
Periventrikuläre Leukomalazie : Untersuchung Cytokin ... · Danksagung Mein Dank gilt Herrn Prof....
129
Periventrikuläre Leukomalazie- Untersuchung Cytokin-induzierter Schädigungen von Oligodendrocyten-Vorläuferzellen und Protektion durch Corticoide Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften der Fakultät für Chemie, Ruhr-Universität Bochum vorgelegt von Dipl.-Chem. Beatrix Feldhaus Bochum, April 2003
Transcript of Periventrikuläre Leukomalazie : Untersuchung Cytokin ... · Danksagung Mein Dank gilt Herrn Prof....

Periventrikuläre Leukomalazie-
Untersuchung Cytokin-induzierter Schädigungen
von Oligodendrocyten-Vorläuferzellen
und Protektion durch Corticoide
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Chemie, Ruhr-Universität Bochum
vorgelegt von
Dipl.-Chem. Beatrix Feldhaus
Bochum, April 2003

Die vorliegende Dissertation wurde von Dezember 1999 bis April 2003 am Lehrstuhl für Molekulare Neurobiochemie der Fakultät für Chemie der Ruhr-Universität Bochum in der Arbeitsgruppe von Prof. Dr. Rolf Heumann angefertigt.
Prüfungskommission:
Prof. Dr. R. Heumann
Prof. Dr. R. Berger
Prof. Dr. C. Wöll

Danksagung
Mein Dank gilt Herrn Prof. Dr. Richard Berger für die Möglichkeit eine interdisziplinäre Dissertation mit einem so interessanten medizinischen Hintergrund anfertigen zu können.
Herrn Professor Dr. Rolf Heumann danke ich für die Möglichkeit, diese Arbeit an seinem Lehrstuhl anfertigen zu können. Zusammen mit Frau PD Dr. Irmgard Dietzel-Meyer haben sie mich durch Dis-kussionen über die Experimente und Ergebnisse immer unterstützt und beraten.
Prof. Dr. Christof Wöll danke ich für seine Bereitschaft als dritter Prüfer zu fungieren.
Herrn Dr. Kai Sven Erdmann danke ich für seine Mühe, mich durch kritische Fragen in der Entwick-lung zu einer erfolgreichen Wissenschaftlerin gefordert und gefördert zu haben. – Und die Kritik der neusten Kinofilme nicht zu vergessen!
Für die Verschönerung des Labor- und vor allem Büroalltages danke ich der ehemaligen und jetzigen Crew des Raumes 169 – Dank ihr gab es immer einen Lichtblick in manchen tristen, weil erfolglosen, Tagen. Danke für die tolle Unterstützung an Jasmin Gerkrath, Sabine Laerbusch, Friederike Thor, Annette Tolle und die „Ehemaligen“ Dr. Kathrin Susen, Dr. Volker Eulenburg und Dr. Matthias Hartmann.
Für die „seelische Unterstützung“ (nach dem Motto: „alle sitzen in einem Boot“, oder „man ist nie alleine“) danke ich meinen weiteren Vorkämpfern bzw. Mitstreitern Dr. Lutz Herrmann, Dr. Eftichia Saccharidou, Dipl.-Biochem. Ina Radtke und Dipl.-Chem. Andreas Gmerek.
Ein extra Dank geht an Annette Tolle für die Sequenzierungen der Konstrukte. Ein lieber Dank auch an Herrn Paikenkamp und Frau Schleicher aus der Zentralen Versuchstierhaltung für die Zucht der Ratten und die gute Zusammenarbeit.
Aus meinem Freundeskreis sei hier besonders meine Schokoladen-Lieferantin Anne erwähnt.
Schließlich möchte ich mich bei meinem Lebensgefährten Jo für seine unendliche Geduld und Unter-stützung bedanken.

Für meine Eltern und Großeltern.

Inhaltsverzeichnis I
INHALTSVERZEICHNIS
Abkürzungen .......................................................................................................................................... V
1 Einleitung...................................................................................................................................... 1
1.1 PERIVENTRIKULÄRE LEUKOMALAZIE - PVL............................................................................. 1 1.1.1 PVL: EINFÜHRUNG ....................................................................................................................... 1 1.1.2 BEDEUTUNG VON INFEKTIONEN IN DER PATHOGENESE EINER PVL............................................. 2 1.1.3 ROLLE VON CYTOKINEN UND OLIGODENDROCYTEN-VORLÄUFERZELLEN.................................. 4 1.2 MYELINISIERUNG UND OLIGODENDROCYTEN ........................................................................... 5 1.2.1 MYELIN: AUFBAU UND FUNKTION ............................................................................................... 5 1.2.2 ENTWICKLUNG VON OLIGODENDROCYTEN .................................................................................. 5 1.2.3 OLIGODENDROCYTEN: REGULATION VON PROLIFERATION UND DIFFERENZIERUNG .................. 7 1.3 CYTOKINE ..................................................................................................................................... 8 1.3.1 EINFÜHRUNG................................................................................................................................. 8 1.3.2 TUMOR-NECROSIS-FACTOR-ALPHA – TNF-α ............................................................................... 9 1.3.2.1 Einführung: TNF-α und seine Rezeptoren.............................................................................. 9 1.3.2.2 Signaltransduktion des TNFR1.............................................................................................. 10 1.3.2.3 Rolle des TNF-α für Myelin und Oligodendrocyten ............................................................. 11 1.3.3 INTERFERON-GAMMA – IFN-γ..................................................................................................... 12 1.3.4 SYNERGISTISCHE BEZIEHUNGEN ZWISCHEN TNF-α UND IFN-γ................................................. 14 1.4 GLUCOCORTICOIDE.................................................................................................................... 15 1.4.1 EINFÜHRUNG............................................................................................................................... 15 1.4.2 KLINISCHE ANWENDUNG............................................................................................................ 16 1.4.3 EINFLUSS AUF MYELIN UND OLIGODENDROCYTEN.................................................................... 17 1.5 ZIELSETZUNG DER ARBEIT ........................................................................................................ 18
2 Materialien .................................................................................................................................. 19
2.1 HERKUNFT DER GERÄTE............................................................................................................ 19 2.2 HERKUNFT VERWENDETER MATERIALIEN UND SUBSTANZEN................................................ 19 2.2.1 CHEMIKALIEN ALLGEMEIN.......................................................................................................... 19 2.2.2 CHEMIKALIEN FÜR DIE ZELLKULTUR.......................................................................................... 20 2.2.3 ENZYME....................................................................................................................................... 20 2.2.4 KITS UND VERBRAUCHSMATERIALIEN ....................................................................................... 21 2.2.5 ANTIKÖRPER ............................................................................................................................... 21 2.2.6 PLASMIDE.................................................................................................................................... 21 2.3 PRIMER........................................................................................................................................ 22 2.4 BAKTERIEN ................................................................................................................................. 22 2.5 GRÖßENSTANDARDS ................................................................................................................... 22

Inhaltsverzeichnis II
2.5.1 PROTEIN-MOLEKULARGEWICHTSSTANDARD ............................................................................. 22 2.5.2 DNA-GRÖßENSTANDARD ........................................................................................................... 22 2.6 VERSUCHSTIERE ......................................................................................................................... 22 2.7 PUFFER UND LÖSUNGEN FÜR MOLEKULARBIOLOGISCHE ARBEITEN..................................... 23 2.8 PUFFER UND LÖSUNGEN FÜR PROTEINCHEMISCHE ARBEITEN............................................... 24 2.9 PUFFER UND LÖSUNGEN FÜR DIE IMMUNCYTOCHEMIE .......................................................... 26 2.10 MEDIEN FÜR DIE BAKTERIENANZUCHT .................................................................................... 26 2.11 LÖSUNGEN FÜR DIE ZELLKULTUR............................................................................................. 27
3 Methoden .................................................................................................................................... 28
3.1 ZELLKULTUR .............................................................................................................................. 28 3.1.1 POLY-D-LYSIN-BESCHICHTUNG VON DECKGLÄSCHEN.............................................................. 28 3.1.2 PRÄPARATION VON PRIMÄREN GLIA-MISCHKULTUREN AUS NEONATALEN RATTEN ................ 28 3.1.3 KULTIVIERUNG VON PRIMÄREN RATTEN-GLIA-MISCHKULTUREN ............................................ 28 3.1.4 ISOLIERUNG VON PRIMÄREN OLIGODENDROCYTEN-VORLÄUFERZELLEN AUS
RATTEN-GLIA-MISCHKULTUREN................................................................................................ 29 3.1.5 KULTIVIERUNG VON PRIMÄREN OLIGODENDROCYTEN-VORLÄUFERZELLEN ............................ 29 3.1.6 FIXIERUNG VON ZELLEN AUF DECKGLÄSCHEN .......................................................................... 30 3.1.7 IMMUNCYTOCHEMIE ................................................................................................................... 30 3.1.8 DETEKTION VON APOPTOTISCHEN ZELLEN (TUNEL-METHODE) .............................................. 30 3.2 PROTEINBIOCHEMISCHE METHODEN ....................................................................................... 32 3.2.1 AUFSCHLUSS VON ZELLEN ZUR GEWINNUNG DER GESAMTPROTEINE....................................... 32 3.2.2 PROTEINBESTIMMUNG MIT BIORAD DC PROTEIN ASSAY.......................................................... 32 3.2.3 DISKONTINUIERLICHE SDS-POLYACRYLAMID GELELEKTROPHORESE (SDS-PAGE) ............... 32 3.2.4 WESTERN-BLOT .......................................................................................................................... 33 3.2.5 PONCEAU-FÄRBUNG VON PROTEINEN ........................................................................................ 34 3.2.6 IMMUNODETEKTION.................................................................................................................... 34 3.2.7 REDETEKTION VON IMMUNOBLOTS („STRIPPEN“)...................................................................... 35 3.3 MOLEKULARGENETISCHE METHODEN..................................................................................... 36 3.3.1 ISOLIERUNG VON GESAMT-RNA ................................................................................................ 36 3.3.2 RT (REVERSE TRANSCRIPTION)-PCR ........................................................................................... 36 3.3.3 SEMI-QUANTITATIVE RT-PCR .................................................................................................... 36 3.3.4 PCR (POLYMERASE CHAIN REACTION)......................................................................................... 37 3.3.5 SOE (SPLICING BY OVERLAP EXTENSION)-PCR............................................................................. 38 3.3.6 GELELEKTROPHORESE VON DNA ............................................................................................... 38 3.3.7 ISOLIERUNG VON DNA-FRAGMENTEN AUS AGAROSEGELSTÜCKEN.......................................... 38 3.3.8 LINEARISIERUNG VON PLASMIDEN ............................................................................................. 38 3.3.9 DEPHOSPHORYLIERUNG VON DNA............................................................................................. 39 3.3.10 PHOSPHORYLIERUNG MIT T4 PHOSPHO-NUKLEOTID-KINASE (PNK).......................................... 39 3.3.11 LIGATION VON DNA ................................................................................................................... 40

Inhaltsverzeichnis III
3.3.12 HERSTELLUNG KOMPETENTER BAKTERIEN NACH DER RBCL2-METHODE ................................. 40 3.3.13 TRANSFORMATION KOMPETENTER BAKTERIEN.......................................................................... 40 3.3.14 PLASMIDPRÄPARATION IM ANALYTISCHEN MAßSTAB................................................................ 41 3.3.15 PLASMIDPRÄPARATION IM PRÄPARATIVEN MAßSTAB................................................................ 41 3.3.16 RESTRIKTION VON DNA ............................................................................................................. 42 3.3.17 IN VITRO-TRANSKRIPTION ........................................................................................................... 43 3.3.18 UV-PHOTOMETRISCHE BESTIMMUNG DER DNA BZW. RNA-KONZENTRATION........................ 43
4 Ergebnisse ................................................................................................................................... 44
4.1 CHARAKTERISIERUNG DER PRIMÄREN OLIGODENDROCYTEN-ZELLKULTUR....................... 44 4.1.1 ENTWICKLUNG DER OLIGODENDROCYTEN-VORLÄUFERZELLEN:
A2B5-IMMUNCYTOCHEMIE ........................................................................................................ 45 4.1.2 ENTWICKLUNG DER OLIGODENDROCYTEN-VORLÄUFERZELLEN:
MOSP-IMMUNCYTOCHEMIE ....................................................................................................... 45 4.1.3 ENTWICKLUNG DER OLIGODENDROCYTEN-VORLÄUFERZELLEN:
EXPRESSION DER PROTEINE MBP, MAG UND CNP................................................................... 48 4.1.4 REINHEIT DER OLIGODENDROCYTEN-VORLÄUFER-ZELLKULTUR:
GFAP-IMMUNCYTOCHEMIE........................................................................................................ 50 4.2 EINFLUSS VON TNF-α AUF DAS ÜBERLEBEN PRIMÄRER OLIGODENDROCYTEN-
VORLÄUFERZELLEN .................................................................................................................. 51 4.3 EINFLUSS VON IFN-γ UND TNF-α AUF PRIMÄRE OLIGODENDROCYTEN-
VORLÄUFERZELLEN .................................................................................................................. 53 4.3.1 EINFLUSS VON IFN-γ UND TNF-α AUF DAS ÜBERLEBEN VON OLIGODENDROCYTEN-
VORLÄUFERZELLEN .................................................................................................................... 54 4.3.2 EINFLUSS VON IFN-γ (100 U/ML) UND TNF-α (100 NG/ML) AUF DIE DIFFERENZIERUNG
VON OLIGODENDROCYTEN-VORLÄUFERZELLEN........................................................................ 58 4.4 SCHÜTZEN CORTICOIDE OLIGODENDROCYTEN-VORLÄUFERZELLEN GEGEN
EINFLÜSSE VON IFN-γ UND TNF-α ? ........................................................................................ 63 4.4.1 GESAMTZELLZAHL UND A2B5-ZELLZAHL ................................................................................. 65 4.4.2 PROTEINEXPRESSION – WESTERN-BLOT-ANALYSEN ................................................................. 69 4.4.2.1 Myelin Basic Protein.............................................................................................................. 70 4.4.2.2 Myelin Associated Glycoprotein............................................................................................ 72 4.4.2.3 2’,3’-Cyclische Nucleotid 3’-Phosphodiesterase ................................................................... 74 4.4.3 RNA-EXPRESSION – SEMIQUANTITATIVE RT-PCR.................................................................... 75 4.4.3.1 Myelin Basic Protein.............................................................................................................. 78 4.4.3.2 Myelin Associated Glycoprotein............................................................................................ 79
5 Diskussion................................................................................................................................... 81
5.1 ETABLIERUNG UND CHARAKTERISIERUNG DER OLIGODENDROCYTEN- VORLÄUFERZELL-KULTUR....................................................................................................... 81

Inhaltsverzeichnis IV
5.1.1 IMMUNCYTOCHEMIE ................................................................................................................... 82 5.1.2 PROTEIN-EXPRESSION................................................................................................................. 82 5.2 EINFLUSS VON TNF-α AUF DAS ÜBERLEBEN VON OLIGODENDROCYTEN-
VORLÄUFERZELLEN .................................................................................................................. 84 5.3 EINFLUSS VON IFN-γ UND TNF-α AUF OLIGODENDROCYTEN-VORLÄUFERZELLEN............. 85 5.3.1 ÜBERLEBEN VON OLIGODENDROCYTEN-VORLÄUFERZELLEN ................................................... 86 5.3.2 DIFFERENZIERUNG VON OLIGODENDROCYTEN-VORLÄUFERZELLEN......................................... 87 5.4 CORTICOIDE: SCHUTZ VOR DURCH IFN-γ UND TNF-α INDUZIERTEM ZELLTOD UND
DIFFERENZIERUNGSSTÖRUNGEN IN OLIGODENDROCYTEN-VORLÄUFERZELLEN?.............. 89 5.4.1 CORTICOIDE: EINFLUSS AUF DAS ÜBERLEBEN VON OLIGODENDROCYTEN-
VORLÄUFERZELLEN .................................................................................................................... 90 5.4.2 CORTICOIDE: EINFLUSS AUF DIE DIFFERENZIERUNG VON OLIGODENDROCYTEN-
VORLÄUFERZELLEN .................................................................................................................... 91 5.5 AUSBLICK .................................................................................................................................... 95
6 Zusammenfassung....................................................................................................................... 97
7 Literatur....................................................................................................................................... 99
8 Anhang...................................................................................................................................... 115
8.1 ABBILDUNGSVERZEICHNIS....................................................................................................... 115 8.2 TABELLENVERZEICHNIS .......................................................................................................... 116 8.3 KONGRESSBEITRÄGE................................................................................................................ 117 8.4 LEBENSLAUF ............................................................................................................................. 118

Abkürzungen V
ABKÜRZUNGEN A Adenin
A Extinktion
Abb. Abbildung
AP-1 Activator Protein 1
APS Ammoniumperoxodisulfat
bFGF Basic Fibroblast Growth Factor
bp Basenpaare
BSA Bovines Serumalbumin
C Corticosteron
C Cytosin
cDNA complementary DNA
cm Zentimeter
CNP 2’, 3’ – Cyclic Nucleotide 3’ - Phosphodiesterase
CNTF Ciliary Neurotrophic Factor
DC Deoxycorticosteron DD Death Domain
DEPC Diethylpyrocarbonat
DEX Dexamethason
DNA Desoxyribonucleotide Acid (Desoxyribonucleinsäure)
DNase Desoxyribonuclease
dNTP 2'-Desoxiribonucleotid-5'-Triphosphat
DMEM Dulbecco’s Modified Eagle’s Medium
DMSO Dimethylsulfoxid
ECL Enhanced Chemiluminescence
EDTA Dinatrium-Ethylendiamintetraacetat
EtBr Ethidiumbromid
FADD Fas-Associated Death Domain protein
FCS Fetal Calf Serum g Erdbeschleunigung (9,81 m/s2)
G Guanin
GAF IFNgamma-Activated Factor
GAS γ−interferon Activation Sequence
GFAP Glial Fibrillary Acidic Protein
GR Glucocorticoid-Rezeptor
GRE Glucocorticoid-receptor Responsive Element

Abkürzungen VI
hsp Heat-Shock-Protein
IFNγ Interferon-γ
IL Interleukin
IRF-1 Interferon-Regulatory-Factor-1
JAK Janus Kinase
kD Kilodalton
LB-Medium Luria-Bertani-Medium
LPS Lipopolysaccharide MAG Myelin Associated Glycoprotein
MBP Myelin Basic Protein
MHC Major Histocompatibility Complex
min Minuten
ml Milliliter
mM Millimolar
MOG Myelin-Oligodendrocyte Glycoprotein MOSP Myelin Oligodendrocyte Specific Protein
MS Multiple Sklerose
µl Mikroliter
NF-κB Nuclear Factor-κB
nm Nanometer
NP-40 Nonidet-P40
OD Optische Dichte
PAGE Polyacrylamidgelelektrophorese
PBS Phosphate Buffered Saline
PCR Polymerase Chain Reaction (Polymerasekettenreaktion)
PDGF Platelet-Derived Growth Factor
PFA Paraformaldehyd
PLP Proteolipid-Protein
PMSF Phenylmethansulfonylfluorid
PNK Phospho-Nukleotid-Kinase
PVL Periventrikuläre Leukomalazie
RIP Receptor-Interacting protein
RNA Ribonucleotide Acid (Ribonucleinsäure)
rpm rounds per minute
RT Raumtemperatur
RT-PCR Reverse Transcription-PCRn
SAP Shrimp Alkaline Phosphatase
SDS Sodium Dodecyl Sulfate (Natriumdodecylsulfat)

Abkürzungen VII
SDS-PAGE SDS-Polyacrylamidgelelektrophorese
SH-2 Src-Homology-2
SET Saline-EDTA-Tris-Puffer SOE-PCR Splicing by Overlapping Extensions PCR
STAT Signal Transducer and Activator ofTtranscription
T3 Triodo-I-Thyronin
TBS Tris Buffered Saline
TE Tris-EDTA-Puffer
TEB Tris-Borat-EDTA-Puffer
TEMED N, N, N’,N’-Tetramethylethyldiamin
TNF(R) Tumor Necrosis Factor (Receptor)
TRADD TNFR-AssociatedDdeath Domain protein
TRAF TNFR-Associated Factor
Tris Trishydroxymethylaminomethan
TUNEL Terminal deoxynucleotidyl transferase-mediated UTP Nick End Labeling
U Enzymeinheit
UV Ultra Violett
v/v Volumen pro Volumen
w/v Weight (Masse) pro Volumen
ZNS Zentralnervensystem

Einleitung 1
1 EINLEITUNG
1.1 Periventrikuläre Leukomalazie - PVL
1.1.1 PVL: Einführung
Trotz vieler Fortschritte in der Perinatalmedizin sind die Ursachen für die Entstehung von frühgeburt-
lichen Hirnschäden noch nicht ausreichend geklärt. Als Anzeichen für eine Hirnschädigung bei Früh-
geborenen gelten spastische motorische Störungen, wobei abhängig von der Größe und Lokalisation
der betroffenen Hirnareale, auch die Sehkraft, das Gehör oder die Sprache beeinträchtigt sein können.
Ein solcher Hirnschaden beeinträchtigt das Leben der betroffenen Kinder, aber auch der Angehörigen,
erheblich. Der Erforschung von präventiven und therapeutischen Maßnahmen kommt deshalb große
Bedeutung zu.
Die typische Schadensläsion bei frühgeborenen Kindern ist die periventrikuläre Leukomalazie (PVL)
und die intraventrikuläre Hirnblutung [Volpe, 1995]. Mit periventrikulärer Leukomalazie, einer sehr
häufigen Art des fetalen Hirnschadens, bezeichnet man eine Schädigung der cerebralen weißen Sub-
stanz. Diese Schädigung wurde zuerst 1867 von Virchow beschrieben und tritt insbesondere bei Früh-
geborenen auf (periventrikulär = um die mit Hirnwasser gefüllten Räume gelegen; Leukomala-
zie = Einweichung bzw. Schädigung der weißen Substanz) [Virchow, 1867; Volpe, 1995; Volpe,
1997].
Man unterschiedet in der PVL zwischen einer fokalen und einer diffusen Komponente [Übersicht in
Volpe, 2001]. Die fokale Komponente ist tief in der weißen Substanz lokalisiert (im Bereich der End-
zonen der langen penetrierenden Arterien) und wird durch sogenannte Nekrose-Herde charakterisiert.
Diese Nekrosen betreffen alle zellulären Elemente und führen später zu Zystenbildungen. Bei der
diffusen Komponente handelt es sich um eine weniger starke, aber zellspezifische Schädigung: In
einem eher großflächigen Bereich (im Gebiet der Endzonen der kurzen penetrierenden Arterien und in
den Gebieten zwischen den langen penetrierenden Arterien) werden dabei Oligodendrocyten-
Vorläuferzellen [siehe Abschnitt 1.2] geschädigt.
Diese Oligodendrocyten-Vorläuferzellen bilden in humanen unreifen Feten die Hauptpopulation an
Zellen der weißen Substanz, weshalb sie wahrscheinlich eine Schlüsselrolle in der Entwicklung einer
PVL spielen.

Einleitung 2
Abbildung 1: Lokalisation der fokalen bzw. diffusen Komponente einer PVL In dieser schematischen Darstellung eines horizontalen Schnittes des Großhirns ist zu erkennen, dass die fokale Komponente der PVL tief in der weißen Substanz lokalisiert ist (im Bereich der Endzonen der langen penetrierenden Arterien), wohingegen die diffuse Komponente in einem großflächigen Bereich vorzufinden ist (im Gebiet der Endzonen der kurzen penetrierenden Arterien und in den Gebieten zwi-schen den langen penetrierenden Arterien) [aus Volpe, 2001].
1.1.2 Bedeutung von Infektionen in der Pathogenese einer PVL
Neben einem hypoxisch-ischämischen Insult [Rosenberg et al., 1989; Hasegawa et al., 1993; Bagen-
holm et al., 1997; Bagenholm et al., 1998; Tan et al., 1999; Taylor et al., 1999] werden in den letzten
Jahren mütterliche und fetale Infektionen sowie Entzündungen mit Cytokin-Freisetzung als eine weite-
re Ursache für eine PVL bei frühgeborenen Kindern diskutiert. So konnte in einer Reihe von klini-
schen und epidemiologischen Studien bereits gezeigt werden, dass entzündliche Reaktionen nicht nur
zu einer Verschlimmerung des cerebralen Schadens nach einer Ischämie führen, sondern dass sie das
Gehirn auch direkt schädigen können:
Nach einer Chorioamnionitis (einer Infektion der Eihäute während einer Schwangerschaft) beispiels-
weise ist das Risiko für eine periventrikuläre Leukomalazie oder eine peri-/intraventrikuläre Hirnblu-
tung bei unreifen Babys signifikant erhöht [Adinolfi, 1993; Perlman et al., 1996; Berger et al., 1997;
Dammann und Leviton, 1997; Grether und Nelson, 1997; Yoon et al., 2000]. Einige Publikationen
deuten darauf hin, dass durch solch eine Chorioamnionitis im Feten entzündliche Reaktionen ausgelöst
werden, die dann zu einem neonatalen Hirnschaden mit anschließender Cerebralparese führen können
[Adinolfi 1993; Perlman et al., 1996; Dammann und Leviton 1998; Leviton et al., 1999].
Folgende Abbildung stellt schematisch dar, wie diese Beziehung zwischen einer mütterlichen Infekti-
on und einem Hirnschaden des Feten erklären werden könnte [Romero und Mazor, 1988]:

Einleitung 3
Abbildung 2: Zusammenhang mütterliche Infektion – kindlicher Hirnschaden An dieser Abbildung lässt sich erklären, wie durch eine aszendierende intrauterine Infektion Hirnschäden im Feten entstehen können: Zum einen können die freigesetzten Prostaglandine (PG) zu vorzeitiger We-hentätigkeit führen, wodurch das Risiko für eine Hirnschädigung erhöht wird. Zum anderen können Cytokine (IL1, TNF) über die Nabelschnur oder das Fruchtwasser in den Feten gelangen, und dort nach Durchdringen der noch unreifen Blut-Hirn-Schranke, das fetale Gehirn schädigen [aus Romero und Ma-zor, 1988]. Die bei einer aufsteigenden intrauterinen Infektion in der Dezidua (der mütterlichen Schicht der Ei-
haut) freigesetzten Cytokine könnten wie in Abbildung 2 dargestellt über die Nabelschnur oder das
Fruchtwasser in die fetale Zirkulation gelangen. Durch die noch unreife Blut-Hirn-Schranke [Adinolfi
1985] könnten sie dann weiter in das fetale Gehirn gelangen und dort verschiedene toxische Reaktio-
nen auslösen. Diese können dann zum Beispiel zu einer periventrikulären Leukomalazie oder einer
periventrikulären Hirnblutung führen.
Diese Annahme wird dadurch gestützt, dass bei Kindern, die eine Cerebralparese (einen frühkindli-
chen Hirnschaden) entwickelten, erhöhte Cytokin-Level im Nabelschnurblut [Yoon et al., 1996], im
Fruchtwasser [Yoon et al., 1997a; Martines et al., 1998] und im Blutkreislauf der Babys [Grether und
Nelson, 1997; Nelson et al., 1998; Grether et al., 1999] nachgewiesen wurden.

Einleitung 4
1.1.3 Rolle von Cytokinen und Oligodendrocyten-Vorläuferzellen
Es gibt immer mehr Hinweise dafür, dass Lipopolysaccharide (LPS), Bestandteile der Zellmembran
gram-negativer Bakterien, welche Astrocyten und Mikroglia anregen, entzündliche Cytokine zu pro-
duzieren, Schäden in der weißen Hirnsubstanz hervorrufen können. Dabei kann von den nachweislich
durch LPS im Gehirn freigesetzten Cytokinen [Gatti und Bartfai, 1993; Hillhouse und Mosley, 1993;
Van Dam et al., 1995; Berger et al., 2000] besonders der Tumor Necrosis Factor alpha (TNF-α) mit
einem Hirnschaden in Verbindung gebracht werden [Barone et al., 1997]. Beispielsweise konnte bei
Ratten-Feten, deren Muttertieren intraperitoneal (in den Bauchraum hinein) LPS injiziert wurde, eine
erhöhte Expression von IL-1β und TNF-α sowie ein leichter Schaden der weißen Substanz beobachtet
werden [Cai et al., 2000]. Ähnliche Ergebnisse wurden bei Versuchen mit durch Bakterien ausgelösten
intrauterinen Infektionen in Kaninchen erzielt [Yoon et al., 1997b].
Das Risiko für eine periventrikuläre Leukomalazie ist genau in der Periode der Embryonalentwicklung
am höchsten, in welcher die aktive Myelin-Bildung noch nicht stattfindet und in welcher Oligodendro-
cyten-Vorläuferzellen proliferieren und beginnen zu Differenzieren [Kinney and Back, 1998; Back et
al., 2001].
Abbildung 3: Diagramm über die Entwicklung der Oligodendrocyten in der humanen weißen Substanz während der zweiten Hälfte der Schwangerschaft: Die Periode der Entwicklung, in der das Risiko für die Entstehung einer PVL sehr hoch ist, fällt mit der Periode der Entwicklung der humanen cerebralen weißen Substanz zusammen, in welcher die Population von späten Oligodendrocyten-Vorläuferzellen dominierend ist (90%, siehe Back et al., 2001). Daraus er-gibt sich, dass Oligodendrocyten-Vorläuferzellen eine wichtige Rolle in der Entstehung von PVL spielen könnten [modifiziert nach Back et al., 2001]. Diese Studien stützen die Theorie, dass Cytokine diese Oligodendrocyten-Vorläuferzellen, sogenannte
O-2A-Vorläuferzellen [siehe 1.2.2.], direkt schädigen könnten [Back und Volpe, 1997; Dammann und
Leviton, 1997]. Ein Verlust der Vorläuferzellen könnte die für eine PVL charakteristische Störung der
Myelinisierung und die Schlüsselrolle dieser Zellpopulation in der Entstehung dieser Krankheit erklä-
ren.

Einleitung 5
1.2 Myelinisierung und Oligodendrocyten
1.2.1 Myelin: Aufbau und Funktion
Im Zentralnervensystem wird Myelin von Oligodendrocyten synthetisiert [Pfeiffer et al., 1993], indem diese ihre Plasmamembran konzentrisch um die Axone von Nervenzellen wickeln. Man unterteilt das Myelin in verschiedene Komponenten [Arroyo and Scherer, 2000]: Das kompakte Myelin bildet den Hauptteil der Myelin-Scheide und ist weitestgehend für die isolierende Eigenschaft verantwortlich. Es enthält unter anderem die Haupt-Myelin-Proteine Proteolipid-Protein (PLP) und Myelin Basic Protein (MBP) [Lees and Brostoff, 1984; Norton and Cammer, 1984; Griffiths et al., 1998]. Die nicht-kompakten Regionen des Myelins sind durch die Proteine 2´3´-Cyclic Nucleotide 3´-Phosphodiesterase (CNP), Myelin-Associated Glycoprotein (MAG), Myelin-Oligodendrocyte Gly-coprotein (MOG) und Connexin-32 (Cx32) gekennzeichnet [Braun et al., 1988; Brunner et al., 1989; Trapp, 1990; Li et al., 1997].
Ein kritischer Faktor in der Myelinisierung des Zentralnervensystems ist die Proliferationsphase von Oligodendrocyten [Übersicht in Wood and Williams, 1984]: Obwohl ein Oligodendrocyt viele ver-schiedene Axone umhüllen kann [Bunge, 1968], muss eine ausreichende Anzahl von Oligodendrocy-ten generiert werden, damit die Axone über ihre ganze Länge mit Myelin umhüllt sind.
Für ein „normales Funktionieren“ des reifen Zentralnervensystems ist das Myelin unbedingt erforder-lich: Die Axone, deren Durchmesser größer als 0,2µm ist, werden von dieser mehrschichtigen isolie-renden Membran umgeben, durch welche sie die benötigte hohe Geschwindigkeit bei der Weiterlei-tung von Signalen aufrechterhalten können. Eine Unterbrechung des Myelins durch beispielsweise eine Verletzung oder Degeneration führt deshalb zu starken funktionellen Defiziten wie zum Beispiel motorischen Störungen.
1.2.2 Entwicklung von Oligodendrocyten
Oligodendrocyten gehören zu der Gruppe der Gliazellen, zu welchen unter anderem Astrocyten und Mikroglia zählen. Gliazellen bilden das Stützgewebe des Zentralnervensystems, sorgen für die „Ernäh-rung“ der Nervenzellen und schützen diese durch ihre Fähigkeit Ionen zu puffern. Oligodendrocyten werden von Neuroepithel-Zellen des Neuralrohres generiert und wandern anschließend aktiv durch das Zentralnervensystem zu ihren Bestimmungsorten [Übersicht in Miller, 1996]. Im adulten Nervensys-tem sind die meisten Oligodendrocyten in der weißen Substanz lokalisiert, wo sie für die Bildung des isolierenden Myelins zuständig sind.
Sie entwickeln sich aus einer Gliavorläuferstufe, den sogenannten O-2A-Vorläuferzellen. Unter ver-schiedenen Zellkulturbedingungen können aus diesen Vorläuferzellen entweder Typ-II-Astrocyten oder reife Oligodendrocyten entstehen [Raff et al., 1983a]. Die Vorläuferzellen können durch die An-tikörper A2B5 [Eisenbarth et al., 1979] und GD3 [Goldman et al.,1986] nachgewiesen werden. Wäh-rend der Differenzierung verändern sich die Zellen morphologisch von bipolaren Zellen zu Zellen mit

Einleitung 6
stark verzweigten Fortsätzen. Dabei können unterschiedliche Entwicklungsstadien mit entsprechenden Antikörpern charakterisiert werden:
Zunächst tritt O4 auf [Sommer und Schachner, 1981], gefolgt von GalC (Nachweis mit O1-Antikörper) und dem gleichzeitigen Verlust von A2B5 / GD3-Immunreaktivität [Raff et al., 1983b]. Wenig später beginnt die CNP-Expression [Kim et al., 1984], dann die MBP- und PLP-Expression [Raff et al., 1979; Kanfer et al., 1989].
Abbildung 4: Entwicklung von Oligodendrocyten-Vorläuferzellen zu reifen Oligodendrocyten mit ent-sprechender Antigen-Expression Die Entwicklung von Oligodendrocyten-Vorläuferzellen (OL Progenitor) zu reifen Oligodendrocyten (Ma-ture-OL) ist mit Veränderungen in der Morphologie, der Antigen-Expression, sowie der Fähigkeit zu migrieren, proliferieren bzw. myelinisieren verbunden. Auf Grund dieser Kenntnisse ist eine Charakteri-sierung der einzelnen Entwicklungsstufen möglich [modifiziert nach Back und Volpe, 1997].
Neben der veränderten Morphologie und der unterschiedlichen Immunreaktivität werden die Entwick-lungsstadien auch durch ihre unterschiedliche migratorische Fähigkeit, proliferative Kapazität und Reaktion auf Wachstumsfaktoren ausgezeichnet [Übersicht in Pfeiffer et al., 1993]. Vor allem letzteres wird in in vitro Studien ausgenutzt: Durch geeignete Wahl von Faktoren können so Proliferation und Differenzierung gesteuert werden.

Einleitung 7
1.2.3 Oligodendrocyten: Regulation von Proliferation und Differenzierung
Es wurde bereits gezeigt, dass eine Reihe von in Nervenzellen produzierten Faktoren die Proliferation und/oder Reifung von Oligodendrocyten regulieren [McMorris and Dubois-Dalcq, 1988; Gard und Pfeiffer, 1993; Barres et al., 1994a].
Einer der am besten charakterisierten Proliferationsfaktoren für A2B5-positive Zellen ist der Platelet-Derived Growth Factor (PDGF) [Noble et al., 1988; Richardson et al., 1988]. Im Zentralnervensystem wird dieser Faktor sowohl von Astrocyten als auch von Neuronen synthetisiert [Yeh et al., 1991]. PDGF alleine kann jedoch nicht zu einer unendlichen Proliferation führen: Nach maximal acht Zelltei-lungen beginnen Oligodendrocyten-Vorläuferzellen in vitro zu differenzieren [Raff et al., 1985; Temple and Raff, 1986; Raff et al., 1988].
Der Fibroblast Growth Factor (FGF) stimuliert zum einen selber die Proliferation von Oligodendrocy-ten-Vorläuferzellen [Eccelston and Silberberg, 1984; Gard und Pfeiffer, 1993; Baron et al., 2000] und
führt zum anderen zu einer erhöhten und anhaltenden Expression des PDGF-Rezeptor-α [McKinnon et
al., 1991], wodurch die Sensitivität der O-2A-Zellen gegenüber PDGF erhöht wird. Dies Kombination aus beiden Faktoren führt so zu einer unbegrenzten Proliferation der Vorläuferzellen [Bogler et al., 1990] und wird deshalb in verschiedenen in vitro Studien zur Induzierung der Proliferation benutzt [Bogler et al., 1990; McKinnon et al. 1990; Huber et al., 1994; Andrews et al., 1998; Back et al., 1998; Cammer 2000].
Der Ciliary Neurotrophic Factor (CNTF) gehört wie PDGF zu der Gruppe der Cytokine, hat aber einen anderen Effekt auf Oligodendrocyten: Er fördert das Überleben und die Differenzierung zu rei-fen Oligodendrocyten.
Es sind aber auch Moleküle bekannt, die außerhalb des Gehirns produziert werden, und ebenfalls an der Entwicklung von Oligodendrocyten beteiligt sind. Thyroid-Hormone zum Beispiel regulieren zum einen die Expression verschiedener Enzyme, welche mit der Myelinisierung des Zentralnervensystems zusammenhängen. Zum anderen regulieren sie die Expression der Hauptbestandteile der Myelinschei-den [Bhat et al., 1979; Almazan et al., 1985]. T3 (Triodo-I-Thyronin) wird deshalb als ein Differenzie-rungsfaktor diskutiert, welcher die Dauer der Proliferationsphase von Oligodendrocyten-Vorläuferzellen kontrolliert und die Differenzierung in reife Oligodendrocyten fördert [Barres et al., 1994b; Baas et al., 1997]. Der Mechanismus dieser Hemmung der Proliferation ist allerdings noch unbekannt.
Die Kombination von T3 mit CNTF und Forskolin wurde 1998 von Back et al. als eine geeignete Zu-sammenstellung von Faktoren zur in vitro Induzierung der Oligodendrocyten-Differenzierung be-schrieben [Back et al., 1998].
Einleitung 8
1.3 Cytokine
1.3.1 Einführung
Als Cytokine bezeichnet man eine große Familie verschiedener Polypeptide, welche im allgemeinen mit Entzündungen, Aktivierung des Immunsystems, Zelldifferenzierung oder Zelltod in Verbindung gebracht werden. Zu ihnen zählen Interleukine (IL), Interferone (IFN), Tumor Necrosis Factor (TNF), Chemokine und Wachstumsfaktoren. In gesundem Gewebe haben sie nur wenig oder gar keine be-kannte Funktion, wohingegen ihre Freisetzung als Reaktion auf Verletzungen, Infektionen oder Ent-
zündungen schnell induziert wird. Man unterscheidet grob zwischen entzündlichen (z.B. IL-1, TNF-α
und IL-6) und anti-entzündlichen Cyktokinen (z.B. IL-1α, IL-10 und TGF-ß), wobei erstere im allge-
meinen neurotoxisch und letztere neuroprotektiv wirken [Allan and Rothwell, 2001].
Die entsprechenden Rezeptoren, Signaltransduktionswege und die Regulierung der Aktivität von Cy-tokinen ist sehr komplex: es sind überlappende, zusammenwirkende und antagonistische Interaktionen
bekannt. Interleukin-1 und TNF-α zum Beispiel führen in humanen fetalen Zellkulturen aus Neuronen
und Gliazellen getrennt nicht zum Zelltod, wohingegen sie zusammen Neurotoxizität auslösen [Chao et al., 1995].
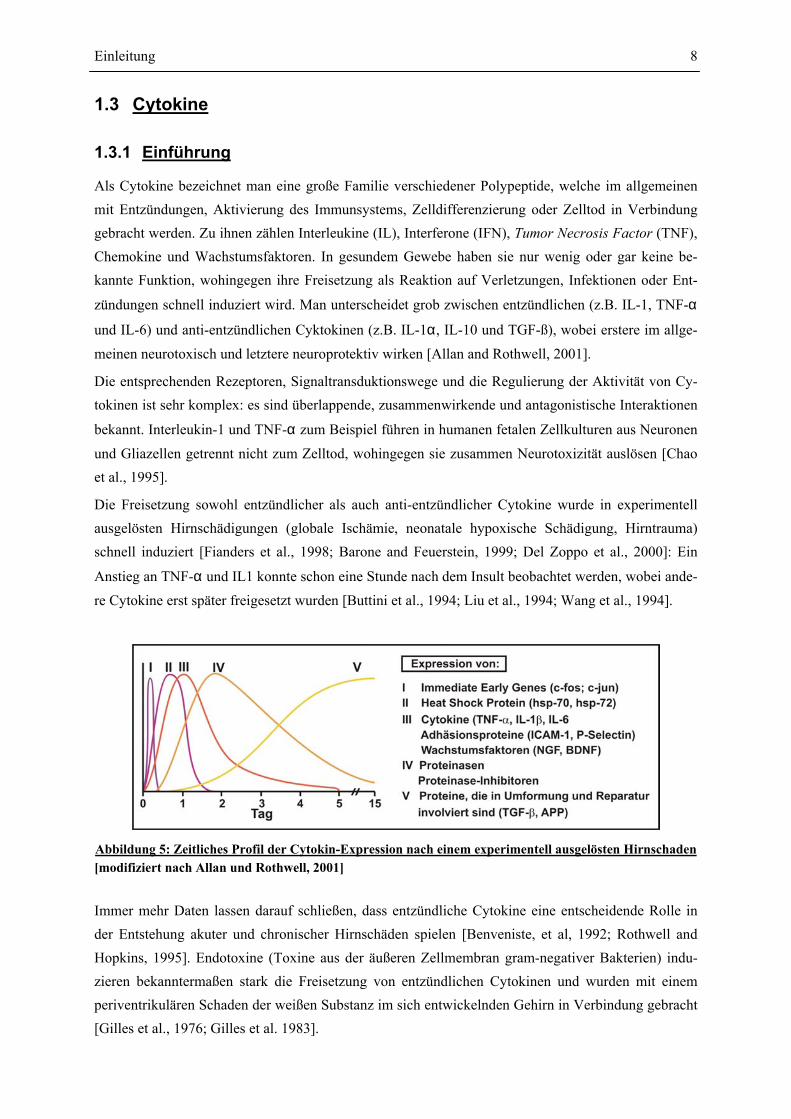
Die Freisetzung sowohl entzündlicher als auch anti-entzündlicher Cytokine wurde in experimentell ausgelösten Hirnschädigungen (globale Ischämie, neonatale hypoxische Schädigung, Hirntrauma) schnell induziert [Fianders et al., 1998; Barone and Feuerstein, 1999; Del Zoppo et al., 2000]: Ein
Anstieg an TNF-α und IL1 konnte schon eine Stunde nach dem Insult beobachtet werden, wobei ande-
re Cytokine erst später freigesetzt wurden [Buttini et al., 1994; Liu et al., 1994; Wang et al., 1994].
Abbildung 5: Zeitliches Profil der Cytokin-Expression nach einem experimentell ausgelösten Hirnschaden [modifiziert nach Allan und Rothwell, 2001]
Immer mehr Daten lassen darauf schließen, dass entzündliche Cytokine eine entscheidende Rolle in der Entstehung akuter und chronischer Hirnschäden spielen [Benveniste, et al, 1992; Rothwell and Hopkins, 1995]. Endotoxine (Toxine aus der äußeren Zellmembran gram-negativer Bakterien) indu-zieren bekanntermaßen stark die Freisetzung von entzündlichen Cytokinen und wurden mit einem periventrikulären Schaden der weißen Substanz im sich entwickelnden Gehirn in Verbindung gebracht [Gilles et al., 1976; Gilles et al. 1983].

Einleitung 9
Allerdings spielen Cytokine auch eine wichtige Rolle in der normalen Entwicklung des Gehirns: Sie werden während der Entwicklung regional und temporal unterschiedlich exprimiert und scheinen so an der Migration und Differenzierung von Zellen beteiligt zu sein [Pousset et al, 1994].
Eine Aufteilung der Cytokine in eine der beiden Gruppen ist aber nicht immer einfach möglich:
TNF-α beispielsweise spielt sowohl in der normalen Entwicklung des Zentralnervensystems eine Rol-
le, wird aber auch als eines der ersten Cytokine nach einer Schädigung des Gehirns freigesetzt [siehe Abb. 5]. Dies spiegelt die Komplexität der großen Familie der Cytokine wieder.
1.3.2 Tumor-Necrosis-Factor-alpha – TNF-α
1.3.2.1 Einführung: TNF-α und seine Rezeptoren
Das Cytokin TNF-α spielt im komplexen Netzwerk der Immunreaktionen eine wichtige Rolle, da es in
die Stimulation des Immunsystems, in Tumorresistenz und in Resistenz gegen Infektionen involviert ist [Aggarwal et al. 1991; Vilcek und Lee, 1991]. Dieses Cytokin wird von vielen verschiedenen Zel-len gebildet, aber zu den wichtigsten TNF-Produzenten gehören aktivierte Makrophagen und Monozy-
ten. Diese setzen besonders nach Stimulation mit Lipopolysacchariden TNF-α frei. [Männel et al.,
1980; Higuchi et al., 1990]. In vielen Schädigungen des Zentralnervensystems (Alzheimer, Multiple
Sklerose, Parkinson) sind erhöhte Level an TNF-α im Hirngewebe und in der cerebrospinal-
Flüssigkeit gefunden worden. Die TNF-α Produktion im Gehirn kann durch viele verschiedene Zellen
nach unterschiedlicher Stimulation erfolgen: beispielsweise setzen Mikroglia und Makrophagen
TNF-α nach einer Ischämie frei [Liu et al., 1994; Uno et al., 1995; Buttini et al., 1996], und Neurone
werden durch Lipopolysaccharide [Breder et al., 1994], eine cerebrale Ischämie [Liu et al., 1994] und Hirnschädigungen [Tchelingerian et al., 1993] stimuliert, diesen Faktor zu produzieren.
TNF-α gehört zu der Familie von Cytokinen, zu der auch Fas (CD95/APO-1), Fas-Ligand (FasL) und
CD40L gehören [Wallach et al., 1999]. Es wird als ein 26 kDa großes, Membran-assoziiertes Protein produziert, welches durch spezifische Metalloproteinasen zu einem 17 kDa großen, löslichen Protein abgespalten wird [Black et al, 1997; Moss et al, 1997]. Beide Formen bilden Trimere aus, um biolo-
gisch aktiv zu werden. TNF-α vermittelt Funktionen, die von der Leitung von Leukozyten zu Entzün-
dungsherden bis hin zur Regulation des Zelltodes durch Proliferation, Zelltoxizität und Apoptose rei-chen [Wallach et al, 1997]. In ischämischen Insulten wurde dieses Cytokin sowohl als neuroprotektiv als auch neurotoxisch beschrieben [Bruce et al., 1996; Rothwell and Luheshi, 1996; Barone et al., 1997].Auch bei der Demyelinisierung und dem Zelltod von Oligodendrocyten scheint es eine ent-scheidende Rolle zu spielen [Selmaj and Raine, 1988]. Diese weite Palette von Aktivitäten wird darauf zurückgeführt, dass zwei Typen des TNF-Rezeptors auf der Oberfläche von fast allen Säugetier-Zellen vorhanden sind: der p55 TNFR (auch TNFR1, CD120a oder TNFRSF1A genannt) und der p75 TNFR (auch TNFR2, CD120b oder TNFRSF1B genannt). Dabei scheinen die meisten biologischen Effekte
des TNF-α durch den TNFR1 vermittelt zu werden [Vandenabeele et al., 1995].

