
Über den Mechanismus der Abscheidung von SiO2-Gallerten in Pflanzenzellen
8
Kleine Mitteilungen Uber den Mechanismus der Abscheidung yon SiO~-Gallerten in Pflanzenzellen Von Hans Pfeiffer, Bremen Mit i Textfigur (Eingegangen den 12. 0ktobcr 1929) 1. Die Verkieselung yon Pflanzenzellen braucht sich nicht immer auf ganze Gewebe zu erstrecken, sondern kann sich auch auf bestimmt Iokali- sierte, oft eigenartig gestaltete Zellen beschr~inken. Als solche habe ich seit langem die yon DVVAL-JovvE (1) entdeckten, sp~iter freilich nut gelegentlich gefundenen (2) Kegelzellen (cellules s fond conique) vieler Cyperaceen untersucht (3). Uber den Verkieselungsvorgang an bestimmt organisierten Zellformen sind bereits einige Ergebnisse (4) vorhanden, nach denen durch eine mit Energieaufwand verbundene Lebenst~tigkeit der Zellen fortgesetzt SiO~ aus benachbarten Gewebeelementen aufgenommen wird, auch wean be- felts eine ges~ttigte LSsung auftritt. Indem in den betr. Zellen kurz vor der Verkieselung keine Plasmolyse bemerkt wird, diese aber in manchen F~llen -- trotz gewisser, von dem Massenverh~ltnis zwischen Cytoplasma und Vakuolen bedingter technischer Schwierigkeiten -- durch SalzlSsungen (durch Na2 SOt am deutlichsten) hervorgerufen werden kann, muff ein Ein- dringungsvermSgen yon SlOe angenommen werden. Well die SiO~-Gallerten schon neben dem erst sp~ter degenerierenden Zellkern vorkommen und wachsen kSnnen, ist zu schliei~en, daft das Cytoplasma yon SiO 2 durch- drungen werden kann und das Sol nicht sogleich zur Ausfiillung zu kommen braucht. Ob es sich bei der Si02-Aufnahme um eine AlkalisilikatlSsung, aus welcher das Si durch Siiuren befreit werden miii~te, oder um geliJste Kieselsiiure handelt, bleibe hier uner~rtet, braucht doch das eine Vorkommen jenes andere nicht auszuschlieflen. Seltener, wie in Elementen der Sekund~ir- rinde yon Licania AUBL. (Moquilea AWL.), wird die Zelle plStzlich ann~hernd ganz yon kSrniger SiO~-Masse erfiillt, welche sich allmi~hlich yon auflen nach innen verdichtet. Sind solcherart die Phi~nomene wiihrend der Abscheidung der Si02-Gallerten bereits weitgehend gekl~rt, so bedarf doch der Mechanis- mus jenes Prozesses erneuter Untersuchung.
-
Upload
hans-pfeiffer -
Category
Documents
-
view
215 -
download
2
Transcript of Über den Mechanismus der Abscheidung von SiO2-Gallerten in Pflanzenzellen

Kleine Mitteilungen Uber den Mechanismus der Abscheidung yon SiO~-Gallerten
in Pflanzenzellen
Von Hans Pfeiffer, Bremen
Mit i Textfigur
(Eingegangen den 12. 0ktobcr 1929)
1. Die Verkieselung yon Pflanzenzellen braucht sich nicht immer auf ganze Gewebe zu erstrecken, sondern kann sich auch auf bestimmt Iokali- sierte, oft eigenartig gestaltete Zellen beschr~inken. Als solche habe ich seit langem die yon DVVAL-JovvE (1) entdeckten, sp~iter freilich nut gelegentlich gefundenen (2) K e g e l z e l l e n (cellules s fond conique) vieler Cyperaceen untersucht (3). Uber den V e r k i e s e l u n g s v o r g a n g an bestimmt organisierten Zellformen sind bereits einige Ergebnisse (4) vorhanden, nach denen durch eine mit Energieaufwand verbundene Lebenst~tigkeit der Zellen fortgesetzt SiO~ aus benachbarten Gewebeelementen aufgenommen wird, auch wean be- felts eine ges~ttigte LSsung auftritt. Indem in den betr. Zellen kurz vor der Verkieselung keine Plasmolyse bemerkt wird, diese aber in manchen F~llen - - trotz gewisser, von dem Massenverh~ltnis zwischen Cytoplasma und Vakuolen bedingter technischer Schwierigkeiten - - durch SalzlSsungen (durch Na2 SO t am deutlichsten) hervorgerufen werden kann, muff ein Ein- dringungsvermSgen yon SlOe angenommen werden. Well die SiO~-Gallerten schon neben dem erst sp~ter degenerierenden Zellkern vorkommen und wachsen kSnnen, ist zu schliei~en, daft das Cytoplasma yon SiO 2 durch- drungen werden kann und das Sol nicht sogleich zur Ausfiillung zu kommen braucht. Ob es sich bei der Si02-Aufnahme um eine AlkalisilikatlSsung, aus welcher das Si durch Siiuren befreit werden miii~te, oder um geliJste Kieselsiiure handelt, bleibe hier uner~rtet, braucht doch das eine Vorkommen jenes andere nicht auszuschlieflen. Seltener, wie in Elementen der Sekund~ir- rinde yon Licania AUBL. (Moquilea AWL.), wird die Zelle plStzlich ann~hernd ganz yon kSrniger SiO~-Masse erfiillt, welche sich allmi~hlich yon auflen nach innen verdichtet. Sind solcherart die Phi~nomene wiihrend der Abscheidung der Si02-Gallerten bereits weitgehend gekl~rt, so bedarf doch der Mech an i s - mus jenes Prozesses erneuter Untersuchung.

