
Biosynthese von Cyanidin in Zellsuspensionskulturen...
5
This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution 4.0 International License. Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschung in Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht: Creative Commons Namensnennung 4.0 Lizenz. BIOSYNTHESE VON CYANIDIN IN ZELLSUSPENSIONSKULTUREN 581 Biosynthese von Cyanidin in Zellsuspensionskulturen von Haplopappus gracilis Biosynthesis of Cyanidin in Cell Suspension Cultures of Haplopappus gracilis HANSJÖRG FRITSCH *, KLAUS HAHLBROCK und H ANS GRISEBACH Lehrstuhl für Biochemie der Pflanzen am Biologischen Institut II der Universität Freiburg i. Br. (Z. Naturforsch. 26 b, 581—585 [1971]; eingegangen am 25. März 1971) Earlier work with buckwheat seedlings showed that dihydrokaempferol (3,5,7,4'-tetrahydroxy- flavanone) (1) is a good precursor for cyanidin and quercetin. Since cell suspension cultures have several advantages for the quantitative comparison of the incorporation of a radioactive precursor into a product, the role of 1 in the biosynthesis of cyanidin was reinvestigated with cell cultures of Haplopappus gracilis. These cultures show good production of cyanidin-glycosides in blue light in a medium containing sucrose, 2,4-D and Fe-EDTA. Maximum anthocyanin production and maximal activity of phenylalanine ammonia-lyase is reached about 60 —70 hrs. after the onset of illumina- tion. The incorporation of 1 and of phenylalanine and 4,2',4',6'-tetrahydroxychalcone into cyanidin was compared under identical conditions after 1, 3 and 6 hrs. feeding time. 1 prooved to be by far the best precursor for cyanidin. The feeding time required was much shorter and the incorpo- ration rates were much higher than in experiments with seedlings. Nach früheren Versuchen über die Biosynthese des Cyamdins beim Buchweizen ( Fagopyrum esculen- tum L.) ist Dihydrokaempferol (1) (3.5.7.4'-Tetra- hydroxyflavanon) eine gute Vorstufe für Cyanidin. Dihydrokaempferol-(G)-[ 3 H] zeigte eine etwa gleichhohe Einbaurate in Cyanidin wie gleichzeitig appliziertes Phenylalanin-[1- 14 C], dagegen war der Einbau von Kaempferol-(G)-[ 3 H] etwa 10-fach ge- ringer Auf Grund dieser Ergebnisse haben wir postuliert, daß 1 eine Zwischenstufe auf dem Bio- syntheseweg vom Chalkon zum Cyanidin ist x . Es erschien jedoch wünschenswert, diese Hypothese durch weitere Versuche zu prüfen. Die mit Buchwei- zenkeimlingen erhaltenen Einbauraten lagen nur in •der Größenordnung von etwa 0,5 Prozent. Auch sonst haben Keimlinge einige Nachteile für den quantitativen Vergleich des Einbaus verschiedener potentieller Vorstufen in ein Biosyntheseprodukt, wie z. B. die Schwierigkeit des sterilen Arbeitens, die Ungewißheit des Stofftransports und die Inhomo- genität des Pflanzenmaterials. Diese Nachteile sollten durch das Arbeiten mit Zellsuspensionen vermieden werden können. Hier ist ein absolut steriles Arbeiten gewährleistet. Außer Sonderdrudeanforderungen an Prof. Dr. H. GRISEBACH, Lehrstuhl für Biochemie der Pflanzen am Biologischen In- stitut II der Universität, D-7800 Freiburg i. Br., Germany, Schänzlestraße 9/11. * Aus der Diplomarbeit von H. FRITSCH, Freiburg 1971. Abkürzungen: 2.4-D = 2.4-Dichlorphenoxyessigsäure, NAA = a-Naphthylessigsäure, Enzyme: L-Phenylalanin ammo- nium-lyase (PAL) E.C. 4.3.1.5. einer Permeation durch die Zellwand und Membran ist für die Vorstufen kein weiterer Stofftransport notwendig, und bei Arbeiten mit Aliquoten aus einer Mutterkultur sind die Ergebnisse streng mit- einander vergleichbar. Für unsere Versuche schienen uns Zellkulturen von Haplopappus gracilis geeignet zu sein. Von Ge- webekulturen dieser Komposite ist bekannt, daß sie im kurzwelligen Bereich des sichtbaren Lichts Cya- nidin-3-glucosid und Cyanidin-3-rutinosid bilden 2 . In der vorliegenden Arbeit haben wir daher den Einbau von 1 in Cyanidin bei Zellkulturen von H. gracilis mit dem Einbau von L-Phenylalanin und 4.2'.4'.6-Tetrahydroxychalkonglucosid nach ver- schiedenen Fütterungszeiten verglichen. Es wird ge- zeigt, daß 1 die beste Vorstufe für Cyanidin ist. Material und Methoden Zellsuspensionskulturen von H. gracilis Eine Suspensionskultur aus dem Sproß von Haplo- pappus gracilis erhielten wir von 0. L. GAMBORG, Saskatoon, Canada. Die Weiterkultivierung erfolgte in dem von GAMBORG angegebenen Medium B5-D 3 , wobei 1 L. PATSCHKE, W . BARZ U. H. GRISEBACH, Z. Naturforsch. 21b, 45 [1966]. 2 R. VON ARDENNE, Z. Naturforsch. 20 b, 186 [1965]; J. B. HARBORNE, Comparative Biochemistry of the Flavonoids, S. 227, Academic Press, London 1967. 3 O. L. GAMBORG, R. A. MILLER U. K. OJIMA, Exp. Cell Res. 50,151 [1968].