Einleitung 10
1.3.2.2 Signaltransduktion des TNFR1
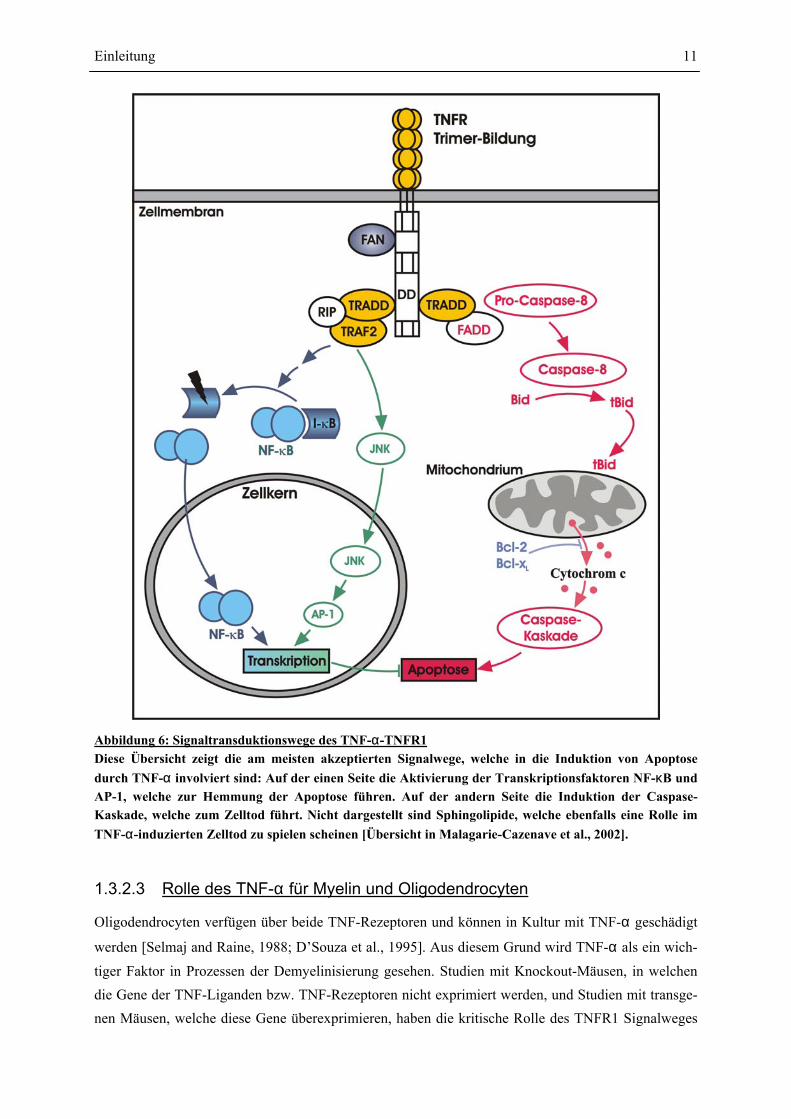
Die Signalwege, die durch TNF-α ausgelöst werden, sind noch nicht vollständig aufgeklärt. Die Bin-
dung von TNF-α führt zu einer Trimerisierung des TNFR1, wodurch dann die verschiedenen Signal-
wege ausgelöst werden. Abbildung 6 [S.11] zeigt eine Übersicht über die Signalwege, welche in die Apoptose (gerichteter Zelltod) involviert sind: Die Trimerisierung führt zu einer Anlagerung von ver-schiedenen Proteinen an die cytoplasmatische Death Domain DD des Rezeptors. Auf der einen Seite führt die Komplexbildung von TRADD (TNFR-Associated Death Domain protein) mit RIP (Receptor Interacting Protein) und TRAF2 (TNFR-Associated Factor 2) zur Aktivierung der zwei Transkripti-
onsfaktoren NF-κB (Nuclear Factor κB) und AP-1 (Activator Protein 1): Die Aktivierung des TNFR1
führt zur Translokalisation des IκB-Kinase-Kompexes an den Rezeptor, wo es dann zur Phosphorylie-
rung von IκB kommt. Damit verbunden wird IκB vom NF-κB Dimer abgelöst, welches dann in den
Zellkern wandern kann. Die Aktivierung des Transkriptionsfaktors AP-1 erfolgt nach Aktivierung der
JNK (c-Jun N-terminal kinase). Diese Aktivierung der Transkriptionsfaktoren NF-κB und AP-1 führt
zur Inhibierung der Apoptose [Karin und Lin, 2002]. Auf der anderen Seite führt die Assoziation von FADD (Fas-Associated Death Domain protein) an TRADD zur Aktivierung der Initiator-Caspase-8. Das aktive Heterodimer Caspase-8 aktiviert dann beispielsweise die Caspase-3. Caspase-8 spaltet zusätzlich das Mitglied der Bcl-2-Familie Bid: die gespaltene Form tBid (truncated Bid) wandert dann zu den Mitochondrien und aktiviert die Freisetzung apoptotischer Faktoren wie Cytochrom c. Zusam-men mit ATP und dem Faktor APAF1 (Apoptotic Protease Activating Factor 1) fördert Cytochrom c die Aktivierung der Caspase-9. Diese wiederum führt dann über die Aktivierung von weiteren Caspa-sen zu Apoptose [Wallach et al., 1999]. Je nachdem, welcher Signalweg überwiegt, wird Apoptose ausgelöst.
Einleitung 11
Abbildung 6: Signaltransduktionswege des TNF-α-TNFR1 Diese Übersicht zeigt die am meisten akzeptierten Signalwege, welche in die Induktion von Apoptose durch TNF-α involviert sind: Auf der einen Seite die Aktivierung der Transkriptionsfaktoren NF-κB und AP-1, welche zur Hemmung der Apoptose führen. Auf der andern Seite die Induktion der Caspase-Kaskade, welche zum Zelltod führt. Nicht dargestellt sind Sphingolipide, welche ebenfalls eine Rolle im TNF-α-induzierten Zelltod zu spielen scheinen [Übersicht in Malagarie-Cazenave et al., 2002].
1.3.2.3 Rolle des TNF-α für Myelin und Oligodendrocyten
Oligodendrocyten verfügen über beide TNF-Rezeptoren und können in Kultur mit TNF-α geschädigt
werden [Selmaj and Raine, 1988; D’Souza et al., 1995]. Aus diesem Grund wird TNF-α als ein wich-
tiger Faktor in Prozessen der Demyelinisierung gesehen. Studien mit Knockout-Mäusen, in welchen die Gene der TNF-Liganden bzw. TNF-Rezeptoren nicht exprimiert werden, und Studien mit transge-nen Mäusen, welche diese Gene überexprimieren, haben die kritische Rolle des TNFR1 Signalweges

Einleitung 12
in der Kontrolle von Demyelinisierung und Entzündung im Zentralnervensystem aufgezeigt [Übersicht in Probert et al., 2000].
In Oligodendrocyten kann TNF-α ein weites Spektrum von Veränderungen auslösen, welche von der
Hemmung des Wachstums der Fortsätze bis hin zu DNA-Fragmentierung und Nekrose reichen [Sel-maj and Raine, 1988; Selmaj et al., 1991; Jenkins and Ikeda 1992; Louis et al., 1993; Mc Larnon et al., 1993; Butt and Jenkins 1994; D’Souza et al., 1995; Soliven and Szuchet 1995; Hisahara et al., 1997].
Es existieren auch wenige Publikationen über Untersuchungen, die sich mit einem Effekt von TNF-α
auf Oligodendrocyten-Vorläuferzellen befassen. Während Andrews und Agresti jeweils nur einen den
Einfluss von IFN-γ auf Oligodendrocyten-Vorläuferzellen potenzierenden Effekt von TNF-α feststel-
len konnten [Agresti et al., 1998; Andrews et al., 1998], beobachtete Cammer auch einen direkten
Einfluss von TNF-α alleine auf die Ausdifferenzierung von angereicherten Oligodendrocyten-
Vorläuferzell-Kulturen [Cammer 2000]. Auch Scurlock und Dawson beobachteten einen direkten
Einfluss von TNF-α auf O2A-Vorläuferzellen: Eine 24stündige Behandlung führte zu 70 % Apoptose
[Scurlock and Dawson, 1999]. Die unterschiedlichen Beobachtungen sind wahrscheinlich auf unter-schiedliche Kultivierungsbedingungen sowie verschiedene Reinheitsgrade der Kulturen zurückzufüh-
ren, so dass eine eindeutige Aussage über die Rolle von TNF-α auf Überleben und Entwicklung von
Oligodendrocyten-Vorläuferzellen schwierig ist.
1.3.3 Interferon-gamma – IFN-γ
Die Interferone sind eine Gruppe von Cytokinen, welche bei der Infektionsabwehr eine wesentliche
Rolle spielen. Sie werden unterteilt in Typ I Interferone (IFN-α, IFN-β, IFN-δ, IFN-ω, IFN-τ) und
Typ II Interferone (IFN-γ). Interferon-γ wird hauptsächlich von T-Zellen und natürlichen Killer-Zellen
produziert [Boehm et al., 1997]. Aber auch Makrophagen [Munder et al., 1998], B-Zellen [Yoshimoto
et al., 1997] und CD8a+ dendritische Zellen [Ohteki et al., 1999] können nach Aktivierung IFN-γ
produzieren.
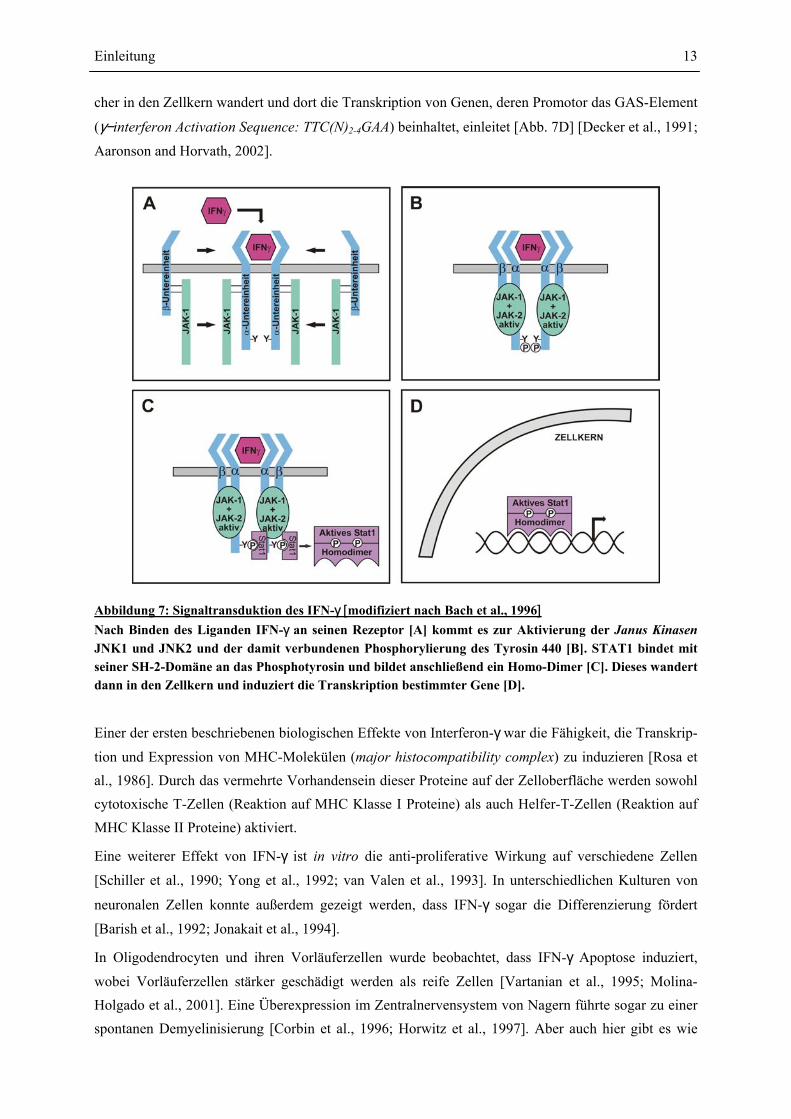
Interferon-γ bindet an einen spezifischen Rezeptor, welcher ubiquitär auf allen kernhaltigen Zellen
exprimiert wird und somit auch auf Oligodendrocyten zu finden ist [Torres et al., 1995]. In unstimu-
lierten Zellen sind die IFN-γ-Rezeptoruntereinheiten α und β mit inaktiven Formen der Proteine JAK1
bzw. JAK2 (Janus Kinase) assoziiert [Kotenko et al., 1995; Sakatsume et al., 1995; Kaplan et al.,
1996]. Der Ligand IFN-γ führt zu einer schnellen Dimerisierung der α-Untereinheiten, wodurch Bin-
dungsstellen für die β-Untereinheiten generiert werden [Fountoulakis et al., 1992; Greenlund et al.,
1993; Marsters et al., 1995]. Die Ligand-Rezeptor-Komplexbildung [Abb. 7A] führt zur Aktivierung der Proteine JAK1 und JAK2 [Müller et al., 1993; Watling et al., 1993]. Die aktivierten Kinasen
phosphorylieren das Tyrosin-440 der α-Untereinheiten, wodurch je eine Bindungsstelle für STAT1
(signal transducer and activator of transcription 1) geschaffen wird [Greenlund et al., 1994; Green-lund et al., 1995] [Abb. 7B]. STAT1 bindet hoch spezifisch mit seiner SH-2-Domäne (Src-Homology-2) an das Phosphotyrosin des Rezeptors. Die Phosphorylierung von STAT1 (Tyrosin 701) führt dann zur Dimerisierung zum sogenannten IFNgamma-Activated Factor (GAF) [Abb. 7C], wel-
Einleitung 13
cher in den Zellkern wandert und dort die Transkription von Genen, deren Promotor das GAS-Element
(γ−interferon Activation Sequence: TTC(N)2-4GAA) beinhaltet, einleitet [Abb. 7D] [Decker et al., 1991;
Aaronson and Horvath, 2002].
Abbildung 7: Signaltransduktion des IFN-γ [modifiziert nach Bach et al., 1996] Nach Binden des Liganden IFN-γ an seinen Rezeptor [A] kommt es zur Aktivierung der Janus Kinasen JNK1 und JNK2 und der damit verbundenen Phosphorylierung des Tyrosin 440 [B]. STAT1 bindet mit seiner SH-2-Domäne an das Phosphotyrosin und bildet anschließend ein Homo-Dimer [C]. Dieses wandert dann in den Zellkern und induziert die Transkription bestimmter Gene [D].
Einer der ersten beschriebenen biologischen Effekte von Interferon-γ war die Fähigkeit, die Transkrip-
tion und Expression von MHC-Molekülen (major histocompatibility complex) zu induzieren [Rosa et al., 1986]. Durch das vermehrte Vorhandensein dieser Proteine auf der Zelloberfläche werden sowohl cytotoxische T-Zellen (Reaktion auf MHC Klasse I Proteine) als auch Helfer-T-Zellen (Reaktion auf MHC Klasse II Proteine) aktiviert.
Eine weiterer Effekt von IFN-γ ist in vitro die anti-proliferative Wirkung auf verschiedene Zellen
[Schiller et al., 1990; Yong et al., 1992; van Valen et al., 1993]. In unterschiedlichen Kulturen von
neuronalen Zellen konnte außerdem gezeigt werden, dass IFN-γ sogar die Differenzierung fördert
[Barish et al., 1992; Jonakait et al., 1994].
In Oligodendrocyten und ihren Vorläuferzellen wurde beobachtet, dass IFN-γ Apoptose induziert,
wobei Vorläuferzellen stärker geschädigt werden als reife Zellen [Vartanian et al., 1995; Molina-Holgado et al., 2001]. Eine Überexpression im Zentralnervensystem von Nagern führte sogar zu einer spontanen Demyelinisierung [Corbin et al., 1996; Horwitz et al., 1997]. Aber auch hier gibt es wie

Einleitung 14
schon beim Cytokin TNF-α konträre Untersuchungen, die keinen cytotoxischen Effekt von IFN-γ
nachweisen konnten [Selmaj et al., 1991; Agresti et al., 1996; Pouly et al., 2000]. Dafür beobachtete die Arbeitsgruppe von Agresti jedoch einen die Proliferation und die Differenzierung hemmenden
Effekt von IFN-γ auf Oligodendrocyten und Oligodendrocyten-Vorläuferzellen [Agresti et al., 1996].
1.3.4 Synergistische Beziehungen zwischen TNF-α und IFN-γ
Die Kombination der Cytokine TNF-α und IFN-γ spielt in einer der bekanntesten Krankheiten mit
gestörter Myelinisierung eine große Rolle: diese Cytokine sind im Gehirn und in der Rückenmarks-flüssigkeit von Patienten mit Multipler Sklerose (MS) stark angereichert, so dass ihnen eine entschei-dende Rolle in der Entstehung dieser Krankheit zugesprochen wird [Raine, 1995; Popko et al., 1997]. Aufgrund der oft beobachteten Degeneration von Oligodendrocyten im Gehirn von MS-Patienten [Ozawa et al., 1994; Lucchinetti and Rodriguez, 1997], wird ein direkter toxischer Effekt dieser Cyto-kine auf diese Zellpopulation diskutiert.
In verschiedenen Systemen konnte die synergistische Wirkung von IFN-γ und TNF-α auf Zellprolife-
ration und Zelldifferenzierung gezeigt werden [Schiller et al., 1990; van Valen et al., 1993; Montaldo et al., 1994; Delvenne et al., 1995]. Auch in CG4-Zellen, einer Oligodendrocyten-Zelllinie, und in
primären Oligodendrocyten-Vorläuferzellen wurde eine synergistische Wirkung von IFN-γ und
TNF-α dokumentiert: Die Arbeitsgruppe um Andrews beobachtete eine Induktion von Apoptose durch
IFN-γ alleine, welche durch TNF-α in einer konzentrationsabhängigen Weise potenziert wurde [And-
rews et al., 1998]. Auch Agresti und Mitarbeiter befassten sich mit dem Einfluss von IFN-γ (alleine
und in Kombination mit TNF-α) auf primäre Oligodendrocyten-Vorläuferzellen: Sie beobachteten
ebenfalls in verschiedenen Versuchen (Zellproliferation; Zelldifferenzierung und Stoffwechselaktivi-
tät) eine Potenzierung des Effektes von IFN-γ durch TNF-α [Agresti et al., 1996].
Die potenzierende Wirkung von TNF-α auf IFN-γ in Oligodendrocyten könnte darauf beruhen, dass in
der Promotor-Region des TNFR1 das GAS-Element vorhanden ist, d.h. dass es durch die Behandlung
mit IFN-γ zu einer erhöhten Expression des TNFα-Rezeptor-1 kommt [Dopp et al., 1997]. Die Sig-
nalweiterleitung beider Cytokine könnte dann zu einer erhöhten Aktivierung des MHC Klasse I Genes führen [Agresti et al., 1998]. In einer Reihe von demyelinisierenden Krankheiten wurde eine anormale Expression dieses Genes beobachtet [Traugott 1987; Traugott and Lebon 1988]. Sogar ein direkter Zusammenhang zwischen Schädigungen von Oligodendrocyten und der Überexpression dieses Genes konnte hergestellt werden [Turnley et al.; 1991; Yoshioka et al., 1991].
Damit ist nicht nur das Finden von Faktoren, die zu einer Unterdrückung der Freisetzung von Cytoki-nen führen, in der Entwicklung von präventiven und therapeutischen Maßnahmen von Bedeutung: Es besteht auch die Möglichkeit, durch Faktoren, deren Signalwege denen der Cytokine entgegenwirken, einen negativen Effekt der Cytokine zu vermeiden. Aufgrund ihrer allgemeinen antientzündlichen Wirkung sind Glucocorticoide für beide Aspekte von großem Interesse.

Einleitung 15
1.4 Glucocorticoide
1.4.1 Einführung
Zu der Gruppe der Glucocorticoide [griechisch: „Gluco“ = Zucker ; „Corticoid“ ≈ Rinde] gehören zum einen Hormone der Nebennierenrinde (Cortisol, Cortison, Corticosteron) und zum anderen struktur-verwandte synthetische Verbindungen (z.B. Dexamethason, Triamcinolon etc.), welche insbesondere den Zuckerstoffwechsel beeinflussen.
Sie wirken allgemein hemmend auf Entzündungen, unterdrücken die Aktivierung des Immunsystems und können sowohl in vitro als auch in vivo Wachstum hemmen [Cupps und Fauci, 1982]. Außerdem
können sie die Transkription vieler Cytokin-Gene, wie beispielsweise IL-1, IL-6, TNF-α und IFNγ,
unterdrücken [Cupps und Fauci, 1982; Culpepper und Lee, 1987; Taniguchi, 1988]. Im Gehirn sind Glucocorticoide für die normale Entwicklung notwendig: Sie sind unter anderem an der Veränderung von Neuron-Neuron- und Neuron-Glia-Wechselwirkungen sowie der Apoptose beteiligt [Übersicht in Matthews, 2000].
Die Wirkung von Corticoiden wird durch intrazelluläre Glucocorticoid-Rezeptoren (GR) bzw. Minera-locorticoid-Rezeptoren (MR) vermittelt, die zuerst in Ratten Thymocyten identifiziert wurden [Munck und Brinck-Johnson, 1968]. Die Corticoid-Rezeptoren gehören zur Familie der Steroid-Hormon-Rezeptoren, welche alle eine homologe DNA-Bindungsdomäne und eine divergierende C-terminale Liganden-Bindungsdomäne besitzen [Evans, 1988; Beato, 1989].
In seiner inaktiven Form liegt der GR in einem Komplex mit Heat-Shock-Proteinen (hsp90 und hsp70) und einem p59 Immunophilin-Molekül im Cytoplasma vor [Pratt, 1993; Guiochon-Mantel et al., 1996]. Mit Bindung des Glucocorticoids an den Rezeptor dissoziiert dieser Komplex und der neu ge-bildete Rezeptor-Ligand-Komplex wandert in den Zellkern. Dort bindet er an die Promotor-Regionen von verschiedenen Genen, welche ein Glucocorticoid-Rezeptor Responsive Element (GRE) tragen und induziert deren Expression [Yamamoto, 1985; Sakai et al., 1988; Drouin et al., 1993]. Neben dieser Induktion der Transkription von Genen, die das spezifisches DNA-Element (GRE) tragen, können
Glucocorticoide auch wirken, indem sie mit anderen Transkriptionsfaktoren wie NF-κB und AP-1
interagieren und so deren Aktivität hemmen [Diamond et al., 1990; Jonat et al., 1990; Yang-Yen et al., 1990; Schule und Evans, 1991; Herrlich, 2001].
Bei der antientzündlichen Wirkung spielt die Unterdrückung der Aktivität des Transkriptionsfaktors
NF-κB wahrscheinlich eine sehr wichtige Rolle. Zum einen kann durch eine Komplexbildung aus NF-
κB, GR und Glucocorticoid die Migration des NF-κB in den Zellkern verhindert werden [Ray und
Prefontaine 1994; Caldenhoven et al., 1995; Scheinman et al., 1995b]. Zum anderen fördern Corti-
costeroide die Synthese des inhibitorischen Proteins des Transkriptionsfaktors NF-κB IκBα [Auphan
et al., 1995; Scheinman et al., 1995a]. Das NF-κB ist ein Heterodimer aus einer p65- (RelA) und einer
p50-Untereinheit [Kawakami et al., 1988; Baeuerle und Baltimore, 1989] und liegt in seiner inaktiven
Form in einem Komplex mit dem IκB-Protein vor [Israel, 1995; Baldwin, 1996]. Viele verschiedene
extrazelluläre Signale führen zu einer Aktivierung des NF-κB, indem sie die Phosphorylierung und

Einleitung 16
Ubiquitinylierung des IκB induzieren [Verma et al., 1995; Finco und Baldwin, 1995; Baeuerle und
Baltimore, 1996]. Dadurch kommt es zu einer Dissoziation des Komplexes, so dass NF-κB in den
Zellkern wandert und dort die Transkription verschiedener Gene (beispielsweise Cytokine und Che-mokine) reguliert [Barnes und Adcock, 1993; Cato und Wade, 1996].
Aufgrund der Tatsache, dass die durch Corticoide ausgelöste vermehrte Synthese des IκBα für eine
Unterdrückung der NF-κB-Aktivität nicht unbedingt erforderlich und sogar nicht unbedingt ausrei-
chend ist [Heck et al., 1996], ist der antientzündliche Mechanismus der Corticosteroide nicht eindeutig geklärt.
Neben der antientzündlichen Wirkung wurde in verschiedenen Zellsystemen auch eine anti-apoptotische Wirkung von Corticosteroiden beobachtet, welche im Falle des Dexamethason besonders stark ausgeprägt ist. Allgemein gehören die Gene der Bcl-2-Familie zu den am stärksten in die Induk-tion und Prävention von Apoptose involvierten Genen [Bergmann et al., 1998; Chao et al., 1998]. Zu dieser Familie gehören pro-apoptotische Gene wie Bcl-xS, Bad, Bax und Bid, und anti-apoptotische Gene wie Bcl-2, Mcl-1 und Bcl-xL [Nagata, 1997]. Bei dem durch Dexamethason induzierten Schutz vor Apoptose zeigte sich, dass je nachdem durch welchen Stimulus die Apoptose eingeleitet wird, die Expression der Gene der Bcl-2-Familie beeinflusst wird [Meßmer et al., 1999; Sasson et al., 2001, 2002], oder aber die anti-apoptotische Wirkung über die Regulation anderer Gene stattfindet [Wen et al., 1997; Evans-Storms und Cidlowski, 2000].
1.4.2 Klinische Anwendung
In der Perinatalmedizin wird das synthetische Glucocorticoid Dexamethason sowohl pränatal als auch postnatal eingesetzt: Bei pränataler Verabreichung wird das Risiko für beispielsweise die Entwicklung eines RDS (respiratory distress syndrom = Atemnotsyndrom des Neugeborenen) gesenkt, postnatal unterstützt Dexamethason die Lungenreifung [Cosmi and DiRenzo, 1989; Kari et al., 1994; Yeh et al., 1997; Omar et al, 2000]. Eine Untersuchung der Folgen für die neurologische Entwicklung nach prä-nataler oder postnataler Dexamethason-Therapie ergab, dass eine pränatale Therapie nicht mit einem verstärkten Vorkommen einer periventrikulären Leukomalazie und auch nicht mit einer verschlechter-ten neurologischen Entwicklung in Verbindung gebracht werden konnte, während eine postnatale Therapie die neurologische Entwicklung der Kinder negativ beeinflusste [LeFlore et al., 2002].
Aufgrund ihrer antientzündlichen und das Immunsystem unterdrückenden Wirkung gehören Corticoi-de weiterhin zu den häufig verabreichten Medikamenten bei der Behandlung von Multipler Sklerose und anderen demyelinisierenden Krankheiten. Es ist jedoch nur wenig und widersprüchliches über den direkten Einfluss auf die Myelin-formenden Zellen (Oligodendrocyten) und auf die Komponenten des Myelins (beispielsweise das Myelin Basic Protein) bekannt [siehe Abschnitt 1.4.3].

Einleitung 17
1.4.3 Einfluss auf Myelin und Oligodendrocyten
Sowohl reife als auch unreife Oligodendrocyten exprimieren die Rezeptoren für Corticoide [Vielkind et al., 1990], wobei verschiedene Corticoide unterschiedlich mit den Rezeptoren interagieren: Corti-costeron kann beide Rezeptoren (GR und MR) aktivieren, während beispielsweise Deoxycorticosteron nur den Typ-I-Rezeptor (MR) und Dexamethason nur den Typ-II-Rezeptor aktiviert (GR). In Kulturen reifer Oligodendrocyten wurden bereits erste Hinweise darauf gefunden, dass diese drei Corticoide
teilweise vor durch TNF-α und IFN-γ induziertem Zelltod schützen. Beide Rezeptoren sind somit in
diesen protektiven Effekt involviert [Melcangi et al., 2000]. Der intrazelluläre Mechanismus, durch welchen Corticoide auf Oligodendrocyten wirken ist jedoch noch nicht aufgeklärt.
Bei in vivo Experimenten wurde überwiegend ein negativer Einfluss von Glucocorticoiden auf die Bildung von Myelin beobachtet: Eine wiederholten Gabe von hohen Dosen Corticosteroiden während der Schwangerschaft führte bei den Feten zu einer signifikante Störung der Myelinisierung [Dunlop et al., 1997; Huang et al., 2001]. Außerdem wurde die Expression der mRNA der Proteine MBP und PLP durch Gabe von Dexamethason in der frühen postnatalen Periode unterdrückt [Tsuneishi et al., 1991]. Auch auf Oligodendrocyten-Vorläuferzellen zeigten Corticosteroide einen Effekt: In mit Corticosteron behandelten adulten Ratten ging die Proliferation von diesen Vorläuferzellen im Hippocampus drama-tisch zurück [Alonso, 2000].
In in vitro Untersuchungen hingegen hatten Glucocorticoide einen die Myelinisierung unterstützenden Effekt: die Expression der Proteine MBP und CNP sowie deren mRNA wird durch Dexamethason gesteigert [Almazan et al., 1986; Kumar et al., 1989; Byravan und Campagnoni, 1994]. Ferner konnte gezeigt werden, dass Glucocorticoide in der Entwicklung von Oligodendrocyten eine wichtige Rolle spielen: Dexamethason beispielsweise verlängert die Dauer eines Zellzyklus dieser Zellen und begüns-tigt so die Differenzierung zu reifen Oligodendrocyten [Barres et al., 1994b].
Diese Unstimmigkeit zwischen in vivo und in vitro Untersuchungen ist wahrscheinlich auf die unter-schiedliche Dosierung zurückzuführen: In weniger häufigen und niedrigeren Dosen als in den in vivo-Studien gegeben, ist ein positiver Effekt von Corticoiden auf die Myelinisierung möglich.

Einleitung 18
1.5 Zielsetzung der Arbeit Um die Rolle von Oligodendrocyten-Vorläuferzellen in der Entstehung einer periventrikulären Leu-komalazie untersuchen zu können, sollte zunächst die Isolierung und Kultivierung von Oligodendrocy-ten-Vorläuferzellen am Lehrstuhl für molekulare Neurobiochemie etabliert werden. Eine wichtige Vorraussetzung für diese Untersuchungen war es, die Zellen gezielt im Vorläuferzell-Stadium halten zu können und sie ebenso gezielt dazu anregen zu können, zu reifen Oligodendrocyten auszudifferen-zieren.
Da in den letzten Jahren Entzündungen und dabei freigesetzte Cytokine immer mehr an Bedeutung in
der Pathogenese einer PVL gewinnen, sollte der Einfluss von Cytokinen wie TNF-α und IFN-γ auf das
Überleben und die weitere Ausdifferenzierung dieser Oligodendrocyten-Vorläuferzellen untersucht und charakterisiert werden.
Bei einem negativer Einfluss der Cytokine auf Oligodendrocyten-Vorläuferzellen, war als nächster Schritt geplant, Reagenzien mit hoher klinischer Relevanz in der Perinatalmedizin (wie beispielsweise Glucocorticoide) auf eine mögliche schützende Funktion hin zu untersuchen.

Materialien 19
2 MATERIALIEN
2.1 Herkunft der Geräte
Megafuge 1.0R Heraeus Sepatech Tischzentrifuge 5415C Eppendorf ELISA-Reader 400 AT SLT-Labinstruments Inkubator (Co2) Heraeus Instruments Mastercycler Gradient Eppendorf Mastercycler Personal Eppendorf Mikroskop Axiovert35 Zeiss Kaltlichtquelle thermoDux Schüttelmaschine ES-W SM1300 Fisher Scientific SDS-PAGE-Apperatur Mini Protean II Biorad Stereolupe Zeiss; Wild Heerbrugg Sterilwerkbank clanLAF Thermomixer 5436 Eppendorf UV-VIS-Densitometer UV-1202 Shimadzu Videodensitometer Biotec Fischer Vortex-Genie II Bender & Hobein AG Warmluftschüttler HAT-Instruments
2.2 Herkunft verwendeter Materialien und Substanzen
2.2.1 Chemikalien allgemein
Acrylamid Roth Agarose Gibco BRL APS Riedel de Häen Ampicillin Appligene Biotin-16-dUTP Roche Bromphenolblau Serva BSA Boehringer Mannheim DMSO Baker dNTP’s Boehringer Mannheim EtBr Serva Immunofluor ICN Biomedicals Magermilchpulver Serva β-Mercaptoethanol Merck NP-40 Fluka NTP’s Boehringer Mannheim Phenol Baker

Materialien 20
Ponceau S Fluka Select Peptone 400 Gibco TEMED Biorad TriPure Roche Tween 20 Serva Alle weiteren nicht näher bezeichneten Chemikalien wurden von den Firmen Appigene, Baker, Fluka, Gibco BRL, Merck, New England Biolabs, Pharmacia, Riedel de Häen, Roth, Serva und Sigma bezo-gen.
2.2.2 Chemikalien für die Zellkultur
B27 (ohne Anti-Oxidantien) Gibco CNTF (rekombinant, Ratte) R&D Systems [557-NT] Corticosteron Sigma [C2505] Deoxycorticosteron Sigma [D6875] Dexamethason Sigma [D8893] DMEM Gibco FCS Gibco bFGF (rekombinant, Human) R&D Systems [233-FB] Forskolin Sigma [F3917] Ham’s F12 Biochrome IFN-γ (rekombinant, Ratte) R&D Systems [585-IF] Neurobasal-Medium (NB) Gibco PDGF-AA (rekombinant, Human) R&D Systems [221-AA] Penicillin/Streptomycin Lösung
(10000 U/ml, 10000 µg/ml) Gibco
TNF−α (rekombinant, Ratte) R&D Systems [510-RT] T3 Sigma
2.2.3 Enzyme
Dnase I, RNase-frei [1 U/µl] Roche Klenow-Enzym, labeling grade [2 U/µl] MBI Fermentas PNK MBI Fermentas Restriktionsenzyme /-Puffer MBI Fermentas RNA-Polymerase [T7/T3] [20 U/µl] MBI Fermentas RNase A Boehringer Mannheimer RNase Inhibitor Roche SAP und Puffer USB T4-DNA-Ligase und Puffer [5 U/µl] MBI Fermentas Terminale Transferase Roche Thermolase [5 U/ml] GL Biotech Vent-DNA-Polymerase New England Biolabs

Materialien 21
2.2.4 Kits und Verbrauchsmaterialien
Affigel 10 Biorad ECL-Detektionskit Amersham Deckgläschen ∅ 12 mm Laborservice Brenzinger Filme Grobo, Agfa, Curix Filterpapier Schleicher & Schüll Nucleobond PC500 Macherey-Nagel Nucleospin RNA II Macherey-Nagel Objektträger Menzel-Glaser Qiagen OneStep RT-PCR Kit Qiagen QIAEX Gel Extraction Kit Qiagen Nitrocellulosemembran BA 85 (0,45µm) Schleicher & Schüll Plastikwaren (Pipettenspitzen, Reaktions-
gefäße, Petrischalen, usw.) Eppendorf, Greiner, Nunc, Sarstedt
TriPure Roche Zellkulturflaschen Nunc
2.2.5 Antikörper
anti-Actin (affinitätsgereinigt) Sigma [A2066] anti-A2B5 Chemicon [MAB312R] anti-Maus-Alexa Fluor™ 488 Molecular Probes anti-CNPase Chemicon [MAB326R] anti-GFAP Sigma [G3893] anti-MAG Chemicon [MAB1567] anti-MBP DPC Biermann [SM1450] anti-MOSP Chemicon [MAB328] anti-Maus-HRP Dianova anti-rabbit-IgG-Peroxidase-Konjugat Sigma [A6154] anti-β-Tubulin Sigma
2.2.6 Plasmide
pBluescript II SK+ Stratagene pcDNA3 Invitrogen

Materialien 22
2.3 Primer Rat MBP-1 5’-GGATGTGATGGCATCACAGAAGAGA-3’ Rat MBP-2 5’-GGTGTACGAGGTGTCACAATGTTCT-3’ MBP-SOE-1 5’-ACAGAGACACGGGCATCCTTGACTCCCGGGGCTCTG-3’ MBP-SOE-2 5’-GAGTCCTTGCCAGAGCCCCGGGAAGTCAAGG-3’ Rat MAG-1 5’-GTCAGCTGCATGGTGCCGGACAACT-3’ Rat MAG-2 5’-CCCGCATCCAAGTCAGCAGCGGTG-3’ MAG-SOE-1 5’-GGATGAAGGCACCTGGGTGCAGGCTGTCAGGC-3’ MAG-SOE-2 5’-GGGGAAGGCAGCCTGACAGCCTGCACCCAG-3'
2.4 Bakterien C600 Genotyp: F- supE44 thi-1 thr-1 leuB6 LacY1 tonA21 Lambda- hsdR- hsdM+
XL2-blue MRF´ Genotyp: ∆(mrcA)183 ∆(mrcCB-hsdSMR-mm)173 endA1 supE44 thi-1 recA1 gyrA96 relA1 lac [F’proAB lacIq Z∆M15 Tn10(tet)r amy Camr]
2.5 Größenstandards
2.5.1 Protein-Molekulargewichtsstandard
SDS-6H Sigma (Banden bei: 205, 116, 97, 66, 45, 29 kD)
SDS-7-Dalton Mark VII L Sigma (Banden bei: 66, 45, 36, 29, 24, 21, 14 kD)
2.5.2 DNA-Größenstandard
BOA-Marker, Tth730-Plasmid mit HinfI und EcoRI geschnitten. (Banden bei: 1400, 800, 517, 396, 265, 191, 88, 78, 65 bp)
2.6 Versuchstiere Die Sprague-Dawley-Ratten wurden in der zentralen Versuchstierhaltung der Medizinischen Fakultät [ZVM] gezüchtet. 1-3-Tage alte Jungtiere wurden zur Gewinnung von Oligodendrocyten-Vorläuferzellen aus dieser Zucht genommen. Eltern-Tiere wurden von den Firmen Harlan-Winkelmann und Charles River bezogen.

Materialien 23
2.7 Puffer und Lösungen für molekularbiologische Arbeiten DEPC-H2O 0,1 % (v/v) DEPC in H2O 1h bei RT rühren und auskochen DNA- Phenol Phenol gepuffert mit TrisHCl, pH 7,9 DNA- Probenpuffer 100 mM EDTA, 20 % (w/v) Ficoll 400 0,025 % (w/v) Bromphenolblau 0,025 % (w/v) Xylencyanol in H2O dNTP Mix 10 mM dATP 10 mM dCTP 10 mM dGTP 10 mM dTTP in H2O EtBr-Stammlösung 10 mg/ml EtBr, in H2O Lösung I 25 mM Tris HCl, pH 7,9 50 mM Glukose 10 mM EDTA in H2O Lösung II 1 % (w/v) SDS 0,2 M NaOH in H2O Lösung III 3 M KAc in H2O, mit Eisessig auf pH 5,5 eingestellt Lithiumchlorid-Stammlösung 5 M LiCl in H2O RF I 100 mM RbCl 50 mM MnCl2 30 mM CH3COOK 10 mM CaCl2 15 % (v/v) Glycerin pH 5,8

Materialien 24
RF II 10 mM MOPS pH 6,8 10 mM RbCl 75 mM CaCl2 15 % (v/v) Glycerin RNase A-Stammlösung 10 mg/ml in H2O TE - Puffer 10 mM Tris HCl, pH 7,9 1 mM EDTA in H2O TEB - Puffer 89 mM Tris Borat, pH 8,0 89 mM Borsäure 2 mM EDTA in H2O TEB- Laufpuffer 0,5 µg/ml EtBr in TEB-Puffer
2.8 Puffer und Lösungen für proteinchemische Arbeiten x % Blockpuffer x % (w/v) Milchpulver in TBS-T Boehringer Lyse Puffer 50 mM Tris HCl pH 7,4 150 mM NaCl 40 mM NaF 5 mM EDTA 5 mM EGTA 1 mM Na3VO3 1 % (v/v) Nonidet P40 0,1 % (w/v) Na-Desoxycholat 0,1 % (w/v) SDS 1 mM PMSF in H2O Ponceau S Lösung 0,2 % (w/v) Ponceau S 3 % (w/v) TCA 3 % (w/v) Sulfosalicylsäure in H2O

Materialien 25
Protein-Transferpuffer 25 mM Tris HCl, pH 8,6 192 mM Glycin 20 % (v/v) Methanol 0,01 % (w/v) SDS in H2O 4x Sammelgel-Puffer 0,5 M Tris HCl, pH 6,8 0,4 % (w/v) SDS in H2O 1x SET 50 mM Tris HCl, pH 7,8 150 mM NaCl 1 mM EDTA in H2O SDS-Elektrophoresepuffer 25 mM Tris HCl, pH 8,6 192 mM Glycin 0,1 % (w/v) SDS in H2O SDS-Probenpuffer (4x) 250 mM Tris HCl, pH 6,8 8 % (w/v) SDS 40 % (v/v) Glycerin 10 % (v/v) β-Mercaptoethanol 0,04 % (w/v) Pyronin Y in H2O Stripping Puffer 68,5 mM Tris, pH 6,8 100 mM β-Mercaptoethanol 2 % (w/v) SDS in H2O TBS (T) 25 mM Tris-HCl 150 mM NaCl pH 7,5 (0,1% (v/v) Tween 20) 4x Trenngel- Puffer 1,5 M Tris HCl, pH 8,8 0,4% (w/v) SDS in H2O

Materialien 26
2.9 Puffer und Lösungen für die Immuncytochemie A2B5-Blockierlösung 5 % BSA 1 % Ziegen-Serum in PBS GFAP-Blockierlösung 1 % BSA in PBS MOSP-Blockierlösung 3 % Ziegen-Serum in PBS Fixierlösung 4 % Paraformaldehyd in PBS, pH 7.4 Permeabilisierungs-Lösung 10 % Succrose 0,5 % Nonidet P40 in PBS
2.10 Medien für die Bakterienanzucht Ampicillin-Stammlösung 50 mg/ml Ampicillin in H2O 100 µg/ml Arbeitskonzentration LB-Platten 15 g/l Select-Agar 100 mg/l Ampicillin in LB-Medium LB-Medium 10 g/l Peptonhydrolysat 5 g/l Hefehydrolysat 5 g/l NaCl ad 1 l mit H2O, pH 7,4

Materialien 27
2.11 Lösungen für die Zellkultur Differenzierungs-Medium (DM) 45 µM T3 5 µM Forskolin 10 ng/ml CNTF 1 x B27 in Neurobasal-Medium Gliamischkultur-Medium 45 % Ham‘s F12 45 % DMEM 5 % 10 % FCS Penicillin [10000 U/ml] Streptomycin [10000 µg/ml] PBS 8,1 mM Na2HPO4 138 mM NaCl 2,7 mM KCl 1,47 mM KH2PO4 Proliferations-Medium (PM) 10 ng/ml PDGF 10 ng/ml bFGF 1 x B27 in Neurobasal-Medium

Methoden 28
3 METHODEN
3.1 Zellkultur
3.1.1 Poly-D-Lysin-Beschichtung von Deckgläschen
Die Kultur der Oligodendrocyten-Vorläuferzellen erfolgte auf Poly-D-Lysin-beschichteten Deckglä-
schen. Dazu wurden die Deckgläschen (∅ 12 mm) zunächst für mehrere Stunden ausgebacken, um
dann über Nacht bei RT in Poly-D-Lysin-Lösung (0,25 mg/ml) inkubiert zu werden. Die Lösung wur-de am nächsten Tag abgenommen und die Deckgläschen dreimal mit sterilem Wasser gewaschen. Nach Trocknen der Deckgläschen im Luftstrom der Sterilbank, wurden diese bis zur weiteren Ver-
wendung bei 4°C gelagert.
3.1.2 Präparation von primären Glia-Mischkulturen aus neonatalen Ratten
[nach Mc Carthy und De Vellis, 1980]
Aus dem Hirn von 1-3 Tage alten Sprague-Dawley-Ratten wurde eine Gliamischkultur präpariert, aus welcher später Oligodendrocyten-Vorläuferzellen isoliert werden sollten.
Nach Dekapierung der Ratten wurde der Schädel geöffnet und das Hirn in gekühltes PBS überführt. Unter dem Mikroskop wurde dann die Hirnhaut entfernt. Die so gereinigten Hirne wurden in einer zweiten mit PBS gefüllten Kulturschale gesammelt. Anschließend wurden diese erst durch ein 125 µm-Sieb und dann durch ein 36 µm-Sieb gesiebt. Die zweite Kulturschale und das zweite Sieb wurden noch einmal mit 10 ml gekühltem PBS gewaschen. Die Waschlösung wurde zur Zellsuspensi-on gegeben.
Nach 10minütiger Zentrifugation bei 900 rpm (Heraeus Megafuge 1.0 R; 153 g) und 4°C wurde der
Überstand verworfen. Das Pellet wurde in Glia-Mischkultur-Medium aufgenommen und auf 75cm2-Flaschen verteilt (1½ Hirne pro Flasche). Das Gesamtvolumen pro Flasche wurde mit Glia-Mischkultur-Medium auf 10 ml eingestellt.
Die Zellen wurden bei 37ºC und 5 % CO2 kultiviert.
3.1.3 Kultivierung von primären Ratten-Glia-Mischkulturen
Nach 3-4 Tagen erfolgte der erste Mediumwechsel. Dazu wurde das alte Medium komplett abgenom-men und durch 10 ml pro Kulturflasche frisches Medium ersetzt.
Anschließend wurde alle zwei Tage das Medium gewechselt, bis nach 7-10 Tagen ein geschlossener Astrocyten-Teppich auf dem Flaschenboden gewachsen war. Zu diesem Zeitpunkt erfolgte dann die Isolierung der Oligodendrocyten-Vorläuferzellen [siehe 3.1.4].

Methoden 29
3.1.4 Isolierung von primären Oligodendrocyten-Vorläuferzellen aus Rat-ten-Glia-Mischkulturen [nach Simpson et al., 1997]
Um die Oligodendrocyten-Vorläuferzellen von den Astrocyten und Mikroglia zu trennen, bedient man sich ihrer unterschiedlichen Affinität: Während die Mikroglia nur locker an den Astrocyten haften, sind die Oligodendrocyten-Vorläuferzellen stärker an diese gebunden. Die Astrocyten selber haften mit einer sehr hohen Affinität an der Oberfläche der Kulturflaschen.
Die Kulturschalen wurden, um zunächst die Mikroglia abzulösen, mit Klebeband auf einem Schüttler im Brutschrank fixiert und für 3 Stunden bei 190 rpm geschüttelt. Der Mikroglia-haltige Überstand wurde abgenommen und verworfen. Um alle Mikroglia zu entfernen, wurden die Flaschen noch ein-mal mit je 5 ml PBS gewaschen. Anschließend wurden 10 ml Glia-Mischkultur-Medium auf die Zel-len gegeben, die Flaschen erneut auf dem Schüttler fixiert und für 16-18 Stunden bei 190 rpm geschüt-telt.
Der Überstand aus dem Schüttelvorgang wurde abgenommen, gleichmäßig auf 15 ml-Tubes verteilt und 5 min bei 1700 rpm (Heraeus Megafuge 1.0 R; 546 g) zentrifugiert. Der Überstand wurde verwor-fen und das Pellet in frischem Glia-Mischkultur-Medium aufgenommen (2 ml pro Kulturflasche). Die Zellsuspension wurde dann auf 10 ml-Kulturschalen verteilt und für 45 min im Brutschrank inkubiert. In dieser Zeit setzten sich Astrocyten auf der Oberfläche der Schale ab, während Oligodendrocyten-Vorläuferzellen im Überstand verblieben. Der Überstand wurde dann abgenommen, die Zellzahl be-stimmt und die Zellen in einer Dichte von 105 Zellen pro Deckglas auf poly-D-Lysin beschichtete Deckgläschen [siehe 3.1.1] ausgesät.
Nach einer Stunde Inkubation hatten sich die Oligodendrocyten-Vorläuferzellen auf den beschichteten Deckgläschen abgesetzt und das Glia-Mischkultur-Medium wurde durch Proliferationsmedium ersetzt. Um das Glia-Mischkultur-Medium vollständig zu entfernen, wurden die Deckgläschen einmal kurz in 100 µl Proliferationsmedium gewaschen.
3.1.5 Kultivierung von primären Oligodendrocyten-Vorläuferzellen
Der erste Mediumwechsel mit Proliferationsmedium erfolgte zeitgleich mit der ersten Behandlung der Zellen 24 Stunden nach Aussaat der Zellen.
Bei einer Kultivierung über den dritten Kulturtag bzw. die Behandlung hinaus wurde dann (nach 72 Stunden) auf Differenzierungsmedium gewechselt. Um das Proliferationsmedium bzw. die eingesetz-ten Faktoren der Behandlung vollständig zu entfernen, wurden die Deckgläschen einmal kurz in 100 µl Differenzierungsmedium gewaschen.
Bis zur Lyse oder Fixierung der Zellen erfolgte alle zwei Tage ein Mediumwechsel mit Differenzie-rungsmedium.

