
Zentrale Regulation des Körpergewichtes - link.springer.com · tisches Polypeptid und Amylin) ......
15
Zentrale Regulation des Körpergewichtes Christian Roth Inhalt 1 Entwicklung in den letzten Jahrzehnten .............................................................. 1 2 Regelkreise der Hunger- und Sättigungsregulation .................................................. 2 2.1 Regulatoren von Hunger und Sättigung aus der Peripherie (Magen-Darm-Trakt und Fettgewebe) . . . 2 2.2 Zentrale Regulatoren und Strukturen der Energiehomöostase .......................................... 5 2.3 Zentrale Kontrolle der Thermogenese über das autonome Nervensystem .............................. 9 3 Adipositas durch hypothalamische Läsionen ......................................................... 10 3.1 Experimentelle Adipositas durch hypothalamische Läsionen im Tiermodell .......................... 10 3.2 Hypothalamische Adipositas beim Menschen ........................................................... 10 4 Genetische Ursachen der Adipositas .................................................................. 11 4.1 Experimentelle Adipositas durch Veränderungen einzelner Gene des zentralen Melanokortinsystems im Tiermodell ............................................................................................. 11 4.2 Humane Adipositasmutationen .......................................................................... 11 4.3 Weitere genetische Faktoren und komplexe Genetik .................................................... 12 5 Einfluss der Darmflora ................................................................................. 14 6 Zusammenfassung ...................................................................................... 14 Literatur ....................................................................................................... 14 1 Entwicklung in den letzten Jahrzehnten In den letzten vier Jahrzehnten konnte man eine dramatische Zunahme der Adipositasrate beobachten. Die Betrachtung von Energiebilanz und Regulation des Körpergewichtes rückt daher zunehmend in den Fokus eines pädiatrischen Endokri- nologen. Das Körpergewicht wird durch zahlreiche äußere und innere Einflussfaktoren sowie periphere und zentrale, meist redundante Regelmechanismen bestimmt. Neben dem individuellen genetischen Hintergrund und dem aktuellen Wandel von soziokulturellen Faktoren und Lifestyle sind Veränderungen der appetitregulierenden Peptide und Regula- tionsstrukturen im zentralen Nervensystem wichtige Fakto- ren für Störungen des Energiegleichgewichtes. Obwohl die Regulation des Körpergewichtes erstaunlich präzise erfolgt, kann es bei längerfristiger Störung der Balance der Energie- zufuhr zu Über- oder Untergewicht kommen. Eine Verände- rung der Homöostase des Energiestoffwechsels kann sowohl durch eine veränderte Nahrungsaufnahme als durch einen veränderten Energieverbrauch verursacht werden, der vom Grundumsatz, der Wärmeproduktion, die je nach Ernäh- rungszustand variiert, der Muskelarbeit und dem Wachstum abhängt. Diese komplexen Regelmechanismen haben sich in der Evolution über einen langen Zeitraum entwickelt, um gegen den Hungertod zu schützen. Durch den modernen Lebensstil kehrt sich der evolutionäre Vorteil in ein höheres Risiko einer adipositasbedingten erhöhten kardiovaskulären Mortalität („thrifty gene hypothesis“) um. C. Roth (*) Division of Endocrinology and Diabetes, Seattle Children’ s Research Institute, Department of Pediatrics, University of Washington, Seattle, Vereinigte Staaten E-Mail: [email protected] # Springer-Verlag GmbH Deutschland, ein Teil von Springer Nature 2018 O. Hiort et al. (Hrsg.), Pädiatrische Endokrinologie und Diabetologie, Springer Reference Medizin, https://doi.org/10.1007/978-3-662-52794-8_18-1 1
Transcript of Zentrale Regulation des Körpergewichtes - link.springer.com · tisches Polypeptid und Amylin) ......

Zentrale Regulation des Körpergewichtes
Christian Roth
Inhalt1 Entwicklung in den letzten Jahrzehnten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
2 Regelkreise der Hunger- und Sättigungsregulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.1 Regulatoren von Hunger und Sättigung aus der Peripherie (Magen-Darm-Trakt und Fettgewebe) . . . 22.2 Zentrale Regulatoren und Strukturen der Energiehomöostase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52.3 Zentrale Kontrolle der Thermogenese über das autonome Nervensystem . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
3 Adipositas durch hypothalamische Läsionen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103.1 Experimentelle Adipositas durch hypothalamische Läsionen im Tiermodell . . . . . . . . . . . . . . . . . . . . . . . . . . 103.2 Hypothalamische Adipositas beim Menschen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
4 Genetische Ursachen der Adipositas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 114.1 Experimentelle Adipositas durch Veränderungen einzelner Gene des zentralen Melanokortinsystems
im Tiermodell . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 114.2 Humane Adipositasmutationen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 114.3 Weitere genetische Faktoren und komplexe Genetik . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
5 Einfluss der Darmflora . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
6 Zusammenfassung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
Literatur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
1 Entwicklung in den letzten Jahrzehnten
In den letzten vier Jahrzehnten konnte man eine dramatischeZunahme der Adipositasrate beobachten. Die Betrachtungvon Energiebilanz und Regulation des Körpergewichtes rücktdaher zunehmend in den Fokus eines pädiatrischen Endokri-nologen. Das Körpergewicht wird durch zahlreiche äußereund innere Einflussfaktoren sowie periphere und zentrale,meist redundante Regelmechanismen bestimmt. Neben demindividuellen genetischen Hintergrund und dem aktuellenWandel von soziokulturellen Faktoren und Lifestyle sind
Veränderungen der appetitregulierenden Peptide und Regula-tionsstrukturen im zentralen Nervensystem wichtige Fakto-ren für Störungen des Energiegleichgewichtes. Obwohl dieRegulation des Körpergewichtes erstaunlich präzise erfolgt,kann es bei längerfristiger Störung der Balance der Energie-zufuhr zu Über- oder Untergewicht kommen. Eine Verände-rung der Homöostase des Energiestoffwechsels kann sowohldurch eine veränderte Nahrungsaufnahme als durch einenveränderten Energieverbrauch verursacht werden, der vomGrundumsatz, der Wärmeproduktion, die je nach Ernäh-rungszustand variiert, der Muskelarbeit und dem Wachstumabhängt. Diese komplexen Regelmechanismen haben sich inder Evolution über einen langen Zeitraum entwickelt, umgegen den Hungertod zu schützen. Durch den modernenLebensstil kehrt sich der evolutionäre Vorteil in ein höheresRisiko einer adipositasbedingten erhöhten kardiovaskulärenMortalität („thrifty gene hypothesis“) um.
C. Roth (*)Division of Endocrinology and Diabetes, Seattle Children’s ResearchInstitute, Department of Pediatrics, University of Washington, Seattle,Vereinigte StaatenE-Mail: [email protected]
# Springer-Verlag GmbH Deutschland, ein Teil von Springer Nature 2018O. Hiort et al. (Hrsg.), Pädiatrische Endokrinologie und Diabetologie, Springer Reference Medizin,https://doi.org/10.1007/978-3-662-52794-8_18-1
1
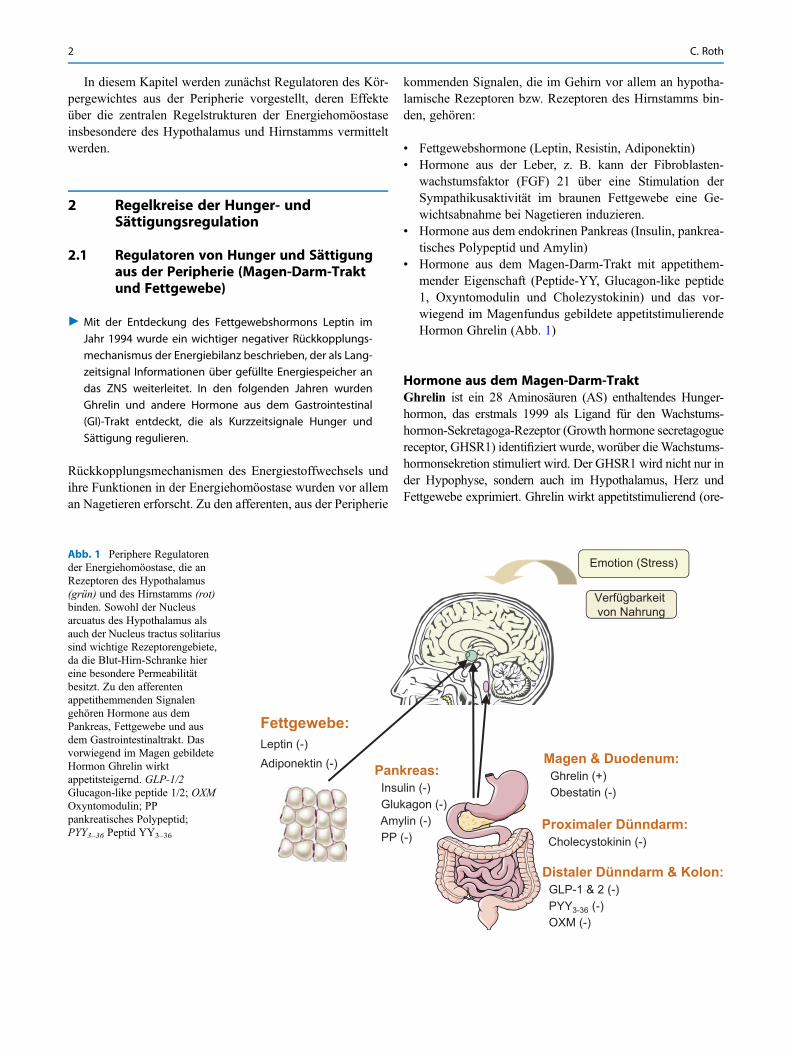
In diesem Kapitel werden zunächst Regulatoren des Kör-pergewichtes aus der Peripherie vorgestellt, deren Effekteüber die zentralen Regelstrukturen der Energiehomöostaseinsbesondere des Hypothalamus und Hirnstamms vermitteltwerden.
2 Regelkreise der Hunger- undSättigungsregulation
2.1 Regulatoren von Hunger und Sättigungaus der Peripherie (Magen-Darm-Traktund Fettgewebe)
" Mit der Entdeckung des Fettgewebshormons Leptin imJahr 1994 wurde ein wichtiger negativer Rückkopplungs-mechanismus der Energiebilanz beschrieben, der als Lang-zeitsignal Informationen über gefüllte Energiespeicher andas ZNS weiterleitet. In den folgenden Jahren wurdenGhrelin und andere Hormone aus dem Gastrointestinal(GI)-Trakt entdeckt, die als Kurzzeitsignale Hunger undSättigung regulieren.
Rückkopplungsmechanismen des Energiestoffwechsels undihre Funktionen in der Energiehomöostase wurden vor alleman Nagetieren erforscht. Zu den afferenten, aus der Peripherie
kommenden Signalen, die im Gehirn vor allem an hypotha-lamische Rezeptoren bzw. Rezeptoren des Hirnstamms bin-den, gehören:
• Fettgewebshormone (Leptin, Resistin, Adiponektin)• Hormone aus der Leber, z. B. kann der Fibroblasten-
wachstumsfaktor (FGF) 21 über eine Stimulation derSympathikusaktivität im braunen Fettgewebe eine Ge-wichtsabnahme bei Nagetieren induzieren.
