
Untersuchungen zur Plastizität der laminären Dendriten im ... · der Struktur sowie der...
141
Aus dem Institut für Zellbiologie und Tierökologie der Tierärztlichen Hochschule Hannover und der Klinik für Hals-Nasen-Ohren-Heilkunde der Medizinischen Hochschule Hannover Untersuchungen zur Plastizität der laminären Dendriten im auditorischen Cortex von akustisch deprivierten sowie chronisch elektrisch intracochleär stimulierten Hauskatzen (Felis domestica) INAUGURAL-DISSERTATION zur Erlangung des Grades einer Doktorin (Dr. med. vet.) durch die Tierärztliche Hochschule Hannover Vorgelegt von Heike Rieger aus Bremen Hannover 2004
-
Upload
nguyenngoc -
Category
Documents
-
view
214 -
download
0
Transcript of Untersuchungen zur Plastizität der laminären Dendriten im ... · der Struktur sowie der...

Aus dem Institut für Zellbiologie und Tierökologie
der Tierärztlichen Hochschule Hannover
und der Klinik für Hals-Nasen-Ohren-Heilkunde
der Medizinischen Hochschule Hannover
Untersuchungen zur Plastizität der laminären Dendriten
im auditorischen Cortex von
akustisch deprivierten sowie chronisch elektrisch intracochleär stimulierten
Hauskatzen (Felis domestica)
INAUGURAL-DISSERTATION
zur Erlangung des Grades einer Doktorin
(Dr. med. vet.)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Heike Rieger
aus Bremen
Hannover 2004

Wissenschaftliche Betreuung: Prof. Dr. rer. nat. G. Bicker
für die Tierärztliche Hochschule Hannover
Prof. Dr. rer. nat. G. Reuter
für die Medizinische Hochschule Hannover
1. Gutachter: Prof. Dr. rer. nat. G. Bicker
2. Gutachter: Prof. Dr. rer. nat. E. Zimmermann
Tag der mündlichen Prüfung: 24.11.2004

PHLQHQ�(OWHUQ��*\QQ\�XQG�)XVVHO�

Abkürzungsverzeichnis
i
Abkürzungsverzeichnis
AI primäres auditorisches Feld
AII sekundäres auditorisches Feld
AAF anteriores auditorisches Feld
AC auditorischer Cortex
AES anteriorer ectosylvanischer Sulcus
AVNC anteriorer ventraler Nucleus cochlearis
CAP zusammengesetztes Aktionspotential des Hörnerven („compound action
potential“)
CF charakteristische Frequenz
CI Cochlea-Implantat
CIS-Strategie “continuous interleaved sampling”-Strategie
dB Dezibel
DNC dorsaler Nucleus cochlearis
DiA 4-Di-16-ASP, Dialkylaminosteryl
DiI DiIC18(3), 1,1´-dioctadecyl-3,3,3´,3´-tetramethylindo-carbocyanine
perchlorate, Dialkylcarbocyanin
DNLL dorsaler Nucleus des lateralen Lemniscus
FAEP frühe akustisch evozierte Potentiale
FEEP frühe elektrisch evozierte Potentiale
FITC Fluoresceine-isothiocyanate
fMRI funktionelle Magnetresonanztomographie
Hz Hertz
IC inferiorer Colliculus
ICC Nucleus centralis des inferioren Colliculus
IHC innere Haarzellen („inner hair cells“)
INLL intermediärer Nucleus des lateralen Lemniscus
KGW Körpergewicht
kHz Kilohertz
LL Nucleus lemnisci lateralis, lateraler Lemniscus
LSO lateraler superiorer Olivenkern
MGB Corpus geniculatum mediale („medial geniculate body”)
MSO medialer superiorer Olivenkern

Abkürzungsverzeichnis
ii
MW Mittelwert
NC Nucleus cochlearis
OHC äußere Haarzellen („outer hair cells“)
p Irrtumswahrscheinlichkeit
PAF posteriores auditorisches Feld
PBS phosphate balanced salt
PEG posteriorer ectosylvanischer Gyrus
PCIT portabler Cochlea-Implantat Tester
PES posteriorer ectosylvanischer Sulcus
PFA Paraformaldehyd
PVNC posteriorer ventraler Nucleus cochlearis
SOC superiorer Olivenkomplex
SPL Lautstärke (“sound pressure level”)
SSS suprasylvanischer Sulcus
RITC Rhodamine-B-isothiocyanate
VAF ventrales auditorisches Feld
VNLL ventraler Nucleus des lateralen Lemniscus
VPAF ventroposteriores auditorisches Feld
VPON ventraler periolivärer Kern
µA Mikroampere

Inhaltsverzeichnis
I
Inhaltsverzeichnis
1. EINLEITUNG 11
2. LITERATURÜBERSICHT 13
2.1 Die Anatomie des Hörorgans 13
2.2 Die Grundlagen des Hörvorgangs 15
2.2.1 Das physiologische Hören 15
2.2.2 Das Hören mit dem Cochlea-Implantat 17
2.3 Die afferente Hörbahn 18
2.4 Der auditorische Cortex 21
2.4.1 Die Lage und makroskopische Anatomie des auditorischen Cortex 21
2.4.2 Der histophysiologische Aufbau des auditorischen Cortex 24
2.4.2.1 Die tonotope Organisation der auditorischen Felder 24
2.4.2.2 Die detaillierte Verteilung der afferenten Projektionen 25
2.4.2.3 Die intra- und intercorticalen Verbindungen
des auditorischen Cortex 28
2.4.3 Die Cytoarchitektur und neuronale Organisation des
auditorischen Cortex 31
2.5 Die Veränderungen im auditorischen System der Katze 38
2.5.1 Die physiologische Entwicklung und Ausreifung des
auditorischen Systems 38
2.5.1.1 Das Hörorgan und die afferente Hörbahn 38
2.5.1.2 Der auditorische Cortex 38
2.5.2 Die plastischen Umstrukturierungen im auditorischen System nach
neonataler akustischer Deprivation 40
2.5.2.1 Das Hörorgan und die afferente Hörbahn 41
2.5.2.2 Der auditorische Cortex 43

Inhaltsverzeichnis
II
2.5.3 Die plastischen Umstrukturierungen im auditorischen System nach
neonataler akustischer Deprivation und anschließender chronischer
elektrischer intracochleärer Stimulation 45
2.5.3.1 Das Hörorgan und die afferente Hörbahn 45
2.5.3.2 Der auditorische Cortex 46
2.6 Die Ziele der Untersuchung 48
3. MATERIAL UND METHODEN 49
3.1 Material 49
3.1.1 Die Versuchstiere 49
3.1.2 Die Sachmaterialien 51
3.1.2.1 Die Pharmaka 51
3.1.2.2 Die Chemikalien 51
3.1.2.3 Die Verbrauchsmaterialien 52
3.1.2.4 Die Gebrauchsgegenstände 52
3.1.2.5 Die technische Ausstattung 53
3.2 Methoden 53
3.2.1 Die experimentelle neonatale Ertaubung der Katzen 53
3.2.2 Die Implantation der elektronischen Innenohrprothesen in neonatal
experimentell ertaubte Katzen 54
3.2.3 Die chronische elektrische intracochleäre Stimulation
der neonatal ertaubten, implantierten Katzen 55
3.2.4 Die histologische Aufbereitung des auditorischen Cortex 56
3.2.5 Die Fluoreszenzfarbstoff-Tracingtechnik 56
3.2.5.1 DiA/DiI als neuronale Tracer 57
3.2.5.2 Die Methode der DiA/DiI-Färbung 57
3.2.6 Die Anfertigung der Gewebeschnitte 59
3.2.7 Die Methode der Kresyl-Violett-Färbung (NISSL-Ersatzfärbung) 61
3.2.8 Die histologisch-morphologische Auswertung des auditorischen Cortex 62
3.2.9 Die statistische Analyse 65

Inhaltsverzeichnis
III
4. ERGEBNISSE 66
4.1 Die Morphometrie der Dendriten der Laminae I bis VI im
auditorischen Cortex 66
4.1.1 Horizontale Dendritenverläufe im AAF, AI und AII 66
4.1.1.1 Hörende Versuchstiergruppe 66
4.1.1.2 Ertaubte Versuchstiergruppe 67
4.1.1.3 Chronisch elektrisch intracochleär stimulierte
Versuchstiergruppe 68
4.1.2 Diagonale Dendritenverläufe im AAF, AI und AII 69
4.1.2.1 Hörende Versuchstiergruppe 70
4.1.2.2 Ertaubte Versuchstiergruppe 71
4.1.2.3 Chronisch elektrisch intracochleär stimulierte
Versuchstiergruppe 72
4.1.3 Vertikale Dendritenverläufe im AAF, AI und AII 73
4.1.3.1 Hörende Versuchstiergruppe 73
4.1.3.2 Ertaubte Versuchstiergruppe 74
4.1.3.3 Chronisch elektrisch intracochleär stimulierte
Versuchstiergruppe 75
4.2 Der direkte Vergleich der Laminae zwischen der hörenden, neonatal
ertaubten sowie chronisch elektrisch intracochleär stimulierten
Versuchstiergruppen im AAF, AI und AII 76
4.3 Eine Zusammenfassung der Ergebnisse 76
5. DISKUSSION 80
5.1 Material und Methoden 80
5.2 Die Morphometrie der Dendriten der Laminae I bis VI im AAF, AI und AII
normal hörender Katzen 85
5.3 Die Auswirkungen einer experimentell induzierten neonatalen akustischen
Deprivation auf die Morphometrie der Dendriten der Laminae I bis VI
im AAF, AI und AII der Katze 88

Inhaltsverzeichnis
IV
5.4 Die Auswirkungen einer chronischen elektrischen intracochleären
Stimulation auf die Morphometrie der Dendriten der Laminae I bis VI
im AAF, AI und AII der Katze 94
5.5 Die Schlussfolgerungen 98
6. ZUSAMMENFASSUNG 100
7. SUMMARY 102
8. LITERATURVERZEICHNIS 104
9. ANHANG 121
9.1 Das Cochlea-Implantat 121
9.2 Die chronische elektrische intracochleäre Stimulation 122
9.3 Die Überprüfung des Hörstatus durch Messung der frühen
akustisch (FAEP) und elektrisch (FEEP) evozierten Potentiale 123
9.4 Die täglichen Körpergewichtszunahmen der Versuchstiere 125
9.5 Die Herstellung der verwendeten Chemikalien-Lösungen 127
9.5.1 PBS-Lösung (Perfusionslösung) 127
9.5.2 Paraformaldehyd-Lösung (Fixationslösung) 127
9.5.3 Agar agar (Verschluss- und Einbettmedium) 128
9.5.4 Kresyl-Violett-Färbelösung (Nissl-Ersatzfärbung) 128
9.5.5 Mowiol (Eindeckelmedium) 128
9.6 Die Fluoreszenz-Farbstoffe DiA und DiI 129
9.6.1 Die Struktur des DiA 129
9.6.2 Das Absoptions- und Emissionsspektrum des DiA 129
9.6.3 Die Struktur des DiI 130
9.6.4 Das Absoptions- und Emissionsspektrum des DiI 130

Inhaltsverzeichnis
V
9.7 Die Daten der Auswertung 131
9.7.1 Das primäre auditorische Feld (AI) 131
9.7.1.1 Hörende Tiere (Prozent) 131
9.7.1.2 Neonatal ertaubte Tiere (Prozent) 131
9.7.1.3 Neonatal ertaubte, anschließend chronisch elektrisch
intracochleär stimulierte Tiere (Prozent) 132
9.7.2 Das sekundäre auditorische Feld (AII) 132
9.7.2.1 Hörende Tiere (Prozent) 132
9.7.2.2 Neonatal ertaubte Tiere (Prozent) 133
9.7.2.3 Neonatal ertaubte, anschließend chronisch elektrisch
intracochleär stimulierte Tiere (Prozent) 133
9.7.3. Das anteriore auditorische Feld (AAF) 134
9.7.3.1 Hörende Tiere (Prozent) 134
9.7.3.2 Neonatal ertaubte Tiere (Prozent) 134
9.7.3.3 Neonatal ertaubte, anschließend chronisch elektrisch
intracochleär stimulierte Tiere (Prozent) 135
10. VERZEICHNIS DER ABBILDUNGEN 136
11. VERZEICHNIS DER TABELLEN 139
12. DANKSAGUNG 141

1. Einleitung
11
1. EINLEITUNG
Das Hören, als im Innenohr stattfindender Transduktionsprozess, ist auf die Funktions-
tüchtigkeit weniger tausend Hörsinneszellen angewiesen. Sind diese inneren Haarzellen
zerstört, kommt es zu einer progressiven Degeneration der in der zentralen auditorischen
Hörbahn nachgeschalteten Nervenzellen (HARDIE und SHEPHERD 1999). Ein
Charakteristikum neuronaler Zellen ist jedoch, dass diese nach ihrer Differenzierung die
Teilungsfähigkeit eingebüßt haben. Zu Beginn der Erforschung neuronaler Vernetzungen und
der Struktur sowie der Entwicklung des Großhirns wurde daher die Fragestellung
aufgeworfen, inwieweit nach Beendigung der embryonalen und neonatalen Entwicklung die
Möglichkeit einer Restrukturierung oder Plastizität innerhalb bereits entwickelter komplexer
neuronaler Strukturen stattfinden kann. Während man früher davon ausging, dass aufgrund
der Charakteristik der zugrundeliegenden zellulären Strukturen nach vollständiger Ausreifung
der cerebralen Strukturen keine Reorganisationsfähigkeit vorliegt, konnten neuere
Untersuchungen (PETITTO et al. 2000; RAUSCHECKER und KORTE 1993; REBILLARD
et al. 1977, 1980) zeigen, dass bereits spezialisierte Gehirnareale nach einem Funktionsverlust
von noch aktiven Bereichen vereinnahmt werden (sogenannte cross-modale Plastizität).
Mit Hilfe einer elektronischen Innenohrprothese (Cochlea-Implantat) können vollständig im
Innenohr ertaubten Patienten Höreindrücke vermittelt werden, die von der Wahrnehmung von
Umgebungsgeräuschen bis zu einem offenen Sprachverständnis des Patienten reichen. Bei
prälingual ertaubten Kindern kann es den Erwerb einer annähernd normalen Vokalisation
ermöglichen (SHEPHERD et al. 1997). Trotz des erfolgreichen Einsatzes des Cochlea-
Implantats bei prä- und postlingual ertaubten Patienten bestehen noch Defizite in Bezug auf
die Auswirkungen einer Taubheit sowie einer chronischen elektrischen Stimulation auf die
Funktion, Plastizität und Reifung der Nervenzellen im auditorischen Cortex.
Verschiedene Arbeiten vergangener Jahre konnten zeigen, dass eine experimentelle
Ertaubung sowie eine chronische elektrische intracochleäre Stimulation Auswirkungen auf
die Physiologie der zentralen auditorischen Hörbahn nach sich zieht (CORDS 1996; KELLER
1997). Auf morphologischer Ebene konnten in diversen Schaltzentren der Hörbahn (HEID et
al. 1998; SNYDER et al. 1990; TERAYAMA et al. 1977; VOGT et al. 1997) sowie dem
auditorischen Cortex (REUTER et al. 2002b; WENKE 1999; WURTH 1999) Veränderungen
im Zellbild beobachtet werden.

1. Einleitung
12
Hinweise darauf, dass eine experimentell erzeugte Taubheit auch funktionelle Auswirkungen
auf den auditorischen Cortex, als höchste Station der zentralen Hörbahn hat, konnten
insbesondere DINSE et al. (1997) und IRVINE et al. (2001) nachweisen. Wie es zu dieser
funktionellen Umstrukturierung kommt und inwieweit dieser eine Veränderung der
neuronalen Verknüpfung in den auditorischen Feldern zugrunde liegt, ist jedoch noch nicht
bekannt.
In der vorliegenden Arbeit sollte untersucht werden, inwieweit durch eine experimentelle
direkte Stimulation des Hörnerven mittels Cochlea-Implantat bei neonatal ertaubten
Individuen eine Plastizität im auditorischen Cortex induziert werden kann. Dazu wurde das
Tiermodell der neonatal ertaubten Hauskatze als Modell für prälingual ertaubte Kinder
verwendet.
Das Ziel der vorliegenden Arbeit ist ein Vergleich der Struktur der Dendriten innerhalb des
auditorischen Cortex normal hörender, neonatal experimentell ertaubter sowie neonatal
experimentell ertaubter, anschließend chronisch elektrisch intracochleär stimulierter Katzen.
Dazu wird mit Hilfe einer Tracingtechnik unter Verwendung von Fluoreszenzfarbstoffen
gearbeitet. Innerhalb des anterioren, primären und sekundären auditorischen Feldes wird jede
der sechs corticalen Schichten betrachtet.
Insbesondere soll in dieser Arbeit der Frage nachgegangen werden, inwieweit es durch eine
neonatale Ertaubung zu einer qualitativen und/oder quantitativen Veränderung der Dendriten
im auditorischen Cortex kommt und ob diese durch den Einsatz eines Cochlea-Implantats
beeinflusst werden kann.

2. Literaturübersicht
13
2. LITERATURÜBERSICHT
2.1 Die Anatomie des Hörorgans
Das Hörorgan besteht aus einer Reihe informationsaufnehmender und informations-
übertragender Elemente. Funktionell und anatomisch besteht es aus drei Abschnitten: dem
äußeren Ohr, dem Mittel- und dem Innenohr (Abb. 1).
Das Außenohr (Auris externa) wird von der Ohrmuschel (Auricula) und dem äußeren
Gehörgang (Meatus acusticus externus) gebildet. Das membranöse Trommelfell (Membrana
tympani) schließt den äußeren Gehörgang ab und bildet die Grenze zum luftgefüllten,
schleimhautausgekleideten Mittelohr (Auris media). Zu den Strukturen des in der Pars
tympanica des Felsenbeins gelegenen Mittelohres gehört die medial vom Trommelfell
gelegene Paukenhöhle (Cavum tympani) mit den drei Gehörknöchelchen (Ossicula auditus).
Von diesen ist der Stiel des Hammers (Malleus) mit dem Trommelfell verwachsen und über
den Amboss (Incus) mit dem Steigbügel (Stapes) verbunden. Die Ossicula bilden ein an
Bändern, Sehnen und Bindegewebe aufgehängtes Hebelsystem zur Schallübertragung auf das
Innenohr (Auris interna).
Abb. 1: Schematische Darstellung der Kranialansicht eines Transversalschnittes durch das rechte Ohr einer Katze (aus HUDSON und HAMILTON 1993)
1 Schädeldach; 2 M. temporalis; 3-6` äußeres Ohr: 3-4 Ohrmuschelknorpel, (3 Scapha, 4 Concha), 5 Halbringförmiger Knorpel, 6 Äußerer Gehörgang (vertikaler Anteil), 6`Äußerer Gehörgang (horizontaler Anteil); 7-12 Mittelohr: 7 Bulla tympanica, 8 Septum bullae, 9 Trommelfell, 10 -12 Knochen des Ohres (10 Hammer, 11 Amboss, 12 Steigbügel); 13 Ohrtrompete (Eustachische Röhre); 14 Pars petrosa der Felsenbeinpyramide; 15-17 Knöchernes Labyrinth: 15 Bogengänge, 16 Vestibulum, 17 Cochlea

2. Literaturübersicht
14
Im Innenohr befindet sich, neben dem Vestibularapparat, das im inneren Gehörgang (Meatus
acusticus internus) gelegene eigentliche Hörorgan, die Hörschnecke (Cochlea). Die Cochlea
ist ein blind endendes schlauchförmiges Organ, das in Form eines Schneckenhauses beim
Menschen in viereinhalb und bei der Katze in zweieinhalb Windungen um die knöcherne
Schneckenspindel (Modiolus) aufgerollt ist. Sie besteht aus drei übereinanderliegenden
Kanälen (Skalen), von denen zwei, die oben gelegene Vorhoftreppe (Scala vestibuli) und die
untere Paukentreppe (Scala tympani), über das Helikotrema miteinander verbunden sind. Die
Membrana vestibularis (Reißnersche Membran) trennt die Scala media (Ductus cochlearis)
von der Scala vestibuli, während die Basilarmembran (Lamina basilaris) die Scala media von
der Scala tympani abgrenzt (Abb. 2). Die Scala vestibuli und die Scala tympani sind mit aus
dem Liquor cerebrospinalis stammender kaliumarmer Perilymphe gefüllt. In der Scala media
befindet sich von der Stria vascularis produzierte kaliumreiche Endolymphe.
Abb. 2: Schematische Darstellung eines Querschnitts durch eine Cochleawindung mit Corti-Organ
(modifiziert nach BLOOM und FAWCETT 1975)
Die zentrale Funktionseinheit der Cochlea ist die häutige Basilarmembran. Sie inseriert am
Labium limbi vestibulare der Lamina spiralis ossea und zieht an die laterale Wand der
Cochlea. Auf ihr ist das Corti-Organ (Organum spirale) mit den in Stützzellen eingebetteten
eigentlichen Sinneszellen des Hörorgans, den Haarzellen, lokalisiert.
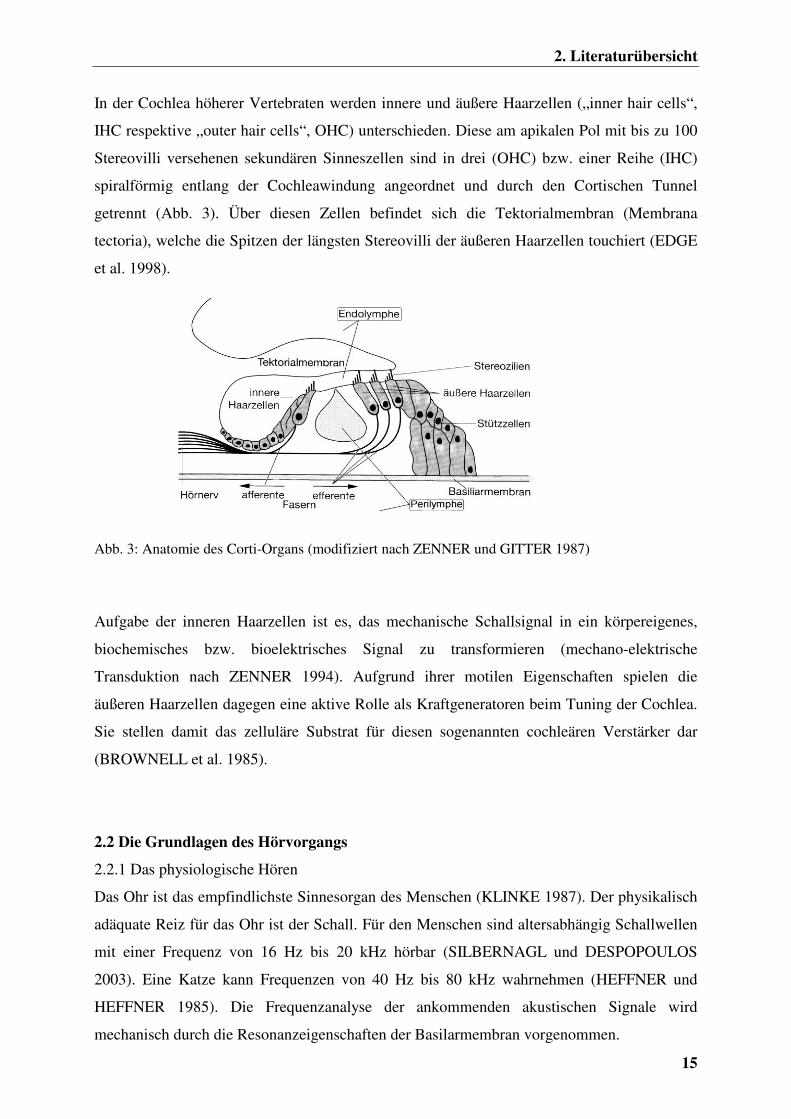
2. Literaturübersicht
15
In der Cochlea höherer Vertebraten werden innere und äußere Haarzellen („inner hair cells“,
IHC respektive „outer hair cells“, OHC) unterschieden. Diese am apikalen Pol mit bis zu 100
Stereovilli versehenen sekundären Sinneszellen sind in drei (OHC) bzw. einer Reihe (IHC)
spiralförmig entlang der Cochleawindung angeordnet und durch den Cortischen Tunnel
getrennt (Abb. 3). Über diesen Zellen befindet sich die Tektorialmembran (Membrana
tectoria), welche die Spitzen der längsten Stereovilli der äußeren Haarzellen touchiert (EDGE
et al. 1998).
Abb. 3: Anatomie des Corti-Organs (modifiziert nach ZENNER und GITTER 1987)
Aufgabe der inneren Haarzellen ist es, das mechanische Schallsignal in ein körpereigenes,
biochemisches bzw. bioelektrisches Signal zu transformieren (mechano-elektrische
Transduktion nach ZENNER 1994). Aufgrund ihrer motilen Eigenschaften spielen die
äußeren Haarzellen dagegen eine aktive Rolle als Kraftgeneratoren beim Tuning der Cochlea.
Sie stellen damit das zelluläre Substrat für diesen sogenannten cochleären Verstärker dar
(BROWNELL et al. 1985).
2.2 Die Grundlagen des Hörvorgangs
2.2.1 Das physiologische Hören
Das Ohr ist das empfindlichste Sinnesorgan des Menschen (KLINKE 1987). Der physikalisch
adäquate Reiz für das Ohr ist der Schall. Für den Menschen sind altersabhängig Schallwellen
mit einer Frequenz von 16 Hz bis 20 kHz hörbar (SILBERNAGL und DESPOPOULOS
2003). Eine Katze kann Frequenzen von 40 Hz bis 80 kHz wahrnehmen (HEFFNER und
HEFFNER 1985). Die Frequenzanalyse der ankommenden akustischen Signale wird
mechanisch durch die Resonanzeigenschaften der Basilarmembran vorgenommen.

2. Literaturübersicht
16
Schallwellen gelangen als Druckschwankungen in der Luft in den äußeren Gehörgang und
treffen auf das Trommelfell. Die per Luftleitung weitergegebene Energie des Schalls führt
zum Schwingen des Trommelfells und damit zu einem Ein- und Auswärtsvibrieren der
Gehörknöchelchenkette (HÜTTENBRINK 1992). Die beweglich im ovalen Fenster der
Cochlea sitzende Fußplatte des Stapes leitet die Schallenergie an die Perilymphe der Scala
vestibuli im Innenohr fort. Diese nicht-kompressible Flüssigkeit drückt in Folge dessen die
Reißnersche Membran in Richtung Scala media sowie das auf der Basilarmembran gelegene
Corti-Organ nach unten. Durch die Auslenkung entsteht eine sich von basal nach apikal
ausbreitende passive Welle der Basilarmembran, so dass die Schallenergie als Wanderwelle
weiterläuft. Die Laufzeit der Wanderwelle vom apikalen bis zum basalen Ende der
Basilarmembran beträgt beim Menschen etwa sechs Millisekunden (KLINKE 1987).
In Abhängigkeit von der Frequenz des auslösenden Schalls kommt es aufgrund der
progressiven Zunahme der Elastizität der Basilarmembran von der Schneckenbasis bis zur
Spitze zu einer maximalen Auslenkung der Wanderwelle an verschiedenen Orten entlang der
cochleären Membranen (VON BÉKÉSY 1928). Nach diesem sogenannten Tonotopieprinzip
finden sich innerhalb der Cochlea die Resonanzstellen der hohen Frequenzen basal, jene der
tiefen Frequenzen sind apikal lokalisiert (Cochleotopie). Die nach dem Frequenz-Ort-
Transformations-Prinzip beobachtete Wanderwelle ist jedoch zu breit, um die scharfe
Frequenzabstimmung des Hörorgans zu erklären. Die theoretischen Arbeiten von KEMP
(1979) zeigten, dass die scharfe Frequenzabstimmung des intakten Hörorgans nur mit Hilfe
eines aktiven, mechanischen Prozesses erreicht werden kann, welcher Energie in die
Wanderwelle einkoppelt.
Die von der Wanderwelle ausgelöste Auslenkung führt zu einer relativen Scherbewegung
zwischen Tektorial- und Basilarmembran und damit zum Abscheren der Stereovilli der
äußeren Haarzellen. Aufgrund der mechanischen Lageveränderung kommt es zur Öffnung
von Ionenkanälen an der Zellapex und zur Auslösung eines Rezeptorpotentials. Es resultieren
aktive, oszillierende Längenänderungen der äußeren Haarzellen, welche zur lokalen
Verstärkung der Wanderwelle führen (REUTER et al. 1991). Dabei weisen die angeregten
Zellen bei einer Hyperpolarisation eine Verlängerung und während einer Depolarisation eine
Verkürzung in einem Bereich von ca. 4 % auf (ASHMORE 1987).

2. Literaturübersicht
17
Als Folge subtektorialer Flüssigkeitsströmungen erfolgt daraufhin die Deflexion der
Stereovilli der inneren Haarzellen (DALLOS 1986), worauf ein Ca2+ -Einstrom am basalen
Zellpol folgt. Im Anschluss wird der Transmitterstoff Glutamat an die afferenten Fasern des
Ganglion spirale cochleae abgegeben (EYBALIN und PUJOL 1987). Nach Überschreiten des
postsynaptischen Schwellenwertes kommt es zur Auslösung von in den Nervenfasern
weitergeleiteten Aktionspotentialen. Da 90 % aller afferenten Neurone mit den inneren
Haarzellen assoziiert sind (SPOENDLIN 1969), wird so die neuronale Weiterleitung des
Höreindrucks über den Hörnerv und die auditiven Kerne im Hirnstamm bis zum auditorischen
Cortex im Temporallappen induziert.
Durch das Tonotopieprinzip und der lokalen Verstärkung der Wanderwelle durch die äußeren
Haarzellen reflektiert die Cochlea die zeitliche Struktur und Intensität ankommender
komplexer Signale und verteilt die Kodierung der spektrotemporalen Komponenten des
Schalls auf die tonotop organisierte Anordnung primärer afferenter Fasern. Jeder inneren
Haarzelle mit den dazugehörigen afferenten Nervenfasern ist somit eine bestimmte optimale
Reizfrequenz zugeordnet, der sogenannten charakteristischen Frequenz (CF). Die Tonotopie,
welche entlang der Cochleawindungen beginnt, setzt sich so entlang der verschiedenen
Strukturen der zentralen Hörbahn bis in den auditorischen Cortex fort (MERZENICH et al.
1982).
Zusätzlich zur Frequenzerkennung ist im Hörnerv eine zweite Kodierungsmöglichkeit
vorhanden (SACHS 1984). Bei der Beschallung des Ohres mit niedrigen Frequenzen treten
die neuronalen Entladungen im Hörnerv bevorzugt zu bestimmten Zeitpunkten innerhalb des
Schwingungszyklus auf (sogenannte frequenzgekoppelte Entladung). Zeitstrukturen im
Schallreiz werden so durch zeitgerecht auftretende Aktionspotentiale im Hörnerven
abgebildet und über die aufsteigende Hörbahn weitergeleitet. Das zentrale Nervensystem ist
in der Lage, das dadurch in den neuronalen Aktionspotentialen entstehende Zeitmuster
auszuwerten und zur Frequenzanalyse zu nutzen (Periodizitätsanalyse).
2.2.2 Das Hören mit dem Cochlea-Implantat
Beim „elektrischen“ Hören mit dem Cochlea-Implantat wird die Funktion der inneren
Haarzellen durch die direkte Aktivierung des Hörnerven mittels eines elektrischen Impulses
der intracochleär platzierten Elektrodenkontakte überbrückt.

2. Literaturübersicht
18
Der Wegfall der mechanischen cochleären Komponente und der damit einhergehende
Zeitgewinn zwischen Schallauftritt und Auslösung von Aktionspotentialen am Hörnerven
spiegelt sich bei der Messung der Hörbahnaktivität in einer Verkürzung der Zeit zwischen
Reizdarbietung und Auftreten von Aktivitäten der auditorischen Hörbahn (sogenannte Latenz)
wieder (CORDS 1996).
Die ortsgebundene Frequenzspezifität wird bei Gebrauch eines Cochlea-Implantats nicht
durch die Auslösung der cochleären mechanischen Prozesse, sondern durch die individuelle
Aktivierung der einzelnen in der Cochlea platzierten Elektrodenkontakte durch geeignete
Stimulationsstrategien erreicht (zur Übersicht siehe BÜCHNER 2002). Die Frequenz-
erkennung des ankommenden Schalls erfolgt hierbei durch den an das Mikrophon
angeschlossenen Sprachprozessor (siehe Anhang 9.1). Dadurch bleibt auch beim Hören mit
einem Cochlea-Implantat über die gesamte auditorische Hörbahn und auf Ebene des
auditorischen Cortex die grundsätzliche cochleäre Tonotopie erhalten (POPELAR et al.
1995).
2.3 Die afferente Hörbahn
Die die inneren und äußeren Haarzellen der Cochlea innervierenden afferenten Nervenfasern
erreichen den auditorischen Cortex im Temporallappen über die aufsteigende Hörbahn. Dabei
divergiert die Information von einem Ort der Cochlea über mehr als 10 Hirnstammkerne und
24 identifizierte Zelltypen und konvergiert schließlich wieder auf ein Frequenzband im
inferioren Colliculus (IRVINE 1992). Bis zum Mittelhirn werden alle Informationen über die
cochleotopen (tonotopen) Verbindungen der sogenannten lemniscalen, zentralen Hörbahn
geschaltet. Ab dem inferioren Colliculus unterscheidet man eine zusätzliche, extralemniscale,
diffuse, nicht-tonotop organisierte Bahn.
Als sekundäre Sinneszellen bilden sowohl die inneren als auch die äußeren Haarzellen keine
eigenen Nervenfortsätze aus. Stattdessen werden sie von den peripheren Dendriten der
Bipolarzellen des Ganglion spirale cochleae, dem in der Schneckenspindel gelegenen Beginn
des Hörnerven, innerviert. Die von den Basalganglien des Ganglion spirale cochleae
ausgehenden afferenten zentralen Fortsätze bilden die Pars cochlearis des Nervus
vestibulocochlearis, dem 8. Gehirnnerv. Sie ziehen durch den inneren Gehörgang im
Felsenbein zum Rautenhirn und projizieren in den jeweiligen Nucleus cochlearis (NC), den
ersten synaptischen Kerngebieten der zentralen Hörbahn (siehe Abb. 4). Der NC jeder Seite
befindet sich oberflächlich nahe der Eintrittsstelle des Hörnerven im Tuberculum acusticum.

2. Literaturübersicht
19
Er besteht aus einem, in der Medulla oblongata gelegenen, dorsalen Nucleus (DNC) sowie
einem ventralen Ganglion, welches aus einem anterioren ventralen (AVNC) und einem
posterioren ventralen (PVNC) Nucleus zusammengesetzt ist (MELCHER et al. 1996). Die
von den äußeren und inneren Haarzellen ausgehenden Axone gabeln sich nach Erreichen des
ipsilateralen Nucleus cochlearis systematisch auf: jede Faser gibt einen aufsteigenden Ast an
den AVNC sowie einen absteigenden Ast ab, welcher im PVNC und DNC endet (LORENTE
DE NÓ 1933). Somit erhält jede Untereinheit des Nucleus cochlearis cochleotope (tonotope)
Informationen vom gesamten Verband der äußeren und inneren Haarzellen. Im DNC kommt
es zudem zur Aufteilung der Hörbahn. Während lediglich 10 % der Fasern ipsilateral weiter
verlaufen, werden etwa 90 % aller afferenten Fasern zur Gegenseite umgeschaltet (HEID et
al. 1997). Vom Nucleus cochlearis aus projizieren die sekundären afferenten Fasern, auch
zweites Neuron genannt, zu Zellgruppen in der Medulla, der Pons und dem Mittelhirn
(AITKIN 1986). Dabei formen sie drei Hauptfaserbündel: die ventrale akustische Stria,
welcher sich das Corpus trapezoideum in der ventralen Pons anschließt, die intermediäre Stria
(Heldsche Stria) und die dorsale akustische Stria (Monakowsche-Stria).
Abb. 4: Schematische Darstellung der afferenten zentralen Hörbahn der linken Cochlea (modifiziert
nach ZENNER 1993)

2. Literaturübersicht
20
Die ventrale Stria erreicht die Hauptzellgruppen des im querverlaufenden Corpus
trapezoideum gelegenen, kontralateralen superioren Olivenkomplexes (SOC). Hierbei handelt
es sich um eine nicht-obligatorische Schaltstelle für alle Fasern der Hörbahn. Die im SOC
entspringenden Fasern verbinden sich mit Projektionen aus dem ipsi- und kontralateralen
Nucleus cochlearis und betreten den ipsilateralen Nucleus lemnisci lateralis (lateraler
Lemniscus, LL). Von dessen dorsalen Nucleus (DNLL) aus steigen sie als drittes Neuron
bilateral sowie vom intermediären (INLL) und ventralen Nucleus (VNLL) vornehmlich
ipsilateral zum rostral gelegenen inferioren Colliculus (IC) der Vierhügelplatte auf (HEID et
al. 1997).
Der IC wird traditionell in einen zentralen, perizentralen und externalen Nucleus unterteilt.
Der Nucleus centralis (ICC) wird als obligatorische Schaltstelle für über den lateralen
Lemniscus übertragene Informationen aller auditorischen Hirnstammkerne angesehen. Dabei
hält er Kontakt mit annähernd 20 Neuronentypen, welche in ungefähr einem duzend Arealen
des Hirnstamms und des Cortex lokalisiert sind. Vom ICC aus werden die tonotopen
Informationen, weiterhin als drittes Neuron, zumeist ungekreuzt über das Brachium colli, dem
im Vorderhirn gelegenen auditorischen Thalamus, übermittelt.
Die hauptsächlich mit dem auditorischen System assoziierten thalamischen Kerne sind das
Corpus geniculatum mediale („medial geniculate body“, MGB) und der laterale Anteil der
posterioren Gruppe der thalamischen Kerne. Diese Nuclei stellen die wichtigsten und letzten
synaptischen Stationen der auditorischen Hörbahn dar. Cytoarchitektonisch lässt sich der
MGB in einen ventralen, einen dorsalen und einen medialen Hauptanteil unterteilen.
Innerhalb dieser Hauptgebiete sind weitere Unterbereiche zu unterscheiden (ANDERSEN et
al. 1980).
Die thalamocorticalen Neurone des lemniscalen, zentralen auditorischen Systems projizieren
als viertes Neuron vor allem aus dem ventralen Anteil über die Hörstrahlung (Radiatio
acustica) in einer Art Punkt- zu Streifen- System zu den tonotop organisierten auditorischen
Rindenfeldern im Temporallappen der Großhirnhemisphären (Henlesche-Querwindung).
Dadurch erhalten das primäre auditorische Feld, das anteriore auditorische Feld sowie das
ventroposteriore (ANDERSEN et al. 1980) und wahrscheinlich auch das posteriore
auditorische Feld (FITZPATRICK et al. 1977) hoch konvergierenden Input von einer
Vielzahl von auditorischen thalamischen Kernen. Die auf diese Weise gebildeten
ausgedehnten Konvergenzen und Divergenzen berücksichtigen auf allen Stufen der
Verarbeitung die von den inneren Haarzellen der Cochlea ausgehende Tonotopie
(MERZENICH et al. 1982).

2. Literaturübersicht
21
Das zweite aufsteigende auditorische System, die extralemniscale, diffuse Hörbahn erhält den
Großteil der afferenten Informationen von außerhalb der zentralen auditorisch-thalamischen
Hörbahn gelegenen Gebieten. Anders als die lemniscale auditorische Hörbahn weist das
extralemniscale System keine oder eine nur gering ausgeprägte tonotope Organisation auf. Es
beinhaltet den externalen und perizentralen Nucleus des inferioren Colliculus, die mediale
Abteilung des Corpus geniculatum mediale und das sekundäre auditorische Feld sowie
weitere sekundäre corticale Bereiche, welche die tonotopen auditorischen Felder ringartig
umgeben. Weiterer „nichtspezifischer“ den auditorischen Cortex erreichender Input entspringt
in der Formatio reticularis des Hirnstamms sowie in den medialen/intralaminären Nuclei des
Thalamus.
Zusätzlich zu dem konvergierenden/divergierenden afferenten auditorischen System existiert
ein eher massives und komplexes System von Bahnen, die als efferente Fasern in den
auditorischen Arealen des Cortex als sogenanntes „Rasmussen-Bündel“ ihren Ursprung
nehmen und bis in den Bereich des Corti-Organs reichen. Dabei durchlaufen sie die
genannten Schaltstationen und werden in der Peripherie als zwei relativ eigenständige Bahnen
unterschieden, welche sich im superioren Olivenkomplex teilen (WARR 1992). Der Hauptteil
der efferenten Fasern erreicht die äußeren Haarzellen und stammt von medial im superioren
Olivenkomplex gelagerten Neuronen. Die inneren Haarzellen erhalten Informationen von
kleinen, unmyelinisierten Neuronen des lateralen superioren Olivenkomplexes.
Das efferente System hat eine inhibitorische Wirkung im Sinne einer selektiven Filterung von
Tönen und Geräuschen. Diese Funktion wird über axodendritische Synapsen zwischen den
afferenten und efferenten Fasern der Haarzellen erreicht (HOTH und LENARZ 1994).
2.4 Der auditorische Cortex
2.4.1 Die Lage und makroskopische Anatomie des auditorischen Cortex
Der auditorische Cortex ist das Zielgebiet der afferenten Neurone der aufsteigenden
lemniscalen und extralemniscalen Hörbahnen. Die Bezeichnung „auditorischer Cortex“
bezieht sich im klassischen Sinne auf die Bereiche des cerebralen Cortex, deren
hauptsächliche afferente Projektionen dem Corpus geniculatum mediale entspringen und
Neurone enthält, die auf akustische Reize reagieren (EHRET 1997).

2. Literaturübersicht
22
Im weiteren Sinnen sind zum auditorischen Cortex auch jene corticalen Areale zu zählen,
welche als „polysensorisch“, „nicht-spezifisch“ oder „assoziativ“ bezeichnet werden.
Anatomisch gehört dieser Bereich zur Facies parietalis des Telencephalon, dem superioren
Bereich des Temporallappens.
Histologische, physiologische und verhaltenskundliche Untersuchungen haben gezeigt, dass
der auditorische Bereich des Neocortex eine komplexe Struktur mit mehreren Teilgebieten ist.
Er ist zusammengesetzt aus einem tonotop organisierten primären auditorischen Feld (AI) und
speziesabhängig von einem oder mehreren weiteren angrenzenden Feldern, welche ebenfalls
eine tonotope Organisation aufweisen können (BRUGGE und REALE 1985).
Die Anzahl der auditorischen Felder nimmt von drei bis vier bei Insektivoren über fünf bis
sieben bei Rodentiern hin zu sechs und mehr als acht bei Carnivoren und Primaten zu (siehe
Tab. 1). Sie scheint abhängig von der auf einer bestimmten Evolutionsstufe erreichten
relativen Größe der corticalen Oberfläche zu sein (MERZENICH und SCHREINER 1992).
Die einzelnen auditorischen Felder können auf der Grundlage ihrer Cytoarchitektur, ihren
Verbindungsstrukturen und ihrer funktionellen Unterschiede differenziert werden.
Tab.1: Auflistung der auditorischen Felder ausgewählter Spezies (modifiziert nach CLAREY et al. 1991)
Der auditorische Cortex des Menschen entspricht den nach BRODMANN (1909)
cytoarchitektonisch eingeteilten Arealen 41, 42 und 22. Nach den cytoarchitektonischen
Untersuchungen von VON ECONOMO und HORN (1930) konnte er in ein primäres
auditorisches Feld (AI) und vier weitere, das AI umgebenen, Areale unterteilt werden.
Das AI, auch Area TC genannt, entspricht dem Brodmann-Areal 41. Es liegt im sylvanischen
Anteil des Gyrus temporalis superior in der Tiefe der Henleschen Querwindung (LIGEOIS-
CHAUVEL et al. 1991). Seine dorsoventrale Ausbreitung beträgt etwa 3 mm, mit einer
Fläche von ca. 900-1600 mm². Es handelt sich um ein primär rezeptives Feld, welches
wichtige thalamische Afferenzen erhält (siehe Kapitel 2.4.2.2).
Spezies Auditorische Felder (zusätzlich zu einem tonotopen primären auditorischen Feld)
Referenzen
Meerschweinchen (Cavia procellus)
Zwei tonotope Felder, drei sekundäre Felder (REDIES et al. 1989)
Hauskatze (Felis domestica)
Drei tonotope Felder, vier sekundäre Felder (REALE und IMIG 1980)
Makake (Macaca mulatta)
Ein rostrolaterales und laterales tonotopes Feld; ein caudomediales sekundäres Feld und ein auditorischer „Gürtel“ mit bis zu drei Untereinheiten
(WOOLSEY und WALZL 1982)

2. Literaturübersicht
23
Dabei scheint es eine Art Verteiler darzustellen, welcher alle einkommenden Informationen
auf die anderen corticalen Felder weiterleitet (PHILIPPS und IRVINE 1982).
Morphologisch ist das primäre auditorische Feld durch eine starke reziproke Verbindung mit
dem ventralen Nucleus des Corpus geniculatum mediale des Thalamus charakterisiert
(MOREL und IMIG 1984). Physiologisch ist es durch Neurone gekennzeichnet, welche stabil
und gut abgestimmt auf Tonhäufungen antworten und in einer Tonotopie angeordnet sind,
welche im wesentlichen den Gradienten der cochleären Frequenzrepräsentation reflektiert
(EHRET 1997). Dabei scheint es zusätzlich, im Vergleich zu den nicht-primären Arealen,
sensibler für Schwankungen innerhalb des Frequenz- und Geräuschlevels zu sein (HALL
2003).
Die nicht-primären Assoziationsareale liegen anterior, posterior und lateral vom AI. Diese
sekundären auditorischen Gebiete entsprechen den Brodmann-Arealen 42 und 22. Sie
schließen sich außen an den Temporallappen an, wo sie die posteriore Hälfte des superioren
temporalen Gyrus bedecken. Das Areal 22 liegt direkt im Planum temporale und ist beim
Menschen an der Sprachfunktion beteiligt. Physiologisch sind die sekundären auditorischen
Felder durch Neurone gekennzeichnet, welche einen breiteren Frequenzantwortbereich
aufweisen. Die Empfindlichkeit dieser Neurone ist dabei im Vergleich zu Neuronen des AI
deutlich geringer (SCHREINER und CYNADER 1984).
Verschiedene Studien haben gezeigt, dass die sekundären auditorischen Bereiche der
humanen Hörrinde in einer Gehirnhälfte, vornehmlich auf der linken, größer sind
(GESCHWIND und LEVITSKY 1968) und bei der physiologischen Sprachverarbeitung
hemisphärische Unterschiede auftreten. So werden in der posterioren Area 22 der linken
Hemisphäre (Wernicke Sprachregion) Sprachsignale verarbeitet, während die
korrespondierende Region der rechten Hemisphäre dazu nicht fähig ist (SELDON 1982). Von
den corticalen Arealen im rechten superioren temporalen Cortex ist vor allem die
Verarbeitung der räumlichen Informationen abhängig. So ist der rechte auditorische Cortex
auf die Verarbeitung der Richtung eintreffender frequenzmodulierter Töne spezialisiert
(BRECHMANN 2003). In diesen Phänomenen liegt die Sprachlateralisation des Menschen
begründet.
Zusätzlich besteht in den Brodmann-Arealen 37, 39 und 40 ein tertiäres Assoziationsgebiet,
welches als Übergangsregion zwischen den sekundären visuellen, auditiven und taktilen bzw.
kinästhetischen Assoziationsgebieten fungiert.

