
Leadership aus neurowissenschaftlicher Sicht
109
Mag. a iur. Stephanie Vorderegger, Bakk.rer.soc.oec. Leadership aus neurowissenschaftlicher Sicht M A S T E R A R B E I T zur Erlangung des akademischen Grades eines Master of Science (MSc) der Studienrichtung Betriebswirtschaft an der Karl-Franzens-Universität Graz Betreuer: Univ. Prof. MMag. Dr. Alfred Gutschelhofer Institut für Unternehmensführung und Entrepreneurship Graz, März 2015
Transcript of Leadership aus neurowissenschaftlicher Sicht

Mag.a iur. Stephanie Vorderegger, Bakk.rer.soc.oec.
Leadership aus neurowissenschaftlicher Sicht
M A S T E R A R B E I T
zur Erlangung des akademischen Grades
eines Master of Science (MSc)
der Studienrichtung Betriebswirtschaft
an der Karl-Franzens-Universität Graz
Betreuer: Univ. Prof. MMag. Dr. Alfred Gutschelhofer
Institut für Unternehmensführung und Entrepreneurship
Graz, März 2015

Ehrenwörtliche Erklärung
Ich erkläre ehrenwörtlich, dass ich die vorliegende Arbeit selbstständig und ohne fremde
Hilfe verfasst, andere als die angegebenen Quellen nicht benutzt und die den Quellen
wörtlich oder inhaltlich entnommenen Stellen als solche kenntlich gemacht habe. Die Ar-
beit wurde bisher in gleicher oder ähnlicher Form keiner anderen inländischen oder aus-
ländischen Prüfungsbehörde vorgelegt und auch noch nicht veröffentlicht. Die vorliegende
Fassung entspricht der eingereichten elektronischen Version.
März 2015 Unterschrift

I
Inhaltsverzeichnis
Inhaltsverzeichnis ............................................................................................................ I
Abbildungsverzeichnis ................................................................................................... V
Tabellenverzeichnis ........................................................................................................ V
Abkürzungsverzeichnis ................................................................................................ VII
A. EINLEITUNG ............................................................................................................. 1
1 Ausgangssituation und Problemstellung ............................................................... 1
2 Forschungsfragen ................................................................................................... 2
3 Ziel und praktische Bedeutung ............................................................................... 2
4 Methodik und Aufbau der Arbeit ............................................................................. 3
B. GRUNDLAGEN DER NEUROWISSENSCHAFTEN UND NEUROLEADERSHIP ..... 5
1 Das Gehirn ................................................................................................................ 5
2 Aufbau und Funktionen des Gehirns ..................................................................... 6
2.1 Großhirn (Telencephalon) ................................................................................. 7
2.1.1 Einteilung der Großhirnrinde .................................................................. 9
2.1.1.1 Okzipitallappen (Hinterhautslappen) ......................................10
2.1.1.2 Temporallappen (Schläfenlappen) ........................................10
2.1.1.3 Parietallappen (Scheitellappen) .............................................11
2.1.1.4 Frontallappen (Stirnlappen) ...................................................12
2.1.1.4.1 Motorkortex.............................................................12
2.1.1.4.2 Präfrontaler Kortex ..................................................13
2.1.1.5 Inselrinde (Insula) ..................................................................14
2.1.1.6 Cingulum (Gyrus cinguli) .......................................................15
2.1.2 Basalganglien .......................................................................................15
2.1.3 Limbisches System ...............................................................................16
2.2 Zwischenhirn (Diencephalon) ...........................................................................17
2.3 Kleinhirn (Cerebellum) .....................................................................................18
2.4 Hirnstamm .......................................................................................................19

II
2.4.1 Mittelhirn (Mesencephalon) ..................................................................19
2.4.2 Brücke (Pons) .......................................................................................20
2.4.3 Verlängertes Mark (Medulla oblongata) ................................................20
2.5 Tabellarischer Überblick...................................................................................20
3 Methoden der Neurowissenschaften .....................................................................23
3.1 Bildgebende Verfahren i.w.S. ...........................................................................23
3.1.1 Elektroenzephalographie (EEG) ...........................................................24
3.1.2 Magnetenzephalographie (MEG) ..........................................................25
3.1.3 Intrakranielle Elektroenzephalographie (iEEG) .....................................25
3.2 Bildgebende Verfahren i.e.S. ...........................................................................26
3.2.1 Positronenemissionstomographie (PET) ...............................................26
3.2.2 Funktionelle Magnetresonanztomographie (fMRT) ...............................27
3.3 Weitere Methoden............................................................................................29
3.3.1 Transkranielle Magnetstimulation (TMS) ..............................................29
3.3.2 Diffusionstensor-Bildgebung (DTI) ........................................................30
3.4 Kritische Würdigung .........................................................................................31
4 (Neuro-)Leadership .................................................................................................33
4.1 Leadership - grundlegende Bedeutung ............................................................33
4.1.1 Führungstheorien .................................................................................34
4.1.1.1 Eigenschaftstheorien .............................................................34
4.1.1.1.1 Attributionstheorie ...................................................35
4.1.1.1.2 Charismatische Führung .........................................35
4.1.1.2 Führungsstiltheorien ..............................................................36
4.1.1.2.1 Eindimensionale Führungsstile ...............................36
4.1.1.2.2 Zweidimensionale Führungsstile .............................38
4.1.1.3 Situative Führungstheorien ...................................................39
4.1.1.4 Transformative vs. Transaktionale Führung ..........................40
4.1.1.4.1 Transaktionale Führung ..........................................40
4.1.1.4.2 Transformative Führung..........................................40
4.2 Neuroleadership ..............................................................................................41
4.2.1 Gehirngerechte Führung nach Hüther ..................................................42
4.2.1.1 Regelmäßig neue Herausforderungen schaffen ....................42
4.2.1.2 Know-How im Unternehmen vernetzen .................................43
4.2.1.3 Eine positive Fehlerkultur schaffen ........................................43
4.2.1.4 Für positive Erfahrungen sorgen ...........................................43

III
4.2.2 SCARF- Modell von Rock .....................................................................44
4.2.2.1 Status (status) .......................................................................44
4.2.2.2 Vorhersehbarkeit (certainty) ..................................................44
4.2.2.3 Autonomie (autonomy) ..........................................................45
4.2.2.4 Soziale Beziehungen (relatedness) .......................................45
4.2.2.5 Gerechtigkeit (fairness) .........................................................45
4.2.3 Neuroleadership nach Elger .................................................................46
4.2.3.1 Das Belohnungssystem als zentrale Schaltstelle...................46
4.2.3.2 Fairness ................................................................................46
4.2.3.3 Beeinflussung der Erwartungen und des Verhaltens durch
Vorinformationen ..................................................................................46
4.2.3.4 Einzigartigkeit jedes Gehirns .................................................47
4.2.3.5 Keine Fakten ohne Emotionen ..............................................47
4.2.3.6 Erfahrungen bestimmen das Verhalten .................................47
4.2.3.7 Eigendynamik von Situationen ..............................................47
4.2.4 Neuroleadership nach Peters/Ghadiri ...................................................48
4.2.4.1 Bindung .................................................................................48
4.2.4.2 Orientierung und Kontrolle ....................................................48
4.2.4.3 Selbstwerterhöhung und Selbstwertschutz ............................48
4.2.4.4 Lustgewinn und Unlustvermeidung........................................49
4.3 Beitrag der Neurowissenschaften für die Führungsforschung ..........................49
4.3.1 Kritische Würdigung .............................................................................51
C. METAANALYSE .......................................................................................................53
1 Charakteristika einer Metaanalyse .........................................................................53
1.1 Schematischer Ablauf ......................................................................................53
1.2 Vor- und Nachteile einer Metaanalyse .............................................................54
2 Zielsetzung und Forschungsfrage .........................................................................55
2.1 Forschungsfragen ............................................................................................55
3 Methode ...................................................................................................................56
3.1 Aufnahmekriterien ............................................................................................56
3.2 Literaturrecherche ............................................................................................57
3.3 Kodierung ........................................................................................................58
3.4 Erhobene Studien ............................................................................................58
4 Datenanalyse ...........................................................................................................61

IV
4.1 Eingesetzte Methoden der Neurowissenschaften ............................................61
4.2 Zeitlicher Verlauf der Studien ...........................................................................62
4.3 Stichprobenanzahl ...........................................................................................63
4.4 Eingesetzte Stimuli ..........................................................................................66
4.5 Beteiligte Hirnregionen bei der Leadershipforschung .......................................70
4.5.1 Neurophysiologische Unterschiede zwischen erfolgreichen und nicht
erfolgreichen Führungskräften .........................................................................70
4.5.2 Soziale Einflussnahme der FK auf den Geführten ................................73
4.5.2.1 Neurophysiologische Auswirkungen von Belohnungen .........73
4.5.2.2 Neurophysiologische Auswirkungen von charismatischen
Führungskräften ...................................................................................75
4.5.2.3 Neurophysiologische Auswirkungen von Beziehungen
zwischen MitarbeiterInnen und Führungskräften...................................76
4.5.3 Neurophysiologische Aktivitäten, die führungsrelevanten Fertigkeiten
zugrunde liegen ...............................................................................................78
4.5.3.1 Entscheidungsfindung ...........................................................78
5 Restriktionen ...........................................................................................................79
6 Diskussion ...............................................................................................................79
6.1.1 Neurophysiologische Unterschiede zwischen erfolgreichen und nicht
erfolgreichen Führungskräften .........................................................................79
6.1.2 Soziale Einflussnahme der Führungskraft auf den Geführten ...............80
6.1.3 Neuropysiologische Aktivitäten, die führungsrelevanten Fertigkeiten
zugrunde liegen ...............................................................................................82
D. ZUSAMMENFASSUNG ............................................................................................83
Ausblick ..........................................................................................................................84
ANHANG .........................................................................................................................85
GLOSSAR .......................................................................................................................89
Literaturverzeichnis .......................................................................................................92

V
Abbildungsverzeichnis
Abbildung 1: Aufbau einer Nervenzelle ............................................................................. 6
Abbildung 2: Aufbau des Gehirns...................................................................................... 7
Abbildung 3: Die zwei Gehirnhälften (Hemisphären) ......................................................... 9
Abbildung 4: Die Lappen des menschlichen Gehirns (Seitenansicht) ................................ 9
Abbildung 5: Einteilung der Gehirnlappen und zugehörige Funktionen (Seitenansicht) ....14
Abbildung 6: Die Insula ....................................................................................................14
Abbildung 7: Gyrus cinguli und seine Funktionsbereiche .................................................15
Abbildung 8: Das limbische System .................................................................................17
Abbildung 9: Hirnstamm und Zwischenhirn ......................................................................19
Abbildung 10: Eindimensionaler Führungsstil nach Tannenbaum/Schmidt ......................37
Abbildung 11: Managerial Grid nach Blake/Mouton .........................................................39
Abbildung 12: Graphische Darstellung der Studienanzahl im Zeitablauf ..........................63
Abbildung 13: Durchschnittliche Stichprobenanzahl im Zeitverlauf ...................................66
Abbildung 14: Relative Häufigkeit der eingesetzten Stimuli ..............................................70
Tabellenverzeichnis
Tabelle 1: Gehirnregionen und deren Funktionen im Überblick ........................................22
Tabelle 2: Übersicht der Methoden der Neurowissenschaften .........................................32
Tabelle 3: Neurowissenschaft und Führungsforschung ....................................................52
Tabelle 4: Alle erhobenen Studien zum Thema Führungsforschung ................................60
Tabelle 5: Absolute und relative Anzahl der verwendeten Methoden ...............................61
Tabelle 6: Zeitlicher Verlauf der erfassten Studien ...........................................................62
Tabelle 7: Stichprobenanzahl der erfassten Studien ........................................................64
Tabelle 8: Durchschnittliche Stichprobenanzahl ...............................................................65
Tabelle 9: Durchschnittliche Stichprobenanzahl im Zeitverlauf .........................................65
Tabelle 10: Stimuli und Studiendesign .............................................................................69
Tabelle 11: Eingesetzte Stimuli ........................................................................................69
Tabelle 12: Aktivierte Gehirnregionen ..............................................................................72
Tabelle 13:Neuropysiologische Auswirkungen von Belohnungen ....................................74
Tabelle 14: Aktivierung von Gehirnregionen, die neurophysiologische Strukturen von
charismatischen Führungskräften untersuchten ...............................................................75
Tabelle 15: Aktivierung von Gehirnregionen, die neurophysiologische Strukturen
zwischenmenschlicher Beziehungen untersuchten ..........................................................77

VI
Tabelle 16: Aktivierte Hirnregionen jener Studie, die das Entscheidungsverhalten
untersuchte ......................................................................................................................78

VII
Abkürzungsverzeichnis
aA anderer Ansicht
ACC anteriores Cingulum
BOLD blood oxygen level dependent
bspw. beispielsweise
d.h. das heißt
DLPFC dorsolateraler präfrontaler Kortex
DMPFC dorsomedialer präfrontaler Kortex
DTI Diffusions-Tensor-Bildgebung
EEG Elektroenzephalographie
et al. und andere
EUR Euro
f folgende
FF Forschungsfrage
FK Führungskraft
fMRT funktionelle Magnetresonanztomographie
grds. grundsätzlich
i.d.R. in der Regel
iEEG Intrakranielle Elektroenzephalographie
IPL inferior Parietallappen
IPS Sulcus intraparietalis
Jg. Jahrgang
Jhd. Jahrhundert
k.A. keine Angabe
MA MitarbeiterInnen
MEG Magnetencephalographie
Mio. Million(en)
Mrd. Milliarde(n)
MRT Magnetresonanztomographie
ms Millisekunde(n)
mwH mit weiteren Hinweisen
NAcc Nucleus accumbens
Nr. Heftnummer
OFC orbifrontaler Kortex
o.V. ohne Verfasser

VIII
PCC posteriores Cingulum
PET Positronenemissionstomographie
SMA supplementär-motorische Rinde
sog. sogenannte
s.o. siehe oben
SQUIDS superconducting quantum interference devices
TMS transkranielle Magnetstimulation
u.a. unter anderem
v.a. vor allem
VLPFC ventrolateraler präfrontaler Kortex
VMPFC ventromedialer präfrontaler Kortex
Vol. Volume (=Jahrgang)
VP virtueller Partner
z.B. zum Beispiel
ZNS Zentrales Nervensystem

A EINLEITUNG
1
1 Ausgangssituation und Problemstellung
Die Neurowissenschaft ist noch ein sehr junges Forschungsfeld, das in den letzten Jahren
einen enormen Aufschwung erlebt hat. Neurowissenschaftliche Studien beschäftigen sich
u.a. mit der Messung der menschlichen Gehirnaktivität während der Lösung von Aufga-
ben. Mit Hilfe bestimmter Methoden kann festgestellt werden, welche Gehirnregionen zur
Lösung bestimmter Aufgaben beitragen. Durch diese Erkenntnisse können in weiterer
Folge Rückschlüsse auf das Denken und Fühlen eines Menschen gezogen werden. Die-
ser Aspekt hat wahrscheinlich auch für den enormen Aufschwung der Neurowissenschaf-
ten und deren Teildisziplinen gesorgt.1
Die Neuroökonomie - als Teildisziplin der Neurowissenschaft - kombiniert neurowissen-
schaftliche Erkenntnisse mit betriebswirtschaftlichen Themenfeldern, wobei sich die Neu-
roökonomie wiederum in verschiedene Spezialbereiche, wie bspw. in Neuromarketing,
Neurofinance, Neuroleadership oder Neuromanagement, aufgliedert.2 Der Zweck der
Neuroökonomie liegt darin, zu erforschen, wie Menschen in unterschiedlichen Situationen
Entscheidungen fällen, wie sie mit Vor- oder Nachteilen umgehen und welches Verhalten
daraus abgeleitet werden kann.3 Das Ziel der Neuroökonomie ist somit die Erfassung af-
fektiver und kognitiver Prozesse in ihrer Gesamtheit, um das menschliche Gehirn als
„Black Box“ besser zu verstehen und somit Zusammenhänge oder theoretische Modelle
empirisch zu fundieren.4 So wurde bspw. mithilfe der Neurowissenschaften im Rahmen
einiger Studien erforscht, dass Emotionen kognitive Prozesse mit steuern, wodurch eine
Abkehr vom homo oeconomicus, der auf Basis von rationalen Überlegungen entscheidet,
gezeichnet wird.5
Neuroleadership, als betriebswirtschaftlicher Teilbereich der Neuroökonomie, versucht
neurowissenschaftliche Erkenntnisse mit bekannten Managementtheorien zu verbinden,
1 Vgl. Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe, J.
(Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.543.
2 Vgl. Peters, T./Ghadiri, A (2011).: Neuroleadership - Grundlagen, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S.13-14. 3 Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern ,
Freiburg, S. 11. 4 Vgl. Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe, J.
(Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.543. 5 Vgl. Reimann, M./Weber, B. (2011): Neuroökonomie - Eine Bestandsaufnahme, in: Reimann, M./Weber, B.
(Hrsg.): Neuroökonomie. Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 7-8; so auch Elger, C.E. (2013): Neuroleadership, Freiburg, S. 31.
A. EINLEITUNG

A EINLEITUNG
2
um im Ergebnis eine gehirngerechtere Führung und damit einhergehend eine bessere
Performance zu erreichen.6
Insbesondere für die Bereiche Neuromarketing sowie für Neurofinance sind bereits un-
zählige Studien und wissenschaftliche Arbeiten veröffentlicht worden. Im Bereich des
Neuroleaderships mangelt es hingegen bis dato an wissenschaftlichen Arbeiten und auch
an durchgeführten Studien.7 Neuroleadership ist noch keine in sich geschlossene Theorie,
sondern befindet sich derzeit in der Entwicklung, wobei davon auszugehen ist, dass die
neurowissenschaftlichen Erkenntnisse auch in diesem Bereich in Zukunft eine veränderte
Sichtweise auf das menschliche Denken und Handeln begründet.8
Überdies stellt sich das Problem vorhandener Studien dahingehend, dass deren Ergeb-
nisse noch nie untereinander verglichen wurden. Diesem Defizit soll durch diese Arbeit
Rechnung getragen werden, indem Anhand einer Metaanalyse alle bisher veröffentlichen
Studien zum Themenfeld „Leadership“ zusammengetragen und systematisiert werden.
2 Forschungsfragen
Im Zusammenhang mit der Problemstellung und der daraus resultierenden Zielsetzung
werden durch die vorliegende Arbeit folgende Forschungsfragen beantwortet:
o FF1: Welche neurowissenschaftliche Methoden können für Aspekte der Führungs-
forschung herangezogen werden und worin liegen deren Vor- und Nachteile?
o FF2: Welchen Beitrag kann die Neurowissenschaft für die Führungsforschung leis-
ten?
o FF3: Welche Hirnregionen sind bei Aspekten der Leadershipforschung beteiligt?
3 Ziel und praktische Bedeutung
Das Ziel dieser Arbeit liegt darin, bisherige Studien zum Themenbereich „Leadership“ zu
erfassen und zu systematisieren. Die Studien sollen insbesondere im Hinblick auf die,
während der Untersuchung, aktivierten Gehirnregionen untersucht und systematisiert
werden. In diesem Zusammenhang wird überprüft, ob die diesbezüglichen Ergebnisse der
einzelnen Studien miteinander übereinstimmen oder eher lose, ohne Verbindung, im
Raum stehen.
6 Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern ,
Freiburg, S. 15. 7 Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlagen, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 5. 8 Vgl. Elger, C.E. (2013), Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern,
Freiburg, S. 15.

A EINLEITUNG
3
Die praktische Bedeutung in dieser Arbeit liegt darin, einzelne, bereits vorhandene Stu-
dien im Bereich des Leaderships zu systematisieren und miteinander zu vergleichen und
im besten Fall Übereinstimmungen hinsichtlich Aktivierungen einzelner Hirnareale festzu-
stellen. Durch diese Arbeit können sich in weiterer Folge Forschungsfelder im Bereich
„Leadership“ herausstellen, die in Zukunft untersucht werden könnten.
4 Methodik und Aufbau der Arbeit
Um einen strukturierten Aufbau zu garantieren, und um ein Verständnis für die teilweise
sehr komplexen und auch bei weitem noch nicht abschließend erforschten9 Prozesse im
Gehirn zu erlangen, ist ein grundlegendes theoretisches Wissen vom Aufbau des Gehirns
nötig, das im ersten Teil der Arbeit vermittelt wird.
Dabei erfolgt eine grobe Unterteilung des Gehirns in die Bereiche Vorderhirn, Kleinhirn
und Hirnstamm, wobei das Großhirn und das Zwischenhirn gemeinsam das Vorderhirn
bilden und der Hirnstamm sowohl das Mittelhirn, den Pons und die Medulla oblongata
umfasst. Neben einer kurzen anatomischen Darstellung dieser Hirnregionen stehen ins-
besondere deren jeweiligen Hauptfunktionen im Mittelpunkt. Abschließend zu diesem Un-
terkapitel werden die wesentlichen Inhalte für das bessere Verständnis tabellarisch aufbe-
reitet.
Neben den verschiedenen Hirnregionen ist es unumgänglich, die wichtigsten Verfahren
der Neurowissenschaften, sowie deren Vor- und Nachteile, vorzustellen. Dies wird vor
allem für das tiefere Verständnis dieser Arbeit sowie für die Beantwortung der ersten For-
schungsfrage essentiell sein. Die neurowissenschaftlichen Methoden können dabei in
bildgebende Verfahren i.w.S. und in bildgebende Verfahren i.e.S. eingeteilt werde. Neben
den klassischen neurowissenschaftlichen Methoden, wie bspw. der Elektroenzepha-
lographie und der funktionellen Magnetresonanztomographie werden mit der transkraniel-
len Magnetstimulation und mit der Diffusions-Tensor-Bildgebung noch zwei weitere, sich
im Vormarsch befindende, Verfahren vorgestellt. Auch dieses Unterkapitel wird zur Ver-
anschaulichung mit einer tabellarischen Aufbereitung der einzelnen vorgestellten neuro-
wissenschaftlichen Methoden und deren Vor- und Nachteile abgeschlossen.
Im Fokus des darauf folgenden Kapitals stehen unterschiedliche Führungsansätze und
die Herstellung einer Verbindung von Leaderhip im betriebswirtschaftlichen Kontext mit
den Neurowissenschaften. In diesem Zusammenhang werden auch der unlängst von Da-
9 Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern,
Freiburg S. 28.

A EINLEITUNG
4
vid Rock konstruierte Begriff des Neuroleaderships10, die Bedeutung der Neurowissen-
schaften für das Leadership und einzelne bereits erforschte Inhalte vermittelt, ehe zum
Abschluss auf drei Bereiche eingegangen wird, in denen die Neurowissenschaft zur Füh-
rungsforschung beitragen kann.
Nach dieser theoretischen Einführung wird im zweiten Teil der Arbeit eine Metaanalyse
durchgeführt. Dabei werden die Ergebnisse der Primärstudien, die nach bestimmten Krite-
rien in die Analyse aufgenommen wurden, erfasst und analysiert. Die durchgeführte Me-
taanalyse entspricht nicht einer Metaanalyse im herkömmlichen Sinn. Aufgrund der gerin-
gen Datenmenge wurde keine Kodierung durchgeführt und können deshalb auch keine
statistischen Effekte nachgewiesen werden. Vielmehr steht im Rahmen dieser Arbeit die
Erfassung und Systematisierung der einzelnen Studien im Vordergrund.
Aufgrund der vielen, für BetriebswirtInnen, nicht geläufigen Fachausdrücke aus der Medi-
zin sowie aus dem psychologischen Bereich, befindet sich am Ende der Arbeit ein Glos-
sar, das alle medizinischen bzw psychologischen Begriffe kurz erläutert.
Zu guter Letzt ist anzumerken, dass für die einzelnen Gehirnareale sowohl die deutschen,
als auch die lateinischen Begriffe parallel verwendet werden.
10
Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern, Freiburg S.14.

5
Dieses einführende Kapitel befasst sich mit den Grundlagen der Neurowissenschaften.
Dabei wird zunächst der anatomische Aufbau des menschlichen Gehirns dargestellt und
auf die, der jeweiligen Gehirnregion zugrunde liegenden, Funktionen eingegangen. Im
Anschluss daran erfolgt eine kurze Darstellung der wichtigsten Verfahren der Neurowis-
senschaften. In diesem Zusammenhang stehen insbesondere die jeweiligen Vor- und
Nachteile jeder Methode im Fokus. Das abschließende Teilkapitel beschäftigt sich sodann
mit (Neuro-)Leadership, wobei zunächst auf den Begriff „Leadership“ sowie auf einzelne
Führungsstiltheorien eingegangen wird, ehe Neuroleadership und in diesem Zusammen-
hang die ersten Ansätze einiger Wissenschaftler vorgestellt werden. Im Anschluss daran
wird aufgezeigt, in welchen Bereichen die Neurowissenschaft zur Führungsforschung bei-
tragen kann. An dieser Stelle ist festzuhalten, dass die Inhalte dieses Kapitels für das tie-
fere Verständnis dieser Arbeit von grundlegender Bedeutung sind.
1 Das Gehirn
Das menschliche Gehirn macht mit ca. 1,5 Kilogramm nur rund drei Prozent der Körper-
masse aus, verbraucht jedoch ungefähr 20 Prozent der gesamten Energie, die der
menschliche Körper für seine Funktionen benötigt.11 Neben dem Rückenmark ist das Ge-
hirn ein Bestandteil des ZNS12 und besteht insgesamt aus 60-100 Mrd. Nervenzellen, den
sog. Neuronen.13
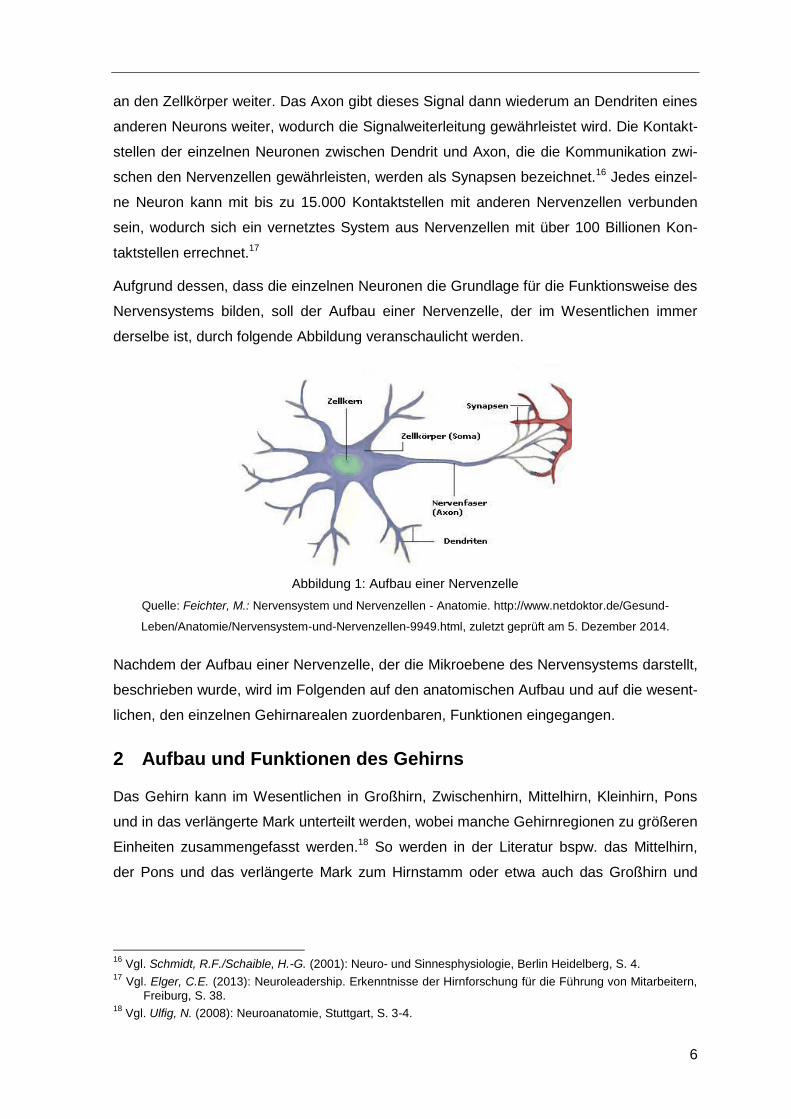
Eine Nervenzelle besteht typischerweise aus dem Zellkörper, den sog. Dendriten und
einem Axon.14 Wie aus Abbildung 1 ersichtlich ist, nimmt der Zellkörper den größten Teil
einer Zelle ein. Der Zellkörper beinhaltet neben dem Zellkern auch weitere Strukturen, die
die grundlegenden Funktionen, wie bspw. die Energieversorgung und den Stoffwechsel,
gewährleisten.15 Vom Zellkörper gehen einige Fortsätze ab, wobei hier zwischen Dendri-
ten und dem Axon unterschieden wird. Die sog. Dendriten, von denen stets mehrere exis-
tieren, nehmen über Neurotransmitter Signale von anderen Neuronen auf und leiten diese
11
Vgl. Elger, C.E. (2013): Neuroleadership. Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern, Freiburg, S. 38.
12 Vgl. Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen. Morphologie, Kernspintomogra-
phie und 3 D-Computerrekonstruktion, Göttingen, S. 7. 13
Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 14.
14 Vgl. Solms, M./Turnbull, O. (2004): Das Gehirn und die innere Welt. Neurowissenschaft und Psychoanaly-
se, Düsseldorf/Zürich, S. 23. 15
Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 14.
B. GRUNDLAGEN DER NEUROWISSENSCHAFTEN UND
NEUROLEADERSHIP
6
an den Zellkörper weiter. Das Axon gibt dieses Signal dann wiederum an Dendriten eines
anderen Neurons weiter, wodurch die Signalweiterleitung gewährleistet wird. Die Kontakt-
stellen der einzelnen Neuronen zwischen Dendrit und Axon, die die Kommunikation zwi-
schen den Nervenzellen gewährleisten, werden als Synapsen bezeichnet.16 Jedes einzel-
ne Neuron kann mit bis zu 15.000 Kontaktstellen mit anderen Nervenzellen verbunden
sein, wodurch sich ein vernetztes System aus Nervenzellen mit über 100 Billionen Kon-
taktstellen errechnet.17
Aufgrund dessen, dass die einzelnen Neuronen die Grundlage für die Funktionsweise des
Nervensystems bilden, soll der Aufbau einer Nervenzelle, der im Wesentlichen immer
derselbe ist, durch folgende Abbildung veranschaulicht werden.
Abbildung 1: Aufbau einer Nervenzelle
Quelle: Feichter, M.: Nervensystem und Nervenzellen - Anatomie. http://www.netdoktor.de/Gesund-
Leben/Anatomie/Nervensystem-und-Nervenzellen-9949.html, zuletzt geprüft am 5. Dezember 2014.
Nachdem der Aufbau einer Nervenzelle, der die Mikroebene des Nervensystems darstellt,
beschrieben wurde, wird im Folgenden auf den anatomischen Aufbau und auf die wesent-
lichen, den einzelnen Gehirnarealen zuordenbaren, Funktionen eingegangen.
2 Aufbau und Funktionen des Gehirns
Das Gehirn kann im Wesentlichen in Großhirn, Zwischenhirn, Mittelhirn, Kleinhirn, Pons
und in das verlängerte Mark unterteilt werden, wobei manche Gehirnregionen zu größeren
Einheiten zusammengefasst werden.18 So werden in der Literatur bspw. das Mittelhirn,
der Pons und das verlängerte Mark zum Hirnstamm oder etwa auch das Großhirn und
16
Vgl. Schmidt, R.F./Schaible, H.-G. (2001): Neuro- und Sinnesphysiologie, Berlin Heidelberg, S. 4. 17
Vgl. Elger, C.E. (2013): Neuroleadership. Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern,
Freiburg, S. 38. 18
Vgl. Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 3-4.

7
das Zwischenhirn zum Vorderhirn (Prosencephalon) zusammengefasst.19 Das Vorderhirn
ist von enormer Bedeutung, zumal es für die Steuerung fast aller Prozesse im ZNS ver-
antwortlich ist.20 Aufgrund dessen erfolgt im Folgenden eine umfassendere Darstellung
dieser beiden Gehirnbereiche, wobei das Hauptaugenmerkt dieses Kapitels auf der
Übermittlung der wesentlichsten Funktionen der einzelnen Gehirnbereiche, und nicht auf
deren umfassender anatomischer Erörterung, liegt. Zuvor soll die nachstehende Abbil-
dung die grobe Einteilung der Gehirnregionen veranschaulichen.
Abbildung 2: Aufbau des Gehirns
Quelle: Pokorna, E., Der menschliche Körper, Teil II. http://www.ftvs.cuni.cz/eknihy/jazyky/ein-
fuhrungindiesportsprache/Texts/5-Korper_II.html, zuletzt geprüft am 4. November 2013.
2.1 Großhirn (Telencephalon)
Mit ca. 85% des gesamten Gewichtes nimmt das Großhirn den größten Teil des mensch-
lichen Gehirns ein.21 Das Großhirn kann aus Vereinfachungsgründen in die Großhirnrinde
(Kortex) und in das Großhirnmark, den Basalganglien, gegliedert werden.22
Entwicklungsgeschichtlich lässt sich das Großhirn in die Urrinde (Archaeokortex), die Alt-
rinde (Palaeokortex) und in die Neurinde (Neokortex) gliedern.23 Als wesentlicher Be-
standteil des Archaeokortex gilt der Hippocampus, der zugleich einen wichtigen Teil des
19
Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 109; eine Einteilung in Vorder-hirn, Mittelhirn und Rautenhirn nehmen u.a. folgende Autoren vor: Carlson, N.R. (2004): Physiologische Psychologie, München, S. 88; siehe dazu auch Rohen, J.W. (2001): Funktionelle Neuroanatomie, Stutt-gart, S.15.
20 Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-
letzt geprüft am 4. November 2013. 21
Vgl. Schandry, R. (2003): Biologische Psychologie, Weinheim, S. 128; Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 145.
22 Vgl. Raab, G./Gernsheimer, O./Schindler, M. (2009): Neuromarketing. Grundlagen-Erkenntnisse-Anwen-
dungen, Wiesbaden, S. 102. 23
Vgl. Raab, G./Gernsheimer, O./Schindler, M. (2009): Neuromarketing. Grundlagen-Erkenntnisse-Anwen-dungen, Wiesbaden, S. 104.

8
limbischen Systems darstellt.24 Der Palaeokortex zählt zu den ältesten Teilen der Groß-
hirnrinde. Sein Zentrum bildet das Riechhirn, das bei Tieren im Vergleich zum Menschen
besser ausgebildet ist. Der Neokortex ist, wie der Name schon vermuten lässt, der ent-
wicklungsgeschichtlich jüngste Teil der Großhirnrinde. Dennoch nimmt er mit insgesamt
rund 90% den größten Teil der Großhirnrinde ein.25
Charakteristisch für die Oberfläche des Kortex ist die starke Faltung. Diese Wölbungen
werden als Gyri (Gehirnwindungen) bezeichnet, die durch Sulci (Furchen) voneinander
getrennt sind.26 Die Großhirnrinde sowie die subkortikalen Kerne im inneren des Gehirns
bilden die graue Substanz, während sich dazwischen Ansammlungen verschiedener Bah-
nen befinden, die als weiße Substanz bezeichnet werden.27
Das Großhirn lässt sich von außen durch eine Furche in zwei Hemisphären unterteilen.
Die linke Hemisphäre besteht dabei anatomisch betrachtet großteils aus vielen kurzen
neuronalen Verbindungen, während in der rechten Hemisphäre längere Verbindungen
auch weiter entfernte Hirnregionen miteinander verknüpfen.28 Beide Hemisphären sind mit
Hilfe eines breiten Bandes von Nervenfasern durch den sog. Balken (corpus callosum)
miteinander verbunden.29 Anatomisch ist besonders auffallend, dass eine besonders enge
Verbindung zwischen der linken Hemisphäre und der rechten Körperhälfte und umgekehrt
besteht. Es wird heutzutage davon ausgegangen, dass beide Gehirnhälften hinsichtlich
ihrer Funktionsweise unterschiedlich spezialisiert sind. So sind in der linken Hemisphäre
vor allem sprachliche und analytische Fähigkeiten lokalisiert, während die rechte Gehirn-
hälfte mehr mit räumlichen Prozessen assoziiert wird.30
24
Zum limbischen System siehe Kapitel 2.3. 25
Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 241. 26
Vgl. Bear, M.F./Connors, B.W./Paradiso, M.A. (2009): Neurowissenschaften. Ein grundlegendes Lehrbuch für Biologie, Medizin und Psychologie, Heidelberg, S. 231; Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 145; Firbas, W./Gruber, H./Mayr, R. (1995): Neuroanatomie, Wien, S. 1.
27 Vgl. Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 145.
28 Vgl. Ahlert, D./Kenning, P. (2006): Neuroökonomik, in: Zeitschrift für Management, Jg. 1, Nr. 1, S. 27.
29 Vgl. Solms, M./Turnbull, O. (2004): Das Gehirn und die innere Welt. Neurowissenschaft und Psychoanaly-
se, Düsseldorf/Zürich, S. 30. 30
Vgl. Anderson, J.R. (2001): Kognitive Psychologie, Heidelberg/Berlin, S. 22-23.

9
Abbildung 3: Die zwei Gehirnhälften (Hemisphären)
Quelle: Bear, M.F./Connors, B.W./Paradiso, M.A. (2009): Neurowissenschaften. Ein grundlegendes Lehrbuch
für Biologie, Medizin und Psychologie, Heidelberg, S. 216.
2.1.1 Einteilung der Großhirnrinde
Sowohl die linke als auch die rechte Gehirnhälfte lassen sich jeweils in Frontallappen,
Parietallappen, Temporallappen, und Okzipitallappen gliedern. Unter dem Frontal- und
Temporallappen verborgen, befindet sich zudem die sog. Insula (Inselrinde), die keinem
der vier Gehirnlappen zugeordnet werden kann. Ebenso wenig stellt der Gyrus cinguli
einen Teil der vier Gerhirnlappen dar.31
Die vier Gehirnlappen werden durch verschiedene Sulci voneinander abgegrenzt. Der
Sulcus lateralis trennt den Frontallappen und den Parietallappen vom Temporallappen.
Der Sulcus centralis trennt den Frontallappen vom Parietallappen und der Sulcus parieto-
occipitalis sorgt für die Trennung von Parietallappen und Okzipitallappen.32
Nachfolgende Abbildung soll die eben ausgeführte Einteilung der Großhirnrinde bildlich
veranschaulichen.
Abbildung 4: Die Lappen des menschlichen Gehirns (Seitenansicht)
Quelle: (Bear, M.F./Connors, B.W./Paradiso, M.A. 2009): Neurowissenschaften. Ein grundlegendes Lehrbuch
für Biologie, Medizin und Psychologie, Heidelberg, S. 217.
31
Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 216. 32
Vgl. Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, Göttingen, S. 14.

10
Funktionell kann das Großhirn und insbesondere der Neokortex in Primärfelder, Sekun-
därfelder und Assoziationsfelder eingeteilt werden. Als Primärfelder werden primäre Endi-
gungsorte bezeichnet, die ihre Sinnesimpulse direkt vom Thalamus erhalten und an das
Bewusstsein weiterleiten. Neben den sensorischen gibt es auch motorische Primärzent-
ren, die den Ursprung für absteigende motorische Bahnen bilden. Sekundärfelder sind
jene Gebiete des Neokortex, die die erhaltenen Sinnesinformationen interpretieren. Ana-
tomisch sind sie mit Primärzentren benachbart und diesen vor- bzw. nachgeschaltet. As-
soziationsfelder bilden eine Art subsidiäre Kategorie; d.h. dazu zählen jene Gebiete des
Neokortex, die weder den Primär- noch den Sekundärfeldern zugeordnet werden kön-
nen.33
Im Folgenden werden die Gehirnlappen einzeln erläutert und auf deren wesentlichen
Funktionen eingegangen, ehe die Insula sowie der Gyrus cinguli, die ebenso zur Groß-
hirnrinde gezählt werden, kurz vorgestellt werden. Im Anschluss daran wird darüber hin-
aus auf die Basalganglien sowie auf das limbische System eingegangen.
2.1.1.1 Okzipitallappen (Hinterhautslappen)
Der Okzipitallappen zählt aufgrund seiner überschaubaren Zugänge und aufgrund seiner
vergleichsweise übersichtlichen Organisation zu den am meisten erforschten Strukturen
im menschlichen Gehirn.34
Im Okzipitallappen befindet sich mit der primären und der sekundären Sehrinde das visu-
elle System. Innerhalb der primären Sehrinde erfolgt eine Bewusstwerdung der visuellen
Signale, während die angrenzende sekundäre Sehrinde diese Signale weiterverarbeitet.35
Somit ist die wesentliche Funktion des Okzipitallappens in der optischen Wahrnehmung
und deren Weiterverarbeitung zu sehen.36
2.1.1.2 Temporallappen (Schläfenlappen)
Nach dem Frontallappen ist der Temporallappen der zweitgrößte Lobi und dient mit vielen
verschiedenen Zentren unterschiedlichen Funktionen.37 Mit dem Gyrus temporalis superi-
33
Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 221-222. 34
Vgl. Leyh, A. (23.09.2011): Der Okzipitallappen, http://dasgehirn.info/entdecken/anatomie/der-occipitallap-
pen/; zuletzt geprüft am 18.September 2014. 35
Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 28.
36 Vgl. Firbas, W./Gruber, H./Mayr, R. (1995): Neuroanatomie, Wien, S. 176.
37 Vgl. Wicht, H. (05.09.2011): Der Temporallappen, http://dasgehirn.info/entdecken/anatomie/der-temporal-
lappen/, zuletzt geprüft am 18.September 2014.

11
or, dem Gyrus temporalis medius und dem Gyrus temporalis inferior zählen drei große
Gehirnwindungen zum Temporallappen.38
Im Temporallappen, der komparabel zum Okzipitallappen ebenso aus einer primären und
einer sekundären Hörrinde besteht, befindet sich das auditive System. Die primäre Hör-
rinde dient dabei der Bewusstwerdung auditiver Signale, während die sekundäre Hörrinde
die akustischen Erinnerungsbilder in weiterer Folge interpretiert. Die sekundäre Hörrinde
wird durch das Erkennen und der Interpretation von Sprache auch „Wernicke-
Sprachzentrum“ genannt, das bei den meisten Menschen in der linken Gehirnhälfte aus-
gebildet ist.39 Neben dem Hören, als die wohl bekannteste Funktion des Temporallap-
pens, dient dieser auch dem Sprechen und Verstehen, sowie dem visuellen Erkennen und
der Gedächtnisbildung.40
2.1.1.3 Parietallappen (Scheitellappen)
Der Parietallappen kann mit der primären und der sekundären somatosensiblen Rinde
einerseits, sowie dem posterioren parietalen Kortex andererseits in zwei größere Bereiche
eingeteilt werden.41 Berührungs-, Temperatur- sowie Schmerzimpulse kommen in der
primären somatosensiblen Rinde, insbesondere im postzentralen Gyrus, zum Bewusst-
sein und werden wiederum von der sekundären somatosensiblen Rinden interpretiert,
indem das Ertastete vom Gehirn erkannt wird.42
Der posteriore parietale Kortex ist für die Raumorientierung entscheidend. Visuelle, vesti-
buläre, auditive und propriozeptive Reize werden hier zusammengetragen und integriert,
um eine räumliche Vorstellung und Orientierung zu ermöglichen.43 Im posterioren Parietal-
lappen befindet sich der Sulcus intraparietalis, der den oberen (superior) Parietallappen
vom unteren (inferior) Parietallappen trennt. Hier befindet sich der „mathematische Kor-
tex“44, da in diesem Bereich numerische Informationen verarbeitet werden.45
38
Vgl. Kahle, W./Frotscher, M. (2009): Taschenatlas Anatomie. Nervensystem und Sinnesorgane, Band 3, Stuttgart, S. 214.
39 Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 242-246 mwH; siehe dazu auch
Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, Göttingen, S. 17. 40
Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-letzt geprüft am 6. September 2014; o.V. ; Wicht (05.09.2011): Der Temporallappen, http://dasgehirn.info/entdecken/anatomie/der-temporal-lappen/, zuletzt geprüft am 18.September 2014; Braus, D.F. (2004): EinBlick ins Gehirn. Moderne Bildgebung in der Psychiatrie, Stuttgart, S. 20f.
41 Vgl. Leyh, A. (09.09.2011): Der Parietallappen, http://dasgehirn.info/entdecken/anatomie/der-parietallap-
pen/, zuletzt geprüft am 18. September 2014. 42
Vgl. Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 154f. 43
Vgl. Leyh, A. (09.09.2011): Der Parietallappen, http://dasgehirn.info/entdecken/anatomie/der-parietallap-pen/, zuletzt geprüft am 18. September 2014; Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 254.
44 Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 254.
45 Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-
letzt geprüft am 6. September 2014.o.V.

12
Der Gyrus angularis gilt als zentrale Schaltstelle zwischen dem „Wernicke-
Sprachzentrum“ und der sekundären Sehrinde und spielt dort eine wesentliche Rolle bei
der Zuordnung visueller Reize zu sprachlichen Begriffen.46
Weitere Bestandteile des Parietallappens sind der temproroparietale Übergang sowie der
Precuneus.47
2.1.1.4 Frontallappen (Stirnlappen)
Der Frontallappen nimmt den gesamten vorderen Teil des Großhirns, bis hin zur Zentral-
furche ein und stellt somit die größte Struktur des menschlichen Gehirns dar.48 Ebenso
wie der Temporallappen enthält der Frontallappen mit dem Gyrus frontalis superior, dem
Gyrus frontalis medius und dem Gyrus frontalis inferior drei große Gehirnwindungen.49
Der Frontallappen, bestehend aus dem Motorkortex, der das frontale Augenfeld sowie das
motorischen Sprachzentrum umfasst, und dem präfrontalen Kortex, ist der Ort des soma-
tomotorischen Systems. 50 Aufgrund der großen Bedeutung des Frontallappens und seiner
höheren geistigen Funktionen werden im Folgenden die einzelnen Hirnregionen, die je-
weils zum Motorkortex bzw. zum präfrontalen Kortex zählen, erläutert.
2.1.1.4.1 Motorkortex
Der Motorkortex umfasst die primäre und sekundäre motorische Rinde, den prämotori-
schen Kortex, das motorische Sprachzentrum, das frontale Augenfeld und schließlich die
supplementär motorische Rinde.51
Die Funktion des Motorkortex liegt in feinmotorischen Bewegungen, die im davor gelege-
nen prämotorischen Kortex initiiert werden. Dabei werden alle Informationen betreffend
der Körpermotorik in der primären motorischen Rinde lokalisiert und in der sekundären
motorischen Rinde dementsprechend interpretiert.52 Das motorische Sprachzentrum,
auch „Broca’sches“ Sprachzentrum genannt, ist für die Produktion von Sprache verant-
wortlich. Während das „Broca’sche“ Sprachzentrum v.a. für Vokabular, Wortbildung und
Grammatik verantwortlich ist, befinden sich im „Wernick’schen“ Sprachzentrum Funktio-
46
Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 235-238 mwH; Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 155.
47 Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-
letzt geprüft am 6. September 2014.o.V. 48
Vgl. Leyh, A. (08.09.2011): Der Frontallappen, http://dasgehirn.info/entdecken/anatomie/der-frontallappen/, zuletzt geprüft am 18. September 2014.
49 Vgl. Kahle, W./Frotscher, M. (2009): Taschenatlas Anatomie. Nervensystem und Sinnesorgane, Band 3,
Stuttgart, S. 214. 50
Vgl. Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, Göttingen, S. 15. 51
Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-letzt geprüft am 6. September 2014.o.V.
52 Vgl. Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, Göttingen, S. 15.

