
Nonenzymatie Formation of Energy Rich Phosphates in the...
6
This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution 4.0 International License. Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschung in Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht: Creative Commons Namensnennung 4.0 Lizenz. Nonenzymatie Formation of Energy Rich Phosphates in the Course of Imidazole Oxidation with Fenton's Reagent* Omer Saygin** Chemical Institute, Veterinary School, D-3000 Hannover, FRG Z. Naturforsch. 85 b, 1009-1014 (1980); received February 29, 1980 Bioids, Oxidative Phosphorylation, Photophosphorylation, Fenton's Reaction, Imidazole Oxidation Oxidation of substituted imidazoles with modified Fenton's reagent (Fe(II), ascorbic acid, EDTA and H2O2) in aqueous solution at pH 5-7 in the presence of inorganic ortho- phosphate gives high yields of an unknown bound phosphate, in the case of 2-methyl- imidazole up to 35% based on inorganic phosphate. The dependence of the yields on reaction parameters and the hydrolytic behaviour of the compound was studied. The products are unstable in alkaline medium and fairly stable in acid solution (half-life period ca. 1 h at 25 °C in I N perchloric acid). Similar bound phosphate was also observed using autoxidation with O2 instead of H2O2. Oxidative phosphorylation and photophospho- rylation are fundamental processes in living organ- isms in which energy is conserved in energy rich organic phosphates serving as standardized energy suppliers in endergonic biochemical reactions. The importance of similar processes providing the thermodynamical driving force for the selforganiza- tion of life-like systems in a prebiological scenario has been stressed by Decker [1-3]. Since thermo- dynamically such systems are dissipative structures, such selforganization should be expected at or near the site where on Earth we have maximal energy dissipation i.e. the dissipation of light energy [4-5]. This rationale suggested a search for an organized production of compounds with high redox- or group potentials through photoredox reactions in an open systems as a thermodynamically feasible source of selforganization. Since in sensitized photoredox reactions intermediates of high reactivity are essen- tial, our first approach was a study of reaction products of high energy radicals in the presence of inorganic phosphate. For this purpose we chose Fenton's reagent, which is one of the most powerful inorganic oxidizing agents, the actual oxidizing species being the hydroxyl radical formed from hydrogen peroxide and ferrous ion [6]: H2O2 + Fe(II)-EDTA = Fe(III)-EDTA + OH- + OH * Bioids I X ; VIII: Ö. Saygin and P. Decker, Z. Natur- forsch. 35b, 727 (1980); I: see ref. [2]. ** Reprint requests to Dr. Ö. Saygin, Tierärztliche Hochschule, Bischofsholer Damm 15, D-3000 Han- nover 1, FRG. 0340-5087/80/0800-1009/$ 01.00/0 In the presence of a suitable reducing agent, here ascorbic acid, Fe(II) is regenerated, AH2 + 2 Fe(III)-EDTA = A + 2 Fe(II)-EDTA + 2 H+ and the system behaves as a model peroxidase system. Using it as a source of energy rich inter- mediates, we looked for any energy rich bound phosphate formation in the course of the oxidation of a substrate in the presence of inorganic ortho- phosphate. Here we report the formation of such products with high yields (about 35% based on inorganic phosphate) using imidazoles as substrate. Materials and Methods Imidazole (p.a.) w r as obtained from Merck, 1-methylimidazole (p.a.) and 4-hydroxymethyl- imidazole-hydrochloride from Riedel de Haen, 2-methylimidazole from Sigma, USA, pyrimidine (98%) from Ega, histamine • 2 HOI (99%) and imi- dazole-4-acetic acid • Na-salt from Serva, isopropyl- acetate from Baker. All other reagents were of reagent degree. Reaction conditions In a typical experiment the complete system contained the following components (//moles in 1 ml of total volume) at pH 4-6 and 0 °C: Na 2 HP0 4 • 2H 2 0 100; FeS0 4 • 7H 2 0 4; ascorbic acid 40; EDTA 20 and substrate 40. The solution was cooled with ice and the reaction was initiated by the addition of 50 ju\ of 30% H2 0 2 . After two minutes the phosphate determination was started. Quantitative determination of bound phosphate 200 /A of the reaction mixture was transfered into a tube containing 1.5 ml HC104 (10%) and 0.3 ml Na 2 Mo0 4 (1.2 M) in the cold (0 °C). Inorganic phos- phate was extracted four times as the molybdate
Transcript of Nonenzymatie Formation of Energy Rich Phosphates in the...