Kleine Mitteihngen 121
2. Es liegt nahe: die SiO~-Abscheidung in kolloidehemischer Hinsieht mit der A u s f l o c k u n g ( K o a g u l a t i o n ) zu vergleichen. Diese ist nfindestens teilweise eine elektrische Erseheinung. Der )Iechanismus der Teilehenaggregation besteht nach der Theorie yon S~OLUCHOWSX~ (5) darin, da~ die Attraktionskr~tfte zwischen den Teilehen nach eingetretener Entladung nicht mehr durch elektrisehe AbstoSungskritfte ausgeglichen werden. In diesem Sinne miil~te also die Si02-Abscheidung einer Entladung der Zell- kolloide parallel gehen und sich als deren Folgewirkung ergeben. Das Auftreten yon SiOs-GaUerten wiirde sohiil den Eintritt eines Neutralteilehenmaximums und eine A n- ni~herung an den I E P der Z e l l a m p h o l y t e begleiten. Die an anderer Stelle in dieser Zeitschrift in den Grundziigen entworfene Arbeitshypothese tiber die Aktivierunff tier Teilungstgtigkeit bis zur Merlstenmtisierun~ yon Dauerzellen (6) miigte mithin zu den teilweise welt zurfickreiehenden Untersuchungsergebnissen fiber die Verkieselung yon Pflanzenzellen (3) in Beziehung gebracht werden kgnnen. Indessen ist die Technik zum Nachweise des IEP yon Protoplasten (7) noeh viel zu sehr in der Entwiekhng begriffen, um eine bequeme Naehpriifung jener Folgerung zu gestatten. Hinzu komrat, da$ zufolge der elektrometrisehen Untersuehung des goagulationsprozesses yon Si02-Solen dutch LASKIN (8) statt der hauptsi~chlich elektrostatischen (entgegengesetzt geladener Ionen oder geladenel: ~olekiile) die andere Wirkungsart "con Elektroiyten (9), ni~mlieh die lyotrope resp. debydratatisierende (mittels Ionen oder Holekiile starken Hydratations- verm(igens), vorzuziehen ist (10). Der Vorgang der Abscheidung" und allmi~hliehen Ver- diehtung yon Si 02-Gallerten macht zwar bereits in Bestatigung dieser Auffassung vSllig den Ein&-uck einer gewghnliehen W a s s e r a b s p a l t u n g s k o n d e n s a t i o n , dutch welche eine Vermindernng der Dispersitiit und LSslichkeit des Sols bis zur irreversiblen Aus- scheidung (11) bewirk~ wird; abet es bedarf doch wohl noch sehlfissiger Grilnde far die Ubertragung der LASKINschen Ansicht. Da die Umsti~ndliehkeit der Methodik die direkte Bestimmung der Distanz gegen den IEP (7) sehr erschwert, ist untersucht worden, ob sich ein anderer Beweis ffir die ~ f i t w i r k u n g d e h y d r a t a t i v e r P r o z e s s e b e 1 d e r S i 02 - A b s c h ei d u n g in Pfianzenzdlen erbringen lagt.
