
vom Anfang bis zum Ende der stationären Phasen, sowie der Stimulationsphasen in das
Datenfile zur späteren Analyse mit aufgenommen.
Das System bot eine Online-Darstellung des Stimulusverlaufs auf dem Computermonitor
(EIZO, 570A) und die zeitgleiche Darstellung der Spikeaktivität in Form von Rasterplots
und PSTH.
2.5 Erstellen des Penetrationsrasters Um eine möglichst hohe Erfolgsrate in der Lokalisation praetectaler richtungsselektiver
Neurone zu erzielen, wurden zwei Methoden kombiniert.
Mit Hilfe der drei Mikromanipulatoren (X-, Y-, Z-Achse) konnte mit der Ableitelektrode
ein Penetrationsraster in 200 µm Schritten im rostralen Bereich des linken Tectum opticum
11

I. Elektrophysiologie 2. Material & Methoden
(TeO) abgefahren werden, um elektrophysiologische Ableitungen im ventral unterhalb des
TeO gelegenen Praetectum durchzuführen.
Als Ursprungskoordinate mit der Position anterior / posterior = 0/0 µm diente der rostrale
Pol des Tectum opticum. Die medio - laterale Position = 0/0 µm war der Torus
longitudinalis, der sich zwischen den beiden Tectum opticum Hälften befindet (Abb. I.
2.2).
TeO
posterior
m/l = 0
lateral a/p > 0
TeO
a/p = 0
m/l > 0 anterior
Abb. I. 2.2: Schematische Darstellung des verwendeten Penetrationsrasters. Dorsalansicht des linken
Tectum opticum (TeO). Abkürzungen: m/l = medial / lateral; a/p = anterior / posterior.
Zusätzlich wurden an jedem Penetrationsort die rezeptiven Feldgrenzen visuell erregbarer
Neuronen des retinotop organisierten TeOs auf der Außenseite der Plexiglashalbkugel
markiert.
In Vorexperimenten wurde festgestellt, dass die Gehirne der untersuchten Fischarten eine
gleichbleibende Größe aufwiesen, so dass sowohl die Koordinaten der
Mikromanipulatoren, als auch die Lage der tectalen rezeptiven Felder zu einer genauen
Lokalisation des praetectalen Areals beitrugen.
12

I. Elektrophysiologie 2. Material & Methoden
2.6 Bestimmung der rezeptiven Felder Die Bestimmung der Grenzen der visuellen rezeptiven Felder wurde mit einem variablen
weißen Lichtpunkt (1° - 25° Sehwinkel) aus einer Handlampe durchgeführt. Änderungen
der Spikeaktivität, die durch wechselnde Lichtintensitäten an den Grenzen der rezeptiven
Felder ausgelöst wurden, wurden am linken und rechten Azimut sowie an der oberen und
unteren Elevation mit einem Filzstift (Edding 3000) an der Außenseite der
Plexiglashalbkugel markiert. Nach Beendigung des Experiments wurden mit Hilfe eines
Laserpointer unterstützten Winkelmessers, dessen Rotationsachse sich exakt an der
Position des rechten, stimulierten Auges befand, die zuvor markierten Feldgrenzen
ausgemessen und protokolliert. Die kartesische Darstellung der Verteilung rezeptiver
Felder im rechten Gesichtsfeld der Versuchstiere wurde mit Sigma Plot© (SPSS Inc.,
Version 6.0) durchgeführt.
2.7 Datenanalyse und Auswertung Die CORTEX - Datenfiles wurden mit programmierten MATLAB (The MathWorks, inc;
Version 5.3.1 (R11.1)) Funktionen quantitativ analysiert.
Die Darstellung der Rasterplots und PSTHs (Peri – Stimulus – Time - Histogram) in 40 ms
bin - Breite erfolgte exakt mit den Triggersignalen zu den Stimulusphasen Anfang-Ende
der Aufzeichnung, Dauer der stationären Phase und Dauer der Bewegungsphasen.
Als Grundlage zur Bestimmung von Richtungsempfindlichkeitskurven (Tuning - Kurven)
richtungsselektiver Zellen wurde die mittlere Spikerate in jeder der acht untersuchten
Stimulusrichtungen über zehn Trials ermittelt. Die Berechnung der Richtungsvektoren
erfolgte durch Vektoraddition mit Hilfe der trigonometrischen Funktionen und der
mittleren Spikerate zur jeweiligen Stimulationsrichtung.
13

I. Elektrophysiologie 2. Material & Methoden
2.7.1 Statistische Verfahren Die Berechnung der statistisch signifikanten Vorzugsrichtung richtungsselektiver
Nervenzellen wurde mit Hilfe der zirkulären Statistik durchgeführt. Dieses Verfahren
beruht auf der Umrechung der mittleren Spikerate pro Stimulationsphase (3000 ms) in die
Frequenz in einer der Stimulusrichtungen (Angabe in Winkelgrad). Diese Umrechnung
wurde für alle acht Stimulusrichtungen und pro Stimulationsphase durchgeführt.
Anschließend wurden die Frequenzdaten zu den entsprechenden Winkeln gruppiert. Durch
Bildung eines Vertrauensintervall zu p = 95 % wurden die Daten mit dem t-Test auf
Signifikanz überprüft. Die abschließende Überprüfung der als signifikant ermittelten Daten
auf ihre Vorzugsrichtung basiert auf dem zirkulären statistischen Verfahren von Rayleigh
Moore (Batschelet, 1981).
Die Verteilungsbestimmung auf signifikante Unterschiede in der Anzahl von
richtungsselektiven Neuronen, die eine horizontale-vertikale bzw. horizontal nach nasal
oder nach temporal verlaufende Stimulusrichtung kodieren, wurde mit dem nicht -
parametrischen χ2-Tests durchgeführt. Das statistische Verfahren beruht auf dem Vergleich
zweier Häufigkeitsverteilungen.
Die Latenzzeitanalyse erfolgte, indem 500 ms vor dem Einsetzen des Bewegungsreizes in
der Vorzugsrichtung die Standardabweichung der mittleren Spontanaktivitätsrate in 10 ms
bin-Breite berechnet wurde. Zum Zeitpunkt der Aktivierung musste die reizgetriebene
Aktivität um die dreifache Standardabweichung über der Spontanaktivitätsrate liegen, um
einen signifikanten Wert (t-Test) zu erreichen, der den Zeitpunkt der Aktivierung
bestimmt.
2.7.2 Histologie und Rekonstruktion des Ableitortes Am Ende der elektrophysiologischen Untersuchungen wurden die Fische mit einer
Überdosis MS-222 (2,5%) eingeschläfert. Anschließend wurden die Tiere mit einer 0,9% -
igen NaCl-Lösung (300 ml), gefolgt von einer 4% -igen Paraformaldehyd - Phosphatpuffer
(0,1 M, pH 7,4) - Lösung mit 20% Sucrose (250 ml) transcardial perfundiert.
Das Gehirn der Fische wurde aus der Schädelkapsel herauspräpariert und die Dura mater
entfernt. Das isolierte Gehirn wurde dann für 24 Stunden in derselben Fixierlösung
14

I. Elektrophysiologie 2. Material & Methoden
belassen. Danach folgte für 12 Stunden ein Gefrierschutz der Gehirne in einer 30 % - igen
Sucrose - Phosphatpuffer (0,1 M, pH 7,4) Lösung.
Die Gehirne wurden in Einbettkammern in Hühnereiweiß-Gelatine-Medium eingebettet,
über Nacht nachfixiert (10 % Paraformaldehyd in Phosphatpuffer mit 30 % Sucrose) und
anschließend in Isopentan bei –70°C schockgefroren. Mit einem Gefriermikrotom (Microm
HM 500 OM) wurden 50 µm dicke Frontalschnitte von rostral nach caudal angefertigt. Die
Schnittserien wurden auf gelatinierte Objektträger aufgezogen und für mehrere Tage in
einen Trockenschrank aufbewahrt. Anschließend erfolgte das Färben der Schnittserien
nach Klüver - Barrea und / oder Nissl und das Eindeckeln mit Depex.
Die Bestimmung der Läsionsorte erfolgte mit dem Lichtmikroskop (Zeiss, Axioskop FS).
Ausgehend von dem ersten Schnitt der Serie, auf dem zweifelsfrei der rostrale Pol des TeO
zu erkennen war, wurden die Schnitte bis zum Läsionsort gezählt. Daraus ergab sich die
Distanz vom rostralen Pol des TeOs zum Läsionsort. Schrumpfungsfaktoren aufgrund der
verwendeten Fixierlösungen wurden nicht berücksichtigt.
15

I. Elektrophysiologie 3. Ergebnisse
3. Ergebnisse
3.1 Lokalisation richtungsselektiver Neurone im Praetectum
von Salmo gairdneri und Esox lucius Nach der Vorgabe des gewählten Penetrationsrasters (Abb. I. 3.1 und I. 2.5) wurden
systematisch elektrophysiologische Ableitungen im Bereich des linken Praetectums von
Salmo gairdneri und Esox lucius durchgeführt. Dabei stand neuronale
Richtungsselektivität im Praetectum, ausgelöst durch die visuelle Stimulation des rechten
Auges mit dem aus dem Planetarium generierten, großflächigen Zufallspunktemuster, als
Suchkriterium im Vordergrund.
Zum Auffinden der richtungsselektiven Neurone wurde ein in seinem Durchmesser
variabler, weißer Lichtpunkt aus einer Handlampe auf die opake Oberfläche der
Plexiglashalbkugel projiziert und damit das rechte Gesichtsfeld kontinuierlich in allen
Raumrichtungen stimuliert und abgesucht.
Nach der Lokalisation visueller Aktivität wurden folgende Charakterisierungen
durchgeführt:
- Lage und Größenbestimmung der rezeptiven Felder mit dem Lichtpunkt aus der
Handlampe.
- ON, OFF, ON/OFF Eigenschaften: getestet durch wechselnde Lichtintensitäten mit der
Handlampe bzw. Raumhelligkeit innerhalb des rezeptiven Felds.
- Richtungsselektivität: getestet durch das sich in acht Stimulusrichtungen bewegende
großflächige Zufallspunktemuster des Planetariums.
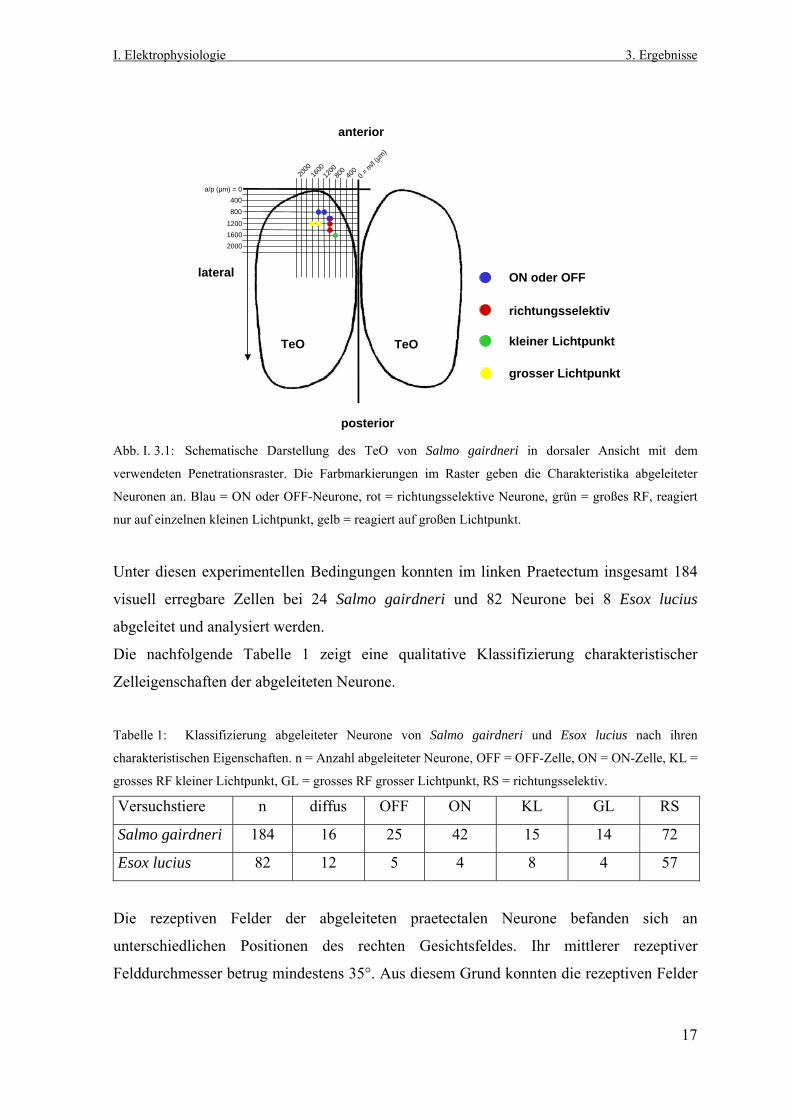
In der Abbildung I. 3.1 ist in dorsaler Ansicht das Penetrationsraster mit der Lage des
Tectum opticum (TeO) von Salmo gairdneri schematisch dargestellt. Die Ableitungen
fanden in der linken Gehirnhemisphäre statt und wurden von dorsal nach ventral durch das
TeO und das darunter befindliche gesamte Praetectum durchgeführt. Die
Farbmarkierungen zeigen die Rasterpositionen mit visueller neuronaler Aktivität. Die
unterschiedlichen Farben kodieren die abgeleiteten Antworteigenschaften.
16
I. Elektrophysiologie 3. Ergebnisse
anterior
Abb. I.
verwen
Neuron
nur auf
Unter
visuel
abgele
Die n
Zellei
Tabelle
charakt
grosses
Versu
Salmo
Esox
Die r
unters
Felddu
a/p (µm) = 0
0 = m
/l (µm
)
posterior
TeO TeO
400
800
120016002000
lateral
400
800
120020
0016
00
richtungsselektiv
ON oder OFF
kleiner Lichtpunkt
grosser Lichtpunkt
3.1: Schematische Darstellung des TeO von Salmo gairdneri in dorsaler Ansicht mit dem
deten Penetrationsraster. Die Farbmarkierungen im Raster geben die Charakteristika abgeleiteter
en an. Blau = ON oder OFF-Neurone, rot = richtungsselektive Neurone, grün = großes RF, reagiert
einzelnen kleinen Lichtpunkt, gelb = reagiert auf großen Lichtpunkt.
diesen experimentellen Bedingungen konnten im linken Praetectum insgesamt 184
l erregbare Zellen bei 24 Salmo gairdneri und 82 Neurone bei 8 Esox lucius
itet und analysiert werden.
achfolgende Tabelle 1 zeigt eine qualitative Klassifizierung charakteristischer
genschaften der abgeleiteten Neurone.
1: Klassifizierung abgeleiteter Neurone von Salmo gairdneri und Esox lucius nach ihren
eristischen Eigenschaften. n = Anzahl abgeleiteter Neurone, OFF = OFF-Zelle, ON = ON-Zelle, KL =
RF kleiner Lichtpunkt, GL = grosses RF grosser Lichtpunkt, RS = richtungsselektiv.
chstiere n diffus OFF ON KL GL RS
gairdneri 184 16 25 42 15 14 72
lucius 82 12 5 4 8 4 57
ezeptiven Felder der abgeleiteten praetectalen Neurone befanden sich an
chiedlichen Positionen des rechten Gesichtsfeldes. Ihr mittlerer rezeptiver
rchmesser betrug mindestens 35°. Aus diesem Grund konnten die rezeptiven Felder
17

I. Elektrophysiologie 3. Ergebnisse
der praetectalen Neurone eindeutig von denen der visuellen Neurone im TeO mit RF
Größen von 2° bis 15° unterschieden werden. Außerdem waren sie anhand der Ableittiefe
eindeutig von visuellen Neuronen des TeO, das eine Schichtdicke von ca. 530 µm aufwies,
zu trennen. Auch sind letztere eindeutig retinotop organisiert.
Als diffus visuell wurden Zellen charakterisiert, die auf Veränderung der Lichtintensität im
Raum reagierten, aber keine eindeutig bestimmbaren rezeptiven Feldgrenzen besaßen und
auch nicht auf Bewegung des Zufallspunktemusters oder auf Bewegung von kleinen und
großen Lichtpunkten aus der Handlampe mit Aktivitätsveränderung reagierten.
Mit einer OFF Eigenschaft konnten 25 Neurone bei Salmo gairdneri und 5 bei Esox lucius
abgeleitet werden. Diese Neurone zeigten ihre höchsten tonischen oder auch phasisch /
tonischen Entladungsraten bei völliger Abdunkelung des Raumes.
Die Anzahl von Neuronen mit einer ON Eigenschaft betrug bei Salmo gairdneri 42 und bei
Esox lucius 4. Ihre maximale tonische oder phasisch/tonische Reaktion erfolgte bei
Belichtung des RFs oder der Plexiglashalbkugel.
Visuelle Neurone mit einem RF, das annähernd dem gesamten rechten Gesichtsfeld
entsprach, und die nur auf einen kleinen (ca. 1° Sehwinkel) bewegten Lichtpunkt mit und
ohne Präsentation des Zufallspunktemuster reagierten, waren etwas seltener (Salmo
gairdneri: n = 14, Esox lucius: n = 8 ).
Außerdem konnten Zellen mit einem großen RF abgeleitet werden, die nur auf
einen großen Lichtpunkt (ca. 5° Sehwinkel) reagierten (Salmo gairdneri: n = 14,
Esox lucius: n = 4).
Die Anzahl dieser klassifizierten Neurone entspricht nicht der tatsächlichen
Ableithäufigkeit. Genauere Angaben sind nicht möglich, da im Verlauf der
elektrophysiologischen Untersuchungen solche Zellen nicht weiter konsequent untersucht
wurden. Die Begründung dafür ist, dass diese Neurone typischerweise im Laufe der
Versuche wiederholt bei verschiedenen Tieren an den gleichen Positionen abgeleitet
wurden und deshalb als Leitstrukturen zum Auffinden des praetectalen Areals mit
richtungsselektiven Neuronen (RS) dienten.
18
I. Elektrophysiologie 3. Ergebnisse
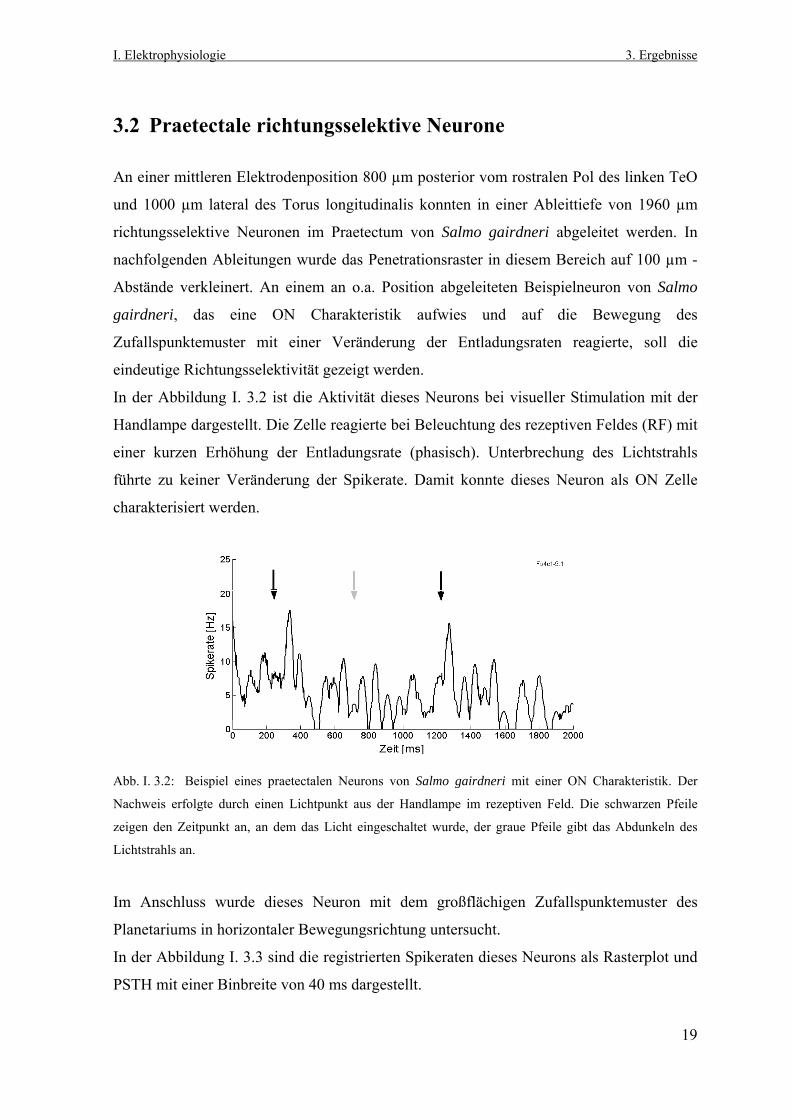
3.2 Praetectale richtungsselektive Neurone An einer mittleren Elektrodenposition 800 µm posterior vom rostralen Pol des linken TeO
und 1000 µm lateral des Torus longitudinalis konnten in einer Ableittiefe von 1960 µm
richtungsselektive Neuronen im Praetectum von Salmo gairdneri abgeleitet werden. In
nachfolgenden Ableitungen wurde das Penetrationsraster in diesem Bereich auf 100 µm -
Abstände verkleinert. An einem an o.a. Position abgeleiteten Beispielneuron von Salmo
gairdneri, das eine ON Charakteristik aufwies und auf die Bewegung des
Zufallspunktemuster mit einer Veränderung der Entladungsraten reagierte, soll die
eindeutige Richtungsselektivität gezeigt werden.
In der Abbildung I. 3.2 ist die Aktivität dieses Neurons bei visueller Stimulation mit der
Handlampe dargestellt. Die Zelle reagierte bei Beleuchtung des rezeptiven Feldes (RF) mit
einer kurzen Erhöhung der Entladungsrate (phasisch). Unterbrechung des Lichtstrahls
führte zu keiner Veränderung der Spikerate. Damit konnte dieses Neuron als ON Zelle
charakterisiert werden.
Abb. I. 3.2: B
Nachweis erf
zeigen den Ze
Lichtstrahls an
Im Anschlu
Planetarium
In der Abbi
PSTH mit e
eispiel eines praetectalen Neurons von Salmo gairdneri mit einer ON Charakteristik. Der
olgte durch einen Lichtpunkt aus der Handlampe im rezeptiven Feld. Die schwarzen Pfeile
itpunkt an, an dem das Licht eingeschaltet wurde, der graue Pfeile gibt das Abdunkeln des
.
ss wurde dieses Neuron mit dem großflächigen Zufallspunktemuster des
s in horizontaler Bewegungsrichtung untersucht.
ldung I. 3.3 sind die registrierten Spikeraten dieses Neurons als Rasterplot und
iner Binbreite von 40 ms dargestellt.
19
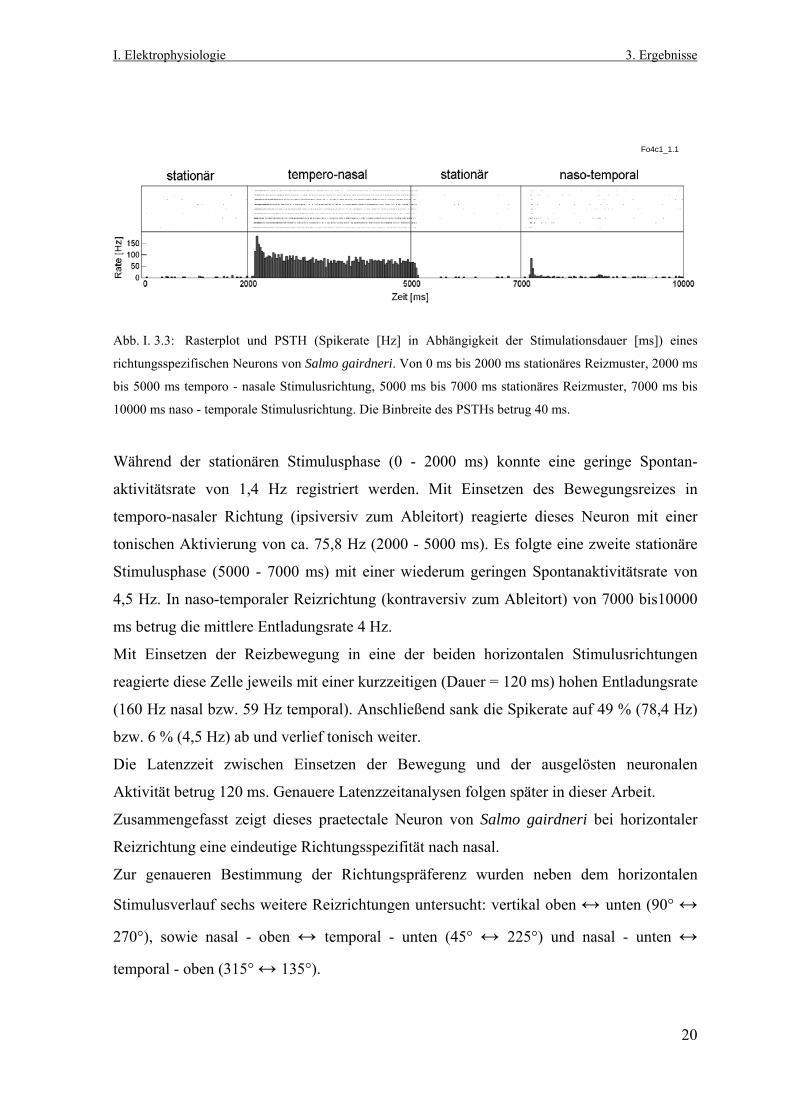
I. Elektrophysiologie 3. Ergebnisse
Fo4c1_1.1
Abb. I. 3.3: Rasterplot und PSTH (Spikerate [Hz] in Abhängigkeit der Stimulationsdauer [ms]) eines
richtungsspezifischen Neurons von Salmo gairdneri. Von 0 ms bis 2000 ms stationäres Reizmuster, 2000 ms
bis 5000 ms temporo - nasale Stimulusrichtung, 5000 ms bis 7000 ms stationäres Reizmuster, 7000 ms bis
10000 ms naso - temporale Stimulusrichtung. Die Binbreite des PSTHs betrug 40 ms.
Während der stationären Stimulusphase (0 - 2000 ms) konnte eine geringe Spontan-
aktivitätsrate von 1,4 Hz registriert werden. Mit Einsetzen des Bewegungsreizes in
temporo-nasaler Richtung (ipsiversiv zum Ableitort) reagierte dieses Neuron mit einer
tonischen Aktivierung von ca. 75,8 Hz (2000 - 5000 ms). Es folgte eine zweite stationäre
Stimulusphase (5000 - 7000 ms) mit einer wiederum geringen Spontanaktivitätsrate von
4,5 Hz. In naso-temporaler Reizrichtung (kontraversiv zum Ableitort) von 7000 bis10000
ms betrug die mittlere Entladungsrate 4 Hz.
Mit Einsetzen der Reizbewegung in eine der beiden horizontalen Stimulusrichtungen
reagierte diese Zelle jeweils mit einer kurzzeitigen (Dauer = 120 ms) hohen Entladungsrate
(160 Hz nasal bzw. 59 Hz temporal). Anschließend sank die Spikerate auf 49 % (78,4 Hz)
bzw. 6 % (4,5 Hz) ab und verlief tonisch weiter.
Die Latenzzeit zwischen Einsetzen der Bewegung und der ausgelösten neuronalen
Aktivität betrug 120 ms. Genauere Latenzzeitanalysen folgen später in dieser Arbeit.
Zusammengefasst zeigt dieses praetectale Neuron von Salmo gairdneri bei horizontaler
Reizrichtung eine eindeutige Richtungsspezifität nach nasal.
Zur genaueren Bestimmung der Richtungspräferenz wurden neben dem horizontalen
Stimulusverlauf sechs weitere Reizrichtungen untersucht: vertikal oben ↔ unten (90° ↔
270°), sowie nasal - oben ↔ temporal - unten (45° ↔ 225°) und nasal - unten ↔
temporal - oben (315° ↔ 135°).
20

I. Elektrophysiologie 3. Ergebnisse
Die neuronale Aktivität unter diesen Stimulationsbedingungen ist in der Abbildung I. 3.4
als Rasterplot und PSTH mit 40 ms Binweite dargestellt.
Abb. I. 3.4: Neuronale Aktivität eines richtungsselektiven Neurons von Salmo gairdneri während visueller
Stimulation mit dem Planetarium als Rasterplot und PSTH mit einer Binweite von 40 ms. Stimulation in den
acht getesteten Stimulusrichtungen mit jeweils zehn Trials pro Stimulationsrichtung und einer
Reizgeschwindigkeit von 10 °/s.
Fo4c1_1.1
Das Zentrum der Abbildung I. 3.4 repräsentiert die Lage des rechten, visuell stimulierten
Auges und die Position des Versuchstieres relativ zur Stimulusrichtung. Die acht
Rasterplots und PSTHs sind nach dem Stimuluszeitverlauf (2000 ms stationäres
Reizmuster, 3000 ms bewegtes Reizmuster) und den acht Stimulationsrichtungen
angeordnet.
Mit dieser Darstellung kann deutlich gezeigt werden, dass horizontale Bewegung des
Zufallspunktemusters nach nasal in einem Bereich von 45° bis 315° die höchsten
Spikeraten auslöst. In entgegengesetzter Reizrichtung (135° bis 225°) nach temporal sind
die Spikeraten signifikant (ANOVA: p < 0,05) niedriger. In den 2000 ms andauernden
stationären Stimulationsphasen sind sie am niedrigsten.
21
I. Elektrophysiologie 3. Ergebnisse
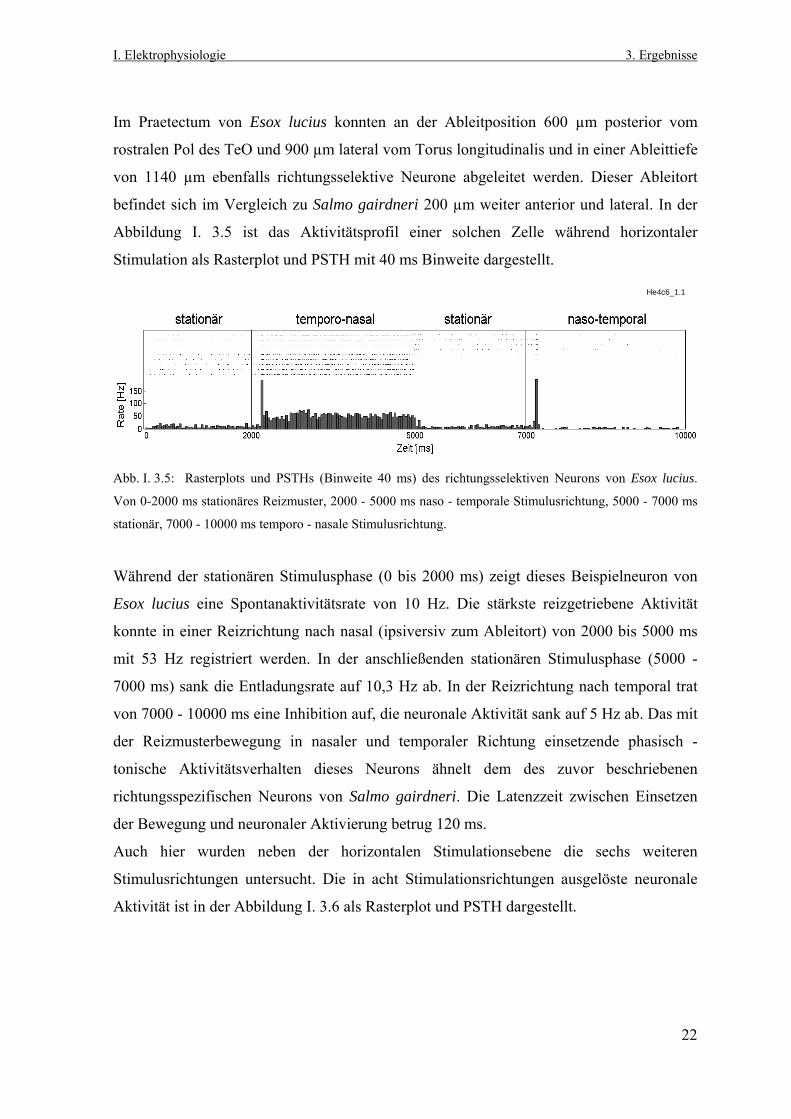
Im Praetectum von Esox lucius konnten an der Ableitposition 600 µm posterior vom
rostralen Pol des TeO und 900 µm lateral vom Torus longitudinalis und in einer Ableittiefe
von 1140 µm ebenfalls richtungsselektive Neurone abgeleitet werden. Dieser Ableitort
befindet sich im Vergleich zu Salmo gairdneri 200 µm weiter anterior und lateral. In der
Abbildung I. 3.5 ist das Aktivitätsprofil einer solchen Zelle während horizontaler
Stimulation als Rasterplot und PSTH mit 40 ms Binweite dargestellt.
He4c6_1.1
Abb. I. 3.5: Rasterplots und PSTHs (Binweite 40 ms) des richtungsselektiven Neurons von Esox lucius.
Von 0-2000 ms stationäres Reizmuster, 2000 - 5000 ms naso - temporale Stimulusrichtung, 5000 - 7000 ms
stationär, 7000 - 10000 ms temporo - nasale Stimulusrichtung.
Während der stationären Stimulusphase (0 bis 2000 ms) zeigt dieses Beispielneuron von
Esox lucius eine Spontanaktivitätsrate von 10 Hz. Die stärkste reizgetriebene Aktivität
konnte in einer Reizrichtung nach nasal (ipsiversiv zum Ableitort) von 2000 bis 5000 ms
mit 53 Hz registriert werden. In der anschließenden stationären Stimulusphase (5000 -
7000 ms) sank die Entladungsrate auf 10,3 Hz ab. In der Reizrichtung nach temporal trat
von 7000 - 10000 ms eine Inhibition auf, die neuronale Aktivität sank auf 5 Hz ab. Das mit
der Reizmusterbewegung in nasaler und temporaler Richtung einsetzende phasisch -
tonische Aktivitätsverhalten dieses Neurons ähnelt dem des zuvor beschriebenen
richtungsspezifischen Neurons von Salmo gairdneri. Die Latenzzeit zwischen Einsetzen
der Bewegung und neuronaler Aktivierung betrug 120 ms.
Auch hier wurden neben der horizontalen Stimulationsebene die sechs weiteren
Stimulusrichtungen untersucht. Die in acht Stimulationsrichtungen ausgelöste neuronale
Aktivität ist in der Abbildung I. 3.6 als Rasterplot und PSTH dargestellt.
22

I. Elektrophysiologie 3. Ergebnisse
He4c6_1.1
Abb. I. 3.6: Neuronale Aktivität einer richtungsselektiven Zelle von Esox lucius mit Rasterplot und PSTH
(Binweite 40 ms) bei acht getesteten Stimulusrichtungen mit jeweils zehn Trials und einer
Reizgeschwindigkeit von 10 °/s. Die vertikalen Linien zu den Zeitpunkten 0 ms, 2000 ms, 5000 ms und 7000
ms geben den Zeitpunkt der Stimulationsänderung an.
Die Darstellung der neuronalen Aktivität in der Abbildung I. 3.6 zeigt, dass diese
praetectale Zelle von Esox lucius eine horizontale, nach nasal verlaufende
Stimulationsrichtung (ipsiversiv zum Ableitort) in einem Bereich von 45° bis 270° mit
einer hohen Spikerate (75Hz) deutlich bevorzugt. In entgegengesetzter Stimulusrichtung,
der Nullrichtung, kann eine Inhibition der neuronalen Aktivität beobachtet werden. Dieses
Funktionsprinzip wird als push-pull bezeichnet. Das phasisch-/-tonische Antwortverhalten
mit Einsetzen und Fortführung der Bewegungsphase ist in allen Reizrichtungen gegeben.
Mit diesen zwei Beispielneuronen kann gezeigt werden, dass im Praetectum beider
Fischarten visuelle, richtungsselektive Zellen existieren, die einen horizontalen nach nasal
verlaufenden visuellen Stimulus (ipsiversiv zum Ableitort) bevorzugen.
Bei beiden Fischarten tritt ein phasisches „movement onset“ Verhalten der
richtungsselektiven Neuronen mit Beginn der Stimulusbewegung auf. Unterschiede zeigen
sich in der Antwortcharakteristik. Die richtungsselektiven Neurone von Salmo gairdneri
zeigen stets geringe bis gar keine Spontanaktivität und reagieren mit Aktivitätserhöhung
23

I. Elektrophysiologie 3. Ergebnisse
im Bereich ihrer Vorzugsrichtung. Die Neurone von Esox lucius sind immer spontanaktiv,
reagieren ebenfalls mit Aktivitätserhöhung auf Reizung in ihrer Vorzugsrichtung, aber mit
Aktivitätserniedrigung auf Reizung in der Nullrichtung (push - pull Prinzip).
Zusammengefasst deuten diese bisherigen Befunde bei beiden Fischarten auf ein
praetectales Areal hin, dass möglicherweise dem NOT und dem nLM der anderen
Vertebratenklassen entspricht.
3.3 Untersuchungen zum Vorkommen weiterer richtungs-
selektiver Neurone In weiteren Experimenten wurde in einem Bereich von ca. 300 µm anterior / posterior und
200 µm medial / lateral relativ zu den ersten Penetrationsorten (siehe I. 3.1) in größeren
Ableittiefen (ca. + 400 µm) Neurone lokalisiert, deren Vorzugsrichtung nicht
ausschließlich ipsiversiv zum Ableitort war.
Zur genauen Bestimmung der Richtungspräferenz wurden Richtungsempfindlichkeits-
kurven (Tuning - Kurven) und die jeweilige Vorzugsrichtung über trigonometrische
Funktionen als Richtungsvektors berechnet.
Der Mittelpunkt der nachfolgend dargestellten Polarplots repräsentiert die Lage des
rechten, visuell stimulierten Auges. Eine Stimulusrichtung nach nasal entspricht 0° in
Polarkoordinaten, eine temporale Reizrichtung 180°, eine vertikale Bewegung nach oben
90°, nach unten 270°, sowie die Bewegungsrichtungen nasal - oben 45°, temporal - unten
225° und temporal - oben 135°, nasal - unten 315°.
Die Tuning-Kurven der richtungsselektiven Neurone ergaben sich aus der Berechnung der
mittleren Spikerate während des 3000 ms andauernden Bewegungsreizes über zehn
Einzeltrials in jeder der acht untersuchten Stimulusrichtungen. Die Radien der Polarplots
geben die mittlere Spikerate pro Sekunde an. Die Position des Richtungsvektors, der die
Vorzugsrichtung des analysierten Neurons anzeigt, wurde mit trigonometrischen
Funktionen berechnet und auf das Radienmaximum normiert. Er repräsentiert nicht die
berechnete Entladungsrate in der Vorzugsrichtung.
Abbildung I. 3.7 A zeigt die Tuning-Kurve mit der Vorzugsrichtung von 353° des oben
beschriebenen, die nasale Reizrichtung (ipsiversiv zum Ableitort) kodierenden Neurons
24

I. Elektrophysiologie 3. Ergebnisse
von Salmo gairdneri (Abb. I. 3.4). Die Abb. I. 3.7 B zeigt ein weiteres richtungsselektives
Neuron mit einer temporalen Vorzugsrichtung (kontraversiv zum Ableitort) von 189°.
BA
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
225°
270°
315°
180°0 20 40 60 80
0
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
temporal
temporal-oben
temporal-unten nasal-unten
nasal
nasal-oben
unten
oben
Abb. I. 3.7: Tuning - Kurve und Richtungsvektor zweier richtungsselektiver Neurone von Salmo gairdneri
in Polarkoordinaten. In A das zuvor beschriebene Neuron (vgl. Abb. I. 3.4), in B ein weiteres Neuron, das
eine horizontale kontraversive Stimulusrichtung kodiert.
Neben Neuronen, die eine ipsiversive (nach nasal) Stimulusrichtung kodieren, konnten
richtungsselektive Neurone mit einer genau entgegengesetzten, also kontraversiven
Vorzugsrichtung (nach temporal) abgeleitet werden.
Das bedeutet, dass in einem räumlich begrenzten Ableitbereich Zellen mit
unterschiedlichen Vorzugsrichtungen vorkommen.
Die Abbildung I. 3.8 zeigt exemplarisch die Tuning - Kurven und Vorzugsrichtungen
weiterer richtungsselektiver Neurone, die spezifisch für eine der sechs weiteren
Stimulusrichtungen sind.
25
I. Elektrophysiologie 3. Ergebnisse
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
B
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
270°
315°
E F
C D
temporal-oben
temporal
nasal-unten
nasal
nasal-oben
unten
oben A
225°temporal-unten
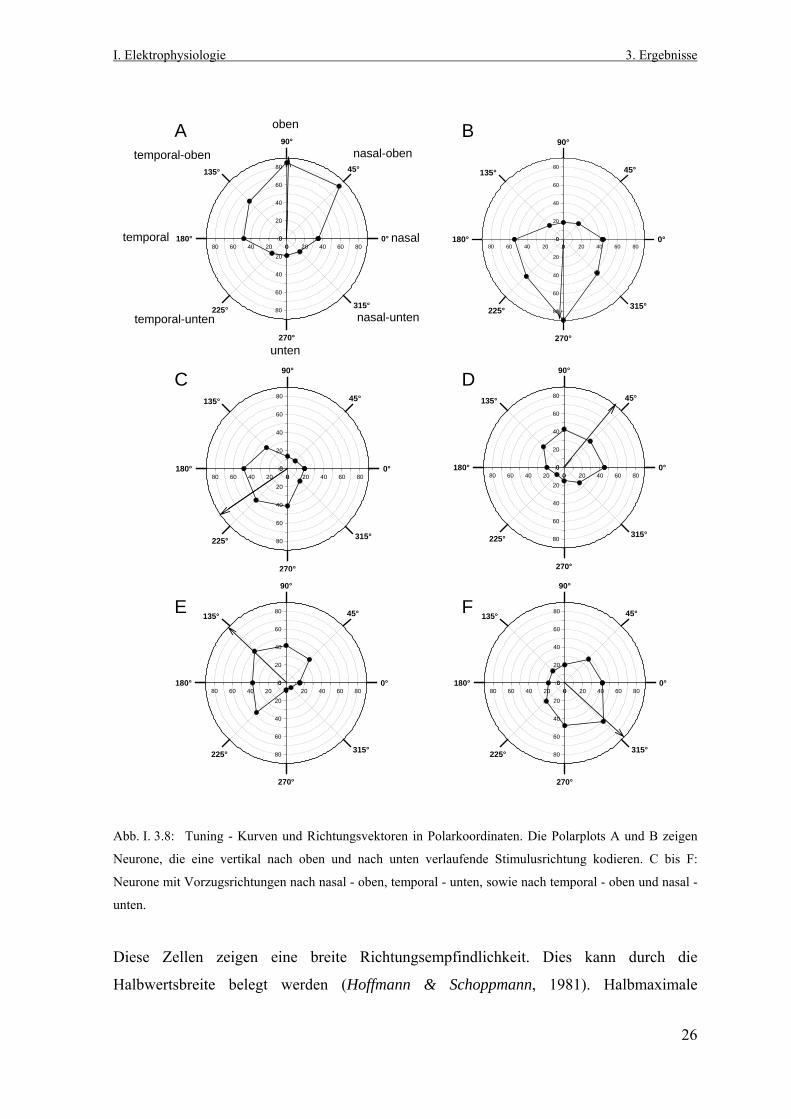
Abb. I. 3.8: Tuning - Kurven und Richtungsvektoren in Polarkoordinaten. Die Polarplots A und B zeigen
Neurone, die eine vertikal nach oben und nach unten verlaufende Stimulusrichtung kodieren. C bis F:
Neurone mit Vorzugsrichtungen nach nasal - oben, temporal - unten, sowie nach temporal - oben und nasal -
unten.
Diese Zellen zeigen eine breite Richtungsempfindlichkeit. Dies kann durch die
Halbwertsbreite belegt werden (Hoffmann & Schoppmann, 1981). Halbmaximale
26

I. Elektrophysiologie 3. Ergebnisse
Entladungsraten werden noch bei Winkelrichtungen, die +/- 45° von der jeweiligen
Vorzugsrichtung abweichen, erreicht.
Im Praetectum von Esox lucius wurden ebenfalls Neurone mit beiden horizontalen
Vorzugsrichtungen abgeleitet, die in der Abbildung I. 3.9 A und B in Polarkoordinaten mit
ihrer Richtungsempfindlichkeitskurve und Richtungsvektor dargestellt sind.
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
A
temporal
temporal-unten nasal-unten
nasal
unten
Bnasal-oben
oben
temporal-oben
Abb. I. 3.9: Tuning - Kurve und Richtungsvektor von richtungsselektiven Neuronen aus dem Praetectum
von Esox lucius. A: ipsiversiv, B: kontraversiv kodierendes Neuron.
Wie bei Salmo gairdneri konnten auch bei Esox lucius in dem untersuchten Areal Neurone
mit vertikalen und schrägen Vorzugsrichtungen abgeleitet werden. Abbildung I. 3.10 zeigt
die Richtungsempfindlichkeitskurven und Richtungsvektoren solcher Neurone.
27

I. Elektrophysiologie 3. Ergebnisse
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
oben
unten
nasal-oben
nasal
nasal-untentemporal-unten
temporal
temporal-oben
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
F
D
E
C
A B
Abb. I. 3.10: Tuning - Kurven und Richtungsvektoren in Polarkoordinaten. Die Polarplots A und B zeigen
Neurone, die eine vertikal nach oben und nach unten verlaufende Stimulusrichtung kodieren. C bis F:
Neurone mit Vorzugsrichtungen nach nasal - oben, temporal - unten, sowie nach temporal - oben und nasal -
unten.
28