• Hormone aus dem endokrinen Pankreas (Insulin, pankrea-tisches Polypeptid und Amylin)
• Hormone aus dem Magen-Darm-Trakt mit appetithem-mender Eigenschaft (Peptide-YY, Glucagon-like peptide1, Oxyntomodulin und Cholezystokinin) und das vor-wiegend im Magenfundus gebildete appetitstimulierendeHormon Ghrelin (Abb. 1)
Hormone aus dem Magen-Darm-TraktGhrelin ist ein 28 Aminosäuren (AS) enthaltendes Hunger-hormon, das erstmals 1999 als Ligand für den Wachstums-hormon-Sekretagoga-Rezeptor (Growth hormone secretagoguereceptor, GHSR1) identifiziert wurde, worüber die Wachstums-hormonsekretion stimuliert wird. Der GHSR1 wird nicht nur inder Hypophyse, sondern auch im Hypothalamus, Herz undFettgewebe exprimiert. Ghrelin wirkt appetitstimulierend (ore-
Emotion (Stress)
Verfügbarkeit von Nahrung
Proximaler Dünndarm:Cholecystokinin (-)
Pankreas:Insulin (-)Glukagon (-)Amylin (-)PP (-)
Magen & Duodenum:Ghrelin (+)Obestatin (-)
Distaler Dünndarm & Kolon:GLP-1 & 2 (-)PYY3-36 (-)OXM (-)
Fettgewebe:Leptin (-)Adiponektin (-)
Abb. 1 Periphere Regulatorender Energiehomöostase, die anRezeptoren des Hypothalamus(grün) und des Hirnstamms (rot)binden. Sowohl der Nucleusarcuatus des Hypothalamus alsauch der Nucleus tractus solitariussind wichtige Rezeptorengebiete,da die Blut-Hirn-Schranke hiereine besondere Permeabilitätbesitzt. Zu den afferentenappetithemmenden Signalengehören Hormone aus demPankreas, Fettgewebe und ausdem Gastrointestinaltrakt. Dasvorwiegend im Magen gebildeteHormon Ghrelin wirktappetitsteigernd. GLP-1/2Glucagon-like peptide 1/2; OXMOxyntomodulin; PPpankreatisches Polypeptid;PYY3–36 Peptid YY3–36
2 C. Roth

xigene Wirkung; Tschop et al. 2000; Kap. ▶ „Fettgewebe alsendokrines Organ“). Es bindet am GHSR im ventromedialenHypothalamus, wodurch Hunger und Nahrungsaufnahmeinduziert werden. Es stimuliert im Nucleus arcuatus dieSekretion von
• Neuropeptid Y, von dem bekannt ist, dass es die Nah-rungsaufnahme steigert, sowie
• AGRP (Agouti-related peptide), wodurch das appetit-hemmmende Melanokortinsystem (Abschn. 2.2) im Hy-pothalamus inhibiert wird.
Beim Fasten ist die endogene Ghrelinsekretion hoch, abersie nimmt innerhalb von Minuten nach Nahrungsaufnahmeab. Über die wachstumshormonstimulierende Wirkung ent-faltet Ghrelin lipolytische Effekte. Darüber hinaus beeinflusstGhrelin auch die mitochondriale Lipidstoffwechselgenex-pression und die Fettverteilung in der Leber und dem Ske-lettmuskel (Barazzoni et al. 2005). Obestatin ist ein Peptid,das gemeinsam mit Ghrelin durch das Ghrelin/Obestatin-Pre-prohormon(GHRL)-Gen kodiert wird, jedoch anorexigenwirkt und die Nahrungsaufnahme drosselt (Zhang et al.2005).
Das Peptid YY3–36 (PYY3–36) ist ein vor Kurzem ent-decktes Sättigungshormon, das in den L-Zellen des Darmesgebildet und nach Nahrungsaufnahme in Abhängigkeit vonder aufgenommenen Nahrungsmenge in den Blutkreislauffreigesetzt wird. Im Blut wird PYY1–36 durch die Dipeptidyl-peptidase IV (DPP-IV) der N-terminale Tyrosin-Prolin-Restabgespaltet, wodurch PYY3–36 entsteht. Im Blut zirkulierenetwa
• 60 % des PYY1–36 (36 Aminosäuren), das an alle vierY-Rezeptoren (Y1, Y2, Y4, Y5) bindet, und
• 40 % des PYY3–36 (34 Aminosäuren), das nach Passierender Blut-Hirn-Schranke spezifisch an hypothalamische
• Y2-Rezeptoren (Y-2R) bindet und hierüber eine anorekti-sche Wirkung entfaltet.
Sowohl bei Nagetieren als auch bei Menschen konntegezeigt werden, dass das PYY3–36 den Appetit hemmt (Bat-terham et al. 2002). Das PYY3–36 bewirkt eine Verminderungdes hypothalamischen mRNA-Spiegels von Neuropeptid Y(NPY) des orexigenen Systems, wodurch der Hungergehemmt wird. Das Glucagon-like peptide 1 (GLP-1) wirdvon intestinalen L-Zellen produziert, aber auch in denA-Zellen des endokrinen Pankreas und in einzelnen Neu-ronen des Hirnstamms exprimiert. Es entsteht durch post-translationale Modifikation aus dem Vorläuferpeptid Prä-proglukagon. Durch Bindung an den GLP-1-Rezeptor(GLP-1R) hemmt GLP-1 die Magenmotilität, wodurch dieNahrungsabsorption verlängert wird. Außerdem entfaltetGLP-1 eine appetithemmende Wirkung und stimuliert die
β-Zellen des Pankreas und damit die Insulinsekretion (In-kretineffekt).
Oxyntomodulin (OXM) wird ebenfalls in den L-Zellendes Darmes gebildet und wirkt über den GLP-1R appetithem-mend.
Cholezystokinin (CCK) ist ein 8 Aminosäuren enthalten-des Darmpeptid, das nach Nahrungsaufnahme von enteren-dokrinen I-Zellen in Duodenum und Jejunum produziertwird. Die Ausschüttung des Hormons wird durch Fettsäurenund Aminosäuren im Nahrungsbrei angeregt. Es bindet anCholezystokinin-1-Rezeptoren (CCK1R) im Nucleus tractussolitarius (NTS) und der Area postrema des Hirnstamms undvermittelt die Beendigung der Nahrungsaufnahme und dieSättigung. Daneben verlangsamt es die Magenentleerung,stimuliert die Pankreassekretion sowie die Kontraktion derglatten Muskulatur der Gallenblasenwand und regt hierdurchdadurch den Gallenfluss an.
Hormone aus dem Pankreas
" Insulin spielt in der Energiehomöostase eine extrem wich-tige Rolle: Insulinrezeptoren werden in verschiedenenhypothalamischen Kerngebieten exprimiert, an die Insulinnach Passieren der Blut-Hirn-Schranke bindet und einezentrale appetithemmende Wirkung auslöst.
Bei Nagetieren wurde nachgewiesen, dass intrazerebroven-trikuläre Insulininjektionen die Nahrungsaufnahme durch dieAktivierung der Insulinrezeptorsubstrat-Phosphatidylinosi-tol-3-OH Kinase (IRS-PI3K) in hypothalamischen ventrome-dialen Neuronen hemmen (Schwartz et al. 2000; Wisse et al.2007). Zahlreiche Insulin-knock-out-Modelle zeigen, dasseine verminderte zentrale Insulinwirkung zu einem adipösenPhänotyp führt.
Das pankreatische Polypeptid (PP) gehört ebenfalls zuden Sättigungshormonen. Es wird in den Pankreasinseln,aber auch im Darm gebildet und gehört zu der gleichenHormongruppe wie PYY und NPY (Neuropeptid-Y). Beistarkem Übergewicht wurden verminderte Serumkonzentra-tionen nachgewiesen, die sich nach effizienter Gewichtsab-nahme normalisierten.
Amylin, das auch „Islet amyloid polypeptide“ (IAPP)genannt wird, ist ein aus 37 Aminosäuren bestehendes Poly-peptid, das zusammen mit Insulin in den β-Zellen des Pan-kreas synthetisiert wird. Amylin ist aber auch Hauptbestand-teil der Amyloidablagerungen in den Langerhans-Inseln desPankreas, die man bei Typ-2-Diabetikern findet. In der Regu-lation des Kohlenhydratstoffwechsels geht man davon aus,dass Amylin und Insulin sich in ihren Wirkungen ergänzen.Amylin hemmt die Glykogensynthese in der Leber und hatblutglukosesteigernde sowie lipolytische Effekte. Es bewirktaber auch als Sättigungspeptid eine Reduktion der Mahlzei-tengröße, Hemmung der Magenentleerung und Stimulation
Zentrale Regulation des Körpergewichtes 3

von Insulin, Glukagon, Pankreasamylase und -lipase. Ratten,denen ein Amylin-Antagonist verabreicht wurde, zeigen einegesteigerte Nahrungsaufnahme.
Fettgewebshormone und AdipozytokineDas Fettgewebe ist nicht nur ein großes Speicherorgan fürEnergie, sondern auch ein sehr aktives endokrines Organ(Kap. ▶ „Fettgewebe als endokrines Organ“). Es bestehtnicht nur aus Adipozyten, sondern auch aus anderen Zellen,wie Fibroblasten, Immunzellen und Endothelzellen, die ver-schiedene Hormone und Zytokine (Adipozytokine) sezernieren,wodurch die metabolischen und immunologischen Funktio-nen beeinflusst werden. Ihre Serumkonzentrationen korrelie-ren mit der Körperfettmasse und spielen in der Pathogenesevon adipositasassoziierten Erkrankungen eine Rolle.
Die im Fettgewebe produzierten proinflammatorischenAdipozytokine wie Leptin, Resistin, Plasminogen-Aktivator-Inhibitor 1, Interleukin-6 (IL-6) und Tumor-Nekrose-Faktorα (TNF-α) führen zu einer Verschlechterung der Insulinwir-kung. Besonders das intraabdominelle (viszerale) Fettge-webe, das bei dem androgenen Fettverteilungstyp vermehrtvorliegt, scheint die Insulinresistenz entscheidend zu beein-flussen. Adipozytokine sind ein mögliches Bindeglied zwi-schen Insulinresistenz und Adipositas (Bornfeldt und Tabas2011; Schrover et al. 2016) (Kap. ▶ „Endokrine Störungenbei Kindern und Jugendlichen mit Adipositas“).
" Adipozytokine führen nicht nur zu Störungen im Glukose-stoffwechsel, sondern auch zu Inflammation und kardio-vaskulären Erkrankungen.
Die Produktion von IL-6 im Fettgewebe führt zur Erhö-hung des C-reaktiven Proteins, das einen Prädiktor und Risi-kofaktor für Arteriosklerose darstellt.