-
Upload
dangkhuong -
Category
Documents
-
view
224 -
download
1
Transcript of Biosynthese von Cyanidin in Zellsuspensionskulturen...

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
BIOSYNTHESE VON CYANIDIN IN ZELLSUSPENSIONSKULTUREN 581
Biosynthese von Cyanidin in Zellsuspensionskulturen von Haplopappus gracilis
Biosynthesis o f Cyanidin in Cell Suspension Cultures of Haplopappus gracilis
HANSJÖRG FRITSCH * , KLAUS HAHLBROCK u n d HANS GRISEBACH
Lehrstuhl für Biochemie der Pflanzen am Biologischen Institut II der Universität Freiburg i. Br.
(Z. Naturforsch. 26 b, 581—585 [1971]; eingegangen am 25. März 1971)
Earlier work with buckwheat seedlings showed that dihydrokaempferol (3,5,7,4'-tetrahydroxy-flavanone) (1) is a good precursor for cyanidin and quercetin. Since cell suspension cultures have several advantages for the quantitative comparison of the incorporation of a radioactive precursor into a product, the role of 1 in the biosynthesis of cyanidin was reinvestigated with cell cultures of Haplopappus gracilis. These cultures show good production of cyanidin-glycosides in blue light in a medium containing sucrose, 2,4-D and Fe-EDTA. Maximum anthocyanin production and maximal activity of phenylalanine ammonia-lyase is reached about 60 —70 hrs. after the onset of illumina-tion. The incorporation of 1 and of phenylalanine and 4,2',4',6'-tetrahydroxychalcone into cyanidin was compared under identical conditions after 1, 3 and 6 hrs. feeding time. 1 prooved to be by far the best precursor for cyanidin. The feeding time required was much shorter and the incorpo-ration rates were much higher than in experiments with seedlings.
Nach früheren Versuchen über die Biosynthese des Cyamdins beim Buchweizen ( F a g o p y r u m esculen-tum L . ) ist Dihydrokaempferol ( 1 ) (3 .5 .7 .4 ' -Tetra-hydroxyf lavanon) eine gute Vorstufe für Cyanidin. D i h y d r o k a e m p f e r o l - ( G ) - [ 3 H ] zeigte eine etwa gleichhohe Einbaurate in Cyanidin wie gleichzeitig appliziertes Phenyla lanin- [1 - 1 4 C] , dagegen war der Einbau von K a e m p f e r o l - ( G ) - [ 3 H ] etwa 10-fach ge-ringer Auf Grund dieser Ergebnisse haben wir postuliert, daß 1 eine Zwischenstufe auf dem Bio-syntheseweg v o m Chalkon zum Cyanidin i s t x . Es erschien jedoch wünschenswert, diese Hypothese durch weitere Versuche zu prüfen. Die mit Buchwei-zenkeimlingen erhaltenen Einbauraten lagen nur in •der Größenordnung von etwa 0 ,5 Prozent. Auch sonst haben Keimlinge einige Nachteile f ü r den quantitativen Vergleich des Einbaus verschiedener potentieller Vorstufen in ein Biosyntheseprodukt, wie z. B. die Schwierigkeit des sterilen Arbeitens, die Ungewißheit des Stofftransports und die Inhomo-genität des Pflanzenmaterials.
Diese Nachteile sollten durch das Arbeiten mit Zellsuspensionen vermieden werden können. Hier ist e in absolut steriles Arbeiten gewährleistet. Außer
Sonderdrudeanforderungen an Prof. Dr. H. GRISEBACH, Lehrstuhl für Biochemie der Pflanzen am Biologischen In-stitut II der Universität, D-7800 Freiburg i. Br., Germany, Schänzlestraße 9/11.
* Aus der Diplomarbeit von H. FRITSCH, Freiburg 1971. Abkürzungen: 2.4-D = 2.4-Dichlorphenoxyessigsäure, NAA
= a-Naphthylessigsäure, Enzyme: L-Phenylalanin ammo-nium-lyase (PAL) E.C. 4.3.1.5.
einer Permeation durch die Zellwand und Membran ist für die Vorstufen kein weiterer Stofftransport notwendig , und bei Arbeiten mit Aliquoten aus einer Mutterkultur sind die Ergebnisse streng mit-einander vergleichbar.