I. Elektrophysiologie 3. Ergebnisse
Die Vorzugsrichtungen entsprechen auch bei Esox lucius den untersuchten acht
Stimulationsrichtungen. Das Tuning der Richtungsempfindlichkeit gleicht dem von Salmo
gairdneri, d.h. halbmaximale Entladungsraten werden noch in einem Winkelbereich +/-
45° um die Vorzugsrichtung erreicht. Auch hier zeigt sich die Inhibition dieser Neurone in
der Nullrichtung, womit das „push - pull“ Prinzip auch für die anderen Richtungen
bestätigt.
Bei beiden untersuchten Spezies liegen in einem begrenzten Areal des Praetectums
richtungsselektive Neurone, die spezifisch auf eine von den acht untersuchten
Stimulusrichtungen reagieren. Diese Zellen weisen ein breites Richtungstuning auf.
3.4 Verteilung der Vorzugsrichtungen aller richtungsselektiven
Neurone Insgesamt wurden 72 richtungsspezifische Neurone bei Salmo gairdneri und 63
richtungsspezifische Neuronen bei Esox lucius abgeleitet. Dabei fiel auf, dass die
Vorzugsrichtungen dieser Zellen sich nicht nur auf eine der acht untersuchten
Stimulusrichtungen beschränkten, sondern unterschiedlich verteilt waren.
Um zu testen, ob alle richtungsselektiven Neurone ausschließlich vom rechten Auge
aktiviert wurden, wurde bei jeder Einzelzelle auch das linke Auge durch eine Veränderung
der Raumhelligkeit und eine direkte Beleuchtung mit der Handlampe gereizt. Eine
Veränderung der Luminanz auf der Retina hätte eine veränderte Entladungsrate des
abgeleiteten praetektalen Neurons bewirken müssen. Keine der untersuchten
richtungsspezifischen Zellen konnte vom linken Auge (ipsilateral) beeinflußt werden. Dies
deutet auf eine totale Überkreuzung des Nervus opticus am Chiasma opticum und keine
Verbindung zwischen den Hirnhälften bei diesen Neuronen hin.
Aufgrund der Vorzugsrichtungsverteilung und des breiten Tunings (Halbwertsbreite > 90°)
wurde mit zirkulärer Statistik die Richtungsselektivität eindeutig bestimmt.
Von den 72 Neuronen der Regenbogenforelle konnten 63 Neurone mit einer statistisch
signifikanten Richtungsselektivität (p < 0,001) ermittelt werden. Von ursprünglich 57
Neuronen des Hechtes sind nach dem zirkulären statistischen Verfahren 45 als signifikant
29

I. Elektrophysiologie 3. Ergebnisse
richtungsselektiv einzuordnen. Ein Vergleich der Vorzugsrichtungen aus den
trigonometrischen Berechnungen mit denen aus der zirkulären Statistik ergab keinen
Unterschied für die eindeutig richtungsselektiven Neurone.
In der Abbildung I. 3.11 sind in Polarkoordinaten die Vorzugsrichtungen der signifikant
richtungsspezifischen Neurone von Salmo gairdneri und Esox lucius dargestellt.
0°
45°
90°
135°
180°
225°
270°
315°
0°
45°
90°
135°
0°
225°
270°
315°
unten
oben
18
temporal-oben
temporal
temporal-unten nasal-unten
nasal
nasal-oben
Esox lucius Salmo gairdneri
Abb. I. 3.11: Verteilung der Vorzugsrichtungen signifikant richtungsspezifischer praetectaler Zellen
(p < 0,001) von Salmo gairdneri (n = 63) und Esox lucius (n = 45) in Polarkoordinaten.
Ob die Vorzugsrichtungen der 63 Neurone von Salmo gairdneri und der 45 Neurone von
Esox lucius im Polarkoordinatensystem gleich verteilt sind, soll im Weiteren untersucht
werden.
Die Lage der acht gewählten Stimulusrichtungen im Polarkoordinatensystem erlaubt eine
Einteilung in jeweils zwei horizontale und vertikale Winkelbereiche: in nasaler Richtung
45° bis 315°, in temporaler Richtung 135° bis 225°, nach oben 45° bis 135° und nach
unten 225° bis 315°.
Die Legitimation zu dieser Einteilung liegt in dem breiten Tuning dieser Neurone und
darin, dass im NOT und nLM der anderen Vertebratenklassen durchaus Neurone
vorkommen, die eine Tuningbreite in diesem Winkelbereich besitzen. Demnach ergeben
sich in der Summe für Salmo gairdneri 39 Neurone, die eine horizontale Stimulusrichtung
30
I. Elektrophysiologie 3. Ergebnisse
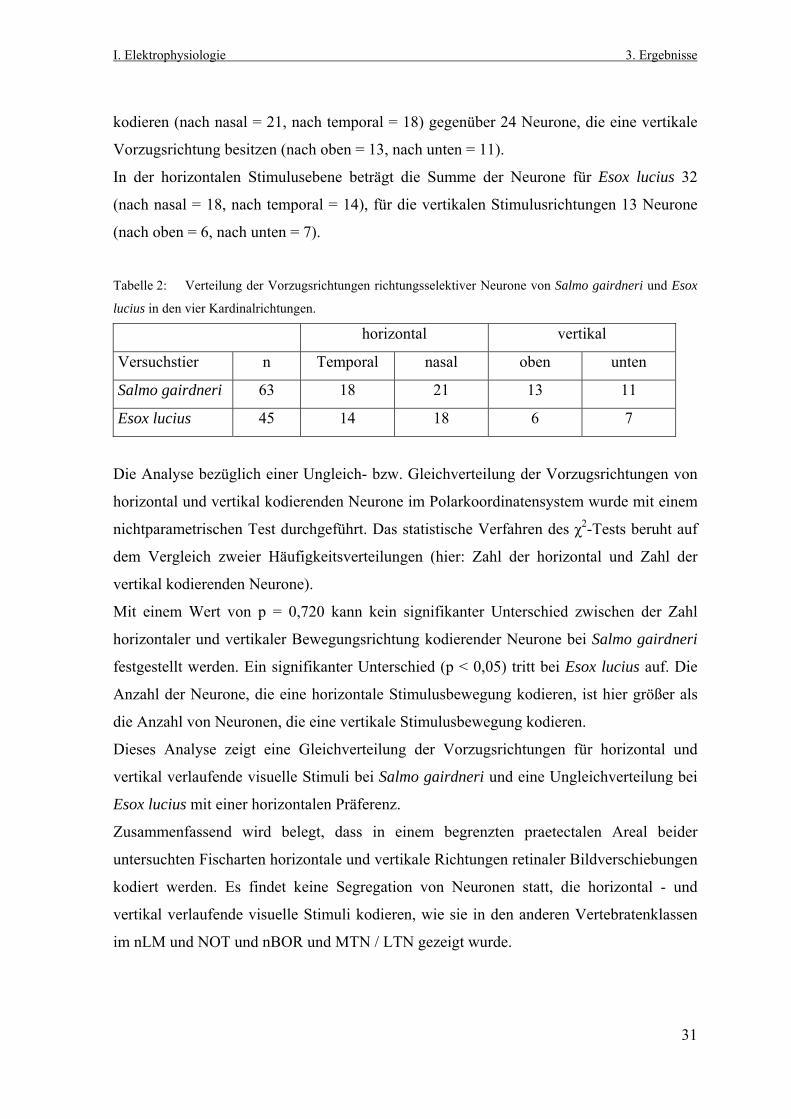
kodieren (nach nasal = 21, nach temporal = 18) gegenüber 24 Neurone, die eine vertikale
Vorzugsrichtung besitzen (nach oben = 13, nach unten = 11).
In der horizontalen Stimulusebene beträgt die Summe der Neurone für Esox lucius 32
(nach nasal = 18, nach temporal = 14), für die vertikalen Stimulusrichtungen 13 Neurone
(nach oben = 6, nach unten = 7).
Tabelle 2: Verteilung der Vorzugsrichtungen richtungsselektiver Neurone von Salmo gairdneri und Esox
lucius in den vier Kardinalrichtungen.
horizontal vertikal
Versuchstier n Temporal nasal oben unten
Salmo gairdneri 63 18 21 13 11
Esox lucius 45 14 18 6 7
Die Analyse bezüglich einer Ungleich- bzw. Gleichverteilung der Vorzugsrichtungen von
horizontal und vertikal kodierenden Neurone im Polarkoordinatensystem wurde mit einem
nichtparametrischen Test durchgeführt. Das statistische Verfahren des χ2-Tests beruht auf
dem Vergleich zweier Häufigkeitsverteilungen (hier: Zahl der horizontal und Zahl der
vertikal kodierenden Neurone).
Mit einem Wert von p = 0,720 kann kein signifikanter Unterschied zwischen der Zahl
horizontaler und vertikaler Bewegungsrichtung kodierender Neurone bei Salmo gairdneri
festgestellt werden. Ein signifikanter Unterschied (p < 0,05) tritt bei Esox lucius auf. Die
Anzahl der Neurone, die eine horizontale Stimulusbewegung kodieren, ist hier größer als
die Anzahl von Neuronen, die eine vertikale Stimulusbewegung kodieren.
Dieses Analyse zeigt eine Gleichverteilung der Vorzugsrichtungen für horizontal und
vertikal verlaufende visuelle Stimuli bei Salmo gairdneri und eine Ungleichverteilung bei
Esox lucius mit einer horizontalen Präferenz.
Zusammenfassend wird belegt, dass in einem begrenzten praetectalen Areal beider
untersuchten Fischarten horizontale und vertikale Richtungen retinaler Bildverschiebungen
kodiert werden. Es findet keine Segregation von Neuronen statt, die horizontal - und
vertikal verlaufende visuelle Stimuli kodieren, wie sie in den anderen Vertebratenklassen
im nLM und NOT und nBOR und MTN / LTN gezeigt wurde.
31

I. Elektrophysiologie 3. Ergebnisse
3.5 Entscheidet die Anzahl richtungsselektiver Neurone über
eine Symmetrie bzw. Asymmetrie des monokularen
horizontalen optokinetischen Reflexes? Mit den bisher gewonnenen Erkenntnissen drängt sich die Frage auf, ob es möglich ist, aus
der Richtungspräferenz der praetectalen richtungsspezifischen Neuronenpopulation beider
Fischarten Rückschlüsse auf eine Asymmetrie des monokularen horizontalen
optokinetischen Reflexes (mhOKR) zu ziehen.
Zur Klärung dieser Fragestellung wurden nur die in der horizontalen Stimulusebene
kodierenden Neuronen beider Fischarten in die Analyse einbezogen. In die Auswertung für
Salmo gairdneri gingen insgesamt 39 Zellen ein, wovon 21 eine nasale und 18 Neuronen
eine temporale Reizrichtung bevorzugten.
Für Esox lucius waren es insgesamt 32 Neuronen, 18 Neuronen kodierten eine nasale
Stimulusrichtung und 14 Neuronen eine temporale Reizrichtung.
In der Abbildung I. 3.12 ist die Verteilung der Neurone von Salmo gairdneri und Esox
lucius in Polarkoordinaten dargestellt.
airdneriSalmo g
0°
45°
90°
135°
180°
225°
270°
315°
0°
45°
90°
135°
180°
225°
270°
315°
temporal-oben
temporal
temporal-unten nasal-unten
nasal
nasal-oben
unten
oben
Esox lucius
Abb. I. 3.12: Verteilung der horizontalen nasalen und temporalen Vorzugsrichtungen praetectaler Zellen von
Salmo gairdneri (n = 39) und Esox lucius (n = 32) in Polarkoordinaten.
32

I. Elektrophysiologie 3. Ergebnisse
Der Test auf eine Gleich- bzw. Ungleichverteilung der Neurone auf die beiden
horizontalen Stimulusrichtungen wurde mit dem χ2-Test durchgeführt.
Das Ergebnis zeigte weder für Salmo gairdneri (p = 0,172) noch für Esox lucius
(p = 0,389) einen signifikanten Unterschied in der Anzahl von Neuronen, die einen in
nasaler oder temporaler Reizrichtung verlaufenen Stimulus kodieren. Damit ist eine
Asymmetrie des OKR aus der Verteilung der Vorzugsrichtung der Neurone nicht
abzuleiten.
3.6 Populationsantwort Eine weitere Analysemöglichkeit zur Charakterisierung des richtungsselektiven Areals in
bezug auf den mhOKR bietet die Untersuchung der Aktivität der gesamten
Neuronenpopulation. Dazu wurde durch Summation der Tuning - Kurven aller signifikant
richtungsselektiven Neuronen die Populationsantwort berechnet. Die Ergebnisse sind für
beide untersuchten Fischarten in der Abbildung I. 3.13 dargestellt.
n = 63
20 40 60 80
20
40
60
80
20406080
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
n = 45
20 40 60 80
20
40
60
80
20406080
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
temporal-oben
temporal
temporal-unten nasal-unten
nasal
nasal-oben
unten
oben
Salmo gairdneri Esox lucius
Abb. I. 3.13: Richtungscharakteristik der Neuronenpopulation von Salmo gairdneri (n = 63) und Esox lucius
(n= 45). Die Radien geben die mittlere Spikerate der Neuronenpopulation an.
33

I. Elektrophysiologie 3. Ergebnisse
Die Richtungscharakteristik der Population zeigt keine Präferenz für eine der vier
Kardinalrichtungen (horizontal: nasal-temporal und vertikal: oben-unten) bei Salmo
gairdneri und bei Esox lucius.
Das deutet auf eine Gleichverteilung der Vorzugsrichtungen im visuellen Raum hin. Eine
Asymmetrie des mhOKRs lässt sich dadurch nicht erklären.
3.7 Weitere Charakterisierung praetectaler richtungsselektiver
Neurone von Salmo gairdneri und Esox lucius Anhand der bisher gewonnenen Ergebnisse kann die Qualität des mhOKR (Klar 1999)
nicht erklärt werden. Aus diesem Grund wurden die signifikant richtungsselektiven
Neuronen von Salmo gairdneri und Esox lucius weiter analysiert. Diese Ergebnisse sollen
zusätzlichen Aufschluss über die Rolle der richtungsselektiven Neurone im
optokinetischen System geben.
3.7.1 Neuronale Aktivität in Abhängigkeit von der Reizgeschwindigkeit Das optokinetische System ist so konzipiert, dass es einer retinalen Bildverschiebung
entgegenwirkt. Dazu muss diesem visuomotorischen System die Richtung, wie auch die
Reizgeschwindigkeit bekannt sein, damit die Augenmuskeln die Augen entsprechend
auslenken können. Die Bestimmung der Bildverschiebungsrichtung kann von den
praetectalen richtungsselektiven Neuronen beider Fischarten geleistet werden. Es stellt sich
nun die Frage, wie diese Zellen auf unterschiedliche Geschwindigkeiten des präsentierten
Zufallspunktemuster reagieren.
Während des elektrophysiologischen Experiments konnte akustisch und mit dem
Aufzeichnungsprogramm CORTEX, das eine Online- Darstellung der Spikeraten
ermöglichte, eine der acht untersuchten Stimulusrichtungen als Vorzugsrichtung sofort
bestimmt werden. In dieser Richtung wurden Geschwindigkeiten des Zufallspunktemusters
von 0 °/s bis 80 °/s in jeweils 10 °/s Schritten bei Salmo gairdneri untersucht. Eine
Reizgeschwindigkeit von 5 °/s wurde bei Esox lucius zusätzlich gemessen, da es sich
34
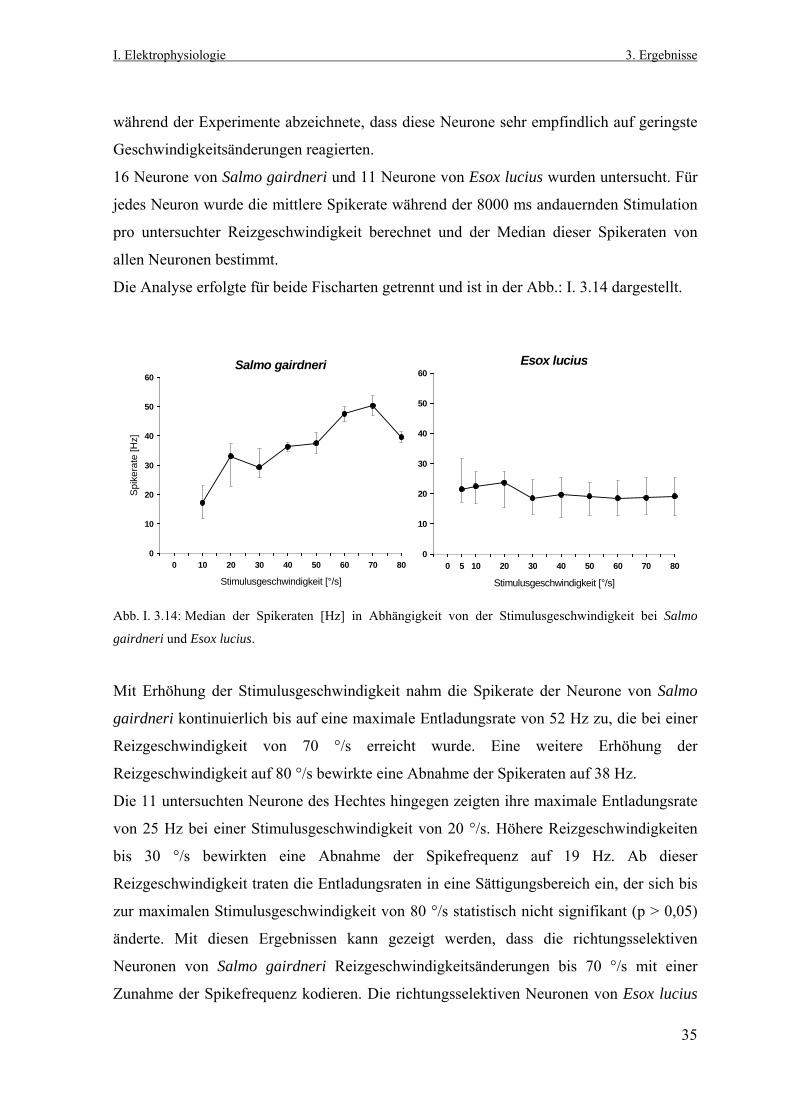
I. Elektrophysiologie 3. Ergebnisse
während der Experimente abzeichnete, dass diese Neurone sehr empfindlich auf geringste
Geschwindigkeitsänderungen reagierten.
16 Neurone von Salmo gairdneri und 11 Neurone von Esox lucius wurden untersucht. Für
jedes Neuron wurde die mittlere Spikerate während der 8000 ms andauernden Stimulation
pro untersuchter Reizgeschwindigkeit berechnet und der Median dieser Spikeraten von
allen Neuronen bestimmt.
Die Analyse erfolgte für beide Fischarten getrennt und ist in der Abb.: I. 3.14 dargestellt.
Esox lucius
Stimulusgeschwindigkeit [°/s]
0 5 10 20 30 40 50 60 70 80
Spi
kera
te [H
z]
0
10
20
30
40
50
60Salmo gairdneri
Stimulusgeschwindigkeit [°/s]
0 10 20 30 40 50 60 70 80
Spi
kera
te [H
z]
0
10
20
30
40
50
60
Abb. I. 3.14: Median der Spikeraten [Hz] in Abhängigkeit von der Stimulusgeschwindigkeit bei Salmo
gairdneri und Esox lucius.
Mit Erhöhung der Stimulusgeschwindigkeit nahm die Spikerate der Neurone von Salmo
gairdneri kontinuierlich bis auf eine maximale Entladungsrate von 52 Hz zu, die bei einer
Reizgeschwindigkeit von 70 °/s erreicht wurde. Eine weitere Erhöhung der
Reizgeschwindigkeit auf 80 °/s bewirkte eine Abnahme der Spikeraten auf 38 Hz.
Die 11 untersuchten Neurone des Hechtes hingegen zeigten ihre maximale Entladungsrate
von 25 Hz bei einer Stimulusgeschwindigkeit von 20 °/s. Höhere Reizgeschwindigkeiten
bis 30 °/s bewirkten eine Abnahme der Spikefrequenz auf 19 Hz. Ab dieser
Reizgeschwindigkeit traten die Entladungsraten in eine Sättigungsbereich ein, der sich bis
zur maximalen Stimulusgeschwindigkeit von 80 °/s statistisch nicht signifikant (p > 0,05)
änderte. Mit diesen Ergebnissen kann gezeigt werden, dass die richtungsselektiven
Neuronen von Salmo gairdneri Reizgeschwindigkeitsänderungen bis 70 °/s mit einer
Zunahme der Spikefrequenz kodieren. Die richtungsselektiven Neuronen von Esox lucius
35
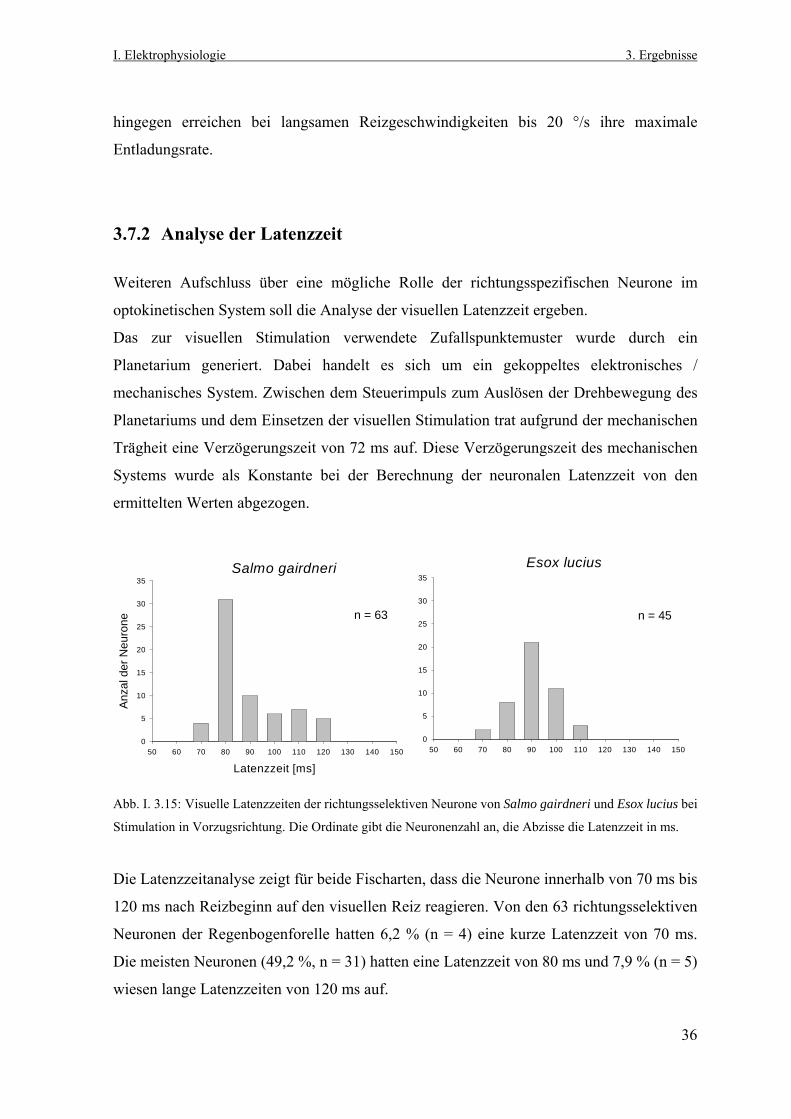
I. Elektrophysiologie 3. Ergebnisse
hingegen erreichen bei langsamen Reizgeschwindigkeiten bis 20 °/s ihre maximale
Entladungsrate.
3.7.2 Analyse der Latenzzeit Weiteren Aufschluss über eine mögliche Rolle der richtungsspezifischen Neurone im
optokinetischen System soll die Analyse der visuellen Latenzzeit ergeben.
Das zur visuellen Stimulation verwendete Zufallspunktemuster wurde durch ein
Planetarium generiert. Dabei handelt es sich um ein gekoppeltes elektronisches /
mechanisches System. Zwischen dem Steuerimpuls zum Auslösen der Drehbewegung des
Planetariums und dem Einsetzen der visuellen Stimulation trat aufgrund der mechanischen
Trägheit eine Verzögerungszeit von 72 ms auf. Diese Verzögerungszeit des mechanischen
Systems wurde als Konstante bei der Berechnung der neuronalen Latenzzeit von den
ermittelten Werten abgezogen.
Abb. I. 3.15: Visuelle Latenzzeiten der richtungsselektiven Neurone von Salmo gairdneri und Esox lucius bei
Stimulation in Vorzugsrichtung. Die Ordinate gibt die Neuronenzahl an, die Abzisse die Latenzzeit in ms.
Esox lucius
50 60 70 80 90 100 110 120 130 140 150
Anza
l der
Neu
rone
0
5
10
15
20
25
30
35Salmo gairdneri
Latenzzeit [ms]50 60 70 80 90 100 110 120 130 140 150
Anz
al d
er
0
5
10
15
20
25
30
35
n = 63 n = 45
Neu
rone
Die Latenzzeitanalyse zeigt für beide Fischarten, dass die Neurone innerhalb von 70 ms bis
120 ms nach Reizbeginn auf den visuellen Reiz reagieren. Von den 63 richtungsselektiven
Neuronen der Regenbogenforelle hatten 6,2 % (n = 4) eine kurze Latenzzeit von 70 ms.
Die meisten Neuronen (49,2 %, n = 31) hatten eine Latenzzeit von 80 ms und 7,9 % (n = 5)
wiesen lange Latenzzeiten von 120 ms auf.
36

I. Elektrophysiologie 3. Ergebnisse
Eine kurze Latenzzeit hatten 4,4 % (n = 2) von insgesamt 45 untersuchten Neuronen des
Hechtes. Die meisten Neurone (46,7 %, n = 21) hatten eine Latenzzeit von 90 ms. Die
längsten Latenzzeiten von 110 ms konnten an 6,7 % (n = 3) bestimmt werden.
Der Median der Latenzzeit beträgt für Salmo gairdneri 80 ms und für Esox lucius 90 ms.
3.7.3 Größe und Verteilung der rezeptiven Felder richtungsselektiver
Neurone
Ein weiteres Kriterium zur Charakterisierung der richtungsselektiven Zellen ist die Größe
der rezeptiven Felder und ihre Lage im Gesichtsfeld.
Klassenunabhängig konnten den richtungsselektiven Neurone des NOT und des nLM stets
großflächige, meist im Bereich des zentralen Gesichtsfelds gelegene rezeptive Felder
zugeordnet werden.
In der vorliegenden Arbeit wurden die Grenzen der rezeptiven Felder mit einem
Lichtpunkt aus der Handlampe bestimmt.
Alle ermittelten rezeptiven Felder der eindeutig richtungsselektiven Neurone hatten eine
Mindestausdehnung im Azimut von 30° und von 25° in der Elevation. Die maximalen
Feldausdehnungen betrugen 135° im Azimut und 90° in der Elevation.
In der Abbildung I. 3.16 sind für die Forelle und in der Abb. I. 3.17 für den Hecht
exemplarisch aus jeweils einer Penetration die ermittelten rezeptiven Felder abgeleiteter
Neurone gezeigt.
37
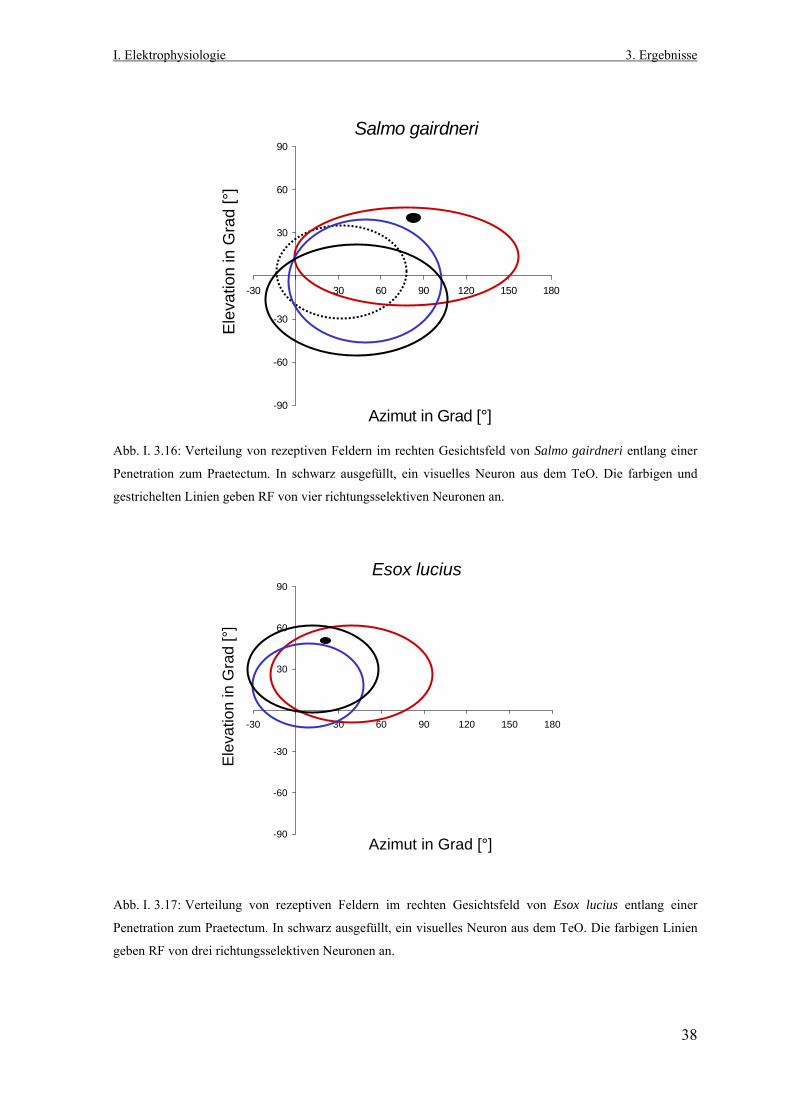
I. Elektrophysiologie 3. Ergebnisse
Salmo gairdneri
Azimut in Grad [°]
-30 30 60 90 120 150 180
Ele
vatio
n in
Gra
d [°
]
-90
-60
-30
30
60
90
Abb. I. 3.16: Verteilung von rezeptiven Feldern im rechten Gesichtsfeld von Salmo gairdneri entlang einer
Penetration zum Praetectum. In schwarz ausgefüllt, ein visuelles Neuron aus dem TeO. Die farbigen und
gestrichelten Linien geben RF von vier richtungsselektiven Neuronen an.
Esox lucius
Azimut in Grad [°]
-30 30 60 90 120 150 180
Ele
vatio
n in
Gra
d [°
]
-90
-60
-30
30
60
90
Abb. I. 3.17: Verteilung von rezeptiven Feldern im rechten Gesichtsfeld von Esox lucius entlang einer
Penetration zum Praetectum. In schwarz ausgefüllt, ein visuelles Neuron aus dem TeO. Die farbigen Linien
geben RF von drei richtungsselektiven Neuronen an.
38

I. Elektrophysiologie 3. Ergebnisse
Die RF der richtungsselektiven Neurone waren bei beiden Fischarten deutlich größer als
die RF der visuellen Neurone aus dem TeO (schwarz ausgefüllt).
Die gezeigten tektalen rezeptiven Felder geben exakt den Penetrationsort an, bei dem die
Ableitung im Praetectum zur Lokalisation richtungsselektiver Neuronen führte:
Salmo gairdneri = 87° Azimut und 42° Elevation.
Esox lucius = 28° Azimut und 48° Elevation.
Die Mittelpunkte der RF praetectaler richtungsselektiver Neurone befinden sich zu 82 % in
einem Bereich des frontalen Gesichtsfeldes zwischen -20° und 45° Azimut bei Salmo
gairdneri und zu 72,3 % zwischen 12° und 54° Azimut bei Esox lucius.
3.8 Rekonstruktion des Ableitortes richtungsselektiver Neurone
im Praetectum von Salmo gairdneri und Esox lucius
3.8.1 Histologische Lokalisation der praetectalen richtungsselektiven
Neurone Die bisherige Charakterisierung der richtungsspezifischen Neurone lässt durchaus die
Vermutung zu, dass diese an der Generierung des OKR beteiligt sind. Nachfolgend soll mit
der histologischen Rekonstruktion das Areal der richtungsspezifischen Neurone im
Praetectum beider Fischarten genauer bestimmt werden. Nach Beendigung der
elektrophysiologischen Experimente wurden Ableitorte richtungsselektiver Neurone durch
eine Mikroläsion markiert. Nach der histologischen Aufarbeitung der Gehirne konnte dann
lichtmikroskopisch die Lage des markierten Areals genauer bestimmt und mit den
Elektrodenpositionen im Ableitschema verglichen werden.
Alle Fischgehirne wurden frontal von rostral nach caudal in 50 µm dicke Schnitte
geschnitten und anschließend Nissl gefärbt.
39

I. Elektrophysiologie 3. Ergebnisse
d
Abb. I .3.18: Sch
der linken Gehir
PG = Praeglom
ventraler Thalam
Abbildung I
Frontalschnitt
diesem Expe
Penetration z
richtungsselek
Tiefe von 190
letzte neurona
Dieses Areal
lateral zum do
Die Abbildu
Frontalschnitt
TeO
PG
L
L
1 mm
Thv
Ve Psp
TeO
PG
L
L
TeO
PG
L
L
1 mm
Thd
Ve Psp
m l
v
warz - weiß Fotografie eines 50 µm dicken, Nissl - gefärbten praetektalen Frontalschnittes
nhemisphäre von Salmo gairdneri. d = dorsal, v = ventral, l =lateral, TeO = Tectum opticum,
erulus, Ve = 3. Ventrikel, Psp = Nucleus praetectalis superficialis parvocellularis, Thd =
us, L = Läsion. Schnitt in frontaler Ansicht.
. 3.18 zeigt eine Schwarz - Weissfotografie eines 50 µm dicken
es von Salmo gairdneri 900 µm caudal vom rostralen Pol des TeOs. In
riment wurden zur dorsalen und ventralen Arealabgrenzung in einer
wei Mikroläsionen an den Ableitorten des ersten und des letzten
tiven Neurons durchgeführt. Damit kann ein dorsaler Arealbeginn in einer
0 µm unterhalb der TeO – Oberfläche gezeigt werden. 500 µm tiefer war die
le Richtungsselektivität bestimmbar.
befindet sich zentral im dorsalen Praetectum, dorsal zum Praeglomerulus und
rsalen Thalamus.
ng I. 3.19 zeigt eine Schwarz-Weissfotografie eines 50 µm dicken
es von Esox lucius mit einer Läsion.
40
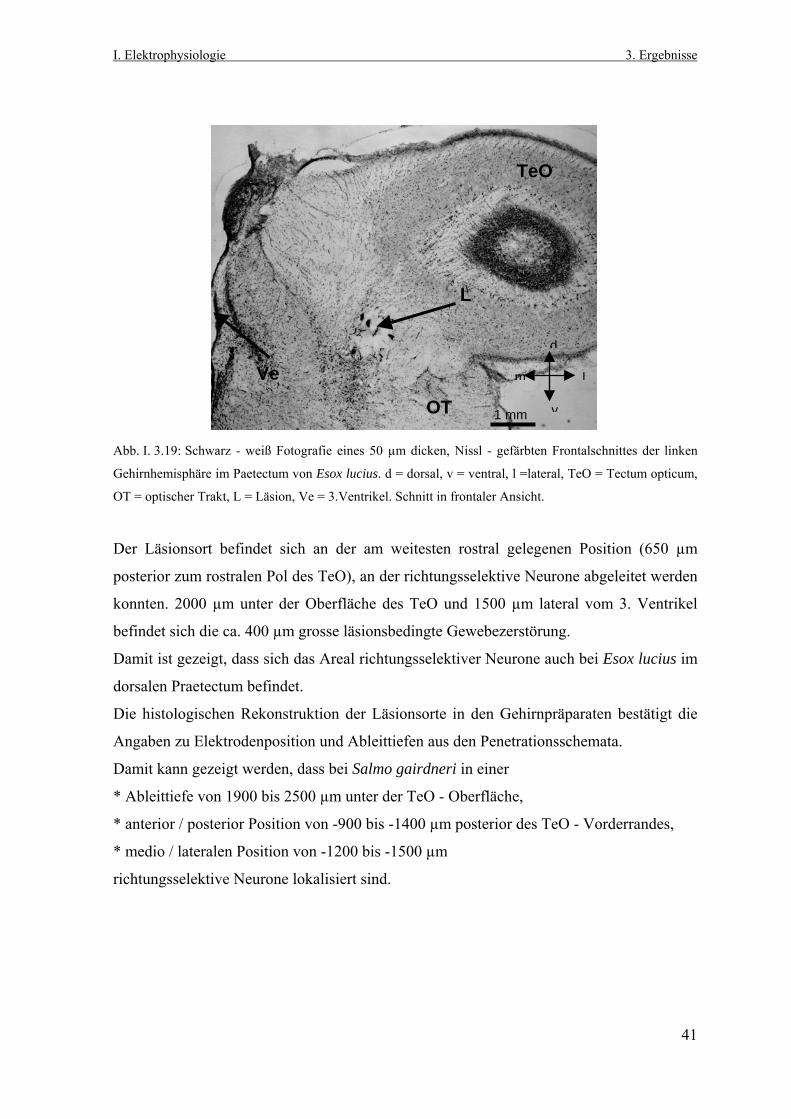
I. Elektrophysiologie 3. Ergebnisse
L
OT
Ve
TeO
1 mm
m l
v
d
Abb. I. 3.19: Schwarz - weiß Fotografie eines 50 µm dicken, Nissl - gefärbten Frontalschnittes der linken
Gehirnhemisphäre im Paetectum von Esox lucius. d = dorsal, v = ventral, l =lateral, TeO = Tectum opticum,
OT = optischer Trakt, L = Läsion, Ve = 3.Ventrikel. Schnitt in frontaler Ansicht.
Der Läsionsort befindet sich an der am weitesten rostral gelegenen Position (650 µm
posterior zum rostralen Pol des TeO), an der richtungsselektive Neurone abgeleitet werden
konnten. 2000 µm unter der Oberfläche des TeO und 1500 µm lateral vom 3. Ventrikel
befindet sich die ca. 400 µm grosse läsionsbedingte Gewebezerstörung.
Damit ist gezeigt, dass sich das Areal richtungsselektiver Neurone auch bei Esox lucius im
dorsalen Praetectum befindet.
Die histologischen Rekonstruktion der Läsionsorte in den Gehirnpräparaten bestätigt die
Angaben zu Elektrodenposition und Ableittiefen aus den Penetrationsschemata.
Damit kann gezeigt werden, dass bei Salmo gairdneri in einer
* Ableittiefe von 1900 bis 2500 µm unter der TeO - Oberfläche,
* anterior / posterior Position von -900 bis -1400 µm posterior des TeO - Vorderrandes,
* medio / lateralen Position von -1200 bis -1500 µm
richtungsselektive Neurone lokalisiert sind.
41

I. Elektrophysiologie 3. Ergebnisse
Bei Esox lucius konnte in einer
* Ableittiefe von 1140 bis 1900 µm unter der TeO - Oberfläche,
* anterior / posterior Position von -500 bis - 1400 µm posterior des TeO - Vorderrandes,
* medio / lateralen Position - 1500 von bis - 1700 µm
das Areal der richtungsselektiven Neurone eingegrenzt werden.
3.8.2 Existiert eine topographische Ordnung? An vier weiteren Salmo gairdneri wurden Experimente mit der Fragestellung nach einer
topographischen Ordnung der richtungsselektiven Neurone innerhalb dieses praetectalen
Areals durchgeführt.
Dazu wurden in jeweils unterschiedlichen rostro - caudalen Bereichen des richtungs-
selektiven Areals im Praetectum die Zellaktivitäten aufgezeichnet, die entlang einer
Penetration während visueller Reizung mit dem Zufallspunktemuster ausgelöst werden
konnten.
Die ermittelten Vorzugsrichtungen wurden horizontalen (nach nasal, nach temporal) und
vertikalen (nach oben, nach unten) Bereichen, entsprechend der vorangegangenen
Untersuchungen, zugeordnet. Anschließend wurden die Ableitpositionen mit einer
Mikroläsion markiert.
Mit der histologischen Rekonstruktion des Ableitortes und der Auswertung der
elektrophysiologisch gewonnenen Daten sollte ein Bezug der Vorzugsrichtung eines
Neurons zum jeweiligen Ableitort hergestellt werden. Abbildung I. 3.20 zeigt von den vier
untersuchten Versuchstieren Frontalschnitte mit Läsionen in unterschiedlichen Bereichen
des Praetectums in einer Anordnung von caudal nach rostral.
42
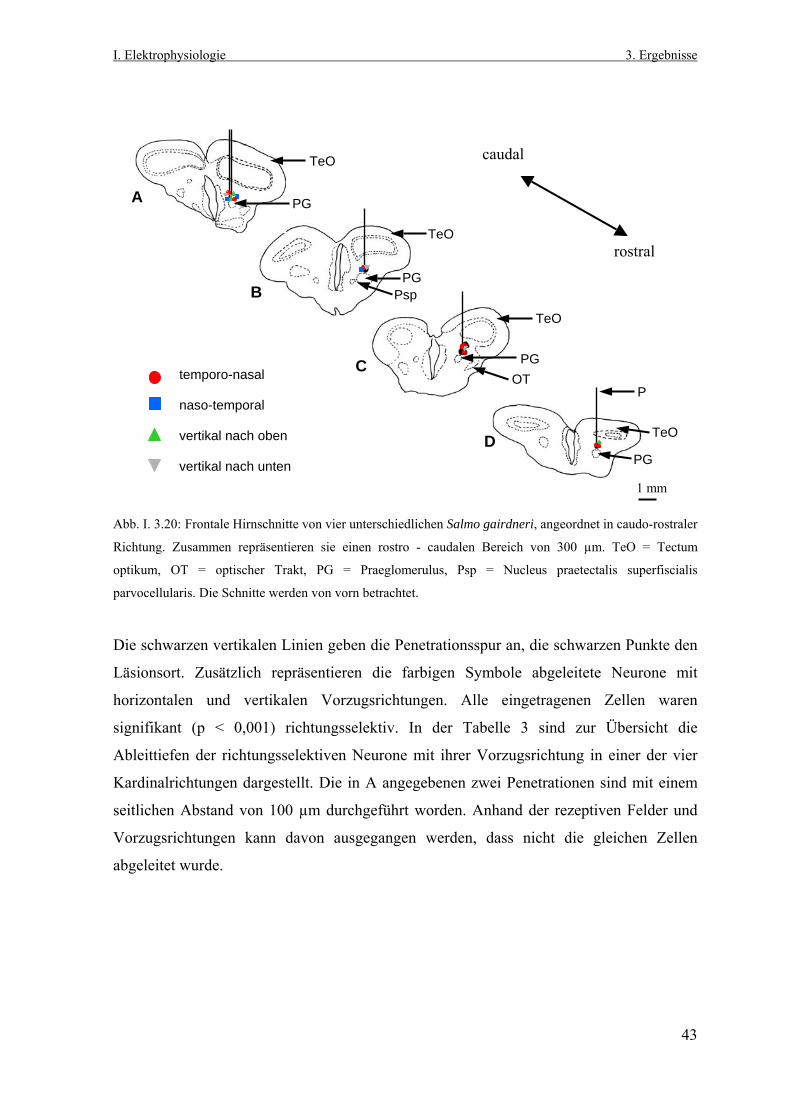
I. Elektrophysiologie 3. Ergebnisse
1 mm
D PG
TeO
C
TeO
OTPG
B Psp
TeO
PG
A
TeO
PG
rostral
caudal
temporo-nasal naso-temporal vertikal nach oben vertikal nach unten
P
Abb. I. 3.20: Frontale Hirnschnitte von vier unterschiedlichen Salmo gairdneri, angeordnet in caudo-rostraler
Richtung. Zusammen repräsentieren sie einen rostro - caudalen Bereich von 300 µm. TeO = Tectum
optikum, OT = optischer Trakt, PG = Praeglomerulus, Psp = Nucleus praetectalis superfiscialis
parvocellularis. Die Schnitte werden von vorn betrachtet.
Die schwarzen vertikalen Linien geben die Penetrationsspur an, die schwarzen Punkte den
Läsionsort. Zusätzlich repräsentieren die farbigen Symbole abgeleitete Neurone mit
horizontalen und vertikalen Vorzugsrichtungen. Alle eingetragenen Zellen waren
signifikant (p < 0,001) richtungsselektiv. In der Tabelle 3 sind zur Übersicht die
Ableittiefen der richtungsselektiven Neurone mit ihrer Vorzugsrichtung in einer der vier
Kardinalrichtungen dargestellt. Die in A angegebenen zwei Penetrationen sind mit einem
seitlichen Abstand von 100 µm durchgeführt worden. Anhand der rezeptiven Felder und
Vorzugsrichtungen kann davon ausgegangen werden, dass nicht die gleichen Zellen
abgeleitet wurde.
43
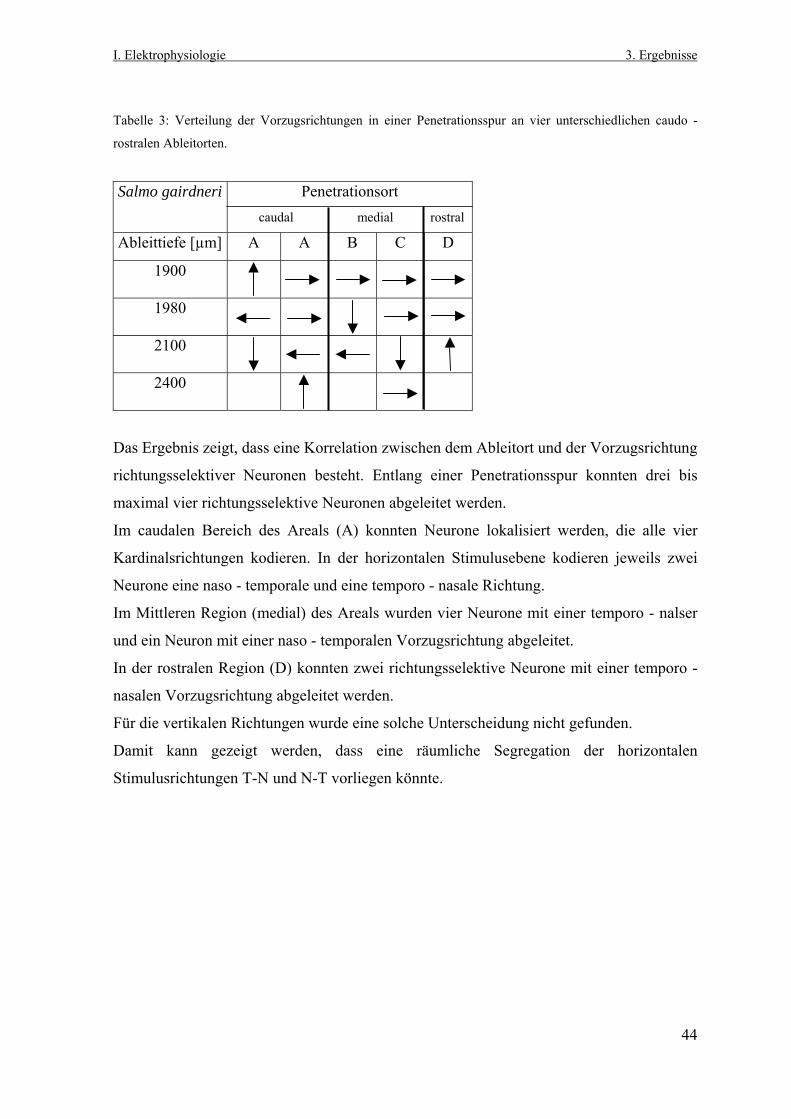
I. Elektrophysiologie 3. Ergebnisse
Tabelle 3: Verteilung der Vorzugsrichtungen in einer Penetrationsspur an vier unterschiedlichen caudo -
rostralen Ableitorten.
Salmo gairdneri Penetrationsort caudal medial rostral
Ableittiefe [µm] A A B C D
1900
1980
2100
2400
Das Ergebnis zeigt, dass eine Korrelation zwischen dem Ableitort und der Vorzugsrichtung
richtungsselektiver Neuronen besteht. Entlang einer Penetrationsspur konnten drei bis
maximal vier richtungsselektive Neuronen abgeleitet werden.
Im caudalen Bereich des Areals (A) konnten Neurone lokalisiert werden, die alle vier
Kardinalsrichtungen kodieren. In der horizontalen Stimulusebene kodieren jeweils zwei
Neurone eine naso - temporale und eine temporo - nasale Richtung.
Im Mittleren Region (medial) des Areals wurden vier Neurone mit einer temporo - nalser
und ein Neuron mit einer naso - temporalen Vorzugsrichtung abgeleitet.
In der rostralen Region (D) konnten zwei richtungsselektive Neurone mit einer temporo -
nasalen Vorzugsrichtung abgeleitet werden.
Für die vertikalen Richtungen wurde eine solche Unterscheidung nicht gefunden.
Damit kann gezeigt werden, dass eine räumliche Segregation der horizontalen
Stimulusrichtungen T-N und N-T vorliegen könnte.
44

