

Abwehr I-II - meduniwien.ac.at · B-Lymphozyten Sekundäre: Endothelzellen der Blutgefäße ... *...
58
Transcript of Abwehr I-II - meduniwien.ac.at · B-Lymphozyten Sekundäre: Endothelzellen der Blutgefäße ... *...

Abwehr I.:angeborene, natürliche Abwehr
Abwehr II.:erworbene, spezifische Abwehr
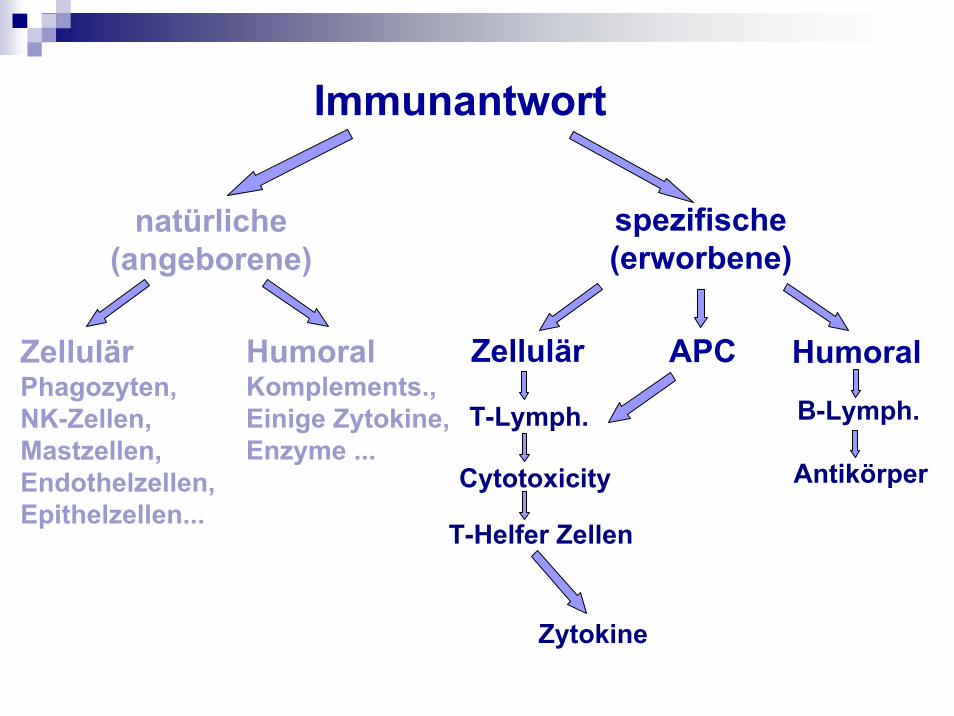
Immunantwort
natürliche(angeborene)
ZellulärPhagozyten, NK-Zellen,Mastzellen, Endothelzellen, Epithelzellen...
HumoralKomplements., Einige Zytokine, Enzyme ...
spezifische(erworbene)
Zellulär HumoralAPC
T-Lymph.
Cytotoxicity
T-Helfer Zellen
Zytokine
B-Lymph.
Antikörper
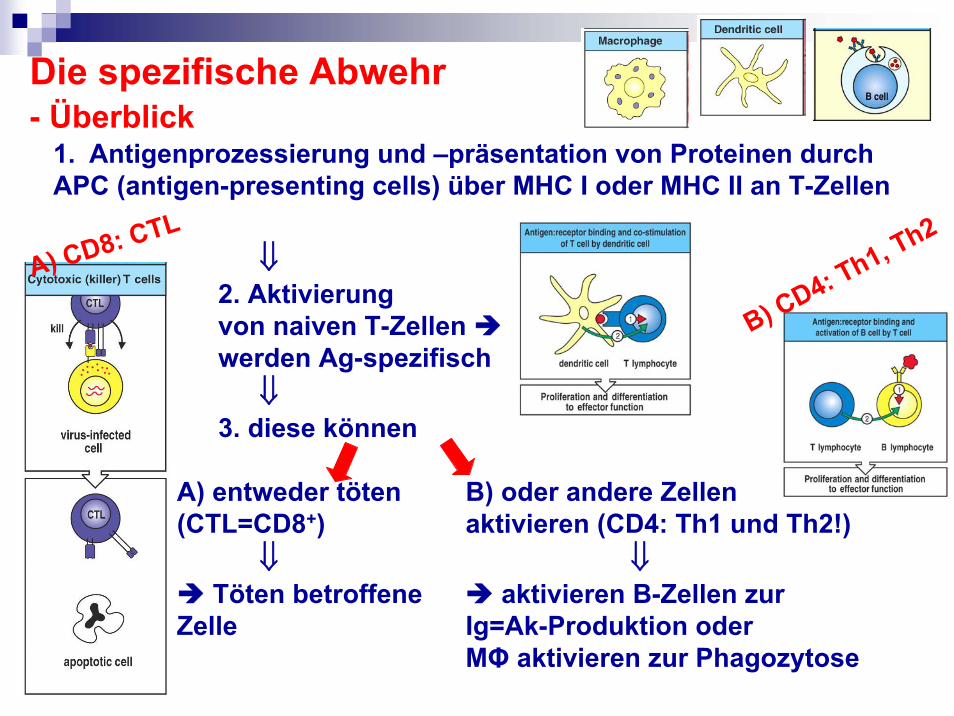
Die spezifische Abwehr- Überblick
1. Antigenprozessierung und –präsentation von Proteinen durchAPC (antigen-presenting cells) über MHC I oder MHC II an T-Zellen
⇓2. Aktivierung von naiven T-Zellen werden Ag-spezifisch
⇓3. diese können
A) entweder töten B) oder andere Zellen (CTL=CD8+) aktivieren (CD4: Th1 und Th2!)
⇓ ⇓Töten betroffene aktivieren B-Zellen zur
Zelle Ig=Ak-Produktion oder MΦ aktivieren zur Phagozytose
A) CD8: CTL
B) CD4: Th1, Th2