Leptin ist ein 167-Aminosäuren-Peptid, das in Adipozytengebildet wird. Die wichtigste Rolle von Leptin besteht darin,Informationen über den Energievorrat bzw. die periphere Ener-giespeicherung in Adipozyten an das Gehirn weiterzuleitenund somit vor dem Hungertod zu schützen. Die im Blutzirkulierenden Leptinspiegel korrelieren mit der Fettgewebs-masse. Die Leptinproduktion wird durch Insulin und Gluko-kortikoide stimuliert. Eine hohe Dichte von Leptinrezeptorenwird in hypothalamischen Kerngebieten, dem Nucleus arcua-tus und im ventromedialen Hypothalamus, gemessen. Leptinbindet an seinen Rezeptor, einen Zytokinrezeptor, der in derSignalweiterleitung die Januskinase 2 (JAK2) aktiviert. Leptinaktiviert außerdem die IRS-PI3K in Neuronen des ventrome-dialen Hypothalamus, wodurch es zu einer zentralen Appetit-hemmung kommt. Leptin vermindert die Nahrungsaufnahmeund erhöht die Aktivität des zentralen Sympatikotonus. Dane-ben ist Leptin an der insulinunabhängigen Regulation desBlutglukosespiegels beteiligt. Die Verabreichung von Leptin
führt zu einem gesteigerten Ruheenergieumsatz, Verminde-rung der Nahrungsaufnahme und Reduktion der Blutglukose-spiegel (Park und Ahima 2015; Schwartz et al. 2013). NiedrigeLeptinspiegel werden bei verminderten Energievorräten ge-messen. Dadurch werden metabolische Prozesse sowie diePubertätsentwicklung gehemmt und der Appetit gesteigert.Gefüllte Energievorräte sind wichtig für Pubertät und Schwan-gerschaft. Es ist bekannt, dass Leptin die pulsatile Gonado-tropin-Releasing-Hormon(GnRH)-Sekretion stimuliert. Leptin-defiziente Menschen haben eine gestörte oder ausbleibendePubertät, was sich durch Leptinsubstitution normalisiert.
" Bei sehr aktiven Leistungssportlerinnen, Balletttänzerin-nen und anorektischen Adoleszentinnen kann es in Folgeder mangelnden Fettmasse zu einer ausbleibenden Puber-tät oder einer primären bzw. sekundären Amenorrhö kom-men. Der Grund hierfür ist ein durch Leptinmangelbedingter hypogonadotroper Hypogonadisms.
Nach der Reduktion der körperlichen Aktivität und demAnstieg des Gewichtes über eine kritische Grenze schreitetdie Pubertät fort oder der menstruelle Zyklus setzt wieder ein.
Adiponektin ist ein Adipozytokin, das ausschließlich imFettgewebe gebildet wird (Kap. ▶ „Fettgewebe als endokri-nes Organ“). Es kann neben seiner appetithemmenden Wir-kung auch antiinflammatorische Effekte entfalten. Es wirktaußerdem einer Insulinresistenz entgegen. Hierdurch wirkt esprotektiv bezüglich der Entwicklung von Diabetes mellitusTyp 2 und kardiovaskulären Erkrankungen. Adiponektinbeeinflusst den Energieverbrauch und die Thermogenese.Es bildet verschieden große Komplexe (High-molecular-weight-, Middle-molecular-weight-, und Low-molecular-weight-Adiponektin), die eine unterschiedliche Assoziationmit Parametern des metabolischen Syndroms aufweisen.Adiponektinrezeptoren (AdipR1 und 2) werden auch imHypothalamus exprimiert. Verabreichung von Adiponektinführt zu einer Verbesserung von Blutglukose und Insulin-resistenz, vermehrter Insulinsekretion sowie einer Reduzie-rung des Körpergewichtes (Bluher und Mantzoros 2015).
" Adiponektinspiegel korrelieren negativ mit einer Insulin-resistenz; niedrige Adiponektinspiegel sind ein starker Prä-diktor für die Entwicklung eines metabolischen Syndromsauch bei Kindern.
Resistin ist ein 12,5-kDa-proinflammatorisches Polypep-tid, das eine Rolle in der Entstehung von Insulinresistenz undDiabetes mellitus Typ 2 zu spielen scheint. In verschiedenenStudien konnte eine Assoziation zwischen Resistinspiegelnund der Ausprägung von Insulinresistenz und Adipositasnachgewiesen werden.
4 C. Roth

Besondere Aspekte der peripheren Regulation
InsulinresistenzInsulin entfaltet im ZNS eine appetithemmende Wirkung.Eine verminderte Insulinwirkung im Hypothalamus, z. B.durch einen Defekt des Insulinrezeptors im Gehirn, führt zueiner Hyperphagie, einer gesteigerten Glukoneogenese undeiner Hypertriglyzeridämie. Auch eine durch die Adipositasbedingte Insulinresistenz kann zu einer gestörten Appetit-hemmung führen. Eine verminderte IRS-PI3K-Aktivität kanndie Effekte von Insulin und Leptin im Gehirn abschwächen.Daneben entstehen ungünstige Effekte in peripheren Gewe-ben, die den Circulus vitiosus der Insulinresistenz in derLeber, im Muskel und im Fettgewebe weiter verstärken. Inβ-Zellen kann eine verminderte IRS-PI3K-Aktivität zurβ-Zell-Erschöpfung und einem Diabetes mellitus Typ 2 füh-ren (Wisse et al. 2007). Eine Reduktion der Hyperinsulinämiedurch eine erfolgreiche Gewichtsreduktion führt auch zueiner erhöhten Leptinsensitivität und damit einer Verbesse-rung der Sättigung (Kap. ▶ „Physiologie und Pathophysio-logie der Insulinsekretion“).
Veränderungen peripherer Hormone desEnergiestoffwechsels bei AdipositasBei Adipositas sind hohe periphere Insulinspiegel Ausdruckeiner Insulinresistenz (s. oben). Die zentrale Insulinresistenzführt zu einer gesteigerten Kalorienaufnahme. Hohe Leptin-spiegel sind zum einen auf die erhöhte Fettmasse, zum ande-ren jedoch auch auf die Entwicklung einer Leptinresistenzzurückzuführen. Sowohl erhöhte Insulin- als auch erhöhteLeptinspiegel können sich bei erfolgreicher Gewichtsreduk-tion normalisieren. Es besteht eine negative Korrelation zwi-schen Ghrelinspiegel und dem Body-Mass-Index (BMI). BeiAdipositas sind Ghrelin-Plasmakonzentrationen vermindert,während sie bei Patienten mit einem Prader-Willi-Syndrommassiv gesteigert sind, was möglicherweise mit der Hyper-phagie dieser Patienten im Zusammenhang steht. Im Gegen-satz zur langsamen Gewichtsreduktion (z. B. über ein Jahr)steigen bei einer kurzfristigen Gewichtsreduktion die Ghre-linspiegel im Blut stark an, wodurch der Hunger stimuliertwird. Dies kann den Erfolg einer gewichtsreduzierendenTherapie gefährden. Patienten mit einer Adipositas habenverminderte Spiegel des Sättigungshormons PYY, das nachNahrungsaufnahme nicht ausreichend hoch und lange anhal-tend ansteigt. Nach effizienter Gewichtabnahme kann eineNormalisierung der PYY-Werte beobachtet werden. DieGLP-1-Sekretion ist bei Adipositas vermindert und zeigteine Normalisierung nach erfolgreicher Gewichtsreduktion.Ebenso sind Adiponektin-Serumspiegel bei Adipositas ver-mindert und steigen nach erfolgter Gewichtsreduktion wiederan (Roth und Reinehr 2010).
Veränderungen von peripheren Hormonen nachMagen-Bypass-OperationBei Patienten mit extremer Adipositas, die nicht an einermonogenen Form der Adipositas leiden, ist eine Magen-Bypass(MB)-Operation die effektivste Methode, Überge-wicht zu reduzieren. Diese kommt nicht allein durch die ver-ringerte Resorption der Nahrung zustande, sondern auch durchVeränderung der Appetitregulation. Es werden nach der Ope-ration deutlich verminderte Ghrelinkonzentrationen im Plasmabei gleichzeitiger Gewichtsabnahme gemessen. Nach Anlageeines Roux-Y-MB wird zusätzlich ein Anstieg der postpran-dialen PYY-, GLP-1- und OXM-Konzentrationen im Plasmagemessen, wodurch das Sättigungsgefühl verstärkt wird. Au-ßerdem steigt die schnelle Insulinsekretion an, was zu einerverbesserten Kontrolle der postprandialen Blutglukose führt.Nach einer MB-Operation kommt es außerdem zu einer ver-minderten Aktivität in Hirnarealen, die zum sog. „Food-reward-System“ (Abschn. 2.2) gehören, einschließlich desmedialen orbitofrontalen Kortex, Amygdala, Putamun, Nu-cleus caudatus und Nucleus accumbens. Dies konnte mittelsfunktioneller Kernspintomografie (fMRT, Abschn. 2.2) nach-gewiesen werden (Scholtz et al. 2014). Nach einer Vagotomiewerden die Hormonsekretion verschiedener gastrointestinalerHormone beeinflusst und dadurch die Vermittlung von Hungerdurch Ghrelin, aber auch die Vermittlung von Sättigung durchGLP-1 und PYYvermindert.
2.2 Zentrale Regulatoren und Strukturen derEnergiehomöostase
Hypothalamische KerngebieteIn den letzten Jahren wurden die zentralen Regulationsstruk-turen, die für die Wahrnehmung von Hunger und Sättigungverantwortlich sind, intensiv erforscht. Insbesondere Kernge-biete des Hypothalamus sind wichtige Regulationszentren.
" Historisch gesehen zählen Nucleus arcuatus (ARC), Nucleusparaventricularis (PVN), Nucleus ventromedialis (VMN) undNucleus dorsomedialis (DMN) zum „Sättigungszentrum“
(Abb. 2).
Diese hypothalamischen Strukturen integrieren afferentehormonelle Signale und Metabolite aus der Körperperipherie(s. unten) und in Neuronen synthetisierte Peptide wie dasNeuropeptid Y, das AGRP, das melanozytenstimulierendeHormon (α-MSH) und das Cocaine-amphetamine-regulatedtranscript (CART). Im ARC werden diese hormonellenSignale in neuronale Signale transformiert, die an denN. paraventricularis (PVN) weitergeleitet werden (Schwartzet al. 2000).
Zentrale Regulation des Körpergewichtes 5

" Der laterale Hypothalamus (LHA) und die perifornikaleRegion (PFA) sind Regionen, die beide appetitsteigerndeNeuropeptide wie Orexin A und B und melaninkonzentrie-rendes Hormon exprimieren und werden daher als „Hun-gerzentrum“ angesehen (Abb. 2; Tab. 1).
Dort lokalisierte Läsionen verursachen temporäre Apha-gie, Adipsie und Gewichtsverlust. Auch Zytokine regulierenhypothalamische Sättigungs- und Hungerzentren. So werdeneine Reihe von Zytokinen, wie der Tumor-Nekrose-Faktor-α,Interleukine und Interferon-γ, mit der bei Tumorpatientenauftretenden Kachexie in Zusammenhang gebracht (DeBoerund Marks 2006). Allerdings kann auch hochkalorische fett-reiche Ernährung eine Inflammation und Dysfunktion desHypothalamus bewirken, die die Entwicklung einer Adiposi-tas möglicherweise begünstigt. Neben den genannten hypo-
thalamischen Neuropeptiden gibt es weitere appetitregulie-rende Rezeptoren. Die Stimulation des Cannabinoidrezeptorsz. B. führt zu einer Appetitsteigerung. Die Applikation einesCannabinoidrezeptorenhemmers bewirkt eine Gewichtsre-duktion.
