
Sox 9-Expression am Haarfollikel – Expressionsverhalten ...
80
Aus der Universitäts-Hautklinik der Albert-Ludwigs-Universität Freiburg i. Br. Sox 9-Expression am Haarfollikel – Expressionsverhalten bezüglich Haarfarbe, Haarergrauen und Haarausfall INAUGURAL-DISSERTATION zur Erlangung des Medizinischen Doktorgrades der Medizinischen Fakultät der Albert-Ludwigs-Universität Freiburg i. Br. Vorgelegt 2013 von Paulina Stoeva geboren in Sofia
Transcript of Sox 9-Expression am Haarfollikel – Expressionsverhalten ...

Aus der Universitäts-Hautklinik
der Albert-Ludwigs-Universität Freiburg i. Br.
Sox 9-Expression am Haarfollikel –
Expressionsverhalten bezüglich Haarfarbe, Haarergrauen
und Haarausfall
INAUGURAL-DISSERTATION
zur
Erlangung des Medizinischen Doktorgrades
der Medizinischen Fakultät
der Albert-Ludwigs-Universität
Freiburg i. Br.
Vorgelegt 2013
von Paulina Stoeva
geboren in Sofia

Dekan: Prof. Dr. Dr. h.c. mult. Hubert E. Blum
Erstgutachter: Prof. Dr. med. Markus Braun-Falco Zweigutachter: Prof. Dr. med. Wilhelm-Bernhard Niebling Jahr der Promotion: 2013

INHALTSVERZEICHNIS: SEITE
1. EINLEITUNG - 5 -
1.1 FUNKTION UND EINTEILUNG DER MENSCHLICHEN HAARE - 5 -
1.2 ANATOMIE DES MENSCHLICHEN TERMINALHAARES - 6 -
1.3 HAARWACHSTUMSZYKLUS, STAMMZELLEN UND ALTERN - 8 -
1.4 MELANOZYTEN UND MELANIN - 11 -
1.5 HAARFARBE - 13 -
1.6 HAARERGRAUEN - 14 -
2. FRAGESTELLUNG - 16 -
3. MATERIAL UND METHODEN - 17 -
3.1 PROBANDEN - 17 -
3.1.1 Gewebeschnitte der Probanden ............................................................................ - 17 -
3.1.2 Probandeninformationen ...................................................................................... - 17 -
3.2. ANFERTIGUNG DER SCHNITTPRÄPARATE - 18 -
3.3 ENTPARAFFINIERUNG - 18 -
3.4 ANTIGENRETRIEVAL-METHODEN - 19 -
3.4.1 Hitzeinaktivierung in Citrat-Puffer im Dampfkocher .......................................... - 19 -
3.4.2 Hitzeinaktivierung in Tris-EDTA-Puffer im Dampfkocher ................................. - 20 -
3.4.3 Permeabilitätserhöhung der Zellwände mit Triton X100 ..................................... - 20 -
3.4.4 Proteolytischer Enzymverdau .............................................................................. - 20 -
3.5 IMMUNHISTOCHEMISCHE FÄRBUNG - 21 -
3.5.1 Färben mit dem TechMate Horizon Färbeautomaten .......................................... - 21 -
3.5.2 Ansetzen der Antikörper ...................................................................................... - 22 -
3.5.3 Beladen der Maschine und Färbevorgang ............................................................ - 22 -
3.5.4 Eindecken der gefärbten Präparate ....................................................................... - 23 -
3.6 MIKROSKOPISCHE AUSWERTUNGEN - 23 -
3.7 PROBANDENGRUPPEN - 24 -
3.8 STATISTISCHE ANALYSE UND AUSWERTEVERFAHREN - 24 -
3.9 MATERIALIEN - 25 -
3.9.1 Antikörper ............................................................................................................ - 25 -
3.9.2 Chemikalien ......................................................................................................... - 25 -

3.9.3 Geräte ................................................................................................................... - 26 -
3.9.4 Sonstiges ............................................................................................................... - 26 -
4. ERGEBNISSE - 27 -
4.1 ETABLIERUNG DER IMMUNHISTOCHEMISCHEN FÄRBUNGEN - 27 -
4.1.1 Testreihen ............................................................................................................. - 27 -
4.1.2 Ergebnisse der Etablierungsversuche ................................................................... - 28 -
4.2 PROBANDENCHARAKTERISIERUNG - 29 -
4.2.1 Allgemeine Angaben zu den Probanden .............................................................. - 29 -
4.2.2 Haarfarben der Probanden .................................................................................... - 31 -
4.3 EXPRESSION VON SOX 9 IM PROBANDENKOLLEKTIV - 34 -
4.4 VERGLEICHENDE ANALYSEN DER SOX 9-EXPRESSION - 38 -
4.4.1 Vergleich der Haarfarben ..................................................................................... - 38 -
4.4.2 Altersabhängigkeit von SOX 9 ............................................................................ - 48 -
4.4.3 SOX 9-Expression abhängig von Alter und Geschlecht ...................................... - 49 -
4.4.4 Differenzierung nach Geschlecht ......................................................................... - 51 -
5. DISKUSSION - 53 -
5.1 DER HAARAPPARAT UND STAMMZELLEN - 54 -
5.1.1 SOX 9 ................................................................................................................... - 56 -
5.1.2 SOX 10 ................................................................................................................. - 56 -
5.1.3 PAX 3 ................................................................................................................... - 56 -
5.1.4 CLIM 2 ................................................................................................................. - 57 -
5.2 SOX 9-EXPRESSION IM HAARFOLLIKEL - 58 -
5.2.1 SOX 9-Expression und Haarfarbe ........................................................................ - 58 -
5.2.2 SOX 9-Expression und Haarergrauen .................................................................. - 60 -
5.2.3 SOX 9-Expression und altersbedingte Alopezie .................................................. - 62 -
5.2.4 SOX 9-Expression in Abhängigkeit vom Alter .................................................... - 62 -
5.2.5 Geschlechtsabhängigkeit der SOX 9-Expression ................................................. - 62 -
5.3 METHODISCHE SCHWÄCHEN - 63 -
5.3.1 Einschränkungen bei den Färbeergebnissen......................................................... - 63 -
5.3.2 Einschränkungen bei der Ergebnisinterpretation ................................................. - 64 -
5.4 AUSBLICK - 66 -
6. ZUSAMMENFASSUNG - 67 -
7. LITERATURVERZEICHNIS - 68 -

8. DANKSAGUNG ERROR! BOOKMARK NOT DEFINED.
9. LEBENSLAUF ERROR! BOOKMARK NOT DEFINED.
10. ANHANG - 79 -
ABKÜRZUNGEN - 79 -

- 5 -
1. Einleitung
„Zweimal färbt sich das Haar; zuerst aus dem Blonden ins Braune,
Bis das Braune sodann silbergediegen sich zeigt.“
(„Weissagungen der Bakis“, Johann Wolfgang von Goethe)
Bereits Goethe beschäftigte sich im Jahre 1827 mit Haarfarben und deren Wandelbarkeit mit
zunehmendem Alter als Symbol für Vergänglichkeit. In diesem Gedicht beschreibt er das
Phänomen, dass Neugeborene und Säuglinge zunächst helleres Kopfhaar haben, welches sich
jedoch im Laufe der Zeit dunkler färbt, um dann nach Jahrzehnten als prototypisches Zeichen
menschlichen Alterns grau zu werden.
1.1 Funktion und Einteilung der menschlichen Haare
Unsere Haare erfüllen vielerlei Funktionen. Das Kopfhaar bietet Schutz gegen
Sonnenstrahlen, die Wimpern und Augenbrauen schützen die Augen vor Schweiß und die
Vibrissae am Naseneingang vor Reizungen der Nasenschleimhaut. Desweiteren spielen Haare
eine Rolle beim Wärmehaushalt, dienen der Reibungsminderung, Berührungsempfindung und
erfüllen eine Signalwirkung.
Beim Menschen unterscheidet man drei verschiedene Haararten. Lanugohaare kommen beim
Feten und unreifen Neugeborenen vor. Vellushaar ist das normale Oberflächenhaar, welches
das menschliche Integument nahezu vollständig bedeckt. Ausnahmen stellen die Handflächen
und Fußsohlen, die Schleimhäute, die Glans penis und das Lippenrot dar. Das Vellushaar ist
normalerweise weniger als 2 cm lang, dünn (< 40 µm), marklos und gewöhnlich farblos. Die
dritte Art, und diejenige, um die es in dieser Arbeit geht, ist das Terminalhaar . Zwischen
80.000 und 120.000 Terminalhaare bedecken das Kapillitium (behaarte Kopfhaut). Sie sind
lang, dick (> 40 µm), pigmentiert und besitzen ein Haarmark. Bereits beim Neugeborenen
sind sie am Kapillitium sowie als Augenbrauen und Wimpern vorhanden. Eine besondere
Form der Terminalhaare stellen die Sexualhaare dar, die in der Pubertät unter
Androgeneinfluss an bestimmten Körperstellen aus Vellushaaren entstehen. Zu ihnen zählen
die Genital- und Achselhaare, sowie die männliche Bart- und Körperbehaarung (Braun-Falco
et al., 2005).

- 6 -
1.2 Anatomie des menschlichen Terminalhaares
a b
Abbildung 1: a) Schema einer Haarwurzel (nach Junqueira and Carneiro, 1991),
b) Haarwurzel im histologischen Schnittpräparat mit Hämatoxylin-Eosin (HE) gefärbt,
Vergrößerung 5 x
Der Haarfollikel lässt sich in Haarschaft (Scapus pili) und Haarwurzel (Radix pili) einteilen.
Er ist aus mehreren konzentrischen Epithelschichten aufgebaut. Die Haarwurzel endet mit
einer Anschwellung, dem Haarbulbus (Abbildung 1, Abbildung 2). Darin befindet sich die
Wachstumszone, die sogenannte Matrix, die die bindegewebige Haarpapille (Papilla pili) mit
den Blutgefäßen enthält. Die Haarmatrixzellen steigen nach der Zellteilung im Bulbus auf und
differenzieren sich zu den Zellen des Haarschafts und der inneren Wurzelscheide. In der
Matrix befinden sich auch die Melanozyten, die ihren Farbstoff Melanin an die Zellen des
wachsenden Haars abgeben.

- 7 -
Abbildung 2: Unterer Teil eines Haarschaftes mit Wulstregion und Bulbus, HE, 5 x
Der Haarschaft liegt im Haarkanal und besteht aus Medulla (Mark), Kortex (Rinde) und
Kutikula (Cuticula pili, Oberhäutchen des Haares) (Abbildung 1). Der innerste Teil, die
Medulla, ist nur bei manchen Terminalhaaren, zum Teil auch nur abschnittsweise,
nachweisbar. Sie enthält schwammartiges Gewebe, das faser- oder kugelartig sein kann,
Cementum und luftgefüllte Hohlräume (Wagner and Joekes, 2007). Die Rinde besteht aus
komplett keratinisierten, axial ausgerichteten Matrixzellen, die die Hauptmasse des Haars
ausmachen. Die äußerste Haarschicht, die Kutikula, wird aus flachen, sich dachziegelartig
überlappenden Zellen gebildet.
Anliegend folgt nun die innere Wurzelscheide (Vagina epithelialis radicularis interna), die
von innen nach außen aus den drei Lagen Kutikula , Huxley-Schicht (Stratum epitheliale
internum [granuliferum]) und Henle-Schicht (Stratum epitheliale externum [pallidum])
aufgebaut ist. Die Kutikula des Haarschafts und die der inneren Wurzelscheide sind
dachziegelartig gegeneinander ausgerichtet, sodass sie lamellenartig ineinander greifen und so
fest verankert sind, dass die innere Wurzelscheide bei Epilation des Haares mit herausgerissen
wird. Die innere Wurzelscheide umgibt die Matrixzellen des Bulbus, schützt und formt den
noch weichen, nach oben steigenden Schaft.
Die äußerste Hülle des Haarfollikels ist die äußere Wurzelscheide (Vagina epithelialis
radicularis externa). Sie geht oberhalb des Infundibulums, der Einmündung der Talgdrüse
(Glandula sebacea), in die Epidermis über. Auf Höhe des Ansatzes des Musculus arrector pili
verdickt sie sich zu einem Wulst (bulge region) (Abbildung 2) und bildet den Sitz der
epithelialen Stammzellen (Abbildung 4 (5)) (Braun-Falco et al., 2005; Tobin D. J., 2011; Wu
et al., 2011).

- 8 -
1.3 Haarwachstumszyklus, Stammzellen und Altern
Im Gegensatz zu allen anderen menschlichen Organen, durchläuft das Haar postnatal
periodisch Zyklen von aktiver Produktion (anagen), Regression (katagen) und Ruhe (telogen).
Dabei stellt der Haarfollikel den Schaftproduktionsort des Haares dar (Abbildung 3).
Während der Anagenphase erreicht der Haarfollikel seine maximale Größe und die
Matrixzellen eine sehr hohe mitotische Aktivität.
Die maximale Haarlänge ist von der Dauer des Anagens, die sich meist auf 3-6 Jahre beläuft,
abhängig und variiert individuell sehr stark. Die Wachstumsgeschwindigkeit beträgt ca. 1 cm
pro Monat oder 0,3 mm pro Tag. Ist die Anagenphase beendet, folgt eine ca. 2-wöchige
Umbauphase, das Katagen. Millionen synchronisierter Apoptosen führen dabei zur
Schrumpfung des Haarfollikels und einer beinahe vollständigen Auflösung der Haarpapille.
Nun schließt sich eine etwa 2-4 Monate dauernde Telogenphase an. Während dieser findet
kein Stoffwechsel statt, was das Haar für schädigende Noxen unempfindlich macht. Das
Abbildung 3: Haarzyklus: Die drei Phasen und ihre jeweilige Dauer (nach Braun-
Falco et al., 2005)
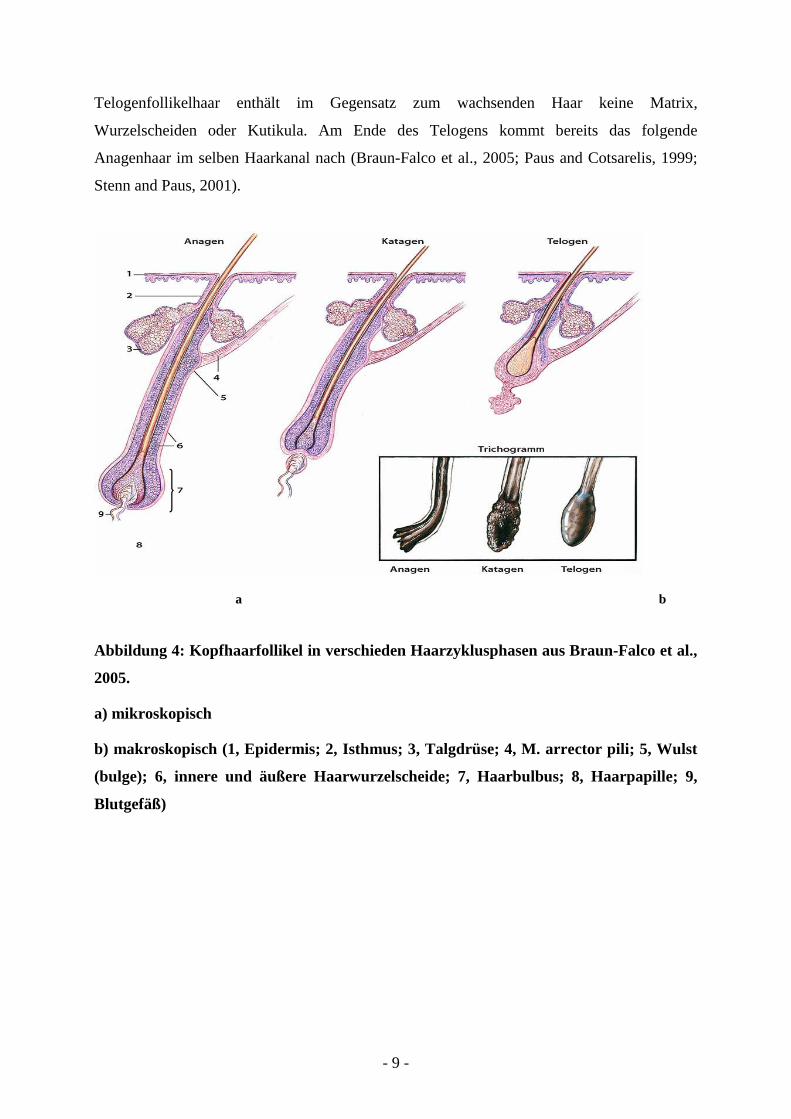
- 9 -
Telogenfollikelhaar enthält im Gegensatz zum wachsenden Haar keine Matrix,
Wurzelscheiden oder Kutikula. Am Ende des Telogens kommt bereits das folgende
Anagenhaar im selben Haarkanal nach (Braun-Falco et al., 2005; Paus and Cotsarelis, 1999;
Stenn and Paus, 2001).
a b
Abbildung 4: Kopfhaarfollikel in verschieden Haarzyklusphasen aus Braun-Falco et al.,
2005.
a) mikroskopisch
b) makroskopisch (1, Epidermis; 2, Isthmus; 3, Talgdrüse; 4, M. arrector pili; 5, Wulst
(bulge); 6, innere und äußere Haarwurzelscheide; 7, Haarbulbus; 8, Haarpapille; 9,
Blutgefäß)

- 10 -
Beim Wachstum eines neuen Haars kommt es zur Stammzellenstimulierung. Diese verlassen
ihre Nische, wandern in Richtung Matrix und beginnen dort zu wachsen und zu einem neuen
Haarschaft zu differenzieren (Oshima et al., 2001). Wichtige Induktoren des Wachstums
scheinen u.a. sonic hedgehog und Wnt/ β-catenin zu sein (Vidal et al., 2005).
Nach Verletzungen oder aufgrund physiologischen Bedarfs liefern Stammzellen Nachschub
an Zellen der betroffenen Gewebe, wie Haut, Knochenmark, Gehirn, Intestinum und
Skelettmuskulatur. Somit erfüllen sie eine Schlüsselrolle bei der Erneuerung und
Aufrechterhaltung adulten Gewebes. Diese Fähigkeit lässt mit zunehmendem Alter allmählich
nach. Im Zuge dessen kommt es, beispielsweise bei Stammzellen des Hämatopoesesystems
oder der Skelettmuskulatur, nicht zu einer signifikant sinkenden Anzahl (Rando T. A., 2006),
sondern lediglich zu Einbußen und Anomalitäten der Regenerierungs- oder
Differenzierungsfähigkeit (Van Zant and Liang, 2003). Ganz im Gegensatz zum
menschlichen Haar, bei dem das Alter zusätzlich ein Sinken der Melanozytenstammzellzahlen
mit sich bringt (Commo et al., 2004; Steingrímsson et al., 2005; Nishimura et al., 2005;
Aubin-Houzelstein et al., 2008). Es kommt dabei zu einer gestörten Melanozytenhomöostase.
Die Stammzellen sind nicht mehr in der Lage, für den nötigen Zellnachschub zu sorgen, was
schließlich als potenzielle Ursache des Haarergrauens zu sehen ist (Sarin and Artandi, 2007).
Zu den möglichen Ursachen des Stammzellenuntergangs zählen oxidativer Stress (Johnson
and Jackson, 1992; Arck et al., 2006), eine gestörte Telomerasefunktion (Chang et al., 2004;
Rudolph et al., 1999), ein nicht ausreichender Apoptoseschutz zum Beispiel durch Bcl 2-
Mangel (Nishimura et al., 2005; Steingrímsson et al., 2005; Arck et al., 2006; Veis et al.,
1993), eine intrinsisch determinierte Lebensspanne und vorbestimmte, limitierte Anzahl von
Zellteilungen (Sharpless and De Pinho, 2004) (s. Kapitel 4).

- 11 -
1.4 Melanozyten und Melanin
Melanozyten sind aus der Neuralleiste abstammende, sekretorisch aktive Zellen, die sich in
der Basalschicht der Epidermis, in der äußeren Wurzelscheide und im Bulbus des
Haarfollikels befinden (Abbildung 1). Ferner sitzen sie in der Uvea des Auges und im
Innenohr.
Beim Übergang des Haarfollikels von telogen zu anagen werden die
Melanozytenstammzellen stimuliert, sodass es während der Anagenphase zu
Melanozytenstammzellproliferation und zur vermehrten Bildung von Melanozytenvorläufern
kommt. Im Laufe der weiteren Entwicklung des Haares wandern die Melanozytenvorläufer
unter Einfluss von c-kit- und Endothelin-Signalen (Barsh G. S., 1996) in den entstehenden
Haarfollikel und teilen sich in zwei Gruppen: differenzierte Melanozyten, die im Haarbulbus
verbleiben und innerhalb ihrer Melanosomen aus Tyrosin das Pigment Melanin synthetisieren,
dieses speichern und an benachbarte Zellen abgeben (Botchkareva et al., 2003), und
Melanozytenstammzellen, die in der Stammzellennische, dem Wulst der äußeren
Wurzelscheide, verbleiben (Abbildung 5) (Nishimura et al., 2005).
Zusammenfassend lässt sich festhalten, dass im reifen Haarfollikel drei anatomisch und
funktionell unterschiedliche Vertreter der Melanozytenzellreihe gleichzeitig vorhanden sind:
Melanozytenstammzellen, Melanozytenvorläufer (hier stellvertretend die Melanoblasten) und
die ausdifferenzierten und für die Melaninsynthese verantwortlichen Melanozyten im Bulbus
(Botchkareva et al., 2001; Tobin and Bystryn, 1996; Kumano et al., 2008). Als
Unterscheidungsmerkmal zwischen diesen verschiedenen Vertretern der
Melanozytenzellreihe gilt u.a. die Enzymproduktion. Nur die reifen Melanozyten produzieren
alle für die Melanogenese wichtigen Enzyme, also auch die Tyrosinase, die Vorläufer
dagegen lediglich Tyrosinase-related protein (Trp 1) und Dopachrom-Tautomerase (Dct)
(Sarin and Artandi, 2007).