Antigen presentierende Zellen Funkzion:Exogene Proteinantigene zu endozytieren, sie zum immunogenen Peptiden zu zersetzen und diese im Komplex mit Histokompatibilitätsantigene der Klasse II MHCII an ihre Oberfläche zu Th-Zellen zu presentieren.
Primäre:MakrophagenDendritische ZellenB-Lymphozyten
Sekundäre:Endothelzellen der BlutgefäßeEpithelzellen im Darm
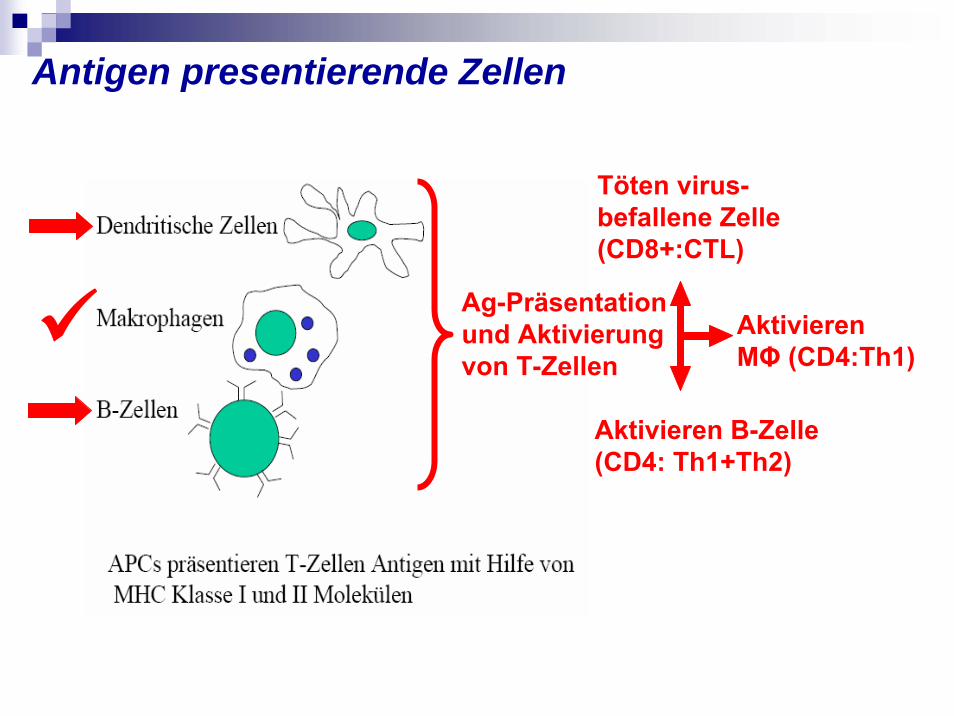
Ag-Präsentation und Aktivierung von T-Zellen
Töten virus-befallene Zelle(CD8+:CTL)
Aktivieren B-Zelle (CD4: Th1+Th2)
Aktivieren MΦ (CD4:Th1)
Antigen presentierende Zellen
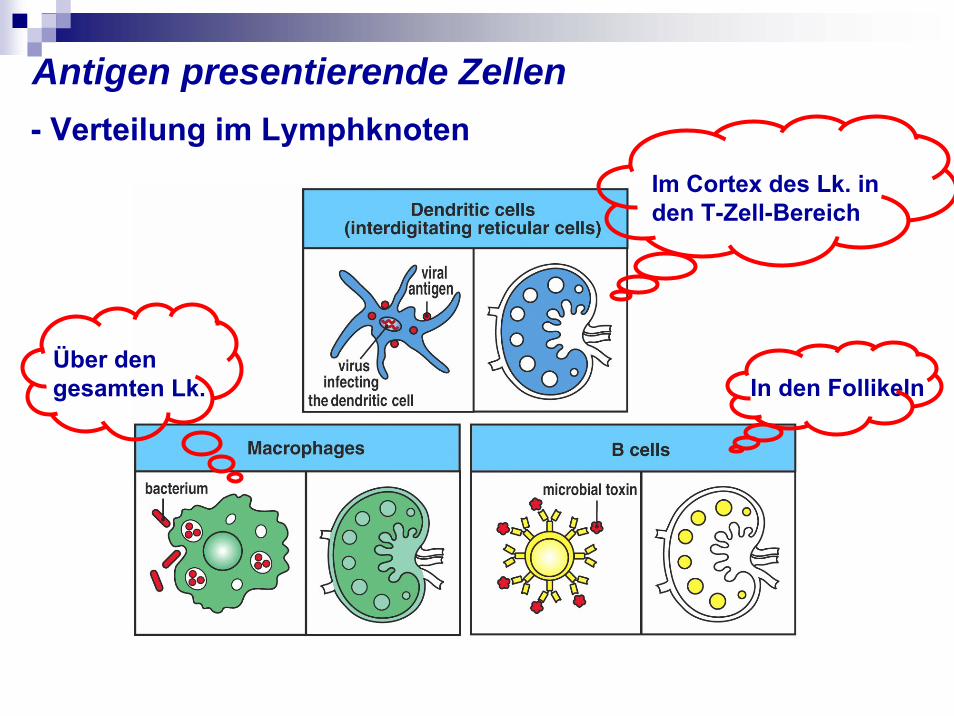
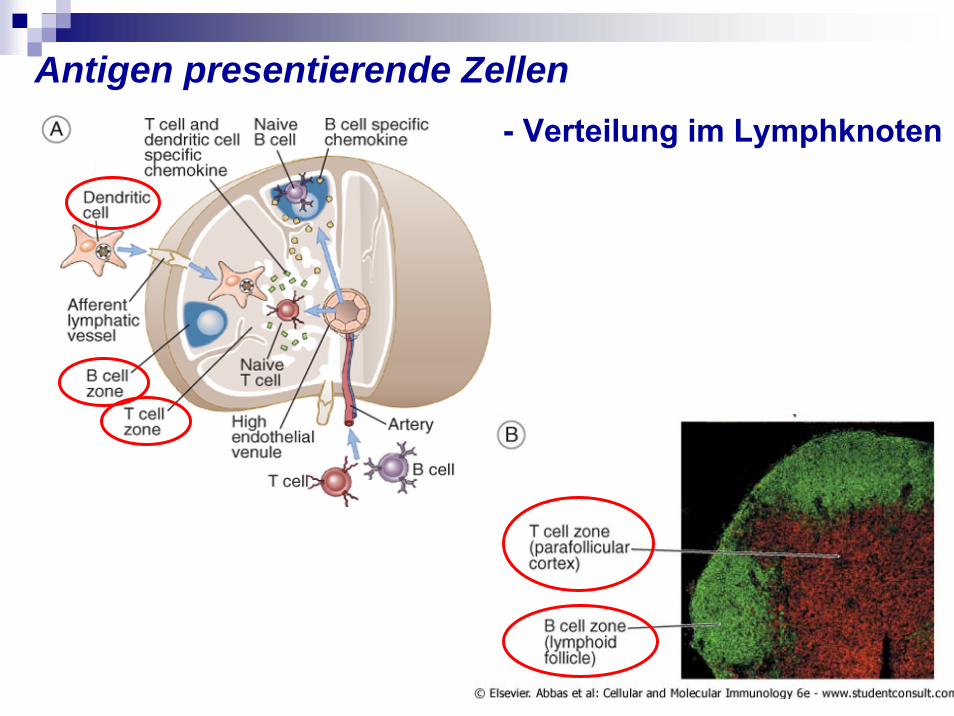
Antigen presentierende Zellen - Verteilung im Lymphknoten
In den Follikeln
Im Cortex des Lk. in den T-Zell-Bereich
Über den gesamten Lk.
Antigen presentierende Zellen - Verteilung im Lymphknoten
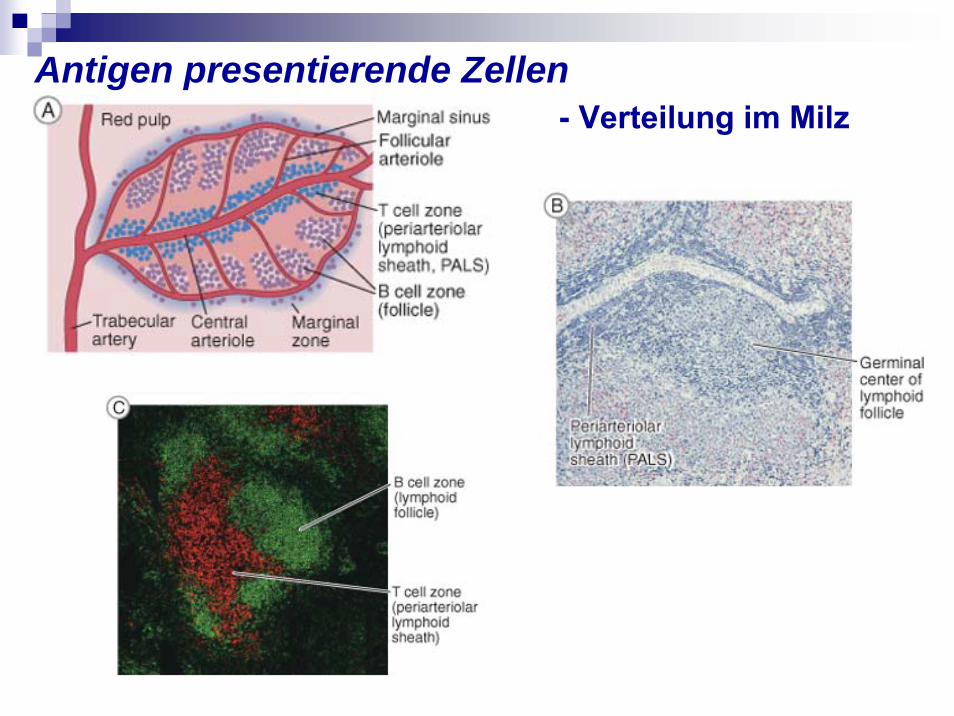
Antigen presentierende Zellen - Verteilung im Milz
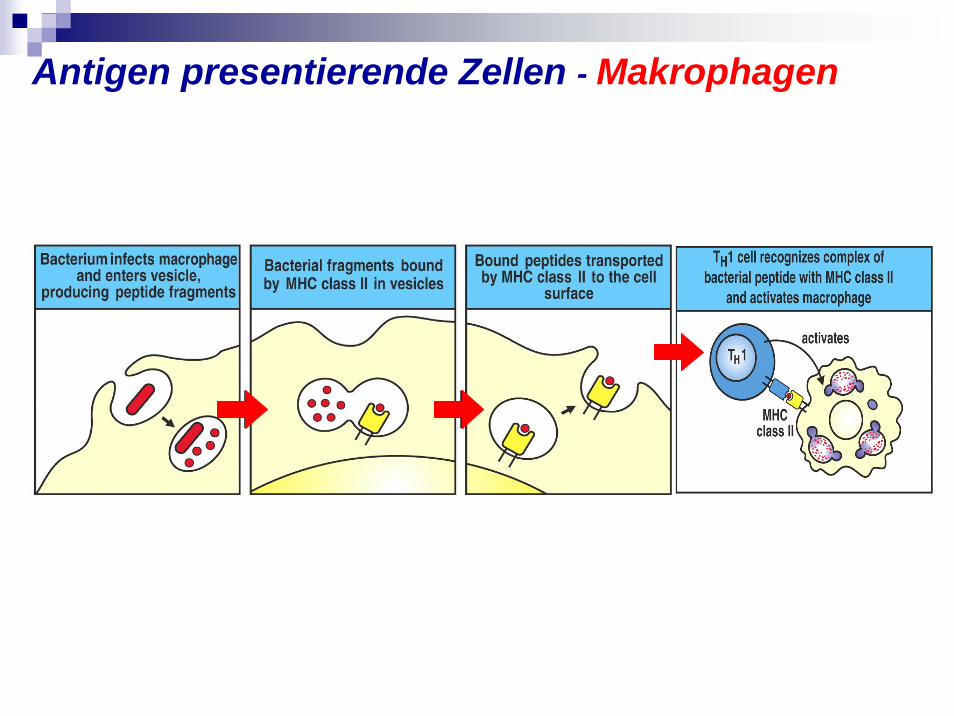
Antigen presentierende Zellen - Makrophagen

Antigen presentierende Zellen - Dendritische Zellenstammen aus der myeloische Linie
niedrigere phagozytäre Kapazität
Antigenpresentation (sind änlich zu Makrophagen)
in alle Gewäben, auser Gehirn
Exprimieren: MHC-Klasse-IMHC-Kalsse-IIMolekül B7 (costimulation)ICAM-1, ICAM-2, LFA-1, LFA-3 (Adhäsionsmoleküle)

