
Environmental · PDF fileEnvironmental Studies Am Herberhaeuser Weinberge 23 D-37075...
68
Environmental Studies Am Herberhaeuser Weinberge 23 D-37075 Goettingen e-mail: [email protected] www.environmental-studies.de ________________________________________________ Radioökologie Abschlussbericht zum Forschungsvorhaben StSch 4206 im Auftrag des Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit „Überprüfung von Ökosystemen nach Tschernobyl hinsichtlich der Strahlenbelastung der Bevölkerung“ Von U. Fielitz Juli 2001 1
Transcript of Environmental · PDF fileEnvironmental Studies Am Herberhaeuser Weinberge 23 D-37075...

Environmental Studies Am Herberhaeuser Weinberge 23 D-37075 Goettingen e-mail: [email protected] www.environmental-studies.de
________________________________________________ Radioökologie
Abschlussbericht zum Forschungsvorhaben StSch 4206
im Auftrag des
Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit
„Überprüfung von Ökosystemen nach Tschernobyl hinsichtlich der Strahlenbelastung der Bevölkerung“
Von U. Fielitz Juli 2001
1

Alle Aktivitätsmessungen im Rahmen dieses Forschungsvorhabens wurden am Isotopenlaboratorium für biologische und medizinische Forschung der Universität Göttingen durchgeführt.
Sämtliche, in diesem Bericht vorkommenden Daten, die sich auf Messungen aus den Jahren 1994 und 1995 beziehen, wurden vom Autor, im Rahmen des Forschungsvorhabens “Radiocäsiumverteilung und –trends bei Wildbret in Bayern”, das von der Bundesanstalt für Fleischforschung, im Auftrag des Bayerischen Staatsministeriums für Landes-entwicklung und Umweltfragen durchgeführt wurde, erhoben.
Alle übrigen Daten wurden, von 1988 bis 1994, in den Forschungsvorhaben StSch 4015 und 4042 „Radioaktivität in Wildtieren“, im Auftrag des Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit gewonnen.
Die Arbeiten wurden mit Mitteln des Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit finanziert.
Der Bericht gibt die Auffassung und Meinung des Auftragnehmers wieder und muss nicht mit der Meinung des Auftraggebers (Bundesministerium für Umwelt, Naturschutz und Reaktorsicherheit) übereinstimmen.
2

Inhaltsverzeichnis Kurzfassung....................................................................................................................
Abstract............................................................................................................................
1 Einleitung.........................................................................................................................
2 Untersuchungsgebiete.....................................................................................................
3 Ziele und Methoden.........................................................................................................
4 Aktivitätsbestimmung.....................................................................................................
5 Cs-137 Aktivität in Waldböden...................................................................................... 5.1 Allgemeines...............................................................................................................5.2 Cs-137 Flächenbelastung der Böden im Untersuchungsgebiet Bodenmais..............5.3 Bodenkontamination der Probeflächen B1 und F1................................................... 5.4 Tiefenverteilung von Cs-137 in Bodenprofilen........................................................ 5.5 Zeitlicher Verlauf von Cs-137 in Böden...................................................................
6 Cs-137 Kontamination in Pflanzen, Beeren und Pilzen...............................................
6.1 Allgemeines..................................................................................................................... 6.2 Voruntersuchungen zur Cs-137 Aktivität von Waldbodenpflanzen......................... 6.3 Cs-137 Aktivität in der Waldbodenvegetation......................................................... 6.4 Zeitverlauf von Cs-137 in Waldbodenpflanzen........................................................ 6.5 Cs-137 in Früchten.................................................................................................... 6.6 Cs-137 in Pilzen.........................................................................................................
7 Cs-137 Aktivität in Wildtieren....................................................................................... 7.1 Cs-137 Aktivität in Wildschweinen.......................................................................... 7.2 Cs-137 Aktivität in Rothirschen............................................................................... 7.3 Cs-137 Aktivität in Rehen.........................................................................................7.4 Cs-137 Aktivität in Kühen........................................................................................ 7.5 Höhe und zeitliche Entwicklung der Cs-137 Kontamination bei Rehen,
Rothirschen und Wildschweinen..............................................................................
8 Cs-137 in verschiedenen Biomedien des Waldes, ein Überblick.................................
9 Transferfaktoren von Cs-137 Boden→Biomedien.......................................................
10 Vorräte von Cs-137 im Waldökosystem der Probefläche B1......................................
11 Zusammenfassende Diskussion......................................................................................
12 Literaturverzeichnis........................................................................................................
Danksagung.....................................................................................................................
3
1
2
3
4
7
8
9 9 10 12 12 21
23
23 24 26 28 36 38
43 43 44 45 49 49
51
52
55
58
62
65

Kurzfassung Seit 1988 wurde, im Rahmen mehrerer Forschungsvorhaben, die Cs-137 Aktivität in rund 7.000 Proben von Boden, Pflanzen, Pilzen und Wildtieren bestimmt, um die Dynamik des Nuklids in Waldökosystemen zu untersuchen. Die Arbeiten wurden hauptsächlich in einem Bergfichtenwald in Bodenmais (Bayern) und einem Eichenwald in Fuhrberg (Nieder-sachsen) durchgeführt, wo ungünstige Bodenparameter und Standortfaktoren relativ hohe Radiocäsium-Aufnahmeraten in Biomedien bedingen. Das Untersuchungsgebiet Boden-mais gehört, mit durchschnittlich 53.690 Bq Cs-137•m-2 (Stand 1. Mai 1986, n=59), zu den durch den Tschernobyl-Fallout am höchsten kontaminierten Gebieten der Bundesrepublik. 1999 wurde ein Buchenwald in Göttingen (Niedersachsen), mit günstigen Standort-faktoren in die Untersuchungen mit einbezogen. Boden- und Pflanzenproben wurden aus-schließlich auf 100x100 m großen Probeflächen entnommen.
Kennzeichnend für die untersuchten Waldökosysteme ist die große Variabilität der Cs-137 Aktivität innerhalb einer Art und zwischen den Arten. In Bodenprofilen aus den drei Untersuchungsgebieten befanden sich im Jahr 2000, durchschnittlich noch 56% des Cs-137 Inventars in den obersten 10 cm und 93% in den obersten 20 cm.
In den untersuchten Pflanzenarten nahm Cs-137 Aktivität von 1987 bis 1994 signifikant ab, wobei sich die Kontamination seit 1995 nur noch wenig veränderte. Bei acht Pflanzen-arten aus zwei der Untersuchungsgebiete betrug die mittlere effektive Halbwertzeit –5,3 Jahre (Minimum –3 Jahre, Maximum –24 Jahre).
Pilze aus dem Untersuchungsgebiet Bodenmais hatten 2000, wie üblich, die höchsten Cs-137 Messwerte: Ziegenlippe und Hallimasch durchschnittlich 400 Bq•kg-1, Pfifferling, Steinpilz und Rotfußröhrling 1.000 Bq•kg-1 und Maronenröhrling 3.030 Bq•kg-1, bezogen auf Frischsubstanz. Mit durchschnittlich 25.660 Bq•kg-1 waren die unterirdisch wachsen-den Hirschtrüffel deutlich höher kontaminiert als alle oberirdisch wachsenden Pilzarten.
Die Werte der aggregierten Transferfaktoren (Tag) für den Übergang von Cs-137 Boden→Pflanze und Boden→Fleisch schwankten um mehrere Größenordnungen, z.B. bei den autotrophen Pflanzenarten, von 0,0001 m2•kg-1 bis 0,41 m2•kg-1. Auf den Dauer-probeflächen B1 und F1 wurden viel höhere Transferfaktoren festgestellt als auf G1. So war der Tag für Wildschweine aus Bodenmais 392mal höher als für Wildschweine aus Göttingen. Diese Unterschiede sind im wesentlichen durch unterschiedliche bodenbürtige Standortparameter der Probeflächen bedingt.
Die Cs-137 Kontamination von Rehen variierte in Bodenmais von 1987 bis 2000 saisonal, mit niedrigen Werten im Frühjahr und deutlich höheren Werten im Herbst. Ähnlich wie bei den Pflanzen, erfolgte bei Rehen, von 1987 bis 1995, eine relativ schnelle Abnahme der Cs-137 Aktivität, während von 1996 bis 2000 keine signifikante Veränderung festzustellen war. Die effektive Halbwertzeit für Cs-137 in Rehwild (n=1.429) beträgt für diesen Zeitraum -6 Jahre. Im Jahr 2000 war der Median der Cs-137 Messwerte von Rehen aus Bodenmais 776 Bq•kg-1, von Wildschweinen 7.890 Bq•kg-1. Bei Wildschweinen ist, von 1987 bis 2000, keine signifikante Abnahme der Kontamination festzustellen.
1

Abstract Since 1988, within the scope of several research projects, in 7,000 samples of soil, plants, mushrooms and game from forest ecosystems, the 137Cs activity concentration was measured, in order to investigate the dynamics of the nuclide. The investigation sites are a spruce mountain forest near the village Bodenmais (Bavaria) and an oak forest close to Fuhrberg (Lower Saxony). In both forests, unfavourable location conditions cause a relativ high transfer of 137Cs into plants and game. At Bodenmais, the mean 137Cs inventory was 53,690 Bq•m-2 (corrected to 1 May 1986, n=59), in Germany this site belongs to the highest contaminated areas by the Chernobyl accident. 1999, a different investigation site, a beech forest close to Göttingen with favourable location conditions and resulting low 137Cs transfers was included in the study. All soil and plant samples were taken exclusively from 100x100 m permanent study plots.
Typifying for the 3 forest sites was the high intra- and interspecies variability of the 137Cs activity concentration. Even 14 years after the Chernobyl-fallout at the 3 investigation sites, the average 137Cs inventory, contained in the top 10 cm of soil was 56% and 93% in the top 20 cm.
From 1987 till 1994, in the leaves of the investigated plant species the 137Cs activity concentration decreased significant, during the following years there was little change. The effective half life of 137Cs varies between –3 years for raspberry and –24 years for the fern Pteridium aquillinum, whereas most of the plant species show half lifes of about -5 years.
In 2000, as usual mushrooms from the Bodenmais investigation site showed the highest 137Cs contaminations: Xerocomus subtomentosus and Armillaria mellea had about 400 Bq•kg-1, Cantharellus cibarius, Boletus edulis and Xerocomus chrysenteron about 1,000 Bq•kg-1, Xerocomus badius 3,030 Bq•kg-1 and Cortinarius hercynicus 6,750 Bq•kg-1, in fresh weight. The 137Cs activity concentration of the underground growing Elaphomyces granulatus had 25,660 Bq Cs-137•kg-1 and exceeded all overground fructifying mush-rooms.
The aggregated transfer factors (Tagg) for soil→plant and soil→flesh varied with several orders of magnitude. Tagg values for soil→autotroph plant species reached from 0,0001 m2•kg-1 to 0,41 m2•kg-1. While at the permanent study plots in Bodenmais and Fuhrberg the Tagg values were of comparable quantity, at Göttingen, they were lower than two orders of magnitude. For example Tagg for Cs-137 in wild boar from Bodenmais was 392 times higher than for wild boar from Göttingen.
From 1987 till 2000, the 137Cs activity in roe-deer from Bodenmais varied according to the seasons, with highest values in autumn, and lowest values in spring. In consequence of the decrease of the 137Cs activity concentration in grazing plants, from 1987 until 1995, the 137Cs contamination in roe deer (n=1.429) declined, but from 1996 till 2000 it stagnated. The effective half-life of Cs-137 in roe deer was –6 years. In 2000, the median of the 137Cs values in roe deer from Bodenmais was 776 Bq•kg-1, for wild boar 7,890 Bq•kg-1. There was no significant change in the 137Cs contaminantion of wild boar, from 1987 till 2000.
2

1 Einleitung Die langfristige Strahlenbelastung der Umwelt durch den Tschernobyl-Fallout wird im wesentlichen durch das Isotop Cäsium-137 verursacht. Während die Radiocäsium-Kontamination in landwirtschaftlich erzeugten Grundnahrungsmitteln pflanzlicher Her-kunft schon seit Jahren wieder auf das Niveau vor dem Reaktorunfall abgesunken ist, werden auch heute noch in einigen Waldgebieten vergleichsweise hohe Cs-137 Gehalte in Beeren, Pilzen und Wildtieren festgestellt. Diese Unterschiede in der Kontamination sind dadurch bedingt, dass der Transfer von Radiocäsium in Pflanzen auf landwirtschaftlich genutzten Böden durch die Fixierung von Cs-Ionen in Tonmineralen, das hohe Nährstoff-angebot und einen hohen pH-Wert, nur gering, auf Waldböden aber, insbesondere im Auflagehumus, relativ hoch ist. Darüber hinaus finden in Waldökosystemen, neben rein physikalischen (Sorption, Fixierung etc.), auch dynamisch biologische Prozesse (Streu-eintrag, Zersetzung etc.) statt, die die Kontamination von Vegetation und Wildtieren in komplexer Weise beeinflussen.
In verschiedenen Forschungsvorhaben wurde bereits von 1987 bis 1998 untersucht, wie sich Radiocäsium in charakteristischen Biomedien aus Waldökosystemen räumlich und zeitlich in Böden, Waldbodenpflanzen, Bäumen, Pilzen und Wildtieren verhält (FIELITZ 1994).
Ziel der vorliegenden Untersuchungen war es, systematisch zu klären, wie hoch die Cs-137 Gehalte, 14 Jahre nach dem Reaktorunfall in Tschernobyl, in den wichtigsten Kompar-timenten verschiedener Waldökosysteme sind. Der überwiegende Teil der Arbeiten wurde in den Wäldern um Bodenmais durchgeführt. Diese sind durch den Tschernobyl-Fallout besonders betroffen, auf der Dauerprobefläche B1 betrug die Cs-137 Flächenaktivität, im Mai 2000, rund 72.000 Bq•m-2. Aufgrund der ungünstigen Standortfaktoren, ergibt sich hier ein relativ hoher Transfer des Nuklids in die Nahrungskette. Die Cs-137 Gehalte von Pflanzen, Beeren, Pilzen und Wildtieren aus den Wäldern von Bodenmais gehören zu den höchsten der Bundesrepublik. Um einen Überblick über die Spannweite der Kontamination und den Transfer von Cs-137 in Biomedien von Waldökosystemen zu erhalten, wurde auch ein Gebiet in Göttingen untersucht. Mit rund 5.000 Bq•m-2 (Stand: Mai 2000) ist die Cs-137 Aktivität des Bodens relativ gering. Die sehr gute Nährstoffversorgung, der hohe pH-Wert und weitere günstige Standortfaktoren führen zu einem geringen Transfer des Nuklids in die Biomasse. Schließlich wurde ein weiteres Waldökosystem in Fuhrberg, Niedersachsen, untersucht das eine Bodenkontamination von 8.050 Bq•m-2 (Stand: Mai 2000) hat, aber schlechte Standortbedingungen, die einen höheren Transfer von Radio-cäsium verursachen.
Die Arbeiten sollen Auskunft darüber geben, wie sich nach dem Reaktorunfall, der Verlauf der Cs-137 Aktivität in Pflanzen und Tieren, in unterschiedlichen Waldökosystemen entwickelt. Aus den mehrjährigen Zeitreihen über die Cs-137 Aktivität in den wichtigsten Biomedien soll die Dynamik des Nuklids ermittelt, sowie effektive Halbwertzeiten und aggregierte Transferfaktoren berechnet werden.
3

2 Untersuchungsgebiete Von 1988 bis 2000 wurden die Untersuchungen, mit kurzen Unterbrechungen, in den Wäldern um die Ortschaften Bodenmais und Fuhrberg durchgeführt. 1999 und 2000 wurde ein Wald bei Göttingen in die Arbeiten mit einbezogen. Die Lage der Untersuchungs-gebiete ist in der Abbildung 1 angegeben.
#
#
#
Fuhrberg
Göttingen
Bodenmais
0 200 400 Kilometer
Abb. 1: Lage der Untersuchungsgebiete
Das Untersuchungsgebiet Bodenmais liegt 30 km nordöstlich von Deggendorf, im Wuchsbezirk "östlicher vorderer Bayerischer Wald" zwischen 550-1.450 m ü. M.. Im Jahresmittel beträgt die Temperatur 5 oC und es fallen, je nach Höhenlage, zwischen 1.140-1.550 mm Niederschlag. Im Winter liegt eine geschlossene Schneedecke von z.T. über 1 Meter Höhe, oft bis April. Das Gebiet ist von zusammenhängenden, submontanen bis montanen Bergmischwäldern, mit Fichte und Buche als Hauptbaumarten, bewachsen. Der vorherrschende Bodentyp ist Braunerde.
Das Untersuchungsgebiet Fuhrberg liegt 15 km nordöstlich von Hannover im "ostniedersächsischen Flachland", im Wuchsbezirk "Süd-Heide", auf 40-80 m ü. M.. Die Hauptbaumarten sind Eiche und Kiefer. Mit durchschnittlich 600 mm Niederschlag pro Jahr ist das Klima hier deutlich trockener als in Bodenmais.
Bei dem Untersuchungsgebiet in Göttingen handelt es sich um einen Kalkbuchenwald, in dem vereinzelt Esche, Spitzahorn, Stieleiche, Traubeneiche und Ulme vorkommen. Im langjährigen Mittel wurden 610 mm Niederschlag gemessen.
4

In allen Untersuchungsgebieten wurden 100 x 100 m große Dauerflächen eingerichtet, die im folgenden als B1 und B2 (Bodenmais), F1 (Fuhrberg) und G1 (Göttingen) bezeichnet werden. Die Probeflächen in Bodenmais und Fuhrberg wurden 1988 ausgewiesen. Bei G1 in Göttingen handelt es sich um eine Probefläche, die seit den 60er Jahren im Rahmen von mehreren nationalen und internationalen Projekten der Waldschadens-Forschung durch die Universität Göttingen untersucht wird. Die wichtigsten Kenndaten der Probeflächen sind in der Tabelle 1 zusammengestellt.
In Bodenmais liegt die Probefläche B1 etwa 920 m ü. M., Hauptbaumart ist Fichte, mit einem Bestandsalter von 120 Jahren. Auf schwach podsoliger Braunerde ist ein 2-8 cm starker, schwach rohhumusartiger Moder ausgebildet. Der pH-Wert beträgt im Auflage-humus 3,2, im Mineralboden (0-5 cm) 3,4.
Die Probefläche B2 befindet sich in 980 m ü. M. und ist mit Fichte (120 jährig), Tanne (120 jährig), Buche (30-110 jährig) und Eberesche (30 jährig) bestockt. Über einer Parabraunerde ist ein 2-8 cm starker, rohhumusartiger Moder ausgebildet. Der pH-Wert beträgt sowohl im Humus als auch im Mineralboden (0-5 cm) 3,3. Die Entfernung zur Fläche B1 beträgt 4 km.
Die Probefläche F1 liegt 41 m ü. M. und ist vorwiegend mit Eiche bestockt (Bestandsalter 130 Jahre). Auf einem Podsol - Gley liegt ein 2–8 cm dicker, schwach rohhumusartiger Moder. Der pH-Wert beträgt im Humus 3,1, im Mineralboden (0-5 cm) 2,9. Es handelt sich um einen grundfeuchten, nur schwach Nährstoff versorgten Standort .
Auf der Probefläche G1 stockt ein 120 jähriger Buchenwald, 400 m ü. M. auf Muschel-kalk, mit vereinzelten Edellaubhölzern. Als Bodentyp hat sich eine Terrafusca Rendzina von durchschnittlich 30 cm, über festem Kalkgestein entwickelt. Die Streu ist nur <1cm stark und wird innerhalb von anderthalb Jahren von bodenwühlenden Tieren in den Mineralboden eingearbeitet. Entsprechend handelt es sich um einen sehr humus- und skelettreichen Ah-Horizont.
Die effektive Kationenaustauschkapazität (AKE), ein Maß für die Nährstoffversorgung, ist auf G1 in den oberen 5 cm des Bodens mit 740 mymol (eq)•g-1 wesentlich höher als auf B1, mit 144 mymol (eq)•g-1, sowie auf B2 mit 138 mymol (eq)•g-1 und F1 mit 51 mymol (eq)•g-1.
Auch die Azidität ist auf G1, in dem Bodenbereich 0-5 cm, mit einem pH-Wert von 6,7, wesentlich höher als auf den anderen Probeflächen, wo die Werte zwischen 2,9 und 3,4 liegen.
Insgesamt führen diese Standortfaktoren zu unterschiedlichen Artenspektren bei Wald-bodenpflanzen und Pilzen. So kommen z.B. auf den Flächen B1 und B2 Heidelbeere und Seegras vor, auf F1 Heidelbeere, Pfeifengras und Adlerfarn, während auf G1 Heidelbeere völlig fehlt und Frauenfarn und Dornfarn nur vereinzelt wachsen.
5

Tab. 1: Charakterisierung der Dauerprobeflächen B1 und B2 in Bodenmais, F1 in Fuhrberg und G1 in Göttingen
Dauerfläche B1 B2 F1 G1
Höhe m ü.M. [m] 920 980 41 400
Niederschlag [mm•a-1] 1.200 1.250 600 610
Bestockung Fichte, (Tanne, Buche) Fichte, Tanne, Buche Eiche Buche
Humusform rohhumusartiger rohhumusartiger rohhumusartiger Mull Moder (0-4 cm) Moder (0-8 cm) Moder (0-4 cm) (0,7 cm)
Bodentyp Podsol - Braunerde Parabraunerde Podsol - Gley Terra fusca Rendzina
pH (CaCl2) 3,4 3,3 2,9 6,7Mineralboden (0-5cm)
Kationenaustausch- K 21; Ca 27; Mg 78; K 31; Ca 45; K 11; Ca 35; Mg 11; kapazität Humus Fe 300; Al 448; Mg 61; Fe 210; Fe 94; Al 89 [mymol(eg)•g-1]
Kationenaustausch- K 3,5; Ca 7,8; Mg 3,8; K 3,0; Ca 10; Mg 4,0; K 1,1; Ca 3,8; Mg 1; K 10,1;Ca 701; kapazität Mineralboden Fe 2,1; Al 117 Fe 8,2; Al 105; Fe 2,2; Al 22; Mg 18,7 0-5 cm [mymol(eg)•g-1]
AKE [mymol(eg)•g-1] 144 138 51 740
Korngrößenverteilung: Mineralb. 0-5 cm [%] Ton 13 5 Schluff 27 43Sand 59 51
Die bodenchemischen Analysen der Flächen B1, B2 und F1 wurden 1989 vom Institut für Bodenkunde und Waldernährung der Universität Göttingen ausgeführt. Die Angaben zu der Fläche G1 stammen aus MEIWES (1984).
6

