
Die Rolle der antimikrobiellen Peptide und des E-Cadherins ... · Die Rolle der antimikrobiellen...
66
Die Rolle der antimikrobiellen Peptide und des E-Cadherins in der Pathogenese der Nekrotisierenden Enterokolitis Der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg zur Erlangung des Doktorgrades Dr. med. vorgelegt von Agnes Kaiser aus Neustadt a.d. Waldnaab
Transcript of Die Rolle der antimikrobiellen Peptide und des E-Cadherins ... · Die Rolle der antimikrobiellen...

Die Rolle der antimikrobiellen Peptide und des E-Cadherins in der
Pathogenese der Nekrotisierenden Enterokolitis
Der Medizinischen Fakultät
der Friedrich-Alexander-Universität
Erlangen-Nürnberg zur
Erlangung des Doktorgrades Dr. med. vorgelegt von Agnes Kaiser
aus Neustadt a.d. Waldnaab

Als Dissertation genehmigt
von der Medizinischen Fakultät
der Friedrich-Alexander-Universität Erlangen-Nürnberg
Tag der mündlichen Prüfung: 12. Mai 2014
Dekan (Vorsitzender des Promotionsorgans): Prof. Dr. med. Dr. h.c.
Jürgen Schüttler
Gutachter: Prof. Dr. Henrik Köhler
Gutachter: Prof. Dr. Wolfgang Rascher

Inhaltsverzeichnis
1. Zusammenfassung 1
1.1. Hintergrund und Ziele der Arbeit 1
1.2. Methoden (Patienten, Material und Untersuchungsmethoden) 1
1.3. Ergebnisse 1
1.4. Schlussfolgerungen 2
2. Summary 3
2.1. Background and Aims of This Thesis 3
2.2. Methodology (subjects, materials and research methods) 3
2.3. Results 3
2.4. Conclusions 4
3. Einleitung 5
3.1. Das Krankheitsbild der Nekrotisierenden Enterokolitis 5
3.1.1. Diagnostik, Therapie, Prognose 5
3.1.2. Epidemiologie 7
3.2. Entstehungsgrundlagen, Modelle der Pathogenese 7
3.2.1. Gastrointestinale Schutzfunktionen 8
3.2.2. Bakterien und Ernährung, zelluläre Transmitter 10
3.2.3. Zelluläre und immunologische Abwehr 13
3.2.4. Andere Risikofaktoren 16
4. Material und Methoden 18
4.1. Patientencharakteristika 18
4.2. Immunhistochemie 21
4.3. TaqMan-PCR 25
5. Ergebnisse 30
5.1. Immunhistochemische Färbungen 30
5.1.1. E-Cadherin 30
5.1.2. α-Defensin 3/5 31
5.1.3. TFF1 34
5.1.4. TFF2 36
5.1.5. LL-37 38
5.1.6. GKN1 40
5.2. PCR der Schnitte 41
6. Diskussion 42

7. Literaturverzeichnis 50
8. Abkürzungsverzeichnis 56
9. Danksagung 59
10. Erklärung 60
11. Lebenslauf 61

1
1. Zusammenfassung
1.1. Hintergrund und Ziele der Arbeit
Die nekrotisierende Enterokolitis (NEC) ist eine der Erkrankungen, die an
Häufigkeit zunimmt, weil sie vornehmlich die frühgeborenen Kinder betrifft.
Welcher entscheidende Mechanismus zur Erkrankung führt, ist heute immer noch
unklar, weshalb Therapie, Prophylaxe und Nachsorge große Herausforderungen
darstellen. Es sei auf diverse Studien von Salzman, Vieten und Schaart (Salzman et
al. 1998, Schaart et al. 2009, Vieten et al. 2005) hingewiesen, die antimikrobielle
Peptide (AMPs) bei NEC-Patienten im Darm untersucht haben und zu teilweise
unterschiedlichen Ergebnissen gekommen sind. Man vermutet mit einer
Verminderung dieser Peptide Parallelen zu den chronisch entzündlichen
Darmerkrankungen. Da die Probandenkollektive für jede der Studien nicht sehr groß
sind, war ein Ziel der vorliegenden Studie, die AMPs α-Defensin 5, TFF1 und
TFF2, LL-37 und das Peptid GKN1 in Expression und Darstellung im Darm (hier
ebenfalls E-Cadherin) zu untersuchen.
1.2. Methoden (Patienten, Material und Untersuchungsmethoden)
Für die Untersuchung griffen wir auf Resektate aus Operationen am Dünndarm
zurück, die in den Jahren von 1998 bis 2009 an der Universitätskinderklinik
Erlangen gesammelt wurden. Für die immunhistologischen Untersuchungen von E-
Cadherin, α-Defensin 5, TFF1, TFF2, LL-37 und GKN1 kamen sieben NEC-
Patienten zur Auswertung. Die drei Kontrollen stammten aus Operationen bei
Atresie oder Mekoniumileus. Um den Verlauf nach NEC zu untersuchen, wurden
fünf Patienten untersucht, die sich einer Stomarückverlagerungsoperation oder einer
Operation bei Stenose unterziehen mussten. Um die Expression des α-Defensin 5,
TFF1, TFF2, LL-37 und GKN1 zu untersuchen, kamen für die TaqMan-PCR vier
NEC-Patienten-, drei Verlauf-NEC-Patienten- und drei Kontroll-Patienten-Proben
zur Auswertung. Die Ergebnisse der PCR wurden nicht quantitativ ausgewertet.
1.3. Ergebnisse
Wir konnten nur geringe Unterschiede zwischen NEC- und Kontrollgruppe in der
immunhistologischen Färbung und der Expression feststellen. Es lassen sich
tendenziell mehr Panethzellen und damit α-Defensin 5, mehr TFF1 und LL-37 bei
der akuten NEC detektieren. In der Verlaufgruppe finden sich intensivere Färbungen

2
von α-Defensin 5 und Expression von TFF2, die sich in den anderen Gruppen nicht
zeigt. Die Expression von TFF1 zeigt sich in allen Gruppen. Vieten et al. kamen hier
zu anderen Ergebnissen (Vieten et al. 2005).
1.4. Schlussfolgerungen
Es zeigen sich Hinweise bei der NEC, die Produktion der AMPs zu steigern,
eventuell läuft diese Reaktion aber zu langsam und zu schwach ab. Möglicherweise
rühren jedoch die deutlicheren Unterschiede in der Verlaufgruppe nur daher, dass
die Patienten dieser Gruppe älter sind als die der anderen. Der Schluss einer
absoluten Verminderung der AMPs, entweder genetisch oder anderweitig bedingt,
kann für den Fall einer akuten NEC nicht gezogen werden. Leider kann bei diesen
Untersuchungen nicht beurteilt werden, wie sich die Verteilung vor Manifestation
der Erkrankung darstellt. Auch lassen sich kleine Unterschiede mit
immunhistologischer Färbung womöglich gar nicht sicher detektieren. Unser
Patientenkollektiv war von der Altersverteilung in den Gruppen NEC und Kontrolle
vergleichbar. Für niedrigere Gestationsalter konnten in anderen Studien verringerte
Mengen an α-Defensinen (Mallow et al. 1996) und β-Defensinen (Richter et al.
2010) nachgewiesen werden. Unklar bleibt, weshalb einzelne reifgeborene Kinder
erkranken.

3
2. Summary
2.1. Background and Aims of This Thesis
Necrotising enterocolitis (NEC) primarily affects premature infants. The definitive
cause of this condition remains obscure, which is why treatment, preventive care
and follow-up pose great challenges. In several studies Salzman, Vieten and Schaart
(Salzman et al. 1998, Schaart et al. 2009, Vieten et al. 2005) analysed antimicrobial
peptids (AMPs) in the intestines of NEC patients, with differing results. It is
assumed that the decrease of these peptides is connected with NEC and chronic
inflammatory bowel diseases. Since all of these studies were conducted on small
groups of test subjects, this thesis is to analyse the expression and representation of
α-defensin 5, TFF1, TFF2, LL-37 AMPs (only representation of α-defensin 3 and E-
cadherin) and the GKN1 peptides in the bowel in another study.
2.2. Methodology (subjects, materials and research methods)
Resections from small intestine, which were collected in the years between 1998
and 2009 at the university’s children’s hospital in Erlangen, were used for research.
Seven NEC patients were examined for the immunohistochemical analysis of E-
cadherin, α-defensin 5, TFF1, TFF2, LL-37 and GKN1. The three control samples
used came from atresia or meconium ileus surgeries because. In order to analyse
post-NEC development five patients which had to undergo ostomy conversion or
stenosis surgery were examined. In order to analyse the expression of α-defensin 5,
TFF1, TFF2, LL-37 and GKN1, samples from four current NEC patients, three
patients developing NEC and three control patients were examined using TaqMan
PCR. The PCR results were not analysed quantitatively.
2.3. Results
No wide differences could be detected between NEC patients and the control group
using immunohistochemical tinting and expression. In acute cases of NEC more
paneth cells and therefore α-defensin 5, more TFF1 and LL-37 may be detected. The
NEC developing group showed stronger tints of α-defensin 5 and expression of
TFF2, which did not occur in the other groups. TFF1 is expressed in all groups.
Here the findings of Vieten et al. differ (Vieten et al. 2005).

4
2.4. Conclusions
A potential for NEC to raise AMP production could be observed, however, this
reaction may occur too slowly. The differences between the groups may be a result
of the patients in the NEC developing group being older than patients in the other
groups. Acute NEC cannot be linked to an absolute drop of AMP levels, neither
genetically nor otherwise. Unfortunately these analyses cannot show AMP
distribution before the manifestation of NEC. It is also possible that small
differences cannot be measured accurately using immunohistochemical tinting. The
patients in the NEC and control groups were of similar ages, which may be another
reason for the lack of measurable differences and the difference in the NEC
developing group. Other studies have proven decreased amounts of α-defensins
(Mallow et al. 1996) and β-defensins (Richter et al. 2010) for younger gestational
ages. It remains uncertain why individual term infants develop the disease.

5
3. Einleitung
Die nekrotisierende Enterokolitis ist eine der Erkrankungen, die nicht trotz, sondern
vielmehr gerade wegen der medizinischen Fortschritte an Häufigkeit zunimmt, weil
sie vornehmlich die frühgeborenen Kinder betrifft. Die Pathogenese ist nicht
vollständig geklärt. Es gibt Puzzleteile und verschiedene Modelle. Eine große
Bedeutung wird den antimikrobiellen Peptiden in der Darmschleimhaut
beigemessen. In unserer Arbeit haben wir α-Defensin 5, die trefoil factors TFF1 und
2, das Cathelicidin LL-37, das Gastrokin GKN1 und das E-Cadherin bei NEC-
Patienten und Kontrollen sowohl färberisch zur Darstellung gebracht als auch ihre
Expression (mit Ausnahme von E-Cadherin) mittels PCR gemessen. Die These war,
dass diese Peptide in der Darmschleimhaut bei akuter NEC verringert sind.
Untersuchungsmaterial waren für diese Peptide Dünndarmschnitte.
3.1. Das Krankheitsbild der Nekrotisierenden Enterokolitis
3.1.1. Diagnostik, Therapie, Prognose
Die NEC ist eine Darmerkrankung, die nur Säuglinge und von diesen besonders
frühgeborene Neugeborene betrifft. Sie trat früher wie heute auf, ja sie ist eine der
Krankheiten, deren Inzidenz zunimmt. Gründe dafür mögen einerseits die bessere
Versorgung frühgeborener Kinder sein, die es heute immer häufiger zu Wege bringt,
dass auch kleinste Kinder, die mit 23 Schwangerschaftswochen geboren werden,
überleben (Ramsay and Santella 2011). Andererseits hat die in den letzten 40 Jahren
verstärkte Forschung auf diesem Gebiet bewirkt, dass heute ein erhöhtes
Augenmerk darauf gelegt wird und Pädiater für diese Erkrankung sensibilisiert sind.
Die Gefahr, diese Erkrankung zu entwickeln, besteht nur ein paar Wochen bis etwa
drei Monate nach Geburt. Wobei jedoch zu beachten ist, dass, je jünger das Kind bei
der Geburt ist, desto länger nach der Geburt die Gefahr besteht, eine NEC zu
entwickeln (González-Rivera et al. 2011). Die Krankheit umfasst in ihrer
Präsentation ein breites Spektrum. Das Krankheitsbild reicht von systemischen
Zeichen wie Apnoen, Bradykardie und geblähtem Abdomen ohne weitere
sonographische oder röntgenologische Zeichen (Bell Stadium I) bis zur
Darmperforation und Sepsis (Bell Stadium III) (siehe Abbildung 1). Erste Zeichen
sind meist Apnoen und Bradykardie. Dann treten die abdominellen Symptome hinzu
wie ein geblähter Bauch, blutige Stühle und Magenreste. In diesem Stadium I wird
zunächst nur von einer suspekten NEC gesprochen. Andere neonatale Erkrankungen

6
müssen ausgeschlossen werden, indem Elektrolyte kontrolliert, infektiöse Ursachen
in Betracht gezogen oder Intoxikationen durch die Mutter ausgeschlossen werden.
Im Bell-Stadium II, der gesicherten NEC, kommen fehlende Darmgeräusche und
eine Abwehrspannung des Abdomens hinzu. Radiologisch oder sonographisch lässt
sich die pathognomonische Pneumatosis intestinalis, die intramurale Luft in der
Darmwand, darstellen oder Luft in den Portalvenen. Es treten schwerere
systemische Beschwerden, eine metabolische Azidose und Thrombozytopenie auf.
Wichtigste und erste Maßnahme zur Therapie ist das Absetzen der oralen Nahrung.
Die Nahrungskarenz entlastet den Darm, in dem sich die Nahrung staut. Eine
Magensonde wird gelegt um Magenreste ableiten zu können. Zusätzlich obligat ist
eine breite antibiotische Therapie, meist bestehend aus einem β-Laktam-
Antibiotikum, einem Aminoglykosid (wie Tobramycin oder Gentamicin) oder
Glykopeptid (wie Vancomycin) und Metronidazol. Im Stadium III ist die Operation
dringend indiziert, kann aber schon im Stadium II notwendig sein. Bei der Operation
zeigt sich ein nekrotischer Darm mit Schleimhautödem, Hämorrhagie und
mukosaler Ulzeration. Die Entscheidung dazu muss individuell gefällt werden (Bell
et al. 1978). Bis zu 50% der NEC-Fälle müssen operiert werden (Ade-Ajayi et al.
1996). Meist wird in einer ersten Operation ein Enterostoma angelegt, das in einer
Folge-Operation nach ein bis zwei Monaten wieder revidiert wird. Komplikationen
der Operation sind das Kurzdarmsyndrom nach ausgedehnten Resektionen,
Strikturen mit notwendiger Reoperation, Platzbauch und intraabdominelle Abszesse
(Lin and Stoll 2006). Die schwere Komplikation der Sepsis bei einer NEC wird mit
einer Inzidenz von 20-40% angegeben (Bhatia 2010).
Stadium I: V. a. NEC milde systemische Zeichen: Apnoe, Bradykardie, Temperaturschwankungen milde intestinale Zeichen: aufgetriebenes Abdomen, Magenreste, blutige Stühle radiologisch unauffällig Stadium II: gesicherte NEC milde bis mäßige systemische Zeichen intestinal zusätzlich fehlende Darmgeräusche, abdomineller Druckschmerz spezifische radiologische Zeichen: Pneumatosis intestinalis oder portalvenöse Luft Laborveränderungen: metabolische Azidose, Thrombozytopenie Stadium III: fortgeschrittene NEC schwere systemische Krankheit: Hypotension intestinal zusätzlich auffallend aufgetriebenes Abdomen, Peritonitis schwere radiologische Zeichen: Pneumoperitoneum zusätzliche Laborveränderungen: auch respiratorische Azidose, DIC Abbildung 1: Stadien der nekrotisierenden Enterokolitis nach Bell (Lin and Stoll 2006)

7
3.1.2. Epidemiologie
In der Literatur sind inzwischen etliche prädisponierende Faktoren beschrieben, die
genaue Ätiologie ist jedoch weiterhin unklar. Die schon erwähnte Frühgeburtlichkeit
spielt mit dem Geburtsgewicht die größte Rolle für die Entwicklung einer NEC und
auch für die Prognose in Bezug auf die Mortalität. Die Inzidenz der Erkrankung
liegt bei 0,5 bis 5 pro 1000 Lebendgeburten (Lin and Stoll 2006). Für besonders
leichte Kinder ist die Inzidenz höher: In einer Fall-Kontroll-Studie des NICHD
(National Institute of Child Health and Human Development) erkrankten 11,5% der
Kinder mit einem Geburtsgewicht von 401-750g, 9,1% mit einem Geburtsgewicht
von 751-1000g, 6,0% mit 1001-1250g, und 3,9% mit 1251-1500g. Insgesamt
erkrankten 7,1% der Kinder mit extrem niedrigem Geburtsgewicht (VLBW) (bis
1500g) (Guillet et al. 2006). Der Einfluss des Gestationsalters geht aus folgenden
Zahlen hervor: Bei Frühgeborenen ist die NEC bis zu zehnmal häufiger, die
Inzidenz liegt hier bei 2 bis 5% (Hunter et al. 2008). Oder anders ausgedrückt: 90%
der Kinder, die an einer NEC erkranken, sind frühgeboren. Die Letalität der NEC
liegt bei 20 bis 50% (Schnabl et al. 2008). Von Kindern mit extrem niedrigem
Geburtsgewicht, die wegen einer NEC oder einer intestinalen Perforation operiert
wurden, starben in einer Studie 49% (Blakely et al. 2005), in einer Studie von
Guthrie et al. lag die Letalität der NEC auf einer Neugeborenen-Intensivstation
(Kinder bis 34 SSW) bei 12%. Die Letalität war höher bei Säuglingen, die sich einer
Operation unterziehen mussten, bei denen also auch die Krankheit fortgeschrittener
und kritischer war (Guthrie et al. 2003). Ein weiterer interessanter Aspekt ist die
höhere Mortalität von männlichen Neugeborenen und afroamerikanischen Kindern,
wobei letztere auch eine höhere Inzidenz der NEC aufweisen (Llanos et al. 2002).
3.2. Entstehungsgrundlagen, Modelle der Pathogenese
Dass vor allem Frühgeborene und untergewichtige Kinder betroffen sind, muss in
der Natur der Erkrankung liegen. Man vermutet, dass die NEC Ausdruck der
inadäquaten Immunabwehr des noch unreifen Darms ist, seiner unreifen Motilität,
Digestion und Kreislaufregulation, der noch nicht abgeschlossenen Besiedelung mit
der späteren kommensalen Keimflora und der fehlenden Fähigkeit, eine
funktionierende Barriere zwischen äußerem und innerem Milieu aufzubauen und zu
erhalten (Lin and Stoll 2006). Die Barriere besteht aus einer Muzinschicht,
Schleimhaut mit Darmepithel und den darin enthaltenen körpereigenen

8
Abwehrsystemen. Es sind verschiedenste antimikrobielle Peptide bekannt, von
denen man weiß, dass sie bei frühgeborenen Kindern noch nicht in den später
vorhandenen Mengen vorliegen (Mallow et al. 1996, Richter et al. 2010, Salzman et
al. 1998). Die Erkrankung ist jedoch auch vereinzelt bei reifen Kindern zu finden,
weshalb man eine genetische Ursache vermutet. Ein Ansatz ist hier die
Genkopienzahl von β-Defensin 2. Entsprechende Untersuchungen haben bei Morbus
Crohn Unterschiede in der Genkopienzahl gezeigt. Auf der anderen Seite wird auch
von überschießenden Abwehrmechanismen gesprochen, die durch eine überstarke
Aktivierung von Entzündungsmediatoren erst die intakte Barriere zerstören, auf
einen Reiz hin gibt also der Darm selbst seine Schutzfunktion auf. Welche dieser
Einflussgrößen der auslösende Faktor ist, ist noch nicht klar (Lin and Stoll 2006),
möglicherweise sind es bei verschiedenen Kindern unterschiedliche. Die These der
Beteiligung von Bakterien an der Entstehung stützt sich auf folgende Beobachtung:
Man hat gesehen, dass die NEC niemals intrauterin auftritt und sehr selten ist bei
Kindern, die nie enteral ernährt wurden (Berseth 2005). Weitere Risikofaktoren sind
einige angeborene Herzfehler oder ein Atemnotsyndrom (Lin and Stoll 2006).
3.2.1. Gastrointestinale Schutzfunktionen
In erster Reihe für die Abwehr pathogener Keime steht der Magen mit seiner
Produktion von Magensäure. Neugeborene weisen noch eine niedrigere
Säuresekretion auf als Erwachsene (Hoy et al. 2000). Der intrauterine
Gastrointestinaltrakt weist zunächst noch keine geregelte Peristaltik auf. Erst mit 34
SSW werden wandernde Kontraktionswellen beobachtet (Berseth 1989). Das
bedeutet, dass vorher durch ungeregelte Kontraktionen die Weiterbeförderung des
Darminhaltes verzögert ist, woraus eine verlängerte Kontaktzeit möglicherweise
pathogener Bakterien mit der Schleimhaut resultiert. Das begünstigt im unreifen
Darm eine Schädigung. Bifidobakterien und Laktobazillen können auf die Motilität
fördernd wirken (Husebye et al. 2001).
Im Kontakt mit der Schleimhaut hat der reife Darm weitere Möglichkeiten der
Abwehr. Er produziert eine Schleimschicht, die eine mechanische Barriere zum
Epithel bildet und somit Schutz vor eindringenden Bakterien und enzymatischem
Eigenverdau ist. Diese Schleimschicht ist im Dünndarm dünner – auf den
Peyer’schen Plaques fehlt sie – und nimmt im Dickdarm zu. Im Rektum nimmt sie
Dicken von 285 µm an. Sie besteht vor allem aus dem Glykoprotein Muzin, das der