- 12 -
a b
Abbildung 5: a) Anagener Haarfollikel mit dem Wulst in der äußeren Wurzelscheide als
Sitz der Melanozytenstammzellen und dem Haarbulbus als Sitz der reifen Melanozyten
(nach Braun-Falco et al., 2005). Stammzellen und Melanozytenvorstufen wandern vom
äußeren Wurzelscheidenwulst in Richtung Bulbus, wo sie dann als ausdifferenzierte
Melanozyten liegen bleiben und sich der Melaninproduktion widmen.
b) grobes Schema der Melanozytendifferenzierung. Ein Teil der Melanozytenstammzellen
(MSZ) differenziert sich zu Melanozytevorstufen, hier stellvertretend Melanoblasten (MB),
und diese schließlich zu Melanin-produzierenden Melanozyten (MZ). Der andere Teil bleibt
als Stammzellenreservoir erhalten.

- 13 -
1.5 Haarfarbe
Die Haarfarbe des Menschen ist komplex polygen determiniert und hängt im Wesentlichen
davon ab, welcher Melanintyp im Haarfollikel produziert wird. Eumelanin kennzeichnet
braunes oder schwarzes Haar, welches bei über 90 % der Weltbevölkerung vorherrscht,
während Phäomelanin bei Blonden und Rothaarigen vorkommt (Tobin D. J., 2008; Ito and
Wakamatsu, 2010 & 2011). Neben Quantität und Qualität des Pigments spielen das Verhältnis
der beiden Pigmentarten zueinander, sowie die Haardicke eine Rolle. So entstehen die vielen
Farbnuancen durch eine Mischung beider Pigmentarten (Ito and Wakamatsu, 2011). Blonde
Haare sind dünn, pigmentarm und enthalten fast kein Mark. Schwarze Haare haben viel
Eumelanin und die größten Melanosomen, die im Haarschaft über Mark und Rinde verteilt
sind. Zahlreiche genetische Faktoren kodieren für Farbmutationen. Auf diese wird im
Diskussions-Kapitel gesondert eingegangen.
Die Anzahl der Melanozyten in der Haut ist bei Individuen unterschiedlicher Hauttypen
gleich, deren melanogene Aktivität variiert jedoch stark. Darunter versteht man die
produzierte Menge und Art des Melanins und die Anzahl und Größe der Melanosomen. Die
Melanogenese beinhaltet die Melaninproduktion und dessen Transfer zu den Keratinozyten
von Haut und Haaren. Beim Haar ist sie streng an das anagene Stadium des Wachstumszyklus
gekoppelt. Schlüsselenzyme der Melanogenese sind die Tyrosinase und die Dopachrom-
Tautomerase.
UV-B-Strahlen stimulieren die Melanozyten direkt oder aber indirekt über parakrine
Stimulatoren. Ferner bewirkt UV-Licht eine direkte DNA-Schädigung, eine sekundäre
Aktivierung der Tyrosinase und somit eine verstärkte Melanogenese. Faktoren, wie ACTH, α-
MSH, α-Melanotropin, Endothelin, TNF-α, u.a., stimulieren das Melanozytenwachstum, die
Dendritenformation und ebenfalls die Melanogenese.
Im Allgemeinen sind follikuläre Melanozyten gegenüber Umwelteinflüssen empfindlicher als
epidermale, was sich im unübersehbaren Vorgang der Canities zeigt. Ursache dafür ist u.a. die
Tatsache, dass das Haarpigment bei der Ausscheidung von Schwermetallen, Chemikalien und
Giften aus dem Körper mitwirkt, indem es diese an sich bindet (Tobin D. J., 2008).
Erkrankungen mit Beteiligung der Melanozyten sind Hypopigmentierungen, wie z.B. der
genetisch bedingte Albinismus oder die erworbene Vitiligo, und Hyperpigmentierungen. Die
häufigste generalisierte Hyperpigmentierung der Haut ist die „Bräune“ als Folge UV-
induzierter Zellaktivierung. An Veränderungen der Haarfarbe unterscheidet man Albinismus,
Poliose, Vitiligo, Canities sowie Heterochromien. Beim Albinismus handelt es sich um eine

- 14 -
rezessiv vererbte Störung der Melaninsynthese, die in weiß-gelblicher Haarfarbe resultiert.
Die Poliose ist eine erworbene Erkrankung, bei der es in den entzündlichen
Kopfhautbereichen fleckförmig zu pigmentlosen Haarbüscheln kommt. Bei Heterochromien
bestehen Unterschiede zwischen Kopf-, Bart- und Körperhaaren (Braun-Falco et al., 2005). In
den Kapiteln 1.6 und 4 wird näher auf die Canities (Haarergrauen) eingegangen.
1.6 Haarergrauen
Eines der sichtbarsten Zeichen menschlichen Alterns ist der Prozess des Haarergrauens
(Canities). Mit zunehmendem Lebensalter kommt es beim Menschen physiologischer Weise
zu verringerter Pigmentproduktion im Haarfollikel und somit zu natürlichem Haarergrauen.
Genauer betrachtet werden die Haare nicht grau, sondern weiß. Dieses Weiß resultiert aus der
Eigenfarbe des Haarkeratins und der, durch Fehlen des Melanins bedingten Reflexion des
Lichts. Das gleichzeitige Vorhandensein pigmentierter und nicht pigmentierter Haare führt
zum optischen Effekt der grauen Haare (Braun-Falco et al., 2005).
Der Beginn des Ergrauens ist genetisch festgelegt und findet bei Kaukasiern im Alter von 34
± 9,6 Jahren, bei Afroamerikanern im Alter von 43,9 ± 10,3 Jahren statt (Tobin and Paus,
2001). Im Alter von 50 Jahren sind bei annähernd 50 % der Menschen 50 % weiße Haare
sichtbar. Üblicherweise werden Barthaare noch vor den Haaren des Kapillitium weiß. Am
Kapillitium sind vorwiegend zuerst die Schläfen betroffen (Braun-Falco et al., 2005).
Rauchen scheint mit einem früheren Haarergrauen einherzugehen (Mosley and Gibbs, 1996).
Tritt das Ergrauen bereits ab dem 20. Lebensjahr auf, spricht man von Canities praecox.
Canities symptomatica kann begleitend bei Malignomen, schweren endokrinologischen
Störungen, Malnutrition (Vitamin-A-, Spurenelement- und Eisenmangel), perniziöser
Anämie, akuten fieberhaften Zuständen, durch Einwirkung von Arzneimitteln, Kosmetika,
Metallen oder auch spontan auftreten (Braun-Falco et al., 2005; Ito and Wakamatsu, 2011).
Das physiologische Haarergrauen geht mit einer abnehmenden Melanozytenfunktion einher
(Sato et al., 1973; Lloyd et al., 1987; Tobin and Paus, 2001). Dabei führt insbesondere eine
verminderte Tyrosinaseaktivität zu zunehmendem Pigmentverlust des Haarschafts. Neben der
reduzierten Pigmentproduktion ist ein eingeschränkter Transport des Melanins zum
wachsenden Haarschaft für den Ergrauungsprozess verantwortlich (Kaplan et al., 2011; Tobin
and Paus, 2001).
Doch nicht nur die Funktionalität der Melanozyten, auch eine altersbedingte Beeinträchtigung
ihrer Homöostase, welche von einer kleinen Menge Stammzellen im äußeren

- 15 -
Wurzelscheidenwulst aufrechterhalten wird, wird für den Ergrauungsprozess verantwortlich
gemacht. Mit anderen Worten, die vorhandenen Melanozytenstammzellen sind zum einen
nicht mehr in der Lage, genügend Melanozyten nachzuliefern (Sarin and Artandi, 2007), zum
anderen sinkt die Anzahl der Melanozytenstammzellen im Haarfollikel mit zunehmendem
senilem Ergrauen der Haare (Nishimura E. K., 2011). Eine Abnahme der Melanozyten sowohl
in der äußeren Wurzelscheide als auch im Haarbulbus wird als verantwortlich für die
Abnahme des Melaningehalts angesehen (Tobin D. J., 2009; Commo et al., 2004). So scheint
folglich neben dem Funktionsverlust auch eine verringerte Melanozyten(stammzellen)anzahl
eine Ursache der Canities darzustellen.
Welche Auslöser führen zu diesen Funktionsverlusten und Zahleneinbußen der Melanozyten
oder deren Vorläufer?
Eine Vielzahl äußerer und innerer Einflüsse, wie UV-Strahlung, Zigarettenrauch,
Entzündungen und psychoemotionaler Stress, generieren oxidativen Stress, dessen
Zusammenhang mit Canities bereits in zahlreichen Publikationen beschrieben worden ist
(Arck et al., 2006; Johnson and Jackson, 1992). Die Melaninsynthese des anagenen
Haarfollikels erzeugt als interner Stressor ebenfalls einen hohen oxidativen Stress und wird
verdächtigt, durch die dabei entstehenden freien Radikale, zu selektivem vorzeitigem Altern
und Apoptose der Haarfollikelmelanozyten und somit zum Altern des Haarbulbus zu führen
(Arck et al., 2006). Johnson und Jackson (1992) zeigten in Versuchen an Mäusen durch
Oxidantien verursachte Defizite innerhalb der Melanozytenzellreihen und bei der
Pigmentation. Spannenderweise kam es bei Albinomäusen nicht zu solch
Melanozytenverlusten, was die These stützt, dass diese durch bei der Melaninsynthese
entstehende toxische oxidative Produkte zustande kommen (Johnson and Jackson, 1992).
Weiterhin weist unpigmentiertes Haar aufgrund der geringeren oxidativen Belastung besseres
Wachstum auf (Arck et al., 2006). Es unterscheidet sich von pigmentiertem Haar durch seine
Beschaffenheit, ist starrer, rauer und anfälliger für äußere Schädigungen, insbesondere durch
UV-Strahlen (Kaplan et al., 2011).

- 16 -
2. Fragestellung
Der Einfluss von Stammzellen innerhalb des Haarfollikels in Bezug auf die Entstehung
unterschiedlicher Haarfarben, den Vorgang des Haarergrauens und des androgenetischen
Haarausfalles sind derzeit nicht vollständig verstanden und eröffnen ein breites Feld an
Untersuchungsmöglichkeiten.
Bei der Haarfarbe spielen Quantität und Qualität der Melanogenese, neben der Melaninart,
eine entscheidende Rolle. Welche Faktoren hierbei auf den Haarfollikel einwirken, und wie
sie das tun, ist derzeit noch offen.
Auch der Vorgang des Haarergrauens ist, obwohl es sich dabei um einen weit verbreiteten,
alterungsbedingten Vorgang handelt, noch nicht vollständig verstanden. Es gibt in der
Literatur die Meinung, dass sowohl die Anzahl als auch die Aktivität der Melanozyten im
Haarfollikel mit dem Alter abnehmen und somit zu Haarergrauen führen (Tobin D. J., 2009;
Commo et al., 2004; Botchkareva et al., 2001; Steingrímsson et al., 2005). Mit sinkender
Haarpigmentierung wird eine Reduktion der Melanozytenstammzellen im Wulst der äußeren
Wurzelscheide postuliert (Steingrímsson et al., 2005). Von pigmentiert über grau hin zu
unpigmentiert sinkt dort die Zellenanzahl (Arck et al., 2006). Sarin und Artandi (2007) sehen
die Ursache des Ergrauens vielmehr im Absinken der Zahlen innerhalb der
Melanozytenzelllinie als in deren Funktionsstörung.
Haarausfall geht mit einem Untergang im Bereich des Stammzellkompartments einher.
Welche zahlentechnischen Unterschiede finden sich bei den Faktoren zwischen Probanden
mit und ohne Haarausfall?
In der vorliegenden Arbeit sollte der Einfluss unterschiedlicher Stammzellmarker auf
Haarpigmentierung, Haarergrauen und Haarverlust anhand der immunhistochemischen
Expression von SOX 9, SOX 10, PAX 3 und CLIM 2 bei einem Kollektiv von Menschen
unterschiedlichen Alters, Geschlechts und unterschiedlicher Haarfarbe evaluiert werden.

- 17 -
3. Material und Methoden
3.1 Probanden
3.1.1 Gewebeschnitte der Probanden
Für diese Arbeit wurden Probanden ausgewählt, die zwischen den Jahren 2005 und 2008 in
der Universitäts-Hautklinik Freiburg am Kopf operiert worden sind, oder die sich einer
Probeentnahme am Kopf unterzogen haben. Die entsprechenden Schnittpräparate wurden
herausgesucht und angesehen. Für die engere Auswahl war im Wesentlichen entscheidend,
dass unabhängig von der vorliegenden Diagnose ein ausreichend großes Gewebestück ohne
Einfluss des pathologischen Prozesses Einblick auf möglichst zahlreiche tangential
geschnittene Haarfollikel erlaubte. Zu kleine Präparate wurden aussortiert.
Schließlich wurden die histologischen Gewebeblöcke der selektierten Probanden
herausgesucht und für weitere Schnitte und immunhistochemische Färbungen verwendet.
3.1.2 Probandeninformationen
Von den selektierten Fällen wurden die Probandenakten herausgesucht und daraus Daten wie
deren Alter, Geschlecht, Diagnose und Haarfarbe entnommen.
79 Probanden im Alter von 1 bis 93 Jahren, davon 35 männlichen und 44 weiblichen
Geschlechts, wurden in die Arbeit eingeschlossen. Sämtliche Präparate entstammen dem
Archiv der Hautklinik Freiburg und sind diagnostisch abgeklärt. Vom harmlosen Naevus,
über Ekzeme und bis hin zu Tumorerkrankungen wie dem malignen Melanom sind
unterschiedlichste Diagnosen vertreten.
Waren Fotos der Operationslokalisation vorhanden, konnte aus diesen die Haarfarbe
entnommen werden. Soweit die Fotodokumentation keine Rückschlüsse auf Haarfarbe und
Haarergrauen erlaubte, wurden die Probanden angefragt nach ihrer Haarfarbe, im Speziellen
der Status des Haarergrauens.

- 18 -
3.2 Anfertigung der Schnittpräparate
Zur Anfertigung immunhistochemischer Schnitte wurden die Paraffinblöcke der ausgewählten
Probanden herausgesucht. Diese wurden mindestens 20 Minuten bei -18 °C gekühlt. Von den
gekühlten Blöcken wurde mittels eines Mikrotoms (Abbildung 6) 4 µm dicke Schnitte
angefertigt, in ein ca. 40 °C warmes Wasserbad gelegt und von dort aus auf beschichtete
Objektträger aufgezogen. Nach Trocknung auf einer Wärmeplatte wurden sie senkrecht in
Aufbewahrungskästen sortiert und solange im Trockenen gelagert, bis sie zum
Entparaffinieren und Färben eingesetzt wurden.
Abbildung 6: Mikrotom
3.3 Entparaffinierung
Die Objektträgerpräparate wurden im Brutschrank 30 Minuten lang bei 60 °C inkubiert und
anschließend entparaffiniert. Für diesen Zweck wurden sie senkrecht auf einen Ständer
verteilt und in eine Xylol-gefüllte Glasküvette gestellt. Nach 15 Minuten wurden sie in zwei
weitere Xylol-gefüllte Küvetten und in einer Alkoholverdünnungsreihe, welche aus Behältern
mit absteigenden Alkoholkonzentrationen (100 %, 100 %, 95 %, 95 %, 80 %, 70 %, 50 %)
bestand, durch mehrfaches Tunken gespült. Daraufhin wurde der Ständer in eine mit Aqua
dest. gefüllte Plastikküvette abgestellt.

- 19 -
3.4 Antigenretrieval-Methoden
Durch die Einbettung in Paraffin kommt es zu einer Konservierung von Gewebe und Zellen.
Diese gewebeschützende Funktion kommt Zustande, indem Proteine durch Koagulation
und/oder chemische Quervernetzung denaturieren. Die Erhaltung der Morphologie bringt
jedoch den Nachteil des Verlustes der Immunreaktivität vieler Antigene mit sich.
Um diese „Antigenmaskierung“ wieder rückgängig zu machen, was für die bessere
immunhistochemische Färbbarkeit notwendig ist, ist eine Demaskierung, auch Epitop-
Retrieval genannt, angezeigt. Hierfür stehen verschiedene Methoden zur Auswahl. In den
folgenden Abschnitten werden die in dieser Arbeit verwendeten Vorgehensweisen erläutert.
3.4.1 Hitzeinaktivierung in Citrat-Puffer im Dampfk ocher
Als Retrieval-Lösung wurde 10 mM Citrat-Puffer mit pH 6,0 verwendet, der jedes Mal vor
Gebrauch frisch hergestellt wurde. Hierfür wurden je 4,5 ml 0,1 mol Zitronensäure und
20,5 ml 0,1 mol Natriumcitrat in einen Glaszylinder gegeben, dieser mit Aqua dest. auf ein
Volumen von 250 ml aufgefüllt und die Flüssigkeit mit einem Magnetrührer gut durchmischt.
Anschließend wurde der pH-Wert gemessen und auf pH 6 eingestellt.
Nun wurde die Pufferlösung in Küvetten gegeben und im Dampfgarer 10 Minuten lang
vorgekocht und danach die entparaffinierten Präparate mit Plastikständer in die Pufferbehälter
gestellt und 20 Minuten lang darin gekocht. Im Anschluss daran wurden die Küvetten in ein
kaltes Wasserbad gestellt. Nach 10 Minuten fand ein Wasserwechsel statt, d.h. das inzwischen
erwärmte Wasser wurde durch frisches kaltes ersetzt und erneut einige Minuten stehen
gelassen. Am Ende wurden die Plastikständer mit den Präparaten in Aqua dest. gestellt.
Bestandteile Citratpuffer (pH 6):
4,5 ml 0,1 Zitronensäure (21.01g in 1000 ml Aqua dest.)
20,5 ml Natriumcitrat (29,41g in 1000 ml Aqua dest.)
ad 250 ml Aqua dest.

- 20 -
3.4.2 Hitzeinaktivierung in Tris-EDTA-Puffer im Dampfkocher
Nach Herstellen der Pufferlösung wurden hier dieselben Schritte des Kochvorgangs an Stelle
von Citrat-Puffer (pH 6) mit Tris-EDTA-Puffer (pH 9) durchgeführt. Ziel des Einsatzes von
Puffern verschiedener pH-Werte ist es, die optimalen Bedingungen für die Immunreaktivität
zu finden.
Bestandteile Tris-EDTA-Puffer (pH 9):
- 5 ml 0,5 mol Tris Puffer pH 7,6
- 0,5 ml 0,5 mol EDTA
- ad 250 ml Aqua dest.
- NaOH (zur Einstellung pH 9)
3.4.3 Permeabilitätserhöhung der Zellwände mit Triton X100
Pro Milliliter verwendeten Antikörperverdünnungsmediums wurden 0,05 g Rinderserum
(Albumin Fraktion V) aufgelöst. Nun wurde langsam 0,3 % Triton X 100 dazugeben. Erst
wenn sich das zähe Triton aufgelöst hatte, wurde der Antikörper hinzupipettiert und nach
sanftem Schütteln in den Färbeautomaten pipettiert. Das Triton als nicht-ionisches Detergenz
hilft dabei, die Membranproteine aus Membranproteinkomplexen zu lösen.
3.4.4 Proteolytischer Enzymverdau
Durch eine Trypsin-Andauung des Gewebes sollte eine Permeabilisierung, und somit eine
verbesserte Immunreaktivität erreicht werden (Boenisch et al., 2003). Hierfür wurden die
Schnitte mit Trypsinlösung (pH 7,8) der Firma DAKO (DAKO S2012) für 15 min bei
Raumtemperatur vorbehandelt.

- 21 -
3.5 Immunhistochemische Färbung
Angewendet wurde die „Zwei-Schritt indirekte Methode“, eine allgemein bekannte
Sandwich-Färbemethode, bei welcher zunächst ein unmarkierter Primärantikörper spezifisch
an das Zielantigen bindet. Im nächsten Schritt bindet ein zweiter enzymmarkierter
Sekundärantikörper an diesen Primärantikörper. Anschließend folgt die Substrat-
Chromogenreaktion, bei der es zu einer roten Färbung kommt (Abbildung 7) (Boenisch et al.,
2003). Die Methode wurde entsprechend der Anleitung der Firma Dako durchgeführt.
3.5.1 Färben mit dem TechMate Horizon Färbeautomaten
Es wurde nach einem Standardprotokoll zur Anfertigung immunhistochemischer Färbungen
(MSAPAL) gearbeitet und diesem entsprechend die TechMate Horizon Färbemaschine
(Abbildung 8) mit Puffern (Dako REAL Wash Buffer 1-4), je sechs Tropfen sekundärem
Antikörper (AB 2), sechs Tropfen Dako Streptavidin Alkaline Phosphatase (AP) und je 400 µl
Dako Hämatoxylin pro Vertiefung befüllt.
Abbildung 7: Prinzip der immunhistochemischen Färbung mit Primär- und Sekundär-
Antikörper (nach Boenisch et al., 2003)

- 22 -
Abbildung 8: TechMate Horizon Färbeautomat
3.5.2 Ansetzen der Antikörper
In Eppendorfreaktionsgefäße wurden jeweils die gewünschte Menge Dako
Antikörperverdünnungsmedium und Antikörper pipettiert und mit einem Vortexer gemischt.
Dem Protokoll folgend, wurden je 300 µl Antikörperverdünnung in die vorgesehenen
Gerätevertiefungen pipettiert. Dabei wurde beachtet, dass sich keine Blasen bilden.
3.5.3 Beladen der Maschine und Färbevorgang
Als nächstes wurde die Maschine mit den Präparaten beladen. Dabei wurden je zwei
Objektträger in einem Spalt befestigt, sodass die Hautschnitte einander zugewandt waren. Bei
ungerader Anzahl wurde ein ungebrauchter Objektträger dagegen gestellt. Somit wurden
jeweils zwischen zwei Objektträgern über Kapillarwirkung die Lösungen in der Maschine
hochgesogen. Die Objektträgerhalter sollten gleichmäßig mit Objektträgern beladen sein.
Bevor der Färbevorgang startet, wurden die Präparate manuell drei Mal in Puffer BUF 2
gespült und auf den Saugkissen abgetupft. Anschließend wurde der beladene
Objektträgerhalter in die Färbemaschine eingesetzt, der Deckel geschlossen und das
Färbeprogramm gestartet.
Nach 90 Minuten wurde Chromogen frisch angesetzt. In einem Eppendorfreaktionsgefäß
wurden die benötigte Menge Dako AP Substratpuffer und Dako Chromogen Rot 1-3 pipettiert
und das Ganze auf dem Vortex geschüttelt. Vorsichtig wurden je 700 µl der hergestellten
Chromogenlösung pro Vertiefung pipettiert, der Deckel erneut geschlossen und die Maschine
weiter den Färbevorgang durchführen gelassen.