3 Ziele und Methoden Voruntersuchungen, die 1987 in verschiedenen Waldökosystemen durchgeführt wurden, ergaben, dass die Cs-137 Gehalte des Bodens und der Pflanzen in jedem Gebiet erheblich variieren. Daraufhin wurden zwei Untersuchungsgebiete, Bodenmais und Fuhrberg, mit unterschiedlich hoher Bodenkontamination und unterschiedlichen Standortparametern, insbesondere bezüglich der Zusammensetzung der Baumarten, ausgewählt. Die Flächen-belastung der Böden wurde in beiden Gebieten durch ein Rasterverfahren ermittelt. Um die Migration von Cs-137 Ionen im Boden und die Dynamik des Nuklids in der pflanzlichen Biomasse zu untersuchen, wurden 1988 in beiden Gebieten Dauerprobeflächen ein-gerichtet. Auf diesen Flächen wurden regelmäßig, in mehrjährigen Abständen Boden-profile entnommen und die Tiefenverteilung von Cs-137 in 2 cm Schichtabständen untersucht. Von 1989 bis 2000 wurden, mit kurzen Unterbrechungen, verschiedene Pflanzenarten mehrmals während der Vegetationsperiode beprobt, sodass langjährige Zeitreihen über den Verlauf der Cs-137 Aktivität in diesen Biomedien vorliegen. In beiden Untersuchungsgebieten führen ungünstige Standortbedingungen zu einem relativ hohen Transfer von Radicäsium in die Nahrungskette. Um die Spannweite von Transferfaktoren zu ermitteln, wurde 1999 und 2000 ein Laubwaldökosystem mit günstigen Standort-bedingungen, bei Göttingen, in die Untersuchungen mit einbezogen.
Boden: Bei jedem Entnahmetermin wurden jeweils Bodenprofilserien, mit einem Bohrstock volumenecht entnommen. Jede Serie bestand aus 10 Bohrkernen, die in 2 cm Schichtfolgen getrennt und jeweils gleiche Schichten zu einer Mischprobe vereinigt wurden. Die Entnahmetiefe betrug 30 cm bis 50 cm, in Abhängigkeit von dem anstehenden Grundgestein. Die Proben wurden in Polyäthylenbeuteln aufbewahrt und bei Standard-bedingungen (105 oC) getrocknet. Anschließend wurden sie durch ein Sieb mit 2 mm Maschenweite gesiebt. Der Feinboden und der auf dem Sieb verbleibende Skelettanteil wurden gewogen, letzterer danach verworfen. Zur Aktivitätsbestimmung wurde der Feinboden pulverisiert und in 500 ml bzw. 120 ml Messbecher eingewogen. Neben der Cs-137 Aktivität wurde auch die K-40 Aktivität bestimmt.
Kraut und Strauchschicht: Da sich die Verteilung der Biomasse in einer Pflanze im Jahresverlauf ändert, wurden ausschließlich Blätter beprobt. Dazu wurden an jedem Entnahmetermin, von jeder untersuchten Pflanzenart, Blattorgane von 20-40 Individuen entnommen und zu einer Mischprobe vereinigt.
Bäume: Von jedem gefällten Baum wurde die gesamte Biomasse entnommen, und nach Stamm (Holz), Rinde (Kambium und alle, weiter außen liegenden Gewebetypen) und Blätter bzw. Nadeln getrennt. Nadeln wurden nach Jahrgängen getrennt und daraus eine Mischprobe gewonnen. Die Probenahme der Blattorgane von Bäumen erfolgte gegen Ende der Vegetationsperiode, wenn die Schwankungen im Mineralstoffgehalt von Blättern und Nadeln am geringsten sind.
Pilze: Von Pilzen wurden nur die oberirdischen Teile, also der Pilzstiel mit Hut entnommen. Jede untersuchte Probe war eine Mischprobe aus mindestens 3-5 Exemplaren, die möglichst vom selben Myzel stammten.
Die Entnahme von Boden- und Vegetationsproben erfolgte ausschließlich auf den Probeflächen.
7

Aufarbeitung der Proben: Die entnommenen Pflanzenteile wurden in Polyäthylenbeutel verpackt und während der meist 2-tägigen Freilandarbeiten kühl gelagert. Im Labor wurde von den Pflanzenproben das Frisch- und Trockengewicht bestimmt, an Wurzeln anhaften-de Bodenpartikel mit Wasser abgespült. Die Trocknung der Proben erfolgte bei 105 °C in Umlufttrockenschränken. Vor der Einwaage in die Messbecher wurden alle Proben in einer Mühle pulverisiert.
Da der Wassergehalt in Pflanzen, Pilzen und Boden im Jahresablauf variiert, wurde die spezifische Cs-137 Aktivität normalerweise auf die Trockensubstanz (TS) berechnet.
Wild: Von Wildtieren wurde ausschließlich reines Muskelfleisch vom Unterarm (Musculus flexor digitorum superficiales/profundus) untersucht. Im Labor wurde das Muskelfleisch vom Knochen abpräpariert, von anhaftenden Sehnen und Fett befreit, homogenisiert und in Messbecher eingewogen (120 ml, 500 ml). Die Aktivitätsangaben bei Wildtieren beziehen sich immer auf Bq•kg-1 Frischsubstanz (FS).
4 Aktivitätsbestimmung Die Aktivitätsmessungen der Proben wurden im Isotopenlaboratorium für biologische und medizinische Forschung der Universität Göttingen durchgeführt. Die Messungen erfolgten an zwei Reinstgermanium-Detektoren. Zusätzlich wurde während des gesamten Untersuchungszeitraums ein Reinstgermanium-Bohrloch-Detektor verwandt.
Gemessen wurden die Aktivitäten der Nuklide Cäsium-134, Cäsium-137 und Kalium-40. Als Messgefäße dienten handelsübliche Plastikbecher. Das Messvolumen betrug, je nach vorhandener Probemenge 500 ml, 180 ml oder 120 ml und bei speziellen Untersuchungen einzelner Pflanzenteile 4 ml. Für die verschiedenen Messvolumina wurde der Wirkungs-grad durch Kalibrierung der Anlage mit einer 8-Nuklide-Standardlösung bestimmt.
Die Aktivitätsmessungen von Cs-137 wurden i.d.R. solange durchgeführt, bis ein statisti-scher Zählratenfehler von ±5 %, für einen Vertrauensbereich von 95 % erreicht war. Für Bodenproben, die aus Tiefen unter 15 cm stammten und teilweise <5 Bq Cs-137 •kg-1 enthielten, bedeutete das Messzeiten >200.000 Sekunden. In dem Aktivitätsbereich 0-2 Bq lag der Zählratenfehler für Cs-137 bei 5-20%. Der statistische Zählratenfehler für K-40 betrug, je nach Aktivitätsgehalt der Proben 5 % bis 10 %.
8

5 Cs-137 Aktivität der Waldböden 5.1 Allgemeines
Die räumliche Verteilung von Radiocäsium im Boden hängt i.w. von dessen Sorptions-eigenschaften und der Wasserbewegung ab. Die Wasserbewegung ergibt sich aus der Niederschlagsmenge, sowie der Anzahl, Größenverteilung und Gestalt der Bodenporen. Die Sorptionseigenschaften werden von dem Anteil und der Zusammensetzung der Ton-fraktion und der organischen Substanz, sowie der Austauschkapazität und dem pH-Wert des Bodens bestimmt. Diese Parameter variieren mit dem Standort, deshalb sind gene-ralisierende Aussagen über die Reaktion von Radiocäsium im Boden kaum möglich (COUGTHRY und THORNE 1983, FRANKE 1982, KÜHN 1982, PAVLOTSKAYA et al. 1967).
Die vertikale Wanderungsgeschwindigkeit von Radiocäsium in ungestörten Böden ist relativ gering (mit Ausnahme von Sandböden und tropischen Lateritböden). Nach überein-stimmenden Ergebnissen vieler Autoren, verbleiben langfristig mindestens 80% der Cs-Aktivität in den oberen 15 cm des Bodens (ANPA 2000, KÜHN 1982, RITCHIE und RUDOLPH 1970, SQUIRE und MIDDLETON 1966, VÖLKEL 1997, WALTON 1963, ZIBOLD et al. 1997).
Die Ursache für diese geringe Tiefenverlagerung ist die Fixierung von Cäsium in den Kristallgittern von Tonmineralen. Tonminerale sind blättchenförmige Gebilde, die aus übereinander liegenden, negativ geladenen Silikatschichten aufgebaut sind. Zwischen diese Schichten werden bei Dreischichttonmineralen Kationen als Ladungsausgleich eingelagert. Cäsium- und Kaliumionen passen nach TAMURA (1963) wegen ihrer Größe und ihrer hohen Polarisierbarkeit besonders gut in diese Zwischenschichten und werden dort fixiert. Aufgrund dieser spezifischen Bindungsplätze sind sie vor Auswaschung in tiefere Boden-schichten geschützt und nur bedingt pflanzenverfügbar (FREDRIKSSON et al. 1958).
In der organischen Substanz der Waldböden erfolgt die Bindung von Kationen adsorptiv an Carboxyl- und Enolgruppen, sowie an phenolische OH-Gruppen (SCHEFFER und SCHACHTSCHNABEL 1984); spezifische Bindungsplätze für Cäsium-Ionen bestehen hier aber nicht (CREMERS et al. 1988). Deshalb ist Radiocäsium in der organischen Substanz gut pflanzenverfügbar.
In Waldökosystemen erfolgt durch abgestorbene Pflanzen- und Baumteile und, in geringerem Maße, auch durch abgestorbene tierische Organismen, ein kontinuierlicher Eintrag auf den Boden. Diese organische Substanz wird von Bodenorganismen zersetzt, humifiziert, teilweise in den Unterboden eingearbeitet und dann mineralisiert. Der Ablauf dieser Umsetzungsprozesse hängt, neben der jährlich produzierten Streumenge und Streu-zusammensetzung, wesentlich von den Standortverhältnissen, also den Lebensbedingungen der Bodenorganismen ab. Bei günstigen Bedingungen wird bei der Streuzersetzung (hoher pH-Wert, gute Wärme- und Feuchte- und Luftverhältnisse, etc.) die organische Substanz weitgehend in den oberen Mineralboden eingearbeitet und umgesetzt, sodass keine oder nur gering mächtige humose Auflagen entstehen. Bei ungünstigen Standortbedingungen (niedriger PH-Wert, Staunässe, geringe Wärme, dauernde Trockenheit, nährstoffarme Böden etc.) ist der Abbau der organischen Substanz verzögert bis stark reduziert, was zur Ausbildung von Humushorizonten führt, die bis zu 30 cm mächtig werden können.
9

Fast alle Waldbodenpflanzen und die meisten Bäume beziehen ihrer Nährstoffe über-wiegend aus den obersten 10 cm des Bodens. Dies ist genau der Bereich, in dem langfristig der Großteil des im Boden vorhandenen Cs-137 Inventars verbleibt und in dem das Nuklid relativ gut pflanzenverfügbar ist.
5.2 Cs-137 Flächenbelastung der Böden im Untersuchungsgebiet Bodenmais durch den Kernwaffen- und Tschernobyl-Fallout
Die Cs-137 Flächenbelastung der Böden im Untersuchungsgebiet Bodenmais wurde 1989 ermittelt. Dazu wurde der Boden an 59 Rasterpunkten bis 20 cm Tiefe beprobt. Um jeden Rasterpunkt wurden, in einem Radius von 50 Metern, 10 Bohrkerne entnommen, jeder nach Humusauflage und Mineralboden getrennt und jede Schichte zu einer Mischprobe vereinigt. In der Abbildung 2 ist die Verteilung der Cs-137 Flächenaktivität der Böden, in Form von Box-Plots dargestellt.
0
20000
40000
60000
80000
100000
140000
160000
Kernwaffen-Cs-137 Tschernobyl-Cs-137Cs-137 gesamt
Cs-137 [Bq/m2]
Abb. 2: Flächenbezogene Cs-137 Aktivität der Böden in Bodenmais (n=59), am 1.5.1986
Der Median der Messwerte war 51.800 Bq•m-2, die mittlere Cs-137 Kontamination der Böden betrug 53.690±26.690 Bq•m-2 (X±SD), reichte jedoch von 12.090 Bq•m-2 bis 146.480 Bq•m-2. Die Daten waren annähernd normal verteilt. Mit diesem Cs-137 Inventar des Bodens gehört das Untersuchungsgebiet Bodenmais zu den höchst belasteten Gebieten der Bundesrepublik. Der größte Teil der Cs-137 Aktivität geht auf den Tschernobyl-Fallout zurück: 1986 wurden durchschnittlich 45.160±25.520 Bq•m-2 auf dem Boden deponiert. Als Minimum wurden 6.740 Bq•m-2, als Maximum 145.130 Bq•m-2 festgestellt, der Median lag bei 44.990 Bq•m-2. Die mittlere Cs-137 Aktivität, die am 1. Mai 1986 infolge des globalen Fallouts noch im Boden vorhanden war, betrug 8.530±4.660 Bq•m-2, der Median lag bei 8.080 Bq•m-2. Die Spannweite der Messwerte war auch hier relativ groß, es wurden Aktivitäten zwischen 1.330 Bq•m-2 und 23.040 Bq•m-2 festgestellt. Die Variationskoeffizienten betrugen für Cs-137 aus den Altlasten 55%, aus dem Tschernobyl-
10

Fallout 57%. Die inhomogene Verteilung der gesamten Cs-137 Aktivität in den Böden ist in Abbildung 3 dargestellt. Mit durchschnittlich 15.320±2.460 Bq Cs-137•m-2 war die Bodenkontamination, im Mai 1986, im Nordwesten des Untersuchungsgebietes (Gebiet 1) am geringsten. Sie nahm in süd- östliche Richtung kontinuierlich zu und erreichte mit 78.940±20.320 Bq•m-2 im Gebiet 4 die höchste durchschnittliche Belastung.
Bodenmais
Regenhütte
Gr. Arber0 1 5 km
40 - 60
0 - 20
20 - 40
> 60
Gebiet 1
Gebiet 2
Gebiet 3
Gebiet 4
Cs-137 Aktivität in kBq/qm
Abb. 3: Bodenkontamination mit Cs-137 im Untersuchungsgebiet Bodenmais, Daten auf den 1.Mai 1986 zerfallskorrigiert (n = 59)
Diese Unterschiede in der Cs-137 Aktivität der Böden sind i.w. durch die kleinräumig unterschiedliche Niederschlagsverteilung am 30.4. und 1.5.1986 bedingt: die Luftmassen-bewegung erfolgte aus süd-östlicher Richtung, Niederschläge gingen größtenteils als Steigungsregen an den oberen Süd-Osthängen nieder, während der Nord-Westen des Untersuchungsgebietes weitgehend trocken blieb. Zwischen dem Cs-137 Gehalt und der Höhenlage der Rasterpunkte konnte weder bei den Alt-, noch bei den Neulasten ein statistisch signifikanter Zusammenhang gefunden werden.
5.3 Bodenkontamination der Probeflächen B1 und F1
Um die Flächenaktivität der Böden der Probeflächen zu bestimmen, wurden 1988 die 100x100 m großen Probeflächen in je 4 Quadranten unterteilt, aus jedem Quadrant 9 Bohrkerne entnommen (Einstichtiefe 20 cm) und zu einer Mischprobe vereinigt. Aus den Messergebnissen der 4 Mischproben wurde die Cs-137 Aktivität pro Quadratmeter, in 0 bis 20 cm Bodentiefe berechnet (Tabelle 2).
11

Tab. 2: Mittlere flächenbezogene Cs-137 Aktivität in Böden der Probeflächen und Variationskoeffizienten (V), am 1.5.1986
Dauerfläche Cs-137Aktivität in 0 - 20 cm Tiefe [Bq•m-2] V [% ]
F1 10.320 5,5
B1 98.450 17 B2 83.370 14
Die Flächenaktivität des Bodens war, entsprechend dem Nord-Süd Gefälle der Cs-137 Kontamination in der Bundesrepublik, auf den Probeflächen B1 mit 98.450 Bq•m-2 und B2 mit 83.370 Bq•m-2 deutlich höher, als auf F1 mit 10.320 Bq•m-2.
5.4 Tiefenverteilung von Cs-137 in Bodenprofilen
Auf den Probeflächen B1 und F1 wurden seit 1989, in mehrjährigen Abständen Bodenprofile entnommen und die vertikale Verteilung von Cs-137 ermittelt. Die letzten Bodenprofile wurden auf den Dauerflächen F1 und G1 im Juni 2000, auf B1 und auf einer Wiese in Bodenmais im September 2000, entnommen. Da in einem Profil aus Bodenmais Unstimmigkeiten bei der Tiefenverteilung der Cs-137 Aktivität auftraten, wurde die Probe-nahme im Mai 2001 wiederholt.
Die Mächtigkeit des Auflagehumus variierte auf den Flächen B1 und F1 zwischen 2 und 8 cm, bei einer mittleren Dicke von 4 cm. Die Ol, Of und Oh Horizonte sind deutlich vonein-ander abgegrenzt und gut zu unterscheiden. Auf der Probefläche G1 liegt als Auflage-humus nur ein Ol-Horizont vor.
Die insgesamt in den Profilen vorhandene Cs-137 Aktivität, also das Gesamtinventar pro Profil, reichte von 5.450 Bq•m-2 auf G1 bis 73.200 Bq•m-2 auf B1.
In der Abbildung 4 sind die Tiefenverteilung der spezifischen Cs-137 Aktivität, die Trockenraumdichte und die flächenbezogene Cs-137 Aktivität von Profil I auf der Probefläche F1, von Juni 2000 dargestellt. Die spezifische Cs-137 Aktivität stieg in den
12

50
40
30
20
10
00 50 100 150 200 250
Cs-137 [Bq/kg] TS
Bode
ntie
fe [c
m]
Waldboden, Profil IProbefläche F1, 6/2000 Humusauflage Mineralboden
50
40
30
20
10
00,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0
Waldboden, Profil IProbefläche F1, 6/2000 Humusauflage Mineralboden
Trockenraumdichte [g/cm3]
Bode
ntie
fe [c
m]
50
40
30
20
10
00 5 10 15 20 25
Waldboden, Profil IProbefläche F1, 6/2000Inventar: 8.840 Bq/m2
Humusauflage Mineralboden
Cs-137 [%/m2/cm]
Bode
ntie
fe [c
m]
Abb. 4: Tiefenverteilung der spezifischen Aktivität (oben), der Trockenraumdichte (Mitte) und der flächenbezogenen Cs-137 Aktivität (unten) in Profil I, Probefläche F1, Juni 2000
13

obersten Zentimetern des Profils kontinuierlich an, erreichte das Maximum in der Tiefenstufe 4-6 cm und nahm dann schnell ab. Dieser Verlauf der Tiefenverteilung ist für viele Waldböden typisch. In Profil I nahm die Aktivität in der Humusauflage von 170 Bq•kg-1 über 214 Bq•kg-1 bis zur ersten Schicht des Ah-Horizontes auf 228 Bq•kg-1 zu. In den darunter liegenden Schichten des Mineralbodens nahm die Aktivität von Kompartiment zu Kompartiment, nahezu modellartig, exponentiell ab. Unterhalb 10 cm Bodentiefe fanden sich immer weniger als 50 Bq•kg-1, unterhalb 32 cm Profiltiefe weniger als 1 Bq•kg-1, unterhalb 42 cm waren etwa 0,2 Bq Cs-137•kg-1 vorhanden.
Die Horizonte der Waldböden haben, aufgrund ihres unterschiedlichen Kompostierungs- und Mineralisierungsgrades, sehr unterschiedliche Dichten. Die Trockenraumdichte ist in der Humusauflage relativ niedrig und steigt mit zunehmender Bodentiefe kontinuierlich an. In Profil I nimmt die Trockenraumdichte von der Humusauflage zum Mineralboden in der Tiefenstufe von 10 bis 12 cm relativ schnell von 0,09 g•cm-3 bis auf 1,35 g•cm-3 zu. Dieser Anstieg ist durch die zunehmende Mineralisierung des Ah-Horizontes bedingt. In den tiefer liegenden Mineralbodenschichten nimmt die Trockenraumdichte nur noch wenig zu, in der Tiefenstufe 30-32 cm beträgt der Wert 1,54 g•cm-3, bei 48-50 cm 1,65 g•cm-3.
Unterschiedliche Standortbedingungen führen bei Waldböden zu unterschiedlichen Trockenraumdichten. Daher ist die Angabe der Cs-137 Aktivität pro Masse Boden (spezi-fische Aktivität), weder geeignet die Tiefenverteilung des Nuklids auf verschiedenen Waldböden miteinander zu vergleichen, noch um Transferfaktoren zu berechnen. Einen realistischeren Vergleich ermöglicht die Angabe der in einem bestimmten Bodenvolumen enthaltenen Cs-137 Aktivität, zumal auch Pflanzenwurzeln Radiocäsium aus einem bestimmten Bodenvolumen aufnehmen.
Deshalb wird die Aktivität von Cs-137 im folgenden auf das Bodenvolumen, pro Quadrat-meter, pro 2 cm Kompartiment bezogen (flächenbezogene Aktivität). Um die Cs-137 Tiefenverteilung in Profilen aus verschiedenen Untersuchungsgebieten mit unterschied-lichem Cs-Inventar vergleichen zu können, wird die Aktivität zusätzlich in Prozent angegeben. Das heißt, dass für jede Schicht, der Cs-137 Anteil an der insgesamt im Boden-profil enthaltenen Cs-137 Aktivität berechnet wird, die Einheit ist dann %•m-2•cm-1.
Die flächenbezogene Aktivität kann nur dann aus der spezifischen Aktivität berechnet werden, wenn das Bodenprofils volumenecht beprobt wurde. Da die Entnahme und Messung von Bodenproben aufwändig ist, macht es Sinn, Cs-137 Messdaten von Tiefen-verteilungen mathematisch zu beschreiben, um zum Beispiel die Aktivität in tiefen, schlecht zugänglichen Bodenschichten abzuschätzen zu können.
In der Abbildung 5 sind die Messwerte der spezifischen Cs-137 Aktivität von Profil I, für die Bodentiefe 8-50 cm, in logarithmischer Skalierung, mit linearem Fit dargestellt. Der Regressionskoeffizient beträgt –0,990 und ist hochsignifikant (P<0.0001). Die Regressionsgerade gibt die ermittelte Verteilung des Nuklids für die Bodentiefe 8-36 cm sehr gut wieder. Für die Regressionsgerade gilt die Beziehung:
lg Cs-137 = 2,4467 – 0,0747 * Bodentiefe [cm]
14