9
Schicht ihre Gelkonsistenz und dem Darm die wichtige Gleitfähigkeit verleiht. Auch
das bakterizide Lysozym ist Bestandteil des Schleims. Sezerniert werden die
Muzine von darmständigen Goblet-Zellen. Diese befinden sich in den Zotten und
Krypten des Dünn- und Dickdarms und produzieren weiterhin den zur Abwehr
wichtigen trefoil factor. Die Sekretion von Muzinen geschieht konstitutiv und kann
u. a. durch inflammatorische Zytokine wie TNFα, IL1, IL6, Neuropeptide und
Hormone gesteigert werden. T-Lymphozyten und lokale Entzündung führten im
Versuch zu einer Veränderung in der Zusammensetzung der Muzine und in der Zahl
der Goblet-Zellen. Bakterien wie Staphylococcus sp., Pseudomonas aeruginosa und
Laktobazillus wirken auf die Schleimproduktion fördernd. Die Muzine sind also die
erste Barriere für Bakterien im Darm und bestimmen die kommensale Keimflora,
die Bakterien wiederum beeinflussen die Expression und Sekretion der Muzine
(Deplancke and Gaskins 2001). Bei frühgeborenen Kindern dürften noch viele
Veränderungen in der Muzinexpression stattfinden (Buisine et al. 1998), auch haben
die Goblet-Zellen noch nicht ihre spätere Reife erlangt (Lin and Stoll 2006). Ein
unterstützender Mechanismus ist die Chlorid-Wassersekretion aus Enterozyten als
Waschfunktion. Der Prozess, der zu dieser Waschfunktion befähigt, ist
kontinuierlich und bei Frühgeborenen noch nicht abgeschlossen (Lebenthal and
Lebenthal 1999).
Ein Protein zur Abwehr, das in dieser Arbeit untersucht wird, ist der schon erwähnte
trefoil factor. Es sind heute drei verschiedene Peptide bekannt, TFF1-3. TFF1, auch
pS2 genannt, und TFF2, oder spasmolytisches Peptid, kommen vor allem im Magen
vor, während der intestinale TFF, oder TFF3, der dominierende TFF der intestinalen
Goblet-Zellen ist (Soriano-Izquierdo et al. 2004). Alle drei werden sie aber in den
muzinbildenden Schleimhäuten wie Gastrointestinaltrakt, den Konjunktiven, dem
Respirationstrakt oder den Speicheldrüsen gebildet. Ihnen kommen wichtige
Aufgaben im Epithelschutz, in der Epithelregeneration nach einer Schädigung durch
die Migration von epithelialen Zellen und in der Verhinderung von Apoptose zu. In
frühen Stadien nach einer Schleimhautverletzung ist ihre Expression erhöht. Ein
Fehlen dieser Produktionssteigerung bei Früh- oder Neugeborenen könnte ein
Faktor in der Pathogenese einer NEC sein. Vieten et al. konnten in einer
Untersuchung ähnlich der unseren im Darm bei akuter NEC keine Expression von
TFF1 und 2, von TFF3 eine verminderte nachweisen (Vieten et al. 2005). Schaart et
al. fanden während der akuten NEC bei epithelialen Schäden in der Zahl verringerte

10
Goblet- und Paneth-Zellen, die zu einem späteren Zeitpunkt nach überstandener
Krankheit wieder normal erhöht war (Schaart et al. 2009). Die fehlende sofortige
Reaktion auf eine Epithelschädigung könnte mitverantwortlich sein für die
Entstehung einer NEC und die Schwere des Krankheitsbildes.
Für den Zusammenhalt der Epithelzellen und die Dichtigkeit der Epithelschicht
zuständig sind die Zonula occludens, bestehend aus tight junctions, und die
Adhärensverbindungen (Zonula adhaerens, Fascia adhaerens, Punctum adhaerens)
und Desmosomen (Macula adhaerens), letztere beide eher für die mechanische
Stärke des Zellverbundes und die Gewährleistung der Kontraktilität des Darms.
Tight junctions bestehen aus den Membranproteinen Occludin und Claudin, welche
vor allem eine Barriere für kleinste Ionen darstellen, aber auch mit dem Aktin-
Zytoskelett in Kontakt stehen. Adhärensverbindungen verbinden vor allem das
Zytoskelett über die Epithelzellen hinweg miteinander, Desmosomen das
Intermediärfilament Keratin, wodurch sie mehr mechanische Funktion haben; dazu
verwenden sie Transmembranproteine. Die wichtigsten sind verschiedene
Cadherine, weswegen wir in unserer Arbeit das E-Cadherin untersuchten. Cadherine
sind Typ1 transmembrane Glykoproteine, die Ca²+-abhängig aneinander binden.
Die Ektodomäne bindet an die Ektodomäne eines Cadherins einer benachbarten
Zelle, die zytoplasmatische Domäne bindet an β-Catenin und darüber indirekt an α-
Catenin, das mit seiner Aktin-bindenden Domäne an Aktin koppelt. Ein anderer
intrazellulärer Partner des Cadherins ist das p120-Catenin, das eine wichtige Rolle
in der Regulation der Stärke der Adhäsion zu spielen scheint und die Endozytose
von Cadherinen verhindert. Es bindet nicht direkt an Aktin, sondern steuert seine
Bewegungen (Niessen and Gottardi 2008). Einen Einfluss auf diese Strukturen
haben verschiedene Mediatoren wie Wachstumsfaktoren und Myosin (Franke 2009,
Niessen and Gottardi 2008). Die chronisch entzündlichen Darmerkrankungen
weisen im entzündeten Darmabschnitt eine Affektion von Adhärensverbindungen
auf. Der betroffene Darmabschnitt zeigt eine verringerte Expression u. a. von E-
Cadherin, die bis zur immunhistochemisch fehlenden Nachweisbarkeit geht (Gassler
et al. 2001).
3.2.2. Bakterien und Ernährung, zelluläre Transmitter
Bei vielen NEC-Fällen werden pathogene Keime gefunden, jedoch gibt es kein
„NEC-Bakterium“ (Peter et al. 1999). Häufigste Keime sind Enterobacteriaceae sp.

11
(E. coli, Salmonellen, Klebsiellen, Enterobacter sp.), es finden sich auch Clostridia
sp., Staphylococcus sp., Viren und Pilze (Bhatia 2010). Man vermutet, dass auch nur
eine Imbalance in der Besiedelung des Darms zu einer NEC führen kann.
Frühgeborene weisen eine andere Keimbesiedelung auf als Reifgeborene. Das mag
dem Umstand geschuldet sein, dass Kaiserschnittgeburten häufiger sind, auf
Intensivstationen bestimmte nosokomiale Keime durch Personal verteilt werden,
häufig Antibiotika gegeben werden und seltener gestillt wird (Hoy et al. 2000). Die
Art der Ernährung hat auf das Gleichgewicht der Darmflora einen großen Einfluss.
Im Mutterleib ist der kindliche Darm noch völlig keimfrei, nach der Geburt wird er
sofort von Bakterien besiedelt. Es ist wichtig, dass sich ein bestimmtes
Gleichgewicht einstellt und dass sich die richtigen Keime ansiedeln. Die richtige
Nahrungsstrategie ist bei Frühgeborenen noch immer umstritten. Zwar zeigen
Kinder, die nie enteral ernährt wurden, sehr selten das Bild einer NEC, jedoch ist
das Risiko dafür bei Beginn der oralen Ernährung erhöht. Fehlende enterale
Nahrung hat die Atrophie des Darms gefördert und nicht seine Reifung. Wenn jetzt
pathogene Keime, auf Intensivstationen vermehrt, in dieses Milieu eindringen, ist
die Entstehung einer NEC wahrscheinlicher. Auf der einen Seite steht also die
Befürchtung, mit oraler Ernährung die Entstehung der NEC zu fördern, auf der
anderen Seite trägt die Nahrung zur Reifung des Darmes bei (Hay 2008). Studien
zeigen eine 3-10fach erhöhte Inzidenz der NEC bei Formula-ernährten Kindern
(Lucas and Cole 1990), wohingegen unter den Frühgeborenen gestillte Kinder
weniger NEC entwickelten (Schanler et al. 1999). Das Stillen gilt als wichtiger
Protektionsfaktor. In der Muttermilch sind sogenannte Prebiotika, nicht verdauliche
Oligosaccharide, enthalten, die die Vermehrung von Bifidobakterien fördern.
Außerdem wirken enthaltene Immunglobuline, aktive Leukozyten, Zytokine,
Laktoferrin und Glykoproteine protektiv (Hunter et al. 2008). In Formulanahrung
wird versucht die Eigenschaften der Muttermilch nachzuahmen, indem sie mit
Prebiotika wie Frukto- und Galakto-Oligosacchariden und Inulin angereichert wird.
Probiotika, das sind Bakterien wie Laktobazillen und Bifidobakterien (z. B.
Infloran®), vermindern das Risiko der Entstehung einer NEC. In einem Cochrane-
Review über 16 randomisierte und quasi-randomisierte kontrollierte Studien traten
signifikant weniger NEC-Fälle bei Frühgeborenen, v. a. bei Kindern < 1500g
Geburtsgewicht auf und es zeigte sich bei den Kindern eine geringere Mortalität. In
Bezug auf die Entstehung einer Sepsis zeigte sich kein Unterschied. In keiner der

12
ausgewerteten Studien fand sich eine systemische Infektion mit dem verabreichten
Bakterium. Die Empfehlung zur Gabe von Probiotika gilt für Kinder > 1000 g
Geburtsgewicht (Alfaleh et al. 2011). Die kommensale Keimflora hat u. a. über
folgende Mechanismen große Bedeutung: Über sogenannte MAMPs (microbial-
associated molecular patterns) signalisieren kommensale Keime wie das
Bifidobakterium den Darmzellen bestimmte Zellvorgänge. MAMPs binden als
Ligand an PRRs (pattern recognition receptors) wie etwa toll-like-Rezeptoren
(TLRs) des Epithels und aktivieren den NF-κB-Signalweg, indem sie den
Transkriptionsfaktor NF-κB (nuclear factor κB) von seinem Inhibitor IκB lösen. NF-
κB tritt in den Zellkern ein und induziert die Transkription von Genen, die wichtig
sind für die Nahrungsaufnahme, die Barrierefunktion, die Angiogenese und die
Reifung (Hooper et al. 2001), und die Transkription proinflammatorischer Zytokine
wie TNFα, IL1β, IL6 und IL8 (Cantó et al. 2009). Ein weiterer Signalweg der TLRs,
der auch in der Transkription dieser Gene mündet, geht über MAPK (mitogen-
activated protein kinase). Eine alternative Aktivierung dieser Kaskaden läuft über
NOD2 (nucleotide-binding oligomerization domain), einen intrazellulären PRR für
Muramyldipeptid (MDP) (Kumar et al. 2009). Dadurch werden die Reaktionen, die
über TLRs getriggert werden, noch verstärkt. Die Vorgänge in der Zelle gibt
folgende Abbildung wieder:
Abbildung 2: Steuerung der antimikrobiellen Peptide (Cantó et al. 2009, Hooper et al. 2001, Kumar et al. 2009)
TAK1
MAPK
NOD2
TLR
IκB NF-κB
Bakterium
MDP
MAMP
inflammatorische Zytokine antimikrobielle Peptide

13
Eine Vorstellung ist, dass Bifidobakterien beispielsweise durch die Stimulation zur
Produktion eines nicht funktionierenden Rezeptors auf diese Weise ganz direkt die
Aufnahme von pathogenen Keimen verhindern (Hunter et al. 2008). Außerdem wird
den Kommensalen eine Rolle in der Inhibition einer überschießenden
Entzündungsantwort und der Apoptose zugeschrieben. Muramyldipeptide,
Bestandteile von gram-positiven und gram-negativen Bakterien, regulieren über
NOD2-Aktivierung die Produktion von proinflammatorischen Zytokinen. Die
Imbalance in der Regulierung der Entzündungsantwort durch die veränderte
Ausschüttung pro- und antiinflammatorischer Substanzen wird als eine Ursache des
Morbus Crohn gesehen, da eine NOD2-Mutation als ein Risikofaktor für die
Entwicklung dieser chronisch-entzündlichen Darmerkrankung gilt (Cantó et al.
2009). NOD2-Mutationen zeigten auch eine verminderte Produktion von α-Defensin
5 und 6. Allgemein waren beim aktivierten Morbus Crohn diese Defensine im Ileum
vermindert, bei Morbus Crohn-Patienten mit NOD2-Mutation war dieses Bild
jedoch noch deutlicher zu sehen (Wehkamp et al. 2004). Studien hierzu konnten bei
NEC-Patienten die typischen Mutationen nicht nachweisen (Zouali et al. 2005).
3.2.3. Zelluläre und immunologische Abwehr
Eine Rolle in der Abwehr spielen Transmitter wie NO und andere
Entzündungsmediatoren wie TNFα, IL1β, IL6, IL8 und IFNγ. Neonatale und unreife
Zellen weisen ein anderes Reaktionsmuster auf (Claud et al. 2004, Shu et al. 1994).
Eine weitere Möglichkeit im angeborenen Immunsystem ist die Abwehr von
Bakterien mit Hilfe von antimikrobiellen Peptiden, die lokal im Darm von Paneth-
Zellen in den Lieberkühn-Krypten des Dünndarms und von Enterozyten gebildet
werden. Das erste entdeckte AMP war das von Alexander Fleming identifizierte
Lysozym, ein antimikrobielles Enzym, das in den Paneth-Zellen produziert wird
(Keshav 2006). Später entdeckte Peptide sind die α-Defensine, β-Defensine und
Cathelicidine (LL-37/hCAP-18). Sie sind eigentliches Untersuchungsziel dieser
Arbeit. Diese zum angeborenen Immunsystem gehörenden Abwehrmechanismen
sind wie die anderen Barrierefaktoren wichtig für die erste Abwehr von jeglichen
Keimen, die Trennung von äußerem und innerem Milieu. Defensine sind kationische
cysteinreiche Peptide mit drei Disulfid-Brücken, die sowohl von Vertebraten,
Invertebraten wie Insekten als auch von Pflanzen gebildet werden. Ihr
Wirkspektrum reicht von Bakterien über Viren bis hin zu Pilzen. Beim Menschen

14
finden sich α- und β-Defensine, die θ-Defensine kommen bei Menschen und
Mäusen nicht vor. Die α-Defensine 1-4 (HNP-1-4) werden in Neutrophilen gebildet,
5 und 6 (HD-5 und -6) sind Produkte der Paneth-Zellen des Dünndarms. Sie werden
konstitutiv exprimiert, gespeichert als Propeptide und durch Trypsin (bei Mäusen
durch MMP7) bei Sekretion aktiviert. Sie sind für die Erhaltung der Homöostase der
Keimflora des Dünndarms verantwortlich (Salzman 2010). Dadurch wirken sie
indirekt auch an der Reifung des erworbenen Immunsystems mit und in der
Modulation von dessen Immunantworten. Sie induzieren Zytokine, der direkte
Wirkmechanismus ist die Abwehr der Pathogene durch Porenbildung in deren
Zellwänden und Membranen (Wiesner and Vilcinskas 2010).
Im Dickdarm kommen die α-Defensine nur in metaplastischen Paneth-Zellen bei
Colitis ulcerosa vor (Hase et al. 2002). Die α-Defensin-Sekretion der Paneth-Zellen
ist reguliert über Acetylcholin, T-Zell-Zytokine und über NOD2 und damit ebenso
wie die Produktion inflammatorischer Zytokine durch bakterielle Stimuli (Keshav
2006). Frühgeborene zeigen eine verminderte Expression der α-Defensine 5 und 6
und haben auch weniger Peptide. In NEC-erkranktem Darm konnten Salzman et al.
eine verstärkte Expression und vermehrt Paneth-Zellen, aber keine adäquat dazu
vermehrten intrazellulären Peptide finden (Salzman et al. 1998). Dies könnte im
Sinne einer Translationsdysfunktion auftreten (Lin and Stoll 2006). Schaart et al.
hingegen fanden bei akuter NEC bei epithelialen Schäden die Paneth-Zellen
vermindert (Schaart et al. 2009). In Versuchen an Mäusen, die HD-5 exprimierten,
zeigte sich eine erhöhte Resistenz gegen Salmonellen-Infektion. Es fand sich auch
eine reduzierte Zahl an Th17-Zellen in der Lamina propria, sozusagen als Antwort
auf bestimmte fehlende Keime. Ein Beispiel dafür, dass Defensine Einfluss auf das
erworbene Immunsystem haben (Salzman 2010).
Die β-Defensine sind breit verteilt über die Schleimhäute. Es sind inzwischen die β-
Defensine 1-4 (hBD-1-4) bekannt. Sie sind chemotaktisch für dendritische Zellen
und T-Zellen (Wiesner and Vilcinskas 2010). hBD-1 wird vornehmlich konstitutiv
exprimiert und ist zu einem kleinen Teil auch induzierbar. hBD-2-4 sind in den
meisten Geweben induzierbar auf entzündliche und pathogene Stimuli hin. Dabei
beschränkt sich die Expression von hBD-2 im Darm auf das Colon (Doss et al.
2010). In anderen Quellen wird es als ubiquitär exprimierbar beschrieben (Hase et
al. 2002). Es wurde in unseren Dünndarmpräparaten nicht bestimmt. Dass
frühgeborene Kinder noch weniger β-Defensin produzieren, konnten Richter et al.