- 23 -
3.5.4 Eindecken der gefärbten Präparate
Sobald die Maschine fertig mit der Färbung war, wurden der Objektträgerhalter entnommen,
die Objektträger auf eine Metallhalterung verteilt und in eine mit 100 % Ethanol gefüllte
Plastikküvette gestellt. Zügig wurde die Metallhalterung je fünf Sekunden in einer Xylol-
Reihe, bestehend aus drei Xylol-enthaltenden Glasküvetten, getaucht. Diese wurden
ursprünglich mit derselben Xylolflüssigkeit befüllt. Da die Objektträger jedoch stets in der
gleichen Reihenfolge in die Küvetten getaucht wurden, wurde in der stets zuerst verwendeten
Xylolflüssigkeit dabei mehr Farbstoff aus der vorangegangenen Färbung gelöst, in der 2.
weniger und in der 3. am wenigsten. Letztere Küvette enthielt somit die klarste Lösung.
Im Anschluss wurden die Objektträger in die Coverslipping-Maschine gestellt, welche die
Deckgläser auf die gefärbten Präparate klebt. Schließlich wurden die Objektträger horizontal
unter einer Abzugshaube gelagert bis die Deckgläser angetrocknet waren.
3.6 Mikroskopische Auswertungen
Es wurden pro Präparat diejenigen drei Haaranschnitte mit den meisten positiv gefärbten
Zellen identifiziert und als horizontal, schräg, travers oder Papille differenziert. Bei 40-facher
Vergrößerung wurden pro Gewebeschnitt die drei Rasterfelder mit der höchsten Anzahl
positiver Zellen ausgezählt. Dieser Vorgang wurde nochmals bei einem zweiten Präparat
desselben Probanden wiederholt. Auf der Evaluation dieser sechs Felder beziehen sich alle
weiteren Berechnungen. Ein Beispiel für die verschiedenen Haaranschnitte und die
Rasterfelder mit positiven Zellen ist in Abbildung 9 gezeigt.
a b c d Abbildung 9: SOX-9 Expression in unterschiedlichen Bereichen des Haarschaftes.
a) horizontal (20 x), b) schräg (40 x), c) travers (40 x), d) Papille (20 x)

- 24 -
3.7 Probandengruppen
Die Probanden wurden anhand unterschiedlicher Merkmale wie Alter, Geschlecht und
Haarfarbe in Gruppen unterteilt. Die Unterteilung erfolgte nach folgenden Gesichtspunkten:
• nach Haarfarben
� Einteilung in drei Farbgruppen: hell, dunkel, grau unter Auszählung aller
Haaranschnittebenen
� Einteilung in neun Farbgruppen: rötlich, blond, dunkelblond, braun,
dunkelbraun, schwarz, grau (beginnend), grau, Alopezie;
hier erfolgte auch eine Differenzierung in unterschiedliche anatomische Areale:
o Auszählung aller Haaranschnittebenen
o nach Haarfarbe unter Ausschluss der Papillenschnitte
o nach Haarfarbe unter isolierter Betrachtung der Papillenschnitte
� nach Haarfarbe unter Streichung bestimmter Mittelwerte pro Proband
• Haarausfall versus nicht-Haarausfall
• graue Haare versus nicht-graue
• nach Altersgruppen: 0-14, 15-40, 41-60, > 61 Jahre
• nach Geschlecht: Frauen versus Männer
3.8 Statistische Analyse und Auswerteverfahren
Für die Berechnung der Mittelwerte, Mediane, Standardabweichungen und
Korrelationskoeffizienten wurde das Programm EXCEL verwendet. Zusätzlich wurden nach
Gruppenbildung mittels GraphPad Prism 5-Programm student’s t-Tests durchgeführt. Als
signifikant wurde ein Unterschied mit p < 0,05 angesehen. Die Tabellen wurden im Word-
Programm, die Grafiken, wie Kreis- oder Balkendiagramme, wurden mithilfe von EXCEL
erstellt. Die Box-Whisker-Plots entstanden durch Einsatz des „QtiPlot“-Programms.
- 25 -
3.9 Materialien
3.9.1 Antikörper Spezies Klonalität Hersteller
SOX 9 (H-90): sc-20095 Kaninchen polyklonal Santa Cruz Biotechnology, Inc.
Santa Cruz, California, USA
PAX 3/7 (H-208): sc-25409 Kaninchen polyklonal Santa Cruz Biotechnology, Inc.
Santa Cruz, California, USA
SOX 10 (N-20): sc-17342 Ziege polyklonal Santa Cruz Biotechnology, Inc.
Santa Cruz, California, USA
CLIM 2 (N-18): sc-11198 Ziege polyklonal Santa Cruz Biotechnology, Inc.
Santa Cruz, California, USA
3.9.2 Chemikalien Bestellnr.: Hersteller:
Albumin Fraktion V (Rind) 8076.4 Carl Roth, Karlsruhe
Triton X 100 93426 Fluka BioChemika, Buchs
Xylol 9713-2 Roth, Karlsruhe
Ethanol
Aqua destillata
Detektionskit Dako, Alkaline Phosphatase/
RED, Rabbit/Mouse K5005 Dako, Hamburg
Antikörperverdünnungsmedium S2022 Dako, Hamburg
Hämatoxylin S2020 Dako, Hamburg
Trypsin S2012 Dako, Hamburg
Pufferkit für TechMate-Färbeautomaten K5006 Dako, Hamburg
Citronensäure-Monohydrat D1.00244 MERCK, Darmstadt
tri-Natriumcitrat-Dihydrat 6448 MERCK, Darmstadt
Target-Retrieval Solution pH 9 S2367 Dako, Hamburg

- 26 -
3.9.3 Geräte
Kühlschrank 4 °C BOSCH, München
Gefrierschrank -18 °C Haier, Bad Homburg
Mikrotom MIKROM HM 360, Walldorf
Wasserbad Medax Nagel KG, Kiel
Wärmeplatte Medax Nagel KG, Kiel
Waage Mettler P160, Giessen
Brutschrank Heraeus, Hanau
Pipetten Eppendorf, Hamburg
Immunostainer TechMate Horizon Dako, Hamburg
Mikroskop Carl Zeiss, Oberkochen
Vortex Heidolph, Kelheim
Magnetrührer IKA Labortechnik, Staufen
Dampfgarer Multigourmet, BRAUN
pH-Meter Seven Easy Mettler Toledo, Giessen
Coverslipping Machine, Automatic Promounter RCM90 Medite, Meisei, CH
3.9.4 Sonstiges
Kapillarspaltobjektträger 100 µm/ 75 µm, S2025/ S2024 Dako, Hamburg
Deckgläser Langenbrinck, Emmendingen
Eindeckmedium Permount, 17986-05 Electron Microscopy Sciences,
Hatfield, USA
Reagenziengefäße für TechMate Horizon, S2039 Dako, Hamburg
Saugkissen für TechMate Horizon, S2043 Dako, Hamburg
Pipettenspitzen Eppendorf, Hamburg
Reaktionsgefäße 1,5 ml Eppendorf, Hamburg
Microtome Blades, Stainless Steel S35, 02.075.00.000 Feather, Osaka, Japan
AxioVision Fotosoftware Carl Zeiss, Oberkochen
Metallhalterungen für Objektträger
Plastikküvetten
Magnetrührstäbchen

- 27 -
4. Ergebnisse
4.1 Etablierung der immunhistochemischen Färbungen
Nach einer Durchsicht der Fachliteratur zu den Stichworten „Haare“ und
„Stammzellfaktoren“ wurde eine Korrelation zu käuflichen Antikörpern erstellt, aus der sich
folgende Antikörper zum immunhistochemischen Nachweis von Stammzellmarker ergab:
SOX 9 (H-90): sc-20095,
SOX 10 (N-20): sc-17342,
PAX 3/ 7 (H-208): sc-25409,
CLIM 2 (N-18): sc-11198.
Soweit vorhanden, wurden methodische Anwendungsbeschreibungen aus der Literatur in die
Etablierungsversuche eingebracht (Kordes and Hagel, 2006; Moriyama et al., 2006).
Die Antikörper wurden in Referenzschnitten von Kopfhautgewebe ausgetestet. Als
Negativkontrolle dienten konsekutive, parallel methodisch mitgeführte Schnittpräparate unter
Auslassung des Primärantikörpers. Als Positivkontrolle dienten Gewebeschnitte von
Malignen Melanomen.
4.1.1 Testreihen
Zuerst wurde im Standardfärbeverfahren ohne Demaskierung eine Verdünnungsreihe der
Antikörper in Citratpuffer pH 6 durchgeführt. Darüber hinaus wurden Verdünnungsreihen
unter Hitzevorbehandlung als Demaskierungsverfahren ausgetestet. Als Pufferlösung für die
Hitzeinaktivierung kam sowohl Citrat- (pH 6), als auch Tris-EDTA-Puffer (pH 9) in
unterschiedlichen Inkubationszeiten bei 100 °C zum Einsatz. Desweiteren wurde eine
Vorbehandlung mit Triton X 100 verwendet, um die Zellmembranen temporär porös zu
machen und somit die Wahrscheinlichkeit einer intrazellulären Färbung zu erhöhen.

- 28 -
Im Konkreten wurden folgende Variablen an den einzelnen Antikörpern ausgetestet:
• SOX 9:
o Antikörperverdünnungsreihe: 1:50, 1:100, 1:200, 1:300, 1:400
o Kochen in Citratpuffer, pH 6
o Kochen in Tris-EDTA-Puffer, pH 9
o Blocken mit 5 % -igem Rinderserum
o 10 min Inkubation in 0,3 % -igem Triton X 100
• SOX 10:
o Antikörperverdünnungsreihe: 1:50, 1:100, 1:200, 1:300, 1:400
o Kochen in Citratpuffer, pH 6
o Kochen in Tris-EDTA-Puffer, pH 9
o mit 5 % -igem Rinderserum blocken
o 10 min Inkubation in 0,3 % -igem Triton X 100
• PAX 3:
o Antikörperverdünnungsreihe: 1:50, 1:100, 1:150, 1:200
o Kochen in Citratpuffer, 10 min/20 min
o Kochen in Tris-EDTA-Puffer, pH 9, 20 min
o mit 5 % -igem Rinderserum blocken
o 10 min Inkubation in 0,3 % -igem Triton X 100
• CLIM 2:
o Antikörperverdünnungsreihe: 1:50, 1:100, 1:200, 1:300, 1:400
o Kochen in Citratpuffer, pH 6
o Kochen in Tris-EDTA-Puffer, pH 9
4.1.2 Ergebnisse der Etablierungsversuche
PAX 3 und CLIM 2
Die Etablierungsversuche der immunhistochemischen Färbung mit den spezifischen
Antikörpern gegen PAX 3 und CLIM 2 erbrachten keinerlei akzeptable Ergebnisse. Trotz
Austesten unterschiedlicher Konzentrationen sowohl von Primär- als auch von

- 29 -
Sekundärantikörpern, sowie unterschiedlicher Demaskierungsmethoden zeigte sich entweder
eine vollständige Negativität oder eine intensive Hintergrundfärbung der Gewebsschnitte,
sodass keine spezifische Auswertung möglich war.
SOX 10
Bei SOX 10 zeigten sich in den Verdünnungsreihen von 1:50 bis 1:400 schwankende und
insgesamt nur sehr geringe Anfärbungen einzelner Zellen. Eine Verlängerung und
Intensivierung der Hitzeinaktivierung führte zu einem Anstieg der Hintergrundsfärbung,
welche sich auch nach Blocken mit Rinderserum nicht unterdrücken ließ. Gleiches geschah
nach Vorbehandlung mit Triton X 100. Eine Zunahme spezifischer Anfärbungen konnte
hierdurch nicht erlangt werden. Aufgrund dieser inkonstanten Anfärbbarkeit wurden weitere
Versuche mit SOX 10 eingestellt.
SOX 9
Bei SOX 9 kristallisierten sich im Laufe der Etablierungsversuche folgende Parameter als
optimal heraus: Primärantikörperverdünnung: 1:100, Demaskieren in Citratpuffer pH 6 über
20 min bei 100 °C und Antikörperinkubation in Inkubationspuffer unter Zusatz von 0,3 %
Triton X 100 und 5 % Rinderserum. Diese Parameter wurden bei den folgenden Versuchen
konstant gehalten.
4.2 Probandencharakterisierung
4.2.1 Allgemeine Angaben zu den Probanden
Anhand der histologischen Datei der Freiburger Universitäts-Hautklinik wurden ca. 250
Probanden ermittelt, die zwischen 2005 und 2008 am Kopf operiert worden waren.
Schnittpräparate der Gewebestücke wurden auf die Beurteilbarkeit von Haarfollikeln
(tangentiale Schnittführung) überprüft. Es wurden nur Präparate ausgewählt, welche
mindestens zehn Haarfollikelanschnitte aufwiesen, welche zudem präferenziell tangential
angeschnitten waren. Dies führte zu einem Ausschluss von ca. 150 Präparaten, welche zu
klein waren.
- 30 -
Von den übrigen ca. 100, ließ sich bei ca. 30 Probanden die Haarfarbe weder anhand von
Fotos in den elektronischen Akten, noch telefonisch eruieren. So blieben zur weiteren
Analyse 79 Probanden, 35 männliche und 44 weibliche Probanden im Alter von 1-93 Jahren.
Das Durchschnittsalter lag bei 47,2 Jahren.
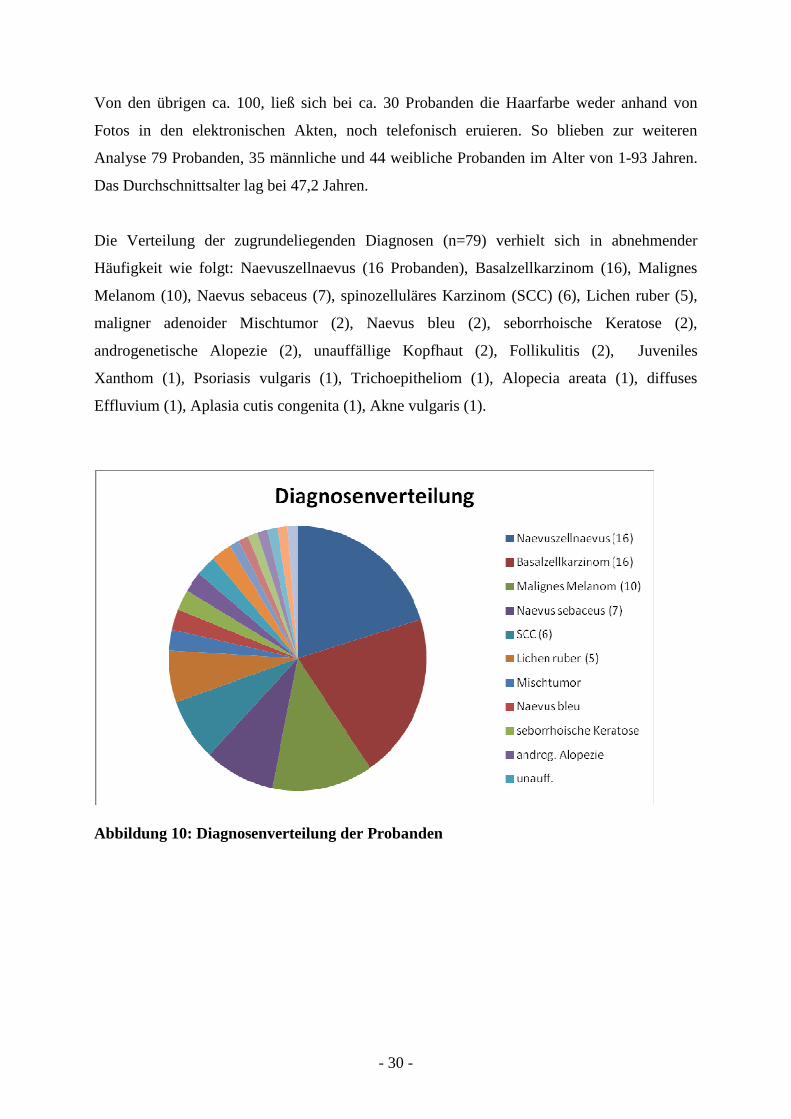
Die Verteilung der zugrundeliegenden Diagnosen (n=79) verhielt sich in abnehmender
Häufigkeit wie folgt: Naevuszellnaevus (16 Probanden), Basalzellkarzinom (16), Malignes
Melanom (10), Naevus sebaceus (7), spinozelluläres Karzinom (SCC) (6), Lichen ruber (5),
maligner adenoider Mischtumor (2), Naevus bleu (2), seborrhoische Keratose (2),
androgenetische Alopezie (2), unauffällige Kopfhaut (2), Follikulitis (2), Juveniles
Xanthom (1), Psoriasis vulgaris (1), Trichoepitheliom (1), Alopecia areata (1), diffuses
Effluvium (1), Aplasia cutis congenita (1), Akne vulgaris (1).
Abbildung 10: Diagnosenverteilung der Probanden

- 31 -
4.2.2 Haarfarben der Probanden
Die Haarfarben der Probanden wurden aus den Probandenakten übernommen. In Fällen, in
denen keine Informationen zur Haarfarbe vorlagen, wurden die Probanden telefonisch befragt.
Die Haarfarben wurden nach zwei unterschiedlichen Schemata unterteilt.
Schema 1:
Einteilung in die drei Farbgruppen hell (rötlich, blond, dunkelblond), dunkel (braun,
dunkelbraun, schwarz), grau (grau beginnend, grau). Hierbei zeigte sich folgende
Fallzahlverteilung:
� hell (n=23)
� dunkel (n=26)
� grau (n=30)
Schema 2:
Einteilung in neun Haarfarben (rötlich, blond, dunkelblond, braun, dunkelbraun, schwarz,
grau (beginnend), grau, Alopezie). In Tabelle 1 sind die einzelnen Zuordnungen aufgeführt.
Tabelle 1: Probanden (Haarfarben, Alter, Geschlecht, Fotobeispiele)
Haarfarbe Alter m/w Fotobeispiele
Rötlich: n=1 blond-rötlich
46 m
Blond: n=8 aschblond (manchmal Strähnchen) aschblond, einzelne grau hellblond hellblond blond blond hell-mittel-blond (Strähnchen blond) mittelblond
67 58 7 22 3 22 28 42
w w m w w w w w
Dunkelblond: n=13 dunkel-blond, vereinzelt grau dunkel-blond, vereinzelt grau dunkel-blond, vereinzelt weiß dunkelblond dunkelblond
66 66 76 35
w w w m
- 32 -
dunkelblond dunkelblond dunkelblond dunkelblond dunkelblond dunkelblond- Kastanie dunkelblond- hellbraun dunkelblond- hellbraun
32 24 37 40 16 9 53 24 27
w w m m m m m m m
Braun: n=13 hellbraun hellbraun mittelbraun mittelbraun mittelbraun braun braun braun braun braun braun braun braun, etwas grau
25 20 18 14 14 20 1 6 1 38 39 12 75
m w w w w w m w m w m w m
Dunkelbraun: n=9 dunkelbraun, einzelne graue dunkelbraun, einzelne graue dunkelbraun dunkelbraun dunkelbraun dunkelbraun dunkelbraun braun, getönt schwarz dunkelbraun
46 50 47 33 28 24 28 38 24
m m m w m m w w w
Schwarz: n=3 dunkelbraun-schwarz schwarz schwarz
10 37 26
m m m
- 33 -
Grau (beginnend): n=8 grau- dunkelbraun 50:50 braun- grau braun- grau braun- grau 50:50 grau- braun grau- braun, braun gefärbt grau- dunkelblond kastanienrot, leicht grau, (gefärbt: kastanienrot)
67 80 80 63 79 55 72 62
w w w w m w m w
Grau: n=16 grau (und dunkelblond) grau (- bräunlich), gefärbt: bräunlich grau, gefärbt: rötlich (früher: dunkelblond) grau (bis mittelblond) grau (- aschblond) grau grau grau grau- meliert/ weiß (früher blond) grau- weiß grau weiß weiß weiß (-grau) grau- weiß weiß- grau
64 57 65 70 72 66 83 69 93 80 76 86 80 91 84 81
m w w m w m m m w m m m m m w w
Alopezie: n=8 hellbraun + Alopezie Alopezie (blond) weiß-grau (+Haarausfall) weiß-grau (+Haarausfall) weiß (wenig) weiß (wenig) weiß, wenig diffuse Alopezie
40 37 75 75 73 73 79 34
w w w w w w w w

- 34 -
4.3 Expression von SOX 9 im Probandenkollektiv
SOX 9-positive Zellen im Bereich der Haarfollikel waren bei Probanden aller Haarfarben zu
erkennen. Diese ließen sich durch ihre rote Kernfärbung klar von anderen Zellen abgrenzen.
Im Folgenden sind Bildbeispiele zur morphologischen Verteilung der SOX 9-Expression von
mehreren Probanden unterschiedlichen Alters, Haarfarbe und Haardichte dargestellt. Die
Auswertungen wurden anhand verschiedener Schnittebenen am Haarfollikel durchgeführt.
a b
c d
Abbildung 11: Sox 9-Expression an vier Haaranschnitten einer 73-jährigen Probandin
mit weißen Haaren. Der Mittelwert von SOX 9-positiven Zellen pro 40x HPF lag bei
20,17.
a) Schräger Anschnitt. 40 x Vergrößerung. 20 SOX 9-positive Zellen
b) Schräger Anschnitt, 40 x Vergrößerung, 29 SOX 9-positive Zellen
c) Traverser Anschnitt, 40 x Vergrößerung, 24 SOX 9-positive Zellen
d) Traverser Anschnitt, 20 x Vergrößerung, 28 SOX 9-positive Zellen (Summe der 40x
Felder 20+8)