Der Fit über die gesamte Profiltiefe liefert eine schlechtere Anpassung, da die Verteilung der Messwerte in den obersten 8 cm des Bodens keiner linearen Funktion folgen. In tieferen Schichten nehmen die Abweichungen zwischen den Werten aus der Gleichung und den Messwerten zu, was offensichtlich durch die sehr geringere Aktivität und den dadurch größeren Messfehler bedingt ist: In der Schichttiefe 32-34 cm betrug die Cs-137 Aktivität 1,1 Bq•kg-1, bei einem Messfehler von 4% (bezogen auf 2 Sigma). Ab der Tiefenstufe 38-40 cm wurden jeweils 0,2 Bq•kg-1 oder weniger Aktivität, mit Messfehlern von 12-20% festgestellt. Daher kann die Aktivität, die tatsächlich im Boden unterhalb 50 cm vorhanden ist, besser über die Regressionsgleichung abgeschätzt werden.
50
40
30
20
10
00,1 1 10 100 400
Cs-137 [Bq/kg] TS
Bode
ntie
fe [c
m]
Waldboden, Profil IProbefläche F1, 6/2000 Humusauflage Mineralboden
Abb. 5: Spezifische Cs-137 Aktivität im Mineralboden, Bodentiefe 8 - 50 cm, mit linearem Fit sowie oberer und unterer Grenze des 95% Konfidenzbereichs, Profil I, F1 2000
Die Unterschiede in der Cs-137 Tiefenverteilung in Profil I, die sich zwischen spezifischer und flächenbezogener Cs-137 Aktivität, insbesondere in den obersten 10 cm des Bodens ergeben, werden durch den Vergleich der oberen mit der unteren Grafik in Abbildung 4 deutlich. Aufgrund der geringeren Trockenraumdichte ist im Vergleich zum Mineralboden (mittlere Grafik), besonders in den Kompartimenten der Humusauflage, die flächen-bezogene Cs-137 Aktivität deutlich niedriger als die massenbezogene Aktivität. Insgesamt befanden sich in dem 50 cm tiefen Profil 8.840 Bq•m-2, wovon in den beiden Kompar-timenten der Humusauflage nur 10,1% (3,5% und 6,6%), in den obersten 10 cm des Bodens 51% des Inventars gebunden waren.
Die Abbildung 6 zeigt die Tiefenverteilung von Cs-137 in dem Profil III von F1, wo die Humusauflage mit 8 cm, mächtiger war, als in Profil I. Das Inventar des Profils betrug 7.250 Bq•m-2, davon befanden sich 39% im Auflagehumus und 54% in den obersten 10
15

cm. Im Gegensatz zu der Verteilung von Cs-137 in allen anderen Bodenprofilen, befindet sich im Profil III das Maximum der flächenbezogenen Aktivität, mit rund 15% in 4 aufeinander folgenden Schichtfolgen. Die Unterschiede in der Tiefenverteilung von Cs-137 in Profil I und Profil III sind beträchtlich, obwohl beide von derselben Probefläche stammen und zeitgleich entnommen wurden. Die Gründe dafür werden in Kapitel 11 diskutiert.
50
40
30
20
10
00 5 10 15 20 25
Waldboden, Profil IIIProbefläche F1, 6/2000Inventar: 7.250 Bq/m2
Humusauflage Mineralboden
Cs-137 [%/m2/cm]
Bode
ntie
fe [c
m]
Abb. 6: Tiefenverteilung der flächenbezogenen Cs-137 Aktivität in Profil III auf F1, Juni 2000
Die flächenbezogene Tiefenverteilung von Cs-137 in 2 Profilen von der Fläche B1 ist in der Abbildung 7 dargestellt. Das obere Bild zeigt die Messergebnisse von Profil IV, das eine durchschnittliche Humusdicke von 6 cm und ein Cs-137 Inventar von 73.200 Bq•m-2 aufwies. Im unteren Bild ist die Tiefenverteilung des Nuklids von Profil V dargestellt. Das Inventar betrug hier 70.650 Bq•m-2, bei einer Humusauflage von 4 cm. Während die Cs-137 Inventare beider Profile gut übereinstimmen (der Variationskoeffizient für die Flächenbelastung von Cs-137 beträgt auf der B1 17%, Kapitel 5.3), gibt es bei der Tiefenverteilung des Nuklids deutliche Unterschiede.
In Profil IV liegt das Maximum der Flächenaktivität deutlich in tieferen Schichten als in Profil V: Bei Profil IV nimmt die Aktivität von der Humusauflage bis zur Tiefenstufe 10 bis 12 cm kontinuierlich zu und erreicht hier, mit rund 20% Anteil am Inventar, das Maximum. Dagegen befindet sich in Profil V die höchste Aktivität mit rund 17%, unmit-telbar unter der Humusauflage im obersten Ah-Horizont, bei 6 bis 8 cm. In der Tiefenstufe 28 bis 30 cm sind in Profil IV noch 1%, in Profil V noch 0,5% des Inventars, das entspricht einer spezifischen Aktivität von 50 bzw. 39 Bq•kg-1. Cs-137 Ionen sind sicherlich auch in noch tiefere Bodenschichten vorgedrungen, allerdings ist eine Profilentnahme unterhalb 30 cm Bodentiefe, aufgrund des anstehenden Gesteins, normalerweise nicht möglich.
16

30
20
10
00 5 10 15 20 25
Cs-137 [%/m2/cm]
Waldboden, Profil IV Probefläche B1, 9/2000Inventar: 73.200 Bq/m2
Humusauflage Mineralboden
Bode
ntie
fe [c
m]
30
20
10
00 5 10 15 20 25
Waldboden, Profil V Probefläche B1, 5/2001Inventar: 70.650 Bq/m2
Humusauflage Mineralboden
Bode
ntie
fe [c
m]
Abb. 7: Tiefenverteilung der flächenbezogenen Cs-137 Aktivität in den Profilen IV (oben) und V (unten) auf B1, September 2000 und Mai 2001
In der Abbildung 8 ist die Verteilung des Nuklids in einem Wiesenboden aus Bodenmais dargestellt. Es handelt sich um eine, unmittelbar am Waldrand der Ortschaft Regenhütte gelegene Wiese, die als Kuhweide genutzt wurde. Das Inventar des Profils hat mit 65.700 Bq Cs-137 •m-2, eine vergleichbar hohe Aktivität wie die Profile IV und V von der Probefläche B1. Die Tiefenverteilung von Cs-137 unterscheidet sich dagegen deutlich. Im Gegensatz zu den Waldbodenprofilen ist die Flächenaktivität des Wiesenbodens in den oberen 10 cm sehr viel höher als in den tiefer liegenden Bodenschichten. Das Maximum
17

der flächenbezogenen Aktivität liegt zu rund 25% in der Schichttiefe 4 bis 6 cm. Da sich sowohl in der Bodenschicht davor als auch in der danach folgenden jeweils 19% des Cs-137 befinden, sind schon 63% des Inventars im Tiefenbereich 2 bis 8 cm vorhanden.
30
20
10
00 5 10 15 20 25
Cs-137 [%/m2/cm]
Bode
ntie
fe
Wiesenboden, Bodenmais 9/2000Inventar 67.740 Bq/m2
Mineralboden
Abb. 8: Tiefenverteilung der flächenbezogene Cs-137 Aktivität in einem Wiesenboden, Bodenmais, September 2000
In der Abbildung 9 sind die spezifische Aktivität, die Trockenraumdichte und die flächen-bezogene Aktivität eines Waldbodenprofils von G1 dargestellt. Die Standortbedingungen unterscheiden sich auf G1 deutlich von B1 und F1, wo als Bodentyp saure Podsole vorliegen. Auf der Fläche G1 liegt als Bodentyp eine Terrafusca-Rendzina, ein flach-gründiger, tonreicher, plastischer, dichter Boden, von nur 25 bis 30 cm Mächtigkeit über Kalkgestein, vor. Der pHCaCl2-Wert beträgt 5,7, in der Tiefenstufe 0-5 cm. Aufgrund der hohen Leistung des Edaphons, besteht die Humusauflage nur als Ol-Horizont mit durch-schnittlich <1 cm Dicke. Daher konnte, um die Probenahme auf allen Flächen gleich durchzuführen, die Humusauflage auf G1 nicht in 2 cm Schritten beprobt werden, sondern wurde komplett erfasst und der darunter befindliche Boden in den üblichen 2 cm Kompar-timenten beprobt.
Die Trockenraumdichte ist, abgesehen von dem geringen Wert des Ol-Horizontes von 0,015 g •cm-3, bereits in den obersten cm des Bodens relativ groß und nimmt von rund 0,6 g •cm-3 (Kompartiment 0-2 cm) auf rund 1 g •cm-3 (Kompartiment 6-8 cm) zu. Deswegen unterscheidet sich die Tiefenverteilung der spezifischen und der flächenbezogenen Aktivität in den obersten vier Kompartimenten wesentlich weniger als in Profil I auf F1 (Abbildung 4).
Im Ol-Horizont war auf G1 der Anteil der spezifischen und der flächenbezogenen Cs-137 Aktivität vernachlässigbar gering, die flächenbezogene Aktivität betrug rund 5 Bq•m-2, das entspricht einem Anteil von nur 0,1% an dem Inventar von 5.450 Bq•m-2. Die flächenbe-
18

30
20
10
0
0 20 40 60 80 100Cs-137 [Bq/kg] TS
Bode
ntie
fe [c
m]
Waldboden, Probefläche G1, 6/2000 Ol Mineralboden
30
20
10
0
0,0 0,2 0,4 0,6 0,8 1,0
Waldboden, Probefläche G1, 6/2000 Ol Mineralboden
Trockenraumdichte [g/cm3]
Bode
ntie
fe [c
m]
30
20
10
0
0 5 10 15 20 25
Waldboden, Probefläche G1, 6/2000Inventar: 5.450 Bq/m2
Ol Mineralboden
Cs-137 [%/m2/cm]
Bode
ntie
fe [c
m]
Abb. 9: Tiefenverteilung der spezifischen Aktivität (oben), der Trockenraumdichte (Mitte) und der flächenbezogenen Cs-137 Aktivität (unten), in einem Waldbodenprofil von der
Probefläche G1, Juni 2000
19

zogene Cs-137 Aktivität ist in der Schichtstufe 2-4 cm am größten und fällt bis 10 cm Tiefe exponentiell ab. In der Bodentiefe 10–20 cm bleibt die Aktivität etwa gleich und nimmt von 20 cm Bodentiefe an weiter ab.
Bei der Tiefenverteilung von Cs-137 fällt auf, dass sich die meiste Aktivität unmittelbar unter der Bodenoberfläche befindet und dann kontinuierlich abnimmt. Dies ist erstaunlich, weil die Biomasse und die Leistung der Bodentiere, insbesondere der Bodenwühler, auf der Fläche sehr hoch ist. Vor allem Regenwürmer, Mistkäfer, aber auch Larven von Schnell- und Maikäfern und letztlich auch Mäuse bewirken die Verlagerung der Streu in tiefere Bodenschichten bzw. die gleichmäßige Durchmischung des Oberbodens. Innerhalb von anderthalb Jahren wird die Streu komplett umgesetzt, beziehungsweise in den Oberboden eingearbeitet. Der jährliche Input an Cs-137 durch die Blätter der Bäume liegt im einstelligen Becquerel-Bereich. Daher wäre, 14 Jahre nach dem Nuklideintrag, eine gleichförmigere Verteilung des Nuklids im Oberboden, mit einem tieferliegenden Aktivi-tätsmaximum zu erwarten gewesen. Da die reale Tiefenverteilung von Cs-137 dem wider-spricht, müssen wohl andere Mechanismen, wie ein aufwärtsgerichteter Transport des Nuklids durch Mikroorganismen oder Pilze, die Migration des Nuklids in dieser humosen Bodenzone wesentlich beeinflussen. Die Ergebnisse von anderen Untersuchungen zu der Dynamik von Cs-137 in der Laubstreu lassen auf einen solchen Mechanismus schließen (FIELITZ 1994, RAFFERTY et al. 1997).
In der Tabelle 3 ist die flächenbezogene Aktivität und der prozentuale Anteil am Gesamtinventar von Cs-137 in verschiedenen Bodenkompartimenten für die untersuchten Profile zusammengestellt.
Tab. 3: Flächenbezogene Cs-137 Aktivität und prozentualer Anteil in den Bodenprofilen
Cs-137 [Bq•m-2] (%)
Boden-kompartiment
B1 Profil IV
B1 Profil V
Bodenmais Wiese
F1 Profil I
F1 Profil III
G1
Humusauflage 12.080 (16) 9.990 (14) 895 (10) 2.835 (39) 5 (0,1)Bodentiefe 0-10 31.840 (43) 42.520 (60) 55.930 (83) 4.520 (51) 3.940 (54) 4.040 (74)Bodentiefe 0-20 67790 (93) 66.590 (94) 65.530 (97) 8.010 (91) 6.611 (91) 5.240 (96)
Profil insgesamt 73.240 70.650 67.740 8.840 7.250 5.450
20

5.5 Zeitlicher Verlauf der Tiefenverteilung von Cs-137 in Böden
Um die vertikale Migration von Cs-137 Ionen im Bodenprofil zu untersuchen, wurde die Tiefenverteilung des Nuklids im Abstand von mehreren Jahren bestimmt. In der Abbildung 10 ist die Verteilung von Cs-137 in Bodenprofilen auf der Fläche B1 und F1, von 1989 bis 2001 dargestellt.
0 10 20 30 40,1
1
10
0
50
1989 1992 1997 2001
Cs-137 [%/cm/qm]
Bodentiefe [cm]
B1
0 10 20 30 40,1
1
10
0
50
1991 1998 2000
F1
Abb. 10: Zeitlicher Verlauf der Tiefenverteilung von Cs-137 in Bodenprofilen der Probeflächen B1 (oben) und F1 (unten)
21

Da die Cs-137 Flächenaktivität auf dieser Probefläche um mehrere Tausend Bq•m-2 schwankt, ist jeweils der prozentuale Cs-137 Anteil pro 2 cm Schicht und pro Quadrat-meter dargestellt. Die Skalierung der Abszisse wurde logarithmisch gewählt.
Jedes Symbol stellt die mittlere prozentuale Verteilung des Nuklids in der entsprechenden Tiefenstufe von jeweils 3 Profilserien dar. In den Jahren 2000 (F1) bzw. 2001 (B1) wurde nur je eine Profilserie berücksichtigt, weswegen der Verlauf der Tiefenverteilung in diesen Jahren weniger gut abgesichert ist als in den Jahren davor. Der Aufwand bei der Entnahme von Bodenprofilen ist erheblich, für die dargestellten Ergebnisse der Probefläche B1 (obere Grafik) wurden insgesamt 1.310 einzelne 2 cm Bodenschichten geschnitten. Auf B1 wurden die Profile 1989 und 1992 bis 22 cm Bodentiefe, 1997 und 2001 bis 30 cm, auf F1 wurde 1991 Boden bis zu einer Tiefe von 28 cm, 1989 bis 40 cm und 2000 bis 50 cm entnommen.
Sowohl die grundlegende Struktur der Tiefenverteilung als auch die Dynamik der Cs-137 Migration ist auf F1 und B1 verblüffend ähnlich. Auf beiden Flächen steigt die Cs-137 Aktivität in jedem Profil zunächst in den ersten 3 Schichtstufen an, erreicht bei 4 bis 6 cm Tiefe das Maximum und nimmt dann im darunter liegenden Boden, von Schichtfolge zu Schichtfolge kontinuierlich ab. Die Bodentiefe 0 bis 6 cm entspricht auf beiden Flächen etwa der Humusauflage, die jeweils zwischen 2 und 8 cm schwankt. Die Verteilung von Cs-137 in der Humusauflage war der Verteilung im Mineralboden grundsätzlich entgegen gesetzt: Zunahme der Aktivität in der Humusauflage mit der Tiefe (Ol→Of→Oh-Horizonte) und Abnahme der Aktivität mit zunehmender Tiefe im Mineralboden.
Die Migration des Nuklids erfolgte mit der Zeit, durch sukzessive Verlagerung von Cs-137-Ionen aus jedem 2 cm Kompartiment, in das darunter liegende. In der Humus-auflage war dabei der Verlust an Aktivität pro Bodenschicht fast immer größer als die Zunahme durch das vorhergehende Kompartiment. Dagegen ist in den aufeinander folgenden Kompartimenten des Mineralbodens die Zunahme der Cs-137 Aktivität jeweils höher als die Abgabe.
Die Schnittstelle in diesem Migrationsprozess liegt bei den Böden von B1 und F1 in der Bodenschicht 6-8 cm, das ist durchschnittlich der obere Bodenbereich des Ah-Horizontes, also der Bereich unterhalb der Humusauflage (auf der Probefläche G1, Abbildung 9, wo nur ein sehr dünner Ol-Horizont vorliegt, ist diese Schnittstelle nicht vorhanden). In dieser Schicht veränderte sich die Flächenaktivität kaum, hier befinden sich auf B1 langfristig 16 bis 17%, auf F1 rund 14% der insgesamt in den Profilen vorhandenen Aktivität. Die Konstanz dieser Werte bedeutet aber nicht, dass an dieser Stelle keine Migration von Cs-137 Ionen erfolgt, vielmehr ist die Dynamik hier am größten, vergleichbar mit der engsten Stelle einer Eieruhr.
22

6 Cs-137 Kontamination in Pflanzen, Beeren und Pilzen 6.1 Allgemeines
Cäsium und Kalium verhalten sich in Pflanzen biochemisch ähnlich, wobei das Stoff-wechselverhalten von Kalium wesentlich besser bekannt ist. Im folgenden wird eine kurze Einführung über das Verhalten beider Elemente in Pflanzen gegeben.
Nach STRASBURGER (1978) ist Kalium das einzige monovalente Kation, das für alle Pflanzen essentiell ist. Bei höheren Pflanzen stellt Kalium oft den höchsten Anteil an allen Makroelementen in der Trockensubstanz (BORESCH 1935). Es wirkt insbesondere als Osmotikum bei der Quellung des Zellplasmas und als Co-Faktor bei Enzymreaktionen. Kalium wird in der Pflanzenzelle nicht in organische Verbindungen eingebaut, es kommt fast ausschließlich im Zellsaft und im Quellungswasser von Plasma und Zellwand vor (SCHMALFUSS 1936). Cäsium kann Kalium nicht in seinen Stoffwechselfunktionen ersetzen. Pflanzen können Radiocäsium auf zwei verschiedenen Wegen aufnehmen: direkt über die Blätter und indirekt über die Wurzeln.
Unmittelbar nach dem Tschernobyl-Fallout war die Cs-137 Aktivität von Waldboden-pflanzen vorwiegend durch Ablagerung des Nuklids auf oberirdische Pflanzenteile bedingt, in den folgenden Jahren dagegen, fast ausschließlich durch die Aufnahme von Cs-137 Ionen über die Wurzeln aus dem Boden. Weitere Wege, wie Cs-137 auf Pflanzengewebe gelangen kann sind a) Einträge über das Traufwasser aus dem Kronenraum der Bäume, b) Einträge über Resuspension und c) Einträge über „rain splash“. Diese Einträge sind im Vergleich zur Aufnahme über die Wurzeln, vernachlässigbar gering (FIELITZ 1994).
Nach AMBERGER (1979) und STRASBURGER (1978) gelangen Cs+- Ionen, zunächst passiv, durch Diffusion aus der Bodenlösung in die Zellwände der Wurzelhaare. Auf dem Weg zu den im Wurzelinneren gelegenen Xylemleitgefäßen müssen sie, spätestens an der Endodermis, in das Cytoplasma übergehen. Dieser Transport durch die Zellwand wird wahrscheinlich durch Carrier vermittelt und ist ein aktiver Prozess. Die Bindungsstellen der Carrier sind kationen- und anionenspezifisch. Chemisch ähnliche Ionen, wie z.B. Cs+ und K+, haben vermutlich gleiche Trägermoleküle und können sich, je nach Konzentration in der Bodenlösung, gegenseitig bei der Aufnahme behindern. An dieser Stelle wird Cäsium, genau wie Kalium, von der Pflanze selektiv, gegen ein Konzentrationsgefälle aufgenommen. Pflanzenzellen reichern K+-Ionen häufig mehr als das 1.000fache der K+-Konzentration in der Bodenlösung an.
Die Cs+Ionen gelangen schließlich in den Transpirationsstrom und damit in oberirdische Organe, wo sie intrazellulär angereichert werden. Da Cäsium, genau wie Kalium, zu den Elementen gehört, die im Xylem und im Phloem mobil sind, kann es, je nach Bedarf, zwischen verschiedenen Organen umverteilt werden (BROWN 1964, LÄUCHLI 1971, STRASBURGER 1978).
23

6.2 Voruntersuchungen zur Cs-137 Aktivität von Waldbodenpflanzen
1988, zu Beginn der Untersuchungen auf den Probeflächen, stellte sich zunächst die Frage, ob die oberirdische Biomasse einer einzelnen Pflanze, oder nur Teile davon, oder aber Mischproben von mehreren Pflanzen einer Art, die aussagefähigsten Ergebnisse liefern würden.
Daher wurde zunächst an verschiedenen Arten untersucht, wie Cs-137 in einzelnen Pflanzenorganen verteilt ist. Es stellte sich heraus, dass die einzelnen Teile einer Pflanze (Wurzel, Stängel, Blätter) immer unterschiedliche Cs-137 Gehalte aufweisen. Als typisches Beispiel sind in der Tabelle 4 die Cs-137 Gehalte verschiedener Teile einer Einzelpflanze von Dornfarn angegeben.. Da sich Cs-137 stoffwechselphysiologisch ähn-lich verhält wie Kalium, ist auch die K-40 Aktivität angegeben.
Tab. 4: Verteilung der Cs-137 Aktivität in verschiedenen Teilen einer Pflanze von Dornfarn auf B1, Probenahme November 1988
Pflanzenteil
Cs-137
[Bq/kg] TS
K-40
[Bq/kg] TS
Wassergehalt
[%]
Blatt 18.900 820 73
Blattanlage 2.860 200 57
Stängel 8.340 530 69
Wurzel 4.420 240 66
Die Blätter enthielten deutlich mehr Cs-137 Aktivität als alle anderen Pflanzenteile. Auffällig war, dass die Blattanlagen, die im Herbst bereits für die kommende Vegetations-periode angelegt sind, verhältnismäßig wenig Cs-137 enthielten. Die Reihenfolge der Cs-137 und K-40 Gehalte in den Pflanzenteilen war gleich, in Pflanzenteilen mit hohen K-40 Aktivitäten befanden sich auch entsprechend mehr Cs-137 Aktivität und umgekehrt.
Die einzelnen Pflanzenteile unterscheiden sich auch in dem Verhältnis von Trocken-gewicht zu Frischgewicht, was die Zusammensetzung aus unterschiedlichen Geweben wiedergibt (Blätter z.B. Photosynthesegewebe und Stängel Festigungsgewebe).
Letztlich unterscheiden sich sogar die einzelnen Blätter von ein und derselben Pflanze im Cs-137 Gehalt, wie Untersuchungen an je 5 Blättern, von 8 verschiedenen Einzelpflanzen Dornfarn, auf der Probefläche B1 belegen. Der durchschnittliche Variationskoeffizient betrug 7,9%, bei einem Messfehler von 1%.
Untersucht wurde auch die Variabilität von Cs-137 innerhalb einer Pflanzenart, wozu auf jeder Probefläche die Aktivität der Blätter von 12 Einzelpflanzen bestimmt wurde. Die Schwankungen waren groß, als Beispiel wurden 1989 auf der Probefläche B2 Variations-koeffizienten von 53% für Wald-Frauenfarn, 53% für Waldhainsimse und 29% für
24

Dornfarn ermittelt. Der Maximalwert bei Wald-Frauenfarn betrug mit 8.800 Bq•kg-1 TS das 12fache des Minimalwertes von 760 Bq Cs-137•kg-1.
Die Aktivitätsmessungen von Einzelpflanzen eignen sich deshalb weder um zeitliche Verläufe von Cs-137 einer Pflanzenart zu untersuchen, noch um einen Überblick über die Cs-137 Kontamination in einem Gebiet zu bekommen, es sei denn, man beprobt größere Kollektive. Aufgrund der Ergebnisse dieser Voruntersuchungen, wurden in den folgenden Jahren bei jeder Probenahme jeweils Blätter von 20 bis 40 Einzelpflanzen einer Art entnommen und zu einer Mischprobe vereinigt.
Der Wassergehalt von Pflanzen ändert sich ebenfalls im Lauf der Vegetationsperiode. In der Abbildung 11 ist der Wassergehalt verschiedener Proben von Dornfarn auf der Probe-fläche B1 und von Pfeifengras auf F1, im Jahresverlauf dargestellt.
Mrz Apr Mai Jun Jul Aug Sep Okt Nov Dez30
40
50
60
70
80
90
100
Dornfarn, B1
Monat
Wassergehalt [%]
Apr Mai Jun Jul Aug Sep Okt Nov Dez30
40
50
60
70
80
90
100
Pfeifengras, F1
Monat
Abb. 11: Wassergehalt in Blättern von Dornfarn auf B1 und Pfeifengras auf F1, 1989
25