15
zeigen. Sie stellten mit Zunahme des Gestationsalters im Stuhl vermehrt β-Defensin
2 fest (Richter et al. 2010).
Das Gen CAMP codiert u. a. das AMP Cathelicidin. Es wird in der Haut vermehrt
bei der Wundheilung gefunden und ist induzierbar. Nach Translation wird
Cathelicidin als Präpropeptid (hCAP-18) gespeichert. Nach Abspaltung von
Cathelin entsteht das aktive antimikrobielle Peptid LL-37. Diese Abspaltung findet
außerhalb der Zelle statt, bei neutrophilen Granulozyten durch die Serinprotease
Proteinase 3 und bei epithelialen Zellen an der Hautoberfläche durch Kallikrein-
Isoenzyme, die LL-37 in die zwei kleineren Peptide RK-31 und KS-30 spalten
(Wiesner and Vilcinskas 2010) (siehe Abbildung 3).
Abbildung 3: LL-37 (Wiesner and Vilcinskas 2010)
Es wird sowohl im Dünn- als auch im Dickdarm exprimiert, im Dünndarm
allerdings deutlich weniger (Hase et al. 2002). Ein weiterer Produktionsort sind
Makrophagen. Eine wichtige Rolle in der Regulation der Expression spielt Vitamin
D. Wie die β-Defensine wirkt LL-37 chemotaktisch für Neutrophile, Monozyten und
T-Zellen. In Monozyten senkt es die Produktion proinflammatorischer Zytokine auf
bakterielle Stimuli. Bei Morbus Crohn im Colon zeigten LL-37 wie hBD-2
verringerte Antworten. Anders bei der Colitis ulcerosa: Hier zeigt sich hBD-2 und
LL-37 im entzündlichen Darm erhöht (Doss et al. 2010). Daher wird die Defizienz
von α-Defensinen im Dünndarm und β-Defensinen im Dickdarm, in der Parallele
Exon 1-3: Signalpeptid, Cathelin-Domäne Exon 4: antimikro- bielle Aktivität
CAMP-Gen
Präpropeptid: Speicherung in zellulären Granula
Propeptid: Freisetzung
extrazelluläre Proteinasen: Cathelin und C-terminales AMP
Cathelin C-terminales antimikrobielles Peptid (LL-37)

16
zum Morbus Crohn, verdächtigt, ursächlich an der Entstehung der NEC mitbeteiligt
zu sein, resultierend aus einer fehlenden Steuerung durch die Zusammensetzung der
Keimflora. Dass die kommensalen Keime wichtig sind für die Aufrechterhaltung der
Schranken, beispielsweise in der Stimulation zur Muzinproduktion, wurde oben
gezeigt (Salzman 2010). Die Peyer-Plaques, die B- und T-Zellen enthalten, sind in
ihrer Zahl und Größe beim Neugeborenen reduziert. Ihre komplette Reifung findet
erst im Laufe der Zeit durch den Kontakt mit Keimen statt (Hunter et al. 2008).
3.2.4. Andere Risikofaktoren
Im Tiermodell der NEC wird die Erkrankung ausgelöst durch
Hypoxie/Ischämie/Asphyxie und Formula-Nahrung (Sodhi et al. 2008). Als Ursache
für eine solche hypoxische Situation kommt beim Säugling beispielsweise ein
offener Ductus arteriosus Botalli (persistierender Ductus arteriosus PDA) in Frage,
der tatsächlich als Risikofaktor für eine NEC identifiziert werden konnte. Im Sinne
eines Steal-Phänomens „entzieht“ der Ductus der Aorta das Blut des systemischen
Kreislaufs. Die Minderperfusion erhöht das Risiko für Schäden wie die
intraventrikuläre Blutung (IVH), transiente renale Insuffizienz und die NEC
(Hamrick and Hansmann 2010). Ein Atemnotsyndrom und perinatale Asphyxie
vermindern direkt den Sauerstoffpartialdruck und wirken damit hypoxisch auf den
Darm. Vor allem bei reifgeborenen Kindern mögen diese zusätzlichen Faktoren von
Bedeutung sein, da hier das Problem der Unreife des Darms weniger gegeben ist
(Lin and Stoll 2006). O’Donovan et al. fanden bei VLBW- und ELBW-Kindern mit
Behandlung eines PDA retrospektiv eine Inzidenz der NEC (mind. Bell-Stadium II)
von 12-14% (O'Donovan et al. 2003). Andere Studien berichten von einer Inzidenz
der NEC bei mit Indometacin behandeltem PDA von 30-35% bei ELBW-Kindern
(Bell-Stadium I eingeschlossen) (Fujii et al. 2002, Grosfeld et al. 1996). Obwohl der
PDA-Verschluss vor der Entwicklung einer NEC schützt, wird Indometacin selbst in
einigen Studien – im Gegensatz zu der von O’Donovan et al. – als eigenständiger
Risikofaktor für spontane intestinale Perforationen beschrieben, möglicherweise
durch die folgende Hyperämie und das Anschwellen des Darms. Besonders die
frühe (innerhalb der ersten 48 Stunden) präventive Behandlung frühgeborener
Kinder – primär zur Verhütung einer IVH, dann bei PDA appliziert – vermindert
zwar die Rate an operationsbedürftigen PDAs, erhöht andererseits aber die Inzidenz

17
intestinaler Perforationen bei NEC im Gegensatz zu späterer Medikamenten-Gabe
bei Indikation. Dieses Phänomen verstärken Steroide (Fujii et al. 2002).

18
4. Material und Methoden
Die Proben entstammen Kollektiven, die zwischen den Jahren 1998 und 2009 an der
Kinder- und Jugendklinik der Universtitätsklinik Erlangen gesammelt wurden. In
dieser Zeit wurde Material in Operationen von an NEC erkrankten Kindern
gewonnen, bei denen Darmresektionen notwendig waren. Die Resektate wurden in
Paraffin eingebettet und somit haltbar gemacht. Ebenso wurde mit Resektaten
verfahren, die nach Abheilung der Krankheit in Stomarückverlagerungsoperationen
oder Stenoseoperationen gewonnen wurden. Diese stellten das Kollektiv der
Verlaufskontrolle nach NEC. Als Kontrolle dienten Resektate aus Operationen bei
Mekoniumileus oder Darmatresie. Nach Sichtung der Patientenakten blieben mit
Erfüllung der Einschlusskriterien (jeweils erste NEC-Operation und klinische
Zeichen der NEC in der NEC-Gruppe, unversehrter Darm bei den Kontrollen,
jeweils betroffener Darmabschnitt Dünndarm) sieben NEC-Patienten, fünf
Verlaufskontroll-Patienten und drei Kontroll-Patienten. Zur Darstellung der
verschiedenen antimikrobiellen Peptide in der Darmschleimhaut wurden
immunhistochemische Schnitte angefertigt. Zu beurteilen waren sowohl Verteilung
als auch Menge der Proteine. Zur weiteren Genauigkeit wurde die Expression der
histologisch gesehenen Proteine mittels TaqMan-RT-PCR untersucht. Hier ergab
sich durch die Art der Konservierung der Proben (Paraffinmaterial) und die
jahrelange Lagerung das Problem, dass in einigen Resektaten keine sauberen
Kurven mehr zu ermitteln waren bzw. keine nennbaren Mengen an
Housekeepinggen-RNA mehr nachzuweisen waren. Daher sind hierbei nur vier
NEC-Patienten, drei Verlauf-NEC-Patienten und die drei Kontroll-Patienten zur
Auswertung gekommen, die deshalb auch nicht quantitativ geschah.
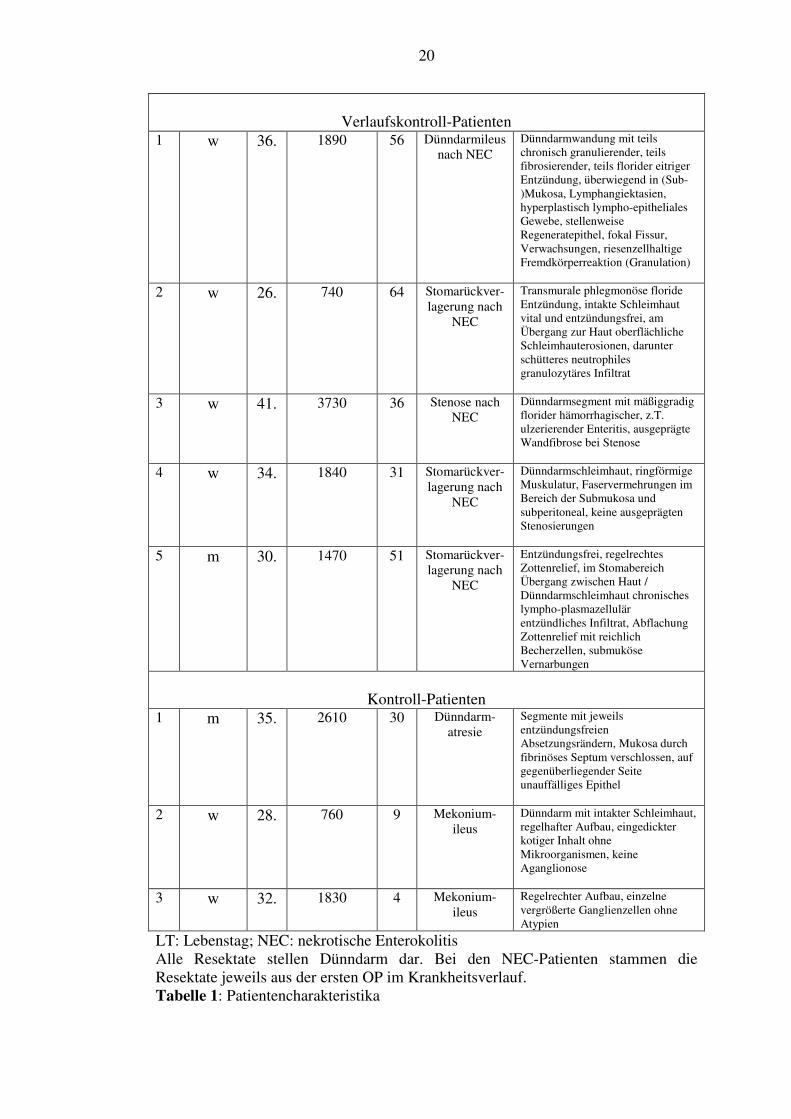
4.1. Patientencharakteristika
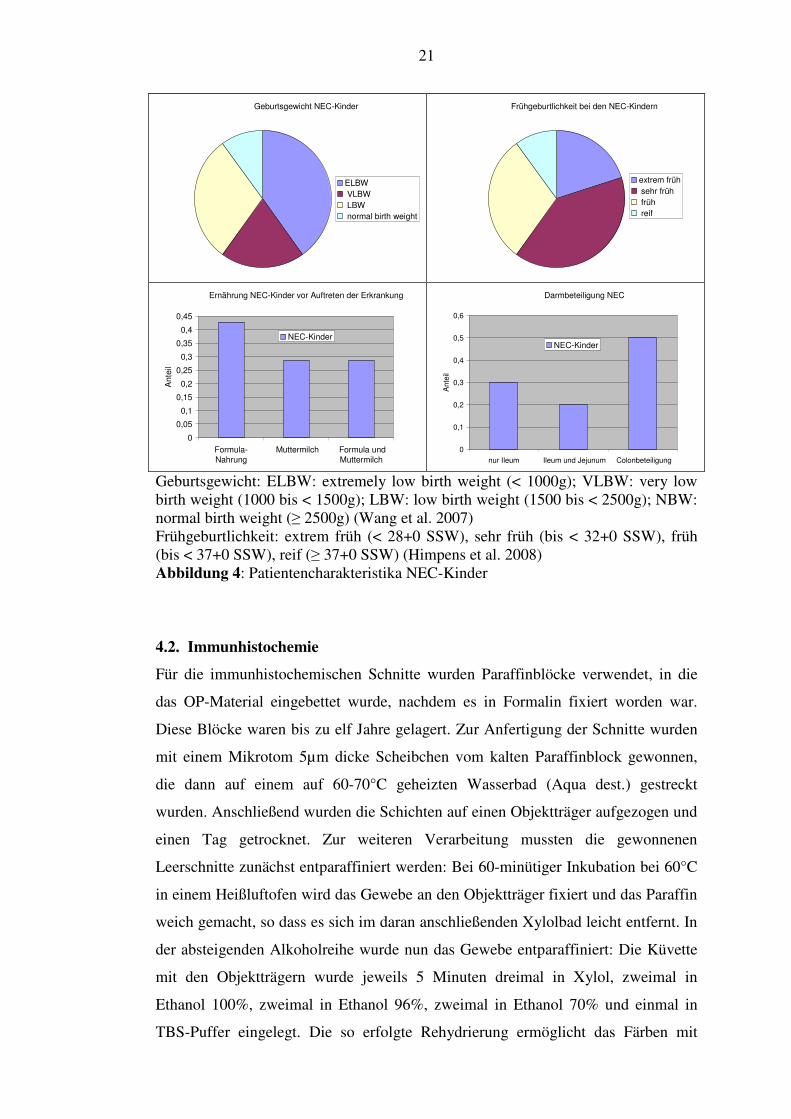
Unsere Patientencharakteristika sind der Tabelle 1 und der Abbildung 4 zu
entnehmen. Es kamen sieben NEC-Patienten zur Auswertung und durchweg nur
Patienten mit mindestens Bell-Stadium II. Mittlerer Wert für das Auftreten der
ersten Symptome war der 5. Lebenstag (4,6. ± 2,2; Bereich von 2. bis 8.), für die
Operation der 10. Lebenstag (10,1. ± 7,2; Bereich von 3. bis 20.). Alle Resektate
sind Dünndarmresektate und stammen aus der ersten Operation im Verlauf der
Krankheit. Zur Verlaufbeurteilung kamen fünf Patienten zur Auswertung, die
Resektate stammen aus der Stomarückverlagerungsoperation oder Operation bei
19
Stenose. Sie fand im Mittel 37 Tage (37,4 ± 11,3; Bereich von 24 bis 52) nach der
ersten NEC-Operation statt. Zwei dieser Patienten kamen schon als NEC-Patienten
zur Auswertung. Die drei Kontrollpatienten wurden wegen eines Mekoniumileus
oder einer Darmatresie im Mittel am 14. Lebenstag (14,3 ± 13,8; Bereich von 4. bis
30.) operiert.
Nr. Geschlecht SSW Geburtsgewicht (g)
Alter bei OP
(LT)
OP Pathologie-Befund
NEC-Patienten
1 w 29. 710 17 NEC Ileum Transmurale und hämorrhagisch durchsetzte Nekrosen mit umschriebener Perforation, dichtere Infiltrate Lymphozyten und Plasmazellen, neutrophile Granulozyten bis an Serosa und Dünndarmabtragung heran
2 w 28. 700 20 NEC Ileum Hämorrhagische Wandnekrosen, Perforation, granulozytäre Infiltrate gesamte Wandung, Epithel mit verstärkter Regeneration, Ulzerationen und Reepithelialisierung, außen subserös Blasenbildung
3 w 41. 3730 4 NEC Dünndarm
-
4 m 30. 730 3 NEC Dünndarm
Weite Strecken und lumenseitig betonte ausgedehnte Nekrosen auf gesamter Darmwandung, Hämorrhagien
5 m 36. 2200 16 NEC Ileum Ulzerationen der Schleimhaut bis Submukosa, stellenweise transmural Einblutung und chronisch-granulierende Entzündung, Ulzerationen bis in Resektionsränder
6 w 34. 1840 7 NEC Ileum Teilweise transmurale Nekrosen, umschriebene Perforation, eitrig-kotige Peritonitis, Oberfläche mit breiten Belägen aus Fibrin und neutrophile Granulozyten
7 w 32. 1210 4 NEC Ileum Ausgeprägte hämorrhagische Infarzierung mit fibrinöser Entzündung, Absetzungsränder entzündungsfrei
20
Verlaufskontroll-Patienten
1 w 36. 1890 56 Dünndarmileus nach NEC
Dünndarmwandung mit teils chronisch granulierender, teils fibrosierender, teils florider eitriger Entzündung, überwiegend in (Sub-)Mukosa, Lymphangiektasien, hyperplastisch lympho-epitheliales Gewebe, stellenweise Regeneratepithel, fokal Fissur, Verwachsungen, riesenzellhaltige Fremdkörperreaktion (Granulation)
2 w 26. 740 64 Stomarückver-lagerung nach
NEC
Transmurale phlegmonöse floride Entzündung, intakte Schleimhaut vital und entzündungsfrei, am Übergang zur Haut oberflächliche Schleimhauterosionen, darunter schütteres neutrophiles granulozytäres Infiltrat
3 w 41. 3730 36 Stenose nach NEC
Dünndarmsegment mit mäßiggradig florider hämorrhagischer, z.T. ulzerierender Enteritis, ausgeprägte Wandfibrose bei Stenose
4 w 34. 1840 31 Stomarückver-lagerung nach
NEC
Dünndarmschleimhaut, ringförmige Muskulatur, Faservermehrungen im Bereich der Submukosa und subperitoneal, keine ausgeprägten Stenosierungen
5 m 30. 1470 51 Stomarückver-lagerung nach
NEC
Entzündungsfrei, regelrechtes Zottenrelief, im Stomabereich Übergang zwischen Haut / Dünndarmschleimhaut chronisches lympho-plasmazellulär entzündliches Infiltrat, Abflachung Zottenrelief mit reichlich Becherzellen, submuköse Vernarbungen
Kontroll-Patienten
1 m 35. 2610 30 Dünndarm-atresie
Segmente mit jeweils entzündungsfreien Absetzungsrändern, Mukosa durch fibrinöses Septum verschlossen, auf gegenüberliegender Seite unauffälliges Epithel
2 w 28. 760 9 Mekonium-ileus
Dünndarm mit intakter Schleimhaut, regelhafter Aufbau, eingedickter kotiger Inhalt ohne Mikroorganismen, keine Aganglionose
3 w 32. 1830 4 Mekonium-ileus
Regelrechter Aufbau, einzelne vergrößerte Ganglienzellen ohne Atypien
LT: Lebenstag; NEC: nekrotische Enterokolitis Alle Resektate stellen Dünndarm dar. Bei den NEC-Patienten stammen die Resektate jeweils aus der ersten OP im Krankheitsverlauf. Tabelle 1: Patientencharakteristika
21
Geburtsgewicht NEC-Kinder
ELBW
VLBW
LBW
normal birth weight
Frühgeburtlichkeit bei den NEC-Kindern
extrem früh
sehr früh
früh
reif
Ernährung NEC-Kinder vor Auftreten der Erkrankung
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
Formula-
Nahrung
Muttermilch Formula und
Muttermilch
Ante
il
NEC-Kinder
Darmbeteiligung NEC
0
0,1
0,2
0,3
0,4
0,5
0,6
nur Ileum Ileum und Jejunum Colonbeteiligung
Ante
il
NEC-Kinder
Geburtsgewicht: ELBW: extremely low birth weight (< 1000g); VLBW: very low birth weight (1000 bis < 1500g); LBW: low birth weight (1500 bis < 2500g); NBW: normal birth weight (≥ 2500g) (Wang et al. 2007) Frühgeburtlichkeit: extrem früh (< 28+0 SSW), sehr früh (bis < 32+0 SSW), früh (bis < 37+0 SSW), reif (≥ 37+0 SSW) (Himpens et al. 2008) Abbildung 4: Patientencharakteristika NEC-Kinder
4.2. Immunhistochemie
Für die immunhistochemischen Schnitte wurden Paraffinblöcke verwendet, in die
das OP-Material eingebettet wurde, nachdem es in Formalin fixiert worden war.
Diese Blöcke waren bis zu elf Jahre gelagert. Zur Anfertigung der Schnitte wurden
mit einem Mikrotom 5µm dicke Scheibchen vom kalten Paraffinblock gewonnen,
die dann auf einem auf 60-70°C geheizten Wasserbad (Aqua dest.) gestreckt
wurden. Anschließend wurden die Schichten auf einen Objektträger aufgezogen und
einen Tag getrocknet. Zur weiteren Verarbeitung mussten die gewonnenen
Leerschnitte zunächst entparaffiniert werden: Bei 60-minütiger Inkubation bei 60°C
in einem Heißluftofen wird das Gewebe an den Objektträger fixiert und das Paraffin
weich gemacht, so dass es sich im daran anschließenden Xylolbad leicht entfernt. In
der absteigenden Alkoholreihe wurde nun das Gewebe entparaffiniert: Die Küvette
mit den Objektträgern wurde jeweils 5 Minuten dreimal in Xylol, zweimal in
Ethanol 100%, zweimal in Ethanol 96%, zweimal in Ethanol 70% und einmal in
TBS-Puffer eingelegt. Die so erfolgte Rehydrierung ermöglicht das Färben mit

22
wässrigen Lösungen. Zuvor muss jedoch eine Antigendemaskierung vorgenommen
werden. Aldehydverkettungen, die durch die Fixierung zustande kommen,
maskieren sonst einige Antigene, so dass sie vom Antikörper nicht erkannt werden
(Flenker 1998). Dazu wurden die Objektträger in der Demaskierungslösung G
(BioLogo BioPrime® Demaskierungslösung G, Konzentrat auf Glykol-Basis) in
einem gewöhnlichen haushaltsgebräuchlichen Steamer gekocht. Der Steamer wurde
mit Aqua dest. gefüllt, die Küvette mit der Demaskierungslösung noch ohne die
Objektträger hineingestellt und 30 Minuten erhitzt. Der Objektträgerhalter mit den
Schnitten wurde direkt aus dem TBS-Puffer in das schon heiße Demaskierungsbad
gebracht. Hier wurden die Schnitte 20 Minuten gekocht. Danach wurden sie in dem
Bad auf Raumtemperatur heruntergekühlt. Nachdem die Objektträger noch zweimal
5 Minuten in einem TBS-Bad gespült worden waren, wurde mit einem Tuch um das
Gewebe herum überschüssige Flüssigkeit abgetupft und das Gewebe mit einem
Fettstift (ImmEdge™ Pen, Vector®) umrandet und wieder in den Puffer überführt.
So wird gewährleistet, dass die folgenden auf das Gewebe zu pipettierenden
Flüssigkeiten auch darauf verbleiben und nicht abfließen. Bevor auf die Schnitte
Antikörper aufgebracht werden können, muss das Gewebe blockiert werden, um
keine unspezifischen Hintergrundfärbungen zu erhalten: Um die endogene
Peroxidase zu zerstören, die sonst wie die später zugeführte Peroxidase reagieren
würde, wurde zunächst Peroxidase-Blocking Solution (Dako® REAL™) auf das
Gewebe gegeben und 5 Minuten in einer Feuchtkammer inkubiert. Dann wurde
überschüssige Flüssigkeit abgeschüttelt und Protein Block (Dako® Serum-free)
aufpipettiert. Auch dieser wurde 5 Minuten inkubiert und dann abgeschüttelt.
Dadurch, dass sich Protein an Kollagen und Bindegewebe bindet, wird auch mit
diesem Schritt die Hintergrundfärbung auf ein Minimum reduziert. Bis jetzt wurde
mit allen Schnitten für alle Färbungen gleich verfahren. Nun wurden die
verschiedenen Primärantikörper verschiedener Spezies und in verschiedenen
Konzentrationen aufgebracht. Für die E-Cadherin-Färbung wurde ein Maus-
Primärantikörper (Abcam Antibody IgG1 Mouse monoclonal HECD-1 to human E-
Cadherin 0,05mg/ml) in Antibodydiluent (Dako® Background Reducing) auf 1:50
verdünnt (1µg/ml). Der TFF1-Antikörper (Sigma-Aldrich Prestige Antibodies® IgG
Rabbit polyklonal to human TFF1 0,4mg/ml) wurde 1:1000 verdünnt aufpipettiert
(0,4µg/ml). Ebenso der TFF2-Antikörper (R&D Systems® Antibody IgG2b Mouse
monoclonal Clone 366508 to human TFF2 0,5mg/ml) in 1:1000-Verdünnung