3. Schon seit ]angerer Zei t is t versucht worden, die Abscheidung in die Zellen e indr ingender Si02-Sole durch U b e r s c h w e m m u n g m i t b a s i s c h e n F a r b s t o f f e n hervorzurufen (12). W i e aus der neuen Monographie KgSTERS (13) ZU ersehen ist, haben zwar schon PFEFFER, OVERTO.~ U. a. bes t immte Fa l lungen in Zellen dutch Behandlung mit ungif t igen Farbs tof fen erzeugt, und aus neuerer Zei t werden von K~STER entsprechende Befunde von KLE~S und SCARTU zit iert . Aber eine spezifisch gelbeschleunigende W i r k u n g basischer Farbs tof fe scheinen doch erst TRAUBE und KOH~ER (14) bewufit angewende t zu haben. PFEIFFERS Beobachtungen (12) stel len also im wesentl ichen eine For t f i ih rung ihrer Versuchsanstel lungen darl) . So betr~chtl ich auch die zahlreichen Angaben fiber das Verhal ten organischer Farbs tof fe zu SiO,, ein-
~) Wi~hrend der Niederschrift sehe ich, was mir bisher entgangen ist (12), da$ in i~hnlieher Weise auch BRESSLAU (15) vorgesehlagen hat, den Plasmaleib yon Infwsorien durch Uberfiirbang mit Opalblau oder Phloxinrhodamin zum G e l a t i n i e r e n zu bringen und so Dauerpr@arate der Tierehen herzustellen, sofern die Konservierung des Kernes unnStig ist.

122 Kleine ]~[itteilungen
ander widersprechen (16), so dare doch mindestens in dem Spezialfalle der Veriinderung yon Si02-Solen pflanzlicher Zellen durch Farbstoffl(~sungen hoher Konzentration eine f r fi h e r e S i- N a c h w eis b a r k e i t als gesichert gelten (12) 1). Die Diskrepanzen zwischen den bisherigen Ergebnissen diirften sich in der Hauptsache aus dem erwiesenen Befunde (18) erkl~tren, daft Adsorptionen jeder Art am Silikagel w e i t g e h e n d von der Basizi t i~t des A d s o r b e n s abh~ngig sind. Mag aueh bei Lis~:I~s Modellversuchen das Resultat von dem Dispersit~tsgrad der Farbstoffe einigermafien unabh~ngig sein, so ist doch die besondere Bedeutung~ welehe ROHLAND~ GLASU~OW U. a. (19) ge- rade dem D i s p e r s i t ~ t s z u s t a n d e zuschreiben~ bei der Behandlung yon Pflanzenzellen mindestens auf dem Umwege fiber unterschiedliches Ein- dringungsvermSgen der Farbstoffteilchen nicht zu iibersehen. So zeigen entsprechende Versuche eine ziemlich gleich gute Verwendbarkeit von Methylen- blau, Neutralrot, Methylgriin und Gentianaviolett, aber ein langsameres Eindringen yon Rhodamin und kaum eine nennenswerte iufnahme yon
Nachtblau in die Kegelzellen~ und in solchen Fi~llen kann auch ein beschleuni- gender Einflufi auf die Bildung der Si02-@allerte night in Betracht kommen.
4. Die grofien Vorteile~ welche die G e w e b e a u f h e l l u n g m i t t e l s P h e n o l (konz. Carbolsiiure) bei der Untersuchung yon Verkieselungen bietet (20), haben den ersten Anlafi zu kombinierten F~irbungsversuchen mit Phenol als Liisungsmittel der Farbstoffe (12;20) gegeben. Phenol ist, wie aus den Ergebnissen zahlreicher Resorptionsversuehe (21) ersichtlich ist, in der wiisserigen LSsung fast nut m o l e k u l a r d i s p e r s verteilt. Fiir unsere Ver- suche sehr gfinstig ist die auffallende Erleichterung, mit welcher in Phenol geISste Farbstoffe aufgenommen werden. Dieser Befund ist erkliirlich, weil ja eine b e s o n d e r s d i f f u s i b l e Fo rma r ~ durch Lipoidl~isliehkeit der Sub- stanz~ welche vielleicht der L(islichkeit in den Lipoiden der Plasmagrenzen parallel geht, gewi~hrleistet wird (22). Dutch die yon jener des gebriiuch- lichen LSsungsmittels (Wasser) stark a b w e i c h e n d e DEK des P h e n o l s werden freilich unter Berficksichtigung des WALDEsschen Gesetzes (23) ganz andere Dissoziationsverhiiltnisse flit die Farbstoffe geschaffen (24). Es ist ni~mlich iiberwiegend mit der A u f n a h m e u n d i s s o z i i e r t e r F a r b s t o i f m o l e - ki i le zu rechnen, sofern die Versuche die Aufnahme des Farbstoffes ergeben. ~brigens kann schon die v e r r i n g e r t e O r i e n t i e r u n g s s t ~ b i l i s a t i o n des D i s p e r s i o n s m i t t e l s bei der Durchtr~nkung mit Phenol (ohne Farbstoff) eine F6rderung der F~llung silicider Hydrogele erkl~ren.