Follikuläre dendritische:• B-Zell Zone• keine Phagozytose• exprimieren keine MHCII• Ag-Ak Komplex einzufangen keine Internalisierung Komplex bleibt auf die Oberfläche B-Zellen erkennen
das Antigen
Antigen presentierende Zellen - Dendritische ZellenFormen
Interdigitierende dendritische Zellen: • T-Zell-Bereich• Stimulatoren der naive T-Zellen
Langerhans-Zellen: in der Epidermis der Haut
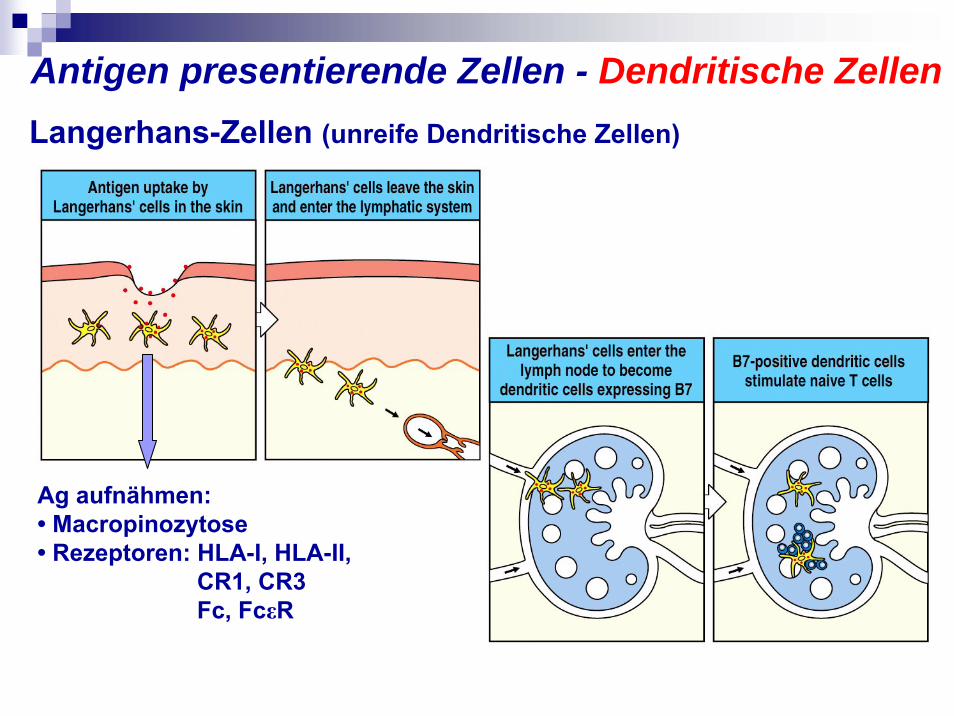
Ag aufnähmen:• Macropinozytose• Rezeptoren: HLA-I, HLA-II,
CR1, CR3Fc, FcεR
Langerhans-Zellen (unreife Dendritische Zellen)
Antigen presentierende Zellen - Dendritische Zellen
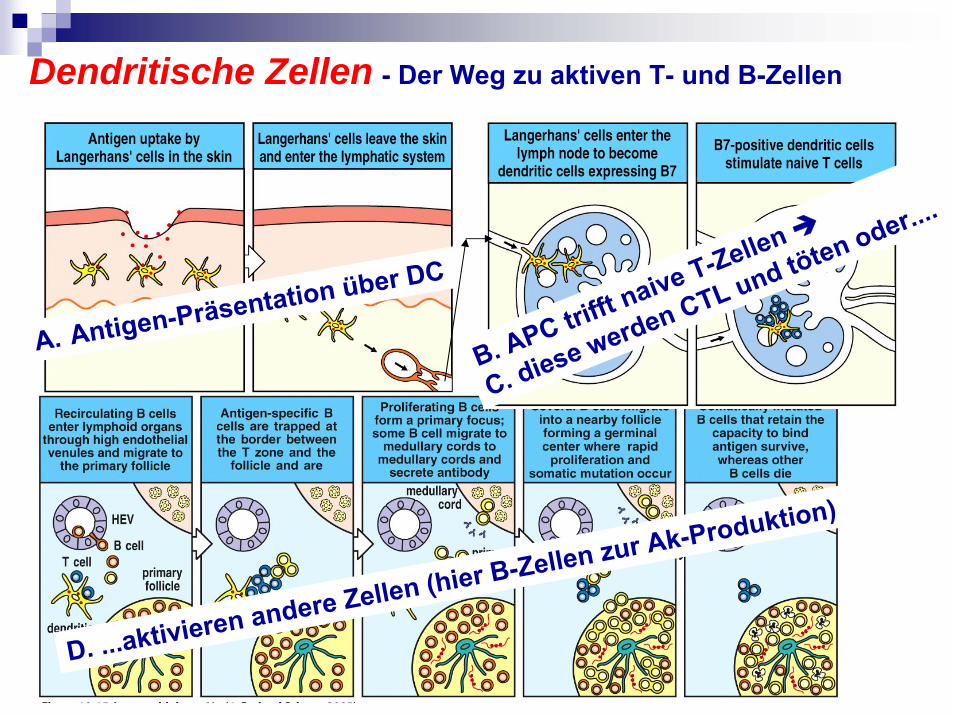
Dendritische Zellen - Der Weg zu aktiven T- und B-Zellen
A. Antigen-Präsentation über DC
D. ...aktivieren andere Zellen (hier B-Zellen zur Ak-Produktion)
B. APC trifft naive T-Zellen
C. diese werden CTL und töten oder....
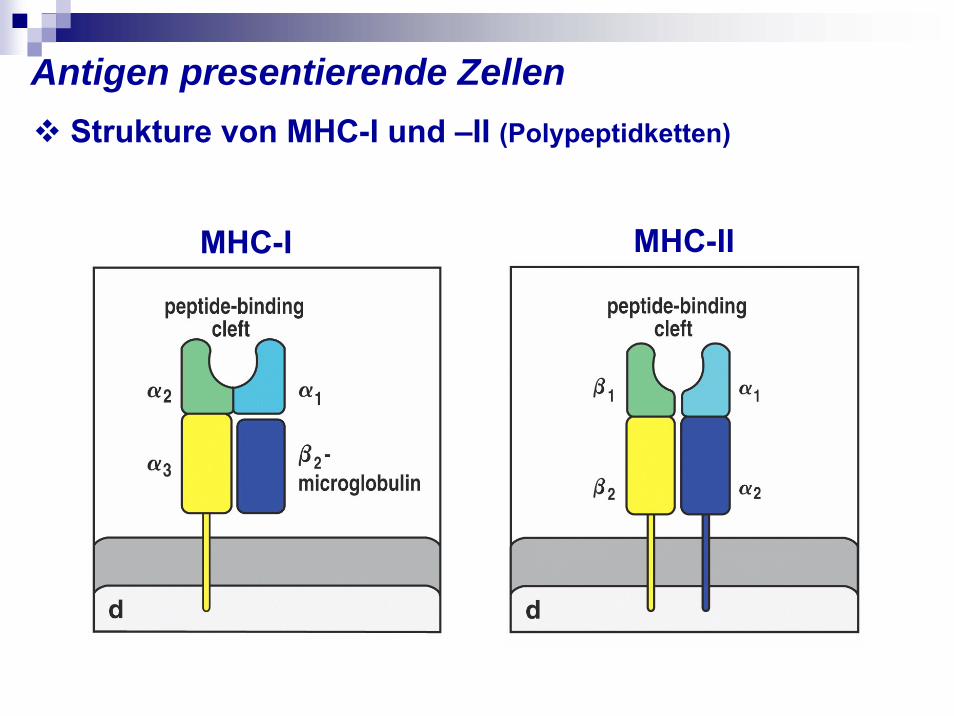
Antigen presentierende Zellen
MHC-I MHC-II
Strukture von MHC-I und –II (Polypeptidketten)
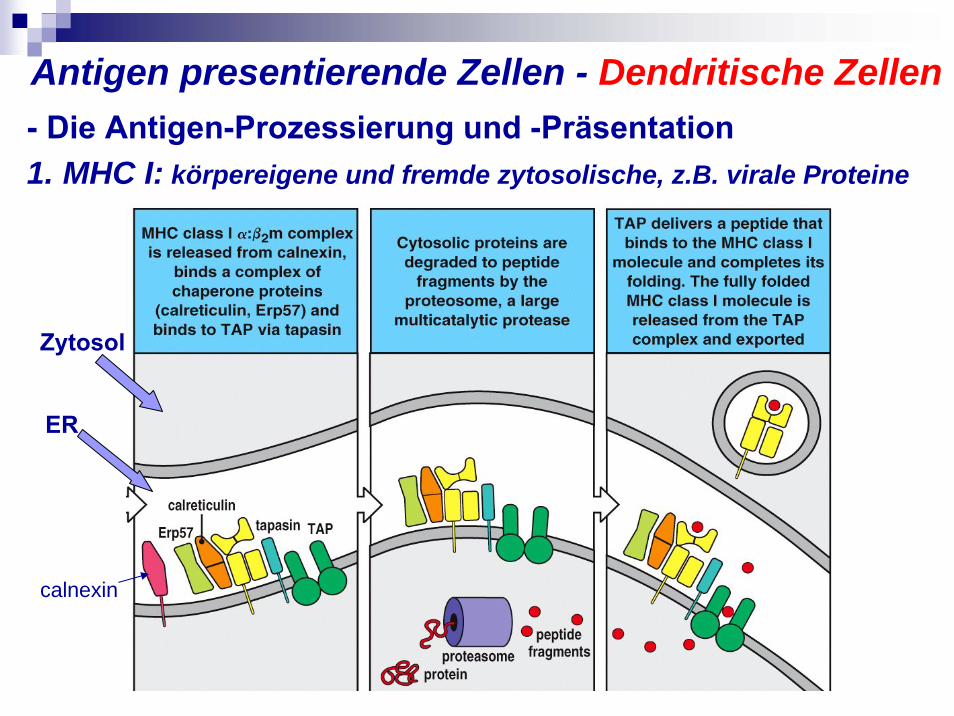
- Die Antigen-Prozessierung und -Präsentation1. MHC I: körpereigene und fremde zytosolische, z.B. virale Proteine
Antigen presentierende Zellen - Dendritische Zellen
ER
Zytosol
calnexin
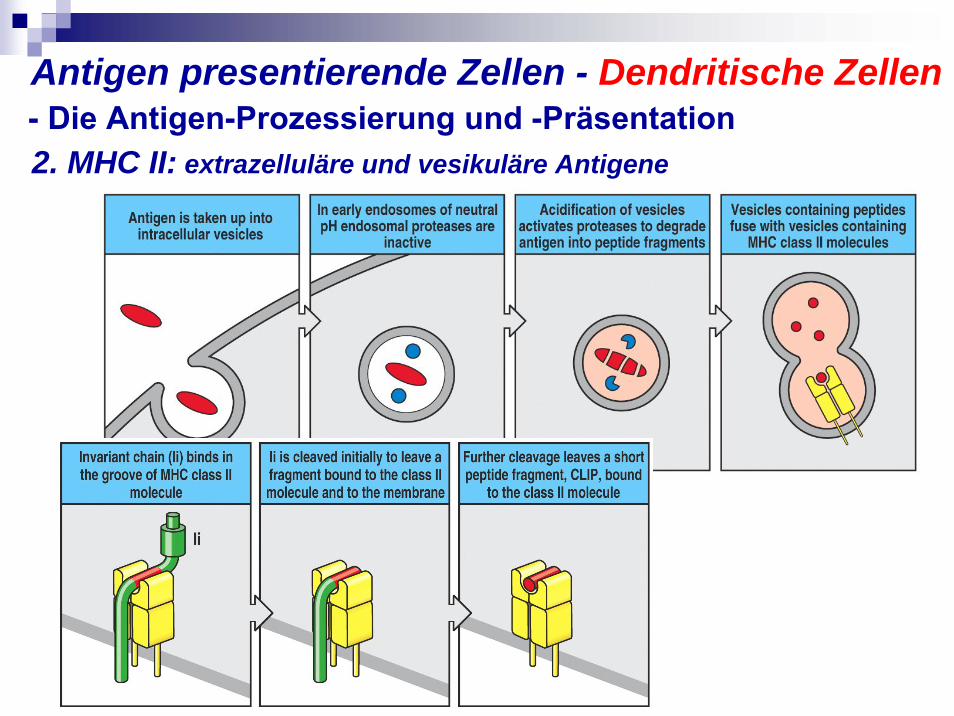
2. MHC II: extrazelluläre und vesikuläre Antigene- Die Antigen-Prozessierung und -PräsentationAntigen presentierende Zellen - Dendritische Zellen
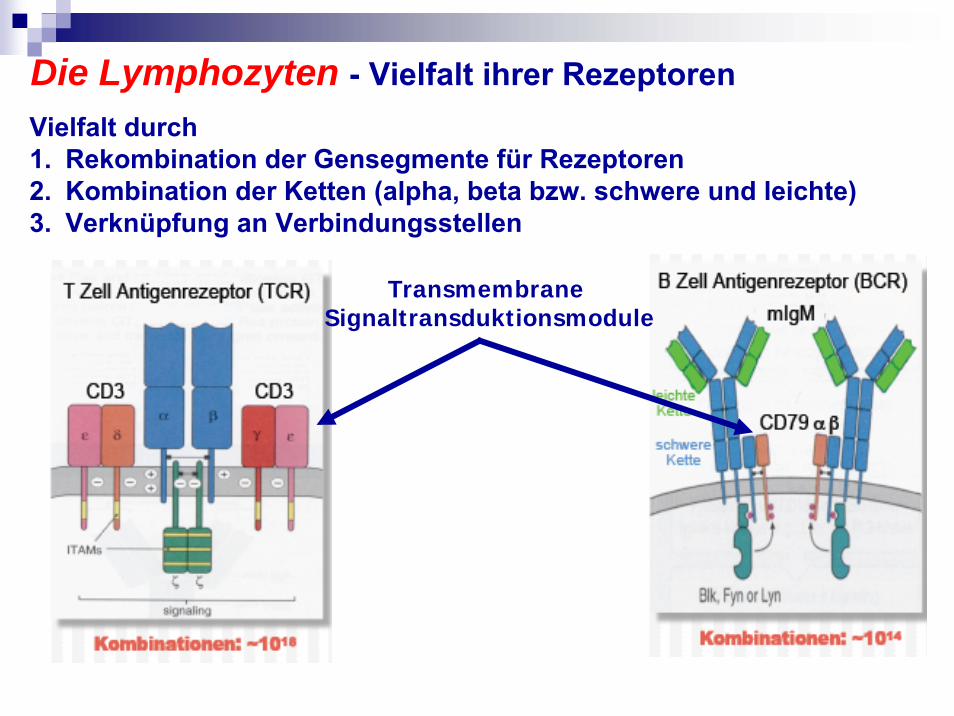
Die Lymphozyten - Vielfalt ihrer Rezeptoren
Transmembrane Signaltransduktionsmodule
Vielfalt durch1. Rekombination der Gensegmente für Rezeptoren2. Kombination der Ketten (alpha, beta bzw. schwere und leichte)3. Verknüpfung an Verbindungsstellen
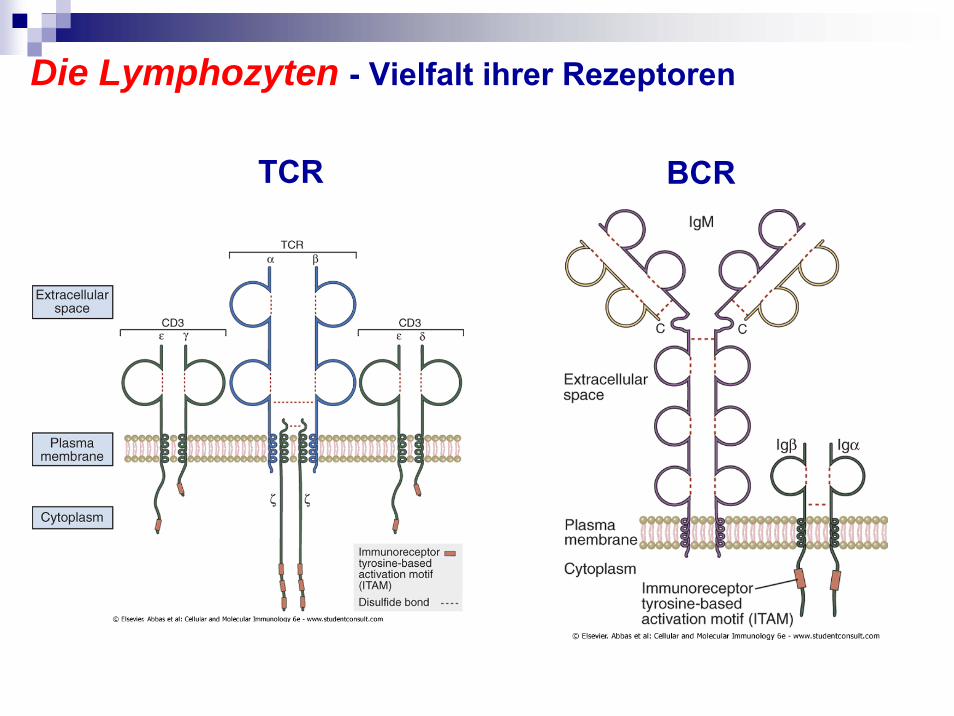
Die Lymphozyten - Vielfalt ihrer Rezeptoren
TCR BCR
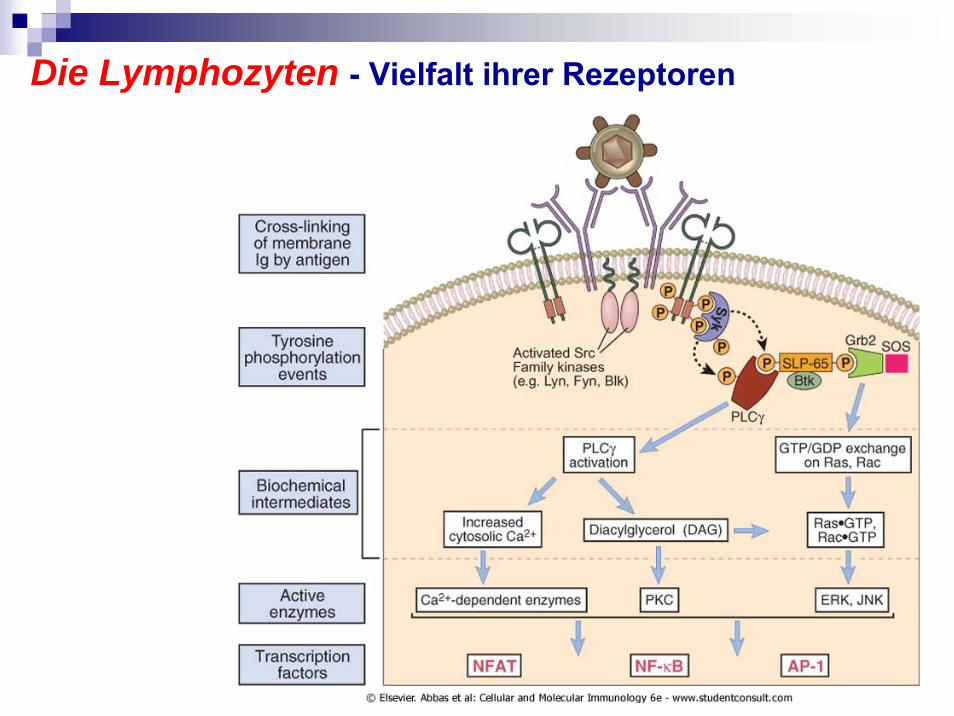
Die Lymphozyten - Vielfalt ihrer Rezeptoren
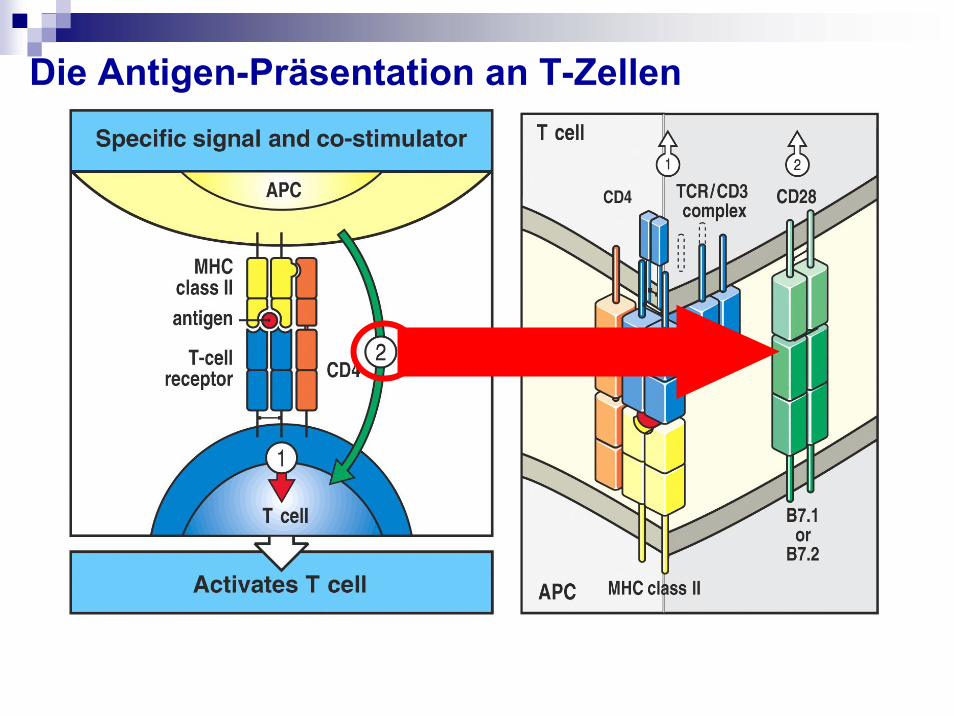
Die Antigen-Präsentation an T-Zellen
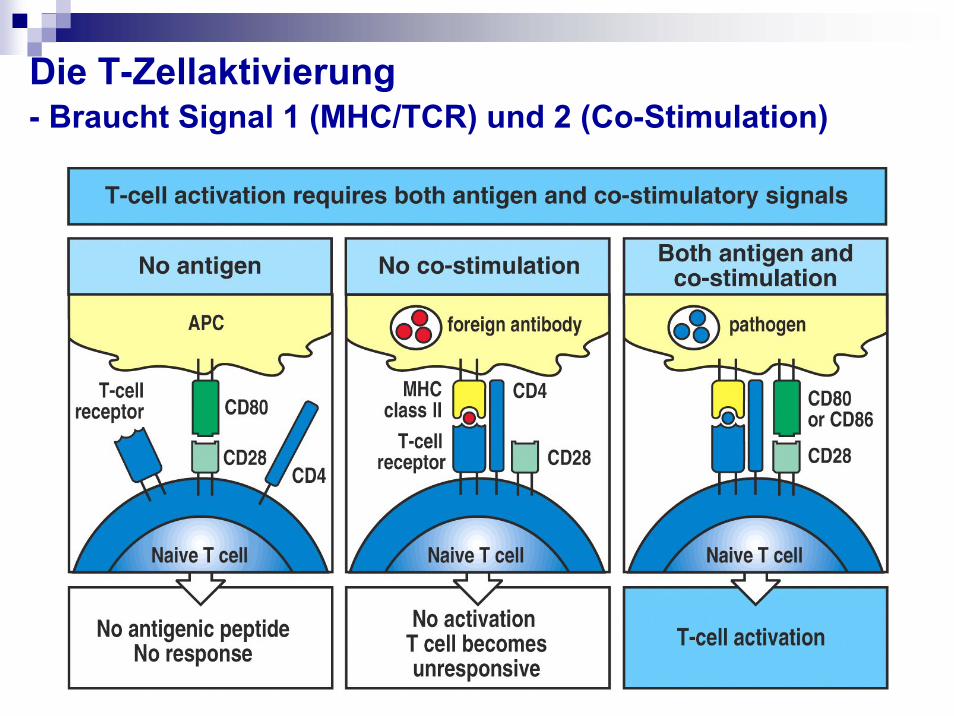
Die T-Zellaktivierung- Braucht Signal 1 (MHC/TCR) und 2 (Co-Stimulation)
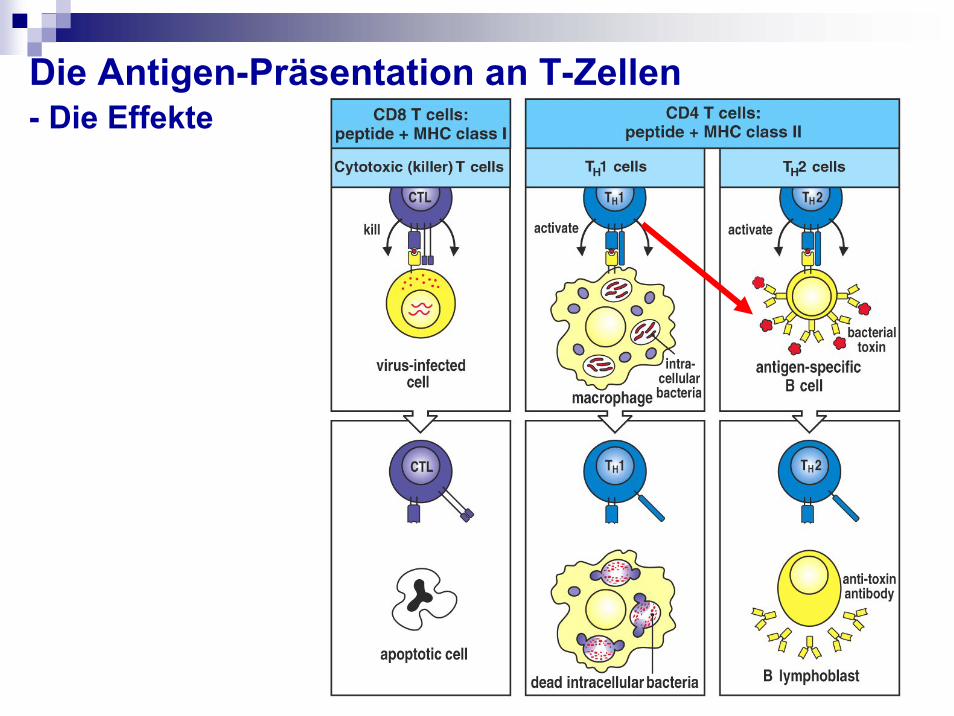
Die Antigen-Präsentation an T-Zellen- Die Effekte
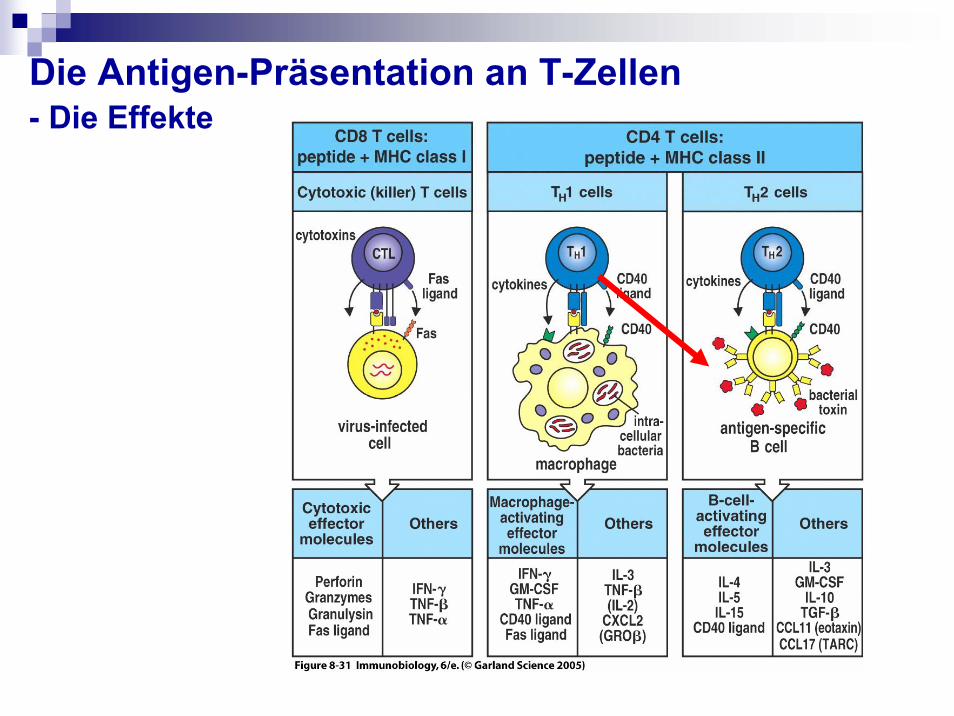
Die Antigen-Präsentation an T-Zellen- Die Effekte
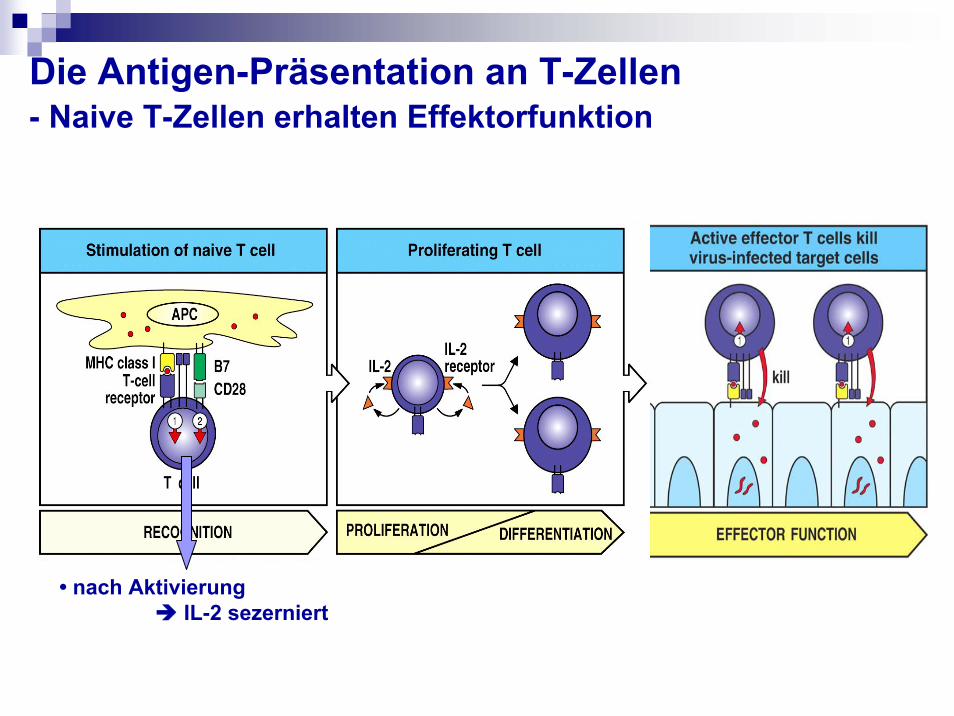
Die Antigen-Präsentation an T-Zellen- Naive T-Zellen erhalten Effektorfunktion
• nach Aktivierung IL-2 sezerniert
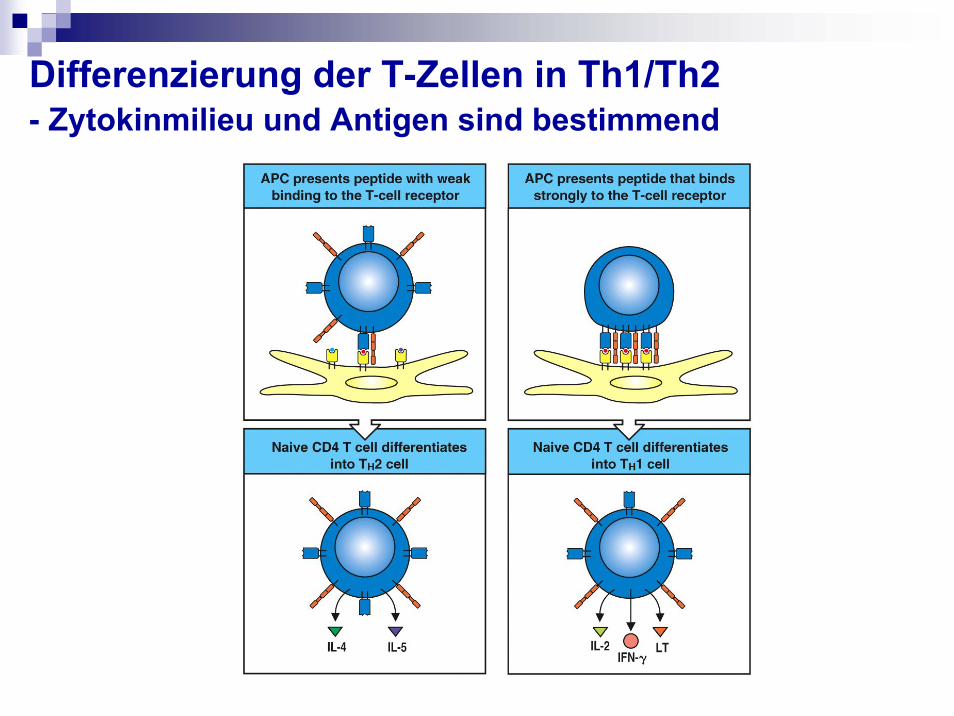
Differenzierung der T-Zellen in Th1/Th2- Zytokinmilieu und Antigen sind bestimmend