23
(0,5µg/ml). Für die LL-37-Färbung wurde der Antikörper (Antibodies-online®
Antibody IgG Rabbit polyclonal to human cAMP 1mg/ml) 1:750 verdünnt
(1,3µg/ml). Die α-Defensine wurden in einer Doppelfärbung gemacht, dazu wurde
α-Defensin 5-Antikörper (Abcam® antibody IgG2b Mouse monoclonal 8C8 to
human alpha Defensin NP5 2,35mg/ml) in einer 1:500-Verdünnung verwendet
(4,7µg/ml), und α-Defensin 3-Antikörper (BioLogo® polyklonal Rabbitserum) in
einer 1:1250-Verdünnung. Bei den Doppelfärbungen wurde genauso verfahren wie
mit den Einzelfärbungen, es konnten beide Antikörper gleichzeitig aufpipettiert
werden, da es sich jeweils um einen Maus- und einen Kaninchen-Antikörper
handelte.
Der Primärantikörper wurde jeweils eine Nacht bei 4°C inkubiert. Am nächsten Tag
wurden die Objektträger dreimal jeweils 5 Minuten in TBS-Puffer gespült um
überschüssige Antikörper zu entfernen. Dann konnte das Gewebe mit dem
Zweitantikörper inkubiert werden. Hierfür wurde ein biotinylierter Goat Anti-Mouse
IgG oder Anti-Rabbit IgG (Vector®) 1:500 verdünnt mit Antibodydiluent
verwendet. Bei der α-Defensin-Doppelfärbung konnten beide Zweitantikörper direkt
nacheinander inkubiert werden, da beide durch eine Färbung dargestellt wurden. Der
Zweitantikörper wurde jeweils 30 Minuten auf dem Gewebe in der Feuchtkammer
inkubiert. Während dieser Zeit wurde der Avidin-Biotin-Peroxidase-Komplex
(Vectastain® Elite ABC Kit) in TBS-Puffer angesetzt. Dafür wurden je zwei
Tropfen Reagens A (Avidin DH solution) und Reagens B (biotinylated enzyme) in
5ml TBS gegeben. Dieser Ansatz wurde im Kühlschrank bei 4°C 30 Minuten stehen
gelassen, um die Vernetzung der Avidin-Moleküle mit den Biotin-Peroxidase-
Molekülen zu gewährleisten. Nach der Zweitantikörper-Inkubation wurden die
Schnitte wiederum dreimal jeweils 5 Minuten in TBS-Puffer gespült, um
überschüssige Antikörper zu entfernen. Daraufhin wurden sie erneut in die
Feuchtkammer gegeben und der vorbereitete Vector ABC Kit aufpipettiert und für
30 Minuten bei Raumtemperatur inkubiert. Hier binden sich Avidin-Biotin-Enzym-
Komlexe an das schon vorhandene Biotin, verstärken somit das Signal und bringen
das gebundene Enzym Peroxidase mit. Die ans Avidin gekoppelte Peroxidase wird
später die Chromogenumwandlung katalysieren. Nach einer weiteren 5-minütigen
Spülung in TBS-Puffer wurden unter mikroskopischer Sicht einige Tropfen frisch
angesetzte Peroxidase-Substrate (Vector® DAB Peroxidase Substrate Kit) -
Wasserstoffperoxid und 3,3’-Diaminobenzidin (DAB) - auf den Objektträger

24
gegeben. Unter Beobachtung der Anfärbung wurde je nach Antigenfärbung 5 bis 30
Sekunden inkubiert, um eine möglichst gute Färbung ohne Hintergrundfärbung zu
erreichen. Das Prinzip dieser Färbemethode ist, durch Biotinkopplung des
Zweitantikörpers und Verstärkung mit Hilfe des Avidin-Biotin-Peroxidase-
Komplexes ein Chromogen sichtbar zu machen. Dies geschieht, indem das
Chromogen DAB und das Substrat, in diesem Fall Wasserstoffperoxid, erst ganz
zum Schluss zugegeben werden. Wasserstoffperoxid setzt dann durch seine
Reaktion mit Peroxidase, die nur am Avidin vorhanden ist, Protonen frei, die das
Chromogen braun werden lassen. So kann der Ort des Antigens genau detektiert und
seine Menge abgeschätzt werden. Die Färbung wurde gestoppt, indem die
Objektträger in ein TBS-Puffer-Bad gestellt wurden. Anschließend wurde kurz in
Aqua dest. gewaschen. Es wurde die Gegenfärbung für 3 Sekunden mit Hämalaun
(Merck® Mayers Hämalaunlösung) durchgeführt und anschließend 7 Minuten unter
laufendem Leitungswasser gebläut. Die lila Gegenfärbung macht die Zellkerne und
das Gewebe sichtbar ohne die braune Färbung zu überdecken. Durch das Bläuen
wird der Farbstoff stabilisiert, sodass er sich im daran anschließenden Alkoholbad
nicht wegwäscht. Analog zur Rehydrierung wurden für die Entwässerung alle
Schnitte in eine aufsteigende Alkoholreihe gegeben. Nach dem dritten Xylolbad
wurden die Objektträger direkt in Entellan (Fluka® Analytical, Eukitt®
Schnelleinschlussmittel) eingedeckt und mit einem Deckglas geschützt.
Für die Negativkontrollen wurde statt des Primärantikörpers jeweils ein Isotyp
desselben verwendet, der nicht gegen spezielle Antigene gerichtet ist. Es sind dies
Immunglobulinfraktionen derselben Tierspezies, aber eines nicht immunisierten
Tieres. Es konnte gezeigt werden, dass keine unspezifischen antikörperbedingten
Färbungen auftreten, sondern nur Antikörper-gebundenes Antigen detektiert wird.
Es wurde verwendet:
Dianova® DLN-05791 IgG1 aus Maus NCG01 0,2mg/ml in 1:200-Verdünnung
(1µg/ml),
Dianova® DLN-05809 IgG2b aus Maus NCG2B.01 0,2mg/ml in 1:42,55-
Verdünnung (4,7µg/ml) und 1:400-Verdünnung (0,5µg/ml) und Dianova® PHA-
70833 Negativkontrollserum aus Kaninchen polyklonal.

25
4.3. TaqMan-PCR
Um die Expression der AMPs zu untersuchen, wurde eine Real-Time-PCR
durchgeführt. Mit dieser Methode kann die mRNA eines bestimmten Gens und
damit seine Expression quantitativ bestimmt werden. Wir haben dazu die übliche
TaqMan-PCR verwendet. Um die RNA per PCR messen zu können, muss sie
zunächst isoliert und dann in einer RT-PCR-Reaktion in kopierte DNA (cDNA)
umgeschrieben werden. Zur RNA-Isolation aus unserem paraffingelagerten Material
wurde der RNeasy° von Qiagen verwendet.
Dazu wurden zunächst acht 5µm-Schnitte pro Paraffinblock gewonnen und in ein
2ml Eppendorfgefäß gegeben. Die folgenden Schritte wurden alle bei
Raumtemperatur durchgeführt. Zur Entparaffinierung wurde 1ml Xylol zupipettiert
und die Probe gevortext. Dann wurde sie 2 Minuten bei 14000 U/min zentrifugiert,
um überstehendes Xylol wieder vom Gewebepellet abpipettieren zu können. Ebenso
wurde mit 100%igem Ethanol verfahren. Dann wurden die Pellets an der Luft
getrocknet, so dass keine Alkoholrückstände mehr vorhanden waren. Es wurden
240µl PKD-Pufferlösung und 10µl Proteinase K zugegeben und durch Vortexen mit
den Zellen des Pellets vermischt, bevor die Eppendorfgefäße bei 55°C und dann bei
80°C jeweils 15 Minuten auf einem Heizblock inkubiert wurden. Dadurch wird die
RNA aus den Zellen freigegeben und durch das Formalin entstandene
Querverbindungen gelöst. Um die noch enthaltene genomische DNA (gDNA) aus
der Probe zu entfernen, wurden 500µl des RBC-Puffers zupipettiert und das ganze
Volumen (750µl) auf eine gDNA Filtersäule gegeben. Die Proben wurden in den
Filtersäulen für 30 Sekunden bei 10000 U/min zentrifugiert und die Säule mit der
gDNA verworfen. Das durchgepresste Eluat wurde mit 1200µl Ethanol 100%
versetzt und durch Pipettieren gut vermischt. Jetzt wurde das ganze Volumen
(1950µl), verteilt auf kleinere Mengen von 700µl, nacheinander auf die RNeasy
MinElute-Säule gegeben und 15 Sekunden bei 10000 U/min zentrifugiert. Das Eluat
wurde jeweils verworfen. Nach dreimaliger Durchführung ist alle Flüssigkeit
entfernt und die RNA in der Säule „gefangen“. Der sich anschließende
Waschvorgang wurde mit RPE-Puffer durchgeführt, der zuvor mit Ethanol versetzt
wurde (11ml Ethanol zu 44ml Puffer-Konzentrat). 500µl dieses Puffers wurden auf
die Säule pipettiert, die dann 2 Minuten bei 10000 U/min zentrifugiert wurde. Das
Eluat wurde jeweils wieder verworfen. Noch einmal wurde 5 Minuten bei 14000
U/min mit geöffnetem Deckel zentrifugiert, um alle Flüssigkeit und Alkohol zu

26
entfernen, und das Eluat verworfen. Die Säule wurde in ein neues 1,5ml-
Sammelröhrchen gestellt und direkt auf die Säulenmembran 24µl RNase-freies
Wasser gebracht. Um die RNA aus der Säule zu lösen, wurde noch einmal 1 Minute
bei 14000 U/min zentrifugiert. Anschließend wurde die Säule verworfen. Die 22µl
Eluat wurden gevortext und auf Eis gelegt bis sie bei 260nm photometrisch auf
ihren RNA-Gehalt bzw. ihre RNA-Konzentration gemessen wurden. Bis zur
weiteren Verarbeitung wurden sie bei -80°C gelagert.
Für die cDNA-Synthese mittels RT-PCR wurde nach gängigem Protokoll
vorgegangen. Dazu wurde die RNA aller Proben in eine Konzentration von
0,125µg/µl gebracht. Mit Hilfe der photometrisch gemessenen RNA-
Konzentrationen konnte genau 1µg RNA einer Probe entnommen werden. Mit
DEPC-Aqua dest. wurde das Volumen von 1µg Gesamt-RNA auf ein
Gesamtvolumen von 8µl aufgefüllt. Da im Doppelansatz vorgegangen wurde, hatten
wir jeweils 2µg in 16µl. Zunächst wurden die benötigten Mengen für alle Proben
mit Excel berechnet und alle Ansätze für die einzelnen Schritte vorbereitet. Wir
verwendeten jeweils 2µl 10x DNase I-Puffer und 2µl RNase-freie DNase I
(RQ1/Promega) für den DNase-Verdau. Dieser wurde 15 Minuten bei
Raumtemperatur durchgeführt und mit 2µl 25mM EDTA Stop-Solution gestoppt. Es
wurde 15 Minuten bei 65°C inkubiert. Der Mastermix A enthielt 1,2µl
Randomprimer, 0,8µl Oligo dT-Primer und 8µl DEPC-Aqua dest. Mit ihm wurde 5
Minuten bei 70°C zur Anlagerung der Primer inkubiert und die Probe anschließend
1 Minute auf Eis gegeben. Für die reverse Transkription wurde der Mastermix B
zupipettiert. Er enthielt 10µl 5x MMLV-Puffer, 2,5µl dNTP-Mix, 1µl RNase-
Inhibitor, 2µl MMLV, und 4,5µl DEPC-Aqua dest. Die reverse Transkription wurde
im Thermoblock 60 Minuten bei 37°C durchgeführt, anschließend wurden die
Proben mit der gewonnenen cDNA auf Eis gegeben und bei -20°C bis zum weiteren
Gebrauch gelagert.
Die quantitative Expressionsbestimmung geschah mittels einer TaqMan-PCR
(Applied Biosystems, 7500 Real Time PCR System). Diese ist ein quantitatives
Echtzeit-PCR-Verfahren. Sie ermöglicht die Detektion von spezifisch synthetisierter
DNA mittels Sonden, die zur Synthese einer bestimmten Nukleotidsequenz und
somit eines speziellen Genprodukts (in unserem Fall in cDNA umgeschriebene
mRNA-Produkte der Gene der AMPs) nötig sind und durch Lichtemission
detektierbar sind. Die Sonden, die also an ein spezielles Gen angepasst sein müssen,

27
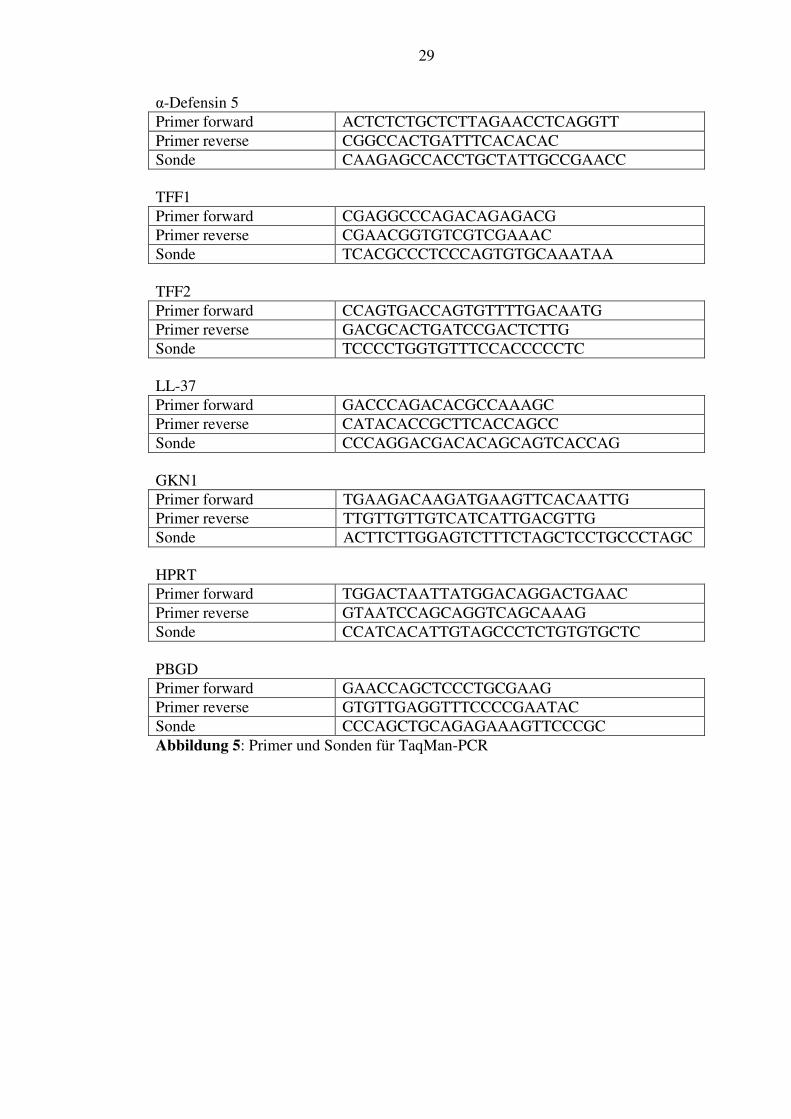
wurden wie die Primer sorgfältig ausgesucht und auf ihre Spezifität mittels
Elekrophoreselauf getestet (siehe Abbildung 5). Um sie und besonders auch ihren
Verbrauch im TaqMan detektieren zu können, sind sie mit zwei verschiedenen
Fluoreszenzfarbstoffen ausgestattet. Am 5’-Ende befindet sich das FAM (6-
Carboxyfluoreszein), am 3’-Ende das TAMRA (Tetramethylrhodamin). FAM ist der
Donorfarbstoff, auch Reporter genannt, TAMRA der Akzeptorfarbstoff oder
Quencher. Durch die Lichtquelle im TaqMan-Gerät wird der Donorfarbstoff
angeregt, der seine Energie aber nicht in Form von Licht abgibt, sondern sie auf den
Akzeptor überträgt (Fluoreszenz-Resonanzenergietransfer bzw. Förster-
Resonanzenergietransfer FRET). Im TaqMan ist dieser Akzeptorfarbstoff nicht
emissiv, er gibt seine Energie nicht in Form von Licht ab, sondern löscht einfach das
Fluoreszenzsignal des Donors (dark quencher). Dies geschieht jedoch nur bei sehr
enger räumlicher Nähe, wie sie bei der intakten Sonde, die nur ein paar Nukleotide
lang ist, gegeben ist. In der TaqMan-PCR macht man sich jetzt die 5’-3’-
Exonuklease-Aktivität der Taq-Polymerase zunutze. Nachdem sie am 3’-Ende eines
Primers angesetzt hat, synthetisiert sie komplementäre DNA und spaltet, sobald sie
auf die ebenfalls gebundene Sonde trifft, diese in ihre einzelnen Nukleotide auf, um
das DNA-Stück bis zum reversen Primer fertigzustellen. Reporter und Quencher
werden so voneinander getrennt und beeinflussen sich gegenseitig nicht mehr. Das
Reportersignal wird nun durch den Quencher nicht mehr unterdrückt und kann als
Fluoreszenz detektiert werden. Die Fluoreszenz steigt direkt proportional mit der
synthetisierten DNA. Gemessen wird die Fluoreszenz am Ende der Elongation eines
jeden PCR-Zyklus. Aus der Menge der Zyklen, die bis zu einer Signalwahrnehmung
nötig sind und damit den Beginn der exponentiellen Phase der DNA-Vermehrung
(Threshold) markieren (Ct-Wert), lässt sich auf die Menge der zu Beginn
vorhandenen cDNA des spezifischen AMPs schließen. Dies geschieht dadurch, dass
der Ct-Wert einer Probe mit denen einer Standardverdünnungsreihe von bekannten
DNA-Konzentrationen, die für jedes Gen mitgeführt wird, verglichen wird. Die
Standardverdünnungsreihe ergibt eine Standardkurve, die jedem Ct-Wert eine
bestimmte Konzentration zuweist. So lässt sich für jede Probe die Konzentration der
Gen-cDNA ablesen. Dabei verhält sich der Ct-Wert indirekt proportional zur DNA-
Menge. Je höher dieser Wert ist, d.h. je mehr Zyklen bis zur Signalwahrnehmung
nötig sind, desto weniger cDNA war zu Beginn in dieser Probe vorhanden. Die
Standardverdünnungsreihe aus 21 verschiedenen Verdünnungen wurde in solchen