1) K~STER (17) vermutet wohl mit Recht bei der yon FR. WEBER konstatierten FSrderung spontaner u bei Helodea-Blattzellen durch Neu- tralrotf~,rloung eine Mitwirkung von Koagulationserscheiuungen~ welche durch den Farb- stoff hervorgerufen werdem

Kleine Mitteilungen 123
Bei der Durchf i ihrung der Versuche (20) hat es sich als zweckma$ig erwiesen~ eine Reihe gleich beschaffener Reagenzgl~ser mit der ~leichen )~[enge kalter PhenollSsunff (gewShnliche Handelsware) zu fiillen and die gleiche ~enge verschiedener Farbstoffe hineinzubrlngen. In die kurz durchschiittelte L(isung werden ganze Versuchspflanzen bezw. unverletzte Sprosse mit der Schnittfl/~che hineingestellt und je nach GrSl3e des Objekts und nach Lebhaftigkeit seiner Transpiration 10 bis 40 Stunden unter trans- pirationsfSrdernde Bedingungen gebracht. Dann saugen die Pflanzen mit dem Trans- pirationsstrome Phenol bezw. gleichzeiti~ die in diesem ffelSsten Farbstoffe nach und nehmen zugleich eine ftir alas Schneiden mit dem Rasierraesser sehr erwiinschte Kon- sistenz an. Die Untersuchung der Schnitte erfolgt in kalter PhenollSsung (Stamml~isung)~ welche man zur ]~[aterialersparnis evtl. mit Wasser verdiinnen kann (his der rStliche Ton der Liisunff verschwunden ist, welchen die am Lichte aufbewahrte StammlSsung trotz Verwendung brauner Gliiser bald anzunehmen pflegt). Weft die Verkieselung der Keffelzellen erfahrungsgem~$ ziemlich plStzlich eintritt, ist es nicht leicht, entsprechende En twiek lungss tad ien helm Mikroskopieren zu fassen (4). Am schnellsten gelangt mad zum Ziele, wenn man am Grunde der jiingsten Internodien jene Zellen durchmustert, welehe spi~ter fiir gewShnlieh die Si02-Ablafferungen ffihren, also Epidermiszellen (,Engzellen") fiber den Festigungselementen der Achse, des Blattes oder der Blattseheide. ~Bei wenigen Objekten, wie bei Cladium Mariseus R. Bx~., fiihrt auch die mikroskopische Untersuehnng der Endodermis der Wurzeln vielfach zum Ziele.
5. Wenn es im Sinne yon LASKI~ (10) richtig ist, dab die Si02-Aus- fallung die Folge einer Dehydratation ist, so mtissen sich die basischen Farb- stoffe je nach ihrem Dehydratationsverm5gen (Hydratationsgrade) ungleich verhalten oder mindestens verschieden geeignet sein. In der beschriebenen Weise sind deshalb V e r s u c h e m i t M e t h y l e n b l a u , R h o d a m i n , N a c h t - b l a u und N e u t r a l r o t angestellt worden. Es hat sich dabei gezeigt, dab bei Verwendung yon Methylenblau-Phenol bereits eine 0,1-prozentige LSsung des Farbstoffes ausreicht, w~hrend die andern Substanzen bei einer solchen Konzentration noch keine siehere Entscheidung zulassen. Dennoch sind sie keineswegs ungeeignet, wie unter Beachtung der yon KI)STER (17) zitierten und gedeuteten Versuchsergebnisse F~. WESERS mindestens ftir Neutralrot zu erwarten ist. Doeh werden wir hierbei eine Konzentration von vielleieht 1% wahlen. Die beiden letzten Farbstoffe scheinen zu der SiO~-Abscheidung ebenfalls yon ungleicher Brauchbarkeit zu sein, indem jene yore Nachtblau einigermaf~en der des Neutralrot gleiehkomlnt, freilich wegen schlechteren EindringungsvermSgens praktisch geringer ist, wiihrend Rhodamin am besten in noch hSheren Konzentrationen zu verwenden ist.