13
nen zum Sprachverständnis. Beide Sprachzentren sind, wie bereits erwähnt, fast bei allen
Menschen, vor allem bei Rechtshändern53, ausschließlich in der linken Gehirnhälfte lokali-
siert.54 Das frontale Augenfeld ist für willkürliche Augenbewegungen zuständig.55
2.1.1.4.2 Präfrontaler Kortex
Der präfrontale Kortex fasst indes alle Areale des Frontallappens, die vor der prämotori-
schen Rinde liegen, zusammen56 und kann grob in den orbitofrontalen, den medialen und
den lateralen Kortex unterteilt werden.57
Der orbifrontale Kortex spielt eine wesentliche Rolle bei der Beurteilung und der Bewer-
tung von Stimuli hinsichtlich der zu erwartenden - positiven oder negativen - Auswirkun-
gen.58 Der mediale präfrontale Kortex kann wiederum in den dorsomedialen präfrontalen
Kortex (DMPFC) und in den ventromedialen präfrontalen Kortex (VMPFC) unterteilt wer-
den, wobei das para-anteriore Cingulum einen Teil des VMPFC darstellt. Der DMPFC
wird hauptsächlich mit sozialer Kognition in Verbindung gebracht, während dem VMPFC
besondere Bedeutung bei der Entscheidungsfindung zukommt. Das para-anteriore Cingu-
lum wird mit der Evaluierung von Belohnungswerten assoziiert. Ebenso wie der mediale
präfrontale Kortex kann der laterale präfrontale Kortex in den dorsolateralen präfrontalen
Kortex (DLPFC) einerseits, und den ventrolateralen präfrontalen Kortex (VLPFC) anderer-
seits, unterteilt werden. Der DLPFC ist insbesondere für Konflikt-Monitoring und für kogni-
tive Kontrolle bekannt, während der VLPFC mit höheren kognitiven Fähigkeiten assoziiert
wird, wodurch dieser Bereich in gewisser Weise das Intelligenzzentrum des Gehirns dar-
stellt.59
Zusammenfassend werden dem präfrontalen Kortex höhere soziale, geistige und psychi-
sche Fähigkeiten des Menschen zugeschrieben.60
Nachfolgende Abbildung soll die eben geschilderte Unterteilung der Großhirnrinde in die
besagten vier Gehirnlappen nochmals veranschaulichen.
53
Vgl. Kahle, W./Frotscher, M. (2009): Taschenatlas Anatomie. Nervensystem und Sinnesorgane, Band 3, Stuttgart, S. 264.
54 Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie.
Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 32 bzw. 34. 55
Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-letzt geprüft am 6. September 2014.o.V.
56 Vgl. Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 153.
57 Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-
letzt geprüft am 6. September 2014.o.V. 58
Vgl. Trepel, M. (2008): Neuroanatomie. Struktur und Funktion, München, S. 250. 59
Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-letzt geprüft am 6. September 2014; Trepel, M. (2008): Neuroanatomie. Struktur und Funktion, Mün-chen, S. 250.o.V.
60 Vgl. Trepel, M.(2011): Neuroanatomie. Struktur und Funktion, München, S. 230.
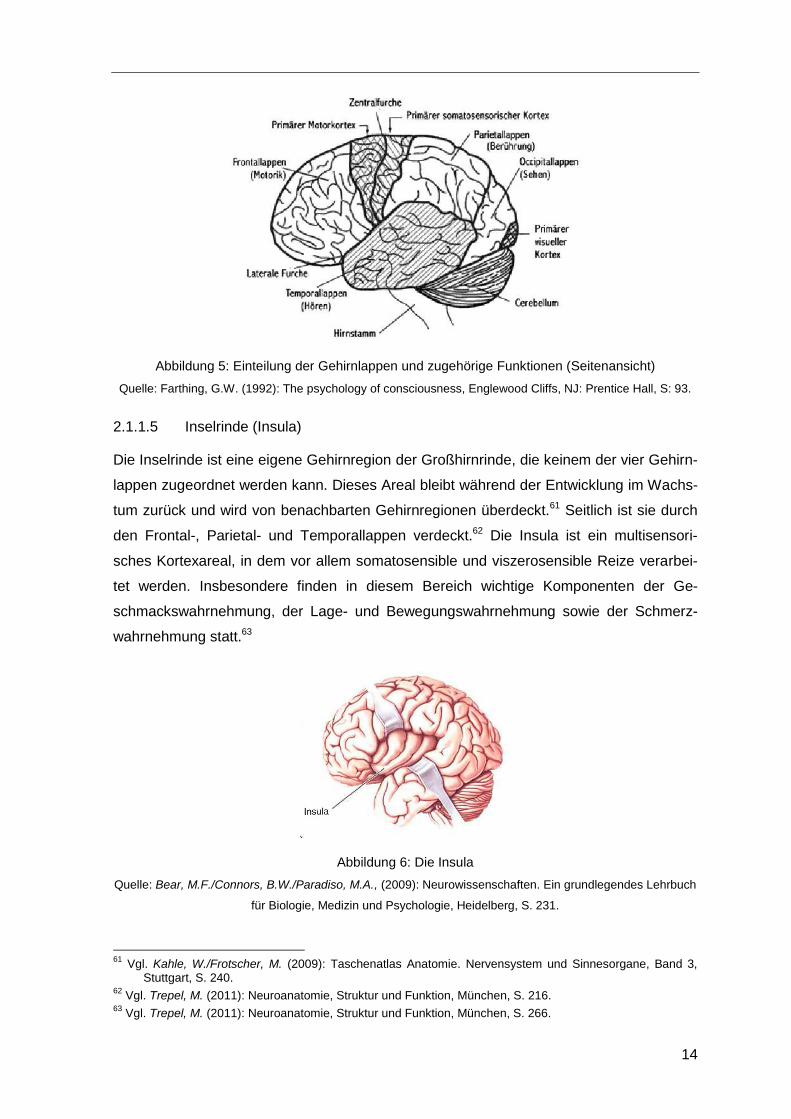
14
Abbildung 5: Einteilung der Gehirnlappen und zugehörige Funktionen (Seitenansicht)
Quelle: Farthing, G.W. (1992): The psychology of consciousness, Englewood Cliffs, NJ: Prentice Hall, S: 93.
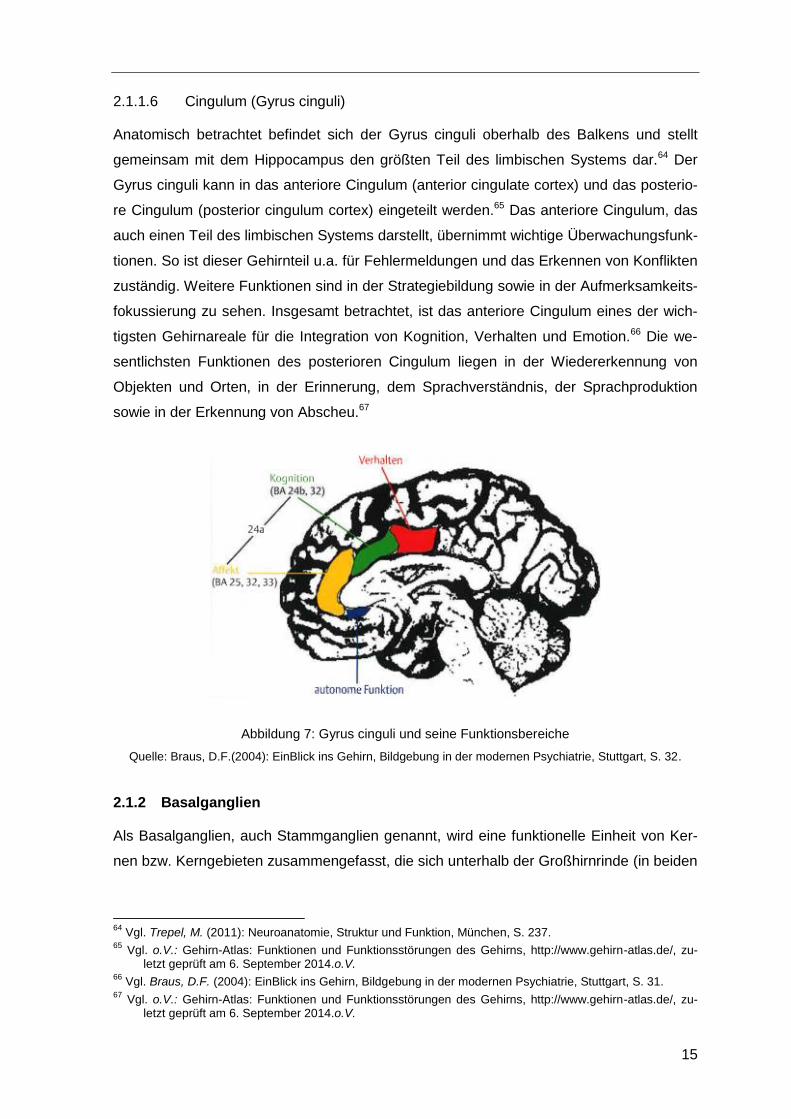
2.1.1.5 Inselrinde (Insula)
Die Inselrinde ist eine eigene Gehirnregion der Großhirnrinde, die keinem der vier Gehirn-
lappen zugeordnet werden kann. Dieses Areal bleibt während der Entwicklung im Wachs-
tum zurück und wird von benachbarten Gehirnregionen überdeckt.61 Seitlich ist sie durch
den Frontal-, Parietal- und Temporallappen verdeckt.62 Die Insula ist ein multisensori-
sches Kortexareal, in dem vor allem somatosensible und viszerosensible Reize verarbei-
tet werden. Insbesondere finden in diesem Bereich wichtige Komponenten der Ge-
schmackswahrnehmung, der Lage- und Bewegungswahrnehmung sowie der Schmerz-
wahrnehmung statt.63
Abbildung 6: Die Insula
Quelle: Bear, M.F./Connors, B.W./Paradiso, M.A., (2009): Neurowissenschaften. Ein grundlegendes Lehrbuch
für Biologie, Medizin und Psychologie, Heidelberg, S. 231.
61
Vgl. Kahle, W./Frotscher, M. (2009): Taschenatlas Anatomie. Nervensystem und Sinnesorgane, Band 3, Stuttgart, S. 240.
62 Vgl. Trepel, M. (2011): Neuroanatomie, Struktur und Funktion, München, S. 216.
63 Vgl. Trepel, M. (2011): Neuroanatomie, Struktur und Funktion, München, S. 266.
15
2.1.1.6 Cingulum (Gyrus cinguli)
Anatomisch betrachtet befindet sich der Gyrus cinguli oberhalb des Balkens und stellt
gemeinsam mit dem Hippocampus den größten Teil des limbischen Systems dar.64 Der
Gyrus cinguli kann in das anteriore Cingulum (anterior cingulate cortex) und das posterio-
re Cingulum (posterior cingulum cortex) eingeteilt werden.65 Das anteriore Cingulum, das
auch einen Teil des limbischen Systems darstellt, übernimmt wichtige Überwachungsfunk-
tionen. So ist dieser Gehirnteil u.a. für Fehlermeldungen und das Erkennen von Konflikten
zuständig. Weitere Funktionen sind in der Strategiebildung sowie in der Aufmerksamkeits-
fokussierung zu sehen. Insgesamt betrachtet, ist das anteriore Cingulum eines der wich-
tigsten Gehirnareale für die Integration von Kognition, Verhalten und Emotion.66 Die we-
sentlichsten Funktionen des posterioren Cingulum liegen in der Wiedererkennung von
Objekten und Orten, in der Erinnerung, dem Sprachverständnis, der Sprachproduktion
sowie in der Erkennung von Abscheu.67
Abbildung 7: Gyrus cinguli und seine Funktionsbereiche
Quelle: Braus, D.F.(2004): EinBlick ins Gehirn, Bildgebung in der modernen Psychiatrie, Stuttgart, S. 32.
2.1.2 Basalganglien
Als Basalganglien, auch Stammganglien genannt, wird eine funktionelle Einheit von Ker-
nen bzw. Kerngebieten zusammengefasst, die sich unterhalb der Großhirnrinde (in beiden
64
Vgl. Trepel, M. (2011): Neuroanatomie, Struktur und Funktion, München, S. 237. 65
Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-letzt geprüft am 6. September 2014.o.V.
66 Vgl. Braus, D.F. (2004): EinBlick ins Gehirn, Bildgebung in der modernen Psychiatrie, Stuttgart, S. 31.
67 Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-
letzt geprüft am 6. September 2014.o.V.

16
Hemisphären) in der weißen Substanz des Gehirns befinden.68 Zu den Kerngebieten zäh-
len neben dem Striatum, das aus dem Putamen und dem Nucleus caudatus besteht, dem
Globus pallidus, dem Nucleus accumbens auch der Nucleus subthalamicus sowie die
Substantia nigra. Die einzelnen Kerne der Basalganglien stehen mittels unzähliger Faser-
bündel in Verbindung zueinander. Primär dienen die Basalganglien der Koordination und
Organisation von Bewegungsabläufen, wobei das Striatum, das seine Afferenzen haupt-
sächlich aus dem Kortex erhält, als Eingangspforte und somit als rezeptiver Teil des Sys-
tems gilt, während die Efferenzen das Basalgangliensystem zumeist über den Globus
pallidus wieder verlassen.69 Das Striatum und der Nucleus accumbens spielen zudem
auch bei Belohnungen eine sehr entscheidende Rolle.70
2.1.3 Limbisches System
Das limbische System, das v.a. aus dem alten Teil der Großhirnrinde - dem Archaeokor-
tex71 - gebildet wird, ist eine Zusammenfassung mehrerer Gehirnstrukturen zu einem Sys-
tem. Im Unterschied zum Neocortex, der im Wesentlichen mit der Verarbeitung von Pro-
zessen, die das Nervensystem betreffen, beschäftigt ist, wird das limbische System vor
allem mit Gedächtnisleistungen72 und Emotionen in Zusammenhang gebracht.73 Da es
sich beim limbischen System um ein theoretisches Konstrukt handelt, werden stets unter-
schiedliche Strukturen als Bestandteile des limbischen Systems zusammengefasst.74
Gemäß Solms/Turnbull zählen der Hypothalamus, der Thalamus, der Hippocampus, der
Gyrus cinguli, die Amygdala, sowie die Faserbahn Fornix (Gewölbebogen) zu den we-
sentlichsten Bestandteilen des limbischen Systems.75
Der Hippocampus ist für die räumliche Orientierung und für die Gedächtnisbildung zu-
ständig.76 Zudem werden im Hippocampus bereits bekannte Situationen bzw. Impulse von
Unbekannten unterschieden.77 Eine zentrale Bedeutung im limbischen System kommt
68
Vgl. CNS Forum Lundbeck Institute, http://de.brainexplorer.org/brain_atlas/Brainatlas_index.shtml, zuletzt geprüft am 10.September 2014.
69 Vgl. Firbas, W./Gruber, H./Mayr, R. (1995): Neuroanatomie, Wien, S. 171; o.V.: Gehirn-Atlas: Funktionen
und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zuletzt geprüft am 6. September 2014.
70 Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-
letzt geprüft am 6. September 2014; Braus, D.F. (2004): EinBlick ins Gehirn, Bildgebung in der moder-
nen Psychiatrie, Stuttgart, S. 34. 71
Vgl. Rohen, J.W. (2001): Funktionelle Neuroanatomie, Stuttgart, S. 160. 72
Vgl. Anderson, J.R. (2001): Kognitive Psychologie, Heidelberg/Berlin, S. 22. 73
Vgl. Rohen, J.W. (2001): Funktionelle Neuroanatomie, Stuttgart, S. 160. 74
Vgl. Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 227. 75
Vgl. Solms, M./Turnbull, O. (2004): Das Gehirn und die innere Welt. Neurowissenschaft und Psychoanaly-se, Düsseldorf/Zürich, S. 31-32.
76 Vgl. Anderson, J.R. (2001): Kognitive Psychologie, Heidelberg/Berlin, S. 22.
77 Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie.
Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 36-37.

17
dem Mandelkern (Amygdala) zu.78 Durch die Aktivierung der lediglich 1-2 cm großen
Amygdala werden bestimmte Affekte, wie Wut, Angst oder Ekel ausgelöst. Des Weiteren
ist die Amygdala der Sitz des emotionalen Gedächtnisses, wobei hier auch emotionales
Lernen stattfindet.79 Insgesamt hat das limbische System einen großen Einfluss auf emo-
tionale und vegetative Parameter, wodurch es eine große Rolle bei Motivation, Lernen
oder auch andere kognitiven Leistungen spielt.80
Abbildung 8: Das limbische System
Quelle: Braus, D.F. (2004): EinBlick ins Gehirn, Bildgebung in der modernen Psychiatrie, Stuttgart, S. 26.
2.2 Zwischenhirn (Diencephalon)
Das Zwischenhirn bildet zusammen mit dem Großhirn das Vorderhirn und ist hauptsäch-
lich in zwei Bereiche, den Thalamus und den Hypothalamus81, gegliedert.82
Der bohnenförmige Thalamus, der mit einer Länge von rund 4 cm einen Teil des Beloh-
nungssystems darstellt83, besteht aus vielen sog. Thalamuskernen, wovon einige in en-
gem Kontakt mit dem Großhirn stehen.84 Diese Kerne leiten alle Sinnesreize, mit Aus-
78
Vgl. Rohen, J.W. (2001): Funktionelle Neuroanatomie, Stuttgart, S. 160. 79
Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 36-37.
80 Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, 220; Ulfig, N. (2008): Neuroana-
tomie, Stuttgart, S. 228. 81
Weitere Gehirnbereiche, die zum Zwischenhirn zählen sind insbesondere der Epithalamus, der Metathala-mus, sowie die Faserbahn Fornix (Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Ge-
hirns, http://www.gehirn-atlas.de/, zuletzt geprüft am 6. September 2014). 82
Vgl. Solms, M./Turnbull, O. (2004): Das Gehirn und die innere Welt. Neurowissenschaft und Psychoanaly-se, Düsseldorf/Zürich, S. 29.
83 Vgl. o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns, http://www.gehirn-atlas.de/, zu-
letzt geprüft am 6. September 2014.o.V. 84
Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 181.

18
nahme des Riechens, zum Kortex oder zu subkortikalen Regionen weiter.85 Dadurch,
dass der Thalamus Informationen sortiert, bevor diese weitergeleitet werden, sprechen
einige Literaturstimmen im Zusammenhang mit dem Thalamus auch vom „Tor des Be-
wusstseins“.86
Die wichtigste Funktion des Thalamus ist die Übertragung von Sinneswahrnehmungen an
den Kortex.87 Eine weitere wichtige Funktion kommt dem Thalamus für das motorische
System zu, indem er Impulse aus den Basalganglien und dem Kleinhirn aufnimmt und
nach deren Verstärkung wieder weiterleitet.88
Der Hypothalamus enthält viele Kerngebiete und gilt funktionell als oberstes Integrations-
zentrum vegetativer Funktionen; d.h. der Hypothalamus ist v.a. für die Koordination von
Atmung, Kreislauf, Nahrungsaufnahme und vieles mehr verantwortlich.89
2.3 Kleinhirn (Cerebellum)
In der hinteren Schädelgrube, hinter dem Pons und der Medulla oblongata befindet sich
das Kleinhirn,90 das morphologisch in zwei Kleinhirnhemisphären und den Kleinhirnwurm
eingeteilt werden kann. Die Oberfläche des Kleinhirns besteht wie jene des Großhirns aus
zahlreichen Furchen, die das Kleinhirn in unterschiedliche Lappen unterteilen.91
Funktionell kann das Kleinhirn in Vestibulocerebellum, Spinocerebellum und Potocerebel-
lum eingeteilt werden. Das Vestibulocerebellum regelt mittels Informationen aus den
Vestibulariskernen das Gleichgewicht sowie die Koordination von Augen- und Körperbe-
wegungen.92 Das Spinocerebellum erhält seine Informationen hauptsächlich vom Rü-
ckenmark und ermöglicht exakte und flexible Bewegungen.93 Das Pontocerebellum erhält
seine Informationen hauptsächlich von den Brückenkernen und dient dazu, Bewegungen
zu planen und harmonisch auszuführen. Zudem steht das Pontocerebellum über den
85
Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 23-24.
86 Vgl. Roth, G. (2003): Fühlen, Denken, Handeln - Wie das Gehirn unser Verhalten steuert, Frankfurt am
Main, S.206. 87
Vgl. CNS Forum Lundbeck Institute : Gehirnatlas, http://de.brainexplorer.org/brain_atlas/Brainatlas_in-dex.shtml, zuletzt geprüft am 6.November 2013.
88 Vgl. Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, Göttingen, S. 24.
89 Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 187- 190 mwH.
90 Vgl. Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, Göttingen, S. 28.
91 Vgl. Ulfig, N. (2008): Neuroanatomie, Stuttgart, S. 117f.
92 Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 170.
93 Vgl. Birbaumer, N./Schmidt, R. F. (2006): Biologische Psychologie, Heidelberg, S. 91-92.
19
Pons in enger Beziehung mit dem Großhirn, wodurch es auch an der Feinregulierung der
Muskeln beteiligt ist.94
Die wichtigste Funktion des Kleinhirns ist die Koordination und Feinabstimmung von Be-
wegungsabläufen. Neben diesen motorischen Funktionen befasst sich das Kleinhirn zu-
dem auch mit kognitiven Funktionen und der Sprache.95
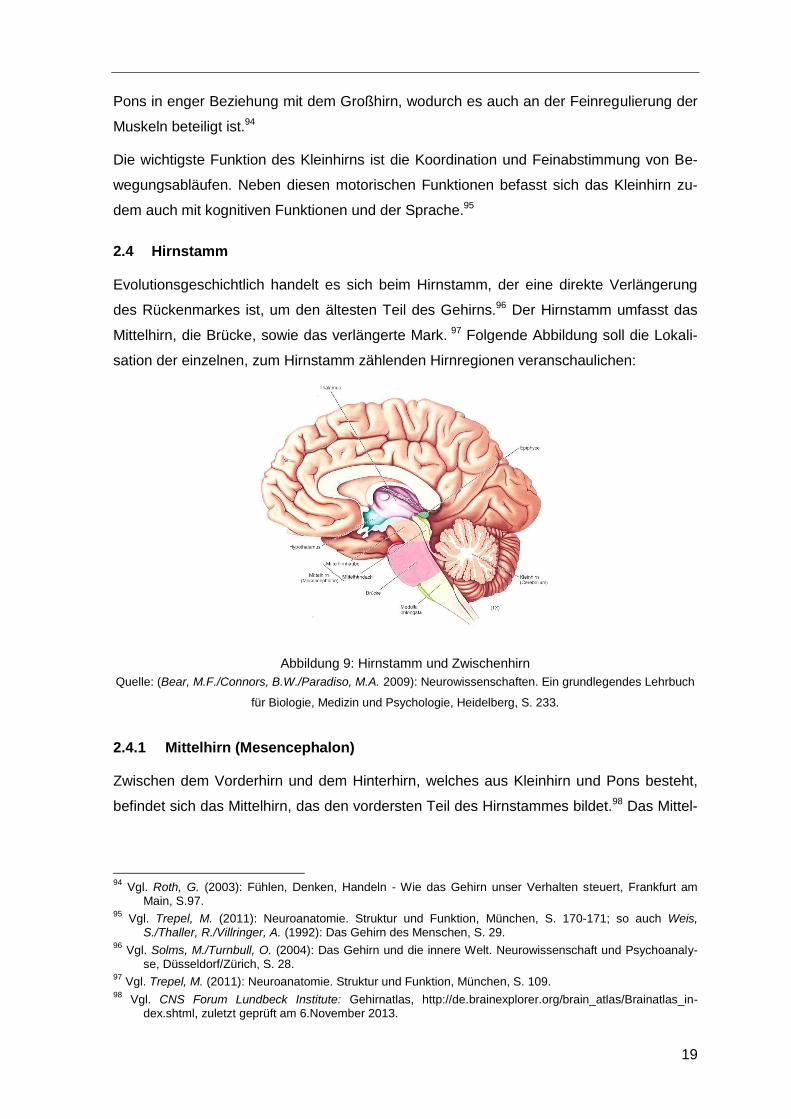
2.4 Hirnstamm
Evolutionsgeschichtlich handelt es sich beim Hirnstamm, der eine direkte Verlängerung
des Rückenmarkes ist, um den ältesten Teil des Gehirns.96 Der Hirnstamm umfasst das
Mittelhirn, die Brücke, sowie das verlängerte Mark. 97 Folgende Abbildung soll die Lokali-
sation der einzelnen, zum Hirnstamm zählenden Hirnregionen veranschaulichen:
Abbildung 9: Hirnstamm und Zwischenhirn
Quelle: (Bear, M.F./Connors, B.W./Paradiso, M.A. 2009): Neurowissenschaften. Ein grundlegendes Lehrbuch
für Biologie, Medizin und Psychologie, Heidelberg, S. 233.
2.4.1 Mittelhirn (Mesencephalon)
Zwischen dem Vorderhirn und dem Hinterhirn, welches aus Kleinhirn und Pons besteht,
befindet sich das Mittelhirn, das den vordersten Teil des Hirnstammes bildet.98 Das Mittel-
94
Vgl. Roth, G. (2003): Fühlen, Denken, Handeln - Wie das Gehirn unser Verhalten steuert, Frankfurt am Main, S.97.
95 Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 170-171; so auch Weis,
S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, S. 29. 96
Vgl. Solms, M./Turnbull, O. (2004): Das Gehirn und die innere Welt. Neurowissenschaft und Psychoanaly-se, Düsseldorf/Zürich, S. 28.
97 Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 109.
98 Vgl. CNS Forum Lundbeck Institute: Gehirnatlas, http://de.brainexplorer.org/brain_atlas/Brainatlas_in-
dex.shtml, zuletzt geprüft am 6.November 2013.

20
hirn ist mit einer Größe von rund 1,5 cm99 der kleinste Gehirnteil dieser Einteilung und
kann in drei Schichten untergliedert werden: Hirnschenkel, Haube und Vierhügelplatte.
Die Vierhügelplatte, oder auch Tectum (Dach) genannt, besteht aus jeweils zwei oberen
und zwei unteren Hügeln und beinhaltet Zellansammlungen, die für das Seh- und Hör-
vermögen essentiell sind.100 Wesentliche Funktionen des Mittelhirns liegen in der Bewe-
gungsregulation, der Körperhaltung, der Kontrolle der Augenfunktion und der Umschal-
tungen von auditiven Reizen.101
2.4.2 Brücke (Pons)
Der rund 2,5 cm lange Pons befindet sich zwischen der Medulla oblongata und dem Mit-
telhirn.102 Der Pons besteht aus dicken Faserbündeln und einigen Kerngebieten. Die Fa-
serbündel verbinden Teile des Großhirns mit dem Kleinhirn, dem Rückenmark und mit
den Hirnnervenkernen.103
Die Funktion der Brücke liegt v.a. in der Weiterleitung vom Kortex ausgehender Impulse
zur Steuerung von Bewegungsabläufen. Zudem ist die Steuerung des Schlaf- und Wach-
rhythmus eine zentrale Aufgabe des Ponses.104
2.4.3 Verlängertes Mark (Medulla oblongata)
Die Medulla oblongata, oder auch verlängertes Mark genannt, ist ca. 2 cm lang und grenzt
anatomisch an den Pons an, wobei eine klare Abgrenzung nach unten zum Rückenmark
nicht möglich ist.105
Das verlängerte Mark übernimmt wichtige Funktionen der vegetativen Steuerung, wie
bspw. der Atmung. Außerdem befinden sich in der Medulla oblongata die Zentren für Re-
flexauslösungen, verursacht durch Schlucken oder auch Husten. Zudem wird hier in ei-
nem Zentrum der Schlaf- und Wachrythmus reguliert.106
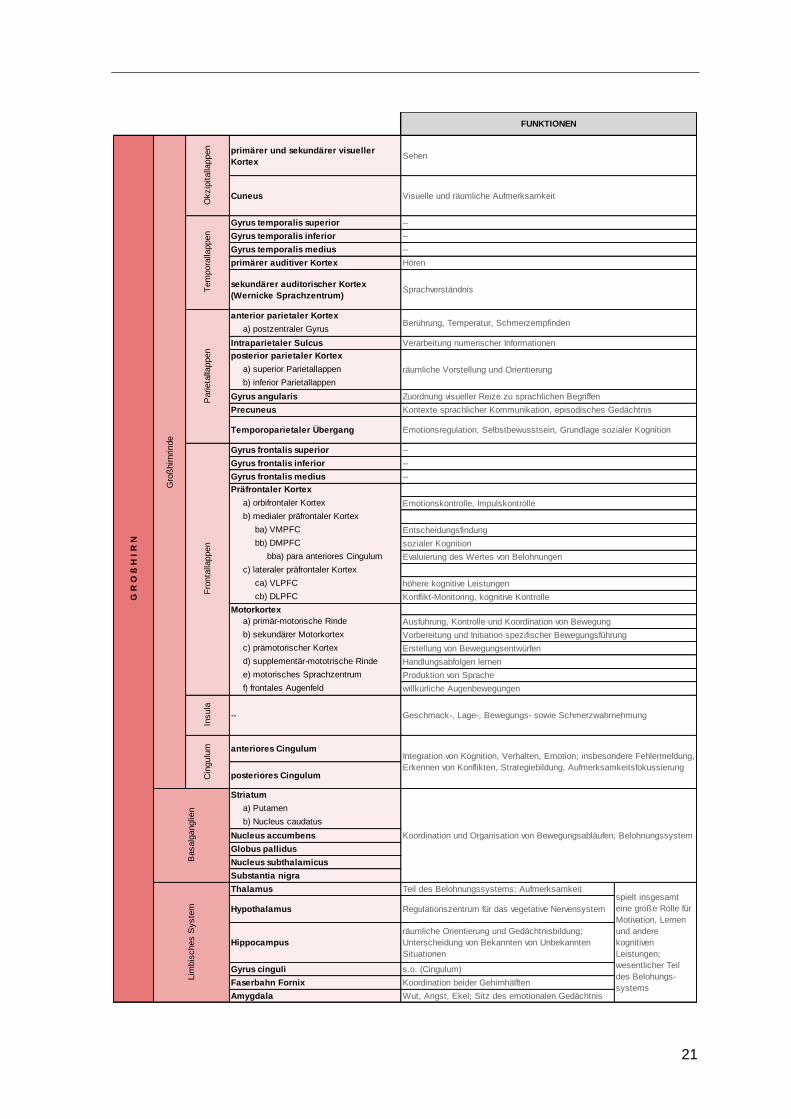
2.5 Tabellarischer Überblick
Abschließend zu diesem Kapitel soll folgende Tabelle die Einteilung des menschlichen
Gehirns sowie deren wesentliche Funktionen nochmals zusammenfassen.
99
Vgl. Trepel, M. (2011): . Struktur und Funktion, München, S.137. 100
Vgl. Thompson, R.F. (2001): Das Gehirn - von der Nervenzelle zur Verhaltenssteuerung, Heidelberg, S.15 101
Vgl. Derouiche, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen - Methoden - Anwendungen, Wiesbaden, S. 24.
102 Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 115.
103 Vgl. Weis, S./Thaller, R./Villringer, A. (1992): Das Gehirn des Menschen, Göttingen, S. 27.
104 Vgl. CNS Forum Lundbeck Institute: Gehirnatlas, http://de.brainexplorer.org/brain_atlas/Brainatlas_in-
dex.shtml, zuletzt geprüft am 6.November 2013. 105
Vgl. Trepel, M. (2011): Neuroanatomie. Struktur und Funktion, München, S. 115. 106
Vgl. Schandry, R. (2003), Biologische Psychologie, Weinheim, S. 109-110.
21
primärer und sekundärer visueller
Kortex
Cuneus
Gyrus temporalis superior
Gyrus temporalis inferior
Gyrus temporalis medius
primärer auditiver Kortex
sekundärer auditorischer Kortex
(Wernicke Sprachzentrum)
anterior parietaler Kortex
a) postzentraler Gyrus
Intraparietaler Sulcus
posterior parietaler Kortex
a) superior Parietallappen
b) inferior Parietallappen
Gyrus angularis
Precuneus
Temporoparietaler Übergang
Gyrus frontalis superior
Gyrus frontalis inferior
Gyrus frontalis medius
Präfrontaler Kortex
a) orbifrontaler Kortex
b) medialer präfrontaler Kortex
ba) VMPFC
bb) DMPFC
bba) para anteriores Cingulum
c) lateraler präfrontaler Kortex
ca) VLPFC
cb) DLPFC
Motorkortex
a) primär-motorische Rinde
b) sekundärer Motorkortex
c) prämotorischer Kortex
d) supplementär-mototrische Rinde
e) motorisches Sprachzentrum
f) frontales Augenfeld
Insula
--
anteriores Cingulum
posteriores Cingulum
Striatum
a) Putamen
b) Nucleus caudatus
Nucleus accumbens
Globus pallidus
Nucleus subthalamicus
Substantia nigra
Thalamus Teil des Belohnungssystems; Aufmerksamkeit
Hypothalamus Regulationszentrum für das vegetative Nervensystem
Hippocampus
räumliche Orientierung und Gedächtnisbildung;
Unterscheidung von Bekannten von Unbekannten
Situationen
Gyrus cinguli s.o. (Cingulum)
Faserbahn Fornix Koordination beider Gehirnhälften
Amygdala Wut, Angst, Ekel; Sitz des emotionalen Gedächtnis
FUNKTIONENG
R O
ß H
I R
N
Gro
ßhirnrinde
Okzip
italla
ppen
Sehen
Visuelle und räumliche AufmerksamkeitT
em
pora
llappen
--
--
--
Hören
Sprachverständnis
Parieta
llappen
Berührung, Temperatur, Schmerzempfinden
Verarbeitung numerischer Informationen
räumliche Vorstellung und Orientierung
Zuordnung visueller Reize zu sprachlichen Begriffen
Kontexte sprachlicher Kommunikation, episodisches Gedächtnis
Emotionsregulation, Selbstbewusstsein, Grundlage sozialer Kognition
Fro
nta
llappen
--
--
--
Erstellung von Bewegungsentwürfen
Handlungsabfolgen lernen
Produktion von Sprache
willkürliche Augenbewegungen
Emotionskontrolle, Impulskontrolle
Entscheidungsfindung
sozialer Kognition
Evaluierung des Wertes von Belohnungen
höhere kognitive Leistungen
Konflikt-Monitoring, kognitive Kontrolle
Ausführung, Kontrolle und Koordination von Bewegung
Vorbereitung und Initiation spezifischer Bewegungsführung
Geschmack-, Lage-, Bewegungs- sowie Schmerzwahrnehmung
Cin
gulu
m
Integration von Kognition, Verhalten, Emotion; insbesondere Fehlermeldung,
Erkennen von Konflikten, Strategiebildung, Aufmerksamkeitsfokussierung
Basalg
anglie
n
Koordination und Organisation von Bewegungsabläufen; Belohnungssystem
Lim
bis
ches S
yste
m
spielt insgesamt
eine große Rolle für
Motivation, Lernen
und andere
kognitiven
Leistungen;
wesentlicher Teil
des Belohungs-
systems

22
KL
EIN
HIR
NM
ITT
EL
HIR
NM
ED
UL
LA
OB
LO
NG
AT
AP
ON
SFUNKTIONEN
Koordination und Feinabstimmung von Bewegungsabläufen; kognitive
Funktion; Sprache
Bewegungsregulation, Körperhaltung, Kontrolle der Augenfunktion und
Umschaltungen von auditiven Reizen
vegetative Steuerung (Atmung); Zentrum für Reflexauslösung (Schlucken,
Husten); Regulierung von Wach- und Schlafrythmus
Steuerung von Schlaf- und Wachrythmus; Weiterleitung vom Kortex
ausgehender Impulse zur Steuerung von Beweungsabläufen
ZW
ISC
HE
NH
IRN
Thalamus Übertragung von Sinneswahrnehmungen an den Kortex
Hypothalamus u.a. Koordination von Atmung, Kreislauf, Nahrungsaufnahme
Tabelle 1: Gehirnregionen und deren Funktionen im Überblick
Quelle: eigene Darstellung

23
3 Methoden der Neurowissenschaften
Dieses Teilkapitel vermittelt Grundlegendes zu den bekanntesten neurowissenschaftli-
chen Methoden. Es wird dabei streng genommen zwischen elektrophysiologischen einer-
seits und bildgebenden Verfahren andererseits unterschieden.107 Die Gemeinsamkeit aller
Verfahren ist der Versuch, auf unterschiedliche Weise, die Aktivität des Gehirns bei der
Durchführung bestimmter kognitiver Aufgaben darzustellen. Durch die Darstellung der
Gehirnaktivität können Rückschlüsse auf die Funktionsweise des Gehirns gezogen wer-
den.108 Mit Hilfe mathematischer Modelle gelang es zunehmend, elektrophysiologische
Verfahren weiterzuentwickeln, wodurch sie heutzutage auch zu den bildgebenden Verfah-
ren gezählt werden können.109 Um dennoch den Unterschied zwischen elektrophysiologi-
schen und bildgebenden Verfahren zu wahren, erfolgt im Folgenden eine Einteilung in
bildgebende Verfahren i.w.S. sowie in bildgebende Verfahren i.e.S. Zudem werden weite-
re Methoden vorgestellt, die sich bis dato noch in der Entwicklung befinden.
Neben der Erörterung unterschiedlicher neurowissenschaftlicher Methoden werden auch
deren Vor- und Nachteile erörtert. Abschließend soll ein tabellarischer Überblick die we-
sentlichen Inhalte zur besseren Übersicht zusammenfassen.
3.1 Bildgebende Verfahren i.w.S.
Zu den bildgebenden Verfahren i.w.S. werden elektrophysiologische Methoden, denen
u.a. die Elektroenzephalographie sowie die Magnetenzephalographie angehören, gezählt.
Mit elektrophysiologischen Verfahren werden neurophysiologischen Vorgänge visualisiert,
wobei dabei aber keine Informationen über anatomischen Strukturen, die den neurophy-
siologischen Vorgängen zugrunde liegen, gewonnen werden können.110 Wie bereits er-
wähnt wurden diese Methoden unter Zuhilfenahme mathematischer Modelle enorm wei-
terentwickelt, wodurch heutzutage auch - wenn auch nicht sehr exakte - anatomische Zu-
ordnungen möglich sind. Daher können die EEG sowie die MEG auch im Sinne eines
bildgebenden Verfahrens eingesetzt werden.111 Aus diesem Grund werden sie in dieser
Arbeit zu den bildgebenden Verfahren i.w.S. gezählt.
107
Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-
schaften, Stuttgart, S. 11 bzw. S. 152. 108
Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 44.
109 Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie.
Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 44; in diesem Sinne auch Jäncke, L. (2005): Me-thoden der Bildgebung in der Psychologie und den kognitiven Neurowissenschaften, Stuttgart, S. 152.
110 Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-
schaften, Stuttgart, S. 11 bzw. S. 152. 111
Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-schaften, Stuttgart, S. 152.

24
Im Folgenden werden die EEG, die MEG sowie die iEEG als bildgebende Verfahren i.w.S.
vorgestellt.
3.1.1 Elektroenzephalographie (EEG)
Durch gezielt, nach einem internationalen Standard, am Kopf angebrachte Elektroden
werden mittels EEG elektrische Aktivitäten in äußeren Hirnregionen gemessen. Die
messbaren elektrischen Aktivitäten entstehen dabei durch die Signalübertragung von
Neuronen, wodurch Neurotransmitter freigesetzt werden.112
Die Messung der elektrischen Aktivität erfolgt im Tausendstelsekundenbereich, womit
durch die EEG eine sehr hohe zeitliche Auflösung gewährleistet ist.113 Neben der hohen
zeitlichen Auflösung kann mittels EEG, im Gegensatz zu den klassischen bildgebenden
Verfahren wie fMRT und PET, sehr preiswert Forschung betrieben werden.114 Ein weiterer
Vorteil dieser Methode ist dahingehend zu sehen, dass die für die Messung benötigten
Elektroden sowie das restliche Equipment sehr leicht transportiert und somit an verschie-
densten Orten flexibel eingesetzt werden können.115 Außerdem misst die EEG direkt die
elektrische Aktivität, während andere (neurowissenschaftliche) Verfahren lediglich indirek-
te Indikatoren der Gehirnaktivität, wie bspw. die Durchblutung, messen.116
Der Nachteil dieser Methode liegt allerdings in ihrer schlechten räumlichen Auflösung, da
über die Elektroden nur Aktivitäten an der Gehirnoberfläche gemessen werden, während
tiefere Hirnregionen nicht lokalisiert werden können. 117 Die ungenaue Lokalisation der
erregten Gehirnareale kann allerdings, wie bereits erwähnt, unter Zuhilfenahme mathema-
tischer Modelle wieder etwas präzisiert werden.118
112
Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 37. 113
Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 45.
114 Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-
schaften, Stuttgart, S. 152; in diesem Sinne auch Dimoka, A. et al (2012): On the use of neurophysiolo-gical tools in IS research: developing a research agenda fpr NeuroIS, in: MIS Quarterly, Vol 36, Appen-dix 6.
115 Vgl. Kenning, P./ Plassmann, H. (2005): NeuroEconomics: An Overview from an Economic Perspective, in:
Brain Research Bulletin, Vol. 67, S. 343-354; so auch Camerer C./Loewenstein G./Prelec D. (2005): Neuroeconomics: How Neuroscience Can Inform Economics, in: Journal of Economic Literature, Vol XLIII, S.12.
116 Vgl. Dimoka, A. et al (2012): On the use of neurophysiological tools in IS research: developing a research
agenda fpr NeuroIS, in: MIS Quarterly, Vol 36, Appendix 6; so auch Camerer C./Loewenstein G./Prelec D. (2005): Neuroeconomics: How Neuroscience Can Inform Economics, in: Journal of Economic Litera-ture, Vol. XLIII, S.12.
117 Vgl. Ahlert, D./Kenning, P. (2006): Neuroökonomie, in: Zeitschrift für Management, S. 24-25.
118 Vgl. Roth, G. (2003): Fühlen, Denken, Handeln - Wie das Gehirn unser Verhalten steuert, Frankfurt am
Main, S. 124.

25
3.1.2 Magnetenzephalographie (MEG)
Eine Weiterentwicklung der EEG ist in der MEG zu sehen.119 Mittels MEG werden magne-
tische Schwankungen gemessen, die durch neuronale Aktivität erzeugt werden. Die Mes-
sung erfolgt dabei i.d.R. mit 100 bis 300 Sensoren, die in einem helmartigen Ganzkopf-
MEG-System integriert sind.120 Da die messbaren Magnetfeldschwankungen sehr
schwach ausgeprägt sind, müssen diese mithilfe heliumgekühlter Sensoren - sogenann-
ten SQUIDS - gemessen werden.121
Genauso wie die EEG verfügt die MEG über eine sehr hohe zeitliche Auflösung. Außer-
dem wird mittels MEG, ebenso wie bei der EEG die neuronale Aktivität auf direktem Wege
gemessen.122 Die räumliche Auflösung ist bei der MEG, im Vergleich zur EEG, deutlich
besser, weil die Magnetfelder nicht von der Leitfähigkeit verschiedener Strukturen im Ge-
hirn beeinflusst werden, wodurch auch eine bessere bzw. präzisere Lokalisierung der
(auch tieferen) Gehirnregionen möglich ist.123
Aufgrund der Messung von magnetischen Schwankungen ist als Nachteil der MEG die
leichte Störanfälligkeit durch verschiedene Störquellen anzusehen, die möglicherweise die
Resultate verfälschen. Um diesen Nachteil abzuwenden, bedarf es stets eines hohen Ab-
schirmungsaufwandes, der mit erheblichen Kosten verbunden ist, wodurch diese Methode
im Vergleich zur EEG wesentlich kostspieliger ist. Die Vorbereitungszeit einer Messung ist
allerdings geringer als bei der EEG, weil keine einzelnen Elektroden am Kopf angebracht
werden müssen.124
3.1.3 Intrakranielle Elektroenzephalographie (iEEG)
Durch die iEEG können einzelnen Nervenzellen und deren Aktionspotentiale abgeleitet
werden125, indem (hauptsächlich bei Tieren) implantierte Elektroden auf die Oberfläche
des Gehirns, oder sogenannte Stabelektronen in tiefere Hirnregionen eingesetzt wer-
den.126 Mithilfe dieser Methode kann nicht nur festgestellt werden, welche Hirnregionen
119
Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 38.
120 Vgl. Büchel, C./Karnath, H.-O/Thier, P. (2012): Methoden der kognitiven Neurowissenschaften, in: Karnath,
H.-O./Thier, P. (Hrsg.): Kognitive Neurowissenschaften, Berlin Heidelberg, S. 25. 121
Vgl. Hermey, G./Mahlke, C./Schwake, M./Sommer, T. (2010): Der Experimentator: Neurowissenschaften, Heidelberg, S. 239.
122 Vgl. Hüsing, B./Jäncke, L./Tag, B. (2006): Impact Assessment of Neuroimaging, Zürich, S. 5.
123 Vgl. Hermey, G./Mahlke, C./Schwake, M./Sommer, T. (2010): Der Experimentator: Neurowissenschaften,
Heidelberg, S. 239. 124
Vgl. Büchel, C./Karnath, H.-O/Thier, P. (2012): Methoden der kognitiven Neurowissenschaften, in: Karnath, H.-O./Thier, P. (Hrsg.): Kognitive Neurowissenschaften, Berlin Heidelberg, S. 25.
125 Vgl. Sog. „Single Neuron Measurement“.
126 Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie.
Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 46.

26
aktiviert sind, sondern es können auch aufschlussreiche Informationen über die Frage
gewonnen werden, warum gewisse Neuronen unter gewissen Umständen erregt oder
gelähmt werden.127
Bei Menschen werden implantierte Elektroden sehr selten und nur aus medizinischen
Gründen für die Ursachendiagnostik eingesetzt. Zudem ist die iEEG aufgrund der Kom-
plexität der Datenauswertung und aufgrund der Notwendigkeit eines chirurgischen Eingrif-
fes (noch) nicht sehr verbreitet.128
3.2 Bildgebende Verfahren i.e.S.
Unter bildgebende Verfahren i.e.S. sind jene neurowissenschaftlichen Methoden zu ver-
stehen, die es ermöglichen, anatomische Strukturen des menschlichen Körpers so präzi-
se wie möglich sichtbar zu machen. Zu den bekanntesten bildgebenden Verfahren i.e.S.
zählen die Positronenemissionstomographie sowie die funktionelle Magnetresonanztomo-
graphie, die beide im Folgenden vorgestellt werden. Weitere bildgebende Verfahren i.e.S.,
auf die in dieser Arbeit nicht näher eingegangen wird, sind etwa die Sonographie, die
Computertomographie sowie die Szintigraphie.129
3.2.1 Positronenemissionstomographie (PET)
Die PET zählt neben der EEG zu den älteren Methoden der Neurowissenschaften. Im
Fokus dieser Methode steht die Messung des Blutflusses (oder des Glukoseverbrau-
ches130), der Aufschluss über die neuronale Aktivität im Gehirn gibt.131 Die Messung mit-
tels PET erfolgt, indem dem/der Probanden/in schwach radioaktive Stoffe intravenös ver-
abreicht werden. Beim Zerfall dieser radioaktiven Stoffe werden Positronen freigesetzt,
deren räumliche Verteilung durch ringförmig um den/die Probanden/in angelegte, Detekto-
ren gemessen werden kann.132 Jene Hirnregionen, die einen überdurchschnittlichen
Stoffwechsel aufweisen, werden durch die verabreichte radioaktive Substanz sichtbar und
können lokalisiert werden.133
127
Vgl. Camerer/Loewenstein G./Prelec D. (2005): Neuroeconomics: How Neuroscience Can Inform Econo-mics, in: Journal of Economic Literature, Vol XLIII, S.15.
128 Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie.
Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 46. 129
Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-schaften, Stuttgart, S. 11.
130 Vgl. Braus, D.F. (2004): EinBlicke ins Gehirn. Moderne Bildgebung in der Psychiatrie, Stuttgart, S. 55.
131 Vgl. Camerer/Loewenstein G./Prelec D. (2005): Neuroeconomics: How Neuroscience Can Inform Econo-
mics, in: Journal of Economic Literature, Vol XLIII, S.12. 132
Vgl. Raab, G./Gernsheimer, O./Schindler, M. (2009): Neuromarketing. Grundlagen-Erkenntnisse-Anwen-dungen, Wiesbaden, S. 184.
133 Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 39.

27
Der Vorteil des PET Verfahrens ist, im Gegensatz zur EEG und MEG, in der besseren
räumlichen Auflösung zu sehen.134 Unter Zuhilfenahme eines Computers ist es möglich,
ein dreidimensionales Bild des Ortes, der Zeit und der Quantität der radioaktiven Zerfälle
zu errechnen und neuronale Aktivitäten zu lokalisieren. Die räumliche Auflösung von PET-
Bildern beträgt, je nach Größe und Anzahl der Detektoren, 3-6 mm.135 Zudem besteht mit
dieser Methode auch die Möglichkeit, tiefere Hirnregionen wie bspw. das Kleinhirn zu
messen bzw. zu lokalisieren.136
Die zeitliche Auflösung des PET-Verfahrens ist jedoch im Vergleich zur EEG bzw. MEG
schlechter.137 Neuronale Aktivitäten werden mittels PET nicht auf direktem Wege, sondern
indirekt über den Blutfluss gemessen.138 Außerdem ist diese neurowissenschaftliche Me-
thode, durch die Verabreichung von - wenn auch nur geringen Mengen - radioaktiven
Substanzen nicht unumstritten, und aus diesem Grund auch auf wenige Versuchsdurch-
läufe begrenzt.139 Zudem muss die zu verabreichende radioaktive Substanz aufgrund der
geringen Halbwertszeit direkt vor Ort mit einem Teilchenbeschleuniger hergestellt werden,
womit diese Methode nicht nur mit einem enormen technischen Aufwand verbunden ist,
sondern zugleich zu den kostspieligsten Verfahren der Neurowissenschaften zählt.140
3.2.2 Funktionelle Magnetresonanztomographie (fMRT)
Die fMRT ist das neueste und zugleich populärste bildgebende Verfahren, das auf zwei
grundlegenden biologischen Phänomenen basiert. Einerseits liegt die Besonderheit des
Hirnstoffwechsels darin, dass in neuronal aktivierten Hirnregionen mehr sauerstoffreiches
Blut transportiert als verbraucht wird. Andererseits wandelt sich der eisenhaltige rote Blut-
farbstoff Hämoglobin nach der Sauerstoffbindung in Oxyhämoglobin um, und weist da-
durch eine andere magnetische Eigenschaft auf. Durch die unterschiedlichen magneti-
schen Eigenschaften von sauerstoffarmen und sauerstoffreichem Blut, die als BOLD-
134
Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-schaften, Stuttgart, S. 136.
135 Vgl. Hermey, G./Mahlke, C./Schwake, M./Sommer, T. (2010): Der Experimentator: Neurowissenschaften,
Heidelberg, S. 229-230. 136
Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-schaften, Stuttgart, S. 136.
137 Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 40. 138
Vgl. Hüsing, B./Jäncke, L./Tag, B. (2006): Impact Assessment of Neuroimaging, Zürich, S. 5. 139
Vgl. Kenning, P./Plassmann, H./Ahlert, D. (2007): Applications of functional magnetic resonance imaging for market research, in: Qualitative Market Research: An International Journal, Vol. 10, Nr. 2, S. 139.
140 Vgl. Birbaumer, N./Schmidt, R. F. (2006): Biologische Psychologie, Heidelberg, S. 486; Braus, D.F. (2004):
EinBlicke ins Gehirn. Moderne Bildgebung in der Psychiatrie, Stuttgart, S. 52.

28
Effekt141 bezeichnet werden, können neuronale Aktivitäten mittels MRT-Scanners gemes-
sen bzw. lokalisiert werden.142
Die räumliche Auflösung der fMRT ist mit ungefähr drei mm3 Voxel sehr präzise und somit
noch besser als die räumliche Auflösung der PET. Auch die zeitliche Auflösung ist im
Vergleich zur PET mit ungefähr zwei Sekunden besser, allerdings schlechter als jene der
EEG sowie der MEG.143 Im Gegensatz zu statischen Röntgenaufnahmen kann mittels
fMRT das arbeitende Gehirn nahezu in Echtzeit dargestellt werden. Es kommt lediglich zu
einer Verzögerung im Ausmaß von 1 bis 6 Sekunden, da die Veränderung der Durchblu-
tung in etwa diese Zeit beansprucht. Hinsichtlich der Auflösung gilt, je stärker der Magnet
ist, desto besser ist die Auflösung144, wobei damit gleichzeitig die Kosten für den To-
mographen ansteigen.145 Nicht nur hinsichtlich der räumlichen und zeitlichen Auflösung ist
die fMRT der PET überlegen, auch die Tatsache, dass die fMRT nach dem derzeitigen
Stand der Wissenschaft ohne jegliche Nebenwirkungen für Versuchspersonen durchge-
führt werden kann, ist als entscheidender Vorteil dieses Verfahrens festzuhalten. So sind
problemlos Versuchswiederholungen und Untersuchungen von Kindern möglich, die Auf-
schluss über die Entwicklung der Gehirnfunktionen ermöglichen sollen.146
Mittels fMRT werden Veränderungen im Blut, sohin keine direkten neuronalen Aktivitäten
gemessen.147 Das BOLD-Signal ist somit nur ein indirektes Maß für neuronale Aktivität.148
Zudem sind die hohen Anschaffungskosten, die sich auf ca. EUR 1-2 Mio. belaufen, sowie
die Betriebs- (EUR 100.000 bis 200.000 jährlich) und Nutzungskosten je Hirnscanner-
Stunde (EUR 750-1.200) ein wesentlicher Nachteil des Verfahrens mittels fMRT.149 Auf-
grund der Schwäche der Signale sind pro Versuchsperson 30-40 Durchgänge erforder-
lich, um statistisch verlässliche Ergebnisse zu erhalten. Außerdem werden für eine Unter-
141
Blood oxygen level dependent. 142
Vgl. Hermey, G./Mahlke, C./Schwake, M./Sommer, T. (2010): Der Experimentator: Neurowissenschaften,
Heidelberg, S. 240. 143
Vgl. Camerer, C. (2007): Neuroeconomics: Using Neuroscience to make Economic Predictions, in: The Economic Journal, Vol. 117, C30.
144 aA Braus, der annimmt, dass stärkere Geräte nicht automatisch bessere Ergebnisse liefern. Vielmehr ist
die zu präferierende Magnetfeldstärke von der jeweiligen Aufgabenstellung sowie von der Qualität der Datenerhebung, insbesondere von den Kenntnissen der auswertenden Personen abhängig (Vgl. Braus, D.F. (2004): EinBlicke ins Gehirn. Moderne Bildgebung in der Psychiatrie, Stuttgart, S. 58).
145 Vgl. Thompson, R.F. (2001): Das Gehirn - von der Nervenzelle zur Verhaltenssteuerung, Heidelberg, S.
426. 146
Vgl. Hermey, G./Mahlke, C./Schwake, M./Sommer, T. (2010): Der Experimentator: Neurowissenschaften, Heidelberg, S. 239.
147 Vgl. Häusel, H.-G. (2008): Methoden der Neuromarketing-Forschung, in: Häusel, H.G. (Hrsg.): Neuromar-
keting. Erkenntnisse der Hirnforschung für Markenführung, Werbung und Verkauf, Planegg/München, S. 213.
148 Vgl. Hermey, G./Mahlke, C./Schwake, M./Sommer, T. (2010): Der Experimentator: Neurowissenschaften,
Heidelberg, S. 241. 149
Vgl. Häusel, G. (2006): Direkt ins Hirn?, in: absatzwirtschaft - Zeitschrift für Marketing, S. 38; Hüsing, B./Jäncke, L./Tag, B. (2006): Impact Assessment of Neuroimaging, Zürich, S. 65.