2. Literaturübersicht
24
Der feline auditorische Cortex lässt sich ebenfalls in mehrere Teilgebiete einteilen. Er
erstreckt sich zwischen dem suprasylvanischen (SSS), anterioren ectosylvanischen (AES) und
posterioren ectosylvanischen Sulcus (PES) (WINER 1992).
Auch bei dieser Spezies ist das primäre auditorische Feld der größte Bereich der Hörrinde. Es
liegt auf dem mittleren ectosylvanischen Gyrus und reicht caudal bis zur caudalen Spitze des
PES sowie in einen kleinen Bereich des posterioren ectosylvanischen Gyrus (PEG) hinein. Im
rostralen Bereich ist die Grenze des AI durch die Spitze des AES gekennzeichnet (BRUGGE
und REALE 1985). Direkt rostral des AI, auf dem suprasylvanischen Gyrus beginnend und
sich auf den anterioren ectosylvanischen Gyrus ausbreitend, ist das anteriore auditorische Feld
(AAF) der Katze gelegen. Auf dem PEG ist caudal des AI das posteriore auditorische Feld
(PAF) und ventrocaudal des AI das ventroposteriore auditorische Feld (VPAF) lokalisiert.
Neben diesen vier tonotop organisierten auditorischen Feldern befindet sich direkt ventral im
Anschluss an das AI zwischen dem AES und dem PES das nicht tonotop aufgebaute
sekundäre auditorische Feld (AII). Dieses breitet sich über mindestens 4 mm nach ventral aus.
Ventrocaudal des AII, längs des mittleren ectosylvanischen Sulcus bis rostral auf den PEG
reichend, liegt das ventrale auditorische Feld (VAF). Auch dieses Areal besitzt eine tonotope
Organisation, deren Frequenzrepräsentation jedoch im Hinblick auf die Repräsentation der
anderen tonotopen Felder keine feste Beziehung aufweist.
Im Bereich des „polysensorischen“ oder „nicht-spezifischen“ Cortex sind bei der Katze drei
weitere auf auditorische Stimuli reagierende Felder gelegen. Sie erhalten neben
konvergierenden auditorischen Informationen der extralemniscalen Hörbahn, visuelle und
somatosensorische Eingänge (REALE und IMIG 1980).
2.4.2 Der histophysiologische Aufbau des auditorischen Cortex
2.4.2.1 Die tonotope Organisation der auditorischen Felder
Wie im Kapitel 2.3 bereits beschrieben, setzt sich die tonotope Organisation der zentralen
auditorischen Hörbahn von der Cochlea über den Hirnstamm bis zum auditorischen Cortex
fort. Durch elektrophysiologische Untersuchungen (Einzelzellableitungen) konnte die
tonotope Organisation in den einzelnen auditorischen Feldern dargestellt werden.
Bei der Katze werden vier tonotope Felder unterschieden (REALE und IMIG 1980): das
primäre auditorische Feld (AI), das anteriore auditorische Feld (AAF), das posteriore
auditorische Feld (PAF) und das ventroposteriore auditorische Feld (VPAF).

2. Literaturübersicht
25
Jedes dieser Felder erhält über die lemniscale Hörbahn afferente Informationen aus dem
Nucleus centralis des inferioren Colliculus sowie der ventralen Abteilung des Corpus
geniculatum mediale (MGB). Dabei wird auf allen Ebenen dieser Verbindung die cochleotope
Tonotopie eingehalten. In den tonotop organisierten Feldern sind Neurone in sogenannten
isofrequenten Bändern angeordnet. Diese beinhalten Nervenzellen mit identischer oder sehr
ähnlicher charakteristischer Frequenz (Frequenz, bei welcher die Reizschwelle einer Zelle am
kleinsten ist, CF) und sind in Streifen in dorsoventraler Richtung (d. h. vertikal zur
Cortexoberfläche) über alle sechs Schichten angeordnet (KRUCKER 1996).
Neben der tonotopen Repräsentation der Schallfrequenz gibt es im auditorischen Cortex
zusätzlich Repräsentationen für die Reizintensität und Tuningschärfe (Breite des zur
Aktivierung des betreffenden Neurons führenden Frequenzbandes) sowie über den binauralen
Vergleich auch eine Repräsentation für die Lokalisation der Schallquelle im Raum (HEIL et
al. 1994; REALE und KETTNER 1986).
Im AI ist eine komplette Repräsentation der Cochlea wiederzufinden. Dabei sind Neurone mit
hoher CF rostral lokalisiert, die Repräsentation der niedrigen Frequenzen aus dem apikalen
Bereich der Cochlea findet caudal innerhalb des AI statt (WOOLSEY und WALZL 1942).
Für den primären auditorischen Cortex des Menschen haben experimentelle Arbeiten von
HOWARD et al. (1996) Hinweise auf eine ebenfalls tonotope Organisation gegeben.
Der Frequenzgradient des AAF stellt sich spiegelbildlich zur Frequenzrepräsentation im AI
dar (WINER 1992). Die aufeinander folgenden Repräsentationsgebiete der niedrigen
Frequenzen liegen anterioventral auf dem anterioren ectosylvanischem Gyrus entlang einer
sogenannten „isofrequenten Kontour“.
Im Gegensatz zu den tonotop organisierten auditorischen Feldern besitzt das sekundäre
auditorische Feld Neurone mit deutlich unschärfer begrenzten charakteristischen Frequenzen
und eine ungenauere Tonotopie. Die relativ breiten Frequenzbereiche verlaufen jedoch
parallel zum Frequenzgradienten des AI (SCHREINER und CYNADER 1984). Diese
Eigenschaften reflektieren die relativ diffusen vom MGB ausgehenden afferenten
Verbindungen.
2.4.2.2 Die detaillierte Verteilung der afferenten Projektionen
Der Neocortex ist ein neuronales Netzwerk mit zahlreichen Zwischenverbindungen, welches
überwiegend durch seine eigene Leistung erregt wird.

2. Literaturübersicht
26
Diese Definition wird durch die Tatsache getragen, dass nur 0,01 bis 0,1 % der Verbindungen
der Pyramidenzellen, als Träger der sensorischen Information, thalamischen Ursprungs sind,
während die übrigen zu corticalen Zellen vor allem in der ipsilateralen Hemisphäre bestehen.
Selbst in der Lamina IV (siehe Kapitel 2.4.3) machen die thalamischen Projektionen nur 15
bis 20 % des gesamten Inputs aus (LEVAY und GILBERT 1976).
Beginnend mit der klassischen Methode der retrograden Degeneration konnte die präzise
Organisation der Verknüpfung der auditorischen thalamischen Kerne mit den verschiedenen
corticalen auditorischen Feldern nicht eindeutig dargestellt werden (SOUSA-PINTO 1973a).
Neuere Untersuchungen mit retrograden und anterograden axonalen Mikroinjektionen
erlauben dagegen neben der detaillierten Darstellung der Verbindungen zusätzlich Aussagen
über die Reziprozität der Projektionen. Die Kombination dieser Tracingtechniken mit elektro-
physiologischen Untersuchungen zeigt, dass jedes der corticalen auditorischen Felder
Afferenzen von mehreren Thalamuskernen erhält. Daneben besteht für jedes Feld ein
gesondertes thalamisches Untergebiet, mit welchem es besonders stark verbunden ist
(ANDERSEN et al. 1980). So wird das primäre auditorische Feld als einziger corticaler
Bereich von einer großen Anzahl thalamocorticaler Projektionen aus dem ovoiden und
besonders aus dem lateralen Anteil der ventralen Subbereiche des MGB gespeist (ROSE und
WOOLSEY 1949). Daneben bestehen Afferenzen vom tiefen dorsalen Anteil des dorsalen
Bereiches, vom nicht-tonotop organisierten medialen Anteil und von Neuronen aus der
lateralen Abteilung der posterioren Gruppe der thalamischen Kerne.
Der hauptsächliche Unterschied der thalamocorticalen Bahnen des primären (AI) und des
anterioren auditorischen Feldes (AAF) bestehen in der Ausprägung der jeweiligen
Verbindungen. Während das AI stärkeren und kontinuierlicheren Input vom ventralen MGB
erhält, verfügt das AAF über eine stärkere Verbindung mit dem lateralen Anteil der
posterioren Gruppe der thalamischen Kerne sowie des tiefen dorsalen Anteils des dorsalen
Bereiches (MOREL und IMIG 1984). Dennoch liefern die Ähnlichkeiten der afferenten
thalamischen Projektionen zwischen dem AI und dem AAF Hinweise auf eine parallele
Verarbeitung von Informationen zwischen diesen beiden Feldern (PHILIPPS und IRVINE
1982), wobei das AAF auf die Verarbeitung von transienten Geräuschen spezialisiert zu sein
scheint (TIAN 1992). Begrenzte Bereiche im AI und AAF erhalten zudem koextensiven Input
von Neuronenbögen, welche den ventralen Anteil des MGB passieren, von Zellkolumnen des
tiefen dorsalen Nucleus des dorsalen MGB, von Neuronen aus dem medialen Anteil des MGB
sowie von Zellen des lateralen Anteils der posterioren Gruppe der thalamischen Kerne.

2. Literaturübersicht
27
Auch senden viele zum AI projizierende Zellen des ventralen MGB Axonkollaterale zum
AAF (MOREL und IMIG 1984). Dabei sind diese Verbindungen sowohl konvergierender als
auch divergierender, isofrequenter Natur (DICKSON und GERSTEIN 1974).
Das posteriore und ventroposteriore auditorische Feld erhalten ausgedehnte afferente
Informationen von mindestens vier Unterbereichen des MGB (IMIG und MOREL 1983).
Das sekundäre auditorische Feld bezieht seine afferenten, nicht tonotopen Informationen von
der über den perizentralen Nucleus des inferioren Colliculus und den medialen Anteil des
MGB geschalteten diffusen, extralemniscalen Hörbahn. Zusätzlich ist es reziprok mit dem
lateralen und dem caudalen Anteil des dorsalen Nucleus des MGB verbunden.
Die laminäre Verteilung der thalamocorticalen Projektionen zeigt innerhalb einer Spezies in
unterschiedlichen corticalen Arealen eine starke Einheitlichkeit. Besonders deutlich zeigt sich
das laminäre Verteilungsmuster im primären auditorischen Feld. Dort erhalten die
pyramidalen Neurone in den Laminae I und III bis VI die Masse des direkten thalamischen
afferenten Inputs (MITANI et al. 1985). Die Afferenzen aus dem medialen Anteil des MGB
ziehen hauptsächlich in die Lamina I und zu einem geringen Anteil in die Lamina IV
(MITANI et al. 1984). Diese Projektionen laufen parallel zu in die Laminae III und IV
ziehenden Verbindungen, sind aber aufgrund der großen Axonendurchmesser und der
monosynaptischen Verbindung langsamer leitend (WINER 1992). Der ventrale Bereich des
MGB projiziert überwiegend in die Lamina IV (LEVAY und GILBERT 1976). Neben diesen
direkten Einflüssen der thalamischen Kerne besteht durch die extensiven intracorticalen
Verbindungsstrukturen der lokal vernetzten Pyramidenzellen ein zusätzlicher indirekter
Einfluss auf die übrigen Laminae. So erhalten die pyramidalen Zellen der Lamina II
zusammen mit den Pyramidenzellen der Lamina III zusätzlich thalamische Eingänge, welche
zuvor über die Lamina IV geschaltet wurden (NIIMI und NAITO 1974). Die Pyramidenzellen
der Laminae II und III senden wiederum Projektionen in ipsilaterale corticocorticale
(WINGUTH und WINER 1986) und comissurale Gebiete (IMIG und BRUGGE 1978).
Weitere Verschaltungen konnten durch intralaminare Messungen von synaptischen Strömen
verifiziert werden. Nach einer direkten thalamocorticalen Aktivierung der Laminae III bis VI
folgten synaptische Aktivitäten in den infragranulären Schichten (KRAL et al. 2000).
Die Gesamtheit dieser Verknüpfungen führt dazu, dass das Projektionsgebiet einer einzelnen
thalamischen Nervenfaser ein Gebiet von mehreren Millimetern und eine Vielzahl von
corticalen Nervenzellen aktiviert.

2. Literaturübersicht
28
Daneben werden durch Projektionen der thalamisch modifizierten pyramidalen Zellen auf
reziprokem Weg der Thalamus, das Mesencephalon und die auditorischen Kerngebiete im
Hirnstamm erreicht.
2.4.2.3 Die intra- und intercorticalen Verbindungen des auditorischen Cortex
Während die thalamischen Kerne untereinander nicht verbunden sind, bestehen im
auditorischen Cortex eine Vielzahl von Interaktionen.
Innerhalb und zwischen den einzelnen Feldern sowie mit anderen funktionellen Arealen
beider Hemisphären und mit subcorticalen Strukturen bestehen zahlreiche Verknüpfungen.
An der Bildung dieser Verbindungen beteiligen sich sowohl pyramidale, als auch nicht-
pyramidale Neurone aus den Laminae II bis IV (WINGUTH und WINER 1986). Die
intrinsischen Verbindungen einzelner corticaler Neurone können als „kurz“, d.h. grob
innerhalb der betroffenen Zellkolumne oder als „lang“, also über die Ausdehnung des
Dendritenbaums hinaus ragend, klassifiziert werden. Dabei können die „langen“
Verbindungsfasern eine Strecke von mehreren Quadratmillimeter bedecken (zur Übersicht
siehe MARTIN 1984).
Der Gesamteindruck der intracorticalen Verbindungen ist, dass die ipsilateralen Felder AI,
AAF, PAF und VPAF topographisch und reziprok miteinander verbunden sind (IMIG und
BRUGGE 1978). Zusätzlich projiziert jedes dieser Felder zu mehreren Arealen des peripheren
auditorischen Gürtels (siehe Tab. 2, 3). Weiterhin ist jedes der vier tonotop organisierten
Felder reziprok und topographisch über das Corpus callosum mit dem jeweilig homologen
Feld der anderen Hemisphäre verbunden. Die Verknüpfungen mit den nicht-tonotopen
Feldern sind dagegen schwächer und divergierender Natur.
AI AAF AII PAF VPAF AI + * * * -
AAF * + ~ * % AII - - + - - PAF - - ~ * -
VPAF - - ~ - *
Termination
Tab. 2: Commissurale Verbindungen der auditorischen Felder (modifiziert nach WINER 1992)
Urs
pru
ng
+: stark * : moderat ~ : schwach -: nicht vorhanden %: unbekannt

2. Literaturübersicht
29
Innerhalb eines auditorischen Feldes können die verschiedenen Verbindungen speziellen
Funktionen zugewiesen werden. So verfügen Regionen des AI, deren Neurone
Summationseffekte bei binauraler Stimulation aufweisen, über viele callosale Verbindungen
(IMIG und BRUGGE 1978), wohingegen Regionen außerhalb der Summationskolumnen
viele ipsilaterale Afferenzen vom AAF und PAF erhalten (IMIG und REALE 1981). Zudem
scheinen Neurone mit gleicher oder sehr ähnlicher charakteristischer Frequenz über die
Feldgrenzen hinweg bevorzugt miteinander verbunden zu sein (CLARKE et al. 1993).
Bei Betrachtung der laminären Verteilung der Verbindungen im primären auditorischen Feld
sind die supragranulären pyramidalen Neurone in erster Linie an der Verbreitung von
Informationen zu anderen corticalen Bereichen beteiligt, während die infragranulären Zellen
die prinzipiellen Elemente für die Übermittlung von Informationen zu subcorticalen
Strukturen darstellen.
In der Lamina I befinden sich die apikalen Verzweigungen der Dendriten von pyramidalen
Neuronen, deren Somata in allen Schichten lokalisiert sind. Zusätzlich beteiligen sich die
laminaeigenen Zellen mit ihren meist parallel zur Cortexoberfläche verlaufenden Dendriten
mit den Endverzweigungen von afferenten Assoziations- und Kommissurenfasern an der
Bildung des intralaminaren feinfaserigen tangentiellen Flechtwerks (Stria laminaris). In den
Laminae II und III befinden sich corticocorticale und callosale pyramidale Zellen. Die
wichtigsten Verbindungen der in der Lamina II lokalisierten Neurone ziehen zu den
angrenzenden sekundären auditorischen Feldern. In der Lamina III befinden sich 75 % der
Gesamtprojektionen der commissuralen Afferenzen (CODE und WINER 1985). Zwei Drittel
dieser Zellen sind Pyramidenzellen, den Rest bilden bipolare und einzelne große multipolare
Zellen.
AI AAF AII PAF VPAF AI - + + * +
AAF + - + + * AII + % - * * PAF + + + - +
VPAF + + + % -
Termination U
rsp
run
g
Tab. 3: Corticocorticale Verbindungen der auditorischen Felder (modifiziert nach WINER 1992)
+: stark * : moderat ~ : schwach -: nicht vorhanden %: unbekannt

2. Literaturübersicht
30
Aus den Laminae III und V stammen zusätzlich jeweils ca. 30 % der Projektionen des AI zum
AII (WINGUTH und WINER 1986) und der Großteil der weiteren corticocorticalen
Verbindungen. Etwa 50 % der Neurone, vor allem kleine Pyramidenzellen und
spindelförmige Zellen, in der Lamina IV sind mit dem Corpus geniculatum mediale (MGB)
verbunden (WONG und KELLY 1981). Zum commissuralen System im primären
auditorischen Feld tragen die Zellen der Lamina IV jedoch wenig bei und projizieren darüber
hinaus auch nicht in die sekundären auditorischen Felder. Sie übernehmen wahrscheinlich
eine lokale Schaltfunktion innerhalb des AI (LEVAY und GILBERT 1976). In der Lamina V
sind große Pyramidenzellen lokalisiert, die zum Striatum und den superioren sowie inferioren
Colliculi projizieren (OJIMA et al. 1992). Die mittelgroßen Pyramidenzellen sind
überwiegend mit dem MGB verbunden. Weitere Verbindungen zu ipsilateralen sekundären
auditorischen Feldern, zum kontralateralen auditorischen Cortex und zum Claustrum zeigen
die Vielfältigkeit der hier entspringenden Verschaltungen. Zudem erhält die Lamina V eine
bedeutende Anzahl von Afferenzen commissuralen und corticocorticalen Ursprungs.
Anhand anatomischer, pharmakologischer und physiologischer Untersuchungen wurde darauf
geschlossen, dass der wichtigste vertikale Informationsfluss innerhalb eines auditorischen
corticalen Feldes durch die corticalen Schichten L-IV:� /-II/III:� /-V:� /-VI verläuft
(BOLZ et al. 1989). Neben diesem vertikalen Informationsfluss besteht eine beträchtliche
horizontale Verbindung, welche Informationen von benachbarten Regionen und spezifischen,
weiter entfernten Bereichen integriert. Wie viele Synapsen innerhalb der einzelnen Schichten
diesem horizontalen Informationsfluss zuzuordnen sind, ist nicht bekannt. In der Lamina IV
jedoch erhalten lediglich 15 bis 20 % der Synapsen thalamische Eingänge (LEVAY und
GILBERT 1976), die meisten Verbindungen scheinen mit intra- und interlaminaren Neuronen
zu bestehen.
Bei den intrinsischen Verbindungen speziell des AI zeigt sich eine dorsoventrale Orientierung
der dendritischen Felder, die mit der Orientierung der isofrequenten Bänder übereinstimmt
(OJIMA et al. 1992). Aufgrund dieser dorsoventralen Ausrichtung der Dendritenbäume sowie
unterschiedlicher thalamocorticaler und corticocorticaler Verbindungen innerhalb des AI der
Katze wird angenommen, dass dieser aus den funktionell unterschiedlichen dorsalen (AId)
und ventralen (AIv) Unterbereichen besteht (SUTTER und SCHREINER 1995). Dabei wird
dem AId vor allem die Unterscheidung und Verarbeitung der räumlichen Informationen
zugerechnet. Die Organisation des AIv deutet auf eine verstärkte Rolle bei der Detektion
sowie der lokalen Verarbeitung räumlicher Informationen hin.

2. Literaturübersicht
31
Weitere Untersuchungen der laminären Verteilung corticocorticaler Verbindungen
offenbarten, dass in der Hierarchie der zentralen Hörbahn das AI sowie das AAF auf einer
gleichen, niedrigeren Ebene liegen als das AII, das PAF und das VPAF (ROUILLER et al.
1991).
2.4.3 Die Cytoarchitektur und neuronale Organisation des auditorischen Cortex
Der Neocortex der Großhirnrinde (Cortex cerebri), zu welchem auch der auditorische Cortex
gehört, zeigt in den meisten Bereichen eine auffallende laminäre, areale und cytochemische
Homogenität (COW und LEIMAN 1970). Er wird deswegen auch als homoitypischer
Isocortex bezeichnet. Dennoch können funktionell verschiedene Regionen oft deutlich anhand
ihrer Cytoarchitektur unterschieden werden (POWELL und MOUNTCASTLE 1959).
Bereits in der ersten Hälfte des 19. Jahrhunderts haben die Studien von VON ECONOMO
und HORN (1930) am menschlichen Cortex die charakteristische sechsschichtige
Grundanordnung der Nervenzellen im Neocortex der Säugetiere offenbart. Die meisten
Untersuchungen über die cytoarchitektonische Struktur und die neuronale Organisation des
auditorischen Cortex sind jedoch bei der Katze vorgenommen worden. Durch Golgi-
Imprägnationen und intrazelluläre Farbstoffinjektionen erhielt man eine Übersicht über
vorhandene Neurone und deren Projektionsgebiete (SOUSA-PINTO 1973b). Nachfolgende
Untersuchungen von MITANI et al. (1985) und WINER (1984 a, b, c) ergänzen diese
Beschreibungen.
Senkrecht zur Hirnoberfläche werden von der Cortexoberfläche zur weißen Substanz die
Molekularschicht (Lamina I; molecularis), die äußere Körnerschicht (Lamina II; granularis
externa), die äußere Pyramidenschicht (Lamina III; pyramidalis externa), die innere
Körnerschicht (Lamina IV; granularis interna), die innere Pyramidenschicht (Lamina V;
pyramidalis interna) und die multiforme Schicht (Lamina VI; multiformis) angetroffen. In
jeder Schicht können, basierend auf Unterschieden in der neuronalen Architektur,
Verknüpfungen mit anderen Gebieten und immunhistochemischen Kriterien, weitere
Unterschichten differenziert werden. Im Bereich des auditorischen Cortex wird diese laminäre
Zellarchitektur durch die Myeloarchitektur der von der Lamina II zu Lamina III zunehmend
dicker werdenden Axonquerschnitte vervollständigt. In den auditorischen Subarealen fällt im
Zusammenhang mit dem allgemeinen Aufbau der einzelnen Laminae eine deutliche granuläre
Heterotypie auf. Der auditorische Cortex wird daher ähnlich den Riech- und Sehfeldern dem
granulären Rindentyp (Coniocortex) zugeordnet.

2. Literaturübersicht
32
Neben der horizontal zur Cortexoberfläche verlaufenden Zellanordnung in Schichten fällt im
gesamten cerebralen Cortex die vertikal zur Oberfläche organisierte Ansammlung von
Nervenzellen auf. Diese Struktureinheiten werden auch als „corticale Kolumnen“ bezeichnet.
Auch diese säulenartige Anordnung erscheint im auditorischen Cortex höher entwickelt als in
anderen Regionen des Neocortex (LORENTE DE NÓ 1949).
Die neuronale Population des cerebralen Cortex besteht aus zwei Hauptkomponenten, welche
anhand der Form ihrer Zellkörper und der Länge ihrer Axone klassifiziert werden. Die
Pyramidenzellen, deren lange Axone in die weiße Substanz eintauchen und meist ihr
Ursprungsfeld verlassen, machen bis zu 85 % aller Neurone im menschlichen Gehirn aus
(BRAITENBERG und SCHÜTZ 1991). Die restlichen Nervenzellen werden als nicht-
pyramidale Zellen bezeichnet und besitzen meist lokal endende Axone. Auf der Basis ihrer
Dendritenbäume werden diese Zellen als multipolar, bipolar oder büschelförmig klassifiziert
(PETERS und JONES 1984). Die morphologischen Variationen dieser zwei Gruppen
corticaler Neurone stehen im Zusammenhang mit ihren jeweiligen funktionellen
Eigenschaften (LUND 1988).
Neben den Unterschieden zu anderen funktionellen Gebieten des Neocortex können auch die
einzelnen Felder des auditorischen Cortex anhand ihrer cytoarchitektonischen Struktur
differenziert werden (ROSE und WOOLSEY 1949).
Ein charakteristisches cytoarchitektonisches Merkmal des primären auditorischen Feldes (AI)
ist die ausgeprägte vertikale Anordnung der dendritischen und axonalen Felder der
pyramidalen Nervenzellen, insbesondere in der Lamina IV. So zeigte die Auswertung der
Dendritenorientierung innerhalb der Laminae IV und V des AI eine deutlich signifikante
(p<0.05) nicht-randomisierte vertikale sowie eine randomisierte tangentiale Verteilung der
basalen Dendriten von pyramidalen Zellen, nicht aber von denen nicht-pyramidaler
Sternzellen (GLASER et al. 1979).
Direkt unter der Pia mater liegt die nur 150 µm breite Molekularschicht, auch Lamina
plexiformis oder Lamina I genannt. Diese Schicht besteht vorwiegend aus Neuropil und
enthält keine Pyramidenzellen sowie nur eine geringe Anzahl nicht-pyramidaler Neurone.

2. Literaturübersicht
33
Das reichhaltige Neuropil zeichnet sich durch eine unter den corticalen Schichten einzigartige
neurochemische Homogenität aus. Anhand der Nervenfaserstruktur kann innerhalb der
Lamina I eine oberflächliche und tiefe Schicht differenziert werden.
Die obere Schicht Ia enthält viele myelinisierte Axone und einige aufsteigende dendritische
Äste. Die tiefe, besonders zellarme und neuropilreiche Schicht Ib hingegen beherbergt
hauptsächlich unmyelinisierte Axone und umfangreichere, aus den tieferen Schichten
aufsteigende, apikale Dendriten (SOUSA-PINTO et al. 1975). Die auffallendsten Neurone in
der Lamina I sind horizontale Zellen, deren marklose Dendriten meist parallel zur
Cortexoberfläche verlaufen. Die mittelgroßen Nervenzellen der Lamina I entsenden ihre
zahlreichen Dendriten in ungeordneter Struktur vertikal in zwei Richtungen. Die wenigen
nicht-pyramidalen Zellen in der Lamina I sind meist spindelförmig und verschieden groß. Sie
besitzen zumeist maximal fünf Dendriten, deren distale Abzweigungen nur wenig entwickelt
sind.
In der äußeren Körnerschicht, oder Lamina II, können sowohl pyramidale als auch nicht-
pyramidale Zellen angetroffen werden, wobei letztere den weitaus größeren Anteil
ausmachen.
Auch diese, sich etwa 150 bis 200 µm unter der weichen Hirnhaut an die Molekularschicht
anschließende Schicht, kann in zwei Unterschichten geteilt werden. Die weniger dichte
oberflächliche Schicht IIa enthält vorwiegend kleinere, zumeist nicht-pyramidale, multipolare
Neurone mit rundem oder blitzförmigem Soma und feinen, meist lokal begrenzten,
ungeordnet verlaufenden nicht-myelinisierten Dendriten. In der unteren Schicht IIb sind vor
allem die lateral und vertikal in andere Schichten projizierenden Pyramidenzellen angesiedelt
(WINGUTH und WINER 1986). Diese sind im Vergleich zu den Pyramidenzellen der
Lamina III kleiner und besitzen weniger gut entwickelte dendritische Felder. Im Vergleich zur
Lamina IIa sind die Dendriten der Lamina IIb jedoch massiver. In Golgi-gefärbten Schnitten
lassen sich von jedem Zelltyp noch verschiedene Größen darstellen. Zu unterscheiden sind
kleine und mittlere Pyramidenzellen, deren apikale Dendriten in die Lamina I projizieren,
bipolare Zellen mit dünnen Dendriten, kleine runde oder spindelförmige multipolare Neurone
mit strahlenförmigen Ausläufern, welche ein enges dendritisches Feld beschreiben, und große
multipolare Zellen, deren gut ausgebildete Fortsätze hoch entwickelt sind. Die nicht-
pyramidalen Körner- oder Sternzellen sind so arrangiert, dass sie als Schaltneurone innerhalb
der vertikalen Zellkolumnen fungieren.

2. Literaturübersicht
34
Die Grenze zwischen der Lamina II und der äußeren Pyramidenschicht, Lamina III, ist durch
das Auftreten der markanten großen Pyramidenzellen bei etwa 400 µm charakterisiert. Mit
300 µm (SOUSA-PINTO 1973b) bzw. 500 µm Dicke (WINER 1992) macht die Lamina III
etwa ein Viertel der Gesamtdicke des primären auditorischen Feldes aus. Die in dieser Schicht
zahlreich vorkommenden pyramidalen Zellen variieren in drei Größen, wobei sich die
charakteristischen großen Pyramidenzellen vor allem in der unteren Teilschicht, Lamina IIIb
befinden. Diese Nervenzellen erhalten Afferenzen aus dem Thalamus, aus ipsilateralen und
intrinsischen corticalen Bereichen und aus Gebieten der kommisuralen Windungen. Ihre
Dendriten bilden ein lokales Netzwerk, welches in die eigene Lamina und die Lamina IV
einstrahlt und durch ausgedehnte afferente Ausläufer das kontralaterale AI innerviert. Einige
projizieren auch in andere ipsilaterale auditorische Felder (CODE und WINER 1985).
Vor allem in der Lamina IIIb sind auch die am zahlreichsten vorkommenden mittleren
Pyramidenzellen lokalisiert. Während die apikalen Ausläufer dieser dicht gepackten Neurone
in die Lamina I projizieren, ziehen die lateral ausgerichteten basilaren Ausläufer in das
laminaeigene Neuropil und demarkieren die schwer erkennbare Grenze zur Lamina IV. Die
Abgrenzung zur Lamina IV ist auch mit Hilfe der Nissl-Färbung nicht eindeutig zu
bestimmen, da sich einige der Pyramidenzellen der Lamina III mit den granulären Zellen der
Lamina IV vermischen.
In der oberflächlichen Schicht IIIa finden sich kleine Zellen mit ovalen Somata und eine
Vielzahl an kleinen Pyramidenzellen, deren apikale Dendritenäste in die Lamina II ziehen.
Die in der Lamina III vorkommenden nicht-pyramidalen Zellen scheinen hauptsächlich
lokalen Einfluss in die Lamina II zu haben. Neben bipolaren Varianten, deren Axon in
Richtung Lamina II orientiert ist, kommen auch kleine multipolare Zellen vor, welche in
genau definierte corticale Gebiete projizieren. Die meisten dieser Neurone sind kleiner als 50
µm und bilden im Gegensatz zu den meist tangential ausgerichteten Dendriten der
Pyramidenzellen wenig vertikal polarisierte, kugelige dendritische Felder. MEYER et al.
(1984) beschrieben zusätzlich nicht-pyramidale Sternzellen, deren meist parallel zu den
Dendriten der Pyramidenzellen verlaufende Ausläufer hauptsächlich lokalen Einfluss
besitzen, und kleine axonfreie Nervenzellen mit feinen Dendriten.
Das Neuropil der Lamina III wird sowohl von feinen Dendriten aus der schichteigenen
Neuroglia, als auch von aufsteigenden Axonen aus der oberflächlichen und tiefen Schicht der
Lamina IV, die Quellen der intracorticalen Verbindungen sind, gebildet.

2. Literaturübersicht
35
Der Beginn der 200 bis 250 µm dünnen inneren Körnerzellschicht, Lamina IV, ist durch
kleine ovale Zellen markiert. Diese Zellschicht ist charakterisiert durch vorwiegend kleine,
dicht gepackte nicht-pyramidale Zellen, die überaus deutliche corticale Kolumnen bilden. Der
säulenartige Aufbau entsteht durch den vertikalen Verlauf der eigenen Perikarya mit ihren
Ausläufern und durch einstrahlende Axone der aufsteigenden intrinsischen Fasern
corticocorticalen und thalamischen Ursprungs.
Von den in dieser Schicht anzutreffenden sechs verschiedenen Nervenzelltypen befinden sich
die kleineren Neurone mit ihrem lokal begrenzten Einfluss zumeist in der oberen Hälfte
(Lamina IVa). Auffällig sind die sogenannten „Double bouquet“ – Zellen („bitufted cells“),
welche zu den höchst entwickelten Zellen dieser Schicht zählen. Sie besitzen die größte
Anzahl an Dendriten, welche pferdeschwanzartig entweder in horizontaler Projektion die
intralaminare Grenze der inneren Körnerschicht markieren, oder lang und stielförmig
ausgebildet in die Lamina III ziehen. In der Lamina IVb befinden sich große, meist
multipolare und büschelförmige Zellen, deren Spine-reiche Ausläufer auch in die Lamina III
reichen.
Die etwa 1000 bis 1100 µm unter der Pia mater beginnende innere Pyramidenzellschicht,
Lamina V, ist wie die Lamina III ungefähr 400 µm dick und enthält eine umfassende
morphologische Ansammlung von pyramidalen und nicht-pyramidalen Zellen. Diese
Nervenzellen sind in weniger dichten vertikalen, strahlenförmigen Fäden angeordnet, welche
nur von dünnen Streifen somafreien Neuropils unterbrochen sind. Die herausragende
Eigenschaft der Lamina V ist das Vorhandensein zahlreicher Pyramidenzellen, von denen
einige die größten Neurone innerhalb des AI sind. Eine signifikante Anzahl dieser Zellen hat
zudem eine seitenverkehrte Orientierung. In sich kann auch die Lamina V in eine obere,
zellarme aber faserreiche Lamina Va und eine untere Teilschicht (Vb) mit zahlreichen
Pyramidenzellen unterschiedlicher Größe differenziert werden.
Insgesamt kommen in der Lamina V sieben unterschiedliche Zelltypen vor, von denen drei zu
den pyramidalen Zellen gezählt werden. Diese Neurone sind relativ groß und besitzen ein
einfaches Axongeflecht sowie eine vielseitige Dendritenanordnung. Bemerkenswert sind die
Riesenpyramidenzellen, auch Betzsche Zellen genannt, deren Perikarya bis zu 100 µm groß
sind. Die Dendriten dieser Zellen haben entweder einen intralaminaren Verlauf oder reichen
vor allem mit ihren apikalen Anteilen in die Laminae I und II. Dort verlaufen sie teilweise
parallel zur Cortexoberfläche und nehmen so Kontakt mit feinen Axonen aus dem Corpus
geniculatum mediale auf (RYUGO und KILLACKEY 1974).

2. Literaturübersicht
36
Sternförmige Pyramidenzellen, auch als „umgekehrte Pyramidenzellen“ bezeichnet, formen
mit ihren Dendriten einen mehr strahlenförmigen Funktionsbereich. Abhängig von den
Klassifizierungskriterien werden diese Zellen entweder wegen ihrer langen apikalen
Dendriten zu den Pyramidenzellen gezählt, oder aber aufgrund der GABAergen Perikarya den
nicht-pyramidalen Zellen zugeordnet (PRIETO et al. 1992).
Zu den verschiedenen in der Lamina V vorkommenden nicht-pyramidalen Neuronen zählen
große und mittlere multipolare Zellen. Die großen multipolaren Nervenzellen besitzen durch
die gesamte Lamina V ziehende, sehr schlanke Dendriten mit wenigen Verzweigungen. Die
dicken nicht-myelinisierten Axone sind lokal weit verzweigt und einige dieser enden bei
vertikaler Projektion wahrscheinlich an den apikalen Dendriten der Pyramidenzellen. Die
mittleren multipolaren Zellen sind zum Großteil in der Lamina Va lokalisiert. Diese Zellen
strahlen mit ihren feinen Dendriten mehr kugelförmig aus. Die ebenfalls dicken nicht-
myelinisierten Axone bilden ein überwiegend lokal begrenztes, laterales Netzwerk, welches
vertikal auf- und absteigende Ausläufer entsendet, deren Ausmaß nicht an das der großen
multipolaren Zellen heranreicht.
Die multiforme Schicht, Lamina VI, beginnt etwa 1500 µm unter der Cortexoberfläche, hat
ebenfalls eine Breite von 400 µm und ist trotz einiger versprengter Nervenzellen deutlich zur
weißen Substanz hin abgegrenzt. Die Lamina VI enthält große, mit myelinisierten Axonen
gefüllte, zellfreie Zonen, welche von dicht und gleichmäßig gelagerten Nervenzellen
akzentuiert werden. Diese Neurone besitzen innerhalb des primären auditorischen Feldes mit
einer Population von mindestens neun verschiedenen Arten die höchste Diversität. Einzigartig
im primären auditorischen Feld und charakteristisch für diese Zellschicht ist das Auftreten
von Riesenmultipolarzellen. Diese Zellen kommen jedoch in nur sehr geringer Anzahl vor.
Sie besitzen einen spindelförmigen Zellkörper, welcher einen Durchmesser von mindestens
50 µm hat. Mit einem dendritischen Feld, welches eine Höhe von 600 µm und eine Breite von
400 µm aufweist, ist eine solche Zelle in der Lage verschiedene Repräsentationsbereiche oder
Isofrequenz- oder Repräsentationsbereiche eines Ohres umfassen (MIDDLEBROOKS et al.
1980). Weiterhin kennzeichnend für die Lamina VI sind horizontale Zellen, welche sowohl
Eigenschaften pyramidaler als auch nicht-pyramidaler Zellen besitzen. Sie entwickeln eine
den bipolaren Nervenzellen ähnliche horizontale Polarisation der Dendriten, welche nicht so
weit verzweigt sind wie jene der Pyramidenzellen.
Die oberflächliche Lamina VIa ist dominiert von Pyramidenzellen, welche vorwiegend eine
vertikale Anordnung zeigen.

2. Literaturübersicht
37
Die Form der kleinen und mittleren Pyramidenzellen unterscheidet sich von deren
Äquilvalenten der Laminae II bis IV durch eine geringere Anzahl und einfachere
Dendritenaufzweigungen, welche eine zellarttypische Vielfältigkeit vermissen lassen. Die
apikalen Dendriten dieser Zellen reichen selten bis in die Lamina III, die Axone projizieren in
die weiße Substanz. Außerdem kommen atypische Pyramidenzellen vor, welche sich mit
enorm langen Axonen bis zur Basis der Lamina IV erstrecken. Auch die sternförmigen
„umgekehrten Pyramidenzellen“ können angetroffen werden. Letztere sind ebenfalls
einfacher aufgebaut als die in der Lamina V vorkommenden Exemplare. In der unteren
Schichthälfte, der Lamina VIb, sind vor allem die nicht-pyramidalen Zellen in einer
heterogenen Anordnung angesiedelt. Obwohl sie in ihrer Anzahl sehr viel geringer sind als in
den anderen Laminae, sind fünf verschiedene Arten ausgeprägt. Kleine und mittlere
Nervenzellen mit einfachen, sternförmigen oder leicht büschelförmigen Fortsätzen bilden ein
eher kleines dendritisches Feld mit einer lateralen Ausbreitung von 150 µm. Des Weiteren
existieren bipolare Nervenzellen mit schlanken Dendriten.
Der histologische Aufbau des sekundären auditorischen Feldes (AII) entspricht im
wesentlichen dem des AI. Charakteristisch für das AII ist das Auftreten von Riesen-
pyramidenzellen in der oberen Hälfte der inneren Pyramidenzellschicht (Lamina Va). Zudem
erscheint die Zelldichte aller sechs Zellschichten insgesamt wesentlich homogener.
Zwischen dem AI und dem anterioren auditorischen Feld (AAF) bestehen ebenfalls keine
großen cytoarchitektonischen Unterschiede. Insgesamt ist die Zelldichte im AAF etwas
geringer als im AI. Zudem sind die Zellkörper der pyramidalen und nicht-pyramidalen Zellen
größer als im primären auditorischen Feld. Besonders in den Laminae IIa und IVa fällt der
Größenunterschied bei den Pyramidenzellen auf. Auch enthält die Lamina IV des AAF
Pyramidenzellen und die Zellarmut der Lamina V ist weniger stark ausgeprägt.
Untersuchungen des humanen auditorischen Cortex zeigen ebenfalls ein Gebiet mit
verschiedenen Schichten, welche im Vergleich mit dem felinen auditorischen Cortex eine
ähnliche Schichtbreite bei ähnlicher Zelldichte aber unterschiedlichen Zellgrößen zeigen. Die
größte Zelldichte in der humanen Hörrinde ist in den Laminae II und IV erkennbar.
Charakteristisch für die Lamina IV ist eine sehr heterogene Population mit verschiedenen
Zellarten. In Studien von ONG und GAREY (1990) konnten mit Hilfe der Golgi-Färbung
morphologische Ähnlichkeiten mit der Zellpopulation der Katze, vor allem bei den nicht-
pyramidalen Zellen, festgestellt werden.

2. Literaturübersicht
38
2.5 Die Veränderungen im auditorischen System der Katze
2.5.1 Die physiologische Entwicklung und Ausreifung des auditorischen Systems
2.5.1.1 Das Hörorgan und die afferente Hörbahn
Während beim Menschen Antworten auf akustische Stimuli schon in der 24.
Schwangerschaftswoche beobachtet werden können (WALSH und MCGEE 1990), entwickelt
sich der Hörsinn bei Katzen erst in den ersten Lebenswochen parallel zur Entwicklung des
äußeren Gehörgangs und der auditorischen Hörbahn (MOORE 1985).
Bei der Katze sind zum Zeitpunkt der Geburt das äußere und mittlere Ohr noch unausgereift.
Während der ersten Lebenswoche ist der äußere Gehörgang geschlossen und das Mittelohr
mit Amnionflüssigkeit gefüllt (WALSH et al. 1986). Zum Ende der ersten Lebenswoche
öffnet sich der äußere Gehörgang und das Trommelfell sowie die noch wenig kalzifizierten
Gehörknöchelchen werden freigelegt. Schließlich wird die Flüssigkeit im Mittelohr resorbiert.
Auch das Innenohr der Katze ist in der ersten Lebenswoche zunächst noch unausgereift
(PUJOL und MARTY 1970). Die anatomische Ausreifung der Cochlea beginnt in der Mitte
der basalen Windung. Nach der Differenzierung der Haarzellen kommt es zu deren
Innervierung durch aussprossende Spiralganglienzellausläufer. Im Anschluss reifen die
Stützzellen des Corti-Organs aus, der Cortische Tunnel öffnet sich und die Basilarmembran
erhält ihre endgültige Beweglichkeit. Anschließend reifen Elemente im apikalen Teil der
Cochlea (EGGERMONT et al. 1991). Die Reifung der mechanischen Elemente der Cochlea
dauert bei der Katze bis zur 3. Lebenswoche an (LINDEMAN et al. 1971).
Die funktionelle Entwicklung der auditorischen Hörbahn findet parallel in den ersten Wochen
nach der Geburt statt. Bei normalhörigen Katzen können ab dem sechsten Lebenstag
akustisch evozierte Hirnstammpotentiale abgeleitet werden (WALSH et al. 1986). Ab dem 30.
Lebenstag ist die auditorische Funktion schließlich voll entwickelt (KELLER 1997).
2.5.1.2 Der auditorische Cortex
Im Gegensatz zu den Strukturen des Ohres und zur zentralen Hörbahn benötigt die
Entwicklung sowohl des felinen als auch des humanen auditorischen Cortex einen weit
ausgedehnteren Zeitraum. So beginnt die laminäre Ausreifung des Cortex beim Menschen
bereits in der 16. Fetalwoche und schreitet bis zum vierten Lebensmonat fort.

2. Literaturübersicht
39
Die vollständige Reifung der sechsschichtigen Cytoarchitektur ist jedoch erst mit dem 3.
Lebensjahr abgeschlossen. Im Alter von 11 bis 12 Jahren ist schließlich die endgültige
axonale Dichte entwickelt (MOORE und GUAN 2001).
Die neuronale Entwicklung und Ausreifung in der Großhirnrinde findet dabei sowohl beim
Menschen (HUTTENLOCHER und DABHOLKAR 1997), als auch bei der Katze (FRIAUF
und SHATZ 1991) von innen nach außen statt. Die Neurone der tiefen corticalen Schichten
entstehen zuerst, während die der äußeren, oberflächlichen Laminae später entstehen
(LUSKIN und SHATZ 1985), um dann zwischen den infragranulären Schichten nach oben zu
wandern (sogenannte Zell-Migration).
Die grundsätzlichen intracorticalen neuronalen Verknüpfungen des felinen auditorischen
Cortex sind bereits bei der Geburt vorhanden. Die Axone bilden jedoch ein dichtes,
kontinuierliches Band, welches sich über den Cortex ausbreitet. Die vertikale Anordnung der
Neurone in Säulen, wie sie im ausgereiften auditorischen Cortex deutlich ausgeprägt ist, ist
bei neonatalen Tieren noch nicht ausgebildet (FENG und BRUGGE 1983). Ungleich dem
Herausbilden der Basisschaltungen im auditorischen Cortex, welche vor dem Beginn des
Hörvermögens entstehen, hält die dendritische Entwicklung lange nach dem Beginn des
Hörvermögens an. Die Initialphase des dendritischen Wachstums ist durch die Produktion
zahlreicher Äste gekennzeichnet. Ihren maximalen Verästelungsgrad erreichen die Neurone
gerade vor Erreichen der Hörfähigkeit. Es folgt eine Phase der Differenzierung, in welcher die
Anzahl der Dendritenäste wieder abnimmt. Dabei reifen proximale Dendriten zuerst aus,
während die terminalen Strukturen lange in unreifem Zustand verbleiben (DeCARLOS et al.
1985).
Für die Funktion des auditorischen Cortex besteht gleich vielen anderen zentralen
sensorischen Bereichen eine kritische Entwicklungsperiode. Diese dehnt sich beim Menschen
bis auf das 7. Lebensjahr aus (SKUSE 1993). Für die Katze existieren widersprüchliche
Literaturangaben, nach denen die kritische Phase bis zur 3. Lebenswoche (REBILLARD et al.
1980) bzw. bis hin zum 6. Lebensmonat (KRAL et al. 2001; KRAL et al. 2002) anhält.
Während dieser kritischen Phase kommt es bei ungestörter Entwicklung mit normaler
akustischer Stimulation zur fortschreitenden Differenzierung, Myelinisierung und Ausbildung
von Synapsen. Wird jedoch in dieser kritischen Periode die normale akustische Entwicklung
gestört, kommt es zu erheblichen plastischen Veränderungen (siehe Kapitel 2.5.2), welche um
so deutlicher ausfallen, je jünger die deprivierten Tiere sind (MOORE 1985).