Die nur geringen Unterschiede in der Brauchbarkeit einiger der basischen Farbstoffe haben bald die Veranlassung gegeben, dat~ das anfangs verfolgte Ziel, in der SiO2-Ablagerung eine Ann~herung an den IEP der Zellampholyte zu erweisen (S. 121), ganz fallen gelassen worden ist. Die yon LASKIN (10) angenommene und d u r c h die F ~ r b u n g s v e r s u c h e n i c h t zu w i d e r l e g e n d e d e h y d r a t a t i s i e r e n d e W i r k u n g b a s i s c h e r F a r b s t o f f e
124 Kleine Mitteflungen
beruht auf der Annahme, dat~ eine infolge der Hydratation auftretende scheinbare KonzentrationserhShung grSfSer ist als die durch Dissoziations- rfiekgang hervorgerufene Abnahme der Teilchenzahl. Indem so die Hydra- ration mit dem LSsungsvorgange eng verbunden ist, mul3 sich, wie Ascm Kn~sI (25) gezeigt hat, die Ltislichkeit einer Substanz (hier des Si 02) dadurch vermindern, dab ihm durch einen andern (hier den Farb-) StoOl das zur Hydratation benStigte Wasser entzogen wird. Die ungleiehe Eignung der verwendeten Farbstoffe zur Ausfiillung yon Si02-Solen mug somit ihren Grund in dem u n t e r s c h i e d l i e h e n D e h y d r a t a t i o n s v e r m S g e n tier in Phenol gel Ssten Farbstoffe haben.
,.q/5o~;m3
~g5
r
o~o3
o, o2
0,oi
0 o,1
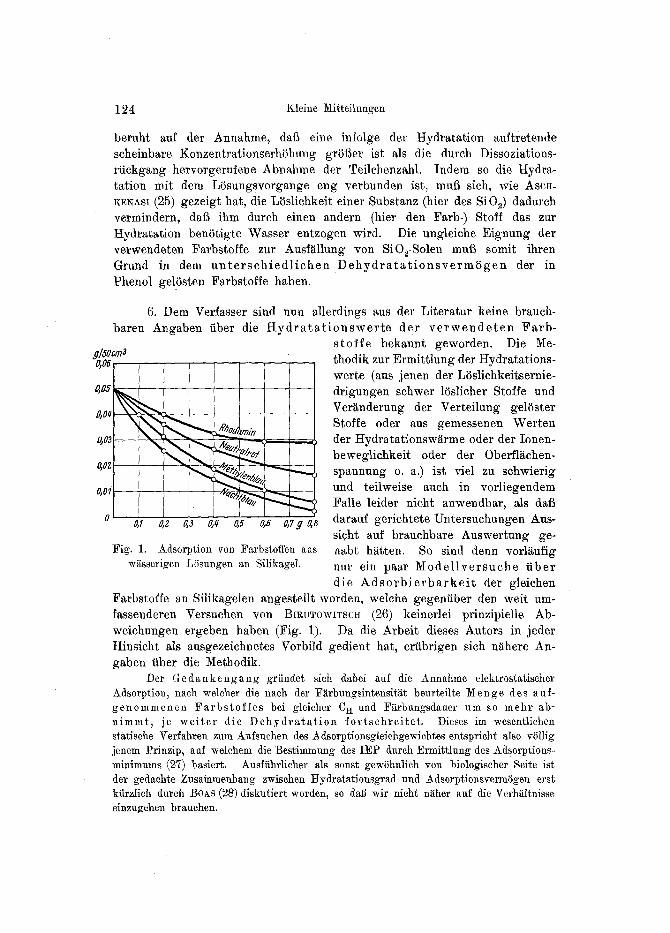
6. Dem Verfasser sind nun allerdings aus der Literatur keine brauch- baren Angaben fiber die t t y d r a t a t i o n s w e r t e der v e r w e n d e t e n Farb-
s t o f f e bekannt geworden. Die Me- thodik zur Ermittlung der Hydrafiations- werte (aus jenen der Li3slichkeitsernie- drigungen schwer 15slicher Stoffe und Veriinderung der Verteilung gelSster
~ ~ ~ m / n Stoffe oder aus gemessenen Werten �9 der Hydratationswiirme oder der Ionen-
beweglichkeit oder der Oberfl~chen- % t ~ , , ,,,~ ~ , ~ " ~ spannung o. a.) ist viel zu schwierig
z ,~ , c ,~ , ,~_ ~ und teilweise auch in vorliegendem | -~ ~ - . ~ Falle leider nicht anwendbar, als dat~
g,z 0,s o,q 0,5 0,8 0,7 g g,s darauf gerichtete Untersuchungen Aus- si.cht auf brauehbare Auswertung ge-
Fig. 1. Adsorption yon Farbstoffen aus nabt h~tten. So sind denn vorlgufig w~sserigen L/isungen an Silikagel. nut ein paar M o d e l l v e r s u c h e f iber
die A d s o r b i e r b a r k e i t der gleichen Farbstoffe an Silikagelen angestellt worden~ welche gegenfiber den weit um- fassenderen Versuchen yon BIRUTOWITSCH (26) keinerlei prinzipielle Ab- weichungen ergeben haben (Fig. 1). Da die Arbeit dieses Autors in jeder Hinsicht als ausgezeichnetes Vorbild gedient hat, eriibrigen sich n~here An- gaben tiber die Methodik.