I. Elektrophysiologie 4. Diskussion
4. Diskussion
4.1 Auswahl der Versuchstiere und die Wahl der Methode Die elektrophysiologische Suche nach dem neuronalen Substrat zur Ausführung der
langsamen Augenfolgephase des hOKNs wurde an zwei Süßwasser - Teleosteern, Salmo
gairdneri und Esox lucius durchgeführt. Die Wahl fiel auf diese echten Knochenfischarten,
da beide Prädatoren sind und in Bächen, Flussoberläufen (Salmo gairdneri) und in Seen
(Esox lucius) beheimatet sind. Die Detektion und das Schlagen von Beute wird von beiden
Fischarten vorwiegend visuell geführt (New et al., 2000). Aus diesem Grund sollten beide
Arten über ein gut entwickeltes visuelles System verfügen.
In meiner Diplomarbeit habe ich neuroanatomische Untersuchungen am visuellen System
und Verhaltensexperimente zum OKN an diesen beiden Fischarten durchgeführt (Polyak,
1968; Bathelt, 1970; Klar, 1999). Die Ergebnisse dieser Arbeiten belegen das gut
entwickelte visuelle System und die Fähigkeit zu horizontalen optokinetischen
Augenbewegungen bei diesen Spezies.
Um die elektrophysiologischen Untersuchungen im Praetectum an beiden Versuchstieren
durchzuführen, musste eine vollständig neue Versuchsapparatur aufgebaut werden. Eine
wichtige Voraussetzung war die homogene visuelle Stimulation des gesamten
Gesichtsfeldes. Das monokulare Gesichtsfeld von Salmo gairdneri hat eine Ausdehnung
von ca. 205° im Azimut und von ca.180° in der Elevation. Annähernd gleich groß ist das
monokulare Gesichtsfeld von Esox lucius (Azimut ca. 225°, Elevation ca. 180°)
(Scheuring, 1920; Kahmann, 1934, 1936). Die Ganzfeldstimulation wurde erreicht, indem
das Versuchtier mit seiner Fixationshalterung und den Mikromanipulatoren samt
Elektrodenvortrieb in der Plexiglashalbkugel um 45° in seiner Körperlängsachse nach
rechts geneigt wurde. Dabei tauchte das zu stimulierende rechte Auge vollständig in das
Wasser. Das obere Drittel des rechten dorsalen Gesichtsfeldes konnte nicht innerhalb des
Wassers stimuliert werden.
Bisherige Untersuchungen zum optokinetischen System der Fische wurden an Tieren in
horizontaler Schwimmlage durchgeführt. (Easter, 1972; Easter & Schmidt, 1977; Kunkel,
1988; Dieringer et al., 1992; Keng & Anastasio, 1997; Easter & Nicola, 1997; Fritsches &
45

I. Elektrophysiologie 4. Diskussion
Marschall, 2002). Diese Untersuchungsmethode birgt Nachteile. Die limitierenden
Faktoren sind die Grenzfläche zwischen den Medien Wasser und Luft (Wasseroberfläche),
sowie die Glaskanten der Aquarien, die eine homogene Ganzfeldreizung stören können.
An der Wasseroberfläche und den Glaskanten der kubusförmigen Aquarien werden die
Lichtstrahlen der optischen Stimulation gebrochen. Dadurch erfährt an dieser Grenzfläche
das Reizmuster diskontinuierliche Geschwindigkeiten.
Um eine möglichst hohe Erfolgsrate der elektrophysiologischen Experimente zu erreichen,
mussten die Ableitorte reproduzierbar sein. Das Fehlen einer stereotaktischen Apparatur
für Fische und eines Gehirnatlanten wurde durch die Tatsache, dass bei den adulten
Versuchstieren eine altersunabhängige Größenkonstanz der Gehirne auftrat, kompensiert.
Aufgrund der Retinotopie des TeOs konnten die RF der ON/OFF Zellen aus dem Stratum
opticum als zusätzliche Orientierungshilfe zur Lokalisation richtungsselektiver Neurone
genutzt werden.
4.2 Elektrophysiologische Ableitungen im Praetectum Mit elektrophysiologischen Untersuchungsmethoden sollte in der vorliegenden Arbeit der
Fragestellung nachgegangen werden, ob es im Praetectum zweier verschiedener Teleostei -
Arten Neurone gibt, die eine Richtungsselektivität für großflächige, horizontal bewegte
visuelle Stimuli besitzen und ob diese Neurone an der Generierung der langsamen
Folgephase der Augen während des horizontalen OKNs beteiligt sein könnten.
Mindestens zwei wichtige funktionelle Kriterien müssen dafür erfüllt sein:
- Richtungsselektivität von Neuronen ist dadurch definiert, dass ein Neuron mit einer
starken, anhaltenden Entladung reagiert, wenn ein Reiz in einer bestimmten Richtung
durch sein rezeptives Feld bewegt wird (Vorzugsrichtung). In entgegengesetzter
Reizrichtung (Nullrichtung) kann nur eine geringe oder keine reizabhängige Aktivität
ausgelöst werden kann (Barlow et al., 1964; Barlow & Levick, 1965).
- Große rezeptive Felder der richtungsselektiven Neurone sind notwendig, um
ausschliesslich auf großflächige Bildverschiebungen reagieren zu können. Störeinflüsse,
wie z.B. die Bewegung von kleinen Objekten, müssen unterdrückt werden, da sie sonst den
46

I. Elektrophysiologie 4. Diskussion
Stabilisierungsvorgang des visuomotorischen Systems erheblich unterbrechen würden
(Howard, 1993).
Diese Charakteristika dienten anfangs als Suchkriterien, mit denen die
elektrophysiologischen Untersuchungen im Praetectum von Salmo gairdneri und Esox
lucius zum Auffinden richtungsselektiver Neurone durchgeführt wurden.
4.2.1 Richtungsselektive Neurone Zu Beginn der elektrophysiologischen Untersuchungen wurden mit Hilfe eines
Penetrationsrasters Ableitungen im Praetectum durchgeführt. Dabei wurden im rostro -
medialen Bereich Zellen lokalisiert, die eine ON- bzw. OFF- Charakteristik und rezeptive
Felder von 6 bis 12° Sehwinkel Durchmesser im frontalen Gesichtsfeld aufwiesen. In
angrenzenden Ableitbereichen konnten Neurone mit grossen RF, die auf bewegte kleine
und grosse Lichtpunkte reagierten lokalisiert, werden. Diese Neurone waren alle nicht
richtungsselektiv. Eine genaue Zuordnung zu bekannten visuellen praetectalen
Kerngebieten gestaltete sich aufgrund widersprüchlicher Neuroanatomie als schwierig
(Finger, & Karten, 1978; Allum et al., 1981; Vanegas & Ito, 1983; Ito & Murakami, 1984;
Fite, 1985; Butler et, al., 1991; Rupp et al., 1996). Diese anfänglichen Befunde wurde in
weiteren Experimenten bestätigt und dienten als zusätzliche Orientierungshilfe zum
Auffinden der richtungsselektiven Neurone.
Die weiteren Ergebnisse belegen eindeutig die Existenz visueller richtungsselektiver
Zellen im Praetectum beider untersuchten Fischarten. Diese wurden zunächst mit
horizontalen Bewegungsreizen identifiziert. Das im Ergebnisteil I. 3.2 als Beispiel
vorgestellte richtungsselektive Neuron von Salmo gairdneri war sehr gering bis gar nicht
spontanaktiv (< 4 Hz). Mit einer nach nasal verlaufenden Stimulationsrichtung konnte zu
Beginn eine kurzzeitige, phasische (40 – 80 ms) Antwort gefolgt von einer über die
verbleibende Stimulationsphase tonischen Reizreaktion auf den präsentierten visuellen
Stimulus ausgelöst werden (Vorzugsrichtung). In einer nach temporal verlaufenden
Stimulusrichtung hingegen entsprachen die Entladungsraten der jeweiligen
Spontanaktivität (Nullrichtung). Die kurzzeitige phasische Komponente mit Einsetzen der
Bewegung trat auch in dieser Reizrichtung auf (Abb. I. 3.4).
47

I. Elektrophysiologie 4. Diskussion
Im Vergleich dazu zeigten die richtungsselektiven Neurone von Esox lucius stets eine
höhere Spontanaktivität (> 10 Hz). In der Vorzugsrichtung nach nasal wurden
erwartungsgemäß die höchsten Entladungsraten registriert. Wie bei Salmo gairdneri folgte
einer kurzzeitigen phasischen Entladung eine tonische Antwort. Dieses „movement onset“-
Verhalten mit Einsetzen des visuellen Bewegungsreizes ist bei beiden Fischarten gleich.
Bei visueller Stimulation in Nullrichtung trat bei Esox im Unterschied zu Salmo gairdneri
eine Inhibition auf (Abb. I. 3.6). Die Arbeitsweise dieses Neurons erfolgt also nach dem
„push-pull“ Prinzip. Beide Aktivierungsformen, rein exzitatorisch (Salmo gairdneri) und
„push-pull“ (Esox lucius), ermöglichen die Detektion einer Stimulusrichtung. Das „push-
pull“ Prinzip ermöglicht zusätzliche Modulationsmöglichkeiten der neuronalen Antwort
für nachgeschaltete Strukturen. Sie sind bei allen Vertebratenklassen in verschiedenen
Gehirnarealen, die der Analyse von Bewegungsrichtung im Raum dienen, zu finden, wie
z. B. im NOT und im nLM (Mammalia: Collewijn, 1975; Hoffmann & Schoppmann, 1975;
Hoffmann, 1985; Kato et al., 1986; Ibbotson, 2001; Amphibia: Katte & Hoffmann, 1980;
Montgomery et al., 1982; Lazar et al., 1983; Reptilia: Ariel, 1997; Ariel & Kogo, 2001;
Aves: Winterson & Brauth, 1985; Wylie & Frost, 1990).
4.2.2 Fanden die ersten Ableitungen tatsächlich im Praetectum statt? Die elektrophysiologischen Ableitungen im Praetectum erfolgten stets nach einer
Penetration durch das Tectum opticum. Für die Teleostei ist das TeO eine wichtige
neuronale Struktur. Sie ist retinotop organisiert, dient der photopischen und skotopischen
Wahrnehmung, der Analyse von Formen und Objekten und ist unter anderem auch an der
Visuomotorik beteiligt (Bathelt, 1970; Guthrie & Sharma, 1979; Douglas & Djamgoz,
1990; Herrero et al., 1998, 1999). Das retinotope Ordnungsprinzip zeigt sich bei den
Neuronen im Stratum opticum und Stratum griseum fibrosum et superficiale des TeOs.
Diese tektalen Zellen erhalten einen direkten Eingang von den Ganglienzellen der
kontralateralen Retina. Bei den meisten Teleosteern findet eine totale Überkreuzung des
Nervus opticus am Chiasma opticum statt. Ausnahmen stellen Osteoglossomorpha und
Clupeomorpha da (Repérant et al., 1982). Bei den Elopomorpha und den Euteleostei, zu
denen Salmo gairdneri und Esox lucius gehören, existieren nur wenige bilaterale
48

I. Elektrophysiologie 4. Diskussion
retinofugale Projektionen zum Gehirn. Die Terminalen der ipsilateralen Projektion liegen
im praeoptischen Areal oder ipsilateralen Bereichen des Thalamus und des TeO.
Allerdings existieren zu diesem Befund widersprüchliche Daten (Sas & Maler, 1986;
Wilms & Fritzsch, 1990, 1992).
Die Repräsentation der visuellen Welt auf der Retina erfährt auf dem Projektionsweg zum
TeO eine Drehung um 180°:
- ventrale Retinabereiche (dorsales Gesichtsfeld) werden im medialen Bereich,
- dorsale Retinabereiche (ventrales Gesichtsfeld) im lateralen Bereich,
- temporale Retinabereiche (nasales Gesichtsfeld) im rostralen Bereich und
- nasale Retinabereiche (temporales Gesichtsfeld) im caudalen Bereich des TeO
repräsentiert.
Die rezeptiven Felder (RF) der tektalen Neurone, sind abhängig von der Exzentrizität,
klein (< 1° Sehwinkel) bis mittelgroß (bis ca. 15° Sehwinkel). Kleine RF befinden sich in
den Bereichen der Retina mit hohen Ganglienzelldichte wie z.B. im horizontalen Streak,
der Area centralis oder der Fovea. Die mittelgroßen bis großen RFs sind mehr in den
peripheren Bereichen der Retina zu finden (Schwassmann, 1968; Schwassmann & Krüger,
1970).
Mit Elektrostimulationen im TeO können schnelle und langsame, konvergente und
divergente Augenbewegungen, sowie unterschiedlich stark ausgeprägte
Schwimmbewegungen und eine Kombination aus Augen- und Rumpfbewegung ausgelöst
werden. Selbst komplexe Verhaltensmuster wie der Fluchtreflex können mit dieser
Methode erzeugt werden (Guthrie & Banks, 1978; Salas et al., 1997; Herrero et al., 1998).
Das Auslösen von Augen- und Rumpfbewegungen ist abhängig von der verwendeten
Stromstärke, der Impulsdauer und dem Stimulationsort im TeO. Der tektale Penetrationsort
befand sich bei Salmo gairdneri und Esox lucius in einem ca. 500 µm x 500 µm großen
Bereich im medio - rostralen TeO. Elektrostimulationen am Goldfisch würden in diesem
Bereich kontraversive Augenbewegungen auslösen (Herrero et al., 1999). Besteht die
Möglichkeit, dass im TeO abgeleitet wurde? Wie kann das Praetectum neurophysiologisch
vom TeO abgegrenzt werden? An dem Ableitort, an dem die richtungsselektiven Neurone
von Salmo gairdneri gefunden wurden, befanden sich die RF im darrüberliegenden TeO in
einem Azimutbereich von 80° bis 90° und einer Elevation von 35° bis 45° bezogen auf die
verlängerte Körperlängsachse.
49

I. Elektrophysiologie 4. Diskussion
Die RF von Esox lucius befanden sich mehr im frontalen Gesichtsfeld, nämlich bei 35° bis
45° Azimut und 20° bis 30° Elevation. Für beide Fischarten konnte eine RF - Größe von
ca. 3,5° Sehwinkel bestimmt werden. Die RF - Größen der richtungsselektiven Zellen von
Salmo gairdneri und Esox lucius hingegen waren mit > 20° Sehwinkel erheblich größer
und unterscheiden sich damit deutlich von den RF des TeO. Sie gleichen in ihrer RF -
Grösse und der Verteilung im Gesichtsfeld den NOT und nLM Neuronen der anderen
Vertebratenklassen (Mammali: Collewijn, 1975; Ballas & Hoffmann, 1985; Aves: Gioanni
et al., 1983; Reptilia: Ariel & Kogo, 2001; Amphibia: Katte & Hoffmann, 1980).
Es ist daher auszuschliessen, dass die Ableitungen der richtungsselektiven Neurone im
TeO erfolgten. Zusätzlich darf nicht ausser Acht gelassen werden, dass sich die optische
Schicht des TeOs in einer Ableittiefe von 250 µm bis 400 µm befand, während die
Ableitungen der richtungsselektiven Neurone in Tiefen von 1600 µm bis 2400 µm
erfolgten. Ein endgültiger Beweis ist das nicht, da das Tectum opticum beim Fisch
halbkreisförmig ist und gerade im rostralen Bereich die optische Schicht kreisförmig
verläuft. Einen genauen Aufschluss über den Ableitort liefert die anatomische
Rekonstruktion mit Hilfe von Mikroläsionen an Ableitstellen richtungsselektiver Neurone.
Die Mikroläsionen befanden sich bei Salmo gairdneri im medio - dorsalen Bereich des
Praetectums. Weiter rostral befanden sich die Läsionen bei Esox lucius. Die Position der
Läsion bei beiden Fischarten war eindeutig nicht im TeO. Damit wird gezeigt, dass die
Ableitungen im Praetectum stattfanden.
Diese ersten Ergebnisse verdeutlichen, dass sich die horizontal richtungsselektiven Zellen
von Salmo gairdneri und Esox lucius im Praetectum befanden und grundsätzlich die
Fähigkeit besitzen, visuelle Stimuli in einer Reizrichtung von temporal nach nasal
(ipsiversiv zum Ableitort) eindeutig zu selektieren. Damit würden diese Neurone von
Salmo gairdneri und Esox lucius die erste und zweite Anforderung zur
Richtungsselektivität sowie zur Größe und Lage der rezeptiven Felder erfüllen.
50

I. Elektrophysiologie 4. Diskussion
4.3 Richtungspräferenz richtungsselektiver Neurone Richtungsselektive Neurone des NOT und nLM, die an der Ausführung der langsamen
Augenfolgebewegung des OKNs, dem OKR beteiligt sind, besitzen keineswegs
ausschließlich horizontale Richtungspräferenzen (Fan et al., 1995). Das
Richtungsverhalten solcher Neurone kann mit Einführung weiterer Stimulationsrichtungen
näher untersucht werden. Die Verwendung von sechs zusätzlichen Stimulusrichtungen
(oben ↔ unten (90°↔270°), sowie nasal - oben ↔ temporal - unten (45°↔225°) und
nasal - unten ↔ temporal - oben (315°↔135°)), ermöglichte die Bestimmung der
Richtungspräferenz dieser Neurone in Form von Tuning - Kurven, sowie den
Vorzugsrichtungsvektoren. Für die gezeigten Beispielzellen beider Fischarten ergab sich
eine horizontale, nach nasal verlaufende Richtungspräferenz in einem Winkelbereich von
45° bis 315°. Aufgrund dieser Vorzugsrichtung (ipsiversiv zum Ableitort) sind diese
Neurone vergleichbar mit richtungsselektiven Zellen im optokinetischen System der
anderen Vertebratenklassen und zwar mit dem nLM der Amphibia, Reptilia, und Aves und
mit dem NOT der Mammalia (Aves: Winterson & Brauth, 1985; Reptilia: Schröder, 1981;
Amphibia: Cochran et al., 1984; Katte & Hoffmann, 1980; Montgomery et al., 1985;
Mammalia: Collewijn, 1975; Hoffmann & Schoppmann, 1975; Mustari & Fuschs, 1990).
In beiden Kerngebieten konnte klassenübergreifend genau diese ipsiversive
Vorzugsrichtung der richtungsselektiven Neurone gezeigt werden, die an der langsamen
horizontalen Augenfolgephase beim horizontalen optokinetischen Reflex (hOKR) beteiligt
sind. Der NOT und der nLM sind die primären visuomotorischen Schaltstationen im
horizontalen optokinetischen System.
Damit stehen die anfänglichen Befunde von Salmo gairdneri und Esox lucius im Einklang
mit den Ergebnissen an anderen Vertebratenklassen und erhöhen die Wahrscheinlichkeit,
dass diese richtungsselektiven Neurone zum primären neuronalen Schaltkreis des OKR der
Knochenfische gehören. Insgesamt konnten 72 Neurone von Salmo gairdneri und 49
Neurone von Esox lucius während der elektrophysiologischen Untersuchungen im
Praetectum abgeleitet werden, die eine Richtungsselektivität aufwiesen. Diese wurden mit
der zuvor angewendeten Auswertungsmethode, der Tuning – Kurven - Berechnung und der
Vorzugsrichtungsbestimmung näher quantifiziert. Die Ergebnisse zeigen, dass nicht nur
ein horizontal ipsiversiv verlaufender visueller Stimulus von diesen richtungsselektiven
51

I. Elektrophysiologie 4. Diskussion
Neuronen beider Fischarten kodiert wird, sondern mindestens alle acht präsentierten
Stimulusrichtungen.
Weiterführend stellte sich die Frage nach dem präzisen Tuning dieser Zellen. Die
signifikanten Ergebnisse aus der zirkulären Statistik (p < 0,001) belegen eindeutig für 63
der ursprünglich 72 Neurone von Salmo gairdneri und 43 von 49 Neurone von Esox lucius,
dass alle Raumrichtungen von diesen richtungsselektiven Neuronen kodiert werden
können. Damit wurde gezeigt, dass innerhalb eines begrenzten praetectalen Areals
richtungsselektive Zellen lokalisiert sind, die horizontale und vertikale
Stimulationsrichtungen kodieren.
Dies ist ein überraschendes Resultat, da in allen bisher untersuchten Vertebratenklassen
eine Segregation der Vorzugsrichtungen richtungsselektiver Neurone, die an der
Ausführung des OKRs beteiligt sind, in verschiedene Kerngebiete des Praetectums und des
AOS nachgewiesen wurde. Bei Amphibia, Reptilia und Aves wurde, wie bereits erwähnt,
der praetectale nLM identifiziert. Die richtungsselektiven Neurone dieses Kerngebietes
kodieren eine ipsiversive Stimulusrichtung, d. h., ein dem rechten Auge präsentiertes
großflächiges Reizmuster, das nach links verläuft, aktiviert die richtungsspezifischen
Zellen im linken nLM. Ein nach rechts bewegtes Reizmuster aktiviert den rechten nLM.
Damit sind die zwei horizontalen Raumrichtungen im linken und rechten nLM
repräsentiert (Winterson & Brauth, 1985; Natal & Britto, 1987; Kogo et al., 1998).
Die Bedeutung des nLM für die Ausführung des OKR dokumentieren Untersuchungen an
Amphibien (Rana spec.) unter vergleichbaren visuellen Stimulationsbedingen, wie sie auch
in dieser Arbeit verwendet wurden (Katte & Hoffmann, 1980; Fite et al., 1989).
Weitere Unterstützung fanden diese Befunde durch Studien, in denen die Auswirkung von
Läsionen auf die langsame Augenfolgephase des hOKR untersucht wurde. Frösche führen
im allgemeinen eher Kopfbewegungen als Augenbewegungen aus. Tiere mit nLM – Läsion
zeigten spontane Kopfbewegungen kontraversiv zum Läsionsort. Auffällige Defizite bei
der Präsentation vertikaler visueller Reize konnten nicht registriert werden (Montgomery et
al., 1982; Lázár et al., 1983). Entsprechende Resultate ergaben elektrophysiologische
Experimente am nLM bei Reptilia (Fan et al., 1995) und Aves ( Gioanni et al.,1983;
Winterson & Brauth, 1985; Wylie & Frost, 1990; Wylie et al., 1996; Fu et al., 1997).
52

I. Elektrophysiologie 4. Diskussion
Der nLM wird aufgrund seiner Lage im Praetectum und der funktionellen Eigenschaften
seiner Neurone als vergleichbare Struktur zum praetectalen NOT der Mammalia
angesehen. Bei Säugern beeinflussen zusätzlich Efferenzen des visuellen Cortex die
Bildverschiebungsneurone des NOT (Hoffmann & Schoppmann, 1975; Pereira et al.,
2000; Hoffmann & Fischer, 2001, Distler et al., 2002). An verschiedenen Säugerspezies
(Kaninchen, Ratte, Frettchen, Katze, Affe) wurden Richtungsselektivität, Richtungstuning
und Aktivitätsprofil der NOT Neurone untersucht (Collewijn, 1975; Hoffmann &
Schoppmann, 1975; Hoffmann, 1985; Kato et al., 1986; Schiff et al., 1990; Soodak &
Simpson, 1988). Die Befunde dieser Studien deuten auf eine große Konstanz dieser
visuomotorischen Struktur hin.
Vertikale optokinetische Augenbewegungen werden über die visuellen Arealen des
akzessorischen optischen Systems (AOS) ausgelöst. Dazu gehören der im Mesencephalon
lokalisierte Nucleus of the basal optic root (nBOR) in den Klassen Amphibien, Reptilien
und Vögeln und der mediale terminale Nukleus (MTN) und der laterale terminale Nukleus
(LTN) der Mammalia (Tan et al., 1993; Van der Togt et al., 1993; Schmidt et al., 1998),
die wie nLM und NOT retinalen Eingang vom kontralateralen Auge erhalten. Im nBOR
gelang es, Zellen zu identifizieren, die selektiv auf vertikale Reizbewegungen reagieren
(Cochran et al., 1984; Katte & Hoffmann, 1980; Lazar et al., 1983). In diesen Arbeiten
konnte weiterhin gezeigt werden, dass die im nBOR lokalisierten, horizontale Bewegungen
kodierenden, richtungsselektiven Neurone keinen bzw. sehr geringen Einfluss auf den
horizontalen OKN haben. Diese Ergebnisse wurden durch Läsionen im nBOR mit
anschliessenden Verhaltensexperimenten bestätigt (Lazar, 1989). Vielmehr scheinen die
horizontal kodierenden Neurone im nBOR einen modulatorischen Einfluss auf die
richtungsselektiven Neurone des nLM zu besitzen (Montgomery et al., 1982; Fite et al.,
1989).
Bei den Reptilien und Vögeln wurden die richtungsselektiven Eigenschaften der nBOR
Neurone bestätigt (Reptilia: Ariel et al., 2001; Fan et al., 1995; Kogo et al., 1998; Bruns &
Wallmann, 1981, Aves: Wylie & Frost, 1990; Wylie, 2001). In der Populationsanalyse
konnten in diesen Studien drei kartesische Richtungen gezeigt werden, die von diesen
Neuronen kodiert werden: vertikal nach oben, vertikal nach unten sowie eine horizontal
kontraversive Richtung. Damit stellt der nBOR eine Mischform der Säugeräquivalente
NOT und MTN / LTN dar. Trotzdem wird der nBOR als eine dem MTN und dem LTN der
53

I. Elektrophysiologie 4. Diskussion
Säuger vergleichbare Struktur angesehen. Innerhalb des MTN / LTN konnten jedoch nur
richtungselektive Zellen für vertikale Reizbewegung lokalisiert werden (Soodak &
Simpson, 1988; Matsuo & Cohen, 1984), die aber sonst in ihren charakteristischen
Eigenschaften mit den richtungsselektiven Neuronen des nBOR vergleichbar sind. Bei den
Säugern tritt eine Trennung der visuellen Informationsverarbeitung für horizontale (NOT)
und vertikale (MTN / LTN) Stimuli im visuomotorischen System auf (Grasse & Cynader,
1988).
Werden die unterschiedlichen Raumrichtungen von einer unterschiedlichen Anzahl
richtungsselektiver Neurone bei den von mir untersuchten Fischarten kodiert? Das
Ergebnis dieser Untersuchung ergab für Salmo gairdneri ein eindeutiges Nein. Mit einem
p = 0,154 konnte mit Hilfe des Chi2-Testes keine signifikante Ungleichverteilung in der
Anzahl horizontal und vertikal kodierender Neurone festgestellt werden.
Für Esox lucius hingegen wurde ein Wert von p < 0,05 ermittelt. Hier kodieren mehr
Neurone die horizontale Stimulusrichtung als die vertikale.
4.4 Praetectale richtungsselektive Neurone und optokinetisches
Verhalten Wie können aus den bisher gewonnenen Erkenntnissen Rückschlüsse auf die
Eigenschaften des monokularen horizontalen optokinetischen Reflexes (mhOKR) gezogen
werden, bzw. kann anhand der Verteilung der Vorzugsrichtungen der praetectalen Neurone
eine Symmetrie bzw. Asymmetrie des mhOKR erklärt werden?
Unter Laborbedingungen kann durch visuelle Ganzgesichtsfeldstimulation der horizontale
optokinetische Nystagmus (hOKN) ausgelöst werden. Dieser besteht aus zwei Phasen in
alternierender Abfolge. Einer langsamen Phase in der die Augen der retinalen
Bildverschiebung folgen (OKR) und einer zweiten Phase mit einer Rückstellsakkade
(schnelle Augenbewegungen), die die langsame Augenfolgephase unterbricht und die
Augen zur Startposition zurückführt. Unter binokularen Stimulationsbedingungen zeigen
die meisten der bis jetzt untersuchten Spezies einen symmetrischen hOKR, d.h. die Güte
bzw. das gain (Quotient der Augenfolgegeschwindigkeit durch die Reizgeschwindigkeit)
54

I. Elektrophysiologie 4. Diskussion
der langsamen Augenfolgephase ist in temporo - nasaler (T-N) und naso - temporaler
(N-T) Richtung gleich. Bei monokularer Stimulation tritt bei vielen untersuchten Tierarten
(z. B. Kanninchen, Maus, Ratte) ein asymmetrischer hOKR auf. Das gain des OKRs ist in
temporo - nasaler Reizrichtung höher als in naso - temporaler. Diese Befunde können mit
den Theorien von Tauber & Adkins (1968) und Van Hof-Van Duin (1976) erklärt werden.
Für einen symmetrischen OKR ist ein grosses binokulares Gesichtsfeld, eine Foveation
und der Dekussationgrad des Nervus opticus relevant. Je mehr Fasern des Nervus opticus
kreuzen umso asymmetrischer wird der OKR.
Fische können optokinetische Augenbewegungen ausführen, die denen der Säuger und der
anderen Vertebratenklassen durchaus vergleichbar sind (Easter, 1972; Kunkel, 1988;
Dieringer et al., 1992; Klar, 1999; Fritschtes & Marshall, 2002).
Entsprechende Untersuchungen an Salmo gairdneri und Esox lucius zum mhOKN wurden
von mir in meiner Diplomarbeit (1999) durchgeführt. In dieser Arbeit konnte gezeigt
werden, dass Salmo gairdneri einen fast symmetrischen mhOKR bei niedrigen
Stimulusgeschwindigkeiten (< 14 °/s) zeigt. Mit zunehmender Reizgeschwindigkeit
(> 14 °/s) stellte sich eine Asymmetrie mit temporo - nasaler Vorzugsrichtung ein. Der
mhOKR von Esox lucius zeigte stets eine Asymmetrie mit naso - temporaler
Vorzugsrichtung.
Diese Verhaltensergebnisse stehen nicht im Einklang mit der oben genannten Theorie. Es
müssen andere Mechanismen greifen, wie z. B. die Anzahl der richtungsselektiven
Neurone die eine T-N oder N-T Richtung kodieren.
Für die Beantwortung der oben angeführten Fragestellung kamen nur richtungsselektive
Zellen in Betracht, die innerhalb von +/- 45° zur horizontalen Stimulationsrichtung eine
nach nasal (ipsiversiv) verlaufende Reizrichtung (45° bis 315°) oder eine nach temporal
(kontraversiv) verlaufende Reizrichtung (135° bis 225°) kodieren. Die Gruppenanalyse des
Chi2-Tests (ipsiversiv versus kontraversiv) ergab keinen signifikanten Unterschied für die
beiden horizontalen Stimulusrichtungen beider Versuchstiere. Dies steht im Einklang mit
den Ergebnissen aus den Verhaltensexperimenten für Salmo gairdneri, aber nicht für Esox
lucius. Eine naso - temporale Vorzugsrichtung des mOKRs kann anhand der Verteilung
richtungsselektiver Neurone nicht gezeigt werden. Von daher ist anzunehmen, dass nicht
nur die Verteilung der Neurone das alleinige Kriterium ist, um eine Symmetrie bzw.
Asymmetrie zu erklären.
55

I. Elektrophysiologie 4. Diskussion
Wenn nicht die Anzahl der Neuronen über eine Vorzugsrichtung des mhOKN entscheidet,
dann vielleicht die Aktivierungsstärke der abgeleiteten Neuronenpopulation. Die
Populationsantwort wurde durch Aufsummierung der Tuning - Kurven untersucht. Die
Tuningkurve zeigt den Richtungsbereich, durch den ein Neuron aktiviert wird. In der
Summe sollten sich daher eventuell bevorzugte Raumrichtungen der Neuronenpopulation
zeigen. Das Ergebnis zeigt für beide untersuchten Fischarten keine eindeutige
Bevorzugung einer Raumrichtung.
Eine Erklärungsmöglichkeit für diesen Befund besteht darin, dass, wie oben erwähnt, nicht
alle praetectalen richtungsselektiven Neurone zu den okulomotorischen Kernen projizieren
und damit auch nicht in ihrer Gesamtanzahl an der Generierung des OKR beteiligt sein
müssen. Mögliche weiterführende Projektionen könnten zum Beispiel zu der dorsalen
Kappe der Oliva inferior, zum Cerebellum, oder zu den Vestibulariskernen ziehen. Diese
Projektionen können dann zur Kalibrierung des vestibulo - okulären Reflex (VOR) genutzt
werden. Die Existenz solcher neuronalen Verbindungen wurde mannigfaltig in
entsprechenden Vertebratenklassen nachgewiesen und wird im zweiten Kapitel dieser
Arbeit detailliert diskutiert.
Eine genaue Klärung dieser Projektionswege an den untersuchten Fischarten kann nur eine
Methode erbringen, in der einzelne richtungsselektive Neurone markiert werden (z.B. mit
Neurobiotin), um die Lage der Terminalen im Gehirn genau bestimmen zu können (Pastor
et al., 1991; Suwa et al., 1999).
4.5 Geschwindigkeitsverhalten der richtungsselektiven Neurone
Bisher wurden nur die Bewegungsrichtungen mit einer konstanten Reizgeschwindigkeit
untersucht. Der verwendete Stimulus beinhaltet zwei Komponenten, die
Bewegungsrichtung und die Geschwindigkeit. Entscheidend für die Ausführung der
langsamen Augenfolgebewegung ist das Geschwindigkeitsverhalten der
richtungsselektiven Zellen, da visuelle Reize auch in natürlicher Umgebung
unterschiedlich schnell sein können. Die Reizgeschwindigkeit wird von
richtungsselektiven Neuronen des optokinetischen Systems über die Spikerate kodiert
(Collewijn, 1975; Hoffmann, 1985; Wylie & Frost, 1990; Winterson & Brauth, 1985).
56

I. Elektrophysiologie 4. Diskussion
Der Median von 16 richtungsselektiven Neuronen bei Salmo gairdneri zeigt ein
kontinuierliches Ansteigen der Spikerate bis zu einem Maximalwert bei einer
Stimulusgeschwindigkeit von 70°/s. Danach nimmt die Entladungsrate der Zellen stetig ab.
Die 11 untersuchten richtungsselektiven Neurone von Esox lucius erreichen ihre maximale
Spikerate bei einer Reizgeschwindigkeit von 20 °/s. Es folgt eine Abnahme der Rate bis
30 °/s Stimulusgeschwindigkeit. Diese Spikerate bleibt bis zur maximal getesteten
Reizgeschwindigkeit von 80°/s konstant.
Mit diesen Ergebnissen können zwei Gruppen von richtungsselektiven Neuronen gebildet
werden: bei Salmo gairdneri richtungsselektive Neurone, die hohe Geschwindigkeiten bis
60°/s kodieren können, und bei Esox lucius richtungsselektive Neurone, die auf geringe
Geschwindigkeitsänderungen mit einer starken neuronalen Aktivität reagieren. Beide
Gruppen sowie ihre Interaktionen wurden auch in den anderen Vertebratenklassen
beschrieben (Kogo et al., 1998; Ariel & Kogo, 2001).
Von besonderer Bedeutung scheint dies bei den niederen Vertebraten im Zusammenspiel
des nLM und nBOR unter Einbeziehung der richtungsselektiven retinalen Ganglienzellen
zu sein. Im nLM finden sich sog. „langsame“ richtungsselektive Neurone, im nBOR
„schnelle“ richtungsselektive Neurone (Wylie & Crowder, 2000. Die Modulation erfolgt
von nBOR Zellen inhibitorisch auf die Neurone des nLM und bezieht sich nicht nur auf
das Geschwindigkeitsverhalten der nLN Neurone sondern auch auf ihr Tuning – Verhalten.
Interessanterweise konnte das bei Esox lucius beschriebene „Push-Pull“ Prinzip an den
nLM und nBOR Neuronen nachgewiesen werden (Ariel & Kogo, 1998, 2001). Im Rahmen
dieser Untersuchungen wurde auch festgestellt, dass die Latenzzeiten der
richtungsselektiven Neurone mit 80 ms relativ lang sind. In diesem Zeitbereich lag auch
der Median der Latenzzeiten von Salmo gairdneri und Esox lucius. Mit diesem Ergebnis
kann kein direkter retinaler Eingang bewiesen werden, ist aber vergleichbar mit
Latenzzeiten anderer Vertebraten (Cochran et al., 1984; Klauer et al., 1990).
4.6 Lage der richtungsselektiven Neurone im Praetectum Die anatomische Rekonstruktion des Ableitortes ergab eine dorsale Position im medialen
Praetectum von Salmo gairdneri und Esox lucius, wobei sich das Areal von Esox lucius
57

I. Elektrophysiologie 4. Diskussion
ca. 300 µm weiter rostral im Praetectum befand. Dieser Ableitort kann mit
neuroanatomischen Untersuchungen an anderen Fischarten (Goldfisch, Zebrafisch,
Flussbarsch), sowie an Salmo gairdneri und an Esox lucius selbst nicht in Einklang
gebracht werden. Diese befanden sich im lateralen Bereich des Praetectums (Finger &
Karten, 1978; Fite, 1985; Butler et al., 1991).
Analog zu den Untersuchungen am optokinetischen System bei Säugern, Reptilien, Vögeln
und Amphibien wurden in der Überordnung Teleostei der Klasse Osteichthyes ebenfalls
zahlreiche Untersuchungen zur neuroanatomischen Identifizierung der beteiligten
Hirnstrukturen durchgeführt. Dies Studien dienten auch dem Versuch einer
Homologisierung von Hirnarealen einschließlich Praetectum und AOS zu anderen
Vertebratenklassen. Hauptsächlich wurden dazu Arten aus der Familie Cyprinidea
(Carassius auratus, Cyprinus carpio) und perciforme Arten untersucht.
Finger und Karten versuchten 1978, das AOS von Carassius auratus und Ictalurus
punctatus näher zu beschreiben und mit dem nLM bzw. nBOR anderer Vertebratenklassen
zu homologisieren. Dabei wurde ein großer Bereich des Praetectums zu einer Region
zusammengefasst, die von ihnen mit Areal P1/P2 bezeichnet wurde. Der Hauptgrund dafür
sind die Projektionen von P1 zum Cerebellum und den okulomotorischen Kernen.
Vanegas und Ito (1983) versuchten, das Areal P1 weiter zu differenzieren, und fanden
heraus, dass sich innerhalb des Areals P1 Zellen mit unterschiedlichen Morphologien
befinden. Dabei ordneten sie eine Neuronenpopulation mit großem Soma am anterioren
Pol von P1 der Area praetectalis zu, von der ebenfalls Projektionen zum Cerebellum und
zu den okulomotorischen Kernen führen. Sie beschreiben diesen Bereich des
Knochenfischgehirns als äquivalent zum akzessorischen optischen System anderer
Vertebraten. In weiteren Untersuchungen wurde diese eindeutige Zuordnung nicht bestätigt
(Sharma, 1972; Springer & Gaffney, 1981).
In neuroanatomischen Arbeiten, die sich auf das Praetectum und das AOS von Salmo
gairdneri und Esox lucius beschränken, ergaben sich erhebliche Unterschiede in der
Positionsangabe der Area praetectalis, sowie den Angaben zur Morphologie und den
efferenten Projektionen (Repérant et al., 1976; Fite, 1985; Butler et al., 1991). Eine genaue
Zuordnung der Kerne des AOS erweist sich als sehr schwierig und wird bis heute
kontrovers diskutiert.
58

I. Elektrophysiologie 4. Diskussion
Dies zeigt, dass mit neuroanatomischen Untersuchungen allein eine exakte Zuordnung von
Zellpopulationen bzw. Kerngebieten äußerst schwierig ist. Daraus begründet sich die
Vorgehensweise, mit elektrophysiologischen Methoden nach den neuronalen Grundlagen
optokinetischer Augenbewegungen im Praetectum und AOS von Salmo gairdneri und
Esox lucius zu suchen.
4.7 Topographische Organisation der Vorzugsrichtungen Die Hypothese einer möglichen topographischen Ordnung in dem dorsalen praetectalen
Areal ist nicht unbegründet. Dieses grundlegende Ordnungsprinzip findet sich in vielen
visuellen Arealen des Gehirns wie z. B. im Tectum opticum (Schwassmann, 1968;
Schwassmann & Krüger, 1970; Guthrie & Banks; 1978, Grover & Sharma, 1979).
Die Untersuchung wurde an vier Salmo gairdneri in vier unterschiedlichen caudo -
rostralen Positionen des richtungsselektiven Zellbereichs durchgeführt. Entlang der
Penetrationsspur wurden alle richtungsselektiven Neuronen analysiert, sowie ihre
Ableittiefe und Vorzugsrichtung bestimmt. Anschließend wurden Ableitort und
Vorzugsrichtung miteinander in Bezug gesetzt.
Das Ergebnis dieser Untersuchung stellt sich wie folgt dar:
Im rostralen Bereich des richtungsselektiven Areals wurden neben vertikalen Richtungen
kodierenden Neuronen nur Zellen mit einer temporo - nasalen Vorzugsrichtung gefunden.
Im mittleren Bereich des richtungsselektiven Areals wurden Neurone mit einer nasalen
oder temporalen Vorzugsrichtung abgeleitet. Im caudalen Bereich überwog die Anzahl von
Neuronen, die eine naso - temporale Reizrichtung bevorzugen.
Diese Befunde deuten auf eine räumliche Trennung von Neuronen unterschiedlicher
Vorzugsrichtung in diesem Kerngebiet hin. Falls der anteriore und der posteriore Teil des
Kerngebietes unterschiedliche Projektionen aufweisen, ließe sich damit möglicherweise
auch die Symmetrie bzw. Asymmetrie des mhOKR erklären.
Weitere Untersuchungen zu diesem Befund werden in II. Verhaltensexperimente dieser
Arbeit vorgestellt.
59