2. Literaturübersicht
40
2.5.2 Die plastischen Umstrukturierungen im auditorischen System nach neonataler
akustischer Deprivation
Unter „Plastizität“ versteht man in Bezug auf das Nervensystem die Verformbarkeit von
neuronalen Verbindungen. Diese können sich auf den Funktionsbereich einzelner
Nervenzellen oder ganzer Neuronenaggregate auswirken.
ELBERT und ROCKSTROH (2004) unterscheiden vier Prinzipien der corticalen Plastizität:
Zum einen führt eine gesteigerte Stimulation eines cerebralen Bereiches zu einer
Vergrößerung der jeweiligen corticalen Repräsentationszonen und kann zusätzlich die
Topographie dieses Areals verändern („Practice makes perfect“). Zum anderen führt eine
Reduktion sowie ein vollständiger Verlust von afferentem Input zu einer Übernahme der
deprivierten Cortexareale durch andere, noch aktivierbare Repräsentationszonen („Use it or
lose it“). Weiterhin führt eine synchrone, für das Verhalten relevante Stimulation von
benachbarten peripheren Rezeptoren zu einer Integration ihrer corticalen Repräsentation
(„Fire together, wire together“). Schließlich finden Veränderungen nur statt, wenn
verhaltensrelevante Aufgaben und deren Übung so intensiv und ausgeprägt sind, dass das
Gehirn deren Verarbeitung auch im Schlaf fortsetzt („You have to dream it to achieve it“).
Von HEBB (1949) wurde postuliert, dass diese Ausbildung und Aufrechterhaltung der
funktionellen cerebralen Organisation auf den Mechanismen synaptischer Interaktionen
beruht.
Übertragen auf die zentrale auditorische Hörbahn und den auditorischen Cortex kann es durch
eine akustische Deprivation zu anhaltenden Veränderungen in der tonotopen Organisation, in
der allgemeinen neuronalen Organisation sowie zu Veränderungen der Dendritenanzahl, der
Dendritenlänge und der Spine- bzw. Synapsenanzahl kommen.
Für experimentelle Arbeiten wird eine akustische Deprivation als Plastizität-auslösendes
Agens entweder durch die lokale oder systemische Applikation von ototoxischen Substanzen
(zumeist Aminoglykosid-Antibiotika) oder durch eine chirurgischen Entfernung der
cochleären Strukturen herbeigeführt. Als im Anschluss beobachtete, für eine Plastizität im
auditorischen System verantwortliche Mechanismen können die schnelle Aufhebung von
Hemmeffekten, sogenannte Demaskierung, oder eine tatsächlich stattfindende Reorganisation
von Zellen mit damit einhergehenden morphologischen Veränderungen fungieren. Diese
können sowohl in subcorticalen Bereichen der auditorischen Hörbahn als auch im
auditorischen Cortex auftreten. Der genaue Mechanismus der neuronalen Plastizität ist jedoch
noch nicht exakt bekannt.

2. Literaturübersicht
41
2.5.2.1 Das Hörorgan und die afferente Hörbahn
Die Haarzellen von Säugetieren unterliegen, im Gegensatz zu denen von Vögeln (zur
Übersicht siehe COTANCHE 1999), keiner spontanen Regeneration und ein Verlust dieser
Rezeptorzellen führt zu einer irreversiblen sensorineuralen Taubheit. In Folge dessen findet
zunächst ein schneller und extensiver Verlust der unmyelinisierten peripheren Dendriten
innerhalb des Corti-Organs statt (TERAYAMA et al. 1977). Diesem folgt eine eher
schrittweise Degeneration der myelinisierten Anteile der peripheren Dendriten innerhalb der
knöchernen Lamina spiralis und der Zellkörper der Spiralganglienzellen (HARDIE und
SHEPHERD 1999). Während die Spiralganglienzellen über die gesamte Lebenszeit einer
retrograden Degeneration unterliegen können, entsteht ein transneuronaler Verlust von
Neuronen des Nucleus cochlearis (NC) und höherer Strukturen fast ausschließlich, wenn es
während der frühen, kritischen Entwicklungsperiode des auditorischen Systems zu
Schädigungen im akustischen System kommt (NORDEEN et al. 1983; TIERNEY et al.
1997). So konnte nach neonataler akustischer Deprivation ein hochsignifikantes Schrumpfen
verschiedener Strukturen im felinen auditorischen Hirnstamm beobachtet werden.
Das Gesamtvolumen des NC verringerte sich beispielsweise um 42 %, die durchschnittliche
Größe sphärischer Zellen um 38 % und die Dichte der sphärischen Zellen im anterioren
ventralen Nucleus des NC nahm um 57 % ab (LUSTIG et al. 1994). Ähnlich Ergebnisse
wurden von HULTCRANTZ et al. (1991) sowie HEID et al. (1998) erarbeitet.
VOGT et al. (1997) konnten nachweisen, dass bei neonatal ertaubten Katzen das
Gesamtvolumen sowie die Zellfläche der zentralen Nuclei der inferioren Colliculi (ICC) im
Vergleich zu normalhörigen Tieren signifikant verringert und die Zelldichte signifikant erhöht
war. Da die Gesamtzellzahl der ICC nicht signifikant verändert war, wurden die
Deprivationseffekte als Atrophie der Zellkörper mit nur geringem Verlust von Zellen und
daraus resultierender erhöhter Zelldichte interpretiert.
In der afferenten Verbindung des Nucleus cochlearis zum inferioren Colliculus (IC) konnten
dagegen zwischen kongenital tauben Katzen und normalhörigen Tieren nur geringe
Unterschiede nachgewiesen werden (HEID et al. 1997). So zeigte sich lediglich eine auffällig
reduzierte nucleotope Organisation innerhalb des kontralateralen posterioren ventralen NC
und des dorsalen Nucleus des lateralen Lemniscus beider Seiten. Das Muster der
Hauptprojektionen zeigte sich trotz kongenitaler auditorischer Deprivation unverändert
(LUSTIG et al. 1994).

2. Literaturübersicht
42
Es sei hier allerdings erwähnt, dass bei kongenital tauben Katzen ein größeres Maß an
auditorischen Afferenzen als bei neonatal ertaubten Katzen bewahrt ist (HEID et al. 1998;
LEAKE et al. 1999).
Untersuchungen an unilateral experimentell auditorisch deafferentierten Versuchstieren haben
hingegen nachgewiesen, dass eine einseitige Cochleaschädigung eine Reorganisation der
afferenten Projektionsmuster nach sich zieht. So sind die vom Nucleus cochlearis zum
kontralateralen inferioren Colliculus aufsteigenden Projektionen von der deafferentierten
Seite signifikant um 30 bis 65 % reduziert (NORDEEN et al. 1983). Weiterhin besteht ein
damit einhergehender Anstieg der Fähigkeit des unbeschädigten Ohres Neurone in den
ipsilateralen inferioren Colliculus (KITZES 1984) und auditorischen Cortex (REALE et al.
1987) zu senden. Auch MOORE (1994) wies nach Zerstörung der Cochlea und langen
Überlebenszeiten eine Erhöhung der Neurone im NC nach, welche Axone an den ipsilateralen
IC senden.
In Einklang mit den morphologischen Auswirkungen einer akustischen Deprivation wird
durch eine Schädigung der Cochlea auch die Neurochemie der zentralen auditorischen
Hörbahn beeinflusst. So ist die Proteinsynthese im NC nach Schädigung oder Inaktivierung
der Cochlea schnell und signifikant herunterreguliert (SIE und RUBEL 1992). Ferner
induziert eine Cochleaschädigung eine komplexe Serie von Veränderungen der Freisetzung,
Bindung und Wiederaufnahme innerhalb der glutamatergen, glycerinergen und GABAergen
Systeme im auditorischen Hirnstamm (MARIANOWSKI et al. 2000; SUNEJA et al. 1998).
Weitere Hinweise für Kompensationsmechanismen innerhalb der zentralen auditorischen
Hörbahn konnten bei physiologischen Versuchen mit Chinchillas gefunden werden. Dort trat
nach lokal begrenzten Cochleaschädigungen, trotz einer dem Verlust der inneren Haarzellen
proportionalen Reduktion der Hörnervaktivität, eine dagegen nur leicht reduzierte Amplitude
des inferioren Colliculus auf. Dieses beruhte zum einen darauf, dass die Abnahme der
Amplitude des zusammengesetzten Aktionspotentials des Hörnerven (CAP) keinen Effekt auf
die Reizschwelle und das Tuning der verbliebenen aktiven Fasern im Hörnerven hatte. Zum
andern kam es innerhalb des IC zu einem schnelleren Anstieg des lokalen Feldpotentials und
einer Erhöhung der maximalen Amplitude von unterhalb der Cochleaschädigung lokalisierten
Frequenzen. Neben der nur geringen Verringerung der Reizantwort des inferioren Colliculus
wurde auf der Ebene des auditorischen Cortex sogar eine über das normale Maß erhöhte
Reaktivität festgestellt (RAGGIO und SCHREINER 1999).

2. Literaturübersicht
43
Ursache für diese Veränderungen ist wahrscheinlich eine Demaskierung von bereits
vorhandenen, aber gehemmten neuronalen Verbindungen. Insbesondere der Verlust von
Inhibitionen aus dem Bereich der Hörschädigung könnte hierbei eine Rolle spielen (SALVI et
al. 2000).
2.5.2.2 Der auditorische Cortex
Weder beim Menschen noch bei anderen Wirbeltieren ist das Gehirn vollständig genetisch
determiniert. Während der Embryonalphase entwickelt sich die cerebrale Mikrostruktur in
Abhängigkeit von den aus der Peripherie aufsteigenden Nervenbahnen. Die Entwicklung der
auf corticaler Ebene nachweisbaren Veränderungen besteht insbesondere in der Aussprossung
oder Rückbildung von thalamocorticalen Afferenzen. Auch postembryonal können im
Bereich des auditorischen Cortex neuronale Neuverknüpfungen auftreten. Die nach Abschluss
der Ontogenese stattfindende Plastizität durch die Zerstörung und Neubildung synaptischer
Verbindungen beschränken sich jedoch auf Bereiche von wenigen Mikrometern und dürfen
nicht als Kompensation für peripher bedingte Taubheit verstanden werden (GILBERT 1993).
Tatsächlich stattfindende funktionelle Veränderungen im Sinne einer Reorganisation im
Bereich des felinen auditorischen Cortex nach einer neonatalen Langzeitertaubung wurden
von DINSE und Mitarbeitern mittels optischer Messungen intrinsischer Signale
nachgewiesen. Als Effekte wurden kleinere aktivierte Flächen und geringere Signalstärken im
Vergleich zu akut ertaubten Versuchstieren gefunden (DINSE et al. 1997). Derartige
Veränderungen lassen sich zudem durch Messung der synaptischen Aktivität darstellen. So
fallen die synaptischen Ströme von Reizantworten mittlerer Latenzen kongenital tauber
Katzen in den Laminae II und III sowie weiterer infragranulärer Schichten im primären
auditorischen Feld signifikant geringer aus als bei hörenden Kontrolltieren (KLINKE et al.
1999; KRAL et al. 2000).
HARRISON et al. (1991) konnten weiterhin an Katzen zeigen, dass sich die tonotope
Organisation des primären auditorischen Feldes nach hochfrequentem bilateralen neonatalen
cochleären Hörverlust durch die Applikation des ototoxischen Aminoglykosid-Antibiotikums
Amikacin umfassend umstrukturiert. Dabei beinhalteten die anterioren Teile des primären
auditorischen Feldes, welche physiologisch Neurone mit hohen charakteristischen Frequenzen
(CF) beherbergen, fast ausschließlich Neurone mit einer speziellen, niedrigeren CF.

2. Literaturübersicht
44
Diese Frequenz entsprach auf Ebene der Cochlea dem Grenzbereich zwischen intakten und
beschädigten Haarzellen. Diese Veränderungen (Übergreifen von periläsionären Frequenzen)
reflektieren einen dynamischen Prozess der Plastizität und sind nicht als passive
Konsequenzen der Schädigung erklärbar (IRVINE et al. 2001).
Die tonotope Abbildung des primären auditorischen Feldes bei neonatal ertaubten Katzen
zeigt im Vergleich zu normalhörenden Tieren generell ein durch irregulär angeordnete
Aktivitätszonen mit mehreren Aktivitätszentren hervorgerufenes diffuses und fleckiges
Erscheinungsbild. Die bei Kontrolltieren gefundenen klaren Aktivitätsveränderungen
aufgrund der Tonotopie der Frequenzabbildung können bei tauben Tieren nicht beobachtet
werden. Neben dieser Verzerrung der corticalen Tonotopie liegen bei tauben Tieren zudem
erhöhte neuronale Schwellenwerte vor (REUTER et al. 2002a).
Cytoarchitektonisch wurde für den felinen auditorischen Cortex gezeigt, dass in der Lamina
III des ipsilateralen und kontralateralen primären auditorischen Feldes neonatal ertaubter
Tiere eine signifikante Reduktion der Gesamtneuronenanzahl und der Anzahl großer
Pyramidenzellen auftritt (REUTER 1997; WENKE 1999). Auch im kontralateralen anterioren
auditorischen Feld kommt es zu einem signifikanten Pyramidenzellverlust in der Lamina III.
Im ebenfalls untersuchten sekundären auditorischen Feld beider Cortexhemisphären zeigten
sich hingegen keine signifikanten Unterschiede (WENKE et al. 1997).
Im Bereich des humanen auditorischen Cortex konnte mittels funktioneller
Magnetresonanztomographie (fMRI) eine Reduktion der corticalen Aktivität bei
hörbeeinträchtigten Personen nachgewiesen werden (FIRSZT et al. 2003). Zudem zeigte sich
bei unilateraler Taubheit eine bilaterale Aktivierung der Hörrinde. Ursache für derartige
zentralauditive Kompensationsmechanismen im Sinne einer Bilateralisation ist auch hier
wahrscheinlich eine Demaskierung vorhandener ipsilateraler Verbindungen (BILECEN et al.
2000).
Hinsichtlich der physiologisch-funktionellen Reorganisation besitzen die übergeordneten
auditorischen Felder, insbesondere das sekundäre auditorische Feld bei Katzen, ebenso wie
die supragranulären Hirnschichten (KACZMAREK et al. 1997) eine größere Plastizität als
das primäre auditorische Feld (WEINBERGER et al. 1984).

2. Literaturübersicht
45
Dahingehend wurde eine corticale Plastizität über die Grenzen des auditorisch-sensorischen
Areals hinweg von REBILLARD et al. (1977, 1980) nachgewiesen. Nach bilateraler
chirurgischer neonataler akustischer Deprivation sowie bei kongenital tauben Katzen konnten
im primären auditorischen Cortex visuell evozierte Potentiale abgeleitet werden. Diese Daten
konnten jedoch in einer späteren Studie nicht bestätigt werden (KRAL et al. 2003).
Umgekehrt konnten RAUSCHECKER und KORTE (1993) an neonatal durch Verschluss der
Augenlider binocular deprivierten Katzen zeigen, dass diese Tiere akustische
Lokalisationsaufgaben besser absolvierten als Kontrolltiere. Anatomisch hatten sich
auditorische Bereiche in Regionen ausgebreitet, die bei sehenden Tieren auf visuelle Reize
reagierten.
In Übereinstimmung mit diesen tierexperimentellen Ergebnissen führt die visuelle
Präsentation von Zeichen bei humanen Patienten nach Langzeitgebrauch der Zeichensprache
zu einer Aktivierung des Sprachsystems in der gleichen Weise, wie es bei hörenden Personen
durch Sprache geschieht (PETITTO et al. 2000). Zusätzlich konnten PARASNIS (1998)
sowie NEVILLE und LAWSON (1987) in ihren Untersuchungen nachweisen, dass taube
Patienten in visuellen und kognitiven Tests besser abschneiden als hörende Personen.
2.5.3 Die plastischen Umstrukturierungen im auditorischen System nach neonataler
akustischer Deprivation und anschließender chronischer elektrischer intracochleärer
Stimulation
Durch in die Scala tympani der Cochlea eingeführte elektronische Reizprothesen, den
Cochlea-Implantaten, kann die Funktion ausgefallener innerer Haarzellen überbrückt werden.
Durch die Verwendung mehrkanaliger Elektroden kann die in der Cochlea vorgegebene
Tonotopie in einem großen Frequenzbereich genutzt werden. Zahlreiche Studien haben die
Auswirkungen der chronischen elektrischen intracochleären Stimulation bei neonatal
ertaubten Tieren untersucht. Dabei zeigte sich, dass Entwicklungsstörungen zumindest
teilweise durch eine ausreichende elektrische Stimulation aufgehoben werden können.
2.5.3.1 Das Hörorgan und die afferente Hörbahn
LUSTIG et al. (1994) fanden bei neonatal ertaubten, unilateral chronisch intracochleär
stimulierten Katzen eine Linderung der durch eine neonatale Ertaubung hervorgerufenen
Schrumpfung der sphärischen Zellen innerhalb des Nucleus cochlearis (NC).

2. Literaturübersicht
46
Gleichzeitig zeigte der Vergleich zwischen ipsi- und kontralateralem NC im Durchschnitt um
6 % signifikant größere Zellen im ipsilateralen NC. Letztere Ergebnisse konnten jedoch von
HULTCRANTZ et al. (1991) nicht reproduziert werden. Das Gesamt-NC-Volumen, sowie
die Dichte der sphärischen Zellen, konnten durch die chronische elektrische Stimulation nicht
beeinflusst werden (LUSTIG et al. 1994). Weiterhin konnte im Vergleich zum
kontralateralen, unstimulierten Ohr dieser Tiere für elektrodennahe Bereiche der stimulierten
Cochlea ein signifikanter Erhalt der Spiralganglienzellen beobachtet werden (LEAKE et al.
1991).
Auf Ebene des inferioren Colliculus konnten nach chronischer elektrischer intracochleärer
Stimulation von neonatal ertaubten Katzen größere Areale innerhalb des zentralen Nucleus
(ICC) aktiviert werden (SNYDER et al. 1990). Zudem zeigte sich die Zellfläche des ipsi- und
kontralateralen ICC im Vergleich zu unstimulierten Tieren signifikant erhöht. In Bezug auf
das Gesamtvolumen und die Gesamtzellzahl der ICC konnte jedoch kein signifikanter
positiver Einfluss einer chronischen elektrischen intracochleären Stimulation nachgewiesen
werden (VOGT et al. 1997).
Der von CORDS (1996) durchgeführte Vergleich der Physiologie der zentralen Hörbahn
mittels früher elektrisch evozierter Potentiale zwischen neonatal ertaubten chronisch
elektrisch intracochleaär stimulierten und implantierten unstimulierten Katzen konnte positive
Einflüsse der Stimulation auf die Ausreifung des auditorischen Systems zeigen.
2.5.3.2 Der auditorische Cortex
Bei humanen Cochlea-Implantat-Trägern korrelieren die durch akustische Reize aktivierten
corticalen Areale mit der Sprachleistung und breiten sich mit Dauer der auditorischen
Erfahrung aus (LEE et al. 2001). Nach einer Cochlea-Implantation durchgeführte Sprach-,
Verständnis- und Verhaltenstests konnten bei prälingual ertaubten Kindern zeigen, dass die
durch eine neonatale Deprivation induzierte fehlerhafte Entwicklung des auditorischen
Systems durch eine frühzeitige Implantat-Versorgung aufgehalten und in eine nahezu normale
Entwicklung gelenkt werden kann. Dabei werden die durch die künstliche Stimulation
ausgelösten Höreindrücke durch ein Hör-Sprachtraining zunehmend mit Bedeutung gefüllt
und Assoziationen mit der eigenen Sprache hergestellt.

2. Literaturübersicht
47
Mittels Optical Recording können die Auswirkungen einer chronischen elektrischen
Stimulation mit Cochlea-Implantat auch am neonatal ertaubten felinen auditorischen Cortex
nachgewiesen werden. Sie zeigten sich als signifikante Erhöhung der Signalstärke,
Verbreiterung der aktivierten corticalen Flächen sowie einer stärkeren Überlappung der
aktivierten Bereiche (DINSE et al. 1997). Daneben findet eine Reorganisation der corticalen
Topographie statt. Ähnlich normalhörigen Tieren konnte eine Aktivitätsverlagerung von
rostral nach caudal innerhalb des primären auditorischen Feldes entsprechend der jeweils
stimulierten Frequenzen registriert werden (REUTER et al. 2002b).
Bei kongenital tauben Katzen konnten durch eine frühzeitige Implantation eines einkanaligen
Cochlear-Implantats ebenfalls Defizite in der Reifung des kontralateralen primären
auditorischen Feldes aufgefangen und sogar degenerative Prozesse aufgehoben werden
(KRAL et al. 2002). Nach chronischer Stimulation mit adäquaten Reizen zeigte sich
insbesondere eine Erhöhung der Amplituden der corticalen Feldpotentiale mittlerer Latenzen,
eine Vergrößerung der kontralateralen antwortgebenden Areale sowie eine Weiterentwicklung
von Antworten langer Latenz. Die beobachteten Aktivitäten ähnelten sowohl in der
Ausprägung, als auch in der Ausbreitung denen akustisch gereizter normalhörender Tiere und
korrelierten positiv mit der Stimulationsdauer. Im ipsilateralen auditorischen Cortex zeigten
sich gleichartige, jedoch im Vergleich geringer ausfallende Aktivitätssteigerungen (KLINKE
et al. 1999; KRAL et al. 2002).
Auch in Bezug auf die laminären synaptischen Aktivitäten im primären auditorischen Feld
konnten positive Auswirkungen einer chronischen elektrischen intracochleären Stimulation
nachgewiesen werden. Im Vergleich zu unstimulierten Tieren wiesen stimulierte kongenital
taube Katzen eine größere synaptische Effizienz auf. Während sich bei den naiven kongenital
tauben Katzen synaptische Ströme der Reizantworten mittlerer Latenzen nahezu auf die
Laminae II bis IV beschränkten, breiteten sie sich bei den stimulierten Tieren, ähnlich
normalhörenden Tieren, auf die Lamina V sowie auf weitere infragranuläre Schichten aus
(KLINKE et al. 1999; KRAL et al. 2000).
Histologisch konnte bei chronisch intracochleär stimulierten neonatal ertaubten Katzen ein
signifikanter relativer Erhalt der Pyramidenzellpopulation in der Lamina III des ispilateralen
und kontralateralen primären auditorischen Feldes nachgewiesen werden.

2. Literaturübersicht
48
Im ipsilateralen auditorischen Cortex fiel dieser Neuronenerhalt erwartungsgemäß geringer
aus (REUTER et al. 2002a). Im anterioren auditorischen Feld konnten die durch eine
neonatale Ertaubung hervorgerufenen Zellverluste durch eine chronische intracochleäre
Stimulation nicht signifikant umgekehrt werden (WENKE 1999).
2.6 Die Ziele der Untersuchung
Im Verlauf der detaillierten Untersuchung der Auswirkungen eines Cochlea-Implantats auf
die zentrale Hörbahn soll mit dieser Arbeit die höchste Ebene der auditorischen Verarbeitung,
der auditorische Cortex, untersucht werden.
Dazu wurde die Struktur der Dendriten in den Laminae I bis VI in den drei wichtigsten
auditorischen Subarealen der Katze, dem primären, sekundären und anterioren auditorischen
Feld untersucht und gegenübergestellt. Dies erfolgte an drei Versuchstiergruppen mit
unterschiedlichem Hörstatus (normal hörend, neonatal ertaubt und neonatal ertaubt,
anschließend chronisch elektrisch intracochleär stimuliert).

3. Material und Methoden
49
3. MATERIAL UND METHODEN
3.1 Material
3.1.1 Die Versuchstiere
Bei dem in dieser Studie untersuchten Material handelt es sich um den auditorischen Cortex
von Hauskatzen (Felis domestica). Die Versuchstiere beiderlei Geschlechts im Alter zwischen
23 und 356 Wochen stammten aus Würfen der Katzenhaltung des Zentralen Tier-
laboratoriums der Medizinischen Hochschule Hannover.
Gehalten wurden die Katzen in Gruppen zu jeweils 5-15 Tieren in 12-24 m2 großen,
vollklimatisierten, fensterlosen Räumen. Die Umweltbedingungen waren mit konstanten 24
°C Raumtemperatur, 40 - 60 % relativer Luftfeuchtigkeit und 12-stündiger täglicher
Beleuchtung für alle Katzen identisch. Die Versuchstiere hatten freien Zugang zu frischem
Wasser sowie frei verkäuflichem Dosen- und Trockenfutter. Der gesamte Bestand wurde
veterinärmedizinisch betreut und hinsichtlich Ekto- und Endoparasiten regelmäßig
kontrolliert. Er war bezüglich des Felinen Leukose Virus (FeLV), des Felinen
Immundefizienz Virus (FIV) und der Toxoplasmose serologisch negativ. Gegen
Katzenschnupfen (Infektiöse Rhinotracheitis) und Katzenseuche (Infektiöse Panleukopenie)
wurde regelmäßig geimpft.
Alle Tiere wurden nach physiologischen Untersuchungen, welche von der Bezirksregierung
Hannover nach Anzeige gemäß §8a des deutschen Tierschutzgesetzes von 1986 unter der
Aktennummer 97/972 genehmigt wurden, dieser Untersuchung zugeführt.
Als Voraussetzung für die Aufnahme in die Untersuchungsgruppen galt ein Lebensalter von
mindestens 30 Tagen, da ab diesem Zeitpunkt von einem vollständig ausgereiften
auditorischen System ausgegangen werden kann (siehe Kapitel 2.5.1.1)
Die Gruppenaufteilung erfolgte nach dem jeweiligen Hörstatus der Tiere: normal hörend,
neonatal experimentell ertaubt und neonatal experimentell ertaubt, anschließend chronisch
elektrisch intracochleär stimuliert. Die Hörfunktion der Katzen wurde vor der Aufnahme in
den Versuch durch Ableitung von frühen akustisch evozierten Potentialen überprüft.
Die Gesamtzahl von 13 Versuchstieren setzt sich aus drei Gruppen zusammen:
1. Hörende Kontrollgruppe (n = 6);
2. Neonatal experimentell ertaubte Versuchsgruppe (n = 4);
3. Neonatal experimentell ertaubte, anschließend chronisch elektrisch intracochleär
stimulierte Versuchsgruppe (n = 3);
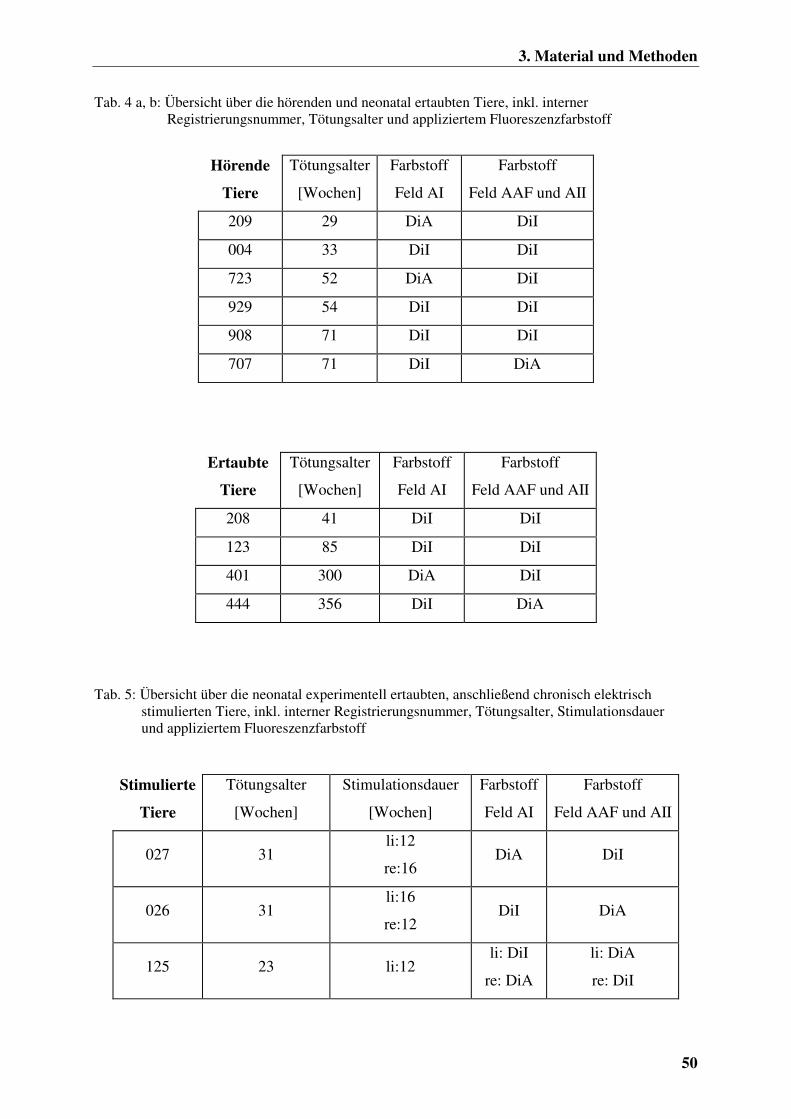
3. Material und Methoden
50
Tab. 4 a, b: Übersicht über die hörenden und neonatal ertaubten Tiere, inkl. interner Registrierungsnummer, Tötungsalter und appliziertem Fluoreszenzfarbstoff
Hörende
Tiere
Tötungsalter
[Wochen]
Farbstoff
Feld AI
Farbstoff
Feld AAF und AII
209 29 DiA DiI
004 33 DiI DiI
723 52 DiA DiI
929 54 DiI DiI
908 71 DiI DiI
707 71 DiI DiA
Tab. 5: Übersicht über die neonatal experimentell ertaubten, anschließend chronisch elektrisch stimulierten Tiere, inkl. interner Registrierungsnummer, Tötungsalter, Stimulationsdauer und appliziertem Fluoreszenzfarbstoff
Ertaubte
Tiere
Tötungsalter
[Wochen]
Farbstoff
Feld AI
Farbstoff
Feld AAF und AII
208 41 DiI DiI
123 85 DiI DiI
401 300 DiA DiI
444 356 DiI DiA
Stimulierte
Tiere
Tötungsalter
[Wochen]
Stimulationsdauer
[Wochen]
Farbstoff
Feld AI
Farbstoff
Feld AAF und AII
027 31 li:12
re:16 DiA DiI
026 31 li:16
re:12 DiI DiA
125 23 li:12 li: DiI
re: DiA
li: DiA
re: DiI

3. Material und Methoden
51
3.1.2 Die Sachmaterialien
3.1.2.1 Die Pharmaka:
a) Atropinsulfat (0,5 mg/ml), Atropinsulfat Braun 0,5 mg; B. Braun Melsungen AG,
Melsungen
b) Benzathin [100 000 I.E.], 24,8 mg Benzylpenicillin-Procain [25 000 I.E.],
156,3 mg Dihydrostreptomycinsulfat [125 mg Dihydrostreptomycin], 1,5 mg Methyl-
4-Penicillin-Streptomycin-Kombination mit Depotwirkung, (je ml 82,6 mg
Benzylpenicillin-hydroxybenzoat, 0,2 mg Propyl-4-hydroxybenzoat, Polysorbat 80,
Natriumcitrat, Saccharose, Lecithin , Wasser für Injektionszwecke), Tardomyocel
comp. III; Bayer Vital GmbH, Leverkusen
c) Carprofen 5% (50 mg/ml) und Benzylalkohol 1% (10 mg/ml), Zenecarp®; C-VET
Limeted, Bury St. Edmunds, England
d) Ketaminhydrochlorid (115,34 mg/ml) und Benzethoniumchlorid (100 µg/ml) in
Natriumchloridlösung, Ketamin
10%; WDT, Garbsen
e) Neomycinsulfat (50 mg/ml) in 0,9 %iger Natriumchloridlösung; Sigma-Aldrich
Chemie GmbH, Steinheim
f) Pentobarbital-Natrium in Natriumchlorid-Lösung (60 mg/ml), Narcoren®; WDT,
Garbsen
g) Prilocainhydrochlorid (20 mg/ml), Xylonest®
2%; AstraZeneca GmbH, Plankstadt
h) Thilo-Tears® Gel; Alcon Pharma (Dr. Thilo) GmbH, Freiburg i. Breisgau
i) Xylazin-Hydrochlorid (23,32 mg/ml) und p-Hydroxybenzoesäuremethylester (1
mg/ml) in Natriumchloridlösung, Rompun
2%; Bayer Vital GmbH, Leverkusen
3.1.2.2 Die Chemikalien:
a) 1 M Essigsäure; Merck KGaA, Darmstadt
b) 1 N Natronlauge; Merck KGaA, Darmstadt
c) Agar agar; Merck KGaA, Darmstadt
d) Cyanacrylat-Klebstoff, Roti-Coll®; Carl Roth GmbH & Co., Karlsruhe
e) Dinatriumydrogenphosphat-Monohydrat (Na2HPO4*2 H2O); Merck KGaA, Darmstadt
f) Entellan; Merck KGaA, Darmstadt
g) Fluoreszenzfarbstoff DiA; Molecular Probes Inc., Eugene, Oregon, USA
h) Fluoreszenzfarbstoff DiI; Molecular Probes Inc., Eugene, Oregon, USA
i) Glycerol; Sigma-Aldrich Chemie GmbH, Steinheim
j) Kresyl-Violett-Farbstoff; Merck KGaA, Darmstadt

3. Material und Methoden
52
k) Mowiol 40-88; Sigma-Aldrich Chemie GmbH, Steinheim
l) Natrium-Acetat; Merck KGaA, Darmstadt
m) Natriumchlorid; Merck KGaA, Darmstadt
n) Natriumdihydrogenphosphat-Monohydrat (NaH2PO4*H2O); Merck KGaA, Darmstadt
o) Paraformaldehyd; Merck KGaA, Darmstadt
p) Tris/HCl-Puffer (0,2 M, pH 8,5); Merck KGaA, Darmstadt
q) Xylol; Merck KGaA, Darmstadt
r) Zinkphosphatzement, Harvard Cement®
, normalhärtend; Hoffmann & Richter
HARVARD Dental GmbH, Berlin
3.1.2.3 Die Verbrauchsmaterialien:
a) Einmal-Becher mit Schraubverschluß, 100 ml; Sarstedt AG & Co., Nümbrecht
b) Intrafix® Air Infusionsset ; B. Braun Melsungen AG, Melsungen
c) Kimwipes Lite; Kimberley-Clark Limeted, Kent, England
d) Rasierklingen normal; Wilkinson sword GmbH, Solingen
e) Serican®-Einmalinjektionskanülen, 19 G x 1¼"; B. Braun Melsungen AG, Melsungen
f) Zentrifugenröhrchen Blue-Max TM, 50 ml; Becton Dickinson GmbH, Heidelberg
3.1.2.4 Die Gebrauchsgegenstände:
a) Aquarell-Pinsel, Gr. 2; Faber Castell Vertrieb GmbH, Stein
b) Chirurgische und anatomische Pinzetten; Aeskulap AG & Co. KG, Tuttlingen
c) Deckgläser (24 x 60 mm); Menzel-Gläser GmbH & Co. KG, Braunschweig
d) Hirnmesser; Aeskulap AG & Co. KG, Tuttlingen
e) hölzerne Vibratomblöcke; Leica, Bensheim
f) Knochenzange; Aeskulap AG & Co. KG, Tuttlingen
g) Objektträger (76 x 26 mm); Menzel-Gläser GmbH & Co. KG, Braunschweig
h) Skalpellgriff inkl. Skalpellklingen Gr. 11 und Gr. 21; Aeskulap AG & Co. KG,
Tuttlingen

3. Material und Methoden
53
3.1.2.5 Die technische Ausstattung:
a) Axioskop; Carl Zeiss, Oberkochen
• Rhodamin-Filter, Filtersatz 15 (BP 546, FT 580, LP 590); Carl Zeiss,
Oberkochen
• Fluorescein Isothiocyanate (FITC) Filter, Filtersatz 10 (BP 450-490, FT 510,
LP 515-565); Carl Zeiss, Oberkochen
• 10x/20 E-Plan Okular; Carl Zeiss, Oberkochen
• 10x/20 E-Plan Okular mit eingraviertem Zählraster; Carl Zeiss, Oberkochen
• 20fach (NA 0,50) Plan-NEOFLUAR Objektiv; Carl Zeiss, Oberkochen
b) Computersoftware:
• Windows NT 4.0® inkl. Excel 2000®; Microsoft GmbH, Unterschleissheim
• SPSS 11.0®; SPSS Inc., Chicago, USA
c) Digitalfotokamera Canon EOS D90; Canon Deutschland GmbH, Krefeld
d) handelsüblicher Computer
e) Leica DMLS Lichtmikroskop; Leica, Bensheim
• 10x/20 C-Plan Okular; Leica, Bensheim
• 10x/20 C-Plan Okular mit eingravierter Messskala; Leica, Bensheim
• 10fach (NA0,10) C-Plan Objektiv; Leica, Bensheim
f) Schwenkarm-Wandmikroskop; Carl Zeiss, Oberkochen
g) Vibratom VT1000S; Leica, Bensheim
3.2 Methoden
Alle Arbeiten wurden durchgehend von einer Person und in konstanter Weise durchgeführt.
3.2.1 Die experimentelle neonatale Ertaubung der Katzen
In Zusammenarbeit mit einer weiteren Arbeitsgruppe der Medizinischen Hochschule
Hannover wurden insgesamt 7 Versuchstiere neonatal experimentell ertaubt.
Dazu wurde eine chronische systemische Applikation des ototoxischen Antibiotikums
Neomycin durchgeführt (CORDS 1996, Übersicht in KRAL et al. 2001). Die Versuchstiere
erhielten vom dritten bis einschließlich 13. Lebenstag eine tägliche subcutane Injektion von
50 mg Neomycin pro kg Körpergewicht (KGW). Zur Überprüfung des Hörstatus wurden am
14. Lebenstag die frühen akustisch evozierten Potentiale (FAEP) abgeleitet (siehe Anhang
9.3).

3. Material und Methoden
54
Konnten bei einer Reizlautstärke von 100 dB SPL noch Antworten der zentralen Hörbahn
registriert werden (Resthörigkeit), wurde die Neomycinapplikation weitere vier Tage
durchgeführt und der Erfolg nochmals mittels FAEP-Ableitung überprüft.
Die vom ersten Lebenstag an für jedes Tier dokumentierten täglichen Körpergewichts-
zunahmen zeigten, dass es durch diese Eingriffe zu keiner Beeinträchtigung des
Allgemeinbefindens kommt (siehe Anhang 9.4). Weiterhin wurden bei keinem der neonatal
ertaubten Tiere Verhaltensauffälligkeiten beobachtet.
3.2.2 Die Implantation der elektronischen Innenohrprothese in neonatal experimentell
ertaubte Katzen
Eines der zuvor neonatal ertaubten Versuchstiere erhielt unilateral, zwei weitere neonatal
ertaubte Tiere erhielten bilateral eine elektronische Innenohrprothese (Cochlea-Implantat). Da
es sich bei der Cochlea-Implantation um einen aufwändigen Eingriff handelt, wurde bei den
beidseitig implantierten Tieren nach dem ersten Eingriff eine Rekonvaleszenzzeit von 4
Wochen eingehalten. Im Verlauf der Implantation wurde nach der sogenannten Soft-Surgery-
Technik nach LEHNHARDT (1993) vorgegangen.
Die nüchternen Tiere erhielten noch in gewohnter Umgebung zunächst Rompun subcutan
(0,1 ml/kg KGW) und nach dem Einsetzen der Wirkung als Narkoseeinleitung Ketamin
intramuskulär (0,1 ml/kg KGW). Die Prämedikation erfolgte mit Atropin (0,1 ml/kg KGW)
und Zenecarp (0,08 ml/kg KGW) subcutan. Als Infektionsprophylaxe wurde vor der
Operation Tardomyocel (0,1 ml/kg KGW) subcutan verabreicht. Zur Aufrechterhaltung der
Narkose wurde Narcoren in individueller Dosierung intravenös verabreicht. Zusätzlich
wurden die Schnittlinien durch eine Infiltrationsanästhesie mit jeweils 0,5 ml des
Lokalanästhetikums Xylonest schmerzunempfindlich gemacht.
Die halbrunden etwa 4 cm langen Hautinzisionen befanden sich retroauriculär. Durch
stumpfes Vorpräparieren wurde unter Schonung der Glandula parotidea die prominent
liegende Bulla tympanica aufgesucht und von ventrolaterocaudal queroval mit einem
Längsdurchmesser von ca. 1 cm eröffnet. Ca. 2 mm apikal des runden Fensters (Fenestra
rotunda erfolgte eine Cochleostomie. Durch diese Öffnung wurde die elektronische
Innenohrprothese in die basale Windung der Scala tympani eingeführt, die Insertionsstelle mit
Bindegewebe abgedeckt und das Implantat endgültig mit Zinkphosphatzement im Inneren der
Bulla tympanica fixiert.

3. Material und Methoden
55
Der die Platindrähte enthaltende Silikonschlauch und die den Elektrodenabschluss
darstellende Steckverbindung des Cochlea-Implantats wurde unter die Musculi
sternooccipitalis et cleidocervicalis hindurchgeleitet und in der Medianen caudal der
Schulterblätter durch eine Hautöffnung wieder an die Körperoberfläche geführt. Die
Fixierung erfolgte subcutan durch eine Tabaksbeutelnaht mit Vicryl (3-0). Die retro-
auriculären Wundhöhlen wurden in der Tiefe durch zwei Sultansche Diagonalhefte mit
Vicryl (3-0) verkleinert. Für die abschließenden zweischichtigen Hautnähte wurde Vicryl (3-
0) und Mersilene (2-0) verwendet.
Während des gesamten Eingriffs lagen die Katzen auf einer auf 38 °C geheizten Wärmematte,
die Augen wurden durch die Tränenersatzflüssigkeit Thilo-Tears-Gel® vor Austrocknung
geschützt.
Um den freiliegenden Anteil der Elektrode zu schützen, wurden den Katzen nach der
Operation kleine Nylonjacken angepasst, welche nur für den Zeitraum der elektrischen
Stimulation durch andere, ebenfalls an die Tiere angepasste, Stimulationsjacken ersetzt
wurden.
3.2.3 Die chronische elektrische intracochleäre Stimulation der neonatal ertaubten,
implantierten Katzen
Mit der elektrischen Stimulation (siehe Anhang 9.2) der mit einem Cochlea-Implantat
versorgten Versuchstiere wurde, um das Einheilen des Implantats abzuwarten, erst 7 Tage
nach der Implantation begonnen. Dafür wurden die Tiere an fünf Tagen der Woche für jeweils
mindestens vier Stunden einzeln in eine geräumige Katzentransportbox verbracht und
beschallt. Dieses Stimulationsschema ist für eine adäquate Reizung des auditorischen Cortex
ausreichend (LUSTIG et al. 1994). Insgesamt ergaben sich Stimulationszeiträume von 84 bis
112 Tagen.
Für die Stimulation wurde die zum Schutz des Cochlea-Implantats von den Tieren getragene
Nylonjacke gegen eine zweite Jackenform mit daran fixierten Implantatempfänger
ausgetauscht. Durch eine Steckverbindung wurde der Kontakt zwischen dem in der Medianen
caudal der Schulterblätter austretenden Elektrodensteckkontakt und dem Implantatempfänger
hergestellt. Die Sprachprozessoren, welche durch ein Kabel mit dem Implantatempfänger in
Verbindung standen, wurden außerhalb der Stimulationsbox abgelegt. Die Beschallung der
Tiere erfolgte durch ein in unmittelbarer Nähe der Transportbox aufgestelltes Radio sowie
durch, gleichzeitig auftretende, Umgebungsgeräusche, welche auch aus Lautäußerungen von
Artgenossen bestanden.

3. Material und Methoden
56
Einmal wöchentlich wurden, analog zu den akustischen, die elektrisch evozierten Hirnstamm-
potentiale abgeleitet. Anhand der dabei für die jeweiligen Elektrodenkonfigurationen
bestimmten Hörschwellen wurde der Sprachprozessor so programmiert, dass die auf Basis der
akustischen Stimuli umgewandelten elektrischen Impulse über der individuellen Hörschwelle
lagen und somit von einer adäquaten Reizung des auditorischen Cortex ausgegangen werden
konnte (LUSTIG et al. 1994).
3.2.4 Die histologische Aufbereitung des auditorischen Cortex
Damit das corticale Nervengewebe in seiner gleichmäßigen Struktur erhalten blieb und um
die für die Fluoreszenzuntersuchungen störenden Erythrozyten zu entfernen, wurden die
Versuchstiere in tiefer Narkose transcardial perfusionsfixiert. Dazu wurde der Thorax der in
Rückenlage fixierten Tiere weiträumig eröffnet, das Herz freigelegt und das Perikard entfernt.
Eine an ein Infusionssystem angeschlossene Einmalinjektionskanüle wurde in den linken
Ventrikel eingeführt. Über eine zweite, in den rechten Ventrikel platzierte Kanüle wurde das
ausgespülte Blut respektive die durchgespülte Perfusionslösung abgelassen. Das Infusions-
system wurde zur Ausspülung des Blutes zunächst mit 1,0 l kalter PBS-Lösung gefüllt. Im
Anschluss wurde ebenfalls über das Infusionssystem mit 1,0 l kalter 4 %iger PFA-Lösung
fixiert. Nach Durchlauf beider Flüssigkeiten wurde der Kopf im atlanto-occipitalen Gelenk
von der Wirbelsäule getrennt und durch vorsichtiges Abtragen der Schädeldecke mit einer
Knochenzange das Gehirn aus dem Schädel herauspräpariert.
Bis zur Applikation der Fluoreszenzfarbstoffe wurden die Gehirne in mit 80 ml 4 %iger PFA-
Lösung gefüllten Einmal-Bechern bei 4 °C in Dunkelheit gelagert. Diese Lagerzeit betrug im
Mittel 75 Wochen.
3.2.5 Die Fluoreszenzfarbstoff-Tracingtechnik
Mittels Tracingtechnik mit Hilfe zweier Fluoreszenzfarbstoffe wurde der prozentuale Anteil
der Dendriten insgesamt im anterioren, primären und sekundären auditorischen Feld des
auditorischen Cortex und deren prozentualer Anteil in den einzelnen corticalen Schichten der
genannten auditorischen Felder bestimmt. Dabei wurden die Dendriten separat nach ihrem
Verlauf relativ zur Cortexoberfläche als horizontal, diagonal oder vertikal ziehend registriert.