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
Nonenzymatie Formation of Energy Rich Phosphates in the Course of Imidazole Oxidation with Fenton's Reagent*
Omer Saygin** Chemical Institute, Veterinary School, D-3000 Hannover, FRG
Z. Naturforsch. 85 b, 1009-1014 (1980); received February 29, 1980
Bioids, Oxidative Phosphorylation, Photophosphorylation, Fenton's Reaction, Imidazole Oxidation Oxidation of substituted imidazoles with modified Fenton's reagent (Fe(II), ascorbic
acid, EDTA and H2O2) in aqueous solution at pH 5 -7 in the presence of inorganic ortho-phosphate gives high yields of an unknown bound phosphate, in the case of 2-methyl-imidazole up to 3 5 % based on inorganic phosphate. The dependence of the yields on reaction parameters and the hydrolytic behaviour of the compound was studied. The products are unstable in alkaline medium and fairly stable in acid solution (half-life period ca. 1 h at 25 °C in I N perchloric acid). Similar bound phosphate was also observed using autoxidation with O2 instead of H2O2.
Oxidative phosphorylation and photophospho-rylation are fundamental processes in living organ-isms in which energy is conserved in energy rich organic phosphates serving as standardized energy suppliers in endergonic biochemical reactions. The importance of similar processes providing the thermodynamical driving force for the selforganiza-tion of life-like systems in a prebiological scenario has been stressed by Decker [1-3]. Since thermo-dynamically such systems are dissipative structures, such selforganization should be expected at or near the site where on Earth we have maximal energy dissipation i.e. the dissipation of light energy [4-5]. This rationale suggested a search for an organized production of compounds with high redox- or group potentials through photoredox reactions in an open systems as a thermodynamically feasible source of selforganization. Since in sensitized photoredox reactions intermediates of high reactivity are essen-tial, our first approach was a study of reaction products of high energy radicals in the presence of inorganic phosphate. For this purpose we chose Fenton's reagent, which is one of the most powerful inorganic oxidizing agents, the actual oxidizing species being the hydroxyl radical formed from hydrogen peroxide and ferrous ion [6]:
H2O2 + Fe(II)-EDTA = Fe(III)-EDTA + OH- + O H
* Bioids I X ; VIII : Ö. Saygin and P. Decker, Z. Natur-forsch. 35b, 727 (1980); I : see ref. [2].
** Reprint requests to Dr. Ö. Saygin, Tierärztliche Hochschule, Bischofsholer Damm 15, D-3000 Han-nover 1, FRG.
0340-5087/80/0800-1009/$ 01.00/0
In the presence of a suitable reducing agent, here ascorbic acid, Fe(II) is regenerated,
AH2 + 2 Fe(III)-EDTA = A + 2 Fe(II)-EDTA + 2 H+
and the system behaves as a model peroxidase system. Using it as a source of energy rich inter-mediates, we looked for any energy rich bound phosphate formation in the course of the oxidation of a substrate in the presence of inorganic ortho-phosphate. Here we report the formation of such products with high yields (about 35% based on inorganic phosphate) using imidazoles as substrate.
Materials and Methods Imidazole (p.a.) wras obtained from Merck,
1-methylimidazole (p.a.) and 4-hydroxymethyl-imidazole-hydrochloride from Riedel de Haen, 2-methylimidazole from Sigma, USA, pyrimidine (98%) from Ega, histamine • 2 HOI (99%) and imi-dazole-4-acetic acid • Na-salt from Serva, isopropyl-acetate from Baker. All other reagents were of reagent degree.