Methoden 30
3.1.6 Fixierung von Zellen auf Deckgläschen
Zunächst wurden die Zellen 3 mal 5 min mit PBS gewaschen. Zur Fixierung wurde 500 µl Fixierlö-sung auf jedes Deckgläschen gegeben und 15 min bei RT inkubiert. Danach wurde erneut 3 mal für 5 min mit PBS gewaschen und die Deckgläschen bis zur weiteren Verwendung in PBS bei 4°C gela-gert.
3.1.7 Immuncytochemie
Die fixierten Zellen wurden zunächst für eine Stunde in entsprechender Blockierlösung [siehe 2.9] inkubiert, um unspezifisches Binden des Erstantikörpers zu vermindern. Anschließend wurden die Zellen für eine Stunde (anti-A2B5, anti-GFAP) bzw. für zwei Stunden bei Raumtemperatur und 16
Stunden bei 4°C (anti-MOSP) in nach Herstellerangaben verdünnter Antikörperlösung inkubiert. Da-
nach wurden die Zellen drei mal 10 min in PBS + 0,5 % Ziegen-Serum gewaschen. Dann erfolgte die Inkubation mit dem entsprechenden 2. Antikörper [siehe Tabelle 1] für eine Stunde bei Raumtempera-tur in einer lichtundurchlässigen Kammer. Nach drei weiteren Waschschritten in PBS + 0,5 % Ziegen-Serum wurden die Deckgläschen mit ImmunoFluor™ auf Objektträgern aufgebracht und nach Aushär-ten unter dem Fluoreszenzmikroskop ausgewertet (25 Sichtfelder pro Deckgläschen).
1. ANTIKÖRPER VERDÜNNUNG 2. ANTIKÖRPER VERDÜNNUNG
anti-A2B5 1:200
anti-GFAP 1:400
anti-MOSP 1:3000
anti-Maus-Alexa Fluor™ 488
1:1000
Tabelle 1: Immuncytochemie - verwendete Antikörper-Konzentrationen
3.1.8 Detektion von apoptotischen Zellen (TUNEL-Methode)
[nach Vaillant et al., 1999]
Für die Detektion von apoptotischen Zellen wurden Zellen auf Deckgläschen verwendet. Nach abge-schlossener Stimulierung wurden diese zunächst fixiert [siehe 3.1.6]. Anschließend erfolgte ein 30 minütiger Permeabilisierungschritt bei RT in einer 0.5 %igen (v/v) Lösung von NP-40 in PBS.
Um eine genaue Charakterisierung des Entwicklungsstandes der apoptotischen Zellen zu erreichen, wurde zunächst eine A2B5-Immuncytochemie [siehe 3.1.7; ohne Eindeckeln] durchgeführt.
Danach wurden die Zellen für 1,5 Stunden bei 37°C mit der TUNEL-Reaktionslösung [10 µl pro
Deckgläschen; siehe Tabelle 2] inkubiert. Nach dreimaligem Waschen mit PBS wurde für 45 min mit
Cy3-gekoppeltem Extravidin (1:400 in PBS) inkubiert. In den nachfolgenden Waschschritten wurde der Waschlösung des ersten Schrittes Hoechst 33258 (1:1000 in PBS) zur Anfärbung der Zellkerne zugefügt.
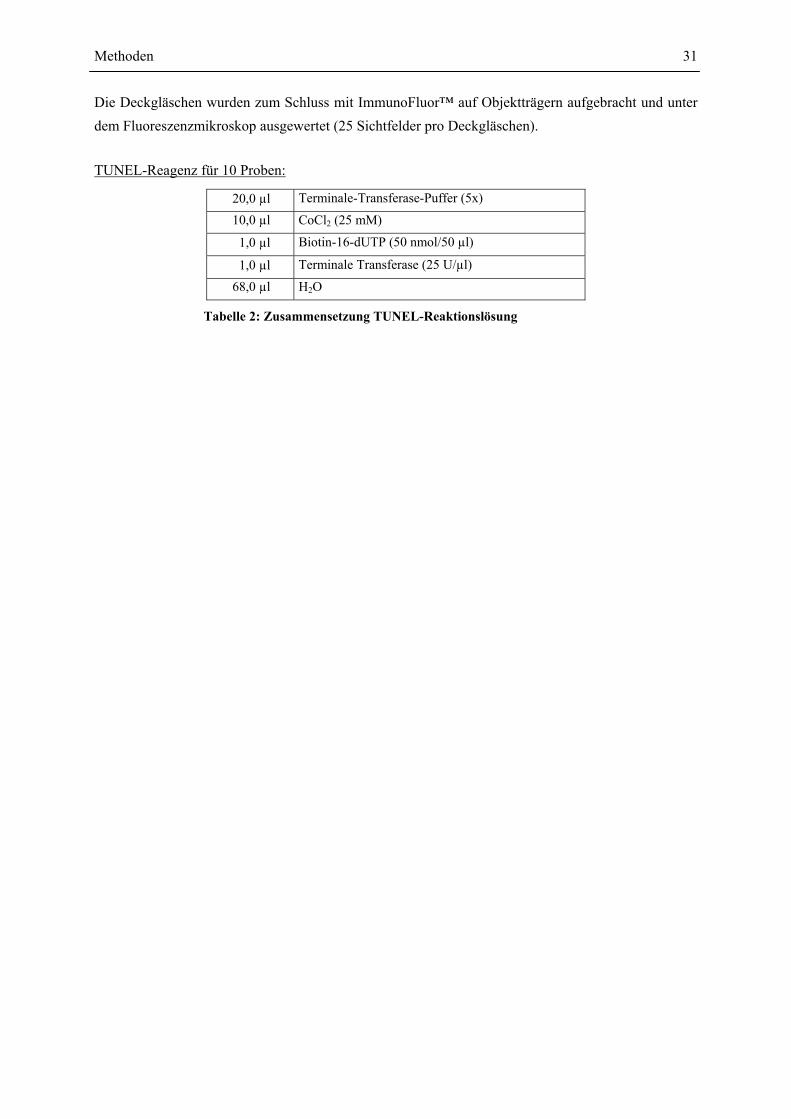
Methoden 31
Die Deckgläschen wurden zum Schluss mit ImmunoFluor™ auf Objektträgern aufgebracht und unter dem Fluoreszenzmikroskop ausgewertet (25 Sichtfelder pro Deckgläschen).
TUNEL-Reagenz für 10 Proben:
20,0 µl Terminale-Transferase-Puffer (5x)
10,0 µl CoCl2 (25 mM)
1,0 µl Biotin-16-dUTP (50 nmol/50 µl)
1,0 µl Terminale Transferase (25 U/µl)
68,0 µl H2O
Tabelle 2: Zusammensetzung TUNEL-Reaktionslösung

Methoden 32
3.2 Proteinbiochemische Methoden
3.2.1 Aufschluss von Zellen zur Gewinnung der Gesamtproteine
Zunächst wurden die zu lysierenden Zellen auf Eis gestellt. Danach wurden sie einmal kurz mit kaltem PBS gewaschen und mit Boehringer-Lysepuffer versetzt (30 µl pro 24-Well-Vertiefung). Nach 5 minütiger Inkubation wurden die Zellen abgeschabt und in ein Reaktionsgefäß überführt. Nach ei-nem Zentrifugationschritt von 5 min bei 14.000 rpm und 4°C (Eppendorf Tischzentrifuge) zum Ent-fernen der Zelldebris, wurde das geklärte Lysat in ein frisches Reaktionsgefäß überführt. Für eine langfristige Lagerung wurden die Lysate bei -20°C eingefroren.
3.2.2 Proteinbestimmung mit BioRad DC Protein Assay
Um zu gewährleisten, dass für die anschließenden Western-Blot-Analysen der Lysate gleiche Prote-inmengen verschiedener Gruppen verwendet wurden, wurde die Proteinkonzentration mit Hilfe des DC-Protein Assay Kits bestimmt:
In eine 96well Mikrotiterplatte wurden für eine Eichgerade 5 µl verschiedener BSA-Lösungen (0,1 µg/µl; 0,25 µg/µl; 0,5 µg/µl; 0,75 µg/µl; 1,0 µg/µl; 1,25 µg/µl und 1,5 µg/µl) und für die Protein-bestimmung je 5 ml der Lysate pipettiert. Dazu wurden je 20 µl der Lösung A‘ (hergestellt aus 1 ml Lösung A und 20 µl Lösung S) und 200 µl der Lösung B gegeben (Lösungen A, S und B sind Bestandteil des Kits). Nach Entfernen von Luftblasen wurde 15 min bei Raumtemperatur inkubiert. Anschließend wurden die Proben im ELISA-Reader bei 620 nm ausgelesen und die Proteinkonzentration anhand der Eichgeraden mit Hilfe von Microsoft Excel® bestimmt.
3.2.3 Diskontinuierliche SDS-Polyacrylamid Gelelektrophorese (SDS-PAGE)
Die Auftrennung von Proteinen erfolgte in einer diskontinuierlichen SDS-Polyacrylamidgelelektrophorese nach der Methode von Laemmli [Laemmli, 1970]. Die Elektrophore-sen wurden in der mini-PROTEAN-Slab Cell-Apparatur der Firma Biorad durchgeführt. Zum Gießen der X %-igen Trenn- bzw. Y %-igen Sammelgele wurden folgende Ansätze (ausreichend für zwei 1,5 mm dicke Minigele) zusammenpipettiert:
X %iges Trenngel (15 ml Ansatz) Y %iges Sammelgel (6 ml Ansatz)
11,25–0,5 x X ml Wasser 4,37-0,2 x Y ml Wasser
3,75 ml Trenngel-Puffer (4 x) 1,5 ml Sammelgel-Puffer (4 x)
0,5 x X ml Acrylamid-Stammlösung 0,2 x Y ml Acrylamid-Stammlösung
150 µl 10 % SDS 60 µl 10 % SDS
100 µl 10 % APS 60 µl 10 % APS
10 µl TEMED 10 µl TEMED

Methoden 33
APS und TEMED - die Chemikalien, welche die chemische Polymerisation einleiten - wurden unmit-telbar vor dem Gießen des Gels zugesetzt.
Nach dem Gießen wurde das Trenngel vorsichtig mit 75 %igem Ethanol überschichtet. Das Ende der Polymerisation zeigt sich nach ca. 10-20 min, durch eine scharfe Grenzlinie zwischen Wasser und Alkohol. Der Überstand wurde abgegossen und weitestgehend entfernt. Nun wurde das Sammelgel gegossen, wobei darauf geachtet wurde, dass sich am Teflonkamm, der die Probentaschen formte, keine Luftblasen bildeten. Die Polymerisation war nach ca. 15 min abgeschlossen. Der Kamm wurde unter Elektrophorese-Puffer vorsichtig herausgezogen.
Proteinproben wurden mit 4 x Laemmli-Probenpuffer versetzt. Vor der Elektrophorese wurden die
Proben zum Denaturieren der Proteine 5 min auf 95°C erhitzt und anschließend im Eisbad gekühlt. Bei
Lysaten, die unter nicht-reduzierenden Bedingungen aufgetrennt werden sollten, enthielt der verwen-
dete Laemmli-Probenpuffer kein β-Mercaptoethanol. Außerdem wurden diese Proben nicht aufge-
kocht. Die Elektrophorese erfolgte bei 100 V, während die Proben das Sammelgel durchwanderten. Danach wurde bei 150 V elektrophoriert, bis der Farbstoff Pyronin Y das Ende des Trenngels erreicht hatte.
3.2.4 Western-Blot
Das Polyacrylamidgel sowie eine entsprechend zurechtgeschnittene Nitrocellulosemembran wurden in Vorbereitung auf den Blot zunächst 10 min in Protein-Transferpuffer inkubiert.
Für den elektrophoretischen Proteintransfer auf die Nitrocellulosemembran wurde unter Transferpuffer ein ‚Sandwich’ zusammengelegt, bestehend aus:
1. 2 dicken Filterpapieren,
2. dem Gel,
3. der Nitrocellulosemembran,
4. dickem Filterpapier und
5. einer Schaumstoffmatte.
Dieses ‚Sandwich’ wurde in die Mini-PROTEAN-Slab-Cell-Apparatur von Biorad eingespannt. Die Nitrocellulosemembran war dabei der Anode, das Gel der Kathode zugewandt.
Das Blotten auf die NC-Membran erfolgte unter Rühren auf dem Magnetrührer entweder für 2 Stun-den bei 100 V oder über Nacht bei 30 V.
Wurde bei 100 V geblottet, kühlte man die Anlage durch ein in den Transferpuffer eintauchendes Kühlaggregat, welches nach einer Stunde durch ein neues ersetzt wurde; Blotten bei 30 V hingegen erfolgte bei Raumtemperatur.

Methoden 34
3.2.5 Ponceau-Färbung von Proteinen
Um den Proteintransfer beim Western-Blot kontrollieren und die Markerbanden auf der Nitrocellulose markieren zu können, wurden die transferierten Proteine reversibel mit Ponceau-S-Lösung angefärbt. Dazu wurde die Membran nach dem Western-Blot 10 min in Ponceau-S-Lösung geschwenkt und an-schließend so lange mit Wasser gewaschen, bis die Proteinbanden deutlich zu erkennen waren. Die Markerbanden wurden markiert und zur Dokumentation wurde zusätzlich eine Kopie der angefärbten Nitrocellulose angefertigt. Dann konnte die Membran durch Waschen in TBST vollständig entfärbt werden.
Die Membranen wurden entweder direkt weiter behandelt oder bis auf weiteres bei 4°C gelagert.
3.2.6 Immunodetektion
Bei der Immunoblot-Analyse wird ein Protein, das in einem SDS-Polyacrylamidgel aus einer komple-xen Proteinprobe aufgetrennt wurde, durch die spezifische Reaktion mit einem mono- oder polyklona-len Antikörpern, der gegen dieses Protein gerichtet ist, detektiert. Die Detektion erfolgt in der Regel nicht unmittelbar, sondern indirekt mit Hilfe eines zweiten Antikörpers, der aus einem anderen Tier gewonnen wurde und gegen die konstante Region des ersten Antikörpers gerichtet ist. Dieser Zweitan-tikörper ist mit einem Enzym gekoppelt, welches mit einem zugesetzten Substrat reagiert, dessen Re-aktionsprodukte ein optisches Signal liefern.
In dieser Arbeit wurde das ECL (Enhanced ChemiLuminescence)-Detektionssystem der Firma Amers-ham verwendet:
Die an den Zweitantikörper gekoppelte Peroxidase katalysiert in Gegenwart von Wasserstoffperoxid die Chemolumineszenzreaktion eines Luminols. Exponiert man den Immunoblot einem lichtempfind-lichen Film, so zeigt sich an den Stellen, an denen sich der Zweitantikörper befindet, eine Schwär-zung.
Zunächst wurde die NC-Membran 1 Stunde mit 5 % Blockpuffer inkubiert. In diesem Schritt sollten unspezifische Bindungsstellen für den Erstantikörper durch Milchproteine abgesättigt werden.
Der Blot wurde 3 mal 10 min mit TBS-T gewaschen und mit dem Erstantikörper (in geeigneter Ver-dünnung, siehe Tabelle 3) 1 Stunde bei Raumtemperatur unter Schütteln inkubiert. Nach gründlichem Waschen (3 mal 10 min in TBS-T schütteln) folgte die Inkubation mit dem zweiten Antikörper, eben-falls für eine Stunde bei Raumtemperatur unter Schütteln. Der Blot wurde erneut gründlich in TBS-T gewaschen (mind. 3 mal 10 min), zwischen zwei Lagen Filterpapier getrocknet und 1 min in frisch zubereiteter ECL-Lösung (1:1 Mischung von Lösung 1 und 2, Fa. Amersham) geschüttelt.
Nachdem überschüssige ECL-Lösung von der NC-Membran abgetropft war, wurde der Blot mit der Proteinseite nach unten luftblasenfrei auf eine durchsichtige Folie aufgelegt und verpackt.
In der Dunkelkammer wurde mit dem Blot ein Film (Hyperfilm-ECL, Fa. Amersham) belichtet, der anschließend in konventioneller Weise entwickelt wurde.
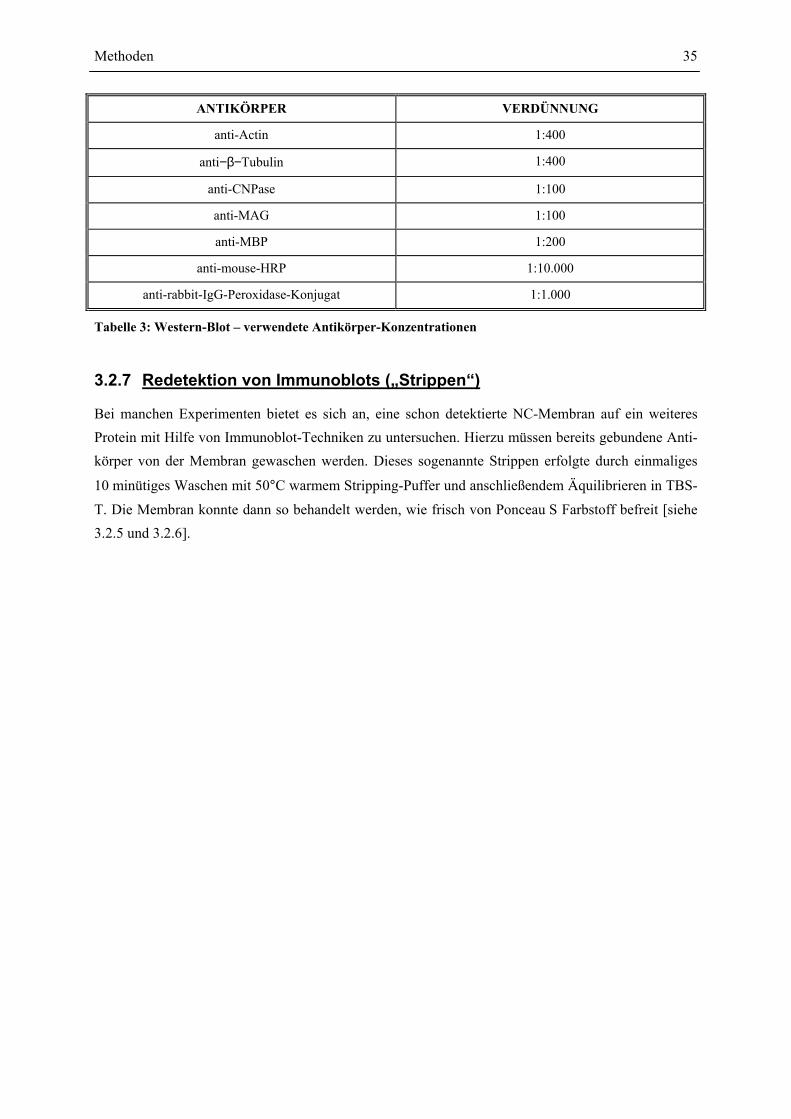
Methoden 35
ANTIKÖRPER VERDÜNNUNG
anti-Actin 1:400
anti−β−Tubulin 1:400
anti-CNPase 1:100
anti-MAG 1:100
anti-MBP 1:200
anti-mouse-HRP 1:10.000
anti-rabbit-IgG-Peroxidase-Konjugat 1:1.000
Tabelle 3: Western-Blot – verwendete Antikörper-Konzentrationen
3.2.7 Redetektion von Immunoblots („Strippen“)
Bei manchen Experimenten bietet es sich an, eine schon detektierte NC-Membran auf ein weiteres Protein mit Hilfe von Immunoblot-Techniken zu untersuchen. Hierzu müssen bereits gebundene Anti-körper von der Membran gewaschen werden. Dieses sogenannte Strippen erfolgte durch einmaliges
10 minütiges Waschen mit 50°C warmem Stripping-Puffer und anschließendem Äquilibrieren in TBS-
T. Die Membran konnte dann so behandelt werden, wie frisch von Ponceau S Farbstoff befreit [siehe 3.2.5 und 3.2.6].

Methoden 36
3.3 Molekulargenetische Methoden
3.3.1 Isolierung von Gesamt-RNA
Die Isolierung von Gesamt-RNA aus frisch präpariertem Ratten-Hirn erfolgte mit Hilfe des Nucle-ospin RNA II Kits der Firma Macherey-Nagel gemäß Herstellerangaben. Die RNA wurde für die Klo-nierung der Positivkontrolle für die semiquantitative RT-PCR verwendet.
Die RNA aus Zellen der Oligodendrocyten-Zellkultur wurde mit Hilfe des TriPure Reagenz der Firma Roche entsprechend der Anleitung des Herstellers isoliert. Die RNA wurde bis zur Verwendung in der
semiquantitativen RT-PCR bei -80° C gelagert.
3.3.2 RT (Reverse Transcription)-PCR
Die RT-PCR (Reverse Transkription mit anschließender PCR-Reaktion) diente zur Amplifizierung von cDNA aus gesamt-RNA. Diese Reaktion wurde mit Hilfe des Qiagen One-Step-RT-PCR-Kit durchgeführt. Entgegen der beigefügten Anleitung wurde dabei je Reaktion ein 25 µl-Ansatz vorberei-tet und anschließend dem RT-PCR-Programm im Mastercycler-Gradient der Firma Eppendorf unter-zogen.
Programm für RT-PCR:
1 T=50°C t=30 min
2 T=95°C t=15 min
3 T=94°C t=1 min
4 T=AT t=1 min 5 T=72°C t=1 min
6 goto 3 rep 25 7 T=72°C t=10 min
8 Hold 4°C
Die Annealing-Temperatur AT wurde zunächst aus den Primersequenzen berechnet und anschließend mit Hilfe des Mastercycler-Gradient experimentell optimiert.
3.3.3 Semi-quantitative RT-PCR
Die semi-quantitative RT-PCR wurde ebenfalls mit dem One-Step-RT-PCR-Kit der Firma Qiagen durchgeführt. Auch hier wurde je Reaktion nur ein 25 µl-Ansatz vorbereitet. Die semi-quantitative RT-PCR unterschied sich in soweit von der RT-PCR, dass neben der zu untersuchenden RNA auch noch ein Standard einer bekannten Konzentration zugefügt wurde. Dadurch ergab sich für jede Reak-tion folgendes Pipettierschema:

Methoden 37
5,0 µl Puffer (10x)
5,0 µl Lösung Q
2,5 µl Primer Mix (100 µM)
1,0 µl dNTP-Mix (10 mM)
5,0 µl RNA-Matrize (50 ng)
5,0 µl Standard-RNA
1,0 µl Polymerase (1 U/µl)
0,2 µl RNase Inhibitor 0,3 µl H2O
Das PCR-Programm entsprach dem unter 3.3.2 beschriebenen.
3.3.4 PCR (Polymerase Chain Reaction)
Die PCR ist eine Technik, mit der unter den richtigen Reaktionsbedingungen gezielt DNA-Abschnitte amplifiziert werden können. Für die Klonierung des Standards für die semiquantitative RT-PCR wur-de in folgendem Ansatz aus dem Plasmid, welches die Posititv-Kontrolle enthielt, je ein Fragment für die spätere SOE-PCR (siehe 3.3.5) amplifiziert:
2,5 µl Puffer (10x)
2,5 µl Primer-Mix (100 µM)
2,5 µl dNTP-Mix (1,25 mM)
5,0 µl MgCl2 (25 mM) 5,0 µl DNA-Matrize
0,2 µl Thermolase (5 U/ml)
7,3 µl H2O
Um RNA auf einen mögliche DNA-Kontamination zu untersuchen, wurde anstelle der DNA-Matrize entsprechend RNA eingesetzt. Die Reaktionsansätze wurden in einem Mastercycler Personal der Fir-ma Eppendorf dem folgenden Programm unterzogen:
1 T=94°C t=5 min
2 T=94°C t=1 min
3 T=AT t=1 min 4 T=72°C t=1 min
5 goto 2 rep 25 6 T=72°C t=10 min
7 Hold 4°C

Methoden 38
3.3.5 SOE (splicing by overlap extension)-PCR [nach Horton R.M. et al., 1988]
DNA-Fragmente, die miteinander verbunden werden sollten, wurden in separaten PCR-Reaktionen amplifiziert, wobei die Primer an den zusammenzufügenden Enden einander teilweise komplementär waren. In einer SOE (splicing by overlap extension)-Reaktion wurden diese Produkte dann gemischt und während der PCR-Reaktion [Programm siehe 3.3.4] miteinander verbunden.
Pipettierschema:
2,5 µl Thermo-Pol-Puffer (10x)
2,5 µl Primer (1.Fragment, N-terminal) (100 µM)
2,5 µl Primer (2.Fragment, C-terminal) (100 µM)
2,5 µl dNTP-Mix (1,25 mM)
2,5 µl Fragment 1 (1 ng/µl)
2,5 µl Fragment 2 (1 ng/µl)
1,0 µl VentTM-Polymerase (1 U/µl)
9,0 µl H2O
3.3.6 Gelelektrophorese von DNA
Die Auftrennung von DNA-Fragmenten erfolgte durch Elektrophorese in 1-2,5 %igen Agarose-Gelen. Dazu wurde die Agarose in einer entsprechenden Menge an TEB-Puffer durch Aufkochen in der Mik-rowelle gelöst und in eine Elektrophorese-Apparatur gegossen. Nach Erkalten der Agarose-Lösung wurde die Apparatur mit TEB-Laufpuffer gefüllt und die DNA-Proben, versetzt mit ¼ Volumen 5 x DNA-Probenpuffer, in die Taschen des Gels geladen. Die Elektrophorese erfolgte dann bei 4,5-6V pro cm Elektrodenabstand.
Das Ethidiumbromid im TEB-Laufpuffer diente als interkalierendes Agens zur Sichtbarmachung der DNA-Banden unter UV-Licht.
3.3.7 Isolierung von DNA-Fragmenten aus Agarosegelstücken
Die Isolierung von DNA-Fragmenten aus Agarose-Gelen erfolgte mit Hilfe des QIAEX Gel Extraktion Kit der Firma Qiagen nach Vorschrift des Herstellers.
3.3.8 Linearisierung von Plasmiden
Für die Linearisierung von Plasmiden wurden 10 µg Plasmid eingesetzt und mit dem entsprechenden Restriktionsenzym/-puffer versetzt. Nach 2 Stunden wurde 1/10 des Ansatzes auf einem 1 %igen Aga-rose-Gel analysiert. Nach einer weiteren Stunde wurde die Linearisierung mittels einer Phenol-Chloroform-Extraktion mit

Methoden 39
1. 100 µl Phenol (mit einem Tropfen Chloroform) 2. 50 µl Phenol und 50 µl Chloroform 3. 100 µl Chloroform
beendet und das linearisierte Plasmid nach Zugabe von 1/5 Vol. LiCl (8 M) sowie dem 3 fachen Vol.
an absolutem Ethanol bei -20ºC (mindestens 1 Stunde) gefällt.
Anschließend wurde die DNA 10 min bei 4ºC und 15800 g sedimentiert, mit 200 µl 70 %igem Ethanol
gewaschen und in 50 µl DEPC-H2O aufgenommen.
3.3.9 Dephosphorylierung von DNA
Mit Restriktionsendonukleasen geöffnete Vektoren werden dephosphoryliert, damit die Ligase keinen einfachen „Ringschluss“ katalysiert.
Dazu wurde folgender Ansatz für 1-1,5 Stunden bei 37ºC in dem von der Herstellerfirma empfohlenen Puffer inkubiert:
Pipettierschema:
X µl linearisiertes Plasmid 3 µl 10x SAP Puffer 0,5 µl SAP (5 U/µl)
ad 30 µl H2O
Die Reaktion wurde dann durch 30 minütige Inkubation bei 65ºC abgestoppt. Anschließend erfolgte eine Phenol-Chloroform-Extraktion mit anschließender Plasmid-Fällung [siehe 3.3.8].
3.3.10 Phosphorylierung mit T4 Phospho-Nukleotid-Kinase (PNK)
Dieses Enzym katalysiert den Transfer des γ-Phosphats von ATP zum 5’-OH-Terminus von DNA.
Diese Reaktion wird benötigt, um mit Hilfe von PCR amplifizierte Fragmente zu phosphorylieren, wenn sie anschließend in blunt-end geschnittene, dephosphorylierte Vektoren einkloniert werden sol-len.
Pipettierschema:
2 µl zu phosphorylierende DNA (2 µg) 1 µl ATP (10 mM)
20 U PNK 5 µl X Puffer
ad 25 µl H2O
Es folgte eine Inkubation von 1,5 Stunden bei 37°C; die Reaktion wurde nach Erhöhen der Temperatur
auf 75°C durch eine 20 minütige Inkubation und anschließendes Schockkühlen auf Eis abgebrochen.

Methoden 40
3.3.11 Ligation von DNA
Das DNA-Fragment und der geöffnete Vektor können sich aneinander lagern und durch eine DNA-Ligase, welche die Bildung einer Phosphodiesterbindung zwischen zwei DNA-Ketten katalysiert, verknüpft werden. Die DNA-Ligase benötigt eine freie OH-Gruppe am 3'-Ende der einen DNA-Kette, eine Phosphatgruppe am 5'-Ende der anderen sowie eine Energiequelle wie ATP.
Für eine erfolgreiche Ligation wurde zunächst auf einem 1 %igen Agarose-Gel das Mengenverhältnis des Vektors zum Fragment abgeschätzt, um dann das Fragment im Überschuss einzusetzen. Für die Reaktion wurden dann folgende Komponenten zusammen gegeben:
Pipettierschema:
x µl Klonierungsvektor y µl DNA-Fragment 1 µl dATP (10 mM) 1 µl 10xLigase-Puffer 1 µl Ligase
ad 10µl H2O.
Nach Mischen wurde dann über Nacht bei 4ºC inkubiert.
3.3.12 Herstellung kompetenter Bakterien nach der RbCl2-Methode
Zunächst wurden in 5 ml LB-Medium mehrere Bakterien-Einzelkolonien einer Glycerin-Stock-
Lösung bei 190 rpm im Warmluftschüttler (37°C) über Nacht inkubiert. Mit 150 µl dieser Bakterien-
suspension wurde dann eine 150 ml Kultur angeimpft und bei 37°C im Warmluftschüttler bis zu einer
optischen Dichte von OD600 = 0,35 – 0,6 (idealerweise OD600 < 0,4) wachsen gelassen.
Das Wachstum der Bakterien wurde durch 5 minütige Inkubation auf Eis abgestoppt. Die Bakterien
wurden dann durch 10 minütige Zentrifugation bei 1700 g / 4°C sedimentiert. Nach vorsichtiger und
vollständiger Resuspendierung in 60 ml RF1-Lösung (4°C, vorgekühlt) wurde 15 min auf Eis inku-
biert und erneut bei 1700 g / 4°C zentrifugiert.
Das Sediment wurde vorsichtig in 12 ml RF2 resuspendiert (4°C, vorgekühlt), aliquotiert und in flüs-
sigem Stickstoff schockgefroren. Die Lagerung der Aliquots erfolgte bei -80°C.
3.3.13 Transformation kompetenter Bakterien
Für jede Transformation wurden 100 µl kompetenter Bakterien für 20 min auf Eis aufgetaut. Damit sich ein Verhältnis Bakterien zu DNA von 2:1 ergab, wurde die DNA-Lösung (100-200 ng Plasmid-DNA oder ein Teil des Ligationsansatzes) mit Wasser auf 50 µl aufgefüllt und zu 100 µl kompetenten Bakterien gegeben.

Methoden 41
Nach einer Inkubation von 30 min bei 0°C wurden die Bakterien einem Hitzeschock von 42 °C (90 s)
unterzogen und im Eisbad für 2 min gekühlt. Nach Zugabe von 500 µl LB-Medium wurde der Trans-
formationsansatz 30 min bei 37°C im Schüttler inkubiert.
Nach Sedimentation der Bakterien bei 320 g/3 min in einer Tischzentrifuge wurden 500 µl des Über-standes wieder abgenommen, das Bakteriensediment resuspendiert und auf LB-Agar-Platten (50 µg/ml Amp, da die zur Transformation eingesetzten Vektoren das Ampicillin-Resistenzgen als Marker ent-
hielten) ausgestrichen. Die Platten wurden bei 37°C inkubiert, bis nach etwa 12-16 Stunden Kolonien
sichtbar wurden.
3.3.14 Plasmidpräparation im analytischen Maßstab
Um nach einer Transformation von Bakterien mit rekombinanten Plasmiden diejenigen Bakterienko-lonien zu identifizieren, die das gewünschte Plasmid tragen, wurden zunächst von einer geeigneten Anzahl willkürlich gewählter Bakterienkolonien 5 ml-Kulturen beimpft.
Dazu wurde mit einer sterilen Pipettenspitze eine vereinzelt gewachsene Bakterienkolonie angesto-chen und in ein Reagenzglas mit 5 ml LB-Medium (100 µg/ml Amp) überführt. Die Kultur wurde
dann im Schüttelinkubator bei 37°C und 190 rpm über Nacht herangezogen.
Die erhaltenen Bakterienkulturen wurden in 1,5 ml Schritten in ein Eppendorf-Reaktionsgefäß über-führt und jeweils 1 min bei 15800 g zentrifugiert. Das erhaltene Bakteriensediment wurde in 200 µl Lösung 1 aufgenommen und durch Zugabe von 450 µl Lösung 2 lysiert. Nach Mischen mit 300 µl eisgekühlter Lösung 3 wurde die ausgefallene chromosomale DNA durch Zentrifugation bei 15800 g für 5 min sedimentiert.
Der Überstand wurde mit dem 0,6 fachen Volumen an Isopropanol versetzt und 5 min bei RT inku-biert. Durch 5 minütige Zentrifugation bei 15800 g und RT wurde das Plasmid-DNA-Sediment mit 1 ml 70 %igem Ethanol gewaschen, kurz unter Vakuum getrocknet und in 100 µl TE-Puffer aufge-nommen.
15 µl dieser Lösung wurden für eine Testrestriktion genutzt [siehe 3.3.16]. Der Rest der Lösung wurde nach Zugabe des 1,5 fachen Volumens an Wasser sowie des 1,5 fachen Volumens an PEG-Lösung für
mindestens 2 Stunden bei -20°C inkubiert. Bei einer positiven Testrestriktion wurde dann bei 15°C
und 20800 g 30 min zentrifugiert, das Sediment wie zuvor beschrieben mit 70 %igem Ethanol gewa-schen und im Vakuum getrocknet. Die aufgereinigte Plasmid-DNA wurde in 20 µl TE-Puffer re-suspendiert und zur Sequenzierung abgegeben.
3.3.15 Plasmidpräparation im präparativen Maßstab
Zunächst wurden mit 100 µl einer 5 ml-Bakterienkultur 100 ml LB-Medium (100 µg/ml Ampicillin) beimpft und über Nacht im Warmluftschüttler (180 U/min) inkubiert.
Die Bakterien wurden in aufeinanderfolgenden Zentrifugationsschritten (je 10 min bei 3023 g und
4°C) in einem 50 ml-RG sedimentiert und in 8 ml Lösung 1 aufgenommen. Nach Zugabe von 16 ml

Methoden 42
Lösung 2 wurde unter gelegentlichem Umschwenken 10 min bei Raumtemperatur inkubiert. Schließ-lich wurde die Lösung mit 12 ml eisgekühlter Lösung 3 versetzt und 15 min auf Eis inkubiert. An-
schließend wurde 10 min bei 3023 g und 4°C zentrifugiert und der Überstand nach Filtration durch
einen Faltenfilter mit 0,6 Volumina Isopropanol versetzt. Es folgte eine Inkubation von mindestens 20 min bei Raumtemperatur, während der die Plasmid-DNA gefällt wurde.
Die DNA wurde mittels Zentrifugation (3032 g / 4°C / 10 min) sedimentiert und in 1,6 ml TE-Puffer
aufgenommen. Die Lösung wurde auf 2,5 M Ammoniumacetat eingestellt und nach 15 minütiger In-
kubation bei 0°C erneut bei 4000 rpm und 4°C zentrifugiert. Die ausgefallene RNA bildete das Sedi-
ment, welches verworfen wurde. Der Überstand wurde mit dem 2,5 fachen Volumen absoluten Etha-
nols gemischt und mindestens 30 min bei -20°C inkubiert.
Die so gefällte Plasmid-DNA wurde sedimentiert (3023 g / 4°C / 10 min), mit 1 ml 70%igem Ethanol
gewaschen und in 0,4 ml TE-Puffer aufgenommen. Anschließend wurde bei 37°C ein einstündiger
RNase-Verdau durch Einstellen auf eine RNase-Konzentration von 50 µg/ml RNase A durchgeführt.
Durch Extraktion mit zweimal 1 Vol. Phenol und 1 Vol. Chloroform und danach einmal mit 1 Vol. Chloroform wurde der Verdau gestoppt. Aus dem wässrigen Extraktionsüberstand wurde durch Zuga-be von 1/5 Volumen Ammoniumacetat-Lösung (10 M) und dem 2,5 fachen Volumen absolutem Etha-
nols die Plasmid-DNA 20 min bei -20°C gefällt.
Die Plasmid-DNA wurde 15 min bei 15800 g und 4°C sedimentiert, mit 200 µl 70 %igem Ethanol
gewaschen und nach Trocknen in 150 µl TE-Puffer aufgenommen. Nach UV-photometrischer Be-stimmung der DNA-Konzentration [siehe 3.3.18] wurde die Lösung auf 1 µg/µl verdünnt und bei
-20°C gelagert.
3.3.16 Restriktion von DNA
Mittels Restriktion von Plasmid-DNA mit den entsprechenden Restriktionsendonukleasen kann ein DNA-Fragment aus dem zirkulären DNA-Strang freigesetzt werden. Durch eine anschließende Agaro-se-Gelelektrophorese kann dann dieses Fragment von der restlichen Plasmid-DNA getrennt werden [siehe 3.3.6].
Die Restriktion erfolgte jeweils mit den gewünschten Restriktionsenzymen nach Angaben des Herstel-lers in den zugehörigen Puffersystemen beim enzymspezifischen Temperaturoptimum.
Um die Anzahl der benötigten Units pro µg Plasmid zu erhalten, benutzt man folgende Formel:
)(Re)()(Re)(
λλ
µ stellenstriktionsPlasmidbpPlasmidstellenstriktionsbp
hgUnits
××=
×

Methoden 43
3.3.17 In vitro-Transkription
Die in vitro-Transkription dient zur Herstellung der Standard-RNA für die semiquantitative RT-PCR. Die entsprechende cDNA ist in ein Plasmid einkloniert, welches mit einem Restriktionsenzym lineari-siert wird, dessen Schnittstelle vom RNA-Polymerase-Promotor aus hinter dem einklonierten DNA-Fragment liegt.
Folgender Reaktionsansatz wurde auf Eis zusammen pipettiert und 90 min bei 37°C inkubiert:
Pipettierschema:
12 µl DEPC-H2O
6 µl Transkriptionspuffer (5x)
5 µl linearisiertes Plasmid
3 µl NTPs (2,5 mM)
1 µl RNase Inhibitor (40 U/µl)
3 µl RNA-Polymerase (20 U/µl)
Nach Zugabe von 1 µl DNAse (RNase-frei; 1 U/µl) wurde 15 min bei 37°C inkubiert. Anschließend
wurden durch eine 45 minütige Minidialyse (Millipore-Filter 0,025 µm) überschüssige NTPs entfernt.
Der Ansatz wurde dann abgenommen, mit DEPC-H2O auf 100 µl aufgefüllt und mit 1 µl RNase Inhi-bitor (40 U/µl) versetzt. Nach Entnahme von je 5 µl für eine Gelanalyse und die Konzentrationsbe-
stimmung wurde die RNA bei -80°C verwahrt.
3.3.18 UV-Photometrische Bestimmung der DNA bzw. RNA-Konzentration
Für die Konzentrationsbestimmung wurden 5 µl DNA bzw. RNA mit 95 µl H2O bzw. DEPC-H2O versetzt und gut durchmischt. Von dieser Lösung wurde dann an einem Photometer die Extinktion bei 260 nm in einer Quarzküvette gemessen. Der Nullausgleich erfolgte für DNA-Messungen mit H2O und für RNA-Messungen mit DEPC-H2O. Bei einer Schichtdicke von 1 cm erhält man mit folgender Formel den Wert der Konzentration:
100020)260(
/ionsfaktorKonzentratnmOD
lgc ××=
µµ
Der Konzentrationsfaktor beträgt dabei für DNA 50 ng/ml und für RNA 40 ng/ml.
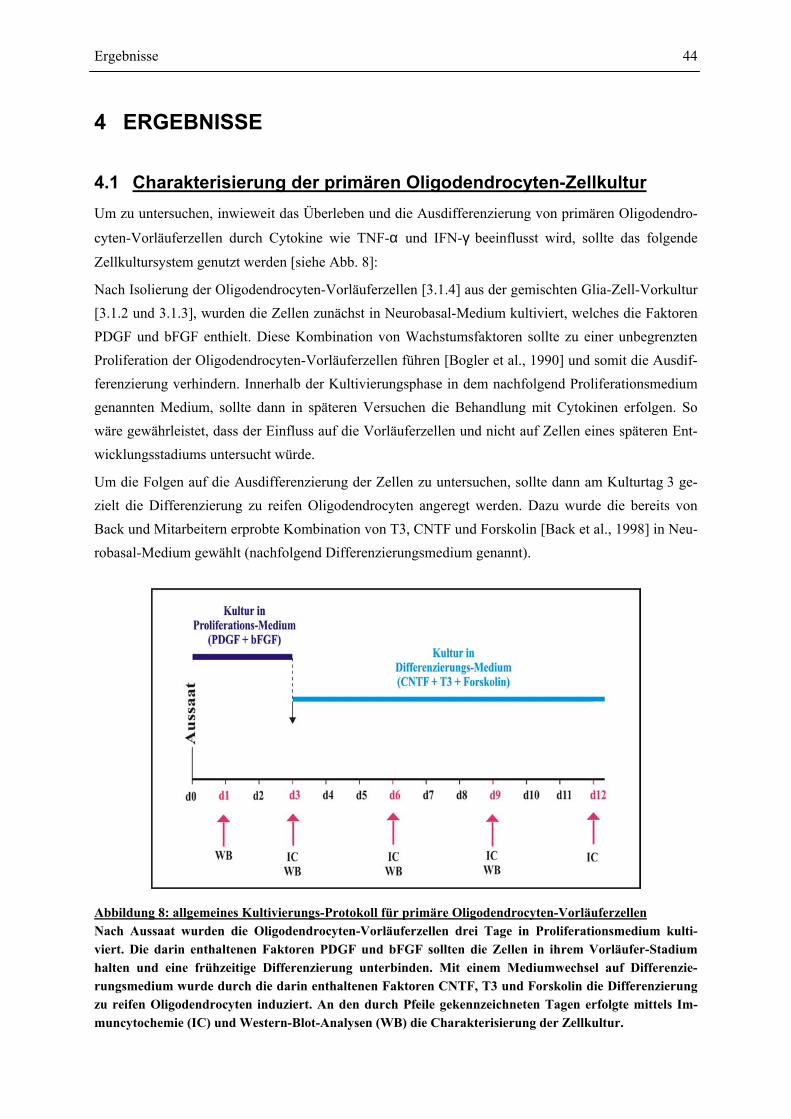
Ergebnisse 44
4 ERGEBNISSE
4.1 Charakterisierung der primären Oligodendrocyten-Zellkultur Um zu untersuchen, inwieweit das Überleben und die Ausdifferenzierung von primären Oligodendro-
cyten-Vorläuferzellen durch Cytokine wie TNF-α und IFN-γ beeinflusst wird, sollte das folgende
Zellkultursystem genutzt werden [siehe Abb. 8]:
Nach Isolierung der Oligodendrocyten-Vorläuferzellen [3.1.4] aus der gemischten Glia-Zell-Vorkultur [3.1.2 und 3.1.3], wurden die Zellen zunächst in Neurobasal-Medium kultiviert, welches die Faktoren PDGF und bFGF enthielt. Diese Kombination von Wachstumsfaktoren sollte zu einer unbegrenzten Proliferation der Oligodendrocyten-Vorläuferzellen führen [Bogler et al., 1990] und somit die Ausdif-ferenzierung verhindern. Innerhalb der Kultivierungsphase in dem nachfolgend Proliferationsmedium genannten Medium, sollte dann in späteren Versuchen die Behandlung mit Cytokinen erfolgen. So wäre gewährleistet, dass der Einfluss auf die Vorläuferzellen und nicht auf Zellen eines späteren Ent-wicklungsstadiums untersucht würde.
Um die Folgen auf die Ausdifferenzierung der Zellen zu untersuchen, sollte dann am Kulturtag 3 ge-zielt die Differenzierung zu reifen Oligodendrocyten angeregt werden. Dazu wurde die bereits von Back und Mitarbeitern erprobte Kombination von T3, CNTF und Forskolin [Back et al., 1998] in Neu-robasal-Medium gewählt (nachfolgend Differenzierungsmedium genannt).
Abbildung 8: allgemeines Kultivierungs-Protokoll für primäre Oligodendrocyten-Vorläuferzellen Nach Aussaat wurden die Oligodendrocyten-Vorläuferzellen drei Tage in Proliferationsmedium kulti-viert. Die darin enthaltenen Faktoren PDGF und bFGF sollten die Zellen in ihrem Vorläufer-Stadium halten und eine frühzeitige Differenzierung unterbinden. Mit einem Mediumwechsel auf Differenzie-rungsmedium wurde durch die darin enthaltenen Faktoren CNTF, T3 und Forskolin die Differenzierung zu reifen Oligodendrocyten induziert. An den durch Pfeile gekennzeichneten Tagen erfolgte mittels Im-muncytochemie (IC) und Western-Blot-Analysen (WB) die Charakterisierung der Zellkultur.

Ergebnisse 45
Bevor eine Untersuchung des Effektes von Cytokinen auf diese Zellkultur möglich war, musste sie zunächst mittels Immuncytochemie [3.1.7] und Western-Blot-Analysen [3.2.4 – 3.2.6] an den in Ab-bildung 8 angegebenen Tagen charakterisiert werden. Außerdem sollte gezeigt werden, dass der Anteil an Astrocyten, welche bekanntermaßen selber Cytokine freisetzen können, vernachlässigbar war.
4.1.1 Entwicklung der Oligodendrocyten-Vorläuferzellen: A2B5-Immuncytochemie
Oligodendrocyten-Vorläuferzellen sind bipolare Zellen, welche in vitro durch Immuncytochemie mit dem monoclonalen Antikörper A2B5 identifiziert werden können [Fredman et al., 1984; Dubois et al., 1990]. Unter den in Abschnitt 4.1. beschriebenen Bedingungen war zu erwarten, dass die Zellen in den ersten drei Tagen positiv mit diesem Antikörper reagieren, während nach dem Wechsel auf Differen-zierungsmedium mit zunehmendem Grad der Differenzierung diese Immunreaktivität abnimmt. Um dies zu untersuchen, wurden die Zellen auf Deckgläschen kultiviert und an den Kulturtagen 3, 6, 9 und 12 mit 4 %iger Paraformaldehyd-Lösung fixiert [3.1.1]. Oligodendrocyten-Vorläuferzellen wurden dann mit dem Antikörper A2B5 detektiert [3.1.7].
Wie in Abbildung 9 [S. 46] zu sehen ist, zeigen die Zellen am Kulturtag 3 die für Oligodendrocyten-Vorläuferzellen typische bipolare Morphologie [siehe Abb. 9A]. Gleichzeitig reagiert der größte Teil der Zellen positiv mit dem Antikörper A2B5 für Oligodendrocyten-Vorläuferzellen [siehe Abb. 9B].
Auszählungen ergaben, dass der Hauptanteil der Zellen A2B5-positiv war [90,4 ± 4,2 %; siehe
Abb. 21]. Ausgelöst durch den Wechsel auf Differenzierungsmedium beginnen die Zellen ihre bipola-re Morphologie zu verlieren und bilden vermehrt Fortsätze aus [siehe Abb. 9C, 9E und 9G]. Dies ist verbunden mit einem Verlust der Immunreaktivität gegen A2B5 [siehe Abb. 9D, 9F und 9H].
4.1.2 Entwicklung der Oligodendrocyten-Vorläuferzellen: MOSP-Immuncytochemie
Oligodendrocyten-Vorläuferzellen, auch O-2A-Vorläuferzellen genannt, können nicht nur zu Oligo-dendrocyten, sondern auch zu Astrocyten differenzieren [Noble und Murray, 1984; Raff et al., 1984; Armstrong et al., 1990; Wren et al., 1992]. Aus diesem Grund wurde mittels Immuncytochemie [3.1.7] gegen das Myelin Oligodendrocyte Specific Protein (MOSP) überprüft, ob die Faktoren T3, CNTF und Forskolin auch im Neurobasal-Medium zur Ausdifferenzierung zu reifen Oligodendrocyten führen [siehe Abb. 10, S.47]. Das MOSP ist ein Oberflächenprotein, welches ausschließlich von Oligo-dendrocyten des Zentralnervensystems exprimiert wird [Dyer et al., 1991]. Die Expression beginnt in dem Stadium der Entwicklung zu reifen Oligodendrocyten, in welchem die Zellen ihre Fortsätze aus-bilden und mit der Bildung von Myelin beginnen [Mu und Dyer, 1994].
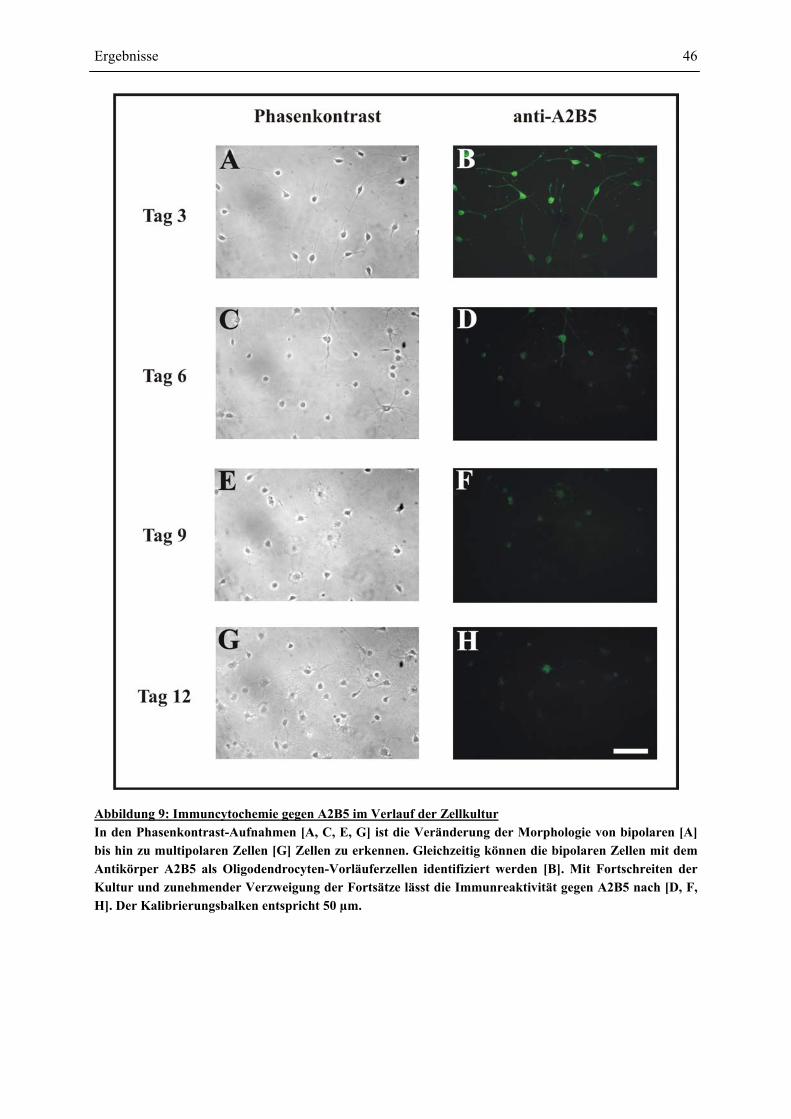
Ergebnisse 46
Abbildung 9: Immuncytochemie gegen A2B5 im Verlauf der Zellkultur In den Phasenkontrast-Aufnahmen [A, C, E, G] ist die Veränderung der Morphologie von bipolaren [A] bis hin zu multipolaren Zellen [G] Zellen zu erkennen. Gleichzeitig können die bipolaren Zellen mit dem Antikörper A2B5 als Oligodendrocyten-Vorläuferzellen identifiziert werden [B]. Mit Fortschreiten der Kultur und zunehmender Verzweigung der Fortsätze lässt die Immunreaktivität gegen A2B5 nach [D, F, H]. Der Kalibrierungsbalken entspricht 50 µm.
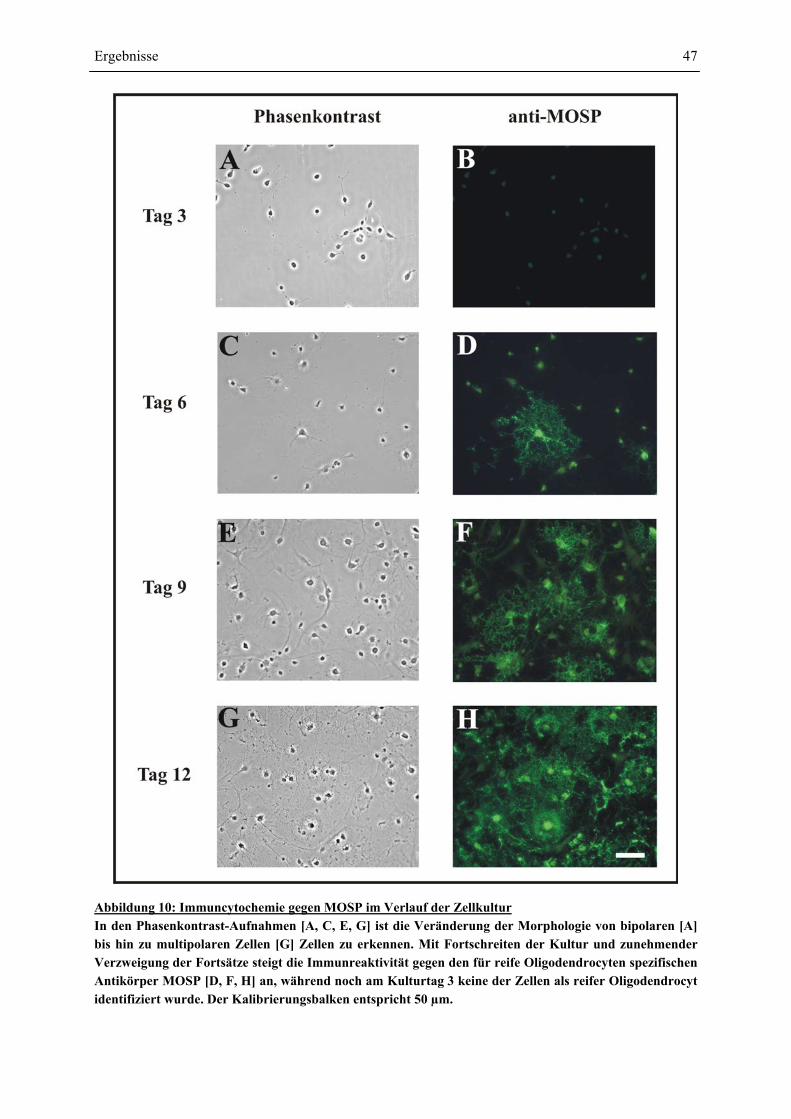
Ergebnisse 47
Abbildung 10: Immuncytochemie gegen MOSP im Verlauf der Zellkultur In den Phasenkontrast-Aufnahmen [A, C, E, G] ist die Veränderung der Morphologie von bipolaren [A] bis hin zu multipolaren Zellen [G] Zellen zu erkennen. Mit Fortschreiten der Kultur und zunehmender Verzweigung der Fortsätze steigt die Immunreaktivität gegen den für reife Oligodendrocyten spezifischen Antikörper MOSP [D, F, H] an, während noch am Kulturtag 3 keine der Zellen als reifer Oligodendrocyt identifiziert wurde. Der Kalibrierungsbalken entspricht 50 µm.

