The Guanine Nucleotide Exchanger Vav2 Interacts with c ...
100
The Guanine Nucleotide Exchanger Vav2 Interacts with c-ErbB-2 and Induces Alveolar Morphogenesis of Mammary Epithelial Cells DISSERTATION zur Erlangung des akademisches Grades doctor rerum naturalium (Dr. rer. nat.) im Fach Biologie eingereicht an der Mathematisch-Naturwissenschaftlichen Fakultät I der Humboldt-Universität zu Berlin von Dipl. Biochem. Silvana Di Cesare geb. am 23.09.63 in La Plata, Argentinien Präsident der Humboldt-Universität zu Berlin Prof. Dr. Jürgen Mlynek Dekan der Mathematisch-Naturwissenschaftlichen Fakultät I Prof. Dr. Bernhard Ronacher Gutachter: 1. Prof. Dr. Harald Saumweber 2. Prof. Dr. Walter Birchmeier 3. Prof. Dr. Franz Theuring Tag der mündlichen Prüfung: 9. November 2001
Transcript of The Guanine Nucleotide Exchanger Vav2 Interacts with c ...

The Guanine Nucleotide Exchanger Vav2 Interacts with c-ErbB-2 and
Induces Alveolar Morphogenesis of Mammary Epithelial Cells
DISSERTATION zur Erlangung des akademisches Grades
doctor rerum naturalium (Dr. rer. nat.)
im Fach Biologie
eingereicht an der
Mathematisch-Naturwissenschaftlichen Fakultät I
der Humboldt-Universität zu Berlin
von
Dipl. Biochem. Silvana Di Cesare
geb. am 23.09.63 in La Plata, Argentinien
Präsident der Humboldt-Universität zu Berlin
Prof. Dr. Jürgen Mlynek
Dekan der Mathematisch-Naturwissenschaftlichen Fakultät I
Prof. Dr. Bernhard Ronacher
Gutachter: 1. Prof. Dr. Harald Saumweber
2. Prof. Dr. Walter Birchmeier
3. Prof. Dr. Franz Theuring
Tag der mündlichen Prüfung: 9. November 2001

i
CONTENTS
1 INTRODUCTION 3 1.1 Development of the Mammary Gland 4
1.1.1 Regulation by Systemic Hormones 4 1.1.2 Local Effects of Multiple Growth Factors 6 1.1.3 The Role of Extracellular Matrix 9
1.2 The ErbB Family of Receptor Tyrosine Kinases 11 1.2.1 ErbB Ligands Contain an EGF-like Domain 12 1.2.2 Generation of Signal Diversity 13 1.2.3 Intracellular Effectors of ErbB Receptors 15 1.2.4 ErbB Signaling in Embryonic Development 16 1.2.5 ErbB Receptors in Mammary Gland Development 18
1.3 Vav Family of Guanine Nucleotide Exchange Factors 23
2 MATERIALS AND METHODS 25 2.1 Amplification of the Hollenberg cDNA Yeast Expression Library 25 2.2 Yeast Plasmids and Strains 25 2.3 Yeast Two-Hybrid Screen 27 2.4 Verification of Interacting Clones 28 2.5 Mutagenesis of the Yeast TprMet-ErbB2 Baits 29 2.6 Mammalian Expression Plasmids 30 2.7 Site-Directed Mutagenesis 30 2.8 Mammalian Cell Lines 31 2.9 Immunoprecipitation and Western Blotting 32 2.10 Matrigel Assays 33 2.11 Light and Electron Microscopy 34 2.12 In Situ Hybridization Analysis 34
3 RESULTS 35 3.1 Yeast Two-Hybrid Screen with ErbB-2 Bait Proteins 35 3.2 Distinct Phosphotyrosine Residues of ErbB-2 Bind to Various Interaction Partners 39 3.3 Vav2 Interacts with Receptor Tyrosine Kinases of the ErbB Subfamily 41 3.4 Vav2 Induces Alveolar Morphogenesis of EpH4 Mammary Epithelial Cells 43 3.5 Vav2 and ErbB-2 Can Both Directly and Indirectly Associate in Mammalian Cells 47 3.6 Vav2 and ErbB-2 Are Associated in Mammary Epithelium During Pregnancy 49 3.7 The Dbl-Homology Domain of Vav2 Is Required for Its Morphogenic Activity in EpH4 Cells 50 3.8 Catalytically Inactive Vav2 Blocks Neuregulin-Mediated Morphogenesis of EpH4 Cells 52

ii
4 DISCUSSION 54 4.1 Modified Yeast Two-Hybrid System: A Powerful Tool to Search for Phosphotyrosine-Interacting
Proteins 54 4.2 New Insights into the Intracellular Signaling Pathways of ErbB-2 55 4.3 Vav2 Couples to Receptor Tyrosine Kinases via Its SH2 Domain 56 4.4 Vav2 Mediates ErbB-2 Signals for Alveolar Morphogenesis of EpH4 Mammary Epithelial Cells 58 4.5 The Morphogenic Activity of Vav2 on Mammary Epithelial Cells May Involve Changes in Actin
Cytoskeleton 62 4.6 Vav2 Is a Specific Effector of Neuregulin Signals for Alveolar Morphogenesis 63 4.7 Possible Molecular Mechanisms Involving Vav2 as Critical Effector of Alveolar Morphogenesis 65 4.8 Conclusions 68
REFERENCES 70
ERKLÄRUNG 95

1
ABSTRACT
The ErbB receptor tyrosine kinases constitute a subfamily of four structurally related
members, the EGF receptor (ErbB-1), ErbB-2, ErbB-3 and ErbB-4. ErbB receptor tyrosine
kinases are critical for embryonic development of central and peripheral neural structures and
heart. In addition, ErbB receptors play an important role in the postnatal development of the
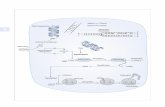
mammary gland. Previous studies showed that activated ErbB-2 receptor induces alveolar
morphogenesis of EpH4 mammary epithelial cells that are cultured on a three-dimensional
matrix (termed Matrigel). However, the downstream signaling proteins that mediate this
biological activity of ErbB-2 were unknown. In this work, Vav2 was identified as a direct
interaction partner of tyrosine-phosphorylated ErbB receptors using the yeast two-hybrid
system. Vav2 is a member of a family of guanine nucleotide exchange factors that induce
cytoskeletal rearrangements, transcriptional alterations, and have oncogenic potential when
activated. To test the ability of Vav2 to mediate morphogenic signals of ErbB-2, EpH4 cells
overexpressing Vav2 protein were cultured on Matrigel. Indeed, Vav2 induces alveolar
morphogenesis of EpH4 cells when activated either by oncogenic mutation or tyrosine
phosphorylation by ErbB-2. The morphogenic activity of Vav2 requires the Dbl homology
domain, which mediates GDP/GTP exchange. Dominant-negative Vav2 specifically blocks
the morphogenic signals of ErbB-2 in EpH4 cells without interfering with ErbB2-induced
mitogenesis. Importantly, Vav2 is co-expressed and interacts with ErbB-2 in the mammary
glands of pregnant mice. Taken together, these results point to Vav2 as a candidate to mediate
ErbB-2 signals for alveolar morphogenesis in vivo, which is a relevant step in the
development of the mammary gland during pregnancy.

2
ZUSAMMENFASSUNG
Die Familie der ErbB-Rezeptor-Tyrosinkinasen besteht aus vier Mitgliedern, dem EGF-
Rezeptor (ErbB-1), ErbB-2, ErbB-3 und ErbB-4. ErbB-Rezeptoren spielen eine wichtige
Rolle bei der Entwicklung des Nervensystems, des Herzens und der Brustdrüsen. Ein Teil
dieser Differenzierungsvorgänge läßt sich in vitro nachvollziehen: so ist zum Beispiel die
Aktivierung des ErbB-2 Rezeptors ausreichend für alveoläre Morphogenese der
Brustdrüsenepithelzellinie EpH4. Intrazelluläre Moleküle, die dieses ErbB2-Signal
übertragen, sind allerdings noch unbekannt. Mit Hilfe eines neuen, modifizierten Hefe-2-
Hybrid-Systems wurde in der vorliegenden Arbeit Vav2 als neuer Interaktionspartner von
ErbB-2 identifiziert. Vav2 assoziiert mit aktiviertem ErbB-2 über eine SH2-Domäne. Die
Interaktion ist direkt und ist von zwei Phosphotyrosinen in ErbB-2 abhängig. Vav2 kann den
GDP/GTP-Austausch bei GTPasen der Rho-Familie vermitteln. Dadurch kann der Umbau des
Zytoskeletts und Veränderungen der Transkription sowie Zelltransformation induziert werden.
In einem dreidimensionalen Zellkultursystem kann aktiviertes Vav2 in EpH4 Zellen die
Bildung von alveolären Zellaggregaten induzieren. In diesen Alveolen umgibt eine Schicht
polarisierter milchproduzierender Zellen ein zentrales Lumen. Diese Vav2-vermittelte
Morphogenese ist abhängig von der katalytischen GDP/GTP-Austausch Aktivität von Vav2.
Katalytisch-inaktives Vav2 kann die morphogenetische Aktivität von ErbB-2 in EpH4-Zellen
verhindern, ohne die mitogene Aktivität von ErbB-2 zu beeinflussen. Vav2 ist mit ErbB-2
coexprimiert und interagiert mit dem Rezeptor in Brustdrüsenzellen schwangerer Mäuse.
Diese Untersuchungen deuten darauf hin, dass Vav2 eine wichtige Funktion bei der durch
ErbB-2 induzierten alveolären Morphogenese der Brustdrüse spielt.

3
1 INTRODUCTION
Epithelium and mesenchyme are two distinct types of tissues that are present in virtually every
organ. Lung, kidney, vascular system and most glands consist of tubular epithelial networks
embedded in mesenchymal tissue. The formation of the first rudiments for these tree-like
structures is specified during embryogenesis; epithelial buds invade the underlying
mesenchyme and ramify to create interconnected tubules, a process known as branching
morphogenesis. Lungs, salivary glands and mammary glands exhibit further developmental
changes, namely alveolar morphogenesis, whereby round hollow alveoli arise from the ductal
tree and differentiate into the functional units of the organ. Mesenchymal-epithelial
interactions are strictly required for patterning morphogenic events (Sawer and Fallon, 1983);
mesenchymal soluble factors bind to their epithelial receptors thus activating several signaling
pathways which lead to local cellular responses like growth, motility, morphogenesis and
differentiation.
The mammary gland is one of the most interesting models to study mesenchymal-epithelial
interactions, as they play critical roles not only in embryonic mammary development but also
in postnatal growth and differentiation of the gland. The mammary gland is a dynamic organ
which has the unique property to undergo main developmental changes after birth. The
complex mechanisms supporting mammary ductal branching and alveolar morphogenesis
have been subject of extensive research, but are still far from being fully unravelled.
This work characterizes Vav2 as a novel intracellular effector of the receptor tyrosine kinase
ErbB-2 that mediates lobulo-alveolar morphogenesis of the mammary gland. The first part of
this Introduction reviews the developmental stages of the mammary gland and their
regulation. The second part concerns the ErbB family of receptor tyrosine kinases and their
role in mammary gland development. The third part describes features of the Vav family of
guanine nucleotide exchange factors that may be useful for interpreting the results.

4
1.1 Development of the Mammary Gland
The development of the murine mammary gland will be here outlined, since the mouse is the
most thoroughly studied mammalian model (reviewed in Sakakura, 1987; Silberstein et al.,
2001). During embryonic life, the mammary anlagen appear between days E10-E12 as an
invagination of epidermal cells into the underlying mesenchyme, the fat pad. The mammary
fat pad differentiates from deeply-placed mesenchymal cells. At day E16, primary ducts
emerge from the epithelial rudiments into the fat pad and undergo an initial round of
branching morphogenesis. Later, the nipples derive from epidermal invagination around the
primary duct. The infant mammary gland consists of a primitive ductal tree that emanates
from the nipple into the proximal fat pad. During puberty (5 to 8 weeks of age), the ducts
elongate and branch further into the fat pad. The growing points for ductal growth are
structures termed end buds, located at the terminal ends of the ducts. These end buds consist
of two distinguishable cell populations: the cap cells, which are the progenitors for
myoepithelium, and the body cells, which give rise to mammary epithelium. During the
estrous cycle, alveolar buds first emerge from the lateral walls of the ducts, then regress and
form again in the next cycle. Alveolar morphogenesis begins early in pregnancy with
extensive budding of alveoli from the ducts; later, alveoli cluster to form lobulo-alveolar
structures that are the secretory units of milk components throughout late pregnancy and
lactation. Following weaning, the glands undergo a remodelling process known as involution
(Lund et al., 1996; Li et al., 1997); during this phase, massive epithelial apoptosis results in
destruction of lobulo-alveolar structures and regression of the gland to the pre-pregnancy
state.
1.1.1 Regulation by Systemic Hormones
The development and biology of the mammary gland is controlled by a complex interplay of
systemic hormones and local growth factors. Hormones are critical regulators of mammary

5
development both in embryonic and postnatal life. Once the embryonic mammary buds are
formed, they stimulate expression of androgen receptors in the surrounding mesenchyme
(Kratochwil, 1986). Testicular androgens from male fetuses induce regression of the epithelial
mammary rudiment at E14, whereas estrogens are apparently not required for prenatal
development (Korach, 1994).
The structure and functionality of the postnatal mammary tissue is orchestrated by hormonal
changes occurring with age, estrous cycle and reproductive status. Pubertal growth and
secretory differentiation of mammary parenchyme is dependent on ovarian steroids (estrogen,
progesterone), prolactin and growth hormone. A role for estrogen and progesterone in ductal
growth has been demonstrated by genetic and tissue recombination studies. Estrogen induces
ductal dichotomous branching via paracrine stimulation of the stromal receptors, whereas the
epithelial receptors are dispensable (Bocchinfuso and Korach, 1997; Cunha et al., 1997).
Conversely, progesterone targets its epithelial receptors at puberty to promote ductal side-
branching (Lydon et al., 1995; Brisken et al., 1998). Genetic ablation of prolactin or its
epithelial receptor revealed that prolactin controls regression of terminal end buds and ductal
side-branching during puberty (Ormandy et al., 1997; Horseman et al., 1997); these effects of
prolactin are indirect and may involve regulation of ovarian production of progesterone
(Brisken et al., 1999). Recent tissue recombination experiments circumvented the sterility of
the abovementioned knockout mice and showed that both progesterone and prolactin act
directly on their epithelial receptors to stimulate lobulo-alveolar development (Brisken et al.,
1998; Brisken et al., 1999). Ectopic expression of growth hormone in transgenic mice leads to
precocious mammary development and epithelial differentiation (Bchini et al., 1991),
indicating that growth hormone promotes functional differentiation of the gland. This effect
may be mediated by insulin-like growth factor-1 (IGF-1), as growth hormone induces
synthesis of IGF-1 in stromal mammary cells (Kleinberg, 1997). Moreover, an essential role
of IGF-1 in end bud formation was shown by the targeted deletion of IGF-1, which resulted in

6
retardation of ductal morphogenesis that could be rescued by exogenous IGF-1 (Ruan and
Kleinberg, 1999).
1.1.2 Local Effects of Multiple Growth Factors
The complexity of signals that control postnatal mammary growth and differentiation is
intriguing. Systemic hormones may synergize with a variety of growth factors that are locally
produced either in the mammary mesenchyme (like HGF/SF, FGF-4, FGF-2, KGF, HRG1α,
TGF-β, insulin-like growth factors) or in the mammary epithelium (Wnts, EGF, TGF-α,
amphiregulin, FGF-1). This section concerns stimulatory and inhibitory growth factors that
are implicated in growth and differentiation of the mammary gland. The function of EGF-like
growth factors and their receptors in mammary development will be addressed separately.
The role of Wnt signals in mammary development begins in embryonic life. Lef-1 null mice
fail to form mammary anlagen (van Genderen et al., 1994). Synergism of Wnt and parathyroid
hormone-related peptide (PTHrP) signals has been shown to be essential for branching
morphogenesis, sexual dimorphism and nipple formation of the mammary gland during
embryogenesis (Wysolmerski et al., 1998; Dunbar and Wysolmerski, 1999; Foley et al.,
2001). In postnatal life, the Wnt pathway is essential for mammary growth and differentiation
during pregnancy. Wnt-4 and progesterone signals trigger interconnected cascades that control
ductal side-branching and alveolar morphogenesis; using genetic and tissue recombination
techniques, Brisken et al. (2000) showed that progesterone impacts nearby epithelial cells to
induce expression of Wnt-4, which in turn synergistically acts in a paracrine fashion to
promote ductal side-branching. Further support for a role of Wnt signaling in mammary
development at pregnancy comes from genetic experiments with β-catenin, a downstream
effector of the Wnt pathway. Mammary glands from virgin mice overexpressing an active β-
catenin mutant show pregnancy-like lobulo-alveolar development and differentiation (Imbert
et al., 2001); however, ductal side-branching is normal in these transgenic mice. These
findings suggest that the canonical Wnt signaling pathway contributes to alveolar