Der Gedankengang grfindet sich dabei auf die Annahme elektrostatischer Adsorption: nach welcher die nach der Fiirbungsintensitiit beurteilte ~Ienge des auf- genommenen Farbs toffes bei ffleicher Ctt und Fih'bunffsdauer um so mehr ab- nimmt~ je welter die Dehydra ta t ion fortschrei tet . Dieses im wesentlichen statische Verfahren zum Aufsuchen des Adsorptionsgleichgewichtes entspricht also v(ifiig jenem Prinzip, auf welchem die Bestimmung des IEP dnrch Ermittlung des Adsorptions- minimums (27) basiert. Ausfiihrlicher als sonst gewShnlich yon biologischer Seite ist der gedachte Zusammenhang zwisehen Hydratationsg-rad und AdsorptionsvermSgeu erst kiirzlich dutch BOAS (28)diskutiert worden~ so dal~ wir nicht n~iher auf die Verh~iltnisse einzugehen brauchen.

Kleine Mitteilungen 125
Wenn die angefiihrten Uberlegungen zutreffen, miissen also die weniger geeigneten Farbstoffe zugleich diejenigen st~rkeren HydratationsvermSgens und jene von Silikagelen besonders gut adsorbierten darstellen. Wie die Figur zeigt, folgen die verwendeten Farbstoffe hinsichtlich ihrer Adsorptions- ~iihigkeit tatsiichlich der fMlenden Reihe:
0,1~ Rhodamin > 0,1~ Neutralrot > 0,1~ Methylenblau > 0,1~ Nachtblau,
d. h. Naehtblau wtirde fiir die Versuche der zweekmi~fiigste Farbstoff sein, wenn es in ausreichender Menge in die Zelle einzudringen vermSchte, M e t h y l e n b l a u und N e u t r a l r o t kommen daher in erster Linie zur Er- zeugung yon SiOe-Gallerten in Pflanzenzellen in Betracht.
Nach diesem Ergebnis kSnnen wir die LASKIN sche Deutung der Si 02-Abscheidung (i0) als Dehydratationsproze$ als gegeben hinnehmen, sofern sich die Farbstoffe auch noch d~rch andere dehydra ta t i s ie rend wirkende Substanzen (Satze) ersetzen lassen. Wirklich sind schon vor mehreren Jahren yon 1)FEIFFER (4) Befunde nach Be- handlung mit Na2S04 (in w~sseriger LSsung) gemaeht worden, die als Best~tigung ge- deutet werden mSgen. Die Schwierigkeiten, welche planm~$igen Versuchen mit Sulfaten, Citraten und Tartraten e~ner- und mit Nitraten nnd Rhodaniden anderseits entgegen- stehen, indem es sich bei den fiir die Si0~-Abscheidung hauptsaehlich in Betracht kommenden Kegelzel]en am sehr plasmareiche, aber zellsaft~rme Elemente handelt, sind noeh nieht geniigend iiberwunden (vielleicht iiberhaupt nicht zu umgehen), am derart ein weiteres G-lied in der Beweisfiihrung zu liefern. Wir miissen deswegen unsere Schliisse vor l~uf ig allein auf die Resultate ~ler Farbstoffversuche gr[inden.
7. Als Grf inde fiir d ie U b e r t r a g b a r k e i t der Befunde LASKINS (10) ~uf die Erscheinungen der Si02-Abscheidung in Pflanzenzellen haben wir aber bereits die fo]genden erkannt. Indem wegen der g e r i n g e n Dis soz i - a t i o n der F a r b s t o f f e in PhenollSsungen weniger Ionen- als haupts~chlich Molekiilwirkungen in Betraeht kommen, aufierden, das Sol in den Pflanzen- zellen bereits d u r c h das in w i i s s e r i g e r LSs~ ~g m o l e k u l a r d i s p e r s e P h e n o l zu r A b s c h e i d u n g g e b r a c h t wird, werden wit uns eher fiir dehydratative als Ladungswirkungen entscheiden wollen. Best~rkt werden wir in diesem Ergebnis, zu dem LASKIN (10) durch exakte 1VIodellversuche ja ebenfalls gekommen ist, durch unsere Beobachtungen tiber die u n t e r s e h i e d - l i c h e E i g n u n g der u n t e r s u c h t e n b a s i s c h e n F a r b s t o f f e , die sich nach ihrem ganzen Verhalten gegeniiber den Pflanzenzellen in die auch nach dem Adsorptionsverhalten und damit nach dem ttydratationsgrade sich ergebende Reihe ordnen. Die friiher yon PFEIFrE~ (4) verSffentlichten Versuche mit d e h y d r a t a t i s i e r e n d w i r k e n d e n S a l z l S s u n g e n widersprechen eben- falls nicht, wenngleich sie noch des besseren Ausbaues in der bezeichneten Richtung bediirfen.