Das Schicksal der naiven T-Zellen im Lymphknoten wird durch 3 Signale(von DC) bestimmt:
1. MHC I-, MHC II-Peptid Liganden Erkennung durch TCR (Signal 1):a. Geringe Interaktion, Selbstpeptide Anergie oder Apoptose oder Tregb. Mittlere Stärke oder Dauer der Interaktion Effektorzellen + central
memory T-Zellenc. Starke, lange Interaktion Effektor-Zellen, die besten werden dann zu
memory Effektorzellen
2. Co-stimulatorische Moleküle (Signal 2): * Aktivierung der T-Zellen(!), Polarisierung, Beeinflussung und Timing
der Zytokinbildung* In Abwesenheit dieser Signale wird die T-Zelle anerg, was zur Toleranz
führen kann (wichtig bei Selbstpeptiden).
3. Zytokine (Signal 3): * Polarisierung der TH-Antwort. Die Art der gebildeten Zytokine kann
vom Subtyp der DCs, von der Art des Ag, der Entzündungsreaktion und dem Mikromilieu abhängen.
Aktivierung der T-Zellen- Zytokinmilieu und Antigen sind bestimmend
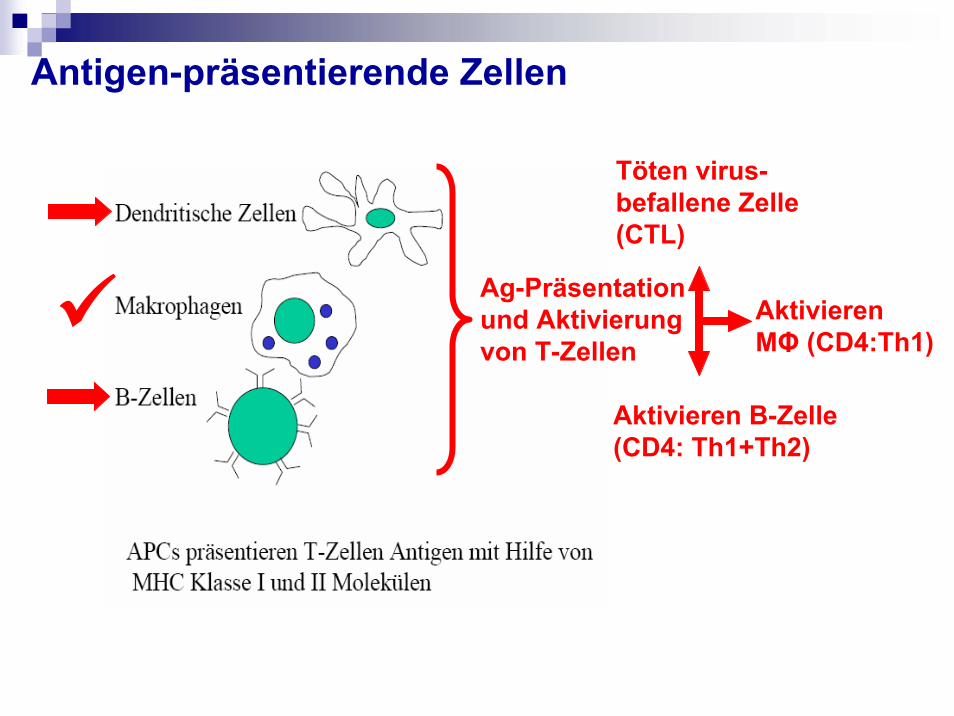
Antigen-präsentierende Zellen
Ag-Präsentation und Aktivierung von T-Zellen
Töten virus-befallene Zelle(CTL)
Aktivieren B-Zelle (CD4: Th1+Th2)
Aktivieren MΦ (CD4:Th1)