7
morphogenesis, while Wnt control of ductal side-branching during pregnancy is not mediated
by β-catenin but by other as yet unknown downstream effectors. Alternative Wnt-induced
effectors are reviewed elsewhere (Hülsken and Birchmeier, 2001).
Previous observations suggest a role for hepatocyte growth factor/scatter factor (HGF/SF) and
its receptor c-Met in ductal growth of the virgin mammary gland. HGF/SF and its receptor c-
Met are coordinately produced in mammary mesenchyme and epithelium, respectively, with
highest expression levels throughout puberty and in adult life until mid-pregnancy (Pepper et
al., 1995; Niranjan et al., 1995; Yang et al., 1995); HGF/SF is expressed at low levels during
late pregnancy and throughout lactation, and expression again increases during involution of
the glands. In support of a role in ductal growth, HGF/SF stimulates branching when human
mammary organoids are cultured on collagen, resembling the ductal elongation events that are
observed in the mammary gland during puberty (Niranjan et al., 1995). Similarly, exogenous
HGF induces extensive ductal branching in organ culture, whereas antisense HGF
oligonucleotides block ductal growth of explanted mouse mammary glands (Yang et al.,
1995). Branching morphogenesis is also observed in organotypic cell culture experiments
whereby primary mammary epithelial cells, or cells from the mammary epithelial cell lines
NMuMG and EpH4 are grown on three-dimensional matrices in the presence of HGF/SF
(Niranjan et al., 1995; Soriano et al., 1995; Niemann et al., 1998). However, direct proof of a
physiological role of HGF/SF signaling in the mammary gland in vivo is still awaited.
Factors that regulate differentiation of cells from the mononuclear phagocytic lineage also
appear to be involved in mammary development. The colony stimulating factor 1 gene (CSF-
1) is disrupted by an inactivating mutation in the recessive osteopetrosis (op) allele;
homozygous op/op mice show a lactational defect due to incomplete mammary ductal side-
branching and lactational failure, despite normal lobulo-alveolar morphogenesis and
expression of milk proteins (Pollard and Hennighausen., 1994). The TNF family member
osteoprotegerin-ligand or osteclast differentiation factor (ODF) synergizes with CSF-1 in

8
osteoclastogenesis (Lacey et al., 1998); absence of ODF or its epithelial receptor RANK
(receptor activator of NFκB) impairs formation of lobulo-alveolar structures during pregnancy
due to enhanced cell death; however, and in contrast to CSF-1 ablation, it does not affect side-
branching (Fata et al., 2000). These results suggest a model whereby CSF-1 regulates ductal
sprouting, and ODF subsequently promotes lobulo-alveolar development and terminal
differentiation.
The formation of the adult mammary gland depends not only on growth stimulation but also
on active inhibition, which prevents infilling the extraglandular mesenchyme. There is ample
evidence that various TGF-β factors reversibly inhibit growth of the mammary end buds
(reviewed in Silberstein, 2001). Overexpression of TGF-β1 in the mammary gland leads to
impaired ductal elongation (Pierce et al., 1993) and absence of alveolar outgrowth and milk
secretion (Jhappan et al., 1993), indicating that TGF-β1 negatively regulates both ductal and
alveolar morphogenesis. Conversely, inhibition of TGF-β1 signaling promotes excessive
ductal branching in mice that overexpress a kinase-deficient TGF-β type II receptor in the
mammary stroma (Gorska et al., 1998; Joseph et al., 1999); this effect may be driven by up-
regulation of HGF/SF expression, thus suggesting that the chronic inhibition of ductal growth
by TGF-β1 results from its down-regulatory effect on the periductal synthesis of HGF/SF.
Transgenic mice and transplantation experiments identified TGFβ-3 as a local factor that is
secreted by alveolar cells upon milk stasis, and initiates apoptosis during the first stage of
involution (Nguyen et al., 2000); the autocrine pro-apoptotic effect of TGFβ-3 involves
activation of Stat3 followed by up-regulation of IGF-binding protein-5 (IGFBP-5), which
sequesters and inactivates the mitogen IGF-1 (Li et al., 1997; Tonner et al., 1997; Chapman et
al., 1999; Nguyen et al., 2000).
The role of fibroblast growth factor (FGF) signaling in the mammary gland is still
controversial; whereas the different members of the FGF family show a temporally and
spatially regulated expression in mammary tissue, genetic experiments suggest that they may

9
have redundant roles in the development of the gland. Expression of endogenous FGF4 and its
receptor, FGFR1, is detected in virgin females, but not during pregnancy and lactation
(Coleman-Knarcik et al., 1994; Chodosh et al., 2000). However, transgenic mice for FGF4
exhibit hyperplastic lobulo-alveolar structures that persist longer after weaning due to
impaired apoptosis, and mice expressing an ectopic dominant-negative form of FGFR1 in
mammary epithelium surprisingly lack a discernible phenotype (Morini et al., 2000). It is
possible that FGF4, like TGFβ factors, plays a role in the control of apoptotic remodelling
during ductal development and involution. In contrast to the transgenics for inactive FGFR1,
mice overexpressing a dominant-negative FGFR2 transgene show an impairment in lobulo-
alveolar development by mid-pregnancy (Jackson et al., 1997); knockout mice for FGF-7, its
putative mammary ligand, show normal mammary development (Guo et al., 1996). Future
research employing combined FGF-factor knockout mice may help to understand the function
of this family in mammary tissue.
To conclude, the mammary development is regulated by multiple growth and differentiation
factors and their intracellular signaling cascades. Loss of a single factor is often not
compensated by the others, indicating that these factors trigger essential interacting or
intersecting signaling cascades. Therefore, integration of these signals and the identification of
new mammogenic factors still represent a major challenge.
1.1.3 The Role of Extracellular Matrix
The inducing effects of embryonic mesenchyme or its postnatal counterpart, termed stroma,
are partially mediated by components of the extracellular matrix (ECM). In the adult gland,
epithelial cells are in direct contact with basal myoepithelial cells and with the ECM structure
known as basement membrane. The basement membrane is a complex and organized three-
dimensional array of laminin, type IV collagen, heparan sulphate, proteoglycans and other
proteins (Timpl, 1996). It is known that cell-ECM interactions influence tissue architecture
through modulation of signaling pathways that affect cell growth, differentiation, survival and

10
morphogenesis (reviewed in Adams and Watt, 1993). The use of reconstituted basement
membranes provides an excellent system to study the essential role of ECM for lactogenic
differentiation in vitro. In organotypic cell culture, synthesis and secretion of milk
components by mammary epithelial cells results from the interrelated effects of hormones,
cell-cell and cell-ECM contacts (reviewed in Lin and Bissell, 1993). Primary mammary
epithelial cells fail to differentiate to a secretory phenotype when cultured on plastic or onto a
thin collagen type I layer (Emerman and Pitelka, 1977; Berdichevsky et al., 1992); however,
these cells undergo alveolar morphogenesis and secrete milk proteins vectorially when
cultured on a reconstituted, laminin-rich matrix from Engelbreth-Holm-Swarm (EHS)
tumours (termed Matrigel; Kleinman et al., 1986; Barcellos-Hoff et al., 1989). Indeed,
adhesion of mammary epithelial cells to laminin is critical for β-casein gene expression and
for activation of Stat5, an essential regulator of milk gene expression (Streuli et al., 1995a;
Streuli et al., 1995b; see also section 2.5 of this Introduction). However, the extent of
morphogenic events in vitro is limited, indicating that additional signals from living stromal
cells are required to support formation of fully-developed ducts or alveoli.
Integrins are cellular receptors for laminin, a major ECM component (Sonnenberg et al.,
1990). Integrins are expressed at the basal membrane of epithelial cells both in mammary
alveolar tissue and in EHS-cultured alveoli (Streuli et al., 1991). A role of integrins as
physiological receptors for ECM was suggested by cell culture studies in which integrin
function was blocked by pan-antibodies (Streuli et al., 1991); such blocking antibodies
prevented the expression of milk proteins by mammary epithelial cells embedded in EHS
matrix, indicating that the ECM induces lactogenic differentiation via integrin signaling.
Genetic studies confirmed that integrins are cellular laminin receptors that control mammary
functional differentiation; targeted expression of a transgene coding for an inactive β1-
integrin subunit impairs lobulo-alveolar development and secretory differentiation during
pregnancy (Faraldo et al., 1998). Moreover, laminin accumulates at the lateral surface of

11
luminal cells in the transgenic glands, indicating that integrin-ECM interactions determine
baso-apical polarity of alveolar cells.
Overall, it is evident that direct cell-ECM contacts contribute to the induction of morphogenic
and lactogenic events in mammary development. Future work will help to fully understand the
cooperative effects of the ECM and other mitogenic factors in growth, morphogenesis and
functional differentiation of the mammary epithelium.
1.2 The ErbB Family of Receptor Tyrosine Kinases
ErbB proteins belong to subclass I of the superfamily of receptor tyrosine kinases. This family
has evolved from a single ligand-receptor pair in Caenorhabditis elegans, lin-3/let-23 (Aroian
et al., 1990). Drosophila melanogaster also express one ErbB receptor, but three activating
ligands and one inhibitor (Freeman, 1998). In vertebrates, the ErbB family comprises four
membrane receptors: the epidermal growth factor receptor (EGF receptor; also termed ErbB-
1, HER1), ErbB-2 (c-Neu, HER2), ErbB-3 (HER3) and ErbB-4 (HER4) (reviewed in
Olayioye et al., 2000). These receptors share a glycosylated extracellular ligand-binding
region with two cysteine-rich domains, a transmembrane stretch, and an intracellular region
that encompasses a tyrosine kinase domain and a C-terminal tail containing the
autophosphorylation sites. ErbB-1, ErbB-2 and ErbB-4 encode ligand-activated tyrosine
kinases, whereas the corresponding ErbB-3 sequence is apparently devoid of kinase activity
(Guy et al., 1994). ErbB receptors show different patterns of expression: ErbB-1 is expressed
by liver parenchymal cells, fibroblasts, keratinocytes and several epithelial tissues, like the
basal layer of the skin (Adamson, 1990; Partanen, 1990); ErbB-2 is expressed in a variety of
tissues including nervous system, connective tissue and secretory epithelium (Kokai et al.,
1987); ErbB-3 is expressed primarily in epithelium of various organs, in peripheral nervous
system and in oligodendrocytes, while ErbB-4 is mostly restricted to central nervous system,
cardiac muscle and glial cells (Pinkas-Kramarski et al., 1997). These differential expression

12
patterns are consistent with specific biological activities during embryonic life (discussed in
Section 2.4 of this Introduction).
The ErbB receptors are activated upon ligand binding, a general mechanism that is shared by
various receptor tyrosine kinases. Ligand binding induces formation of receptor dimers; this
key step allows each receptor subunit to cross-phosphorylate tyrosine residues in the
activation loop of the kinase domain of its partner, thus enhancing the catalytic activity
(Hubbard et al., 1998). Following activation of the kinases, specific tyrosine residues on the
C-terminal tail of the receptors become autophosphorylated; these phosphotyrosine residues
are docking sites for intracellular signaling molecules that couple the receptors to signal
transduction cascades, thus ultimately resulting in specific cellular responses. The hallmark of
the ErbB family is the formation of both homo- and heterodimers following ligand binding.
Moreover, each ligand induces the formation of preferential dimers in tissues where more than
two ErbB receptors are expressed, leading to signal diversification (see below). The cellular
routing of each receptor after ligand binding also differs for each family member: ErbB-1
undergoes rapid internalization and targets EGF for lysosomal degradation, whereas the other
ErbBs are slowly internalized and are recycled back to the cell surface without significant
degradation of the endocytosed ligand (Baulida et al., 1996; Pinkas-Kramarski et al., 1996;
Lenferink et al., 1998).
1.2.1 ErbB Ligands Contain an EGF-like Domain
ErbB receptors are activated upon binding ligands that are known as EGF-related growth
factors. These factors are mostly produced as transmembrane precursors, which can be
proteolytically cleaved, thus releasing the extracellular, biologically active region. ErbB
ligands are characterized by the presence of a conserved EGF-like domain of 35-50 amino
acids that is essential for receptor binding. Six cystein residues form three disulfide bonds that
hold together the characteristic three-loop structure of this motif. EGF-like ligands can be
grouped according to their receptor-binding affinity: EGF, amphiregulin and transforming

13
growth factor-α (TGF-α) specifically bind to ErbB-1; betacellulin, heparin-binding EGF (HB-
EGF) and epiregulin bind both ErbB-1 and ErbB-4; neuregulins are a complex family of
proteins that include NRG-1 and NRG-2, which bind to ErbB-3 and ErbB-4, and the recently
described NRG-3 and NRG-4, which bind to ErbB-4. So far, no direct ligand for ErbB-2 has
been described (Peles et al., 1993; Tzahar et al., 1994).
NRG-1 (also termed Neu differentiation factor) was first isolated from medium of Ras-
transformed rat fibroblasts, and the human counterpart Heregulin was detected in medium of
breast cancer cells (reviewed in Peles and Yarden, 1993). Two neuronal factors, termed glial
growth factor (GGF) and acetylcholine receptor-inducing activity (ARIA) are alternatively
spliced variants of NRG-1. Four different nrg genes code for the neuregulin isoforms and their
related variants (Holmes et al., 1992; Wen et al., 1992; Falls et al., 1993; Marchionni et al.,
1993; Carraway et al., 1997; Chang et al., 1997; Harari et al., 1999). NRG-1 shows a wide
expression pattern during embryogenesis, being detectable in the central nervous system and
in ventricular endothelium (Meyer et al., 1995). NRG-2 is also expressed in embryonic neural
tissue and heart but otherwise is largely non-overlapping with NRG-1 expression (Carraway et
al., 1997; Chang et al., 1997). NRG-3 expression is restricted to developing and adult nervous
tissue (Zhang et al., 1997); NRG-4 is highly expressed in adult pancreatic tissue and weakly
in muscle, but no data exist about embryonic expression (Harari et al., 1999).
1.2.2 Generation of Signal Diversity
It has been proposed that EGF-like ligands are bivalent; in case of NRG-1, an N-terminally
located high affinity site with narrow specificity first binds a direct receptor (ErbB-3 or ErbB-
4), and then a second C-terminal site recruits a co-receptor with lower affinity but broader
specificity, ErbB-2 usually being the preferred one (Tzahar et al., 1997). Such a mechanism
may account for the diversity of receptor dimers that are observed with a single ligand, as well
as for the activation of the orphan receptor ErbB-2 in response to different EGF-like growth
factors (Karunagaran et al., 1996; Graus-Porta et al., 1997). Evidence for the existence of all