28
Schritten gewählt, dass jede Verdünnung noch einmal auf das Doppelte verdünnt
wurde (1:1, 1:2, 1:4, 1:8, …, 1:33554432). Verwendet wurden dafür Kryo-Magen-
und -Dünndarm-Proben und mit Qiagenkit gewonnene Blut-cDNA. Ein Ansatz für
die TaqMan-PCR enthielt 2,5µl 10x Taq-Puffer II, 1,75µl ROX Passive, 1µl dNTP-
Mix (UTP) 5mM, 2µl MgCl2 4mM, 0,13µl AmpliTaq gold (5U/µl), 0,25µl
AmpERaseUNg, 7,37µl H2O, 2,5µl Sonde taq (2µM), 2,5µl Primer forward (3µM),
2,5µl Primer reverse (3µM) und 2,5µl des bei der reversen Transkrition gewonnenen
Templates. Nachdem eine Background-Messung mit der leeren Platte durchgeführt
wurde, wurde 2 Minuten bei 50°C inkubiert, dann zur Denaturierung 10 Minuten bei
95°C. Dann wurden 40 Zyklen durchgeführt: 15 Sekunden 94°C zur Denaturierung,
60 Sekunden 64°C zur Strangsynthese. An jedem Zyklusende nach dem Annealing
wurde die Fluoreszenz gemessen. Um die Güte der Proben zu prüfen, wurden
Housekeeping-Gene für jede gemessen. Housekeeping-Gene sind konstitutiv
exprimierte, nicht regulierte Gene, die gewebsunspezifisch sind. So konnten die
gefundenen AMP-Gen-Mengen immer auf die Housekeeping-Gen-Mengen dieser
Probe bezogen werden. Um auch hier ausschließen zu können, dass für dieses
Housekeeping-Gen individuelle Expressionsmuster vorliegen, wurden zwei
verschiedene Housekeeping-Gene (Hypoxanthinphosphoribosyltransferase HPRT
und Porphobilinogendeaminase PBGD) gemessen und die AMP-Gene auch auf
beide bezogen. Es ergab sich jedoch innerindividuell kein Unterschied in der
Expression. Ausgewertet wurde mit dem Programm 7500 System Software,
Grafiken wurden mit Graph Pad Prism 4 erstellt.
29
α-Defensin 5 Primer forward ACTCTCTGCTCTTAGAACCTCAGGTT Primer reverse CGGCCACTGATTTCACACAC Sonde CAAGAGCCACCTGCTATTGCCGAACC TFF1 Primer forward CGAGGCCCAGACAGAGACG Primer reverse CGAACGGTGTCGTCGAAAC Sonde TCACGCCCTCCCAGTGTGCAAATAA TFF2 Primer forward CCAGTGACCAGTGTTTTGACAATG Primer reverse GACGCACTGATCCGACTCTTG Sonde TCCCCTGGTGTTTCCACCCCCTC LL-37 Primer forward GACCCAGACACGCCAAAGC Primer reverse CATACACCGCTTCACCAGCC Sonde CCCAGGACGACACAGCAGTCACCAG GKN1 Primer forward TGAAGACAAGATGAAGTTCACAATTG Primer reverse TTGTTGTTGTCATCATTGACGTTG Sonde ACTTCTTGGAGTCTTTCTAGCTCCTGCCCTAGC HPRT Primer forward TGGACTAATTATGGACAGGACTGAAC Primer reverse GTAATCCAGCAGGTCAGCAAAG Sonde CCATCACATTGTAGCCCTCTGTGTGCTC PBGD Primer forward GAACCAGCTCCCTGCGAAG Primer reverse GTGTTGAGGTTTCCCCGAATAC Sonde CCCAGCTGCAGAGAAAGTTCCCGC Abbildung 5: Primer und Sonden für TaqMan-PCR
30
5. Ergebnisse
5.1. Immunhistochemische Färbungen
5.1.1. E-Cadherin
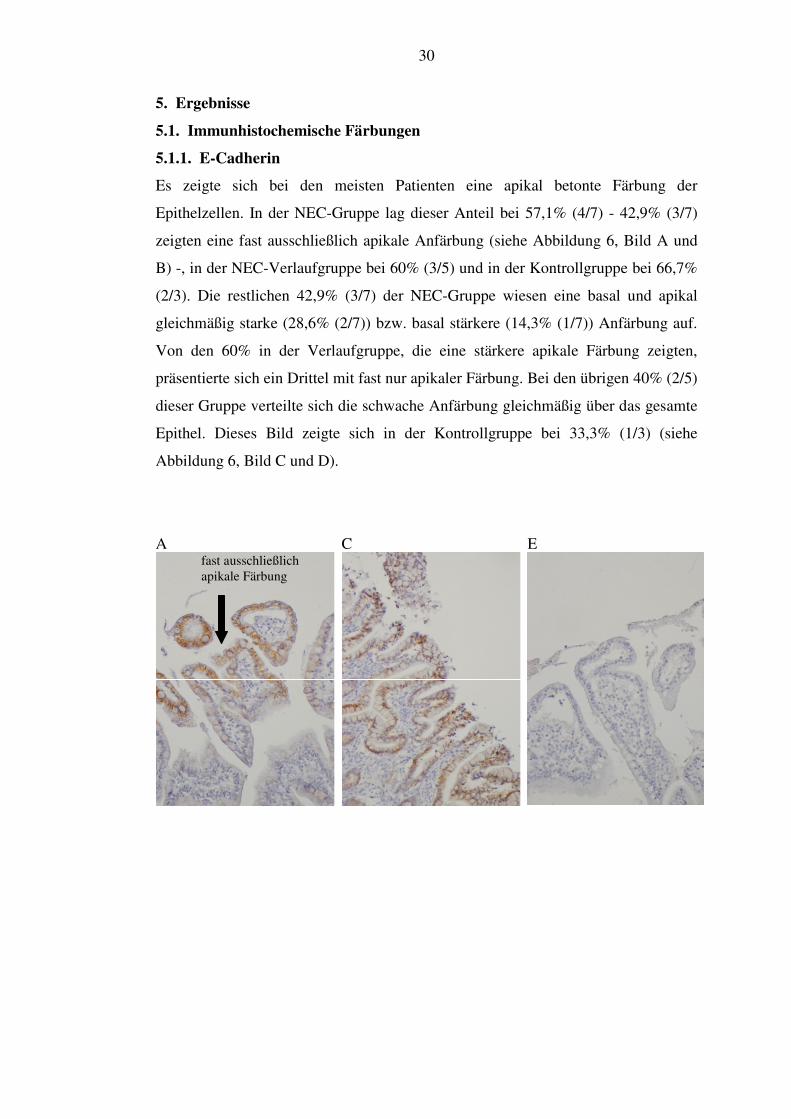
Es zeigte sich bei den meisten Patienten eine apikal betonte Färbung der
Epithelzellen. In der NEC-Gruppe lag dieser Anteil bei 57,1% (4/7) - 42,9% (3/7)
zeigten eine fast ausschließlich apikale Anfärbung (siehe Abbildung 6, Bild A und
B) -, in der NEC-Verlaufgruppe bei 60% (3/5) und in der Kontrollgruppe bei 66,7%
(2/3). Die restlichen 42,9% (3/7) der NEC-Gruppe wiesen eine basal und apikal
gleichmäßig starke (28,6% (2/7)) bzw. basal stärkere (14,3% (1/7)) Anfärbung auf.
Von den 60% in der Verlaufgruppe, die eine stärkere apikale Färbung zeigten,
präsentierte sich ein Drittel mit fast nur apikaler Färbung. Bei den übrigen 40% (2/5)
dieser Gruppe verteilte sich die schwache Anfärbung gleichmäßig über das gesamte
Epithel. Dieses Bild zeigte sich in der Kontrollgruppe bei 33,3% (1/3) (siehe
Abbildung 6, Bild C und D).
A C E
fast ausschließlich apikale Färbung
31
B D F
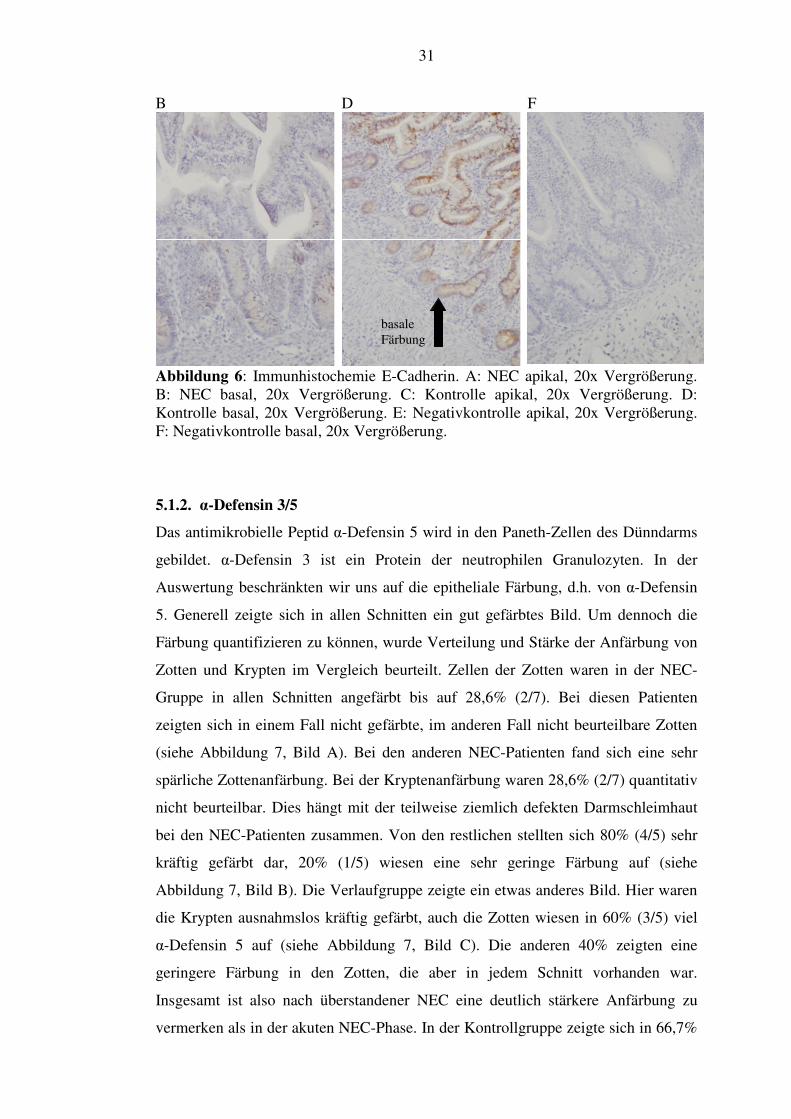
Abbildung 6: Immunhistochemie E-Cadherin. A: NEC apikal, 20x Vergrößerung. B: NEC basal, 20x Vergrößerung. C: Kontrolle apikal, 20x Vergrößerung. D: Kontrolle basal, 20x Vergrößerung. E: Negativkontrolle apikal, 20x Vergrößerung. F: Negativkontrolle basal, 20x Vergrößerung.
5.1.2. α-Defensin 3/5
Das antimikrobielle Peptid α-Defensin 5 wird in den Paneth-Zellen des Dünndarms
gebildet. α-Defensin 3 ist ein Protein der neutrophilen Granulozyten. In der
Auswertung beschränkten wir uns auf die epitheliale Färbung, d.h. von α-Defensin
5. Generell zeigte sich in allen Schnitten ein gut gefärbtes Bild. Um dennoch die
Färbung quantifizieren zu können, wurde Verteilung und Stärke der Anfärbung von
Zotten und Krypten im Vergleich beurteilt. Zellen der Zotten waren in der NEC-
Gruppe in allen Schnitten angefärbt bis auf 28,6% (2/7). Bei diesen Patienten
zeigten sich in einem Fall nicht gefärbte, im anderen Fall nicht beurteilbare Zotten
(siehe Abbildung 7, Bild A). Bei den anderen NEC-Patienten fand sich eine sehr
spärliche Zottenanfärbung. Bei der Kryptenanfärbung waren 28,6% (2/7) quantitativ
nicht beurteilbar. Dies hängt mit der teilweise ziemlich defekten Darmschleimhaut
bei den NEC-Patienten zusammen. Von den restlichen stellten sich 80% (4/5) sehr
kräftig gefärbt dar, 20% (1/5) wiesen eine sehr geringe Färbung auf (siehe
Abbildung 7, Bild B). Die Verlaufgruppe zeigte ein etwas anderes Bild. Hier waren
die Krypten ausnahmslos kräftig gefärbt, auch die Zotten wiesen in 60% (3/5) viel
α-Defensin 5 auf (siehe Abbildung 7, Bild C). Die anderen 40% zeigten eine
geringere Färbung in den Zotten, die aber in jedem Schnitt vorhanden war.
Insgesamt ist also nach überstandener NEC eine deutlich stärkere Anfärbung zu
vermerken als in der akuten NEC-Phase. In der Kontrollgruppe zeigte sich in 66,7%
basale Färbung
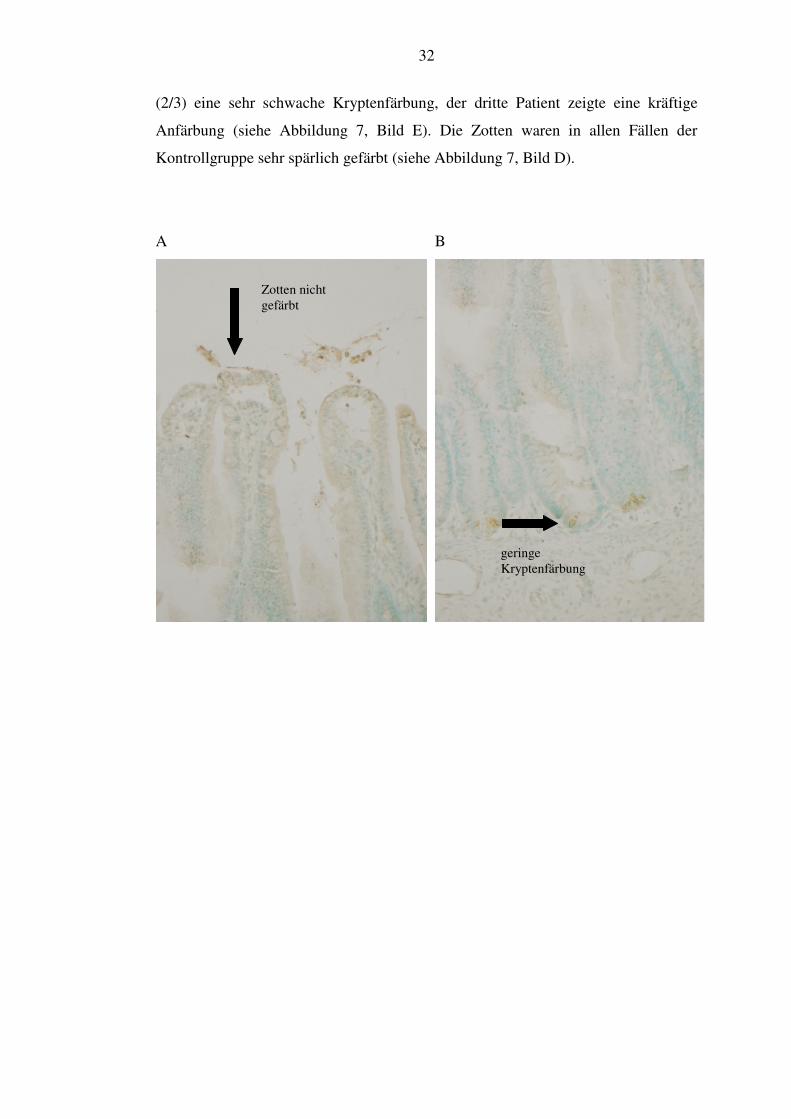
32
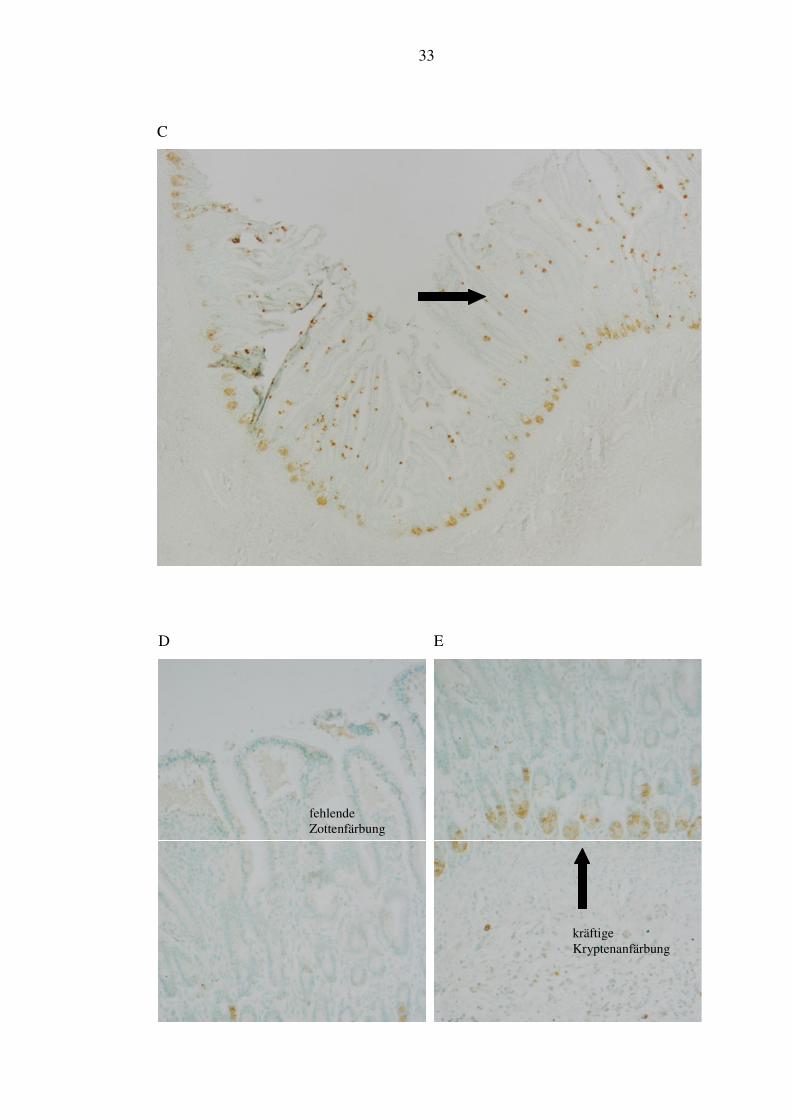
(2/3) eine sehr schwache Kryptenfärbung, der dritte Patient zeigte eine kräftige
Anfärbung (siehe Abbildung 7, Bild E). Die Zotten waren in allen Fällen der
Kontrollgruppe sehr spärlich gefärbt (siehe Abbildung 7, Bild D).
A B
Zotten nicht gefärbt
geringe Kryptenfärbung
33
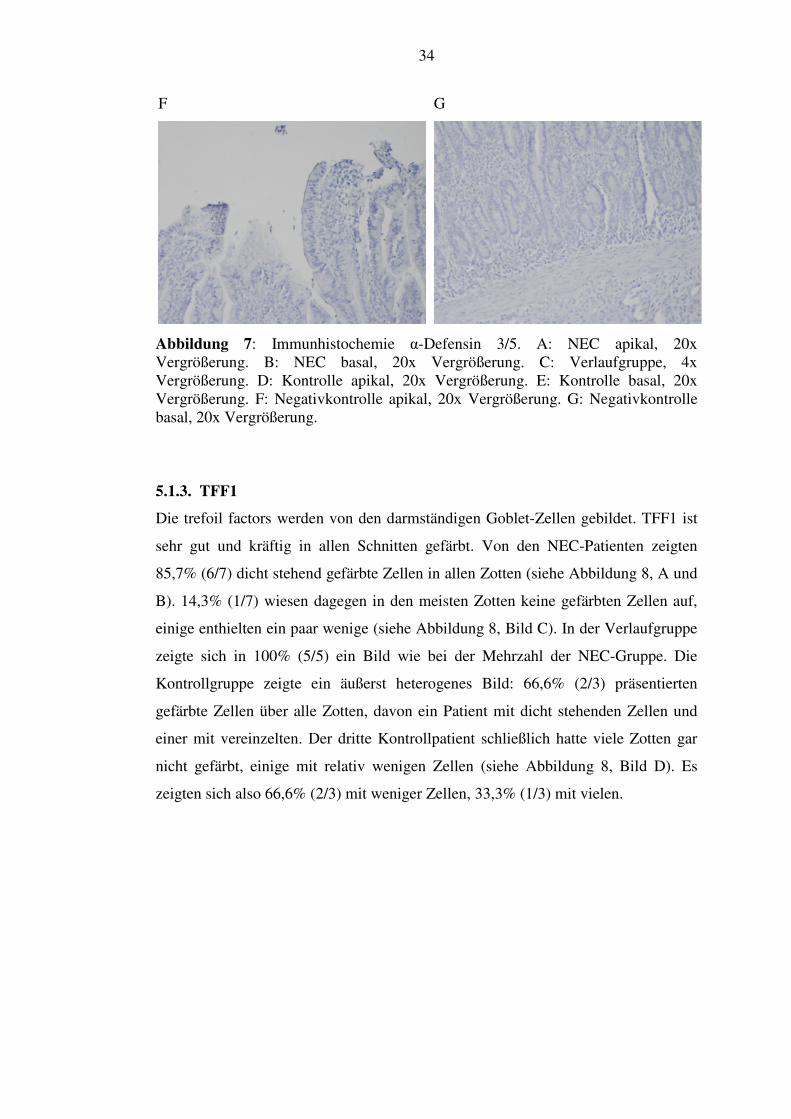
C
D
E
fehlende Zottenfärbung
kräftige Kryptenanfärbung
34
F G
Abbildung 7: Immunhistochemie α-Defensin 3/5. A: NEC apikal, 20x Vergrößerung. B: NEC basal, 20x Vergrößerung. C: Verlaufgruppe, 4x Vergrößerung. D: Kontrolle apikal, 20x Vergrößerung. E: Kontrolle basal, 20x Vergrößerung. F: Negativkontrolle apikal, 20x Vergrößerung. G: Negativkontrolle basal, 20x Vergrößerung.
5.1.3. TFF1
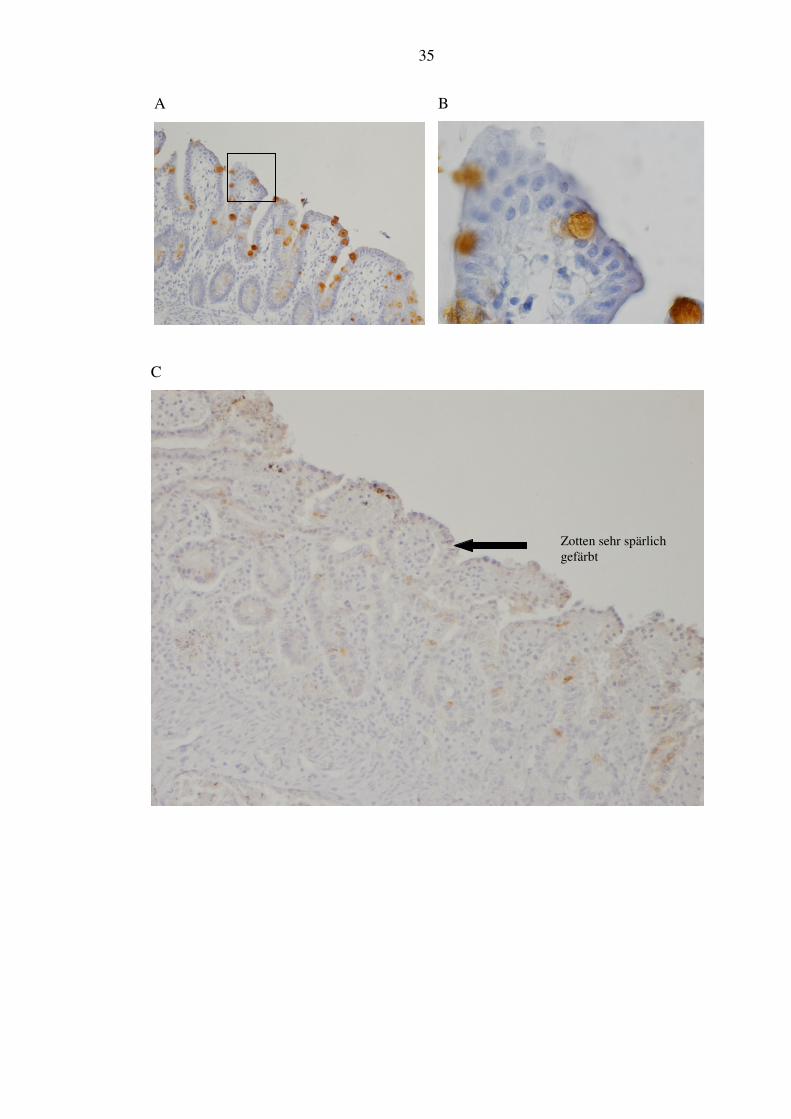
Die trefoil factors werden von den darmständigen Goblet-Zellen gebildet. TFF1 ist
sehr gut und kräftig in allen Schnitten gefärbt. Von den NEC-Patienten zeigten
85,7% (6/7) dicht stehend gefärbte Zellen in allen Zotten (siehe Abbildung 8, A und
B). 14,3% (1/7) wiesen dagegen in den meisten Zotten keine gefärbten Zellen auf,
einige enthielten ein paar wenige (siehe Abbildung 8, Bild C). In der Verlaufgruppe
zeigte sich in 100% (5/5) ein Bild wie bei der Mehrzahl der NEC-Gruppe. Die
Kontrollgruppe zeigte ein äußerst heterogenes Bild: 66,6% (2/3) präsentierten
gefärbte Zellen über alle Zotten, davon ein Patient mit dicht stehenden Zellen und
einer mit vereinzelten. Der dritte Kontrollpatient schließlich hatte viele Zotten gar
nicht gefärbt, einige mit relativ wenigen Zellen (siehe Abbildung 8, Bild D). Es
zeigten sich also 66,6% (2/3) mit weniger Zellen, 33,3% (1/3) mit vielen.
35
A
B
C
Zotten sehr spärlich gefärbt