29
suchung idealerweise zumindest 20 ProbandInnen benötigt, wodurch sich aufsummiert
die Untersuchungskosten samt Vorbereitungen und Datenauswertung auf rund EUR
30.000 belaufen.150 Neben dem Kostenfaktor ist ein Nachteil dieser Methode dahingehend
zu sehen, dass nicht geklärt werden kann, ob der erhöhte Sauerstoffgehalt im Blut auf
aktivierte oder gehemmte Nervenzellen zurückzuführen ist.151 Weitere Nachteile einer
fMRT Untersuchung sind in der Einschränkung auf einfache Untersuchungsdesigns sowie
in der äußerst komplexen Datenauswertung zu sehen.152
3.3 Weitere Methoden
Neben den bekanntesten und am weitesten verbreiteten bildgebenden Verfahren, wozu
v.a. die EEG, die MEG, sowie die PET und die fMRT zählen, existieren auch andere neu-
rowissenschaftliche Methoden, die es ermöglichen sollen, die biologischen Grundlagen
menschlichen Verhaltens besser zu verstehen.153 Zu diesen Methoden zählen u.a. die
transkranielle Magnetstimulation sowie die Diffusiontensor-Bildgebung, die im Folgenden
in aller Kürze vorgestellt werden.
3.3.1 Transkranielle Magnetstimulation (TMS)
Mit Hilfe der transkraniellen Magnetstimulation können Gehirnregionen entweder stimuliert
oder gehemmt werden.154 Dies erfolgt, indem der unter der Kopfhaut angebrachten Mag-
netspule liegende, Kortex mit wechselnden Magnetfeldern mit niedrigen oder hohen Fre-
quenzen stimuliert wird.155 Die Erfahrung hat gezeigt, dass bei Frequenzen unter einem
Hertz die darunter liegenden Hirnregionen gehemmt werden, während bei Frequenzen
über fünf Hertz regelmäßig von einer Stimulierung der Hirnareale auszugehen ist.156 Der
genaue Wirkungsmechanismus ist allerdings bis dato noch nicht erforscht. Der Zweck
dieser Methode liegt v.a. darin, krankheitsbedingte Ausfälle von Gehirnregionen nachzu-
ahmen, damit in weiterer Folge die Notwendigkeit dieser Regionen in Bezug auf ihre kog-
nitiven Funktionen untersucht werden kann.
150
Vgl. Häusel, H.-G. (2006): Direkt ins Hirn?, in: absatzwirtschaft - Zeitschrift für Marketing, S. 38. 151
Vgl. Schubert, S. (2004): Dem Kunden ins Gehirn geschaut, in: Psychologie heute, S.38-41. 152
Vgl. Häusel, H.-G. (2008): Methoden der Neuromarketing-Forschung, in: Häusel, H.G. (Hrsg.): Neuromar-
keting. Erkenntnisse der Hirnforschung für Markenführung, Werbung und Verkauf, Planegg/München, S. 215-218 mwH.
153 Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie.
Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 51. 154
Vgl. Camerer, C.(2007): Neuroeconomics: Using Neuroscience to make Economic Predictions, in: The Economic Journal, Vol. 117, C30.
155 Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie.
Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 52-53. 156
Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-schaften, Stuttgart, S. 203.

30
Allerdings können mittels TMS nur oberflächlich liegende Gehirnregionen untersucht wer-
den, da die Eindringungstiefe des Magnetfeldes effektiv nur wenige Zentimeter beträgt.157
Bei epileptischen PatientInnen soll TMS vermieden werden, da diese Methode Krampfan-
fälle hervorrufen kann.158
3.3.2 Diffusionstensor-Bildgebung (DTI)
Eine Weiterentwicklung der MRT ist in der Diffusionstensor-Bildgebung zu sehen, mit der
Einblicke in feinstrukturelle Auffälligkeiten neuronaler Netzwerkverbindungen gewonnen
werden können.159 Vereinfacht ausgedrückt werden mittels DTI Informationen über die
Lage der neuronalen Bahnen in der weißen Gehirnsubstanz gewonnen. Konkret werden
dabei Diffusionsbewegungen von Wassermolekülen in Flüssigkeiten gemessen, wobei die
Diffusion der Moleküle mit diesem Verfahren in allen Richtungen erfasst werden kann.160
Nach mehreren Durchgängen kann für jedes Voxel festgestellt werden, wie stark sich die
Zellflüssigkeit in die verschiedenen untersuchten Richtungen bewegt, wobei diese Werte
als dreidimensionaler Tensor ausgedrückt werden. Daraus kann die durchschnittliche Dif-
fusität, die in den Hohlräumen des Hirns hoch ist, weil die Wassermoleküle dort selten auf
Barrieren stoßen, abgeleitet werden. 161
Die Methode der DTI ermöglicht es, anatomische Faserverbindungen in der weißen Sub-
stanz zu visualisieren, wodurch auch Rückschlüsse auf den Verlauf neuronaler Entwick-
lungen im Gehirn gezogen werden können.162 Zudem verhilft die Methode der DTI dazu,
Verknüpfungen zwischen den einzelnen Gehirnregionen aufzuklären und neurologische
Erkrankungen besser zu diagnostizieren, da der Aufbau sowie die Dichte dieser Faser-
bahnen für etliche Leistungen unseres Gehirns essentiell sind.
Nachteile dieser Methode sind zum einen die hohe Störanfälligkeit, bspw. bei minimaler
Bewegung der Versuchspersonen, sowie zum anderen die relativ grobe Auflösung mit
157
Vgl. Weber, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.): Neuroökonomie. Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 52-53.
158 Vgl. Jäncke, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven Neurowissen-
schaften, Stuttgart, S. 205. 159
Vgl. Braus, D.F./Tost, H./Hirsch, J.G./Gass, A. (2001): Diffusions-Tensor-Bildgebung (DTI) und funktionelle Magnetresonanztomographie (fMRT) erweitern das Methodenspektrum in der psychiatrischen For-schung, in: Der Nervenarzt, Vol. 72, Nr. 5, S. 385.
160 Vgl. Hermey, G./Mahlke, C./Schwake, M./Sommer, T. (2010): Der Experimentator: Neurowissenschaften,
Heidelberg, S. 223-224. 161
Vgl. Goebel, R./Zimmermann, J. (2011): Pfadfinder durch die weiße Substanz, in: Gehirn und Geist, Nr. 6, S. 64-65.
162 Vgl. Büchel, C./Karnath, H.-O/Thier, P. (2012): Methoden der kognitiven Neurowissenschaften, in: Karnath,
H.-O./Thier, P. (Hrsg.): Kognitive Neurowissenschaften, Berlin Heidelberg, S. 21; Braus, D.F. (2004): EinBlicke ins Gehirn. Moderne Bildgebung in der Psychiatrie, Stuttgart, S. 61.

31
einer Voxelgröße von mehreren mm3, wodurch feinere Fasern nicht aufgelöst werden
können.163
3.4 Kritische Würdigung
Wie aus diesem Kapitel hervorgeht, sind für Aspekte der Führungsforschung vor allem die
fMRT sowie die EEG als neurowissenschaftliche Methoden von Relevanz. Neben diesen
zwei, wohl populärsten, Methoden stellt die Neurowissenschaft mit der PET, der MEG, der
iEEG, der TMS und der DTI weitere Verfahren zur Verfügung, die für die Klärung von füh-
rungsrelevanten Fragestellungen herangezogen werden können.
Jede einzelne Methode weist unterschiedliche Vor- und Nachteile auf.164 Abhängig von
der jeweiligen Forschungsfrage ist die eine oder andere Methode zu präferieren. Werden
z.B. nur oberflächliche Gehirnregionen untersucht, und kann von einer exakten Lokalisati-
on der einzelnen Areale abgesehen werden, ist wohl die EEG im Hinblick auf Kosten, ört-
liche Flexibilität, Vorbereitungszeit und Datenauswertung die am besten geeignetste Me-
thode. Wird hingegen das Ziel verfolgt, eine möglichst genaue Lokalisierung der erregten
Gehirnregionen zu erreichen, ist die Messung (indirekter) neuronaler Aktivität mittels
fMRT erstrebenswert. Allerdings muss bedacht werden, dass bei dieser Methode hohe
Kosten anfallen und die Datenauswertung aufgrund der Komplexität Fachpersonal erfor-
dert.165 Neben dem gewünschten Resultat können auch verschiedene Studiensdesigns,
die Anzahl der ProbandInnen und ähnliches für die Auswahl der neurowissenschaftlichen
Methode ausschlaggebend sein.
Zusammenfassend soll folgende Tabelle einen Überblick über die populärsten Methoden
der Neurowissenschaften und deren Vor- und Nachteile geben. Weil jede einzelne Me-
thode, wie bereits erwähnt, sowohl Vor-, als auch Nachteile aufweist, wird in der Literatur
vielfach die Ansicht vertreten, verschiedene Methoden zu kombinieren.166
163
Vgl. Goebel, R./Zimmermann, J. (2011): Pfadfinder durch die weiße Substanz, in: Gehirn und Geist, Nr. 6,
S. 64-67. 164
Vgl dazu Tabelle 2. 165
Siehe dazu Kapitel B.2. 166
Vgl. Raab, G./Gernsheimer, O./Schindler, M. (2009): Neuromarketing. Grundlagen-Erkenntnisse-Anwen-dungen, Wiesbaden, S. 192; so auch Goebel, R./Zimmermann, J. (2011): Pfadfinder durch die weiße Substanz, in: Gehirn und Geist, Nr. 6, S. 67.

32
Methode Vorgehensweise
+ zeitliche Auflösung
+ Messung direkter neuronaler Aktivität
+ geringe Kosten
+ örtliche Flexibilität
~ mäßige Vorbereitungszeit
- räumliche Auflösung
-tiefere Hirnregionen können nicht
lokalisiert werden
+ zeitliche Auflösung
+ Messung direkter neuronaler Aktivität
+ geringe Vorbereitungszeit
~Lokalisation tieferer Hirnregionen
möglich
- örtliche Flexibilität
~ räumliche Auflösung
- Störanfälligkeit
~ relativ hohe Kosten
~/+ räumliche Auflösung
+Lokalisation tieferer Hirnregionen
möglich
-Messung indirekter neuronaler
Aktivität
- hohe Vorbereitungszeit
- örtlich unflexibel
- zeitliche Auflösung
-Verabreichung einer radioaktiven
Substanz
- hohe Kosten
- komplexe Datenauswertung
+Lokalisation tieferer Hirnregionen
möglich
+ räumliche Auflösung
~ mäßige Vorbereitungszeit
~ zeitliche Auflösung
- örtlich unflexibel
-Messung indirekter neuronaler
Aktivität
- hohe Kosten
- komplexe Datenauswertung
Vor- und Nachteile
fMRTMessung der Stoffwechselaktivität
des Gehirns mittels fMRT-Scanner
Bil
dg
eb
en
de V
erf
ah
ren
i.w
.S.
PET
Messung des Blutflusses mittels
Detektoren nach Verabreichung einer
radioaktiven Substanz
Bil
dg
eb
en
de V
erf
ah
ren
i.e
.S.
Messung elektrischer Aktivität mit
am Kopf angebrachten ElektrodenEEG
MEGMessung magnetischer
Schwankungen mittels Sensoren
Tabelle 2: Übersicht der Methoden der Neurowissenschaften
In Anlehnung an: Raab, G./Gernsheimer, O./Schindler, M. (2009): Neuromarketing. Grundlagen - Erkenntnis-
se - Anwendungen, Wiesbaden, S. 193; Kenning, P./Plassmann, H./Ahlert, D. (2007): Applications of functio-
nal magnetic resonance imaging for market research, in: Qualitative Market Research: An International Jour-
nal, Vol. 10, Nr. 2, S.138.

33
4 (Neuro-)Leadership
Dieses Teilkapitel beschäftigt sich zunächst mit der grundlegenden Bedeutung des Begrif-
fes „Leadership“, einer Teildisziplin der Betriebswirtschaftslehre. In diesem Zusammen-
hang wird auf die wesentlichsten Führungsstile eingegangen, die sich im Zeitverlauf stetig
veränderten bzw. weiterentwickelten, ehe im darauffolgenden Kapitel eine Verbindung
zwischen Leadership im betriebswirtschaftlichen Sinn und den Neurowissenschaften her-
gestellt wird. Im Zusammenhang mit Neuroleadership werden vier Forschungsansätze
einer gehirngerechten Führung näher vorgestellt. Abschließend zu diesem Kapitel werden
jene Bereiche aufgezeigt, in denen die Neurowissenschaft zur Führungsforschung beitra-
gen kann.
4.1 Leadership - grundlegende Bedeutung
John Kotter hat Ender der 1980er Jahre im Zusammenhang mit seinen Arbeiten zu Ver-
änderungsprozessen in Unternehmen167 maßgeblich zur Entwicklung des Leadership-
Ansatzes beigetragen.168 Vor allem in den letzten Jahren nahm das Interesse an Lea-
dership immer mehr zu, was vor allem unzählige Publikationen rund um dieses Thema
untermauern.169
Die Suche nach einer einheitlichen Definition des Begriffes „Leadership“ - zu Deutsch
„Führung“ - gestaltet sich jedoch als äußerst schwierig. Vielmehr finden sich nahezu so
viele unterschiedliche Definitionen zu diesem Begriff wie es Menschen gab, die versuch-
ten, eine geeignete Begriffsbeschreibung zu finden.170 Die Problematik der Unmengen an
unterschiedlichen Definitionen zum Thema Führung ist wohl auf die unterschiedlichen
wissenschaftlichen Ansätze rund um den Begriff zurückzuführen.171
In dieser Arbeit wird unter Leadership ein Prozess172 verstanden, womit ein Individuum
eine Gruppe von Individuen beeinflusst, um ein bestimmtes Ziel zu erreichen.173
Leadership ist von Management zu unterscheiden, zumal Leadership weit über die Beein-
flussung von Mitarbeitern hinausgeht. Den Unterschied zwischen Leadership und Mana-
167
Vgl. dazu Kotter, J. (1991): Leadership lässt sich lernen, in: Harward Business manager, Jg. 13, Nr. 1, S.35-43; Kotter, J. (1988): The Leadership Factor, New York mwH.
168 Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 654; so auch Hegele-Raih, C. (2004):
Was ist Leadership?, in: Harvard Business Manager, http://www.harvardbusinessma-nager.de/heft/artikel/a-620896.html, zuletzt geprüft am 22. November 2013.
169 Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 654.
170 Vgl. Northouse, P.G. (2013): Leadership. Theory and Practice, California, S. 2.
171 Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 655.
172 Ein Prozess impliziert, dass der Leader seine „Followers“ beeinflusst, als auch umgekehrt.
173 Vgl. Northouse, P.G. (2013): Leadership. Theory and Practice, California, S. 5; so ähnlich auch z.B. Stähle,
W. (1999): Management, München, S. 328.

34
gement hat Peter Drucker wie folgt zusammengefasst: „Management is doing things right;
leadership is doing the right things.“ In diesem Sinne umfasst Leadership die Entwicklung
von Visionen und Strategien, die wiederum bei den MitarbeiterInnen einen Sinn stiften
und zur Identifikation mit gemeinsamen Zielen führen. Management hingegen ist vor allem
für die Umsetzung von Strategien sowie für die Lösung etwaig auftretender Probleme zu-
ständig. Während beim Management Planung und Kontrolle die dominierende Rolle ein-
nehmen, steht beim Leadership die Personalführung im Vordergrund.174
Die Frage, welche Komponenten für eine gute Führung ausschlaggebend sind, beschäf-
tigt die Betriebswirtschaftslehre bereits seit langer Zeit. Alle Versuche, eine allgemein gül-
tige Führungstheorie zu entwickeln sind bis dato fehlgeschlagen.175 Im Folgenden werden
die wesentlichsten Führungstheorien erläutert.
4.1.1 Führungstheorien
Führungstheorien sind theoretische Aussagensysteme, die erklären sollen, wie Führung
auf die Geführten wirkt und welche Faktoren für gute Führung ausschlaggebend sind.
Obwohl bis dato keine Führungstheorie entwickelt wurde, die Allgemeingültigkeit erlangte
(d.h. immer zum Erfolg führt), lassen sich, historisch betrachtet, gewisse Phasen in der
Entwicklung der einzelnen Führungstheorien identifizieren, die jeweils durch besondere
Merkmale gekennzeichnet sind.176 Im Sinne dieser Histologie werden im Folgenden chro-
nologisch die zum jeweiligen Zeitpunkt vorherrschenden Führungstheorien vorgestellt.
4.1.1.1 Eigenschaftstheorien
Die Eigenschaftstheorien dominierten in den ersten Jahrzenten des 20. Jhd. und sind da-
her zu den ältesten Erklärungsansätzen der Führung zu zählen.177 Nach diesen Theorien
ist der Führungserfolg primär von den persönlichen Eigenschaften der Führungsperson
abhängig. Dabei wurde angenommen, dass diese, für den Führungserfolg erforderlichen,
Eigenschaften angeboren sind oder bereits in einem sehr frühen Entwicklungsstadium
erworben wurden.178 Um die für den Erfolg ausschlaggebenden Eigenschaften zu identifi-
zieren, die eine Führungskraft gegenüber den Geführten auszeichnet, wurden zahlreiche
empirische Studien durchgeführt.179 Beispiele für Eigenschaften, die Führungskräften zu-
174
Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 655. 175
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 358. 176
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 358. 177
Vgl. Schaufelbühl, K./Hugentobler, W./Blattner, M. (Hrsg.) (2007): Betriebswirtschaftslehre für Bachelor, Zürich, S. 889; Jung, H.(2010): Allgemeine Betriebswirtschaftslehre, München, S. 216; Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 359.
178 Vgl. Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis,
Wiesbaden, S. 259. 179
Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 644.

35
geschrieben wurden sind einerseits physische Merkmale wie Größe und Gesundheit, an-
dererseits psychische Merkmale wie Intelligenz, Willensstärke, Entschlusskraft und
Selbstvertrauen.180
Die Eigenschaftstheorie ist aufgrund ihrer Inkonsistenz, Widersprüchlichkeit181 und Unein-
heitlichkeit der Ergebnisse als weitgehend gescheitert anzusehen, zumal Stogdill182 124
Studien zu diesem Thema auf ihre Gemeinsamkeiten empirisch analysierte und nur fünf
Eigenschaften identifizieren konnte, die in mehr als 15 Studien bei Führungskräften (im
Vergleich zu den Geführten), häufiger auftraten.183
4.1.1.1.1 Attributionstheorie
Obwohl die Eigenschaftstheorie heftig kritisiert wurde, wurden dennoch immer wieder
Versuche unternommen, den Fokus auf persönliche Merkmale zu legen. Einen solchen
Versuch stellt die eigenschaftsorientierte Attributionstheorie dar, die auf Calder zurück-
geht und in den 70er Jahren des 20. Jhd. zum ersten Mal erwähnt wurde. Gemäß dieser
Theorie sind nicht objektiv messbare Eigenschaften von Führungspersonen für den Füh-
rungserfolg entscheidend. Vielmehr ist es essentiell, dass die Geführten diese persönli-
chen Merkmale auch subjektiv wahrnehmen und der Führungskraft zuschreiben. Die we-
sentliche Stärke der eigenschaftsorientierten Attributionstheorie ist die Fokussierung auf
den/die Geführten. Allerdings blieb auch diese Theorie nicht ohne Kritik, zumal es nicht
gelang, konsistente Eigenschaftsmuster zu identifizieren, die einen wesentlichen Einfluss
auf den Führungserfolg haben.184
4.1.1.1.2 Charismatische Führung
In jüngerer Zeit wurden die eigenschaftstheoretischen Führungsansätze durch die erneute
Entdeckung der charismatischen Führung in gewisser Weise wiederbelebt. Bei diesem
Ansatz wird versucht zu ermitteln, welche persönlichen Merkmale und Verhaltensweisen
charismatische Führungskräfte, wie bspw. J.F. Kennedy, Steve Ballmer oder Jack Welch,
im Vergleich zu weniger charismatischen Führungskräften aufweisen. Diese Studien führ-
ten - wie der eigenschaftstheoretische Ansatz - auch nicht zum gewünschten Erfolg; d.h.
180
Vgl. Jung, H. (2010): Allgemeine Betriebswirtschaftslehre, München, S. 216; Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis, Wiesbaden, S. 259.
181 Widersprüche ergaben sich daraus, dass bestimmte Eigenschaften in manchen Studien vermehrt bei FÜh-
rungskräfte auftraten, in anderen Studien jedoch bei den Geführten (Vgl. Mann, R.D. (1959): A review of the relationships between personality and leadership and popularity, in: Psychological Bulletin, S. 241-270.
182 Vgl. Stogdill, R.M. (1948): Personal factors associated with leadership: A survey of the literature, in: Jour-
nal of Psychology, 25. Jg., S. 35-71. 183
Vgl. Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis, Wiesbaden, S. 259.
184 Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 3361,
363.

36
es konnten keine gemeinsamen persönlichen Merkmale der charismatischen Führungs-
kräfte identifiziert werden. Indessen wird Charisma Führungspersonen aufgrund eines
bestimmten Verhaltens attribuiert. 185
4.1.1.2 Führungsstiltheorien
Mit der Erkenntnis, dass die Qualität der Führung nicht nur anhand von persönlichen Ei-
genschaften, sondern auch anhand des Verhaltens der Führungsperson gegenüber den
MitarbeiterInnen beurteilt werden kann, sind in den 30er Jahren des 20. Jhd. die Füh-
rungsstiltheorien entstanden.186
Unter Führungsstil wird ein andauerndes persönliches Verhaltensmuster einer Führungs-
person verstanden, durch welches sich dessen individuelle Grundhaltung ausdrückt.187
Demnach liegt den Führungsstiltheorien die Annahme zugrunde, dass sich bestimmte
Führungsstile identifizieren lassen, die mehr oder weniger zum Führungserfolg beitragen.
Im Unterschied zu den eigenschaftsorientierten Führungsansätzen werden nunmehr be-
stimmte Verhaltensmuster als Messgröße für den Führungserfolg herangezogen;188d.h.
der jeweilige Führungsstil ist die ausschlaggebende Determinante für die Einstellung der
MitarbeiterInnen gegenüber der Organisation, für die Aufgabenzuweisung und bestimmt
etwa, wie Entscheidungen getroffen werden.189
Abhängig von der Anzahl der Beurteilungskriterien können die Führungsstiltheorien in
eindimensionale, zweidimensionale oder auch mehrdimensionale Ansätze eingeteilt wer-
de.190 Die eindimensionalen sowie die zweidimensionalen Führungsstile werden im Fol-
genden vorgestellt.
4.1.1.2.1 Eindimensionale Führungsstile
Charakteristisch für eindimensionale Führungsstile ist, dass die Systematisierung der
Führungsstile anhand eines einzigen Beurteilungskriteriums erfolgt. Die zu beurteilenden
Führungsstile stellen Endpunkte dar, zwischen denen es aber noch einige Abstufungen
gibt. Eine stärkere Ausprägung in Richtung eines Führungsverhaltens bedeutet somit eine
schwächere Ausprägung des anderen Stils.191
185
Vgl. Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis, Wiesbaden, S. 261; Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Hei-delberg, S. 363.
186 Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 364.
187 Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 645.
188 Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 364f.
189 Vgl. Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis,
Wiesbaden, S. 275; Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 645. 190
Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 645. 191
Vgl. Jung, H. (2010): Allgemeine Betriebswirtschaftslehre, München, S. 221.

37
Ausgangspunkt für das Konzept der eindimensionalen Führungsstile war eine Reihe von
Experimenten von Lewin, in denen er untersuchte, welchen Einfluss ein autoritäres bzw.
ein demokratisches Führungsverhalten auf Produktivität, Arbeitsklima und auf die Kreativi-
tät hatte. Dem Ergebnis zufolge war der demokratische Führungsstil dem autoritären in
allen Belangen überlegen.192
Enorme Popularität erlangte in weiterer Folge die Kontinuumtheorie von Tannen-
baum/Schmidt193. Das zentrale Beurteilungskriterium dieser Theorie ist das Ausmaß an
Entscheidungsspielraum, das den MitarbeiterInnen bei Entscheidungen zugestanden
wird.194 Im Unterschied zu Lewin, der lediglich ein autoritäres mit einem demokratischen
Führungsverhalten verglich, gehen Tannenbaum/Schmidt davon aus, dass zwischen dem
autoritären und dem kooperativen Führungsstil noch fünf weitere Führungsstile zu finden
sind, die je nach Entscheidungsfreiheit der MitarbeiterInnen abgestuft sind.195 Folgende
Abbildung soll eben Ausgeführtes veranschaulichen:
Abbildung 10: Eindimensionaler Führungsstil nach Tannenbaum/Schmidt
Quelle: Tannenbaum, R./Schmidt, W. (1958): How to choose a leadership pattern, in: Harward Business Re-
view, 36. Jg., S. 97; Schaufelbühl, K./Hugentobler, W./Blattner, M. (Hrsg.) (2007): Betriebswirtschaftslehre für
Bachelor, Zürich, S. 899.
Die Autoren gehen nicht davon aus, dass einer der sieben Führungsstile immer der erfolg-
reichste ist. Vielmehr ist der richtige Führungsstil von den Charakteristika des/der Vorge-
setzten, der MitarbeiterInnen sowie von der jeweiligen Situation abhängig. Der Schlüssel
192
Vgl. Schaufelbühl, K./Hugentobler, W./Blattner, M. (Hrsg.) (2007): Betriebswirtschaftslehre für Bachelor,
Zürich, S. 898. 193
Vgl. Tannenbaum, R./Schmidt, W. (1958): How to choose a leadership pattern, in: Harward Business Re-view, 36. Jg., S.95-102.
194 Vgl. Schaufelbühl, K./Hugentobler, W./Blattner, M. (Hrsg.) (2007): Betriebswirtschaftslehre für Bachelor,
Zürich, S. 898. 195
Vgl. Jung, H. (2010): Allgemeine Betriebswirtschaftslehre, München, S. 221.

38
zum Erfolg liegt demnach in der Flexibilität des Führungsverhaltens, da je nach Zusam-
mensetzung der Charakteristika ein unterschiedlicher Führungsstil erforderlich ist.196
Kritik erfuhr die Kontinuumtheorie vor allem deswegen, weil sie mit der Entscheidungspar-
tizipation der MitarbeiterInnen nur ein Verhaltensmerkmal der Führung berücksichtigte.197
4.1.1.2.2 Zweidimensionale Führungsstile
Im Vergleich zu den eindimensionalen Führungsstiltheorien waren die zweidimensionalen
Theorien zeitlich später angesiedelt.198 Zweidimensional ist ein Führungsstil dann, wenn
zur Systematisierung zwei voneinander unabhängige Verhaltensdimensionen herangezo-
gen werden. Der Führungsstil ergibt sich sodann aus der Kombination der unterschiedli-
chen Ausprägungsgrade der jeweiligen Dimension.199
Die zweidimensionalen Führungsstiltheorien gehen auf zwei Forschergruppen der Ohio-
State University und der Michigan University zurück, die mit der MitarbeiterInnen- und der
Aufgabenorientierung zwei Hauptdimensionen des Führungsverhaltens identifizierten.
Aufbauend auf diesen Forschungsarbeiten entwickelten Blake und Mouton200 das sog.
Managerial Grid - ein allgemeines Schema für Führungsverhalten. Bei diesem zweidi-
mensionalen Verhaltensgitter ist jede Hauptdimension in neun verschiedene Ausprä-
gungsgrade unterteilt. Insgesamt ergeben sich aus den unterschiedlichen Kombinationen
der Ausprägungsgrade 81 verschiedene Führungsstile, wobei die Autoren nur auf fünf
Kombinationen näher eingehen.201 Folgende Abbildung soll das Verhaltensgitter, sowie
die fünf wesentlichen Führungsstile, sowohl graphisch als auch inhaltlich veranschauli-
chen.
196
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 367. 197
Vgl. Jung, H. (2010): Allgemeine Betriebswirtschaftslehre, München, S. 222. 198
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 365. 199
Vgl. Schaufelbühl, K./Hugentobler, W./Blattner, M. (Hrsg.) (2007): Betriebswirtschaftslehre für Bachelor, Zürich, S. 899; Jung, H.(2010): Allgemeine Betriebswirtschaftslehre, München, S. 223.
200 Vgl. Blake, R.R./Mouton, J. (1964): The Managerial Grid. The Key to Leadership Excellence, Houston.
201 Vgl. Jung, H. (2010): Allgemeine Betriebswirtschaftslehre, München, S. 223f.

39
Abbildung 11: Managerial Grid nach Blake/Mouton
Quelle: Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis,
Wiesbaden, S. 278.
Der 9.9 Führungsstil wird von den Autoren als Idealstil angesehen, während der Stil 1.1
als zu theoretisch, der 9.1. Führungsstil als zu pessimistisch, das Verhalten nach 1.9 als
zu idealistisch und der Stil Nummer 5.5 als zu kompromissartig gilt.202
Auch diese Führungsstiltheorie blieb nicht ohne Kritik. Vor allem die Empfehlung der Auto-
ren, den Führungsstil 9.9 als universell anwendbar zu erklären, stößt in der Literatur auf
Unverständnis.203 Auch empirische Untersuchungen konnte die Vorteilhaftigkeit der Theo-
rie 9.9 nicht generell nachweisen.204
4.1.1.3 Situative Führungstheorien
Eine erneute Weiterentwicklung in der Führungsforschung ist in den situativen Führungs-
theorien zu erblicken, die in den 60er Jahren des 20. Jhd. entwickelt wurden.205 Den Situ-
ationstheorien liegt die Annahme zugrunde, dass der Führungserfolg nicht ausschließlich
aufgrund von persönlichen Merkmalen oder aufgrund eines bestimmten Verhaltens ab-
hängt, sondern vielmehr von der jeweiligen Situation.206 Wesentliche Aufgabe einer erfolg-
reichen Führungskraft ist somit, die jeweilige Situation in Hinblick auf die Aufgabe, sowie
der Gruppenbeziehung zu analysieren, und das Führungsverhalten flexibel anzupas-
sen.207
202
Vgl. Jung, H.(2010): Allgemeine Betriebswirtschaftslehre, München, S. 225. 203
Vgl. Jung, H.(2010): Allgemeine Betriebswirtschaftslehre, München, S. 225 204
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 372. 205
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 358. 206
Vgl. Dillerup, R./Stoi, R. (2013): Unternehmensführung, München, S. 644, 648. 207
Vgl. Jung, H.(2010): Allgemeine Betriebswirtschaftslehre, München, S. 217.

40
Eine der bekanntesten Situationstheorien sind die Kontingenztheorie von Fiedler208 sowie
die situative Führungstheorie von Hersey und Blanchard209, auf die in dieser Arbeit aller-
dings nicht eingegangen wird.
Beide Theorien wurden erheblich kritisiert und konnten empirisch nur sehr eingeschränkt
bestätigt werden. Dies liegt vor allem an Messproblemen, sowie an konzeptionellen
Schwächen der Modelle. Ein weiterer Kritikpunkt ist in der unzureichenden Berücksichti-
gung des Einflusses des Führungsstils auf unterschiedliche Situationen zu erblicken.210
4.1.1.4 Transformative vs. Transaktionale Führung
Jüngst findet die Differenzierung zwischen transformativer und transaktionaler Führung in
der Führungslehre großen Anklang, wobei davon ausgegangen wird, dass sich beide An-
sätze nicht gegenseitig ausschließen, sondern aufeinander aufbauen. So ist in manchen
Situationen transaktionale Führung völlig angemessen. Darauf aufbauend kann transfor-
mative Führung jedoch einen höheren Einsatzwillen der MitarbeiterInnen hervorbringen.211
Aus diesem Grund werden im Folgenden die wesentlichen Merkmale von transaktionaler
Führung einerseits und transformativer Führung andererseits vorgestellt.
4.1.1.4.1 Transaktionale Führung
Grundlegend für transaktionale Führung ist das Austauschprinzip; d.h. die Führungsper-
son klärt ihre MitarbeiterInnen darüber auf, welche Anforderungen und Erwartungen an
sie gestellt werden. Gleichzeitig soll den Geführten kommuniziert werden, welche Anreize
sie im Gegenzug für ihre Leistungen erhalten, wobei individuelle Wünsche, Erwartungen
und Befürchtungen eines/r jeden MitarbeitersIn berücksichtigt werden müssen. Sohin be-
steht bei der transaktionalen Führung die wesentliche Aufgabe der Führungsperson darin,
den Austauschprozess von Leistungen und Anreizen zu steuern.212
4.1.1.4.2 Transformative Führung
Im Fokus der transformativen Führung steht die Veränderung der Einstellung, der Wün-
sche sowie der Vorstellungen der MitarbeiterInnen. Eine Führungsperson soll gemäß die-
sem Führungsstil tiefgründige Werte und Überzeugungen vermitteln, die Geführten mitzu-
208
Siehe dazu Fiedler, F.E. (1967): A theory of leadership effectivness, New York, mwH. 209
Siehe dazu Hersey P./Blanchard K.H. (1977): Management of organizational behavior, Englewood Cliffs,
mwH. 210
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 376, 379.
211 Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 382.
212 Vgl. Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis,
Wiesbaden, S. 277.

41
reißen und dazu anzuregen, Dinge völlig neu zu sehen.213 Dieser Führungsstil weist große
Ähnlichkeit zum charismatischen Führungsstil auf.214
Wenig überraschend bestätigen empirische Studien, dass der transformative Führungsstil
dem transaktionalen Führungsverhalten überlegen ist. So führt der transformative Stil in
etwa zu einer niedrigeren Fluktuation und gleichzeitig zu einer höheren Produktivität, wo-
bei dieses Führungsverhalten, im Gegensatz zum transaktionalen Stil, wesentlich schwie-
riger zu verwirklichen ist.215
Zusammenfassend kann festgehalten werden, dass sich im Zeitablauf unterschiedliche
Führungstheorien entwickelten und vertreten wurden, wobei keine einzige davon Allge-
meingültigkeit erlangen konnte. Um diesem Ziel in Zukunft eventuell ein Stück näher zu
kommen wird versucht, auf neurowissenschaftliche Erkenntnisse zurückzugreifen. Aus
diesem Grund wird in der Folge der eine Verbindung zwischen Leadership (im betriebs-
wirtschaftlichen Sinn) und den Neurowissenschaften hergestellt. In diesem Zusammen-
hang wird auch auf den status quo der Forschung eingegangen.
4.2 Neuroleadership
Die Wirksamkeit der traditionellen Methoden zur Führungskräfteentwicklung wurden
jüngst stark hinterfragt. Nicht zuletzt aufgrund des Fortschritts durch die Neurowissen-
schaften, sprechen sich viele Forscher dafür aus, über die traditionellen Methoden zur
Bewertung von Führungskräften, zu denen u.a. die Befragung zählt, hinauszugehen. Im
Speziellen soll dabei die Neurowissenschaft Einblicke in das menschliche Gehirn und so
Aufschluss über eine effektive und wirksame Führung, sowie über deren Bewertung und
ihre Entwicklung geben.216 Für die Führungsforschung hat dabei der neurowissenschaftli-
che Teilbereich der sozialen kognitiven Neurowissenschaften die größte Anwendbarkeit.
Die sozial-kognitiven Neurowissenschaften sind ein aufstrebendes, interdisziplinäres Feld,
das versucht, menschliche Interaktionen im Schnittpunkt von sozialen, kognitiven und
neuronalen Sphären zu verstehen. Die sozial-kognitiven Neurowissenschaften sollen ins-
besondere Einblicke darüber gewähren, inwiefern das menschliche Gehirn Führungskräf-
te in vielen Aspekten hinsichtlich Verhalten und Kognition unterstützen kann.217
213
Vgl. Hungenberg, H./Wulf, T. (2011): Grundlagen der Unternehmensführung, Berlin Heidelberg, S. 383. 214
Vgl. Schreyögg, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Studium und Praxis, Wiesbaden, S. 277.
215 Vgl. Robbins, S.P. (2006): Organisation der Unternehmung, München, S. 388.
216 Vgl. Waldman, D.A./Balthazard, P.A./Peterson, S.J. (2011): Leadership and Neuroscience: Can We Revo-
lutionize the Way that Inspirational Leaders Are Identified and Developed?, in: Academy of Management Perspectives, Vol. 25, S. 60.
217 Vgl. Ochsner, K.N/Liebermann, M.D. (2001): The emergence of social cognitive neuroscience, in: Ameri-
can Psychologist, Vol. 56, S.717-734.

42
Für Führungskräfte sind auch gute, harmonische Beziehungen zu ihrem Umfeld, allen
voran zu den MitarbeiterInnen, essentiell. Dabei ist anzunehmen, dass die wechselseitige
Beziehung zwischen Führungskraft und MitarbeiterIn umso harmonischer ist, je inspirie-
render und mitfühlender die Führungskraft agiert. Vice versa schlägt sich eine geringe
Empathie von Führungskräften oftmals in verringerter Rendite nieder und führt zu einer
Dissonanz hinsichtlich der Beziehung zu den MitarbeiterInnen. Auch für dieses Problem-
feld kann die Neurowissenschaft überzeugende Einblicke und überraschende Beobach-
tungen liefern.218
Der Grund dafür, dass die Erforschung einer gehirngerechten Führung im Vergleich zu
anderen Disziplinen der Neuroökonomie wenig fortgeschritten ist, liegt wohl darin, dass
der vielschichtige betriebliche Alltag schwer durch neurowissenschaftliche Methoden ab-
gebildet werden kann. Im Unterschied zu Marketingentscheidungen und Finanztransakti-
onen, wo viele bildgebende Methoden ideal angewendet werden können, kann das viel-
seitige Führungsverhalten und die Reaktionen der MitarbeiterInnen sowie die unterschied-
lichen organisatorischen Möglichkeiten kaum mit Hilfe von neurowissenschaftlichen Me-
thoden abgebildet werden. Vielmehr geht es darum, Erkenntnisse der Hirnforschung für
die MitarbeiterInnenführung zu systematisieren und ein Studiendesign zu entwickeln.219
Basierend auf Erkenntnissen der Hirnforschung gibt es bis dato bereits Ansätze von Neu-
ro- und WirtschaftswissenschaftlerInnen, die Handlungsempfehlungen zur Führung von
MitarbeiterInnen abgeben. Vier dieser Ansätze werden im Folgenden in aller Kürze vorge-
stellt, ehe abschließend diskutiert wird, welchen Beitrag die Neurowissenschaft für die
Führungsforschung leisten kann. Bei der Vorstellung der Ansätze wird auch auf die, den
Handlungsempfehlungen zugrunde liegenden Erkenntnisse aus der Hirnforschung, einge-
gangen.
4.2.1 Gehirngerechte Führung nach Hüther
Hüther entwickelte auf Basis neurowissenschaftlicher Erkenntnisse folgende Handlungs-
empfehlungen für ManagerInnen bzgl. einer effizienten und erfolgreichen Gestaltung des
Umfeldes für MitarbeiterInnen in einem Unternehmen.
4.2.1.1 Regelmäßig neue Herausforderungen schaffen
Das menschliche Gehirn ist aus neurobiologischer Sichtweise nicht dazu geschaffen,
Routinen abzuarbeiten. Vielmehr benötigt das Gehirn permanent neue Herausforderun-
218
Vgl. Boyatzis , R.E.: Neuroscience and the link between inspirational leadership and resonant relations-hips, in: Ivey Business Journal, January/February 2012.
219 Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 14; siehe dazu auch Peters, T./Ghadiri, A. (2012): Mitarbeiter führen heißt Gehirne führen, in: Personalmanager, S. 67.

43
gen, wodurch kreatives Problemlösen optimiert wird. Durch eine neue Herausforderung
wird eine emotionale Erregung im Gehirn ausgelöst, die beruhigt wird, indem das Gehirn
nach einer Lösung sucht. Im Ergebnis bleibt das Denken beweglich.
Führungskräfte sollten demnach MitarbeiterInnen in regelmäßigen Abständen vor neue
Herausforderungen, bspw. durch „Job Rotation“, stellen, damit sich das Gehirn nicht an
Routineabläufen gewöhnt.220
4.2.1.2 Know-How im Unternehmen vernetzen
Werden im Gehirn viele neuronale Netzwerke gleichzeitig aktiviert, funktioniert die kreati-
ve Lösungssuche am Besten, indem bereits vorhandenes Wissen auf eine neue Art und
Weise miteinander verbunden wird.
Für Führungskräfte bedeutet dies, dass Umstände geschaffen werden sollen, die es Mit-
arbeiterInnen mit unterschiedlichem Know-How ermöglichen, sich auszutauschen bzw
zusammenzuarbeiten. Konkrete Maßnahmen, um kreative Lösungen zu fördern wären
bspw. abteilungsübergreifende Teams oder Großgruppenkonferenzen.221
4.2.1.3 Eine positive Fehlerkultur schaffen
Verspürt ein Mensch Angst, so wird im Gehirn ein Notfallprogramm ausgelöst, dass ent-
weder Angriff, Flucht oder Erstarrung hervorruft. Kreatives Denken wird im Angstzustand
nicht nur gehemmt, sondern ist völlig unmöglich.
Aus diesem Grund sind Führungskräfte dazu angehalten eine positive Fehlerkultur im
Unternehmen zu installieren, damit die MitarbeiterInnen so wenig Druck und Versagens-
angst wie möglich verspüren. Unter einer positiven Fehlerkultur ist eine Kultur zu verste-
hen, in der Fehler nicht sanktioniert werden, sondern als Chance gesehen werden, um
daraus zu lernen.222
4.2.1.4 Für positive Erfahrungen sorgen
Durch das Aneinanderkoppeln sämtlicher Netzwerke im Gehirn, die gleichzeitig aktiviert
werden, verbindet das Gehirn bestimmte Emotionen mit bestimmten Situationen, Gerü-
chen, oder Personen.
Damit das Zugehörigkeitsgefühl und die Leistungsbereitschaft der MitarbeiterInnen stei-
gen, soll die Führungskraft versuchen, die Beziehung zu den MitarbeiterInnen durch Lob,
220
Vgl. Hüther, G. (2009): Wie gehirngerechte Führung funktioniert -Neurobiologie für Manager, in: manager-Seminare, Nr: 130, S.30-34.
221 Vgl. Hüther, G. (2009): Wie gehirngerechte Führung funktioniert -Neurobiologie für Manager, in: manager-
Seminare, Nr: 130, S.30-34. 222
Vgl. Hüther, G. (2009): Wie gehirngerechte Führung funktioniert -Neurobiologie für Manager, in: manager-Seminare, Nr: 130, S.30-34.

44
Interesse an deren Person, oder durch Rat in schwierigen Situationen, auszubauen. Da-
durch erfolgt im Gehirn der MitarbeiterInnen eine Verknüpfung positiver Erfahrungen mit
dementsprechenden Emotionen.223
4.2.2 SCARF- Modell von Rock
Das SCARF-Modell von Rock basiert auf der Erkenntnis, dass das Gehirn stets versucht,
erfolgreiche Situationen anzustreben, um eine Aktivierung des Belohnungssystems her-
vorzurufen. Rock identifizierte dabei folgende fünf Treiber, die für eine gute Zusammenar-
beit am Arbeitsplatz sorgen sollen224:
4.2.2.1 Status (status)
Unter Status wird die relative Wichtigkeit, Rangfolge und Alter unter den MitarbeiterInnen
verstanden. Fühlt sich ein/e MitarbeiterIn besser als ein/e andere/r äußert sich dies durch
die Aktivierung des Belohnungssystems. Der Status einer Person kann durch alltägliche
Situationen, etwa durch Lob, positiv, oder durch Kritik, negativ, beeinflusst werden. Be-
reits Ratschläge oder Hilfestellungen eines/r KollegIn können den Status bedrohen.
Ebenso bedrohlich wirken sich Leistungsbeurteilungen durch den/die Vorgesetze/n aus.
Neben Beförderungen kann der Status von MitarbeiterInnen auch durch positives Feed-
back, insbesondere durch öffentliche Anerkennung positiv beeinflusst werden.225
4.2.2.2 Vorhersehbarkeit (certainty)
Um die nahe Zukunft vorherzusehen, will das Gehirn stets gewisse augenblicklich auftre-
tende Muster erkennen. Dies ist nur möglich, wenn die auftretenden Muster dem Gehirn
bekannt sind. Kann das Gehirn Wahrnehmungen nicht vorhersehen, so benötigt es viel
mehr Energie um diese Situationen zu verarbeiten, als in gewohnten, vorhersehbaren
Situationen.
Aus diesem Grund lohnt es sich für Führungskräfte, Vorhersehbarkeit in Hinblick auf viele
tägliche Situationen zu kreieren. Bspw. wird Vorhersehbarkeit dadurch erzeugt, dass Mit-
arbeiterInnen im Fall einer Restrukturierung mitgeteilt wird, zu welchem Zeitpunkt sie
mehr Informationen über die anstehende Restrukturierung erfahren. Des Weiteren ist es
hilfreich, Projekte in mehrere Teilprojekte zu untergliedern sowie Businesspläne, Strate-
223
Vgl. Hüther, G. (2009): Wie gehirngerechte Führung funktioniert -Neurobiologie für Manager, in: manager-Seminare, Nr: 130, S.30-34.
224 Vgl. Peters, T./Ghadiri, A. (2012): Mitarbeiter führen heißt Gehirne führen, in Personalmanagement, S. 67.
225 Vgl. Rock, D. (2008): SCARF: A Brain-Based Model For Collaborating With And Influencing Others, in:
NeuroLeadership journal, Vol. 1, S. 3-4.

45
gien, Organisationsstrukturen und ähnliches so klar und transparent wie möglich zu kom-
munizieren.226
4.2.2.3 Autonomie (autonomy)
Unter Autonomie versteht Rock die Möglichkeit der MitarbeiterInnen ihr Umfeld zu kontrol-
lieren und zu beeinflussen. Wird den MitarbeiterInnen alles bis ins kleinste Detail vorge-
schrieben, sind diese einer Stresssituation ausgesetzt, weil eine Reduzierung der Auto-
nomie als Gefährdung empfunden wird.
Es ist sehr schwierig, eine erkennbare Autonomie in einem Unternehmen herzustellen.
Aber bereits die subtile Wahrnehmung von Autonomie, wie bspw. die freie Arbeitsplatz-,
sowie Arbeitszeit- und Arbeitsablaufgestaltung, kann das Belohnungssystem aktivieren.227
4.2.2.4 Soziale Beziehungen (relatedness)
Soziale Beziehungen bedingen zu entscheiden, ob jemand zu einer Gruppe gehört oder
nicht, ob jemand Freund oder Feind ist. Durch die Zugehörigkeit zu einer Gruppe wird der
Botenstoff Oxytocin vermehrt ausgeschüttet, der gleichzeitig stabilere Beziehungen be-
wirkt. Soziale Beziehungen hängen sehr eng mit Vertrauen zusammen: Je größer das
Vertrauen, desto besser und enger ist die Zusammenarbeit und damit die Kommunikation
der Beteiligten.
Soziale Beziehungen zwischen MitarbeiterInnen können bspw. durch Videokonferenzen
und kleineren Teams, gefördert werden.228
4.2.2.5 Gerechtigkeit (fairness)
Unabhängig von anderen Faktoren belegen unterschiedliche Studien, dass Ungerechtig-
keit durch Aktivierung emotionaler Gehirnregionen einen Abwehrmechanismus auslöst,
um Gefahren zu vermeiden.
Bestehende Ungerechtigkeiten im betrieblichen Kontext können reduziert werden, indem
Transparenz, Kommunikation und Mitwirkung der MitarbeiterInnen im Unternehmen ver-
bessert werden. Beispielsweise sollen klare Erwartungen in jeder Situation kommuniziert
werden und einzelnen Teams die Möglichkeit gegeben werden, eigene Regeln aufzustel-
len.229
226
Vgl. Rock, D. (2008): SCARF: A Brain-Based Model For Collaborating With And Influencing Others, in: NeuroLeadership journal, Vol. 1, S. 4-5.
227 Vgl. Rock, D. (2008): SCARF: A Brain-Based Model For Collaborating With And Influencing Others, in:
NeuroLeadership journal, Vol. 1, S. 5. 228
Vgl. Rock, D. (2008): SCARF: A Brain-Based Model For Collaborating With And Influencing Others, in: NeuroLeadership journal, Vol. 1, S. 5-6.
229 Vgl. Rock, D. (2008) SCARF: A Brain-Based Model For Collaborating With And Influencing Others, in:
NeuroLeadership journal, Vol. 1, S. 6.

46
4.2.3 Neuroleadership nach Elger
Die Basis für den Ansatz von Elger bilden vier Gehirnsysteme. Dazu zählen das Beloh-
nungssystem, das emotionalem System, das Gedächtnis- und das Entscheidungssystem.
Durch die Übertragung dieser Gehirnsysteme auf den betrieblichen Kontext leitet Elger
folgende sieben Grundregeln des Neuroleaderships ab:
4.2.3.1 Das Belohnungssystem als zentrale Schaltstelle
Die Aktivierung des Belohnungssystems wirkt sich motivierend auf die MitarbeiterInnen
aus, weswegen auf dieses besonderes Augenmerkt gelegt werden sollte. Durch das Be-
lohnungssystem wird ein starkes Wohlbefinden erzeugt, das auch bei dauernder Stimulie-
rung nicht in Gewöhnung umschwenkt. Wesentliche Faktoren, um das Belohnungssystem
zu aktivieren sind bspw. das Arbeitsklima, das Ambiente am Arbeitsplatz sowie die Ar-
beitsplatzgestaltung.230
4.2.3.2 Fairness
Das Gehirn als soziales Organ strebt nach Fairness. Fühlt sich ein Mensch als ungerecht
behandelt, so legt dieser ein Verhalten an den Tag, das die Gerechtigkeit wieder herstel-
len soll. In diesem Zusammenhang wird vom altruistischen Bestrafen gesprochen. Um
altruistisches Bestrafen, zu minimieren, soll dem Verhandlungspartner stets ein positives
Feedback gegeben werden, um so sein Belohnungssystem zu aktivieren.231
4.2.3.3 Beeinflussung der Erwartungen und des Verhaltens durch Vorinformationen
Vom Gehirn werden ständig Vorhersagen über bestimmte Situationen getroffen und in
weiterer Folge versucht, diese zu bestätigen. Durch Vorinformationen, positiv sowie nega-
tiv, werden die Vorhersagen beeinflusst, wobei sich positive Vorinformationen weniger
schnell verbreiten und kürzer in Erinnerung bleiben als Negative. Aus diesem Grund sind
ManagerInnen gut beraten, Imagepflege zu betreiben, denn jede noch so kleine Äußerung
oder Gerücht kann die Erwartungen und das Verhalten der MitarbeiterInnen erheblich
beeinflussen. Eine positive Vorinformation führt zur Aktivierung des Belohnungssystems,
womit anstehende Verhandlungen maßgeblich erleichtert werden.232
230
Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern, Freiburg, S. 168-170.
231 Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern,
Freiburg, S. 170. 232
Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern, Freiburg, S. 171-172.

