
TECHNISCHE UNIVERSITÄT MÜNCHEN II. Medizinische Klinik und ... · TECHNISCHE UNIVERSITÄT...
107
TECHNISCHE UNIVERSITÄT MÜNCHEN II. Medizinische Klinik und Poliklinik des Klinikums rechts der Isar Die Bedeutung von Socs3 im in-vivo-Modell der pankreatischen Karzinogenese Thuy Trang Phan
Transcript of TECHNISCHE UNIVERSITÄT MÜNCHEN II. Medizinische Klinik und ... · TECHNISCHE UNIVERSITÄT...

TECHNISCHE UNIVERSITÄT MÜNCHEN
II. Medizinische Klinik und Poliklinik
des Klinikums rechts der Isar
Die Bedeutung von Socs3 im in-vivo-Modell der pankreatischen Karzinogenese
Thuy Trang Phan

TECHNISCHE UNIVERSITÄT MÜNCHEN
II. Medizinische Klinik und Poliklinik
des Klinikums rechts der Isar
(Direktor: Univ.-Prof. Dr. Roland M. Schmid)
Die Bedeutung von Socs3 im in-vivo-Modell der pankreatischen Karzinogenese
Thuy Trang Phan
Vollständiger Abdruck der von der Fakultät für Medizin
der Technischen Universität München
zur Erlangung des akademischen Grades eines
Doktors der Medizin
genehmigten Dissertation.
Vorsitzender: Univ.-Prof. Dr. Ernst J. Rummeny
Prüfer der Dissertation: 1. Priv.-Doz. Dr. Hana Algül
2. Univ.-Prof. Dr. Helmut Friess
Die Dissertation wurde am 10.07.2014 bei der Technischen Universität München
eingereicht und durch die Fakultät für Medizin am 15.04.2015 angenommen.

Meinen Eltern gewidmet

INHALTSVERZEICHNIS
INHALTSVERZEICHNIS
I. ABKÜRZUNGEN ......................................................................................................... 3
II. EINLEITUNG ............................................................................................................... 5
1. Das Pankreaskarzinom ................................................................................................ 5
1.1. Epidemiologie ................................................................................................................. 5
1.2. Molekularbiologische und –genetische Hintergründe .................................................... 6
2. Das murine Pankreaskarzinommodell ..................................................................... 11
2.1. Das Cre/loxP-Rekombinationssystem .......................................................................... 12
2.2. Das KrasG12D-Mausmodell ........................................................................................... 13
3. Suppressors of cytokine signaling (SOCS) ............................................................... 16
3.1. Die Struktur von Socs3 ................................................................................................. 16
3.2. Socs3 als Zytokin-Signaltransduktionsinhibitor ........................................................... 17
4. Zielsetzung der Arbeit ................................................................................................ 21
III. MATERIAL UND METHODEN .............................................................................. 22
1. Material ....................................................................................................................... 22
1.1. Chemikalien .................................................................................................................. 22
1.2. Geräte und Hilfsmittel .................................................................................................. 24
1.3. Puffer und Lösungen .................................................................................................... 25
1.4. Gele ............................................................................................................................... 27
1.5. Antikörper ..................................................................................................................... 28
1.5.1. Antikörper für Immunhistochemie (IHC).................................................................. 28
1.5.2. Antikörper für Western Blot (WB) ............................................................................ 28
1.6. Primer für Genotypisierung .......................................................................................... 29
1.7. Mäuse ........................................................................................................................... 29
2. Methoden ..................................................................................................................... 30
2.1. Versuchstiere ................................................................................................................ 30
2.1.1. Behandlung mit BrdU................................................................................................ 30
2.1.2. Gewebeentnahme in vivo-Präparation und Materialgewinnung ............................... 30
2.2. Molekularbiologische Methoden .................................................................................. 31
2.2.1. DNA-Extraktion aus der Mausschwanzspitze ........................................................... 31
2.2.2. Polymerasekettenreaktion (polymerase chain reaction, PCR)................................... 31
2.2.3. Agarose-Gelelektrophorese ....................................................................................... 32
2.3. Detektion und Quantifizierung der Gen-Transkription ................................................ 33
2.3.1. Isolation von mRNA aus murinem Pankreasgewebe ................................................ 33
2.3.2. cDNA-Synthese ......................................................................................................... 33
2.3.3. Quantitative real-time PCR (qRT-PCR) .................................................................... 33
2.3.4. Reaktionsansatz und Reaktionsbedingungen ............................................................. 34
2.3.5. Quantifizierung der Genexpression ........................................................................... 35
2.4. Proteinchemische Methoden......................................................................................... 35
2.4.1. Proteinisolierung/Proteingewinnung aus Mauspankreasgewebe ............................... 35
2.4.2. Proteinkonzentrationsbestimmung nach Bradford .................................................... 35
2.4.3. Western Blot Analyse ................................................................................................ 36 2.5. Histochemische Methoden ........................................................................................... 38
2.5.1. Immunhistochemie .................................................................................................... 38
2.5.2. Hämatoxylin-Eosin-Färbung (H.E.-Färbung) ............................................................ 39
IV. ERGEBNISSE ............................................................................................................. 41

II. EINLEITUNG
2
1. Pankreasspezifische Inaktivierung von Socs3 in der Maus .................................... 41 1.1. Generierung einer pankreasspezifisch Socs3-defizienten Mauslinie Socs3
Δpanc unter Anwendung des Cre/loxP-Rekombinationssystems ..................................................... 41
1.2. Morphologische Charakterisierung des exokrinen und endokrinen Pankreaskompartiments von Soc3
Δpanc-Mäusen ........................................................... 44
2. Nachweis der Aktivierung des Stat3/Socs3-Signalweges im KrasG12D-Mausmodell
...................................................................................................................................... 46
2.1. Nachweis der pankreatischen Aktivierung von p-Stat3 im KrasG12D-Mausmodell ...... 46
2.2. Expression des Stat3-abhängigen endogenen Inhibitors Socs3 im KrasG12D-
Mausmodell .................................................................................................................. 47
2.3. Aktivierung der an der Jak2/Stat3-Signaltransduktion beteiligten Proteine in KrasG12D-
Mäusen ......................................................................................................................... 49
3. Homozygote Socs3-Deletion führt zu einer konstitutiven Aktivierung des Stat3-Signalweges im Kras
G12D-Mausmodell ...................................................................... 51
3.1. Generierung der pankreasspezifisch Socs3-defizienten Tumormodellmaus Kras
G12D;Socs3Δpanc ....................................................................................................... 51
3.2. Proteinbiochemische und morphologische Charakterisierung von KrasG12D
;Socs3Δpanc-
Mäusen ......................................................................................................................... 51
3.2.1. Proteinbiochemischer Nachweis verstärkter Stat3-Phosphorylierung und Expression Stat3-abhängiger Proteine ............................................................................................ 51
3.2.2. Konstitutive Aktivierung des onkogenen K-Ras-Proteins in KrasG12D
;Socs3Δpanc-
Mäusen ......................................................................................................................... 53
3.2.3. Gewichts- und Pankreasanalyse bei KrasG12D
;Socs3Δpanc-Mäusen ............................... 54
4. Einfluss der Inaktivierung des endogenen Inhibitorproteins Socs3 auf die PanIN-Progression im Kras
G12D-Mausmodell ....................................................................... 56
4.1. Beschleunigung der PanIN-Progression in KrasG12D
;Socs3Δpanc-Mäusen .................... 56
4.1.1. Morphologische und quantitative Charakterisierung der PanIN-Läsionen ............... 56
V. DISKUSSION ............................................................................................................. 66
1. Rolle von Socs3 für die pankreatische Karzinogenese im KrasG12D-Mausmodell . 66
2. Einfluss von Socs3 auf die Apoptose und Proliferation im KrasG12D-Mausmodell
...................................................................................................................................... 68 3. Entzündliche Prozesse fördern die Initiierung und Progression der
pankreatischen Vorläuferläsionen zu duktalem Pankreaskarzinom .................... 71
4. Rolle von Socs3 bei der Pankreasfibrosierung/Desmoplasie in KrasG12D-
Tumormäusen ............................................................................................................. 75
5. Die Bedeutung von Socs3 in Transdifferenzierungsprozessen im Pankreas der Kras
G12D-Tumormäuse ................................................................................................ 78
6. Ausblick ....................................................................................................................... 81
VI. ZUSAMMENFASSUNG ............................................................................................ 82
VII. LITERATURVERZEICHNIS .................................................................................. 85
VIII. ABBILDUNGSVERZEICHNIS ................................................................................ 99
IX. TABELLENVERZEICHNIS .................................................................................. 101
X. DANKSAGUNG ....................................................................................................... 102
XI. ERKLÄRUNG .......................................................................................................... 104

II. EINLEITUNG
3
I. ABKÜRZUNGEN
A Ampere
Abb. Abbildung
APS Ammoniumperoxiddisulfat
Beta-HCG Humanes Choriongonadotropin
Bcl-2 B-cell-lymphoma-2
Bcl-XL B-cell lymphoma-extra large
bp Basenpaare
BrdU 5-Brom-2-desoxyuridin
BSA Bovines Serumalbumin
c Konzentration
CA Carbohydrate Antigen
CaCl2 Kalziumchlorid
CEA Carcino-Embryonales Antigen
CK-19 Cytokeratin-19
Cox-2 Cyclooxygenase-2
Cre cyclization recombination
DAB Diaminobenzidin
dH2O einfach destilliertes Wasser
ddH2O zweifach destilliertes Wasser
DNA desoxyribonucleic acid (Desoxyribonukleinsäure)
DTT Dithiothreitol
EGF epidermal growth factor
EDTA Ethylendiamintetraacetat
F Flox
FAMMM familial atypical mole-malignant melanoma
GAP GTPase aktivierendes Protein
GDP Guanosindiphosphat
gp130 Glykoprotein 130
GTP Guanosintriphosphat
h Stunde(n)
HCL Hydrogenchlorid
H.E. Hämatoxylin-Eosin
HEPES 2-(4-(2-Hydroxyethyl)-1-piperazinyl)-
ethansulfonsäure
H2O2 Wasserstoffperoxid
IHC Immunhistochemie
IL Interleukin
Jak Januskinase
KH2PO4 Kaliumdihydrogenphosphat
KCl Kaliumchlorid
kD Kilodalton

II. EINLEITUNG
4
l Liter
loxP locus of x-over of P1
LSL lox-STOP-lox
M Molar
Mcl-1 myeloid leukemia cell differentiation protein 1
Min. Minute (n)
MAPK mitogen-activated protein kinase
Muc5 Mucin 5
NaCl Natriumchlorid
Na2HPO4 Natriumbicarbonat
NaOH Natriumhydroxid
p Phospho
PanIN pankreatische intraepitheliale Neoplasie
PBS phosphate-buffered saline
PCR polymerase chain reaction
(Polymerasekettenreaktion)
PDAC pancreatic ductal adenocacinoma
PFA Paraformaldehyd
PMSF Phenylmethylsulfonylfluorid
PSC pancreatic stellate cell
RNA ribonucleid acid (Ribonukleinsäure)
RNAse Ribinuklease
rpm rounds per minute (Umdrehungen pro Minute)
RT Raumtemperatur
RTK Rezeptortyrosinkinase
SDS-PAGE Sodium Dodecylsulfat-
Polyacrylamidgelelektrophorese
SH2 Src-homology 2
SOCS suppressor of cytokine signaling
STAT signal transducer and activator of transcription
(human)
Stat signal transducer and activator of transcription
(murin)
TAE Tris-Acetat-EDTA
TBS Tris-buffered saline
TEMED N,N,N´,N´-Tetramethylethylendiamin
TGFß transforming growth factor-beta
Tris Tris(hydroxymethyl)-aminomethan
UV Ultraviolett
V Volt
WB Western Blot

II. EINLEITUNG
5
II. EINLEITUNG
1. Das Pankreaskarzinom
1.1. Epidemiologie
Das Pankreaskarzinom umfasst eine heterogene Gruppe von pankreatischen Malignomen. Im
Jahr 2011 liegt die Anzahl der Neuerkrankungen in den USA bei 44.030, 37.660 Patienten
verstarben am Pankreaskarzinom. Seit 1998 wird eine stetige Zunahme der Inzidenz dieser
aggressiven Tumorerkrankung verzeichnet. Somit stellt das duktale Adenokarzinom des
Pankreas eine fast immer fatal verlaufende Erkrankung dar, die sowohl bei Männern als auch
bei Frauen an neunter Stelle der geschätzten malignen Neuerkrankungen und an vierter Stelle
der tumorbedingten Todesfälle steht (American Cancer Society 2011). Mit einer medianen
Überlebenszeit von etwa drei bis fünf Monaten und einer 5-Jahresüberlebensrate von weniger
als 5% hat das Pankreaskarzinom eine düstere Prognose (Hezel et al., 2006).
Das hochaggressive Ausbreitungsmuster des Pankreaskarzinoms in Form frühzeitiger
Metastasierung in lymphatische und periphere Organe trägt zusätzlich zur infausten Prognose
dieser Tumorerkrankung bei. In der Tat weisen etwa 80% der Patienten bei der
Diagnosestellung Fernmetastasen auf (Yeo et al., 2002b). Darüber hinaus ist die frühzeitige
Infiltration und Ausbreitung der Karzinomzellen entlang der intra- und extrapankreatischen
Nerven charakteristisch für das PDA (Pour et al., 1991; Okusaka et al., 2001; Hirai et al.,
2002). Aufgrund der weitgehenden Resistenz des fortgeschrittenen Karzinoms gegenüber
einem breiten Spektrum an konventioneller Chemo- und Radiotherapie bzw.
Radiochemotherapie (Lionetto et al., 1995), stellt die chirurgische Resektion die einzige
kurative Therapieoption dar, wobei weniger als 20% der Tumore überhaupt resezierbar sind
(Yeo et al., 1995; Rosewitz et al., 1997). Nur etwa 20% der chirurgisch therapierten Patienten
überleben die ersten drei Jahre nach der Operation (Shaib et al., 2007).
Das mittlere Erkrankungsalter bei der Diagnosestellung liegt bei 70 Jahren. Nur etwa 10% der
Patienten entwickeln diesen Tumor vor dem 50. Lebensjahr. Somit stellt dieses
Karzinomleiden eine Erkrankung des hohen Lebensalters dar. Neben dem Alter konnte das
Rauchen als führender vermeidbarer Risikofaktor in der Pankreaskarzinogenese deklariert
werden. 25% der Karzinome lassen sich auf den Zigarettenkonsum zurückführen. Als weitere
denkbare Risikofaktoren werden eine fett- und fleischreiche, ballaststoffarme Ernährung,
Übergewicht, eine chronische Pankreatitis und ein lange bestehender Diabetes Mellitus
beschrieben (Lowenfels et al., 2006). 5-10% aller Pankreastumore sind mit einer familiären
Prädisposition assoziiert und zeigen somit eine familiäre Häufung. Patienten mit positiver

II. EINLEITUNG
6
Familienanamnese haben gegenüber der Normalbevölkerung ein zweifach erhöhtes Risiko an
Bauchspeicheldrüsenkrebs zu erkranken (Schenk et al., 2001).
In den meisten Fällen gehäuften Auftretens maligner Neoplasien des Pankreas ist die
genetische Ursache nicht klar definiert. Bei den familiären Tumorprädispositionssyndromen,
die einen kleinen Teil der hereditären Karzinome ausmachen, sind jedoch autosomal-
dominant vererbte Keimbahnmutationen bestimmter Gene für die hohe Tumorpenetranz
verantwortlich (Lynch et al., 1996). Folgende Tumorprädispositionssyndrome sind mit dem
Pankreaskarzinom assoziiert (Hong et al., 2011):
- Das familiäre Mamma- und Ovarialkarzinom mit der BRCA2 (breast cancer gene 2)-
Mutation;
- das Peutz-Jeghers-Syndrom mit dem mutierten Tumorsuppressorgen STK11
(Serin/Threonin Kinase 11);
- das FAMMM (familial atypical mole-malignant melanome)-Syndrom, das mit einer
Mutation des Genlokus CDKN24 (cyclin-dependent kinase inhibitor 2A) einhergeht;
- die hereditäre Pankreatitis, bei der das kationische Trypsinogen-Gen PRSS1 (Protease,
Serin1) betroffen ist;
- darüber hinaus das hereditäre non-polypöse kolorektale Karzinom (HNPCC)-Syndrom
mit Keimzellmutationen der DNA Reparatur-Gene hMSH1, hMSH2, etc (Aarnio et al.,
1995)
- und die familiäre adenomatöse Polyposis (FAP) mit funktioneller Inaktivierung des
Genprodukts des APC-Gens (Su et al., 2000).
Untersuchungen haben allerdings gezeigt, dass die mit diesen Syndromen assoziierten
Genveränderungen besonders in fortgeschrittenen Tumorstadien zu finden sind, weniger in
prämalignen Läsionen. Infolge dessen geht man davon aus, dass diese Genmutationen eher
die Tumorprogression der Präkanzerose und weniger die Tumorinitiierung fördern (Sunil et
al., 2003; Hezel et al., 2006)
1.2. Molekularbiologische und –genetische Hintergründe
Der häufigste histologische Subtyp des Pankreaskarzinoms entwickelt sich in über 85% der
Fälle aus Pankreasgangzellen und wird als pankreatisches duktales Adenokarzinom (PDA)
bezeichnet. Etwa 15-20% der Neoplasien wie zystische Tumoren (seröses oligozystisches
Adenom, muzinöse nichtneoplastische Zyste), solid-pseudopapilläre Neoplasien,
Azinuszellzystadenome, Azinuszellkarzinome, Plattenepithelkarzinome und Lymphome
entwickeln sich aus anderen Zellen des exokrinen Pankreas. Endokrine Tumoren wie das
II. EINLEITUNG
7
Insulinom, Gastrinom und Glukagonom treten selten auf (Mulkeen et al., 2006).
In diesem Abschnitt der Arbeit soll besonders auf das PDA und dessen Vorläuferläsionen
eingegangen werden. Begriffe wie Pankreaskarzinom, Pankreastumor und duktales
Adenokarzinom werden in diesem Zusammenhang synonym verwendet.
In den letzten Jahren haben klinische, histopathologische und molekulargenetische
Untersuchungen drei wichtige Typen von Vorläuferläsionen identifiziert, die sich über einen
schrittweisen Transformationsprozess zum duktalen Adenokarzinom entwickeln können. Zu
diesen Vorstufen gehören zum einen die muzinös-zystischen Neoplasien (MCN), die
intraduktalen papillär-muzinösen Neoplasien (IPMN) und zum anderen die pankreatisch
intraepithelialen Neoplasien (PanINs) (Brugge et al., 2004; Maitra et al., 2005).
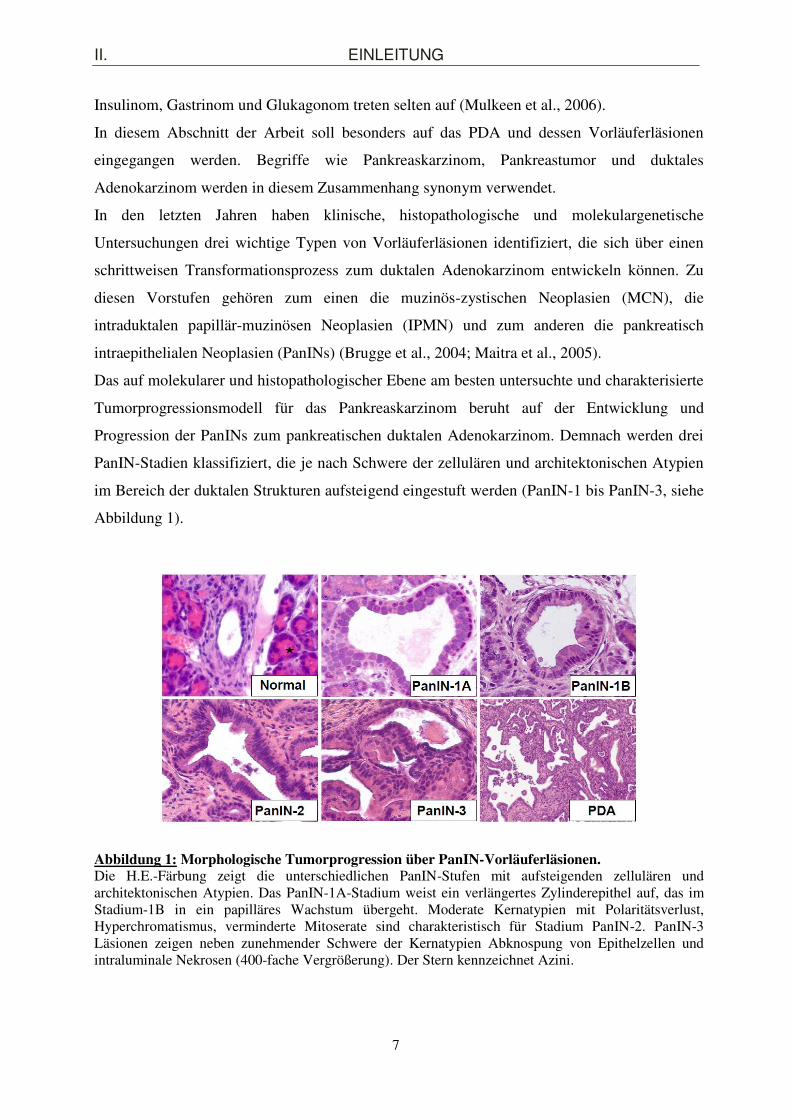
Das auf molekularer und histopathologischer Ebene am besten untersuchte und charakterisierte
Tumorprogressionsmodell für das Pankreaskarzinom beruht auf der Entwicklung und
Progression der PanINs zum pankreatischen duktalen Adenokarzinom. Demnach werden drei
PanIN-Stadien klassifiziert, die je nach Schwere der zellulären und architektonischen Atypien
im Bereich der duktalen Strukturen aufsteigend eingestuft werden (PanIN-1 bis PanIN-3, siehe
Abbildung 1).
Abbildung 1: Morphologische Tumorprogression über PanIN-Vorläuferläsionen. Die H.E.-Färbung zeigt die unterschiedlichen PanIN-Stufen mit aufsteigenden zellulären und architektonischen Atypien. Das PanIN-1A-Stadium weist ein verlängertes Zylinderepithel auf, das im Stadium-1B in ein papilläres Wachstum übergeht. Moderate Kernatypien mit Polaritätsverlust, Hyperchromatismus, verminderte Mitoserate sind charakteristisch für Stadium PanIN-2. PanIN-3 Läsionen zeigen neben zunehmender Schwere der Kernatypien Abknospung von Epithelzellen und intraluminale Nekrosen (400-fache Vergrößerung). Der Stern kennzeichnet Azini.

II. EINLEITUNG
8
PanIN-1A und PanIN-1B sind durch eine Zellkörperverlängerung und eine vermehrte
Schleimproduktion gekennzeichnet. Die im PanIN-1A-Stadium noch vorhandene flache
epitheliale Architektur des Pankreasganges geht im Stadium-1B in ein papilläres
Wachstumsmuster über. Erstaunlicherweise lassen sich diese frühesten Vorläuferformen in
40% aller adulten Pancreata nachweisen, ohne dass ein malignes Geschehen vorliegt. Das
PanIN-2-Stadium weist moderate Kernatypien in Form von Polaritätsverlust,
Hyperchromatismus, nukleärer Vergrößerung und verminderter Mitoserate auf, die im
Stadium-3 an Schwere zunehmen. Zusätzlich beobachtet man bei diesem terminalen
Läsionsgrad weit in das Ganglumen ragende epitheliale Zellknospen sowie luminale Nekrosen
als Zeichen von Zelluntergang und –abstoßung. PanIN-Läsionen der Stufe 3 findet man in der
Regel in weniger als 5% der gesunden Organe, während 30-50% aller invasiven Karzinome
diese typischen schweren Veränderungen aufzeigen.
Pankreasadenokarzinome weisen darüber hinaus ein an das Tumorgewebe angrenzendes
reaktives desmoplastisches Stroma auf. Dieses „Tumor-Microenvironment“ setzt sich aus einer
Vielzahl unterschiedlicher Zellen, extrazellulären Matrixproteinen und neugebildeten Gefäßen
zusammen, die das Wachstum und die Progression des PDAC fördern. Infiltrierende
inflammatorische Zellen und Makrophagen setzen Chemokine und Zytokine frei, die wiederum
eine reaktive Aktivierung von Fibroblasten und pankreatische Sternzellen (pancreatic stellate
cells, PSC) bewirken. Fibroblasten und aktivierte PSCs produzieren daraufhin Fibronektin und
Kollagen, was die Fibrosierung des Pankreasparenchyms verstärkt und das Tumorwachstum
begünstigt (Algül et al., 2007b).
Molekulare Untersuchungen haben gezeigt, dass die morphologischen Auffälligkeiten
einzelner PanIN-Progressionsstufen bis zum invasiven Karzinom bestimmte genetische
Veränderungen aufweisen, die entsprechend ihrer Anzahl und Schwere mit dem Grad der
Läsion korrelieren (siehe Abbildung 2). Wie bei anderen Tumorentitäten kommt in diesem
Zusammenhang Onkogenen und Tumorsuppressorgenen eine besondere Bedeutung zu
(Sakorafas et al., 2001).

II. EINLEITUNG
9
Abbildung 2: Morphologisches und genetisches Progressionsmodell des duktalen Pankreasadenokarzinoms (Hruban et al., 2000).
Die Kras-Punktmutation am Kodon 12 ist die wohl bedeutendste mit der Tumorgenese
assoziierte Mutation eines Onkogens. Initiale PanIN-Läsionen und Pankreasadenokarzinome
weisen eine Mutationsrate von bis zu 44% bzw. 100% auf (Shibata et al., 1990; Magee et al.,
2001). Das K-Ras-Protein stellt das Genprodukt von Kras dar und gehört zu den kleinen GTP-
bindenden Proteinen der Ras-Familie. Diese auch als G-Proteine genannten Moleküle sind an
der Innenseite der Plasmamembran lokalisiert und werden nach Bindung von Zytokinen,
Hormonen, Wachstumsfaktoren an die extrazelluläre Domäne der G-Protein-gekoppelten
Transmembranrezeptoren aktiviert. Hierbei kommt es zur Umwandlung von inaktivem, GDP-
gebundenen K-Ras in die aktive, GTP-assoziierte Form. Über nachgeschaltete intrazelluläre
Signaltransduktionswege wie zum Beispiel die Raf-mitogen-activated protein kinase (Raf-
MAP-Kinase)- oder die Phosphoinositol-3-Kinase-Signalkaskade wird der extrazelluläre
Stimulus in den Nucleus übertragen, wo eine Aktivierung weiterer Transkriptionsfaktoren
initiiert wird. Somit übernehmen die R-Proteine eine bedeutende Funktion in der
Zellproliferation, Differenzierung und Apoptose (Friday et al., 2008).
Bei Kras-Mutationen handelt es sich meist um eine Punktmutation eines einzelnen Nukleotids,
wodurch ein Aminosäureaustausch (Glycin wird durch Valin, Arginin oder Aspartat ersetzt)
stattfindet und infolge dessen ein Genprodukt mit Verlust der intrinsischen katalytischen
Eigenschaft entsteht. Aufgrund der Unfähigkeit zur Hydrolyse des gebundenen GTP bleibt das
„falsche“ G-Protein konstitutiv aktiv, sodass durch permanente Stimulierung der Downstream-
Kaskade kontinuierlich Signale an den Zellkern vermittelt werden, die zu dereguliertem
unkontrollierbaren Zellwachstum, Zellteilung und Differenzierung führen (Downward 2003).

II. EINLEITUNG
10
Molekulare Analysen haben eine signifikant verstärkte Amplifikation von Her2/neu mit
permanenter Aktivierung der entsprechenden Signaltransduktion und erhöhte EGF-
Konzentrationen beim PDA nachgewiesen. Bereits in frühen PanIN-Stadien ist die
Überexpression vorzufinden, sodass sie offenbar neben der Kras-Mutation zu den frühzeitig im
Ablauf der Tumorprogression auftretenden genetischen Veränderungen zählt (Lei et al., 1995;
Tsiambas et al., 2006; Hudis 2007).
Neben den Onkogenen werden die Tumorsuppressorgene als zweithäufigste mutierte
Genklasse im Pankreasadenokarzinom betrachtet. Mutationen an den jeweiligen Genloki
bewirken entweder eine Funktionsreduktion oder sogar einen vollständigen Funktionsverlust
der entsprechenden Genprodukte.
Zu den wichtigsten Vertretern der Tumorsuppressorgene gehören INK4A (inhibitor of cyclin
dependent kinase 4A) und ARF (Alternative Reading Frame), die sich beide auf der gleichen
Region 9p21 des Chromosoms 9 befinden. Das auch als CDKN2A (cyclin dependent kinase
inhibitor 2A) bezeichnete INK4A kodiert für das tumorsuppressive Protein p16, dessen
Inaktivierung in etwa 98% aller sporadischen Pankreaskarzinome auftritt (Schutte et al., 1997).
Über eine inhibitorische Bindung von p16 an die Cyklin-abhängigen Kinasen Cdk4 und Cdk6,
wodurch die funktionelle Phosphorylierung des Retinoblastom-Proteins verhindert wird,
kommt es zur Hemmung der Progression des Zellzyklus vor dem Eintritt in die S-Phase. Aus
der INK4A-Mutation resultiert konsequenterweise eine unkontrollierte Zellproliferation durch
Störungen in der Zellzyklusregulation. Der zweite vom gleichen Genlokus kodierte
Tumorsuppressor ist das Transkript der ARF-Gensequenz und wird als p19 bezeichnet. Die
Aufgabe dieses Proteins besteht in der Hemmung der MDM2-abhängigen Ubiquitinierung und
somit der Verhinderung des proteasomalen Abbaus von p53. Konsequenterweise führt die
Loss-of-function-Mutation des betroffenen Genabschnitts zur Reduktion des p19-Spiegels und
zum Abfall des p53-Levels (Wilentz et al., 1998; Lowe et al., 2003).
Das auf dem Chromosom 17 ansässige Tumorsuppressorgen TP53 kodiert einen weiteren für
die Kontrolle der Zellzyklus-Progression wichtigen „Wächter des Genoms“ p53. TP53 ist das
am häufigsten mutierte Gen in malignen Tumoren des Menschen. Beim Pankreaskarzinom
beträgt die Mutationsrate 50 bis 75%, wobei diese genetische Veränderung zumeist in
fortgeschrittenen PanIN-3-Läsionen vorzufinden ist (Talar-Wojnarowska et al., 2006). Das
Protein p53 stimuliert als Transkriptionsfaktor bei Replikationsstress mit Akkumulation von
gravierenden DNA-Schäden eine Reihe von Genen, die einen Zellzyklusarrest einleiten. Bei
Funktionsverlust des Proteins durch homozygote Mutation kommt es zum Ausfall der
tumorsuppressiven Eigenschaft des Genprodukts und das Wachstum von Zellen mit

II. EINLEITUNG
11
prokarzinogenen chromosomalen Aberrationen wird gefördert (Sherr 2004; Efeyan et al.,
2007).
Ein weiterer an der späteren Pankreastumorgenese beteiligter Tumorsuppressor ist SMAD4
bzw. DPC4 (deleted in pancreatic cancer locus 4), dessen Genprodukt SMAD4 bzw. DPC4
eine besondere Bedeutung in der Transforming Growth Faktor-ß (TGF-ß)-Signalkaskade
besitzt. Über dieses Messenger-Protein löst TGF-ß die Transkription spezieller Zielgene mit
antiproliferativer Wirkung aus. Infolgedessen kommt es bereits in der G1-Phase des Zellzyklus
zu einem Arrest, wodurch das Wachstum und die Differenzierung epithelialer Zellen zum
Stillstand kommen. Läsionen in diesem Genlokus führen zu unkontrollierter Proliferation
duktaler Zellen im Pankreas (Bardeesy et al. 2002; Miyaki et al., 2003). Ebenso sind
genetische Veränderungen des BRCA2-Gens in fortgeschrittenen Stadien der
Tumorprogression beschrieben worden. Eine Inaktivierung bedeutet Dysregulation der DNA-
Reparaturprozesse und daraus resultierende zelluläre Entartung (Hahn et al., 2003; Couch et
al., 2007).
2. Das murine Pankreaskarzinommodell
Trotz des zunehmenden Verständnisses der Signale und Mechanismen der Kanzerogenese
gehört das duktale Pankreasadenokarzinom zu den sehr schwer therapierbaren
Tumorerkrankungen. Deshalb hat sich die Pankreaskarzinom-Forschung zum Ziel gesetzt, die
humane Erkrankung und ihre Charakteristika in möglichst identischen präklinischen
Modellorganismen widerzuspiegeln und somit eine effizientere Therapie zu entwickeln.
Erstaunlicherweise lassen sich die molekularbiologischen und –genetischen Veränderungen
des humanen Pankreaskarzinoms und dessen Vorstufen in der Maus exakt nachbilden. Auf der
Grundlage dieser Erkenntnis wurden in den letzten Jahren einige Mausmodelle entwickelt, bei
denen mittels moderner molekularer Technik die Aktivierung von relevanten Onkogenen bzw.
das Ausschalten von Tumorsuppressorgenen ermöglicht wird (Hruban et al., 2006; Olive et al.,
2006). Zur Generierung genetisch gezielt modifizierter Mäuse bietet sich idealerweise das
Cre/loxP-Rekombinationssystem an, welches zur Etablierung des in dieser Arbeit verwendeten
KrasG12D-Mausmodells essentiell war. Demzufolge werden im Folgenden sowohl die Cre/loxP-
Technologie als auch das KrasG12D-Mausmodell ausführlich beschrieben.

II. EINLEITUNG
12
2.1. Das Cre/loxP-Rekombinationssystem
Die Rekombination ist definiert als ein durch spezifische Enzyme (Rekombinasen)
katalysierter Prozess der Spaltung und Neuverknüpfung der DNA. Auf diesem Prinzip
basierend ermöglicht die Cre/loxP-Technik eine gezielte Entfernung von DNA-Sequenzen in
relevanten Gewebe- oder Zellarten, ohne dass andere davon betroffen sind. Ein in der
molekulargenetischen Forschung häufig verwendetes Rekombinationsverfahren beruht auf der
Aktivität der Cre (cyclization recombination)-Rekombinase des Bakteriophagen P1. Dieses 38
kD schwere, in allen Organismen vorkommende Protein katalysiert die ortsspezifische
Rekombination zwischen zwei angrenzenden locus of x-over of P1 (loxP)-
Erkennungssequenzen. Das loxP-Motiv besteht aus einer 8 bp Spacer-Region, die wiederum
von zwei, jeweils 13 bp langen, invertierten Wiederholungen (inverted repeats) flankiert wird.
Die invers repetitiven Komponenten dienen der Erkennung und DNA-Bindung von Cre. Die
zwei eingebauten loxP-Sequenzen ermöglichen eine effiziente Exzision des loxP-flankierten
(„gefloxten“) DNA-Abschnitts. Das herausgeschnittene DNA-Segment wird als zirkuläres
Rekombinationsprodukt intrazellulär abgebaut. Lediglich verbleibt eine einzelne loxP-Sequenz
im modifizierten Genmaterial. Mittlerweile ist das Cre/loxP-Rekombinationssystem als eine
gängige molekulargenetische Methode zur Herstellung gewebespezifischer Knockout-Mäuse
etabliert. Zur Generierung dieser transgenen Tiere werden zwei genetisch veränderte
Mauslinien benötigt (siehe Abbildung 3).
Abbildung 3: Gewebespezifische Deletion von Mausgenen mittels Cre/loxP-Technologie. Der grüne Pfeil kennzeichnet den Vorgang der Transkription. In Zelltypen mit aktiver Cre-Rekombinase in der Cre/loxP-Maus ist die Genfunktion durch Exzision zerstört.

II. EINLEITUNG
13
Die Mauslinie 1 der F0-Generation trägt den gefloxten Genabschnitt. Hierzu werden die
gleichgerichteten loxP-Sequenzen vor und nach dem betroffenen Allel in die flankierenden
Intronbereiche integriert, somit bleibt das Zielexon funktionsfähig. Die Mauslinie 2 der
Parentalgeneration exprimiert die Cre-Rekombinase selektiv in bestimmten Geweben bzw.
Zelltypen. Zur Generierung der Cre-Maus wird ein Cre-Transgen in das Genom eingebracht.
Die Auswahl des vorgeschalteten Promotors, unter dessen Kontrolle das Cre-Transgen gestellt
wird, bestimmt über Zeitpunkt und Gewebespezifität der Cre-Expression. Daher wird die Cre-
Rekombinase nur von denjenigen Zellen gebildet, die auch über den entsprechenden Promotor
verfügen (Claudine 2004).
Nach Verpaarung der parentalen Cre-exprimierenden und gefloxten Mauslinien geht eine
Maus (Mauslinie 3) in der F1-Generation hervor, die beide genetische Veränderungen in ihrem
Erbgut trägt. Cre-Rekombinase exprimierende Zelltypen weisen eine Exzision des loxP-
markierten Zielexons und daraus resultierend eine Deletion des definierten Gens auf. Im
Gegensatz dazu bleibt in allen anderen Geweben ohne Cre-Transkription die entsprechende
Genfunktion unbeeinflusst. Somit lässt sich mithilfe der Cre/loxP-basierenden Methode ein
gewebespezifischer Gen-Knockout in der Mausgenetik etablieren.
Eine weitere Bedeutung gewinnt das Cre/loxP-System in der Induktion gewebespezifischer
Mutationen über die Verwendung loxP-flankierter STOP-Kassetten. Diese Art der
Genommodifikation wurde im KrasG12D-Mausmodell zur Herstellung onkogener Kras-
Mutationen angewendet und wird im folgenden Abschnitt detailliert beschrieben.
2.2. Das Kras
G12D-Mausmodell
Mit der Entwicklung des LSL-KrasG12D Mausmodells für das PDA gelang dem Forscherteam
unter David Tuveson und Tyler Jacks die Etablierung eines der bedeutendsten murinen
pankreatischen Karzinommodelle, welches auch in dieser Arbeit eine wesentliche Stellung
einnimmt. Grundidee dieses Tiermodells ist die vom endogenen Kras-Locus ausgehende
heterozygote pankreasspezifische Aktivierung des mutierten Kras(G12D)-Onkogens. Über die
Transfektion eines Plasmids in einen Mausstamm wurde das Kras-Allel so verändert, sodass
dieses ein modifiziertes Exon 1 mit einer Guanin-zu-Adenin-Transition auf dem Kodon 12
(G12D) und folglich einen Glycin-Aspartat-Aminosäureaustausch im Genprodukt aufweist
(Jackson et al., 2001). Unter Anwendung des bereits beschriebenen Cre/loxP-
Rekombinationssystems wird die gezielte Expression des veränderten Kras-Allels spezifisch in
Pankreaszellen erreicht. Hierzu wird dem das Kodon 12 enthaltende Exon 1 eine Lox-STOP-
Lox (LSL)-Sequenz vorgeschaltet (siehe Abbildung 4). Dieser LSL-Konstrukt verhindert die

II. EINLEITUNG
14
Transkription des veränderten Strukturgens und somit die Überexpression des onkogenen K-
Ras-Proteins in der LSL-KrasG12D-Mauslinie. Um eine Kras
G12D-Aktivierung selektiv im
Pankreas vorzunehmen, wird die LSL-KrasG12D-Maus mit einer Mauslinie gekreuzt, in der die
Cre-Rekombinase unter pankreassspezifischem Promotor Pdx-1 oder als Knock-in heterozygot
durch Entfernung des Exons 1 in einen Ptf1a-Lokus eingebaut exprimiert wird. Die daraus
resultierende Mauslinie bezeichnet man als Ptf1a-Creex1 bzw. Pdx1-Cre (Hingorani et al.,
2003a; Nakkai et al., 2007).
Pdx1 kodiert für den Transkriptionsfaktor Pankreatischer und duodenaler Homebox Faktor 1
(Pdx1), der auch als Insulin-Promotor-Faktor 1 bezeichnet und ab dem Entwicklungstag 8,5
produziert wird. Eine homozygote Deletion von Pdx1 endet letal in der Embryonalperiode. Das
von Ptf1a-Genbereich kodierte Ptf1a-Protein, welches auch als p48 genannt wird, stellt die
Untereinheit des heterotrimerischen Pankreastranskriptionsfaktorkomplex 1 dar (Rose et al.,
2001). P48 wird ab Tag 9,5 der Embryonalentwicklung exprimiert und ist gemeinsam mit Pdx1
für die pankreasspezifische Differenzierung der Zellen verantwortlich. Das Vorhandensein
beider Faktoren ist essenziell für die morphologische und funktionelle Entwicklung der
Vorläuferzellen zu Pankreasgewebszellen, wobei erwachsene Mäuse Pdx1 zur Bildung der
Inselzellen und p48 zur Initiierung der Azinuszelldifferenzierung benötigen (Rose et al., 2001;
Beres et al., 2006). Demzufolge führt die vollständige Deletion der p48-Gene zu einer
Agenesie des exokrinen Pankreas und konsequenterweise zum Tod der Mäuse früh postnatal
(Krapp et al., 1998; Sellik et al., 2004).
Nach Kreuzung der transgenen Mauslinien LSL-KrasG12D und Ptf1-Cre
ex1 bzw. Pdx1-Cre geht
nun eine Mausmutante LSL-Kras+/G12D;Ptf1a-Cre
ex1 bzw. LSL-Kras+/G12D ;Pdx1-Cre hervor,
die eine Heterozygotie für das KrasG12D-Allel aufweist. Aufgrund der Ptf1a- bzw. Pdx1-Cre-
Rekombinase assoziierten Exzision der loxP-flankierten STOP-Sequenz in diesem auch als
KrasG12D bezeichneten Tumormausmodell entfällt die transkriptionsinhierende Funktion der
STOP-Kassette, was wiederum zur deregulierten hochfrequenten Transkription des mutierten
Exon 1 führt (siehe Abbildung 4).