36
D E
F
Abbildung 8: Immunhistochemie TFF1. A: NEC, 20x Vergrößerung. B: NEC, 100x Vergrößerung aus A. C: NEC, 20x Vergrößerung. D: Kontrolle, 20x Vergrößerung. E: Negativkontrolle apikal (Rabbitserum), 20x Vergrößerung. F: Negativkontrolle basal (Rabbitserum), 20x Vergrößerung.
5.1.4. TFF2
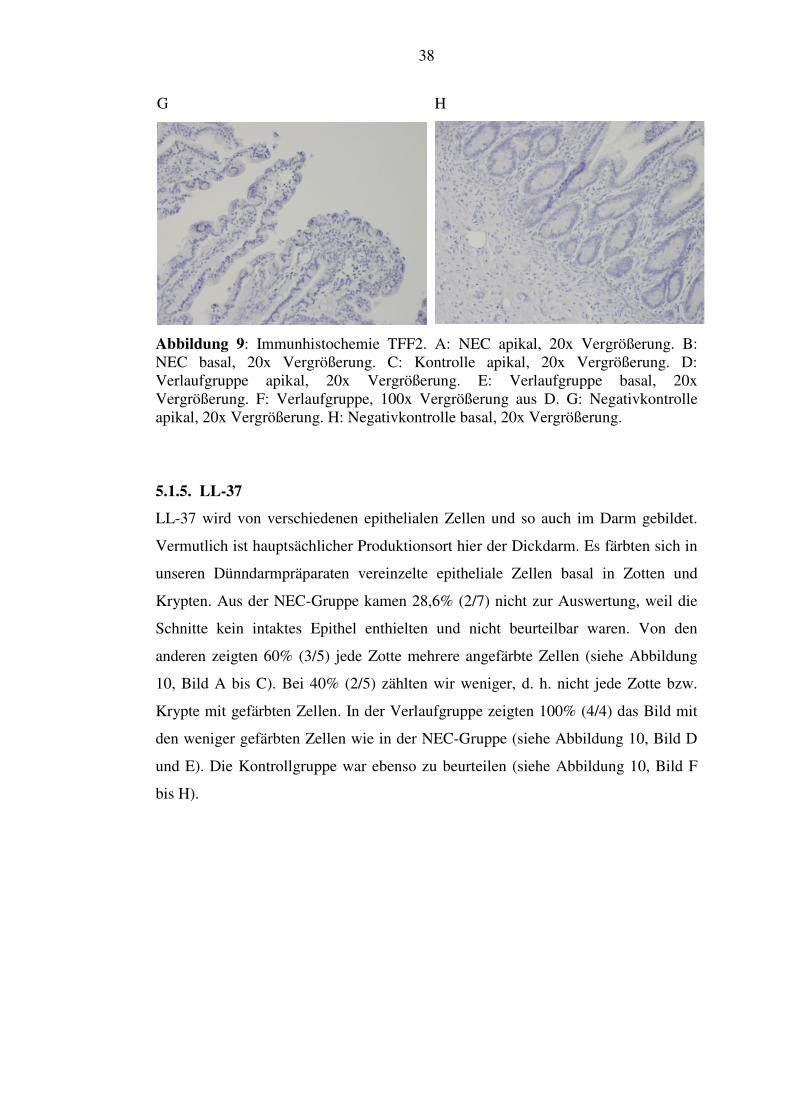
Dagegen war TFF2 in den meisten Schnitten nicht nachzuweisen. Aus der NEC-
Gruppe zeigte nur ein Präparat ein paar gefärbte Zellen apikal, die restlichen 85,7%
(6/7) waren nicht gefärbt (siehe Abbildung 9, Bild A und B). Ebenso in den anderen
Gruppen: Die Verlaufgruppe hatte ein Präparat mit einer ähnlichen Färbung
aufzuweisen (Abbildung 9, Bild D bis F), die Kontrollgruppe ebenfalls. Es zeigten
sich 80% (4/5) der Verlaufgruppe ungefärbt und 66,6% (2/3) der Kontrollgruppe
(siehe Abbildung 9, Bild C).
einzelne Zotten mit wenig gefärbten Zellen

37
A B
C D
E F
apikal ein paar gefärbte Zellen
TFF2-Färbung
basal keine gefärbten Zellen
38
G H
Abbildung 9: Immunhistochemie TFF2. A: NEC apikal, 20x Vergrößerung. B: NEC basal, 20x Vergrößerung. C: Kontrolle apikal, 20x Vergrößerung. D: Verlaufgruppe apikal, 20x Vergrößerung. E: Verlaufgruppe basal, 20x Vergrößerung. F: Verlaufgruppe, 100x Vergrößerung aus D. G: Negativkontrolle apikal, 20x Vergrößerung. H: Negativkontrolle basal, 20x Vergrößerung.
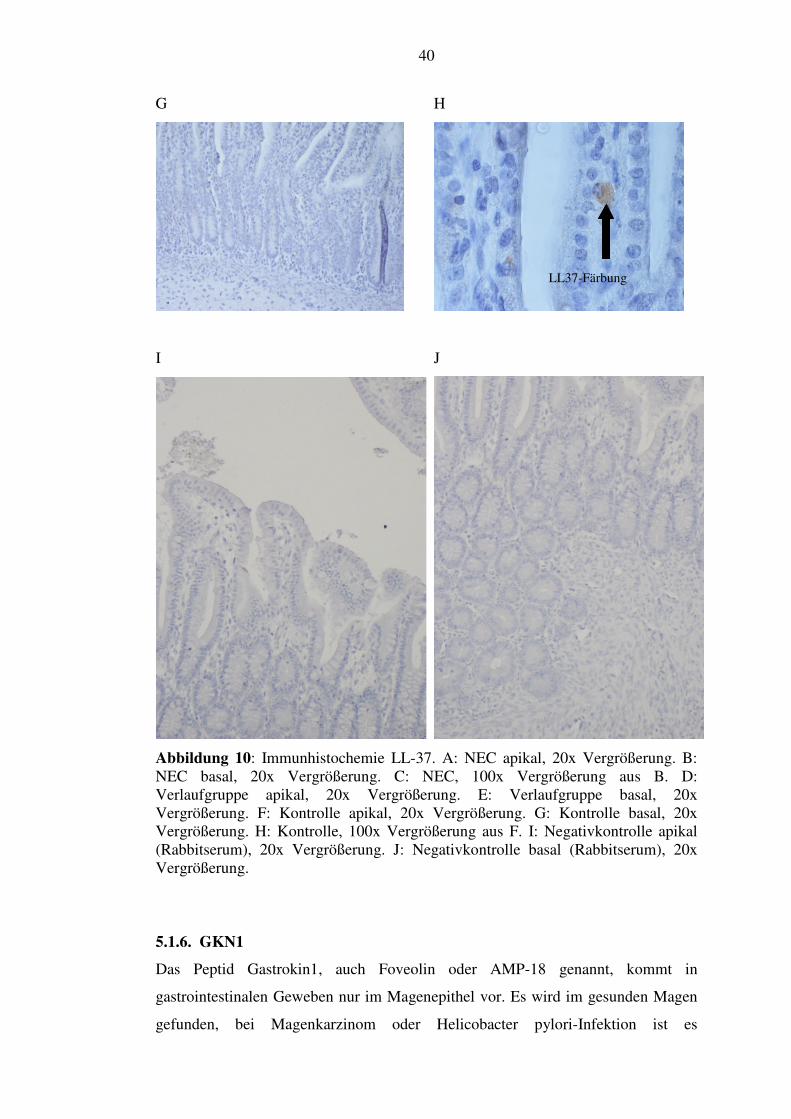
5.1.5. LL-37
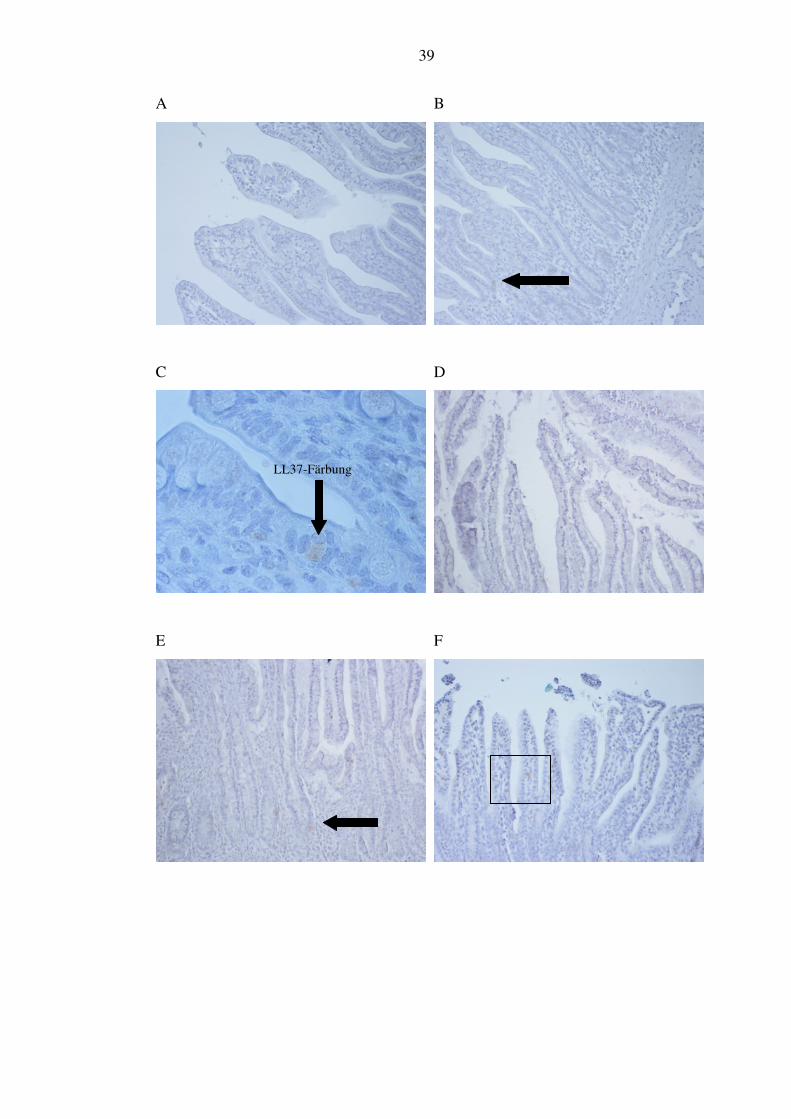
LL-37 wird von verschiedenen epithelialen Zellen und so auch im Darm gebildet.
Vermutlich ist hauptsächlicher Produktionsort hier der Dickdarm. Es färbten sich in
unseren Dünndarmpräparaten vereinzelte epitheliale Zellen basal in Zotten und
Krypten. Aus der NEC-Gruppe kamen 28,6% (2/7) nicht zur Auswertung, weil die
Schnitte kein intaktes Epithel enthielten und nicht beurteilbar waren. Von den
anderen zeigten 60% (3/5) jede Zotte mehrere angefärbte Zellen (siehe Abbildung
10, Bild A bis C). Bei 40% (2/5) zählten wir weniger, d. h. nicht jede Zotte bzw.
Krypte mit gefärbten Zellen. In der Verlaufgruppe zeigten 100% (4/4) das Bild mit
den weniger gefärbten Zellen wie in der NEC-Gruppe (siehe Abbildung 10, Bild D
und E). Die Kontrollgruppe war ebenso zu beurteilen (siehe Abbildung 10, Bild F
bis H).
39
A B
C D
E F
LL37-Färbung
40
G
H
I J
Abbildung 10: Immunhistochemie LL-37. A: NEC apikal, 20x Vergrößerung. B: NEC basal, 20x Vergrößerung. C: NEC, 100x Vergrößerung aus B. D: Verlaufgruppe apikal, 20x Vergrößerung. E: Verlaufgruppe basal, 20x Vergrößerung. F: Kontrolle apikal, 20x Vergrößerung. G: Kontrolle basal, 20x Vergrößerung. H: Kontrolle, 100x Vergrößerung aus F. I: Negativkontrolle apikal (Rabbitserum), 20x Vergrößerung. J: Negativkontrolle basal (Rabbitserum), 20x Vergrößerung.
5.1.6. GKN1
Das Peptid Gastrokin1, auch Foveolin oder AMP-18 genannt, kommt in
gastrointestinalen Geweben nur im Magenepithel vor. Es wird im gesunden Magen
gefunden, bei Magenkarzinom oder Helicobacter pylori-Infektion ist es
LL37-Färbung
41
herunterreguliert. Studien konnten eine schnellere Wundheilung bei Vorhandensein
des Proteins feststellen (Rippa et al. 2010). Alle Schnitte waren für dieses Peptid
negativ (keine Abbildung).
5.2. PCR der Schnitte
Alle Proben zeigten eine Expression des Panethzell-Defensins α-Defensin 5 (siehe
Abbildung 11, Bild A), von TFF1 (siehe Abbildung 11, Bild B) und von
Cathelicidin (siehe Abbildung 11, Bild D). In keiner Probe der Kontrollgruppe
konnte eine Expression von TFF2 nachgewiesen werden. Dagegen zeigten sich alle
Proben der Verlaufkontrolle für cDNA von TFF2 positiv. In der NEC-Gruppe war
eine Probe positiv (siehe Abbildung 11, Bild C). Analog zur immunhistochemischen
Peptiddarstellung zeigte sich in allen Proben keine Expression von GKN1 (keine
Abbildung).
A
B
C
D
Abbildung 11: PCR cDNA. Mittelwert und Standardabweichung. A: α-Defensin 5. B: TFF1. C: TFF2. D: LL-37.