47
4.2.3.4 Einzigartigkeit jedes Gehirns
Durch die Plastizität sowie den komplexen Aufbau und die äußerst komplizierte Funkti-
onsweise ist jedes Gehirn einzigartig; d.h. jedes Gehirn ist unterschiedlich vernetzt. Zwar
gibt es Hirnregionen, in denen bei allen Menschen vergleichbare Prozesse stattfinden,
dennoch kann der Feinaufbau des Gehirns bei verschiedenen Menschen sehr unter-
schiedlich sein. Für Führungskräfte ist es essentiell, die Denkweise ihrer MitarbeiterInnen,
die auf unterschiedlichen Wahrnehmungen und Erinnerungen beruht, zu verstehen. Um
diese Denkweise verstehen zu können, bedarf es einer gewissen Erfahrung, die nicht in
Vorträgen oder Seminaren erlernt werden kann.233
4.2.3.5 Keine Fakten ohne Emotionen
Das Gehirn bewertet jede eingehende oder weitergeleitete Information und ordnet ihr eine
Emotion zu. Durch die unbewusste Zuordnung dieser Emotionen wird das daraus resultie-
rende Verhalten von Menschen beeinflusst. Dies lässt die Schlussfolgerung zu, dass es
keine Fakten ohne emotionale Komponenten gibt. Auf der anderen Seite können Emotio-
nen einzelne Fakten schaffen, man denke bspw. an den Placebo-Effekt, wobei hier das
Verhältnis zwischen Arzt/Ärztin und PatientIn einen wesentlichen Einfluss auf den Erfolg
hat. Ebenso verhält es sich zwischen der Führungskraft und den MitarbeiterInnen.234
4.2.3.6 Erfahrungen bestimmen das Verhalten
Erfahrene MitarbeiterInnen können ähnliche Situationen besser bewältigen als Unerfahre-
ne, weil Unerfahrenheit zumeist Stress verursacht, womit in weiterer Folge das Gehirn
blockiert wird. Außerdem spielt auch die emotionale Komponente eine Rolle, weil unerfah-
rene MitarbeiterInnen stärkere Emotionen zeigen, während routinierte MitarbeiterInnen
ihre Emotionen besser regulieren können.235
4.2.3.7 Eigendynamik von Situationen
Das Verhalten eines Menschen ist situationsbezogen, d.h. je nachdem in welcher Situati-
on sich ein Mensch befindet, kann er sich sehr unterschiedlich verhalten. Dieses situati-
onsbezogene Verhalten beruht auf emotionalen Bewertungen und auf der Reaktion des
Belohnungssystems. Bereits im Unterbewusstsein befindliche Aggressionen können eine
gewisse Eigendynamik entwickeln und vom/von der VerhalndlungspartnerIn gespürt wer-
233
Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern, Freiburg, S. 172-173.
234 Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern,
Freiburg, S. 173-174. 235
Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern, Freiburg, S. 174-175.

48
den. Für Führungskräfte ist es daher essentiell, ein Gespür für diese Situationen zu entwi-
ckeln und Situationen so gut wie möglich zu planen.236
4.2.4 Neuroleadership nach Peters/Ghadiri
Im Sinne des Neuroleaderships nach Peters/Ghadiri soll eine Führungskraft die Arbeits-
umwelt für die MitarbeiterInnen so gestalten, dass die vier Grundbedürfnisse des Men-
schen erfüllt werden. Im Ergebnis wird dadurch eine gehirngerechte Umgebung für
den/die MitarbeiterIn geschaffen. Peters/Ghadiri versuchen in ihrem Ansatz nicht primär
neurowissenschaftliche Aspekte, sondern vor allem Aspekte der Personal- und Organisa-
tionslehre stärker zu berücksichtigen.237 Es handelt sich im Einzelnen um folgende vier
Grundbedürfnisse:
4.2.4.1 Bindung
Unter Bindung ist das Bedürfnis eines Menschen zu verstehen, eine Bindung zu einer
Bezugsperson zu entwickeln. Dieses Grundbedürfnis ist sehr von den Erfahrungen in der
frühen Kindheit abhängig, wodurch auch das spätere Verhalten geprägt wird. Vor allem in
der Zeit der Einstellung sind für MitarbeiterInnen Bezugspersonen, bspw. in Form eines
Mentors, sehr wichtig, um Stresssituationen zu vermeiden.238
4.2.4.2 Orientierung und Kontrolle
Jeder Mensch hat das Bedürfnis, seine Umgebung durch bestimmte Handlungsweisen zu
beeinflussen bzw. einen, nach Möglichkeit großen, Handlungsspielraum zu erhalten. In
engem Zusammenhang damit stehen die Einschätzungen bzw. das Wissen über be-
stimmte Situationen. Ein fehlender Überblick oder fehlende Informationen über bestimmte
Situationen lösen bei MitarbeiterInnen im limbischen System eine Stressreaktion aus, die
sich negativ auf deren Zufriedenheit auswirkt. Aus diesem Grund ist es für ManagerInnen
ratsam, MitarbeiterInnen nicht nur einen Handlungsspielraum und Autonomie einzuräu-
men, sondern auch die Zielvorstellungen klar und transparent zu kommunizieren.239
4.2.4.3 Selbstwerterhöhung und Selbstwertschutz
Menschen, die sich selbst als Individuum wahrnehmen und reflexiv denken, sind bestrebt,
ihren Selbstwert, durch bestimmte Handlungen zu erhöhen oder zu schützen. Bei diesem
Bedürfnis handelt es sich um ein spezifisches menschliches Grundbedürfnis, dass auf-
236
Vgl. Elger, C.E. (2013): Neuroleadership, Erkenntnisse der Hirnforschung für die Führung von Mitarbeitern,
Freiburg, S. 175-176. 237
Vgl. Peters, T./Ghadiri, A. (2012): Mitarbeiter führen heißt Gehirne führen, in: Personalmanager, S. 68. 238
Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 73-74.
239 Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 75-76.

49
grund der Komplexität weniger erforscht ist. Menschen mit geringerem Selbstwertgefühl
versuchen nur selten eine Selbstwerterhöhung zu erreichen, weil sie schmerzhafte Erfah-
rungen in der Vergangenheit zu einer passiven Haltung veranlassen. Diese MitarbeiterIn-
nen kompensieren zumeist das nicht befriedigte Grundbedürfnis der Selbstwerterhöhung
mit anderen Grundbedürfnissen. Um eine Selbstwerterhöhung zu erreichen sind Projekt-
erfolge, aber auch die Anerkennung der Abteilung sowie der Vergleich zu den Mitarbeite-
rInnen förderlich.240
4.2.4.4 Lustgewinn und Unlustvermeidung
Der Mensch ist stets danach bestrebt, angenehme Situationen zu erleben und unange-
nehme Zustände zu vermeiden. Durch diese Bestrebung werden konkrete Erfahrungen
und Umstände im Unterbewusstsein automatisch in die Kategorien „gut“ und „schlecht“
eingeteilt. Um negative Erfahrungen zu vermeiden, können Führungskräfte unter anderem
darauf achten, Teams nach Kriterien zusammenzustellen, die ein positives Gruppenklima
ermöglichen. Empfindet ein/e MitarbeiterIn Teamarbeiten als positiv, wird er/sie auch in
Zukunft mit Freude an einer Teamarbeit teilnehmen.241
Zusammenfassend ist festzuhalten, dass die unterschiedlichen Ansätze der Wissen-
schaftlerInnen gewisse Parallelen bzw. Gemeinsamkeiten aufweisen. Eine Führungskraft
soll bestrebt sein, das Belohnungszentrum der MitarbeiterInnen zu aktivieren, indem
bspw. Stresssituationen vermieden werden. Zur Aktivierung des Belohnungssystems tra-
gen in etwa eine gute Beziehung zwischen Führungskraft und MitarbeiterIn, ein großer
Handlungsspielraum des/der MitarbeiterIn sowie Feedback bei.
Nachdem nun die wesentlichen Ansätze der obgenannten WissenschaftlerInnen, die auf
aktuellen Erkenntnissen der Hirnforschung basieren, vorgestellt wurden, werden in Folge
Bereiche identifiziert, in denen die Neurowissenschaft zur Führungsforschung beitragen
kann.
4.3 Beitrag der Neurowissenschaften für die Führungsforschung
Nach der Durchsicht von (hauptsächlich) fMRI und EEG- Studien, die im Zeitraum von
2009 bis 2014 zum Themenbereich Leadership veröffentlicht wurden242, können grund-
sätzlich drei Bereiche identifiziert werden, in denen die Neurowissenschaft zur Führungs-
forschung beitragen kann.
240
Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 76-77.
241 Vgl. Peters, T./Ghadiri, A. (2011): Neuroleadership - Grundlage, Konzepte, Beispiele. Erkenntnisse der
Neurowissenschaften für die Mitarbeiterführung, Wiesbaden, S. 77-78. 242
Siehe dazu Kapitel C.

50
Zunächst können Studien der Neurowissenschaften zur Klärung beitragen, ob sich erfolg-
reiche von nicht erfolgreichen Führungskräften neurophysiologisch unterscheiden bzw.
aufklären, wie sich eine etwaige Unterscheidung manifestiert.243 Diesbezüglich wurden für
die vorliegende Arbeit innerhalb des besagten Zeitraumes insgesamt sechs Studien244,
die Reaktionen von Geführten auf erfolgreiche bzw. weniger erfolgreiche Führungsperso-
nen verglichen, untersucht.245
Einen weiteren Beitrag kann die Neurowissenschaft leisten, indem sie Aufschluss darüber
gibt, welche neurophysiologischen Auswirkungen die soziale Einflussnahme durch Füh-
rungspersonen bei den MitarbeiterInnen hat, die bei diesen im Idealfall eine Verhaltens-
oder Einstellungsänderung hervorruft.246 Die soziale Einflussnahme und somit die Ände-
rung des Verhaltens oder der Einstellung des Geführten kann auf unterschiedliche Weise
erfolgen. Beispielsweise durch eine charismatische Führungskraft247, durch die wahrge-
nommene Qualität der Beziehung zwischen MitarbeiterInnen und der Führungsperson248
sowie auch durch Belohnung.249
243
Vgl. Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe, J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff.
244 Vgl. Cloutier, J./Ambady, N./Meagher, T./Gabrieli, J.D.E. (2012): The neural substrates of person percepti-
on: Spontaneus use of financial and moral status knowledge, in: Neuropsychologia, Vol. 50, Nr. 9, S. 2371-2376; Chiao, J.Y./ Harada, T./Oby, E.R./Li, Z./Parrish, T./Bridge, D.J. (2009): Neural representati-ons of social status hierarchy in human inferior parietal cortex, in: Neuropsychologia, Vol. 47, Nr. 2, S. 354-363; Cloutier, J./Kelley, W.M./Heatherton, T.F. (2011): The influence of perceptual an knowledge-
based familiarity on the neural substrates of face perception, in: Social Neuroscience, Vol. 6, Nr. 1, S. 63-75; Rule, N.O./Moran, J.M./Freeman, J.B./Whitfield-Gabrieli, S./Gabrieli, J.D.E./Ambady, N. (2011): Face value: Amygdala response reflects the validity of first impressions, in: NeuroImage, Vol. 54, S. 734-741; Doallo, S./Raymond, J.E./Shapiro, K.L./Kiss, M./Eimer, M./Nobre, A.C. (2012): Response inhibition
results in the emotional devaluation of faces: neural correlates as revealed by fMRI, in: Social Neuros-cience, Vol. 7, S. 649-659; Chiao J.Y./Adams, R.B./Tse, P.U./Lowenthal L./Richeson, J.A./Ambady, N. (2009): Knowing Who’s Boss: fMRI and ERP Investigations of Social Dominance Perception, in: Group Processes Intergroup Relations, Vol. 11, Nr. 2, S. 201-214; Rule, N.O./Freeman, J.B./Moran, J.M./Gabrieli J.D.E./Adams Jr., R.B./Ambady, N. (2010): Voting behavior is reflected in amygdala res-ponse across cultures, in: Social Cognitive and Affective Neuroscience, Vol. 5, S. 349-355.
245 Detailliertere Informationen zum Studiendesign/ -ablauf sowie zu den Stimuli siehe Kapitel C.
246 Vgl. Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe, J.
(Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff. 247
Vgl. Schjoedt, U./Stodkilde-Jorgensen, H./Geertz, A.W./Lund, T.E./Roepstorff, A. (2011): The power of charisma - perceived charisma inhibits the frontal executive network of believers in intercessory prayer, in: Social Cognitive and Affective Neuroscience, Vol. 6, Nr. 1, S. 119-127.
248 Vgl. Boyatzis, R.E./Passarelli, A.M./Koenig, K./Lowe, M./Mathew, B./Stoller, J.K./Phillips, M. (2012): Ex-
amination of the neural substrates activated in memories of experiences with resonant and dissonant le-aders, in: The Leadership Quarterly, S. 259-272; Kang, Y./Williams, L.E./Clark, M.S./Gray, J.R./Bargh, J.A. (2011): Physical temperature effects on trust behavior: the role of insula, in: Social Cognitive and Af-fective Neuroscience, Vol. 6, Nr. 4, S. 507–515; Fairhurst, M.R./Janata, P./Keller, P.E. (2013): Being and Feeling in Sync with an Adaptive Virtual Partner: Brain Mechanisms Underlying Dynamic Cooperativity, in: Cerebral Cortex, Vol. 23, Nr. 11, S. 2592–2600; Fairhurst, M.R./Janata, P./Keller, P.E. (2014): Lea-ding the follower: An fMRI investigation of dynamic cooperativity and leader-follower strategies in syn-chronization with an adaptive virtual partner, in: NeuroImage, S. 688–697.
249 Vgl. Spreckelmeyer, K.N./Krach, S./Kohls, G./Rademacher, L./Irmak, A./Konrad, K./Kircher, T./Gründer, G.
(2009): Anticipation of monetary and social reward differently activates mesolimbic brain structures in men and women, in: Social Cognitive and Affective Neuroscience, Vol. 4, Nr. 2, S. 158–165; Lin, A./Adolphs, R./Rangel, A. (2012): Social and monetary reward learning engage overlapping neural sub-strates, in Social Cognitive and Affective Neuroscience, Vol. 7, Nr. 3, S. 274–281.

51
Der dritte Bereich, in welchem die Neurowissenschaft einen wesentlichen Beitrag zur Füh-
rungsforschung leisten kann, ist die Aufklärung, welche neurophysiologischen Prozesse
führungsrelevanten Fähigkeiten zugrunde liegen.250 Eine wesentliche führungsrelevante
Fertigkeit ist die Entscheidungsfindung251, die für Führungskräfte eine zentrale Aufgabe
darstellt. 252
Abschließend soll folgende Tabelle die drei wesentlichen Bereiche, in denen die Neuro-
wissenschaft einen wesentlichen Beitrag für die Führungsforschung leisten kann, veran-
schaulichen. Auf diese Einteilung wird auch bei der Auswertung der Ergebnisse der ein-
zelnen Studien nochmals zurückgegriffen.
4.3.1 Kritische Würdigung
Wie aus diesem Kapitel hervorgeht, kann die Neurowissenschaft in unterschiedlichen Be-
reichen zur Führungsforschung beitragen. So kann die Neurowissenschaft Einblicke dar-
über geben, wie sich erfolgreiche von weniger erfolgreichen Führungskräften neurophy-
siologisch unterscheiden. Außerdem bietet die Neurowissenschaft die Möglichkeit zu un-
tersuchen, wie sich bestimmte bewussten Einflussnahmen von Führungskräften bei den
MitarbeiterInnen neurophysiologisch auswirken; d.h. welche Gehirnsysteme durch z.B.
charismatisches Verhalten, oder durch eine in Aussicht gestellte Belohnung aktiviert oder
gehemmt werden. Der dritte Bereich, in dem die Neurowissenschaft einen wesentlichen
Beitrag zur Führungsforschung beitragen kann, ist die Klärung, welche neurophysiologi-
sche Aktivität führungsrelevanten Fertigkeiten, wie bspw. der Entscheidungsfindung,
zugrunde liegt.253
Die folgende Tabelle soll die drei unterschiedlichen Bereiche, in denen die Neurowissen-
schaft zur Führungsforschung beitragen veranschaulichen.
250
Vgl. Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe, J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff.
251 Vgl. Caspers, S./ Heim, S./Lucas, M.G./Stephan, E./Fischer, L./Amunts, K./Zilles, K. (2012): Dissociated
Neural Processing for Decisions in Managers and Non-Managers, in: PLOS one, Vol. 7, Nr. 8. 252
Vgl. Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe, J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff.
253 Für Details siehe Kapitel B.4.3.

52
1. Unterscheiden sich erfolgreiche von nicht erfolgreichen Führungskräften neurophysiologisch?
2. Welche neurophysiologische Auswirkungen hat die bewusste soziale Einflussnahme durch
Führungspersonen bei MitarbeiterInnen?
(i) Qualität der Beziehung zwischen MitarbeiterIn und Führungsperson
(ii) charismatische Führungskraft
(iii) Belohnung
3. Welche neurophysiologische Aktivitäten liegen führungsrelevanten Fähigkeiten zu Grunde?
(i) Entscheidungsfindung
Bereiche, in denen die Neurowissenschaft zur Führungsforschung beitragen kann
Tabelle 3: Bereiche, in denen die Neurowissenschaft zur Führungsforschung beitragen kann
Quelle: eigene Darstellung

53
Dieses Kapitel beschäftigt sich mit einer Metaanalyse und stellt sohin den empirischen
Teil der Masterarbeit dar. Ehe diesbezüglich auf die Forschungsfrage sowie auf die Me-
thodik eingegangen wird, werden Charakteristika, Zielsetzung und der schematische Ab-
lauf einer Metaanalyse vorgestellt. Im Anschluss daran werden die erfassten Studien in
Hinblick auf den zeitlichen Verlauf, der Stichprobenanzahl sowie der eingesetzten Stimuli
ausgewertet. Der Fokus dieses Kapitels soll neben der Methodik hauptsächlich auf der
Darstellung der aktivierten Hirnregionen liegen. Abschließend werden die aus der Meta-
analyse gewonnenen Ergebnisse diskutiert.
1 Charakteristika einer Metaanalyse
Charakteristisch für eine Metaanalyse ist die Zusammenfassung der Ergebnisse empiri-
scher Untersuchungen zu einem bestimmten, gemeinsamen Thema, um so einen Über-
blick über den aktuellen Forschungsstand zu dieser Thematik zu gewinnen.254 Das Ziel
der Metaanalyse ist die Generalisierung von mehreren in einem bestimmten Forschungs-
bereich vorhandenen Studien.255 Aufgrund der Untersuchung und Darstellung einzelner
empirischer Studien, unter Zugrundelegung eines (gemeinsamen) Forschungsinteresses,
handelt es sich bei einer Metaanalyse um eine Sekundäranalyse.256
1.1 Schematischer Ablauf
Der prototypische Ablauf einer Metastudie besteht aus fünf Stufen. Zunächst ist die For-
schungsfrage zu konkretisieren, ehe im zweiten Schritt relevante Daten für die Beantwor-
tung der Forschungsfrage gesammelt werden. Danach werden diese Daten, nach vorge-
nommener Kodierung und Bewertung, analysiert. Am Ende einer Metaanalyse werden die
gewonnenen Erkenntnisse präsentiert und interpretiert.257
In der vorliegenden Arbeit wird dieser schematische Ablauf allerdings nicht eingehalten.
Aufgrund der geringen Anzahl vorhandener Studien ist eine statistische Auswertung nicht
möglich. Daher geht es in dieser Arbeit vorwiegend darum, die Studien zu erfassen und
254
Vgl. Bortz, J./Döring, N. (2006): Forschungsmethoden und Evaluation für Human- und Sozialwissenschaft-ler, Berlin, S. 672.
255 Vgl. Lueginger, E./Renger, R. (2013): Das weite Feld der Metaanalyse. Sekundär-, literatur- und metaana-
lytische Verfahren im Vergleich, in: kommunikation.medien, Nr. 2, S. 17. 256
Vgl. Lueginger, E./Renger, R. (2013): Das weite Feld der Metaanalyse. Sekundär-, literatur- und metaana-lytische Verfahren im Vergleich, in: kommunikation.medien, Nr. 2, S. 3, wobei die Autoren streng ge-nommen zwischen einer Sekundäranalyse und einer Metaanalyse differenzieren.
257 Vgl. Eisend, M.(2004): Metaanalyse - Einführung und kritische Diskussion, in: Betriebswirtschaftliche Rei-
he, Nr. 8; http://www.econstor.eu/bitstream/10419/49908/1/668827637.pdf; zuletzt geprüft am 25. Sep-tember 2014, S. 6.
C. METAANALYSE

54
zu systematisieren. Eine Kodierung258, wie im schematischen Ablauf dargestellt, findet
sohin in der vorliegenden Arbeit nicht statt. Auch werden keine abhängigen und unabhän-
gigen Variablen definiert. Die Datenanalyse erfolgt ebenso wie die Kodierung nicht einer
klassischen Metaanalyse, da es mangels Kodierung nicht möglich ist, statistische Effekte
nachzuweisen bzw. zu widerlegen.
1.2 Vor- und Nachteile einer Metaanalyse
Als wesentlicher Vorteil der Metaanalyse ist die Zusammenfassung mehrerer Studien zu
einem Gesamten zu nennen, wodurch dem Ergebnis eine höhere (statistische) Relevanz
zukommt als den einzelnen Studien. Ein weiterer Vorzug der Metaanalyse ist der genaue
Aufbau bzw. die Struktur, womit jeder Schritt nachvollziehbar und somit auch die Ergeb-
nisse überprüfbar sind.259
Neben den Vorzügen bleibt die Metaanalyse nicht ohne Kritik. In der Literatur zur Meta-
analyse werden neben den hohen Forschungsaufwand260 übereinstimmend vier wesentli-
che Argumente als die wesentlichsten Kritikpunkte an der Metaanalyse angeführt.261 Auch
als „Äpfel-Birnen-Argument“ bekannt, wird kritisiert, dass durch Metaanalysen nicht ver-
gleichbare Untersuchungen integriert werden. Ein Problem der Vergleichbarkeit kann ins-
besondere dann entstehen, wenn Studien vermengt werden, die sich hinsichtlich der Ei-
genschaften von Untersuchungspersonen in den Stichproben oder durch Auswertungs-
methoden unterscheiden.262 Ein weiterer, immer wieder angeführter Nachteil einer Meta-
analyse ist dahingehend zu erblicken, dass methodisch gute Arbeiten von methodisch
schlechten Arbeiten bei der Integration nicht unterschieden werden. Dieses Argument ist
auch unter dem Namen „garbage in - garbage out“ bekannt.263Neben den bereits ange-
führten Kritikpunkten ist als weiteres Gegenargument hinsichtlich einer Metaanalyse anzu-
führen, dass die Selektivität von WissenschaftlerInnen nicht dem wahren Forschungs-
stand entspricht, zumal überwiegend nur signifikante Ergebnisse in eine Metaanalyse
aufgenommen werden. Darüber hinaus werden in einer Metaanalyse abhängige Daten mit
258
Darunter versteht man die Einteilung bestimmter Parameter in abhängigen und unabhängigen Variablen, um eine statistische Auswertung zu ermöglichen.
259 Vgl. Lipsey, M.W./Wilson, D.B. (2001): Practical Meta-Analysis, Thousands Oaks, S. 5f.
260 Vgl. Lueginger, E./Renger, R. (2013): Das weite Feld der Metaanalyse. Sekundär-, literatur- und metaana-
lytische Verfahren im Vergleich, in: kommunikation.medien, Nr. 2, S 21. 261
Vgl. Drinkmann, A. (1990): Methodenkritische Untersuchungen zur Metaanalyse, Weinheim, S. 20ff; Hun-ter, J.E./Schmidt, F.L. (1990): Methods of Meta-Analysis. Correcting Error and Bias in Research Fin-dings, Newbury Park, S. 506ff; Glass, G.V./McGaw, B./Smith, M.L. (1981): Meta-Analysis in Social Re-search, Beverly Hills, S. 217ff.
262 Vgl. Hunter, J.E./Schmidt, F.L. (1990): Methods of Meta-Analysis. Correcting Error and Bias in Research
Findings, Newbury Park, S. 506ff. 263
Vgl. Glass, G.V./McGaw, B./Smith, M.L. (1981): Meta-Analysis in Social Research, Beverly Hills, S. 217ff.

55
einbezogen und wie unabhängige Daten behandelt, was als weiteres Problem einer Me-
tastudie zu sehen ist.264
2 Zielsetzung und Forschungsfrage
Das Ziel der vorliegenden Arbeit besteht darin, einen Überblick über den Forschungs-
stand der Neurowissenschaften zum Themenfeld „Leadership“ zu geben, wobei anhand
einer Metaanalyse untersucht werden soll, ob die einzelnen empirischen Ergebnisse sich
ergänzen, konsistent oder widersprüchlich sind.
Da sich die vorliegenden Studien mit unterschiedlichen Forschungszielen befassten, und
teilweise widersprüchliche Ergebnisse lieferten, ist es naheliegend, anhand einer Meta-
analyse zu untersuchen, inwieweit die Ergebnisse übereinstimmen. Außerdem ist die Me-
taanalyse als Sekundäranalyse das geeignete Instrument, um die gegenständlichen Stu-
dien zu einem Ganzen zusammenzutragen und die Ergebnisse in einem gemeinsamen
Kontext zu analysieren. In diesem Zusammenhang können mit der Metaanalyse auch
etwaige Forschungslücken aufgedeckt werden.
2.1 Forschungsfragen
Um das Ziel der vorliegenden Arbeit erfüllen zu können, wurden einige empirische Stu-
dien der Neurowissenschaften herangezogen, die sich mit Fragestellungen beschäftigten,
die für die Leadershipforschung relevant sind. Die Hauptforschungsfrage lautet, wie be-
reits in der Einleitung erwähnt, wie folgt:
FF: Welche Hirnregionen sind bei Aspekten der Leadershipforschung beteiligt?
Da „Leadership“ ein sehr breites Themenfeld ist, musste diesbezüglich eine weitere Un-
terteilung vorgenommen werden, da andernfalls nicht vergleichbare Studien integriert
werden265, die zu einer Verfälschung der Ergebnisse führen würde. Die eingangs erwähn-
te Forschungsfrage wird sohin in weitere Forschungsfragen unterteilt, wobei die Untertei-
lung korrespondierend zu Kapitel B.4.3 erfolgt. Die Unterforschungsfragen lauten wie
folgt:
FF a) Welche Hirnregionen sind bei Geführten beim Anblick von erfolgreichen im
Vergleich zu weniger erfolgreichen Führungspersonen beteiligt?
264
Vgl. Drinkmann, A. (1990): Methodenkritische Untersuchungen zur Metaanalyse, Weinheim, S. 20ff; Hun-ter, J.E./Schmidt, F.L. (1990): Methods of Meta-Analysis. Correcting Error and Bias in Research Fin-dings, Newbury Park, S. 506ff; Glass, G.V./McGaw, B./Smith, M.L. (1981): Meta-Analysis in Social Re-
search, Beverly Hills, S. 217ff. 265
Siehe „Apfel-Birnen-Argument“, Kapitel C.1.2.

56
FF b) Welche Hirnregionen sind bei einer bewussten sozialen Einflussnahme
der Führungsperson auf die MitarbeiterInnen, insbesondere in Form ei-
ner
ba) Belohnung
bb) charismatischen Führungskraft
bc) qualitativen Beziehung zwischen den/r Geführten und der Füh-
rungskraft,
beteiligt?
FF c) Welche Hirnregionen spielen eine Rolle im Hinblick auf führungsrele-
vante Fertigkeiten?
3 Methode
Nachdem einleitend zu diesem Kapitel kurz die Grundzüge einer Metaanalyse erklärt und
neuerlich auf die Zielsetzung der vorliegenden Arbeit sowie auf die konkret zu beantwor-
tenden Forschungsfragen eingegangen wurde, spielt nun die methodische Vorgehens-
weise die zentrale Rolle. Zunächst werden jene Kriterien, die zur Aufnahme der Studien
führten, vorgestellt, ehe darauf eingegangen wird, wie und v.a. auch in welchen Daten-
banken die Literaturrecherche durchgeführt wurde. Abschließend zu diesem methodi-
schen Teil wird noch kurz auf die Kodierung eingegangen, die in der vorliegenden Arbeit
aufgrund mangelnder Studienanzahl keine Rolle spielen wird.
3.1 Aufnahmekriterien
Zur Durchführung der gegenständlichen Metaanalyse wurden empirische Studien, die
anhand neurowissenschaftlicher Methoden Aspekte der Leadershipforschung untersuch-
ten, aufgenommen. Zu den Aspekten der Leadershipforschung zählen insbesondere die
in Kapitel B.4.3 aufgezählten Bereiche. Allerdings wurden nur jene Studien ins Set aufge-
nommen, die eine Untersuchung mittels fMRT oder mittels EEG durchführten. Dies des-
halb, weil die fMRT u.a. aufgrund der sehr guten räumlichen Auflösung zu den populärs-
ten neurowissenschaftlichen Methoden zählt266 und dementsprechend eine höhere Anzahl
an Studien mit dieser Methoden vorliegen. Die Miteinbeziehung von EEG Studien ist dar-
auf zurückzuführen, dass diese Methode vergleichsweise sehr einfach durchgeführt wer-
den kann, weshalb auch etliche Studien, die mit dieser Methode arbeiten, vorliegen.
Die Studien wurden zeitlich so eingegrenzt, dass nur jene berücksichtigt wurden, die zwi-
schen den Jahren 2009 und 2014 veröffentlicht wurden. Außerdem wurden nur jene empi-
266
Vgl. dazu Kapitel B.3.2.2.

57
rischen Forschungsarbeiten berücksichtigt, bei denen die Primärliteratur zugänglich war.
Studien, die in Zeitungsartikel publiziert wurden, bei denen die Primärquelle unauffindbar
war, mussten demnach ausgeschieden werden.267 Außerdem konnten auch noch nicht
publizierte neurowissenschaftliche Experimente, die Aspekte der Führungsforschung be-
handelten, mangels Zitierfähigkeit nicht berücksichtigt werden.268 Aufgrund bis dato kaum
vorhandener Studien, in denen Führungspersonen als ProbandInnen herangezogen wur-
den, wurden alle Studien, unabhängig von den Professionen der Untersuchungspersonen,
mit einbezogen.
3.2 Literaturrecherche
Die Literaturrecherche lief in mehreren Schritten ab. In der ersten Phase wurden diverse
Datenbanken, insbesondere „EBSCO“, „wiso“ und „Google Scholar“ mit Schlüsselwörtern
wie „fMRT“ bzw. „fMRI“, „EEG“, zusammen mit Wörtern wie „trust“, „charisma“, „decision
making“, „face perception“, jeweils in Verbindung mit dem Wort „Leadership“ auf brauch-
bare Studien durchforstet. Die daraufhin ausgeworfenen wissenschaftlichen Artikel wur-
den sodann aufgrund der Kurzzusammenfassung, sofern dies als passend erschien, in
die nähere Auswahl mit einbezogen. Jene Artikel, die auch nach Durchsicht der Kurzzu-
sammenfassung vielversprechend erschienen, wurden zu den potentiellen Studien, die für
die gegenständliche Metaanalyse herangezogen werden konnten, gezählt.
Parallel zur Literaturrecherche wurden jene Artikel, die potentiell für die Analyse herange-
zogen werden konnten auf eventuelle weitere, für die gegenständliche Metaanalyse rele-
vante Studien durchsucht. Auf diese Art und Weise konnten auch einige Studien, die bei
der Recherche über die obgenannten Datenbanken nicht ausfindig gemacht werden konn-
ten, in die Analyse mit einbezogen werden.
In die gegenständliche Analyse wurden nur Studien mit einbezogen, die allgemein, bezie-
hungsweise mittels oben genannter Datenbanken der Universität Graz kostenfrei zugäng-
lich waren. Im Umkehrschluss bedeutet dies, dass Studien, auf die der Campus der Uni-
versität keinen kostenfreien Zugang hatte, in die Metaanalyse grds. nicht einbezogen
werden. Davon ausgenommen war lediglich die Studie von Boyatzis et al.269, die das Insi-
tut für Entrepreneurship und Unternehmensführung für diese Arbeit zur Verfügung stellte.
267
z.B. Boyatzis, R.E/Jack, A./Cesaro, R./Passarelli, A./Khawaja, M. (2010): Coaching with Compassion. An fMRI Study of Coaching to the Positive or Negative Emotional Attractor, presented at the Annual Mee-ting of the Academy of Management, Montreal.
268 Studie des Institutes für Unternehmensführung und Entrepreneurship an der Karl-Franzens-Universität
Graz zum Thema „Was motiviert mich im Berufsleben“ (noch nicht publiziert). 269
Vgl. Boyatzis, R.E./Passarelli, A.M./Koenig, K./Lowe, M./Mathew, B./Stoller, J.K./Phillips, M. (2012): Ex-
amination of the neural substrates activated in memories of experience with resonant and dissonant lea-ders, in: The Leadership Quarterly, Jg. 23, S. 259-272.
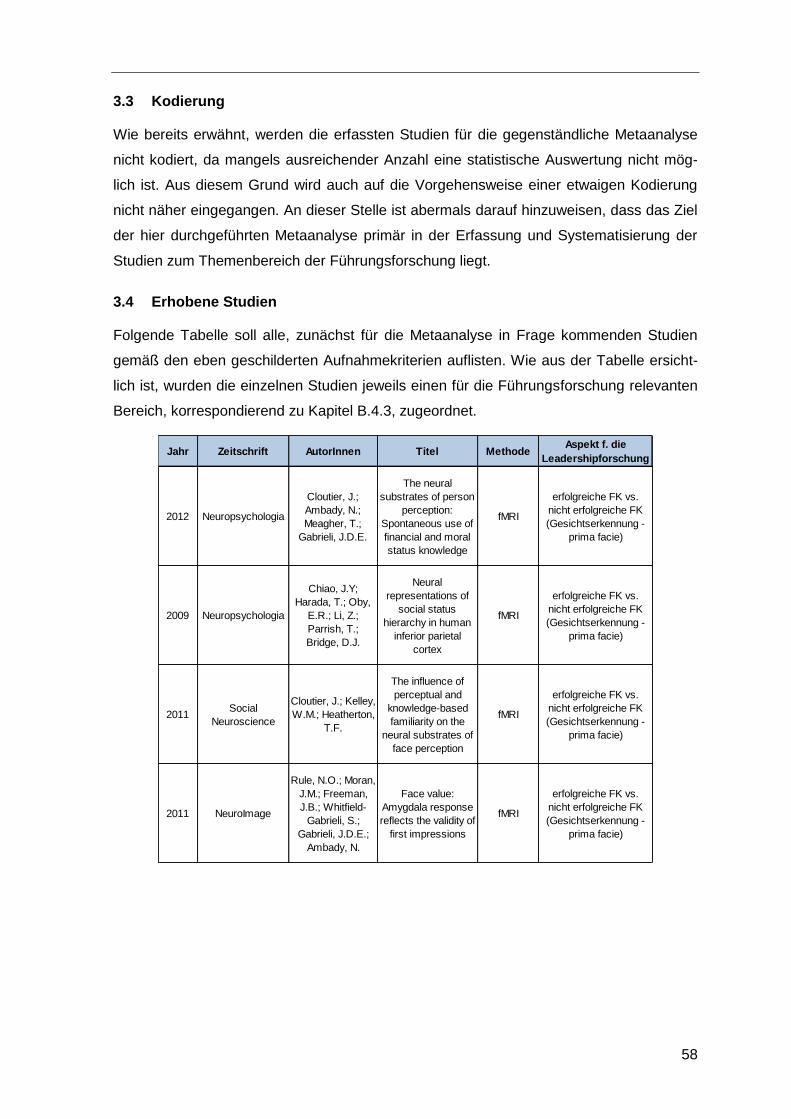
58
3.3 Kodierung
Wie bereits erwähnt, werden die erfassten Studien für die gegenständliche Metaanalyse
nicht kodiert, da mangels ausreichender Anzahl eine statistische Auswertung nicht mög-
lich ist. Aus diesem Grund wird auch auf die Vorgehensweise einer etwaigen Kodierung
nicht näher eingegangen. An dieser Stelle ist abermals darauf hinzuweisen, dass das Ziel
der hier durchgeführten Metaanalyse primär in der Erfassung und Systematisierung der
Studien zum Themenbereich der Führungsforschung liegt.
3.4 Erhobene Studien
Folgende Tabelle soll alle, zunächst für die Metaanalyse in Frage kommenden Studien
gemäß den eben geschilderten Aufnahmekriterien auflisten. Wie aus der Tabelle ersicht-
lich ist, wurden die einzelnen Studien jeweils einen für die Führungsforschung relevanten
Bereich, korrespondierend zu Kapitel B.4.3, zugeordnet.
Jahr Zeitschrift AutorInnen Titel MethodeAspekt f. die
Leadershipforschung
2012 Neuropsychologia
Cloutier, J.;
Ambady, N.;
Meagher, T.;
Gabrieli, J.D.E.
The neural
substrates of person
perception:
Spontaneous use of
financial and moral
status knowledge
fMRI
erfolgreiche FK vs.
nicht erfolgreiche FK
(Gesichtserkennung -
prima facie)
2009 Neuropsychologia
Chiao, J.Y;
Harada, T.; Oby,
E.R.; Li, Z.;
Parrish, T.;
Bridge, D.J.
Neural
representations of
social status
hierarchy in human
inferior parietal
cortex
fMRI
erfolgreiche FK vs.
nicht erfolgreiche FK
(Gesichtserkennung -
prima facie)
2011Social
Neuroscience
Cloutier, J.; Kelley,
W.M.; Heatherton,
T.F.
The influence of
perceptual and
knowledge-based
familiarity on the
neural substrates of
face perception
fMRI
erfolgreiche FK vs.
nicht erfolgreiche FK
(Gesichtserkennung -
prima facie)
2011 NeuroImage
Rule, N.O.; Moran,
J.M.; Freeman,
J.B.; Whitfield-
Gabrieli, S.;
Gabrieli, J.D.E.;
Ambady, N.
Face value:
Amygdala response
reflects the validity of
first impressions
fMRI
erfolgreiche FK vs.
nicht erfolgreiche FK
(Gesichtserkennung -
prima facie)
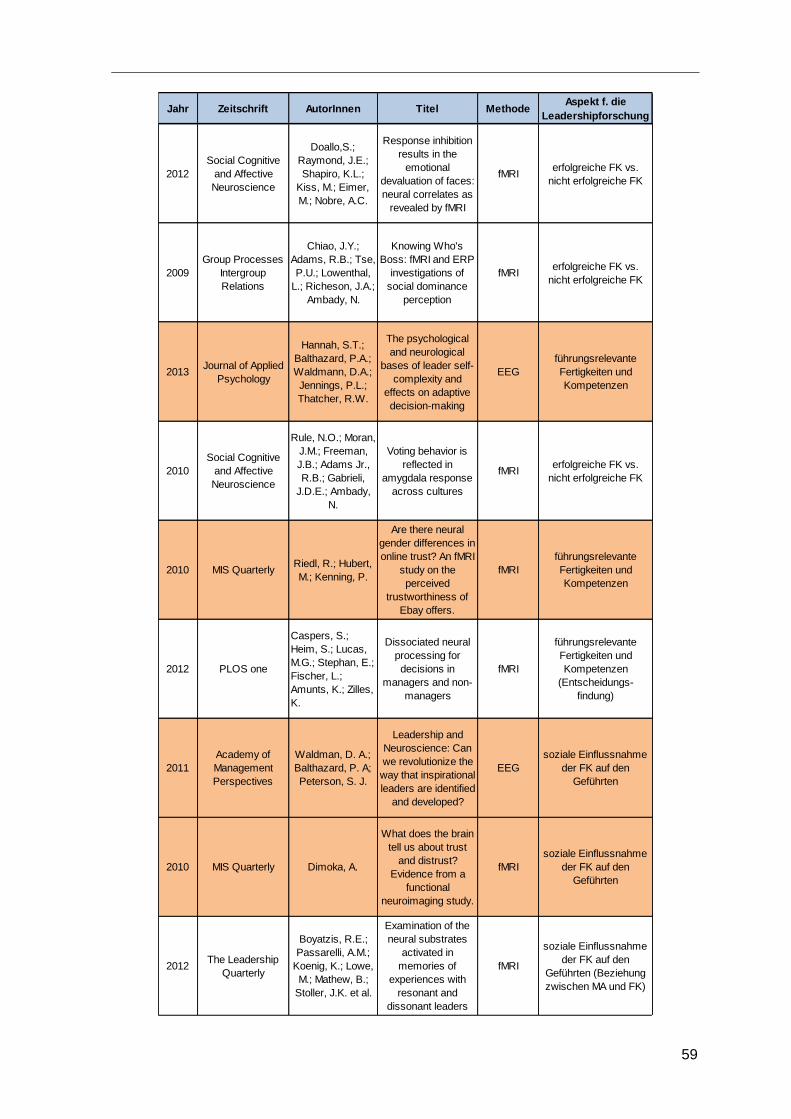
59
Jahr Zeitschrift AutorInnen Titel MethodeAspekt f. die
Leadershipforschung
2012
Social Cognitive
and Affective
Neuroscience
Doallo,S.;
Raymond, J.E.;
Shapiro, K.L.;
Kiss, M.; Eimer,
M.; Nobre, A.C.
Response inhibition
results in the
emotional
devaluation of faces:
neural correlates as
revealed by fMRI
fMRIerfolgreiche FK vs.
nicht erfolgreiche FK
2009
Group Processes
Intergroup
Relations
Chiao, J.Y.;
Adams, R.B.; Tse,
P.U.; Lowenthal,
L.; Richeson, J.A.;
Ambady, N.
Knowing Who's
Boss: fMRI and ERP
investigations of
social dominance
perception
fMRIerfolgreiche FK vs.
nicht erfolgreiche FK
2013Journal of Applied
Psychology
Hannah, S.T.;
Balthazard, P.A.;
Waldmann, D.A.;
Jennings, P.L.;
Thatcher, R.W.
The psychological
and neurological
bases of leader self-
complexity and
effects on adaptive
decision-making
EEG
führungsrelevante
Fertigkeiten und
Kompetenzen
2010
Social Cognitive
and Affective
Neuroscience
Rule, N.O.; Moran,
J.M.; Freeman,
J.B.; Adams Jr.,
R.B.; Gabrieli,
J.D.E.; Ambady,
N.
Voting behavior is
reflected in
amygdala response
across cultures
fMRIerfolgreiche FK vs.
nicht erfolgreiche FK
2010 MIS QuarterlyRiedl, R.; Hubert,
M.; Kenning, P.
Are there neural
gender differences in
online trust? An fMRI
study on the
perceived
trustworthiness of
Ebay offers.
fMRI
führungsrelevante
Fertigkeiten und
Kompetenzen
2012 PLOS one
Caspers, S.;
Heim, S.; Lucas,
M.G.; Stephan, E.;
Fischer, L.;
Amunts, K.; Zilles,
K.
Dissociated neural
processing for
decisions in
managers and non-
managers
fMRI
führungsrelevante
Fertigkeiten und
Kompetenzen
(Entscheidungs-
findung)
2011
Academy of
Management
Perspectives
Waldman, D. A.;
Balthazard, P. A;
Peterson, S. J.
Leadership and
Neuroscience: Can
we revolutionize the
way that inspirational
leaders are identified
and developed?
EEG
soziale Einflussnahme
der FK auf den
Geführten
2010 MIS Quarterly Dimoka, A.
What does the brain
tell us about trust
and distrust?
Evidence from a
functional
neuroimaging study.
fMRI
soziale Einflussnahme
der FK auf den
Geführten
2012The Leadership
Quarterly
Boyatzis, R.E.;
Passarelli, A.M.;
Koenig, K.; Lowe,
M.; Mathew, B.;
Stoller, J.K. et al.
Examination of the
neural substrates
activated in
memories of
experiences with
resonant and
dissonant leaders
fMRI
soziale Einflussnahme
der FK auf den
Geführten (Beziehung
zwischen MA und FK)
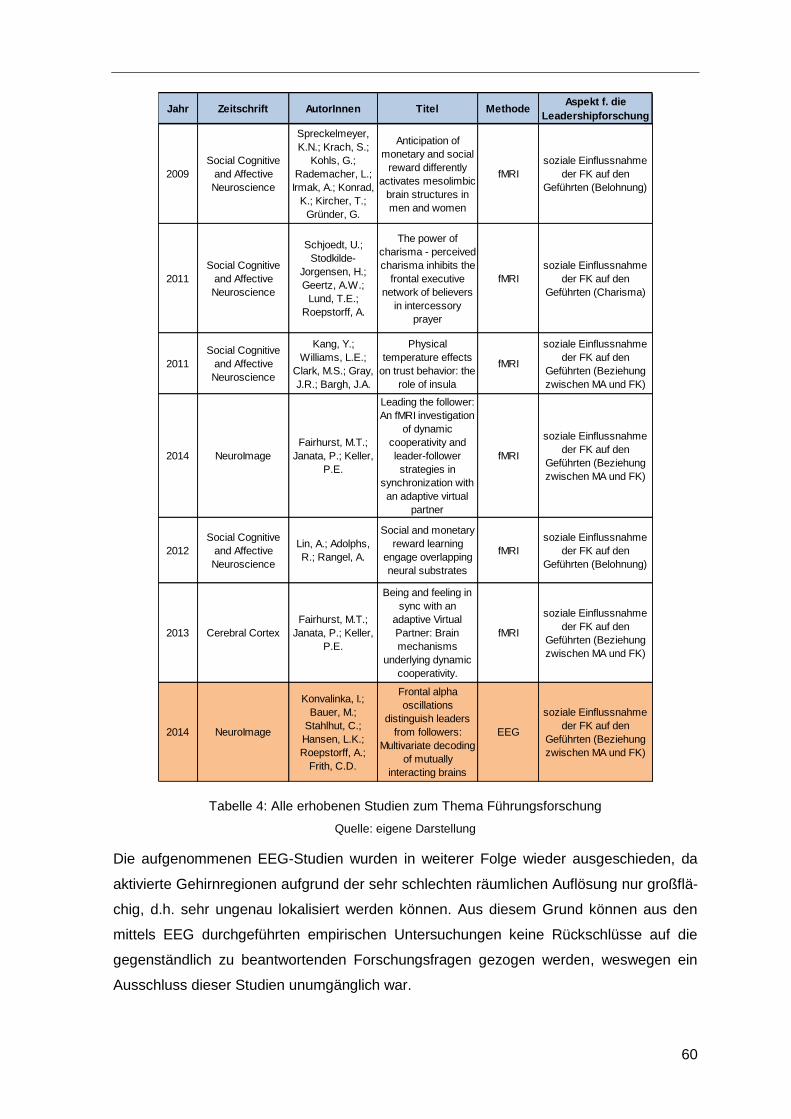
60
Jahr Zeitschrift AutorInnen Titel MethodeAspekt f. die
Leadershipforschung
2009
Social Cognitive
and Affective
Neuroscience
Spreckelmeyer,
K.N.; Krach, S.;
Kohls, G.;
Rademacher, L.;
Irmak, A.; Konrad,
K.; Kircher, T.;
Gründer, G.
Anticipation of
monetary and social
reward differently
activates mesolimbic
brain structures in
men and women
fMRI
soziale Einflussnahme
der FK auf den
Geführten (Belohnung)
2011
Social Cognitive
and Affective
Neuroscience
Schjoedt, U.;
Stodkilde-
Jorgensen, H.;
Geertz, A.W.;
Lund, T.E.;
Roepstorff, A.
The power of
charisma - perceived
charisma inhibits the
frontal executive
network of believers
in intercessory
prayer
fMRI
soziale Einflussnahme
der FK auf den
Geführten (Charisma)
2011
Social Cognitive
and Affective
Neuroscience
Kang, Y.;
Williams, L.E.;
Clark, M.S.; Gray,
J.R.; Bargh, J.A.
Physical
temperature effects
on trust behavior: the
role of insula
fMRI
soziale Einflussnahme
der FK auf den
Geführten (Beziehung
zwischen MA und FK)
2014 NeuroImage
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Leading the follower:
An fMRI investigation
of dynamic
cooperativity and
leader-follower
strategies in
synchronization with
an adaptive virtual
partner
fMRI
soziale Einflussnahme
der FK auf den
Geführten (Beziehung
zwischen MA und FK)
2012
Social Cognitive
and Affective
Neuroscience
Lin, A.; Adolphs,
R.; Rangel, A.
Social and monetary
reward learning
engage overlapping
neural substrates
fMRI
soziale Einflussnahme
der FK auf den
Geführten (Belohnung)
2013 Cerebral Cortex
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Being and feeling in
sync with an
adaptive Virtual
Partner: Brain
mechanisms
underlying dynamic
cooperativity.
fMRI
soziale Einflussnahme
der FK auf den
Geführten (Beziehung
zwischen MA und FK)
2014 NeuroImage
Konvalinka, I.;
Bauer, M.;
Stahlhut, C.;
Hansen, L.K.;
Roepstorff, A.;
Frith, C.D.
Frontal alpha
oscillations
distinguish leaders
from followers:
Multivariate decoding
of mutually
interacting brains
EEG
soziale Einflussnahme
der FK auf den
Geführten (Beziehung
zwischen MA und FK)
Tabelle 4: Alle erhobenen Studien zum Thema Führungsforschung
Quelle: eigene Darstellung
Die aufgenommenen EEG-Studien wurden in weiterer Folge wieder ausgeschieden, da
aktivierte Gehirnregionen aufgrund der sehr schlechten räumlichen Auflösung nur großflä-
chig, d.h. sehr ungenau lokalisiert werden können. Aus diesem Grund können aus den
mittels EEG durchgeführten empirischen Untersuchungen keine Rückschlüsse auf die
gegenständlich zu beantwortenden Forschungsfragen gezogen werden, weswegen ein
Ausschluss dieser Studien unumgänglich war.