Reaction conditions In a typical experiment the complete system
contained the following components (//moles in 1 ml of total volume) at pH 4-6 and 0 °C: Na2HP04 • 2H 2 0 100; FeS04 • 7H 2 0 4; ascorbic acid 40; EDTA 20 and substrate 40. The solution was cooled with ice and the reaction was initiated by the addition of 50 ju\ of 30% H202 . After two minutes the phosphate determination was started.
Quantitative determination of bound phosphate 200 /A of the reaction mixture was transfered into
a tube containing 1.5 ml HC104 (10%) and 0.3 ml Na2Mo04 (1.2 M) in the cold (0 °C). Inorganic phos-phate was extracted four times as the molybdate

1010 O. Saygin • Nonenzymatic Formation of Energy Rieh Phosphates
complex using 4 ml portions of isopropyl actate. Fast phase separation was enforced by centrifuga-tion for 1 min. Under these conditions even labile phosphates remain mainly unhydrolysed in the aqueous phase [7]. The fourth extraction was usually free of inorganic phosphate. Bound phosphate in the aqueous phase was then hydrolyzed by heating to 100 °C for ten minutes. After cooling to room temperature any newly formed phosphate was obtained (to 99% [7]) by an additional extraction. For a quantitative determination 0.1 ml of freshly prepared 0.5% SnCL • 2H2O in methanol was added to 2 ml of the organic phase and the extinction of molybdene blue was determined at 691 nm using an Eppendorf photometer. Since the bound phosphates obtained in our system were almost immediately hydrolyzed at pH values greater than 9, in later experiments we- omitted the heating step for hydrolysis; instead after the fourth extraction step, we increased for 30 sec the pH of the aqueous phase to pH 12-14 using NaOH, and acidified the solution again to pH 1 with HCIO4.
This method allows the detection of 0 .1% of bound phosphate in the presence of inorganic phos-phate.
Column chromatography
A column (3 X 25 cm) with Dowex 1 x 8 (200 to 400 mesh) anion exchange resin (Serva) was trans-formed into the acetate form by washing with 2 M NaOH and 2 M CH3COOH. After equilibration with 0.05 M CH3COOH 3.4 ml of a reaction mixture of 1.5 mmoles Na2HP04 • 2H 2 0 , 2.5 mmole 2-methyl-imidazole, 0.26 mmoles EDTA, 0.13 mmoles FeS04 • 7H 2 0, 1.2 mmoles ascorbic acid, 1.2 ml H2O2 (30%) at pH 5.7 was applied to the column and washed with 0.05 M CH3COOH at 2 °C. Frac-tions of 5 ml were collected and assayed for bound phosphate.
Results Chosing imidazole as the substrate, in dilute
solutions we observed the formation of a bound phosphate with a yield of 0 .9% (based on the inorganic phosphate). Control experiments (Table I) show that omission of phosphate, imidazole, FeEDTA complex, or H2O2, suppressed bound phos-phate formation. In contrast, omission of ascorbic acid only reduced the yield to 0 .2% as expected regarding the role of ascorbic acid as a reductant of the Fe(III), and doubling the initial Fe(II) concen-tration increased the yield again to 0 .5%.
EDTA was also not essential for the formation of bound phosphate since it could be replaced by oxalic acid or diethylene-triamine 1,1,4,7,7 penta-acetic acid. Replacing Fe(II) by Co(II), Mn(II),
Table I. Bound phosphate formation: The complete system contained the following components (/imol/ml). A : inorganic phosphate 100, imidazole 40, Fe(II) 4, E D T A 20, ascorbic acid 40, H 2 0 2 1700. B : inorganic phosphate 300, imidazole 800, Fe(II) 25, EDTA 50, ascorbic acid 250, H 2 0 2 2000. Reaction conditions were as described in the methods section. Yield ( % ) is based on inorganic phosphate.