" Der VMN und der DMN sind an der Regulation von Sym-patikotonus, Herzfrequenz, Blutdruck und Thermoregu-lation beteiligt und sind daher für den Energieverbrauchwichtige Kerngebiete. Beide sind mit anderen für dieRegulation des Energiegleichgewichtes wichtigen Kernge-bieten (PVN, ARC, LHA, NTS) eng verschaltet.
Area postrema/Nucleus tractus solitariiNeben dem Hypothalamus spielen Rezeptoren in der Nucleus-tractus-solitarii-/Area-postrema(NTS/AP)-Region des Hirn-
ARCPOMCCART
NPYAGRP
VMNBDNFGABA
III.Ventr.
PVNOxytocin
CRFTRH
LHA, PFOrexinMCH
Periphere Peptide und Nährstoffe
+/-
DMNNPY,
GABA
AgRPNPYa-MSH
a-MSHNPYAgRP
a-MSHNTS,AP
Hirnstamm und Rückenmark: Regulation peripherer Gewebe über autonomes Nervensystem (Leber, Herz, Muskel, etc)
OxytocinCRF
Höhere kortikale Regionen-Orbitofrontaler KortexMittelhirn/limbisches System-Nucleus accumbens-Corpus striatum-Ventrotegmentale Region
Hypophyse:EndokrineReaktion
Afferenter Vagus& GI-Peptide (-)
Medialer Hypothalamus
Lateraler Hypothalamus
Hirnstamm
Ghrelin (+) Leptin (-)PYY3-36 (-) Insulin (-)GLP-1 (-)PP (-)
Abb. 2 Zentrale Regulation von Hunger und Sättigung. Medial gele-gene hypothalamische Kerngebiete (Nucleus arcuatus, PVN, VMN,DMN) gehören zum sog. Sättigungszentrum. Projektionen von diesenNeuronengebieten und deren Neuropeptide wirken appetithemmend(α-MSH, Oxytocin, TRH, CRH) oder appetitsteigernd (AGRP, NPY).Im lateralen Hypothalamus werden Neuropeptide gebildet, die denHunger stimulieren (MCH sowie Orexin A und B). Das Energiegleich-gewicht wird auch über leptinsensitive Verbindungen zwischen Hypo-thalamus und Hirnstamm (Nucleus tractus solitarius, Area postrema)beeinflusst. Die hypothalamischen Kerngebiete beeinflussen das Appe-titverhalten auch über Projektionen in höher gelegene Hirnareale imKortex, wie dem orbitofrontalen Kortex, sowie im Mittelhirn gelegenenlimbischen Strukturen, die die Nahrungsaufnahme im Zusammenhangmit Erkennen, Belohnungsverhalten und Gefühlen bringen. Läsionen
der medialen Kerngebiete resultieren in Hyperphagie, Adipositas undeinem verminderten Energieverbrauch, während Läsionen des lateralenHypothalamus hingegen zu Hypophagie und Gewichtsverlust bei gestei-gertem Energieverbrauch führen. AGRPAgouti-related peptide; APAreapostrema; ARCNucleus arcuatus; BDNF Brain-derived neutrophic factor;CART Cocaine-amphetamine-regulated transcript; CRF Corticotropine-releasing factor; DMN Nucleus dorsomedialis; GABA γ-Aminobutter-säure; GLP-1 Glucagon-like peptide 1; LHA lateraler Hypothalamus;MCH melaninkonzentrierendes Hormon; α-MSH melanozytenstimulie-rendes Hormon; NPY Neuropeptid Y; NTS Nucleus tractus solitarius;PF perifornikale Region; POMC Proopiomelanokortin; PP pankreati-sches Polypeptid; PVN Nucleus paraventricularis; PYY3-36 PeptidYY3–36; TRH Thyreoliberin; VMN Nucleus ventromedialis
6 C. Roth

stamms eine zentrale Rolle für Bindung und Signalweiterlei-tung der aus der Körperperipherie im ZNS eintreffenden Pep-tide. Sowohl die Eminentia mediana des Hypothalamus alsauch die NTS/AP-Region gehören zu den Zirkumventrikular-Organen, die außerhalb der Blut-Hirn-Schranke liegen unddadurch leicht von im Blut zirkulierenden Hormonen erreichtwerden können. Diese Region ist eng mit hypothalamischenKerngebieten verschaltet.
Food-Reward-SystemDer Hypothalamus und der Hirnstamm gehören zu den pri-mären Zentren für die Regulation des Körpergewichtes.Jedoch wird die Nahrungsaufnahme auch durch Kognition,Gefühle und Belohnungsverhalten beeinflusst. Heute weißman, dass die oben beschriebenen Regelkreise der Appetit-regulation entscheidend von höheren Zentren, die zum sog.Reward-System gehören, beeinflusst werden.
Hierzu gehören insbesondere Regionen des limbischenSystems, aber auch bestimmte Kortexareale des Gehirns.Die Aktivität dieser Hirnareale wird möglicherweise auchdurch Insulin, Leptin und Ghrelin beeinflusst. Das Corpusstriatum ist eine wesentliche Komponente des Reward-Systems. Hierbei sind v. a. Projektionen dopaminerger Neu-ronen des Mittelhirns in der Area tegmentalis ventralis (VTA)und der Substantia nigra zum Corpus striatum einschließlichder spezialisierten Subregion, dem Nucleus accumbens,bedeutsam (Everitt et al. 1999).
" Motivation und Belohnung spielen beim Reward-Systemeine wichtige Rolle. Dies konnte sowohl bei der Abhängig-keit von Drogen als auch bezüglich des Ernährungsverhal-tens in Abhängigkeit von sozialen Faktoren und Umge-bungsfaktoren gezeigt werden.
Wahrnehmung und Erwartung von nahrungsabhängigerBelohnung (Reward) tragen zur Entwicklung von Über-gewicht eine Rolle. Übergewichtige Kinder und Erwachsenefinden oft kalorienreiche (zucker- und fettreiche) Nahrungmehr ansprechend als Normalgewichtige. Durch Aktivierungdes limbischen Dopaminsystems werden durch Nahrungs-reize Belohnungsverhalten, Glücksgefühle und schließlichMotivation zum Essen getriggert (Cornier 2011; Balleineet al. 2007).
Reward- und Appetitkonditionierung sind wichtige psy-chologische Mechanismen. Führen Situationen oder Lebens-phasen zu einem Reward-Mangel, so kann es kompensato-risch zu erhöhter Nahrungsaufnahme kommen (Frustessen).Das Prinzip der Appetitkonditionierung wird auch in derNahrungsmittelwerbung genutzt und ist möglicherweise be-deutsam für das Konsumverhalten und die rasante Zunahmeder Adipositasprävalenz in den westlichen Industrienationenund anderen Ländern mit steigendem Lebensstandard. SowohlLeptin- als auch Insulinrezeptoren werden in diesen Hirnarea-len exprimiert. Aus Untersuchungen an Nagetieren weiß man,dass Lernen und Belohnungsverhalten durch Schwankungender Insulin- und Leptinkonzentrationen beeinflusst werdenkönnen (Figlewicz et al. 2007).
Mediatoren der EnergiehomöostaseZum anorexigenen System gehören das Proopiomelanoakor-tin (POMC) und das kokainamphetaminregulierte Peptid(CART, Cocaine-amphetamine-regulated transcript), wäh-rend zum orexigenen Mechanismus das Neuropeptid Y (NPY)und das Agouti-related peptide (AGRP) zählen. Diese beidenPrinzipien konkurrieren um Bindung an Melanokortinrezep-toren (MC3R und MC4R) im PVN und dem lateralen Hypo-thalamus. Das α-MSH induziert eine Appetithemmung durchdie Bindung an Melanokortinrezeptoren (PVN, LHA, NTS).
Orexigenes SystemNPYund AGRP werden von verschiedenen Neuronen in dergleichen hypothalamischen Region, dem Nucleus arcuatus,kolokalisiert. Das AGRP ist ein endogener Melanokortinan-tagonist. Er bewirkt eine Stimulation der Nahrungsaufnahme.Das NPY ist das primäre orexigene Signal. Fasten undGewichtsverlust induzieren eine erhöhte NPY-Expressionim Nucleus arcuatus, wodurch der Hunger gesteigert wird,während das Sättigungshormon PYY3–36 nach Bindung andie Y2-Rezeptoren und Leptin die NPY-RNA-Spiegel redu-zieren (Batterham et al. 2002). Das NPY hat aber nochweitere Funktionen: Neben der Appetitsteigerung ist es ander der Regulation des Pubertätsbeginns beteiligt. Das mela-ninkonzentrierende Hormon (MCH) ist ein Peptid, das inder Zona incerta und dem lateralen Hypothalamus exprimiertwird. Die MCH-Neuronen bilden synaptische Kontakte zumGroßhirn und dem kortikolimbischen Reward-System, wo-
Tab. 1 Zentrale Faktoren der Appetitregulation und Energiehomö-ostase
Zentrale Appetitregulation
Appetithemmend Appetitsteigernd
POMC, α-MSH Neuropeptid Y
CART AGRP
CRH MCH
Serotonin Opioide
Dopamin Orexin A und B
TRH GABA
Oxytocin
FGF-21
Neurotensin
AGRP Agouti-related peptide; CART Cocaine-amphetamine-regulatedtranscript; CRH Kortikotropin-Releasing-Hormon; FGF-21 Fibroblastgrowth factor 21; MCH melaninkonzentrierendes Hormon; MSH mela-nozytenstimulierendes Hormon; POMC Proopiomelanokortin; TRHThyreoliberin
Zentrale Regulation des Körpergewichtes 7

rüber sie die Nahrungsaufnahme beeinflussen. MCH-kno-ckout-Mäuse haben eine verminderte Nahrungsaufnahmeund sind schlank, während Mäuse mit einer MCH-Überex-pression Übergewicht und eine Insulinresistenz entwickeln.Leptindefiziente homozygote Obese-(ob/ob-)knock-out-Mäu-se (Abschn. 4.1) sind stark übergewichtig und haben einehohe Expression des MCH (Shimada et al. 1998). Orexin Aund B sind 33- und 28-Aminosäuren-Peptide, die im LHAdie NPY-Freisetzung stimulieren, wodurch sich ihre orexige-ne Wirkung erklärt. Orexin-knock-out-Mäuse weisen eineNarkolepsie, Hypophagie und Übergewicht auf (Chou et al.2001).
" Orexine stimulieren den zentralen Sympathikotonus, denEnergieverbrauch und die Wachheit. Sie sind jedoch auchwichtig für das Lernen und die hedonische Reward-Funk-tion.
Endocannabinoide (EC) stimulieren die Nahrungsauf-nahme über CB1-Rezeptoren, die in Kortikotropin-releasing-Hormon (CRH), Neuronen des PVN, in CART-Neuronen desVMN und in MCH/Orexineuronen des LHA und der PFAexprimiert werden. Erhöhte EC-Spiegel finden sich beimMenschen z. B. unter Glukokortikoidtherapie und bei leptin-defizienten ob/ob-Mäusen (Malcher-Lopes et al. 2006).Durch Leptinverabreichung fallen die EC-Spiegel ab, wasden Einfluss von EC auf die zentrale Regulation der Energie-balance unterstreicht.