Die B-Lymphozyten- Prinzip der B-Zellfunktion: Antigenerkennung führt zu monospezifischer Antikörperproduktion
Knochenmarkwird Isotyp und spezifität determiniert von B-Zellennegative Selektion
unreife B-Zellen IgM und IgD an die Oberfläche
Treffen mit Antigen in sekundäre lymphatische Organen
Plasmazellen
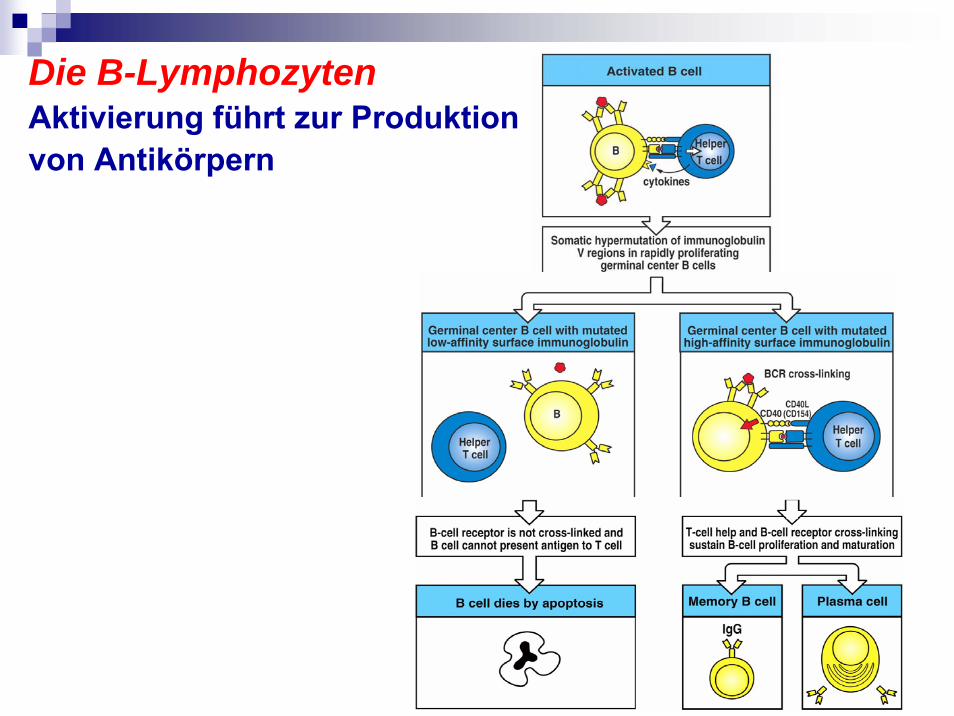
Die B-LymphozytenAktivierung führt zur Produktion von Antikörpern
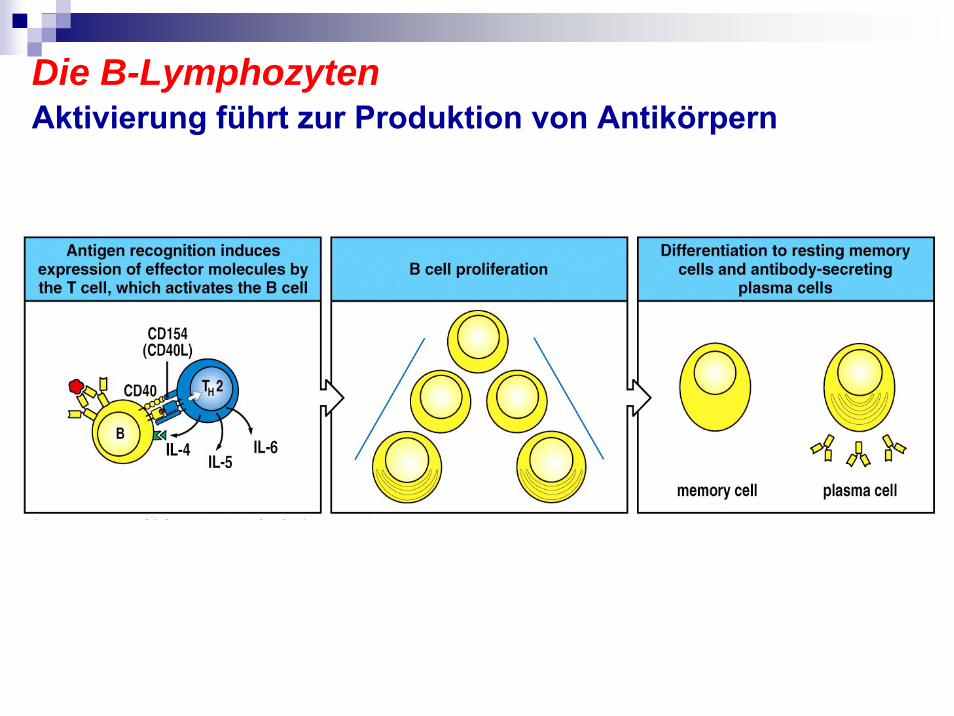
Die B-LymphozytenAktivierung führt zur Produktion von Antikörpern
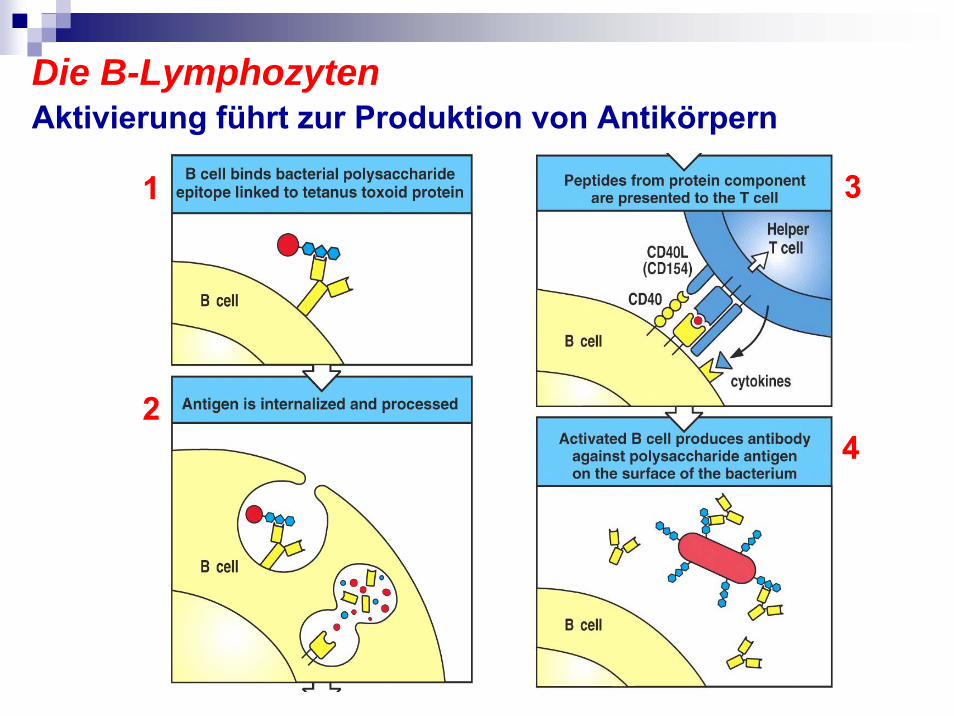
1
2
3
Die B-LymphozytenAktivierung führt zur Produktion von Antikörpern
4