II. Verhaltensuntersuchungen 1. Einleitung
II. Verhaltensexperimente zum visuomotorischen
System von Salmo gairdneri
1. Einleitung Die elektrophysiologischen Experimente zur Identifizierung des neuronalen Substrats des
OKR an Salmo gairdneri und Esox lucius haben eindeutig gezeigt, dass bei den
Salmoniden neben horizontal verlaufenden, visuellen Stimuli auch vertikale Reize von den
richtungsselektiven Neuronen im dorsalen Praetectum kodiert werden.
Bei den Säugern werden horizontal bewegte optokinetische Stimuli im NOT, vertikal
bewegte Reize dagegen im LTN und MTN verarbeitet (Grasse & Cynander, 1988; Mustari
& Fuchs, 1989).
Auch Vögel, Reptilien und Amphibien besitzen zwei neuronale Strukturen für die
Verarbeitung verschiedener Stimulusrichtungen, nämlich den praetectalen Nucleus
lentiformis mesencephali (nLM), der eine horizontale ipsiversive Reizrichtung kodiert und
den Nucleus of the basal optic root (nBOR). Der nBOR beinhaltet richtungsselektive
Neuronenpopulationen, die neben vertikal verlaufenden Stimuli auch horizontale
kontraversive Reizrichtungen kodieren (Reptilia: Ariel, 1997; Amphibia: Katte &
Hoffmann, 1980; Aves: Wylie & Forst, 1990).
In Verhaltensexperimenten mit vertikal bewegten visuellen Reizmustern wurde bei allen
bisher untersuchten Tierarten eine Asymmetrie in der Güte der langsamen Phase des
vertikalen OKR nachgewiesen (Mammalia: Matsuo & Cohen, 1984; Bohmer & Baloh,
1990; Borel & Lacour, 1992; Fuchs, 1989).
Bei Menschen, Affen und Katzen wird eine aufwärts gerichtete Stimulusrichtung besser
kompensiert (Grasse & Cynander, 1988; Fuchs, 1989), während z.B. beim Huhn
abwärtsverlaufende Reize besser kompensiert werden (Wallman & Velez, 1985).
In der Überordnung Teleostei wurden bisher keine Verhaltensuntersuchungen zum
vertikalen OKR durchgeführt, obgleich dieser bei Bewohnern einer dreidimensionalen
Unterwasserwelt nicht unbedeutend seien dürfte. Eine vertikale Stimulationsrichtung
würde einer Rollbewegung über die Körperlängsachse des Fisches entsprechen.
60

II. Verhaltensuntersuchungen 1. Einleitung
Eine weitere Möglichkeit, einer retinalen Bildverschiebung entgegenzuwirken, wird von
dem vestibulo - okulären System bereitgestellt. Es dient der Detektion von translatorischen
Beschleunigungen und Drehbeschleunigungen des Kopfes bzw. des Körpers Ausgehend
von den drei Bogengängen im Innenohr, dem Canalis semicircularis lateralis, C.s. anterior
und dem C.s. posterior werden über nur drei synaptische Verbindungen die Augenmuskeln
angesteuert. Die Projektion führt von den Bogengängen ausgehend über den VIII.
Gehirnnerv (Nervus statoacusticus) zu den Vestibulariskernen, von dort weiter zu den
Augenmuskelkernen und anschließend zu den Augenmuskeln. Dieses System ist bei allen
Vertebraten inklusive der Teleostei (Graf et al., 1997, 2001) nachgewiesen.
Das vestibulo - okuläre und das optokinetische System interagieren miteinander, um eine
Optimierung der Bildstabilisierung auf der Retina bei unterschiedlichen Reizeinflüssen zu
erzielen. Der visuelle Weg führt, ausgehend von den Kernen des AOS, zur ipsilateralen
dorsalen Kappe der Oliva inferior und von dort über die Kletterfasern zum Cerebellum. Er
dient der Kalibrierung des vestibulo - okulären Reflexes (Ito et al., 1974; Ito, 1982). Ein
weiterer Pfad führt vom AOS zum Nucleus praepositus hypoglossi, der den visuellen
Eingang für die Vestibulariskerne liefert. Dies sind die zentralnervösen Bereiche, in denen
beide sensorischen Informationen, retinale und vestibuläre, miteinander verrechnet werden.
Untersuchungen am vestibulo - okulären System echter Knochenfische wurden
hauptsächlich am Goldfisch (Carassius auratus) mit und ohne visuelle Stimulation
ausschließlich in horizontaler Richtung durchgeführt (Schairer & Bennett, 1986; Pastor et
al., 1992; Graf et al., 1997; Mc Elligott et al., 1998). In diesen Verhaltensuntersuchungen
konnte stets eine Verbesserung des gains kompensatorischer Augenbewegungen des VOR
durch visuelle Stimulation gezeigt werden (Mc Elligott, 1988).
Über die Anordnung der Bogengänge werden alle Raumrichtungen kodiert. Das Ziel dieses
Teils der Arbeit ist die Untersuchung vertikaler Augenbewegungen an Salmo gairdneri, da
auch vertikale Bewegungen kodierende richtungsselektive Neuronen im Praetectum
identifiziert wurden.
Bei diesem komplexen sensomotorischen System drängt sich außderdem die Frage auf,
welche Folgen eine Teilzerstörung des praetectalen, richtungsselektiven Areals hat.
61

II. Verhaltensuntersuchungen 1. Einleitung
Mit unilateralen reversiblen Inaktivierungen oder Läsionen des NOT bzw. nLM konnten in
den unterschiedlichen Vertebratenklassen Veränderungen im optokinetischen Verhalten
aufgezeigt werden. Typischerweise zeigten Versuchstiere mit einer solchen Inaktivierung
Spontannystagmus, wobei die langsame Folgephase vom Läsionsort weggerichtet ist
(Mammalia: Hoffmann & Fischer; 2001). Bei Fröschen können zusätzlich Kopfnystagmen
beobachtet werden (Katte & Hoffmann, 1980; Fite, 1985). Bei visueller Stimulation sind
die Augenfolgebewegungen in ihrer Geschwindigkeit stark beeinträchtigt. Vergleichbare
Experimente wurden bei Knochenfischen bislang nicht durchgeführt.
Daraus ergeben sich folgende Fragestellungen, die an der Regenbogenforelle (Salmo
gairdneri) untersucht wurden.
1. Kann ein vertikaler OKR, der einer Rollbewegung um die Körperlängsachse
kompensiert, bei Salmo gairdneri ausgelöst werden?
2. Wie wirken sich unterschiedliche visuelle Reizbedingungen auf die kompensatorischen
Augenbewegungen des vestibulo - okulären Reflexes in vertikaler Stimulusrichtung
aus?
3. Welche Defizite beim mhOKR sind nach unilateraler Zerstörung des dorsalen
praetectalen Areals richtungsselektiver Zellen feststellbar?
62

II. Verhaltensuntersuchungen 2. Material & Methoden
2. Material und Methoden
2.1 Versuchstiere Die Verhaltensuntersuchungen wurden an insgesamt 17 Salmo gairdneri beiderlei
Geschlechts durchgeführt. Ihr Körpergewicht lag zwischen 180 bis 350 g. Die Herkunft der
Tiere und die Hälterungsbedingungen entsprechen den Angaben aus dem ersten Teil dieser
Arbeit. Die Untersuchungen zum vertikalen OKR wurden an drei Salmo gairdneri
durchgeführt. Ihr Alter betrug 15 bis 23 Monate. An acht weiteren Fischen in einem Alter
von 9 bis 14 Monaten fanden die Experimente zum vestibulo-okulären Reflex (VOR) statt.
Weitere sechs Tiere wurden in den Läsionsexperimenten untersucht.
2.2 Verhaltensexperimente
2.2.1 Kälte - Immobilisierung und Fixation Die Fische wurden in einen 15 Liter fassenden Styroporbehälter, der mit 4°C kaltem
Wasser gefüllt war, eingesetzt. Aufgrund der Poikilothermie trat die Kälteimmobilisierung
von Salmo gairdneri nach 30 bis 50 Minuten ein. Um Verletzungen der Fischschleimhaut
zu vermeiden, wurden die Fische anschließend bis auf die Kopf- und Kiemenregion mit
einem feuchten Zellulosepapier ummantelt und in eine Plexiglas - Fixationshalterung
eingesetzt.
Diese bestand aus einer Grundplatte (30 cm x 10 cm), an deren vorderem Ende sich eine
Maulhalterung mit einem integrierten Tubus befand. Die Maulhalterung diente der
Vermeidung von Kopfbewegungen während des Experiments. Über den Tubus wurde
sauerstoffangereichertes und temperiertes Wasser (12°C) mit einem Durchflussvolumen
von 300 ml pro Minute in das Maul der Fische zugeführt. Zwei bewegliche
Seitenhalterungen waren innenseitig mit Schaumstoff ausgekleidet und dienten der
Fixierung des Rumpfs, so dass die Fische in der Halterung bewegungsunfähig blieben.
63

II. Verhaltensuntersuchungen 2. Material & Methoden
2.2.2 Versuchsaufbau für die Verhaltensexperimente zum
optokinetischen und vestibulo - okulären Reflex Bei der Messapparatur zur Erfassung optokinetischer und vestibulo - okulärer
Augenpositionsänderungen handelte es sich um ein elektrisches Wechselmagnetfeld, das in
Anlehnung an Robinson (1963) nach dem Prinzip der Erzeugung einer Induktionsspannung
arbeitet. Im Inneren eines Kubus waren zwei Feldspulenpaare orthogonal zueinander
angeordnet und erzeugten ein annähernd homogenes, horizontales Wechselmagnetfeld mit
einer Frequenz von 35 kHz und ein vertikales Wechselmagnetfeld mit einer Frequenz von
17,5 kHz. Mit dieser Versuchsanordnung wird das Prinzip der magnetischen Induktion
genutzt, indem eine Messspule (Augenmessspule) in das Wechselmagnetfeldes eingebracht
wird, in dereine Induktionswechselspannung UInd(t) erzeugt wird.
dttBdBAAn
Ind tU )]([)),(cos()( •Φ••=
In der Gleichung steht B(t) für die mit der Trägerfrequenz modulierende, zeitabhängige
Größe des magnetischen Wechselfeldes, A(t) ist die Fläche der Spule, n die Zahl der
Spulenwindungen. Φ(A,B) bezeichnet den Winkel zwischen den Feldlinien des
Magnetfeldes und der Flächennormalen der Spule.
Zwei Spulensysteme unterschiedlicher Größe wurden verwendet.
A. Großes Spulensysteme mit den Maßen 80 x 80 x 80 cm (vOKR und mhOKR)
In den Kubus des Wechselmagnetfeldes wurde die opake Plexiglashalbbkugel (siehe
elektrophysiologisches Experiment) eingesetzt. Das Versuchstier befand sich in der
Fixationshalterung mit dem rechten Auge zentriert in der wassergefüllten Halbkugel.
Das in einem Abstand von 10 cm über dem Kopf zentrierte Planetarium projizierte ein
Zufallspunktemuster durch das Wasser auf die opake innere Oberfläche der
Plexiglashalbkugel. Zwischen dem Kopf des Versuchstieres und dem Planetarium
wurde eine Sichtblende eingesetzt, um visuelle Irritationen zu vermeiden.
In das gleiche Wechselmagnetfeld konnte auch eine Reiztrommel mit einem
Durchmesser von 70 cm und einer Höhe von 30 cm eingesetzt werden. Innenseitig war
diese mit einem schwarz-weißen Zufallspunktemuster ausgekleidet. Der Innenraum
wurde mit einer Glühbirne künstlich ausgeleuchtet. Das Versuchstier befand sich in
64

II. Verhaltensuntersuchungen 2. Material & Methoden
seiner Fixationshalterung, mit den Augen zentriert in der Mitte der Apparatur innerhalb
eines durchsichtigen wassergefüllten Plexiglasbeckens (Klar 1999).
B. Kleines Spulensystem mit den Maßen 30 x 30 x 30 cm (VOR)
Der Kubus des Spulensystems war über eine zentrierte Achse vertikal und horizontal
drehbar. Über eine durch einen Elektromotor angetriebene Exzenterscheibe, die mit
dem Magnetfeld verbunden war, konnten vertikale und horizontale sinusoidale
Drehbewegungen mit Auslenkungsamplituden von +/- 10° bei einer Frequenz von 0,1
Hz ausgeführt werden. Die Versuchstiere befanden sich kopfzentriert innerhalb des
Wechselmagnetfelds in einer mit der Drehachse verbundenen Fixationshalterung in
einer vollständig wassergefüllten Glasglocke. Um die Glasglocke herum befand sich
ein Zylinder, der innenseitig mit einem schwarz-weißen Zufallspunktemuster
ausgekleidet war und mit Kunstlicht beleuchtet werden konnte.
Der Aufbau wurde vervollständigt durch eine externe Wasseraufbereitungsanlage
bestehend aus einem Kryostaten, der eine gleichbleibende Wassertemperatur (12° C)
erzeugte, einer Membranpumpe zur Sauerstoffanreicherung des Wassers und einer
Wasserpumpe, die das aufbereitete Wasser wieder mit einer Flussrate von 300 ml/min über
den Tubus in das Maul der Versuchstiere zurückführte.
HFS2
VFS1
VFS2
HFS1
BA
Abb. II. 2.1: Schematische Darstellung der zwei Versuchsapparaturen zur Erfassung von Augenbewegungen.
In A ist das Wechselmagnetfeld zur OKR - Stimulation, in B das Wechselmagnetfeld zur VOR - Stimulation
dargestellt. HFS1 und HFS2 = horizontale Feldspulen, VFS1 und VFS2 = vertikale Feldspulen, PI =
Plexiglashalbkugel, P = Planitarium.
65

II. Verhaltensuntersuchungen 2. Material & Methoden
2.2.3 Reizmodalitäten
2.2.3.1 Vertikaler optokinetischer Reflex
Diese Verhaltensexperimente fanden in der Plexiglashalbkugel mit dem durch das
Planetarium generierten Zufallspunktemuster im großen Wechselmagnetfeld statt.
Untersucht wurden monokulare, horizontale optokinetische Augenbewegungen
(Kontrollexperiment) und monokulare / binokulare, vertikale optokinetische
Augenbewegungen. Die Stimulusgeschwindigkeiten betrugen 5, 10, 15, 20, 25 und 30 °/s.
Horizontal wurde in temporo - nasaler (T-N) und naso - temporaler (N-T) Richtung
stimuliert, die vertikale Reizung erfolgte nach oben und unten mit den gleichen
Reizgeschwindigkeiten.
2.2.3.2 Kompensatorische Augenbewegungen des vestibulo - okulären Reflexes
Die sinusförmige Drehbewegungen des Magnetfeldkubus mit dem Fisch in seiner
Fixationshalterung hatten in der vertikalen und horizontalen Stimulationsebene eine
Amplitude von +/- 10° bei einer Frequenz von 0,1 Hz.
Drei Stimulationsparadigmen wurden in beiden Stimulationsebenen untersucht:
A) Das Reizmuster drehte sich mit exakt der gleichen Geschwindigkeit in die gleiche
Richtung wie das Versuchstier, so dass keine Relativbewegung zwischen
Fischhalterung und Reiztrommel auftrat.
B) Das Reizmuster blieb stationär im Raum, während das Versuchstier sinusoidal
gedreht wurde.
C) Das Reizmuster drehte mit exakt der gleichen Geschwindigkeit gegen die
sinusoidale Drehrichtung des Versuchstieres, so dass die relative Bewegung von
Fisch und Reiztrommel eine Amplitude von 20° hatte.
Zum Vergleich wurden Experimente im Dunkeln mit Drehbewegungen in vertikaler und
horizontaler Stimulationsebene durchgeführt.
2.2.3.3 Monokularer horizontaler optokinetischer Reflex im Läsionsexperiment
Mit der Reiztrommel wurde der mhOKR untersucht. Das rechte Auge wurde mit einem
schwarz-weißen Zufallspunktemuster stimuliert. Dabei wurden Reizgeschwindigkeiten von
4, 9, 14, 18, 24, 28 und 32 °/s in naso - temporaler (N-T) und temporo - nasaler (T-N)
66

II. Verhaltensuntersuchungen 2. Material & Methoden
Richtung verwendet. Die Positionen des visuell stimulierten und des abgedeckten Auges
wurden aufgezeichnet.
Die Aufzeichnungsdauer in allen Verhaltensexperimenten betrug 30 Sekunden in der
steady – state - Phase der Stimulation. Die Abfolge der Geschwindigkeiten war stets
zufällig gewählt.
2.2.4 Messdatenerfassung, Analyse und Auswertung Den Versuchstieren wurde irritationsfrei eine Augenmessspule (∅ = 2 mm, 40 Windungen,
Eigenbau) mit einer parallelen Ausrichtung zu den horizontalen bzw. vertikalen
Magnetfeldlinien auf das Augenlid geklebt (Sekundenkleber, Patex).
Augenpositionsänderungen bewirkten eine Veränderungen der Induktionsspannung in der
Augenspule. Die Induktionsspannungsänderung wurde abgegriffen und in zwei LOCK - IN
Verstärker für die horizontale und die vertikale Komponente geleitet (Princeton Applied
Resarch, Model 128 A). Mit Hilfe der Referenzsignale der magnetischen Wechselfelder
wurde die Amplitude des Trägersignals aus dem induzierten Augenspulensignal
elektronisch bestimmt. Die unmodulierten Augenpositionssignale wurden dann über einen
parallel geschalteten Zweikanal-Kontrolloszillographen zu einem Computer (MAX DATA
386) mit einer A/D Wandlerkarte weitergeleitet. Mit der Computersoftware EYEMOVE
wurden die horizontalen und vertikalen Augenpositionssignale mit einer Rate von 100 Hz
aufgezeichnet. Die graphische Online-Darstellung der beiden Augenpositionssignale
erfolgte auf einem Monitor als Spannung (V) gegen die Zeit (ms).
Alle verwendeten Augenspulen wurden entsprechend der Versuchsbedingungen der
Verhaltensexperimente für das rechte und das linke Auge kalibriert. Dazu wurden die
Augenspulen an die exakte Augenposition der Versuchstiere innerhalb der
Wechselmagnetfeld - Messapparaturen gebracht. Für die OKR - Kalibrierungen wurden
die Augenspulen an einem Goniometer angebracht und nacheinander exakt in eine 10°
nasale - und 10° temporale horizontale Richtung, sowie in vertikaler Richtung 10° nach
oben und 10° nach unten gedreht. Zur Erstellung der VOR Kalibrierungssignale wurde die
67

II. Verhaltensuntersuchungen 2. Material & Methoden
Augenspule an der Fixationshalterung befestigt und der Kubus in vertikaler und
horizontaler Richtung gedreht. Diese Kalibrationsmessungen wurden mit EYEMOVE
aufgezeichnet und dienten als Berechnungsgrundlage der Augenpositionssignale.
2.2.4.1 Vertikaler und horizontaler optokinetischer Reflex
Eine in MATLAB (The MathWorks, Inc., Version 5.3.1 (R11.1)) programmierte
Analysesoftware errechnete aus den aufgezeichneten Augenpositionssignalen
entsprechende Geschwindigkeitssignale. Sakkadische Augenbewegungen zeichnen sich
durch ihre hohen Geschwindigkeiten aus. Sie konnten durch eine variable
Geschwindigkeitsschwelle detektiert und eliminiert werden. Durch Integration wurde das
bearbeitete Augengeschwindigkeitssignal ohne die Sakkaden zurück in Positionssignale
transformiert. Über die Kalibrationsmessung der Augenspulen (+/- 10° Auslenkung)
erfolgte die Umrechnung der Augenpositionssignale in Grad (°).
Mit einer zeitlich seriellen Bearbeitung der Datenfiles wurden die Anfangs- und
Endzeitpunkte der Augenfolgebewegungen bestimmt, sowie die Anfangs- und Endwerte
der Augenpositionsamplituden (Grad). Damit konnten die Geschwindigkeiten aller
langsamen Augefolgephasen innerhalb der 30 s Aufzeichnungen berechnet und deren
Median ermittelt werden. Aus dem Geschwindigkeitsmedian und der entsprechenden
Reizgeschwindigkeit wurde durch Quotientenbildung das gain berechnet.
Weiterhin erstellte das Programm Daten über die Anzahl der Augenfolgebewegungen
innerhalb der 30 s Aufzeichnungen und den Median der Amplituden dieser
Augenfolgebewegungen.
2.2.4.2 Vestibulo-okulärer Reflex
Die quantitative Berechnung des gains der kompensatorischen Augenbewegungen während
des VORs wurde gleichfalls mit einer in MATLAB programmierten Routine durchgeführt.
Im ersten Schritt der Analyse wurde aus der ersten Ableitung der sinusförmigen
Augenpositionssignale das Geschwindigkeitssignal berechnet (d.h. ein Cosinus). Durch
eine anschließende schnelle Fourier-Transformation (FFT) wurden die
Geschwindigkeitssignale auf ihre Energie, Frequenzspektrum und Phasenverschiebung hin
untersucht.
68

II. Verhaltensuntersuchungen 2. Material & Methoden
Das periodische Leistungssignal sP(t) nimmt in einem endlichen Zeitintervall (0s bis 30s
Aufzeichnungsdauer) hohe Werte an. Damit hat das Signal eine ∞ hohe Energie E.
∫∞
∞−
∞⇒= dttsE P2)(
In der Spektralanalyse (die Signalaufzeichnungsrate betrug 100 Hz) wurde ein Bereich von
0 Hz bis 50 Hz in 0,1 Hz Schritten analysiert Die maximale Energiedichte trat bei 0,1 Hz
auf. Damit trat keine Frequenzverschiebung gegenüber der Stimulationsfrequenz auf.
Aufgetretene Seitenenergien +/- 0,1 Hz sind auf ein Signalrauschen zurückzuführen. Der
sakkadische Anteil befindet sich wegen seines hohen Geschwindigkeitsanteils in einem
Frequenzbereich > 50 Hz.
Die Energie des Maximums bei 0,1 Hz wurde in die Amplitude der zugehörigen
Schwingung umgerechnet (Quadratwurzel) und dann mit der Stimulusgeschwindigkeit
zum gain verrechnet.
Durch das Kontrollexperiment mit den paralysierten Versuchstieren wurde der apparative
Fehler des Messsystems bestimmt. Dieser Wert wurde von dem VOR - gain der
Augenbewegungen abgezogen. Die Normierung des gains erfolgte durch die Werte der
Kalibrierungsdatei, die einem gain - Wert von 1 entsprechen.
2.2.5 Statistische Verfahren zur Signifikanzbestimmung
2.2.5.1 Vestibulo - okulärer Reflex
Für die Ergebnisse der einzelnen Tiere wurde aus fünf Einzeltrials (je 30 s) eines
Stimulationsparadigmas der Mittelwert errechnet. Die Mittelwerte der jeweiligen
untersuchten Stimulationsparadigmen wurden mit dem parametrischen t-Test auf
signifikante Unterschiede überprüft. In die Gruppenanalyse gingen die Mittelwerte aller
untersuchten Tiere zu einem Stimulationsparadigma ein. Anschließend wurde mit Hilfe der
Ein – Wege - ANOVA (p < 0,05) auf signifikante Unterschiede der untersuchten
Stimulationsparadigmen einer Versuchsreihe getestet.
69

II. Verhaltensuntersuchungen 2. Material & Methoden
2.2.5.2 Läsionsexperiment
Veränderungen des gains der Augenfolgebewegungen in Bezug auf die
Stimulationsrichtungen wurden auf signifikante Unterschiede mit der Einwege - ANOVA
(p < 0,05) untersucht. Gleiches gilt für die Anzahl und Amplitudengrößen der
Augenfolgebewegungen.
Die statistischen Analysen erfolgten mit der Computersoftware SigmaStat© (SPSS Inc.,
Version 2.03).
2.3 Elektrophysiologisches Läsionsexpriment
2.3.1 Fischanästhesie und Präparation Die Anästhesie und Präparation der sechs untersuchten Salmo gairdneri entsprachen der in
I. 2.3 beschriebenen Vorgehensweise. Ein Unterschied bestand in dem nicht vollständigen
Entfernen des Haut - Muskelgewebes im Kopfbereich. Es blieb über den Zeitraum des
elektrophysiologischen Experiments an der lateralen Seite erhalten.
Im Anschluss an die elektrophysiologische Lokalisation des praetectalen Areals und die
Läsion wurde die Schädelöffnung wieder verschlossen. Ein genau auf die Öffnung
angepasstes steriles Stück Paraplast wurde mit Gewebekleber (Braun) am Knochen plan
und möglichst wasserdicht aufgeklebt. Anschließend wurde das zu Versuchsbeginn
erhaltene Haut-Muskelgewebe wieder in die Wundöffnung eingesetzt. Der vollständige
und wasserabweisende Verschluss der Wunde erfolgte abschließend mit Gewebekleber.
Die Wunde wurde mit Lokalanästhetikum Xylonest® versorgt. Anschließend erfolgte das
Ausleiten der MS-222 Narkose. Die unterstützende Beatmung wurde erst eingestellt,
nachdem der Fisch selbständig und gleichmäßig Kiemenbewegungen ausführte. Dies
geschah ca. 3 - 4 Stunden nach dem Wundverschluss.
70

II. Verhaltensuntersuchungen 2. Material & Methoden
2.3.2 Versuchaufbau und Versuchsdurchführung Der Versuchsaufbau zur Lokalisation praetectaler richtungsselektiver Neurone und der
visuelle Stimulationsmodus entsprach den in I. 2. 4 beschriebenen Bedingungen.
Die elektrolytischen Mikroläsionen erfolgten mit den glasisolierten Wolframdraht-
Ableitelektroden. Über eine netzunabhängige Stromversorgung (A-M Systems, Model
2100) wurde 30 Sekunden lang ein Strom von 10 µA (Elektrodenspitze positiv) erzeugt.
2.3.3 Bestimmung der neuronalen Richtungscharakteristik Die Auswertung der Tuning - Kurven richtungsselektiver Zellen und die statistische
Berechnung der Richtungsvektoren erfolgte mit MATLAB (I. 2.7).
2.3.4 Histologische Lokalisation des Läsionsortes Die Fische wurden mit einer Überdosis MS-222 (5%) eingeschläfert. Danach wurden die
Tiere mit 4% Formol in 0,1 M Phosphatpuffer transcardial perfundiert. Von den Gehirnen
wurden 50 µm dicke Gefrierschnitte angefertigt. Nach der Nissl - Färbung wurden die
Läsionsorte mit einem Zeiss Mikroskop (AXIOSKOP) lokalisiert und mit einer Zeiss
Digitalkamera (Axiocam color, 412-312) fotografiert.
71
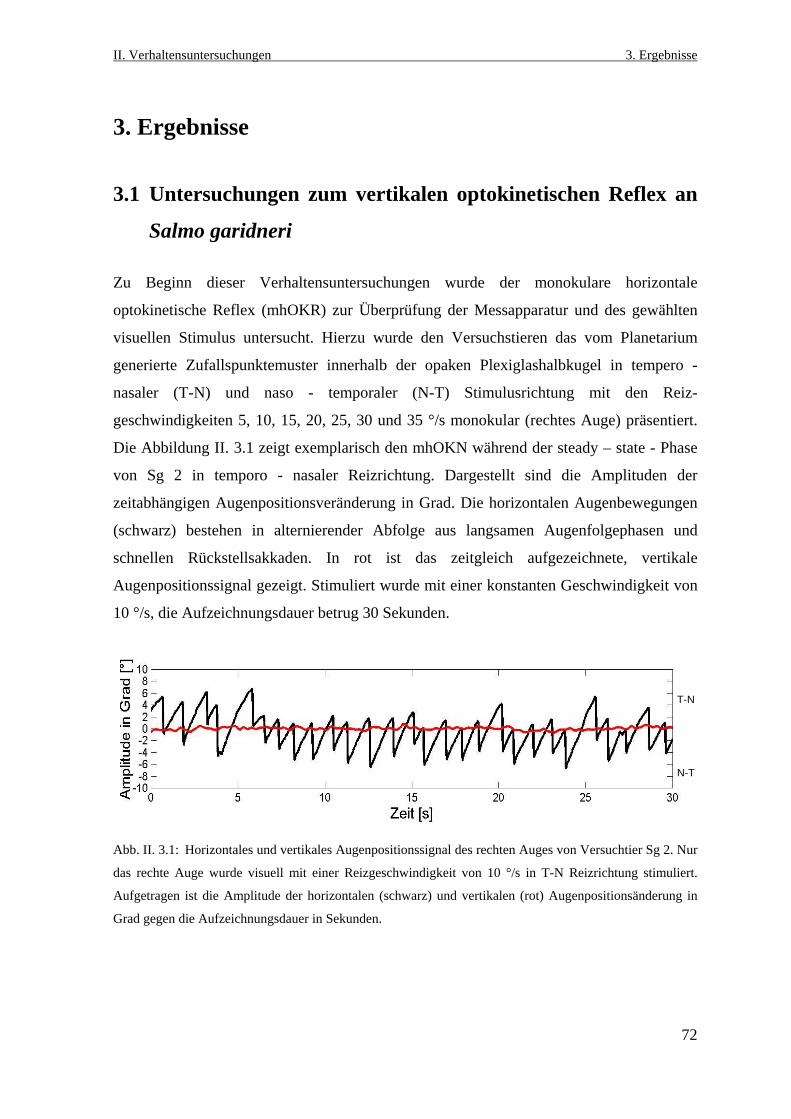
II. Verhaltensuntersuchungen 3. Ergebnisse
3. Ergebnisse
3.1 Untersuchungen zum vertikalen optokinetischen Reflex an
Salmo garidneri Zu Beginn dieser Verhaltensuntersuchungen wurde der monokulare horizontale
optokinetische Reflex (mhOKR) zur Überprüfung der Messapparatur und des gewählten
visuellen Stimulus untersucht. Hierzu wurde den Versuchstieren das vom Planetarium
generierte Zufallspunktemuster innerhalb der opaken Plexiglashalbkugel in tempero -
nasaler (T-N) und naso - temporaler (N-T) Stimulusrichtung mit den Reiz-
geschwindigkeiten 5, 10, 15, 20, 25, 30 und 35 °/s monokular (rechtes Auge) präsentiert.
Die Abbildung II. 3.1 zeigt exemplarisch den mhOKN während der steady – state - Phase
von Sg 2 in temporo - nasaler Reizrichtung. Dargestellt sind die Amplituden der
zeitabhängigen Augenpositionsveränderung in Grad. Die horizontalen Augenbewegungen
(schwarz) bestehen in alternierender Abfolge aus langsamen Augenfolgephasen und
schnellen Rückstellsakkaden. In rot ist das zeitgleich aufgezeichnete, vertikale
Augenpositionssignal gezeigt. Stimuliert wurde mit einer konstanten Geschwindigkeit von
10 °/s, die Aufzeichnungsdauer betrug 30 Sekunden.
T-N N-T
Abb. II. 3.1: Horizontales und vertikales Augenpositionssignal des rechten Auges von Versuchtier Sg 2. Nur
das rechte Auge wurde visuell mit einer Reizgeschwindigkeit von 10 °/s in T-N Reizrichtung stimuliert.
Aufgetragen ist die Amplitude der horizontalen (schwarz) und vertikalen (rot) Augenpositionsänderung in
Grad gegen die Aufzeichnungsdauer in Sekunden.
72
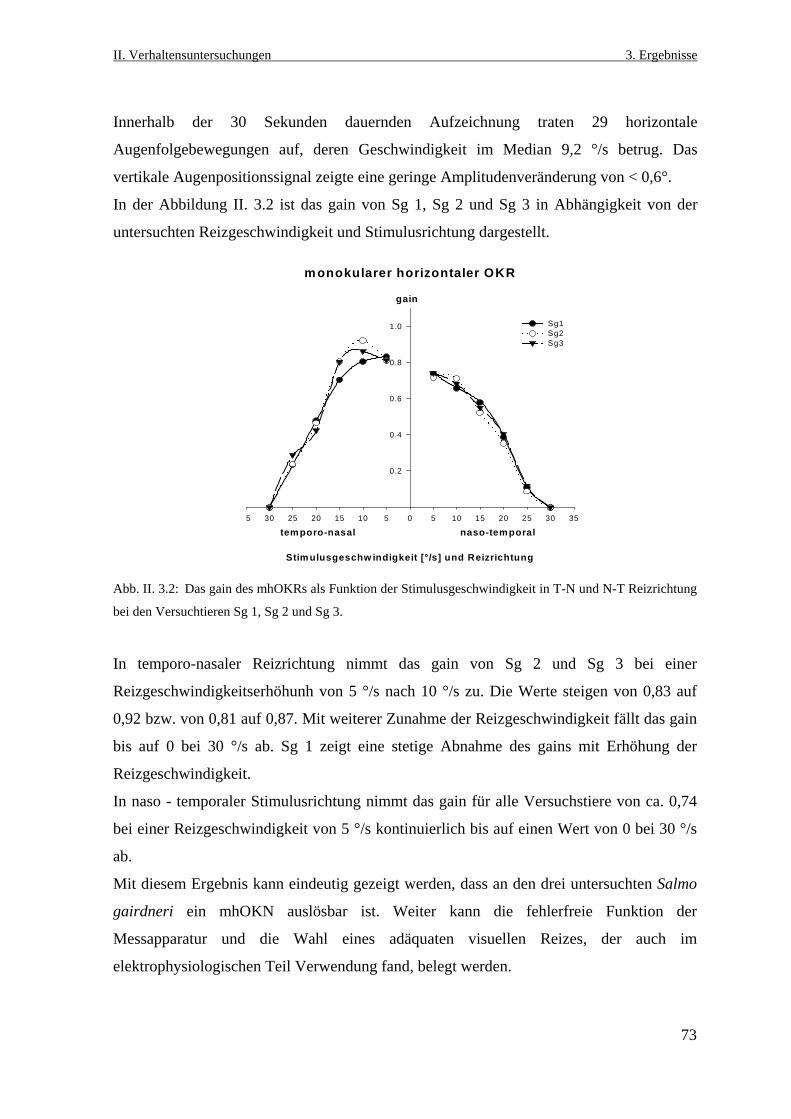
II. Verhaltensuntersuchungen 3. Ergebnisse
Innerhalb der 30 Sekunden dauernden Aufzeichnung traten 29 horizontale
Augenfolgebewegungen auf, deren Geschwindigkeit im Median 9,2 °/s betrug. Das
vertikale Augenpositionssignal zeigte eine geringe Amplitudenveränderung von < 0,6°.
In der Abbildung II. 3.2 ist das gain von Sg 1, Sg 2 und Sg 3 in Abhängigkeit von der
untersuchten Reizgeschwindigkeit und Stimulusrichtung dargestellt.
monokularer horizontaler OKR
Stimulusgeschw indigkeit [°/s] und Reizrichtung
35 30 25 20 15 10 5 0 5 10 15 20 25 30 35
0.2
0.4
0.6
0.8
1.0 Sg1Sg2Sg3
gain
naso-temporaltemporo-nasal
Abb. II. 3.2: Das gain des mhOKRs als Funktion der Stimulusgeschwindigkeit in T-N und N-T Reizrichtung
bei den Versuchtieren Sg 1, Sg 2 und Sg 3.
In temporo-nasaler Reizrichtung nimmt das gain von Sg 2 und Sg 3 bei einer
Reizgeschwindigkeitserhöhunh von 5 °/s nach 10 °/s zu. Die Werte steigen von 0,83 auf
0,92 bzw. von 0,81 auf 0,87. Mit weiterer Zunahme der Reizgeschwindigkeit fällt das gain
bis auf 0 bei 30 °/s ab. Sg 1 zeigt eine stetige Abnahme des gains mit Erhöhung der
Reizgeschwindigkeit.
In naso - temporaler Stimulusrichtung nimmt das gain für alle Versuchstiere von ca. 0,74
bei einer Reizgeschwindigkeit von 5 °/s kontinuierlich bis auf einen Wert von 0 bei 30 °/s
ab.
Mit diesem Ergebnis kann eindeutig gezeigt werden, dass an den drei untersuchten Salmo
gairdneri ein mhOKN auslösbar ist. Weiter kann die fehlerfreie Funktion der
Messapparatur und die Wahl eines adäquaten visuellen Reizes, der auch im
elektrophysiologischen Teil Verwendung fand, belegt werden.
73
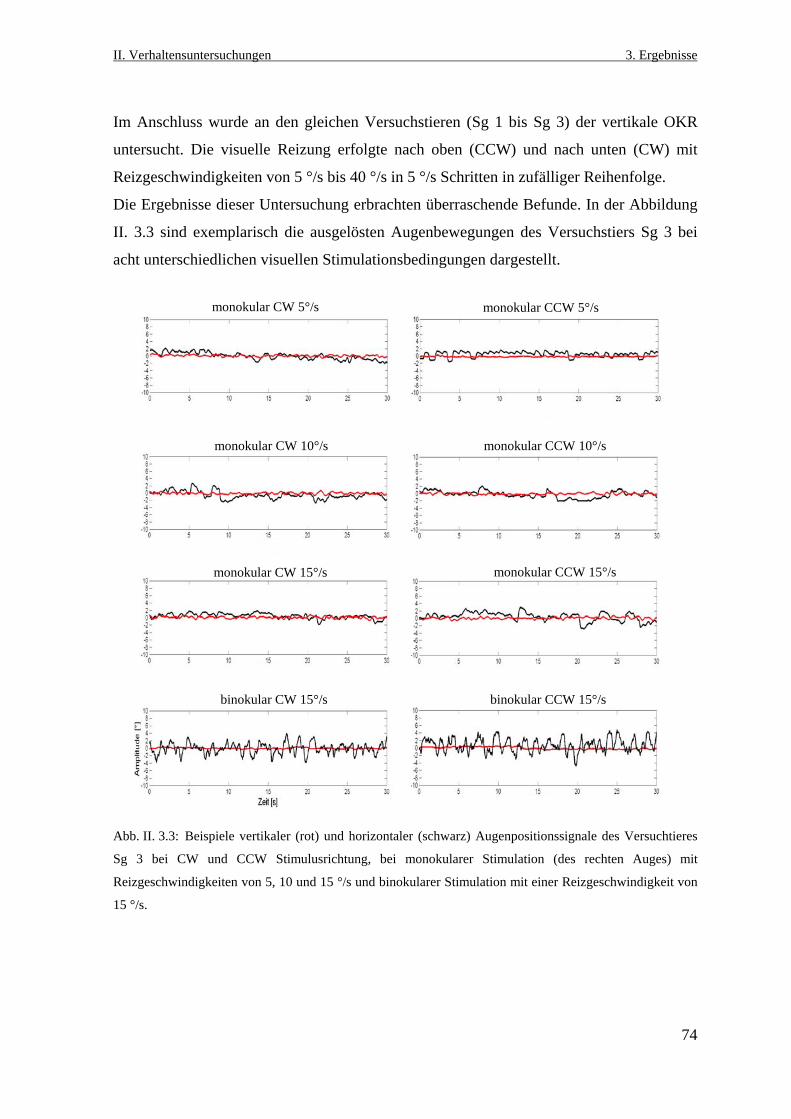
II. Verhaltensuntersuchungen 3. Ergebnisse
Im Anschluss wurde an den gleichen Versuchstieren (Sg 1 bis Sg 3) der vertikale OKR
untersucht. Die visuelle Reizung erfolgte nach oben (CCW) und nach unten (CW) mit
Reizgeschwindigkeiten von 5 °/s bis 40 °/s in 5 °/s Schritten in zufälliger Reihenfolge.
Die Ergebnisse dieser Untersuchung erbrachten überraschende Befunde. In der Abbildung
II. 3.3 sind exemplarisch die ausgelösten Augenbewegungen des Versuchstiers Sg 3 bei
acht unterschiedlichen visuellen Stimulationsbedingungen dargestellt.
binokular CW 15°/s
monokular CW 5°/s monokular CCW 5°/s
monokular CW 10°/s
monokular CW 15°/s
binokular CCW 15°/s
monokular CCW 15°/s
monokular CCW 10°/s
Abb. II. 3.3: Beispiele vertikaler (rot) und horizontaler (schwarz) Augenpositionssignale des Versuchtieres
Sg 3 bei CW und CCW Stimulusrichtung, bei monokularer Stimulation (des rechten Auges) mit
Reizgeschwindigkeiten von 5, 10 und 15 °/s und binokularer Stimulation mit einer Reizgeschwindigkeit von
15 °/s.
74

II. Verhaltensuntersuchungen 3. Ergebnisse
Gezeigt werden drei Reizgeschwindigkeiten (5 °/s, 10 °/s, und 15 °/s) bei monokularer
Reizung in beiden Reizrichtungen. Die horizontalen (schwarz) und vertikalen (rot)
Augenpositionssignale in Grad sind gegen den Zeitverlauf in Sekunden aufgetragen.
Zusätzlich werden zwei binokuare Stimulationen in CW und CCW Richtung gezeigt.
Die visuellen Stimulationen mit 5 °/s, 10 °/s und 15 °/s Reizgeschwindigkeit führen in
beiden vertikalen Stimulusrichtungen zu keinem für den optokinetischen Reflex typischen
Sägezahnmuster, wie er in Abb. II. 3.1 für den mhOKR gezeigt wurde.
Das vertikale Augenpositionssignal des rechten Auges verändert im Mittel seine Lage um
1,4° ohne langsame Augenfolgephase und Rückstellsakkaden, wie es für nystagmische
Augenbewegungen typisch wäre.
In den horizontalen Augenpositionssignalen können Lageveränderungen des Auges mit
Amplituden von 2,3 bis 3,4° bei vertikaler Stimulation registriert werden, die durch
sakkadische Augenpositionsänderungen erreicht werden.
In einem weiteren Beispiel (Abb. II. 3.3 letzte Zeile) werden Augenpositionssignale
gezeigt, die bei binokularer Stimulation mit einer Reizgeschwindigkeit von 15°/s in CW
und CCW Stimulusrichtung beobachtet wurden. Auch in diesen Beispielen können keine
vertikalen nystagmischen Augenbewegungen registriert werden. Der Anteil vertikaler
Augenpositionsänderungen ist vergleichbar mit den zuvor gezeigten monokularen
Beispielen.
Die auftretenden horizontalen Positionsveränderungen des Auges mit Amplituden bis zu
7,8° in CW und 8,2° in CCW Stimulusrichtung gleichen spontanen Augenbewegungen mit
einem hohen sakkadischen Anteil.
Diese Ergebnisse belegen, dass Salmo gairdneri unter den hier verwendeten
experimentellen Bedingungen keinen vertikalen OKN ausführen kann.
75

II. Verhaltensuntersuchungen 3. Ergebnisse
3.2 Visueller Einfluss auf die kompensatorischen Augen -
bewegungen beim vestibulo - okulären Reflex Da kein vertikaler OKR ausgelöst werden kann, führt das Ergebnis aus dem
elektrophysiologischen Teil dieser Arbeit, in dem gezeigt werden konnte, dass die
praetektalen richtungsselektiven Neurone alle Raumrichtungen kodieren, zu folgender
neuer Hypothese: Die visuelle Information der richtungsselektiven Zellen im dorsalen
Praetectum, die einen vertikale retinale Bildverschiebung kodieren, wird zur Verbesserung
kompensatorischer Augenbewegungen beim vestibulo - okulären Reflex (VOR) genutzt.
3.2.1 Der vertikale vestibulo-okuläre Reflex An acht Salmo gairdneri (Sg 4 bis Sg 11) wurden Verhaltensuntersuchungen durchgeführt,
um qualitativ und quantitativ bestimmen zu können, inwieweit das visuelle System einen
Beitrag zur Verbesserung der vestibulär ausgelösten vertikalen kompensatorischen
Augenbewegungen leisten kann.
Dazu wurden drei Stimulationsparadigmen gewählt, in denen die Versuchstiere eine
sinusförmige Drehbewegung erfahren, die einer Rollbewegung um die Körperlängsachse
entsprechen:
A: Das visuelle Stimulusmuster dreht sich exakt mit der vertikalen sinusförmigen
Drehbewegung um die Körperlängsachse des Fisches mit.
B: Das visuelle Stimulusmuster bleibt stationär im Raum, der Fisch erfährt eine
vertikale sinusförmige Drehbewegung um seine Körperlängsachse.
C: Das visuelle Stimulusmuster dreht sich exakt mit der gleichen Geschwindigkeit
entgegen der vertikalen sinusförmigen Drehbewegung des Fisches.
Die Arbeitshypothese lautet:
Wenn richtungsselektive Zellen aus der praetectalen Neuronenpopulation mit vertikaler
Vorzugsrichtung Einfluss auf die Güte vestibulo - okulärer kompensatorischer Augen-
bewegungen haben, sollte sich dies in den Werten des VOR - gains widerspiegeln.
76
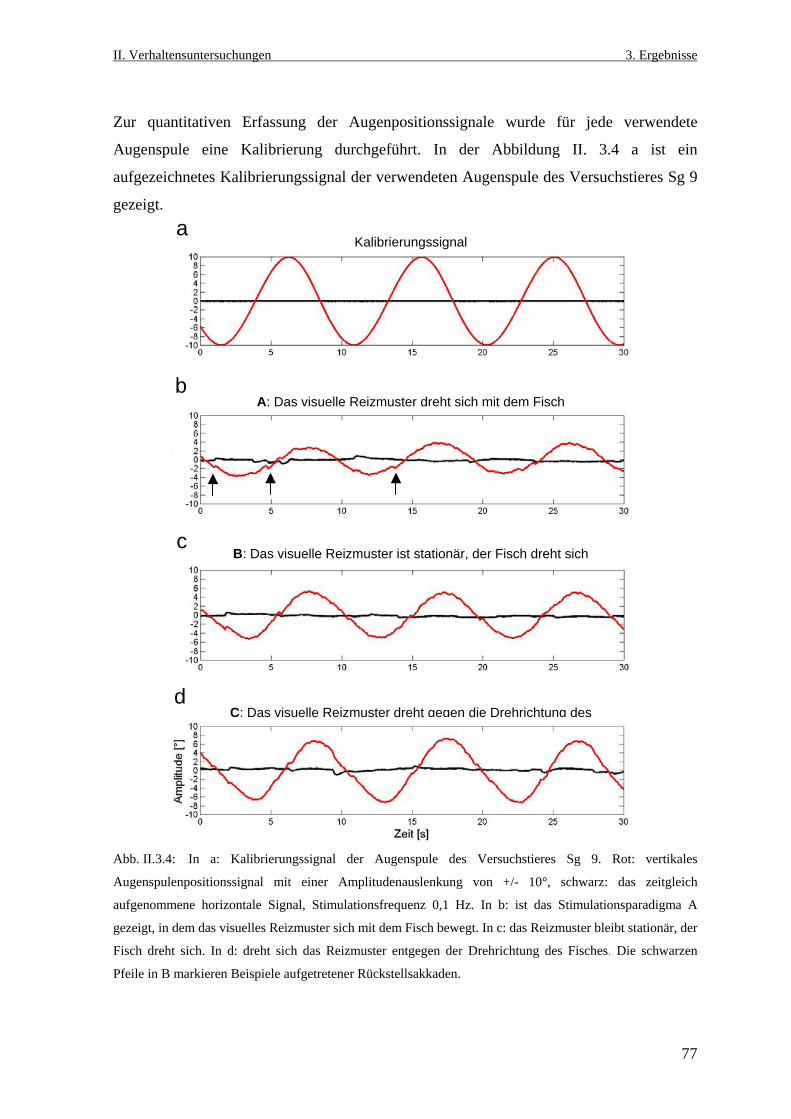
II. Verhaltensuntersuchungen 3. Ergebnisse
Zur quantitativen Erfassung der Augenpositionssignale wurde für jede verwendete
Augenspule eine Kalibrierung durchgeführt. In der Abbildung II. 3.4 a ist ein
aufgezeichnetes Kalibrierungssignal der verwendeten Augenspule des Versuchstieres Sg 9
gezeigt.
Abb. II.3.4: In a: Kalibrierungssignal der Augenspule des Versuchstieres Sg 9. Rot: vertikales
Augenspulenpositionssignal mit einer Amplitudenauslenkung von +/- 10°, schwarz: das zeitgleich
aufgenommene horizontale Signal, Stimulationsfrequenz 0,1 Hz. In b: ist das Stimulationsparadigma A
gezeigt, in dem das visuelles Reizmuster sich mit dem Fisch bewegt. In c: das Reizmuster bleibt stationär, der
Fisch dreht sich. In d: dreht sich das Reizmuster entgegen der Drehrichtung des Fisches. Die schwarzen
Pfeile in B markieren Beispiele aufgetretener Rückstellsakkaden.
b
c B: Das visuelle Reizmuster ist stationär, der Fisch dreht sich
d C: Das visuelle Reizmuster dreht gegen die Drehrichtung des
A: Das visuelle Reizmuster dreht sich mit dem Fisch
Kalibrierungssignal a
77