II. EINLEITUNG
15
Abbildung 4: Systematik der Generierung des Kras
G12D-Tumormausmodells. Grüne Pfeile beschreiben die Transkription der Strukturgene. Roter Balken kennzeichnet die transkriptionsinhibierende Wirkung der STOP-Kassette (modifiziert nach Hingorani et al 2003).
Mithilfe der LSL-KrasG12D Mauslinie ist die selektive endogene Expression des onkogenen K-
Ras-Proteins in Ptf1a bzw. Pdx1 positiven und Cre-Rekombinase positiven
Pankreasgewebszellen möglich. Somit kann die Tumorentwicklung pankreasspezifisch initiiert
und die Entstehung des humanen Pankreaskarzinoms exakt in der Maus rekapituliert werden.
Die genetisch veränderten KrasG12D-Mäuse zeigen daraufhin alle Stadien der PanIN-Läsionen,
die analog zum humanen PDA in ihrer Anzahl und Schwere mit dem Alter der Mäuse
zunehmen. In 10% der Fälle und nach einer Latenz von etwa neun Monaten entwickeln sich
auch invasive und metastasierende Karzinome. Der Nachweis der Kras-Mutation bei allen
PanIN-Stufen in den Progenitorzellen bestärkt dessen Bedeutung in der Induktion und
Progression der Pankreaskarzinogenese. Bei LSL-KrasG12D;Pdx1-Cre-Mäusen treten zusätzlich
andere Tumoren mit variierender Penetranz auf, wie die intestinale Hyperplasie des
Magenepithels, monokutane Papillome und hyperplastische Polypen des Duodenums. Die
simultane Existenz dieser Malignome beweist die embryonale Expression von Pdx1 in
endothelialen Vorläuferzellen (Rose et al., 2001). Ausgehend von diesem etablierten KrasG12D-
Tumormausmodell sind verschiedene komplexe Tiermodellsysteme entwickelt worden, die
additiv zu dem mutierten Kras-Allel andere genetische Veränderungen aufzeigen. Damit lässt
sich der Einfluss einzelner Genmutationen auf die Karzinomentwicklung und –progression
genauer analysieren und mögliche Ansatzpunkte für ein gezieltes medizinisches Eingreifen
finden.

II. EINLEITUNG
16
3. Suppressors of cytokine signaling (SOCS)
Die Mitglieder der SOCS-Familie stellen wichtige Feedback-Inhibitoren des Zytokin-
Signaltransduktionswegs dar. Mittlerweile sind acht SOCS-Proteine (CIS, Socs1-Socs7)
beschrieben worden, die auf ein Zytokinsignal gewebeabhängig exprimiert werden. Von diesen
auch als STAT-induzierte STAT-Inhibitoren (SSI) bezeichneten negativen Regulatoren der
Zytokin-Signalkaskade sind die Proteine Socs1, Socs2 und Socs3 besonders gut charakterisiert
und nehmen eine wichtige Stellung in der Interleukin-6 (IL-6) Signalkaskade ein. Im
Gegensatz dazu sind Socs4 bis Socs7 bislang noch wenig erforscht (Krebs et al., 2000).
Aufgrund der tragenden Rolle von Socs3 in dieser Arbeit, soll im Folgenden besonders auf
diesen inhibitorischen Regulationsfaktor eingegangen werden.
3.1. Die Struktur von Socs3
Socs3 stellt ein aus 225 Aminosäuren (AS) bestehendes Molekül dar, das aus unterschiedlichen
Domänen zusammengesetzt ist (vergleiche Abbildung 5). Die einzelnen Abschnitte erfüllen im
Rahmen der Signaltransduktion bestimmte Funktionen, die in ihrer Gesamtheit für den
negativen Feedback-Mechanismus des Socs3-Proteins von großer Bedeutung sind. Die N-
terminale Kinase-Inhibitor-Region (KIR) bindet als Pseudosubstrat an das aktive Zentrum von
Janus-Kinasen (Jak), wodurch das Andocken nativer Substrate blockiert wird. Eine zentrale
Stellung nimmt die SH2 (Scr-homology-2)-Domäne im Socs3-Molekül ein. Diese bildet die
eigentliche Kinase-Bindungsstelle, die eine hohe Affinität zu phosphorylierten Tyrosinresten
im Aktivierungsloop der Jaks aufweist. Unterstützt wird die SH2-Sequenz von den beiden
flankierenden N-ESS (extended SH2 subdomain) und C-ESS-Domänen, die strukturell die
Assoziation und die Affinität von SH2 an die Tyrosinmotive erhöht. Am C-terminalen Ende
der SH2-Domäne liegt eine PEST-Insertion zwischen zwei konservierten Sekundär-
Strukturelementen. Untersuchungen haben gezeigt, dass dieses PEST-Motiv als wichtiger
Regulator der Proteinstabilität fungiert und seine Entfernung weder zu einer
Strukturveränderung führt noch die Funktion von Socs3 beeinträchtigt. Jedoch zeigen PEST-
Knockout Mäuse eine Zunahme der Socs3-Halbwertszeit durch Beeinflussung des Proteasom-
vermittelten Abbaumechanismus (Babon et al., 2006). Die für die hemmende Wirkung von
Socs3 ebenfalls wichtige SOCS-Box-Region ist C-terminal lokalisiert (siehe Abbildung 5).

II. EINLEITUNG
17
Abbildung 5: Schematische Darstellung der Socs3-Architektur. Die Socs3-Struktur besteht aus einem kleinen N-Terminus, einer Kinase-Inhibitor-Region (KIR), einer SH2-Domäne und einer C-terminalen SOCS-Box. Die zentrale SH2-Domäne setzt sich aus 141 Aminosäuren (AS) zusammen und wird durch ein aus 35 AS bestehendes PEST-Motiv unterbrochen. N-terminal bzw. C-terminal wird die SH2-Domäne durch eine N-ESS- bzw. C-ESS-Sequenz flankiert (modifiziert nach Babon et al. 2006).
Durch Rekrutierung von Elongin B, Elongin C, Cullin 2 und das Ringfinger-Protein Rbx1 an
der SOCS-Box bildet sich ein E3-Ubiquitin-Ligase-Komplex (siehe Abbildung 6), in dem
Socs3 als Adaptormolekül die Ubiquitination und Degradation von bestimmten an der
Signaltransduktion beteiligten Proteinen fördert (Kamura et al., 1998; Kamura et al. 2004).
Abbildung 6: Socs3 als Trägermolekül des E3-Ubiquitin-Ligase-Komplexes (modifiziert nach Krebs et al. 2000).
3.2. Socs3 als Zytokin-Signaltransduktionsinhibitor
Die intrazelluläre Socs3-Proteinkonzentration ist unter normalen Umständen relativ gering.
Für die Aktivierung des auf Chromosom 11 liegenden Socs3-Gens (entspricht dem humanen
Socs3-Gen auf Chromosom 17) ist die Interaktion von Zytokinen besonders der Interleukin
(IL)-6-Familie, Wachstumsfaktoren wie den Insulin-like growth factor oder Hormonen mit
den spezifischen Zelloberflächenrezeptoren essentiell. Innerhalb der Untergruppen zeigen die
Zytokine ähnliche Molekül- und Rezeptorstrukturen. Stellvertretend soll an dieser Stelle der
IL-6-Rezeptor und dessen anschließende Signaltransduktionswege näher erläutert werden.
Der IL-6-Rezeptor verfügt über zwei Untereinheiten. Durch Bindung an das extrazelluläre
Zytokin-bindende Modul, das auch als Glykoprotein (gp)180 bekannt ist, übt IL-6 seine
biologische Aktivität aus. Intrazellulär bilden die ubiquitär exprimierten gp130-
Rezeptoruntereinheiten die signalvermittelte Struktur durch Aktivierung der Signalkaskade

II. EINLEITUNG
18
und stellen gleichzeitig Ziel verschiedener Rückkopplungsmechanismen dar. Weitere
Charakteristika des IL-6-Rezeptors sind die fehlende intrinsische katalytische
Tyrosinkinaseaktivität und die Verankerung in der Plasmamembran über eine singuläre
Transmembrandomäne. Bei Interaktion des Liganden IL-6 mit der extrazellulären
Rezeptorkomponente kommt es zunächst zur Aktivierung rezeptorassoziierter, intrazellulärer
Janus-Kinasen (Jak) durch Autophosphorylierung. Daraufhin phosphorylieren die aktivierten
Jaks die zytosolischen Zytokinrezeptoranteile am Tyrosinrest 759, der als Bindungsstelle
sowohl für diverse Transkriptionsfaktoren wie Stat3 als auch Inhibitoren wie Socs3-Moleküle
fungiert. Die Rezeptor-Ligand-Interaktion setzt somit den Jak/Stat-Signalweg ebenso wie die
PI3K- und die Ras/Raf/MAPK-Signalkaskade in Gang.
Die als Monomere im Zytoplasma lokalisierten Stat3-Moleküle binden an die gp130-
Untereinheit und werden anschließend Jak-vermittelt am Tyrosin 705 phosphoryliert. Die
aktivierten phospho(p)-Stat3-Elemente lagern sich dann zu Homo-oder Heterodimeren
zusammen, die in den Zellkern translozieren und dort an Enhancer-Sequenzen in Promotoren
von IL-6-Zielgenen binden und folglich deren Transkriptionsfrequenz positiv modulieren
(siehe Abbildung 7). In vielen malignen Tumoren kommt es durch eine Hyperaktivität der
Rezeptor-Tyrosinkinasen zu einer konstitutiven Phosphorylierung und Aktivierung von Stat3
und folglich zu einer Überexpression der an verschiedenen physiologischen Prozessen wie
Proliferation, Apoptose, Angiogense, Inflammation beteiligten Zielgene. Dies führt zur
Dysregulation des Zellzyklus und der Zellfunktion, was wiederum das Wachstum und
Überleben transformierter Zellen begünstigt und die Karzinomentwicklung fördert.
Abbildung 7: Die Mechanismen der Aktivierung der IL-6/Jak/Stat3-Signalkaskade und dessen Feedback-Inhibition durch Socs3.

II. EINLEITUNG
19
Die Aktivierung der Ras/Raf/MAPK-Kaskade setzt die Bildung eines Molekülkomplexes
bestehend aus SHP2 (SH2-Domäne enthaltende Phosphatase 2), Grb2 (growth factor receptor
bound protein 2), SOS (son of sevenless) und Gab1 (GRB2-associated binding protein 1)
voraus. Diese Proteineinheit bindet über SHP2 an die phosphorylierte Y759-Domäne und
vermittelt die Umwandlung des GDP-Ras in die energiereiche GTP-gebundene Form. Im
weiteren Verlauf kommt es nacheinander zur Aktivierung von Raf (rat fibrosarcoma), MEK
(Mitogen-activated protein kinase kinase) und MAPK (Mitogen-activated protein kinase),
welche anschließend intranukleär Transkriptionsfaktoren phosphoryliert und auf diese Weise
die Transkription von verschiedenen Ras-Zielgenen initiiert. Die Genprodukte beeinflussen
Wachstum, Proliferation, Zelldifferenzierung und Apoptose (Lim et al., 2005 Friday et al.,
2008).
Abbildung 8: Die Aktivierung der Ras/Raf/MAPK- und PI3K/AKT–Signalkaskade durch IL-6 und dessen negative Regulation durch Socs3.
Ein weiterer mit der IL-6-Signaltransduktion assoziierter Signalweg verläuft über die
Phosphatidylinositol-3-Kinase (PI3K). Obwohl diesem eine große Bedeutung in der
Regulation mannigfaltiger Zellprozesse zugesprochen wird, ist die Komplexität der
Signalkaskade noch nicht vollständig verstanden. Nach Aktivierung durch Bindung an die
pY759-Einheit oder durch das aktive G-Protein Ras ist PI3K in der Lage, bestimmte
Membranlipide wie PIP2 (Phosphatidylinositol-4,5-Bisphosphat) in PIP3
(Phosphatidylinositol-3,4,5-Trisphosphat) zu phosphorylieren. Als second-messenger
rekrutiert PIP3 die Proteinkinase B (PKB; AKT) an die Plasmamembran, wo die
Phosphorylierung und Aktivierung dieses Enzyms stattfindet. In ihrer aktiven Form
beeinflusst AKT eine Reihe von zellulären Prozessen wie die Hemmung der Apoptose durch
Inhibition pro-apoptotischer Proteine der Bcl-Familie, die Stimulation der Proteintranslation

II. EINLEITUNG
20
an den Ribosomen durch Aktivierung von mTOR (mammalian Target of Rapamycin) und die
Förderung der Proliferation durch Inaktivierung des proliferationshemmenden Forkhead-Box-
Protein O3 (FOXO), welches die Zellteilung und Zellwachstum supprimiert. Alle genannten
Abläufe fördern letztendlich bei konstitutiver Aktivierung der Signalkaskade die
Tumorinitiierung und –progression (Manning et al. 2007; Assinder et al., 2009). Als
Antagonist von PI3K wird die Phosphatase PTEN (Phosphatase and Tensin homolog)
beschrieben, die PIP3 zu PIP2 dephosphoryliert und somit folglich die Aktivierung von AKT
verhindert. Eine Loss-of-function Mutation von PTEN resultiert daher in einer anhaltenden
Funktion der Proteinkinase B und führt letztlich zu unkontrolliertem Wachstum und
Proliferation der Zelle (Dubrovska et al., 2008).
Um einer Dauerstimulation entgegen zu wirken wird die IL-6-Wirkung über verschiedene
Mechanismen reguliert. In diesem Zusammenhang kommt insbesondere der Feedback-
Inhibition durch Socs3-Proteine als negative Rückkopplung eine große Bedeutung zu. Nach
Zytokin- bzw. Stat-induzierter Expression von Socs3 (Lang et al., 2003), bindet dieses
Genprodukt mit der SH2-Domäne direkt an das Phosphotyrosin 759 der zytoplasmatischen
gp130-Domäne und verhindert dadurch die Rekrutierung und Aktivierung nativer an der
Signaltransduktion beteiligter Substrate wie Stat3 (vergleiche Abbildung 7). Die Affinität zu
Jaks ist jedoch im Gegensatz zu Socs1 relativ gering, sodass höhere Konzentrationen für die
gleiche Hemmwirkung benötigt werden (Babon et al., 2006). Darüber hinaus fördert Socs3
den proteasomalen Abbau der Signal-Proteine durch Bildung des E3-Ubiquitin-Ligase-
Komplexes. Es kommt schließlich zur Terminierung der IL-6-Signaltransduktion. Die starke
Induktion des endogenen Inhibitors legt eine regulierende Funktion des Socs3 in der
pankreatischen Onkogenese nahe. Das Fehlen von Socs3-Molekülen durch homozygote
Deletion des entsprechenden Gens resultiert in konstitutiver IL-6-Signalvermittlung und führt
infolgedessen zu ungehindertem Wachstum und Entwicklung maligner Prozesse.

II. EINLEITUNG
21
4. Zielsetzung der Arbeit
Das duktale Adenokarzinom des Pankreas ist eine hochmaligne Tumorerkrankung des
Menschen mit einer Mortalitätsrate von 100% und einer frustranen Prognose (Lowensfels et
al., 2006). Trotz der zunehmenden Kenntnisse aus intensiven Forschungsuntersuchungen über
die histopathologischen und molekulargenetischen Ursachen stellt die Prognose eines
Pankreaskarzinoms für den heutigen Stand der Therapie immer noch eine große
Herausforderung dar. Mithilfe neu entwickelter Tumormausmodelle versucht man die
Mechanismen der Tumorentwicklung und- progression nachzuvollziehen und somit
Rückschlüsse auf das humane Pankreaskarzinom zu ziehen.
Die KrasG12D-Mutation spielt in der Tumorinitiierung und Tumorprogression eine tragende
Rolle. Untersuchungen haben diese genetische Veränderung sowohl in den PanIN-
Vorläuferläsionen als auch in fortgeschrittenen metastasierten und invasiven Tumorstadien
nachgewiesen. Andere aktivierende und hemmende molekulare Mechanismen im Rahmen der
PanIN-Progression zu invasiven duktalen Adenokarzinomen sind bisher weitgehend
unbekannt und sind Gegenstand gegenwärtiger Forschungsarbeiten.
Der suppressor of cytokine signaling 3 (Socs3) ist als negativer Regulator zahlreicher
Signaltransduktionswege besonders der IL-6/Jak/Stat3-Signalkaskade beschrieben worden
(Lang et al. 2003). Obwohl für Socs3 eine zunehmende Bedeutung in der Karzinogenese
postuliert wird (Ogata et al., 2006), ist dessen Funktion und Wirkungsweise in der
pankreatischen Onkogenese bisher nur unzureichend untersucht.
Die vorliegende Arbeit beschäftigt sich mit der Rolle bzw. dem Einfluss von Socs3 auf die
durch die KrasG12D-Mutation induzierte murine Pankreaskarzinogenese. Im Rahmen der
durchgeführten Untersuchungen werden dabei folgende wichtige Fragestellungen formuliert:
- Lässt sich in PanIN-Läsionen von LSL-KrasG12D;Ptf1a-Cre
ex1 (Kras
G12D)-Mäusen eine
verstärkte Expression und folglich eine erhöhte Konzentration von Socs3 nachweisen?
- Welche Konsequenz hat dann eine homozygote Deletion des Socs3-Gens mit
konsekutiver Socs3-Inaktivierung für die Entwicklung und Progression von PanINs in
KrasG12D-Mäusen?
- Was bedeutet dieser Genverlust für das Wachstum und Entwicklung der genetisch
veränderten Tiere?

III. MATERIAL UND METHODEN
22
III. MATERIAL UND METHODEN
1. Material
1.1. Chemikalien
Chemikalie Name und Firma
ABC-Lösung ABC Elite Kit Vector, Burlingame, USA
Acrylamid Rotiphorese Gel 30 (Roth,Karlsruhe)
Antigen Unmasking Solution H-330 Vector Labs, Burlingame, USA
APS Sigma, Steinheim
Avidin Avidin/Biotin Blocking, Vector Labs, Burlingame, USA
Biorad Protein Assay Biorad, München
Biotin Avidin/Biotin Blocking, Vector Labs, Burlingame, USA
Bradford (5x) Protein Assay Dye Reagent Concentrate # 500-0006, Bio-Rad
Bromphenolblau Bromphenol blue sodium salt (Sigma, Steinheim)
BrdU Sigma, Steinheim
BSA Cohn V fraction (Sigma, Steinheim)
ddH2O Aqua ad injectabilia (Diaco,Naila)
dH2O Aqua Delta Select Spüllösung (Delta Select, München)
DAB DAB Substrate kit for Peroxidase, Vector Laboratories
DirectPCR® Lysis Reagent Peqlab, Erlangen
DMEM Gibco
DNA-Größenstandard DNA-Leiter-Mix (Peqlab, Erlangen)
ECL Amersham ECL Western Blotting Detection Reagents (GE Healthcare, München)
EDTA Sigma, Steinheim
Eindeckmedium (Mounting Medium) Pertex, Medite
Eosin Croma, Münster
Ethanol Apotheke MRI
Ethidiumbromid Sigma, Steinheim
Goat Serum Sigma, Steinheim
Hämalaun Merck, Darmstadt
Hämatoxylin Merck, Darmstadt
HCl Merck, Darmstadt

III. MATERIAL UND METHODEN
23
HEPES Sigma, Steinheim
Histoclear Roti-Histol (Roth, Karlsruhe)
30%H2O2 Merck, Darmstadt
Isofluran Forene (Abbott, Wiesbaden)
LE Agarose Biozym Scientific, Oldendorf
β-Mercaptoethanol Sigma, Steinheim
Magermilchpulver Sigma, Steinheim
Methanol Roth, Karlsruhe
NP-40 Roche, Mannheim
PBS-Pulver Instamed PBS Dulbecco (Biochrom AG, Berlin),
Pertex-Eindeckmedium Medite, Burgdorf
PFA Apotheke MRI
Phosphatase Inhibitor Cocktail I Sigma, Steinheim
Phosphatase Inhibitor Cocktail II Sigma, Steinheim
Protease-Inhibitor Cocktail Sigma, Steinheim
Proteinase K Roche, Mannheim
Protein-Größenstandard Precision plus protein™ standard (Biorad, München)
Rabbit serum Sigma, Steinheim
Ras Activation Assay Kit Biomal, Hamburg
RedTaq ReadyMix Sigma, Steinheim
RNAseZap Sigma, Steinheim
SDS SDS Ultra pure (Roth, Karlsruhe)
Streptavidin/Biotin block Vector Laboratories, Orton Southgate
TEMED Sigma, Steinheim
Tris-HCL Roth, Karlsruhe
Tris-Base Roth, Karlsruhe
Tween-20 Roth, Karlsruhe
Tabelle 1: Chemikalien.

III. MATERIAL UND METHODEN
24
1.2. Geräte und Hilfsmittel
Geräte Name und Firma Agarosegel-Bildwandler Molecular Imager Gel Doc XR System (Bio-
rad, München) Agarosegelkammer Sub-Cell GT (Biorad, München)
Automatischer Filmprozessor Amersham Hyperprocessor (GE Healthcare)
Computer Hardware Apple
Deckgläser #1 H868, 24x60 mm
Elektrophoreseapparatur Mini-PROTEAN Tetracell (Biorad, München)
Entwicklermaschine Amersham Hyperprocessor (GE, Healthcare, München)
Eppis Eppendorfer
Falcons Falcon®, BD, Franklin Lakes, USA
Feinwaage Analytic Sartorius, Göttingen
Filmkassette BAS Cassette 2025 (Fujifilm, Kleve)
Heizplatte Thermomixer compact (Eppendorf, Hamburg)
Histologiekassetten Bio-Cassettes 07-7130, Bio-Optica, Mailand,lt.
Homogenisator Diax 900 (Heidolph, Schwabach)
Hyperfilm Amersham Hyperfilm ECL (GE Healthcare, München)
Kamera ApoTome, Zeiss, Oberkochen
Magnetrührer MR 3001 (Heidolph, Schwabach)
Microtome HM 355, Microm GmbH
Mikroskope Axio Imager A1 (Zeiss, München) Axiover 40 CFL (Zeiss, München) MZ75 (Leica, Wetzlar)
Mikrowelle Siemens, München
Neubauer®-Zählkammer Marienfeld
Objektträger Superfrost®Plus, Menzel-Gläser, Braunschweig Thermo Scientific, Schwerte
Parafilm Parafilm®PM-992, pechiney plastic packaging, Brampton, USA
pH-Meter Inolab pH 720 (WTW, Weilheim)
Pipetten Eppendorf Research, Hamburg
Pipettenspitzen SafeSeal-Tips® Professional, (Biozym Oldendorf)
Power supply PowerPac basic, Bio-Rad

III. MATERIAL UND METHODEN
25
Scanner Canon
Schwenktisch L40 (Labinco, Breda, Niederlande)
Slides-Folien Thermo scientific
Spektrophotometer 2001 (Anthos, Krefeld)
Stromquelle Power Pac Basic (Biorad, München)
Thermoblock/Schüttler Thermomixer compact (Eppendorf, Hamburg)
Thermocycler Mastercycler (Eppendorf, Hamburg)
Transferapparat Mini Trans-Blot Cellfer (Biorad, München)
Transfermembran Immobilon-P Transfer Membrane (Millipore, Schwalbach)
Vortexer Reax top (Heidolph, Schwabach)
Waage BP 2100 S (Sartorius Göttingen)
Well-Platte Microtest TM 96 Zellkulturplatte (Becton Dickinson, Heidelberg)
Whatman-Papier Biometra, Göttingen
WinRead Anthos
Zentrifuge Centrifuge 5415R (Eppendorf, Hamburg)
Tabelle 2: Geräte und Hilfsmittel.
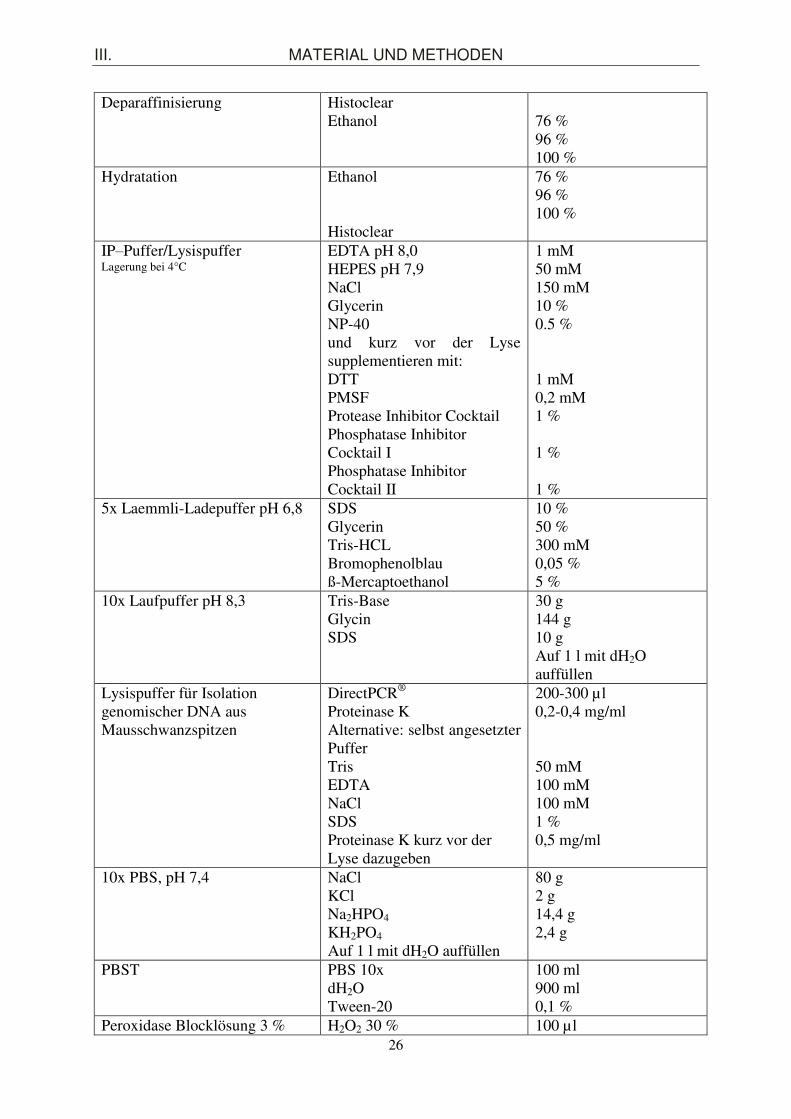
1.3. Puffer und Lösungen
Puffer/Lösung Zusammensetzung Menge ABC-Lösung Pufferlösung
Reagenz A
Reagenz B
2,5 ml 1 Tropfen
1 Tropfen
Antigendemaskierungslösung pH 8,0
1 mM EDTA oder Citratpuffer
Blocklösung (5 %) für IHC Serum Avidin Pufferlösung (PBS/PBS-T, TBS/TBS-T)
50 µl 4 Tropfen 945 µl
Blocking-Puffer (5 %) für WB Skim milk TBS/T oder PBS/T
10 g 200 ml 200 ml
BrdU Stock-Lösung 50 µg/µl BrdU dH2O
500 ml 10 ml mit NaOH alkalisieren bis BrdU vollständig aufgelöst ist, steril filtrieren und Stocklösung bei -20°C aufbewahren
DAB-Lösung dH2O Buffer Stock-Lösung DAB-Lösung H2O2-Lösung
2,5 ml 1 Tropfen 1 Tropfen 1 Tropfen
III. MATERIAL UND METHODEN
26
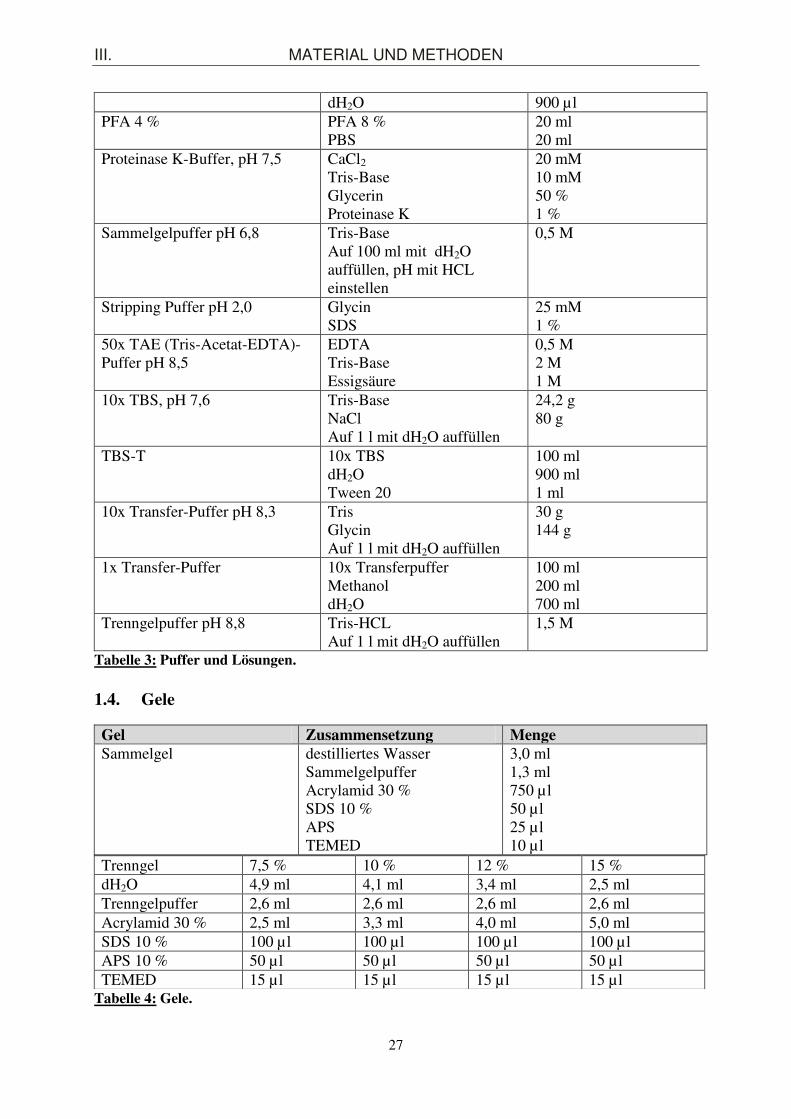
Deparaffinisierung Histoclear Ethanol
76 % 96 % 100 %
Hydratation Ethanol Histoclear
76 % 96 % 100 %
IP–Puffer/Lysispuffer Lagerung bei 4°C
EDTA pH 8,0 HEPES pH 7,9 NaCl Glycerin NP-40 und kurz vor der Lyse supplementieren mit: DTT PMSF Protease Inhibitor Cocktail Phosphatase Inhibitor Cocktail I Phosphatase Inhibitor Cocktail II
1 mM 50 mM 150 mM 10 % 0.5 % 1 mM 0,2 mM 1 % 1 % 1 %
5x Laemmli-Ladepuffer pH 6,8 SDS Glycerin Tris-HCL Bromophenolblau ß-Mercaptoethanol
10 % 50 % 300 mM 0,05 % 5 %
10x Laufpuffer pH 8,3 Tris-Base Glycin SDS
30 g 144 g 10 g Auf 1 l mit dH2O auffüllen
Lysispuffer für Isolation genomischer DNA aus Mausschwanzspitzen
DirectPCR®
Proteinase K Alternative: selbst angesetzter Puffer Tris EDTA NaCl SDS Proteinase K kurz vor der Lyse dazugeben
200-300 µl 0,2-0,4 mg/ml 50 mM 100 mM 100 mM 1 % 0,5 mg/ml
10x PBS, pH 7,4 NaCl KCl Na2HPO4
KH2PO4
Auf 1 l mit dH2O auffüllen
80 g 2 g 14,4 g 2,4 g
PBST PBS 10x dH2O Tween-20
100 ml 900 ml 0,1 %
Peroxidase Blocklösung 3 % H2O2 30 % 100 µl
III. MATERIAL UND METHODEN
27
dH2O 900 µl PFA 4 % PFA 8 %
PBS 20 ml 20 ml
Proteinase K-Buffer, pH 7,5 CaCl2
Tris-Base Glycerin Proteinase K
20 mM 10 mM 50 % 1 %
Sammelgelpuffer pH 6,8 Tris-Base Auf 100 ml mit dH2O auffüllen, pH mit HCL einstellen
0,5 M
Stripping Puffer pH 2,0 Glycin SDS
25 mM 1 %
50x TAE (Tris-Acetat-EDTA)-Puffer pH 8,5
EDTA Tris-Base Essigsäure
0,5 M 2 M 1 M
10x TBS, pH 7,6 Tris-Base NaCl Auf 1 l mit dH2O auffüllen
24,2 g 80 g
TBS-T 10x TBS dH2O Tween 20
100 ml 900 ml 1 ml
10x Transfer-Puffer pH 8,3 Tris Glycin Auf 1 l mit dH2O auffüllen
30 g 144 g
1x Transfer-Puffer 10x Transferpuffer Methanol dH2O
100 ml 200 ml 700 ml
Trenngelpuffer pH 8,8 Tris-HCL Auf 1 l mit dH2O auffüllen
1,5 M
Tabelle 3: Puffer und Lösungen.
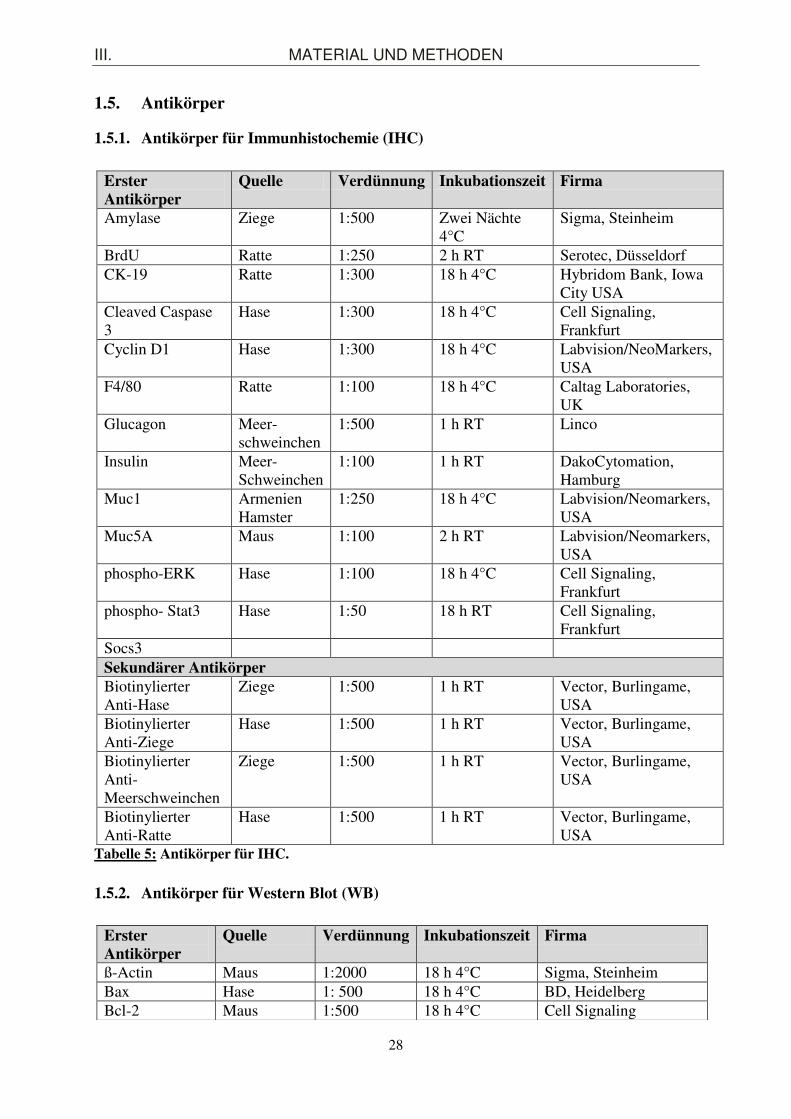
1.4. Gele
Gel Zusammensetzung Menge Sammelgel destilliertes Wasser
Sammelgelpuffer Acrylamid 30 % SDS 10 % APS TEMED
3,0 ml 1,3 ml 750 µl 50 µl 25 µl 10 µl
Tabelle 4: Gele.
Trenngel 7,5 % 10 % 12 % 15 % dH2O 4,9 ml 4,1 ml 3,4 ml 2,5 ml Trenngelpuffer 2,6 ml 2,6 ml 2,6 ml 2,6 ml Acrylamid 30 % 2,5 ml 3,3 ml 4,0 ml 5,0 ml SDS 10 % 100 µl 100 µl 100 µl 100 µl APS 10 % 50 µl 50 µl 50 µl 50 µl TEMED 15 µl 15 µl 15 µl 15 µl
III. MATERIAL UND METHODEN
28
1.5. Antikörper
1.5.1. Antikörper für Immunhistochemie (IHC)
Tabelle 5: Antikörper für IHC.
1.5.2. Antikörper für Western Blot (WB)
Erster Antikörper
Quelle Verdünnung Inkubationszeit Firma
Amylase Ziege 1:500 Zwei Nächte 4°C
Sigma, Steinheim
BrdU Ratte 1:250 2 h RT Serotec, Düsseldorf CK-19 Ratte 1:300 18 h 4°C Hybridom Bank, Iowa
City USA Cleaved Caspase 3
Hase 1:300 18 h 4°C Cell Signaling, Frankfurt
Cyclin D1 Hase 1:300 18 h 4°C Labvision/NeoMarkers, USA
F4/80 Ratte 1:100 18 h 4°C Caltag Laboratories, UK
Glucagon Meer- schweinchen
1:500 1 h RT Linco
Insulin Meer- Schweinchen
1:100 1 h RT DakoCytomation, Hamburg
Muc1 Armenien Hamster
1:250 18 h 4°C Labvision/Neomarkers, USA
Muc5A Maus 1:100 2 h RT Labvision/Neomarkers, USA
phospho-ERK Hase 1:100 18 h 4°C Cell Signaling, Frankfurt
phospho- Stat3 Hase 1:50 18 h RT Cell Signaling, Frankfurt
Socs3 Sekundärer Antikörper Biotinylierter Anti-Hase
Ziege 1:500 1 h RT Vector, Burlingame, USA
Biotinylierter Anti-Ziege
Hase 1:500 1 h RT Vector, Burlingame, USA
Biotinylierter Anti-Meerschweinchen
Ziege 1:500 1 h RT Vector, Burlingame, USA
Biotinylierter Anti-Ratte
Hase 1:500 1 h RT Vector, Burlingame, USA
Erster Antikörper
Quelle Verdünnung Inkubationszeit Firma
ß-Actin Maus 1:2000 18 h 4°C Sigma, Steinheim Bax Hase 1: 500 18 h 4°C BD, Heidelberg Bcl-2 Maus 1:500 18 h 4°C Cell Signaling
III. MATERIAL UND METHODEN
29
Tabelle 6: Antikörper für Western Blot.
1.6. Primer für Genotypisierung
Gen Primername Oligonukleotidsequenz (5ʼ- 3ʼ) Socs3 Socs3 loxpA
Socs3 loxpB -GCGGGCAGGGGAAGAGACTGTCTGGGGTTG- -GGCGCACGGAGCCAGCGTGGATCTGCG-
Ptf1a-cre Cre487 p48as1642
-GTCCAATTTACTGACCGTACACCAA- -CCTCGAAGGCGTCGTTGATGGACTGCA-
Kras Kras 3 Kras 5
-CCATGGCTTGAGTAAGTCTGCG- -CGCAGACTGTAGAGCAGCG-
Tabelle 7: Primer.
1.7. Mäuse
Maus Kurzbezeichnung Referenz LSL-Kras
G12D (Jackson et al., 2001) Ptf1a-Cre
ex1 (Nakhai et al., 2007) Socs3
Flox/Flox Socs3
F/F (Okada, Nakamura 2006)
LSL-KrasG12D;Ptf1a-Cre
ex1 KrasG12D Eigene Kreuzung
Socs3F/F;Ptf1a-Cre
ex1 Socs3Δpanc Eigene Kreuzung
LSL-KrasG12D;Socs3
F/F;Ptf1a-Creex1 Kras
G12D;Socs3Δpanc Eigene Kreuzung
Tabelle 8: Mäuse.
Frankfurt Bcl-XL Hase 1:500 18 h 4°C Cell Signaling,
Frankfurt Cox-2 Ziege 1:200 18 h 4°C Caymann, Ann Arbor,
USA Cyclin D1 Maus 1:500 18 h 4°C Santa Cruz, Heidelberg HSP70 Hase 1:500 18 h 4°C Cell Signaling,
Frankfurt Mcl-1 Hase 1:500 18 h 4°C Epitomics, Burlingame,
USA p21 Maus 1:500 18 h 4°C BD Pharmingen,
Heidelberg p53 Maus 1:500 18 h 4°C Santa Cruz, Heidelberg PCNA Maus 1:500 18 h 4°C Santa Cruz, Heidelberg phospho-ERK Hase 1:500 18 h 4°C Cell Signaling,
Frankfurt Phospho-Jak 2 Hase 1:500 18 h 4°C Cell Signaling,
Frankfurt Phospho-Stat3Y705
Hase 1:500 18 h RT Cell Signaling, Frankfurt
Stat3 Hase 1:500 18 h RT Santa Cruz, Heidelberg Survivin Maus 1:500 18 h RT Santa Cruz, Heidelberg Sekundärer Antikörper Anti-Hase IgG Ziege 1:2000 1 h RT Promega, Mannheim Anti-Maus IgG Schaf 1:2000 1 h RT GE Healthcare,
München

III. MATERIAL UND METHODEN
30
2. Methoden
2.1. Versuchstiere
2.1.1. Behandlung mit BrdU
5-Brom-2-desoxyuridin (BrdU) ist ein Thymidinanalogon, welches von proliferierenden
Zellen in vitalen Geweben aufgenommen werden kann. In phosphorylierter Form wird BrdU
anstelle des Nukleotids Desoxythymidintriphosphat (dTTP) in die neu synthetisierte DNA
(Desoxyribonukleinsäure) aufgenommen. Mithilfe spezifischer Antikörper gegen BrdU kann
eine stattgefundene DNA Synthese immunhistochemisch nachgewiesen werden. Damit dient
die Behandlung mit 5-Brom-2-desoxyuridin der labordiagnostischen Markierung bzw. dem
Nachweis proliferierender Zellen.
Für die Detektion und Quantifizierung der proliferierenden Zellen wird die BrdU Stock-
Lösung 1:10 mit 0,9 % NaCl auf eine Endkonzentration von 2 mg/ml verdünnt. Die Mäuse
wurden zwei Stunden vor Präparation intraperitoneal mit 10 µl/g Körpergewicht (50 µg
BrdU/g Körpergewicht Maus) injiziert.
2.1.2. Gewebeentnahme in vivo-Präparation und Materialgewinnung
Zur Gewebegewinnung wurden die zu untersuchenden Tiere zwei Stunden nach der BrdU-
Injektion mit einer Überdosis Isofluran euthanasiert und das Gewicht bestimmt.
Nach Desinfektion des Abdomens der Mäuse mit 70 % Alkohol, Freilegung der Bauch- und
Thoraxhöhle erfolgte zuerst die Inspektion auf ggf. vorhandenen Aszites, Blut, Metastasen,
vergrößerte Lymphknoten etc.. Anschließend wurden Pankreas, Milz, Leber, Lunge und
Duodenum entnommen. Nach Bestimmung des Pankreasgewichts wurden Gewebestückchen
aus verschiedenen Pankreasanteilen (Caput, Corpus und Cauda) zur Protein- und RNA-
Bestimmung reseziert und in kleinen Eppis in Flüssigstickstoff schockgefroren. Das restliche
Pankreasgewebe und die anderen entnommenen Präparate wurden in 4 % Paraformaldehyd
(PFA) über Nacht fixiert. Am nächsten Tag wurde die Paraformaldehydlösung durch
phosphatgepufferte Salzlösung (phosphate buffered saline, PBS) ersetzt. Zur weiteren
Verarbeitung wurden die Proben ins Institut für Pathologie der Technischen Universität
München geschickt, wo sie schließlich in Paraffin eingebettet wurden und somit für die
Immunhistochemie zur Verfügung standen.
Zur Nachgenotypisierung (PCR) wurden die Schwanzspitzen der verwendeten Mäuse getrennt
und bei -20°C bis zur Anwendung gelagert. Alter, Geschlecht, Gewicht und Genotyp der
Tiere sowie die bei der Resektion beobachteten Auffälligkeiten wurden notiert.