System Yield of bound phosphate A B (/imol/ml) ( % ) (pmol/ml) ( % )
Complete system 0.9 Imidazole omitted 0.0 Phosphate omitted 0.0 Fe(II)-EDTA omitted 0.0 H2O2 omitted 0.0 Ascorbic acid omitted 0.2 Ascorbic acid omitted and initial Fe(II)-EDTA concen-tration doubled 0.5
E D T A replaced by equimolar
0.8 oxalic acid ethylene -triamine 1,1,4,7,7 penta acetic acid 0
Fe(II) replaced by equimolar
Mn(II) 0.0 Co(II) 0.0 V(IV) 0.0 Cu(II) and E D T A omitted 0.9
Imidazole replaced by equimolar
1 -Methylimidazole 2 -Methylimidazole H y droxymethyl -imidazole Histamine Pyrimidine
0.4 1.4
0.3 0.2
0.2
Imidazole replaced by 2 -methylimidazole and 1 h bubbling with air instead of H 2 0 2 0.7
0.9 23.2
0.2
8.0
0.5
0.8
0.9
0.9
0.4 1.4
0.3 0.2
0.2
0.7
3.2 95.0
1 . 1 32.7
V(IV) gave negative results; however, Cu(II) in the absence of EDTA (the complex with EDTA is not reduced by ascorbic acid) yielded the same amount of bound phosphate as Fe(II).
Replacing imidazole by other substrates, 1-methylimidazole, 2-methylimidazole, 4-hydroxy-

1011 O. Saygin • Nonenzymatic Formation of Energy Rieh Phosphates
BOUND PHOSPHATE
M
1 2 3 TIME [ h ]
Fig. 1. Hydrolysis of bound phosphate(s): p H and temperature were adjusted to different values after two minutes of reaction time. The initial yield after this period was taken as 1 0 0 % . Reaction conditions: A (see Table I).
methylimidazole, histamine and pyrimidine gave positive results, 2-methylimidazole giving by far the best yields (Table I) ; the following substrates showed no detectable bound phosphate formation: imidazole-4-acrylic acid, histidine, adenine, xan-thine, guanine, uracil, pyrazole, pyrazine, 1,2,4 triazole, acetonitril, imidazolidinthion, acetamide, cyanamid, dicyanamid, N,N dimethylformamid, oxamide, ethyleneurea.
Studying the time course of the reaction showed that bound phosphate attained a maximum im-mediately after the addition of hydrogen peroxide; then after a reaction period of maximally two minutes it is hydrolysed more or less rapidly, depending on the conditions. In order to charac-terize the hydrolytic behaviour, we adjusted the temperature and the pH of the solution to different values after two minutes of reaction time; Fig. 1 shows that both, increased temperature and pH, increase the rate of hydrolysis. The determination of the kinetic parameters from the initial slopes of the curves gave an observed rate constant of k = 1.7 • 10-4 sec-1 at 25 °C and pH = 0.1 and an activation energy of 20 kcal/mol between 25 and 35 °C for a first order rate of hydrolysis. The shape of the curve at pH 4.5 suggests the existence of at least two different phosphate species.
BOUND PHOSPHATE j mol / ml J
Fig. 2. Effect of p H on bound phosphate forma-tion: Substrates:
I = 2-methylimidazole; I I = imidazole;
I I I = 4-hydroxymethyl-imidazole;
I V = 1-methylimidazole; V = pyrimidine.
Reaction conditions: A (see Table I).