Anorexigenes SystemDas POMC wird in verschiedenen Geweben und Neuronenenzymatisch gespalten. Nach Bindung an Leptinrezeptoren imARC wird durch Leptin die Bildung von α-MSH induziert,welches eine anorektische Wirkung nach Bindung an Melano-kortinrezeptoren im PVN und LHA entfaltet. Das CART isthierbei ein hypothalamisches Neuropeptid, das durch Leptininduziert und durch Fasten reduziert wird. Wird das endogeneCART durch einen Antagonisten blockiert, kommt es zurerhöhten Kalorienaufnahme. Leptin bewirkt auch eine Stimu-lation vonOxytocin in dem parvozellulären Bereich des PVN,welches die Nahrungsaufnahme nach Bindung an Rezeptorenin Neuronen des limbischen Systems und des NTS im Hirn-stamm hemmt (Blevins und Baskin 2015).
" Melanokortinrezeptoren im PVN und LHA modulierenanorexigene und orexigene Informationen und steuernden efferenten Sympathikotonus im VMN, wodurch dieEnergiespeicherung und der Energieverbrauch gesteuertwerden (Abschn. 2.3).
Andere MediatorenNoradrenalin und Serotonin sind ebenfalls Regulatoren derGewichtsbalance. Noradrenalinneurone regulieren über Sy-napsen mit VMH-Neuronen die Nahrungsaufnahme, wobeidie Wirkung nicht einheitlich ist. Intrahypothalamische No-radrenalininfusionen wirken über zentrale α2- und β-adrener-ge Rezeptoren appetitsteigernd, während zentrale Applika-tionen von α1-Agonisten den Appetit hemmen. Serotoninwirkt anorexigen. Serotonin(5-HT2C)-Rezeptor-Agonisteninduzieren im Hypothalamus eine Sättigung, während Sero-toninantagonisten die Nahrungsaufnahme stimulieren.
Weitere Aspekte der Energiehomöostase
Funktionelle MagnetresonanztomografieDie Anwendung der funktionellen Magnetresonanztomogra-fie (fMRT) als bildgebendes Verfahren eröffnet einen nicht-invasiven Zugang zu den Vorgängen im Gehirn mit hoherräumlicher Auflösung. Die fMRT ist eine einzigartige Me-thode, um die Aktivierung des menschlichen Gehirns durchbestimmte Stimuli in vivo nachzuweisen. Durch Messungeines sauerstoffabhängigen Signals wird indirekt die lokaleneuronale Stoffwechselaktivität des Gehirns gemessen, umRückschlüsse auf aktivierte Hirnareale in Abhängigkeit inne-rer und äußerer Stimuli zu erhalten. Mithilfe dieser Technikgelingt es, eine akute Aktivierung von verschiedenen Regio-nen des Gehirns, die bei der Regulation des Appetits beteiligtsind, nach dem Ansehen von Nahrungsfotografien nachzu-weisen. Hierzu gehören:
• Präfrontaler Kortex• Orbitofrontaler Kortex• Amygdala• Insula• Ventrales (Nucleus accumbens) und dorsales Striatum• Hypothalamus• Hirnstammregionen
Mittels fMRT lässt sich die Interaktion zwischen zirkulie-renden Peptiden wie Leptin, PYY, Ghrelin, GLP-1 und denHirnregionen, die an der Regulation der Nahrungsaufnahmebeteiligt sind, nachweisen. Bei leptindefizienten Patienten,die sich Bilder hochkalorischer Nahrungsmittel ansahen,wurde eine gesteigerte Aktivität im Nucleus accumbens undim Putamen nachgewiesen (Farooqi et al. 2007).
Gustatorische SignaleDie Wahrnehmung von gustatorischen Signalen ist für dieAppetitregulation und Ausschüttung appetitregulierender Hor-mone wichtig. Inzwischen sind die Genfamilien der in derMundhöhle lokalisierten Geschmacksrezeptoren für bitter,süß, sauer, salzig und umami (für Aminosäuren wie Glutamat)bekannt. Interessanterweise werden Süß-Rezeptoren auch im
8 C. Roth

Darmepithel exprimiert, wodurch möglicherweise gastrointes-tinale Hormone durch die Wahrnehmung „süß“ stimuliert wer-den können (Sternini et al. 2008).
Im Darm sind Geschmacksrezeptoren in Nachbarschaft zuden enteroendokrinen Zellen des Darmes (z. B. L- oderI-Zellen) lokalisiert. Enteroendokrine Zellen sind das größteendokrine Organ des Körpers und beeinflussen durch dieGI-Hormone die Motilität des GI-Traktes und auch Hungerund Sättigung. Daneben bilden enteroendokrine Zellen auchdirekte Verbindungen mit Neuronen, wodurch ebenfalls dieFunktion des GI-Traktes beeinflusst wird (Latorre et al.2016).
2.3 Zentrale Kontrolle der Thermogeneseüber das autonome Nervensystem
Die Kerngebiete des VMN, DMN und PVN gelten als zen-trale Integrationsstellen von Sättigungskreisläufen, als Regu-lationszentrum der hypothalamischen Hormonproduktion so-wie des autonomen Nervensystems bzw. der sympathischenRegulation des Energieverbrauchs.
Diese Kerngebiete senden außerdem efferente Projektio-nen zum dorsalen motorischen Kern des N. vagus (DMV),um die efferente Vagusaktivität zu beeinflussen.
Für die Regulation des Tag-Nacht-Rhythmus ist derNucleus suprachiasmaticus wichtig. Er liegt oberhalb desChiasma opticum und erhält über den retinohypothalami-schen Trakt Lichtinformationen aus dem Auge und hat Ver-bindungen zur Glandula pinealis, worüber die Melatoninse-kretion beeinflusst wird. Er ist eine „biologische Uhr“ undregelt biologische Rhythmen, im Zusammenhang mit Tagund Nacht sowie Essen und Fasten.
Sympathisches NervensystemEine Erhöhung der sympathischen Aktivität und die Vermin-derung der Nahrungsaufnahme durch Noradrenalin werdenüber β2- und β3-Rezeptoren im Gehirn reguliert. Leptinstimuliert über Neurone in Hypothalamus und Hirnstammdie Lipolyse im braunen Fettgewebe sowie die Thermoge-nese und die Bewegung, was alles zu erhöhtem Energiever-brauch führt. Insulin und Leptin bewirken über den PVN eineStimulation von Thyreoliberin (TRH) und über den VMNund den DMN eine Erhöhung der zentralen Sympathikusak-tivität. Durch die erhöhte Sympathikusaktivität wird auch dasthyreoideastimulierende Hormon (TSH) sowie die Lipolyseüber β3-adrenerge Rezeptoren in den Adipozyten stimuliert,während im Skelettmuskel über β2-adrenerge Rezeptoren derEnergieverbrauch gesteigert wird. Die Anregung des β3-adrenergen Rezeptors in den Adipozyten führt zur Stimula-tion des zyklischen Adenosinmonophosphats (cAMP) undhierdurch zur Aktivierung der Proteinkinase A, was dieExpression von PPARγ-Koaktivator 1α stimuliert (PGC-1α).
PGC-1α ist ein wichtiges Bindeglied zur Wärmeproduktionaus dem Adenosintriphosphat (ATP), indem es die Expres-sion der Uncoupling proteins UCP1 und -2 anregt. UCPreduzieren den Protonengradienten an der inneren Mitochon-drienmembran, weshalb ATP für die Wärmeproduktion ver-braucht und somit gespeicherte Energie in Wärmeenergieumgeleitet wird. UCP1 ist ein Protein an der inneren Mito-chondrienmembran, das den Protonenfluss zur ATP-Produk-tion entkoppelt und somit gespeicherte Energie in Form vonWärmeenergie im braunen Fettgewebe verpuffen lässt. UCP2wird in den meisten Geweben exprimiert, während UCP3vorwiegend im Skelettmuskel vorkommt (Lowell und Spie-gelman 2000).
VagussystemOrexigene und anorexigene Informationen aus den Hypotha-lamuskernen werden zum DMV weitergeleitet, wodurch dasefferente Vagussystem – als Gegenspieler zum Sympathikus –die Energiespeicherung und Drosselung des Energiever-brauchs einschließlich der Verminderung der Herzfrequenzinduziert, während es die Darmperistaltik stimuliert. Inzwi-schen weiß man über retrograde Anfärbung, dass Vagusfasernaus dem DMV kommend bis zum Fettgewebe ziehen und hierdie Aufnahme von Glukose und freien Fettsäuren und somiteine Energiespeicherung stimulieren. Der Vagus stimuliertaber auch die Freisetzung von GLP-1 und Insulin. Nebendiesem efferenten System spielt der afferente N. vagus, dereine primäre neuronale Verbindung zwischen dem Verdau-ungsapparat und dem Gehirn darstellt, auch bei der Vermitt-lung von Hunger eine Rolle. Zahlreiche Rezeptoren fürGI-Hormone werden im afferenten N. vagus exprimiert, wiez. B. Y-2R, GLP-1R, GHSR1, und CCK1R, wodurch auchHunger und Sättigung vermittelt werden. Das afferente Vagus-system leitet außerdem Informationen über die mechanischeSpannung des Magens und Zwölffingerdarms zum NTS.
Veränderungen bei GewichtsabnahmeBei einer Gewichtsabnahme infolge kalorischer Restriktionkommt es zur Verminderung von Leptin und dem aktivenSchildrüsenhormon T3 (Trijodthyronin) bei gleichzeitigerZunahme des inaktiven Schildrüsenhormons Reverse-T3.Hierdurch finden eine Reihe von Stoffwechseladaptionenstatt, um Energieverbrauch zu reduzieren. Durch Reduktiondes Sympathikotonus werden Thermogenese, Fettoxidation,körperliche Aktivität, Blutdruck und Herzfrequenz reduziert.Durch den hierdurch reduzierten Grundumsatz wird einerweiteren Gewichtsabnahme entgegengesteuert. Zusätzlichkommt es durch die abfallenden Leptin- und ansteigendenGhrelinspiegel zu einem ausgeprägten Hungergefühl und derMotivation zu essen, was eine Gewichtszunahme begünstigt.Hierbei wird oft eine unerwünschte und schnelle Gewichts-zunahme nach einer Reduktionsdiät erreicht, wodurch das
Zentrale Regulation des Körpergewichtes 9

neue Endgewicht oft höher ist als das Ausgangsgewicht vorder kalorischen Restriktion (Jo-Jo-Effekt).
3 Adipositas durch hypothalamischeLäsionen
3.1 Experimentelle Adipositas durchhypothalamische Läsionen im Tiermodell
Es sind verschiedene Tiermodelle einer induzierten Adiposi-tas nach experimentellen ZNS-Läsionen beschrieben worden.Läsionen des medialen Hypothalamus, insbesondere desVMN, führen zu Hyperphagie, persistierender Körperge-wichtszunahme und durch eine Reduktion des zentralenSympathikotonus sowie des β3-adrenergen Tonus zu einerAbnahme der Thermogenese im braunen Fettgewebe (Bray2000). Dieses Syndrom kann durch eine Vagotomie unterhalbdes Zwerchfells abgeschwächt werden. Umgekehrt führt einesympathische Denervierung im Tierversuch ähnlich wie eineLäsion im VMN zu einem vergleichbaren Gewichtsanstieg.Dies unterstreicht die Bedeutung des autonomen Nervensys-tems an der Manifestation der hypothalamischen Adipositas.