III. MATERIAL UND METHODEN
31
Alle Experimente wurden nach den Richtlinien des lokalen Tierschutzkommitees
durchgeführt.
2.2. Molekularbiologische Methoden
2.2.1. DNA-Extraktion aus der Mausschwanzspitze
Die DNA-Extraktion ist Voraussetzung für die Analyse des entsprechenden DNA-Abschnitts
mittels Polymerasekettenreaktion (PCR), die wiederum für die Bestimmung des Genotyps der
Versuchstiere essentiell ist.
Genomische DNA aus Mausschwänzen wurde mithilfe des DirectPCR®Lysis Reagent
(peqlab) isoliert. Die Isolation erfolgte nach dem Protokoll DirectPCR®Lysis Reagent.
Die ca. 0,5-1 cm großen Mäuseschwänze wurden mit 200-250 µl DirectPCR®Lysis Reagent,
komplettiert mit 0,2-0,3 mg/ml Proteinkinase K, in Reaktionsgefäßen bei 55 °C über Nacht in
einem Thermoschüttler bis zur kompletten Lyse des Gewebes inkubiert. Am nächsten Tag
wurden die Proben zur Inaktivierung der Proteinase K bei 85 °C für 45 Min. erhitzt und
anschließend für zehn Sekunden zentrifugiert. Der Überstand wurde so für die nachfolgende
PCR gewonnen und das Pellet verworfen. Für die anschließende Genotypisierung wurde 1 µl
pro 25 µl PCR-Reaktion eingesetzt.
2.2.2. Polymerasekettenreaktion (polymerase chain reaction, PCR)
Die PCR diente der in vitro erfolgten enzymatischen Amplifikation der zu untersuchenden
Genabschnitte (spezifische DNA-Sequenzen) der verwendeten Versuchstiere. Mithilfe einer
hitzestabilen DNA-Polymerase und zwei definierter Oligonukleotidprimer, die den zu
amplifizierenden DNA-Abschnitt am 5ʼ- und 3ʼ-Ende flankieren, lässt sich die spezifische
DNA-Sequenz polymerisieren und somit exponentiell vervielfältigen. Die PCR ermöglicht
somit den Nachweis kleinster DNA-Mengen.
Zur Gewinnung einer ausreichenden DNA-Menge ist die PCR in drei Schritte unterteilt, die
sich in 12-50 genau definierten Zyklen wiederholen:
1. Denaturierung des DNA-Doppelstranges
2. Anlagerung (Annealing) der spezifischen Primer
3. Verlängerung (Elongation) der einsträngigen DNA durch die DNA-Polymerase

III. MATERIAL UND METHODEN
32
Die PCR-Proben mit jeweils 25 µl wurden wie folgt angesetzt:
Reagenz Menge dH2O 10,5 µl Primer 1 0,5 µl Primer 2 0,5 µl (oder PrimerMix) 1 µl RedTag Ready Mix 12,5 µl DNA 1 µl
Folgende Primer-Sequenzen wurden für die Genotypisierung verwendet: siehe Tabelle 7
Alle Ansätze wurden noch einmal kurz herunter zentrifugiert, bevor die PCR in einem
automatischen Thermocycler unter folgenden Bedingungen durchgeführt wurde:
1. Initiale Denaturierung 5 Min. 95°C
2. Denaturierung 30 Sek. 95°C
Annealing der Primer 30 Sek. 60°C 40 Zyklen
Elongation 90 Sek. 72°C
3. Terminale Elongation 10 Min. 72°C
Die erhaltenen DNA-Amplifikate können dann direkt mithilfe der Agarose-Gelelektrophorese
analysiert und der Erfolg der DNA-Amplifikation überprüft werden.
2.2.3. Agarose-Gelelektrophorese
Mittels der Agarose-Gelelektrophorese ist eine Größentrennung der DNA-Stränge und durch
Vergleich mit Strängen bekannter Größe eine DNA-Größenbestimmung möglich. Zur
horizontalen Auftrennung der Moleküle wird ein aus Agarosepolymeren vernetztes Gel
hergestellt, dessen Konzentration je nach Fragmentgröße zwischen 1 % und 3 % liegt. Der
Molekularsiebeffekt des Gels dient dabei als Trennprinzip. Die Agarose wurde in 1x TAE-
Puffer in der Mikrowelle fünf Minuten bis zum Kochen erwärmt. Anschließend wurde pro 10
ml Gellösung 1 µl Ethidiumbromid substituiert. Das flüssige Gel wurde dann blasenfrei in die
entsprechende Gelkammer gegossen und ein Gelkamm eingesetzt. Nach Verfestigung des
Gels erfolgte die Beladung der Geltaschen mit jeweils 15 µl der in der PCR gewonnenen
DNA-Amplifikate und dem Größenstandard, der der Größenbestimmung der Fragmente
diente. Aufgrund der negativen Ladung wanderten die DNA-Moleküle nach Anlegen einer
elektrischen Spannung von 120V von der Kathode zur Anode, wobei die
Wanderungsgeschwindigkeit der DNA-Moleküle durch die Gelmatrix von ihrer Größe
abhängig war. Je kleiner das Molekül, desto schneller bewegte es sich. Die Trennschärfe wird
ebenfalls von der Gelkonzentration, der angelegten Spannung und der Laufzeit beeinflusst.

III. MATERIAL UND METHODEN
33
Zur Visualisierung der aufgetrennten DNA dient die Interkalation der Ethidiumbromid-
Moleküle zwischen den Basen der DNA. Durch die so verstärkte fluoreszierende Eigenschaft
von Ethidiumbromid im ultravioletten Licht kann die daran gebundene DNA sichtbar gemacht
werden. Das Ergebnis wurde mit einer LCD-Kamera festgehalten und die Größenanalyse
mithilfe des Gel Doc XR Systems durchgeführt.
2.3. Detektion und Quantifizierung der Gen-Transkription
2.3.1. Isolation von mRNA aus murinem Pankreasgewebe
Zur Gewinnung von RNA wurde zunächst das Pankreasgewebe entnommen und in RLT-
Puffer/1 % (v/v) β-Mercaptoethanol überführt. Anschließend erfolgte die Zerkleinerung
mithilfe eines Homogenisators DIAX 900. Die Extraktion der RNA aus den Homogenisaten
wurde mit dem RNeasy®Mini Kit nach den Protokollangaben des Herstellers durchgeführt.
Dieses Kit enthält mit Silicagel gefüllte Säulen, über welche die RNA aufgereinigt wird. Die
RNA-Konzentration wurde photometrisch unter Zuhilfenahme eines Eppendorf
BioPhotometers gemessen. Nicht angewendete Lysate wurden in flüssigem Stickstoff
schockgefroren und bei -80 °C gelagert.
2.3.2. cDNA-Synthese
Die Umschreibung von RNA in cDNA erfolgte durch reverse Transkription gemäß
Herstellerprotokollen. Hierzu wurden 5 µg RNA nach Substitution von 1 µl Oligo(dT)-Primer
und 1 µl SuperScriptTM II Reverse Transkriptase in cDNA enzymatisch umgewandelt. Bis zur
weiteren Anwendung wurden die cDNA-Produkte bei -20 °C gelagert.
2.3.3. Quantitative real-time PCR (qRT-PCR)
Die Reverse Transkriptase-Polymerase-Kettenreaktion (RT-PCR) stellte die Kombination aus
zwei molekulargenetischen Methoden, der Reversen Transkriptase (RT) und der Polymerase-
Kettenreaktion (PCR), dar, die dem Nachweis von RNA aus Genexpression spezifischer Gene
in Zellen, Geweben etc. dienten. Via qRT-PCR konnte die Produktsynthese in Echtzeit
verfolgt werden. Hierzu wurde die DNA mit einem interkalierenden Fluoreszenzfarbstoff
markiert, der am Ende eines jeden Zyklus optisch angeregt wurde, sodass die
Fluoreszenzemission anschließend gemessen werden konnte. Hierbei korrelierte die Zunahme
der Fluoreszenz mit der Zunahme der DNA-Konzentration. Der häufig, auch in dieser Arbeit
verwendete Fluoreszenzfarbstoff ist SYBRTMGreen I, welcher sich an doppelsträngige DNA
anlagerte. DNA-Einzelstränge und einzelsträngige Oligonukleotid-Primer wurden von
SYBRTMGreen I mit einer geringeren Affinität gebunden, sodass diese die Messung

III. MATERIAL UND METHODEN
34
behinderte. Um die Messung unspezifischer Fluoreszenz durch so genannte Primerdimere zu
minimieren, wurde die Detektionstemperatur angehoben. Da Primerdimere einen geringeren
Schmelzpunkt als spezifische PCR-Produkte besaßen, konnten sie bei höheren Temperaturen
nicht mehr detektiert werden. Deswegen erfolgte die Registrierung der Fluoreszenz erst nach
einer kurzen Inkubation bei 80 °C, anstatt schon nach der Elongation bei 72 °C. Der Nachteil
der geringen Spezifität konnte durch Optimierung des Primerdesigns und der gewählten
Reaktionsbedingungen ausgeglichen werden. Desweiteren ließ sich die Bildung spezifischer
PCR-Produkte anhand der Fragmentlänge nach abgelaufener PCR mithilfe einer
Schmelzkurvenanalyse überprüfen. Hierbei wurde die doppelsträngige DNA bei
kontinuierlicher Temperaturerhöhung von 70 °C auf 90 °C aufgeschmolzen. An dem für das
Fragment definierten Schmelzpunkt zerfiel der DNA-Doppelstrang in Einzelstränge, wodurch
eine Abnahme der Fluoreszenz der Probe registriert wurde. Aufgrund der höheren
Schmelztemperatur der spezifischen doppelsträngigen PCR-Produkte gegenüber
unspezifischen Primerdimeren, widerspiegelte die Höhe des Peaks der Schmelzkurve
annähernd die Fragment-Konzentration.
2.3.4. Reaktionsansatz und Reaktionsbedingungen
Das für die qRT-PCR verwendete Reaktionsgemisch setzte sich zusammen aus SYBR Green
PCR Master Mix 2x, 100 ng cDNA und je 900 nM sense- und antisense-Primer in einem
Reaktionsvolumen von 25 µl. Die anschließende PCR fand in einem ABI-PRISM 7700
Sequenz-Detektions-System unter folgenden Reaktionsbedingungen statt:
1. Initiale Denaturierung 3 Min. 94°C
2. Inkubation 30 Sek. 94°C
3. Inkubation 30 Sek. 60°C 40 Zyklen
4. Inkubation 30 Sek. 72°C
3. Inkubation 3 Sek. 80°C
Nach dem letzten Inkubationsschritt eines jeden PCR-Zyklus erfolgte die
Fluoreszenzmessung. Zur Bestimmung der Schmelzkurven wurde die Temperatur in 0,5°C-
Schritten von 70 bis 90°C angehoben und jeweils für drei Sekunden gehalten und
währenddessen die Fluoreszenz registriert.

III. MATERIAL UND METHODEN
35
2.3.5. Quantifizierung der Genexpression
Die Analyse der Expressionsniveaus der Gene erfolgte entweder durch absolute
Quantifizierung anhand einer gegebenen Kalibrierkurve oder durch relative Quantifizierung
mit Einbezug eines oder mehrerer Referenzgene. Diese auch als „housekeeping“-Gene
bezeichneten Referenzgene wurden als Ladekontrolle eingesetzt, da deren Expressionsniveau
in allen Proben gleichbleibend hoch ist. Häufig wird das Haushaltsgen Cyclophilin eingesetzt.
Als Maß für die Quantifizierung der DNA-Startmenge wurde der sogenannte Ct-(Cycle
Threshold für Schwellenwert-Zyklus) bzw. CP-(Crossing Point)-Wert verwendet, welcher die
Anzahl der nötigen PCR-Zyklen zum Erreichen eines konstant definierten Fluoreszenzniveaus
beschrieb. Unter optimalen Bedingungen und bei 100 %iger Effizienz kommt es zur
Verdopplung der DNA-Menge und analog dazu auch der Fluoreszenzemission nach jedem
PCR-Zyklus. Zur Berechnung des Expressionsniveaus wurde zunächst für jede untersuchte
Probe die Differenz zwischen dem CP-Wert des Referenzgens und des zu untersuchenden
Gens ermittelt: ΔCP = CP-Zielgen – CP-Referenzgen und anschließend in die Gleichung 2-ΔCP
eingesetzt.
2.4. Proteinchemische Methoden
2.4.1. Proteinisolierung/Proteingewinnung aus Mauspankreasgewebe
Zur Proteingewinnung aus Mauspankreasgewebe wurde zunächst das nach der Resektion der
Mäuse schockgefronene Pankreasgewebe auf Eis aufgetaut und danach mit einem Skalpell
zerkleinert. Um die intrazellulären Proteine zu isolieren, wurde das Gewebe mit je 600 µl IP-
Puffer/Lysispuffer resuspendiert und homogenisiert. Nachdem die Proben für 30 Min. auf Eis
gelagert wurden, erfolgte abschließend eine zehnminütige Zentrifugation bei 4°C und 13.200
rpm. Der dabei gewonnene proteinhaltige Überstand wurde für die weitere Diagnostik
abgenommen, während das Pellet verworfen wird.
2.4.2. Proteinkonzentrationsbestimmung nach Bradford
Die Proteinkonzentrationsbestimmung nach Bradford (Bradford, 1976) erfolgte mit dem Bio-
Rad Protein-Assay. Hierbei handelt es sich um eine photometrische Methode, bei der der
Triphenylmethanfarbstoff Coomassie-Brillant-Blau G-250 zur Komplexbildung mit Proteinen
eingesetzt wird. Je nach Proteinkonzentration ist das Maß der Proteinbindung an den Farbstoff
unterschiedlich, was wiederum zu unterschiedlichen Farbveränderungen führt.
Coomassie-Brillant-Blau G-250 hat in ungebundener Form ein Absorptionsmaximum bei 465
nm, das in der gebundenen Form auf 595 nm erhöht wird. Die Absorptionsverschiebung ist

III. MATERIAL UND METHODEN
36
photometrisch messbar und stellt somit ein Maß für die Proteinkonzentration der verwendeten
Probe dar.
Zunächst wird der Bio Rad Protein-Assay im Verhältnis 1:5 mit dH2O verdünnt.
Anschließend wurde auf einer 96-Well-Platte 10 µl der Probe oder 10 µl des Standards mit
250 µl 1x Bradford-Lösung gemischt. Als Standardprotein diente BSA (c=1 mg/ml), welches
in Form einer Standardreihe (0, 1, 2, 4, 6, 8 mg/ml) eingesetzt wurde. Die Proteinlösungen
(aus 2.3.1.) wurden im Verhältnis 1:5 mit dH2O verdünnt, bevor sie in die Wells pipettiert
wurden. Anschließend wurde die Extinktion der Proben mithilfe des Spektrophotometers bei
595 nm gemessen und die Proteinkonzentration anhand der Kalibrierkurve aus den
Konzentrationen der Standards berechnet.
2.4.3. Western Blot Analyse
Unter Western Blot versteht man die Übertragung von Proteinen von einem Elektrophorese-
Gel auf eine Membran, um diese immunologisch zu detektieren bzw. nachzuweisen. Bei
dieser Methode wird zunächst das Proteingemisch der zu untersuchenden Proben mittels
Gelelektrophorese nach Ladung und Größe aufgetrennt und anschließend auf eine Membran
transferiert. Durch das Aufbringen spezifischer Primär- und Sekundärantikörper auf die
Membran erfolgt der Proteinnachweis.
SDS-Polyacrylamidgelelektrophorese (SDS-PAGE)
Die SDS-Polyacrylamidgelelektrophorese (SDS-PAGE) dient der Auftrennung der Proteine
der eingesetzten Proben nach ihrer Größe und Ladung. Dazu wurde das Mini-PROTEAN 3-
Gelelektrophorese-System (BioRad) verwendet. Der Ausgangspunkt der Proteinauftrennung
ist die Überführung der Tertiär- und Sekundärstruktur der Proteine in die Primärstruktur durch
Erhitzung der Proben mit einem SDS-Überschuss auf 95 °C. Dadurch erfolgt die Aufspaltung
der Wasserstoffbrücken und Streckung der Moleküle. SDS als anionisches Detergenz
maskiert die Eigenladung der Proteine, wodurch Mizellen mit konstanter negativer Ladung
entstehen. Die Aufspaltung der Schwefelbrücken zwischen Cysteinen wurde durch die
Substitution von ß-Mercaptoethanol unterstützt. Basis für das Prinzip der SDS-PAGE ist das
Polyacrylamid-Gel, dessen Gelmatrix aus einem Trenngel und Sammelgel besteht und somit
eine diskontinuierliche Elektrophorese gewährleistet. Das weitporige Sammelgel als
Molekularsieb verhinderte die Aggregation der Proteine beim Eintritt in das Gel, während das
engporige Trenngel die Trennung der Proteine ermöglichte. Die variierenden Konzentrationen
des Trenngels (7,5; 10; 12; 15 %; siehe Tabelle 4) mit unterschiedlichem Anteil an Acrylamid
waren umgekehrt proportional zur untersuchten Proteingröße und bestimmten die

III. MATERIAL UND METHODEN
37
Trenneigenschaften des Gels. Durch die Wahl der richtigen Gelkonzentration sollte eine
optimale Bandenschärfe im untersuchten Molekulargewichtsbereich erreicht werden. Das
Trenngel-Gemisch wird mit Ammoniumpersulfat, einem Radikalstarter und TEMED, einem
Polymerisierungskatalysator, versetzt und zügig zwischen zwei abgedichtete, durch einen
Abstandhalter (Spacer) voneinander getrennte Glasplatten blasenfrei gegossen. Der Abstand
zwischen den Glasplatten, die sauber und fettfrei sein müssen, beträgt 1,5 mm. Das Trenngel
wurde nach oben hin mit dH2O überschichtet, welches zum einen eine Glättung der
Geloberfläche ermöglichte und zum anderen eine Störung der Gelpolymerisation durch den
Luftsauerstoff verhinderte. Nach Verfestigung des Gels wurde dH2O abgegossen und das
Sammelgel etwa einen Zentimeter hoch über das Trenngel pipettiert. Ein spezieller Kamm
wurde in das Sammelgel eingesteckt, um Taschen zu erhalten. Diese wurden anschließend mit
den Proben nach definierten Proteinmengen gefüllt. Eine Geltasche wurde mit dem
Längenstandard belegt. Nach Auffüllen der Elektrophorese-Apparatur mit Laufpuffer erfolgte
der Gellauf bei einer Anfangsspannung von 70 V im Sammelgel und 90-120 V (je nach
Gelkonzentration) im Trenngel.
Proteintransfer
Die aufgetrennten Proteine wurden mithilfe eines vertikalen Tank-Blot Systems (Mini Trans-
Blot) auf eine Nitrozellulose-Membran transferiert. Die Nitrozellulose-Membran wurde in
Methanol 100 % bei Raumtemperatur (RT) für eine Minute aktiviert. Anschließend wurden
das zu blottende Gel, die Membran und die Filterpapiere in Transferpuffer äquilibriert und
danach als Sandwich luftblasenfrei in eine Blotkassette gelegt. Die Kassette wurde in eine
Blotkammer (Mini Blot Transfer CellTM) mit eiskaltem Transferpuffer überführt und der
Transfer bei 350 mA für 1-2 Std. durchgeführt. Die Transferbedingungen mussten dabei auf
die Molekülgröße des gewünschten Proteins angepasst werden, da verschieden große Proteine
unterschiedliche Wanderungsgeschwindigkeiten aufzeigten.
Proteindetektion
Die geblottete Membran wurde nach dem Transfer kurz bei RT mit einer Pufferlösung
gewaschen und anschließend eine Stunde bei RT im Blocking-Puffer unspezifisch geblockt.
Der Nachweis der auf die Nitrocellulosemembran geblotteten Proteine erfolgte durch
Immunodetektion mit spezifischen Antikörpern. Die Inkubation mit dem entsprechenden
Primärantikörper (siehe Tabelle 6) und der Pufferlösung erfolgte in einer eingeschweißten
Folie bei 4 °C über Nacht. Am nächsten Tag wurde die Membran zunächst 5x10 Min. mit der
Pufferlösung gewaschen, um danach eine Stunde bei RT auf einem Schüttler mit

III. MATERIAL UND METHODEN
38
peroxidasegekoppeltem Sekundärantikörper (siehe Tabelle 6) inkubiert zu werden. Nach
erneutem Waschgang für 5x10 Min. mit der entsprechenden Pufferlösung erfolgte die
Proteindetektion mit dem Amersham ECLTM Western Blotting Detektion Reagents auf
Amersham HyperfilmTMECL, welcher mithilfe des Amersham Automatic Film-Prozessor
entwickelt wurde.
Wiederverwendung der Membran (Membran-Stripping)
Wenn verschiedene Proteine auf einer Membran sequenziell detektiert werden sollten, wurde
diese zunächst mit Pufferlösung gewaschen, danach in Stripping-Pufferlösung für 30 Min.
gestrippt. Nach erneutem Waschgang wurde die Membran für eine Stunde in Blockierlösung
5 % geschwenkt, bevor diese mit dem nächsten Primärantikörper inkubiert wurde.
2.5. Histochemische Methoden
2.5.1. Immunhistochemie
Das Prinzip der Immunhistochemie beruht auf der Affinität von Antikörpern zu einem
bestimmten Gewebeantigen als Antigen-Antikörper-Reaktion. Die immunhistochemischen
Techniken ermöglichen den hochspezifischen Nachweis von Proteinen und Strukturen, gegen
die Antikörper gebildet werden können. Mithilfe von markierten Antikörpern können diese
detektiert und somit sichtbar gemacht werden. Je nach Art und Lokalisation des zu
untersuchenden Proteins bzw. der Art und Beschaffenheit des zu untersuchenden Gewebes ist
die Wahl der richtigen Methode aus einer Vielzahl an Methoden der immunhistochemischen
Färbung zu treffen.
Das Ergebnis und die Qualität der Färbung sind von mehreren Variablen abhängig: Zum einen
beeinflussen die Fixierung des Materials, die Einbettungsmethode und die
Vorbehandlungsmethoden die Gewebequalität, zum anderen ist die Antigen-Antikörper-
Reaktion von der optimalen Temperatur, der Antikörperkonzentration und der Inkubationszeit
abhängig. Die Immunhistochemie wurde in dieser Arbeit an Paraffingewebeschnitten
durchgeführt.
Herstellung von Paraffinschnitten
Unmittelbar nach der Resektion der Mäuse werden die entnommenen Gewebeproben in 4 %
gepuffertem Paraformaldehyd über Nacht fixiert, um die Autolyse bzw. Heterolyse zu
verhindern. Am nächsten Tag wurde das Fixierungsmittel ausgewaschen und das
Gewebematerial in heißem Paraffinwachs getränkt, das bei Abkühlung erstarrt. Das Gewebe
erhielt dadurch Stabilität und eine gleichmäßige Konsistenz, was die Herstellung dünner und

III. MATERIAL UND METHODEN
39
gleichmäßiger Schnitte ermöglichte. Paraffinschnitte ließen sich mittels Schlittenmikrotomen
anfertigen. Die 3,5 µm dicken Gewebeschnitte wurden zunächst in ein Warmwasserbad
übertragen, in dem sie gestreckt und von Falten befreit wurden. Danach wurden sie auf
beschichtete Objektträger aufgezogen und über Nacht bei RT getrocknet. Zur Färbung wurden
die Schnitte zuerst in Xylol (dreimal fünf Minuten) entparaffiniert und dann durch eine
absteigende Alkoholreihe (jeweils drei Minuten in 100 %, 96 % und 76 % Alkohol) bis zum
dH2O hydratisiert. Die Vorbehandlung der Gewebeschnitte mit EDTA- oder mit Citrat-
Pufferlösung (Antigen Unmasking Solution) in der Mikrowelle für zehn Minuten diente der
Demaskierung der Antigene. Danach wurden die Schnitte 15 Min. bei RT abgekühlt, mit
dH2O und/ohne Waschpuffer gewaschen und anschließend zur Blockierung der endogenen
Peroxidaseaktivität 15 Min. in frisch angesetzter 3 % H2O2-Lösung inkubiert. Nach jedem
Schritt wurden die Gewebeschnitte mit dH2O und 3x5 Min. mit Waschpuffer (PBS bzw.
PBS/T oder TBS bzw. TBS/T) gewaschen. Die darauffolgende immunhistochemische
Färbung wurde nach der (Strept)Avidin-Biotin-Komplex-Methode (ABC-Methode)
durchgeführt.
Streptavidin-Biotin-Komplex-Methode
Die Streptavidin-Biotin-Methode als indirekte Nachweismethode nutzt die starke Affinität
von Avidin bzw. Streptavidin für Biotin zur Bildung von Komplexen aus enzymmarkierten
Avidin-Biotin-Komplexen mit biotinylierten Sekundärantikörpern. Dabei bindet das
Glykoprotein Avidin bzw. Streptavidin an den Sekundärantikörper. Bei der Färbung werden
nacheinander Primärantikörper (siehe Tabelle 5), biotinylierter Sekundärantikörper (siehe
Tabelle 5) und Avidin-Biotin-Enzymkomplex-Substrat-Chromogenlösung appliziert, wobei
die Meerrettichperoxidase am häufigsten als Enzym verwendet wird. Die Funktion der
Peroxidase liegt in der Elektronenübertragung von Chromogen DAB (3,3-Diaminobenzidin)
auf das zugegebene Substrat Wasserstoffperoxid, das dadurch zu Wasser reduziert wird. Im
Rahmen dieser Redoxreaktion fällt DAB zu einem unlöslichen, bräunlichen Endprodukt aus,
das eine signalverstärkende Wirkung hat.
2.5.2. Hämatoxylin-Eosin-Färbung (H.E.-Färbung)
Die H.E.-Färbung ist eine der Routinefärbemethoden in der Histologie und dient durch die
Anfärbung verschiedener Strukturen eines Gewebeschnittes der morphologischen
Untersuchung. Diese klassische Übersichtsfärbung wird typischerweise vor den
immunhistochemischen Färbungen durchgeführt und setzt sich aus zwei Einzelfärbungen

IV. ERGEBNISSE
40
zusammen, mithilfe derer physiologische aber pathologische Aspekte von Zellen,
Zellbestandteilen und Extrazellularmatrix untersucht werden können.
Hämatoxylin
Hämatoxylin ist ein Inhaltstoff aus dem Blauholzbaum und wird als natürlicher Farbstoff
verwendet. Die färbende Eigenschaft entwickelt das Hämatoxylin durch dessen Aufbereitung
zu Hämalaun. Hämalaun färbt saure bzw. basophile Strukturen wie Zellkerne mit der darin
enthaltenen DNA, Mitochondrien, das raue endoplasmatische Retikulum (rER),
Kollagenfasern und Elastin blau.
Eosin
Eosin ist ein synthetischer Farbstoff und färbt alle basischen (eosinophilen) bzw. azidophilen
Strukturen rot, insbesondere die Zytoplasmaproteine.
Die Färbung
Zur Färbung wurden zunächst die Paraffinschnitte (siehe 2.4.1.) 2x5 Min. in Histoclear
(Xylol) entparaffiniert und dann in einer absteigenden Alkoholreihe (2x100 %, 2x96 % und
2x76 %) bis zum dH2O hydratisiert. Die Gewebeschnitte wurden anschließend für 10 Min.
mit Hämalaun gefärbt. Nach der Hämalaun-Färbung erschienen die Zellkerne aufgrund des
niedrigen pH-Wertes der Färbelösung rötlich-braun. Der Farbton schlug sich nach Spülen der
Schnitte in laufendem Leitungswasser in blauviolett um. Danach erfolgte die Gegenfärbung
mit Eosin für 5 Min., das Waschen in 96 % Ethanol und Isopropanol für 25 Sek. Bevor die
Schnitte mit Pertex blasenfrei eingedeckt wurden, erfolgte die Behandlung mit Histoclear für
2x3 Min..

IV. ERGEBNISSE
41
IV. ERGEBNISSE
1. Pankreasspezifische Inaktivierung von Socs3 in der Maus
1.1. Generierung einer pankreasspezifisch Socs3-defizienten Mauslinie Socs3
Δpanc unter Anwendung des Cre/loxP-Rekombinationssystems
Socs3 scheint für die Karzinogenese vieler maligner Tumoren eine besondere Rolle zu spielen
(Ogata et al. 2006). Als negativer Regulator des Jak/Stat3-Signalweges ist dieses Molekül
bereits in verschiedenen Arbeiten beschrieben worden. Um die Bedeutung des Socs3-Proteins
in der pankreatischen Onkogenese zu evaluieren, generierten wir zunächst eine Mauslinie, die
eine pankreasspezifische Inaktivierung des Socs3-Gens und somit eine Socs3-Defizienz
aufweist. Davon ausgehend untersuchten wir in einem weiteren Schritt den Effekt der Socs3-
Deletion auf die PanIN-Entwicklung in KrasG12D-Tumormäusen.
Socs3 wird nach Zytokin-Rezeptor-Interaktion über die Aktivierung des Transkriptionsfaktors
p-Stat3 durch Janus-Kinasen vermehrt exprimiert. Da wir eine Inaktivierung von Socs3 und
den daraus resultierenden Wegfall des negativen Feedback-Mechanismus auf den Jak/Stat3-
Signalkaskade nur in murinen Pankreasgewebszellen anstrebten, wurde das im Abschnitt
I.2.1. beschriebene Cre/loxP-Rekombinationssystem angewendet (siehe Abbildung 9).
Abbildung 9: Schematische Darstellung der Generierung der Socs3Δpanc-Mauslinie.
Nach Kreuzung der SocsF/F-Maus mit der Ptf1a-Cre
ex1-Maus entsteht die Socs3Δpanc-Mauslinie durch
Exzision des Exons 2 mithilfe der Cre-Rekombinase.
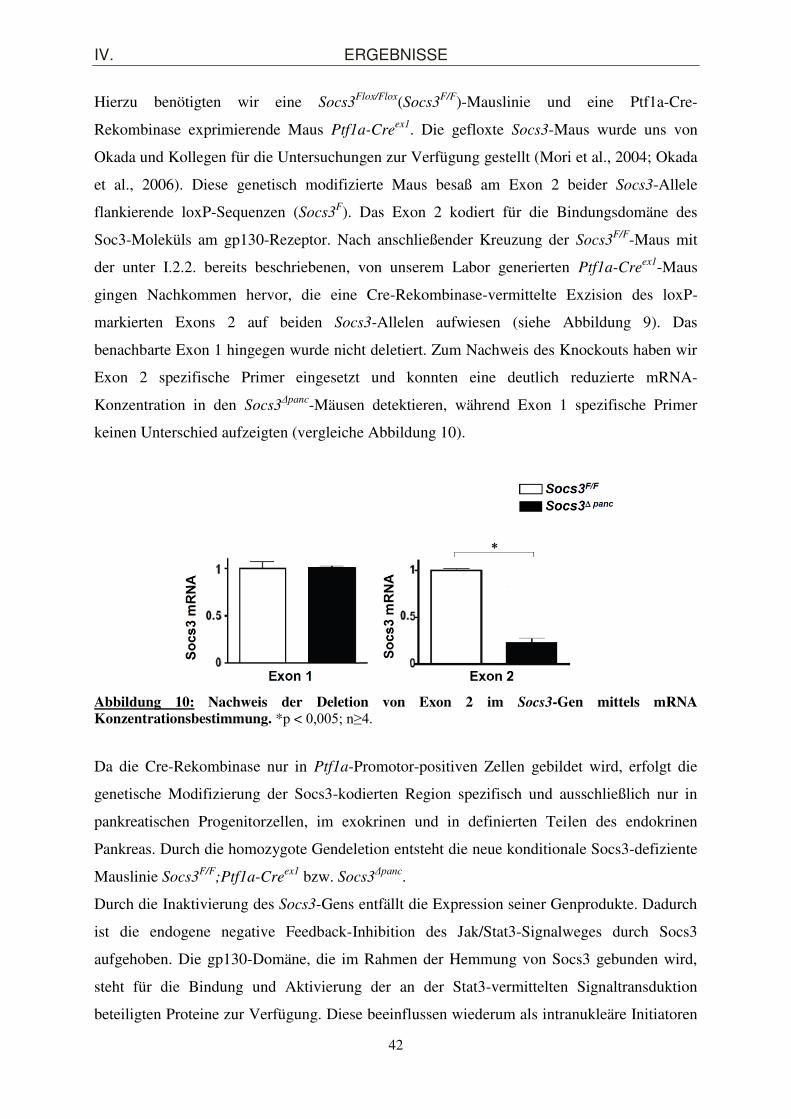
IV. ERGEBNISSE
42
Hierzu benötigten wir eine Socs3Flox/Flox(Socs3
F/F)-Mauslinie und eine Ptf1a-Cre-
Rekombinase exprimierende Maus Ptf1a-Creex1. Die gefloxte Socs3-Maus wurde uns von
Okada und Kollegen für die Untersuchungen zur Verfügung gestellt (Mori et al., 2004; Okada
et al., 2006). Diese genetisch modifizierte Maus besaß am Exon 2 beider Socs3-Allele
flankierende loxP-Sequenzen (Socs3F). Das Exon 2 kodiert für die Bindungsdomäne des
Soc3-Moleküls am gp130-Rezeptor. Nach anschließender Kreuzung der Socs3F/F-Maus mit
der unter I.2.2. bereits beschriebenen, von unserem Labor generierten Ptf1a-Creex1-Maus
gingen Nachkommen hervor, die eine Cre-Rekombinase-vermittelte Exzision des loxP-
markierten Exons 2 auf beiden Socs3-Allelen aufwiesen (siehe Abbildung 9). Das
benachbarte Exon 1 hingegen wurde nicht deletiert. Zum Nachweis des Knockouts haben wir
Exon 2 spezifische Primer eingesetzt und konnten eine deutlich reduzierte mRNA-
Konzentration in den Socs3Δpanc-Mäusen detektieren, während Exon 1 spezifische Primer
keinen Unterschied aufzeigten (vergleiche Abbildung 10).
Abbildung 10: Nachweis der Deletion von Exon 2 im Socs3-Gen mittels mRNA Konzentrationsbestimmung. *p < 0,005; n≥4.
Da die Cre-Rekombinase nur in Ptf1a-Promotor-positiven Zellen gebildet wird, erfolgt die
genetische Modifizierung der Socs3-kodierten Region spezifisch und ausschließlich nur in
pankreatischen Progenitorzellen, im exokrinen und in definierten Teilen des endokrinen
Pankreas. Durch die homozygote Gendeletion entsteht die neue konditionale Socs3-defiziente
Mauslinie Socs3F/F
;Ptf1a-Creex1 bzw. Socs3
Δpanc.
Durch die Inaktivierung des Socs3-Gens entfällt die Expression seiner Genprodukte. Dadurch
ist die endogene negative Feedback-Inhibition des Jak/Stat3-Signalweges durch Socs3
aufgehoben. Die gp130-Domäne, die im Rahmen der Hemmung von Socs3 gebunden wird,
steht für die Bindung und Aktivierung der an der Stat3-vermittelten Signaltransduktion
beteiligten Proteine zur Verfügung. Diese beeinflussen wiederum als intranukleäre Initiatoren

