
Übungen Zellbiologie und Physiologie - uni-muenster.de · Woche 1 Chromatographie AG Hippler 1....
62
Übungen Zellbiologie und Physiologie Sommersemester 2014 Primär- wurzel Spross- meristem Aleuron- schicht Samen- schale GA Scutellum Endosperm (Stärke u.a. Reservestoffe) Zucker -Amylasen Embryo
Transcript of Übungen Zellbiologie und Physiologie - uni-muenster.de · Woche 1 Chromatographie AG Hippler 1....

1
Übungen Zellbiologie und Physiologie
Sommersemester 2014
Primär- wurzel
Spross- meristem Aleuron-
schicht
Samen- schale
GA
Scutellum
Endosperm (Stärke u.a. Reservestoffe)
Zucker
-Amylasen Embryo

Kur
spla
n fü
r di
e Ü
bung
en Z
ellb
iolo
gie
und
Phys
iolo
gie
Som
mer
sem
este
r 20
14
12 G
rupp
en m
it je
wei
ls 4
-5 S
tudi
eren
den,
14:
00-1
8:00
Uhr
(Mitt
woc
hs 1
4:15
– 1
8:15
), H
P141
, 10
Ver
such
swoc
hen
Der
Kur
s fin
det a
uf G
rund
der
hoh
en T
eiln
ehm
erza
hl d
iese
s Jah
r au
ch a
m F
reita
g st
att
Vor
besp
rech
ung:
07.
04.1
4, 1
3:15
Uhr
, HS
SP7
Vor
lesu
ng: L
abor
met
hode
n de
r B
iolo
gie,
Mon
tag
13:0
0 –
14:0
0 U
hr B
egin
n 14
.04.
14 H
S SP
7 K
ursb
egin
n: 1
4.04
.14,
HP
141
(Ach
tung
: die
ers
te V
ersu
chsw
oche
wird
auf
Gru
nd d
er O
ster
tage
get
eilt)
W
oche
1
14.0
4. –
16.
04.
24.0
4. –
25.
04
Hip
pler
C
hrom
atog
raph
ie
Woc
he 2
28
.04.
– 0
2.05
. Pa
ul /
Zeis
M
etab
olis
mus
W
oche
3
05.0
5. –
09.
05.
Web
er
Mus
kelp
hysi
olog
ie
Woc
he 4
12
.05.
– 1
6.05
. K
läm
bt /
Lam
mel
El
ektro
phor
ese
Woc
he 5
19
.05.
– 2
3.05
. Pa
ul /
Boi
ani
Atm
ung
Woc
he 6
26
.05.
– 3
0.05
. K
udla
/ B
öhm
er
Pfla
nzen
pigm
ente
W
oche
7
02.0
6. –
06.
06.
Kud
la /
Bat
istic
En
zym
e Pf
ings
ten
09.0
6. –
13.
06.
Woc
he 8
16
.06.
– 2
1.06
. vo
n Sc
haew
en /
Rip
s En
twic
klun
g un
d B
eweg
ung
W
oche
9
23.0
6. –
27.
06.
Klä
mbt
/ La
mm
el
Mol
ekul
arbi
olog
ie I
Woc
he 1
030
.06.
– 0
4.07
. K
udla
/ W
einl
M
olek
ular
biol
ogie
II

- 1 -
- 2 -

- 3 -
- 4 -

- 5 -
- 6 -

- 7 -
- 8 -

- 9 -
- 10
-

Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
1. A
llgem
eine
Bem
erku
ngen
Chr
omat
ogra
phis
che
Ver
fahr
en d
iene
n in
ers
ter L
inie
zur
Tre
nnun
g vo
n Su
bsta
nzge
mis
chen
.
Die
s be
ginn
t bei
ein
fach
en k
lein
eren
Sub
stan
zen
wie
Am
inos
äure
n, N
ucle
otid
en u
nd Z
ucke
rn
und
ende
t be
i de
n en
tspr
eche
nden
M
akro
mol
ekül
en.
Ver
wen
det
wird
de
r B
egrif
f
Chr
omat
ogra
phie
sei
t ein
er B
esch
reib
ung
von
Tsw
ett i
m J
ahre
190
3, d
em e
ine
Tren
nung
von
Bla
ttpig
men
ten
an e
inem
fest
en A
dsor
ptio
nsm
ater
ial g
elan
g.
Eine
r de
r be
deut
ends
ten
Anw
endu
ngsb
erei
che
in d
er B
ioch
emie
ist
die
Iso
lieru
ng v
on
Prot
eine
n au
s ein
em S
ubst
anzg
emis
ch. P
rote
ine,
als
Mak
rom
olek
üle,
zei
chne
n si
ch d
urch
ein
e
Rei
he u
nter
schi
edlic
her E
igen
scha
ften
wie
Lad
ung,
hyd
roph
obes
bzw
. hyd
roph
iles
Ver
halte
n,
Grö
ße,
biol
ogis
che
Funk
tion
usw
. au
s, di
e si
ch a
us d
er P
rimär
sequ
enz
erge
ben
und
dam
it
spez
ifisc
h fü
r das
jew
eilig
e Pr
otei
n si
nd.
Allg
emei
n gi
lt fü
r ch
rom
atog
raph
isch
e V
erfa
hren
, daß
die
Tre
nnun
g de
r Su
bsta
nzge
mis
che
imm
er a
ufgr
und
der
Wec
hsel
wirk
unge
n de
r en
tspr
eche
nden
Sub
stan
z m
it ei
ner
mob
ilen
(zum
eist
flü
ssig
en)
Phas
e un
d ei
ner
stat
ionä
ren
(zum
eist
fes
ten)
Pha
se e
rfol
gt.
Je n
ach
verw
ende
tem
C
hrom
atog
raph
iem
ater
ial
unte
rsch
eide
t m
an
grun
dsät
zlic
h fo
lgen
de
chro
mat
ogra
phis
che
Ver
fahr
en:
Ver
teilu
ngsc
hrom
atog
raph
ie
Aff
initä
tsch
rom
atog
raph
ie
Grö
ßena
ussc
hluß
chro
mat
ogra
phie
Ione
naus
taus
chch
rom
atog
raph
ie
Gas
chro
mat
ogra
phie
(auc
h ei
ne V
erte
ilung
schr
omat
ogra
phie
, jed
och
oft g
eson
dert
aufg
efüh
rt)
2. V
erte
ilung
schr
omat
ogra
phie
Bei
di
esem
V
erfa
hren
w
ird
das
unte
rsch
iedl
iche
Lö
sung
sver
halte
n bz
w.
der
hydr
ophi
le/h
ydro
phob
e C
hara
kter
der
ein
zeln
en K
ompo
nent
en e
ines
Sto
ffge
mis
ches
in e
inem
Zwei
phas
ensy
stem
gen
utzt
. In
der
Reg
el u
nter
sche
iden
sic
h be
ide
Phas
en d
urch
ihr
e
unte
rsch
iedl
iche
Pol
aritä
t. A
ls m
obile
Pha
se d
ient
hie
r hä
ufig
ein
unp
olar
es L
aufm
ittel
wie
Chl
orof
orm
, al
s st
atio
näre
Pha
se d
ient
zum
eist
das
pol
are
Lösu
ngsm
ittel
Was
ser,
das
an
Kie
selg
el o
der
Cel
lulo
se „
imm
obili
sier
t“ is
t. D
iese
kön
nen
in e
iner
dün
nen
Schi
cht a
uf G
las
oder
Alu
min
ium
auf
getra
gen
(Dün
nsch
icht
chro
mat
ogra
phie
(D
C))
ode
r ab
er i
n ei
n R
ohr
eing
efül
lt w
erde
n (S
äule
nchr
omat
ogra
phie
). A
n di
esem
pol
aren
Mat
eria
l der
stat
ionä
ren
Phas
e
hafte
t ei
n dü
nner
Was
serf
ilm, d
er d
ann
die
eige
ntlic
he s
tatio
näre
Pha
se d
arst
ellt.
Zw
isch
en
dies
en
beid
en
Phas
en
(unp
olar
es
Lauf
mitt
el
/ po
lare
H
ydra
thül
le)
stel
lt si
ch
ein
- 11
-
Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Ver
teilu
ngsg
leic
hgew
icht
in A
bhän
gigk
eit v
om p
olar
en b
zw. u
npol
aren
Cha
rakt
er d
er S
ubst
anz
ein.
Je
unpo
lare
r ein
e Su
bsta
nz is
t, de
sto
stär
ker l
iegt
die
ses
Gle
ichg
ewic
ht a
uf d
er S
eite
des
unpo
lare
n La
ufm
ittel
s. D
er T
renn
effe
kt i
m F
alle
ein
er D
C,
bei
der
die
besc
hich
tete
Pla
tte
senk
rech
t in
das
Lau
fmitt
el g
este
llt w
ird,
beru
ht n
un d
arau
f, da
ß di
e m
obile
Pha
se s
ich
glei
chm
äßig
dur
ch K
apill
arkr
äfte
von
unt
en n
ach
oben
bew
egt u
nd e
ine
in d
iese
m u
npol
aren
Lauf
mitt
el g
ut l
öslic
he (
unpo
lare
) Su
bsta
nz m
it de
m L
aufm
ittel
bes
ser
und
dam
it sc
hnel
ler
mitw
ande
rt al
s ei
ne s
chle
cht l
öslic
he (p
olar
e). D
ie re
lativ
e W
ande
rung
sges
chw
indi
gkei
t ein
er
Subs
tanz
läßt
sich
dur
ch ih
ren
soge
nann
ten
Rf-W
ert c
hara
kter
isie
ren.
Wan
deru
ngst
reck
e Su
bsta
nz
Rf
=
--
----
----
----
----
----
----
----
----
----
----
-
Wan
deru
ngss
treck
e La
ufm
ittel
Die
ser i
st u
.a. a
bhän
gig
von
der W
ahl d
es L
aufm
ittel
s, de
r ver
wen
dete
n st
atio
näre
n Ph
ase
und
der T
empe
ratu
r. R
f-Wer
te v
ersc
hied
ener
Läu
fe si
nd d
aher
nur
bed
ingt
ver
glei
chba
r. A
us d
iese
m
Gru
nd lä
ßt m
an -
wen
n im
mer
mög
lich
- gle
ichz
eitig
bek
annt
e V
ergl
eich
ssub
stan
zen
mitl
aufe
n
Das
bis
her
besc
hrie
bene
Ver
fahr
en t
renn
t nu
r in
ein
e La
ufric
htun
g. D
urch
Dre
hen
der
Chr
omat
ogra
phie
plat
te u
m 9
0° k
ann
aber
ein
e be
reits
vor
getre
nnte
Pro
be n
ochm
als
in e
ine
ande
re L
aufr
icht
ung
getre
nnt
wer
den.
Der
Vor
teil
liegt
hie
r in
der
Mög
lichk
eit,
zwei
vers
chie
dene
Lau
fmitt
el w
ähle
n zu
kön
nen,
um
Sub
stan
zen
aus k
ompl
exen
Gem
isch
en a
nhan
d
zwei
er u
nter
schi
edlic
her P
aram
eter
zu
trenn
en. D
iese
Var
iant
e w
ird a
uch
als z
wei
dim
ensi
onal
e
DC
bez
eich
net.
Eine
wei
tere
Var
iant
e be
steh
t in
der U
mke
hrun
g de
r Pol
aritä
ten
der m
obile
n
und
der
stat
ionä
ren
Phas
e. D
iese
s V
erfa
hren
wird
als
rev
erse
d ph
ase
(rp)
-Chr
omat
ogra
phie
beze
ichn
et.
- 12
-

Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
3. A
ffin
itäts
chro
mat
ogra
phie
Die
se
Tech
nik
beru
ht
auf
der
Nut
zung
sp
ezifi
sche
r
Wec
hsel
wirk
unge
n zw
isch
en M
olek
ülen
, die
sich
auc
h in
ihre
r
biol
ogis
chen
Fun
ktio
n w
ider
spie
geln
.
Solc
he
spez
ifisc
hen
Wec
hsel
wirk
unge
n bi
lden
z.
B.
die
Gru
ndla
ge
für
die
Bild
ung
von
Ant
igen
/Ant
ikör
per-
,
Hor
mon
/Rez
epto
r-, u
nd E
nzym
/Sub
stra
t-Kom
plex
en
Die
Ide
e de
s V
erfa
hren
s is
t, da
ß ei
ner
der
Partn
er s
olch
er
Kom
plex
e ko
vale
nt a
n ei
ne s
tatio
näre
Mat
rix g
ebun
den
wird
,
und
dies
er d
ann
in d
er L
age
ist,
aus
eine
r mob
ilen
Phas
e (z
.B.
eine
r in
eine
r Puf
fers
ubst
anz
gelö
sten
Pro
benm
isch
ung)
sein
en
ents
prec
hend
en P
artn
er e
xtre
m sp
ezifi
sch
zu b
inde
n un
d da
mit
aus
dem
Su
bsta
nzge
mis
ch
zu
entfe
rnen
. D
urch
di
e
ansc
hlie
ßend
e El
utio
n ka
nn s
o de
r zu
vor
gebu
nden
e Pa
rtner
hoch
rein
gew
onne
n w
erde
n.
Als
G
rund
lage
fü
r di
e K
ompl
exbi
ldun
g de
r be
iden
Pa
rtner
di
enen
ni
cht-k
oval
ente
Wec
hsel
wirk
unge
n w
ie H
-Brü
cken
, hy
drop
hobe
Wec
hsel
wirk
unge
n, v
an d
er W
aals
-Krä
fte
usw
. Die
Bild
ung
dies
er K
ompl
exe
ist e
ine
Gle
ichg
ewic
htsr
eakt
ion,
wod
urch
ein
e re
vers
ible
Bin
dung
und
Elu
tion
mög
lich
wird
. A
ls M
atrix
kön
nen
vers
chie
dene
Zuc
kerp
olym
ere
(Cel
lulo
se,
Aga
rose
, D
extra
n),
Poly
ster
olde
rivat
e od
er P
olya
mid
gele
ver
wen
det
wer
den.
Vor
auss
etzu
ng is
t, da
ß ...
ausr
eich
end
funk
tione
lle G
rupp
en v
orha
nden
sin
d, a
n di
e de
r „K
ompl
expa
rtner
“ im
mob
ilisi
ert
wer
den
kann
;
chem
isch
e St
abili
tät
unte
r de
n ve
rwen
dete
n B
edin
gung
en z
ur L
igan
dbin
dung
, El
utio
n un
d
Reg
ener
atio
n ge
wäh
rleis
tet i
st;
das v
erw
ende
te M
ater
ial e
ine
mög
lichs
t ger
inge
Eig
enad
sorp
tion
von
Mak
rom
olek
ülen
bes
itzt;
gute
Flie
ßeig
ensc
hafte
n be
i gle
ichz
eitig
hoh
er P
oros
itat g
ewah
rt bl
eibe
n.
Abb
. 1: P
rinzi
p de
r Aff
initä
ts-
chro
mat
ogra
phie
- 13
-
Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Um
W
echs
elw
irkun
gen
mit
der
Mat
rix
(hyd
roph
obe
Wec
hsel
wirk
unge
n, s
teris
che
Beh
inde
rung
en d
er o
ft se
hr
groß
en
Kom
plex
e w
ie
z.B
. A
ntig
en/A
ntik
örpe
r)
zu
min
imie
ren
oder
die
spe
zifis
che
Bin
dung
an
den
Liga
nden
(kov
alen
t ge
bund
ener
Kom
plex
partn
er)
über
haup
t er
st z
u
erm
öglic
hen,
wer
den
oft s
ogen
annt
e Sp
acer
mol
ekül
e (z
.B.
Hex
amet
hyle
ndia
min
NH
2-(C
H2)
6-N
H2)
ein
geba
ut.
Nac
h de
r Pro
bena
uftra
gung
soll
die
zu re
inig
ende
Sub
stan
z (z
.B. e
in P
rote
in) a
n de
n Li
gand
en
bind
en.
Die
se B
indu
ng e
rfol
gt a
ufgr
und
eine
s si
ch e
inst
elle
nden
Gle
ichg
ewic
htes
. D
iese
(rev
ersi
ble)
Bin
dung
ber
uht i
n je
dem
Fal
l auf
der
Ein
stel
lung
ein
es G
leic
hgew
icht
es. B
ei d
er
Prob
enau
ftrag
ung
müs
sen
desh
alb
die
Bed
ingu
ngen
(z
.B.
pH-W
ert,
Salz
konz
entra
tion,
Tem
pera
tur)
so
gew
ählt
wer
den,
daß
die
ses
Gle
ichg
ewic
ht m
öglic
hst w
eit a
uf d
er S
eite
des
Kom
plex
es l
iegt
. Nur
dan
n ge
lingt
es,
die
zu r
eini
gend
e Su
bsta
nz i
nner
halb
ein
er s
char
fen
Ban
de a
uf d
er S
äule
zu
foku
ssie
ren.
Zur
Elu
tion
kann
man
sic
h gr
unds
ätzl
ich
zwei
er
vers
chie
dene
r Ver
fahr
en b
edie
nen:
Was
chen
mit
eine
r Sub
stan
z, d
eren
Aff
initä
t zum
imm
obili
sier
ten
Kom
plex
partn
er g
röße
r ist
, als
die
Aff
initä
t der
zu
isol
iere
nden
Sub
stan
z;
Erni
edrig
ung
der
Aff
initä
t de
r zu
iso
liere
nden
Sub
stan
z zu
m i
mm
obili
sier
ten
Kom
plex
partn
er
durc
h Ä
nder
ung
der B
indu
ngsb
edin
gung
en (w
ie p
H-W
ert,
Tem
pera
tur o
der I
onen
stär
ke).
Die
Bed
ingu
ngen
sow
ohl f
ür d
ie B
indu
ng a
ls a
uch
für d
ie E
lutio
n si
nd s
o zu
wäh
len,
daß
der
Kom
plex
zwar
gel
öst,
aber
die
zu is
olie
rend
e Sub
stan
z (z.
B. e
in P
rote
in) d
abei
nic
ht d
enat
urie
rt
bzw
. and
erw
eitig
ges
chäd
igt w
ird.
4. G
röße
naus
schl
ußch
rom
atog
raph
ie
Die
ses
auch
Gel
filtra
tion
oder
Gel
perm
eatio
n ge
nann
te V
erfa
hren
tren
nt S
ubst
anze
n au
fgru
nd
ihre
r un
ters
chie
dlic
hen
Grö
ße.
Der
gen
aue
Mec
hani
smus
ist
bis
heu
te n
icht
im
Det
ail
vers
tand
en, j
edoc
h gi
bt e
s ein
e se
hr g
ute
Mod
ellv
orst
ellu
ng ü
ber d
en T
renn
vorg
ang.
Abb
. 2: S
pace
r erm
öglic
hen
die
Bin
dung
eine
s grö
ßere
n Pr
otei
nkom
plex
es
- 14
-

Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Eine
Gel
filtra
tions
säul
e be
steh
t au
s ei
nem
sch
wam
mar
tigen
Gel
mit
Pore
n m
it de
finie
rtem
Grö
ßenb
erei
ch. M
olek
üle,
die
so
groß
sin
d, d
aß s
ie n
icht
in d
iese
Por
en d
iffun
dier
en k
önne
n,
wer
den
auf
kürz
este
m W
eg u
nd d
esha
lb
sehr
schn
ell e
luie
rt. M
olek
üle,
die
so k
lein
sind
, daß
sie
gut i
n di
e Po
ren
des M
ater
ials
diff
undi
eren
kön
nen,
lege
n be
i der
Elu
tion
eine
se
hr
lang
e St
reck
e zu
rück
un
d
elui
eren
ent
spre
chen
d sp
ät.
Mitt
elgr
oße
Mol
ekül
e, d
ie i
n ei
nige
, ab
er n
icht
alle
Pore
n di
ffun
dier
en
könn
en,
elui
eren
dem
zufo
lge z
wis
chen
den
gan
z gro
ßen
und
den
ganz
kl
eine
n M
olek
ülen
. D
ie
Tren
nsch
ärfe
ein
er s
olch
en S
äule
nim
mt
mit
ihre
r Län
gezu
.
Der
Ein
satz
bere
ich
eine
s so
lche
n G
els
häng
t zu
dem
von
der
mol
ekul
aren
Aus
schl
uß-
und
Eins
chlu
ßgre
nze
ab. D
iese
Wer
te h
änge
n vo
n de
r Grö
ßenv
erte
ilung
der
Por
en im
Gel
mat
eria
l
(der
Mat
rix) a
b. D
ie g
rößt
en P
oren
def
inie
ren
die
Aus
schl
ußgr
enze
, alle
Mol
ekül
e, d
ie z
u gr
oß
sind
, um
sel
bst
in d
iese
Por
en z
u di
ffun
dier
en,
elui
eren
gem
eins
am f
rüh
von
der
Säul
e
(„V
orla
uf“)
. Die
kle
inst
en P
oren
def
inie
ren
ents
prec
hend
die
Ein
schl
ußgr
enze
, alle
Mol
ekül
e,
die
so k
lein
sin
d, d
aß s
ie s
elbs
t in
die
se P
oren
diff
undi
eren
kön
nen,
elu
iere
n eb
enfa
lls
gem
eins
am s
pät
von
der
Säul
e. N
ur M
olek
üle,
der
en G
röße
n zw
isch
en A
ussc
hluß
- un
d
Eins
chlu
ßgre
nze
der
Mat
rix l
iege
n, k
önne
n m
it di
esem
Säu
lenm
ater
ial
gem
äß i
hrer
Grö
ße
getre
nnt w
erde
n. In
der
Pra
xis
char
akte
risie
rt m
an e
ine
solc
he S
äule
dur
ch d
rei v
ersc
hied
ene
Vol
umin
a:
1. d
as G
esam
tvol
umen
der
Säu
le V
t
2. d
as V
olum
en w
elch
es d
as G
el e
inni
mm
t Vx
3. d
as V
olum
en d
er u
mge
bend
en L
ösun
g V
0
V0
wird
auc
h al
s A
ussc
hluß
volu
men
bez
eich
net
und
nim
mt
in
der
Reg
el
ca.
35
%
des
Ges
amtv
olum
ens e
in. D
as A
ussc
hluß
volu
men
ist d
ie M
enge
an E
luat
, die
übe
r die
Säu
le fl
ieße
n
Abb
. 3 :
Tren
nung
von
kle
inen
und
gro
ßen
Mol
ekül
en m
it de
r
Gel
filtra
tions
-Chr
omat
ogra
phie
.
Die
Mol
ekül
e si
nd s
chem
atis
ch a
ls k
lein
e un
d gr
oße
schw
arze
Kre
ise
wie
derg
egeb
en.
Abb
. 4: C
hara
keris
tisch
e G
röße
n ei
ner G
elfil
tratio
nssä
ule
- 15
-
Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
muß
, um
Mol
ekül
e je
nsei
ts d
er m
olek
ular
en A
ussc
hluß
gren
ze z
u el
uier
en. E
in e
ntsp
rech
ende
r
Zusa
mm
enha
ng b
este
ht z
wis
chen
dem
Ges
amtv
olum
en u
nd e
r Ein
schl
ußgr
enze
.
Das
Elu
tions
volu
men
(Ve)
eine
r Sub
stan
z is
t die
Men
ge a
n El
uat,
die
zwis
chen
dem
Auf
gebe
n
der
Subs
tanz
und
ihre
m A
ustre
ten
über
die
Säu
le f
ließt
. Pra
ktis
ch w
ird V
e als
das
Vol
umen
best
imm
t, be
i dem
die
hal
bmax
imal
e Pe
akhö
he (
erm
ittel
t übe
r B
rech
ungs
inde
x, b
iolo
gisc
he
Akt
ivitä
t usw
.) er
reic
ht is
t.
Mit
den
ange
gebe
nen
Grö
ßen
kann
dan
n au
f dre
i ver
schi
eden
e A
rten
der R
ückh
alte
grad
ein
er
Subs
tanz
bes
chrie
ben
wer
den.
das
rela
tive
Elut
ions
volu
men
(REV
)
Ve
REV
=
----
----
-
V0
die
Ret
entio
ns-K
onst
ante
(rez
ipro
ker W
ert v
on R
EV)
V
0
R =
--
----
--
V
e
der
Ver
teilu
ngsk
oeff
izie
nt K
d ode
r Kav
Ve -
V0
Kav
=
----
----
---
Vx
Am
häu
figst
en v
erw
ende
t wird
der
Ver
teilu
ngsk
oeff
izie
nt.
Abb
. 5 :
Abh
ängi
gkei
t der
Kav
-Wer
te
vers
chie
dene
r Pro
tein
e vo
n de
ren
Mol
ekul
arge
wic
hten
exe
mpl
aris
ch
für e
in b
estim
mte
s Gel
mat
eria
l
(Sep
hade
x G
200
)
Aus
schl
ußgr
enze
:
Ve =
V0
Kav
= 0
Eins
chlu
ßgre
nze:
Ve =
Vt
Kav
= 1
- 16
-

Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
5. lo
nena
usta
usch
- Chr
omat
ogra
phie
Die
Ion
enau
stau
sch-
Chr
omat
ogra
phie
ist
ein
e be
sond
ere,
seh
r w
eit
verb
reite
te F
orm
der
Aff
initä
tsch
rom
atog
raph
ie, b
ei d
er re
lativ
uns
pezi
fisch
e io
nisc
he W
echs
elw
irkun
gen
zwis
chen
den
zu tr
enne
nden
Sub
stan
zen
und
eine
r (p
ositi
v od
er n
egat
iv)
gela
dene
n M
atrix
aus
genu
tzt
wer
den.
Mit
dies
er A
rt de
r Chr
omat
ogra
phie
kön
nen
som
it nu
r gel
aden
e Su
bsta
nzen
get
renn
t
wer
den.
Bei
der
Ion
enau
stau
sch-
Chr
omat
ogra
phie
bef
inde
n si
ch a
n ei
ner
poly
mer
en M
atrix
gela
dene
fu
nktio
nelle
G
rupp
en.
An
dies
e ge
lade
nen
Gru
ppen
si
nd
reve
rsib
el
Geg
enio
nen
gebu
nden
. M
an
unte
rsch
eide
t gr
unds
ätzl
ich
zwis
chen
K
atio
nen-
un
d
Ani
onen
aust
ausc
hern
. B
eim
Kat
ione
naus
taus
cher
sin
d di
e
funk
tione
llen
Gru
ppen
der
Mat
rix n
egat
iv g
elad
en u
nd
bind
en
dahe
r (p
ositi
v ge
lade
ne)
Kat
ione
n,
beim
Ani
onen
aust
ausc
her
sind
die
Lad
ungs
verh
ältn
isse
gen
au
entg
egen
gese
tzt.
Solc
he f
unkt
ione
llen
Gru
ppen
kön
nen
im
Falle
ein
es K
atio
nena
usta
usch
ers
Car
boxy
l-Gru
ppen
(z.
B.
CM
-Sep
hade
x),
im
Falle
ei
nes
Ani
onen
aust
ausc
hers
Am
inog
rupp
en (
z.B
. D
EAE-
Cel
lulo
se)
sein
. D
ie M
atrix
solc
her
Aus
taus
cher
bes
teht
wie
im
Fal
le d
er A
ffin
itäts
-Chr
omat
ogra
phie
aus
Zuc
kern
(Sep
haro
se, C
ellu
lose
usw
.), P
olys
tyro
len
oder
Pol
yam
iden
.
Nac
h Pr
oben
auftr
ag v
erdr
änge
n nu
n di
e zu
tren
nend
en g
elad
enen
Mol
ekül
e (I
onen
) der
Pro
be
die
reve
rsib
el
ange
lage
rten
Geg
enio
nen
der f
unkt
ione
llen
Gru
ppen
und
bind
en s
o an
die
Mat
rix.
Die
ser
„Ion
enau
stau
sch“
fin
det
natü
rlich
nur
stat
t, w
enn
die
zu tr
enne
nde
Subs
tanz
fest
er a
n di
e fu
nktio
nelle
Gru
ppe
der
Mat
rix b
inde
t als
das
Geg
enio
n. A
uch
hier
ha
ndel
t es
si
ch
um
eine
Gle
ichg
ewic
htsr
eakt
ion.
D
ie
Bed
ingu
ngen
be
im
Prob
enau
ftrag
müs
sen
also
so
gew
ählt
wer
den,
daß
das
Gle
ichg
ewic
ht w
eit
auf
der
Seite
der „
Bin
dung
“ lie
gt.
Abb
. 6: T
ypis
che
funk
tione
lle G
rupp
en
an A
nion
enau
stau
sche
rn (o
ben)
und
Kat
ione
naus
taus
cher
n (u
nten
)
- 17
-
Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Bei
der
ans
chlie
ßend
en E
lutio
n m
acht
man
sich
zun
utze
, daß
die
Bin
dung
sstä
rke
nich
t nur
von
der f
unkt
ione
llen
Gru
ppe
der M
atrix
und
dem
geb
unde
nen
Mol
ekül
abh
ängi
g is
t, so
nder
n au
ch
durc
h di
e U
mge
bung
sbed
ingu
ngen
bee
influ
ßt w
erde
n ka
nn.
So i
st d
ie B
indu
ngss
tärk
e u.
a.
abhä
ngig
von
der
Ione
nstä
rke
der m
obile
n Ph
ase.
Die
lone
nstä
rke
eine
r Lös
ung
ist w
ie fo
lgt d
efin
iert:
25,0
ii
zc
c i
ist d
abei
die
mol
are
Kon
zent
ratio
n un
d z i
die
Ladu
ng d
er S
ubst
anz
i.
Dad
urch
ist
es
mög
lich,
mit
eine
m a
ufst
eige
nden
Kon
zent
ratio
nsgr
adie
nten
ein
es I
ons
die
gebu
nden
e Su
bsta
nz
wie
der
zu
elui
eren
, da
be
i ge
nüge
nd
hohe
n lo
nens
tärk
en
das
Gle
ichg
ewic
ht d
er B
indu
ngsr
eakt
ion
auf
die
Seite
der
Dis
sozi
atio
n ve
rlage
rt w
ird:
das
zur
Elut
ion
benu
tzte
Ion
konk
urrie
rt m
it de
r zu
trenn
ende
n Su
bsta
nz u
m d
ie fu
nktio
nelle
n G
rupp
en
der M
atrix
. Die
Bin
dung
sstä
rke
eine
r Sub
stan
z zu
den
funk
tione
llen
Gru
ppen
ein
er M
atrix
ist
unte
r ge
gebe
nen
Bed
ingu
ngen
ein
e ch
arak
teris
tisch
e St
offk
onst
ante
. D
emzu
folg
e ka
nn m
it
eine
m G
radi
ente
n de
s Elu
tions
-Ion
s ein
e Tr
ennu
ng e
ines
Sub
stan
zgem
isch
es e
rfol
gen,
da
jede
s
gebu
nden
e M
olek
ül im
Idea
lfall
bei e
iner
spe
zifis
chen
lone
nstä
rke
im E
luen
ten
von
der S
äule
gelö
st w
ird.
Obi
ge B
etra
chtu
ngen
gel
ten
nich
t nu
r fü
r kl
eine
Mol
ekül
e m
it ei
ner p
ositi
ven
oder
neg
ativ
en L
adun
g,
sond
ern
auch
fü
r gr
oße
Mol
ekül
e m
it vi
elen
Ladu
ngen
. Pr
otei
ne
z.B
. kö
nnen
an
ih
ren
Seite
ngru
ppen
so
woh
l po
sitiv
e al
s au
ch
nega
tive
Ladu
ngen
trag
en, s
ie z
eich
nen
sich
abe
r im
mer
dur
ch
eine
eind
eutig
e Net
tola
dung
aus.
Im F
alle
der
Pro
tein
e
ist
dies
e N
etto
ladu
ng
vom
pH
-Wer
tdes
Lösu
ngsp
uffe
rs
abhä
ngig
. O
berh
alb
des
Isoe
lekt
risch
en P
unkt
es (p
I ode
r IEP
) lie
gt e
in P
rote
in
mit
nega
tiver
Net
tola
dung
vor
, dar
unte
r mit
posi
tiver
Net
tola
dung
. Am
Isoe
lekt
risch
en P
unkt
glei
chen
sich
die
pos
itive
n un
d ne
gativ
en L
adun
gen
aus,
die
Net
tola
dung
ist n
ull.
Abb
. 8: E
influ
ss d
es p
H-W
erte
s auf
die
Net
tola
dung
eine
s Pro
tein
s
- 18
-

Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Die
s rüh
rt vo
n de
r unt
ersc
hied
liche
n Io
nisi
erba
rkei
t der
ein
zeln
en A
min
osäu
ren
eine
s Pro
tein
s
her,
die
je n
ach
dem
pH
-Wer
tder
Um
gebu
ng n
eutra
l, po
sitiv
ode
r neg
ativ
gel
aden
sein
kön
nen.
Die
Lad
ung
der
auf
der
Obe
rflä
che
des
Prot
eins
exp
onie
rten
Am
inos
äure
n ad
dier
en s
ich
zur
effe
ktiv
en N
etto
ladu
ng d
es P
rote
ins.
Wäh
lt m
an e
inen
gee
igne
ten
pH-W
ertla
ssen
sic
h al
so
Prot
eine
sow
ohl a
n ei
nen
Kat
ione
n-, a
ls a
uch
an e
inen
Ani
onen
aust
ausc
her b
inde
n.
Eine
wei
tere
Anw
endu
ng f
inde
t di
eses
Phä
nom
en b
ei d
er T
renn
ung
von
Prot
eine
n m
it
sign
ifika
nt u
nter
schi
edlic
hen
Isoe
lekt
risch
en P
unkt
en, S
o lä
ßt s
ich
ein
pH-W
ertfi
nden
, bei
dem
das
eine
Pro
tein
ein
e ne
gativ
e N
etto
ladu
ng, d
as a
nder
e ab
er e
ine
posi
tive
Net
tola
dung
bes
itzt.
Unt
er d
iese
n B
edin
gung
en b
inde
t das
neg
ativ
gel
aden
e Pr
otei
n an
ein
en A
nion
enau
stau
sche
r,
das
posi
tiv g
elad
ene
Prot
ein
dage
gen
nich
t meh
r. Eb
enso
läßt
sic
h üb
er e
ine
Ver
ände
rung
des
pH-W
erte
s ein
an
den
Ione
naus
taus
cher
geb
unde
nes P
rote
in w
iede
r elu
iere
n.
6. G
asch
rom
atog
raph
ie
Die
G
asch
rom
atog
raph
ie
oder
G
as/F
lüss
igke
its-C
hrom
atog
raph
ie
ist
eine
ex
trem
leis
tung
sfäh
ige A
rt de
r Ver
teilu
ngsc
hrom
atog
raph
ie. I
hre N
acht
eile
bes
tehe
n da
rin, d
aß si
ch ih
r
Eins
atzg
ebie
t auf
Sub
stan
zen
mit
rela
tiv h
ohem
Dam
pfdr
uck
besc
hrän
kt, d
.h. a
uf S
ubst
anze
n
die
leic
ht v
erda
mpf
bar s
ind,
und
daß
sie
nur
im a
naly
tisch
en, n
icht
im p
räpa
rativ
en M
aßst
ab
durc
hfüh
rbar
ist.
Das
Prin
zip
beru
ht a
uf d
er V
erte
ilung
ein
es S
toff
es z
wis
chen
ein
er st
atio
näre
n Fl
üssi
gkei
t und
eine
r m
obile
n G
asph
ase.
Hie
rfür
wird
als
sta
tionä
re P
hase
ein
e sc
hwer
ver
dam
pfba
re
- 19
-
Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Flüs
sigk
eit a
n di
e In
nenw
and
eine
r meh
rere
Met
er la
ngen
Kap
illar
e ad
sorb
iert.
Die
se K
apill
are
wird
von
eine
m in
erte
n G
as d
urch
strö
mt,
wel
ches
die
mob
ile P
hase
dar
stel
lt. G
ebrä
uchl
ich
sind
hier
für N
2, H
e od
er H
2. D
as z
u tre
nnen
de S
ubst
anzg
emis
ch w
ird im
Inje
ktor
so
star
k er
hitz
t,
daß
es v
erda
mpf
t, un
d da
nn m
it de
m in
erte
n G
as d
urch
die
Kap
illar
e ge
leite
t. Es
ste
llt s
ich,
anal
og z
ur D
C,
ein
Ver
teilu
ngsg
leic
hgew
icht
zw
isch
en s
tatio
näre
r flü
ssig
er u
nd m
obile
r
gasf
örm
iger
Pha
se e
in, d
esse
n R
esul
tat e
ine
unte
rsch
iedl
iche
Wan
deru
ngsg
esch
win
digk
eit d
er
vers
chie
dene
n Su
bsta
nzen
in
der
Kap
illar
e is
t. A
ls D
etek
tore
n di
enen
unt
er a
nder
em
Flam
men
ioni
satio
nsde
tekt
oren
(F
ID)
oder
M
asse
nspe
ktro
met
er
(MS)
am
A
usga
ng
der
Kap
illar
e.
7. B
emer
kung
en z
um H
PLC
ode
r FP
LC
Ver
fahr
en
Bei
die
sen
Ver
fahr
en h
ande
lt es
sich
nic
ht u
m ei
gene
, spe
ziel
le C
hrom
atog
raph
iear
ten,
sond
ern
ledi
glic
h um
app
arat
iv o
ptim
ierte
For
men
der
obe
n be
schr
iebe
nen
Chr
omat
ogra
phie
met
hode
n.
Die
Tre
nnsc
härf
e be
ispi
elsw
eise
bei
ein
er G
elfil
tratio
n ni
mm
t mit
der F
einh
eit d
es M
ater
ials
und
der S
äule
nlän
ge z
u. G
leic
hzei
tig h
ierm
it si
nken
bei
her
köm
mlic
her A
nwen
dung
abe
r auc
h
die
Dur
chflu
ßrat
en, s
o da
ß de
r erf
orde
rlich
e Ze
itauf
wan
d so
gro
ß w
ird, d
aß e
ine
Tren
nung
bei
der
ents
prec
hend
gro
ßen
Tren
nsch
ärfe
pra
ktis
ch n
icht
meh
r m
öglic
h is
t. B
ei e
iner
HPL
C (
=
high
pres
sure
liqu
id ch
rom
atog
raph
y)-A
nlag
e wird
nun
die
Dur
chflu
ßrat
e mitt
els h
oher
Drü
cke
aufr
echt
erh
alte
n. D
ies
erm
öglic
ht e
ine
Tren
nung
in
eine
m r
elat
iv k
urze
n Ze
itrau
m b
ei
glei
chze
itig
hohe
r Tr
enns
chär
fe.
Prin
zipi
ell
aber
is
t an
ei
ner
HPL
C-A
nlag
e je
des
Säul
ench
rom
atog
raph
ie-V
erfa
hren
anw
endb
ar.
Jedo
ch s
telle
n di
e ho
hen
Drü
cke
extre
me
Anf
orde
rung
en a
n da
s Mat
eria
l.
Bei
eine
r FPL
C (=
fast
prot
ein
liqui
d ch
rom
atog
raph
y) si
nd d
ie V
erhä
ltnis
se se
hr äh
nlic
h. A
uch
hier
wird
mitt
els P
umpe
n di
e D
urch
flußr
ate
kons
tant
geh
alte
n, je
doch
wer
den
bei w
eite
m n
icht
so
hohe
D
rück
e au
fgeb
aut.
Die
FP
LC
wird
da
her
auch
of
t M
PLC
od
er
Mitt
eldr
uck-
Chr
omat
ogra
phie
gen
annt
. Typ
isch
erw
eise
die
nt d
ie F
PLC
der
Auf
rein
igun
g vo
n
Prot
eine
n in
w
äßrig
en
Elue
nten
, di
e H
PLC
eh
er
der
Auf
trenn
ung
nied
erm
olek
ular
er
Subs
tanz
en, m
eist
in o
rgan
isch
en L
ösun
gsm
ittel
n.
- 20
-

Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Ver
such
sdur
chfü
hrun
g
Das
Prin
zip
der
Ione
naus
taus
chch
rom
atog
raph
ie s
oll
am B
eisp
iel
des
Häm
oglo
bins
gez
eigt
wer
den.
Häm
oglo
bin
ist d
as S
auer
stof
f-üb
ertra
gend
e Pr
otei
n im
Blu
tkre
isla
uf. E
s be
steh
t aus
vier
Unt
erei
nhei
ten,
die
jew
eils
ein
kov
alen
t geb
unde
nes E
isen
prot
opor
phyr
in (=
Häm
grup
pe)
trage
n. H
ierd
urch
ers
chei
nt e
s br
äunl
ich
gefä
rbt u
nd is
t dah
er s
chon
mit
bloß
em A
uge
gut z
u
erke
nnen
.
Zunä
chst
sol
len
die
äuße
ren
Bed
ingu
ngen
– d
ie g
ünst
igst
en p
H-W
erte
für
Bin
dung
bzw
.
Elu
tion
des P
rote
ins u
nd d
ie B
indu
ngsk
apaz
ität d
er S
äule
- be
stim
mt u
nd o
ptim
iert
wer
den,
um e
inen
erf
olgr
eich
en A
nsat
z zu
r Rei
nigu
ng d
es H
ämog
lobi
ns a
us e
inem
Gem
isch
zu
finde
n.
Die
se V
orve
rsuc
he f
inde
n in
Epp
endo
rfre
aktio
nsge
fäße
n im
sog
enan
nten
bat
ch-V
erfa
hren
stat
t. D
ie g
ewon
nene
n Er
kenn
tnis
se s
olle
n da
nn g
enut
zt w
erde
n, u
m d
as H
ämog
lobi
nüb
er
Säul
en m
it K
atio
nena
usta
usch
er-M
ater
ial
aus
eine
m P
rote
inge
mis
chzu
iso
liere
n. D
as
verw
ende
te P
rote
inge
mis
ch e
nthä
lt H
ämog
lobi
n un
d da
s rö
tlich
gef
ärbt
e C
ytoc
hrom
c (
ein
Prot
ein
der
Atm
ungs
kette
, w
elch
es
eben
falls
ei
ne
Häm
grup
pe
trägt
) in
gl
eich
en
Kon
zent
ratio
nen.
Ben
ötig
te G
erät
e:
„Sp
eckb
rettr
otor
“, T
isch
zent
rifug
e, E
ppen
dorf
gefä
ße m
it Sä
ulen
mat
eria
l, Ep
pend
orfp
ipet
ten
(200
-100
0μl,
20-
200μ
l),
Phot
omet
er,
Küv
ette
n,
pH-M
eter
, ge
pack
te
Säul
en
mit
Car
boxy
met
hylc
ellu
lose
-Sep
hade
x
(Kat
ione
naus
taus
cher
-Mat
eria
l), R
eage
nzgl
äser
, Pas
teur
pipe
tten,
Gum
mis
auge
r
Ben
ötig
te C
hem
ikal
ien:
100
ml 2
0 m
M T
ris-P
uffe
r (Tr
is-(
hydr
oxym
ethy
l)-am
inom
etha
n)
100
ml 2
0 m
M M
alei
nsäu
re-P
uffe
r (ci
s-B
uten
disä
ure)
Säul
enm
ater
ial C
arbo
xym
ethy
lcel
lulo
se-S
epha
dex
vorg
equo
llen
Häm
oglo
bin-
Lösu
ng (5
mg/
ml)
Häm
oglo
bin/
Cyt
ochr
om c
-Gem
isch
(je
5 m
g/m
l)
Kal
ium
chlo
rid (M
olek
ular
gew
icht
74,
55 g
/mol
)
10 μ
l Asc
orbi
nsäu
re (1
00 m
M)
- 21
-
Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Vor
vers
uch
1: B
estim
mun
g de
r ge
eign
eten
pH
-Wer
te fü
r B
indu
ng u
nd E
lutio
n
Um
zu
unte
rsuc
hen,
bei
wel
chem
pH
-Wer
t das
Häm
oglo
bin
am S
äule
nmat
eria
l bin
det,
soll
zunä
chst
ein
e pH
-Rei
he m
it de
n pH
-Wer
ten
5 bi
s 9 h
erge
stel
lt w
erde
n. E
in P
uffe
r, de
r aus
den
beid
en K
ompo
nent
en T
ris u
nd M
alei
nsäu
re h
erge
stel
lt w
ird, h
at e
inen
Puf
ferb
erei
ch v
on e
twa
pH 5
bis
pH
9. E
s w
ird d
er T
ris-P
uffe
r vor
gele
gt u
nd m
it de
m M
alei
nsäu
re-P
uffe
r titr
iert,
bis
er ei
nen
der g
ewün
scht
en p
H-W
erte
erre
icht
hat
. Auf
die
se W
eise
wer
den
alle
ben
ötig
ten
Puff
er
herg
este
llt. V
on je
dem
pH
-Wer
t wer
den
ca. 4
0 m
l in
eine
m 5
0 m
l Fal
con-
Tube
abg
efül
lt.
Fünf
Epp
endo
rfge
fäße
n m
it Sä
ulen
mat
eria
l (V
olum
en =
25 μl
) wer
den
mit
je 1
ml P
uffe
r ein
es
best
imm
ten
pH-W
erte
s ve
rseh
en u
nd 5
Min
uten
lang
am S
peck
bret
troto
r äq
uilib
riert.
Dan
n
wer
den
die
Eppe
ndor
fgef
äße
kurz
zen
trifu
gier
t (1
min
, 13
.000
rpm
), da
mit
sich
das
Säul
enm
ater
ial
am B
oden
abs
etzt
. D
er P
uffe
r w
ird a
bpip
ettie
rt (A
ber
Vor
sich
t! D
as
Säul
enm
ater
ial n
icht
mit
abpi
pett
iere
n!),
durc
h fr
isch
en e
rset
zt u
nd d
ie Ä
quili
brie
rung
ein
zwei
tes M
al d
urch
gefü
hrt.
Die
Gef
äße
wer
den
nach
ern
eute
r A
bnah
me
des
Übe
rsta
ndes
mit
jew
eils
10 μl
der
rei
nen
Häm
oglo
bin-
Lösu
ng b
esch
ickt
und
mit
dem
ent
spre
chen
den
Puff
er a
uf 1
ml
aufg
efül
lt un
d
wie
derg
ut d
urch
mis
cht.
Die
Ein
stel
lung
des
Bin
dung
sgle
ichg
ewic
htes
erfo
lgt 5
Min
uten
lang
am S
peck
bret
troto
r. N
ach
dem
Abz
entri
fugi
eren
des
Säu
lenm
ater
ials
wird
der
Übe
rsta
nd
abge
nom
men
, in
Küv
ette
n pi
petti
ert u
nd p
hoto
met
risch
bei
410
nm
die
Ext
inkt
ion
gem
esse
n.
Aus
den
Erg
ebni
ssen
wird
bes
timm
t, w
elch
e pH
-Wer
te fü
r die
vol
lstä
ndig
e re
vers
ible
Bin
dung
des H
ämog
lobi
ns a
n da
s Säu
lenm
ater
ial u
nd d
esse
n an
schl
ieße
nder
Elu
tion
am b
este
n ge
eign
et
sind
.
Vor
vers
uch
2: E
rmitt
lung
der
Bin
dung
skap
azitä
t der
Säu
le
Um
bei
ein
er A
ufre
inig
ung
eine
Übe
rladu
ng d
er S
äule
zu
verm
eide
n, so
llte,
falls
mög
lich,
auc
h
die
Auf
nahm
ekap
azitä
t der
Säu
le ü
berp
rüft
wer
den.
Es
wer
den,
wie
obe
n be
schr
iebe
n, e
rneu
t
sieb
en E
ppen
dorf
gefä
ße m
it Sä
ulen
mat
eria
l zw
eim
al ä
quili
brie
rt. H
ierb
ei w
ird d
er P
uffe
r m
it
dem
gee
igne
tste
n pH
-Wer
t für
die
Bin
dung
ver
wen
det.
Der
Puf
fer w
ird a
bpip
ettie
rt, d
ie 2
5 μl
Säu
lenm
ater
ial m
it je
5 /
10 /
25 /
50 /
75 /
100 μl
der
Häm
oglo
bin-
Lösu
ng v
erse
tzt u
nd a
uf 1
ml m
it de
m g
eeig
nete
n Pu
ffer
auf
gefü
llt. D
ann
wird
5
Min
uten
lang
am
Spe
ckbr
ettro
tor i
nkub
iert.
Nac
h er
neut
er Z
entri
fuga
tion
wird
der
Übe
rsta
nd
in K
üvet
ten
pipe
ttier
t und
pho
tom
etris
ch b
ei 4
10 n
m d
er re
lativ
e Häm
oglo
bin-
Geh
alt b
estim
mt.
- 22
-

Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Aus
der
Men
ge d
es v
erw
ende
ten
Säul
enm
ater
ials
und
der
erm
ittel
ten
Bin
dung
skap
azitä
t sol
l
bere
chne
t wer
den,
wie
viel
Pro
be a
uf d
ie v
orge
ferti
gten
Säu
len
(Vol
umen
: 1 m
l) th
eore
tisch
aufg
etra
gen
wer
den
könn
te, o
hne
sie
zu ü
berla
den.
Säul
ench
rom
atog
raph
ie: I
solie
rung
des
Häm
oglo
bins
aus
ein
em P
rote
inge
mis
ch
Tip
:zur
Bes
chle
unig
ung
der
Flus
srat
e ka
nn m
it ei
nem
Gum
mis
auge
r ei
n le
icht
er D
ruck
auf
die
Säul
e au
sgeü
bt w
erde
n; v
gl. F
PLC
-Ver
fahr
en.
Übe
rste
hend
e Fl
üssi
gkei
t w
ird s
o w
eit
abge
lass
en, d
ass
die
Säul
e ge
rade
nic
ht t
rock
enfä
llt.
Ans
chlie
ßend
wird
die
Säu
le m
it ei
nem
Säu
lenv
olum
en (
= 1
ml)
des
Puff
ers
mit
dem
zuv
or
erm
ittel
ten
optim
alen
pH
-Wer
t fü
r di
e H
ämog
lobi
nbin
dung
äqu
ilibr
iert.
Ist
der
Puf
fer
durc
hgel
aufe
n, w
erde
n 20
0 μl
des
Pro
tein
gem
isch
es a
ufge
trage
n un
d ei
nsin
ken
gela
ssen
.
Prot
eine
m
it ei
nem
pI
kl
eine
r al
s de
r ak
tuel
le
pH-W
ert
wer
den
mit
eine
m
halb
en
Säul
envo
lum
en P
uffe
r (0,
5 m
l) el
uier
t und
als
Vor
lauf
ver
wor
fen.
Im z
wei
ten
Schr
itt w
ird d
as H
ämog
lobi
n el
uier
t. D
azu
wer
den
eini
ge M
illili
ter
desj
enig
en
Puff
ers
auf d
ie S
äule
geg
eben
, des
sen
pH-W
ert a
ls o
ptim
al fü
r die
Elu
tion
des
Häm
oglo
bins
best
imm
t w
urde
(s.
oben
). D
er D
urch
lauf
kan
n so
lan
ge v
erw
orfe
n w
erde
n, b
is d
ie f
arbi
ge
Ban
de d
as E
nde
der S
äule
err
eich
t, ab
dan
n m
uss d
as E
luat
auf
gefa
ngen
wer
den.
Prot
eine
mit
eine
m p
I grö
ßer a
ls d
er E
lutio
nspu
ffer
ver
blei
ben
wei
terh
in a
uf d
er S
äule
. Die
se
könn
en d
urch
Puf
fer
mit
noch
höh
eren
pH
-Wer
ten
(Em
pfin
dlic
hkei
t de
s Sä
ulen
mat
eria
ls
beac
hten
!) o
der
hohe
n Io
nens
tärk
en e
luie
rt w
erde
n. H
ier
soll
letz
tere
Met
hode
ang
ewan
dt
wer
den.
Daz
u w
erde
n 5
ml d
es fü
r die
Elu
tion
des H
ämog
lobi
n ve
rwen
dete
n Pu
ffer
s auf
1,3
4 M
an K
Cl e
inge
stel
lt (W
ievi
el K
Cl m
uß a
bgew
ogen
wer
den?
)und
das
ver
blie
bene
Pro
tein
mit
eini
ge M
illili
ter d
es sa
lzha
ltige
n Pu
ffer
s von
der
Säu
le v
erdr
ängt
. Das
Elu
at w
ird a
ufge
fang
en.
Von
be
iden
Fr
aktio
nen
wird
je
wei
ls
ein
Spek
trum
(
=300
-600
nm
) am
Ph
otom
eter
aufg
enom
men
und
mit
Ref
eren
zspe
ktre
n ve
rglic
hen.
Nac
hdem
die
Spe
ktre
n au
fgen
omm
en
wur
den,
wird
zu
dem
Elu
at d
er le
tzen
Fra
ktio
n 10
μl e
iner
100
mM
Asc
orbi
nsäu
re-L
ösun
g in
die
Küv
ette
pip
ettie
rt, d
urch
mis
cht u
nd fü
r 2 m
in b
ei R
aum
tem
pera
tur i
nkub
iert.
Ans
chlie
ßend
wird
aber
mal
s ein
Spe
ktru
m (
=300
-600
nm
) am
Pho
tom
eter
aufg
enom
men
und
mit
dem
zuvo
r
aufg
enom
men
en S
pekt
rum
ver
glic
hen.
Lite
ratu
r C
oope
r T G
. Bio
chem
isch
e A
rbei
tsm
etho
den,
1. A
ufl.,
de
Gry
ter B
erlin
New
Yor
k 19
80
Kle
ber H
.-P.;
Schl
ee D
.; Sc
höpp
W.:
Bio
chem
isch
es P
rakt
ikum
, 5. A
ufl.,
Gus
tav
Fisc
her V
erla
g Je
na S
tuttg
art L
übec
k U
lm
1997
Stry
er L
., B
ioch
emie
, Spe
ktru
m V
erla
g 19
96
- 23
-
Woc
he 1
Chr
omat
ogra
phie
A
G H
ippl
er
Frag
en z
ur V
orbe
reitu
ng:
1.Fo
lgen
de d
rei S
ubst
anzg
emis
che
sind
chr
omat
ogra
phis
ch a
ufzu
rein
igen
:
a)Ei
ne a
us P
flanz
en e
xtra
hier
te M
isch
ung
aus
nied
erm
olek
ular
en P
heno
len,
von
den
en d
er B
lattf
arbs
toff
-
Car
otin
abg
etre
nnt w
erde
n so
ll.
b)Ei
n G
emis
ch z
wei
er S
trukt
urpr
otei
ne d
er p
flanz
liche
n Ze
llwan
d: e
inem
Gly
cin-
reic
hen
Prot
ein
(GR
P;
MW
=15,
2 kD
a; p
I=5,
78)
und
eine
m P
rolin
-rei
chen
Pro
tein
(PR
P; M
W=6
0,3
kDa,
pI=
5,81
), w
obei
bei
de
Prot
eine
von
eina
nder
get
renn
t wer
den
solle
n.
c)Ei
n G
emis
ch v
on z
wei
Iso
form
en d
er P
heny
lala
nin-
Am
mon
ium
-Lya
se (
PAL)
: PA
L I
(MW
=332
kD
a,
pI=4
,57)
und
PA
L II
(M
W=3
29 k
Da,
pI=
8,24
), w
obei
wie
deru
m b
eide
Pro
tein
e in
mög
lichs
t rei
ner
Form
gew
onne
n w
erde
n so
llen.
Ihne
n st
ehen
folg
ende
Chr
omat
ogra
phie
med
ien
zur V
erfü
gung
:
I.Ei
ne G
elfil
tratio
nssä
ule
mit
eine
r Aus
schl
ußgr
enze
von
50
kDa
und
eine
r Ein
schl
ußgr
enze
von
8 k
Da;
II.
Eine
Gel
filtra
tions
säul
e m
it ei
ner A
ussc
hluß
gren
ze v
on 3
00 k
Da
und
eine
r Ein
schl
ußgr
enze
von
70
kDa;
III.
Eine
Kie
selg
el-D
C-P
latte
, wah
lwei
se m
it 50
% E
than
ol o
der e
inem
Chl
orof
orm
/Tol
uol-G
emis
ch (1
:1) a
ls
Lauf
mitt
el;
IV.
Eine
mit
Prot
onen
abg
esät
tigte
Kat
ione
naus
taus
cher
säul
e, w
obei
als
Auf
tragu
ngsp
uffe
r ein
Puf
fers
yste
m
mit
eine
m p
H-W
ert v
on 6
,0 g
enut
zt w
erde
n so
ll.
V.
Eine
mit
OH
- abg
esät
tigte
Ani
onen
aust
ausc
hers
äule
, wob
ei a
ls A
uftra
gung
spuf
fer e
in P
uffe
rsys
tem
mit
eine
m p
H-W
ert v
on 3
,0 b
is 4
,0 g
enut
zt w
erde
n ka
nn.
VI.
Eine
abg
esät
tigte
Con
cana
valin
A -
Säu
le (
Con
A i
st e
in L
ectin
, d.
h. e
s is
t in
der
Lag
e, s
pezi
fisch
Zuck
erre
ste
zu b
inde
n),
wob
ei a
ls A
uftra
gung
spuf
fer
ein
Puff
ersy
stem
mit
eine
m p
H-W
ert
von
7,0
genu
tzt w
erde
n so
ll.
Wel
ches
C
hrom
atog
raph
iesy
stem
bi
etet
si
ch
für
wel
ches
Pr
oble
m
an?
Beg
ründ
en
Sie
(kur
z!)
Ihre
Ents
chei
dung
en! W
arum
sind
ein
ige
der a
ufge
führ
ten
Syst
eme
hier
gar
nic
ht a
nwen
dbar
?
2.Ei
ne e
luie
rte F
rakt
ion
enth
ält n
ahez
u re
ines
Cyt
ochr
om c
. Sie
mes
sen
nach
zeh
nfac
her V
erdü
nnun
g be
i 550
nm e
ine
Extin
ktio
n vo
n 0,
185.
Wie
gro
ß is
t die
Kon
zent
ratio
n vo
n C
ytoc
hrom
c in
der
Fra
ktio
n? W
ähle
n Si
e
zur D
arst
ellu
ng d
es E
rgeb
nis e
ine
sinn
volle
Ein
heit!
(Sch
icht
dick
e de
r Küv
ette
= 1
cm
; 55
0nm
= 2
9,5
l m
ol-
1 c
m-1
)
3.Er
läut
ern
Sie
kurz
das
Prin
zip
der
Aff
initä
tsch
rom
atog
raph
ie.
Wel
che
natü
rlich
en P
roze
sse
wer
den
nach
geah
mt b
zw. a
usge
nutz
t? W
elch
e K
räfte
bin
den
die
zu is
olie
rend
en M
olek
üle
an d
ie im
mob
ilisi
erte
n
Kom
plex
partn
er?
4.In
form
iere
n Si
e si
ch ü
ber
die
Subs
tanz
en H
ämog
lobi
n, C
ytoc
hrom
c u
nd A
scor
bins
äure
(Fu
nktio
nale
Gru
ppen
, son
stig
e Ei
gens
chaf
ten)
.
5.Fe
rtige
n S
ie e
in A
blau
fdia
gram
m fü
r die
Ver
such
e an
.
- 24
-

Woc
he 2
M
etab
olitb
estim
mun
g A
G P
aul/Z
eis
Phot
omet
risc
he M
etab
olitb
estim
mun
g am
Bei
spie
l des
ana
erob
en
Gly
koly
se-E
ndpr
oduk
tes L
akta
t
Biol
ogis
cher
Hin
terg
rund
L-
Lakt
at, d
as A
nion
der
Milc
hsäu
re, f
ällt
in v
iele
n tie
risch
en O
rgan
ism
en a
ls S
toff
wec
hsel
-pr
oduk
t an
, w
enn
die
Ver
sorg
ung
der
Gew
ebe
mit
Saue
rsto
ff n
icht
gew
ährle
iste
t is
t. D
er
Bed
arf
an e
nerg
iere
iche
n Ph
osph
atve
rbin
dung
en (
ATP
) ka
nn d
ann
nich
t m
ehr
über
die
ox
idat
ive
Phos
phor
ylie
rung
ge
deck
t w
erde
n,
da
ohne
Sa
uers
toff
al
s En
dakz
epto
r de
r El
ektro
nent
rans
port
der A
tmun
gske
tte z
um E
rlieg
en k
omm
t. N
ur a
naer
obe
Stof
fwec
hsel
weg
e (A
naer
obio
se)
wie
die
Gly
koly
se s
tehe
n un
ter
dies
en B
edin
gung
en z
ur V
erfü
gung
. Die
hie
r “e
ntst
ehen
den”
Ele
ktro
nenp
aare
(N
AD
H +
H+ )
wer
den
dabe
i ni
cht
meh
r au
f Sa
uers
toff
, so
nder
n au
f or
gani
sche
Sub
stan
zen
(Met
abol
ite)
über
trage
n, z
um B
eisp
iel
auf
Pyru
vat,
das
Endp
rodu
kt d
er G
lyko
lyse
, das
dad
urch
zu
Lakt
at r
eduz
iert
wird
, wäh
rend
NA
DH
zu
NA
D
oxid
iert
wird
. Dam
it w
ird v
erhi
nder
t, da
ss s
ich
der
Pool
an
aufn
ahm
eber
eite
n R
eduk
tions
-äq
uiva
lent
en e
rsch
öpft.
M
an u
nter
sche
idet
zw
isch
en f
unkt
ions
bedi
ngte
r un
d bi
otop
bedi
ngte
r A
naer
obio
se. I
m e
rste
n Fa
ll üb
erst
eigt
der
Sau
erst
offb
edar
f de
s ak
tiven
Gew
ebes
die
Sau
erst
offv
erso
rgun
g, z
um
Bei
spie
l bei
exz
essi
ver M
uske
ltätig
keit.
So
kann
es
bei h
oher
kör
perli
cher
Lei
stun
g zu
ein
em
Man
gel a
n en
ergi
erei
chen
Pho
spha
tver
bind
unge
n au
ch b
ei A
nwes
enhe
it vo
n O
2 kom
men
, der
nu
r du
rch
zusä
tzlic
he E
nerg
iege
win
nung
übe
r A
naer
obio
se (
nebe
n de
r sa
uers
toff
limiti
erte
n ox
idat
iven
Pho
spho
rylie
rung
) ged
eckt
wer
den
kann
. Fun
ktio
nsbe
ding
te A
naer
obio
se k
ann
nur
über
kur
ze Z
eit a
ufre
chte
rhal
ten
wer
den
(z.B
. Flu
chtre
aktio
n, E
ndph
ase
eine
r Beu
teja
gd o
.ä.).
B
ei d
er b
ioto
pbed
ingt
en A
naer
obio
se v
erhi
nder
t dag
egen
Sau
erst
offm
ange
l in
der U
mge
bung
ei
nen
aero
ben
Stof
fwec
hsel
. Le
bens
räum
e m
it w
echs
elnd
em S
auer
stof
fgeh
alt
sind
z.B
. H
abita
te im
Küs
tenb
erei
ch, i
n st
ehen
den
Gew
ässe
rn o
der
im B
oden
. In
dies
em F
all k
önne
n an
aero
be S
toff
wec
hsel
weg
e au
ch ü
ber
läng
ere
Zeit
eine
Ver
sorg
ung
der
Gew
ebe
mit
ener
gier
eich
en P
hosp
hatv
erbi
ndun
gen
aufr
echt
erha
lten.
Fu
nktio
nsbe
ding
te A
naer
obio
se u
nter
sche
idet
sic
h vo
n de
r bio
topb
edin
gten
Ana
erob
iose
als
o hi
nsic
htlic
h de
r D
ynam
ik,
aber
auc
h hi
nsic
htlic
h de
r B
etei
ligun
g vo
n sy
stem
isch
en u
nd
met
abol
isch
en
Leis
tung
en
(z.B
. V
entil
atio
ns-
und
Perf
usio
nsan
pass
ung,
“m
etab
olic
de
pres
sion
”, s
pezi
fisch
e an
aero
be S
toff
wec
hsel
weg
e).
So k
önne
n ne
ben
der
„Milc
hsäu
re-
gäru
ng“
wei
tere
Sto
ffw
echs
elw
ege
an d
er a
naer
oben
Ene
rgie
vers
orgu
ng b
etei
ligt s
ein
und
zu
ande
ren
Stof
fwec
hsel
prod
ukte
n fü
hren
(Et
hano
l, O
pine
etc
.) In
vie
len
Fälle
n is
t ab
er e
ine
erhö
hte
Lakt
atko
nzen
tratio
n im
Gew
ebe
bzw
. de
n K
örpe
rflü
ssig
keite
n In
dika
tor
für
eine
n an
aero
ben
Stof
fwec
hsel
. M
etho
disc
her H
inte
rgru
nd
Vor
auss
etzu
ng fü
r die
enz
ymat
isch
e B
estim
mun
g ei
ner M
etab
olitk
onze
ntra
tion
in e
iner
Pro
be
ist
eine
spe
zifis
che
Rea
ktio
n, d
ie p
hoto
met
risch
ver
folg
t w
erde
n ka
nn. D
ie U
mse
tzun
g de
s M
etab
olite
n al
s Su
bstra
t ei
ner
Enzy
mre
aktio
n fü
hrt
zur
Änd
erun
g de
r op
tisch
en
Eige
nsch
afte
n de
r Lös
ung,
d.h
. die
Lic
htab
sorp
tion
durc
h di
e Pr
obe
wird
ver
ände
rt.
- 25
-
Woc
he 2
M
etab
olitb
estim
mun
g A
G P
aul/Z
eis
Die
opt
isch
en E
igen
scha
ften
eine
r Lös
ung
bei e
iner
bes
timm
ten
Wel
lenl
änge
b
esch
reib
t das
La
mbe
rt-B
eer´
sche
Ges
etz:
Dur
ch U
mfo
rmun
g er
gibt
sich
:
Der
Log
arith
mus
von
I 0/I
wird
als
Ext
inkt
ion
E b
ezei
chne
t. G
leic
hbed
eute
nd w
ird a
uch
der
Beg
riff
“Abs
orpt
ion”
ver
wen
det
oder
von
der
“op
tisch
en
Dic
hte”
de
r Lö
sung
ge
spro
chen
. D
ie
linea
re
Abh
ängi
gkei
t de
r Ex
tinkt
ion
von
der
Kon
zent
ratio
n de
s Chr
omop
hors
ist n
ur in
ver
dünn
ten
Lösu
ngen
geg
eben
. Im
Fal
le d
es L
akta
ttest
s ka
nn d
ie B
ildun
g vo
n Py
ruva
t und
NA
DH
(Chr
omop
hor,
s.u.)
durc
h di
e La
ktat
dehy
drog
enas
e (L
DH
) –
also
di
e U
mke
hr
der
Lakt
atbi
ldun
gsre
aktio
n im
St
offw
echs
el
- zu
r en
zym
atis
chen
La
ktat
best
imm
ung
hera
ngez
ogen
w
erde
n,
da
die
Extin
ktio
nsän
deru
ng p
ropo
rtion
al z
ur K
onze
ntra
tion
des L
akta
ts e
rfol
gt.
LD
H
La
ktat
+ N
AD
+ --
----
----
-
Py
ruva
t + N
AD
H +
H+
In e
inem
gee
igne
ten
Puff
ersy
stem
wird
das
Lak
tat
der
Prob
e du
rch
Zuga
be v
on N
AD
und
LD
H
zu
Pyru
vat
umge
setz
t, w
obei
N
AD
H
in
äqui
mol
aren
M
enge
n en
tste
ht.
Das
A
bsor
ptio
nspe
ktru
m d
er b
eide
n N
ukle
otid
e un
ters
chei
det
sich
im
Wel
lenl
änge
nber
eich
von
34
0 nm
: hie
r abs
orbi
ert N
AD
nic
ht, w
ähre
nd N
AD
H e
in E
xtin
ktio
nsm
axim
um a
ufw
eist
( =
6,
3 L
* m
mol
-1 *
cm
-1).
Anh
and
der o
ptis
chen
Dic
hte
bei 3
40 n
m k
ann
also
die
enz
ymat
isch
um
gese
tzte
Met
abol
itmen
ge q
uant
ifizi
ert w
erde
n. D
ie U
mse
tzun
g vo
n N
AD
/NA
DH
in e
iner
R
edox
reak
tion
stel
lt ei
n al
lgem
eine
s Pr
inzi
p en
zym
atis
cher
Ana
lyse
dar
, das
dur
ch g
eeig
nete
W
ahl e
ines
Enz
ymsy
stem
s nah
ezu
belie
bige
Met
abol
ite q
uant
itativ
erf
asse
n ka
nn.
I =
Inte
nsitä
t des
dur
chtre
tend
en L
icht
s I 0
= In
tens
ität d
es e
inge
stra
hlte
n Li
chts
c
= K
onze
ntra
tion
des C
hrom
opho
rs
= m
olar
er E
xtin
ktio
nsko
effiz
ient
des
Chr
omop
hors
d
= W
eglä
nge
des L
icht
s dur
ch d
ie P
robe
CO
O-
|
HO
–C–H
L
akta
t
|
C
H3
CO
O-
|
C=O
Pyr
uvat
|
C
H3
- 26
-

Woc
he 2
M
etab
olitb
estim
mun
g A
G P
aul/Z
eis
Zu
näch
st k
ann
mit
eine
r St
anda
rdlö
sung
bek
annt
er L
akta
tkon
zent
ratio
n di
e Fu
nktio
n de
s en
zym
atis
chen
Te
stsy
stem
s un
d di
e lin
eare
K
onze
ntra
tions
abhä
ngig
keit
der
Rea
ktio
n üb
erpr
üft
und
eine
Eic
hger
ade
erst
ellt
wer
den.
Die
Ext
inkt
ions
ände
rung
nac
h Zu
gabe
un
ters
chie
dlic
her
Lakt
atm
enge
n w
ird
dazu
um
ei
nen
Leer
wer
t ko
rrig
iert,
de
r di
e Ex
tinkt
ions
ände
rung
ber
ücks
icht
igt,
die
bere
its o
hne
Zuga
be d
er P
robe
(zu
m B
eisp
iel d
urch
di
e A
bsor
ptio
n de
s En
zym
s o.
ä.)
erfo
lgt.
Dan
n ka
nn m
it H
ilfe
der
Eich
gera
den
die
Kon
zent
ratio
n de
r Pr
obe
unbe
kann
ter
(zu
best
imm
ende
r) L
akta
tkon
zent
ratio
n an
hand
der
er
folg
ten
Extin
ktio
nsän
deru
ng b
erec
hnet
wer
den.
M
ater
ial:
- 20
Kra
bben
(Car
cinu
s mae
nas)
- F
angn
etz
- Epp
endo
rfpi
pette
n - 1
ml-S
pritz
en u
nd K
anül
en (0
,4 m
m)
- Epp
endo
rfze
ntrif
uge
- Hei
zblo
ck
- Gly
cin-
Hyd
razi
n-Pu
ffer
pH
9,0
(0,9
mol
/l G
lyci
n; 0
,75
mol
/l H
ydra
zin.
Vor
sich
t: G
iftig
) - N
AD
+ -Lös
ung
40 m
mol
/l *
- Lak
tat-S
tand
ard
5 m
mol
/l *
- LD
H-L
ösun
g 2,
5 m
g/m
l *
Die
mit
* ge
kenn
zeic
hnet
en L
ösun
gen
sind
auf
Eis
auf
zube
wah
ren.
Dur
chfü
hrun
g:In
die
sem
Kur
s so
ll di
e La
ktat
konz
entra
tion
in d
er H
ämol
ymph
e ru
hend
er u
nd z
ur A
ktiv
ität
vera
nlas
ster
Tie
re b
estim
mt
und
mite
inan
der
verg
liche
n w
erde
n. D
azu
wird
Stra
ndkr
abbe
n (C
arci
nus m
aena
s) o
hne
und
nach
loko
mot
oris
cher
Akt
ivitä
t (fu
nktio
nsbe
ding
te A
naer
obio
se)
eine
Häm
olym
phpr
obe
entn
omm
en.
Die
Ent
nahm
e er
folg
t m
it ei
ner
Sprit
ze a
us e
inem
Abs
orpt
ions
spek
tren
von
NA
DH
(off
ene
Kre
ise)
und
N
AD
+ (gef
üllte
Kre
ise)
. D
er E
xtin
ktio
nsun
ters
chie
d be
i 340
nm
erm
öglic
ht d
ie
Kon
zent
ratio
nsbe
stim
mun
g im
enz
ymat
isch
en T
est.
(Kar
lson
, P:
Kur
zes
Lehr
buch
der
Bio
chem
ie,
Thie
me-
Ver
lag,
Stu
ttgar
t)
- 27
-
Woc
he 2
M
etab
olitb
estim
mun
g A
G P
aul/Z
eis
Gel
enkh
äutc
hen
am A
nsat
z de
r C
oxa
(Dem
onst
ratio
n du
rch
die
Bet
reue
r). D
en T
iere
n w
ird
Häm
olym
phe
entn
omm
en (
Kon
trollw
ert),
dan
ach
wer
den
sie
mit
Hilf
e ei
nes
Stab
es z
u m
axim
aler
Akt
ivitä
t ve
ranl
asst
. N
ach
der
Akt
ivitä
t w
ird d
em T
ier
erne
ut H
ämol
ymph
e en
tnom
men
und
der
Lak
tatg
ehal
t bes
timm
t. V
on d
en e
inze
lnen
Kur
sgru
ppen
wird
der
Eff
ekt
von
unte
rsch
iedl
iche
r Akt
ivitä
tsda
uer n
ach
folg
ende
m S
chem
a un
ters
ucht
: D
ie H
ämol
ymph
prob
en w
erde
n 10
min
hitz
eden
atur
iert,
10
min
bei
130
00 rp
m z
entri
fugi
ert
und
der Ü
bers
tand
mit
Hilf
e de
s enz
ymat
isch
en T
ests
(sie
he u
nten
) ana
lysi
ert.
La
ktat
-Tes
t: D
urch
führ
ung:
Phot
omet
risch
e M
essu
ng b
ei 3
40 n
m.
Der
N
ulla
bgle
ich
des
Phot
omet
ers
erfo
lgt
mit
eine
r pu
ffer
gefü
llten
K
üvet
te,
dana
ch
Eins
tellu
ngen
des
Pho
tom
eter
s nic
ht m
ehr v
erän
dern
. D
ie E
xtin
ktio
n v
on L
eerw
erte
n, S
tand
ardl
ösun
gen
und
Prob
en w
ird v
or d
er e
nzym
atis
chen
R
eakt
ion
best
imm
t, da
nn e
rfol
gt d
er S
tart
der
Enzy
mre
aktio
n du
rch
Zuga
be v
on L
DH
und
na
ch
Abl
auf
eine
r ei
nstü
ndig
en
Inku
batio
n be
i R
aum
tem
pera
tur
die
Bes
timm
ung
der
Extin
ktio
n na
ch a
bges
chlo
ssen
er e
nzym
atis
cher
Um
setz
ung.
V
erm
eide
n Si
e di
e B
erüh
rung
der
Spi
tzen
von
Rüh
rspa
teln
und
Pip
ette
nspi
tzen
mit
den
Fing
ern,
um
Kon
tam
inat
ione
n m
it pr
oben
frem
dem
Lak
tat z
u ve
rhin
dern
.
1. L
eerw
ert
Bes
timm
ung
der E
xtin
ktio
nsän
deru
ng, d
ie o
hne
umge
setz
tes
Subs
trat a
llein
dur
ch d
ie Z
ugab
e de
s En
zym
s (L
DH
) er
folg
t (N
ullw
ert
der
Eich
gera
den)
. In
zw
ei
Eppe
ndor
fgef
äße
(Dop
pelb
estim
mun
g) w
ird in
der
ang
egeb
enen
Rei
henf
olge
pip
ettie
rt:
Puff
er:
860 μl
N
AD
+ : 20
μl
H2O
: 10
0 μl
Gru
ppe
Akt
ives
Tie
r: H
ämol
ymph
entn
ahm
e na
ch
1 / 2
1
min
3
/ 4
2 m
in
5 / 6
3
min
7
/ 8
4 m
in
9 / 1
0 6
min
11
/ 12
8
min
- 28
-

Woc
he 2
M
etab
olitb
estim
mun
g A
G P
aul/Z
eis
Dur
chm
isch
en S
ie d
ie S
ubst
anze
n (V
orte
x),
brin
gen
Sie
die
Lösu
ng i
n ei
ne H
albm
ikro
-K
üvet
te u
nd st
elle
n Si
e di
ese
in d
en S
trahl
enga
ng. N
otie
ren
Sie
die
Extin
ktio
n. P
ipet
tiere
n Si
e di
e Lö
sung
in
das
Eppe
ndor
fgef
äß z
urüc
k. F
ügen
Sie
20 μl
LD
H z
u un
d m
isch
en S
ie d
ie
Lösu
ng g
ut d
urch
(V
orte
x).
Lass
en S
ie d
ie R
eakt
ion
1h a
blau
fen
und
pipe
ttier
en S
ie d
ie
Lösu
ng i
n di
e K
üvet
te.
Bes
timm
en S
ie d
en E
ndw
ert
der
Extin
ktio
n im
Pho
tom
eter
. D
er
Mitt
elw
ert d
er E
xtin
ktio
nsän
deru
ng b
eide
r L
eerp
robe
n w
ird a
ls L
eerw
ert v
on a
llen
E –
Wer
ten
abge
zoge
n.
2. S
tand
ards
B
estim
mun
g de
r Ex
tinkt
ions
ände
rung
, di
e du
rch
beka
nnte
Sub
stra
tmen
gen
herv
orge
rufe
n w
ird. F
ühre
n Si
e di
e ob
en b
esch
riebe
ne M
essu
ng m
it de
m S
tand
ard
(Lak
tatlö
sung
bek
annt
er
Kon
zent
ratio
n)
durc
h.
Pipe
ttier
en
Sie
dazu
je
weils
zw
eim
al
(Dop
pelb
estim
mun
g)
in
Eppe
ndor
fgef
äße:
St
anda
rd
I II
II
I
Puff
er:
860 μl
86
0 μl
86
0 μl
NA
D+ :
20 μ
l 20
μl
20 μ
l St
anda
rd (5
mm
ol/l
Lakt
at)
10 μ
l 20
μl
30 μ
l W
asse
r 90
μl
80 μ
l 70
μl
Dur
chm
isch
en S
ie d
ie S
ubst
anze
n un
d be
stim
men
Sie
jew
eils
in e
iner
Küv
ette
die
Ext
inkt
ion.
N
ach
dem
Um
fülle
n in
ein
Epp
endo
rfge
fäß
20 μ
l LD
H z
ufüg
en u
nd g
ut d
urch
mis
chen
. La
ssen
Sie
die
Rea
ktio
n je
wei
ls 1
h ab
lauf
en u
nd n
otie
ren
Sie
die
Endw
erte
der
Ext
inkt
ion.
3. P
robe
n B
estim
mun
g de
r Ex
tinkt
ions
ände
rung
dur
ch e
ine
Lösu
ng m
it un
beka
nnte
r (z
u m
esse
nder
) Su
bstra
tkon
zent
ratio
n. F
ür d
ie M
essu
ng d
er P
robe
pip
ettie
ren
Sie
in p
aral
lele
n A
nsät
zen
jewe
ils zw
eim
al (D
oppe
lbes
timm
ung)
in E
ppen
dorf
gefä
ße:
Puff
er:
860 μl
86
0 μl
NA
D+ :
20 μ
l 20
μl
Prob
e 10
μl
20 μ
l W
asse
r 90
μl
80 μ
l D
urch
mis
chen
Sie
die
Sub
stan
zen,
pip
ettie
ren
Sie
sie
in e
ine
Küv
ette
und
brin
gen
Sie
dies
e in
de
n St
rahl
enga
ng. N
otie
ren
Sie
die
Extin
ktio
n. N
ach
dem
Um
fülle
n in
das
Epp
endo
rfge
fäß
20 μ
l LD
H z
ufüg
en u
nd m
isch
en. L
asse
n Si
e di
e R
eakt
ion
1h a
blau
fen
und
notie
ren
Sie
die
- 29
-
Woc
he 2
M
etab
olitb
estim
mun
g A
G P
aul/Z
eis
Endw
erte
. Fal
ls S
ie e
ine
sehr
nie
drig
e od
er s
ehr
hohe
Ext
inkt
ion
erha
lten,
ver
ände
rn S
ie d
ie
Prob
emen
ge. M
esse
n Si
e di
e Ex
tinkt
ions
ände
rung
der
Pro
ben
I (R
uhe)
und
II (A
ktiv
ität)
. Er
stel
lung
der
Eic
hger
aden
: Er
mitt
eln
Sie
für
die
gem
esse
nen
Stan
dard
lösu
ngen
die
Ver
ände
rung
in d
er E
xtin
ktio
n (
E)
durc
h di
e En
zym
reak
tion,
inde
m S
ie d
en A
nfan
gsw
ert v
om E
ndw
ert a
bzie
hen.
Von
dem
so
erha
ltene
n W
ert
für
E zi
ehen
Sie
den
E-
Wer
t fü
r de
n Le
erw
ert
ab.
Die
res
ultie
rend
en
Wer
te tr
agen
Sie
nac
h B
estim
mun
g de
s M
ittel
wer
tes
gege
n di
e K
onze
ntra
tion
auf.
Der
Lee
r-w
ert i
st T
eil d
er G
erad
en (N
ullp
unkt
). D
ie
E-W
erte
für d
ie P
robe
n w
erde
n ge
naus
o er
mitt
elt (
Leer
wer
t abz
iehe
n!).
Nun
kön
nen
Sie
aus
der
mit
Hilf
e de
r St
anda
rdm
essu
ngen
sel
bst e
rste
llten
Eic
hger
aden
die
Lak
tatk
onze
ntra
-tio
n in
den
Pro
ben
bere
chne
n. G
eben
Sie
den
Mitt
elw
ert
der
Lakt
atko
nzen
tratio
n in
der
H
ämol
ymph
e de
r Tie
re je
wei
ls fü
r die
Mes
sung
en in
Ruh
e (I
) und
nac
h A
ktiv
ität (
II) a
n.
Mög
liche
Feh
lerq
uelle
n:
- Pro
be is
t nic
ht g
enug
dur
chge
mis
cht -
-> K
üvet
te m
it Pa
rafil
m v
ersc
hlie
ßen,
schü
tteln
- L
uftb
lase
n in
der
Küv
ette
-->
mit
dem
Fin
ger v
orsi
chtig
geg
en d
ie K
üvet
te k
lopf
en o
der m
it R
ührs
pate
l Luf
tbla
sen
entfe
rnen
Au
swer
tung
:V
ergl
eich
en S
ie d
ie L
akta
tkon
zent
ratio
nen
in d
er H
ämol
ymph
e vo
n R
uhet
iere
n un
d ak
tiven
Ti
eren
. N
otie
ren
Sie
die
Mes
swer
te a
ller
Gru
ppen
. W
elch
en E
influ
ss h
at d
ie D
auer
der
A
ktiv
ität
auf
die
Lakt
atko
nzen
tratio
n? F
ertig
en S
ie e
in D
iagr
amm
an,
in
dem
Sie
die
A
bhän
gigk
eit d
er L
akta
tkon
zent
ratio
n in
der
Häm
olym
phe
von
der A
ktiv
itäts
daue
r dar
stel
len.
Pr
otok
oll:
1. E
inle
itung
: Zie
l des
Ver
such
s 2.
Erg
ebni
sse:
-
Tabe
lle d
er O
rigin
al-M
essw
erte
(eig
ene
Gru
ppe)
-
Eich
gera
de (D
iagr
amm
und
Ger
aden
funk
tion,
eig
ene
Gru
ppe)
-
Ber
echn
ete
Lakt
atko
nzen
tratio
nen
der H
ämol
ymph
prob
en (e
igen
e G
rupp
e)
-Ta
belle
der
Erg
ebni
sse
alle
r Gru
ppen
-
Gra
fisch
e D
arst
ellu
ng d
es Z
eitv
erla
ufs (
Abh
ängi
gkei
t von
CLa
ktat v
on d
er A
ktiv
itäts
daue
r)
-B
esch
reib
ung
der E
rgeb
niss
e in
Tex
tfor
m
3. D
isku
ssio
n
- 30
-

Woc
he 2
M
etab
olitb
estim
mun
g A
G P
aul/Z
eis
Frag
en zu
m V
erst
ändn
is:
-W
elch
e St
offw
echs
elw
ege
führ
en b
ei a
usre
iche
nder
Sau
erst
offv
erso
rgun
g zu
r B
ildun
g vo
n A
TP, w
elch
e un
ter S
auer
stof
fman
gel?
-
Was
ver
steh
t m
an u
nter
fun
ktio
nelle
r A
naer
obio
se,
was
unt
er b
ioto
pbed
ingt
er A
naer
o-bi
ose?
-
Wel
che
Rea
ktio
n ka
taly
sier
t die
Lak
tatd
ehyd
roge
nase
in d
er Z
elle
? -
Was
ist d
ie E
xtin
ktio
n ei
ner L
ösun
g un
d vo
n w
elch
en F
akto
ren
wird
sie
best
imm
t?
-D
ie M
alat
dehy
drog
enas
e (M
DH
) kat
alys
iert
folg
ende
Rea
ktio
n:
M
DH
M
alat
+ N
AD
+ --
----
----
O
xala
ceta
t + N
AD
H +
H+
Wel
che
Kom
pone
nten
mus
s ei
n en
zym
atis
ches
Tes
tsys
tem
ent
halte
n, m
it de
ssen
Hilf
e di
e O
xala
ceta
tkon
zent
ratio
n ei
ner
Prob
e be
stim
mt
wer
den
kann
? W
elch
e Ä
nder
ung
der
Extin
ktio
n er
war
ten
Sie
bei e
iner
Pro
be m
it ei
nem
Oxa
lace
tatg
ehal
t, vo
n 0,
1 m
mol
/l (in
de
r Küv
ette
). (Z
u-/A
bnah
me,
Zah
lenw
ert d
er E
xtin
ktio
nsän
deru
ng)?
Li
tera
tur:
B
ergm
eyer
, Han
s Ulri
ch: G
rund
lage
n de
r enz
ymat
isch
en A
naly
se. V
erla
g C
hem
ie W
einh
eim
. K
arls
on, P
eter
: Kur
zes L
ehrb
uch
der B
ioch
emie
. Thi
eme
Ver
lag,
Stu
ttgar
t. Lo
ttspe
ich,
F.,
Zorb
as, H
: Bio
anal
ytik
. Spe
ktru
m V
erla
g H
eide
lber
g, B
erlin
. Pa
ul, R
üdge
r: Ph
ysio
logi
e de
r Tie
re. S
yste
me
und
Stof
fwec
hsel
. Thi
eme
Ver
lag,
Stu
ttgar
t.
- 31
-- 3
2 -
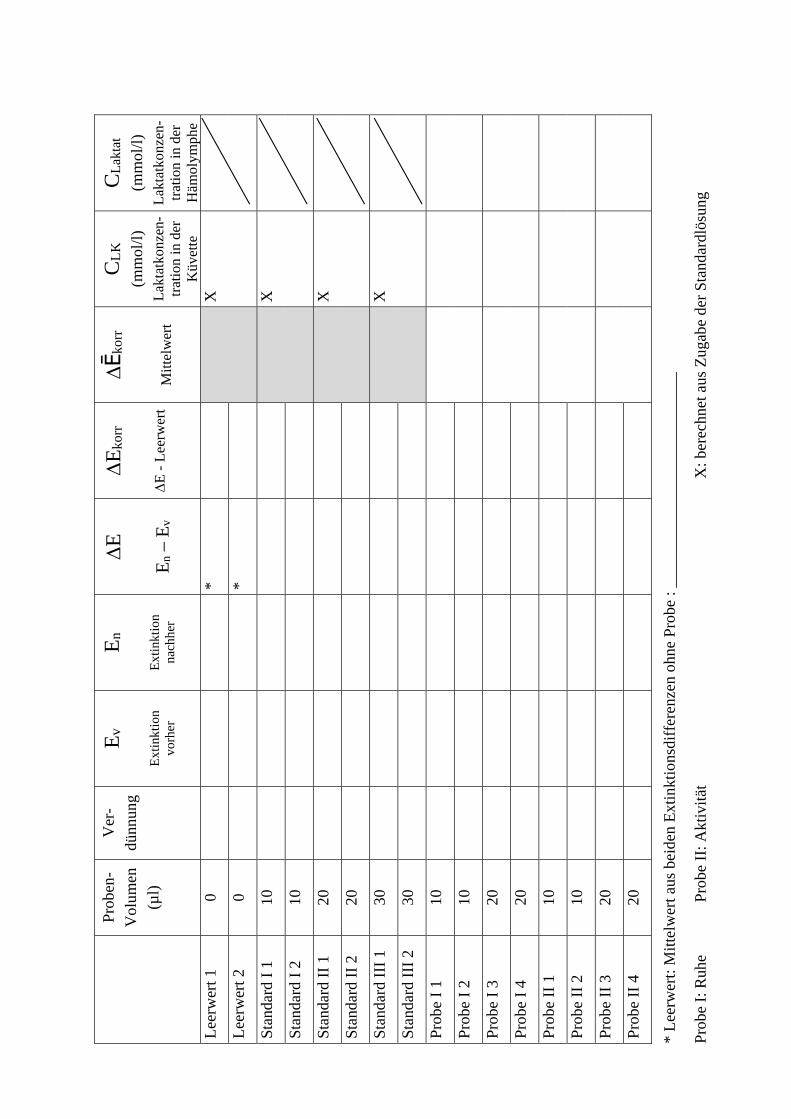
Pr
oben
- V
olum
en
(µl)
Ver
- dü
nnun
g E v
Extin
ktio
n vo
rher
E n
Ex
tinkt
ion
nach
her
E
E n
– E
v
Eko
rr
E
- Le
erw
ert
Ēko
rr
M
ittel
wer
t
CLK
(m
mol
/l)
Lakt
atko
nzen
-tra
tion
in d
er
Küv
ette
CLa
ktat
(m
mol
/l)
Lakt
atko
nzen
-tra
tion
in d
er
Häm
olym
phe
Leer
wer
t 1
0
*
X
Leer
wer
t 2
0
*
Stan
dard
I 1
10
X
Stan
dard
I 2
10
Stan
dard
II 1
20
X
Stan
dard
II 2
20
Stan
dard
III 1
30
X
Stan
dard
III 2
30
Prob
e I 1
10
Prob
e I 2
10
Prob
e I 3
20
Prob
e I 4
20
Prob
e II
1
10
Prob
e II
2
10
Prob
e II
3
20
Prob
e II
4
20
* Le
erw
ert:
Mitt
elw
ert a
us b
eide
n Ex
tinkt
ions
diffe
renz
en o
hne
Prob
e : _
____
____
____
____
____
____
__
Prob
e I:
Ruh
e
Prob
e II
: Akt
ivitä
t
X
: ber
echn
et a
us Z
ugab
e de
r Sta
ndar
dlös
ung

Woc
he 3
Met
abol
ism
usA
G W
eber
Mec
hani
smen
der
Mus
kelk
ontr
aktio
n
Them
en, d
ie z
ur V
orbe
reitu
ng g
ehör
en:
-M
uske
ltype
n -A
ufba
u de
s Ske
lettm
uske
ls, m
olek
ular
er M
echa
nism
us d
er M
uske
lkon
trakt
ion,
B
edeu
tung
von
ATP
und
Ca2+
-S
teue
rung
der
Mus
kelk
ontra
ktio
n
Lite
ratu
r:
Silv
erth
orn:
„Ph
ysio
logi
e“, P
ears
on S
tudi
um
K
apite
l 12:
„M
usku
latu
r“
Cam
pbel
l et a
l.: „
Bio
logi
e“, S
pekt
rum
Ver
lag,
200
9
Kap
itel 4
9: „
Sens
orik
und
Mot
orik
“ H
ickm
an e
t al.:
„Zo
olog
ie“,
Pea
rson
Stu
dium
, 200
8
Kap
itel 2
9: „
Hal
t, Sc
hutz
, Bew
egun
g“
Allg
emei
nes
Die
Fäh
igke
it zu
r B
eweg
ung
ist e
in K
ennz
eich
en d
er T
iere
und
erm
öglic
ht ih
nen
nich
t nur
N
ahru
ng z
u er
lang
en, s
onde
rn a
uch
Gef
ahre
n ab
zuw
ehre
n un
d Pa
rtner
zu
finde
n. U
m s
ich
fortz
ubew
egen
, müs
sen
die
Tier
e - u
nabh
ängi
g vo
n de
r Art
der B
eweg
ung
- aus
reic
hend
Kra
ft ge
gen
die
Um
wel
t aus
üben
, um
Rei
bung
skrä
fte u
nd S
chw
erkr
aft z
u üb
erw
inde
n. D
iese
Kra
ft w
ird
durc
h en
ergi
ever
brau
chen
de
zellu
läre
Pr
ozes
se
erze
ugt
(->
Bio
ener
getik
). D
en
unte
rsch
iedl
iche
n A
rten
der
Bew
egun
g lie
gen
Mec
hani
smen
zug
rund
e, d
ie a
llen
Tier
en
gem
eins
am s
ind.
Auf
der
zel
lulä
ren
Eben
e be
ruht
jeg
liche
Bew
egun
g au
f ko
ntra
ktile
n Sy
stem
en u
nd m
olek
ular
en M
otor
en.
Eine
Mus
kelk
ontra
ktio
n al
lein
e fü
hrt
jedo
ch n
icht
un
mitt
elba
r zu
ein
er B
eweg
ung
des
Tier
es:
alle
Bew
egun
gsar
ten
basi
eren
ent
sche
iden
d da
rauf
, das
s M
uske
ln g
egen
ein
fest
es (H
ydro
,- Ex
o- o
der E
ndo-
) Ske
lett
arbe
iten
könn
en. D
a M
uske
ln s
ich
nur
kont
rahi
eren
kön
nen,
müs
sen
sie
in a
ntag
onis
tisch
wirk
ende
n Pa
aren
am
Sk
elet
t ans
etze
n um
ein
zeln
e K
örpe
rteile
hin
- und
her
bew
egen
zu
könn
en.
Der
Ske
lettm
uske
l de
r W
irbel
tiere
ist
an
Kno
chen
ver
anke
rt un
d er
mög
licht
, di
ese
zu
bew
egen
. Er b
este
ht a
us la
ngen
Mus
kelfa
serb
ünde
ln (
0,1
– 1
mm
), di
e si
ch m
eist
übe
r die
ge
sam
te L
änge
des
Mus
kels
ers
treck
en. E
in B
ünde
l bes
teht
aus
meh
rere
n M
uske
lfase
rn (
10
– 90
μm
). D
ie M
uske
lfase
r ist
ein
e ei
nzel
ne, m
ehrk
erni
ge Z
elle
, die
aus
der
Fus
ion
meh
rere
r em
bryo
nale
r Ze
llen
gebi
ldet
wird
. Je
de M
uske
lfase
r be
steh
t au
s ru
nd 1
000
long
itudi
nal
ange
ordn
eten
Myo
fibril
len.
Jed
e M
yofib
rille
ist
dur
ch Z
-Sch
eibe
n in
ca.
2,5
μm
lan
ge
‚Fäc
her’
(Sa
rkom
ere)
unt
erte
ilt. D
ie S
arko
mer
e si
nd d
ie k
ontra
ktile
n Ei
nhei
ten
des
Mus
kels
. Si
e be
steh
en a
us z
wei
Arte
n vo
n M
yofil
amen
ten:
Dün
ne F
ilam
ente
bes
tehe
n au
s je
zw
ei
Act
inst
räng
en u
nd T
ropo
myo
sin-
Strä
ngen
. Dic
ke F
ilam
ente
wer
den
aus
zahl
reic
hen
Myo
sin
Mol
ekül
en g
ebild
et.
Eine
Mus
kelk
ontra
ktio
n er
folg
t da
nn,
wen
n di
e Fi
lam
ente
ane
inan
der
vorb
eigl
eite
n un
d da
bei d
ie S
arko
mer
e ve
rkür
zen.
D
as M
yosi
nmol
ekül
bes
itzt e
inen
zw
eige
teilt
en K
opf,
der d
ie A
TPas
e-A
ktiv
ität e
nthä
lt. E
r ist
ge
lenk
artig
mit
eine
m H
alss
tück
ver
bund
en (
Kop
f +
Hal
s =
schw
eres
Mer
omyo
sin)
, an
das
sich
, w
iede
rum
ge
lenk
artig
ve
rbun
den,
da
s Sc
hwan
zstü
ck
ansc
hlie
ßt
(=
leic
htes
M
erom
yosi
n). E
in M
yosi
nfila
men
t bes
teht
aus
ca.
150
-360
sol
cher
Mol
ekül
e, d
ie b
ünde
larti
g zu
sam
men
gefa
sst s
ind.
Die
gel
enka
rtige
Bew
eglic
hkei
t des
Kop
f-H
als-
Stüc
kes e
rmög
licht
die
re
vers
ible
Bin
dung
des
Myo
sins
mit
dem
Akt
in (
der
Akt
in-M
yosi
n-K
ompl
ex).
Akt
in is
t ein
gl
obul
äres
Pro
tein
(G
-Akt
in),
von
dem
jew
eils
~40
0 ei
ne p
erls
chnu
rarti
ge K
ette
bild
en (
F-A
ktin
). Je
wei
ls z
wei
mite
inan
der
verd
rillte
Akt
in-K
ette
n bi
lden
ein
Akt
infil
amen
t. D
as
eben
falls
fade
nför
mig
e Tr
opom
yosi
n w
inde
t sic
h um
das
Akt
infil
amen
t, w
obei
ca.
alle
40
nm
ein
Trop
onin
mol
ekül
ang
ehef
tet
ist.
Wäh
rend
der
Kon
trakt
ion
legt
sic
h de
r Tr
opom
yosi
n-
- 33
-
Woc
he 3
Met
abol
ism
usA
G W
eber
Fade
n in
die
Rin
ne z
wis
chen
zw
ei A
ktin
-F-K
ette
n un
d gi
bt d
eren
Bin
dung
sste
llen
für
das
Myo
sin
frei
. Als
mol
ekul
arer
Sch
alte
r wirk
t dab
ei d
as C
a2+-s
ensi
tive
Trop
onin
. D
er K
ontra
ktio
nsm
echa
nism
us d
es W
irbel
tierm
uske
ls i
st h
eute
bis
in
sein
e m
olek
ular
en
Det
ails
bek
annt
. Der
ers
te S
chrit
t bei
der
Akt
ivie
rung
ein
es M
uske
ls is
t die
Dep
olar
isat
ion
der
Mus
kelz
elle
. A
us d
em S
arko
plas
mat
isch
en R
etik
ulum
wird
Ca2+
ins
Cyt
osol
frei
gese
tzt.
Die
C
a2+K
onze
ntra
tion
in d
er M
uske
lzel
le e
rhöh
t da
bei
sich
inn
erha
lb v
on 2
0 m
s um
das
ca.
50
0fac
he.
Ca2+
verb
inde
t si
ch
mit
Trop
onin
, w
odur
ch
Trop
omyo
sin
sein
e he
mm
ende
W
irkun
g au
f di
e A
ktin
-Myo
sin-
Ver
bind
ung
verli
ert.
Die
ene
rgie
liefe
rnde
Rea
ktio
n lä
uft a
m
Kop
f des
Myo
sinm
olek
üls
ab, w
elch
er A
TP b
inde
n un
d in
AD
P +
P i h
ydro
lisie
ren
kann
. Ein
Te
il de
r H
ydro
lisie
rung
sene
rgie
übe
rträg
t si
ch a
uf d
as M
yosi
n, w
odur
ch d
iese
s in
ein
e en
ergi
erei
che
Kon
form
atio
n üb
erfü
hrt
wird
. D
er a
nger
egte
Myo
sink
opf
glei
cht
nun
eine
m
gesp
annt
en H
ebel
und
bin
det a
n ei
ne s
pezi
fisch
e B
indu
ngss
telle
am
Act
in u
nd b
ildet
so
eine
Q
uerb
rück
e zw
isch
en A
ctin
und
Myo
sin
(Act
omyo
sin-
Kom
plex
). Je
tzt w
ird d
ie g
espe
iche
rte
Ener
gie
frei
gese
tzt u
nd d
er M
yosi
nkop
f geh
t wie
der i
n di
e en
ergi
eärm
ere
Kon
form
atio
n üb
er,
wod
urch
sic
h de
r W
inke
l änd
ert,
unte
r de
m d
er K
opf
an d
en S
chw
anz
gebu
nden
ist.
Dur
ch
dies
es U
mkl
appe
n w
ird e
ine
Kra
ft au
f da
s A
ctin
filam
ent
ausg
eübt
und
die
ses
wird
in
Ric
htun
g de
s Sa
rkom
er-Z
entru
ms
gezo
gen.
Die
Bin
dung
zw
isch
en M
yosi
n un
d A
ctin
wird
w
iede
r ge
löst
, w
enn
ein
Mol
ekül
ATP
am
Myo
sink
opf
bind
et (
‚Wei
chm
ache
rwirk
ung
des
ATP
’) u
nd d
er Z
yklu
s ka
nn v
on n
euem
beg
inne
n: d
er fr
eie
Kop
f kan
n da
s ne
ue A
TP s
palte
n,
die
ener
gier
eich
e, g
espa
nnte
Kon
form
atio
n ei
nneh
men
und
an
die
Bin
dung
sste
lle e
ines
an
dere
n A
ctin
mol
ekül
s an
dock
en. J
eder
der
ca.
350
– 4
00 K
öpfe
ein
es d
icke
n Fi
lam
ents
kan
n 5
Que
rbrü
cken
pro
Sek
unde
bild
en u
nd lö
sen,
so d
ass d
ie F
ilam
ente
kon
tinui
erlic
h an
eina
nder
vo
rbei
gezo
gen
wer
den.
Ei
n ei
nzel
ner
Gle
itzyk
lus
führ
t ‚n
ur’
zu e
iner
Mus
kelv
erkü
rzun
g um
ca.
1 %
. Fü
r ei
ne
norm
ale
Mus
kelv
erkü
rzun
g (m
ax. 3
0 %
) is
t des
halb
ein
oft
wie
derh
olte
r A
blau
f de
s Zy
klus
no
twen
dig:
Bin
dung
der
Köp
fe –
Abk
nick
en u
nd G
leite
n –
Lösu
ng d
er B
indu
ng –
Bin
dung
an
eine
r nac
hfol
gend
en S
telle
des
Fila
men
ts.
- 34
-