- 35 -
Zunächst sehen wir exemplarisch vier unterschiedliche Haarfollikel einer Probandin mit
weißen Haaren und einer insgesamt hohen SOX 9-Expression (Abbildung 11). In zwei
schrägen Anschnitten wurden pro ausgezähltes Rasterfeld die Zellanzahl 20 und 29 für
SOX 9-positive Zellen ermittelt (Abbildung 11 a, b). Die beiden traversen Anschnitte liefern
24 und 28 SOX 9-positive Zellen (Abbildung 11 c, d). Abbildung 11 d zeigt, im Gegensatz zu
den drei Haarfollikeln in 40-facher Vergrößerung (Abbildung 11 a, b, c), eine 20-fache
Vergrößerung mit insgesamt 28 positiv angefärbten Zellen. Da für die Auswertung jedoch
deren Anzahl im 40-er Feld gilt, zählen wir hier zwei 40x Felder mit je 20 und
8 SOX 9-positiven Zellen.
Um einen besseren Eindruck über die Verteilung der SOX 9-positiven Zellen am Haarfollikel
zu erhalten, sind vier Probanden mit unterschiedlichen Haarfarben und hoher SOX 9-
Expression beispielhaft dargestellt (Abbildung 12). Unabhängig von der Anschnittart, also ob
travers, horizontal oder schräg, sind die SOX 9-positiven Zellen regelmäßig entlang der
inneren Wurzelscheide verteilt. Zunächst sehen wir in einem schrägen Haarschaftanschnitt
einer hellblonden Probandin 23 SOX 9-positive Zellen (Abbildung 12 a). Im transversen
Anschnitt zeigt das Haar eines 46-jährigen Mannes mit dunkelbrauner Haarfarbe 27
(Abbildung 12 b), ein horizontaler, leicht schräger Anschnitt beim Haar eines 3-jährigen,
blonden Mädchens 38 SOX 9-positive Zellen (Abbildung 12 c). Ein weiterer schräger
Haaranschnitt zeigt 17 SOX 9-positive Zellen bei einem dunkelblonden Probanden
(Abbildung 12 d).
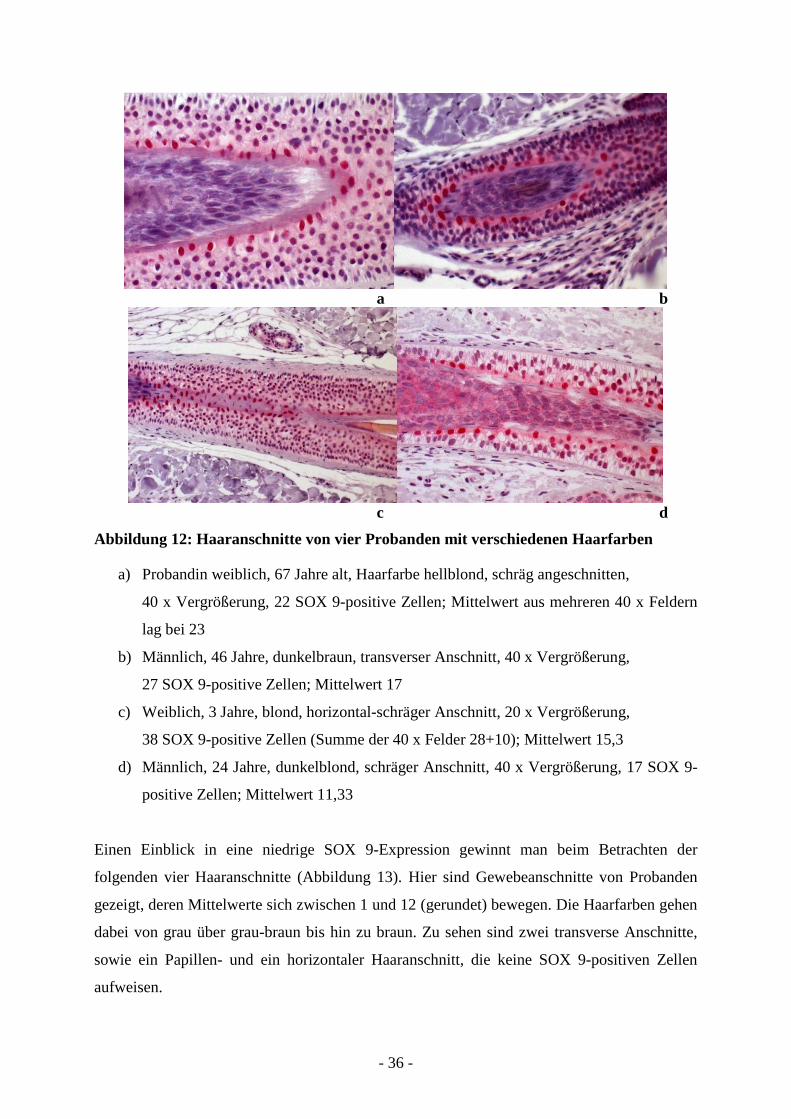
- 36 -
a b
c d
Abbildung 12: Haaranschnitte von vier Probanden mit verschiedenen Haarfarben
a) Probandin weiblich, 67 Jahre alt, Haarfarbe hellblond, schräg angeschnitten,
40 x Vergrößerung, 22 SOX 9-positive Zellen; Mittelwert aus mehreren 40 x Feldern
lag bei 23
b) Männlich, 46 Jahre, dunkelbraun, transverser Anschnitt, 40 x Vergrößerung,
27 SOX 9-positive Zellen; Mittelwert 17
c) Weiblich, 3 Jahre, blond, horizontal-schräger Anschnitt, 20 x Vergrößerung,
38 SOX 9-positive Zellen (Summe der 40 x Felder 28+10); Mittelwert 15,3
d) Männlich, 24 Jahre, dunkelblond, schräger Anschnitt, 40 x Vergrößerung, 17 SOX 9-
positive Zellen; Mittelwert 11,33
Einen Einblick in eine niedrige SOX 9-Expression gewinnt man beim Betrachten der
folgenden vier Haaranschnitte (Abbildung 13). Hier sind Gewebeanschnitte von Probanden
gezeigt, deren Mittelwerte sich zwischen 1 und 12 (gerundet) bewegen. Die Haarfarben gehen
dabei von grau über grau-braun bis hin zu braun. Zu sehen sind zwei transverse Anschnitte,
sowie ein Papillen- und ein horizontaler Haaranschnitt, die keine SOX 9-positiven Zellen
aufweisen.
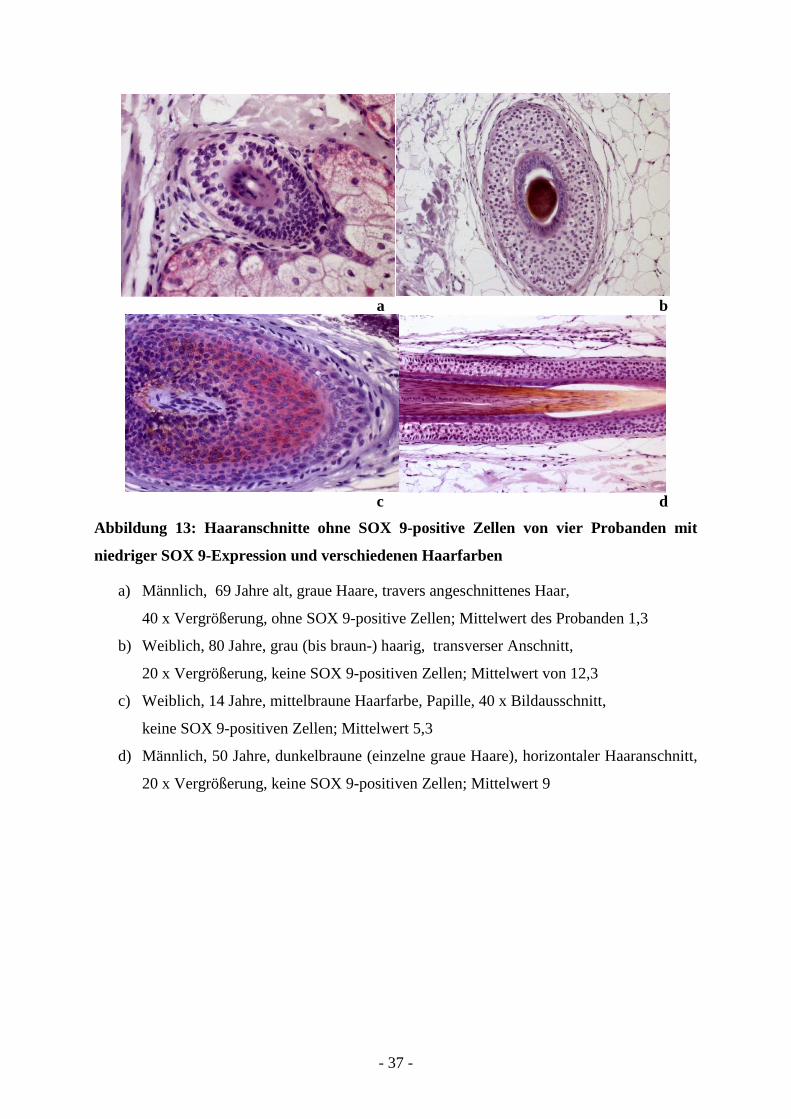
- 37 -
a b
c d
Abbildung 13: Haaranschnitte ohne SOX 9-positive Zellen von vier Probanden mit
niedriger SOX 9-Expression und verschiedenen Haarfarben
a) Männlich, 69 Jahre alt, graue Haare, travers angeschnittenes Haar,
40 x Vergrößerung, ohne SOX 9-positive Zellen; Mittelwert des Probanden 1,3
b) Weiblich, 80 Jahre, grau (bis braun-) haarig, transverser Anschnitt,
20 x Vergrößerung, keine SOX 9-positiven Zellen; Mittelwert von 12,3
c) Weiblich, 14 Jahre, mittelbraune Haarfarbe, Papille, 40 x Bildausschnitt,
keine SOX 9-positiven Zellen; Mittelwert 5,3
d) Männlich, 50 Jahre, dunkelbraune (einzelne graue Haare), horizontaler Haaranschnitt,
20 x Vergrößerung, keine SOX 9-positiven Zellen; Mittelwert 9
- 38 -
4.4 Vergleichende Analysen der SOX 9-Expression
4.4.1 Vergleich der Haarfarben
Im Folgenden wurden die unterschiedlichen Gruppen (Schema 1 und 2, Kapitel 4.2.2) in ihrer
SOX 9-Expression miteinander verglichen und statistisch ausgewertet. Der Mittelwert eines
jedes Probanden ergab sich dabei aus den sechs 40x Rasterfeldern mit der höchsten Anzahl
positiver SOX 9-positiver Zellen. Nach Gruppenbildung wurde aus den Mittelwerten der
dazugehörigen Probanden ein Gruppenmittelwert gebildet.
4.4.1.1 Vergleich von hellem, dunklem und grauem Haar (Schema 1)
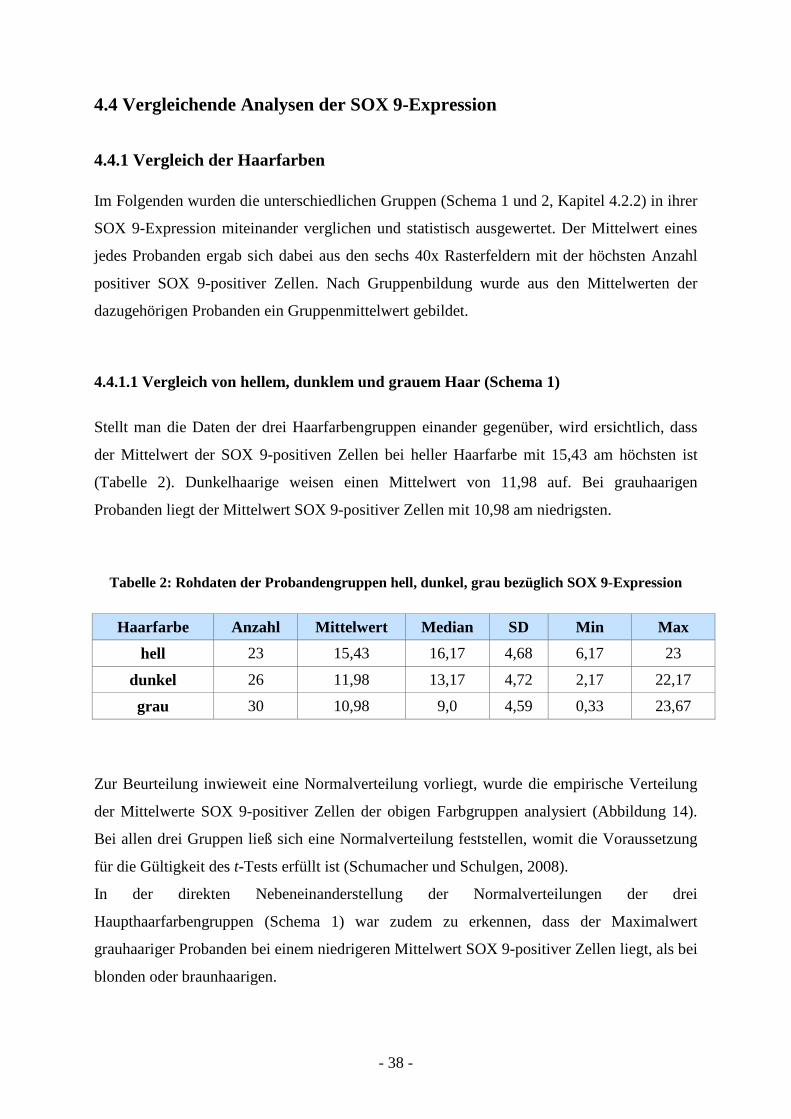
Stellt man die Daten der drei Haarfarbengruppen einander gegenüber, wird ersichtlich, dass
der Mittelwert der SOX 9-positiven Zellen bei heller Haarfarbe mit 15,43 am höchsten ist
(Tabelle 2). Dunkelhaarige weisen einen Mittelwert von 11,98 auf. Bei grauhaarigen
Probanden liegt der Mittelwert SOX 9-positiver Zellen mit 10,98 am niedrigsten.
Tabelle 2: Rohdaten der Probandengruppen hell, dunkel, grau bezüglich SOX 9-Expression
Haarfarbe Anzahl Mittelwert Median SD Min Max
hell 23 15,43 16,17 4,68 6,17 23
dunkel 26 11,98 13,17 4,72 2,17 22,17
grau 30 10,98 9,0 4,59 0,33 23,67
Zur Beurteilung inwieweit eine Normalverteilung vorliegt, wurde die empirische Verteilung
der Mittelwerte SOX 9-positiver Zellen der obigen Farbgruppen analysiert (Abbildung 14).
Bei allen drei Gruppen ließ sich eine Normalverteilung feststellen, womit die Voraussetzung
für die Gültigkeit des t-Tests erfüllt ist (Schumacher und Schulgen, 2008).
In der direkten Nebeneinanderstellung der Normalverteilungen der drei
Haupthaarfarbengruppen (Schema 1) war zudem zu erkennen, dass der Maximalwert
grauhaariger Probanden bei einem niedrigeren Mittelwert SOX 9-positiver Zellen liegt, als bei
blonden oder braunhaarigen.

- 39 -
Abbildung 14: Verteilung der Mittelwerte SOX 9-positiver Zellen („hell ■“, „dunkel ■“
und „grau ■“ im Vergleich)
Die oben tabellarisch aufgeführten Rohdaten der SOX 9-positiven Zellen wurden als Box-
Whisker-Plot visuell dargestellt (Abbildung 15). Diese Diagrammart erlaubt gleichzeitig die
Visualisierung von Mittelwerten (□), Medianen (──), Perzentilen (95 %- Quantil ┬,
5 %- Quantil ┴), Minima (unteres x) und Maxima (oberes x). Die Grenzen der farbigen
Boxen markieren das 25 %- und 75 %-Quantil der Verteilung, die Boxen unterteilende,
waagerechte Gerade das Median. Die antennenförmigen Schwänze, Whiskers genannt,
reichen vom 5 %- bis zum 95 %-Quantil der empirischen Verteilung.
Im vorliegenden Gruppenvergleich fiel ein Absinken der Zellzahlen von „hell“ nach „dunkel“
zu „grau“ auf (Abbildung 15). So verhielten sich sowohl Mittelwerte, Mediane und Minima,
als auch die 5 %- und 25 %-Quantilwerte absteigend. Bei den Maxima, den 75 %- und 95 %-
Quantilen ließ sich dieser Trend jedoch nicht feststellen. Innerhalb der Probandengruppe
„grau“ ließ sich anhand der langen Whiskers und der weit auseinander liegenden Minimal-
und Maximalwerte eine relativ große Schwankungsbreite der SOX 9-positiven Zellzahlen
erkennen.
Verteilung der Mittelwerte SOX9-positiver
Zellen
0123456789
10 11 12
0 5 10 15 20 25 30Mittelwert
Häufigkeit
helldunkel grau

- 40 -
Abbildung 15: Box-Whisker-Plot zur Darstellung der Mittelwerte SOX 9-pos. Zellen
von „hell ■“, „dunkel ■“ und „grau ■“ im Vergleich
Der Vergleich der SOX 9-Expression bei hellen und dunklen Haarfarben zeigte signifikante
Unterschiede bezüglich der Mittelwerte. Die statistische Berechnung der Daten zeigt
signifikante Unterschiede zwischen „hell“ und „dunkel“ (p=0,02), sowie zwischen „hell“ und
„grau“ (p=0,01). Keine Signifikanz lag vor zwischen den Kohortengruppen „dunkelhaarig“
und „grau“ (p=0,54) (Tabelle 3).
Tabelle 3: t-Test (p-Wert) für den Vergleich der Gruppen "hell" , "dunkel" und "grau"
Haarfarbenvergleich (Mittelwerte) p-Wert
hell (15,43) > dunkel (11,98) p=0,02
hell (15,43) > grau (10,98) p=0,01
dunkel (11,98) > grau (10,98) p=0,54
- 41 -
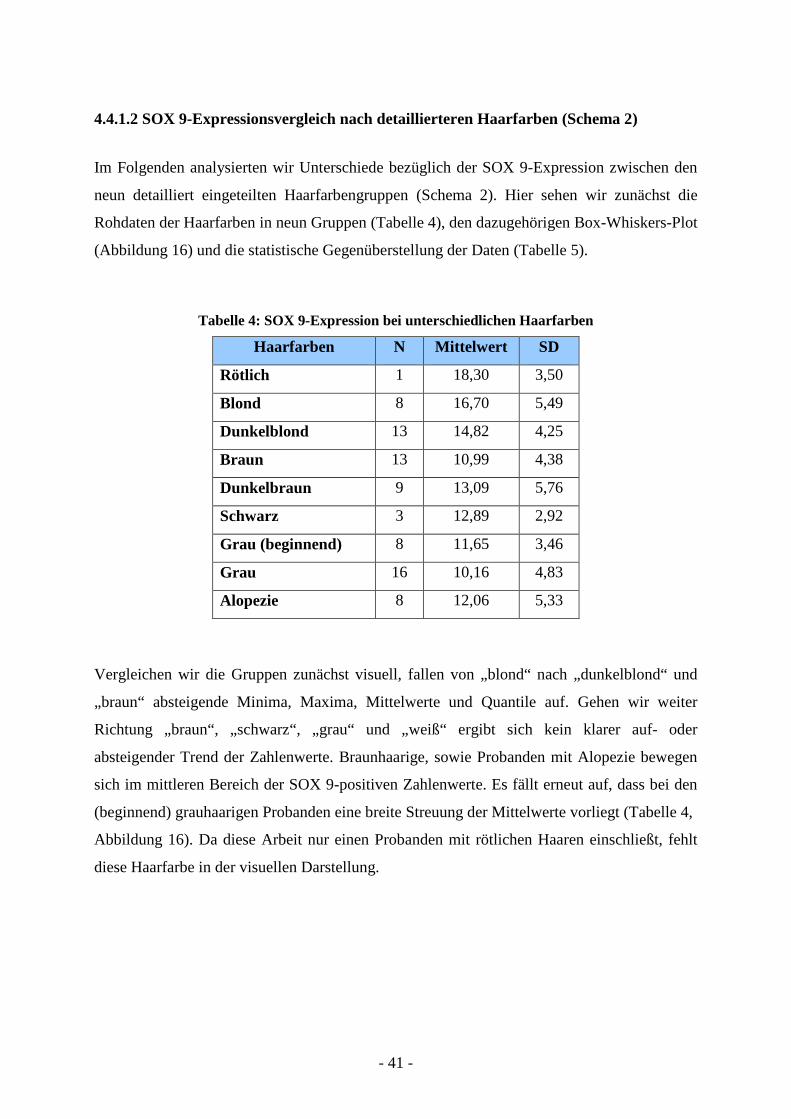
4.4.1.2 SOX 9-Expressionsvergleich nach detaillierteren Haarfarben (Schema 2)
Im Folgenden analysierten wir Unterschiede bezüglich der SOX 9-Expression zwischen den
neun detailliert eingeteilten Haarfarbengruppen (Schema 2). Hier sehen wir zunächst die
Rohdaten der Haarfarben in neun Gruppen (Tabelle 4), den dazugehörigen Box-Whiskers-Plot
(Abbildung 16) und die statistische Gegenüberstellung der Daten (Tabelle 5).
Tabelle 4: SOX 9-Expression bei unterschiedlichen Haarfarben
Haarfarben N Mittelwert SD
Rötlich 1 18,30 3,50
Blond 8 16,70 5,49
Dunkelblond 13 14,82 4,25
Braun 13 10,99 4,38
Dunkelbraun 9 13,09 5,76
Schwarz 3 12,89 2,92
Grau (beginnend) 8 11,65 3,46
Grau 16 10,16 4,83
Alopezie 8 12,06 5,33
Vergleichen wir die Gruppen zunächst visuell, fallen von „blond“ nach „dunkelblond“ und
„braun“ absteigende Minima, Maxima, Mittelwerte und Quantile auf. Gehen wir weiter
Richtung „braun“, „schwarz“, „grau“ und „weiß“ ergibt sich kein klarer auf- oder
absteigender Trend der Zahlenwerte. Braunhaarige, sowie Probanden mit Alopezie bewegen
sich im mittleren Bereich der SOX 9-positiven Zahlenwerte. Es fällt erneut auf, dass bei den
(beginnend) grauhaarigen Probanden eine breite Streuung der Mittelwerte vorliegt (Tabelle 4,
Abbildung 16). Da diese Arbeit nur einen Probanden mit rötlichen Haaren einschließt, fehlt
diese Haarfarbe in der visuellen Darstellung.
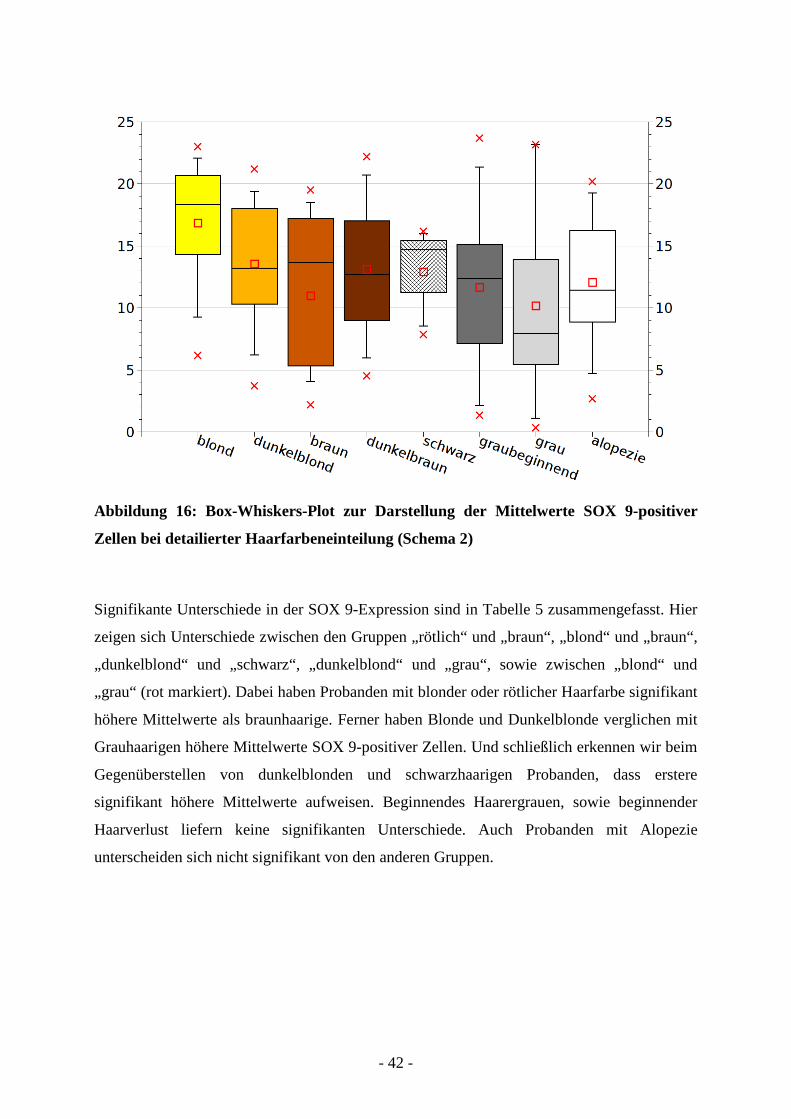
- 42 -
Abbildung 16: Box-Whiskers-Plot zur Darstellung der Mittelwerte SOX 9-positiver
Zellen bei detailierter Haarfarbeneinteilung (Schema 2)
Signifikante Unterschiede in der SOX 9-Expression sind in Tabelle 5 zusammengefasst. Hier
zeigen sich Unterschiede zwischen den Gruppen „rötlich“ und „braun“, „blond“ und „braun“,
„dunkelblond“ und „schwarz“, „dunkelblond“ und „grau“, sowie zwischen „blond“ und
„grau“ (rot markiert). Dabei haben Probanden mit blonder oder rötlicher Haarfarbe signifikant
höhere Mittelwerte als braunhaarige. Ferner haben Blonde und Dunkelblonde verglichen mit
Grauhaarigen höhere Mittelwerte SOX 9-positiver Zellen. Und schließlich erkennen wir beim
Gegenüberstellen von dunkelblonden und schwarzhaarigen Probanden, dass erstere
signifikant höhere Mittelwerte aufweisen. Beginnendes Haarergrauen, sowie beginnender
Haarverlust liefern keine signifikanten Unterschiede. Auch Probanden mit Alopezie
unterscheiden sich nicht signifikant von den anderen Gruppen.