3.2 Hypothalamische Adipositas beimMenschen
Gerade bei Patienten mit hypophysären und hypothalamischenErkrankungen stellt die Adipositas ein erhebliches Problemdar. Manche Patienten verspüren oft in Phasen der dynami-schen Gewichtszunahme einen ungebremsten Drang zu essen.Nach Operationen im Bereich des Hypothalamus kommt esinsbesondere innerhalb des ersten postoperativen Jahres oft zugesteigertemAppetit und zu schneller Gewichtszunahme, wes-halb diese Periode für die Prävention der hypothalamischenAdipositas besonders wichtig ist (Roth et al. 2015). Eineverminderte Sekretion hypophysärer Hormone (GH, TSH,LH, FSH) kann zur Entwicklung der Adipositas beitragen,weshalb eine konsequente Hormonsubstitution zu gewährleis-ten ist. Aber selbst bei optimaler Hormonsubstitution persis-tiert das Problem der gestörten Appetitregulation bei vielenPatienten. Bei der hypothalamischen Adipositas liegt meisteine Schädigung mehrerer medial gelegener hypothalamischerStrukturen vor, wodurch mehrere redundante Systeme kom-pensatorischer Mechanismen verloren gehen und folglich diepostprandiale Sättigung schwerwiegend gestört ist. Dies kannbei manchen Patienten zur Hyperphagie mit unkontrollierterAufnahme großer Nahrungsmengen und/oder ständigemEssen führen.
" Dieses durch organische Läsionen relevanter ZNS-Kernge-biete verursachte unkontrollierte Essverhalten wurde schonoft als psychisches Fehlverhalten missinterpretiert.
Aber selbst diejenigen Patienten, die ihren Appetit gutkontrollieren und eine kalorienreduzierte Diät einnehmen,können an Gewicht zunehmen, da die Leptinresistenz zueiner verminderten körperlichen Aktivität und einem gedros-selten Energieverbrauch führt.
Patienten mit einem Kraniopharyngeom sind häufig auf-fallend träge und bei gestörtem Sättigungsgefühl extrem adi-pös. Als Folge des Tumors, der Operation oder der Bestrah-lung ist von einer gestörten Leptinwirkung und von einemMangel an α-MSH und Oxytocin im Bereich des Hypo-thalamus auszugehen (Roth 2015). Einige Patienten leidenunter extremer Tagesmüdigkeit und haben eine verminderteBewegungsaktivität. Die Zerstörung hypothalamischer Kern-gebiete durch ein Kraniopharyngeom führt wahrscheinlich zueiner Verminderung der Bewegungsaktivität infolge einesverminderten Sympathikotonus und folglich zum verminder-ten Energieverbrauch sowie zur Unterbrechung der Regel-kreise afferenter appetitregulierender Peptide. Außerdemführen Läsionen des VMN zu einer Desinhibition des effe-renten Vagotonus mit konsekutiver Stimulation der pankrea-tischen β-Zellen und postprandialer Hyperinsulinämie, wasebenfalls zur Adipositas führen kann (Kap. ▶ „Physiologieund Pathophysiologie der Insulinsekretion“). Risikofaktorenfür eine schnelle postoperative Gewichtszunahme sind: inva-siver transkranialer Operationsweg gegenüber endoskopi-schem transnasalem Zugang, Anstieg des BMI und Abnahmeder Wachstumsrate schon vor der Diagnose bzw. Operation,großer hypothalamischer Defekt, der bis zum posteriorenHypothalamus reicht und die wichtigen Kerngebiete ARC,PVN, VMN und DMNmiteinschließt. Die Entwicklung einesDiabetes insipidus gilt als endokriner Risikomarker für dieEntwicklung einer hypothalamischen Adipositas (Roth et al.2015; Muller 2016).
Eine Adipositas kann unter folgenden Risikofaktoren beiverschiedenen Hirntumoren auftreten:
Risikofaktoren bei verschiedenen Hirntumoren für dieEntwicklung einer Adipositas• Hypothalamische Tumorlage• Affektion hypothalamischer Gewebe• Ausmaß der Operation• Hypothalamische Endokrinopathie• Hypothalamische Bestrahlung mit über 50 Gy
10 C. Roth

4 Genetische Ursachen der Adipositas
4.1 Experimentelle Adipositas durchVeränderungen einzelner Gene deszentralen Melanokortinsystems imTiermodell
An Mäusen gelang es, bestimmte Adipositasformen mono-genen Defekten zuzuordnen. So bewirkt die Obese(ob)-Mutation eine Leptindefizienz, während die sog. Diabetes(db)-Maus ein mutiertes Gen für den Leptinrezeptor besitzt.Bei beiden Mutationen ist eine extreme Adipositas die Folge,die bei der ob-Maus durch die Verabreichung von Leptinverhindert werden kann, nicht aber bei der db-Maus. Aufder appetithemmenden Seite wird POMC enzymatisch u. a.in das α-MSH und das adrenokortikotrope Hormon (ACTH)gespalten, ein Vorgang, der durch die Bindung von Leptin anvorwiegend im Nucleus arcuatus lokalisierte Leptinrezepto-ren stimuliert wird. Das α-MSH wirkt nach Bindung an diehypothalamischen Melanokortin-4(MC4)-Rezeptoren appe-tithemmend. Mutationen im POMC-Gen führen entspre-chend zu einer endokrinen Störung mit Adipositas, roterFellfarbe und Störung der adrenalen Steroidgenese. Sind dieMelanokortinrezeptoren durch überexprimierte Antagonisten(Agouti-Protein bzw. AGRP) inhibiert, resultiert darauszusätzlich zur Adipositas eine gelbe Haarpigmentierung(Ay-Maus). Eine Mutation des hypothalamischen MC4-Re-zeptors bewirkt eine isolierte Adipositas bei Hyperphagie,aber auch Mutationen des MC3-Rezeptors (MC3-R) führeninfolge eines verminderten Grundumsatzes zur Adipositas.
4.2 Humane Adipositasmutationen
Diese aus monogenen Mausmodellen gewonnenen Erkennt-nisse sind bzgl. einiger Störungen auf den Menschen übertrag-bar (s. auch Kap. ▶ „Angeborene Schilddrüsenerkrankungenbei Neugeborenen und Kleinkindern“). Bei Menschen sindmonogene Formen der Adipositas bekannt wie Leptindefizienz,Leptinrezeptordefizienz, inaktives Leptin, oder Gendefekte fürPOMC, PPARγ, Prohormon-Convertase-1 (Abschn. 4.3),jedoch sind weltweit bisher nur eine kleine Anzahl von Men-schen mit Gendefekten identifiziert worden.
Bei einem POMC-Gendefekt kommt es zu einer Störungdes Splicings, wodurch die Entstehung der Peptide α-MSHund ACTH aus dem gemeinsamen Vorläufer gestört ist. Diesführt im Gehirn zu einer Störung der Sättigung durch man-gelnde Aktivierung von MC4-Rezeptoren und in der Periphe-rie zu einer hellen Hautpigmentierung und roten Haaren durcheine verminderte Aktivierung von MC1-Rezeptoren sowie zueinem ACTH-Mangel-bedingten Hypokortisolismus (Krudeet al. 2003). Beim Prohormon-Convertase-1-Mangel ist die
Spaltung von Preprohormonen in die aktiven Hormone wiePOMC in ACTH und α-MSH oder Proinsulin in Insulin ver-mindert. Diese Störung ist ebenfalls extrem rar und führt zurfrühmanifesten massiven Adipositas, ACTH-Mangel und Hy-perproinsulinämie (Farooqi und O’Rahilly 2006).
Häufiger treten Mutationen des MC4-Rezeptors auf, dieautosomal-dominant vererbt werden. In bisherigen Studienkonnten bei 3–6 % der Patienten mit einem Body-Mass-Index (BMI) über 40 kg/m2 Körperoberfläche (KOF) Muta-tionen im MC4-Rezeptorgen nachgewiesen werden(Farooqiund O’Rahilly 2006). Neuere Familienuntersuchungen erga-ben, dass bei erwachsenen Mutationsträgern der BMI um4 kg/m2 KOF (Männer) bzw. 9,5 kg/m2 KOF (Frauen) höherist als bei nichtbetroffenen Familienmitgliedern (Dempfleet al. 2004). Allerdings gibt es auchMC4R-Polymorphismen,die negativ mit einem erhöhten Körpergewicht assoziiertsind. In einer kürzlich erschienenen Studie bei 899 Kindernmit Übergewicht oder Adipositas konnten bei 1,6 % Varian-ten im MC4R-Gen und dessen Promotorbereich nachgewie-sen werden, die zu einer verminderten Funktion des MC4Rführen (Vollbach et al. 2017). Kürzlich wurden bei Menschenmit Adipositas-Varianten im MRAP2-Gen (Melanocortinreceptor acessory protein 2) nachgewiesen. Man geht davonaus, dass diese Varianten die MC4R-Funktion beeinflussen.Heterozygote MRAP2-Genvarianten können beim Menschenzu früher Manifestation einer extremen Adipositas führen(Schonnop et al. 2016; Asai et al. 2013).
Mit Adipositas assoziierte monogene Adipositassyndro-me und Kandidatengene sind in Tab. 2 mit Angaben aus derOMIM-Datenbank (Online Mendelian Inheritance in Man)aufgelistet.
Mit Adipositas assoziiert sind auch genetische Syndrome(Tab. 3). Man unterscheidet Formen der Adipositas mit Entwick-lungsverzörungen (BBS, Borjeson-Forsman-Lehman, Cohen,Fragiles X, Pseudohypoparathreoidismus, SIM1-Defizienz,BDNF/TrkB-Defizienz) und ohne Entwicklungsverzörungen(Alström, Leptin, Leptinrezeptor, MC4R, POMC, und PCSK1-Defizienz). Bei Cohen-Syndrom und Alström-Syndrom liegenZilienfunktionsstörungen (Ziliopathien) vor (Styne et al. 2017).Ein weiteres Syndrom, das mit extremer „Early-onset-Adi-positas“ assoziiert ist, ist das ROHHAD/ROHHADNET-Syndrom (Rapid-onset obesity, hypothalamic dysfunction,hypoventilation, and autonomic dysregulation; neural cresttumor). Durch die schnelle und sehr ausgeprägte Gewichts-zunahme sowie eine schwere Atemstörung, die schon ab demAlter vor 1–2 Jahren beginnen kann, kann diese Erkrankungzu frühzeitiger Morbidität und Mortalität führen. Die Hor-monstörungen sind vielfältig und umfassen u. a. eine zentraleHypothyreose, Wachstumshormonmangel, Nebennierenrin-deninsuffizienz, Diabetes insipidus, Pubertätsentwicklungs-störungen und Hyperprolaktinämie. Weitere Auffälligkeitensind autonome Dysregulation, Verhaltens- und Entwick-
Zentrale Regulation des Körpergewichtes 11
lungsstörungen sowie neuroendokrine Tumoren. Die Genetikist bei diesem Syndrom noch weitgehend unklar.
4.3 Weitere genetische Faktoren undkomplexe Genetik
Zwillingsstudien legen nahe, dass 40–70 % der Veranlagungder Adipositas vererbt wird, mit einer Konkordanz von0,7–0,9 bei eineiigen gegenüber 0,35–0,45 bei zweieiigenZwillingen. Die Determinierung der interindividuellen Varia-tion der Körperfettmasse ist genetisch komplex und man gehtvon einer polygenen Vererbung aus.