42
6. Diskussion
Ein Unterschied in der Intaktheit der Epithelbarriere zwischen den Gruppen zeigte
sich in unserer Untersuchung nicht. Die mehrheitlichen Schnitte in der NEC-Gruppe
wiesen Stellen geringerer Färbung von E-Cadherin auf im apikal-basalen Vergleich,
nur ein geringer Teil hingegen zeigte durchgehend starke Färbungen. Die
Darstellung in der Kontrollgruppe ist ähnlich der NEC-Gruppe. Stellenweise nicht
intaktes Epithel zeigt sich ebenfalls in beiden Gruppen. In der Verlaufgruppe zeigten
sich alle Patienten stellenweise oder global mit einer recht schwachen Färbung.
Zusammenfassend konnte jedenfalls bei den NEC-Kindern im Vergleich mit der
Verlauf- und Kontrollgruppe kein E-Cadherin-Mangel festgestellt werden, der
gegenüber pathogenen Faktoren wie Keimen eine mechanische Abwehrschwäche
darstellen und folglich zu einer überschießenden Immun- und Entzündungsantwort
führen würde. Es sei allerdings auf die kleine Untersuchungsgruppe für die
Kontrolle hingewiesen.
Hanby et al. sehen in E-Cadherin nicht nur einen Marker für die Epithelbarriere,
sondern schließen aus Menge und Verteilung auf die Aktivität der Regeneration. Sie
stellen die These auf, dass bei Wundheilungsprozessen des Epithels E-Cadherin
stellenweise verringert ist und die Epithelzellen so die notwendige Mobilität
erhalten, andererseits aber auch Inseln mit vermehrtem E-Cadherin vorhanden sind,
um die Position von deren Zellen zu halten. Dazu untersuchten sie neben einer
gesunden Kontrollgruppe ileales und kolonisches Epithel bei der chronisch
entzündlichen Darmerkrankung Morbus Crohn, gastroduodenale peptische Ulzera
und ein in-vitro-Modell mit einer Epithelzelllinie des kolorektalen Karzinoms. Sie
sahen in gesundem Epithel durchgehend starke Färbungen, bei der chronisch
entzündlichen Darmerkrankung an entzündeten Stellen Krypten und apikale Zellen
stark gefärbt, die im mittleren Bereich dazwischenliegenden Zellen schwach bis gar
nicht. Ebenso bei Gastritis. Bei Ulzera waren die Färbungen der apikalen Zellen und
vor allem der Krypten meist schwach, mit Säulen stärkerer Färbung dazwischen.
Hanby et al. erklären die fehlenden oder schwachen Färbungen bei Ulzera mit der
Regeneration der Zellen und ihrer notwendigen Mobilität aufgrund der
Epithelerneuerung, die bei Entzündung mit dem erhöhten Zellumsatz. In den stärker
gefärbten Stellen bei Ulzera sehen sie kürzlich erneuertes Epithel als Ausdruck der
Chronizität (Hanby et al. 1996). Analog dazu sehen wir vielleicht in der schwachen
Färbung der Verlaufgruppe einen frühen Wundheilungsprozess in der Situation der

43
Stomarückverlagerung und sowohl in der NEC-Gruppe als auch in der
Kontrollgruppe generell regenerative oder entzündliche und, vor allem in der NEC-
Gruppe, auch ulzeröse Prozesse.
Insgesamt ist nach überstandener NEC eine deutlich stärkere Anfärbung von α-
Defensin 5 zu vermerken als in der akuten NEC-Phase bzw. in der Kontrolle. Im
Vergleich der NEC- mit der Kontrollgruppe lässt sich andeutungsweise eine
vermehrte Färbung der Krypten bei akuter NEC feststellen. Vermindert konnten wir
die Panethzellen bzw. deren Produkte bei NEC im Vergleich mit der Kontrolle nicht
finden. Salzman et al. stellten in einer Untersuchung ähnlich der unseren α-Defensin
5 immunhistologisch und die Expression von α-Defensin 5 und 6 mittels mRNA-
Messung dar. Sie untersuchten dazu sechs Ileum- und Jejunum-Schnitte von
operiertem Darm von NEC-Kindern, fünf Dünndarm-Kontrollschnitte von Kindern
mit Darmatresie und Mekoniumileus kamen zur Auswertung. Die NEC-Kinder
waren mit 25 bis 31 SSW alle jünger als die Kontrollkinder, die in den SSW 35 bis
40 geboren wurden. Das Lebensalter der NEC-Kinder war 5 bis 45 Tage, die
Kontrollkinder wurden am ersten oder zweiten Lebenstag operiert. In einer weiteren
Untersuchung zeigte ein Fetus der SSW 24 ein geringeres Level an α-Defensin 5,
sowohl mRNA als auch Peptid, als reifgeborene Kinder oder Erwachsene. Die NEC-
Kinder zeigten eine vermehrte Expression von α-Defensin 5 und 6 und mehr Paneth-
Zellen als die Kontrollgruppe. Die Peptide waren aber nicht gleichermaßen erhöht
und in NEC-Gruppe und Kontrolle gleich. Physiologisch müsste die Expression
erniedrigt sein bei den NEC-Kindern aufgrund ihres geringeren Gestationsalters,
Salzman et al. folgern deshalb, die NEC induziere die Expression und erhöhe auch
die Paneth-Zellzahl. Sie deuten diese Befunde als Auswirkung der NEC und, was
auch denkbar wäre, nicht als Ursache für sie. Dass sie nicht das höhere postnatale
Alter der NEC-Kinder widerspiegeln, sehen sie darin bestätigt, dass die
Veränderungen über die verschiedenen Zeitpunkte der Operation bei den einzelnen
NEC-Patienten (5. bis 45. Lebenstag) sehr konsistent auftreten. Sie folgern, dass
durch die inflammatorischen Vorgänge bei der NEC die α-Defensine 5 und 6
vermehrt exprimiert und sezerniert werden, weshalb die intrazellulären Peptide sich
auch nicht erhöht darstellen (Salzman et al. 1998). Die vermehrte Anfärbung von α-
Defensin 5 in der NEC- im Vergleich mit der Kontrollgruppe, sei es durch mehr
Paneth-Zellen oder stärkere Färbung der einzelnen Zelle, können wir in unserer
Untersuchung auch tendenziell sehen. Aufgrund des Alters der Proben und der

44
resultierenden Anfälligkeit der Werte einer PCR haben wir unsere
Expressionsuntersuchung nicht quantitativ ausgewertet. Sieht man sich die Werte im
Vergleich an, lässt sich aber eine Tendenz zu der von Salzman et al. gefundenen
gesteigerten Expression in der akuten NEC-Phase noch nicht beobachten, erst in der
Verlaufgruppe. Problematisch sind bei Salzman et al. die deutlich geringeren
Gestationswochen der NEC-Gruppe, jedoch sehen sie darin, vermutlich auch zu
Recht, noch eine Bestätigung für die gefundene verstärkte Expression bei NEC.
Trotz des relativ gleichen Gestationsalters unserer Gruppen NEC und Kontrolle,
also der geringen Gestationsalter der Kontrollkinder, sehen wir keinen starken
Unterschied im Sinne einer geringeren Färbung bei der Kontrolle. Dennoch können
wir die These von Salzman et al. unterstützen, dass die NEC oder die Prozesse, die
zur NEC führen, eine vermehrte Produktion von α-Defensin 5 hervorrufen. Die
vermehrte Anfärbung in der Verlaufgruppe kann einerseits eine Steigerung der im
akuten Stadium schon und im abgeheilten noch zu sehenden Vermehrung von α-
Defensin 5 – als Abwehrreaktion und Bestandteil der NEC – bedeuten, oder aber
auch nur die natürliche Zunahme dieses Peptids mit dem Alter zeigen.
Schaart et al. untersuchten in den Resektionsrändern von Darmpräparaten bei NEC
u. a. das Goblet-Zell-Peptid TFF3 und die Paneth-Zellen mit Hilfe von deren
Lysozym-Produktion. Sie verwendeten dazu sowohl Dünn- als auch Dickdarm aus
21 NEC-Operationen, 17 Stomarückverlagerungen und 9 Kontrollen. Auch hier
wies die NEC-Gruppe wie bei Salzman et al. ein geringeres Gestationsalter als die
Kontrollgruppe auf (25 bis 39 SSW, Median 32 vs. 35 bis 40 SSW, Median 39). Sie
weisen darauf hin, dass TFF3 ab der 12. SSW exprimiert wird und sich Lysozym in
der SSW 20 findet. Für die Paneth-Zellen treffen sie die Feststellung, dass sie nur an
der Kryptenbasis zu finden sind. Bei akuter NEC stellen sie sowohl eine schwächere
Färbung der einzelnen Zelle fest als auch bei schweren Zellschäden eine verringerte
Zahl an Zellen. Zum Zeitpunkt der Stomarückverlagerung sehen sie vermehrt
Paneth-Zellen (Schaart et al. 2009). Im Gegensatz dazu sehen wir Paneth-Zellen
auch in den Zotten, kräftig in den meisten Schnitten der Verlaufgruppe. Es mag sein,
dass Paneth-Zellen das Lysozym nur in den basalen Zellen exprimieren im
Gegensatz zum α-Defensin. Auch sehen wir ihr Produkt α-Defensin 5 in unseren
NEC-Schnitten kräftig gefärbt, wohingegen die Kontrollgruppe in der Mehrzahl eine
schwache Färbung zeigt. Diese Ergebnisse müssen nicht zwingend im Widerspruch
stehen, da Schaart et al. mit Lysozym ein anderes Produkt der Paneth-Zellen

45
untersuchen. Wir ziehen jedoch die Frühgeburtlichkeit der NEC-Kinder als
mögliche Ursache der geringeren Lysozym-Färbung in Betracht. Diesen
Altersaspekt in ihrem Untersuchungsmodell lassen Schaart et al. außer Acht und
schließen aus ihren Ergebnissen nur auf die insuffiziente Reaktion auf einen Angriff
des Epithels. Wegen der kleinen Kollektive lässt sich nur eine Tendenz in der
Auswertung der Befunde von α-Defensin 5 erkennen, die aber die Ergebnisse der
Untersuchung von Salzman et al. bestätigt und eine vermehrte Peptidproduktion
durch die Paneth-Zellen bei akuter NEC postuliert.
In der Expression lassen sich bei TFF1 keine Unterschiede in den Gruppen
feststellen. In der Immunhistochemie zeigt die Mehrzahl der Kontrollen weniger
gefärbte Zellen, die Mehrheit der NEC-Gruppe zeigt viele Zellen und die
Verlaufgruppe schließlich ausnahmslos viele Zellen. Einen Mangel an TFF1 weist
der Großteil der NEC-Patienten gegenüber den Kontrollen in unserer Untersuchung
also nicht auf und TFF1 ist in allen Schnitten deutlich angefärbt. Vieten et al. haben
Expression und Immunhistochemie von TFF1-3 getestet bei zwölf NEC-Patienten,
acht Verlaufkontrollen, 20 Kontrollen und 15 Fetuskontrollen. Die NEC-Gruppe
war durchschnittlich deutlich jünger als die Kontrollgruppe, die Reifgeborene
repräsentierte. Die Feten entstammten der 11. bis 13. SSW. Durch die Untersuchung
der Feten stellten sie dar, dass diese schon das gleiche Peptidprofil wie Erwachsene
aufweisen und somit das Alter keine Rolle spielt. Für TFF3 fanden sie in den
Kontrollen die höchsten Werte, für TFF1 und 2 in der Verlaufgruppe, in der
regeneratives Epithel die stärkste Expression und Färbung zeigte. Allerdings waren
die Färbungen und auch die Expression für TFF1 jeweils nur schwach. Zwei NEC-
Proben zeigten bei ihnen schwache Färbung. TFF1 und 2 stellen sie als
normalerweise vom gesunden Darm nicht produziertes Peptid dar, nur TFF3 ist
konstitutiv exprimiert. Für TFF3 postulieren sie daher, dass bei NEC die notwendige
Hochregulierung fehlt, bei TFF1 und 2 die Ingangsetzung der Expression. Grund für
die Verringerung von TFF3 bei NEC sehen sie sowohl in dem Verlust von
Gobletzellen als auch dem Verlust von deren Produktion, da sie auch leere
Gobletzellen sehen (Vieten et al. 2005).
Schaart et al. haben in ihrer oben genannten Untersuchung TFF3 als Marker für die
Goblet-Zellen benutzt. Sie sehen in nicht so stark geschädigtem Epithel bei akuter
NEC eine normale bis erhöhte Goblet-Zellzahl und erhaltene TFF3-Produktion bei
allen Zellen. Sie vermuten jedoch, dass die Expression insuffizient ist im Zustand

46
der NEC (Schaart et al. 2009). Wir haben TFF3, den typischeren TFF des Darms,
nicht untersucht. Jedoch sehen wir für TFF1 viel stärkere Färbungen als Vieten et al.
Es zeigt sich wie bei ihnen in NEC- und Verlaufgruppe deutlicher als in den
Kontrollen. Die Aussage, dass die Expression von TFF1 bei NEC fehlt, können wir
nicht bestätigen. Allerdings lässt sich eine Tendenz zur Hochregulierung in unseren
Expressionen auch nicht finden, von einer Probe abgesehen. Ein Kritikpunkt an der
Untersuchung von Vieten et al. ist die Nichtunterscheidung von Dünn- und
Dickdarm. Es ist nicht nachvollziehbar, in welchen Schnitten sie Färbungen fanden
und in welchen nicht. Möglicherweise liegt auch hierin der Grund für die
unterschiedlichen Ergebnisse in Bezug auf TFF1. Wir finden keinen Mangel an
TFF1 gegenüber den Kontrollen bei akuter NEC, tendenziell eher eine Steigerung
der Produktion bzw. vermehrt Gobletzellen wie auch Schaart et al. Das Ergebnis ist
somit vergleichbar mit den Ergebnissen bei α-Defensin 5.
Aus der Färbung von TFF2 lassen sich keine Schlüsse ziehen. Die Ergebnisse
stimmen insofern mit denen von Vieten et al. überein, dass der Darm keine oder nur
eine schwache Färbung für dieses Peptid aufweist (Vieten et al. 2005). Anders als
TFF1 scheint TFF2 nur nach einer Schleimhautschädigung exprimiert zu werden
und sich im gesunden Darm nicht zu finden. In der Expression zeigte sich, mit einer
Ausnahme in der NEC-Gruppe, nur die Verlaufgruppe positiv. In der Proteinfärbung
war es dort in den meisten Schnitten (noch) nicht nachzuweisen. Allerdings ließ sich
auch in der Kontrollgruppe bei einem Schnitt färberisch Peptid nachweisen, ohne
dass es in der PCR gemessen werden konnte. Wir können also die Expression von
TFF2 in einer Probe der NEC-Gruppe messen, färberisch können wir es in einer
Probe der Kontrollgruppe nachweisen. Kontroll- und NEC-Gruppe unterscheiden
sich, alle Ergebnisse in Bezug auf TFF2 vergleichend, in unserer Untersuchung
nicht, die NEC-Gruppe weist keinen Mangel an TFF2 auf gegenüber der
Kontrollgruppe. Das Ergebnis zeigt aber auch, dass zum Zeitpunkt der Operation
der NEC noch keine sichtbare Steigerung der TFF2-Expression mit dem Ziel der
Regeneration und Wundheilung stattgefunden hat. Dass wir es färberisch sowohl in
der NEC- als auch in der Verlaufgruppe nicht messen können, kann auch auf eine
Translationsstörung bei der NEC zurückzuführen sein. Vieten et al. konnten zeigen,
dass die Produktion der TFFs schon im ersten Trimenon beim Feten stattfindet wie
beim Erwachsenen (Vieten et al. 2005). Möglicherweise läuft aber der Prozess, der

47
die Produktion als Reaktion auf eine Schädigung in Gang kommen lässt, bei
Frühgeborenen noch zu langsam ab und prädisponiert deshalb zur NEC.
Für LL-37 konnten keine gruppenspezifischen Unterschiede beobachtet werden,
evtl. lässt sich die Tendenz festhalten, dass es in der NEC-Gruppe färberisch etwas
deutlicher nachzuweisen war und sich in der Verlaufgruppe vermehrt exprimiert
darstellte. Es zeigt sich generell sehr schwach gefärbt. LL-37 gilt als ein Peptid der
differenzierten Zelle, weshalb es sich apikal im gesunden Darm des Erwachsenen
detektieren lässt. Es ist beschrieben, dass es sich im Dünndarm schwächer zeigt als
im Dickdarm. Es soll anders reguliert sein als andere AMPs wie die Defensine.
Beispielsweise wird es unabhängig von einer Keimflora exprimiert (Hase et al.
2002). Erniedrigt ist dieses Peptid nicht nur beim Morbus Crohn (Doss et al. 2010),
sondern auch bei einer Shigellen-Infektion im Darm (Schauber et al. 2003).
Möglicherweise sind Cathelicidine bei Neugeborenen tatsächlich noch verringert,
als NEC-spezifisch vermindert stellt es sich bei uns nicht dar.
Es war ein Ziel der Arbeit, eine parallele Untersuchung der AMPs an jeweils
denselben Präparaten durchzuführen. Hierbei haben wir in den verschiedenen
Färbungen der AMPs unterschiedliche Ergebnisse gefunden. Während bei akuter
NEC das α-Defensin 5, TFF1 und, sehr leicht, LL-37 gegenüber den Kontrollen
ansatzweise erhöht war, ließ sich eine Steigerung der Produktion von TFF2 nicht
feststellen. Wir folgern daraus, dass sie jeweils unterschiedlich reguliert werden.
Ein Problem der Untersuchung der NEC sind die jeweils kleinen Fallzahlen
einzelner Studien, die die Aussagekraft der Ergebnisse einschränken. Ein weiterer
kritischer Punkt ist die Subjektivität der Auswertung der immunhistochemischen
Bilder, die wir durch Auszählung und Berücksichtigung der Verteilung der
angefärbten Peptide möglichst minimiert haben. Jedoch zeigt sich in unserer
Auswertung weitgehende Übereinstimmung mit ähnlichen Untersuchungen der
Literatur. Wir finden die Kapazität des erkrankten Darms, auf die Erkrankung zu
reagieren, in der Darstellung der Bilder der Verlaufkontrolle bestätigt. Diese
gefundenen Reaktionen mögen jedoch zu langsam erfolgen, möglicherweise
spiegelt, wie oben schon erwähnt, dieses Ergebnis auch nur die normale Reifung des
Darms wider: Unsere Kontrollgruppe und NEC-Gruppe sind von der
Altersverteilung in etwa gleich (Mittelwerte NEC-Gruppe: Alter bei OP 10
Lebenstage, Geburt in 32,9. SSW. Mittelwerte Kontrollgruppe: Alter bei OP 14
Lebenstage, Geburt in 31,7. SSW), die Verlaufgruppe ist mit einem Mittelwert von

48
56 Lebenstagen, Geburt in 35. SSW deutlich älter. Kontroll- und NEC-Gruppe
zeigen sich, auch aufgrund der kleinen Kollektive, bis auf Tendenzen in der Färbung
von α-Defensin 5, TFF1 und LL-37 weitgehend gleich. Eine Verringerung von α-
Defensin 5 – sei es als Resultat einer pathologischen Keimflora oder als Ursache für
eine solche, oder als Ausdruck einer Rezeptormutation – ist vermutlich nicht die
einzige Ursache für das Entstehen der NEC. Bei Frühgeborenen mag es eine Rolle
spielen, da die Paneth-Zellen und α-Defensine bei ihnen verringert sind, wie Mallow
et al. gezeigt haben (Mallow et al. 1996). Die Produktionssteigerung, die wir bei
akuter NEC sehen, ist in Bezug auf die Schwere der Erkrankung möglicherweise
noch nicht adäquat und α-Defensin 5 relativ verringert. Möglicherweise ist aber
auch eine vermehrte Produktion von α-Defensin 5 ein begünstigender Faktor als
pathologische Reaktion in der Kaskade der Induktion und Expression. Denkbar ist
nämlich, dass die Veränderungen, die wir sehen, nicht nur durch die NEC verursacht
sind – die AMPs waren verringert aufgrund der Frühgeburtlichkeit, was zur NEC
geführt hat; in ihrem Zuge wurde die Produktion im möglichen Rahmen, aber
insuffizient gesteigert –, sondern auch Prozesse sind, die zur NEC beigetragen
haben – durch AMPs wird eine Entzündungskaskade in Gang gesetzt.
Möglicherweise stimmt das Gleichgewicht in der Reaktion der verschiedenen AMPs
nicht. Die gleichzeitige Erhöhung von TFF2 fehlt beispielsweise. Eine verringerte
Reaktion in der Produktion von TFF spielt wahrscheinlich eine Rolle in der
Pathogenese der NEC. Die Tendenzen bleiben in weiteren Studien zu bestätigen.
Vermutlich spielen viele Faktoren mit hinein: die inflammatorische Antwort der
Zytokine, dieser Weg wird beispielsweise anders reguliert als die Produktion der
Cathelicidine (Hase et al. 2002), Neutrophile in hypoxischen Situationen spielen
eine Rolle, weil sie zu einem verminderten oxidativen Burst führen (Carr 2000), die
tight junctions sind in Bezug auf Zusammensetzung und Funktion bei
Frühgeborenen noch unreif (Hunter et al. 2008), Hypoxie und Ischämie führen
ebenfalls zur Barrierestörung und Einwanderung von Bakterien (Sodhi et al. 2008).
Festzuhalten bleibt, α-Defensin 5, TFF1 und LL-37 sind, absolut gesehen, nicht
vermindert im akuten Stadium der NEC im Vergleich zu den gleichaltrigen
Kontrollen, sie sind aber auch nicht deutlich erhöht. Es ist daher wahrscheinlich,
dass diese Reaktion zu langsam und möglicherweise auch zu wenig aggressiv
stattfindet. Das ist aus unseren Ergebnissen zu dem anderen AMP TFF2 zu lesen.
Dieses reagiert eventuell langsamer als das α-Defensin 5, TFF1 und LL-37, bei

49
denen wir die Erhöhung zum Zeitpunkt der Operation schon andeutungsweise
sehen. Nur durch eine multifaktorielle Genese kann erklärt werden, weshalb einige
reifgeborene Kinder erkranken oder andererseits nicht alle frühgeborenen Kinder
erkranken.