Für unsere Versuche schienen uns Zellkulturen von Haplopappus gracilis geeignet zu sein. V o n Ge-webekulturen dieser Komposite ist bekannt, daß sie im kurzwelligen Bereich des sichtbaren Lichts Cya-nidin-3-glucosid und Cyanidin-3-rutinosid b i l d e n 2 .
In der vorl iegenden Arbeit haben wir daher den Einbau von 1 in Cyanidin bei Zellkulturen von H. gracilis mit dem Einbau von L-Phenylalanin und 4 .2 ' . 4 ' . 6 -Tetrahydroxychalkong lucos id nach ver-schiedenen Fütterungszeiten verglichen. Es wird ge-zeigt, daß 1 die beste Vorstufe für Cyanidin ist.
Material und Methoden
Zellsuspensionskulturen von H. gracilis
Eine Suspensionskultur aus dem Sproß von Haplo-pappus gracilis erhielten wir von 0 . L. GAMBORG, Saskatoon, Canada. Die Weiterkultivierung erfolgte in dem von GAMBORG angegebenen Medium B5-D 3, wobei
1 L . PATSCHKE, W . BARZ U. H. GRISEBACH, Z. Naturforsch. 21b , 45 [1966].
2 R. VON ARDENNE, Z. Naturforsch. 20 b, 186 [1965]; J. B. HARBORNE, Comparative Biochemistry of the Flavonoids, S. 227, Academic Press, London 1967.
3 O. L. GAMBORG, R . A . MILLER U. K . OJIMA, Exp. Cell Res. 50 ,151 [1968].

5 8 2 H.-J. FRITSCH, K. HAHLBROCK UND H. GRISEBACH
als Eisenquelle ein Fe-EDTA-Komplex 4 verwendet wurde. Die Kultur wurde im Dunkeln durchgeführt bei 25 — 28° auf einem Rotationsschüttler (New Bruns-wick Scientific Co.) bei 120 Upm in 200-ml - E r l e n -m e y e r - Kolben, die jeweils ca. 40 ml Zellsuspension enthielten. Tochterkulturen wurden alle 7 Tage ange-legt, indem aus einer Mutterkultur ca. 5 ml Zellsuspen-sion mit einer Pipette in 40 ml frisches Medium über-impft wurden.
Quantitative CyanidinbeStimmung in den Kulturen
Ein Aliquot der Zellen wurde mit kaltem Methanol/ H 2 0 ( 9 : 1 ) , das 0,1% konz. HCl enthielt, versetzt und in einem Homogenisator nach Potter und Elvehjem aufgeschlossen. Nach Zentrifugation in der Kühlzentri-fuge (Sorvall GC2-B, 2700 g, 10 min) wurde im Über-stand bei 535 nm der Cyanidingehalt nach einer Eich-kurve bestimmt.
Isolierung und Reinigung von Cyanidin
Lösungsmittel: L M 1 M e 0 H / H 2 0 ( 9 : 1 ) mit 1 m l konz. HCl / / ; LM 2 MeOH/konz. HCl (94 : 6 ) .
Laufmittel für Papierchromatographie: Forestal Eisessig/H20/konz. HCl (30 : 10 : 3 ) ; B E W Butanol/ Eisessig/H20 ( 4 : 1 : 1 ) .
Nach Extraktion des Zellmaterials (3 — 4 g) mit etwa 100 ml LM 1 ohne Aufschluß über Nacht wurden die Zellen abgesaugt und das Filtrat am Rotationsver-dampfer (25 — 30° ) zur Entfernung von Methanol ein-gedampft. Beim Einengen verklebte das Pigment mit zähen Lipoidklumpen und war dann kaum mehr zu extrahieren. Deshalb wurde der klumpige Rückstand nochmals mit Methanol aufgelöst und wieder einge-dampft. Der sich nun bildende feinverteilte Nieder-schlag am Kolbenrand konnte nach Zusatz von H 2 0 leicht mit Essigester extrahiert werden. Die wäßrige Phase wurde auf eine Duolite-C-25-Säule (Fa. Roth, Karlsruhe) gegeben und mit 0,5 l 1-proz. HCl, 11 H 2 0 und mindestens 2 l Methanol gespült. Nach Extraktion des gereinigten Pigments mit etwa 250 ml LM 2 wurde wieder eingedampft und der Rückstand zur Hydrolyse (90° , 2 h) mit ca. 30 ml 10-proz. HCl versetzt. Wegen der hohen Lichtempfindlichkeit des Cyanidins wurden die Hydrolyse und alle folgenden Arbeitsschritte im Dunkeln durchgeführt. Nach Ausschtüteln des Cyani-dins aus der stark salzsauren Lösung mit i-Amylalkohol und Abziehen des Alkohols am Rotationsverdampfer wurde der Rückstand 'mit LM 1 aufgenommen, auf 25 ml aufgefüllt und die Konzentration durch Messung des Extinktionsmaximums bei ca. 542 nm bestimmt. Für spätere Ausbeutebestimmungen (Einbauraten) diente dieser Wert als 100 Prozent. Die Lösung wurde wieder eingedampft, mit Methanol aufgenommen und für die Papierchromatographie als Band auf Whatman-3MM-Papier aufgetragen.