3. Material und Methoden
57
Die Ergebnisse der hörenden Kontrolltiergruppe wurden mit den Ergebnissen der neonatal
experimentell ertaubten Versuchstiere sowie mit den neonatal ertaubten, anschließend
chronisch elektrisch intracochleär stimulierten Tieren verglichen.
3.2.5.1 DiA/DiI als neuronale Tracer
Für das Tracing der Dendriten wurde die Markierung mit den Fluoreszenzfarbstoffen DiA (4-
Di-16-ASP) und DiI (DiIC18(3) bzw. 1,1´-dioctadecyl-3,3,3´,3´-tetramethylindo-carbocyanine
perchlorate) verwendet.
Diese Farbstoffe sind lipophile Substanzen, welche auch die retrograde Färbung von
Neuronen in vitro ermöglichen (HONIG 1993; HONIG und HUME 1989).
An formaldehydfixierten Geweben können aufgrund der guten Haftung und der Möglichkeit
die Applikation unter mikroskopischer Kontrolle durchzuführen, DiA/DiI-Kristalle direkt auf
die Cortexoberfläche aufgebracht werden. Hierbei handelt es sich um das Mittel der Wahl um
mittels fokaler Applikation selektiv Nervenfasern anzufärben (KÖBBERT et al. 2000).
Die Farbstoffe werden mit ihren hydrophoben, fettlöslichen Hydrocarbonketten parallel in die
äußere Lipidfraktion der Plasmamembran aller mit dem Kristall in Berührung kommender
Dendriten eingefügt und breiten sich ähnlich nativer Lipide durch passive laterale Diffusion
frei entlang des Konzentrationsgefälles aus, wobei die hydrophilen Anteile der Membran als
Diffusionsbarriere wirken. Als Konsequenz werden Zellen in ihrer Gesamtheit, inklusive ihrer
feinen Ausläufer (Abb. 10, 11), angefärbt (GODEMENT et al. 1987; KÖBBERT et al. 2000).
Die Diffusion von DiI in paraformaldehydfixiertem Gewebe erfolgt mit Geschwindigkeiten
von ca. 0,2 Millimetern pro Tag (LUKAS et al. 1998). GALUSKE et al. (2000) konnten im
humanen cerebralen Cortex angefärbte neuronale Fasern im Abstand von bis zu 7 Millimetern
zur Applikationsstelle nachweisen.
3.2.5.2 Die Methode der DiA/DiI- Färbung
Zur Applikation der Fluoreszenzfarbstoffe auf dem anterioren, primären und sekundären
auditorischen Feld wurden die Gehirne dem Fixans entnommen und Farbstoffkristalle in die
Cortexoberfläche inseriert. Die Identifizierung der einzelnen auditorischen Subareale erfolgte
dabei anhand der von BRUGGE und REALE (1985) angefertigten Abbildungen des
Katzencortex.
Stereomikroskopisch wurden zunächst bei 112,5facher Vergrößerung 500 bis 800 µm große
Kristalle der Fluoreszenzfarbstoffe DiA und DiI selektiert.

3. Material und Methoden
58
Geringe Unterschiede in der Größe der applizierten Kristalle sind dabei unvermeidbar, führen
aber lediglich zu einer größeren Anzahl potentiell auswertbarer Gewebeschnitte. Durch die
für die Auswertung herangezogene Berechnung der prozentualen Anteile der angefärbten
Dendriten wurden diese Unterschiede wieder ausgeglichen.
Der zu färbende Cortex wurde unter einem Abzug mit Hilfe von Kimwipes vorsichtig von
Paraformaldehydrückständen befreit. Anschließend wurde die Pia mater durch leichten Zug
mit Hilfe einer chirurgischen Pinzette an den in dieser verlaufenden Blutgefäßen von der
Cortexoberfläche abgehoben und entfernt. Um die Diffusion der Farbstoffkristalle störende
hydrophile Flüssigkeit zu entfernen, wurde die Oberfläche des auditorischen Cortex
anschließend nochmals mittels Kimwipes trockengetupft. Mit einer Skalpellklinge der Größe
11 wurde schließlich unter mikrosko-pischer Kontrolle jeweils ein vorselektierter
Fluoreszenzfarbstoffkristall aufgenommen und auf die Oberfläche des zu färbenden
auditorischen Feldes aufgetragen (Abb. 6). Um die Diffusion des Farbstoffs zu gewährleisten
wurde dabei während der Applikation eine minimale Läsion in die Cortexoberfläche gesetzt.
An jedem Cortex wurde jeweils das anteriore (AAF), primäre (AI) sowie das sekundäre
auditorische Feld (AII) der rechten und linken Hemisphäre mit einem Farbkristall belegt
(Abb. 5). Zu Beginn dieser Studie war eine 3-dimensionale Rekonstruktion der kompletten
Dendritenbäume angefärbter Neurone mittels Confokalem Laser-Scanning-Mikroskop ange-
dacht. Um eventuelle Querverschaltungen zwischen den betrachteten Subarealen zu
identifizieren wurden daher jeweils das AAF und das AII mit einem und das AI mit dem
jeweils anderen Fluoreszenzfarbstoff markiert (Tab. 4 a und b, Tab. 5, Abb. 7).
Cerebellum
AI AAF AII
Abb. 5: Lateralansicht eines freipräparierten felinen Gehirns. AI: Primäres auditorisches Feld, AII: Sekundäres auditorisches Feld, AAF: Anteriores auditorisches Feld, sss: suprasylvanischer Sulcus, aes: anteriorer ectosylvanischer Sulcus, pes: posteriorer ectosylvanischer Sulcus
sss
aes pes
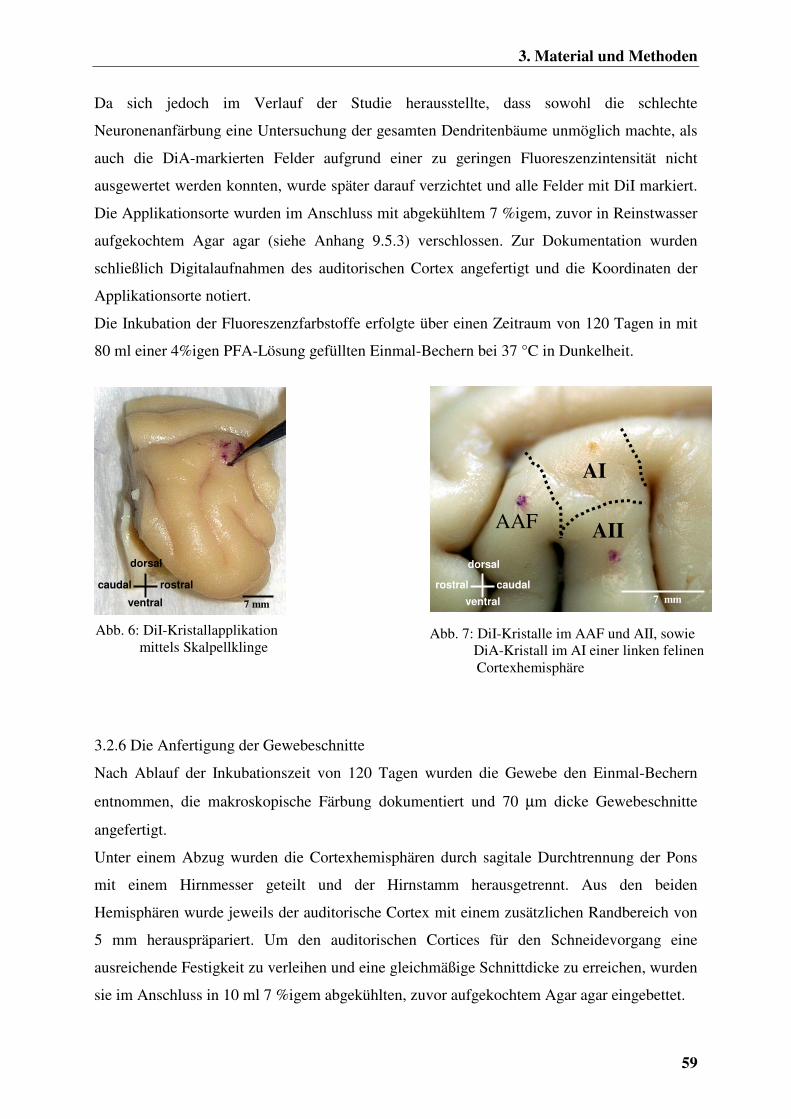
3. Material und Methoden
59
Da sich jedoch im Verlauf der Studie herausstellte, dass sowohl die schlechte
Neuronenanfärbung eine Untersuchung der gesamten Dendritenbäume unmöglich machte, als
auch die DiA-markierten Felder aufgrund einer zu geringen Fluoreszenzintensität nicht
ausgewertet werden konnten, wurde später darauf verzichtet und alle Felder mit DiI markiert.
Die Applikationsorte wurden im Anschluss mit abgekühltem 7 %igem, zuvor in Reinstwasser
aufgekochtem Agar agar (siehe Anhang 9.5.3) verschlossen. Zur Dokumentation wurden
schließlich Digitalaufnahmen des auditorischen Cortex angefertigt und die Koordinaten der
Applikationsorte notiert.
Die Inkubation der Fluoreszenzfarbstoffe erfolgte über einen Zeitraum von 120 Tagen in mit
80 ml einer 4%igen PFA-Lösung gefüllten Einmal-Bechern bei 37 °C in Dunkelheit.
3.2.6 Die Anfertigung der Gewebeschnitte
Nach Ablauf der Inkubationszeit von 120 Tagen wurden die Gewebe den Einmal-Bechern
entnommen, die makroskopische Färbung dokumentiert und 70 µm dicke Gewebeschnitte
angefertigt.
Unter einem Abzug wurden die Cortexhemisphären durch sagitale Durchtrennung der Pons
mit einem Hirnmesser geteilt und der Hirnstamm herausgetrennt. Aus den beiden
Hemisphären wurde jeweils der auditorische Cortex mit einem zusätzlichen Randbereich von
5 mm herauspräpariert. Um den auditorischen Cortices für den Schneidevorgang eine
ausreichende Festigkeit zu verleihen und eine gleichmäßige Schnittdicke zu erreichen, wurden
sie im Anschluss in 10 ml 7 %igem abgekühlten, zuvor aufgekochtem Agar agar eingebettet.
Abb. 6: DiI-Kristallapplikation mittels Skalpellklinge
Abb. 7: DiI-Kristalle im AAF und AII, sowie DiA-Kristall im AI einer linken felinen
Cortexhemisphäre
AAF
AI
AII
7 mm 7 mm
rostral caudal
ventral
dorsal
rostral
dorsal
caudal
ventral

3. Material und Methoden
60
Nach einer Aushärtezeit von mindestens einer Stunde wurden die entstandenen rechteckigen
Agar agar-Blöcke in Form geschnitten und mit einem Cyanacrylat-Klebstoff auf hölzernen
Vibratomblöcken befestigt (Abb. 8).
Beim Zurechtschneiden der Agarblöcke wurden, um ein Herausbrechen des Untersuchungs-
gewebes während des Schneidevorganges zu verhindern, in allen Richtungen des Raumes
mindestens 5 mm Agar agar um das Gewebe herum stehen gelassen.
Der Vibratomblock wurde nun in die Halteeinrichtung des Vibratoms eingespannt und 70 µm
dicke sagitale Serienschnitte angefertigt. Dabei wurde die Winkeleinstellung auf 15° und die
Geschwindigkeit sowie die Frequenz auf Stufe 4 eingestellt. Statt der vom Hersteller
empfohlenen Klingen wurden halbierte Rasierklingen verwendet, da diese sich als
kostengünstiger und präparatschonender erwiesen. Für jede Cortexhemisphäre wurde eine
frische Klinge eingesetzt. Da durch eine Austrocknung des untersuchten Gewebes eine starke
Reduzierung der DiA/DiI-Färbung auftritt (GODEMENT et al. 1987), wurde die
Schneideschale des Vibratoms mit kaltem Reinstwasser gefüllt und dieses regelmäßig
erneuert.
Während des Schneidevorganges wurde anhand der makroskopischen Cortexarchitektur
(siehe Kapitel 2.4.1) eine Identifikation des primären, sekundären und anterioren
auditorischen Feldes vorgenommen und die jeweiligen Schnitte entsprechend beschriftet.
Die ersten zwei Schnitte jedes auditorischen Feldes wurden mittels Pinsel einzeln auf
beschichtete Objektträger gezogen und für die Kresyl-Violett-Färbung verwendet (siehe
Kapitel 3.2.7). Alle weiteren Schnitte wurden zu zweit auf unbeschichtete Objektträger
verbracht, nass mit frisch aufgetautem Mowiol eingedeckelt und bis zur Auswertung bei 4 °C
in Dunkelheit gelagert.
Abb. 8: In Agar agar eingebetteter, auf hölzernem Vibratomblock aufgeklebter rechter auditorischer Cortex. Das anteriore und sekundäre auditorische Feld sind mit DiI markiert. Auf das primäre auditorische Feld ist ein DiA-Kristall aufgetragen.
17,5 mm
17,5 mm

3. Material und Methoden
61
3.2.7 Die Methode der Kresyl-Violett-Färbung (NISSL-Ersatzfärbung)
Mit Hilfe der NISSL-Färbungen werden die Zellkerne und Tigroidschollen von Neuronen
sichtbar gemacht. Es kommt dabei zu einer elektropolaren Anlagerung des basischen
Farbstoffs an die sauren Gruppen der DNS. Bei der verwendeten NISSL-Ersatzfärbung
(Schnell-NISSL-Methode) wurde der Oxazin-Farbstoff Kresyl-Violett verwendet. Die Zell-
kerne und NISSL-Substanz werden violett, die Neurone selbst ganz schwach blau und die
restlichen Strukturen farblos dargestellt. Aufgrund der für die einzelnen Laminae
charakteristisch stark ausgeprägte Anfärbung sowie durch die Identifikation von Lamina-
charakteristika wie dem Auftreten spezieller Zelltypen, der Zellanordnung sowie dem
Verhältnis von Zellen und Neuropil kann eine Schichtdickenbestimmung durchgeführt
werden (ROSE 1949, WINER 1984 a, b, c).
Für jedes auditorische Feld wurde die Dicke der einzelnen Zellschichten an den ersten,
einzeln aufgezogenen Schnitten bestimmt und die Ergebnisse mit den Literaturdaten
verglichen. Im Bereich der mit einem Farbkristall versehenen Gebiete wurde, um einen
Datenverlust zu vermeiden, mit Hilfe des zur Auszählung verwendeten Zählrasters zur
direkten Ausmessung der Schichtdicken nach den Literaturangaben übergegangen.
Die einzeln auf beschichtete Objektträger aufgezogenen Schnitte wurden über Nacht auf einer
auf 37 °C geheizten Wärmeplatte luftgetrocknet.
Anschließend wurde durch eine absteigende Alkoholreihe entfettet,
96 %iges Ethanol 15 min
Isopropanol 10 min
70 %iges Ethanol 10 min
gespült,
Reinstwasser 10 min
und schließlich gefärbt und differenziert
Acetatpuffer (pH 4,6) 10 min
Kresyl-Violett-Färbelösung 1-2 min
Reinstwasser 1 min
70 %iges Ethanol 3-5 min
70 %iges Ethanol 5 min.
96 %iges Ethanol 8-10 min
96 %iges Ethanol 10 min.
Die Durchführung der letztgenannten Schritte erfolgte unter lichtmikroskopischer Kontrolle.

3. Material und Methoden
62
Abschließend wurden die Schnitte weiter entwässert,
Isopropanol 5 min
Xylol 15 min
nass mit Entellan eingedeckelt
und zum Ausdampfen mindesten 24 Stunden unter einem Abzug belassen.
3.2.8 Die histologisch-morphologische Auswertung des auditorischen Cortex
Zum Abgleich der vertikalen Ausbreitung der sechs corticalen Schichten jedes betrachteten
auditorischen Subareals mit den Literaturangaben wurden zunächst die Kresyl-Violett
gefärbten Schnitte mit 40facher Vergrößerung durch ein Okular mit integrierter Messskala
lichtmikroskopisch betrachtet und die Dicke der Laminae I bis VI ausgemessen.
Die eigentliche Auswertung der angefärbten Dendriten erfolgte fluoreszenzmikroskopisch mit
Hilfe eines modifizierten Axioskops. Die mit DiI gefärbten auditorischen Areale wurden unter
Grünanregung (Rhodaminfilter, Filtersatz No. 15) betrachtet. Dabei erscheint DiI in einem
leuchtenden Rotorange auf einem optimalerweise schwarzen Hintergrund. Trotz einer bei den
hier untersuchten Gewebeproben stark ausgeprägten Hintergrundfluoreszenz konnten die
angefärbten Dendriten in ausreichender Intensität dargestellt werden. In den DiA-gefärbten
Bereichen erschienen die markierten neuronalen Strukturen unter dem Filtersatz No. 10
(FITC-Filter) gelbgrün fluoreszierend. In ihrer Intensität war die DiA-Färbung jedoch so
gering ausgeprägt, dass eine Auswertung aller mit diesem Farbstoff angefärbten auditorischen
Felder nicht möglich war.
Aus allen für jedes betrachtete auditorische Subareal angefertigten Schnitten (siehe Kapitel
3.2.6) wurde zunächst fluoreszenzmikroskopisch eine Schnittserie identifiziert, in welcher in
jedem Schnitt einzeln identifizierbare angefärbte Dendriten vorlagen und die Hintergrund-
fluoreszenz eine detaillierte Auswertung erlaubte. Innerhalb dieser Serien wurden nach
randomisiertem Beginn insgesamt 5 Schnitte ausgewertet. Am jeweils ersten dieser
ausgewählten Schnitte wurde eine Färbezone ausgewählt, in welcher zwischen 15 und 40
einzeln verlaufende Dendriten innerhalb des definierten Zählrasters aufzufinden waren (Abb.
9). Unter zu Hilfenahme anatomisch markanter Punkte wie der Rundung der Cortexoberfläche
und anhand von Blutgefäßen wurde dieses Gebiet in jedem der aufeinander folgenden
Schnitte identifiziert.
Anschließend wurden nach den Literaturangaben die einzelnen Schichtausmaße festgelegt.
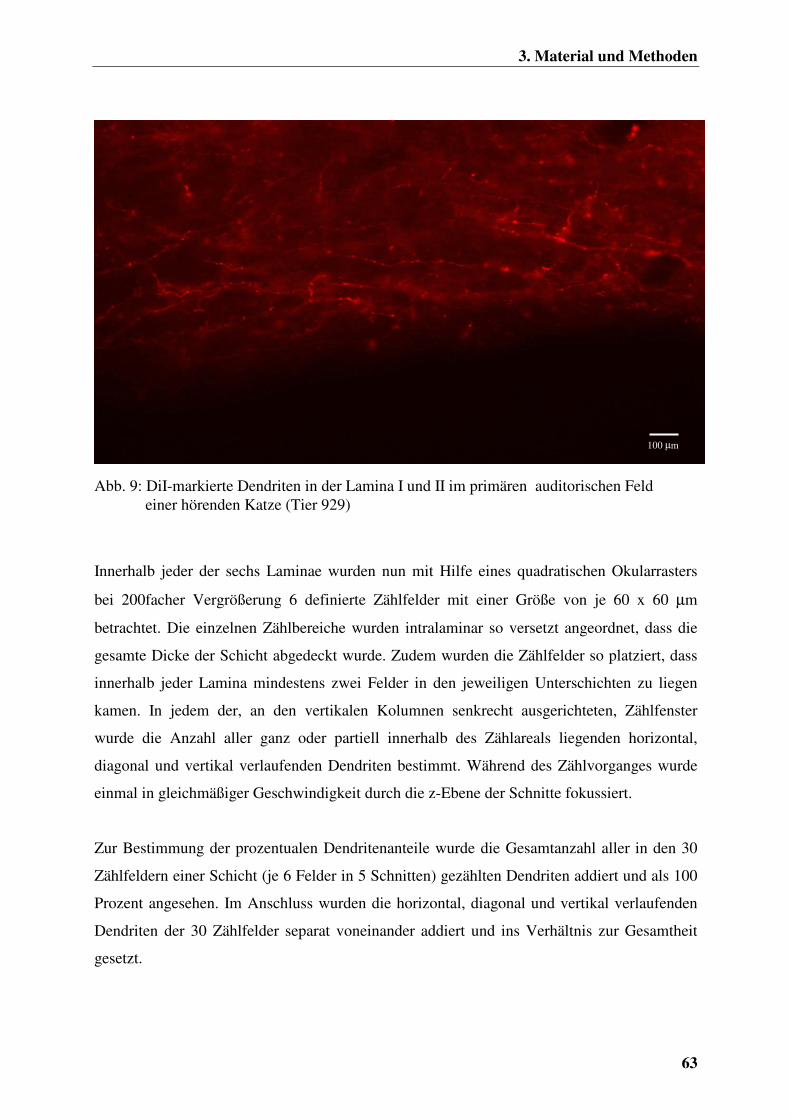
3. Material und Methoden
63
Innerhalb jeder der sechs Laminae wurden nun mit Hilfe eines quadratischen Okularrasters
bei 200facher Vergrößerung 6 definierte Zählfelder mit einer Größe von je 60 x 60 µm
betrachtet. Die einzelnen Zählbereiche wurden intralaminar so versetzt angeordnet, dass die
gesamte Dicke der Schicht abgedeckt wurde. Zudem wurden die Zählfelder so platziert, dass
innerhalb jeder Lamina mindestens zwei Felder in den jeweiligen Unterschichten zu liegen
kamen. In jedem der, an den vertikalen Kolumnen senkrecht ausgerichteten, Zählfenster
wurde die Anzahl aller ganz oder partiell innerhalb des Zählareals liegenden horizontal,
diagonal und vertikal verlaufenden Dendriten bestimmt. Während des Zählvorganges wurde
einmal in gleichmäßiger Geschwindigkeit durch die z-Ebene der Schnitte fokussiert.
Zur Bestimmung der prozentualen Dendritenanteile wurde die Gesamtanzahl aller in den 30
Zählfeldern einer Schicht (je 6 Felder in 5 Schnitten) gezählten Dendriten addiert und als 100
Prozent angesehen. Im Anschluss wurden die horizontal, diagonal und vertikal verlaufenden
Dendriten der 30 Zählfelder separat voneinander addiert und ins Verhältnis zur Gesamtheit
gesetzt.
Abb. 9: DiI-markierte Dendriten in der Lamina I und II im primären auditorischen Feld einer hörenden Katze (Tier 929)
100 µm
100 µm
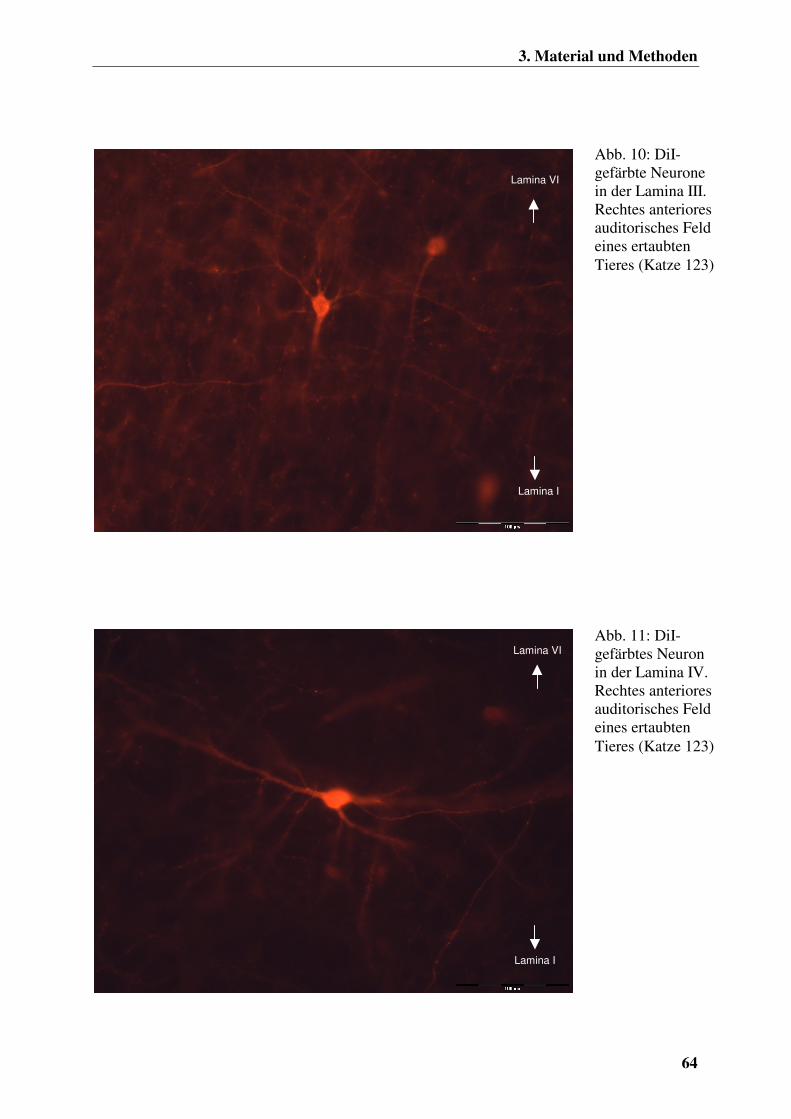
3. Material und Methoden
64
Abb. 10: DiI-gefärbte Neurone in der Lamina III. Rechtes anteriores auditorisches Feld eines ertaubten Tieres (Katze 123)
Abb. 11: DiI-gefärbtes Neuron in der Lamina IV. Rechtes anteriores auditorisches Feld eines ertaubten Tieres (Katze 123)
Lamina I
Lamina VI
Lamina VI
Lamina I
Lamina VI

3. Material und Methoden
65
3.2.9 Die statistische Analyse
Die erhobenen Daten wurden mit dem Tabellenkalkulationsprogramm Excel 2000 zur
Ermittlung der Mittelwerte, Standardabweichungen und Prozentzahlen weiterverarbeitet.
Die statistische Analyse der erhobenen Daten diente der Fragestellung, inwieweit sich eine
experimentelle neonatale Ertaubung und eine chronische elektrische intracochleäre
Stimulation mittels Cochlea-Implantat auf die laminäre Dendritenstruktur auswirkt. Dazu
wurde mittels GraphPad Prism Version 3.02 eine Prüfung auf Normalverteilung nach
Kolmogorov und Smirnov durchgeführt. Die statistische Untersuchung der hörenden und
neonatal experimentell ertaubten Versuchsgruppen erfolgte mittels one-way ANOVA mit
einem Signifikanzniveau von 95%. Anschließend wurde mittels Post Hoc Test nach
Bonferroni ein paarweiser Datenvergleich vorgenommen. Aus allen automatisch
durchgeführten Datenvergleichen wurden die relevanten Datensätze extrahiert und
interpretiert. Bei der statistischen Untersuchung der Daten der chronisch elektrisch
intracochleär stimulierten Tiere wurde aufgrund einer zu geringen Stichprobenanzahl auf den
Kruskal-Wallis-Test mit anschließendem Post Hoc Test nach Dunn ausgewichen.
Bei der statistischen Auswertung der zu lediglich zwei Gruppen zusammengefassten Werte
der diagonalen Dendritenverläufe wurde nach der Überprüfung auf Normalverteilung nach
Kolmogorov und Smirnov der t-Test mit unabhängigen Variablen angewendet.