Bei beiden Arten nimmt der Wassergehalt im Laufe der Vegetationsperiode ab, die Unterschiede zwischen Frühjahr und Herbst sind dabei beträchtlich. Bei Dornfarn geht der Wassergehalt von April bis Oktober von rund 90% auf 70% zurück. Ein Teil der Popu-lation bleibt auch im Winter grün, was an den höheren Messwerten Anfang Dezember zu sehen ist. Die zur selben Zeit absterbenden Blätter haben deutlich niedrigere Wasser-gehalte. Bei Pfeifengras beträgt der Wassergehalt während der Vegetationsperiode rund 70%. Da die Blätter im Herbst abzusterben beginnen, nimmt auch der Wassergehalt kontinuierlich ab.
Um die Messdaten von Proben mit unterschiedlicher jahreszeitlicher Herkunft besser vergleichen zu können, wird die spezifische Cs-137 Aktivität von Pflanzen im folgenden auf das Trockengewicht (TS) bezogen. Die Ausnahme sind Untersuchungen über die Ursachen der Kontamination von Wildtieren, weil diese die Nahrungspflanzen als Frisch-substanz aufnehmen.
6.3 Cs-137 Aktivität in der Waldbodenvegetation
Die Ergebnisse der Untersuchungen von 1998 bis 2000 zeigen, dass sich die einzelnen Pflanzenarten, auf jeder Probefläche, in den mittleren Cs-137 Gehalten der Blätter unterschieden, wobei die Reihenfolge der Kontamination auf B1, B2 und F1 gleich oder zumindest ähnlich ist. Grundsätzlich waren Blätter von Farnen durchschnittlich am höchsten, von Himbeere und Brombeere am geringsten belastet. In der Abbildung 12 sind die Cs-137 Aktivitäten verschiedener Pflanzenarten auf B2 und F1 dargestellt.
Die Ordinaten sind unterschiedlich skaliert, weil die Pflanzenarten auf B2 wesentlich höhere Aktivitätswerte haben als auf F1.
Typischerweise gehören die Farne zu den am höchsten kontaminierten autotrophen Pflanzenarten. Auf B2 lagen die Messwerte von Dornfarn (Dryopteris carthusiana) zwischen 3.950 Bq•kg-1 und 12.690 Bq•kg-1, bei einem Mittelwert von 7.560 Bq•kg-1. Dagegen enthielten Himbeerblätter (Rubus idaeus) durchschnittlich nur 473 Bq•kg-1, Brombeerblätter (Rubus fruticosus) 403 Bq•kg-1, bei Maximalwerten von 1.290 Bq•kg-1 bzw. 1.045 Bq•kg-1. Seegras (Carex brizoides) und Wald-Frauenfarn (Athyrium filix femina) waren mit durchschnittlich 2.240 Bq•kg-1 und 2.340 Bq•kg-1 ähnlich hoch kontaminiert.
Auf F1 ist das Artenspektrum etwas anders als auf B2. Typischerweise enthielten auf F1 Brombeere und Himbeere mit durchschnittlichen 153 Bq•kg-1 und 181 Bq•kg-1 erneut die geringsten Cs-137 Aktivitäten. Es folgten Pfeifengras (Molinia coerulea) und Heidelbeere (Vaccinium myrtillus) mit durchschnittlich 341 und 349 Bq•kg-1 und schließlich, wie gewohnt in der Reihenfolge, Wald-Frauenfarn mit 755 Bq•kg-1 und Dornfarn mit 1.060 Bq•kg-1. Mit deutlichem Abstand war Adlerfarn (Pteridium aquillinum), mit durch-schnittlich 1.750 Bq•kg-1, die am höchsten kontaminierte Pflanzenart. Die mittlere Cs-137 Aktivität der drei Farnarten unterschied sich hochsignifikant von allen anderen unter-suchten Spezies (P<0.001).
26

Himbe
ere
Brombe
ere
Wald
hains
imse
Seegra
s
Wald
-Frau
enfar
n
Heidelb
eere
Männl.
Dorn
farn
Dornfar
n --
100
1000
10000B2
30000
Cs-137 [Bq/kg] TS
Brombe
ere
Himbe
ere
Pfeifen
gras
Heidelb
eere
Wald
-Frau
enfar
n
Dornfar
n
Adlerfa
rn --
100
1000
F1
3000
Abb. 12: Spezifische Cs-137 Aktivität in Blättern verschiedener Pflanzenarten auf B2 (n =je 13) und F1 (n= je 9), von 1998 bis 2000. Mittelwerte und Minimal- bzw.
Maximalwerte.
27

6.4 Zeitverlauf von Cs-137 in Waldbodenpflanzen
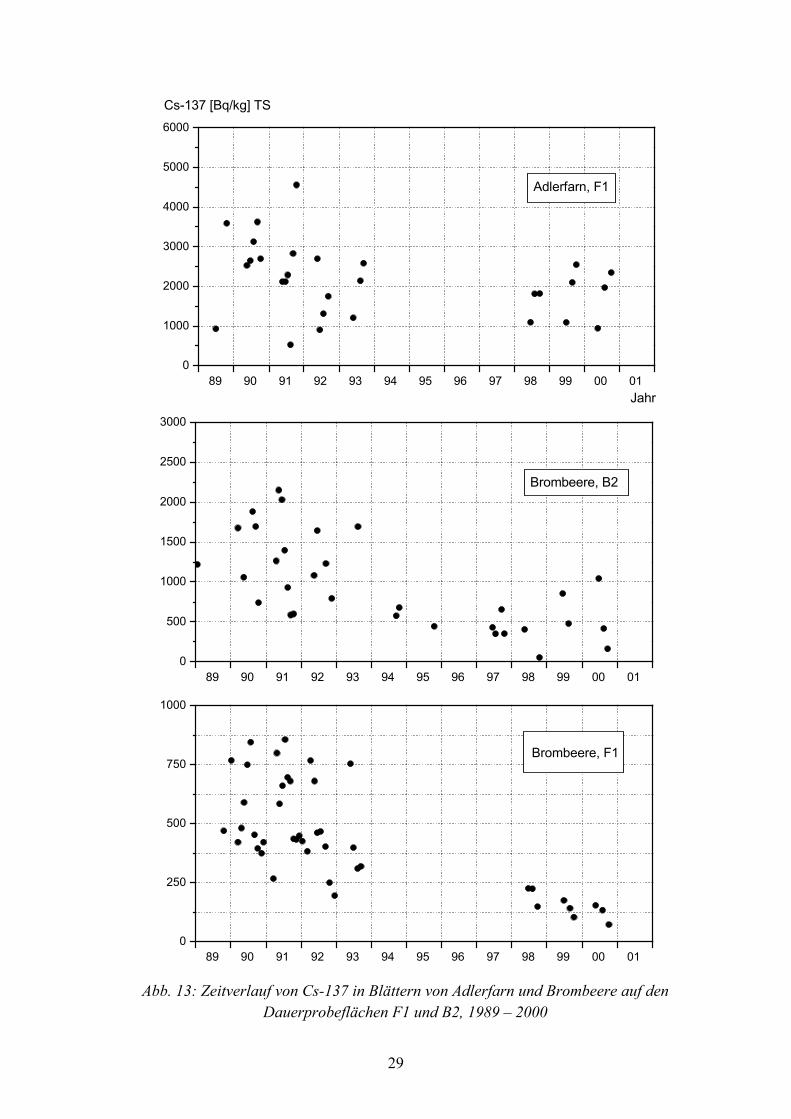
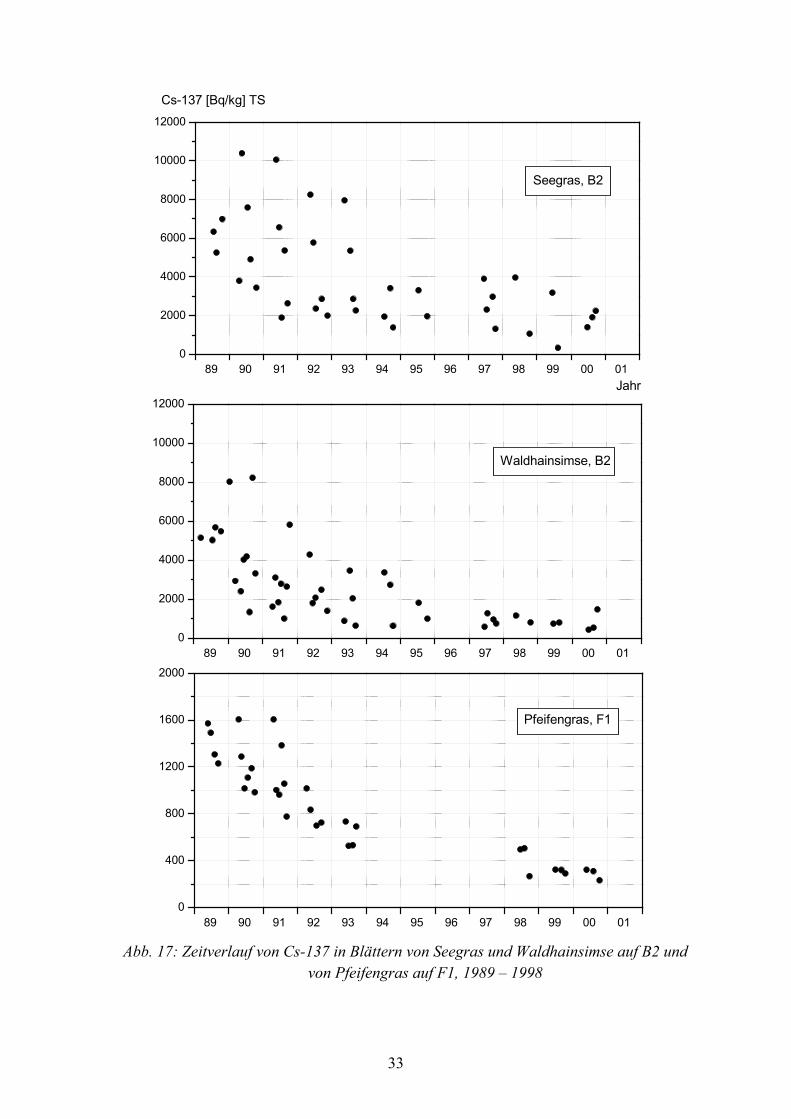
In den Abbildungen 13 bis 18 ist der zeitliche Verlauf von Cs-137 in Blättern, bezogen auf Trockensubstanz, in verschiedenen Pflanzenarten auf B1, B2 und F1, von 1989–2000 dargestellt. Bei jedem Symbolpunkt handelt es sich um den Messwert einer Mischprobe aus 20 bis 40 Einzelpflanzen einer Art.
Von den Flächen B1 und B2 liegen von 1989 bis 2000, mit Ausnahme von 1996, aus jedem Jahr Messwerte vor, während sich die Daten von der Fläche F1 auf die Jahre 1989-1993 und 1998 bis 2000 beschränken. Diese unterschiedliche Datengrundlage von den 3 Probeflächen ist bei der Beurteilung von Langzeittrends sowie beim Vergleich der effektiven Halbwertzeiten von Cs-137 in Pflanzenarten zu beachten.
In den Blättern vieler Pflanzenarten variierten die Cs-137 Gehalte während des Unter-suchungszeitraumes saisonal. Die größten Aktivitätsunterschiede innerhalb der Vegeta-tionsperioden kamen bei Adlerfarn auf F1 vor (Abbildung 13), der in jedem Jahr eine Zunahme der Cs-137 Gehalte vom Frühjahr zum Herbst hin hatte. So wurden im Juni 1999 und 2000 rund 1.000 Bq•kg-1, im Oktober dagegen rund 2.500 Bq•kg-1 festgestellt. Auch in Dornfarn auf B2 und F1 (Abbildung 14) und in Wald-Frauenfarn auf B2 (Abbildung 18) erfolgte eine, wenn auch nur geringe Zunahme der Aktivität während der Vegetations-periode.
Im Jahresverlauf abnehmende Cs-137 Gehalte zeigten die Blätter von Brombeere auf B2 und F1 (Abbildung 13), von Himbeere auf allen 3 Probeflächen (Abbildung 16) sowie von Pfeifengras auf F1 (Abbildung 17).
Mit Ausnahme von Adlerfarn, waren die jahreszeitlichen Schwankungen bei allen Pflanzenarten ab 1997 geringer, als von 1987 bis 1994, was besonders bei Dornfarn, Heidelbeere, Himbeere, Waldhainsimse, Pfeifengras und Wald-Frauenfarn auf F1 offen-sichtlich wird.
Bei allen untersuchten Pflanzenarten nahm die Cs-137 Aktivität von 1987 bis 2000 deutlich ab, nur bei Adlerfarn war der Rückgang weniger ausgeprägt. So betrug 1989 die mittlere Cs-137 Kontamination von Dornfarnblättern auf B1 56.600 Bq•kg-1, im Jahr 2000 dagegen nur 10.210 Bq•kg-1, auf F1 nahm die Aktivität durchschnittlich von 4.500 Bq•kg-1 auf 860 Bq•kg-1 ab.
Besonders gravierend war auf allen Probeflächen der Rückgang der Aktivität in Himbeerblättern, von 1989 bis 2000 nahmen die mittleren Cs-137 Gehalte auf B1 von 4.500 Bq•kg-1 bis 1.010 Bq•kg-1, auf B2 von 3.200 Bq•kg-1 bis 610 Bq•kg-1 und auf F1 von 940 Bq•kg-1 bis 150 Bq•kg-1 ab.
Aus radioökologischer Sicht ist es von besonderem Interesse zu wissen, in welcher Zeit die Cs-137 Kontamination von Produkten aus einem definierten Waldökosystem um die Hälfte abnimmt. Dieser Zeitraum wird als effektive Halbwertzeit (Teff) bezeichnet. Die effektive Halbwertzeit berücksichtigt alle Faktoren, wie radioaktiven Zerfall, Veränderung der Kontamination durch Massenvermehrung bei Pflanzen, Veränderung in der biologischen Verfügbarkeit des Nuklids im Boden usw., die eine Abnahme der Kontamination bedingen.
28
89 90 91 92 93 94 95 96 97 98 99 00 010
1000
2000
3000
4000
5000
6000
Adlerfarn, F1
Jahr
Cs-137 [Bq/kg] TS
89 90 91 92 93 94 95 96 97 98 99 00 010
500
1000
1500
2000
2500
3000
Brombeere, B2
89 90 91 92 93 94 95 96 97 98 99 00 010
250
500
750
1000
Brombeere, F1
Abb. 13: Zeitverlauf von Cs-137 in Blättern von Adlerfarn und Brombeere auf den Dauerprobeflächen F1 und B2, 1989 – 2000
29

89 90 91 92 93 94 95 96 97 98 99 00 010
20000
40000
60000
80000
100000
Dornfarn, B1
Jahr
Cs-137 [Bq/kg] TS
89 90 91 92 93 94 95 96 97 98 99 00 010
10000
20000
30000
40000
50000
Dornfarn, B2
89 90 91 92 93 94 95 96 97 98 99 00 010
1000
2000
3000
4000
5000
6000
Dornfarn, F1
Abb. 14: Zeitverlauf von Cs-137 in Blättern von Dornfarn auf den Dauerprobeflächen B1, B2, und F1, 1989 – 2000
30

89 90 91 92 93 94 95 96 97 98 99 00 010
4000
8000
12000
16000
20000
24000
Heidelbeere, B1
Jahr
Cs-137 [Bq/kg] TS
89 90 91 92 93 94 95 96 97 98 99 00 010
2000
4000
6000
8000
10000
12000
14000
16000
Heidelbeere, B2
89 90 91 92 93 94 95 96 97 98 99 00 010
400
800
1200
1600
2000
Heidelbeere, F1
Abb. 15: Zeitverlauf von Cs-137 in Blättern von Heidelbeere auf den Dauerprobeflächen B1, B2, und F1, 1989 – 2000
31

89 90 91 92 93 94 95 96 97 98 99 00 010
2000
4000
6000
8000
10000
12000
Himbeere, B1
Jahr
Cs-137 [Bq/kg] TS
89 90 91 92 93 94 95 96 97 98 99 00 010
1000
2000
3000
4000
5000
6000
Himbeere, B2
89 90 91 92 93 94 95 96 97 98 99 00 010
400
800
1200
1600
Himbeere, F1
J h
Abb. 16: Zeitverlauf von Cs-137 in Blättern von Himbeere auf den Dauerprobeflächen B1, B2, und F1, 1989 – 2000
32
89 90 91 92 93 94 95 96 97 98 99 00 010
2000
4000
6000
8000
10000
12000
Seegras, B2
Jahr
Cs-137 [Bq/kg] TS
89 90 91 92 93 94 95 96 97 98 99 00 010
2000
4000
6000
8000
10000
12000
Waldhainsimse, B2
89 90 91 92 93 94 95 96 97 98 99 00 010
400
800
1200
1600
2000
Pfeifengras, F1
Abb. 17: Zeitverlauf von Cs-137 in Blättern von Seegras und Waldhainsimse auf B2 und von Pfeifengras auf F1, 1989 – 1998
33

89 90 91 92 93 94 95 96 97 98 99 00 010
4000
8000
12000
16000
20000
Wald-Frauenfarn, B1
Jahr
Cs-137 [Bq/kg] TS
89 90 91 92 93 94 95 96 97 98 99 00 010
2000
4000
6000
8000
10000
Wald-Frauenfarn, B2
89 90 91 92 93 94 95 96 97 98 99 00 010
1000
2000
3000
4000
5000
6000
Wald-Frauenfarn, F1
Abb. 18: Zeitverlauf von Cs-137 in Blättern von Wald-Frauenfarn auf den Dauerprobeflächen B1, B2, und F1, 1989 – 2000
34

Nach dem Zerfallsgesetz gilt für die Abnahme der spezifischen Aktivität:
C/C0 = exp (-λ eff ∗ t)
wobei sich die Ausscheidungskonstante λeff aus der physikalischen Zerfallskonstanten, der biologischen Ausscheidungskonstanten und der Ausscheidungskonstanten für Massen-vermehrung zusammensetzt.
In vorliegender Arbeit wurde die effektive Halbwertzeit berechnet, indem durch die aktuellen, logarithmierten Cs-137 Messdaten eine lineare Regression gelegt und der Steigungskoeffizient (λ eff) der Regressionsgeraden berechnet wurde (z.B. rote Gerade in Abbildung 24). In der Tabelle 5 sind die effektiven Halbwertzeiten von Cs-137 für verschiedene Pflanzenarten auf den 3 Probeflächen angegeben.
Tab. 5: Effektive Halbwertzeiten (Teff) von Waldbodenpflanzen auf den Probeflächen B1, B2 und F1, von 1989 bis 2000
Spezies Teff [Jahre] Teff [Jahre]
B1 B2 F1 Mittelwert
Adlerfarn 23,8 23,8
Dornfarn 4,7 6,2 5,2 5,4
Brombeere 4,8 4,6 4,7
Heidelbeere 7,9 7,2 6,0 7,0
Himbeere 3,1 2,9 4,7 3,6
Pfeifengras 4,8 4,8
Seegras 5,3 5,3
Wald-Frauenfarn 8,4 7,3 5,9 7,2
Waldhainsimse 4,0 4,0
Die längste effektive Halbwertzeit von Cs-137 hat Adlerfarn mit 23,8 Jahren. Für die anderen Pflanzenarten liegen die Werte zwischen 2,9 und 8,4 Jahren.
Die Halbwertzeiten bei den einzelnen Pflanzenarten stimmen auf den 3 Probeflächen relativ gut überein. Himbeere hat immer die kürzeste effektive Halbwertzeit, mit einem Mittelwert von 3,6 Jahren. Blätter von Wald-Frauenfarn und Heidelbeere haben immer längere Halbwertzeiten, von durchschnittlich 7,2 bzw. 7,0 Jahren.
Die effektiven Halbwertzeiten wurden für den Zeitraum 1989 bis 2000 berechnet. Die Cs-137 Aktivitäten nahmen allerdings, wie aus den Abbildungen 13 bis 18 hervorgeht, in den Pflanzen nicht gleichförmig ab. Vielmehr erfolgte bei allen untersuchten Arten (Ausnahme: Adlerfarn), von 1989 bis etwa 1994, ein relativ schneller Rückgang der Aktivität. In der nachfolgenden Phase, von 1995 bis 2000, zeigten einige Arten eine deut-liche langsamere Abnahme (z.B. Brombeere und Himbeere auf F1, Dornfarn auf B1 und B2), keine Veränderung (z.B. Heidelbeere auf B1 und B2, Waldhainsimse auf B2) oder
35