4 Vgl. O. L. GAMBORG, Canad. J. Biochem. 44, 791 [1966]. 5 F. KALBERER U. J. RUTSCHMANN, Helv. chim. Acta 44,
1956 [1961]. 6 L . M . BLAKELY u. F . C . STEWARD, A m e r . J. B o t . 4 8 , 3 5 1
[1961].
Bei einer Bandlänge von 20 cm konnte etwa 1 mg Cyanidin aufgetragen werden. Nach dem 1. Lauf (Lauf-mittel: Forestal, Z?/ = 0,5) wurde die Cyanidin-Bande ausgeschnitten, mit LM 1 extrahiert, auf 25 ml aufge-füllt und der Pigmentgehalt bestimmt. 5 ml dieser Lö-sung wurden für die Radioaktivitätsmessung (Schönin-ger-Verbrennung) eingesetzt. Der Rest wurde einer 2. Papierchromatographie auf Whatman-3-MM (Lauf-mittel: BEW, Rf = 0,7) unterworfen. Die Bestimmung der spezifischen Aktivität erfolgte wie oben. Eine dritte Papierchromatographie mit dem Eluat wurde wieder mit dem Forestal-Laufmittel durchgeführt.
Aufschluß der Zellen für die PAL-BeStimmung
1 g Zellen (Frischgewicht) wurden bei 4 ° mit 0,5 g Aktivkohle und 3 ml 0,1 M Boratpuffer (pH 8,8) im Mörser mit 0,5 g Quarzsand verrieben. Nach Zentrifu-gieren (20 min, 45000 g) wurde der klare Überstand zum Test verwendet.
Radioaktivitätsmessung
Alle Radioaktivitätsmessungen erfolgten in Beckman-Szintillationszählern LS-100 oder LS-230. Wäßrige Lö-sungen wurden im Dioxan-Cocktail (5 g PPO, 100 g Naphthalin, 11 Dioxan), mit Toluol mischbare Lösun-gen in Toluol-Cocktail (5 g PPO, 11 Toluol) gemessen. Die Zählausbeute wurde jeweils durch Zugabe eines internen Standards (Toluol-Standardlösung von New England Nuclear, Boston) bestimmt. Der statistische Zählfehler war immer 1 Prozent.
S c h ö n i n g e r - Verbrennung
Die S c h ö n i n g e r -Verbrennung des Cyanidins wurde nach KALBERER und RUTSCHMANN5 durchge-führt. Als Absorptionslösung für C 0 2 bzw. HTO diente Phenyläthylamin in Methanol (Fluka AG, Buchs, va-kuumdestilliert, 54 ml auf 450 ml Methanol).
Ergebnisse und Diskussion
Anthocyanbildung in Zellsuspensionskulturen von Haplopappus gracilis
Über Anthocyanbi ldung in Kalluskulturen von H. gracilis wurde früher ber i chtet 6 ' 7 . Anhaltspunkte für die günstigsten Kulturbedingungen gab weiter-hin die Doktorarbeit von LACKMANN8. Für die Anthocyanbi ldung in Zellsuspensionskulturen wur-den fo lgende Medien verglichen: I. Med ium B-5 nach GAMBORG3 ohne Aminosäuren, 5 1 0 - 6 M 2 .4 -D, 28 mg/Z Sequestrene 138 Fe ( G e i g y ) .