14
ten possible homo- and heterodimers of ErbB proteins has been reported, including the ErbB-
2 homodimer that is stabilized by oncogenic mutation or overexpression (Riese et al., 1995;
Tzahar et al., 1996). This network of inter-receptor interactions displays a strict hierarchy
rather than a random pattern (Tzahar et al., 1996). In fact, ErbB-2 is the preferred
heterodimerization partner for all other ErbB family members. A driving force for the
preferential binding to ErbB-2 could be that heterodimers containing ErbB-2 have a very low
rate of ligand dissociation compared to other receptor pairs; this property of ErbB-2 can
significantly prolong cell stimulation by every ErbB ligand (Graus-Porta et al., 1995;
Karunagaran et al., 1996). In the absence of ErbB-2, NRG-1 induces formation of other
heterodimers like ErbB-1/ErbB-3 and ErbB-1/ErbB-4 heterodimers, which explains the
inhibition in trans of EGF binding to ErbB-1 when NRG-1 is present (Karunagaran et al.,
1995). The existence of several ligands, together with their distinct ability to stabilize
preferential homo- and heterodimeric receptor pairs, points to the existence of a hierarchical
network of ligand-stimulated receptor dimerization events within the ErbB family (Pinkas-
Kramarski et al., 1996).
Autophosphorylation of tyrosine residues may also be influenced by the heterodimer
combination (Olayioye et al., 1998); in this way, each receptor has the ability to interact with
distinct sets of intracellular signaling proteins thus increasing the functional versatility of the
ErbB family. There are four potential mechanisms which may account for ligand-induced
differential phosphorylation of the receptors (reviewed in Sweeney and Carraway, 2000). One
ligand may induce receptor dimerization and phosphorylation of a particular subset of tyrosine
residues. Binding to a different ligand could influence site usage for phosphorylation by
promoting the association of the dimeric receptor complex with other cellular proteins like
kinases, phosphatases or even cell surface molecules, which may mediate phosphorylation or
dephosphorylation of specific sites. Alternatively, this second ligand could stimulate the
assembly of oligomeric receptor complexes, or induce a different conformation of the receptor
pair. Taken together, combinatorial dimerization and ligand-induced diversification of

15
signaling appear to confer ErbB receptors the potential to give rise to a broad range of cellular
responses; moreover, because each receptor is unique in terms of catalytic activity, cellular
routing and transmodulation, the resulting network allows fine tuning and stringent control of
cellular functions.
1.2.3 Intracellular Effectors of ErbB Receptors
As for other receptor tyrosine kinases, ligand-induced autophosphorylation of ErbB receptors
on specific tyrosine residues creates docking sites for cytoplasmic signaling proteins
containing Src-homology2 (SH2) or phosphotyrosine-binding (PTB) domains (reviewed in
Pawson, 1995). The binding specificity of these proteins is determined by the amino acid
sequences adjacent to the phosphorylated tyrosines; amino acids located N-terminally
determine the binding of specific PTB domains, and amino acids that are C-terminally located
select SH2 domains. All ErbB family members, including the C. elegans and D. melanogaster
homologs Let23 and DER, couple via Shc and/or Grb-2 to the mitogen-activated protein
kinase (MAPK) pathway (Pinkas-Kramarski et al., 1996). However, certain intracellular
proteins are preferential substrates of specific ErbB receptors. ErbB-3 is a preferred activator
of p85 subunit of phosphatidylinositol-3-kinase (PI-3-K) due to the multiple specific binding
motifs present in the ErbB-3 C-terminal tail, which are virtually absent in case of ErbB-2
(Fedi et al., 1994; Prigent et al., 1994). Similarly, the negative regulator c-Cbl and
phospholipase Cγ1 (PLCγ1) couple to both ErbB-1 and ErbB-2 but not to ErbB-3 or ErbB-4
(Fazioli et al., 1991; Fedi et al., 1994; Cohen et al., 1996; Levkowitz et al., 1998; Klapper et
al., 2000; Levkowitz et al., 2000). Olayioye et al. (2000) offers an excellent review of the
present knowledge about the intracellular mediators of ErbB signals. As this work aimed to
find novel substrates for the ErbB-2 receptor, its known downstream effectors will be
described in detail.
ErbB-2 contains 5 putative autophosphorylation sites in its C-terminal tail, termed here Y1
(the most N-terminal tyrosine residue) to Y5 (the most C-terminal one; Hazan et al., 1989;

16
Segatto et al., 1990; Akiyama et al., 1991). It has been described that Shc binds to Y4 through
its PTB domain (Segatto et al., 1993; Ricci et al., 1995; Dankort et al., 1997). Grb-2 directly
binds to Y2 via its SH2 domain, and indirectly via Shc (Ricci et al., 1995; Dankort et al.,
1997). Chk binds to Y5 (Zrihan-Licht et al., 1998). Grb-7, c-Src, Ras-GTPase activating
protein (Ras-GAP) and the abovementioned PLCγ1 also interact with ErbB-2, though the
binding sites are unclear (Fazioli et al., 1991; Jallal et al., 1992; Stein et al., 1994;
Muthuswamy and Muller, 1995b).
The functional role of the ErbB-2 autophosphorylation sites in receptor-mediated
transformation has been assessed by mutational analysis of the rodent constitutively active
ErbB-2, termed Neu (Dankort et al., 1997). Absence of all major autophosphorylation sites of
Neu dramatically decreases transforming activity upon overexpression in fibroblasts. The C-
terminal tyrosine residues Y2 to Y5 can independently mediate transformation, since they
fully restore transforming activity when individually added back to the inactive receptor. In
contrast, the first tyrosine residue Y1 may not be involved in receptor-mediated
transformation, as the resulting add-back mutant lacks transforming potential; moreover, Y1
may represent a negative regulatory site, since it supresses transforming activity when restored
in combination with any other single tyrosine residue. Recent studies show that the
functionally-redundant add-back mutants containing tyrosines Y2, Y4 and Y5 activate Ras to
induce transformation, whereas the add-back mutant containing tyrosine Y3 operates
independently of Ras to activate MAPK (Dankort et al., 2001).
1.2.4 ErbB Signaling in Embryonic Development
The importance of ErbB signaling in development was revealed by genetic studies in mice.
Null mutations of individual ErbB receptor loci are embryonic lethal. Loss of erbB-1 leads to
embryonic or perinatal lethality depending on the genetic background of the host (Miettinen et
al., 1995; Sibilia et al., 1995; Threadgill et al., 1995; Sibilia et al., 1998); the mice display
abnormalities in multiple organs including brain, skin, lung and gastrointestinal tract. Mice

17
that show spontaneous or targeted mutation of TGFα, one of the various ErbB-1 ligands,
show only part of the phenotype observed in erbB-1 null mice, like impaired development of
the eyes and hair follicles (Luetteke et al., 1993; Mann et al., 1993); this partial overlap
suggests that each ErbB-1 ligand may play a distinct developmental role. The information
gained by targeted mutation of erbB-2, erbB-3 and erbB-4 receptors and their ligand NRG-1
clearly demonstrates that distinct receptor heterodimers are essential for different
developmental events. ErbB-2 -/- mice die at midgestation (E10.5) due to malformation of
myocardial trabeculae in the heart ventricle (Lee et al., 1995), a phenotype that is shared by
the NRG-1 (Meyer et al., 1995) and the erbB-4 null mice (Gassmann et al., 1995). These
results are consistent with the view that NRG-1, which is expressed in endothelial cells of the
endocardium (Meyer and Birchmeier, 1994), is required for activation of myocardial ErbB-
2/ErbB-4 heterodimers to promote trabecular formation in the developing heart. In contrast to
ErbB-2 and ErbB-4, ErbB-3 is not expressed in myocardium but in mesenchyme of the pre-
valvular endocardial cushions. Accordingly, erbB-3 null mice die at day E13.5 displaying
normal heart trabeculation but defective valve formation (Erickson et al., 1997; Riethmacher
et al., 1997).
In addition to cardiac abnormalities, erbB-3 -/- mice show a generalized neural crest defect
that affects both central and peripheral nervous structures. ErbB-3 mutant mice fail to form
cranial sensory ganglia due to impaired migration of neurons from the hindbrain neural crest
(Erickson et al., 1997; Riethmacher et al., 1997). This phenotype is also observed in mice
lacking erbB-2 or NRG-1 (Lee et al., 95; Meyer et al., 1995); in contrast, erbB-4 deficient
mice do not exhibit deficient cellularity of cranial ganglia but rather the innervation of these
ganglia is disrupted, thus suggesting a unique role for ErbB-4 (Gassmann et al., 1995). In the
peripheral nervous system, erbB-3 -/- mice completely lack Schwann cell precursors and
Schwann cells that normally accompany nerves (Erickson et al., 1997; Riethmacher et al.,
1997). In addition, degenerative motor and sensory neurons are found in the dorsal root
ganglia (Erickson et al., 1997; Riethmacher et al., 1997), and migration of sympathogenic

18
neural crest cells is also impaired in erbB-3 mutants (Britsch et al., 1998); similar defects
have been observed in NRG-1 and erbB-2 deficient mice (Meyer et al., 1995; Kramer et al.,
1996; Britsch et al., 1998). It is evident from these studies that ErbB-2/ErbB-3 heterodimers
transmit NRG-1 signals for neural crest cells to migrate. Recently, the early mortality of erbB-
2 null mice has been rescued by myocardial expression of an erbB-2 transgene (Morris et al.,
1999; Woldeyesus et al., 1999); erbB-2 rescued mice show striking similarities with the erbB-
3 null mice, thus confirming the role of ErbB-2/ErbB-3 complexes in the development of the
peripheral nervous system. Data on genetic ablation of the other NRG genes are largely
missing. Mice carrying a targeted mutation of the NRG-3 gene, however, do not show an overt
phenotype, but may deserve a more detailed analysis (T. Müller and C. Birchmeier,
unpublished data). So far, the differential embryonic expression of the various neuregulin
proteins and their distinct biological properties in vitro suggest different physiological
functions (Carraway et al., 1997; Chang et al., 1997; Crovello et al., 1998). Together, these
observations show the critical function of ErbB receptors and their ligands during
embryogenesis. Moreover, the above genetic studies define distinct developmental roles for
certain receptor combinations and are therefore strong support for the occurrence of signal
diversification in vivo.
1.2.5 ErbB Receptors in Mammary Gland Development
There is considerable evidence that ErbB signaling has important roles in both normal and
neoplastic growth of the mammary gland. All four ErbB receptors are found in mammary
tissue (Schroeder et al., 1998). In prepubescent mammary gland, ErbB-1 and ErbB-2 are
widely expressed in epithelium, stroma and mesenchymal fat, with ErbB-1 levels being
particularly high in stromal cells. In the mature gland, ErbB-3 and ErbB-4 are also detected;
just at this stage, ErbB-1 and ErbB-2 are differentially located: ErbB-1 is present in stroma
and adipose compartments, while ErbB-2 is prominent in epithelium. During pregnancy, the
four ErbB receptors are coordinately expressed in mammary epithelium; ErbB-1 and ErbB-2
are found at high levels in the alveolar epithelium throughout pregnancy, whereas ErbB-3 and

19
ErbB-4 levels increase preferentially in the ductal epithelium later in pregnancy. During
lactation, receptor levels are low. ErbB-1 and ErbB-2 expression markedly increases during
involution, while ErbB-3 expression declines and ErbB-4 expression is not detectable.
Despite expression of all ErbB receptors during puberty, only ErbB-1 and ErbB-2 are
tyrosine-phosphorylated while ductal growth occurs (Sebastian et al., 1998). ErbB-3 and
ErbB-4 seem to be present in a non-phosphorylated, inactive state at this stage, suggesting that
none of them is relevant for ductal morphogenesis. Endogenous phosphorylation of all four
receptors is detected during late pregnancy and lactation, with increasing levels of tyrosine-
phosphorylated ErbB-1, ErbB-2 and ErbB-4 later at lactation. Information of the
phosphorylation state of the ErbB receptors during involution is missing.
Like the receptors, the six EGFR ligands and NRG-1 are differentially expressed in mammary
tissue (Schroeder et al., 1998). TGFα, betacellulin and heparin-binding EGF (HB-EGF)
transcripts are found in prepubescent mammary gland and through mid-pregnancy, drop
markedly during late pregnancy and lactation and again increase at involution. Amphiregulin
and epiregulin transcripts appear in mature virgin glands and early in pregnancy, respectively;
transcript levels of both factors decline later in pregnancy, remain low throughout lactation
and involution, and reappear as the gland again resembles the mature virgin state. EGF
transcripts are present at low levels in the virgin gland, and they dramatically increase during
late pregnancy and lactation and return to low levels during involution. The NRG-1α isoform
is found in mammary mesenchyme and shows a strongly regulated pattern of expression
(Yang et al., 1995). NRG-1 is present at low levels in the virgin gland. At mid-pregnancy,
NRG-1 exhibits a sudden concentration peak and then rapidly decreases to basal levels, which
are constant later in pregnancy, throughout lactation and involution.
Several lines of evidence support a role of ErbB signaling in the development of the
mammary gland. In early studies using mice, pellets containing EGF-like factors were

20
surgically implanted into the mammary glands to allow the slow local release of these factors.
Implants of EGF, TGFα and NRG-1α stimulate ductal side-branching and lobulo-alveolar
morphogenesis in virgin glands (Vonderhaar et al., 1987; Jones et al., 1996). EGF- or TGFα-
induced alveoli lack secretory activity; in contrast, alveoli that derive from NRG-1α treatment
are differentiated into secretory structures, which accumulate secreted milk proteins in their
luminal compartment. These observations indicate that all factors can promote formation of
alveoli but only NRG-1α stimulates their terminal differentiation. In other experiments, EGF-
like growth factors were directly injected into the mammary glands to study their ability to
induce tyrosine phosphorylation of ErbB receptors. Treatment of prepubescent glands with
EGF stimulates tyrosine phosphorylation of stromal ErbB-1 (the EGF receptor) and ErbB-2
(Schroeder et al., 1998; Sebastian et al., 1998); as above mentioned, endogenous
phosphorylation of ErbB-1 and ErbB-2 is observed in mammary tissue during puberty
(Sebastian et al., 1998). Therefore, it is likely that locally-produced EGF induces at puberty
the formation of active ErbB-1/ErbB-2 heterodimers, which may be essential for mammary
ductal growth. Exogenous EGF induces phosphorylation of ErbB-1 and ErbB-2 at pregnancy,
despite all ErbB receptors being present; in contrast, administration of NRG-1β results in
trans-phosphorylation of all ErbB receptors, clearly indicating that neuregulin can induce the
formation of combinatorial receptor complexes at pregnancy. Together with the pregnancy-
restricted expression of neuregulin in mammary mesenchyme (Yang et al., 1995), these
findings strongly suggest that NRG-1 may play a major role in the mammary gland during
pregnancy to promote alveolar morphogenesis via trans-activation of ErbB heterodimeric
receptors.
Convincing evidence of the physiological role of ErbB signals in mammary development has
been supplied by genetic studies in mice; moreover, the phenotypes of knockout or transgenic
mice support the differential roles of the ErbB receptors and their ligands that could be
expected from the abovementioned stimulation studies, and from expression and activation
patterns in vivo. Mammary glands from mice carrying a targeted mutation of the amphiregulin

21
gene show impaired ductal growth, whereas TGFα/EGF double null mice show normal
mammary development at this stage (Luetteke et al., 1999). Amphiregulin null mammary
glands are competent for lobuloalveolar differentiation; however, additional loss of TGFα
and/or EGF severely compromises lactogenesis. Coherent with these findings, expression of a
TGFα transgene in mammary tissue induces precocious alveolar development in virgin
females, alveolar hyperplasia during pregnancy and delayed involution (Matsui et al., 1990;
Sandgren et al., 1995). Together, these results suggest distinct functions of the various ErbB-1
ligands in the mammary gland: amphiregulin may be critical for ductal growth during puberty,
while TGFα or EGF may be involved in alveolar differentiation during pregnancy. ErbB-1 is
activated in virgin tissue; mammary expression of a transgene encoding a dominant-negative
ErbB-1 receptor (the EGF receptor) inhibits ductal branching in the glands of virgin mice thus
showing a role for ErbB-1 signaling in pubertal mammary development (Xie et al., 1997);
similarly, female waved-2 mice, which carry a spontaneous inactivating mutation of the erbB-
1 gene, display impaired glandular development (Fowler et al., 1995). Transplantation and
tissue recombination experiments further support a role for ErbB-1 in ductal morphogenesis.
Mammary gland grafts from neonatal erbB-1 -/- mice fail to undergo ductal growth (Sebastian
et al., 1998); however, they develop lobulo-alveolar structures when stimulated by prolactin,
indicating that the EGF receptor is essential for ductal branching but not for alveolar
morphogenesis (Wiesen et al., 1999). Moreover, tissue recombinants revealed that wild-type
fat pad supports outgrowth of erbB-1 -/- epithelium whereas the -/- fat pad does not, thus
clearly showing the relevant role of stromal ErbB-1 in mammary ductal growth.
A role for NRG-1 in mammary gland development during pregnancy is substantiated by its
restricted expression and pan-activating effect on ErbB receptors at this stage (see above).
Indeed, mammary glands from mice that lack NRG-1α fail to undergo lobulo-alveolar
morphogenesis at pregnancy (Li et al., submitted); NRG-1α is the isoform that is normally
expressed in the glands. In contrast, NRG-1β null mice die during embryogenesis; the NRG-
1β isoform accounts for the cardiac and neural crest phenotypes that were described in section