Ergebnisse 48
Um zu untersuchen, ob die Differenzierung der Oligodendrocyten-Vorläuferzellen unter den angege-benen Bedingungen induziert werden konnte, wurden die Zellen auf Deckgläschen kultiviert und an den Kulturtagen 3, 6, 9 und 12 mit 4 %iger Paraformaldehyd-Lösung fixiert [3.1.1]. Reife Oligo-dendrocyten wurden dann mit dem Antikörper MOSP detektiert [3.1.7].
Durch die Immuncytochemie gegen das Myelin Oligodendrocyte Specific Protein konnte gezeigt wer-den, dass die Oligodendrocyten-Vorläuferzellen, welche selber nicht positiv mit diesem Antikörper reagieren [siehe Abb. 10B], im Laufe der Kultivierung in Differenzierungsmedium zu reifen Oligo-dendrocyten ausdifferenzierten. Zum einen bildeten sie immer mehr Fortsätze aus, bis sie die für reife Oligodendrocyten typische bäumchenartige Struktur aufwiesen [siehe Abb. 10C, 10E und 10G]. Zum anderen zeigten die Zellen eine immer stärker werdende Immunreaktivität gegen den MOSP-Antikörper [siehe Abb. 10D, 10F und 10H]. Die Induktion der Ausdifferenzierung zu reifen Oligo-dendrocyten war somit in Neurobasal-Medium, welches die Faktoren T3, CNTF und Forskolin enthält, möglich.
4.1.3 Entwicklung der Oligodendrocyten-Vorläuferzellen: Expression der Proteine MBP, MAG und CNP
Eine weitere Methode, um die Entwicklung der Oligodendrocyten-Vorläuferzellen zu reifen Oligo-dendrocyten zu untersuchen, ist die Western-Blot-Analyse von Zell-Lysaten unterschiedlicher Kulti-vierungstage. Dazu wurden die Zellen an den Kulturtagen 3, 6, 9 und 12 lysiert. Je 10 µg des Lysates wurden für die Untersuchung der Proteine MBP und CNP auf einem 12 %igem SDS-Polyacrylamidgel aufgetrennt [3.2.3]. Für das Protein MAG wurde ein 8 %iges SDS-Polyacrylamidgel unter nicht-reduzierenden Bedingungen für die Auftrennung der Proteine des Lysates gewählt. Nach dem Transfer auf eine Nitrocellulosemembran wurden die Proteine mit den entsprechenden Antikörpern nachgewie-sen [3.2.4 – 3.2.6].
Es wurden die drei Proteine MBP, CNP und MAG gewählt, da ihre Expression charakteristisch für reife Oligodendrocyten ist: Das MBP gehört mit einem Anteil von 30% zu einem der Hauptbestandtei-le des Myelins des Zentralnervensystems [Mirsky et al., 1980]. Das CNP gehört zu den Proteinen, welche recht früh von sich entwickelnden Oligodendrocyten exprimiert werden [siehe Abb. 4; Vogel und Thompson, 1988] und das MAG, welches nur einen geringen Anteil im Gesamtprotein des Myelin ausmacht, spielt eine wichtige Rolle in der Interaktion zwischen Gliazellen und Axonen [Quarles et al., 1992].
Wie in Abbildung 11 [S.49] zu sehen ist, wurde die Expression der Myelin-spezifischen Proteine durch die Faktoren im Differenzierungsmedium erfolgreich induziert: An den Kulturtagen 1 und 3 war keines der drei Proteine nachweisbar, während sie mit Fortschreiten der Kultur immer stärker expri-miert wurden. Um zu zeigen, dass alle Proben gleich viel Protein enthielten, und dieser Effekt nicht auf unterschiedliche Proteinmengen zurückzuführen ist, wurde der Western-Blot mit einem anti-Tubulin Antikörper redetektiert.
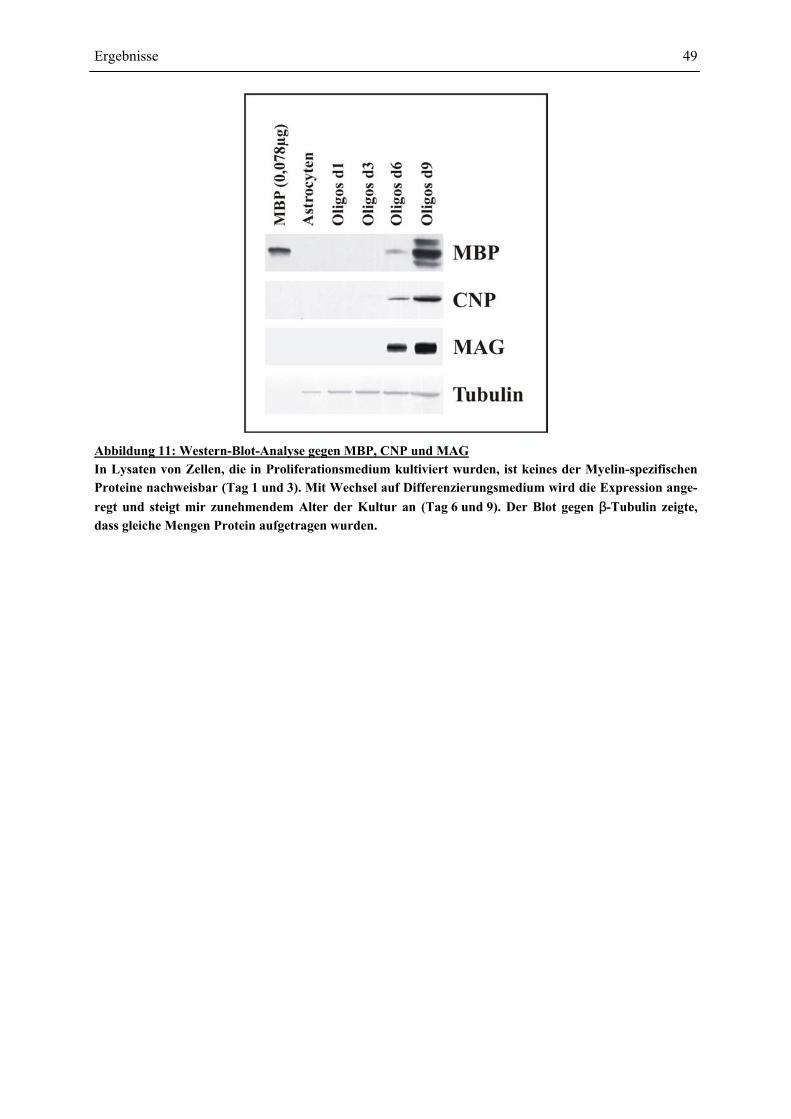
Ergebnisse 49
Abbildung 11: Western-Blot-Analyse gegen MBP, CNP und MAG In Lysaten von Zellen, die in Proliferationsmedium kultiviert wurden, ist keines der Myelin-spezifischen Proteine nachweisbar (Tag 1 und 3). Mit Wechsel auf Differenzierungsmedium wird die Expression ange-regt und steigt mir zunehmendem Alter der Kultur an (Tag 6 und 9). Der Blot gegen β-Tubulin zeigte, dass gleiche Mengen Protein aufgetragen wurden.

Ergebnisse 50
4.1.4 Reinheit der Oligodendrocyten-Vorläufer-Zellkultur: GFAP-Immuncytochemie
Der Anteil an Astrocyten sollte in der Zellkultur so gering wie möglich gehalten werden, da diese Zellen Hormone und Cytokine produzieren können, welche die biologische Aktivität von beispiels-
weise TNF-α beeinflussen können [Rothwell und Relton, 1993; Hopkins und Rothwell, 1995]. Effekte
von TNF-α auf die Zellkultur könnten somit bei einem hohen Anteil von Astrocyten auch durch Pro-
dukte dieser Zellen verändert werden.
Um zu überprüfen, wie hoch der Anteil an Astrocyten in dieser Kultur war, wurden die Zellen auf Deckgläschen kultiviert und mit 4 %iger Paraformaldehyd-Lösung fixiert [3.1.1]. Anschließend wurde eine Immuncytochemie mit dem Astrocyten-spezifischen Antikörper GFAP durchgeführt [3.1.7].
Abbildung 12: Immuncytochemie gegen GFAP Die anti-GFAP-Immuncytochemie zeigt, dass sich nur ein sehr geringer Anteil von Astrocyten in der Kul-tur befinden (B, gekennzeichnet durch Pfeile). Der Kalibrierungsbalken entspricht 50 µm
Abbildung 12 zeigt exemplarisch, dass der Anteil an GFAP-positiven Zellen, also Astrocyten, sehr gering war. Zusammen mit der Tatsache, dass der Anteil an A2B5-positiven Zellen gut 90 % der Zel-len ausmachte [siehe 4.1.1 und Abbildung 21] konnte davon ausgegangen werden, dass ein durch Cytokine erzeugter Effekt auf die Oligodendrocyten-Vorläuferzellen nicht auf Produkte von Astrocy-ten zurückzuführen wäre.
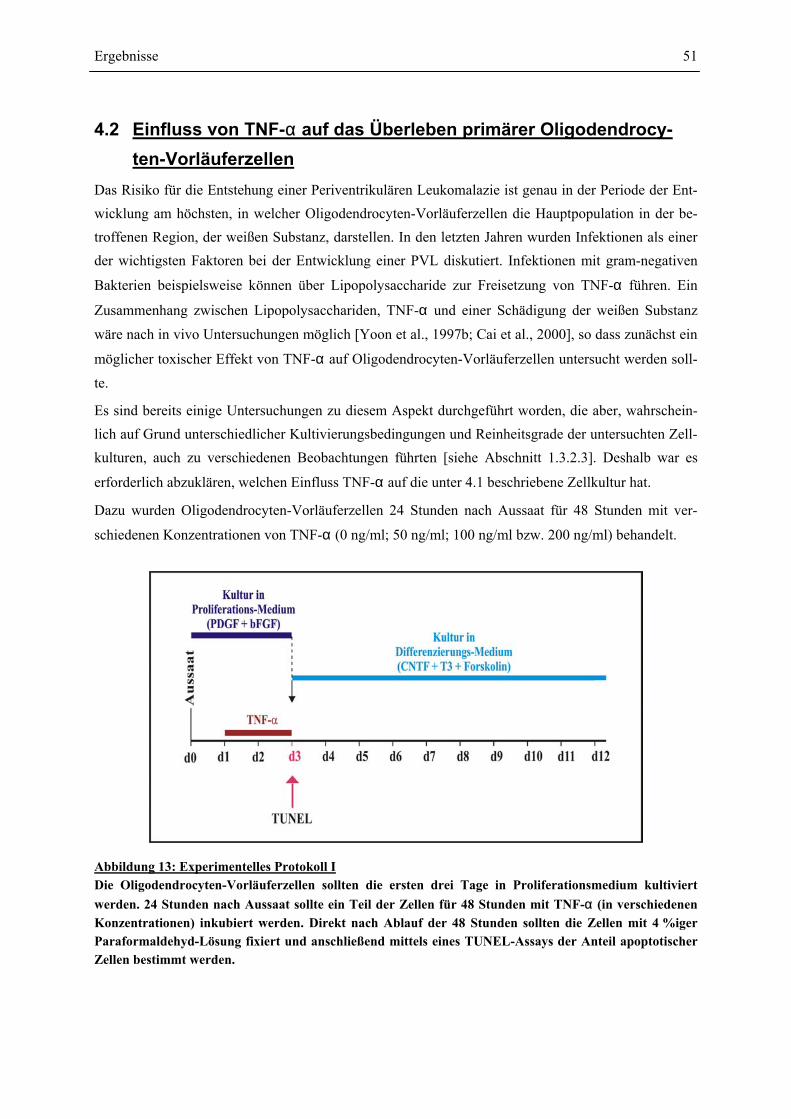
Ergebnisse 51
4.2 Einfluss von TNF-α auf das Überleben primärer Oligodendrocy-ten-Vorläuferzellen
Das Risiko für die Entstehung einer Periventrikulären Leukomalazie ist genau in der Periode der Ent-wicklung am höchsten, in welcher Oligodendrocyten-Vorläuferzellen die Hauptpopulation in der be-troffenen Region, der weißen Substanz, darstellen. In den letzten Jahren wurden Infektionen als einer der wichtigsten Faktoren bei der Entwicklung einer PVL diskutiert. Infektionen mit gram-negativen
Bakterien beispielsweise können über Lipopolysaccharide zur Freisetzung von TNF-α führen. Ein
Zusammenhang zwischen Lipopolysacchariden, TNF-α und einer Schädigung der weißen Substanz
wäre nach in vivo Untersuchungen möglich [Yoon et al., 1997b; Cai et al., 2000], so dass zunächst ein
möglicher toxischer Effekt von TNF-α auf Oligodendrocyten-Vorläuferzellen untersucht werden soll-
te.
Es sind bereits einige Untersuchungen zu diesem Aspekt durchgeführt worden, die aber, wahrschein-lich auf Grund unterschiedlicher Kultivierungsbedingungen und Reinheitsgrade der untersuchten Zell-kulturen, auch zu verschiedenen Beobachtungen führten [siehe Abschnitt 1.3.2.3]. Deshalb war es
erforderlich abzuklären, welchen Einfluss TNF-α auf die unter 4.1 beschriebene Zellkultur hat.
Dazu wurden Oligodendrocyten-Vorläuferzellen 24 Stunden nach Aussaat für 48 Stunden mit ver-
schiedenen Konzentrationen von TNF-α (0 ng/ml; 50 ng/ml; 100 ng/ml bzw. 200 ng/ml) behandelt.
Abbildung 13: Experimentelles Protokoll I Die Oligodendrocyten-Vorläuferzellen sollten die ersten drei Tage in Proliferationsmedium kultiviert werden. 24 Stunden nach Aussaat sollte ein Teil der Zellen für 48 Stunden mit TNF-α (in verschiedenen Konzentrationen) inkubiert werden. Direkt nach Ablauf der 48 Stunden sollten die Zellen mit 4 %iger Paraformaldehyd-Lösung fixiert und anschließend mittels eines TUNEL-Assays der Anteil apoptotischer Zellen bestimmt werden.
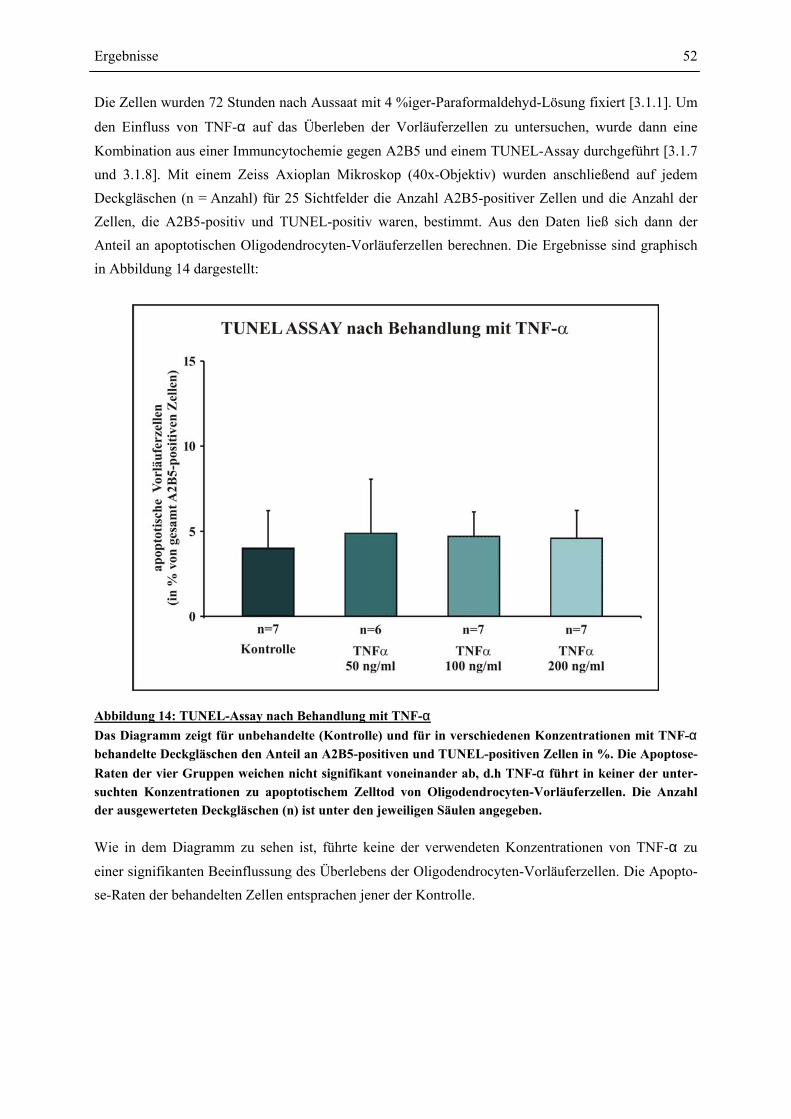
Ergebnisse 52
Die Zellen wurden 72 Stunden nach Aussaat mit 4 %iger-Paraformaldehyd-Lösung fixiert [3.1.1]. Um
den Einfluss von TNF-α auf das Überleben der Vorläuferzellen zu untersuchen, wurde dann eine
Kombination aus einer Immuncytochemie gegen A2B5 und einem TUNEL-Assay durchgeführt [3.1.7 und 3.1.8]. Mit einem Zeiss Axioplan Mikroskop (40x-Objektiv) wurden anschließend auf jedem Deckgläschen (n = Anzahl) für 25 Sichtfelder die Anzahl A2B5-positiver Zellen und die Anzahl der Zellen, die A2B5-positiv und TUNEL-positiv waren, bestimmt. Aus den Daten ließ sich dann der Anteil an apoptotischen Oligodendrocyten-Vorläuferzellen berechnen. Die Ergebnisse sind graphisch in Abbildung 14 dargestellt:
Abbildung 14: TUNEL-Assay nach Behandlung mit TNF-α Das Diagramm zeigt für unbehandelte (Kontrolle) und für in verschiedenen Konzentrationen mit TNF-α behandelte Deckgläschen den Anteil an A2B5-positiven und TUNEL-positiven Zellen in %. Die Apoptose-Raten der vier Gruppen weichen nicht signifikant voneinander ab, d.h TNF-α führt in keiner der unter-suchten Konzentrationen zu apoptotischem Zelltod von Oligodendrocyten-Vorläuferzellen. Die Anzahl der ausgewerteten Deckgläschen (n) ist unter den jeweiligen Säulen angegeben.
Wie in dem Diagramm zu sehen ist, führte keine der verwendeten Konzentrationen von TNF-α zu
einer signifikanten Beeinflussung des Überlebens der Oligodendrocyten-Vorläuferzellen. Die Apopto-se-Raten der behandelten Zellen entsprachen jener der Kontrolle.
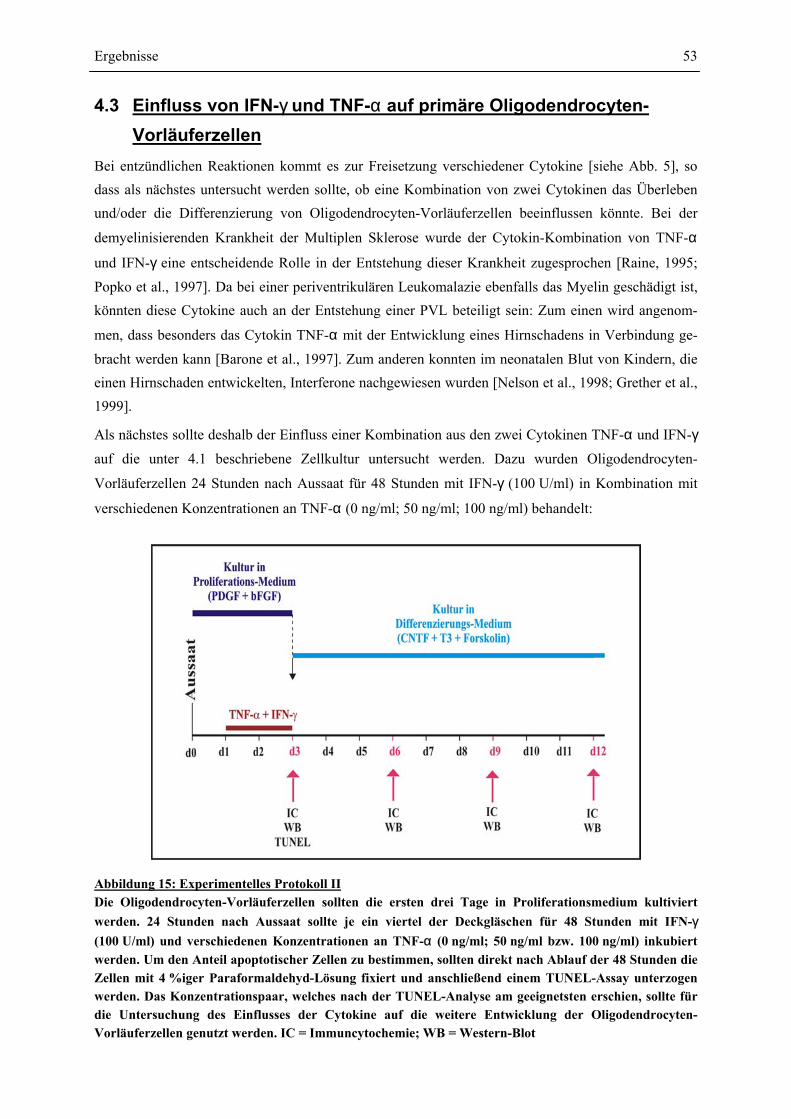
Ergebnisse 53
4.3 Einfluss von IFN-γ und TNF-α auf primäre Oligodendrocyten-Vorläuferzellen
Bei entzündlichen Reaktionen kommt es zur Freisetzung verschiedener Cytokine [siehe Abb. 5], so dass als nächstes untersucht werden sollte, ob eine Kombination von zwei Cytokinen das Überleben und/oder die Differenzierung von Oligodendrocyten-Vorläuferzellen beeinflussen könnte. Bei der
demyelinisierenden Krankheit der Multiplen Sklerose wurde der Cytokin-Kombination von TNF-α
und IFN-γ eine entscheidende Rolle in der Entstehung dieser Krankheit zugesprochen [Raine, 1995;
Popko et al., 1997]. Da bei einer periventrikulären Leukomalazie ebenfalls das Myelin geschädigt ist, könnten diese Cytokine auch an der Entstehung einer PVL beteiligt sein: Zum einen wird angenom-
men, dass besonders das Cytokin TNF-α mit der Entwicklung eines Hirnschadens in Verbindung ge-
bracht werden kann [Barone et al., 1997]. Zum anderen konnten im neonatalen Blut von Kindern, die einen Hirnschaden entwickelten, Interferone nachgewiesen wurden [Nelson et al., 1998; Grether et al., 1999].
Als nächstes sollte deshalb der Einfluss einer Kombination aus den zwei Cytokinen TNF-α und IFN-γ
auf die unter 4.1 beschriebene Zellkultur untersucht werden. Dazu wurden Oligodendrocyten-
Vorläuferzellen 24 Stunden nach Aussaat für 48 Stunden mit IFN-γ (100 U/ml) in Kombination mit
verschiedenen Konzentrationen an TNF-α (0 ng/ml; 50 ng/ml; 100 ng/ml) behandelt:
Abbildung 15: Experimentelles Protokoll II Die Oligodendrocyten-Vorläuferzellen sollten die ersten drei Tage in Proliferationsmedium kultiviert werden. 24 Stunden nach Aussaat sollte je ein viertel der Deckgläschen für 48 Stunden mit IFN-γ (100 U/ml) und verschiedenen Konzentrationen an TNF-α (0 ng/ml; 50 ng/ml bzw. 100 ng/ml) inkubiert werden. Um den Anteil apoptotischer Zellen zu bestimmen, sollten direkt nach Ablauf der 48 Stunden die Zellen mit 4 %iger Paraformaldehyd-Lösung fixiert und anschließend einem TUNEL-Assay unterzogen werden. Das Konzentrationspaar, welches nach der TUNEL-Analyse am geeignetsten erschien, sollte für die Untersuchung des Einflusses der Cytokine auf die weitere Entwicklung der Oligodendrocyten-Vorläuferzellen genutzt werden. IC = Immuncytochemie; WB = Western-Blot

Ergebnisse 54
Um den Einfluss von den verschiedenen Kombinationen an IFN-γ und TNF-α auf das Überleben der
Vorläuferzellen zu untersuchen, sollte am Kulturtag 3 mittels eines TUNEL-Assays der Anteil an apoptotischen Vorläuferzellen bestimmt werden. Dieser TUNEL-Assay diente gleichzeitig dazu, ein geeignetes Konzentrationspaar zu finden, welches zum einen signifikant das Überleben der Zellen beeinflusste, und zum anderen eine Untersuchung des Differenzierungsverhaltens der überlebenden Zellen ermöglichte.
An den im experimentellen Protokoll II angegebenen Tagen sollte anschließend der Effekt der Cytoki-ne auf die weitere Entwicklung der Zellkultur untersucht werden. Dazu wurde die Gesamtzellzahl bestimmt, die Immuncytochemie gegen die Oberflächenantigene A2B5 und MOSP sowie Western-Blot-Analysen der Myelin-spezifischen Proteine MBP, MAG und CNP durchgeführt.
4.3.1 Einfluss von IFN-γ und TNF-α auf das Überleben von Oligodendrocy-ten-Vorläuferzellen
Zunächst sollte geklärt werden, ob IFN-γ alleine und/oder in Kombination mit TNF-α in Oligodendro-
cyten-Vorläuferzellen Apoptose induzieren kann. Dazu wurden die Zellen entsprechend dem experi-mentellen Protokoll in Abbildung 15 für 48 Stunden mit verschiedenen Konzentrationspaaren an
IFN-γ und TNF-α (0/0; 100/0; 100/50; 100/100) inkubiert. Anschließend wurden die Zellen in 4 %iger
Paraformaldehyd-Lösung fixiert [3.1.1] und einer Kombination aus A2B5-Immuncytochemie und TUNEL-Assay unterzogen [3.1.7 und 3.1.8].
Die Auswertung des TUNEL-Assays erfolgte anhand einer in Abbildung 16 [S.54] gezeigten Abfolge von Mikroskop-Aufnahmen. Die A2B5-Färbung [siehe Abb. 16B] diente zur Bestimmung der Anzahl an Oligodendrocyten-Vorläuferzellen. Apoptotische Zellen wurden anhand der TUNEL-Färbung [sie-he Abb. 16D] bestimmt. Durch einen Vergleich der beiden Färbungen war es möglich apoptotische Oligodendrocyten-Vorläuferzellen zu identifizieren. Die Hoechst-Färbung in Abbildung 16C diente dazu, sicher zu gehen, dass es sich bei der TUNEL-Färbung um die Färbung der Zellkerne und nicht um unspezifische Anfärbungen von Zelltrümmern handelt.
Die quantitative Auswertung aus drei unabhängigen Versuchen ist in Abbildung 17 dargestellt:
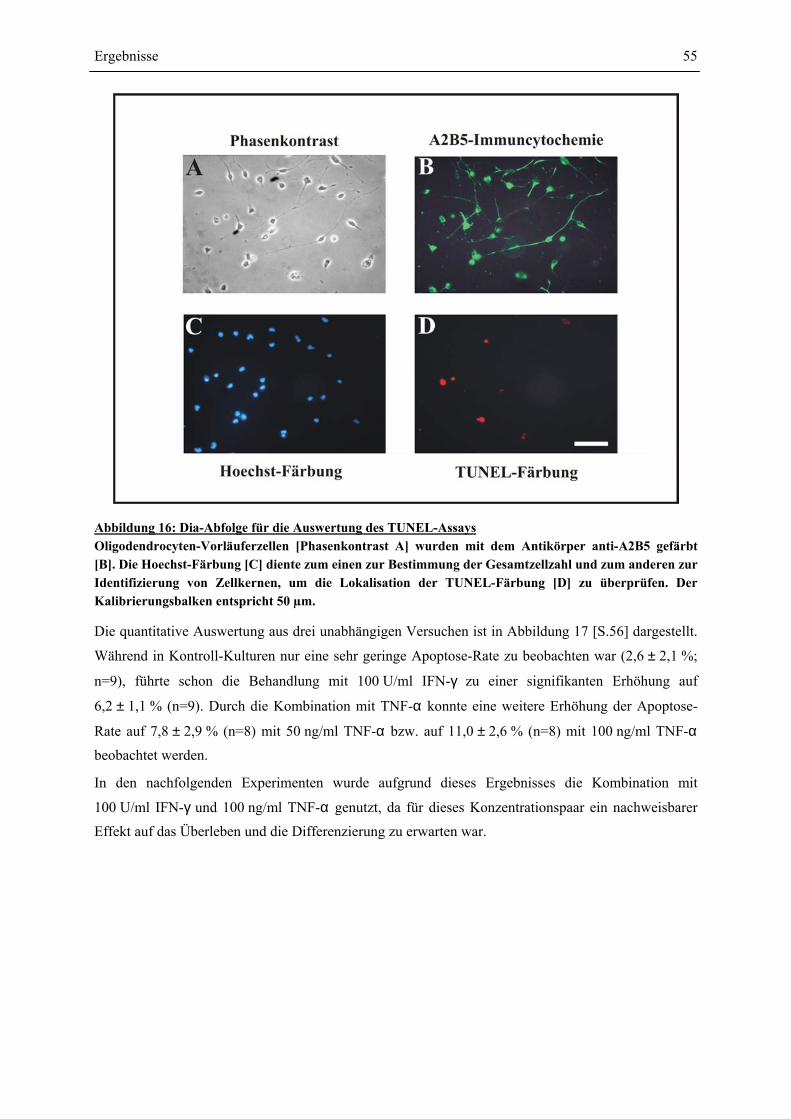
Ergebnisse 55
Abbildung 16: Dia-Abfolge für die Auswertung des TUNEL-Assays Oligodendrocyten-Vorläuferzellen [Phasenkontrast A] wurden mit dem Antikörper anti-A2B5 gefärbt [B]. Die Hoechst-Färbung [C] diente zum einen zur Bestimmung der Gesamtzellzahl und zum anderen zur Identifizierung von Zellkernen, um die Lokalisation der TUNEL-Färbung [D] zu überprüfen. Der Kalibrierungsbalken entspricht 50 µm.
Die quantitative Auswertung aus drei unabhängigen Versuchen ist in Abbildung 17 [S.56] dargestellt.
Während in Kontroll-Kulturen nur eine sehr geringe Apoptose-Rate zu beobachten war (2,6 ± 2,1 %;
n=9), führte schon die Behandlung mit 100 U/ml IFN-γ zu einer signifikanten Erhöhung auf
6,2 ± 1,1 % (n=9). Durch die Kombination mit TNF-α konnte eine weitere Erhöhung der Apoptose-
Rate auf 7,8 ± 2,9 % (n=8) mit 50 ng/ml TNF-α bzw. auf 11,0 ± 2,6 % (n=8) mit 100 ng/ml TNF-α
beobachtet werden.
In den nachfolgenden Experimenten wurde aufgrund dieses Ergebnisses die Kombination mit
100 U/ml IFN-γ und 100 ng/ml TNF-α genutzt, da für dieses Konzentrationspaar ein nachweisbarer
Effekt auf das Überleben und die Differenzierung zu erwarten war.
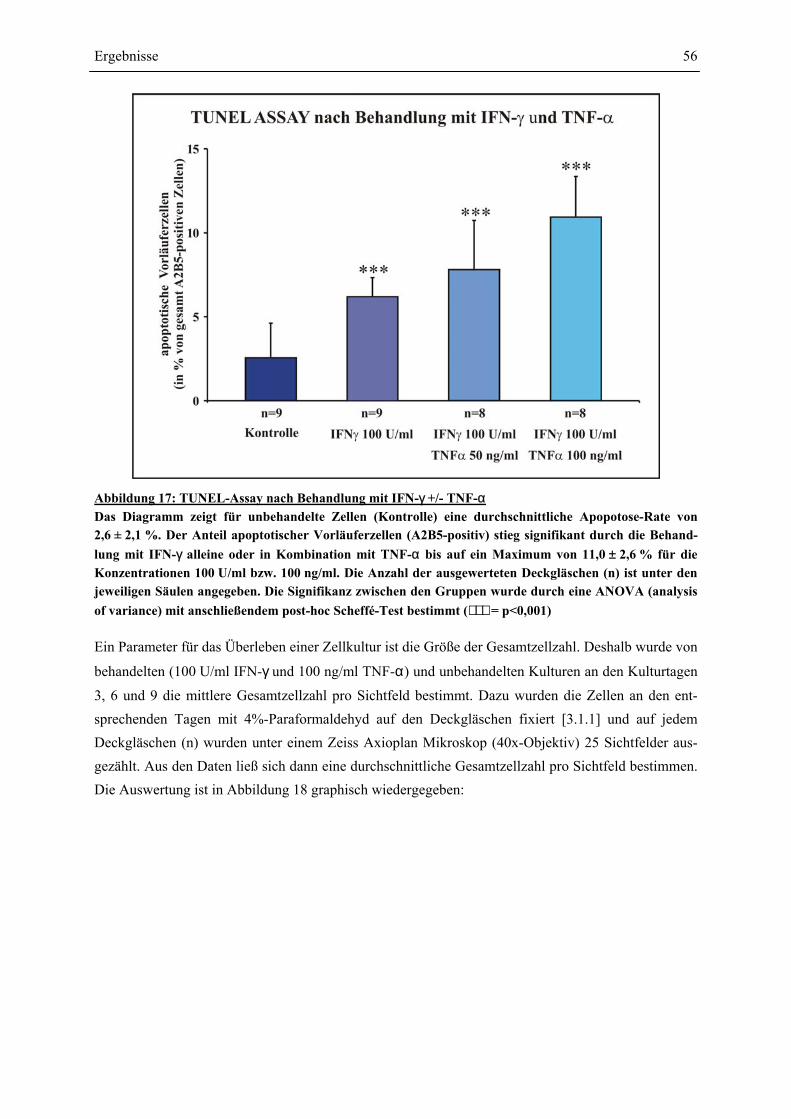
Ergebnisse 56
Abbildung 17: TUNEL-Assay nach Behandlung mit IFN-γ +/- TNF-α Das Diagramm zeigt für unbehandelte Zellen (Kontrolle) eine durchschnittliche Apopotose-Rate von 2,6 ± 2,1 %. Der Anteil apoptotischer Vorläuferzellen (A2B5-positiv) stieg signifikant durch die Behand-lung mit IFN-γ alleine oder in Kombination mit TNF-α bis auf ein Maximum von 11,0 ± 2,6 % für die Konzentrationen 100 U/ml bzw. 100 ng/ml. Die Anzahl der ausgewerteten Deckgläschen (n) ist unter den jeweiligen Säulen angegeben. Die Signifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem post-hoc Scheffé-Test bestimmt (∗∗∗ = p<0,001)
Ein Parameter für das Überleben einer Zellkultur ist die Größe der Gesamtzellzahl. Deshalb wurde von
behandelten (100 U/ml IFN-γ und 100 ng/ml TNF-α) und unbehandelten Kulturen an den Kulturtagen
3, 6 und 9 die mittlere Gesamtzellzahl pro Sichtfeld bestimmt. Dazu wurden die Zellen an den ent-sprechenden Tagen mit 4%-Paraformaldehyd auf den Deckgläschen fixiert [3.1.1] und auf jedem Deckgläschen (n) wurden unter einem Zeiss Axioplan Mikroskop (40x-Objektiv) 25 Sichtfelder aus-gezählt. Aus den Daten ließ sich dann eine durchschnittliche Gesamtzellzahl pro Sichtfeld bestimmen. Die Auswertung ist in Abbildung 18 graphisch wiedergegeben:
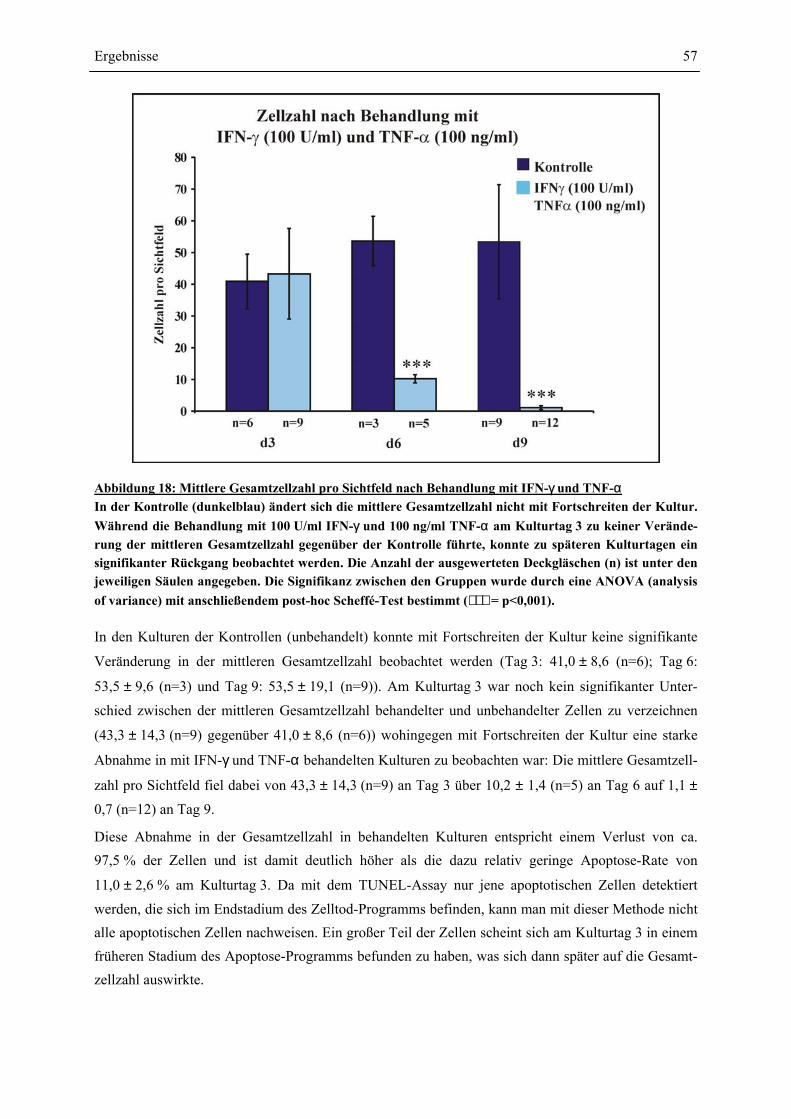
Ergebnisse 57
Abbildung 18: Mittlere Gesamtzellzahl pro Sichtfeld nach Behandlung mit IFN-γ und TNF-α In der Kontrolle (dunkelblau) ändert sich die mittlere Gesamtzellzahl nicht mit Fortschreiten der Kultur. Während die Behandlung mit 100 U/ml IFN-γ und 100 ng/ml TNF-α am Kulturtag 3 zu keiner Verände-rung der mittleren Gesamtzellzahl gegenüber der Kontrolle führte, konnte zu späteren Kulturtagen ein signifikanter Rückgang beobachtet werden. Die Anzahl der ausgewerteten Deckgläschen (n) ist unter den jeweiligen Säulen angegeben. Die Signifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem post-hoc Scheffé-Test bestimmt (∗∗∗ = p<0,001).
In den Kulturen der Kontrollen (unbehandelt) konnte mit Fortschreiten der Kultur keine signifikante
Veränderung in der mittleren Gesamtzellzahl beobachtet werden (Tag 3: 41,0 ± 8,6 (n=6); Tag 6:
53,5 ± 9,6 (n=3) und Tag 9: 53,5 ± 19,1 (n=9)). Am Kulturtag 3 war noch kein signifikanter Unter-
schied zwischen der mittleren Gesamtzellzahl behandelter und unbehandelter Zellen zu verzeichnen
(43,3 ± 14,3 (n=9) gegenüber 41,0 ± 8,6 (n=6)) wohingegen mit Fortschreiten der Kultur eine starke
Abnahme in mit IFN-γ und TNF-α behandelten Kulturen zu beobachten war: Die mittlere Gesamtzell-
zahl pro Sichtfeld fiel dabei von 43,3 ± 14,3 (n=9) an Tag 3 über 10,2 ± 1,4 (n=5) an Tag 6 auf 1,1 ±
0,7 (n=12) an Tag 9.
Diese Abnahme in der Gesamtzellzahl in behandelten Kulturen entspricht einem Verlust von ca. 97,5 % der Zellen und ist damit deutlich höher als die dazu relativ geringe Apoptose-Rate von
11,0 ± 2,6 % am Kulturtag 3. Da mit dem TUNEL-Assay nur jene apoptotischen Zellen detektiert
werden, die sich im Endstadium des Zelltod-Programms befinden, kann man mit dieser Methode nicht alle apoptotischen Zellen nachweisen. Ein großer Teil der Zellen scheint sich am Kulturtag 3 in einem früheren Stadium des Apoptose-Programms befunden zu haben, was sich dann später auf die Gesamt-zellzahl auswirkte.
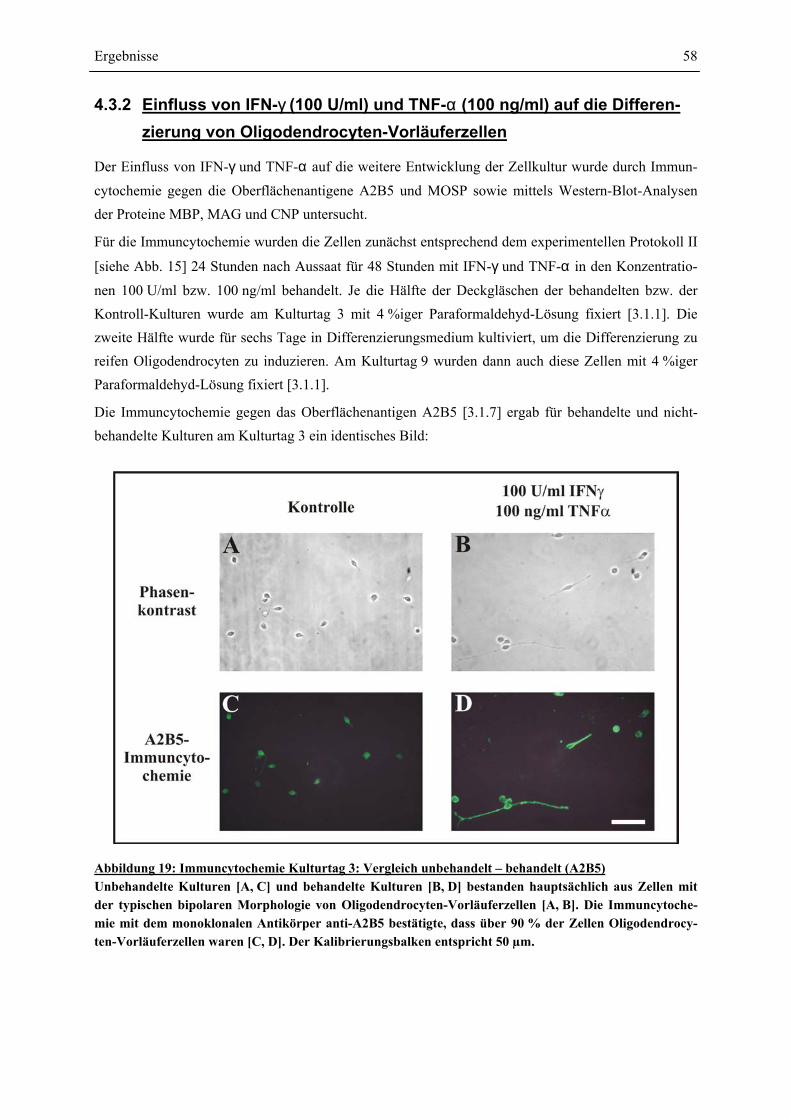
Ergebnisse 58
4.3.2 Einfluss von IFN-γ (100 U/ml) und TNF-α (100 ng/ml) auf die Differen-zierung von Oligodendrocyten-Vorläuferzellen
Der Einfluss von IFN-γ und TNF-α auf die weitere Entwicklung der Zellkultur wurde durch Immun-
cytochemie gegen die Oberflächenantigene A2B5 und MOSP sowie mittels Western-Blot-Analysen der Proteine MBP, MAG und CNP untersucht.
Für die Immuncytochemie wurden die Zellen zunächst entsprechend dem experimentellen Protokoll II
[siehe Abb. 15] 24 Stunden nach Aussaat für 48 Stunden mit IFN-γ und TNF-α in den Konzentratio-
nen 100 U/ml bzw. 100 ng/ml behandelt. Je die Hälfte der Deckgläschen der behandelten bzw. der Kontroll-Kulturen wurde am Kulturtag 3 mit 4 %iger Paraformaldehyd-Lösung fixiert [3.1.1]. Die zweite Hälfte wurde für sechs Tage in Differenzierungsmedium kultiviert, um die Differenzierung zu reifen Oligodendrocyten zu induzieren. Am Kulturtag 9 wurden dann auch diese Zellen mit 4 %iger Paraformaldehyd-Lösung fixiert [3.1.1].
Die Immuncytochemie gegen das Oberflächenantigen A2B5 [3.1.7] ergab für behandelte und nicht-behandelte Kulturen am Kulturtag 3 ein identisches Bild:
Abbildung 19: Immuncytochemie Kulturtag 3: Vergleich unbehandelt – behandelt (A2B5) Unbehandelte Kulturen [A, C] und behandelte Kulturen [B, D] bestanden hauptsächlich aus Zellen mit der typischen bipolaren Morphologie von Oligodendrocyten-Vorläuferzellen [A, B]. Die Immuncytoche-mie mit dem monoklonalen Antikörper anti-A2B5 bestätigte, dass über 90 % der Zellen Oligodendrocy-ten-Vorläuferzellen waren [C, D]. Der Kalibrierungsbalken entspricht 50 µm.
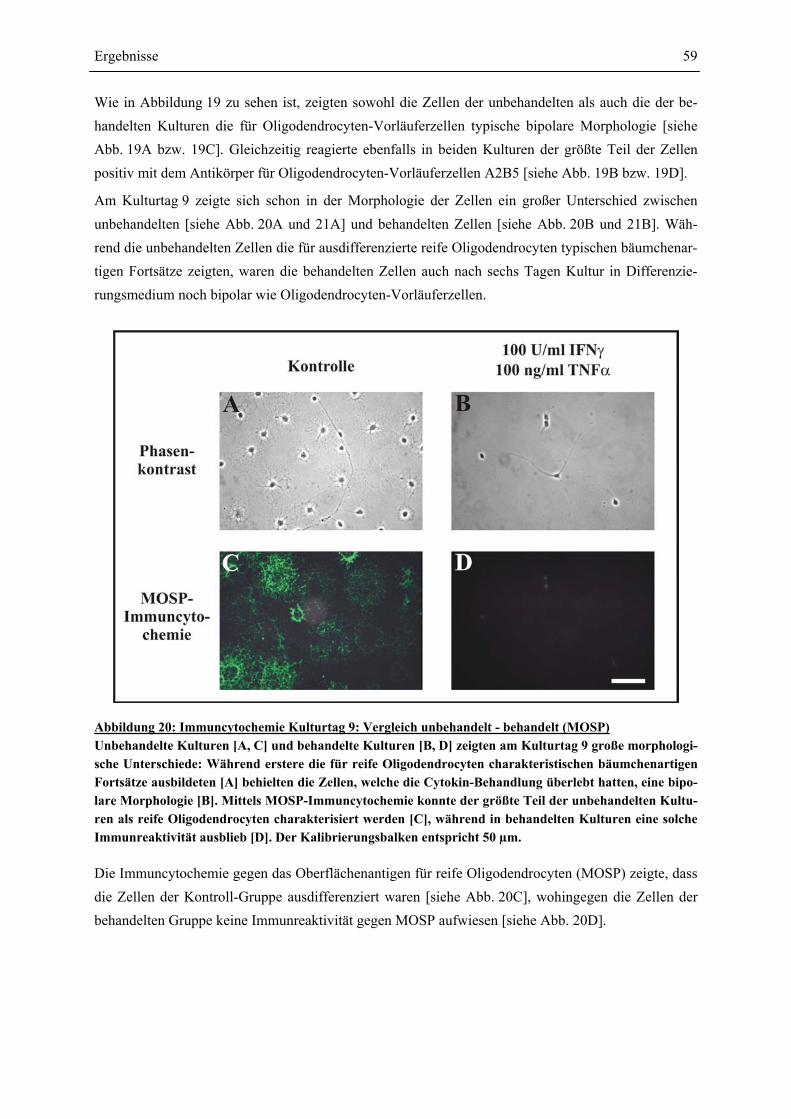
Ergebnisse 59
Wie in Abbildung 19 zu sehen ist, zeigten sowohl die Zellen der unbehandelten als auch die der be-handelten Kulturen die für Oligodendrocyten-Vorläuferzellen typische bipolare Morphologie [siehe Abb. 19A bzw. 19C]. Gleichzeitig reagierte ebenfalls in beiden Kulturen der größte Teil der Zellen positiv mit dem Antikörper für Oligodendrocyten-Vorläuferzellen A2B5 [siehe Abb. 19B bzw. 19D].
Am Kulturtag 9 zeigte sich schon in der Morphologie der Zellen ein großer Unterschied zwischen unbehandelten [siehe Abb. 20A und 21A] und behandelten Zellen [siehe Abb. 20B und 21B]. Wäh-rend die unbehandelten Zellen die für ausdifferenzierte reife Oligodendrocyten typischen bäumchenar-tigen Fortsätze zeigten, waren die behandelten Zellen auch nach sechs Tagen Kultur in Differenzie-rungsmedium noch bipolar wie Oligodendrocyten-Vorläuferzellen.
Abbildung 20: Immuncytochemie Kulturtag 9: Vergleich unbehandelt - behandelt (MOSP) Unbehandelte Kulturen [A, C] und behandelte Kulturen [B, D] zeigten am Kulturtag 9 große morphologi-sche Unterschiede: Während erstere die für reife Oligodendrocyten charakteristischen bäumchenartigen Fortsätze ausbildeten [A] behielten die Zellen, welche die Cytokin-Behandlung überlebt hatten, eine bipo-lare Morphologie [B]. Mittels MOSP-Immuncytochemie konnte der größte Teil der unbehandelten Kultu-ren als reife Oligodendrocyten charakterisiert werden [C], während in behandelten Kulturen eine solche Immunreaktivität ausblieb [D]. Der Kalibrierungsbalken entspricht 50 µm.
Die Immuncytochemie gegen das Oberflächenantigen für reife Oligodendrocyten (MOSP) zeigte, dass die Zellen der Kontroll-Gruppe ausdifferenziert waren [siehe Abb. 20C], wohingegen die Zellen der behandelten Gruppe keine Immunreaktivität gegen MOSP aufwiesen [siehe Abb. 20D].