7 J. REINERT, H . CLAUSS U. R . VON ARDENNE, Naturwissen Schäften 51, 8 7 [ 1 9 6 4 ] .
8 I. LACKMANN, Dissertation, Freie Universität Berlin 1968.
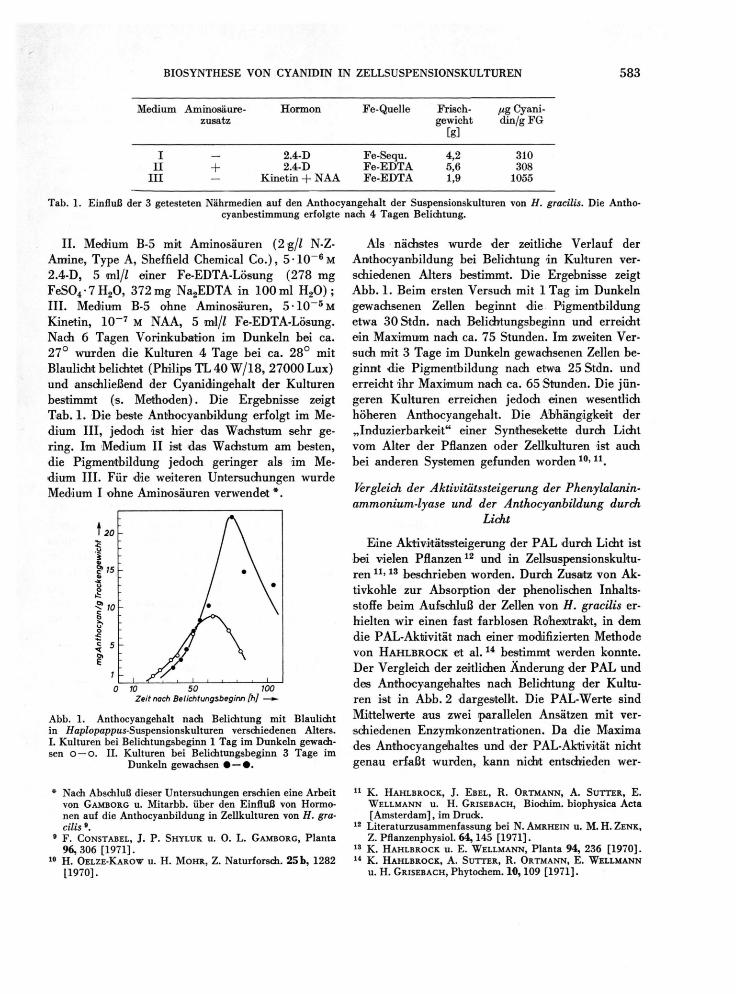
BIOSYNTHESE VON CYANIDIN IN ZELLSUSPENSIONSKULTUREN 583
Medium Aminosäure-zusatz
Hormon Fe-Quelle Frisch-gewicht
[g]
/ig Cyani-din/gFG
I 2.4-D Fe-Sequ. 4,2 310 II + 2.4-D Fe-EDTA 5,6 308
III Kinetin + N A A Fe-EDTA 1,9 1055
Tab. 1. Einfluß der 3 getesteten Nährmedien auf den Anthocyangehalt der Suspensionskulturen von H. gracilis. Die Antho-cyanbestimmung erfolgte nach 4 Tagen Belichtung.
II . Med ium B-5 mit Aminosäuren (2 g/Z N-Z-Amine , Type A , Sheffield Chemical C o . ) , 5 - 1 0 ~ 6 M 2.4 -D, 5 ml/Z einer Fe -EDTA-Lösung ( 2 7 8 m g F e S 0 4 • 7 H 2 0 , 372 mg N a 2 E D T A in 1 0 0 ml H 2 0 ) ; III . Med ium B-5 ohne Aminosäuren, 5 - 1 0 _ 5 M Kinetin, 1 0 - 7 M N A A , 5 ml/Z Fe -EDTA-Lösung . Nach 6 Tagen Vorinkubation im Dunkeln bei ca. 2 7 ° wurden die Kulturen 4 Tage bei ca. 2 8 ° mit Blaulicht belichtet (Phil ips T L 4 0 W / 1 8 , 2 7 0 0 0 L u x ) und anschließend der Cyanidingehalt der Kulturen bestimmt (s. Methoden ) . Die Ergebnisse zeigt Tab . 1. D ie beste Anthocyanbi ldung er fo lgt im Me-dium III , jedoch ist hier das Wachstum sehr ge-ring. I m Medium II ist das Wachstum am besten, die Pigmentbildung jedoch geringer als i m Me-d ium III . Für die weiteren Untersuchungen wurde Medium I ohne Aminosäuren verwendet * .
t 20
t i 0) g>75
7 0 10 50 100
Zeit nach Belichtungsbeginn [h] —»-
Abb. 1. Anthocyangehalt nach Belichtung mit Blaulicht in Haplopappus-Suspensionskulturen verschiedenen Alters. I. Kulturen bei Belichtungsbeginn 1 Tag im Dunkeln gewach-sen o —o. II. Kulturen bei Belichtungsbeginn 3 Tage im
Dunkeln gewachsen • — • .
* Nach Abschluß dieser Untersuchungen erschien eine Arbeit von GAMBORG U. Mitarbb. über den Einfluß von Hormo-nen auf die Anthocyanbildung in Zellkulturen von H. gra-cilis 9.
9 F . CONSTABEL, J. P . SHYLUK U. O . L . GAMBORG, P lanta 9 6 , 3 0 6 [ 1 9 7 1 ] .
1 0 H . OELZE-KAROW U. H . MOHR, Z . Naturforsch. 2 5 b , 1 2 8 2 [ 1 9 7 0 ] .
Als nächstes wurde der zeitliche Verlauf der Anthocyanbi ldung bei Belichtung in Kulturen ver-schiedenen Alters bestimmt. Die Ergebnisse zeigt A b b . 1. Be im ersten Versuch mit 1 Tag im Dunkeln gewachsenen Zellen beginnt die Pigmentbi ldung etwa 3 0 Stdn. nach Belichtungsbeginn und erreicht ein M a x i m u m nach ca. 7 5 Stunden. Im zweiten Ver-such mit 3 Tage im Dunkeln gewachsenen Zellen be-ginnt die Pigmentbi ldung nach etwa 25 Stdn. und erreicht ihr Max imum nach ca. 65 Stunden. Die jün-geren Kulturen erreichen jedoch einen wesentlich höheren Anthocyangehalt . D ie Abhängigkeit der „ Induz ierbarke i t " einer Synthesekette durch Licht v o m Alter der Pflanzen oder Zellkulturen ist auch bei anderen Systemen gefunden worden 1 0 ' 1 1 .