1012 O. Saygin • Nonenzymatic Formation of Energy Rieh Phosphates
BOUND PHOSPHATE J/umol/ml]
5 0
BOUNO PHOSPHATE £ pmot / ml J
XX) 2 0 0 3 0 0 iOO 500 ORTHOPHOSPHATE
[/u mol/ml J 2 0 0 4 0 0 6 0 0 8 0 0 1000 2-METHYUMDAZOLE
[jumol / ml]
60
3 0
10
BOUND PHOSPHATE
Aunol/ ml]
(C)
20 30 A0 SO Fe«I)[/umol / ml]
BOUND PHOSPHATE
£ pmol / ml ]
(D)
2 0 0 4 0 0 6 0 0 8 0 0 1 0 0 0 ASCORBIC ACTD [firwl/frt]
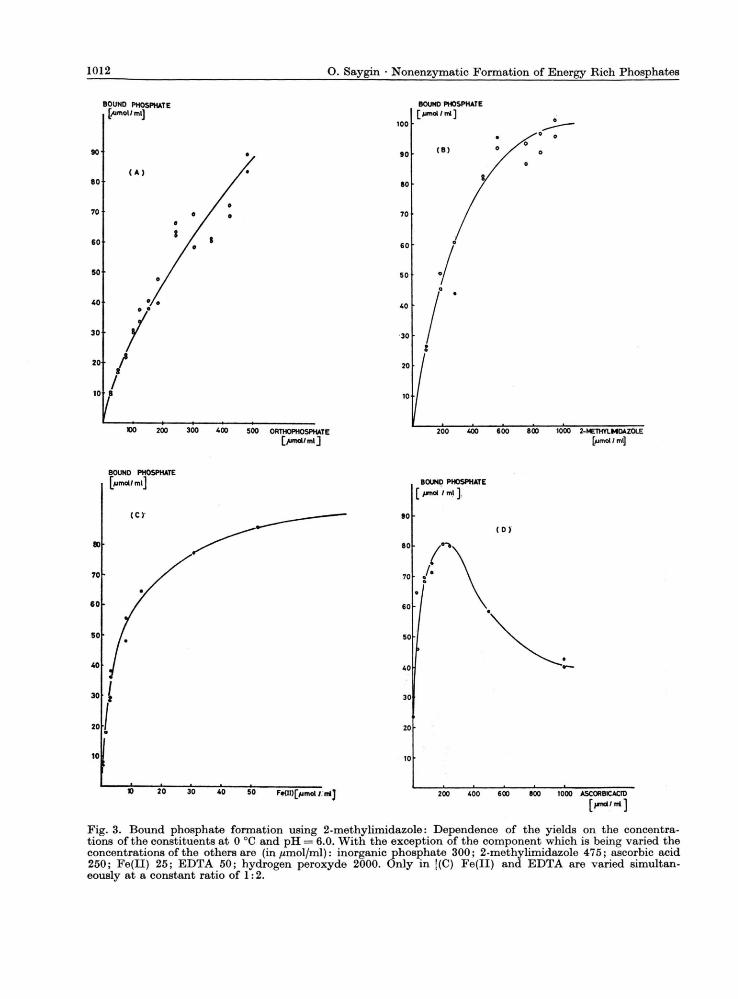
Fig. 3. Bound phosphate formation using 2-methylimidazole: Dependence of the yields on the concentra-tions of the constituents at 0 °C and pH = 6.0. With the exception of the component which is being varied the concentrations of the others are (in jumol/ml): inorganic phosphate 300; 2-methylimidazole 475; ascorbic acid 250; Fe(II) 25; EDTA 50; hydrogen peroxyde 2000. Only in '(C) Fe(II) and EDTA are varied simultan-eously at a constant ratio of 1:2.

1013 O. Saygin • Nonenzymatic Formation of Energy Rieh Phosphates
Fig. 2 shows the dependence of the bound phos-phate formation on the pH of the solution. As dependent on the pH of the solution at the begin-ning, we obtained characteristic maxima for each substrate.
Since 2-methylimidazole gave the greatest yields at pH 6 we used this substrate and higher concentra-tions of reagents for a study of the dependence of the yields on the various parameters. Fig. 3 shows the yields obtained from 2-methylimidazole as functions of the concentration of the components. In the case of inorganic phosphate and 2-methyl-imidazole as variables we observed linear depend-ence in small concentrations and saturation at higher concentrations. With Fe(II)-EDTA as variable, saturation was reached at very low concentrations, indicating the catalytic role of iron. Only in the case of ascorbic acid a maximum was observed. To understand the decrease of the yield at higher concentrations, in additional experiments we added ethanol or £erf-butanol (250 //moles per ml) and observed also a decrease of the yield (50%). Since alcohols are known radical trapping agents, the observed decrease of the yield at higher concentra-tions of ascorbic acid can be explained by trapping the OH radicals.
In other experiments we tried to replace hydrogen peroxide by O2. Using 2-methylimidazole as sub-strate, after one hour of bubbling air through the solution (whithout H2O2) easily detectable amounts of bound phosphate were obtained (Table I).