IV. ERGEBNISSE
43
die Transkription bestimmter Zielgene und steuern somit Prozesse der Proliferation,
Apoptosis und Inflammation.
Um den Effekt der Ptf1a-Cre-assoziierten DNA-Mutation im Bereich des Socs3-Lokus zu
evaluieren, wurde der Genotyp der aus der Kreuzung von Socs3F/F- und Ptf1a-Cre
ex1-Mäusen
hervorgehenden Nachkommen mittels Polymerasekettenreaktion (PCR) bestimmt. Das hierfür
benötigte genetische Material gewannen wir aus der Schwanzspitze der Mäuse. Anschließend
führten wir die PCR wie unter II.2.2. beschrieben durch. Dabei verwendeten wir die Socs3-
Primer A und B, die eine Abgrenzung des Wildtyp-Allels von am Exon 2 gefloxtem Socs3-
Allel ermöglichten. Zusätzlich dienten zwei Primer Cre487 und p48as1642 dem Nachweis des
Ptf1a-Cre-Transgens auf der DNA (vergleiche Tabelle 8).
Die PCR-Ergebnisse in der Abbildung 9 zeigen bezüglich des Genotyps drei mögliche
Kombinationen, wobei alle Nachkommen homozygot für das Ptf1a-Cre-Transgen sind.
Socs3+/+
;Ptf1a-Creex1-Mäuse weisen eine deutliche Bande für das Socs3-Wildtyp-Allel auf,
d.h. sie zeigen keine mutative Veränderung im Bereich dieses Genlokus und sind deshalb
homozygot für das entsprechende Allel. Anders besitzen Socs3+/Δ
;Ptf1a-Creex1-Mäuse ein
Allel entsprechend dem Wildtyp-Genotyp und ein loxP-flankiertes Exon 2 auf dem zweiten
Allel. Die für uns relevante und interessante Kombination waren die auch als Socs3Δ/Δ
;Ptf1a-
Creex1
bzw. Socs3Δpanc bezeichneten Mäuse. Die pankreasspezifische Cre-Rekombinase-
vermittelte, homozygote Deletion des Exons 2 auf beiden Allelen des Socs3-Gens lässt sich
nur in diesen Mäusen initiieren, sodass diese die Grundlage unserer weiteren Untersuchungen
darstellen.
Abbildung 11: PCR-Analysen der möglichen Genotypen aus der Kreuzung von Socs3
F/F-und Ptf1a-Cre
ex1-Mäusen. loxP=gefloxtes Allel; wt=Wildtyp-Allel.
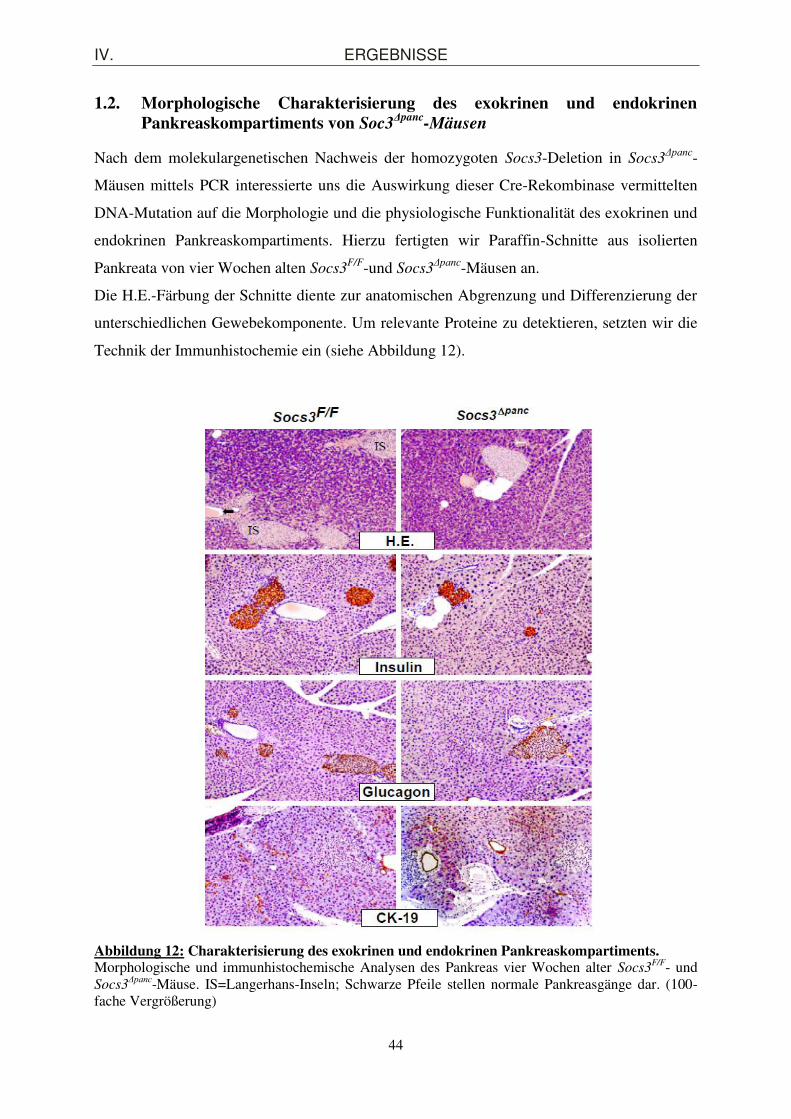
IV. ERGEBNISSE
44
1.2. Morphologische Charakterisierung des exokrinen und endokrinen Pankreaskompartiments von Soc3
Δpanc-Mäusen
Nach dem molekulargenetischen Nachweis der homozygoten Socs3-Deletion in Socs3Δpanc-
Mäusen mittels PCR interessierte uns die Auswirkung dieser Cre-Rekombinase vermittelten
DNA-Mutation auf die Morphologie und die physiologische Funktionalität des exokrinen und
endokrinen Pankreaskompartiments. Hierzu fertigten wir Paraffin-Schnitte aus isolierten
Pankreata von vier Wochen alten Socs3F/F-und Socs3
Δpanc-Mäusen an.
Die H.E.-Färbung der Schnitte diente zur anatomischen Abgrenzung und Differenzierung der
unterschiedlichen Gewebekomponente. Um relevante Proteine zu detektieren, setzten wir die
Technik der Immunhistochemie ein (siehe Abbildung 12).
Abbildung 12: Charakterisierung des exokrinen und endokrinen Pankreaskompartiments. Morphologische und immunhistochemische Analysen des Pankreas vier Wochen alter Socs3
F/F- und Socs3
Δpanc-Mäuse. IS=Langerhans-Inseln; Schwarze Pfeile stellen normale Pankreasgänge dar. (100-fache Vergrößerung)

IV. ERGEBNISSE
45
Die in Abbildung 12 veranschaulichte H.E.-Übersichtsfärbung verdeutlicht, dass es keinen
Unterschied in der Pankreasmorphologie zwischen der Socs3Δpanc-Maus und der Socs3
F/F-
Kontrollmaus besteht. In beiden Darstellungen erkennen wir die Pankreas-typische
Gewebestruktur zusammengesetzt aus Lobuli, welche von Bindegewebssepten voneinander
abgegrenzt sind. Da dieses Organ hauptsächlich aus serös-exokrinen Drüsen besteht, ist in
beiden Schnittpräparaten eine große Dichte an basophilen Azinuszellen und hellen
zentroazinären Zellen deutlich erkennbar. Zwischen den Läppchen verlaufen
Ausführungsgänge, die sich im Ductus pancreaticus vereinigen. Vereinzelt erkennt man
scharf abgrenzbare Langerhans-Inseln, welche die endokrine Funktion des Organs erfüllen.
Aus diesem unauffälligen Befund beider Präparate lässt sich schließen, dass die homozygote
Socs3-Deletion in der Socs3Δpanc-Linie die Morphologie des exokrinen und endokrinen
Pankreaskompartiments nicht beeinträchtigt.
Zur Evaluation der Funktion der endokrinen Zellgruppen detektierten wir mittels
immunhistochemischer Färbung Insulin und Glucagon unter Nutzung der Spezifität
markierter Primärantikörper. Auch hier konnten wir keinen Unterschied in der
Hormonproduktion feststellen. Es lassen sich im Vergleich bei beiden Mauslinien sowohl
Insulin, welches von den zentral liegenden dominierenden β-Zellen produziert wird, als auch
Glucagon, das von den randständig lokalisierten α-Zellen sezerniert wird, nachweisen. Somit
bleibt die endokrine Funktion der Langerhans-Inselzellen von der Socs3-Mutation
unbeeinflusst.
Zusätzlich hatten wir das Pankreasgewebe beider exemplarisch dargestellten Tiere
immunhistochemisch nach Cytokeratin-19 (CK-19) detektiert. CK-19 ist ein zytoskelettales
Protein, welches von einschichtigem Epithel und luminalen Drüsenzellen zur Stabilisierung
und Formgebung produziert wird. Hierbei zeigen sich sowohl bei der Socs3Δpanc-Maus als
auch der Kontrollmaus Socs3F/F eine annähernd gleiche CK-19 Expression.
Desweiteren interessierte uns die Gewichtsentwicklung der Tiere aus beiden Mauslinien.
Hierzu ermittelten wir ab der dritten bis ca. 20. Lebenswoche wöchentlich das Gewicht von
zehn Socs3-Knockoutmäusen und 15 SocsF/F-Mäusen. Die gemessenen Gewichte wurden
tabellarisch aufgelistet, der Mittelwert und die Standardabweichung daraus berechnet und in
Abbildung 13 graphisch dargestellt. Auch hier konnten wir keine gravierenden Unterschiede
zwischen den Knockout- und den Kontrollmäusen feststellen. Zwar zeigen die Socs3Δpanc-
Tiere in der Verlaufsbeobachtung gegenüber gleichaltrigen Vergleichstieren leicht höhere
Mittelwerte, diese gleichen sich aber ab der 18. Lebenswoche den Werten der SocsF/F-Mäuse
an und bleiben im Verlauf konstant.
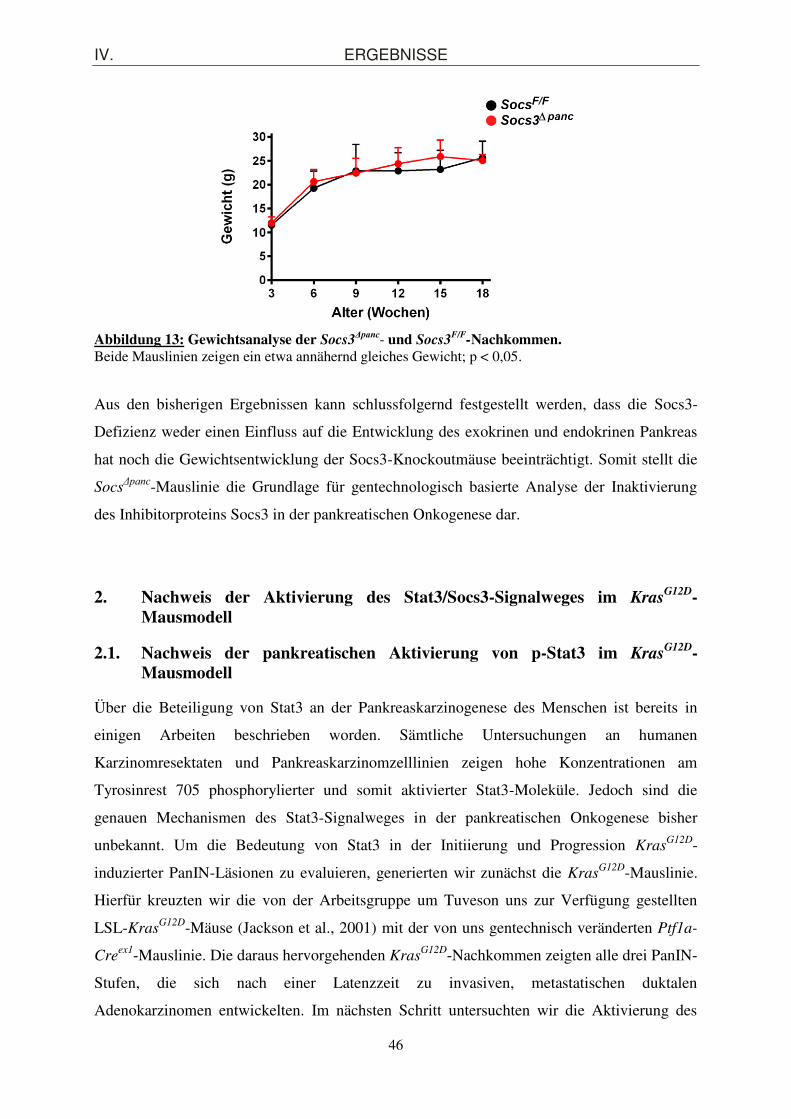
IV. ERGEBNISSE
46
Abbildung 13: Gewichtsanalyse der Socs3Δpanc- und Socs3
F/F-Nachkommen.
Beide Mauslinien zeigen ein etwa annähernd gleiches Gewicht; p < 0,05.
Aus den bisherigen Ergebnissen kann schlussfolgernd festgestellt werden, dass die Socs3-
Defizienz weder einen Einfluss auf die Entwicklung des exokrinen und endokrinen Pankreas
hat noch die Gewichtsentwicklung der Socs3-Knockoutmäuse beeinträchtigt. Somit stellt die
SocsΔpanc-Mauslinie die Grundlage für gentechnologisch basierte Analyse der Inaktivierung
des Inhibitorproteins Socs3 in der pankreatischen Onkogenese dar.
2. Nachweis der Aktivierung des Stat3/Socs3-Signalweges im KrasG12D-
Mausmodell
2.1. Nachweis der pankreatischen Aktivierung von p-Stat3 im KrasG12D-
Mausmodell
Über die Beteiligung von Stat3 an der Pankreaskarzinogenese des Menschen ist bereits in
einigen Arbeiten beschrieben worden. Sämtliche Untersuchungen an humanen
Karzinomresektaten und Pankreaskarzinomzelllinien zeigen hohe Konzentrationen am
Tyrosinrest 705 phosphorylierter und somit aktivierter Stat3-Moleküle. Jedoch sind die
genauen Mechanismen des Stat3-Signalweges in der pankreatischen Onkogenese bisher
unbekannt. Um die Bedeutung von Stat3 in der Initiierung und Progression KrasG12D-
induzierter PanIN-Läsionen zu evaluieren, generierten wir zunächst die KrasG12D-Mauslinie.
Hierfür kreuzten wir die von der Arbeitsgruppe um Tuveson uns zur Verfügung gestellten
LSL-KrasG12D-Mäuse (Jackson et al., 2001) mit der von uns gentechnisch veränderten Ptf1a-
Creex1-Mauslinie. Die daraus hervorgehenden Kras
G12D-Nachkommen zeigten alle drei PanIN-
Stufen, die sich nach einer Latenzzeit zu invasiven, metastatischen duktalen
Adenokarzinomen entwickelten. Im nächsten Schritt untersuchten wir die Aktivierung des

IV. ERGEBNISSE
47
Stat3-Signalweges in den Pankreata dieser Tumormodellmäuse. Mittels der unter II.2.3.
beschriebenen Western Blot-Methode wiesen wir die Expression von Stat3-Molekülen nach,
die dann durch Phosphorylierung am Tyrosin 705 zu p-Stat3 aktiviert werden. In der
Abbildung 14 sind die Ergebnisse dargestellt.
Der Nachweis von Stat3 bzw. p-Stat3 in den präneoplastischen Läsionen des Pankreas führt
zu der Hypothese, dass Stat3 an der pankreatischen Onkogenese im KrasG12D-Mausmodell
beteiligt sein könnte.
Abbildung 14: Proteinbiochemischer Nachweis der Expression von Stat3 und p-Stat3 in Pankreata neun Wochen alter Kras
G12D-Mäuse.
2.2. Expression des Stat3-abhängigen endogenen Inhibitors Socs3 im Kras
G12D-Mausmodell
Nachdem wir die Stat3-Expression und dessen Phosphorylierung zu p-Stat3 in der KrasG12D-
Maus mittels Western Blot analysiert haben, überprüften wir anschließend die Aktivierung
von Socs3. Das Socs3-Gen mit Stat3-responsiven Elementen in ihren Promotoren gehört zu
den Zielgenen dieser Moleküle. Das Genprodukt Socs3 reguliert in einer negativen
Rückkopplung (negatives feedback) die Stat3-Signalvermittlung und verhindert somit eine
konstitutive Aktivierung der Signalkaskade.
Durch Bindung der Stat3-Dimeren an die vorgeschaltete Promotorregion kommt es zur
Induktion der Socs3-Transkription und zur Bildung der mRNA, einer einsträngigen messenger
Ribonukleinsäure, die als Matrize der Proteinbiosynthese dient. Konsequenterweise führt die
vermehrte Stat3-Aktivierung in der KrasG12D-Tumormauslinie zu einer erhöhten
Transkriptionsfrequenz des Socs3-Genlokus mit gesteigerter Synthese der mRNA. Abbildung
15 veranschaulicht die von uns mittels quantitativer Real-Time-PCR (quantitative Echtzeit-
PCR) gemessene relative Socs3-mRNA in Pankreata von KrasG12D-Mäusen im Vergleich zu
den Kontroll-LSL-KrasG12D-Tieren, bei denen die vorgeschaltete gefloxte STOP-Kassette die
Transkription des onkogenen Kras verhindert.

IV. ERGEBNISSE
48
Abbildung 15: Quantitative Real-Time-PCR Analysen der vermehrten Expression von Socs3-mRNA in neun Wochen alten Kras
G12D-Mäusen. Mittelwert ± Standardabweichung (n ≥ 5), *p < 0.005
Die Ergebnisse zeigen für die Mauslinie KrasG12D eine deutlich erhöhte mRNA-Expression
des endogenen Inhibitors Socs3 gegenüber der der Kontrolltiere. Konsequenterweise führt die
gesteigerte mRNA-Bildung zu einer verstärkten Socs3-Proteinbiosynthese an den Ribosomen.
Im Folgenden überprüften wir die Expression von Socs3 in der KrasG12D-Mauslinie auf
histochemischer Ebene. Wie Abbildung 16 demonstriert, konnten wir immunhistochemisch
mithilfe eines gegen Socs3 gerichteten Primärantikörpers dessen vermehrte Bildung in Azini-
Zellen und PanIN-Läsionen neun Wochen alter KrasG12D-Mäuse nachweisen. Somit legt die
starke Induktion des endogenen Inhibitors eine regulierende Funktion des Socs3 in der
pankreatischen Onkogenese nahe.
Abbildung 16: Immunhistochemische Detektion von Socs3 in Zellen der Azini (schwarze Pfeile) und in den PanIN-Läsionen (weiße Pfeile) neun Wochen alter Kras
G12D-Mäuse.

IV. ERGEBNISSE
49
2.3. Aktivierung der an der Jak2/Stat3-Signaltransduktion beteiligten Proteine in Kras
G12D-Mäusen
Neben der Induktion des endogenen Inhibitors Socs3 regulieren phosphorylierte Stat3-Dimere
intranukleär die Transkription bestimmter Gene, die an der Steuerung von Proliferation,
Wachstum, Apoptose und Inflammation beteiligt sind. Der Nachweis der exprimierten
Genprodukte lässt auf die Aktivierung des Jak2/Stat3-Signaltransduktionsweges
rückschließen. Hierzu setzen wir erneut die Methode der Immunhistochemie ein und
detektieren die relevanten Proteine. Das zu färbende Pankreasgewebe stammte von 13
Wochen alten KrasG12D-Mäusen und offenbart histologisch PanIN-Vorläuferläsionen (siehe
Abbildung 17). Charakteristisch für die pankreatischen intraepithelialen Neoplasien in diesem
Wochenalter ist die Zunahme der Anzahl der Ausführungsgänge, eine Verlängerung des
epithelialen Zellkörpers der Ducti und eine erhöhte zytoplasmatische Muzinproduktion. Hinzu
geht das normalerweise flach dominierende Ausführungsgang-Epithel in ein papilläres
Wachstumsmuster über. Diese auf Abbildung 17 aufgeführten Eigenschaften weisen auf
PanIN-Stufen 1A bis 1B hin, die zusätzlich ein umgebendes desmoplasmatisches Stroma
aufweisen. Sowohl in den Azinuszellen als auch in den Pankreasgangzellen innerhalb der
PanIN-Läsionen lässt sich das phosphorylierte Jak2-Protein (p-Jak2) nachweisen. Das aktive
p-Jak2 phosphoryliert das Stat3-Monomer am Tyrosinrest Y705 zu p-Stat3 (Babon et al.,
2006). Dementsprechend konnten wir, wie Abbildung 17 veranschaulicht, sowohl in den
Azini als auch in den duktalen Zellen eine erhöhte p-Stat3-Konzentration detektieren.
Darüber hinaus ist es uns gelungen, die Expression Stat3-abhängiger Proteine in den
präneoplastischen Läsionen und in den Azinuszellen 13 Wochen alter KrasG12D-Mäuse zu
demonstrieren. Hierzu zählen Cyclin D1, Survivin, die antiapoptisch wirkenden und zur
Proteinfamilie der Bcl-2 (B-cell lymphoma-2) gehörigen Mitglieder Mcl-1 (Myloid cell
lymphoma-1) und Bcl-XL. Zusätzlich ließen sich andere Moleküle bei unseren
immunhistochemischen Untersuchungen detektieren, wie die Matrix Metalloproteinase-2
(MMP-2), welche im Abbau der extrazellulären Matrix involviert ist und somit für den
Prozess der Metastasierung eine entscheidende Rolle spielt und das proinflammatorische
Protein Cox-2 (Cyclooxygenase-2). Auch die Expression des von uns nachgewiesenen
Hitzeschockproteins-70 (heat shock protein 70, HSP-70) ist Stat3-abhängig und wird durch
dessen Aktivierung gesteigert. HSP-70 stabilisiert in Situationen des zellulären Stresses
andere Eiweißstrukturen, um diese vor Denaturierung und Abbau zu schützen. Andererseits
unterstützt HSP-70 die proteasomale Degradation funktionsunfähiger Proteinmoleküle.
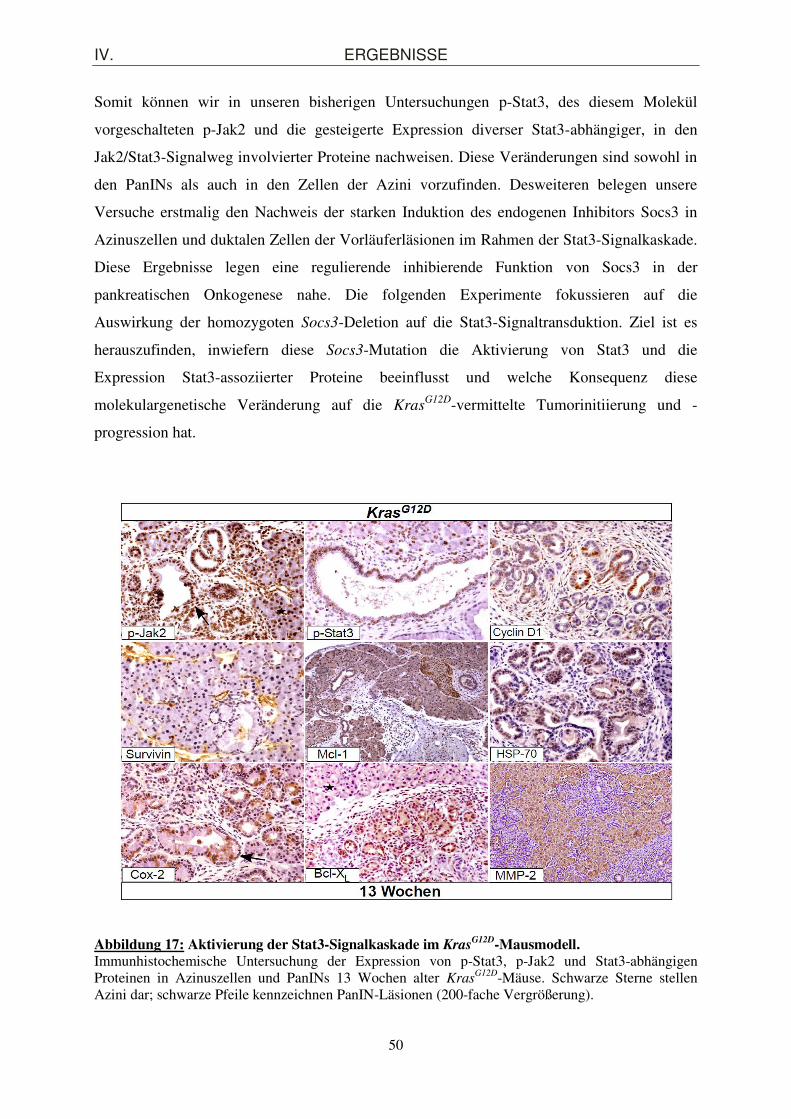
IV. ERGEBNISSE
50
Somit können wir in unseren bisherigen Untersuchungen p-Stat3, des diesem Molekül
vorgeschalteten p-Jak2 und die gesteigerte Expression diverser Stat3-abhängiger, in den
Jak2/Stat3-Signalweg involvierter Proteine nachweisen. Diese Veränderungen sind sowohl in
den PanINs als auch in den Zellen der Azini vorzufinden. Desweiteren belegen unsere
Versuche erstmalig den Nachweis der starken Induktion des endogenen Inhibitors Socs3 in
Azinuszellen und duktalen Zellen der Vorläuferläsionen im Rahmen der Stat3-Signalkaskade.
Diese Ergebnisse legen eine regulierende inhibierende Funktion von Socs3 in der
pankreatischen Onkogenese nahe. Die folgenden Experimente fokussieren auf die
Auswirkung der homozygoten Socs3-Deletion auf die Stat3-Signaltransduktion. Ziel ist es
herauszufinden, inwiefern diese Socs3-Mutation die Aktivierung von Stat3 und die
Expression Stat3-assoziierter Proteine beeinflusst und welche Konsequenz diese
molekulargenetische Veränderung auf die KrasG12D-vermittelte Tumorinitiierung und -
progression hat.
Abbildung 17: Aktivierung der Stat3-Signalkaskade im Kras
G12D-Mausmodell. Immunhistochemische Untersuchung der Expression von p-Stat3, p-Jak2 und Stat3-abhängigen Proteinen in Azinuszellen und PanINs 13 Wochen alter Kras
G12D-Mäuse. Schwarze Sterne stellen Azini dar; schwarze Pfeile kennzeichnen PanIN-Läsionen (200-fache Vergrößerung).

IV. ERGEBNISSE
51
3. Homozygote Socs3-Deletion führt zu einer konstitutiven Aktivierung des Stat3-Signalweges im Kras
G12D-Mausmodell
3.1. Generierung der pankreasspezifisch Socs3-defizienten Tumormodellmaus Kras
G12D;Socs3Δpanc
Um der Bedeutung von Socs3 auf die KrasG12D-vermittelte Tumorinitiierung und -progression
genauer nachzugehen, generierten wir zunächst eine Socs3-defiziente KrasG12D-Mauslinie.
Hierzu benötigten wir erneut zwei Mauslinien: die bereits unter I.2.2. beschriebene LSL-
KrasG12D;Ptf1a-Cre
ex1 (KrasG12D)-Tumormaus und die unter III.1.1. charakterisierte
Socs3F/F
;Ptf1a-Creex1 (Socs3
Δpanc)-Mauslinie. Nachdem wir beide Mauslinien miteinander
gekreuzt haben, gingen Nachkommen hervor, die einen Genotyp LSL-KrasG12D
;Socs3F/F
;
Ptf1a-Creex1 aufwiesen. Diese neu entstandene auch als Kras
G12D;Socs3
Δpanc bezeichnete
Tumormauslinie besaß sowohl das aktive pankreasspezifische Onkogen KrasG12D, welches die
Entwicklung und Progression der PanINs förderte, als auch die Ptf1a-Cre-vermittelte
homozygote Deletion des Exons 2 des Socs3-Gens, die zu einer fehlenden Expression von
Socs3 führte. Somit haben wir mit der Etablierung des neuen KrasG12D
;Socs3Δpanc-
Tumormausmodells die Basis für die daran anschließenden Versuche geschaffen. Hierin galt
es die Charakteristika der von uns generierten KrasG12D
;Socs3Δpanc-Maus mit den der nicht
Socs3-defizienten KrasG12D-Kontrollmaus zu vergleichen und so die Bedeutung bzw. die
Relevanz des Inhibitorproteins Socs3 auf die KrasG12D-vermittelte Entwicklung
pankreatischer präneoplastischer Läsionen und die Stat3-Phosphorylierung zu untersuchen.
3.2. Proteinbiochemische und morphologische Charakterisierung von Kras
G12D;Socs3
Δpanc-Mäusen
3.2.1. Proteinbiochemischer Nachweis verstärkter Stat3-Phosphorylierung und Expression Stat3-abhängiger Proteine
Der Tyrosinrest 759 an der zytosolischen gp130-Untereinheit stellt im phosphorylierten
Zustand die Andockstelle für die Stat3-Monomere und Socs3-Moleküle dar. Gebunden an
diesem aktiven Phosphotyrosin werden die Stat3-Proteine anschließend Jak2-vermittelt am
Tyrosin 705 zu p-Stat3 umgewandelt. Das phosphorylierte Tyrosin 759 ist zugleich
Bindungsort von dem endogenen Inhibitor Socs3, an dem dieser den inhibierenden Feedback-
Mechanismus auf den Stat3-Signaltransduktionsweg ausübt (Lang et al., 2000).
Ausgehend von diesen Vorkenntnissen untersuchten wir im folgenden Schritt die Auswirkung
der pankreasspezifischen Socs3-Defizienz auf die Induktion von p-Stat3 und den Stat3-
abhängigen Proteinen in KrasG12D
;Socs3Δpanc-Mäusen. Hierzu verglichen wir die Western

IV. ERGEBNISSE
52
Blot-Analysen der Socs3-defizienten KrasG12D-Maus mit den der Kras
G12D-Kontrollmaus. Die
von uns im Rahmen der Experimente verwendeten Antikörper sind in Tabelle 7 aufgelistet.
Zunächst überprüften wir proteinbiochemisch die Expression von Stat3, p-Stat3 und p-Jak2 in
beiden Mauslinien. Wie Abbildung 18 demonstriert, exprimieren sowohl vier Wochen alte
KrasG12D
;Socs3Δpanc-Mäuse als auch gleich alte Kras
G12D-Kontrolltiere Stat3 und p-Stat3.
Jedoch weist das Pankreasgewebe der KrasG12D
;Socs3Δpanc-Mäuse eine frühzeitige und
prolongierte Stat3-Phosphorylierung auf, was an dem hohen Proteinlevel von p-Stat3 zu
erkennen ist. Auch die Phosphorylierung der dem Stat3-Molekül vorgeschalteten Jak2 ist
ebenso frühzeitig nachweisbar.
Desweiteren interessierte uns die Auswirkung der Socs3-Defizienz auf die Expression Stat3-
abhängiger Proteine. Wie die Western Blot-Analysen in der Abbildung 18 veranschaulichen,
führt die frühzeitige und prolongierte Aktivierung von p-Stat3 ebenfalls zu einer gesteigerten
Expression diverser Stat3-Zielproteine in den exemplarisch dargestellten vier Wochen alten
KrasG12D
;Socs3Δpanc-Mäusen im Vergleich zu den Kras
G12D-Kontrolltumormäusen. Für Bcl-
XL, Mcl-1, Cox-2, HSP-70 und Cyclin D1 weisen die Socs3-deletierten KrasG12D-Mäuse ein
höheres Proteinlevel auf. ß-actin wurde hier als Ladekontrolle eingesetzt.
A B
Abbildung 18: Proteinbiochemischer Nachweis der Socs3-Defizienz in Kras
G12D;Socs3
Δpanc-Mäusen. A: Nachweis der frühzeitigen und prolongierten Stat3-Phosphorylierung im Pankreasgewebe vier Wochen alter Socs3-defizienter Kras
G12D-Mäuse. B: Verstärkte Expression Stat3-abhängiger Proteine in Western Blot-Analysen vier Wochen alter Kras
G12D;Socs3
Δpanc-Mäuse.

IV. ERGEBNISSE
53
3.2.2. Konstitutive Aktivierung des onkogenen K-Ras-Proteins in Kras
G12D;Socs3
Δpanc-Mäusen
Das G-Protein K-Ras bindet im aktiven Zustand Guanosintriphosphat (GTP). Durch die
intrinsische Hydrolyseaktivität wird das GTP-gebundene K-Ras bei Terminierung des
Ligandensignals in die inaktive GDP-assoziierte Form umgewandelt. Bei der KrasG12D-
Mutation liegt eine ungenügende Überführung von GTP-korreliertem K-Ras in die inaktive
GDP-gebundene Konfiguration, wodurch eine konstitutive, wachstumsfördernde K-Ras-
Proteinaktivität entsteht (Shibata et al., 1990). Da in den Socs3-defizienten KrasG12D-Mäusen
sowohl eine KrasG12D-Mutation als auch eine Socs3-Deletion vorliegt, überprüften wir
mithilfe des Ras-Aktivierungsassays mit dem Ras Activation Assay Kit, ob die Inaktivierung
von Socs3 einen Einfluss auf die Ausprägung der G12D-Mutation ausübt. Das Prinzip dieser
eingesetzten Methode beruht auf der hochspezifischen Bindung der Ras-bindenden Domäne
(RBD) des im Kit enthaltenen Proteins Raf-1 (Ras effector kinase) an aktives GTP-
korreliertes Ras-Protein. Nach Herstellung der Proteinlysate aus Pankreasgewebe beider
Mauslinien wurden diese jeweils mit dem Raf-1 RBD Agarose enthaltenden Reaktionsreagenz
versetzt. Anschließend ließ sich der entstandene Raf-RBD/GTP-Ras Komplex im Western
Blot mit einem konjugierten Ras-spezifischen Sekundärantikörper detektieren. Die Western
Blot-Ergebnisse des durchgeführten Ras Pull-Down Assays in der Abbildung 19 zeigen für
beide vier Wochen alte KrasG12D- und Kras
G12D;Socs3
Δpanc-Mauslinien eine annähernd gleich
starke Expression von RBD- und damit von GTP-assoziiertem Ras-Protein im Pankreas. Aus
unseren gewonnenen Daten kann demnach geschlossen werden, dass die pankreasspezifische
Inaktivierung des Socs3-Gens keinen Einfluss auf die Ras-Aktivität bzw. auf die Expression
des konstitutiv aktiven K-Ras-Proteins ausübt. Beide genetische Veränderungen existieren
nebeneinander und scheinen synergistisch die Progression von pankreatischen
intraepithelialen Neoplasien zum duktalen Adenokarzinom zu verstärken.
Abbildung 19: Nachweis der Expression konstitutiv aktiver K-Ras-Proteine. In beiden Mauslinien liegt eine unveränderte Aktivierung des onkogenen K-Ras-Proteins vor.
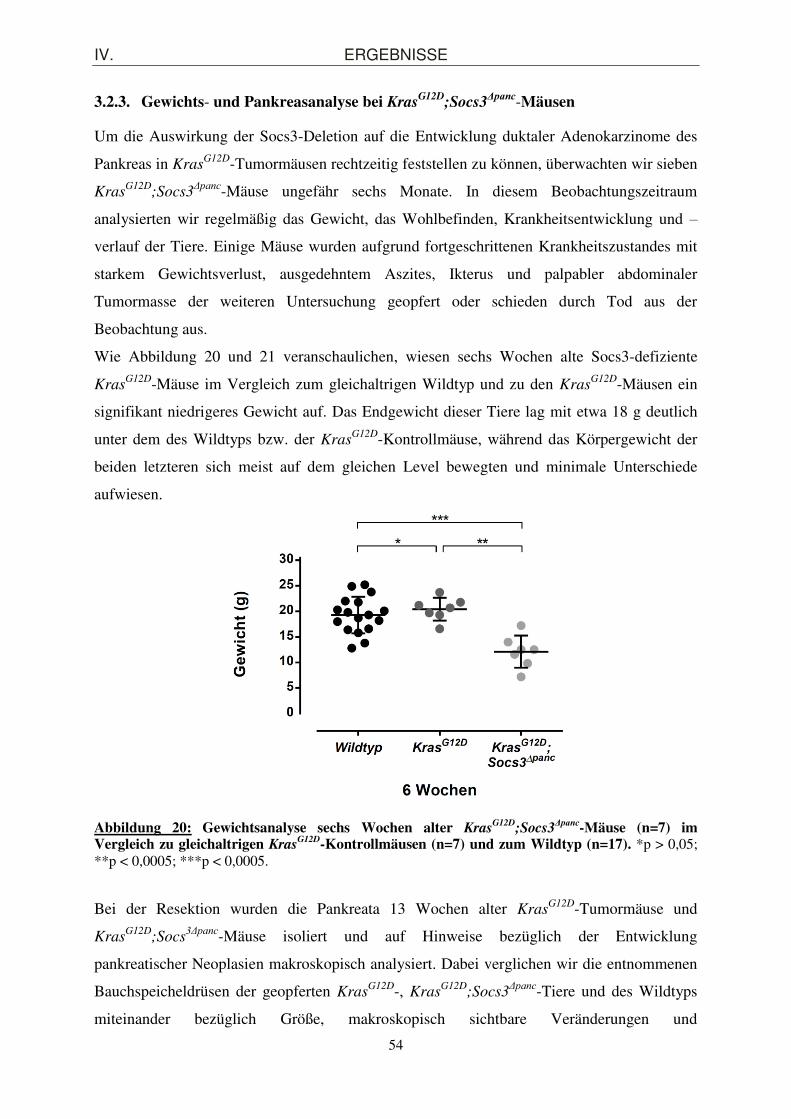
IV. ERGEBNISSE
54
3.2.3. Gewichts- und Pankreasanalyse bei KrasG12D
;Socs3Δpanc-Mäusen
Um die Auswirkung der Socs3-Deletion auf die Entwicklung duktaler Adenokarzinome des
Pankreas in KrasG12D-Tumormäusen rechtzeitig feststellen zu können, überwachten wir sieben
KrasG12D
;Socs3Δpanc-Mäuse ungefähr sechs Monate. In diesem Beobachtungszeitraum
analysierten wir regelmäßig das Gewicht, das Wohlbefinden, Krankheitsentwicklung und –
verlauf der Tiere. Einige Mäuse wurden aufgrund fortgeschrittenen Krankheitszustandes mit
starkem Gewichtsverlust, ausgedehntem Aszites, Ikterus und palpabler abdominaler
Tumormasse der weiteren Untersuchung geopfert oder schieden durch Tod aus der
Beobachtung aus.
Wie Abbildung 20 und 21 veranschaulichen, wiesen sechs Wochen alte Socs3-defiziente
KrasG12D-Mäuse im Vergleich zum gleichaltrigen Wildtyp und zu den Kras
G12D-Mäusen ein
signifikant niedrigeres Gewicht auf. Das Endgewicht dieser Tiere lag mit etwa 18 g deutlich
unter dem des Wildtyps bzw. der KrasG12D-Kontrollmäuse, während das Körpergewicht der
beiden letzteren sich meist auf dem gleichen Level bewegten und minimale Unterschiede
aufwiesen.
Abbildung 20: Gewichtsanalyse sechs Wochen alter KrasG12D
;Socs3Δpanc-Mäuse (n=7) im
Vergleich zu gleichaltrigen KrasG12D-Kontrollmäusen (n=7) und zum Wildtyp (n=17). *p > 0,05;
**p < 0,0005; ***p < 0,0005.
Bei der Resektion wurden die Pankreata 13 Wochen alter KrasG12D-Tumormäuse und
KrasG12D
;Socs3Δpanc-Mäuse isoliert und auf Hinweise bezüglich der Entwicklung
pankreatischer Neoplasien makroskopisch analysiert. Dabei verglichen wir die entnommenen
Bauchspeicheldrüsen der geopferten KrasG12D-, Kras
G12D;Socs3
Δpanc-Tiere und des Wildtyps
miteinander bezüglich Größe, makroskopisch sichtbare Veränderungen und

IV. ERGEBNISSE
55
Gewebebeschaffenheit. Die Abbildung 21 demonstriert ein im Vergleich zum Wildtyp stark
vergrößertes Pankreas der KrasG12D-Tumormaus. Zusätzlich lassen sich Lobulierungen und
Indurationen im Organgewebe nachweisen.
Die zusätzliche Socs3-Defizienz in KrasG12D-Tumormäusen bewirkt jedoch eine starke
Abnahme des Organgewichts (siehe Abbildung 21).
Abbildung 21: Analyse des makroskopischen Pankreasaspekts. Größe, Form und Konsistenz der Pankreata 13 Wochen alter Kras
G12D; Socs3
Δpanc-Mäuse und KrasG12D-Kontrollmäuse im Vergleich
zum Wildtyp. Zum Größenvergleich ist aus jeder Mauslinie ein Tier exemplarisch dargestellt.
Abbildung 21 demonstriert ein im Vergleich zum Wildtyp bis auf ein Fünftel der Organmasse
geschrumpfte Bauchspeicheldrüse mit diffus nodulärer und lobulierter Parenchymstruktur.
Die ausgedehnte Fibrosierung des sonst weichen Gewebes lässt das Organ derb, knotig und
formlos erscheinen. Diese makroskopisch sichtbaren Veränderungen an den Pankreata der
von uns generierten KrasG12D
;Socs3Δpanc-Mauslinie deuten auf vorhandene präneoplastische
und/oder neoplastische Prozesse hin, die sich scheinbar in weiter fortgeschrittenen Stadien
befinden als die der gleichaltrigen Mäuse mit singulärer KrasG12D-Mutation.
Im Beobachtungszeitraum von sechs Monaten entwickelten 28,57% (2/7) der
KrasG12D
;Socs3Δpanc-Mäuse ein Pankreasadenokarzinom, während in der Kras
G12D-Kohorte
keine Karzinomentwicklung zu beobachten war. Die Tumorlatenz von etwa 63 Tagen bei den
KrasG12D
;Socs3Δpanc-Versuchstieren war somit geringer im Vergleich zu den Kras
G12D-
Kontrolltieren.
Aus unseren bisher gewonnen Daten können wir damit annehmen, dass die
pankreasspezifische Deletion des Socs3-Gens und die dadurch aufgehobene negative
Rückkopplung auf den Stat3-Signalweg einen maßgeblichen Effekt auf die Tumorprogression
in der KrasG12D-induzierten pankreatischen Onkogenese haben könnte. Die im Folgenden
durchgeführten Experimente sollen unsere angenommene Hypothese belegen.