II. Verhaltensuntersuchungen 3. Ergebnisse
Die Kalibrierung erfolgte mit der stationären Augenmessspule exakt an der genauen
Position des gemessenen rechten Auges der Fische mit einer vertikalen sinusoidalen
Auslenkung von +/- 10° des Wechselmagnetfeldes um die Augenspule mit einer
Stimulationsfrequenz von 0,1 Hz und einer Aufzeichnungsdauer von 30 s. Anhand dieser
Kalibrierung war es möglich, aus den aufgezeichneten Augenpositionssignale der
Versuchstiere das VOR gain quantitativ zu bestimmen.
Weiter in der Abbildung II. 3.4 sind am Beispiel der Regenbogenforelle Sg 9 die
Augenpositionssignale für die drei Stimulationsparadigmen A, B und C dargestellt. In rot
ist jeweils das vertikale, in schwarz das horizontale Augenpositionssignal zu sehen.
Folgende qualitative Feststellungen können anhand dieser drei Beispiele von Sg 9 (Abb. II.
3.4 b bis d) getroffen werden:
1. In den Stimulationsparadigmen A, B, C können vertikale vestibulo - okuläre
kompensatorische Augenbewegungen ausgelöst werden, deren Amplituden von A nach C
zunehmen und deren maximale Amplitude nicht die der Kalibrierung von +/- 10° in a
erreichen.
2. Während der Ausführung vertikaler kompensatorischer Augenbewegungen werden von
dem Versuchstier unterschiedlich häufig Sakkaden entgegen der Drehrichtung ausgeführt
(Pfeile in Abb. II. 3.4 c).
3. Das horizontale Augenpositionssignal bleibt in seiner Lage relativ konstant. Es treten
vereinzelt horizontale Sakkanden, gefolgt von sehr langsamen Augenbewegungen zum
Ausgangspunkt, auf.
78
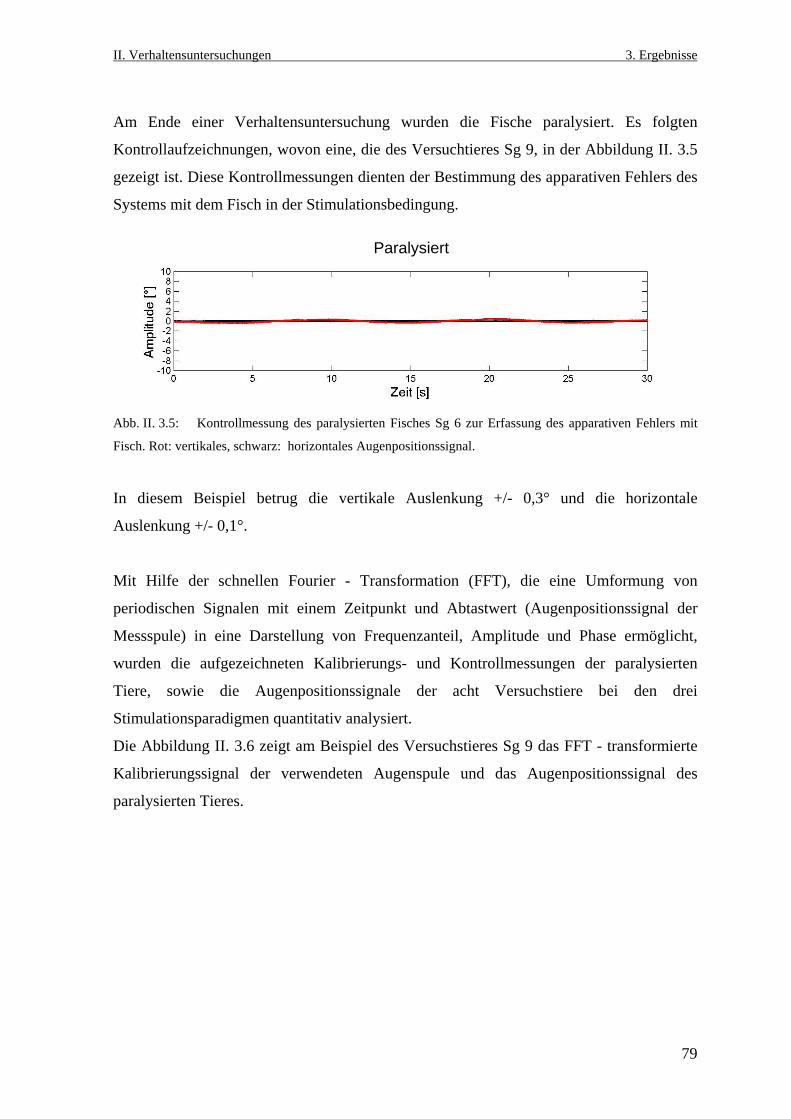
II. Verhaltensuntersuchungen 3. Ergebnisse
Am Ende einer Verhaltensuntersuchung wurden die Fische paralysiert. Es folgten
Kontrollaufzeichnungen, wovon eine, die des Versuchtieres Sg 9, in der Abbildung II. 3.5
gezeigt ist. Diese Kontrollmessungen dienten der Bestimmung des apparativen Fehlers des
Systems mit dem Fisch in der Stimulationsbedingung.
Paralysiert
Abb. II. 3.5: Kontrollmessung des paralysierten Fisches Sg 6 zur Erfassung des apparativen Fehlers mit
Fisch. Rot: vertikales, schwarz: horizontales Augenpositionssignal.
In diesem Beispiel betrug die vertikale Auslenkung +/- 0,3° und die horizontale
Auslenkung +/- 0,1°.
Mit Hilfe der schnellen Fourier - Transformation (FFT), die eine Umformung von
periodischen Signalen mit einem Zeitpunkt und Abtastwert (Augenpositionssignal der
Messspule) in eine Darstellung von Frequenzanteil, Amplitude und Phase ermöglicht,
wurden die aufgezeichneten Kalibrierungs- und Kontrollmessungen der paralysierten
Tiere, sowie die Augenpositionssignale der acht Versuchstiere bei den drei
Stimulationsparadigmen quantitativ analysiert.
Die Abbildung II. 3.6 zeigt am Beispiel des Versuchstieres Sg 9 das FFT - transformierte
Kalibrierungssignal der verwendeten Augenspule und das Augenpositionssignal des
paralysierten Tieres.
79
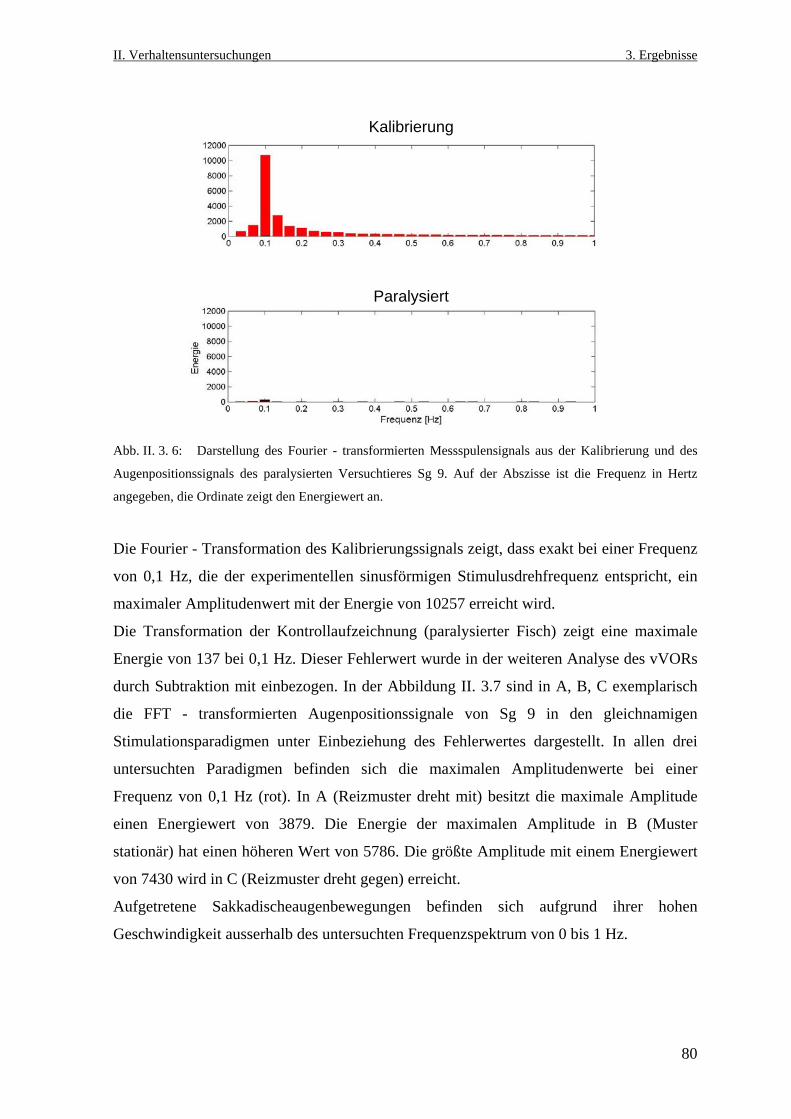
II. Verhaltensuntersuchungen 3. Ergebnisse
Paralysiert
Kalibrierung
Abb. II. 3. 6: Darstellung des Fourier - transformierten Messspulensignals aus der Kalibrierung und des
Augenpositionssignals des paralysierten Versuchtieres Sg 9. Auf der Abszisse ist die Frequenz in Hertz
angegeben, die Ordinate zeigt den Energiewert an.
Die Fourier - Transformation des Kalibrierungssignals zeigt, dass exakt bei einer Frequenz
von 0,1 Hz, die der experimentellen sinusförmigen Stimulusdrehfrequenz entspricht, ein
maximaler Amplitudenwert mit der Energie von 10257 erreicht wird.
Die Transformation der Kontrollaufzeichnung (paralysierter Fisch) zeigt eine maximale
Energie von 137 bei 0,1 Hz. Dieser Fehlerwert wurde in der weiteren Analyse des vVORs
durch Subtraktion mit einbezogen. In der Abbildung II. 3.7 sind in A, B, C exemplarisch
die FFT - transformierten Augenpositionssignale von Sg 9 in den gleichnamigen
Stimulationsparadigmen unter Einbeziehung des Fehlerwertes dargestellt. In allen drei
untersuchten Paradigmen befinden sich die maximalen Amplitudenwerte bei einer
Frequenz von 0,1 Hz (rot). In A (Reizmuster dreht mit) besitzt die maximale Amplitude
einen Energiewert von 3879. Die Energie der maximalen Amplitude in B (Muster
stationär) hat einen höheren Wert von 5786. Die größte Amplitude mit einem Energiewert
von 7430 wird in C (Reizmuster dreht gegen) erreicht.
Aufgetretene Sakkadischeaugenbewegungen befinden sich aufgrund ihrer hohen
Geschwindigkeit ausserhalb des untersuchten Frequenzspektrum von 0 bis 1 Hz.
80
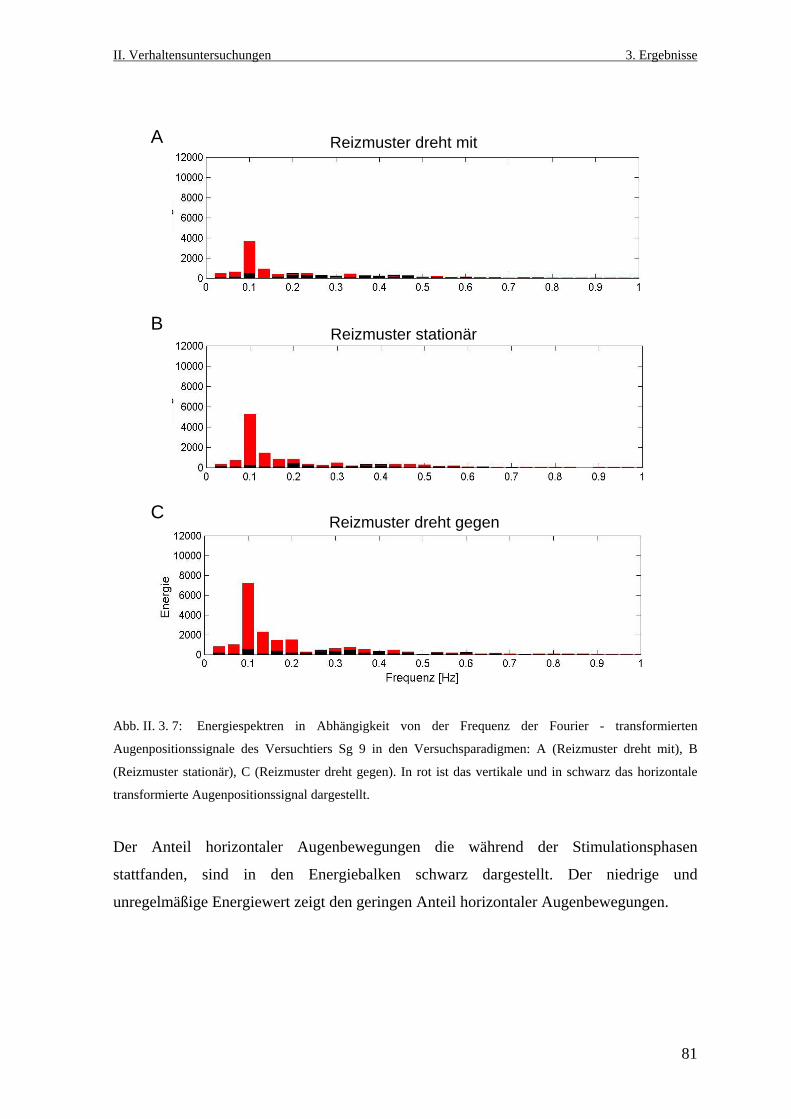
II. Verhaltensuntersuchungen 3. Ergebnisse
Reizmuster dreht gegen
Reizmuster stationär
Reizmuster dreht mit
C
B
A
Abb. II. 3. 7: Energiespektren in Abhängigkeit von der Frequenz der Fourier - transformierten
Augenpositionssignale des Versuchtiers Sg 9 in den Versuchsparadigmen: A (Reizmuster dreht mit), B
(Reizmuster stationär), C (Reizmuster dreht gegen). In rot ist das vertikale und in schwarz das horizontale
transformierte Augenpositionssignal dargestellt.
Der Anteil horizontaler Augenbewegungen die während der Stimulationsphasen
stattfanden, sind in den Energiebalken schwarz dargestellt. Der niedrige und
unregelmäßige Energiewert zeigt den geringen Anteil horizontaler Augenbewegungen.
81
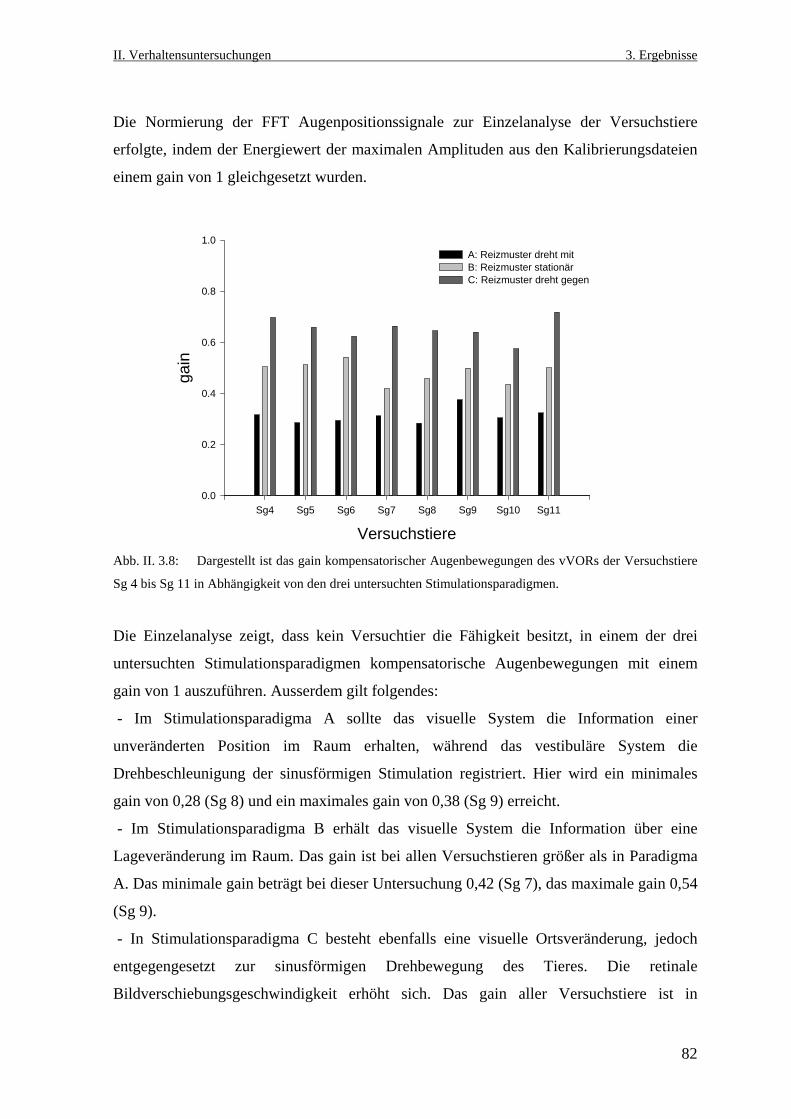
II. Verhaltensuntersuchungen 3. Ergebnisse
Die Normierung der FFT Augenpositionssignale zur Einzelanalyse der Versuchstiere
erfolgte, indem der Energiewert der maximalen Amplituden aus den Kalibrierungsdateien
einem gain von 1 gleichgesetzt wurden.
VersuchstiereSg4 Sg5 Sg6 Sg7 Sg8 Sg9 Sg10 Sg11
gain
0.0
0.2
0.4
0.6
0.8
1.0A: Reizmuster dreht mitB: Reizmuster stationärC: Reizmuster dreht gegen
Abb. II. 3.8: Dargestellt ist das gain kompensatorischer Augenbewegungen des vVORs der Versuchstiere
Sg 4 bis Sg 11 in Abhängigkeit von den drei untersuchten Stimulationsparadigmen.
Die Einzelanalyse zeigt, dass kein Versuchtier die Fähigkeit besitzt, in einem der drei
untersuchten Stimulationsparadigmen kompensatorische Augenbewegungen mit einem
gain von 1 auszuführen. Ausserdem gilt folgendes:
- Im Stimulationsparadigma A sollte das visuelle System die Information einer
unveränderten Position im Raum erhalten, während das vestibuläre System die
Drehbeschleunigung der sinusförmigen Stimulation registriert. Hier wird ein minimales
gain von 0,28 (Sg 8) und ein maximales gain von 0,38 (Sg 9) erreicht.
- Im Stimulationsparadigma B erhält das visuelle System die Information über eine
Lageveränderung im Raum. Das gain ist bei allen Versuchstieren größer als in Paradigma
A. Das minimale gain beträgt bei dieser Untersuchung 0,42 (Sg 7), das maximale gain 0,54
(Sg 9).
- In Stimulationsparadigma C besteht ebenfalls eine visuelle Ortsveränderung, jedoch
entgegengesetzt zur sinusförmigen Drehbewegung des Tieres. Die retinale
Bildverschiebungsgeschwindigkeit erhöht sich. Das gain aller Versuchstiere ist in
82
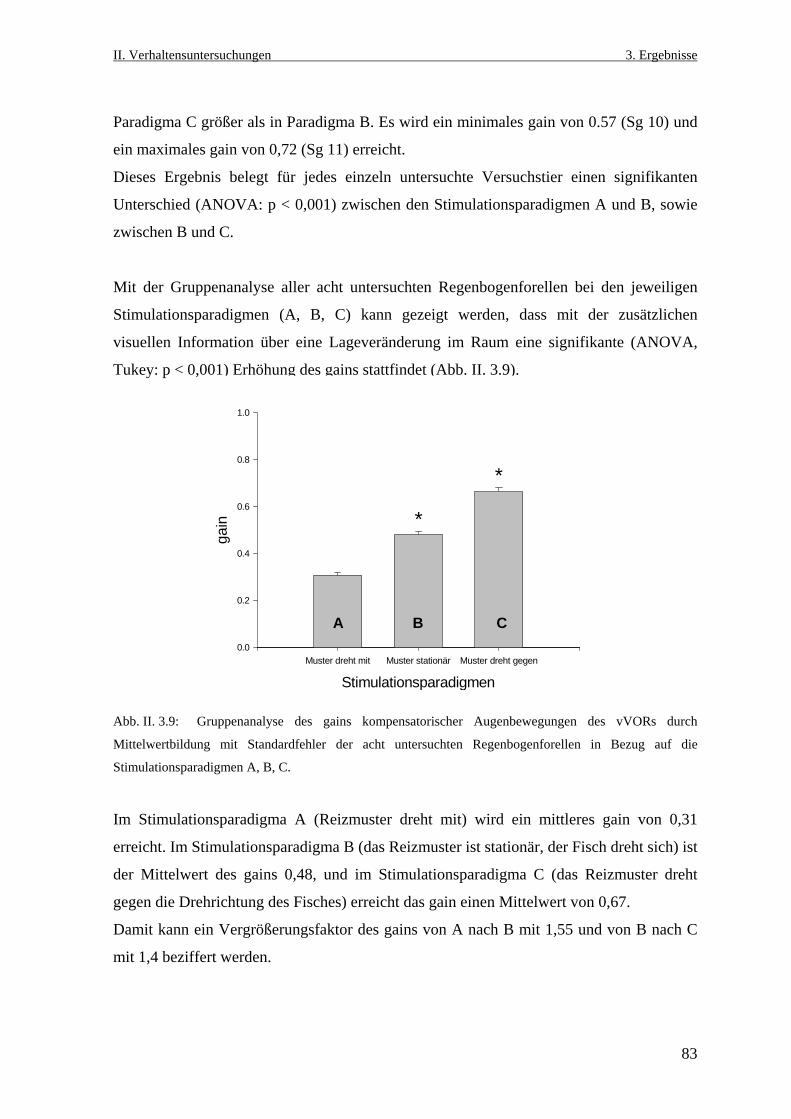
II. Verhaltensuntersuchungen 3. Ergebnisse
Paradigma C größer als in Paradigma B. Es wird ein minimales gain von 0.57 (Sg 10) und
ein maximales gain von 0,72 (Sg 11) erreicht.
Dieses Ergebnis belegt für jedes einzeln untersuchte Versuchstier einen signifikanten
Unterschied (ANOVA: p < 0,001) zwischen den Stimulationsparadigmen A und B, sowie
zwischen B und C.
Mit der Gruppenanalyse aller acht untersuchten Regenbogenforellen bei den jeweiligen
Stimulationsparadigmen (A, B, C) kann gezeigt werden, dass mit der zusätzlichen
visuellen Information über eine Lageveränderung im Raum eine signifikante (ANOVA,
Tukey: p < 0,001) Erhöhung des gains stattfindet (Abb. II. 3.9).
StimulationsparadigmenMuster dreht mit Muster stationär Muster dreht gegen
gain
0.0
0.2
0.4
0.6
0.8
1.0
*
*
A B C
Abb. II. 3.9: Gruppenanalyse des gains kompensatorischer Augenbewegungen des vVORs durch
Mittelwertbildung mit Standardfehler der acht untersuchten Regenbogenforellen in Bezug auf die
Stimulationsparadigmen A, B, C.
Im Stimulationsparadigma A (Reizmuster dreht mit) wird ein mittleres gain von 0,31
erreicht. Im Stimulationsparadigma B (das Reizmuster ist stationär, der Fisch dreht sich) ist
der Mittelwert des gains 0,48, und im Stimulationsparadigma C (das Reizmuster dreht
gegen die Drehrichtung des Fisches) erreicht das gain einen Mittelwert von 0,67.
Damit kann ein Vergrößerungsfaktor des gains von A nach B mit 1,55 und von B nach C
mit 1,4 beziffert werden.
83
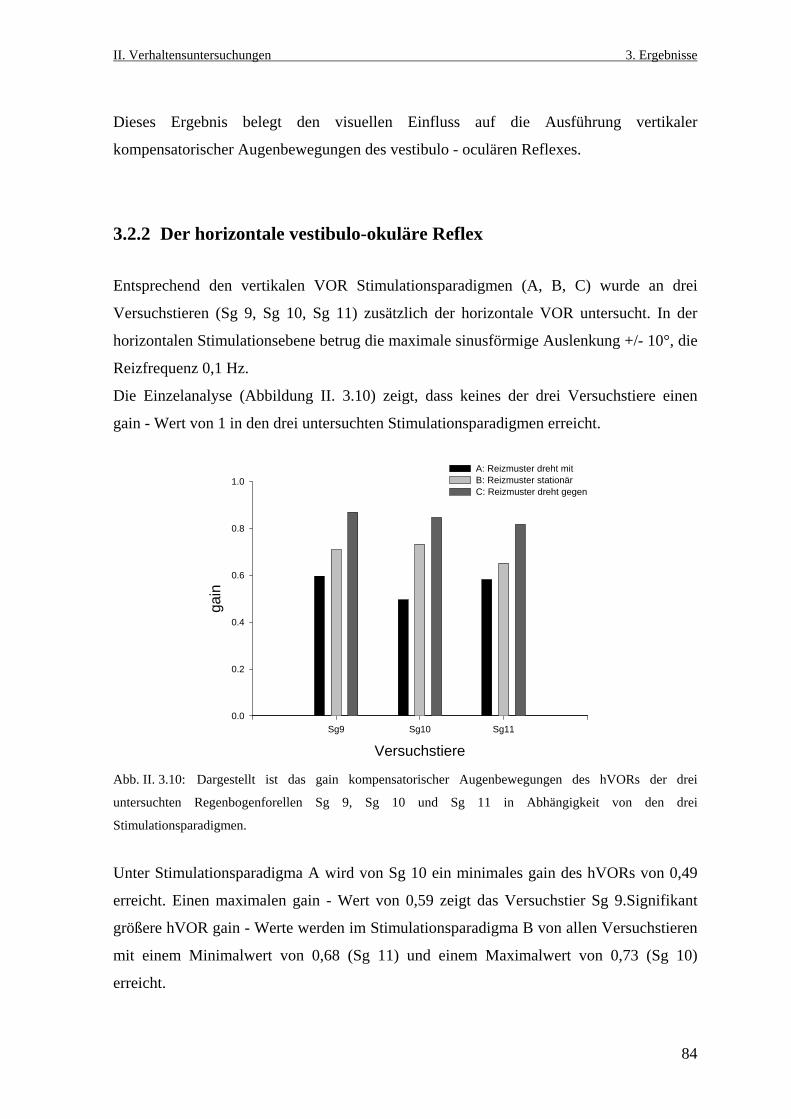
II. Verhaltensuntersuchungen 3. Ergebnisse
Dieses Ergebnis belegt den visuellen Einfluss auf die Ausführung vertikaler
kompensatorischer Augenbewegungen des vestibulo - oculären Reflexes.
3.2.2 Der horizontale vestibulo-okuläre Reflex
Entsprechend den vertikalen VOR Stimulationsparadigmen (A, B, C) wurde an drei
Versuchstieren (Sg 9, Sg 10, Sg 11) zusätzlich der horizontale VOR untersucht. In der
horizontalen Stimulationsebene betrug die maximale sinusförmige Auslenkung +/- 10°, die
Reizfrequenz 0,1 Hz.
Die Einzelanalyse (Abbildung II. 3.10) zeigt, dass keines der drei Versuchstiere einen
gain - Wert von 1 in den drei untersuchten Stimulationsparadigmen erreicht.
VersuchstiereSg9 Sg10 Sg11
gain
0.0
0.2
0.4
0.6
0.8
1.0A: Reizmuster dreht mitB: Reizmuster stationärC: Reizmuster dreht gegen
Abb. II. 3.10: Dargestellt ist das gain kompensatorischer Augenbewegungen des hVORs der drei
untersuchten Regenbogenforellen Sg 9, Sg 10 und Sg 11 in Abhängigkeit von den drei
Stimulationsparadigmen.
Unter Stimulationsparadigma A wird von Sg 10 ein minimales gain des hVORs von 0,49
erreicht. Einen maximalen gain - Wert von 0,59 zeigt das Versuchstier Sg 9.Signifikant
größere hVOR gain - Werte werden im Stimulationsparadigma B von allen Versuchstieren
mit einem Minimalwert von 0,68 (Sg 11) und einem Maximalwert von 0,73 (Sg 10)
erreicht.
84
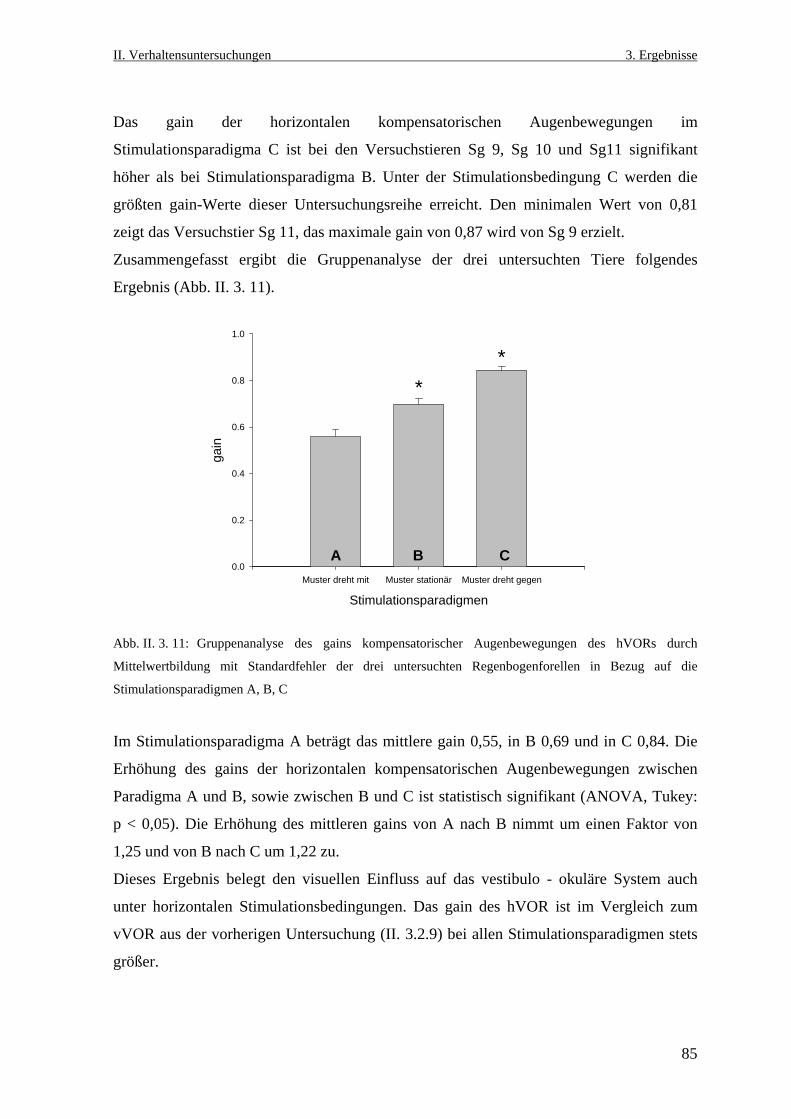
II. Verhaltensuntersuchungen 3. Ergebnisse
Das gain der horizontalen kompensatorischen Augenbewegungen im
Stimulationsparadigma C ist bei den Versuchstieren Sg 9, Sg 10 und Sg11 signifikant
höher als bei Stimulationsparadigma B. Unter der Stimulationsbedingung C werden die
größten gain-Werte dieser Untersuchungsreihe erreicht. Den minimalen Wert von 0,81
zeigt das Versuchstier Sg 11, das maximale gain von 0,87 wird von Sg 9 erzielt.
Zusammengefasst ergibt die Gruppenanalyse der drei untersuchten Tiere folgendes
Ergebnis (Abb. II. 3. 11).
Stimulationsparadigmen
Muster dreht mit Muster stationär Muster dreht gegen
gain
0.0
0.2
0.4
0.6
0.8
1.0
**
A B C
Abb. II. 3. 11: Gruppenanalyse des gains kompensatorischer Augenbewegungen des hVORs durch
Mittelwertbildung mit Standardfehler der drei untersuchten Regenbogenforellen in Bezug auf die
Stimulationsparadigmen A, B, C
Im Stimulationsparadigma A beträgt das mittlere gain 0,55, in B 0,69 und in C 0,84. Die
Erhöhung des gains der horizontalen kompensatorischen Augenbewegungen zwischen
Paradigma A und B, sowie zwischen B und C ist statistisch signifikant (ANOVA, Tukey:
p < 0,05). Die Erhöhung des mittleren gains von A nach B nimmt um einen Faktor von
1,25 und von B nach C um 1,22 zu.
Dieses Ergebnis belegt den visuellen Einfluss auf das vestibulo - okuläre System auch
unter horizontalen Stimulationsbedingungen. Das gain des hVOR ist im Vergleich zum
vVOR aus der vorherigen Untersuchung (II. 3.2.9) bei allen Stimulationsparadigmen stets
größer.
85
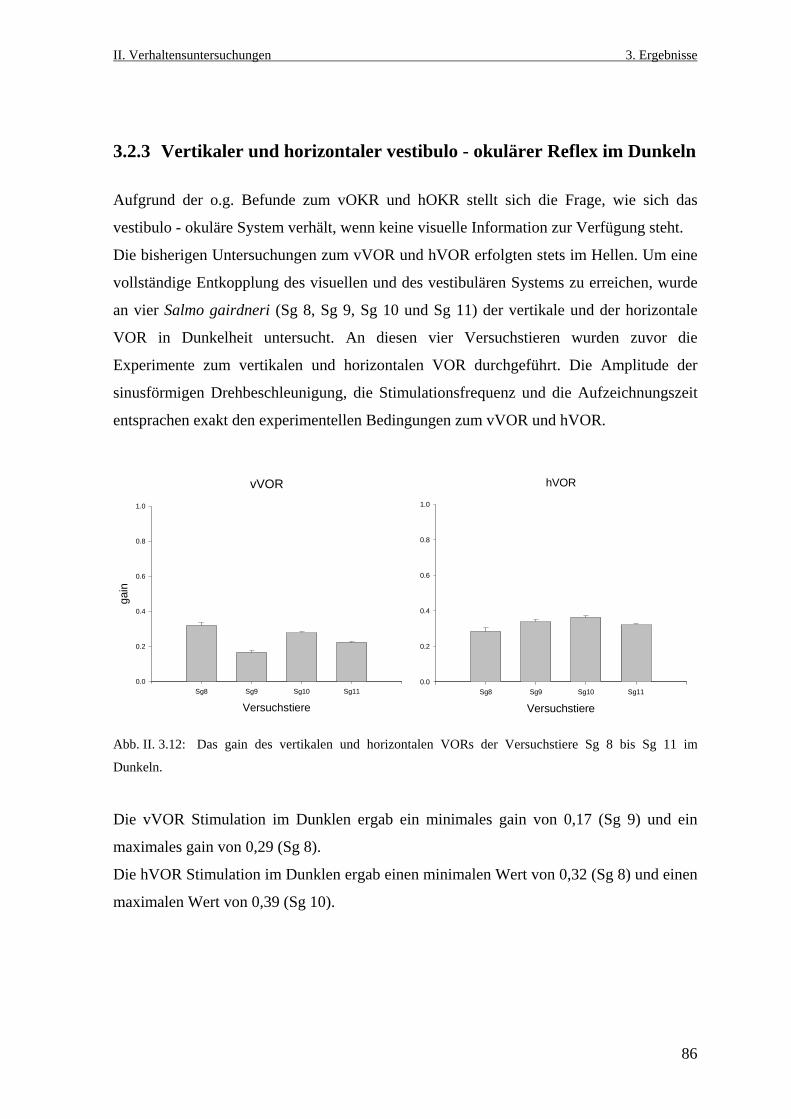
II. Verhaltensuntersuchungen 3. Ergebnisse
3.2.3 Vertikaler und horizontaler vestibulo - okulärer Reflex im Dunkeln Aufgrund der o.g. Befunde zum vOKR und hOKR stellt sich die Frage, wie sich das
vestibulo - okuläre System verhält, wenn keine visuelle Information zur Verfügung steht.
Die bisherigen Untersuchungen zum vVOR und hVOR erfolgten stets im Hellen. Um eine
vollständige Entkopplung des visuellen und des vestibulären Systems zu erreichen, wurde
an vier Salmo gairdneri (Sg 8, Sg 9, Sg 10 und Sg 11) der vertikale und der horizontale
VOR in Dunkelheit untersucht. An diesen vier Versuchstieren wurden zuvor die
Experimente zum vertikalen und horizontalen VOR durchgeführt. Die Amplitude der
sinusförmigen Drehbeschleunigung, die Stimulationsfrequenz und die Aufzeichnungszeit
entsprachen exakt den experimentellen Bedingungen zum vVOR und hVOR.
hVOR
VersuchstiereSg8 Sg9 Sg10 Sg11
gain
0.0
0.2
0.4
0.6
0.8
1.0
vVOR
VersuchstiereSg8 Sg9 Sg10 Sg11
0.0
0.2
0.4
0.6
0.8
1.0
gain
Abb. II. 3.12: Das gain des vertikalen und horizontalen VORs der Versuchstiere Sg 8 bis Sg 11 im
Dunkeln.
Die vVOR Stimulation im Dunklen ergab ein minimales gain von 0,17 (Sg 9) und ein
maximales gain von 0,29 (Sg 8).
Die hVOR Stimulation im Dunklen ergab einen minimalen Wert von 0,32 (Sg 8) und einen
maximalen Wert von 0,39 (Sg 10).
86
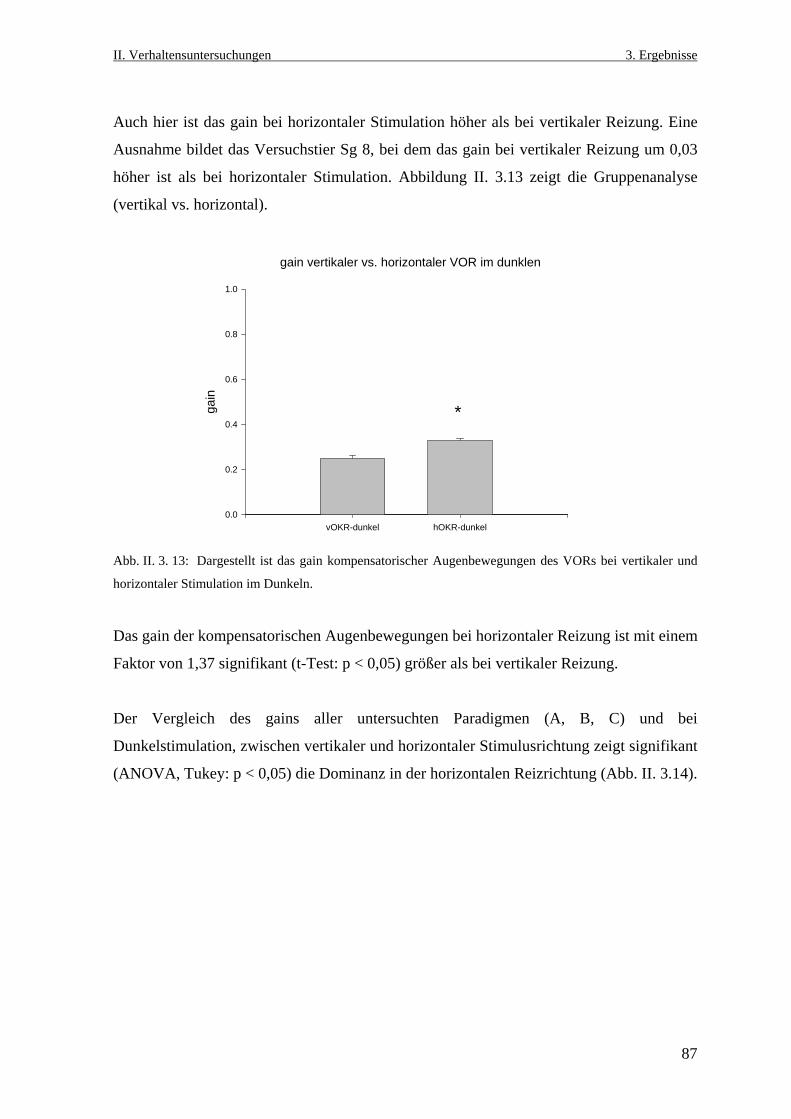
II. Verhaltensuntersuchungen 3. Ergebnisse
Auch hier ist das gain bei horizontaler Stimulation höher als bei vertikaler Reizung. Eine
Ausnahme bildet das Versuchstier Sg 8, bei dem das gain bei vertikaler Reizung um 0,03
höher ist als bei horizontaler Stimulation. Abbildung II. 3.13 zeigt die Gruppenanalyse
(vertikal vs. horizontal).
vOKR-dunkel hOKR-dunkel
gain
0.0
0.2
0.4
0.6
0.8
1.0
gain vertikaler vs. horizontaler VOR im dunklen
*
Abb. II. 3. 13: Dargestellt ist das gain kompensatorischer Augenbewegungen des VORs bei vertikaler und
horizontaler Stimulation im Dunkeln.
Das gain der kompensatorischen Augenbewegungen bei horizontaler Reizung ist mit einem
Faktor von 1,37 signifikant (t-Test: p < 0,05) größer als bei vertikaler Reizung.
Der Vergleich des gains aller untersuchten Paradigmen (A, B, C) und bei
Dunkelstimulation, zwischen vertikaler und horizontaler Stimulusrichtung zeigt signifikant
(ANOVA, Tukey: p < 0,05) die Dominanz in der horizontalen Reizrichtung (Abb. II. 3.14).
87
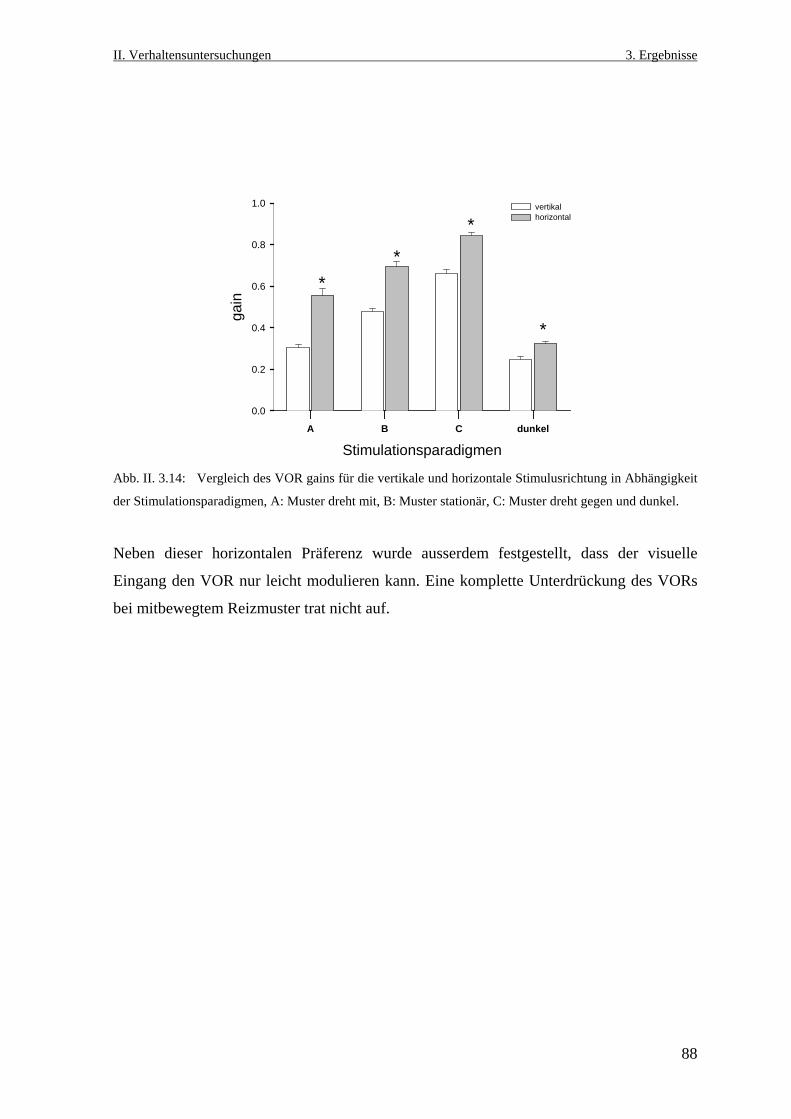
II. Verhaltensuntersuchungen 3. Ergebnisse
Abb. II. 3.14: Ver
der Stimulationspar
Neben dieser h
Eingang den VO
bei mitbewegtem
StimulationsparadigmenA B C dunkel
gain
0.0
0.2
0.4
0.6
0.8
1.0 vertikal horizontal
* *
*
*
gleich des VOR gains für die vertikale und horizontale Stimulusrichtung in Abhängigkeit
adigmen, A: Muster dreht mit, B: Muster stationär, C: Muster dreht gegen und dunkel.
orizontalen Präferenz wurde ausserdem festgestellt, dass der visuelle
R nur leicht modulieren kann. Eine komplette Unterdrückung des VORs
Reizmuster trat nicht auf.
88