eine Zunahme der Cs-137 Aktivität (z.B. Adlerfarn auf F1, Himbeere auf B1). Diese Trendverläufe sind, aufgrund der kurzen Zeitspanne und der geringen Veränderungen der Cs-137 Aktivität, statistisch nicht signifikant.
Auf der Fläche F1 fällt der hohe Wert der Teff von 23,8 Jahren bei Adlerfarn auf, für den Zeitraum 1989 bis 2000. Seit 1998 nehmen die Cs-137 Gehalte der Blätter wieder zu, die Steigung durch die logarithmierten Werte hat ein positives Vorzeichen.
6.5 Cs-137 in Früchten
Proben von Früchten wurden auf den Probeflächen B1 und F1 entnommen, wobei jeweils Beeren von 20 Einzelpflanzen zu einer Mischprobe vereinigt wurden. In der Tabelle 6 sind die Ergebnisse von 1999 bis 2000 zusammengefasst.
Tab. 6: Mittlere Cs-137 Aktivitäten in der Frischsubstanz von Früchten auf den Probeflächen B1 und F1, 1999 -2000
Früchte Cs-137 [Bq/kg] FS
n B1 n F1
Heidelbeeren 14 436 12 26 Brombeeren 3 180 5 13 Himbeeren 3 130 3 2 Eicheln 7 136 Roggen* 3 2
* von landwirtschaftlicher Nutzfläche bei F1
Auf beiden Probeflächen nahm die Cs-137 Aktivität in der Reihenfolge Himbeeren→Brombeeren→Heidelbeeren zu.
Wie die Blätter der Waldbodenpflanzen, unterschieden sich auch die Früchte der Heidelbeere in ihrem Cs-137 Gehalt auf allen Probeflächen, an jedem Entnahmetermin erheblich. Zum Beispiel enthielten im August 1999, auf B1 entnommene Mischproben zwischen 170 und 770 Bq•kg-1 (FS). Von der Fläche F1 wurden von 1998 bis 2000, ins-gesamt 12 Mischproben Heidelbeeren gemessen. Die mittlere Cs-137 Aktivität, bezogen auf Frischsubstanz, betrug 26 Bq•kg-1, als Minimalwert wurden 2, als Maximalwert 52 Bq•kg-1 ermittelt.
In der Abbildung 19 ist der Aktivitätsverlauf in Heidelbeeren auf B1, von 1989 bis 2000, in Frischsubstanz dargestellt. Das Verhältnis Frischsubstanz : Trockensubstanz beträgt etwa 10 : 1. Von 1989 bis 2000 nahm die Cs-137 Aktivität von Heidelbeeren signifikant ab (P<0,0001), die effektive Halbwertzeit betrug 6,2 Jahre. Die mittlere Cs-137 Aktivität der Heidelbeeren betrug 1989 noch 1.040 Bq•kg-1 (FS). Dagegen enthielten die im Juli 2000
36
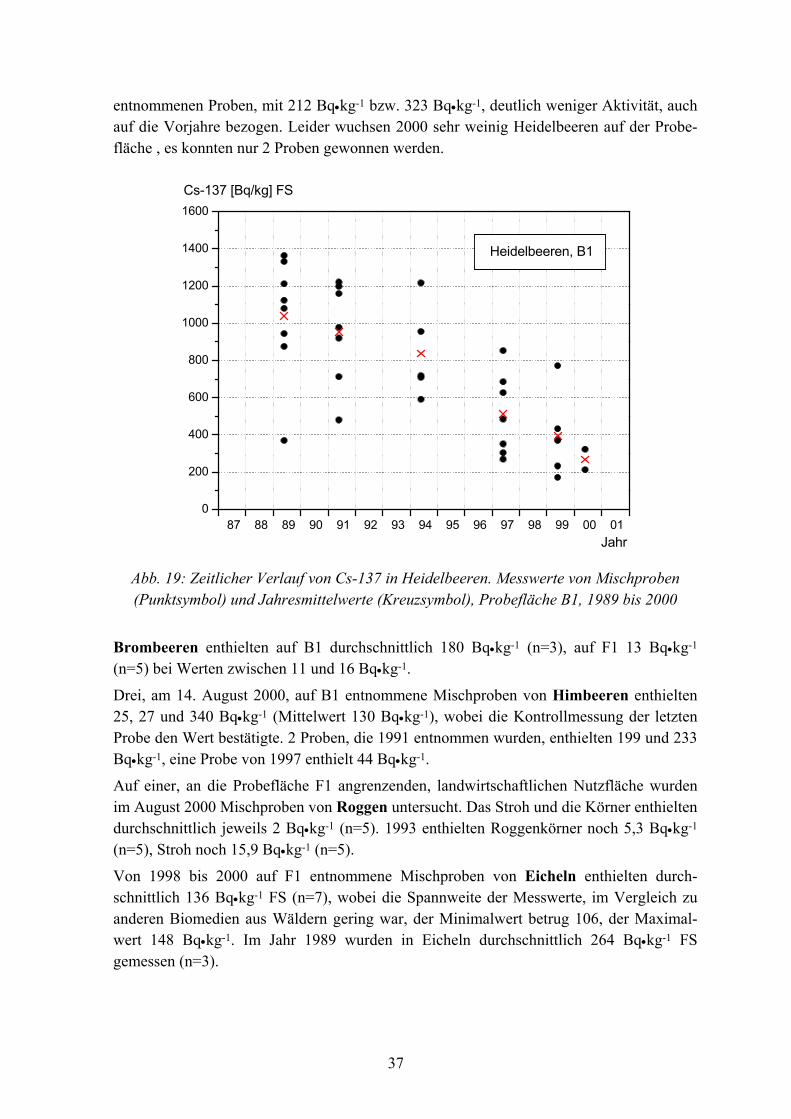
entnommenen Proben, mit 212 Bq•kg-1 bzw. 323 Bq•kg-1, deutlich weniger Aktivität, auch auf die Vorjahre bezogen. Leider wuchsen 2000 sehr weinig Heidelbeeren auf der Probe-fläche , es konnten nur 2 Proben gewonnen werden.
87 88 89 90 91 92 93 94 95 96 97 98 99 00 010
200
400
600
800
1000
1200
1400
1600
Heidelbeeren, B1
Cs-137 [Bq/kg] FS
Jahr
Abb. 19: Zeitlicher Verlauf von Cs-137 in Heidelbeeren. Messwerte von Mischproben (Punktsymbol) und Jahresmittelwerte (Kreuzsymbol), Probefläche B1, 1989 bis 2000
Brombeeren enthielten auf B1 durchschnittlich 180 Bq•kg-1 (n=3), auf F1 13 Bq•kg-1 (n=5) bei Werten zwischen 11 und 16 Bq•kg-1.
Drei, am 14. August 2000, auf B1 entnommene Mischproben von Himbeeren enthielten 25, 27 und 340 Bq•kg-1 (Mittelwert 130 Bq•kg-1), wobei die Kontrollmessung der letzten Probe den Wert bestätigte. 2 Proben, die 1991 entnommen wurden, enthielten 199 und 233 Bq•kg-1, eine Probe von 1997 enthielt 44 Bq•kg-1.
Auf einer, an die Probefläche F1 angrenzenden, landwirtschaftlichen Nutzfläche wurden im August 2000 Mischproben von Roggen untersucht. Das Stroh und die Körner enthielten durchschnittlich jeweils 2 Bq•kg-1 (n=5). 1993 enthielten Roggenkörner noch 5,3 Bq•kg-1 (n=5), Stroh noch 15,9 Bq•kg-1 (n=5).
Von 1998 bis 2000 auf F1 entnommene Mischproben von Eicheln enthielten durch-schnittlich 136 Bq•kg-1 FS (n=7), wobei die Spannweite der Messwerte, im Vergleich zu anderen Biomedien aus Wäldern gering war, der Minimalwert betrug 106, der Maximal-wert 148 Bq•kg-1. Im Jahr 1989 wurden in Eicheln durchschnittlich 264 Bq•kg-1 FS gemessen (n=3).
37

6.6 Cs-137 in Pilzen
Da Pilze auf den Probeflächen während des gesamten Untersuchungszeitraums sehr unter-schiedlich fruktifizierten, konnten von 1989 bis 2000 nur in größeren Abständen Pilze von B1 und F1 entnommen werden. Zum Beispiel wurden 1994, 1997 und 1998 an den Probe-nahmeterminen nur sehr wenig oberirdisch wachsende Fruchtkörper gefunden. Dagegen kam es 2000, in allen 3 Untersuchungsgebieten zu einer „Pilzschwemme“. Dieses seltene Massenvorkommen von Pilzen, insbesondere in Bodenmais, bot die Möglichkeit, die innerartliche Verteilung der Cs-137 Aktivität bei Maronenröhrlingen und Steinpilzen genauer zu untersuchen. In der Abbildung 20 ist die Cs-137 Aktivität verschiedener Pilzarten von der Probefläche B1 und Umgebung dargestellt.
Ziegen
lippe
Hallim
asch
Pfifferl
ing
Steinp
ilz
Rotfußrö
hrlin
g
Graue
r Wulst
ling
Marone
Kartoffe
lbovis
t
Dickfuß
Hirsch
trüffe
l
100
1.000
10.000
100.000
855405824373
Cs-137 [Bq/kg] FS
Abb. 20: Cs-137 Aktivität in Pilzen, Probefläche B1 und Umgebung, Herbst 2000. Säulen = Mittelwerte, untere bzw. obere Whisker = Minimal- bzw. Maximalwerte
Durchschnittlich waren Ziegenlippe (Xerocomus subtomentosus), mit 390 Bq•kg-1 und Hallimasch (Armillaria mellea) mit 400 Bq•kg-1 in der Frischsubstanz, die am geringsten kontaminierten Speisepilze. Pfifferling (Cantharellus cibarius), Steinpilz (Boletus edulis) und Rotfußröhrling (Xerocomus chrysenteron) hatten Aktivitäten von durchschnittlich 1.000 Bq•kg-1, im mittleren Kontaminationsbereich. Im Durchschnitt deutlich höhere Cs-137 Gehalte hatten Grauer Wulstling (Amanita spissa) und Maronenröhrling (Xerocomus badius), mit 1.700 bzw. 3.030 Bq•kg-1. Viel höher kontaminiert war Nadelwald-Dickfuß (Cortinarius hercynicus) mit 6.750 Bq•kg-1. Die Cs-137 Aktivität der unterirdisch wachsen-
38

den Hirschtrüffel (Elaphomyces granulatus)übertraf, mit durchschnittlich 25.660 Bq•kg-1,die aller oberirdisch wachsenden Pilzarten um ein Vielfaches.
Die Ergebnisse zeigen, dass bei den Pilzarten, wie bei den autotrophen Pflanzen auch, die Cs-137 Aktivität artspezifisch variiert. Auch die innerartliche Variabilität war bei Pilzen groß: Die Cs-137 Gehalte schwankten jeweils um das 5- bis 10fache. In der Tabelle 7 sind die Daten für Maronenröhrlinge und Steinpilze, von 2 Probenahmen auf B1 und Um-gebung, im Herbst 2000, zusammengestellt.
Tab. 7: Statistische Daten von Maronenröhrlingen und Steinpilzen, Probefläche B1 und Umgebung, Probenahme 14.8. und 23.9. 2000
Kenngröße
Maronenröhrling
Cs-137 [Bq•kg-1] FS
Steinpilz
Cs-137 [Bq•kg-1] FS
n 40 24
Minimum 323 306
Maximum 12.110 1.830
Arithmetischer Mittelwert 3.030 1.000
Geometrischer Mittelwert 2.210 900
Standardabweichung 2.510 425
Median 1.890 890
Variationskoeffizient 83% 42%
Die Verteilung der Messwerte war bei beiden Pilzarten rechtsschief. Die Mittelwerte lagen über den geometrischen Mittelwerten und diese über den Median-Werten. Bei Maronen-röhrlingen reichten die Werte zum Beispiel von 323 Bq•kg-1 bis 12.110 Bq•kg-1, bei Stein-pilzen von 306 Bq•kg-1 bis 1.830 Bq•kg-1. Der Variationskoeffizient war bei Maronenröhr-lingen mit 83%, fast doppelt so groß wie bei Steinpilzen mit 42%, allerdings war der Probenumfang bei Maronenröhrlingen auch größer.
Um den zeitlichen Verlauf der Cs-137 Aktivität in den Fruchtkörpern von Pilzen genau zu ermitteln, ist es notwendig, die Aktivität auf Trockensubstanz zu beziehen. Der Grund dafür liegt in der Stoffwechselphysiologie: Pilze haben, im Gegensatz zu Pflanzen, keine äußeren Abschlussgewebe, die effektiv vor Verdunstung schützen. Dadurch ist die spezifische Cs-137 Aktivität im Frischgewicht maßgeblich von der Bodenfeuchte und der Witterung abhängig. Der zeitliche Verlauf der Cs-137 Aktivität in Maronenröhrlingen und Steinpilzen von B1 ist in den Abbildungen 21, bezogen auf Trockensubstanz, dargestellt. Die Variabilität der Messwerte je Probenahme ist bei beiden Pilzarten erheblich. Bei Maronenröhrlingen und Steinpilzen ist während des gesamten Untersuchungszeitraums keine signifikante Änderung der Cs-137 Aktivität festzustellen. Tendenziell steigt bei
39
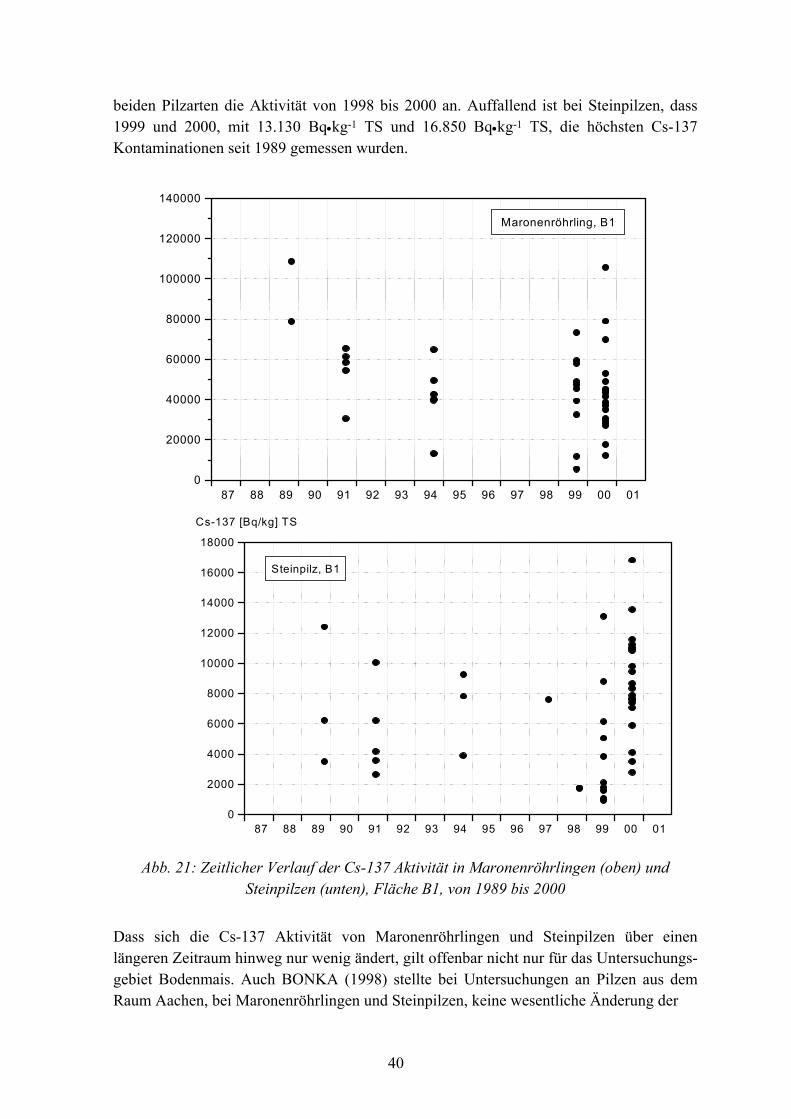
beiden Pilzarten die Aktivität von 1998 bis 2000 an. Auffallend ist bei Steinpilzen, dass 1999 und 2000, mit 13.130 Bq•kg-1 TS und 16.850 Bq•kg-1 TS, die höchsten Cs-137 Kontaminationen seit 1989 gemessen wurden.
87 88 89 90 91 92 93 94 95 96 97 98 99 00 010
20000
40000
60000
80000
100000
120000
140000
Maronenröhrling, B1
87 88 89 90 91 92 93 94 95 96 97 98 99 00 010
2000
4000
6000
8000
10000
12000
14000
16000
18000
Steinpilz, B1
Cs-137 [Bq/kg] TS
Abb. 21: Zeitlicher Verlauf der Cs-137 Aktivität in Maronenröhrlingen (oben) und Steinpilzen (unten), Fläche B1, von 1989 bis 2000
Dass sich die Cs-137 Aktivität von Maronenröhrlingen und Steinpilzen über einen längeren Zeitraum hinweg nur wenig ändert, gilt offenbar nicht nur für das Untersuchungs-gebiet Bodenmais. Auch BONKA (1998) stellte bei Untersuchungen an Pilzen aus dem Raum Aachen, bei Maronenröhrlingen und Steinpilzen, keine wesentliche Änderung der
40

Cs-137 Aktivität, von 1989 bis 1996 fest. GOUSSIOS und NIESSL (1997) dokumentierten den zeitlichen Verlauf der Jahresmittelwerte der spezifischen Cs-137 Aktivität in wild-wachsenden Speisepilzen aus ganz Bayern, von 1986 bis 1996. Während bei Maronen-röhrlingen eine Abnahme der Kontamination festzustellen war, stiegen die Cs-137 Gehalte bei Steinpilzen tendenziell an.
In der Abbildung 22 sind die Messergebnisse der Pilze, die 2000 auf F1 in Fuhrberg ent-nommen wurden, dargestellt. Genau wie in Bodenmais unterschieden sich auch hier die einzelnen Arten in ihren mittleren Cs-137 Gehalten, die innerartliche Variabilität war ebenfalls hoch. Die Cs-137 Messwerte von 4 Proben Violetter Lacktrichterling (Laccaria amethysta), bezogen auf Frischsubstanz, reichten von 57 bis 430 Bq•kg-1, bei einem Mittel-wert von 170 Bq•kg-1. Bei 12 Proben Maronenröhrlingen wurden Werte zwischen 154 und 1.020 Bq Cs-137•kg-1 ermittelt.
Apfeltä
ublin
g
Grauer
Wuls
tling
Violett
er La
cktric
hterlin
g
Sandrö
hrling
Kartoff
elbov
ist
Rotfuß
röhrlin
g
Marone
0
100
200
300
400
1000
1100
3 1235435
Cs-137 [Bq/kg] FS
Abb. 22: Cs-137 Aktivität in Pilzen, Probefläche F1 und Umgebung, Herbst 2000. Säulen = Mittelwerte, untere bzw. obere Whisker = Minimal- bzw. Maximalwerte
Die Reihenfolge der Cs-137 Gehalte in verschiedenen Pilzarten war auf F1 ähnlich wie auf der Probefläche B1. Am niedrigsten war die Aktivität von Parasol (Macrolepiota procera) mit 5 Bq•kg-1. Auch Pfifferlinge enthielten mit 97 Bq•kg-1 erneut relativ wenig Cs-137. Da von diesen beiden Arten jeweils nur eine Probe untersucht werden konnte, sind die Mess-ergebnisse in der Abbildung 22 nicht dargestellt. Durchschnittlich enthielten Apfeltäubling (Russula paludosa) mit 89 Bq•kg-1 die niedrigste Cs-137 Aktivität. Deutlich höhere mitt-lere Aktivitätswerte hatten mit 309 Bq•kg-1 Kartoffelbovisten, mit 354 Bq•kg-1 Maronen-
41

röhrlinge und mit 359 Bq•kg-1 Rotfußröhrlinge. Aus den Vorjahren liegen von den Pilz-arten nicht genügend Messwerte vor, um Trends über den zeitlichen Verlauf der Cs-137 Aktivität zu untersuchen.
Auf der Probefläche G1, in Göttingen ist das Artenspektrum von Pilzen, aufgrund der Bodenverhältnisse und insbesondere wegen des hohen pH-Wertes, anders als auf B1 und F1. Bei mehreren Pilzarten lag die Aktivität, bezogen auf Trockensubstanz, unter der Nachweisgrenze von 0,5 Bq•kg-1, etwa bei Birnenstäubling (Lycoperdon pyriforme), Fleischiger Anis-Champignon (Agaricus silvicola) und Grauer Wulstling. Auch Röhrlinge enthielten in der Frischsubstanz nur relativ wenig Cs-137 Aktiviät: Maronenröhrlinge hatten durchschnittlich 8,8 Bq•kg-1 (n=6), Rotfußröhrlinge 6,4 Bq•kg-1 (n=1). Höhere Aktivitäten hatten je eine Probe Grüner Speisetäubling (Russula heterophylla) 28,4 Bq•kg-1 und Mehlstiel-Täubling (Russula farinipes) 178 Bq•kg-1.
Bezogen auf alle 3 Untersuchungsgebiete, wurde 2000 die geringste Cs-137 Aktivität mit mit <0,5 Bq•kg-1 in der Frischsubstanz, in Birnenstäubling, Dünnfleischiger Anis-Champignon und Grauer Wulstling auf der Probefläche G1 ermittelt, die höchste mit 34.870 Bq•kg-1 in Hirschtrüffeln, auf der Probefläche B1.
Diese Werte sind wahrscheinlich die Minimal- bzw. Maximalwerte von Cs-137 in Pilzen in Bundesrepublik im Jahr 2000, die Aktivität variierte damit über 5 Größenordnungen. Die Abbildung 23 verdeutlicht die Spannweite der Cs-137 Messwerte bei dem Speisepilz Maronenröhrling.
1
10
100
1000
10000
61240
Maronenröhrling
GöttingenFuhrbergBodenmais
Cs-137 [Bq/kg] FS
Abb. 23: Cs-137 Aktivität von Maronenröhrlingen in den Untersuchungsgebieten, 2000
42

7 Cs-137 Aktivität in Wildtieren 7.1 Cs-137 Aktivität in Wildschweinen
Das Probenaufkommen von Wildschweinen aus Bodenmais ist seit 1994 sehr gering, was hauptsächlich mit der geänderten Jagdstrategie des zuständigen Forstamtes Bodenmais zusammenhängt. Wildschweine sind in den geschlossenen Waldgebieten des Unter-suchungsgebietes nur sehr schwer zu bejagen und wurden bisher an Lockfütterungen (Kirrungen) angefüttert und dort erlegt. Seit 1994 werden Wildschweine, aus ökologischen Gründen, nicht mehr angefüttert, mit der Folge, dass nur noch vereinzelt Tiere erlegt werden. In der Abbildung 24 ist der Zeitverlauf der Cs-137 Aktivität in Wildschweinen aus dem Untersuchungsgebiet Bodenmais und einem angrenzenden Jagdrevier in Zwiesel dargestellt. Die Bodenkontamination ist in beiden Gebieten ähnlich hoch.
01 01 198601 01 198701 01 198801 01 198901 01 199001 01 199101 01 199201 01 199301 01 199401 01 199501 01 199601 01 199701 01 199801 01 199901 01 200001 01 200101 01 2002
1000
10000
50000
1986
Wildschwein
2000199819961994199219901988
Cs-137 [Bq/kg] FS
Jahr
Abb. 24: Zeitverlauf von Cs-137 in Wildschweinen aus Bodenmais und Zwiesel, mit Regressionsgerade, 1987 bis 2001 (n=103)
Die Cs-137 Kontamination der Wildschweine stagnierte während des Untersuchungs-zeitraum auf hohem Niveau. So war die durchschnittliche Cs-137 Aktivität von Wild-schweinen, im ersten Halbjahr 2001, mit 8.990 Bq•kg-1 (n=7) in der Frischsubstanz, noch höher, als im Gesamtjahr 1987 mit 7.240 Bq•kg-1 (n=14). Allerdings muss berücksichtigt werden, dass 4 der Proben von 2001 aus dem Frühjahr stammten, wenn Wildschweine die höchsten Radiocäsiumwerte im Jahresablauf aufweisen. Der Verlauf der Regressions-geraden zeigt, dass die Kontamination der Tiere ansteigt. Die Korrelation ist mit R= 0,238 nur schwach, aber signifikant (P=0,0154). Aufgrund der diskontinuierlichen Verteilung der Messwerte über den Untersuchungszeitraum und nur wenigen Proben aus den letzten Jahren, sollten die Ergebnisse nur als Trend gesehen werden. Aus der Steigung der
43

Geraden ergibt sich eine positive effektive Halbwertzeit von 10,4 Jahren, für Cs-137 in Wildschweinen.
Zu einer vergleichbaren Aussage kommt auch HECHT (2000), der die Cs-137 Konta-mination von Wildschweinen langfristig in ganz Bayern untersuchte. Der Autor stellte in den Jahren nach dem Tschernobyl-Fallout eine immer geringer werdende Abnahme der Cs-137 Aktivität in Wildschweinen fest, dann eine Zeit lang keine Veränderung der Messwerte und seit 1995, trendmäßig, eine erneute Aktivitätszunahme. Für den Zeitraum 1986–1996 ergab sich aus den logarithmierten Messwerten eine (positive) effektive Halb-wertzeit von 62 Jahren (HECHT 1997).
Aus dem Untersuchungsgebiet Göttingen wurden im Jahr 2000 auch Proben von Wild-schweinen untersucht. Die Werte reichten von 1,5 bis 3,1 Bq Cs-137•kg-1 (n=4), bei einem Durchschnittswert von 2,2 Bq•kg-1. Diese Aktivitäten dürften zu den niedrigsten Cs-137 Werten zählen, bei Wildschweinen aus der Bundesrepublik gemessen wurden, während die im Raum Bodenmais festgestellten Messwerte zu den höchsten zählen. Die Kontami-nation von Wildschweinen variiert damit um fast 4 Größenordnungen.
7.2 Cs-137 Aktivität in Rothirschen
In der Abbildung 25 ist der Zeitverlauf der Cs-137 Aktivität in Rothirschen aus dem Untersuchungsgebiet Bodenmais und einem angrenzenden Jagdrevier in Zwiesel, aus dem auch die Wildschweinproben stammten, dargestellt.
01 01 198601 01 198701 01 198801 01 198901 01 199001 01 199101 01 199201 01 199301 01 199401 01 199501 01 199601 01 199701 01 199801 01 199901 01 200001 01 200101 01 2002
10
100
1.000
1986
Rothirsch
2000199819961994199219901988
Cs-137 [Bq/kg] FS
Jahr
Abb. 25: Zeitverlauf von Cs-137 in Rothirschen aus Bodenmais und Zwiesel, mit Regressionsgerade, 1987 bis 2000 (n=116)
44