4. Ergebnisse
66
4. ERGEBNISSE
4.1 Die Morphometrie der Dendriten der Laminae I bis VI im auditorischen Cortex
Die in die Untersuchung eingegangenen n-Zahlen ergeben sich aus der Addition der mit DiI
gefärbten rechten und linken Cortexhälften, abzüglich einzelner Hemisphären, welche auf-
grund einer zu großen Hintergrundfluoreszenz nicht ausgewertet werden konnten (siehe
Anhang 9.7). Die Zusammenlegung der Cortexhemisphären ist möglich, da bei allen drei
Versuchsgruppen kein signifikanter interhemisphärischer Unterschied besteht (nicht
dargestellt).
4.1.1 Horizontale Dendritenverläufe im AAF, AI und AII
Bei der Betrachtung des prozentualen Anteils aller horizontal verlaufenden Dendriten an den
insgesamt in den 30 Zählfeldern vorhandenen Dendriten innerhalb jeder der sechs aus-
gewerteten Laminae fällt über die Gesamtdicke der auditorischen Felder eine ausgeprägte
Systematik auf. Nach dieser horizontalen „Schichten-Hierarchie“ nimmt der prozentuale
Anteil der horizontalen Dendritenverläufe mit zunehmender Cortextiefe ab (Abb. 12, 13, 14).
Im folgenden wurde intraareal statistisch überprüft, inwieweit die Lamina I im Vergleich zu
den Laminae II bis VI dominant ist.
4.1.1.1 Hörende Versuchstiergruppe
Die prozentualen Anteile der horizontal verlaufenden Dendriten der hörenden Tiere an den
Laminae I bis VI des anterioren (AAF), primären (AI) und sekundären auditorischen Feldes
(AII) sind in der Tabelle 6 aufgeführt.
Der statistische Vergleich der Lamina I mit den Laminae II bis VI innerhalb der drei
auditorischen Felder ergibt jeweils einen signifikanten Unterschied mit p<0.001 (Abb. 12)
und beweist damit die angenommene Dominanz der Lamina I.
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=9) 42±6 27±4 19±3 18±5 17±4 16±4 AI (n=7) 50±5 23±4 21±3 18±4 20±5 24±3 AII (n=10) 52±6 24±9 25±8 18±6 16±6 17±8
Tab. 6: Prozentualer Anteil der Dendriten (horizontaler Verlauf; MW±SD)
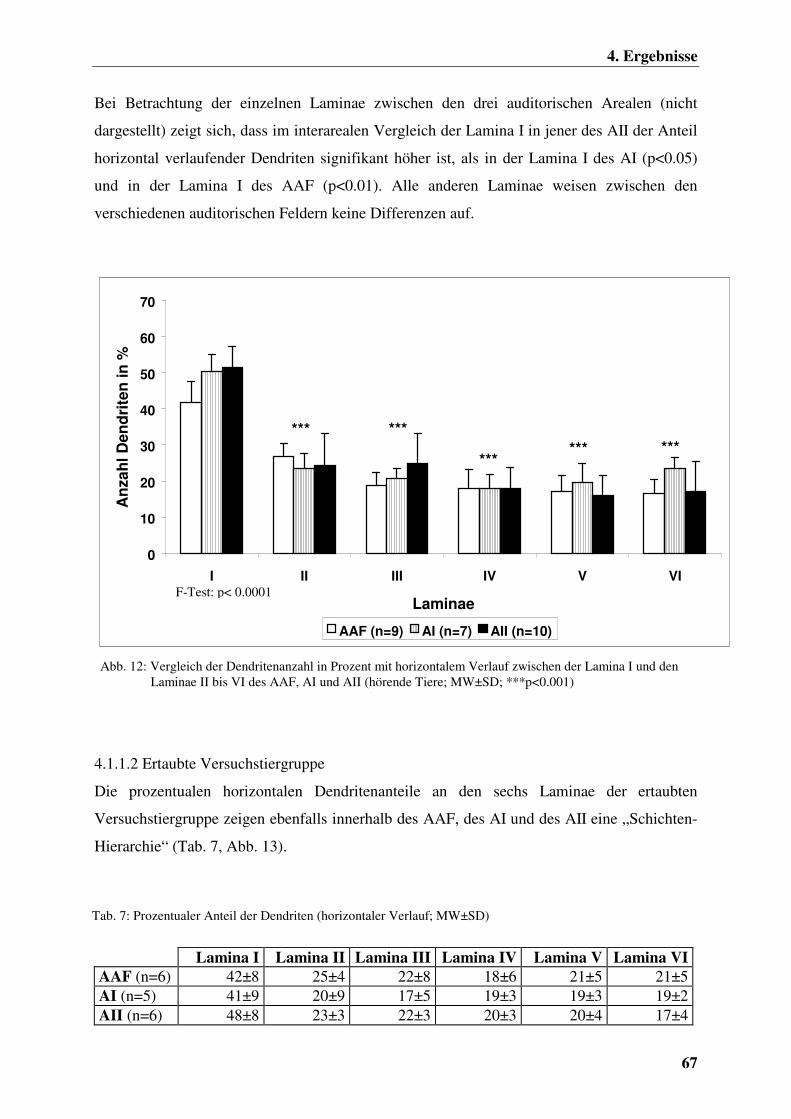
4. Ergebnisse
67
Bei Betrachtung der einzelnen Laminae zwischen den drei auditorischen Arealen (nicht
dargestellt) zeigt sich, dass im interarealen Vergleich der Lamina I in jener des AII der Anteil
horizontal verlaufender Dendriten signifikant höher ist, als in der Lamina I des AI (p<0.05)
und in der Lamina I des AAF (p<0.01). Alle anderen Laminae weisen zwischen den
verschiedenen auditorischen Feldern keine Differenzen auf.
4.1.1.2 Ertaubte Versuchstiergruppe
Die prozentualen horizontalen Dendritenanteile an den sechs Laminae der ertaubten
Versuchstiergruppe zeigen ebenfalls innerhalb des AAF, des AI und des AII eine „Schichten-
Hierarchie“ (Tab. 7, Abb. 13).
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=6) 42±8 25±4 22±8 18±6 21±5 21±5 AI (n=5) 41±9 20±9 17±5 19±3 19±3 19±2 AII (n=6) 48±8 23±3 22±3 20±3 20±4 17±4
0
10
20
30
40
50
60
70
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=9) AI (n=7) AII (n=10)
*** ***
****** ***
F-Test: p< 0.0001
Abb. 12: Vergleich der Dendritenanzahl in Prozent mit horizontalem Verlauf zwischen der Lamina I und den Laminae II bis VI des AAF, AI und AII (hörende Tiere; MW±SD; ***p<0.001)
Tab. 7: Prozentualer Anteil der Dendriten (horizontaler Verlauf; MW±SD)
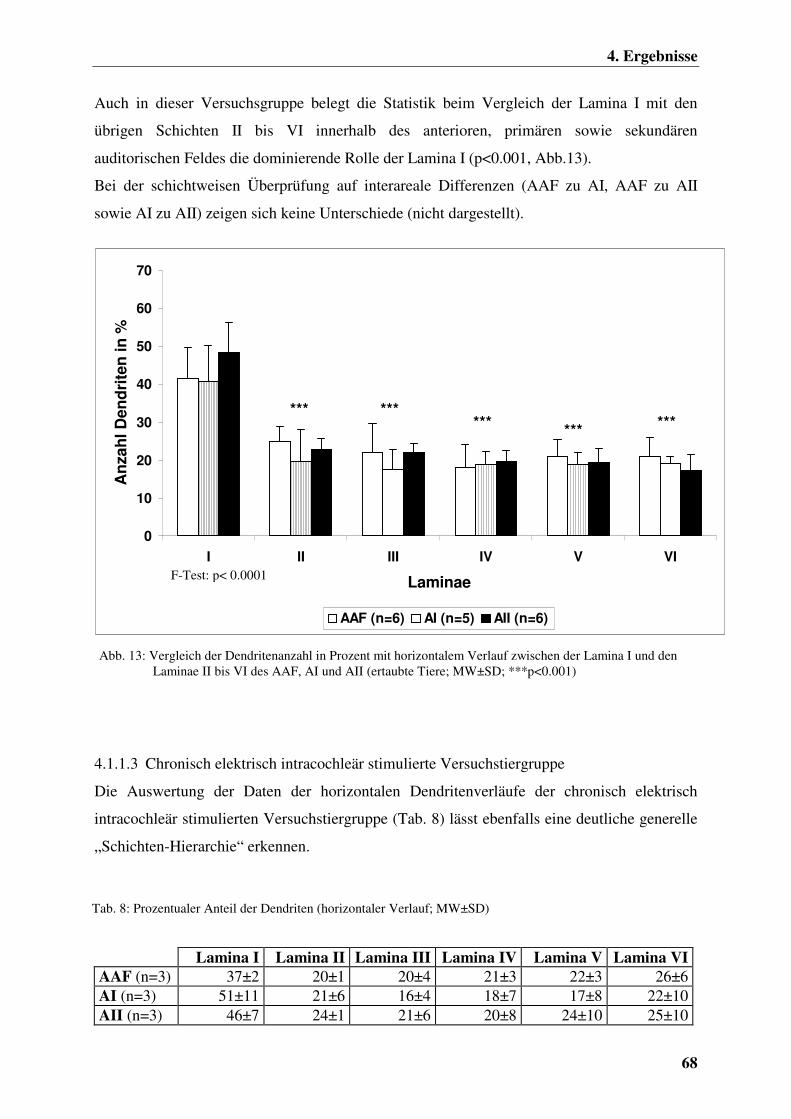
4. Ergebnisse
68
Auch in dieser Versuchsgruppe belegt die Statistik beim Vergleich der Lamina I mit den
übrigen Schichten II bis VI innerhalb des anterioren, primären sowie sekundären
auditorischen Feldes die dominierende Rolle der Lamina I (p<0.001, Abb.13).
Bei der schichtweisen Überprüfung auf interareale Differenzen (AAF zu AI, AAF zu AII
sowie AI zu AII) zeigen sich keine Unterschiede (nicht dargestellt).
4.1.1.3 Chronisch elektrisch intracochleär stimulierte Versuchstiergruppe
Die Auswertung der Daten der horizontalen Dendritenverläufe der chronisch elektrisch
intracochleär stimulierten Versuchstiergruppe (Tab. 8) lässt ebenfalls eine deutliche generelle
„Schichten-Hierarchie“ erkennen.
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=3) 37±2 20±1 20±4 21±3 22±3 26±6 AI (n=3) 51±11 21±6 16±4 18±7 17±8 22±10 AII (n=3) 46±7 24±1 21±6 20±8 24±10 25±10
0
10
20
30
40
50
60
70
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=6) AI (n=5) AII (n=6)
*** ******
******
F-Test: p< 0.0001
Abb. 13: Vergleich der Dendritenanzahl in Prozent mit horizontalem Verlauf zwischen der Lamina I und den Laminae II bis VI des AAF, AI und AII (ertaubte Tiere; MW±SD; ***p<0.001)
Tab. 8: Prozentualer Anteil der Dendriten (horizontaler Verlauf; MW±SD)
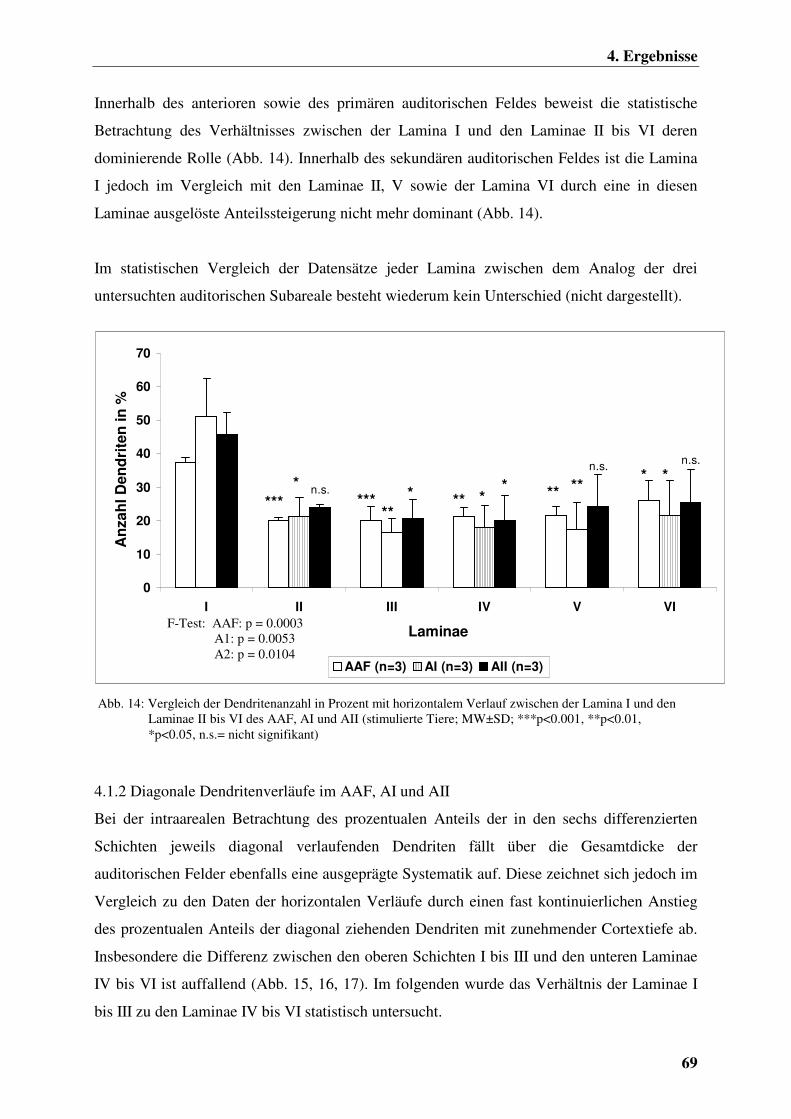
4. Ergebnisse
69
Innerhalb des anterioren sowie des primären auditorischen Feldes beweist die statistische
Betrachtung des Verhältnisses zwischen der Lamina I und den Laminae II bis VI deren
dominierende Rolle (Abb. 14). Innerhalb des sekundären auditorischen Feldes ist die Lamina
I jedoch im Vergleich mit den Laminae II, V sowie der Lamina VI durch eine in diesen
Laminae ausgelöste Anteilssteigerung nicht mehr dominant (Abb. 14).
Im statistischen Vergleich der Datensätze jeder Lamina zwischen dem Analog der drei
untersuchten auditorischen Subareale besteht wiederum kein Unterschied (nicht dargestellt).
4.1.2 Diagonale Dendritenverläufe im AAF, AI und AII
Bei der intraarealen Betrachtung des prozentualen Anteils der in den sechs differenzierten
Schichten jeweils diagonal verlaufenden Dendriten fällt über die Gesamtdicke der
auditorischen Felder ebenfalls eine ausgeprägte Systematik auf. Diese zeichnet sich jedoch im
Vergleich zu den Daten der horizontalen Verläufe durch einen fast kontinuierlichen Anstieg
des prozentualen Anteils der diagonal ziehenden Dendriten mit zunehmender Cortextiefe ab.
Insbesondere die Differenz zwischen den oberen Schichten I bis III und den unteren Laminae
IV bis VI ist auffallend (Abb. 15, 16, 17). Im folgenden wurde das Verhältnis der Laminae I
bis III zu den Laminae IV bis VI statistisch untersucht.
F-Test: AAF: p = 0.0003 A1: p = 0.0053 A2: p = 0.0104
0
10
20
30
40
50
60
70
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=3) AI (n=3) AII (n=3)
*** ****
**
*
**** **
*
**
* n.s.
n.s. n.s.
Abb. 14: Vergleich der Dendritenanzahl in Prozent mit horizontalem Verlauf zwischen der Lamina I und den Laminae II bis VI des AAF, AI und AII (stimulierte Tiere; MW±SD; ***p<0.001, **p<0.01, *p<0.05, n.s.= nicht signifikant)
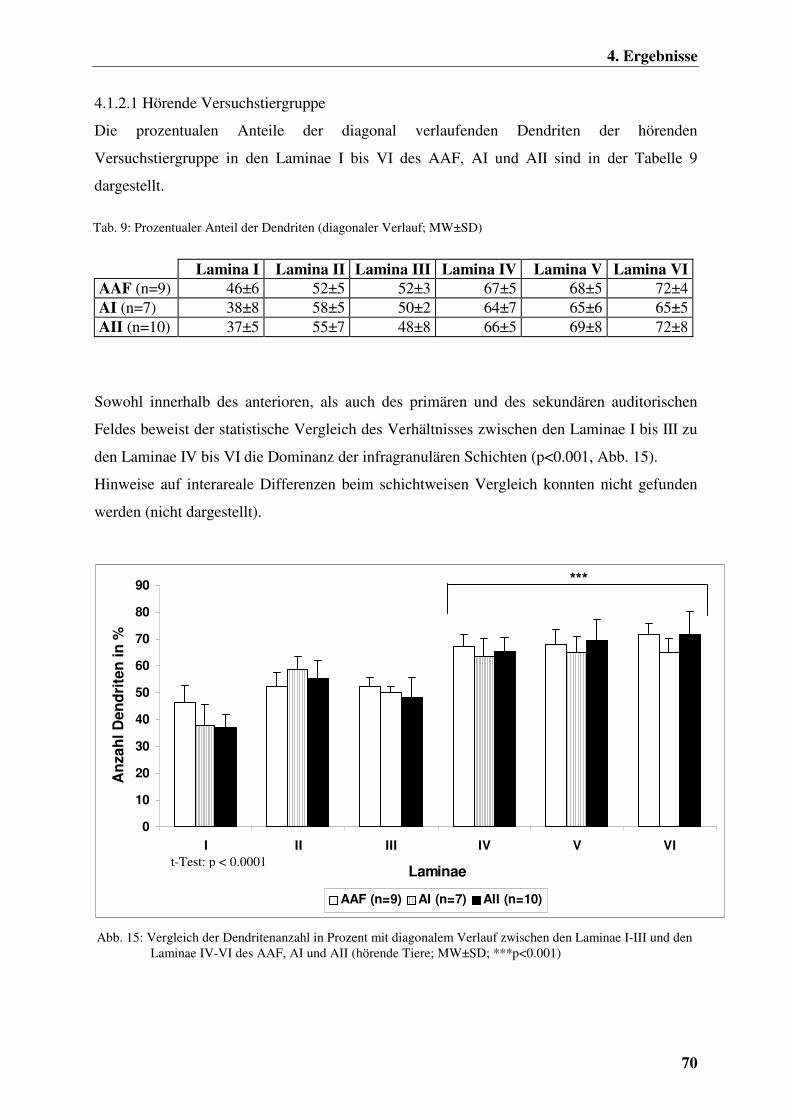
4. Ergebnisse
70
4.1.2.1 Hörende Versuchstiergruppe
Die prozentualen Anteile der diagonal verlaufenden Dendriten der hörenden
Versuchstiergruppe in den Laminae I bis VI des AAF, AI und AII sind in der Tabelle 9
dargestellt.
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=9) 46±6 52±5 52±3 67±5 68±5 72±4 AI (n=7) 38±8 58±5 50±2 64±7 65±6 65±5 AII (n=10) 37±5 55±7 48±8 66±5 69±8 72±8
Sowohl innerhalb des anterioren, als auch des primären und des sekundären auditorischen
Feldes beweist der statistische Vergleich des Verhältnisses zwischen den Laminae I bis III zu
den Laminae IV bis VI die Dominanz der infragranulären Schichten (p<0.001, Abb. 15).
Hinweise auf interareale Differenzen beim schichtweisen Vergleich konnten nicht gefunden
werden (nicht dargestellt).
0
10
20
30
40
50
60
70
80
90
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=9) AI (n=7) AII (n=10)
***
t-Test: p < 0.0001
Tab. 9: Prozentualer Anteil der Dendriten (diagonaler Verlauf; MW±SD)
Abb. 15: Vergleich der Dendritenanzahl in Prozent mit diagonalem Verlauf zwischen den Laminae I-III und den Laminae IV-VI des AAF, AI und AII (hörende Tiere; MW±SD; ***p<0.001)
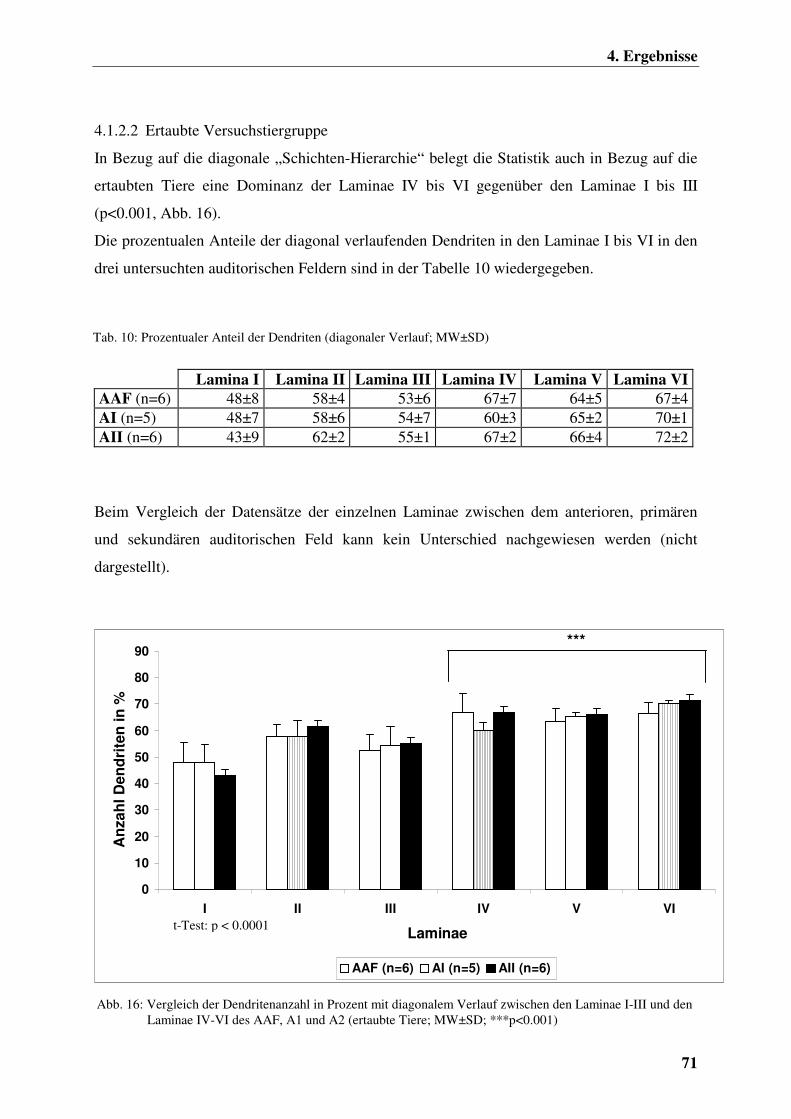
4. Ergebnisse
71
4.1.2.2 Ertaubte Versuchstiergruppe
In Bezug auf die diagonale „Schichten-Hierarchie“ belegt die Statistik auch in Bezug auf die
ertaubten Tiere eine Dominanz der Laminae IV bis VI gegenüber den Laminae I bis III
(p<0.001, Abb. 16).
Die prozentualen Anteile der diagonal verlaufenden Dendriten in den Laminae I bis VI in den
drei untersuchten auditorischen Feldern sind in der Tabelle 10 wiedergegeben.
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=6) 48±8 58±4 53±6 67±7 64±5 67±4 AI (n=5) 48±7 58±6 54±7 60±3 65±2 70±1 AII (n=6) 43±9 62±2 55±1 67±2 66±4 72±2
Beim Vergleich der Datensätze der einzelnen Laminae zwischen dem anterioren, primären
und sekundären auditorischen Feld kann kein Unterschied nachgewiesen werden (nicht
dargestellt).
0
10
20
30
40
50
60
70
80
90
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=6) AI (n=5) AII (n=6)
***
t-Test: p < 0.0001
Tab. 10: Prozentualer Anteil der Dendriten (diagonaler Verlauf; MW±SD)
Abb. 16: Vergleich der Dendritenanzahl in Prozent mit diagonalem Verlauf zwischen den Laminae I-III und den Laminae IV-VI des AAF, A1 und A2 (ertaubte Tiere; MW±SD; ***p<0.001)
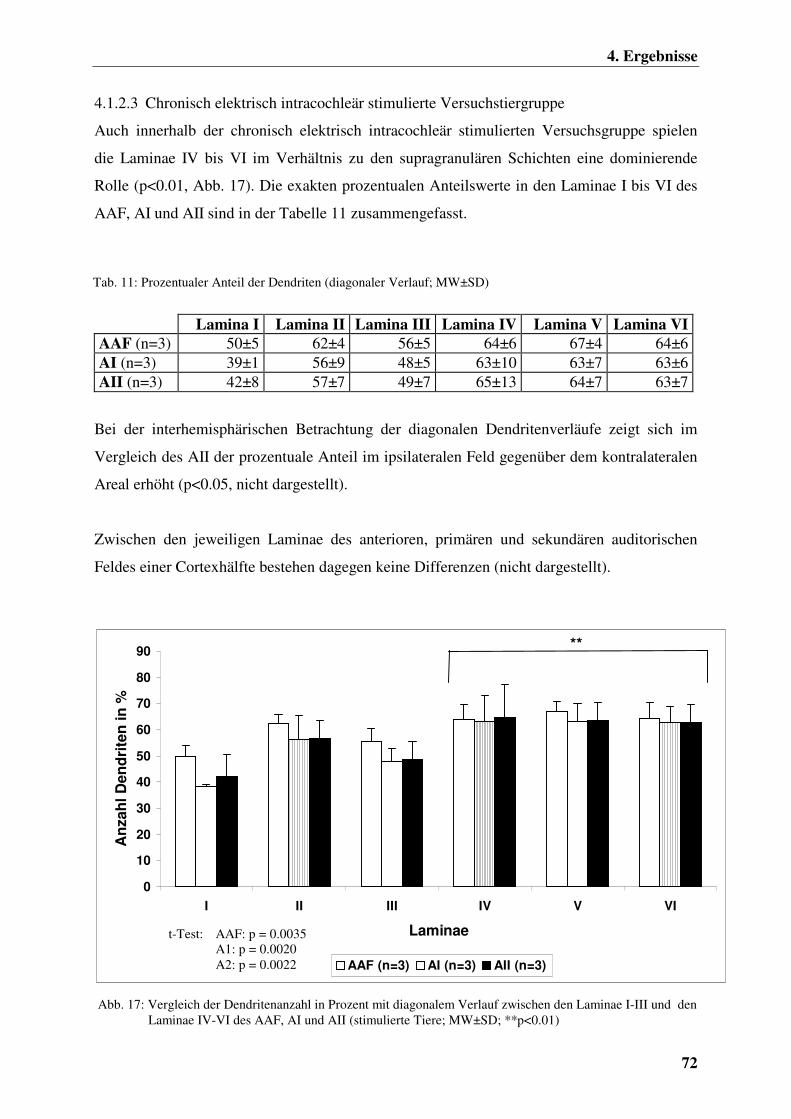
4. Ergebnisse
72
4.1.2.3 Chronisch elektrisch intracochleär stimulierte Versuchstiergruppe
Auch innerhalb der chronisch elektrisch intracochleär stimulierten Versuchsgruppe spielen
die Laminae IV bis VI im Verhältnis zu den supragranulären Schichten eine dominierende
Rolle (p<0.01, Abb. 17). Die exakten prozentualen Anteilswerte in den Laminae I bis VI des
AAF, AI und AII sind in der Tabelle 11 zusammengefasst.
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=3) 50±5 62±4 56±5 64±6 67±4 64±6 AI (n=3) 39±1 56±9 48±5 63±10 63±7 63±6 AII (n=3) 42±8 57±7 49±7 65±13 64±7 63±7
Bei der interhemisphärischen Betrachtung der diagonalen Dendritenverläufe zeigt sich im
Vergleich des AII der prozentuale Anteil im ipsilateralen Feld gegenüber dem kontralateralen
Areal erhöht (p<0.05, nicht dargestellt).
Zwischen den jeweiligen Laminae des anterioren, primären und sekundären auditorischen
Feldes einer Cortexhälfte bestehen dagegen keine Differenzen (nicht dargestellt).
0
10
20
30
40
50
60
70
80
90
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=3) AI (n=3) AII (n=3)
**
t-Test: AAF: p = 0.0035 A1: p = 0.0020 A2: p = 0.0022
Abb. 17: Vergleich der Dendritenanzahl in Prozent mit diagonalem Verlauf zwischen den Laminae I-III und den Laminae IV-VI des AAF, AI und AII (stimulierte Tiere; MW±SD; **p<0.01)
Tab. 11: Prozentualer Anteil der Dendriten (diagonaler Verlauf; MW±SD)
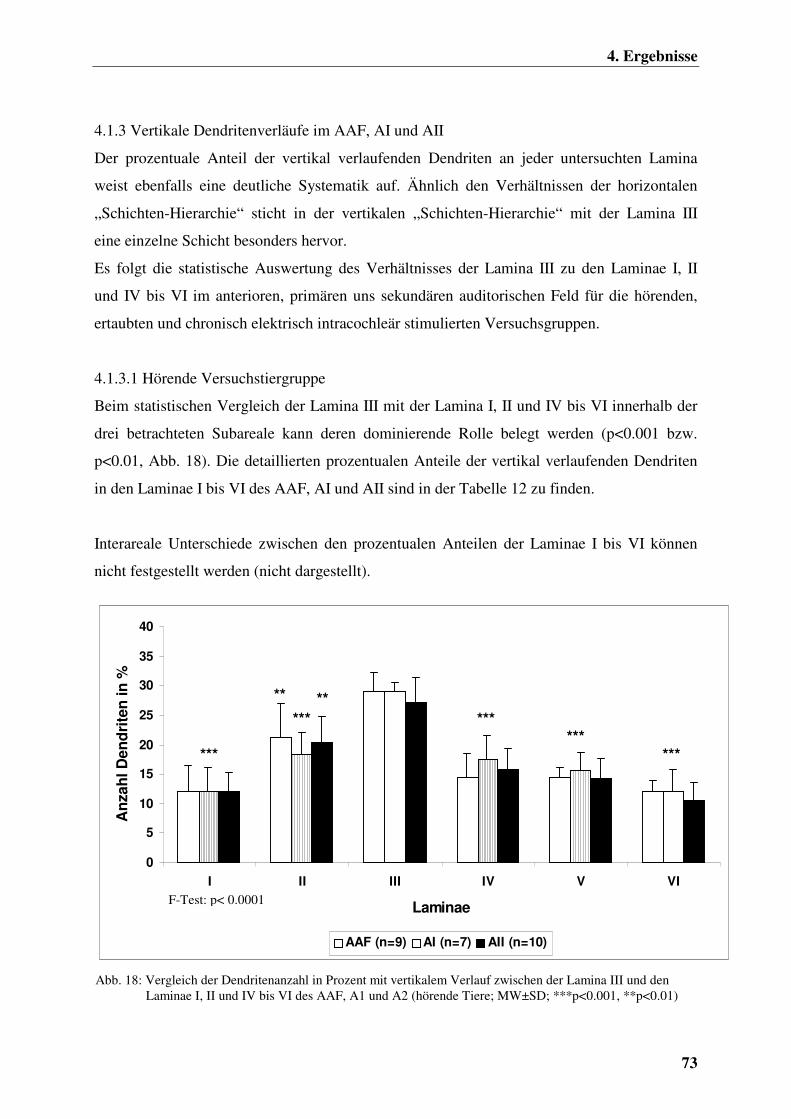
4. Ergebnisse
73
4.1.3 Vertikale Dendritenverläufe im AAF, AI und AII
Der prozentuale Anteil der vertikal verlaufenden Dendriten an jeder untersuchten Lamina
weist ebenfalls eine deutliche Systematik auf. Ähnlich den Verhältnissen der horizontalen
„Schichten-Hierarchie“ sticht in der vertikalen „Schichten-Hierarchie“ mit der Lamina III
eine einzelne Schicht besonders hervor.
Es folgt die statistische Auswertung des Verhältnisses der Lamina III zu den Laminae I, II
und IV bis VI im anterioren, primären uns sekundären auditorischen Feld für die hörenden,
ertaubten und chronisch elektrisch intracochleär stimulierten Versuchsgruppen.
4.1.3.1 Hörende Versuchstiergruppe
Beim statistischen Vergleich der Lamina III mit der Lamina I, II und IV bis VI innerhalb der
drei betrachteten Subareale kann deren dominierende Rolle belegt werden (p<0.001 bzw.
p<0.01, Abb. 18). Die detaillierten prozentualen Anteile der vertikal verlaufenden Dendriten
in den Laminae I bis VI des AAF, AI und AII sind in der Tabelle 12 zu finden.
Interareale Unterschiede zwischen den prozentualen Anteilen der Laminae I bis VI können
nicht festgestellt werden (nicht dargestellt).
0
5
10
15
20
25
30
35
40
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=9) AI (n=7) AII (n=10)
***
******
***
***
** **
F-Test: p< 0.0001
Abb. 18: Vergleich der Dendritenanzahl in Prozent mit vertikalem Verlauf zwischen der Lamina III und den Laminae I, II und IV bis VI des AAF, A1 und A2 (hörende Tiere; MW±SD; ***p<0.001, **p<0.01)
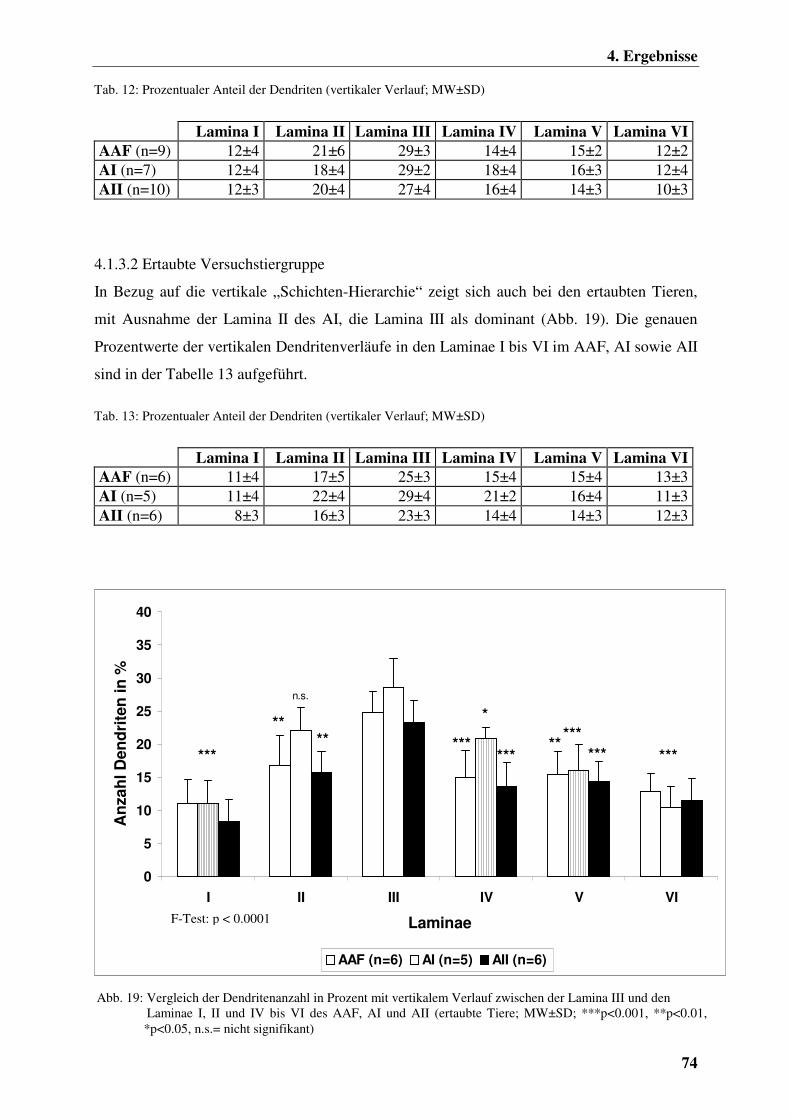
4. Ergebnisse
74
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=9) 12±4 21±6 29±3 14±4 15±2 12±2 AI (n=7) 12±4 18±4 29±2 18±4 16±3 12±4 AII (n=10) 12±3 20±4 27±4 16±4 14±3 10±3
4.1.3.2 Ertaubte Versuchstiergruppe
In Bezug auf die vertikale „Schichten-Hierarchie“ zeigt sich auch bei den ertaubten Tieren,
mit Ausnahme der Lamina II des AI, die Lamina III als dominant (Abb. 19). Die genauen
Prozentwerte der vertikalen Dendritenverläufe in den Laminae I bis VI im AAF, AI sowie AII
sind in der Tabelle 13 aufgeführt.
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=6) 11±4 17±5 25±3 15±4 15±4 13±3 AI (n=5) 11±4 22±4 29±4 21±2 16±4 11±3 AII (n=6) 8±3 16±3 23±3 14±4 14±3 12±3
0
5
10
15
20
25
30
35
40
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=6) AI (n=5) AII (n=6)
***
**
*** ***
*** ** **
*
n.s.
*** ***
F-Test: p < 0.0001
Tab. 13: Prozentualer Anteil der Dendriten (vertikaler Verlauf; MW±SD)
Abb. 19: Vergleich der Dendritenanzahl in Prozent mit vertikalem Verlauf zwischen der Lamina III und den Laminae I, II und IV bis VI des AAF, AI und AII (ertaubte Tiere; MW±SD; ***p<0.001, **p<0.01, *p<0.05, n.s.= nicht signifikant)
Tab. 12: Prozentualer Anteil der Dendriten (vertikaler Verlauf; MW±SD)
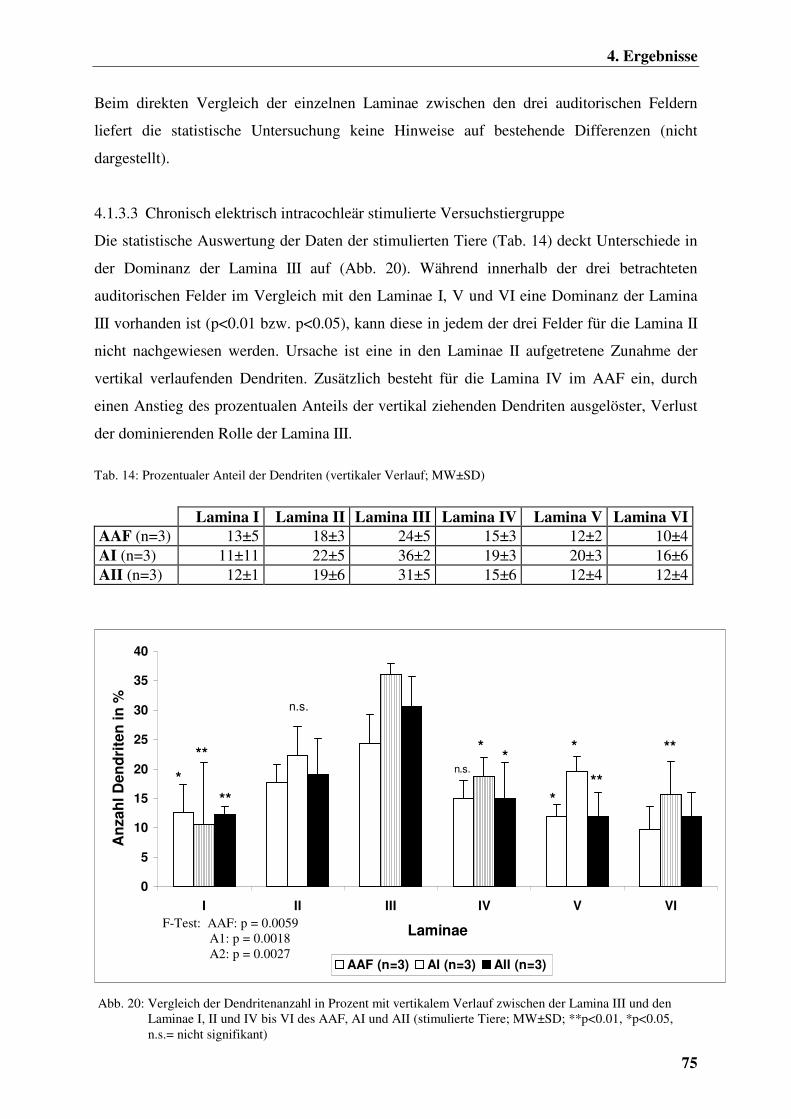
4. Ergebnisse
75
Beim direkten Vergleich der einzelnen Laminae zwischen den drei auditorischen Feldern
liefert die statistische Untersuchung keine Hinweise auf bestehende Differenzen (nicht
dargestellt).
4.1.3.3 Chronisch elektrisch intracochleär stimulierte Versuchstiergruppe
Die statistische Auswertung der Daten der stimulierten Tiere (Tab. 14) deckt Unterschiede in
der Dominanz der Lamina III auf (Abb. 20). Während innerhalb der drei betrachteten
auditorischen Felder im Vergleich mit den Laminae I, V und VI eine Dominanz der Lamina
III vorhanden ist (p<0.01 bzw. p<0.05), kann diese in jedem der drei Felder für die Lamina II
nicht nachgewiesen werden. Ursache ist eine in den Laminae II aufgetretene Zunahme der
vertikal verlaufenden Dendriten. Zusätzlich besteht für die Lamina IV im AAF ein, durch
einen Anstieg des prozentualen Anteils der vertikal ziehenden Dendriten ausgelöster, Verlust
der dominierenden Rolle der Lamina III.
Lamina I Lamina II Lamina III Lamina IV Lamina V Lamina VI AAF (n=3) 13±5 18±3 24±5 15±3 12±2 10±4 AI (n=3) 11±11 22±5 36±2 19±3 20±3 16±6 AII (n=3) 12±1 19±6 31±5 15±6 12±4 12±4
0
5
10
15
20
25
30
35
40
I II III IV V VI
Laminae
An
zah
l D
en
dri
ten
in
%
AAF (n=3) AI (n=3) AII (n=3)
* ** ** *
** **
*
*
n.s.
n.s.
*
F-Test: AAF: p = 0.0059 A1: p = 0.0018 A2: p = 0.0027
Tab. 14: Prozentualer Anteil der Dendriten (vertikaler Verlauf; MW±SD)
Abb. 20: Vergleich der Dendritenanzahl in Prozent mit vertikalem Verlauf zwischen der Lamina III und den Laminae I, II und IV bis VI des AAF, AI und AII (stimulierte Tiere; MW±SD; **p<0.01, *p<0.05, n.s.= nicht signifikant)

4. Ergebnisse
76
Interareale Unterschiede des prozentualen Anteils der vertikal verlaufenden Dendriten an den
Laminae I bis VI konnten nicht nachgewiesen werden (nicht dargestellt).
4.2 Der direkte Vergleich der Laminae zwischen der hörenden, neonatal ertaubten sowie
chronisch elektrisch intracochleär stimulierten Versuchstiergruppen im AAF, AI
und AII
Neben der Auswertung der Unterschiede zwischen den „Schichten-Hierarchien“ der drei
Versuchsgruppen mit unterschiedlichem Hörstatus wurde ein direkter statistischer Vergleich
jeder einzelnen der sechs betrachteten Laminae jedes auditorischen Feldes mit der jeweilig
analogen Schicht zwischen den drei Tiergruppen durchgeführt (nicht dargestellt).
Die Auswertung der Daten der prozentualen Anteile der horizontalen, diagonalen und
vertikalen Dendritenverläufe der einzelnen Schichten ergibt sowohl innerhalb des anterioren,
wie auch des primären und des sekundären auditorischen Feldes beim Vergleich zwischen den
hörenden und ertaubten Tieren keine signifikanten Unterschiede. Damit ist ein Einfluss der
neonatalen experimentellen Ertaubung auf die laminäre Dendritenstruktur beim direkten
Schichtvergleich nicht nachzuweisen.
Auch bei Betrachtung der horizontal, diagonal und vertikal verlaufenden Dendritendatensätze
der hörenden Versuchsgruppe im Vergleich zu den Prozentzahlen der chronisch elektrisch
intracochleär stimulierten Versuchsgruppe zeigen sich beim Vergleich der analogen Schichten
innerhalb des AAF, AI und des AII keine Einflüsse einer Stimulation mittels Cochlea-
Implantat.
Der ebenfalls durchgeführte Test auf signifikante schichtbezogene Unterschiede der
laminären prozentualen Dendritenanteile zwischen der ertaubten und chronisch elektrisch
intracochleär stimulierten Gruppen weist sowohl in Bezug auf die drei untersuchten
Verlaufsrichtungen als auch in Bezug auf die drei ausgewerteten auditorischen Areale kein
positives Ergebnis auf.
4.3 Eine Zusammenfassung der Ergebnisse
Im Hinblick auf die prozentualen Anteile der horizontal verlaufenden Dendriten über die
Gesamtdicke des anterioren, primären und sekundären auditorischen Feldes ist eine
hochsignifikant (p<0.001) von der Lamina I dominierte „Schichten-Hierarchie“ bei den
hörenden Tieren der Kontrollgruppe zu erkennen.

4. Ergebnisse
77
Die prozentualen horizontalen Dendritenanteile an den sechs Laminae der neonatal
experimentell ertaubten Versuchstiergruppe entsprechen innerhalb des AAF, des AI und des
AII in ihrer Systematik den Ergebnissen der normal hörenden Kontrolltiergruppe.
Die Auswertung der Daten der horizontalen Dendritenverläufe der chronisch elektrisch
intracochleär stimulierten Versuchstiergruppe macht hinsichtlich der „Schichten-Hierarchie“
Unterschiede zu der hörenden und neonatal ertaubten Gruppe deutlich. Innerhalb des
anterioren sowie des primären auditorischen Feldes fällt bei der statistischen Betrachtung
außer deutlich geringer ausfallenden Signifikanzniveaus insbesondere im AI kein Unterschied
in der Dominanz der Lamina I auf. Für die Verhältnisse der Lamina I zu den Laminae II, V
und VI im sekundären auditorischen Feld besteht jedoch keine signifikante dominierende
Rolle der Lamina I und offenbart damit durch die Versorgung mit einem Cochlea-Implantat
aufgetretene Veränderungen innerhalb der „Schichten-Hierarchie“.
In Bezug auf die diagonal ziehenden Dendritenanteile jeder einzelnen Lamina besteht bei der
hörenden Kontrollgruppe eine „Schichten-Hierarchie“, welche durch signifikant über die
Laminae I bis III dominierende Laminae IV bis VI gekennzeichnet ist.
Gleich den Verhältnissen zwischen den ertaubten und hörenden Versuchsgruppen in Bezug
auf die horizontale Systematik zeigt sich auch bei Betrachtung der diagonalen „Schichten-
Hierarchie“ im AAF, AI und AII kein Einfluss der neonatalen Ertaubung mittels chronischer
subcutaner Neomycinapplikation.
Im Gegensatz zum Vergleich der horizontalen “Schichten-Hierarchie“ zeigt sich bezüglich
der diagonalen Systematik keine Veränderung aufgrund einer chronischen elektrischen
intracochleären Stimulation.
Bei Betrachtung der laminaspezifischen vertikalen Prozentzahlen fällt bei den hörenden
Tieren eine „Schichten-Hierarchie“ mit einer signifikant stärker ausgeprägten Lamina III auf.
Hier zeigt sich erstmals eine Veränderung nach neonataler pharmakologischer Ertaubung.
Beim statistischen Vergleich der Lamina III mit der Lamina II im primären auditorischen Feld
der neonatal ertaubten Versuchstiere ist das Verhältnis nicht signifikant verschieden und
unterscheidet sich damit deutlich von dem Schichtenverhältnis der hörenden
Kontrolltiergruppe.
Bei der statistischen Auswertung der Daten der chronisch elektrisch intracochleär stimulierten
Versuchsgruppe im anterioren, primären und sekundären auditorischen Feld fallen die nach
der Intervention aufgetretenen Veränderungen sehr stark aus.

4. Ergebnisse
78
Während für die Laminae I, V und VI im AAF, AI und AII wie bei den vorausgegangenen
Gruppen eine Dominanz der Lamina III vorliegt, kann im Verhältnis zur Lamina II innerhalb
des anterioren sowie des primären und sekundären auditorischen Feldes keine dominierende
Lamina III nachgewiesen werden. Ebenso ist im AAF die Lamina III nicht dominant
gegenüber der Lamina IV.
Der statistische Vergleich der getrennt voneinander ausgewerteten rechten und linken
Cortexhälfte offenbarte bei den hörenden und ertaubten Versuchstieren in keinem Fall
signifikante interhemisphärische Unterschiede. Im Hinblick auf die chronisch elektrisch
stimulierten Tiere zeigte sich jedoch bei den diagonal verlaufenden Dendriten ein signifikant
gegenüber dem kontralateralen Areal erhöhter prozentualer Anteil im ipsilateralen Cortex des
sekundären auditorischen Feldes (p<0.05).
Bei der Durchführung des direkten Vergleiches zwischen den einzelnen Laminae der drei
auditorischen Felder mit ihren Analoga ist in der hörenden Kontrollgruppe ein signifikant
höherer prozentualer Anteil horizontal verlaufender Dendriten in der Lamina I des AII
gegenüber der Lamina I im AI (p<0.05) und im AAF (p<0.01) vorhanden
Des weiteren bestehen jedoch sowohl innerhalb, als auch zwischen den Tiergruppen mit
unterschiedlichen Hörstatus bei der schichtweisen Überprüfung auf interareale Differenzen
keine signifikanten Unterschiede. Damit können Einflüsse der neonatalen experimentellen
Ertaubung und der chronischen elektrischen intracochleären Stimulation auf die prozentuale
Dendritenstruktur bei einem direkten Schichtenvergleich zwischen den Versuchsgruppen
nicht nachgewiesen werden.

4. Ergebnisse
79
Lamina AAF AI AII AAF AI AII AAF AI AII
Experimentell ertaubte Tiere Chronisch stimulierte Tiere
Tab. 15: Zusammenfassung der prozentualen Dendritenverläufe in den auditorischen Subarealen der drei Versuchstiergruppen (MW)
I
II
III
IV
V
VI
horizontal
diagonal
vertikal
Hörende Tiere

5. Diskussion
80
5. DISKUSSION
5.1 Material und Methoden
Zur Untersuchung von Einflüssen einer prälingualen Ertaubung und einer chronischen
elektrischen intracochleären Stimulation mittels Cochlea-Implantat besteht in der
Medizinischen Hochschule Hannover ein seit langem etabliertes Tiermodell. Als Versuchstier
wurde die Katze gewählt, da die auditorische Hörbahn dieser Spezies bereits am 30.
Lebenstag ausgereift ist (KELLER 1997). Zudem ähneln sich neben der Funktion und
Entwicklung (FULLERTON et al. 1987) auch die Morphologie der auditorischen Systeme
von Mensch und Katze sehr stark (ONG und GAREY 1990). Dies ist als Vorraussetzung für
die Übertragbarkeit der Ergebnisse zwischen den beiden Arten von Vorteil. Weiterhin handelt
es sich bei dem felinen auditorischen Cortex um einen der bestuntersuchten auditorischen
Bereiche unter den Säugetieren (CLAREY et al. 1991).
Alle aus der Katzenzucht der Medizinischen Hochschule Hannover stammenden Versuchs-
tiere waren gesund und optimal ernährt. Einflüsse auf die Neuronenpopulation durch
Fehlernährung und sonstige, z. B. neurologische Erkrankungen, waren auszuschließen. Um
die prälinguale Taubheit von Kindern zu simulieren, wurde bei sieben Katzen die normale,
auf akustische Reize angewiesene Entwicklung des auditorischen Systems ab dem ersten
Lebenstag durch die chronische systemische Gabe des ototoxischen Aminoglycosid-
Antibiotikums Neomycin verhindert.
Die Ertaubung mittels Neomycin führt durch eine irreversible Blockade von extra- und
intrazellulären Rezeptoren zu einer selektiven Zerstörung der cochleären Haarzellen
(ZENNER und SCHACHT 1986) und lässt die Mehrheit der für die Weiterleitung von
elektrischen Signalen essentiellen Spiralganglienzellen primär intakt (LUSTIG et al. 1994),
obgleich sie mit zunehmender Ertaubungszeit einer langsamen sekundären Degeneration
unterlaufen (LEAKE und HRADEK 1988). LEAKE et al. (1999) fanden nach
pharmakologischer Ertaubung eine Reduktion der Spiralganglienzellen innerhalb von 6
Monaten um 50 % und nach drei Jahren um 90 %.
Für eine adäquate Aktivierung des auditorischen Cortex durch eine elektrische
Innenohrprothese ist diese Restzahl jedoch ausreichend (REUTER et al. 2002a).

5. Diskussion
81
Arbeiten von POPELAR et al. (1995) an mit einem Cochlea-Implantat versorgten, nicht
experimentell ertaubten Katzen haben zudem gezeigt, dass es durch ein Cochlea-Implantat zu
einer tonotopen Aktivierung des auditorischen Cortex kommt und die zeitlichen
Eigenschaften der aktivierten Neurone denen akustisch stimulierter Zellen gleichen.
Die erfolgreiche Ertaubung der Versuchstiere wurde durch ein Fehlen neuronaler Antworten
während der Messung der frühen akustisch evozierten Potentiale (FAEP) bei Darbietung des
akustischen Click-Reizes mit einer technisch maximal möglichen Lautstärke von 100 dB SPL
definiert (siehe Anhang 9.3, Abb.22). Die hörenden Kontrolltiere wiesen bereits bei einer
Lautstärke von 30 dB SPL deutliche FAEPs auf (siehe Anhang 9.3, Abb. 21).
Damit im Versuchsaufbau die Simulation einer chronischen elektrischen intracochleären
Stimulation den Verhältnissen am Menschen möglichst gleicht, wurden den Versuchstieren
aus der Humanmedizin stammende, lediglich in der Länge der Platindrähte angepasste,
Innenohrelektroden implantiert. Die ordnungsgemäße Funktion des Cochlea-Implantats wurde
durch eine wöchentlich erfolgte Messung der elektrisch evozierten Hirnstammpotentiale
überprüft.
Zum Zeitpunkt der Materialentnahme für diese Untersuchung wiesen alle Tiere ein
Lebensalter von mindestens 23 Wochen auf und verfügten somit über ein vollständig
ausgereiftes auditorisches System (KELLER 1997).
Mit der Anfertigung von Vibratomschnitten und der Kresyl-Violett-Färbung wurde mit
etablierten histologischen Methoden gearbeitet.
Im Zusammenhang mit der DiI-Tracertechnik gewährleisten Vibratomschnitte von nativem
Material gegenüber Kryostatschnitten von tiefgefrorenen Strukturen eine bessere Gewebe-
und Färbequalität (KÖBBERT et al. 2000; LUKAS et al. 1998).Trotz der hier durchgeführten
Einbettung des nativen Cortexmaterials in Agar agar konnte die geringe strukturelle
Eigenstabilität des Gewebes nicht ausreichend verstärkt werden, so dass die histologische
Schnittdicke, um einen Verlust einzelner Schnitte zu verhindern, auf 70 µm festgelegt werden
musste.
Bei der histologischen Aufarbeitung des Untersuchungsmaterials mit Hilfe der DiI-
Tracingtechnik wurde eine seit vielen Jahren erprobte Methode durchgeführt. Diese
ermöglicht eine postmortale Anfärbung von Neuronen und damit die Untersuchung von
bereits vorfixiertem Cortexmaterial.

5. Diskussion
82
Bei der Applikation von DiI-Kristallen kommt es zur Anfärbung aller in direktem Kontakt
zum Kristall stehenden Neuronenausläufer. Eine unspezifische Färbung weiterer Fasern ist
aufgrund der ausgeprägten Hydrophobie des Farbstoffs und der damit praktisch nicht
stattfindenden Verbreitung im Interzellularraum nicht zu erwarten.
DiI tritt jedoch aus gefärbten Neuronen heraus, wenn diese beim Schneidevorgang zerstört
worden sind. Für retrograd angefärbte Dendriten und bei ausreichend dicken Gewebeschnitten
stellt sich dieses Problem jedoch äußerst gering dar (HONIG und HUME 1989).
Ein weiterer Vorteil dieser Methode besteht in der Stabilität der Färbung. Da in fixiertem
Gewebe keine Stoffwechselaktivitäten wie Endo- oder Exozytose stattfinden, bleibt eine
Membrananfärbung wie sie durch DiI-Kristallapplikation geschieht, insbesondere bei
Aufbewahrung im Kühlschrank, auch über längere Lagerungszeiten bestehen (HONIG 1993).
KÖBBERT et al. (2000) fanden eine für über ein Jahr bestehenbleibende Färbung und eine
nur sehr gering ausfallende Verblassung der Fluoreszenzintensität über die Zeit sowie nach
durchgeführter Illumination.
Die Auflösung feiner Prozesse im fixierten Gewebe durch Carbocyanine wie dem DiI ist
gleich der in lebendem Gewebe und besser als durch Golgi-Methoden und anderen
intraaxonal transportierten Tracer-Substanzen (GODEMENT et al. 1987). Bei höherer
Vergrößerung können, an komplett mit DiI angefüllten Neuronen, alle morphologischen
Details dargestellt werden (GODEMENT et al. 1987; KÖBBERT et al. 2000).
Für die komplette Anfärbung von Neuronen inklusive der Somata ist jedoch die Zeitspanne
zwischen Todeszeitpunkt bzw. dem Beginn der Fixation und der Farbstoffapplikation
ausschlaggebend (KÖBBERT et al. 2000; LUKAS et al. 1998). Diese ist möglichst gering zu
halten. Die Auswertung des hier vorliegenden Materials sowie die Ergebnisse von WURTH
(1999) deuten darauf hin, dass für die Untersuchung des felinen auditorischen Cortex eine
postmortale Fixationszeit vor Applikation des DiI-Kristalls von zwei Tagen optimal ist.
Da diese Zeitspanne bei der Mehrzahl der für die hier durchgeführte Untersuchung
vorliegenden Gewebe weit überschritten wurde, war eine detaillierte Auswertung der
Dendritenmorphologie einzelner selektiv angefärbter Neurone aufgrund fehlender oder
unvollständiger Anfärbung der Zellen nicht möglich.
Bei der Auswertung der für diese Studie vorliegenden Gewebe wurde deshalb die Gewichtung
auf die deskriptive Darstellung und prozentuale Bestimmung der Verlaufsrichtung der
Dendriten in den Laminae I bis VI des anterioren, primären und sekundären auditorischen
Feldes gelegt.

5. Diskussion
83
Hierzu wurden die durch die Markierung mit DiI sichtbaren Dendriten nach ihrem Verlauf
relativ zur Cortexoberfläche der Gruppe „horizontal“ (parallel), „diagonal“ (schräg) oder
„vertikal“ (senkrecht) verlaufend zugeteilt. Dabei wurde für die horizontal verlaufenden
Dendriten eine Bedeutung bei der intralaminaren Verbindung sowie für die Verknüpfung
unterschiedlicher auditorischer Areale postuliert. Der senkrechte Verlauf eines Dendriten
weist auf seine Rolle in der Verbreitung von Informationen zwischen unterschiedlichen
Schichten innerhalb eines auditorischen Feldes hin. Die diagonalen Dendritenverläufe
nehmen eine Zwischenstellung ein. Ihnen steht wahrscheinlich eine Bedeutung bei der lokalen
Quervernetzung von Neuronen zu.
Die Bestimmung der zur eigentlichen Datenerhebung benötigten Anzahl histologischer
Schnitte und Zählfelder pro Schnitt wurde in Anlehnung an die hierarchische Proben-Strategie
für die stereologische Auswertung nach GUPTA et al. (1993) vorgenommen. In einem
solchen hierarchischen System werden Probenvarianzen und Messfehler in die biologischen
Variationen zwischen Individuen eingeblendet (GRIFFITH 1993).
Um zwischen den Unterschichten jeder Lamina auftretende Unregelmäßigkeiten zu
eliminieren, wurden die Zählfelder so platziert, dass innerhalb jeder Lamina mindestens zwei
Felder in den jeweiligen Unterschichten zu liegen kamen. Weiterhin wurde, um Zählfehler im
Sinne von Doppelzählungen zu verhindern, während der Zählung einmal in gleichmäßiger
Geschwindigkeit durch die z-Ebene der Schnitte fokussiert und dabei alle Dendriten gezählt,
welche ganz oder partiell innerhalb des Zählareals lagen (SCHNEIDER 1978).
Obwohl bei der Auswahl der in jedem Schnitt ausgewerteten Fläche mit den Kriterien von
einer minimalen bzw. maximalen Anzahl von 15 bis 40 einzeln identifizierbaren Dendriten
Schwankungen in der Anzahl der ausgezählten Dendriten minimiert wurden, konnte die
Auswertung aufgrund der Färbemethode nicht auf die absolut gezählte Dendritenanzahl
bezogen werden. Dies liegt zum einen in der Applikation von Farbstoffkristallen und zum
anderen in der Individualität der Untersuchungsgewebe begründet. In Hinblick auf die
Kristallapplikation kam es trotz Vorselektion der Farbstoffkristalle zu geringen Größen-
schwankungen. Da die Anzahl der angefärbten Dendriten jedoch mit dem Ausmaß des
Kristalls positiv korrelieren dürfte, könnte dieses unter Umständen zu falsch hohen oder
falsch geringen Dendritenzählungen führen.

5. Diskussion
84
In Bezug auf die Individualität der Gewebe spielt vor allem die unterschiedlich lange
Verweildauer im Fixans vor der Anfärbung eine herausragende Rolle. Im Verlauf der Studie
konnte festgestellt werden, dass eine lange Vorfärbefixationszeit positiv mit einer hohen
Hintergrundfluoreszenz korreliert. Dadurch musste bei außerordentlich lange fixierten
Präparaten die Auszählung in Regionen mit relativ stärkerer Dendritenanfärbung und damit
einhergehenden höheren Dendritenanzahlen verlegt werden.
Dieses Problem tritt sowohl beim Vergleich unterschiedlicher Felder eines Cortex, als auch
beim Vergleich der einzelnen Individuen auf. Um diesen methodischen Fehler zu eliminieren,
wurde daher für alle Betrachtungen lediglich das prozentuale Verhältnis der horizontal,
diagonal und vertikal verlaufenden Dendriten zueinander verwendet.
Neben der Ausarbeitung und Auswertung der „Schichten-Hierarchie“ für die Daten der
horizontal, diagonal und vertikal verlaufenden Dendriten innerhalb des anterioren, primären
und sekundären auditorischen Feldes für jede Versuchsgruppe und dem Vergleich dieser
Systematiken zwischen den Versuchsgruppen wurden ebenfalls die einzelnen analogen
Schichten der drei untersuchten auditorischen Areale innerhalb der Versuchstiergruppen
untersucht (siehe Kapitel 4.1). Im zweiten Schritt wurde ein direkter Vergleich jeder
einzelnen der sechs corticalen Schichten des AAF, AI und AII mit der analogen Schicht der
anderen Versuchsgruppen verglichen (siehe Kapitel 4.2).
Der Vorteil der letztlich durchgeführten Methode liegt, neben der Aufarbeitung von bereits
vorhandenem, fixiertem Gewebe, vor allem in der Berücksichtigung aller mit dem Farbstoff
in Berührung stehenden und damit angefärbten Dendriten und gibt damit einem Einblick über
die Gesamtheit der vorhandenen Zellausläufer. Dies steht im Gegensatz zu anderen
Methodiken mit intrazellular angefärbten Neuronen, bei welchen automatisch zumindest eine
Selektion in Hinblick auf die Vollständigkeit der intrazellularen Färbung erfolgt.
Die viel zu langen postmortalen Fixierungszeiten der verwendeten Gewebe vor der DiI-
Applikation führten jedoch zu einer begrenzten Aussagekraft der Dendritenstruktur, da eine
Zuordnung der angefärbten Dendriten zu den Zellsomata nicht möglich war. Aus diesem
Grund war es nicht möglich eine Differenzierung von intralaminaren, intraarealen sowie
intracorticalen Ausläufern im Gegensatz zu in anderen Schichten, auditorischen Feldern oder
subcorticalen Strukturen lokalisierten Zellen vorzunehmen. Weiterhin konnte eine 3-
dimensionale Darstellung der Dendritenbäume einzelner Neurone mittels Confokalem-Laser-
Scanning Mikroskop nicht wie beabsichtigt realisiert werden.

5. Diskussion
85
Zudem ist bei der Verwendung von bereits fixiertem Gewebe eine Überprüfung der Funktion
einzelner Zellen nicht mehr möglich. Eine Aussage über die Funktionalität der angefärbten
Dendriten war aufgrund fehlender elektrophysiologischer Daten wie z.B. von Einzelzell-
ableitungen daher ebenfalls nicht möglich.
5.2 Die Morphometrie der Dendriten der Laminae I bis VI im AAF, AI und AII normal
hörender Katzen
Mit dieser Studie wurde untersucht, inwieweit auf der höchsten Ebene der auditorischen
Informationsverarbeitung, dem auditorischen Cortex, durch eine neonatale Ertaubung sowie
eine anschließende chronische elektrische intracochleäre Stimulation eine Plastizität der
laminären Dendriten hervorgerufen wird. Grundlage für eine Veränderung wären die bereits
nachgewiesenen Veränderungen innerhalb der zentralen auditorischen Hörbahn (HEID et al.
1998; SNYDER et al. 1990; TERAYAMA et al. 1977; VOGT et al. 1997), welche sich
aufgrund der neuronalen Verschaltungen in die cerebralen auditorischen Subareale fortsetzen
könnten. Bei den wichtigsten, von einer qualitativen und/oder quantitativen Veränderung
betroffenen afferenten Bahnen, handelt es sich um die Afferenzen aus den thalamischen
Kernen der zentralen auditorischen Bahn, welche an den pyramidalen Neuronen der Laminae
I und III bis VI enden (MITANI et al. 1985). Dieser direkte thalamische Einfluss wird durch
die extensiven intracorticalen Verbindungsstrukturen der lokal vernetzten Pyramidenzellen
auf die übrigen Laminae übertragen (siehe Kapitel 2.4.2.2). Durch die zusätzlich vorhandenen
interarealen (commissuralen) und intercorticalen Verbindungen des auditorischen Cortex
(siehe Kapitel 2.4.2.3), welche vor allem in den Laminae II und III enden, besteht ein
zusätzlicher Informationsaustausch zwischen den verschiedenen Subarealen einer sowie der
rechten und linken Cortexhemisphäre.
Die Untersuchung einer von diesen Bahnen auf den auditorischen Cortex ausgebreiteten
Plastizität wurde durch die Bestimmung der prozentualen Anteile der, relativ zur Cortex-
oberfläche betrachtet, horizontal, diagonal und vertikal verlaufenden Dendriten in den
Laminae I bis VI der auditorischen Felder durchgeführt.
Bei der Betrachtung des prozentualen Anteils der in die drei Verlaufsrichtungen horizontal,
diagonal oder vertikal verlaufend eingeteilten Dendriten innerhalb jeder der sechs
ausgewerteten Laminae fällt über die Gesamtdicke der auditorischen Felder eine ausgeprägte
Systematik („Schichten-Hierarchie“) auf.