22
2.4 of this Introduction (C. Birchmeier, unpublished data). These results definitely
demonstrate that NRG-1α is the naturally occurring isoform in mammary tissue, where it
functions during pregnancy as a critical growth factor that promotes alveolar morphogenesis
and secretory differentiation. Further insights into the mechanisms of neuregulin-induced
morphogenesis have been gained from genetic studies with ErbB receptors. Expression of
dominant-negative forms of erbB-2 and erbB-4 in the mammary gland of transgenic mice
revealed a physiological role for both receptors in lobulo-alveolar development at late
pregnancy and lactation (Jones et al., 1999; Jones and Stern, 1999). Moreover, there is genetic
and biochemical evidence that Stat5a mediates morphogenic signals of ErbB-4 in the
mammary gland during lactation (Liu et al., 1997; Jones et al., 1999). According to this data,
it can be speculated that NRG-1α induces alveolar morphogenesis via signaling pathways that
involve trans-activation of ErbB-2 and ErbB-4 and transcriptional regulation by Stat5a.
As already mentioned in Section 1.3 of this Introduction, the physiological processes that
prepare the mammary gland for lactogenesis can be mimicked in vitro using organ culture and
organotypic assays. By these means, our group has extensively contributed to understand the
function of neuregulin in the mammary gland. The pioneering work of Yang et al. (1995) has
shown that NRG promotes lobulo-alveolar differentiation of mammary gland explants in
organ culture. The same morphogenic effect of neuregulin has been observed by Niemann et
al. (1998) in organotypic cell culture. Treatment with neuregulin induces EpH4 mammary
epithelial cells to form alveoli-like structures when cultured on a Matrigel reconstituted
matrix; moreover, alveolar cells functionally differentiate and secrete milk components into a
luminal compartment, thus reproducing in vitro the physiological responses of mammary
epithelium to neuregulin. In addition, these studies show that activation of exogenous ErbB-2
receptor tyrosine kinase is sufficient for EpH4 cells to undergo alveolar morphogenesis in the
Matrigel system. The present work contributes to understand the intracellular mechanisms
underlying the morphogenic events of neuregulin-stimulated EpH4 cells in organotypic

23
culture, for it identifies Vav2 as an essential mediator of ErbB-2 specific signals leading to
alveolar morphogenesis.
1.3 Vav Family of Guanine Nucleotide Exchange Factors
Vav proteins constitute a family of structurally related guanine nucleotide exchange factors
that are involved in signaling pathways leading to cytoskeletal rearrangements and changes in
gene expression (reviewed in Bustelo, 2000). Vav, the first member of the family, was
identified as an oncogene encoding a constitutively active protein which lacks 67 amino acids
at the N-terminus and induces transformation of NIH 3T3 fibroblasts (Katzav et al., 1989;
Katzav et al., 1991). Vav expression and function is restricted to the hematopoietic system,
and T and B lymphocytes derived from vav -/- mice reveal defects in antigen-receptor induced
proliferation (Fischer et al., 1995; Tarakhovsky et al., 1995; Zhang et al., 1995; Fischer et al.,
1998). The recently discovered vav2 and vav3 are also expressed in tissues of non-
hematopoietic origin, among them epithelia (Schuebel et al., 1996; Movilla et al., 1999).
Transient expression of N-terminally truncated Vav2 and Vav3 proteins in fibroblasts results
in morphological changes such as membrane ruffling and formation of lamellipodia, and N-
terminally truncated Vav2 is transforming in fibroblasts (Schuebel et al., 1996; Schuebel et
al., 1998; Movilla et al., 1999). Vav2 has been implicated in cell-mediated killing by
cytotoxic lymphocytes and in cellular responses following activation of B cell receptor and
CD19 (Billadeau et al., 2000; Doody et al., 2000; Doody et al., 2001; Tedford et al., 2001).
Vav and Vav3 can enhance NFκB-dependent transcription after TCR engagement (Moores et
al., 2000). No biological function has as yet been assigned to Vav2 and Vav3 in epithelial
tissues. Vav proteins share many different structural domains, among them a Dbl-homology
domain that is responsible for their GDP/GTP exchange activity, and an SH2 domain that may
link them to receptor tyrosine kinases (Bustelo, 2000). Indeed, tyrosine phosphorylation is
required for the activation of full-size but not of N-terminally truncated forms of Vav proteins
(Bustelo, 2000). Recently, crystallographic analysis revealed that the N-terminus of Vav acts

24
as an autoinhibitory site, and that phosphorylation of one specific tyrosine residue within this
region relieves autoinhibition (Aghazadeh et al., 2000).

25
2 MATERIALS AND METHODS
Standard protocols for various techniques in molecular biology, like preparation and analysis
of DNA and RNA, enzymatic manipulation, transfection of DNA into mammalian cells,
protein analysis and setup of polymease chain reaction (PCR) were performed according to
Current Protocols in Molecular Biology (Ausubel et al., 1994).
2.1 Amplification of the Hollenberg cDNA Yeast Expression Library
The Hollenberg mouse embryo cDNA library (Behrens et al., 1996) was amplified to gain
enough plasmid for yeast transformation. E.coli library culture was thawed on ice, and 103 and
106 dilutions were made in LB-ampicillin selective medium. One µl of the 103 dilution, and
50-100 µl of the 106 dilution was mixed with LB-ampicillin medium, spread onto LB-
ampicillin plates and incubated at 37°C overnight. On the next day, colonies were counted to
calculate the library titer (number of colony forming units per ml of bacterial suspension). A
volume of this suspension containing twice as much cells as the number of independent
clones of the library (5x106) was diluted in LB-ampicillin, plated at 200,000 colonies per 15-
cm dish (nearly confluent) and incubated at 37°C overnight. Colonies were scraped into 1.5-
liter selective medium; this suspension was incubated at 37°C with shaking for a further 2 h,
bacteria were pelleted and large-scale plasmid isolation was performed using Megaplasmid
columns 2500 (Qiagen).
2.2 Yeast Plasmids and Strains
Bait proteins for the yeast two-hybrid system are encoded in the pBTM116 vector (Bartel and
Fields, 1995). This vector carries the TRP1 gene and has a polylinker downstream of LexA
coding sequences. The E. coli repressor LexA consists of two domains: a C-terminal domain,
which is responsible for dimerization prior to DNA binding, and an N-terminal domain
responsible for specific binding to a palindromic operator containing the CTGTNNNN
consensus half-site. The Hollenberg library is inserted in the VP16 vector, which carries the

26
LEU2 gene and contains a nuclear localization signal-VP16 activation domain sequence
upstream of the multiple cloning site. Both pBTM116 and VP16 yeast expression vectors bear
a bacterial origin of replication and an ampicillin resistance gene, which allow plasmid
amplification in bacteria.
To generate TprMet-ErbB2 yeast baits, a cDNA fragment encoding amino acids 1005-1260 of
rodent ErbB-2 harbouring a Y1253F mutation was inserted into the Not I/Sal I sites of pSK+
(Stratagene) This region stretches over Y1028 (Y1), Y1144 (Y2), 1201 (Y3) and 1226/7 (Y4)
of ErbB-2. A cDNA sequence coding for dimerization and kinase domains of TprMet (amino
acids 1-481; Park 86 cell) was generated by PCR; this fragment, flanked by 5´ Not I/EcoR I
and 3´ Not I sites, was inserted into the Not I site at 5´ end of ErbB-2 in pSK+, thus rendering
the hybrid cDNA for the TprMet-ErbB2(Y1-4) bait. The complete cDNA sequence was
finally subcloned downstream of LexA into EcoR I/Sal I sites of pBTM116 yeast expression
vector. A new TprMet hybrid protein was constructed to include Y1253 (Y5) of ErbB-2 in the
bait. An ErbB-2 sequence encoding amino acids 1197-1260 was fused to TprMet as above to
generate the TprMet-ErbB2(Y3-5) bait; thus, Y1201 (Y3), 1226/1227 (Y4) and 1253 (Y5) of
ErbB-2 were present in this bait. A BTM-TprMet-∆tail control plasmid lacking the Met C-
terminal multiple docking site was constructed by inserting a PCR fragment encoding amino
acids 1-479 of TprMet into the EcoR I site of pBTM116. To generate kinase-deficient bait
proteins, the wild type sequences of TprMet were excised by EcoR I/Not I and replaced by a
PCR fragment containing the inactivating mutation K243A (Rodrigues and Park, 1993).
Both TprMet-ErbB2(Y1-4) and TprMet-ErbB2(Y3-5) bait proteins were used to screen the
Hollenberg cDNA library from E10.5 mouse embryos. Saccharomyces cerevisiae strain L40
was used as host. This yeast strain carries two reporter genes, HIS3 and lacZ, whose
expression is driven by minimal GAL1 promoters fused to multimerized LexA binding sites;
therefore, yeast expressing LexA activators can be detected as histidine prototrophs or by
measurable β-galactosidase activity. A pBTM116-lamin plasmid was used as a further control

27
to eliminate false positive clones. Reagents and methods for yeast two-hybrid analyses were
adapted from MATCHMAKERTM Handbook (PT1265-1, Clontech).
2.3 Yeast Two-Hybrid Screen
Bait plasmids and library were sequentially introduced into host L40 yeast strain to improve
the efficiency of transformation. Briefly, yeast was initially transformed with bait plasmid in a
small-scale procedure, and then library screens were performed (Fields and Song, 1989). To
prepare competent yeast, a single colony of L40 yeast was inoculated into 20 ml of YPD (10
g/l yeast extract, Difco; 20 g/l peptone, Difco; 2% dextrose) medium and incubated at 30°C
overnight. On the next day, culture was diluted 10-fold and further incubated until OD600 was
0.5. Cells were pelleted, washed with water and resuspended in 1.5 ml of sterile 1X TE/LiAc
(10 mM Tris-HCl pH 7.5, 1 mM EDTA, 100 mM lithium acetate). Competent L40 yeast was
then transformed with bait plasmid; 0.5 µg of plasmid DNA was mixed with 50 µg of salmon
sperm carrier DNA, 50 µl of competent yeast and 300 µl PEG/TE/LiAc (40% polyethylene
glycol, 1X TE/LiAc), vortexed and incubated at 30°C for 30 min. DMSO was added to 10%,
and heat-shock transformation was performed at 42°C for 15 min. Mixture was chilled on ice,
cells were pelleted, resuspended in 250 µl water and spread onto selection agar plates lacking
tryptophan. Colonies appear after 2 or 3 days. For the library screen, one single colony of the
bait transformants was grown to obtain 200 ml of a saturated cell suspension (OD600 greater
than 1) in medium without tryptophan. This culture was added to 800 ml YPD medium to
prepare 20 ml of competent yeast as above. Yeast suspension was incubated at room
temperature for 10 min, and 10 mg carrier DNA, 250 µg library DNA and 140 ml
PEG/TE/LiAc was addded. Mixture was incubated at 30°C for 30 min, 17.6 ml of DMSO was
added, and heat-shock transformation was performed as above. Co-transformed yeast cells
were resuspended in 1 liter YPD and incubated at 30°C for 1 h. Cells were pelleted,
resuspended in selection medium lacking tryptophan and leucine and further incubated at
30°C for 8 h. Finally, cells were resuspended in 10 ml water and 200 µl of a dilution series
was plated onto 50 selection agar plates without tryptophan, leucine and histidine in the

28
presence of 20 mM 3-aminotriazole and incubated at 30°C. Double transformants that express
interacting proteins rendered colonies within 8-10 days, which were re-plated onto fresh
selection agar plates for further analysis.
2.4 Verification of Interacting Clones
Interaction between bait and preys was confirmed by β-galactosidase activity filter assay and
by re-transfection of bait and prey plasmids back into yeast. For filter assay, colonies from
selection plates (without tryptophan and leucine) were replica-transferred onto a Whatman
Nr.1 filter. The replica filter was submerged in liquid nitrogen to permeabilize cells, and then
allowed to thaw. The filter was placed (colonies facing up) onto another filter presoaked in Z
buffer/X-gal solution (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, pH
7.0, 0.27% β-mercaptoethanol and 0.334 g/l X-gal) and incubated at 30°C. Colonies
expressing β-galactosidase appeared blue within 1-12 h.
For yeast re-transformation, library plasmids encoding interacting proteins were isolated from
individual positive colonies. To remove the bait plasmid from double transformant yeast,
colonies were inoculated into medium lacking leucine and cultured at 30°C for 1 day. Growth
in the absence of tryptophan selection allows survival of yeast segregants without bait
plasmid, which is randomly lost. To isolate the library plasmid from yeast segregants, cells
from 1 ml of the above culture were pelleted, lysed in 200 µl of lysis buffer (2% Triton X-100,
1% SDS, 100 mM NaCl, 1 mM EDTA, 10 mM Tris pH 8.0) and disrupted in the presence of
200 µl phenol/chloroform/isoamyl alcohol (25:24:1) and 0.3 g of acid-washed glass beads by
vortexing for 2 min; suspension was clarified by centrifugation and plasmid was recovered
from the supernatant by standard ethanol precipitation. Library plasmid was amplified in E.
coli HB101 strain, which is leucine auxotroph due to a leuB mutation. HB101 cells were
electroporated with the isolated library plasmid and plated onto selection agar plates without
leucine; therefore, transformants can be selected by complementation with the yeast LEU2
gene from the VP16 library plasmid. Bait and prey plasmids were re-transformed into yeast

29
according to the small-scale transformation protocol previously described. True positive
interacting clones grew again on selective agar plates without tryptophan, leucine and
histidine and were positive in β-galactosidase assays.
2.5 Mutagenesis of the Yeast TprMet-ErbB2 Baits
Mutational analysis of the bait proteins was performed to characterize the ErbB-2 binding
sites for each interacting protein found in the screen. Deletion mutants bearing single or
tandem tyrosine residues of the ErbB-2 multiple docking region were generated by standard
PCR. The PCR fragments were flanked by 5´ Not I and 3´ Sal I sites, and were used to replace
the ErbB-2 wild type sequence of the hybrid TprMet-ErbB2 baits. The ErbB-2 deletion
mutants fused to TprMet were: Y1 (amino acids 1014-1081) includes Y1028; Y1-2 (amino
acids 1014-1160) includes Y1028 and Y1144; Y2 (amino acids 1138-1160) includes Y1144;
Y2-3 (amino acids 1138-1221) includes Y1144 and Y1201; Y2F-3 is a variant of Y2-3 in
which Y2 was mutated to F using a commercial kit (Clontech); Y3 (amino acids 1194-1221)
includes Y1201; Y3-4 (amino acids 1194-1244) includes Y1201 and Y1226/1227; Y4 (amino
acids 1220-1244) includes Y1226/1227; Y4-5 (amino acids 1220-1260) includes Y1226/1227
and Y1253; Y5 (amino acids 1248-1260) includes Y1253.
Expression of the various TprMet-ErbB2 proteins in the yeast was checked by Western blot
analysis using anti-LexA antibodies (Clontech). L40 yeast transformants were grown in
selective medium without tryptophan at 30°C. During exponential growth phase, OD600 was
measured for 1 ml suspension; cells were pelleted from an aliquot corresponding to OD600=
0.5 and resuspended in 100 µl 2X denaturing buffer for SDS-PAGE (100 mM Tris base, pH
6.8, 20% glycerol, 4% SDS, 2% β-mercaptoethanol, 0.2% bromphenol blue). Twenty µl of
this suspension was loaded per slot onto an 8% polyacrylamide gel, proteins were resolved,
blotted in a PVDF membrane and Western Blotting was performed according to standard
protocols.