II. Verhaltensuntersuchungen 3. Ergebnisse
3.3 Optokinetische Augenbewegungen vor und nach Läsion des
dorsalen Praetectums Mit Hilfe elektrolytischer Mikroläsionen wurde untersucht, ob die im
elektrophysiologischen Teil dieser Arbeit beschriebenen praetectalen richtungsselektiven
Neurone von Salmo gairdneri an der Generierung der langsamen Augenfolgephase des
hOKRs essentiell beteiligt sind und somit das neuronale Substrat für optokinetische
Augenbewegungen in der Überordnung Teleostei stellen können.
Zu Beginn dieser Experimentreihe wurden sechs Salmo gairdneri (Sg 12 bis Sg 17) auf die
Auslösbarkeit ihres mhOKRs hin untersucht. Auch diese sechs Versuchstiere konnten vor
dem Läsionsexperiment einen mhOKR bei Reizgeschwindigkeiten von 4 °/s, 9 °/s, 14 °/s
und 18 °/s in beiden Stimulusrichtungen (T-N und N-T) ausführen (siehe Abb. II. 3.15).
3.3.1 Elektrophysiologisches Läsionsexperiment Zur Lokalisation des praetectalen Areals richtungsselektiver Neurone wurden
elektrophysiologische Ableitungen an den zuvor in den Verhaltenstests untersuchten, sechs
Regenbogenforellen durchgeführt. An je einem Ableitort eindeutig richtungsselektiver
Neurone erfolgten bei fünf Fischen (Sg 12 bis Sg 16) elektrolytische Mikroläsionen. Das
sechste Versuchstier (Sg 17) diente als Kontrolle. An ihm wurden nur
elektrophysiologische Ableitungen im peripheren Bereich des richtungsselektiven Areals
in vergleichbaren Ableittiefen durchgeführt.
In der Abbildung II. 3.15 sind in Polarkoordinaten die Tuning-Kurven und
Vorzugsrichtungen signifikant richtungsselektiver Neurone gezeigt, die vor der Läsion bei
den fünf Versuchstieren abgeleitet wurden. Der Mittelpunkt des Polarplots repräsentiert die
Lage des rechten, stimulierten Auges. Die Radien geben die Spikerate pro Sekunde an.
89

II. Verhaltensuntersuchungen 3. Ergebnisse
Abb. II. 3.15: Tuning - Kurven und Richtungsvektoren fünf richtungsselektiver Neurone der Versuchstiere
Sg 12 bis Sg 16 vor der Läsion. Die Radien geben die Aktivität in Spikes / Sekunde an.
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
Sg 15
Sg 14
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
270°
315°
nasal-unten
temporal-oben
temporal
unten
oben
nasal
nasal-oben
Sg 12
225°
temporal-unten
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
0 20 40 60 800
20
40
60
80
0204060800
20
40
60
80
0°
45°
90°
135°
180°
225°
270°
315°
Sg 16
Sg 14
Sg 13
Im Praetectum der fünf Versuchstiere konnten vor der Läsion folgende richtungsselektive
Neurone abgeleitet werden:
- Sg 12 nasal, 9,5°
- Sg 13 unten, 261°
- Sg 14 nasal - oben, 38° und nasal - unten, 309°
- Sg 15 temporal - oben, 121°
- Sg 16 temporal, 169°
Nach erfolgter Läsion konnte während des Zurückziehens der Ableitelektrode bei keinem
der fünf Versuchstiere neuronale Aktivität innerhalb eines Bereiches von 200 – 350 µm
oberhalb des Läsionsortes registriert werden. Die genaue histologische Rekonstruktion der
Läsionsorte erfolgte nach Beendigung der anschließenden Verhaltensexperimente zum
mhOKR.
90

II. Verhaltensuntersuchungen 3. Ergebnisse
3.3.2 Verhaltensbeobachtung nach Läsion des dorsalen Praetectums Im Anschluss an das Läsionsexperiment und nach eindeutig abgeklungener
Narkosewirkung konnten folgende Verhaltensbeobachtungen im Hälterungsbecken der
getrennt gehaltenen Versuchstiere gemacht werden.
Alle fünf Fische mit einem lädiertem Praetectum zeigten eine starke Einschränkung in
ihrer Mobilität. Sie vermieden es, im Becken frei umher zu schwimmen, wie es für Salmo
gairdneri charakteristisch wäre. Vielmehr verweilten sie häufig mit einer leichten Neigung
um ihre Körperlängsachse nach links auf ihre Brustflossen gestützt am Boden des
Hälterungsbeckens. In den wenigen beobachteten Aktivitätsphasen wurden überwiegend
Schwimmrichtungen entgegen dem Uhrzeigersinn (ipsiversiv zur Läsion) registriert. Mit
einem weißen Stock (∅ = 2 cm), der im frontalen Gesichtsfeld horizontal bewegt wurde,
konnten die Fische zum Schwimmen animiert werden. Die bevorzugte Schwimmrichtung
entgegen dem Uhrzeigersinn behielten alle fünf Versuchstiere aber bei.
Ein völlig anderes Verhalten zeigte der Kontrollfisch im gleichen postoperativen Zeitraum.
Eine Einschränkung der Mobilität wurde nicht beobachtet. Der Fisch schwamm stetig im
Hälterungsbecken umher. Ebenso war kein Unterschied zwischen den Schwimmrichtungen
entgegen dem Uhrzeigersinn (ipsiversiv zum Ableitort) oder im Uhrzeigersinn
(kontraversiv zum Ableitort) festzustellen. Dieses Kontrolltier zeigte ein arttypisches
Aktivitätsverhalten.
3.3.3 Der monokulare horizontale optokinetische Reflex nach der Läsion Im Anschluss an die elektrolytische Läsion durften sich die Fische acht bis maximal zwölf
Stunden erholen, um zu gewährleisten, dass der Einfluss der verwendeten Narkotika
vollständig abgeklungen war.
Danach wurde den Versuchstieren wie vor der Läsion der visuelle Stimulus monokular,
rechtes Auge sehend, bei den gleichen Geschwindigkeiten und in den gleichen
Stimulusrichtungen (T-N und N-T) präsentiert.
91
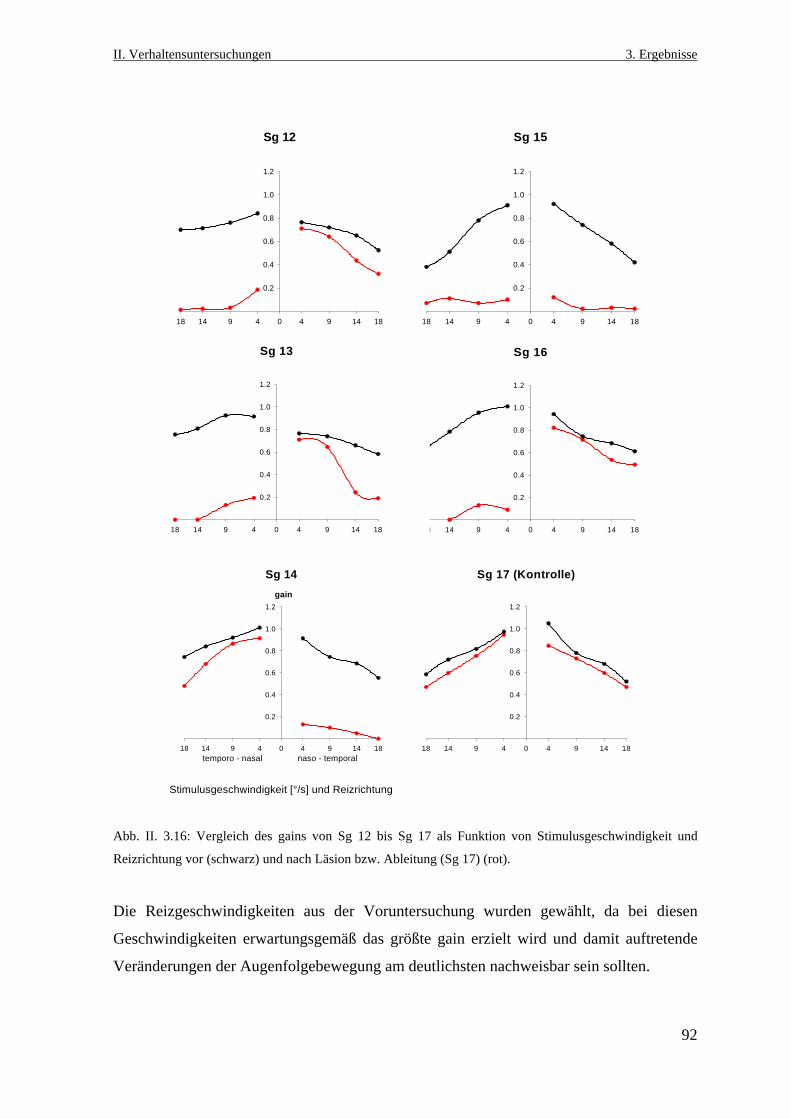
II. Verhaltensuntersuchungen 3. Ergebnisse
Sg 12
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 15
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 16
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 17 (Kontrolle)
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 13
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 14
Stimulusgeschwindigkeit [°/s] und Reizrichtung
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
naso - temporaltemporo - nasal
gain
Sg 12
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 15
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 16
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 17 (Kontrolle)
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 13
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
Sg 14
Stimulusgeschwindigkeit [°/s] und Reizrichtung
18 14 9 4 0 4 9 14 18
0.2
0.4
0.6
0.8
1.0
1.2
naso - temporaltemporo - nasal
gain
Abb. II. 3.16: Vergleich des gains von Sg 12 bis Sg 17 als Funktion von Stimulusgeschwindigkeit und
Reizrichtung vor (schwarz) und nach Läsion bzw. Ableitung (Sg 17) (rot).
Die Reizgeschwindigkeiten aus der Voruntersuchung wurden gewählt, da bei diesen
Geschwindigkeiten erwartungsgemäß das größte gain erzielt wird und damit auftretende
Veränderungen der Augenfolgebewegung am deutlichsten nachweisbar sein sollten.
92

II. Verhaltensuntersuchungen 3. Ergebnisse
In der Abbildung II. 3.16 ist für jedes Versuchstier (Sg 12 bis Sg 17) das gain als Funktion
der Stimulusgeschwindigkeit und Reizrichtung vor (schwarz) und nach (rot) Läsion bzw.
Ableitung des linken Praetectums gezeigt. Die Versuchstiere Sg 12, Sg 13 und Sg 16
zeigen im Vergleich zur Voruntersuchung eine deutliche Abnahme der Geschwindigkeit
der Augenfolgereaktion des mhOKRs in temporo - nasaler Stimulusrichtung (ipsiversiv).
Schon bei 4 °/s Reizgeschwindigkeit reduziert sich das gain um 80 % auf einen Wert von
< 0,2. Bei höheren Reizgeschwindigkeiten nimmt das gain weiter ab, so dass ab einer
Reizgeschwindigkeit von 14 °/s die gain - Werte unterhalb von 0,05 lagen. Das gain in
naso - temporaler Stimulusrichtung ist weniger stark reduziert als in temporo - nasaler
Stimulusrichtung bei gleichen Stimulusgeschwindigkeiten. Es verringert sich für alle drei
Tiere im Vergleich zur Voruntersuchung um ca. 30 %.
Das Versuchstier Sg 14 zeigt bei einer Reizgeschwindigkeit von 4 °/s eine starke
Reduktion des gains um 87 % in naso - temporaler Stimulusrichtung. Ab einer Reiz-
geschwindigkeit von 14 °/s war das gain kleiner als 0,05. Kaum beeinträchtigt hingegen ist
die temporo - nasale Stimulusrichtung, bei der das gain um 10 % (4 °/s) bis 23 % (18 °/s)
abnahm.
Eine Verringerung des gains in beiden Stimulusrichtungen im Vergleich zur
Voruntersuchung tritt bei Versuchstier Sg 15 auf. Hier kommt es zu einer Reduktion um
82 % in temporo - nasaler und um 91 % in naso - temporaler Stimulusrichtung.
Das Kontrolltier Sg 17 (sham - Operation) zeigte nach dem Ableitexperiment über den
gesamten untersuchten Geschwindigkeitsbereich und für beiden Stimulusrichtungen ein
gain, das ähnlich (Reduktion zwischen 2 % und 8 %) zu den vor dem Experiment
gemessenen Werten war. Mit dieser Kontrolle kann belegt werden, dass die Operation und
die Penetrationen mit der Mikroelektrode durch das Tectum opticum nicht zu
Beeinträchtigungen der langsamen Augenfolgephase des mhOKR führen.
Diese Ergebnisse zeigen, dass die irreversible Inaktivierung des Areals richtungsselektiver
Neurone im Praetectum Auswirkungen auf die Geschwindigkeit der langsamen
Augenfolgephase des mhOKRs hat. Zusätzlich wird deutlich, dass die Reduktion des gains
nicht auf eine der untersuchten Stimulusrichtungen beschränkt sein muss.
93

II. Verhaltensuntersuchungen 3. Ergebnisse
3.3.4 Analyse der Augenfolgebewegungen während der
Stimulationsphasen Als Augenfolgebewegungsereignisse werden Veränderungen, die während der
monokularen visuellen Stimulationsphasen innerhalb der 30 s Aufzeichnungsdauer
registriert wurden, definiert. Dazu gehören die Anzahl der Augenfolgebewegungen, also
die Frequenz, sowie die Amplituden der Augenpositionsveränderung des abgedeckten
linken und des stimulierten rechten Auges vor und nach der Läsion.
3.3.4.1 Anzahl der Augenfolgebewegungen
In der Abbildung II. 3.17 ist von allen Versuchstieren die absolute Anzahl aufgetretener
Augenfolgephasen (ohne Berücksichtigung ihrer Folgegeschwindigkeit) pro 30 Sekunden
Aufzeichnungsdauer gezeigt. Das Versuchstier Sg 12 steht aufgrund der vergleichbaren
Reduktion des gains in temporo-nasaler Reizrichtung repräsentativ auch für die
Regenbogenforellen Sg 13 und Sg 16.
Bei keinem der Tiere zeigte sich vor der praetectalen Läsion ein signifikanter Unterschied
in der Anzahl der Augenfolgebewegungen des linken und des rechten Auges. Signifikante
Unterschiede treten erst nach der Läsion bei den Tieren Sg 12 bis Sg 16 auf.
Die Anzahl der Augenfolgebewegungen in N-T Stimulusrichtung von Versuchstier Sg 12
ist nach der Läsion für das linke Auge im Mittel um 14,8%, für das rechte Auge um 18,6
% gegenüber dem präoperativen Werten reduziert. In T-N Stimulusrichtung tritt dagegen
eine signifikante Reduktion um 66,7 % für das linke und um 74,5% für das rechte Auge
gegenüber der Anzahl von Augenfolgebewegungen beim intakten Tier auf (p < 0,05).
94
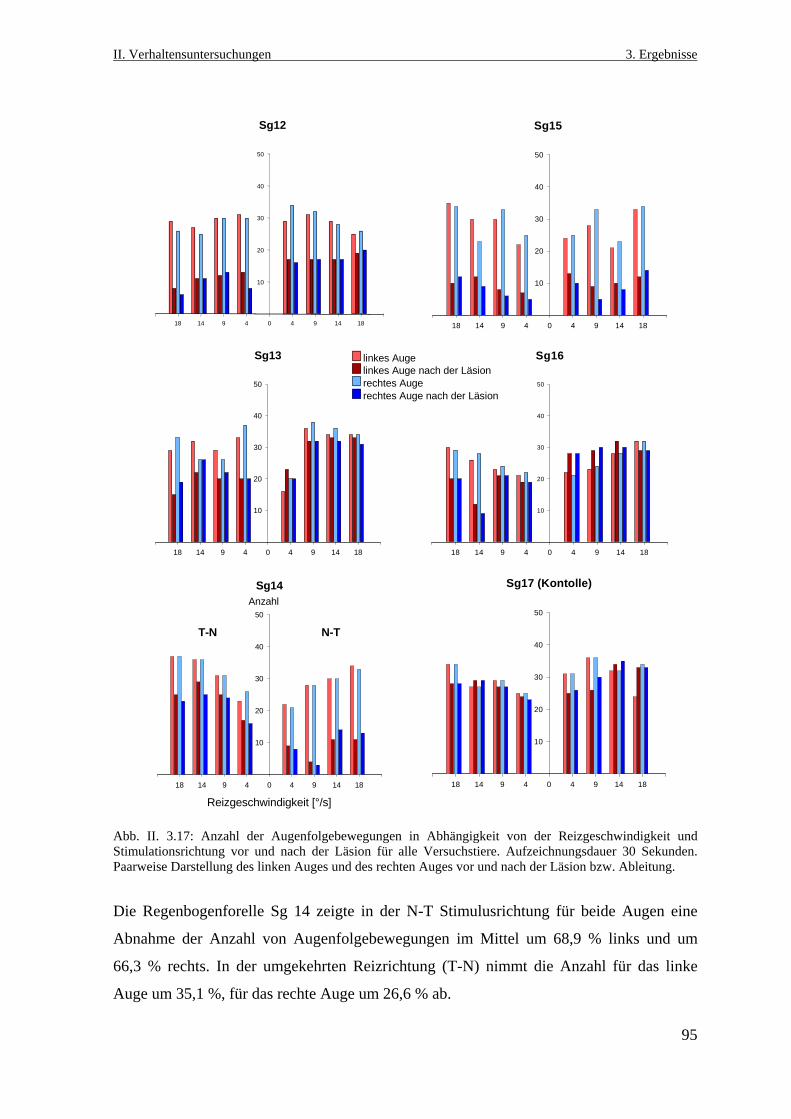
II. Verhaltensuntersuchungen 3. Ergebnisse
Sg12 Sg15
AbbStimPaa
Die
Ab
66,
Au
18 14 9 4 0 4 9 14 18
10
20
30
40
50
18 14 9 4 0 4 9 14 18
10
20
30
40
50
Sg13
18 14 9 4 0 4 9 14 18
10
20
30
40
50
Sg14
18 14 9 4 0 4 9 14 18
10
20
30
40
50
N-TT-N
Reizgeschwindigkeit [°/s]
Anzahl
Sg16
18 14 9 4 0 4 9 14 18
10
20
30
40
50
Sg17 (Kontolle)
18 14 9 4 0 4 9 14 18
10
20
30
40
50
linkes Augelinkes Auge nach der Läsionrechtes Augerechtes Auge nach der Läsion
. II. 3.17: Anzahl der Augenfolgebewegungen in Abhängigkeit von der Reizgeschwindigkeit und ulationsrichtung vor und nach der Läsion für alle Versuchstiere. Aufzeichnungsdauer 30 Sekunden.
rweise Darstellung des linken Auges und des rechten Auges vor und nach der Läsion bzw. Ableitung.
Regenbogenforelle Sg 14 zeigte in der N-T Stimulusrichtung für beide Augen eine
nahme der Anzahl von Augenfolgebewegungen im Mittel um 68,9 % links und um
3 % rechts. In der umgekehrten Reizrichtung (T-N) nimmt die Anzahl für das linke
ge um 35,1 %, für das rechte Auge um 26,6 % ab.
95

II. Verhaltensuntersuchungen 3. Ergebnisse
Eine deutliche, läsionsbedingte Herabsetzung der Anzahl der Augenfolgebewegungen in
beiden Stimulusrichtungen wird von Versuchstier Sg 15 gezeigt. In der N-T
Stimulusrichtung nimmt die Anzahl um 71,8 % für das linke Auge und um 67,2 % für das
rechte Auge ab, in der T-N Reizrichtung um 57,4 % für das linke Auge und um 67,2 % für
das rechte Auge.
Das Kontrolltier Sg 17 zeigt eine Abnahme in der Anzahl von Augenfolgephasen nach
dem Ableitexperiment um 10,9 % für das linke und um 14,9 % für das rechte Auge in T-N
Reizrichtung und um 11,2 % (linkes Auge) bzw. 17,8 % (rechtes Auge) in N-T Richtung.
Die Unterschiede in der Anzahl der Bewegungen der beiden Augen zeigen die
Entkopplung des linken und des rechten Auges.
3.3.4.2 Amplituden der Augenfolgebewegungen
Die Bestimmung der Amplituden der Augenfolgebewegungen in Abhängigkeit von den
untersuchten Reizgeschwindigkeiten und Stimulusrichtungen vor und nach Läsion des
Praetectums soll weiteren Aufschluss über den Reduktionsfaktor des gains der
Augenfolgebewegung geben.
Hierzu wurde die Augenposition zu Beginn und am Ende jeder aufgetretenen langsamen
Augenfolgephase innerhalb der 30 Sekunden Stimulusphase bestimmt und deren Median
berechnet. Wie in der Untersuchung zuvor repräsentiert das Versuchstier Sg 12 auch die
Ergebnisse von Sg 13 und Sg 16.
Vor der Läsion des Praetectums lag bei allen Versuchstieren, unabhängig von Reizrichtung
und Geschwindigkeit, der Median der Augenfolgebewegungsamplituden zwischen 3,2°
(Sg 12) und 7,9° (Sg 15).
96
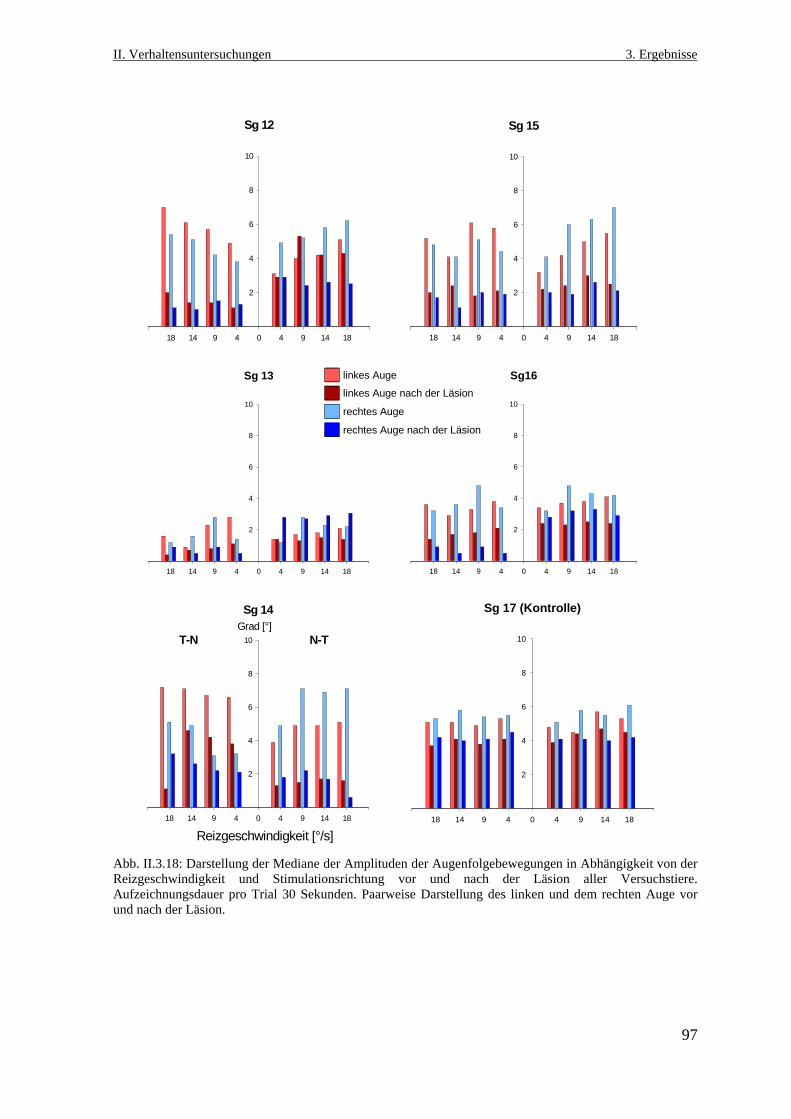
II. Verhaltensuntersuchungen 3. Ergebnisse
ARAu
bb. II.3.18: Darstellung der Mediane der Amplituden der Augenfolgebewegungen in Abhängigkeit von der eizgeschwindigkeit und Stimulationsrichtung vor und nach der Läsion aller Versuchstiere. ufzeichnungsdauer pro Trial 30 Sekunden. Paarweise Darstellung des linken und dem rechten Auge vor nd nach der Läsion.
Sg 12
18 14 9 4 0 4 9 14 18
2
4
6
8
10
Sg 13
18 14 9 4 0 4 9 14 18
2
4
6
8
10
Sg 15
18 14 9 4 0 4 9 14 18
2
4
6
8
10
Sg16
18 14 9 4 0 4 9 14 18
2
4
6
8
10
linkes Auge
linkes Auge nach der Läsion
rechtes Auge
rechtes Auge nach der Läsion
Sg 14
18 14 9 4 0 4 9 14 18
2
4
6
8
10 N-TT-N
Reizgeschwindigkeit [°/s]
Grad [°]
Sg 17 (Kontrolle)
18 14 9 4 0 4 9 14 18
2
4
6
8
10
97

II. Verhaltensuntersuchungen 3. Ergebnisse
Nach dem Läsionsexperiment sind bei allen Versuchtieren die Amplituden der
Augenfolgebewegung beider Augen signifikant (ANOVA: p < 0,05) verringert (Abb.
III.3.18).
Das Tier Sg 12, wie auch Sg 13 und Sg 16, zeigt signifikant kleinere Amplituden der
Folgebewegung des rechten und des linken Auges in T-N im Vergleich zur N-T Reizung.
In N-T Reizrichtung findet sich eine signifikante Abnahme der Amplituden bei
Versuchstier Sg 14.
Das Versuchstier Sg 15 zeigt nach Läsion in beiden Stimulusrichtungen geringere
Amplituden der Augenfolgebewegung.
Das Kontrolltier Sg 17 zeigt nach dem Ableitexperiment eine geringe Abnahme in der
Amplitude der Augenfolgebewegungen. Es besteht kein signifikanter Unterschied in der
Amplitude zwischen der T-N und N-T Stimulusrichtung.
3.3.4.3 Geschwindigkeit der Augenfolgebewegung
Mit dem direkten Vergleich der drei Parameter Augenfolgegeschwindigkeit, Amplitude der
Augenfolgebewegung und Frequenz der Augenfolgebewegungen soll das durch die Läsion
ausgelöste Defizit näher charakterisiert werden.
In Tabelle 3 sind diese Werte für alle untersuchten Versuchstiere nach den
Stimulusrichtungen und Geschwindigkeiten aufgeschlüsselt. Zusätzlich wurde das Produkt
aus Amplitude und Frequenz der Augenfolgebewegung berechnet. Diese Werte stimmen
erwartungsgemäss grösstenteils mit den gemessenen Geschwindigkeiten der
Augenfolgebewegungen überein.
98
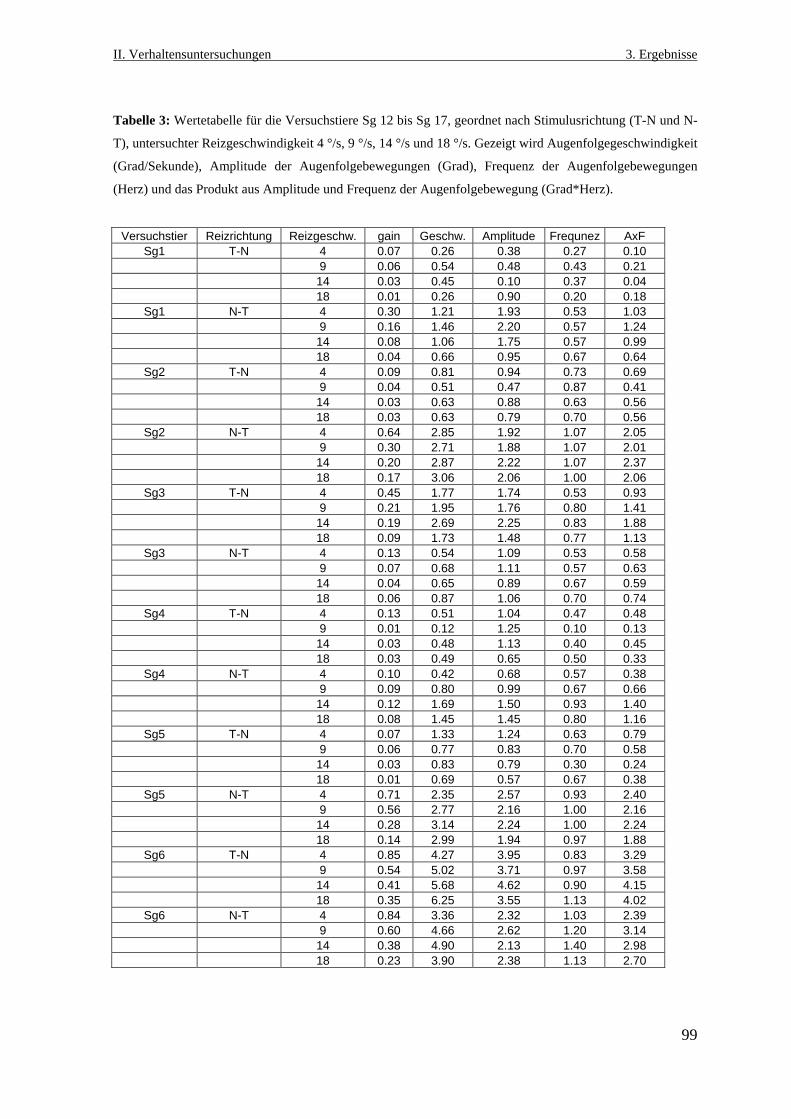
II. Verhaltensuntersuchungen 3. Ergebnisse
Tabelle 3: Wertetabelle für die Versuchstiere Sg 12 bis Sg 17, geordnet nach Stimulusrichtung (T-N und N-
T), untersuchter Reizgeschwindigkeit 4 °/s, 9 °/s, 14 °/s und 18 °/s. Gezeigt wird Augenfolgegeschwindigkeit
(Grad/Sekunde), Amplitude der Augenfolgebewegungen (Grad), Frequenz der Augenfolgebewegungen
(Herz) und das Produkt aus Amplitude und Frequenz der Augenfolgebewegung (Grad*Herz).
Versuchstier Reizrichtung Reizgeschw. gain Geschw. Amplitude Frequnez AxF Sg1 T-N 4 0.07 0.26 0.38 0.27 0.10
9 0.06 0.54 0.48 0.43 0.21 14 0.03 0.45 0.10 0.37 0.04 18 0.01 0.26 0.90 0.20 0.18
Sg1 N-T 4 0.30 1.21 1.93 0.53 1.03 9 0.16 1.46 2.20 0.57 1.24 14 0.08 1.06 1.75 0.57 0.99 18 0.04 0.66 0.95 0.67 0.64
Sg2 T-N 4 0.09 0.81 0.94 0.73 0.69 9 0.04 0.51 0.47 0.87 0.41 14 0.03 0.63 0.88 0.63 0.56 18 0.03 0.63 0.79 0.70 0.56
Sg2 N-T 4 0.64 2.85 1.92 1.07 2.05 9 0.30 2.71 1.88 1.07 2.01 14 0.20 2.87 2.22 1.07 2.37 18 0.17 3.06 2.06 1.00 2.06
Sg3 T-N 4 0.45 1.77 1.74 0.53 0.93 9 0.21 1.95 1.76 0.80 1.41 14 0.19 2.69 2.25 0.83 1.88 18 0.09 1.73 1.48 0.77 1.13
Sg3 N-T 4 0.13 0.54 1.09 0.53 0.58 9 0.07 0.68 1.11 0.57 0.63 14 0.04 0.65 0.89 0.67 0.59 18 0.06 0.87 1.06 0.70 0.74
Sg4 T-N 4 0.13 0.51 1.04 0.47 0.48 9 0.01 0.12 1.25 0.10 0.13 14 0.03 0.48 1.13 0.40 0.45 18 0.03 0.49 0.65 0.50 0.33
Sg4 N-T 4 0.10 0.42 0.68 0.57 0.38 9 0.09 0.80 0.99 0.67 0.66 14 0.12 1.69 1.50 0.93 1.40 18 0.08 1.45 1.45 0.80 1.16
Sg5 T-N 4 0.07 1.33 1.24 0.63 0.79 9 0.06 0.77 0.83 0.70 0.58 14 0.03 0.83 0.79 0.30 0.24 18 0.01 0.69 0.57 0.67 0.38
Sg5 N-T 4 0.71 2.35 2.57 0.93 2.40 9 0.56 2.77 2.16 1.00 2.16 14 0.28 3.14 2.24 1.00 2.24 18 0.14 2.99 1.94 0.97 1.88
Sg6 T-N 4 0.85 4.27 3.95 0.83 3.29 9 0.54 5.02 3.71 0.97 3.58 14 0.41 5.68 4.62 0.90 4.15 18 0.35 6.25 3.55 1.13 4.02
Sg6 N-T 4 0.84 3.36 2.32 1.03 2.39 9 0.60 4.66 2.62 1.20 3.14 14 0.38 4.90 2.13 1.40 2.98 18 0.23 3.90 2.38 1.13 2.70
99
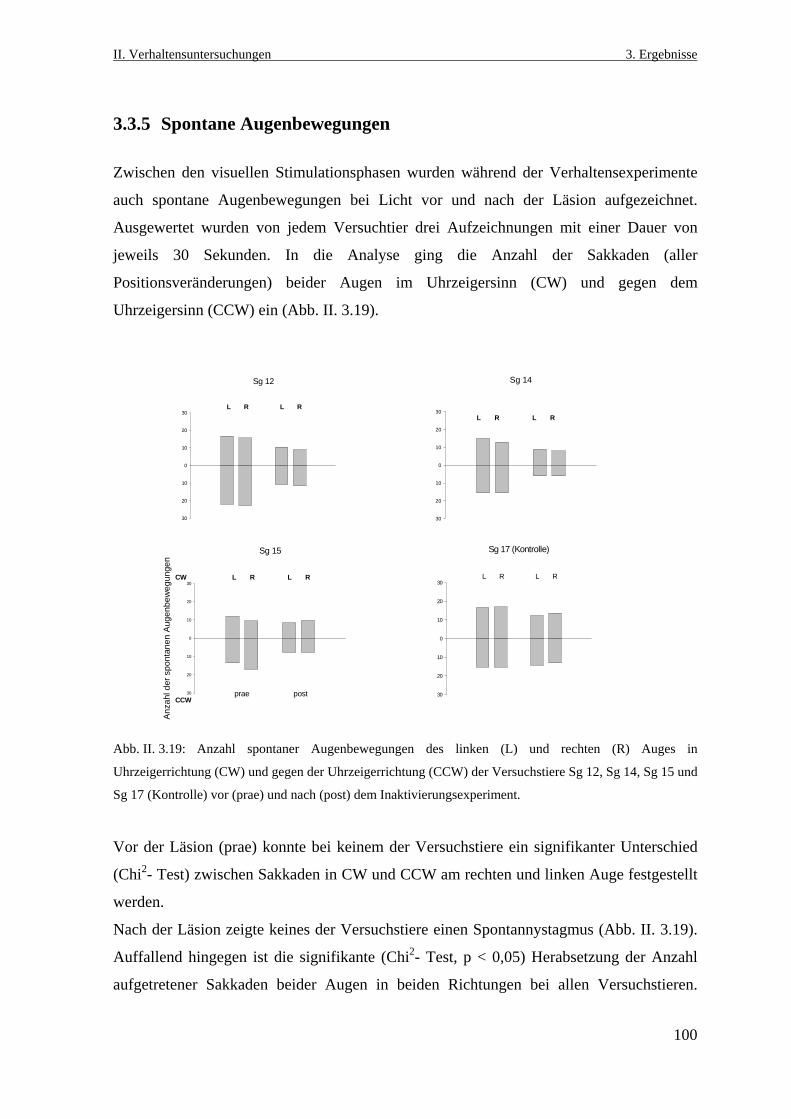
II. Verhaltensuntersuchungen 3. Ergebnisse
3.3.5 Spontane Augenbewegungen Zwischen den visuellen Stimulationsphasen wurden während der Verhaltensexperimente
auch spontane Augenbewegungen bei Licht vor und nach der Läsion aufgezeichnet.
Ausgewertet wurden von jedem Versuchtier drei Aufzeichnungen mit einer Dauer von
jeweils 30 Sekunden. In die Analyse ging die Anzahl der Sakkaden (aller
Positionsveränderungen) beider Augen im Uhrzeigersinn (CW) und gegen dem
Uhrzeigersinn (CCW) ein (Abb. II. 3.19).
Sg 12
30
20
10
0
10
20
30 L L R R
Sg 15
Anza
hl d
er s
pont
anen
Aug
enbe
weg
unge
n
30
20
10
0
10
20
30 L L R
prae post
RCW
CCW
Sg 14
30
20
10
0
10
20
30 L L R R
Sg 17 (Kontrolle)
30
20
10
0
10
20
30 L L R R
Abb. II. 3.19: Anzahl spontaner Augenbewegungen des linken (L) und rechten (R) Auges in
Uhrzeigerrichtung (CW) und gegen der Uhrzeigerrichtung (CCW) der Versuchstiere Sg 12, Sg 14, Sg 15 und
Sg 17 (Kontrolle) vor (prae) und nach (post) dem Inaktivierungsexperiment.
Vor der Läsion (prae) konnte bei keinem der Versuchstiere ein signifikanter Unterschied
(Chi2- Test) zwischen Sakkaden in CW und CCW am rechten und linken Auge festgestellt
werden.
Nach der Läsion zeigte keines der Versuchstiere einen Spontannystagmus (Abb. II. 3.19).
Auffallend hingegen ist die signifikante (Chi2- Test, p < 0,05) Herabsetzung der Anzahl
aufgetretener Sakkaden beider Augen in beiden Richtungen bei allen Versuchstieren.
100

II. Verhaltensuntersuchungen 3. Ergebnisse
Unterschiede in der Anzahl von Sakkaden für das rechte und linke Auge in CW bzw. CCW
Richtung können nicht gezeigt werden.
3.3.6 Histologische Lokalisation des Läsionsortes Nach Abschluss der Experimente wurden die Versuchstiere eingeschläfert, transcardial
perfundiert, die Gehirne entnommen und histologisch aufgearbeitet. In den 50 µm dicken
Nissl-gefärben Frontalschnitten konnte anschließend der Läsionsort näher identifiziert
werden.
Im Praetectum der Versuchstiere Sg 12, Sg 13, Sg 16 lagen die Läsionen ca. 900 µm
posterior vom rostralen Pol des Tectum opticum und ca. 1900 µm unter der tectalen
Oberfläche. Bei Sg 15 lag die Läsion ca. 1150 µm hinter dem rostralen Pol des TeO und
2250 µm unter seiner Oberfläche.
Mit einem Abstand von 1400 µm zum TeO – Pol und einer Tiefe von 2600 µm lag der
Läsionsort von Sg 14 am weitesten posterior.
Abbildung II. 3.20 zeigt Mikrofotografien der frontalen Gehirnschnitte von den
Versuchstieren Sg 12 (A), Sg 15 (B), Sg 14 (C) und dem Kontrolltier Sg 17 (D bis F) in
rostro-caudaler Richtung. Die Distanz zwischen den Läsionsorten in Abb. II. 3.20 A
(Sg 12, Sg 13, Sg 16) und C (Sg14) beträgt ca. 350 µm, die mittlere Tiefe unter der
tektalen Oberfläche 2350µm. Mit diesen Koordinaten kann die Lage des Areals
richtungsselektiver Neuronen im Praetectum der Regenbogenforelle beschrieben werden.
101
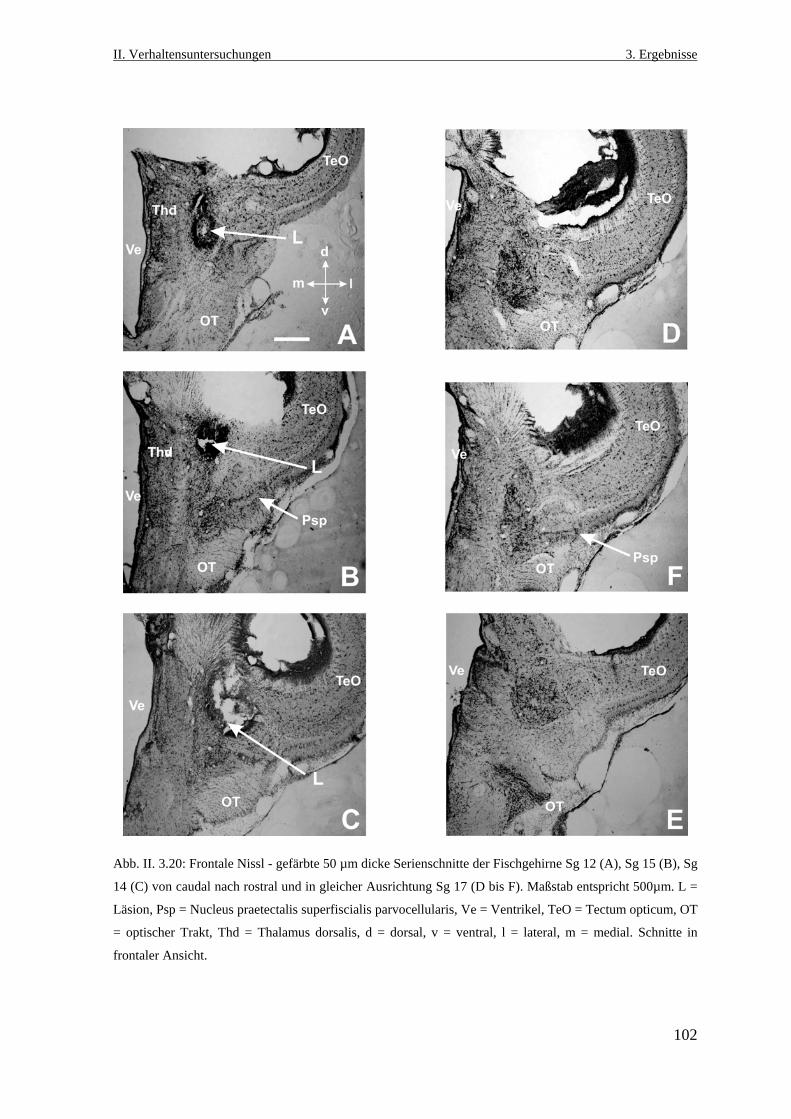
II. Verhaltensuntersuchungen 3. Ergebnisse
Abb. II. 3.20: Frontale Nissl - gefärbte 50 µm dicke Serienschnitte der Fischgehirne Sg 12 (A), Sg 15 (B), Sg
14 (C) von caudal nach rostral und in gleicher Ausrichtung Sg 17 (D bis F). Maßstab entspricht 500µm. L =
Läsion, Psp = Nucleus praetectalis superfiscialis parvocellularis, Ve = Ventrikel, TeO = Tectum opticum, OT
= optischer Trakt, Thd = Thalamus dorsalis, d = dorsal, v = ventral, l = lateral, m = medial. Schnitte in
frontaler Ansicht.
102

II. Verhaltensuntersuchungen 3. Ergebnisse
Die Schnittserie D bis F des Kontrolltieres Sg 17 in der Abb. II. 3.20 zeigt einen
vergleichbaren praetectalen Bereich wie A, B und C. Diese Schnittserie veranschaulicht,
dass in Folge des Ableitsexperiments keine sichtbaren Gewebeschädigungen im
Praetectum aufgetreten sind.
Eine Korrelation zwischen den Vorzugsrichtungen der abgeleiteten Neurone (Abb. II.
3.14) und den aufgetretenen Defiziten bei den optokinetischen Augenbewegungen beider
untersuchten Stimulusrichtungen kann belegt werden.
Läsionen im
- caudalen Bereich (Sg 12, Vorzugsrichtung nasal; Sg 13, Vorzugrichtung unten;
Sg16, Vorzugsrichtung temporal) bewirken eine Einschränkung in N-T
Stimulusrichtung,
- im mittleren Bereich des Areals (Sg 15, Vorzugsrichtung temporal - oben)
bewirken eine Einschränkung in beiden Stimulusrichtungen,
- rostralen Bereich (Sg 14, Vorzugsrichtung nasal) bewirken eine Einschränkung in
N-T Stimulusrichtung.
Mit den Untersuchungsergebnissen des Kontrolltieres Sg 17 kann weiter belegt werden,
dass nicht die Penetrationen mit der Mikroelektrode, die immer durch das TeO geführt
haben und dabei zu Verletzungen des tektalen Gewebes führten, diese Defizite in der
Ausführung des OKR hervorrufen.
103