5. Diskussion
86
In Bezug auf die horizontal ziehenden Dendriten nimmt der prozentuale Anteil bei einer
überaus prominenten Lamina I mit zunehmender Cortextiefe ab. Die deskriptiven
Auffälligkeiten in der Verteilung der horizontal verlaufenden Dendriten über die sechs
corticalen Schichten des anterioren, primären und sekundären auditorischen Feldes werden
durch die statistische Auswertung bestätigt. Danach bestehen zwischen der Lamina I und den
übrigen Laminae statistisch hochsignifikante (p<0.001) Unterschiede (siehe Kapitel 4.1.1.1,
Abb. 12).
Bestätigt wird die Dominanz der Lamina I durch vorangegangene Studien (ROSE 1949),
welche gezeigt haben, dass sich die in der Lamina I lokalisierten Zellen mit ihren meist
parallel zur Cortexoberfläche verlaufenden Dendriten an der Bildung des intralaminaren
feinfaserigen tangentiellen Flechtwerks (Stria laminaris) beteiligen. Zudem verlaufen die
Dendriten der auffallendsten Neurone der Lamina I zumeist parallel zur Cortexoberfläche.
Bei der intraarealen Betrachtung des prozentualen Anteils der in den sechs differenzierten
Schichten jeweils diagonal verlaufenden Dendriten zeichnet sich eine von der horizonatalen
Systematik deutlich unterscheidende „Schichten-Hierarchie“ ab. Diese ist durch einen fast
kontinuierlichen Anstieg des prozentualen Anteils der diagonal ziehenden Dendriten mit
zunehmender Cortextiefe charakterisiert. Insbesondere die Differenz zwischen den oberen
Schichten I bis III und den unteren Laminae IV bis VI ist auffallend (siehe Kapitel 4.1.2.1,
Abb. 15). Nach dieser diagonalen „Schichten-Hierarchie“ wurde das Verhältnis der Laminae I
bis III zu den Laminae IV bis VI auf einen statistisch signifikanten Unterschied überprüft und
eine hochsignifikante (p<0.001) Dominanz der infragranulären Schichten nachgewiesen.
Deskriptiv ist in allen ausgewerteten auditorischen Feldern eine zwischen den Laminae
unterschiedliche Morphologie der diagonal verlaufenden Dendriten auffällig. Während in den
Laminae I bis III die diagonalen Dendriten kurze Strecken zurücklegen und damit
höchstwahrscheinlich Interaktionen zwischen relativ dicht benachbarten Neuronen bewirken,
weisen die diagonal verlaufenden Dendriten der Laminae IV bis VI deutlich längere Verläufe
auf und ziehen vornehmlich in die weiße Substanz. Eventuell stellen letztere Strukturen
Verbindungen zu den subcorticalen Schaltstellen der lemniscalen und extralemniscalen
Hörbahn dar.

5. Diskussion
87
Auch in Bezug auf den prozentualen Anteil der vertikal verlaufenden Dendriten innerhalb
jeder Lamina ist über alle sechs Schichten eine deutliche Systematik zu erkennen. Ähnlich
den Verhältnissen der horizontalen „Schichten-Hierarchie“ sticht in der vertikalen Systematik
mit der Lamina III eine einzelne Lamina hervor. Die herausragende Rolle der Lamina III in
der deskriptiven Betrachtung konnte durch die vergleichende statistische Auswertung als
hochsignifikante (p<0.001) Dominanz der Lamina III gegenüber den übrigen Schichten
bestätigt werden (siehe Kapitel 4.1.3.1, Abb. 18).
Da in der Lamina III die überwiegende Anzahl der commissuralen Verbindungen ankommen
(CODE und WINER 1985), stellt dies unter Umständen ein Zeichen für die weitere
intraareale Verteilung der commissuralen Eingänge auf die übrigen Laminae dar. Zudem
erhält die Lamina IV Afferenzen aus dem ventralen MGB (LEVAY und GILBERT 1976) und
projiziert direkt in die Lamina II und III (NIIMI und NAITO 1974), was eine hohe Anzahl
von vertikal verlaufenden Dendriten nötig erscheinen lässt. Weiterhin sind in der Lamina IIb
die unter anderem stark vertikal in die unteren Schichten projizierenden Pyramidenzellen
angesiedelt (WINGUTH und WINER 1986). In der Lamina III sind damit intralaminare und
von den Laminae II und IV ausgesandte vertikale Dendriten anzutreffen und führen somit zu
der herausragenden vertikalen Dendritenanzahl.
Ein signifikanter Unterschied zwischen der rechten und linken Cortexhemisphäre bei den
hörenden Versuchstieren konnte in keinem Fall dargestellt werden und ist nach den
Ergebnissen bisheriger Studien auch nicht zu erwarten (WENKE 1999; WURTH 1999).
Unterschiede zwischen den analogen Laminae des AAF, AI und AII in Bezug auf die
prozentuale Dendritenstruktur konnten hier, mit Ausnahme der Lamina I, ebenfalls nicht
dargestellt werden.
In vorhergegangenen histologischen Studien wurde zwischen dem anterioren, primären und
sekundärem auditorischen Feld eine unterschiedliche Anzahl von Pyramidenzellen in der
Lamina III beobachtet (WENKE 1999). Da in der vorliegenden Arbeit jedoch nicht die
absolute Zahl der Dendriten, sondern lediglich das prozentuale Verhältnis der von den
vorhandenen Neuronen ausgehenden, angefärbten Dendriten zueinander beurteilt wurde,
stehen sich diese Erkenntnisse nicht widersprüchlich gegenüber. Vielmehr macht dies
deutlich, dass auch in den auditorischen Feldern mit geringerer Nervenzellzahl als dem AI,
dem AAF und AII, die prinzipielle Dendritenstruktur erhalten ist.

5. Diskussion
88
5.3 Die Auswirkungen einer experimentell induzierten neonatalen akustischen
Deprivation auf die Morphometrie der Dendriten der Laminae I bis VI im AAF, AI
und AII der Katze
Verschiedene Arbeiten deuten für den auditorischen Cortex des Menschen und der Katze, wie
auch für andere sensorische Bereich des Cortex beschrieben, auf eine kritische
Entwicklungsperiode hin (KRAL et al. 2001; KRAL et al. 2002; REBILLARD et al. 1980;
SKUSE 1993). Eine frühe auditorische Deprivation führt auf verschiedenen Stufen der
zentralen Hörbahn zu der Induktion von Atrophien (HULTCRANTZ et al. 1991; LUSTIG et
al. 1994; TIERNEY et al. 1997; VOGT et al. 1997) und einer Plastizität des sich
entwickelnden auditorischen Systems (MARIANOWSKI et al. 2000; SUNEJA et al. 1998). Je
früher die Störung der normalen akustischen Entwicklung stattfindet, desto größer ist der
negative Effekt auf die Entwicklung des auditorischen Systems und damit einhergehend
deutlicher die strukturellen und funktionellen Veränderungen (MOORE 1985).
Mit dieser Studie konnte der Einfluss einer neonatalen experimentellen Ertaubung mittels
chronischer subcutaner Applikation eines ototoxischen Aminoglycosid-Antibiotikums auf die
„Schichten-Hierarchie“ der vertikal verlaufenden Dendriten des primären auditorischen
Feldes nachgewiesen werden. Eine laminaspezifische Veränderung der einzelnen
prozentualen Anteile der Dendritenverlaufsrichtungen aufgrund der pharmakologischen
Ertaubung mittels Neomycin konnte beim Vergleich mit den anderen Versuchsgruppen
hingegen nicht nachgewiesen werden.
Die prozentualen horizontalen Dendritenanteile an den Laminae I bis VI der ertaubten
Versuchstiergruppe entsprechen innerhalb des anterioren, primären und sekundären
auditorischen Feldes in ihrer Systematik den Ergebnissen der hörenden Versuchstiergruppe.
Die Übereinstimmung der dominanten Lamina I bezüglich der „Schichten-Hierarchie“ zeigt
sich auch bei der vergleichenden statistischen Betrachtung des Verhältnisses der Lamina I zu
den übrigen Schichten in einem zwischen den Gruppen identischen Signifikanzniveau von
p<0.001.
Gleich den Verhältnissen zwischen der ertaubten und hörenden Versuchsgruppe in Bezug auf
die horizontale „Schichten-Hierarchie“ zeigt sich auch bei Betrachtung der diagonalen
Systematik kein Unterschied zwischen den Tieren mit hörendem oder neonatal ertaubten
Hörstatus.

5. Diskussion
89
In Bezug auf die Struktur der prozentualen Anteile der vertikalen Dendritenverläufe über die
sechs betrachteten Laminae zeigt sich dagegen ein deutlicher Unterschied zur hörenden
Kontrollgruppe. So ist innerhalb des primären auditorischen Feldes die Lamina III im
Verhältnis zur Lamina II nicht dominant. Bei Betrachtung der Verhältnisse innerhalb des
anterioren und des sekundären auditorischen Feldes besteht gleich den Ergebnissen der
hörenden Tiere für jedes betrachtete Verhältnis eine Dominanz der Lamina III, wenn auch die
Höhe der Signifikanzniveaus geringfügige Differenzen aufweist.
Bei Betrachtung der Gesamtheit der mit der hier verwendeten Methodik untersuchten
Systematiken zeigt sich eine insgesamt sehr geringe Veränderung der laminären
Dendritenstruktur aufgrund der neonatalen experimentellen Ertaubung. Ursächlich kann
hierfür zum einen die geringe Sensitivität der verwendeten Methodik sein. Mit dem zur
Verfügung stehenden Material konnten weitere Anzeichen einer stattgefundenen Plastizität
wie eine Verlängerung oder Verkürzung der Dendriten, eine Zu- oder Abnahme der absoluten
Dendritenanzahl oder Veränderungen in Bezug auf die Spine- bzw. Synapsenanzahl nicht
untersucht werden.
Zum anderen könnte eine mögliche Erklärung für die minimal ausfallenden Veränderungen in
der Tatsache liegen, dass nicht durch einen primär andersartigen „Input“ eine Plastizität
induziert wurde, sondern lediglich ein Wegfall des afferenten Informationsflusses
stattgefunden hat.
Ein weitaus größer ausfallender Unterschied zwischen den Gruppen der hörenden und
ertaubten Tiere wäre einerseits denkbar, da die Ausreifung der dendritischen Strukturen bei
neonatalen Tieren noch nicht abgeschlossen ist und damit nicht das Muster des ausgereiften
auditorischen Cortex aufweist (FENG und BRUGGE 1983). Den neonatal experimentell
ertaubten Versuchstieren dieser Studie wurde durch die Intervention die physiologische
Entwicklung des Hörsystems auf Basis einer akustischen Stimulation vorenthalten, so dass bei
den untersuchten Tieren dieser Versuchsgruppe zumindest theoretisch eine Dendritenstruktur,
welche mit neonatalen auditorisch unreifen Katzen vergleichbar ist vorliegen dürfte.
Andererseits konnte insbesondere in einer von WALSH et al. (1986) durchgeführten Studie
Hinweise auf eine, durch spontane Neuronenaktivitäten in subcorticalen Bereichen
hervorgerufene, geringe Stimulation des auditorischen Cortex auch ohne akustischen Input
gefunden werden.

5. Diskussion
90
Dieses Phänomen könnte für die Entwicklung der prinzipiellen Dendritenstruktur
verantwortlich sein und damit den hier gefundenen nur geringen Einfluss einer neonatalen
Ertaubung erklären.
Eine vollständige Umstrukturierung im Sinne der Bildung neuartiger Verknüpfungen ist
weiterhin vor dem Hintergrund unwahrscheinlich, dass die prinzipiellen intracorticalen
Verbindungen bereits zur Geburt vorliegen und nur in der Ausprägung einer gewissen
Variabilität unterliegen (FENG und BRUGGE 1983).
Während auf Ebene der zentralen auditorischen Hörbahn durch eine neonatale Ertaubung sehr
ausgeprägte strukturelle Veränderungen bewirkt werden (siehe Kapitel 2.5.2.1), treten nach
den hier präsentierten Ergebnissen innerhalb des auditorischen Cortex nur relativ gering
ausfallende Veränderungen auf. Ursache hierfür und damit eine Erklärung für die hier
gefundenen geringen Unterschiede könnte weiterhin im Grad der Deafferentierung liegen.
Während subcorticale Strukturen mit einer akustischen Deprivation den Hauptanteil ihrer
afferenten Verbindungen verlieren, kommt es innerhalb des auditorischen Cortex durch die
parallele Verknüpfung mit verschiedenen Untereinheiten der auditorischen Hirnstamm-
strukturen sowie der interarealen Verknüpfungen zu einem, relativ gesehen, weitaus
geringeren Verlust der totalen afferenten Innervation (REALE et al. 1987).
In der von WENKE (1999) durchgeführten Untersuchung zum Einfluss einer neonatalen
Ertaubung auf die Gesamt- und Pyramidenzellzahl in der Lamina III des felinen anterioren,
primären und sekundären Feldes zeigten sich hingegen deutliche Auswirkungen der
ausgebliebenen auditorischen Entwicklung. Bei neonatal ertaubten Tieren war im AI eine um
29 % signifikant erniedrigte Gesamtzellzahl vorhanden. Die Pyramidenzellzahl reduzierte
sich in diesem Areal sogar um 87 %. Innerhalb des AAF trat ein signifikanter
Pyramidenzellverlust von 28 % auf. Die Gesamtzellzahl war nicht signifikant verändert. Im
AII kam es durch eine neonatale Ertaubung zu keiner signifikanten Veränderung der
Zellzahlen.
Diese Ergebnisse unterstreichen zum einen die hier gefundenen nicht stattgefundenen
Veränderungen in Bezug auf das sekundäre auditorische Feld, lassen auf der anderen Seite
jedoch einen größeren als den hier nachgewiesenen Unterschied zwischen den hörenden und
ertaubten Tieren im Bereich des anterioren und primären auditorischen Feldes erwarten.

5. Diskussion
91
Allerdings ist es möglich, dass hier der gleiche Effekt eingetroffen ist, welcher bei den
hörenden Kontrolltieren in Bezug auf die unterschiedlichen auditorischen Felder beschrieben
wurde. Danach würde eine in den absoluten Anzahlen der Neurone eventuell vorliegende
Differenz durch die in dieser Studie angewandte prozentuale Auswertung hinsichtlich der
Verlaufsrichtungen der angefärbten Dendriten diesen Unterschied nicht aufzeigt werden
können.
Nach Zerstörung der Cochlea kam es jedoch bei verschiedenen Tiermodellen im Bereich des
auditorischen Hirnstamms wiederum auch zu Umstrukturierungen wie einer Axonen-
sprossung, neuem Wachstum von Nervenzellen (BILAK et al. 1997) und sogar zur Bildung
neuer Synapsen (BENSON et al. 1997). Diese Veränderungen im subcorticalen Bereich
könnten sich prinzipiell zusätzlich auf den auditorischen Cortex auswirken und hier zu einem
Erhalt der Dendritenstruktur führen.
Im Gegensatz dazu konnte jedoch auch gezeigt werden, dass Neurone im primären
auditorischen Feld von naiven kongenital tauben Katzen im Vergleich zu normalhörenden
Kontrolltieren eine insgesamt geringere Verzweigungs-häufigkeit der Dendriten aufweisen
(WURTH et al. 1999). Es zeigte sich eine signifikant reduzierte Anzahl primärer und
sekundärer basaler Dendriten. Gleichzeitig waren die gemessenen Dendritenspannweiten in
der Lamina II und IV der kongenital tauben Tiere signifikant reduziert.
Allerdings wurde in dieser Studie lediglich eine Cortexhemisphäre einer kongenital tauben
Katze sowie nur zwei auditorische Cortices einer normalhörenden Katze ausgewertet.
Weiterhin ist nicht bekannt, inwieweit sich die corticalen auditorischen Dendritenstrukturen
per se zwischen neonatal ertaubten und kongenital tauben Katzen unterscheiden.
Die Ergebnisse jener Untersuchung sind daher einerseits mit Vorsicht zu betrachten,
andererseits scheinen Ergebnisse von KLINKE et al. (1999) die Studie jedoch zu
untermauern, da bei den kongenital tauben Tieren auch geringer ausfallende synaptische
Ströme gemessen wurden.
Nach unilateraler neonataler Deprivation von Kaninchen konnten MCMULLEN et al. (1988)
in den Laminae III und IV des kontralateralen auditorischen Cortex ebenfalls signifikante
Veränderungen der Dendritenstruktur nachweisen. In dieser Studie zeigte sich jedoch, dass
die Dendriten nicht-pyramidaler Neurone der einseitig ertaubten Tiere eine Verlängerung um
durchschnittlich 27 % relativ zu denen normalhörender Geschwistertiere aufweisen.

5. Diskussion
92
Die maximale Verlängerung wurde dabei in tangentialer Ausrichtung sowie in Richtung der
weißen Substanz gefunden. Zusätzlich konnte ein abnormales Dendritenwachstum in Form
von zum Zellsoma rekurrierenden Dendriten dargestellt werden. Hinweise auf eine Reduktion
der Dendritenanzahl wurden nicht gefunden.
Am gleichen Versuchsmodell konnten MCMULLEN und GLASER (1988) eine Reduktion
der Spines an den basalen Dendriten der pyramidalen Neurone der Laminae III und IV des
kontralateralen auditorischen Cortex der unilateral deprivierten Tiere um durchschnittlich 38
% nachweisen. Dieses stellt einen eindeutigen Hinweis auf einen Verlust von Synapsen nach
einer Deprivation dar. Hinsichtlich der Somataquerschnittsfläche, der Gesamtzahl der
Dendritenäste und der basalen Dendriten wie auch der Gesamtlänge der basalen Dendriten
wurden wiederum keine Unterschiede zu normalhörenden Geschwistertieren gefunden.
Bei Betrachtung dieser Ergebnisse muss jedoch auf die methodischen Unterschiede
hingewiesen werden. Bei einseitigen Ertaubungen kann theoretisch über die intakte
ipsilaterale afferente Bahn eine Veränderung auf Ebene des auditorischen Cortex induziert
werden. Dieser Einfluss ist jedoch aufgrund der grundsätzlichen Verschaltung der afferenten
Bahnen als eher gering einzuschätzen, da der Anteil aus der ipsilateralen Cochlea lediglich 30
% beträgt (IRVINE et al. 1996).
Mittels Optical Recording konnten von REUTER et al. (2002a) die funktionellen Aus-
wirkungen einer neonatalen Ertaubung verifiziert werden. Bei den experimentell ertaubten
Katzen dieser Studie war zur Auslösung von neuronaler Aktivität im AI eine im Mittel
doppelt so hohe Stimulationsenergie wie bei hörenden Kontrolltieren nötig. Gleichzeitig
zeigte sich die bei den hörenden Tieren deutliche tonotope Darstellung der
Stimulationsfrequenzen bei den ertaubten Tieren stark verzerrt. Aktivierte Areale erschienen
dabei als kleine Inseln, welche von Bereichen unterschiedlicher Sensitivität umgeben waren.
In Bezug auf die durch angrenzende Stimulationsfrequenzen aktivierten corticalen Areale und
der registrierten „Überlappungszone“ zwischen diesen zwei Frequenzen zeigten sich deutliche
signifikante Unterschiede. Während bei den hörenden Tiere nur geringe Überlappungszonen
(26 %) aufgezeichnet werden konnten, wiesen die ertaubten Tiere weitaus größere (40 %) und
kompaktere Überlappungszonen auf.

5. Diskussion
93
Neben den deutlichen Unterschieden zwischen den hörenden und ertaubten Tieren
hinsichtlich der Funktionalität des AI zeigen diese Daten jedoch auch, dass trotz einer
Ertaubung und einem damit einhergehenden Fehlen von akustischen Sensationen und einer
darauf beruhenden nur rudimentären Entwicklung der auditorischen Hörbahn sowie des
auditorischen Cortex, letzterer durch adäquate Reize stimuliert werden kann. Dies bedeutet
weiterhin, dass selbst bei neonatal ertaubten Tieren die Grundstruktur der afferenten Hörbahn
und der corticalen Neuronenverschaltungen, also die Dendritenstruktur, prinzipiell vorhanden
ist.
Ähnliche Ergebnisse an neonatal ertaubten Katzen konnten von RAGGIO und SCHREINER
(1999) dargestellt werden. Es zeigte sich hier ebenfalls, dass der auditorische Cortex ohne
afferenten Informationsfluss unterentwickelt bleibt, zum anderen jedoch auch eine
rudimentäre, aber dennoch tontotope, aktivierbare Struktur im primären auditorischen Feld
ausgebildet ist. Letztere Ergebnisse lassen große Unterschiede bezüglich der Dendriten-
struktur bei der neonatal ertaubten Katze unwahrscheinlich erscheinen.
Signifikante Unterschiede zwischen den beiden separat ausgewerteten Cortexhälften in der
Gruppe der neonatal ertaubten Tiere konnten in dieser Studie nicht nachgewiesen werden.
Auch die histologische Untersuchung von WENKE (1999) ergab für neonatal ertaubte Katzen
keinen signifikanten interhemisphärischen Unterschied.
Innerhalb der ertaubten Versuchsgruppe konnte beim Vergleich der erhobenen Daten für die
Laminae I bis VI zwischen den untersuchten auditorischen Feldern kein signifikanter
Unterschied und damit kein Einfluss der neonatalen Deafferentierung nachgewiesen werden.
Die Ursache hierfür ist wahrscheinlich darin begründet, dass durch die nicht-selektive
Deprivation auf der niedrigsten Stufe der auditorischen Hörbahn durch die Schädigung der
Hörsinneszellen mittels Aminoglycosid-Antibiotikum alle der hier untersuchten auditorischen
Felder die Auswirkungen der Deprivation in gleicher Ausprägung erfahren.
Bei dem direkten Vergleich der prozentualen Dendritenanteile der analogen Laminae der
ertaubten und der hörenden Versuchsgruppe zeigten sich ebenfalls keine signifikanten
Unterschiede.

5. Diskussion
94
5.4 Die Auswirkungen einer chronischen elektrischen intracochleären Stimulation auf
die Morphometrie der Dendriten der Laminae I bis VI im AAF, AI und AII der
Katze
Elektrische Innenohrprothesen, auch Cochlea-Implantat (CI) genannt, sind in der Lage die
Funktion der sensorischen Zellen im Innenohr zu ersetzen, so dass die Verarbeitung von
akustischen Reizen ermöglicht wird. Im Gegensatz zur normalen akustischen Stimulation,
welche sich auf die Erregung weniger Frequenzbereiche des Hörnerven beschränkt (6 bis 8),
werden durch die direkte elektrische Stimulation mit einem CI alle Frequenzbereiche des
Hörnerven aktiviert. Eine Veränderung der sensorischen Afferenzen in Folge einer
elektrischen Stimulation konnte bereits nachgewiesen werden (siehe Kapitel 2.5.3.1), so dass
auch eine Plastizität auf corticaler Ebene erwartet werden kann.
Mit dieser Studie konnte der Einfluss einer chronischen elektrischen intracochleären
Stimulation mittels Cochlea-Implantat auf die „Schichten-Hierarchie“ der horizontal
verlaufenden Dendriten des sekundären auditorischen Feldes sowie der vertikal verlaufenden
Dendriten in allen drei betrachteten auditorischen Subarealen nachgewiesen werden. Eine
laminaspezifische Veränderung der einzelnen prozentualen Anteile der Dendriten-
verlaufsrichtungen aufgrund der Versorgung mit einem CI konnte beim Vergleich mit den
anderen Versuchsgruppen hingegen nicht nachgewiesen werden.
Die Auswertung der Daten der horizontalen Dendritenverläufe der chronisch elektrisch
intracochleär stimulierten Tiere macht hinsichtlich der „Schichten-Hierarchie“ aufgrund
deutlicher Unterschiede zu der hörenden und ertaubten Versuchsgruppe eine differenzierte
Betrachtung der drei auditorischen Felder notwendig.
Innerhalb des anterioren sowie des primären auditorischen Feldes zeigt sich bei der
statistischen Betrachtung der Dendritenverhältnisse über die Laminae kein herausragender
Einfluss der Stimulation mittels Cochlea-Implantat, wenngleich sich die Signifikanzniveaus,
insbesondere des AI, deutlich unterscheiden. Hinsichtlich des sekundären auditorischen
Feldes besteht jedoch im Verhältnis zur Lamina II, V und Lamina VI keine Dominanz der
Lamina I. Damit wird ein deutlicher Einfluss der chronischen elektrischen intracochleären
Stimulation deutlich.

5. Diskussion
95
Im Gegensatz zu der horizontalen Systematik unterscheidet sich die elektrisch stimulierte
Versuchsgruppe bezüglich der diagonalen „Schichten-Hierarchie“ lediglich durch ein geringer
ausfallendes Signifikanzniveau (p<0.01) von der hörenden und der ertaubten Versuchsgruppe.
Bei der statistischen Auswertung der prozentualen Anteile der vertikalen Dendritenverläufe
fällt ein sehr deutlicher Einfluss der CI-Versorgung auf. Während für die Laminae I, V und VI
wie bei den vorausgegangenen Versuchsgruppen eine Dominanz der Lamina III vorhanden
ist, kann für die Lamina II innerhalb des anterioren, primären und sekundären auditorischen
Feldes keine dominierende Rolle der Lamina III nachgewiesen werden. Ebenso weist die
Lamina IV im AAF keinen signifikanten Unterschied zur Lamina III auf.
In der Gruppe der chronisch elektrisch intracochleär stimulierten Tiere scheint bei
Betrachtung der vorliegenden Ergebnisse eine strukturelle Veränderung im Sinne einer
Plastizität als Folge der Stimulation mit einem Cochlea-Implantat und der damit
einhergehenden Veränderung des „akustischen Inputs“ stattgefunden zu haben.
Eine funktionelle Reorganisation im auditorischen Cortex chronisch elektrisch intracochleär
stimulierter Katzen konnte auch durch die Messung intrinsischer corticaler Signale
nachgewiesen werden (REUTER et al. 2002a). So war zur Aktivierung der corticalen
Aktivität bei diesen Tieren eine signifikant geringere Stimulationsintensität als bei den
normalhörenden Tieren notwendig. Weiterhin hat bei den chronisch elektrisch intracochleär
stimulierten Tieren eine substantielle Reorganisation der Tonotopie stattgefunden. Im
Vergleich zu normalhörenden Tieren wurde bei dieser Versuchsgruppe durch Stimulation
mehrerer Elektrodenpaare eine signifikant größere corticale Fläche aktiviert. Dies lässt auf
einen Verlust der repräsentativen Selektivität schließen. Dennoch konnte auch bei den
chronisch elektrisch intracochleär stimulierten Tieren die tonotope Verlagerung der
aktivierten Cortexareale von caudal nach rostral in Abhängigkeit der Stimulusfrequenz
beobachtet werden (REUTER et al. 2002b).
Diese Ergebnisse zeigen neben einer funktionellen Rehabilitation im auditorischen Cortex
nach Versorgung mit einem Cochlea-Implantat jedoch auch, dass auch nach chronischer
elektrischer intracochleärer Stimulation nach einer Ertaubung eine substantielle Unreife in der
corticalen Aktivierung bestehen bleibt.

5. Diskussion
96
In diesem Sinne konnten auch PONTON und EGGERMONT (2001) zeigen, dass bei
Erwachsenen und Kindern, welche nach einer kritischen Taubheitsdauer mit einem CI
versorgt werden, die corticalen auditorisch evozierten Potentiale nicht denen normalhörender
Personen entsprechen. Auch die Reizantworten von chronisch intracochleär stimulierten
Katzen weisen auf eine nicht vollständig ausgereifte Funktion der Axone in den superficialen
Schichten des auditorischen Cortex hin (KLINKE et al. 1999).
Die deutlichen Veränderungen in der „Schichten-Hierarchie“ in Bezug auf die horizontal und
vertikal verlaufenden Dendriten können dadurch erklärt werden, dass für die Ausbildung
dieser Verbindungen Einflüsse aus anderen Strukturen relevant sind und sich dort
stattgefundene Veränderungen in der Systematik der hier untersuchten auditorischen Felder
wiederspiegeln. Im Falle der horizontal verlaufenden Dendriten handelt es sich dabei
wahrscheinlich vor allem um Interaktionen mit anderen corticalen Bereichen, während die
vertikalen Dendritenverläufe höchstwahrscheinlich die Veränderungen der thalamocorticalen
Bahnen reflektieren.
Der vergleichsweise gering ausfallende Unterschied in der diagonalen Hierarchie ist unter
Umständen darauf zurückzuführen, dass die diagonal verlaufenden Dendriten
höchstwahrscheinlich einen eher lokalen Einfluss haben und daher nicht in dem Maße wie die
horizontal und vertikal verlaufenden Dendriten von externen Strukturen wie der afferenten
Hörbahn und den anderen auditorischen Feldern beeinflusst werden.
Histologisch zeigte sich für die Lamina III der Einfluss einer chronischen elektrischen
intracochleären Stimulation im ipsilateralen und kontralateralen AI in einer signifikant über
der Anzahl einer neonatal ertaubten Tiergruppe liegenden Gesamt- und Pyramidenzellzahl
(WENKE 1999). In Hinblick auf die Daten der dort verwendeten hörenden Versuchsgruppe
zeigte sich jedoch in einer signifikant niedrigeren Gesamt- und Pyramidenzellzahl der
stimulierten Tiere, dass eine chronische elektrische Stimulation den aufgetretenen Zellverlust
sowohl ipsilateral als auch kontralateral nicht vollständig ausgleichen kann. Die bei der
stimulierten Gruppe gefundenen Zellzahlen entsprechen circa 89 % (Gesamtzellzahl) bzw. 65
% (Pyramidenzellzahl) den bei hörenden Kontrolltieren gefundenen Zahlen (REUTER et al.
2002a). Im sekundären auditorischen Feld konnte in dieser Studie durch eine chronische
elektrische intracochleäre Stimulation keine signifikante Veränderung der Zellzahlen
gegenüber der ertaubten Versuchsgruppe erreicht werden. Auch innerhalb des AAF konnte
der aufgetretene Pyramidenzellverlust nicht durch die Stimulation aufgehoben werden.

5. Diskussion
97
Bei Betrachtung der horizontalen Systematik zwischen der hörenden und stimulierten
Versuchsgruppe dieser Studie fällt dagegen in den hier präsentierten Daten eine ausgeprägtere
Differenz in Bezug auf das sekundäre auditorische Feld auf. Dies steht im Einklang mit
vorangegangenen Studien, welche in den übergeordneten auditorischen Feldern, insbesondere
dem AII der Katze, eine größere Plastizität als dem primären auditorischen Feld nachwiesen
(WEINBERGER et al. 1984).
Im Vergleich der ipsi- und kontralateralen auditorischen Cortices zeigte sich in der
histologischen Studie von WENKE (1999) in der Lamina III im primären auditorischen
Cortex ein signifikanter Unterschied. Während das ipsilaterale primäre auditorische Feld eine
um 20 % erniedrigte Gesamtzellzahl aufwies, war diese im kontralateralen AI um lediglich 12
% reduziert. In Bezug auf die Pyramidenzellzahl fand sich ipsilateral eine signifikant zur
kontralateralen Seite verschiedene Reduzierung von 64 % bzw. 35 %.
In Bezug auf die Dendritenstruktur zeigt sich in der vorliegenden Studie mit Ausnahme der
diagonalen Verläufe im sekundären auditorischen Feld kein Unterschied zwischen dem
ipsilateral und kontralateral stimuliertem Cortex. In diesem Fall liegt allerdings im
ipsilateralen AII ein signifikant gegenüber dem kontralateralen Areal erhöhter prozentualer
Anteil der diagonal verlaufenden Dendriten vor (p<0.05). Dies steht im Gegensatz zu den
oben genannten Ergebnissen sowie den durch die Anatomie der Hörbahn begründeten zu
erwartenden ausgeprägteren Einflüssen einer cochleären Stimulation auf den kontralateralen
auditorischen Cortex. Auf der anderen Seite zeigte sich bei einer unilateralen Taubheit beim
Menschen eine bilaterale Aktivierung der Hörrinde. Ursache für derartige zentralauditive
Kompensationsmechanismen im Sinne einer Bilateralisation ist wahrscheinlich eine
Demaskierung vorhandener ipsilateraler Verbindungen (BILECEN et al. 2000).
Das Auftreten eines signifikanten interhemisphärischen Unterschiedes im AII spricht
wiederum für seine übergeordnete Fähigkeit zur Plastizität (WEINBERGER et al. 1984). Zum
anderen werden die Veränderungen in Bezug auf die Systematik der diagonal verlaufenden
Dendriten, wenn man von einer eher lokalen Vernetzungsfunktion dieser Strukturen ausgeht,
von den funktionellen Veränderungen hinsichtlich der veränderten Größe der aktivierten
corticalen Areale durch die Ergebnisse von REUTER et al. (2002b) unterstützt.
Innerhalb der stimulierten Versuchstiergruppe konnte beim Vergleich der erhobenen Daten
für die Laminae I bis VI zwischen den untersuchten auditorischen Feldern kein signifikanter
Unterschied nachgewiesen werden.

5. Diskussion
98
Die Ursache hierfür ist wahrscheinlich darin begründet, dass durch die Aktivierung der
auditorischen Hörbahn auf niedrigster Stufe alle der hier untersuchten auditorischen Felder
die Auswirkungen der Plastizitäten in gleicher Ausprägung erfahren.
Bei dem direkten Vergleich der prozentualen Dendritenanteile der analogen Laminae der
chronisch elektrisch intracochleär stimulierten Tiere mit denen der ertaubten und der
hörenden Versuchsgruppe zeigen sich ebenfalls keine signifikanten Unterschiede.
5.5 Die Schlussfolgerungen
Ziel der vorliegenden Untersuchung war es, die Auswirkungen einer akustischen Deprivation
und einer chronischen elektrischen intracochleären Stimulation auf die Dendritenstruktur im
auditorischen Cortex der Hauskatze zu untersuchen. Dazu wurde das an der Medizinischen
Hochschule Hannover seit langem etablierte Tiermodell der Katze verwendet.
Für die Arbeit standen drei Versuchsgruppen zur Verfügung. Neben den normalhörenden
Tieren der Kontrollgruppe wurden die Tiere der beiden weiteren Gruppen durch die
systemische Applikation eines Aminoglykosid-Antibiotikums neonatal ertaubt. Zwei dieser
Tiere wurden bilateral, ein weiteres ertaubtes Tier unilateral mit einem Cochlea-Implantat
versorgt und bildeten damit die Gruppe der chronisch elektrisch intracochleär stimulierten
Tiere.
Zahlreiche Untersuchungen haben bereits gezeigt, dass es bei der Katze in Folge einer
neonatalen Ertaubung und einer chronischen elektrischen Stimulation zu Veränderungen auf
zellulärer Ebene der auditorischen Hörbahn (CORDS 1996; HEID et al. 1998; KELLER
1997; SNYDER et al. 1990; TERAYAMA et al. 1977) sowie des auditorischen Cortex
kommt (REUTER et al. 2002b; WENKE 1999; WURTH 1999).
Die Auswertung der Dendritenstruktur erfolgte in der vorliegenden Studie durch die
Berechnung der prozentualen Anteile der horizontal, diagonal oder vertikal verlaufenden
Dendriten in den Laminae I bis VI des anterioren (AAF), primären (AI) und sekundären (AII)
auditorischen Feldes des rechten und linken auditorischen Cortex. Dazu wurde die
Gesamtanzahl aller in 30 Zählfeldern einer Schicht gezählten Dendriten addiert und als 100
Prozent angesehen. Im Anschluss wurden die horizontal, diagonal und vertikal verlaufenden
Dendriten der 30 Zählfelder separat voneinander addiert und ins Verhältnis zur Gesamtheit
gesetzt.

5. Diskussion
99
An der Kontrollgruppe konnte gezeigt werden, dass für die jeweiligen Verlaufsrichtungen
unterschiedliche intraareale Systematiken („Schichten-Hierarchien“) vorliegen, welche sich
zwischen den drei untersuchten Feldern nicht unterscheiden. Im Vergleich der „Schichten-
Hierarchien“ für die horizontal, diagonal und vertikal verlaufenden Dendriten der Kontroll-
tiere mit den Systematiken der neonatal ertaubten Tieren zeigten sich nur sehr geringe
Veränderungen. Eine Plastizität als Folge der akustischen Deprivation scheint sich dabei auf
die vertikal verlaufenden Dendriten innerhalb des AI zu beschränken. Bei Betrachtung der
Ergebnisse der chronisch elektrisch intracochleär stimulierten Versuchsgruppe zeigen sich
sowohl hinsichtlich der horizontal verlaufenden Dendriten innerhalb des AII sowie
insbesondere in Bezug auf die vertikalen Dendritenanteile im AAF, AI und AII Hinweise auf
reorganisatorische Prozesse.
Die zwischen dem Mensch und der Katze bestehenden Parallelen in der Funktion und
Entwicklung (FULLERTON et al. 1987) sowie der Morphologie (ONG und GAREY 1990)
des auditorischen Systems machen es wahrscheinlich, dass die hier gezeigten Änderungen der
Dendritensystematik auch bei mit einem Cochlea-Implantat versorgten Patienten auftreten.
Daraus kann gefolgert werden, dass die hier nachgewiesenen Veränderungen die Grundlage
für die auf der Ebene des auditorischen Cortex stattfindende Plastizität darstellen, welche
beim Cochlea-Implantat-Nutzer zur Vermittlung von Höreindrücken bis hin zum offenen
Sprachverständnis führt.
Um einen genaueren Einblick in die Auswirkungen und die Funktion der hier gezeigten
Plastizität zu erhalten ist jedoch ein Ausbau der hier verwendeten Methode zwingend
notwendig. Zum einen kann die Einhaltung kürzerer postmortaler Fixationszeiten von
optimalen zwei Tagen eine Auswertung der kompletten Dendritenbäume einzelner Neurone
ermöglichen. Neben einer quantitativen Auswertung, bezogen auf die Anzahl und
Ausdehnung der Dendriten pro Neuron, wäre damit bei weiterhin verbleibender Verwendung
von 70 µm dicken Vibratomschnitten auch eine 3-dimensionale Rekonstruktion der
Dendritenbäume mit Hilfe eines Confokalen-Laser-Scanning Mikroskops möglich. Die
Grundlagen für eine derartige Auswertung wurden im Rahmen dieser Untersuchung
erarbeitet. Sinnvoll wäre zum anderen eine Kombination der postmortalen Zellfärbung mit
zuvor durchgeführten Messungen der physiologischen Zellfunktion im Sinne einer Ableitung
intrinsischer corticaler Signale. Auch eine Überprüfung der Funktionalität mittels
immunhistochemischer Methoden könnte angewendet werden. So wäre eine Bestimmung der
Anzahl sowie eine Darstellung der Verteilung von GABA-Rezeptoren vorstellbar.

6. Zusammenfassung
100
6. ZUSAMMENFASSUNG
Untersuchungen zur Plastizität der laminären Dendriten im auditorischen Cortex von
akustisch deprivierten sowie chronisch elektrisch intracochleär stimulierten
Hauskatzen (Felis domestica)
Heike Rieger
Cochlea-Implantate (CI) sind in der Lage Menschen, deren Taubheit auf einem Funktions-
verlust der Haarzellen beruht, durch eine direkte elektrische Stimulation des Hörnerven zu
Hörsensationen bis hin zum offenen Sprachverständnis zu verhelfen. Sie bieten damit bei
vollständig ertaubten Patienten die einzige Möglichkeit der Rehabilitation.
Im Gegensatz zum rapide voranschreitenden technischen Fortschritt in der Entwicklung
leistungsfähigerer Innenohrprothesen, schreiten die physiologischen Untersuchungen der
Einflüsse einer chronischen elektrischen intracochleären Stimulation auf die Strukturen der
Hörrinde nur langsam voran. Weitergehende Untersuchungen über die Auswirkung einer
akustischen Deprivation und einer Stimulation mittels CI auf die neuronale Organisation im
auditorischen Cortex sind daher dringend erforderlich. Diese Auswirkungen werden in der
vorliegenden Studie anhand einer Auswertung der Dendritenverläufe in den Laminae I bis VI
des anterioren (AAF), primären (AI) und sekundären (AII) auditorischen Feldes dargestellt.
Die Fragestellung dieser Arbeit lautet, inwieweit es bei einer neonatalen Ertaubung sowie
einer anschließenden chronischen elektrischen intracochleären Stimulation zu plastischen
Veränderungen in den auditorischen Subarealen kommt.
Sieben Katzen wurden für diese Studie durch eine tägliche subcutane Injektion des
ototoxischen Aminoglycosid-Antibiotikums Neomycin ertaubt. Der Erfolg der Ertaubung
wurde mittels Hirnstammaudiometrie nach akustischer Stimulation (Click-Reiz, 100 dB SPL)
kontrolliert. Zwei der neonatal ertaubten Tiere wurden im Alter von 11 bis 12 Wochen
bilateral, ein weiteres Tier unilateral mit humanen Cochlea-Implantaten versorgt. Über einen
Zeitraum von 84 bis 112 Tagen erfolgte eine tägliche mindestens vierstündige Stimulation der
Tiere nach der „continuous interleaved sampling“-Strategie 2 dB über der individuellen
Hörschwelle. Als Kontrollgruppe dienten 6 normal hörende Tiere.
Alle Versuchstiere wurden in Allgemeinanästhesie transcardial perfundiert und der
auditorische Cortex präpariert. Nach Applikation von 500 bis 800 µm großen Kristallen des
Carbocyanin-Fluoreszenzfarbstoffs DiI auf das AAF, AI und AII beider Hemisphären wurden
die Gewebe für 120 Tage in 4 %igem Paraformaldehyd bei 37 °C und Dunkelheit inkubiert.

6. Zusammenfassung
101
Im Anschluss an die Anfertigung von 70 µm dicken Vibratomschnitten wurden für jedes der
Subareale in der Lamina I bis VI jeweils 30 Zählfelder (je 6 in 5 Schnitten) mit einer Größe
von 60 x 60 µm betrachtet. Gezählt wurden alle Dendriten, welche ganz oder partiell
innerhalb des Zählareals lagen. Diese wurden nach ihrer relativ zur Cortexoberfläche
gesehenen Verlaufsrichtung als horizontal, diagonal oder vertikal ziehend eingeordnet.
Die statistische Auswertung erfolgte nach Überprüfung auf Normalverteilung mittels one-way
ANOVA mit einem Signifikanzniveau von 95 %. Anschließend erfolgte mittels Post Hoc Test
nach Bonferroni ein paarweiser Datenvergleich. In Einzelfällen wurde auf den Kruskal-
Wallis-Test mit anschließendem Post Hoc Test nach Dunn ausgewichen. Die Auswertung der
Ergebnisse der diagonalen Datenverläufe erfolgte nach Prüfung auf Normalverteilung mittels
t-Test mit unabhängigen Variablen.
Bei der hörenden Kontrollgruppe konnte sowohl für die horizontal und diagonal als auch für
die vertikal verlaufenden Dendriten eine voneinander differierende Systematik über die sechs
Cortexschichten beobachtet werden. Diese Systematiken waren prinzipiell auch in der
neonatal ertaubten sowie der chronisch elektrisch intracochleär stimulierten Versuchsgruppe
vorhanden. Es zeigten sich jedoch für die ertaubte Tiergruppe bei Betrachtung der vertikalen
Dendritenverläufe im AI Hinweise auf eine stattgefundene Plastizität. Im Verhältnis zur
Lamina II trat ein durch einen Anstieg des prozentualen Anteils der vertikal ziehenden
Dendriten in dieser Schicht ausgelöster Verlust der signifikant dominierenden Rolle der
Lamina III auf. In der Gruppe der stimulierten Tiere zeigten sich im Vergleich zur ertaubten
Versuchsgruppe stärker ausgeprägte Unterschiede zur hörenden Kontrollgruppe. In Bezug auf
die vertikal ziehenden Dendriten kam es in der Lamina II aller drei auditorischen Felder sowie
in der Lamina IV des AAF zu einem einem Verlust der signifikanten Dominanz der Lamina
III. Zusätzlich traten bei den horizontalen Dendritenverläufen in den Laminae II, V und VI im
AII ebenfalls ein Dominanzverlust der Lamina I auf. Auch hierbei handelt es sich um
deutliche, auf eine plastische Veränderung hindeutende, Abweichungen zur Kontrollgruppe.
Mit dieser Studie konnte auf Ebene des auditorischen Cortex eine plastische Veränderung der
Dendritensystematik über die Laminae I bis VI nach pharmakologisch induzierter Cochlea-
schädigung sowie nachgeschalteter chronischer elektrischer intracochleärer Stimulation nach-
gewiesen werden. Aufgrund der Parallelen zwischen dem Hörsystemen von Mensch und
Katze (FULLERTON et al. 1987; ONG und GAREY 1990) ist eine Übertragung der
Versuchsergebnisse auf den Menschen möglich. Inwieweit die hier gezeigten histo-
morphologischen Veränderungen an eine Funktionsänderung des auditorischen Cortex
gekoppelt sind, bedarf weiterer Untersuchungen.

7. Summary
102
7. SUMMARY
Investigation into laminar dendritic plasticity in the auditory cortex of
acoustically deprivied and chronic electrically intracochlear stimulated cats
(Felis domestica)
Heike Rieger
Cochlear implants (CI) are in the position to achieve hearing sensations up to open speech
understanding in patients, whose deafness is caused by a functional loss of hair cells. By
directly stimulating the auditory nerve the missing hair cell activity is reconciled. This is the
only method for rehabilitating profoundly deaf subjects.
In contrast to the rapidly progressing technical improvement of efficient cochlear implants,
the investigation into the physiological influences of chronic electric intracochlear stimulation
to the central auditory pathway and the auditory cortex is progressing slowly. Therefore more
extensive studies on the consequence of acoustic deprivation and chronic stimulation by CI on
the neuronal organization of auditory fields are essentially required.
These possible effects are examined by means of a descriptive and semiquantitative study of
the pattern of dendrites across Laminae I to VI of the anterior, primary and secondary
auditory field. To verify indications for a reorganisation and stimulation-depending effects of
plasticity is in the centre of attention.
To this end, an animal model already established at the ENT department of the Medical
School of Hanover was used. Seven cats received a daily subcutaneous injection of the
ototoxic aminoglycoside antibiotic Neomycin for a period of 13 to 17 days, starting at the first
day of life. The result of the deafening procedure was controlled by brainstem audiometry. In
two neonatal deafened cats multichannel electrodes were implanted into the scala tympani of
the cochlea bilaterally, in one other animal unilaterally at the age of 11 to 12 weeks.
Subsequently, these cats were stimulated with environmental sounds by the “continuous
interleaved sampling” strategy over a period of 84 to 112 days 2 dB above hearing threshold.
Six normal hearing cats made up the control group. All animals were transcardially perfused
under general anaesthesia and the brains were removed. After application of 500 to 800 µm
large crystals of the carbocyanine fluorescent dye DiI onto the anterior, primary and
secondary auditory field of the right and left cortex, the examinated tissue was incubated in
parafomaldehyde for 120 days at 37 °C in absence of light.