Ergebnisse 60
Abbildung 21: Immuncytochemie Kulturtag 9: Vergleich unbehandelt - behandelt (A2B5) Auch in dieser Abbildung werden die morphologischen Unterschiede zwischen unbehandelten Kulturen [A, C] und behandelten Kulturen [B, D] deutlich. Die A2B5-Immuncytochemie zeigte, dass unbehandelte Kulturen diese Immunreaktivität aufgrund der erfolgten Differenzierung verloren hatten [B], während in behandelten Kulturen ein sehr großer Teil der Zellen A2B5-positiv war [D]. Der Kalibrierungsbalken entspricht 50 µm.
Statt dessen zeigten die mit IFN-γ und TNF-α behandelten Zellen am Kulturtag 9 einen sehr hohen
Anteil an A2B5-positiven Zellen [siehe Abb. 21D], während in Zellen der Kontrolle fast keine Im-munreaktivität gegen dieses Oberflächenantigen nachgewiesen wurde [siehe Abb. 21C].
Die quantitative Auswertung der A2B5-Immunreaktivität bestätigte den visuellen Eindruck eines er-
höhten Anteils an undifferenzierten Zellen in mit IFN-γ und TNF-α behandelten Kulturen [siehe
Abb. 22]. An den Kulturtagen 3 und 9 wurden dazu Zellen mit 4 %iger Paraformaldehyd-Lösung fi-xiert [3.1.1] und anschließend mit dem Antikörper anti-A2B5 angefärbt [3.1.7]. Unter einem Zeiss Axioplan Mikroskop (40x-Objektiv) wurde dann aus 25 Sichtfeldern pro Deckglas die Gesamtzellzahl und die Anzahl an A2B5-positiven (grün-fluoreszierenden) Zellen bestimmt. Aus dem Verhältnis von A2B5-positiven Zellen zur Gesamtzellzahl ließ sich dann der Anteil A2B5-positiver Zellen errechnen. Abbildung 22 zeigt die graphische Auswertung:
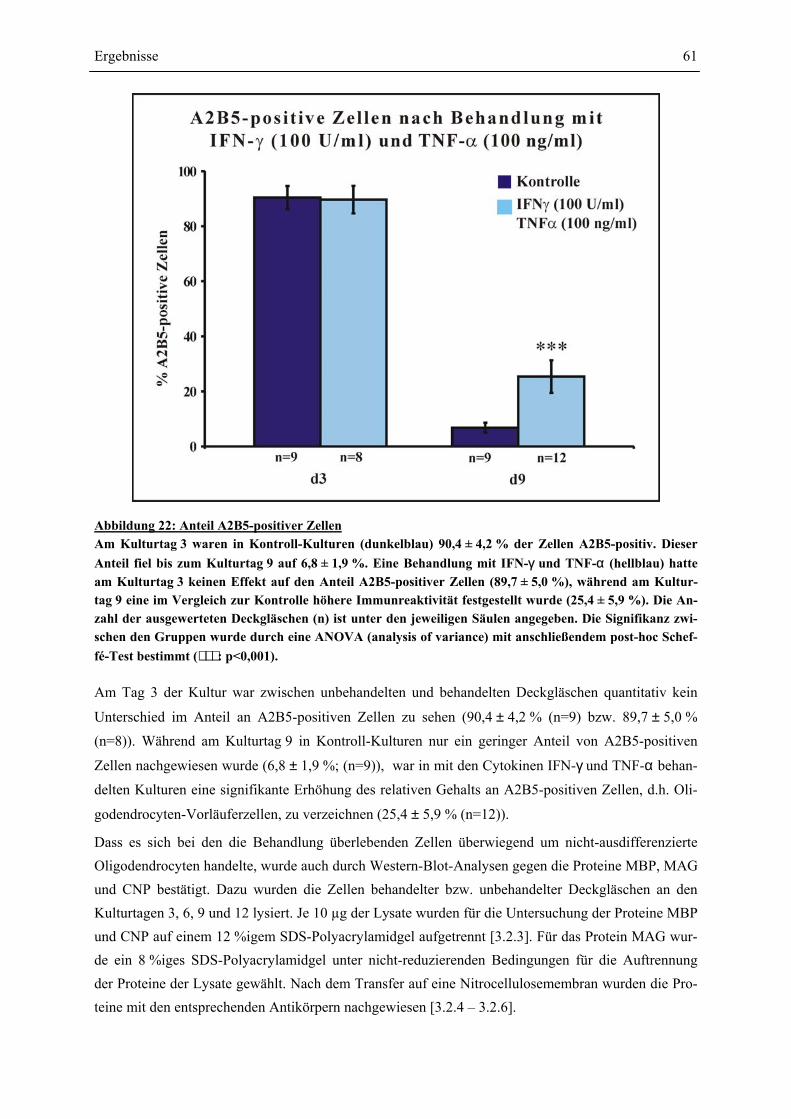
Ergebnisse 61
Abbildung 22: Anteil A2B5-positiver Zellen Am Kulturtag 3 waren in Kontroll-Kulturen (dunkelblau) 90,4 ± 4,2 % der Zellen A2B5-positiv. Dieser Anteil fiel bis zum Kulturtag 9 auf 6,8 ± 1,9 %. Eine Behandlung mit IFN-γ und TNF-α (hellblau) hatte am Kulturtag 3 keinen Effekt auf den Anteil A2B5-positiver Zellen (89,7 ± 5,0 %), während am Kultur-tag 9 eine im Vergleich zur Kontrolle höhere Immunreaktivität festgestellt wurde (25,4 ± 5,9 %). Die An-zahl der ausgewerteten Deckgläschen (n) ist unter den jeweiligen Säulen angegeben. Die Signifikanz zwi-schen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem post-hoc Schef-fé-Test bestimmt (∗∗ ∗ : p<0,001).
Am Tag 3 der Kultur war zwischen unbehandelten und behandelten Deckgläschen quantitativ kein
Unterschied im Anteil an A2B5-positiven Zellen zu sehen (90,4 ± 4,2 % (n=9) bzw. 89,7 ± 5,0 %
(n=8)). Während am Kulturtag 9 in Kontroll-Kulturen nur ein geringer Anteil von A2B5-positiven
Zellen nachgewiesen wurde (6,8 ± 1,9 %; (n=9)), war in mit den Cytokinen IFN-γ und TNF-α behan-
delten Kulturen eine signifikante Erhöhung des relativen Gehalts an A2B5-positiven Zellen, d.h. Oli-
godendrocyten-Vorläuferzellen, zu verzeichnen (25,4 ± 5,9 % (n=12)).
Dass es sich bei den die Behandlung überlebenden Zellen überwiegend um nicht-ausdifferenzierte Oligodendrocyten handelte, wurde auch durch Western-Blot-Analysen gegen die Proteine MBP, MAG und CNP bestätigt. Dazu wurden die Zellen behandelter bzw. unbehandelter Deckgläschen an den Kulturtagen 3, 6, 9 und 12 lysiert. Je 10 µg der Lysate wurden für die Untersuchung der Proteine MBP und CNP auf einem 12 %igem SDS-Polyacrylamidgel aufgetrennt [3.2.3]. Für das Protein MAG wur-de ein 8 %iges SDS-Polyacrylamidgel unter nicht-reduzierenden Bedingungen für die Auftrennung der Proteine der Lysate gewählt. Nach dem Transfer auf eine Nitrocellulosemembran wurden die Pro-teine mit den entsprechenden Antikörpern nachgewiesen [3.2.4 – 3.2.6].

Ergebnisse 62
Abbildung 23: Western-Blot-Analyse gegen MBP, MAG und CNP An den Kulturtagen 9 und 12 war in den Lysaten der Kontroll-Kulturen eine starke Expression von MBP, MAG und CNP nachzuweisen, während in den Lysaten behandelter Zellen an keinem der untersuchten Tage eine positive Reaktion mit den entsprechenden Antikörpern zu sehen war. Der Blot gegen Actin zeigte, dass gleiche Mengen Protein aufgetragen wurden.
In Abbildung 23 ist zu sehen, dass in den Zellen der Kontroll-Gruppe die Expression der Myelin-spezifischen Proteine durch die Faktoren im Differenzierungsmedium erfolgreich angeregt wurde: Während am Kulturtag 3 keines der drei Proteine nachweisbar war, wurden sie nach Wechsel auf Dif-ferenzierungsmedium mit Fortschreiten der Kultur immer stärker exprimiert. Die Lysate der behandel-ten Zellen hingegen zeigten an keinem der untersuchten Tage ein positives Signal, d.h. die Zellen differenzierten nicht aus. Um zu zeigen, dass alle Proben gleich viel Protein enthielten, und dieser Effekt nicht auf unterschiedliche Proteinmengen zurückzuführen ist, wurde der Western-Blot mit ei-nem anti-Actin Antikörper redetektiert.

Ergebnisse 63
4.4 Schützen Corticoide Oligodendrocyten-Vorläuferzellen gegen Ein-flüsse von IFN-γ und TNF-α ?
Corticoide werden aufgrund ihrer antientzündlichen Wirkung häufig in der Behandlung von Multipler Sklerose und anderen demyelinisierenden Krankheiten eingesetzt und finden auch in der Perinatalme-dizin sowohl pränatal als auch postnatal Anwendung. Aufgrund dieser klinischen Relevanz lag es nahe zu untersuchen, ob Corticoide eine Rolle im Schutz vor einer diffusen periventrikulären Leukomalazie spielen könnten, indem sie Oligodendrocyten-Vorläuferzellen vor den Folgen einer Infektion, d.h. vor den Einflüssen der dabei freigesetzten Cytokine, schützen. Für reife Oligodendrocyten gibt es bereits erste Hinweise darauf, dass die Corticoide Aldosteron, Corticosteron, Deoxycorticosteron und Dexa-methason teilweise vor durch Cytokinen induziertem Zelltod schützen [Melcangi et al., 2000].
Die in den bisher gezeigten Experimenten genutzte Konzentration an IFN-γ und TNF-α von 100 U/ml
bzw. 100 ng/ml hatte einen so starken Effekt auf das Überleben und die Differenzierung der Oligo-dendrocyten-Vorläuferzellen, dass diese ungeeignet für das Finden protektiver Reagenzien war. Des-halb musste zunächst eine Konzentration gefunden werden, welche gegenüber der Kontrolle einen erkennbaren, aber deutlich schwächeren Effekt als die Kombination 100/100 zeigte. Dazu wurden Oligodendrocyten-Vorläuferzellen entsprechend dem experimentellem Protokoll in Abbildung 15 für
48 Stunden mit verschiedenen Konzentrationspaaren an IFN-γ und TNF-α (100/10; 50/10; 25/10;
10/10) inkubiert. Anschließend wurde das Medium auf Differenzierungsmedium gewechselt und die Zellen bis zum 9. Kulturtag kultiviert. Am Kulturtag 9 wurden die Zellen lysiert und je 10 µg der Ly-sate wurden für die Untersuchung des Proteins MBP auf einem 12 %igem SDS-Polyacrylamidgel aufgetrennt [3.2.3]. Für das Protein MAG wurde ein 8 %iges SDS-Polyacrylamidgel unter nicht-reduzierenden Bedingungen für die Auftrennung der Proteine der Lysate gewählt. Nach dem Transfer auf eine Nitrocellulosemembran wurden die Proteine mit den entsprechenden Antikörpern nachgewie-sen [3.2.4 – 3.2.6].

Ergebnisse 64
Abbildung 24: Western-Blot-Analyse zur Konzentrationsfindung In den Lysaten behandelter Zellen war für alle Konzentrationspaare eine gegenüber der Kontrolle ver-minderte Expression des MBP nachweisbar. Das Protein MAG hingegen wurde außer in der Kontrolle nur in mit 10 U/ml IFN-γ und 10 ng/ml TNF-α behandelten Kulturen detektiert. Der Blot gegen Actin zeigte, dass gleiche Mengen Protein aufgetragen wurden.
Abbildung 24 zeigt je einen für drei unabhängige Experimente repräsentativen Blot. Das Konzentrati-
onspaar mit 10 U/ml IFN-γ und 10 ng/ml TNF-α wurde aufgrund dieses Ergebnisses für die weiteren
Versuche genutzt: Die Cytokine führten in diesen Konzentrationen zu einer gegenüber der Kontrolle deutlich verminderten Expression des Myelin Basic Proteins und des Myelin Associated Glycoprotein, wobei die Expression aber noch nachweisbar blieb.
Um einen möglichen schützenden Effekt von Glucocorticoiden auf mit IFN-γ und TNF-α behandelte
Oligodendrocyten-Vorläuferzellen zu untersuchen, wurde das in Abbildung 25 [S.64] dargestellte experimentelle Protokoll entworfen.
Die Oligodendrocyten-Vorläuferzellen wurden 24 Stunden nach Aussaat für 48 Stunden mit IFN-γ
(10 U/ml) und TNF-α (10 ng/ml) behandelt. Die Corticoide Corticosteron (C), Deoxycorticosteron
(DC) und Dexamethason (DEX) wurden in der Konzentration von 1 µM je einem viertel der mit Cyto-kinen behandelten bzw. unbehandelten Deckgläschen im gleichen Zeitraum zugesetzt. Insgesamt 72 Stunden nach Aussaat erfolgte dann ein Mediumwechsel auf Differenzierungsmedium. Die Zellen wurden bis zum Kulturtag 9 in diesem Medium kultiviert. Am Kulturtag 9 wurden die Zellen für wei-tere Experimente fixiert (Immuncytochemie; Bestimmung der Gesamtzellzahl), lysiert (Western-Blot-Analysen) oder für RNA-Untersuchungen (RT-PCR) geerntet.
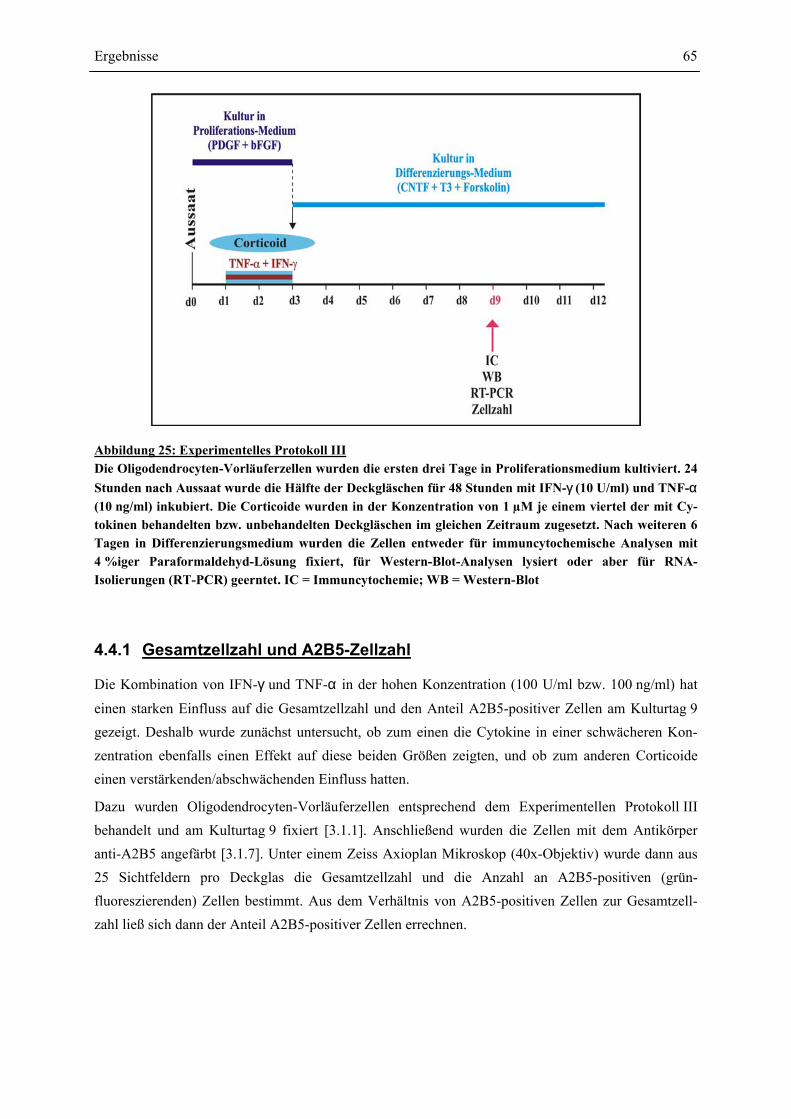
Ergebnisse 65
Abbildung 25: Experimentelles Protokoll III Die Oligodendrocyten-Vorläuferzellen wurden die ersten drei Tage in Proliferationsmedium kultiviert. 24 Stunden nach Aussaat wurde die Hälfte der Deckgläschen für 48 Stunden mit IFN-γ (10 U/ml) und TNF-α (10 ng/ml) inkubiert. Die Corticoide wurden in der Konzentration von 1 µM je einem viertel der mit Cy-tokinen behandelten bzw. unbehandelten Deckgläschen im gleichen Zeitraum zugesetzt. Nach weiteren 6 Tagen in Differenzierungsmedium wurden die Zellen entweder für immuncytochemische Analysen mit 4 %iger Paraformaldehyd-Lösung fixiert, für Western-Blot-Analysen lysiert oder aber für RNA-Isolierungen (RT-PCR) geerntet. IC = Immuncytochemie; WB = Western-Blot
4.4.1 Gesamtzellzahl und A2B5-Zellzahl
Die Kombination von IFN-γ und TNF-α in der hohen Konzentration (100 U/ml bzw. 100 ng/ml) hat
einen starken Einfluss auf die Gesamtzellzahl und den Anteil A2B5-positiver Zellen am Kulturtag 9 gezeigt. Deshalb wurde zunächst untersucht, ob zum einen die Cytokine in einer schwächeren Kon-zentration ebenfalls einen Effekt auf diese beiden Größen zeigten, und ob zum anderen Corticoide einen verstärkenden/abschwächenden Einfluss hatten.
Dazu wurden Oligodendrocyten-Vorläuferzellen entsprechend dem Experimentellen Protokoll III behandelt und am Kulturtag 9 fixiert [3.1.1]. Anschließend wurden die Zellen mit dem Antikörper anti-A2B5 angefärbt [3.1.7]. Unter einem Zeiss Axioplan Mikroskop (40x-Objektiv) wurde dann aus 25 Sichtfeldern pro Deckglas die Gesamtzellzahl und die Anzahl an A2B5-positiven (grün-fluoreszierenden) Zellen bestimmt. Aus dem Verhältnis von A2B5-positiven Zellen zur Gesamtzell-zahl ließ sich dann der Anteil A2B5-positiver Zellen errechnen.

Ergebnisse 66
Abbildung 26: A2B5-Immuncytochemie nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9) Nicht mit Cytokinen behandelte Kulturen [A-H] zeigten im Phasenkontrast die für reife Oligodendrocyten charakteristischen bäumchenartigen Fortsätze [A-D] und keine Immunreaktivität gegen den Antikörper A2B5 [E-H]. Eine 48-stündige Behandlung mit Corticoiden [B-D; F-H] schien zu einer erhöhten Gesamt-zellzahl zu führen. Nur mit IFN-γ und TNF-α behandelte Kulturen [I, M] waren im Vergleich zu Kon-trollkulturen [A-H] in ihrer Gesamtzellzahl reduziert [I]. Diese Kulturen bestanden zum großen Teil aus bipolaren Zellen mit einer starken A2B5-Immunreaktivität [M]. Nach einer gleichzeitigen Behandlung mit Corticoiden [J-L; N-P] war eine erhöhte Gesamtzellzahl mit einer für reife Oligodendrocyten typi-schen Morphologie zu beobachten [J-L]. Damit einher ging der Verlust der A2B5-Immunreaktivität. Der Kalibrierungsbalken entspricht 50 µm.
In Abbildung 26 sind Mikroskop-Aufnahmen zu sehen, welche bei einer von vier dieser Versuchsrei-hen aufgenommen wurden. Diese Bilder deuten darauf hin, dass Corticoide sowohl in Kontroll- als
auch in mit IFN-γ und TNF-α behandelten Kulturen zu einer Erhöhung der mittleren Gesamtzellzahl
führten. Die Behandlung mit Corticoiden hatte auch auf die Morphologie der Zellen einen starken Einfluss: Mit Corticoiden und Cytokinen behandelte Zellen zeigten deutlich die Morphologie von ausdifferenzierten Oligodendrocyten, wohingegen nur mit Cytokinen behandelte Kulturen zu einem großen Teil aus bipolaren Zellen bestanden. Dieser Eindruck wird durch die unterschiedliche A2B5-Immunreaktivität bestätigt [M-P]. Die Morphologie und A2B5-Immunreaktivität von mit Corticoiden und Cytokinen behandelten Zellen scheint annähernd identisch mit der von Kontroll-Kulturen zu sein.
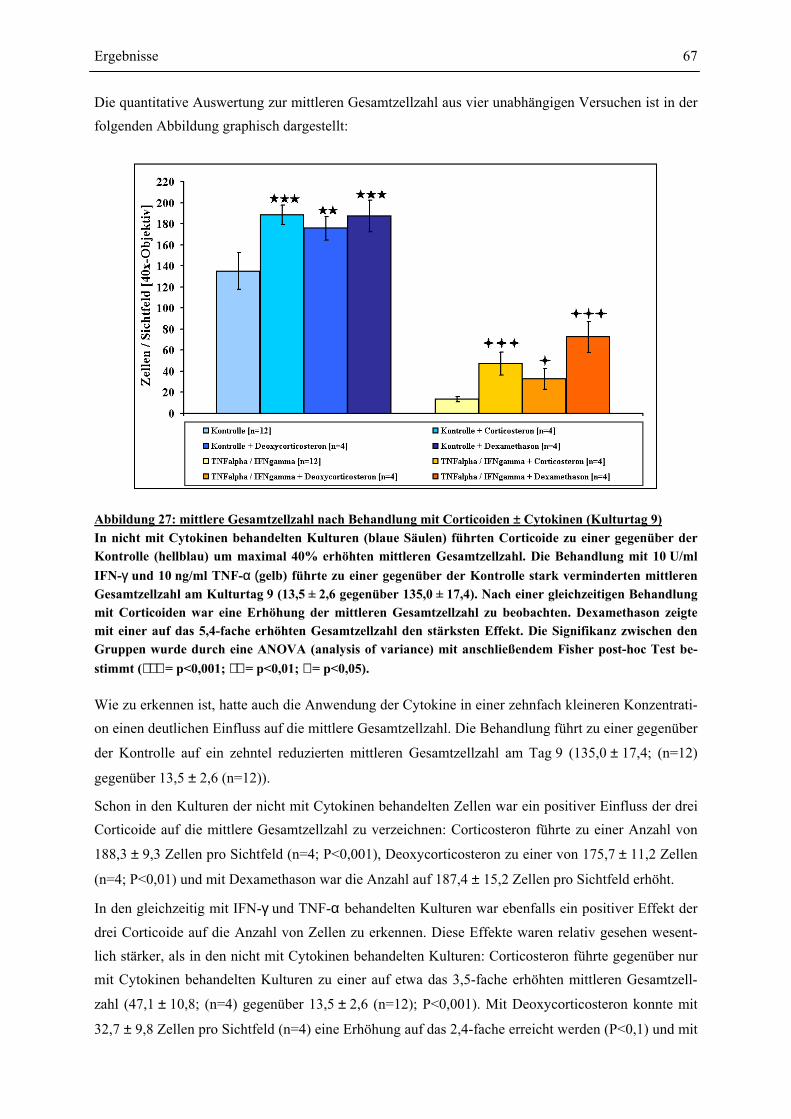
Ergebnisse 67
Die quantitative Auswertung zur mittleren Gesamtzellzahl aus vier unabhängigen Versuchen ist in der folgenden Abbildung graphisch dargestellt:
Abbildung 27: mittlere Gesamtzellzahl nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9) In nicht mit Cytokinen behandelten Kulturen (blaue Säulen) führten Corticoide zu einer gegenüber der Kontrolle (hellblau) um maximal 40% erhöhten mittleren Gesamtzellzahl. Die Behandlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α (gelb) führte zu einer gegenüber der Kontrolle stark verminderten mittleren Gesamtzellzahl am Kulturtag 9 (13,5 ± 2,6 gegenüber 135,0 ± 17,4). Nach einer gleichzeitigen Behandlung mit Corticoiden war eine Erhöhung der mittleren Gesamtzellzahl zu beobachten. Dexamethason zeigte mit einer auf das 5,4-fache erhöhten Gesamtzellzahl den stärksten Effekt. Die Signifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem Fisher post-hoc Test be-stimmt (∗∗ ∗ = p<0,001; ∗∗ = p<0,01; ∗ = p<0,05).
Wie zu erkennen ist, hatte auch die Anwendung der Cytokine in einer zehnfach kleineren Konzentrati-on einen deutlichen Einfluss auf die mittlere Gesamtzellzahl. Die Behandlung führt zu einer gegenüber
der Kontrolle auf ein zehntel reduzierten mittleren Gesamtzellzahl am Tag 9 (135,0 ± 17,4; (n=12)
gegenüber 13,5 ± 2,6 (n=12)).
Schon in den Kulturen der nicht mit Cytokinen behandelten Zellen war ein positiver Einfluss der drei Corticoide auf die mittlere Gesamtzellzahl zu verzeichnen: Corticosteron führte zu einer Anzahl von
188,3 ± 9,3 Zellen pro Sichtfeld (n=4; P<0,001), Deoxycorticosteron zu einer von 175,7 ± 11,2 Zellen
(n=4; P<0,01) und mit Dexamethason war die Anzahl auf 187,4 ± 15,2 Zellen pro Sichtfeld erhöht.
In den gleichzeitig mit IFN-γ und TNF-α behandelten Kulturen war ebenfalls ein positiver Effekt der
drei Corticoide auf die Anzahl von Zellen zu erkennen. Diese Effekte waren relativ gesehen wesent-lich stärker, als in den nicht mit Cytokinen behandelten Kulturen: Corticosteron führte gegenüber nur mit Cytokinen behandelten Kulturen zu einer auf etwa das 3,5-fache erhöhten mittleren Gesamtzell-
zahl (47,1 ± 10,8; (n=4) gegenüber 13,5 ± 2,6 (n=12); P<0,001). Mit Deoxycorticosteron konnte mit
32,7 ± 9,8 Zellen pro Sichtfeld (n=4) eine Erhöhung auf das 2,4-fache erreicht werden (P<0,1) und mit
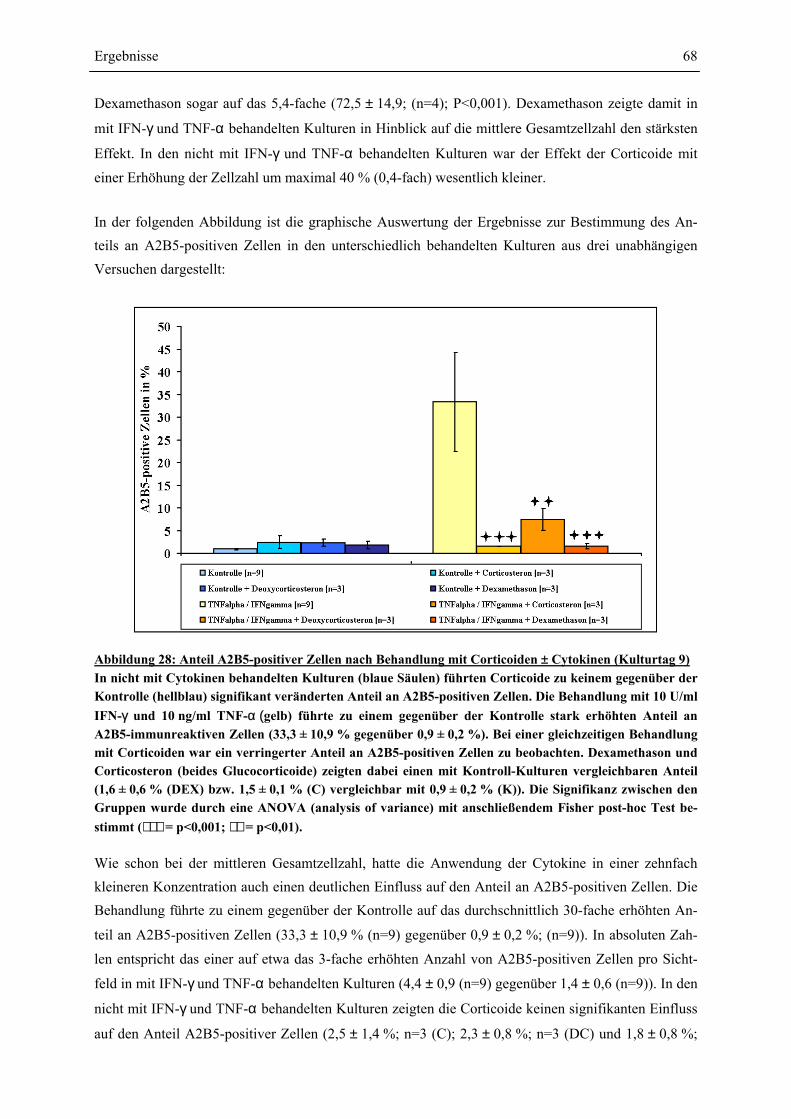
Ergebnisse 68
Dexamethason sogar auf das 5,4-fache (72,5 ± 14,9; (n=4); P<0,001). Dexamethason zeigte damit in
mit IFN-γ und TNF-α behandelten Kulturen in Hinblick auf die mittlere Gesamtzellzahl den stärksten
Effekt. In den nicht mit IFN-γ und TNF-α behandelten Kulturen war der Effekt der Corticoide mit
einer Erhöhung der Zellzahl um maximal 40 % (0,4-fach) wesentlich kleiner.
In der folgenden Abbildung ist die graphische Auswertung der Ergebnisse zur Bestimmung des An-teils an A2B5-positiven Zellen in den unterschiedlich behandelten Kulturen aus drei unabhängigen Versuchen dargestellt:
Abbildung 28: Anteil A2B5-positiver Zellen nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9) In nicht mit Cytokinen behandelten Kulturen (blaue Säulen) führten Corticoide zu keinem gegenüber der Kontrolle (hellblau) signifikant veränderten Anteil an A2B5-positiven Zellen. Die Behandlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α (gelb) führte zu einem gegenüber der Kontrolle stark erhöhten Anteil an A2B5-immunreaktiven Zellen (33,3 ± 10,9 % gegenüber 0,9 ± 0,2 %). Bei einer gleichzeitigen Behandlung mit Corticoiden war ein verringerter Anteil an A2B5-positiven Zellen zu beobachten. Dexamethason und Corticosteron (beides Glucocorticoide) zeigten dabei einen mit Kontroll-Kulturen vergleichbaren Anteil (1,6 ± 0,6 % (DEX) bzw. 1,5 ± 0,1 % (C) vergleichbar mit 0,9 ± 0,2 % (K)). Die Signifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem Fisher post-hoc Test be-stimmt (∗∗ ∗ = p<0,001; ∗∗ = p<0,01).
Wie schon bei der mittleren Gesamtzellzahl, hatte die Anwendung der Cytokine in einer zehnfach kleineren Konzentration auch einen deutlichen Einfluss auf den Anteil an A2B5-positiven Zellen. Die Behandlung führte zu einem gegenüber der Kontrolle auf das durchschnittlich 30-fache erhöhten An-
teil an A2B5-positiven Zellen (33,3 ± 10,9 % (n=9) gegenüber 0,9 ± 0,2 %; (n=9)). In absoluten Zah-
len entspricht das einer auf etwa das 3-fache erhöhten Anzahl von A2B5-positiven Zellen pro Sicht-
feld in mit IFN-γ und TNF-α behandelten Kulturen (4,4 ± 0,9 (n=9) gegenüber 1,4 ± 0,6 (n=9)). In den
nicht mit IFN-γ und TNF-α behandelten Kulturen zeigten die Corticoide keinen signifikanten Einfluss
auf den Anteil A2B5-positiver Zellen (2,5 ± 1,4 %; n=3 (C); 2,3 ± 0,8 %; n=3 (DC) und 1,8 ± 0,8 %;

Ergebnisse 69
n=3 (DEX) gegenüber 0,9 ± 0,2 %; n=9 (Kontrolle)). In mit den Cytokinen behandelten Kulturen hin-
gegen zeigten erneut alle drei Corticoide einen starken Effekt: Sie führten zu einem signifikant gerin-geren Anteil an Oligodendrocyten-Vorläuferzellen. Corticosteron und Dexamethason zeigten dabei
erneut den stärksten Effekt (1,5 ± 0,1 %; n=3 (C) bzw. 1,6 ± 0,6 %; n=3 (DEX); jeweils P<0,001 im
Vergleich zu nur mit IFN-γ und TNF-α behandelten Kulturen). Beide Glucocorticoide führten damit
zu einem mit Kontroll-Kulturen vergleichbaren Anteil an A2B5-positiven Zellen. Deoxycorticosteron hingegen zeigte einen etwas schwächeren Effekt (7,4 ± 2,4 %; n=3; P<0,01). Auch ein Vergleich der absoluten Zahlen A2B5-positiver Zellen spiegelt deutlich den Einfluss der Corticoide auf die Diffe-renzierung der die Cytokin-Behandlung überlebenden Zellen wieder: 0,76 ± 1,12 (C); 2,2 ± 1,9 (DC); und 1,16 ± 1,6 (DEX).
Aus diesen Daten lässt sich schließen, dass Corticoide dem die Differenzierung von Oligodendrocy-
ten-Vorläuferzellen unterdrückenden Effekt von IFN-γ und TNF-α entgegenwirken können. Dies soll-
te durch Western-Blot-Analysen und semiquantitative RT-PCR weiter untersucht werden.
4.4.2 Proteinexpression – Western-Blot-Analysen
In vorangegangenen Experimenten wurde beobachtet, dass die Expression von Myelin-spezifischen
Proteinen am Kulturtag 9 durch IFN-γ und TNF-α im Vergleich zu Kontroll-Kulturen herabgesetzt ist
[siehe Abb. 23 und Abb. 24]. Um zu untersuchen, ob Corticoide diesem Effekt entgegen wirken, und somit einen positiven Einfluss auf die Differenzierung der die Behandlung überlebenden Zellen zei-gen, wurden Western-Blot-Analysen gegen die Proteine MBP, MAG und CNP durchgeführt.
Dazu wurden Oligodendrocyten-Vorläuferzellen entsprechend dem experimentellen Protokoll III be-handelt. Am Kulturtag 9 wurden die Zellen lysiert und je 10 µg der Lysate wurden für die Untersu-chung der Proteine MBP und CNP auf einem 12 %igem SDS-Polyacrylamidgel aufgetrennt [3.2.3]. Für das Protein MAG wurde ein 8 %iges SDS-Polyacrylamidgel unter nicht-reduzierenden Bedingun-gen für die Auftrennung der Proteine der Lysate gewählt. Nach dem Transfer auf eine Nitrocellulose-membran wurden die Proteine mit den entsprechenden Antikörpern nachgewiesen [3.2.4 – 3.2.6]. Um sicher zu gehen, dass alle Proben gleich viel Protein enthielten, wurden die Western-Blot mit einem anti-Actin Antikörper redetektiert.
Anschließend erfolgte die densitometrische Auswertung der Western-Blots: Mit dem Programm TINA 9.0 wurden die Intensitäten der einzelnen Antikörper-Banden bestimmt. Die Intensitäten der Antikör-per-Banden der Proteine MBP, MAG und CNP wurden gegen die des entsprechenden Actin-Blots normiert. Jeder Blot enthielt folgende acht Proben:
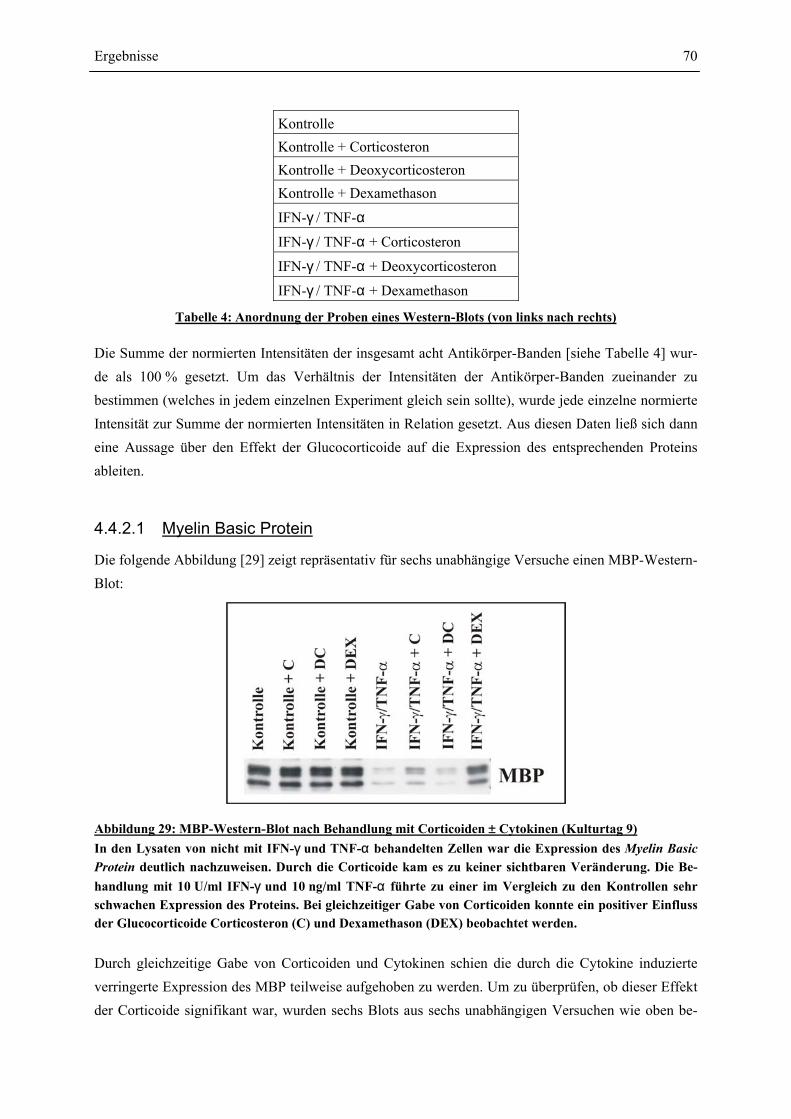
Ergebnisse 70
Kontrolle Kontrolle + Corticosteron Kontrolle + Deoxycorticosteron Kontrolle + Dexamethason
IFN-γ / TNF-α
IFN-γ / TNF-α + Corticosteron
IFN-γ / TNF-α + Deoxycorticosteron
IFN-γ / TNF-α + Dexamethason
Tabelle 4: Anordnung der Proben eines Western-Blots (von links nach rechts)
Die Summe der normierten Intensitäten der insgesamt acht Antikörper-Banden [siehe Tabelle 4] wur-de als 100 % gesetzt. Um das Verhältnis der Intensitäten der Antikörper-Banden zueinander zu bestimmen (welches in jedem einzelnen Experiment gleich sein sollte), wurde jede einzelne normierte Intensität zur Summe der normierten Intensitäten in Relation gesetzt. Aus diesen Daten ließ sich dann eine Aussage über den Effekt der Glucocorticoide auf die Expression des entsprechenden Proteins ableiten.
4.4.2.1 Myelin Basic Protein
Die folgende Abbildung [29] zeigt repräsentativ für sechs unabhängige Versuche einen MBP-Western-Blot:
Abbildung 29: MBP-Western-Blot nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9) In den Lysaten von nicht mit IFN-γ und TNF-α behandelten Zellen war die Expression des Myelin Basic Protein deutlich nachzuweisen. Durch die Corticoide kam es zu keiner sichtbaren Veränderung. Die Be-handlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α führte zu einer im Vergleich zu den Kontrollen sehr schwachen Expression des Proteins. Bei gleichzeitiger Gabe von Corticoiden konnte ein positiver Einfluss der Glucocorticoide Corticosteron (C) und Dexamethason (DEX) beobachtet werden.
Durch gleichzeitige Gabe von Corticoiden und Cytokinen schien die durch die Cytokine induzierte verringerte Expression des MBP teilweise aufgehoben zu werden. Um zu überprüfen, ob dieser Effekt der Corticoide signifikant war, wurden sechs Blots aus sechs unabhängigen Versuchen wie oben be-
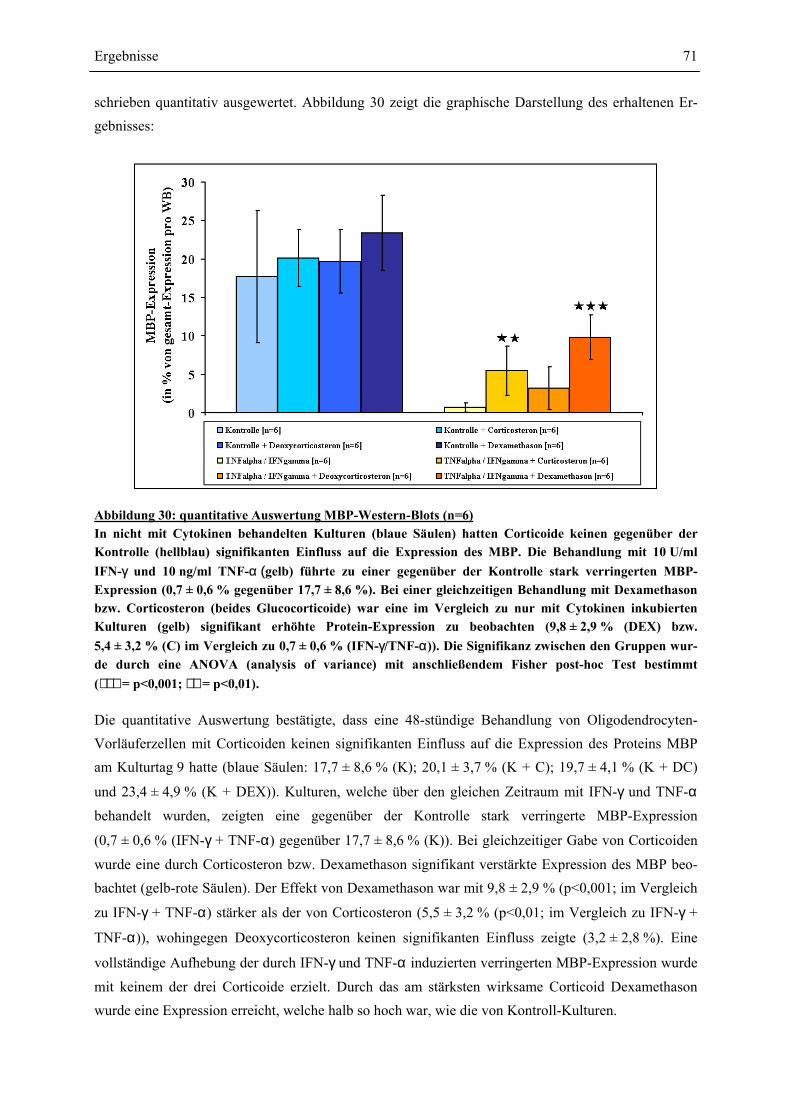
Ergebnisse 71
schrieben quantitativ ausgewertet. Abbildung 30 zeigt die graphische Darstellung des erhaltenen Er-gebnisses:
Abbildung 30: quantitative Auswertung MBP-Western-Blots (n=6) In nicht mit Cytokinen behandelten Kulturen (blaue Säulen) hatten Corticoide keinen gegenüber der Kontrolle (hellblau) signifikanten Einfluss auf die Expression des MBP. Die Behandlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α (gelb) führte zu einer gegenüber der Kontrolle stark verringerten MBP-Expression (0,7 ± 0,6 % gegenüber 17,7 ± 8,6 %). Bei einer gleichzeitigen Behandlung mit Dexamethason bzw. Corticosteron (beides Glucocorticoide) war eine im Vergleich zu nur mit Cytokinen inkubierten Kulturen (gelb) signifikant erhöhte Protein-Expression zu beobachten (9,8 ± 2,9 % (DEX) bzw. 5,4 ± 3,2 % (C) im Vergleich zu 0,7 ± 0,6 % (IFN-γ/TNF-α)). Die Signifikanz zwischen den Gruppen wur-de durch eine ANOVA (analysis of variance) mit anschließendem Fisher post-hoc Test bestimmt (∗∗ ∗ = p<0,001; ∗∗ = p<0,01).
Die quantitative Auswertung bestätigte, dass eine 48-stündige Behandlung von Oligodendrocyten-Vorläuferzellen mit Corticoiden keinen signifikanten Einfluss auf die Expression des Proteins MBP am Kulturtag 9 hatte (blaue Säulen: 17,7 ± 8,6 % (K); 20,1 ± 3,7 % (K + C); 19,7 ± 4,1 % (K + DC)
und 23,4 ± 4,9 % (K + DEX)). Kulturen, welche über den gleichen Zeitraum mit IFN-γ und TNF-α
behandelt wurden, zeigten eine gegenüber der Kontrolle stark verringerte MBP-Expression
(0,7 ± 0,6 % (IFN-γ + TNF-α) gegenüber 17,7 ± 8,6 % (K)). Bei gleichzeitiger Gabe von Corticoiden
wurde eine durch Corticosteron bzw. Dexamethason signifikant verstärkte Expression des MBP beo-bachtet (gelb-rote Säulen). Der Effekt von Dexamethason war mit 9,8 ± 2,9 % (p<0,001; im Vergleich
zu IFN-γ + TNF-α) stärker als der von Corticosteron (5,5 ± 3,2 % (p<0,01; im Vergleich zu IFN-γ +
TNF-α)), wohingegen Deoxycorticosteron keinen signifikanten Einfluss zeigte (3,2 ± 2,8 %). Eine
vollständige Aufhebung der durch IFN-γ und TNF-α induzierten verringerten MBP-Expression wurde
mit keinem der drei Corticoide erzielt. Durch das am stärksten wirksame Corticoid Dexamethason wurde eine Expression erreicht, welche halb so hoch war, wie die von Kontroll-Kulturen.

Ergebnisse 72
4.4.2.2 Myelin Associated Glycoprotein
Repräsentativ für vier unabhängige Versuche ist in der folgenden Abbildung ein MAG-Western-Blot abgebildet:
Abbildung 31: MAG-Western-Blot nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9) In den Lysaten von nicht mit IFN-γ und TNF-α behandelten Zellen war die Expression des Myelin Associ-ated Glycoprotein deutlich nachweisbar. Durch die Corticoide kam es zu keiner sichtbaren Veränderung. Die Behandlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α führte zu einer im Vergleich zu den Kontrollen sehr schwachen Expression des Proteins. Bei gleichzeitiger Gabe von Corticoiden konnte ein positiver Einfluß des Glucocorticoids Dexamethason (DEX) beobachtet werden.
Die durch die Cytokine induzierte verringerte Expression des MAG schien durch die gleichzeitige Gabe von Dexamethason teilweise aufgehoben zu werden. Um zu überprüfen, ob dieser Effekt signifi-kant war, wurden vier Blots aus vier unabhängigen Versuchen wie oben beschrieben quantitativ aus-gewertet. Das Ergebnis ist in Abbildung 32 [S.73] graphische dargestellt.
Die quantitative Auswertung ergab, dass eine 48-stündige Behandlung von Oligodendrocyten-
Vorläuferzellen mit Corticoiden weder in gleichzeitig mit IFN-γ und TNF-α behandelten Kulturen
(gelb-rote Säulen) noch in nur mit Corticoiden behandelten Kulturen (blaue Säulen) einen signifikan-ten Einfluss auf die Expression des Proteins MAG am Kulturtag 9 hatte (gelb-rote Säulen: 0,9 ± 0,9 %
(IFN-γ/TNF-α); 4,3 ± 3,0 % (IFN-γ/TNF-α + C); 0,3 ± 0,2 % (IFN-γ/TNF-α + DC) und 5,8 ± 4,4 %
(IFN-γ/TNF-α + DEX); blaue Säulen: 18,4 ± 4,9 % (K); 22,7 ± 3,8 % (K + C); 19,3 ± 2,4 % (K + DC)
und 27,2 ± 5,9 % (K + DEX)).
Kulturen, welche nur mit IFN-γ und TNF-α behandelt wurden, zeigten eine gegenüber der Kontrolle
stark verringerte MAG-Expression (0,9 ± 0,9 % (IFN-γ + TNF-α) gegenüber 18,4 ± 4,9 % (K)). Mit
keinem der drei Corticoide konnte dieser Effekt der Cytokine aufgehoben oder zumindest abge-schwächt werden.
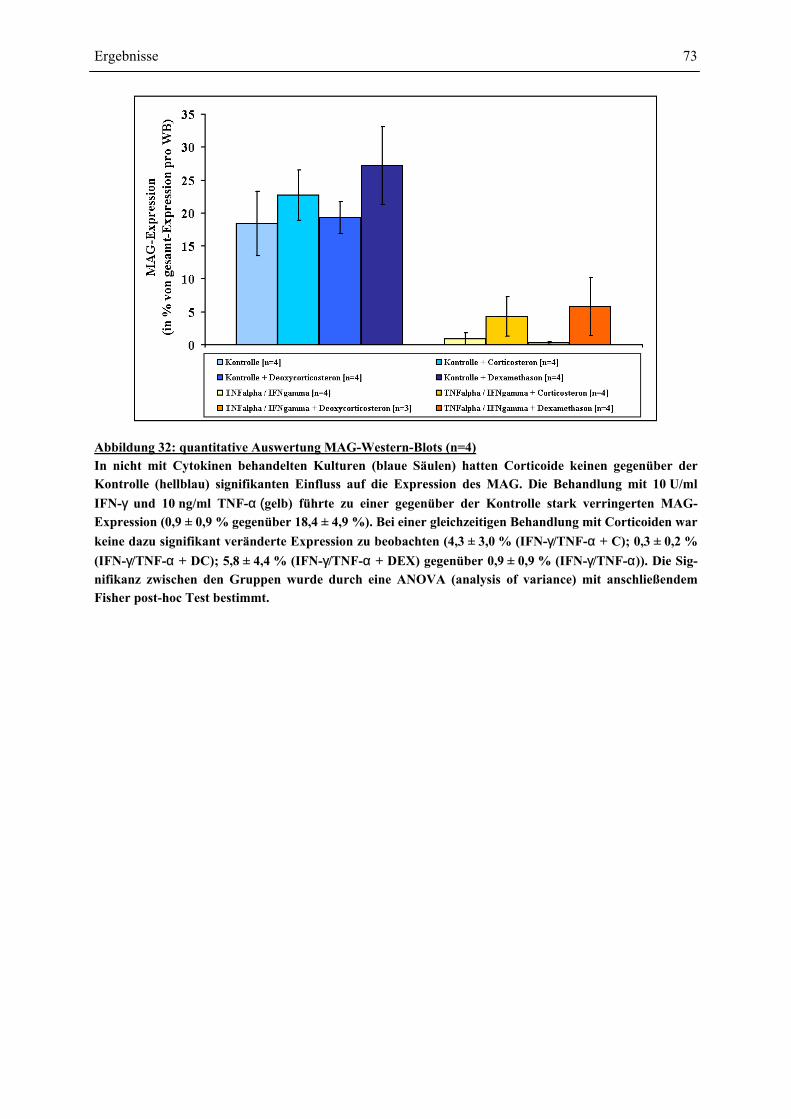
Ergebnisse 73
Abbildung 32: quantitative Auswertung MAG-Western-Blots (n=4) In nicht mit Cytokinen behandelten Kulturen (blaue Säulen) hatten Corticoide keinen gegenüber der Kontrolle (hellblau) signifikanten Einfluss auf die Expression des MAG. Die Behandlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α (gelb) führte zu einer gegenüber der Kontrolle stark verringerten MAG-Expression (0,9 ± 0,9 % gegenüber 18,4 ± 4,9 %). Bei einer gleichzeitigen Behandlung mit Corticoiden war keine dazu signifikant veränderte Expression zu beobachten (4,3 ± 3,0 % (IFN-γ/TNF-α + C); 0,3 ± 0,2 % (IFN-γ/TNF-α + DC); 5,8 ± 4,4 % (IFN-γ/TNF-α + DEX) gegenüber 0,9 ± 0,9 % (IFN-γ/TNF-α)). Die Sig-nifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem Fisher post-hoc Test bestimmt.