61
Außerdem wurden die Studie von Dimoka, A. (2010)270 sowie die Studie von Riedl,
R./Hubert, M./Kenning, P. (2010)271 von der weiteren Auswertung ausgeschlossen, zumal
diese Studiendesigns neurophysiologische Auswirkungen untersuchten, die dem be-
triebswirtschaftlichen Teilbereich „Marketing“ zuzuordnen sind.
Insgesamt wurden sohin, nach Abzug der in der Tabelle rot hinterlegten EEG-Studien und
der beiden Studien, die dem Themenbereich „Marketing“ zuzuordnen sind, 15 Studien für
die gegenständliche Metaanalyse erfasst, die in weiterer Folge analysiert werden.
4 Datenanalyse
Dieses Teilkapitel beschäftigt sich nun mit der Analyse der Daten der erfassten Studien.
Zunächst wird der zeitliche Verlauf der Studienanzahl untersucht. Im Anschluss daran
erfolgt die Berechnung der durchschnittlichen Stichprobenanzahl der erfassten fMRT Stu-
dien. Des Weiteren wird die durchschnittliche Stichprobenanzahl im zeitlichen Verlauf
analysiert. Hierbei soll beantwortet werden, ob im Zeitablauf die Anzahl der Studienteil-
nehmerInnen gestiegen ist. Danach wird untersucht, welche Stimuli am häufigsten einge-
setzt wurden, ehe im Abschluss dieses Kapitels - je nach Aspekt für die Leadershipfor-
schung aufgegliedert - die Studien im Hinblick auf die jeweils aktivierten Hirnregionen
ausgewertet werden.
4.1 Eingesetzte Methoden der Neurowissenschaften
Im ersten Schritt wurden alle272, zunächst erfassten Studien auf die jeweilige zur Anwen-
dung gekommene neurowissenschaftliche Methode untersucht.273 Tabelle 5 spiegelt die
absoluten sowie relativen Zahlen der verwendeten neurowissenschaftlichen Methoden
wieder.
fMRT 17 85%
EEG 3 15%
20 100%
Tabelle 5: Absolute und relative Anzahl der verwendeten Methoden
Quelle: eigene Darstellung
270
Vgl. Dimoka, A. (2010): What does the brain tell us about trust and distrust? Evidece from a functional neuroimaging study, in: MIS Quarterly, Vol. 34, Nr.2, S. 373–396.
271 Vgl. Riedl, R./Hubert, M./Kenning, P. (2010): Are there neural gender differences in online trust? An fMRI
study on the perceived trustworthiness of ebay offers, in: MIS Quarterly, Vol. 34, Nr. 2, S. 397–428. 272
Für diese Analyse wurden auch die ausgeschlossenen Studien mit einbezogen. 273
Siehe dazu Tabelle 4.
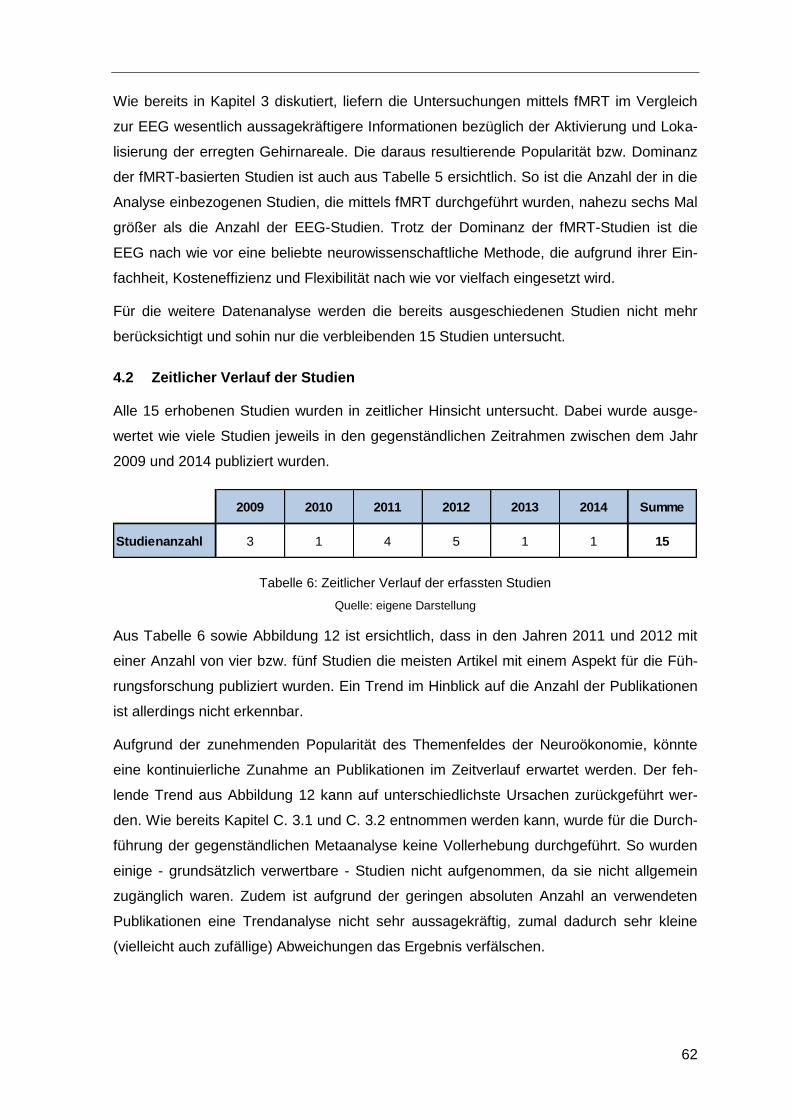
62
Wie bereits in Kapitel 3 diskutiert, liefern die Untersuchungen mittels fMRT im Vergleich
zur EEG wesentlich aussagekräftigere Informationen bezüglich der Aktivierung und Loka-
lisierung der erregten Gehirnareale. Die daraus resultierende Popularität bzw. Dominanz
der fMRT-basierten Studien ist auch aus Tabelle 5 ersichtlich. So ist die Anzahl der in die
Analyse einbezogenen Studien, die mittels fMRT durchgeführt wurden, nahezu sechs Mal
größer als die Anzahl der EEG-Studien. Trotz der Dominanz der fMRT-Studien ist die
EEG nach wie vor eine beliebte neurowissenschaftliche Methode, die aufgrund ihrer Ein-
fachheit, Kosteneffizienz und Flexibilität nach wie vor vielfach eingesetzt wird.
Für die weitere Datenanalyse werden die bereits ausgeschiedenen Studien nicht mehr
berücksichtigt und sohin nur die verbleibenden 15 Studien untersucht.
4.2 Zeitlicher Verlauf der Studien
Alle 15 erhobenen Studien wurden in zeitlicher Hinsicht untersucht. Dabei wurde ausge-
wertet wie viele Studien jeweils in den gegenständlichen Zeitrahmen zwischen dem Jahr
2009 und 2014 publiziert wurden.
2009 2010 2011 2012 2013 2014 Summe
Studienanzahl 3 1 4 5 1 1 15
Tabelle 6: Zeitlicher Verlauf der erfassten Studien
Quelle: eigene Darstellung
Aus Tabelle 6 sowie Abbildung 12 ist ersichtlich, dass in den Jahren 2011 und 2012 mit
einer Anzahl von vier bzw. fünf Studien die meisten Artikel mit einem Aspekt für die Füh-
rungsforschung publiziert wurden. Ein Trend im Hinblick auf die Anzahl der Publikationen
ist allerdings nicht erkennbar.
Aufgrund der zunehmenden Popularität des Themenfeldes der Neuroökonomie, könnte
eine kontinuierliche Zunahme an Publikationen im Zeitverlauf erwartet werden. Der feh-
lende Trend aus Abbildung 12 kann auf unterschiedlichste Ursachen zurückgeführt wer-
den. Wie bereits Kapitel C. 3.1 und C. 3.2 entnommen werden kann, wurde für die Durch-
führung der gegenständlichen Metaanalyse keine Vollerhebung durchgeführt. So wurden
einige - grundsätzlich verwertbare - Studien nicht aufgenommen, da sie nicht allgemein
zugänglich waren. Zudem ist aufgrund der geringen absoluten Anzahl an verwendeten
Publikationen eine Trendanalyse nicht sehr aussagekräftig, zumal dadurch sehr kleine
(vielleicht auch zufällige) Abweichungen das Ergebnis verfälschen.
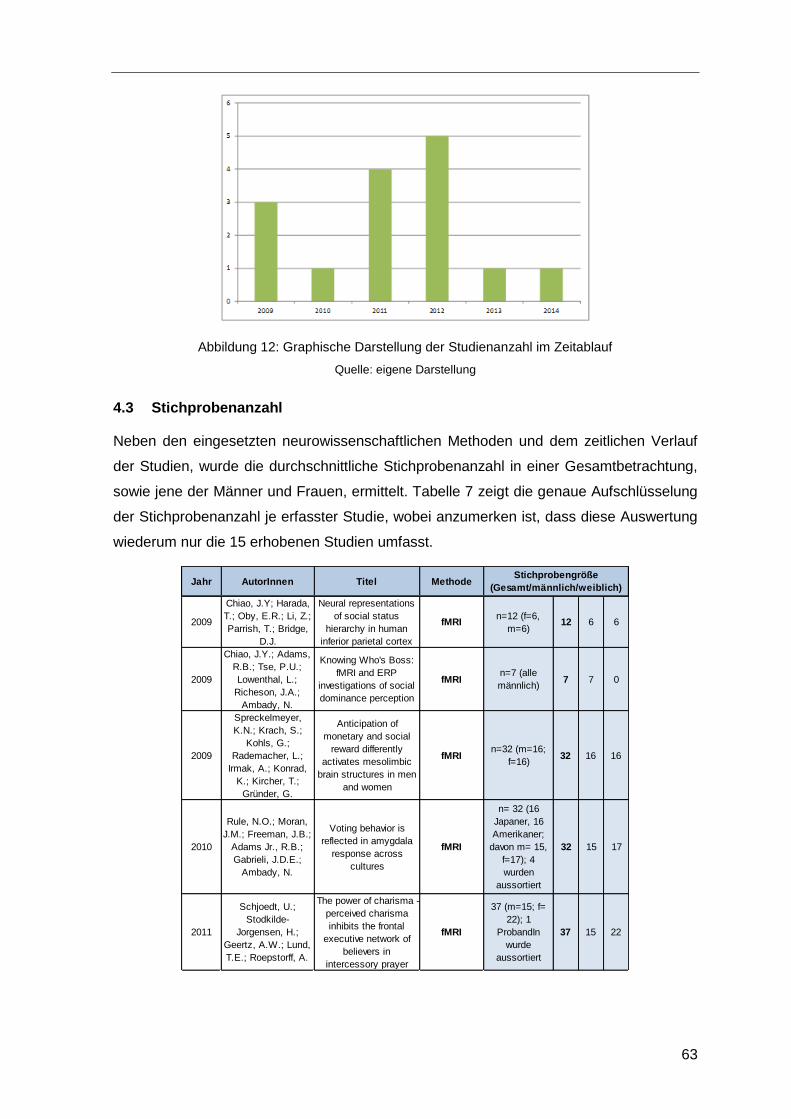
63
Abbildung 12: Graphische Darstellung der Studienanzahl im Zeitablauf
Quelle: eigene Darstellung
4.3 Stichprobenanzahl
Neben den eingesetzten neurowissenschaftlichen Methoden und dem zeitlichen Verlauf
der Studien, wurde die durchschnittliche Stichprobenanzahl in einer Gesamtbetrachtung,
sowie jene der Männer und Frauen, ermittelt. Tabelle 7 zeigt die genaue Aufschlüsselung
der Stichprobenanzahl je erfasster Studie, wobei anzumerken ist, dass diese Auswertung
wiederum nur die 15 erhobenen Studien umfasst.
Jahr AutorInnen Titel Methode
2009
Chiao, J.Y; Harada,
T.; Oby, E.R.; Li, Z.;
Parrish, T.; Bridge,
D.J.
Neural representations
of social status
hierarchy in human
inferior parietal cortex
fMRIn=12 (f=6,
m=6)12 6 6
2009
Chiao, J.Y.; Adams,
R.B.; Tse, P.U.;
Lowenthal, L.;
Richeson, J.A.;
Ambady, N.
Knowing Who's Boss:
fMRI and ERP
investigations of social
dominance perception
fMRIn=7 (alle
männlich)7 7 0
2009
Spreckelmeyer,
K.N.; Krach, S.;
Kohls, G.;
Rademacher, L.;
Irmak, A.; Konrad,
K.; Kircher, T.;
Gründer, G.
Anticipation of
monetary and social
reward differently
activates mesolimbic
brain structures in men
and women
fMRIn=32 (m=16;
f=16)32 16 16
2010
Rule, N.O.; Moran,
J.M.; Freeman, J.B.;
Adams Jr., R.B.;
Gabrieli, J.D.E.;
Ambady, N.
Voting behavior is
reflected in amygdala
response across
cultures
fMRI
n= 32 (16
Japaner, 16
Amerikaner;
davon m= 15,
f=17); 4
wurden
aussortiert
32 15 17
2011
Schjoedt, U.;
Stodkilde-
Jorgensen, H.;
Geertz, A.W.; Lund,
T.E.; Roepstorff, A.
The power of charisma -
perceived charisma
inhibits the frontal
executive network of
believers in
intercessory prayer
fMRI
37 (m=15; f=
22); 1
ProbandIn
wurde
aussortiert
37 15 22
Stichprobengröße
(Gesamt/männlich/weiblich)
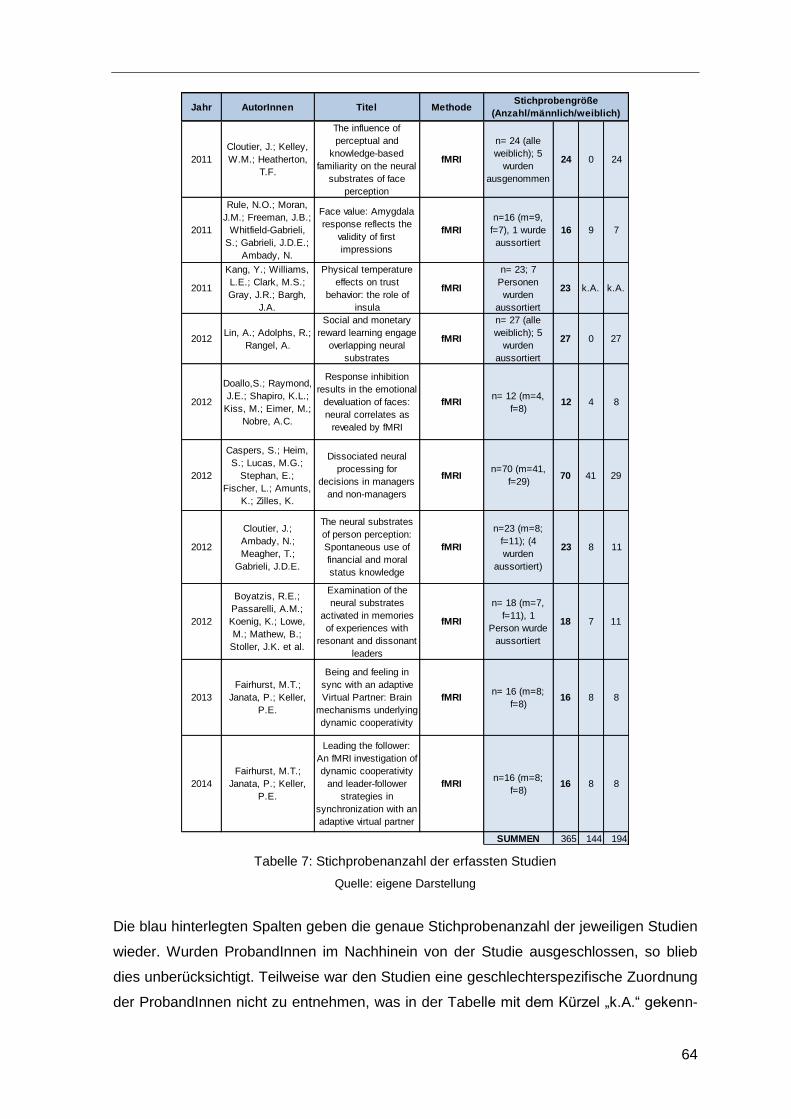
64
Jahr AutorInnen Titel Methode
2011
Cloutier, J.; Kelley,
W.M.; Heatherton,
T.F.
The influence of
perceptual and
knowledge-based
familiarity on the neural
substrates of face
perception
fMRI
n= 24 (alle
weiblich); 5
wurden
ausgenommen
24 0 24
2011
Rule, N.O.; Moran,
J.M.; Freeman, J.B.;
Whitfield-Gabrieli,
S.; Gabrieli, J.D.E.;
Ambady, N.
Face value: Amygdala
response reflects the
validity of first
impressions
fMRI
n=16 (m=9,
f=7), 1 wurde
aussortiert
16 9 7
2011
Kang, Y.; Williams,
L.E.; Clark, M.S.;
Gray, J.R.; Bargh,
J.A.
Physical temperature
effects on trust
behavior: the role of
insula
fMRI
n= 23; 7
Personen
wurden
aussortiert
23 k.A. k.A.
2012Lin, A.; Adolphs, R.;
Rangel, A.
Social and monetary
reward learning engage
overlapping neural
substrates
fMRI
n= 27 (alle
weiblich); 5
wurden
aussortiert
27 0 27
2012
Doallo,S.; Raymond,
J.E.; Shapiro, K.L.;
Kiss, M.; Eimer, M.;
Nobre, A.C.
Response inhibition
results in the emotional
devaluation of faces:
neural correlates as
revealed by fMRI
fMRIn= 12 (m=4,
f=8)12 4 8
2012
Caspers, S.; Heim,
S.; Lucas, M.G.;
Stephan, E.;
Fischer, L.; Amunts,
K.; Zilles, K.
Dissociated neural
processing for
decisions in managers
and non-managers
fMRIn=70 (m=41,
f=29)70 41 29
2012
Cloutier, J.;
Ambady, N.;
Meagher, T.;
Gabrieli, J.D.E.
The neural substrates
of person perception:
Spontaneous use of
financial and moral
status knowledge
fMRI
n=23 (m=8;
f=11); (4
wurden
aussortiert)
23 8 11
2012
Boyatzis, R.E.;
Passarelli, A.M.;
Koenig, K.; Lowe,
M.; Mathew, B.;
Stoller, J.K. et al.
Examination of the
neural substrates
activated in memories
of experiences with
resonant and dissonant
leaders
fMRI
n= 18 (m=7,
f=11), 1
Person wurde
aussortiert
18 7 11
2013
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Being and feeling in
sync with an adaptive
Virtual Partner: Brain
mechanisms underlying
dynamic cooperativity
fMRIn= 16 (m=8;
f=8)16 8 8
2014
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Leading the follower:
An fMRI investigation of
dynamic cooperativity
and leader-follower
strategies in
synchronization with an
adaptive virtual partner
fMRIn=16 (m=8;
f=8)16 8 8
SUMMEN 365 144 194
Stichprobengröße
(Anzahl/männlich/weiblich)
Tabelle 7: Stichprobenanzahl der erfassten Studien
Quelle: eigene Darstellung
Die blau hinterlegten Spalten geben die genaue Stichprobenanzahl der jeweiligen Studien
wieder. Wurden ProbandInnen im Nachhinein von der Studie ausgeschlossen, so blieb
dies unberücksichtigt. Teilweise war den Studien eine geschlechterspezifische Zuordnung
der ProbandInnen nicht zu entnehmen, was in der Tabelle mit dem Kürzel „k.A.“ gekenn-
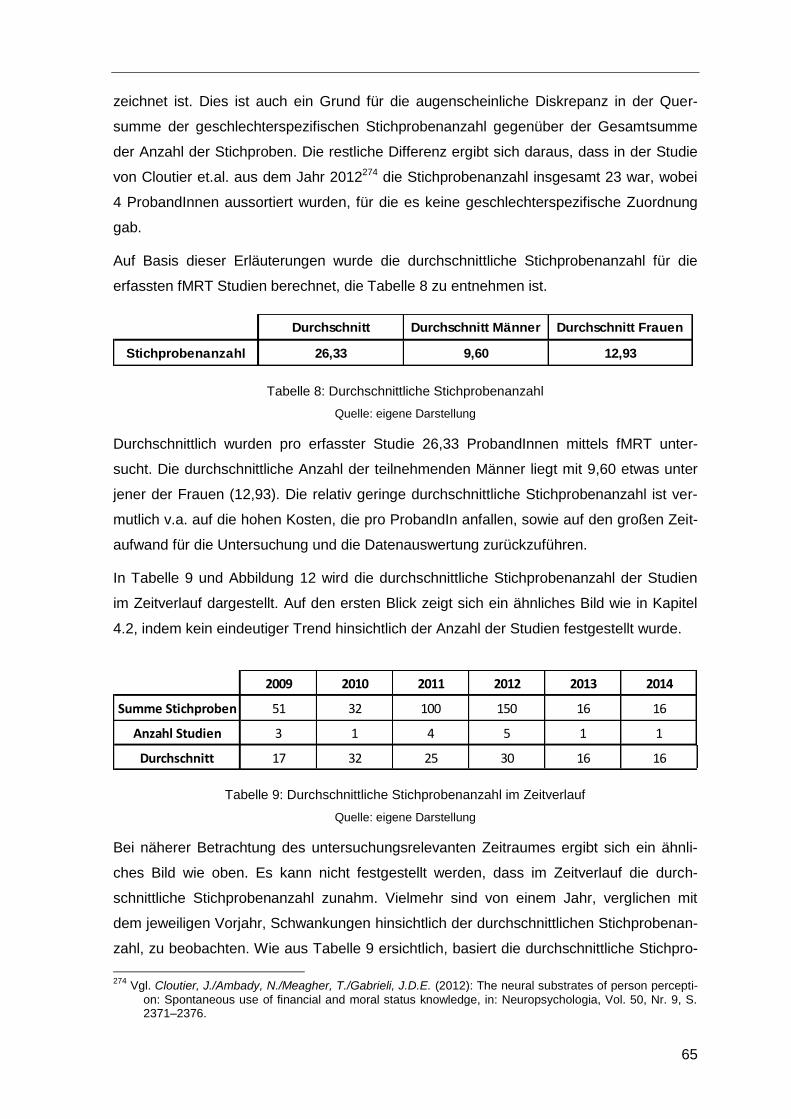
65
zeichnet ist. Dies ist auch ein Grund für die augenscheinliche Diskrepanz in der Quer-
summe der geschlechterspezifischen Stichprobenanzahl gegenüber der Gesamtsumme
der Anzahl der Stichproben. Die restliche Differenz ergibt sich daraus, dass in der Studie
von Cloutier et.al. aus dem Jahr 2012274 die Stichprobenanzahl insgesamt 23 war, wobei
4 ProbandInnen aussortiert wurden, für die es keine geschlechterspezifische Zuordnung
gab.
Auf Basis dieser Erläuterungen wurde die durchschnittliche Stichprobenanzahl für die
erfassten fMRT Studien berechnet, die Tabelle 8 zu entnehmen ist.
Durchschnitt Durchschnitt Männer Durchschnitt Frauen
Stichprobenanzahl 26,33 9,60 12,93
Tabelle 8: Durchschnittliche Stichprobenanzahl
Quelle: eigene Darstellung
Durchschnittlich wurden pro erfasster Studie 26,33 ProbandInnen mittels fMRT unter-
sucht. Die durchschnittliche Anzahl der teilnehmenden Männer liegt mit 9,60 etwas unter
jener der Frauen (12,93). Die relativ geringe durchschnittliche Stichprobenanzahl ist ver-
mutlich v.a. auf die hohen Kosten, die pro ProbandIn anfallen, sowie auf den großen Zeit-
aufwand für die Untersuchung und die Datenauswertung zurückzuführen.
In Tabelle 9 und Abbildung 12 wird die durchschnittliche Stichprobenanzahl der Studien
im Zeitverlauf dargestellt. Auf den ersten Blick zeigt sich ein ähnliches Bild wie in Kapitel
4.2, indem kein eindeutiger Trend hinsichtlich der Anzahl der Studien festgestellt wurde.
2009 2010 2011 2012 2013 2014
Summe Stichproben 51 32 100 150 16 16
Anzahl Studien 3 1 4 5 1 1
Durchschnitt 17 32 25 30 16 16
Tabelle 9: Durchschnittliche Stichprobenanzahl im Zeitverlauf
Quelle: eigene Darstellung
Bei näherer Betrachtung des untersuchungsrelevanten Zeitraumes ergibt sich ein ähnli-
ches Bild wie oben. Es kann nicht festgestellt werden, dass im Zeitverlauf die durch-
schnittliche Stichprobenanzahl zunahm. Vielmehr sind von einem Jahr, verglichen mit
dem jeweiligen Vorjahr, Schwankungen hinsichtlich der durchschnittlichen Stichprobenan-
zahl, zu beobachten. Wie aus Tabelle 9 ersichtlich, basiert die durchschnittliche Stichpro-
274
Vgl. Cloutier, J./Ambady, N./Meagher, T./Gabrieli, J.D.E. (2012): The neural substrates of person percepti-
on: Spontaneous use of financial and moral status knowledge, in: Neuropsychologia, Vol. 50, Nr. 9, S. 2371–2376.

66
benanzahl der Studien aus den Jahren 2013 und 2014 jeweils auf einer einzigen Studie.
Dies ist insbesondere darauf zurückzuführen, dass für die gegenständliche Analyse nur
Studien, die bis zum Stichtag mit 20. Juni 2014 publiziert wurden, mit einbezogen wurden.
Außerdem ist abermals darauf hinzuweisen, dass aufgrund der erwähnten Aufnahmekrite-
rien keine Vollerhebung durchgeführt wurde. Daher kann auch für diese Jahre keine aus-
sagekräftige Bewertung der durchschnittlichen Anzahl der Stichproben pro fMRT-Studie
vorgenommen werden. Die Entwicklung der durchschnittlichen Stichprobenanzahl wird
durch folgende Tabelle graphisch veranschaulicht.
17
32
25
30
16 16
0
5
10
15
20
25
30
35
2009 2010 2011 2012 2013 2014
Abbildung 13: Graphische Darstellung der durchschnittlichen Stichprobenanzahl im Zeitverlauf
Quelle: eigene Darstellung
4.4 Eingesetzte Stimuli
Die in die Analyse mit einbezogenen Studien werden in diesem Kapitel im Hinblick auf die
eingesetzten Stimuli untersucht. Tabelle 10 gibt einen ersten Überblick über den Typus
der eingesetzten Reize sowie über den Ablauf der Studien selbst. Die Zielsetzungen bzw.
Hypothesen der jeweiligen Studien sind dem Anhang zu entnehmen.
Jahr AutorInnen Titel
2009
Chiao, J.Y; Harada,
T.; Oby, E.R.; Li, Z.;
Parrish, T.; Bridge,
D.J.
Neural representations
of social status
hierarchy in human
inferior parietal cortex
Die Stimuli bestanden aus 96 grauen Fotos, wovon 24 Fotos arabische Ziffern
zwischen 33 und 99 zeigten, 24 Fotos stellten Statusabzeichen von NROTC
dar, 24 Fotos lichteten verschiedene NROTC Offiziere ab und die restlichen 24
Fotos zeigten verschiedene Autos der Marke "Toyota". Den ProbandInnen wurde
jeweils ein Bild (Zahl, Statusabzeichen, Offizier oder Toyota) gezeigt, dass sie
in weiterer Folge bewerten mussten, und zwar dahingehend, ob das Bild im
Hinblick auf einen gewählten Mittelwert besser, schlechter oder gleich gut ist.
Jede Runde (insgesamt 4) bestand aus 3 Blöcken zu je 3 Typen, die
hinsichtlich der semantischen Entfernung variierten. Jede Runde inkludierte
auch einen Kontrollaufgabe, in welcher die TeilnehmerInnen einen Knopf drücken
mussten, sobald die Farbe des gezeigten Kreuzes von weiß auf rot wechselte.
Fotos/Bilder
Stimuli und Design
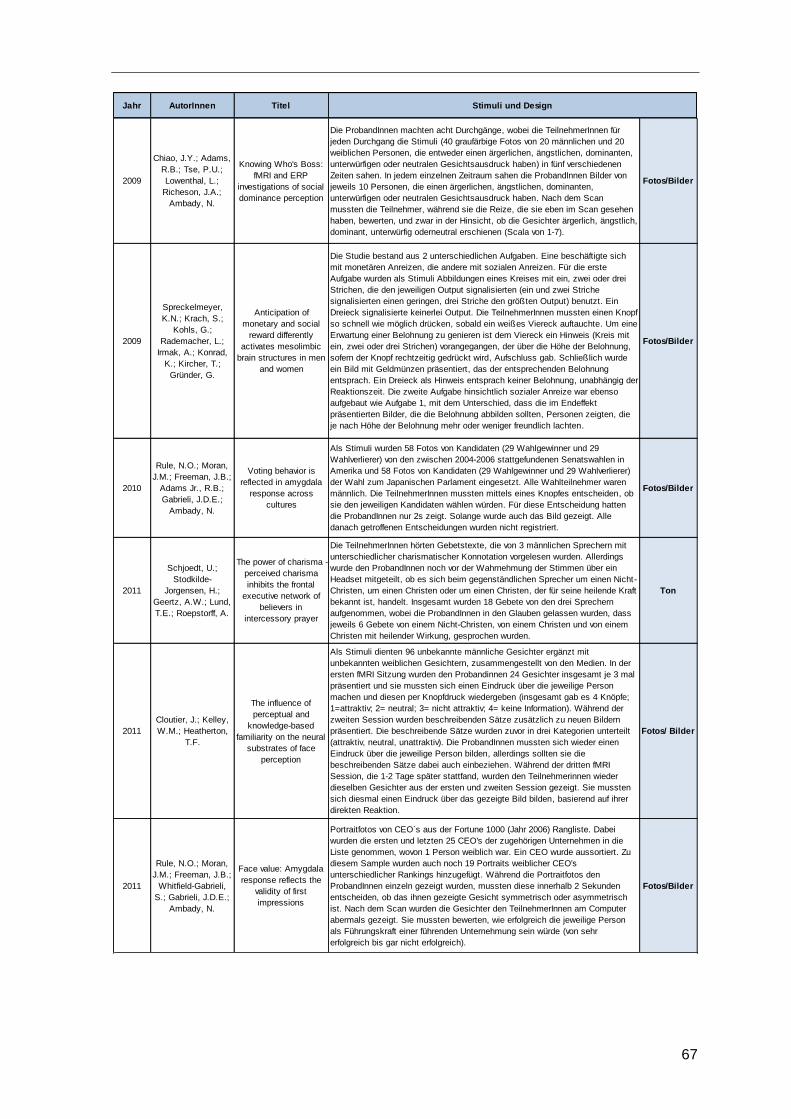
67
Jahr AutorInnen Titel
2009
Chiao, J.Y.; Adams,
R.B.; Tse, P.U.;
Lowenthal, L.;
Richeson, J.A.;
Ambady, N.
Knowing Who's Boss:
fMRI and ERP
investigations of social
dominance perception
Die ProbandInnen machten acht Durchgänge, wobei die TeilnehmerInnen für
jeden Durchgang die Stimuli (40 graufärbige Fotos von 20 männlichen und 20
weiblichen Personen, die entweder einen ärgerlichen, ängstlichen, dominanten,
unterwürfigen oder neutralen Gesichtsausdruck haben) in fünf verschiedenen
Zeiten sahen. In jedem einzelnen Zeitraum sahen die ProbandInnen Bilder von
jeweils 10 Personen, die einen ärgerlichen, ängstlichen, dominanten,
unterwürfigen oder neutralen Gesichtsausdruck haben. Nach dem Scan
mussten die Teilnehmer, während sie die Reize, die sie eben im Scan gesehen
haben, bewerten, und zwar in der Hinsicht, ob die Gesichter ärgerlich, ängstlich,
dominant, unterwürfig oderneutral erschienen (Scala von 1-7).
Fotos/Bilder
2009
Spreckelmeyer,
K.N.; Krach, S.;
Kohls, G.;
Rademacher, L.;
Irmak, A.; Konrad,
K.; Kircher, T.;
Gründer, G.
Anticipation of
monetary and social
reward differently
activates mesolimbic
brain structures in men
and women
Die Studie bestand aus 2 unterschiedlichen Aufgaben. Eine beschäftigte sich
mit monetären Anreizen, die andere mit sozialen Anreizen. Für die erste
Aufgabe wurden als Stimuli Abbildungen eines Kreises mit ein, zwei oder drei
Strichen, die den jeweiligen Output signalisierten (ein und zwei Striche
signalisierten einen geringen, drei Striche den größten Output) benutzt. Ein
Dreieck signalisierte keinerlei Output. Die TeilnehmerInnen mussten einen Knopf
so schnell wie möglich drücken, sobald ein weißes Viereck auftauchte. Um eine
Erwartung einer Belohnung zu genieren ist dem Viereck ein Hinweis (Kreis mit
ein, zwei oder drei Strichen) vorangegangen, der über die Höhe der Belohnung,
sofern der Knopf rechtzeitig gedrückt wird, Aufschluss gab. Schließlich wurde
ein Bild mit Geldmünzen präsentiert, das der entsprechenden Belohnung
entsprach. Ein Dreieck als Hinweis entsprach keiner Belohnung, unabhängig der
Reaktionszeit. Die zweite Aufgabe hinsichtlich sozialer Anreize war ebenso
aufgebaut wie Aufgabe 1, mit dem Unterschied, dass die im Endeffekt
präsentierten Bilder, die die Belohnung abbilden sollten, Personen zeigten, die
je nach Höhe der Belohnung mehr oder weniger freundlich lachten.
Fotos/Bilder
2010
Rule, N.O.; Moran,
J.M.; Freeman, J.B.;
Adams Jr., R.B.;
Gabrieli, J.D.E.;
Ambady, N.
Voting behavior is
reflected in amygdala
response across
cultures
Als Stimuli wurden 58 Fotos von Kandidaten (29 Wahlgewinner und 29
Wahlverlierer) von den zwischen 2004-2006 stattgefundenen Senatswahlen in
Amerika und 58 Fotos von Kandidaten (29 Wahlgewinner und 29 Wahlverlierer)
der Wahl zum Japanischen Parlament eingesetzt. Alle Wahlteilnehmer waren
männlich. Die TeilnehmerInnen mussten mittels eines Knopfes entscheiden, ob
sie den jeweiligen Kandidaten wählen würden. Für diese Entscheidung hatten
die ProbandInnen nur 2s zeigt. Solange wurde auch das Bild gezeigt. Alle
danach getroffenen Entscheidungen wurden nicht registriert.
Fotos/Bilder
2011
Schjoedt, U.;
Stodkilde-
Jorgensen, H.;
Geertz, A.W.; Lund,
T.E.; Roepstorff, A.
The power of charisma -
perceived charisma
inhibits the frontal
executive network of
believers in
intercessory prayer
Die TeilnehmerInnen hörten Gebetstexte, die von 3 männlichen Sprechern mit
unterschiedlicher charismatischer Konnotation vorgelesen wurden. Allerdings
wurde den ProbandInnen noch vor der Wahrnehmung der Stimmen über ein
Headset mitgeteilt, ob es sich beim gegenständlichen Sprecher um einen Nicht-
Christen, um einen Christen oder um einen Christen, der für seine heilende Kraft
bekannt ist, handelt. Insgesamt wurden 18 Gebete von den drei Sprechern
aufgenommen, wobei die ProbandInnen in den Glauben gelassen wurden, dass
jeweils 6 Gebete von einem Nicht-Christen, von einem Christen und von einem
Christen mit heilender Wirkung, gesprochen wurden.
Ton
2011
Cloutier, J.; Kelley,
W.M.; Heatherton,
T.F.
The influence of
perceptual and
knowledge-based
familiarity on the neural
substrates of face
perception
Als Stimuli dienten 96 unbekannte männliche Gesichter ergänzt mit
unbekannten weiblichen Gesichtern, zusammengestellt von den Medien. In der
ersten fMRI Sitzung wurden den Probandinnen 24 Gesichter insgesamt je 3 mal
präsentiert und sie mussten sich einen Eindruck über die jeweilige Person
machen und diesen per Knopfdruck wiedergeben (insgesamt gab es 4 Knöpfe;
1=attraktiv; 2= neutral; 3= nicht attraktiv; 4= keine Information). Während der
zweiten Session wurden beschreibenden Sätze zusätzlich zu neuen Bildern
präsentiert. Die beschreibende Sätze wurden zuvor in drei Kategorien unterteilt
(attraktiv, neutral, unattraktiv). Die ProbandInnen mussten sich wieder einen
Eindruck über die jeweilige Person bilden, allerdings sollten sie die
beschreibenden Sätze dabei auch einbeziehen. Während der dritten fMRI
Session, die 1-2 Tage später stattfand, wurden den Teilnehmerinnen wieder
dieselben Gesichter aus der ersten und zweiten Session gezeigt. Sie mussten
sich diesmal einen Eindruck über das gezeigte Bild bilden, basierend auf ihrer
direkten Reaktion.
Fotos/ Bilder
2011
Rule, N.O.; Moran,
J.M.; Freeman, J.B.;
Whitfield-Gabrieli,
S.; Gabrieli, J.D.E.;
Ambady, N.
Face value: Amygdala
response reflects the
validity of first
impressions
Portraitfotos von CEO`s aus der Fortune 1000 (Jahr 2006) Rangliste. Dabei
wurden die ersten und letzten 25 CEO's der zugehörigen Unternehmen in die
Liste genommen, wovon 1 Person weiblich war. Ein CEO wurde aussortiert. Zu
diesem Sample wurden auch noch 19 Portraits weiblicher CEO's
unterschiedlicher Rankings hinzugefügt. Während die Portraitfotos den
ProbandInnen einzeln gezeigt wurden, mussten diese innerhalb 2 Sekunden
entscheiden, ob das ihnen gezeigte Gesicht symmetrisch oder asymmetrisch
ist. Nach dem Scan wurden die Gesichter den TeilnehmerInnen am Computer
abermals gezeigt. Sie mussten bewerten, wie erfolgreich die jeweilige Person
als Führungskraft einer führenden Unternehmung sein würde (von sehr
erfolgreich bis gar nicht erfolgreich).
Fotos/Bilder
Stimuli und Design
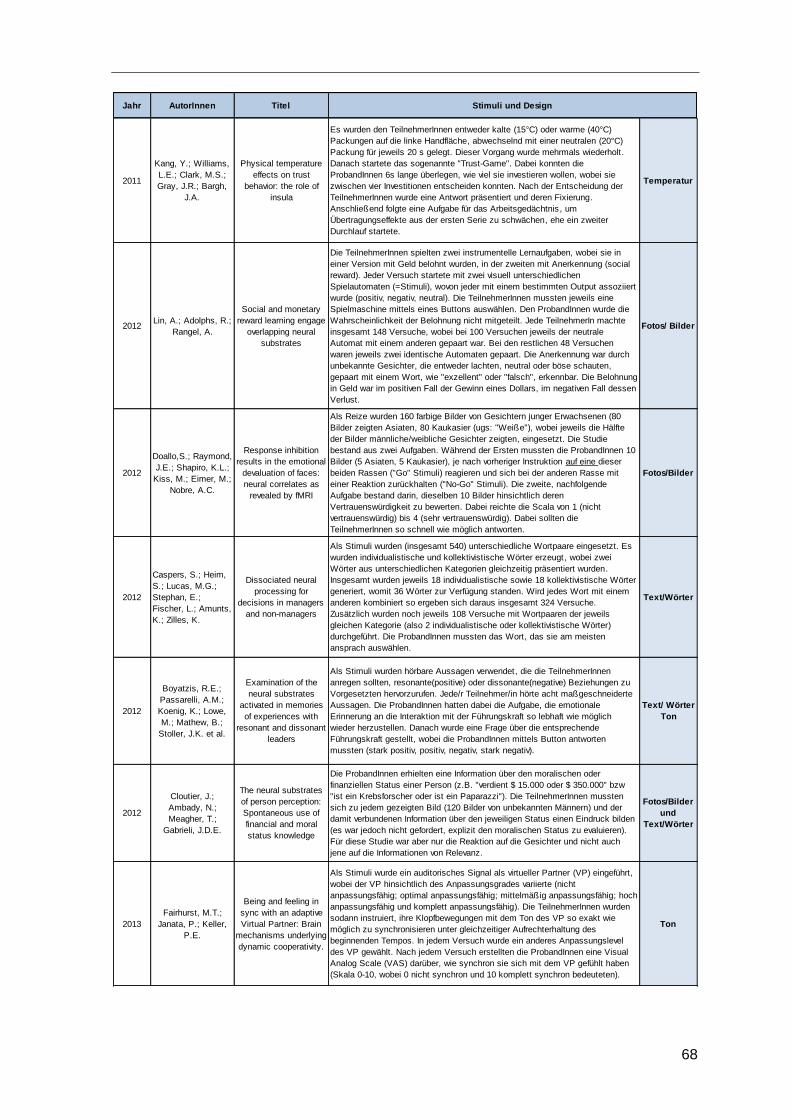
68
Jahr AutorInnen Titel
2011
Kang, Y.; Williams,
L.E.; Clark, M.S.;
Gray, J.R.; Bargh,
J.A.
Physical temperature
effects on trust
behavior: the role of
insula
Es wurden den TeilnehmerInnen entweder kalte (15°C) oder warme (40°C)
Packungen auf die linke Handfläche, abwechselnd mit einer neutralen (20°C)
Packung für jeweils 20 s gelegt. Dieser Vorgang wurde mehrmals wiederholt.
Danach startete das sogenannte "Trust-Game". Dabei konnten die
ProbandInnen 6s lange überlegen, wie viel sie investieren wollen, wobei sie
zwischen vier Investitionen entscheiden konnten. Nach der Entscheidung der
TeilnehmerInnen wurde eine Antwort präsentiert und deren Fixierung.
Anschließend folgte eine Aufgabe für das Arbeitsgedächtnis, um
Übertragungseffekte aus der ersten Serie zu schwächen, ehe ein zweiter
Durchlauf startete.
Temperatur
2012Lin, A.; Adolphs, R.;
Rangel, A.
Social and monetary
reward learning engage
overlapping neural
substrates
Die TeilnehmerInnen spielten zwei instrumentelle Lernaufgaben, wobei sie in
einer Version mit Geld belohnt wurden, in der zweiten mit Anerkennung (social
reward). Jeder Versuch startete mit zwei visuell unterschiedlichen
Spielautomaten (=Stimuli), wovon jeder mit einem bestimmten Output assoziiert
wurde (positiv, negativ, neutral). Die TeilnehmerInnen mussten jeweils eine
Spielmaschine mittels eines Buttons auswählen. Den ProbandInnen wurde die
Wahrscheinlichkeit der Belohnung nicht mitgeteilt. Jede TeilnehmerIn machte
insgesamt 148 Versuche, wobei bei 100 Versuchen jeweils der neutrale
Automat mit einem anderen gepaart war. Bei den restlichen 48 Versuchen
waren jeweils zwei identische Automaten gepaart. Die Anerkennung war durch
unbekannte Gesichter, die entweder lachten, neutral oder böse schauten,
gepaart mit einem Wort, wie "exzellent" oder "falsch", erkennbar. Die Belohnung
in Geld war im positiven Fall der Gewinn eines Dollars, im negativen Fall dessen
Verlust.
Fotos/ Bilder
2012
Doallo,S.; Raymond,
J.E.; Shapiro, K.L.;
Kiss, M.; Eimer, M.;
Nobre, A.C.
Response inhibition
results in the emotional
devaluation of faces:
neural correlates as
revealed by fMRI
Als Reize wurden 160 farbige Bilder von Gesichtern junger Erwachsenen (80
Bilder zeigten Asiaten, 80 Kaukasier (ugs: "Weiße"), wobei jeweils die Hälfte
der Bilder männliche/weibliche Gesichter zeigten, eingesetzt. Die Studie
bestand aus zwei Aufgaben. Während der Ersten mussten die ProbandInnen 10
Bilder (5 Asiaten, 5 Kaukasier), je nach vorheriger Instruktion auf eine dieser
beiden Rassen ("Go" Stimuli) reagieren und sich bei der anderen Rasse mit
einer Reaktion zurückhalten ("No-Go" Stimuli). Die zweite, nachfolgende
Aufgabe bestand darin, dieselben 10 Bilder hinsichtlich deren
Vertrauenswürdigkeit zu bewerten. Dabei reichte die Scala von 1 (nicht
vertrauenswürdig) bis 4 (sehr vertrauenswürdig). Dabei sollten die
TeilnehmerInnen so schnell wie möglich antworten.
Fotos/Bilder
2012
Caspers, S.; Heim,
S.; Lucas, M.G.;
Stephan, E.;
Fischer, L.; Amunts,
K.; Zilles, K.
Dissociated neural
processing for
decisions in managers
and non-managers
Als Stimuli wurden (insgesamt 540) unterschiedliche Wortpaare eingesetzt. Es
wurden individualistische und kollektivistische Wörter erzeugt, wobei zwei
Wörter aus unterschiedlichen Kategorien gleichzeitig präsentiert wurden.
Insgesamt wurden jeweils 18 individualistische sowie 18 kollektivistische Wörter
generiert, womit 36 Wörter zur Verfügung standen. Wird jedes Wort mit einem
anderen kombiniert so ergeben sich daraus insgesamt 324 Versuche.
Zusätzlich wurden noch jeweils 108 Versuche mit Wortpaaren der jeweils
gleichen Kategorie (also 2 individualistische oder kollektivistische Wörter)
durchgeführt. Die ProbandInnen mussten das Wort, das sie am meisten
ansprach auswählen.
Text/Wörter
2012
Boyatzis, R.E.;
Passarelli, A.M.;
Koenig, K.; Lowe,
M.; Mathew, B.;
Stoller, J.K. et al.
Examination of the
neural substrates
activated in memories
of experiences with
resonant and dissonant
leaders
Als Stimuli wurden hörbare Aussagen verwendet, die die TeilnehmerInnen
anregen sollten, resonante(positive) oder dissonante(negative) Beziehungen zu
Vorgesetzten hervorzurufen. Jede/r Teilnehmer/in hörte acht maßgeschneiderte
Aussagen. Die ProbandInnen hatten dabei die Aufgabe, die emotionale
Erinnerung an die Interaktion mit der Führungskraft so lebhaft wie möglich
wieder herzustellen. Danach wurde eine Frage über die entsprechende
Führungskraft gestellt, wobei die ProbandInnen mittels Button antworten
mussten (stark positiv, positiv, negativ, stark negativ).
Text/ Wörter
Ton
2012
Cloutier, J.;
Ambady, N.;
Meagher, T.;
Gabrieli, J.D.E.
The neural substrates
of person perception:
Spontaneous use of
financial and moral
status knowledge
Die ProbandInnen erhielten eine Information über den moralischen oder
finanziellen Status einer Person (z.B. "verdient $ 15.000 oder $ 350.000" bzw
"ist ein Krebsforscher oder ist ein Paparazzi"). Die TeilnehmerInnen mussten
sich zu jedem gezeigten Bild (120 Bilder von unbekannten Männern) und der
damit verbundenen Information über den jeweiligen Status einen Eindruck bilden
(es war jedoch nicht gefordert, explizit den moralischen Status zu evaluieren).
Für diese Studie war aber nur die Reaktion auf die Gesichter und nicht auch
jene auf die Informationen von Relevanz.
Fotos/Bilder
und
Text/Wörter
2013
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Being and feeling in
sync with an adaptive
Virtual Partner: Brain
mechanisms underlying
dynamic cooperativity.
Als Stimuli wurde ein auditorisches Signal als virtueller Partner (VP) eingeführt,
wobei der VP hinsichtlich des Anpassungsgrades variierte (nicht
anpassungsfähig; optimal anpassungsfähig; mittelmäßig anpassungsfähig; hoch
anpassungsfähig und komplett anpassungsfähig). Die TeilnehmerInnen wurden
sodann instruiert, ihre Klopfbewegungen mit dem Ton des VP so exakt wie
möglich zu synchronisieren unter gleichzeitiger Aufrechterhaltung des
beginnenden Tempos. In jedem Versuch wurde ein anderes Anpassungslevel
des VP gewählt. Nach jedem Versuch erstellten die ProbandInnen eine Visual
Analog Scale (VAS) darüber, wie synchron sie sich mit dem VP gefühlt haben
(Skala 0-10, wobei 0 nicht synchron und 10 komplett synchron bedeuteten).
Ton
Stimuli und Design
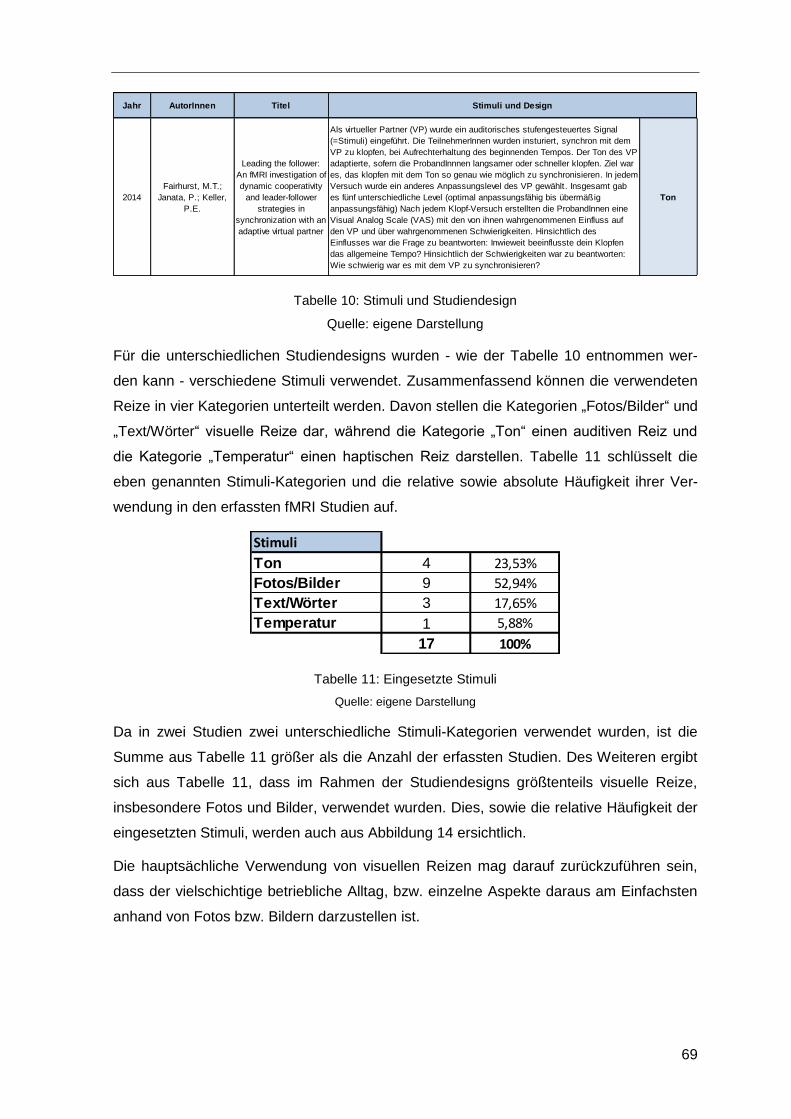
69
Jahr AutorInnen Titel
2014
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Leading the follower:
An fMRI investigation of
dynamic cooperativity
and leader-follower
strategies in
synchronization with an
adaptive virtual partner
Als virtueller Partner (VP) wurde ein auditorisches stufengesteuertes Signal
(=Stimuli) eingeführt. Die TeilnehmerInnen wurden insturiert, synchron mit dem
VP zu klopfen, bei Aufrechterhaltung des beginnenden Tempos. Der Ton des VP
adaptierte, sofern die ProbandInnnen langsamer oder schneller klopfen. Ziel war
es, das klopfen mit dem Ton so genau wie möglich zu synchronisieren. In jedem
Versuch wurde ein anderes Anpassungslevel des VP gewählt. Insgesamt gab
es fünf unterschiedliche Level (optimal anpassungsfähig bis übermäßig
anpassungsfähig) Nach jedem Klopf-Versuch erstellten die ProbandInnen eine
Visual Analog Scale (VAS) mit den von ihnen wahrgenommenen Einfluss auf
den VP und über wahrgenommenen Schwierigkeiten. Hinsichtlich des
Einflusses war die Frage zu beantworten: Inwieweit beeinflusste dein Klopfen
das allgemeine Tempo? Hinsichtlich der Schwierigkeiten war zu beantworten:
Wie schwierig war es mit dem VP zu synchronisieren?
Ton
Stimuli und Design
Tabelle 10: Stimuli und Studiendesign
Quelle: eigene Darstellung
Für die unterschiedlichen Studiendesigns wurden - wie der Tabelle 10 entnommen wer-
den kann - verschiedene Stimuli verwendet. Zusammenfassend können die verwendeten
Reize in vier Kategorien unterteilt werden. Davon stellen die Kategorien „Fotos/Bilder“ und
„Text/Wörter“ visuelle Reize dar, während die Kategorie „Ton“ einen auditiven Reiz und
die Kategorie „Temperatur“ einen haptischen Reiz darstellen. Tabelle 11 schlüsselt die
eben genannten Stimuli-Kategorien und die relative sowie absolute Häufigkeit ihrer Ver-
wendung in den erfassten fMRI Studien auf.
Stimuli
Ton 4 23,53%
Fotos/Bilder 9 52,94%
Text/Wörter 3 17,65%
Temperatur 1 5,88%
17 100%
Tabelle 11: Eingesetzte Stimuli
Quelle: eigene Darstellung
Da in zwei Studien zwei unterschiedliche Stimuli-Kategorien verwendet wurden, ist die
Summe aus Tabelle 11 größer als die Anzahl der erfassten Studien. Des Weiteren ergibt
sich aus Tabelle 11, dass im Rahmen der Studiendesigns größtenteils visuelle Reize,
insbesondere Fotos und Bilder, verwendet wurden. Dies, sowie die relative Häufigkeit der
eingesetzten Stimuli, werden auch aus Abbildung 14 ersichtlich.
Die hauptsächliche Verwendung von visuellen Reizen mag darauf zurückzuführen sein,
dass der vielschichtige betriebliche Alltag, bzw. einzelne Aspekte daraus am Einfachsten
anhand von Fotos bzw. Bildern darzustellen ist.