Vergleich der Aktivitätssteigerung der Phenylalanin-ammonium-lyase und der Anthocyanbildung durch
Licht
Eine Aktivitätssteigerung der P A L durch Licht ist bei vielen P f lanzen 1 2 und in Zellsuspensionskultu-ren n ' 1 3 beschrieben worden . Durch Zusatz von Ak-tivkohle zur Absorpt ion der phenolischen Inhalts-stoffe be im Aufschluß der Zellen von H. gracilis er-hielten wir einen fast farblosen Rohextrakt, in d e m die PAL-Aktivität nach einer modif izierten Methode von HAHLBROCK et a l . 1 4 bestimmt werden konnte. Der Vergleich der zeitlichen Änderung der P A L und des Anthocyangehaltes nach Belichtung der Kultu-ren ist in A b b . 2 dargestellt. Die PAL-Werte sind Mittelwerte aus zwei parallelen Ansätzen mit ver-schiedenen Enzymkonzentrationen. Da die Max ima des Anthocyangehaltes und der PAL-Aktivität nicht genau erfaßt wurden, kann nicht entschieden wer-
1 1 K . HAHLBROCK, J. EBEL, R . ORTMANN, A . SUTTER, E . WELLMANN U. H. GRISEBACH, Biochim. biophysica Acta [Amsterdam], im Drude.
1 2 L i teraturzusammenfassung bei N . AMRHEIN U. M . H . ZENK, Z . Pf lanzenphysiol . 6 4 , 1 4 5 [ 1 9 7 1 ] .
1 3 K . HAHLBROCK U. E . WELLMANN, Planta 9 4 , 2 3 6 [ 1 9 7 0 ] . 1 4 K . HAHLBROCK, A . SUTTER, R . ORTMANN, E . WELLMANN
u. H . GRISEBACH, Phytochem. 1 0 , 1 0 9 [ 1 9 7 1 ] .
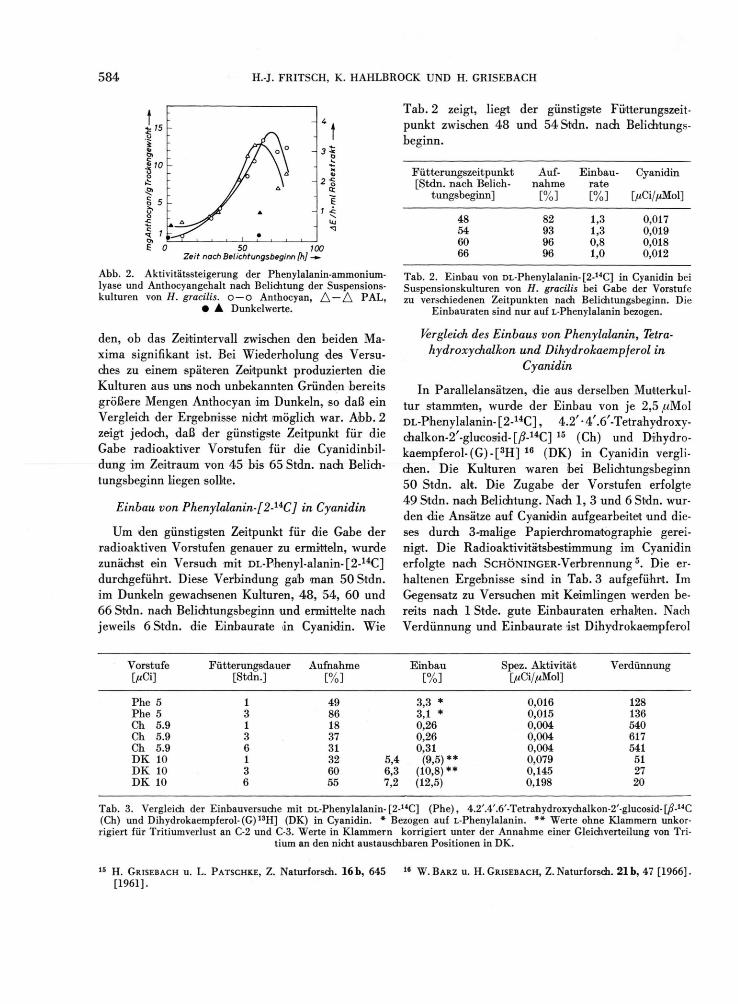
5 8 4 H.-J. FRITSCH, K. HAHLBROCK UND H. GRISEBACH
o> E 0 50 100
Zeit nach Belichtungsbeginn [hj
Abb. 2. Aktivitätssteigerung der Phenylalanin-ammonium-lyase und Anthocyangehalt nach Belichtung der Suspensions-kulturen von H. gracilis. O — O Anthocyan, A — A PAL,
• A Dunkelwerte.