Ergebnisse 74
4.4.2.3 2’,3’-Cyclische Nucleotid 3’-Phosphodiesterase
Der Einfluss auf die Expression dieses Proteins durch eine Behandlung mit Corticoiden mit bzw. ohne
IFN-γ und TNF-α ist in Form eines für sechs Versuche repräsentativen Western-Blots in der folgenden
Abbildung gezeigt:
Abbildung 33: CNP-Western-Blot nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9) In den Lysaten von nicht mit IFN-γ und TNF-α behandelten Zellen war die Expression der 2’,3’-Cyclischen Nucleotid 3’-Phosphodiesterase deutlich nachweisbar. Durch die Corticoide kam es zu keiner sichtbaren Verstärkung. Die Behandlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α führte zu einer im Ver-änderung zu den Kontrollen sehr schwachen Expression des Proteins. Bei gleichzeitiger Gabe von Corti-coiden konnte ein positiver Einfluss der Glucocorticoide Corticosteron (C) und Dexamethason (DEX) beobachtet werden.
Durch gleichzeitige Gabe von Corticoiden und Cytokinen schien die durch die Cytokine induzierte verringerte Expression des CNP zum Teil aufgehoben zu werden. Um zu überprüfen, ob dieser Effekt der Corticoide signifikant war, wurden sechs Blots aus sechs unabhängigen Versuchen wie oben be-schrieben quantitativ ausgewertet. Abbildung 34 [S.75] zeigt die graphische Darstellung des Ergebnis-ses.
Die quantitative Auswertung bestätigte, dass eine 48-stündige Behandlung von Oligodendrocyten-Vorläuferzellen mit Corticoiden keinen signifikanten Einfluss auf die Expression des Enzyms CNP hatte (blaue Säulen: 14,2 ± 8,5 % (K); 15,1 ± 3,9 % (K + C); 17,6 ± 4,9 % (K + DC) und 16,6 ± 4,6 %
(K + DEX)). Kulturen, welche über den gleichen Zeitraum mit IFN-γ und TNF-α behandelt wurden,
zeigten eine gegenüber der Kontrolle verringerte CNP-Expression (4,1 ± 3,4 % (IFN-γ + TNF-α) ge-
genüber 17,7 ± 8,6 % (K)). Bei gleichzeitiger Gabe von Corticoiden wurde eine durch Corticosteron bzw. Dexamethason signifikant verstärkte Expression des CNP beobachtet (gelb-rote Säulen:
11,0 ± 5,5 % (IFN-γ/TNF-α + C) bzw. 12,4 ± 4,3 % (IFN-γ/TNF-α + DEX) gegenüber 4,1 ± 3,4
(IFN-γ/TNF-α) mit jeweils p<0,05), wohingegen Deoxycorticosteron keinen signifikanten Effekt zeig-
te. Eine vollständige Aufhebung der durch IFN-γ und TNF-α induzierten verringerten CNP-
Expression wurde dabei mit keinem der beiden Glucocorticoide erzielt. Die mit Corticosteron bzw. Dexamethason erreichte Expression entsprach nur ca. 75 % bzw. ca. 85 % der Expression von Kon-troll-Kulturen.
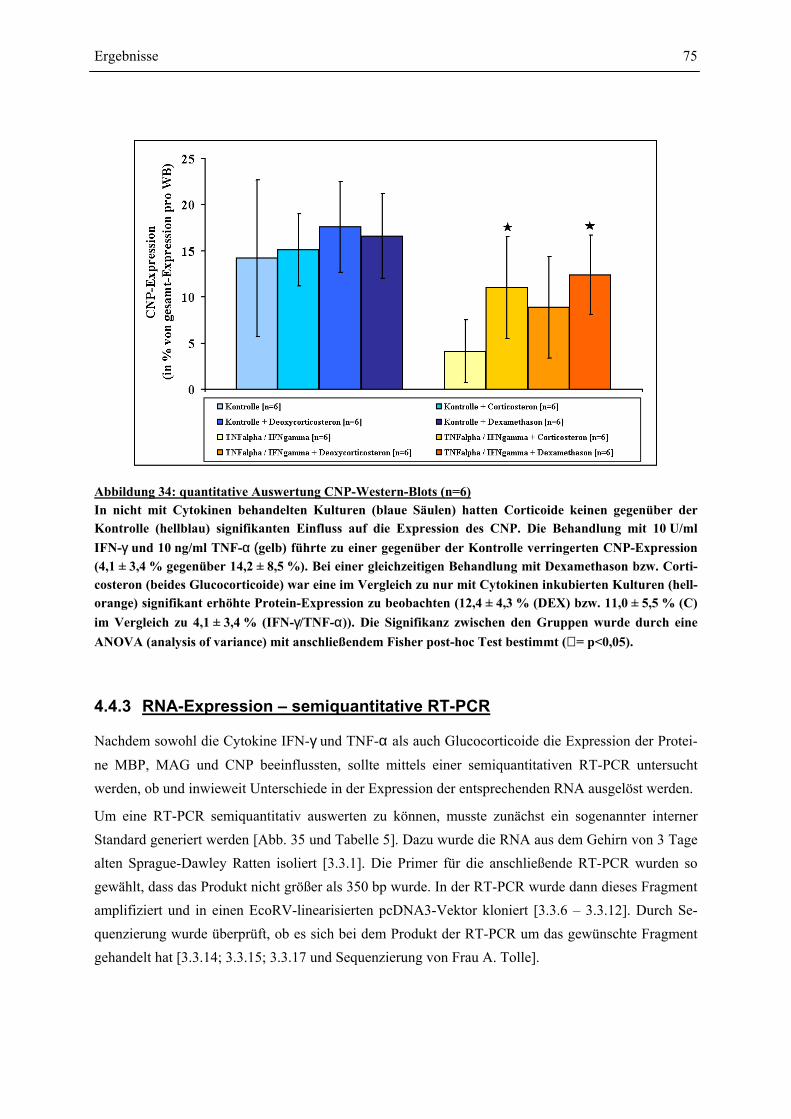
Ergebnisse 75
Abbildung 34: quantitative Auswertung CNP-Western-Blots (n=6) In nicht mit Cytokinen behandelten Kulturen (blaue Säulen) hatten Corticoide keinen gegenüber der Kontrolle (hellblau) signifikanten Einfluss auf die Expression des CNP. Die Behandlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α (gelb) führte zu einer gegenüber der Kontrolle verringerten CNP-Expression (4,1 ± 3,4 % gegenüber 14,2 ± 8,5 %). Bei einer gleichzeitigen Behandlung mit Dexamethason bzw. Corti-costeron (beides Glucocorticoide) war eine im Vergleich zu nur mit Cytokinen inkubierten Kulturen (hell-orange) signifikant erhöhte Protein-Expression zu beobachten (12,4 ± 4,3 % (DEX) bzw. 11,0 ± 5,5 % (C) im Vergleich zu 4,1 ± 3,4 % (IFN-γ/TNF-α)). Die Signifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem Fisher post-hoc Test bestimmt (∗ = p<0,05).
4.4.3 RNA-Expression – semiquantitative RT-PCR
Nachdem sowohl die Cytokine IFN-γ und TNF-α als auch Glucocorticoide die Expression der Protei-
ne MBP, MAG und CNP beeinflussten, sollte mittels einer semiquantitativen RT-PCR untersucht werden, ob und inwieweit Unterschiede in der Expression der entsprechenden RNA ausgelöst werden.
Um eine RT-PCR semiquantitativ auswerten zu können, musste zunächst ein sogenannter interner Standard generiert werden [Abb. 35 und Tabelle 5]. Dazu wurde die RNA aus dem Gehirn von 3 Tage alten Sprague-Dawley Ratten isoliert [3.3.1]. Die Primer für die anschließende RT-PCR wurden so gewählt, dass das Produkt nicht größer als 350 bp wurde. In der RT-PCR wurde dann dieses Fragment amplifiziert und in einen EcoRV-linearisierten pcDNA3-Vektor kloniert [3.3.6 – 3.3.12]. Durch Se-quenzierung wurde überprüft, ob es sich bei dem Produkt der RT-PCR um das gewünschte Fragment gehandelt hat [3.3.14; 3.3.15; 3.3.17 und Sequenzierung von Frau A. Tolle].

Ergebnisse 76
Abbildung 35: Klonierungs-Strategie Positiv-Kontrolle und Interner Standard Zunächst wurde aus Ratten-Hirn Gesamt-RNA isoliert. Aus dieser RNA wurde dann mittels RT-PCR (1. PCR, Primer siehe Tabelle 5) ein Fragment der relevanten mRNA von ca. 300-350 bp amplifiziert. Die benutzten Primer sollten auch bei der späteren semiquantitativen RT-PCT eingesetzt werden, so dass es sich bei dem erhaltenen Fragment um die Positiv-Kontrolle handelte. Nach Klonierung in einen EcoRV geschnittenen Vektor und Überprüfung der Sequenz wurden aus dieser Positiv-Kontrolle in zwei getrenn-ten PCR-Reaktionen das 1. SOE- und das 2. SOE-Fragment amplifiziert (2. und 3. PCR, Primer siehe Tabelle 5). In der letzten PCR wurden die dabei erhaltenen Fragmente als Matrize eingesetzt und mitein-ander verbunden. Bei dem Produkt handelte es sich um den internen Standard, welcher sich um 50 bp von der Positiv-Kontrolle unterschied. Nach Klonierung und Sequenzüberprüfung konnte aus dem Vek-tor der interne Standard mittels in vitro-Transkription generiert werden.
Ein interner Standard sollte so gewählt werden, dass für dessen Amplifizierung die selben Primer verwendet werden, wie für die zu untersuchende RNA. Außerdem sollte der Längenunterschied zwi-schen dem Standard und dem Fragment der RT-PCR nicht größer als 50 bp sein und die Sequenzen sollten sich so wenig wie möglich voneinander unterscheiden. Mit diesen Vorraussetzungen ist eine Vergleichbarkeit der beiden Fragmente am besten gesichert.
Im nächsten Schritt sollte deshalb ein Stück von ca. 50 bp aus dem zuvor klonierten RT-PCR Frag-ment herauskloniert werden. Dazu wurden in je einer PCR die Fragmente für eine anschließende SOE-PCR amplifiziert [siehe Tabelle 5]. Mittels der SOE-PCR wurden die beiden Fragmente zusammenge-fügt und dann in einen EcoRV-linearisierten pcDNA3-Vektor kloniert [3.3.6 – 3.3.12]. Durch Sequen-zierung wurde überprüft, ob es sich bei dem Produkt der SOE-PCR um das gewünschte Fragment gehandelt hatte [3.3.14; 3.3.15; 3.3.17 und Sequenzierung von Frau A. Tolle].
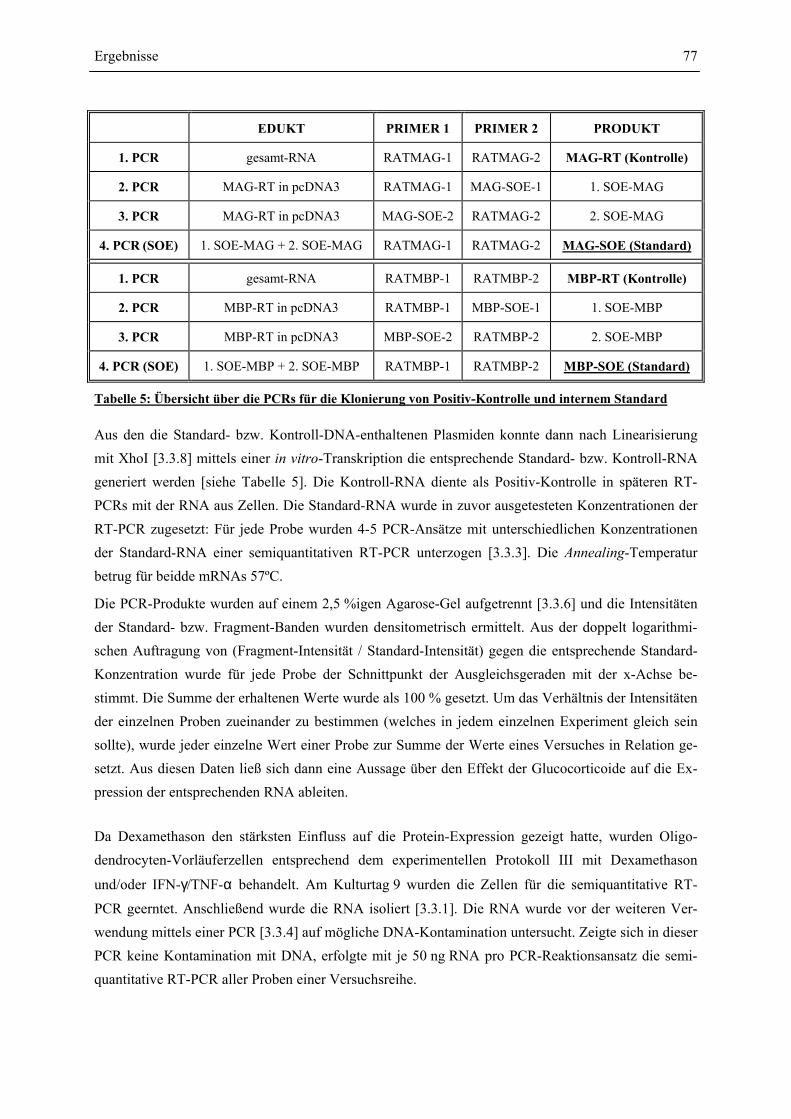
Ergebnisse 77
EDUKT PRIMER 1 PRIMER 2 PRODUKT
1. PCR gesamt-RNA RATMAG-1 RATMAG-2 MAG-RT (Kontrolle)
2. PCR MAG-RT in pcDNA3 RATMAG-1 MAG-SOE-1 1. SOE-MAG
3. PCR MAG-RT in pcDNA3 MAG-SOE-2 RATMAG-2 2. SOE-MAG
4. PCR (SOE) 1. SOE-MAG + 2. SOE-MAG RATMAG-1 RATMAG-2 MAG-SOE (Standard)
1. PCR gesamt-RNA RATMBP-1 RATMBP-2 MBP-RT (Kontrolle)
2. PCR MBP-RT in pcDNA3 RATMBP-1 MBP-SOE-1 1. SOE-MBP
3. PCR MBP-RT in pcDNA3 MBP-SOE-2 RATMBP-2 2. SOE-MBP
4. PCR (SOE) 1. SOE-MBP + 2. SOE-MBP RATMBP-1 RATMBP-2 MBP-SOE (Standard)
Tabelle 5: Übersicht über die PCRs für die Klonierung von Positiv-Kontrolle und internem Standard
Aus den die Standard- bzw. Kontroll-DNA-enthaltenen Plasmiden konnte dann nach Linearisierung mit XhoI [3.3.8] mittels einer in vitro-Transkription die entsprechende Standard- bzw. Kontroll-RNA generiert werden [siehe Tabelle 5]. Die Kontroll-RNA diente als Positiv-Kontrolle in späteren RT-PCRs mit der RNA aus Zellen. Die Standard-RNA wurde in zuvor ausgetesteten Konzentrationen der RT-PCR zugesetzt: Für jede Probe wurden 4-5 PCR-Ansätze mit unterschiedlichen Konzentrationen der Standard-RNA einer semiquantitativen RT-PCR unterzogen [3.3.3]. Die Annealing-Temperatur betrug für beidde mRNAs 57ºC.
Die PCR-Produkte wurden auf einem 2,5 %igen Agarose-Gel aufgetrennt [3.3.6] und die Intensitäten der Standard- bzw. Fragment-Banden wurden densitometrisch ermittelt. Aus der doppelt logarithmi-schen Auftragung von (Fragment-Intensität / Standard-Intensität) gegen die entsprechende Standard-Konzentration wurde für jede Probe der Schnittpunkt der Ausgleichsgeraden mit der x-Achse be-stimmt. Die Summe der erhaltenen Werte wurde als 100 % gesetzt. Um das Verhältnis der Intensitäten der einzelnen Proben zueinander zu bestimmen (welches in jedem einzelnen Experiment gleich sein sollte), wurde jeder einzelne Wert einer Probe zur Summe der Werte eines Versuches in Relation ge-setzt. Aus diesen Daten ließ sich dann eine Aussage über den Effekt der Glucocorticoide auf die Ex-pression der entsprechenden RNA ableiten.
Da Dexamethason den stärksten Einfluss auf die Protein-Expression gezeigt hatte, wurden Oligo-dendrocyten-Vorläuferzellen entsprechend dem experimentellen Protokoll III mit Dexamethason
und/oder IFN-γ/TNF-α behandelt. Am Kulturtag 9 wurden die Zellen für die semiquantitative RT-
PCR geerntet. Anschließend wurde die RNA isoliert [3.3.1]. Die RNA wurde vor der weiteren Ver-wendung mittels einer PCR [3.3.4] auf mögliche DNA-Kontamination untersucht. Zeigte sich in dieser PCR keine Kontamination mit DNA, erfolgte mit je 50 ng RNA pro PCR-Reaktionsansatz die semi-quantitative RT-PCR aller Proben einer Versuchsreihe.
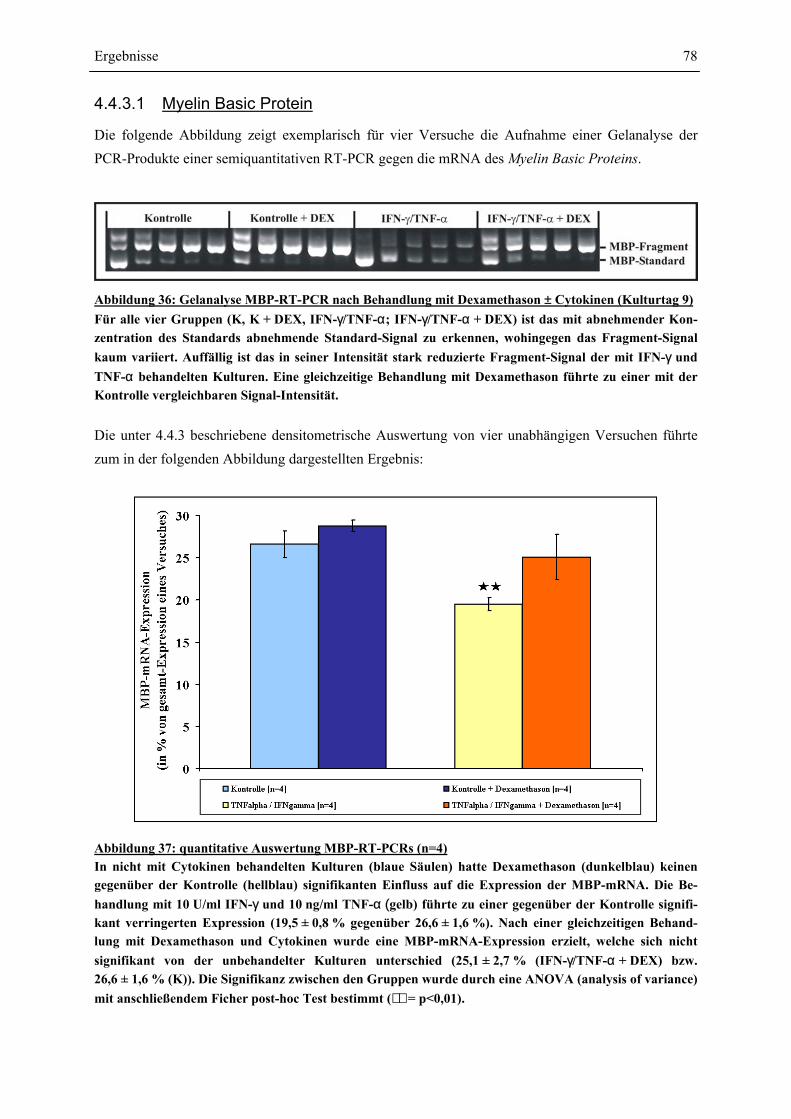
Ergebnisse 78
4.4.3.1 Myelin Basic Protein
Die folgende Abbildung zeigt exemplarisch für vier Versuche die Aufnahme einer Gelanalyse der PCR-Produkte einer semiquantitativen RT-PCR gegen die mRNA des Myelin Basic Proteins.
Abbildung 36: Gelanalyse MBP-RT-PCR nach Behandlung mit Dexamethason ± Cytokinen (Kulturtag 9) Für alle vier Gruppen (K, K + DEX, IFN-γ/TNF-α; IFN-γ/TNF-α + DEX) ist das mit abnehmender Kon-zentration des Standards abnehmende Standard-Signal zu erkennen, wohingegen das Fragment-Signal kaum variiert. Auffällig ist das in seiner Intensität stark reduzierte Fragment-Signal der mit IFN-γ und TNF-α behandelten Kulturen. Eine gleichzeitige Behandlung mit Dexamethason führte zu einer mit der Kontrolle vergleichbaren Signal-Intensität.
Die unter 4.4.3 beschriebene densitometrische Auswertung von vier unabhängigen Versuchen führte zum in der folgenden Abbildung dargestellten Ergebnis:
Abbildung 37: quantitative Auswertung MBP-RT-PCRs (n=4) In nicht mit Cytokinen behandelten Kulturen (blaue Säulen) hatte Dexamethason (dunkelblau) keinen gegenüber der Kontrolle (hellblau) signifikanten Einfluss auf die Expression der MBP-mRNA. Die Be-handlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α (gelb) führte zu einer gegenüber der Kontrolle signifi-kant verringerten Expression (19,5 ± 0,8 % gegenüber 26,6 ± 1,6 %). Nach einer gleichzeitigen Behand-lung mit Dexamethason und Cytokinen wurde eine MBP-mRNA-Expression erzielt, welche sich nicht signifikant von der unbehandelter Kulturen unterschied (25,1 ± 2,7 % (IFN-γ/TNF-α + DEX) bzw. 26,6 ± 1,6 % (K)). Die Signifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem Ficher post-hoc Test bestimmt (∗∗ = p<0,01).

Ergebnisse 79
Die quantitative Auswertung ergab, dass eine 48-stündige Behandlung von Oligodendrocyten-Vorläuferzellen mit Dexamethason keinen signifikanten Einfluss auf die Expression der mRNA des Myelin Basic Protein hatte (blaue Säulen: 26,6 ± 1,6 % (K) und 28,8 ± 0,7 % (K + DEX)). Kulturen,
welche über den gleichen Zeitraum mit IFN-γ und TNF-α behandelt wurden, zeigten eine gegenüber
der Kontrolle signifikant verringerte MBP-mRNA-Expression (19,5 ± 0,8 % (IFN-γ + TNF-α) gegen-
über 26,6 ± 1,6 % (K); p<0,01). Die gleichzeitige Gabe von Dexamethason führte am Kulturtag 9 zu einer mit unbehandelten Kontrollen vergleichbaren mRNA-Expression des MBP. Auf RNA-Ebene
induziert Dexamethason demnach eine Aufhebung des durch IFN-γ und TNF-α ausgelösten Effektes
(25,1 ± 2,7 % (IFN-γ/TNF-α + DEX) entsprechend 26,6 ± 1,6 (K)).
4.4.3.2 Myelin Associated Glycoprotein
Die Gelanalyse der PCR-Produkte einer semiquantitativen RT-PCR gegen die mRNA des Myelin Associated Glycoprotein ist in Abbildung 38 gezeigt.
Abbildung 38: Gelanalyse MAG-RT-PCR nach Behandlung mit Dexamethason ± Cytokinen (Kulturtag 9) Für alle vier Gruppen (K, K + DEX, IFN-γ/TNF-α; IFN-γ/TNF-α + DEX) ist das mit abnehmender Kon-zentration des Standards abnehmende Standard-Signal zu erkennen, wohingegen das Fragment-Signal kaum variiert. Auffällig ist das in seiner Intensität stark reduzierte Fragment-Signal der mit IFN-γ und TNF-α behandelten Kulturen. Eine gleichzeitige Behandlung mit Dexamethason führte zu einer im Ver-gleich dazu erhöhten Signal-Intensität.
Die semiquantitative Auswertung der RT-PCRs aus drei unabhängigen Versuchen führte zu dem in Abbildung 39 [S.80] graphisch dargestellten Ergebnis.
Die Auswertung der MAG-RT-PCR ergab entsprechend der MBP-RT-PCR-Auswertung, dass eine 48-stündige Behandlung von Oligodendrocyten-Vorläuferzellen mit Dexamethason keinen signifikanten Einfluss auf die Expression der mRNA dieses Myelin-spezifischen Proteins hatte (blaue Säulen: 25,7 ± 2,8 % (K) und 31,4 ± 3,0 % (K + DEX)). Kulturen, welche über den gleichen Zeitraum mit
IFN-γ und TNF-α behandelt wurden, zeigten eine gegenüber der Kontrolle signifikant verringerte
MAG-mRNA-Expression (17,8 ± 3,4 % (IFN-γ + TNF-α) gegenüber 25,7 ± 2,8 % (K); p<0,01). Auch
beim MAG führte die gleichzeitiger Gabe von Dexamethason zu einer am Kulturtag 9 mit unbehandel-ten Kontrollen vergleichbaren mRNA-Expression des MAG. Entgegen der Ergebnisse auf Protein-
Level wurde durch Dexamethason eine Aufhebung des durch IFN-γ und TNF-α ausgelösten Effektes
erzielt (26,1 ± 3,8 % (IFN-γ/TNF-α + DEX) entsprechend 25,7 ± 2,8 (K)).
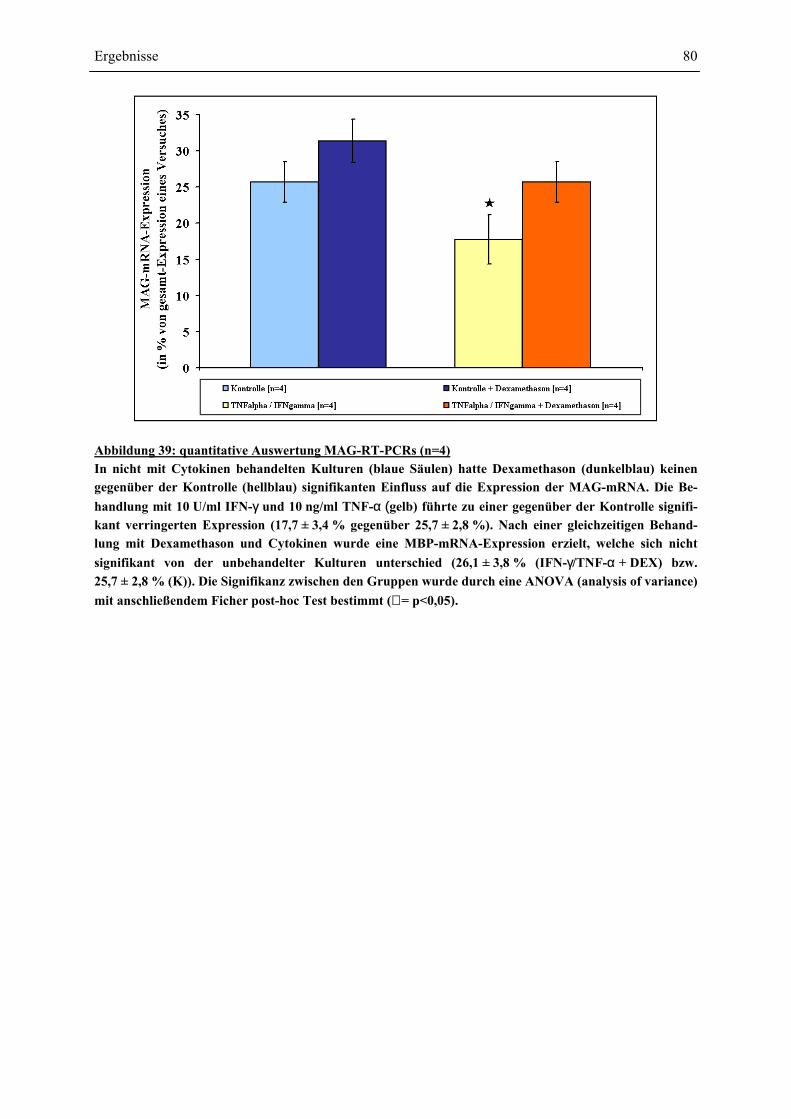
Ergebnisse 80
Abbildung 39: quantitative Auswertung MAG-RT-PCRs (n=4) In nicht mit Cytokinen behandelten Kulturen (blaue Säulen) hatte Dexamethason (dunkelblau) keinen gegenüber der Kontrolle (hellblau) signifikanten Einfluss auf die Expression der MAG-mRNA. Die Be-handlung mit 10 U/ml IFN-γ und 10 ng/ml TNF-α (gelb) führte zu einer gegenüber der Kontrolle signifi-kant verringerten Expression (17,7 ± 3,4 % gegenüber 25,7 ± 2,8 %). Nach einer gleichzeitigen Behand-lung mit Dexamethason und Cytokinen wurde eine MBP-mRNA-Expression erzielt, welche sich nicht signifikant von der unbehandelter Kulturen unterschied (26,1 ± 3,8 % (IFN-γ/TNF-α + DEX) bzw. 25,7 ± 2,8 % (K)). Die Signifikanz zwischen den Gruppen wurde durch eine ANOVA (analysis of variance) mit anschließendem Ficher post-hoc Test bestimmt (∗ = p<0,05).

Diskussion 81
5 DISKUSSION Oligodendrocyten-Vorläuferzellen wird eine wichtige Rolle in der Entstehung einer periventrikulären Leukomalazie zugesprochen. Sie bilden in dem Zeitraum der Entwicklung der humanen cerebralen weißen Substanz, in welchem das Risiko für die Entstehung einer PVL nachweislich sehr hoch ist, die dort dominierende Zellpopulation [Back et al., 2001].
Es werden verschiedene Faktoren diskutiert, welche zu der für eine diffuse PVL charakteristischen Störung der Myelinisierung führen können [Übersicht in Volpe, 2001]. In den letzten Jahren wird dabei mütterlichen und fetalen Infektionen eine wachsende Bedeutung in der Entstehung von frühge-burtlichen Hirnschädigungen zugeschrieben. Neben der durch Infektionen ausgelösten Kreislaufdysre-gulation, welche dann zu einer Hirnschädigung führen kann [Garnier et al., 2001], lassen immer mehr Studien den Schluss zu, dass ein direkter toxischer Effekt, von beispielsweise während einer Infektion freigesetzten Cytokinen, auf Oligodendrocyten-Vorläuferzellen ein weiterer möglicher Mechanismus in der Entstehung von Hirnschäden sein könnte.
Ein Auslösen des Zelltod-Programms von Oligodendrocyten-Vorläuferzellen oder eine Störung der Differenzierung zu reifen Oligodendrocyten sind mögliche Mechanismen, durch welche Cytokine eine Störung der Myelinisierung in der weißen Substanz von betroffenen Kindern induzieren könnten. Im Rahmen dieser Arbeit wurde deshalb, um die These eines direkten toxischen Effektes von Cytokinen auf Oligodendrocyten-Vorläuferzellen zu untersuchen, die Charakterisierung der Zellkultur und der Folgen einer Cytokin-Behandlung auf diese beiden Mechanismen hin vorgenommen.
5.1 Etablierung und Charakterisierung der Oligodendrocyten-Vorläuferzell-Kultur
Ziel der vorliegenden Arbeit war es, die Rolle von Oligodendrocyten im Vorläuferzell-Stadium in der Entstehung einer periventrikulären Leukomalazie tiefergehend zu untersuchen. Dazu musste zunächst eine Zellkultur von Oligodendrocyten-Vorläuferzellen etabliert werden, welche so wenig wie möglich mit Astrocyten verunreinigt war. Dies war erforderlich, da Astrocyten Hormone und Cytokine produ-
zieren können, welche die biologische Aktivität von beispielsweise TNF-α beeinflussen können
[Rothwell und Relton, 1993; Hopkins und Rothwell, 1995]. Bei einer zu starken Kontamination mit Astrocyten hätten dann die Ergebnisse der Experimente nicht direkt auf Oligodendrocyten-Vorläuferzellen bezogen werden können.
Die Kultivierungsbedingungen wurden so gewählt, dass die Zellen zum einen gezielt im Vorläuferzell-Stadium gehalten werden können und zum anderen ebenso gezielt dazu angeregt werden können, zu reifen Oligodendrocyten auszudifferenzieren. Die Kombination von PDGF und bFGF wurde zur Un-terdrückung der Differenzierung gewählt, so dass die Zellen im Vorläufer-Zell-Stadium verweilten sollten [Bogler et al., 1990]. Um die Differenzierung zu reifen Oligodendrocyten anzuregen, wurde die Kombination von T3, CNTF und Forskolin verwendet [Back et al., 1998].

Diskussion 82
Unter den in Abbildung 8 angegebenen allgemeinen Kultivierungsbedingungen war zu erwarten, dass die Zellen in den ersten drei Tagen morphologisch und immuncytochemisch als Oligodendrocyten-Vorläuferzellen charakterisiert werden können, während nach dem Mediumwechsel auf Differenzie-rungsmedium die Zellen immer mehr Eigenschaften (Morphologie, Immuncytochemie, Protein-Expression) von reifen Oligodendrocyten zeigen sollten.
5.1.1 Immuncytochemie
Oligodendrocyten-Vorläuferzellen können durch Immuncytochemie mit dem monoklonalen Antikör-per A2B5 charakterisiert werden [Fredman et al., 1984; Dubois et al., 1990], wohingegen reife Oligo-dendrocyten immuncytochemisch unter anderem mit einem monoklonalen Antikörper, welcher gegen das Myelin Oligodendrocyte Specific Protein gerichtet ist, identifiziert werden können. Dieses Protein ist ein Oberflächenprotein, welches von Oligodendrocyten des ZNS exprimiert wird, sobald die Zellen beginnen, Myelin zu bilden [Dyer et al., 1991; Mu und Dyer 1994].
Die nach einem Protokoll von Simpson aufgereinigten Zellen zeigten in den ersten drei Tagen eine hohe Immunreaktivität gegen den Antikörper A2B5 [siehe Abb. 9] [Simpson et al., 1997]. Auszählun-gen der A2B5-Immuncytochemie ergaben, dass die Kulturen am Tag 3 aus durchschnittlich 90 % A2B5-positiven Zellen, d.h. Oligodendrocyten-Vorläuferzellen, bestanden [siehe Abb. 21]. Die sehr geringe Immunreaktivität gegen den Astrocyten-spezifischen monoklonalen Antikörper GFAP [siehe Abb. 12] zeigt ebenfalls, dass es sich bei der etablierten Zellkultur um eine Kultur von hoher Reinheit handelte.
Durch die Immuncytochemie gegen MOSP konnte gezeigt werden, dass die Oligodendrocyten-Vorläuferzellen durch dem Wechsel auf Differenzierungsmedium [Back et al., 1998] erfolgreich dazu angeregt wurden auszudifferenzieren [siehe Abb. 10]: Sowohl die Anzahl von Zellen, welche die Morphologie reifer Oligodendrocyten zeigten, nahm zu, als auch die Anzahl MOSP-positiver Zellen.
5.1.2 Protein-Expression
Um zu untersuchen, ob Cytokine direkt zu einer Störung der Differenzierung von Oligodendrocyten-Vorläuferzellen führen können, sollte neben den morphologischen und immuncytochemischen Be-trachtungen, auch der Einfluss auf die Proteinexpression einiger Myelin-spezifischer Proteine charak-terisiert werden. Die folgenden Proteine wurden aus den folgenden Gründen für diese Charakterisie-rung ausgewählt:
Das Myelin Basic Protein gehört neben dem Proteolipid-Protein zu den Haupt-Proteinen des zentralen (30 %) und des peripheren (18 %) Nervensystems. Diese Proteine werden in der Entwicklung von Oligodendrocyten erst im letzten Stadium, im Stadium der reifen Oligodendrocyten, exprimiert, wes-halb sie auch zur Charakterisierung dieses Zellstadiums dienen [siehe Abb. 4]. Es existieren verschie-dene Isoformen des MBP, die in der Größe von 14 bis 21 kDa variieren. Der in dieser Arbeit verwen-dete Antikörper erkennt hauptsächlich die 21 kDa-Form.

Diskussion 83
Das Myelin Associated Glycoprotein macht sowohl im Zentralen als auch im peripheren Nervensystem quantitativ nur einen sehr geringen Anteil am Gesamt-Protein aus. Trotzdem ist es vermutlich gerade in der Entwicklung von demyelinisierenden Krankheiten von großer Bedeutung: Bei Patienten mit Multipler Sklerose war dieses Protein gegenüber anderen Myelin-spezifischen Proteinen am Rande von sich entwickelnden Plaques am stärksten reduziert [Quarles et al., 1992]. Aus diesem Grunde wurde auch seine Expression im Rahmen dieser Arbeit untersucht.
Die CNPase wird früher in der Entwicklung von Oligodendrocyten exprimiert als MBP und MAG: Die Expression beginnt bereits im Stadium der unreifen Oligodendrocyten [siehe Abb. 4]. In Mäusen führte eine Überexpression dieses Enzyms zu einer in der Entwicklung früher einsetzenden Expression der Proteine MBP und CNP, zu einer erhöhten Anzahl verlängerter Fortsätze der Oligodendrocyten und zu einem ungewöhnlich hohen Gehalt an Myelin [Gravel et al., 1996]. Aufgrund dieser Ergebnis-se nimmt man an, dass die CNPase eine große Rolle in der Regulation der Myelinisierung von Axonen spielt. Das Enzym war somit auch im Rahmen dieser Arbeit von großem Interesse.
Da die Expression dieser drei Myelin-spezifischen Proteine erst mit Fortschreiten der Entwicklung von Oligodendrocyten einsetzt und dann ansteigt [Übersicht in Back und Volpe, 1997; Abb. 4], war sie auch dazu geeignet, die Zellkultur zu charakterisieren. Western-Blot-Analysen zeigten, dass die Ex-pression der Proteine MBP und CNP mit Fortschreiten der Kultur zunahm, woraus man schließen kann, dass die Entwicklung zu reifen Oligodendrocyten unter den verwendeten Kultur-Bedingungen erfolgreich angeregt werden konnte. Gleichzeitig war in der Phase der Kultivierung in Proliferations-medium keine Expression dieser Proteine nachweisbar (Tag 1 und Tag 3) [siehe Abb. 11].
Zusammenfassend kann man sagen, dass durch die gewählten Kultur-Bedingungen die Vorrausset-zungen für eine gezielte Untersuchung des Einflusses von Cytokinen auf das Überleben und die Diffe-renzierung von Oligodendrocyten-Vorläuferzellen erfüllt waren.

Diskussion 84
5.2 Einfluss von TNF-α auf das Überleben von Oligodendrocyten-Vorläuferzellen
Als Ursache für eine periventrikuläre Leukomalazie bei frühgeborenen Kindern gewinnen in den letz-ten Jahren mütterliche und fetale Infektionen an Bedeutung. Beispielsweise konnten Lipopolysaccha-ride, Bestandteile der Zellmembran gram-negativer Bakterien, nach intraperitonealer Injektion in die Muttertiere direkt mit einem Schaden der weißen Hirnsubstanz der Rattenfeten in Verbindung ge-
bracht werden [Cai et al., 2000]. Gleichzeitig wurde eine erhöhte Expression von IL-1β und TNF-α
beobachtet. Folgende in vivo-Studien zeigten ebenfalls eine erhöhte Expression von TNF-α und einen
auch Oligodendrocyten betreffenden Hirnschaden: Nach einer Ligatur beider Halsschlagadern von Ratten wurde unter anderem ein Schaden der weißen Substanz, sowie mittels RT-PCR eine erhöhte
Expression von TNF-α und seinem Rezeptor TNFR-1 nachgewiesen [Masumura et al., 2001]. Auch
die Arbeitsgruppe um Lee hatte schon eine Verbindung zwischen dem apoptotischen Zelltod von Oli-
godendrocyten und einem erhöhten Level an TNF-α beobachtet [Lee et al., 2000]. In transgenen Mäu-
sen, welche entweder eine ZNS-spezifische Expression von TNF-α aufwiesen, oder eine Astrocyten-
spezifische Expression des membrangebundenen TNF-α, wurden ebenfalls Oligodendrocyten- und
Myelin-Schädigungen bzw. eine selektive Apoptose von Oligodendrocyten vermittelt [Akassoglou et
al. 1998]. Da TNF-α demnach nicht nur in Kombination mit IL-1β [Cai et al., 2000] sondern auch
einzeln zu Schädigungen von Myelin und Myelin-spezifischen Zellen führt [Akassoglou et al., 1998; Lee et al., 2000; Masumura et al., 2001], kann man zusammenfassend von einer entscheidenden Rolle dieses Cytokines in der Entwicklung von Hirnschäden ausgehen.
In der Entwicklung einer PVL sind Oligodendrocyten-Vorläuferzellen von großer Bedeutung, da sie im Zeitraum der Entwicklung der humanen cerebralen weißen Substanz, in dem das Risiko für die Entstehung einer PVL nachweislich sehr hoch ist, die dort dominierende Zellpopulation bilden [Back et al., 2001]. Aus diesen Gründen wurde im Rahmen dieser Arbeit mittels TUNEL-Analysen ein mög-
licher toxischer Effekt von TNF-α auf Oligodendrocyten-Vorläuferzellen untersucht. Dabei konnte in
drei unabhängigen Versuchen mit keiner der untersuchten Konzentrationen von TNF-α signifikant
Apoptose induziert werden [siehe Abb. 14].
In der Literatur waren zu diesem Zeitpunkt divergierende Ergebnisse über den Einfluss von TNF-α auf
mit Oligodendrocyten-Vorläuferzellen angereicherte Kulturen bekannt: Während Merrill, Agresti und Andrews keinen Effekt nachweisen konnten, zeigte Cammer zumindest einen Langzeiteffekt auf die Ausdifferenzierung der Zellkultur und Scurlock und Dawson beobachteten sogar bis zu 70 % Apopto-se sowie eine Aktivierung der Caspase-3 [Merrill, 1991; Agresti et al., 1996; Andrews et al., 1998; Scurlock und Dawson, 1999; Cammer, 2000]. Die Untersuchungen hinsichtlich einer Apoptose-Induktion wurden sowohl von Scurlock und Dawson als auch von Agresti und Andrews mittels eines MTT-Assays untersucht; ein TUNEL-Assay hierzu wurde in primären Zellkulturen bisher nur in einer gemischten Glia-Zellkultur durchgeführt [Ye und D’Ercole, 1999].

Diskussion 85
Auch in Experimenten mit reifen Oligodendrocyten sind unterschiedliche Effekte von TNF-α bekannt:
Während in einigen Studien eine direkte Zellschädigung durch TNF-α beobachtet wurde [Selmaj et
al., 1991; Louis et al., 1993; D’Souza et al., 1995; Wilt et al., 1995; Hisahara et al., 1997], konnte dies in anderen Studien nicht bestätigt werden [Merrill, 1991; Soliven et al., 1991; McLarnon et al., 1993; Andrews et al., 1998]. Allgemein werden diese Differenzen auf die verschiedensten Gründe zurückge-führt: Die untersuchten Zellen unterscheiden sich beispielsweise in ihrer Herkunft (Art, Rasse), in ihrer Reinheit (angereicherte Vorläuferzellen, Astrocyten- und Mikroglia-Kontaminationen) und in ihrer Kultivierung (verwendete Seren, Wachstumsfaktoren, Zelldichte).
Das im Rahmen dieser Arbeit gewählte Kultivierungsprotokoll unterschied sich in mindestens einem dieser Punkte von den bisher untersuchten Kulturen: Agresti und Andrews beispielsweise kultivierten ihre Oligodendrocyten-Vorläuferzellen durchgehend in einem Medium, welches T3 enthielt. Dieser Faktor ist für seine die Differenzierung fördernde Fähigkeit bekannt [Bhat et al., 1979; Almazan et al., 1985]. Eine Induktion der Differenzierung während der Cytokin-Behandlung war für die Fragestellung dieser Arbeit unerwünscht, so dass auf dieses bekannte Kultivierungsprotokoll nicht zurückgegriffen werden konnte. Die Studie, welche als einzige ebenfalls ein serumfreies Medium mit den Faktoren PDGF und bFGF nutzte, enthielt keine Angaben über den Reinheitsgrad der Zellkultur [Cammer,
2000], so dass eine Untersuchung der eigenen Kultur auf einen direkten toxischen Effekt von TNF-α
erforderlich war.
Nachdem die TUNEL-Analysen ergaben, dass TNF-α unter den gewählten Kulturbedingungen keinen
toxischen Effekt auf Oligodendrocyten-Vorläuferzellen hatte, wurden keine weiteren Experimente mit
diesem Cytokin alleine durchgeführt. Dieses Ergebnis weißt darauf hin, dass TNF-α einzeln nicht
ausreichend ist, um die schweren Schädigungen der weißen Substanz in einer PVL zu induzieren.
5.3 Einfluss von IFN-γ und TNF-α auf Oligodendrocyten-Vorläuferzellen
Neben TNF-α könnten auch noch weitere Cytokine mit der Entwicklung einer periventrikulären Leu-
komalazie in Verbindung gebracht werden. So wurden im neonatalen Blut von Kindern, welche einen
Hirnschaden entwickelten, Interferone nachgewiesen. Es ist bekannt, dass Interferon-γ einen schädli-
chen Einfluss auf Myelin hat: Eine Behandlung von Patienten mit Multipler Sklerose führte zu einer Verschlimmerung der Krankheit [Panitch et al., 1987]. Auch in Tierexperimenten konnte dieser Effekt beobachtet werden: In transgenen Mäusen, welche dieses Cytokin überexprimierten wurden Abnorma-litäten des Myelins festgestellt [Corbin et al., 1996; Horwitz et al., 1997].
Aus diesen Gründen wurde im Rahmen dieser Arbeit als nächstes geprüft, ob IFN-γ (alleine oder in
Kombination mit TNF-α) zu einer Schädigung von Oligodendrocyten-Vorläuferzellen führen kann,
welche in ihrem Ausmaß die schweren Schädigungen der weißen Substanz einer PVL erklären könnte.

Diskussion 86
5.3.1 Überleben von Oligodendrocyten-Vorläuferzellen
Die TUNEL-Assay-Analyse von drei unabhängigen Zellkulturen zeigte, dass IFN-γ in einer Konzent-
ration von 100 U/ml über einen Behandlungszeitraum von 48 Stunden signifikant zu einer gegenüber Kontroll-Kulturen erhöhten Apoptose von Oligodendrocyten-Vorläuferzellen führt [siehe Abb. 17].
Das Cytokin TNF-α, welches alleine zu keiner erhöhten Apoptose-Rate führte [siehe Abb. 14], ver-
stärkte den Effekt von IFN-γ bis zu einer Apoptose-Rate von 11.0 ± 2.6 %.
In einer Arbeit von Baerwald und Popko wurde bereits ein ebenfalls durch TUNEL-Assay-Analysen
ermittelter apoptotischer Effekt von IFN-γ auf Oligodendrocyten-Vorläuferzellen (A2B5-positiv) be-
schrieben [Baerwald und Popko, 1998]. Die Werte der Apoptose in unbehandelten Kulturen und in mit
100 U/ml IFN-γ behandelten Kulturen der eigenen Arbeit liegen in der gleichen Größenordnung wie
die von Baerwald und Popko ermittelten. Diese benutzten jedoch ein Medium ohne die eine Differen-zierung unterdrückenden Faktoren PDGF und bFGF, auf welche in der vorliegenden Arbeit nicht ver-
zichtet werden konnte. Da in einer Studie beobachtet wurde, dass IFN-γ der proliferativen Wirkung
jedes einzelnen Faktoren entgegen wirkte [Agresti et al., 1996], war es erforderlich, den Einfluss von
TNF-α und IFN-γ auf Oligodendrocyten-Vorläuferzellen, die in einem diese Faktoren enthaltenen
Medium kultiviert wurden, zu bestimmen. Außerdem zeigten Untersuchungen von Agresti (LDH-
Assay und Propidium-Iodid-Färbung) keinen cytotoxischen Effekt von IFN-γ mit/ohne TNF-α auf
Oligodendrocyten-Vorläuferzellen [Agresti et al., 1996]. Auf die TUNEL-Analyse der eigenen Zell-kultur konnte aus diesen Gründen in dieser Arbeit nicht verzichtet werden.
Der verstärkende Effekt von TNF-α auf die durch IFN-γ induzierten Wirkungen ist zum einen aus
anderen Zellsystemen [Montaldo et al., 1994; Delvenne et al., 1995; Suk et al., 2001a; Suk et al., 2001b], aber auch aus Experimenten mit Oligodendrocyten und deren Vorläuferzellen bekannt
[Agresti et al., 1996; Andrews et al., 1998]. Der Synergismus von IFN-γ und TNF-α kann durch ver-
schiedene molekulare Mechanismen erklärt werden. Zum einen wurde gezeigt, dass IFN-γ die Expres-
sion des TNF-α-Rezeptors hochregulieren kann, so dass die Zellen sensitiver für einen durch TNF-α
vermittelten Zelltod werden [Dopp et al., 1997]. Zum anderen induziert IFN-γ auch die Expression des
Interferon-Regulatory-Factor-1 (IRF-1), welches bekannt dafür ist, die durch TNF-α induzierte NF-
κB-Aktivität zu hemmen. Dies führt zu einer Verschiebung des Verhältnisses von pro- zu anti-
apoptotischen Signalwegen zugunsten der pro-apoptotischen [Suk et al., 2001a].
In dieser Arbeit wurde durch den synergistischen Effekt von IFN-γ und TNF-α eine Apoptose-Rate
von 11.0 ± 2.6 % erreicht. Die Kombination von 100 U/ml IFN-γ und 100 ng/ml TNF-α führte zu
dieser Apoptose-Rate. Diese Konzentrationen wurden für die weiteren Untersuchungen des Überle-bens und der Differenzierung der Oligodendrocyten-Vorläuferzellen gewählt.
Die Untersuchungen der mittleren Gesamtzellzahl von behandelten Deckgläschen im Vergleich zu unbehandelten zeigten, dass es während der Behandlung zu keiner Veränderung dieser Größe kam. Erst nach Entfernen der Faktoren und Induzierung der Differenzierung entwickelten sich Unterschiede in der mittleren Gesamtzellzahl zwischen den beiden Gruppen. Während in den Kontrollen mit Fort-

Diskussion 87
schreiten der Kultur keine signifikanten Änderungen nachweisbar waren, kam es bis zum Kulturtag 9 durch die Behandlung zu einem Verlust von ca. 97,5 % der Zellen. Dieser große Verlust in der Ge-samtzellzahl war nach der Apoptose-Rate von 11,0 ± 2,6 % am Kulturtag 3 nicht zu erwarten gewesen. Dies kann aber dadurch erklärt werden, dass mit dem TUNEL-Assay nur die Zellen detektiert werden, die sich im Endstadium des Zelltod-Programms befinden. Ein großer Teil der Zellen war wahrschein-lich in einem früheren Stadium des Apoptose-Programms, was sich erst auf die Gesamtzellzahl späte-rer Kultivierungszeitpunkte auswirkte.
5.3.2 Differenzierung von Oligodendrocyten-Vorläuferzellen
Alle Experimente, die sich mit dem Einfluss von IFN-γ und TNF-α auf die Differenzierung der die
Behandlung überlebenden Zellen beschäftigten, zeigten, dass diese Zellen keine Anzeichen für eine Ausdifferenzierung zu reifen Oligodendrocyten vorwiesen. Zum einen blieb die morphologische Ent-wicklung von bipolaren Zellen zu Zellen mit bäumchenartigen Fortsätzen aus [Phasenkontrast B in Abb. 20 und 21]. Gleichzeitig blieben behandelte Zellen A2B5-positiv [siehe Abb. 21D] und zeigten weder eine Expression des für reife Oligodendrocyten spezifischen Oberflächenantigens MOSP [siehe Abb. 20D], noch eine Expression der drei Myelin-spezifischen Proteine MAG, MBP und CNP [siehe Abb. 23].
Für diese Ergebnisse gibt es zwei mögliche Erklärungen: Zum einen kann durch die Behandlung mit den Cytokinen eine Selektion einer Population von Oligodendrocyten-Vorläuferzellen, welche von sich aus nicht in der Lage sind auszudifferenzieren, stattgefunden haben. Zum anderen wäre eine akti-ve Beeinflussung der Differenzierung möglich, wofür es auch schon in der Literatur erste Hinweise gibt:
Die Arbeitsgruppe um Agresti beispielsweise zeigte nach 48-stündiger Behandlung ebenfalls eine Zunahme von Oligodendrocyten-Vorläuferzellen (LB1-positiv) und gleichzeitig eine Abnahme von reifen Oligodendrocyten (Galc-positiv) [Agresti et al., 1996]. Und auch Baerwald und Popko beobach-
teten bei einer durchgehenden Behandlung mit IFN-γ eine Abnahme der Galc-Immunreaktivität
[Baerwald und Popko, 1998]. Die Effekte dieser beiden Studien sind aber wesentlich schwächer, als die in der vorliegenden Arbeit; bei Agresti waren sie sogar reversibel. Der Unterschied in der Stärke des Einflusses auf die Differenzierung ist vermutlich durch die unterschiedlichen Kultivierungsbedin-gungen zu erklären: Agresti und Baerwald kultivierten ihre Zellen jeweils durchgehend in einem Me-dium. Schon während der Behandlung mit den Cytokinen war dem Medium in beiden Fällen mindes-tens ein Faktor beigesetzt, welcher die Differenzierung von Oligodendrocyten-Vorläuferzellen indu-zieren kann. So waren bei Agresti am Kulturtag 3 nur noch etwa 50 % der Zellen Oligodendrocyten-Vorläuferzellen, während es in der vorliegenden Arbeit noch 90 % waren [siehe Abb. 22]. So können bei Agresti die beobachteten Effekte nicht unbedingt nur auf Oligodendrocyten-Vorläuferzellen bezo-gen werden, sondern vielmehr auf sich entwickelnde Oligodendrocyten, die sich in einer der Entwick-lungsstufen zwischen Vorläufer- und reifen Zellen befinden.