Seit mehr als 10 Jahren wurden einige genomweite Asso-ziationsstudien (GWAS) durchgeführt, um genetische Fak-toren der Adipositas zu identifizieren. Inzwischen sind vieleVarianten („single nucleotide polymorphisms“) bekannt, dieeinen mehr oder weniger starken Einfluss auf das Körperge-wicht haben. Einige dieser Varianten betreffen auch Gene desLeptin-Melanokortin-Systems. In verschiedenen Kopplungs-analysen wurde von über 150 Genloci („quantitative traitloci“), die sich in mehreren Studien replizieren ließen und
einen Einfluss auf das Körpergewicht haben, berichtet(Lu und Loos 2013). Als Kandidatengen gilt z. B. das FTO-Gen (Fat mass and obesity-associated). Homozygote Trägerder Risikoallele sind durchschnittlich 3 kg schwerer undhaben ein 1,7-fach erhöhtes Adipositasrisiko verglichen mitIndividuen, die das Risikoallel nicht tragen (Frayling et al.2007). Bei vielen dieser Gene ist noch nicht bekannt, wie sichdie Genvarianten auf den Energiestoffwechsel auf zellulärerund molekularer Ebene auswirken. Es ist jedoch festzuhalten,dass einzelne dieser Genvarianten, die zwar in großen gene-tischen Studien zu statistisch signifikanten Ergebnissen führ-ten, das Körpergewicht eines Individuums gering beeinflus-sen, wenn es nicht zur additiven Wirkung verschiedenerEffekte kommt.
" Durch die Verfügbarkeit moderner DNA-Chiptechnologiekonnte im Rahmen von genomweiten Assoziationsstudienin den letzten Jahren eine Reihe von Genvariantenbeschrieben werden, die mit einem erhöhten Adipositas-risiko assoziiert sind.
Tab. 2 Monogene Adipositassyndrome und Kandidatengene
Defizienz, GenOMIM-Nr.
Locus/Chromosom/Vererbung Klinische Charakteristika
Brain-derivedneurotrophic factor(BDNF)
113505/612469
11p14.1Haploinsuffizienz
Manifestation in früher Kindheit, Hyperphagie, kognitive Störung,Hyperaktivität, Konzentrationsschwäche, gestörtes Kurzzeitgedächtnis,verminderte SchmerzwahrnehmungAssoziation mit WAGRO-Syndrom (Willms-Tumor, Aniridie, urogenitaleAuffälligkeiten, mentale Retardierung, Adipositas)
Leptin, LEP, OB-Gen 164160 7p32.1AR
Adipositasmanifestation im Säuglingsalter, extreme Adipositas mit Hyperphagieund vermindertem Sympathikotonus, hypogonadotroper Hypogonadismus,Hyperinsulinämie, reduzierte Schilddrüsenhormonspiegel und T-Zell-Defekt(Zahl und Funktion) mit rekurrierenden Infektionen. Sehr niedrige Leptinspiegel,Leptininjektionen als kausale Therapie möglich
Leptinrezeptor 601007 1p31.3AR
Adipositasmanifestation im Säuglingsalter, mit Hyperphagie und vermindertemSympathikotonus, hypogonadotropem Hypogonadismus. Lineares Wachstumvermindert, Wachstumshormonmangel, Hypothyreose. ErhöhteLeptinkonzentrationen, Leptintherapie nicht wirksam
Melanocortin-4-Rezeptor(MC4R)
155541 18q21.32AD
Manifestation in früher Kindheit, Hyperphagie mit autonomer Dysfunktion,Sprachentwicklungsverzörung, Verhaltensstörungen, beschleunigtes Wachstum,Hyperinsulinämie, erhöhte Muskelmasse und Knochendichte. Herzfrequenz undBlutdruck sind relativ niedrig (niedriger Sympathikotonus)
Prohormone convertase1 (PCSK1)
162150 5q15AR
Manifestation in früher Kindheit, intestinale Absorptionsstörung, schwereDiarrhö bei kleinen Säuglingen, moderate Hyperphagie, hypogonadotroperHypogonadismus, Hypokortisolismus infolge ACTH-Defizienz, postprandialeHypoglykämie, niedrige Insulinspiegel, Diabetes mellitus, Diabetes insipidus,Hypothyreose
Proopiomelanocortin(POMC)
609734 2p23.3ARHaploinsuffizienz
Frühe Adipositasmanifestation, extreme Adipositas mit Hyperphagie,cholestatischer Ikterus oder Nebennierenkrise bei Neugeborenen (ACTH-/Kortisol-Defizienz), blasses Hautkolorit, rote Haare
Single-minded 1 (SIM1) 603128 6q16.3Haploinsuffizienz
Manifestation in früher Kindheit, Hyperphagie, beschleunigtes Wachstum(Funktion „downstream“ von MC4R), Phänotyp „Prader-Willi-like“
Neurotrophic tyrosinereceptor kinase (NTRK2)
613886 9q21.33Haploinsuffizienz
Manifestation in früher Kindheit, Hyperphagie, mentale Retardierung
AD autosomal-dominant; AR autosomal-rezessiv
12 C. Roth
Des Weiteren konnten inzwischen epigenetische Mecha-nismen nachgewiesen werden, die die Entwicklung des Kör-pergewichtes beeinflussen. Dies ist schon seit Langembekannt im Zusammenhang mit extremer intrauteriner Wachs-tumsretardierung bzw. Mangelgeburt durch mangelnde Nah-rungsaufnahme der Mutter, z. B. in Kriegsjahren. Manchedieser Nachkommen weisen eine überschießende Gewichts-zunahme und frühe Entwicklung einer Adipositas und einesmetabolischen Syndroms auf. Kürzlich konnte jedoch auchein erhöhtes Adipositasrisiko bei Menschen mit verändertem
Methylierungsmuster im POMC-Gen gezeigt werden (Kuh-nen et al. 2016).
" Cave Auch wenn das Körpergewicht entscheidend durchgenetische Faktoren prädisponiert wird, ist das Problemder hohen Adipostasprävalenz nicht auf Mutationen ein-zelner Gene zurückzuführen und ein Mutationsscreeningist bis auf gezielte wissenschaftliche Fragestellungen der-zeit nicht zu empfehlen.
Tab. 3 Syndromale Erkrankungen, die häufig mit einer Adipositas assoziiert sind
SyndromOMIMNr. Gen/Chromosom Klinische Charakteristika
Alström 203800 ALMS2p13
Frühe Adipositas, Hyperphagie, Retinitis pigmentosa,sensorineuraler Hörverlust, extreme Insulinresistenz, Acanthosisnigricans, Typ-2-Diabetes mellitus, Hyperlipidämie, dilatativeKardiomyopathie, progressive Nieren-, Leber- undLungendysfunktion, Nierenversagen, bei JungenHypogonadismus
Bardet-Biedl (BBS) 209900 Multiple (BBS1-15) 16q21,+15q22-q23 u. a.
Frühe stammbetonte Adipositas, Retinitis pigmentosa, ulnare/fibulare Polydaktylie, Nierenmalformationen undNierenversagen, Leberfibrose, Diabetes mellitus, Herzanomalien,Hypogenitalismus und Urogenitalfehlbildungen, oft mentaleRetardierung, verminderte körperliche Aktivität
Beckwith-Wiedemann 130650 H19, KCNQ10T1, CDKN1C11p15.5-p15.4NSD1 5q35
Adipositas mit allgemeinem Riesenwuchs, Hemihypertrophie,grobe Gesichtszüge, Makroglossie, Kerbenohren,Viszeromegalie, Bauchwanddefekte, Hypoglykämie, erhöhteRate an Tumoren
Borjeson-Forssman-Lehman Xq26-27 PHF6Xq26-27
Moderate stammbetonte Adipositas, meist erst im Schulalter,große fleischige Ohrmuscheln, Hypotonie,Entwicklungsverzögerung, Hypogonadismus, Gynäkomastie
Cohen 216550 COH18q22-q23
Moderate stammbetonte Adipositas, chorioretinale Dystrophie,wellenförmige Augenlider, hoher Nasenrücken, dysplastischeOhren, offener Mund, kurzes Philtrum, Mikrozephalie,Hypotonie, überstreckbare Gelenke, motorischeSchwerfälligkeit, Kleinwuchs, verzögerte Pubertät, leichte bisausgeprägte Retardierung
Carpenter 201000 RAB236p11.2
Moderate stammbetonte Adipositas in später Kindheit,Akrozephalie, evtl. temporal ausladend, flacher Nasenrücken,Exophthalmie, hoher Gaumen, dorsalwärts rotierte Ohren, kurzeHände, Polydaktylie, Syndaktylie, Genu valgum, Herzfehler,Hypogonadismus
Fragiles-X 300624 FMR1-GenXq27.3
Frühe Adipositas, Makrozephalie, prominente Backenknochen,große Ohren, hohe Stimme, Makroorchidismus, Autismus,Hochwuchs, mesenchymale Schwäche, mentale Retardierung
Prader-Willi 176270 Paternale 15q11-q13Deletion, oder maternaleuniparentale Disomie
Extreme Adipositas, mandelförmige Augen, zeltförmiger Mund,hoher Gaumen, Kleinwuchs, kleine Hände und Füße. Hypotonie,Trinkschwäche als Säugling gefolgt von gesteigertem Appetit.Typ-2-Diabetes mellitus, Hypogonadismus, Schlafstörungen,Stimmungslabilität, Zwanghaftigkeit, leichte/mäßigeRetardierung
PseudohypoparathyroidismusTyp 1a
103580 GNAS120q13.2
Meist moderate Adipositas, rundes Gesicht, gedrungenerKleinwuchs, verkürzte Metacarpalia, Endorganresistenzverschiedener Hormone möglich (GHRH, TSH, PTH, α-MSH),Hypogonadismus, evtl. mentale Retardierung, Hypokalzämie,Hyperphosphatämie, ektope Verkalkungen
Smith-Margenis 617074 RAI117p11.2
Frühmanifeste Adipositas, gestörter Tag-Nacht-Rhythmus,Schlafstörungen, gestörte Melatoninsekretion,Verhaltensauffälligkeiten, Entwicklungsretardierung
Zentrale Regulation des Körpergewichtes 13

5 Einfluss der Darmflora
In den letzten Jahren wurde die Besiedelung des Darmes mitverschiedenen Darmbakterien im Zusammenhang mit Ge-wichtsregulation und Entstehung von Adipositas und Dia-betes intensiv erforscht. Die Zusammensetzung der Darm-flora hängt von vielen verschiedenen Einflüssen, u. a. auchUmweltfaktoren und Ernährungsgewohnheiten, ab und un-terscheidet sich erheblich zwischen normalgewichtigen undübergewichtigen Personen. Durch eine veränderte Darmflora(Dysbiose) kann es zu erhöhter Bildung von Entzündungs-mediatoren, Entzündung der Darmschleimhaut, Beeinflus-sung der Sekretion von Sättigungshormonen aus dem Darm,erhöhter Darmpermeabilität, Penetrieren von Darmbakterienund deren Stoffwechselprodukten in den Systemkreislauf,systemischer Entzündung, Steatohepatitis, chronischer syste-mischer Entzündung („low grade inflammation“), gestörterSättigung und möglicherweise auch Begünstigung einer kar-diovaskulären Erkrankung kommen (Henao-Mejia et al.2012).