30
2.6 Mammalian Expression Plasmids
Full-length cDNA of Vav2 (∆29 isoform; Schuebel et al., 1996) was isolated from a λgt11
E17.5 mouse embryo cDNA library, using a cDNA fragment of Vav2 from a clone found in
the yeast two-hybrid screen. To generate Flag-tagged Vav2 proteins, the corresponding cDNA
sequences were inserted in frame in the Not I site downstream of a Flag epitope tag in pcDNA
3.1(+) (Invitrogen). The cDNAs of ∆N-Vav2 (encoding amino acids 189-839 of full-
sizeVav2) and ∆N-Vav2-∆C (encoding amino acids 189-595) were amplified by PCR using
full-size Vav2 as template. To generate ∆N-Vav2-dblmut and ∆N-Vav2-∆C∆PH, the
sequences encoding amino acids 336-342 and 383-507, respectively, were further deleted by
overlapping PCR. To generate the cDNA of TrkErbB2-∆tail, an Nco I/Not I fragment
encoding the ErbB-2 C-terminal tail (amino acids 1006-1260) was removed from a
pSK+/TrkErbB2 plasmid (Sachs et al., 1996) and replaced by a triple HA epitope tag flanked
by Nco I and compatible EagI sites; then, the sequence coding for HA-tagged TrkErbB2-∆tail
was excised EcoR I/Eag I and subcloned into pcDNA 3.1(+) using EcoR I/Not I sites. The
cDNA of HA-tagged TrkErbB2-Vav2 was constructed by inserting the full-size sequence of
Vav2 (encoding residues 2-839) into the Eag I site downstream from triple HA. To generate
the kinase-deficient mutant TrkErbB2-Kin-, a 951 bp-cDNA fragment containing the
inactivating K758A mutation was excised by Nco I/Mro I from the pLSV-K758A plasmid
(Ben-Levy et al., 1994) and inserted into pUC118-TrkErbB2 (Sachs et al., 1996) in place of
the wild-type sequence; the cDNA of TrkErbB2-Kin- was excised by Acc65 I/Spe I from
pUC118 and inserted into Acc65 I/Xba I sites of pcDNA 3.1(+). In TrkErbB2mut, Y1028 and
Y1201 of ErbB-2 C-terminal tail were mutated to phenylalanine using a commercial kit
(Clontech).
2.7 Site-Directed Mutagenesis
The TransformerTM Site-Directed Mutagenesis kit (Clontech) was used to generate Trk-ErbB2
variants in which single or pairs of tyrosine residues of the ErbB-2 C-terminal tail were

31
mutated to phenylalanine. Two oligonucleotide primers were used to introduce a base change
in double-stranded DNA. One primer (referred to as the mutagenic primer) introduced the
desired mutation. The second primer (referred to as the selection primer) disrupted a unique
restriction site to facilitate the ultimate selection of the mutated plasmid. A unique Sca I of
pUC118-Trk-ErbB2 was mutated to a Hpa I site with the selection primer 5'
CAGAATGACTTGGTTAACTACTCACCAGTC 3' (highlighted is the mutated base).
Mutagenic primers were Y2F (5' CCCCAGCCCGAGTTTGTGAACCAA 3'), Y3F (5' GGAG
AACCCTGAATTCTTAGTACCGAGAGAAGGC 3') and Y4F (5' GCCCAGCCTTTGACA
ACCTCTTTTTCTGGGACCAG 3'). The two primers were simultaneously annealed to one
DNA strand of the target plasmid. After standard elongation, ligation and a primary selection
by digestion with Sca I and Hpa I, the mixture of wild-type and mutated plasmid was
transformed into BMH 71-18 mutS E. coli strain, which contains a DNA mismatch repair
deficiency mutation. Plasmid DNA was prepared from the mixed bacterial population and
isolated DNA was again digested by Sca I and Hpa I. Mutated DNA was resistant to digestion
with Sca I but sensitive to Hpa I, whereas parental DNA was linearized by Sca I. A new
transformation of E. coli with the Sca I restriction mixture rendered high recovery of the
mutated plasmid. Protocols were carried out according to Clontech's user manual. Mutated
Trk-ErbB-2 DNAs were digested by Acc65 I/Spe I from pUC118 and inserted into the Kpn
I/Xba I sites of pcDNA3.1(+). A TrkErbB2 variant without both Vav2 binding sites (termed
TrkErbB2mut) was generated by Y1F mutation of pcDNA-TrkErbB2-Y3F, using a unique
Nde I site for the selection primer (5' TACATCAAGTGTATCCTCTGCCAAGTACGCCCC
CTA 3') and a Y1F mutagenic primer (5' CTGGTAGACGCTGAAGAGTTTCTGGTGCCCC
AGC 3').
2.8 Mammalian Cell Lines
Human embryo kidney 293T cells were used for transient transfections. For morphogenesis
assays on matrigel, EpH4 mouse mammary epithelial cells were used. EpH4 cells (Lopez-
Barahona et al., 1995) are a clonal epithelial derivative of IM-2 mouse mammary gland

32
epithelial cells, which were originally isolated from mammary tissue of a mid-pregnant
mouse (Reichmann et al., 1989). In our lab, the variant EpH4 K6 was subcloned (Niemann et
al., 1998), which exhibits a pronounced morphogenic potential on matrigel. The K6 subclone
was used in this work.
2.9 Immunoprecipitation and Western Blotting
3x106 293T cells were plated per 15-cm dish and co-transfected with 4 µg Flag-tagged Vav2
and 200 ng TrkErbB2, TrkErbB2-Kin- or TrkErbB2mut expression vectors by
calcium/phosphate precipitation. Twelve hours after transfection, cells were starved in serum-
free DMEM medium for 36 h. Prior to lysis, cells were stimulated with 50 ng/ml NGF
(Promega) in the presence of 1 µM phenylarsine oxide at 37°C for 10 min, and then washed
with ice-cold PBS (Sachs et al., 1996). Cells were harvested in 800 µl lysis buffer
supplemented with phosphatase and protease inhibitors (50 mM HEPES, 50 mM NaCl, 1.5
mM MgCl2, 1 mM EGTA, 10% glycerol, 1% Triton X-100, pH 7.5; inhibitors were added at
the following end concentrations: 10 mM NaF, 1 mM Na orthovanadate, 10 mM Na
pyrophosphate, 1 mM PMSF, 0.5 µg/ml aprotinin) and incubated on ice for 15 min. Lysates
were clarified by centrifugation at 14,000 rpm for 10 min at 4°C. For immunoprecipitations,
300 µl cell lysate was mixed with an equal volume of HNTG buffer (20 mM HEPES, 150
mM NaCl, 10% glycerol, 0.1% Triton X-100, pH 7.5; inhibitors were used as for lysis buffer)
and incubated with primary antibodies at 4°C overnight. Immunoprecipitates were
resuspended in 70 µl denaturing loading buffer and heated at 95°C for 3 min. Proteins were
resolved by SDS-PAGE, and transferred to PVDF membranes for 1 h at 100 mAmp per blot
using a Trans-Blot SD semi-dry electroblotter (BioRad). Membranes were blocked with 5%
low-fat milk powder in PBS at 25°C for 3 h, or with 0.5% Tween 20 and 10% FCS in PBS for
anti-phosphotyrosine primary antibodies. Membranes were incubated with primary antibodies
in blocking solution at 4°C overnight. Following 3 washes with PBS containing either 5%
milk or 0.05% Tween (for anti-phosphotyrosine Western blots), membranes were incubated
with peroxidase-conjugated secondary antibodies at 25°C for 45 min. After thorough washing,

33
proteins were detected by enhanced chemiluminescence (Amersham). For protein interaction
studies in mammary epithelial cells, EpH4 cells were serum-starved for 1 day, stimulated with
2 ng/ml neuregulin-β1 (R&D Systems) at 37°C for 10 min and lysed as for 293T cells.
Immunoprecipitation followed by SDS-PAGE and Western blotting was performed as above.
For detection of Vav2/ErbB-2 complexes in mammary tissue, mammary glands from 17.5-day
pregnant mice were solubilized in lysis buffer and incubated with anti-Vav2 antibodies at 4°C
for 7 h. Co-immunoprecipitated ErbB-2 was detected as described above.
Antibodies used for immunoprecipitation and Western blotting were: anti-Flag (affinity beads,
Sigma), anti-Flag Octapeptide (Zymed), anti-Vav2 (DPH, Calbiochem), anti-ErbB-2 (Ab-8,
NeoMarkers) and anti-PY antibodies (PY20, Transduction Laboratories). Peroxidase-
conjugated secondary antibodies were: goat anti-rabbit (Calbiochem), goat anti-mouse
(Jackson ImmunoResearch) and rabbit anti-sheep, Upstate Biotechnology). Protein G
Sepharose 4 FAST FLOW (Amersham Pharmacia Biotech) was used for immuno-
precipitation.
2.10 Matrigel Assays
EpH4 cells were transfected with Flag-tagged ∆N-Vav2, Flag-tagged ∆N-Vav2 mutants, HA-
tagged TrkErbB2-Vav2 or TrkErbB2-∆tail expression vectors by standard calcium/phosphate
method and selected for neomycin resistance in the presence of 800 µg/ml G418. Individual
clones stably expressing the exogenous proteins were expanded and tested for morphogenesis
on Matrigel (basement membrane from Engelbreth-Holm-Swarm murine tumor, Sigma;
Niemann et al., 1998). 24-well plates were cooled on ice, and each well was coated with 70
µl Matrigel solution. Plates were incubated at 37°C for 30-60 min until Matrigel solidified.
EpH4 cells were plated dropwise as a suspension containing approximately 300-500 cell
clusters/ml in DMEM supplemented with 10% FCS and the following hormones: bovine
prolactin (at 3 µg/ml; Sigma), hydrocortisone (at 1 µg/ml; Merck) and insulin (at 5 µg/ml;
Sigma). After one day of culture at 37°C, medium was replaced by serum-free DMEM

34
containing hormones. Neuregulin-β1 (at 1 ng/ml) or NGF (at 100 ng/ml) were added to
medium, which was changed daily. Assays were terminated after 5 days of culture. For
branching morphogenesis assays, 24-well plates were coated with a 5 mm-thick layer of
collagen; then, matrigel was added on top and cells were plated as above. Assays were
performed in the presence of DMEM medium supplemented with 10% FCS, hormones and
HGF/SF (at 10 units/ml).
2.11 Light and Electron Microscopy
For light and electron microscopy studies, matrigel specimens were fixed with 2.5 %
glutaraldehyde in 0.1 M phosphate buffer and 0.18 M sucrose at 4°C for 2 days, postfixed
with 1% OsO4 in 0.1 M phosphate buffer for 2 h, dehydrated in a graded ethanol series and
embedded in Poly/BedR 812 (Polysciences, Inc). For light microscopy, semithin section (1
µm) were stained with toluidine blue and analyzed in a Zeiss Axioplan II imaging microscope.
Ultrathin sections (70 nm) were contrasted with 2% uranyl acetate and lead citrate (Merck)
and analyzed in a Philips EM 400T electron microscope.
2.12 In Situ Hybridization Analysis
For in situ hybridization (Hülsken et al., 2001), sections of third thoracic glands of 16-day
pregnant mice were used. Digoxygenin-labelled RNA probes were synthesized with T3 or T7
RNA polymerase using DIG RNA kit (Boehringer Mannheim). Probes were: neuregulin,
nucleotides 391-1458 (Yang et al., 1995); ErbB-2, nucleotides 1746-3780; Vav2, nucleotides
1-2232 plus 504 nucleotides 5´ from start codon. Pyrogenin was used for red counterstaining.

35
3 RESULTS
3.1 Yeast Two-Hybrid Screen with ErbB-2 Bait Proteins
Previous studies implicate the receptor tyrosine kinase ErbB-2 in lobulo-alveolar development
of the mammary gland (Niemann et al., 1998; Jones and Stern, 1999). In search for new
substrates of ErbB-2 that mediate its function in mammary alveolar morphogenesis, yeast
two-hybrid screens were performed with ErbB-2 baits (Fields and Song, 1989). The classical
baits for receptor tyrosine kinases are hybrid proteins, which consist of an E.coli LexA DNA-
binding and dimerization domain followed by the intracellular region of the receptor tyrosine
kinase (O'Neill et al., 1994; Weidner et al., 1996). However, baits spanning the full-length
cytoplasmic region of the ErbB-2 receptor were highly unstable in the yeast (my own
observation; data not shown). Therefore, modified yeast baits were conceived on the basis of
previous studies with chimeric receptors; when C-terminal sequences of c-Met are fused to
the kinase of TrkA (the nerve growth factor receptor), the hybrid receptor elicits Met-specific
morphogenic responses upon activation with NGF (Sachs et al., 1996). Thus, the C-terminal
docking region of a receptor is sufficient to determine its signaling specificity; this region
contains tyrosine residues that, when phosphorylated, become docking sites for receptor-
specific downstream effectors. In view of these results, chimeric ErbB-2 bait proteins were
generated: the tyrosine kinase TprMet was fused to LexA, and the C-terminal tail of Met was
replaced by the C-terminal tail of ErbB-2 (Figure 1). TprMet was chosen as heterologous bait
kinase since it is strongly active when expressed in the yeast (my own observation; data not
shown). TprMet is the human oncogenic counterpart of c-Met (Park et al., 1986). Tpr
(translocated promoter region) codes for two putative dimerization domains, and is a movable
DNA element that has spontaneously translocated into a genomic region upstream of the Met
kinase; the resulting TprMet protein dimerizes in a ligand-independent fashion through its
newly-acquired Tpr sequences, and is therefore a cytoplasmic, constitutively active tyrosine
kinase protein with the biological activity of c-Met. It is therefore likely that the presence of a
C-terminal tail of ErbB-2 in the modified TprMet-ErbB2 baits bestows ErbB-2 signaling

36
properties on the chimeric proteins (see below). Two ErbB-2 baits, TprMet-ErbB2(Y1-4) and
TprMet-ErbB2(Y3-5), were generated to cover the five tyrosine residues that are putative
autophosphorylation sites of ErbB-2 (Fig. 1).
Figure 1. Structure of the chimeric ErbB-2 baits used in yeast two-hybrid screens.
The baits consist of a LexA DNA-binding and dimerization region, followed by the kinase domain of TprMet;
the C-terminal tail of TprMet is substituted for sequences of the ErbB-2 multiple docking region. The TprMet-
ErbB2(Y1-4) bait stretches over tyrosines Y1028 (for simplicity Y1 in the scheme), Y1144 (Y2), Y1201 (Y3)
and Y1226/7 (Y4) of ErbB-2, with Y1253 (Y5) being mutated to F; the TprMet-ErbB2(Y3-5) bait includes
tyrosines Y1201 (Y3), Y1226/7 (Y4) and Y1253 (Y5).
TprMet-ErbB2(Y1-4) contains the four N-terminal tyrosines of c-ErbB-2 (Y1 to Y4); TprMet-
ErbB2(Y3-5) includes the three most C-terminal tyrosines (Y3 to Y5). Both baits were
efficiently expressed in the yeast and were constitutively phosphorylated on tyrosine residues
(data not shown). In preliminary tests, these baits exhibited ErbB-2 selectivity for binding
downstream signaling molecules: TprMet-ErbB2(Y1-4) interacted with Grb-2 and Shc, while
TprMet-ErbB2(Y3-5) interacted with Shc (data not shown); it has already been described that
Grb-2 and Shc directly interact with phosphorylated Y2 and Y4 residues of ErbB-2,
respectively (Dankort et al., 1997).
Both chimeric ErbB-2 baits were used to screen the Hollenberg library (see Materials and
Methods). The library consists of mouse E10.5 embryonic cDNAs that are fused to the VP16
activation domain, and has extensively been used in our group to find novel interaction

37
partners of receptor tyrosine kinases (Weidner et al., 1996; Grimm et al., 2001). In several
independent screens, clones encoding the SH2 domains of known and novel interaction
partners of ErbB-2 were isolated. Table 1 summarizes the results of screens for each ErbB-2
bait protein. Fig. 2 indicates the amino acid region encoded in the interacting clones. Src and
PLCγ1 have previously been described as substrates of c-ErbB-2 (Fazioli et al., 1991;
Muthuswamy and Muller, 1995b); their isolation in the yeast screen corroborates ErbB-2
binding specificity of the chimeric baits. Vav2, Nck and Grb10 were here identified as novel
interaction partners of the ErbB-2 receptor.
Table 1. Interaction partners of ErbB-2 isolated in independent yeast two-hybrid
screens using different baits.
Clone TprMet-ErbB2(Y1-4) TprMet-ErbB2(Y3-5)
Vav2 2 1
PLCγ1 6 7
Nck - 1
Src - 2
Grb10 - 34
Numbers indicate frequency of each isolated interaction partner, including overlapping
clones.
Interaction of the prey proteins with the baits was confirmed by co-transfection into yeast
cells, followed by analysis of yeast growth and β-galactosidase activity assays (data not
shown). Sequence comparison analyses revealed that the interacting regions spanned the SH2
domain of each ErbB-2 partner (Fig. 2), strongly suggesting that interaction involved
phosphotyrosine residues of the baits. This hypothesis was confirmed by using kinase-
defective baits; a point mutation of lysine 243 to alanine (K243A) was introduced in the ATP-

38
binding site of the TprMet kinase, which completely disrupts the catalytic activity (Rodrigues
and Park, 1993). The inactivating mutation of the bait kinases abolished interaction,
confirming that the isolated SH2 domains bound to phosphotyrosine residues (Fig. 3, middle
panel; compare to interaction with wild-type bait, upper panel). Furthermore, deletion of the
ErbB-2 C-terminal tail also impaired binding (Fig. 3, lower panel), clearly indicating that the
clones interacted with phosphotyrosines that were located on the ErbB-2 tail and not within
the TprMet sequences of the baits.
Figure 2. Interaction partners of ErbB-2 in the yeast system are SH2 domains.