- 43 -
Tabelle 5: Signifikante Unterschiede der SOX 9-Expression der unterschiedlichen Haarfarben
Blond Dunkel-blond
Braun Dunkel-braun
Schwarz Grau (beginn.)
Grau Alopezie
Rötlich p=0,78 p=0,2 p=0,04 p=0,15 p=0,10 p=0,16 p=0,07 p=0,09
Blond p=0,24 p=0,01 p=0,12 p=0,23 p=0,08 p=0,02 p=0,07
Dunkel-blond
p=0,06 p=0,42 p=0,5 p=0,22 p=0,049 p=0,22
Braun p=0,39 p=0,59 p=0,82 p=0,73 p=0,67
Dunkel-braun
p=0,96 p=0,65 p=0,30 p=0,71
Schwarz p=0,79 p=0,53 p=0,83 Grau
(beginnend) p=0,63 p=0,90
Grau p=0,52
4.4.1.3 Vergleich der Haarfarben unter verschiedenen Aspekten
A) SOX 9-Expression entlang des Haarschaftes (ohne Papillenanschnitte)
Die Papille stellt die Hauptwachstumszone dar, während Stammzellen ihre Stammzellnische
innerhalb des Haarschaftes haben. Um einen Unterschied zwischen Proliferationszone und
Stammzellreservoir zu erkennen ist es sinnvoll, das Haar in Stamm und Papille zu unterteilen
und diese, die SOX 9-positiven Zellen betreffend, getrennt zu betrachten (Tabelle 6,
Tabelle 8).
Tabelle 6: SOX 9-Expression entlang des Haarschaftes ohne Papillenanschnitte
bei Probanden unterschiedlicher Haarfarben
Haarfarben N Mittelwert SD
Rötlich 1 18,30 3,50
Blond 8 16,70 5,47
Dunkelblond 13 14,23 4,34
Braun 13 10,90 4,45
Dunkelbraun 9 12,44 5,86
Schwarz 3 12,67 3,20
Grau (beginnend) 8 12,09 3,19
Grau 16 10,25 4,64
Alopezie 8 12,20 5,44

- 44 -
Wird die SOX 9-Expression entlang der Haarschaftanschnitte isoliert betrachtet, d.h. unter
Aussparen aller Mittelwerte von Papillenanschnitten, zeigten sich Signifikanzen bei der
Gegenüberstellung von „blond“ und „braun“, sowie von „blond“ und „grau“ (rot markiert)
(Tabelle 7). Diese unterscheiden sich nicht, von den bisher gewonnenen Ergebnissen ohne
diese anatomisch getrennte Betrachtung.
Tabelle 7: Vergleich der Haarfarben ohne Papillenanschnitte (p-Werte)
Blond Dunkel-blond
Braun Dunkel-braun
Schwarz Grau
(beginn.) Grau Alopezie
Rötlich p=0,70 p=0,17 p=0,08 p=0,18 p=0,17 p=0,31 p=0,15 p=0,19
Blond p=0,23 p=0,03 p=0,13 p=0,28 p=0,19 p=0,04 p=0,13
Dunkel-blond p=0,09 p=0,38 p=0,55 p=0,41 p=0,09 p=0,35
Braun p=0,52 p=0,61 p=0,68 p=0,80 p=0,61
Dunkel-braun p=0,95 p=0,91 p=0,44 p=0,93
Schwarz p=0,91 p=0,60 p=0,90
Grau (beginnend) p=0,58 p=0,97
Grau p=0,52
B) SOX 9-Expression in der Papillenregion
Nicht alle Probanden dieser Arbeit wiesen Papillenanschnitte auf, sodass hier die Rohdaten
von nur 38 Probanden verwendet werden konnten (Tabelle 8). Teilweise fand sich nur ein
Papillenanschnitt pro Proband.

- 45 -
Tabelle 8: Nur Papillenanschnitte, Einteilung nach Haarfarben,
n= 38 Probanden (51 Papillenanschnitte)
Haarfarben N Mittelwert SD
Rötlich 1 16,0 0
Blond 2 13,0 5,67
Dunkelblond 7 13,71 6,58
Braun 3 15,33 2,08
Dunkelbraun 8 14,94 7,74
Schwarz 1 17,5 0,71
Grau (beginnend) 6 12,92 5,14
Grau 8 9,63 4,11
Alopezie 2 16 8,49
Beim isolierten Studieren der SOX 9-Expression der Papillenanschnitte, fand sich lediglich
eine signifikant höhere Anzahl SOX 9-positiver Zellen bei schwarzhaarigen gegenüber
grauhaarigen Probanden (rot markiert) (Tabelle 9). Die übrigen Gegenüberstellungen der
Haarfarben lieferten keine signifikanten Ergebnisse, wobei die statistische Relevanz bei den
niedrigen Probandenzahlen in Frage zu stellen ist, da sich nur bei 38 Probanden
Papillenanschnitte fanden, teilweise sogar nur einer pro Proband.
Tabelle 9: Vergleich der Haarfarben (p-Werte), nur Papillenanschnitte
Blond Dunkel-blond
Braun Dunkel-braun
Schwarz Grau
(beginn.) Grau Alopezie
Rötlich p=0,53 p=0,65 p=0,70 p=0,86 p=0,10 p=0,45 p=0,07 p=1
Blond p=0,89 p=0,54 p=0,75 p=0,38 p=0,99 p=0,35 p=0,72
Dunkel-blond p=0,70 p=0,75 p=0,46 p=0,81 p=0,17 p=0,69
Braun p=0,93 p=0,27 p=0,47 p=0,05 p=0,90
Dunkel-braun p=0,67 p=0,59 p=0,11 p=0,87
Schwarz p=0,28 p=0,03 p=0,83
Grau (beginnend) p=0,21 p=0,54
Grau p=0,14

- 46 -
Betrachtet man die Papillenregion separat, zeigte sich lediglich ein signifikanter Unterschied
in der SOX 9-Expression zwischen schwarzem und grauem Haar (Tabelle 9). Die bei
Auswertung des gesamten Haares signifikanten Unterschiede zwischen Blond/Braun und
Blond/Grau konnten in der Papillenregion nicht aufgezeigt werden und umgekehrt. Es gilt
jedoch hervorzuheben, dass die Anzahl der zur Untersuchung gekommenen Papillenregionen
möglicherweise zu niedrig war, um hier kleinere Expressionsunterschiede signifikant
aufzeigen zu können.
C) Signifikanzberechung unter Streichung von Ausreißern
Bei den Rohdaten pro Proband wurden die zwei auffälligsten „Ausreißer“, d.h. die zwei am
weitesten vom Mittelwert abweichenden Werte SOX 9-positiver Zellen, für die weitere
Berechnung gestrichen. Die statistischen Auswertungen wurden mit den verbleibenden vier
Mittelwerten pro Proband durchgeführt (Daten nicht aufgeführt). Dabei konnten die bereits
gewonnenen Ergebnisse bestätigt werden.
Es zeigte sich unter Streichung der niedrigsten zwei Mittelwerte pro Proband eine signifikante
Unterscheidung zwischen „blond“ und „braun“, sowie zwischen „blond“ und „grau“. Wieder
wiesen blonde Probanden höhere SOX 9-positive Mittelwerte auf.
Wurden jeweils der niedrigste und höchste Mittelwert SOX 9-positiver Zellen pro Proband
weggelassen, erkannten wir signifikante Unterschiede zwischen den Haarfarben „rötlich“ und
„grau“, „blond“ und „grau“, „dunkelblond“ und „grau“, sowie „blond“ und „braun“ im Sinne
der bereits gewonnenen Ergebnisse.
Zusammengefasst stellen sich keine statistisch relevanten Unterschiede ein, ob die
zugrundeliegenden Berechnungen auf allen sechs Rohwerten oder den am engsten
zusammenliegenden vier Rohwerten unter Streichung von zwei Ausreißern basieren. Dies
bekräftigt die Verwendung aller Rohwerte bei der Auswertung.
- 47 -
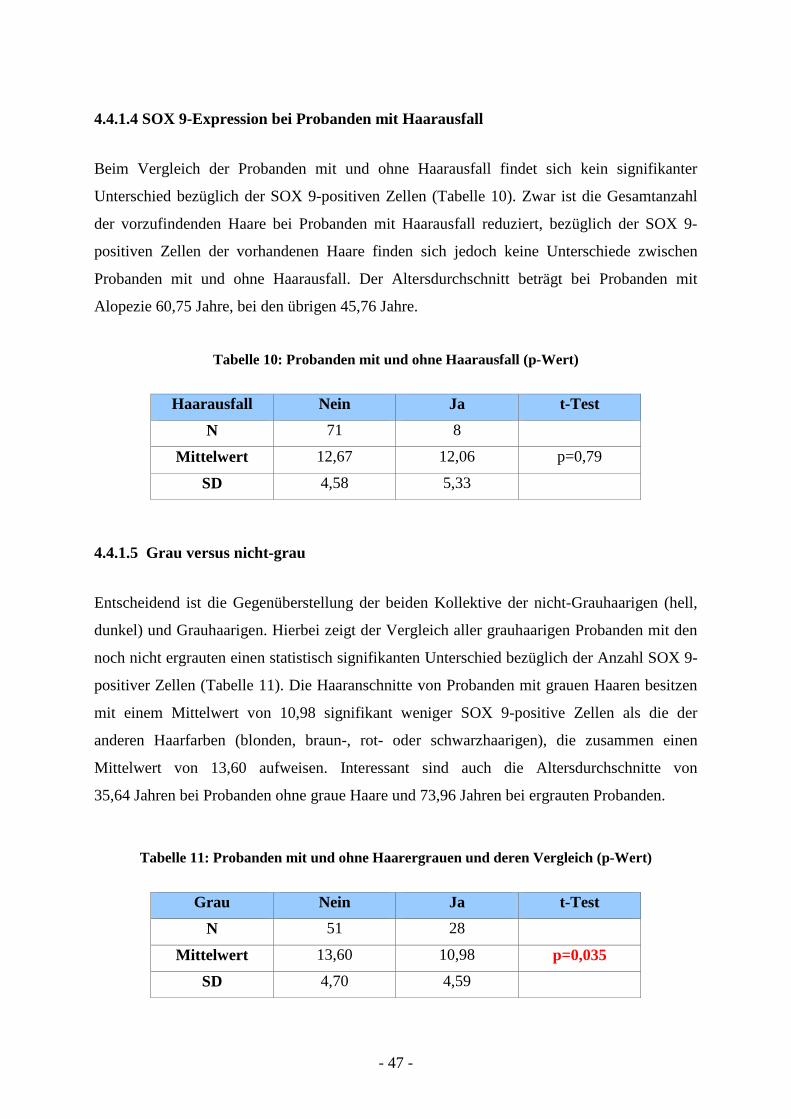
4.4.1.4 SOX 9-Expression bei Probanden mit Haarausfall
Beim Vergleich der Probanden mit und ohne Haarausfall findet sich kein signifikanter
Unterschied bezüglich der SOX 9-positiven Zellen (Tabelle 10). Zwar ist die Gesamtanzahl
der vorzufindenden Haare bei Probanden mit Haarausfall reduziert, bezüglich der SOX 9-
positiven Zellen der vorhandenen Haare finden sich jedoch keine Unterschiede zwischen
Probanden mit und ohne Haarausfall. Der Altersdurchschnitt beträgt bei Probanden mit
Alopezie 60,75 Jahre, bei den übrigen 45,76 Jahre.
Tabelle 10: Probanden mit und ohne Haarausfall (p-Wert)
Haarausfall Nein Ja t-Test
N 71 8 Mittelwert 12,67 12,06 p=0,79
SD 4,58 5,33
4.4.1.5 Grau versus nicht-grau
Entscheidend ist die Gegenüberstellung der beiden Kollektive der nicht-Grauhaarigen (hell,
dunkel) und Grauhaarigen. Hierbei zeigt der Vergleich aller grauhaarigen Probanden mit den
noch nicht ergrauten einen statistisch signifikanten Unterschied bezüglich der Anzahl SOX 9-
positiver Zellen (Tabelle 11). Die Haaranschnitte von Probanden mit grauen Haaren besitzen
mit einem Mittelwert von 10,98 signifikant weniger SOX 9-positive Zellen als die der
anderen Haarfarben (blonden, braun-, rot- oder schwarzhaarigen), die zusammen einen
Mittelwert von 13,60 aufweisen. Interessant sind auch die Altersdurchschnitte von
35,64 Jahren bei Probanden ohne graue Haare und 73,96 Jahren bei ergrauten Probanden.
Tabelle 11: Probanden mit und ohne Haarergrauen und deren Vergleich (p-Wert)
Grau Nein Ja t-Test
N 51 28 Mittelwert 13,60 10,98 p=0,035
SD 4,70 4,59
- 48 -
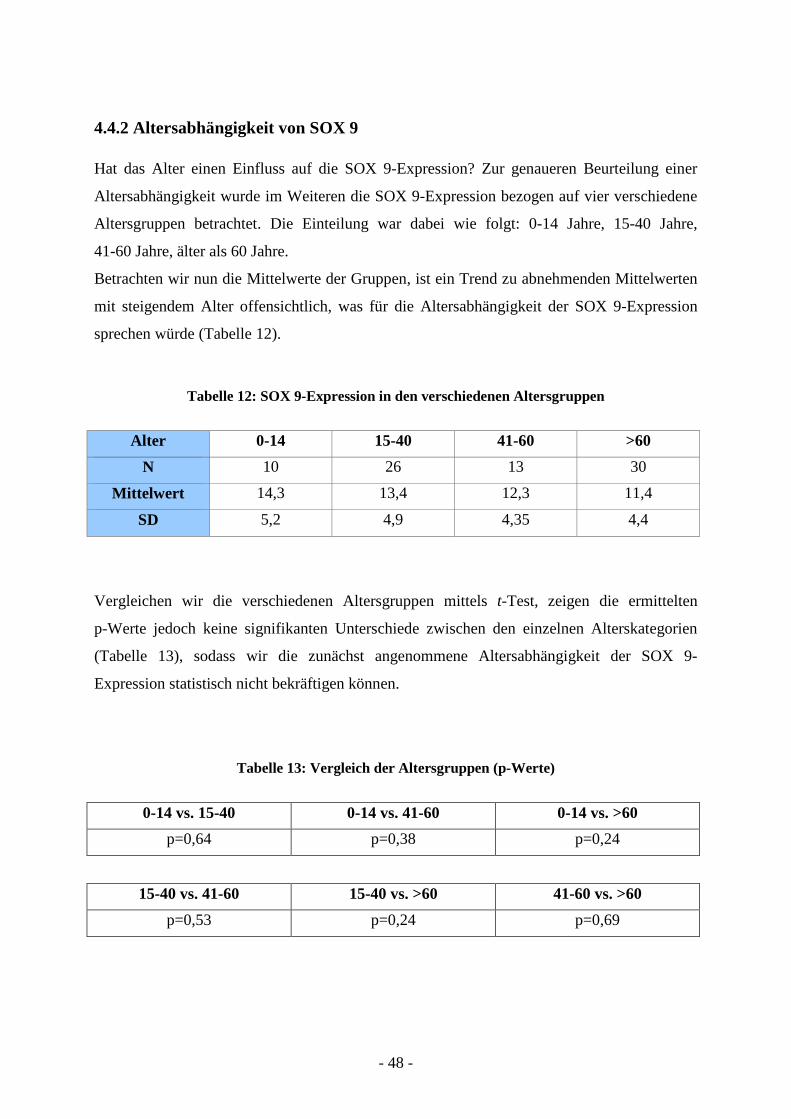
4.4.2 Altersabhängigkeit von SOX 9
Hat das Alter einen Einfluss auf die SOX 9-Expression? Zur genaueren Beurteilung einer
Altersabhängigkeit wurde im Weiteren die SOX 9-Expression bezogen auf vier verschiedene
Altersgruppen betrachtet. Die Einteilung war dabei wie folgt: 0-14 Jahre, 15-40 Jahre,
41-60 Jahre, älter als 60 Jahre.
Betrachten wir nun die Mittelwerte der Gruppen, ist ein Trend zu abnehmenden Mittelwerten
mit steigendem Alter offensichtlich, was für die Altersabhängigkeit der SOX 9-Expression
sprechen würde (Tabelle 12).
Tabelle 12: SOX 9-Expression in den verschiedenen Altersgruppen
Alter 0-14 15-40 41-60 >60
N 10 26 13 30
Mittelwert 14,3 13,4 12,3 11,4
SD 5,2 4,9 4,35 4,4
Vergleichen wir die verschiedenen Altersgruppen mittels t-Test, zeigen die ermittelten
p-Werte jedoch keine signifikanten Unterschiede zwischen den einzelnen Alterskategorien
(Tabelle 13), sodass wir die zunächst angenommene Altersabhängigkeit der SOX 9-
Expression statistisch nicht bekräftigen können.
Tabelle 13: Vergleich der Altersgruppen (p-Werte)
0-14 vs. 15-40 0-14 vs. 41-60 0-14 vs. >60
p=0,64 p=0,38 p=0,24
15-40 vs. 41-60 15-40 vs. >60 41-60 vs. >60
p=0,53 p=0,24 p=0,69
- 49 -
4.4.3 SOX 9-Expression abhängig von Alter und Geschlecht
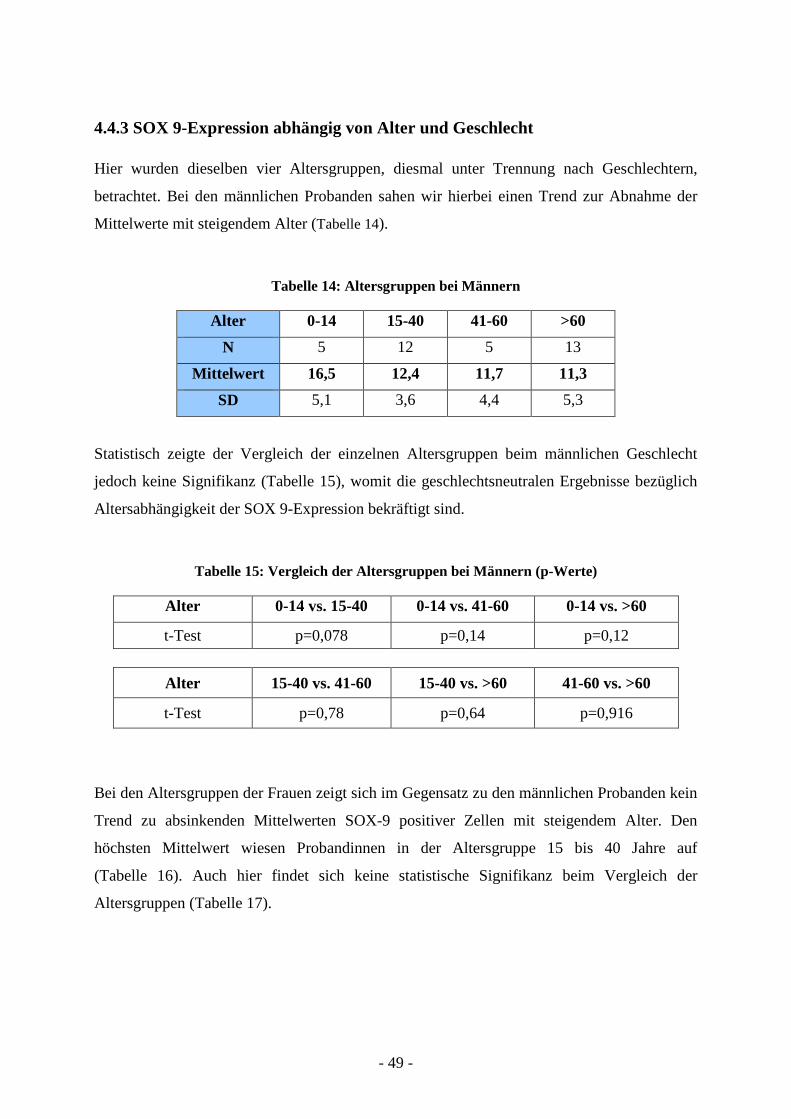
Hier wurden dieselben vier Altersgruppen, diesmal unter Trennung nach Geschlechtern,
betrachtet. Bei den männlichen Probanden sahen wir hierbei einen Trend zur Abnahme der
Mittelwerte mit steigendem Alter (Tabelle 14).
Tabelle 14: Altersgruppen bei Männern
Alter 0-14 15-40 41-60 >60
N 5 12 5 13
Mittelwert 16,5 12,4 11,7 11,3
SD 5,1 3,6 4,4 5,3
Statistisch zeigte der Vergleich der einzelnen Altersgruppen beim männlichen Geschlecht
jedoch keine Signifikanz (Tabelle 15), womit die geschlechtsneutralen Ergebnisse bezüglich
Altersabhängigkeit der SOX 9-Expression bekräftigt sind.
Tabelle 15: Vergleich der Altersgruppen bei Männern (p-Werte)
Alter 0-14 vs. 15-40 0-14 vs. 41-60 0-14 vs. >60
t-Test p=0,078 p=0,14 p=0,12
Alter 15-40 vs. 41-60 15-40 vs. >60 41-60 vs. >60
t-Test p=0,78 p=0,64 p=0,916
Bei den Altersgruppen der Frauen zeigt sich im Gegensatz zu den männlichen Probanden kein
Trend zu absinkenden Mittelwerten SOX-9 positiver Zellen mit steigendem Alter. Den
höchsten Mittelwert wiesen Probandinnen in der Altersgruppe 15 bis 40 Jahre auf
(Tabelle 16). Auch hier findet sich keine statistische Signifikanz beim Vergleich der
Altersgruppen (Tabelle 17).