Das Staatliche Forstamt Bodenmais ist rotwildfreies Gebiet, das heißt, Rothirsche werden nicht geduldet, wenn Tiere zuwandern, müssen sie abgeschossen werden. Entsprechend gering ist das Probenaufkommen für Untersuchungen.
Die Cs-137 Aktivität von Rothirschen nahm während des Untersuchungszeitraumes deutlich ab. Während die Proben 1987 durchschnittlich 1.510 Bq•kg-1 (n=32) enthielten, lagen die Werte, seit 1995, immer unter 1.000 Bq•kg-1. Aus der Regressionsgeraden errechnet sich die effektive Halbwertzeit von -5,7 Jahren, die Abnahme der Kontamination ist hochsignifikant (P<0.0001).
Eine Abnahme der Cs-137 Kontamination bei Rotwild ermittelte auch HECHT (1997), bei Untersuchungen, von 1986-1996, in Bayern, die effektive Halbwertzeit betrug –11,4 Jahre.
7.3 Cs-137 Aktivität in Rehen
Im Muskelfleisch von Rehen variierte die Cs-137 Aktivität jedes Jahr ausgesprochen saisonal, mit niedrigen Werten im Frühjahr und deutlich höheren Werten im Herbst. Als Beispiel ist der Verlauf der Kontamination von 2000/2001 in Abbildung 26 dargestellt.
Je nach Vegetationszustand und Zeitpunkt des Wintereinbruchs, fressen sich Rehe, von Ende September bis November ihren “Winterspeck” an, nehmen also wesentlich mehr Nahrung auf als sonst, hauptsächlich Blätter von Farnen, Himbeere, Brombeere, Heidelbeere und Pilze. Dornfarn und einige Pilzarten sind gerade dann relativ hoch mit Cs-137 belastet, also erreicht auch die Kontamination von Rehwild, durchschnittlich im Oktober/November ihren höchsten Stand.
Mit dem ersten Schneefall geht der Nahrungsbedarf der Rehe schlagartig zurück und sinkt, aufgrund des nun verminderten Stoffwechsels während des Winters bis fast auf die Hälfte der Menge ab, die im Oktober aufgenommen wird (ELLENBERG 1978).
Da die biologische Halbwertzeit von Cs-137 in Rehen, mit ca. 10 Tagen relativ klein ist, nimmt auch die Kontamination der Tiere, mit Beginn des Schneefalls, schnell ab. Dieser Zusammenhang wird aus der Abbildung 26 deutlich: Anfang Dezember 2000 fiel im Untersuchungsgebiet Bodenmais Schnee, der zu einer festen Schneedecke wurde. Als Folge davon nahm die Cs-137 Aktivität der Rehe schnell ab, von durchschnittlich 3.200 Bq•kg-1 im November, auf nur noch 259 Bq•kg-1 im Januar.
Im Winter liegt in Bodenmais i.d.R. mehr als 1 Meter Schnee. Rehe nehmen dann den Großteil ihrer Nahrung an Winterfütterungen auf, wo nur gering kontaminiertes Futter angeboten wird. Zusätzlich fressen sie noch geringe Mengen Knospen von Sträuchern und jungen Bäumen, die aus dem Schnee herausragen, bzw. andere Waldpflanzen, die sie frei scharren. Knospen sind i.d.R. geringer belastet als die Blätter der meisten Waldpflanzen während der Vegetationsperiode. In Folge dessen bleiben die Cs-137 Gehalte im Muskel-fleisch der Rehe den ganzen Winter über niedrig. Im Frühjahr beginnt mit der Aufnahme der ersten, relativ Cs-137 haltigen Waldbodenpflanzen, im Cs-Stoffwechsel von Rehwild ein, dem Winter entgegengesetzter Zyklus: Der Cs-137 Input durch die Nahrung nimmt zu, wodurch das Nuklid in der Muskulatur angereichert wird.
45

A M J J A S O N D J F M A M
0
1000
2000
3000
4000
5000
geschlosseneSchneedeckeReh
Monat
Cs-137 [Bq/kg] FS
Abb. 26: Cs-137 Aktivität in Rehen aus Bodenmais, Mai 2000 bis Mai 2001 (n=73)
Dieses saisonale Muster der Kontamination zeigte sich in jedem Jahr. In der Abbildung 27 ist der Verlauf der Cs-137 Aktivität in Rehwild aus Bodenmais, von 1987 bis 2001, anhand der Monatsmediane gezeigt. Die Unterschiede zwischen der minimalen Kontamination im Frühjahr und der maximalen im Herbst sind erheblich und betragen eine Größenordnung.
Von 1987 bis 2001 nahm die Cs-137 Aktivität in Rehen ab. Der höchste Medianwert wurde im Oktober 1987, mit 7.040 Bq•kg-1 festgestellt. 1993 betrug der höchste Monats-median im Oktober 3.810 Bq•kg-1, 1999 im September noch 1.220 Bq•kg-1. 2000 erfolgte ein deutlicher Anstieg der Kontamination, mit 3.370 Bq•kg-1 wurde im November 2000 der höchste monatliche Median seit 1993 festgestellt.
Für den gesamten Untersuchungszeitraum, von 1987-2000, ergibt sich für Cs-137 in Rehen (n= 1.429) eine effektive Halbwertzeit von –6 Jahren. (P<0.0001). Dieser Wert entsteht aus der Kombination von zwei Phasen mit sehr unterschiedlicher Dynamik: In der ersten Phase, von 1987 bis 1995, erfolgt ein relativ schneller Rückgang der Aktivität, mit einer Halbwertzeit von -4,6 Jahren (n=1.091, P<0.0001), während von 1996 bis 2000, mit einer Halbwertzeit von -89 Jahren (n=338, P=0.8454) eine deutliche Verlangsamung eintrat. Obwohl dieser Wert nicht signifikant ist und durch die ausnehmend hohe Kontamination der Rehe im Jahr 2000 besonders beeinflusst wird, besteht seit 1996 der Trend zu größeren, effektiven Halbwertzeiten. Zur gleichen Trendaussage kommt auch HECHT (1997), bei Untersuchungen an Rehen im Bayerischen Wald; er ermittelte eine effektive Halbwertzeit von -12,7 Jahren für Rehe aus ganz Bayern, von 1986-1996. ZIBOLD et al. (2001) geben für Rehe aus Fichtenbeständen in Oberschwaben, von 1987-1998, effektive Halbwertzeiten von 3,5 Jahren und für Rehe aus Moorgebieten 17 Jahre an.
46
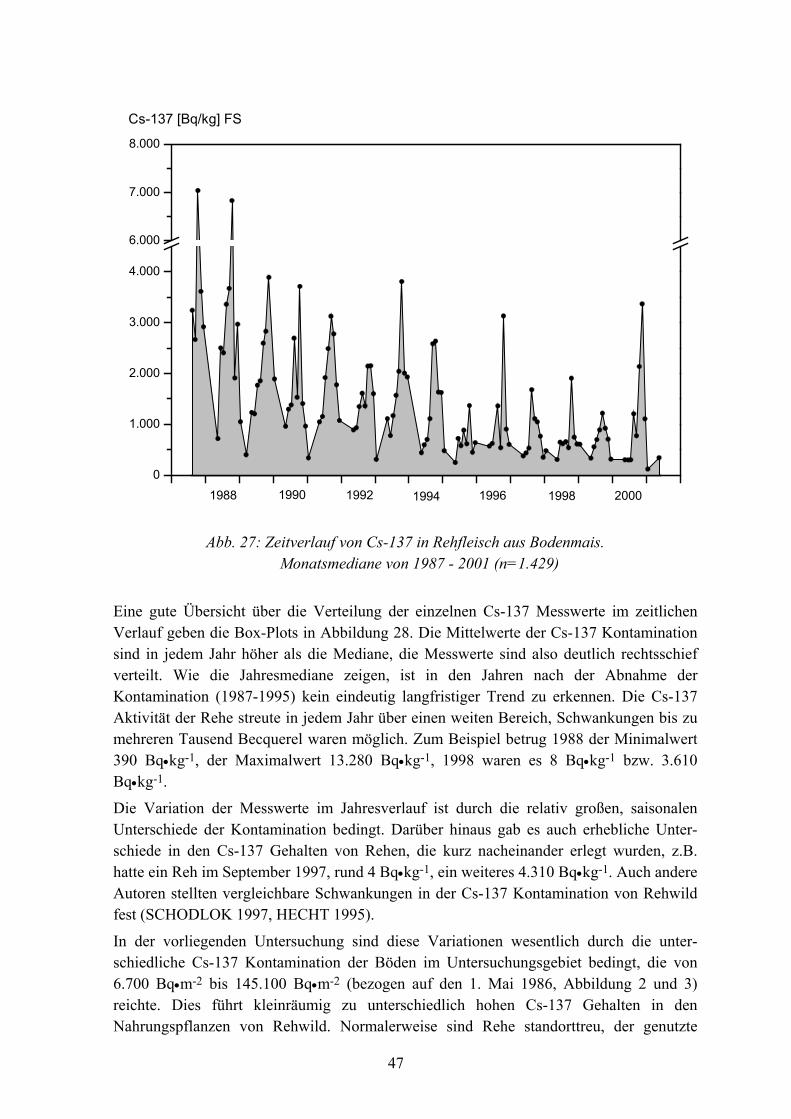
0
1.000
2.000
3.000
4.000
6.000
7.000
8.000
2000
Cs-137 [Bq/kg] FS
199819961994 1992 1990 1988
Abb. 27: Zeitverlauf von Cs-137 in Rehfleisch aus Bodenmais. Monatsmediane von 1987 - 2001 (n=1.429)
Eine gute Übersicht über die Verteilung der einzelnen Cs-137 Messwerte im zeitlichen Verlauf geben die Box-Plots in Abbildung 28. Die Mittelwerte der Cs-137 Kontamination sind in jedem Jahr höher als die Mediane, die Messwerte sind also deutlich rechtsschief verteilt. Wie die Jahresmediane zeigen, ist in den Jahren nach der Abnahme der Kontamination (1987-1995) kein eindeutig langfristiger Trend zu erkennen. Die Cs-137 Aktivität der Rehe streute in jedem Jahr über einen weiten Bereich, Schwankungen bis zu mehreren Tausend Becquerel waren möglich. Zum Beispiel betrug 1988 der Minimalwert 390 Bq•kg-1, der Maximalwert 13.280 Bq•kg-1, 1998 waren es 8 Bq•kg-1 bzw. 3.610 Bq•kg-1.
Die Variation der Messwerte im Jahresverlauf ist durch die relativ großen, saisonalen Unterschiede der Kontamination bedingt. Darüber hinaus gab es auch erhebliche Unter-schiede in den Cs-137 Gehalten von Rehen, die kurz nacheinander erlegt wurden, z.B. hatte ein Reh im September 1997, rund 4 Bq•kg-1, ein weiteres 4.310 Bq•kg-1. Auch andere Autoren stellten vergleichbare Schwankungen in der Cs-137 Kontamination von Rehwild fest (SCHODLOK 1997, HECHT 1995).
In der vorliegenden Untersuchung sind diese Variationen wesentlich durch die unter-schiedliche Cs-137 Kontamination der Böden im Untersuchungsgebiet bedingt, die von 6.700 Bq•m-2 bis 145.100 Bq•m-2 (bezogen auf den 1. Mai 1986, Abbildung 2 und 3) reichte. Dies führt kleinräumig zu unterschiedlich hohen Cs-137 Gehalten in den Nahrungspflanzen von Rehwild. Normalerweise sind Rehe standorttreu, der genutzte
47
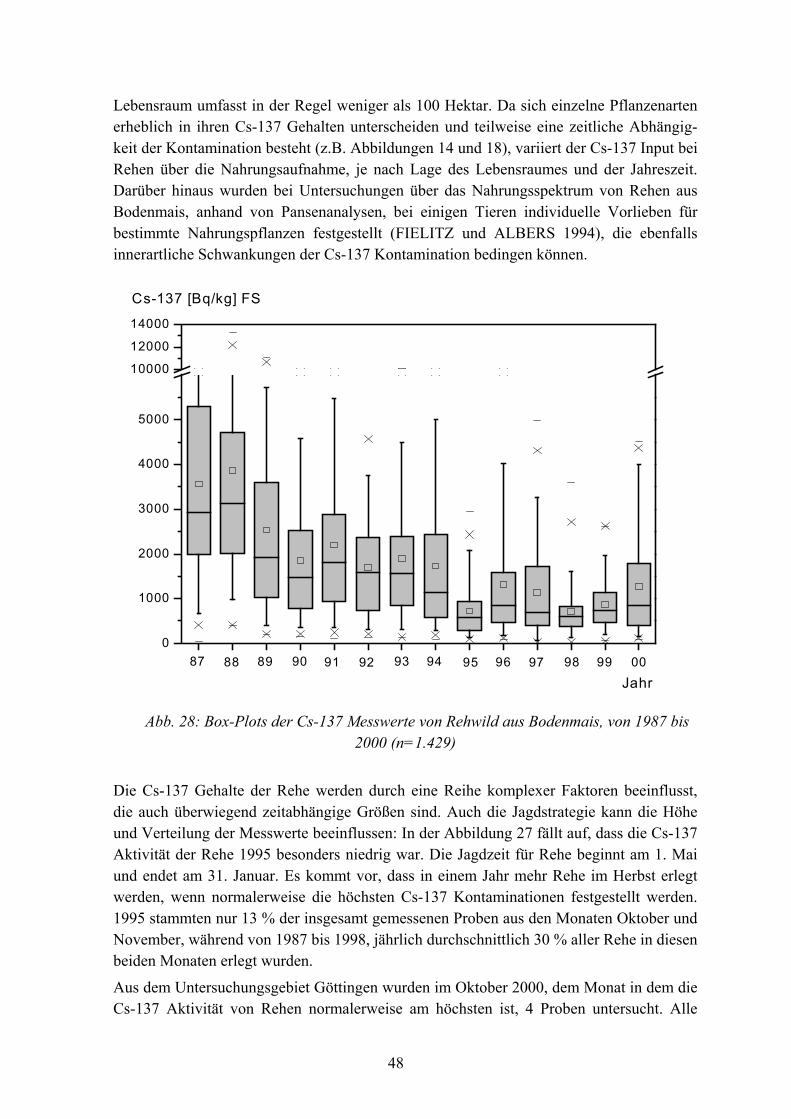
Lebensraum umfasst in der Regel weniger als 100 Hektar. Da sich einzelne Pflanzenarten erheblich in ihren Cs-137 Gehalten unterscheiden und teilweise eine zeitliche Abhängig-keit der Kontamination besteht (z.B. Abbildungen 14 und 18), variiert der Cs-137 Input bei Rehen über die Nahrungsaufnahme, je nach Lage des Lebensraumes und der Jahreszeit. Darüber hinaus wurden bei Untersuchungen über das Nahrungsspektrum von Rehen aus Bodenmais, anhand von Pansenanalysen, bei einigen Tieren individuelle Vorlieben für bestimmte Nahrungspflanzen festgestellt (FIELITZ und ALBERS 1994), die ebenfalls innerartliche Schwankungen der Cs-137 Kontamination bedingen können.
0
1000
2000
3000
4000
5000
10000
12000
14000
99 00989796959493929190898887
Jahr
Cs-137 [Bq/kg] FS
Abb. 28: Box-Plots der Cs-137 Messwerte von Rehwild aus Bodenmais, von 1987 bis 2000 (n=1.429)
Die Cs-137 Gehalte der Rehe werden durch eine Reihe komplexer Faktoren beeinflusst, die auch überwiegend zeitabhängige Größen sind. Auch die Jagdstrategie kann die Höhe und Verteilung der Messwerte beeinflussen: In der Abbildung 27 fällt auf, dass die Cs-137 Aktivität der Rehe 1995 besonders niedrig war. Die Jagdzeit für Rehe beginnt am 1. Mai und endet am 31. Januar. Es kommt vor, dass in einem Jahr mehr Rehe im Herbst erlegt werden, wenn normalerweise die höchsten Cs-137 Kontaminationen festgestellt werden. 1995 stammten nur 13 % der insgesamt gemessenen Proben aus den Monaten Oktober und November, während von 1987 bis 1998, jährlich durchschnittlich 30 % aller Rehe in diesen beiden Monaten erlegt wurden.
Aus dem Untersuchungsgebiet Göttingen wurden im Oktober 2000, dem Monat in dem die Cs-137 Aktivität von Rehen normalerweise am höchsten ist, 4 Proben untersucht. Alle
48

Proben enthielten weniger als 0,5 Bq•kg-1. Die Cs-137 Kontamination von Rehen, wie auch von Wildschweinen, variiert innerhalb der Bundesrepublik um 4 Größenordnungen.
7.4 Cs-137 Aktivität in Kühen
In der Abbildung 8 ist die Tiefenverteilung von Cs-137 in einer Wiese, im Unter-suchungsgebiet Bodenmais, dargestellt. Die Wiese wurde 2000 als Weide für zwei Kühe genutzt. Die Tiere wurden im November 2000, im Alter von 11/2 Jahren geschlachtet. Die Cs-137 Gehalte im Muskelfleisch betrugen 2 und 39 Bq•kg-1.
7.5 Höhe und zeitliche Entwicklung der Cs-137 Kontamination bei Rehen, Rothirschen und Wildschweinen
Die Höhe und die zeitliche Entwicklung der Cs-137 Kontamination unterscheidet sich bei Rehen, Rothirschen und Wildschweinen aus dem Untersuchungsgebiet Bodenmais deutlich. In der Abbildung 24 sind die Regressionsgeraden, die sich aus den logarithmisch skalierten Messwerten für jede der drei Wildarten ableiten lassen, dargestellt.
Die Cs-137 Aktivitäten von Rehen und Rothirschen nahmen von 1986 bis 2000 ab, bei Wildschweinen nahmen sie zu. Dies ist ein Trendaussage, denn die Anzahl der unter-suchten Rothirsche und Wildschweine ist mit 116 bzw. 103 relativ gering, im Vergleich zu 1.429 untersuchten Rehen. HECHT (1997) kommt zu ähnlichen Trendverläufen bei den 3 Wildarten. Wie kommt es zu dieser unterschiedlichen Entwicklung in der Cs-137 Kontamination?
Reh und Rothirsch sind Pflanzenfresser, die sich von oberirdischer, grüner Biomasse ernähren. Die Cs-137 Aktivität nimmt seit 1986 in der pflanzlichen Biomasse ab, ent-sprechend geht auch die Aktivität im Muskelfleisch der Tiere zurück. Die Abnahme verläuft bei beiden Wiederkäuerarten etwa parallel, mit einer effektiven Halbwertzeit von rund 6 Jahren, wobei Rehe durchschnittlich immer höher kontaminiert waren als Rothirsche. Dies hängt damit zusammen, dass mit zunehmender Körpermasse der Transfer Futter → Fleisch geringer wird und Rothirsche andere Nahrungspflanzen bevorzugen als Rehe. So ist z.B. der Anteil von Gras im Nahrungsspektrum von Rotwild deutlich höher als bei Rehen. Und im selben Lebensraum enthält Gras durchschnittlich weniger Cs-137 als Blätter von Heidelbeeren, Farnen und anderen Waldbodenpflanzen, die von Rehen bevorzugt aufgenommen werden. Dadurch ist die mit der Nahrung aufgenommene Cs-137 Aktivität, bezogen auf die Körpermasse, bei Rehen höher als bei Rothirschen.
Dagegen waren Rehe und Wildschweine in Bodenmais, unmittelbar nach dem Tscher-nobyl-Fallout 1986, im Durchschnitt etwa gleich hoch kontaminiert. In den folgenden Jahren nahm die Cs-137 Aktivität bei Rehen ab, bei Wildschweinen zu. Diese entgegen gesetzte Entwicklung ist durch die Wechselwirkung mehrerer Faktoren verursacht. Wildschweine sind Allesfresser mit einem breiten Nahrungsspektrum, die, je nach Ange- bot, auf unterschiedlichste Nahrungsressourcen zugreifen können. Der wesentliche Unterschied zu den Pflanzenfressern besteht darin, dass Wildschweine pflanzliche und
49

tierische Nahrung auch direkt aus dem Boden wühlen und diesen als Begleitstoff mit aufnehmen.
100
1000
10000
1986
2000199819961994199219901988
Cs-137 [Bq/kg] FS
Jahr
Rothirsch, Teff= - 5,7 a
Wildschwein, Teff= + 10,4 a
Reh, Teff= - 6,0 a
Abb. 28: Regressionsgeraden zu den Cs-137 Messwerten von Rehen, Rothirschen und Wildschweinen, Untersuchungsgebiet Bodenmais, 1986-2000
Als „Delikatesse“ fressen sie Hirschtrüffel, die sie aus der Humusauflage wühlen. Hirsch-trüffel sind kleine, knollige, braune Pilze, etwa 1-2 cm breit und 2-5 cm lang, die nestartig zwischen der Humusauflage und dem Mineralboden wachsen. Diese Pilzart hat, mit durchschnittlich 25.660 Bq•kg-1,die höchsten Cs-137 Aktivitäten in der Frischsubstanz, die bisher in einem biologischen Medium aus dem Untersuchungsgebiet Bodenmais gemessen wurden (Abbildung 20). Im Vergleich dazu enthielt Dornfarn, die am höchsten kontaminierte Pflanzenart, die von Rehen gefressen wird, im Jahr 2000 rund 3.069 Bq•kg-1 und Maronenröhrlinge 3.030 Bq•kg-1, bezogen auf Frischsubstanz. Wichtig ist hier der Bezug der Cs-137 Aktivität auf Frischsubstanz, denn in der Trockensubstanz haben Pilze oft deutlich höhere Aktivitäten als Pflanzen. Aber Tiere fressen das Zeug nun mal grün und nicht getrocknet.
Wieso aber waren Wildschweine und Rehe 1986 etwa gleich hoch kontaminiert?
Weil Wildschweine, mit einer Resorptionsrate von 90 bis 100% (MOORE und COMAR 1962, VOIGT et al. 1989), deutlich besser Cs-137 resorbieren als die Wiederkäuer, mit nur 50 bis 80% (HOEK 1976, ILIN und MOSKALEV 1957, MC CLELLAN et al. 1962, SANSOM 1966, VOIGT et al. 1989). Und weil die Cs-137 Aktivität in der von Wild-schweinen aus dem Boden aufgenommenen, pflanzlichen Biomasse vermutlich geringer war, als in der oberirdisch wachsenden Nahrung von Rehen, denn Cs-137 befand sich hauptsächlich in den obersten Zentimetern des Bodens. Darüber hinaus war ein beträcht-
50

licher Teil des Nuklids noch in der oberirdischen Biomasse der Bäume adsorbiert und gelangte erst nach und nach auf den Waldboden. Nach BUNZL et al. (1989) wurden in einem 100-jährigen Fichtenwald im Bayerischen Wald, 60 % der insgesamt zugeführten Cs-134 Aktivität, zunächst im Kronenraum festgehalten. Mit der Zeit gelangte ein Teil, durch Auswaschung, Windeinwirkung und Streufall auf den Waldboden. Dieser Prozess war auch 600 Tage nach dem Fallout noch nicht vollständig abgeschlossen.
Deswegen war 1986 und 1987 die oberirdische pflanzliche Biomasse höher kontaminiert als die unterirdische. In den folgenden Jahren änderte sich dieses Verhältnis, Cs-137 wanderte langsam einige Zentimeter tiefer in den Boden (Abbildung 10). Darüber hinaus fanden biotische und physikalische Umlagerungs- und Bindungsprozesse statt, die die Verfügbarkeit des Nuklids veränderten. Wahrscheinlich führten diese Faktoren dazu, dass trotz des radioaktiven Zerfalls, der die Cs-137 Aktivität, von 1986 bis 2000 um rund 30% verminderte, Hirschtrüffel 2000 ähnlich hoch kontaminiert waren wie 1986. Im Gegensatz dazu ist 2000 die Cs-137 Kontamination aller Waldbodenpflanzen deutlich geringer als 1986 (Abbildungen 13 bis 18).
8 Cs-137 in verschiedenen Biomedien des Waldes, ein Überblick Unabhängig von den Untersuchungsgebieten, ist die Variabilität der Cs-137 Aktivität in einzelnen Kompartimenten des Waldes sehr ausgeprägt. Einen Überblick gibt die Abbildung 29, wo die Cs-137 Gehalte, bezogen auf Frischsubstanz, in verschiedenen Biomedien der Dauerprobefläche B1, als Box-Plots dargestellt sind. Die Gruppe „Holz“ umfasst Messwerte von Buche, Fichte und Tanne, die Gruppe „Pflanzen“ umfasst Blätter der Spezies Dornfarn, Wald-Frauenfarn, Heidelbeere, Himbeere, Seegras und Wald-hainsimse. Bei der Gruppe „Pilze“ wurden nur die Speisepilze Grauer Wulstling, Halli-masch, Pfifferling, Marone, Rotfußröhrling, Steinpilz und Maronenröhrling berücksichtigt. Ausgewertet wurden die Messwerte von Proben, die von 1997 bis 2000, auf der Dauerprobefläche B1 entnommen wurden. Bei den Rehen handelt es sich um Proben von Tieren, die auf Flächen in der Umgebung von B1, mit vergleichbarer Bodenkontamination, erlegt wurden.
Die Cs-137 Aktivität in Holz ist deutlich niedriger als in anderen Biomedien. Der Mittel-wert von 128 Bq•kg-1 unterscheidet sich signifikant von allen anderen Mittelwerten (P<0.01). Zwischen den Mittelwerten von Heidelbeeren 436 Bq•kg-1, Rehen 1.007 Bq•kg-1, Pflanzen 1.250 und Speisepilzen 1.450 Bq•kg-1 bestehen dagegen keine signifi-kanten Unterschiede. Der Minimalwert der Cs-137 Aktivität war 4 Bq•kg-1 in einer Probe Fichtenholz, der Maximalwert 12.112 Bq•kg-1 in Maronenröhrlingen.
Die Mittelwerte der Cs-137 Aktivität von Heidelbeeren, Pflanzen, Rehen und Pilzen liegen deutlich über den Medianen, ein Hinweis für die rechtsschiefe Verteilung der Messwerte in
51