39
The overall structure of the interacting proteins is schematically shown, relevant domains are highlighted. The
amino acid stretch encoded in the isolated clones is indicated by the bars below the protein structures,
overlapping clones are shown. AD: acidic domain; CH: calponin-homology domain; DH: Dbl-homology domain;
PH: pleckstrin-homology domain; PR: proline-rich region; SH2: Src-homology-2 domain; SH3: Src-homology-3
domain; ZF: zinc finger.
Figure 3. The SH2 domains of substrates isolated in yeast two-hybrid screens interact with
phosphotyrosine residues of the ErbB-2 C-terminal tail.
Preys were co-transfected into yeast together with wild-type TprMet-ErbB2(Y3-5) (TprMet-ErbB2 for
simplicity), kinase-defective (TprMet-ErbB2-Kin-) or C-terminally deleted (TprMet-∆tail) baits (schematic
structures are on the right), and interaction was tested in a yeast colony-growth assay on selection plates. The
various preys interacted with the wild-type TprMet-ErbB2 bait, but not with the kinase-defective or the deletion
mutant without ErbB-2 C-terminal tail.
3.2 Distinct Phosphotyrosine Residues of ErbB-2 Bind to Various Interaction Partners
The phosphotyrosine residues of ErbB-2 that are direct binding sites for its interaction
partners were mapped by mutational analysis of the bait proteins. Sequences of the C-terminal
tail of ErbB-2 containing single tyrosines or pairs of consecutive tyrosine residues were fused
to TprMet to generate bait deletion mutants (see Materials and Methods); these new baits
were tested with the library clones for interaction in the yeast, which was evaluated by colony-
growth on selection plates and β-galactosidase activity. Grb-2 and Shc were included in these

40
experiments as controls. The results of yeast colony-growth and β-galactosidase assays are
summarized in Table 2. Nck interacted with baits containing Y3 (Y1201) and Grb10 with
baits containing Y4 (Y1226/7); Src required simultaneously both phosphorylated Y3 and Y4
(Y1201 and Y1226/7); PLCγ1 interacted with every tyrosine-phosphorylated bait mutant (data
not shown). Vav2 directly bound to phosphorylated Y1 (Y1028) and Y3 (Y1201) of ErbB-2
(see also Fig. 4). Interestingly, both tyrosines that bound Vav2 are located within a pYLVP
motif, which apparently constitutes the consensus binding sequence for the SH2 domain of
Vav2. These results again validate the reliability of this modified yeast two-hybrid approach
to identify phosphotyrosine binding proteins and to map their direct binding motifs.
Table 2. Characterization of specific binding sites of ErbB-2 for its
interaction partners.
Bait* Vav2 Nck Src Grb10 Grb2 Shc
Y1 + - - - - -
Y1-2 + - - - + -
Y2 - - - - + -
Y2-3 + + - - + -
Y3 + + - - - -
Y3-4 + + + + - +
Y3-5§ + + - + - +
Y4-5 - - - + - +
Y4 - - - + - +
Y5 - - - - - +
Plus (+) indicates interaction, minus (-) indicates no interaction.

41
*Baits are TprMet-ErbB2 deletion mutants; numbers indicate single or pairs
of tyrosine residues in the C-terminal tail of ErbB-2.
§TprMet-ErbB2(Y3-5) bait was included as internal control.
Figure 4. Vav2 directly interacts with phosphotyrosines Y1 and Y3 of c-ErbB-2.
The full-size cDNA of Vav2 was isolated from a phage library (see Materials and Methods) and inserted in VP16
yeast vector. To characterize the Vav2 binding sites on c-ErbB-2, the full-length Vav2 prey protein was tested
with various ErbB-2 bait mutants for interaction in the yeast. The results from colony-growth (left panel) and β-
galactosidase assays (right panel) are shown. ErbB-2 bait mutants are those from Table 2; additionally, Y2 and
Y3 are mutated to phenylalanine in the TprMet-ErbB2(Y2F-3) and TprMet-ErbB2(Y2-3F) baits, respectively.
TprMet-∆tail lacks an ErbB-2 C-terminal tail. Kin denotes kinase-deficient bait mutants. Hybrid TprMet bait
proteins for the other ErbB receptors (EGF receptor, ErbB-3 and ErbB-4) were simultaneously tested (see
below).
3.3 Vav2 Interacts with Receptor Tyrosine Kinases of the ErbB Subfamily
Next, the receptor specificity of each ErbB-2 interaction partner for other RTKs was tested in
the yeast system. Src, Nck, PLCγ1 and Grb10 interacted with various receptor tyrosine kinases
from unrelated subfamilies (Table 3). In contrast, Vav2 appears to preferentially bind
receptors of the ErbB subfamily; Vav2 bound to baits encoding TprMet fusion proteins with

42
the C-terminal tails of EGF receptor, ErbB-3 and ErbB-4 (Table 3 and Fig. 4); it additionally
interacted with a constitutively active PDGFβ receptor (Table 3).
Table 3. Interaction of the various ErbB-2 partners with different receptor tyrosine
kinases.
Receptor* PLCγγγγ1 Src Nck Grb10 Vav2
TprMet-ErbB2 + + + + +
TprMet-EGFR + + + + +
TprMet-ErbB3 - + + + +
TprMet-ErbB4 - - - + +
TprMet-∆tail - - - - -
Met + + + + -
Insulin Receptor + - + + -
Ret + + - + -
Kit - - - - -
KGF Receptor - - - - -
Ros + + - + -
Sea + - - - -
PDGFβ Receptor + + + + +
Plus (+) indicates interaction.
*Baits are LexA-receptor fusion proteins (Weidner et al., 1996). Baits containing TprMet
sequences are indicated. EGFR: epidermal growth factor receptor; KGF: keratinocyte growth
factor; PDGF: platelet-derived growth factor.

43
Together, these results point to Vav2 as a putative candidate to mediate ErbB signals in
mammary gland development. First, Vav2 specifically interacts with all the ErbB receptors;
second, all ErbB receptors are essential at different stages of mammary development (see
section 2.5 of the Introduction). Lastly, activated Vav2 induces cytoskeletal rearrangements,
which are a critical step for morphogenic events. Therefore, the potential of Vav2 to mediate
ErbB-2 signals in alveolar morphogenesis was next tested in organotypic cell culture
experiments.
3.4 Vav2 Induces Alveolar Morphogenesis of EpH4 Mammary Epithelial Cells
Activation of endogenous or ectopic ErbB-2 receptor instructs EpH4 mammary epithelial cells
to form functional alveoli-like structures when these cells are cultured on EHS matrix (termed
Matrigel; Niemann et al., 1998). To examine whether Vav2 mediates the morphogenic effect
of ErbB-2 in organotypic culture, EpH4 mammary epithelial cells were stably transfected with
a cDNA encoding N-terminally truncated Vav2, the oncogenic, constitutively active form of
Vav2 (termed ∆N-Vav2; structures are shown in Fig. 5; see Schuebel et al., 1996); afterwards,
transfectants were cultured on Matrigel to test their ability to undergo alveolar morphogenesis.
∆N-Vav2 was preferred to full-size Vav2 for the Matrigel assay in view of its functional
properties: ∆N-Vav2 is constitutively active as guanine nucleotide exchange factor and
induces cytoskeletal rearrangements when overexpressed in fibroblasts (Schuebel et al.,
1996); in contrast, full-size Vav2 requires tyrosine phosphorylation to elicit such responses
(Schuebel et al., 1998). Remarkably, overexpression of ∆N-Vav2 induced the arrangement of
the transfected EpH4 cells into large alveolar structures within two days of culture on Matrigel
(Fig. 5B, compare with control in A). The alveoli consisted of monolayers of tightly
associated cells facing a lumen (Fig. 5D). Ultrastructural studies revealed that the alveolar
cells were polarized; microvilli were scattered at the luminal side, and tight junctions were
observed at apical cell-cell borders (Fig. 6B, C). Polarization of alveolar cells in the mammary
gland during pregnancy and lactation has been described (Nemanic et al., 1971). Control
44
EpH4 cells formed small solid aggregates, and cells were non-polarized and were only loosely
connected (Fig. 6A).
Figure 5. Constitutively active Vav2 induces alveolar morphogenesis of EpH4 mammary epithelial cells.
(B) Cells expressing ∆N-Vav2 (see schematic structures below) form large alveoli-like structures on matrigel.
(D) Alveolar cells are facing a lumen (asterisk), as observed in semithin sections. (A, C) Mock-transfected
controls form small aggregates. Bar: 50 µm. CH: calponin-homology domain; AD: acidic domain; DH: Dbl-
homology domain; PH: pleckstrin-homology domain; ZF: zinc finger.

45
Figure 6. Constitutively active Vav2 induces polarization of alveolar EpH4 mammary epithelial cells.
(B, C) Microvilli (arrows) are present at the luminal side (asterisk) of alveolar EpH4 cells expressing ∆N-Vav2;
apical tight junctions (arrowheads) are revealed by electron-microscopy of ultrathin sections. (A) Mock-
transfected controls form small clumps of loosely associated cells. Bars: (A, B) 5 µm; (C) 0.5 µm. ECM:
extracellular matrix.
Full-size Vav2 is inactive as GDP/GTP exchanger in a non-phosphorylated state (Schuebel et
al., 1998). Not surprisingly, EpH4 cells overexpressing full-size Vav2 did not produce alveoli
(data not shown), suggesting that full-size Vav2 may also require tyrosine-phosphorylation to
become morphogenic. It has previously been shown that C-terminal fusion of a substrate to its
receptor results in efficient tyrosine phosphorylation of the substrate (Schaeper et al., 2000);
moreover, the insulin receptor substrate sequences are fused to the insulin receptor in D.
melanogaster (Yenush et al., 1996). Therefore, the fusion protein TrkErbB2-Vav2 was
generated by replacing the C-terminal tail of ErbB-2 with full-length Vav2 in a TrkErbB2
receptor (Sachs et al., 1996; structures are shown in Fig. 7). The resulting protein lacks the
docking sites of ErbB-2 and hence could only signal through the coupled substrate when
activated by nerve growth factor (NGF). Indeed, EpH4 cells expressing TrkErbB2-Vav2
formed large alveoli following stimulation with NGF (Fig. 7C, D). In the absence of NGF or
in transfectants lacking Vav2 in the fusion protein, respectively, none or only rudimentary
structures were seen (Fig. 7A, B and E, F). Taken together, these findings demonstrate that
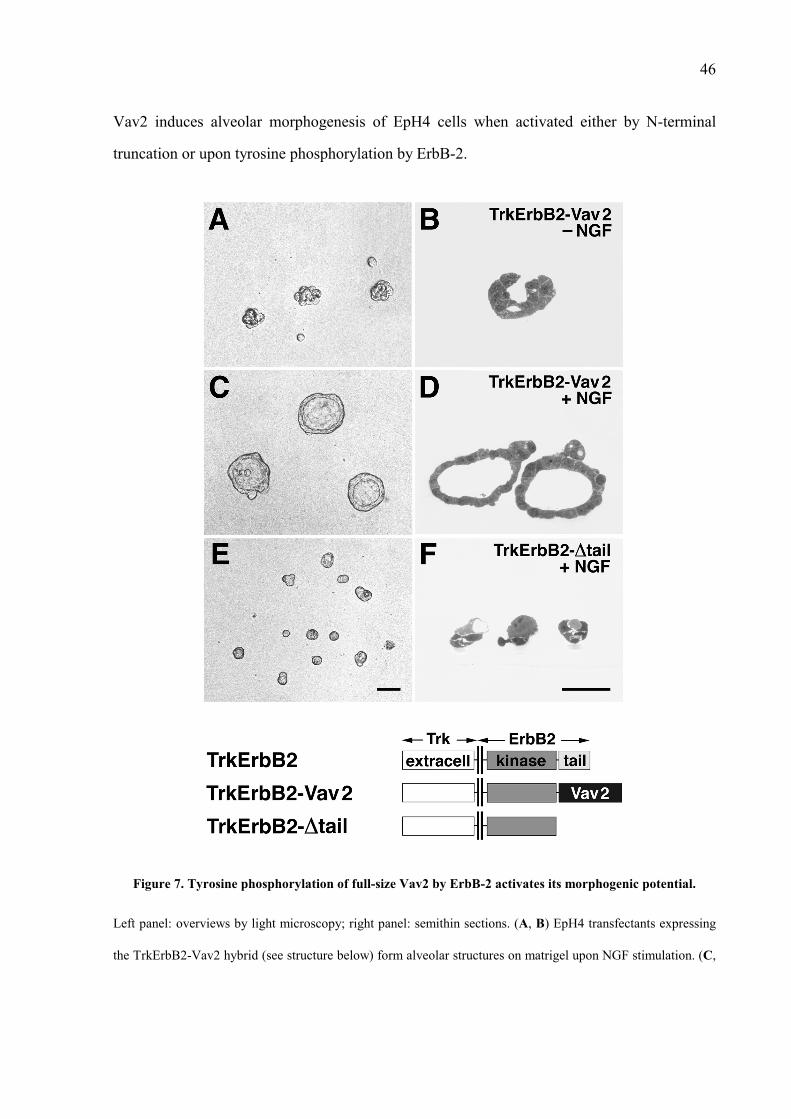
46
Vav2 induces alveolar morphogenesis of EpH4 cells when activated either by N-terminal
truncation or upon tyrosine phosphorylation by ErbB-2.
Figure 7. Tyrosine phosphorylation of full-size Vav2 by ErbB-2 activates its morphogenic potential.
Left panel: overviews by light microscopy; right panel: semithin sections. (A, B) EpH4 transfectants expressing
the TrkErbB2-Vav2 hybrid (see structure below) form alveolar structures on matrigel upon NGF stimulation. (C,