IV. ERGEBNISSE
56
4. Einfluss der Inaktivierung des endogenen Inhibitorproteins Socs3 auf die PanIN-Progression im Kras
G12D-Mausmodell
4.1. Beschleunigung der PanIN-Progression in KrasG12D
;Socs3Δpanc-Mäusen
4.1.1. Morphologische und quantitative Charakterisierung der PanIN-Läsionen
In der vergleichenden Analyse der PanIN-Entwicklung in Pankreata von KrasG12D- und
KrasG12D
;Socs3Δpanc-Mäusen im folgenden Abschnitt möchten wir den Einfluss der aus der
Inaktivierung des Inbibitorproteins Socs3 fehlenden Hemmung des Stat3-Signalweges auf die
alterskorrelierte PanIN-Progression in Socs3-defizienten KrasG12D-Mäusen näher beleuchten.
Zunächst untersuchten wir anhand H.E.-gefärbter Gewebefeinschnitte vier, neun, 13 und 18
Wochen alter KrasG12D-Kontrollmäuse und Kras
G12D;Socs3
Δpanc-Mäuse die
Pankreasmorphologie im Vergleich. Die Ergebnisse dieser Übersichtsfärbung sind in zehn-
und 100-facher Vergrößerung in der Abbildung 22 dargestellt.
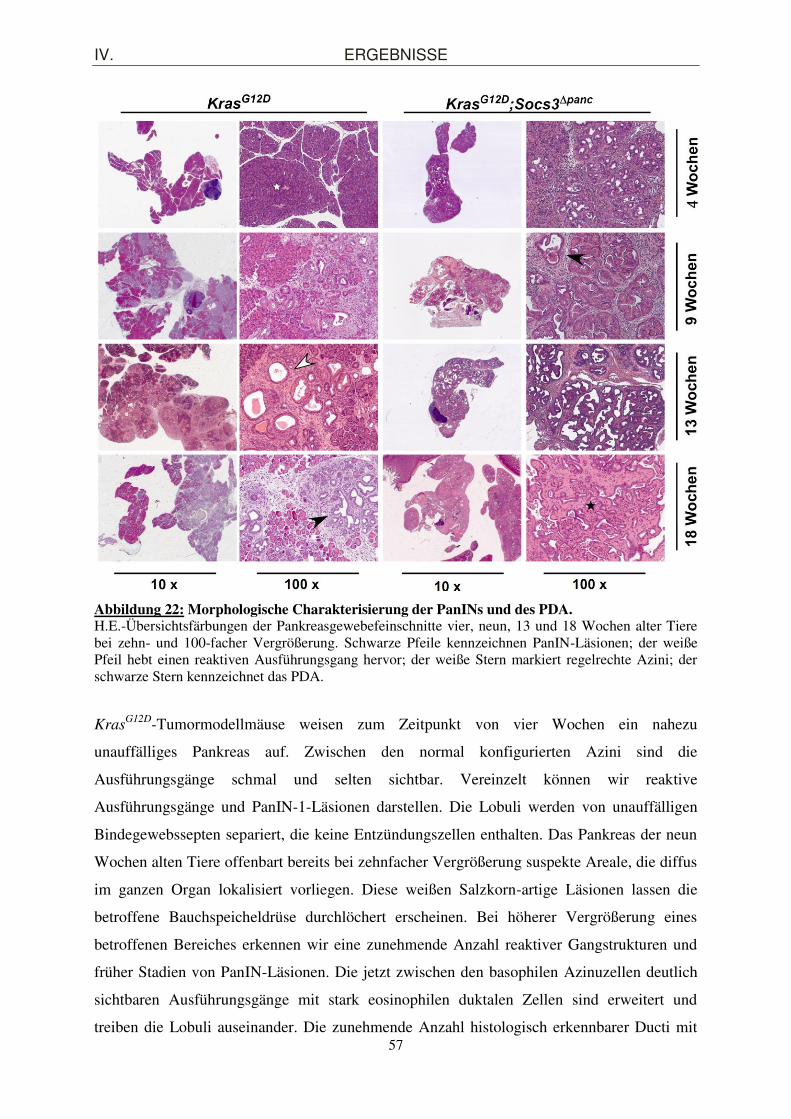
IV. ERGEBNISSE
57
Abbildung 22: Morphologische Charakterisierung der PanINs und des PDA. H.E.-Übersichtsfärbungen der Pankreasgewebefeinschnitte vier, neun, 13 und 18 Wochen alter Tiere bei zehn- und 100-facher Vergrößerung. Schwarze Pfeile kennzeichnen PanIN-Läsionen; der weiße Pfeil hebt einen reaktiven Ausführungsgang hervor; der weiße Stern markiert regelrechte Azini; der schwarze Stern kennzeichnet das PDA.
KrasG12D-Tumormodellmäuse weisen zum Zeitpunkt von vier Wochen ein nahezu
unauffälliges Pankreas auf. Zwischen den normal konfigurierten Azini sind die
Ausführungsgänge schmal und selten sichtbar. Vereinzelt können wir reaktive
Ausführungsgänge und PanIN-1-Läsionen darstellen. Die Lobuli werden von unauffälligen
Bindegewebssepten separiert, die keine Entzündungszellen enthalten. Das Pankreas der neun
Wochen alten Tiere offenbart bereits bei zehnfacher Vergrößerung suspekte Areale, die diffus
im ganzen Organ lokalisiert vorliegen. Diese weißen Salzkorn-artige Läsionen lassen die
betroffene Bauchspeicheldrüse durchlöchert erscheinen. Bei höherer Vergrößerung eines
betroffenen Bereiches erkennen wir eine zunehmende Anzahl reaktiver Gangstrukturen und
früher Stadien von PanIN-Läsionen. Die jetzt zwischen den basophilen Azinuzellen deutlich
sichtbaren Ausführungsgänge mit stark eosinophilen duktalen Zellen sind erweitert und
treiben die Lobuli auseinander. Die zunehmende Anzahl histologisch erkennbarer Ducti mit

IV. ERGEBNISSE
58
Formanomalie lässt auf einen neoplastischen duktalen Prozess hindeuten. Zudem weist das
vermehrte desmoplastische Stroma auf eine verstärkte Entzündungsreaktion mit fibrotischer
Begleitreaktion hin. Mit zunehmendem Alter nimmt sowohl die Dichte als auch die Schwere
der PanIN-Läsionen zu. Die zehnfache Vergrößerung des Pankreas der 13 Wochen alten
KrasG12D-Kontrollmaus veranschaulicht, dass in diesem Stadium ein dominierender Organteil
von den PanINs eingenommen ist und die zunehmende Ausdehnung dieser eine Verdrängung
der exokrinen Azini bedeutet. Bei 100-facher Vergrößerung erkennen wir im Vergleich zur
neun Wochen alten Maus Areale mit übermäßig dilatierten Gangstrukturen, die statt dem
normal flachen Epithel ein papilläres Wachstumsmuster offenbaren. Zusätzlich bewirkt die
raumfordernde Expansion des umgebenden desmoplastischen Stromas ein Auseinanderdriften
der Azini. Um die 18. Lebenswoche zeigt die 10-fach vergrößerte Pankreasaufnahme der
Mäuse mit KrasG12D-Mutation eine weitere Progression der PanIN-Dichte und des
Schweregrades der Neoplasien. Wie Abbildung 22 verdeutlicht, erstrecken sich
präneoplastische Läsionen und das Stroma über fast das ganze Organ mit daraus
resultierendem deutlichem Rückgang der Azinuszellen. In der höheren Vergrößerung
erkennen wir zusätzlich intraluminale Pankreasgangnekrosen als Charakteristikum der PanINs
im Stadium 3 und atrophierte Azinuszellen. Die exemplarisch dargestellten Pankreata der
KrasG12D-Mäuse unterschiedlichen Alters zeigen somit die für diese Tumormauslinie typische
zeit- bzw. altersabhängige PanIN-Progression.
Die Socs3-Defizienz in KrasG12D
-Mäusen lässt in den H.E.-gefärbten Gewebefeinschnitten
eine Beschleunigung der PanIN-Progression erkennen. Im Vergleich zu gleichaltrigen
KrasG12D-Mäusen ist das Pankreas dieser Mäuse interessanterweise schon nach vier Wochen
morphologisch sehr stark umgewandelt. Bereits bei 10-facher Vergrößerung demaskieren sich
duktale Vorläuferläsionen aufgrund starker Gangdilatation, die annähernd die Hälfte des
Organs eingenommen haben. Während Pankreata vier Wochen alter KrasG12D-Mäuse nahezu
unauffällig erscheinen, fallen bei näherer histologischer Betrachtung der Gewebestruktur
gleichaltriger KrasG12D
;Socs3Δpanc-Mäuse ausgedehnte fibrotisch umgebaute heterogene
Areale auf. Neben normalen Azinis und reaktiven Gängen sind ausgeprägte hochgradig
veränderte Ausführungsgänge in Form von PanIN-1- bis PanIN-3-Läsionen zu erkennen. Die
schon in der vierten Lebenswoche eingetretenen Veränderungen der Gangstruktur deuten auf
ein im Vergleich zu der KrasG12D-Tumormauslinie sehr früh einsetzendes neoplastisches
duktales Geschehen mit ausgedehnter fibrotischer Begleitreaktion hin.
Um das Ausmaß der PanIN-Progression bei pankreasspezifischer Inaktivierung von Socs3
noch genauer beurteilen zu können, führten wir anschließend einen quantitativ analytischen
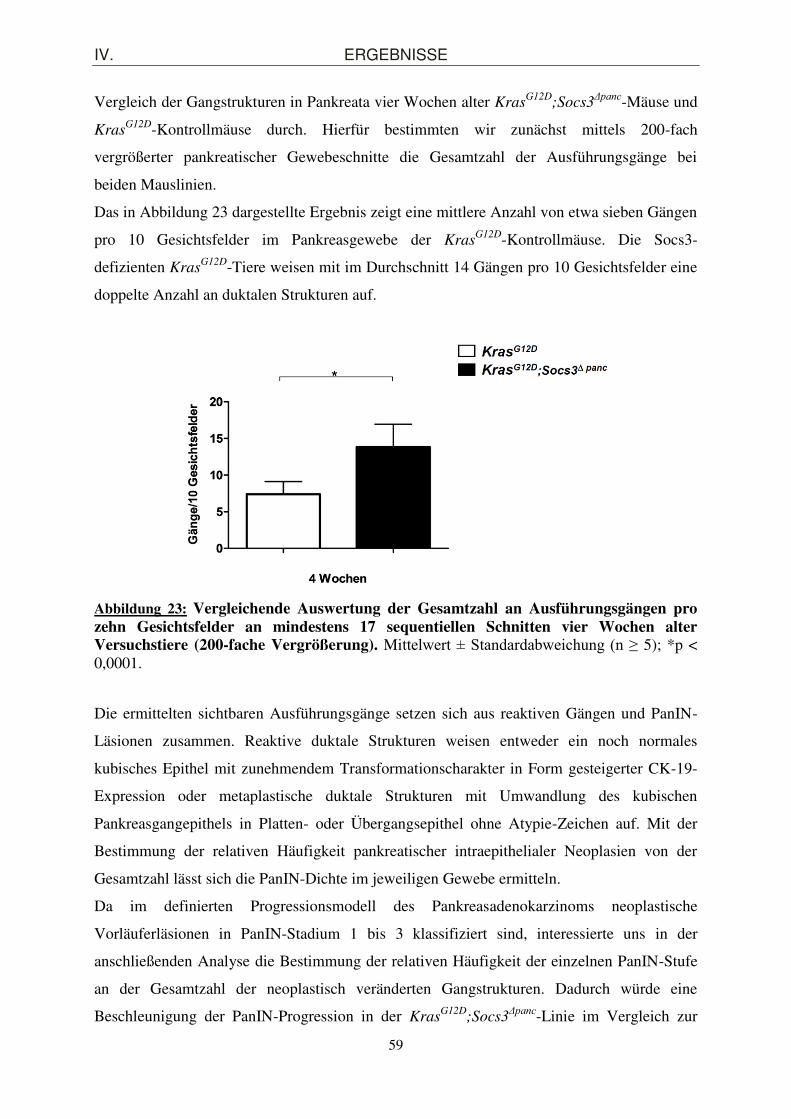
IV. ERGEBNISSE
59
Vergleich der Gangstrukturen in Pankreata vier Wochen alter KrasG12D
;Socs3Δpanc-Mäuse und
KrasG12D-Kontrollmäuse durch. Hierfür bestimmten wir zunächst mittels 200-fach
vergrößerter pankreatischer Gewebeschnitte die Gesamtzahl der Ausführungsgänge bei
beiden Mauslinien.
Das in Abbildung 23 dargestellte Ergebnis zeigt eine mittlere Anzahl von etwa sieben Gängen
pro 10 Gesichtsfelder im Pankreasgewebe der KrasG12D-Kontrollmäuse. Die Socs3-
defizienten KrasG12D-Tiere weisen mit im Durchschnitt 14 Gängen pro 10 Gesichtsfelder eine
doppelte Anzahl an duktalen Strukturen auf.
Abbildung 23: Vergleichende Auswertung der Gesamtzahl an Ausführungsgängen pro zehn Gesichtsfelder an mindestens 17 sequentiellen Schnitten vier Wochen alter Versuchstiere (200-fache Vergrößerung). Mittelwert ± Standardabweichung (n ≥ 5); *p < 0,0001.
Die ermittelten sichtbaren Ausführungsgänge setzen sich aus reaktiven Gängen und PanIN-
Läsionen zusammen. Reaktive duktale Strukturen weisen entweder ein noch normales
kubisches Epithel mit zunehmendem Transformationscharakter in Form gesteigerter CK-19-
Expression oder metaplastische duktale Strukturen mit Umwandlung des kubischen
Pankreasgangepithels in Platten- oder Übergangsepithel ohne Atypie-Zeichen auf. Mit der
Bestimmung der relativen Häufigkeit pankreatischer intraepithelialer Neoplasien von der
Gesamtzahl lässt sich die PanIN-Dichte im jeweiligen Gewebe ermitteln.
Da im definierten Progressionsmodell des Pankreasadenokarzinoms neoplastische
Vorläuferläsionen in PanIN-Stadium 1 bis 3 klassifiziert sind, interessierte uns in der
anschließenden Analyse die Bestimmung der relativen Häufigkeit der einzelnen PanIN-Stufe
an der Gesamtzahl der neoplastisch veränderten Gangstrukturen. Dadurch würde eine
Beschleunigung der PanIN-Progression in der KrasG12D
;Socs3Δpanc-Linie im Vergleich zur
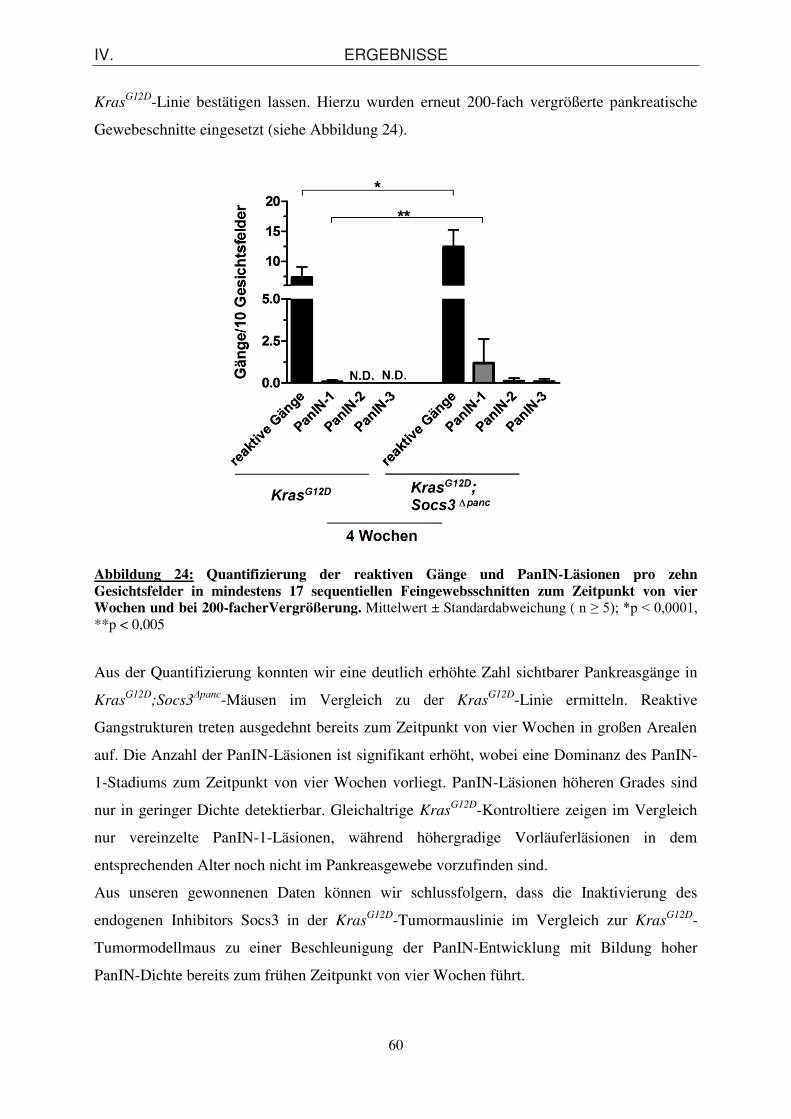
IV. ERGEBNISSE
60
KrasG12D-Linie bestätigen lassen. Hierzu wurden erneut 200-fach vergrößerte pankreatische
Gewebeschnitte eingesetzt (siehe Abbildung 24).
Abbildung 24: Quantifizierung der reaktiven Gänge und PanIN-Läsionen pro zehn Gesichtsfelder in mindestens 17 sequentiellen Feingewebsschnitten zum Zeitpunkt von vier Wochen und bei 200-facherVergrößerung. Mittelwert ± Standardabweichung ( n ≥ 5); *p < 0,0001, **p < 0,005
Aus der Quantifizierung konnten wir eine deutlich erhöhte Zahl sichtbarer Pankreasgänge in
KrasG12D
;Socs3Δpanc-Mäusen im Vergleich zu der Kras
G12D-Linie ermitteln. Reaktive
Gangstrukturen treten ausgedehnt bereits zum Zeitpunkt von vier Wochen in großen Arealen
auf. Die Anzahl der PanIN-Läsionen ist signifikant erhöht, wobei eine Dominanz des PanIN-
1-Stadiums zum Zeitpunkt von vier Wochen vorliegt. PanIN-Läsionen höheren Grades sind
nur in geringer Dichte detektierbar. Gleichaltrige KrasG12D-Kontroltiere zeigen im Vergleich
nur vereinzelte PanIN-1-Läsionen, während höhergradige Vorläuferläsionen in dem
entsprechenden Alter noch nicht im Pankreasgewebe vorzufinden sind.
Aus unseren gewonnenen Daten können wir schlussfolgern, dass die Inaktivierung des
endogenen Inhibitors Socs3 in der KrasG12D-Tumormauslinie im Vergleich zur Kras
G12D-
Tumormodellmaus zu einer Beschleunigung der PanIN-Entwicklung mit Bildung hoher
PanIN-Dichte bereits zum frühen Zeitpunkt von vier Wochen führt.
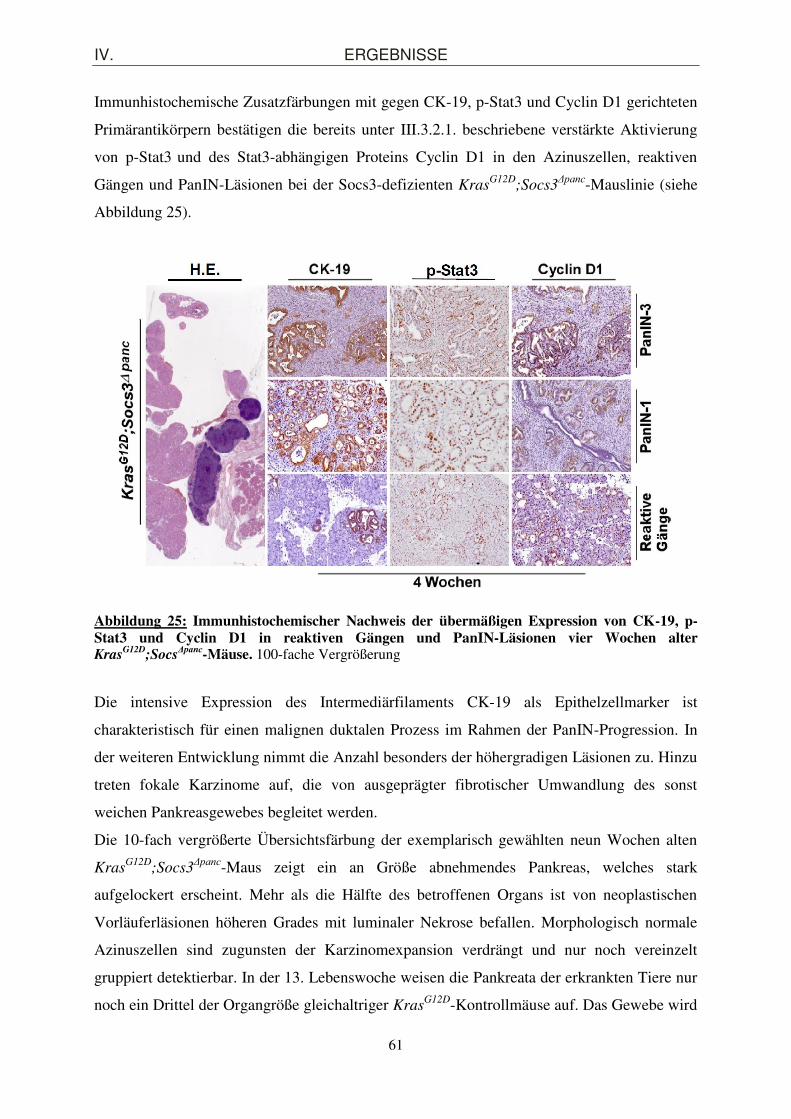
IV. ERGEBNISSE
61
Immunhistochemische Zusatzfärbungen mit gegen CK-19, p-Stat3 und Cyclin D1 gerichteten
Primärantikörpern bestätigen die bereits unter III.3.2.1. beschriebene verstärkte Aktivierung
von p-Stat3 und des Stat3-abhängigen Proteins Cyclin D1 in den Azinuszellen, reaktiven
Gängen und PanIN-Läsionen bei der Socs3-defizienten KrasG12D
;Socs3Δpanc-Mauslinie (siehe
Abbildung 25).
Abbildung 25: Immunhistochemischer Nachweis der übermäßigen Expression von CK-19, p-Stat3 und Cyclin D1 in reaktiven Gängen und PanIN-Läsionen vier Wochen alter Kras
G12D;Socs
Δpanc-Mäuse. 100-fache Vergrößerung
Die intensive Expression des Intermediärfilaments CK-19 als Epithelzellmarker ist
charakteristisch für einen malignen duktalen Prozess im Rahmen der PanIN-Progression. In
der weiteren Entwicklung nimmt die Anzahl besonders der höhergradigen Läsionen zu. Hinzu
treten fokale Karzinome auf, die von ausgeprägter fibrotischer Umwandlung des sonst
weichen Pankreasgewebes begleitet werden.
Die 10-fach vergrößerte Übersichtsfärbung der exemplarisch gewählten neun Wochen alten
KrasG12D
;Socs3Δpanc-Maus zeigt ein an Größe abnehmendes Pankreas, welches stark
aufgelockert erscheint. Mehr als die Hälfte des betroffenen Organs ist von neoplastischen
Vorläuferläsionen höheren Grades mit luminaler Nekrose befallen. Morphologisch normale
Azinuszellen sind zugunsten der Karzinomexpansion verdrängt und nur noch vereinzelt
gruppiert detektierbar. In der 13. Lebenswoche weisen die Pankreata der erkrankten Tiere nur
noch ein Drittel der Organgröße gleichaltriger KrasG12D-Kontrollmäuse auf. Das Gewebe wird
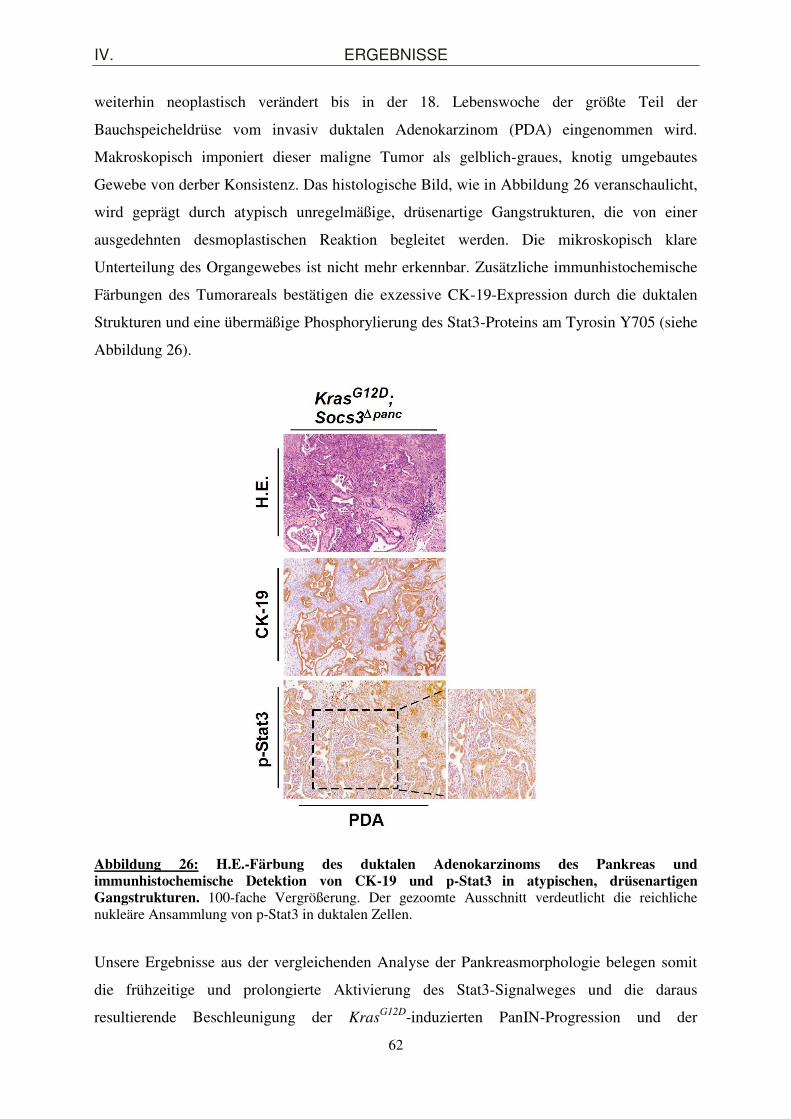
IV. ERGEBNISSE
62
weiterhin neoplastisch verändert bis in der 18. Lebenswoche der größte Teil der
Bauchspeicheldrüse vom invasiv duktalen Adenokarzinom (PDA) eingenommen wird.
Makroskopisch imponiert dieser maligne Tumor als gelblich-graues, knotig umgebautes
Gewebe von derber Konsistenz. Das histologische Bild, wie in Abbildung 26 veranschaulicht,
wird geprägt durch atypisch unregelmäßige, drüsenartige Gangstrukturen, die von einer
ausgedehnten desmoplastischen Reaktion begleitet werden. Die mikroskopisch klare
Unterteilung des Organgewebes ist nicht mehr erkennbar. Zusätzliche immunhistochemische
Färbungen des Tumorareals bestätigen die exzessive CK-19-Expression durch die duktalen
Strukturen und eine übermäßige Phosphorylierung des Stat3-Proteins am Tyrosin Y705 (siehe
Abbildung 26).
Abbildung 26: H.E.-Färbung des duktalen Adenokarzinoms des Pankreas und immunhistochemische Detektion von CK-19 und p-Stat3 in atypischen, drüsenartigen Gangstrukturen. 100-fache Vergrößerung. Der gezoomte Ausschnitt verdeutlicht die reichliche nukleäre Ansammlung von p-Stat3 in duktalen Zellen.
Unsere Ergebnisse aus der vergleichenden Analyse der Pankreasmorphologie belegen somit
die frühzeitige und prolongierte Aktivierung des Stat3-Signalweges und die daraus
resultierende Beschleunigung der KrasG12D-induzierten PanIN-Progression und der
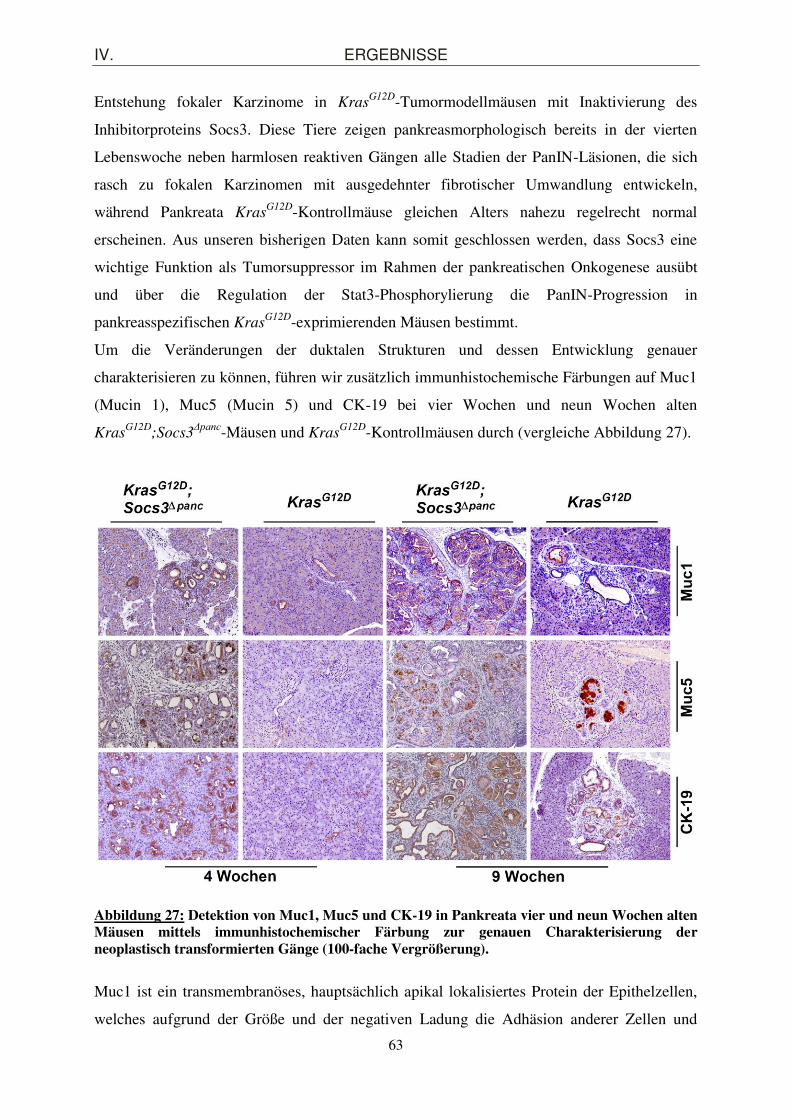
IV. ERGEBNISSE
63
Entstehung fokaler Karzinome in KrasG12D-Tumormodellmäusen mit Inaktivierung des
Inhibitorproteins Socs3. Diese Tiere zeigen pankreasmorphologisch bereits in der vierten
Lebenswoche neben harmlosen reaktiven Gängen alle Stadien der PanIN-Läsionen, die sich
rasch zu fokalen Karzinomen mit ausgedehnter fibrotischer Umwandlung entwickeln,
während Pankreata KrasG12D-Kontrollmäuse gleichen Alters nahezu regelrecht normal
erscheinen. Aus unseren bisherigen Daten kann somit geschlossen werden, dass Socs3 eine
wichtige Funktion als Tumorsuppressor im Rahmen der pankreatischen Onkogenese ausübt
und über die Regulation der Stat3-Phosphorylierung die PanIN-Progression in
pankreasspezifischen KrasG12D-exprimierenden Mäusen bestimmt.
Um die Veränderungen der duktalen Strukturen und dessen Entwicklung genauer
charakterisieren zu können, führen wir zusätzlich immunhistochemische Färbungen auf Muc1
(Mucin 1), Muc5 (Mucin 5) und CK-19 bei vier Wochen und neun Wochen alten
KrasG12D
;Socs3Δpanc-Mäusen und Kras
G12D-Kontrollmäusen durch (vergleiche Abbildung 27).
Abbildung 27: Detektion von Muc1, Muc5 und CK-19 in Pankreata vier und neun Wochen alten Mäusen mittels immunhistochemischer Färbung zur genauen Charakterisierung der neoplastisch transformierten Gänge (100-fache Vergrößerung).
Muc1 ist ein transmembranöses, hauptsächlich apikal lokalisiertes Protein der Epithelzellen,
welches aufgrund der Größe und der negativen Ladung die Adhäsion anderer Zellen und

IV. ERGEBNISSE
64
Mikroorganismen verhindert. Bei zahlreichen Adenokarzinomen oder anderen epithelialen
Tumoren kommt es zu einer Überexpression dieses Moleküls. Durch Polaritätsverlust der
Zelle bei fortgeschrittenen Tumoren wird Muc1 vermehrt auch basolateral gebildet, was eine
verminderte Zelladhäsion hervorruft. Die Abnahme des Zellzusammenhalts stellt somit die
Voraussetzung der Metastasenbildung bei invasiven Karzinomen dar. Muc5 gehört ebenfalls
zu den Glykoproteinen, die jedoch von Schleimhäuten als sekretorische Mucine abgesondert
werden. Eine verstärkte Expression dieses Makromoleküls liegt in PanIN-Läsionen und
Adenokarzinomen vor. Wie die in Abbildung 27 dargestellten Immunhistochemie-Bilder
veranschaulichen, lassen sich zahlreiche sowohl Muc1- als auch Muc5-exprimierende
Ausführungsgänge und PanINs in vier Wochen alten KrasG12D
;Socs3Δpanc-Mäusen
nachweisen, die in der neunten Lebenswoche an Dichte und Schweregrad zunehmen. Im
Vergleich dazu sind im Pankreasgewebe der exemplarisch gewählten KrasG12D-Kontrollmaus
die Mucin-positiven Gänge und intraepithelialen Neoplasien erst zum Zeitpunkt von neun
Wochen detektierbar, während vier Wochen alte Tiere nur vereinzelt Muc1 in den reaktiven
Gängen produzieren. Auch die Anzahl der CK-19-exprimierenden Gangstrukturen und
PanINs in Pankreata der KrasG12D-Linie ist deutlich geringer als im Gewebe gleichaltriger
Socs3-defizienten KrasG12D-Mäuse.
Somit unterstützen die gewonnenen Daten einer frühzeitigen und intensiven Expression
PanIN-spezifischer Markerproteine Muc1, Muc5 und CK-19 in der Bauchspeicheldrüse von
KrasG12D
;Socs3Δpanc-Mäusen die Aussage: Die Deletion von Socs3 in der Kras
G12D;Socs3
Δpanc-
Mauslinie führt zu einer deutlichen Beschleunigung der Progression KrasG12D-induzierter
PanINs mit frühem Auftreten von invasiv duktalen Pankreasadenokarzinomen.
5. Zusammenfassung der Ergebnisse
In Bezug auf die unter I.4. formulierte Zielsetzung können die Ergebnisse der im Rahmen
dieser Arbeit durchgeführten Untersuchungen wie folgt zusammengefasst werden:
1) Im LSL-KrasG12D
;Ptf1a-Creex1(Kras
G12D)-Tumormausmodell konnten wir in den
PanIN-Läsionen eine Aktivierung des Stat3-Signaltransduktionsweges nachweisen.
Das Homodimer aus zwei aktivierten p-Stat3 fungiert als intranukleärer
Transkriptionsfaktor und reguliert die Expression diverser Stat3-Zielproteine. Socs3
als wichtiger Tumorsuppressor lässt sich in KrasG12D-Mäusen in hoher Konzentration
detektieren und reguliert den IL6/Stat3-Signalweg über negative Rückkopplung.