50
7. Literaturverzeichnis
1. Ade-Ajayi N, Kiely E, Drake D, Wheeler R, Spitz L. Resection and primary
anastomosis in necrotizing enterocolitis. J R Soc Med. 1996;89(7):385-388.
2. Alfaleh K, Anabrees J, Bassler D, Al-Kharfi T. Probiotics for prevention of
necrotizing enterocolitis in preterm infants. Cochrane Database Syst Rev.
2011.
3. Bell M, Ternberg J, Feigin R, Keating J, Marshall R, Barton L, Brotherton T.
Neonatal necrotizing enterocolitis. Therapeutic decisions based upon clinical
staging. Ann Surg. 1978;187(1):1-7.
4. Berseth C. Feeding strategies and necrotizing enterocolitis. Curr Opin
Pediatr. 2005;17(2):170-173.
5. Berseth C. Gestational evolution of small intestine motility in preterm and
term born infants. J Pediatr. 1989;115(4):646-651.
6. Bhatia J. Strategies to prevent necrotising enterocolitis. Chin Med J (Engl).
2010;123(20):2759-2765.
7. Blakely M, Lally K, McDonald S, Brown R, Barnhart D, Ricketts R,
Thompson W, Scherer L, Klein M, Letton R, Chwals W, Touloukian R,
Kurkchubasche A, Skinner M, Moss R, Hilfiker M. Postoperative outcomes
of extremely low birth-weight infants with necrotizing enterocolitis or
isolated intestinal perforation: a prospective cohort study by the NICHD
Neonatal Research Network. Ann Surg. 2005;241(6):984-994.
8. Buisine M, Devisme L, Savidge T, Gespach C, Gosselin B, Porchet N,
Aubert J. Mucin gene expression in human embryonic and fetal intestine.
Gut. 1998;43(4):519-524.
9. Cantó E, Moga E, Ricart E, Garcia-Bosch O, Garcia-Planella E, Juarez C,
Vidal S. MDP-Induced selective tolerance to TLR4 ligands: impairment in
NOD2 mutant Crohn's disease patients. Inflamm Bowel Dis.
2009;15(11):1686-1696.
10. Carr R. Neutrophil production and function in newborn infants. Br J
Haematol. 2000;110(1):18-28.
11. Claud E, Lu L, Anton P, Savidge T, Walker W, Cherayil B.
Developmentally regulated IkappaB expression in intestinal epithelium and

51
susceptibility to flagellin-induced inflammation. Proc Natl Acad Sci U S A.
2004;101(19):7404-7408.
12. Deplancke B, Gaskins H. Microbial modulation of innate defense: goblet
cells and the intestinal mucus layer. Am J Clin Nutr. 2001;73(6):1131S-
1141S.
13. Doss M, White M, Tecle T, Hartshorn K. Human defensins and LL-37 in
mucosal immunity. J Leukoc Biol. 2010;87(1):79-92.
14. Flenker H. Basiswissen Histologie und Zytologie. Arbeitsbuch für MTA und
BTA. 2 ed: Stein KH; 1998.
15. Franke W. Discovering the molecular components of intercellular junctions
– a historical view. Cold Spring Harb Perspect Biol. 2009;1(3):a003061.
16. Fujii A, Brown E, Mirochnick M, O'Brien S, Kaufman G. Neonatal
necrotizing enterocolitis with intestinal perforation in extremely premature
infants receiving early indomethacin treatment for patent ductus arteriosus. J
Perinatol. 2002;22(7):535-540.
17. Gassler N, Rohr C, Schneider A, Kartenbeck J, Bach A, Obermüller N, Otto
H, Autschbach F. Inflammatory bowel disease is associated with changes of
enterocytic junctions. Am J Physiol Gastrointest Liver Physiol.
2001;281(1):G216-228.
18. González-Rivera R, Culverhouse R, Hamvas A, Tarr P, Warner B. The age
of necrotizing enterocolitis onset: an application of Sartwell's incubation
period model. J Perinatol. 2011:Epub ahead of print.
19. Grosfeld J, Chaet M, Molinari F, Engle W, Engum S, West K, Rescorla F,
Scherer Lr. Increased risk of necrotizing enterocolitis in premature infants
with patent ductus arteriosus treated with indomethacin. Ann Surg.
1996;224(3):350-357.
20. Guillet R, Stoll B, Cotton C, Gantz M, McDonald S, Poole W, Phelps D.
Association of H2-blocker therapy and higher incidence of necrotizing
enterocolitis in very low birth weight infants. Pediatrics. 2006;117(2):e137-
142.
21. Guthrie S, Gordon P, Thomas V, Thorp J, Peabody J, Clark R. Necrotizing
enterocolitis among neonates in the United States. J Perinatol.
2003;23(4):278-285.

52
22. Hamrick S, Hansmann G. Patent ductus arteriosus of the preterm infant.
Pediatrics. 2010;125(5):1020-1030.
23. Hanby A, Chinery R, Poulsom R, Playford R, Pignatelli M. Downregulation
of E-cadherin in the reparative epithelium of the human gastrointestinal tract.
Am J Pathol. 1996;148(3):723-729.
24. Hase K, Eckmann L, Leopard J, Varki N, Kagnoff M. Cell differentiation is
a key determinant of cathelicidin LL-37/human cationic antimicrobial
protein 18 expression by human colon epithelium. Infect Immun.
2002;70(2):953-963.
25. Hay WJ. Strategies for feeding the preterm infant. Neonatology.
2008;94(4):245-254.
26. Himpens E, Van den Broeck C, Ostra A, Calders P, Vanhaesebrouck P.
Prevalence, type, distribution, and severity of cerebral palsy in relation to
gestational age: a meta-analytic review. Dev Med Child Neurol.
2008;50(5):334-340.
27. Hooper L, Wong M, Thelin A, Hansson L, Falk P, Gordon J. Molecular
analysis of commensal host-microbial relationships in the intestine. Science.
2001;291(5505):881-884.
28. Hoy C, Wood C, Hawkey P, Puntis J. Duodenal microflora in very-low-
birth-weight neonates and relation to necrotizing enterocolitis. J Clin
Microbiol. 2000;38(12):4539-4547.
29. Hunter C, Upperman J, Ford H, Camerini V. Understanding the
susceptibility of the premature infant to necrotizing enterocolitis (NEC).
Pediatric Research. 2008;63(2):117-123.
30. Husebye E, Hellström P, Sundler F, Chen J, Midtvedt T. Influence of
microbial species on small intestinal myoelectric activity and transit in germ-
free rats. Am J Physiol Gastrointest Liver Physiol. 2001;280(3):G368-380.
31. Keshav S. Paneth cells: leukocyte-like mediators of innate immunity in the
intestine. J Leukoc Biol. 2006;80(3):500-508.
32. Kumar H, Kawai T, Akira S. Pathogen recognition in the innate immune
response. Biochem J. 2009;420(1):1-16.
33. Lebenthal A, Lebenthal E. The ontogeny of the small intestinal epithelium.
JPEN J Parenter Enteral Nutr. 1999;23(5 Suppl):S3-6.
34. Lin P, Stoll B. Necrotising enterocolitis. Lancet. 2006;368(9543):1271-1283.

53
35. Llanos A, Moss M, Pinzòn M, Dye T, Sinkin R, Kendig J. Epidemiology of
neonatal necrotising enterocolitis: a population-based study. Paediatr
Perinat Epidemiol. 2002;16(4):342-349.
36. Lucas A, Cole T. Breast milk and neonatal necrotising enterocolitis. Lancet.
1990;336(8730):1519-1523.
37. Mallow E, Harris A, Salzman N, Russell J, DeBerardinis R, Ruchelli E,
Bevins C. Human enteric defensins. Gene structure and developmental
expression. J Biol Chem. 1996;271(8):4038-4045.
38. Niessen C, Gottardi C. Molecular components of the adherens junction.
Biochim Biophys Acta. 2008;1778(3):562-571.
39. O'Donovan D, Baetiong A, Adams K, Chen A, Smith E, Adams J, Weisman
L. Necrotizing enterocolitis and gastrointestinal complications after
indomethacin therapy and surgical ligation in premature infants with patent
ductus arteriosus. J Perinatol. 2003;23(4):286-290.
40. Peter C, Feuerhahn M, Bohnhorst B, Schlaud M, Ziesing S, von der Hardt H,
Poets C. Necrotising enterocolitis: is there a relationship to specific
pathogens? Eur J Pediatr. 1999;158(1):67-70.
41. Ramsay S, Santella R. The definition of life: a survey of obstetricians and
neonatologists in new york city hospitals regarding extremely premature
births. Matern Child Health J. 2011;15(4):446-452.
42. Richter M, Topf H, Gröschl M, Fröhlich T, Tzschoppe A, Wenzl T, Köhler
H. Influence of gestational age, cesarean section, and type of feeding on
fecal human beta-defensin 2 and tumor necrosis factor-alpha. J Pediatr
Gastroenterol Nutr. 2010;51(1):103-105.
43. Rippa E, La Monica G, Allocca R, Romano M, De Palma M, Arcari P.
Overexpression of gastrokine 1 in gastric cancer cells induces fas-mediated
apoptosis. J Cell Physiol. 2010(Epub ahead of print).
44. Salzman N. Paneth cell defensins and the regulation of the microbiome:
Détente at mucosal surfaces. Gut Microbes. 2010;1(6):401-406.
45. Salzman N, Polin R, Harris M, Ruchelli E, Hebra A, Zirin-Butler S, Jawad
A, Martin Porter E, Bevins C. Enteric defensin expression in necrotizing
enterocolitis. Pediatr Res. 1998;44(1):20-26.
46. Schaart M, de Bruijn A, Bouwman D, de Krijger R, van Goudoever J,
Tibboel D, Renes I. Epithelial functions of the residual bowel after surgery

54
for necrotising enterocolitis in human infants. J Pediatr Gastroenterol Nutr.
2009;49(1):31-41.
47. Schanler R, Shulman R, Lau C. Feeding strategies for premature infants:
beneficial outcomes of feeding fortified human milk versus preterm formula.
Pediatrics. 1999;103(6 Pt 1):1150-1157.
48. Schauber J, Svanholm C, Termén S, Iffland K, Menzel T, Scheppach W,
Melcher R, Agerberth B, Lührs H, Gudmundsson G. Expression of the
cathelicidin LL-37 is modulated by short chain fatty acids in colonocytes:
relevance of signalling pathways. Gut. 2003;52(5):735-741.
49. Schnabl K, Van Aerde J, Thomson A, Clandinin M. Necrotizing
enterocolitis: a multifactorial disease with no cure. World J Gastroenterol.
2008;14(14):2142-2161.
50. Shu U, Demeure C, Byun D, Podlaski F, Stern A, Delespesse G. Interleukin
12 exerts a differential effect on the maturation of neonatal and adult human
CD45R0- CD4 T cells. J Clin Invest. 1994;94(4):1352-1358.
51. Sodhi C, Richardson W, Gribar S, Hackam D. The development of animal
models for the study of necrotizing enterocolitis. Dis Model Mech. 2008;1(2-
3):94-98.
52. Soriano-Izquierdo A, Gironella M, Massaguer A, May F, Salas A, Sans M,
Poulsom R, Thim L, Piqué J, Panés J. Trefoil peptide TFF2 treatment
reduces VCAM-1 expression and leukocyte recruitment in experimental
intestinal inflammation. J Leukoc Biol. 2004;75(2):214-223.
53. Vieten D, Corfield A, Carroll D, Ramani P, Spicer R. Impaired mucosal
regeneration in neonatal necrotising enterocolitis. Pediatr Surg Int.
2005;21(3):153-160.
54. Wang C, Shoji H, Sato H, Nagata S, Ohtsuka Y, Shimizu T, Yamashiro Y.
Effects of oral administration of bifidobacterium breve on fecal lactic acid
and short-chain fatty acids in low birth weight infants. J Pediatr
Gastroenterol Nutr. 2007;44(2):252-257.
55. Wehkamp J, Harder J, Weichenthal M, Schwab M, Schäffeler E, Schlee M,
Herrlinger K, Stallmach A, Noack F, Fritz P, Schröder J, Bevins C,
Fellermann K, Stange E. NOD2 (CARD15) mutations in Crohn´s disease are
associated with diminished mucosal alpha-defensin expression. Gut.
2004;53(11):1658-1664.

55
56. Wiesner J, Vilcinskas A. Antimicrobial peptides: the ancient arm of the
immune system. Virulence. 2010;1(5):440-464.
57. Zouali H, Bonnard A, De Lagausie D, Farnoux C, Aigrain Y, Cezard J,
Peuchmaur M, Hugot J, Berrebi D. CARD15/NOD2 is not a predisposing
factor for necrotizing enterocolitis. Dig Dis Sci. 2005;50(9):1684-1687.

56
8. Abkürzungsverzeichnis
AMP antimikrobielles Peptid
APC Antigen-präsentierende Zellen
Aqua dest. Aqua destillata
cDNA komplementäre Desoxyribonukleinsäure
CAMP Cathelicidin antimikrobielles Peptid
DAB 3,3’-Diaminobenzidin
DEPC-Aqua dest. Diethylpyrocarbonat-Aqua destillata
DIC disseminierte intravasale Gerinnung
DNA Desoxyribonukleinsäure
dNTP Desoxyribonukleosidtriphosphat
EDTA Ethylendiamintetraessigsäure
ELBW extremely low birth weiht
ELISA enzyme-linked immunosorbent assay
FAM 6-Carboxyfluorescein
FRET Förster/Fluoreszenz-Resonanzenergietransfer
GALT gut associated lymphoid tissue
gDNA genomische Desoxyribonukleinsäure
GKN1 Gastrokin 1
hBD-2 humanes β-Defensin 2
hCAP-18 humanes kationisches antimikrobielles Protein 18
HD-5, HD-6 humanes Defensin 5 und 6
HLHS hypoplastisches Linksherz-Syndrom
HNP-1-4 humane Neutrophilen-Peptide 1-4
HPRT Hypoxanthin-Guanin-Phosphoribosyltransferase
IFNγ Interferon γ
IgA, IgG Immunglobuline A und G
IκB Inhibitor von κB
IL1, 1β, 4, 6, 8, 12, 18 Interleukine 1, 1β, 4, 6, 8, 12, 18
iNOS induzierbare NO-Synthase
IVH intraventrikuläre Hämorrhagie
KS-30 alternativ prozessiertes Derivat von LL-37
LBW low birth weight

57
LL-37 antimikrobiell wirksames C-terminales CAMP-Gen
Produkt aus 37 Aminosäuren
LPS Lipopolysaccharid
MAMP microbe-associated molecular pattern
MAPK Mitogen-aktivierte Proteinkinase
MDP Muramyldipeptid
MEF minimal enteral feeding
MLPA multiplex ligation-dependent probe amplification
MMLV-Puffer Puffer zur reversen Transkription
MMP7 Matrixmetalloproteinase 7
mRNA messenger RNA
NBW normal birth weight
NEC nekrotisierende Enterokolitis
NFκB nuclear factor κB
NICHD National Institute of Child Health and Human
Development
NK-Zelle natürliche Killerzelle
NO nitric oxide (Stickstoffmonoxid)
NOD2 Nucleotide-binding oligomerization domain-
containing protein 2
NOS NO-Synthase
OEIS-Syndrom kaudales Fehlbildungssyndrom
ONOOˉ Peroxynitrit
OP Operation
PAF Plättchen-aktivierender Faktor
Pank. an. Pankreas anulare
PBGD Porphobilinogen-Desaminase
PBMC peripheral blood mononuclear cells
PCR Polymerase Kettenreaktion
PDA persistierender Ductus arteriosus Botalli
PKD-Puffer Puffer zur Lösung von Formaldehydbrücken zwischen
Nukleinsäuren
PRR pattern recognition receptor
PRT paralogue ratio test

58
pS2 anderer Name für TFF1
RK-31 alternativ prozessiertes Derivat von LL-37
RNA Ribonukleinsäure
ROX Farbstoff als passive Referenz
RPE-Puffer Waschpuffer
RT-PCR reverse Transkriptase-Polymerase Kettenreaktion
sp. species
SSW Schwangerschaftswoche
TAK1 transforming growth factor β-aktivierte Kinase 1
TAMRA Tetramethylrhodamin
TaqMan-RT-PCR TaqMan-Real Time-Polmerase Kettenreaktion
TBS-Puffer Tris-Salz-Puffer
TFF trefoil factor
TLR toll-like receptor
TNFα Tumornekrosefaktor α
VACTERL-Assoziation komplexes Fehlbildungssyndrom
VLBW very low birth weight
Wnt/Tcf-4 Signalweg

59
9. Danksagung
Ich möchte mich bedanken bei meinem Doktorvater Prof. Dr. Henrik Köhler für das
in mich gesetzte Vertrauen und die Betreuung meiner Arbeit. Nachdem Prof. Dr.
Köhler in die Schweiz gerufen worden war, bedanke ich mich ganz besonders bei
Aline Rückel, die es übernommen hat, jederzeit vor Ort ansprechbar zu sein und ein
offenes Ohr für meine Probleme zu haben. Vielen Dank für deine kritischen und
konstruktiven Anmerkungen!
Mein großer Dank gilt Elke Licha, die später Elke Schirmer wurde, aus dem
Forschungslabor der Kinderklinik Erlangen. Sie hat mich in alle Methoden
eingeführt, immer Zeit und Rat für mich gehabt. Ich danke dir!
Und allen anderen aus dem Laborteam: Krissy, Kathrin, Gudrun, Ida, Yvonne, Julia,
Sabine, Manu, Manfred, Tina, Beate, Annika, Nico und Ulla.
An das Humangenetische Institut der Universitätsklinik Erlangen ergeht mein Dank
für das Überlassen der Ergebnisse der Messung der Genkopienzahl von β-Defensin
2, auch wenn diese schließlich in der Arbeit nicht verwendet wurden. Vielen Dank
an Frau PD Dr. Ulrike Hüffmeier!
Ich danke Herrn Dr. Fröhlich, der nach dem Umzug des Forschungslabors in die alte
Dermatologie meine Immunhistochemie-Schnitte aufbewahrt hat.
Meinem Bruder Jeremias und meiner Schwester Hanna danke ich für das
Korrekturlesen meiner Arbeit. Vielen Dank an Sabine Neumann für das
Korrekturlesen der englischen Zusammenfassung!
Schlussendlich möchte ich mich bei meiner ganzen Familie für die Ermöglichung
meines Studiums, für das Interesse und die Unterstützung bedanken.

60
10. Erklärung
Hiermit erkläre ich, dass die vorliegende Dissertation von mir in allen Teilen
selbstständig angefertigt wurde und keine anderen als die angegebenen Quellen und
Hilfsmittel benutzt wurden. Darüber hinaus erkläre ich, dass die Dissertationsschrift
weder vollständig, noch teilweise einer anderen Fakultät mit dem Ziel vorgelegt
wurde, einen akademischen Grad zu erwerben.

61
11. Lebenslauf
Persönliche Daten:
Name Kaiser
Vornamen Agnes Gabriele
Geburtsdatum 25.05.1986
Geburtsort Neustadt a.d. Waldnaab
Familienstand ledig
Religionszugehörigkeit evangelisch
Eltern Hanns-Friedrich Kaiser, Kantor
Friederike Kaiser, Hausfrau
Geschwister Dorothea Kaiser, Ausbildung Geigenbau
Hanna Kaiser, Studentin der Biologie
Jeremias Kaiser, Student der Elektrotechnik,
Elektronik und Informationstechnik
Schulausbildung:
1992 – 1996 Clausnitzer-Grundschule in Weiden i.d. Opf.
1996 – 2005 Humanistisches Augustinus-Gymnasium Weiden
i.d. Opf., Abschluss mit dem Abitur
Hochschulausbildung:
April 2006 – Juni 2012 Studium der Humanmedizin an der Friedrich-
Alexander-Universität Erlangen
01.04.2008 Erster Abschnitt der Ärztlichen Prüfung
Februar 2011 – Januar 2012 Praktisches Jahr:
1. Tertial: Chirurgische Klinik,
Universitätsklinikum Erlangen
2. Tertial: Kinder- und Jugendklinik,
Universitätsklinikum Erlangen
3. Tertial: Medizinische Kliniken 1 und 2,
Universitätsklinikum Erlangen
21.06.2012 Zweiter Abschnitt der Ärztlichen Prüfung

62
Mai 2010 – heute Doktorarbeit bei Prof. Dr. H. Köhler,
Universitätskinderklinik Erlangen, Thema: „Die
Rolle der antimikrobiellen Peptide und des E-
Cadherins in der Pathogenese der
Nekrotisierenden Enterokolitis“
Pflegepraktikum:
Januar – März 2006 Neurologische Klinik, Klinikum Weiden i. d. Opf.;
Dr. med. M. Angerer
Famulaturen:
August 2008 Dr. med. H. Vielhaber, Klinik für Kinderheilkunde
und Jugendmedizin, Klinikum Weiden
September 2008 Prof. Dr. med. Dr. rer. nat. U. Eysholdt,
Phoniatrische und Pädaudiologische Abteilung,
Universitätsklinikum Erlangen
Februar – März 2009 Dr. med. H. Beyer, Kardiologische Intensivstation,
Waldkrankenhaus Erlangen
Juli 2009 Dr. med. D. Kramer, Praktische Ärztin
Naturheilverfahren, Dechsendorf
September 2009 Prof. Dr. med. K.-H. Dietl, Kinik für Allgemein-,
Thorax- und Viszeralchirurgie, Klinikum Weiden
März 2010 Dr. med. T. Ebert, Kinder- und Jugendarztpraxis,
Veitsbronn
September 2010 Dr. med. J. Altmeppen, Klinik für Anästhesie,
operative Intensivmedizin und Notfallmedizin,
Klinikum Weiden
Beruflicher Werdegang:
September 2012 Assistenzärztin in der Klinik für Kinder und
Jugendmedizin des Klinikum Coburg



![4.21 Peptide und Proteine 4.22 Eigenschaften und Nachweis ... · molekül abgespalten, die Reaktion ist daher eine Kondensationsreaktion [B2]. ... Röntgenstrukturanalysen zeigen,](https://static.fdokument.com/doc/165x107/5d67735e88c993f41f8b8ff2/421-peptide-und-proteine-422-eigenschaften-und-nachweis-molekuel-abgespalten.jpg)