7. Summary
103
Following cutting 70 µm thick sections by a vibratome 30 counting areas (5 sections each
with 6 areas) with dimensions of 60 to 60 µm were examined in the laminae I to VI from each
of the studied auditory fields of both cerebral hemispheres. All dendrites, which laid totally or
partially inside these counting areas were counted. Counted dendrites were classified as
horizontal, diagonal or vertical by their course in relation to the cortical surface.
For statistical evaluation the Goodness-of-fit test and a one-way test of variance with a 95 %
level of significance were performed. At the same time a paired data comparison by post hoc
test of Bonferroni was carried out. All relevant data sheets were extracted and interpreted. In
individual cases it had been necessary to switch to the Kruskal-Wallis test with adjacent post
hoc test of Dunn. The evaluation of the diagonal dendrites, which were summarized to merely
two data sheets, was performed after testing normal distribution by a t-test with independent
variables.
In the normal hearing control group a different system across the six cortical layers for the
horizontal as well as for the diagonal and vertical dendrites could be recognized. These
patterns were also present in the deafened and the stimulated group. On closer examination of
the vertical dendrites indications for a reorganisation could be verified. Inside lamina II of the
primary auditory field a loss of significant dominance of lamina III by an increase of the
percentage of vertical dendrites could be observed.
In the group of the chronic electric intracochlear stimulated cats the differences to the
normally hearing cats were even greater. In case of the vertical dendrites an increase could be
seen inside lamina II of all three auditory fields and inside lamina IV of the anterior auditory
field. This caused a loss of significant dominance of lamina III. Additional the horizontal
dendrites inside laminae II, V and VI of the secondary auditory field increased causing a loss
of dominance of lamina I. This has to be considered as a clear indication of plasticity.
These results showed a plasticity of the system of dendrites across the six laminae in the
auditory cortex after pharmacologically induced cochlear damage and chronic electrical
intracochlear stimulation.

8. Literaturverzeichnis
104
8. LITERATURVERZEICHNIS
AITKIN, L. M. (1986): The Auditory Midbrain. Structure and Function in the Central Auditory Pathway. Humana Press, Clifton, New Jersey
ANDERSEN, R. A., P. L. KNIGHT u. M. M. MERZENICH (1980): The thalamocortical and corticothalamic connections of AI, AII, and the anterior auditory field (AAF) in the cat: evidence for two largely segregated systems of connections. J. Comp. Neurol., 194, S. 663-701
ASHMORE, J. F. (1987): A fast motile response in guinea-pig outer hair cells: The cellular basis of the cochlear amplifier. J. Physiol., 388 , S. 323-347
BENSON, C. G., J. S. GROSS, S. K. SUNEJA u. S. J. POTASHNER (1997): Synaptophysin immunoreactivity in the cochlear nucleus after unilateral cochlear or ossicular removal. Synapse, 25, S. 243-257
BILAK, M., J. KIM, S. J. POTASHNER, B. A. BOHNE u. D. K. MOREST (1997): New growth of axons in the cochlear nucleus of adult chinchillas after acoustic trauma. Exp. Neurol., 147, S. 256-268
BILECEN, D., E. SEIFRITZ, E. W. RADÜ, N. SCHMID, S. WETZEL, R. PROBST u. K. SCHEFFLER (2000): Cortical reorganization after acute unilateral hearing loss traced by fMRI. Neurology, 54, S. 765-767
BLOOM, W. u. D. W. FAWCETT (1975): A Textbook of Histology. 10. Auflg. Saunders, Philadelphia, USA
BOLZ, J., C. D. GILBERT u. T. N. WIESEL (1989): Pharmacological analysis of cortical circuitry. Trends Neurosci., 12, S. 292-296
BRAITENBERG, V. u. A. SCHÜTZ (1991): Anatomy of the cortex. Springer-Verlag, Berlin BRECHMANN, A. (2003): A functional parcellation of the auditory cortex in humans using fMRI. 26th annual Midwinter Research Meeting, Daytona Beach, Florida, USA, S. 135
BRODMANN, K. (1909): Vergleichende Lokalisationslehre der Großhirnrinde in ihren Prinzipien dargestellt auf Grund des Zellaufbaus. Neudruck 1925 Bath-Verlag, Leipzig

8. Literaturverzeichnis
105
BROWNELL, W., C. BADER, D. BERTRAND u. Y. DE RIBEAUPIERRE (1985): Evoked mechanical responses of isolated cochlear outer hair cells. Science, 227, S. 194-196
BRUGGE, J. F. u. R. A. REALE (1985): Auditory cortex. in: A. PETERS u. E. G. E. JONES (Hrsg.) Cerebral Cortex, Vol. 4: Association and Auditory Cortices, Plenum Press, New York, S. 229-271
BÜCHNER, A. (2002): Entwicklung und Evaluation einer signalabhängigen "n aus m" -Sprachverarbeitungsstrategie für Cochlea-Implantate. Dissertation, Medizinische Hochschule Hannover
CHOW, K. L. u. A. LEIMAN (1970): The structural and functional organization of the neocortex. Neurosci. Res. Prog. Summ., 5, S. 153-220
CLAREY, J. C., P. BARONE u. T. J. IMIG (1991): Physiology of Thalamus and Cortex. in: A. N. POPPER u. R. R. FAY (Hrsg.) The Mammalian Auditory Pathway: Neurophysiology, 2. Auflg. Springer-Verlag, New York, Berlin, Heidelberg, S. 232-334
CLARKE, S. C., F. DE RIBAUPIERRE, E. M. ROUILLER u. Y. DE RIBAUPIERRE (1993): Several neuronal and axonal types form long intrinsic connections in the cat primary auditory cortical field. Anat. Embryol., 188, S. 117-138
CODE, R. A. u. J. A. WINER (1985): Commisural neurons in layer III of cat primary auditory cortex (AI): pyramidal and non-pyramidal cell input. Hear. Res., 23, S. 205-222
CORDS, S. M. (1996): Einfluß elektrischer intracochleärer Stimulation auf die Latenzen elektrisch evozierter auditorischer Hirnstammpotentiale bei der neonatal ertaubten Katze. Dissertation, Medizinische Hochschule Hannover/Tierärztliche Hochschule Hannover
COTANCHE, D. (1999): Structural recovery from sound and aminoglycoside damage in the avian cochlea. Audiol. Neurootol., 4, S. 271-285
DALLOS, P. (1986): Neurobiology of cochlear inner and outer hair cells: Intracellular recordings. Hear. Res., 22, S. 185-198
DeCARLOS, J. A., L. LOPEZ-MASCARAQUE u. F. VALVERDE (1985): Development, morphology and topography of chandelier cells in the auditory cortex of the cat. Dev. Brain Res., 22, S. 293-300

8. Literaturverzeichnis
106
DIAMOND, D. M. u. N. M. WEINBERGER (1984): Physiological plasticity of single neurons in auditory cortex of the cat during acquisition of the pupillary conditioned response: II. Secondary field (AII). Behav. Neurosci., 98, S. 189-210
DICKSON, J. W. u. G. L. GERSTEIN (1974): Interactions between neurons in the auditory cortex of the cat. J. Neurophysiol., 37, S. 1239-1261
DINSE, H. R., G. REUTER, S. M. CORDS, B. GODDE, T. HILGER u. T. LENARZ (1997): Optical imaging of cat auditory cortical organization following electrical stimulation of a multi-channel cochlear implant: differential effects of acute and chronic stimulation. Am. J. Otol., 18, S. 17-18
EDGE, R. M., B. N. EVANS, M. PEARCE, C.-P. RICHTER, X. HU u. P. DALLOS (1998): Morphology of the unfixed cochlea. Hear. Res., 124, S. 1-16
EGGERMONT, J. J., C. W. PONTON, S. G. COUPLAND u. R. WINKELAAR (1991): Frequency dependent maturation of the cochlea and brainstem evoked potentials. Acta Otolarngol. (Stockh.), 111, S. 220-224
EHRET, G. (1997): The auditory cortex. J. Comp. Physiol.[A], 181, S. 547-557
ELBERT, T. u. B. ROCKSTROH (2004): Reorganization of Human Cerebral Cortex: The Range of Changes following Use and Injury. Neuroscientist, 10, S. 129-141
EYBALIN, M. u. R. PUJOL (1987): Choline acetyltransferase (ChAT) immunelectron microscopy distinguishes at least three types of efferent synapses in the organ of Corti. Exp. Brain Res., 65, S. 679-684
FENG, J. Z. u. J. F. BRUGGE (1983): Postnatal development of auditory callosal connections in the kitten. J. Comp. Neurol., 214, S. 416-426
FIRSZT, J. B., W. GAGGL, P. A. WACKYM, J. L. ULMER, u. R. W. PROST (2003): Hemispheric activation in human auditory cortex of normal hearing and hearing loss subjects. 26th annual Midwinter Research Meeting, Daytona Beach, Florida, USA, S. 229
FITZPATRICK, K. A., T. J. IMIG u. R. A. REALE (1977): Thalamic projections to the posterior auditory field in cat. Brain Res., 15, S. 305-340
FRIAUF, E. u. C. J. SHATZ (1991): Changing patterns of synaptic input to subplate and cortical plate during development of visual cortex. J. Neurophysiol., 66, S. 2059-2071

8. Literaturverzeichnis
107
FULLERTON, B. C., R. A. LEVINE, H. L. HOSFORD DUNN u. N. Y. KIANG (1987): Comparsion of cat and human brainstem auditory evoked potentials. Electroencephalogr. Clin. Neuropyhsiol., 66, S. 547-570
GALUSKE, R. A., W. SCHLOTE, H. BRATZKE u. W. SINGER (2000): Interhemispheric asymetries of the modular structure in human temporal cortex. Science, 289, S. 1946-1949
GESCHWIND, N. u. W. LEVITSKY (1968): Human brain: left-right asymetries in temporal speech region. Science, 161, S. 186-187
GILBERT, C. D. (1993): Rapid dynamic changes in adult cerebral cortex. Curr. Opin. Neurobiol., 3, S. 100-103
GLASER, E. M., H. VAN DER LOOS u. M. GISSLER (1979): Tangential Orientation and Spatial Order in Dendrites of Cat Auditory Cortex: A Computer Microscope Study of Golgi-impregnated Material. Exp. Brain Res., 36, S. 411-431 GODEMENT, P., J. VANSELOW, S. THANOS u. F. BONHOEFFER (1987): A study in developing visual systems with a new method of staining neurones and their processes in fixed tissue. Development, 101, S. 697-713
GRIFFITH, G. (1993): Quantitative Aspects of Immunocytochemistry. in: Fine Structure Immunocytochemistry, Springer-Verlag, S. 371-411
GUPTA, M., T. M. MAYHEW, K. S. BEDI, A. K. SHARMA u. F. H. WHITE (1993): Inter-animal variation and its influence on the overall precision of morphometric estimates based on nested sampling designs. J. Microsc., 131, S. 147-154
HALL, D. (2003): Functional subdivisions within human auditory cortex: An overview. 26th annual Midwinter Research Meeting, Daytona Beach, Florida, USA, S. 133
HARDIE, N. A. u. R. K. SHEPHERD (1999): Sensorineural hearing loss during development: Morphological and physiological response of the cochlea and auditory brainstem. Hear. Res., 128, S. 147-165
HARRISON, R. V., A. NAGASAWA, D. W. SMITH, S. STANTON u. R. J. MOUNT (1991): Reorganization of auditory cortex after neonatal high frequency cochlear hearing loss. Hear. Res., 54, S. 11-19
HEBB, O. D. (1949): The organization of behaviour. A neurophysiological theory. Wiley, New Jork

8. Literaturverzeichnis
108
HEFFNER, R. S. und H. E. HEFFNER (1985): Hearing range of the domestic cat. Hear. Res., 19, S. 85-88 HEID, S., T. K. JÄHN-SIEBERT, R. KLINKE, R. HARTMANN u. G. LANGNER (1997): Afferent projection patterns in the auditory brainstem in normal and congenitally deaf white cats. Hear. Res., 110, S. 191-199
HEID, S., R. HARTMANN u. R. KLINKE (1998): A model for prelingual deafness, the congenitally deaf white cat - population statistics and degenerative changes. Hear. Res., 115, S. 101-112
HEIL, P., R. RAJAN u. D. R. F. IRVINE (1994): Topographic representations of tone intensity along the isofrequency axis of cat primary auditor cortex. Hear. Res., 76, S. 188-202
HONIG, M. G. u. R. I. HUME (1989): DiI and DiO: versatile fluorescent dyes for neuronal labeling and pathway tracing. Trends Neurosci., 12, S. 333-341
HONIG, M. G. (1993): DiI Labeling. in: Neuroscience Protocols, Elsevier Science Publishers, S. 1-20
HOTH, S. u. T. LENARZ (1994): Elektrische Reaktionsaudiometrie. Springer-Verlag, Berlin, Heidelberg
HOWARD, M. A., I. O. VOLKOV, P. J. ABBAS, H. DAMASIO, M. C. OLLENDIECK u. M. A. GRANNER (1996): A chronic microelectrode investigation of the tonotopic organization of the human auditory cortex. Brain Res., 724, S. 260-264
HUDSON, L. S. u. W. P. HAMILTON (1993): Atlas of feline anatomy for veterinarians. Saunders Company, Philadelphia, London
HULTCRANTZ, M., R. L. SNYDER, S. J. REBSCHER u. P. A. LEAKE (1991): Effects of neonatal deafening and chronic intracochlear electrical stimulation on the cochlear nucleus in cats. Hear. Res., 54, S. 272-280
HUTTENLOCHER, P. R. u. A. S. DABHOLKAR (1997): Regional differences in synaptogenesis in human cerebral cortex. J. Comp. Neurol., 387, S. 167-178

8. Literaturverzeichnis
109
HÜTTENBRINK, K.-B. (1992): Die Mechanik und Funktion des Mittelohres. Teil 1: Die Ossikelkette und die Mittelohrmuskeln. Laryngol. Rhinol. Otol., 71, S. 545-551
IMIG, T. J. u. J. F. BRUGGE (1978): Sources and terminations of callosal axons related to binaural and frequency maps in primary auditory cortex of the cat. J. Comp. Neurol., 182, S. 637-660
IMIG, T. J. u. R. A. REALE (1981): Ipsilateral cortico-cortical projections related to binaural columns in cat primary auditory cortex. J. Comp. Neurol., 203, S. 1-14
IMIG, T. J. u. A. MOREL (1983): Organization of the thalamocortical auditory system in the cat. Annu. Rev. Neurosci., 6, S. 95-120
IRVINE, D. R. F. (1992): Physiology of the auditory brainstem. in: R. R. FAY u. A. N. POPPER (Hrsg.) The Physiology of the mammalian auditory CNS, Springer-Verlag, Berlin, Heidelberg, New York, S. 153-231
IRVINE, D. R. F., R. RAJAN u. L. AITKIN (1996): Sensitivity to interaural intensity differences of neurons in primary auditory cortex of the cat. I. Types of sensitivity and effects of variations in sound pressure level. J. Neurophysiol., 75, S. 75-96
IRVINE, D. R. F., R. RAJAN u. M. BROWN (2001): Injury- and Use-Related Plasticity in Adult Auditory Cortex. Audiol. Neurootol., 6, S. 192-195
KACZMAREK, L., M. KOSSUT u. J. SKANGIEL-KRAMSKA (1997): Glutamate receptors in cortical plasticity: Molecular and cellular biology. Physiol. Rev., 77, S. 217-255
KELLER, P. (1997): Untersuchungen zur Entwicklung der frühen akustisch evozierten Potentiale (FAEP) bei der Katze für den Einsatz in der Grundlagenforschung und zur klinischen Anwendung. Dissertation, Tierärztliche Hochschule Hannover
KEMP, D. T. (1979): Evidence of mechanical nonlinearity and frequency selective wave amplification in the cochlea. Arch. Otorhinolaryngol., 224, S. 37-45
KITZES, L. M. (1984): Some physiological consequences of neonatal cochlear destruction in the inferior colliculus of the gerbil, Meriones unguiculatus. Brain Res., 306, S. 171-178

8. Literaturverzeichnis
110
KLINKE, R. (1987): Die Verarbeitung von Schallreizen im Innenohr - eine Übersicht über neuere Forschungsergebnisse. HNO, 35, S. 139-148
KLINKE, R. u. R. HARTMANN (1997): Basic neurophysiology of cochlear implants. Am. J. Otol., 18, S. 7-10
KLINKE, R., A. KRAL, S. HEID, J. TILLEIN u. R. HARTMANN (1999): Recruitment of the Auditory Cortex in Congenitally Deaf Cats by Long-Term Cochlear Electrostimulation. Science, 285, S. 1729-1733
KÖBBERT, C., R. APPS, I. BECHMANN, J. L. LANCIEGO, J. MEY u. S. THANOS (2000): Current concepts in neuroanatomical tracing. Prog. Neurobiol., 62, S. 327-351
KRAL, A., R. HARTMANN, J. TILLEIN, S. HEID u. R. KLINKE (2000): Congenital Auditory Deprivation Reduces Synaptic Activity within the Auditory Cortex in a Layer-specific Manner. Cereb. Cortex, 10, S. 714-726
KRAL, A., R. HARTMANN, J. TILLEIN, S. HEID u. R. KLINKE (2001): Auditory developmental plasticity in cats: A sensitive period of 6 months. Assoc. Res. Otolaryngol., 24, S. 128
KRAL, A., R. HARTMANN, J. TILLEIN, S. HEID u. R. KLINKE (2002): Hearing after Congenital Deafness: Central Auditory Plasticity and Sensory Deprivation. Cereb. Cortex, 12, S. 797-807
KRAL, A., J. H. SCHRÖDER, R. KLINKE u. A. K. ENGEL (2003): Absence of cross-modal reorganization in the primary auditory cortex of congenitally deaf cats. Exp. Brain Res., 153, S. 605-613
KRUCKER, T. (1996): Contribution to the pharmacology and electrophysiology of the primary auditory cortex in the cat. Dissertation, Sandoz Forschungsinstitut Bern LEAKE, P. A. u. G. T. HRADEK (1988): Cochlear pathology of long term neomycin induced deafness in cats. Hear. Res., 33, S. 11-34
LEAKE, P. A., G. T. HRADEK, S. J. REBSCHER u. R. L. SNYDER (1991): Chronic intracochlear electrical stimulation induces selective survival of spiral ganglion neurons in neonatally deafened cats. Hear. Res., 54, S. 251-271

8. Literaturverzeichnis
111
LEAKE, P. A., G. T. HRADEK u. R. L. SYNDER (1999): Chronic electrical stimulation by a cochlear implant promotes survival of spiral ganglion neurons after neonatal deafness. J. Comp. Neurol., 412, S. 543-562
LEE, D. S., J. S. LEE, S. H. OH, S.-K. KIM, J.-W. KIM, J.-K. CHUNG, M. C. LEE u. C. S. KIM (2001): Deafness - Cross-modal plasticity and cochlear implants. Nature, 409, S. 149-150
LEHNHARDT, E. (1993): Intracochleäre Platzierung der Cochlear-Implant-Elektroden in soft surgery technique. HNO, 41, S. 356-359
LEVAY, S. u. C. D. GILBERT (1976): Laminar patterns of geniculocortical projection in the cat. Brain Res., 175, S. 233-245
LIGEOIS-CHAUVEL, C., A. MUSOLINO u. P. CHAUVEL (1991): Localization of the primary auditory cortex in man. Brain, 114, S. 139-153
LINDEMAN, H. H., H. W. ADES, G. BREDBERG u. H. ENGSTROM (1971): The sensory hairs and the tectorial membrane in the development of the cat´s organ of Corti. A scanning electron microscopic study. Acta Otolaryngol., 72, S. 229-242
LOMAX, M. I., T. W. GONG, Y. CHO, L. HUANG, S. H. OH, H. J. ADLER, Y. RAPHAEL u. R. A. ALTSCHULER (2001): Differential Gene Expression Following Noise Trauma in Birds and Mammals. Noise Health, 3, S. 19-35
LORENTE DE NÓ, R. (1933): Anatomy of the eigth nerve. III. General plan of structure of the primary cochlear nuclei. Laryngoscope, 43, S. 327-350
LORENTE DE NÓ (1949): Cerebral Cortex: architecture, intracortical connections, motor projections. in: J. F. FULTON (Hrsg.) The Physiology of the Nervous System, Oxford Univ. Press, London, S. 288-330
LUKAS, J.-R., M. AIGNER, M. DENK, H. HEINZL, M. BURIAN u. R. MAYR (1998): Carbocyanine postmortem neuronal tracing. Influence of different parameters on tracing distance and combination with immunocytochemistry. J. Histochem. Cytochem., 46, S. 901-910
LUND, J. S. (1988): Anatomical organization of macaque monkey striate cortex. Annu. Rev. Neurosci., 11, S. 253-288

8. Literaturverzeichnis
112
LUSKIN, M. B. u. C. J. SHATZ (1985): Neurogeneses of the cat´s primary visual cortex. J. Comp. Neurol., 242, S. 611-631
LUSTIG, L. R., P. A. LEAKE, R. L. SNYDER u. S. J. REBSCHER (1994): Changes in cat cochlear nucleus following neonatal deafening and chronic intracochlear electrical stimulation. Hear. Res., 74, S. 29-37
MARIANOWSKI, R., W.-H. LIAO, T. VAN DEN ABBEELE, P. FILLIT, P. HERMAN, B. FRACHET u. P. TRAN BA HUY (2000): Expression of NMDA, AMPA and GABAA receptor subunit mRNAs in the rat auditory brainstem. I. Influence of early auditory deprivation. Hear. Res., 150, S. 1-11
MARTIN, K. A. C. (1984): Neuronal circuits in cat striate cortex. in: E. G. JONES u. A. PETERS (Hrsg.) Cerebral Cortex, Functional properties of cortical cells, 2. Auflg. Plenum Press, New York, London, S. 241-284
MCMULLEN, N. T. u. E. M. GLASER (1988): Auditory cortical responses to neonatal deafening: pyramidal neuron spine loss without changes in growth or orientation. Exp. Brain Res., 72, S. 195-200
MCMULLEN, N. T., B. GOLDBERGER, C. M. SUTER u. E. M. GLASER (1988): Neonatal Deafening Alters Nonpyramidal Dendrite Orientation in Auditory Cortex: A Computer Microscope Study in the Rabbit. J. Comp. Neurol., 267, S. 92-106
MCMULLEN, N. T., C. B. SMELSER u. R. K. DE VENECIA (1994): A Quantitative Analysis of Parvalbumin Neurons in Rabbit Auditory Neocortex. J. Comp. Neurol., 349, S. 493-511
MELCHER, J. R., J. J. GUINAN, I. M. KNUDSEN u. N. Y. KIANG (1996): Generators of the brainstem auditory evoked potential in cat II. Correlating lesion sites with waveform changes. Hear. Res., 93, S. 28-51
MERZENICH, M. M. u. C. E. SCHREINER (1992): Mammalian auditory cortex – some comparative observations. in: D. F. R. WEBSTER (Hrsg.) The evolutionary biology of hearing., Springer, Berlin, Heidelberg, New York, S. 673-689
MERZENICH, M., S. A. COLWELL u. R. A. ANERSEN (1982): Auditory forebrain organization. in: C. ED. WOOSLEY (Hrsg.) Cortical Sensory Organization. Vol. 3: Multiple Auditory Areas, Humana Press, Clifton, New Jersey, S. 43-57

8. Literaturverzeichnis
113
MEYER, G., A. CASTANEYRA-PERDOMO u. R. FERRES-TORRES (1984): A type of apparently axonless granule cell in the cat auditory cortex. A Golgi study. Neurosciene, 33, S. 489-498
MIDDLEBROOKS, J. C., R. W. DYKES u. M. M. MERZENICH (1980): Binaural response-specific banks in primary auditory cortex (AI) of the cat: topographic organization orthogonal to isofrequency contours. Brain Res., 181, S. 31-48
MITANI, A., K. ITOH, S. NOMURA, M. KANEKO u. N. MIZUNO (1984): Thalamocortical projections to layer I of the primary auditory cortex in the cat: a horseradish peroxidase study. Brain Res., 310, S. 347-350
MITANI, A., M. SHIMOKOUCHI, K. ITOH, S. NOMURA, M. KUDO u. N. MIZUNO (1985): Morphology and laminar organization of electrophysiologically identified neurons in the primary auditory cortex in the cat. J. Comp. Neurol., 235, S. 430-447
MOORE, D. R. (1985): Postnatal development of the mammalian central auditory system and the neural consequences of auditory deprivation. Acta Otolaryngol. Suppl., 421, S. 19-30
MOORE, D. R. (1994): Auditory brainstem of the ferret - long survival following cochlear removal progressively changes projections from the cochlear nucleus to the inferior colliculus. J. Comp. Neurol., 339, S. 301-310
MOORE, J. K. u. Y.-L. GUAN (2001): Cytoarchitectural and axonal maturation in human auditory cortex. JARO, 2, S. 297-311
MOREL, A. u. T. J. IMIG (1984): Neurons in the tonotopic thalamus of the cat are topographically organized with respect to their target fields in auditory cortex. Soc. Neurosci. Abstr., 10, S. 244
NEVILLE, H. J. u. D. LAWSON (1987): Attention to central and peripheral visual space in a movement detection task. III. Seperate effects of auditory deprivation and acquisition of a visual language. Brain Res., 405, S. 284-294
NIIMI, K. u. F. NAITO (1974): Cortical projections of the medial geniculate body in the cat. Exp. Brain Res., 19, S. 326-342
NORDEEN, K. W., H. P. KILLACKEY u. L. M. KITZES (1983): Ascending projections to the inferior colliculus following unilateral cochlear ablation in the neonatal gerbil, Meriones unguiculatus. J. Comp. Neurol., 214, S. 144-153

8. Literaturverzeichnis
114
OJIMA, H., C. N. HONDA u. E. G. JONES (1992): Characteristics of Intracellularly Injected Infragranular Pyramidal Neurons in Cat Primary Auditory Cortex. Cereb. Cortex, 2, S. 197-216
ONG, W. J. u. L. J. GAREY (1990): Neuronal archirecture of the human temporal cortex and of the mustache bat. Anat. Embryol., 181, S. 351-364
PARASNIS, I. (1998): Cognitive diversity in deaf people: Implication for communication and education. Scand. Audiol. Suppl., 49, S. 109-115
PETERS, A. u. E. G. JONES (1984): Classification of cortical neurons. in: A. PETERS u. E. G. JONES (Hrsg.) Cerebral cortex, Cellular components of the cerebral cortex, 1. Auflg. Plenum Press, New York, London, S. 107-121
PETITTO, L. A., R. J. ZATORRE, K. GAUNA, E. J. NIKELSKI, D. DOSTIE u. A. C. EVANS (2000): Speech-like cerebral activity in profoundly deaf people processing signed language: Implications for neural basis of human language. Proc. Natl. Acad. Sci. USA, 97, S. 13961-13966
PHILIPPS, D. P. u. D. R. F. IRVINE (1982): Properties of single neurons in the anterior auditory field (AAF) of cat cerebral cortex. Brain Res., 248, S. 237-244
PONTON, C. W. u. J. J. EGGERMONT (2001): Of Kittens and Kids: Altered Cortical Maturation following Profound Deafness and Cochlear Implant Use. Audiol. Neurootol., 6, S. 363-380
POPELAR, J., R. HARTMANN, J. SYKA u. R. KLINKE (1995): Middle latency responses to acoustical and electrical stimulation of the cochlea in cats. Hear. Res., 92, S. 63-77
POWELL, T. P. S. u. V. B. MOUNTCASTLE (1959): Some aspects of the functional organization of the postcentral gyrus of the monkey: a correlation of findings obtained in a single unit analysis with architecture. Bull. Johns Hopkins Hosp., 105, S. 133-162
PRIETO, J. J., B. A. PETERSON u. J. A. WINER (1994): Morphology and spatial distribution of GABAergic neurons in cat primary auditory cortex (AI). J. Comp. Neurol., 344, S. 349-382
PUJOL, R. u. R. MARTY (1970): Postnatal maturation in the cochlea of the cat. J. Comp. Neurol., 139, S. 115-120

8. Literaturverzeichnis
115
RAGGIO, M. W. u. C. E. SCHREINER (1999): Neuronal responses in cat primary auditory cortex to electrical cochlear stimulation III. Activation patterns in short- and long-term deafness. J. Neurophysiol., 82, S. 3506-3526
RAUSCHECKER, J. P. u. M. KORTE (1993): Auditory compensation for early blindness in cerebral cortex. J. Neurosci., 13, S. 4538-4548
REALE, R. A. u. T. J. IMIG (1980): Tonotopic organization in auditory cortex of the cat. J. Comp. Neurol., 192, S. 265-291
REALE, R. A. u. R. E. KETTNER (1986): Topography of binaural organization in primary auditory cortex of the cat: Effects of changing interaural intensity. J. Neurophysiol., 56, S. 663-682
REALE, R. A., J. F. BRUGGE u. J. C. K. CHAN (1987): Maps of auditory cortex in cats reared after unilateral cochlear ablation in the neonatal period. Brain Res., 431, S. 281-290
REBILLARD, G., E. CARLIER, M. REBILLARD u. R. PUJOL (1977): Enhancement of visual responses on the primary auditory cortex of the cat after an early destruction of cochlear receptors. Brain Res., 129, S. 162-164
REBILLARD, G., M. REBILLARD u. R. PUJOL (1980): Factors effecting the recording of visual-evoked potentials from the deaf cat primary auditory cortex (AI). Brain Res., 188, S. 252-254
REDIES, H., J. F. BRUGGE u. J. C. K. CHAN (1989): Functional subdivisions in the auditory cortex of the guinea pig. J. Comp. Neurol., 282, S. 473-488
REUTER, G. (1997): Untersuchungen zur Aufhebung von neonataler akustischer Deprivation durch das Cochlea Implantat: Sinnesersatz bei gehörlosen Kindern. Habilitation, Medizinische Hochschule Hannover
REUTER, G., A. H. GITTER u. H. P. ZENNER (1991):
Morphologischer Nachweis schneller Bewegungen äußerer Haarzellen des Innenohres als Ursprung des cochleären Verstärkungsprozesses. Verh. Anat. Ges., 83, S. 583-584
REUTER, G., B. GODDE, S. M. CORDS, T. HILGER, T. LENARZ, u. H. R. DINSE (2002a): Functional and structural reorganization of auditory cortex following long-term cochlear implant stimulation. 7th European Workshop on Cochlear Implants, Auditory Brainstem Implants and Implantable Hearing Aids, Hannover

8. Literaturverzeichnis
116
REUTER, G., S. M. CORDS, T. HILGER, B. GODDE, H. R. DINSE, u. T. LENARZ (2002b): Reorganization of the deafened auditory cortex after chronical electrical intracochlear multi-channel stimulation revealed by optical imaging. 7th European Workshop on Cochlear Implants, Auditory Brainstem Implants and Implantable Hearing Aids, Hannover
ROSE, J. E. (1949): The cellular structure of the auditory region of the cat. J. Comp. Neurol., 19, S. 409-439
ROSE, J. E. u. C. N. WOOLSEY (1949): Structure and relations of thalamic connections, cellular structure and evocable electrical activity in the auditory region of the cat. J. Comp. Neurol., 91, S. 441-466
ROUILLER, E. M., G. M. SIMM, A. E. P. VILLA, Y. DE RIBAUPIERRE u. F. DE RIBAUPIERRE (1991): Auditory corticocortical interconnections in the cat: evidence for parallel and hierarchical arrangement of the auditory cortical areas. Exp. Brain Res., 86, S. 483-505
RYUGO, D. K. u. H. P. KILLACKEY (1974): Differential telencephalic projections of the medial and ventral divisions of the medial geniculate body of the rat. Brain Res., 82, S. 173-177
SACHS, M. B. (1984): Neural coding of Complete Sounds. Ann. Rev. Physiol., 46, S. 261-273
SALVI, R. J., J. WANG u. D. DING (2000): Auditory plasticity and hyperactivity following cochlear damage. Hear. Res., 147, S. 261-274
SCHNEIDER, T. (1978): The influence of counting rules on the Number and on the size distribution of fibres. Ann. Occup. Hyg., 21, S. 341-350
SCHREINER, C. E. u. M. S. CYNADER (1984): Basic functional organization of second auditory cortical field (AII) of the cat. J. Neurophysiol., 51, S. 1284-1305
SELDON, H. L. (1982): Structure of Human Auditory Cortex. III. statistical Analysis of Dendritic Trees. Brain Res., 249, S. 211-221
SHEPHERD, R. K., R. HARTMANN, S. HEID, N. A. HARDIE u. R. KLINKE (1997): The central auditory system and auditory deprivation: experience with cochlear implants in the congenitally deaf. Acta Oto-Laryngol. Suppl., 532, S. 28-33

8. Literaturverzeichnis
117
SIE, K. C. u. E. W. RUBEL (1992): Rapid changes in protein synthesis and cell size in the cochlear nucleus following eighth nerve activity blockade or cochlear ablation. J. Comp. Neurol., 320, S. 501-508
SILBERNAGL, S. u. A. DESPOPOULOS (2003): Taschenatlas der Physiologie. 5. Auflg. Thieme-Verlag, Stuttgart, S. 310-370
SKUSE, D. H. (1993): Extreme deprivation in early childhood. in: D. BISHOP u. K. MOGFORD (Hrsg.) Language Development in Exceptional Circumstances, Erlbaum, Hillsdale, S. 29-46
SNYDER, R. L., S. J. REBSCHER, K. CAO, P. A. LEAKE u. K. KELLY (1990): Chronic intracochlear electrical stimulation in the neonatally deafened cat. I. Expansions of central representations. Hear. Res., 50, S. 7-34
SOUSA-PINTO, A. (1973a): Cortical projections of the medial geniculate body in the cat. Adv. Anat. Embryol. Cell. Biol., 48, S. 1-42
SOUSA-PINTO, A. (1973b): The structure of the first auditory cortex (AI) in the cat. I. Light microscopic observations on its structure. Arch. Ital. Biol., 111, S. 112-137
SOUSA-PINTO, A., M. M. PAULA-BARBOSSA u. M. D. C. MATOS (1975): A Golgi and electron microscopic study of nerve cells in layer 1 of the cat auditory cortex. Brain Res., 95, S. 443-458
SPOENDLIN, H. (1969): Innervation patterns of the organ of Corti of the cat. Acta Oto-Laryngol., 67, S. 239-254
SUNEJA, S. K., C. G. BENSON u. S. J. POTASHNER (1998): Glycerine receptors in adult guinea pig brain stem auditory nuclei: Regulation after unilateral cochlear ablation. Exp. Neurol., 154, S. 473-488
SUTTER, M. L. u. C. E. SCHREINER (1995): Topography of intensity tuning in cat primary auditory cortex: single-neuron versus multiple-neuron recordings. J. Neurophysiol., 73, S. 190-204
TERAYAMA, Y., Y. KANEKO, K. KAWAMOTO u. N. SAKAI (1977): Ultrastructural changes of the nerve elements following disruption of the organ of Corti. I. Nerve elements in the organ of Corti. Acta Otolaryngol., 83, S. 291-302

8. Literaturverzeichnis
118
TIAN, B. (1992): A neuroethological approach to the study of cat non-primary auditory cortex. Dissertation, Universität Tübingen
TIERNEY, T. S., F. A. RUSSEL u. D. R. MOORE (1997): Susceptibility of developing cochlear nucleus neurons to deafferentation-induced death abruptly ends just before the onset of hearing. J. Comp. Neurol., 378, S. 295-306
VOGT, P., S. M. CORDS, P. KELLER, T. LENARZ, u. G. REUTER (1997): Chronic electrical intracochlear multichannel stimulation effects the auditory pathway in neonatally deafened kittens: III. Inferior Colliculus. 25th Göttingen Neurobiology Conference
VON BÉKÉSY, G. (1928): Zur Theorie des Hörens: Die Schwingungsform der Basilarmembran. Physik in unserer Zeit, 29, S. 793-810
VON ECONOMO, C. u. L. HORN (1930): Über Windungsrelief, Masse und Rindenarchitektonik der Supratemporalfläche, ihre individuellen und ihre Seitenunterschiede. Z. Ges. Neurol. Psychol., 130, S. 687-759
WALSH, E. J., J. MCGEE u. E. JAVEL (1986): Development of auditory-evoked potentials in the cat. I. Onset of response and development of sensitivity. J. Acoust. Soc. Am., 79, S. 712-724
WALSH, E. J. u. J. MCGEE (1990): Development of auditory coding in the central nervous system: implications for in utero hearing. Seminars in Perinatology, 14, S. 281-293
WARR, W. (1992): Efferent pathways. in: A. N. POPPER u. R. R. FAY (Hrsg.) The Mammalian Auditory Pathway: Neuroanatomy, Springer-Verlag, New York
WEINBERGER, N. M., W. HOPKINS u. D. M. DIAMOND (1984): Physiological plasticity of single neurons in auditory cortex of the cat during acquisition of the pupillary conditioned response: I. Primary Field (AI). Behav. Neurosci., 98, S. 171-188
WENKE, K. A., S. M. CORDS, P. ERFURT, T. LENARZ, u. G. REUTER (1997): Chronic electrical intracochlear multichannel stimulation effects the auditory pathway in neonatally deafened kittens: IV. Auditory Cortex. 25th Göttingen Neurobiology Conference

8. Literaturverzeichnis
119
WENKE, K. A. (1999): Morphologische Veränderungen im auditorischen Cortex der Katze nach neonataler Ertaubung und chronischer intracochleärer Stimulation. Dissertation, Medizinische Hochschule Hannover/Tierärztliche Hochschule Hannover
WILSON, B. S., C. C. FINLEY, D. T. LAWSON, R. D. WOLFORD, D. K. EDDINGTON u. W. M. RABINOWITZ (1991): Better speech recognition with cochlear implants. Nature, 352, S. 236-238
WINER, J. A. (1984a): The non-pyramidal neurons in layer III of cat primary auditory cortex (AI). J. Comp. Neurol., 229, S. 512-530
WINER, J. A. (1984b): The pyramidal cells in layer III of cat primary auditory cortex (AI). J. Comp. Neurol., 229, S. 476-496
WINER, J. A. (1984c): Anatomy of the layer IV in cat primary auditory cortex (AI). J. Comp. Neurol., 224, S. 535-567
WINER, J. A. (1992): The functional architecture of the medial geniculate body and the primary auditory cortex. in: D. F. R. WEBSTER (Hrsg.) The mammalian auditory pathways: Neuroanatomy, Springer-Verlag, Berlin, Heidelberg, New York, S. 222-409
WINGUTH, S. D. u. J. A. WINER (1986): Corticocortical connections of cat primary auditory cortex (AI): laminar organization and identification of supragranular neurons projecting to area AII. J. Comp. Neurol., 248, S. 36-56
WONG, D. u. J. P. KELLY (1981): Differentially projecting cells in individual layers of the auditory cortex: a double-labeling study. Brain Res., 230, S. 362-366
WOOLSEY, C. N. u. E. M. WALZL (1942): Topical projection of nerve fibres from local regions of the cochlea to the cerebral cortex of the cat. Bull. Johns Hopkins Hosp., 71, S. 315-344
WURTH, N. N. (1999): Vergleichende Morphologie der Neurone im primären auditorischen Cortex der kongenital gehörlosen weißen Katze. Dissertation, Johann Wolfgang Goethe-Universität Frankfurt am Main
WURTH, N. N., S. HEID, A. KRAL, u. R. KLINKE (1999): Morphology of neurons in the primary auditory Cortex (AI) in normally and congenitally deaf cats - a study of DiI labeled cells. 27th Göttingen Neurobiology Conference, S. 318

8. Literaturverzeichnis
120
ZENNER, H. P. u. J. SCHACHT (1986): Hörverlust durch Aminoglykosid-Antibiotika: Angriff am Membranbaustein PIP2 in äußeren Haarzellen als Wirkungsmechanismus. HNO, 34, S. 417-423
ZENNER, H. P. u. A. H. GITTER (1987): Die Schallverarbeitung des Ohres. Physik in unserer Zeit, 4, S. 97-105
ZENNER, H. P. (1993): Hören. in: R. F. SCHMIT (Hrsg.) Neuro- und Sinnesphysiologie, Springer-Verlag, Berlin, Heidelberg, S. 305-328
ZENNER, H. P. (1994): Physiologische und biochemische Grundlagen des normalen und gestörten Gehörs. in: H. H. NAUMANN, J. HELMS, C. HERBERHOLD u. E. H. KASTENBAUER (Hrsg.) Oto-Laryngologie in Klinik und Praxis, Thieme-Verlag, Stuttgart, S. 81-231

9. Anhang
121
9. ANHANG
9.1 Das Cochlea-Implantat
Cochlea-Implantate sind elektrische Innenohrprothesen, welche bei Vorliegen eines Ausfalls
des Corti-Organs dessen Funktion ersetzen. Durch die Versorgung mit einem Cochlea-
Implantat ist es möglich Personen, deren Hörverlust auf eine sensorische, von den Haarzellen
ausgehende Schallempfindungsschwerhörigkeit bei intakter zentraler Hörbahn beruht, zu
einem offenen Sprachverständnis zu verhelfen. Dabei werden adäquate Schallinformationen
über die elektrischen Kontakte des Cochlea-Implantats als elektrische Reize direkt an den
Nervus vestibulocochlearis weitergeleitet (KLINKE und HARTMANN 1997).
Das im Rahmen dieser Untersuchung verwendete Cochlea-Implantat bestand aus einer
Stimulationselektrode, einem Sprachprozessor mit Mikrophon und externem Implantat-
empfänger sowie einer Programmiereinheit. Alle genutzten Bestandteile werden auch klinisch
am humanen Patienten eingesetzt.
Bei der Stimulationselektrode handelte es sich um eine Silikon-ummantelte mehrkanalige
elektrische Innenohrprothese der Firma Cochlear Ltd. (Sydney, Australien). Die an der Spitze
der Prothese lokalisierten Elektrodenkontakte werden durch 6 bis 8 Plättchen oder Ringe aus
Platin-Iridium dargestellt, ein die Verbindung zum Implantatempfänger herstellender
Steckkontakt bildet den Elektrodenabschluss. Dieser Steckkontakt und die Elektroden-
kontakte sind durch Platindrähte verbunden.
Der Sprachprozessor, Modell MMT-5202, der Firma Advanced Bionics (Sylmar, Californien)
wandelt die über das Mikrophon eingehenden akustischen Signale in einen elektrischen Code
um. Dieser wird an den Implantatempfänger weitergeleitet, dekodiert, in elektrische Signale
umgewandelt und der Mehrkanalelektrode mit den verschiedenen Elektrodenkontakten
zugeführt. Die sich anschließende Steckverbindung besteht aus mehreren kunststoff-
ummantelten Kupferdrähten, welche jeweils einen Ausgang mit einem Elektrodenkontakt der
Elektrode verbinden.
Das Mikrophon befindet sich im sogenannten Kopfstück, welches mit dem Implantat-
empfänger über einen Magneten verbunden ist.
Aufgrund der anatomischen Verhältnisse bei der Katze konnte der Implantatempfänger in
diesen Untersuchungen nicht wie beim Menschen retroauriculär implantiert werden, sondern
musste extern in einem Kunststoffgehäuse an einer dem Tier angepassten Nylonjacke platziert
werden.

9. Anhang
122
Die Mehrkanalelektrode und der externe Implantatempfänger standen über eine Steck-
verbindung in Kontakt. Das an den Implantatempfänger angeschlossene Mikrophon und der
Sprachprozessor verfügten über eine zusätzliche, zweiteilige Kabelverbindung, so dass
Informationen einerseits vom Mikrophon zum Sprachprozessor, andererseits auch vom
Prozessor zum Implantatempfänger geleitet werden konnten.
Zur Programmierung des Sprachprozessors wurde ein Personal Computer (386er Prozessor)
mit Interface und integrierter Software der Firma Advanced Bionics genutzt. Eine direkte
Verbindung von Prozessor und Interface ermöglichte die Datenübertragung von der
Rechnereinheit zum Sprachprozessor. Um den Prozessor neu programmieren zu können,
wurden bereits vorhandene Parameter mit Hilfe einer Ultraviolett-Lichtbrücke gelöscht.
9.2 Die chronische elektrische intracochleäre Stimulation
Für die chronische elektrische Stimulation der drei mit einem Cochlea-Implantat versehenen
Versuchstiere wurde eine Beschallung durch ein in unmittelbarer Nähe der Tiere aufgestelltes
Radio sowie durch Umgebungsgeräusche vorgenommen.
Zur Codierung der akustischen Signale wurde die „continuous interleaved sampling“ (CIS)
Stimulationsstrategie eingesetzt (WILSON et al. 1991). Dabei werden die über das Mikrofon
eingehenden Signale verstärkt, Hochpass-gefiltert, komprimiert und nach Digitalisierung in
Form von pulsatilen Wellen, nahezu simultan auf vier den entsprechenden Elektrodenpaaren
zugeordneten Frequenzbändern weitergeleitet. Durch die unterschiedlichen Elektroden-
konfigurationen ist eine Aufteilung des Frequenzbereichs von 350 bis 5500 Hz auf vier
verschiedene Kanäle möglich. Innerhalb eines jeden Frequenzbandes wurde das Signal nach
Durchlaufen eines Gleichrichters und nach Tiefpass-Filterung logarithmisch komprimiert und
als Radiosignal über ein Kabel an den Implantatempfänger weitergegeben. Das vom
Implantatempfänger dekodierte Signal wurde schließlich über eine entsprechende
Steckverbindung in die Elektrode eingespeist.
Die Phasenlänge des zur Stimulation verwendeten biphasischen Rechteckimpulses betrug 75
�P� PLW� HLQHU� )UHTXHQ]� YRQ� ����� +]�� ,P� ELSRODUHQ� 0RGXV� ZXUGHQ� GLUHNW� EHQDFKEDUWH�Elektrodenkontakte gegeneinander stimuliert. Andere Stimulationskombinationen ergaben
sich nur dann, wenn das Ausweichen auf eine andere Kombination aufgrund des Defekts
eines oder mehrerer Elektrodenkontakte notwendig wurde.