IV. ERGEBNISSE
65
2) Socs3-defiziente KrasG12D-Mäuse weisen im Verlauf der Beobachtung ein im
Vergleich zu KrasG12D-Kontrolltieren deutlich reduziertes Körper- und
Pankreasgewicht auf.
3) Mithilfe des Cre/loxP-Rekombinationssystems gelang es uns die Generierung der
LSL-KrasG12D
;Socs3F/F
;Ptf1a-Creex1(Kras
G12D;Socs3
Δpanc)-Mauslinie. Diese genetisch
veränderten Tiere weisen eine pankreasspezifische Deletion des Exons 2 des Socs3-
Gens auf. Die auf diesem Wege erreichte Inaktivierung des endogenen
Inhibitorproteins Socs3 im Pankreas führt zu einer frühzeitigen und prolongierten
Jak2-vermittelten Stat3-Phosphorylierung am Tyrosinrest Y705 (p-Stat3), die bereits
zum Zeitpunkt von vier Wochen detektierbar ist. Sowohl Morphologie und
Physiologie des endokrinen und exokrinen Pankreas als auch die PanIN-induzierte
konstitutive Aktivierung des onkogenen KrasG12D bleiben von der Socs3-Deletion
unbeeinflusst. Histomorphologische und proteinbiochemische Untersuchungen des
Pankreas der KrasG12D
;Socs3Δpanc-Mäuse im Vergleich zu Kras
G12D-Kontrollmäusen
belegen jedoch die beschleunigte Entwicklung und Progression der altersabhängigen
KrasG12D-induzierten PanIN-Läsionen schon nach der vierten Lebenswoche.
4) Im Beobachtungszeitraum von sechs Monaten entwickelten 28,57% (2/7) der
KrasG12D
;Socs3Δpanc-Mäuse ein Pankreasadenokarzinom, während in der Kras
G12D-
Kohorte keine Karzinomentwicklung zu beobachten war. Die Tumorlatenz von etwa
63 Tagen bei den KrasG12D
;Socs3Δpanc-Versuchstieren war somit kürzer im Vergleich
zu den KrasG12D-Kontrolltieren. Das Pankreasgewebe ist schon zum Zeitpunkt von
vier Wochen morphologisch stark umgewandelt. Es finden sich heterogene Areale mit
normalen Azinis, PanIN-Läsionen unterschiedlichen Grades mit ausgedehnter
desmoplasmatischer Stromareaktion vor.
5) Aus den gewonnenen Ergebnissen lässt sich schlussfolgern, dass die Inaktivierung des
endogenen Tumorsuppressors Socs3 die Progression präneoplastischer Läsionen im
murinen KrasG12D-Tumormodell beschleunigt. Socs3 übt somit durch negative
Rückkopplung eine tragende Funktion im Akzelerationsprozess der PanINs aus und
bestimmt über die Stat3-Phosphorylierung die Regulation der PanIN-Entwicklung und
–Progression zu invasiv duktalem Adenokarzinom.

V. DISKUSSION
66
V. DISKUSSION
1. Rolle von Socs3 für die pankreatische Karzinogenese im Kras
G12D-Mausmodell
Obwohl Socs3 bereits in einigen humanen Tumorzelllinien und –geweben als ein wichtiger
Tumorsuppressor beschrieben wurde, blieb dessen genaue Rolle und Funktion in der
pankreatischen Karzinogenese bisher unbeleuchtet.
In einigen Studien wurde über eine endogene DNA-Hypermethylierung als epigenetischer
Inaktivierungsmechanismus bestimmter Socs-Gene im Rahmen der Karzinogenese spekuliert.
He und Kollegen konnten in humanen NSCLC (Nichtkleinzelliges Karzinom)-Zellinien,
Mesotheliom-Zelllinien und operativ gewonnenem Lungenkarzinomgewebe eine im
Vergleich zu den normalen Kontrollzelllinien supprimierte Expression des Socs3-Transkripts
feststellen. Die Socs3-Defizienz in den Tumorzellen resultierte in einer Hyperaktivität des
Jak/Stat-Signaltransduktionsweges mit proliferativer und onkogener Eigenschaft. Eine
genetische Wiederherstellung der Socs3-Funktion induzierte hingegen durch Inhibition der
Stat3-Aktivität Apoptose und Suppression des Zellwachstums.
Interessanterweise wurden hypermethylierte CpG-Inseln des Socs3-Gens in allen
Tumorzelllinien gefunden, während normale Gewebszellen mit regelrechter Socs3-Expression
keine Veränderung der entsprechenden Genareale zeigten (He et al., 2003).
CpG-Inseln haben ihre Bedeutung in der Regulation der Genexpression und stellen somit
einen Mechanismus der epigenetischen Genregulation dar (Antequera 2003). Die
Methylierung der CpG-Inseln eines Gens hat die Konsequenz, dass dieses Gen nicht mehr
transkribiert wird, was unter anderem in der Tumorentstehung durch das Abschalten
relevanter Tumorsuppressorgene eine große Rolle spielt. Untersuchungen mittels
molekulargenetischer Analysen an ösophagealem Plattenepithelkarzinom und Barrett-
Adenokarzinom (Tischoff et al., 2007), humanem Kopf- und Hals-Plattenepithelkarzinom und
deren dysplastischen Vorläuferläsionen (Weber et al., 2005), hepatozellulären Karzinomen
(HCC) und deren präneoplastischen Läsionen (Niwa et al., 2005) offenbarten ebenfalls eine
Hypermethylierung der CpG-Inseln enthaltenden Socs3-Promotorregion. Auch hier kam es
als Folge der Socs3-Defizienz zu einer konstitutiven Aktivierung des Stat3-Signalweges. Im
Kolon bietet die endogene Induktion der Socs3-Expression demzufolge einen protektiven
Mechanismus gegenüber Verletzung-induzierter Kryptenhyperplasie und Entzündungs-
assoziierter Karzinogenese durch Hemmung der IL-6/Stat3-Signalkaskade (Rigby et al.,
2007).

V. DISKUSSION
67
In den meisten Karzinomzellen der Ovarien und der Mamma kommt es zu einer Inaktivierung
des Socs1- und Socs2-Gens durch CpG-Hypermethylierung in der Promotorregion, wodurch
diese Gene nicht mehr transkribiert werden. Im Gegensatz zu den Ergebnissen von He und
Kollegen war die Promotorregion des Socs3-Gens in den beiden genannten Tumorentitäten
nicht hypermethyliert und das Genprodukt dementsprechend intrazellulär in hoher
Konzentration detektierbar. Darüber hinaus wurde eine erhöhte Stat3-Aktivität in den
betroffenen Tumorzellen registriert, die auf die fehlende Expression des Socs1-Moleküls mit
wachstums- und proliferationsinhibierender Wirkung zurück geführt wurde (Yoshikawa et
al., 2001; Lee et al., 2006). Die konstitutive Stat3-Aktivierung ist mit einer Überexpression
von Socs3 assoziiert und führte zur Zunahme der Proliferationsrate der mamillären
Epithelzellen. Die gewonnenen Ergebnisse ließen Sutherland und Kollegen annehmen, dass
das endogene Inhibitorprotein Socs3 in dieser Zellart keine Tumorsuppressor-Aktivität erfüllt,
was wiederum dessen vielseitige teilweise unerforschte Funktion in verschiedenen
Karzinomarten verdeutlicht (Sutherland et al., 2004).
Drei Jahre nach der Veröffentlichung der Arbeit von Sutherland und Kollegen, wurde die
Studie von der Arbeitsgruppe um Evans publik, in der sie entgegen Sutherlands Aussagen
eine Zunahme der Expression von Socs2 bis Socs7 postulierten. Im Vergleich zu den
normalen mamillären Epithelzellen reagierten die Tumorzelllinien trotz hypermethylierter
CpG-Inseln adäquat auf Zytokin- bzw. Wachstumsfaktor-Signale mit erhöhter Expression von
Socs1 und Socs3, wodurch der Mechanismus der Socs-vermittelten Feedback-Inhibition nicht
gestört wurde (Evans et al., 2007). In Übereinstimmung mit den Ergebnissen von der
Arbeitsgruppe um Sutherland, konnten sie belegen, dass trotz Zunahme des Socs3-Levels in
Mammakarzinomzellen keine effiziente Feedback-Inhibition auf konstitutive Stat3-
Aktivierung erfolgte. Socs3 fungierte stattdessen als Tumorpromotor, welcher die IFN-
α/Stat1-vermittelte Wachstumsinhibition hemmte.
Die intrazelluläre Stat3-Aktivierung im Rahmen der pankreatischen Onkogenese scheint
ebenfalls unter der Socs3-Kontrolle zu stehen. In unseren Versuchen konnten wir erstmalig
zeigen, dass in KrasG12D-Tumormäusen eine erhöhte Socs-3 Expression vorliegt. Um den
Einfluss von Socs3 auf die Pankreaskarzinogenese im KrasG12D-transgenen Mausmodell zu
untersuchen, generierten wir eine weitere Mauslinie mit pankreasspezifischer Socs3-Deletion.
Eine Inaktivierung des Socs3-Gens führte demnach zu einer prolongierten und verstärkten
Stat3-Phosphorylierung, was wiederum die Beschleunigung der PanIN-Progression und die
Entstehung von Pankreaskarzinomen förderte.

V. DISKUSSION
68
Im Einklang mit den aufgeführten Daten konnten wir in unserer Arbeit zeigen, dass innerhalb
des Beobachtungszeitraums von sechs Monaten 28,57% (2/7) der KrasG12D
;Socs3Δpanc-Mäuse
ein Pankreasadenokarzinom entwickelten, während in der KrasG12D-Kohorte keine
Karzinomentwicklung zu beobachten war. Die Tumorlatenz von etwa 63 Tagen bei den
KrasG12D
;Socs3Δpanc-Versuchstieren war somit kürzer im Vergleich zu den Kras
G12D-
Kontrolltieren.
2. Einfluss von Socs3 auf die Apoptose und Proliferation im Kras
G12D-Mausmodell
Proliferation und Apoptose sind zellautonome Effekte, die das Gleichgewicht zwischen
Zellwachstum und –eliminierung gewährleisten. Ein Ungleichgewicht dieser teilweise
komplexen Mechanismen kann eine unkontrollierte Proliferation begünstigen und die
Tumorentstehung fördern. Während in frühen und späten PanIN-Stadien die Apoptose als ein
seltenes Ereignis anzusehen ist, stellt die Proliferation einen dominierenden Mechanismus in
der Tumorprogression dar (Luttges et al., 2003).
Stat3 ist bereits in vielen Untersuchungen als einen wichtigen Regulator des programmierten
Zelltods und des Zellzyklus beschrieben worden (Konnikova et al., 2003; Lin et al., 2005;
Xihong et al., 2008). Aktivierte p-Stat3 Homo- oder Heterodimere aktivieren als
Transkriptionsfaktoren zahlreiche Zielgene. U.a. werden Proteine wie Bcl-XL, Mcl-1,
Survivin und Cyclin D1 vermehrt gebildet. Bcl-XL und Mcl-1 gelten als anti-apoptotische
Signalmoleküle, die durch verstärkte Expression zum Überleben transformierter Zellen in
einigen humanen Tumorerkrankungen beiträgt (Krajewska et al., 1996; Okaro et al., 2001; Ma
et al., 2004; Bhattacharya et al., 2005; Yu et al., 2009; Placzek et al., 2010).
Untersuchungen an humanen Pankreastumorzelllinien haben gezeigt, dass eine Inaktivierung
von Stat3 in einer Abnahme der Transkriptionsrate von Bcl-XL und Mcl-1 resultierte und
folglich die Apoptose induzierte (Huang et al., 2010). In unseren weiterführenden
Experimenten konnten wir zum ersten Mal nachweisen, dass die homozygote Deletion des
negativen Regulators Socs3 in vier Wochen alten KrasG12D-Tumormäusen eine frühe und
prolongierte Stat3-Aktivierung hervorruft und somit eine im Vergleich zu gleichaltrigen
KrasG12D-Tumormäusen stärkere Expression der anti-apoptotischen Proteine Bcl-XL und Mcl-
1 verursacht.
Zusätzlich zu den o.g. Ergebnissen konnten wir proteinbiochemisch eine stärkere Cyclin-D1
Expression in Socs3-defizienten KrasG12D-Tumormäusen im Vergleich zu den Kras
G12D-
Kontrollmäusen nachweisen. Cyclin D1 ist ein proliferationsförderndes, intranukleär

V. DISKUSSION
69
lokalisiertes Protein, welches die G1/S-Progression innerhalb des Zellzyklus fördert und somit
die proliferative Aktivität einer Zelle steuert (Baldin et al., 1993). Die Überexpression von
Cyclin D1 in KrasG12D
;Socs3Δpanc-Mäusen kennzeichnete somit eine Verkürzung des G1/S-
Phasenübergangs, die wiederum die Progression transformierter Zellen innerhalb des
Zellzyklus beschleunigt und die Entwicklung von Karzinomen wesentlich beeinflusst. In
anderen Studien konnte in Übereinstimmung mit unseren Resultaten ebenfalls eine
Überexpression von Cyclin D1 in humanen Pankreaskarzinomzellen nachgewiesen werden,
die mit einer schlechten Prognose assoziiert ist (Gansauge et al., 1997). Die Bedeutung von
Cyclin D1 in der Tumorgenese nicht nur im Pankreasadenokarzinom, sondern auch in
zahlreichen anderen Tumorarten wird auch in anderen Arbeiten betont (Musgrove et al.,
2011).
In unseren Versuchen konnten wir erstmalig zeigen, dass die übermäßige Stat3 Expression in
vier Wochen alten KrasG12D-Mäusen eine starke Aktivierung der genannten Proteine zur
Folge hat. Dieses Ergebnis verdeutlicht die Proliferation als dominierender Prozess in der
PanIN-Progression. Die zusätzliche, homozygote Deletion des Socs3-Gens in diesen
Tumormäusen führt zu einer im Vergleich zu gleichaltrigen KrasG12D-Kontrollmäusen
verstärkten und prolongierten Aktivierung von Stat3 und der Stat3-assoziierten Proteine durch
Wegfall des negativen Inhibitors. Die dadurch hervorgerufene, persistierende Stat3 Aktivität
und die vermehrte Bildung anti-apoptotischer und proliferationsfördernder Moleküle
verursachten eine Beschleunigung der PanIN-Progression zu pankreatischem duktalem
Adenokarzinom.
Im murinen Modell der Hepatitis-assoziierten Hepatokarzinogenese stieg nach Injektion des
Karzinogens DEN bzw. con A die IL-6 Serumkonzentration sowohl im Wildtyp als auch in
den Soc3-Knockouts ohne signifikanten Unterschied dramatisch an. Jedoch war in den Socs3-
defizienten Tieren nach 24 Stunden eine anhaltende und stärkere Stat3-Phosphorylierung im
Vergleich zu den Wildtypen detektierbar. In diesem Zusammenhang nahm die
Transkriptionsfrequenz der Stat3-Zielgene Bcl-XL, Mcl-1, Cyclin D1 zu. Die homozygote
Deletion des Socs3-Gens in den Leberparenchymzellen resultierte in einer verstärkten,
prolongierten Stat3-Aktivierung und beschleunigte infolgedessen die Entwicklung des HCC
(Ogata et al., 2006).
Ein weiteres Ergebnis unserer experimentellen Untersuchungen stellte die
immunhistochemiche Detektion des Enzyms Cyclooxygenase 2 (Cox-2) in präneoplastischen
pankreatischen Läsionen dar. Cox-2 gehört zu den Zielgenen von Stat3 und wird bei dessen
Aktivierung verstärkt transkribiert (Yu et al., 2008). Cox-2-Synthese wird erst bei

V. DISKUSSION
70
Entzündungen bzw. Verletzung des Gewebes induziert und fördert anschließend die
Produktion von pro-inflammatorischen Prostaglandinen, die chronische Entzündungen
unterhalten. Inflammatorische Prozesse sind bedeutende epigenetische Faktoren, die
maßgeblich zur Entstehung und Progression zahlreicher Tumoren führen (Shacter et al.,
2002). Die PanIN-Progression und die Tumorentwicklung in der Pankreaskarzinogenese
scheinen auch entzündungsassoziiert zu sein (Raimondi et al., 2010). Da Cox2 ein Zielgen
von Stat3 darstellt und dessen Genprodukt pro-inflammatorische Wirkung besitzt, kann davon
ausgegangen werden, dass Stat3 maßgeblich als Regulator tumor-assoziierter Entzündung
fungiert. Mittels immunhistochemischer Färbungen konnten wir in PanINs der KrasG12D-
Tumormäuse Cox-2 nachweisen. Unsere Western-Blot Analysen demonstrierten eine bereits
in der vierten Lebenswoche verstärkte und prolongierte Expression von Cox-2 in Socs3-
defizienten KrasG12D-Mäusen im Vergleich zu den gleichaltrigen Kras
G12D-Kontrolltieren.
Diese Ergebnisse deuten darauf hin, dass die homozygote Deletion von Socs3 die Stat3-
vermittelte Cox-2-Synthese verstärkt, die wiederum inflammatorische Mechanismen bereits in
der vierten Lebenswoche aktivieren und so eine tumor-assoziierte Entzündung unterhalten,
die die PanIN-Progression und Karzinomentwicklung beschleunigen. Weiterhin begünstigt
das durch Cox-2 gebildete Prostaglandin E2 in vielen Tumorarten die Bildung des
Angiogenese-Faktors VEGF, das wiederum das Tumorwachstum fördert. Demzufolge wird
vermutet, dass Cox-2 eine wesentliche Rolle in der Tumorgenese spielen könnte (Simmons et
al., 2004).
In KrasG12D-induzierten PanINs war uns ebenfalls der proteinbiochemische Nachweis von
HSP-70 möglich. HSP-70 stellt ein wichtiger Bestandteil des Chaperonsystems dar, das
ubiquitär in eukaryontischen Zellen vorkommt. Die wichtigsten Funktionen von HSP-70
liegen in der richtigen Faltung und Aktivierung vieler Proteine. Zusätzlich ermöglicht dieses
Molekül über anti-apoptotische Mechanismen das Überleben und Wachstum von Tumorzellen
(Hatfield et al., 2012). HSP-70 wird reichlich in malignen Tumoren unterschiedlichen
Ursprungs gebildet und seine Expression korreliert mit erhöhter Zellproliferation, schlechter
Differenzierung, Lymphknotenmetastasen und schlechtem Therapie-Outcome (Ciocca et al.,
1993; Vargas-Roig et al., 1997; Jäättelä, 1999). Das Gen HSP-70 gehört ebenfalls zu den
Zielgenen von Stat3. Analog zur verstärkten und prolongierten Aktivierung von Stat3 in vier
Wochen alten, Socs3-defizienten KrasG12D-Tumormäusen konnten wir mittels Western Blot-
Methode ebenfalls eine im Vergleich zu den Kontrolltieren verstärkte HSP-70 Bildung
nachweisen. Unsere Ergebnisse deuten darauf hin, dass HSP-70 als zytoprotektives Molekül

V. DISKUSSION
71
für das Überleben und Wachstum transformierter Azinuszellen im Rahmen der
Tumorprogression von großer Bedeutung ist.
Die Rolle von Socs3 im Rahmen der pankreatischen Karzinogenese ist bisher kaum
beleuchtet worden. In einigen humanen Tumorzelllinien wurde die Bedeutung von Socs3 als
negativer Regulator in der Inhibition der IL-6/Jak2/Stat3-Signalkaskade und der daraus
resultierenden Reduktion zellulärer Proliferation, Induktion von Apoptose und Zellzyklus-
Arrest hervorgehoben. Demzufolge wird diesem Molekül ein Anti-Tumoreffekt
zugeschrieben (Iwahori et al., 2011).
In unseren experimentellen Untersuchungen konnten wir erstmals die Expression von Socs3
im murinen in-vivo KrasG12D-Modell belegen. Demzufolge scheint dieses Molekül in der
Regelung zellulärer Prozesse wie Apoptose, Proliferation, Zelldifferenzierung nicht nur unter
physiologischen Bedingungen sondern auch im Rahmen der pankreatischen Karzinogenese
eine wichtige Stellung einzunehmen. Neben Socs3 konnten wir mittels
immunhistochemischer Methode auch andere Stat3-Zielproteine nachweisen, die in ihrer
Gesamtheit zu kontinuierlicher Proliferation, apoptotischer Resistenz,
Angiogenesestimulation, zellulärer Invasivität und Metastasierung sowie tumor-assoziierter
Inflammation führen. Durch die zusätzliche homozygote Deletion von Socs3 in KrasG12D-
Tumormäusen entfällt dessen Inhibition auf den Stat3-Signalweg, wodurch diese Zielgene
bereits in frühen Lebenswochen vermehrt transkribiert werden und tumor-fördernde
Mechanismen aktivieren.
3. Entzündliche Prozesse fördern die Initiierung und Progression der pankreatischen Vorläuferläsionen zu duktalem Pankreaskarzinom
Inflammation ist definiert als eine lokale oder systemische Reaktion eines Organismus auf
einen internen oder externen Fremdstoff oder ein Antigen mit dem Ziel der Entfernung dieser
potentiell schädigenden Reize, um Voraussetzungen für anschließende
Gewebereparaturprozesse zu schaffen. Damit ist sie Ausdruck der Immunreaktion.
Nach Gewebsverletzung wandern Entzündungszellen wie Mastzellen, Makrophagen,
Granulozyten, Lymphozyten aber auch Fibroblasten in die geschädigte Geweberegion und
aktivieren Reparaturprozesse mit Angiogenese, Koordination der Zellmigration und
Wiederherstellung der extrazellulären Matrix. In der Regel ist die Inflammation ein
selbstlimitierender Prozess aufgrund der balancierten Verhältnisse zwischen initial
freigesetzten, pro-inflammatorischen Botenstoffen und anti-inflammatorischen

V. DISKUSSION
72
Signalmolekülen. Eine Dysregulation dieser Mechanismen kann zu einer chronischen
Entzündung des betroffenen Gewebes führen mit der Gefahr der Tumorentwicklung (Barton,
2008; Medzhitov, 2008; Raimondi et al., 2010).
Das Pankreas als Digestionsorgan enthält in den Zellen der exokrinen Drüsen viele
Enzymvorstufen, die bei Schädigung des Gewebes freigesetzt und aktiviert werden. Es
entsteht eine akute Pankreatitis. Rezidivierende Pankeatitiden führen letztendlich zur
Entstehung einer chronischen Entzündung mit irreversiblen Veränderungen des
Organgewebes, was wiederum der Ausgangspunkt für die Entstehung eines
Pankreaskarzinoms sein kann (Raimondi et al., 2010).
In den letzten Jahren wurde im Rahmen zahlreicher Pankreatitis-Forschungen ein
Zusammenhang zwischen chronischer Pankreatitis und pankreatischer Karzinogenese
postuliert (Lowenfels et al., 1993; Malka et al., 2002).
Kras-Mäuse mit einer chronischen Pankreatitis wie z.B. durch Caerulein (ein
Cholezystokinin-Analogon) induziert entwickelten bereits im fünften Lebensmonat
ausgeprägte Atrophie der Azini und zunehmende Fibrosierung im Vergleich zu den
Kontrolltieren. Desweiteren zeigte sich in den Pankreata der Kras-Tiere eine deutliche azinär-
duktale Metaplasie sowie multifokale metaplastische Gänge mit atypischen Kernstrukturen
und luminalen Veränderungen, die auf prä-maligne Vorgänge hindeuten. Die Hälfte der Kras-
Mäuse entwickelten bereits in frühem Lebensalter von fünf Monaten frühe PanIN-Läsionen
im Vergleich zu gleichaltrigen Kontrolltieren. Die Schwere der Läsionen nahm mit
zunehmendem Alter der Tiere zu, bis schließlich ein Drittel der Kras-Tumormäuse invasive
duktale Adenokarzinome im achten Lebensmonat entwickelten. Im Vergleich dazu waren in
den Organen der ebenfalls mit Caerulein behandelten Kontrolltieren weder PanINs noch
invasive Karzinome nachweisbar. Somit stellt die chronische Pankreatitis ein wesentlicher
Faktor dar, der zur Beschleunigung der Kras-induzierten PanIN-Progression führt (Guerra et
al., 2007).
In Entzündungsprozessen wurden dem inflammatorisch-potenten Zytokin IL-6 und dessen
Signalkaskade über Stat3/Socs3 eine besondere Bedeutung zugesprochen (Hodge et al.,
2005). Die Aktivierung von Stat3 sowohl in humanen Pankreasadenokarzinomen (Scholz et
al., 2003; Hutzen et al., 2009; Glienke et al., 2010; Lin et al., 2010b) als auch in den PanIN-
Läsionen der Tumormäuse (Lesina et al., 2011; Fukuda et al., 2011) lässt vermuten, dass
entweder zell-autonome Mechanismen oder autokrine bzw. parakrine Stimulation der Zellen
für diesen Effekt verantwortlich sind. Unsere immunhistochemischen Ergebnisse offenbarten
ausgedehnte Jak2- und Stat3-Aktivierung sowohl in den duktalen als auch infiltrierenden und

V. DISKUSSION
73
Azini-Zellen. Die Jak2-Phosphorylierung wies möglicherweise auf eine parakrine oder
autokrine, weniger auf eine zell-autonome Aktivierung der Signalkaskade hin (Heinrich et al.,
1998; Bravo et al., 2000). Bei den Liganden handelt es sich dabei um Zytokine wie IL-6, IL-
11, IL-27, aber auch LIF (leukemia inhibitor factor) und OSM (Oncostatin M). Die intensive,
ubiquitäre Jak2/Stat3-Aktivierung im KrasG12D-Tumormausmodell lässt vermuten, dass
gp130-Rezeptorliganden in der pankreatischen Onkogenese eine besondere Stellung
einnehmen könnten. In einigen klinischen Studien konnte gezeigt werden, dass eine
Korrelation zwischen der Höhe der IL-6-Serumkonzentration und dem Outcome bzw. der
Prognose der an Pankreaskarzinom erkrankten Patienten besteht (Ebrahimi et al., 2004; Talar-
Wojnarowska et al., 2009).
In zahlreichen Untersuchungen konnten Azinuszellen als epitheliale Quellen von Zytokinen
wie IL-6 und IL-11 in einer mit Caerulein induzierten Pankreatitis nachgewiesen werden
(Grady et al., 1997; Blinman et al., 2000; Bhatia et al., 2002). Die Expression von IL-6 und
IL-11 in den Azini von Stat3-defizienten KrasG12D-Mäusen war eindeutig niedriger als in den
Azini der KrasG12D-Kontrolltiere (Fukada et al., 2011). Diese Ergebnisse und die Kenntnis,
dass sowohl IL-6, IL-11, als auch IL-6Rα und gp130 Zielgene von Stat3 sind (Ernst et al.,
2008; Toth et al., 2011), lassen schlussfolgern, dass Stat3 möglicherweise selbst durch
Steigerung der IL-6 bzw. IL-11 Expression und über einen autokrinen Mechanismus die
Jak2/Stat3-Signalkaskade im murinen KrasG12D-Modell des Pankreaskarzinoms aktiviert
(Fukada et al., 2011).
Im Einklang mit den Ergebnissen der Arbeitsgruppe um Fukuda konnten wir in unseren
Versuchen ebenfalls eine höhere IL-6 mRNA-Konzentration in KrasG12D-Tumormäusen als in
den LSL-KrasG12D-Kontrollmäusen messen. Zusätzlich ist unserer Arbeitsgruppe gelungen, in
isolierten Tumorzelllinien, die im Gegensatz zum Primärtumor oder Metastasen keine bzw.
geringfügige Stat3-Aktivierung aufzeigen, nach IL-6 Stimulation eine starke Stat3-
Phosphorylierung zu induzieren (Lesina et al., 2011). Somit konnten wir zeigen, dass neben
der autokrinen Zellstimulation ein parakriner Mechanismus der Stat3-Aktivierung existiert.
Ein typisches, morphologisches Merkmal des duktalen Pankreaskarzinoms ist die reaktive,
stromale Desmoplasie angrenzend an die PanINs und das Tumorgewebe. Dieses
desmoplastische Stroma stellt ein für die Karzinomentwicklung komplexes „Tumor-
Microenvironment“ dar, das sich aus extrazellulärer Matrix, myofibroblastischen,
pankreatischen Sternzellen und v.a. inflammatorischen Zellen und Makrophagen
zusammensetzt. Untersuchungen haben gezeigt, dass Entzündungszellen insbesondere
Makrophagen für die Synthese von Chemokinen und inflammatorischen Zytokinen wie IL-6

V. DISKUSSION
74
verantwortlich sind (Algül et al., 2007b; Erkan et al., 2008). Weiterführende Experimente
unserer Arbeitsgruppe konnten tatsächlich infiltrierende Makrophagen als wichtigste
Produzenten von IL-6 identifizieren, das zur Aktivierung der Stat3/Socs3-Signalkaskade in
den Tumorvorläufer- bzw. Tumorzellen führt (Lesina et al., 2011). In Zusammenschau der
Forschungsergebnisse sowohl unserer Arbeitsgruppe als auch von Fukada und Kollegen kann
geschlussfolgert werden, dass eine parakrine Stimulation der transformierten Zellen durch IL-
6-Freisetzung aus den Makrophagen die intrazelluläre Jak2/Stat3/Socs3-Signalkaskade
aktiviert. Der Traskriptionsfaktor Stat3 fördert in seiner aktiven Form wiederum die Bildung
von IL-6 und IL-11, was zusätzlich als autokriner Mechanismus die Aktivität des Stat3-
Signalweges aufrechterhält.
Einige Analysen haben gezeigt, dass Stat3 sowohl in Tumorzellen als auch in
Entzündungszellen wie Makrophagen, natürlichen Killerzellen und neutrophilen
Granulozyten im „Tumor-Microenvironment“ konstitutiv aktiviert ist. Durch diese
Dauerstimulation verhindert Stat3 zum einen die Expression vieler für die Immunantwort
gegen die transformierten Zellen wichtiger Botenstoffe und zum anderen fungiert dieser
Transkriptionsfaktor als Aktivator diverser für die Immunsuppression bedeutender Gene u.a.
auch IL-6. Somit unterdrückt die konstitutive Aktivierung der Stat3-Signalkaskade bei Stat3-
Defizienz die antitumoröse Immunantwort, was die Bedeutung dieses Signalweges sowohl in
der Immunmodulation als auch der Tumorentwicklung unterstreicht (Yu et al., 2007).
Die IL-6/Jak2/Stat3-Signalkaskade ist ein ubiquitär vorkommender Signalweg, der bereits im
Rahmen entzündungsassoziierter Onkogenese in anderen Organen wie der Lunge (Li et al.,
2007) und dem Kolon (Bollrath et al., 2009; Grivennikov et al., 2009) beschrieben wurde.
Die komplexe Bedeutung von Socs3 im Rahmen akuter und chronischer Inflammation wurde
in einigen Studien untersucht. Demnach kam es nach Stimulation von isolierten,
pankreatischen Azinus-Zellen mit Lipopolysaccharide (LPS), Tumornekrosefaktor (TNF)-α,
IL-6 oder IL-1ß zu einer erhöhten mRNA-Synthese sowohl von Stat3 als auch Socs3.
Die Induktion des endogenen, negativen Regulators im Pankreas könnte für die Hemmung der
Synthese pro-inflammatorischer Gene eine wichtige Rolle spielen. Die Socs3 Hoch-
Regulation in Azinuszellen inhibiert möglicherweise die weitere Zytokin-Induktion und
hemmt somit die Prolongation und Chronifizierung entzündlicher Prozesse (Vona-Davis et
al., 2005).
In thermisch behandelten Hepatozyten konnte ebenfalls eine deutlich erhöhte Stat3-
Expression und –Aktivierung sowie eine gesteigerte Socs3-Synthese beobachtet werden.
Nicht-Leberparenchymzellen wie Kupffer-Zellen wurden als Quelle inflammatorischer

V. DISKUSSION
75
Zytokine wie IL-6, IL-1 und TNF-α identifiziert. Somit stellten diese wichtigen Zellen der
Socs3-Aktivierung in Rahmen immunologischer Prozesse dar. Das erhöhte Socs3-Proteinlevel
ging mit einer abnehmenden Stat3-Phosphorylierung einher. Die Stimulation primär isolierter
Hepatozyten mit IL-6 führte ebenfalls zu einer Zunahme der Socs3-Konzentration und
Abnahme von Stat3-Aktivität im Vergleich zu nicht mit IL-6 stimulierten Zellen. Daraus kann
geschlussfolgert werden, dass Socs3 die Stat3-Aktivierung hemmte und somit die IL6-
Signaltransduktion terminierte (Kong et al., 2002).
Im Rahmen chronisch entzündlicher Darmerkrankungen limitiert die Socs3-Überexpression
die Aktivierung der IL-6/Stat3-Signalkaskade in den betroffenen Darmzellen und verhindert
eine Hyperproliferation der Darmkrypten und die entzündungs-assoziierte Karzinogenese
(Suzuki et al., 2001; Rigby et al., 2007). In Übereinstimmung zu unseren Ergebnissen führt
eine Socs3-Deletion zur Hyperaktivierung der Stat3-Signalkaskade, was wiederum die
chronische Entzündung und die damit assoziierte Tumorinitiierung und –progression fördert.
Einige Studien haben gezeigt, dass eine intrazelluläre Proteintherapie mit Socs3 das Ausmaß
entzündlicher Prozesse und Apoptose vermindert. Die auf diesem Wege erfolgte Auffüllung
der endogenen Socs3-Reserve bei Defizienz terminierte die Zytokin-Signaltransduktion wie
z.B. durch IL-6 und verhindert somit Apoptose und hämorrhagische Nekrosen im Rahmen
einer akuten Entzündung der Leber (Jo et al., 2005).
Socs3 scheint wichtige Mechanismen im Rahmen von Entzündungsprozessen zu regulieren.
Um die Bedeutung dieses Moleküls für die entzündungsassoziierte Tumorgenese genauer
beschreiben zu können sind weiterführende Untersuchungen notwendig.
4. Rolle von Socs3 bei der Pankreasfibrosierung/Desmoplasie in Kras
G12D-Tumormäusen
Ein bedeutendes, histomorphologisches Kerncharakteristikum des duktalen
Pankreasadenokarzinoms ist die Bildung einer ausgeprägten Desmoplasie. Diese Veränderung
ist gekennzeichnet durch eine massive Fibrosierung des Pankreasgewebes, was als Folge einer
gestörten Homöostase der extrazellulären Matrix (ECM) darstellt. Unter normalen
physiologischen Umständen wird die Organarchitektur durch ein Gleichgewicht zwischen
Synthese und Degradation der ECM aufrechterhalten. Demzufolge führt eine Imbalance
zugunsten der vermehrten Bildung zu einer unphysiologischen Fibrosierung des Gewebes
(Algül et al., 2007; Pandol et al., 2009). Untersuchungen haben gezeigt, dass die Aktivität der

V. DISKUSSION
76
Desmoplasie einen prognostischen Wert bei Patienten mit Pankreaskarzinom besitzt, wobei
eine hohe Aktivität mit einer schlechteren Prognose assoziiert ist (Erkan et al., 2008).
Zu den wichtigsten zellulären Bestandteilen des desmoplasmatischen Stromas zählen neben
inflammatorischen Zellen und Makrophagen v.a. die pankreatischen Sternzellen (pancreatic
stellate cells, PSC). Die PSCs sind Äquivalente zu den hepatischen Sternzellen (auch als Ito
Zellen oder HSCs genannt) und sind in ihrem aktivierten Zustand für die Produktion von
Proteinen der ECM wie Kollagene Typ I und III sowie Fibronektin verantwortlich. Im
normalen, gesunden Pankreasgewebe sind ruhende, gering proliferierende PSCs in den
periazinären und interlobulären Organbereichen lokalisiert und synthetisieren kaum ECM.
Eine parakrine Aktivierung erfolgt bei Entzündungsprozessen oder im Rahmen tumoröser
Erkrankungen durch infiltrierende Zellen wie mononukleare Zellen, die v.a. TGF-ß1, PDGF,
TGF-α sowie Zytokine IL-1, IL-6 und Sauerstoffradikale freisetzen (Algül et al., 2007;
Shimizu, 2008). Die aktivierten PSCs veränderten ihre Morphologie zu proliferierenden
myofibroblastenähnlichen Zellen, die neben den o.g. Proteinen α-smooth muscle actin (α-
SMA), Matrix-Metalloproteasen (MMPs) und deren Inhibitoren exprimieren. Das
Vorkommen von α-SMA beschränkt sich normalerweise auf vaskuläre, glatte Muskelzellen
und ist im pankeatischen Gewebe sehr gering. Vielmehr ist dieses Protein als intrazelluläres,
in Mikrofilamentenbündel lokalisiertes Molekül in Myofibroblasten, glatten Muskelzellen und
Myoepithelzellen zu finden (Gugliotta et al., 1988; Desmouliere et al,. 1993).
MMPs und ihre spezifische Gewebeinhibitoren (TIMPs) werden sowohl von PSCs als auch
von Tumorzellen gebildet. MMPs dienen der Degradation der ECM durch Abbau der
Matrixproteine sowie der Basalmembran und tragen somit zur Metastasierung von
Tumorzellen bei. TIMP-1 und TIMP-2 inhibieren dagegen die MMPs und verhindern damit
den Abbau der ECM (Algül et al., 2007).
PSCs werden nicht nur parakrin stimuliert. Der Nachweis der Expression des TGF-ß1
Rezeptors an der Zelloberfläche und des Signalmoleküls TGF-ß1 zeigt, dass die Aktivität der
PSCs, einst durch exogene Faktoren getriggert, durch autokrine Stimulationsmechanismen
auch bei Abwesenheit exogener Noxen aufrechterhalten wird.
Trotz zahlreicher Studien zur Entstehung einer Desmoplasie im Rahmen diverser
Pankreaserkrankungen sind die zugrundeliegenden Mechanismen zur dessen Entstehung und
Rolle in der pankreatischen Karzinogenese sehr komplex und bisher noch nicht ausreichend
geklärt. Jedoch ist die Akkumulation von gering durchblutetem Stroma ein typisches
Merkmal für PanIN-Läsionen, wobei die Ausprägung mit dem Grad der PanIN-Stufen
zunimmt (Erkan et al., 2012).

V. DISKUSSION
77
In unseren Untersuchungen wiesen die Pankreata Socs3-defizienter KrasG12D-Tumormäuse
morphologisch ebenfalls ausgedehnte Fibrosierungsareale auf. Die Ergebnisse demonstrierten
ein im Vergleich zum Wildtyp bis auf ein Fünftel der Organmasse geschrumpfte
Bauchspeicheldrüse mit diffus nodulärer und lobulierter Parenchymstruktur. Die ausgedehnte
Fibrosierung des sonst weichen Gewebes lässt das Organ derb, knotig und formlos
erscheinen. Diese makroskopisch sichtbaren Veränderungen an den Pankreata der von uns
generierten KrasG12D
;Socs3Δpanc-Mauslinie deuten auf vorhandene fibrotische Prozesse hin,
die sich scheinbar in weiter fortgeschrittenen Stadien befinden als die der gleichaltrigen
Mäuse mit singulärer KrasG12D-Mutation. Während Pankreata vier Wochen alter Kras
G12D-
Mäuse nahezu unauffällig erscheinen, fallen bei näherer histologischer Betrachtung der
Gewebestruktur gleichaltriger KrasG12D
;Socs3Δpanc-Mäuse ausgedehnte fibrotisch umgebaute
heterogene Areale auf. Dieses desmoplasmatische Stroma ist besonders um PanIN-Läsionen
detektierbar. Die Fibrosierungsprozesse scheinen durch die Socs3-Deletion beschleunigt zu
sein. In Anlehnung an die Arbeit von Ogata und Kollegen könnte die Erklärung für unsere
Ergebnisse darin liegen, dass bei chronischen Entzündungsprozessen, Einwirkung von
chemischen Noxen oder Apoptoseprozessen eine dynamische Kaskade von Reaktionen
hervorgerufen wird, bei der Entzündungszellen wie Makrophagen aktiviert werden, die
anschließend eine Reihe von Zytokinen wie IL-6 und Wachstumsfaktoren produzieren und
freisetzen, wodurch ruhende PSCs zu proliferierenden Myofibroblasten aktiviert werden. Die
zunehmende Transkription des Stat3-Zielgens TGF-ß1 in den Makrophagen aber
wahrscheinlich auch in den transformierten Zellen verursacht eine parakrine Stimulation der
PSCs zu proliferierenden Myofibroblasten, die dann vermehrt ECM-Proteinen wie Kollagene
und Fibronektin freisetzen. Somit stellt TGF-ß1 ein wichtiges Signalmolekül für
Fibrosierungsprozesse dar (Saxena et al., 2002; Ogata et al., 2006). Diese Matrixproteine
fördern in ihrer Gesamtheit die Pankreasfibrosierung. Durch autokrine
Stimulationsmechanismen kann die Aktivierung der PSCs trotz Wegfall der Stimuli
aufrechterhalten werden.
In KrasG12D
;Socs3Δpanc-Mäusen kommt es durch die Socs3-Deletion zu einem Ausfall der
negativen Rückkopplung der Stat3-Signalkaskade, wodurch es möglicherweise zu einer
verstärkten TGF-ß1 Expression und zu einer Beschleunigung der Fibrosierungsprozesse
kommt. Um die Komplexität und die genauen, grundlegenden Mechanismen der
Pankreasfibrosierung genauer verstehen zu können, sind weiterführende Untersuchungen
erforderlich.