Woc
he 3
Met
abol
ism
usA
G W
eber
Ver
such
V
iele
ele
men
tare
Sch
ritte
des
mol
ekul
aren
Kon
trakt
ions
mec
hani
smus
kön
nen
an i
solie
rten
Myo
fibril
len
("Fi
brill
enm
odel
l")
dem
onst
riert
wer
den.
Für
die
sen
Ver
such
ben
utze
n w
ir ha
ndel
sübl
iche
s H
ackf
leis
ch (
som
it ka
nn d
iese
r V
ersu
ch g
gf.
auch
gut
in
der
Schu
le
durc
hgef
ührt
wer
den)
. Sie
sol
len
am F
ibril
lenm
odel
l zei
gen,
das
s di
e M
uske
lkon
trakt
ion
von
Cal
cium
ione
n ( C
a2+)
und
ATP
abh
ängt
und
das
s da
bei
ATP
ges
palte
n w
ird (
ATP
ase-
Akt
ivitä
t). D
ie A
TP-S
paltu
ng s
oll
quan
tifiz
iert
wer
den.
Im
ein
zeln
en w
erde
n di
e Fi
brill
en
folg
ende
n B
edin
gung
en u
nter
wor
fen:
-Z
ugab
e vo
n A
TP
-Zug
abe
von
Ca2+
-Zug
abe
von
EGTA
(2
-Am
inoe
thyl
-Gly
colä
ther
-N,N
,N',N
'-Tet
raes
sigs
äure
), ei
nes
Che
latb
ildne
rs,
der
Ca2+
kom
plex
iert
(„w
egfä
ngt“
). K
ontra
ktio
nen
sind
an
eine
r de
utlic
hen
Vol
umen
abna
hme,
sc
hnel
lere
n Se
dim
enta
tion
der
Fibr
illen
un
d/od
er
eine
s si
ch
zusa
mm
enzi
ehen
den
Gel
zylin
ders
aus
den
Fib
rille
n er
kenn
bar.
Das
Aus
maß
der
ATP
-Sp
altu
ng w
ird ü
ber d
as fr
eige
setz
te a
norg
anis
che
Phos
phat
bes
timm
t. M
ater
ial
5 g
Rin
derh
ackf
leis
ch;
1.2
mol
/l Tr
ichl
ores
sigs
äure
(TC
A);
0.2
mol
/l A
TP i
n Pu
ffer
* (*
Pu
ffer
= 4
0 m
mol
/l Im
idaz
ol, 8
0 m
mol
/l K
Cl,
5 m
mol
/l M
gCl 2
, pH
7.2
), 0.
002
mol
/l C
aCl 2
in P
uffe
r*; 0
.4 m
ol/l
EGTA
in P
uffe
r*
Phos
phat
test
:Lö
sung
1: A
mm
oniu
mva
nada
t-Sal
pete
rsäu
reLö
sung
2: A
mm
oniu
mm
olyb
dat-S
chw
efel
säur
eLö
sung
3: P
hosp
hats
tand
ard
(200
μm
ol/l)
Dur
chfü
hrun
g 1.
H
erst
elle
n de
r Fib
rille
nsus
pens
ion
Das
Hac
kfle
isch
wird
mit
ca.
45 m
l ei
skal
tem
Puf
fer
vers
etzt
und
mit
dem
Ultr
a-Tu
rrax
ho
mog
enis
iert
(3 x
20
sec)
, um
die
Myo
fibril
len
frei
zuse
tzen
. Das
Hom
ogen
at w
ird d
urch
Pe
rlong
aze
filtri
ert
und
ansc
hlie
ßend
in
eine
m g
ewog
enen
Zen
trifu
genb
eche
r (F
alco
ntub
e)
bei 3
000U
pM fü
r 10
min
zen
trifu
gier
t. D
er Ü
bers
tand
(lös
liche
Pro
tein
e u.
a.) w
ird v
erw
orfe
n,
das
Myo
fibril
len-
Sedi
men
t w
ird z
um A
usw
asch
en l
öslic
her
Prot
eine
mit
45 m
l ei
skal
tem
Puff
er
vers
etzt
un
d m
it de
m
Ultr
a-Tu
rrax
se
hr
kurz
re
susp
endi
ert.
Nac
h er
neut
er
Zent
rifug
atio
n (w
ie o
ben)
wird
das
Myo
fibril
len-
Sedi
men
t im
20-
fach
en V
olum
en s
eine
s G
ewic
hts i
n Pu
ffer
auf
geno
mm
en u
nd se
hr k
urz
resu
spen
dier
t.
- 35
-
Woc
he 3
Met
abol
ism
usA
G W
eber
2.V
ersu
chsa
nsät
zeIn
den
Ans
ätze
n 1
- 4 w
erde
n je
wei
ls 2
ml v
erdü
nnte
Fib
rille
nsus
pens
ion
(FS)
, im
Ans
atz
5w
erde
n 2
ml
Puffe
r an
stel
le v
on F
ibril
lens
uspe
nsio
n un
d 10
0 μl
der
ang
egeb
enen
Zus
ätze
zu
sam
men
pip
ettie
rt (in
Rea
genz
gläs
ern!
!).
Dan
n w
ird
mit
Puff
er
auf
ein
Endv
olum
en
von
2.2
ml
aufg
efül
lt.
Vor
dem
Ans
etze
n de
r V
ersu
che
müs
sen
Sie
sich
ein
Pip
ettie
rsch
ema
über
lege
n, d
enn
die
Kon
trakt
ion
star
tet m
it de
r Zug
abe
von
ATP
!
Ans
atz
1:
FS +
CaC
l 2A
nsat
z 2:
FS
+ A
TP
Ans
atz
3:
FS +
CaC
l 2 +
ATP
A
nsat
z 4:
FS
+ E
GTA
+ A
TP
Ans
atz
5:
Puff
er +
ATP
Sie
solle
n di
e V
orgä
nge
ca. 1
5 m
in la
ng b
eoba
chte
n un
d si
ch a
lle V
erän
deru
ngen
not
iere
n.
3.Ph
otom
etris
che
Phos
phat
best
imm
ung:
Zu
r B
estim
mun
g de
r in
den
jew
eilig
en V
ersu
chen
fre
iges
etzt
en P
hosp
hatm
enge
wer
den
die
Fibr
illen
wie
der r
esus
pend
iert.
100
μl d
er S
uspe
nsio
n w
erde
n en
tnom
men
und
in e
inem
1,5
-m
l-Rea
ktio
nsge
fäß
mit
1 m
l TC
A (
Tric
hlor
essi
gsäu
re,
1,2
mol
/l) v
erse
tzt.
Die
dad
urch
au
sgef
ällte
n Pr
otei
ne
wer
den
abze
ntrif
ugie
rt (1
m
in,
Eppe
ndor
f-Ze
ntrif
uge)
. Zu
r Ph
osph
atbe
stim
mun
g w
erde
n D
oppe
lbes
timm
unge
n ge
mac
ht:
In
2-m
l-Rea
ktio
nsge
fäße
n w
erde
n fü
r di
e R
eage
nzie
nlee
rwer
te j
e 40
0 μl
TC
A v
orge
legt
, für
die
Sta
ndar
ds j
e 40
0 μl
ei
ner
200 μm
ol/l
Phos
phat
-Lös
ung,
für
die
Tes
tans
ätze
1-5
je
400 μl
der
jew
eilig
en
Übe
rstä
nde
nach
TC
A-F
ällu
ng.
Zu a
llen
Prob
en w
erde
n je
wei
ls 4
00 μ
l Lö
sung
1 u
nd 4
00 μ
l Lö
sung
2 g
egeb
en u
nd g
ut
gem
isch
t. N
ach
10 m
in p
hoto
met
risch
e B
estim
mun
g de
r Ext
inkt
ion
bei 4
05 n
m.
Das
Pho
tom
eter
wird
vom
Bet
reue
r in
Bet
rieb
geno
mm
en u
nd d
emon
strie
rt.
Prot
okol
l
Das
Pro
toko
ll so
ll en
thal
ten
(neb
en d
en ü
blic
hen
Abs
chni
tten)
: - P
ipet
tiers
chem
a fü
r die
ver
schi
eden
en F
ibril
lena
nsät
ze
- Ein
e Ta
belle
zum
Kon
trakt
ions
verh
alte
n in
den
ver
schi
eden
en F
ibril
lena
nsät
zen
- Die
Mes
sdat
en d
er P
hosp
hatb
estim
mun
g
- Die
Ber
echn
ung
der u
mge
setz
ten
ATP
-Men
gen
- Pr
ozen
t der
um
gese
tzte
n A
TP-M
enge
n be
zoge
n au
f de
n Fi
brill
enan
satz
mit
dem
höc
hste
n U
msa
tz
- 36
-
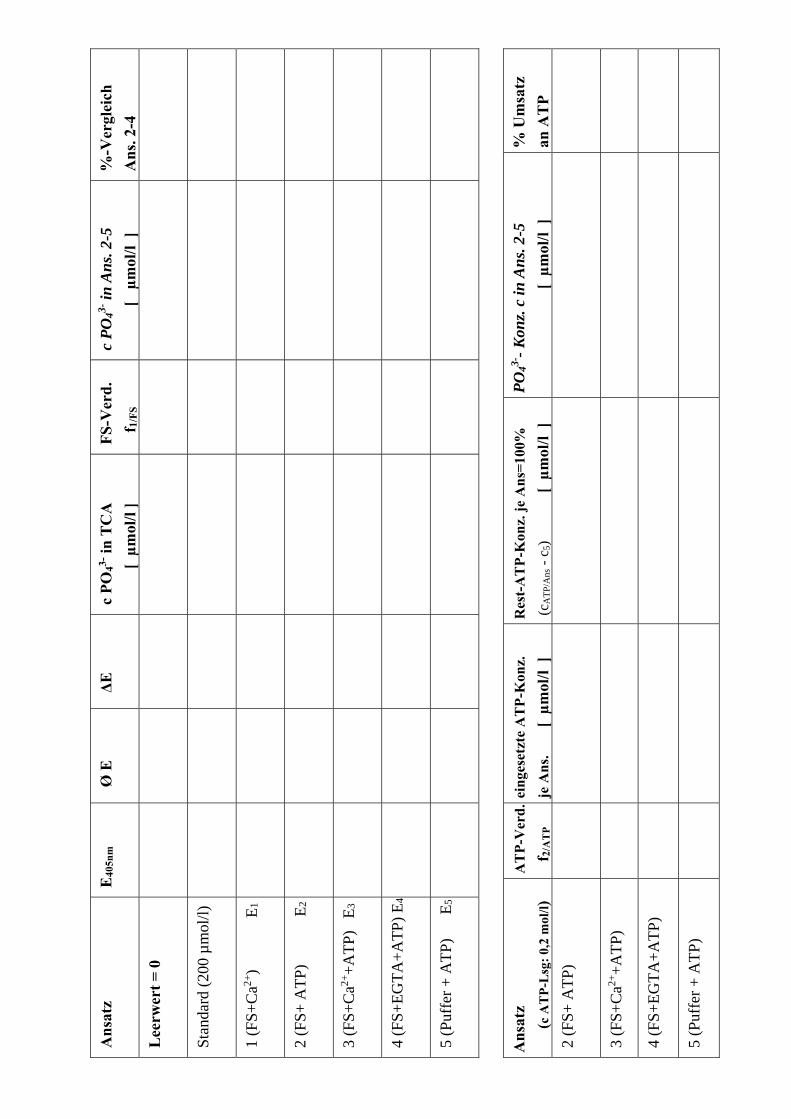
Ans
atz
E40
5nm
Ø
E
ΔE
c PO
43- in
TC
A
[
µmol
/l ]
FS-V
erd.
f 1/F
S c
PO43-
in A
ns. 2
-5
[
µm
ol/l
] %
-Ver
glei
ch
Ans
. 2-4
L
eerw
ert =
0
Stan
dard
(200
µm
ol/l)
1 (F
S+C
a2+)
E
1
2 (F
S+ A
TP)
E
2
3 (F
S+C
a2++A
TP)
E3
4
(FS+
EGTA
+ATP
) E4
5 (P
uffe
r + A
TP)
E
5
A
nsat
z
(c
AT
P-Ls
g: 0
,2 m
ol/l)
A
TP-
Ver
d.
f 2
/AT
P ei
nges
etzt
e A
TP-
Kon
z.
je A
ns.
[
µm
ol/l
] R
est-
AT
P-K
onz.
je A
ns=1
00%
(c
ATP
/Ans
- c 5
)
[ µ
mol
/l ]
PO43-
- Kon
z. c
in A
ns. 2
-5
[ µ
mol
/l ]
% U
msa
tz
an A
TP
2 (F
S+ A
TP)
3 (F
S+C
a2++A
TP)
4 (F
S+EG
TA+A
TP)
5 (P
uffe
r + A
TP)

Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Gru
ndla
gen
Wen
n ge
lade
ne T
eilc
hen
bzw
. Mol
ekül
e in
ein
ele
ktris
ches
Fel
d ge
brac
ht w
erde
n, w
ird a
uf d
ie
Mol
ekül
e, e
ntsp
rech
end
ihre
r N
etto
ladu
ng (
q) u
nd d
er v
erw
ende
ten
Feld
stär
ke (
E v),
eine
besc
hleu
nige
nde K
raft
(Fe)
ausg
ewirk
t (F e
= q
* E
), di
e zu
eine
r Wan
deru
ng d
er M
olek
üle f
ührt.
Die
in ei
ner E
lekt
roph
ores
e ver
wen
dete
Fel
dstä
rke e
rgib
t sic
h au
s der
ange
legt
en S
pann
ung
(U)
und
der
Dis
tanz
der
Ele
ktro
den
(l) (
E v =
U/l)
. D
ie W
ande
rung
sric
htun
g de
s M
olek
üls
ist
abhä
ngig
von
sei
ner
Ladu
ng (
posi
tiv o
der
nega
tiv).
Die
Ges
amtla
dung
ist
das
Pro
dukt
der
Ladu
ngsz
ahl (
z) u
nd d
er E
lem
enta
rladu
ng (e
) (q
= z
*e).
Die
erz
eugt
e B
eweg
ung
des M
olek
üls
wird
dur
ch d
ie V
isko
sitä
t des
Med
ium
s bzw
. die
Por
engr
öße d
er v
erw
ende
ten
Mat
rix g
ebre
mst
.
Die
Rei
bung
seig
ensc
hafte
n de
s Med
ium
s / d
er M
atrix
wer
den
durc
h de
n R
eibu
ngsk
oeff
izie
nten
(fc)
bes
chrie
ben.
Die
wirk
ende
Rei
bung
skra
ft (F
fr) i
st d
as P
rodu
kt d
es R
eibu
ngsk
oeff
izie
nten
und
der W
ande
rung
sges
chw
indi
gkei
t des
Mol
ekül
s (v)
(Ffr =
f c*
v).
Das
Ver
hältn
is d
er b
eweg
ende
n K
raft
(Fe)
und
der
bre
mse
nden
Kra
ft (F
fr)
bew
irkt
eine
kons
tant
e B
eweg
ung
des M
olek
üls i
m e
lekt
risch
en F
eld
(v =
q *
E/ f
c.).
Die
Bew
egun
g vo
n kl
eine
n ku
gelfö
rmig
en K
örpe
rn i
n ei
ner
Flüs
sigk
eit
wird
dur
ch d
as
Stok
e´sc
he G
eset
z be
schr
iebe
n:
W
= 6
rv
Wob
ei W
der
Rei
bung
swid
erst
and,
v d
ie B
eweg
ungs
gesc
hwin
digk
eit d
er K
ugel
, V
isko
sitä
t
der F
lüss
igke
it un
d r d
er R
adiu
s der
Kug
el is
t.
Die
Mob
ilitä
t ( v
on M
olek
ülen
kan
n be
schr
iebe
n w
erde
n du
rch
die
Gle
ichu
ng:
= z
* e
/ 6
r
Aus
die
ser
Gle
ichu
ng w
ird e
rsic
htlic
h, d
ass
die
Mob
ilitä
t ein
es M
olek
üls,
das
sich
in e
inem
elek
trisc
hen
Feld
dur
ch e
in M
ediu
m o
der e
ine
Gel
mat
rix b
eweg
t, ab
häng
t von
sei
ner L
adun
g
und
Form
. Die
obe
n ge
zeig
te F
orm
el b
esch
reib
t die
Mob
ilitä
t ein
es k
ugel
förm
igen
Kör
pers
, da
Prot
eine
abe
r nic
ht d
iese
r ide
alis
ierte
n Fo
rm e
ntsp
rech
en, g
ilt d
iese
For
mel
für P
rote
ine
nur m
it
Kor
rekt
urfa
ktor
en. D
ie T
renn
ung
von
Mol
ekül
en d
urch
Ele
ktro
phor
ese
ist d
amit
abhä
ngig
von
den
unte
rsch
iedl
iche
n La
dung
s- u
nd R
eibu
ngse
igen
scha
ften
der M
olek
üle.
Das
bed
eute
t, da
ss
Mol
ekül
e, d
ie s
ich
in d
iese
n Ei
gens
chaf
ten
star
k un
ters
chei
den,
gut
von
eina
nder
get
renn
t
wer
den
könn
en u
nd so
lche
mit
ähnl
iche
n Ei
gens
chaf
ten
schl
echt
er z
u tre
nnen
sind
. Die
Län
ge
der
Lauf
stre
cke
wäh
rend
der
Ele
ktro
phor
ese
ist b
ei A
uftre
nnun
g vo
n Pr
otei
nen
eben
falls
ein
Fakt
or,
da M
olek
üle
mit
ähnl
iche
n M
obili
täts
eige
nsch
afte
n er
st b
ei l
ange
n La
ufdi
stan
zen
deut
lich
vone
inan
der g
etre
nnt w
erde
n kö
nnen
.
Elek
troph
oret
isch
e A
uftre
nnun
gen
könn
en in
flü
ssig
en M
edie
n du
rchg
efüh
rt w
erde
n, je
doch
sind
di
e en
tste
hend
en Z
onen
, in
den
en s
ich
die
Mol
ekül
e s
amm
eln,
anf
ällig
(D
iffus
ion,
- 37
-
Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Bew
egun
gen)
. Dah
er w
erde
n m
eist
fes
te S
ubst
anze
n al
s M
atric
es f
ür d
ie e
lekt
roph
oret
isch
e
Auf
trenn
ung
genu
tzt.
Es
könn
en
z.B
. Pa
pier
, Ze
llulo
se-A
ceta
t, St
ärke
, A
garo
se
oder
Poly
acry
lam
id in
Ver
bind
ung
mit
vers
chie
dene
n El
ektro
phor
esep
uffe
rn v
erw
ende
t wer
den.
Bei
der V
erw
endu
ng v
on G
elm
atric
es fü
r die
ele
ktro
phor
etis
che
Auf
trenn
ung
kom
mt z
u de
n ob
en
besc
hrie
bene
n M
obili
täts
para
met
ern
(Lad
ung
und
Rei
bung
seig
ensc
hafte
n) d
ie G
röße
der
zu
trenn
ende
n M
olek
üle
vers
tärk
t als
Tre
nnkr
iteriu
m h
inzu
. Bei
der
Auf
trenn
ung
in fe
sten
Med
ien
ist d
ie P
oren
größ
e der
ver
wen
dete
n M
atrix
für d
en L
aufw
ider
stan
d de
r zu
trenn
ende
n M
olek
üle
maß
gebl
ich.
Gro
ße M
olek
üle
wer
den
unte
r di
esen
Bed
ingu
ngen
stä
rker
geb
rem
st a
ls k
lein
e
Mol
ekül
e, so
das
s ein
Mol
ekul
arsi
ebef
fekt
bei
der
Auf
trenn
ung
in G
elen
hin
zuko
mm
t.
Von
den
obe
n ge
nann
ten
fest
en E
lekt
roph
ores
emed
ien
sind
Aga
rose
und
Pol
yacr
ylam
id d
ie a
m
wei
test
ver
brei
tete
n. A
garo
se is
t ein
Pol
ysac
char
id, d
as s
ich
nach
Auf
koch
en b
eim
Abk
ühle
n
zu F
aser
n zu
sam
men
lage
rt un
d G
ele
mit
rela
tiv g
roße
n Po
reng
röße
n bi
ldet
. A
garo
sege
le
wer
den
häuf
ig v
erw
ende
t be
i de
r A
uftre
nnun
g vo
n N
ukle
insä
uren
, kö
nnen
abe
r au
ch z
ur
Auf
trenn
ung
von
groß
en P
rote
inen
, Pro
tein
kom
plex
en o
der
Nuk
lein
säur
e/Pr
otei
nkom
plex
en
verw
ende
t wer
den.
Für
die
Auf
trenn
ung
kom
plex
er P
rote
inge
mis
che
ist P
olya
cryl
amid
das
häu
figst
ver
wen
dete
Gel
med
ium
. Gel
e au
s Po
lyac
ryla
mid
wer
den
durc
h ra
dika
lisch
e Po
lym
eris
atio
n er
zeug
t. D
ie
Pore
ngrö
ße d
er p
olym
eris
ierte
n A
cryl
amid
gele
ist d
urch
das
leic
ht z
u va
riier
ende
Ver
hältn
is
der
verw
ende
ten
Rea
genz
ien
gut
zu b
eein
fluss
en,
was
die
Gen
erie
rung
von
Gel
en m
it
geei
gnet
en
Tren
neig
ensc
hafte
n fü
r M
olek
üle
unte
rsch
iedl
iche
r G
röße
er
laub
t.
Poly
acry
lam
idge
le k
önne
n üb
er w
eite
pH
-Ber
eich
e ei
nges
etzt
wer
den,
sin
d tra
nspa
rent
und
chem
isch
wei
tgeh
end
iner
t. D
adur
ch la
ssen
sic
h Pr
otei
nban
den
durc
h ei
ne F
ärbu
ng w
ie z
.B.
Coo
mas
sie-
Bla
u im
Gel
nac
hwei
sen.
Die
von
uns
im P
rakt
ikum
ver
wen
dete
n Po
lyac
ryla
mid
gele
sind
flac
he G
ele,
die
sich
zw
isch
en
zwei
Gla
spla
tten
befin
den.
Zum
Anf
ertig
en e
ines
Pol
yacr
ylam
idge
ls w
ird e
ine
Mis
chun
g au
s
Acr
ylam
id, N
,N´-
Met
hyle
nbis
acry
lam
id, N
, N, N
´, N
´– T
etra
met
hyl-e
thyl
endi
amin
(TEM
ED)
und
Am
mon
ium
pero
xodi
sulfa
t (A
PS)
in w
ässr
iger
Puf
ferlö
sung
gem
isch
t und
zw
isch
en d
ie
abge
dich
tete
n G
lasp
latte
n ge
goss
en.
Dur
ch
die
Mis
chun
g de
r K
ompo
nent
en
wird
di
e
Poly
mer
isat
ions
reak
tion
gest
arte
t. B
ei
der
radi
kalis
chen
Po
lym
eris
atio
n di
ent
Am
mon
ium
pero
xodi
sulfa
t al
s R
adik
alst
arte
r un
d TE
MED
kat
alys
iert
die
Ents
tehu
ng f
reie
r
Rad
ikal
e au
s A
mm
oniu
mpe
roxo
disu
lfat.
Aus
den
Mon
omer
en d
es A
cryl
amid
wer
den
linea
re
Poly
mer
e ge
bild
et,
die
durc
h da
s ei
nges
etzt
e N
, N
´-M
ethy
lenb
isac
ryla
mid
que
r ve
rnet
zt
wer
den.
Die
Por
engr
öße
der
so e
rzeu
gten
Gel
e hä
ngt a
b vo
n de
m V
erhä
ltnis
von
Acr
ylam
id
und
N,N
´-M
ethy
lenb
isac
ryla
mid
(C
-Wer
t )
und
von
der
Men
ge d
iese
s G
emis
ches
, da
s zu
r
- 38
-

Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Rea
ktio
nsm
isch
ung
zuge
gebe
n w
ird (T
-Wer
t). D
er T
-Wer
t bes
chre
ibt d
ie M
enge
der
insg
esam
t
eing
eset
zten
Mon
omer
e (A
cryl
amid
und
N,N
´-M
ethy
lenb
isac
ryla
mid
) in
Gew
icht
spro
zent
pro
Vol
umen
(%
w/v
) un
d er
rech
net
sich
aus
fol
gend
er F
orm
el:
T =
Acr
ylam
id (
g) +
N,N
´-
Met
hyle
nbis
acry
lam
id (
g)*
100
/ 10
0 m
l. D
er C
-Wer
t gi
bt d
as V
erhä
ltnis
von
N,N
´-
Met
hyle
nbis
acry
lam
id z
u A
cryl
amid
im G
emis
ch in
Gew
icht
spro
zent
(%
w/w
) an
und
wird
bere
chne
t mit
der F
orm
el: C
= N
,N´-
Met
hyle
nbis
acry
lam
id (g
) *
100
/ A
cryl
amid
(g) +
N,N
´-
Met
hyle
nbis
acry
lam
id (
g).
Bei
kon
stan
tem
C-W
ert
und
stei
gend
em T
-Wer
t si
nkt
die
Pore
ngrö
ße. B
ei h
ohen
und
nie
drig
en C
-Wer
ten
nim
mt d
ie P
oren
größ
e zu
und
ein
Min
imum
liegt
bei
C =
5%
. Die
Acr
ylam
idko
nzen
tratio
n, u
nd d
amit
die
erze
ugte
Por
engr
öße
des
Gel
s,
beei
nflu
sst d
en T
renn
bere
ich
der P
rote
ine.
Wir
verw
ende
n hi
er e
in M
onom
erge
mis
ch a
us 3
0%
Acr
ylam
id u
nd 0
,8%
N,N
´-M
ethy
lenb
isac
ryla
mid
. Um
ein
e op
timal
e A
uftre
nnun
g de
r Pro
tein
e
vers
chie
dene
r G
röße
n zu
err
eich
en, s
ollte
die
gee
igne
te K
onze
ntra
tion
Acr
ylam
id v
erw
ende
t
wer
den.
Die
Acr
ylam
idko
nzen
tratio
n fü
r di
e A
uftre
nnun
g vo
n Pr
otei
nen
vers
chie
dene
r
Grö
ßenb
erei
che
ist i
n de
r Tab
elle
auf
gefü
hrt.
Prot
eing
röße
(kD
a)
% fi
nale
Acr
ylam
id K
onze
ntra
tion
40-2
05
5%
25-2
05
7,5%
15-2
05
10%
15-6
6 12
,5%
15-4
5 15
%
Elek
troph
ores
en
könn
en
unte
rteilt
w
erde
n in
ko
ntin
uier
liche
un
d di
skon
tinui
erlic
he
Elek
troph
ores
en.
In
kont
inui
erlic
hen
Gel
elek
troph
ores
en
wer
den
dies
elbe
n Pu
ffer
m
it
kons
tant
em p
H-W
ert i
m G
el, d
en E
lekt
rode
nkam
mer
n un
d Pr
oben
ver
wen
det,
wen
n au
ch b
ei
spez
ielle
n A
pplik
atio
nen
in u
nter
schi
edlic
hen
Kon
zent
ratio
nen.
Im G
egen
satz
daz
u kö
nnen
bei
disk
ontin
uier
liche
n El
ektro
phor
esen
unt
ersc
hied
liche
Puf
ferk
onze
ntra
tione
n, p
H-W
erte
und
Pore
ngrö
ßen
inne
rhal
b de
s Gel
s und
der
Ele
ktro
denk
amm
ern
verw
ende
t wer
den.
Bei
der
hie
r ve
rwen
dete
n di
skon
tinui
erlic
hen
Gel
elek
troph
ores
e (L
aem
mli
1970
) w
ird i
m
unte
ren
Ber
eich
der
Gla
spla
tten
zunä
chst
das
Tre
nnge
l ge
goss
en.
Im T
renn
gel
wird
die
Puff
erko
nzen
tratio
n vo
n 0,
375m
ol/L
Tris
-Cl
(pH
8,8
), 0,
1% S
DS
eing
eset
zt.
Übe
r de
m
Tren
ngel
wird
spä
ter e
in S
amm
elge
l geg
osse
n m
it ei
ner P
uffe
rkon
zent
ratio
n vo
n 0,
125m
ol/L
Tris
-Cl (
pH 6
.8),
0,1%
SD
S. D
er v
erw
ende
te E
lekt
rode
npuf
fer i
st z
usam
men
gese
tzt a
us 2
5mM
Tris
(p
H
8,3)
, 0,
192M
G
lyci
n,
0,1%
SD
S.
Es
wer
den
hier
al
so
unte
rsch
iedl
iche
Puff
erko
nzen
tratio
nen
und
pH-W
erte
in T
renn
- und
Sam
mel
-Gel
ver
wen
det.
- 39
-
Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Zusä
tzlic
h zu
den
dis
kont
inui
erlic
hen
pH-W
erte
n w
ird im
Sam
mel
gel e
ine
höhe
re P
oren
größ
e
verw
ende
t. D
ie P
robe
n w
erde
n da
nn in
Gel
tasc
hen
auf d
as S
amm
elge
l auf
getra
gen.
Dur
ch d
en
nied
rigen
pH
-Wer
t im
Sam
mel
gel
und
die
Ver
wen
dung
von
Gly
cin
(Fol
ge-I
onen
) im
Elek
trode
npuf
fer
und
Cl- -I
onen
(L
eit-I
onen
) im
Tr
enn-
/Sam
mel
gel-P
uffe
r w
ird
ein
Sam
mel
effe
kt d
er P
rote
ine
in d
er P
robe
erz
eugt
. Die
Pro
tein
e sa
mm
eln
sich
zu
eine
r sch
mal
en
Ban
de z
wis
chen
Lei
t- un
d Fo
lge-
Ione
n an
. Bei
m Ü
berg
ang
der P
rote
ine
vom
Sam
mel
gel i
ns
Tren
ngel
erh
öht s
ich
der
Lauf
wid
erst
and
aufg
rund
der
ger
inge
ren
Pore
ngrö
ße im
Tre
nn-G
el
und
der p
H-W
ert i
st im
Tre
nnge
l höh
er a
ls im
Sam
mel
gel.
Die
Gly
cin-
Ione
n üb
erho
len
weg
en
ihre
r ger
inge
ren
Mol
ekül
größ
e un
d de
s ve
ränd
erte
n pH
-Wer
tes
die
Prot
eine
, die
sic
h zu
ein
er
schm
alen
Ban
de a
n de
r G
renz
e zu
m T
renn
gel
sam
mel
n. N
un e
rfol
gt d
ie A
uftre
nnun
g de
r
Prot
eine
in e
inem
hom
ogen
en P
uffe
r im
Tre
nnge
l.
Prot
eine
si
nd
aus
Am
inos
äure
n zu
sam
men
gese
tzt,
die
sich
be
zügl
ich
ihre
r
Ladu
ngse
igen
scha
ften
in
den
Seite
nres
ten
zum
Te
il st
ark
unte
rsch
eide
n.
Da
die
elek
troph
oret
isch
e M
obili
tät
von
Prot
eine
n au
ch v
on d
er L
adun
g ab
häng
t, sp
ielt
nebe
n de
r
Mol
ekül
größ
e eb
enfa
lls d
ie A
min
osäu
rese
quen
z ei
ne R
olle
. Is
t ei
ne P
rote
inau
ftren
nung
unab
häng
ig ih
rer E
igen
ladu
ng e
rwün
scht
, z.B
. zur
Mol
ekul
arge
wic
htsb
estim
mun
g, w
ird e
ine
Poly
acry
lam
id-G
elel
ektro
phor
ese
in A
nwes
enhe
it vo
n SD
S (e
ngl.
Sodi
um-D
odec
ylsu
lfate
Na+ [C
H3-
(CH
2)10
-CH
2OSO
3- ]),
eine
s an
ioni
sche
n D
eter
gens
, du
rchg
efüh
rt. S
DS
bind
et a
n
hydr
opho
be R
egio
nen
des
Prot
eins
und
ent
falte
t es
. M
ultim
ere
Prot
eine
dis
sozi
iere
n na
ch
Erhi
tzen
in S
DS
halti
ger L
ösun
g (<
95°
C) i
n ih
re U
nter
einh
eite
n. M
it de
r Bin
dung
von
ca.
1,4
g
SDS
/ g P
rote
in w
erde
n st
arke
neg
ativ
e La
dung
en e
inge
führ
t, de
ren
Sum
me
prop
ortio
nal z
ur
Prot
einm
asse
ist
. D
isul
fidbr
ücke
n w
erde
n du
rch
Zuga
be r
eduz
iere
nder
Sub
stan
zen,
wie
-
Mer
capt
oeth
anol
(HO
-CH
2-C
H2-
SH) o
der D
ithio
thre
itol (
DTT
, HS-
CH
2-C
HO
H-C
HO
H-C
H2-
SH)
gelö
st.
Die
Bew
eglic
hkei
t de
r Pr
otei
ne d
urch
die
Tre
nnge
lmat
rix i
st u
nter
die
sen
Bed
ingu
ngen
übe
r w
eite
Ber
eich
e lin
ear
zu d
en L
ogar
ithm
en i
hrer
Mol
ekul
arge
wic
hte.
Dad
urch
ist e
s m
öglic
h, d
as re
lativ
e M
olek
ular
gew
icht
ein
es P
rote
ins
durc
h V
ergl
eich
mit
der
elek
troph
oret
isch
en M
obili
tät v
on P
rote
inen
bek
annt
en M
olek
ular
gew
icht
s (G
röße
nmar
ker)
zu
erm
ittel
n.
Für
die
Bes
timm
ung
des
Mol
ekul
arge
wic
hts
von
Prot
eine
n w
ird
eine
Gel
elek
troph
ores
e du
rchg
efüh
rt, b
ei d
er a
uf e
inem
Gel
die
Pro
be d
es P
rote
ins
und
ein
Grö
ßenm
arke
r in
un
ters
chie
dlic
hen
Spur
en
nebe
nein
ande
r au
fget
renn
t w
erde
n.
Nac
h
Aus
mes
sen
der
Wan
deru
ngss
treck
en
der
einz
elne
n G
röße
nmar
kerb
ande
n na
ch
der
Elek
troph
ores
e ka
nn e
ine
Eich
gera
de e
rste
llt w
erde
n.
- 40
-

Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Anf
ertig
en v
on A
cryl
amid
gele
n fü
r di
e SD
S-PA
GE
D
ie G
ele
für
den
Kur
s w
erde
n ge
stel
lt. D
er A
ufba
u fü
r da
s G
ieße
n de
r G
ele
wird
im K
urs
geze
igt w
erde
n. D
a di
e R
eakt
ions
mis
chun
g fü
r die
Pol
ymer
isat
ion
der A
cryl
amid
gele
zun
ächs
t
flüss
ig is
t, m
uss v
or d
em B
efül
len
der A
ppar
atur
sich
erge
stel
lt w
erde
n, d
ass d
iese
dic
ht is
t. D
ies
kann
get
este
t wer
den,
inde
m m
an E
than
ol z
wis
chen
die
Gla
spla
tten
gibt
und
beo
bach
tet,
ob d
er
ober
e Fl
üssi
gkei
tssp
iege
l abs
inkt
. Wen
n da
s de
r Fa
ll is
t, m
uss
die
App
arat
ur n
eu a
ufge
baut
wer
den.
Wen
n di
e G
ele
in d
en G
ießs
tänd
en d
icht
sin
d, k
ann
die
Rea
ktio
nsm
isch
ung
zusa
mm
enge
setz
t w
erde
n (e
ntsp
rech
end
der
unte
n be
schr
iebe
nen
Rez
eptu
r).
Bei
de
r
Anf
ertig
ung
der G
ele
ist d
arau
f zu
acht
en, d
ass A
cryl
amid
in fl
üssi
ger
Form
gift
ig is
t! A
lso
wer
den
alle
Arb
eite
n m
it de
r nö
tigen
Sor
gfal
t, un
ter
Ver
wen
dung
gee
igne
ter
pers
önlic
her
Schu
tzau
srüs
tung
, unt
er e
inem
Abz
ug d
urch
gefü
hrt.
Die
Res
te d
er R
eakt
ions
mis
chun
g w
erde
n
vor
der
Ents
orgu
ng
poly
mer
isie
rt.
Alle
ko
ntam
inie
rten
Abf
älle
w
erde
n in
di
e da
für
vorg
eseh
enen
Son
dera
bfal
lbeh
älte
r geg
eben
.
Ver
wen
dete
Lös
unge
n zu
r A
nfer
tigun
g ei
nes A
cryl
amid
-Gel
s
Acr
ylam
id S
tam
mlö
sung
:
30%
(w/v
) Acr
ylam
id
0,
8% (w
/v) N
,N´-
Met
hyle
nbis
acry
lam
id
SDS
10%
(w/v
)
A
mm
oniu
mpe
roxo
disu
lfat:
10
% (w
/v)
TE
ME
D
pr
o an
alys
is 9
9%
Sam
mel
gelp
uffe
r:
0,5M
Tris
-Cl p
H 6
,8
Tre
nnge
lpuf
fer:
1,
5M T
ris-C
l pH
8,8
E
lekt
rode
npuf
fer:
0,
025M
Tris
, 0,1
92M
Gly
cin,
0,1
% S
DS
pH 8
.3
Zus
amm
ense
tzun
g de
r R
eakt
ions
mis
chun
g fü
r da
s Tre
nnge
l D
ie R
eakt
ions
mis
chun
g fü
r zw
ei G
ele
eine
r Kon
zent
ratio
n vo
n 10
% w
ird b
este
ht a
us:
Tren
ngel
puff
er
7,5m
ldd
H2O
12
ml
SDS
10%
0,
3ml
Acr
ylam
id-S
tam
mlö
sung
10m
l TE
MED
10l
APS
10%
15
0l
Das
Tre
nnge
l wird
bis
zu
eine
r Höh
e vo
n et
wa
6 cm
zw
isch
en d
ie v
orbe
reite
ten
Plat
ten
gefü
llt,
vors
icht
ig m
it w
enig
Isop
ropa
nol ü
bers
chic
htet
und
die
Pol
ymer
isat
ion
wird
abg
ewar
tet.
- 41
-
Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Zus
amm
ense
tzun
g de
r R
eakt
ions
mis
chun
g fü
r da
s Sam
mel
gel
Die
Rea
ktio
nsm
isch
ung
für
zwei
Gel
e m
it ei
ner
Sam
mel
gelk
onze
ntra
tion
von
4% w
ird
zusa
mm
enge
setz
t aus
:
Sam
mel
gelp
uffe
r 2,
5ml
ddH
2O
6ml
SDS
10%
0,
1ml
Acr
ylam
id-S
tam
mlö
sung
1,
3ml
TEM
ED5
lA
PS 1
0%50
l
Vor
der
Zug
abe
der
Sam
mel
gellö
sung
mus
s da
s Is
opro
pano
l en
tfern
t w
erde
n, m
it de
m d
as
Tren
ngel
übe
rsch
icht
et w
urde
. N
achd
em d
ie S
amm
elge
llösu
ng z
ugeg
eben
wur
de,
wird
der
Prob
enka
mm
in d
ie S
amm
elge
llösu
ng zw
isch
en d
ie G
lasp
latte
n ge
stec
kt. W
enn
das S
amm
elge
l
ausg
ehär
tet i
st, w
ird d
er P
robe
nkam
m e
ntfe
rnt u
nd d
as G
el w
ird m
it Sp
acer
n un
d G
lasp
latte
n
in d
ie E
lekt
roph
ores
ekam
mer
übe
rfüh
rt. D
ie P
ositi
on d
er P
robe
ntas
chen
und
die
Gre
nze
von
Sam
mel
- un
d Tr
enn-
Gel
wer
den
mar
kier
t. V
or d
em B
elad
en d
er G
elta
sche
n m
it de
n Pr
oben
wird
der
Ele
ktro
denp
uffe
r in
die
Lau
fapp
arat
ur g
efül
lt. B
evor
die
Pro
ben
eing
efül
lt w
erde
n,
bitte
dar
auf a
chte
n, d
ass
das
Gel
luftb
lase
nfre
i an
beid
en E
nden
mit
dem
Ele
ktro
denp
uffe
r in
Kon
takt
ste
ht. N
achd
em d
ie P
robe
n in
die
Pro
bent
asch
en e
inge
brac
ht w
orde
n si
nd k
ann,
die
Elek
troph
ores
e ge
star
tet w
erde
n.
Prot
einp
robe
n fü
r di
e SD
S-Pa
ge
Bei
der
ele
ktro
phor
etis
chen
Auf
trenn
ung
von
kom
plex
en P
rote
inge
mis
chen
, w
ie s
ie d
urch
eine
n A
ufsc
hluß
von
Zel
len
oder
von
Ges
amto
rgan
ism
en e
ntst
ehen
, bi
lden
sic
h be
i de
r
Auf
trenn
ung
eine
Vie
lzah
l vo
n B
ande
n, d
ie d
ie g
roße
Men
ge u
nter
schi
edlic
her
Prot
eine
wid
ersp
iege
ln,
die
von
Zelle
n od
er O
rgan
ism
en p
rodu
zier
t w
erde
n. E
ine
groß
e Za
hl v
on
Prot
eine
n is
t in
alle
n Ze
llen
vorh
ande
n, d
a si
e fü
r den
Sto
ffw
echs
el o
der d
ie Z
ells
trukt
ur a
ller
Zelle
n es
sent
iell
sind
. Den
noch
unt
ersc
heid
et si
ch d
er S
atz
von
Prot
eine
n in
nerh
alb
der Z
elle
n
eine
s O
rgan
ism
us
oder
ve
rsch
iede
ner
Entw
ickl
ungs
stad
ien,
da
ve
rsch
iede
ne
Zellt
ypen
unte
rsch
iedl
iche
Fun
ktio
nen
besi
tzen
und
dah
er P
rote
ine
synt
hetis
iere
n, d
ie f
ür d
ie z
u
erfü
llend
e Fu
nktio
n be
nötig
t wer
den.
Die
Dar
stel
lung
der
diff
eren
tiell
expr
imie
rten
Prot
eine
erfo
rder
t ent
wed
er h
ocha
uflö
send
e Tre
nnsy
stem
e (2D
-Ele
ktro
phor
ese)
und
/ode
r ein
e vor
herig
e
Frak
tioni
erun
g.
Unt
er g
ünst
igen
Bed
ingu
ngen
kan
n ei
n un
ters
chie
dlic
her
Prot
einb
esta
nd j
edoc
h au
ch i
n
Ges
amte
xtra
kten
da
rges
tellt
w
erde
n.
Als
B
eisp
iel
eine
r na
chw
eisb
aren
di
ffer
entie
llen
- 42
-

Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Prot
eine
xpre
ssio
n di
enen
für
das
Pra
ktik
um D
otte
rpro
tein
e vo
n D
roso
phila
mel
anog
aste
r.
Dot
terp
rote
ine
wer
den
von
den
Mut
terti
eren
in
die
Ooc
yten
ein
gela
gert
und
als
Nah
rung
s-
liefe
rant
für d
ie E
ntw
ickl
ung
der E
mbr
yone
n ge
nutz
t. D
ie H
aupt
kom
pone
nte
des D
otte
rs w
ird
von
drei
Pol
ypep
tiden
ges
tellt
, die
als
Vite
lloge
nin
1-3
beze
ichn
et w
erde
n. D
ie G
ene,
die
die
se
Prot
eine
kod
iere
n un
d al
s YP
(1-3
) (en
gl. D
otte
r = y
olk)
bez
eich
net w
erde
n, li
egen
auf
dem
X-
Chr
omos
om. D
ie V
itello
geni
ne w
erde
n na
ch p
osttr
ansl
atio
nale
n M
odifi
katio
nen
in d
er O
ocyt
e
eing
elag
ert.
Prob
envo
rber
eitu
ngfü
r di
e SD
S -P
AG
E
Es w
erde
n Ep
pend
orfm
ikro
reak
tions
gefä
ße g
este
llt,
in d
enen
sic
h ad
ulte
Wei
bche
n, a
dulte
Män
nche
n un
d Em
bryo
nen
in je
wei
ls 5
0l P
BS/
2%SD
S in
ein
gefr
oren
em Z
usta
nd b
efin
den.
Mit
eine
m H
omog
enis
atio
nspi
stel
l wer
den
die
Prob
en m
echa
nisc
h au
f Eis
zer
klei
nert,
mit
50l
2xPr
oben
puff
er v
erse
tzt,
für 5
Min
uten
in e
inem
Was
serb
ad (>
95°C
) erh
itzt,
auf E
is a
bgek
ühlt
und
für 5
Min
uten
mit
13.0
00 rp
m in
ein
er T
isch
zent
rifug
e ze
ntrif
ugie
rt. D
ann
wer
den
je 1
0l
für d
en G
ella
uf in
die
Pro
bent
asch
en d
es G
els ü
berf
ührt.
Die
se P
robe
n un
d w
eite
re v
orbe
reite
te
Prob
en m
it Pr
otei
nen
vers
chie
dene
n M
olek
ular
gew
icht
s für
die
Ele
ktro
phor
ese w
erde
n ge
stel
lt.
Lös
unge
nPB
S/2%
SDS
0,13
M N
aCl
2x
Prob
enpu
ffer
0,
125M
Tris
-Cl (
pH 6
,8)
7mM
Na 2
HPO
4
4%SD
S
3m
M N
aH2P
O4
20
% G
lyce
rol (
v/v)
2%
SD
S (w
/v)
0,
2M D
TT
(pH
7,6
)
0,
02%
Bro
mph
enol
Bla
u (w
/v)
- 43
-
Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Ver
wen
dete
Pro
tein
mar
ker:
Rot
iR-M
ark-
Stan
dard
(Fir
ma
Car
l Rot
h)
Prot
ein
MW
(kD
a)
Kon
z.
(mg/
ml)
Myo
sin,
Rin
d 21
2 0,
2
ß-G
alak
tosi
dase
, E.c
oli
118
0,2
Seru
mal
bum
in, R
ind
glyc
. 66
0,
2
Ova
lbum
in, H
uhn
glyc
. 43
0,
27
Car
bona
nhyd
rase
29
0,
13
Tryp
sin
Inhi
bito
r, So
ja
20
0,2
Lyso
zym
, Huh
n 14
,5
0,2
Nac
hdem
alle
Pro
ben
und
der G
röße
nmar
ker i
n di
e Pr
oben
tasc
hen
des
Gel
s pi
petti
ert w
urde
n,
wer
den
die
Elek
trode
n au
f di
e G
ella
ufap
para
tur
gest
eckt
und
in d
er r
icht
igen
Pol
ung
in d
ie
Span
nung
sque
lle g
este
ckt.
Die
Kab
el d
er E
lekt
rode
n un
d di
e St
eckp
lätz
e de
r Spa
nnun
gsqu
elle
sind
far
big
mar
kier
t. D
ie K
abel
müs
sen
in d
ie S
teck
plät
ze m
it de
rsel
ben
Farb
e ei
nges
teck
t
wer
den.
Wäh
rend
der
Wan
deru
ng d
er P
robe
n im
Sam
mel
gel w
ird e
ine
Span
nung
von
80-
90 V
ange
legt
. Nac
h Ü
bertr
itt d
er P
robe
n in
s Tre
nnge
l wird
die
Spa
nnun
g bi
s auf
120
-130
V e
rhöh
t.
Die
Ele
ktro
phor
ese
daue
rt ca
. 2 h
. Wen
n di
e B
rom
phen
olbl
au-F
ront
kur
z vo
r de
m E
nde
des
Tren
ngel
s ste
ht w
ird d
ie E
lekt
roph
ores
e be
ende
t.
Prot
einf
ärbu
ngD
as G
el w
ird a
us d
er G
ella
ufap
para
tur g
enom
men
. Die
Gla
spla
tten
wer
den
mit
eine
m S
pace
r
vors
icht
ig v
onei
nand
er g
elös
t und
die
obe
re P
latte
wird
vor
sich
tig an
geho
ben.
Die
Orie
ntie
rung
des
Gel
s w
ird m
arki
ert,
inde
m e
ine
Ecke
abg
esch
nitte
n w
ird. D
ann
wird
das
Gel
vor
sich
tig
abge
hobe
n un
d in
ein
e Sc
hale
mit
Färb
elös
ung
gele
gt. D
as G
el w
ird a
uf e
inem
Tau
mel
schü
ttler
in e
twa
20-3
0 m
l Fä
rbel
ösun
g in
kubi
ert
bis
die
Ban
den
deut
lich
sich
tbar
sin
d (c
a.10
-20
Min
uten
). D
anac
h w
ird
die
Färb
elös
ung
in
die
dafü
r vo
rges
ehen
en
Sam
mel
behä
lter
abge
goss
en.
Die
Hin
terg
rund
färb
ung
des
Gel
s ka
nn d
urch
Was
chen
mit
Entfä
rbel
ösun
g
redu
zier
t wer
den.
Färb
elös
ung
Rot
i®-B
lue
quic
k E
ntfä
rbel
ösun
g
20%
Eth
anol
- 44
-

Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Aus
wer
tung
Fe
rtige
n Si
e fü
r Ih
r G
el e
ine
Eich
gera
de u
nter
Ver
wen
dung
des
Pro
tein
größ
enm
arke
rs a
n.
Hie
rzu
wer
den
die W
ande
rung
sdis
tanz
en d
er ei
nzel
nen
Grö
ßenm
arke
rban
den
auf d
em G
el n
ach
der
Elek
troph
ores
e au
sgem
esse
n (L
aufr
icht
ung
beac
hten
!!!)
und
anh
and
der
beka
nnte
n
Mol
ekul
arge
wic
hte
wird
ein
e G
erad
e er
stel
lt. H
ierz
u w
erde
n au
f der
Ord
inat
e di
e lo
g 10 W
erte
der
Mol
ekul
arge
wic
hte
der
Grö
ßenm
arke
rpro
tein
e in
Kilo
dalto
n (k
Da)
und
auf
der
Abs
ziss
e
die W
ande
rung
sdis
tanz
en d
er je
wei
ligen
Pro
tein
e auf
getra
gen.
Ans
telle
der
Wan
deru
ngss
treck
e
der
Prot
einb
ande
n ka
nn
auch
de
r R
f-W
ert
gege
n de
n Lo
garit
hmus
(lo
g 10)
de
s
Mol
ekul
arge
wic
hts
aufg
etra
gen
wer
den.
Der
Rf-
Wer
t erm
ittel
t sic
h au
s de
m Q
uotie
nten
der
Lauf
stre
cke
der j
ewei
ligen
Pro
tein
e / L
aufs
treck
e de
r Mar
kerf
ront
. Um
das
Mol
ekul
arge
wic
ht
eine
s Pr
otei
ns z
u be
stim
men
, wird
die
Wan
deru
ngss
treck
e ge
mes
sen
und
der
Wer
t au
f de
r
Ger
aden
(e
rste
llt
aus
den
Lauf
dist
anze
n un
d be
kann
ten
Mol
ekul
arge
wic
hten
de
r
Mar
kerp
rote
ine)
ein
getra
gen.
Das
kor
resp
ondi
eren
de (
log 1
0) M
olek
ular
gew
icht
kan
n da
nn in
der G
rafik
auf
der
Mol
ekul
arge
wic
htsa
chse
abg
eles
en, o
der r
echn
eris
ch e
rmitt
elt w
erde
n.
Ver
glei
chen
Si
e di
e B
ande
nmus
ter
der
Prob
en.
Find
en
sie
Unt
ersc
hied
e im
Prot
einb
ande
nmus
ter
der
Dro
soph
ila-P
robe
n vo
n W
eibc
hen,
Män
nche
n un
d Em
bryo
nen?
Bes
timm
en S
ie d
as M
olek
ular
gew
icht
des
/der
unt
ersc
hied
lich
expr
imie
rten
Prot
eins
/e.
Ver
glei
chen
Sie
die
Ban
denm
uste
r der
Pro
ben,
die
Ihne
n zu
r Ver
fügu
ng g
este
llt w
urde
n un
d
best
imm
en
Sie
die
Mol
ekul
arge
wic
hte
der
Prot
einb
ande
n.
Wie
ve
ränd
ern
sich
di
e
Prot
einb
ande
n, w
enn
unte
rsch
iedl
iche
Pro
tein
men
gen
in ih
nen
vorh
ande
n si
nd?
Auf
tren
nung
von
Ser
umpr
otei
nen
auf C
ellu
lose
acet
atfo
lie
Der
hie
r ver
wen
dete
Lau
fpuf
fer u
nd d
ie m
etha
nolh
altig
en L
ösun
gen
sind
ges
undh
eits
schä
dlic
h
oder
gift
ig, d
aher
Exp
ositi
on v
erm
eide
n un
d ge
eign
ete S
chut
zkle
idun
g un
d H
ands
chuh
e tra
gen.
Die
Fär
bung
und
Ent
färb
ung
wird
unt
er e
inem
Abz
ug d
urch
gefü
hrt.
Dur
chfü
hrun
g de
r Se
rum
auft
renn
ung
Cel
lulo
seac
etat
stre
ifen
wer
den
in L
aufp
uffe
r vo
rinku
bier
t. V
or d
em A
uftra
gen
der
Prob
e
wer
den
die
Cel
lulo
seac
etat
stre
ifen
auf
3MM
-Pap
ier
(Wha
tman
) fü
r 1-
2 M
inut
en a
n de
r Lu
ft
getro
ckne
t. D
ann
wer
den
5-10μl
nat
ives
Ser
um (
Zieg
e) m
it 2μ
l Far
blös
ung
gem
isch
t und
in
eine
m S
treife
n qu
er z
ur L
ängs
richt
ung
in d
er M
itte
des
Cel
lulo
seac
etat
stre
ifens
auf
getra
gen.
Nac
h ku
rzem
Ant
rock
nen
wer
den
die
Cel
lulo
seac
etat
stre
ifen
mit
der g
leic
hen
Orie
ntie
rung
in
die
Lauf
kam
mer
n ge
legt
. Die
End
en d
er C
ellu
lose
acet
atst
reife
n m
üsse
n m
it de
m L
aufp
uffe
r in
Kon
takt
sein
. Die
Ele
ktro
phor
ese
wird
mit
200V
für 6
0-90
Min
uten
dur
chge
führ
t. Zu
m F
ärbe
n
der
Prot
einb
ande
n w
erde
n di
e C
ellu
lose
acet
atst
reife
n fü
r 5
Min
uten
in
Am
idos
chw
arz-
- 45
-
Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Färb
elös
ung
gele
gt u
nd v
orsi
chtig
ges
chüt
telt
(Unt
er A
bzug
arb
eite
n).
Zur
Entfe
rnun
g de
r
Hin
terg
rund
färb
ung
wer
den
die
Cel
lulo
seac
etat
stre
ifen
in E
ntfä
rbe-
Lösu
ng g
eleg
t und
leic
ht
gesc
hütte
lt. D
ie E
ntfä
rbe-
Lösu
ng w
ird m
ehre
re M
ale
gew
echs
elt,
bis
die
Hin
terg
rund
färb
ung
vers
chw
inde
t. D
ie F
ärbe
lösu
ng w
ird w
iede
r ver
wen
det u
nd d
ie b
enut
zte
Entfä
rbe-
Lösu
ng w
ird
in S
amm
elbe
hälte
r für
Lös
emitt
elab
fälle
geg
eben
.
Lau
fpuf
fer
0,
067M
Nat
rium
-Bar
bita
l
0,01
3M B
arbi
tal
pH
8,6
Am
idos
chw
arz-
Färb
elös
ung
10g
Am
idos
chw
arz
50m
l Ess
igsä
ure
400m
l Met
hano
l
400m
l ddH
2O
Ent
färb
e-L
ösun
g
50
ml E
ssig
säur
e
600m
l Met
hano
l
200m
l ddH
2O
Farb
lösu
ng
0,
25%
w/v
Bro
mph
enol
blau
0,
25%
w/v
Xyl
enec
yano
l FF
30%
v/v
Gly
cerin
In w
ievi
ele
Ban
den
wird
das
Ser
um a
ufge
trenn
t? W
ie is
t die
Tre
nnsc
härf
e de
r Ban
den?
Ver
glei
chen
Sie
die
Erg
ebni
sse
(z.B
. Zah
l der
Pro
tein
band
en) m
it de
n La
ufsp
uren
, in
dene
n
Seru
m in
der
SD
S-PA
GE
aufg
etre
nnt w
urde
.
Auf
tren
nung
und
Nac
hwei
s von
Glu
cose
-6-P
hosp
hat-
Deh
ydro
gena
se
Als
B
eisp
iel
für
eine
Pr
otei
ntre
nnun
g m
it A
garo
sege
len
wer
den
wir
im
Kur
s ei
nen
Prot
eine
xtra
kt o
der
Enzy
mlö
sung
en a
uftre
nnen
und
die
Enz
ymak
tivitä
t de
r G
luco
se-6
-
Phos
phat
Deh
ydro
gena
se (G
6PD
) mit
eine
r Far
brea
ktio
n na
chw
eise
n. D
ie A
uftre
nnun
g er
folg
t
hier
in
ei
nem
A
garo
sege
l (F
orbe
s et
.al.
1991
). D
as
Ziel
di
eser
A
uftre
nnun
g is
t di
e
Unt
ersc
heid
ung
von
Var
iant
en d
er G
luco
se-6
-Pho
spha
t Deh
ydro
gena
se, d
ie si
ch u
nter
nat
iven
Bed
ingu
ngen
in
der
elek
troph
oret
isch
en M
obili
tät
unte
rsch
eide
n. D
a da
s A
nfer
tigen
von
Aga
rose
gele
n w
enig
er a
ufw
endi
g is
t al
s vo
n A
cryl
amid
gele
n, k
ann
bei
ausr
eich
end
groß
en
Mob
ilitä
tsun
ters
chie
den
auch
in A
garo
sege
len
eine
Aus
sage
übe
r Pro
tein
mer
kmal
e an
hand
der
Lauf
dist
anzu
nter
schi
ede
gem
acht
wer
den.
Die
(na
tive)
Pro
tein
prob
e w
ird f
ür d
en V
ersu
ch z
ur V
erfü
gung
ges
tellt
. D
ie G
ele
wer
den
vorb
erei
tet
und
müs
sen
vor
dem
A
uftra
gen
der
Prob
e 5
Min
uten
m
it 10
0V
in
der
Elek
troph
ores
ekam
mer
äqu
ilibr
iert
wer
den.
Dan
n w
erde
n je
5μl
der
Pro
ben
mit
2μl F
arbl
ösun
g
- 46
-

Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
(s. o
.) in
die
Gel
tasc
hen
pipe
ttier
t. D
ie G
elta
sche
n w
erde
n au
f der
Sei
te d
er K
atho
de p
latz
iert
und
die
Elek
troph
ores
e w
ird g
esta
rtet.
Nac
h de
m G
ella
uf w
ird d
as G
el a
uf e
in m
it Fä
rbel
ösun
g
geträ
nkte
s Fi
lterp
apie
r ge
legt
und
bei
37°
C i
n ei
ner
feuc
hten
Kam
mer
ink
ubie
rt, b
is d
ie
Farb
reak
tion
sich
tbar
wird
.
Lau
fpuf
fer:
13,6
mM
Tris
pH
9,1
15
mM
Bor
säur
e
4m
M E
DTA
Gel
:
0,
9% A
garo
se in
Lau
fpuf
fer
0,13
mM
NA
DP
G6P
D-F
ärbe
lösu
ng:
4mM
MgC
l 2
10
mM
Glu
kose
-6-P
hosp
hat
0,19
6mM
Phe
nazi
n m
etho
sulfa
t (PM
S)
12m
M 3
-(4,
5-di
-met
hylth
iazo
l-2-y
l)-2,
5-di
phen
ylte
trazo
lium
bro
mid
Aus
wer
tung
:
Ver
glei
chen
Sie
die
Lau
fspu
ren
mit
den
vers
chie
dene
n Pr
oben
nac
h de
r Fär
bung
. Ist
ein
Unt
ersc
hied
zw
isch
en d
en L
aufw
eite
n de
r jew
eilig
en P
robe
n zu
erk
enne
n?
Lite
ratu
r-B
artle
tt R
.C. R
apid
Cel
lulo
se A
ceta
t Ele
ctro
phor
esis
. I. S
erum
Pro
tein
s. C
linic
al C
hem
istry
V
ol.9
No3
:317
-324
(196
3)
-Bre
nnan
M.D
., W
eine
r A.J.
, Gor
alsk
i T.J.
, Mah
owal
d A
.P.,
The
folli
cle
cells
are
a m
ajor
si
te o
f vite
lloge
nin
synt
hesi
s in
Dro
soph
ila m
elan
ogas
ter.
Dev
. Bio
l. 89
(198
2) 2
25-2
36
- For
bes J
., St
eytle
r J.G
, van
Hee
rden
R. A
garo
se g
el e
lect
roph
ores
is o
f glu
cose
-6-p
hosp
hate
-de
hydr
ogen
ase
isoe
nzym
es. C
linic
a C
him
ica
Act
a 19
9:27
9-28
2 (1
991)
-K
uhn
R. &
Hof
fste
tter-
Kuh
n S.
Cap
illar
y El
ectro
phor
esis
: Prin
cipl
es a
nd P
ract
ice.
Spr
inge
r V
erla
g B
erlin
Hei
delb
erg
(199
3)
-Lae
mm
li U
.K. C
leav
age
of st
ruct
ural
pro
tein
s dur
ing
asse
mbl
y of
the
head
of B
acte
rioph
age
T4. N
atur
e 22
7:68
0-68
5 (1
970)
-L
otts
peic
h F.
& Z
orba
s H. B
ioan
alyt
ik. S
pekt
rum
Aka
dem
isch
er V
erla
g H
eide
lber
g (1
998)
-R
ickw
ood
D.,
Ham
esB
. D. H
rsg.
Gel
Ele
ctro
phor
esis
of P
rote
ins.
Prac
tical
App
roac
h Se
ries,
Oxf
ord
Uni
vers
ity P
ress
Oxf
ord
(199
2)
-Rus
sell
J. H
. Mill
ipor
e te
chni
ques
in p
hysi
olog
y an
d bi
oche
mis
try. E
lect
roph
oret
ic
tech
niqu
es. I
n K
erku
t G.A
. (Ed
.) Ex
perim
ents
in p
hysi
olog
y an
d bi
oche
mis
try V
ol. 6
A
cade
mic
Pre
ss L
ondo
n an
d N
ew Y
ork
(197
3)
-Stry
er L
. Bio
chem
ie. S
pekt
rum
Aka
dem
isch
er V
erla
g H
eide
lber
g (1
990)
-W
este
rmei
er R
. Ele
ctro
phor
esis
in P
ract
ice,
A G
uide
to M
etho
ds a
nd A
pplic
atio
ns o
f DN
A
and
Prot
ein
Sepa
ratio
ns. F
ourth
, rev
ised
and
enl
arge
d Ed
ition
. Wile
y-V
CH
Ver
lag
Wei
nhei
m (2
005)
- 47
-
Woc
he 4
Prot
ein-
Ele
ktro
phor
ese
AG
Klä
mbt
/Lam
mel
Frag
en z
ur V
orbe
reitu
ng
Wel
cher
Fak
tor h
at k
eine
n Ei
nflu
ß au
f die
Wan
deru
ngsg
esch
win
digk
eit v
on T
eilc
hen
in e
inem
el
ektri
sche
n Fe
ld?
V
isko
sitä
t des
Med
ium
s
Ladu
ng d
es T
eilc
hens
Form
des
Tei
lche
ns
O
rient
ieru
ng d
es e
lekt
risch
en F
elde
s
Wel
che
Kom
pone
nten
bes
timm
en d
ie P
oren
göße
von
Acr
ylam
id-G
elen
?
A
cryl
amid
TEM
ED
A
mm
oniu
mpe
roxo
disu
lfat
N
, N´-
Met
hyle
nbis
acry
lam
id
SD
S
Wel
che
Kom
pone
nte
ist f
ür d
ie Q
uerv
erne
tzun
g in
Acr
ylam
id-G
elen
not
wen
dig?
A
mm
oniu
mpe
roxo
disu
lfat
N
, N´-
Met
hyle
nbis
acry
lam
id
TEM
ED
Acr
ylam
id
SD
S
Wel
che
Kom
pone
nten
wer
den
als
Rad
ikal
star
ter
bei
der
Erze
ugun
g vo
n A
cryl
amid
-Gel
en
verw
ende
t?
N
, N´-
Met
hyle
nbis
acry
lam
id
A
cryl
amid
SDS
Tr
is
Am
mon
ium
pero
xodi
sulfa
t
Wel
che
Bez
iehu
ng b
este
ht z
wis
chen
der
Lau
fdis
tanz
von
Pro
tein
en b
ei e
iner
SD
S Pa
ge u
nd
ihre
m M
olek
ular
gew
icht
?
War
um si
nd S
DS
und
DTT
im P
robe
npuf
fer e
ntha
lten?
Wel
che
Mat
rices
wer
den
für d
ie v
ersc
hied
enen
Ele
ktro
phor
esen
ver
wen
det?
Bei
wel
chen
Ele
ktro
phor
esen
sind
die
auf
getre
nnte
n Pr
otei
ne in
nat
ivem
Zus
tand
?
- 48
-

Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
1. A
llgem
eine
Bem
erku
ngen
Res
piro
met
risch
e Mes
sung
en w
erde
n in
der
Tie
rphy
siol
ogie
eing
eset
zt, u
m In
form
atio
nen
über
den
phys
iolo
gisc
hen
Stat
us e
ines
Tie
res
zu g
ewin
nen.
Oftm
als
liefe
rn s
olch
e M
essu
ngen
auc
h
wic
htig
e H
inw
eise
üb
er
die
Anp
assu
ngsf
ähig
keit
von
Tier
en
an
verä
nder
te
Um
wel
tbed
ingu
ngen
. Je
nac
hdem
, ob
es
sich
um
Luf
tatm
er o
der
Was
sera
tmer
han
delt,
kom
men
unt
ersc
hied
liche
Mes
sprin
zipi
en u
nd B
erec
hnun
gsw
ege
zur
Anw
endu
ng. I
n di
esem
Kur
sabs
chni
tt so
ll m
it H
ilfe
resp
irom
etris
cher
Met
hode
n di
e Te
mpe
ratu
rabh
ängi
gkei
t de
r
Stof
fwec
hsel
rate
ein
es h
omöo
ther
men
Luf
tatm
ers
und
eine
s po
ikilo
ther
men
Was
sera
tmer
s
unte
rsuc
ht w
erde
n.
2. M
etho
disc
he G
rund
lage
n 2.
1.
Mes
sung
de
r O
2-V
erbr
auch
srat
e ei
nes
Luf
tatm
ers
in
eine
m
gesc
hlos
sene
n
Res
piro
met
er
Prin
zipi
ell
könn
en
Gas
wec
hsel
mes
sung
en
in
eine
m
gesc
hlos
sene
n Sy
stem
un
ter
Kon
stan
thal
tung
des
Dru
cks
oder
unt
er K
onst
anth
altu
ng d
es V
olum
ens
durc
hgef
ührt
wer
den.
Im K
ursv
ersu
ch s
oll
der
O2-
Ver
brau
ch e
ines
Luf
tatm
ers
bei
kons
tant
em D
ruck
bes
timm
t
wer
den.
Hie
rzu
wird
das
in A
bb. 1
dar
gest
ellte
Res
piro
met
er v
erw
ende
t. N
ach
Auf
setz
en d
es
Kam
mer
deck
els
und
Ver
schl
ieße
n de
s D
reiw
egeh
ahns
kom
mt
es v
orüb
erge
hend
zu
eine
m
Dru
ckab
fall
in d
er K
amm
er, d
a de
r Sa
uers
toff
ver
atm
et u
nd d
as a
bgeg
eben
e K
ohle
ndio
xid
durc
h K
OH
geb
unde
n w
ird.
Nac
h Er
reic
hen
eine
r be
stim
mte
n D
ruck
diff
eren
z w
ird d
as
Kam
mer
volu
men
übe
r ein
e Sp
ritze
ver
ringe
rt un
d di
e D
ruck
diff
eren
z au
f Nul
l zur
ückg
efüh
rt.
Das
"ve
rbra
ucht
e" S
pritz
envo
lum
en V
(ml)
ents
pric
ht d
er M
enge
an
verb
rauc
htem
Sau
erst
off.
Die
gew
onne
nen
Mes
swer
te m
üsse
n nu
n no
ch a
uf S
tand
ardb
edin
gung
en (
STPD
: st
anda
rd
tem
pera
ture
and
pres
sure
, dry
air)
um
gere
chne
t wer
den.
Nac
h de
m a
llgem
eine
n G
asge
setz
gilt
:
0
0
0
PV
PV
TT
(1
)
wob
ei P
der
Dru
ck (
kPa)
, V
das
verb
rauc
hte
O2-
Vol
umen
(m
l) un
d T
die
abso
lute
Mes
stem
pera
tur
T (in
Kel
vin!
) is
t. P 0
, V0
und
T 0 s
ind
fest
e G
röße
n un
d be
zieh
en s
ich
auf
Stan
dard
bedi
ngun
gen
(sie
he T
ab. 1
). D
er D
ruck
P m
uss
aus
der D
iffer
enz
von
Luftd
ruck
PL
(mB
ar)
und
Was
serd
ampf
druc
k P W
(kP
a) b
erec
hnet
wer
den
(P=P
L–P W
; Ei
nhei
ten
vorh
er
umre
chne
n!).
Der
Was
serd
ampf
druc
k P W
ent
spric
ht d
em P
artia
ldru
ck d
es W
asse
rs, d
er si
ch b
ei
100
%ig
er L
uftfe
ucht
e ei
nste
llt. E
r ist
abh
ängi
g vo
n de
r Ver
such
stem
pera
tur u
nd k
ann
aus T
ab.
3 en
tnom
men
wer
den.
Nac
h U
mst
ellu
ng v
on G
l. (1
) ka
nn d
as v
erbr
auch
te O
2-V
olum
en
(kor
rigie
rt au
f STP
D) w
ie fo
lgt b
erec
hnet
wer
den:
- 49
-
Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
0
00
PV
TV
PT
(2
)
Die
Sau
erst
offv
erbr
auch
srat
e V
(ml h
–1) e
rgib
t sic
h na
ch D
ivis
ion
durc
h di
e M
essd
auer
t (
h):
t
VV
0
(3)
Um
die
O2-
Ver
brau
chsr
ate
vers
chie
den
groß
er T
iere
ver
glei
chen
zu
könn
en, w
ird s
chlie
ßlic
h
noch
ein
e N
orm
ieru
ng a
uf G
ram
m K
örpe
rmas
se v
orge
nom
men
. Die
se so
gen
annt
e sp
ezifi
sche
O2-
Ver
brau
chsr
ate
spez
V (m
l h–1
g–1
) erh
ält m
an n
ach:
mV
Vsp
ez
(4)
Tab.
1. P
aram
eter
zur
Ber
echn
ung
der O
2-V
erbr
auch
srat
e ei
nes L
ufta
tmer
s
Sy
mbo
l W
ert/E
inhe
it B
esch
reib
ung
Bem
erku
ngen
m
g
Kör
perm
asse
des
Tie
res
P kP
a D
ruck
unt
er V
ersu
chsb
edin
gung
en
P 0
101.
3 kP
a St
anda
rddr
uck
unte
r Ver
such
sbed
ingu
ngen
P L
m
bar
Luftd
ruck
10
mba
r = 1
kPa
P W
kP
a W
asse
rdam
pfdr
uck
sieh
e Ta
b. 3
T
K
Ver
such
stem
pera
tur
0 °C
= 2
73.1
5 K
T 0
27
3.15
K
Stan
dard
tem
pera
tur
V m
l ve
rbra
ucht
es O
2-V
olum
en (g
emes
sen)
V 0
m
l ve
rbra
ucht
es O
2-V
olum
en (k
orrig
iert
auf S
TPD
) V
m
l h–1
O
2-V
erbr
auch
srat
e sp
ezV
m
l h–1
g–1
sp
ezifi
sche
O2-
Ver
brau
chsr
ate
t h
Mes
sdau
er
- 50
-

Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
Abb.
1.��
��e�
�t�s
��e�
���
ste�
����
�e��
es��
es��
��ss
e�e�
��es
p���
�et
e�s�z
����
est��
���
���e
s����
Ve��
����
�s��
����
��t�
t�e�
����
���
��e�
��es
��e�
��es
��e�
���e
t�s��
��e�
�e��
��ss
����
e��
�t�e�
�e�
��
���
��s�
��e�
s���
��
���s
����
����s
�T�e
�����
����
���e�
�pe�
����
e�te
����
�s��
e���
�e��
�ese
tzt��
���
e��e
���
��t�
�t��
�t��e
���t
ze��
e���
��
���s
����
z���
e��
e��e
����
����
��st
��t�
��t�
����
e��
Te�
pe��
t����
� ���
��s��
ess�
e���
����e
��e�
�te�
pe��
e���
�e��
��s
se��
����
e�se
��t��
2.2.
M
essu
ng
der
O2-
Ver
brau
chsr
ate
eine
s W
asse
ratm
ers
in
eine
m
gesc
hlos
sene
n
Res
piro
met
er
Die
Sau
erst
offv
erbr
auch
srat
e ei
nes
Was
sera
tmer
s ka
nn in
ein
em g
esch
loss
enen
�es
piro
met
er
mitt
els e
iner
Sau
erst
offe
lekt
rode
gem
esse
n �
erde
n. D
as M
essp
rinzi
p be
ruht
auf
der
�ed
uktio
n
von
Saue
rsto
ff a
n de
r (ne
gativ
gel
aden
en) P
latin
-Kat
hode
�
2
22
2O
2�O
2e�
O2O
�.
�us
dem
geb
ildet
en �
2O2 e
ntst
ehen
nac
h �
ufna
hme
von
z�ei
�ei
tere
n �l
ektro
nen
z�ei
O�
– -
�one
n. �
n de
r �ef
eren
z-Si
lber
-�no
de �
ird g
leic
hzei
tig S
ilber
unt
er �
bgab
e ei
nes �
lekt
rons
zu
�g�
o�id
iert.
D
ie
�ed
uktio
n vo
n Sa
uers
toff
f�
hrt
zu
eine
m
Stro
mflu
ss�
�el
cher
de
r
Saue
rsto
ffko
nzen
tratio
n in
der
�es
piro
met
erka
mm
er p
ropo
rtion
al is
t.
Vor
�ede
r Mes
sung
ist d
ie S
auer
stof
fele
ktro
de m
it gu
t bel
�fte
tem
Was
ser (
100
� L
ufts
�ttig
ung)
und
O2-
frei
em W
asse
r (0
� L
ufts
�ttig
ung)
zu
kalib
riere
n. �
ach
�ins
etze
n de
s Ti
eres
in
bel�
ftete
m W
asse
r kom
mt e
s au
fgru
nd d
er �
tmun
gsak
tivit�
t zu
eine
m �
bfal
l des
O2-
�eh
alts
in d
er �
espi
rom
eter
kam
mer
. �s �
ird d
ie �
eitd
auer
t (
h) b
is z
um �
rrei
chen
ein
er b
estim
mte
n
- 5�
-
Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
Lufts
�ttig
ungs
diff
eren
z �
(�)
gem
esse
n.
�us
di
esen
be
iden
�
r��e
n ka
nn
die
Saue
rsto
ffve
rbra
uchs
rate
des
Tie
res
� (n
mol
h–1
) �ie
folg
t ber
echn
et �
erde
n�
t
��
��
100
(5
)
Die
�
r��e
�
(n
mol
) st
ellt
die
in
bel�
ftete
m
Was
ser
(100
�
Lufts
�ttig
ung)
ge
l�st
e
Saue
rsto
ffm
enge
da
r. Si
e �
ird
aus
dem
�l
�ssi
gkei
tsvo
lum
en
V (l)
� de
m
L�sl
ichk
eits
koef
fizie
nten
von
O2
in W
asse
r (�
mol
l–1 k
Pa–1
) un
d de
m O
2-Pa
rtial
druc
k P O
2
(kPa
) err
echn
et�
2
O�
VP
. (�
)
Der
O
2-Pa
rtial
druc
k er
gibt
si
ch
aus
der
Diff
eren
z vo
n Lu
ftdru
ck
P L
(m�
ar)
und
Was
serd
ampf
druc
k P W
(P=P
L–P W
� �in
heite
n vo
rher
um
rech
nen�
)� �
obei
der
�ak
tor 0
.20�
den
frak
tione
llen
�nt
eil v
on O
2 in
der L
uft �
ider
spie
gelt�
W
LO
20�
.02
PP
P.
(7)
Tab.
2. P
aram
eter
zur
�er
echn
ung
des O
2-V
erbr
auch
s ein
es W
asse
ratm
ers
S�m
bol
�
inhe
it �
esch
reib
ung
�em
erku
ngen
�
nm
ol
Saue
rsto
ffm
enge
�
nm
ol h
–1
O2-
Ver
brau
chsr
ate
2OP
kP
a Sa
uers
toff
parti
aldr
uck
P L
mba
r Lu
ftdru
ck
10 m
bar =
1 k
Pa
P W
kPa
Was
serd
ampf
druc
k si
ehe
Tab.
3
V l
�l�s
sigk
eits
volu
men
si
ehe
Ver
such
sanl
eitu
ng
�m
ol l–1
kPa
–1
L�sl
ichk
eits
koef
fizie
nt v
on O
2 in
Was
ser
sieh
e Ta
b. 3
�
�
Lufts
�ttig
ungs
diff
eren
z t
h M
essd
auer
3.
U
nter
such
unge
n zu
r T
empe
ratu
rabh
ängi
gkei
t de
r St
offw
echs
elra
te
eine
s
hom
öoth
erm
en L
ufta
tmer
s und
ein
es p
oiki
loth
erm
en W
asse
ratm
ers
Die
Tem
pera
tur b
eein
fluss
t ein
e V
ielz
ahl v
on p
h�si
olog
isch
en P
roze
ssen
. �in
e �r
h�hu
ng d
er
Tem
pera
tur f
�hrt
dabe
i mei
st z
u ei
ner �
esch
leun
igun
g di
eser
Vor
g�ng
e. S
o st
eigt
zum
�ei
spie
l
die
Sto
ff�
echs
elra
te e
ines
Tie
res
bei e
iner
�rh
�hun
g de
r K�r
perte
mpe
ratu
r um
10
°C in
der
�eg
el
um
das
2–3-
fach
e.
Ver
�nde
rung
en
der
�m
gebu
ngst
empe
ratu
r k�
nnen
�e
doch
unte
rsch
iedl
iche
�
us�
irkun
gen
auf
K�r
perte
mpe
ratu
r un
d St
off�
echs
elra
te
habe
n.
�ei
- 5�
-

Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
p���
���t�
e��
e�
Tier
en
pass
t si
ch
die
K�r
perte
mpe
ratu
r m
ehr
oder
�
enig
er
der
�m
gebu
ngst
empe
ratu
r an
. St
eige
nde
�m
gebu
ngst
empe
ratu
ren
f�hr
en s
omit
auch
zu
eine
r
�una
hme
der
Stof
f�ec
hsel
rate
. �m
�eg
ensa
tz d
azu
sind
��
���
t�e��
e Ti
ere
(V�g
el u
nd
S�ug
er)
bef�
higt
� ih
re
K�r
perte
mpe
ratu
r in
nerh
alb
eine
s be
stim
mte
n �
erei
ches
de
r
�m
gebu
ngst
empe
ratu
r ko
nsta
nt z
u ha
lten
(The
rmor
egul
atio
n).
Die
Sto
ff�
echs
elra
te v
on
hom
�oth
erm
en
Tier
en
zeig
t da
bei
eine
ga
nz
ande
re
�bh
�ngi
gkei
t vo
n de
r
�m
gebu
ngst
empe
ratu
r. �n
nerh
alb
eine
r so
gena
nnte
n th
erm
isch
neu
trale
n �o
ne� e
inem
�en
ige
�ra
d br
eite
n �
erei
ch d
er �
mge
bung
stem
pera
tur�
ist i
hre
Stof
f�ec
hsel
rate
min
imal
. Obe
rhal
b
und
unte
rhal
b de
r th
erm
isch
ne
utra
len
�one
ni
mm
t di
e St
off�
echs
elin
tens
it�t
zu�
da
W�r
mea
bgab
emec
hani
smen
(S
ch�
itzen
� �
eche
ln)
bz�
. W
�rm
epro
dukt
ions
mec
hani
smen
(Mus
kelz
itter
n� z
itter
frei
e Th
erm
ogen
ese)
akt
ivie
rt �
erde
n. D
ie S
toff
�ec
hsel
rate
ein
es T
iere
s
kann
indi
rekt
dur
ch M
essu
ng d
es S
auer
stof
fver
brau
chs (
�es
piro
met
rie) b
estim
mt �
erde
n.
Ver
such
I:
�es
timm
en S
ie d
ie s
pezi
fisch
e Sa
uers
toff
verb
rauc
hsra
te
spez
V (
ml h
–1 g
–1)
eine
r
Mau
s bei
�m
gebu
ngst
empe
ratu
ren
von
5 °C
� �au
mte
mpe
ratu
r und
30
°C.
Tier
e �
Mat
eria
l�
Mau
s (�
�s��
�s��
��s)
� �es
piro
met
erge
f��
mit
Dru
ckse
nsor
und
Com
pute
r� 20
�-ig
e K
O�
-
L�su
ng� t
empe
rierte
Was
serb
ecke
n (5
°C� �
aum
tem
pera
tur�
30 °C
)
Dur
chf�
hrun
g un
d �
us�
ertu
ng�
�un�
chst
mus
s ein
e �i
chun
g de
s Dru
ckse
nsor
s f�r
�ede
Mes
stem
pera
tur d
urch
gef�
hrt �
erde
n.
Daz
u �
ird d
as �
ef��
(si
ehe
�bb
. 1)
vers
chlo
ssen
und
der
Dre
i�eg
ehah
n so
ein
gest
ellt�
das
s
eine
Ver
bind
ung
z�is
chen
Kam
mer
und
Spr
itze�
nic
ht a
ber z
ur �
u�en
luft
best
eht.
Dan
n �
ird
stuf
en�
eise
der
�nt
erdr
uck
erh�
ht�
inde
m d
er S
tem
pel
der
Sprit
ze i
n Sc
hritt
en v
on 2
ml
aufg
ezog
en �
ird. D
ie er
halte
nen
Mes
s�er
te �
erde
n no
tiert
und
es �
ird ei
ne �
ichg
erad
e ers
tellt
.
�ur
Mes
sung
de
s Sa
uers
toff
verb
rauc
hs
das
�es
piro
met
erge
f��
mit
KO
�-g
etr�
nkte
m
�ilte
rpap
ier
(CO
2-�
bsor
bens
) be
f�lle
n un
d Tr
ennb
oden
ein
setz
en� d
amit
das
Tier
nic
ht m
it
KO
� (s
tark
�tz
end�
) in
Ver
bind
ung
kom
mt.
�ns
chlie
�end
kan
n da
s Ti
er in
die
Mes
skam
mer
eing
eset
zt u
nd d
er D
ecke
l der
�es
piro
met
erka
mm
er h
alb
vers
chlo
ssen
�er
den.
Ste
mpe
l der
�n�e
ktio
nssp
ritze
auf
die
10
ml-M
arke
brin
gen.
Der
Dre
i�eg
ehah
n is
t so
einz
uste
llen�
das
s ein
e
Ver
bind
ung
zur
�u�
enlu
ft er
halte
n bl
eibt
. M
essk
amm
er 1
0 m
in t
empe
riere
n. �
ach
dies
er
Vor
bere
itung
spha
se d
en D
rei�
egeh
ahn
so v
erst
elle
n� d
ass
die
Ver
bind
ung
zur
�u�
enlu
ft
- 5�
-
Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
unte
rbro
chen
�ird
� �e
doch
die
Ver
bind
ung
z�is
chen
�n�
ektio
nssp
ritze
und
Mes
skam
mer
erha
lten
blei
bt. S
topp
uhr
star
ten�
Das
Tie
r ve
rbra
ucht
Sau
erst
off
und
gibt
Koh
lend
io�i
d ab
�
�el
ches
dur
ch K
O�
abs
orbi
ert
�ird
. �n
folg
edes
sen
ents
teht
im
�es
piro
met
erge
f��
ein
�nt
erdr
uck�
der
ein
Ma�
f�r d
ie M
enge
des
ver
atm
eten
Sau
erst
offs
ist.
Der
in d
er M
essk
amm
er
sich
aus
bild
ende
�nt
erdr
uck
�ird
mit
�ilf
e de
s Dru
ckm
essg
er�t
es �
ber�
acht
.
�m V
ersu
ch �
ird d
ie �
eitd
auer
gem
esse
n� d
ie f
�r d
as �
rrei
chen
ein
es �
nter
druc
kes
ben�
tigt
�ird
� der
ein
em �
asvo
lum
en v
on 5
ml e
ntsp
richt
. �st
die
ser W
ert e
rrei
cht�
�ird
�be
r die
Spr
itze
das
Vol
umen
ve
rkle
iner
t (�
kom
prim
iert�
)� bi
s de
r �
nter
druc
k be
seiti
gt
ist.
��r
�ede
�m
gebu
ngst
empe
ratu
r si
nd M
essz
eit
und
verb
rauc
htes
Spr
itzen
volu
men
zu
prot
okol
liere
n
(min
dest
ens
3 M
essu
ngen
). D
as K
�rpe
rge�
icht
des
Tie
res
�ird
am
�nd
e de
s V
ersu
ches
best
imm
t. �
us d
en M
essd
aten
�ird
die
spez
ifisc
he S
auer
stof
fver
brau
chsr
ate
spez
V (m
l h–1
g–1
)
f�r �
ede
�m
gebu
ngst
empe
ratu
r ber
echn
et u
nd in
ein
em D
iagr
amm
dar
gest
ellt.
�us
den
Dat
en
alle
r �ru
ppen
�er
den
f�r �
ede
Ver
such
stem
pera
tur m
ittle
re V
erbr
auch
srat
en b
erec
hnet
und
es
�ird
mit
�ilf
e de
s �-
Test
s un
d t-T
ests
gep
r�ft�
ob
sich
die
mitt
lere
n V
erbr
auch
srat
en b
ei d
en
vers
chie
dene
n Te
mpe
ratu
ren
stat
istis
ch u
nter
sche
iden
.
Ver
such
II:
�es
timm
en S
ie d
ie S
auer
stof
fver
brau
chsr
ate
� (
nmol
h–1
) be
im g
ro�e
n
Was
serf
loh
bei 1
5 °C
und
25
°C.
Tier
e �
Mat
eria
l�
Was
serf
loh
(��p
����
��
����
� C
rust
acea
)� �
espi
rom
eter
gef�
� m
it O
2-�l
ektro
de
und
�eg
istri
erge
r�t
(Sch
reib
er)�
��h
rfis
ch�
Mag
netr�
hrer
� Th
erm
osta
t� be
l�fte
tes
Med
ium
�
�at
rium
dith
ioni
t (�
a 2S 2
O�)
� Pla
stik
pipe
tten�
Kam
mer
volu
men
auf
1.5
ml e
inst
elle
n
Dur
chf�
hrun
g un
d �
us�
ertu
ng�
Die
O2-
�lek
trode
ist �
e���e
�e��
�es
ste�
pe��
t��
mit
tem
perie
rtem
� bel
�fte
tem
Was
ser
(100
�
Lufts
�ttig
ung)
und
mit
Was
ser
unte
r �u
satz
von
ein
igen
Kris
talle
n �
atriu
mdi
thio
nit
(0�
Lufts
�ttig
ung)
zu
kalib
riere
n. �
atriu
mdi
thio
nit e
ntfe
rnt d
en S
auer
stof
f im
Was
ser �
ie fo
lgt�
2
2�
22
�3
�a
SO
O�
O�
a�SO
�a�
SO.
Die
Kal
ibra
tion
ist a
uf d
em S
chre
iber
papi
er z
u no
tiere
n. �
ach
meh
rmal
igem
Sp�
len
�ird
die
Kam
mer
mit
bel�
ftete
m W
asse
r be
f�llt
� de
r W
asse
rflo
h ei
nges
etzt
und
die
Kam
mer
so
vers
chlo
ssen
� das
s ke
ine
Luftb
lase
n vo
rhan
den
sind
. Die
�bn
ahm
e im
O2-
�eh
alt �
ird d
urch
- 5�
-

Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
den
Schr
eibe
r auf
geze
ichn
et. �
ei c
a. 7
0 �
Luf
ts�t
tigun
g �
ird d
ie T
ierm
essu
ng b
eend
et. �
alls
eine
�bn
ahm
e au
f 70
� n
icht
inne
rhal
b vo
n 30
min
m�g
lich
ist�
�ird
nac
h �
bspr
ache
mit
dem
Kur
slei
ter
die
Mes
sung
nac
h 30
min
bee
ndet
. �ac
h �n
tnah
me
des
Tier
es u
nd �
uff�
llen
des
Kam
mer
med
ium
s �
ird
die
Kam
mer
er
neut
ve
rsch
loss
en
und
eine
Le
er�
ertm
essu
ng
durc
hgef
�hrt�
um
sp�t
er d
en �
igen
verb
rauc
h de
r O2-
�lek
trode
und
der
�ak
terie
n zu
erm
ittel
n.
�us
dem
zei
tlich
en �
bfal
l de
s Sa
uers
toff
geha
ltes
in d
er K
amm
er l
asse
n si
ch a
nhan
d de
r
�le
ichu
ngen
5–7
die
O2-
Ver
brau
chsr
aten
f�r
die
Tie
r- u
nd d
ie L
eer�
ertm
essu
ng b
erec
hnen
.
Die
O2-
Ver
brau
chsr
ate
des
Tier
es �
(nm
ol h
–1)
erh�
lt m
an� i
ndem
man
den
Wer
t au
s de
r
Leer
�er
tmes
sung
von
dem
der
Tie
rmes
sung
sub
trahi
ert.
�us
den
Dat
en a
ller
�ru
ppen
des
Kur
ses
�er
den
f�r d
ie b
eide
n V
ersu
chst
empe
ratu
ren
die
mitt
lere
n V
erbr
auch
srat
en b
erec
hnet
.
Pr�f
en S
ie m
it �
ilfe
des
�-Te
sts
und
t-Tes
ts� o
b si
ch d
ie m
ittle
ren
Ver
brau
chsr
aten
bei
den
beid
en V
ersu
chst
empe
ratu
ren
stat
istis
ch u
nter
sche
iden
.
4. L
itera
tur
Bau
er, H
. C.,
Hof
er, R
., K
napp
, W. u
nd M
oser
, H. (
1�7�
). �o
olog
isch
e ��
perim
ente
.
M�n
chen
� Deu
tsch
er T
asch
enbu
chve
rlag.
Eck
ert,
R. (
2000
). Ti
erph
�sio
logi
e. S
tuttg
art�
Thie
me.
Eck
ert,
M. u
nd H
erte
l, W
. (1�
�3).
Prak
tikum
der
Tie
rph�
siol
ogie
. �en
a� S
tuttg
art�
�us
tav
�isc
her V
erla
g.
Em
mer
ich,
H. (
1��0
). St
off�
echs
elph
�sio
logi
sche
s Pra
ktik
um. S
tuttg
art�
�e�
�or
k� �
eorg
Thie
me
Ver
lag.
Nac
htig
all,
W. (
1�72
). �o
oph�
siol
ogis
cher
�ru
ndku
rs. �
rbei
tsb�
cher
�io
logi
e� T
asch
ente
�t
�. W
einh
eim
��er
gstr.
� Ver
lag
Che
mie
.
5. F
rage
n un
d A
ufga
ben
zur
theo
retis
chen
Vor
bere
itung
Was
bes
agt d
ie a
llgem
eine
�as
glei
chun
g�
Was
ver
steh
t man
unt
er d
em P
artia
ldru
ck e
ines
�as
es�
Was
ver
steh
t man
unt
er d
em L
�slic
hkei
ts- o
. �bs
orpt
ions
koef
fizie
nten
ein
es �
ases
�
�rkl
�ren
Sie
die
ph�
siol
ogis
chen
�eg
riffe
hom
�oth
erm
� poi
kilo
ther
m� h
eter
othe
rm� e
ndot
herm
�
ekto
ther
m�
Was
ver
steh
t man
unt
er z
itter
frei
er T
herm
ogen
ese�
- 55
-
Woc
he 5
Res
piro
met
rie
AG
Pau
l/Boi
ani
6. A
nhan
g
T���
����
�sse
����
p���
����
�P����
����
�s���
��e�
t�����
����
��e�
st��
�����
����
����
��ee
���s
se���
����
����
���e
�t���
���e
��Te�
pe��
t���
T
(°C
) P �
(kPa
) (�
mol
l–1 k
Pa–1
)
T (°
C)
P � (k
Pa)
(�m
ol l–1
kPa
–1)
S���
asse
r M
eer�
asse
r S�
��as
ser
Mee
r�as
ser
0.5
0.�3
3 21
.�
1�.7
20.5
2.�1
113
.�
11.0
1.
0 0.
�57
21.1
1�
.521
.02.
��7
13.�
11
.0
1.5
0.��
1 20
.�
1�.3
21.5
2.5�
�13
.�
10.�
2.
0 0.
705
20.5
1�
.122
.02.
���
13.2
10
.�
2.5
0.73
1 20
.3
15.�
22.5
2.72
513
.1
10.7
3.
0 0.
75�
20.0
15
.�23
.02.
�0�
13.0
10
.�
3.5
0.7�
5 1�
.7
15.5
23.5
2.��
512
.�
10.5
�.
0 0.
�13
1�.�
15
.�2�
.02.
���
12.�
10
.�
�.5
0.��
2 1�
.2
15.2
2�.5
3.07
�12
.7
10.�
5.
0 0.
�72
1�.0
15
.025
.03.
1�7
12.5
10
.3
5.5
0.�0
3 1�
.�
1�.�
25.5
3.2�
312
.5
10.2
�.
0 0.
�35
1�.5
1�
.72�
.03.
3�1
12.�
10
.1
�.5
0.��
� 1�
.3
1�.�
2�.5
3.��
212
.2
10.1
7.
0 1.
002
1�.1
1�
.327
.03.
5�5
12.2
10
.0
7.5
1.03
� 17
.�
1�.2
27.5
3.�7
112
.1
�.�
�.0
1.07
2 17
.�
1�.0
2�.0
3.7�
012
.0
�.�
�.5
1.11
0 17
.�
13.�
2�.5
3.��
111
.�
�.�
�.0
1.1�
� 17
.3
13.7
2�.0
�.00
�11
.�
�.7
�.5
1.1�
7 17
.0
13.7
2�.5
�.12
311
.7
�.�
10.0
1.
22�
1�.�
13
.530
.0�.
2�3
11.�
�.
5 10
.5
1.2�
� 1�
.7
13.�
30.5
�.3�
�11
.�
�.5
11.0
1.
312
1�.�
13
.231
.0�.
��2
11.5
�.
� 11
.5
1.35
7 1�
.3
13.1
31.5
�.�2
211
.3
�.3
12.0
1.
�02
1�.1
13
.032
.0�.
755
11.3
�.
2 12
.5
1.��
� 15
.�
12.�
32.5
�.��
111
.2
�.2
13.0
1.
��7
15.�
12
.733
.05.
030
11.1
�.
2 13
.5
1.5�
7 15
.�
12.�
33.5
5.17
311
.0
�.1
1�.0
1.
5��
15.5
12
.53�
.05.
31�
11.0
�.
0 1�
.5
1.�5
1 15
.2
12.3
3�.5
5.��
�10
.�
�.�
15.0
1.
705
15.1
12
.235
.05.
�23
10.�
�.
� 15
.5
1.7�
1 1�
.�
12.1
35.5
5.7�
010
.7
�.�
1�.0
1.
�1�
1�.�
12
.03�
.05.
��1
10.7
�.
� 1�
.5
1.�7
7 1�
.�
11.�
3�.5
�.10
�10
.7
�.7
17.0
1.
�37
1�.5
11
.�37
.0�.
275
10.�
�.
� 17
.5
2.00
0 1�
.�
11.�
37.5
�.��
�10
.5
�.�
1�.0
2.
0��
1�.3
11
.�3�
.0�.
�25
10.�
�.
� 1�
.5
2.12
� 1�
.1
11.5
3�.5
�.�0
�10
.�
�.5
1�.0
2.
1�7
1�.0
11
.33�
.0�.
��2
10.3
�.
� 1�
.5
2.2�
7 13
.�
11.3
3�.5
7.1�
110
.2
�.�
20.0
2.
33�
13.7
11
.2�0
.07.
37�
10.2
�.
3
- 5�
-