47
D) Non-stimulated transfectants and (E, F) cells expressing a TrkErbB2 protein without the C-terminal tail of
ErbB-2 (TrkErbB2-∆tail) lack morphogenic activity. Bars: 50 µm.
3.5 Vav2 and ErbB-2 Can Both Directly and Indirectly Associate in Mammalian Cells
The interaction between Vav2 and ErbB-2 was also studied in cultured mammalian cells.
Human epithelial kidney 293T cells were transiently co-transfected with Vav2 and wild-type
or mutated TrkErbB2 receptors, and interaction after stimulation with NGF was examined by
Western blotting. Exogenous Vav2 co-immunoprecipitated with a TrkErbB2 hybrid receptor
in lysates of transfected 293T cells, but not with kinase-deficient receptor TrkErbB2-Kin-
(Fig. 8A, upper panel). These results indicate that association of Vav2 with ErbB-2 requires
tyrosine phosphorylation of the receptor, as previously observed in the yeast system.
Surprisingly, Vav2 still interacted with TrkErbB2mut, which lacks the Vav2 binding sites of
ErbB-2 (Y1028 and Y1201) identified in the yeast two-hybrid system. This suggests that
Vav2 may bind directly and indirectly to ErbB-2. However, interaction of Vav2 with
TrkErbB2 or TrkErbB2mut depends on activation of the hybrid receptors by NGF (Fig. 8A,
upper panel). Increased tyrosine phosphorylation of Vav2 was observed upon binding to
ErbB-2 proteins that contain an activated kinase (Fig. 8A, middle panel).
Interaction between Vav2 and ErbB-2 was also tested in EpH4 cells that are cultured under
standard conditions. Subconfluent EpH4 cells were treated shortly with neuregulin-1 (see
Materials and Methods) to activate the endogenous ErbB-2 receptor (Niemann et al., 1998)
and then lysed. Immunoprecipitation of endogenous Vav2 from cell lysates followed by
Western blotting revealed that Vav2 associated with ErbB-2 in lysates of neuregulin-treated
EpH4 cells (Fig. 8B, upper left panel). In addition, increased tyrosine phosphorylation of
ErbB-2 and Vav2 was observed in lysates of stimulated EpH4 cells (Fig. 8B, upper right
panel). These findings indicate that interaction between Vav2 and ErbB-2 occur in EpH4 cells
after activation of ErbB-2 by neuregulin. Moreover, activated ErbB-2 may further
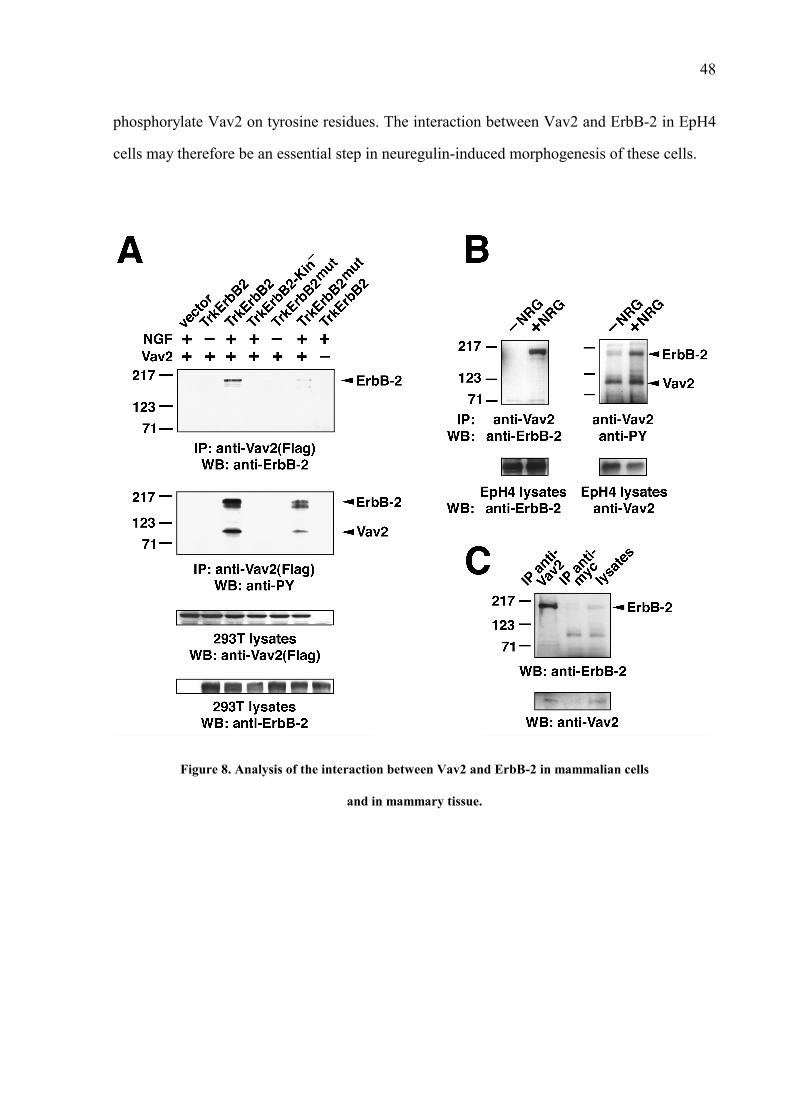
48
phosphorylate Vav2 on tyrosine residues. The interaction between Vav2 and ErbB-2 in EpH4
cells may therefore be an essential step in neuregulin-induced morphogenesis of these cells.
Figure 8. Analysis of the interaction between Vav2 and ErbB-2 in mammalian cells
and in mammary tissue.

49
(A) Co-immunoprecipitation of Vav2 with various TrkErbB2 receptors in transfected 293T cells. (Top) Vav2 co-
immunoprecipitates with TrkErbB2 and TrkErbB2mut, when these hybrid receptors are activated by NGF. Vav2
does not associate with kinase-defective TrkErbB2-Kin-. (Middle) Binding of Vav2 to activated TrkErbB-2
receptors results in increased tyrosine phosphorylation of Vav2. (Bottom) Control immunoblots of total cell
lysates. Molecular weight markers on the left are in kDa. (B) Interaction between Vav2 and ErbB-2 in EpH4
mammary epithelial cells. (Top) Vav2 co-immunoprecipitates with ErbB-2 in lysates of neuregulin-treated cells
(left panel); association of Vav2 with activated ErbB-2 results in increased tyrosine phosphorylation of Vav2
(right panel). (Bottom) Control immunoblots of total cell lysates. (C) In vivo association between Vav2 and
ErbB-2 in mammary glands during pregnancy. Vav2 co-immunoprecipitates with ErbB-2 in mammary gland
lysates from pregnant mice. Anti-c-Myc antibody was used as non-specific control; Vav2 immuno-precipitation
is shown below.
3.6 Vav2 and ErbB-2 Are Associated in Mammary Epithelium During Pregnancy
Interaction between Vav2 and ErbB-2 was also studied in mammary glands from pregnant
mice. Lysis of all mammary glands from a female mouse at day 17.5 of pregnancy was
followed by immunoprecipitation of endogenous Vav2 and Western blotting as above. Indeed,
Vav2 co-immunoprecipitated with ErbB-2 from the mammary lysates (Fig. 8C), suggesting
the presence of endogenous complexes between Vav2 and ErbB-2 in mammary tissue during
pregnancy.
Cellular localization of Vav2 and ErbB-2 was studied in mammary tissue from pregnant mice
by in situ hybridization. Importantly, Vav2 and ErbB-2 are co-expressed in the epithelial cell
layer that lines alveoli and ducts of mammary glands during pregnancy (Fig. 9; Schroeder et
al., 1998). The co-localization of Vav2 and ErbB-2 in mammary epithelium supports the
presence of interaction in vivo during pregnancy, as above suggested by biochemical analysis
(see Fig. 8C); moreover, it rules out the possibility of association during the immuno-
precipitation of Vav2. Taken together, these observations indicate that ErbB-2 and Vav2 can
functionally associate in the mammary gland while lobulo-alveolar development takes place.

50
Figure 9. Vav2 and ErbB-2 are co-expressed in mammary alveolar epithelium during pregnancy.
(Top) Vav2 and ErbB-2 co-localize in alveolar (black arrowheads) and ductal (arrows) epithelia of the mammary
gland at late pregnancy. (Bottom) Neuregulin (NRG) is located in the mesenchyme surrounding alveoli and ducts
(white arrows); a control section (control) was hybridized with a Vav2 sense probe. Bar: 100 µm.
3.7 The Dbl-Homology Domain of Vav2 Is Required for Its Morphogenic Activity in
EpH4 Cells
The structural domains of Vav2 confer the protein two interesting features: first, Vav2 acts as
a GDP/GTP exchange factor for Rho/Rac GTPases via its catalytic Dbl-homology domain;
second, Vav2 has the capability to participate in signal transduction cascades by binding
further downstream proteins via its C-terminal SH2 and SH3 domains (Bustelo, 2000). To
define the functional role of the various domains of Vav2 in alveolar morphogenesis,
mutational analyses of constitutively active Vav2 were performed. ∆N-Vav2-∆C encodes an
active protein which lacks SH2 and both SH3 domains (structures are shown in Fig. 10). ∆N-
Vav2-dblmut lacks the GTP/GDP exchange activity due to a deletion of seven conserved
amino acids within the Dbl homology domain (336LLLKELL342), which has been reported
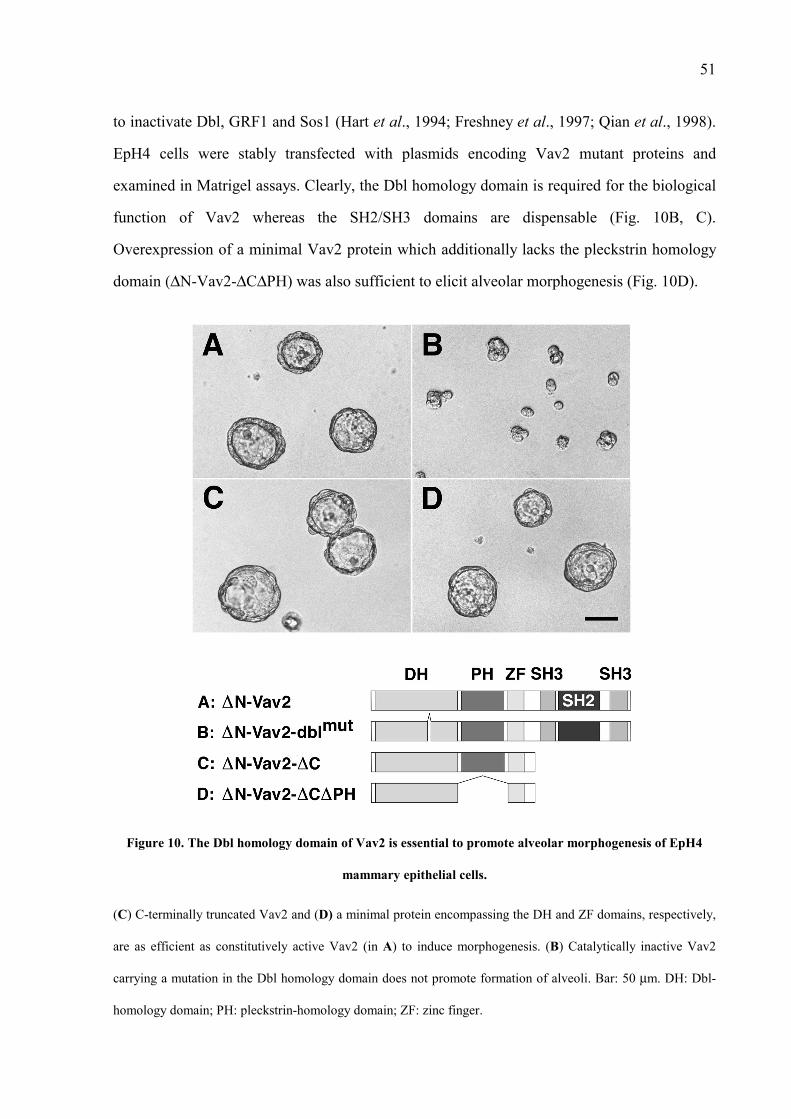
51
to inactivate Dbl, GRF1 and Sos1 (Hart et al., 1994; Freshney et al., 1997; Qian et al., 1998).
EpH4 cells were stably transfected with plasmids encoding Vav2 mutant proteins and
examined in Matrigel assays. Clearly, the Dbl homology domain is required for the biological
function of Vav2 whereas the SH2/SH3 domains are dispensable (Fig. 10B, C).
Overexpression of a minimal Vav2 protein which additionally lacks the pleckstrin homology
domain (∆N-Vav2-∆C∆PH) was also sufficient to elicit alveolar morphogenesis (Fig. 10D).
Figure 10. The Dbl homology domain of Vav2 is essential to promote alveolar morphogenesis of EpH4
mammary epithelial cells.
(C) C-terminally truncated Vav2 and (D) a minimal protein encompassing the DH and ZF domains, respectively,
are as efficient as constitutively active Vav2 (in A) to induce morphogenesis. (B) Catalytically inactive Vav2
carrying a mutation in the Dbl homology domain does not promote formation of alveoli. Bar: 50 µm. DH: Dbl-
homology domain; PH: pleckstrin-homology domain; ZF: zinc finger.

52
3.8 Catalytically Inactive Vav2 Blocks Neuregulin-Mediated Morphogenesis of EpH4
Cells
EpH4 cells that express catalytically inactive Vav2 (∆N-Vav2-dblmut) were tested for
alveolar morphogenesis following neuregulin treatment. Importantly, ∆N-Vav2-dblmut
interferred with the formation of alveoli when cells were treated with neuregulin: instead of
large hollow structures (Fig. 11B, C), cell aggregates without lumina were observed (Fig. 11E,
F), indicating that alveolar morphogenesis is prevented while cell growth still occurs. Only
small cell groups were formed in absence of neuregulin treatment (Fig. 11A, D).
Figure 11. Catalytically inactive Vav2 harbouring a mutation in the Dbl homology domain blocks the
morphogenic signals of neuregulin in EpH4 mammary epithelial cells.
(E, F) EpH4 cells expressing ∆N-Vav2-dblmut (see structure below) do not form hollow alveoli but rather large
cell aggregates upon stimulation with neuregulin. (B, C) Mock-transfected cells form alveoli-like structures in

53
response to neuregulin, as previously described (Niemann et al., 1998). (A, D) Non-stimulated controls. Bars: 50
µm. DH: Dbl-homology domain; PH: pleckstrin-homology domain; ZF: zinc finger.
Interestingly, ∆N-Vav2-dblmut did not interfere with tubular branching when EpH4 cells were
treated with hepatocyte growth factor/scatter factor (Fig. 12D, compare to control cells in B;
Niemann et al., 1998). Thus, catalytically inactive Vav2 blocks neuregulin-specific signals for
alveolar morphogenesis of EpH4 cells, while it does not affect cellular responses to other
morphogenic stimuli.
Figure 12. Catalytically inactive Vav2 does not interfere with branching morphogenesis
of EpH4 mammary epithelial cells.
(D) EpH4 cells expressing ∆N-Vav2-dblmut (see structure in Fig. 11) form tubular structures upon stimulation
with HGF/SF. (B) Mock-transfected cells exhibit branching morphogenesis in response to HGF/SF, as previously
described (Niemann et al., 1998). (A, C) Non-stimulated controls. Bars: 50 µm. DH: Dbl-homology domain; PH:
pleckstrin-homology domain; ZF: zinc finger.