70
23,53%
52,94%
17,65%
5,88%
Ton
Fotos/Bilder
Text/Wörter
Temperatur
Abbildung 14: Relative Häufigkeit der eingesetzten Stimuli
Quelle: eigene Darstellung
Nachdem nun die Datengrundlage, d.h. die verwendeten Studien, eingehend analysiert
wurden, wird im restlichen Kapitel auf die bei den einzelnen Studien beteiligten Hirnregio-
nen eingegangen.
4.5 Beteiligte Hirnregionen bei der Leadershipforschung
Dieses Kapitel beschäftigt sich konkret mit den Ergebnissen der einzelnen Studien, näm-
lich damit, welche Hirnregionen bei den einzelnen zugrunde liegenden Studiendesigns
messbar beteiligt waren. Es werden sohin die einzelnen, jeweils beteiligten Hirnregionen
tabellarisch aufgelistet, ehe im Anschluss an dieses Kapitel diskutiert wird, welche aussa-
gen hinsichtlich der Studienergebnisse getroffen werden können.
Die Ergebnisse der Studien werden korrespondierend zu Kapitel B 4.3., präsentiert.
4.5.1 Neurophysiologische Unterschiede zwischen erfolgreichen und nicht erfolg-
reichen Führungskräften
Zunächst werden die Ergebnisse jener Studien tabellarisch aufgelistet, die zur Klärung
beitragen können, wie sich erfolgreiche von weniger erfolgreichen Führungskräften neu-
rophysiologisch unterscheiden. Zu dieser Kategorie wurden jene Studien gezählt, bei de-
nen ProbandInnen laut Studiendesign auf Bilder von bestimmten (erfolgreichen, oder
auch weniger erfolgreichen) Personen bzw. Statussymbolen reagierten. In manchen Stu-
dien erhielten die ProbandInnen neben den Bildern von Personen zusätzlich bestimmte

71
Informationen (z.B. über den finanziellen oder moralischen Status von Personen275 oder
beschreibende Sätze276) zu den gezeigten Bildern.
Tabelle 12 soll die Ergebnisse der Studien, die einen Beitrag dazu leisten können, erfolg-
reiche von weniger erfolgreichen Führungskräften zu unterscheiden, veranschaulichen.
In der Tabelle 12 sind jene Hirnregionen eingetragen, bei denen eine signifikante Aktivie-
rung in den jeweiligen Studien festgestellt werden konnte. Aus der Tabelle geht zudem
hervor, ob die Erregung der Hirnregion in der rechten, linken oder in beiden Hemisphären
gemessen werden konnte. Für den Fall, dass den Studien nicht entnommen werden konn-
te, ob die rechte, linke oder beide Seiten des jeweiligen Gehirnareales während der Stu-
die aktiviert war, wurde dies in der Tabelle mit einem „x“ dargestellt. Leere Zellen zeigen
keine Aktivierung der jeweiligen Hirnregion in Bezug auf die jeweilige Studie an.
Im Vergleich zu Tabelle 1 sind in Tabelle 12 nicht alle Hirnregionen aufgelistet. Es wurden
diejenigen Regionen ausgeblendet, bei denen keine Aktivierung gemessen werden konn-
te.
Aus Tabelle 12 geht hervor, dass die in Hinblick auf ihr Studiendesign vergleichbaren
Studien, unterschiedliche Gehirnaktivitäten nachwiesen. Dies mag auch darauf zurückzu-
führen sein, dass die untersuchten Studiendesigns nicht völlig ident waren. Immerhin
konnte in vier von sieben untersuchten Studien jeweils eine Aktivierung des superior tem-
poral Sulcus, des Gyrus temporalis superior, des Gyrus frontalis medius, des posterioren
Cingulum sowie der Amygdala und des Kleinhirns festgestellt werden. Dies deutet darauf-
hin, dass diese Hirnregionen bei der Unterscheidung erfolgreicher von weniger erfolgrei-
chen Führungspersonen eine wesentliche Rolle einnehmen. Allerdings ist darauf hinzu-
weisen, dass - mit Ausnahme des Kleinhirns - die jeweiligen Gehirnregionen in den unter-
schiedlichen Studien in verschiedenen Gehirnhälften erregt waren.277
Hervorzuheben ist des Weiteren der Gyrus praecentralis, bei dem in insgesamt drei Stu-
dien jeweils eine Aktivierung in derselben Hemisphäre festgestellt werden konnte. Auch
hier liegt die Vermutung nahe, dass dieses Areal für die Klärung, wie sich erfolgreiche von
weniger erfolgreichen Führungspersonen unterscheiden, bedeutend ist.278
Zudem waren auch die Amygdala, die Insula sowie der Gyrus fusiform bei insgesamt 3
Studien erregt, jedoch nicht immer in derselben Gehirnhälfte.
275
Vgl. Cloutier, J./Ambady, N./Meagher, T./Gabrieli, J.D.E. (2012): The neural substrates of person percepti-
on: Spontaneus use of financial and moral status knowledge, in: Neuropsychologia, Vol. 50 (9), S. 2371-2376.
276Vgl. Cloutier, J./Kelley, W.M./Heatherton, T.F. (2011): The influence of perceptual and knowledge-based
familiarity on the neural substrates of face perception, in: Social Neuroscience, Vol. 6 (1), S. 63-75. 277
Vgl. dazu Tabelle 12. 278
Vgl. dazu Tabelle 12.
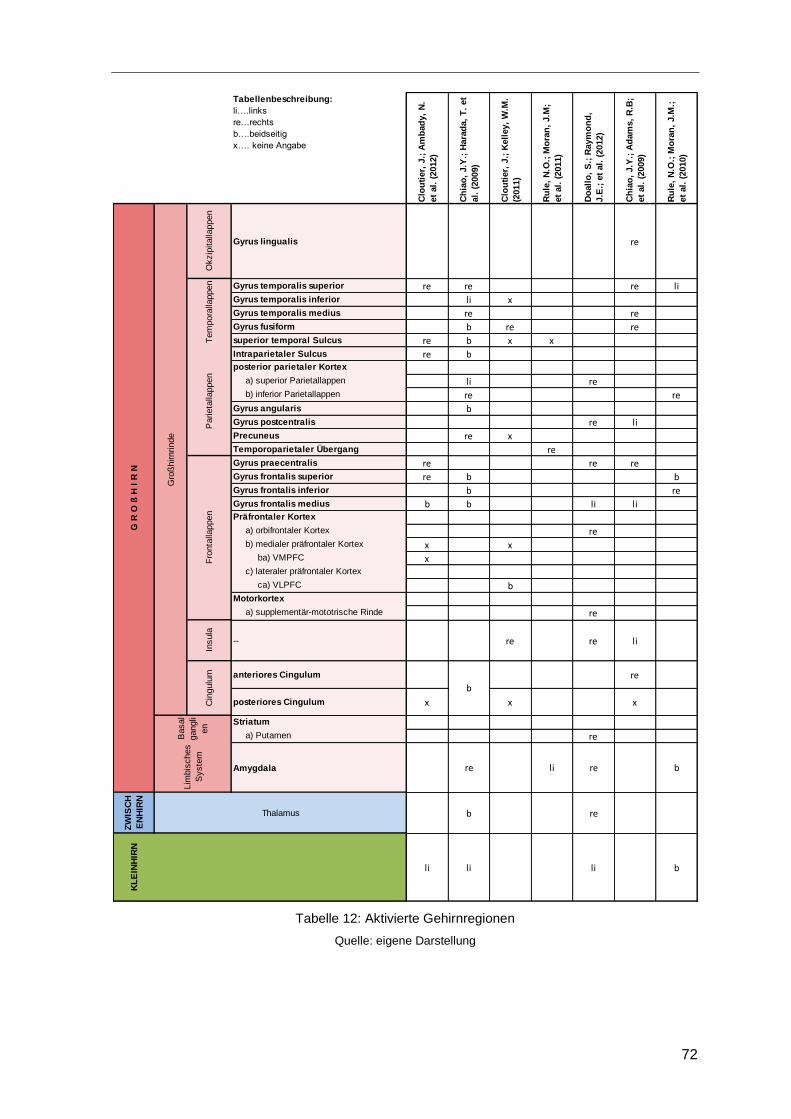
72
Tabellenbeschreibung:
li….links
re…rechts
b….beidseitig
x…. keine Angabe
Clo
uti
er,
J.;
Am
bad
y,
N.
et
al.
(2012)
Ch
iao
, J.Y
.; H
ara
da,
T.
et
al.
(2009)
Clo
uti
er,
J.;
Kell
ey,
W.M
.
(2011)
Ru
le,
N.O
.; M
ora
n,
J.M
;
et
al.
(2011)
Do
all
o,
S.;
Raym
on
d,
J.E
.; e
t al.
(2012)
Ch
iao
, J.Y
.; A
dam
s,
R.B
;
et
al.
(2009)
Ru
le,
N.O
.; M
ora
n,
J.M
.;
et
al.
(2010)
Gyrus lingualis re
Gyrus temporalis superior re re re li
Gyrus temporalis inferior li x
Gyrus temporalis medius re re
Gyrus fusiform b re re
superior temporal Sulcus re b x x
Intraparietaler Sulcus re bposterior parietaler Kortex
a) superior Parietallappen li reb) inferior Parietallappen re re
Gyrus angularis b
Gyrus postcentralis re li
Precuneus re x
Temporoparietaler Übergang re
Gyrus praecentralis re re re
Gyrus frontalis superior re b b
Gyrus frontalis inferior b re
Gyrus frontalis medius b b li liPräfrontaler Kortex
a) orbifrontaler Kortex reb) medialer präfrontaler Kortex x x
ba) VMPFC xc) lateraler präfrontaler Kortex
ca) VLPFC bMotorkortex
a) supplementär-mototrische Rinde re
Insula
-- re re li
anteriores Cingulum re
posteriores Cingulum x x x
Striatum
a) Putamen re
Amygdala re li re b
ZW
ISC
H
EN
HIR
N
b re
KL
EIN
HIR
N
li li li b
G R
O ß
H I
R N
Gro
ßhirnrinde
Okzip
italla
ppen
Tem
pora
llappen
Parieta
llappen
Fro
nta
llappen
Cin
gulu
m
b
Basal
gangli
en
Lim
bis
ches
Syste
m
Thalamus
Tabelle 12: Aktivierte Gehirnregionen
Quelle: eigene Darstellung

73
4.5.2 Soziale Einflussnahme der FK auf den Geführten
In diesem Kapitel werden die Ergebnisse jener Studien präsentiert, die Aufschluss dar-
über geben, welche neurophysiologischen Auswirkungen die soziale Einflussnahme durch
Führungspersonen bei den Geführten hat. Durch die soziale Einflussnahme der Füh-
rungsperson werden im Idealfall beim/bei der Geführten Verhaltens- oder Einstellungsän-
derungen hervorgerufen.279 Da die soziale Einflussnahme und somit die Änderung des
Verhaltens oder der Einstellung des/der Geführten auf unterschiedliche Weise erfolgen
kann, werden die Ergebnisse der Studien nach den jeweiligen Möglichkeiten der Einfluss-
nahme gegliedert. Zunächst werden die Ergebnisse dargestellt, die die neurophysiologi-
schen Auswirkungen von Belohnungen untersuchten. Im Anschluss daran werden jene
Ergebnisse präsentiert, die sich mit den Auswirkungen bei den Geführten durch eine cha-
rismatische Führungskraft beschäftigten, ehe zu guter Letzt die verbleibenden Studien,
die sich mit der wahrgenommene Qualität der Beziehung zwischen MitarbeiterInnen und
der Führungsperson beschäftigten, dargestellt werden.
4.5.2.1 Neurophysiologische Auswirkungen von Belohnungen (bei Geführten)
Belohnung zählt auch zu den Formen sozialer Einflussnahme der Führungskraft auf die
Geführten. In insgesamt zwei Studien280 wurden die neurophysiologischen Auswirkungen
von finanzieller Belohnung im Vergleich zur sozialen Anerkennung als Belohnungsform
untersucht. Die finanzielle Belohnung wurde bildlich jeweils mit Geldmünzen dargestellt
und die soziale Anerkennung anhand lächelnde oder traurige Gesichter.
Wie aus Tabelle 13 ersichtlich ist, wurde in beiden Studien im primären und sekundären
visuellen Kortex, im Precuneus, sowie im Nucleus caudatus und Putamen eine erhöhte
Aktivierung nachgewiesen.281 Hervorzuheben ist, dass in der Studie von Lin, A.; Adolphs,
R. et al. (2012) nur Frauen teilnahmen und das keine genaue Lokalisierung der jeweiligen
Hirnregionen - also ob die rechte, linke oder beide Hemisphären erregt waren - vorge-
nommen wurde.
Des Weiteren muss angemerkt werden, dass die Ergebnisse in Hinblick auf neurophysio-
logische Auswirkungen von Anreizen nicht repräsentativ sind, da nur zwei Studien dies-
bezüglich erhoben wurden. Außerdem wurde in der Studie von Spreckelmeyer, K.N;
279
Vgl. bereits Kapitel B.4.3. sowie Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissen-schaftlicher Sicht, in: Felfe, J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff.S. 8; 12ff.
280 Vgl. Lin, A./Adolphs, R./Rangel, A. (2012): Social and monetary reward learning engage overlapping neural
substrates, in: Social Cognitive and Affective Neuroscience, Vol. 7, Nr. 3, S. 274–281; Spreckelmeyer, K.N./Krach, S./Kohls, G./Rademacher, L./Irmak, A./Konrad, K./Kircher, T./Gründer, G. (2009): Anticipati-on of monetary and social reward differently activates mesolimbic brain structures in men and women, in: Social Cognitive and Affective Neuroscience, Vol. 4, Nr. 2, S. 158–165.
281 Vgl. dazu Tabelle 13.
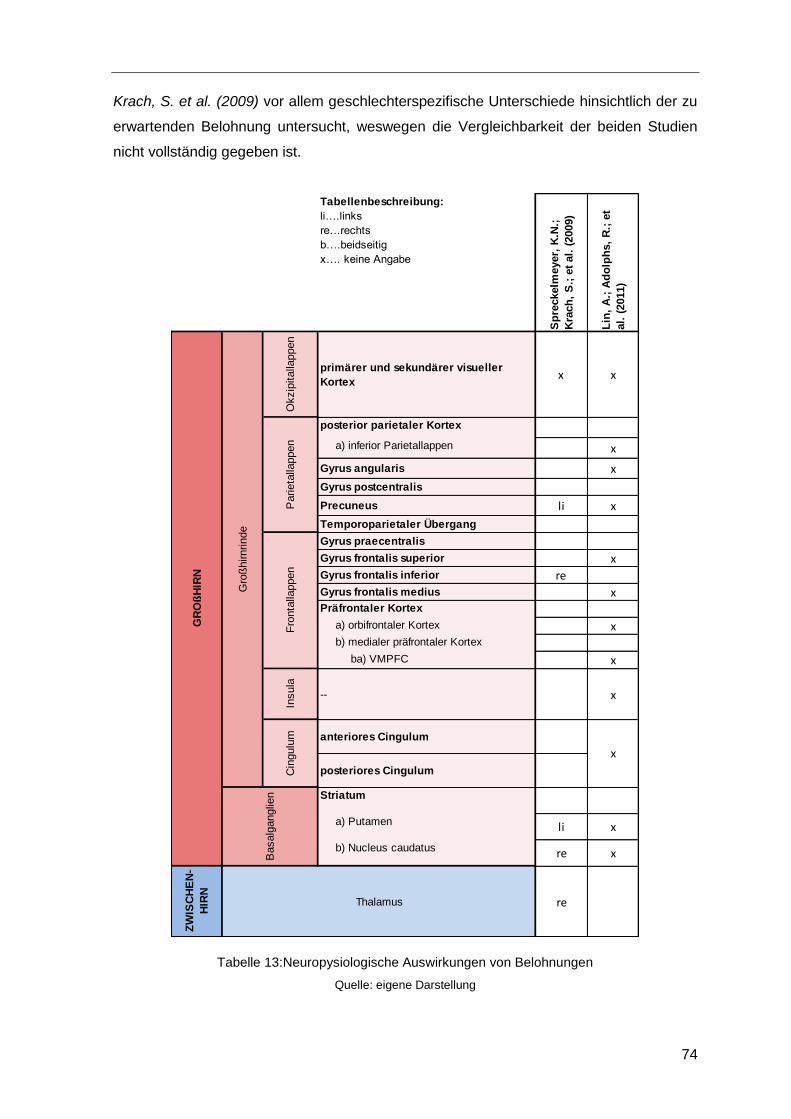
74
Krach, S. et al. (2009) vor allem geschlechterspezifische Unterschiede hinsichtlich der zu
erwartenden Belohnung untersucht, weswegen die Vergleichbarkeit der beiden Studien
nicht vollständig gegeben ist.
Tabellenbeschreibung:
li….links
re…rechts
b….beidseitig
x…. keine Angabe
Sp
reckelm
eyer,
K.N
.;
Kra
ch
, S
.; e
t al.
(2009)
Lin
, A
.; A
do
lph
s,
R.;
et
al.
(2011)
Okzip
italla
ppen
primärer und sekundärer visueller
Kortexx x
posterior parietaler Kortex
a) inferior Parietallappen x
Gyrus angularis x
Gyrus postcentralis
Precuneus li x
Temporoparietaler Übergang
Gyrus praecentralis
Gyrus frontalis superior x
Gyrus frontalis inferior re
Gyrus frontalis medius xPräfrontaler Kortex
a) orbifrontaler Kortex xb) medialer präfrontaler Kortex
ba) VMPFC x
Insula
-- x
anteriores Cingulum
posteriores Cingulum
Striatum
a) Putamen li x
b) Nucleus caudatus re x
ZW
ISC
HE
N-
HIR
N
re
x
Basalg
anglie
n
Thalamus
GR
Oß
HIR
N
Gro
ßhirnrinde
Parieta
llappen
Fro
nta
llappen
Cin
gulu
m
Tabelle 13:Neuropysiologische Auswirkungen von Belohnungen
Quelle: eigene Darstellung

75
4.5.2.2 Neurophysiologische Auswirkungen von charismatischen Führungskräften (bei
Geführten)
Neben Belohnungen lösen auch vermeintlich charismatische Führungskräfte bei Geführ-
ten bestimmte neurophysiologische Strukturen aus. Dies haben Schjoed, U.; Stodkilde,
Jorgensen, H. et al. (2011) in ihrer Studie nachgewiesen.
Der Tabelle 14 sind alle Hirnareale zu entnehmen, die während der Untersuchung eine
erhöhte Aktivierung zeigten. An dieser Stelle ist darauf hinzuweisen, dass aufgrund einer
einzigen erfassten Studie keine weitergehende Analyse hinsichtlich der Konsistenz des
Ergebnisses durchgeführt werden kann.
Tabellenbeschreibung:
li….links
re…rechts
b….beidseitig
x…. keine Angabe
Sch
joed
, U
.; S
tod
kil
de-
Jo
rgen
sen
, H
.; e
t al.
(2011)
Tem
pora
llappen
Gyrus temporalis inferior x
Parieta
llappen
Precuneus x
Gyrus praecentralis xPräfrontaler Kortex x
a) medialer präfrontaler Kortex xb) lateraler präfrontaler Kortex
ba) DLPFC x
Insula
-- x
Cin
gulu
m
anteriores Cingulum x
Amygdala x
ME
DU
LL
A
OB
LO
NG
AT
A
x
Lim
bis
ches
Syste
m
GR
Oß
HIR
N
Gro
ßhirnrinde
Fro
nta
llappen
Tabelle 14: Aktivierung von Gehirnregionen, die neurophysiologische Strukturen von charismati-schen Führungskräften untersuchten
Quelle. eigene Darstellung

76
4.5.2.3 Neurophysiologische Auswirkungen von Beziehungen zwischen MitarbeiterIn-
nen und Führungskräften (bei Geführten)
MitarbeiterInnen können nicht nur durch Belohnungen oder durch charismatisches Verhal-
ten beeinflusst werden, sondern auch durch eine gute Austauschbeziehung zwischen
Ihnen und der Führungskraft. Diesbezüglich werden in diesem Kapitel insgesamt vier Stu-
dien aufgelistet, die neurophysiologische Auswirkungen von zwischenmenschlichen Be-
ziehungen untersuchten. In der Studie von Boyatzis, R.E; Pasarelli A.M. et al. (2012)282
mussten sich MitarbeiterInnen an Situationen erinnern, in denen sich die Führungskraft
ihnen gegenüber unterstützend bzw. hinderlich verhalten hat. In der Studie von Kang, Y.;
Williams, L.E. (2011)283 wurde zwischenmenschliches Vertrauen anhand unterschiedlicher
Temperaturen simuliert und untersucht. Die Studien von Fairhurst, M.; Janata, P. et al.
(2013 bzw. 2014)284 untersuchten neuronale Korrelate von Kooperation, wobei in diesen
Studien „Leader“ von „Followern“ differenziert werden konnten.
Wie aus Tabelle 15 ersichtlich ist, konnte in allen vier Studien eine signifikante Aktivierung
der Insula - zumeist beidseitig - festgestellt werden. In drei der vier erhobenen Studien ist
eine erhöhte Erregung des inferioren Parietallappens, des Gyrus frontalis inferior, des
anterioren Cingulum sowie des Thalamus ersichtlich. Diese Areale scheinen bei der Mit-
arbeiterInnen-Vorgesetzten Beziehung eine wesentliche Rolle einzunehmen.285
282
Vgl. Boyatzis, R.E./Passarelli, A.M./Koenig, K./Lowe, M./Mathew, B./Stoller, J.K./Phillips, M. (2012): Ex-
amination of the neural substrates activated in memories of experiences with resonant and dissonant le-aders, in: The Leadership Quarterly, Jg. 23, S. 259–272.
283 Vgl. Kang, Y./Williams, L.E./Clark, M.S./Gray, J.R./Bargh, J.A. (2011): Physical temperature effects on trust
behavior: the role of insula, in: Social Cognitive and Affective Neuroscience, Vol. 6, Nr. 4, S. 507–515. 284
Vgl. Fairhurst, M.R./Janata, P./Keller, P.E. (2013): Being and Feeling in Sync with an Adaptive Virtual Part-ner: Brain Mechanisms Underlying Dynamic Cooperativity, in: Cerebral Cortex, Vol. 23, Nr.11, S. 2592–2600; Fairhurst, M.R./Janata, P./Keller, P.E. (2014): Leading the follower: An fMRI investigation of dy-namic cooperativity and leader-follower strategies in synchronization with an adaptive virtual partner, in: NeuroImage, S. 688–697.
285 Siehe dazu auch Kapital C.6.
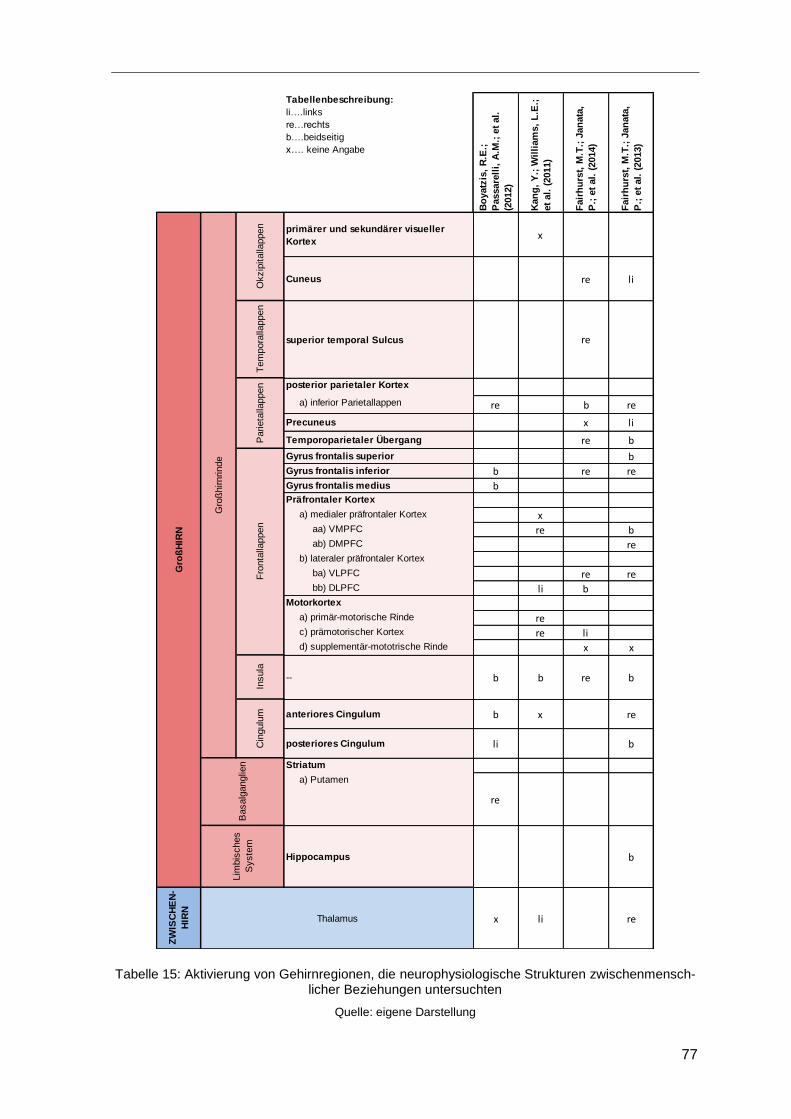
77
Tabellenbeschreibung:
li….links
re…rechts
b….beidseitig
x…. keine Angabe
Bo
yatz
is,
R.E
.;
Passare
lli,
A.M
.; e
t al.
(2012)
Kan
g,
Y.;
Wil
liam
s,
L.E
.;
et
al.
(2011)
Fair
hu
rst,
M.T
.; J
an
ata
,
P.;
et
al.
(2014)
Fair
hu
rst,
M.T
.; J
an
ata
,
P.;
et
al.
(2013)
primärer und sekundärer visueller
Kortexx
Cuneus re li
Tem
pora
llappen
superior temporal Sulcus re
posterior parietaler Kortex
a) inferior Parietallappen re b re
Precuneus x li
Temporoparietaler Übergang re b
Gyrus frontalis superior b
Gyrus frontalis inferior b re re
Gyrus frontalis medius bPräfrontaler Kortex
a) medialer präfrontaler Kortex xaa) VMPFC re bab) DMPFC re
b) lateraler präfrontaler Kortex
ba) VLPFC re rebb) DLPFC li b
Motorkortex
a) primär-motorische Rinde rec) prämotorischer Kortex re lid) supplementär-mototrische Rinde x x
Insula
-- b b re b
anteriores Cingulum b x re
posteriores Cingulum li b
Striatum
a) Putamen
re
Hippocampus b
ZW
ISC
HE
N-
HIR
N
x li re
Gro
ßH
IRN
Gro
ßhirnrinde
Okzip
italla
ppen
Parieta
llappen
Fro
nta
llappen
Cin
gulu
m
Basalg
anglie
nLim
bis
ches
Syste
m
Thalamus
Tabelle 15: Aktivierung von Gehirnregionen, die neurophysiologische Strukturen zwischenmensch-licher Beziehungen untersuchten
Quelle: eigene Darstellung

78
4.5.3 Neurophysiologische Aktivitäten, die führungsrelevanten Fertigkeiten
zugrunde liegen
Der dritte Bereich, in welchem die Neurowissenschaft einen wesentlichen Beitrag zur Füh-
rungsforschung leisten kann, ist die Aufklärung, welche neurophysiologischen Prozesse
grundlegend den führungsrelevanten Fähigkeiten zugrunde liegen.286 Zu diesem Bereich
zählt insbesondere die Entscheidungsfindung, die für Führungskräfte eine zentrale Aufga-
be darstellt.287
4.5.3.1 Entscheidungsfindung
Zur Entscheidungsfindung wurde für den besagten Zeitraum nur eine fMRT-Studie erho-
ben. Ziel dieser Studie war es, den Entscheidungsprozess zwischen Managern und Nicht-
Managern zu untersuchen. Dementsprechend waren die TeilnehmerInnen der Studien
zum Teil Führungskräfte, zum anderen Teil Menschen ohne Führungserfahrung.
Tabellenbeschreibung:
li….links
re…rechts
b….beidseitig
x…. keine Angabe
Casp
ers
, S
.; H
eim
, S
.; e
t
al.
(2012)
Okzip
italla
ppen
primärer und sekundärer visueller
Kortexx
primärer auditorischer Kortex x
sekundärer auditorischer Kortex
(Wernicke Sprachzentrum)x
posterior parietaler Kortex
a) superior Parietallappen
x
Präfrontaler Kortex
a) lateraler präfrontaler Kortex
aa) DLPFC x
Insula
-- x
Cin
gulu
m
anteriores Cingulum x
Striatum
a) Nucleus caudatusx
Basalg
anglie
n
GR
Oß
HIR
N
Gro
ßhirnrinde
Tem
pora
llappen
Parieta
llappen
Fro
nta
llappen
Tabelle 16: Aktivierte Hirnregionen jener Studie, die das Entscheidungsverhalten untersuchte
Quelle: eigene Darstellung
286
Vgl. Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe, J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff.
287 Vgl. Bass, B.M./Bass, R. (2009): The Bass bandbook of leadership: Theory, research, and managerial
applications, New York.

79
Auch diesbezüglich ist darauf hinzuweisen, dass aufgrund einer einzigen erfassten Studie
keine weitergehende Analyse im Rahmen dieser Arbeit durchgeführt werden kann.
5 Restriktionen
Wie bereits erwähnt, wurden nur jene Studien erhoben, die kostenfrei mittels Datenban-
ken der Universität Graz zugänglich waren. Außerdem wurden nur jene Studien einbezo-
gen, die bis zum Stichtag mit 20. Juni 2014 veröffentlich wurden. Darüber hinaus wurden
nur fMRT sowie EEG Studien erhoben, wobei die EEG Studien in weiterer Folge aufgrund
mangelnder Aussagekraft für die vorliegende Arbeit wieder aussortiert wurden. Von den
übrig gebliebene 17 fMRT Studien wurden zwei Studien aussortiert, da sich diese Studien
mit Untersuchungsdesign beschäftigen, die dem Marketing zugeordnet werden können
und daher für die vorliegende Arbeit nicht von Relevanz sind. Insgesamt wurden sohin 15
Studien erhoben und hinsichtlich der aktivierten Hirnregionen untersucht. Anzumerken ist,
dass alle Hirnregionen, bei denen eine signifikante Erregung gemessen werden konnte,
erfasst wurden. Es wurde dabei nicht hinsichtlich der Stärke der jeweiligen Erregung diffe-
renziert.
Aufgrund der geringen Anzahl erhobener Studien (n=15) ist eine statistische Auswertung
nicht möglich. Das vorrangige Ziel dieser Arbeit besteht daher nicht in der Bestimmung
von abhängigen und unabhängigen Variablen (=Kodierung) und deren statistischen Aus-
wertung, sondern in der Erfassung und Systematisierung der einzelnen Studien.
6 Diskussion
In diesem abschließenden Kapital werden die Ergebnisse aus Kapitel C. 4.5 diskutiert.
Dabei wird wiederum korrespondierend zu Kapitel B. 4.3. und Kapitel C.4.5. vorgegangen;
d.h. die Ergebnisse werden nach den unterschiedlichen Bereichen, die zur Führungsfor-
schung beitragen können, geordnet erörtert.
6.1.1 Neurophysiologische Unterschiede zwischen erfolgreichen und nicht erfolg-
reichen Führungskräften
Wie aus Tabelle 12 ersichtlich ist, waren bei jenen Studien, die neurophysiologische Un-
terschiede von erfolgreiche und weniger erfolgreichen Führungskräften untersuchten, fol-
genden Hirnarealen erregt: Gyrus lingualis, Gyrus temporalis superior, Gyrus temporalis
inferior, Gyrus temporalis medius, Gyrus fusiform, superior temporal Sulcus, superior Pa-
rietallappen, inferior Parietallappen, Gyrus angularis, Gyrus postcentralis, Precuneus,
temporoparietaler Übergang, Gyrus praecentralis, Gyrus frontalis superior, Gyrus frontalis
inferior, Gyrus frontalis medius, OFC, medialer präfrontaler Kortex, VMPFC, VLPFC,

80
supplementär-motorische Rinde, anteriores und posteriores Cingulum, Putamen, Amyg-
dala, Thalamus und im Kleinhirn. Die Aktivierung war jeweils entweder in der rechten,
linken oder in beiden Hemisphären nachweisbar.288
Wie aus dieser langen Aufzählung ersichtlich ist, waren insgesamt sehr viele, verschiede-
ne Gehirnareale in den einzelnen Studien erregt. Auffallend ist, dass die einzelnen Stu-
dien auf den ersten Blick uneinheitliche Ergebnisse liefern. Dies mag wahrscheinlich auch
darauf zurückzuführen sein, dass die einzelnen Studiendesigns - wenn auch nur gering -
unterschiedlich sind. Allerdings konnte in der Mehrheit der untersuchten Studien eine Ak-
tivierung des des superior temporal Sulcus, des Gyrus temporalis superior, des Gyrus
frontalis medius, des posterioren Cingulum sowie der Amygdala und des Kleinhirns fest-
gestellt werden.
Erfolgreiche Führungskräfte lösten bei den ProbandInnen eine signifikant höhere Aktivie-
rung der Amygdala aus. Die Amygdala ist wesentlicher Bestandteil des limbischen Sys-
tems und dafür bekannt, bei emotionaler Bewertung von emotionalen und gesellschaftli-
chen Reizen involviert zu sein.289 Auch der superior temporale Gyrus spielt für die Beurtei-
lung, ob eine Führungskraft mehr oder weniger erfolgreich ist, eine wesentliche Rolle.290
Eine Aktivierung des superior temporalen Gyrus wurde bereits beim Anblick dominanter
Gesichter nachgewiesen.291 Der superior temporale Gyrus und die Amygdala sind eng
vernetzt, womit die Annahme nahe liegt, dass eine Aktivierung beider Hirnregionen durch
den Anblick dominanter Gesichter ausgelöst wird.292 Neben der Amygdala und dem supe-
rior temporalen Gyrus, ist beim Anblick dominanter Gesichter auch die Insula aktiviert.293
6.1.2 Soziale Einflussnahme der Führungskraft auf den Geführten
Wie aus der Arbeit hervorging, kann soziale Einflussnahme auf ganz unterschiedliche
Weise erfolgen. In dieser Arbeit wurden Studien, die sich mit der neurophysiologischen
Auswirkung von Belohnungen, charismatischen Führungskräften und Beziehungen zwi-
schen Führungskräften und MitarbeiterInnen beschäftigen, untersucht.
288
Für Details siehe Tabelle 12. 289
Vgl. CNS Forum Lundbeck Institute: Gehirnatlas, http://de.brainexplorer.org/brain_atlas/Brainatlas_in-
dex.shtml, zuletzt geprüft am 6.November 2013. 290
Vgl. Rule, N.O./Moran, J.M./Freeman, J.B./Whitfield-Gabrieli, S./Gabrieli, J.D.E./Ambady, N. (2011): Face value: Amygdala response reflects the validity of first impressions, in: NeuroImage, Jg. 54, Nr. 1, S. 734-741.
291 Vgl. Chiao J.Y./Adams, R.B./Tse, P.U./Lowenthal L./Richeson, J.A./Ambady, N. (2009): Knowing Who's
Boss: fMRI and ERP Investigations of Social Dominance Perception, in: Group Processes & Intergroup Relations, Jg. 11, Nr. 2, S. 201-214.
292 So auch Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe,
J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff. 293
Vgl. dazu Tabelle 12.

81
Hinsichtlich der zwei Studien, die sich mit den neuropysiologischen Auswirkungen von
Belohnungen beschäftigten, konnte eine Aktivierung des primären und sekundären visuel-
len Kortex, des inferioren Parietallappen, des Gyrus angularis, des Precuneus, des Gyrus
frontalis superior, des Gyrus frontalis inferior, des Gyrus frontalis medius, des OFC sowie
des VMPFC, des anterioren und posterioren Cingulum, des Putamen und des Nucleus
caudatus sowie des Thalamus festgestellt werden.294
Eine Aktivierung des Nucleus caudatus, des Putamen sowie des Precuneus konnte in
beiden Studien nachgewiesen werden. Allerdings ist darauf hinzuweisen, dass die beiden
Studien nicht vollständig vergleichbar sind, zumal Spreckelmeyer et.al. geschlechterspezi-
fische Differenzen untersuchten, während an der Studie von Lin et.al. nur Frauen teilnah-
men.
Hinsichtlich der Studie von Schjoed et. al., die sich mit den neurophysiologischen Auswir-
kungen von charismatischen Führungskräften bei Geführten beschäftigte, wurde eine Er-
regung in folgenden Gehirnarealen nachgewiesen: Gyrus temporalis inferior, Precuneus,
Gyrus praecentralis, präfrontaler Kortex (medialer präfrontaler Kortex und DLPFC), ante-
riores Cingulum, Amygdala und Medulla oblongata. Ob es Übereinstimmungen mit ande-
ren Ergebnissen gibt, kann mangels weiterer Studien nicht festgestellt werden.295
Insgesamt vier Studien untersuchten, inwiefern sich eine gute Beziehung zwischen Mitar-
beiterInnen und Führungskräften bei Geführten neuropysiologisch auswirkt. Insgesamt
konnten Aktivierungen im primären und sekundären visuellen Kortex, im Cuneus, im su-
perior temporal Sulcus, im inferioren Parietallappen, im Precuneus, im temproparietalen
Übergang, im Gyrus frontalis superior, im Gyrus frontalis inferior, im Gyrus frontalis medi-
us, im medialen präfrontalen Kortex (insbesondere im VMPFC und DMPFC), im lateralen
präfrontalen Kortex (insbesondere im DLPFC), in der primär-motorischen Rinde, im prä-
motorischen Kortex, sowie in der supplementär motorischen Rinde, in der Insula, dem
anterioren und posterioren Cingulum, im Putamen, im Hippocampus und auch im Thala-
mus, nachgewiesen werden.296
In allen vier Studien konnte eine signifikante Aktivierung der Insula festgestellt werden.
Des Weiteren konnte in drei der vier Studien eine erhöhte Erregung des inferioren Parie-
tallappens, des Gyrus frontalis inferior, des anterioren Cingulum, sowie des Thalamus
nachgewiesen werden. Aufgrund der mehrheitlichen Übereinstimmung scheinen diese
Areale bei der Beziehung zwischen MitarbeiterInen und Vorgesetzten eine entscheidende
Rolle einzunehmen.
294
Vgl dazu Tabelle 13. 295
Vgl. dazu Tabelle 14. 296
Vgl. dazu Tabelle 15.

82
6.1.3 Neuropysiologische Aktivitäten, die führungsrelevanten Fertigkeiten
zugrunde liegen
Die letzte ausgewertete Studie von Caspers et. al. beschäftigte sich mit neurophysiologi-
schen Strukturen, die der Entscheidungsfindung, als eine wesentliche führungsrelevante
Fertigkeit, zugrunde liegen. Dabei konnte eine Aktivierung folgender Areale festgestellt
werden: primärer und sekundärer visueller Kortex, primärer und sekundärer auditiver Kor-
tex, superior Parietallappen, DLPFC, Insula, anteriores Cingulum, und Nucleus caudatus.
Mangels einer zweiten Studie zu diesem Themenbereich können keine Vergleiche hin-
sichtlich der Konsistenz bzw. Übereinstimmung der Ergebnisse gezogen werden.
Alles in allem verdeutlichen die Ergebnisse der vorliegenden Arbeit, dass der For-
schungsbereich des „Neuroleaderships“ noch in den Fußstapfen steckt. Um eine Überein-
stimmungen herauszufinden, bedarf es mehreren Studien, um diese sodann auch auf
statistische Effekte untersuchen zu können. Aufgrund der wenig vorhandenen Studien
lassen sich kaum Übereinstimmungen hinsichtlich der erregten Areale finden. Dies mag
vor allem darauf zurückzuführen sein, dass die unterschiedlichen Studiendesigns - obwohl
eine Unterteilung vorgenommen wurde - alles in allem zu verschieden und daher nicht
wirklich vergleichbar sind. Daher gibt es in allen identifizierten Bereichen noch enormen
Forschungsbedarf, bis es möglich sein wird, allgemeine, übereinstimmende Aussagen zu
liefern.

83
Zusammenfassend ist festzuhalten, dass das Gehirn in das Großhirn, das Zwischenhirn,
das Kleinhirn sowie in den Hirnstamm unterteilt werden kann. Jedem dieser Hirnareale
liegen bestimmte Funktionen zugrunde, wobei darauf hinzuweisen ist, dass der Großteil
des Gehirns bzw. dessen Funktionsweise noch gänzlich unerforscht ist.297
Für Aspekte der Führungsforschung sind vor allem die fMRT sowie die EEG als neuro-
wissenschaftliche Methoden von Relevanz. Neben diesen zwei, derzeit wohl populärsten
Methoden, stellt die Neurowissenschaft mit der PET, MEG, iEEG, TMS und DTI weitere
Verfahren zur Verfügung, die für die Klärung von führungsrelevanten Fragestellungen
herangezogen werden können.
Jede einzelne Methode weist unterschiedliche Vor- und Nachteile auf.298 Abhängig von
der jeweiligen Forschungsfrage, ist die eine oder andere Methode besser geeignet. Wer-
den z.B. nur oberflächliche Gehirnregionen untersucht, und kann von einer exakten Loka-
lisierung der einzelnen Areale abgesehen werden, ist wohl die EEG im Hinblick auf Kos-
ten, örtliche Flexibilität, Vorbereitungszeit und Datenauswertung die am besten geeignete
Methode. Wird hingegen das Ziel verfolgt, eine möglichst genaue Lokalisierung der erreg-
ten Gehirnregionen zu erreichen, ist die Messung (indirekter) neuronaler Aktivität mittels
fMRT erstrebenswert. Allerdings müssen bei dieser Methode auch die hohen Kosten und
die komplexe Datenauswertung bedacht werden..299
Wie aus dieser Arbeit hervorgeht, kann die Neurowissenschaft in unterschiedlichen Berei-
chen zur Führungsforschung beitragen. So kann die Neurowissenschaft Einblicke darüber
geben, wie sich erfolgreiche von weniger erfolgreichen Führungskräften neurophysiolo-
gisch unterscheiden. Außerdem bietet die Neurowissenschaft Möglichkeiten, zu untersu-
chen, wie sich bestimmte bewussten Einflussnahmen von Führungskräften auf Mitarbeite-
rInnen neurophysiologisch auswirken; d.h. welche Gehirnsysteme durch z.B. charismati-
sches Verhalten, oder durch eine in Aussicht gestellte Belohnung aktiviert oder gehemmt
werden. Der dritte Bereich, in dem die Neurowissenschaft einen wesentlichen Beitrag zur
Führungsforschung leisten kann, ist in der Klärung, welche neurophysiologische Aktivitä-
ten führungsrelevanten Fertigkeiten, wie z.B. der Entscheidungsfindung, zugrunde lie-
gen.300
297
Vgl. Tabelle 1. 298
Vgl dazu Tabelle 2. 299
Siehe dazu Kapitel B.3. 300
Für Details siehe Kapitel B.4.3.
D. ZUSAMMENFASSUNG

84
Mittels einer Metaanalyse wurden in der vorliegenden Arbeit insgesamt 15 Studien, die
(teilweise indirekt) Aspekte der Führungsforschung untersuchten, im Hinblick auf die akti-
vierten Hirnregionen ausgewertet. Die Auswertung erfolgte dabei korrespondierend zu
Kapitel B.4.3. d.h. es wurden vergleichbare Studien zusammengefasst und deren Ergeb-
nisse, aufgeschlüsselt nach aktivierten Hirnregionen, tabellarisch dargestellt. Wie aus den
jeweiligen Tabellen ersichtlich ist, lieferten die einzelnen Studien durchaus unterschiedli-
che Ergebnisse. Ein Grund dafür mögen die nicht völlig identischen Untersuchungsde-
signs sein. Außerdem muss an dieser Stelle erwähnt werden, dass das Ziel dieser Arbeit
in der Systematisierung und Erfassung der aktivierten Hirnregionen lag, und sohin nicht
auf die unterschiedlich stark ausgeprägten Erregungen der jeweiligen Areale eingegangen
wurde. Welche Gehirnregionen für die einzelnen Bereiche in den jeweiligen Studien kon-
kret aktiviert waren ist den Tabellen 12-15 zu entnehmen.
Ausblick
Die Untersuchung führungsrelevanter Fragestellungen anhand von neurowissenschaftli-
chen Methoden ist ein noch sehr junges Forschungsgebiet. Dies wurde durch die sehr
uneinheitlichen Ergebnisse der einzelnen Studien untermauert. Dennoch liefern erste
Studien klare Ergebnisse über neuroanatomische Strukturen und Netzwerke, die bei füh-
rungsrelevanten Aspekten beteiligt sind. So werden erfolgreiche Führungskräfte im Ver-
gleich zu weniger erfolgreichen Führungskräften neurophysiologisch unterschiedlich
wahrgenommen. Außerdem können Führungskräfte durch die bewusste soziale Einfluss-
nahme bestimmte neurophysiologische Strukturen und Prozesse bei ihren MitarbeiterIn-
nen aktivieren. Schließlich lassen sich auch führungsrelevante Fertigkeiten neurophysio-
logisch abbilden.301
Wie jedoch die Ergebnisse aus Kapitel C.4.5. zeigen, sind die einzelnen Ergebnisse der
Studien nicht deckungsgleich, sondern stehen eher noch sehr lose im Raum. Damit wird
deutlich, dass sich die Leadershipforschung noch in den Fußstapfen befindet. Aus diesem
Grund ist es unabdingbar, in Zukunft weitere Studien zu Aspekten der Führungsforschung
zu untersuchen, um dem ursprünglichen Ziel, eine allgemein gültige, d.h. in jedem Um-
weltzustand anwendbare, erfolgreiche Führungstheorie zu entwerfen, ein bisschen näher
zu kommen.302
301
So auch Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe,
J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff.
302 So auch Bergner, S./Rybnicek, R. (2015): Führungsforschung aus neurowissenschaftlicher Sicht, in: Felfe,
J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen, S.552ff.