Die B-LymphozytenTreffen in Lymphorganen die spezifischen T-Zellen
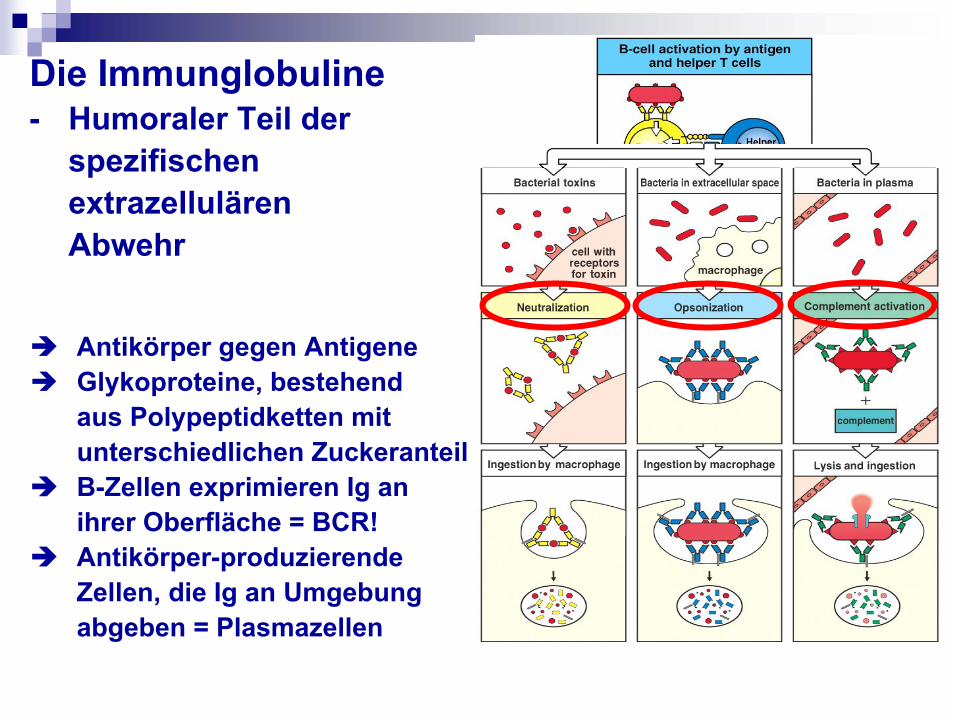
Die Immunglobuline- Humoraler Teil der
spezifischen extrazellulären Abwehr
Antikörper gegen AntigeneGlykoproteine, bestehend aus Polypeptidketten mit unterschiedlichen Zuckeranteil B-Zellen exprimieren Ig an ihrer Oberfläche = BCR!Antikörper-produzierende Zellen, die Ig an Umgebung abgeben = Plasmazellen

Funktionen
Antikörper neutralisierenbinden an andere Zelltypen aktivieren das KomplementAntigene binden (jedes Immunglobulin kann dabei nur an ein bestimmtes Antigen binden an die spezifischeantigene Determinante = Epitop)Valenz: mindestens 2, IgA = 4, IgM = 10
Die Funktionen der Immunglobuline

Bindung eines Antikörpers an ein Antigen: oft keine unmittelbaren Auswirkungen auf das Antigen erst, wenn weitere Effektor-Funktionen ins Spiel kommen (abhängig von der Bindung des Antikörpers an sein Antigen und Art der Antikörper-Moleküle)Neutralisation: Pathogen und/oder seine Produkte binden und sie damit zu neutralisieren (bakterielle Toxine, Oberfläche von Viren oder Bakterien)Bindung an verschiedene Zellarten: Phagozyten, Lymphozyten, Dendritische Zellen, Thrombozyten, Mastzellen und Basophile tragen an ihrer Oberfläche Rezeptoren (Fc Rezeptoren) Phagozytose wesentlich besser
Die Funktionen der Immunglobuline

binden auch an derartige Rezeptoren auf denThrophoblastzellen der Plazenta mütterlichen Ig durch die Plazenta in den fetalen Kreislauf schützen das Ungeborene vor InfektionenKomplementbindung und – aktivierung: Antikörper können durch Bindung an bestimmte Komplementkomponenten zur Aktivierung des Komplementsystems führen Lyse der beladenen Mikroorganismen, dabei Freisetzung von biologisch aktiven Mediatorsubstanzen und zur Beladung der Erreger mit bestimmten Komplementkomponenten noch bessere Opsonisierung
Die Funktionen der Immunglobuline

Monomereuniversiellste Immungloblinklasse (kann die meisten Funktionen der Immunglobuline ausüben)häufigstes Immunglobulin im Serum: bis 75% der Antikörperhäufigstes und wichtigstes Immunglobulin im Extravaskulärraum (Gewebe)einzige Ig Klasse, die Plazentaschranke überqueren kann(Subklassen können unterschiedlich gut übertreten, IgG2 am schlechtesten)Komplementbindung ist unterschiedlich ausgeprägt, je nach Subklasse (IgG4 bindet das Komplement nicht)Makrophagen, Monozyten und manche Lymphozyten haben Rezeptoren für Fc-Region von IgG (Subklassen binden unterschiedlich gut: IgG2 und IgG4 z.B. gar nicht) IgG ist somit ein gutes Opsonin
IgG: Der universale Antikörper

Pentamerejeweils alle schweren und leichten Ketten ident, die Valenz beträgt maximal 10, Aufbau des Pentamers mit J-Kette, die durch Disulfidbrücken an 2 schwere Ketten gebunden istIgM wird als erstes vom Fetus produziertauch das erste Immunglobulin, das von einer naiven B-Zelle produziert wird, wenn sie durch ein Antigen aktiviert wirdAufgrund seiner pentameren Struktur kann IgM das Komplement besonders gut bindenIgM kann gut agglutinieren: Mikroorganismen und Antikörper verklumpen miteinander, die Keime werden eliminiert
IgM: Der erste Antikörper

im Serum als Monomer, in Sekreten als Dimerauch hier ist eine J-Kette mit den beiden schweren (alpha) Ketten verbundenIgA ist der wichtigste Antikörper in Sekreten wie Tränen, Speichel, Kolostrum, Schleim besonders wichtig zur Ausbildung der lokalen (mukosalen) ImmunantwortIn Sekreten ist IgA mit einem weiteren Protein zusammen, demsekretorischen Stück für den Transport durch die Epithelzellen notwendig (wird von den Epithelzellen produziert und an ihrerbasalen Seite exprimiert) sekretorisches Stück beschützt das sezernierte IgA davor, in den Sekreten verdaut zu werdenIgA bindet an CD89 (FcαRI-hochaffine Rezeptor) auf eosinoplilen und neutrophilen Granulozyten, Monozyten, MakrophagenAntigenerkennung durch zwei IgA Zellen degranulieren
IgA: Der sezernierte Antikörper

nur als MonomereIgD nur in sehr geringen Mengen im SerumBedeutung ist unklarzusammen mit IgM ist als frühe Klasse on der B-Zellen koexprimiertIgD wird auf der Oberfläche von reifen naiven B-Zellengefunden und fungiert dort als Antigenrezeptor
IgD: Der unbekannte Antikörper
IgD:Mission unknown

nur MonomereIgE ist das seltenste Immunglobulin im Serumauch ohne Antigen bindet es an Oberfläche von Basophilen und Mastzellen an Fc Rezeptoren für IgE (FcεRI+II)an allergischen Reaktionen beteiligt: bindet ein Antigen an zwei oberflächengebundene IgE-Moleküle gleichzeitig Quervernetzung der Fc Rezeptoren der Mastzelle
Freisetzung von pharmakologisch aktiven Substanzen, die die klinische Symptomatik der Allergie hervorrufenwichtige Rolle bei der Abwehr von parasitären Wurmerkrankungengute Opsonine: IgE-Moleküle binden an Antigene auf den Parasiten docken sie an Fc Rezeptoren von Eosinophilen an Abtöten der Parasiten
IgE: Der allergische Antikörper