- 50 -
Tabelle 16: Altersgruppen bei Frauen
Alter 0-14 15-40 41-60 >60
N 5 14 8 17
Mittelwert 12,2 14,3 12,7 11,5
SD 5,2 6,1 4,3 3,7
Tabelle 17: Vergleich der Altersgruppen bei Frauen (p-Werte)
Alter 0-14 vs. 15-40 0-14 vs. 41-60 0-14 vs. >60
t-Test p=0,47 p=0,87 p=0,87
Alter 15-40 vs. 41-60 15-40 vs. >60 41-60 vs. >60
t-Test p=0,53 p=0,26 p=0,70
Auch eine Gegenüberstellung beider Geschlechter innerhalb der jeweiligen Altersgruppen
zeigt im t-Test keine signifikanten Unterschiede (Tabelle 18).
Tabelle 18: Vergleich der Geschlechter innerhalb der Altersgruppen
Alter 0-14 15-40 41-60 >60
t-Test p=0,19 p=0,34 p=0,76 p=0,93
So können wir festhalten, dass zwar ein gewisser Trend zu sinkenden SOX 9-positiven
Mittelwerten mit steigendem Alter zu erkennen ist, dieser jedoch mittels statistischer
Untersuchungen, auch nach Hinzuziehen der Variable „Geschlecht“, zu keinen signifikanten
Ergebnissen führt.
- 51 -
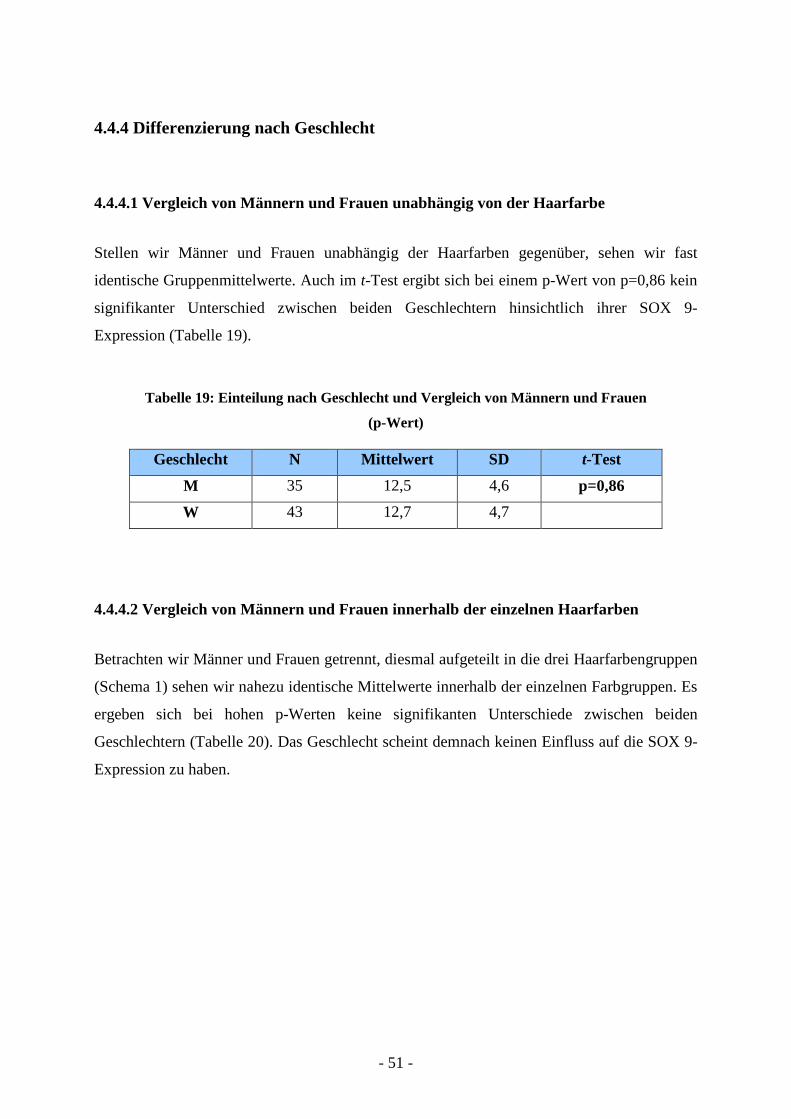
4.4.4 Differenzierung nach Geschlecht
4.4.4.1 Vergleich von Männern und Frauen unabhängig von der Haarfarbe
Stellen wir Männer und Frauen unabhängig der Haarfarben gegenüber, sehen wir fast
identische Gruppenmittelwerte. Auch im t-Test ergibt sich bei einem p-Wert von p=0,86 kein
signifikanter Unterschied zwischen beiden Geschlechtern hinsichtlich ihrer SOX 9-
Expression (Tabelle 19).
Tabelle 19: Einteilung nach Geschlecht und Vergleich von Männern und Frauen
(p-Wert)
Geschlecht N Mittelwert SD t-Test
M 35 12,5 4,6 p=0,86
W 43 12,7 4,7
4.4.4.2 Vergleich von Männern und Frauen innerhalb der einzelnen Haarfarben
Betrachten wir Männer und Frauen getrennt, diesmal aufgeteilt in die drei Haarfarbengruppen
(Schema 1) sehen wir nahezu identische Mittelwerte innerhalb der einzelnen Farbgruppen. Es
ergeben sich bei hohen p-Werten keine signifikanten Unterschiede zwischen beiden
Geschlechtern (Tabelle 20). Das Geschlecht scheint demnach keinen Einfluss auf die SOX 9-
Expression zu haben.

- 52 -
Tabelle 20: a) Hell-, b) dunkel- und c) grauhaarige Probanden nach Geschlecht getrennt
a
Geschlecht Anzahl Mittelwerte SD t-Test w 13 16,03 4,72 p=0,49 m 10 14,63 4,62
23
b
Geschlecht Anzahl Mittelwerte SD t-Test w 13 11,85 5,73 p=0,89 m 13 12,13 3,72
26
c
Geschlecht Anzahl Mittelwerte SD t-Test w 18 10,94 4,04 p=0,97 m 12 11,04 5,41
30

- 53 -
5. Diskussion
Die Kopfhaare sind eines der wichtigsten äußeren Erscheinungsmerkmale eines jeden
Individuums. Sie unterscheiden sich in ihrer Farbe, ihrer Festigkeit, ihrer Neigung eher glatt,
gewellt oder gelockt zu sein. Mit zunehmendem Alter mischen sich die farbigen Haare mit
weißen Haaren, bis man über ein meliertes Zwischenstadium zunehmend weiße Kopfhaare
bekommt. Parallel verläuft während des Alterungsprozesses der langsam zunehmende Verlust
der Haare, welcher in erster Linie beim Mann zur Glatzenbildung bis hin zum vollständigen
Haarverlust führen kann. Individuelle wie generelle Faktoren herauszufinden, welche die
Entstehung unterschiedlicher Haarfarben, sowie Faktoren, welche das Haarergrauen und den
primär genetisch bedingten Haarverlust beeinflussen, sind Gegenstand langjähriger
Forschung. Neben genetischen Faktoren hat die Stammzellforschung in den letzten ein bis
zwei Jahrzehnten zunehmend Bedeutung in dieser Fragestellung erhalten.
Ziel der vorliegenden Arbeit war es auf relativ einfache methodische Weise, sprich unter
Verwendung immunhistochemischer Methoden, screeningartig herauszufinden, welche
Stammzellmarker auf die Entstehung der Haarfarbe, sowie auf Haarergrauen und Haarausfall
Einfluss nehmen könnten.
Dieses Screening wurde mit einer Literaturrecherche zu den Themen „Stammzellen und
Haarfarbe, Haarergrauen und Haarausfall“ begonnen. Aus der Vielzahl der sich hieraus
ergebenden Faktoren bzw. Moleküle wurden jene ausgewählt, welche immunhistochemisch
detektierbar waren und für die kommerziell erhältliche Antikörper zur Verfügung standen.
Dies führte zu einer ersten Selektion folgender Moleküle: SOX 9, SOX 10, PAX 3 und
CLIM 2.
Die Probandenauswahl erfolgte anhand bereits vorhandener Schnittpräparate aus dem Archiv
der dermatologischen Universitätsklinik Freiburg. Es wurde in einem umschriebenen
Zeitraum nach Präparaten gesucht, welche aus der Kopfhaut stammten. Dabei war es von
Wichtigkeit, dass mehrere Haarfollikel möglichst tangential zu deren Verlaufsrichtung im
histologischen Präparat getroffen und ohne pathologischen Prozess waren. Insgesamt konnten
79 Probanden in die Studie eingeschlossen werden. Die Haarfarbe der Probanden, sowie das
Vorhandensein einer androgenetischen Alopezie wurden aus der
Patientenphotodokumentation übernommen oder telefonisch bei den Probanden erfragt.

- 54 -
Hieraus ergab sich eine Zuordnung der Probanden in unterschiedliche Gruppen entsprechend
ihrer Haarfarben bzw. Haardichte.
Es folgte die Etablierungsphase der immunhistochemischen Färbemethoden von SOX 9,
SOX 10, PAX 3 und CLIM 2 an einzelnen Gewebeschnitten. Bei den Etablierungsversuchen
wurden unterschiedliche Konzentrationen der Primär- und Sekundärantikörper in
ansteigenden Konzentrationsreihen ausgetestet. Zudem wurden unterschiedliche
Demaskierungsverfahren, wie Vorkochen in Citrat- und Tris-EDTA-Puffer, zur Optimierung
der Färbeergebnisse eingesetzt. Bei den Versuchsreihen zeigte sich, dass die Verwendung der
Antikörper gegen SOX 10, PAX 3 und CLIM 2 keine spezifischen Resultate erbrachte –
entweder kam es zu einer überschießenden Backgroundfärbung (PAX3/7 und CLIM 2) oder
zu so inkonstanten Färbeergebnissen (SOX 10), dass man die Zuverlässigkeit der Aussage
anzweifeln musste. Ausnahme war SOX 9 – hier zeigten die Vorversuche reproduzierbare und
spezifische Ergebnisse. Aus diesem Grunde wurden in der weiteren Arbeit ausschließlich
Färbungen mit Antikörpern gegen SOX 9 durchgeführt.
Im Vergleich der SOX 9-Expression bei Probanden mit unterschiedlichen Haarfarben bzw.
Haardichte zeigten sich folgende Ergebnisse:
Blonde Probanden unterschieden sich signifikant aufgrund einer erhöhten SOX 9-Expression
sowohl von braun-, als auch von grauhaarigen Individuen. Im Vergleich von Grauhaarigen zu
Nicht-Grauhaarigen zeigte sich ebenfalls eine höhere SOX 9-Expression bei nicht-
grauhaarigen als bei grauhaarigen. Mit zunehmendem Alter zeigte sich ein Trend zu einer
kontinuierlichen Abnahme von SOX 9, dieser erreichte jedoch keine statistische Signifikanz.
Geschlechtsspezifische Unterschiede konnten nicht aufgezeigt werden. Die acht untersuchten
Individuen mit Alopezie zeigten keine signifikante Reduktion von SOX 9.
5.1 Der Haarapparat und Stammzellen
In den vergangenen zwei Jahrzehnten wurden die funktionellen Abläufe der
Haarmorphogenese, Einflussfaktoren auf den Haarzyklus, das Phänomen des Haarverlustes,
wie des Haarergrauens in zahlreichen wissenschaftlich höchst anspruchsvollen Arbeiten
untersucht. Hierzu zählten unterschiedlichste Knockout-Mausmodelle, in denen unter
anderem der Einfluss von Genen wie Ragged/Opossum, Tabby, Waved 2, Fuzzy, Hairless,
Angora, Nude, Balding oder Lethal spotting beleuchtet und in einer Übersichtsarbeit von
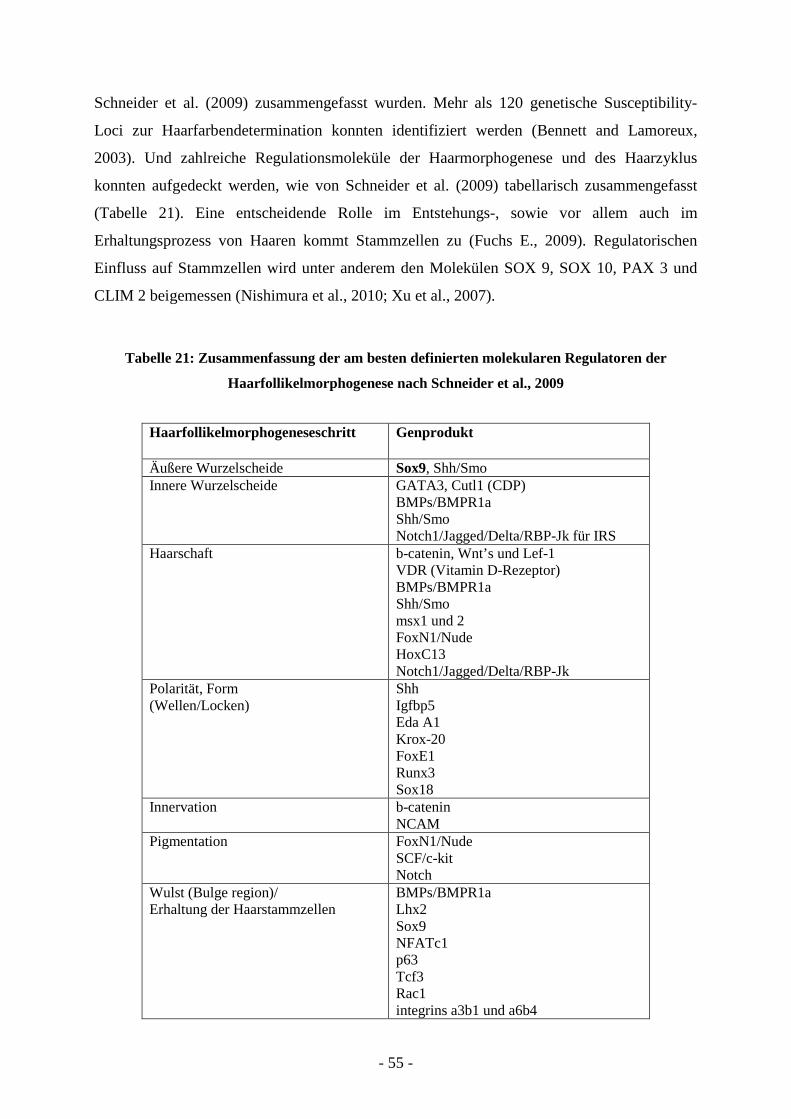
- 55 -
Schneider et al. (2009) zusammengefasst wurden. Mehr als 120 genetische Susceptibility-
Loci zur Haarfarbendetermination konnten identifiziert werden (Bennett and Lamoreux,
2003). Und zahlreiche Regulationsmoleküle der Haarmorphogenese und des Haarzyklus
konnten aufgedeckt werden, wie von Schneider et al. (2009) tabellarisch zusammengefasst
(Tabelle 21). Eine entscheidende Rolle im Entstehungs-, sowie vor allem auch im
Erhaltungsprozess von Haaren kommt Stammzellen zu (Fuchs E., 2009). Regulatorischen
Einfluss auf Stammzellen wird unter anderem den Molekülen SOX 9, SOX 10, PAX 3 und
CLIM 2 beigemessen (Nishimura et al., 2010; Xu et al., 2007).
Tabelle 21: Zusammenfassung der am besten definierten molekularen Regulatoren der
Haarfollikelmorphogenese nach Schneider et al., 2009
Haarfollikelmorphogeneseschritt Genprodukt
Äußere Wurzelscheide Sox9, Shh/Smo Innere Wurzelscheide GATA3, Cutl1 (CDP)
BMPs/BMPR1a Shh/Smo Notch1/Jagged/Delta/RBP-Jk für IRS
Haarschaft b-catenin, Wnt’s und Lef-1 VDR (Vitamin D-Rezeptor) BMPs/BMPR1a Shh/Smo msx1 und 2 FoxN1/Nude HoxC13 Notch1/Jagged/Delta/RBP-Jk
Polarität, Form (Wellen/Locken)
Shh Igfbp5 Eda A1 Krox-20 FoxE1 Runx3 Sox18
Innervation b-catenin NCAM
Pigmentation FoxN1/Nude SCF/c-kit Notch
Wulst (Bulge region)/ Erhaltung der Haarstammzellen
BMPs/BMPR1a Lhx2 Sox9 NFATc1 p63 Tcf3 Rac1 integrins a3b1 und a6b4

- 56 -
5.1.1 SOX 9
SOX 9 ist essentiell für die Entwicklung und Differenzierung zahlreicher Zellreihen, unter
anderem dieser, die embryologisch gesehen von Neuralleiste (Cheung et al., 2005) und
Neuralrohr (Stolt et al., 2003) abstammen. Aus der Neuralleiste entwickeln sich insbesondere
Gewebe des Nervensystems und der Haut und Hautanhangsgebilde, zu denen auch die Haare
zählen. Für diese Arbeit von besonderem Interesse ist, dass SOX 9 in der äußeren
Wurzelscheide des Haarfollikels exprimiert und in allen Stadien des Haarzyklus nachweisbar
ist (Vidal et al., 2005). Die fundamentale Bedeutung von SOX 9 für den Haarapparat zeigt
sich in SOX 9-Knockout-Mäusen, bei denen es zu einem vollständigen Verlust der Haare und
des Haarfollikel-Stammzellkompartiments kommt (Cook et al., 2005; Vidal et al., 2005).
Außerdem ist es bei der Genese von Tumoren epidermalen Ursprungs, nämlich des
semimalignen Basalzellkarzinoms und der benignen Trichoepitheliome und Trichilemmome
(Vidal et al., 2008), sowie bei der embryologischen Entwicklung des Nagelbettes und der
ekkrinen Schweissdrüsen plantar (Krahl and Sellheyer, 2010) involviert.
5.1.2 SOX 10
SOX 10 reguliert über die Expression des Endothelin-Rezeptor-B (EDNRB)-Gens die
terminale Migration von Melanoblasten und anderen Melanozytenvorläuferzellen (Yokoyama
et al., 2006; Cook et al., 2005). Mutationen von SOX 10-kodierenden Genen ergeben
Syndrome, die mit Hautpigmentationsabnormitäten assoziiert sind, wie das Waardenburg
Syndrom und das „Yemenite Deaf-Blind Hypopigmentation“ Syndrom (Pingault et al., 2010).
Ferner wird SOX 10 von relativ differenzierten Tumoren neuroektodermalen Ursprungs, wie
malignen Melanomen und Gliomen, produziert (Gershon et al., 2005; Ferletta et al., 2007).
5.1.3 PAX 3
PAX 3 ist essentiell für die Differenzierung von Melanozytenstammzellen, da es die
melanogene Kaskade initiiert (Lang et al., 2005; Nishimura et al., 2005; Steingrímsson et al.,
2005). Als Schlüsselmolekül aktiviert es einerseits synergistisch zu SOX 10 direkt die
Expression von Mitf und konkurriert andererseits um die Bindung an einen für die Dct-
Expression erforderlichen Enhancer und hemmt somit die Mitf-/ SOX 10-vermittelte
Aktivierung. Dct kodiert für das Enzym Dopachrom-Tautomerase, das einen wichtigen