85
ANHANG
Jahr Zeitschrift AutorInnen Titel Hypothese Zielsetzung MethodeStichproben-
größeStimuli und Design
Typus der
ProbandInnenAnmerkungen Ergebnisse Diskussion Kategorie
2012 Neuropsychologia
Cloutier, J.;
Ambady, N.;
Meagher, T.;
Gabrieli, J.D.E.
The neural substrates
of person perception:
Spontaneous use of
financial and moral
status knowledge
Ziel: Identifizierung von
neuronalen Reaktionen auf
soziale Zielobjekte anhand
zwei unterschiedlicher
'Status-Dimensionen'
(finanzieller Status vs.
moralischer Status).
fMRI
n=23 (m=8;
f=11); (4
wurden
aussortiert)
Die ProbandInnen erhielten eine Information über den
moralischen oder finanziellen Status einer Person (z.B.
"verdient $ 15.000 oder $ 350.000" bzw "ist ein
Krebsforscher oder ist ein Paparazzi"). Die
TeilnehmerInnen mussten sich zu jedem gezeigten Bild
(120 Bilder von unbekannten Männern) und der damit
verbundenen Information über den jeweiligen Status
einen Eindruck bilden (es war jedoch nicht gefordert,
explizit den moralischen Status zu valuieren). Für diese
Studie war aber nur die Reaktion auf die Gesichter und
nicht auch jene auf die Informationen von Relevanz.
TeilnehmerInnen aus
einer städtischen
Universitäts-
gemeinschaft
Diese Studie zeigt die
Einbindung von verschiedenen
Gehirnregionen während einer
spontanen Bewertung von
sozialen Zielobjekten als eine
Funktion von deren sozialen
Status.
Es wurde herausgefunden, dass eine Aktivierung des intraparietal Sulcus (IPS) empfindlich
auf den finanziellen Status eines Individuums reagiert, während eine Aktivierung des
ventromedialen präfontalen Cortex auf den moralischen Status empfindlich reagiert. Die
Aktivierung des rechten IPS war für jene Gesichter, die mit einem niedrigem finanziellen
Status gepaart waren, höher als für diejenigen, die mit einem hohen finanziellen Status
assoziiert wurden. Genauso konnte eine höhere Aktivierung des mittleren frontales Gyrus,
des linken Cerebellum, des rechten superior temporal Sulcus, des rechten superior frontal
Gyrus, des linken mittleren frontalen Gyrus sowie des posterioren cingularen Gyrus
beobachtet werden, wobei die Aktivierung für Gesichter mit niedrigerem finanziellen Status
größer war. Aktivierungen im VMPFC sowie des rechten superior temporalen Gyrus und des
medialen präfrontalen Kortex waren für Gesichter, die mit hohem moralischem Status
gepaart wurden, im Vergleich mit jenen Gesichtern mit niedrigem moralischem Status,
höher. Es konnte weder eine unterschiedliche Aktivierung des IPS zwischen Gesichtern,
gepaart mit hohem und niedrigem moralischen Status sowie auch keine unterschiedliche
Aktivierung des VMPFC hinsichtlich Gesichtern, die mit hohen oder niedrigen finanziellen
Status gepaart wurden, ausfindig gemacht werden.
Die ProbandInnen haben entgegen früheren Studien die
Information über den Status einer Person spontan den
jeweiligen Gesicht zugeschrieben. Es ist denkbar, dass die
erhöhte Aktivierung des IPS hinsichtlich Gesichter mit
niedrigerem finanziellen Status auf den spontanen Vergleich
mit dem eigenen (im Durchschnitt niedrigen) finanziellen
Status zurückzuführen ist. Die höhere Aktivierung des
VMPFC bei Gesichtern, die mit einem höherem moralischen
Status assoziiert wurden ist kongruent mit dessen
Beteiligung hinsichtlich der moralischen Urteilskraft. Dieses
Resultat unterstützt auch die Hypothese, dass der VMPFC
die affektive Beurteilung der Wahrnehmung eines Status
unterstützt. Dies indiziiert zudem, dass die TeilnehmerInnen
den Gesichtern mit höheren moralischen Status einen
größeren Wert beimessen.
erfolgreiche FK vs.
nicht erfolgreiche FK
2009 Neuropsychologia
Chiao, J.Y; Harada,
T.; Oby, E.R.; Li, Z.;
Parrish, T.; Bridge,
D.J.
Neural representations
of social status
hierarchy in human
inferior parietal cortex
Ziel: untersuchen, in
welchem Ausmaß
Entscheidungen über
sozialen Status, Hierarchie,
und Nummern (im Vergleich
zueinander) Subregionen
des inferior parietalen Kortex
erfassen bzw. aktivieren.
fMRIn=12 (f=6,
m=6)
Die Stimuli bestanden aus 96 grauen Fotos, wovon 24
Fotos arabische Ziffern zwischen 33 und 99 zeigten, 24
Fotos stellten Statusabzeichen von NROTC dar, 24
Fotos lichteten verschiedene NROTC Offiziere ab und
die restlichen 24 Fotos zeigten verschiedene Autos der
Marke "Toyota". Den ProbandInnen wurde jeweils ein
Bild (Zahl, Statusabzeichen, Offizier oder Toyota)
gezeigt, das sie in weiterer Folge bewerten mussten,
und zwar dahingehend, ob das Bild im Hinblick auf
einen gewählten Mittelwert besser, schlechter oder
gleich gut ist. Jede Runde (insgesamt 4) bestand aus 3
Blöcken zu je 3 Typen, die hinsichtlich der
semantischen Entfernung variierten. Jede Runde
inkludierte auch eine Kontrollaufgabe, in welcher die
TeilnehmerInnen einen Knopf drücken mussten, sobald
die Farbe des gezeigten Kreuzes von weiß auf rot
wechselte.
Naval Reserve Officer
Training Corp
(angehende
Marineoffiziere)
Die Reaktionszeit bei Bildern,
nahe dem Mittelwert war
signifikant langsamer als bei
Bildern weit entfernt vom
Mittelwert. Quer durch alle vier
Bereiche war die Exaktheit im
Gesamten sehr hoch.
In allen vier Bild-Bereichen konnte eine erhöhte beidseitige Aktivierung des IPS festgestellt
werden (im Vergleich zur Kontrollaufgabe). Der Vergleich von Ziffern zeigte zudem eine
Aktivierung im linken posterior superior Parietallappen, sowie im linken medialen frontalen
Gyrus und im rechten inferior frontalen Gyrus. Ähnlich des Ziffern-Vergleichs konnte bei den
restlichen Status-Symbolen jeweils eine zweiseitige Aktivierung des inferior frontal Gyrus
festgestellt werden. Uniform-Status-Vergleiche in Relation zu Auto-Status-Vergleiche
zeigten eine höhere Aktivierung im rechten mittleren frontalen Gyrus. Des Weiteren konnte
ein "Distanz-Effekt" beobachtet werden, wonach neurale Aktivität innerhalb des beidseitigen
IPS für den Mittelwert naheliegende Nummern und sozialen Status' steigt (im Verhältnis zu
Entfernten). Zusätzliche Gehirnregionen innerhalb des frontalen, parietalen und temporalen
Kortex offenbarten ebenso eine stärkere Aktivierung für, den Mittelwert näher liegende Werte
und Symbole, im Vergleich zu Entfernten. Für den numerischen Vergleich waren dies:
bilateral Precuneus, der rechte supramarginal Gyrus, der rechte mittlere und superior
temporal Gyrus, der rechte inferior, der mittlere und der superior frontale Gyrus sowie der
linke cingulare Gyrus. Für die Uniform-Vergleiche: rechter superior und inferior frontal Gyrus,
im rechten mittleren und superior temporal Gyrus. Für die Gesichts-Vergleiche: bilateral
Precuneus, rechter mittlerer temporal Gyrus, linker superior temporal Gyrus, linker cingulare
Gyrus. Die Auto-Vergleiche: linker Precuneus, bilateral posterior superior Parietallappen,
bilateral mittlerer frontale Gyrus, bilateral Thalamus, linker inferior temporal Gyrus und
rechter superior temporal Gyrus. Diese Ergebnisse über die Vergleiche von sozialen Status'
sowie von Nummern ergänzen unterschiedliche sowie überlappende neuronale
Vorstellungen innerhalb des menschlichen inferior parietalen Kortex.
Sowohl sozialer Status als auch numerische Vergleiche
aktivierten unterschiedliche sowie überlappende
Gehirnregionen des inferioren parietalen Kortex. Die
Resultate dieser Studie zeigen ein neues Ausmaß der
neuralen Darstellung im inferioren parietalen Kortex, die nicht
nur für wahrgenommenen und kognitive Bereiche, sondern
auch für Bereiche der sozialen Kenntnis, wie insbesondere
die soziale Status-Hierarchie, gilt. Übereinstimmend mit
älteren Studien sind im Hinblick auf soziale Vergleiche ein
Netzwerk von Gehirnregionen, innerhalb des
occipitotemporalen und frontalen Kortex beteiligt.
erfolgreiche FK vs.
nicht erfolgreiche FK
2011 Social Neuroscience
Cloutier, J.; Kelley,
W.M.; Heatherton,
T.F.
The influence of
perceptual and
knowledge-based
familiarity on the neural
substrates of face
perception
Ziel: das neuronale Stubstrat
von Vertrautheit im Hinblick
auf das Gesicht und Wissen
über die Person zu
untersuchen. Hypothese:
verschiedene erweiterte
Gehirnnetzwerke
differenzieren hinsichtlich
der Wahnehmung von (a)
neuen Gesichtern (b) neuen
Gesichtern mit einer
personenbezogenen
Beschreibung (c) vertrauten
Gesichtern und (d)
vertrauten Gesichtern mit
einer erlernten
Beschreibung.
fMRI
n= 24 (alle
weiblich); 5
wurden
aussortiert
Als Stimuli dienten 96 unbekannte männliche Gesichter
ergänzt mit unbekannten männlichen Gesichtern,
zusammengestellt von den Medien. In der ersten fMRI
Sitzung wurden den ProbandInnen 24 Gesichter
insgesamt je 3 mal präsentiert und sie mussten sich
einen Eindruck über die jeweilige Person machen und
diesen per Knopfdruck wiedergeben (insgesamt gab es
4 Knöpfe; 1=attraktiv; 2= neutral; 3= nicht attraktiv; 4=
keine Information). Während der zweiten Session
wurden beschreibende Sätze zusätzlich zu neuen
Bildern präsentiert. Die beschreibende Sätze wurden
zuvor in drei Kategorien unterteilt (attraktiv, neutral,
unattraktiv). Die ProbandInnen mussten sich wieder
einen Eindruck über die jeweilige Person bilden,
allerdings sollten sie die beschreibenden Sätze dabei
auch einbeziehen. Während der dritten fMRI Session,
die 1-2 Tage später stattfand, wurden den
Teilnehmerinnen wieder die selben Gesichter aus der
ersten und zweiten Session gezeigt. Sie mussten
diesmal einen Eindruck über das gezeigte Bild bilden,
basierend auf ihrer direkten Reaktion.
Teilnehmerinnen
wurden vom lokalen
Darthmouth College
rekrutiert.
--
Der mittlere präfrontale Kortex (MPFC) zeigte eine erhöhte Aktivierung hinsichtlich neuer
Gesichter (a) und Gesichter für die eine persönliche Beschreibung erhältlich ist (b).
Statistische Auswertungen ergaben zudem, dass die Aktivierung für Gesichter mit einer
Beschreibung höher war als für Gesichter ohne Beschreibung. Vertraute Gesichter mit einer
Beschreibung (d) führten wiederum zu einer erhöhten Aktivierung des MPFC als vertraute
Gesichter ohne einer Beschreibung (c). Einige Gehirnregionen zeigten eine erhöhte
Aktivierung als die Gesichter mit einer Beschreibung gepaart wurden: so der mediale
präfrontale Kortex; der ventrale laterale präfrontale Kortex beidseitig; der
Precuneus/posterior cingulare Gyrus und die rechte Insula. Der Precuneus/posteriores
Cingulum war vorzugsweise aktiviert bei der Meinungsbildung von wahrgenommenen
vertrauten Gesichtern im Vergleich zu neuen Gesichtern. Wenn über neue Gesichter, im
Vergleich zu vertrauten Gesichtern ein Eindruck gebildet wird, sind viele Gehirnregionen
aktiviert. Vorzugsweise wurde eine Aktivierung von neuen Gesichtern im rechten
fusiform/inferior temporalen Gyrus beobachtet. Bei der Meinungsbildung von vertrauten
Gesichtern gepaart mit einer Beschreibung zeigten einige Gehirnregionen eine erhöhte
Aktivierung: mediale präfrontaler Kortex; der Precuneus, der linke ventrolateraler präfrontaler
Kortex; und der posterior superior temporal Sulcus (STS). Dies sind alles Gehirnregionen,
die mit der sozialen Kognition assoziiert werden. Bei der Meinungsbildung von vertrauten
Gesichtern verglichen mit vertrauten Gesichtern mit Beschreibung waren vor allem
Aktivierungen im rechten fusiform Gyrus beobachtbar.
Diese Studie zeigt, dass der MPFC vorzugsweise aktiviert
ist, wenn Wissen über die Person vorhanden ist. Außerdem
werden durch das Vorhandensein von Wissen über eine
Person viele Regionen des sogenannten "sozialen Gehirns"
aktiviert. Die gegenständliche Studie bestätigt zudem die
EInbindung des Precuneus im Hinblick auf wahrgenommene
Vertrautheit.
erfolgreiche FK vs.
nicht erfolgreiche FK
2011 Neuroimage
Rule, N.O.; Moran,
J.M.; Freeman, J.B.;
Whitfield-Gabrieli,
S.; Gabrieli, J.D.E.;
Ambady, N.
Face value: Amygdala
response reflects the
validity of first
impressions
Hypothese: wenn die
Gesichter der
Führungskräfte
wahrgenommen werden,
wird die Amygdala auf die
tatsächliche Leadership-
Fähigkeit reagieren. Diese
Reaktion entspricht der
Erregung, die durch
subjektive Eindrücke der
Leadership-Qualität und
objektiven Indikatoren des
Erfolges einer Führungskraft
vorhergesagt wird.
fMRI
n=16 (m=9,
f=7), 1 wurde
aussortiert
Als Reize dienten Portraitfotos von CEO`s aus der
Fortune 1000 (Jahr 2006) Rangliste. Dabei wurden die
ersten und letzten 25 CEO's der zugehörigen
Unternehmen in die Liste genommen, wovon eine
Person weiblich war. Ein CEO wurde aussortiert. Zu
diesem Sample wurden auch noch 19 Portraits
weiblicher CEO's unterschiedlicher Rankings
hinzugefügt. Während die Portraitfotos den
ProbandInnen einzeln gezeigt wurden, mussten diese
innerhalb von 2 Sekunden entscheiden, ob das ihnen
gezeigte Gesicht symmetrisch oder asymmetrisch ist.
Nach dem Scan wurden die Gesichter den
TeilnehmerInnen am Computer abermals gezeigt. Sie
mussten bewerten, wie erfolgreich die jeweilige Person
als Führungskraft einer führenden Unternehmung sein
würde (von sehr erfolgreich bis gar nicht erfolgreich).
StudentInnen
Größere Aktivierung der
Amygdala wurde mit einer
besseren Führungsqualität
assoziiert und mit größeren
Reingewinnen des
Unternehmens, in dem der
jeweilige CEO arbeitete.
Bei den meisten TeilnehmerInnen konnte eine positive Korrelation zwischen der Aktivierung
der linken Amygdala und dem Reingewinn eines Unternehmens beobachtet werden (mehr
Reaktion, wenn ein Unternehmen mehr Umsätze erwirtschaftete). Auch die Aktivierung im
rechten temporoparietal Übergang und in der linken superior temporal Sulcus zeigt eine
Korrelation mit dem Reingewinn eines Unternehmens. Außerdem konnte eine signifikante
positive Korrelation zwischen der Aktivierung der linken Amygdala und der
wahrgenommenen Führungsqualität festgestellt werden. Die Aktivierung der linken
Amygdala erhöht sich, wenn das Unternehmen des CEO mehr Reingewinn macht und wenn
die Leadership-Fähigkeit besser eingeschätzt wird. Die Gesichter jener erfolgreicheren
Führungskräfte bewirken somit eine höhere Aktivierung in der linken Amygdala. Eine
Analyse des gesamten Gehirn zeigte neben einer erhöhten Aktivierung der linken Amygdala
zudem auch eine höhere Aktivierung im superior temporalen Sulcus und des
temporoparietalen Überganges. Diese Regionen werden oftmals als "theory of mind"
bezeichnet.
Die Attraktivität des Gesichtes zeigte bei der Wahrnehmung
der CEO's keinen Zusammenhang mit der Aktivierung der
Amygdala. Auch Ratings bezüglich emotionalen Ausdruck
oder die Entscheidungen über die Symmetrie des Gesichtes
zeigten einen signifikanten Zusammenhang mit der
Aktivierung der Amygdala. Diese Ergebnisse sind nicht
überraschend, weil bereits in früheren Studien
herausgefunden wurde, dass die Amygdala hinsichtlich der
Bewertung von emotionalen und gesellschaftlichen Reizen
eine Rolle spielt. Auch ist bekannt, dass die Amygdala umso
stärker reagiert, desto mehr Reize erregen. In der
Leadershipforschung wird deshalb vorgeschlagen, dass
CEO's jene Personen sein sollen, die Dominanz und Macht
ausstrahlen.
erfolgreiche FK vs.
nicht erfolgreiche FK
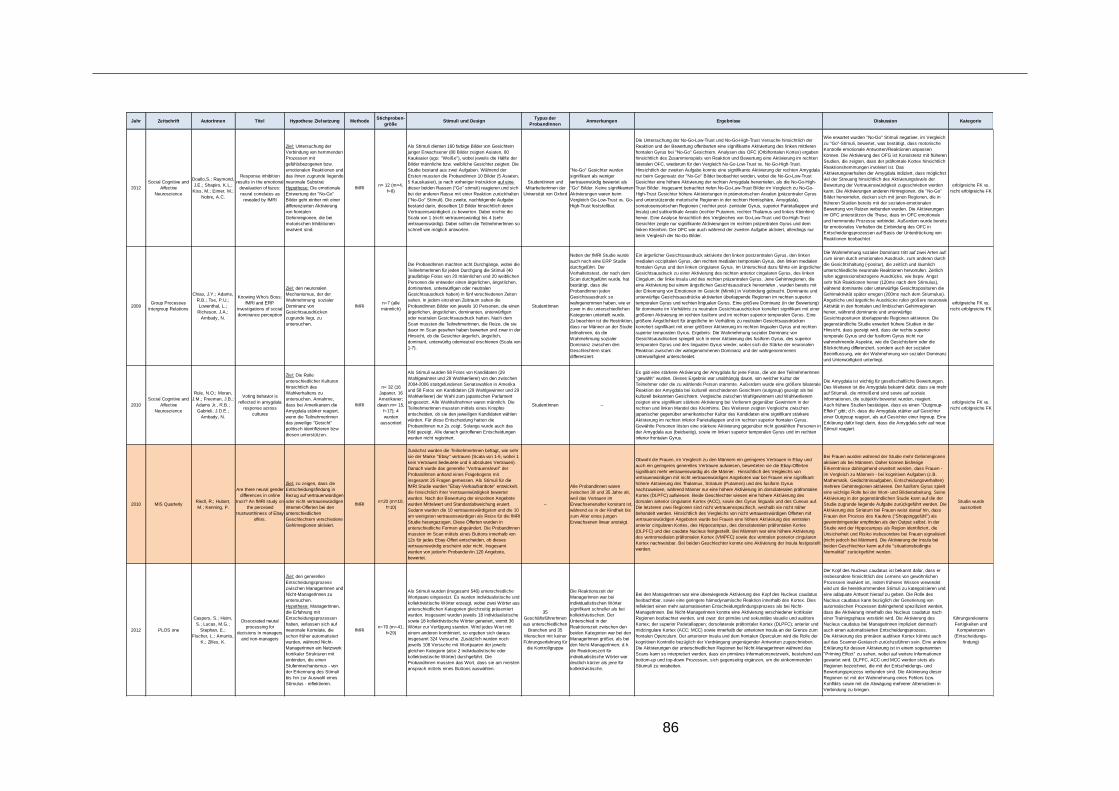
86
Jahr Zeitschrift AutorInnen Titel Hypothese Zielsetzung MethodeStichproben-
größeStimuli und Design
Typus der
ProbandInnenAnmerkungen Ergebnisse Diskussion Kategorie
2012
Social Cognitive and
Affective
Neuroscience
Doallo,S.; Raymond,
J.E.; Shapiro, K.L.;
Kiss, M.; Eimer, M.;
Nobre, A.C.
Response inhibition
results in the emotional
devaluation of faces:
neural correlates as
revealed by fMRI
Ziel: Untersuchung der
Verbindung von hemmenden
Prozessen mit
gefühlsbezogenen bzw.
emotionalen Reaktionen und
das ihnen zugrunde liegende
neuronale Substrat
Hypothese: Die emotionale
Entwertung der "No-Go"
Bilder geht einher mit einer
differenzierten Aktivierung
von frontalen
Gehirnregionen, die bei
motorischen Inhibitionen
involviert sind.
fMRIn= 12 (m=4,
f=8)
Als Stimuli dienten 160 farbige Bilder von Gesichtern
junger Erwachsener (80 Bilder zeigten Asiaten, 80
Kaukasier (ugs: "Weiße"), wobei jeweils die Hälfte der
Bilder männliche bzw. weibliche Gesichter zeigten. Die
Studie bestand aus zwei Aufgaben. Während der
Ersten mussten die ProbandInnen 10 Bilder (5 Asiaten,
5 Kauskasier), je nach vorheriger Instruktion auf eine
dieser beiden Rassen ("Go" stimuli) reagieren und sich
bei der anderen Rasse mit einer Reaktion zurückhalten
("No-Go" Stimuli). Die zweite, nachfolgende Aufgabe
bestand darin, dieselben 10 Bilder hinsichtlich deren
Vertrauenswürdigkeit zu bewerten. Dabei reichte die
Scala von 1 (nicht vertrauenswürdig) bis 4 (sehr
vertrauenswürdig). Dabei sollten die TeilnehmerInnen so
schnell wie möglich antworten.
StudentInnen und
MitarbeiterInnen der
Universität von Oxford
"No-Go" Gesichter wurden
signifikant als weniger
vertrauenwürdig bewertet als
"Go" Bilder. Keine signifikanten
Aktivierungen waren beim
Vergleich Go-Low-Trust vs. Go-
High-Trust feststellbar.
Die Untersuchung der No-Go-Low-Trust und No-Go-High-Trust Versuche hinsichtlich der
Reaktion und der Bewertung offenbarten eine signifikante Aktiviertung des linken mittleren
frontalen Gyrus bei "No-Go" Gesichtern. Analysen des OFC (Orbifrontalen Kortex) ergaben
hinsichtlich des Zusammenspiels von Reaktion und Bewertung eine Aktivierung im rechten
lateralen OFC, wiederum für den Vergleich No-Go-Low-Trust vs. No-Go-High-Trust.
Hinsichtlich der zweiten Aufgabe konnte eine signifikante Aktivierung der rechten Amygdala
nur beim Gegensatz der "No-Go" Bilder beobachtet werden, wobei die No-Go-Low-Trust
Gesichter eine höhere Aktivierung der rechten Amygdala hervorriefen, als die No-Go-High-
Trust Bilder. Insgesamt betrachtet riefen No-Go-Low-Trust Bilder im Vergleich zu No-Go-
High-Trust Gesichter höhere Aktiviertungen in prämotorischen Arealen (präzentraler Gyrus
und unterstützende motorische Regionen in der rechten Hemisphäre, Amygdala),
somatosensorischen Regionen ( rechter post- zentraler Gyrus, superior Parietallappen und
Insula) und subkortikale Areale (rechter Putamen, rechter Thalamus und linkes Kleinhirn)
hervor. Eine Analyse hinsichtlich des Vergleiches von Go-Low-Trust und Go-High-Trust
Gesichter zeigte nur signifikante Aktivierungen im rechten präzentralen Gyrus und dem
linken Kleinhirn. Der OFC war auch während der zweiten Aufgabe aktiviert, allerdings nur
beim Vergleich der No-Go Bilder.
Wie erwartet wurden "No-Go" Stimuli negativer, im Vergleich
zu "Go"-Stimuli, bewertet, was bestätigt, dass motorische
Kontrolle emotionale Antworten/Reaktionen anpassen
können. Die Aktivierung des OFG ist Konsistenz mit früheren
Studien, die zeigten, dass der präfrontale Kortex hinsichtlich
Reaktionshemmungen involviert ist. Das
Aktivierungsverhalten der Amygdala indiziert, dass möglichst
viel der Streuung hinsichtlich des Aktivierungslevels der
Bewertung der Vertrauenswürdigkeit zugeschrieben werden
kann. Die Aktivierungen anderen Hirnregionen, die "No-Go"
Bilder hervorriefen, decken sich mit jenen Regionen, die in
früheren Studien bereits mit der sozialen-emotionalen
Bewertung von Reizen verbunden wurden. Die Aktivierungen
im OFC unterstützen die These, dass im OFC emotionale
und hemmende Prozesse verbindet. Außerdem wurde bereits
für emotionales Verhalten die Einbindung des OFC in
Entscheidungsprozessen auf Basis der Unterdrückung von
Reaktionen beobachtet.
erfolgreiche FK vs.
nicht erfolgreiche FK
2009Group Processes
Intergroup Relations
Chiao, J.Y.; Adams,
R.B.; Tse, P.U.;
Lowenthal, L.;
Richeson, J.A.;
Ambady, N.
Knowing Who's Boss:
fMRI and ERP
investigations of social
dominance perception
Ziel: den neuronalen
Mechanismus, der der
Wahrnehmung sozialer
Dominanz von
Gesichtsausdrücken
zugrunde liegt, zu
untersuchen.
fMRIn=7 (alle
männlich)
Die ProbandInnen machten acht Durchgänge, wobei die
TeilnehmerInnen für jeden Durchgang die Stimuli (40
graufärbige Fotos von 20 männlichen und 20 weiblichen
Personen die entweder einen ärgerlichen, ängstlichen,
dominanten, unterwürfigen oder neutralen
Gesichtsausdruck haben) in fünf verschiedenen Zeiten
sahen. In jedem einzelnen Zeitraum sahen die
ProbandInnen Bilder von jeweils 10 Personen, die einen
ärgerlichen, ängstlichen, dominanten, unterwürfigen
oder neutralen Gesichtsausdruck hatten. Nach dem
Scan mussten die TeilnehmerInnen, die Reize, die sie
davor im Scan gesehen haben bewerten und zwar in der
Hinsicht, ob die Gesichter ärgerlich, ängstlich,
dominant, unterwürfig oderneutral erschienen (Scala von
1-7).
StudentInnen
Neben der fMRI Studie wurde
auch noch eine ERP Studie
durchgeführt. Der
Verhaltenstest, der nach dem
Scan durchgeführt wurde, hat
bestätigt, dass die
ProbandInnen jeden
Gesichtsausdruck so
wahrgenommen haben, wie er
zuvor in die unterschiedlichen
Kategorien unterteilt wurde.
Zu beachten ist die Restriktion,
dass nur Männer an der Studie
teilnahmen, da die
Wahrnehmung sozialer
Dominanz zwischen den
Geschlechtern stark
differenziert.
Ein ärgerlicher Gesichtsausdruck aktivierte den linken postzentralen Gyrus, den linken
medialen occipitalen Gyrus, den rechten medialen temporalen Gyrus, den linken medialen
frontalen Gyrus und den linken cingularen Gyrus. Im Unterschied dazu führte ein ängstlicher
Gesichtsausdruck zu einer Aktivierung des rechten anterior cingularen Gyrus, des linken
Cingulum, der linke Insula und des rechten präzentralen Gyrus. Jene Gehirnregionen, die
eine Aktivierung bei einem ängstlichen Gesichtsausdruck hervorriefen , wurden bereits mit
der Erkennung von Emotionen im Gesicht (Mimik) in Verbindung gebracht. Dominante und
unterwürfige Gesichtsausdrücke aktivierten überlappende Regionen im rechten superior
temporalen Gyrus und rechten lingualen Gyrus. Eine größere Dominanz (in der Bewertung)
für dominante im Verhältnis zu neutralen Gesichtsausdrücken korreliert signifikant mit einer
größeren Aktivierung im rechten fusiform und im rechten superior temporalen Gyrus. Eine
größere Ängstlichkeit für ängstliche im Verhältnis zu neutralen Gesichtsausdrücken
korreliert signifikant mit einer größerer Aktivierung im rechten lingualen Gyrus und rechten
superior temporalen Gyrus. Ergebnis: Die Wahrnehmung sozialer Dominanz von
Gesichtsausdrücken spiegelt sich in einer Aktivierung des fusiform Gyrus, des superior
temporalen Gyrus und des lingualen Gyrus wieder, wobei sich die Stärke der neuronalen
Reaktion zwischen der wahrgenommenen Dominanz und der wahrgenommenen
Unterwürfigkeit unterscheidet.
Die Wahrnehmung sozialer Dominanz tritt auf zwei Arten auf:
zum einen durch emotionalen Ausdruck, zum anderen durch
die Gesichtshaltung (-positur), die zeitlich und räumlich
unterschliedliche neuronale Reaktionen hervorrufen. Zeitlich
rufen aggressionsbezogene Ausdrücke, wie bspw. Angst
sehr früh Reaktionen hervor (120ms nach dem Stimulus),
während dominante oder unterwürfige Gesichtsposituren die
Gehirnaktivität später erregen (200ms nach dem Stiumulus).
Ängstliche und ärgerliche Ausdrücke rufen größere neuronale
Aktivität in den frontalen und limbischen Gehirnregionen
hervor, während dominante und unterwürfige
Gesichtsposituren überlappende Regionen aktivieren. Die
gegenständliche Studie erweitert frühere Studien in der
Hinsicht, dass gezeigt wird, dass der rechte superior
temporale Gyrus und der fusiform Gyrus nicht nur
wahrnehmende Aspekte, wie die Gesichtsform oder die
Blickrichtung differenziert, sondern auch der sozialen
Beeinflussung, wie der Wahrnehmung von sozialer Dominanz
und Unterwürfigkeit unterliegt.
erfolgreiche FK vs.
nicht erfolgreiche FK
2010
Social Cognitive and
Affective
Neuroscience
Rule, N.O.; Moran,
J.M.; Freeman, J.B.;
Adams Jr., R.B.;
Gabrieli, J.D.E.;
Ambady, N.
Voting behavior is
reflected in amygdala
response across
cultures
Ziel: Die Rolle
unterschiedlicher Kulturen
hinsichtlich des
Wahlverhaltens zu
untersuchen. Annahme,
dass bei Amerikanern die
Amygdala stärker reagiert,
wenn die TeilnehmerInnen
das jeweilige "Gesicht"
politisch identifizieren bzw
diesen unterstützen.
fMRI
n= 32 (16
Japaner, 16
Amerikaner;
davon m= 15,
f=17); 4
wurden
aussortiert
Als Stimuli wurden 58 Fotos von Kandidaten (29
Wahlgewinner und 29 Wahlverlierer) von den zwischen
2004-2006 stattgefundenen Senatswahlen in Amerika
und 58 Fotos von Kandidaten (29 Wahlgewinner und 29
Wahlverlierer) der Wahl zum japanischen Parlament
eingesetzt. Alle Wahlteilnehmer waren männlich. Die
TeilnehmerInnen mussten mittels eines Knopfes
entscheiden, ob sie den jeweiligen Kandidaten wählen
würden. Für diese Entscheidung hatten die
ProbandInnen nur 2s zeigt. Solange wurde auch das
Bild gezeigt. Alle danach getroffenen Entscheidungen
wurden nicht registriert.
StudentInnen --
Es gab eine stärkere Aktivierung der Amygdala für jene Fotos, die von den TeilnehmerInnen
"gewählt" wurden. Dieses Ergebnis war unabhängig davon, von welcher Kultur der
Teilnehmer oder die zu wählende Person stammte. Außerdem wurde eine größere bilaterale
Reaktion der Amygdala bei kulturell verschiedenen Gesichtern (outgroup) gezeigt als bei
kulturell bekannten Gesichtern. Vergleiche zwischen Wahlgewinnern und Wahlverlierern
zeigten eine signifikant stärkere Aktivierung bei Verlierern gegenüber Gewinnern in der
rechten und linken Mandel des Kleinhirns. Des Weiteren zeigten Vergleiche zwischen
japanischer gegenüber amerikanischer Kultur des Kandidaten eine signifikant stärkere
Aktivierung im rechten inferior Parietallappen und im rechten superior frontalen Gyrus.
Gewählte Personen lösten eine stärkere Aktivierung gegenüber nicht gewählten Personen in
der Amygdala aus (beidseitig), sowie im linken superior temporalen Gyrus und im rechten
inferior frontalen Gyrus.
Die Amygdala ist wichtig für gesellschaftliche Bewertungen.
Des Weiteren ist die Amygdala bekannt dafür, dass sie mehr
auf Stiumuli, die mitreißend sind sowie auf soziale
Informationen, die subjektiv bewertet wurden, reagiert.
Auch frühere Studien bestätigen, dass es einen "Outgroup-
Effekt" gibt; d.h. dass die Amygdala stärker auf Gesichter
einer Outgroup reagiert, als auf Gesichter einer Ingroup. Eine
Erklärung dafür liegt darin, dass die Amygdala sehr auf neue
Stimuli reagiert.
erfolgreiche FK vs.
nicht erfolgreiche FK
2010 MIS QuarterlyRiedl, R.; Hubert,
M.; Kenning, P.
Are there neural gender
differences in online
trust? An fMRI study on
the perceived
trustworthiness of Ebay
offers.
Ziel: zu zeigen, dass die
Entscheidungsfindung in
Bezug auf vertrauenwürdigen
oder nicht vertrauenwürdigen
Internet-Offerten bei den
unterschiedlichen
Geschlechtern verschiedene
Gehirnregionen aktiviert.
fMRIn=20 (m=10,
f=10)
Zunächst wurden die TeilnehmerInnen befragt, wie sehr
sie der Marke "Ebay" vertrauen (Scala von 1-5, wobei 1
kein Vertrauen bedeutete und 5 absolutes Vertrauen).
Danach wurde das generelle "Vertrauenslevel" der
ProbandInnen anhand eines Fragebogens mit
insgesamt 25 Fragen gemessen. Als Stimuli für die
fMRI Studie wurden "Ebay-Verkaufsanbote" entwickelt,
die hinsichtlich ihrer Vertrauenwürdigkeit bewertet
wurden. Nach der Bewertung der einzelnen Angebote
wurden Mittelwert und Standardabweichung eruiert.
Sodann wurden die 10 vertrauenswürdigsten und die 10
am wenigsten vertrauenswürdigen als Reize für die fMRI
Studie herangezogen. Diese Offerten wurden in
unterschiedliche Formen abgeändert. Die ProbandInnen
mussten im Scan mittels eines Buttons innerhalb von
12s für jedes Ebay-Offert entscheiden, ob dieses
vertrauenswürdig erscheint oder nicht. Insgesamt
wurden von jeder/m Probanden/in 120 Angebote,
bewertet.
--
Alle ProbandInnen waren
zwischen 30 und 35 Jahre alt,
weil das Vertauen im
Erwachsenenalter konstant ist,
während es in der Kindheit bis
zum Alter eines jungen
Erwachsenen linear ansteigt.
Obwohl die Frauen, im Vergleich zu den Männern ein geringeres Vertrauen in Ebay und
auch ein geringeres generelles Vertrauen aufwiesen, bewerteten sie die Ebay-Offerten
signifikant mehr vertrauenswürdig als die Männer. Hinsichtlich des Vergleichs von
vertrauenwürdigen mit nicht vertrauenwürdigen Angeboten war bei Frauen eine signifikant
höhere Aktivierung des Thalamus, Striatum (Putamen) und des fusiform Gyrus
nachzuweisen, während Männer nur eine höhere Aktivierung im dorsolateralen präfrontalen
Kortex (DLPFC) aufwiesen. Beide Geschlechter wiesen eine höhere Aktivierung des
dorsalen anterior cingularen Kortex (ACC), sowie des Gyrus lingualis und des Cuneus auf.
Die letzteren zwei Regionen sind nicht vertrauensspezifisch, weshalb sie nicht näher
behandelt werden. Hinsichtlich des Vergleichs von nicht vertauenswürdigen Offerten mit
vertrauenswürdigen Angeboten wurde bei Frauen eine höhere Aktivierung des ventralen
anterior cingularen Kortex, des Hippocampus, des dorsolateralen präfrontalen Kortex
(DLPFC) und des caudate Nucleus festgestellt. Bei Männern war eine höhere Aktivierung
des ventromedialen präfrontalen Kortex (VMPFC) sowie des ventralen posterior cingularen
Kortex nachweisbar. Bei beiden Geschlechter konnte eine Aktivierung der Insula festgestellt
werden.
Bei Frauen wurden während der Studie mehr Gehirnregionen
aktiviert als bei Männern. Daher können bisherige
Erkenntnisse dahingehend erweitert werden, dass Frauen -
im Vergleich zu Männern - bei kognitiven Aufgaben (z.B.
Mathematik, Gedächtnisaufgaben, Entscheidungsverhalten)
mehrere Gehirnregionen aktivieren. Der fusiform Gyrus spielt
eine wichtige Rolle bei der Wort- und Bildverarbeitung. Seine
Aktivierung in der gegenständlichen Studie kann auf die der
Studie zugrunde liegende Aufgabe zurückgeführt werden. Die
Aktivierung des Striatum bei Frauen weist darauf hin, dass
Frauen den Prozess des Kaufens ("Shoppinggefühl") als
gewinnbringender empfinden als den Output selbst. In der
Studie wird der Hippocampus als Region identifiziert, die
Unsicherheit und Risiko insbesondere bei Frauen signalisiert
(nicht jedoch bei Männern). Die Aktivierung der Insula bei
beiden Geschlechter kann auf die "situationsbedingte
Normalität" zurückgeführt werden.
Studie wurde
aussortiert
2012 PLOS one
Caspers, S.; Heim,
S.; Lucas, M.G.;
Stephan, E.;
Fischer, L.; Amunts,
K.; Zilles, K.
Dissociated neural
processing for
decisions in managers
and non-managers
Ziel: den generellen
Entscheidungsprozess
zwischen ManagerInnen und
Nicht-ManagerInnen zu
untersuchen.
Hypothese: ManagerInnen,
die Erfahrung mit
Entscheidungsprozessen
haben, verlassen sich auf
neuronale Korrelate, die
schon früher automatisiert
wurden, während Nicht-
ManagerInnen ein Netzwerk
kortikaler Strukturen mit
einbinden, die einen
Stufenmechanismus - von
der Erkennung des Stimuli
bis hin zur Auswahl eines
Stimulus - reflektieren.
fMRIn=70 (m=41,
f=29)
Als Stimuli wurden (insgesamt 540) unterschiedliche
Wortpaare eingesetzt. Es wurden individualistische und
kollektivistische Wörter erzeugt, wobei zwei Wörter aus
unterschiedlichen Kategorien gleichzeitig präsentiert
wurden. Insgesamt wurden jeweils 18 individualistische
sowie 18 kollektivistische Wörter generiert, womit 36
Wörter zur Verfügung standen. Wird jedes Wort mit
einem anderen kombiniert, so ergeben sich daraus
insgesamt 324 Versuche. Zusätzlich wurden noch
jeweils 108 Versuche mit Wortpaaren der jeweils
gleichen Kategorie (also 2 individualistische oder
kollektivistische Wörter) durchgeführt. Die
ProbandInnen mussten das Wort, dass sie am meisten
ansprach mittels eines Buttons auswählen.
35
GeschäftsführerInnen
aus unterschiedlichen
Branchen und 35
Menschen mit keiner
Führungserfahrung für
die Kontrollgruppe
Die Reaktionszeit der
ManagerInnen war bei
individualistischen Wörter
signifikant schneller als bei
kollektivistischen. Der
Unterschied in der
Reaktionszeit zwischen den
beiden Kategorien war bei den
ManagerInnen größer, als bei
den Nicht-ManagerInnen; d.h.
die Reaktionszeit für
individualistische Wörter war
deutlich kürzer als jene für
kollektivistische.
Bei den ManagerInnen war eine überwiegende Aktivierung des Kopf des Nucleus caudatus
beobachtbar, sowie eine geringere hämodynamische Reaktion innerhalb des Kortex. Dies
reflektiert einen mehr automatisierten Entscheidungsfindungsprozess als bei Nicht-
ManagerInnen. Bei Nicht-ManagerInnen konnte eine Aktivierung verschiedener kortikaler
Regionen beobachtet werden, und zwar: der primäre und sekundäre visuelle und auditore
Kortex; der superior Parietallappen; dorsolaterale präfrontaler Kortex (DLPFC); anterior und
midcingulare Kortex (ACC; MCC) sowie innerhalb der anterioren Insula an der Grenze zum
frontalen Operculum. Der anterioren Insula und dem frontalen Operculum wird die Rolle der
kognitiven Kontrolle bezüglich der Verdrängung ungenügender Antworten zugeschrieben.
Die Aktivierungen der unterschiedlichen Regionen bei Nicht-ManagerInnen während des
Scans kann so interpretiert werden, dass ein primäres Informationsnetzwerk, bestehend aus
bottom-up und top-down Prozessen, sich gegenseitig ergänzen, um die einkommenden
Stiumuli zu verabeiten.
Der Kopf des Nucleus caudatus ist bekannt dafür, dass er
insbesondere hinsichtlich des Lernens von gewöhnlichen
Prozessen involviert ist, indem früheres Wissen verwendet
wird um die hereinkommenden Stimuli zu kategorisieren und
eine adäquate Antwort hierauf zu geben. Die Rolle des
Nucleus caudatus kann bezüglich der Generierung von
automatischen Prozessen dahingehend spezifiziert werden,
dass die Aktivierung innerhalb des Nucleus caudatus nach
einer Trainingsphase verstärkt wird. Die Aktivierung des
Nucleus caudatus bei ManagerInnen impliziert demnach
auch einen automatisierten Entscheidungsprozess.
Die Aktivierung des primären auditiven Kortex könnte auch
auf das Scanner-Geräusch zurüchzuführen sein. Eine andere
Erklärung für dessen Aktivierung ist in einem sogenannten
"Priming Effect" zu sehen, wobei auf weitere Informationen
gewartet wird. DLPFC, ACC und MCC werden stets als
Regionen bezeichnet, die mit der Entscheidungs- und
Bewertungsprozess verbunden sind. Die Aktivierung dieser
Regionen ist mit der Wahrnehmung eines Fehlers bzw.
Konflikts sowie mit die Abwägung mehrerer Alternativen in
Verbindung zu bringen.
führungsrelevante
Fertigkeiten und
Kompetenzen
(Entscheidungs-
findung)
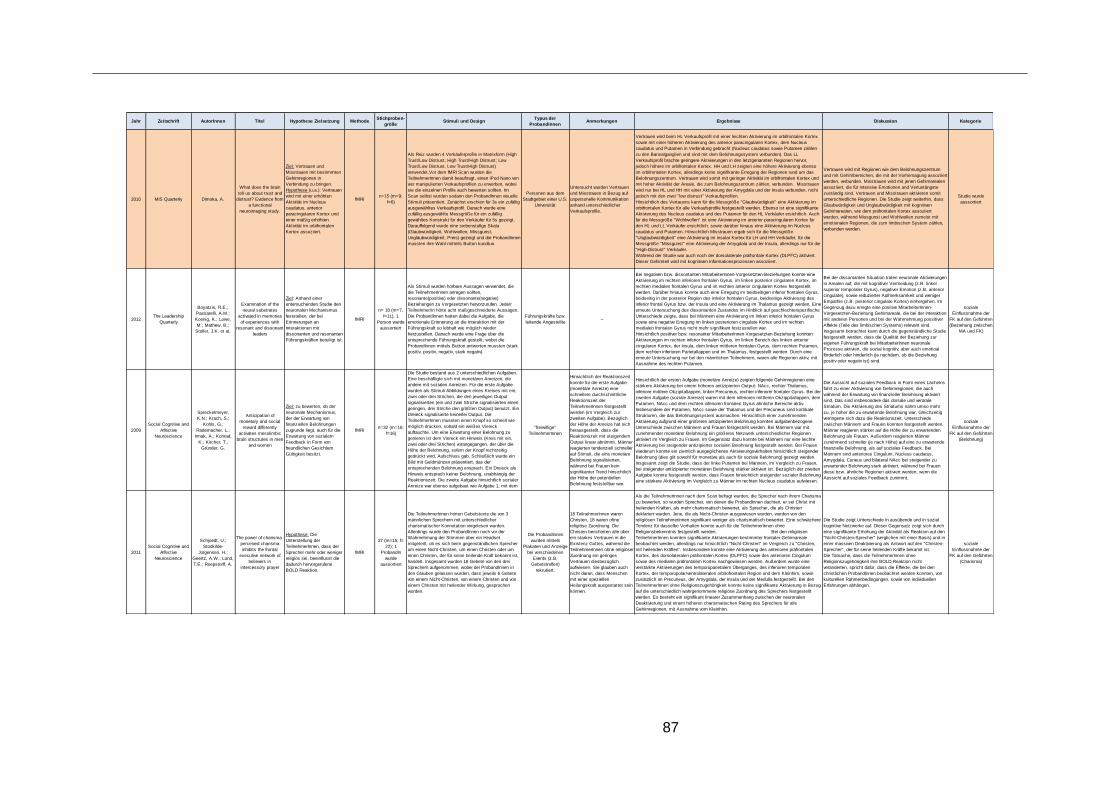
87
Jahr Zeitschrift AutorInnen Titel Hypothese Zielsetzung MethodeStichproben-
größeStimuli und Design
Typus der
ProbandInnenAnmerkungen Ergebnisse Diskussion Kategorie
2010 MIS Quarterly Dimoka, A.
What does the brain
tell us about trust and
distrust? Evidence from
a functional
neuroimaging study.
Ziel: Vertrauen und
Misstrauen mit bestimmten
Gehirnregionen in
Verbindung zu bringen.
Hypothese (u.a.): Vertrauen
wird mit einer erhöhten
Aktivität im Nucleus
caudatus, anterior
paracingularen Kortex und
einer mäßig erhöhten
Aktivität im orbifrontalen
Kortex assoziiert.
fMRIn=15 (m=9;
f=6)
Als Reiz wurden 4 Verkäuferprofile in Matrixform (High
Trust/Low Distrust; High Trust/High Distrust; Low
Trust/Low Distrust, Low Trust/High Distrust)
verwendet.Vor dem fMRI Scan wurden die
TeilnehmerInnen damit beauftragt, einen iPod Nano von
vier manipulierten Verkaufsprofilen zu erwerben, wobei
sie die einzelnen Profile auch bewerten sollten. Im
Scan selbst wurden sodann den ProbandInnen visuelle
Stimuli präsentiert. Zunächst erschien für 3s ein zufällig
ausgewähltes Verkaufsprofil. Danach wurde eine
zufällig ausgewählte Messgröße für ein zufällig
gewähltes Konstrukt für den Verkäufer für 5s gezeigt.
Darauffolgend wurde eine siebenstufige Skala
(Glaubwürdigkeit, Wohlwollen, Missgunst,
Ungläubwürdigkeit, Preis) gezeigt und die ProbandInnen
mussten ihre Wahl mittells Button kundtun.
Personen aus dem
Stadtgebiet einer U.S.
Universität
Untersucht wurden Vertrauen
und Misstrauen in Bezug auf
unpersonelle Kommunikation
anhand unterschiedlicher
Verkaufsprofile.
Vertrauen wird beim HL Verkaufsprofil mit einer leichten Aktivierung im orbifrontalen Kortex
sowie mit einer höheren Aktivierung des anterior paracingularen Kortex, dem Nucleus
caudatus und Putamen in Verbindung gebracht (Nucleus caudatus sowie Putamen zählen
zu den Bansalganglien und sind mit dem Belohnungssystem verbunden). Das LL
Verkaufsprofil brachte geringere Aktivierungen in den letztgenannten Regionen hervor,
jedoch höhere im orbifrontalen Kortex. HH und LH zeigten eine höhere Aktivierung ebenso
im orbifrontalen Kortex, allerdings keine signifikante Erregung der Regionen rund um das
Belohnungszentrum. Vertrauen wird somit mit geringer Aktivität im orbifrontalen Kortex und
mit hoher Aktivität der Areale, die zum Belohnungszentrum zählen, verbunden. Misstrauen
wird nur bei HL und HH mit einer Aktivierung der Amygdala und der Insula verbunden, nicht
jedoch mit den zwei "low distrust" Verkaufsprofilen.
Hinsichtlich des Vertauens kann für die Messgröße "Glaubwürdigkeit" eine Aktivierung im
orbifrontalen Kortex für alle Verkaufsprofile festgestellt werden. Ebenso ist eine signifikante
Aktivierung des Nucleus caudatus und des Putamen für den HL Verkäufer ersichtlich. Auch
für die Messgröße "Wohlwollen" ist eine Aktivierung im anterior paracingularen Kortex für
den HL und LL Verkäufer ersichtlich, sowie darüber hinaus eine Aktivierung im Nucleus
caudatus und Putamen. Hinsichtlich Misstrauen ergab sich für die Messgröße
"Unglaubwürdigkeit" eine Aktivierung im insular Kortex für LH und HH Verkäufer, für die
Messgröße "Missgunst" eine Aktivierung der Amygdala und der Insula, allerdings nur für die
"High-Distrust" Verkäufer.
Während der Studie war auch noch der dorsolaterale präfrontale Kortex (DLPFC) aktiviert.
Dieser Gehirnteil wird mit kognitiven Informationsprozessen assoziiert.
Vertrauen wird mit Regionen wie dem Belohnungszentrum
und mit Gehirnbereichen, die mit der Vorhersagung assoziiert
werden, verbunden. Misstrauen wird mit jenen Gehirnarealen
assoziiert, die für intensive Emotionen and Verlustängste
zuständig sind. Vertrauen und Misstrauen aktivieren somit
unterschiedliche Regionen. Die Studie zeigt weiterhin, dass
Glaubwürdigkeit und Unglaubwürdigkeit mit kognitiven
Gehirnarealen, wie dem präfrontalen Kortex assoziiert
werden, während Missgunst und Wohlwollen zumeist mit
emotionalen Regionen, die zum limbischen System zählen,
verbunden werden.
Studie wurde
aussortiert
2012The Leadership
Quarterly
Boyatzis, R.E.;
Passarelli, A.M.;
Koenig, K.; Lowe,
M.; Mathew, B.;
Stoller, J.K. et al.
Examination of the
neural substrates
activated in memories
of experiences with
resonant and dissonant
leaders
Ziel: Anhand einer
untersuchenden Studie den
neuronalen Mechanismus
feststellen, der bei
Erinnerungen an
Interaktionen mit
dissonanten und resonanten
Führungskräften beteiligt ist.
fMRI
n= 18 (m=7,
f=11), 1
Person wurde
aussortiert
Als Stimuli wurden hörbare Aussagen verwendet, die
die TeilnehmerInnen anregen sollten,
resonante(positive) oder dissonante(negative)
Beziehungen zu Vorgesetzten hervorzurufen. Jede/r
Teilnehmer/in hörte acht maßgeschneiderte Aussagen.
Die ProbandInnen hatten dabei die Aufgabe, die
emotionale Erinnerung an die Interaktion mit der
Führungskraft so lebhaft wie möglich wieder
herzustellen. Danach wurde eine Frage über die
entsprechende Führungskraft gestellt, wobei die
ProbandInnen mittels Button antworten mussten (stark
positiv, positiv, negativ, stark negativ).
Führungskräfte bzw.
leitende Angestellte--
Bei negativen bzw. dissontanten MitarbeiterInnen-Vorgesetzten-Beziehungen konnte eine
Aktivierung im rechten inferioren frontalen Gyrus, im linken posterior cingularen Kortex, im
rechten medialen frontalen Gyrus und im rechten anterior cingularen Kortex festgestellt
werden. Darüber hinaus konnte auch eine Erregung im beidseitigen inferior frontalen Gyrus,
beidseitig in der posterior Region des inferior frontalen Gyrus, beidseitige Aktivierung des
inferior frontal Gyrus bzw. der Insula und eine Aktivierung im Thalamus gezeigt werden. Eine
erneute Untersuchung des dissontanten Zustandes im Hinblick auf geschlechterspezifische
Unterschiede zeigte, dass bei Männern eine Aktivierung im linken inferior frontalen Gyrus
sowie eine negative Erregung im linken posterioren cingulate Kortex und im rechten
medialen frontalen Gyrus nicht mehr signifikant festzustellen war.
Hinsichtlich positiver bzw. resonanter MitarbeiterInnen-Vorgesetzten-Beziehung konnten
Aktivierungen im rechten inferior frontalen Gyrus, im linken Bereich des linken anterior
cingularen Kortex, der Insula, dem linken mittleren frontalen Gyrus, dem rechten Putamen,
dem rechten inferioren Parietallappen und im Thalamus, festgestellt werden. Durch eine
erneute Untersuchung nur bei den männlichen Teilnehmern, waren alle Regionen aktiv, mit
Ausnahme des rechten Putamen.
Bei der dissontanten Situation traten neuronale Aktivierungen
in Arealen auf, die mit kognitiver Vermeidung (z.B. linker
superior temporaler Gyrus), negativer Emotion (z.B. anterior
cingulate), sowie reduzierter Aufmerksamkeit und weniger
Empathie (z.B. posterior cingulate Kortex) einhergehen. Im
Gegenzug dazu erregte eine positive MitarbeiterInnen-
Vorgesetzten-Beziehung Gehirnareale, die bei der Interaktion
mit anderen Personen und bei der Wahrnehmung possitiver
Affekte (Teile des limbischen Systems) relevant sind.
Insgesamt betrachtet kann durch die gegenständliche Studie
festgestellt werden, dass die Qualität der Beziehung zur
eigenen Führungskraft bei MitarbeiterInnen neuronale
Prozesse aktiviert, die sozial kognitiv, aber auch emotioal
förderlich oder hinderlich (je nachdem, ob die Beziehung
positiv oder negativ ist) sind.
soziale
Einflussnahme der
FK auf den Geführten
(Beziehung zwischen
MA und FK)
2009
Social Cognitive and
Affective
Neuroscience
Spreckelmeyer,
K.N.; Krach, S.;
Kohls, G.;
Rademacher, L.;
Irmak, A.; Konrad,
K.; Kircher, T.;
Gründer, G.
Anticipation of
monetary and social
reward differently
activates mesolimbic
brain structures in men
and women
Ziel: zu bewerten, ob der
neuronale Mechanismus,
der der Erwartung von
finanziellen Belohnungen
zugrunde liegt, auch für die
Erwartung von sozialem
Feedback in Form von
freundlichen Gesichtern
Gültigkeit besitzt.
fMRIn=32 (m=16;
f=16)
Die Studie bestand aus 2 unterschiedlichen Aufgaben.
Eine beschäftigte sich mit monetären Anreizen, die
andere mit sozialen Anreizen. Für die erste Aufgabe
wurden als Stimuli Abbildungen eines Kreises mit ein,
zwei oder drei Strichen, die den jeweiligen Output
signalisierten (ein und zwei Striche signalisierten einen
geringen, drei Striche den größten Output) benutzt. Ein
Dreieck signalisierte keinerlei Output. Die
TeilnehmerInnen mussten einen Knopf so schnell wie
möglich drücken, sobald ein weißes Viereck
auftauchte. Um eine Erwartung einer Belohnung zu
genieren ist dem Viereck ein Hinweis (Kreis mit ein,
zwei oder drei Strichen) vorangegangen, der über die
Höhe der Belohnung, sofern der Knopf rechtzeitig
gedrückt wird, Aufschluss gab. Schließlich wurde ein
Bild mit Geldmünzen präsentiert, das der
entsprechenden Belohnung ensprach. Ein Dreieck als
Hinweis entsprach keiner Belohnung, unabhängig der
Reaktionszeit. Die zweite Aufgabe hinsichtlich sozialer
Anreize war ebenso aufgebaut wie Aufgabe 1, mit dem
Unterschied, dass die im Endeffekt präsentierten Bilder,
"freiwillige"
TeilnehmerInnen
Hinsichtlich der Reaktionszeit
konnte für die erste Aufgabe
(monetäre Anreize) eine
schnellere durchschnittliche
Reaktionszeit der
TeilnehmerInnen festgestellt
werden (im Vergleich zur
zweiten Aufgabe). Bezüglich
der Höhe der Anreize hat sich
herausgestellt, dass die
Reaktionszeit mit steigendem
Output linear abnimmt. Männer
reagierten tendenziell schneller
auf Stimuli, die eine monetäre
Belohnung signalisierten,
während bei Frauen kein
signifikanter Trend hinsichtlich
der Höhe der potentiellen
Belohnung feststellbar war.
Hinsichtlich der ersten Aufgabe (monetäre Anreize) zeigten folgende Gehirnregionen eine
stärkere Aktivierung bei einem höheren antizipierten Output: NAcc, rechter Thalamus,
inferiore mittlere Okzipitallappen, linker Precuneus, rechter inferiorer frontaler Gyrus. Bei der
zweiten Aufgabe (soziale Anreize) waren mit dem inferioren mittleren Okzippitallappen, dem
Putamen, NAcc und dem rechten inferioren frontalen Gyrus ähnliche Bereiche aktiv.
Insbesondere der Putamen, NAcc sowie der Thalamus und der Precuneus sind kortikale
Strukturen, die das Belohnungssystem ausmachen. Hinsichtlich einer zunehmenden
Aktivierung aufgrund einer größeren antizipierten Belohnung konnten aufgabenbezogene
Unterschiede zwischen Männern und Frauen festgestellt werden. Bei Männern war mit
zunehmender monetärer Belohnung ein größeres Netzwerk unterschiedlicher Regionen
aktiviert im Vergleich zu Frauen. Im Gegensatz dazu konnte bei Männern nur eine leichte
Aktivierung bei steigender antizipierter sozialen Belohnung festgestellt werden. Bei Frauen
wiederum konnte ein ziemlich ausgeglichenes Aktivierungsverhalten hinsichtlich steigender
Belohnung (dies gilt sowohl für monetäre als auch für soziale Belohnung) gezeigt werden.
Insgesamt zeigt die Studie, dass der linke Putamen bei Männern, im Vergleich zu Frauen,
bei steigender antizipierter monetären Belohnung stärker aktiviert ist. Bezüglich der zweiten
Aufgabe konnte festgestellt werden, dass Frauen hinsichtlich steigender sozialer Belohnung
eine stärkere Aktivierung im Vergleich zu Männer im rechten Nucleus caudatus aufwiesen.
Die Aussicht auf soziales Feedback in Form eines Lächelns
führt zu einer Aktivierung von Gehirnregionen, die auch
während der Erwartung von finanzieller Belohnung aktiviert
sind. Das sind insbesondere das dorsale und ventrale
Striatum. Die Aktivierung des Striatums nahm umso mehr
zu, je höher die zu erwartende Belohnung war. Gleichzeitig
verringerte sich dazu die Reaktionszeit. Unterschiede
zwischen Männern und Frauen konnten festgestellt werden.
Männer reagieren stärker auf die Höhe der zu erwartenden
Belohnung als Frauen. Außerdem reagierten Männer
zunehmend schneller (je nach Höhe) auf eine zu erwartende
finanzielle Belohnung, als auf soziales Feedback. Bei
Männern sind anteriores Cingulum, Nucleus caudatus,
Amygdala, Cuneus und bilateral NAcc bei steigender zu
erwartender Belohnung stark aktiviert, während bei Frauen
diese bzw. ähnliche Regionen aktiviert werden, wenn die
Aussicht auf soziales Feedback zunimmt.
soziale
Einflussnahme der
FK auf den Geführten
(Belohnung)
2011
Social Cognitive and
Affective
Neuroscience
Schjoedt, U.;
Stodkilde-
Jorgensen, H.;
Geertz, A.W.; Lund,
T.E.; Roepstorff, A.
The power of charisma -
perceived charisma
inhibits the frontal
executive network of
believers in
intercessory prayer
Hypothese: Die
Unterstellung der
TeilnehmerInnen, dass der
Sprecher mehr oder weniger
religiös sei, beeinflusst die
dadurch hervorgerufene
BOLD Reaktion.
fMRI
37 (m=15; f=
22); 1
ProbandIn
wurde
aussortiert
Die TeilnehmerInnen hörten Gebetstexte die von 3
männlichen Sprechern mit unterschiedlicher
charismatischer Konnotation vorgelesen wurden.
Allerdings wurde den ProbandInnen noch vor der
Wahrnehmung der Stimmen über ein Headset
mitgeteilt, ob es sich beim gegenständlichen Sprecher
um einen Nicht-Christen, um einen Christen oder um
einen Christen, der für seine heilende Kraft bekannt ist,
handelt. Insgesamt wurden 18 Gebete von den drei
Sprechern aufgenommen, wobei dei ProbandInnen in
den Glauben gelassen wurden, dass jeweils 6 Gebete
von einem Nicht-Christen, von einem Christen und von
einem Christen mit heilender Wirkung, gesprochen
wurden.
Die ProbandInnen
wurden mittels
Plakaten und Anzeige
bei verschiedenen
Events (z.B.
Gebetstreffen)
rekrutiert.
18 TeilnehmerInnen waren
Christen, 18 waren ohne
religiöse Zuordnung. Die
Christen berichteten alle über
ein starkes Vertrauen in die
Existenz Gottes, während die
TeilnehmerInnen ohne religiöser
Zuordnung ein geringes
Vertrauen diesbezüglich
aufwiesen. Sie glauben auch
nicht daran, dass Menschen
mit einer speziellen
Heilungskraft ausgestattet sein
können.
Als die TeilnehmerInnen nach dem Scan befragt wurden, die Sprecher nach ihrem Charisma
zu bewerten, so wurden Sprecher, von denen die ProbandInnen dachten, er sei Christ mit
heilenden Kräften, als mehr charismatisch bewertet, als Sprecher, die als Christen
deklariert wurden. Jene, die als Nicht-Christen ausgewiesen wurden, wurden von den
religiösen TeilnehmerInnen signifikant weniger als charismatisch bewertet. Eine schwächere
Tendenz für dasselbe Verhalten konnte auch für die TeilnehmerInnen ohne
Religionsbekenntnis festgestellt werden. Bei den religiösen
TeilnehmerInnen konnten signifikante Aktivierungen bestimmter frontaler Gehirnareale
beobachtet werden, allerdings nur hinsichtlich "Nicht-Christen" im Vergleich zu "Christen,
mit heilenden Kräften". Insbesondere konnte eine Aktivierung des anterioren präfrontalen
Kortex, des dorsolateralen präfrontalen Kortex (DLPFC) sowie des anterioren Cingulum
sowie des medialen präfrontalem Kortex nachgewiesen werden. Außerdem wurde eine
verstärkte Aktivierungen des temporoparietalem Überganges, des inferioren temporalen
Kortex, der temporopolar/ventrolateralen orbitofrontalen Region und dem Kleinhirn, sowie
zusätzlich im Precuneus, der Amygdala, der Insula und der Medulla festgestellt. Bei den
TeilnehmerInnen ohne Religionszugehörigkeit konnte keine signifikante Aktivierung in Bezug
auf die unterschiedlich wahrgenommene religiöse Zuordnung des Sprechers festgestellt
werden. Es besteht ein signifikant linearer Zusammenhang zwischen der neuronalen
Deaktivierung und einem höheren charismatischen Rating des Sprechers für alle
Gehirnregionen, mit Ausnahme vom Kleinhirn.
Die Studie zeigt Unterschiede in ausübende und in sozial
kognitive Netzwerke auf. Dieser Gegensatz zeigt sich durch
eine signifikante Erhöhung der Aktivität als Reaktion auf den
"Nicht-Christen-Sprecher" (verglichen mit einer Basis) und in
einer massiven Deaktivierung als Antwort auf den "Christen-
Sprecher", der für seine heilenden Kräfte bekannt ist.
Die Tatsache, dass die TeilnehmerInnen ohne
Religionszugehörigkeit ihre BOLD Reaktion nicht
veränderten, spricht dafür, dass die Effekte, die bei den
christlichen ProbandInnen beobachtet werden konnten, von
kulturellen Rahmenbedingungen, sowie von individuellen
Erfahrungen abhängen.
soziale
Einflussnahme der
FK auf den Geführten
(Charisma)
88
Jahr Zeitschrift AutorInnen Titel Hypothese Zielsetzung MethodeStichproben-
größeStimuli und Design
Typus der
ProbandInnenAnmerkungen Ergebnisse Diskussion Kategorie
2011
Social Cognitive and
Affective
Neuroscience
Kang, Y.; Williams,
L.E.; Clark, M.S.;
Gray, J.R.; Bargh,
J.A.
Physical temperature
effects on trust
behavior: the role of
insula
Ziel: die Rolle der Insula im
Hinblick auf den "Temperatur-
Vertrauen-Effekt" zu
untersuchen. Hypothese:
Kälte (Wärme) führt zu
geringeren (größeren)
Ausdruck
zwischenmenschlichen
Vertrauens und der Effekt
der unterschiedlichen
Temperaturen spiegelt sich
in der Aktivität der Insula
wieder.
fMRI
n= 23; 7
Personen
wurden
aussortiert
Es wurden den TeilnehmerInnen entweder kalte (15°C)
oder warme (40°C) Packungen auf die linke Handfläche,
abwechselnd mit einer neutralen (20°C) Packung für
jeweils 20s gelegt. Dieser Vorgang wurde mehrmals
wiederholt. Danach startete das sogenannte "Trust-
Game". Dabei konnten die ProbandInnen 6s lange
überlegen wie viel sie investieren wollen, wobei sie
zwischen vier Investitionen entscheiden konnten. Nach
der Entscheidung der TeilnehmerInnen wurde eine
Antwort präsentiert und deren Fixierung. Anschließend
folgte eine Aufgabe für das Arbeitsgedächtnis, um
Übertragungseffekte aus der ersten Serie zu
schwächen, ehe ein zweiter Durchlauf startete.
--
Es konnte kein Effekt
hinsichtlich der investierten
Menge in Bezug auf die
unterschiedlichen
Temperaturen festgestellt
werden. Ergebnis: Kälte führt
zu geringeren Vertrauen, im
Vergleich zu Wärme.
Sowohl kalte als auch warme Temperaturen riefen eine größere Aktivierung im rechten
primären somatosensorischen Kortex im Vergleich zu neutralen Temperaturen hervor.
Weiters wurde festgestellt, dass mit kalten Temperaturen eine größere Aktivierung der
bilateralen Insula und des bilateralen zentralen und parietal opercularen Kortex einher geht.
Werden die Gehirnregionen, die sowohl bei kalten, als auch bei warmen Temperaturen
empfindlich waren, untersucht, so geht hervor, dass Kälte größere Aktivierung in der Nähe
von posterior insular-opercularen Regionen hervorrief als Wärme. Regionen nahe des
bilateral insular-opercularen Kortex, dem rechtem primären somatosensorischen Zentrum
und des Temporallappens waren mehr aktiv während Kälteempfinden. Wärme hingegen
führte zu größerer Aktivierung im posterioren cingulären Kortex (PCC), sowie in der
inferioren medialen frontalen Region. Gehirnregionen, die eine größere Aktivierung während
der Entscheidungsphase des "Trust-Games" nach Temperatur-Manipulation aufwiesen sind
der okzipitale poles, der anteriore cingulare Kortex, der linke Thalamus sowie der
dorsolaterale präfrontale Kortex (DLPFC). Die linke anterior Insula war signifikant mehr
aktiviert während dem "Trust Game" für jene Sitzungen, denen kalte Temperaturen
vorangegangen sind. Hinsichtlich des direkten Vergleichs von kalter und warmer Temperatur-
Manipulation war festzustellen, dass mit einer kalten Temperatur eine größere Aktivierung
der bilateralen anterior Insula und des zentralen Operculum einherging. Zusätzlich waren
auch der rechte VMPFC, der rechte primäre somatosensorische Kortex, der rechte
prämotorische Kortex und der rechte primäre motorische Kortex auch aktivierter in der
Entscheidungsphase nach einer Temperatur-Manipulation durch Kälte. Die Aktivierung in der
Entscheidungsphase war größer nach Manipulation durch Kälte als durch Wärme.
Die linke anteriore Insel sowie das Operculum wurden bereits
in früheren Studien als Regionen, die eine große Rolle in
Temperatur-Prozessen spielen, identifiziert. Sie funktionieren
als eine Vermittlungsregion, wo innere Sinnenausdrücke in
Emotionen umgewandelt werden. Auch die Aktivierung von
weiteren Regionen nahe der Insula und des ACC während der
Entscheidungsfindung ist ebenso bereits gut dokumentiert.
Ganz allgemein gesprochen unterstützt diese Studie die
These, dass Kälte zu einem geringeren Vertrauen führt, im
Vergleich zu Wärme. Temperaturunterschiede führen nicht
nur zu unterschiedlichem Verhalten, sondern auch zu
unterschiedliche neuronale Aktivierung.
soziale
Einflussnahme der
FK auf den Geführten
(Beziehung zwischen
MA und FK)
2014 NeuroImage
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Leading the follower:
An fMRI investigation of
dynamic cooperativity
and leader-follower
strategies in
synchronization with an
adaptive virtual partner
Ziel: Untersuchung der
neuronalen Korrelate von
Führung.
fMRIn=16 (m=8;
f=8)
Als virtueller Partner (VP) wurde ein auditorisches
stufengesteuertes Signal (=Stimuli) eingeführt. Die
TeilnehmerInnen wurden insturiert, synchron mit dem
VP zu klopfen, bei Aufrechterhaltung des beginnenden
Tempos. Der Ton des VP adaptierte, sofern die
ProbandInnnen langsamer oder schneller klopfen. Ziel
war es, das Klopfen mit dem Ton so genau wie möglich
zu synchronisieren. In jedem Versuch wurde ein
anderes Anpassungslevel des VP gewählt. Insgesamt
gab es fünf unterschiedliche Level (optimal
anpassungsfähig bis übermäßig anpassungsfähig).
Nach jedem Klopf-Versuch erstellten die ProbandInnen
eine Visual Analog Scale (VAS) mit den von ihnen
wahrgenommenen Einfluss auf den VP und über
wahrgenommenen Schwierigkeiten. Hinsichtlich des
Einflusses war die Frage zu beantworten: Inwieweit
beeinflusste dein Klopfen das allgemeine Tempo?
Hinsichtlich der Schwierigkeiten war zu beantworten:
Wie schwierig war es mit dem VP zu synchronisieren?
Freiwillige
TeilnehmerInnen
Als "Leader" wurden diejenigen
bezeichnet, die Versuche, bei
denen sie größeren Einfluss
verspürten, als weniger
schwierig bezeichneten,
während "Followers" diejenigen
waren, die Versuche mit mehr
Einfluss als schwieriger
bezeichneten.
Die wesentlichsten Auswirkungen der VP Anpassbarkeit zeigten eine Aktivierung der
rechten lateralen Strukturen, insbesondere der anterior Insula, des ventrolateralen
präfrontalen Kortex (VLPFC) des inferioren frontalen Gyrus, des superioren temporalen
Gyrus und des inferioren Parietallappen. Die rechte frontale Aktivierung von inferior frontalen
Gyrus und der anterior Insula war größer, als die ProbandInnen mit einen optimal
anpassungsfähigen VP (im Vergleich zu einem übermäßig anpassungsfähigen VP) klopften.
Die Haupteffekte der Gruppen (Leader vs. Follower) zeigten eine Aktivierung des
dorsolateralen präfrontalen Kortex (DLPFC), der anterior Insula, des inferioren frontalen
Gyrus, des linken prämotorischen Kortex, des rechten midcingulate, der Pre SMA, des
Precuneus und des rechten Cuneus sowie des linken IPL. Die Aktivierung war bei Leadern
größer als bei Followern. Werden Leader und Followershinsichtlich der zwei Levels von VP
Anpassbarkeit verglichen, so kann kein signifikanter Unterschied hinsichtlich des optimalen
Anpassungslevels festgestellt werden, aber eine größere Aktivierung der sensomotorischen
Region des Precuneus bei Leadern, und zwar bei klopfen mit einem übermäßig adaptiven
VP.
Mit dieser Studie konnte gezeigt werden, dass Führung mit
einer Aktivierung des rechten lateralen Gehirnbereiches, der
in kognitiven Kontrollprozessen sowie in selbstbezogenen
Prozessen involiert ist korreliert. "Leader" waren hinsichtlich
der Anpassungsfähigkeit stabiler als "Follower".
Einhergehend mit anderen Studien konnte gezeigt werden,
dass "Leader" umso effizienter führen und umso mehr
Kontrolle über die Geführten wahrnehmen, wenn ein sicheres
Level von sozialem Austausch existiert. Die laterale
Aktivierung geht auch mit früheren Studien einher, die
herausgefunden haben, dass die Aktivierung in der rechten
Hemisphäre größer ist, wenn man selbst nachgemacht wird.
Zusätzlich wurde durch diese Studie herausgefunden, dass
die Aktivierung des inferior frontalen Gyrus sowie des IPL mit
subjektiven Bewertungen von wahrgenommenen Einfluss
positiv korreliert.
soziale
Einflussnahme der
FK auf den Geführten
(Beziehung zwischen
MA und FK)
2012
Social Cognitive and
Affective
Neuroscience
Lin, A.; Adolphs, R.;
Rangel, A.
Social and monetary
reward learning engage
overlapping neural
substrates
Ziel: herauszufinden, ob
dieselben Gehirnregionen im
Hinblick auf
belohnungsbezogenes
Lernen für soziale sowie für
monetäre Belohnung
aktiviert werden, oder ob
unterschiedliche
Gehirnregionen in Bezug auf
soziale Belohnung einerseits
und monteäre Belohnung
andererseits erregt werden.
fMRI
n= 27 (alle
weiblich); 5
wurden
aussortiert
Die Teilnehmerinnen spielten zwei instrumentelle
Lernaufgaben, wobei sie in einer Version mit Geld
belohnt wurden, in der Zweiten mit Anerkennung (social
reward). Jeder Versuch startete mit zwei visuell
unterschiedlichen Spielautomaten (=Stimuli), wovon
jeder mit einem bestimmten Output assoziiert wurde
(positiv, negativ, neutral). Die Teilnehmerinnen mussten
jeweils eine Spielmaschine mittels eines Buttons
auswählen. Den Probandinnen wurde die
Wahrscheinlichkeit der Belohnung nicht mitgeteilt. Jede
Teilnehmerin machte insgesamt 148 Versuche, wobei
bei 100 Versuchen jeweils der neutrale Automat mit
einem anderen gepaart war. Bei den restlichen 48
Versuchen waren jeweils zwei identische Automaten
gepaart. Die Anerkennung war durch unbekannte
Gesichter, die entweder lachten, neutral oder böse
schauten, gepaart mit einem Wort, wie "exzellent" oder
"falsch", erkennbar. Die Belohnung in Geld war im
positiven Fall der Gewinn eines Dollars, im negativen
Fall dessen Verlust.
Teilnehmerinnen
stammen aus der
Gemeinschaft des
Californischen Institute
of Technology
Die Probandinnen lernten
zuverlässig jenen
Spielautomaten zu wählen, der
die größte Wahrscheinlichkeit
eines positiven Outputs
(innerhalb weniger Versuche,
sowohl für Geld als auch für
Anerkennung), aufwies. Die
Probandinnen lernten das Nicht-
Wählen des negativen
Spielautomaten langsamer, als
das Wählen des positiven
Automatens.
In der Version mit der Geldentlohnung konnte herausgefunden werden, dass eine
Aktivierung des VMPFC mit den Wert des Stimulus korreliert. So auch das mid-cingulum,
der superior frontale Gyrus sowie der angulare Gyrus. Außerdem korreliert in der monetären
(sozialen) Version das Vorhersagefehler-Signal mit einer Aktivierung im Caudate (Striatum)
und Putamen. Zudem korreliert die Höhe des Outputs mit einer Aktivierung im VMPFC, der
Insula,des okzipitalen Kortex, des cingularen Gyrus und des superior frontalen Gyrus. In der
Version mit Anerkennung als Belohnung wurde ebenso festgestellt, dass eine Aktivierung
des VMPFC mit dem Stimulus Wert korreliert. Außerdem korreliert in dieser Version die
Höhe des Outputs mit einer Aktivierung des VMPFC.
Durch diese Studie wurde - übereinstimmend mit älteren
Studien - herausgefunden, dass überlappende
Gehirnregionen des VMPFC und des ventralen Striatum
Wertsignale sowohl für monetäre, als auch für soziale
Belohnungen kodieren.
soziale
Einflussnahme der
FK auf den Geführten
(Belohnung)
2013 Cerebral Cortex
Fairhurst, M.T.;
Janata, P.; Keller,
P.E.
Being and feeling in
sync with an adaptive
Virtual Partner: Brain
mechanisms underlying
dynamic cooperativity.
Ziel: das neuronale Substrat
von Kooperativität in einer
Aufgabe zu untersuchen
bzw. neuronalen Korrelate
identifizieren, die das Gefühl
synchron zu sein
ausdrücken.
fMRIn= 16 (m=8;
f=8)
Als Stimuli wurde ein auditorisches Signal als virtueller
Partner (VP) eingeführt, wobei der VP hinsichtlich des
Anpassungsgrades variierte (nicht anpassungsfähig;
optimal anpassungsfähig; mittelmäßig
anpassungsfähig; hoch anpassungsfähig und komplett
anpassungsfähig). Die TeilnehmerInnen wurden sodann
insturiert, ihre Klopfbewegungen mit dem Ton des VP
so exakt wie möglich zu synchronisieren unter
gleichzeitiger Aufrechterhaltung des beginnenden
Tempos. In jedem Versuch wurde ein anderes
Anpassungslevel des VP gewählt. Nach jedem Versuch
erstellten die ProbandInnen eine Visual Analog Scale
(VAS) darüber, wie synchron sie sich mit dem VP
gefühlt haben (Skala 0-10, wobei 0 nicht synchron und
10 komplett synchron bedeuteten).
Freiwillige
TeilnehmerInnen
Die Synchronisierung wurde
am Besten durch einen VP, der
eine geringe
Anpassungfähigkeit aufweist,
unterstützt. Ein übermäßiger
anpassungsfähiger VP, und
somit ein unseriöser VP,
erschwerte die Aufgabe. Die
Synchronisierung mit einem
übermäßig (α > 0,75)
anpassungsfähigen VP wurde
als schwieriger empfunden als
die Synchronisierung mit einem
optimal anpassungsfähigen
VP.
Die erste Analyse identifizierte jene Bereiche im Gehirn, die als Funktion der objektiven
Messgröße hinsichtlich des Gefühls, mit dem VP synchron zu sein, aktiviert waren. Diese
Regressionsanalyse identifzierte Gehirnregionen in denen die Aktivierung als Funktion der
Standardabweichung der Asynchronität je nach Anpassungsgrad des VP variiert.
Synchronität, also geringe Standardabweichung der Asychronität, korreliert mit einer
erhöhten Aktivierung in folgenden Regionen: VMPFC, ergänzende motorische Regionen,
Hippocampus, primärer somatosensorischer Kortex, posterior Cingulum und Precuneus. Im
Gegensatz dazu korreliert eine Asynchronität (=hohe Standardabweichung für
Asynchronität) mit einer erhöhten Aktivierung eines umfangreichen Netzwerkes: anterior
Insula, inferior und superior frontal Gyri, ventrolateral präfrontaler Kortex (VLPFC) und der
inferiore Parietallappen. Diese Regressionsanalyse wurde wiederholt während der
Steuerung der wahrgenommenen Schwierigkeit. Wieder konnten zwei unterschiedliche
Aktivierungsverhalten festgestellt werden. Eine Aktivierung der anterioren Insula, des
superioren frontalen Gyrus, des inferioren frontalen Gyrus und des DMPFC korreliert mit
dem Gefühl assynchron zu sein, während eine Aktivierung des Hippocampus, sowie des
Posterior Cingulate und des Precuneus mit einer Synchronität assoziiert wird.
Anhand einer weiteren Regressionsanalyse wurden jene Gehirnregionen identifiziert, dessen
Aktivität mit den subjektiven Bewertungen hinsichtlich der Schwierigkeit der Aufgabe
korrelierte. Steigerungen der wahrgenommener Schwierigkeit synchron zu sein, korrelierte
mit größerer Aktivierung im rechten inferior frontalen Gyrus, in der rechten anterior Insula, im
posterior DMPFC, beidseitig im VLPFC, im superior frontal Gyrus. Außerdem war eine
inferior parietale Aktivität in der Region des temporoparietalen Überganges festzustellen.
Eine Abnahme von wahrgenommener Schwierigkeit der Aufgabe aktivierte
SMA/midcingulate, den VMPFC, inferior Parietallappen sowie den Hippocampus.
Durch die Variierung der Anpassungsfähigkeit des VP konnte
begleitend eine Aktivierung von zwei unterschiedlichen
Gehirnnetzwerken festgestellt werden. Eines dieser
Netzwerke wird primär mit selbstreferentielle, soziale und
emotionale Prozesse assoziiert, während die andere Region
mit kognitiver Kontrolle verbunden wird.
Die Aktivierung des SMA und S1/M1 (sensomotorische
Region) in Verbindung mit dem mittleren frontalen-parietalen
Netzwerk könnte eine kritische Verbindung zwischen der
Handlungskomponente der zweiteiligen Interaktion und der
sozio-affektiven Komponente der Kooperation aufzeigen. Die
relevante Verbindung hierzu könnte der PCu sein, da dieser
anatomisch und funktionell zu beiden, dem SMA und dem
auditorischen Kortex im posterior superior temporalen Gyrus
verbunden ist. Der PCu ist bekannt dafür, dass er externe
und eigene Informationen integriert. Aktivierungen innerhalb
des VMPFC wurden in früheren Studien immer dann
beobachtet, wenn Individuen Objekte im Vergleich zu ihnen
selbst beurteilten, andere bewerteten oder bei einer
Lernreihe. Die Regionen rund um die rechte anterior Insula
und dem angrenzenden inferior frontal Gyrus sind während
kooperative und kompetitive Spiele miteingebunden.
soziale
Einflussnahme der
FK auf den Geführten
(Beziehung zwischen
MA und FK)