1
10
100
1000
10000
e
30000
7266911412
PilzeRehPflanzenHeidelbeerenHolz
Cs-137 [Bq/kg] FS
Abb. 29: Cs-137 Aktivität in der Frischsubstanz verschiedener Kompartimente, Bodenmais, Dauerprobefläche B1, von 1997 – 2000
der Grundgesamtheit. Auffällig ist die ausgeprägte Variation der Aktivität innerhalb jeder Gruppe, was durch a) saisonale Kontaminationsschwankungen, wie sie besonders bei Rehwild und in geringerem Maße auch bei Pflanzen vorkommen und b) kleinräumige Unterschiede in der Flächenbelastung der Böden sowie der Standortparameter, wie z.B. Dicke der Humusauflage etc., bedingt sein kann.
9 Transferfaktoren von Cs-137 Boden → Biomedien Aufgrund der unterschiedlichen Bodenkontamination der Dauerflächen, lässt sich die Radiocäsiumaufnahme der untersuchten Pflanzenarten am besten anhand der Trans-ferfaktoren vergleichen. Nach der Richtlinie zu § 45 der Strahlenschutzverordnung (BMI 1979) ist der Transferfaktor als das Verhältnis der spezifischen Aktivität eines Nuklids in der Pflanze zu der spezifischen Aktivität in der Wurzelzone des Bodens definiert:
Aktivität in der Pflanze (in Bq• kg-1 Frischsubstanz) TPf = Aktivität im Boden (in Bq• kg-1 Trockensubstanz)
Dieses Konzept ist für die Berechnung von Transferfaktoren auf landwirtschaftlich genutzten Böden ausgelegt, die durch das Pflügen eine vergleichsweise homogene Trockenraumdichte haben und wo Radiocäsium relativ gleichmäßig, über eine Pflug-schartiefe von 30 cm verteilt ist. Für Waldböden ist die Berechnung von Transferfaktoren
52

nach dieser Gleichung nicht geeignet, weil, sowohl die vertikale Verteilung von Radio-cäsium, als auch die Trockenraumdichte in den obersten 15 cm äußerst inhomogen ist. Da sich Pflanzenarten in ihrem Wassergehalt unterscheiden, werden TPf häufig mit der spezifischen Aktivität in Trockensubstanz berechnet.
Um bei der Berechnung der Transferfaktoren Unterschiede in den Bodendichten und der Tiefenverteilung von Radiocäsium zu umgehen, wurden sie in dieser Arbeit als aggregierte Transferfaktoren (Tag) berechnet. Für den Übergang des Nuklids vom Boden in die Pflanze, ist er definiert als:
Cs-137 Aktivität pro kg Pflanzentrockensubstanz [Bq• kg-1] Tag = Cs-137 Inventar im Trockenboden [Bq• m-2], 0 - 10 cm Bodentiefe
Der Bezug des aggregierten Transferfaktors auf die Cs-137 Aktivität in dem Boden-kompartiment 0 bis 10 cm berücksichtigt den Nährstoffeinzugsbereich der meisten Waldbodenpflanzen und Bäume.
Der aggregierten Transferfaktor Boden→ Fleisch wird berechnet, indem die spezifische Cs-137 Aktivität in der Frischsubstanz von Fleisch durch das Cs-137 Inventar in 0-10 cm Bodentiefe pro Quadratmeter, dividiert wird.
Die Transferfaktoren sind in Tabelle 8 angegeben. Bei der pflanzlichen und tierischen Biomasse wurden, für jede Art, die Mittelwerte der Cs-137 Aktivität von 1999 bis 2000, zur Berechnung verwendet. Die flächenbezogene Cs-137 Aktivität des Bodens wurde aus den zuletzt entnommenen Profilen berechnet (Mittelwerte).
Die Werte der Transferfaktoren für Cs-137 in autotrophen Waldpflanzenarten reichten von 0,0001 m2•kg-1 bis 0,41 m2•kg-1. Farne hatten auf allen Probeflächen die größten Tag. Auf B1 war der Tag Wert für Dornfarn, mit 0,38 m2•kg-1, rund 24 höher als für Himbeere, die mit 0,016 m2•kg-1 den niedrigsten Wert hatte.
Die Werte der aggregierten Transferfaktoren waren für Früchte von Brombeere, Heidelbeere und Himbeere etwa gleich hoch wie für Blätter. Der arithmetische Mittelwert für die Tag der 3 Beerenarten auf B1 betrug 0,067 m2•kg-1, auf F1 betrug er 0,032 m2•kg-1.
Bei Pilzen war die Spannweite der Tag Werte noch größer als bei den Pflanzenarten und reichte von <0,0001 bis 3,3 m2•kg-1. Besonders hohe Tag Werte wurden für Dickfuß, Milchling und Hirschtrüffel, mit 3,3 bzw. 2,1 und 1,9 m2•kg-1 auf B1 ermittelt, während sie für Maronenröhrlinge mit 0,77 m2•kg-1, deutlich niedriger waren. Überwiegend niedrige Aufnahmeraten für Cs-137 wurden bei Pilzen auf G1 festgestellt, z.B. bei Maronen-röhrlingen 0,03 m2•kg-1. Nur der Tag für Mehlstiel-Täubling fiel mit 0,47 m2•kg-1 aus dem Rahmen.
Auch bei den untersuchten Tierarten aus Bodenmais gab es große Schwankungen bei den Tag Werten für Cs-137 Boden→Fleisch. Der, mit Abstand niedrigste Tag Wert wurde bei Kühen, mit 0,0006 m2•kg-1, der höchste bei Wildschweinen, mit 0,21 m2•kg-1 ermittelt,
53

Tab. 8: Transferfaktoren für Cs-137 Boden → Biomedien, Probeflächen B1, G1 und F1, Zeitraum 1999-2000
Tag• 10-2 [m2•kg-1 ] Biomedium B1 F1 G1
Pflanzen (Blätter) Adlerfarn 41,4 Brombeere 3,9 3,6 Dornfarn 38,1 25,0 0,64 Heidelbeere 14,3 8,3 Himbeere 1,6 4,4 0,01 Pfeifengras 8,1 Seegras 0,01 Wald-Frauenfarn 12,3 17,9 0,11 Früchte Brombeeren 4,8 3,1 Heidelbeeren 11,7 6,2 Himbeeren 3,5 0,5 Pilze Anis-Champignon <0,01 Dickfuß 330,2 Grauer Wulstling 49,2 45,7 Hallimasch 8,8 Hirschtrüffel 191,5 Kartoffelbovist 70,5 97,8 Maronenröhrling 76,7 120,1 3,0 Mehlstiel-Täubling Milchling 211,6 46,8 Rotfußröhrling 39,9 101,1 2,0 Pfifferling 27,6 Sandröhrling 72,9 Steinpilz 26,1 Ziegenlippe 13,2 Holz Fichte 0,4 Fleisch Kuh* 0,06 Reh 2,7 <0,006 Rothirsch 1,0 Wildschwein 21,2 0,05
*= Kuh von der untersuchten Wiese, s. Abbildung 8
54

während die Werte für Reh und Rothirsch mit 0,027 bzw. 0,01 m2•kg-1 im mittleren Bereich lagen.
Während auf B1 und F1 die Transferfaktoren ähnlich hoch waren, wurden für G1 um Größenordnungen kleinere Werte festgestellt. Zum Beispiel war der Tag bei Wild-schweinen aus Bodenmais 392mal höher als in Göttingen. Der Tag von Dornfarn auf B1 war mit 0,38 m2•kg-1, 60 mal höher als auf G1 mit 0,0064 m2•kg-1.
Die geometrischen Mittelwerte der aggregierten Transferfaktoren für alle untersuchten Pilzarten waren auf B1 und F1 mit 0,54 bzw. 0,83 m2•kg-1, deutlich höher als für alle Pflanzenarten, mit 0,084 bzw. 0,088 m2•kg-1. Allerdings sind, aufgrund der unter-schiedlichen Verhältnisse von Trockensubstanz zu Frischsubstanz, bei autotrophen Pflanzen und Pilzen (Kapitel 6.2) die Unterschiede bei den Tag Werten, bezogen auf Frischsubstanz, niedriger.
Die in der Literatur angegebenen aggregierten Transferfaktoren schwanken über einen weiten Bereich. Grundsätzlich besteht das Problem, dass die Tag von verschiedenen Autoren unterschiedlich berechnet werden: Die flächenbezogene Cs-137 Aktivität des Bodens wird entweder auf die organischen Horizonte (STEINER und WIRTH 1997), die obersten 10, 15 oder 20 cm des Bodens, oder aber auf das Gesamtinventar bezogen. Zum Beispiel liegen, nach BERESFORD et al. (1999), die Werte für aggregierte Transfer-faktoren von Pilzen, die in den Ländern des Zusammenschlusses von unabhängigen Staaten (CIS) untersucht wurden, zwischen 0,0000712 und 10,08 m2•kg-1.
10 Vorräte von Cs-137 im Waldökosystem der Probefläche B1 In den voran gegangenen Kapiteln wurde die Verteilung von Cs-137 im Boden, in der pflanzlichen Biomasse und in Wildtieren dargestellt. Aus diesen Daten wurde der Vorrat des Nuklids in dem Waldökosystem der Probefläche B1 errechnet (Tabelle 8).
Dazu wurde das untersuchte Waldökosystem in die Kompartimente Humusauflage (0-5 cm), Mineralboden (5-30 cm), Pilze, Waldbodenvegetation, Baumbestand und Rehwild aufgeteilt. Die Cs-137 Vorräte wurden durch Multiplikation der spezifischen Cs-137 Aktivität der Kompartimente mit ihrer Biomasse, bezogen auf das Frischgewicht, pro Quadratmeter, berechnet.
Auf B1 wurde die Hauptbaumart Fichte, stellvertretend für das Systemkompartiment „Baumbestand“ ausgewählt. Für den Fichtenbestand wurde die Biomassenverteilung nach (ELLENBERG et al. 1986) angenommen und, mit Hilfe der Ertragstafel für Holz und Rinde, nach (SCHOBER 1979), auf 120 Jahre Bestandsalter hochgerechnet. Die Biomasse der Systemkompartimente Waldbodenvegetation und Pilze wurde, über grobe Bestands-dichte- und Biomassenbestimmung der Arten abgeschätzt. Zusätzlich wurden Angaben über die Biomassenverteilung der Waldbodenvegetation von SCHMIDT (1992) verwendet. Die Dichte von Rehwild beträgt in Bodenmais ca. 3 Stück pro 100 Hektar, das entspricht einer Biomasse von 6•10-5 kg•m-2.
55
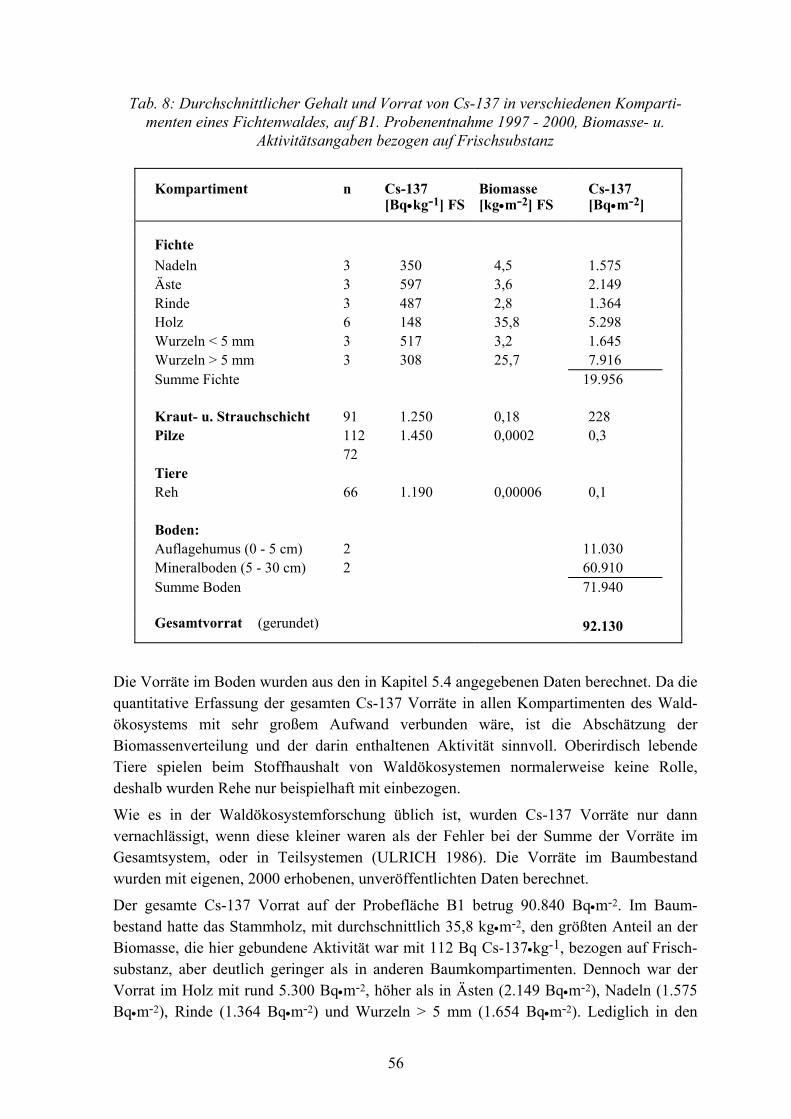
Tab. 8: Durchschnittlicher Gehalt und Vorrat von Cs-137 in verschiedenen Komparti-menten eines Fichtenwaldes, auf B1. Probenentnahme 1997 - 2000, Biomasse- u.
Aktivitätsangaben bezogen auf Frischsubstanz
Kompartiment n Cs-137 Biomasse Cs-137 [Bq•kg-1] FS [kg•m-2] FS [Bq•m-2]
Fichte Nadeln 3 350 4,5 1.575 Äste 3 597 3,6 2.149 Rinde 3 487 2,8 1.364 Holz 6 148 35,8 5.298 Wurzeln < 5 mm 3 517 3,2 1.645 Wurzeln > 5 mm 3 308 25,7 7.916 Summe Fichte 19.956 Kraut- u. Strauchschicht 91 1.250 0,18 228 Pilze 112 1.450 0,0002 0,3 72 Tiere Reh 66 1.190 0,00006 0,1 Boden: Auflagehumus (0 - 5 cm) 2 11.030 Mineralboden (5 - 30 cm) 2 60.910 Summe Boden 71.940 Gesamtvorrat (gerundet) 92.130
Die Vorräte im Boden wurden aus den in Kapitel 5.4 angegebenen Daten berechnet. Da die quantitative Erfassung der gesamten Cs-137 Vorräte in allen Kompartimenten des Wald-ökosystems mit sehr großem Aufwand verbunden wäre, ist die Abschätzung der Biomassenverteilung und der darin enthaltenen Aktivität sinnvoll. Oberirdisch lebende Tiere spielen beim Stoffhaushalt von Waldökosystemen normalerweise keine Rolle, deshalb wurden Rehe nur beispielhaft mit einbezogen.
Wie es in der Waldökosystemforschung üblich ist, wurden Cs-137 Vorräte nur dann vernachlässigt, wenn diese kleiner waren als der Fehler bei der Summe der Vorräte im Gesamtsystem, oder in Teilsystemen (ULRICH 1986). Die Vorräte im Baumbestand wurden mit eigenen, 2000 erhobenen, unveröffentlichten Daten berechnet.
Der gesamte Cs-137 Vorrat auf der Probefläche B1 betrug 90.840 Bq•m-2. Im Baum-bestand hatte das Stammholz, mit durchschnittlich 35,8 kg•m-2, den größten Anteil an der Biomasse, die hier gebundene Aktivität war mit 112 Bq Cs-137•kg-1, bezogen auf Frisch-substanz, aber deutlich geringer als in anderen Baumkompartimenten. Dennoch war der Vorrat im Holz mit rund 5.300 Bq•m-2, höher als in Ästen (2.149 Bq•m-2), Nadeln (1.575 Bq•m-2), Rinde (1.364 Bq•m-2) und Wurzeln > 5 mm (1.654 Bq•m-2). Lediglich in den
56

Wurzeln < 5 mm des Baumbestandes war der Vorrat mit 7.916 Bq•m-2,noch größer. Insgesamt waren im Fichtenbestand 18.667 Bq•m-2 gebunden.
Die mittlere spezifische Cs-137 Aktivität der Kraut- und Strauchschicht betrug durch-schnittlich 1.250 Bq•kg-1 (FS). Berücksichtigt wurden die Messwerte von 91 Proben von 8 verschiedenen Pflanzenarten, wobei die Proben überwiegend von den Arten, Brombeere, Dornfarn, Heidelbeere, Himbeere und Wald-Frauenfarn stammten. Aufgrund der geringen Biomasse von 0,18 kg•m-2, waren in der Waldbodenvegetationen nur 228 Bq•m-2 gebunden, rund 46 mal weniger als im oberirdischen Fichtenbestand.
Die mittlere spezifische Cs-137 Aktivität von Pilzen betrug 1.450 Bq•kg-1. Da die Bio-masse, mit 0,0002 kg•m-2 sehr gering war, betrug der Vorrat nur 0,3 Bq Cs-137•m-2. Rehe waren 2000 durchschnittlich mit 1.190 Bq•kg-1 kontaminiert. Die Biomasse der untersuchten Tiere auf der Probefläche B1 war so gering, dass der hier gebundene Cs-137 Vorrat vernachlässigbar ist. In Rehfleisch waren durchschnittlich nur etwa 0,1 Bq Cs-137 •m-2 enthalten.
Für den Boden sind keine spezifischen Aktivitäten und Biomassen angegeben, da Radio-cäsium in diesem Systemkompartiment nicht homogen verteilt ist (Kapitel 5.3). Insgesamt waren hier 71.940 Bq Cs-137•m-2 gebunden, 3,5mal mehr als in der oberirdischen Bio-masse. 11.030 Bq•m-2 befanden sich im 5 cm mächtigen Auflagehumus, 60.910 Bq•m-2 im Mineralboden bis 30 cm Bodentiefe. Bis zu dieser Beprobungstiefe wurde das im Boden vorhandene Cs-137 nahezu vollständig erfasst (Abbildung 7).
Auflagehumus 12 %
Mineralboden 67 %
0 - 5 cm
5 – 30 cm
10 %
0, 3 %
Wurzeln 10, 5 %
Abb. 30: Vorrat von Cs-137 in einem Fichtenbestand auf B1, in % vom Gesamtvorrat = 92.130 Bq•m-2, Probenahme 1998-2000
57