- 57 -
Reaktionsschritt bei der Melaninentstehung katalysiert und führt über Verdrängung von
PAX 3 zur Anregung von Melanozytenstammzellen (Lang et al., 2005; Nishimura et al.,
2005; Steingrímsson et al., 2005; Prince et al., 2001; Smit et al., 2000; Vachtenheim and
Novotná, 1999). In Analogie zu SOX 10 führen Mutationen in PAX 3 ebenfalls zum
Depigmentations-Syndrom Waardenburg. PAX 3 und sein Homolog PAX 7 werden
desweiteren von schlecht differenzierten und malignen Tumoren neuroektodermalen
Ursprungs produziert. In Studien konnten die Isoformen PAX 3 a-e in Hautmelanomzelllinien
nachgewiesen werden (Gershon et al., 2005; Parker et al., 2004).
5.1.4 CLIM 2
CLIM 2 wird hauptsächlich von Gewebe mit mehrschichtigem Epithel exprimiert, das in der
Lage ist sich selbst zu erneuern, wie beispielsweise Epidermis, Haarfollikel und Kornea
(Sugihara et al., 1998). Als Co-Faktor von LIM-Domänen ist es an der Erhaltung von
Haarfollikelstammzellen im Wulstbereich der äußeren Wurzelscheide verantwortlich (Xu et
al., 2007). Mäuse mit dominant negativer CLIM-Expression zeigten eine progressive
Haarfollikeldysfunktion mit Haarausfall (Xu et al., 2007).
Wie bereits erwähnt, stammt der überaus größte Teil der wissenschaftlichen Erkenntnisse zum
Haarapparat und zum Haarwachstum aus (transgenen) Mausstudien oder unter Verwendung
aufwendiger molekularer Methoden von humanem Gewebe, dann meist von nur wenigen
Probanden.
Daher war die Zielsetzung in der vorliegenden Arbeit, die Bedeutung von Stammzellfaktoren
auf relativ einfache methodische Weise, nämlich unter Verwendung immunhistochemischer
Anfärbungen von humanem Gewebe, quasi als Screeningmethode zu evaluieren. Ein
Entscheidungsmerkmal zugunsten der Faktoren SOX 9, SOX 10, PAX 3 und CLIM 2 war das
Vorhandensein käuflich erhältlicher Antikörper. Obwohl eine Anwendbarkeit in Formalin-
fixiertem und Paraffin-eingebettetem Gewebe nur für SOX 9 in der Literatur nachlesbar war,
wollten wir für die weiteren Antikörper zumindest einen Nachweisversuch in Formalin-
fixiertem Paraffin-eingebettetem Gewebe durchführen. Trotz Austesten unterschiedlicher
Konzentrationsreihen und Demaskierungsverfahren blieben die Versuchsreihen unter
Anwendung der Antikörper gegen SOX 10, PAX 3 und CLIM 2 ohne spezifischen Nachweis
der entsprechenden Faktoren. Bei SOX 10 ließ sich zwar eine Positivität von Zellen im

- 58 -
Bereich des Haarfollikels nachweisen, doch war dieser Nachweis in
Bestätigungsuntersuchungen so inkonstant, dass auch für SOX 10 eine zuverlässige
Anwendung des Antikörpers in Frage gestellt werden musste. Aufgrund der fehlenden
Spezifität für die Antikörper gegen PAX 3 und CLIM 2, sowie der inkonstanten
Färbeergebnisse des SOX 10-Antikörpers wurden weiterreichende Untersuchungen mit diesen
Antikörpern eingestellt.
Die SOX 9-Antikörper waren dagegen spezifisch und zeigten wiederholt kongruente
Ergebnisse, sodass sich die weitere Studie auf die Expression von SOX 9 im Haarfollikel
fokussiert.
5.2 SOX 9-Expression im Haarfollikel
In der vorliegenden Studie wurde die Expression von SOX 9 an den Haarfollikeln von 79
nicht verwandten Probanden immunhistochemisch bestimmt. Expressionsunterschiede
wurden hinsichtlich der Haarfarbe der Probanden, des Ergrauens, des Vorhandenseins einer
Alopezie, sowie der Alters- und Geschlechtsunterschiede statistisch miteinander verglichen.
Als positiv wurde eine Anfärbung des Zellkernes von Zellen innerhalb des Haarfollikels
gewertet, ähnlich den Ergebnissen aus der Arbeit von Krahl und Sellheyer (2010). Pro
Proband wurden zwei voneinander unabhängig gefärbte Präparate mikroskopisch ausgewertet.
Von jedem Präparat wurden die drei 40x-Gesichtsfelder mit der höchsten Anzahl SOX 9-
positiver Zellen ausgezählt. Der Mittelwert aus der Gesamtheit der sechs Gesichtsfelder
diente als Vergleichszahl zwischen einzelnen Probanden.
5.2.1 SOX 9-Expression und Haarfarbe
In einer ersten Analyse wurden die unterschiedlichen Haarfarben in helle und dunkle
aufgeteilt. Hierbei zeigte sich ein signifikanter Expressionsunterschied zwischen hellen und
dunklen Haaren, wobei die hellen Haare eine höhere SOX 9-Expression aufwiesen.

- 59 -
Bei einer Unterteilung der Haarfarben in die Gruppen rötlich, blond, dunkelblond, braun,
dunkelbraun und schwarz zeigte sich eine signifikant höhere Expression von SOX 9 bei
Blonden gegenüber Braunhaarigen, bei Rötlichen gegenüber Braunhaarigen, sowie
Dunkelblonden gegenüber Schwarzhaarigen.
Tendenziell zeigten somit Hellhaarige eine höhere SOX 9-Expression als Dunkelhaarige.
Interessanterweise ergab eine Pubmedrecherche mit den Suchworten „hair color“ and
„SOX 9“ keine einzige Publikation und eine Suche mit „hair color“ and „SOX“ nur eine
einzige Arbeit über SOX 10 bei Waardenburg Syndrom (Hwang et al., 2009). Somit geben
diese immunhistochemischen Expressionsdaten erste Hinweise auf einen Einfluss von SOX 9
auf die Haarfarbe, wobei weiterreichende Untersuchungen zum Einen eine Bestätigung der
Daten, zum Anderen detaillierter funktionelle Zusammenhänge liefern müssen.
Bislang sind klare Vorstellungen zu Einflussfaktoren auf die Haar- und auch auf die
Pigmentierungsfarbe der Haut eher rar. Im Fokus stehen die sogenannten „Pigmentation
Genes“, von denen ca. 150 unterschiedliche bekannt sind, wobei MC1R (Melanocortin-1-
Rezeptor), ASIP (agouti-signaling protein), Mitf (Mikrophthalmie-assoziierter
Transkriptionsfaktor) und Enzyme der Melanogenese (Tyrosinase, Trp1 (Tyrosinase-related
protein 1), Dct (Dopachrom-Tautomerase)) im Vordergrund des Interesses stehen (Ito and
Wakamatsu, 2010; Harris et al., 2010; Millar et al., 1995). Beispielsweise wurden in Genom-
weiten Assoziationsstudien über 80 Single-Nucleotide-Polymorphismen (SNP) in MC1R,
sowie in TYP, TYRP1, ASIP und anderen mit Einfluss auf die Pigmentierung gefunden
(Sturm R. A., 2009; Han et al., 2008; Ito and Wakamatsu, 2010).
Chemische Untersuchungen am Haarfollikel von Ito und Wakamatsu (2010) zeigten
Unterschiede im Verhältnis von Eumelanin (braun bis schwarz) zu Pheomelanin (blond bis
rötlich) bei unterschiedlich gefärbten Haaren. Hierbei scheint auch der pH-Wert innerhalb des
Melanosoms eine wichtige Rolle zu spielen, der bei hellen Haaren eher im saueren Milieu, bei
dunklen im neutralen Bereich liegt.

- 60 -
5.2.2 SOX 9-Expression und Haarergrauen
Ein Vergleich der SOX 9-Expression von Nicht-Grauhaarigen im Vergleich zu Grauhaarigen
zeigte eine signifikant niedrigere Expression von SOX 9 bei Grauhaarigen. Hierbei fällt ein
tendenzieller Abfall der SOX 9-Expression von Hellhaarigen zu Dunkelhaarigen und weiter
zu Grauhaarigen auf, wobei ersterer im Einzelvergleich der Gruppen zueinander signifikant
ist, zweiterer nicht.
SOX 9 ist nicht nur in der äußeren Haarwurzelscheide nachweisbar, sondern hat auch
wesentlichen Einfluss auf die Melanogenese (Harris et al., 2010). Die Expression von SOX 9
steigt mit der Differenzierung vom Melanoblasten zum Melanozyten an, wohingegen
diejenige von SOX 10 abnimmt. Nach UV-Exposition steigt die Expression von SOX 9 in
Melanozyten an und aktiviert über Mitf, Dct und Tyr die Melaninproduktion (Passeron et al.,
2007). Somit könnte SOX 9 indirekt über seine aktivierende Regulation auf die Melanozyten
und die Melaninsynthese Einfluss auf den Grad der Pigmentierung von Haaren nehmen,
wobei jedoch derzeit keine Daten von SOX 9 zum Agingprozess des Haares und seiner
Pigmentierung vorliegen.
Bislang aus mehreren Studien bekannt ist die Tatsache, dass mit abnehmender
Haarpigmentierung auch die Anzahl der Melanozyten abnimmt (Tobin D. J., 2009; Commo et
al., 2004). Tobin (2009) spricht davon, dass graue Haare weniger, weiße sogar gar keine
Melanozyten enthalten. Commo et al. (2004) berichten von nur wenigen im Bulbus
vorhandenen Melanozyten bei grauhaarigen Probanden, die aber noch in der Lage sind,
Tyrosinase und Trp 1 und somit Melanin zu produzieren. Im Unterschied dazu sind bei
weißen Haaren im Bulbus keine Melanozyten mehr auffindbar. Zudem besitzt pigmentiertes
Haar ruhende Melanozyten in der äußeren Wurzelscheide. Dieselben inaktiven, nicht-
Melanin-produzierenden Melanozyten finden sich auch bei Grau- bzw. Weißhaarigen, jedoch
in niedrigerer Anzahl (Horikawa et al., 1996; Takada et al., 1992). Andere wissenschaftliche
Arbeiten gehen von Funktionseinbußen der Melanozyten als Grund des Haarergrauens aus,
anstatt oder zusätzlich zu ihrem zahlenmäßigen Untergang. Vermutlich als Folge einer
eingeschränkten Tyrosinaseaktivität zeigte graues Haar eine Abnahme der Menge an
Melaninkörnchen (Sato et al., 1973; Lloyd et al., 1987). Tobin und Paus (2001) machen neben
einer verringerten Tyrosinasefunktion, suboptimale Melanozyten-Kortexkeratinozyten-

- 61 -
Interaktionen und eine fehlerhafte Migration der Melanozyten vom Reservoir in die obere
äußere Wurzelscheide für Pigmentverdünnung und Haarergrauen verantwortlich. Es ist
denkbar, dass solche funktionseingeschränkten Melanozyten ein anderes Färbeverhalten
aufweisen als normal funktionsfähige und wir deshalb bei ergrauten Haaren weniger SOX 9-
Posivität feststellten.
Darüber hinaus scheinen Melanozytenvorstufen mit zunehmendem Ergrauen der Haare an
Zahl abzunehmen. Mit zunehmendem Alter kommt es zu stressinduzierten Veränderungen der
pigmentproduzierenden, reparierenden und antioxidativ wirksamen Enzyme, Co- und
Wachstumsfaktoren, wie Bcl 2 und die c-Kit. Der oxidative Stress führt zu Schädigung und
zum Zusammenbrechen der Redoxkapazitäten der Haarfollikelmelanozyten und so kommt es
folglich mit sinkender Haarpigmentation zur Reduktion der Melanozytenstammzellen (MSZ)
im Wulst der äußeren Wurzelscheide (Arck et al., 2006). Das Fehlen von frisch
differenzierten Melanozyten, die in den Haarbulbus einwandern können, ist Folge dieses
Stammzellenmangels (Steingrímsson et al., 2005). In Anlehnung an Stammzellen anderer
Gewebe wird vermutet, dass die Melanozytenstammzellen im Alter zwar teilweise noch
vorhanden sind, jedoch veränderte Eigenschaften besitzen (Commo et al., 2004).
SOX 9 spielt eine Schlüsselrolle bei der Erhaltung von Melanozytenstammzellen (MSZ) und
Melanozytenvorläuferzellen (Cook et al., 2005; Vidal et al., 2005) und ist essentiell für deren
Differenzierung (Nowak et al., 2008). So lässt sich die hier gezeigte reduzierte SOX 9-
Expression bei grauhaarigen Probanden auch auf eine herabgesetzte Anzahl von
Melanozytenstammzellen bei grauen Haaren zurückführen, was als eine Ursachen
menschlichen Haarergrauens gilt (Nishimura E. K., 2011).
Im „ergrauenden Mausmodel“ zeigte sich mit jedem Haarzyklus eine graduelle Abnahme der
Melanozytenstammzellen. Dieses Verhalten fand sich auch beim Menschen. Hier wurden
mehr Melanoblasten in den Bulbi der pigmentierten Follikel als in denen der ergrauten
gefunden. Daraus lässt sich folgern, dass Haarergrauen einen Defekt in der
Melanozytenstammzellerhaltung beinhaltet und in einem mit dem Alter voranschreitenden
stufenweisen Verlust der pigmentierten Tochterzellen resultiert (Nishimura et al., 2005;
Vachtenheim and Novotná, 1999). Wichtig für das Überleben der Melanozytenvorstufen und
MSZ sind neben SOX 9 der Notch-Signalweg (Moriyama et al., 2006; Schouwey et al., 2006),
sowie die Faktoren Mitf und Bcl 2. Letztere zwei sorgen für eine Abnahme der MSZ (Veis et
al., 1993; Nishimura et al., 2005).

- 62 -
5.2.3 SOX 9-Expression und altersbedingte Alopezie
Neben Haarergrauen ist Haarausfall (Alopezie) eines der Kennzeichen menschlichen Alterns.
Studien zeigen, dass mit zunehmendem Alter die Kopfhaardichte abnimmt. Bezogen auf den
Wachstumszyklus des Haares bedeutet dies eine verkürzte Anagenphase und eine längere
Telogenphase. Die normalerweise auf der Kopfhaut vorherrschenden Terminalhaare werden
lokal durch feines Vellushaar ersetzt. Eine wichtige Rolle dabei spielt die graduelle Abnahme
von Haarfollikelstammzellen (Leirós et al., 2012; Al-Refu K., 2012). Bei Männern zeigt sich
hauptsächlich androgenbedingter Haarausfall, der vor allem genetisch bedingt ist. Im
Gegensatz dazu scheinen bei Frauen eher Umwelteinflüsse eine Rolle für das Entwickeln der
Alopezie zu spielen (Messenger A. G., 2011).
Im hier untersuchten Klientel waren acht Probanden mit Alopezie. Bei diesen war keine
signifikante Abnahme in der SOX 9-Expression feststellbar. Zufälligerweise handelte es sich
ausschließlich um Frauen, sodass eine Interpretation dieser sowieso schon niedrigen Anzahl
an Probanden nur unter größtem Vorbehalt zu sehen ist.
5.2.4 SOX 9-Expression in Abhängigkeit vom Alter
Obwohl keine signifikanten SOX 9-Expressionsunterschiede beim Vergleich von Probanden
mit und ohne Alopezie gefunden wurden, wäre eine Abnahme der SOX 9-Expression mit
zunehmendem Alter ein Kriterium für androgenetische Alopezie und Canities. Eine
Unterteilung unserer Probanden in vier Altersgruppen und deren Vergleich erbrachte keine
konkreten signifikanten Expressionsunterschiede von SOX 9, zeigte aber einen absteigenden
Trend der SOX 9-Expression mit zunehmendem Alter.
5.2.5 Geschlechtsabhängigkeit der SOX 9-Expression
Ein Geschlechtsvergleich der SOX 9-Expression durch alle Altersgruppen zeigte im
vorliegenden Klientel keinerlei Unterschiede.

- 63 -
Geschlechtsspezifische Untersuchungen bezüglich Haarergrauens zeigten ebenfalls keine
Unterscheide zwischen Männern und Frauen die Ausprägung oder das Alter, in dem
Haarergrauen auftritt betreffend (Trueb R. M., 2005; Lapeere et al., 2008; Keogh and Walsh,
1965; Cline D. J., 1988). Jo et al. (2011) zeigen geschlechtsspezifisch lediglich eine
Verschiedenheit in der Verteilung der ergrauenden Haare. So beginnt das Ergrauen bei Frauen
vor allem frontal, während Männer temporal und occipital zuerst ergrauen.
5.3 Methodische Schwächen
5.3.1 Einschränkungen bei den Färbeergebnissen
Von den verwendeten Antikörpern gelangen die Färbungen nur mit dem Antikörper SOX 9,
sodass sich sämtliche Ergebnisse dieser Arbeit auf mittels SOX 9-Antikörper angefärbte
Zellen des Haares beziehen. Doch warum haben die Färbungen mit SOX 10-, PAX 3- und
CLIM 2-Antikörpern nicht funktioniert? Für das Misslingen der Färbungen mit den übrigen,
oben genannten Antikörpern kommen mehrere Ursachen in Frage.
Konservierung
Eine Überlegung bezüglich der misslungenen Färbungen mit den anderen Antikörpern bezieht
sich auf die Dauer und Art der Konservierung. Die Idee dabei ist, dass zum einen durch eine
jahrelange Konservierung in Paraffin ein Teil der Gewebeeigenschaften verloren geht.
Zum anderen ist möglicherweise die Paraffinart nicht kompatibel zur Färbemethode und den
ausgewählten Antikörpern. In einigen Studien werden die Untersuchungen anstelle von
Paraffin- an Gefrierschnitten durchgeführt. In diesem Fall kommt es sofort nach Entnahme
aus der Körperregion zum Einsatz flüssigen Stickstoffs (Arck et al., 2006; Moriyama et al.,
2006). An diesen Arbeiten konnten wir uns aufgrund der grundsätzlich anderen Methodik
nicht orientieren.
Demaskierung
Eine weitere Ursache für das Misslingen der Färbungen ist, dass wir aufgrund der
verwendeten Demaskierungsmethoden keine spezifischen Ergebnisse erzielen konnten. So
war entweder zu viel Backgroundfärbung zu sehen oder es war nichts angefärbt, zum Beispiel

- 64 -
weil spezifische Antigenstrukturen nicht freigelegt wurden und die Antikörper somit nicht an
ihre Zielstruktur andocken konnten. Ein längeres Vorkochen beispielsweise hatte jedoch zur
Folge, dass die Präparate verkocht und für weitere Färbungen nicht verwertbar waren.
Antikörper
Möglicherweise waren die eingesetzten Primär- oder Sekundärantikörperkonzentrationen
nicht richtig gewählt, was zu einer zu hohen Hintergrundfärbung führen kann.
Gehen wir davon aus, dass es sich bei den SOX 9-positiven Zellen um weit differenzierte
Zellen der Melanozytenzellreihe handeln könnte, ist eine mögliche Erklärung für das
Nichtanfärben mittels SOX 10-Antikörper, dass es mit zunehmender
Melanozytendifferenzierung zu steigendem SOX 9- und gleichzeitig abfallendem SOX 10-
Gehalt kommt (Cook et al., 2005; Gershon et al., 2005).
5.3.2 Einschränkungen bei der Ergebnisinterpretation
Subjektivität der Haarfarbe
Die Gruppenzugehörigkeit einzelner Probanden war zum Teil nicht eindeutig beurteilbar,
insbesondere bei den telefonisch nach ihrer Haarfarbe befragten Probanden, welche ca. die
Hälfte des Kollektivs ausmachen. Die Haarfarbe könnte dabei subjektiv eingeschätzt werden
und es würde beispielsweise der dunkelblonde Proband in die Gruppe „braun“ gelangen. Ein
ähnliches Problem ergibt sich auch bei der Einschätzung des Grauanteils. Hat der Proband nur
vereinzelt graue Haare oder ist schon die Hälfte oder mehr ergraut? Oder kennt der Proband
womöglich das Ausmaß des eigenen Ergrauens gar nicht, weil er sich schon seit Jahren die
Haare färbt?
Desweiteren taucht die Problematik der „grauen Haare“ auf. Diese sind nicht ausnahmslos
grau, sondern oft „meliert“, das heißt der Proband hat gleichzeitig neben grauen und weißen
Haaren auch Haare seiner Ursprungshaarfarbe. Da bei der Auswertung der histologischen
Präparate pro Proband die sechs Haare mit den meisten SOX 9-positiven Zellen ausgewählt

- 65 -
wurden, ist davon auszugehen, dass diese, auch bei Probanden, die größtenteils graue
beziehungsweise weiße Haare besitzen, eher die pigmentierten als die grauen waren. So
lassen sich auch die Ergebnisse der Minima und Maxima erklären. Die Maxima erreichen bei
allen drei Gruppen annähernd dieselben Werte, da, wie eben erörtert, auch pigmentierte Haare
in die Auswertung einbezogen wurden. Die Minima hingegen unterscheiden sich deutlich. Da
vermutlich hier tatsächlich auch graue und weiße Haare erfasst sind, kommen Haare mit
ausschließlich negativem Färbeergebnis, sprich ohne SOX 9-positive Zellen, in die
Berechnungen mit hinein und es ergeben sich Werte von annähernd Null als Minima.
Statistik
Die relativ hohen Standardabweichungen sprechen dafür, dass bei den einzelnen Probanden
von Haar zu Haar hohe Schwankungen der SOX 9-positiven Zellzahlen vorkommen
(Vgl. Kapitel 4. Ergebnisse). In der vorliegenden Arbeit müssen einige Einschränkungen bei
den Aussagen der Ergebnisse gemacht werden. Zum einen ist die Anzahl der Probanden mit
n = 79 relativ gering, was die Wahrscheinlichkeit für statistische Fehler 1. oder 2. Art erhöht.
Zum anderen wären Färbungen mit mehreren Antikörpern anstatt nur mit einem
aussagekräftiger gewesen. Größere Datensammlungen, sowie weitere
Antikörperetablierungsversuche und -testreihen hätten allerdings den Rahmen der
Doktorarbeit gesprengt.