Woc
he 6
Pf
lanz
enpi
gmen
te
AG
Kud
la/B
öhm
er
Iden
tifiz
ieru
ng u
nd C
hara
kter
isie
rung
von
Gen
en d
er P
igm
ents
ynth
ese
in
Bras
sica
rapa
Ler
nzei
le:
Am
End
e di
eses
Kur
stag
es so
llten
Sie
Fol
gend
es g
eler
nt h
aben
:
Ver
glei
ch
von
anth
ocya
ninl
osen
, ge
lbgr
ünen
M
utan
ten
(���
� ��
�)
mit
Wild
typp
flanz
en
(AN
L/Y
GR
) M
essu
ng v
on A
ntho
cyan
und
Chl
orop
hyllk
onze
ntra
tione
n in
Blä
ttern
M
essu
ng v
on T
rans
krip
tions
fakt
orak
tivitä
t mitt
els d
er T
abak
-1-H
ybrid
Met
hode
Ein
leitu
ng:
Die
Rol
le d
er C
hlor
ophy
lle u
nd a
nder
er T
etra
pyrr
ole
Jähr
lich
wer
den
durc
h C
hlor
ophy
lle,
der
men
gen-
mäß
ig g
rößt
en G
rupp
e de
r Tet
rapy
rrol
e, sc
hätz
ungs
-w
eise
1025
J an
Sonn
enen
ergi
e ab
sorb
iert
und
für d
ie
Ber
eits
tellu
ng v
on c
hem
isch
er E
nerg
ie (
ATP
) un
d R
eduk
tions
äqui
vale
nten
(N
AD
PH)
nutz
bar
gem
acht
. Daz
u st
eht e
ine
Chl
orop
hyllm
enge
von
ca.
2,
4 x
108
t zu
r V
erfü
gung
. D
ie
jähr
lich
bios
ynth
etis
ierte
Ges
amtm
enge
wird
auf
1,2
x 1
010 t
gesc
hätz
t. C
hlor
ophy
lle si
nd c
yclis
che
Tetra
pyrr
ole,
d.
h.,
vier
Pyr
rolri
nge
sind
übe
r sog
. Met
hinb
rück
en
mite
inan
der
zu e
inem
Mak
rocy
clus
ver
knüp
ft, i
n de
ssen
Ze
ntru
m
sich
ei
n M
agne
sium
-Ato
m
als
Zent
rala
tom
bef
inde
t (A
bbild
ung
1).
Das
Pyr
rol a
ls G
rund
baus
tein
des
Mak
rocy
clus
stel
lt ei
nen
fünf
glie
drig
en R
ing
aus
vier
C-u
nd e
inem
N-
Ato
m
dar,
der
in
den
einz
elne
n Te
trapy
rrol
en
unte
rsch
iedl
ich
subs
titui
ert
ist.
Die
H
äme,
eine
w
eite
re w
icht
ige
Gru
ppe
von
Tetra
pyrr
olen
, bes
itzen
al
s Ze
ntra
lato
m E
isen
. B
eisp
iele
für
die
se G
rupp
e si
nd d
er r
ote
Blu
tfarb
stof
f H
ämog
lobi
n un
d da
s M
yogl
obin
, die
Cyt
ochr
ome
der
Atm
ungs
- un
d de
r ph
otos
ynth
etis
chen
El
ektro
nent
rans
portk
ette
, so
wie
di
e pr
osth
etis
che
Gru
ppe
der
Kat
alas
e un
d Pe
roxi
dase
. Siro
häm
ist a
ls C
o-fa
ktor
in d
en E
nzym
en N
itrit-
Red
ukta
se u
nd S
ulfit
redu
ktas
e en
thal
ten.
D
as in
met
hano
gene
n B
akte
rien
vork
omm
ende
Tet
rapy
rrol
F43
0 en
thäl
t Nic
kel a
ls Z
entra
lato
m. C
obal
t is
t in
das
Cor
rin-G
erüs
t des
Vita
min
s B
12 ei
ngeb
aut.
Neb
en d
en c
yclis
chen
Tet
rapy
rrol
en g
ibt e
s au
ch
offe
nket
tige
Tetra
pyrr
ole,
z .
B.
die
Phyc
obili
nePh
ycoe
ryth
rinun
d Ph
ycoc
yani
nin
den
Rot
alge
n,
Cry
ptop
hyce
en u
nd d
en C
yano
bakt
erie
n. A
uch
der C
hrom
opho
r des
Phy
toch
rom
sis
t ein
off
enke
ttige
s Te
trapy
rrol
, ebe
nso
wie
die
Gal
lenf
arbs
toff
e (B
iline
). D
ie F
unkt
ion
der C
hlor
ophy
lle u
nd P
hyco
bilin
e be
steh
t in
der A
bsor
ptio
n vo
n Li
cht u
nd d
er W
eite
rgab
e de
r Ene
rgie
an
die
Rea
ktio
nsze
ntre
n.
Abbildun
g1Strukturen
vonßCarotin
,Luteinun
dCh
loroph
ylla
+b(nachHe
ldtP
flanzen
bioche
mie,Spe
ktrum,
Akadem
ische
rVerlag,He
idelbe
rg)
- 5�
-
Woc
he 6
Pf
lanz
enpi
gmen
te
AG
Kud
la/B
öhm
er
Chl
orop
hyll
Anr
egun
g un
d A
bsor
ptio
nssp
ektr
um
Lich
tqua
nten
wer
den
von
Chl
oro-
phyl
l a u
nd b
in d
en „
Ant
enne
n“ d
er
Phot
osys
tem
e ab
sorb
iert.
Es
en
t-st
ehen
ele
ktro
nisc
h an
gere
gte
Zu-
stän
de,
Chl
*.
Dab
ei
sind
zw
ei
Anr
egun
gszu
stän
de
mög
lich:
D
er
ener
gier
eich
ere
Anr
egun
gszu
stan
d („
2. A
nreg
ungs
zust
and“
) wird
dur
ch
Abs
orpt
ion
eine
s en
ergi
erei
chen
bl
auen
Lic
htqu
ants
err
eich
t. Er
ent
-sp
richt
der
bla
uen
Abs
orpt
ions
band
e de
r Chl
orop
hylle
(s. E
nerg
iesc
hem
a re
chts
). D
iese
r Z
usta
nd i
st j
edoc
h se
hr u
nsta
bil
und
zerf
ällt
inne
rhal
b vo
n 10
-12 s
in e
inen
ene
rgie
ärm
eren
A
nreg
ungs
zust
and,
de
ssen
En
ergi
egeh
alt
dem
ein
es r
oten
Lic
htqu
ants
ent
spric
ht.
Die
Ene
rgie
diff
eren
z zw
isch
en d
en b
eide
n A
nreg
ungs
zust
ände
n w
ird d
urch
Mol
ekül
vibr
atio
nen
als t
herm
isch
e En
ergi
e ab
gege
ben.
Der
nie
drig
ere
Anr
egun
gszu
stan
d („
1. A
nreg
ungs
zust
and“
) w
ird d
urch
Abs
orpt
ion
von
rote
n Li
chtq
uant
en e
rrei
cht
(rot
e A
bsor
ptio
nsba
nde
der C
hlor
ophy
lle) u
nd is
t etw
as st
abile
r (Le
bens
daue
r 10-9
s).
Nur
die
Ene
rgie
des
„st
abile
ren“
1. A
nreg
ungs
zust
ande
s ka
nn p
hoto
synt
hetis
ch v
erw
erte
t wer
den.
Die
zu
sätz
liche
Ene
rgie
des
bla
uen
Lich
tqua
nts
geht
ver
lore
n. A
ufgr
und
der e
ngen
exi
toni
sche
n K
oppl
ung
der
Chl
orop
hylle
„sp
ringe
n“ d
ie 1
. A
nreg
ungs
zust
ände
inn
erha
lb d
er A
nten
ne v
on C
hlor
ophy
ll zu
C
hlor
ophy
ll, w
erde
n sc
hlie
ßlic
h zu
m R
eakt
ions
zent
rum
des
jew
eilig
en P
hoto
syst
ems
trans
ferie
rt un
d lö
sen
dort
die
Frei
setz
ung
eine
s El
ektro
ns a
us. E
rst i
n de
n R
eakt
ions
zent
ren
von
PS II
und
PS
I fin
det
som
it La
dung
stre
nnun
g un
d da
mit
die
Abg
abe
eine
s El
ektro
ns s
tatt.
Chl
orop
hyll
a un
d b
bild
en
zusa
mm
en m
it de
n C
arot
inoi
den
den
sog.
Lic
htsa
mm
elko
mpl
ex (
engl
.: Li
ght
Har
vest
ing
Com
plex
(L
HC
)). I
nner
halb
die
ses
Kom
plex
es w
ird d
ie E
nerg
ie s
trahl
ungs
los
und
ohne
Ele
ktro
nenü
bertr
agun
g du
rch
sog.
För
ster
trans
fer o
der E
xcito
nenw
ande
rung
wei
terg
egeb
en, e
bens
o w
ie in
der
Cor
e-A
nten
ne,
die
aus C
hlor
ophy
ll a
und
Car
otin
oide
n be
steh
t. N
ur in
den
Rea
ktio
nsze
ntre
n vo
n PS
II, P
680,
und
PS I,
P 7
00, f
inde
t die
Prim
ärre
aktio
n de
r Pho
tosy
nthe
se st
att,
die
wie
folg
t for
mul
iert
wer
den
kann
:
Chl
a (P
680)
+ h
v
Chl
a*
Chl
a+ +
e-
Der
gle
iche
Pro
zess
find
et a
m P
700 d
es P
S I s
tatt.
Das
Chl
a+ -R
adik
al d
es P
680 w
ird d
urch
die
Ele
ktro
nen,
di
e de
r w
asse
rspa
ltend
e M
anga
n-K
ompl
ex l
iefe
rt, w
iede
r re
duzi
ert,
also
reg
ener
iert.
Ver
einf
acht
fo
rmul
iert:
2 H
20
4 H
+ + O
2 +
4 e-
P 700
wird
von
den
Ele
ktro
nen
rege
nerie
rt, d
ie ü
ber d
ie E
lekt
rone
ntra
nspo
rtket
te v
on P
S II
kom
men
. Die
El
ektro
nen,
die
P70
0 abg
ibt,
redu
zier
en le
tzte
ndlic
h da
nn N
AD
P+ zu
NA
DPH
2.
Chl
orop
hyll
Fluo
resz
enz
Ein
durc
h Li
cht a
nger
egte
s Pi
gmen
t kan
n en
twed
er u
nter
Wär
mea
bgab
e od
er d
urch
Abs
trahl
ung
eine
s Li
chtq
uant
s in
den
Gru
ndzu
stan
d zu
rück
kehr
en. D
eakt
ivie
rung
dur
ch L
icht
abst
rahl
ung
beze
ichn
et m
an
als
Fluo
resz
enz.
Auf
grun
d de
r be
sond
eren
Mol
ekül
stru
ktur
des
Chl
orop
hylls
(„s
tarr
es“
ringf
örm
iges
Te
trapy
rrol
) wer
den
durc
h de
n 1.
Anr
egun
gszu
stan
d w
enig
„th
erm
isch
e“ V
orgä
nge
im M
olek
ül (z
.B.
Abbildun
g2Energieschem
aCh
loroph
yllanregun
g
- 5�
-

Woc
he 6
Pf
lanz
enpi
gmen
te
AG
Kud
la/B
öhm
er
Mol
ekül
vibr
atio
n) a
usge
löst
. D
aher
kan
n ei
n Te
il de
r vo
n C
hlor
ophy
llen
abso
rbie
rten
Lich
tene
rgie
w
iede
r in
For
m v
on F
luor
esze
nz a
bges
trahl
t w
erde
n. D
a de
nnoc
h st
ets
etw
as E
nerg
ie d
urch
in
tram
olek
ular
e V
orgä
nge
verlo
ren
geht
, ist
der
Ene
rgie
geha
lt de
s em
ittie
rten
Lich
tqua
nts g
erin
gfüg
ig,
aber
nic
ht w
esen
tlich
nie
drig
er a
ls d
er u
rspr
üngl
iche
Ene
rgie
geha
lt de
s Anr
egun
gszu
stan
ds. A
us d
iese
m
Gru
nd
ist
die
Chl
orop
hyll
Fluo
resz
enz
imm
er
dunk
elro
t, w
ähre
nd
der
Ener
gieg
ehal
t de
s 1.
A
nreg
ungs
zust
and
dem
ein
es h
ellro
ten
Lich
tqua
nts e
ntsp
richt
(s���
�e��
�es�
�e�
����
e�).
Wen
ngle
ich
nur
ein
rela
tiv g
erin
ger
Teil
der
eing
estra
hlte
n Li
chte
nerg
ie w
iede
r al
s Fl
uore
szen
z ab
gest
rahl
t w
ird,
so i
st d
ie F
luor
esze
nz a
n is
olie
rten
Chl
orop
hylle
n do
ch s
ehr
gut
sich
tbar
. In
den
C
hlor
opla
sten
sel
bst,
z.B
. im
Bla
ttgew
ebe
ist d
ie C
hlor
ophy
ll Fl
uore
szen
z w
esen
tlich
sch
wäc
her.
Nur
1–
3 %
der
ein
gest
rahl
ten
Lich
tene
rgie
wird
wie
der
als
Fluo
resz
enz
emitt
iert.
Phy
siol
ogis
ch i
st d
ie
Chl
orop
hyll
Fluo
resz
enz
dam
it be
deut
ungs
los.
Jedo
ch h
at s
ie e
rheb
liche
Bed
eutu
ng in
der
For
schu
ng
und
dem
opt
isch
en „
mon
itorin
g“ d
er P
flanz
ende
cke
in B
ioto
pen.
Das
hän
gt d
amit
zusa
mm
en, d
ass
die
„Flu
ores
zenz
ausb
eute
“ vo
n ph
otos
ynth
etis
chem
Gew
ebe
eine
n zw
ar k
ompl
exen
, inz
wis
chen
abe
r gu
t ve
rsta
nden
en Z
usam
men
hang
mit
der
phot
osyn
thet
isch
en L
eist
ung
aufw
eist
. So
läss
t si
ch z
.B. e
ine
Schw
ächu
ng d
er P
hoto
synt
hese
leis
tung
dur
ch S
tress
gut
sich
tbar
mac
hen.
Stru
ktur
und
Fun
ktio
nen
der
Car
otin
oide
D
ie C
arot
inoi
de s
telle
n ei
ne w
icht
ige
und
groß
e U
nter
grup
pe d
er Is
opre
noid
e da
r. D
iese
r Obe
rbeg
riff
leite
t si
ch v
om G
rund
baus
tein
alle
r Is
opre
noid
e, d
em I
sopr
en,
ab.
Die
in
der
Bio
synt
hese
akt
iven
Fo
rmen
des
Iso
pren
s si
nd d
ie C
5-Kör
per
lsop
ente
nylp
yrop
hosp
hat (
IPP)
und
sei
n Is
omer
, Dim
ethy
l-al
lyl-P
P (P
reny
l-PP)
. Zw
ei I
PP w
erde
n zu
m M
onot
erpe
n G
eran
yl-P
P zu
sam
men
gela
gert,
wei
tere
A
dditi
on v
on I
PP f
ührt
der
Rei
he n
ach
zum
Far
nesy
l-PP
und
dann
zum
Ger
anyl
gera
nyl-P
P, d
em
Aus
gang
spro
dukt
für
den
Phy
tolre
st d
es C
hlor
ophy
lls a
und
b. Z
wei
Ger
anyl
gera
nyl-P
P sc
hlie
ßlic
h bi
lden
das
Tet
rate
rpen
(4 T
erpe
ne a
us 8
lsop
rene
n, C
40) P
hyto
en, d
as ü
ber v
ersc
hied
ene
Zwis
chen
stuf
en
zu
-und
ß-C
arot
in u
mge
wan
delt
wird
. D
iese
Ver
bind
unge
n be
steh
en n
ur a
us K
ohle
nsto
ff u
nd
Was
sers
toff
und
bild
en d
ie G
rupp
e de
r C
arot
ine
(Abb
ildun
g 1)
. W
erde
n Sa
uers
toff
grup
pen,
z.B
. A
lhoh
ol-,
Ethe
r-,
Ald
ehyd
-, K
eto-
, C
arbo
xyl-u
nd a
uch
Epox
y-G
rupp
en,
in d
ie C
arot
ine
eing
efüh
rt,
gela
ngt m
an z
u de
n X
anth
ophy
llen.
Dur
ch d
ie E
lekt
rone
gativ
ität d
er S
auer
stof
f Fun
ktio
nen
erha
lten
die
Xan
thop
hylle
ei
nen
etw
as
pola
rere
n C
hara
kter
al
s da
s ex
trem
lip
ophi
le
�-C
arot
in.
Wic
htig
e X
anth
ophy
lle s
ind
Lute
in, V
iola
xant
hin
und
Zeax
anth
in (
Abb
ildun
g 1)
. Car
otin
e un
d X
anth
ophy
lle
bild
en z
usam
men
die
Obe
rgru
ppe
der C
arot
inoi
de.
Car
otin
oide
flu
ores
zier
en n
icht
, obg
leic
h si
e si
chtb
ares
Lic
ht s
ehr
gut a
bsor
bier
en. D
as h
ängt
dam
it zu
sam
men
, da
ss i
n la
ngge
stre
ckte
n C
arot
inoi
den
(40
C-A
tom
e, 2
0 ko
njug
ierte
Dop
pel-b
indu
ngen
) du
rch
Lich
tabs
orpt
ion
Mol
ekül
vibr
atio
nen
ausg
elös
t w
erde
n, d
urch
wel
che
die
Anr
egun
gsen
ergi
e te
ilwei
se th
erm
isch
dea
ktiv
iert
wird
. Dah
er s
ind
Car
otin
oide
als
pho
tosy
nthe
tisch
e A
nten
nenp
igm
ente
("
akze
ssor
isch
e Pi
gmen
te")
w
eit
wen
iger
ge
eign
et
als
Chl
orop
hylle
. Fu
nktio
nell
erfü
llen
die
Car
otin
oide
den
noch
meh
rere
wic
htig
e A
ufga
ben.
So
sind
sie
, wie
ber
eits
erw
ähnt
, zu
eine
m g
ewis
sen
Um
fang
an
der L
icht
abso
rptio
n de
r Pho
tosy
nthe
se b
etei
ligt u
nd s
chlie
ßen
dabe
i zu
eine
m T
eil d
ie s
og.
"Grü
nlüc
ke"
der
Chl
orop
hylla
bsor
ptio
n. E
ine
wei
tere
wic
htig
e Fu
nktio
n de
r C
arot
inoi
de i
st i
hre
Bet
eilig
ung
am s
trukt
urel
len
Auf
bau
vor
alle
m v
on P
hoto
syst
em I
I. O
hne
Car
otin
oide
gib
t es
kei
n fu
nktio
nsfä
hige
s PS
II.
Eine
drit
te u
nd w
ohl w
icht
igst
e Fu
nktio
n is
t der
Sch
utz
der
Chl
orop
hylle
vor
Ph
otoo
xida
tion.
Dur
ch L
icht
ang
ereg
tes
Chl
orop
hyll
kann
Sau
erst
off i
n di
e se
hr a
ggre
ssiv
e Fo
rm d
es
sog.
Sin
glet
t-Sau
erst
offs
übe
rfüh
ren,
der
dan
n da
s Chl
orop
hyll
irrev
ersi
bel o
xidi
ert.
Car
otin
oide
hab
en
die
Fähi
gkei
t, Si
ngle
tt-Sa
uers
toff
zu
��e�
��e�
�d.h
. sei
ne E
nerg
ie z
u üb
erne
hmen
, ohn
e da
bei s
elbs
t ze
rstö
rt zu
wer
den.
Auc
h kö
nnen
sie
Chl
orop
hylle
, die
ihre
Ene
rgie
nic
ht so
fort
über
trage
n kö
nnen
, �e�
e���
t�e�e
�(a
breg
en)
und
dam
it di
e B
ildun
g vo
n Si
ngle
tt-Sa
uers
toff
von
vor
n he
rein
ver
hind
ern.
- 5�
-
Woc
he 6
Pf
lanz
enpi
gmen
te
AG
Kud
la/B
öhm
er
Schl
ießl
ich
fung
iere
n di
e C
arot
inoi
de a
uch
noch
als
gel
be u
nd o
rang
efar
bene
bis
bra
une
Blü
ten-
und
Fruc
htfa
rbst
offe
.
Stru
ktur
und
Fun
ktio
n de
r A
ntho
cyan
e A
ntho
cyan
idin
e un
d A
ntho
cyan
e si
nd fü
r die
Rot
-Vio
lettf
ärbu
ng v
on P
flanz
enor
gane
n (B
lätte
r, B
lüte
n,
Spro
sse)
ver
antw
ortli
ch. I
m G
egen
satz
zu
den
Chl
orop
hylle
n un
d C
arot
inoi
den
sind
sie
was
serlö
slic
h un
d lie
gen
dahe
r in
den
Vak
uole
n vo
r.
Abs
orpt
ions
spek
tros
kopi
e
Spek
trosk
opis
che
Met
hode
n w
ie z
. B
. die
heu
te a
nzuw
ende
nde
Abs
orpt
ions
spek
trosk
opie
sin
d zu
r Id
entif
izie
rung
und
Qua
ntifi
zier
ung
von
(unb
ekan
nten
) Sub
stan
zen
heut
e ni
cht m
ehr w
egzu
denk
en. S
ie
beru
ht a
uf d
em P
rinzi
p de
r Mes
sung
der
Lic
htab
sorp
tion
beim
Dur
chtri
tt ei
nes L
icht
stra
hles
dur
ch e
ine
Prob
e be
i ein
er o
der
meh
rere
n W
elle
nlän
gen.
Ver
schi
eden
e Su
bsta
nzen
zei
gen
bei u
nter
schi
edlic
hen
Wel
lenl
änge
n un
ters
chie
dlic
he
Abs
orpt
ione
n vo
n Li
cht.
Die
au
f di
ese
Wei
se
gew
onne
nen
Abs
orpt
ions
spek
tren
sind
oft
char
akte
ristis
ch f
ür d
ie j
ewei
ligen
Rei
nsub
stan
zen.
Mis
st m
an d
ie
Abs
orpt
ion
eine
r Lös
ung
in d
er e
ine
rein
e Su
bsta
nz in
ein
em in
die
sem
Ber
eich
nic
ht a
bsor
bier
ende
n Lö
sung
smitt
el g
elös
t is
t, so
fin
det
man
inn
erha
lb e
ines
gew
isse
n K
onze
ntra
tions
bere
iche
s ei
nen
quas
iline
aren
Zus
amm
enha
ng z
wis
chen
der
Abs
orpt
ion
und
der
Kon
zent
ratio
n ei
ner
Lösu
ng. D
iese
r Zu
sam
men
hang
wird
dur
ch d
as L
ambe
rt-B
eer's
che
Ges
etz
besc
hrie
ben:
� =
·c·d
Dab
ei i
st �
= E
xtin
ktio
n,
= m
olar
er E
xtin
ktio
nsko
effiz
ient
, c =
Kon
zent
ratio
n in
Mol
·L-1
und
d =
Sc
hich
tdic
ke d
er L
ösun
g in
cm
. Ext
inkt
ion
beze
ichn
et d
en d
ekad
isch
en L
ogar
ithm
us d
er in
ein
e Pr
obe
eing
estra
hlte
n Li
chtin
tens
ität I
0 im
Ver
hältn
is z
u de
r nac
h de
r Pro
be g
emes
sene
n In
tens
ität I
:
Dur
ch d
as L
ambe
rt-B
eer’
sche
Ges
etz
wird
al
so e
ine
e-Fu
nktio
n lin
earis
iert.
Des
halb
da
rf b
ei d
esse
n A
nwen
dung
nur
der
unt
ere,
qu
asi-l
inea
re B
erei
ch d
er F
unkt
ion
benu
tzt
wer
den
(es
gilt
also
nu
r fü
r ge
ringe
K
onze
ntra
tione
n,
für
quan
titat
ive
Bes
timm
unge
n so
llte
E 0
,8 se
in!)
.
Pigm
entk
onze
ntra
tione
n kö
nnen
mit
Hilf
e ei
nes
Spek
troph
otom
eter
s un
d de
m B
eer-
Lam
berts
chen
G
eset
zes
unte
r Zu
hilfe
-na
hme
des
Extin
ktio
nsko
effiz
ient
en
für
Chl
orop
hyll
bere
chne
t w
erde
n. U
m d
ie
Kon
zent
ratio
nen
von
Chl
orop
hyll,
Car
o-tin
oide
n un
d A
ntho
cyan
en z
u m
esse
n, s
ind
vers
chie
dene
B
erec
hnun
gen
besc
hrie
ben
wor
den,
die
meh
r ode
r wen
iger
akk
urat
arb
eite
n [1
-4].
Sim
s un
d G
amon
nut
zten
dab
ei z
ur E
xtra
ktio
n de
r Chl
orop
hylle
ein
e M
isch
ung
aus
Ace
ton
und
Tris
Puff
er (8
0:20
Vol
; pH
=7.8
) und
ver
öffe
ntlic
hten
G
leic
hung
en, w
elch
e de
n Ei
nflu
ss v
on A
ntho
cyan
en b
erüc
ksic
htig
en:
Abbildun
g3Extin
ktionskoeffizienten
fürC
hlorop
hyll,Carotin
oide
undAn
thocyane
inAb
hängigkeitvonde
rWellenlänge.
- �0
-

Woc
he 6
Pf
lanz
enpi
gmen
te
AG
Kud
la/B
öhm
er
Mes
sung
en in
Ace
ton/
Tris
-Puf
fer:
Chl
orop
hyll
a (μ
mol
ml-1
) = 0
.013
73 A
663
- 0.0
0089
7 A
537
- 0.0
0304
6 A
647
Chl
orop
hyll
b (μ
mol
ml-1
) = 0
.024
05 A
647
- 0.0
0430
5 A
537
- 0.0
0550
7 A
663
Car
otin
oide
(μm
ol m
l-1) =
(A47
0 - (
17.1
x (C
hl a
+ C
hl b
) - 9
.479
x a
ntho
cyan
in))
/119
.26
Mes
sung
en in
saur
em M
etha
nol:
Ant
hocy
ane
(μm
ol m
l-1) =
0.0
3333
3 A
529
– 0.
0075
99 A
650
Tab
ak-1
-Hyb
rid
Die
Met
hode
des
Tab
ak-1
-Hyb
rid b
asie
rt au
f de
r U
nter
such
ung
von
Prot
ein-
DN
A-I
nter
-ak
tione
n,
ähnl
ich
dem
H
efe-
1-H
ybrid
w
elch
er d
ie B
indu
ng e
ines
Tra
nskr
iptio
nsf-
akto
rs a
n ei
ne D
NA
-Seq
uenz
nac
hwei
st. D
er
Vor
teil
des T
abak
-1-H
ybrid
bes
teht
neb
en d
er
Nut
zung
ein
es p
flanz
liche
n O
rgan
ism
us i
n de
r B
estim
mun
g de
r Tr
ansk
riptio
nsfa
ktor
-ak
tivitä
t.
Daf
ür w
ird d
ie z
u un
ters
uche
nde
Prom
otor
-se
quen
z vo
r ein
Rep
orte
rgen
(hie
r Luc
ifera
se)
fusi
onie
rt un
d m
it H
ilfe
der
Agr
obak
teriu
m-
verm
ittel
ten
Tran
sfor
mat
ion
in
Taba
k ex
prim
iert
(Abb
ildun
g 4
A).
Die
Pro
mot
or-
aktiv
ität k
ann
über
die
Luc
ifera
seex
pres
sion
er
mitt
elt
wer
den.
D
afür
w
erde
n di
e tra
nsfo
rmie
rten
Taba
kblä
tter
mit
Luci
ferin
in
filtri
ert
und
die
Men
ge
an
Che
milu
min
esze
nz g
emes
sen.
Die
zu
anal
y-si
eren
den
Tran
skrip
tions
fakt
oren
wer
den
co-
expr
imie
rt um
die
Änd
erun
g de
r Rep
orte
rgen
-ak
tivitä
t im
V
ergl
eich
zu
er
mitt
eln
(Abb
ildun
g 4
B).
Dad
urch
wird
die
Akt
ivitä
t de
s Tr
ansk
riptio
nsfa
ktor
s (a
ktiv
iere
nd/ r
epri-
mie
rend
) auf
den
zu
unte
rsuc
hend
en P
rom
oter
be
stim
mt.
Bra
ssic
a ra
pa
Ziel
die
ses
Ver
such
stag
es is
t zw
eier
lei.
Zum
ein
en w
erde
n Si
e ��
�ss�
����
�p�
als
schn
ell w
achs
ende
n M
odel
lorg
anis
mus
nut
zen
um d
esse
n Pi
gmen
tzus
amm
ense
tzun
gen
zu s
tudi
eren
und
in v
ersc
hied
enen
G
enot
ypen
zu
quan
tifiz
iere
n. Z
um a
nder
en le
rnen
Sie
die
Met
hode
des
Tab
ak-1
-Hyb
rid a
ls M
etho
de z
ur
Mes
sung
von
Tra
nskr
iptio
nsfa
ktor
aktiv
itäte
n ke
nnen
. Wild
typ
���s
s���
���p
� Pf
lanz
en h
aben
vio
lette
St
iele
und
grü
ne B
lätte
r. D
ie V
iole
ttfär
bung
erg
ibt s
ich
aus d
er P
rodu
ktio
n vo
n A
ntho
cyan
en.
Abbildun
g4Tabak1Hy
brid
ASche
matisc
heDa
rstellung
des
verw
ende
ten
Prom
otor
Repo
rtergen
Konstrukt.
BDe
tektion
derLucife
raseaktivitä
tmittelse
ines
Imagers(bildlich).
- ��
-
Woc
he 6
Pf
lanz
enpi
gmen
te
AG
Kud
la/B
öhm
er
Ver
such
:N
ach
eine
r che
mis
chen
Mut
agen
ese
mit
Ethy
lmet
hans
ulfo
nat
(EM
S) w
urde
ein
e M
utan
te id
entif
izie
rt,
die
grün
e St
iele
(��
�) un
d ge
lbgr
üne
Blä
tter
(���
) be
sitz
t. Fo
lgen
de F
rage
stel
lung
en e
rgeb
en s
ich
aus
dem
Phä
noty
pen
dies
er M
utan
te:
1.H
at d
iese
Mut
ante
(���
) ger
inge
re K
onze
ntra
tione
n vo
n A
ntho
cyan
en a
ls W
ildty
ppfla
nzen
(AN
L)?
2.H
at d
iese
gel
bgrü
ne (�
��) M
utan
te g
erin
gere
Kon
zent
ratio
nen
an C
hlor
ophy
ll al
s W
ildty
ppfla
nzen
(Y
GR
)?
3.H
at d
ie g
elbg
rüne
Mut
ante
(��
�) h
öher
e K
onze
ntra
tione
n vo
n C
arot
enoi
den
als
Wild
typp
flanz
en
(YG
R)?
4.
Hab
en d
ie N
achk
omm
en (
F1)
von
Kre
uzun
gen
von
Wild
typp
flanz
en (
AN
L/Y
GR
) m
it di
eser
M
utan
te (�
��/�
��) e
ine
mitt
lere
Kon
zent
ratio
n de
r zu
unte
rsuc
hend
en P
igm
ente
? 5.
Wie
wird
die
Exp
ress
ion
des
YG
R G
ens
durc
h di
e Tr
ansk
riptio
nsfa
ktor
en Y
GI1
, YG
I2 u
nd Y
GI3
re
gulie
rt?
Mes
sung
von
Car
oten
oid
und
Chl
orop
hyll-
Kon
zent
ratio
nen
Wir
verw
ende
n 10
Tag
e al
te �
��ss
����
��p�
Pfla
nzen
zur
Ana
lyse
der
Pig
men
tzus
amm
ense
tzun
g. M
it ei
nem
Kor
kboh
rer d
er G
össe
#5
wer
den
2 B
latts
chei
ben
aus
den
Blä
ttern
der
���
ss��
����
p� P
flanz
en
entn
omm
en.
Bes
timm
en s
ie d
en D
urch
mes
ser
der
Bla
ttsch
eibe
n (L
inea
l) un
d be
rech
nen
sie
die
Bla
ttflä
che!
Geb
en s
ie j
ewei
ls 1
Bla
ttsch
eibe
in
ein
1,5
mL
Rea
ktio
nsge
fäß
zusa
mm
en m
it 40
0 μL
A
ceto
n/Tr
is P
uffe
r (80
:20
Vol
; pH
=7.8
) und
mör
sern
sie
die
se m
it H
ilfe
des
Pist
ills.
Tran
sfer
iere
n Si
e di
e Su
spen
sion
in e
in 1
0 m
L R
eakt
ions
gefä
ß (F
alco
n). W
asch
en si
e da
s 1,5
mL
Rea
ktio
nsge
fäß
und
das
Pist
ill m
it 1
mL
Ace
ton/
Tris
und
geb
en s
ie d
ie S
uspe
nsio
n eb
enfa
lls in
das
15
mL
Rea
ktio
nsge
fäß.
Fü
llen
sie
ansc
hlie
ßend
die
Sus
pens
ion
im R
eakt
ions
gefä
ß au
f 6 m
L m
it A
ceto
n/Tr
is P
uffe
r (80
:20
Vol
; pH
=7.8
) auf
.
Mes
sen
Sie
nun
die
Abs
orpt
ion
des
Übe
rsta
ndes
bei
den
Wel
lenl
änge
n 47
0, 5
37, 6
47, u
nd 6
63 n
m
(not
iere
n si
e di
e M
essw
erte
in T
abel
le 1
). M
esse
n Si
e au
ch d
ie Ü
bers
tänd
e in
den
Rea
ktio
nsge
fäße
n ih
rer T
isch
nach
barn
bis
sie
5 M
essw
erte
pro
Gen
otyp
vor
liege
n ha
ben.
Mes
sung
der
Ant
ocya
nkon
zent
ratio
nen
Wei
l di
e G
leic
hung
en f
ür d
ie B
erec
hnun
g de
r A
ntho
cyan
-Kon
zent
ratio
nen
die
oben
bes
chrie
ben
wur
den,
nic
ht b
eson
ders
akk
urat
sin
d, w
erde
n w
ir ei
ne m
odifi
zier
te M
etho
de v
erw
ende
n. E
ntne
hmen
si
e w
iede
rum
mit
Hilf
e ei
nes
Kor
kboh
rers
der
Gös
se #
5 2
Bla
ttsch
eibe
n au
s de
n B
lätte
rn d
er �
��ss
����
��p�
Pfla
nzen
. B
estim
men
sie
den
Dur
chm
esse
r de
r B
latts
chei
ben
(Lin
eal)
und
bere
chne
n si
e di
e B
lattf
läch
e. G
eben
sie
die
Bla
ttsch
eibe
n in
ein
1,5
mL
Rea
ktio
nsge
fäß,
die
smal
zus
amm
en m
it 40
0μl
saur
em M
etha
nol
(Met
hano
l/HC
l/Was
ser;
90:1
:1),
und
mör
sern
sie
die
se m
it H
ilfe
des
Pist
ills.
Tran
sfer
iere
n Si
e di
e Su
spen
sion
in
ein
15 m
L R
eakt
ions
gefä
ß (F
alco
n).
Was
chen
sie
das
1,5
mL
Rea
ktio
nsge
fäß
und
das
Pist
ill m
it 1
mL
saur
em M
etha
nol u
nd g
eben
sie
die
Sus
pens
ion
eben
falls
in
das
Rea
ktio
nsge
fäß.
Fül
len
sie
ansc
hlie
ßend
die
Sus
pens
ion
im R
eakt
ions
gefä
ß au
f 6 m
L m
it sa
urem
M
etha
nol a
uf. M
esse
n Si
e nu
n di
e A
bsor
ptio
n de
s Übe
rsta
ndes
bei
den
Wel
lenl
änge
n 52
9 un
d 65
0 nm
. B
erec
hnen
sie
dan
n di
e G
esam
tchl
orop
hyll-
, C
hlor
ophy
ll A
-, C
hlor
ophy
ll B
- un
d C
arot
inoi
d-K
onze
ntra
tione
n (T
abel
len
2-5)
.
- ��
-

Woc
he 6
Pf
lanz
enpi
gmen
te
AG
Kud
la/B
öhm
er
Mes
sung
der
tran
skri
ptio
nelle
n R
egul
atio
n vo
n Y
GR
dur
ch Y
GI1
, YG
I2 u
nd Y
GI3
�
���t
����
��e�
t���
����
Pfla
nzen
wur
den
bere
its m
ittel
s Agr
obak
teriu
m-I
nfilt
ratio
n tra
nsfo
rmie
rt. D
abei
w
urde
das
Pro
mot
or-L
ucife
rase
-Kon
stru
kt Y
GR
alle
in s
owie
mit
den
Tran
skrip
tions
fakt
oren
YG
I1,
YG
I2 u
nd Y
GI3
co-
infil
trier
t. In
jizie
ren
sie
mit
Hilf
e ei
ner 1
mL
Sprit
ze (o
hne
Kan
üle)
etw
a 1
mL
eine
r 10
0 μM
Luc
iferin
Lös
ung
in je
wei
ls 1
Bla
tt de
r ber
eits
tran
sfor
mie
rten
���
�t��
����
e�t�
����
�� P
flanz
en.
Ach
ten
Sie
dara
uf,
dass
das
ges
amte
Bla
tt m
it Lö
sung
dur
chträ
nkt
ist.
Nac
h ei
ner
10 m
inüt
igen
In
kuba
tion
bei R
aum
tem
pera
tur w
ird d
ie C
hem
ilum
ines
zenz
der
vie
r tra
nsfo
rmie
rten
Blä
tter m
it H
ilfe
eine
s Im
ager
s auf
geno
mm
en (U
SB-S
tick
mitb
ringe
n).
Dis
kuss
ion
Das
Pro
toko
ll so
llte
die
gew
onne
n R
esul
tate
wie
folg
t zus
amm
enfa
ssen
:
1.K
ompl
ett a
usge
füllt
e Ta
belle
n 1-
5.
2.Ei
n Sä
ulen
diag
ram
m m
it de
n M
essu
ngen
für C
hlor
ophy
ll-, C
arot
inoi
d- u
nd A
ntho
cyan
konz
en-
tratio
nen
für d
ie d
rei G
enot
ypen
. 3.
Eine
bes
chrif
tete
Abb
ildun
g, d
es T
abak
-1-H
ybrid
s (C
hem
ilum
ines
zenz
bild
).
Im W
eite
ren
sollt
en fo
lgen
de F
rage
n im
Pro
toko
ll be
antw
orte
t wer
den:
1.W
arum
sind
die
���
����
Pfla
nzen
gel
bgrü
n?
2.W
ie is
t die
Pig
men
tpro
dukt
ion
in d
en F
1 H
ybrid
en b
eein
fluss
t?
3.W
ie w
ird Y
GR
tran
skrip
tione
ll vo
n Y
GI1
, YG
I2 u
nd Y
GI3
regu
liert?
Lite
ratu
r:
1.
Porr
a, R
.J. (2
002)
. The
cheq
uere
d hi
stor
y of
the d
evel
opm
ent a
nd u
se o
f sim
ulta
neou
s equ
atio
ns
for t
he a
ccur
ate
dete
rmin
atio
n of
chl
orop
hylls
a a
nd b
. Pho
tosy
nth
Res
��,
149
-156
. 2.
Li
chte
ntha
ler,
H.,
and
Wel
lbur
n,
A.
(198
3).
Det
erm
inat
ions
of
to
tal
caro
teno
ids
and
chlo
roph
ylls
a a
nd b
of l
eaf e
xtra
cts i
n di
ffer
ent s
olve
nts.
Bio
chem
ical
Soc
iety
Tra
nsac
tions
��,
59
1 - 5
92.
3.
Arn
on,
D.I.
(1
949)
. C
OPP
ER
ENZY
MES
IN
IS
OLA
TED
C
HLO
RO
PLA
STS.
PO
LYPH
ENO
LOX
IDA
SE IN
BET
A V
ULG
AR
IS. P
lant
Phy
siol
. ��,
1-1
5.
4.
Sim
s, D
., an
d G
amon
, J. (
2002
). R
elat
ions
hip
betw
een
pigm
ent c
onte
nt a
nd sp
ectra
l ref
lect
ance
ac
ross
a w
ide
rang
e of
spe
cies
, lea
f st
ruct
ures
and
dev
elop
men
tal s
tage
s. R
emot
e Se
nsin
g of
En
viro
nmen
t ��,
337
-354
.
- ��
-- �
� -

A
nhan
g Ta
belle
1 C
hlor
ophy
ll Pi
gmen
tdat
en fü
r Wild
typ
und
Mut
ante
AN
L/Y
GR
F1
Hyb
ride
anl/y
gr
Wel
lenl
änge
(nm
) 1
2 3
4 5
1 2
3 4
5 1
2 3
4 5
470
537
647
663
Tabe
lle 2
Ant
hocy
an-D
aten
für W
ildty
ppfla
nzen
und
Mut
ante
AN
L/Y
GR
F1
Hyb
ride
anl/y
gr
Wel
lenl
änge
(nm
) 1
2 3
4 5
1 2
3 4
5 1
2 3
4 5
529
650
Tabe
lle 3
Ant
hocy
an-K
onze
ntra
tione
n fü
r Wild
typp
flanz
en u
nd M
utan
te. W
erte
wer
den
bere
chne
t nac
h Si
ms
und
Gam
on [4
]. Ko
nzen
tratio
nen
in d
er P
flanz
en w
erde
nor
mal
isie
rt fü
r da
s Vo
lum
en d
er S
uspe
nsio
n (6
mL)
und
die
Flä
che
der
Bla
ttsch
eibe
(D
urch
mes
ser=
____
____
cm;
Fläc
he=_
____
___m
2 )
A
NL/
YG
R
F1 H
ybrid
e an
l/ygr
1 2
3 4
5 1
2 3
4 5
1 2
3 4
5 A
ntho
cyan
in
µmol
ml-1
Ant
hocy
anin
µm
ol m
-2

Ta
belle
4 C
hlor
ophy
ll un
d C
arot
inoi
d-K
onze
ntra
tione
n fü
r W
ildty
ppfla
nzen
und
Mut
ante
. Wer
te w
erde
n be
rech
net n
ach
Sim
s un
d G
amon
[4]
. K
onze
ntra
tione
n in
der
Pfla
nzen
wer
de n
orm
alis
iert
für
das
Vol
umen
der
Sus
pens
ion
(6 m
L) u
nd d
ie F
läch
e de
r B
latts
chei
be (Ø
=___
____
__; F
läch
e=__
____
__m
2 )
A
NL/
YG
R
F1 H
ybrid
e an
l/ygr
1 2
3 4
5 1
2 3
4 5
1 2
3 4
5 C
hlor
ophy
ll a
(µm
ol m
l-1)
Chl
orop
hyll
b (µ
mol
ml-1
)
Car
otin
oide
(µ
mol
ml-1
)
Chl
orop
hyll
a (µ
mol
m-2
)
Chl
orop
hyll
b (µ
mol
m-2
)
Car
otin
oide
(µ
mol
m-2
)
Ges
amt-
Chl
orop
hyll
(µm
ol
m-2
)
Chl
orop
hyll
a/b
Ver
hältn
is
Tabe
lle 5
Zus
amm
enfa
ssun
g de
r Chl
orop
hyll
Kon
zent
ratio
nen
in W
ildty
ppfla
nzen
und
Mut
ante
AN
L/YG
R
F1 H
ybrid
e an
l/ygr
C
hlor
ophy
ll a
(µm
ol m
-2)
C
hlor
ophy
ll b
(µm
ol m
-2)
C
arot
inoi
de (µ
mol
m-2
)
Ges
amt-C
hlor
ophy
ll (µ
mol
m-2
)
chl a
/b V
erhä
ltnis

Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Ver
such
1: E
nzym
kine
tik
Subs
trat
sätt
igun
g un
d M
icha
elis
kons
tant
e de
r L
acta
t-D
ehyd
roge
nase
Ver
such
szie
l: M
it H
ilfe
des
Opt
isch
en T
ests
sol
l die
kat
alyt
isch
e A
ktiv
ität (
Um
satz
pro
Zei
t) ei
nes
Enzy
ms
quan
titat
iv
erfa
sst
und
die
Abh
ängi
gkei
t de
r A
ktiv
ität
von
der
Subs
tratk
onze
ntra
tion
(Sub
stra
tsät
tigun
gsku
rve)
bes
timm
t wer
den.
Die
Bez
iehu
ng zw
isch
en S
ubst
rat u
nd E
nzym
wird
du
rch
die
Mic
hael
is-M
ente
n-Th
eorie
bes
chrie
ben.
The
oret
isch
er H
inte
rgru
nd
Die
kata
lytis
che
Akt
ivitä
t ein
es E
nzym
s ist
die
Um
satz
rate
bei
def
inie
rter S
ubst
ratk
onze
ntra
-tio
n (z
.B. s
ättig
ende
Kon
zent
ratio
n) u
nd d
efin
ierte
r Tem
pera
tur.
Die
inte
rnat
iona
le S
I–Ei
nhei
t fü
r die
kat
alyt
isch
e A
ktiv
ität i
st d
as K
atal
(kat
). 1
kat l
iegt
vor
, wen
n 1
Mol
Sub
stra
t pro
sec
um
gese
tzt w
ird. D
ie E
inhe
it lä
sst s
ich
auch
auf
ein
e be
stim
mte
Men
ge a
n En
zym
mol
ekül
en
bezi
ehen
: 1 k
at =
�·m
ol E
nzym
, wob
ei �
ein
Prop
ortio
nalit
ätsf
akto
r ist
, der
von
der
Um
satz
rate
ab
häng
t. D
ie fr
üher
übl
iche
Enz
ymei
nhei
t U (u
nit)
ents
pric
ht d
em U
msa
tz v
on 1
m
ol m
in-1
.
Die
Ges
chw
indi
gkei
t der
enzy
mat
isch
en K
atal
yse e
ines
Enz
yms w
ird im
Wes
entli
chen
dur
ch
die
Tem
pera
tur u
nd d
ie K
onze
ntra
tion
des S
ubst
rate
s bee
influ
sst.
Die
Um
satz
rate
ein
es E
nzym
s st
eigt
exp
onen
tiell
mit
der
Tem
pera
tur.
Im E
xpon
entia
lfakt
or
(„St
eilh
eit“
der
Exp
onen
tialfu
nktio
n) s
pieg
elt
sich
die
Akt
ivie
rung
sene
rgie
der
Rea
ktio
n (A
rrhe
nius
bezi
ehun
g). I
n de
r Pr
axis
wird
die
Tem
pera
tura
bhän
gigk
eit h
äufig
dur
ch d
en Q
10ge
kenn
zeic
hnet
. D
iese
r gi
bt d
en F
akto
r an
, um
den
die
bet
reff
ende
Rea
ktio
n be
i ei
ner
Tem
pera
ture
rhöh
ung
um 1
0o C g
este
iger
t wird
. Für
vie
le E
nzym
reak
tione
n lie
gt d
er Q
10 b
ei 2
bi
s 3.
Im P
rakt
ikum
wird
die
Abh
ängi
gkei
t de
r G
esch
win
digk
eit
eine
r En
zym
reak
tion
von
der
Subs
tratk
onze
ntra
tion
unte
rsuc
ht.
Mit
Blic
k au
f di
e A
rrhe
nius
bezi
ehun
g m
uss
dabe
i di
e
Rea
ktio
nste
mpe
ratu
r kon
stan
t geh
alte
n w
erde
n.
Subs
trat
abhä
ngig
keit
der
Enz
ymak
tivitä
t (M
icha
elis
-Men
ten
Bez
iehu
ng)
Eine
enz
ymat
isch
e R
eakt
ion
kata
lysi
ert i
m e
infa
chst
en F
all d
ie U
mw
andl
ung
eine
s Su
bstra
tes
S in
ein
Pro
dukt
P:
���s
t��t
(S)
��
P����
�t�(P
)
Zur B
estim
mun
g de
r Ges
chw
indi
gkei
t die
ser R
eakt
ion
im E
xper
imen
t wird
die
Abn
ahm
e de
s Su
bstra
tes o
der d
ie Z
unah
me
des P
rodu
ktes
mit
der Z
eit b
eoba
chte
t. A
nfan
gsge
schw
indi
gkei
t:
Man
be
stim
mt
die
Rea
ktio
nsge
schw
indi
gkei
t (�
) ei
ner
enzy
mat
isch
en R
eakt
ion
expe
rimen
tell
in e
iner
Mes
sküv
ette
, die
ein
en M
essp
uffe
r, so
wie
das
En
zym
und
sei
n Su
bstra
t in
def
inie
rter
Kon
zent
ratio
n en
thäl
t. D
ie R
eakt
ion
wird
ent
wed
er
durc
h Zu
gabe
des
Enz
yms
(bei
anw
esen
dem
Sub
stra
t od
er d
urch
Zug
abe
des
Subs
trats
bei
an
wes
ende
m E
nzym
) ge
star
tet.
Man
beo
bach
tet,
dass
� n
icht
kon
stan
t bl
eibt
, so
nder
n m
it fo
rtsch
reite
nder
Sub
stra
tabn
ahm
e (-
c S/
t) un
d Pr
oduk
tzun
ahm
e (+
c P/
t) be
stän
dig
abni
mm
t (s
. Abb
.). N
eben
der
beg
inne
nden
Rüc
krea
ktio
n un
d de
r „Pr
oduk
them
mun
g“ d
es E
nzym
s spi
elt
dabe
i vor
alle
m d
ie a
bneh
men
de V
erfü
gbar
keit
von
S ei
ne R
olle
.
- �5
-
Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Es
ist
dahe
r w
icht
ig,
im
Expe
rimen
t de
n St
art
der
Rea
ktio
n zu
er
fass
en
und
die
����
��s�
es��
���
����
e�t��
o zu
best
imm
en.
Subs
trat
sätt
igun
gsku
rve:
Die
Abh
ängi
gkei
t der
Anf
angs
gesc
hwin
digk
eit �
o[m
ol L
-1 s-1
]von
der S
ubst
ratk
onze
ntra
tion
c S [m
ol L
-1]
(bei
kon
stan
ter T
empe
ratu
r und
Enz
ymko
nzen
tratio
n)
folg
t in
der R
egel
ein
er h
yper
bolis
chen
Fun
ktio
n. D
iese
���
st��
ts�t
t����
�s��
��e
läss
t sic
h in
3
Ber
eich
e ze
rlege
n: B
ei se
hr n
iedr
igen
Sub
stra
tkon
zent
ratio
nen
stei
gt �
o ann
äher
nd li
near
mit
c S, d
.h.,
die R
ate i
st v
olls
tänd
ig v
on d
er S
ubst
ratk
onze
ntra
tion
best
imm
t (�e
��t��
����
����
����
).B
ei s
ättig
ende
r Sub
stra
tkon
zent
ratio
n is
t die
Enz
ymak
tivitä
t max
imal
(�o u
nabh
ängi
g vo
n c S
;�e
��t��
����
��
����
��).
Die
be
iden
G
renz
bere
iche
w
erde
n vo
n ei
nem
��
pe��
���s
��e�
��e
����
�s�e
�e��
� ve
rbun
den.
Nac
h de
r The
orie
von
MIC
HA
ELIS
und
MEN
TEN
folg
t die
Sät
tigun
gsku
rve
der F
unkt
ion:
��
�
��
��
�m
ax0
Dah
er la
ssen
sich
aus
der
Sub
stra
tsät
tigun
gsku
rve
wic
htig
e ki
netis
che
Para
met
er e
iner
En
zym
reak
tion
able
iten,
vor
alle
m �
max
und
KM
.
Anf
angs
gesc
hwin
digk
eit
v 0 ge
gen
Sub
stra
tkon
zent
rati
onbe
i ko
nsta
nter
Enz
ymko
nzen
trat
ion
[Sub
stra
t]
v 0
Lin
ewea
ver-
Bur
k pl
ot:
1/v 0
gege
n 1/
[Sub
stra
t]
1/[S
ubst
rat]
1/v 0
� ���
����
� ���
��
��� �
��
��
���
����
���
Co
l 1 v
s C
ol 2
Pro
du
kte
ntw
ick
lun
g a
lsF
un
ktio
n d
er Z
eit
Zei
t
Pro
du
kt]
��
���
�ss
te��
��
���
�
- ��
-

Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Max
imal
gesc
hwin
digk
eit
v max
: En
zym
gesc
hwin
digk
eit
v be
i sä
ttige
nder
Su
bstra
t-ko
nzen
tratio
n c S
;
� max
nim
mt p
ropo
rtion
al m
it de
r Enz
ymko
nzen
tratio
n zu
; bei
kon
stan
ter E
nzym
konz
entra
tion
nim
mt �
max
exp
onen
tiell
mit
der T
empe
ratu
r zu
(Arr
heni
usbe
zieh
ung,
s. o
ben)
;
Mic
hael
isko
nsta
nte
KM
: Su
bstra
tkon
zent
ratio
n be
i ha
lbm
axim
aler
R
eakt
ions
-ge
schw
indi
gkei
t (v
= v m
ax /
2).
KM
ist e
in n
umer
isch
es M
aß fü
r die
Aff
initä
t des
Enz
yms
zu s
eine
m S
ubst
rat.
Je n
iedr
iger
der
K
M W
ert,
dest
o hö
her i
st d
ie A
ffin
ität z
um S
ubst
rat.
Für v
ersc
hied
ene
Enzy
me
varii
ert d
er K
M
Wer
t zw
isch
en e
twa
10-2
und
10-6
mol
/l.
Enz
ym-S
ubst
rat-
Kom
plex
: Zu
r en
zym
atis
chen
Kat
alys
e is
t di
e B
ildun
g ei
nes
Kom
plex
es
zwis
chen
frei
em E
nzym
(E) u
nd S
ubst
rat (
S) er
ford
erlic
h. D
ie U
mse
tzun
g de
s Sub
stra
tes e
rfol
gt
im K
ompl
ex, w
elch
er d
ann
sehr
schn
ell i
n fr
eies
Enz
ym u
nd P
rodu
kt z
erfä
llt:
P�
�P��
��
Mic
hael
is-M
ente
n-T
heor
ie:
Sie
besa
gt,
dass
di
e G
esch
win
digk
eit
v o
der
gesa
mte
n En
zym
reak
tion
in e
rste
r Näh
erun
g vo
n de
r rel
ativ
en K
onze
ntra
tion
des a
ktiv
en K
ompl
exes
��
best
imm
t ist
:
v o =
�[��
]
Wob
ei�
eine
Rea
ktio
nsko
nsta
nte
für
die
kata
lytis
che
Zerf
alls
reak
tion
[��]
����
�P is
t und
si
ch fü
r jed
es E
nzym
unt
ersc
heid
et. D
ie E
nzym
reak
tion
läss
t sic
h in
dre
i Tei
lsch
ritte
zer
lege
n:
1. S
chrit
t: D
ie S
ubst
ratb
indu
ng a
n da
s En
zym
(��
���
���
�) is
t rev
ersi
bel u
nd s
chne
ll. 2
. Sc
hritt
: Im
ES
– K
ompl
ex w
ird d
as S
ubst
rat i
n da
s Pro
dukt
P u
mge
wan
delt:
��
��P.
Die
ser
Schr
itt is
t ebe
nfal
ls se
hr sc
hnel
l und
reve
rsib
el. 3
. Sch
ritt:
Das
Gle
ichg
ewic
ht d
er F
olge
reak
tion
(„ka
taly
tisch
er Z
erfa
ll“: �
P��
���P
) lie
gt s
ehr w
eit a
uf d
er re
chte
n Se
ite, d
.h.,
dies
er le
tzte
Sc
hritt
ist p
rakt
isch
irre
vers
ibel
. Jed
och
ist d
iese
Rea
ktio
n de
utlic
h la
ngsa
mer
als
die
übr
igen
R
eakt
ione
n un
d be
gren
zt so
mit
kine
tisch
die
ges
amte
Enz
ymre
aktio
n, d
aher
vo
=�[
��].
Bei
seh
r nie
drig
en S
ubst
ratk
onze
ntra
tione
n st
eigt
vo l
inea
r mit
[��]
. Mit
stei
gend
em c
s nim
mt
� ozu
. Bei
Sub
stra
tsät
tigun
g st
eigt
�o
nich
t wei
ter
(vm
ax).
Bei
�m
ax li
egen
ann
äher
nd a
lle, b
ei
Hal
bsät
tigun
g (v
max
/ 2
= K
M) d
ie H
älfte
der
Enz
ymm
olek
üle
als �
� K
ompl
ex v
or.
Die
wic
htig
en k
inet
isch
en G
röße
n K
M u
nd v
max
las
sen
sich
lei
cht
aus
der
LIN
EWEA
VER
-B
UR
K-T
rans
form
atio
n de
r Sub
stra
tsät
tigun
gsfu
nktio
n ab
leite
n (d
oppe
lt-re
zipr
oke A
uftra
gung
, 1/
c S g
egen
1/�
o). D
ie h
yper
bolis
che
Sätti
gung
sfun
ktio
n er
gibt
ein
e G
erad
e m
it de
r St
eigu
ng
KM
/�m
ax. A
us d
en A
bszi
ssen
- und
Ord
inat
ensc
hnitt
punk
ten
dies
er F
unkt
ion
könn
en K
M u
nd � m
ax
best
imm
t wer
den
(s���
�����
���)
.
Bei
allo
ster
isch
en E
nzym
en m
it „k
oope
rativ
er“
Subs
tratb
indu
ng i
st d
ie S
ubst
ratb
indu
ng
wes
entli
ch k
ompl
exer
. D
ie S
ättig
ungs
funk
tion
hat
dann
ein
e si
gmoi
de F
orm
und
die
Li
new
eave
r-B
urk-
Tran
sfor
mat
ion
ist n
icht
meh
r lin
ear.
- ��
-
Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Die
Lac
tat-
Deh
ydro
gena
se (L
DH
) kat
alys
iert
die R
eduk
tion
von
Pyru
vat z
u La
ctat
und
ist d
as
Enzy
m d
er M
ilchs
äure
gäru
ng. I
n de
m V
ersu
ch w
ird d
ie A
bhän
gigk
eit d
er E
nzym
aktiv
ität v
on
der S
ubst
ratk
onze
ntra
tion
best
imm
t. D
ie „
Anf
angs
gesc
hwin
digk
eit“
vo w
ird b
ei v
ersc
hied
enen
La
ctat
konz
entra
tione
n an
hand
des
opt
isch
en T
ests
gem
esse
n: d
ie Z
unah
me
von
NA
DH
mit
der
Zeit
wird
als
Abs
orpt
ions
zuna
hme
bei
340
nm g
emes
sen.
Die
Erg
ebni
sse
wer
den
als
Subs
trats
ättig
ungs
kurv
e au
fget
rage
n,
aus
der
Line
wea
ver-
Bur
k-Tr
ansf
orm
atio
n w
ird
die
Mic
hael
isko
nsta
nte
(KM
) und
die
max
imal
e A
ktiv
ität (
v max
) bes
timm
t.
Für
den
Ver
such
wird
ein
kom
mer
ziel
l er
hältl
iche
s ge
rein
igte
s En
zym
ver
wen
det.
Das
G
leic
hgew
icht
der
LD
H R
eakt
ion
liegt
auf
der
link
en S
eite
(Gle
ichg
ewic
htsk
onst
ante
� =
2,9
· 10
-12 m
ol/l)
.
L-(+
)-La
ctat
+ N
AD
+ P
yruv
at +
NA
DH
+ H
+
Um
de
nnoc
h ei
ne
optim
ale
Oxi
datio
n vo
n La
ctat
zu
er
reic
hen,
w
ird
das
chem
isch
e G
leic
hgew
icht
auf
die
rec
hte
Seite
, al
so z
ugun
sten
der
Pro
dukt
e Py
ruva
t un
d N
AD
H
vers
chob
en, i
ndem
1.
di
e K
onze
ntra
tion
des C
o-Su
bstra
tes N
AD
+ sehr
hoc
h ge
setz
t wird
, 2.
da
s Pro
dukt
Pyr
uvat
che
mis
ch d
urch
Bild
ung
eine
s Hyd
razo
ns
(Kom
plex
bild
ung)
aus
dem
Gle
ichg
ewic
ht g
ezog
en w
ird.
Die
vol
lstä
ndig
e G
leic
hung
laut
et d
aher
:
L-(+
)-La
ctat
+ N
AD
+ + H
ydra
zin
Pyr
uvat
-hyd
razo
n +
NA
DH
+ H
+
Die
Gle
ichg
ewic
htsk
onst
ante
die
ser R
eakt
ion
liegt
bei
ca.
7·
102 m
ol/l.
Ent
spre
chen
d de
m p
H
Opt
imum
der
LD
H w
ird d
ie M
essu
ng b
ei p
H 9
.0 d
urch
gefü
hrt.
Exp
erim
ente
ller
Tei
l
Ben
ötig
te R
eage
nzie
n:
N
AD
(M
G 6
63,4
g/m
ol)
G
lyci
n (z
ur P
uffe
rung
; MG
75,
07 g
/mol
)
L-(+
)-La
ctat
(Na-
Salz
; MG
112
,1 g
/mol
)
Hyd
razi
nhyd
roch
lorid
(MG
50,
06 g
/mol
)
L-LD
H (5
mg/
ml;
550
units
/mg)
= L
acta
t-Deh
ydro
gena
se (E
nzym
)
Stam
mlö
sung
en (v
orha
nden
):
R
eakt
ions
puff
er: 0
,4 M
Hyd
razi
n, 0
,5 M
Gly
zin,
pH
9.0
NA
D, 3
0 m
g/m
l (be
rech
nen
Sie
die
Mol
aritä
t, vo
r Kur
sbeg
inn!
)
LDH
, 0,
01 m
g/m
l
L-(+
)-La
ctat
500
mg/
ml
(ber
echn
en S
ie d
ie M
olar
ität,
vor K
ursb
egin
n!)
Prak
tisch
e T
ipps
für
das e
xper
imen
telle
Arb
eite
n
Mik
rolit
erpi
pett
en
gibt
es i
n ve
rsch
iede
nen
Ver
sion
en, e
ntw
eder
mit
fest
ein
gest
ellte
n od
er m
it ve
rste
llbar
em
Vol
umen
(z.B
. 10-
100 μl
).
- ��
-

Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Pi
pett
ensp
itzen
:B
lau:
für V
olum
ina
bis 1
000 μl
G
elb:
für V
olum
ina
bis 2
00 μ
l W
eiß:
für V
olum
ina
bis 1
0 μl
B
edie
nung
von
aut
omat
isch
en M
ikro
liter
pipe
tten
Zu d
en P
ipet
ten
gibt
es
Kun
stst
offs
pitz
en
(gel
b, b
lau
und
wei
ß), d
ie je
wei
ls n
ur a
uf d
ie
ents
prec
hend
en P
ipet
ten
pass
en. D
ie P
ipet
ten
habe
n m
eist
dr
ei
Dru
ckpu
nkte
: de
r er
ste
Dru
ckpu
nkt
ist
derje
nige
, vo
n de
m l
angs
am
(!!!
) au
fges
augt
wird
; be
im E
ntle
eren
der
Sp
itze
wird
der
Kol
ben
lang
sam
bis
zu
dies
em
erst
en D
ruck
punk
t gef
ührt;
zur
vol
lstä
ndig
en
Entle
erun
g (d
ies i
st d
as g
ewün
scht
e Vol
umen
) de
r Spi
tze
wird
nun
vol
lend
s bi
s zu
m z
wei
ten
Dru
ckpu
nkt
durc
hged
rück
t. B
eim
dr
itten
D
ruck
punk
t wird
zum
inde
st b
ei d
en M
odel
len
von
Eppe
ndor
f die
Spi
tze
abge
wor
fen.
Bei
den
ver
stel
lbar
en P
ipet
ten
(bei
Epp
endo
rf-P
ipet
ten
Her
ausz
iehe
n de
s Bed
ienu
ngsk
nopf
es u
nd v
orsi
chtig
es D
rehe
n) w
ird d
as e
inge
stel
lte V
olum
en
in μ
l ang
ezei
gt.
Ach
tung
! Pip
ette
n nu
r bis
zum
Max
imal
volu
men
auf
dreh
en. N
iem
als g
egen
ein
en W
ider
stan
d w
eite
rdre
hen!
Pip
ette
n ni
emal
s ohn
e Pi
pette
nspi
tze
verw
ende
n.
Her
stel
len
eine
r L
acta
t–K
onze
ntra
tions
reih
e:A
us d
er L
acta
t-Sta
mm
lösu
ng w
ird e
ine
Ver
dünn
ungs
reih
e he
rges
tellt
. Pi
petti
eren
Sie
in
num
mer
ierte
Röh
rche
n (N
r.1-9
) na
ch d
em f
olge
nden
Sch
ema
und
bere
chne
n Si
e vo
r Ku
rsbe
ginn
die
Mol
aritä
t.
Nr.
Lact
at
Stam
mlö
sung
m
l
H2O ml
Err
echn
ete
Kon
zent
ratio
n im
R
öhrc
hen
(mm
ol/l)
Err
echn
ete
Kon
zent
ratio
n in
de
r K
üvet
te
(mm
ol/l)
1
0,5
--
2 0,
2 0,
2
3
0,2
0,3
4 0,
1 0,
4
5
0,1
0,9
6 0,
1 1,
4
7
0,1
2,4
8 0,
1 4,
9
9
--
1,0
- ��
-
Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Vor
bere
itung
:
Phot
omet
er u
nd a
nges
chlo
ssen
en M
essc
ompu
ter a
nste
llen.
Pr
ogra
mm
zur
Auf
nahm
e vo
n En
zym
kine
tiken
star
ten.
D
ie E
inw
eisu
ng e
rfol
gt v
om A
ssis
tent
en.
Mac
hen
Sie
sich
mit
dem
Abl
auf a
m M
essc
ompu
ter v
ertra
ut in
dem
Sie
ein
e Te
stm
essu
ng (a
n N
r. 1)
dur
chfü
hren
.B
enöt
igte
Lös
unge
n un
d Pi
pette
n be
reits
telle
n.
Mes
sabl
auf:
Pipe
ttier
en S
ie in
die
Mes
sküv
ette
(seh
r gen
au!)
:
1,25
ml R
eakt
ions
puff
er (H
ands
chuh
e!)
+
0,1
ml N
AD
Sta
mm
lösu
ng
+ 50
μl L
acta
t au
s der
Kon
zent
ratio
nsre
ihe
(Sta
rten
Sie
mit
Nr.1
)
Ans
atz
mit
eine
m R
ührs
pate
l gut
mis
chen
, Küv
ette
in d
as P
hoto
met
er st
elle
n un
d de
n R
efer
enzw
ert a
ufne
hmen
+ 50
μl (
sehr
gen
au) L
-LD
H z
upip
ettie
ren
Erne
ut m
it de
m R
ührs
pate
l gut
und
rasc
h m
isch
en!
Phot
omet
erde
ckel
schl
ieße
n un
d R
egis
trier
ung
am C
ompu
ter s
ofor
t sta
rten.
Rei
nige
n Si
e di
e be
nutz
te K
üvet
te in
tens
iv m
it H
2O u
m E
nzym
rest
e fü
r die
näc
hste
Mes
sung
ko
mpl
ett z
u en
tfern
en. D
iese
r Flü
ssig
abfa
ll w
ird g
eson
dert
gesa
mm
elt!
Wie
derh
olen
Sie
die
sen
Abl
auf f
ür a
lle L
acta
tkon
zent
ratio
nen.
- �0
-

Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Aus
wer
tung
:
Die
Aus
wer
tung
wird
wäh
rend
des
Pra
ktik
ums m
it M
icro
soft
Exce
l aus
gefü
hrt.
Aus
der
Extin
ktio
nszu
nahm
e pr
o Ze
it w
ird d
ie R
eakt
ions
gesc
hwin
digk
eit
v o b
erec
hnet
(mol
NA
DH
, bzw
. Lac
tat *
sec-1
* m
g En
zym
-1).
Die
s erf
olgt
mit
Hilf
e des
Lam
bert-
Bee
rsch
en-
Ges
etze
s (
NA
DH
, 340
nm
= 6
,3 L
* m
mol
-1 *
cm-1
). D
abei
ist
dar
auf
zu a
chte
n, d
ass
nur
die
Stei
gung
de
s lin
eare
n A
nfan
gsbe
reic
hs
zur
Aus
wer
tung
he
rang
ezog
en
wird
(„
Anf
angs
gesc
hwin
digk
eit“
vo;
sieh
e Ei
nfüh
rung
obe
n).
Jede
r Sub
stra
tkon
zent
ratio
n c S
(mol
/l) k
ann
dann
ein
Wer
t für
vo z
ugeo
rdne
t wer
den.
Tra
gen
Sie
in e
inem
Dia
gram
m v
o geg
en c
S auf
(Su
bstra
tsät
tigun
gsfu
nktio
n; s
. Abb
ildun
g). I
n ei
nem
zw
eite
n D
iagr
amm
trag
en S
ie 1
/vo g
egen
1/c
S auf
(Lin
ewea
ver-
Bur
kD
arst
ellu
ng;
s. A
bb.).
In
dies
er D
arst
ellu
ng s
ollte
n di
e M
essp
unkt
e au
f ei
ner
Ger
aden
lie
gen
(Lin
ewea
ver-
Bur
k D
arst
ellu
ng; s
. Abb
.). In
die
ser D
arst
ellu
ng s
ollte
n di
e M
essp
unkt
e au
f ein
er G
erad
en li
egen
. A
us d
en S
chni
ttpun
kten
der
extra
polie
rten
Aus
glei
chsg
erad
en m
it O
rdin
ate u
nd A
bszi
sse l
asse
n si
ch d
ie W
erte
für v
max
(mol
Lac
tat *
sec-1
* m
g En
zym
-1) u
nd K
M fü
r Lac
tat (
mol
* L
-1) a
blei
ten
Abb
ildun
g: S
ubst
ratfu
nktio
n de
r Lac
tat D
ehyd
roge
nase
- 71
-
Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Ver
such
2: P
erox
idas
eIs
olie
rung
, Tre
nnun
g un
d hi
stoc
hem
isch
er N
achw
eis p
flanz
liche
r Pe
roxi
dase
-Iso
enzy
me
Ver
such
szie
l:
In d
iese
m V
ersu
chst
eil s
olle
n pf
lanz
liche
Iso
enzy
me
am B
eisp
iel d
er P
erox
idas
enzu
näch
stau
s pf
lanz
liche
n G
eweb
en i
solie
rt, ü
ber
eine
nat
ive
Gel
-Ele
ktro
phor
ese
aufg
etre
nnt
und
ansc
hlie
ßend
im
Gel
mitt
els
eine
r sp
ezifi
sche
n Fä
rber
eakt
ion
sich
tbar
gem
acht
wer
den
(Ver
such
2a)
. Ein
dire
kter
Nac
hwei
s de
r Pe
roxi
dase
n in
pfla
nzlic
hen
Gew
eben
erf
olgt
dur
ch
eine
wei
tere
Fär
bere
aktio
n (V
ersu
ch 2
b).
The
oret
isch
er H
inte
rgru
nd
Isoe
nzym
e si
nd E
nzym
e, d
ie i
dent
isch
e ch
emis
che
Rea
ktio
nen
kata
lysi
eren
, si
ch a
ber
in
wei
tere
n Ei
gens
chaf
ten
(z.B
. Mol
mas
se, i
soel
ektri
sche
r Pun
kt, L
okal
isat
ion
im G
eweb
e et
c.)
vone
inan
der u
nter
sche
iden
. Iso
enzy
me
könn
en z
.B. P
rodu
kte
vers
chie
dene
r Gen
e se
in, w
elch
e si
ch in
der
Prim
ärst
rukt
ur (A
min
osäu
rese
quen
z de
s Pr
otei
ns) u
nter
sche
iden
. Neb
en g
enet
isch
be
ding
ten
Isoe
nzym
en k
önne
n ab
er a
uch
post
trans
latio
nale
Mod
ifika
tione
n ei
nes
Prot
eins
zu
meh
rere
n En
zym
varia
nten
füh
ren.
Da
die
gena
ue E
ntst
ehun
g vo
n m
ultip
len
Form
en e
ines
En
zym
s im
Ein
zelfa
ll hä
ufig
nic
ht le
icht
zu
klär
en is
t, ve
rwen
det m
an d
en I
soen
zym
begr
iff
mei
st „
oper
atio
nal“
, d.
h. f
ür a
lle f
unkt
ione
ll äh
nlic
hen
Enzy
me
eine
s O
rgan
ism
us (
glei
che
Nac
hwei
srea
ktio
n).
Pero
xida
sen
sind
ubi
quitä
r in
Pfla
nzen
, Tie
ren
und
Mik
roor
gani
smen
ver
brei
tet.
Die
Meh
rzah
l de
r Pe
roxi
dase
n si
nd
eise
nhal
tige
Porp
hyrin
-Gly
copr
otei
ne
(„H
ämin
-Enz
yme“
), an
dere
hi
ngeg
en
besi
tzen
ei
n C
ytoc
hrom
e-C
ty
pisc
hes-
, ei
n Se
len-
od
er
Van
adiu
m-Z
entru
m.
Pero
xida
sen
benö
tigen
als
Sub
stra
t ent
wed
er H
2O2 o
der O
2, um
ein
e V
ielz
ahl v
on R
eduk
tand
en
oxid
iere
n zu
kön
nen.
Es w
urde
n bi
sher
ber
eits
übe
r 200
mög
liche
Sub
stra
te id
entif
izie
rt.
Pero
xida
sen
RH
2 + H
2O2
2 H
2O +
R
Pfla
nzlic
he P
erox
idas
en d
er K
lass
e II
I (P
OX
, EC
1.1
1.1.
7; E
C =
enz
yme
com
mis
sion
) si
nd
Häm
-hal
tige
Gly
copr
otei
ne u
nd k
omm
en s
owoh
l int
ra-,
als
auch
ext
raze
llulä
r vor
und
kön
nen
aufg
rund
ihre
s is
oele
ktris
chen
Pun
ktes
(3,
5 -
10)
als
neut
ral,
saue
r od
er b
asis
ch k
lass
ifizi
ert
wer
den.
Höh
ere P
flanz
en b
esitz
en in
der
Reg
el ei
ne V
ielz
ahl P
erox
idas
e-Is
oenz
ymen
, die
dur
ch
eine
Mul
tigen
-Fam
ilie
codi
ert
wer
den.
Bis
zu
100
vers
chie
dene
Iso
enzy
me
wur
den
z.B
. in
Arab
idop
sise
ntde
ckt.
Die
Fu
nktio
nen
von
Pero
xida
sen
sind
vi
elfä
ltig;
si
e sp
iele
n ei
ne
Rol
le
bei
der
Bio
synt
hese
/Abb
au v
on L
igni
n in
der
Zel
lwan
d, d
er V
erkn
üpfu
ng v
on Z
ellw
andp
rote
inen
, dem
„E
ntgi
ften“
von
H2O
2, de
r Pro
dukt
ion
von
antim
ikro
biel
len
Rad
ikal
en, d
er W
undh
eilu
ng, d
er
Oxi
datio
n vo
n to
xisc
hen
Red
ukta
nden
, de
m A
uxin
-Kat
abol
ism
us u
nd d
er A
bweh
r ge
gen
Path
ogen
e un
d In
sekt
en.
- 72
-

Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Bes
onde
rs g
ut s
ind
die
Funk
tione
n de
r pf
lanz
liche
n Pe
roxi
dase
n be
i de
r Li
gnifi
zier
ung
(rad
ikal
isch
e Po
lym
eris
atio
n de
r Li
gnin
mon
omer
e) u
nter
such
t w
orde
n. Z
ellw
andg
ebun
dene
Pe
roxi
dase
n la
ssen
sich
dah
er h
isto
chem
isch
in g
roße
n M
enge
n im
ent
steh
ende
n X
ylem
in d
er
Näh
e de
r K
ambi
en
nach
wei
sen.
D
a ei
ne
vers
tärk
te
Lign
ifizi
erun
g hä
ufig
au
ch
bei
Abw
ehrr
eakt
ione
n un
d V
erw
undu
ngen
ein
e R
olle
spi
elt,
ist
es n
icht
ver
wun
derli
ch,
dass
Pa
thog
enbe
fall,
Stre
ss u
nd V
erle
tzun
g zu
r Ind
uktio
n vo
n Pe
roxi
dase
n fü
hren
.
Im �
nter
schi
ed z
u tie
risch
en e
ntha
lten
pfla
nzlic
he P
erox
idas
en v
iele
Koh
lenh
ydra
te
(
max
. 25
� d
er M
olek
ülm
asse
). D
ies
schü
tzt
sie
vor
dest
abili
sier
ende
n Ei
nflü
ssen
, w
ie z
.B.
Prot
eoly
se. A
ufgr
und
der
leic
hten
Zug
ängl
ichk
eit
und
hohe
n St
abili
tät
wer
den
Pero
xida
sen
viel
fach
als
Hilf
senz
yme
in b
ioch
emis
chen
ode
r im
mun
olog
isch
en A
naly
seve
rfah
ren
(z.B
. EL
ISA
-Tes
t, W
este
rn-B
lots
, Sch
wan
gers
chaf
tste
st e
tc.)
eing
eset
zt.
Da
die
Ele
ktro
phor
ese
in V
ersu
chsw
oche
5 a
usfü
hrlic
h be
hand
elt w
ird, s
oll d
as P
rinzi
p di
eser
M
etho
de
an
dies
er
Stel
le
nur
kurz
be
schr
iebe
n w
erde
n:
Die
El
ektro
phor
ese
ist
ein
bioc
hem
isch
es
Tren
nver
fahr
en,
das
auf
unte
rsch
iedl
iche
n ph
ysik
alis
ch-c
hem
isch
en
Eige
nsch
afte
n vo
n M
akro
mol
ekül
en b
eruh
t. M
ittel
s El
ektro
phor
ese
lass
en s
ich
Gem
isch
e ge
lade
ner S
ubst
anze
n (z
.B. P
rote
ine)
tren
nen.
Dab
ei h
ängt
die
Wan
deru
ngsg
esch
win
digk
eit d
er
Subs
tanz
en v
on d
er F
elds
tärk
e un
d vo
n G
röße
, Ges
talt
und
Ladu
ng d
er T
eilc
hen
ab. B
ei d
er
Prot
eint
renn
ung
durc
h ei
ne P
olya
cryl
amid
-Gel
-Ele
ktro
phor
ese
�PA
GE
� w
ird a
ls M
ediu
m
ein
Poly
acry
lam
idge
l ver
wen
det.
Poly
acry
lam
id (
sieh
e A
bbild
ung)
ist
ein
Pol
ymer
, he
rges
tellt
dur
ch d
ie V
erke
ttung
von
A
cryl
amid
-Ein
heite
n un
d de
ren
�ue
rver
netz
ung
über
N,N
�-M
ethy
lenb
isac
ryla
mid
.
D
ie
Poly
mer
isat
ion
erfo
lgt d
urch
den
Rad
ikal
star
ter A
mm
oniu
mpe
rsul
fat.
Zur S
tabi
lisie
rung
d
er
so
ents
tehe
nden
fr
eien
R
adik
ale
wird
de
m
Gem
isch
Te
tram
ethy
lend
iam
in
(T
EMED
) zu
gese
tzt.
Die
Po
reng
röße
de
s G
els
wird
du
rch
die
Acr
ylam
id-
und
N,N
�-M
ethy
lenb
isac
ryla
mid
-Kon
zent
ratio
n be
stim
mt.
Abb
ildun
g. H
erst
ellu
ng v
on
Poly
acry
lam
id d
urch
V
erne
tzun
g vo
n A
cryl
amid
un
dN
,N�M
ethy
lenb
isac
ryla
mid
. A
ls R
adik
alst
arte
r wird
A
mm
oniu
mpe
rsul
fat
zuge
setz
t. ( D
. Voe
t � J.
G.
Voe
t, B
ioch
emis
try, W
iley,
N
ew �
ork,
1��
5).
Die
PA
GE
kann
entw
eder
unt
er n
ativ
en o
der u
nter
den
atur
iere
nden
Bed
ingu
ngen
erfo
lgen
. Ein
e na
tive P
AG
E tr
ennt
Pro
tein
e in
eine
m g
egen
über
dem
in��
i�o-
Zust
and
unve
ränd
erte
n Zu
stan
d.
Enzy
me
beha
lten
ihre
kat
alyt
isch
e A
ktiv
ität,
Olig
omer
e w
erde
n (m
öglic
hst)
erha
lten.
- 7�
-
Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
Im K
urs w
ird e
ine
Ver
tikal
gelk
amm
er n
ach
Stud
ier v
erw
ende
t (si
ehe
unte
n). D
ie G
ele
wer
den
in d
iese
r Ver
such
swoc
he g
este
llt u
nd w
urde
n, w
ie in
Ver
such
swoc
he 5
bes
chrie
ben,
geg
osse
n.
Abb
ildun
g. V
ertik
alge
lkam
mer
nac
h St
udie
r
Akt
ivit�
tsf�
r�un
gen
im G
el o
der
im G
eweb
e si
nd n
eben
den
ber
eits
ken
neng
eler
nten
ge
kopp
elte
n En
zym
test
s ei
ne
wei
tere
M
etho
de
der
Enzy
mbe
stim
mun
g.
Mitt
els
dies
er
soge
nann
ten
„kol
orim
etri
sche
n E
nzym
test
s“ l
asse
n si
ch E
nzym
e od
er a
uch
Met
abol
ite
kolo
rimet
risch
sow
ohl
qual
itativ
, al
s au
ch q
uant
itativ
nac
hwei
sen.
Kol
orim
etris
che
Test
s w
erde
n in
der
Ind
ustri
e un
d de
r M
ediz
in r
outin
emäß
ig e
inge
setz
t z.B
. zur
Bes
timm
ung
des
Blu
tzuc
kers
pieg
els u
nd z
ur B
estim
mun
g vo
n Pe
stiz
iden
im G
rund
was
ser.
Exp
erim
ente
ller
Tei
l
Ver
such
2a:
Isol
ieru
ng v
on P
erox
idas
e Is
oenz
ymen
, Auf
tren
nung
�
�er
nativ
e E
lekt
roph
ores
e un
d ��
r�un
g im
Gel
Zum
Nac
hwei
s de
r Pe
roxi
dase
Akt
ivitä
t ve
rwen
det
man
sta
tt de
r w
enig
abs
orbi
eren
den
natü
rlich
en P
heno
le a
rtifiz
ielle
Akz
epto
ren,
die
bei
ihre
r enz
ymat
isch
en �
mw
andl
ung
eine
m
star
ken
Farb
wec
hsel
unt
erlie
gen.
Im v
orlie
gend
en T
est v
erm
ittel
t die
Per
oxid
ase
die
Oxi
datio
n de
s in
der r
eduz
ierte
n Fo
rm fa
rblo
sen
Was
sers
toff
dona
tors
4-C
hlor
-1-n
apht
hol z
um o
xidi
erte
n,
blau
schw
arz
gefä
rbte
n 4-
Chl
or-1
-nap
htho
l.
Pfla
nzen
mat
eria
l: �
�e 2�
� m
g�
T
omat
e:
gro�
e B
l�tt
er
Ta�
ak:
gro�
e B
l�tt
er, k
lein
e B
l�tt
er, W
urze
l
Ger
�te:
N
2-B
ehäl
ter,
Eisk
iste
, Waa
ge, R
asie
rklin
ge, B
rettc
hen,
Stö
ßel,
Eppe
ndor
fgef
äße,
vor
gekü
hlte
Epp
endo
rftis
chze
ntrif
uge,
Sch
utzb
rille
, Sca
nner
��s
unge
n/�
hem
ikal
ien:
Auf
schl
ussp
uffe
r
0,1
M K
H2P
O4/
M K
2HPO
4, pH
7,0
- 7�
-

Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
3 m
M E
DTA
0,
5� (w
/v) P
olyc
lar A
T
5 m
M D
ithio
eryt
hrito
l, D
TE
nativ
es G
el (1
0�ig
)
4,�
ml A
. bid
est
3,0
ml 1
,5 M
Tris
/HC
l, pH
�,�
4,0
ml 3
0� A
cryl
amid
(fer
tig a
nges
etzt
, gift
ig!!
�10
0 �l
10�
Am
mon
ium
pers
ulfa
t
10
�l T
EMED
Elek
troph
ores
elau
fpuf
fer
0,1�
M G
lyci
n 0,
0125
M T
ris, p
H �
,�
Prob
enpu
ffer
3 m
l Gly
cerin
1
Spat
elsp
itze
Bro
mph
enol
blau
7
ml A
. bid
est
Inku
batio
nspu
ffer
50 m
M K
2HPO
4/KH
2PO
4, p
H 7
,350
0 m
M N
aCl
Färb
elös
ung
20
ml I
nkub
atio
nspu
ffer
5
ml M
eOH
3,75
mg
4-C
hlor
-1-n
apht
hol
(Han
dsch
uhe�
)
H2O
2-Lö
sung
(30�
ig)
(ätze
nd!!
!)
Auf
ar�e
itung
der
Pro
�en
Je
200
mg
fris
ch g
eern
tete
s Pfla
nzen
mat
eria
l (W
aage
) wur
den
in e
in b
esch
rifte
tes 1
,5
ml E
ppen
dorf
gefä
ß ge
gebe
n un
d m
it H
ilfe
in fl
üssi
gen
Stic
ksto
ff g
efro
ren
(�ch
utz�
rille
���)
G
efro
rene
Pro
be m
it ei
nem
Stö
ßel f
ein
pulv
eris
iere
n,an
schl
ieße
nd 4
00 �
l Auf
schl
ussp
uffe
r daz
ugeb
en
Pr
oben
15
min
bei
140
00 rp
m u
nd 4
oC
zent
rifug
iere
n
die
klar
en �
bers
tänd
e m
ittel
s Pip
ette
in 1
,5 m
l Epp
endo
rfge
fäße
übe
rfüh
ren
Sollt
en si
ch n
och
Zellb
ruch
stüc
ke e
tc. i
m �
bers
tand
bef
inde
n er
neut
zen
trifu
gier
en
(s.o
.)
10 �
l aus
dem
�be
rsta
nd in
ein
neu
es E
ppi ü
berf
ühre
n
Alle
4 P
robe
n m
it �e
2,5
�l P
robe
npuf
fer v
erse
tzen
Prob
en b
is z
ur E
lekt
roph
ores
e au
f Eis
auf
bew
ahre
n
Vor
�ere
itung
und
�ur
chf�
hrun
g de
r E
lekt
roph
ores
e
M
arki
eren
Sie
die
Tas
chen
böde
n au
f der
Gla
spla
tte m
it ei
nem
Edd
ing
Pr
oben
-Kam
m v
orsi
chtig
nac
h ob
en a
us d
em G
el z
iehe
n
D
ie G
lasp
latte
n in
die
Ele
ktro
phor
esea
ppar
atur
ein
setz
en
Ei
nmal
den
Dec
kel a
ufse
tzen
, um
zu
test
en, d
ass e
r ric
htig
sitz
t
- 7�
-
Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
V
orra
tsbe
hälte
r (ob
en u
nd u
nten
) mit
Lauf
puff
er b
efül
len;
dab
ei d
arau
f ach
ten,
das
s sic
h
kein
e Lu
ftbla
sen
an d
er u
nter
en G
elka
nte
befin
den
D
as G
esam
tvol
umen
der
Pro
ben
mit
eine
r Pip
ette
in d
ie T
asch
en fü
llen
St
rom
stär
ke v
on 3
0 M
illi-A
mpe
re p
ro G
el e
inst
elle
n un
d de
n La
uf st
arte
n
(Lau
fzei
t ca.
1h)
Anf
�r�e
n de
r Pe
roxi
dase
-Ban
den
im G
el
G
lass
chei
ben
aus E
lekt
roph
ores
e-A
ppar
atur
ent
fern
en u
nd v
orsi
chtig
auf
hebe
ln
Gel
ent
nehm
en u
nd in
ein
er P
last
iksc
hale
in 3
0 m
l Ink
ubat
ions
puff
er 5
min
inku
bier
en
Puff
er a
bgie
ßen
und
die
Färb
elös
ung
(Han
dsch
uhe�
) auf
s Gel
gie
ßen
10
0 �l
30�
iges
H2O
2 in
die
Färb
elös
ung
pipe
ttier
en (�
chut
z�ri
lle�
) und
vor
sicht
ig
sc
hwen
ken
Fä
rbun
g de
r Ban
den
wird
nac
h ca
. 15
min
sich
tbar
Aus
wer
tung
:
Das
Gel
wird
zur
Aus
wer
tung
ein
gesc
annt
. R
este
der
Fär
belö
sung
wer
den
geso
nder
t ges
amm
elt�
Ver
such
2�:
His
tolo
gisc
her
Nac
hwei
s von
Per
oxid
ase
Enzy
me,
bzw
. Iso
enzy
me
sind
nic
ht g
leic
hmäß
ig im
ges
amte
n G
eweb
e ei
ner P
flanz
e ve
rteilt
, so
nder
n ze
igen
spe
zifis
che
Mus
ter,
die
unte
r an
dere
m m
it H
ilfe
von
hist
oche
mis
chen
N
achw
eism
etho
den
sich
tbar
ge
mac
ht
wer
den
könn
en.
Im
Folg
ende
n w
ird
eine
N
achw
eism
etho
de fü
r in
der Z
ellw
and
loka
lisie
rte P
erox
idas
en b
esch
riebe
n un
d du
rchg
efüh
rt.
Als
Sub
stra
t w
ird f
arbl
oses
3,3
�,5,5
�-Te
tram
ethy
lben
zidi
n (T
MB
) ve
rwen
det,
das
durc
h di
e R
eakt
ion
der P
erox
idas
e in
ein
tief
blau
gef
ärbt
es R
adik
alio
n üb
erge
ht (s
iehe
Abb
.) D
ie B
lau-
Färb
ung
ist ü
ber e
inig
e St
unde
n st
abil,
ble
icht
dan
ach
�edo
ch a
us.
Abb
ildun
g: B
ildun
g de
s bla
ugef
ärbt
en 3
,3�,5
,5�-
Tetra
-met
hylb
enzi
din-
Rad
ikal
ions
aus
3,
3�,5
,5�-
Tetra
met
hylb
enzi
din
(TM
B) d
urch
kat
alyt
isch
e W
irkun
g de
r Per
oxid
ase
�nac
h G
ALL
ATI
�PR
AC
HT,
1��
5�
- 7�
-

Woc
he 7
Enz
yme
AG
Kud
la/B
atis
tic
�ur
chf�
hrun
g:TM
B-F
ärbe
lösu
ng:
50 m
l 0,1
M N
aAce
tat,
pH 4
,5
5 m
g Te
tram
ethy
lben
zidi
n in
2,5
ml E
tOH
gel
öst,
ad 1
00 m
l A.b
ides
t
Bev
or S
ie d
ie S
chni
tte a
nfer
tigen
, ber
eite
n Si
e bi
tte i
n Pe
trisc
hale
n fo
lgen
de L
ösun
gen
vor
(Han
dsch
uhe�
):Pr
obe
1: 5
ml H
2OPr
obe
2: 5
ml T
MB
Pr
obe
3: 5
ml T
MB
, Sch
nitte
in d
er L
ösun
g 10
min
inku
bier
en, d
anac
h 15
�l H
2O2
zuge
ben
Prob
e 4:
2,5
ml T
MB
, 2,5
ml A
scor
bat,
Schn
itte
in d
er L
ösun
g 10
min
inku
bier
en, d
anac
h
15
�l H
2O2
zuge
ben
Ferti
gen
Sie
ansc
hlie
ßend
von
fris
chen
Tom
aten
stän
geln
mit
eine
r R
asie
rklin
ge d
ünne
�
uers
chni
tte a
n. D
ie S
chni
tte w
erde
n di
rekt
in d
ie v
orbe
reite
ten
Lösu
ngen
geg
eben
, dam
it di
e �
uers
chni
tte n
icht
an
der L
uft l
iege
n un
d au
stro
ckne
n.
Dok
umen
tiere
n Si
e ih
re B
eoba
chtu
ngen
in e
iner
sche
mat
isch
en Z
eich
nung
und
mac
hen
Sie
ein
Foto
dur
chs M
ikro
skop
� Mar
kier
en S
ie d
ie b
lau
gefä
rbte
n B
erei
che�
Pr
obe
1 (W
asse
rkon
trolle
) so
llte
farb
los
blei
ben.
War
um i
st u
nter
�m
stän
den
�edo
ch e
ine
leic
hte S
chw
arzb
raun
färb
ung
sich
tbar
� In
Prob
e 2 w
ird d
en P
erox
idas
en n
ur d
er F
arbs
toff
TM
B
als S
ubst
rat z
ur V
erfü
gung
ges
tellt
. War
um w
ird a
uch
hier
scho
n ei
ne B
lauf
ärbu
ng si
chtb
ar�
In
Prob
e 3
wer
den
der F
arbs
toff
und
H2O
2 als
Sub
stra
t hin
zu g
egeb
en, s
o da
ss d
ie P
erox
idas
en im
G
eweb
e lo
kalis
iert
wer
den
könn
en. D
isku
tiere
n Si
e ih
re B
eoba
chtu
ng u
nter
Ber
ücks
icht
igun
g de
r Fun
ktio
n de
r Per
oxid
asen
. Was
beo
bach
ten
Sie
in P
robe
4 (A
scor
bat �
Far
bsto
ff�
H2O
2)�
R
este
der
Fär
belö
sung
wer
den
geso
nder
t ges
amm
elt�
- 77
-- 7
� -

Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
A�
Indu
ktio
n de
r -A
myl
ase-
�ynt
hese
dur
ch G
i��e
relli
ns�u
re in
Get
reid
e-k�
rner
n Pf
lanz
enho
rmon
e (A
uxin
e, C
ytok
inin
e, G
ibbe
relli
ne, u
.a.)
habe
n m
ultip
le W
irksa
mke
it; s
ie
führ
en z
u ei
ner V
erän
deru
ng d
er g
enet
isch
en A
ktiv
ität d
er b
etro
ffen
en Z
elle
n. Z
ur E
rklä
rung
di
eses
Phä
nom
ens
geht
man
dav
on a
us, d
ass
es im
Gen
om n
eben
akt
iven
und
inak
tiven
auc
h po
tent
iell
aktiv
e un
d in
aktiv
e G
ene
gibt
. Die
se p
oten
tiell
aktiv
en o
der i
nakt
iven
Gen
e kö
nnen
z.
. du
rch
Hor
mon
e ak
tivie
rt od
er i
nakt
ivie
rt w
erde
n. D
abei
wirk
t da
s H
orm
on n
ur a
ls
Aus
löse
r, w
as d
ie m
ultip
le W
irksa
mke
it er
klär
t. D
irekt
e Fo
lge
der H
orm
onw
irkun
g m
uss d
ann
die
Neu
synt
hese
ode
r der
Syn
thes
esto
p vo
n „m
esse
nger
“ (m
)RN
A s
ein,
was
die
Neu
synt
hese
od
er d
ie H
emm
ung
der B
iosy
nthe
se v
on P
rote
inen
zur
Fol
ge h
at.
Die
Wirk
ung
eine
s Ph
ytoh
orm
ons
läss
t sic
h in
ein
druc
ksvo
ller
Wei
se a
n de
r Sa
men
keim
ung
von
Get
reid
e de
mon
strie
ren.
Die
Frü
chte
der
Grä
ser (
Kar
yops
en) b
este
hen
aus d
em to
ten,
abe
r st
ärke
reic
hen
Endo
sper
m, d
as v
on le
bend
en Z
elle
n (A
leur
onsc
hich
t) um
gebe
n is
t und
an
eine
r Se
ite a
n da
s Scu
tellu
m d
es E
mbr
yos g
renz
t. B
ei d
er K
eim
ung
wird
die
Stä
rke
gesp
alte
n; a
us d
er d
abei
frei
gese
tzte
n G
luco
se g
ewin
nt d
er
Embr
yo d
ie z
ur K
eim
ung
und
zum
(anf
ängl
ich)
het
erot
roph
en W
achs
tum
not
wen
dige
Ene
rgie
so
wie
sein
e B
aust
offe
. Die
Kei
mun
g w
ird a
lso
erst
dur
ch S
tärk
espa
ltung
mög
lich!
Hie
rfür
sind
be
sond
ere
Enzy
me:
Am
ylas
en (
= H
ydro
lase
n) n
otw
endi
g, d
ie v
or d
er K
eim
ung
kein
en
nenn
ensw
erte
n A
bbau
der
Stä
rke
veru
rsac
hen.
Der
Sam
en e
nthä
lt -A
myl
asen
ber
eits
im u
n-ge
keim
ten
Zust
and:
Die
se E
xoen
zym
e alle
in k
önne
n na
ch d
em Q
uelle
n di
e Stä
rkek
örne
r jed
och
nur b
is z
um G
renz
dext
rin h
ydro
lysi
eren
. Dag
egen
wer
den
-Am
ylas
en, d
ie d
ie S
tärk
ekör
ner
volls
tänd
ig a
uflö
sen
könn
en, e
rst n
ach
Que
llung
in d
er A
leur
onsc
hich
t neu
syn
thet
isie
rt. D
ies
wird
vom
(ge
quol
lene
n) E
mbr
yo a
usge
löst
(in
duzi
ert),
und
zw
ar d
urch
Fre
iset
zung
des
Ph
ytoh
orm
ons G
ibbe
relli
nsäu
re (G
A3)
.
Indo
l-Ess
igs�
ure
�IE
�, e
in A
uxin
� G
i��e
relli
ns�u
re �G
A�,
ein
Gi�
�ere
llin�
D
ie In
dukt
ion
der
-Am
ylas
e-Sy
nthe
se in
Get
reid
ekör
nern
ist f
ür G
ibbe
relli
nsäu
re s
pezi
fisch
un
d ka
nn a
ls ä
ußer
st e
mpf
indl
iche
r Bio
test
ben
utzt
wer
den.
Neb
en A
myl
asen
wird
dur
ch d
ie
Sekr
etio
n di
eses
Hor
mon
s au
ch d
ie S
ynth
ese
von
Prot
ease
n un
d R
ibon
ukle
asen
indu
zier
t. D
ie
neu
synt
hetis
ierte
n En
zym
e w
erde
n vo
n de
n Ze
llen
der A
leur
onsc
hich
t in
das t
ote
Endo
sper
m-
gew
ebe
sekr
etie
rt (d
ie m
RN
A w
ird a
m „
raue
n“ E
R tr
ansl
atie
rt un
d di
e En
zym
e na
ch F
altu
ng
über
den
Gol
gi-A
ppar
at z
ur P
lasm
amem
bran
tra
nspo
rtier
t), u
m d
as g
espe
iche
rte R
eser
ve-
mat
eria
l zu
mob
ilisi
eren
(s. n
ächs
te A
bb.).
CH3O
CH2
COO
HH
O
O
OH
NHCH
2—
CO
OH
- 7�
-
Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
A
bb. B
eisp
iel:
�t�r
kehy
drol
yse
Mob
ilisi
erun
g vo
n R
eser
vem
ater
ial i
m E
ndos
perm
-ge
web
e vo
n G
etre
idek
örne
rn n
ach
Indu
ktio
n vo
n hy
drol
ytis
chen
Enz
ymen
(
-Am
ylas
en) d
urch
Gib
bere
llins
äure
(GA
). D
ie A
bbau
prod
ukte
der
Res
erve
stof
fe w
erde
n al
s M
alz
beze
ichn
et. D
a G
ibbe
relli
nsäu
re a
lso
(indi
rekt
) die
Mal
zbild
ung
indu
zier
t, gi
bt m
an in
der
Bra
uere
i heu
te z
usät
zlic
h di
eses
Hor
mon
de
n ke
imen
den
Ger
sten
körn
ern
zu, u
m d
ie M
alza
usbe
ute
zu e
rhöh
en.
Der
Nac
hwei
s der
Indu
ktio
n de
r -A
myl
ase-
Synt
hese
dur
ch d
as P
flanz
enho
rmon
Gib
bere
llin-
säur
e in
Wei
zenk
örne
rn b
asie
rt au
f fol
gend
en Ü
berle
gung
en:
a) E
mbr
yofr
eie
Kor
nhäl
ften
könn
en b
ei Q
uellu
ng k
eine
Gib
bere
llins
äure
zur
Ind
uktio
n de
r -A
myl
ase-
Synt
hese
frei
setz
en. K
ünst
liche
Zuf
uhr v
on G
ibbe
relli
nsäu
re z
u so
lche
n K
orn-
hälft
en s
ollte
abe
r zu
eine
r Ind
uktio
n de
r -A
myl
ase-
Synt
hese
führ
en, u
nd z
war
gen
au s
o w
ie d
ies i
n em
bryo
halti
gen
Kör
nern
ode
r Kor
nhäl
ften
gesc
hieh
t. b)
Es
ist z
u be
wei
sen,
das
s -A
myl
asen
neu
syn
thet
isie
rt w
erde
n. D
afür
wird
ein
Kon
troll-
ansa
tz z
usät
zlic
h m
it ei
nem
Pro
tein
synt
hese
-Inh
ibito
r ink
ubie
rt (z
.B. A
ctid
ion
oder
Cyc
lo-
hexi
mid
). V
ersu
chsd
urch
f�hr
ung:
�de
r V
ersu
ch w
ird
in 2
Tei
len
durc
hgef
�hrt
�
Der
ers
te T
eil,
d.h.
die
��e
ri�is
a�io
n� �
�e���
n� �
nd �n
��ba
�ion
der
�ei
zen�
�rne
r, w
ird d
urch
di
e B
etre
uer d
urch
gefü
hrt.
Sie
erha
lten
für d
en z
wei
ten
Teil,
der
von
Ihne
n du
rchz
ufüh
ren
ist,
die v
orbe
hand
elte
n un
d ei
ngef
rore
nen
Kor
nhäl
ften
und
begi
nnen
mit
der E
xtra
ktio
n (S
.3 u
nten
).
- ��
-

Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
�� T
eil:
��ur
chf�
hrun
g du
rch
die
Bet
reue
r�
Für j
e 4
Arb
eits
grup
pen
wer
den
je 3
0 W
eize
nkör
ner m
it ei
ner R
asie
rklin
ge a
uf e
iner
Gla
spla
tte
äqua
toria
l in
zwei
Häl
ften
zerte
ilt, s
o da
ss m
an e
ine
embr
yoha
ltige
und
ein
e em
bryo
frei
e H
älfte
er
hält.
Je
wei
ls 1
0 em
bryo
�a��i
�e u
nd 3
0 em
bryo
�rei
e K
ornh
älfte
n fü
r: A
nsat
z 1)
10
em
bryo
�a��i
�e K
ornh
älfte
n
2)
10
em
bryo
�rei
e
Kor
nhäl
ften
3)
10
"
"
4)
10
"
"
wer
den
in b
esch
rifte
te 1
00 m
l Bec
herg
läse
r gef
üllt.
Je
der A
nsat
z wird
kur
z zur
Obe
rflä
chen
ster
ilisa
tion
in 1
2% N
atriu
mhy
poch
lorid
geb
adet
. Dan
n w
ird d
ie N
atriu
mhy
poch
lorid
-Lös
ung
abde
kant
iert
und
jede
r Ans
atz
3mal
mit
jew
eils
ca.
50
ml
Aqu
a bi
dest
. ge
was
chen
. N
ach
Abd
ekan
tiere
n de
r le
tzte
n W
asch
was
serp
ortio
n w
ird j
eder
A
nsat
z no
ch m
it ca
. 20
ml d
er e
ntsp
rech
ende
n In
kuba
tions
lösu
ng g
ewas
chen
: A
nsat
z 1)
A
qua
bide
st.
2)
A
qua
bide
st.
3)
10
-6 M
GA
3-Lö
sung
(99
ml A
qua
bide
st. +
1 m
l 10-4
M G
A3-
Lösu
ng)
4)
10-6
M G
A3-
Lösu
ng (9
8 m
l Aqu
a bi
dest
. + 1
ml 1
0-4 M
GA
3-Lö
sung
) plu
s 1 m
l C
hx. (
Cyc
lohe
xim
id-L
ösun
g, 2
mg/
ml)
Dan
n w
erde
n di
e A
nsät
ze in
ca.
50
ml d
er e
ntsp
rech
ende
n In
kuba
tions
lösu
ng b
ei 4
°C im
Küh
l-sc
hran
k üb
er N
acht
geq
uolle
n.
Am
näc
hste
n Ta
g w
ird d
ie Q
uellu
ngsf
lüss
igke
it vo
n de
n K
ornh
älfte
n ab
deka
ntie
rt. V
ier P
VC
-Pe
trisc
hale
n w
erde
n m
it 8-
lagi
gem
Zel
lsto
ff a
usge
legt
und
mit
so v
iel
der
jew
eils
fris
ch
ange
setz
ten
Inku
batio
nslö
sung
get
ränk
t, da
ss d
ie F
lüss
igke
it ge
rade
noc
h in
den
tief
sten
Del
len
des
Papi
ers
frei
ste
ht (
das
Papi
er s
oll
also
seh
r na
ss,
aber
nic
ht ü
berf
lute
t se
in).
Die
10
Kor
nhäl
ften
von
jede
m d
er 4
Ans
ätze
wer
den
dann
so in
die
Pet
risch
alen
ein
gebr
acht
, das
s die
Sc
hnitt
fläch
en n
ach
oben
in d
ie L
uft r
agen
. D
ie v
ersc
hlos
sene
n Pe
trisc
hale
n w
erde
n nu
n bi
s zu
m n
ächs
ten
Tag
bei 3
0°C
im B
ruts
chra
nk
inku
bier
t und
ans
chlie
ßend
bei
-20°
C a
ufbe
wah
rt.
2� T
eil:
�Eig
ene
�ur
chf�
hrun
g - V
ersu
chst
ag �
� E
xtra
ktio
n de
r K
ornh
�lft
en:
�Her
stel
len
des R
ohex
trak
ts�
Gru
ppen
1, 5
, 6 u
nd 1
0 ex
trahi
eren
Roh
extra
kt (R
E) a
us A
nsat
z 1
(em
bryo
�a��i
�e K
ornh
älfte
n in
Aqu
a bi
dest
. ink
ubie
rt), G
rupp
en 2
und
7 e
xtra
hier
en R
E au
s A
nsat
z 2
(em
bryo
�rei
e K
orn-
hälft
en in
Aqu
a bi
dest
.), G
rupp
en 3
und
8 e
xtra
hier
en R
E au
s A
nsat
z 3
(em
bryo
�rei
e K
orn-
hälft
en in
GA
3-Lö
sung
), G
rupp
en 4
und
9 e
xtra
hier
en R
E au
s A
nsat
z 4
(em
bryo
�rei
e K
orn-
hälft
en i
n G
A3-
Lösu
ng p
lus
Cyc
lohe
xim
id).
A�
HT
�N
G:
�ie
RE
m�s
sen
�eim
n�c
hste
n V
ersu
chss
chri
tt �
Am
ylas
e-N
achw
eis�
zw
isch
en d
en G
rupp
en s
o au
sget
ausc
ht w
erde
n,
dass
�ede
r G
rupp
e al
le �
ver
schi
eden
en R
E z
ur V
erf�
gung
steh
en�
- �1
-
Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
Die
10
Kor
nhäl
ften
eine
s A
nsat
zes
wer
den
in e
inem
Mör
ser
mit
etw
as S
and
und
6 m
l des
t. W
asse
r zu
ein
em m
öglic
hst h
omog
enen
Bre
i zer
riebe
n. D
as M
örse
rn w
ird m
it 3
ml W
asse
r be
gonn
en u
nd e
rst n
ach
wei
tgeh
ende
r Ze
rstö
rung
der
Kor
nstru
ktur
(un
d Fi
ltrie
ren,
s.u
.) m
it w
eite
ren
3 m
l Was
ser
nach
gesp
ült.
Das
Hom
ogen
at w
ird d
urch
4-f
ache
n M
ull f
iltrie
rt (M
ull
ausw
inde
n!)
und
unlö
slic
he Z
ellb
esta
ndte
ile 1
0 m
in l
ang
in d
er T
isch
zent
rifug
e be
i m
ax.
Ges
chw
indi
gkei
t abg
etre
nnt.
Der
Übe
rsta
nd w
ird a
ls je
wei
liger
Roh
extra
kt (
RE)
bez
eich
net
und
für d
ie w
eite
ren
Unt
ersu
chun
gen
verw
ende
t. N
achw
eis d
er A
myl
asen
dur
ch R
eakt
ion
im �
t�rk
e-A
gar:
�Ver
such
stag
� u
nd 2
� 2
stär
keha
ltige
Aga
rpla
tten
(2%
Aga
r, 1%
lösl
iche
Stä
rke,
0,1
% C
aCl 2
in P
VC
-Pet
risch
alen
) w
erde
n ge
stel
lt (b
itte
mit
Gru
ppen
num
mer
und
Dat
um b
esch
rifte
n!).
Mit
Hilf
e ei
ner
blau
en
Pipe
ttens
pitz
e w
erde
n 7
glei
chm
äßig
ang
eord
nete
Löc
her a
us d
er A
gars
chic
ht a
usge
stan
zt u
nd
auf
der
Rüc
ksei
te d
er P
etris
chal
e nu
mm
erie
rt. I
n je
des
Loch
wird
nun
der
Roh
extra
kt (
bitte
zw
isch
en d
en G
rupp
en so
aus
taus
chen
, das
s am
End
e al
le 4
ver
schi
eden
en R
ohex
trakt
e in
jede
r G
rupp
e vo
rhan
den
sind
) sow
ie a
uthe
ntis
che
-Am
ylas
e bz
w.
-Am
ylas
e (w
ird g
este
llt) n
ach
dem
folg
ende
n Sc
hem
a pi
petti
ert (
Löch
er ra
ndvo
ll pi
petti
eren
, je
ca. 5
00 μ
l):
in L
och
1
RE
vom
Ans
atz
1)
2
"
"
"
2)
3
"
"
"
3)
4
"
"
"
4)
5
-A
myl
ase
je 1
μl k
äufli
ches
Enz
ym (R
oche
) in
10 m
l Was
ser
6
-A
myl
ase
(= p
ositi
ve K
ontro
llen)
, fer
tig!
7
Aqu
a bi
dest
. (=
neg
ativ
e K
ontro
lle)
Bei
de P
latte
n w
erde
n na
ch d
em V
erde
ckel
n im
Bru
tsch
rank
bei
30°
C i
nkub
iert
(bitt
e se
hr
vors
icht
ig t
rans
porti
eren
!). N
un k
önne
n di
e im
Roh
extra
kt g
elös
ten
Am
ylas
en i
n di
e A
gar-
schi
cht h
inei
ndiff
undi
eren
und
die
dar
in g
elös
te S
tärk
e ab
baue
n. N
ach
3 h
30 m
in (E
nde
des
Kur
stag
es) u
nd n
ach
20 S
td. (
am n
ächs
ten
Kur
stag
) wird
je e
ine
Plat
te a
us d
em B
ruts
chra
nk
geno
mm
en u
nd a
ngef
ärbt
. A
usw
ertu
ng:
�Ver
such
stag
� u
nd 2
� D
ie A
garp
latte
wird
mit
ca. 2
0 m
l ver
dünn
ter (
1:4)
Jod
-Jod
-Kal
ium
(JJK
)-Lö
sung
übe
rgos
sen
und
ca. 1
min
steh
en g
elas
sen.
Dan
n w
ird d
ie JJ
K-L
ösun
g in
ein
e A
bfal
lflas
che
deka
ntie
rt, d
ie
Aga
robe
rflä
che
mit
Leitu
ngsw
asse
r abg
espü
lt un
d di
e Pl
atte
geg
en h
elle
s Lic
ht b
etra
chte
t und
da
s Er
gebn
is d
okum
entie
rt (m
öglic
hst d
urch
Fot
ogra
fiere
n, D
igita
lkam
era
mitb
ringe
n!).
Noc
h im
Aga
r ver
blie
bene
Stä
rke
wird
dur
ch Jo
d-Ei
nlag
erun
g (S
tärk
espi
rale
) ang
efär
bt, w
obei
nic
ht
gesp
alte
ne S
tärk
e du
nkel
viol
ett
und
-Gre
nzde
xtrin
(A
bbau
prod
ukt
von
-Am
ylas
en)
rosa
er
sche
int.
Da
Jod
sich
nur
in
Stär
ke b
zw.
das
-Gre
nzde
xtrin
ein
lage
rn k
ann,
ers
chei
nen
Ber
eich
e m
it vo
llstä
ndig
abg
ebau
ter S
tärk
e w
eiß
(Hin
wei
s auf
-A
myl
ase-
Akt
ivitä
t).
Das
ein
gela
gerte
Jod
sub
limie
rt re
lativ
sch
nell,
dah
er is
t es
wic
htig
, das
s ze
itnah
foto
graf
iert
wird
ode
r alle
Far
bnua
ncen
gle
ich
nach
dem
Anf
ärbe
n ta
bella
risch
not
iert
wer
den!
Dab
ei s
oll
auch
auf
das
Aus
maß
und
die
Far
bqua
lität
der
Ent
färb
ungs
zone
n be
urte
ilt w
erde
n.
�r��
�ren
Sie
Ihre
Erg
ebni
sse!
- �2
-

Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
B� �
osis
-Eff
ekt-
Kur
ve d
er IE
�-W
irku
ng a
uf d
as �
trec
kung
swac
hstu
m
Hor
mon
e ko
mm
en in
der
Pfla
nze
nur i
n se
hr g
erin
gen
Men
gen
vor.
Für d
en d
irekt
en N
achw
eis
und
vor
alle
m f
ür d
ie q
uant
itativ
e B
estim
mun
g de
r Ph
ytoh
orm
one
steh
en n
ur ä
ußer
st
aufw
endi
ge V
erfa
hren
zur
Ver
fügu
ng (z
.B. r
adio
aktiv
mar
kier
te H
orm
one,
Rad
ioim
mun
test
). Zu
r qu
antit
ativ
en B
estim
mun
g ve
rwen
det m
an a
ber
auch
den
sog
. „B
iote
st“.
Des
sen
Prin
zip
best
eht d
arin
, das
s man
an
lebe
nden
, auf
das
Hor
mon
ans
prec
hend
en O
bjek
ten
eine
spez
ifisc
he
Rea
ktio
n m
isst
. Die
Stä
rke
der
Rea
ktio
n w
ird d
ann
mit
eine
r Ei
chku
rve
(Rea
ktio
n au
f da
s en
tspr
eche
nde
Hor
mon
bei
bek
annt
en K
onze
ntra
tione
n) v
ergl
iche
n. E
in s
olch
er B
iote
st b
iete
t no
ch e
ine
wei
tere
Unt
ersu
chun
gsm
öglic
hkei
t: M
an k
ann
dam
it pr
üfen
, ob
syn
thet
isch
he
rges
tellt
e St
offe
die
gle
iche
Rea
ktio
n he
rvor
rufe
n w
ie d
as p
flanz
enei
gene
Hor
mon
. Fü
r ein
en B
iote
st v
erw
ende
t man
sol
che
Pfla
nzen
teile
, die
auf
das
zu
unte
rsuc
hend
e H
orm
on
star
k re
agie
ren.
Bei
den
Aux
inen
(wie
z. B
. Ind
oles
sigs
äure
) ist
das
kla
ssis
che
und
auch
heu
te
noch
mei
st v
erw
ende
te O
bjek
t di
e K
oleo
ptile
(K
eim
sche
ide)
von
Grä
sern
(vo
rwie
gend
A�
ena,
Haf
er) i
m e
tiolie
rten
Zust
and
(d.h
. im
Dun
keln
ang
ezog
en).
In K
oleo
ptile
n w
ird IE
S in
de
r Spi
tze
(api
kal)
prod
uzie
rt, w
ande
rt na
ch u
nten
(bas
ipet
al) u
nd lö
st d
adur
ch in
Stre
ckun
gs-
zone
n st
arke
s Wac
hstu
m a
us. E
ntfe
rnt m
an d
ie S
pitz
e, w
ird d
as ü
brig
e K
oleo
ptilg
eweb
e w
egen
M
ange
l an
eige
ner I
ES fü
r von
auß
en z
ugeg
eben
e IE
S se
hr e
mpf
indl
ich.
In
die
sem
Ver
such
wird
die
Kon
zent
ratio
nsab
häng
igke
it de
s Stre
ckun
gsw
achs
tum
s der
Haf
er-
kole
optil
en v
on e
xoge
n zu
gefü
hrte
r IES
gem
esse
n (D
osis
-Eff
ekt-K
urve
der
IES
Wirk
ung
auf
das S
treck
ungs
wac
hstu
m).
Ver
such
sdur
chf�
hrun
g: In
7 E
rlenm
eyer
kolb
en s
olle
n si
ch je
9 m
l ein
er 1
0-3, 1
0-4, 1
0-5, 1
0-6, 1
0-7, 1
0-8 u
nd 0
mol
aren
IE
S-Lö
sung
bef
inde
n. D
azu
wird
ein
e l0
-3 m
olar
e IE
S-Lö
sung
folg
ende
rmaß
en h
erge
stel
lt:
17,5
2 m
g IE
S w
erde
n in
wen
igen
Tro
pfen
1 N
NaO
H g
elös
t, un
d ca
. 50
– 60
ml A
qua
bide
st.
zuge
gebe
n. D
ie L
ösun
g w
ird m
it 1
N H
CI
auf
eine
n pH
-Wer
t um
5 e
inge
stel
lt un
d da
nn
quan
titat
iv in
ein
en 1
00 m
l Mes
skol
ben
über
führ
t (d.
h. 3
mal
mit
ca. 5
ml A
qua
bide
st. n
ach-
spül
en).
Ans
chlie
ßend
wird
mit
Aqu
a bi
dest
. au
f 10
0 m
l au
fgef
üllt.
(D
er V
ergl
eich
bark
eit
halb
er u
nd a
us G
ründ
en d
er Z
eite
rspa
rnis
wird
die
10-3
M IE
S-Lö
sung
ges
tellt
). H
erst
elle
n de
r ln
ku�a
tions
med
ien
f�r
die
Kol
eopt
ilen:
In e
inen
Erle
nmey
erko
lben
wer
den
9 m
l Aqu
a bi
dest
. pip
ettie
rt (K
ontro
llans
atz
ohne
IES)
. Aus
de
r 10
-3 m
olar
en I
ES-L
ösun
g w
erde
n di
e an
dere
n IE
S-Lö
sung
en d
urch
Ver
dünn
en i
n 10
er
Schr
itten
her
gest
ellt.
Daz
u w
erde
n 10
ml d
er 1
0-3 m
olar
en L
ösun
g in
ein
en E
rlenm
eyer
kolb
en
pipe
ttier
t. H
ierv
on w
ird 1
ml i
n ei
nen
zwei
ten
Erle
nmey
er p
ipet
tiert
und
9 m
l Aqu
a bi
dest
. da
zuge
gebe
n. N
ach
grün
dlic
hem
Dur
chm
isch
en w
ird a
us d
iese
r 10
-4 m
olar
en I
ES-L
ösun
g w
iede
r 1 m
l ent
nom
men
(usw
.).
Her
stel
len
der
Kol
eopt
ilseg
men
te:
Etio
lierte
Haf
erko
leop
tilen
wer
den
aus
der
Anz
ucht
scha
le e
ntno
mm
en (
Kol
eopt
ile n
icht
dr
ücke
n S
chäd
igun
g!)
und
das
Prim
ärbl
att
entfe
rnt.
Mit
eine
m S
chne
idea
ppar
at w
erde
n m
ehre
re K
oleo
ptile
n gl
eich
zeitg
um
2 m
m d
ekap
itier
t und
5 m
m la
nge S
tück
e gew
onne
n. D
iese
K
oleo
ptilz
ylin
der w
erde
n so
fort
mith
ilfe
eine
r Pin
zette
in d
ie E
rlenm
eyer
mit
der I
ES-L
ösun
g ge
gebe
n, d
.h. j
e 15
Seg
men
te p
ro L
ösun
g. D
ie E
rlenm
eyer
wer
den
mit
Alu
folie
abg
edec
kt u
nd
für c
a. 2
4 St
unde
n du
nkel
ges
tellt
. Es m
uss d
arau
f gea
chte
t wer
den,
das
s alle
Kol
eopt
ilzyl
inde
r in
der
Lös
ung
schw
imm
en u
nd n
icht
am
Ran
d de
s G
efäß
es h
afte
n! D
anac
h w
ird d
er V
ersu
ch
vom
Vor
tag
ausg
ewer
tet:
Die
Län
gen
der
Kol
eopt
ilseg
men
te w
erde
n au
f M
illim
eter
papi
er
ausg
emes
sen
(auf
0,5
mm
gen
au) u
nd m
it de
n A
usga
ngsw
erte
n ve
rrec
hnet
.
- ��
-
Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
�tat
istik
: A
ufgr
und
der S
treuu
ng d
es b
iolo
gisc
hen
Mat
eria
ls u
nd w
egen
der
Feh
lerm
öglic
hkei
ten
bei d
en
Mes
sung
en w
erde
n fo
lgen
de s
tatis
tisch
e R
echn
unge
n fü
r di
e Zu
wac
hslä
ngen
(En
dlän
ge d
er
Kol
eopt
ilseg
men
te m
inus
Anf
angs
läng
e) d
urch
gefü
hrt.
a) D
ie E
inze
lwer
te x
i der
jew
eilig
en IE
S-K
onz.
wer
den
zum
�itt
elw
ert (
MW
) ver
rech
net:
n
ist d
ie A
nzah
l der
gem
esse
nen
Wer
te, h
ier a
lso
n =
10.
b) D
er M
ittel
wer
t sag
t jed
och
nich
ts d
arüb
er a
us, w
ie st
ark
die
Ein
zelw
erte
um
ihn
heru
m
stre
uen,
z.B
.
1.
Mes
srei
he:
2. M
essr
eihe
:
x 1i
19
x 2i
10
20
20
21
30
= 6
0
= 60
MW
1 =
20
MW
2 =
20
n-
1 =
1 n-
1 =
10
Bei
de M
essr
eihe
n er
gebe
n de
n gl
eich
en M
ittel
wer
t, ob
woh
l die
ers
te v
iel w
enig
er st
reut
als
die
zw
eite
. A
ls M
aß f
ür d
ie S
treuu
ng n
imm
t m
an d
ie S
umm
e de
r D
iffer
enze
n zw
isch
en d
en
einz
elne
n M
essw
erte
n un
d de
m M
ittel
wer
t: (
x i
��
). W
egen
der
unt
ersc
hied
liche
n V
or-
zeic
hen
kann
die
se S
umm
e ab
er N
ull e
rgeb
en (s
iehe
obi
ges B
eisp
iel).
Dah
er w
ird d
ie D
iffer
enz
x i
MW
vor
der
Sum
men
bild
ung
quad
riert,
so
dass
nur
noc
h po
sitiv
e W
erte
vor
kom
men
. Als
St
anda
rdab
wei
chun
g be
zeic
hnet
man
folg
ende
n A
usdr
uck:
Ei
n w
eite
res M
aß fü
r die
Stre
uung
ist d
er �
mitt
lere
�eh
ler
des �
ittel
wer
tes�
(auc
h al
s St
anda
rdab
wei
chun
g de
s Mitt
elw
erte
s M
W b
ezei
chne
t)
n��
�i
1)
(2
1n
��
��
in
)1(
)(
2
nn
��
�n�
mi
- ��
-

Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
� un
d m
kan
n m
an fü
r die
ein
zeln
en IE
S-K
onze
ntra
tione
n m
it H
ilfe
der S
tatis
tikfu
nktio
n vo
n Ta
sche
nrec
hner
n be
rech
nen
und
als
± -W
ert z
um M
ittel
wer
t pro
toko
llier
en. A
ls g
raph
isch
es
Maß
für
die
Stre
uung
wird
m a
ls s
enkr
echt
er S
trich
nac
h ob
en (
+) u
nd n
ach
unte
n (-
) vo
m
Mitt
elw
ert a
us d
arge
stel
lt. D
amit
erhä
lt m
an e
in g
raph
isch
es (q
ualit
ativ
es) M
aß fü
r die
Fra
ge
nach
der
Irrtu
msw
ahrs
chei
nlic
hkei
t: Is
t der
Unt
ersc
hied
zw
isch
en z
wei
Mitt
elw
erte
n au
f Gru
nd
vers
chie
dene
r Ver
such
sbed
ingu
ngen
ent
stan
den
oder
ist d
er U
nter
schi
ed re
in z
ufäl
lig?
Eine
wei
tere
Mög
lichk
eit d
ie Ir
rtum
swah
rsch
einl
ichk
eit z
wis
chen
zw
ei M
ittel
wer
ten
zu p
rüfe
n is
t de
r so
gena
nnte
t-T
est.
Die
ser
wur
de z
uers
t 19
08 v
on W
.S. G
osse
t be
schr
iebe
n, d
er a
ls
Che
mik
er u
nd M
athe
mat
iker
in
der
Qua
lität
skon
trolle
der
Gui
nnes
s-B
raue
rei
in D
ublin
ar
beite
te. G
osse
t dur
fte s
eine
Erg
ebni
sse
nich
t unt
er s
eine
m N
amen
pub
lizie
ren.
Er
benu
tzte
da
s Pse
udon
ym "
Stud
ent"
. Aus
die
sem
Gru
nd h
eißt
der
Tes
t auc
h �t
uden
t-t-
Tes
t. A
usw
ertu
ng:
Zur
Aus
wer
tung
des
Ver
such
es w
erde
n di
e M
ittel
wer
te d
er Z
uwac
hslä
ngen
(m
m)
in e
inem
D
iagr
amm
geg
en d
ie e
ntsp
rech
ende
n IE
S-K
onze
ntra
tione
n (M
) au
fget
rage
n. F
ür d
ie M
ittel
-w
erte
der
höc
hste
n IE
S-K
onze
ntra
tion
(l0-3
M) u
nd d
er M
olar
ität d
er g
rößt
en L
änge
nzun
ahm
e w
erde
n di
e Wer
te v
on m
ber
echn
et u
nd in
die
Gra
phik
eing
etra
gen.
Um
zu p
rüfe
n, o
b de
r Unt
er-
schi
ed z
wis
chen
die
sen
Wer
ten
real
ode
r zuf
alls
bedi
ngt i
st, w
ird d
er t-
Test
zum
Ver
glei
ch d
er
beid
en M
ittel
wer
te d
urch
gefü
hrt.
Daz
u w
erde
n di
e en
tspr
eche
nden
Wer
te d
er S
tand
ard-
abw
eich
ung
� un
d de
s Mitt
elw
erte
s ��
in d
ie o
bige
For
mel
ein
gese
tzt.
A
us d
er b
eilie
gend
en t
-Tab
elle
wird
dan
n en
tnom
men
, w
elch
er P
roze
ntsa
tz
an
Irrtu
ms-
wah
rsch
einl
ichk
eit (
Irrtu
m h
eißt
zuf
ällig
e V
ersc
hied
enhe
it) d
em e
rmitt
elte
n t-W
ert e
ntsp
richt
. D
ie Z
ahl d
er F
reih
eits
grad
e f (
1. S
palte
) leg
t die
Zei
le in
der
Tab
elle
fest
, in
der d
er g
efun
dene
t-W
ert a
ufge
such
t wer
den
mus
s (f
= n
1 +
n 2 -2
). A
us d
er u
nter
sten
Zei
le d
er T
abel
le k
ann
die
ents
prec
hend
e Ir
rtum
swah
rsch
einl
ichk
eit
(ev
entu
ell d
urch
Ext
rapo
liere
n zw
isch
en d
en a
n-ge
gebe
nen
Wer
ten)
abg
eles
en w
erde
n.
)1
1 (2
)(
)(
21
21
22
22
11
21
nn
nn
��
��
��
��
��
�i
i
)1
1 (2
)1(
)1(
21
21
22 2
12 1
21
nn
nn
n�
n�
��
��
- ��
-
Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
t - V
ER
TE
I��
NG
��t
uden
t-t-
Tes
t�
f:
1
1,00
0 2,
41
6,31
12
,70
31,8
2 63
,70
318,
30
637,
00
2
0,81
6 1,
60
2,29
4,
30
6,97
9,
92
22,3
3 31
,60
3
0,76
5 1,
42
2,35
3,
18
4,54
5,
84
10,2
2 12
,90
4
0,74
1 1,
34
2,13
2,
78
3,75
4,
60
7,17
8,
61
5
0,72
7 1,
30
2,01
2,
57
3,37
4,
03
5,89
6,
86
6
0,71
8 1,
27
1,94
2,
45
3,14
3,
71
5,21
5,
96
7
0,71
1 1,
25
1,89
2,
36
3,00
3,
50
4,79
5,
40
8
0,70
6 1,
24
1,86
2,
31
2,90
3,
36
4,50
5,
04
9
0,70
3 1,
23
1,83
2,
26
2,82
3,
25
4,30
4,
75
10
0,70
0 1,
22
1,81
2,
23
2,76
3,
17
4,14
4,
59
11
0,69
7 1,
21
1,80
2,
20
2,72
3,
11
4,03
4,
44
12
0,69
5 1,
21
1,78
2,
18
2,68
3,
05
3,93
4,
32
13
0,69
4 1,
20
1,77
2,
16
2,65
3,
01
3,85
4,
22
14
0,69
2 1,
20
1,76
2,
14
2,62
2,
98
3,79
4,
14
15
0,69
1 1,
20
1,75
2,
13
2,60
2,
95
3,73
4,
07
16
0,69
0 1,
19
1,75
2,
12
2,58
2,
92
3,69
4,
01
17
0,68
9 1,
19
1,74
2,
11
2,57
2,
90
3,65
3,
96
18
0,68
8 1,
19
1,73
2,
10
2,55
2,
88
3,61
3,
92
19
0,68
8 1,
19
1,73
2,
09
2,54
2,
86
3,58
3,
88
20
0,68
7 1,
18
1,73
2,
09
2,53
2,
85
3,55
3,
85
21
0,68
6 1,
18
1,72
2,
08
2,52
2,
83
3,53
3,
82
22
0,68
6 1,
18
1,72
2,
07
2,21
2,
82
3,51
3,
79
23
0,68
5 1,
18
1,71
2,
07
2,50
2,
81
3,49
3,
77
24
0,68
5 1,
18
1,71
2,
06
2,49
2,
80
3,47
3,
74
25
0,68
4 1,
18
1,71
2,
06
2,49
2,
79
3,45
3,
72
26
0,68
4 1,
18
1,71
2,
06
2,48
2,
78
3,44
3,
71
27
0,68
4 1,
18
1,71
2,
05
2,47
2,
77
3,42
3,
69
28
0,68
3 1,
17
1,70
2,
05
2,47
2,
76
3,41
3,
67
29
0,68
3 1,
17
1,70
2,
05
2,46
2,
76
3,40
3,
66
30
0,68
3 1,
17
1,70
2,
04
2,46
2,
75
3,39
3,
65
40
0,68
1 1,
17
1,68
2,
02
2,42
2,
70
3,31
3,
55
60
0,67
9 1,
16
1,67
2,
00
2,39
2,
66
3,23
3,
46
120
0,
677
1,16
1,
66
1,98
2,
36
2,62
3,
17
3,37
121
0,67
4 1,
15
1,64
1,
96
2,33
2,
58
3,09
3,
29
:
50
25
10
5 2
1 0,
2 0,
1 =
Irrtu
msw
ahrs
chei
nlic
hkei
t für
zw
eise
itige
Fra
gest
ellu
ng (i
n %
) f =
Fre
ihei
tsgr
ad d
er t-
Ver
teilu
ng, f
= n
1 +
n 2 -
2 al
tern
ativ
: http
://m
edw
eb.u
ni-m
uens
ter.d
e/in
stitu
te/im
ib/le
hre/
skrip
te/b
iom
athe
/utte
st.h
tml
- ��
-
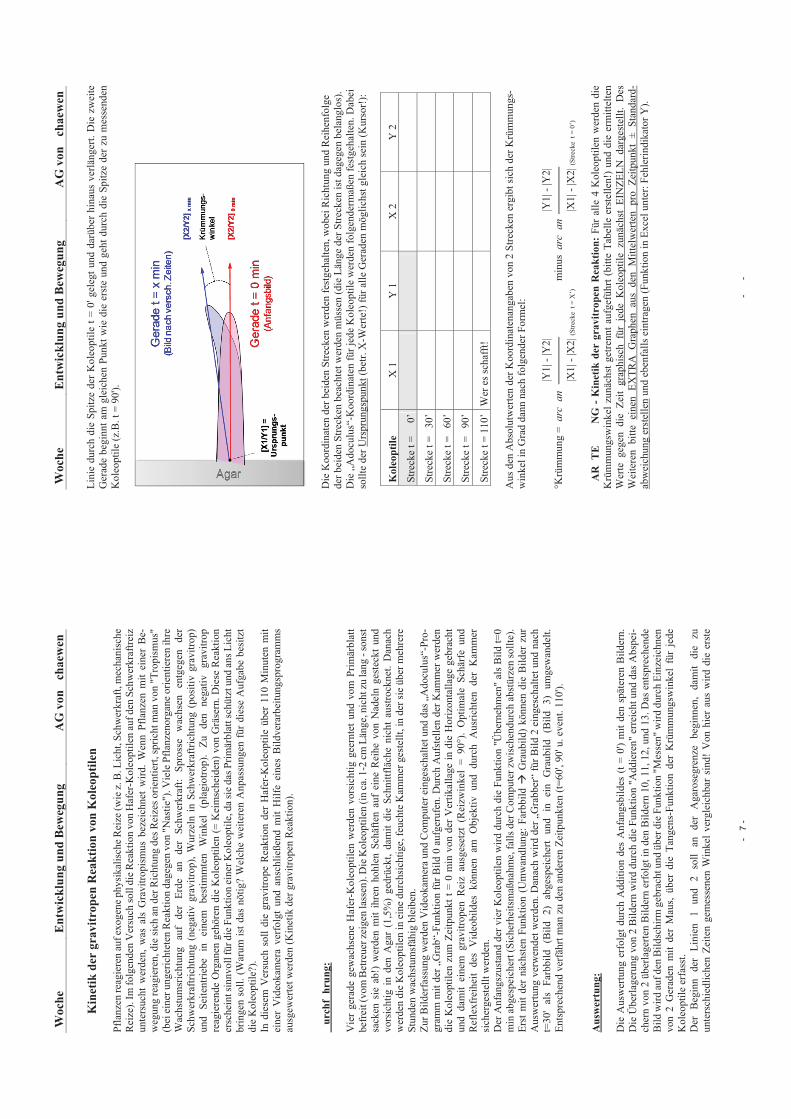
Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
��
Kin
etik
der
gra
vitr
open
Rea
ktio
n vo
n K
oleo
ptile
n Pf
lanz
en re
agie
ren
auf e
xoge
ne p
hysi
kalis
che R
eize
(wie
z. B
. Lic
ht, S
chw
erkr
aft,
mec
hani
sche
R
eize
). Im
folg
ende
n V
ersu
ch so
ll di
e Rea
ktio
n vo
n H
afer
-Kol
eopt
ilen
auf d
en S
chw
erkr
aftre
iz
unte
rsuc
ht w
erde
n, w
as a
ls G
ravi
tropi
smus
bez
eich
net
wird
. W
enn
Pfla
nzen
mit
eine
r B
e-w
egun
g re
agie
ren,
die
sich
an
der R
icht
ung
des R
eize
s orie
ntie
rt, sp
richt
man
von
"Tro
pism
us"
(bei
ein
er u
nger
icht
eten
Rea
ktio
n da
gege
n vo
n "N
astie
"). V
iele
Pfla
nzen
orga
ne o
rient
iere
n ih
re
Wac
hstu
msr
icht
ung
auf
der
Erde
an
der
Schw
erkr
aft:
Spro
sse
wac
hsen
ent
gege
n de
r Sc
hwer
kraf
trich
tung
(ne
gativ
gra
vitro
p), W
urze
ln in
Sch
wer
kraf
trich
tung
(po
sitiv
gra
vitro
p)
und
Seite
ntrie
be i
n ei
nem
bes
timm
ten
Win
kel
(pla
giot
rop)
. Zu
den
neg
ativ
gra
vitro
p re
agie
rend
e O
rgan
en g
ehör
en d
ie K
oleo
ptile
n (=
Kei
msc
heid
en) v
on G
räse
rn. D
iese
Rea
ktio
n er
sche
int s
innv
oll f
ür d
ie F
unkt
ion
eine
r Kol
eopt
ile, d
a sie
das
Prim
ärbl
att s
chüt
zt u
nd an
s Lic
ht
brin
gen
soll.
(War
um is
t das
nöt
ig?
Wel
che
wei
tere
n A
npas
sung
en fü
r die
se A
ufga
be b
esitz
t di
e K
oleo
ptile
?).
In d
iese
m V
ersu
ch s
oll d
ie g
ravi
trope
Rea
ktio
n de
r H
afer
-Kol
eopt
ile ü
ber
110
Min
uten
mit
eine
r V
ideo
kam
era
verf
olgt
und
ans
chlie
ßend
mit
Hilf
e ei
nes
Bild
vera
rbei
tung
spro
gram
ms
ausg
ewer
tet w
erde
n (K
inet
ik d
er g
ravi
trope
n R
eakt
ion)
. �
urch
f�hr
ung:
Vie
r ge
rade
gew
achs
ene
Haf
er-K
oleo
ptile
n w
erde
n vo
rsic
htig
gee
rnte
t und
vom
Prim
ärbl
att
befr
eit (
vom
Bet
reue
r zei
gen
lass
en).
Die
Kol
eopt
ilen
(in ca
. 1-2
cm L
änge
, nic
ht zu
lang
- so
nst
sack
en s
ie a
b!)
wer
den
mit
ihre
n ho
hlen
Sch
äfte
n au
f ei
ne R
eihe
von
Nad
eln
gest
eckt
und
vo
rsic
htig
in
den
Aga
r (1
,5%
) ge
drüc
kt, d
amit
die
Schn
ittflä
che
nich
t au
stro
ckne
t. D
anac
h w
erde
n di
e K
oleo
ptile
n in
ein
e du
rchs
icht
ige,
feuc
hte
Kam
mer
ges
tellt
, in
der s
ie ü
ber m
ehre
re
Stun
den
wac
hstu
msf
ähig
ble
iben
. Zu
r Bild
erfa
ssun
g w
erde
n V
ideo
kam
era
und
Com
pute
r ein
gesc
halte
t und
das
„A
docu
lus“
-Pro
-gr
amm
mit
der „
Gra
b“-F
unkt
ion
für B
ild 0
auf
geru
fen.
Dur
ch A
ufst
elle
n de
r Kam
mer
wer
den
die
Kol
eopt
ilen
zum
Zei
tpun
kt t
= 0
min
von
der
Ver
tikal
lage
in d
ie H
oriz
onta
llage
geb
rach
t un
d da
mit
eine
m g
ravi
trope
n R
eiz
ausg
eset
zt (
Rei
zwin
kel
= 90
°).
Opt
imal
e Sc
härf
e un
d R
efle
xfre
ihei
t de
s V
ideo
bild
es k
önne
n am
Obj
ektiv
und
dur
ch A
usric
hten
der
Kam
mer
si
cher
gest
ellt
wer
den.
D
er A
nfan
gszu
stan
d de
r vie
r Kol
eopt
ilen
wird
dur
ch d
ie F
unkt
ion
"Übe
rneh
men
" al
s Bild
t=0
min
abg
espe
iche
rt (S
iche
rhei
tsm
aßna
hme,
falls
der
Com
pute
r zw
isch
endu
rch
abst
ürze
n so
llte)
. Er
st m
it de
r nä
chst
en F
unkt
ion
(Um
wan
dlun
g: F
arbb
ild
Gra
ubild
) kö
nnen
die
Bild
er z
ur
Aus
wer
tung
ver
wen
det w
erde
n. D
anac
h w
ird d
er „
Gra
bber
“ fü
r Bild
2 e
inge
scha
ltet u
nd n
ach
t=30
' al
s Fa
rbbi
ld (
Bild
2)
abge
spei
cher
t un
d in
ein
Gra
ubild
(B
ild 3
) um
gew
ande
lt.
Ents
prec
hend
ver
fähr
t man
zu
den
ande
ren
Zeitp
unkt
en (t
=60'
, 90'
u. e
vent
. 110
').
Aus
wer
tung
: D
ie A
usw
ertu
ng e
rfol
gt d
urch
Add
ition
des
Anf
angs
bild
es (
t = 0
') m
it de
n sp
äter
en B
ilder
n.
Die
Übe
rlage
rung
von
2 B
ilder
n w
ird d
urch
die
Fun
ktio
n "A
ddie
ren"
err
eich
t und
das
Abs
pei-
cher
n vo
n 2
über
lage
rten
Bild
ern
erfo
lgt i
n de
n B
ilder
n 10
, 11,
12,
und
13.
Das
ent
spre
chen
de
Bild
wird
auf d
en B
ildsc
hirm
geb
rach
t und
übe
r die
Fun
ktio
n "M
esse
n" w
ird d
urch
Ein
zeic
hnen
vo
n 2
Ger
aden
mit
der
Mau
s, üb
er d
ie T
ange
ns-F
unkt
ion
der
Krü
mm
ungs
win
kel
für
jede
K
oleo
ptile
erf
asst
. D
er B
egin
n de
r Li
nien
1 u
nd 2
sol
l an
der
Aga
rose
gren
ze b
egin
nen,
dam
it di
e zu
un
ters
chie
dlic
hen
Zeite
n ge
mes
sene
n W
inke
l ver
glei
chba
r si
nd!
Von
hie
r au
s w
ird d
ie e
rste
- �7
-
Woc
he �
E
ntw
ickl
ung
und
Bew
egun
gA
G v
on �
chae
wen
Lini
e du
rch
die
Spitz
e de
r Kol
eopt
ile t
= 0'
gel
egt u
nd d
arüb
er h
inau
s ve
rläng
ert.
Die
zw
eite
G
erad
e be
ginn
t am
gle
iche
n Pu
nkt w
ie d
ie e
rste
und
geh
t dur
ch d
ie S
pitz
e de
r zu
mes
send
en
Kol
eopt
ile (z
.B. t
= 9
0').
D
ie K
oord
inat
en d
er b
eide
n St
reck
en w
erde
n fe
stge
halte
n, w
obei
Ric
htun
g un
d R
eihe
nfol
ge
der b
eide
n St
reck
en b
each
tet w
erde
n m
üsse
n (d
ie L
änge
der
Stre
cken
ist d
ageg
en b
elan
glos
). D
ie „
Ado
culu
s“-K
oord
inat
en fü
r jed
e K
oleo
ptile
wer
den
folg
ende
rmaß
en fe
stge
halte
n. D
abei
so
llte
der U
rspr
ungs
punk
t (be
tr. X
-Wer
te!)
für a
lle G
erad
en m
öglic
hst g
leic
h se
in (K
urso
r!):
Kol
eopt
ile �
� X
1
Y 1
X
2
Y 2
Stre
cke
t =
0’
Stre
cke
t =
30’
Stre
cke
t =
60’
Stre
cke
t =
90’
Stre
cke
t = 1
10’
Wer
es s
chaf
ft!
A
us d
en A
bsol
utw
erte
n de
r Koo
rdin
aten
anga
ben
von
2 St
reck
en e
rgib
t sic
h de
r Krü
mm
ungs
-w
inke
l in
Gra
d da
nn n
ach
folg
ende
r For
mel
:
|Y
1| -
|Y2|
|Y1|
- |Y
2|
°Krü
mm
ung
= a
rc �a
n
min
us a
rc �a
n
|X
1| -
|X2|
(Stre
cke
t =
X’)
|X1|
- |X
2| (S
treck
e t
= 0’
) �
AR
�TE
��
�N
G -
Kin
etik
der
gra
vitr
open
Rea
ktio
n: F
ür a
lle 4
Kol
eopt
ilen
wer
den
die
Krü
mm
ungs
win
kel z
unäc
hst g
etre
nnt a
ufge
führ
t (bi
tte T
abel
le e
rste
llen!
) un
d di
e er
mitt
elte
n W
erte
geg
en d
ie Z
eit
grap
hisc
h fü
r je
de K
oleo
ptile
zun
ächs
t EI
NZE
LN d
arge
stel
lt. D
es
Wei
tere
n bi
tte e
inen
EX
TRA
Gra
phen
aus
den
Mitt
elw
erte
n pr
o Ze
itpun
kt ±
Sta
ndar
d-ab
wei
chun
g er
stel
len
und
eben
falls
ein
trage
n (F
unkt
ion
in E
xcel
unt
er: F
ehle
rindi
kato
r Y).
- ��
-

- 89
-- 9
0 -

- 9�
-- 9
� -

- 9�
-- 9
� -

- 9�
-- 9
� -

- 9�
-- 9
8 -

- 99
-- �
00 -

Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
Plas
mid
isol
ieru
ng, R
estr
iktio
n &
Tra
nsfo
rmat
ion
The
orie
D
er h
eutig
e V
ersu
chst
ag d
es P
rakt
ikum
s be
schä
ftige
n si
ch m
it de
n gr
undl
egen
den
Met
hode
n de
r Mol
ekul
arge
netik
, die
heu
te in
pra
ktis
ch a
llen
Ber
eich
en d
er B
iolo
gie
eing
eset
zt w
erde
n.
Heu
te w
erde
n Si
e Pl
asm
id-D
NA
aus
Bak
terie
n is
olie
ren,
rei
nige
n un
d zu
r K
ontro
lle e
inen
R
estri
ktio
nsve
rdau
dur
chfü
hren
. Si
e w
erde
n ko
mpe
tent
e B
akte
rien
mit
dem
ger
eini
gten
Pl
asm
id
trans
form
iere
n un
d w
erde
n de
n Tr
ansf
orm
atio
nser
folg
au
f se
lekt
iven
M
edie
n üb
erpr
üfen
.
Plas
mid
ePl
asm
ide s
ind
extra
-chr
omos
omal
e, in
der
Reg
el ri
ngfö
rmig
e (zi
rkul
äre)
dop
pels
träng
ige D
NA
-M
olek
üle,
die
in
viel
en M
ikro
orga
nism
en h
äufig
auc
h na
türli
ch v
orko
mm
en.
Sie
besi
tzen
G
röße
n zw
isch
en 2
und
200
kb u
nd d
iene
n vi
elfä
ltige
n bi
olog
isch
en F
unkt
ione
n.
Die
in d
er M
olek
ular
gene
tik v
erw
ende
ten
Plas
mid
e (a
uch
als
Vek
tore
n be
zeic
hnet
, da
sie
zur
Übe
rtrag
ung
zu k
loni
eren
der G
ene
in B
akte
rien
dien
en) s
tam
men
nur
von
ein
er k
lein
en Z
ahl
bakt
erie
ller
Plas
mid
e ab
, di
e üb
er g
ezie
lte D
NA
-Rek
ombi
natio
n fü
r di
e ve
rsch
iede
nen
Anw
endu
ngen
opt
imie
rt w
urde
n. A
lle d
iese
Pla
smid
e (i.
d.R
. 2 –
15k
b) tr
agen
ein
Gen
für e
ine
Ant
ibio
tika-
Res
iste
nz, d
amit
Plas
mid
-hal
tige,
tran
sgen
e B
akte
rien
im G
egen
satz
zu
Plas
mid
-lo
sen
Wild
typ-
Bak
terie
n au
f A
ntib
iotik
a-ha
ltige
m
Sele
ktio
nsm
ediu
m
wac
hsen
kö
nnen
. D
aneb
en en
thal
ten
alle
Pla
smid
e ein
en R
eplik
atio
nsur
spru
ng (O
RI:
„orig
in o
f rep
licat
ion“
), de
r si
e zu
aut
onom
er R
eplik
atio
n im
Bak
teriu
m b
efäh
igt.
Der
OR
I ent
sche
idet
übe
r die
Kop
ienz
ahl
des
Plas
mid
s pr
o B
akte
rienz
elle
(u
nd
dam
it üb
er
die
Plas
mid
ausb
eute
pr
o M
illili
ter
Kul
turm
ediu
m):
man
unt
ersc
heid
et lo
w-c
opy-
Plas
mid
e (<
20 K
opie
n je
Zel
le) u
nd h
igh-
copy
-Pl
asm
ide
(>20
Kop
ien
je Z
elle
). V
iele
mod
erne
Lab
or-P
lasm
ide
enth
alte
n de
n C
olE1
-Orig
in,
der
zu e
iner
Kop
ienz
ahl
von
meh
rere
n hu
nder
t Pl
asm
iden
pro
Bak
terie
nzel
le f
ührt.
Zur
A
ufna
hme
zu k
loni
eren
der
Gen
e tra
gen
die
Labo
rpla
smid
e m
eist
ein
en P
olyl
inke
r (m
ultip
le
clon
ing
site
, m
cs)
– ei
ne
kurz
e sy
nthe
tisch
e D
NA
-Seq
uenz
, di
e an
eina
nder
gere
iht
Erke
nnun
gsse
quen
zen
zahl
reic
her
gäng
iger
und
sel
tene
r R
estri
ktio
nsen
zym
e en
thäl
t. D
er
Poly
linke
r wird
häu
fig a
uf b
eide
n Se
iten
flank
iert
von
unte
rsch
iedl
iche
n Pr
omot
oren
(z.B
. den
Pr
omot
oren
T3
und
T7, d
ie v
on d
en R
NA
-Pol
ymer
asen
des
T3-
bzw
. T7-
Phag
en s
pezi
fisch
er
kann
t wer
den)
, so
dass
das
klo
nier
te G
en u
nabh
ängi
g vo
n se
iner
Orie
ntie
rung
tran
skrib
iert
wer
den
kann
. G
ängi
ge P
lasm
ide
sind
pB
R32
2 (1
5-20
Kop
ien)
, pU
C (
500-
700
Kop
ien)
, pB
lues
crip
t (30
0-50
0 K
opie
n) u
nd p
GEM
(300
-400
Kop
ien)
.
E. c
oli
Esch
eric
hia
coli
ist e
in m
eist
har
mlo
ses,
Gra
m-n
egat
ives
Dar
mba
kter
ium
. Pra
ktis
ch a
lle im
La
bor v
erw
ende
ten
E.-c
oli-S
täm
me
gehe
n au
f den
Sic
herh
eits
stam
m K
-12
zurü
ck. V
on K
-12
abge
leite
te S
täm
me,
die
sich
gut
für d
ie K
loni
erun
g vo
n Pl
asm
iden
eign
en, s
ind
z.B
. DH
1, D
H5
und
XL1
-Blu
e. B
eson
ders
häu
fig v
erw
ende
t w
ird d
er S
tam
m X
L1-B
lue,
der
zw
ar e
twas
la
ngsa
mer
wäc
hst,
aber
den
Vor
teil
hat,
reko
mbi
natio
nsde
fizie
nt (
recA
- ) zu
sei
n, s
o da
ss
uner
wün
scht
e R
ekom
bina
tione
n de
r DN
A, d
ie so
nst v
orko
mm
en k
önnt
en, n
icht
auf
trete
n.
E. c
oli
ist
ein
sehr
gen
ügsa
mes
Bak
teriu
m,
das
in e
inem
flü
ssig
en o
der
auf
eine
m f
este
n M
inim
alm
ediu
m a
us G
luko
se (
C-
und
Ener
gieq
uelle
) un
d ei
nige
n Sa
lzen
(u.
a. N
-Que
lle)
- �0�
-
Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
wac
hsen
kan
n. U
m e
in s
chne
llere
s W
achs
tum
zu
erre
iche
n, v
erw
ende
t man
jedo
ch m
eist
ein
ko
mpl
exer
es M
ediu
m, z
.B. L
B (L
uria
-Ber
tani
, mit
Hef
eext
rakt
, Bac
totry
pton
und
NaC
l) od
er
TB (t
errif
ic b
roth
). Tr
ansg
ene
Bak
terie
n en
thal
ten
in d
er R
egel
ein
Pla
smid
, das
neb
en d
em z
u kl
onie
rend
en G
en a
ls M
arke
r ei
n G
en f
ür e
ine
Ant
ibio
tika-
Res
iste
nz t
rägt
. In
die
sem
Fal
l w
erde
n di
e B
akte
rien
in o
der a
uf e
inem
Ant
ibio
tika-
halti
gen
Med
ium
ang
ezog
en, s
o da
ss n
ur
trans
gene
Bak
terie
n, n
icht
abe
r B
akte
rien
des
Wild
typs
wac
hsen
kön
nen.
Vie
le A
ntib
iotik
a,
z.B
. das
häu
fig v
erw
ende
te A
mpi
cilli
n, s
ind
tem
pera
ture
mpf
indl
ich,
so
dass
sie
dem
Med
ium
er
st n
ach
dem
Aut
okla
vier
en u
nd A
bküh
len
zuge
setz
t wer
den
dürf
en.
Um
ein
gut
es u
nd g
leic
hmäß
iges
Wac
hstu
m z
u er
ziel
en, w
erde
n di
e B
akte
rien,
aus
gehe
nd v
on
eine
r ein
zeln
en K
olon
ie a
uf e
iner
Aga
rpla
tte, z
unäc
hst i
n ei
ner V
orku
ltur i
n flü
ssig
em M
ediu
m
über
Nac
ht v
erm
ehrt
(ÜN
-Kul
tur)
, be
vor
die
eige
ntlic
he H
aupt
kultu
r in
der
gew
ünsc
hten
M
enge
an
Flüs
sigm
ediu
m a
us d
iese
r V
orku
ltur
ange
impf
t wird
. Je
nach
Pla
smid
beda
rf u
nd
dem
ents
prec
hend
em V
olum
en d
er K
ultu
r unt
ersc
heid
et m
an z
wis
chen
Min
i-Prä
p(ar
atio
n) (2
-10
ml),
Mid
i-Prä
p (2
5-10
0ml)
und
Max
i-Prä
p (>
100m
l).
Plas
mid
-Iso
lieru
ng u
nd R
eini
gung
Fü
r vie
le m
olek
ular
gene
tisch
e A
rbei
ten
ist d
ie Q
ualit
ät d
er e
inge
setz
ten
DN
A (o
der R
NA
) von
en
tsch
eide
nder
Wic
htig
keit.
Kon
zent
ratio
n au
f so
rgfä
ltige
s un
d sa
uber
es A
rbei
ten
bei
der
Isol
ieru
ng u
nd R
eini
gung
der
Nuk
lein
säur
en is
t dah
er n
icht
nur
bes
onde
rs w
icht
ig, s
onde
rn
ersp
art h
äufig
auc
h vi
el F
rust
ratio
n be
i den
folg
ende
n A
rbei
tssc
hritt
en. I
nsbe
sond
ere
ist a
uf d
ie
Ver
mei
dung
von
Kon
tam
inat
ione
n du
rch
DN
Ase
n (b
zw. R
NA
sen)
zu
acht
en.
Der
ers
te S
chrit
t der
Pla
smid
isol
ieru
ng is
t das
Auf
brec
hen
der B
akte
rienz
elle
n. D
ies g
esch
ieht
m
eist
übe
r ei
ne a
lkal
isch
e Ly
se. F
ür d
ie a
lkal
isch
e Ly
se, d
ie a
uch
den
mei
sten
kom
mer
ziel
l er
hältl
iche
n K
its z
ur P
lasm
idis
olat
ion
zugr
unde
lieg
t, w
erde
n di
e B
akte
rien
pelle
tiert
und
nach
En
tfern
en d
es M
ediu
ms
in E
DTA
-hal
tigem
Puf
fer r
esus
pend
iert.
Das
ED
TA k
ompl
exie
rt di
e zw
eiw
ertig
en K
atio
nen
(Ca2+
, Mg2+
, and
ere
Schw
erm
etal
le) a
us d
er b
akte
rielle
n Ze
llwan
d un
d de
stab
ilisi
ert
dies
e. D
aneb
en e
nthä
lt de
r Pu
ffer
RN
Ase
n, u
m u
nerw
ünsc
hte
RN
A M
olek
üle
abzu
baue
n. D
ie a
nsch
ließe
nde
Inku
batio
n in
SD
S (N
atriu
m-D
odec
ylsu
lfat)
und
NaO
H
dena
turie
rt di
e Ze
llmem
bran
und
mac
ht d
ie D
NA
des
Bak
teriu
ms
zugä
nglic
h. N
aOH
de
natu
riert
die
in d
er B
akte
rienz
elle
vor
hand
ene
DN
A u
nd P
rote
ine.
Ein
e vo
llstä
ndig
e Ly
se
und
Den
atur
ieru
ng is
t ent
sche
iden
d fü
r die
Qua
ntitä
t und
die
Qua
lität
der
isol
ierte
n Pl
asm
id-
DN
A. A
nsch
ließe
nd w
ird d
ie a
lkal
isch
e Ly
se d
urch
den
Zus
atz
von
saur
em K
aliu
mac
etat
-Pu
ffer
ges
topp
t. H
ierb
ei b
ilden
sich
Kom
plex
e au
s unl
öslic
hem
Kal
ium
-Dod
ecyl
sulfa
t und
den
de
natu
rierte
n Pr
otei
nen
aus
den
Bak
terie
nzel
len.
Die
chr
omos
omal
e D
NA
ble
ibt
mit
den
Prot
eink
ompl
exen
ass
oziie
rt un
d is
t sch
wer
er a
ls d
ie P
lasm
id-D
NA
und
kan
n zu
sam
men
mit
den
Zellt
rüm
mer
n (D
ebris
) abz
entri
fugi
ert w
erde
n. D
ie k
lein
eren
Pla
smid
e re
natu
riere
n un
ter
den
neut
rale
n B
edin
gung
en u
nd b
leib
en im
Übe
rsta
nd.
Die
Pla
smid
e kö
nnen
aus
den
Übe
rsta
nd d
er a
lkal
isch
en L
yse
durc
h Zu
gabe
von
eis
kalte
m
Etha
nol
oder
Iso
prop
anol
gef
ällt
und
ansc
hlie
ßend
abz
entri
fugi
ert
wer
den.
DN
A i
st i
n de
r G
egen
war
t mon
oval
ente
r Kat
ione
n in
Eth
anol
unl
öslic
h. Z
unäc
hst w
ird d
er P
lasm
id-h
altig
en
Lösu
ng N
atriu
mac
etat
, ans
chlie
ßend
100
%ig
er E
than
ol (
2,5
– 3
fach
es V
olum
en)
zuge
setz
t. D
ie F
ällu
ng e
rfol
gt d
ann
mei
st b
ei 4
°C, u
m d
ie A
usbe
ute
zu v
erbe
sser
n. A
nsch
ließe
nd w
ird d
ie
präz
ipiti
erte
D
NA
du
rch
Zent
rifug
atio
n au
s de
r Lö
sung
ab
getre
nnt.
Je
nach
V
erw
endu
ngsz
wec
k m
üsse
n di
e Pl
asm
ide
jedo
ch n
och
wei
ter
aufg
erei
nigt
bzw
. en
tsal
zt
- �0�
-

Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
wer
den.
Der
let
zte
Schr
itt d
er P
lasm
idis
olie
rung
ist
dah
er d
ie R
eini
gung
der
DN
A d
urch
W
asch
en m
it 70
%ig
em E
than
ol.
Das
noc
h m
itgef
ällte
Sal
z lö
st s
ich
im W
asse
rant
eil
der
Was
chlö
sung
und
wird
nac
h Ze
ntrif
ugat
ion
verw
orfe
n. D
ie W
asch
schr
itte
wer
den
in d
er R
egel
1
– 2
Mal
wie
derh
olt u
nd d
as v
erbl
eibe
nde
DN
A-P
elle
t vor
sich
tig g
etro
ckne
t und
ans
chlie
ßend
in
Was
ser o
der P
uffe
r auf
geno
mm
en u
nd b
ei -2
0°C
gel
ager
t.
Qua
ntitä
t und
Qua
lität
der
erh
alte
nen
Plas
mid
-DN
A k
önne
n ph
otom
etris
ch b
estim
mt w
erde
n.
Die
arom
atis
chen
Rin
ge d
er B
asen
in d
en N
ukle
insä
uren
abso
rbie
ren
UV
-Lic
ht b
ei 2
60nm
: ein
e Lö
sung
von
50μ
g/m
l dop
pels
träng
iger
DN
A (d
sDN
A) e
rgib
t (be
i ein
er S
chic
htdi
cke
von
1cm
) ei
ne o
ptis
che
Dic
hte
(OD
) vo
n 1
(ein
zels
träng
ige
ssD
NA
: 40μ
g/m
l, ss
RN
A: 3
3μg/
ml).
Zur
Ü
berp
rüfu
ng d
er R
einh
eit d
er P
lasm
id-D
NA
mis
st m
an m
eist
noc
h di
e A
bsor
ptio
n be
i 280
nm
(Abs
orpt
ions
max
imum
der
aro
mat
isch
en A
min
osäu
ren
in P
rote
inen
): re
ine
DN
A h
at e
inen
O
D26
0/OD
280-
Wer
t von
1,8
(rei
ne R
NA
ein
en v
on 2
,0).
Ver
unre
inig
unge
n de
r Nuk
lein
säur
en
mit
Prot
eine
n fü
hren
zu
sign
ifika
nt n
iedr
iger
en W
erte
n.
Res
trik
tions
enzy
me
Res
trikt
ions
enzy
me
sind
En
donu
klea
sen,
di
e be
stim
mte
B
asen
sequ
enze
n in
ei
ner
dopp
elst
räng
igen
DN
A e
rken
nen
und
beid
e St
räng
e in
ein
er sp
ezifi
sche
n W
eise
öff
nen,
inde
m
sie
Phop
shod
iest
erbi
ndun
gen
des
Zuck
er/P
hosp
hat-R
ückg
rats
de
r D
NA
hy
drol
ysie
ren.
R
estri
ktio
nsen
zym
e sin
d ur
sprü
nglic
h Te
il de
r bak
terie
llen
Abw
ehr:
sie d
iene
n de
r Ver
nich
tung
vo
n au
fgen
omm
ener
Fre
md-
DN
A (z
.B. e
inge
drun
gene
r Pha
gen-
DN
A).
Eige
ne D
NA
wird
bei
de
r Sy
nthe
se d
urch
spe
zifis
che
Mod
ifika
tione
n (M
ethy
lieru
ng v
on A
deni
n od
er C
ytos
in)
an
den
betre
ffen
den
Res
trikt
ions
schn
ittst
elle
n vo
r de
m
Abb
au
durc
h di
e ei
gene
n R
estri
ktio
nsen
zym
e ge
schü
tzt.
In
der
Mol
ekul
arge
netik
w
erde
n R
estri
ktio
nsen
zym
e he
ute
aus
zahl
reic
hen
Bak
terie
n ve
rwen
det (
z.B
. Eco
RI a
us E
sche
rich
ia c
oli;
Bam
HI a
us B
acill
us a
myl
oliq
uefa
cien
s), d
ie s
ich
in d
er j
ewei
ligen
spe
zifis
chen
Erk
ennu
ngss
eque
nz u
nter
sche
iden
(da
bei
wer
den
nur
so
gena
nnte
Typ
-II-
Res
trikt
ions
enzy
me
eing
eset
zt,
die
im G
egen
satz
zu
Typ-
I- u
nd T
yp-I
II-
Res
trikt
ions
enzy
men
kei
ne e
igen
e M
ethy
lieru
ngsa
ktiv
ität a
ufw
eise
n un
d di
e in
nerh
alb
(ode
r se
lten
in
der
unm
ittel
bare
n N
achb
arsc
haft)
ih
rer
Erke
nnun
gsse
quen
z sc
hnei
den)
. D
ie
gäng
igst
en R
estri
ktio
nsen
zym
e er
kenn
en m
eist
ein
e pa
lindr
omis
che
Nuk
leot
idse
quen
z au
s 4 –
8
Bas
en.
Von
gr
oßer
B
edeu
tung
fü
r di
e M
olek
ular
gene
tik
ist
die
Tats
ache
, da
ss
eini
ge
Res
trikt
ions
enzy
me
eine
n gl
atte
n Sc
hnitt
dur
ch b
eide
Strä
nge
der
DN
A s
etze
n un
d so
mit
Frag
men
te m
it st
umpf
en E
nden
(blu
nt e
nds)
lief
ern,
wäh
rend
and
ere
Res
trikt
ions
enzy
me
eine
n ve
rset
zten
Sch
nitt
durc
h de
n D
oppe
lstra
ng f
ühre
n, s
o da
ss F
ragm
ente
mit
über
steh
ende
n 5'
- od
er 3
'-End
en (s
ticky
end
s) e
ntst
ehen
. Die
se ü
bers
tehe
nden
End
en s
ind
beso
nder
s vo
rteilh
aft,
da m
it ih
rer H
ilfe
die
Liga
tion
von
DN
A-F
ragm
ente
n w
esen
tlich
erle
icht
ert w
ird. W
ird z
.B. e
in
Gen
mit
Hilf
e ei
nes
best
imm
ten
Res
trikt
ions
enzy
ms
aus
dem
Gen
om a
usge
schn
itten
(da
für
müs
sen
auf d
er D
NA
in n
icht
zu
groß
er E
ntfe
rnun
g vo
r (up
stre
am) u
nd h
inte
r (do
wns
trea
m)
dem
G
en
die
ents
prec
hend
en
Erke
nnun
gsse
quen
zen
liege
n,
wäh
rend
im
G
en
dies
e Er
kenn
ungs
sequ
enz
nich
t vor
kom
men
dar
f), s
o ka
nn d
iese
s R
estri
ktio
nsfr
agm
ent d
ann
rela
tiv
leic
ht in
eine
n V
ekto
r ein
gebr
acht
wer
den,
der
mit
dem
gle
iche
n R
estri
ktio
nsen
zym
ges
chni
tten
wur
de (d
afür
mus
s der
Vek
tor d
ie e
ntsp
rech
ende
Erk
ennu
ngss
eque
nz im
Pol
ylin
ker e
ntha
lten,
je
doch
an
kein
er a
nder
en S
telle
). B
eide
DN
A-M
olek
üle,
das
Res
trikt
ions
frag
men
t, w
elch
es d
as
- �0�
-
Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
Gen
ent
hält
und
der
linea
risie
rte V
ekto
r w
eise
n je
tzt
die
glei
chen
und
auf
grun
d de
s pa
lindr
omis
chen
Cha
rakt
ers
der E
rken
nung
sseq
uenz
kom
plem
entä
re ü
bers
tehe
nde
Ende
n au
f. D
iese
kön
nen
hybr
idis
iere
n un
d so
mit
ein
wie
deru
m z
irkul
äres
Pla
smid
aus
der
urs
prün
glic
hen
Vek
tors
eque
nz u
nd d
em zu
klo
nier
ende
n G
en b
ilden
. Der
so b
erei
ts st
abili
sier
te R
ing
wird
dan
n ab
schl
ieße
nd
durc
h di
e A
ktiv
ität
eine
r Li
gase
, di
e di
e ve
rble
iben
den,
ve
rset
zten
Ei
nzel
stra
ngbr
üche
dur
ch K
nüpf
en v
on P
hosp
hodi
este
rbin
dung
en v
erbi
ndet
, fe
rtig
gest
ellt
(rek
ombi
nant
e D
NA
). D
ie L
igat
ion
zwei
er F
ragm
ente
mit
glat
ten
Ende
n is
t wei
t sch
wie
riger
, da
es
ke
ine
stab
ilisi
eren
den
Hyb
ridis
ieru
ngsm
öglic
hkei
ten
zwis
chen
de
n be
iden
zu
ve
rbin
dend
en D
NA
-Mol
ekül
en g
ibt.
Um
die
sen
Effe
kt z
u ko
mpe
nsie
ren,
wer
den
Liga
tione
n vo
n Fr
agm
ente
n m
it gl
atte
n En
den
bei e
iner
Tem
pera
tur v
on 1
5°C
dur
chge
führ
t. H
ierb
ei is
t die
th
erm
isch
e B
eweg
ungs
ener
gie
der
Ende
n w
esen
tlich
ger
inge
r un
d di
e Ef
fizie
nz d
er L
igat
ion
wird
deu
tlich
erh
öht.
Die
Klo
nier
ung
unte
r zu
Hilf
enah
me
nur
eine
s R
estri
ktio
nsen
zym
s ha
t ab
er a
uch
bei
über
häng
ende
n En
den
den
Nac
htei
l, da
ss d
as z
u kl
onie
rend
e G
en i
n zw
ei v
ersc
hied
enen
O
rient
ieru
ngen
in d
en V
ekto
r lig
iert
wer
den
kann
. Die
ses P
robl
em k
ann
durc
h de
n Ei
nsat
z vo
n zw
ei v
ersc
hied
enen
Res
trikt
ions
enzy
men
um
gang
en w
erde
n, d
ie u
nter
schi
edlic
he ü
bers
tehe
nde
Ende
n er
zeug
en (
in d
iese
m F
all m
uss
das
eine
Res
trikt
ions
enzy
m v
or d
em G
en, d
as a
nder
e hi
nter
dem
Gen
schn
eide
n, u
nd d
ie E
rken
nung
sseq
uenz
en fü
r bei
de R
estri
ktio
nsen
zym
e müs
sen
im P
olyl
inke
r vo
rhan
den
sein
). Pr
oble
mat
isch
kan
n be
i die
sem
Vor
gehe
n di
e Ta
tsac
he s
ein,
da
ss v
ersc
hied
ene R
estri
ktio
nsen
zym
e unt
ersc
hied
liche
Bed
ingu
ngen
für d
ie o
ptim
ale A
ktiv
ität
benö
tigen
: z.B
. Puf
fers
ubst
anz
(mei
st T
ris),
Salz
e (m
eist
MgC
l 2, N
aCl o
der
KC
l), p
H-W
ert
(mei
st p
H 7
,5-8
), Te
mpe
ratu
r (m
eist
37°
C).
Die
Akt
ivitä
t der
Res
trikt
ions
enzy
me
unte
r de
n je
wei
ligen
Bed
ingu
ngen
wird
in E
inhe
iten
(Uni
ts)
ange
gebe
n: 1
Uni
t spa
ltet 1
μg
DN
A (
als
Stan
dard
wird
DN
A d
es B
akte
rioph
agen
Lam
bda
verw
ende
t) in
1 S
tund
e vo
llstä
ndig
.
Aga
rose
-Gel
elek
trop
hore
seU
m v
ersc
hied
enen
, bei
ein
em R
estri
ktio
nsve
rdau
ent
steh
ende
n, D
NA
-Fra
gmen
te v
onei
nand
er
trenn
en z
u kö
nnen
nut
zt m
an d
ie c
hem
isch
en E
igen
scha
ften
der D
NA
aus
: alle
DN
A-M
olek
üle
trage
n au
fgru
nd i
hres
Zuc
ker/P
hosp
hat-R
ückg
rats
zah
lreic
he n
egat
ive
Ladu
ngen
, wob
ei d
ie
Ladu
ngsd
icht
e alle
r DN
A-M
olek
üle i
st g
leic
h. D
emzu
folg
e ist
die
elek
troph
oret
isch
e Mob
ilitä
t, al
so d
ie W
ande
rung
sges
chw
indi
gkei
t in
eine
m e
lekt
risch
en F
eld,
für
alle
DN
A-M
olek
üle
in
frei
er L
ösun
g gl
eich
gro
ß. F
inde
t die
Ele
ktro
phor
ese
dage
gen
in e
iner
fes
ten
Mat
rix s
tatt,
so
wer
den
die
DN
A-M
olek
üle
je n
ach
ihre
r G
röße
unt
ersc
hied
lich
star
k zu
rück
geha
lten:
kle
ine
DN
A-F
ragm
ente
wan
dern
schn
elle
r dur
ch d
ie M
atrix
als
gro
ße D
NA
-Fra
gmen
te. E
ntsc
heid
end
ist,
dass
zwei
gle
ich
groß
e DN
A-F
ragm
ente
, una
bhän
gig
von
ihre
r Nuk
leot
idse
quen
z, au
fgru
nd
ihre
r gl
eich
en L
adun
gsdi
chte
gle
ich
schn
ell
wan
dern
(K
raft
= M
asse
x B
esch
leun
igun
g).
Nat
ürlic
h hä
ngt
die
Wan
deru
ngsg
esch
win
digk
eit
alle
r D
NA
-Fra
gmen
te
nebe
n de
r K
onze
ntra
tion
der M
atrix
und
dam
it de
r Por
engr
öße
zusä
tzlic
h vo
n de
r ang
eleg
ten
Span
nung
, de
r Art
des L
aufp
uffe
rs (i
.d.R
. Tris
/Ace
tat,
TAE)
und
geg
eben
enfa
lls d
er A
nwes
enhe
it w
eite
rer
Subs
tanz
en (z
.B. E
thid
ium
brom
id, S
alz)
ab.
Je n
ach
Grö
ße d
er z
u tre
nnen
den
DN
A-F
ragm
ente
wäh
lt m
an a
ls M
atrix
Aga
rose
(rel
ativ
gro
ße
Pore
n, d
aher
für
rel
ativ
gro
ße D
NA
-Fra
gmen
te g
eeig
net)
oder
Pol
yacr
ylam
id (
rela
tiv k
lein
e Po
ren,
dah
er b
eson
ders
für r
elat
iv k
lein
e D
NA
-Fra
gmen
te g
eeig
net).
Pol
yacr
ylam
idge
le e
igne
n si
ch n
ur f
ür d
ie T
renn
ung
von
DN
A-F
ragm
ente
n m
it G
röße
n zw
isch
en e
twa
5 un
d 1.
000
Bas
enpa
aren
. Die
Tre
nnsc
härf
e so
lche
r Gel
e is
t bei
ent
spre
chen
der G
ellä
nge
alle
rdin
gs e
norm
,
- �0�
-

Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
so d
ass
Frag
men
te, d
eren
Län
ge s
ich
nur u
m e
in B
asen
paar
unt
ersc
heid
et, a
ufge
trenn
t wer
den
könn
en. A
garo
se-K
onze
ntra
tione
n vo
n 0,
5 bi
s 2,
0% (
Gew
icht
spro
zent
, wei
ght/v
olum
e, w
/v)
erge
ben
Gel
e, d
ie fü
r die
Auf
trenn
ung
von
linea
ren
DN
A-F
ragm
ente
n m
it Lä
ngen
von
60
kb
bis h
inun
ter z
u et
wa
100
bp g
eeig
net s
ind.
Wäh
rend
line
are
DN
A-F
ragm
ente
im A
garo
se-G
el n
ur a
ufgr
und
ihre
r Grö
ße g
etre
nnt w
erde
n,
ist d
ie S
ituat
ion
für z
irkul
äre
Plas
mid
e ko
mpl
izie
rter.
Zirk
ulär
e Pl
asm
ide
wei
sen
in d
er R
egel
ve
rsch
iede
ne K
onfo
rmat
ione
n au
f: si
e lie
gen
sow
ohl a
ls o
ffen
es zi
rkul
äres
Mol
ekül
vor
, wei
sen
aber
auc
h ve
rsch
iede
ne G
rade
der
Übe
rdre
hung
(Su
perh
elix
, sup
er c
oil)
auf.
Die
kom
pakt
e su
perh
elik
ale
Form
ei
nes
zirk
ulär
en
Plas
mid
s w
ande
rt sc
hnel
ler
als
das
glei
ch
groß
e lin
earis
ierte
Pla
smid
. D
ie s
perr
ige
offe
ne F
orm
des
zirk
ulär
en P
lasm
ids
(ohn
e Su
perh
elix
) da
gege
n w
ande
rt hi
ngeg
en d
eutli
che
lang
sam
er a
ls d
ie b
eide
n an
dere
n Fo
rmen
. Is
t ei
n R
estri
ktio
nsve
rdau
unv
olls
tänd
ig, s
o si
nd d
ie v
ersc
hied
enen
Kon
form
atio
nen
des
Plas
mid
s im
A
garo
sege
l na
chw
eis-
und
unt
ersc
heid
bar.
Die
se T
atsa
che
kann
man
nut
zen,
um
mitt
els
Gel
elek
troph
ores
e de
n Er
folg
ei
ner
Line
aris
ieru
ng
zu
über
prüf
en.
Supe
rhel
ikal
e K
onfo
rmat
ione
n vo
n Pl
asm
iden
sin
d fü
r Res
tiktio
nsen
zym
e au
f Gru
nd d
er S
pira
lisie
rung
nur
sc
hwer
zug
ängl
ich
und
wer
den
desh
alb
schl
echt
er g
esch
nitte
n un
d si
nd o
ft im
Gel
noc
h si
chtb
ar.
Zur B
elad
ung
eine
s Aga
rose
gels
wer
den
die
DN
A-P
robe
n in
ein
em L
adep
uffe
r, de
r aus
ein
em
TE-P
uffe
r, G
lyce
rin u
nd B
rom
phen
olbl
au b
este
ht a
ufge
nom
men
. Dur
ch d
as G
lyce
rin w
ird d
ie
Dic
hte
des P
uffe
r-D
NA
-Gem
isch
s sow
eit e
rhöh
t, da
ss d
ie L
ösun
g si
cher
er in
die
Pro
bent
asch
e de
s G
els
gela
den
wer
den
kann
. Bro
mph
enol
blau
die
nt a
ls f
arbi
gen
Mar
ker,
der
etw
a gl
eich
sc
hnel
l wie
ein
300b
p gr
oßes
DN
A-F
ragm
ent w
ande
rt un
d so
der
For
tsch
ritt d
er E
lekt
roph
ores
e vi
suel
l nac
hver
folg
t wer
den
kann
und
der
Gel
lauf
rech
tzei
tig b
eend
et w
erde
n ka
nn. P
aral
lel m
it de
n au
fzut
renn
ende
n Pr
oben
wird
im
mer
ein
en L
änge
nsta
ndar
d (M
arke
r) a
uf d
as G
el
aufg
etra
gen,
der
Fra
gmen
te m
it ge
nau
defin
ierte
n G
röße
n en
thäl
t. R
elat
iv z
u di
esem
Mar
ker
läss
t si
ch d
ann
die
Grö
ße d
er g
etre
nnte
n D
NA
-Fra
gmen
te l
eich
t be
stim
men
. Geb
räuc
hlic
he
Mar
ker
sind
die
100
bp-L
eite
r, di
e 1k
b-Le
iter,
die
Ban
den
in 1
00bp
- bz
w. 1
000b
p-Sc
hritt
en
enth
alte
n.
Nac
h A
bsch
luss
der
Gel
elek
troph
ores
e m
üsse
n di
e D
NA
-Fra
gmen
te z
ur L
änge
naus
wer
tung
si
chtb
ar g
emac
ht w
erde
n. D
ies
gelin
gt m
it Et
hidi
umbr
omid
(3,
8-D
iam
ino-
5-et
hyl-6
-phe
nyl-
phen
anth
ridin
ium
brom
id),
das
aufg
rund
se
iner
pl
anar
en
Stru
ktur
in
di
e D
NA
-Hel
ix
inte
rkal
iere
n, si
ch a
lso
zwis
chen
die
Bas
en d
er D
NA
schi
eben
kan
n. D
er in
terk
alie
rte F
arbs
toff
flu
ores
zier
t in
Ver
bind
ung
mit
der D
NA
bei
ein
er W
elle
nlän
ge v
on 5
90nm
, wen
n er
mit
UV
-Li
cht (
254
– 36
6nm
) ang
ereg
t wird
. Bei
m U
mga
ng m
it Et
hidi
umbr
omid
ist d
esha
lb V
orsi
cht
gebo
ten:
zum
ein
en i
st E
thid
ium
brom
id a
ufgr
und
sein
er i
nter
kalie
rend
en E
igen
scha
ften
ein
pote
ntie
lles
Mut
agen
, zu
m a
nder
en k
ann
das
zur
Det
ektio
n ve
rwen
dete
UV
-Lic
ht b
ei
unge
schü
tzte
n A
ugen
sc
hnel
l ei
ne
äuße
rst
schm
erzh
afte
B
inde
haut
entz
ündu
ng
(Sch
neeb
lindh
eit)
veru
rsac
hen.
Ethi
dium
brom
id w
ird m
eist
ber
eits
in d
as A
garo
se-G
el e
inpo
lym
eris
iert,
kan
n da
s Gel
nac
h de
r El
ektro
phor
ese
in e
inem
Eth
idiu
mbr
omid
bad
gefä
rbt w
erde
n. M
it Et
hidi
umbr
omid
lass
en si
ch
auf d
em U
V-L
euch
ttisc
h (T
rans
illum
inat
or) e
twa 1
0 –
20ng
dsD
NA
in ei
ner B
ande
nac
hwei
sen.
- �0�
-
Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
Tra
nsfo
rmat
ion
von
Bak
teri
en
Als
Tra
nsfo
rmat
ion
beze
ichn
et d
ie V
erän
deru
ng d
es G
enot
yps e
iner
Zel
le d
urch
die
Auf
nahm
e fr
emde
r, na
ckte
r DN
A a
us d
er U
mge
bung
. Tra
nsdu
ktio
n be
zeic
hnet
die
Übe
rtrag
ung
der D
NA
(o
der R
NA
) ein
es P
hage
n od
er V
irus
in e
ine
prok
aryo
ntis
che
oder
euk
aryo
ntis
che
Zelle
(als
o ei
ne F
orm
der
Inf
ektio
n); K
onju
gatio
n is
t die
dire
kte
Übe
rtrag
ung
der
Kop
ie e
ines
Tei
ls d
er
DN
A e
iner
Bak
terie
nzel
le in
ein
e an
dere
. Vie
le B
akte
rien
besi
tzen
ein
en s
olch
en s
pezi
fisch
en
Auf
nahm
emec
hani
smus
für
DN
A a
us d
er U
mge
bung
, w
obei
sie
ihr
Res
trikt
ions
enzy
m-
basi
erte
s „Im
mun
syst
em“
vor a
rtfre
mde
r DN
A sc
hütz
t.
Das
im L
abor
vie
lfach
ein
gese
tzte
Bak
teriu
m E
. col
i ver
fügt
jedo
ch n
atür
liche
rwei
se n
icht
übe
r ei
nen
solc
hen
Mec
hani
smus
, so
das
s di
ese
Bak
terie
n zu
näch
st f
ür d
ie T
rans
form
atio
n m
it fr
emde
r DN
A v
orbe
reite
nt, d
.h. k
ompe
tent
gem
acht
wer
den
müs
sen.
Die
s ge
schi
eht i
n ei
nem
em
piris
ch o
ptim
ierte
n Pr
ozes
s, de
r di
e In
kuba
tion
der
Bak
terie
n in
ein
er C
a2+-L
ösun
g,
wie
derh
olte
Ink
ubat
ion
auf
Eis
und
Zent
rifug
atio
nssc
hritt
e be
inha
ltet.
Kom
pete
nte
E.-c
oli-
Zelle
n kö
nnen
dan
n be
i -80
°C ti
efge
fror
en w
erde
n un
d be
halte
n ih
re K
ompe
tenz
auc
h na
ch
dem
Auf
taue
n fü
r ei
ne g
ewis
se Z
eit b
ei. D
ie e
igen
tlich
e Tr
ansf
orm
atio
n de
r B
akte
rien
kann
da
nn n
ach
vers
chie
dene
n V
erfa
hren
erf
olge
n, b
eson
ders
ein
fach
ist
die
Cal
cium
chlo
rid-
Met
hode
, bes
onde
rs e
ffiz
ient
die
Ele
ktro
pora
tion.
Bei
der
Cal
cium
chlo
rid-M
etho
de w
ird d
ie
DN
A m
it C
aCl 2
auf
der
Obe
rflä
che
der
kom
pete
nten
Bak
terie
nzel
len
präz
ipiti
ert.
Von
hie
r ge
lang
t di
e D
NA
en
twed
er
dire
kt
durc
h di
e M
embr
an,
dere
n In
tegr
ität
durc
h di
e C
alci
umch
lorid
kris
talle
ges
tört
wird
, od
er d
urch
End
ocyt
ose
ins
Inne
re d
er Z
elle
. B
ei d
er
Elek
tropo
ratio
n w
erde
n di
e ko
mpe
tent
en B
akte
rienz
elle
n in
ein
er D
NA
-Lös
ung
in e
in
elek
trisc
hes
Feld
geb
rach
t un
d ku
rzen
Pul
sen
hohe
r Fe
ldst
ärke
aus
gese
tzt.
Dab
ei w
erde
n ku
rzze
itig
Pore
n in
der
Zel
lmem
bran
erz
eugt
zu
wer
den,
dur
ch d
ie M
akro
mol
ekül
e un
d se
lbst
ga
nze
Plas
mid
e in
die
Zel
le g
elan
gen
könn
en. N
ach
der
Tran
sfor
mat
ion,
als
o de
r A
ufna
hme
der
Plas
mid
-DN
A,
benö
tigen
die
jet
zt t
rans
gene
n Ze
llen
zunä
chst
ein
e Er
holu
ngsp
hase
in
flüss
igen
Kul
turm
ediu
m, b
evor
sie
in A
ntib
iotik
a-ha
ltige
Sel
ektio
nsm
edie
n um
gese
tzt w
erde
n kö
nnen
. In
dies
er E
rhol
ungs
zeit
kom
mt e
s zu
r Exp
ress
ion
des
auf d
em P
lasm
id b
efin
dlic
hen
Ant
ibio
tika-
Res
iste
nzge
ns, w
elch
es d
ie tr
ansg
enen
Zel
len
(und
nur
die
se) i
n di
e La
ge v
erse
tzt,
auf d
em A
ntib
iotik
a-ha
ltige
n Se
lekt
ions
med
ium
zu
wac
hsen
.
Für d
ie S
elek
tion
von
E.co
li w
ird o
ft da
s A
ntib
iotik
a A
mpi
cilli
n ei
nges
etzt
. Am
pici
llin
gehö
rt zu
r K
lass
e de
r -L
acta
m-A
ntib
iotik
a un
d he
mm
t in
den
Bak
terie
n da
s En
zym
Ala
nin-
Tran
spep
tidas
e da
ss b
ei d
er Z
ellte
ilung
und
der
dam
it ve
rbun
dene
n Ze
llwan
dneu
synt
hese
be
teili
gt i
st.
Wei
tere
wic
htig
e A
ntib
iotik
a in
der
Mol
ekul
arbi
olog
ie s
ind
Kan
amyc
in u
nd
Tetra
cycl
in w
elch
e di
e ba
kter
ielle
Pro
tein
bios
ynth
ese
durc
h B
lock
ieru
ng d
er ri
boso
mal
en 3
0S
Unt
erei
nhei
t hem
men
.
Bla
u/W
eiß-
Sele
ktio
n vo
n T
rans
form
ande
n A
ntib
iotik
a-ha
ltige
Med
ien
erla
uben
zw
ar d
ie S
elek
tion
der w
enig
en tr
ansf
orm
ierte
n, P
lasm
id-
halti
gen
Zelle
n au
s de
r V
ielz
ahl
der
untra
nsfo
rmie
rten
Wild
typ-
Bak
terie
n. S
ie e
rlaub
en
alle
rdin
gs n
icht
die
Unt
ersc
heid
ung
von
trans
form
ierte
Bak
terie
n, d
ie d
as P
lasm
id m
it de
m z
u kl
onie
rend
en G
en (I
nser
t) en
thal
ten,
von
sol
chen
tran
sfor
mie
rten
Bak
terie
n, d
ie le
digl
ich
das
Plas
mid
ohn
e In
sert
trage
n (b
ei d
er L
igat
ion
kann
sich
, wen
n m
it nu
r ein
em R
estri
ktio
nsen
zym
ge
arbe
itet w
urde
, das
line
aris
ierte
Pla
smid
auc
h w
iede
r zum
Rin
g sc
hlie
ßen,
ohn
e da
ss d
as z
u kl
onie
rend
e G
en e
inge
baut
wur
de).
Die
se U
nter
sche
idun
g w
ird d
urch
die
Ver
wen
dung
des
Pl
asm
ids p
Blu
escr
ipt (
pKS)
ode
r ver
wan
dter
Vek
tore
n er
mög
licht
.
- �0�
-

Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
Solc
he V
ekto
ren,
die
ein
e so
gen
annt
e B
lau/
Wei
ß-Se
lekt
ion
erla
uben
, ent
halte
n ne
ben
dem
A
ntib
iotik
a-R
esis
tenz
gen
noch
den
5‘-
Teil
des l
acZ-
Mar
kerg
ens
(-F
ragm
ent),
das
für d
as N
-te
rmin
ale
Frag
men
t der
-G
alak
tosi
dase
cod
iert.
Der
C-te
rmin
ale
Ber
eich
des
lacZ
-Gen
s (
-Fr
agm
ent)
wird
hin
gege
n im
Gen
om d
er v
erw
ende
ten
E.co
li Ze
llen
kodi
ert.
Bei
de l
acZ-
Gen
frag
men
te st
ehen
unt
er d
er K
ontro
lle e
ines
indu
zier
bare
n Pr
omot
ors,
der n
ur in
Geg
enw
art
der
Indu
ktor
mol
ekül
en
Lact
ose
oder
IP
TG
(Iso
prop
ylth
ioga
lakt
osid
, ei
nem
La
ctos
e-A
nalo
gon)
, akt
ivie
rt w
ird. I
m G
egen
satz
zu
Lact
ose
kann
IPTG
jedo
ch v
on d
en B
akte
rien
nich
t ab
geba
ut w
erde
n un
d ak
tivie
rt di
e G
enex
pres
sion
der
lacZ
-Gen
-Fra
gmen
te d
auer
haft.
Hat
das
B
akte
rium
nu
n be
i de
r Tr
ansf
orm
atio
n ei
n Pl
asm
id
das
für
das
-Fra
gmen
t ko
dier
t au
fgen
omm
en so
wer
den
auf I
PTG
-hal
tigem
Med
ium
bei
de la
cZ-G
en-F
ragm
ente
aktiv
iert,
und
di
e be
iden
Pr
otei
nunt
erei
nhei
ten
un
d
der
-Gal
akto
sida
se
wer
den
gebi
ldet
un
d re
kons
titui
eren
zu
eine
m f
unkt
ione
llen
Prot
ein.
Ent
hält
das
Med
ium
dar
über
hin
aus
das
farb
lose
, abe
r ch
rom
ogen
e X
-Gal
(Su
bstra
t 5-B
rom
o-4-
chlo
ro-3
-indo
lyl-
-D-G
alak
tosi
d), s
o w
ird d
ie c
hrom
opho
re G
rupp
e vo
n de
r -G
alak
tosi
dase
vom
Res
t abg
espa
lten
und
bild
et u
nter
Sa
uers
toff
einw
irkun
g ei
nen
chro
mop
hore
s D
imer
: K
olon
ien
eine
s en
tspr
eche
nden
E.-c
oli-
Klo
ns fä
rben
sich
bla
u.
In p
Blu
escr
ipt P
lasm
iden
ist d
ie „
mul
tiple
clo
ning
site
“ ku
rz h
inte
r dem
Sta
rtcod
on A
TG d
es
lacZ
-Gen
-Fra
gmen
ts ei
ngef
ügt w
orde
n so
das
s den
noch
eine
funk
tions
fähi
ges
-Gal
akto
sida
se-
Frag
men
t geb
ildet
wer
den
kann
(de
r Po
lylin
ker
ist s
o ko
nstru
iert,
das
s er
das
Les
eras
ter
des
Gen
s nic
ht v
ersc
hieb
t; es
wer
den
nur e
inig
e zu
sätz
liche
, in
der u
rspr
üngl
iche
n -G
alak
tosi
dase
ni
cht v
orha
nden
e A
min
osäu
ren
eing
ebau
t, di
e ab
er d
ie T
ertiä
tstru
ktur
des
Pro
tein
s un
d da
mit
die
Akt
ivitä
t des
Enz
yms n
icht
wes
entli
ch b
eein
fluss
en).
Träg
t der
Vek
tor j
edoc
h ei
n In
sert,
ist
also
ein
DN
A-F
ragm
ent i
n de
n Po
ylin
ker
ligie
rt w
orde
n, s
o un
terb
richt
die
ses
Inse
rt in
den
m
eist
en F
älle
n da
s la
cZ-G
en u
nd e
s w
ird k
eine
akt
ive
-Gal
akto
sida
se m
ehr
gebi
ldet
(z.
B.
durc
h V
ersc
hieb
en d
es L
eser
aste
rs, d
urch
Ein
füge
n ei
nes S
topp
codo
ns o
der d
urch
Ver
ände
rung
de
r Ter
tiärs
trukt
ur d
es g
ebild
eten
Fus
ions
prot
eins
). E.
-col
i-Klo
ne m
it In
sert-
halti
gem
Pla
smid
er
sche
inen
dah
er au
f der
IPTG
/X-G
al-P
latte
wei
ß. D
as la
cZ-G
en is
t eig
entli
ch T
eil d
es L
akto
se-
Ope
rons
von
E. c
oli.
Für d
as B
lau/
Wei
ß-Sc
reen
ing
dürf
en d
aher
nur
E.-c
oli-S
täm
me v
erw
ende
t w
erde
n, d
ie k
ein
endo
gene
s Lak
tose
-Ope
ron
meh
r tra
gen.
- �0�
-
Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
Prax
is V
ersu
chsü
bers
icht
Als
bio
logi
sche
s A
usga
ngsm
ater
ial w
erde
n dr
ei I
hnen
unb
ekan
nte
bakt
erie
lle „
Übe
r-N
acht
“-
Kul
ture
n (Ü
N) b
erei
tges
tellt
. Ihr
e Auf
gabe
ist d
ie Is
olat
ion
der P
lasm
ide a
us d
en zu
r Ver
fügu
ng
gest
ellte
n K
ultu
ren
und
anha
nd d
er E
rgeb
niss
e des
Res
trikt
ions
verd
aus u
nd d
er T
rans
form
atio
n
von
E.co
li Ze
llen
die
enth
alte
nen
Kon
stru
kte
zu id
entif
izie
ren.
Fol
gend
e K
onst
rukt
e gi
lt es
zu
best
imm
en:
1.pB
lues
crip
t (30
00bp
) + c
DN
A G
en 1
(600
bp)
2.pB
lues
crip
t (30
00bp
) + c
DN
A G
en 2
(120
0bp)
3.pB
lues
crip
t (30
00bp
)
Jede
Gru
ppe
wäh
lt zw
ei d
er d
rei
unbe
kann
ten
Prob
en z
u A
naly
se a
us.
Zur
volls
tänd
igen
Aus
wer
tung
wer
den
dann
auc
h di
e Er
gebn
isse
der
Nac
hbar
grup
pe h
eran
gezo
gen.
Dur
chfü
hrun
g:
Die
Bak
terie
n w
erde
n ge
ernt
et u
nd d
ie P
lasm
id-D
NA
nac
h de
r M
etho
de v
on B
irnbo
im u
nd
Dol
y im
kle
inen
Maß
stab
(„M
ini-P
räp“
) iso
liert.
Ein
Tei
l der
DN
A w
ird im
Ans
chlu
ss m
ittel
s
Res
trikt
ions
-End
onuk
leas
en g
espa
lten
und
ansc
hlie
ßend
mit
nich
t ges
palte
ner P
lasm
id-D
NA
in
eine
m
Aga
rose
gel
verg
liche
n.
Die
ge
won
nene
n Pl
asm
ide
wer
den
durc
h ch
emis
che
Tran
sfor
mat
ion
wie
der
in B
akte
rien
eing
ebra
cht.
Für
die
ansc
hlie
ßend
e Se
lekt
ion
und
Kla
ssifi
zier
ung
durc
h B
lau/
Wei
ß-Se
lekt
ion
wird
ein
Tei
l de
r tra
nsfo
rmie
rten
Bak
terie
n au
f
antib
iotik
ahal
tigem
IPT
G/X
-Gal
-Aga
r au
spla
ttier
t. A
lle R
eakt
ions
gefä
ße k
lar
und
eind
eutig
besc
hrift
en u
nd d
ie e
inze
lnen
Exp
erim
ente
nac
hvol
lzie
hen
zu k
önne
n
1.Pl
asm
id-I
solie
rung
aus
Bak
teri
en (n
ach
Bir
nboi
m u
nd D
oly)
2ml-R
eakt
ions
gefä
ße v
orbe
reite
n un
d m
it K
ultu
rnum
mer
und
Gru
ppe
besc
hrift
en
2ml
eine
r Ü
N-K
ultu
r in
jew
eils
ein
vor
bere
itete
s R
eakt
ions
gefä
ß üb
erfü
hren
und
1 M
inut
e be
i m
axim
aler
Dre
hzah
l in
der
Tis
chze
ntrif
uge
zent
rifug
iere
n um
die
Bak
terie
n zu
pel
letie
ren
Den
Übe
rsta
nd d
urch
Abk
ippe
n in
den
Aut
okla
vier
beut
el v
erw
erfe
n, v
erbl
eibe
ndes
Med
ium
mit
eine
m Z
ells
toff
auf
nehm
en.
Das
Pel
let d
urch
vor
texe
n in
200μl
BIB
DO
1 vo
llstä
ndig
resu
spen
dier
en u
nd 5
Min
uten
bei R
aum
tem
pera
tur i
nkub
iere
n
400μ
l BIB
DO
2 zu
gebe
n un
d vo
rsic
htig
mis
chen
bis
die
Lös
ung
klar
wird
und
kur
z au
f
Eis i
nkub
iere
n
- �08
-

Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
300μ
l BIB
DO
3 zu
gebe
n, m
isch
en b
is e
in w
eiße
r Nie
ders
chla
g en
tste
ht u
nd k
urz
auf E
is
inku
bier
en.
Lösu
ng 5
Min
uten
bei
4°C
und
max
imal
er D
rehz
ahl z
entri
fugi
eren
In d
er Z
wis
chen
zeit
neue
2m
l-Rea
ktio
nsge
fäß
vorb
erei
ten
und
besc
hrift
en
Vor
sich
tig 5
00μl
des
Übe
rsta
ndes
abn
ehm
en u
nd in
die
vor
bere
itete
n R
eakt
ions
gefä
ße
über
führ
en u
nd s
ofor
t 2,
5 V
olum
en e
iska
lten
100%
igen
Eth
anol
zug
eben
und
gut
mis
chen
Lösu
ng b
ei m
axim
aler
Dre
hzah
l für
15
Min
uten
bei
4°C
zen
trifu
gier
en
Übe
rsta
nd
abki
ppen
un
d 1m
l 70
%ig
en
Etha
nol
zuge
ben,
5
Min
uten
be
i 4°
C
zent
rifug
iere
n
Übe
rsta
nd a
bkip
pen
und
1ml 7
0%ig
en E
than
ol z
ugeb
en, 5
Min
uten
zen
trifu
gier
en b
ei
4°C
zen
trifu
gier
en
Jetz
t Ü
bers
tand
vo
rsic
htig
ab
kipp
en
und
verb
leib
ende
n Et
hano
l du
rch
kurz
es
Zent
rifug
iere
n sa
mm
eln
und
vors
icht
ig m
it ei
ner 2
00μl
-Pip
ette
abn
ehm
en
Pelle
t bei
37°
C o
ffen
im H
eizb
lock
troc
knen
und
dan
n in
50μ
l Was
ser a
ufne
hmen
2.R
estr
iktio
nsve
rdau
Mitt
els
Res
trikt
ions
endo
nukl
ease
n ka
nn D
NA
an
spez
ifisc
hen
Erke
nnun
gsse
quen
zen
gezi
elt
gesc
hnitt
en w
erde
n. D
ie in
Ver
such
stei
l 1 is
olie
rten
Plas
mid
e w
erde
n nu
n m
it je
wei
ls e
inem
oder
zw
ei
Res
trikt
ions
enzy
men
ve
rdau
t, um
m
öglic
he
Inse
rts
aus
dem
Pl
asm
id
hera
uszu
schn
eide
n.
1,5m
l-Rea
ktio
nsge
fäße
bes
chrif
ten
und
Was
ser u
nd R
eakt
ions
puff
er v
orle
gen.
Hie
rbei
Vol
umen
für E
inze
l- un
d D
oppe
lver
dau
beac
hten
.
Plas
mid
e au
s Ver
such
1 z
ugeb
en u
nd A
nsat
z au
f Eis
vor
kühl
en
Enzy
me
nach
Anw
eisu
ngen
des
Bet
reue
rs z
ugeb
en
Ans
ätze
für c
a. 1
h be
i 37°
C in
kubi
eren
- �09
-
Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
Ei
nzel
verd
au
Dop
pelv
erda
u W
asse
r 12
,5μl
12μl
10
x-Pu
ffer
2μ
l 2μ
l Pl
asm
id-L
ösun
g 5μ
l 5μ
l En
zym
1
0,5μ
l 0,
5μl
Enzy
m 2
--
0,
5μl
Ges
amtv
olum
en
20μl
20μl
3.
Gel
elek
trop
hore
se
3.1
Her
stellu
ng d
es A
garo
sege
ls
Für d
ie sp
äter
e A
uftre
nnun
g de
r Res
trikt
ions
prod
ukte
mus
s ein
Aga
rose
gel g
egos
sen
wer
den.
1% (w
/v) A
garo
se in
50m
l TA
E-Pu
ffer
geb
en u
nd in
der
Mik
row
elle
auf
koch
en b
is d
ie
Aga
rose
vol
lstä
ndig
gel
öst i
st.
Aga
rose
ans
chlie
ßend
auf
ein
em R
ühre
r abk
ühle
n la
ssen
.
Gel
kam
mer
, Käm
me
und
Schl
itten
vor
bere
iten
In d
ie h
andw
arm
e Lö
sung
0,1μl
Eth
idiu
mbr
omid
pro
ml G
el p
ipet
tiere
n, k
urz
Rüh
ren
und
das G
el in
den
Gel
schl
itten
gie
ßen.
Gel
circ
a 30
Min
uten
aus
härte
n la
ssen
.
3.2
Gel
vor
bere
iten
und
Auftr
agen
der
Pro
ben
Nac
h de
m A
ushä
rten
des
Gel
s: K
ämm
e zi
ehen
und
in
Lauf
richt
ung
in d
ie K
amm
er
eins
etzt
en
Kam
mer
bis
zur
Fül
llini
en m
it 1x
TA
E-Pu
ffer
fülle
n
Die
Pla
smid
e m
üsse
n zu
m A
uftra
gen
auf d
as G
el m
it D
NA
-Lad
epuf
fer v
erse
tzt w
erde
n, u
m si
e
durc
h di
e da
nn h
öher
e D
icht
e in
die
Tas
chen
des
Gel
s lad
en z
u kö
nnen
:
Ver
daut
e Pl
asm
ide
mit
2μl L
adep
uffe
rver
setz
en, k
urz
mis
chen
und
abz
entri
fugi
eren
20μl
der
jew
eilig
en P
lasm
ide
in je
de T
asch
e la
den
Je K
amm
3μl
kB
-Mar
ker l
aden
Gel
lauf
für c
a. 1
Stu
nde
bei 8
0 –
100V
. Dab
ei is
t auf
die
rich
tige
Polu
ng d
er E
lekt
rode
n
zu a
chte
n.
Nac
h A
bsch
luss
des
Lau
fs w
ird d
as G
el in
der
Gel
-Dok
umen
tatio
nsan
lage
unt
er U
V-
Lich
t bet
rach
tet u
nd d
okum
entie
rt.
Bitt
e ei
nen
USB
-Stic
k zu
m T
rans
fer
der
Bild
er m
itbri
ngen
- ��0
-

Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
4.T
rans
form
atio
n vo
n B
akte
rien
Zur
Tran
sfor
mat
ion
wer
den
chem
isch
kom
pete
nte
E. c
oli
Zelle
n ei
nges
etzt
die
mit
CaC
l-
Met
hode
her
gest
ellt
wur
den.
Um
ein
e ho
he T
rans
form
atio
nsra
te z
u er
reic
hen,
müs
sen
die
Zelle
n im
mer
auf
Eis
geh
alte
n w
erde
n, d
a si
e be
i hö
here
n Te
mpe
ratu
ren
ihre
Kom
pete
nz
schn
ell v
erlie
ren.
Vor
gehe
n:
Che
mis
ch k
ompe
tent
en Z
elle
n la
ngsa
m a
uf E
is a
ufta
uen
lass
en.
Eppi
s für
die
Tra
nsfo
rmat
ions
ansä
tze
vorb
erei
ten
und
vork
ühle
n.
100μ
l Zel
len
vorle
gen
und
mit
je 5μl
der
vor
her i
solie
rten
Plas
mid
e m
isch
en.
30 m
in a
uf E
is in
kubi
eren
.
Ans
ätze
im W
asse
rbad
für 1
min
bei
42°
C in
kubi
eren
.
1ml a
ntib
iotik
umfr
eies
LB
-Med
ium
zug
eben
.
Zelle
n fü
r 30
– 60
min
bei
37°
C im
Sch
üttle
r ink
ubie
ren.
5.Se
lekt
ion
der
Bak
teri
en
Um
Tra
nsfo
rman
den
mit
inse
rthal
tigen
Vek
tore
n vo
n so
lche
n oh
ne I
nser
t un
ters
chei
den
zu
könn
en, b
edie
nt m
an s
ich
der B
lau/
wei
ß-Se
lekt
ion.
Die
Aga
r-Pl
atte
n, a
uf d
enen
die
Bak
terie
n
ausp
latti
ert w
erde
n, e
ntha
lten
desh
alb
nebe
n A
mpi
cilli
n no
ch X
-Gal
als
Lak
tose
-Ana
logo
n un
d
IPTG
als
Indu
ktor
des
-G
alak
tosi
dase
-Gen
s.
Vor
gehe
n:
Sele
ktio
nspl
atte
n au
f dem
Bod
en b
esch
rifte
n
Bun
senb
renn
er a
nmac
hen
und
Drig
alsk
y-Sp
atel
kur
z ab
flam
men
.
200μ
l Tra
nsfo
rmat
ions
ansa
tz a
uf d
ie P
latte
n pi
petti
eren
Ans
atz m
it de
m S
pate
l gle
ichm
äßig
ver
teile
n un
d so
lang
e wei
ter v
erte
ilen,
bis
die
Pla
tte
trock
en is
t.
Plat
ten
bei 3
7°C
im W
ärm
esch
rank
übe
r Nac
ht k
opfü
ber i
nkub
iere
n.
Plat
ten
am n
ächs
ten
Tag
doku
men
tiere
n un
d au
swer
ten.
Hie
rfür
ein
e K
amer
a od
er H
andy
mitb
ring
en, u
m d
ie P
latt
en fo
togr
afie
ren
zu k
önne
n
- ���
-
Woc
he 1
0 M
olek
ular
biol
ogie
II
AG
Kud
la/W
einl
K
arte
und
MC
S de
s ver
wen
dete
n pB
lues
crip
t-Vek
tors
Frag
en im
Pro
toko
ll zu
bea
ntw
orte
n:
1.Er
läut
ern
Sie
die
Funk
tion
der s
chem
atis
ch d
arge
stel
lten
Elem
ente
des
obe
n ab
gebi
ldet
en
Klo
nier
ungs
vekt
ors p
Blu
escr
ipt.
2.B
eant
wor
ten
Sie
folg
ende
n Fr
agen
un
d ve
rwen
den
Sie
hier
für
nach
M
öglic
hkei
t
Abb
ildun
gen:
Was
ist d
ie u
rspr
üngl
iche
Fun
ktio
n de
s lac
Z G
enes
?
Wie
ist d
as la
cZ G
ene
im G
enom
von
E. c
oli o
rgan
isie
rt?
Wel
che
Stru
ktur
hat
die
-G
alak
tosi
dase
?
Wel
che
Funk
tion
habe
n G
alak
tosi
dase
n im
Men
sche
n?
Bitt
e ge
ben
Sie
auf d
em P
roto
koll
bitt
e ei
ne M
ail-A
dres
se a
n un
ter
der
wir
Sie
erre
iche
n kö
nnen
, um
Ihn
en d
ie B
ewer
tung
mitt
eile
n zu
kön
nen.
Die
Rüc
kgab
e de
r Pr
otok
olle
erf
olgt
nac
h A
bspr
ache
mit
dem
Bet
reue
r.
- ���
-


