Der besseren Übersicht wegen sind in der Abbildung 30 die Vorräte von Cs-137 in den verschiedenen Systemkompartimenten, prozentual auf den Gesamtvorrat bezogen, grafisch dargestellt.
In dem untersuchten Fichtenbestand waren 79,2 %, also der überwiegende Anteil des insgesamt vorhandenen Cs-137, im Boden festgelegt. In der Biomasse der Fichtenwurzeln befanden sich 10,5 %. Insgesamt waren 89,7% der in dem Waldökosystem vorhandenen Cs-137 Aktivität unterirdisch festgelegt. In der oberirdischen pflanzlichen Biomasse befanden sich 10,3%, davon 10% im Fichtenbestand und nur 0,3 % in der Kraut- und Strauchschicht.
11 Zusammenfassende Diskussion Bei allen Untersuchungen über das Verhalten von Cs-137 in der Umwelt besteht das Problem der extremen Schwankungen der spezifischen Aktivität in der zu betrachtenden Grundgesamtheit.
Cs-137 wurde bei dem Tschernobyl-Fallout im April 1986, bedingt durch kleinräumige Unterschiede bei der Verteilung der Niederschläge, in der Bundesrepublik sehr ungleich deponiert. Diese inhomogene Verteilung tritt in der Regel unabhängig von der Größe des zu betrachteten Systems auf. Zum Beispiel ist die Variabilität der Flächenkontamination von Böden in der Bundesrepublik auf jeder Betrachtungsebene sehr ausgeprägt:
national → regional → lokal→ Standort → Kleinstandort → Mikrobereich
Der Variationskoeffizient beträgt für die Cs-137 Aktivität in den Böden der Bundesrepublik 126% (berechnet aus den Daten des BfS zur „Bodenkontamination der Bundesrepublik Deutschland durch Cs-137 infolge des Reaktorunfalls in Tschernobyl“, n=3.448). Für das Untersuchungsgebiet Bodenmais wurde an 59 Rasterpunkten, über eine Fläche von 7.500 ha, ein Variationskoeffizient von 50% ermittelt. Auf einer 100x100 m großen Alpenweide im Nationalpark Berchtesgarden ermittelten BUNZL et al. (2000) Unterschiede in der Cs-137 Flächenkontamination des Bodens um eine Größenordnung. Der Minimal-und Maximalwert unterschieden sich um den Faktor 40.
Generell sind in Waldökosystemen, im Vergleich zu landwirtschaftlichen Flächen, die Schwankungen der Cs-137 Aktivität in einzelnen Biomedien wesentlich höher. Der Grund dafür sind systeminterne Besonderheiten, die z.B. in Waldböden die hohe Variabilität bei der horizontalen und vertikalen Verteilung von Cs-137, auch auf kleiner Fläche bedingen. Die Unterschiede bei der horizontalen Verteilung der Flächenaktivität sind weitgehend durch den heterogenen Nuklideintrag aus der Kronentraufe bedingt: das Radiocäsium aus dem Tschernobyl-Fallout gelangte Ende April bzw. Anfang Mai 1986, mit großen Regenmengen zunächst auf die Baumkronen. Aufgrund der natürlichen Abweichungen in der Benadelungs- und Astdichte, passierten die Niederschläge den Kronenraum nicht gleichförmig, was eine inhomogene Verteilung des Nuklids auf dem Waldboden zur Folge hatte. Insbesondere wurde bei Buchen, aus dem Kronenraum ausgewaschenes Cs-137 mit dem Stammablauf in stammnahe Bodenbereiche transportiert. FÖRSTER et al. (1991) ermittelten in Einzelfällen bis zu 30mal höhere Cs-137 Gehalte in Böden am Stammfuß
58

von Buchen als an weiter entfernten Stellen; es bestanden hochsignifikante Unterschiede in der Cs-137 Aktivität von stammnahen und stammfernen Waldböden.
Im Untersuchungsgebiet Bodenmais bestanden auf der 100x100 m großen Probefläche B1 erhebliche Unterschiede in der Flächenkontamination des Bodens: drei, 1989 entnommene Profile enthielten 65.800 Bq•m-2, 74.900 Bq•m-2 und 122.500 Bq•m-2. Auch in der Tiefen-verteilung von Cs-137 unterschieden sich die gleichzeitig auf einer Probefläche ent-nommen Profile deutlich, wie die Abbildung 31 zeigt.
30
20
10
00,1 1 10 50
Waldboden, B13 ProfileHerbst 1997
B
oden
tiefe
[cm
]
Cs-137 [%/cm/qm]
Abb. 31: Verteilung von Cs-137 in 3 Bodenprofilen von B1, Mittel- , Minimal und Maximalwerte in % vom Gesamtinventar, Oktober 1997
Die Ursache hierfür sind Schwankungen der Standortbedingungen auf der Probefläche: Die Dicke der Humusauflage variierte zwischen 2 und 8 cm. Der Wassereintrag in den Boden ist durch Stammablauf viel höher als durch die Kronentraufe. Die Temperatur in der Humusauflage, ein wichtiger Parameter für die Leistung von Mikroorganismen und Pilzen, variiert kleinräumig, in Abhängigkeit von Mikrorelief und Sonnen- durchlass. Steine, abgestorbene Wurzeln, etc. sind inhomogen im Waldboden verteilt und führen zu unter-schiedlichen Sorptionsbedingungen. Ähnlich große Spannweiten in der vertikalen Verteilung von Cs-137 in jeder Schichtfolge, bei gleichzeitig entnommenen Profilen wurden auch auf der Probefläche F1 festgestellt. Aussagefähige Ergebnisse zur lang-fristigen Dynamik von Cs-137 in Waldböden sind daher nur durch die Entnahme von mehreren Bodenprofilen und die entsprechende statistische Auswertung zu gewinnen.
Auch 15 Jahre nach dem Tschernobyl-Fallout befanden sich auf den Probeflächen in den 3 Untersuchungsgebieten durchschnittlich noch 56,4% (Minimum 43%, Maximum 74%) der flächenbezogenen Cs-137 Aktivität in den obersten 10 cm und 93% (Minimum 91%, Maximum 97%) in den obersten 20 cm der untersuchten Waldböden. Cs-137 Ionen sind bisher nur in sehr geringem Maß in Bodenschichten tiefer als 20 cm vorgedrungen. In
59

dieser Tiefe liegen B-Horizonte vor, die i.d.R. einen hohen Anteil Mineralboden auf-weisen, in dessen Kristallgittern die Cs-137 Ionen effektiv fixiert werden. Dagegen wanderten Cs-137 Ionen aus den obersten 10 cm zwar sehr langsam, aber kontinuierlich in den Tiefenbereich 10 bis 20 cm.
Während längst bekannt ist, dass die sehr langsame vertikale Wanderung des Nuklids im Mineralboden durch Fixierungsprozesse an Tonminerale (und anderen Bodenbestand-teilen) verursacht ist, konnte weltweit bis heute nicht geklärt werden, wieso sich Cs-Ionen langfristig in der Humusauflage aufhalten. Eigentlich sollten Cs-Ionen, genau wie die bio-chemisch ähnlich reagierenden K-40 Ionen, aus der Humusauflage relativ schnell aus-gewaschen werden und in tiefere Bodenschichten gelangen. Als mögliche Erklärung werden verschiedene Hypothesen aufgestellt. So vermuten LANGER et al. (1997) die Bindung von Cs-137 Ionen als Komplex an organische Trägermaterialien, die von Boden-organismen in der Humusschicht erzeugt werden. OLSSEN et al. (1990) weisen auf die Bedeutung des Pilzmyzels als Speicher von Radiocäsium im Boden hin. Nach Unter-suchungen von BRÜCKMANN (1992), könnte der mikrobiellen Biomasse eine wichtige Funktion beim Transport und als Speicher von Radiocäsium in der Humusauflage zukommen. Die tatsächlichen Abläufe der Dynamik von Cs-137 in den Auflagehorizonten und im obersten Mineralboden sind bisher nicht geklärt.
Die oben erwähnte Inhomogenität von Cs-137 im Boden und bei den Standortbedingungen verursachen auf allen Probeflächen große Schwankungen der spezifischen Cs-137 Aktivität in Pflanzen und Pilzen (z.B. Abbildung 29). Was die radioökologische Inter-pretation von Messdaten aus Waldökosystemen erheblich erschwert und zeigt, dass Messwerte von Einzelproben einer Art nur wenig aussagefähig sind.
Die Bedeutung der Bodenverhältnisse und auch der Standortparameter für die Aufnahme-raten von Cs-137 in die Pflanze wird bei dem Vergleich der aggregierten Transferfaktoren deutlich. Durchschnittlich waren die Tag Werte sämtlicher Pflanzenarten auf B1 und F1 rund 120 mal größer als auf G1.
Die deutlich geringere Cs-137 Aufnahme der Pflanzen auf G1 ist, im Vergleich zu den Probeflächen B1 und F1 wahrscheinlich durch die wesentlich bessere Nährstoffversorgung (höhere Gesamaustauschkapazität, höhere Kaliumaustauschkapazität) und den höheren pH Wert bedingt (Tabelle 1). Die Austauschkapazität ist auf G1, mit 740 mymol(eg)•g-1 wesentlich günstig als mit 144 bzw. 51 mymol(eg)•g-1 auf B1 und F1. Gleiches gilt für die Kalium-Austauschkapazität, mit 10,1 mymol(eg)•g-1 auf G1 und nur 3,5 bzw 1,1 mymol(eg)•g-1 auf B1 und F1. Da die Cs-137 Ionen mit den Kalium Ionen an den Carriern der Durchlassgewebe von Wurzelzellen konkurrieren, bedingt die bessere Pflanzen-verfügbarkeit von Kalium eine reduzierte Aufnahme der Cs-137 Ionen. Ferner befindet sich das Maximum der Cs-137 Aktivität auf G1 zwar in den obersten 4 cm des Ober-bodens, es handelt sich aber nicht, wie auf B1 und F1 um Auflagehumus, sondern bereits um den Ah-Horizont. Ein Teil des Cs-137 ist also schon im Mineralboden fixiert und nur schlecht pflanzenverfügbar.
Auf den Probeflächen der Untersuchungsgebiete Bodenmais und Fuhrberg nahm die Cs-137 Kontamination in Boden, Waldbodenpflanzen, Bäumen und Beeren, von 1987 bis 2000 ab. Gleichzeitig in Bodenmais, auf 7.500 ha durchgeführte Messungen an Rehen,
60

ergaben ebenfalls einen signifikanten Rückgang der Cs-137 Aktivität. Dieser Rückgang erfolgte deutlich schneller, als nur durch den physikalischen Zerfall des Nuklids, wird also auch durch physikalische und biologische Fixations- und Umwandlungsprozesse des Nuklids verursacht. Die Abnahme der Cs-137 Gehalte in den Ökosystemkompartimenten erfolgte nicht gleichförmig, sondern zeigte eine Phase mit relativ schnellem (1988 bis 1993) und eine Phase mit deutlich langsamerem Rückgang (von 1994 bis 2000).
Ein zeitlich ähnlicher Aktivitätsverlauf wurde auch für Waldökosysteme mit anderen Standortfaktoren ermittelt. So finden ZIBOLD et al. (2001) bei Untersuchungen in Ober-schwaben (Baden-Württemberg) ebenfalls eine relativ schnelle Abnahme der Cs-137 Aktivität bei Waldbodenpflanzen, nach dem Tschernobyl-Fallout bis 1991 und wenig Änderung des Aktivitätsniveaus in den folgenden Jahren. Auch bei Untersuchungen im Rahmen eines in mehreren Ländern durchgeführten Forschungsvorhabens der Euro-päischen Gemeinschaft wurde eine deutliche verringerte Aktivitätsabnahme von Cs-137 in Pflanzen, im Vergleich zu der Zeit vor 1994 ANPA (2000) festgestellt.
Die in vorliegender Untersuchung ermittelte, zweiphasige Dynamik von Cs-137 stimmt in Bodenmais bei Pflanzen und Rehen weitgehend überein. Die parallele Entwicklung ist logisch, weil Rehe Cs-137 ausschließlich über die pflanzliche Nahrung aufnehmen. Bei Wildschweinen wird die weitere Entwicklung der Cs-137 Kontamination wesentlich von der Cs-137 Aktivität in Hirschtrüffeln abhängen. Über das Verhalten des Nuklids in diesen Pilzen ist bisher kaum etwas bekannt.
Grundsätzlich gilt zwar, dass die ermittelten Ergebnisse nur für das untersuchte Ökosystem gelten. Es zeigt sich aber, dass in den 450 km auseinander liegenden, unterschiedlichen Waldökosystemen „Fichtenwald“ in Bodenmais und „Eichenwald“ in Fuhrberg, bei unter-schiedlichen Cs-137 Boden-Inventaren, die Werte der aggregierten Transferfaktoren für Cs-137 bei den untersuchten Pflanzenarten gut übereinstimmen. Offenbar führen ähnliche Bodenparameter in beiden Gebieten, langfristig zu einer vergleichbaren, relativ hohen Aufnahmerate von Radiocäsium. Auch die schon diskutierte zeitliche Entwicklung der Cs-137 Aktivität in Pflanzen verläuft in beiden Untersuchungsgebieten gleich.
Daraus folgt, dass ähnliche Bodentypen mit ähnlichen Bodeneigenschaften, unabhängig vom Vegetationsbestand, gleiche Muster in der Dynamik von Cs-137 hervorbringen. Obwohl die Kontamination der Nahrungskette wesentlich von den Cs-137 Gehalten der Pflanzen und Pilze abhängt, kommt den obersten 10 cm des Bodens die eigentliche Schlüsselrolle zu.
Die Untersuchungen in Göttingen zeigen, dass günstige Bodenverhältnisse, auch bei Flächenbelastungen von rund 5.500 Bq•m-2 dazu führen, dass die spezifische Cs-137 Aktivität von Rehen und einigen Pilzarten unter der Nachweisgrenze von 0,5 Bq•kg-1 liegen kann. Dagegen muss in Waldgebieten mit ungünstigen Bodenverhältnissen und mittleren bis hohen Cs-137 Flächenbelastungen langfristig mit erhöhter Cs-137 Kontamination der pflanzlichen und tierischen Biomasse gerechnet werden.
61

12 Literaturverzeichnis Albers B.P., Steindl H., Schimmack W., Bunzl K.; 2000: Soil-to-plant and plant-to-cow’s
milk transfer of radiocaesium in alpine pastures: significance of seasonal variability. Chemosphere 41: 717-723.
Amberger A., 1979: Pflanzenernährung. Eugen Ulmer Verlag, Stuttgart. ANPA 2000: SEMINAT. Long-term dynamics of radionuclides in semi-natural environ-
ments: derivation of parameters and modelling. Final report 1996-1999, European Commission-Nuclear Fission Safety Programme.
Beresford N. A., Howard B. J., Barnett C. L., Voigt G., Semiochkina N., Ratnikov A., Travnikov I., Gillet A. G., Mehli H., Skuterrud L., 1999: Reducing the consum-ption of Cs-137 via forest fungi provision of „self-help“ advice. In: Linkov I. and Schell W.R. (eds.): Contaminated forests: 359-368. NATO Science Series. Series 2: Environmental Security-Vol. 58.
BMI Bundesministerium des Inneren, 1979: Allgemeine Berechnungsgrundlage für die Strahlenexposition bei radioaktiven Ableitungen mit der Abluft oder in Oberflächengewässern (Richtlinie zu § 45 StrlSchv). Gemeinsames Ministerialblatt Nr.21: 369-436.
Bonka H., 1998: Radionuklidtransport nach dem Kernreaktorunfall in Tschernobyl und daraus ableitbare radioökologische Parameter. Schriftenreihe Reaktorsicherheit und Strahlenschutz. BMU-1998-507.
Boresch K., 1935: Gehalt der Pflanzen an Mineralstoffen. Tab. Biol. Per. 4: 485-522. Brown D. N., 1964: Caesium in liriodendron and other woody species: organic bounding
sites. Science 143: 368-369. Brückmann A., 1992: Radioaktives Cäsium nach dem Tschernobyl-Reaktorunfall in Wald-
ökosystemen. Dissertation, Universität Göttingen, Institut für Bodenkunde und Waldernährung.
Bunzel K., Albers B.P., Schimack W., Belli M., Ciuffo L., Menegon S.; 2000: Examination of a relationship between Cs-137 concentrations in soil and plants from alpine pastures. Journal of Environmental Radioactivity 48: 145-158.
Bunzl K., 1996: GSF-Jahresbericht 1995, Neuherberg: 107-110. Bunzel K., Schimack W., Kreutzer K., Schierl R., 1989: Interception and retention of
chernobyl-derived Cs-134, Cs-137 and Ru-106 in a spruce stand. The Science of the Total Environment 78: 77-87.
Cougthry P. J., Thorne M. C., 1983: Radionuclide distribution and transport in terrestrial and aquatic ecosystems. Vol.I.: Caesium. Balkema, Rotterdam: 321-424.
Cremers A., Elsen A., De Preter P., Maes A., 1988: Quantitaive analysis of radiocesium retention in soil. Nature 335: 247-249.
Ellenberg H., Mayer R., Schauermann J. (eds.), 1986: Ökosystemforschung: Ergebnisse des Sollingprojekts 19866-1986. Ulmer Verlag, Stuttgart.
Ellenberg H., 1978: Zur Populationsökologie des Rehes (Capreolus capreolus L., Cervidae) in Mitteleuropa. Spixiania 2.
Fielitz U., Albers U., 1996: Nahrungspektrum von Rehen aus dem Bayerischen Wald. Z. Jagdwiss: 195-202.
Fielitz U., 1994: Radioaktivität in Wildtieren. BMU 1994, 408.
62

Förster H., Schimmack W., Kreutzer K.E., 1991: Die horizontale Verteilung von Radiocäsium im Waldboden unter Fichte und Buche. Z. Pflanzernähr. Bodenk. 154: 87-92
Franke B., 1982: Transfer radioaktiver Stoffe aus dem Boden in Pflanzen. In: Arbeits-gemeinschaft für Umweltfragen (Hrsg.): Das Umweltgespräch. Tagungsprotokoll: Radioökologiesymposium vom 15/16 Okt. 1981, Univ. Stuttgart: 152-183.
Fredriksson L., Eriksson B., Rasmuson B., Gahne Bo., Edvarson D., Löw K., 1958: Studies on soil-plant-animal interrelationsships with respect to fission products. Part A: Plant uptake of Sr-90 and Cs-137 from soils. Geneva Conference Paper P/177: 449-470.
Goussios K., Niessl A., 1998: Allgemeines zur Strahlenschutzvorsorge. In: Honikel K.O., Hecht H. (Hersg.): Radiocäsium in Wald und Wild. 2. Veranstaltung, Kulmbach, 10/11.06.1997. Bundesanstalt für Fleischforschung, Kulmbach: 189-213.
Hecht H., 2000: Radiocäsiumgehalte des Schwarzwildbretes speziell im Gebiet des Bayerischen Waldes. Abschlußbericht zum Forschungsvorhaben. Bundesanstalt für Fleischforschung, Kulmbach.
Hecht H., 1997: Radiocäsiumverteilung und –trends bei Wildbret in Bayern. Abschluss-bericht zum Forschungsvorhaben im Auftrag des Bayerischen Staatsministeriums für Landesentwicklung und Umweltfragen. Bundesanstalt für Fleischforschung, Kulmbach.
Hoek van den J., 1976: Cesium metabolism in sheep and influence of orally ingested bentonite on cesium adsorption and metabolism. Z. Tierphysiol., Tierernähr. u. Futtermittelkunde 37: 315-321.
Ilin D. I., Moskalev Y. I., 1957: On the metabolism of cesium, strontium and a mixture of ß-emitters in cow. J. Nuclear Energy II, 5: 413-420.
Kühn W., 1982: Ausbreitung radioaktiver Stoffe im Boden. In: Arbeitsgemeinschaft für Umweltfragen (Hrsg.): Das Umweltgespräch. Tagungsprotokoll: Radioökologie-symposium vom 15/16 Okt. 1981, Univ. Stuttgart: 76-99.
Langer R., Rieth K.-H., Müller M., Lindner G.; 1997: Indizien für die Beteiligung von organischen Trägerstoffen beim Pflanzentransfer von Cäsium-Radionukliden in Nadelwald-Böden. In: Honikel K.O., Hecht H. (Hersg.): Radiocäsium in Wald und Wild. 2. Veranstaltung, Kulmbach 10/11.06.1997. Bundesanstalt für Fleisch-forschung, Kulmbach: 25-33.
Läuchli A., Pflüger R., 1978: Potassium transport through plant cell membranes and metabolic role of potassium in plants. Proc. 11th Congr. Int. Potash. Inst. Bern. 11: 111-163
Mc Clellan R.O., Mc Kenney J.R., Bustad L.K., 1962: Metabolism and dosimetry of Cs-137 in male sheep. AEC research and development report HW-72511. Hanfort Atomic Products Operation. Richland, Washington.
Meiwes K.J., 1984: Arbeitsbericht zur bodenkundlichen Grundaufnahme, zum chemischen Bodenzustand und zur chemischen Bodeninventur in einem Buchen-waldökosystem auf Kalkgrundgestein im Projekt der DFG Nr. BE 713/8-1. Abteilung für Bodenkunde und Waldernährung der Universität Göttingen.
Moore M., Comar C. L., 1962: Absorption of caesium-137 from the gastro-intestinal- tract of the rat. Int. J. Rad. Biol. 5 (3): 247-254.
63

Olssen, R.A., Joner E., Bakken L.R., 1990: Soil fungi and the fate of radiocesium in soil ecosystems – a discussion of possible mechnisms involved in the radiocesium accumulation in fungi, and the role of fungi as a Cs-sink in the soil. In Desmet, G., Nassimbeni P., and Belli M. (eds.): Transfer of radionuclides in natural ecosystems. Elsevier, 657 – 663.
Pavlotskaya, F.I., et al. 1967: On the mobility of strontium and some other components of global fallout in soils and their accumulation in plants. in: Aberg B.; Hungate, F.P. (eds.): Radiological concentration processes. (Proceedings of an international symposium held in Stockholm, April 25-29, 1966). Pergamon Press, Oxford:: 25-32
Rafferty B., Dawson D., Kliashtorin A.; 1997: Decomposition in two pine forests: the mobilisation of Cs-137 and K from forest litter. Soil Biol. Biochem. 29: 1673-1681.
Ritchie J.C., Rudolph W.K., 1970: Distribution of fallout and natural gamma radionuclides in litter, humus and surface mineral soil layers under natural vegetation in the Great Smoky Mountains, North Carolinia-Tennessee. Health Phys. 18: 479-489.
Sansom B. F., 1966: The metabolism of cesium in dairy cows. J. Agric. Sci. 66: 389-393. Scheffer F., Schachtschnabel P., 1984: Lehrbuch der Bodenkunde. F. Enke Verlag.
Stuttgart Schober R., 1979: Ertragstafeln wichtiger Baumarten. Sauerländer-Verlag, Frankfurt/M. Schmalfuss K., 1936: Das Kalium. Eine Studie zum Kationenproblem im Stoffwechsel und
bei der Ernährung der Pflanze. Naturwissenschaft und Landwirtschaft Heft 19. Schmidt G.: schriftliche Mitteilung. Schodlock H. J., 1997: Darstellung von Forschungsergebnissen aus der Radioökologie
sowie Erfahrungsbericht aus einer zehnjährigen Messtätigkeit bei der Überwachung der Radiocäsiumkontamination von Reh- und Schwarzwild. In: Honikel K.O., Hecht H. (Hersg.): Radiocäsium in Wald und Wild. 2. Veranstaltung, Kulmbach 10/11.06.1997. Bundesanstalt für Fleischforschung, Kulmbach: 165-188.
Squire H.M., Middleton L.J., 1966: Behavior of Cs-137 in soils and pastures. A long term experiment. Radiation Botany 6: 413-423.
Steiner M., Wirth E., 1997: Die Migration von Radiocäsium in Waldböden – Konsequenzen für die zukünftige radioaktive Belastung von Pilzen und Beeren-pflanzen. In: Honikel K.O., Hecht H. (Hersg.): Radiocäsium in Wald und Wild. 2. Veranstaltung, Kulmbach 10/11.06.1997. Bundesanstalt für Fleischforschung, Kulmbach: 1-11.
Strasburger, E., 1978: Lehrbuch der Botanik. Gustav Fischer Verlag. Stuttgart. Tamura T., 1963: Cesium sorption reactions as indicator of clay mineral structures.
International clay conference. Proceedings of the conference held at Stockholm, Sweden, August 12-16.
Ulrich B. Mayer R., Matzner E., 1986: Vorräte und Flüsse der chemischen Elemente. In: Ellenberg H., Mayer R., Schauermann J. (eds.): Ökosystemforschung: Ergebnisse des Sollingprojekts 1986-1986. Ulmer Verlag, Stuttgart: 375-415
Voigt G., Pröhl G., Müller H., Bauer T., Lindner J.P., Probstmeier G., Röhrmoser G., 1989: Determination of the transfer of cesium and jodine from feed into domestic animals. The Science of Total Environment. 85: 329-338.
64

Völkl J., 1997: “Modelle zur Cäsiumfixierung in Böden radioaktiv hoch kontaminierter Gebiete Ost-Bayerns” im Auftrag des Bayerischen Staatsministeriums für Landes-entwicklung und Umweltfragen. Universität Regensburg, AG Landschafts-ökologie und Bodenkunde, März 1997.
Walton A., 1963: The distribution of radioactivity in soils from weapon tests. J. Geophys. Res. 68 (5):1485-1496.
Zibold G., Drissner J., Kaminski S., Klemt E., Miller R., 2001: Time-depency of the radiocaesium contamination of roe deer: measurement and modelling. J. Environ. Radioactivity. 55: 5-27.
Zibold G., Drissner J., Klemt E., Konopleva A.V., Konoplev A.V., Miller R.,: 1997: Biologische Verfügbarkeit von Cäsium-Radionukliden in Waldgebieten des nördlichen und südlichen Voralpenlandes. In: Honikel K.O., Hecht H. (Hersg.): Radiocäsium in Wald und Wild. 2. Veranstaltung, Kulmbach 10/11.06.1997.
Danksagung
Für die Aktivitätsmessungen der Proben am Isotopenlaboratorium für biologische und medizinische Forschung der Universität Göttingen, von 1997 bis 2001, bedanke ich mich herzlich bei Frau U. Wittig und Herrn J. Schikowski.
Die auf Seite 60 angegebenen Daten zur „Bodenkontamination der Bundesrepublik Deutschland durch Cs-137 infolge des Reaktorunfalls in Tschernobyl“ , wurden dem Autor freundlicherweise vom BfS, Fachgebiet ST 2.4 zur Verfügung gestellt.
Die Daten über die Cs-137 Aktivität in Rehwild aus Bodenmais von 1996 wurden dem Autor freundlicherweise von Herrn Dr. H. Hecht, Bundesanstalt für Fleischforschung (Kulmbach) zur Verfügung gestellt.
65


