V. DISKUSSION
78
5. Die Bedeutung von Socs3 in Transdifferenzierungsprozessen im Pankreas der Kras
G12D-Tumormäuse
Die häufigste bisher bekannte und auch histologisch am besten untersuchte Vorläuferläsion
des Pankreaskarzioms ist die PanIN, die einen duktalen Phänotyp besitzt sowie einen duktalen
Ursprung aufweist. Untersuchungen an genetisch veränderten Mausmodellen haben jedoch
gezeigt, dass durch azinär-duktale Metaplasie tubuläre Komplexe entstehen, die als flache,
intermediäre Strukturen eine weitere, alternative Vorläuferläsion des PDAC bilden können.
Die Metaplasie ist definiert als Umwandlung einer differenzierten Gewebe- bzw. Zellart in
Anpassung auf die vorherrschende Umgebung. Rezidivierende Reizzustände durch
chemische, entzündliche und mechanische Stimuli rufen demzufolge eine meist qualitative
Änderung des Gewebes bzw. der Zellen hervor. Zum Beispiel entsteht durch chronische
Reflux-Ösophagitis aus dem Plattenepithel des unteren Ösophagusbereichs ein
magenschleimhautähnliches Zylinderepithel des Barrett-Ösophagus. In der Regel stellt die
Metaplasie einen reversiblen Vorgang dar, der bei Beendigung des einwirkenden Reizes
eintritt. Bleibt der Stimulus jedoch persistent, kann sich die Metaplasie zu einer Dysplasie
entwickeln (Lowenfels et al., 2000; Hruban et al., 2001; Fléjou, 2005).
Azinär-duktale Metaplasie (ADM) wurde bereits bei der Caerulein-induzierten Pankreatitis im
Kras-Mausmodell beobachtet (Guerra et al., 2007). Es ist jedoch unklar, ob die ADM primär
aus expandierten, duktalen Zellen und durch anschließenden Austausch der azinären Zellen
oder durch direkte Reprogrammierung azinärer Zellen in Zellen mit duktalen Eigenschaften
entwickelt, wobei azinären Zellen bei der PanIN-Entwicklung eine bedeutende Rolle
zugesprochen werden. In KrasG12D-exprimierenden Mäusen treten azinär-duktale Metaplasien
entweder zusammen mit PanIN-Läsionen auf oder sie sind diesen vorangestellt. Es wird somit
postuliert, dass die onkogene Kras-Mutation die ADM, die PanIN-Entwicklung und
letztendlich die Entstehung des Pankreasadenokarzinoms induziert (Kopp et al., 2012).
Die metaplastischen Strukturen enthalten sowohl azinäre Zellen mit Zymogengranula als auch
Übergangszellen mit duktalen Merkmalen wie muzinöses Zytoplasma, CK-19 Expression und
Sox9-Bildung (Zhu et al., 2007; Morris et al., 2010). Wie im humanen Pankreaskarzinom
entwickelten sich in den Pankreata der KrasG12D
;Pdx1-Cre- und LSL-KrasG12D
;p48Cre-
Mäuse drei verschiedene Zonen von Zellen, die aus PanINs, ADM und normalen Azinus-
Zellen bestehen. Aufgrund des frühen Auftretens der ADM im Rahmen der humanen,
pankreatischen Karzinogenese ist ihre Bedeutung im Rahmen der Tumorprogression bisher
nur wenig erforscht. Im in-vivo-Mausmodell jedoch zeigte sich bereits bei vier Wochen alten
LSL-KrasG12D
;p48Cre-Tumormäusen ausgedehnte azinäre Metaplasie, die aufgetriebene,

V. DISKUSSION
79
offene Lumina besitzen und muzin-produzierende Gang-ähnliche Zellen enthalten (Zhu et al.,
2007). Der Nachweis sowohl von Amylase (spezifischer Marker von Azinus-Zellen) als auch
von K19 (CK-19, spezifischer Marker duktaler Zellen) bestätigte das Vorliegen einer
metaplastischen Veränderung. Weiterführende Immunoblot-Untersuchungen offenbarten eine
Abnahme der azinären Genprodukte, während die Bildung duktaler Genprodukte anstieg.
Diese Ergebnisse der Arbeitsgruppe um Zhu demonstrierten deutlich die Kras-induzierte
Veränderung der Azinus- zu duktalen Zellen in KrasG12D
;p48Cre-Mäusen. Die
Zellproliferation innerhalb der metaplastischen Strukturen ist sehr hoch, wobei sich die Zellen
erst nach Umwandlung in Gang-ähnliche Zellen aktiv teilen.
In der vorliegenden Arbeit konnten wir in Übereinstimmung mit den Ergebnissen von Zhu
und Kollegen bereits in der vierten Lebenswoche azinär-duktale Metaplasie und vereinzelte,
frühe PanIN-Läsionen detektieren. Mit steigendem Alter der Tiere nahm die Anzahl
histologisch erkennbarer Ducti mit Formanomalie zu, die auf einen metaplastischen duktalen
Prozess hindeutete. Die positive immunhistochemische Detektion von duktalen Markern wie
CK-19 und Mucinen sowie die morphologisch ausgeweiteten, offenen Lumina der tubulären
Strukturen deuteten ebenfalls auf eine azinär-duktale Metaplasie hin.
In vielen Untersuchungen versuchten Forscher herauszufinden, welche molekulargenetische
Mechanismen für die metaplastische Veränderungen verantwortlich sind und inwiefern sich
diese von den der PanIN-Vorläuferläsionen unterscheiden. Dabei konnten sie feststellen, dass
viele in den metaplastischen Komplexen aktivierten Signalkaskaden auch in frühen PanIN-
Vorläufern vorzufinden waren (Kopp et al., 2012). Unter anderem werden dem Notch- (Zhu
et al., 2007), EGF- und dem Hedgehog-Signaltransduktionsweg eine große Bedeutung
zugeschrieben (Means et al., 2005; Pasca di Magliano et al., 2006; De La O et al., 2008;
Fendrich et al., 2008).
Ein weiterer für die ADM- bzw. PanIN-Entwicklung relevanter Signalweg stellt die Stat3-
Signalkaskade dar. Die Akkumulation von ADM und PanIN-Vorläuferläsionen in Pdx1-
Cre;LSL-KrasG12D-Mäusen veranlasste zur Überprüfung der Rolle von Stat3 in der
Entwicklung dieser für die pankreatische Karzinogenese wichtigen Veränderungen. Die
Pankreata der Stat3-defizienten KrasG12D-Mäuse wiesen im Vergleich zu den Pdx1-Cre;LSL-
KrasG12D
;Stat3F/+-Kontrolltieren zum größten Teil ein normales Pankreasparenchym mit
vereinzelten, sporadisch auftretenden ADM und PanINs auf. Das Organgewebe der
Kontrollmäuse zeigte hingegen ausgedehnte azinär-duktale Metaplasien und PanIN-
Formationen als Antwort auf eine exzessive Stat3-Aktivierung zu p-Stat3. Somit konnte
geschlussfolgert werden, dass die Stat3-Deletion die Kras-induzierte ADM- und PanIN-

V. DISKUSSION
80
Bildung bzw. -Progression zwar nicht verhindert, diese aber deutlich vermindert. Somit stellt
der Stat3-Signalweg bzw. die IL-6-Signaltransduktion mögliche, therapeutische Ziele in der
Behandlung des pankreatischen Adenokarzinoms dar (Corcoran et al., 2011).
Eine Stat3-Überexpression im Rahmen der azinär-duktalen Differenzierung wurde ebenfalls
in der genetisch generierten Ptf1a-Cre;CAG-CAT-PDX1-Mauslinie der Arbeitsgruppe um
Miyatsuka beobachtet. Neben der persistierenden Pdx1-Synthese, was die Induktion von
ADM verursacht, wurde von einer verstärkten Stat3-Aktivierung in den metaplastisch
veränderten, duktalen Einheiten des Organparenchyms berichtet. Die Ergebnisse von
Miyatsuka und Kollegen verdeutlichten die essenzielle Bedeutung der Hoch-Regulation von
Pdx-1 und der konstitutiven Stat3-Aktivierung für die Formation tubulärer Komplexe. Als
Antwort auf die Stat3-Hyperaktivierung in Ptf1a-Cre;CAG-CAT-PDX1-Mäusen fanden sie
eine Überexpression von Socs3. Dieses Resultat legte dem negativen Regulator Socs3 eine
regulierende Funktion auf den Stat3-Signalweg im Rahmen der ADM nahe (Miyatsuka et al.,
2006).
Unsere immunhistochemische Schnittfärbungen von KrasG12D-Mäusen bestätigen die
Ergebnisse der oben beschriebenen Arbeiten, dass die metaplastischen Veränderungen
vermehrt Stat3 exprimieren. Diese Ergebnisse deuten darauf hin, dass die verstärkte
Aktivierung von Stat3 in diesem Mausmodell ein frühes Ereignis bei der Entwicklung von
metaplastischen Strukturen darstellt. Im Rahmen weiterführender Untersuchungen
überprüften wir die Abhängigkeit der Reprogrammierung von Azini in duktale Zellen in vivo
von Socs3. Auch hier fanden wir Socs3 als Antwort auf die verstärkte Stat3-Aktivierung in
den metaplastischen Pankreasstrukturen der KrasG12D-Mäuse überexprimiert vor. Somit legt
die starke Induktion des endogenen Inhibitors eine regulierende Funktion des Socs3 in der
azinär-duktalen Metaplasie nahe. Eine Socs3-Deletion in der von uns generierten
KrasG12D
;Socs3Δpanc-Mauslinie mit früher und verstärkter Stat3-Expression resultiert in einer
Beschleunigung der ADM bzw. PanIN-Progression in den entsprechenden Organen. Im
Vergleich zu gleichaltrigen KrasG12D-Kontrollmäusen sind metaplastisch duktale
Veränderungen bereits in der vierten Lebenswoche ausgedehnt im Pankreasgewebe der
KrasG12D
;Socs3Δpanc-Tumormäuse vorzufinden, während die Geweben der Kontrollmäuse
nahezu normal erscheinen. Immunhistochemische Untersuchungen auf CK-19, Mucin 1 und
Mucin 5 dienten zur genauen Charakterisierung der ADM und der Bestätigung der
Beschleunigung der ADM in Socs3-deletierten Mäusen. Bereits im Alter von vier Wochen
entwickeln KrasG12D
;Socs3Δpanc-Tumormäuse metaplastische Gänge mit ausgedehnter CK-19,
Muc1 und Muc5-Expression, während gleichaltrige KrasG12D-Kontrollmäuse ein nahezu

V. DISKUSSION
81
unauffälliges Organ aufweisen. Diese azinär-duktale Metaplasie könnte die Entstehung von
pankreatischen Neoplasien hervorrufen, sodass weiterführende Untersuchungen benötigt
werden.
6. Ausblick
Über die Inaktivierung von Socs3 in humanen Karzinomen und Tumorzelllinien ist in einigen
Arbeiten berichtet worden (He et al., 2003; Sutherland et al., 2004; Weber et al., 2005; Niwa
et al., 2005). Die Bedeutung von Socs3 im murinen in-vivo-Modell des Pankreaskarzinoms
blieb jedoch bisher unbeleuchtet. In unseren Untersuchungen konnten wir zeigen, dass die
KrasG12D-Mauslinie zeit- und altersabhängig alle PanIN-Stufen entwickelten, die den
humanen pankreatischen Vorläuferläsionen entsprechen. Zudem ließ sich eine starke Stat3-
Phosphorylierung am Tyrosinrest Y705 in den KrasG12D-Tumormäusen nachweisen. Socs3
wird Stat3-abhängig exprimiert und fungiert in den PanIN-Läsionen als essentieller Regulator
der Aktivierung und Inhibition von Stat3 und bestimmt somit über die zeitabhängige
Progression der PanINs im KrasG12D-Tumormausmodell.
Aus unseren Ergebnissen können wir damit annehmen, dass auch in humanen
Pankreaskarzinomen die Ausschaltung der Socs3-Aktivität eine Beschleunigung der malignen
Transformation und Progression bedeutet.
In einigen durchgeführten experimentellen Untersuchungen der letzten Jahre versuchten
Forscher die Socs3-Effizienz auf molekulargenetischer Ebene wiederherzustellen. In humanen
Tumorzelllinien wurde die häufig vorzufindende Methylierung und folglich Inaktivierung des
Socs3-Gens durch die Behandlung mit der demethylierenden Verbindung 5-aza-2-
deoxycytidin (5-AZA-DC; Sutherland et al., 2004; Weber et al., 2005; Niwa et al., 2005) oder
Socs3-Gen tragender viraler (CMV, Adenoviren) Vektoren (Lin et al., 2010; Rossa et al.,
2012) aufgehoben. Die dadurch ermöglichte Reaktivierung der Socs3-Expression reduzierte
die Proliferationsrate und induzierte die Apoptose von humanen Tumorzellen. Ebenfalls kam
es zu einer Down-Regulierung der Stat3-Phosphorylierung sowie der Stat3-abhängigen
Zielproteine, die diverse Mechanismen des Zellwachstums und der Zelldifferenzierung, der
Apoptose und Inflammation steuerten (He et al., 2003; Weber et al., 2005; Iwahori et al.,
2011). Diese Ergebnisse demonstrierten, dass das endogene Inhibitorprotein Socs3 einen
potenziell tumorsuppressiven Effekt besitzt und dessen Inaktivierung eine Beschleunigung der
Tumorprogression bedeutet. Bisher ist der Einfluss von Socs3 auf die Initiierung und
Progression von präneoplastischen Läsionen des Pankreas zu duktalen Adenokarzinomen
kaum beleuchtet worden. Mit den gewonnenen Daten gelang unserer Arbeitsgruppe erstmalig

VI. ZUSAMMENFASSUNG
82
der Nachweis, dass Socs3 die Rolle eines essentiellen Regulators der zeitabhängigen
Progression der PanINs in der KrasG12D-initiirenden Onkogenese des Pankreas einnimmt und
die Inaktivierung dieses endogenen Inhibitors letztlich in einer malignen Entartung duktaler
Zellen endet. Diese Erkenntnisse über die Funktion und Mechanismen des Stat3/Socs3-
Signalweges sollen den Ausgangspunkt für weitere sowohl experimentelle als auch klinische
Untersuchungen zur Validierung eines spezifischen und vielversprechenden Targets in der
Therapie des hochaggressiven Pankreasadenokarzinoms darstellen.
VI. ZUSAMMENFASSUNG
Das Pankreaskarzinom steht an vierter Stelle der tumorbedingten Todesfälle. Mit einer
medianen Überlebenszeit von etwa drei bis fünf Monaten und einer 5-Jahresüberlebensrate
von weniger als 5% hat das Pankreaskarzinom eine düstere Prognose (Hezel et al., 2006).
Wesentliche Ursachen für das schlechte Outcome dieser Tumorerkrankung sind die späte
Diagnosestellung, frühe Metastasierung und mangelnde Therapieoptionen (Lionetto et al.,
1995; Yeo et al., 2002b; Kleef et al., 2007a).
Für die Entstehung des duktalen Pankreasadenokarzinoms wurde auf histopathologischer und
molekulargenetischer Ebene ein spezifisches Tumorprogressionsmodell entwickelt.
Demzufolge entsteht der invasive Tumor aus definierten, präneoplastischen
Vorläuferläsionen, den sogenannten pankreatischen intraepithelialen Neoplasien (PanINs), die
wiederum in drei Stadien eingeteilt sind. Mit steigendem Schweregrad dieser Läsionen
nehmen die architektonischen und zellulären Veränderungen der duktalen Strukturen
ebenfalls zu. Molekulargenetische Untersuchungen haben gezeigt, dass die einzelnen PanIN-
Progressionsstufen bestimmte genetische Veränderungen in Form von Mutationen der
Onkogene und Tumorsuppressorgene aufweisen (Hruban et al., 2000).
Mithilfe des Cre/loxP-Rekombinationssystems gelang David Tuveson und Tyler Jacks die
Etablierung eines Mausmodells, in dem die pankreasspezifische KrasG12D-Mutation am Exon1
des entsprechenden Gens vorliegt. In diesem Tumormausmodell konnte das komplette PanIN-
Spektrum rekapituliert werden, da die genetisch veränderten Mäuse alters- und zeitabhängig
alle Stadien dieser Vorläuferläsionen aufzeigen (Jackson et al., 2001). Ausgehend von diesem
etablierten KrasG12D-Tumormausmodell sind verschiedene komplexe Tiermodellsysteme
entwickelt worden, die additiv zu dem mutierten Kras-Allel andere genetische Veränderungen
aufzeigen. Damit lässt sich der Einfluss einzelner Genmutationen auf die

VI. ZUSAMMENFASSUNG
83
Karzinomentwicklung und –progression genauer analysieren und mögliche Ansatzpunkte für
ein gezieltes medizinisches Eingreifen finden.
Das Socs3-Molekül hat intrazellulär eine auf den IL6/Jak2/Stat3-Signalweg inhibierende
Funktion. Stat3-vermittelt exprimiert, moduliert dieses Inhibitorprotein in einer Feedback-
Schleife die Stat3-Phosphorylierung und somit die Stat3-abhängige Expression von
bestimmten Zielgenen, die Prozesse der Proliferation, Apoptose, Angiogenese und
Inflammation steuerten. In zahlreichen humanen Tumorgeweben bzw. -zelllinien konnte
bereits eine Inaktivierung von Socs3 wie z.B. durch eine Hypermethylierung der
Promotorregion und folglich eine Hemmung der Socs3-Transkription als epigenetische
Veränderung nachgewiesen werden (Sutherland et al., 2004; Weber et al., 2005; Niwa et al.,
2005). Die Auswirkung einer Socs3-Defizienz auf die Progression der Vorläuferläsionen zu
duktalen Adenokarzinomen in der pankreatischen Onkogenese blieb bisher jedoch
unbeleuchtet.
In dieser Arbeit haben wir zunächst die Expression von Stat3 und Stat3-abhängigen Proteinen
v.a. Socs3 in KrasG12D-Tumormäusen auf proteinbio- und immunhistochemischer Ebene
bestätigt. Daraus konnte geschlussfolgert werden, dass Socs3 eine balancierende Funktion auf
die Stat3-Phosphorylierung ausübt.
Um die Auswirkung einer Socs3-Defizienz auf die PanIN-Progression genau beurteilen zu
können, generierten wir in einem weiteren Schritt mithilfe der Cre/loxP-Technologie eine
KrasG12D
;Socs3Δpanc-Mauslinie, die eine Deletion des für die Bindungsdomäne am gp130-
Rezeptor kodierenden Exons 2 im Socs3-Gen aufweisen. Durch diese genetische Mutation ist
die Bindung an den intrazellulären Rezeptorbestandteil und folglich die Feedback-Hemmung
der Stat3-Phosphorylierung nicht mehr gewährleistet.
Socs3-defiziente KrasG12D-Mäuse weisen keine Beeinträchtigung in der morphologischen und
funktionellen Entwicklung des exokrinen und endokrinen Pankreas auf. Ebenso wird die
Ausprägung der KrasG12D-Mutation durch die Socs3-Deletion nicht beeinflusst. Diese
Mauslinie ermöglicht somit die Untersuchung des Inhibitorproteins Socs3 auf die
pankreatische Karzinogenese und die Stat3-Phosphorylierung.
Unsere morphologische und biochemische Forschungsergebnisse demonstrieren für
KrasG12D
;Socs3Δpanc-Mäuse im Vergleich zu den Kras
G12D-Kontrolltieren eine frühzeitige und
prolongierte Stat3-Phosphorylierung, die bereits nach vier Wochen postpartum detektierbar
ist, sowie eine Beschleunigung der KrasG12D-initiierenden PanIN-Progression.
Interessanterweise ist das Pankreas der Socs3-defizienten KrasG12D-Mäuse schon zum
Zeitpunkt von vier Wochen morphologisch sehr stark umgewandelt. In dem fibrotisch

84
umgebauten Organ sind heterogne Areale detektierbar, die neben normalen Azinis ausgeprägt
hochgradig veränderte Gänge in Form von PanINs aufweisen.
Das endogene Inhibitorprotein Socs3 übt somit die Rolle eines Tumorsuppressors in der
Onkogenese des Pankreas aus und bestimmt über die Regulation der Stat3-Phosphorylierung
die PanIN-Progression. Unsere Ergebnisse dienen dazu, einen neuen vielversprechenden
Ansatz in der Therapie und Prophylaxe des Pankreaskarzinoms zu definieren und zu
etablieren, weshalb weitere experimentelle und klinische Studien angeschlossen werden
sollten.

VII. LITERATURVERZEICHNIS
85
VII. LITERATURVERZEICHNIS
Aarnio M., Mecklin J.P., Aaltonen L.A., Nystrom-Lahti M., Jarvinen, H.J. (1995). Life-time risk of different cancers in hereditary non-polyposis colorectal cancer (HNPCC) syndrome. Int. J. Cancer 64, 430-433.
Abell K., Bilancio A., Clarkson R.W.E., Tiffen P.G., Altaparmakov A.I., Burdon T.G., Asano
T., Vanhaesebroeck B., Watson C.J. (2005). Stat3-induced apoptosis requires a molecular switch in PI(3)K subunit composition. Nature Cell Biology 7, 392–398.
Adler G., Seufferlein T. Bischoff S.C., Brambs H.J., Feuerbach S., Grabenbauer G., Hahn S., Heinemann V., Hohenberger W., Langrehr J.M., Lutz M.P., Micke O., Neuhaus H., Neuhaus
P., Oettle H., Schlag P.M., Schmid R., Schmiegel W., Schlottmann K., Werner J., Wiedenmann
B., Kopp I. (2007). S3- Guidelines ˝Exocrine pancreatic cancer˝ 2007. Z Gastroenterol 45(6), 487-523.
Ahmad N.A., Lewis J.D., Ginsberg G.G., Rosato E.F., Morris J.B., Kochman M.L. (2000).
Endoscopic ultrasound in preoperative staging of pancreatic cancer. Gastrointest Endosc 52,
463-8.
Alexander W.S., Starr R., Metcalf D., Nicholson S.E., Farley A., Elefanty A.G., Brysha M., Kile B.T., Richardson R., Baca M., Zhang J.-G., Willson T.A., Viney E.M., Sprigg N.S., Rakar S., Corbin J., Mifsud S., DiRago L., Cary D., Nicola N.A., Hilton D.J. (1999). Suppressors of cytokine signaling (SOCS): negative regulators of signal transduction. Journal of Leukocyte Biology 66(4), 588-592.
Algül H., Treiber M:, Lesina M., Schmid R.M. (2007b). Mechanisms of Disease: chronic inflammation and cancer in the pancreas-a potential role for pancreatic stellate cells? Nat Clin Pract Gastroenterol Hepatol 4, 454-462.
American Cancer Society 2011.
Antequera F. (2003). Structure, function and evolution of CpG island promoters. Cell Mol Life Sci 60(8), 1647-58.
Assinder S.J., Dong Q., Kovacevic Z., Richardson D.R. (2009). The TGF-β, PI3K/Akt and PTEN pathways: established and proposed biochemical integration in prostate cancer. Biochem J 417, 411–421.
Babon J.J., McManus E.J., Yao S., DeSouza D.P., Mielke L.A., Sprigg N.S., Willson T.A., Hilton D.J., Nicola N.A., Baca M., Nicholson S.E., Norton R.S. (2006). The Structure of SOCS3 Reveals the Basis of the Extended SH2 Domain Function and Identifies an Unstructured Insertion That Regulates Stability. Molecular Cell 22, 205–216.
Baldin V., Lukas J., Marcote M.J., Pagano M., Draetta G. (1993). Cyclin D1 is a nuclear protein required for cell cycle progression in G1. Genes Dev 7(5), 812-21.
Bardeesy N., DePinho R.A. (2002). Pancreatic cancer biology and genetics. Nat Rev Cancer 2, 897-909.
Barton G.M. (2008). A calculated response: control of inflammation by the innate immune system. J Clin Invest 118(2), 413-420.

VII. LITERATURVERZEICHNIS
86
Beres T.M., Masui T., Swift G.H., Shi L., Henke R.M., MacDonald R.J. (2006). PTF1 is an Organ-Specific and Notch-Independent Basic Helix-Loop-Helix complex containing the mammalian suppressor of Hairless (RBP-J) or its paralogue, RBP-L. Mol Cell Biol 26(1), 117-130.
Bhatia M., Brady M., Kang Y.K., Costello E., Newton D.J., Christmas S.E., Neoptolemos J.P., Slavin J. (2002). MCP-1 but not CINC synthesis is increased in rat pancreatic acini in response to cerulein hyperstimulation. Am J Physiol Gastrointest Liver Physiol 282(1), G77-85.
Bhattacharya S., Ray R.M., Johnson L.R. (2005). STAT3-mediated transcription of Bcl-2, Mcl-1 and c-IAP2 prevents apoptosis in polyamine-depleted cells. Biochem J 392, 335–344.
Bissell D.M., Wang S.S., Jarnagin W.R., Roll F.J. (1995). Cell-specific expression of transforming growth factor-ß in rat liver: evidence for autocrine regulation of hepatocyte proliferation. J Clin Invest 96, 447-455.
Blinman T.A., Gukovsky I., Mouria M., Zaninovic V., Livingston E., Pandol S.J., Gukovskaya A.S. (2000). Activation of pancreatic acinar cells on isolation from tissue: cytokine upregulation via p38 MAP kinase. Am J Physiol Cell Physiol 279, C1993–2003.
Bollrath J., Phesse T.J., von Burstin V.A., Putoczki T., Bennecke M., Bateman T., Nebelsiek T., Lundgren-May T., Canli O., Schwitalla S., Matthews V., Schmid R.M., Kirchner T., Arkan M.C., Ernst M., Greten F.R. (2009). gp130-mediated Stat3 activation in enterocytes regulates cell survival and cell-cycle progression during colitis-associated tumorigenesis. Cancer Cell 15(2), 91-102.
Bravo J., Heath J.K. (2000). Receptor recognition by gp130 cytokines. EMBO J 19, 2399.
Brembeck F.H., Schreiber F.S., Deramaudt T.B., Craig L., Rhoades B., Swain G.; Grippo P., Stoffers D.A., Silberg D.G., Rustgi A.K. (2003). The mutant K-ras oncogene causes pancreatic periductal lymphocytic infiltration and gastric mucous neck cell hyperplasia in transgenic mice. Cancer Res 63, 2005-2009.
Bromberg, J. (2002). Stat proteins and oncogenesis. J Clin Invest 109, 1139-1142.
Brugge W.R., Lauwers G.Y., Sahani D., Fernandez-del Castillo C., Warshaw A.L. (2004). Cystic neoplasms of the pancreas. N Engl J Med 351, 1218-1226.
Burris H.A., Moore M.J., Andersen J., Green M.R., Rothenberg M.L., Modiano M.R., Cripps M.C., Portenoy R.K., Storniolo A.M., Tarassoff P., Nelson R., Dorr F.A., Stephens C.D., Von Hoff D.D. (1997) Improvements in survival and clinical benefit with gemcitabine as first-line therapy for patients with advanced pancreas cancer: a randomized trial. J Clin Oncol 15, 2403-13.
Cappell M.S. (2005). The pathophysiology, clinicalpresentation, and diagnosis of colon cancer and adenomatous polyps. Med Clin N Am 89, 1-42.
Ciocca D.R., Clark G.M., Tandon A.K., Fuqua S.A.W., Welch W.J., McGuire W.L. (1993). Heat shock protein hsp70 in patients with axillary lymph node-negative breast cancer: Prognostic Implications. JNCI J Natl Cancer Inst 85(7), 570-574.
Claudine H. (2004). Methods in Nutrition Science: Cre/loxP System for Generating Tissue-specific Knockout Mouse Models. Nutrition Reviews 62(6), 243-246.

VII. LITERATURVERZEICHNIS
87
Corcoran R.B., Contino G., Deshpande V., Tzatsos A., Conrad C., Benes C.H. Levy D.E., Settleman J., Engelman J.A., Bardeesy N. (2011). Stat3 plays a critical role in Kras-induced pancreatic tumorigenesis. Cancer Res 71, 5020-5029.
Couch F.J., Johnson M.R., Rabe K.G., Brune K., de Andrade M., Goggins M., Rothenmund H., Gallinger S., Klein A., Petersen G.M., Hruban R.H.(2007). Cancer Epidemiol Biomarkers Prev 16, 342-346.
Croker B.A., Krebs D.L., Zhang J.-G., Wormald S., Willson T.A., Stanley E.G., Robb L., Greenhalgh D.J., Förster I., Clausen B.E.,Nicola N.A., Metcalf D., Hilton D.J. , Roberts A.W., Alexander W.S. (2003). SOCS3 negatively regulates IL-6 signaling in vivo. Nature Immunology 4, 540-545.
Dang C.V., O'Donnell K.A., Zeller K.I., Nguyen T., Osthus R.C., Li F. (2006). The c-Myc target gene network. Semin Cancer Biol 16(4), 253-64.
De La O J.-P., Emerson L.L., Goodman J.L., Froebe S.C., Illum B.E., Curtis A.B., Murtaugh L.C. (2008). Notch and Kras reprogram pancreatic acinar cells to ductal intraepithelial neoplasia. PNAS 105(48), 18907-18912.
Desmouliere A., Geinoz A., Gabbiani F., Gabbiani G. (1993). Transforming growthfactor-beta 1 induces alpha-smooth muscle acin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. JBC 122(1), 103-111.
Domagk D., Wessling J., Reimer P., Hertel L., Poremba C., Senninger N., Heinecke A., Domschke W., Menzel J. (2004). Endoscopic retrograde cholangiopancreatography, intraductal ultrasonography, and magnetic resonance cholangiopancreatography in bile duct strictures: a prospective comparison of imaging diagnostics with histopathological correlation. Am J Gastroenterol 99, 1684-9.
Downward J. (2003). Targeting RAS signalling pathways in cancer therapy. Nature Reviews Cancer 3, 11-22.
Dubrovska A., Kim S., Salamone R.J., Walker J.R., Maira S.-M., García-Echeverría C., Schultz P.G., Reddy V.A. (2008). The role of PTEN/Akt/PI3K signaling in the maintenance and viability of prostate cancer stem-like cell populations. PNAS 106(1), 268-273.
Ebrahimi B., Tucker S.L., Li D., Abbruzzese J.L., Kurzrock R. (2004). Cytokines in pancreatic carcinoma: correlation with phenotypic characteristics and prognosis. Cancer 101(12), 2727-36.
Efeyan A., Serrano M. (2007). p53: Guardian of the Genome and Policeman of the Oncogenes. Cell Cycle 6(9), 1006-1010.
Emanuelli B., Peraldi P., Filloux C., Chavey C., Freidinger K., Hilton D.J., Hotamisligil G.S., Van Obberghen E. (2001). SOCS-3 inhibits insulin signaling and is up-regulated in response to tumor necrosis factor-α in the adipose tissue of obese mice. J Biol Chem 276, 47944-47949.
Emanuelli B., Peraldi P., Filloux C., Sawka-Verhelle D., Hilton D., Van Obberghen E. (2000). SOCS-3 is an insulin-induced negative regulator of insulin signaling. J Biol Chem 275, 15985-15991.

VII. LITERATURVERZEICHNIS
88
Erkan M., Hausmann S., Michalski C.W., Fingerie A.A., Dobritz M., Kleeff J., Friess H. (2012). The role of stroma in pancreatic cancer: diagnostic and therapeutic implications. Nat Rev Gastroenterol and Hepatol 9, 454-467.
Erkan M., Michalski C.W., Rieder S., Reiser-Erkan C., Abiatari I., Kolb A., Giese N.A., Esposito I., Friess H., Kleef J. ( 2008). The activated stroma index is a novel and independent prognostic marker in pancreatic ductal adenocacinoma. Clin Gastroenterol Hepatol 6, 1155-1161.
Ernst M., Najdovska M., Grai D., Lundgren-May T., Buchert M., Tye H., Matthews V.B., Armes J., Bhathal P.S., Hughes N.R., Marcusson E.G., Karras J.G., Na S., Sedgwick J.D., Hertzog P.J., Jenkins B.J. (2008). STAT3 and STAT1 mediate IL-11-dependent and inflammation-associated gastric tumorigenesis in gp130 receptor mutant mice. J Clin Invest 118(5), 1727-1738.
Evans M.K., Yu C.-R., Lohani A., Mahdi R.M., Liu X., Trzeciak A.R. Egwuagu C.E. (2007). Expression of SOCS1 and SOCS3 genes is differentially regulated in breast cancer cells in response to proinflammatory cytokine and growth factor signals. Oncogene 26, 1941-1948.
Fendrich V., Esni F., Garay M.V., Feldmann G., Habbe N., Jensen J.N., Dor Y., Stoffers D., Jensen J., Leach S.D., Maitra A. (2008). Hedgehog signaling is required for effective regeneration of exocrine pancreas. Gastroenterology 135(2), 621-31.
Ferrara N. (2002). VEGF and the quest for tumour angiogenesis factors. Nature Reviews Cancer 2, 795-803.
Fléjou J.F. (2005). Barrett’s oesophagus: from metaplasia to dysplasia and cancer. Gut 54, i6-i12.
Friday B.B., Adjei A.A. (2008). Advances in Targeting the Ras/Raf/MEK/Erk Mitogen-Activated Protein Kinase Cascade with MEK Inhibitors for Cancer Therapy. Clin Cancer Res 14, 342.
Friess H., Kleeff J., Fischer L., Muller M., Buchler M.W. (2003). Chirurgische Standardtherapie beim Pankreaskarzinom. Chirurg 74, 183-190.
Fukuda A., Wang S.C., Morris J.P., Folias A.E., Liou A., Kim G.E., Akira S., Boucher K.M., Firpo M.A., Mulvihill S.J., Hebrok M. (2011). Stat3 and MMP7 contribute to pancreatic ductal adenocarcinoma initiation and progression. Cancer Cell 19(4), 441-55.
Gansauge S., Gansauge F., Ramadani M., Stobbe H., Rau B., Harada N, Beger H.G. (1997). Overexpression of Cyclin Dl in Human Pancreatic Carcinoma is associated with poor prognosis. Cancer Research 57, 1634-1637.
Glienke W., Maute L., Wicht J., Bergmann L. (2010). Curcumin inhibits constitutive STAT3 phosphorylation in human pancreatic cancer cell lines and downregulation of survivin/BIRC5 gene expression. Cancer Invest 28(2), 166-71.
Grady T., Liang P., Ernst S.A., Logsdon C.D. (1997). Chemokine gene expression in rat pancreatic acinar cells is an early event associated with acute pancreatitis. Gastroenterology 113, 1966-1975.
Grivennikov S., Karin E., Terzic J., Mucida D., Yu G.Y., Vallabhapurapu S., Scheller J, Rose-John S., Cheroutre H., Eckmann L., Karin M. (2009). IL-6 and Stat3 are required for

VII. LITERATURVERZEICHNIS
89
survival of intestinal epithelial cells and development of colitis-associated cancer. Cancer Cell 15(2), 103-13.
Guerra C., Schuhmacher A.J., Canamero M., Grippo P.J., Verdaguer L., Perez-Gallego L., Dubus P., Sandgren E.P., Barbacid M. (2007). Chronic pancreatitis is essential for induction of pancreatic ductal adenocarcinoma by K-Ras oncogenes in adult mice. Cancer Cell 11(3), 291-302.
Gugliotta P., Sapino A., Macri L., Skalli O., Gabbiani G., Bussolati G. (1988). Specific demonstration of myoepithelial cells by anti-alpha smooth muscle actin antibody. J Histochem Cytochem 36(6), 659-663.
Hahn S.A., Greenhalf B., Ellis I., Sina-Frey M., Rieder H., Korte B., Gerdes B., Kress R., Ziegler A., Raeburn J.A., Campra D., Grützmann R., Rehder H., Rothmund M., Schmiegel W., Neoptolemos J.P.,Bartsch D.K. (2003). BRCA2 Germline Mutations in Familial Pancreatic Carcinoma. JNCI J Natl Cancer Inst 95(3), 214-221.
Hamilton K., Lund, Kay P., Galanko, J.; Sandler R. Keku T. (2010). Suppressor of cytokine signaling 3 (SOCS3) is not an independent biomarker of colorectal adenoma risk. BMC Research Notes 1, 144.
Hatfield M.P.D., Lovas S. (2012). Role of Hsp70 in cancer growth and survival. Protein & Peptide Letters 19, 616-624.
He B., You L., Uematsu K., Zang K., Xu Z., Lee A.Y., Costello J.F., McCormick F., Jablons D.M. (2003). SOCS-3 is frequently silenced by hypermethylation and suppresses cell growth in human lung cancer. Proc Natl Acad Sci U S A 100(24), 14133-8.
Heinrich P.C., Behrmann I., Müller-Newen G., Schaper F., Graeve L. (1998). IL-6-type cytokine signalling through the gp130/JAK/STAT pathway. Biochem J 334, 297.
Hezel A.F., KimmelmannA.C., Stanger B.Z., Bardeesy N., Depinho R.A. (2006). Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev 20, 1218-49.
Hingorani S.R., Petricoin III E.F., Maitra A., Rajapakse V., King C., Jacobetz M.A., Ross S., Conrads T.P., Veenstra T.D., Hitt B.A., Kawaguchi Y., Johann D., Liotta L.A., Crawford H.C., Putt M.E., Jacks T., Wright C.V.E., Hruban R., Lowy A.M., Tuveson D.A. (2003a). Preinvasive and invasive ductal pancreatic cancer and its early detection in the mouse. Cancer Cell 4, 437-450.
Hirai I., Kimura W., Ozawa K., Kudo S., Suto K., Kuzu H., Fuse A.(2002). Perineural invasion in pancreatic cancer. Pancreas 24, 15-25.
Hodge D.R., Hurt E.M., Farrar W.L. (2005). The role of IL-6 and STAT3 in inflammation and cancer. Eur J Cancer 41(16), 2502-12.
Hong S., Park J., Hruban R., Goggins M. (2011). Molecular Signatures of Pancreatic Cancer. Archives of Pathology & Laboratory Medicine 135(6), 716-727.
Horton K.M. (2002). Multidetector CT and three-dimensional imaging of the pancreas: state of the art. J Gastrointest Surg 6, 126-128.
House M.G., Choti M.A. (2005). Palliative therapy for pancreatic/biliary cancer. The Surgical Clinics of North America 85(2), 359-371.