II. Verhaltensuntersuchungen 4. Diskussion
4. Diskussion
4.1 Der vertikale optokinetische Reflex bei Salmo gairdneri Die Verhaltensuntersuchungen zum vertikalen optokinetischen Reflex (vOKR) an Salmo
gairdneri wurden aufgrund der Ergebnisse der elektrophysiologischen Experimente
durchgeführt. Danach existiert in einem begrenzten Areal im dorsalen Praetectum eine
Population von Neuronen, von denen jedes durch Bewegung großflächiger visueller
Stimuli in eine bestimmte Richtung aktiviert wird. In dieser Population wurden damit auch
richtungsselektive Zellen lokalisiert, die einen vertikalen visuellen Stimulus kodieren.
Diese visuelle Bewegung entspricht einer Rollbewegung um die Körperlängsachse.
Aufgrund der charakteristischen Antworteigenschaften und der Richtungsselektivität für
vertikal verlaufende Stimuli erfüllen diese Neurone die Grundvoraussetzung, den OKR in
vertikaler Richtung auszulösen. Die Untersuchungsergebnisse an Salmo gairdneri zum
vertikalen optokinetischen Reflex (vOKR) haben unerwartete Resultate ergeben.
Entgegen aller Erwartungen war es bei keinem der drei untersuchten Versuchstiere
möglich, vertikale Augenbewegungen auszulösen, die einem optokinetischen Nystagmus,
d.h. einer alternierenden Abfolge von langsamen Augenfolgebewegungen und
Rückstellsakkaden, entsprachen. Eine Analyse der OKR - Symmetrie war deshalb nicht
möglich.
Unter der monokularen Stimulationsbedingung während der steady-state-Phase wichen die
vertikalen Augenpositionen nur unregelmäßig und selten von ihrer horizontalen Lage mit
Amplituden von +/- 0,8° ab. Vereinzelt konnten horizontale Augenpositionsveränderungen
registriert werden, deren Amplituden bis zu 3,4° betrugen und nicht mit den vertikalen
Bewegungsrichtungen korrelierten.
Binokulare Stimulation löste maximal fünf vertikale Augenpositionsveränderungen (im
Mittel +/- 0,7°) aus. Diese folgten nicht dem Sägezahnmuster eines optokinetischen
Nystagmus. Eine Korrelation zwischen Augenbewegungsrichtung und
Stimulationsrichtung war nicht gegeben. Die zeitgleichen Aufzeichnungen der
horizontalen Augenpositionsveränderungen ergaben eine Zunahme der Amplituden von
4,9° im Vergleich zur monokularen Stimulation. Horizontale und vertikale
Augenbewegungen waren wiederum nicht gekoppelt.
104

II. Verhaltensuntersuchungen 4. Diskussion
4.1.2 Gründe für das Ausbleiben des vertikalen optokinetischen Reflexes
4.1.2.1 Anordnung der extraokulären Muskeln
Untersuchungen an den extraokulären Muskeln der Teleostei haben gezeigt, dass zwei
schräg verlaufende Muskeln (M. obliquus inferior, M. obliquus superior) nasal zum
anterioren Myodom verlaufen. Vier gerade Muskeln, nämlich der M. rectus inferior, M.
rectus superior, M. rectus posterior und M. rectus anterior ziehen in das posteriore
Myodom, das an der ventralen Schädelbasis anliegt. Trotz dieses gegenüber den anderen
Vertebratenklassen unterschiedlichen Verlaufes der Augenmuskeln befähigen diese sechs
äußeren Augenmuskeln die Teleostei, ihre Augen in alle Raumrichtungen zu bewegen.
Horizontale Augenbewegungen in temporale Richtung werden durch die Kontraktion des
M. rectus posterior ausgeführt, in nasale Richtung durch Kontraktion des M. rectus
anterior. Vertikale Augenbewegungen nach oben erfolgen durch die Kontraktion des M.
obliquus superior und M. rectus superior. Nach unten gerichtete Augenbewegungen
werden durch den M. obliquus inferior und den M. rectus inferior bewirkt (Starck, 1982;
Band 3; Bauchot et al., 1989).
Graf und McGurck (1985) haben unter dem Aspekt der vergleichenden Kinematik die
extraokulären Muskeln des Goldfisches (Carassius auratus) mit denen anderer Vertebraten
mit lateralständigen Augen verglichen. In ihrer Studie zeigten sie, dass trotz veränderter
Ansätze der Augenmuskeln am Bulbus (dorso - ventral abgeflacht) und veränderten
Verlaufes im Schädelbereich (siehe oben) die kinematische Charakteristik der Goldfisch-
Augenmuskulatur qualitativ mit anderen Tieren (z.B. Kaninchen, Ratte) mit lateralen
Augen im Einklang steht (Ezure & Graf, 1984; Simpson & Graf, 1981). Institutseigene
Untersuchungen an Salmo gairdneri ergaben vergleichbare Befunde über Ansatz und
Verlauf der Augenmuskeln im Schädel. Unterschiede zum Goldfisch konnten in der Länge
des M. rectus posterior und des M. rectus anterior aufgezeigt werden. Diese sind bei Salmo
gairdneri im Verhältnis zum Goldfisch um ca. 1/3 länger und verlaufen bis zum
posterioren Ende des posterioren Myodoms, wo sie über eine kurze Sehne am
Schädelknochen ansetzen. Die Stärke und Länge der Augenmuskeln zur Ausführung
vertikaler Augenbewegungen sind bei Regenbogenforelle und Goldfisch annähernd gleich.
Demnach besitzt Salmo gairdneri die anatomische Grundlage für vertikale
105

II. Verhaltensuntersuchungen 4. Diskussion
Augenbewegungen (Ergebnisse in dieser Arbeit nicht gezeigt), die sie auch unter VOR-
Stimulation zeigt.
Zusammengefasst weisen diese Ergebnisse auf eine Dominanz horizontaler
Augenbewegungen hin. Unterstützung findet diese Annahme in der Beobachtung
spontaner Augenbewegungen. Diese wurden ausschließlich in der horizontalen
Bewegungsrichtung ausgeführt. Das bedeutet aber nicht, dass vertikale Augenbewegungen
nicht möglich sind, wie die VOR - Ergebnisse belegen.
4.1.2.2 Die Wahl des adäquaten visuellen Reizes
Das Auslösen des OKRs ist abhängig von der Wahl des visuellen Stimulus. Wichtige
Reizparameter sind:
- die Wahl des Reizmusters (z. B. schwarz - weißes Streifenmuster, schwarz-weißes
Zufallspunktemuster, weiße Lichtpunkte, die durch ein Planetarium erzeugt
werden)
- die Größe des Reizmusters (von >1° Sehwinkel bis 20° Sehwinkel)
- Luminanz und Kontrastverhältnisse
Diese Parameter müssen an das zu untersuchende Versuchstier mit seinen individuellen
optischen Eigenschaften optimal angepasst sein (Schor & Narayan 1981; Flandrin et al.,
1990).
Unter Verwendung zweier unterschiedlicher Reizmuster konnten Maioli und Precht (1984)
in Untersuchungen zum OKR an der Katze zeigen, dass das gain der Augenfolgebewegung
mit einem Zufallspunktemuster größer ist als mit einem Streifenmuster. Allerdings galt
dies nur für höhere Reizgeschwindigkeiten. Eine Erklärung dieses Befundes liegt in der
niedrigeren räumlichen Frequenz des Zufallspunktemusters gegenüber dem verwendeten
Streifenmuster.
Ein nicht optimales Reizmuster kann zu großen Fehleinschätzungen führen. Ein Beispiel
dafür ist der optokinetische Reflex von Albinoratten, der mit einem Streifenmuster nicht
ausgelöst werden kann (Precht & Cazin, 1979). Wählt man allerdings andere
Reizparameter (veränderte Lichtintensitäten, Zufallspunktemuster statt Streifen), so läßt
sich ein OKR auch bei albinotischen Ratten auslösen (Sirkin et al., 1985).
106

II. Verhaltensuntersuchungen 4. Diskussion
In dem in der vorliegenden Arbeit durchgeführtem Verhaltensexperiment zum
monokularen horizontalen optokinetischen Reflex (mhOKR), das der Überprüfung der
Messmethode und des verwendeten visuellen Stimulus diente, konnte eindeutig gezeigt
werden, dass das vom Planetarium erzeugte Zufallspunktemuster einen adäquaten Stimulus
darstellt, um bei Salmo gairdneri monokulare, horizontale optokinetische
Augenbewegungen auszulösen. Entsprechend sollte man mit diesem Reizmuster auch
vertikale Augenbewegungen auslösen können. Dieses Kontrollexperiment bestätigt
außerdem die Ergebnisse zum mhOKR an Salmo gairdneri aus meiner Diplomarbeit. In
diesen Verhaltensexperimenten wurde mit einer Reiztrommel, die innenseitig mit einem
schwarz-weißen Zufallspunktemuster ausgekleidet war, unter künstlicher Beleuchtung
visuell stimuliert.
Die Untersuchungsergebnisse zum vOKR gelten nur für Salmo gairdneri. Vergleichsdaten
aus der Literatur an Salmo gairdneri oder an anderen Knochenfischenarten liegen nicht
vor. Bisher noch unveröffentlichte Beobachtungen zum optokinetischen System an
Serranus cabrilla zeigen jedoch, dass dieser Knochenfisch durchaus die Fähigkeit besitzt,
einen vertikalen OKR auszuführen (Ergebnisse sind in dieser Arbeit nicht gezeigt).
4.1.2.3 Anpassung an das Habitat
Hinweise für eine Anpassung des optokinetischen Systems an das Habitat liefert die Studie
von Dieringer, Graf und Reichenberger (1992), die hOKR und hVOR an unterschiedlichen
Süßwasser- und marinen Knochenfischarten aus den Familien Cyprinidae, Percidae,
Osteoglossidae, Scorpaenidae, Batrachoididae, Siluridae und Characidae durchführten.
Ihre Befunde zeigen einen Bezug zwischen der Besiedlung verschiedener Habitate und der
Visuomotorik beim OKR und VOR und damit verbunden die große Variabilität, die in der
Überordnung Teleostei existiert. Untersuchungen des optokinetischen- und vestibulo-
okulären Reflexes bei Goldfisch (Carassius auratus) und Austernfisch (Opsanus tau)
zeigen zwei unterschiedliche Funktionsprinzipien. Während der Goldfisch bei beiden
Stimulationen (OKR und VOR) stets regelmäßige Augenbewegungen ausführte, waren die
Augenbewegungen des Austernfisches unregelmäßig und hatten stets ein niedrigeres gain.
Auch der optokinetische Nachnystagmus hielt am Goldfisch mit 10 bis 12 s fünf Mal
107

II. Verhaltensuntersuchungen 4. Diskussion
länger an. Dies weist auf das Fehlen eines neuronalen Geschwindigkeitsspeichers im
optokinetischen Systems des Austernfisches hin.
Der Goldfisch ist, wie alle Cypriniden, ein aktiver Schwimmer. Dadurch bedingte
Ortsveränderungen der Umwelt auf der Retina werden durch Augenbewegungen
kompensiert. Im Gegensatz dazu ist der Austernfisch in seinem Lebensraum relativ
ortskonstant. Die Anpassung besteht aus einer perfekten Tarnung. Unbeweglich lauert er
am Boden auf Beutefische, die er bei zu nahem Vorbeischwimmen schlägt. Er vermeidet
ein ununterbrochenes Umherschwimmen und damit Augenbewegungen (Dieringer et al.,
1992).
Salmo gairdneri wird den Strömungsfischen zugeordnet. Ihr bevorzugtes Habitat sind
Flussoberläufe oder Flüsse sowie strömungsreiche Seen. Eine Hypothese über das
Ausbleiben des vOKR könnte darin begründet sein, dass die anatomische Lage der
Schwimmblase den Fisch aufrecht im Wasser hält. Zusätzlich stabilisiert sich der Körper
des Tieres selbst, durch das ständige Schwimmen gegen die Strömungrichtung des
Wasseres. Weitere Strömungsschwankungen des Wassers, die eine instabile Schwimmlage
bewirken können, werden durch Gleichgewichtsorgan und das Seitenlinienorgan detektiert
und zentralnervöse weiter verarbeitet (Bleckmann et al., 1989). Dadurch wird eine
aufrechte Schwimmlage des Fisches im Wasser ermöglicht und ein Rollen um die
Körperlängsachse vermieden. Aktive Schwimmrichtungsveränderungen erfolgen in der
horizintalen oder in „Pitch“ – Richtung, das einer vertikalen Aufwärts- bzw. Abwärts-
Schwimmrichtung entspricht und zur Bildstabilisierung Torsionsbewegung des Auges zur
Folge hat. Die auftretenden retinale Bildverschiebung werden über den optokinetischen
Reflex (OKR) und vestibulo - okulären Reflex (VOR) kompensiert.
4.2 Der visuelle Einfluss auf das vestibulo-okuläre System
4.2.1 Der vertikale vestibulo-okuläre Reflex Die Ergebnisse der vOKR – Studie führten zu folgender Hypothese:
Wenn die vertikale Bewegungen kodierenden richtungsselektiven Neurone im Praetectum
keinen Beitrag zum vOKR leisten, dann könnte ihre visuelle Information über eine
108

II. Verhaltensuntersuchungen 4. Diskussion
vertikale retinale Bildverschiebung zur Kalibrierung des gains der kompensatorischen
Augenbewegungen des vestibulo - okulären Systems genutzt werden, das im Idealfall den
Wert von 1 haben sollte.
Verhaltensstudien, die sich mit dem vestibulo - okulären System befassten, haben bei allen
bisher untersuchten Tieren (inklusive der Teleostei) gezeigt, dass zusätzliche visuelle
Information Auswirkungen auf das gain der kompensatorischen Augenbewegungen des
VOR hat. Mit visueller Positionsinformation im Raum erreicht das gain der
kompensatorischen Augenbewegungen einen Wert von Eins. Beim Fehlen der visuellen
Referenz ist das gain kleiner als Eins. Die Kopplung des optokinetischen und des
vestibulo - okulären Systems wurde zudem anatomisch und elektrophysiologisch nach-
gewiesen ( Ito, 1972, 1982; Dichgans et al., 1973; Precht & Anderson 1979; Cazin et al.,
1980, 1984; Stahl & Simpsion, 1995; Watanabe et al., 1993; Raphan & Cohen, 2002).
Daraus ergab sich die Wahl der drei verwendeten Stimulationsparadigmen:
A. Das visuelle Reizmuster dreht sich exakt mit der sinusförmigen Drehrichtung der
vestibulären Stimulation.
B. Das visuelle Reizmuster bleibt stationär in der Umwelt, das Tier erfährt die
sinusförmige vestibuläre Stimulation.
C. Das visuelle Reizmuster dreht sich mit exakt der gleichen Geschwindigkeit wie das
Tier, aber in entgegengesetzter Richtung.
Bei einer absoluten visuellen Dominanz auf das gain des VOR ergibt sich im
Stimulationsparadigma A ein Erwartungswert von 0, da keine retinale Bildverschiebung
auftritt und somit die kompensatorischen Augenbewegungen des vestibulo - okulären
Systems unterdrückt werden sollten.
Im Stimulationsparadigma B wäre der gain - Erwartungswert 1. Die stationäre Umwelt im
Raum löst die optokinetischen Mechanismen aus, die einer retinalen Bildverschiebung
entgegenwirken und es greifen die vestibulo - okulären Reflexleistungen.
Das dritte Stimulationsparadigma führt zu einer Verdopplung der visuellen
Reizgeschwindigkeit. Die visuell ausgelösten kompensatorischen Augenbewegungen
führen zu einem Erwartungswert des gains von 2. Das Ergebnis aus der Gruppenanalyse
109
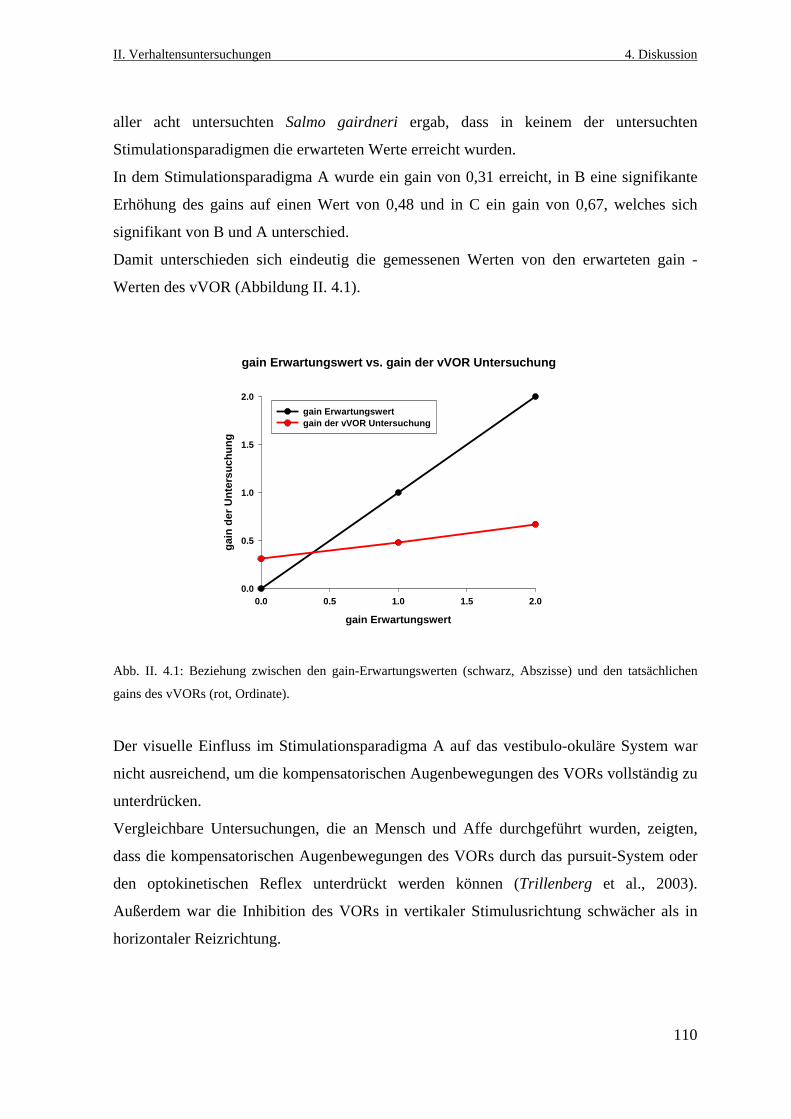
II. Verhaltensuntersuchungen 4. Diskussion
aller acht untersuchten Salmo gairdneri ergab, dass in keinem der untersuchten
Stimulationsparadigmen die erwarteten Werte erreicht wurden.
In dem Stimulationsparadigma A wurde ein gain von 0,31 erreicht, in B eine signifikante
Erhöhung des gains auf einen Wert von 0,48 und in C ein gain von 0,67, welches sich
signifikant von B und A unterschied.
Damit unterschieden sich eindeutig die gemessenen Werten von den erwarteten gain -
Werten des vVOR (Abbildung II. 4.1).
gain Erwartungswert vs. gain der vVOR Untersuchung
gain Erwartungswert
0.0 0.5 1.0 1.5 2.0
gain
der
Unt
ersu
chun
g
0.0
0.5
1.0
1.5
2.0gain Erwartungswertgain der vVOR Untersuchung
Abb. II. 4.1: Beziehung zwischen den gain-Erwartungswerten (schwarz, Abszisse) und den tatsächlichen
gains des vVORs (rot, Ordinate).
Der visuelle Einfluss im Stimulationsparadigma A auf das vestibulo-okuläre System war
nicht ausreichend, um die kompensatorischen Augenbewegungen des VORs vollständig zu
unterdrücken.
Vergleichbare Untersuchungen, die an Mensch und Affe durchgeführt wurden, zeigten,
dass die kompensatorischen Augenbewegungen des VORs durch das pursuit-System oder
den optokinetischen Reflex unterdrückt werden können (Trillenberg et al., 2003).
Außerdem war die Inhibition des VORs in vertikaler Stimulusrichtung schwächer als in
horizontaler Reizrichtung.
110

II. Verhaltensuntersuchungen 4. Diskussion
Um den visuellen Einfluss auf das vestibulo - okuläre System näher zu charakterisieren,
wurde an drei der acht untersuchten Tiere der vertikale VOR auch bei Dunkelheit
untersucht. Der Erwartungswert des gains sollte dabei größer sein als im
Stimulationsparadigma A und kleiner als in B (McElligott, 1997; Baker et al., 1998).
Die Untersuchung zeigte, dass das gain mit 0,22 niedriger war als im
Stimulationsparadigma A. Dieses unerwartete Ergebnis deutet auf einen zusätzlichen
unspezifischen visuellen Einfluss (aufmerksamkeitssteigernd) auf das vestibulo - okuläre
System der Regenbogenforelle hin.
In dem Stimulationsparadigma B erfuhren die Versuchstiere eine vestibuläre Stimulation.
Gleichzeitig trat aufgrund des stationären Reizmusters eine vertikale Bildverschiebung auf
der Retina auf. Die signifikante Erhöhung des gains im Vergleich zum
Stimulationsparadigma A ist ein eindeutiger Beleg für den visuellen Einfluss auf das
vestibulo- okuläre System. Unterstützung fand dieser Befund im Stimulationsparadigma C.
Durch eine Phasenverschiebung um 180° zwischen der vestibulären und der visuellen
Stimulation bei gleichbleibender Reizfrequenz wurde eine Verdoppelung der visuellen
Stimulusgeschwindigkeit erreicht. In der Gruppenanalyse erreichte das gain im Vergleich
zu den Paradigmen B und A den höchsten, wenn auch nicht den doppelten Wert von B, der
theoretisch zu erwarten gewesen wäre.
Der Verstärkungsfaktor des gemessenen VOR - gains aus der Gruppenanalyse in
Abhängigkeit von den Stimulationsparadigmen A - B und B - C liegt bei Werten von 1,55
und 1,4. Diese Werte geben den Modulationsfaktor an, den das visuelle System auf die
Ausführung kompensatorischer Augenbewegungen beim VOR tatsächlich ausübt.
Es bleibt die Frage, warum das gain in den Stimulationsparadigmen B und C nicht die zu
erwartenden Werte von 1 und 2 annahm. Die Ursache könnte in der Ausrichtung des
Bogengangsystems relativ zur Zugrichtung der Augenmuskeln begründet sein (Simpson &
Graf, 1985). Der Canalis semicircularis anterior steht je nach Spezies in einem Winkel von
45° bis 55° zur Körperlängsachse und in einem Winkel von 90° zum C.s. posterior.
Bezogen auf den vVOR bedeutet dies, dass mit der vertikalen sinusförmigen
Drehbeschleunigung, die einer Rollbewegung des Fisches um die Körperlängsachse
entspricht, die Bogengänge C.s. anterior und C.s. posterior stimuliert werden. Vertikale
Augenbewegungen werden vom M. obliquus superior und M. rectus superior nach oben
und vom M. obliquus inferior und M. rectus inferior nach unten ausgeführt. Es findet also
111

II. Verhaltensuntersuchungen 4. Diskussion
eine „Mischstimulation“ zweier Bogengänge und zweier Muskelpaare statt. Eine optimale
Stimulusrichtung für je einen einzelnen Bogengang wäre (in Bogengangkoordinaten) links
- anterior / rechts - posterior (LARP) und rechts-anterior / links-posterior (RALP); dies
entspräche auch der Zugrichtung der Augenmuskeln (Simpson & Graf, 1981).
4.2.2 Der horizontale vestibulo-okuläre Reflex An drei der acht untersuchten Salmo gairdneri wurde unter den Stimulationsparadigmen A,
B und C auch das gain des horizontalen VORs mit der Erwartung untersucht, dass das gain
aufgrund der Lageübereinstimmung des horizontalen Bogenganges mit der horizontalen
Zugrichtung der Augenmuskeln größer ist als in der vertikalen Stimulationsrichtung und
sich entsprechen der theoretischen Voraussage verhält.
Das Ergebnis der Gruppenanalyse stellte sich wie folgt dar. In dem Stimulationsparadigma
A wurde von den Versuchstieren ein gain von 0,55 erreicht. In B war das gain signifikant
auf 0,69, in C auf 0,84 erhöht. Der Vergrösserungsfaktor kann von Stimulationsparadigma
A nach B mit 1,25 und von B nach C 1,22 beziffert werden. Auch in der horizontalen
Stimulationsebene wurden die gain Erwartungswerte also nicht erreicht. Die
Modulationsfaktoren sind annähernd gleich für die vertikale und horizontale
Stimulationsrichtung.
Im hVOR - Dunkelexperiment, das an drei der acht untersuchten Tiere durchgeführt
wurde, stellte sich wie zuvor das niedrigste gain von 0.33 ein. Der Vergleich zwischen dem
vVOR und dem hVOR zeigt, dass das gain der kompensatorischen Augenbewegungen in
der horizontalen Stimulationsebene in allen Paradigmen signifikant größer war.
Dieser Befund spricht für eine sehr enge Beziehung zwischen der horizontalen
Zugrichtung der Augenmuskeln (M. rectus posterior, M. rectus anterior) und der Lage des
horizontalen Bogenganges Canalis semicircularis lateralis, und für eine suboptimale
Anordnung der entsprechenden Strukturen in der vertikalen Roll-Situation.
112

II. Verhaltensuntersuchungen 4. Diskussion
4.2.3 Die Kopplung des optokinetischen und des vestibulo - okulären
Systems Bei den Amnioten sowie den Amphibien wurde eine visuelle Projektion des AOS zur
Oliva inferior nachgewiesen (Mammalia: Giolli et al., 1984; Simpson, 1984; Aves: Brauth
et al., 1978; Wylie, 2001; Reptilia: Reiner & Karten, 1978; Amphibia: Montogomery et al.,
1981, 1982, 1985). Von dort wird die visuelle Information über die Kletterfasern zu den
Purkinjezellen des Vestibulocerebellums geleitet (Ito, 1982; Magnin et al., 1983, 1989;
DeZeeuw et al., 1994, 1995), die ihrerseits den inhibitorischen Eingang auf die
vestibulären Neurone stellen (Ito, 1972; Miles & Lisberger, 1981; Lisberger & Sejnowski,
1992).
Im Vestibulocerebellum findet die Kalibrierung des VORs statt. Dazu dienen die
neuronalen Informationen über die Differenz zwischen Augenbewegung (OKR) und
Reizbewegung (VOR) (Graf et al., 1988; DeZeeuw et al., 1994, 1995).
Eine entsprechende Projektion wurde bei den Teleostei bisher nicht nachgewiesen. Durch
Injektion anterograder Tracer in das AOS (P1/P2: Finger und Karten, 1978; Nucleus
praetectalis ventralis: Luiten; 1986; nucleus praetectalis: Grover und Sharma, 1981; Butler
et. al., 1991) wurde beim Goldfisch unter Umgehung des Projektionsweges zur IO und
Kernen des Mesencephalons eine direkte Projektionen in das Vestibulocerebellum
identifiziert.
Pastor et al. (1994) gelang es, beim Goldfisch die Projektionen der Purkinjezellen im
Vestibulocerebellum zu den Vestibulariskernen nachzuweisen. Durch Elektrostimulation
der Vestibulariskerne konnten sie Augenbewegungen auslösen. Eine Modulierung der
simple – spike - Aktivität von Purkinjezellen durch alleinige VOR - Stimulation und durch
alleinige Augenbewegungen wurden ebenfalls nachgewiesen. Im Vestibulocerebellum
kann also das gain der kompensatorischen Augenbewegungen des VORs durch das
optokinetische und vestibuläre System moduliert werden. Unterstützt werden diese
Ergebnisse durch Adaptationsexperimente zum VOR von McElligott (1998), Michnovicz
und Bennet (1987) und Pastor et al. (1994), die unter dem Aspekt cerebellaerer Plastizität
und des Lernens durchgeführt wurden.
Eine zweite visuelle Projektion, des AOS der Säuger führt zum Nucleus praepositus
hypoglossi. Dieses Kerngebiet liefert den visuellen Eingang für die Vestibulariskerne.
113

II. Verhaltensuntersuchungen 4. Diskussion
Durch Integration der visuellen und der vestibulären Information wird das
Geschwindigkeitssignal der Augen gebildet (McCrea & Baker, 1985; Delgardo-Garcia et
al., 1989). Auch diese Projektion wurde bei Teleostei bisher nicht nachgewiesen. Ein
vergleichbares Areal im Hirnstamm des Goldfisches wurde von Pastor et al. (1994) als
Area II bezeichnet. Eine Inaktivierung der Area II mit Lidocain führte zu einer Reduktion
des gains von VOR und OKR.
Bei den Säugern existiert ein drittes Areal, der Nucleus dorsolateralis pontis. Er erhält
ebenfalls Eingang vom AOS und stellt über die Moosfasern eine Verbindung zum
Kleinhirn her.
Welche Projektionen gehen vom dorsalen praetectalen Areal richtungsselektiver Zellen bei
Salmo gairdneri und Esox lucius aus? Bei den Teleostei sind bisher zwei Wege bekannt,
der direkte Pfad in das Vestibulocerebellum und der Pfad über die Area II. Ein relativ
geringes Gewicht würde ich der direkten Projektion in das Vestibulocerebellum zuordnen.
In Inaktivierungsexperimenten konnten nur geringe Veränderungen des gains gezeigt
werden (Michnovicz & Bennet, 1987; Pastor et al., 1994; DeZeeuw et al., 1995). Der
massive Einfluss einer Area II Inaktivierung auf das gain von OKR und VOR weist auf die
wichtige Rolle der Projektion des Praetectums zur Area II hin (Pastor et al., 1994;
McElligott et al., 1998). Da funktionelle Untersuchungen zu den vom Praetektum
ausgehenden Projektionen derzeit nicht vorliegen, kann die Frage nach der relativen
Bedeutung der verschiedenen Projektionen hier nicht abschließend geklärt werden.
114

II. Verhaltensuntersuchungen 4. Diskussion
4.3 Auswirkungen einer unilateralen praetectalen Läsion auf
den optokinetischen Reflex Durch die Läsion des dorsalen Praetectums wurde die Bedeutung der richtungsselektiven
Neurone von Salmo gairdneri für den horizontalen optokinetischen Reflex untersucht.
Welche Voraussagen können getroffen werden?
Eine für Säuger formulierte Modellvorstellung geht davon aus, dass der linke und der
rechte NOT jeweils vom kontralateralen Auge richtungsselektiven Eingang erhalten. Wenn
keine visuelle Stimulation erfolgt, besitzen beide NOT - Neuronenpopulationen ein
ausgeglichenes Aktivitätsniveau und die Augen bleiben auf ihrer Position. Bei visueller
Stimulation kodiert der linke NOT Bewegungsrichtungen nach links und der rechte NOT
Bewegungsrichtungen nach rechts. Bei Reizung nach links wird die neuronale Aktivität im
linken NOT erhöht, im rechten NOT aber abgesenkt. Es resultiert eine Augenbewegung
nach links.
Wenn nun dieses Gleichgewichtssystem durch reversible Inaktivierung oder Läsion eines
NOTs gestört wird, löst dies aufgrund des künstlich hervorgerufenen
Aktivitätsungleichgewichts langsame Augenfolgebewegungen zur intakten, normal aktiven
Seite hin aus. Rückstellsakkaden erfolgen zur inaktivierten Seite. Die Folge ist ein
Spontannystagmus.
Genau dieses Erscheinungsbild zeigen reversible Inaktivierungen und Läsionsexperimente
in verschiedenen Vertebratenklassen (Amphibia: Montgomery et al., 1982; Aves &
Reptilia: Fite et al., 1979; Reptilia: Fan et al., 1995; Ariel & Kogo, 2001; Kogo et al.,
1998; Aves: Wylie & Frost, 1990).
Welche Auswirkungen haben praetectale Läsionen bei der Regenbogenforelle?
4.3.1 Die Voruntersuchung zum mhOKR und das Läsionsexperiment
Mit der Voruntersuchung zum mhOKR sollte sichergestellt werden, dass bei allen sechs
Salmo gairdneri, die in dieser Studie untersucht wurden, vor dem Eingriff ein mhOKR
auslösbar war.
115

II. Verhaltensuntersuchungen 4. Diskussion
Die Läsion an den fünf Tieren erfolgten an möglichst unterschiedlichen rostro-caudalen
Bereichen des dorsalen praetektalen Areals. Der Grund dafür ist der Befund, dass Neurone
mit unterschiedlichen Vorzugsrichtungen an unterschiedlichen anterior - posterior
Positionen im dorsalen Praetectum liegen. Damit unterscheidet sich dieses praetectale
Areal der Teleostei vom nLM der Amphibia, Reptilia und Aves sowie vom NOT der
Mammalia, in denen ipsiversive horizontale Augenfolgebewegungen kodiert werden
(Amphibia: Katte & Hoffmann, 1980; Fite, 1985; Montgomery et al., 1985; Reptilia:
Schröder, 1981; Mammalia: Collewijn, 1975; Hoffmann & Schoppmann, 1975; Hoffmann
& Fischer, 2001). Das sechste Versuchstier diente als Kontrolle ohne Praetectumläsion.
Dieses Experiment sollte Aufschluss darüber geben, ob Penetrationen mit der
Ableitelektrode zu Gewebeverletzungen im Tectum opticum (TeO) bzw. in benachbarten
Fasertrakten führen, die Veränderungen der Augenbewegungen zur Folge haben. Das TeO
ist ein multisensorisches Areal mit u. a. visuomotorischer Funktion. Elektrostimulation im
TeO führen zu Augen- und/oder Rumpfbewegungen (Herrero et al., 1998). Ausgedehnte
Schädigungen des TeO könnten damit zu Defiziten bei den Augenbewegungen führen.
4.3.2 Verhaltensbeobachtung nach der praetectalen Läsion Nach dem vollständigen Abklingen der Narkosewirkung wurde das Verhalten der
Versuchstiere in ihren Aquarien beobachtet. Alle fünf Salmoniden mit lädiertem linken
Praetectum zeigten eine starke Einschränkung in ihrem aktiven Schwimmverhalten, waren
bodenorientiert mit einer leichten Körperneigung zum Läsionsort. Während der wenigen
Aktivitätsphasen wurden fast ausschließlich Schwimmrichtungen zur Läsionsseite
(ipsiversiv) beobachtet. Das sechste Versuchstier (Kontrolle) zeigte ein normales
Aktivitätsverhalten und keine Präferenz in einer Schwimmrichtung. Dieser Befund
unterstützt die Theorie des Aktivitätsgleichgewichtes im praetectalen Areal, da beim
schwimmen schnelle Bewegungen zur inaktiven Seite ausgeführt werden. Die spontanen
Augenbewegungen wurde am linken abgedeckten Auge und am rechten stimulierten Auge
vor und nach dem Läsionsexperiment analysiert. Eine Bevorzugung einer
Sakkadenrichtung konnte vor der Läsion nicht festgestellt werden. Nach der Läsion nahm
die Sakkadenhäufigkeit bei allen Tieren signifikant ab. In keinem Fall konnte aber eine
116

II. Verhaltensuntersuchungen 4. Diskussion
Bevorzugung einer Sakkadenrichtung an beiden Augen festgestellt werden. Ein
Spontannystagmus blieb aus.
Der Spontannystagmus ist ein eindeutiges Indiz für eine Zerstörung oder Inaktivierung des
NOTs oder nLM. Wenn die Folgestrukturen im Gehirn keine Aktivität von einem NOT
bzw. nLM erhalten, wird das gesamte optokinetische System in einen
Ungleichgewichtszustand versetzt.
Zusammengefasst sprechen diese ersten Ergebnisse gegen einen Mechanismus des
Aktivitätsgleichgewichts des linken und rechten praetectalen richtungsselektiven Areals im
optokinetischen System von Salmo gairdnerii weil kein Spontannystagmus auftrat.
4.3.3 Der mhOKN nach der Inaktivierung Welche weiteren Folgen können im Zusammenhang mit der praetectalen Läsion
festgestellt werden?
Von den sechs Versuchstieren zeigten fünf Tiere eine signifikante Herabsetzung des gains.
Dabei war auffällig, dass die Tiere Sg 12, Sg 13 und Sg 16 gegenüber den praeoperativen
Messungen eine signifikante Reduktion des gains in temporo - nasaler Reizrichtung
zeigten, Sg14 in naso - temporaler Reizrichtung und Sg 15 in beiden Stimulusrichtungen.
Das Kontrolltier (Sg 17) zeigte ein zum Vorexperiment vergleichbares gain über alle
untersuchten Reizgeschwindigkeiten. Eine unilaterale Läsion des Praetectums wirkt sich
also auf das gain der langsamen Augenfolgephase während des OKR in einer der
horizontalen Stimulusrichtungen aus. Darin unterscheidet sich das praetectale Areal der
Regenbogenforelle vom NOT und nLM der anderen bisher untersuchten
Vertebratenklassen. Bei diesen bewirkt eine unilaterale Läsion oder pharmakologische
Inaktivierung eine Reduktion des gains der langsamen Augenfolgephase nur in Richtung
der inaktivierten Seite.
117

II. Verhaltensuntersuchungen 4. Diskussion
4.3.4 Welche Anteile des horizontalen optokinetischen Reflexes sind
während der visuellen Stimulationsphasen betroffen? In der bisherigen Überlegung wurde die Abnahme der Güte der
Augenfolgegeschwindigkeit betrachtet. In der weiterführenden Analyse wurde die
Frequenz und die Amplitude der Augenfolgebewegung untersucht. An allen fünf
Versuchstieren mit Praetectumläsion sank für das stimulierte Auge in der jeweiligen
betroffenen Stimulusrichtung die Frequenz der Augenfolgebewegungen signifikant um ca.
72 % und für das nicht stimulierte Auge um 68 %. Der Grund für die unterschiedliche
Reduktion liegt in der Entkopplung der Augen (Fritsche & Marshall, 2002). Auch die
Amplituden der Augenfolgebewegungen zeigen eine signifikante Verringerung um ca.
42 %. Die Abnahme der Frequenz kann damit erklärt werden, dass bei der Zerstörung des
neuronalen Gewebes nicht ausschließlich richtungsselektive Neurone betroffen waren. Es
ist davon auszugehen, dass Nervenfasern mitbetroffen waren. Diese könnten z. B.
Informationen für sakkadische Augenbewegungen, z. B. vom TeO tragen.
Damit kann die Hypothese gestützt werden, dass die praetectalen richtungsselektiven
Zellen die langsame Augenfolgephase des OKR steuern und somit Teil des neuronalen
Substrats des OKRs sind. Dieser Befund ergänzt die Arbeit von Rösner und Baier (2003),
die Laser - Ablationen des GFP-markierten TeO an Zebrafischmutanten durchführten. In
dieser Studie wurden Ausfälle im OKR und der optomotorischen Reaktion (OMR) nach
Zerstörung einer Hälfte des TeOs untersucht. Entgegen bisheriger Erkenntnisse am
Goldfisch trat hier kein Defizit in der OMR auf (Herrero, 1996). Beim OKR dagegen kam
es zu einer signifikanten Reduktion der Anzahl und der Amplitude der sakkadischen
Augenbewegungen. Unbeeinträchtigt blieb der Geschwindigkeitsbereich der langsamen
Augenfolgephase im OKR. Das neuronale Substrat zur visuellen Auslösung langsamer
Augenfolgebewegungen während des OKRs ist also nicht im TeO lokalisiert.
Die histologische Lokalisation des Läsionsortes innerhalb des Areals der
richtungsselektiven Neurone steht im Zusammenhang mit der Herabsetzung des gains und
118

II. Verhaltensuntersuchungen 4. Diskussion
der beeinträchtigten Richtung der Augenfolgephase.
- Eine caudale Läsion bewirkt eine signifikante Reduktion des gains in temporo -
nasaler Reizrichtung.
- Eine Läsion im zentralen Bereich des Kerngebietes senkt das gain in beiden
Stimulusrichtungen.
- Eine rostrale Läsion bewirkt eine signifikante Reduktion in naso - temporaler
Reizrichtung.
Dieser Befund korrespondiert zur möglichen topographischen Ordnung, die im
elektrophysiologischen Teil dieser Studie (I. 4.10) gezeigt wurde. Damit unterscheidet sich
das Areal deutlich von den Befunden der anderen Vertebratenklasen, in dem nur eine
horizontale Richtung kodiert wird.
4.3.5 Das Modell des optokinetischen Schaltkreises für echte
Knochenfische Anhand den gesamten Untersuchungsergebnisse der vorliegenden Studie, die im 1. Kapitel
an Salmo gairdneri und Esox lucius und im 2. Kapitel an Salmo gairdneri gewonnen
wurden, schlage ich folgenden optokinetischen Schaltkreis bei den Teleostei vor:
Eine temporo - nasale oder naso - temporale Bildverschiebung wird von der Retina
detektiert und kodiert. Diese visuelle Richtungsinformation wird über der Nervus opticus
auf die kontralaterale Seite in das Praetectum projiziert und zwar auf die
richtungsselektiven Zellen im dorsalen praetectalen Areal. Diese Neurone verarbeiten
Richtung und Geschwindigkeit der retinalen Bildverschiebung und leiten diese Information
weiter an vier Kerngebiete.
1. Die Area II befindet sich im Hirnstamm. Sie erhält vom AOS Information über retinale
Bildverschiebungsgeschwindigkeit und bildet den Geschwindigkeitsspeicher für
optokinetische Augenbewegungen. 2. Die Area I mit ihrer Verbindung zu den
Vestibulariskernen erhält vom AOS visuelle Information, die das gain des VOR optimiert.
3. Der direkte Projektionsweg vom AOS zum Vestibulocerebellum dient der Kalibrierung
des VORs (Pastor et al., 1997). 4. Vom AOS erfolgt außerdem eine direkte Projektion zu
den Augenmuskelkernen. Die richtungsselektiven Neurone im Praetectum beider
119

II. Verhaltensuntersuchungen 4. Diskussion
untersuchten Fischarten zeigen die charakteristischen Eigenschaften von Neurone aus dem
AOS anderer Vertebraten.
Diese Projektionswege sind schematisch in der Abbildung II. 4.2 gezeigt.
dpA dpA
AIIAI
Ve
Ce
Amk
Abb. II. 4.2: Modell des neuronalen Schaltkreises des optokinetischen Systems mit dem vestibulo - okulären
System bei Teleostei. dpA = dorsales praetectales Areal, AI und AII = Area I und Area II, Ve = vestibularis
Kerne, Ce = Cerebellum, Amk = Augenmuskelkerne
Die in der Abb. 4.2 dargestellten durchgehenden schwarzen Pfeile, geben bekannte
Projektionswege im Fischgehirn an. Die gestrichelten Pfeile geben Projektionen an, die
von richtungsselektiven Neuronen des dorsalen praetectalen Areals, zu bekannten
Strukturen im optokinetischen Schaltkreis ausgehen. Diese sind zur Zeit nicht bestätigt
bzw. untersucht.
120

III. Zusammenfassung _____________________________________________________
III. Zusammenfassung Die vorliegende Studie untersucht bei zwei echten Knochenfischen, der Regenbogenforelle
(Salmo gairdneri) und dem Hecht (Esox lucius) die Lokalisation von visuellen Neuronen,
die an der Ausführung kompensatorischer Augenbewegungen zur Blickstabilisierung
beteiligt sind. Die Arbeit gliedert sich in einen elektrophysiologischen und in einen
verhaltensphysiologischen Teil zum optokinetischen und zum vestibulo - okulären Reflex.
I. Elektrophysiologische Experimente 1. In den elektrophysiologischen Experimenten gelang der Nachweis richtungsselektiver
Neurone im Praetectum. Insgesamt konnten 72 Neuronen bei Salmo gairdneri und 45
Neuronen bei Esox lucius abgeleitet werden.
2. Diese Neurone unterscheiden sich in ihrer Aktivität bei den beiden Fischarten. Die
richtungsselektiven Neurone von Salmo gairdneri sind wenig oder nicht spontanaktiv
(< 4 Hz), reagieren tonisch auf Stimulation in ihrer Vorzugsrichtung und zeigen keine
Inhibition bei Stimulation in Nullrichtung.
Die richtungsselektiven Neuron von Esox lucius sind spontanaktiv (> 10 Hz), reagieren mit
Aktivitätserhöhung auf Stimulation in Vorzugsrichtung und inhibitorisch mit
Aktivitätserniedrigung in Nullrichtung.
Die Neurone beider Fischarten zeigen außerdem eine starke phasische Aktivierung bei
Einsetzen der Stimulusbewegung in alle Richtungen.
3. Die Vorzugsrichtungen der praetectalen richtungsselektiven Neurone repräsentieren bei
beiden Fischarten alle Raumrichtungen. Das Richtungstuning ist breit (50% der max.
Spikefrequenz bleibt bei +/- 45° Abweichung von der Vorzugsrichtung erhalten).
4. Die Häufigkeit richtungsselektiver Neuronen mit horizontaler bzw. vertikaler
Vorzugsrichtung ist bei Salmo gairdneri gleich. Bei Esox lucius kodieren signifikant mehr
Neurone horizontale als vertikale Stimulusrichtungen (p < 0,05).
5. Bei beiden Fischarten wird die temporo - nasale und naso - temporale Reizrichtung von
der gleichen Anzahl von Neuronen kodiert.
6. Die richtungsselektiven Neurone von Salmo gairdneri kodieren Geschwindigkeiten bis
70 °/s mit einer Erhöhung der Entladungsrate. Die Neurone von Esox lucius erreichen ihre
maximale Aktivität bei 20 °/s.
121