6 Zusammenfassung
Die wesentliche Regulation von Hunger und Sättigunggeschieht über Ausschüttung von Peptidhormonen aus derKörperperipherie (vor allem Fettgewebe, Pankreas, GI-Trakt), die an hypothalamische Rezeptoren binden. Kernge-biete des Hypothalamus und des Hirnstamms sind wichtigeRegulationsstrukturen und Integrationszentren für die Wei-terleitung an höhere Hirnzentren und das limbische Systemsowie für die Umsetzung hormoneller peripherer Signale inneuronale Signale. Vom ZNS werden efferente Signale überdas autonome Nervensystem an die Körperperipherie vermit-telt. Läsionen medial gelegener hypothalamischer Strukturenkönnen zu ungebremstem Appetit und zur therapierefraktärenAdipositas führen. Nur wenige monogene Erkrankungen, diemeist einen für die Appetitregulation wichtigen Botenstoffoder Rezeptor des Leptin-Melanokortin-Systems betreffen,sind bisher bekannt.
Literatur
Asai M, Ramachandrappa S, Joachim M, Shen Y, Zhang R,Nuthalapati N, Ramanathan V, Strochlic DE, Ferket P, Linhart K,Ho C, Novoselova TV, Garg S, Ridderstrale M, Marcus C, Hirsch-horn JN, Keogh JM, O’Rahilly S, Chan LF, Clark AJ, Farooqi IS,Majzoub JA (2013) Loss of function of the melanocortin 2 receptoraccessory protein 2 is associated with mammalian obesity. Science341:275–278
Balleine BW, Delgado MR, Hikosaka O (2007) The role of the dorsalstriatum in reward and decision-making. J Neurosci 27:8161–8165
Barazzoni R, Bosutti A, Stebel M, Cattin MR, Roder E, Visintin L,Cattin L, Biolo G, Zanetti M, Guarnieri G (2005) Ghrelin regulatesmitochondrial-lipid metabolism gene expression and tissue fat dis-tribution in liver and skeletal muscle. Am J Physiol EndocrinolMetab 288:E228–E235
Batterham RL, Cowley MA, Small CJ, Herzog H, Cohen MA, DakinCL, Wren AM, Brynes AE, Low MJ, Ghatei MA, Cone RD, BloomSR (2002) Gut hormone PYY(3-36) physiologically inhibits foodintake. Nature 418:650–654
Blevins JE, Baskin DG (2015) Translational and therapeutic potential ofoxytocin as an anti-obesity strategy: insights from rodents, nonhu-man primates and humans. Physiol Behav 152(Pt B):438–449
Bluher M, Mantzoros CS (2015) From leptin to other adipokines inhealth and disease: facts and expectations at the beginning of the 21stcentury. Metabolism 64:131–145
Bornfeldt KE, Tabas I (2011) Insulin resistance, hyperglycemia, andatherosclerosis. Cell Metab 14:575–585
Bray GA (2000) Reciprocal relation of food intake and sympatheticactivity: experimental observations and clinical implications. Int JObes Relat Metab Disord 24(Suppl 2):8–17
Chou TC, Lee CE, Lu J, Elmquist JK, Hara J, Willie JT, Beuckmann CT,Chemelli RM, Sakurai T, Yanagisawa M, Saper CB, Scammell TE(2001) Orexin (hypocretin) neurons contain dynorphin. J Neurosci21:RC168
Cornier MA (2011) Is your brain to blame for weight regain? PhysiolBehav 104:608–612
DeBoer MD, Marks DL (2006) Therapy insight: use of melanocortinantagonists in the treatment of cachexia in chronic disease. Nat ClinPract Endocrinol Metab 2:459–466
Dempfle A, Hinney A, Heinzel-Gutenbrunner M, Raab M, Geller F,Gudermann T, Schafer H, Hebebrand J (2004) Large quantitativeeffect of melanocortin-4 receptor gene mutations on body massindex. J Med Genet 41:795–800
Everitt BJ, Parkinson JA, Olmstead MC, Arroyo M, Robledo P, RobbinsTW (1999) Associative processes in addiction and reward. The roleof amygdala-ventral striatal subsystems. Ann N Y Acad Sci877:412–438
Farooqi S, O’Rahilly S (2006) Genetics of obesity in humans. EndocrRev 27:710–718
Farooqi IS, Bullmore E, Keogh J, Gillard J, O’Rahilly S, Fletcher PC(2007) Leptin regulates striatal regions and human eating behavior.Science 317:1355
Figlewicz DP, MacDonald Naleid A, Sipols AJ (2007) Modulation offood reward by adiposity signals. Physiol Behav 91:473–478
Frayling TM, Timpson NJ, Weedon MN, Zeggini E, Freathy RM,Lindgren CM, Perry JR, Elliott KS, Lango H, Rayner NW,Shields B, Harries LW, Barrett JC, Ellard S, Groves CJ, Knight B,Patch AM, Ness AR, Ebrahim S, Lawlor DA, Ring SM,Ben-Shlomo Y, Jarvelin MR, Sovio U, Bennett AJ, Melzer D,Ferrucci L, Loos RJ, Barroso I, Wareham NJ, Karpe F, Owen KR,Cardon LR, Walker M, Hitman GA, Palmer CN, Doney AS, MorrisAD, Smith GD, Hattersley AT, McCarthy MI (2007) A commonvariant in the FTO gene is associated with body mass index andpredisposes to childhood and adult obesity. Science 316:889–894
Henao-Mejia J, Elinav E, Jin C, Hao L, Mehal WZ, Strowig T, ThaissCA, Kau AL, Eisenbarth SC, Jurczak MJ, Camporez JP, ShulmanGI, Gordon JI, Hoffman HM, Flavell RA (2012) Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity.Nature 482:179–185
Krude H, Biebermann H, Schnabel D, Tansek MZ, Theunissen P, MullisPE, Gruters A (2003) Obesity due to proopiomelanocortin defici-ency: three new cases and treatment trials with thyroid hormone andACTH4-10. J Clin Endocrinol Metab 88:4633–4640
Kuhnen P, Handke D, Waterland RA, Hennig BJ, Silver M, Fulford AJ,Dominguez-Salas P, Moore SE, Prentice AM, Spranger J, Hinney A,Hebebrand J, Heppner FL, Walzer L, Grotzinger C, Gromoll J,
14 C. Roth

Wiegand S, Gruters A, Krude H (2016) Interindividual variation inDNA methylation at a putative POMC metastable epiallele is asso-ciated with obesity. Cell Metab 24:502–509
Latorre R, Sternini C, De Giorgio R, Greenwood-Van Meerveld B(2016) Enteroendocrine cells: a review of their role in brain-gutcommunication. Neurogastroenterol Motil 28:620–630
Lowell BB, Spiegelman BM (2000) Towards a molecular understandingof adaptive thermogenesis. Nature 404:652–660
Lu Y, Loos RJ (2013) Obesity genomics: assessing the transferability ofsusceptibility loci across diverse populations. Genome Med 5:55
Malcher-Lopes R, Di S, Marcheselli VS, Weng FJ, Stuart CT, BazanNG, Tasker JG (2006) Opposing crosstalk between leptin and glu-cocorticoids rapidly modulates synaptic excitation via endocannabi-noid release. J Neurosci 26:6643–6650
Muller HL (2016) Craniopharyngioma and hypothalamic injury: latestinsights into consequent eating disorders and obesity. Curr OpinEndocrinol Diabetes Obes 23:81–89
Park HK, Ahima RS (2015) Physiology of leptin: energy homeostasis,neuroendocrine function and metabolism. Metabolism 64:24–34
Roth CL (2015) Hypothalamic obesity in craniopharyngioma patients:disturbed energy homeostasis related to extent of hypothalamicdamage and its implication for obesity intervention. J Clin Med4:1774–1797
Roth CL, Reinehr T (2010) Roles of gastrointestinal and adipose tissuepeptides in childhood obesity and changes after weight loss due tolifestyle intervention. Arch Pediatr Adolesc Med 164:131–138
Roth CL, Eslamy H, Werny D, Elfers C, Shaffer ML, Pihoker C,Ojemann J, Dobyns WB (2015) Semiquantitative analysis of hypo-thalamic damage on MRI predicts risk for hypothalamic obesity.Obesity (Silver Spring) 23:1226–1233
Scholtz S, Miras AD, Chhina N, Prechtl CG, Sleeth ML, Daud NM,Ismail NA, Durighel G, Ahmed AR, Olbers T, Vincent RP,Alaghband-Zadeh J, Ghatei MA, Waldman AD, Frost GS, Bell JD,le Roux CW, Goldstone AP (2014) Obese patients after gastric
bypass surgery have lower brain-hedonic responses to food thanafter gastric banding. Gut 63:891–902
Schonnop L, Kleinau G, Herrfurth N, Volckmar AL, Cetindag C,Muller A, Peters T, Herpertz S, Antel J, Hebebrand J, Biebermann H,Hinney A (2016) Decreased melanocortin-4 receptor function con-ferred by an infrequent variant at the human melanocortin receptoraccessory protein 2 gene. Obesity (Silver Spring) 24:1976–1982
Schrover IM, Spiering W, Leiner T, Visseren FL (2016) Adipose tissuedysfunction: clinical relevance and diagnostic possibilities. HormMetab Res 48:213–225
Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, Baskin DG (2000)Central nervous system control of food intake. Nature 404:661–671
Schwartz MW, Seeley RJ, Tschop MH, Woods SC, Morton GJ, MyersMG, D'Alessio D (2013) Cooperation between brain and islet inglucose homeostasis and diabetes. Nature 503:59–66
Shimada M, Tritos NA, Lowell BB, Flier JS, Maratos-Flier E (1998)Mice lacking melanin-concentrating hormone are hypophagic andlean. Nature 396:670–674
Sternini C, Anselmi L, Rozengurt E (2008) Enteroendocrine cells: a siteof ‚taste‘ in gastrointestinal chemosensing. Curr Opin EndocrinolDiabetes Obes 15:73–78
Styne DM, Arslanian SA, Connor EL, Farooqi IS, Murad MH, Silver-stein JH, Yanovski JA (2017) Pediatric obesity-assessment, treat-ment, and prevention: an endocrine society clinical practice guide-line. J Clin Endocrinol Metab 102:709–757
Tschop M, Smiley DL, Heiman ML (2000) Ghrelin induces adiposity inrodents. Nature 407:908–913
Vollbach H, Brandt S, Lahr G, Denzer C, von Schnurbein J, DebatinKM, Wabitsch M (2017) Prevalence and phenotypic characterizationof MC4R variants in a large pediatric cohort. Int J Obes 41:13–22
Wisse BE, Kim F, Schwartz MW (2007) Physiology. an integrative viewof obesity. Science 318:928–929
Zhang JV, Ren PG, Avsian-Kretchmer O, Luo CW, Rauch R, Klein C,Hsueh AJ (2005) Obestatin, a peptide encoded by the ghrelin gene,opposes ghrelin’s effects on food intake. Science 310:996–999
Zentrale Regulation des Körpergewichtes 15


