Wenn wir unter bestimmten Bedingungen (Uberschwemmung mit den in Phenol gelSsten basischen Farbstoffen) die Abscheidung yon Si02-Gallerten

126 Klcine Mitteilungen
ais notwendige Wi rkung hervorrufen und also eine untrennbare Verbindung der physiko-chemischen Erscheinungen mit gewissen Zellfunktionen kon- statieren, so stehen wir damit erst am Anfang unseres Forschens. W i t miissen uns nun bemiihen (29), die e i g e n t l i c h e N a t u r t ie r p h y s i k o - c h e m i s c h e n V e r ~ n d e r u n g e n zu analysieren. Trotz gewalt iger Schwierig- keiten, welche in der Organisat ion des Untersuchungsmater ials begr i indet liegen, darf aber wohl in dem vorliegenden Falle dieser wei tere Schrit t als gegliickt angesehen werden, der uns die Abscheidung yon Si02-Gebflden in pflanzlichen Zellen als D e h y d r a t a t i o n d e r an Z e l l m e m b r a n e n a d s o r - b i e r t e n T e i l c h e n v o n s i l i c i d e n H y d r o s o l e n (11) deuten l~tgt.
Literaturanmerkungen
1. J. DUVAL-JOUYE, Bull. Soc. bot. France 20, 1, 1873; )[dm. Aead. so. )fontpellier 8, 227, 1873.
2. S. KAPH~.tIN, Beih. Bot. Ctb. (I) 18, 233, 1905; O. WERNER, Biol. gener. 4, 403, 1928. 31 ]~. ~PFEIFFER, Ber. Dtseh. bot. Ges. 89, 353, 1921; Beih. Bot. Ctb. (I) 88, 401, 1921;
(II) 89, 436, 1923; (I) 44, 90, 98, 1927; Bot. Arch. 12, 446, 460, 1925 u. a. 4. i~[. FROHNMEYER, Bibl. Bet. 86, Stuttgart (E. Schweizerbart) 1914, S. 12; tt. PFEIFFER,
Bet. Dtsch. bot. CTes. 43, [26], 1925; 47, [78], 1929; FR. NETOLITZKY, Die Kiesel- kSrper, in K. LINSBAUER, Handb. d. Pfl.-Anat., (I), 1 C, I I I /1 a, Berlin (Ge- briider Borntraeger) 1929, S. 4.
5. N. v. SMOLUCnOWSKI, KolI.-Ztsehr. 18, 194, 1916; 21, 98, 1917; Ztsehr. physik. Chem. 92, 129, 1917; R. HOBER, Physikalischc Chemie der Zelle und der Gewebc, 5. Aufl., Leipzig (W. Engelmann) 1924, S. 233; H. 13EEIFFER, Elektrizititt und Eiweil~e, Dresden u. Leipzig (Th. Steinkopff) 1929, S. 77; H. BECrInOLD, Die Kolloide in Biologie und Medizin, 5. Aaft., Dresden u. Leipzig (Th. Steinkopff) 1929, S. 95.
6. H. PFEIFEER, Protoplasma 6, 377, 416, 1929. 7. H. 1)FEIFFER, Der isoelektrische Punkt (IEP) yon Protoplasten und seine Ermittlung,
in E. ABDERHALDEN, tIandb, d. biol. Arbeitsmethod. (u 2/2, S. 1563--15967 1929, 8. ]~. LASKIN, Ztsehr. physik. Chem. 116, 97, 1925i 181, 313; 188, 203; 18~, 387, 1928. 9. SH. DOKAN, Koll.-Ztschr. 84, 155, 1924.