III. Zusammenfassung _____________________________________________________
7. Der Median der visuellen Latenzzeit beträgt für Salmo gairdneri 80 ms, für Esox lucius
90 ms.
8. Die rezeptiven Felder erreichen bei beiden Spezies im Azimut eine Ausdehnung bis zu
130°, in der Elevation bis zu 64°. Sie befinden sich vornehmlich im frontalen Gesichtsfeld
und sind deutlich größer als rezeptive Felder des Tectum opticum.
9. Die Ableitorte richtungsselektiver Neurone liegen im dorsalen Praetectum.
10. In diesem Areal besteht eine topographische Ordnung. Im rostralen Bereich liegen vor
allem Neurone, die nach nasal verlaufende Bewegungen kodieren, im caudalen Bereich
Neurone, die nach temporal verlaufende Bewegungen.
11. Diese Antworteigenschaften gleichen denen von Neuronen bei anderen Vertebraten,
die in den neuronalen Schaltkreis für den OKR eingebunden sind.
II. Verhaltensexperimente 1. Der vertikale optokinetische Reflex wurde an der Regenbogenforelle untersucht, um die
Funktion richtungsselektiver Neurone mit vertikaler Vorzugsrichtung zu ergründen. Diese
Richtung tritt auf bei einer Roll - Bewegung des Fisches um die Körperlängsachse auf und
bewirkt eine vertikale Bildverschiebung auf der Retina. Bei keinem der Tiere konnte ein
vertikaler optokinetischer Reflex ausgelöst werden. Dagegen traten während der
Stimulationsphasen zahlreiche horizontale Augenbewegungen auf.
2. Die visuelle vertikale Richtungsinformation führte zu einer Verbesserung des gains
kompensatorischer Augenbewegungen beim vertikalen VOR. Drei unterschiedliche
visuelle Stimulationsparadigmen wurden getestet. Das Reizmuster bewegte sich exakt mit
dem Fisch mit (1), das Reizmuster bleibt gegenüber der Drehbewegung der Fisches
stationär im Raum (2) und das Reizmuster bewegt sich mit der gleichen
Stimulationsgeschwindigkeit gegen die Drehrichtung des Fisches (3). Das gain der
kompensatorischen Augenbewegungen nahm signifikant von Stimulationsparadigma 1 zu
2 und von 2 zu 3 zu. Damit ist der visuelle Einfluss auf das vestibuläre System belegt. Eine
erwartete Aufhebung des VOR in Paradigma 1 fand nicht statt. Der visuelle Einfluss auf
das vestibuläre System ist bei der Forelle nicht stark genug.
3. Entsprechende Ergebnisse ergaben sich für den horizontalen VOR. Der Vergleich des
gains vertikaler und horizontaler kompensatorischer Augenbewegungen zeigte eine
122

III. Zusammenfassung _____________________________________________________
signifikante Dominanz in der horizontalen Stimulusebene. Dies kann mit der Zugebene der
beteiligten Augenmuskeln und der Anordnung der drei Bogengänge des
Gleichgewichtorgans im Raum erklärt werden.
4. Eine unilaterale Läsion des Areals richtungsselektiver Neurone im Praetectum führt zu
einer signifikanten Verschlechterung des gains des monokularen horizontalen
optokinetischen Reflexes. Eine praetectale Läsion im rostralen Bereich führt zu einer
Beeinträchtigung in naso - temporaler Richtung, eine Läsion im caudalen Bereich in
temporo - nasaler Richtung. Im mittleren Bereich des Areals sind beide Richtungen
betroffen.
5. Diese Verhaltensuntersuchungen weisen darauf hin, dass die im dorsalen Praetectum
kodierte Richtungsinformation über vertikale Stimulusbewegungen keinen bzw. einen
geringen Einfluss auf den vertikalen optokinetischen Reflex hat, sondern zur Modulation
vertikaler kompensatorischer Augenbewegungen beim vestibulo - okulären Reflex genutzt
wird. Die visuelle Information über horizontale Bewegungen ist für das optokinetische und
das vestibulo - okuläre System relevant.
123

III. Zusammenfassung _____________________________________________________
Während dieser Studie wurden vorläufige Ergebnisse an folgenden Stellen publiziert:
Klar M. & K.-P. Hoffmann, 2000. Different asymmetries of optokinetic eye-movements in
rainbow trout and pike. Zoology-Analysis of Complex Systems-Formerly. Zoologische
Jahrbücher, Supplement III, p. 68.
Klar, M &. Hoffmann,K.-P., 2001. Are direction-selective neurons in the pretectum of the
rainbow trout (Salmo gairdneri) the neuronal substrate for OKN generation? In: 3rd
European Conference on comparative neurobiology: Modern view on brain homologies
(III ECCN). Abtsr. 126.
Klar. M & K.-P. Hoffmann. 2001. Direction-selective visual neurons in the pretectum of
the rainbow trout (Salmo gairdneri). In: Proceedings of the 27thGöttingen Neurobiology
Conference. 566.
Klar M. & K.-P. Hoffmann, 2002. Visual direction-selective neurons in the pretectum of
the rainbow trout. Brain Res. Bul. Vol. 57. 431-433.
Klar M. & K.-P. Hoffmann, 2003. How are the rainbow trout pretectal direction-selective
neurons involved in the optokinetic reflex? In: Proceedings of the 28thGöttingen
Neurobiology Conference. 236.
124

IV. Literaturverzeichnis
IV. Literaturverzeichnis
Allum, J. H., Graf, W., Dichgans J. & Schmidt, C. L., 1976. Visual-vestibular interactions
in the vestibular nuclei of the goldfish. Exp. Brain Res. 26, 463-85.
Ariel, M., 1997. Open loop optokinetic responses of the turtle. Vision Res. 37, 925-933.
Ariel, M. & Kogo, N., 2001. Direction tuning of inhibitory inputs to the turtle accessory
optic system. J. Neurophysiol. 86, 2919-2930.
Ashton, J. A., Milleret, C. & Donaldson, I. M., 1989. Effects of afferent signals from the
extraocular muscles upon units in the cerebellum, vestibular nuclear complex and
oculomotor nucleus of the trout. Neuroscience 31, 529-541.
Baker, R., Gilland, E., Green, A., Straker, H. & Suwa, H., 1998. Rhombomeric
organization of horizontal optokinetic and vestibulo-ocular reflexes in goldfish. Soc.
Neurosci. Abstr. 24, 1411.
Ballas, I. & Hoffamnn, K. P., 1985. A correlation between receptive field properties and
morphological structures in the pretectum of the cat. J. Comp. Neurol. 238, 417-428.
Barlow, H. B., Hill, R. M. & Levick, W. R., 1964. Retinal ganglion cells responding
selectively to direction and speed of image motion in the rabbit. J. Physiol. 173, 377-407.
Barlow, H. B. & Levick, W. R., 1965. The mechanism of directionally selective units in
rabbit's retina. J. Physiol. 178, 477-504.
Bathelt, D., 1970. Experimentelle und vergleichende morphologische Untersuchungen am
visuellen System von Teleostiern. Zool. Jb. 87, 402-470.
Batschelet, E., 1981. Circular Statistics in Biology. Academic Press.
125

IV. Literaturverzeichnis
Bauchot, R., Thomot, A. & Bauchot, M. L., 1989. The eye muscles and their innervation in
Chaetodon trifasciatus (Pisces, Teleostei, Chaetodontidae). Enviro. Biol. of Fishes. 25,
221-233.
Bleckmann, H., Tittel, G. & Blübaum-Grönau, E., 1989. Lateral line system of surface
feeding fish: anatomy, physiology and behavior. In the mechanosensory lateral line:
Neurobiology and Evolution. 501-526.
Bohmer, A. & Baloh, R. W., 1990. Vertical optokinetic nystagmus and optokinetic
afternystagmus in humans. J. Vestib. Res. 1, 309-315.
Borel, L. & Lacour, M., 1992. Functional coupling of the stabilizing eye and head reflexes
during horizontal and vertical linear motion in the cat. Exp. Brain Res. 91, 191-206.
Brauth, S. E., 1977. Direct accessory optic projections to the vestibulo-cerebellum: a
possible channel for oculomotor control systems. Exp. Brain Res. 28, 73-84.
Brauth, S. E., Ferfguson, J. L. & Kitt, C. A., 1978. Prosencephalic pathways related to the
paleostriatum of the pigeon (Columba livia). Brain Res. 147, 205-221.
Bruns, S. & Wallmann, J., 1981. Relation of single unit properties to the oculomotor
function of the nucleus of the basal optic root. Exp. Brain. Res. 42, 171-80.
Burr, D. C., Morrone, M. C. & Ross, J., 1994. Selective suppression of the magnocellular
visual pathway during saccadic eyemovements. Nature 371, 511-513.
Butler, A. B., Wullimann, M. F. & Northcutt, R.G., 1991. Comparative cytoarchitectonic
analysis of some visual pretectal nuclei in teleosts. Brain Behav. Evol. 38, 92-144.
Cazin, L., Precht, W. & Lannou, J., 1980. Pathways mediating optokinetic responses of
vestibular nucleus neurons in the rat. Pflügers Arch. 384, 19-29.
126

IV. Literaturverzeichnis
Cazin, L., Magnin, M. Lannou, J., 1982. Non-cerebellar visual afferents to the vestibular
nuclei involving the prepositus hypoglossal complex: an autoradiographic study in the rat.
Exp. Brain Res. 48, 309-313.
Cazin, L., Lannou, J. & Precht, W., 1984. An electrophysiological study of pathways
mediating optokinetic responses to the vestibular nucleus in the rat. Exp. Brain Res.
54, 337-348.
Cochran, S. L., Dieringer, N. & Precht, W., 1984. Basic optokinetic- ocular reflex
pathways in the frog. J. Neurosci. 4, 43-57.
Cohen, B., Schiff, D. & Buettner, J., 1990. Contribution of the nucleus of the optic tract to
optokinetic nystagmus and optokinetic afternystagmus in the monkey: clinical
implications. Res. Publ. Assoc. Res. Nerv Ment. Dis. 67, 233-255.
Collewijn, H., 1975. Oculomotor areas in the rabbits midbrain and pretectum. J. Neurobiol.
6, 3-22.
Collewijn, H., 1975. Direction-selective units in the rabbit’s nucleus of the optic tract.
Brain Res. 100, 489-508.
Cook, J. E. & Podugolnikowa, T. A., 2001. Evidence for spatial regularity among retinal
ganglion cells that project to the accessory optic system in a frog, a reptile, a bird, and a
mammal. Vis. Neurosci. 18, 289-297.
Delgardo-Garcia, J. M., Vidal, P. P., Gomez, C. & Berthoz, A. A., 1989.
Neurophysiological study of prepositus hypoglossi neurons projecting to oculomotor and
preoculomotor nuclei in alert cat. Neurosci. 29, 291-307.
DeZeeuw, C.I., Wylie, D. R., DiGiorgi, P. L. & Simpson, J. L., 1994. Projections of
individual Purkinje cells of identified zones in the flocculus to the vestibular and cerebellar
nuclei in the rabbit. J. Comp. Neurol. 349, 428-447.
127

IV. Literaturverzeichnis
DeZeeuw, C. I. & Berrebi, A. S., 1995. Postsynaptic targets of Purkinje cell terminals in
the cerebellar and vestibular nuclei of the rat.. Eur. J. Neurosci. 7, 2322-2333.
DeZeeuw, C. I., Wylie, D. R., Stahl, J. S. & Simpson, J. L., 1995. Phase relations of
Purkinje cells in the rabbit flocculus during compensatory eye movements. J.
Neurophysiol. 74, 2051-2064.
Dichgans, J., Schmidt, C. L. & Graf, W., 1973. Visual input improves the speedometer
function of the vestibular nuclei in the Goldfish. Exp. Brain Res. 18, 319-322.
Dieringer, N., Reichenberger, I. & Graf, W., 1992. Differences in optokinetic and
vestibular ocular reflex performance in teleosts and their relationship to different life
styles. Brain Behav. Evol. 39, 289-304.
Distler, C., Mustari, M. J. & Hoffmann, K. P., 2002. Cortical projections to the nucleus of
the optic tract and dorsal terminal nucleus and to the dorsolateral pontine nucleus in
macaques: a dual retrograde tracing study. J. Comp. Neurol. 444, 144-158.
Douglas, R. H. & Dkamgoz, M. B. A., (Eds.), 1990. The visual system of fish. Chapman
and Hall, London.
Easter, S. S., 1972. Pursuit eye movements in the goldfish. Vision Res. 12, 673-688.
Easter, S. S. & Schmidt, J. T., 1977. Reversed visuomotor behavior mediated by induced
ipsilateral retinal projections in goldfish. J. Neurophysiol. 40, 1245-1254.
Easter, S. S. & Nicola G. N., 1997. The development of eye movements in the zebrafish
(Danio rerio). Dev. Psychobiol. 31, 267-276.
Ezure, K. & Graf, W., 1984. A quantitative analysis of the spatial organization of the
vestibulo-ocular reflex in lateral- and frontal-eyed animals—I. Orientation of semicircular
canals and extraocular muscles. Neuroscience. 12 , 85-93.
128

IV. Literaturverzeichnis
Fan, T. X., Weber, A. E., Pickard, G. E., Faber, K. M. & Ariel, M., 1995. Visual responses
and connectivity in the turtle pretectum. J. Neurophysiol. 73, 2507-2521.
Finger, T. E. & Karten, H. J., 1978. The accessory optic system in teleosts. Brain Res.153,
144-149.
Fite, K. V., Reiner, A. & Hunt, S. P., 1979. Optokinetic nystagmus and the accessory optic
system of pigeon and turtle. Brain Behav. Evol. 16, 192-202.
Fite, K. V., 1985. Pretectal and accessory-optic visual nuclei of fish, amphibia and reptiles:
theme and variations. Brain Behav. Evol. 26, 71-91.
Fite, K. V. & Hayden, D., 1985. The accessory optic system and temporal correlates of
visuomotor orientation. Brain Behav. Evol. 27, 48-56.
Fite, K. V., Kwei- Levy, C. & Bengston, L., 1989. Neurophysiological investigation of the
pretectal nucleus lentiformis mesencephali in Rana pipiens. Brain Behav. Evol. 34, 164-
170.
Flandrin, J. M., Courjon, J. H., Magnin, M. & Arzi, M., 1990. Horizontal optokinetic
responses under stroboscopic illumination in cat, monkey and man. Exp. Brain Res. 81,
59-69.
Fritsches, K. A. & Marshall, N. J., 2002. Independent and conjugate eye movements
during optokinesis in teleost fish. J. Exp. Biol. 205, 1241-1252.
Fuchs, A. F., Kaneko, C. R. S. & Scudder, C. A., 1985. Brainstem control of saccadic eye
movements. Ann. Rev. Neurosci. 8, 307-337.
Fuchs, A.F., Mustari, M. J., Robinson, F. R. & Kaneko, C. R. S., 1992. Visual signals in
the nucleus of the optic tract and their brainstem destinations. Ann. N. Y. Acad. Sci. 656,
266-276.
129

IV. Literaturverzeichnis
Gioanni, H., Rey, J., Villalobos, J., Richard, D. & Dalbera, A., 1983. Optokinetic
nystagmus in the pigeon (Columba livia). II. Role of the pretectal nucleus of the accessory
optic system (AOS). Exp. Brain. Res. 50, 237-247.
Giolli, R. A., Blanks, R. H. & Torigoe, Y., 1984. Pretectal and brain stem projections of
the medial terminal nucleus of the accessory optic system of the rabbit and rat as studied
by anterograde and retrograde neuronal tracing methods. J. Comp. Neurol. 227, 228-251.
Goltz, M., 1998. Quantitative Analyse des optokinetischen und vestibulären Reflexes beim
Hecht (Esox lucius). Diplomarbeit, Ruhr-Universität-Bochum.
Graf, W. & McGurk, J. F., 1985. Peripheral and central oculomotor organization in the
goldfish, Carassius auratus. J. Comp. Neurol. 239, 391-401.
Graf, W., 1988. Motion detection in physical space and its peripheral and central
representation. Ann. N. Y. Acad. Sci. 545, 154-169.
Graf, W. & Baker, R., 1990. Neuronal adaptation accompanying metamorphosis in the
flatfish. J. Neurobiol. 21, 1136-1152.
Graf, W., Spencer, R., Baker, H. & Baker, R., 1997. Excitatory and inhibitory vestibular
pathways to the extraocular motor nuclei in goldfish. J. Neurophysiol. 77, 2765-2779.
Graf, W., Spencer, R., Baker, H. & Baker, R., 2001 Vestibuloocular reflex of the adult
flatfish. III. A species-specific reciprocal pattern of excitation and inhibition. J.
Neurophysiol. 86, 1376-1388.
Grasse, K. L. & Cynader, M. S., 1988. The effect of visual cortex lesions on vertical
optokinetic nystagmus in the cat. Brain Res. 455, 385-390.
Grover, B. G. & Sharma, S. C., 1979 Tectal projections in the goldfish (Carassius
auratus): A degeneration Study. J. Comp. Neurol. 184, 435-454.
130

IV. Literaturverzeichnis
Grover B. G. & Sharma, S. C., 1981. Organization of extrinsic tectal connections in
goldfish (Caraccius auratus). J. Comp. Neurol. 196(3). 471-88.
Guthrie, D. M. & Banks, J. R., 1978. The receptive field structure of visual cells from the
optic tectum of the freshwater perch (Perca Fluviatilis). Brain Res. 141, 211-225.
Guthrie, D. M. & Sharma, S. C., 1979. Tectal prejections in the goldfisch (Carassius
auratus). J. Comp. Neurol. 184(3), 435-454.
Herrero, L., Rodriguez, F., Salas, C. & Torres, B., 1998. Tail and eye movements evoked
by electrical microstimulation of the optic tectum in goldfish. Exp. Brain Res. 120, 291-
305.
Herrero, L., Perez, P., Nunez Abades, P., Hardy, O. & Torres, B., 1999. Tectotectal
connectivity in goldfish. J. Comp. Neurol. 411, 455-471.
Hoffmann, K. P. & Schoppmann, A., 1975. Retinal input to direction selective cells in the
nucleus tractus optici of the cat. Brain Res. 99, 359-366.
Hoffmann, K. P., 1983. Effects of early monocular deprivation on visual input to cat
nucleus of the optic tract. Exp. Brain Res. 51, 236-246
Hoffmann, K. P., 1985. Direction selective cells in the nucleus of the optic tract in the
squirrel monkey. Neurosci. Abstr. 11, 1009.
Hoffmann, K. P., 1986. Visual inputs relevant for the optokinetic nystagmus in mammals.
Brain Res. 64, 75-84.
Hoffmann, K. P. & Fischer, W. H., 2001. Directional effect of inactivation of the nucleus
of the optic tract on optokinetic nystagmus in the cat. Vision Res. 41, 3389-3398.
131

IV. Literaturverzeichnis
Hoffmann, K. P., Bremmer, F., Thiele, A. & Distler, C., 2002. Directional asymmetry of
neurons in cortical areas MT and MST projecting to the NOT- DTN in macaques.
J. Neurophysiol. 87, 2113-2123.
Howard, I. P., 1993. The optokinetic system. In: Sharpe, J. A. & Barber, H. O. (Eds.), The
vestibulo- ocular reflex and vertigo. Raven Press, New York, pp. 163-184.
Ibbotson, M. R., Clifford, C. W. G. & Mark, R. F., 1998. Adaptation to visual motion in
directional neurons of the nucleus of the optic tract. J. Neurophysiol. 79, 1481-1493.
Ibbotson, M. R. & Clifford, C. W. G., 2001. Interactions between on and off signals in
directional motion detectors feeding the NOT of the wallaby. J. Neurophysiol. 86,
1134-1141.
Ibbotson, M. R. & Price, N. S. C., 2001. Spatiotemporal tuning of directional neurons in
mammalian and avian pretectum: a comparison of physiological properties. J.
Neurophysiol. 86, 2621-2624.
Ibbotson, M. R., Marotte, L. R. & Mark, R. F., 2002. Investigations into the source of
binocular input to the nucleus of the optic tract in an australian marsupial, the wallaby
Macropus eugenii. Exp. Brain Res. 147, 80-88.
Ito, M., 1972. Neural design of the cerebellar motor control system. Brain Res. 40, 81-84.
Ito, M., Shiida, T., Yagi, N. & Yamamoto, M., 1974. The cerebellar modification of
rabbit’s horizontal vestibulo-ocular reflex induced by sustained head rotation combined
with visual stimulation. Proc. Japan Acad. 50, 85-89.
Ito, M., 1982. Cerebellar control of the vestibulo-ocular reflex- around the flocculus
hypothesis. Ann. Rev. Neurosci. 5, 275-296.
132

IV. Literaturverzeichnis
Ito, H. & Murakami, T., 1984. Retinal ganglion cells in two teleost species, Sebasticus
marmoratus and Navodon modestus. J. Comp. Neurol. 229, 80-96.
Ito, H., Vanegas, H., Murakami, T. & Morita, Y., 1984. Diameters and terminal patterns of
retinofugal axons in their target areas: a HRP study in two teleosts (Sebastiscus and
Navodon). J. Comp. Neurol. 230, 179-197.
Ito, M., 1993. Synaptic plasticity in the cerebellar cortex and its role in motor learning.
Can. J. Neurol. Sci. 20, 70-74.
Jardon, B. & Bonaventure, N., 1995. Are retinal or mesencephalic dopaminergic systems
involved in monocular optokinetic nystagmus asymmetry in the frog? Vision Res. 35,
381-388.
Kahmann, H., 1934. Über das Vorkommen einer Fovea centralis im Knochenfischauge.
Zool. Anz. 106, 49-56.
Kahmann, H., 1936. Über das foveale Sehen der Wirbeltiere. I. Über die Fovea centralis
und die Fovea lateralis bei einigen Wirbeltieren. Arch. Opthalmol. 135, 265-276.
Kato, I., Harada, K., Hasegawa, T., Igarashi, T., Koike, Y. & Kawasaki, T., 1986. Role of
the nucleus of the optic tract in monkeys in relation to optokinetic nystagmus. Brain Res.
364, 12-22.
Katte, O. & Hoffmann, K. P., 1980. Direction specific neurons in the pretectum of the frog
(Rana esculenta). J. Comp. Physiol. 140, 53-57.
Keng, M. J. & Aanstasio, T. J., 1997. The horizontal optokinetic response of the goldfish.
Brain Behav. Evol. 49, 214-229.
Klar, M., 1999. Zusammnehang zwischen Ausbildung einer Area centralis in der Retina
und Augenbewegungen bei Knochenfischen. Diplomarbeit, Ruhr- Universität Bochum.
133

IV. Literaturverzeichnis
Klar, M. & Hoffmann, K. P., 2002. Visual direction- selective neurons in the pretectum of
the rainbow trout. Brain Res. Bull. 57, 431-433.
Klauer, S., Sengpiel, F. & Hoffmann, K. P., 1990. Visual response properties and afferents
of nucleus of the optic tract in the ferret. Exp. Brain Res. 83, 178-189.
Kogo, N., McGartland Rubio, D. & Ariel, M., 1998. Direction tuning of individual retinal
inputs to the turtle accessory optic system. J. Neurosci. 18, 2673-2684.
Kunkel, A. W., 1988. Untersuchung zum monokularen optokinetischen Reflex bei Fischen.
Diplomarbeit, Universität Ulm.
Lazar, G., Alkonyi, B. & Toth, P., 1983. Re- investigation of the role of the accessory optic
system and pretectum in the horizontal optokinetic head nystagmus of the frog. Lesion
experiments. Acta Biol. Hung. 34, 385-393.
Lazar, G., 1989. Altering the direction of optokinetic head nystagmus: a lesion study and a
hypothetical model. Exp. Brain Res. 77, 193-200.
Lemire, M. & Reperant, J., 1976. Radioautoradiographic analysis of primary visual
projections in the trout, Salmo irideus Gibb (comparison with other fresh water
Teleosteans). C. R. Acad. Sci. Hebd. Seances Acad. Sci. D. 283, 951-954.
Lisberger, S. G. & Sejnowski, T. J., 1992. Motor learning in a recurrent network model
based on the vestibulo-ocular reflex. Nature 360, 159-161.
Luiten, P. G., 1981. Afferent and efferent connection of the optic tectum in the carp
(Cyprinus caprio L.). Brain Res. 220, 51-64.
Maekawa, K., Takeda, T. & Kimura, M., 1981. Neural activity of nucleus reticalaris
tegmenti pontis- the origin of visual mossy fiber afferents to the cerebellar flocculus of
rabbits. Brain Res. 210, 17-30.
134

IV. Literaturverzeichnis
Magnin, M., Courjon, J. H. & Flandrin, J. M., 1983. Possible visual pathways to the cat
vestibular nuclei involving the nucleus prepositus hypoglossi. Exp. Brain Res. 51, 298-303.
Magnin M., Kennedy, H. & Hoffmann, K. P., 1989. A double labeling investigation of the
pretectal visuo-vestibular pathways. Vis. Neurosci. 3, 53-58.
Maioli, C. & Precht, W., 1984. The horizontal optokinetic nystagmus in the cat. Exp. Brain
Res. 55, 494-506.
Martin, J., Kogo, N. & Ariel, M., 1998. Morphology of basal optic tract terminals in the
turtle, Pseudemys scripta elegans. J. Comp. Neurol. 393, 267-283.
Matsuo, V. & Cohen, B., 1984. Vertical optokinetic nystagmus and vestibular nystagmus
in the monkey: up-down asymmetry and effects of gravity. Exp. Brain Res. 53, 197-216.
McCrea, R. A. & Baker, R., 1985. Anatomical connections of the nucleus prepositus of the
cat. J. Comp. Neurol. 237, 377-407.
McElligott, J. G. & Freedman, W., 1988. Vestibulo-ocular reflex adaptation in cats before
and after depletion of norepinephrine. Exp. Brain Res. 69, 509-521.
McElligott, J. G., Weiser, M. & Baker, R., 1995. Effect of temperature on the normal and
adapted vestibulo-ocular reflex in the goldfish. J. Neurophysiol. 74, 1463-1472.
McElligott, J. G., 1997. Effect of light and dark on retention of an adapted vestibulo-ocular
reflex (VOR) gain change. Soc. Neurosci. Abstr. 23, 509.
McElligott, J. G., Beeton, P. & Polk, J., 1998. Effect of cerebellar inactivation by lidocaine
microdialysis on the vestibuloocular reflex in goldfish. J. Neurophysiol. 79, 1286-1294.
Michnovicz, J. J. & Bennett, M. V., 1987. Effects of rapid cerebellectomy on adaptive gain
control of the vestibulo-ocular reflex in alert goldfish. Exp. Brain Res. 66, 287-294.
135

IV. Literaturverzeichnis
Miles, F. A. & Lisberger, S. G., 1981. Plasticity in the vestibulo-ocular reflex: a new
hypothesis. Annu. Rev. Neurosci. 4, 273-299.
Miles, F. A. & Wallmann, J., 1993. In: Reviews of oculomotor research: visual motion and
ist role in the stabilization of gaze. Elsevier, Amsterdam.
Montgomery, N., Fite, K. V. & Bengston, L., 1981. The accessory optic system of Rana
pipiens: neuroanatomical connections and intrinsic organization. J. Comp. Neurol. 203,
595-612.
Montgomery, N., Fite, K. V., Taylor, M. & Bengston, L., 1982. Neural correlates of
optokinetic nystagmus in the mesencephalon of Rana pipiens: a functional analysis. Brain
Behav. Evol. 21, 137-150.
Montgomery, N. M., Fite, K. V. & Grigonis, A. M., 1985. The pretectal nucleus
lentiformis mesencephali of Rana pipiens. J. Comp. Neurol. 234, 264-275.
Mustari, M. J. & Fuchs, A. F., 1989. Response properties of single units in the lateral
terminal nucleus of the accessory optic system in the behaving primate. J. Neurophysiol.
61, 1207-1220.
Mustari, M. J. & Fuchs, A. F., 1990. Discharge patterns of neurons in the pretectal nucleus
of optic tract (NOT) in behaving primate. J. Neurophysiol. 64. 77-90..
Mustari, M. J., Fuchs, A. F., Kaneko, C. R. S. & Robinson, F. R., 1994. Anatomical
connections of the primate pretectal nucleus of the optic tract. J. Comp. Neurol. 349,
111-128.
Natal, C.L. & Britto, L. R., 1987. The pretectal nucleus of the optic tract modulates the
direction selectivity of accessory optic neurons in rats. Brain Res. 419, 320-323.
136

IV. Literaturverzeichnis
New, J. G., A. Fewkes, L. A. & Khan, A. N., 2001. Strike feeding behavior in the
muskellunge, Esox masquinongy: contributions of the lateral line and visual sensory
systems. J. Exp. Biol. 204, 1207-1221.
Pastor, A. M., Torres, B., Delgado-Garcia, J. M. & Baker, R., 1991. Discharge
characteristics of medial rectus and abducens motoneurons in the goldfish. J.
Neurophysiol. 66, 2125-2140.
Pastor, A. M., de la Cruz, R. R. & Baker, R., 1992. Characterization and adaptive
modification of the goldfish vestibuloocular reflex by sinusoidal and velocity step
vestibular stimulation. J. Neurophysiol. 68, 2003-2015.
Pastor, A. M., de la Cruz, R. R. & Baker, R., 1994. Cerebellar role in adaptation of the
goldfish vestibuloocular reflex. J. Neurophysiol. 72, 1383-1394.
Pastor, A. M., de la Cruz, R. R. & Baker, R., 1997. Characterization of purkinje clls in the
Goldfish cerebellum during eye movement and adaptive modification of the vestibulo-
ocular reflex. Prog. Brain Res. 114, 359-81.
Pastor, A. M., de la Cruz, R. R. & Baker, R., 1997. Eye position and eye velocity integrator
reside in separate brainstem nuclei. Proc. Natl. Acad. Sci. USA. 91, 807-11.
Pereira, A., Volchan, E., Vargas, C. D., Penetra, L. & Rocha- Miranda, C. E., 2000.
Cortical and subcortical influences on the nucleus of the optic tract of the opossum.
Neuroscience 95, 953-963.
Polyak, S., 1968. The vertebrat visual system (Hrsg). Elsevier, Amsterdam. Precht, W. & Anderson, J. H., 1979. Effects of cerebellectomy on the cat’s vertical
vestibuloocular reflex. Pflügers Arch. 382, 51-55.
Precht, W. & Cazin, L., 1979. Functional deficits in the optokinetic system of albino rats.
Exp. Brain Res., 37: 183-186
137

IV. Literaturverzeichnis
Raphan, T., Matsuo, V. & Cohen, B., 1979. Velocity storage in the vestibulo-ocular reflex
arc (VOR). Exp. Brain Res. 35, 229-248.
Raphan, T. & Cohen, B., 2002. The vestibulo-ocular reflex in three dimensions. Exp. Brain
Res. 145, 1-27.
Reiner, A. & Karten, H. J., 1978. A bisynaptic retinocerebellar pathway in the turtle. Brain
Res. 150, 163-169.
Reperant, J., Lemire, M., Miceli, D. & Peyrichoux, J., 1976. A radioautographic study of
the visual system in fresh water teleosts following intraocular injection of tritiated fucose
and proline. Brain Res. 118, 123-131.
Reperant, J., Vesselkin, N. P., Ermakova, T. V., Rustamov, E. K., Rio, J. P., Palatnikov,
G. K., Peyrichoux, J. & Kasimov, R. V., 1982. The retinofugal pathways in a primitive
actinopterygian, the chondrostean Acipenser guldenstadti. An experimental study using
degeneration, radioautographic and HRP methods. Brain Res. 251, 1-23.
Roeser, T. & Baier, H., 2003. Visuomotor behaviors in larval zebrafish after GFP-guided
laser ablation of the optic tectum. J. Neurosci. 23, 3726-3734.
Romer, A. S. , Parsons, T. S.,1991. Vergleichende Anatomie der Wirbeltiere. Verlag Paul
Parey, Hamburg und Berlin.
Rupp, B., Wullimann, M.F. & Reichert, H., 1996. The zebrafish brain: a neuroanatomical
comparsion with the goldfish. Anat. Embryol. (Berl). 194, 187-203.
Salas, C., Herrero, L., Rodriguez, F. & Torres, B., 1997. Tectal codification of eye
movements in goldfish studied by electrical microstimulation. Neuroscience 78, 271-288.
Sas, E. & Maler, L., 1986. Retinofugal projections in a weakly electric gymnotid fish
(Apteronotus leptorhynchus). Neuroscience. 18 (1), 247-59.
138

IV. Literaturverzeichnis
Schairer, J. O. & Bennett, M. V., 1986. Changes in the gain of the vestibulo-ocular reflex
induced by combined visual and vestibular stimulation in goldfish. Brain Res. 373,
164-176.
Schairer, J. O. & Bennett, M. V., 1986. Changes in the gain of the vestibulo-ocular reflex
induced by sinusoidal visual stimulation in goldfish. Brain Res. 373, 177-181.
Scheuring, L., 1920. Beobachtungen und Betrachtungen über die Beziehung der Augen
zum Nahrungserwerb bei Fischen. Zool. Jb. 38, 113-137.
Schiff, D., Cohen, B., Buttner- Ennever, J. & Matsuo, V., 1990. Effects of lesions of the
nucleus of the optic tract on optokinetic nystagmus and after- nystagmus in the monkey.
Exp. Brain Res. 79, 225-239.
Schmidt, M., Zhang, H. Y. & Hoffmann, K. P., 1993. OKN-related neurons in the rat
nucleus of the optic tract and dorsal terminal nucleus of the accessory optic system receive
a direct cortical input. J. Comp. Neurol. 330, 147-157.
Schmidt, M., van der Togt, C., Wahle, P. & Hoffmann, K. P., 1998. Characterization of a
directional selective inhibitory input from the medial terminal nucleus to the pretectal
nuclear complex in the rat. Eur. J. Neurosci. 10, 1533-1543.
Schor, C. & Narayan, V., 1981. The influence of field size upon the spatial frequency
response of optokinetic nystagmus. Vision Res. 21, 985-994.
Schröder, D. M., 1981. Retinal afferents and efferents of an infrared sensitive snake,
Crotalus viridis. J. Morphol. 170, 29-42.
Starck, D., 1982. Vergleichende Anatomie der Wirbeltiere. Band 3. Springer-Verlag.
Berlin, Heidelberg, New York.
139

IV. Literaturverzeichnis
Schwassmann, H. O., 1968. Visual projection upon the optic tectum in foveate marine
teleosts. Vision Res. 8, 1337-1348.
Schwassmann, H. O. & Krüger, L., 1970. Organization of the visual projection upon the
optic tectum of some freshwater fish. J. Comp. Neurol. 124, 113-126.
Sharma, S. C., 1972. The retinal projections in the goldfish: an experimental study. Brain
Res. 39, 213-223.
Simpson, J. I. & Graf, W., 1981. Eye-muscle geometry and compensatory eye movements
in lateral-eyed and frontal-eyed animals. Ann. N. Y. Acad. Sci. 374, 20-30.
Simpson, J. I., 1984, The accessory optic system. Rev. Annu Rev Neurosci. 7, 14-41.
Simpson, J. I. & Graf, W., 1985. The selection of reference frames by nature and its
investigators. Rev. Oculomot. Res.1, 3-16.
Sirkin, D. W., Precht, W. & Courjon, J.H., 1984. Initial, rapid phase of recovery from
unilateral vestibular lesion in rat not dependent on survial of central portion of vestibular
nerve. Brain Res. 302, 245-56.
Sirkin, D. W., Hess B. J. & Precht, W., 1985. Optokinetic nystagmus in albino rats
depends on stimulus pattern. Exp. Brain Res. 61 (1), 218-21.
Soodak, R. E. & Simpson, J. I., 1988. The accessory optic system of rabbit. I. Basic visual
response properties. J. Neurophysiol. 60, 2037-2054.
Springer, A. D., Easter, S. S. & Agrano, B. W., 1977. The role of the optic tectum in
various visually mediated behaviors of goldfish. Brain Res. 128, 393-404.
Springer, A. D. & Gaffney, J. S., 1981. Retinal projections in the goldfish: a study using
cobaltous-lysine. J. Comp. Neurol. 203, 401-424.
140

IV. Literaturverzeichnis
Springer, A. D. & Mednick, A. S., 1985. Retinofugal and retinopetal projections in the
cichlid fish Astronotus ocellatus. J. Comp. Neurol. 236, 179-196.
Springer, A.D. & Mednick, A. S., 1985. Topography of the retinal projection to the
superficial pretectal parvicellular nucleus of goldfish: A cobaltous-lysine study. J. Comp.
Neurol. 237, 239-250.
Stahl, J. S. & Simpson, J. L., 1995. Dynamics of rabbit vestibular nucleus neurons and the
influence of the flocculus. J. Neurophysiol. 73, 1396-1413.
Stahl, J. S. & Simpson, J. L., 1995. Dynamics of abducens nucleus neurons in the awake
rabbit. J. Neurophysiol. 73, 1383-1395.
Suwa, H., Gilland, E. &. Baker, R., 1999. Otolith ocular reflex function of the tangential
nucleus in teleost fish. Ann N. Y. Acad. Sci. 871, 1-14.
Tan, H. S., van der Stehen , J., Simpson, J. I. & Collewijn, H., 1993. Three-dimensional
organization of optokinetic response in the rabbit. J. Neurophysiol. 69, 303-317.
Tauber, E. S. & Atkin, A., 1968. Optomotor responses to monocular stimulation: relation
to visual system organization. Science 160, 1365-1367.
Trillenberg, P., Shelhamer, D., Roberts, D. C. & Zee, D. S., 2003. Cross-axis adaptation of
torsional components in the yaw-axis vestibulo.ocular reflex. Exp. Brain Res. 148,
158-165.
Van der Togt, C., van der Want, J. J. L. & Schmidt, M., 1993. Segregation of direction
selective neurons and synaptic organization of inhibitory intranuclear connections in the
medial terminal nucleus of the rat: an electrophysiological and immunoelectron
microscopical study. J. Comp. Neurol. 338, 175-192.
141

IV. Literaturverzeichnis
Van Hof-Van Duin, J., 1976. Early and permanent effects of monocular depriviation on
pattern discrimination and visuomotor behavior in cats. Brain Res. 111. 261-276.
Vanegas, H. & Ito, H., 1983. Morphological aspects of the teleostean visual system:
a review. Brain Res. Rev. 6, 117-137.
Wallmann, J. & Velez, J., 1985. Directional asymmetries of optokinetic nystagmus:
developmental changes and relation to the accessory optic system and to the vestibular
system. J. Neurosci. 5, 317-329.
Watanabe, S., Kato, I., Sato, S. & Norita, M., 1993. Direct projection from the nucleus of
the optic tract to the medial vestibular nucleus in the cat. Neurosci. Res. 17, 325-329.
Wilm, C. & Fritzsch, B., 1990. Ipsilateral tectinofugal projections in a percomorph bony
fish: Their experimental induction, specificity and maintenance. Brain Behav. Evol. 36,
271-299.
Wilm, C. & Fritzsch, B., 1992. Evidence for driving role of ingrowing axons for the
shifting of older retinal terminals in the tectum of fisch. J. Neurobiol. 23, 149-162.
Winterson, B. J. & Brauth, S. E., 1985. Direction- selective single units in the nucleus
lentiformis mesencephali of the pigeon (Columba livia). Exp. Brain Res. 60, 215-226.
Wylie, D. R. & Frost, B. J., 1990. The visual response properties of neurons in the nucleus
of the basal optic root of the pigeon: a quantitative analysis. Exp. Brain Res. 82, 327-336.
Wylie, D. R. W. & Frost, B. J., 1996. The pigeon optokinetic system: visual input in
extraocular muscle coordinates. Vis. Neurosci. 13, 945-953.
Wylie, D. R. W. & Crowder, N. A., 2000. Spatiotemporal properties of fast and slow
neurons in the pretectal nucleus lentiformis mesencephali in pigeons. J. Neurophysiol. 84,
2529-2540.
142

IV. Literaturverzeichnis
Wylie, D. R. W., 2000. Binocular neurons in the nucleus lentiformis mesencephali in
pigeons: responses to translational and rotational optic flowfields. Neurosci. Lett. 291,
9-12.
Wylie, D. R. W., 2001. Projections from the nucleus of the basal optic root and nucleus
lentiformis mesencephali to the inferior olive in pigeons (Columba livia). J. Comp. Neurol.
429, 502-513.
143

V. Abkürzungen
V. Abkürzungen
ANOVA Analysis of variance
AOS akzessorisch optisches System
a/p anterior-posterior
ccw gegen den Uhrzeigersinn (eng. counterclockwise)
cw im Uhrzeigersinn (eng. clockwiese)
hOKN horizontaler optokinetischer Nystagmus
hOKR horizontaler optokinetischer Reflex
hVOR horizontaler vestibulo-okulärer Reflex
i.m. intramusculaer
IO Oliva inferior
LTN laterale terminale Nukleus
m/l medio- lateral
MTN mediale terminale Nukleus
nBOR Nucleus of the basal optic root
nLM Nucleus lentiformis mesencephali
NOT Nucleus tractus optici
NPH Nucleus praepositus hypoglossi
N-T naso-temporal
OT optischer Trakt
Psp Nucleus praetectalis superfiscialis parvocellularis
PSTH Peri-Stimulus-Time-Histogram
RF rezeptives Feld
TeO Tectum opticum
Thd Thalamus dorsalis
T-N temporo-nasal
VOR vestibulo-okulärer Reflex
vVOR vertikaler vestibulo-okulärer Reflex
144

Curriculum vitae
Persönliche Daten
Name: Matthias Klar Geburtsdatum: 02. August 1962 Geburtsort: Dortmund Familienstand: ledig Eltern: Ruth-Eva Klar, geborene Greinert Dipl. Ing. Helmut Klar Schulausbildung 1968- 1973 Suitbertus-Grundschule in Dortmund 1973- 1974 Leibnitz-Gymnasium in Dortmund 1974- 1980 Albertus-Magnus-Realschule in Dortmund 1980- 1981 Dr. Zimmermannsche-Wirtschaftsschule in Koblenz 1988- 1991 Abendgymnasium in Dortmund Ausbildung 1982- 1986 Auszubildender als Radio-Fernseh-Techniker Anstellungen 1986- 1987 Computer-Service-Techniker bei Computer Reschke GmbH 1987- 1989 Leiter der Computer-Werkstatt und Service-Techniker bei CASDO Computer Steuer in Dortmund Akademische Ausbildung 1991- 1997 Studium der Biologie, Ruhr-Universität Bochum 1997- 1999 Diplom am Lehrstuhl für Allgemeine Zoologie und
Neurobiologie, Ruhr-Universität Bochum; Leiter: Prof. Dr. K. P. Hoffmann Thema der Arbeit: Zusammenhang zwischen Ausbildung einer Area centralis in der Retina und Augenbewegungen bei Knochenfischen.
Seit 1999 Doktorand am Lehrstuhl für Allgemeine Zoologie und Neurobiologie, Ruhr-Universität Bochum

Danksagung An dieser Stelle möchte ich mich bei all denjenigen bedanken, die mir während meiner
Promotion mit Rat und Tat zur Seite standen.
Insbesondere gilt mein Dank meinem Doktorvater Prof. Dr. K.-P. Hoffmann, nicht nur für
die Bereitstellung dieses interessanten Themas und die Freiheit bei der Ausgestaltung
meiner Arbeit, sondern auch dafür, dass er sich stets für mich Zeit nahm und mir mit
seinem außerordentlichen Wissens- und Erfahrungsschatz zur Seite stand. Nicht zuletzt sei
hier seine Geduld erwähnt.
Frau P.D. Dr. C. Distler danke ich für ihre wertvollen Ratschläge auf dem Gebiet der
Histologie, Anatomie und ihrer Hilfsbereitschaft.
Herrn H.-J. Korbmacher bin ich nicht nur für seine Mikroelektroden zu großem Dank
verpflichtet. Nicht vergessen möchte ich Frau M. Möllmann, die mir in der Histologie zur
Seite stand. Herrn Dr. L. Lünenburger danke ich für seine inspirierende Diskussions- und
seine Hilfsbereitschaft, Herrn Dr. B. Krekelberg für seine Hilfe in Matlab und Herrn W.
Junke für seine Unterstützung bei Computerproblemen.
Eewähnen möchte ich auch Herrn E. Reuter, Herrn G. Tiney, Herrn H. Marquart, Herrn B.
Thyner, sowie Herrn S. Dobers, die mir bei elektrischen und mechanischen Problemen eine
große Hilfe waren. Weiterhin schulde ich den Tierpflegern Herrn V. Rostek und Frau M.
Schmidt für die gewissenhafte Versorgung der Fische Dank.
Nicht zuletzt danke ich meinen „ alten“ und „neuen“ Zimmergenossen Frau Dr. A.
Schlack, Dr. M. Kubischik, Frau vet .med. M. Grewing, Frau Dipl.-Biol.W. Blaszcyk und
Herrn C. Schönberger für ein stetig gutes Arbeitsklima und so manche Lacher.
Abschließend möchte ich mich besonders bei meinen Eltern bedanken, die mich auch in
schwierigen Zeiten stets unterstützt haben.


