- 66 -
5.4 Ausblick
In der vorliegenden Arbeit wurde die SOX 9-Expression in Probandengruppen verschiedener
Haarfarben, sowie von Gruppen nach dem Vorhandensein von Haarergrauen oder Alopezie
eingeteilt und verglichen.
Aufgrund des relativ kleinen Probandenkollektivs sind einige Aussagen leider wenig
repräsentativ. Beispielsweise sind lediglich acht Probanden mit Alopezie in die Studie
eingeschlossen, welche auch noch zufällig alle weiblichen Geschlechts sind. Auch bei der
Einteilung in die detaillierteren Haarfarbengruppen ergeben sich geringe Probandenzahlen.
Neben einem größeren Probandenpool sollten zukünftige Studien zu diesem Thema
objektivere Methoden zur Haarfarbeneinstufung und Einteilung in die Gruppen verwenden.
Wo liegt beispielsweise die Grenze zwischen blond und dunkelblond oder ab welchem Anteil
grauer Haare zählt der Proband zu den Grauhaarigen?
Es gibt bislang wenige Publikationen, die sich mit den Unterschieden zwischen den
verschiedenen Haarfarben auf der Ebene der Stammzellmarker beschäftigen. Hierfür wäre
neben SOX 9 der zuverlässige Einsatz anderer Stammzellfaktoren hilfreich, sei es in
immunhistochemischen Färbungen oder anderen experimentellen Methoden.

- 67 -
6. Zusammenfassung
Die Haare gehören mit ihrer vielfältigen Farbigkeit, Form und vielseitigen Gestaltbarkeit zu
den wichtigsten äußeren Erkennungsmerkmalen der Menschen. Im Alter kommt es
zunehmend zu Haarergrauen, sowie vornehmlich bei Männern auch zu Haarverlust. Schon
seit Jahrzehnten interessiert sich die Forschung für Faktoren, welche die verschiedenen
Haarfarben und den Prozess des Haarergrauens und –verlusts beeinflussen.
Ziel der vorliegenden Arbeit war es, anhand immunhistochemischer Färbungen an
Kopfhautpräparaten den Einfluss von Stammzellmarkern auf die Entstehung der Haarfarbe,
sowie bei den Prozessen des Haarergrauens und Haarverlusts herauszufinden. Basierend auf
Literaturrecherche zu diesen drei Themen suchten wir Stammzellmarker heraus, die sich für
immunhistochemische Färbungen eigneten und deren zugehörige Antikörper käuflich zu
erwerben waren. Die Auswahl führte zu den Faktoren SOX 9, SOX 10, PAX 3 und CLIM 2.
Die Selektion der 79 in die Studie eingeschlossenen Probanden erfolgte anhand bereits
vorhandener Schnittpräparate. Aus dem Archiv der dermatologischen Universitätsklinik
Freiburg wurden der Kopfhaut entstammende Präparate aus einem bestimmten Zeitraum
herausgesucht. Voraussetzung war, dass mehrere Haarfollikel möglichst tangential zu deren
Verlaufsrichtung im histologischen Präparat getroffen waren. Informationen wie Haarfarbe
und Vorhandensein einer Alopezie wurden aus der Photodokumentation übernommen oder
telefonisch bei den Probanden erfragt.
Während einer Etablierungsphase immunhistochemischer Färbemethoden mit den SOX 9-,
SOX 10-, PAX 3- und CLIM 2-Antikörpern wurden verschiedene Konzentrationen der
Primär- und Sekundärantikörper, sowie unterschiedliche Demaskierungsverfahren
ausprobiert. Lediglich die Färbungen mit SOX 9-Antikörpern ergaben reproduzierbare und
spezifische Resultate, sodass sämtliche weiteren Färbungen dieser Arbeit nur mit SOX 9-
Antikörpern durchgeführt wurden.
Beim Vergleich der unterschiedlichen, nach Haarfarbe und –dichte eingeteilten Gruppen
bezüglich ihrer SOX 9-Expression zeigten hellhaarige Probanden signifikant höhere Zahlen
SOX 9-positiver Zellen im Vergleich zu dunkel- und grauhaarigen. Auch wiesen nicht-
Grauhaarige eine signifikant höhere SOX 9-Positivität als Grauhaarige auf. Zwar zeigt sich
mit zunehmendem Alter eine Tendenz zur sinkenden SOX 9-Expression, statistisch war diese
jedoch nicht von Signifikanz. Zudem schien die SOX 9-Expression unabhängig von
Geschlecht und Haarausfall zu sein. Die Ergebnisse der vorliegenden Arbeit können die
wichtige Rolle von SOX 9 in Bezug auf Haarfarbe und Haarergrauen somit bestätigen.

- 68 -
7. Literaturverzeichnis
A
Al-Refu K. (2012) Stem Cell and Alopecia: A Review to Pathogenesis. Br J Dermatol. j.
1365-2133
Arck P. C., Overall R., Spatz K., Liezman C., Handjiski B., Klapp B. F., Birch-Machin M. A.,
Peters E. M. J. (2006) Towards a “free radical theory of graying”: melanocyte apoptosis in the
aging human hair follicle is an indicator of oxidative stress induced tissue damage. The
FASEB Journal 20: 908-920
Aubin-Houzelstein G., Djian-Zaouche J., Panthier J. J. (2008) Melanocyte stem cells in
adults. J Soc Biol. 202(1): 25-32
B
Barsh G. S. (1996) The genetics of pigmentation: From fancy genes to complex traits. Trends
in Genetics 12: 299-305
Bennett D. C., Lamoreux M. L. (2003) The color loci of mice – a genetic century. Pigment
Cell Res. 16: 333-344
Boenisch T., Farmilo A. J., Stead R. H., Key M., Welcher R., Harvey R., Atwood K. N.,
Tscherkassy M., Henne C. (2003) Immunchemische Färbemethoden, Handbuch, 3. Aufl.
DakoCytomation Corp. Carpinteria, CA, USA
Botchkareva N. V., Botchkarev V. A., Gilchrest B. A. (2003) Fate of melanocytes during
development of the hair follicle pigmentary unit. J Invest Dermatol Symp Proc 8: 76-79
Botchkareva N. V., Khlgatian M., Longley B. J., Botchkarev V. A., Gilchrest B. A. (2001)
SCF/c-kit signaling is required for cyclic regeneration of the hair pigmentation unit. The
FASEB Journal 15: 645-658
Braun-Falco O., Plewig G., Wolff H. H., Burgdorf W. H. C., Landthaler M. (2005)
Dermatologie und Venerologie, 5. Aufl. Springer, Heidelberg

- 69 -
C
Chang S., Multani A. S., Cabrera N. G., Naylor M. L., Laud P., Lombard D., et al. (2004)
Essential role of limiting telomeres in the pathogenesis of Werner syndrome. Nature Genetics
36: 877-882
Cheung M., Chaboissier M. C., Mynett A., Hirst E., Schedl A., Briscoe J. (2005) The
transcriptional control of trunk neural crest induction, survival, and delamination. Dev Cell 8:
179-192
Cline D. J. (1988) Changes in hair color. Dermatol Clin, 6: 295-303
Commo S., Gaillard O., Bernard B. A. (2004) Human hair graying is linked to a specific
depletion of hair follicle melanocytes affecting both the bulb and the outer root sheath. Fr J
Dermatol 150(3): 435-443
Cook A. L., Smith A. G., Smit D. J., Leonard J. H., Sturm R. A. (2005) Co-expression of
SOX 9 and SOX 10 during melanocytic differentiation in vitro. Exp Cell Res 308(1): 222-235
F
Ferletta M., Uhrbom L., Olofsson T., Pontén F., Westermark B. (2007) Sox 10 has a broad
expression pattern in gliomas and enhances platelet-derived growth factor-B-induced
gliomagenesis. Mol Cancer Res 5(9): 891-897
Fuchs E. (2009) The Tortoise and the Hair: Slow-Cycling Cells in the Stem Cell Race. Cell
137: 811-819
G
Gershon T. R., Oppenheimer O., Chin S. S., Gerald W.L. (2005) Temporally Regulated
Neural Crest Transcription Factors Distinguish Neuroectodermal Tumors of Varying
Malignancy and Differentiation. Neoplasia 7(6): 575-584
Goethe J. W. (1827) Gedichte (Ausgabe letzter Hand. 1827), Weissagungen der Bakis, 22.
Bd. 1-4: Gedichte, Cotta, Stuttgart und Tübingen

- 70 -
H
Han J., Kraft P., Nan H., Guo Q., Chen C., Qureshi A., Hankinson S. E., Hu F. B., Duffy D.
L., Zhao Z. Z., Martin N. G., Montgomery G. W., Hayward N. K, Thomas G., Hoover R. N.,
Chanock S., Hunter D. J. (2008) A Genome-Wide Association Study Identifies Novel Alleles
Associated with Hair Color and Skin Pigmentation. PLoS Genet 4(5): e1000074
Harris M. L., Baxter L. L., Loftus S. K., Pavan W. J. (2010) Sox proteins in melanocyte
development and melanoma. Pigment Cell Melanoma Res. 23(4): 496-513
Horikawa T., Norris D. A., Johnson T. W. et al. (1996) DOPA-negative melanocytes in the
outer root sheath of human hair follicles express premelanosomal antigens but not a
melanosomal antigen or the melanosome-associated glycoproteins tyrosinase, TRP-1 and
TRP-2. J Invest Dermatol 106: 28-35
Hwang K. C., Cho S. K., Lee S. H., Park J. Y., Kwon D. N., Choi Y. J., Park C., Kim J. H.,
Park K. K., Hwang S., Park S. B., Kim J. H. (2009) Depigmentation of skin and hair color in
the somatic cell cloned pig. Dev Dyn 238(7): 1701-8
I
Ito S., Wakamatsu K. (2010) Human hair melanins: what we have learned and have not
learned from mouse coat color pigmentation. Pigment Cell Melanoma Res. 24(1): 63-74
Ito S., Wakamatsu K. (2011) Diversity of human hair pigmentation as studied by chemical
analysis of eumelanin and pheomelanin. JEADV 25: 1369-1380.
J
Jo S. J., Paik S. H., Choi J. W., Lee J. H., Cho S., Kim K. H., Eun H. C., Kwon O. S. (2011)
Hair Graying Pattern Depends on Gender, Onset Age and Smoking Habits, Acta Derm
Venereol 92: 160-161
Johnson R., Jackson I. J. (1992) Light is a dominant mouse mutation resulting in premature
cell death. Nature Genetics 1: 226-229
Junqueira L. C., Carneiro J. (1991) Histologie, 3. Aufl., Springer, Berlin

- 71 -
K
Kaplan P. D., Polefka T., Grove G., Daly S., Jumbelic L., Harper D., Nori M., Evans T.,
Ramaprasad R., Bianchini R. (2011) Grey hair: clinical investigation into changes in hair
fibres with loss of pigmentation in a photoprotected population. International Journal of
Cosmetic Science 33: 171-182
Keogh E. V., Walsh R. J. (1965) Rate of greying of human hair. Nature 207: 877-878
Kordes U., Hagel C. (2006) Expression of SOX9 and SOX10 in central neuroepithelial tumor.
J Neurooncol 80: 151-155
Krahl D., Sellheyer K. (2010 a) Sox9, more than a marker of the outer root sheath:
spatiotemporal expression pattern during human cutaneous embryogenesis. J Cutan Pathol 27:
350-356
Kumano K., Masuda S., Sata M., Saito T., Lee S.Y., Sakata-Yanagimoto M., Tomita T.,
Iwatsubo T., Natsugari H., Kurokawa M., Ogawa S., Chiba S. (2008) Both Notch1 and
Notch2 contribute to the regulation of melanocyte homeostasis. Pigment Cell Melanoma Res.
21 (1): 70-78
L
Lang D., Lu M. M., Huang L., Engleka K. A., Zhang M., Chu E. Y., Lipner S., Skoultchi A.,
Millar S. E., Epstein J. A. (2005) Pax3 functions at a nodal point in melanocyte stem cell
differentiation. Nature 433: 884-887
Lapeere H., Boone B., Schepper S. D., Verhaeghe E. (2008) Hypomelanoses and
hypermelanoses. Fitzpatrick’s dermatology in general medicine, 7th Edition McGraw-Hill
(622-640), New York City
Leirós G. J., Attorresi A. I., Balan͂á M. E. (2012) Hair follicle stam cell differentiation is
inhibited through cross-talk between Wnt/β-catenin and androgen signaling in dermal papilla
cells from patients with androgenetic alopecia. Br J Dermatol 166 (5): 1035-42

- 72 -
Lloyd T., Garry F. L., Manders E. K. et al. (1987) The effect of age and hair colour on human
hair bulb tyrosinase activity. Br J Dermatol 116: 485-489
M
Messenger A. G. (2011) Hair through the female life cycle. Br Jour Dermatol 165(3): 2-6
Millar S. E., Miller M. W., Stevens M. E., Barsh G. S. (1995) Expression and transgenic
studies of the mouse agouti gene provide insight into the mechanisms by which mammalian
coat color patterns are generated. Development 121: 3223–3232
Moriyama M., Osawa M., Mak S. S., Ohtsuka T., Yamamoto N., Han H., Delmas V.,
Kageyama R., Beermann F., Larue L., Nishikawa S. I. (2006) Notch signaling via Hes1
transcription factor maintains survival of melanoblasts and melanocyte stem cells. The
Journal of Cell Biology 173(3): 333–339
Mosley J.G., Gibbs A.C. (1996) Premature grey hair and hair loss among smokers. A new
opportunity for health education? BMJ 313: 1616
N
Nishimura E. K. (2011) Melanocyte stem cells: a melanocyte reservoir in hair follicles for
hair and skin pigmentation. Pigment Cell Melanoma Re 24: 401-410
Nishimura E. K., Suzuki M., Igras V., Du J., Lonning S., Miyachi Y., Roes J., Beermann F.,
Fisher D. E. (2010) Key roles for transforming growth factor beta in melanocyte stemm cell
maintenance. Cell Stem Cell 6(2): 130-140
Nishimura E. K., Granter S. R., Fisher D. E. (2005) Mechanism of Hair Graying: Incomplete
Melanocyte Stem Cell Maintenance in the Niche. Science 307: 720-723
Nowak J. A., Polak L., Pasolli H. A., Fuchs E. (2008) Hair Follicle Stem Cells are Specified
and Function in Early Skin Morphogenesis. Cell Stemm Cell 3(1): 33-43

- 73 -
O
Oshima H., Rochat A., Kedzia C., Kobayashi K., Barrandon Y. (2001) Morphogenesis and
renewal of hair follicles from adult multipotent stem cells. Cell 104: 233-245
P
Parker C. J, Shawcross S. G., Li H., Wang Q. Y., Herrington C.S., Kumar S., MacKie R. M.,
Prime W., Rennie I. G., Sisley K., Kumar P. (2004) Expression of PAX3 alternatively spliced
transcripts and identification of two new isoforms in human tumors of neural crest origin. Int
J Cancer 108: 314-320
Passeron T., Valencia J. C., Bertolotto C., Hoashi T., Le Pape E., Takahashi K., Ballotti R.,
Hearing V. J. (2007) Sox9 is a key player in ultraviolet B-induced melanocyte differentiation
and pigmentation. Proc Natl Acad Sci USA 104(35): 13984-9
Paus R., Cotsarelis G. (1999) The biology of hair follicles. N Engl J Med 341: 491-497
Pingault V., Ente D., Dastot-Le Moal F., Goossens M., Marlin S., Bondurand N. (2010)
Review and update of mutations causing Waardenburg syndrome. Hum Mutat 31(4): 391-406
Prince S., Illing N., Kidson S. H. (2001) SV-40 large T antigen reversibly inhibits expression
of tyrosinase, TRP-1, TRP-2 and Mitf, but not Pax-3, in conditionally immortalized mouse
melanocytes. Cell Biol Int 25(1): 91-102
R
Rando T. A. (2006) Stem cells, ageing and the quest for immortality. Nature 441: 1080-1086
Rudolph K. L., Chang S., Lee H. W., Blasco M., Gottlieb G. J., Greider C., et al. (1999)
Longevity, stress response, and cancer in aging telomerase-deficient mice. Cell 96: 701-712
S
Sarin K. Y., Artandi S. E. (2007) Aging, Graying and Loss of Melanocyte Stem Cells. Stem
Cell Rev 3(3): 212-217

- 74 -
Sato K, Kukita A., Jimbow K. (1973) Electron microscopic studies of dendritic cells in the
human grey and white hair matrix during anagen. Pigment Cell 1: 20-26
Schneider M. R., Schmidt-Ullrich R., Paus R. (2009) The Hair Follicle as a Dynamic
Miniorgan. Current Biology 19: 132-142
Schouwey K., Delmas V., Larue L., Zimber-Strobl U., Strobl L. J., Radtke F., Beermann F.
(2006) Notch1 and Notch2 receptors influence progressive hair graying in a dose-dependent
manner. Dev Dyn 236(1): 282-289
Schumacher M., Schulgen G. (2008) Methodik klinischer Studien, 3. Aufl., Springer, Berlin,
Heidelberg
Sharpless N. E., De Pinho R. A. (2004) Telomeres, stem cells, senescence and cancer. J Clin
Invest. 113(2): 160-8
Smit D. J., Smith A. G., Parsons P. G., Muscat G. E. O., Sturm R. A. (2000) Domains of Brn-
2 that mediate homodimerization and interaction with general and melanocytic transcription
factors. Eur J Biochem 267: 6413-6422
Steingrímsson E., Copeland N. G., Jenkins N. A. (2005) Melanocyte Stem Cell Maintenance
and Hair Graying. Cell 121: 9-12
Stenn K. S., Paus R. (2001) Controls of hair follicle cycling. Physiol Rev 81: 449-494
Stolt C. C., Lommes P., Sock E., Chaboissier M. C., Schedl A., Wegner M. (2003) The Sox9
transcription factor determines glial fate choice in the developing spinal cord. Genes Dev 17:
1677-1689
Sturm R. A. (2009) Molecular genetics of human pigmentation diversity. Hum Mol Genet
18(1): R9-R17

- 75 -
Sugihara T. M., Bach I., Kioussi C., Rosenfeld M. G., Andersen B. (1998) Mouse deformed
epidermal autoregulatory factor 1 recruits a LIM domain factor, LMO-4, and CLIM
coregulators. Proc Natl Acad Sci USA 95: 15418-15423
T
Takada K., Sugiyama K., Yamamoto I. et al. (1992) Presence of quiescent melanocytes within
the outer root sheath in senile white hair. J Invest Dermatol 99: 629-633
Tobin D. J. (2011) The cell biology of human hair follicle pigmentation. Pigment Cell
Melanoma Res. 24 (1): 75-88
Tobin D. J. (2009) Aging of the Hair Follicle Pigmentation System. Int J Trichology 1(2): 83-
93
Tobin D. J. (2008) Human hair pigmentation – biological aspects. International Journal of
Cosmetic Science 30: 233-257
Tobin D.J., Bystryn J. C. (1996) Different populations of melanocytes are present in hair
follicles and epidermis. Pigment Cell Research 9: 304-310
Tobin D. J., Paus R. (2001) Graying: gerontobiology of the hair follicle pigmentary unit. Exp
Gerontol 36(1): 29-54
Trueb R. M., (2005) Aging of hair. J Cosmet Dermatol 4: 60-72
V
Vachtenheim J., Novotná H. (1999) Expression of genes for microphthalmia isoforms, Pax3
and MSG1, in human melanomas. Cell Mol Biol (Noisy-le-grand) 45(7): 1075-1082
Van Zant G., Liang Y. (2003) The role of stem cells in aging. Experimental Hematology 31:
659-672

- 76 -
Veis D. J., Sorenson C. M., Shutter J. R., Korsmeyer S. J. (1993) Bcl-2-deficient mice
demonstrate fulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented hair. Cell
75(2): 229-240
Vidal V. P. I., Chaboissier M. C., Lützkendorf S., Cotsarelis G., Mill P., Hui C. C., Ortonne
N., Ortonne J. P., Schedl A. (2005) Sox9 Is Essential for Outer Root Sheath Differentiation
and the Formation of the Hair Stem Cell Compartment. Current Biology 15: 1340-1351
Vidal V. P. I., Ortonne N., Schedl A. (2008) SOX9 expression is a general marker of vasal
cell carcinoma and adnexal-related neoplasms. J Cutan Pathol 15: 373-379
W
Wagner R., Joekes I. (2007) Hair medulla morphology and mechanical properties. J Cosmet
Sci. 58(4): 359-368
Wu X., Shen Q. T., Oristian D. S., Lu C.P., Zheng Q., Wang H. W., Fuchs E. (2011) Skin
Stem Cells Orchestrate Directional Migration by Regulating Microtubule-ACF7 Connections
through GSK3β. Cell 144 (3): 341-352
X
Xu S., Mannik J., Kudryavtseva E., Lin K. K., Flanagan L. A., Spencer J., Soto A., Wang N.,
Lu Z., Yu Z., Monuki E. S., Andersen B. (2007) Co-factors of LIM domains (Clims/Ldb/Nli)
regulate corneal homeostasis and maintenance of hair follicle stem cells. Developmental
Biology 312: 484-500
Y
Yokoyama S., Takeda K., Shibahara S. (2006) SOX10, in combination with Sp1, regulates the
endothelin receptor type B gene in human melanocyte lineage cells. FEBS Journal 273: 1805-
1820

- 77 -
An dieser Stelle ist im Original die Danksagung.

- 78 -
An dieser Stelle ist im Original der Lebenslauf.

- 79 -
10. Anhang
Abkürzungen
Abb. Abbildung
AB Antibody
ACTH Adrenocorticotropes Hormon
AP Alkaline Phophatase
Aqua dest. Aqua destillata
ASIP agouti-signaling protein
CLIM Co-Faktor von LIM-Domänen
Dct Dopachrom-Tautomerase
HE Hämatoxylin-Eosin
HMG high-mobility group
LIM L in11, Isl-1 und Mec-3
m männlich
Mitf Mikrophthalmie-assoziierter Transkriptionsfaktor
Mitfvit Mitfvitiligo
MB Melanoblasten
MC1R Melanocortin-1-Rezeptor
MSH Melanozyten-stimulierendes Hormon
MSZ Melanozytenstammzellen
MZ Melanozyten
s siehe
PAX Paired-box
SD Standardabweichung
SOX SRY-type HMG box
SRY sex-determining region on Y chromosome
TNF α Tumornekrosefaktor α
Trp Tyrosinase-related protein
vgl. vergleiche
VK Variationskoeffizienten
w weiblich


