Die Funktionen der Immunglobuline
+++ dominierend++ weniger wichtig+ sehr selten* Opsonin bei bestimmten Fc-Rezeptor-Allotyp
(50% der weißen Bevölkerung)

Die Verteilung der Immunglobuline

Toleranzmechanismen
Zentrale Toleranz
Antikörper-Moleküle und T-Zellen-Rezeptoren können jedes andere Molekül erkennen, ob fremd oder körpereigen, sogar künstlich synthetisierte das Immunsystem ist bezüglich seiner Eigenschaft Antigene zu erkennen vollständigToleranzmechanismen gegenüber „selbst“ sowie „fremd-ungefährlich“von entscheidender Bedeutung
Im Thymus: mTEC-Ag erkannt Apoptose =klonale Deletion, Elimination
Periphere Toleranz
• klonale Anergie• mangelnde Kommunikation
= Ignoranz
• klonale Anergie• klonale Ignoranz
T-Zellen B-Zellen
Im KM: Stromazelleklonale Deletion=
Elimination oder neuer Rezeptor
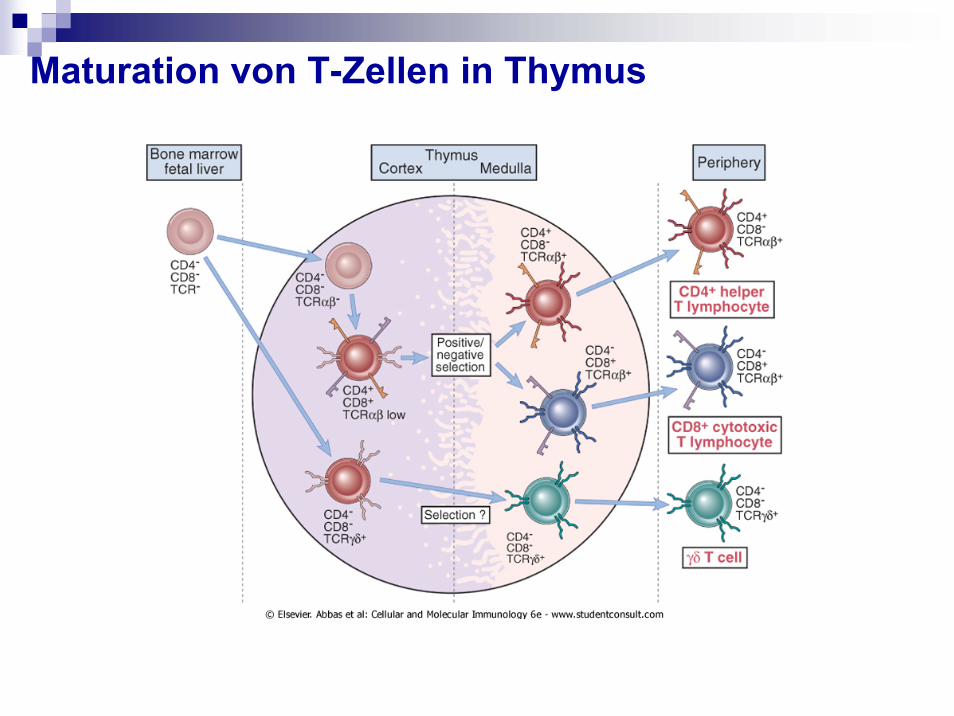
Maturation von T-Zellen in Thymus
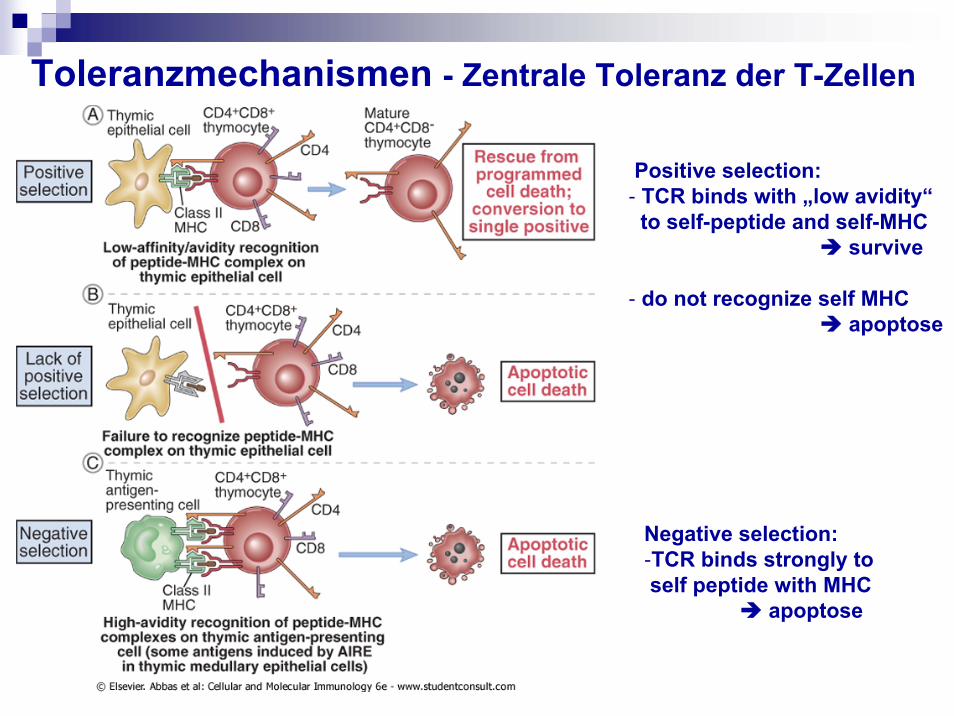
Toleranzmechanismen - Zentrale Toleranz der T-Zellen
Positive selection:- TCR binds with „low avidity“
to self-peptide and self-MHC survive
- do not recognize self MHC apoptose
Negative selection:-TCR binds strongly to self peptide with MHC
apoptose
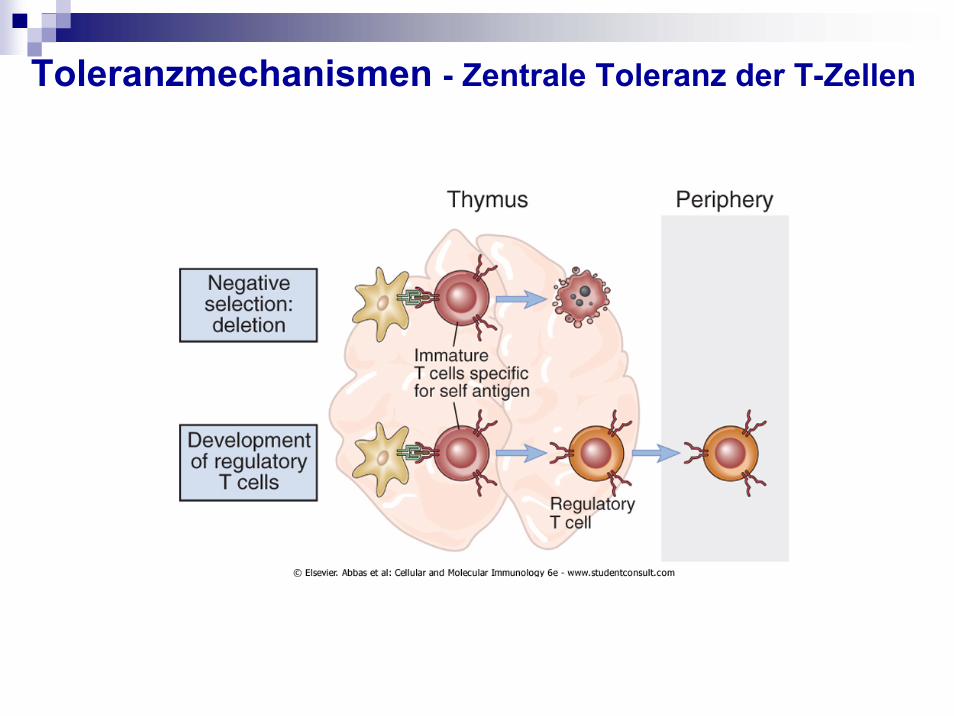
Toleranzmechanismen - Zentrale Toleranz der T-Zellen

Toleranzmechanismen - Periphere Toleranz der T-Zellen
1. Apoptose = klonale Elimination, Deletion2. Anergy = funktionelle Nicht-Reaktivität3. Aktive Suppression durch regulatorische T-Zellen4. Mangelnde Kommunikation = klonale Ignoranz:
isolierte Moleküle in ZNS und Augen; fehlende MHC II-Moleküle; Antigen unterhalb Schwellenwert exprimiertspezifische T-Zellen weder aktiviert noch deletiert

1. Apoptose
Induktion von Activation-induced cell death (AICD) von T-Zellen: nach Restimulierung von kürzlich aktivierten CD4+
T-Zellen kommt es zur Induktion von Fas-Ligand (FasL) und Fasbei Bindung dieser beiden (auf Zelle selbst) oder durch nahe
T-Zellen Apoptose
Verstärkt durch IL-2, also wenn viele T-Zellen aktiviert werden Deletion-induzierte Toleranz
(FasL auch von HIV induziert, FasL auch auf Tumorzellenexprimiert)
Toleranzmechanismen - Periphere Toleranz der T-Zellen
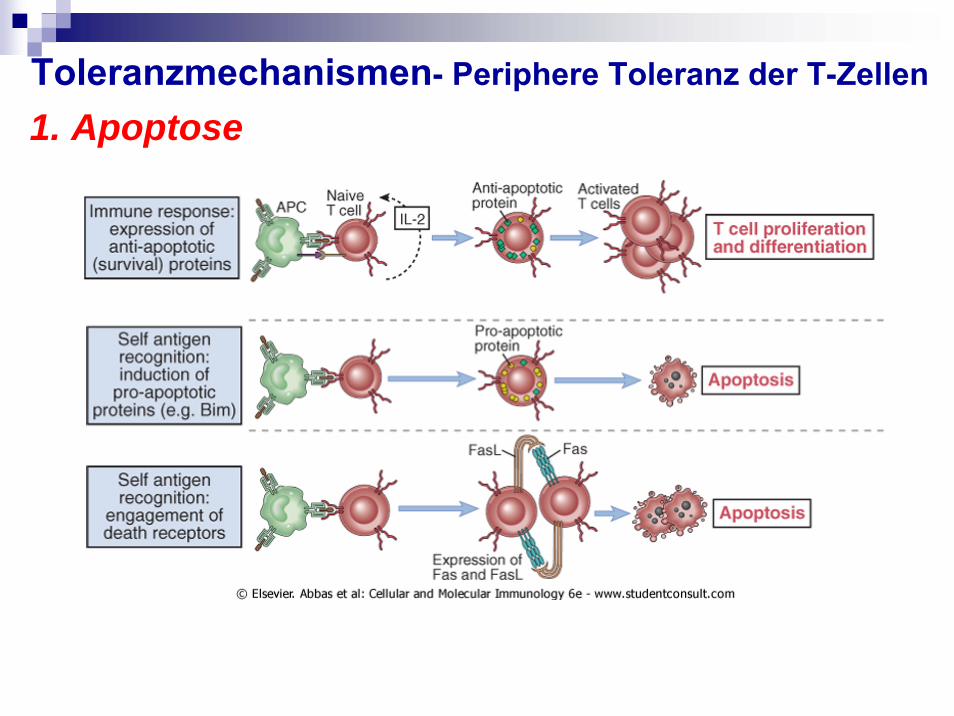
Toleranzmechanismen- Periphere Toleranz der T-Zellen1. Apoptose