Diskussion 88
Diese Arbeit ist zudem die erste, in welcher durch Kultivierungsbedingungen die Vorläuferzell-Phase von der Differenzierungs-Phase der Oligodendrocyten getrennt wurde. In einigen Studien konnte be-reits gezeigt werden, dass Oligodendrocyten-Vorläuferzellen gegenüber proinflammatorischen Stimuli sowie oxidativem Stress anfälliger sind, als reife Oligodendrocyten [Andrews et al., 1998; Back et al., 1998; Baerwald und Popko, 1998; Molina-Holgado et al., 2001]. Diese höhere Anfälligkeit verbunden mit der gelungenen Unterbindung der Differenzierung könnte die Diskrepanz zwischen den Arbeiten erklären.
Die Tatsache, dass im Rahmen dieser Arbeit keine Expression der Myelin-spezifischen Proteine in behandelten Zellen nachweisbar war, spricht dafür, dass durch die Cytokin-Behandlung eine Selektion einer Population von Oligodendrocyten-Vorläuferzellen induziert worden sein könnte, welche von sich aus nicht in der Lage sind auszudifferenzieren. Da die absolute Anzahl an A2B5-positiven Zellen auf unbehandelten Deckgläschen größer war, als die absolute Gesamtzellzahl auf behandelten Deck-
gläschen, kann dies für die Konzentrationen 100 U/ml IFN-γ und 100 ng/ml TNF-α nicht ausgeschlos-
sen werden. Aber zumindest für eine Cytokin-Behandlung in der Konzentration 10 U/ml IFN-γ bzw.
10 ng/ml TNF-α kann eine Selektion als Wirk-Mechanismus ausgeschlossen werden. In dieser Kon-
zentration war die absolute Anzahl von A2B5-positiven Zellen auf behandelten Deckgläschen mit 4,4 ± 0,9 Zellen gegenüber 1,4 ± 0,6 Zellen auf unbehandelten Deckgläschen erhöht. Bei einem selek-tiven Effekt wäre eine identische Anzahl zu erwarten gewesen.
Insgesamt wurden Oligodendrocyten-Vorläuferzellen durch die Cytokine IFN-γ (100 U/ml) und
TNF-α (100 ng/ml) in ihrem Überleben und der Fähigkeit zu Differenzieren stark beeinträchtigt. Dies
stützt die Theorie, dass sie auch in der Entwicklung einer PVL eine wichtige Rolle spielen könnten. Von großem Interesse war deshalb nun das Charakterisieren von protektiven Reagenzien, welche all-gemein klinisch Anwendung finden.

Diskussion 89
5.4 Corticoide: Schutz vor durch IFN-γ und TNF-α induziertem Zelltod und Differenzierungsstörungen in Oligodendrocyten-Vorläuferzellen?
Neben ihrer normalen Funktion in der Regulation der Entwicklung von Oligodendrocyten und anderen Zellen des ZNS [Bohn and Friedrich, 1982; Meyer, 1985; Kumar et al. 1989; Poduslo et al., 1990; Vielkind et al., 1990], haben Glucocorticoide aufgrund ihrer anti-entzündlichen und das Immunsystem beeinflussenden Wirkung [Leung and Munck, 1975] auch in verschiedenen Bereichen der Medizin an Bedeutung gewonnen. Sie haben sich als die stärksten und effektivsten Reagenzien bei der Behand-lung chronischer entzündlicher Krankheiten, wie der Multiplen Sklerose, herausgestellt, und werden auch bei der Unterstützung der Lungenreifung von Frühgeborenen eingesetzt [Cosmi und DiRenzo, 1989; Kari et al., 1994; Yeh et al., 1997; Omar et al, 2000]. Die Risiken und Nebenwirkungen der Corticoide sind weitestgehend erforscht. So haben beispielsweise Untersuchungen von Kindern, wel-chen pränatal einmalig Corticoid verabreicht wurde, gezeigt, dass hieraus keine neurologischen und kognitiven Veränderungen resultierten [MacArthur et al., 1982; Collaborative group on antenatal steroid therapy, 1984; Smolder-de-Hass et al., 1990].
Die Mechanismen der antientzündlichen Wirkung von Glucocorticoiden sind noch nicht eindeutig
geklärt. Es ist jedoch bekannt, dass sie die Transkription verschiedener Cytokine (IFN-γ, TNF-α, IL-1,
IL-2, IL-6) unterdrücken [Cupps und Fauci, 1982; Culpepper und Lee, 1987; Taniguchi, 1988]. Nach bereits erfolgter Freisetzung von Cytokinen, konnte in verschiedenen Zellsystemen weiterhin eine anti-apoptotische Wirkung der Glucocorticoide beobachtet werden [Kull et al., 1988; Pagliacci et al., 1993; Cox et al., 1995; Cox and Austin, 1997; Wen et al., 1997]. Unter anderem gibt es dabei auch für
Kulturen reifer Oligodendrocyten erste Hinweise auf eine schützende Funktion vor durch IFN-γ und
TNF-α induziertem Zelltod [Melcangi et al., 2000].
Im Rahmen dieser Arbeit sollte untersucht werden, ob ein protektiver Effekt auch in Oligodendrocy-ten-Vorläuferzellen aktiviert wird und wie sich die Corticoid-Behandlung auf die weitere Differenzie-rung der Zellen auswirkt. Die bis zu diesem Zeitpunkt im Rahmen dieser Arbeit eingesetzten Konzent-
rationen an IFN-γ und TNF-α hatten für den Nachweis eines Einflusses der Corticoide einen zu star-
ken Effekt gezeigt. Aus diesem Grunde wurde zunächst mittels Western-Blot-Analysen das Konzent-
rationspaar mit 10 U/ml IFN-γ und 10 ng/ml TNF-α bestimmt: Nach Behandlung mit dieser Kombina-
tion war am Kulturtag 9 die Expression der Proteine MBP und MAG nachweisbar und unterschied sich sichtbar von der Expression in Kontroll-Zellen [siehe Abb. 24]. Die Cytokine beeinflussten dem-nach auch in dieser geringeren Konzentration die Differenzierung der Oligodendrocyten-Vorläuferzellen, während noch genügend Spielraum gegeben war, einen positiven wie einen negativen Einfluss von Corticoiden nachweisen zu können.

Diskussion 90
5.4.1 Corticoide: Einfluss auf das Überleben von Oligodendrocyten-Vorläuferzellen
Die Glucocorticoide Corticosteron und Dexamethason sowie das Mineralcorticoid Deoxycorticosteron
führten sowohl in nicht mit Cytokinen behandelten Kulturen, als auch in gleichzeitig mit IFN-γ und
TNF-α behandelten Kulturen zu einer zur jeweiligen Kontrolle signifikanten Erhöhung der mittleren
Gesamtzellzahl. In nicht behandelten Kulturen war der Effekt mit einer Erhöhung der Zellzahl um ca. 40 % wesentlich kleiner als in mit Cytokinen behandelten Kulturen Dort wurde eine Erhöhung auf das 2,4-fache für Deoxycorticosteron, auf das 3,5-fache für Corticosteron und auf das 5,4-fache für Dexa-methason beobachtet.
Diese großen Unterschiede zwischen den beiden Gruppen lassen den Schluss zu, dass die drei geteste-ten Corticoide neben einem allgemeinen Effekt auf Oligodendrocyten-Vorläuferzellen (40 % mehr
überlebende Zellen) auch noch einen schützenden Effekt vor der Cytokin-Kombination IFN-γ und
TNF-α haben. Dabei zeigte Dexamethason von den drei getesteten Corticoiden den stärksten Effekt,
gefolgt von Corticosteron und mit dem schwächsten Ergebnis Deoxycorticosteron. Diese Reihenfolge in der Effektivität ist sehr wahrscheinlich auf die ungleichen Verhältnisse zwischen Gluco- und Mine-ralcorticoidaktivität zurückzuführen. Während Deoxycorticosteron keine Glucocorticoidaktivität auf-weist und beim Corticosteron eine schwache Glucocorticoidaktivität vorhanden ist, wirkt Dexametha-son nur über den Glucocorticoid-Rezeptor [Pschyrembel, 1998].
In einigen Zellsystemen konnte eine durch IFN-γ bzw. TNF-α induzierte gesteigerte Expression des
Glucocorticoid-Rezeptors beobachtet werden: IFN-γ führte in Makrophagen zu einer erhöhten Anzahl
an GR-Rezeptoren [Salkowski und Vogel, 1992] und TNF-α hatte auf L929-Fibroblasten einen die
Rezeptoranzahl steigernden Effekt [Costas et al., 1996]. Dieser Effekt konnte nicht in allen Zellen beobachtet werden [Betancur et al., 1995; Costas et al., 1996], so dass mittels RT-PCR- und Western-
Blot-Analysen geklärt werden muss, ob die Cytokine IFN-γ und TNF-α auch in Oligodendrocyten-
Vorläuferzellen zu einer gesteigerten Expression des Glucocorticoid-Rezeptors führen. In einem sol-chen Falle könnte der großen Unterschied zwischen dem Einfluss der Corticoide auf unbehandelte und auf mit Cytokinen behandelte Kulturen ansatzweise erklärt werden.
In der Studie von Melcangi, in welcher erste Hinweise auf einen protektiven Effekt der Corticoide Aldosteron, Deoxycorticosteron (beides reine Mineralcorticoide), Dexamethason (reines Glucocorti-
coid) und Corticosteron (sowohl Gluco- als auch Mineralcorticoid) vor durch IFN-γ und TNF-α indu-
zierter Apoptose reifer Oligodendrocyten (> 80 % MBP-positive Zellen) beobachtet wurden, sind keine Unterschiede in der Effektivität der einzelnen Corticoide zu erkennen gewesen [Melcangi et al., 2000]. Melcangi folgerte daraus, dass beide Rezeptoren in den protektiven Effekt der Corticoide in-volviert sind, während die Ergebnisse dieser Arbeit auf eine dominierende Rolle des Typ II Rezeptors hindeuten [Rezeptoren siehe Abschnitt 1.4]. Dieser Unterschied könnte auch auf die unterschiedliche Entwicklungsphase der untersuchten Zellen, und damit eine mögliche ungleiche Expression der beiden Rezeptoren, zurückzuführen sein. In diesem Falle wäre aber auch in nicht mit Cytokinen behandelten Kulturen ein stärkerer Unterschied zwischen den drei getesteten Corticoiden wahrscheinlich gewesen,

Diskussion 91
es sei denn, durch die Cytokine würde in Oligodendrocyten-Vorläuferzellen eine Verschiebung des Verhältnisses der Expression der Rezeptoren zueinander induziert werden. Eine genaue Klärung könn-ten möglicherweise Untersuchungen der Rezeptor-Expression in den verschieden behandelten und unterschiedlich weit entwickelten Zellen bringen.
5.4.2 Corticoide: Einfluss auf die Differenzierung von Oligodendrocyten-Vorläuferzellen
Um zu untersuchen, ob die Corticoide einen Einfluss auf die weitere Differenzierung der die Behand-
lung mit IFN-γ und TNF-α überlebenden Zellen hatten, wurden sowohl immuncytochemische Analy-
sen des Oberflächenantigens A2B5 als auch Western-Blot-Analysen gegen die Proteine MBP, MAG und CNP durchgeführt.
Die Immuncytochemie zeigte zunächst, dass die Cytokine auch in der schwächeren Konzentration
(10 U/ml IFN-γ; 10 ng/ml TNF-α) einen deutlichen Einfluss auf den Anteil an A2B5-positiven Zellen
am Kulturtag 9 hatte. Der gegenüber der Kontrolle auf das durchschnittlich 30-fache erhöhte Anteil an A2B5-positiven Zellen auf behandelten Deckgläschen entsprach einer 3-fach erhöhten absoluten An-
zahl. Aus diesen absoluten Zahlen kann man schließen, dass die Cytokine IFN-γ und TNF-α nicht nur
zur Selektion einer Teilpopulation von Oligodendrocyten-Vorläuferzellen führte, welche nicht in der Lage waren auszudifferenzieren, sondern dass deren Differenzierung zu reifen Oligodendrocyten auch aktiv beeinflusst wurde.
Während die Corticoide in Kontroll-Kulturen keinen Einfluss auf den Anteil A2B5-positiver Zellen zeigten, führten sie bei mit Cytokinen behandelten Deckgläschen zu einem signifikant geringeren Anteil an Oligodendrocyten-Vorläuferzellen. Das reine Mineralcorticoid Deoxycorticosteron zeigte dabei erneut den schwächsten Effekt, wohingegen Corticosteron und Dexamethason sogar zu einem mit Kontroll-Kulturen vergleichbaren Anteil an Oligodendrocyten-Vorläuferzellen führten.
Ein Vergleich der absoluten Zahlen A2B5-positiver Zellen zeigte ebenfalls deutlich, dass Corticoste-
ron und Dexamethason die mit IFN-γ und TNF-α behandelten Kulturen am stärksten beeinflussten.
Der durch die Cytokine ausgelöste Defekt der Differenzierung war bei gleichzeitiger Gabe eines der beiden Glucocorticoide mittels A2B5-Immuncytochemie nicht mehr nachweisbar. Dieses Ergebnis
gibt erste Hinweise darauf, dass die durch IFN-γ und TNF-α induzierte Differenzierungsstörung durch
gleichzeitige Gabe von Corticoiden aufgehoben oder zumindest geschwächt werden kann.
Eine Untersuchung der Expression der unter anderem für verschiedene Entwicklungsstufen von Oli-godendrocyten spezifischen und für eine Myelinisierung notwendigen Proteine MAG, MBP und CNP sollte genauere Aussagen hierzu möglich machen.
Verschiedene Studien hatten sich bereits mit dem Einfluss unterschiedlicher Corticoide auf die Myeli-nisierung und die Expression der dazu erforderlichen Proteine beschäftigt. Dabei wurden überwiegend reife Oligodendrocyten untersucht, und die Ergebnisse reichen von negativen bis hin zu positiven Effekten.

Diskussion 92
In in vivo Experimenten mit Ratten wurden allgemeine Beeinträchtigungen der Entwicklung des Ge-hirns durch Corticosteroide beobachtet. Neben einem reduzierten Gewicht des Gehirns [Gumbinas et al., 1973; Velazquez und Romano, 1987; Benesova und Pavlik, 1989; Uno et al., 1990] wurden außer-dem Veränderungen in der Synthese von DNA, RNA und Proteinen nachgewiesen [Velazquez und Romano, 1987; Benesova und Pavlik, 1989]. Auch Beeinträchtigungen der Myelinisierung bzw. der Expression Myelin-spezifischer mRNA wurde in einigen in vivo Studien beobachtet [Tsuneishi et al., 1991; Dunlop et al., 1997; Huang et al., 2001]. In diesen Versuchen wurden jedoch mehrfach sehr hohe Dosen an Corticosteroiden gegeben, wie sie in der Perinatalmedizin nicht zur Anwendung kom-men würden. In dieser Arbeit wurde entsprechend nur eine geringe Konzentration an Corticoiden (1 µM) eingesetzt.
In vitro-Studien hingegen zeigten überwiegend einen positiven Effekt auf die Expression Myelin-spezifischer Proteine. Während in einigen Studien nur nach Langzeit-Behandlungen ein die Expressi-on des Proteins MBP fördernder Effekt beobachtet wurde [Ved et al., 1989; Byravan und Campagno-ni, 1994] konnte Kumar bereits nach 12 bzw. 24 Stunden Inkubation mit Hydrocortison mittels Im-muncytochemie eine erhöhte Expression nachweisen [Kumar et al., 1989]. Auch für Dexamethason konnten positive Effekte auf die Myelinisierung beobachtet werden: In gemischten Kulturen aus Neu-ronen und Schwann´schen Zellen, den Myelin-formenden Zellen des peripheren Nervensystems, wur-de durch Dexamethason die Synthese-Rate des Myelins erhöht [Chan et al., 1998]. In Zellkultur konn-te ferner gezeigt werden, dass während der Gliazell-Differenzierung die Expression der CNPase durch Dexamethason erhöht wurde [Almazan et al., 1986]. Ob und wie die Expression des nur zu einem geringen Teil am Myelin beteiligten Proteins MAG durch Corticoide beeinflusst wird, ist außer im Rahmen dieser Arbeit bisher nicht untersucht worden.
Diese beschriebenen Studien unterscheiden sich in zwei wichtigen Punkten von der vorliegenden Ar-beit: Zum einen wurde keine gleichzeitige Behandlung mit Cytokinen durchgeführt und zum anderen sind reife Oligodendrocyten und nicht Oligodendrocyten-Vorläuferzellen untersucht worden. Sowohl aus in vivo [Dunlop et al., 1997] als auch in vitro Studien [Kumar et al., 1989] gibt es jedoch Hinweise darauf, dass der Zeitpunkt der Behandlung mit Corticoiden entscheidend für die Entwicklung eines Effektes sein kann.
Die Ergebnisse der eigenen Western-Blot-Analysen [siehe Abb. 29-34] zeigten, dass in Kontroll-Kulturen (blaue Säulen) am Kulturtag 9 die Expression der drei Proteine MAG, MBP und CNP durch eine 48-stündige Behandlung mit einem der drei untersuchten Corticoide nicht signifikant gegenüber der entsprechenden Kontrolle verändert war. In Oligodendrocyten-Vorläuferzellen scheinen Corticoide demnach im allgemeinen keine zu einem so späten Zeitpunkt nach Abschluss der Behandlung noch nachweisbaren Effekte auf die Expression der drei untersuchten Proteine zu haben.
In mit IFN-γ und TNF-α behandelten Kulturen (gelb-rote Säulen) konnte für das Protein MAG für
keines der untersuchten Corticoide eine gegenüber nur mit Cytokinen behandelten Zellen signifikante Veränderung nachgewiesen werden. Die Expression von CNP bzw. MBP hingegen wurde durch die
gleichzeitige Gabe von Corticosteron bzw. Dexamethason gegenüber nur mit IFN-γ und TNF-α be-

Diskussion 93
handelten Kulturen erhöht. Dass diese beiden Glucocorticoide in unbehandelten Kulturen keinen Ef-fekt zeigten, könnte durch eine schon für andere Zellen beobachtete Erhöhung der Glucocorticoid-Rezeptor-Expression durch Cytokine erklärt werden [Salkowski und Vogel, 1992; Costas et al., 1996].
Die Theorie einer solchen Wirkung von IFN-γ und/oder TNF-α auf die GR-Expression in Oligo-
dendrocyten-Vorläuferzellen müsste noch mittels RNA- und Protein-Expressions-Analysen bestätigt werden.
Für die Expression des CNP konnte durch die gleichzeitige Behandlung mit einem der beiden Gluco-corticoide annähernd das Level von unbehandelten Kontroll-Kulturen erreicht werden. Allerdings ist der Effekt der Cytokine auf die Expression dieses Proteins im Vergleich zu MAG und MBP schon deutlich schwächer gewesen. Corticosteron und Dexamethason beeinflussten auch die Expression des Myelin Basic Protein nachweislich signifikant. Sie führten zwar nicht zu mit Kontroll-Kulturen ver-gleichbaren Protein-Leveln, zeigten aber im Vergleich zu nur mit Cytokinen behandelten Kulturen
einen sehr starken Effekt. Bei einem Vergleich von nur mit IFN-γ und TNF-α behandelten Kulturen
und zusätzlich mit diesen Glucocorticoiden behandelten Kulturen ist der Effekt auf die Expression des MBP stärker, als auf die des CNP.
Dieser Unterschied könnte auf das in der Entwicklung von Oligodendrocyten unterschiedlich frühe Einsetzen der Expression der beiden Proteine zurückzuführen sein: Während CNP schon im Stadium unreifer Oligodendrocyten exprimiert wird, ist MBP erst im Stadium reifer Zellen nachweisbar [siehe Abb. 4]. Auch der verschieden starke Effekt der Cytokin-Behandlung auf die Proteinexpressionen (ca. 95 % Reduktion gegenüber der Kontrolle für MBP; ca. 65 % Reduktion gegenüber der Kontrolle für CNP) könnte so erklärt werden.
Die unterschiedliche starke Wirkung von Dexamethason auf die CNP- und MBP-Expression in mit Cytokinen behandelten Kulturen könnte auch dadurch erklärt werden, dass die Translation von MBP durch Dexamethason erhöht werden kann [Verdi und Campagnoni, 1990]. Ein Ausbleiben dieses Ef-fektes in Kontroll-Kulturen wiederum könnte man mit einer zu geringen Anzahl an Glucocorticoid-Rezeptoren erklären. Deren Expression kann durch Cytokine erhöht werden [Salkowski und Vogel, 1992; Costas et al., 1996], so dass Dexamethason dann die Translation verstärken kann. Diese Theorie bedarf jedoch noch klärender Experimente.
Die gleichzeitige Gabe der Cytokine und Corticosteron bzw. Dexamethason führt zu einer verstärkten
Expression der Proteine MBP und CNP und damit zu einer Abschwächung des durch IFN-γ und
TNF-α ausgelösten Defektes der Differenzierung. Die Ergebnisse der A2B5-Immuncytochemie hatten
noch den Schluss zugelassen, dass die beiden Glucocorticoide vollständig vor diesem Effekt schützen könnten. Durch die Western-Blot-Analysen konnte dies nicht bestätigt werden, zumal kein signifikan-ter Effekt auf die Expression des MAG beobachtet wurde. Glucocorticoide schützen demnach teilwei-
se vor durch IFN-γ und TNF-α induzierten Schädigungen von Oligodendrocyten-Vorläuferzellen.
Durch Untersuchungen der mRNA-Expression sollte geklärt werden, ob der positive Effekt der Glu-cocorticoide durch eine verstärkte Translation oder schon eine gesteigerte Transkription vermittelt wird.

Diskussion 94
Da Dexamethason den stärksten Effekt zeigte, sollte der Einfluss dieses Glucocorticoids auf die mRNA-Expression mittels semiquantitativer RT-PCR bestimmt werden. Untersucht wurden die mRNAs der Proteine MBP (relativ stärkster Effekt auf Proteinexpression) und MAG (gar kein Effekt auf Proteinexpression).
Für reife Oligodendrocyten war in Zellkultur-Experimenten bereits eine durch Hydrocortison erhöhte Expression der mRNA der Proteine MBP (2-fach erhöht gegenüber der Kontrolle) und PLP (3-fach erhöht gegenüber der Kontrolle) beobachtet worden [Kumar et al., 1989]. Diese Untersuchungen fan-den jedoch direkt nach Abschluss der Behandlung statt, wohingegen in dieser Arbeit auf einen Lang-zeit-Effekt hin untersucht werden sollte.
In eigenen Versuchen war die mRNA-Expression beider Proteine in nur mit Dexamethason behandel-ten Zellen nach 9 Tagen Kultivierung nicht signifikant gegenüber der Kontrolle verändert. Da diese semiquantitative RT-PCR nicht direkt nach Behandlung der Oligodendrocyten-Vorläuferzellen durch-geführt wurde, ist ein direkter Vergleich mit den Ergebnissen der Arbeit von Kumar, und damit ein Vergleich reifer Zellen mit Vorläuferzellen, nicht möglich.
In mit IFN-γ und TNF-α behandelten Zellen war eine auf 70 % (für MAG) bzw. 75 % (für MBP) ver-
ringerte Expression der entsprechenden mRNA auch 6 Tage nach Abschluss der Behandlung noch zu beobachten. Eine gleichzeitige Gabe von Dexamethason führte dazu, dass die mRNA-Level von MBP und MAG zum Zeitpunkt der Untersuchung denen der unbehandelten Kontrollen entsprachen.
Obwohl die mRNA-Level von mit Cytokinen und Dexamethason behandelten Zellen denen der unbe-handelten Kontrollen gleichkamen, war trotzdem ein Unterschied in der Protein-Expression der beiden
Gruppen für MBP und MAG zu beobachten. Dieser Unterschied wurde auch für nur mit IFN-γ und
TNF-α behandelte Kulturen beobachtet: Während die Protein-Expression nur ca. 5 % der Expression
in Kontroll-Kulturen betrug, war die mRNA auf nur 70 % (für MAG) bzw. 75 % (für MBP) verringert. Diese Differenzen zwischen Protein- und RNA-Expression könnten auf eine allgemein durch Apopto-se-induzierende Reagenzien ausgelöste Hemmung der Proteinsynthese zurückzuführen sein. In MCF-
7-Brustkrebs-Zellen beispielsweise hemmt TNF-α die gesamte Translation über einen Caspase-
abhängigen Mechanismus [Jeffrey et al., 2002]. Es ist beschrieben, dass Glucocorticoide durch eine
verstärkte Expression von Genen der Bcl-2-Familie gegen TNF-α−induzierte Apoptose wirken können
[Meßmer et el., 2000; Sasson et al.,2001; Sasson et al., 2002]. In diesem Falle würde der pro-apoptotische Signalweg nach der Aktivierung der Caspase-8 gehemmt werden und die Caspase-abhängige Inhibierung der Proteinsynthese könnte durch Corticoide nicht vollständig aufgehoben werden. Diese Erklärung der Differenzen zwischen RNA- und Protein-Expression müsste durch ge-nauere Untersuchungen der mRNA-Expression der verschiedenen pro- und anti-apoptotischen Gene bestätigt werden. Eine solche Untersuchung ist beispielsweise mit Hilfe von cDNA-Expressions-Arrays möglich.
In verschiedenen Experimenten wurde beobachtet, dass auch Glucocorticoide die Translation sowohl negativ [Kunz et al., 1996; Newton et al., 1998; Gewert et al., 1999; Grosset et al., 1999;] als auch positiv beeinflussen können [Verdi und Campagnoni, 1990; Colson et al., 1992; Sarantos et al., 1994; McGraw et al., 1995; Ambühl et al., 1999]. Unter anderem konnte in einem Zell-freien System der

Diskussion 95
positive Effekt von Dexamethason auf die Translation des Myelin Basic Protein nachgewiesen werden (2-3fach erhöht). Da Dexamethason jedoch in unbehandelten Kulturen nicht zu einer erhöhten Expres-
sion des MBP führte, sind die Unterschiede zwischen RNA- und Protein-Expression in mit IFN-γ und
TNF-α behandelten Kulturen auf diese Cytokine zurückzuführen. In Kontroll-Kulturen ist die Anzahl
an Glucocorticoid-Rezeptoren vielleicht zu gering, um einen Effekt auf die Protein-Expression zu zeigen. Die Behandlung mit Cytokinen könnte die Anzahl der GR jedoch erhöhen und so einen die Translation steigernden Effekt möglich machen [Salkowski und Vogel, 1992; Costas et al., 1996].
5.5 Ausblick Die Ursachen für die Entstehung von frühgeburtlichen Hirnschäden, insbesondere der periventrikulä-ren Leukomalazie sind noch nicht ausreichend geklärt. Bei der diffusen Komponente der PVL sind Oligodendrocyten-Vorläuferzellen die Zellen, die durch verschiedene Faktoren wie oxidativen Stress, erhöhte Level an exogenem Glutamat und auch die Freisetzung von Cytokinen geschädigt werden. Eine Schädigung dieser Zellen kann zu einer unzureichenden Myelinisierung wichtiger motorischer Nervenfasern und anschließend zu spastischen Lähmungen führen.
In dieser Arbeit wurde ein Zellkultursystem mit angereicherten Kulturen von Oligodendrocyten-
Vorläuferzellen etabliert, und ein direkter Einfluss der Cytokine IFN-γ und TNF-α auf das Überleben
und die weitere Differenzierung dieser Zellen nachgewiesen. Diese Ergebnisse stützen die Theorie, dass nach einer Infektion freigesetzte Cytokine direkt das unreife Gehirn eines Ungeborenen schädi-gen können, indem sie die für eine erfolgreiche Myelinisierung wichtige Zellpopulation der Oligo-dendrocyten-Vorläuferzellen in ihrer Größe reduzieren und ihre Fähigkeit, zu reifen Oligodendrocyten auszudifferenzieren, beeinträchtigen.
Die Signalwege der einzelnen Cytokine sind bereits bekannt [Übersicht in Malagarie-Cazenave et al.,
2002 bzw. Bach et al., 1996] und auch der Synergismus von IFN-γ und TNF-α ist in Ansätzen er-
forscht [Agresti et al., 1998]. Eine Untersuchung, ob und inwieweit die Apoptose durch Inhibitoren der einzelnen Signalwege gehemmt werden kann, ist dabei von großem Interesse. Anhand des etablier-ten Zellkultur-Systems könnten beispielsweise Caspase-Inhibitoren und JAK-Inhibitoren in dieser Hinsicht getestet werden.
Bei der Vermeidung von frühgeburtlichen Hirnschäden ist oft nur eine präventive Medikation möglich und erforderlich. Es sind nur wenige Medikamente bzw. Substanzen klinisch getestet, welche geringe Nebenwirkungen zeigen. Dies ist besonders aufgrund der allgemeinen Anfälligkeit von Frühgeborenen von großer Bedeutung. Daher richtet sich die Forschung hauptsächlich auf Reagenzien mit breitflächi-ger Wirkung und geringen Nebenwirkungen bzw. abschätzbaren Risiken.
In dieser Arbeit wurden deshalb Corticoide auf einen möglichen protektiven Effekt hin untersucht:
Corticosteron und Dexamethason führten zu einem signifikanten Schutz vor den durch IFN-γ und
TNF-α induzierten Effekten der Apoptose und der Beeinträchtigung der Differenzierung von Oligo-
dendrocyten-Vorläuferzellen. Die dabei beobachtete Diskrepanz zwischen den Effekten auf die Prote-in- und die mRNA-Expression eröffnet einen neuen Bereich für weitere Experimente: Mit Hilfe von

Diskussion 96
cDNA-Expressions-Arrays könnten neue Hinweise auf die pro-apoptotische Wirkung der Cytokine und auf die anti-apoptotische Wirkung der Glucocorticoide in Oligodendrocyten-Vorläuferzellen ge-wonnen werden. Die Hemmung der Apoptose durch Corticoide wurde schon für einige wenige Zell-systeme beobachtet, unter anderem spielen dabei die Proteine der Bcl-2 Familie eine entscheidende Rolle [Meßmer et al., 2000; Sasson et al., 2001; 2002]. Welche Gene in die anti-apoptotische Wirkung der Corticoide in dem in dieser Arbeit genutzten System involviert sind, könnte in diesen Experimen-ten geklärt werden.
Die Glucocorticoide zeigten nur in mit Cytokinen behandelten Kulturen einen signifikanten Effekt auf die Proteinexpression von MBP und CNP. Ob dies, wie für andere Zellen beschrieben, auf eine durch Cytokine induzierte erhöhte Expression des Glucocorticoid-Rezeptors zurückzuführen ist, könnte Gegenstand weiterer Experimente sein.
Die Corticoide schützen in diesem Zellsystem nicht vollständig vor durch IFN-γ und TNF-α induzier-
ten Schädigungen. Vor allem der ausbleibende Effekt auf die Expression des Proteins MAG könnte, da es sich um ein für Glia-Axon-Wechselwirkungen wichtiges Protein handelt, in in vivo Experimenten zu weniger positiven Ergebnissen führen. Eine Umsetzung der Ergebnisse dieser Arbeit in ein in vivo Modell ist auch deshalb ein weiterer möglicher Schritt.
Mit Hilfe des etablierten Zellkultur-Systems kann man zum einen Corticoide mit anderen Reagenzien kombinieren und deren Wirkung auf das Überleben und die Differenzierung der Zellen charakterisie-ren. Zum anderen können aber auch andere Faktoren, wie beispielsweise Östrogen, welches neuropro-tektiv wirken kann [Übersicht in Behl, 2002], mit diesem System getestet werden.

Zusammenfassung 97
6 ZUSAMMENFASSUNG Die periventrikuläre Leukomalazie (PVL) ist eine typische Schadensläsion der weißen Hirnsubstanz von frühgeborenen Kindern. In den letzten Jahren gewinnen mütterliche und fetale Infektionen immer mehr an Bedeutung für die Entwicklung eines solchen Schadens. Bei der PVL werden die Myelin-bildenden Oligodendrocyten und deren Vorläuferzellen geschädigt. Deshalb sollte in dieser Arbeit untersucht werden, ob Cytokine, die bei einer Infektion freigesetzt werde, einen direkten toxischen Effekt auf Oligodendrocyten-Vorläuferzellen haben oder die Differenzierung zu reifen Oligodendrocy-ten beeinflussen können. Als Reagenzien, welche vor einer solchen Beeinflussung schützen, sollten Corticoide getestet werden. Sie sind für ihre antientzündliche Wirkung bekannt und können aufgrund ihrer bekannten Nebenwirkungen auch zur Behandlung von Frühgeborenen verwendet werden.
Da die Cytokine TNF-α (Tumor Necrosis Factor) und IFN-γ (Interferon-γ ) aufgrund experimenteller
Befunde mit einer Entwicklung eines Hirnschadens in Verbindung gebracht werden können, wurden sie für diese Untersuchungen gewählt. Die Kultur von Oligodendrocyten-Vorläuferzellen wurde so etabliert, dass eine Behandlung der Zellen mit Cytokinen im Vorläuferzell-Stadium und eine gezielte Induktion der Differenzierung möglich waren.
Durch TUNEL-Assay-Analysen wurde gezeigt, dass TNF-α alleine keinen toxischen Effekt auf Oli-
godendrocyten-Vorläuferzellen ausübte. Das Cytokin IFN-γ (100 U/ml) jedoch führte zu einer signifi-
kant erhöhten Apoptose-Rate in diesen Zellen (6,1 ± 1,1 %). TNF-α (100 ng/ml) führte zu einer weite-
ren Erhöhung der Apoptose-Rate auf 11,0 ± 2,6 %. Während direkt nach der Behandlung kein Unter-schied in der mittleren Gesamtzellzahl zwischen behandelten und unbehandelten Kulturen zu ver-zeichnen war, wurde bei längerer Kultivierung in Differenzierungsmedium eine gegenüber Kontroll-Kulturen stark verminderte Zellzahl beobachtet. Bei den diese Cytokinexposition überlebenden Zellen konnten keine für die Ausdifferenzierung spezifischen Oberflächenantigene (MOSP) nachgewiesen werden. Daneben blieb im Gegensatz zur Kontrollgruppe die Expression der Myelin-spezifischen Proteine MBP, MAG und CNP aus. Stattdessen zeigte ein sehr großer Anteil der Zellen eine Immun-reaktivität gegen das für Vorläuferzellen spezifische Oberflächenantigen A2B5.
Auch bei einer Behandlung mit den Cytokinen in einer niedrigeren Konzentration (10 U/ml IFN-γ und
10 ng/ml TNF-α) wurde am Kulturtag 9 eine dramatische Abnahme der Gesamtzellzahl mit einem
hohen Anteil an A2B5-positiven Zellen beobachtet. Außerdem war nur eine schwache Expression der Proteine MBP, MAG und CNP nachzuweisen und die mRNA für MBP und MAG war gegenüber der Kontrolle reduziert.
Als nächstes wurde getestet, ob Corticoide vor diesen durch Cytokine induzierten Störungen des Über-lebens und der Differenzierung von Oligodendrocyten-Vorläuferzellen schützen können. Die Gesamt-zellzahl am Kulturtag 9 konnte durch die gleichzeitige Applikation von Corticoiden (Corticosteron, Deoxycorticosteron, Dexamethason; je 1 µM) signifikant gesteigert werden. Die überlebenden Zellen waren, wie durch A2B5-Immuncytochemie gezeigt, nahezu alle ausdifferenziert. Die mRNA-Level für MBP und MAG konnten durch Dexamethason bis auf das Niveau von Kontroll-Kulturen angehoben werden. Auch eine erhöhte Expression der Proteine MBP und CNP wurde nach Behandlung mit den

Zusammenfassung 98
Glucocorticoiden Corticosteron bzw. Dexamethason beobachtet. Dabei reichte die Erhöhung aber für keine der untersuchten Kombinationen an das entsprechende Protein-Level in Kontroll-Kulturen her-
an. Die durch IFN-γ und TNF-α induzierten Störungen können demnach durch Corticosteron und
Dexamethason nur teilweise behoben werden.
In nicht mit Cytokinen behandelten Zellen führten die Corticoide zu einer um 40 % erhöhten Gesamt-zellzahl. Auf die Protein- und mRNA-Expression der untersuchten Proteine bzw. mRNAs hatten sie in diesen Kulturen keinen Einfluss. Diese Unterschiede zwischen Cytokin-behandelten und unbehandel-
ten Kulturen, sowie die Diskrepanz in der Wirkung auf Protein- und mRNA-Level in mit IFN-γ und
TNF-α behandelten Zellen bieten Ansatzpunkte für weiterführende Forschungen.

Literatur 99
7 LITERATUR
Aaronson DS and Horvath CM. (2002) A Road Map for those who don´t know JAK-STAT. Science 296:1653-1655
Adinolfi M. (1985) The development of the human blood-CSF-brain barrier. Dev Med Child Neurol 27:532-537
Adinolfi M. (1993) Infectious diseases in pregnancy, cytokines and neurological impairment: an hypothe-sis. Dev Med Child Neurol 35:549-53
Aggarwal B, Vilcek J, Eds. (1991) Tumor necrosis factors: structure, function and mechanism. Marcel Dekker Publishers, New York
Agresti C, Dúrso D, Levi G. (1996) Reversible inhibitory effects of interferon-γ and tumour necrosis fac-tor-α on oligodendroglial lineage cell proliferation and differentiation in vitro. Eur J Neurosci 8:1106-1116
Agresti C, Bernardo A, Del Russo N, Marziali G, Battistini A, Aloisi F, Levi G, Coccia M. (1998) Syner-gistic stimulation of MHC class I and IRF-1 gene expression by IFN-γ and TNF-α in oligodendrocytes. Eur J Neurosci 10:2975-2983
Akassoglou K, Bauer J, Kassiotis G, Pasparakis M, Lassmann H, Kollias G, Probert L. (1998) Oligoden-drocyte apoptosis and primary demyelination induced by local TNF/p55TNF receptor signalling in the central nervous system of transgenic mice. Am J Pathology 153:801-813
Allan SM and Rothwell NJ. (2001) Cytokines and acute neurodegeneration. Nature reviews Neurosci 2:734-744
Almazan G, Honegger P, Matthieu JM. (1985) Triiodothyronine stimulation of oligodendroglial differen-tiation and myelination. A developmental study. Dev Neurosci 7:45-54
Almazan G, Honegger P, Du Pasquier P, Matthieu JM. (1986) Dexamethasone stimulates the biochemical differentiation of fetal forebrain cells in reaggregating cultures. Dev Neurosci 8:14-23
Aloisi F, Agresti C, D'Urso D, Levi G. (1988) Differentiation of bipotential glial precursors into oligoden-drocytes is promoted by interaction with type-1 astrocytes in cerebellar cultures. Proc Natl Acad Sci USA 85:6167-6171
Alonso G. (2000) Prolonged corticosterone treatment of adult rats inhibits the proliferation of oligoden-drocyte progenitors present throughout white and gray matter regions of the brain. Glia 31:219-231
Ambühl PM, Yang X, Peng Y, Preisig PA, Moe OW, Alpern RJ. (1999) Glucocorticoids enhance acid activation of the Na+/H+ exchanger 3 (NHE3). J Clin Invest 103:429-435
Andrews T, Zhang P, Bhat NR. (1998) TNFα potentiates IFNγ-induced cell death in oligodendrocyte progenitors. J Neurosci Res 54:574-583
Armstrong RC, Harvath L, Dubois-Dalcq ME. (1990) Type 1 astrocytes and oligodendrocyte-type 2 astro-cyte glial progenitors migrate towards distinct molecules. J Neurosci Res 27:400-407
Arroyo EJ and Scherer SS. (2000) On the molecular architecture of myelinated fibers. Histochem Cell Biol 113:1-18
Auphan N, Di Donato, JA, Rosette C, Helmberg A, Karin M. (1995) Immunosupression by glucocorti-coids: inhibition of IκB synthesis. Science 270:286-290

Literatur 100
Baas D, Bourbeau D,. Sarlieve LL, Ittel ME, Dussault JH, Puymirat J. (1997) Oligodendrocyte maturation and progenitor cell proliferation are independently regulated by thyroid hormone. Glia 19:324-332
Bach EA, Tanner JW, Marsters S, Ashkenazi A, Aguet M, Shaw AS, Schreiber RD. (1996) Ligand-induced assembly and activation of the gamma interferon receptor in intact cells. Mol Cell Biol 16:3214-3221
Back SA and Volpe JJ. (1997) Cellular and molecular pathogenesis of periventricular white matter injury. Ment Retard Dev 3:96-107
Back SA, Gan X, Li Y, Rosenberg PA, Volpe JJ. (1998) Maturation-dependent vulnerability of oligoden-drocytes to oxidative stress-induced death caused by glutathione depletion. J Neurosci 18:6241-6253
Back SA, Luo NL, Borenstein NS, Levine JM, Volpe JJ, Kinney HC. (2001) Late oligodendrocyte pro-genitors coincide with the developmental window of vulnerability for human perinatal white matter in-jury. J Neurosci 21:1302-12
Baerwald KD and Popko B. (1998) Developing and mature oligodendrocytes respond differently to the immune cytokine interferon-gamma. J Neurosci Res 52:230-239
Baeuerle PA, Baltimore D. (1989) A 65-kD subunit of active NF-κB is required for inhibition of NF-κB by IκB. Genes Dev 3:1689-1698
Baeuerle PA, Baltimore D. (1996) NF-κB: Ten years later. Cell 87:13-20
Bagenholm R, Nilsson UA, Kjellmer I. (1997) Formation of the free radicals in hypoxic ischemic brain damage in the neonatal rat, assessed by an endogenous spin trap and lipid peroxidation. Brain Res 773:132-138
Bagenholm R, Nilsson A, Gotborg CW, Kjellmer I. (1998) Free radicals are formed in the brain of fetal sheep during reperfusion after cerebral ischemia. Pediatr Res 43:271-275
Baldwin AS Jr. (1996) The NF-kappa B and I kappa B proteins: new discoveries and insights. Annu Rev Immunol 14:649-683
Barnes PJ and Adcock I. (1993) Anti-inflammatory actions of steroids: molecular mechanisms. Trends Pharmacol Sci 14:436-441
Baron W, Metz B, Bansal R, Hoekstra D, de Vries H. (2000) PDGF and FGF-2 signaling in oligodendro-cyte progenitor cells: regulation of proliferation and differentiation by multiple intracellular signalling pathways. Mol Cell Neurosci 15:314-329
Barone FC, Arvin B, White RF, Miller A, Webb CL, Willette RN, Lysko PG, Feuerstein GZ. (1997) Tu-mor necrosis factor-α: a mediator of focal ischemic brain injury. Stroke 28:1233-1244
Barone FC and Feuerstein GZ. (1999) Inflammatory mediators and stroke: new opportunities for novel therapeutics. J Cereb Blood Flow Metab 19:819-834
Barres BA, Raff MC, Gaese F, Bartke I, Dechand G, Barde YA. (1994a) A crucial role for neurotrophin-3 in oligodendrocyte development. Nature 367:371-375
Barres BA, Lazar MA, and Raff MC. (1994b) A novel role for thyroid hormone, glucocorticoids and reti-noic acid in timing oligodendrocyte development. Development 120:1097-1108
Barish ME, Mansdorf NB, Raissdana SS. (1991) γ-Interferon promotes differentiation of cultured cortical and hippocampal neurons. Dev Biol 144:412-423
Beato M. (1989) Gene regulation by steroid hormones. Cell 56:335-344

Literatur 101
Behl C. (2002) Oestrogen as a neuroprotective hormone. Nature Reviews 3:433-442
Benesova O and Pavlik A. (1989) Perinatal treatment with glucocorticoids and the risk of maldevelopment of the brain. Neuropharmacol 28:89-97
Benveniste EN. (1992) Inflammatory cytokines within the central nervous system: source, function, and mechanism of action. Am J Physiol 263:C1-C16
Berger R, Bender S, Sefkow S, Klingmüller V, Künzel W, Jensen A. (1997) Peri/ intraventricular haemor-rhage: a cranial ultrasound study on 5286 neonates. Eur J Obstet Gynecol 75:191-203
Berger R, Garnier Y, Pfeiffer D, Jensen A. (2000) Lipopolysaccharides do not alter energy metabolism and protein synthesis in an in vitro model of fetal cerebral ischemia. Pediatric Research 48:531-535
Bergmann A; Agapite J, Steller H. (1998) Mechanisms and control of preogrammed cell death in inverte-brates. Oncogene 17:3215-3223
Betancur C, Borrell J, Guaza C. (1995) Cytokine regulation of corticosteroid receptors in the rat hippo-campus: effects of interleukin-1, interleukin-6, tumor necrosis factor and lipopolysaccharide. Neuroen-docrinology 62:47-54
Bhat NR, Sarlieve LL, Subba Rao G, Pieringer RA. (1979) Investigations on myelination in vitro. Regula-tion by thyroid hormone in cultures of dissociated brain cells from embryonic mice. J Biol Chem 254:9342-9344
Black RA, Rauch CT, Kozlosky CJ, Peschon JJ, Slack JL, Wolfson MF, Castner BJ, Stocking KL, Reddy P, Srinivasan S, Nelson N, Boiani N, Schooley KA, Gerhart M, Davis R, Fitzner JN, Johnson RS, Pax-ton RJ, March CJ, Cerretti DP. (1997) A metalloproteinase disintegrin that releases tumour-necrosis factor-alpha from cells. Nature 385:729-733
Boehm U, Klamp T, Groot M, and Howard JC. (1997) Cellular responses to interferongamma. Annu Rev Immunol 15:749-795
Bogler O, Wren D, Barnett SC, Land H, Noble M. (1990) Cooperation between two growth factors pro-mote extended self renewal and inhibits differentiation of oligodendrocyte-type-2 astrocyte (O-2A) progenitor cells. Proc Natl Acad Sci USA 87:6368-6372
Bohn MC and Friedrich VL Jr. (1982) Recovery of myelination in rat optic nerve after developmental retardation by cortisol. J Neurosci 2:1292-1298
Braun PE, Sandillon F, Edwards A, Matthieu JM, Privat A. (1988) Immunocytochemical localization by electron microscopy of 2´3´-cyclic nucleotide 3´-phosphodiesterase in developing oligodendrocytes of normal and mutant brain. J Neurosci 8:3057-3066
Breder CD, Hazulea C, Ghayur T, Klug C, Huginin M, Yasuda K, Teng M, Saper CB. (1994) Regional induction of tumor necrosis factor α expression in the mouse brain after systemic lipopolysaccharide administration. Proc Natl Acad Sci USA 91:11393-11397
Bruce AJ, Bowling W, Kindy MS, Peschon J, Kraemer PJ, Carpenter MK, Holtsberg FW, Mattson MP. (1996) Altered neuronal and microglial responses to excitotoxic and ischemic brain injury in mice lack-ing TNF receptors. Nat Med 2:788-794
Brunner C, Lassmann H, Waehneldt TV, Matthieu JM, Linington C. (1989) Differential ultrastructural localization of myelin basic protein, myelin/oligodendroglial glycoprotein, and 2´3´-cyclic nucleotide 3´-phosphodiesterase in the CNS of adult rats. J Neurochem 52:296-304
Bunge RP. (1968) Glial cells and the central myelin sheath. Physiol Rev 48:197-251

Literatur 102
Butt AM and Jenkins HG. (1994) Morphological changes in oligodendrocytes in the intact mouse optic nerve following intravitreal injection of tumour necrosis factor. J Nueroimmunol 51:27-33
Buttini M, Sauter A, Boddeke HWGM. (1994) Induction of interleukin-1β mRNA after focal cerebral ischaemia in the rat. Brain Res Mol Brain Res 23:126-124
Buttini M, Appel K, Sauter A, Gebicke-Haerter PJ, Boddeke HWGM. (1996) Expression of tumor necro-sis factor alpha after focal cerebral ischemia in the rat. Neusoscience 71:1-16
Byravan S and Campagnoni AT. (1994) Serum factors and hydrocortisone influence the synthesis of mye-lin basic proteins in mouse brain primary culture. Int J Dev Neurosci 12:343-351
Cai Z, Pan Z, Pang Y, Evans OB, Rhodes PG. (2000) Cytokine induction in fetal rat brains and brain in-jury in neonata rats after maternal lipopolysaccharide administration. Pediatr Res 47:64-72
Caldenhoven E, Liden J, Wissink S, van de Stolpe A, Raaijmakers J, Koenderman L, Okret S, Gustafsson J-Å, van der Saag PT. (1995) Negative cross-talk between Rel A and the Glucocorticoid receptor: a possible mechanism for the anti-inflammatory action of glucocorticoids. Mol Endocrinol 9:401-412
Cammer W. (2000) Effects of TNFalpha on immature and mature Oligodendrocytes and their progenitros in vitro. Brain Res 864:213-219
Cato ACB and Wade E. (1996) Molecular mechanisms of anti-inflammatory action of glucocorticoids. BioEssays 18:371-378
Chan JR, Phillips LJ, Glaser M. (1998) Glucocorticoids and progesterins signal the initiation and enhance the rate of myelin formation. Proc Natl Acad Sci USA 95:10459-10465
Chao CC, Hu S, Ehrlich L, Peterson PK. (1995) Interleukin-I and tumor necrosis factor-α synergistically mediate neurotoxicity: incolvement of nitric oxide and of N-methyl-D-aspartate receptors. Brain Behav Immun 9:355-365
Chao DT and Korsmeyer SJ. (1998) BCL-2 family: regulators of cell death. Annu Rev Immunol 16:395-419
Collaborative group on antenatal steroid therapy. (1984) Effects of antenatal dexamethasone administra-tion in the infant: long term follow-up. Pediatrics 104:259-267
Colson P, Ibarondo J, Devilliers G, Balestre MN, Duvoid A, Guillon G. (1992) Upregulation of Via vaso-pressin receptors by glucocorticoids. Am J Physiol Endocrinol Metab 263:E1054-E1062
Corbin JG, Kelly D, Rath EM, Baerwals KD, Suzuki K, Popko B. (1996) Targeted CNS expression of interferon-gamma in transgenic mice leads to hypomyelination, reactive gliosis, and abnormal cerebel-lar development. Mol Cell Neurosci 7:354-370
Cosmi EV and DiRenzo GC. (1989) Prevention and treatment of fetal lung immaturity. Fetal Ther 1:52-62
Costas M, Trapp T, Pereda MP, Sauer J, Rupprecht R, Nahmod VE, Reul JM, Holdboer F, Arzt E. (1996) Molecular and functional evidence for in vitro cytokine enhancement of human and murine target cell sensitivity to glucocorticoids. TNF-alpha priming increases glucocorticoid inhibition of TNF-alpha-induced cytotoxicity/apoptosis. J Clin Invest 98:1409-1416
Cox G. (1995) Glucocorticoid treatment inhibits apoptosis in human neutrophils. Separation of survival and activation outcomes. J Immunol 154:4719-4725
Cox G and Austin RC. (1997) Dexamethasone-induced suppression of apoptosis in human neutrophils requires continuous stimulation of new protein synthesis. J Leukoc Biol 61:224-230
Culpepper J and Lee F. (1987) Glucocorticoid regulation of lymphokine production by murine T lympho-cytes. Lymphokines 13:275-289