9. Anhang
123
9.3 Die Überprüfung des Hörstatus durch Messung der frühen akustisch (FAEP) und
elektrisch (FEEP) evozierten Potentiale
Zur Überprüfung der medikamentösen Ertaubung der neonatalen Versuchstiere wurden am
14. und bei einem Resthörvermögen (Antwort der auditorischen Hörbahn auf Click-Reize bei
100 dB SPL) zusätzlich am 19. Lebenstag die frühen akustisch evozierten Potentiale (FAEP)
abgeleitet (siehe Abb.22). Dabei handelt es sich um durch Schallreize ausgelöste und infolge
der auditorischen Reizverarbeitung und -wahrnehmung entstehende elektrische Spannungen,
welche an der Körperoberfläche registriert werden können (HOTH und LENARZ 1994). Vor
Aufnahme in die Kontrollgruppe wurde diese physiologische Untersuchung ebenfalls an den
hörenden Tieren durchgeführt (siehe Abb. 21).
Über einen Luftleiterkopfhörer wurden hierzu den narkotisieren Tieren, durch direkte
Positionierung des Endstückes mit den dazugehörigen Dichtungsstöpseln im äußeren
Gehörgang, akustische Click-Reize mit definierten Lautstärken (0 bis 100 DB SPL) und einer
Reizfrequenz von 20 Hz zugeführt. Diese biphasischen Rechteckimpulse führen durch eine
kurze Reizdauer (150 µs) und der damit verbundenen schnellen Änderung des Schalldruckes
zu einer synchronen Stimulation einer großen Anzahl von Haarzellen. Zur Detektion der
physiologischen Spannungsänderungen wurden Feinnadel-Ableitelektroden jeweils subcutan
an der Stirn (gemeinsamer Plus-Pol), am rechten und linken Mastoid (Minus-Pol) und am
Nacken (Erdung) platziert.
Bei den chronisch elektrisch intracochleär stimulierten Tieren wurden analog zu den
akustischen, die frühen elektrisch evozierten Potentiale (FEEP) gemessen (siehe Abb. 23).
Dabei erfolgt die Reizgebung nicht über akustische Signale, sondern über das intracochleär
platzierte Cochlea-Implantat.
Mit Hilfe eines Messsystems der Firma ZLE-Systemtechnik GmbH (München), bestehend
aus einer Rechnereinheit, einem Signalgenerator mit integrierter Stromquelle und einem
DATA-Acquisition-System mit integriertem Verstärker wurden die FAEPs und FEEPs
abgeleitet, verstärkt, Bandpass-gefiltert, digitalisiert, gemittelt, aufgezeichnet und auf dem
Monitor als Messkurven dargestellt. Um Spannungsschwankungen durch andere, im gleichen
Stromnetz betriebene Großgeräte auszuschließen, wurde ein Spannungskonstanthalter
zwischen die Messapparatur und das lokale Stromnetz geschaltet. Er gewährleistete einen
Überspannungsschutz und eine Batteriepufferung.
9. Anhang
124
Abb. 21: FAEP-Kurven einer hörenden Katze (ipsilaterale Ableitungen vom rechten (re) und linken
(li) Ohr bei 10 bis 80 dB SPL)
Abb. 22: FAEP-Kurven einer experimentell neonatal ertaubten Katze (ipsilaterale Ableitungen vom rechten (re) und linken (li) Ohr bei 50 bis 100 dB SPL)
9. Anhang
125
Abb. 23: FEEP-Kurven einer chronisch elektrisch intracochleär stimulierten Katze (ipsilaterale
Ableitung vom linken (li) Ohr bei 180 bis 400 µA)
9.4. Die täglichen Körpergewichtszunahmen der Versuchstiere
Um eventuell auftretende Störungen des Allgemeinbefindens bei den neonatal experimentell
ertaubten Katzen aufzudecken, wurden alle Versuchstiere in den ersten 14 Lebenstagen
täglich gewogen und die Körpergewichtszunahmen dokumentiert.
Tab. 16a, b: Übersicht über die täglichen Körpergewichtszunahmen in Gramm [g] der hörenden
Versuchstiere inkl. interner Registrierungsnummer, Lebenstag und Geburtsgewicht [g]
Lebenstag 209 004 723 929 908 707
0 94 126 88 82 90 130
1 +14 +26 +0 +6 +20 +20
2 +12 +21 +11 +12 +12 +4
3 +8 +17 +9 +2 +20 +7
4 +16 +33 nicht bestimmt
+1 +10 +21
5 +13 +23 nicht bestimmt
+26 +21 +25
6 +15 +10 124 -2 +11 +9
7 +15 +5 +0 +11 +19 nicht bestimmt
9. Anhang
126
Tab. 17: Übersicht über die täglichen Körpergewichtszunahmen in Gramm [g] der neonatal
experimentell ertaubten Versuchstiere inkl. interner Registrierungsnummer, Lebenstag und Geburtsgewicht [g]
Lebenstag 209 004 723 929 908 707
8 +11 +21 +23 +11 +8 nicht bestimmt
9 +22 +16 +14 +18 +25 269
10 +7 +23 +18 +14 +11 +20
11 +18 +21 nicht bestimmt
+10 +17 +10
12 +12 +18 nicht bestimmt
+1 +7 +22
13 +16 nicht bestimmt
nicht bestimmt
+29 +17 nicht bestimmt
14 +16 nicht bestimmt
nicht bestimmt
+10 nicht bestimmt
nicht bestimmt
Lebenstag 208 123 401 444 027 026 125
0 122 148 90 109 125 103 160
1 +11 +19 +9 +12 +15 +15 +19
2 +18 +29 +9 +20 +27 +21 +25
3 +16 +16 +16 +13 +32 +23 +17
4 +25 +18 +20 +19 +24 +23 +16
5 +14 +23 +13 +5 +15 +25 +29
6 +19 +10 +23 +5 +16 +12 +12
7 +7 +22 +11 +5 +21 +22 +17
8 +18 +9 +10 +21 +8 +17 +5
9 +11 +18 +7 +10 +15 +10 +22
10 +21 +10 +11 +29 +24 +20 +17
11 +9 +17 +12 +13 nicht bestimmt
nicht bestimmt
+18
12 nicht bestimmt
+11 +9 nicht bestimmt
nicht bestimmt
nicht bestimmt
+14
13 +11 +19 +8 nicht bestimmt
nicht bestimmt
nicht bestimmt
+26
14 nicht bestimmt
nicht bestimmt
nicht bestimmt
nicht bestimmt
nicht bestimmt
nicht bestimmt
nicht bestimmt

9. Anhang
127
9.5 Die Herstellung der verwendeten Chemikalien-Lösungen
9.5.1 PBS-Lösung (Perfusionslösung)
Ansatz der Phosphatpuffer-Stammlösung (0,2 M, pH 7,4):
1. Na2HPO4 x 2 H2O 35,61 g in
Reinstwasser 1000 ml lösen.
2. NaH2PO4 x H2O 27,60 g in
Reinstwasser 1000 ml lösen.
Mischung der Phosphatpuffer-Gebrauchslösung (0,1 M, pH 7,4):
1. Lösung 405 ml mit
2. Lösung 95 ml mit
Reinstwasser 500 ml mischen.
Lagerung bei 4 °C in Dunkelheit.
9.5.2 Paraformaldehyd-Lösung (Fixationslösung)
Für die Herstellung der Fixierungslösung wurde, um beim Lösen des Paraformaldehyds nicht
die Pufferkapazität zu verbrauchen, der Phosphatpuffer doppelt so konzentriert wie benötigt
angesetzt und das Paraformaldehyd zunächst in Reinstwasser gelöst.
Ansatz der Paraformaldehyd-Stammlösung (8 %ig):
Paraformaldehyd 80,0 g in
Reinstwasser 1000 ml lösen, unter Rühren auf 60 °C
Erwärmen,
1 N NaOH 2-5 Tropfen zur Klärung der Lösung dazugeben,
Lösung auf Zimmertemperatur abkühlen
lassen.
Mischung der Paraformaldehyd-Gebrauchslösung (4 %ig in PBS, pH 7,4):
Paraformaldehyd-Stammlösung 1000 ml in
0,2 M Phosphatpuffer 1000 ml lösen, filtrieren,
auf pH 7,4 einstellen.
Lagerung bei 4 °C in Dunkelheit.

9. Anhang
128
9.5.3 Agar agar (Verschluss- und Einbettmedium)
Agar agar 7,0 g mit
Reinstwasser 100 ml versetzen, unter Rühren aufkochen.
Zum Einbetten des Gewebes erkalten lassen.
9.5.4 Kresyl-Violett-Färbelösung (Nissl-Ersatzfärbung)
Ansatz des Acetatpuffers (2,721 %iges Na-Acetat in 1,201 %iger Essigsäure):
1. Na-Acetat 2,721 g in
Reinstwasser 100 ml lösen.
2. 1 M Essigsäure 12,01 ml in
Reinstwasser 1000 ml lösen.
1 Teil Lösung 1. mit 4 Teilen Lösung 2. mischen und bei pH 3,8 – 4,0 einstellen.
Ansatz der Kresyl-Violett-Färbelösung (1 %ig):
Kresyl-Violett 1,0 g in
Acetatpuffer 100 ml lösen, filtrieren.
9.5.5 Mowiol (Eindeckelmedium)
Hierbei handelt es sich um ein nicht eigenfluoreszentes Eindeckelmedium.
97-99 %iges Glycerol 6,0 g in ein 50 ml Zentrifugenröhrchen
einwiegen,
Mowiol 2,4 g dazu geben,
Reinstwasser 7 ml dazu geben,
Zentrifugenröhrchen im Wasserbad
auf 60 °C erwärmen, dabei mehrfach
umrühren,
Tris/HCl 0,2 M, pH 8,5 13 ml dazu geben, gut vermischen,
bei 10.000 U/min für 5 min
zentrifugieren.
Lagerung bei -20 °C in Dunkelheit. 12 Stunden vor dem Gebrauch schonend auftauen. Kein
erneutes Einfrieren möglich.
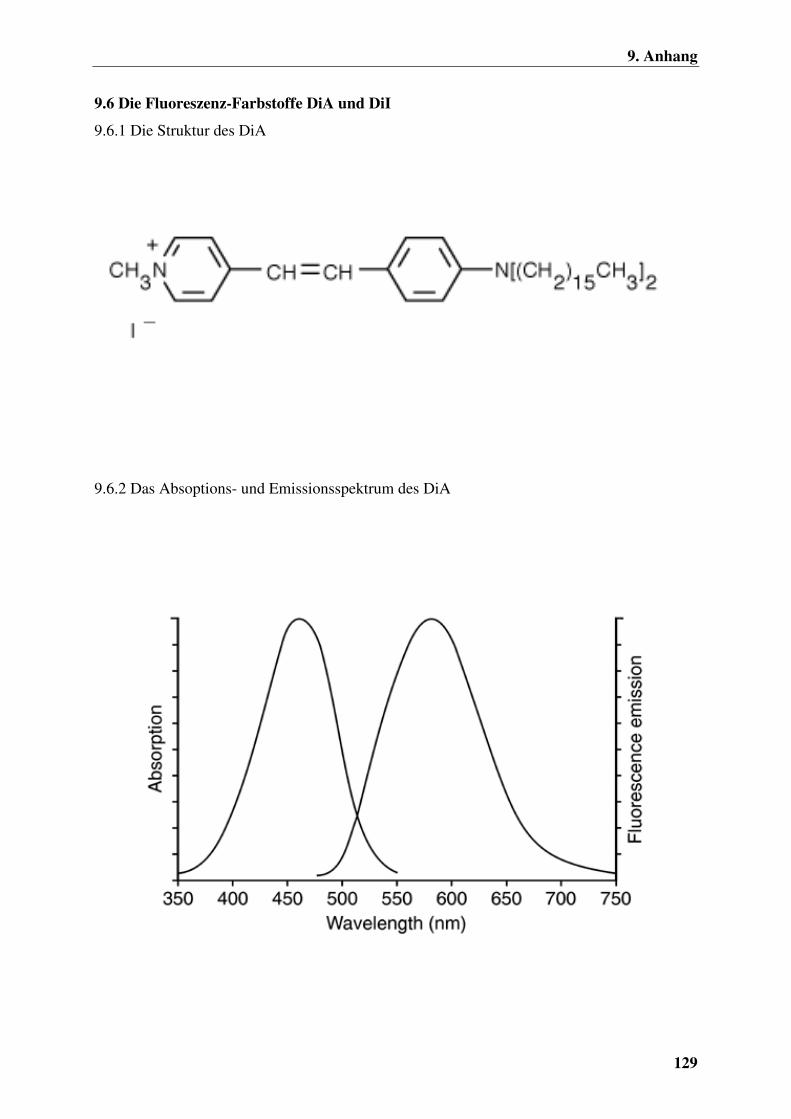
9. Anhang
129
9.6 Die Fluoreszenz-Farbstoffe DiA und DiI
9.6.1 Die Struktur des DiA
9.6.2 Das Absoptions- und Emissionsspektrum des DiA
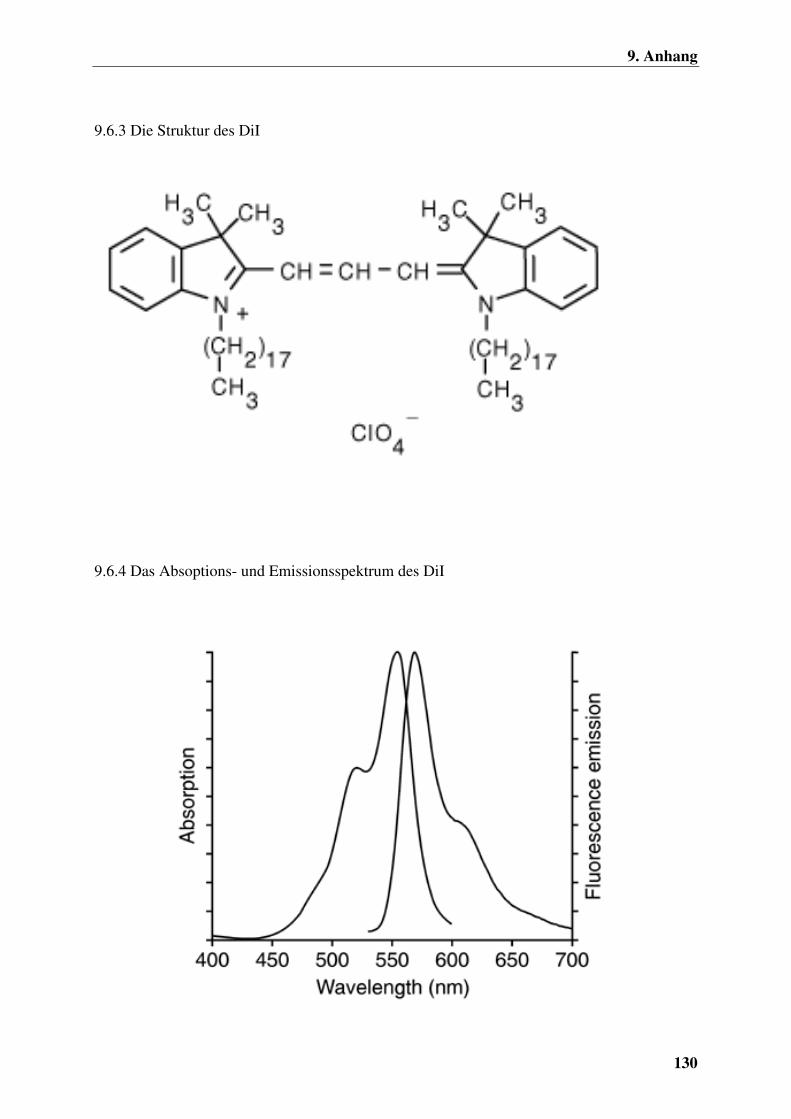
9. Anhang
130
9.6.3 Die Struktur des DiI
9.6.4 Das Absoptions- und Emissionsspektrum des DiI
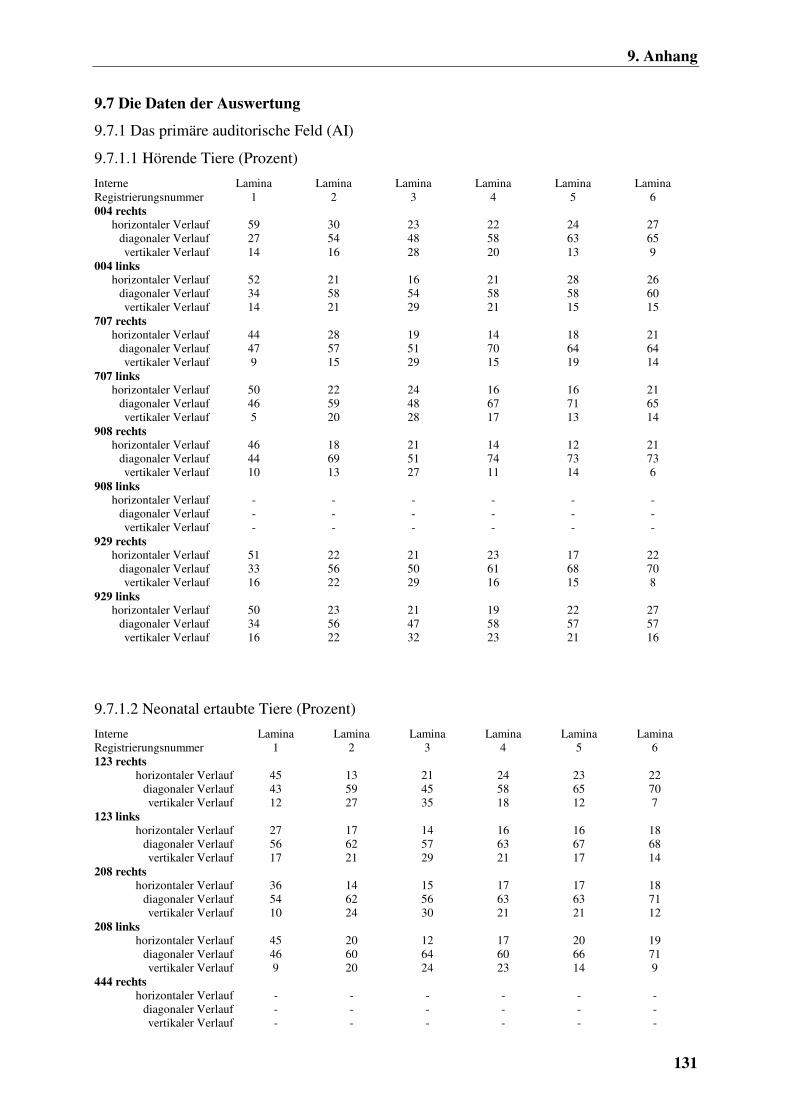
9. Anhang
131
9.7 Die Daten der Auswertung
9.7.1 Das primäre auditorische Feld (AI)
9.7.1.1 Hörende Tiere (Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
004 rechts horizontaler Verlauf 59 30 23 22 24 27
diagonaler Verlauf 27 54 48 58 63 65 vertikaler Verlauf 14 16 28 20 13 9
004 links horizontaler Verlauf 52 21 16 21 28 26
diagonaler Verlauf 34 58 54 58 58 60 vertikaler Verlauf 14 21 29 21 15 15
707 rechts horizontaler Verlauf 44 28 19 14 18 21
diagonaler Verlauf 47 57 51 70 64 64 vertikaler Verlauf 9 15 29 15 19 14
707 links horizontaler Verlauf 50 22 24 16 16 21
diagonaler Verlauf 46 59 48 67 71 65 vertikaler Verlauf 5 20 28 17 13 14
908 rechts horizontaler Verlauf 46 18 21 14 12 21
diagonaler Verlauf 44 69 51 74 73 73 vertikaler Verlauf 10 13 27 11 14 6
908 links horizontaler Verlauf - - - - - -
diagonaler Verlauf - - - - - - vertikaler Verlauf - - - - - -
929 rechts horizontaler Verlauf 51 22 21 23 17 22
diagonaler Verlauf 33 56 50 61 68 70 vertikaler Verlauf 16 22 29 16 15 8
929 links horizontaler Verlauf 50 23 21 19 22 27
diagonaler Verlauf 34 56 47 58 57 57 vertikaler Verlauf 16 22 32 23 21 16
9.7.1.2 Neonatal ertaubte Tiere (Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
123 rechts horizontaler Verlauf 45 13 21 24 23 22
diagonaler Verlauf 43 59 45 58 65 70 vertikaler Verlauf 12 27 35 18 12 7
123 links horizontaler Verlauf 27 17 14 16 16 18
diagonaler Verlauf 56 62 57 63 67 68 vertikaler Verlauf 17 21 29 21 17 14
208 rechts horizontaler Verlauf 36 14 15 17 17 18
diagonaler Verlauf 54 62 56 63 63 71 vertikaler Verlauf 10 24 30 21 21 12
208 links horizontaler Verlauf 45 20 12 17 20 19
diagonaler Verlauf 46 60 64 60 66 71 vertikaler Verlauf 9 20 24 23 14 9
444 rechts horizontaler Verlauf - - - - - -
diagonaler Verlauf - - - - - - vertikaler Verlauf - - - - - -

9. Anhang
132
444 links horizontaler Verlauf 51 34 25 21 - -
diagonaler Verlauf 41 48 50 57 - - vertikaler Verlauf 8 18 25 21 - -
9.7.1.3 Neonatal ertaubte, anschließend chronisch elektrisch intracochleär stimulierte Tiere
(Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
026 rechts horizontaler Verlauf - 15 13 11 8 10
diagonaler Verlauf - 65 52 74 71 68 vertikaler Verlauf - 20 34 15 22 22
026 links horizontaler Verlauf 59 24 21 19 22 30
diagonaler Verlauf 38 57 42 61 61 56 vertikaler Verlauf 3 19 38 20 17 14
125 rechts
horizontaler Verlauf - - - - - - diagonaler Verlauf - - - - - - vertikaler Verlauf - - - - - -
125 links horizontaler Verlauf 43 25 15 24 22 25
diagonaler Verlauf 39 47 49 55 58 64 vertikaler Verlauf 18 28 36 21 20 11
9.7.2 Das sekundäre auditorische Feld (AII)
9.7.2.1 Hörende Tiere (Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
004 rechts horizontaler Verlauf 48 12 15 10 8 9
diagonaler Verlauf 40 65 63 73 84 85 vertikaler Verlauf 12 23 22 16 8 6
004 links horizontaler Verlauf 40 23 17 15 15 16
diagonaler Verlauf 43 53 52 63 72 74 vertikaler Verlauf 18 24 31 21 13 10
209 rechts horizontaler Verlauf 48 18 23 26 22 37
diagonaler Verlauf 39 59 50 57 64 57 vertikaler Verlauf 12 23 28 17 14 6
209 links horizontaler Verlauf 53 22 28 27 22 23
diagonaler Verlauf 42 65 43 62 62 67 vertikaler Verlauf 5 13 29 11 16 10
723 rechts horizontaler Verlauf 56 34 29 23 19 11
diagonaler Verlauf 35 51 49 66 65 75 vertikaler Verlauf 9 15 22 10 16 13
723 links horizontaler Verlauf 53 32 29 17 13 14
diagonaler Verlauf 34 49 42 63 70 72 vertikaler Verlauf 13 19 29 19 16 14
908 rechts horizontaler Verlauf 54 16 18 16 15 15
diagonaler Verlauf 37 58 54 71 72 75 vertikaler Verlauf 6 26 28 14 13 9
908 links horizontaler Verlauf 48 18 18 12 14 16
diagonaler Verlauf 40 57 46 69 65 71 vertikaler Verlauf 12 25 35 19 20 13
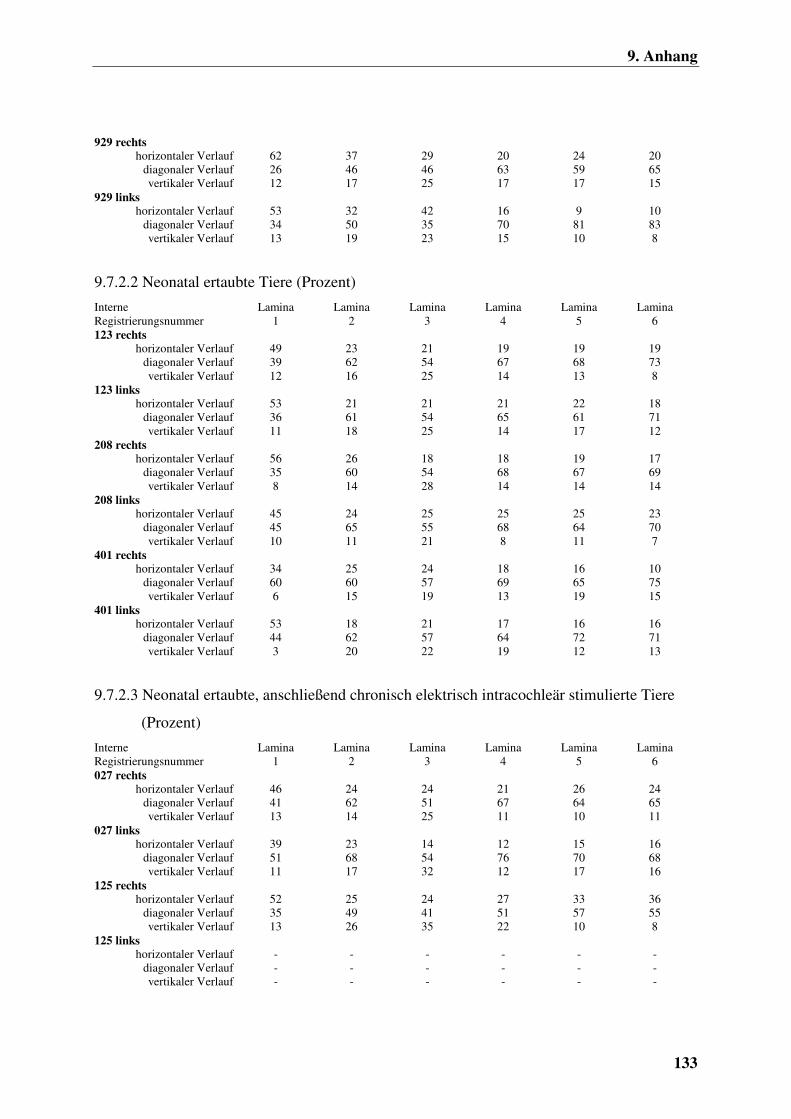
9. Anhang
133
929 rechts
horizontaler Verlauf 62 37 29 20 24 20 diagonaler Verlauf 26 46 46 63 59 65 vertikaler Verlauf 12 17 25 17 17 15
929 links horizontaler Verlauf 53 32 42 16 9 10
diagonaler Verlauf 34 50 35 70 81 83 vertikaler Verlauf 13 19 23 15 10 8
9.7.2.2 Neonatal ertaubte Tiere (Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
123 rechts horizontaler Verlauf 49 23 21 19 19 19
diagonaler Verlauf 39 62 54 67 68 73 vertikaler Verlauf 12 16 25 14 13 8
123 links horizontaler Verlauf 53 21 21 21 22 18
diagonaler Verlauf 36 61 54 65 61 71 vertikaler Verlauf 11 18 25 14 17 12
208 rechts horizontaler Verlauf 56 26 18 18 19 17
diagonaler Verlauf 35 60 54 68 67 69 vertikaler Verlauf 8 14 28 14 14 14
208 links horizontaler Verlauf 45 24 25 25 25 23
diagonaler Verlauf 45 65 55 68 64 70 vertikaler Verlauf 10 11 21 8 11 7
401 rechts horizontaler Verlauf 34 25 24 18 16 10
diagonaler Verlauf 60 60 57 69 65 75 vertikaler Verlauf 6 15 19 13 19 15
401 links horizontaler Verlauf 53 18 21 17 16 16
diagonaler Verlauf 44 62 57 64 72 71 vertikaler Verlauf 3 20 22 19 12 13
9.7.2.3 Neonatal ertaubte, anschließend chronisch elektrisch intracochleär stimulierte Tiere
(Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
027 rechts horizontaler Verlauf 46 24 24 21 26 24
diagonaler Verlauf 41 62 51 67 64 65 vertikaler Verlauf 13 14 25 11 10 11
027 links horizontaler Verlauf 39 23 14 12 15 16
diagonaler Verlauf 51 68 54 76 70 68 vertikaler Verlauf 11 17 32 12 17 16
125 rechts horizontaler Verlauf 52 25 24 27 33 36
diagonaler Verlauf 35 49 41 51 57 55 vertikaler Verlauf 13 26 35 22 10 8
125 links horizontaler Verlauf - - - - - -
diagonaler Verlauf - - - - - - vertikaler Verlauf - - - - - -
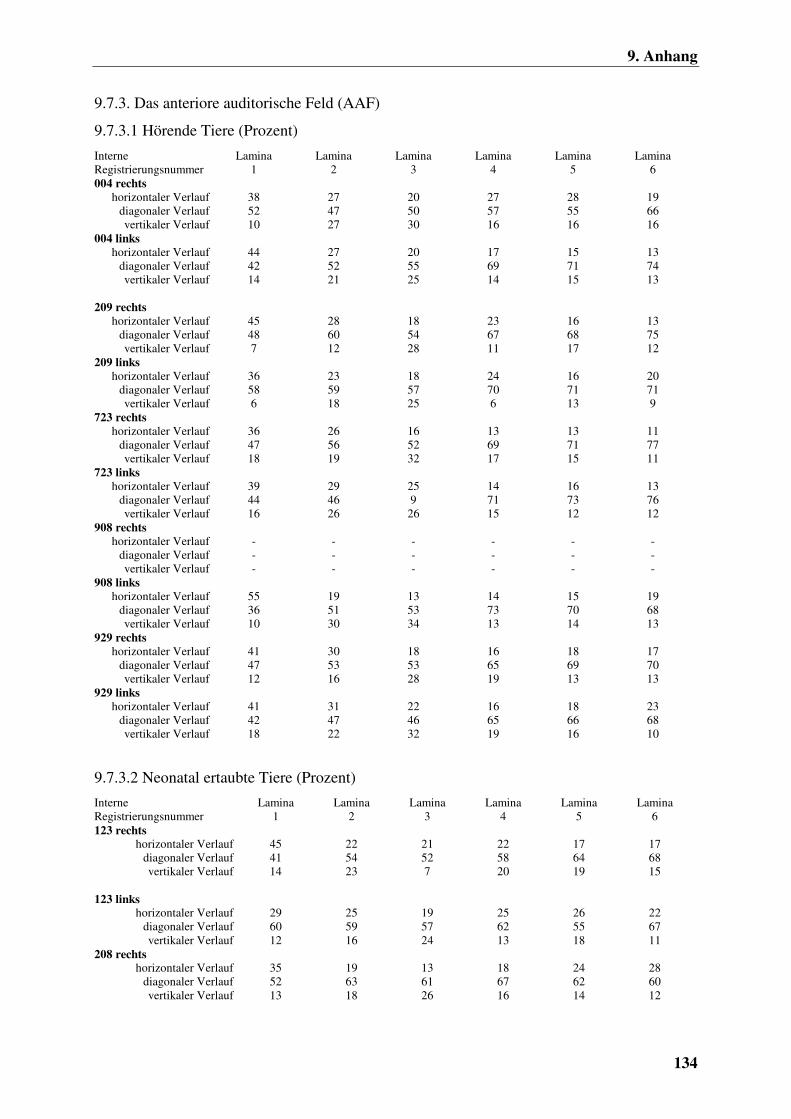
9. Anhang
134
9.7.3. Das anteriore auditorische Feld (AAF)
9.7.3.1 Hörende Tiere (Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
004 rechts horizontaler Verlauf 38 27 20 27 28 19
diagonaler Verlauf 52 47 50 57 55 66 vertikaler Verlauf 10 27 30 16 16 16
004 links horizontaler Verlauf 44 27 20 17 15 13
diagonaler Verlauf 42 52 55 69 71 74 vertikaler Verlauf 14 21 25 14 15 13
209 rechts
horizontaler Verlauf 45 28 18 23 16 13 diagonaler Verlauf 48 60 54 67 68 75 vertikaler Verlauf 7 12 28 11 17 12
209 links horizontaler Verlauf 36 23 18 24 16 20
diagonaler Verlauf 58 59 57 70 71 71 vertikaler Verlauf 6 18 25 6 13 9
723 rechts horizontaler Verlauf 36 26 16 13 13 11
diagonaler Verlauf 47 56 52 69 71 77 vertikaler Verlauf 18 19 32 17 15 11
723 links horizontaler Verlauf 39 29 25 14 16 13
diagonaler Verlauf 44 46 9 71 73 76 vertikaler Verlauf 16 26 26 15 12 12
908 rechts horizontaler Verlauf - - - - - -
diagonaler Verlauf - - - - - - vertikaler Verlauf - - - - - -
908 links horizontaler Verlauf 55 19 13 14 15 19
diagonaler Verlauf 36 51 53 73 70 68 vertikaler Verlauf 10 30 34 13 14 13
929 rechts horizontaler Verlauf 41 30 18 16 18 17
diagonaler Verlauf 47 53 53 65 69 70 vertikaler Verlauf 12 16 28 19 13 13
929 links horizontaler Verlauf 41 31 22 16 18 23
diagonaler Verlauf 42 47 46 65 66 68 vertikaler Verlauf 18 22 32 19 16 10
9.7.3.2 Neonatal ertaubte Tiere (Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
123 rechts horizontaler Verlauf 45 22 21 22 17 17
diagonaler Verlauf 41 54 52 58 64 68 vertikaler Verlauf 14 23 7 20 19 15
123 links
horizontaler Verlauf 29 25 19 25 26 22 diagonaler Verlauf 60 59 57 62 55 67 vertikaler Verlauf 12 16 24 13 18 11
208 rechts horizontaler Verlauf 35 19 13 18 24 28
diagonaler Verlauf 52 63 61 67 62 60 vertikaler Verlauf 13 18 26 16 14 12
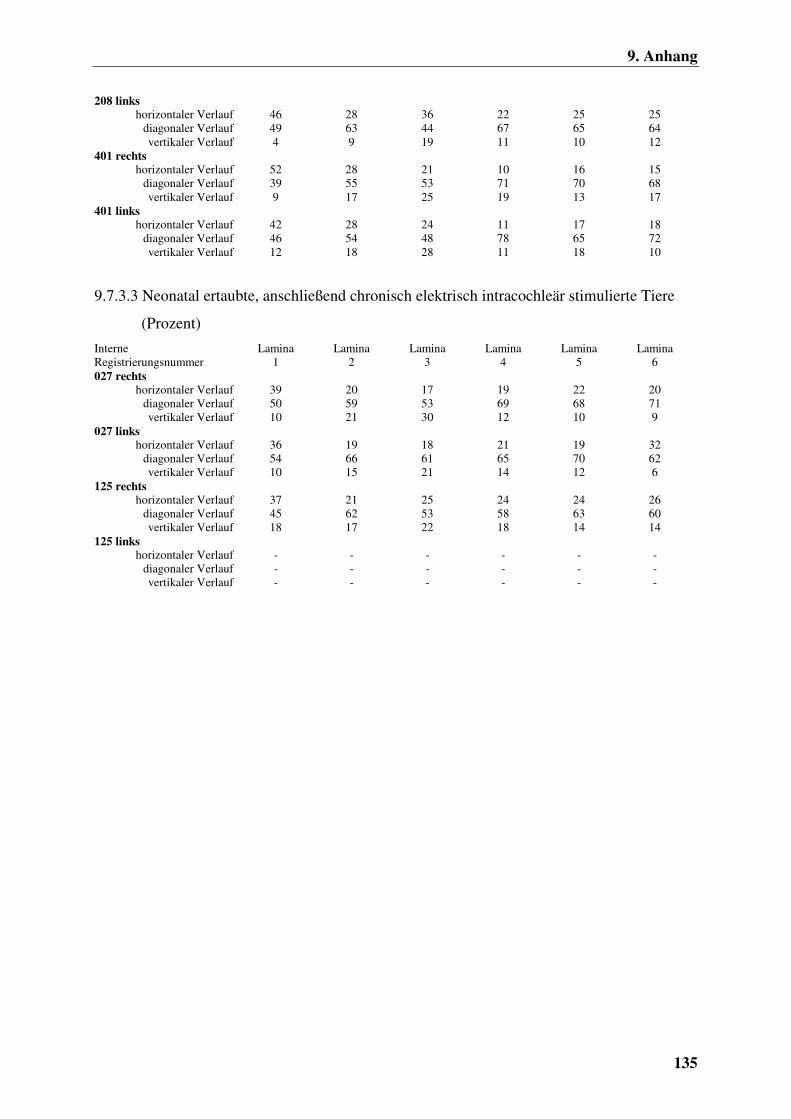
9. Anhang
135
208 links horizontaler Verlauf 46 28 36 22 25 25
diagonaler Verlauf 49 63 44 67 65 64 vertikaler Verlauf 4 9 19 11 10 12
401 rechts horizontaler Verlauf 52 28 21 10 16 15
diagonaler Verlauf 39 55 53 71 70 68 vertikaler Verlauf 9 17 25 19 13 17
401 links horizontaler Verlauf 42 28 24 11 17 18
diagonaler Verlauf 46 54 48 78 65 72 vertikaler Verlauf 12 18 28 11 18 10
9.7.3.3 Neonatal ertaubte, anschließend chronisch elektrisch intracochleär stimulierte Tiere
(Prozent)
Interne Registrierungsnummer
Lamina 1
Lamina 2
Lamina 3
Lamina 4
Lamina 5
Lamina 6
027 rechts horizontaler Verlauf 39 20 17 19 22 20
diagonaler Verlauf 50 59 53 69 68 71 vertikaler Verlauf 10 21 30 12 10 9
027 links horizontaler Verlauf 36 19 18 21 19 32
diagonaler Verlauf 54 66 61 65 70 62 vertikaler Verlauf 10 15 21 14 12 6
125 rechts horizontaler Verlauf 37 21 25 24 24 26
diagonaler Verlauf 45 62 53 58 63 60 vertikaler Verlauf 18 17 22 18 14 14
125 links horizontaler Verlauf - - - - - -
diagonaler Verlauf - - - - - - vertikaler Verlauf - - - - - -

10. Verzeichnis der Abbildungen
136
10. VERZEICHNIS DER ABBILDUNGEN
Abb. 1: Schematische Darstellung der Kranialansicht eines Transversalschnittes
durch das rechte Ohr einer Katze
(modifiziert nach HUDSON und HAMILTON 1993) 13
Abb. 2: Schematische Darstellung eines Querschnitts durch eine Cochlea-
windung mit Corti-Organ
(modifiziert nach BLOOM und FAWCETT 1975) 14
Abb. 3: Anatomie des Corti-Organs
(modifiziert nach ZENNER und GITTER 1987) 15
Abb. 4: Schematische Darstellung der afferenten zentralen Hörbahn der linken
Cochlea (modifiziert nach ZENNER 1993) 19
Abb. 5: Lateralansicht eines freipräparierten felinen Gehirns.
AI: Primäres auditorisches Feld, AII: Sekundäres auditorisches Feld,
AAF: Anteriores auditorisches Feld, sss: suprasylvanischer Sulcus,
aes: anteriorer ectosylvanischer Sulcus, pes: posteriorer ecto-
sylvanischer Sulcus 58
Abb. 6: DiI-Kristallapplikation mittels Skalpellklinge 59
Abb. 7: DiI-Kristalle im AAF und AII, sowie DiA-Kristall im AI einer linken felinen Cortexhemisphäre 59
Abb. 8: In Agar agar eingebetteter, auf hölzernem Vibratomblock aufgeklebter
rechter auditorischer Cortex. Das anteriore und sekundäre
auditorische Feld sind mit DiI markiert. Auf das primäre auditorische
Feld ist ein DiA-Kristall aufgetragen. 60
Abb. 9: DiI-markierte Dendriten in der Lamina I und II im primären
auditorischen Feld einer hörenden Katze (Tier 929) 63

10. Verzeichnis der Abbildungen
137
Abb. 10: DiI-gefärbte Neurone in der Lamina III. Rechtes anteriores
auditorisches Feld eines ertaubten Tieres (Katze 123) 64
Abb. 11: DiI-gefärbtes Neuron in der Lamina IV. Rechtes anteriores
auditorisches Feld eines ertaubten Tieres (Katze 123) 64
Abb. 12: Vergleich der Dendritenanzahl in Prozent mit horizontalem Verlauf
zwischen der Lamina I und den Laminae II bis VI des AAF, AI und
AII (hörende Tiere; MW±SD; ***p<0.0001) 67
Abb. 13: Vergleich der Dendritenanzahl in Prozent mit horizontalem Verlauf
zwischen der Lamina I und den Laminae II bis VI des AAF, AI und
AII (ertaubte Tiere; MW±SD; ***p<0.001) 68
Abb. 14: Vergleich der Dendritenanzahl in Prozent mit horizontalem Verlauf
zwischen der Lamina I und den Laminae II bis VI des AAF, AI und
AII (stimulierte Tiere; MW±SD; ***p<0.001, **p<0.01, *p<0.05,
n.s.= nicht signifikant) 69
Abb. 15: Vergleich der Dendritenanzahl in Prozent mit diagonalem Verlauf
zwischen den Laminae I-III und den Laminae IV-VI des AAF, AI und
AII (hörende Tiere; MW±SD; ***p<0.0001) 70
Abb. 16: Vergleich der Dendritenanzahl in Prozent mit diagonalem Verlauf
zwischen den Laminae I-III und den Laminae IV-VI des AAF, AI und
AII (ertaubte Tiere; MW±SD; ***p<0.001) 71
Abb. 17: Vergleich der Dendritenanzahl in Prozent mit diagonalem Verlauf
zwischen den Laminae I-III und den Laminae IV-VI des AAF, AI und
AII (stimulierte Tiere; MW±SD; **p<0.01) 72
Abb. 18: Vergleich der Dendritenanzahl in Prozent mit vertikalem Verlauf
zwischen der Lamina III und den Laminae I, II und IV bis VI des AAF,
AI und AII (hörende Tiere; MW±SD; ***p<0.001, **p<0.01) 73

10. Verzeichnis der Abbildungen
138
Abb. 19: Vergleich der Dendritenanzahl in Prozent mit vertikalem Verlauf
zwischen der Lamina III und den Laminae I, II und IV bis VI des AAF,
AI und AII (ertaubte Tiere; MW±SD; ***p<0.001,**p<0.01, *p<0.05,
n.s.= nicht signifikant) 74
Abb. 20: Vergleich der Dendritenanzahl in Prozent mit vertikalem Verlauf
zwischen der Lamina III und den Laminae I, II und IV bis VI des AAF,
AI und AII (stimulierte Tiere; MW±SD; **p<0.01, *p<0.05,
n.s.= nicht signifikant) 75
Abb. 21: FAEP-Kurven einer hörenden Katze (ipsilaterale Ableitungen vom
rechten (re) und linken (li) Ohr bei 10 bis 80 dB SPL) 125
Abb. 22: FAEP-Kurven einer experimentell neonatal ertaubten Katze
(ipsilaterale Ableitungen vom rechten (re) und linken (li) Ohr bei 10
bis 80 dB SPL) 125
Abb. 23: FEEP-Kurven einer chronisch intracochleär stimulierten Katze
(ipsilaterale Ableitung vom linken (li) Ohr bei 180 bis 400 µA) 126

11. Verzeichnis der Tabellen
139
11. VERZEICHNIS DER TABELLEN
Tab. 1: Auflistung der auditorischen Felder ausgewählter Spezies (modifiziert
nach CLAREY et al. 1991) 22
Tab. 2: Commissurale Verbindungen der auditorischen Felder (modifiziert
nach WINER 1992) 28
Tab. 3: Corticocorticale Verbindungen der auditorischen Felder (modifiziert
nach WINER 1992) 29
Tab. 4 a, b: Übersicht über die hörenden und neonatal ertaubten Tiere, inkl. interner
Registrierungsnummer, Tötungsalter und appliziertem
Fluoreszenzfarbstoff 50
Tab. 5: Übersicht über die neonatal experimentell ertaubten, anschließend
chronisch elektrisch intracochleär stimulierten Versuchstiere, inkl.
interner Registrierungsnummer, Tötungsalter, Stimulationsdauer
und appliziertem Fluoreszenzfarbstoff 50
Tab. 6: Prozentualer Anteil der Dendriten (horizontaler Verlauf; MW±SD) 66
Tab. 7: Prozentualer Anteil der Dendriten (horizontaler Verlauf; MW±SD) 67
Tab. 8: Prozentualer Anteil der Dendriten (horizontaler Verlauf; MW±SD) 68
Tab. 9: Prozentualer Anteil der Dendriten (diagonaler Verlauf; MW±SD) 70
Tab. 10: Prozentualer Anteil der Dendriten (diagonaler Verlauf; MW±SD) 71
Tab. 11: Prozentualer Anteil der Dendriten (diagonaler Verlauf; MW±SD) 72
Tab. 12: Prozentualer Anteil der Dendriten (vertikaler Verlauf; MW±SD) 74

11. Verzeichnis der Tabellen
140
Tab. 13: Prozentualer Anteil der Dendriten (vertikaler Verlauf; MW±SD) 74
Tab. 14: Prozentualer Anteil der Dendriten (vertikaler Verlauf; MW±SD) 75
Tab. 15: Zusammenfassung der prozentualen Dendritenverläufe in den
auditorischen Subarealen der drei Versuchstiergruppen (MW) 79
Tab.16a, b: Übersicht über die täglichen Körpergewichtszunahmen in Gramm [g]
der hörenden Versuchstiere inkl. interner Registrierungsnummer,
Lebenstag und Geburtsgewicht [g] 126
Tab.17: Übersicht über die täglichen Körpergewichtszunahmen in Gramm [g]
der neonatal experimentell ertaubten Versuchstiere inkl. interner
Registrierungsnummer, Lebenstag und Geburtsgewicht [g] 127

12. Danksagung
141
12. DANKSAGUNG
Herrn Prof. Dr. Günter Reuter danke ich herzlich für die Überlassung des äußerst
interessanten Dissertationsthemas.
Herrn Prof. Dr. Gerd Bicker gilt mein besonderer Dank für die sehr freundliche und
unkomplizierte Betreuung der Arbeit.
Mein ganz besonderer Dank gilt Herrn Jörg Bornemann für seine Unterstützung, seine stets
konstruktive Kritik und sein Verständnis insbesondere in der Phase der Korrektur.
Frau Anja Schlinkert und Frau Dina Wilkens danke ich für die kollegiale und
freundschaftliche Zusammenarbeit.
Herrn Peter Erfurt danke ich für die Anfertigung der digitalen Fotographien.
Herrn Bernhard Vaske danke ich für die Beratung der statistischen Auswertung.
Meinen Eltern danke ich dafür, dass sie immer an ein gutes Ende geglaubt haben.


