den, ob das Zeitintervall zwischen den beiden Ma-xima signifikant ist. Bei Wiederholung des Versu-ches zu einem späteren Zeitpunkt produzierten die Kulturen aus uns noch unbekannten Gründen bereits größere Mengen Anthocyan im Dunkeln, so daß ein Vergleich der Ergebnisse nicht mögl ich war. A b b . 2 zeigt jedoch, daß der günstigste Zeitpunkt für die Gabe radioaktiver Vorstufen für die Cyanidinbil -dung im Zeitraum von 4 5 bis 65 Stdn. nach Belich-tungsbeginn liegen sollte.
Einbau von Phenylalanin-[2-uC] in Cyanidin
U m den günstigsten Zeitpunkt für die Gabe der radioaktiven Vorstufen genauer zu ermitteln, wurde zunächst ein Versuch mit DL -Phenyl-alanin-[2-1 4C] durchgeführt. Diese Verbindung gab man 5 0 Stdn. im Dunkeln gewachsenen Kulturen, 4 8 , 54 , 6 0 und 6 6 Stdn. nach Belichtungsbeginn und ermittelte nach jeweils 6 Stdn. die Einbaurate in Cyanidin. W i e
Tab . 2 zeigt, liegt der günstigste Fütterungszeit-punkt zwischen 4 8 und 54 Stdn. nach Belichtungs-beginn.
Fütterungszeitpunkt Auf- Einbau- Cyanidin [Stdn. nach Belich- nahme rate
tungsbeginn] [%] [%] [^Ci/^Mol]
48 82 1,3 0,017 54 93 1,3 0,019 60 96 0,8 0,018 66 96 1,0 0,012
Tab. 2. Einbau von DL-Phenylalanin-[2-14C] in Cyanidin bei Suspensionskulturen von H. gracilis bei Gabe der Vorstufe zu verschiedenen Zeitpunkten nach Belichtungsbeginn. Die
Einbauraten sind nur auf L-Phenylalanin bezogen.
Vergleich des Einbaus von Phenylalanin, Tetra-hydroxychalkon und Dihydrokaempferol in
Cyanidin
In Parallelansätzen, die aus derselben Mutterkul-tur stammten, wurde der Einbau von je 2 ,5 ^ M o l DL-Phenylalanin- [ 2 - 1 4 C ] , 4 .2 ' • 4 ' .6 ' -Tetrahydroxy-chalkon-2 ' -g lucosid- [ / 5- 1 4 C] 15 (Ch) und Dihydro -k a e m p f e r o l - ( G ) - [ 3 H ] 1 6 ( D K ) in Cyanidin vergli-chen. Die Kulturen waren bei Belichtungsbeginn 5 0 Stdn. alt. Die Zugabe der Vorstufen erfolgte 4 9 Stdn. nach Belichtung. Nach 1, 3 und 6 Stdn. wur-den die Ansätze auf Cyanidin aufgearbeitet und die-ses durch 3-malige Papierchromatographie gerei-nigt. D ie Radioaktivitätsbestimmung im Cyanidin erfolgte nach SCHÖNINGER-Verbrennung5. Die er-haltenen Ergebnisse s ind in Tab . 3 aufgeführt. Im Gegensatz zu Versuchen mit Keimlingen werden be-reits nach 1 Stde. gute Einbauraten erhalten. Nach Verdünnung und Einbaurate ist Dihydrokaempferol
Vorstufe Fütterungsdauer Aufnahme Einbau Spez. Aktivität Verdünnung l>Ci] [Stdn.] [%] [%] [^Ci/^uMol]
Phe 5 1 49 3,3 * 0,016 128 Phe 5 3 86 3,1 * 0,015 136 Ch 5.9 1 18 0,26 0,004 540 Ch 5.9 3 37 0,26 0,004 617 Ch 5.9 6 31 0,31 0,004 541 D K 10 1 32 5,4 (9 ,5)** 0,079 51 D K 10 3 60 6,3 (10,8)** 0,145 27 D K 10 6 55 7,2 (12,5) 0,198 20
Tab. 3. Vergleich der Einbauversuche mit DL-Phenylalanin-[2-14C] (Phe), 4.2'.4'.6'-Tetrahydroxychalkon-2'-glucosid-[/?-14C (Ch) und Dihydrokaempferol- (G) 13H] (DK) in Cyanidin. * Bezogen auf L-Phenylalanin. ** Werte ohne Klammern unkor-rigiert für Tritiumverlust an C-2 und C-3. Werte in Klammern korrigiert unter der Annahme einer Gleichverteilung von Tri-
tium an den nicht austauschbaren Positionen in DK.
15 H. GRISEBACH U. L. PATSCHKE, Z. Naturforsch. 16 b, 645 16 W. BARZ U. H. GRISEBACH, Z. Naturforsch. 21 b, 47 [1966]. [1961].