VII. LITERATURVERZEICHNIS
90
Hruban R.H., Adsay N.V., Albores-Saavedra J., Anver M.R., Biankin A.V., Boivin G.P., Furth E.E., Furukawa T., Klein A., Klimstra D.S., Kloppel G., Lauwers G.Y., Longnecker D.S., Luttges J., Maitra A., Offerhaus G.J., Pérez-Gallego L., Redston M., Tuveson D.A. (2006). Pathology of genetically engineered mouse models of pancreatic exocrine cancer: consensus report and recommendations. Cancer Res 66, 95-106.
Hruban R.H., Goggins M., Parsons J., Kern S.E. (2000). Progression Model for Pancreatic Cancer. Clin Cancer Res 6, 2969.
Hruban R.H., Takaori K., Klimstra D.S., Adsay N.V., Albores-Saavedra J., Biankin A.V., Biankin S.A., Compton C., Fukushima N., Furukawa T., Goggins M., Kato Y., Kloppel G., Longnecker D.S., Luttges J., Maitra A., Offerhaus G.J., Shimizu M., Yonezawa S. (2004). An illustrated consensus on the classification of pancreatic intraepithelial neoplasia and intraductal papillary mucinous neoplasms. Am J of Surg Pathol 28(8), 977-987.
Huang S., Sinicrope F.A. (2010). Sorafenib inhibits STAT3 activation to enhance TRAIL-mediated apoptosis in human pancreatic cancer cells. Mol Cancer Ther 9(3), 742-750.
Hudis C.A. (2007). Trastuzumab- Mechanism of Action and Use in Clinical Practice. N Engl J Med 357, 39-51.
Hutzen B., Friedman L., Sobo M., Lin L., Cen L., De Angelis S., Yamakoshi H., Shibata H., Iwabuchi Y., Lin J. (2009). Curcumin analogue GO-Y030 inhibits STAT3 activity and cell growth in breast and pancreatic carcinomas. Int J Oncol 35, 867-872.
Iwahori K., Serada S., Fujimoto M., Nomura S., Osaki T., Lee C.M., Mizuguchi H., Takahashi T., Ripley B., Okumura M., Kawase I., Kishimoto T., Naka T. (2011). Overexpression of SOCS3 exhibits preclinical antitumor activity against malignant pleural mesothelioma. Int J Cancer 129(4), 1005-17.
Jackson E.L., Willis N., Mercer K., Bronson R.T., Crowley D., Montoya R., Jacks T., Tuveson D.A. (2001). Analysis of lung tumor initiation and progression using conditional expression of oncogenic K-ras. Genes Dev 15, 3243-3248.
Jäättelä M. (1999). Heat shock proteins as cellular lifeguards. Ann Med 31(4), 261-71.
Jo D., Liu D., Yao S., Collins R.D., Hawiger J. (2005). Intracellular protein therapy with SOCS3 inhibits inflammation and apoptosis. Nat Med 11(8), 892-8.
Kamura T., Maenaka K., Kotoshiba S., Matsumoto M., Kohda D., Conaway R.C., Conaway J.W., Nakayama K.I. (2004). VHL‐box and SOCS‐box domains determine binding specificity for Cul2‐Rbx1 and Cul5‐ Rbx2 modules of ubiquitin ligases. Genes Dev 18, 3055-3065.
Kamura T., Sato S., Haque D., Liu L., Kaelin W.G. Jr., Conaway R.C., Conaway J.W. (1998). The Elongin BC complex interacts with the conserved SOCS‐box motif present in members of the SOCS, ras. WD‐40 repeat, and ankyrin repeat families. Genes Dev 12, 3872‐3881.
Kim J.E., Lee K.T., Lee J.K., Paik S.W., Rhee J.C., Choi K.W. (2004). Clinical usefulness of carbohydrate antigen 19-9 as a screening test for pancreatic cancer in an asymptomatic population. J Gastroenterol Hepatol 19, 182-186.
Kim S., Keku T.O., Martin C., GalankoJ., Woosley J.T., Schroeder J.C., Satia J.A., Halabi S.,
Sandler R.S. (2008). Circulating Levels of Inflammatory Cytokines and Risk of Colorectal
Adenomas. Cancer Res 68, 322-328.

VII. LITERATURVERZEICHNIS
91
Kile B.T., Schulman B.A., Alexander W.S., Nicola N.A., Martin H.M., Hilton D.J. (2002). The SOCS box: a tale of destruction and degradation. Trends Biochem Sci 27(5), 235-41.
Kleeff J., Beckhove P., Esposito I., Herzig S., Huber P.E., Lohr J.M., Friess H. (2007a). Pancreatic cancer microenvironment. Int J Cancer 121, 699-705.
Klingler A., Klocker J., Springer P., Kober F., Glaser K. (2000). Combined laparoscopy and laparoscopic ultrasonography in the oncologic diagnostics of pancreas and liver. European Surgery 32(5), 228-232.
Kong F., Guo X., Noel J.G., Wells D.A., Lovell G.J., Ogle C.K. (2002). Thermal injury-induced increases of hepatocyte SOCS3 lead to decreases in STAT3. Shock 18(4), 374-9.
Konnikova L., Kotecki M., Kruger M.M., Cochran B.H. (2003). Knockdown of STAT3 expression by RNAi induces apoptosis in astrocytoma cells. BMC Cancer 3, 23.
Kopp J.L., von Figura G., Mayes E., Liu F.-F., Dubois C.L., Morris J.P., Pan F.C., Akiyama H., Wright C.V.E., Jensen K., Hebrok M., Sander M. (2012). Identification of Sox9-dependent acinar-to-ductal reprogramming as the principal mechanism for initiation of pancreatic ductal adenocarcinoma. Cancer Cell 22, 737-750.
Kos C.H. (2004). Cre/loxP System for Generating Tissue-specific Knockout Mouse Models. Nutrition Reviews 62(6), 243-246.
Krajewska M., Krajewski S., Epstein J.I., Shabaik A., Sauvageot J., Song K., Kitada S, Reed J.C. (1996). Immunohistochemical analysis of bcl-2, bax, bcl-X, and mcl-1 expression in prostate cancers. Am J Pathol 148(5), 1567-76.
Krapp A., Knofler M., Ledermann B., Burki K., Berney C., Zoerkler N., Hagebuchle O., Wellauer P.K. (1998). The bHLH protein PTF1-p48 is essential for the formation of the exocrine and the correct spatial organization of the endocrine pancreas. Genes Dev 12, 3752-3763.
Krebs D.L., Hilton D.J. (2000). SOCS: physiological suppressors of cytokine signaling. J Cell Sci 113, 2813-2819.
Lang R., Pauleau A.-L., Parganas E., Takahashi Y., Mages J., Ihle J.N., Rutschman R., Murray P.J. (2003). SOCS3 regulates the plasticity of gp130 signaling. Nature Immunology 4(6), 546-550.
Lin L., Hutzen B., Zuo M., Ball S., Deangelis S., Foust E., Pandit B., Ihnat M.A., Shenoy S.S., Kulp S., Li P.K., Li C., Fuchs J., Lin J. (2010b). Novel STAT3 phosphorylation inhibitors exhibit potent growth-suppressive activity in pancreatic and breast cancer cells. Cancer Res 70(6), 2445-54.
Lee T.L., Yeh J., Van Waes C., Chen Z. (2006). Epigenetic modification of SOCS-1 differentially regulates STAT3 activation in response to interleukin-6 receptor and epidermal growth factor receptor signaling through JAK and/or MEK in head and neck squamous cell carcinomas. Mol Cancer Ther 5(1), 8-19.
Lei S., Appert H.E., Nakata B., Domenico D.R.,Kim K., Howard J.M. (1995). Overexpression of HER2/neu Oncogene in Pancreatic Cancer Correlates with Shortened Survival. Int J Pancreatol 17(1), 15-21.

VII. LITERATURVERZEICHNIS
92
Lesina M., Kurkowski M.U., Ludes K., Rose-John S., Treiber M., Klöppel G., Yoshimura A., Reindl W., Sipos B., Akira S., Schmid R.M., Algül H. (2011). Stat3/Socs3 activation by IL-6 transsignaling promotes progression of pancreatic intraepithelial neoplasia and development of pancreatic cancer. Cancer Cell 19(4), 456-69.
Li Y., Du H., Qin Y., Roberts J., Cummings O.W., Yan C. (2007). Activation of the signal transducers and activators of the transcription 3 pathway in alveolar epithelial cells induces inflammation and adenocarcinomas in mouse lung. Cancer Res 67, 8494-8503.
Lim K.-H., Counte C.M. (2005). Reduction in the requirement of oncogenic Ras signaling to activation of PI3K/AKT pathway during tumor maintenance. Cancer Cell 8, 381-392.
Lin Y.-C., Lin C.-K., Tsai Y.-H., Weng H.-H.; Li Y.-C., Liang Y., Chen J.-K., Jablons D.M., Yang C.-T. (2010). Adenovirus-mediated SOCS3 gene transfer inhibits the growth and enhances the radiosensitivity of human non-small cell lung cancer cells. Oncology reports 24(6), 1605-1612
Lionetto R., Pugliese V., Bruzzi P., Rosso R. (1995). No Standard treatment is available for advanced pancreatic Cancer. Eur J Cancer 31A, 882-887.
Lin Q., Lai R., Chirieac L.R., Li C., Thomazy V.A., Grammatikakis I., Rassidakis G.Z., Zhang W., Fujio Y., Kunisada K., Hamilton S.R.,Amin H.M. (2005). Constitutive activation of JAK3/STAT3 in colon carcinoma tumors and cell lines: inhibition of JAK3/STAT3 signaling induces apoptosis and cell cycle arrest of colon carcinoma cells. Am J Pathol 167(4), 969-80.
Lowe S.W., Sherr C.J. (2003). Tumor suppression by Ink4a-Arf: progress and puzzles. Curr Opin Genet Dev 13(1), 77-83.
Lowenfels A. B., Maisonneuve P. (2006). Epidemiology and risk factors for pancreatic cancer. Best Pract Res Clin Gastroenterol 20, 197-209.
Lowenfels A.B., Maisonneuve P., Cavallini G., Ammann R.W., Lankisch P.G., Andersen J.R., Dimagno E.P., Andren-Sandberg A., Domellof L. and the International Pancreatitis Study Group (1993). Pancreatitis and the Risk of Pancreatic Cancer. N Engl J Med 328, 1433-1437.
Lu Y., Fukuyama S., Yoshida R., Kobayashi T., Saeki K., Shiraishi H., Yoshimura A., Takaesu G. (2006). Loss of SOCS3 Gene Expression Converts STAT3 Function from Anti-apoptotic to Pro-apoptotic. The Journal of Biological Chemistry 281(48), 36683-36690.
Lüttges J., Neumann S., Jesenofsky R., Borries V., Löhr M., Klöppel G. (2003). Lack of apoptosis in PanIN-1 and PanIN-2 lesions associated with pancreatic ductal adenocarcinoma is not dependent on K-ras status. Pancreas 27(3), e57-62.
Lynch H.T., Smyrk T., Kern S.E., Hruban R.H., Lightdale C.J., Lemon S.J., Lynch J.F., Fusaro L.R., Fusaro R.M., Ghadirian P. (1996). Familial pancreatic cancer: a review. Semin Oncol 23, 251-275.
Ma X.T.,Wang S., Ye Y.J., Du R.Y., Cui Z.R., Somsouk M. (2004). Constitutive activation of Stat3 signaling pathway in human colorectal carcinoma. World J Gastroenterol 10(11), 1569-73.

VII. LITERATURVERZEICHNIS
93
Magee C.J., Greenhalf W., Howes N. Ghaneh P., Neoptolemos J.P. (2001). Molecular pathogenesis of pancreatic ductal adenocarcinoma and clinical implications. Surg Oncol 10, 1–23.
Maitra A., Fukushima N., Takaori K., Hruban R.H. (2005). Precursors to invasive pancreatic cancer. Adv Anat Pathol 12, 81-91.
Malka D., Hammel P., Maire F., Rufat P., Madeira I., Pessione F., Lévy P., Ruszniewski P. (2002). Risk of pancreatic adenocarcinoma in chronic pancreatitis. Gut 51, 849-852.
Manning B.D. and Cantley L.C. (2007). AKT/PKB Signaling: Navigating Downstream. Cell 129(7), 1261-1274.
Means A.L., Meszoely I.M., Suzuki K., Miyamoto Y., Rustgi A.K., Coffey R.J. Jr, Wright C.V., Stoffers D.A., Leach S.D. (2005). Pancreatic epithelial plasticity mediated by acinar cell transdifferentiation and generation of nestin-positive intermediates. Development 132(16), 3767-76.
Medzhitov R. (2008). Origin and physiological roles of inflammation. Nature 454, 428-435.
Menssen A., Hermeking H. (2002). Characterization of the c-MYC-regulated transcriptome by SAGE: Identification and analysis of c-MYC target genes. Proc Natl Acad Sci U S A. 99(9), 6274-6279.
Miyaki M., Kuroki T. (2003). Role of Smad4 (DPC4) inactivation in human cancer. Biochemical and Biophysical Research Communications 306(4), 799-804.
Miyatsuka T., Kaneto H., Shiraiwa T., Matsuoka T.A., Yamamoto K., Kato K., Nakamura Y., Akira S., Takeda K., Kajimoto Y., Yamasaki Y.,Sandgren E.P., Kawaguchi Y., Wright C.V., Fujitani Y. (2006). Persistent expression of PDX-1 in the pancreas causes acinar-to-ductal metaplasia through Stat3 activation. Genes Dev 20(11), 1435-40.
Mori H., Hanada R., Hanada T., Aki D., Mashima R., Nishinakamura H., Torisu T., Chien K. R., Yasukawa H., Yoshimura A. (2004). Socs3 deficiency in the brain elevates leptin sensitivity and confers resistance to diet-induced obesity. Nature Medicine 10, 739-743.
Morris J.P., Cano D.A., Sekine S., Wang S.C., Hebrok M. (2010). β-catenin blocks Kras-dependent reprogramming of acini into pancreatic cancer precursor lesions in mice. J Clin Invest 120(2), 508-20.
Mulkeen A.L., Yoo P.S., Cha C. (2006). Less common neoplasms of the pancreas. World J Gastroenterol 12(20), 3180-3185.
Musgrove E.A., Caldon C.E., Barraclough J., Stone A., Sutherland R.L. (2011). Cyclin D as a therapeutic target in cancer. Nature Reviews Cancer 11, 558-572.
Nakhai H., Sel S., Favor J., Mendoza-Torres L., Paulsen F., Duncker G.I., Schmid R.M. (2007). Ptf1a is essentialnfor the differentiation of GABAergic and glycinergic amacrine cells and horizontal cells in the mouse retina. Development 134, 1151-1160.
Nazli O., Bozdag A.D., Tansug T., Kir R., Kaymak E. (2000). The diagnostic importance of CEA and CA 19-9 for the early diagnosis of pancreatic carcinoma. Hepatogastroenterology 47, 1750-2.

VII. LITERATURVERZEICHNIS
94
Niwa Y., Kanda H., Shikauchi Y., Saiura A., Matsubara K., Kitagawa T., Yamamoto J., Kubo T., Yoshikawa H. (2005). Methylation silencing of SOCS-3 promotes cell growth and migration by enhancing JAK/STAT and FAK signalings in human hepatocellular carcinoma. Oncogene 24(42), 6406-17.
Oettle H., Post S., Neuhaus P., Gellert K., Langrehr J., Ridwelski K., Harald Schramm H., Fahlke J., Zuelke C., Burkart C., Gutberlet K., Kettner E., Schmalenberg H., Weigang-Koehler K., Bechstein W.-O., Niedergethmann M., Schmidt-Wolf I., Roll L.; Doerken B., Riess H. (2007). Adjuvant Chemotherapy with Gemcitabine vs Observation in Patients Undergoing Curative-Intent Resection of Pancreatic CancerA Randomized Controlled Trial. JAMA 297(3), 267-277.
Ogata H., Chinen T., Yoshida T., Kinjyo I., Takaesu G., Shiraishi H., Iida M., Kobayashi T. (2006). Loss of Socs3 in the liver promotes fibrosis by enhancing Stat3-mediated TGF-ß1 production. Oncogene 25, 2520-2530.
Ogata H., Kobayashi T., Chinen T., Takaki H., Sanada T., Minoda Y., Koga K., Takaesu H., Maehara Y., Iida M., Yoshimura A. (2006). Deletion of the SOCS3 Gene in Liver Parenchymal Cells Promotes Hepatitis-Induced Hepatocarcinogenesis. Gastroenterol 131, 179-193.
Okada S., Nakamura M., Katoh H., Miyao T., Shimazaki T., Ishii K., Yamane J., Yoshimura
A., Iwamoto Y., Toyama Y., Hideyuki Okano H. (2006). Conditional ablation of Stat3 or Socs3 discloses a dual role for reactive astrocytes after spinal cord injury. Nature Medicine 12, 829-834.
Okaro A.C., Deery A.R., Hutchins R.R., Davidson B.R. (2001). The expression of antiapoptotic proteins Bcl-2, Bcl-XL, and Mcl-1 in benign, dysplastic, and malignant biliary epithelium. J Clin Pathol 54, 927-932.
Okusaka T., Okada S., Ueno H., Ikeda M., Shimada K., Yamamoto J., Kosuge T., Yamasaki S., Fukushima N., Sakamoto M. (2001). Abdominal pain in patients with resectable pancreatic cancer with reference to clinicopathologic findings. Pancreas 22, 279-84.
Olive K.P., Tuveson D.A. (2006). The Use of Targeted Mouse Models for Preclinical Testing of Novel Cancer Therapeutics. Clin Cancer Res 12, 5277.
Pandol S., Edderkaoui M., Gukovsky I., Lugea A., Gukovskaya A. (2009). Desmoplasia of pancreatic ductal adenocarcinoma. Clin Gastroenterol Hepatol 7(11), 44-47.
Pasca di Magliano M., Sekine S., Ermilov A., Ferris J., Dlugosz A.A., Hebrok M. (2006). Hedgehog/Ras interactions regulate early stages of pancreatic cancer. Genes Dev 20(22), 3161-73.
Pensa S., Regis G., Boselli D., Novelli F., Poli V. (2009). STAT1 and STAT3 in Tumorigenesis: Two Sides of the Same Coin? Madame Curie Bioscience Database (Internet).
Placzek W.J., Wei J., Kitada S., Zhai D., Reed J.C., Pellecchia M. (2010). A survey of the anti-apoptotic Bcl-2 subfamily expression in cancer types provides a platform to predict the efficacy of Bcl-2 antagonists in cancer therapy. Cell Death and Disease 1, e40.
Pour P.M., Egami H., Takiyama Y. (1991). Patterns of growth and metastases of induced pancreatic cancer in relation to the prognosis and its clinical implications. Gastroenterology 100, 529-36.

VII. LITERATURVERZEICHNIS
95
Raimondi S., Lowenfels A.B., Morselli-Labate A.M., Maisonneuve P., Pezzilli R. (2010). Pancreatic cancer in chronic pancreatitis; aetiology, incidence, and early detection. Best Pract Res Clin Gastroenterol 24(3), 349-58.
Riehle K.J., Campbell J.S., McMahan R.S., Johnson M.M., Beyer R.P., Bammler T.K., Fausto N. (2007). Regulation of liver regeneration and hepatocarcinogenesis by suppressor of cytokine signaling 3. JEM 205(1), 91-103.
Rigby R.J., Simmons J.G., Greenhalgh C.J., Alexander W.S., Lund P.K. (2007). Suppressor of cytokine signaling 3 (SOCS3) limits damage-induced crypt hyperproliferation and inflammation-associated tumorigenesis in the colon. Oncogene 26, 4833-4841.
Roberts A.W., Robb L., Rakar S., Hartley L., Cluse L., Nicola N.A., Metcalf D., Hilton D.J., Alexander W.S. (2001). Placental defects and embryonic lethality in mice lacking suppressor of cytokine signaling 3. PNAS 98(16), 9324-9329.
Rose S.D., Swift G.H., Peyton M. J., Hammer R.E., McDonald R.J. (2001). The role of PTF1-P48 in pancreatic acinar gene expression .J Biol Chem 276, 44018-44026.
Rosewicz S., Wiedenmann B. (1997). Pancreatic carcinoma. Lancet 349(9050), 485-9.
Rösch T., Dittler H.J., Strobel K., Meining A., Schusdziarra V., Lorenz R., Allescher H.-D., Kassem A.M., Gerhardt P., Siewert J.-R., Höfler H., Classen M.( 2000). Endoscopic ultrasound criteria for vascular invasion in the staging of cancer of the head of the pancreas: a blind reevaluation of videotapes. Gastrointest Endosc 52, 469-77.
Rossa C., Sommer G., Spolidorio L.C., Rosenzweig S.A., Watson D.K., Kirkwood K.L. (2012). Loss of Expression and Function of SOCS3 Is an Early Event in HNSCC: Altered Subcellular Localization as a Possible Mechanism Involved in Proliferation, Migration and Invasion. PLoS ONE 7(9), e45197.
Rui L., Yuan M., Frantz D., Shoelson S., White, M.F. (2002). SOCS-1 and SOCS-3 block insulin signaling by ubiquitin-mediated degradation of IRS1 and IRS2. J Bio Chem 277, 42394-42398.
Saad E.D., Machado M.C.,Wajsbrot D., Abramoff R., Hoff P.M., Tabacof J., Katz A., Simon S.D., Gansl S.C. (2002). Pretreatment Ca 19-9 level as a prognostic factor in patients with advanced pancreatic cancer treated with gemcitabine. Int J Gastrointest Cancer 32(1), 35-41.
Sakorafas G.H., Tsiotos G.G.(2001). Molecular biology of pancreatic cancer: potential clinical implications. BioDrugs 15, 439-52.
Saxena N.K., Ikeda K., Rockey K.I., Friedmann S.L., Anania F.A. (2002). Leptin in hepatic fibrosis: Evidence for increased collagen production in stellate cells and lean littermates of ob/ob mice. Hepatology 35(4), 762-771.
Schenk M., Schwartz A.G., O’Neal E., Kinnard M., Greenson J.K., Fryzek J.P., Ying G.S., Garabrant D.H. (2001). Familial risk of pancreatic cancer. J Natl Cancer Inst 93, 640-644.
Schindeler A. ,Kolind M., Little D.G. (2013). Cellular transitions and tissue engineering. Cell Reprogram 15(2), 101-6.
Scholz A., Heinze S., Detjen K.M., Peters M., Welzel M., Hauff P., Schirner M., Wiedenmann B., Rosewicz S. (2003). Activated signal transducer and activator of

VII. LITERATURVERZEICHNIS
96
transcription 3 (STAT3) supports the malignant phenotype of human pancreatic cancer. Gastroenterology 125(3), 891-905.
Schutte M., Hruban R.H., Geradts J., Maynard R., Hilgers W., Rabindran S.K., Moskaluk C.A., Hahn S.A., Schwarte-Waldhoff I., Schmiegel W., Baylin S.B., Kern S.E., Herman J.G. (1997). Abrogation of the Rb/p16 Tumor-suppressive Pathway in Virtually All Pancreatic Carcinomas. Cancer Research 57, 3126-313.
Sellik G.S., Barker K.T., Stolte-Dijkstra I., Fleischmann C., Coleman R.J., Garrett C., Gloyn A.L., Edghill E.L., Hattersley A.T., Wellauer P.K., Goodwin G., Houlston R.S. (2004). Mutations in PTF1A cause pancreatic and cerebellar agenesis. Nat Genet 36, 1301-1305.
Senn J.J., Klover P.J., Nowak I.A., Zimmers T.A., Koniaris L.G., Furlanetto R.W. Mooney R.A. (2003). Suppressor of cytokine signaling-3 (SOCS-3), a potential mediator of interleukin-6-dependent insulin resistance in hepatocytes. J Biol Chem 278, 13740-13746.
Shacter E., Weitzman S.A. (2002). Chronic inflammation and cancer. Oncology (Williston Park) 16(2), 217-26.
Shaib Y., Davila J., Naumann C., El-Serag H. (2007). The impact of curative intent surgery on the survival of pancreatic cancer patients: a U.S. Population based study. Am J Gastroenterol 102, 1377-82.
Sherr C.J. (2004). Principles of Tumor Suppression. Cell 116(2), 235-246.
Shibata D., Capella G., Perucho M. (1990). Mutational activation of the c-K-ras gene in human pancreatic carcinoma. Baillière's Clinic Gastroenterol 4(1), 151-169.
Shimizu K. (2008). Mechanism of pancreatic fibrosis and applications to the treatment of chronic pancreatitis. J Gastroenterol 43, 823-832.
Simmons D.L., Botting R.M., Hla T. (2004). Cyclooxygenase Isozymes: The Biology of Prostaglandin Synthesis and Inhibition. Pharmacol Rev 56, 387-437.
Song M.M., Shuai K. (1998). The Suppressor of Cytokine Signaling (SOCS) 1 and SOCS3 but Not SOCS2 Proteins Inhibit Interferon-mediated Antiviral and Antiproliferative Activities. Journal of Biological Chemistry 273, 35056-35062.
Starr R., Willson T.A., Viney E.M., Murray L.J.L., Rayner J.R., Jenkins B.J., Gonda T.J., Alexander W.S., Metcalf D., Nicola N.A., Hilton D.J. (1997). A family of cytokine inducible inhibitors of signalling. Nature 387, 917-921.
Stocken D.D., Büchler M.W., Dervenis C., Bassi C., Jeekel H., Klinkenbijl J.H.G., Bakkevold K.E., Takada T., Amano H., Neoptolemos J.P. (2005). Meta-analysis of randomised adjuvant therapy trials for pancreatic cancer. British Journal of Cancer 92, 1372-1381.
Su L.K., Steinbach G., Sawyer J.C., Hindi M., Ward P.A., Lynch P.M. (2000). Genomic rearrangements of the APC tumor-suppressor gene in familial adenomatous polyposis. Hum Genet 106, 101-107.
Sutherland K.D., Lindeman G.J., Choong D.Y.H., Wittlin S., Brentzell L., Phillips W., Campbell I.G., Visvader J.E. (2004). Differential hypermethylation of SOCS genes in ovarian and breast carcinomas. Oncogene 23, 7726-7733.

VII. LITERATURVERZEICHNIS
97
Suzuki A., Hanada T., Mitsuyama K., Yoshida T., Kamizono S., Hoshino T., Kubo M., Yamashita A., Okabe M., Takeda K., Akira S., Matsumoto S., Toyonaga A., Sata M., Yoshimura A. (2001). CIS3/SOCS3/SSI3 plays a negative regulatory role in STAT3 activation and intestinal inflammation. J Exp Med 193(4), 471-81.
Takahashi Y., Carpino1 N., Cross J.C., Torres M., Parganas E., Ihle J.N. (2003). SOCS3: an essential regulator of LIF receptor signaling in trophoblast giant cell differentiation. The EMBO Journal 22(3), 372-384.
Talar-Wojnarowska R., Gasiorowska A., Smolarz B., Romanowicz-Makowska H., Kulig A., Malecka-Panas E. (2009). Clinical significance of interleukin-6 (IL-6) gene polymorphism and IL-6 serum level in pancreatic adenocarcinoma and chronic pancreatitis. Dig Dis Sci. 54(3), 683-9.
Talar-Wojnarowska R., Malecka-Panas E. (2006). Molecular pathogenesis of pancreatic adenocarcinoma: potential clinical implications. Med Sci Monit 12, 186-193.
Testini M., Gurrado A., Lissidini G., Venezia P., Greco L., Piccinni G. (2010). Management of mucinous cystic neoplasms of the pancreas. World J Gastroenterol 16(45), 5682-5692.
Tischoff I., Hengge U.R., Vieth M., Ell C., Stolte M., Weber A., Schmidt W.E., Tannapfel A. (2007). Methylation of SOCS‐3 and SOCS‐1 in the carcinogenesis of Barrett's adenocarcinoma. Gut. 56(8), 1047-1053.
Torisu T., Sato N., Yoshiga D., Kobayashi T., Yoshioka T., Mori H., Iida M., Yoshimura A. (2007). The dual function of hepatic SOCS3 in insulin resistance in vivo. Genes to Cells 12, 143-154.
Toth K.G., McKay B.R., De Lisio M., Little J.P., Tarnopolsky M.A., Parise G. (2011). IL-6 Induced STAT3 Signalling Is Associated with the Proliferation of Human Muscle Satellite Cells Following Acute Muscle Damage. PLoS ONE 6(3), 1-12.
Tsiambas E., Karameris A., Dervenis C., Lazaris A.C., Giannakou N., Gerontopoulos K., Patsouris E. (2006). HER2/neu Expression and Gene Alterations in Pancreatic Ductal Adenocarcinoma: A Comparative Immunohistochemistry and Chromogenic in Situ Hybridization Study Based on Tissue Microarrays and Computerized Image Analysis. JOP J Pancreas 7(3), 283-294.
Ueki. K., Kondo T., Kahn, C.R. (2004a). Suppressor of cytokine signaling 1 (SOCS-1) and SOCS-3 cause insulin resistance through inhibition of tyrosine phosphorylation of insulin receptor substrate proteins by discrete mechanisms. Mol Cell Biol 24, 5434-5446.
Ueki, K., Kondo T., Tseng Y.H., Kahn, C.R. (2004b). Central role of suppressors of cytokine signaling proteins in hepatic steatosis, insulin resistance, and the metabolic syndrome in the mouse. Pro Natl Acad Sci USA 101, 10422-10427.
Vargas-Roig L.M., Fanelli M.A., López L.A., Gago F.E., Tello O., Aznar J.C., Ciocca D.R. (1997). Heat shock proteins and cell proliferation in human breast cancer biopsy samples. Cancer Detect Prev 21(5), 441-51.
Velasco J.M., Rossi H., Hieken T.J., Fernandez M. (2000). Laparoscopic ultrasound enhances diagnostic laparoscopy in the staging of intra-abdominal neoplasms. Am Surg 66(4), 407-411.

VII. LITERATURVERZEICHNIS
98
Vona-Davis L.C., Frankenberry K.A., Waheed U., Peterson E., McFadden D.W. (2005). Expression of STAT3 and SOCS3 in pancreatic acinar cells. J Surg Res 127(1), 14-20.
Weber A., Hengge U.R., Bardenheuer W., Tischoff I., Sommerer F., Markwarth A., Dietz A., Wittekind C., Tannapfel A. (2005). SOCS-3 is frequently methylated in head and neck squamous cell carcinoma and its precursor lesions and causes growth inhibition. Oncogene 24, 6699-6708.
White G.E., Cotterill A., Addley M.R., Soilleux E.J., Greaves D.R. (2011). Suppressor of cytokine signalling protein SOCS3 expression is increased at sites of acute and chronic inflammation. J Mol Histol 42(2), 137-51.
Wilentz R.E., Geradts J., Maynard R., Offerhaus G.J.A., Kang M., Goggins M., Yeo C.J., Kern S.E., Hruban R.H. (1998). Inactivation of the p16 (INK4A) Tumor-suppressor Gene in Pancreatic Duct Lesions: Loss of Intranuclear Expression. Cancer Research 5, 4740-4744.
Xiong H., Zhang Z.G., Tian X.Q., Sun D.F., Liang Q.C., Zhang Y.J., Lu R., Chen Y.X., Fang J.Y. Inhibition of JAK1, 2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces tumor cell invasion in colorectal cancer cells. 2008. Neoplasia, 10(3), 287-97.
Xu Q., Briggs J., Park S., Niu G., Kortylewski M., Zhang S., Gritsko T., Turkson J., KayH., Semenza G.L., Cheng J.Q., Jove R., Yu H. (2005). Targeting Stat3 blocks both HIF-1 and VEGF expression induced by multiple oncogenic growth signaling pathways. Oncogene 24, 5552-5560.
Yeo C.J., Cameron J.L., Lillemoe K.D., Sitzmann J.V., Hruban R.H., Goodman S.N., Dooley W.C., Coleman J., Pitt H.A. (1995). Pancreaticoduodenectomy for cancer of the head of the pancreas. 201 patients. Ann Surg 221, 721-31.
Yeo T.P., Hruban R.H., Leach S.D., Wilentz R.E., Sohn T.A., Kern S.E., Iacobuzio-Donahue C.A., Maitra A., Goggins M., Canto M.I., Abrams R.A., Laheru D., Jaffee E.M., Hidalgo M., Yeo C.J. (2002b). Pancreatic cancer. Curr Probl Cancer 26, 176-275.
Yoshikawa H., Matsubara K., Qian G.S., Jackson P., Groopman J.D., Manning J.E., Harris C.C., Herman J.G.(2001). SOCS-1, a negative regulator of the JAK/STAT pathway, is silenced by methylation in human hepatocellular carcinoma and shows growth-suppression activity. Nat Genet 28(1), 29-35.
Yu H., Kortylewski M., Pardoll D. (2007). Crosstalk between cancer and immune cells: role of Stat3 in the tumor microenvironment. Nat Rev Immunol 7(1), 41-51.
Yu H., Pardoll D., Jove R. (2009). STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer 9, 798-809.
Yu L.-J., Wu M.-L., Li H., Chen X.-Y., Wang Q., Sun Y., Kong Q.-K., Liu J. (2008). Inhibition of STAT3 Expression and Signaling in Resveratrol-Differentiated Medulloblastoma Cells. Neoplasia 10(7), 736-744.
Zhu L., Shi G., Schmidt C.M., Hruban R.H., Konieczny S.F. (2007). Acinar cells contribute
to the molecular heterogeneity of pancreatic intraepithelial neoplasia. Am J Pathol 171(1), 263-73.

VIII. ABBILDUNGSVERZEICHNIS
99
VIII. ABBILDUNGSVERZEICHNIS
Abbildung 1: Morphologische Tumorprogression über PanIN-Vorläuferläsionen ................... 8
Abbildung 2: Morphologisches und genetisches Progressionsmodell des duktalen Pankreaskarzinoms. .................................................................................................................. 10
Abbildung 3: Gewebespezifische Deletion von Mausgenen mittels Cre/loxP-Technologie. .. 13
Abbildung 4: Systematik der Generierung des KrasG12D-Tumormausmodells. ....................... 16
Abbildung 5: Schematische Darstellung der SOCS3-Architektur. .......................................... 18
Abbildung 6: SOCS3 als Trägermolekül des E3-Ubiquitin-Ligase-Komplexes ..................... 18
Abbildung 7: Die Mechanismen der Aktivierung der IL-6/Jak/Stat3-Signalkaskade und dessen Feedback-Inhibition durch SOCS3-Moleküle .......................................................................... 19
Abbildung 8: Die Aktivierung der Ras/Raf/MAPK- und PI3K/AKT-Signalkaskade durch IL-6 und dessen negative Regulation durch Socs3........................................................................... 20
Abbildung 9: Schematische Darstellung der Generierung der Socs3Δpanc-Mauslinie. ............. 42
Abbildung 10: Nachweis der Deletion von Exon 2 im Socs3-Gen mittels mRNA Konzentrationsbestimmung ...................................................................................................... 43
Abbildung 11: PCR-Analysen der möglichen Genotypen aus der Kreuzung von Socs3F/F-und
Ptf1a-Creex1-Mäusen. ............................................................................................................... 44
Abbildung 12: Charakterisierung des exokrinen und endokrinen Pankreaskompartiments. ... 45
Abbildung 13: Gewichtsanalyse der Socs3Δpanc- und Socs3
F/F-Nachkommen. ........................ 47
Abbildung 14: Proteinbiochemischer Nachweis der Expression von Stat3 und p-Stat3 in Pankreata neun Wochen alter Kras
G12D-Mäuse ........................................................................ 48
Abbildung 15: Quantitative Real-Time-PCR Analysen der vermehrten Expression von Socs3-mRNA in neunWochen alten Kras
G12D-Mäusen. ..................................................................... 49
Abbildung 16: Immunhistochemische Detektion von Socs3 in Zellen der Azini (schwarze Pfeile) und in den PanIN-Läsionen (weiße Pfeile) neun Wochen alter Kras
G12D-Mäuse. ....... 49
Abbildung 17: Aktivierung der Stat3-Signalkaskade im KrasG12D-Mausmodell. .................... 51
Abbildung 18: Proteinbiochemischer Nachweis der Socs3-Defizienz in KrasG12D
;Socs3Δpanc-
Mäusen. .................................................................................................................................... 53
Abbildung 19: Nachweis der Expression konstitutiv aktiver K-Ras-Proteine. ........................ 54
Abbildung 20: Gewichtsanalyse sechs Wochen alter KrasG12D
;Socs3Δpanc-Mäuse (n=7) im
Vergleich zu gleichaltrigen KrasG12D-Kontrollmäusen (n=7) und zum Wildtyp (n=17). ........ 55
Abbildung 21: Analyse des makroskopischen Pankreasaspekts. ............................................. 56

100
Abbildung 22: Morphologische Charakterisierung der PanINs und des PDA ......................... 58
Abbildung 23: Vergleichende Auswertung der Gesamtzahl an Ausführungsgängen pro zehn Gesichtsfelder an mindestens 17 sequentiellen Schnitten vier Wochen alter Versuchstiere (200-fache Vergrößerung) ........................................................................................................ 60
Abbildung 24: Quantifizierung der reaktiven Gänge und PanIN-Läsionen pro zehn Gesichtsfelder in mindestens 17 sequentiellen Feingewebsschnitten zum Zeitpunkt von vier Wochen und bei 200-facher Vergrößerung .............................................................................. 61
Abbildung 25: Immunhistochemischer Nachweis der übermäßigen Expression von CK-19, p-Stat3 und Cyclin D1 in reaktiven Gängen und PanIN-Läsionen vier Wochen alter Kras
G12D;Socs
Δpanc-Mäuse. 100-fache Vergrößerung .............................................................................................................. 62
Abbildung 26: H.E.-Färbung des duktalen Adenokarzinoms des Pankreas und immunhistochemische Detektion von CK-19 und p-Stat3 in atypischen, drüsenartigen Gangstrukturen. .............................. 63
Abbildung 27: Detektion von Muc1, Muc5 und CK-19 in Pankreata vier und neun Wochen alten Mäusen mittels immunhistochemischer Färbung zur genauen Charakterisierung der neoplastisch transformierten Gänge. 100-fache Vergrößerung. .......................................................................... 64

IX. TABELLENVERZEICHNIS
101
IX. TABELLENVERZEICHNIS Tabelle 1: Chemikalien ............................................................................................................ 24
Tabelle 2: Geräte und Hilfsmittel ............................................................................................. 26
Tabelle 3: Puffer und Lösungen ............................................................................................... 28
Tabelle 4: Gele ......................................................................................................................... 28
Tabelle 5: Antikörper für IHC .................................................................................................. 29
Tabelle 6: Antikörper für Western Blot ................................................................................... 30
Tabelle 7: Primer ...................................................................................................................... 30
Tabelle 8: Mäuse ...................................................................................................................... 30

X. DANKSAGUNG
102
X. DANKSAGUNG
Die Dissertation wurde an der II. Medizinischen Klinik und Poliklinik der Medizinischen
Fakultät der Technischen Universität durchgeführt.
An dieser Stelle bedanke ich mich bei all den Menschen, die am Zustandekommen dieser
Dissertation beteiligt waren:
Mein ganz besonderer Dank geht an meinen Doktorvater Herrn Priv.-Doz. Dr. Hana Algül,
der mich herzlich in sein Forschungsteam aufgenommen und mir das überaus interessante und
anspruchsvolle Thema dieser Arbeit zur Verfügung gestellt hat. Durch ihn ergab es für mich
die Möglichkeit in einer hochprofessionellen Arbeitsgruppe tätig sein zu dürfen und auf
fachlich höchstem Niveau Einblicke in die Grundlagenforschung zu erhalten. Desweiteren
möchte ich mich bei meinem Doktorvater für seine Betreuung, Geduld, Motivation und
konzeptionelle Unterstützung bei der Fertigstellung der Dissertation bedanken. Trotz seines
weitreichenden Engagements sowohl im klinischen als auch wissenschaftlichen Bereich stand
er mir zu jeder Zeit mit Rat und Tat zur Seite.
Vielen herzlichen Dank!
Dem Direktor der II. Medizinischen Klinik und Poliklinik Herrn Prof. Dr. med. R.M. Schmid
danke ich für die Möglichkeit, in seiner Klinik promovieren zu dürfen.
Bei Frau Marina Lesina und Frau Karen Dlubatz möchte ich mich besonders für die
hochkompetente Einführung in die wissenschaftlich-experimentelle Arbeit des
Gastroenterologischen Forschungslabors und ihre unerschöpfliche Geduld bedanken. Mit
ihrer Hilfe war es mir möglich enorme praktische Kenntnisse und Erfahrungen zu gewinnen.
Ebenso möchte ich mich bei all den wissenschaftlichen Mitarbeitern des
Gastroenterologischen Forschungslabors für ihre Hilfsbereitschaft bedanken. Auch meinen
Mitdoktoranden danke ich für ihre Unterstützung bei manchen Experimenten.
Meinen Eltern Herrn Phan Nhan Hoa und Frau Chu Thi Minh Hao und meiner Schwester
Phan Ngoc Trang danke ich für ihre Fürsorge, Liebe und Unterstützung. Ohne sie wären mein
Medizinstudium und diese Promotion nicht möglich und ihnen möchte ich diese Arbeit
widmen.
Vielen Dank, liebe Eltern und Schwester!
Bei meinem Freund Nico Brehm möchte ich mich für die tatkräftige Unterstützung nicht nur
bei der Fertigstellung der Dissertation sondern auch bei zahlreichen Prüfungen bedanken. In

103
vielen schlaflosen Nächten war er die Person, die immer an meiner Seite stand und mich
immer wieder psychisch und physisch aufbaut.
Vielen, vielen Dank Nico!
Mein letzter Dank geht an Herrn Andreas Kaiser, der diese Arbeit für mich Korrektur las.
Herzlichen Dank für die ausgezeichneten und wertvollen Hinweise!

XI. ERKLÄRUNG
104
XI. ERKLÄRUNG
Ich erkläre hiermit, dass ich die vorliegende Arbeit selbstständig und unter Verwendung der
angegebenen Hilfsmittel, persönlichen Mitteilungen und Quellen angefertigt habe.
München, den 23.03.2014


