10. :E. LASKIN, Koll.-Ztsehr. 45, 131, 135, 1928. 11. u H. ~VISLICENUS, Koll.-Ztsehr. 27, 209, 217, 1920; P. KLASON, Bet. Dtsch,
chem. Ges. 58, 1864, 1920. 12. ]:[. PFEIFFER, Verb. Abt. exper. Zellforsch. X. Internat. Zool.-Kongr. Budapest (Arch.
exper. Zellforsch. 61, 418, 428, 1928. - - u auch R. Zs igmondy . Kolloid- chemie, 3. Aufl., Leipzig (0. Spamer) 1920, S. 212.
13. E. K(~STER, Pathologie der Pflanzenzelle, 1 (Protoplasma-Nonogr. 3), Berlin (Gebriider Borntraeger) 1929, S. 118, 155.
14. J. TRAUBE und F. KOHLER, Internat. Ztsehr. physik-ehem. Biol. 2, 197, 202, 1915, 15. ]E. BRESSLAU, Arch. Protistenkd. 43, 467, 1921.

Kleine ~[itteilungen 12 7
16. H. RHEINBOLDT und ]~. WEDEKIND, Kolloidehem. Bei]a. 17, 115: 118, 132, 161, 1923; H. PFEIFFER, a. a . O. 429.
17. ]~. K~ISTER, a. a. 0.: S. 32: 118. 18. D.P. ~-RETTIE nod ]:~. J. WILLIAI~IS, Journ, Amer. chem. Soc. 50: 668, 1928. 19. P. ROHLAND, Ztschr. anorg. Chem. 89: 164: 1914; )f. GLASUNOW, Ztschr. wiss.
1VIikrosk. 44, 9:19271 H. PFEIFFER: Protoplasma 8: 261, 273: 1929. 20. I-t. PFEIFFER, Arch. exper. Zellforsch. 6: 418, 430: 1928; Ber. Dtseh. hot. Ges. 47,
[78], 1929. 21. H. SCHADE, Die physikalische Chemie in der inneren Medizin: 3. Aufl, Dresden u.
Leipzig (Th. Steinkopff) 1923, S. 115. 25. H. ]~EYER und R. ~OTTLIEB, Die experimentelle Pharmakologie: Wien u. Leipzig
(Urban u. 8ehwarzenberg) 1910: 8. 147. 23. P. WALDEN: Ztsehr. physik. Chem. 70: 584, 1910. 24. R. :KELLER: Bioehem. Ztschr. 115: 134: 1921; Die Elektrizitat in der Zelle, 2. Aufi.
hIahrisch-Ostrau (Kittl) 1925, 8. 18; E. A. HAFNER, Ergebn. Physiol. 24, 566, 1926; O. BLt~, Physik. Ztsehr. 27, 226~ 1926; Jos. GICKLHORN: Protoplasma 1, 124: 1926; Kolloidehem. Beih. 28, 328, 1929; KELLER und GlCt~HORN, Methoden der Bioelektrostatik, in E. ABDERItALDEN, Handb. d. biol. Arbeitsmethod. (V), 2: 1189, 1226, 1928; H. PFEIFFER: E]ektrizitat und EiweiBe, Dresden u. Leipzig (Th. Steinkopff) 1929, S. 48, 131 u. a.; Protoplasma 8, 261, 277, 1929; R. FORTY, Kolloidehem. Beih. 28, 314, 320 und 322: 325: 1929; ]Kethoden zur Bestimmung der elektrischen Struktur kolloider 8toffe, insbesondere der Biokolloide, in ]~. •BDERHALDEN, Handb. d. biol. Arbeitsmethod. (III), B, 775: 836, 854: 1929.
25. S. ASCHKENASI: Ztschr. Elektroehem. 28, 58: 106, 1922. 26. ST. BIRUTOWlTSC~, Koll.-Ztschr. 44, 239, 1928. 27. H. PFEIFFER: in E. ABDERttALDEN, Eandb. d. biol. Arbeitsmethod. (V), 2/2, 1563,
1571: 1929. 28. FR. BOAS: Die Pflanze als ko]loides System: Freising-Miinchen (F. P. Datterer) 1928,
S. 21, 29:106 u. a. -- Vgl. auch A. FRUMKIN: Ztschr. physiol. Chem. 109, 84, 1924. 29. L.V. ]~EILBRUNN, Colloid Symposium ~onogr. 8: 135, 150, 1925; The colloid
chemistry of protoplasm (Protop]asma-Monogr. 1),Berlin (Gebriider Borntraeger) 1928~ 8. 8; tI. PFEIFFER: Elektrizit~t und Eiweil~e, Dresden u. Leipzig (Th. Steinkopff} 1929, S. 2: 127.


