2. Anergy
Geringe Interaktion von DC und T-ZellenGeringe Menge an co-stimulatorischen Molekülen und MHC MolekülenGeringe Affinität des TCR zum MHC/Peptid KomplexAntigen-Erkennung parallel mit CTLA-4 statt CD28 (auf T-Zellen) und B7 (auf DC)
Toleranzmechanismen - Periphere Toleranz der T-Zellen
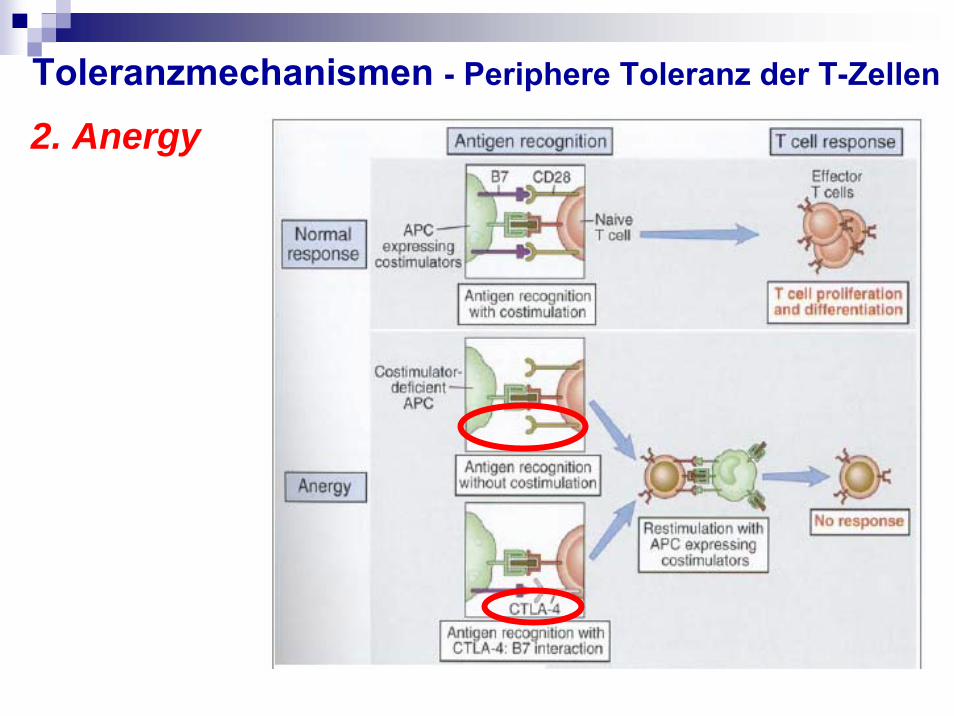
Toleranzmechanismen - Periphere Toleranz der T-Zellen
2. Anergy

3.Regulatorische T-Zellen
Exprimieren IL-2 Rezeptor-α (CD25)Sind generiert nach „self-antigen recognition” in Thymus aber kann auch in peripherisches LymphknotenGenerierung ist abhängig von IL-2, TGF-βCharakterisiert durch die Produktion von
immunsuppressiven Faktoren: IL-10, TGF-βLymphozyten, Makrophagen sind gehemmt
T-reg werden gebildet, wenn mit DC interagierendeT-Zellen wiederholt hohen Mengen an IL-10 ausgesetzt
werdenIL-10 wird von einigen DC-Subtypen produziertT-reg unterdrücken TH1 und TH2-Antworten
Toleranzmechanismen - Periphere Toleranz der T-Zellen
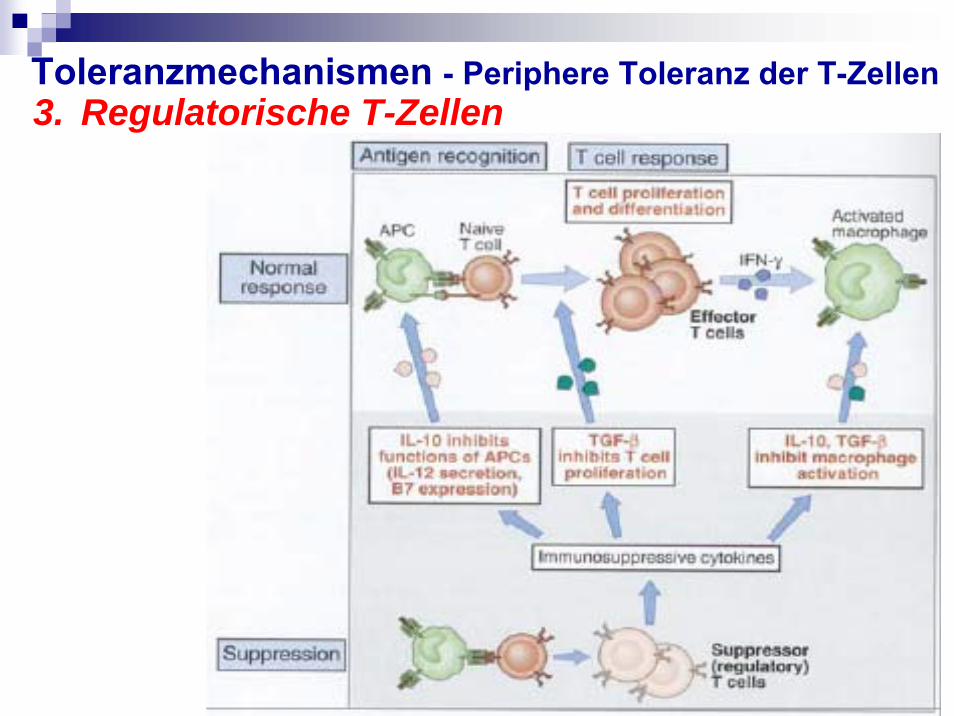
Toleranzmechanismen - Periphere Toleranz der T-Zellen3. Regulatorische T-Zellen
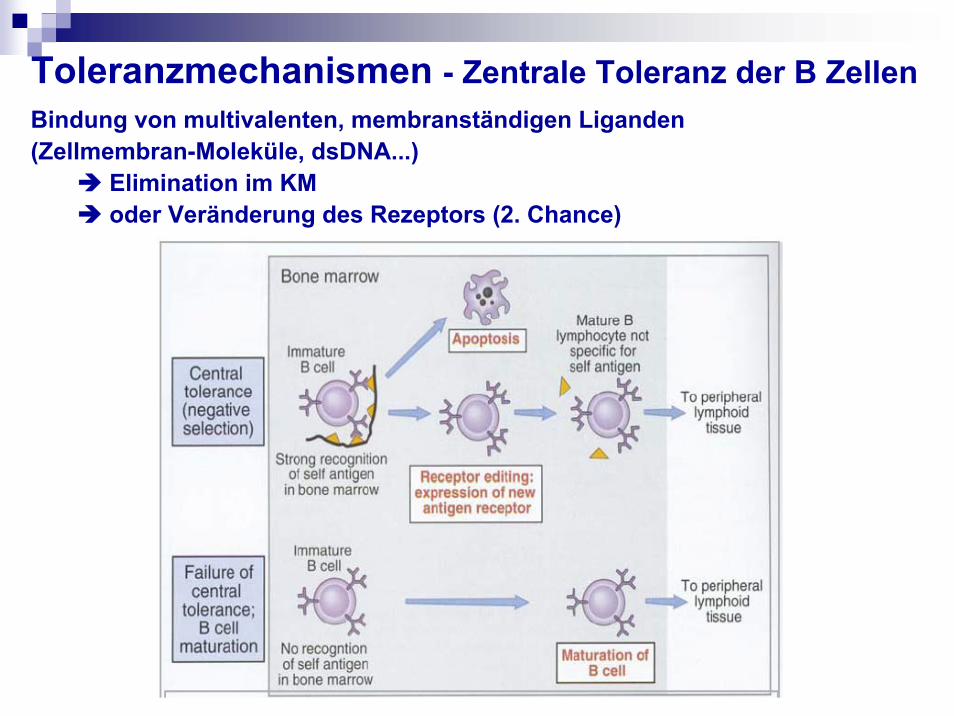
Toleranzmechanismen - Zentrale Toleranz der B ZellenBindung von multivalenten, membranständigen Liganden(Zellmembran-Moleküle, dsDNA...)
Elimination im KM oder Veränderung des Rezeptors (2. Chance)
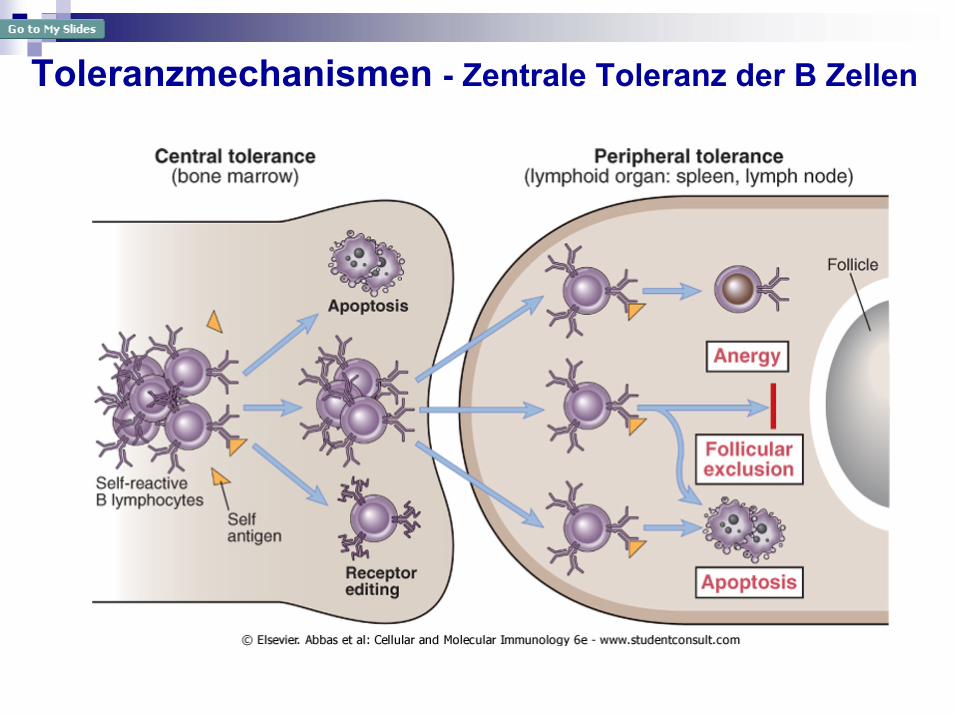
Toleranzmechanismen - Zentrale Toleranz der B Zellen

Toleranzmechanismen - Zusammenfassung


