89
GLOSSAR
Afferenz: Zuflüsse von Information zu einer definierten Struktur.
Anterior: Anatomische Bezeichnung für eine bestimmte Lage. Richtung Nase gelegen.
Arbeitsgedächtnis: Die vorübergehende Speicherung von Informationen, die in ihrer
Kapazität begrenzt ist und bei der ständige Wiederholung notwendig ist.
Axon: Teil einer Nervenzelle, das auf die Leitung von Nervenimpulsen spezialisiert ist.
Balken: Teil des Großhirns. Besteht aus Axonen und verbindet die beiden Gehirnhälften.
Basalganglien: Eine Zellgruppenreihe im Großhirn. Umfasst den Nucleaus caudatus, das
Putamen, das Pallidum und den Nucleus subthalamicus.
Broca-Zentrum: Bereich des Frontallappens (Großhirn).
Brücke: Teil des Hirnstammes
Brückenkerne: Ein Bündel von Neuronen, das als Umschaltstation Informationen aus der
Großhirnrinde an die Kleinhirnrinde weiterleitet.
Dendrit: Teil einer Zelle, der auf die Aufnahme von synaptisch übertragenen Informatio-
nen durch andere Neuronen spezialisiert ist.
Diffusion: Die temperaturabhängige Bewegung von Molekülen von Bereichen mit hohen
Konzentrationen zu Bereichen mit niedriger Konzentration.
Efferenz: Weiterleitung von Informationen von einer definierten Struktur; das Gegenteil
von Afferenz.
Elektroenzephalogramm: Messung elektrischer Aktivität des Gehirns mittels an der
Kopfhaut angebrachten Elektroden.
Fornix: Faserbündel
Frontallappen: Teil des Großhirns
Graue Substanz: Eine Bezeichnung für eine Ansammlung von Nervenzellen im ZNS.

90
Großhirn: Besteht grds. aus der Großhirnrinde und den Basalganglien.
Großhirnrinde: Die Schicht grauer Substanz, direkt unter der Oberfläche des Gehirns.
Gyrus: Eine Wölbung, die zwischen sich zwischen den Furchen des Großhirns befindet.
Hertz: Einheit mit der die Schallfrequenz in Schwingungen pro Sekunde angegeben wird.
Hippocampus: Teil des Temporallappens, der vermutlich eine entscheidende Rolle beim
Lernen und der Gedächtnisbildung spielt.
Hirnstamm: Umfasst Mittelhirn, Brücke und das verlängerte Rückenmark.
Hypothalamus: Teil des Zwischenhirns, der vor allem bei der Kontrolle des vegetativen
Nervensystems beteiligt ist.
Kleinhirn: Teil des Gehirn, der ein wichtiges Zentrum für die Kontrolle von Bewegungen
ist.
Limbisches System: Bestimmte Hirnregionen, die anatomisch miteinander verbunden
sind und denen vermutlich besondere Bedeutung an der Verarbeitung von Emotionen, am
Lernen und an der Gedächtnisbildung zukommt.
Medial: Anatomische Bezeichnung für eine bestimmte Lage: Richtung Mittellinie gesehen.
Medulla oblongata: Teil des Hirnstammen.
Mittelhirn: Besteht aus Mittelhirndach und Mittelhirnhaube.
Motorkortex: Teil des präfrontalen Kortex, der direkt an der Kontrolle von willkürlichen
Bewegungen beteiligt ist.
Neuron: einzelne Nervenzelle, die durch Synapsenübertragung Informationen weiterleitet.
Neurotransmitter: Eine chemische Substanz, die nach einer Stimulierung freigesetzt wird
und postsynaptische Rezeptoren aktiviert.
Nucleus caudatus: Teil der Basalganglien, der an der Kontrolle der Motorik beteiligt ist.
Okzipitallappen: Teil der Großhirnrinde.
Parietallappen: Teil der Großhirnrinde.

91
Posterior: Anatomische Bezeichnung für eine bestimmte Lage; Richtung Schwanz.
Präfrontaler Kortex: Teil des Frontallappens.
Prämotorisches Areal: Teil des Frontallappens, der an der Kontrolle willkürlicher Bewe-
gungen beteiligt ist.
Propriozeptiv: Wahrnehmungen aus dem eigenen Körper vermittelnd (bspw. aus Mus-
keln)
Putamen: Teil der Basalganglien, der an der motorischen Kontrolle beteiligt ist.
Striatum: besteht aus Nucleus caudatus und Putamen und ist an der Initiation von willkür-
lichen Bewegungen des Körpers beteiligt und spielt eine Rolle beim prozeduralen Ge-
dächtnis.
Synapse: Kontaktbereich, in dem ein Neuron eine Information an ein anderes Neuron
überträgt.
Temporallappen: Teil der Großhirnrinde.
Thalamus: Teil des Zwischenhirns, der sehr stark mit dem Neokortex verknüpft ist.
Vestibulariskern: Kern in der Medulla oblongata.
Vorderhirn: Dazu zählen das Großhirn sowie das Zwischenhirn.
Voxel: Gitterpunkt in einem dreidimensionalen Gitter.
Weiße Substanz: Ein Bündel von Nervenzellen, das sich direkt unterhalb der Großhirn-
rinde befindet.
Wernicke Areal: Region im Temporallappen
Zentrales Nervensystem: Das Gehirn und das Rückenmark.
Zwischenhirn: besteht u.a. aus Thalamus und Hypothalamus.

92
Literaturverzeichnis
AHLERT, D./KENNING, P. (2006): Neuroökonomik, in: Zeitschrift für Management, Nr. 1,
S. 24–47.
ANDERSON, J. R. (2001): Kognitive Psychologie. 3. Aufl., Heidelberg, Berlin.
BASS, B.M./BASS, R. (2009): The Bass bandbook of leadership: Theory, research, and
managerial applications, New York.
BEAR, M. F./CONNORS, B. W./PARADISO, M. A. (2009): Neurowissenschaften. Ein
grundlegendes Lehrbuch für Biologie, Medizin und Psychologie. 3. Aufl., Heidelberg.
BERGNER, S./RYBNICEK, R. (2015): Führungsforschung aus neurowissenschaftlicher
Sicht, in: Felfe, J. (Hrsg.): Trends in der psychologischen Führungsforschung, Göttingen,
543ff
BIRBAUMER, N./SCHMIDT, R. F. (2006): Biologische Psychologie. 6. Aufl., Heidelberg.
BORTZ, J./DÖRING, N. (2006): Forschungsmethoden und Evaluation für Human- und
Sozialwissenschaftler, Berlin.
BOYATZIS, R.: Neurscience and the link between inspirational leadership and resonant
relationsships, in: Ivey Business Journal, January/February 2012.
BOYATZIS, R.E./PASSARELLI, A.M./KOENIG, K./LOWE, M./MATHEW, B./STOLLER,
J.K./PHILLIPS, M. (2012): Examination of the neural substrates activated in memories of
experiences with resonant and dissonant leaders, in: The Leadership Quaterly, Jg. 23,
S. 259–272.
BRAUS, D. F. (2004): EinBlick ins Gehirn. Bildgebung in der modernen Psychiatrie, Stutt-
gart.
BRAUS, D.F./TOST, H./HIRSCH, J.G./GASS, A. (2001): Diffusions-Tensor-Bildgebung
(DTI) und funktionelle Magnetresonanztomographie (fMRI) erweitern das Methodenspekt-
rum in der psychiatrischen Forschung, in: Der Nervenarzt, Vol. 72, Nr. 5, S. 384–390.

93
BÜCHEL, C./KARNATH, H.-O/THIER, P. (2012): Methoden der kognitiven Neurowissen-
schaften, in: Karnath, H.-O./ Thier, P. (Hrsg.): Kognitive Neurowissenschaften, Berlin Hei-
delberg, S. 9–34.
CAMERER, C. (2007): Neuroeconomics: Using Neuroscience to make economic predicti-
ons, in: The Economic Journal, S. C26-C42.
CAMERER, C./LOEWENSTEIN G./PRELEC D. (2005): Neuroeconomics: How Neuros-
cience Can Inform Economics, in: Journal of Economic Literature, XLIII, S. 9–64.
CARLSON, N. R. (2004): Physiologische Psychologie. 8. Aufl., München u.a.
CASPERS, S./ HEIM, S./LUCAS, M.G./STEPHAN, E./FISCHER, L./AMUNTS,
K./ZILLES, K. (2012): Dissociated Neural Processing for Decisions in Managers and Non-
Managers, in: PLOS One, Vol. 7, Nr. 8.
CHIAO J.Y./ADAMS, R.B./TSE, P.U./LOEWENTHAL L./RICHESON, J.A./AMBADY, N.
(2009): Knowing Who's Boss: fMRI and ERP Investigations of Social Dominance Percep-
tion, in: Group Processes Intergroup Relations, Vol. 11, Nr. 2, S. 201–214.
CHIAO, J.Y./ HARADA, T./OBY, E.R./LI, Z./PARRISH, T./BRIDGE, D.J. (2009): Neural
representations of social status hierarchy in human inferior parietal cortex, in: Neuropsy-
chologia, Vol. 47, Nr. 2, S. 354–363.
CLOUTIER, J./AMBADY, N./MEAGHER, T./GABRIELI, J.D.E. (2012): The neural sub-
strates of person perception: Spontaneous use of financial and moral status knowledge,
in: Neuropsychologia, Vol. 50, Nr. 9, S. 2371–2376.
CLOUTIER, J./KELLEY, W.M./HEATHERTON, T.F. (2011): The influence of perceptual
and knowledge-based familiarity on the neural substrates of face perception, in: Social
Neuroscience, Vol. 6, Nr. 1, S. 63–75.
CNS FORUM LUNDBECK INSTITUTE: Gehirnatlas.
http://de.brainexplorer.org/brain_atlas/Brainatlas_index.shtml.
DEROUICHE, A. (2011): Eine kleine Neuroanatomie, in: Reimann, M./Weber, B. (Hrsg.):
Neuroökonomie. Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 11–39.
DILLERUP, R./Stoi, R. (2013): Unternehmensführung. 4. Aufl., München.

94
DIMOKA, A. (2010): What does the brain tell us about trust and distrust? Evidece from a
functional neuroimaging study, in: MIS Quarterly, Vol. 34, Nr. 2, S. 373–396.
DIMOKA, A. et al (2012): On the use of neurophysiological tools in IS research: develo-
ping a research agenda for NeuroIS, in: MIS Quarterly, Vol. 36, S. 679–702.
DOALLO, S./RAYMOND, J.E./SHAPIRO, K.L./KISS, M./EIMER, M./NOBRE, A.C.
(2012): Response inhibition results in the emotional devaluation of faces: neural correlates
as revealed by fMRI, in: Social Neuroscience, Vol. 7, S. 649–659.
DRINKMANN, A. (1990): Methodenkritische Untersuchungen zur Metaanalyse, Wein-
heim.
DRUMM, H. J. (2008): Personalwirtschaft. 6. Aufl., Berlin Heidelberg.
EISEND, M.: Metaanalyse - Einführung und kritische Diskussion, in: : Betriebswirtschaftli-
che Reihe Nr.8/2004, S. 1–40.
ELGER, C. E. (2013): Neuroleadership. Erkenntnisse der Hirnforschung für die Führung
von Mitarbeitern. 2. Aufl., Freiburg.
FAIRHURST, M.R./JANATA, P./KELLER, P.E. (2013): Being and Feeling in Sync with an
Adaptive Virtual Partner: Brain Mechanisms Underlying Dynamic Cooperativity, in: Ce-
rebral Cortex, Vol. 23, Nr. 11, S. 2592–2600.
FAIRHURST M.R./JANATA, P./KELLER, P.E. (2014): Leading the follower: An fMRI in-
vestigation of dynamic cooperativity and leader-follower strategies in synchronization with
an adaptive virtual partner, in: NeuroImage, S. 688–697.
FARTHING, G. W. (1992): The psychology of consciousness.
FEICHTER, M.: Nervensystem und Nervenzellen - Anatomie.
http://www.netdoktor.de/Gesund-Leben/Anatomie/Nervensystem-und-Nervenzellen-
9949.html.
FIEDLER, F. E. (1967): A theory of leadership effectivness, New York.
FIRBAS, W./GRUBER, H./MAYR, R. (1995): Neuroanatomie. 2. Aufl., Wien.

95
GLASS, G. V./MCGAW B./SMITH, M. L. (1981): Meta-Analysis in Social Research, Be-
verly Hills.
GOEBEL, R./ZIMMERMANN, J. (2011): Pfadfinder durch die weiße Substanz, in: Gehirn
und Geist, Nr. 6, S. 64–68.
HÄUSEL, H.-G. (2006): Direkt ins Hirn?, in: absatzwirtschaft-Zeitschrift für Marketing, Vol.
9, S. 36–40.
HÄUSEL, H.-G. (2008): Methoden der Neuromarketing-Forschung, in: Häusel, H.-G.
(Hrsg.): Neuromarketing. Erkenntnisse der Hirnforschung für Markenführung, Werbung
und Verkauf, Planegg/München.
HEGELE-RAIH (2004): Was ist Leadership?
http://www.harvardbusinessmanager.de/heft/artikel/a-620896.html.
HERMEY, G./MAHLKE, C./SCHWAKE, M./SOMMER, T. (2010): Der Experimentator:
Neurowissenschaften, Heidelberg.
HERSEY P./BLANCHARD K.H. (1977): Management of Organizational Behavior - Utili-
zing Human Resources. 3. Aufl., Englewood Cliffs.
HUNGENBERG, H./WULF, T. (2011): Grundlagen der Unternehmensführung. 4. Aufl.,
Berlin Heidelberg.
HUNTER, J. E./SCHMIDT, F. L. (1990): Methods of Meta-Analysis. Correcting Error and
Bias in Research Findings, Newbury Park.
HÜSING, B./JÄNCKE, L./TAG, B. (2006): Impact Assessment of Neuroimaging, Zürich.
HÜTHER, G. (2009): Wie gehirngerechte Führung funktioniert - Neurobiologie für Mana-
ger, in: managerSeminare, Nr. 130, S. 30–34.
JÄNCKE, L. (2005): Methoden der Bildgebung in der Psychologie und den kognitiven
Neurowissenschaften, Stuttgart.
JUNG, H. (2010): Allgemeine Betriebswirtschaftslehre. 12. Aufl., München.

96
KAHLE, W./FROTSCHER, M. (2009): Taschenatlas Anatomie. Nervensystem und Sin-
nesorgane. 10. Aufl., Stuttgart.
KANG, Y./WILLIAMS, L.E./CLARK, M.S./GRAY, J.R./BARGH, J.A. (2011): Physical
temperature effects on trust behavior: the role of insula, in: Social Cognitive and Affective
Neuroscience, Vol. 6, Nr. 4, S. 507–515.
KENNING, P./ PLASSMANN, H. (2005): NeuroEconomics: An Overview from an Econo-
mic Perspective, in: Brain Research Bulletin, Vol. 67, S. 343–354.
KENNING, P./PLASSMANN, H./AHLERT, D. (2007): Applications of functional magnetic
resonance imaging for market research, in: Qualitative Market Research: An International
Journal, Vol. 10, Nr. 2, S. 135–152.
KOTTER, J. (1988): The Leadership Factor, New York.
KOTTER, J. (1991): Leadership lässt sich lernen, in: Harvard Business manager, 13. Jg.,
Nr., S. 35–43.
LEYH, A. (08.09.2011): Der Frontallappen. http://dasgehirn.info/entdecken/anatomie/der-
frontallappen/.
LEYH, A. (09.09.2011): Der Parietallappen. http://dasgehirn.info/entdecken/anatomie/der-
parietallappen/.
LEYH, A. (23.09.2011): Der Okzipitallappen.
http://dasgehirn.info/entdecken/anatomie/der-occipitallappen/.
LIN, A./ADOLPHS, R./RANGEL, A. (2012): Social and monetary reward learning engage
overlapping neural substrates, in: Social Cognitive and Affective Neuroscience, Vol. 7,
Nr. 3, S. 274–281.
LIPSEY, M. W./Wilson, D. B. (2001): Practical Meta-Analysis, Thousands Oaks.
LUEGINGER, E./RENGER, R. (2013): Das weite Feld der Metaanalyse. Sekundär-, litera-
tur- und metaanalytische Verfahren im Vergleich, in: kommunikation.medien, Nr. 2, S. 1–
31.

97
MANN, R. D. (1959): A review of the relationships between personality and leadership
and popularity, in: Psychological Bulletin, S. 241–270.
NORTHOUSE, P. G. (2013): Leadership. Theory and Practice. 6. Aufl., California.
o.V.: Gehirn-Atlas: Funktionen und Funktionsstörungen des Gehirns. http://www.gehirn-
atlas.de.
OCHSNER, K.N/LIEBERMANN, M.D. (2001): The emergence of social cognitive neuros-
cience, in: American Psychologist, Vol. 56, S. 717–734.
PETERS, T./GHADIRI, A. (2011): Neuroleadership - Grundlagen, Konzepte, Beispiele.
Erkenntnisse der Neurowissenschaften für die Mitarbeiterführung, Wiesbaden.
PETERS, T./GHADIRI, A. (2012): Mitarbeiter führen heißt Gehirne führen, in: Personal-
manager, S. 67–69.
POKORNA, E.: Der menschliche Körper, Teil II.
http://www.ftvs.cuni.cz/eknihy/jazyky/einfuhrungindiesportsprache/Texts/5-Korper_II.html.
RAAB, G./GERNSHEIMER, O./SCHINDLER, M. (2009): Neuromarketing. Grundlagen-
Erkenntnisse-Anwendungen. 2. Aufl., Wiesbaden.
RIEDL, R./HUBERT, M./KENNING, P. (2010): Are there neural gender differences in on-
line trust? An fMRI study on the perceived trustworthiness of ebay offers, in: MIS Quarter-
ly, Vol. 34, Nr. 2, S. 397–428.
ROBBINS, S. P. (2006): Organisation der Unternehmung. 9. Aufl., München.
ROCK, D. (2008): SCARF: A Brain-Based Model for Collaborating With and Influencing
Other, in: NeuroLeadership, Vol. 1, S. 44–52.
ROHEN, J. W. (2001): Funktionelle Neuroanatomie. 6. Aufl., Stuttgart.
ROTH, G. (2003): Fühlen, Denken, Handeln - Wie das Gehirn unser Verhalten steuert,
Frankfurt am Main.

98
RULE, N.O./FREEMAN, J.B./MORAN, J.M./GABRIELI, J.D.E./ADAMS, Jr.,
R.B./AMBADY, N. (2010): Voting behavior is reflected in amygdala response across cul-
tures, in: Social Cognitive and Affective Neuroscience, Vol. 5, S. 349–355.
RULE, N.O./MORAN, J.M./FREEMAN, J.B./WHITFIELD-GABRIELI, S./GABRIELI,
J.D.E./AMBADY, N. (2011): Face value: Amygdala response reflects the validity of first
impressions, in: NeuroImage, Vol. 54, S. 734–741.
SCHANDRY, R. (2003): Biologische Psychologie. 3. Aufl., Weinheim.
SCHAUFFELBÜHLK./HUGENTOBLER, W./BLATTNER, M. . (2007): Betriebswirtschafs-
lehre für Bachelor, Zürich.
SCHJOED, U./STODKILDE-JORGENSEN, H./GEERTZ, A.W./LUND,
T.E./ROEPSTORFF, A. (2011): The power of charisma - perceived charisma inhibits the
frontal executive network of believers in intercessory prayer, in: Social Cognitive and Af-
fective Neuroscience, Vol. 6, Nr. 1, S. 119–127.
SCHMIDT, R.F./SCHAIBLE, H.-G. (Hrsg.) (2001): Neuro- und Sinnesphysiologie. 4. Aufl.,
Berlin Heidelberg.
SCHREYÖGG, G./Koch, J. (2010): Grundlagen des Managements. Basiswissen für Stu-
dium und Praxis. 2. Aufl., Wiesbaden.
SCHUBERT, S. (2004): Dem Kunden ins Gehirn geschaut, in: Psychologie heute, S. 38–
41.
SOLMS, M./TURNBULL, O. (2004): Das Gehirn und die innere Welt. Neurowissenschaft
und Psychoanalyse, Düsseldorf/Zürich.
SPRECKELMEYER, K.N./KRACH, S./KOHLS, G./RADEMACHER, L./IRMAK,
A./KONRAD, K./KIRCHER, T./GRÜNDER, G. (2009): Anticipation of monetary and social
reward differently activates mesolimbic brain structures in men and women, in: Social
Cognitive and Affective Neuroscience, Vol. 4, Nr. 2, S. 158–165.
STOGDILL, R. M. (1948): Personal factors associated with leadership: A survey of the
literature, in: Journal of Psychology, 25. Jg., S. 35–71.

99
TANNENBAUM, R./SCHMIDT, W. (1958): How to choose a leadership pattern, in: Har-
ward Business Review, 36. Jg., 95-102.
THOMPSON, R. F. (2001): Das Gehirn - von der Nervenzelle zur Verhaltenssteuerung.
3. Aufl., Heidelberg.
TREPEL, M. (2011): Neuroanatomie: Struktur und Funktion. 5. Aufl., München.
ULFIG, N. (2008): Neuroanatomie, Stuttgart.
WALDMAN, D.A./BALTHAZARD, P.A./PETERSON, S.J. (2011): Leadership and Neu-
roscience: Can We Revolutionize the Way That Inspirational Leaders Are Identified and
Developed?, in: Academy of Management Perspectives, Vol. 25, S. 60–74.
WEBER, B. (2011): Methoden der Neuroökonomie, in: Reimann, M./Weber, B. (Hrsg.):
Neuroökonomie. Grundlagen-Methoden-Anwendungen, Wiesbaden, S. 41–55.
WEIS, S./THALLER, R./VILLRINGER, A. (1992): Das Gehirn des Menschen. Morpholo-
gie, Kernspintomographie und 3 D-Computerrekonstruktion, Göttingen/Toronto/Zürich.
WICHT, H. (05.09.2011): Der Temporallappen.
http://dasgehirn.info/entdecken/anatomie/der-temporallappen/.


