By chromatography of the reaction mixture on ion exchange columns two fractions of bound phos-phate were obtained between elution volumes of 40-80 and 125-200 ml; inorganic phosphate remains absorbed on the column under these conditions. Work on isolation and identification of the phos-phorylated product(s) is in progress.
Discussion At this point little can be said about the mecha-
nism of this unusual condensation reaction, except
that energy rich radicals are probably involved in the trapping of inorganic phosphate. This may open a new aspect on biochemical processes like oxidative phosphorylation and photophosphorylation. Indeed, the molecular mechanism of both stays uncleared since the work of Lohmann on ATP [8]. Radical mechanisms in relation to these processes have been stressed rather exceptionally [9]. Usually two-electron processes have been considered in phos-phorylation reactions [10] and general interest is presently focused on vectorial processes [11] in heterogenic systems, rather than on homogeneous reactions. Conspicuously enough, one electron proc-esses are involved in the successful model systems of Wieland and Bäuerlein [12].
Only few cases of photophosphorylation models have been reported [9, 13]. However, such systems intrinsically imply the idea of one electron processes. Therefore, it appears significant that preliminary experiments with photosensitized oxidation of 2-methylimidazole under similar experimental con-ditions indeed gave rise [15] to bound phosphate formation. These results apparently justify the use of radical reactions as an experimentally more convenient model of photoreactions in the course of our "Hannover program" [3, 4, 14] - the experi-mental search for the first pre biological "Bioids" [1-3], selforganizing systems which were capable to acquire and conserve the first bits of evolutionary information represented as steady states stabilized by distinct chemical feedback structures. Indeed, apart of feedback reactions, energy acquisition and transformation must be considered as the most important element of such mechanisms.
Note added in proof: A crystalline product, Schmp. 130 °C, has been isolated from the fraction eluted at 125-200 ml. Elemental analysis and NMR data are compatible with a O-monophosphoric acid ester of 4,5-dihydroxy-2-methyl-zl2-imidazoline [15].
[1] P. Decker 1974, Evolution in offenen Systemen: prize essay submitted to the Bavarian Academy of Sciences; reprints (170pp.) available from Documentation Center on the Origin of Life, Universitätsbibliothek, Ulm, FRG. ; c/. P. Decker, Nachr. Chem. Techn. 23, 167 (1975).
[2] P. Decker and A. Speidel, Z. Naturforsch. 2 7 b , 257 (1972). Abstract in: FEBS, Abstracts of
Communications 6th Meeting Madrid, April 1969, Publ. by the Spanish Biochemical Soc., Madrid 1969, Abstract No. 435, p. 146.
[3] P. Decker and W . Heidmann in H. Nöda (ed.): Origin of Life, Proc. 2. ISSOL Mtg. and 5. ICOL Mtg. Kyoto 1977. Japan Sei. Soc. Press, Tokyo 1978 (JSSP No. 01312-1104), pp. 617-624.
[4] P. Decker in D. C. Walker (ed.): Origins of

1014 O. Saygin • Nonenzymatic Formation of Energy Rieh Phosphates
Optical Activity in Nature, Elsevier, Amsterdam 1979, pp. 109-124.
[5] P. Decker and Ö. Saygin, Z. Naturforsch. 34c, 649 (1979).
[6] C. Walling, Acc. Chem. Res. 8, 125 (1975). [7] B. E. Wahler and A. Wollenberger, Biochem. Z.
329, 508 (1958). [8] K . Lohmann, Naturwissenschaften 17, 624 (1929). [9] N. V. Goncharova and V. B. Evstigneef, Biokhi-
mia 42, 963 (1977).
[10] V . M. Clark, D. W . Hutchinson, A. J. Kirby, and S .G . Warren, Angew. Chem. 76, 704 (1964).
[11] P. Mitchell, Eur. J. Biochem. 95, 1 (1979). [12] T. Wieland and E. Bäuerlein, Naturwissenschaf-
ten 54, 80 (1967). [13] Shu-I Tu and Jui H. Wang, Biochemistry 9, 4505
(1970). [14] P. Decker, Ann. New York Acad. Sei. 316, 236
(1979). [15] P. Decker and 0 . Saygin, in preparation.


