Literatur 103
Cupps TR and Fauci AS. (1982) Corticosteroid-mediated immunoregulation in man. Immunol Rev 65:133-155
Dammann O and Leviton A. (1997) Maternal intrauterine infection, cytokines, and brain damage in the preterm newborn. A Review. Pedriatr Res 42:1-8
Dammann O and Leviton A. (1998) Infection remote from the brain, neonatal white matter damage, and cerebral palsy in the preterm infant. Semin Pediatr Neurol 5:190-201
Decker T, Lew DJ, Mirkovitch J, Darnell JE Jr. (1991) Cytoplasmic activation of GAF, an IFN-gamma-regulated DNA-binding factor. EMBO J 10:927-932
Delvenne P, Al-Saleh W, Gilles C, Thiry A, Boniver J. (1995) Inhibition of growth of normal and human papilloma virus-transformed keratinocytes in monolayer and organotypic cultures by interferon-γ and tumor necrosis factor-α. Am J Pathol 146:589-598
Del Zoppo G, Ginis I, Hallenbeck JM, Iadecola C, Wang X, Feuerstein GZ. (2000) Inflammation and stroke: putataive role for cytokines, adhesion molecules and iNOS in brain response to ischemia. Brain Pathol 10:95-112
Diamond ML, Miner JN, Yoshinaga SK, Yamamoto KR. (1990) Transcription factor interactions selectors of positive or negative regulation from a single DNA. Science 249:1266-1272
Dopp JM, Mackenzie-Graham A, Otero GC, Merrill JE. (1997) Differential expression, cytokine modula-tion, and specific functions of type-1 and type-2 tumor necrosis factor receptors in rat glia. J Neuroim-munol 75:104-112
Drouin J, Sun YL, Chamberland M, Gauthier Y, De Lean A, Nemer M, Schmidt TJ. (1993) Novel gluco-corticoid receptor complex with DNA element of the hormone-repressed POMC gene. EMBO J 12:145-156
D’Souza S, Alinauskas K, McCrea E, Goodyer C, Antel JP. (1995) Differential susceptibility of human CNS-derived populations to TNF-α-dependent and independent immune-mediated injury. J Neurosci 15:7293-7300
Dubois C, Manuguerra JC, Hauttecoeur B, Maze J. (1990) Monoclonal antibody A2B5, which detects cell surface antigens, binds to ganglioside GT3 (II3 (NeuAc)3LacCer) and to its 9-O-acetylated derivative. J Biol Chem 265:2797-2803
Dunlop SA, Archer MA, Quinlivan JA, Beazley LD, Newnham JP. (1997) Repeated prenatal corticoster-oids delay myelination in the ovine central nervous system. J Maternal-Fetal Med 6:309-313
Dyer CA, Hickey WF, Geisert EE Jr. (1991) Myelin/oligodendrocyte-specific protein: a novel surface membrane protein that associates with microtubules. J Neurosci Res 28:607-613
Eccelston PA and Silberberg DH. (1984) Fibroblast growth factor is a mitogen for oligodendrocytes in vitro. Brain Res 210:315-318
Eisenbarth GS, Walsh FS, Nierenberg M. (1979) Monoclonal antibody to a plasma membrane antigen of neurons. Proc Natl Acad Sci USA 76:4913-4917
Evans RM. (1988) The steroid and thyroid hormone receptor superfamily. Science 240:889
Evans-Storms RB und Cidlowski JA. (2000) Delineation of an antiapoptotic action of glucocorticoids in hepatoma cells: the role of nuclear factor-κB. Endocrinol 141:1854-1862
Fianders KC, Ren RF, Lippa CF. (1998) Transforming growth factor-βs in neurodegenerative disease. Prog Neurobiol 54:71-85

Literatur 104
Finco TS, Baldwin AS. (1995) Mechanistic aspects of NF-kappa B regulation: the emerging role of phos-phorylation and proteolysis. Immunity 3:263-272
Fountoulakis M, Zulauf M, Lustig A, Garotta G. (1992) Stoichiometry of interaction between interferon-γ and its receptor. Eur J Biochem 208:781–787
Fredman P, Magnani JL, Nirenberg M, Ginsburg V. (1984) Monoclonal antibody A2B5 reacts with many gangliosides in neuronal tissue. Arch Biochem Biophys 233:661-666
Gambinas M, Oda M, Huttenlocher P. (1973) The effects of corticosteroids on myelination of the develop-ing rat brain. Biol Neonate 22:355-366
Gard AL and Pfeiffer SE. (1993) Glial cell mitogens bFGF and PDGF differentially regulate development of O4+GalC- oligodendrocyte progenitors. Dev Biol 159:618-630
Garnier Y, Coumans A, Jensen A, Berger R, Hasaart THM. (2001) Endotoxemia severely affects circula-tion during normoxia and asphyxia in immature fetal sheep. J Soc Gynecol Investig 8:134-142
Gatti S and Bartfai T. (1993) Induction of tumor necrosis factor-alpha mRNA in the brain after peripheral endotoxin treatment: comparison with interleukin-1 family and interleukin-6. Brain Res 624:291-294
Gewert K, Svensson U, Andersson K, Holst E, Sundler R. (1999) Dexamethasone differentially regulates cytokine transcription and translation in macrophages responding to bacteria or okadaic acid. Cell Sig-nal 11:665-670
Gilles FH, Leviton A, Kerr CS. (1976) Susceptibility of the neonatal feline telencephalic white matter to lipopolysaccharide. J Neurol Sci 27:183-191
Gilles FH, Leviton A, Dooling EC. (1983) The developing human brain: growth and epidemiologic neuro-pathology. Boston: John Wrigth, Inc.
Goldman JE, Geier SS, Hirano M. (1986) Differentiation of astrocytes and oligodendrocytes from germi-nal matrix cells in primary cultures. J Neurosci 6:52-60
Gravel M, Peterson J, Yong VW, Kottis V, Trapp B, Braun PE. (1996) Overexpression of 2’,3’-cyclic nucleotide 3’-phosphodiesterase in transgenic mice alters oligodendrocyte development and produces aberrant myelination. Mol Cell Neurosci 7:453-466
Greenlund AC, Schreiber RD, Goeddel DV, Pennica D. (1993) Interferon-γ induces receptor dimerization in solution and on cells. J Biol Chem 268:18103–18110
Greenlund AC, Farrar MA, Viviano BL, Schreiber RD. (1994) Ligand induced IFNγ receptor phosphorylation couples the receptor to its signal transduction system (p91). EMBO J 13:1591–1600
Greenlund AC, Morales MO, Viviano BL, Yan H, Krolewski J, Schreiber RD. (1995). Stat recruitment by tyrosine-phosphorylated cytokine receptors: an ordered reversible affinity-driven process. Immunity 2:677–687
Grether JK and Nelson KB. (1997) Maternal infection and cerebral palsy in infants of normal birth weight. JAMA 278:207-11
Grether JK, Nelson KB, Dambrosia JM, Phillips TM. (1999) Interferons and cerebral pulsy. J Pediatr 134:324-332
Griffiths IR, Klugmann M, Anderson TJ, Thomson C, Vouyiouklis D, Nave KA. (1998) Current concepts of PLP and its role in the nervous system. Microsc Res Tech 41:344-358
Grosset C, Taupin JL, Lemercier C, Moreau JF, Reiffers J, Ripoche J. (1999) Leukaemia inhibitory factor expression is inhibited by glucocorticoids through post transcriptional mechanisms. Cytokine 11:29-36

Literatur 105
Guiochon-Mantel A, Delabre K, Lescop P, Milgrom E. (1996) Intracellular traffic of steroid hormone receptors. J Steroid Biochem Mol Biol 56:3-9
Hasegawa K, Yoshioka H, Sawada T, Nishikawa H. (1993) Direct measurement for free radicals in the neonatal mouse brain subjected to hypoxia: an electron spin resonance spectroscopic study. Brain Res 607:161-166
Heck S, Bender K, Kullmann M, Göttlicher M, Herrlich P, Cato ACB. (1997) IκBα-independent down-regulation of NF-κB activity ba glucocorticoid receptor. EMBO J 16:4698-4707
Herrlich P. (2001) Cross-talk between Glucocorticoid receptors and AP-1. Oncogene 20:2465-2475
Higuchi M, Higashi N, Taki H, and Osawa T. (1990) Cytolytic mechanism of activated macrophages. Tumor necrosis factor and L-arginine dependent mechanisms act synergistically as the major cytolytic mechanisms of activated macrophages. J Immunol 144:1425-1431
Hillhouse EW and Mosley K. (1993) Peripheral endotoxin induces hypothalamic immunoreactive inter-leukin-1 beta in the rat. Br J Pharmacol 109:289-290
Hisahara S, Shoji S, Okano H, Miura M. (1997) ICE/CED-3 family executes oligodendrocyte apoptosis by tumor necrosis factor. J Neurochem 69:10-20
Hopkins SJ and Rothwell NJ. (1995) Cytokines and the nervous system. I: Expression and recognition. Trends Neurosci 18:83-88
Horwitz MS, Evans CF, McGavern DB, Rodriguez M, Oldstone MB. (1997) Primary demyelination in transgenic mice expressing interferon-gamma. Nat Med 3:1037-1041
Huang WL, Harper CG, Evans SF, Newnham JP, Dunlop SA. (2001) Repeated prenatal corticosteroid administration delays myelination of the corpus callosum in fetal sheep. Int J Devl Neuroscience 19:415-425
Huber LA, Madison DL, Simons K, Pfeiffer SE. (1994) Myelin membrane biogenesis by oligodendro-cytes: developmental regulation of low molecular weight GTP-binding proteins. FEBS Lett 347:273-278
Israel A. (1995) A role of phosphorylation and degradation in the control of NF-kappa B activity. Trends Genet 11:203-205
Jeffrey IW, Bushell M, Tilleray VJ, Morley S, Clemens MJ. (2002) Inhibition of protein synthesis in apop-tosis: differential requirements by the tumor necrosis factor alpha family and a DNA-damaging agent for caspases and the double-stranded RNA-dependent protein kinase. Cancer Res 62:2272-2280
Jenkins HG and Ikeda H. (1992) Tumor necrosis factor causes an increase in axonal transport of protein and demyelination in the mouse optic nerve. J Neurol Sci 108:99-104
Jonakait GM, Wei R, Sheng ZL, Hart RP, Ni I. (1994) Interferon-γ promotes cholinergic differentiation of embryonic septal nuclei and adjacent basal forebrain. Neuron 12:1149-1159
Jonat C, Rahmsdorf HJ, Park KK, Cato AC, Gebel S, Ponta H, Herrlich P. (1990) Antitumor promotion and antiinflammation: downregulation of AP-1 (Fos/Jun) activity by Glucocorticoid hormone. Cell 62:1189-1204
Kanfer J, Parenty M, Goujat-Zalc C, Mmonge M, Bernier L, Campagnoni AT, Dautigny A, Zalc B. (1989) Developmental exopression of myelin proteolipid, basic protein, and 2’,3’-cyclic nucleotide 3’-phosphodiesterase transcripts in different rat brain regions. J Mol Neurosci 1:39-46
Kaplan DK, Greenlund AC, Tanner JW, Shaw AS, Schreiber RD. (1996) Identification of an interferon-γ receptor α chain sequence required for JAK-1 binding. J Biol Chem 271:9–12

Literatur 106
Kari MA, Hallman M, Eronen M, Terano K, Virtanen M, Koivisto M, Ikonen RS. (1994) Prenatal dexa-methasone treatment in conjunction with rescue therapy of human surfactant: a randomized placebo-controlled multicenter study. Pediatrics 93:730-736
Karin M and Lin A. (2002) NF-kappaB at the crossroads of life and death. Nat Immunol 3:221-227
Kawakami K, Scheidereit C, Roeder RG. (1988) Identification and purification of a human immunoglobu-lin-enhancer-binding protein (NF-κB) that activates transcription from a human immunodeficiency vi-rus type 1 promoter in vitro. Procl Natl Acad Sci USA 85:4700-4704
Kim SU, McMorris FA, Sprinkle TJ. (1984) Imunofluorescence demonstration of 2’,3’-cyclic-nucleotide 3’-phosphodiesterase in cultured oligodendrocytes of mouse, rat, calf and human. Brain Res 300:195-199
Kinney HC and Back SA. (1998) Human oligodendroglial development: relationship to periventricular leukomalacia. Semin Pediat .Neurol 5:180-189
Kotenko S, Izotova L, Pollack B, Mariano T, Donnelly R, Muthuku-maran G, Cook J, Garotta G, Silven-noinen O, Ihle J, Pestka S. (1995) Interaction between the components of the interferon gamma recep-tor complex. J Biol Chem 270:20915–20921
Kull FC. (1988) Reduction in tumor necrosis factor receptor affinity and cytotoxicity by glucocorticoids. Biochem Biophys Res Commun 153:402-409
Kumar S, Cole R, Chiappelli F, De Vellis J. (1989) Differential regulation of oligodendrocyte markers by glucocorticoids: Post-transcriptional regulation of both proteolipid protein and myelin basic protein and transcriptional regulation of glycerol phosphate dehydrogenase. Proc Natl Acad Sci USA 86:6807-6811
Kunz D, Walker G, Eberhardt W, Pfeilschifter J. (1996) Molecular mechanisms of dexamethasone inhibi-tion of nitric oxide synthase expression in interleukin 1 beta stimulated mesangial cells: evidence for the involvement of transcriptional and post-transcriptional regulation. Proc Natl Acad Sci USA 93:255-259
Laemmli UK. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685
Lee YB, Yune TY, Baik SY, Shin YH, Du S, Rhim H, Lee EB, Kim YC, Shin ML, Markelonis GJ, Oh TH. (2000) Role of tumor necrosis factor-alpha in neuromal and glial apoptosis after spinal cord injury. Exp Neurol 166:190-195
Lees MB, Brostoff SW. (1984) Proteins of myelin. In: Merell P, editor. Myelin. New York: Plenum Press, 197-217
LeFlore JL, Salhab WA, Broyles RS, Engle WD. (2002) Association of antenatal and postnatal dexa-methasone exposure with outcomes in extremely low birth weight neonates. Pediatrics 110:275-279
Leung K and Munck A. (1975) Peripheral actions of glucocorticoids.Annu Rev Physiol 37:245-272
Leviton A, Paneth N, Reuss L, et al. (1999) Maternal infection, fetal inflammatory response, and brain damage in very low birth weight infants. Pediatr Res 46:566-75
Li J, Hertzberg EL, Nagy JI. (1997) Connexin32 in oligodendrocytes and association with myelinated fibers in mouse and rat brain. J Comp Neurol 379:571-591
Liu T, Clark RK, McDonnell PC, Young PR, White RF, Barone FC, Feuerstein GZ. (1994) Tumor necro-sis factor−α in ischemic neurons. Stroke 25:1481-1488
Louis JC, Magal E, Takayama S, Varon S. (1993) CNTF protection of oligodendrocytes against natural and tumor necrosis factor-induced death. Science 259:689-692

Literatur 107
Lucchinette CF and Rodriguez M. (1997) The controversy surrounding the pathogenesis of the multiple sclerosis lesion. Mayo Clin Proc 72:665-678
MacArther BA, Howie RN, Dezoete JA. (1982) School progress and cognitive development of 6-year old children whose mothers were treated antenatally with betamethasone. Pediatrics 70:99-105
Männel DN, Moore RN, Mergenhagen SE. (1990) Macrophages as a source of tumoricidal activity (tu-mor-necrotizing factor). Infec Imm 30:523-530
Malagarie-Cazenave S, Andrieu-Abadie N, Ségui B, Gouazé V, Tardy C, Cuvillier O, Levade T. (2002) Sphingolipid signalling: molecular basis and role in TNF−α−induced cell death. Exp Rev mol med 20 December, http://www.expertreviews.org/0200546X
Marsters S, Pennica D, Bach E, Schreiber RD, Ashkenazi A. (1995) Interferon γ signals via a high-affinity multisubunit receptor complex that con-tains two types of polypeptide chain. Pro. Natl Acad Sc. USA 92:5401–5405
Martinez E, Figueroa R, Garry D, Visintainer P, Patel K, Verma U, Sehgal PB, Tejani N. (1998) Elevated amniotic fluid interleukin-6 as a predictor of neonatal periventricular leukomalacia and intraventricular hemorrhage. J Matern Fetal Investig 8:101-107
Masumura M, Hata R, Nagai Y, Sawada T. (2001) Oligodendroglial cell death with DNA fragmentation in the white matter under chronic cerebral hypoperfusion: comparison between normotensive and sponta-neously hypertensive rats. Neurosci Res 39:401-412
Matthews SG. (2000) Antenatal glucocorticoids and programming of the developing CNS. Pediatr Res 47:291-300
McCarthy KD and De Vellis J (1980) Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J Cell Biol 85:890-902
McGraw DW, Chai SE, Hiller FC, Cornett LE. (1995) Regulation of the beta 2-adrenergic receptor and its mRNA in the rat lung by dexamethasone. Exp Lung Res 21:535-546
McKinnon RD, Toshimitsu M, Dubois-Dalcq M, Aaronson S. (1990) FGF modulates the PDGF-driven pathway of oligodendrocyte development. Neuron 5:603-614
McKinnon RD, Matsui T, Aranda M, Dubois-Dalcq M. (1991) A role for fibroblast growth factor in oli-godendrocyte development. Dev Biol 152:363-372
McLarnon JG, Michikawa M, Kim SU. (1993) Effects of Tumor necrosis factor on inward potassium current and cell morphology in cultured human oligodendrocytes. Glia 9:120-126
McMorris FA and Dubois-Dalcq M. (1988) Insulin-like growth factor I promotes cell proliferation and oligodendroglial commitment in rat glial progenitor cells developing in vitro. J Neurosci Res 21:199-209
Melcangi RC, Cavarretta I, Magnaghi V, Ciusani E, Salmaggi E. (2000) Corticosteroids protect oligoden-drocytes from cytokine-induced cell death. Neuroreport 11:3969-3972
Merrill (1991) Effects of interleukin-1 and tumor necrosis factor-α on astrocytes, microglia, oligodendro-cytes and glial precursors in vitro. Dev Neurosci 13:130-137
Meßmer UK, Winkel G, Briner VA, Pfeilschifter J. (2000) Suppression of apoptosis by glucocorticoids in glomerular endothelial cells: effects on proapoptotic pathways. British J Pharmacol 129:1673-1683
Meyer WT. (1985) Biochemical effects of corticosteroids for fetal maturation on perinatal outcomes. Physiol Rev 65:946-1020
Miller RH. (1996) Oligodendrocyte origins. Trends in Neurosci 19:92-96

Literatur 108
Mirsky R, Winter J, Abney ER, Pruss RM, Gavrilovic J, Raff MC. (1980) Myelin-specific proteins and glycolipids in rat Schwann cells and oligodendrocytes in culture. Cell Biol 84:483-494
Molina-Holgado E, Vela JM, Arévalo-Matrín A, Guaza C. (2001) LPS/IFN−γ cytotoxicity in oligodendro-glial cells: role of nitric oxide and protection by the anti-inflammatory cytokine IL-10. Eur J Neurosci. 13:493-502
Montaldo PG, Carbone R, Corrias MV, Ferrarix PC, Puzoni M. (1994) Synergistic differentiation-promoting activity of interferon−γ and tumor necrosis factor−α: Role of receptor regulation on human neuroblasts. JNCI 86:1694-1701
Moss ML, Jin SL, Milla ME, Bicket DM, Burkhart W, Carter HL, Chen WJ, Clay WC, Didsbury JR, Has-sler D, Hoffmann CP, Kost IA, Mitchell J, Moyer M, Pahel G, Rocque W, Overton LK, Schoenen F, Seaton T, Su JL, Warner J, Willard D, Becherer D. (1997) Cloning of a disintegrin metalloproteinase that processes precursor tumour-necrosis factor-alpha. Nature 385:733-736
Mu QQ and Dyer C (1994) Developmental expression of MOSP in cultured oligodendrocytes. Neurochem Res 19:1033-1038
Müller M, Briscoe J, Laxton C, Guschin D, Ziemiecki A, Silvennoinen O, Harpur AG, Barbier G, Wit-thuhn BA, Schindler C, Pellegrini S, Wilks AF, Ihle JN, Stark GR, Kerr IM. (1993) The protein tyro-sine kinase JAK1 complements a mutant cell line defective in the interferon-alpha/beta and -gamma signal transduction pathways. Nature 366:129–135
Munck A, Brinck-Johnsen T. (1968) Specific and nonspecific physicochemical interactions of glucocorti-coids and related steroids with rat thymus cells in vitro. J Biol Chem 243:5556-5565
Munder M, Mallo M, Eichmann K, Modolell M. (1998) Murine macrophages secrete interferon gamma upon combined stimulation with interleukin (IL)-12 and IL-18: A novel pathway of autocrine macro-phage activation. J Exp Med 187:2103-2108
Nagata S. (1997) Apoptosis by death factor. Cell 88:355-365
Nelson KB, Dambrosia JM, Grether JK. (1998) Neonatal cytokines and coagulation factors in children with cerebral palsy. Ann Neurol 44:665-75
Newton R, Seybold J, Kuitert LM, Bergmann M, Barnes PJ.(1998) Repression of cyclooxygenase-2 and prostaglandin E2 release by dexamethasone occurs by transcriptional and post-transcriptional mecha-nisms involving loss oj polyadenylated mRNA. J Biol Chem 273:32312-32321
Noble M and Murray K (1984) Purified astrocytes promote the in vitro division of a bipotential glial pro-genitor cell. EMBO J 3:2243-2247
Noble M, Murray K, Stroobant P, Waterfield MD, Riddle P. (1988) Platelet-derived growth factor pro-motes division and motility and inhibits premature differentiation of the oligodendrocyte/type-2 astro-cyte progenitor cell. Nature 333:560-562
Norton WT and Cammer W. (1984) Isolation and characterization of myelin. In: Morell P, editor. Myelin. New York: Plenum Press, 147-195
Ohteki T, Fukao T, Suzue K, Maki C, Ito M, Nakamura M, Koyasu S. (1999) Interleukin 12-dependent interferon gamma production by CD8alpha+ lymphoid dendritic cells. J Exp Med 189:1981-1986
Omar SA, DeCristofaro JD, Agarwal BJ, La Gamma EF. (2000) Effect of prenatal steroids an potassium balance in extremely low birth weight neonates. Pediatrics 106:561-567
Ozawa K, Suchanek G, Breitschopf H, Brück W, Budka H, Jellinger K, Lassmann H. (1994) Patterns of oligodendroglia pathology in multiple sclerosis. Brain 117:1311-1322

Literatur 109
Pagliacci MC, Migliorati G, Smacchia M, Grignani F, Riccardi C, Nicoletti I. (1993) Cellular stress and glucocorticoid hormones protect L929 mouse fibroblasts from tumor necrosis factor alpha cytotoxicity. J Endocrinol Invest 16:591-599
Panitch HS, Hirsch RL, Schindler J, Johnson KP. (1987) Treatment of multiple sclerosis with gamma interferon: exacerbation associated with activation of the immune system. Neurology 37:1097-1102
Perlman JM, Risser R, Broyles RS (1996) Bilateral cystic periventricular leukomalacia in the premature infant: Associated risk factors. Pediatrics 97:822-827
Pfeiffer SE, Warrington AE, Bansal R. (1993) The oligodendrocyte and its many cellular processes. Trends Cell Biol 3:191-197
Poduslo SE, Pak CH, Miller K. (1990) Hydrocortisone induction during oligodendroglial differentiation. Neurosci Lett 113:84-88
Popko B, Corbin JG, Baerwald KD, Dupree J, Garcia AM. (1997) The effects of interferon−γ on the nerv-ous system. Mol Neurobiol 14:19-35
Pouly S, Becher B, Blain M, Antel JP. (2000) Interferon−γ modulates human oligodendrocyte susceptibil-ity to fas-mediated apoptosis. J Neuropath Exp Neurology 59:280-286
Pousett F. (1994) Developmental expression of cytokine genes in the cortex and hippocampus of the rat central nervous system. Dev Brain Res 81:143-146
Pratt WB. (1993) The role of heat shock proteins in regulating the function, folding and trafficking of the glucocorticoid receptor. J Biol Chem 268:21455-21458
Probert L, Eugster HP, Akassoglou K, Bauer J, Frei K, Lassmann H, Fontana A. (2000) TNFR1 signalling is critical for the development of demyelination and the limitation of T-cell responses during immune-mediated CNS disease. Brain 123:2005-2019
Pschyrembel Klinisches Wörterbuch (1998) 258. neu bearbeitete Auflage. De Gruyter, Berlin
Quarles R, Colman D, Salzer J, Trapp B. (1992) Myelin-associated glycoprotein: structure-function rela-tionships and involvement in neurological diseases. In Myelin: Biology and chemistry (Martensen R., ed.), CRC, Boca Raton, FL, 4413-4448
Raff MC, Fields KL, Hakoori SI, Mirsky R, Pruss RM, Winter J. (1979) Cell-type specific markers for distinguishing and studying neurons and the major classes of glial cells in culture. Brain Res 174:283-308
Raff MC, Miller RH, Noble M. (1983a) A glial progenitor cell that develops in vitro into an astrocyte or an oligodendrocyte depending on culture medium. Nature 303:389-396
Raff MC, Miller RH, Noble M. (1983b). Glial cell lineages in the rat optic nerve. Cold Spring Harb Symp Quant Biol 48 Pt2:569-572
Raff MC, Williams BP, Miller RH. (1984) The in vitro differentiation of a bipotential glial progenitor cell. EMBO J 3:1857-1864
Raff MC, Abney ER, Fok-Seang J. (1985) Reconstitution of a developmental clock in vitro: a critical role for astrocytes in the timing of oligodendrocyte differentiation. Cell 42:61-69
Raff MC, Lillien LE, Richardson WD, Burne JF, Noble MD. (1988) Plateled-derived growth factor from astrocytes drives the clock that times oligodendrocyte development in culture. Nature 333:562-565
Raine CS (1995) Multiple sclerosis: TNF revisited, with promise. Nature Med 1:211-214

Literatur 110
Ray A and Prefontaine KE (1994) Physical association and antagonism between the p65 subunit of tran-scription factor NF−κB and the Glucocorticoid receptor. Proc Natl Acad Sci USA 91:752-756
Richardson WD, Pringle N, Mosley MJ, Westermark B, and Dubois-Dalcq M. (1988) A role for platelet-derived growth factor in normal gliogenesis in the central nervous system. Cell 53:309-319
Romero R and Mazor M. (1988) Infection and preterm labor: pathways for intrauterine infections. Clin Obstet Gynecol 31:558
Rosa FM, Cochet MM, Fellous M. (1986) Interferon and major histocompatibility complex genes: a model to analyse eukaryotic gene regulation? Interferon 7:47-87
Rosenberg AA, Murdangh E, White CW. (1989) The role of oxygen free radicals in postasphyxia cerebral hypoperfusion in newborn lambs. Pediatr Res 26:215-219
Rothwell NJ and Hopkins SJ. (1995) Cytokines and the nervous system II: Actions and mechanisms of action. Trends Neurosci 18:130-136
Rothwell NJ and Luheshi GN. (1996) Brain TNF: damage limitation or damage reputation? Nat Med 2:746-747
Rothwell NJ and Relton JK. (1993) Involvement of cytokines in acute neurodegeneration in the CNS. Neurosci Biobehav Rev 17:217-227
Sakai DD, Helms S, Carlstedt Duke J, Gustafsson JA, Rottman FM, Yamamoto KR. (1988) Hormone-mediated repression: a negative glucocorticoid response element from bovine prolactin gene. Genes Dev 2:1144-1154
Salkowski CA and Vogel SN. (1992) IFN-gamma mediates increased Glucocorticoid receptor expression in murine macrophages. J Immunol 148:2770-2777
Sarantos P, Chakrabarti R, Copeland EM, Souba WW. (1994) Dexamethasone increases jejunal glutamine synthetase expression via translational regulation. Am J Surg 167:8-13
Sasson R, Tajima K, Amsterdam A. (2001) Glucocorticoids protect against apoptosis induced by serum deprivation, cyclic adenosine 3’,5’-monophosphate and p53 activation in immortalized human granu-losa cells: involvement of Bcl-2. Endocrinology 142:802-811
Sasson R, Winder N, Kees S, Amsterdam A. (2002) Induction of apoptosis in granulosa cells by TNFα and its attenuation by glucocorticoids involves modulation of Bcl2. Biochem Biophys Res Commun 294:51-59
Scheinman RI, Cogswell PC, Lofquist AK, Baldwin AS Jr. (1995a) Role of transcriptional activation of IκB−α in mediation of immunosuppression by glucocorticoids. Science 270:283-286
Scheinman RI, Gualberto A, Jewell CM, Cidlowski JA, Baldwin AS, Jr. (1995b) Characterisation of mechanisms involved in transrepression of NF-kappa B by activated glucocorticoid receptors. Mol Cell Biol 15:943-953
Schiller JH, Storer B, Bittner G, Horisberger MA. (1990) Characterization of the synergistic antiprolifera-tive effects of interferon−γ and tumor necrosis factor−α on human colon carcinoma cell line. J Inter-feron Res 10:129-139
Schule R and Evans RM. (1991) Retinoic acid is a negative regulator of AP-1-responsive genes Proc Natl Acad Sci USA 88:6092-6096
Scurlock B and Dawson G. (1999) Differential responses of oligodendrocytes to tumor necrosis factor and other pro-apoptotic agents: Role of ceramide in apoptosis. J Neurosci Res 55:514-522

Literatur 111
Selmaj K and Raine CS. (1988) Tumor necrosis factor mediates myelin and oligodendrocyte damage in vitro. Ann Neurol 23:339-346
Selmaj K, Raine CS, Farooq M, Norton WT, Brosnan CF. (1991) Cytokine cytotoxicity against oligoden-drocytes: apoptosis induced by lymphotoxin. J Immunol 147:1522-1529
Sheehan KCF, Calderon J, Schreiber RD. (1988) Generation and characterization of monoclonal antibod-ies specific for the human IFN-gamma receptor. J Immunol 140:4231–4237
Simpson PB, Mehotra S, Lange GD, Russell JT (1997) High density distribution of endoplasmic reticulum proteins and mitochondria at specialized Ca2+ release sites in oligodendrocyte processes. J Biol Chem 272:22654-22661
Smolder-de-Hass H, Neuvel J, Schmand B, Treffers PE, Koppe JG, Hocks J. (1990) Physical development and medical history of children who were treated antenatally with corticosteroids to prevent respiratory distress syndrome: a 10- to 12-year follow-up. Pediatrics 86:65-70
Soliven B, Szuchet S, Nelson DJ. (1991) Tumor necrosis factor inhibits K+ current expression in cultured oligodendrocytes. J Memb Biol 124:127-137
Soliven B and Szuchet S. (1995) Signal transduction pathways in oligodendrocytes: role of tumor necrosis factor−α. Int J Dev Neurosci 13:351-367
Suk K, Kim S, Kim YH, Kim KA, Chang I, Yagita H, Shong M, Lee MS. (2001a) IFN-γ/TNF-α syner-gism as the final effector in autoimmune diabetes: A key role for STAT/IFN regulatory factor-1 path-way in pancreatic β cell death. J Immunol 166:4481-4489
Suk K, Chang I, Kim YH, Kim S, Kim JY, Kim H, Lee, MS. (2001b) Interferon γ (IFNγ) and tumor necro-sis factor α synergism in ME-180 cervical cancer cell apoptosis and necrosis. J Biol Chem 16:13153-13159
Sommer I and Schachner M. (1981) Monoclonal antibodies (O1 to O4) to oligodendrocyte cell surfaces: an immunocytological study in the central nervous system. Dev Biol 83:311-327
Tan S, Zhou F, Nielsen VG, Wang ZW, Gladson CL, Parks DA. (1999) Increased injury following inter-mittent fetal hypoxia-reoxygenation is associated with increased free radical production in fetal rabbit brain. J Neuropathol Exp Neurol 58:972-981
Taniguchi T. (1988) Regulation of cytokine gene expression. Annu Rev Immunol 6:439-464
Taylor DL, Edwards AD, Mehmet H. (1999) Oxidative metabolism, apoptosis and perinatal brain injury. Brain Pathol 9:93-117
Tchelingerian JL, Quinonero J, Booss J, Jacque C. (1993) Localization of TNFα and IL-1α immunoreac-tivities in striatal neurons after surgical injury to the hippocampus. Neuron 1:213-224
Temple S and Raff MC. (1986) Clonal analysis of oligodendrocyte development in culture: evidence for a developmental clock that counts cell divisions. Cell 44:773-779
Torres C, Aranguez I, Rubio N. (1995) Expression of interferon−γ receptors on murine oligodendrocytes and its regulation by cytokines and mitogens. Immunol 86:250-255
Trapp BD. (1990) Myelin-associated glycoprotein: location and potential function. Ann NY Acad Sci 605:29-43
Traugott U. (1987) Multiple sclerosis: Relevance of class I and class II MHC-expressing cells to lesion development. J Neuroimmunol 16:283-302

Literatur 112
Traugott U and Lebon P. (1988) Multiple sclerosis: Involvement of interferon in lesion pathogenesis. Ann Neurol 24:243-251
Tsuneishi S, Takada S, Motoike T, Ohashi T, Sano K, Nakamura H. (1991) Effects of dexamethasone on the expression of myelin basic protein, proteolipid protein, and glial fibrillary acidic protein genes in developing rat brain. Brain Res Dev Brain Res 61:117-123
Turnley AM, Morahan G, Okano H, Bernard O, Mikoshiba K, Allison J, Bartlett PF, Miller JFAP. (1991) Dysmyelination in transgenic mice resulting from expression of class I histocompatibility molecules in oligodendrocytes. Nature 353:566-569
Uno H, Lohmiller L, Thieme C, Kemnitz JW, Engle MJ, Toeker EB, Farrell PM. (1990) Brain damage induced by prenatal exposure to dexamethasone in fetal rhesus macaques: I. Hippocampus. Dev Brain Res 53:157-167
Uno H, Matsuyama T, Tagaya M, Hata R, Akita H, Nakamura H, Sugita M. (1995) Cerebral ischemia triggers early production of tumor necrosis factor-α by microglia. J Cereb Blood Flow Metab 15:18
Vaillant AR, Mazzoni I, Tudan C, Boudreau M, Kaplan DR, Miller FD. (1999) Depolarization and neuro-trophins converge on the phosphatidylinositol 3-kinase-Akt pathway to synergistically regulate neu-ronal survival. J Cell Biol 146:955-966
Van Dam AM, Bauer J, Tilders FJ, Berkenbosch F. (1995) Endotoxin-induced appearance of immunoreac-tive interleukin-1 beta in ramified microglia in rat brain: a light and electron microscopic study. Neuro-science 65:815-26
Vandenabeele P, Declercq W, Beyaert R, Fiers W. (1995) Two tumour necrosis factor receptors: structure and function. Trends Cell Biol 5:392-399
Van Valen F, Winkelmann W, Burdach S, Gobel U, Jürgens H. (1993) Interferon−γ and tumor necrosis factor−α induce a synergistic antiproliferative response in human Ewing´s sarcoma cells in vitro. J Cancer Res Clin Oncol 119:615-621
Vartanian T, Li Y, Zhao M, Stefansson K. (1995) Interferon−γ−induced oligodendrocyte death: Implica-tion for the pathogenesis of multiple sclerosis. Mol Med 1:732-743
Ved HS, Gustow E, Pieringer RA. (1989) Effect of hydrocortisone on myelin basic protein in developing primary brain cultures. Neurosci Lett 99:203-207
Velazquez PN and Romano MC. (1987) Corticosterone therapy during gestation: Effects on the develop-ment of the rat cerebellum. Int J Dev Neurosci 5:189-194
Verdi JM and Campagnoni AT. (1990) Translation regulation by steroids. Identification of a steroid modu-latory element in the 5’-untranslated region of the myelin basic protein messenger RNA. J Biol Chem 265:20314-20320
Verma IM, Stevenson JK, Schwarz EM, Antwerp DV, Miyamoto S. (1995) Rel/NF−κB/IκB family: inti-mate tales of association and dissociation. Genes Dev 9:2723-2735
Vielkind U, Walencewicz A, Levine JM, Bohn MC. (1990) Type II glucocorticoid receptors are expressed in oligodendrocytes and astrocytes. J Neurosci Res 27:360-373
Vilcek J and Lee TH. (1991) Tumor necrosis factor. New sights into the molecular mechanism of its mul-tiple actions. J Biol Chem 266:7313-7316
Virchow R. (1867) Zur pathologischen Anatomie des Gehirns I. Congenitale Encephalitis und Myelitis. Virchow Arch Pathol Anat 38:129

Literatur 113
Vogel US and Thompson RJ. (1988) Molecular structure, localization and possible functions of the myelin associated enzyme 2’,3’-cyclic nucleotide phosphodiesterase. J Neurochem 50:1667-1677
Volpe JJ. (1995) Neurology of the newborn. Philadelphia: Saunders
Volpe JJ. (1997) Brain injury in the premature infant: neuropathology, clinical aspects and pathogenesis. MRDD Research Reviews 3:3-12
Volpe JJ. (2001) Neurobiology of periventricular leukomalacia in the premature infant. Pediatr Res 50:553-562
Wallach D, Boldwin M, Varfolomeev E, Beyaert R, Vandenbeele P, Fiers W. (1997) Cell death induction by receptors of the TNF family: towards a molecular understanding. FEBS Lett 410:96-106
Wallach D, Varfolomeev EE, Malinin NL, Goltsev YV, Kovalenko AV, Boldin MP. (1999) Tumor necro-sis factor receptor and Fas signalling mechanisms. Annu Rev Immunol 17:331-367
Wang X, Yue TL, Barone FC, White RF, Gagnon RC, Feuerstein GZ. (1994) Concomitant cortical expres-sion of TNF−α and IL-1β mRNAs follow early response gene expression in transient focal ischemi. Mol Chem Neuropathol 23:103-114
Watling D, Guschin D, Müller M, Silvennoinen O, Witthuhn BA, Quelle FW, Rogers NC, Schindler C, Stark GR, Ihle JN, Kerr IM. (1993) Complementation by the protein tyrosine kinase JAK2 of a mutant cell line defective in the interferon-gamma signal transduction pathway. Nature 366:166–170
Wen LP, Madani K, Fahrni JA, Duncan SR, Rosen GD. (1997) Dexamethasone inhibits lung epithelial cell apoptosis induced by IFN−γ and Fas. Am J Physiol 273:L921-L929
Wilt SG, Milward E, Zhou JM, Nagasato K, Patton H, Rusten R, Griffin DE, O’Conner M, Dubois-Dalq M. (1995) In vitro evidence for a dual role of tumor necrosis factor-α in human immunodeficiency vi-rus type I encephalopathy. Ann Neurol 37:381-394
Wood PM and Williams AK (1984) Oligodendrocyte proliferation and CNS myelination in cultures con-taining dissociated embryonic neuroglia and dorsal root ganglion neurons. Dev Brain Res 12:225-241
Wren D, Wolswijk G, Noble M (1992) In vitro analysis of the origin and maintenance of O-2A adult pro-genitor cells. J Cell Biol 116:167-176
Yamamoto KR. (1985) Steroid receptor regulated transcription of specific genes and gene network. Annu Rev Genet 19:209
Yang-Yen HF, Chambard JC, Sun YL, Smeal T, Schmidt TJ, Drouin J, Karin M. (1990) Transcriptional interference between c-jun and the glucocorticoid receptor: mutual inhibitin of DNA binding due to di-rect protein-protein interactions. Cell 62:1205-1215
Ye P und D’Ercole AJ. (1999) insulin-like growth factor I protects oligodendrocytes from tumor necrosis factor-α-induced injury. Endocrinology 140:3063-3072
Yeh HJ, Ruit KG, Wang YX, Parks WC, Snider WD, Deuel TF. (1991) PDGF A-chain gene is expressed by mammalian neurons during development and in maturity. Cell 64:209-216
Yeh TF, Lin YJ, Hsieh WS, Lin HC, Lin CH, Chen JY, Kao HA, Chien CH. (1997) Early postnatal dexa-methasone therapy for the prevention of chronic lung disease in preterm infants with respiratory dis-tress syndrome: a multicenter clinical trial. Pediatrics 100:E3
Yong VW, Tejada-Berges T, Goodyer CG, Antel JP, Yong FP. (1992) Differential proliferative response of human and mouse astrocytes to gamma-interferon. Glia 6:269-280

Literatur 114
Yoon BH, Romero R, Yang SH, Jun JK, Kim IO, Choi JH, Syn HC. (1996) Interleukin-6 concentrations in umbilical cord plasma are elevated in neonates with white matter lesions associated with periventriku-lar leukomalacia. Am J Obstet Gynecol 177:1433-1440
Yoon BH, Jun JK, Romero R, Park KH, Gomez R, Choi JH, Kim IO. (1997a) Amniotic fluid inflamma-tory cytokines (interleukin-6, interleukin-1β, and tumor necrosis factor-α), neonatal brain white matter lesions, and cerebral palsy. Am J Obstet Gynecol 177:19-26
Yoon BH, Kim CJ, Romero R, Jun JK, Park KH, Choi ST, Chi JG. (1997b) Experimentally induced in-trauterine infection causes fetal brain white matter lesions in rabbits. Am J Obstet Gynecol 177:797-802
Yoon BH, Romero R, Park JS, Kim CJ, Kim SH, Choi JH, Han TR. (2000) Fetal exposure to an intra-amniotic inflammation and the development of cerebral palsy at the age of three years. Am J Obstet Gynecol 182:675-81
Yoshimoto T., Okamura H, Tagawa YI, Iwakura Y, Nakanishi K. (1997) Interleukin 18 together with interleukin 12 inhibits IgE production by induction of interferon-gamma production from activated B cells. Proc Natl Acad Sci USA 94:3948-3953
Yoshioka T, Feigenbaum L, Jay G. (1991) Transgenic mouse model for central nervous system demyelina-tion. Mol Cell Biol 11:5479-5486

Anhang 115
8 ANHANG
8.1 Abbildungsverzeichnis
Abbildung 1: Lokalisation der fokalen bzw. diffusen Komponente einer PVL ...................................... 2 Abbildung 2: Zusammenhang mütterliche Infektion – kindlicher Hirnschaden ..................................... 3 Abbildung 3: Diagramm über die Entwicklung der Oligodendrocyten in der humanen weißen
Substanz während der zweiten Hälfte der Schwangerschaft:........................................... 4 Abbildung 4: Entwicklung von Oligodendrocyten-Vorläuferzellen zu reifen Oligodendrocyten mit
entsprechender Antigen-Expression................................................................................. 6 Abbildung 5: Zeitliches Profil der Cytokin-Expression nach einem experimentell ausgelösten
Hirnschaden...................................................................................................................... 8 Abbildung 6: Signaltransduktionswege des TNF-α-TNFR1 ................................................................ 11 Abbildung 7: Signaltransduktion des IFN-γ [modifiziert nach Bach et al., 1996] ................................ 13 Abbildung 8: allgemeines Kultivierungs-Protokoll für primäre Oligodendrocyten-Vorläuferzellen ... 44 Abbildung 9: Immuncytochemie gegen A2B5 im Verlauf der Zellkultur ............................................ 46 Abbildung 10: Immuncytochemie gegen MOSP im Verlauf der Zellkultur ......................................... 47 Abbildung 11: Western-Blot-Analyse gegen MBP, CNP und MAG.................................................... 49 Abbildung 12: Immuncytochemie gegen GFAP ................................................................................... 50 Abbildung 13: Experimentelles Protokoll I........................................................................................... 51 Abbildung 14: TUNEL-Assay nach Behandlung mit TNF-α ............................................................... 52 Abbildung 15: Experimentelles Protokoll II ......................................................................................... 53 Abbildung 16: Dia-Abfolge für die Auswertung des TUNEL-Assays.................................................. 55 Abbildung 17: TUNEL-Assay nach Behandlung mit IFN-γ +/- TNF-α ............................................... 56 Abbildung 18: Mittlere Gesamtzellzahl pro Sichtfeld nach Behandlung mit IFN-γ und TNF-α .......... 57 Abbildung 19: Immuncytochemie Kulturtag 3: Vergleich unbehandelt – behandelt (A2B5)............... 58 Abbildung 20: Immuncytochemie Kulturtag 9: Vergleich unbehandelt - behandelt (MOSP) .............. 59 Abbildung 21: Immuncytochemie Kulturtag 9: Vergleich unbehandelt - behandelt (A2B5) ............... 60 Abbildung 22: Anteil A2B5-positiver Zellen........................................................................................ 61 Abbildung 23: Western-Blot-Analyse gegen MBP, MAG und CNP.................................................... 62 Abbildung 24: Western-Blot-Analyse zur Konzentrationsfindung ....................................................... 64 Abbildung 25: Experimentelles Protokoll III ........................................................................................ 65 Abbildung 26: A2B5-Immuncytochemie nach Behandlung mit Corticoiden ± Cytokinen
(Kulturtag 9)................................................................................................................... 66 Abbildung 27: mittlere Gesamtzellzahl nach Behandlung mit Corticoiden ± Cytokinen
(Kulturtag 9)................................................................................................................... 67 Abbildung 28: Anteil A2B5-positiver Zellen nach Behandlung mit Corticoiden ± Cytokinen
(Kulturtag 9)................................................................................................................... 68

Anhang 116
Abbildung 29: MBP-Western-Blot nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9)...... 70 Abbildung 30: quantitative Auswertung MBP-Western-Blots (n=6).................................................... 71 Abbildung 31: MAG-Western-Blot nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9) ..... 72 Abbildung 32: quantitative Auswertung MAG-Western-Blots (n=4)................................................... 73 Abbildung 33: CNP-Western-Blot nach Behandlung mit Corticoiden ± Cytokinen (Kulturtag 9)....... 74 Abbildung 34: quantitative Auswertung CNP-Western-Blots (n=6) .................................................... 75 Abbildung 35: Klonierungs-Strategie Positiv-Kontrolle und Interner Standard ................................... 76 Abbildung 36: Gelanalyse MBP-RT-PCR nach Behandlung mit Dexamethason ± Cytokinen
(Kulturtag 9)................................................................................................................... 78 Abbildung 37: quantitative Auswertung MBP-RT-PCRs (n=4) ........................................................... 78 Abbildung 38: Gelanalyse MAG-RT-PCR nach Behandlung mit Dexamethason ± Cytokinen
(Kulturtag 9)................................................................................................................... 79 Abbildung 39: quantitative Auswertung MAG-RT-PCRs (n=4) .......................................................... 80
8.2 Tabellenverzeichnis Tabelle 1: Immuncytochemie - verwendete Antikörper-Konzentrationen............................................ 30 Tabelle 2: Zusammensetzung TUNEL-Reaktionslösung ...................................................................... 31 Tabelle 3: Western-Blot – verwendete Antikörper-Konzentrationen.................................................... 35 Tabelle 4: Anordnung der Proben eines Western-Blots (von links nach rechts)................................... 70 Tabelle 5: Übersicht über die PCRs für die Klonierung von positiv-Kontrolle und internem
Standard ................................................................................................................................. 77

Anhang 117
8.3 Kongressbeiträge
Production, expression and molecular evolution of a recombinant antibody against the intracellular signalling protein Ras.
Gronemeyer T., Feldhaus B., Damoiseaux R., Herrmann L., Kuhlmann J., and Heumann R.
52nd MOSBACHER KOLLOQUIUM, S10 (2001)
Oligodendrocyten-Vorläuferstufen: Einfluss von TNF-α und IFN-γ.
Feldhaus B., Dietzel-Meyer I., Jensen A., Heumann R., Berger R.
5. Statuskonferenz zu Forschungsprojekten der Kinderneurologie, Berlin 2002
TNF-alpha und IFN-gamma beeinträchtigen das Überleben und die Differenzierung von Oligodendro-cyten-Vorläuferstufen.
Feldhaus B., Dietzel-Meyer I., Jensen A., Heumann R., Berger R.
201. Tagung der NRWGGG (Niederrheinisch-Westfälischen Gesellschaft für Gynäkologie und Ge-burtshilfe), Detmold 2002
Oligodendrocyten-Vorläuferstufen: Einfluss von TNFα und IFNγ auf Überleben und Differenzierung.
Feldhaus B., Dietzel-Meyer I., Jensen A., Heumann R., Berger R.
54. Kongress der DGGG (Deutsche Gesellschaft für Gynäkologie und Geburtshilfe), Düsseldorf 2002
Archives of Gynecology and Obstetrics; 267 S.119 (Suppl.1) 2002
Einfluss der Cytokine TNF-α und IFN-γ auf Überleben und Differenzierung von Oligodendrocyten-
Vorläuferstufen.
Feldhaus B.
13. Jahrestagung der Arbeitsgemeinschaft ‚Neonatologie und pädiatrische Intensivmedizin Ruhrge-biet’, Bochum 2002

Anhang 118
8.4 Lebenslauf
PERSÖNLICHE DATEN
Name: Beatrix Feldhaus Geburtsdatum: 19. Juni 1975 Geburtsort: Neuss
SCHULAUSBILDUNG
1981 – 1985 Katholische Grundschule, Hochneukirch 1985 – 1994 Franz-Meyers-Gymnasium, Mönchengladbach-
Giesenkirchen; Abschluss: Allgemeine Hochschulreife
HOCHSCHULAUSBILDUNG
1994 Beginn des Studiums der Chemie an der Ruhr-Universität Bochum
März 1997 Vordiplom
Juni/Juli 1999 Diplom am Lehrstuhl für molekulare Neurobiochemie (Prof. Dr.
R. Heumann) mit dem Thema: „Regulation der Neurotrophine
und deren Rezeptoren durch neuronale p21-ras-Aktivität“
Juli 1999 - wissenschaftliche Hilfskraft am Lehrstuhl für molekulare Oktober 1999 Neurobiochemie (RUB)
Dezember 1999 - Anfertigung der vorliegenden Dissertation am Lehrstuhl für April 2003 molekulare Neurobiochemie (Prof. Dr. R. Heumann) unter
Betreuung von Herrn Prof. Dr. R. Berger (Universitäts-
Frauenklinik, Knappschaftskrankenhaus Bochum)


