585 BENZOQUINONE PENETRATION AND PPHOTOREDUCTION
( 1 ) die weitaus beste Vorstufe für Cyanidin. Da der prozentuale Anteil von Trit ium an C-2 und C-3 von 1 unbekannt ist und dieses Tritium be im Ubergang in Cyanidin verloren gehen muß, läßt sich eine ge-naue Einbaurate von 1 nicht angeben. Sie muß aber auf jeden Fall höher liegen als die gemessenen Werte.
Im Gegensatz zu den früheren Ergebnissen mit Keimlingen ist der Unterschied des Einbaus von 1 zu Phenylalanin signifikant besser, so daß 1 als echte Zwischenstufe der Cyanidinbiosynthese ange-sehen werden kann. Mögliche Reaktionswege für die Umwandlung von 1 in Cyanidin sind bereits früher diskutiert worden 1 7 .
17 H. GRISEBACH, Biosynthetic Patterns in Microorganisms and Higher Plants, S. 13, J. Wiley, New York 1967.
Auffa l lend ist der im Vergleich zu früheren Er-gebnissen 18 relativ schlechte Einbau des Chalkons. Dies könnte daran liegen, daß das Chalkonglucosid und nicht das freie Chalkon eingesetzt wurde. Die A u f n a h m e des Chalkons in die Zellen war ebenfalls schlecht. Versuche über die Umwandlung von Di-hydrokaempfero l bzw. Dihydroquercetin in Cyani-din in zellfreien Extrakten aus H. gracilis sind im Gange.
Dr. 0 . L. GAMBORG, Saskatoon, Canada, danken wir für die Überlassung einer Suspensionskultur von H. gracilis, und Prof. J. REINERT, Berlin, für die Disser-tation von I. LACKMANN. Die Arbeit wurde durch die Deutsche Forschungsgemeinschaft unterstützt (SFB 4 6 ) .
18 L. PATSCHKE u. H. GRISEBACH, Z . Naturforsch. 2 0 b , 1039 [1965].
On the Mechanism of Benzoquinone Penetration and Photoreduction by Whole Cells
H . GIMMLER * a n d M . A V R O N
Biochemistry Department, The Weizmann Institute of Science, Rehovot, Israel
(Z. Naturforsch. 26 b, 585—588 [1971] ; received February 3, 1971)
Short time treatment of intact Porphyridium cruentum cells with benzoquinone results in changes of the cell membranes, which lead to a higher permeability. This increased permeability allows the measurements of photosynthetic electron transport reactions with various electron donors, ac. ceptors and mediators, which cannot enter untreated cells. The capacity of benzoquinone to act as a H i l l - reagent in vivo is interpreted as due to a double action of this compound: changing the permeability of the cells by reacting with the cell membrane coupled with the ability of the un-reacted molecules to serve as electron acceptors.
Investigation of the rate of electron transport in intact cells have been hampered b y the lade of per-meant electron acceptors. C 0 2 , the natural electron acceptor cannot be used since its assimilation and the accompanying 0 2 evolution depend on A T P formation. Thus, uncouplers which inhibit A T P -formation, inhibit C0 2 -assimilation, but not neces-sarily electron flow.
The one exception, observed many years ago , was benzoquinone, which was shown to serve as a H i l l - reagent in whole cells 1 - 5 . Most of the other electron acceptors, successfully employed in experi-
Reprints request to M. AVRON, Weizmann Institute of Science, Rehovoth, Israel.
* On leave from the Institute of Botany, University of Würz-burg, Germany.
ments with chloroplasts, failed in inducing oxygen evolution in whole cells. It was therefore concluded that only benzoquinone could enter the inner com-partments of the cells. The reason f o r this facilitated entrance was not known.
Recently, PARK and collaborators 6 - 9 reported a method, by which membranes of whole cells became permeable to certain electron acceptors, donors and mediators . It involved a short time fixation of the b io log ica l material with low concentrations of alde-hydes. Aldehydes act as cross linking agents in b io -logical m a t e r i a l 1 0 - 1 3 and this was believed to lead
Abbreviations: DAD = diaminodurene = 2,3,5,6-tetramethyl-p-phenylenediamine. DPIP = 2,6-dichlorophenolindophe-nol, Diquat (D.Q.) = 2,2-ethylene-2,2-dipyridilium dibro-mide, DCMU = 3,4-dichlorophenyl-l,l-dimethylurea, FCCP = carbonyl cyanide p-trifluoro-phenylhydrazone. PMS = phenazinmethosulfate.
![Blütenöle – Chemische Analyse, Biosynthese und ... · Cucurbitaceen wurden Ketofettsäuren vermutet, deren Strukturen jedoch nicht gesichert bestimmt werden konnten [4].](https://static.fdokument.com/doc/165x107/5ba0cc7a09d3f2497e8be451/bluetenoele-chemische-analyse-biosynthese-und-cucurbitaceen-wurden.jpg)

