54
4 DISCUSSION
Previous results from our and other groups suggest that neuregulin activates the receptor c-
ErbB-2 to promote lobulo-alveolar development of the mammary gland (Yang et al., 1995;
Jones et al., 1996; Niemann et al., 1998; Jones and Stern, 1999). However, little was known
about the intracellular effectors that mediate this morphogenic effect. Here, a modified yeast
two-hybrid screen was developed to search for new substrates of ErbB-2 that are involved in
alveolar morphogenesis of the mammary gland. The guanine nucleotide exchange factor Vav2
was identified among other proteins as a novel interaction partner of ErbB-2. A full
characterization of the ErbB-2 docking sites for these interacting proteins was performed in
the yeast system, thus adding complexity to the previous knowledge on intracellular effectors
of ErbB-2. Next, the potential function of Vav2 in ErbB2-induced morphogenesis was tested
in EpH4 mammary gland epithelial cells that were cultured on a reconstituted basement
membrane (Matrigel). These studies show two main points: first, Vav2 indeed promotes
alveoli-like growth and reorganization of EpH4 cells, either when constitutively active or
upon activation by ErbB-2; second, co-expression of Vav2 and ErbB-2 in mammary alveolar
epithelium is observed in pregnant mice. These findings suggest a functional association
between Vav2 and ErbB-2 during alveolar morphogenesis in vivo. Together, this work
supports a model for neuregulin-induced mammary alveolar morphogenesis in vivo, in which
Vav2 is a physiological target downstream of ErbB-2 that plays a crucial role in transduction
of signals for lobulo-alveolar morphogenesis.
4.1 Modified Yeast Two-Hybrid System: A Powerful Tool to Search for
Phosphotyrosine-Interacting Proteins
The commonly-used yeast two-hybrid baits to search for interaction partners of receptor
tyrosine kinases are hybrid proteins, which consist of a LexA DNA-binding and dimerization
sequence fused to the intracellular region of the receptors (O'Neill et al., 1994; Weidner et al.,
1996; Grimm et al., 2001). Bait molecules dimerize via LexA when expressed in yeast cells,

55
thus enabling the activation of receptor tyrosine kinases without requirement of a ligand. This
approach could not be used in case of the receptor ErbB-2, since the resulting hybrid proteins
were rapidly degraded in the yeast. Therefore, modified yeast baits were generated: C-terminal
sequences of ErbB-2 containing the autophosphorylation sites were fused to an oncogenic
TprMet kinase in order to obtain constitutively active TprMet-ErbB2 baits. TprMet encodes
the intracellular region of c-Met in frame with a leucin-zipper dimerization domain derived
from a different locus (Park et al., 1986). The heterologous TprMet kinase efficiently
phosphorylates tyrosine residues of ErbB-2, thus creating docking sites for signaling proteins
with SH2 and PTB domains. By this means, known and novel interacting proteins for ErbB-2
were identified in a cDNA library screen in yeast.
This modified yeast two-hybrid system has proven to be useful for standard search of proteins
that interact with peptides and proteins containing phosphorylated tyrosine residues; even
unrelated protein sequences become tyrosine-phosphorylated when fused to the TprMet kinase
and can be used as yeast baits. For example, coupling Gab1 to TprMet enabled the analysis of
the interaction between phosphorylated Gab1 and the SH2 domains of PI-3-K, PLCγ, Shp2
and CRKL (Schaeper et al., 2000). Thus, the new method presented here offers an easy
alternative for yeast screens as opposed to a tribrid approach, which includes an additional
kinase (Licitra and Liu, 1996). The main advantage of the here described modified two-hybrid
over a tribrid system is that only two selectable markers instead of three markers are
necessary; therefore, the efficiency of yeast transformation is improved, and protein
expression levels in the yeast are higher than those of the tribrid method.
4.2 New Insights into the Intracellular Signaling Pathways of ErbB-2
In independent cDNA library screens with tyrosine phosphorylated ErbB-2 baits, novel
ErbB2-interacting proteins like Nck, Grb-10 and Vav2 were identified, along with the known
partners Src and PLCγ1 (Muthuswamy and Muller, 1995b; Fazioli et al., 1991). These
proteins belong to the group of multiadaptor signaling molecules; the most conspicuous

56
feature of such proteins is the presence of several specialized regions that allow protein-
protein interactions (reviewed in Pawson and Scott, 1997). The ErbB-2 partners identified in
the present workt contain SH2 domains; indeed, these SH2 domains were always present in
the interacting clones and therefore mediate the association with phosphotyrosine residues of
ErbB-2. The direct binding sites of ErbB-2 for these proteins were mapped by mutational
analysis of the baits in the yeast system. So far, tyrosines Y2 and Y4 of the ErbB-2
multidocking site were known to directly bind Grb-2 and Shc, respectively (Ricci et al., 1995;
Dankort et al., 1997). Here, direct binding of Vav2 to Y1 and Y3, Grb-10 to Y4, Nck to Y3
and Src to complexed Y3-Y4 were characterized. The overlap of Vav2, Nck and Src for
binding to Y3 suggests that these proteins may represent alternative effectors of ErbB-2.
Alternatively, these proteins may assemble to form a hierarchy of different protein
multicomplexes; any of such supramolecular complexes may lead to a unique outcoming
signal, regardless of the protein-protein interaction pattern within the complexes. The
formation of functionally redundant protein alignments between Src, Nck and Vav2 (possibly
together with some other proteins) is indeed conceivable. Preliminary data indicate that Vav2
can directly bind Src or Nck; using the modified yeast two-hybrid method, a TprMet-Vav2
fusion bait was recently tested with several SH2 domains for interaction in the yeast. Indeed,
tyrosine-phosphorylated Vav2 interacted with the SH2 domains of Src and Nck but not with
that of Grb2 (data not shown). A mechanism whereby Vav2 is ultimately recruited and
activated via different protein-protein associations may account for the direct and indirect
binding of Vav2 to ErbB-2 (see also below).
4.3 Vav2 Couples to Receptor Tyrosine Kinases via Its SH2 Domain
Direct association of the SH2 domain of Vav2 with ErbB-2 involves recognition of the motif
pYLVP, present at Y1 (Y1028) and Y3 (Y1201). Using an entirely different selection
procedure, the sequence pYXEP (where X is L, M or E) has been identified to bind the SH2
domain of Vav (Songyang et al., 1994). The SH2 domains of Vav and Vav2 share 55%
identity (Schuebel et al., 1996), and it is therefore possible that they have distinct binding

57
preferences. From all the ErbB-2 partners that were found in the yeast screens, only Vav2
showed a particular affinity for binding to the various ErbB receptors. Vav2 also interacts
with the PDGFβ receptor the yeast system (Table 3). In fact, recent biochemical and mass
spectrometry analyses have shown that all three Vav proteins can form complexes with EGF
or PDGFβ receptors and become phosphorylated on tyrosine residues following stimulation
with EGF or PDGF (Bustelo et al., 1992; Margolis et al., 1992; Lopez-Lago et al., 2000;
Moores et al., 2000; Pandey et al., 2000). However, it was not clear from these studies
whether Vav proteins couple directly to the receptors or indirectly via binding to adaptor
proteins. The results from yeast two-hybrid analyses that are here presented do not rule out the
possibility of indirect interaction (see below); however, they provide the first line of evidence
that indeed Vav2 can directly bind to receptor tyrosine kinases via its SH2 domain, a
mechanism that may be shared by the other Vav proteins.
The above identified pYLVP consensus motif for binding to Vav2 is conserved in the C-
terminal region of ErB-4 (Y1022) but not in the EGF receptor, ErbB-3 or the
PDGFβ receptor; however, the rather similar sequences pYLIP in the EGF receptor (at
Y1012), pYLMP in ErbB-3 (at Y1159) and pYIIP in PDGFβ receptor (at Y1021) may be
predicted as Vav2 binding sites. This suggests that positions 1+ and 2+ of the consensus motif
for Vav2 are flexible but, in contrast to the pYXEP binding motif of Vav, they may selectively
be occupied by amino acids with a non-polar side chain.
The examination of ErbB-2/Vav2 interactions in mammalian cells rendered an unexpected
result: in 293T cells, a mutant ErbB-2 receptor that lacks both Y1 and Y3, the direct docking
sites for Vav2 identified in the yeast system, still binds Vav2. This indicates that, in addition
to direct binding, Vav2 may also interact indirectly with ErbB-2 via a further adaptor protein.
Similarly, direct and indirect binding has been reported for recruitment of the adaptor Gab1 to
the receptor tyrosine kinase c-Met (Fixman et al., 1995; Holgado-Madruga et al., 1996;
Weidner et al., 1996; Bardelli et al., 1997; Nguyen et al., 1997; Schaeper et al., 2000). ErbB-2

58
binds to Grb2, Shc, the Csk-homologous kinase, PLCγ1, Nck, Grb10 and Src (Fazioli et al.,
1991; Ricci et al., 1995; Muthuswamy and Muller, 1995; Dankort et al., 1997; Zrihan-Licht
et al., 1998; this work), which may indirectly recruit Vav2 to the receptor. Sequence analysis
of Vav2 reveals the presence of putative consensus motifs for binding the SH2 domains of
Src, Lck (a Src-related tyrosine kinase) and Shc (Songyang et al., 1994). Ongoing studies
suggest that Vav2 may directly interact with the SH2 domains of Src and Nck.
4.4 Vav2 Mediates ErbB-2 Signals for Alveolar Morphogenesis of EpH4 Mammary
Epithelial Cells
Recent work has shown that normal development of the mammary gland during pregnancy to
prepare lactogenesis can be mimicked in vitro. The reorganization of mammary epithelial
cells to form lobulo-alveolar structures is markedly influenced by the matrix on which these
cells are cultured. Barcellos-Hoff et al. (1989) demonstrated that primary mammary epithelial
cells form functional, albeit rudimentary, alveoli-like structures when cultured on a
reconstituted three-dimensional matrix, in the presence of lactogenic hormones and absence of
serum. Within the first days of culture, cells remodel the exogenous basement membrane and
form matrix-ensheated aggregates, which subsequently cavitate; by day 6 of culture, cells are
reorganized into hollow spheres composed of morphologically polarized cells facing a lumen.
These cells are functionally differentiated and secrete milk proteins vectorially into the
luminal compartment. A reconstituted basement membrane alone, however, does not account
for the complex epithelial-mesenchymal interactions that are linked to mammary
development. The spontaneously immortalized, non tumorigenic mouse mammary cell line
IM-2 consists of both epithelial and fibroblastic cell populations (Reichmann et al., 1989); it
has been shown that the fibroblastic cells render the epithelial cells competent to undergo
cytoskeletal rearrangements and to functionally differentiate on Matrigel. The physiological
signals from the mammary mesenchyme can alternatively be supplied by addition of growth
factors to mammary epithelial cells in organotypic culture. Previous studies pointed to
neuregulin as a mesenchymal growth factor that stimulates alveolar morphogenesis of

59
mammary glands in organ culture (Yang et al., 1995). In view of these results, Niemann et al.
(1998) used EpH4 mammary cells (an epithelial subclone derived from the abovementioned
IM-2 cell line) to test the effect of neuregulin in a Matrigel system; EpH4 cells indeed form
large alveoli-like structures when cultured on Matrigel in the presence of neuregulin.
Additionally, biochemical studies revealed tyrosine phosphorylation of endogenous ErbB-2 in
EpH4 cell lysates following neuregulin treatment (Fig. 8). Formation of similar alveolar
structures is observed when EpH4 transfectants that stably overexpress a TrkErbB2 chimeric
receptor are cultured on Matrigel in the presence of nerve growth factor (Niemann et al.,
1998). These results indicate that activation of the overexpressed ErbB-2 receptor is sufficient
to elicit alveolar morphogenesis in organotypic cultures of mammary epithelial cells. Together
with the mesenchymal localization of neuregulin transcripts and the epithelial expression of
ErbB-2 in mammary tissue at mid-pregnancy, these findings strongly suggest that stromal
neuregulin activates epithelial ErbB-2 to induce alveolar morphogenesis of the mammary
gland in vivo. Moreover, this model of neuregulin signaling supports the increasing evidence
of the role of epithelial–mesenchymal interactions in postnatal growth and differentiation of
the mammary gland (reviewed in Cunha and Hom, 1996; Robinson et al., 1999; Silberstein,
2001).
To understand the molecular mechanisms of neuregulin/ErbB signaling in mammary alveolar
morphogenesis, this work aimed at the identification of intracellular effectors of ErbB-2 that
mediate these morphogenic effects. In yeast two-hybrid screens with ErbB-2 baits, Vav2 was
identified as a novel partner of ErbB-2. Vav2 belongs to a family of guanine nucleotide
exchange factors for small GTPases of the Rho superfamily, like Rho, Rac and Cdc42
(reviewed in Bustelo, 2000). Vav proteins contain a common array of domains, which include
an N-terminal regulatory acidic region, a catalytic Dbl-homology domain and several C-
terminal SH2 and SH3 domains (see Fig. 2). Unlike Vav, Vav2 expression is not restricted to
the hematopoietic system, but is also found in several epithelia, for example in the mammary
gland epithelium (Fig. 9). Though recent reports involve Vav2 in immune responses of certain

60
hematopoietic lineages (Billadeau et al., 2000; Doody et al., 2000), a function of Vav2 in
epithelial tissues is also possible. Upon phosphorylation of a single regulatory tyrosine residue
in the N-terminus, Vav proteins gain catalytic activity towards Rho GTPases, thus leading to
changes in the actin cytoskeleton and gene transcription (reviewed in Bustelo, 2000). Vav2
associates not only with ErbB-2 in the yeast system, but also with all other ErbB receptors
(Fig. 4), which are known to be involved at different stages of mammary development (Xie et
al., 1997; Jones and Stern, 1999; Jones et al., 1999). The capability of Vav2 to induce
cytoskeletal reorganization, together with its high affinity for ErbB receptors, favored the
choice of Vav2 as candidate to mediate morphogenic signals of ErbB-2.
The morphogenic potential of Vav2 was here tested in a Matrigel assay. It was known that
truncation of the N-terminal region of Vav proteins eliminates an autoinhibitory loop and
exposes the catalytic site to small GTPases, thus enhancing GDP/GTP exchange (Aghazadeh
et al., 2000). Therefore, EpH4 mammary epithelial cells were stably transfected with a cDNA
encoding an N-truncated, constitutively active Vav2 protein (Schuebel et al., 1996), and
transfectants were cultured on Matrigel under serum-free conditions and in the absence of
growth factors. In this organotypic system, catalytically active Vav2 was sufficient to induce
formation of large alveolar structures that resembled those that are observed following
activation of ErbB-2 by neuregulin (see Niemann et al., 1998). Alveoli consisted of a
monolayer of polarized epithelial cells, which enclose a luminal compartment by means of
apical tight junctions. These results represent the first evidence of a biological response
elicited by overexpression of active Vav2 in epithelial cells. Constitutively active but not
wild-type Vav2 elicited these morphogenic events in EpH4 cells, indicating that GDP/GTP
exchanger activity, and therefore cytoskeletal reorganization, may be essential in these
processes. In line with these findings, it has been reported that truncated forms of all Vav
family members have a deregulated activity as guanine nucleotide exchange factors in vitro,
are highly transforming in focus formation assays, and elicit changes in cell morphology upon
transient transfection into NIH 3T3 fibroblasts (Bustelo, 2000 and references therein).

61
Activation of an overexpressed TrkErbB2-Vav2 fusion protein by nerve growth factor also
induced alveolar morphogenesis of EpH4 transfectants. This finding suggests that the
activated ErbB-2 kinase of the fusion protein efficiently phosphorylated wild-type Vav2.
Phosphorylation may occur on the putative regulatory tyrosine Y172 of Vav2; thus, the
autoinhibitory loop of wild-type Vav2 is disrupted and therefore, phosphorylated full-size
Vav2 can now behave as its oncogenic truncated counterpart to elicit morphogenesis.
Biochemical studies revealed association between Vav2 and ErbB-2 in lysates from EpH4
cells following neuregulin treatment (Fig. 8B, left panel). Activation of ErbB-2 was observed
after stimulation with neuregulin as an increase in phosphotyrosine content, despite high basal
phosphorylation levels (Fig. 8B, right panel). Importantly, endogenous complexes between
Vav2 and ErbB2 were found in lysates of mammary glands from pregnant mice (Fig. 8C). In
addition, in situ hybridization studies showed spatial co-localization of Vav2 and ErbB-2 in
mammary alveolar epithelium (Fig. 9), which allows the physical interaction between the
receptor and its putative effector in the mammary gland during pregnancy.
Taken together, Vav2 can induce alveolar morphogenesis of EpH4 cells, both in its
constitutively active state or when activated by ErbB-2. This places Vav2 as a downstream
effector of ErbB-2 for this biological response. In both cases, alveolar structures were similar
to those that are formed upon neuregulin treatment or activation of ectopic ErbB-2 (Niemann
et al., 1998); along with the functional link between neuregulin and ErbB-2, and with the
physical association of ErbB-2 and Vav2 in mammary tissue, these findings support a relevant
role of Vav2 as a downstream effector of neuregulin/ErbB-2 signals for alveolar
morphogenesis of the mammary gland in vivo. Furthermore, they provide clear evidence of the
as yet suggested function of Vav2 in signaling of receptor tyrosine kinases that lead to a
concrete biological response.

62
4.5 The Morphogenic Activity of Vav2 on Mammary Epithelial Cells May Involve
Changes in Actin Cytoskeleton
Mutational analysis revealed that a functional Dbl-homology domain is necessary and
sufficient for Vav2 to elicit alveolar morphogenesis. The Dbl-homology domain enables Vav2
to function as guanine nucleotide exchange factor towards small GTPases of the Rho family
with an as yet unclear specificity (Schuebel et al., 1998; Abe et al., 2000). Disruption of this
domain in a mutant Vav2 protein completely abolished its ability to promote alveolar
morphogenesis of EpH4 mammary epithelial cells. Moreover, the isolated Dbl-homology
domain of Vav2 still retained morphogenic properties, suggesting that the GDP/GTP
exchange activity is critical in this process, whereas the SH2/SH3 adaptor domains are
dispensable. Similarly, integrity of the Dbl-homology domain in the absence of SH2/SH3
modules is required by Vav3 to promote formation of lamellipodia and membrane ruffling in
transfected NIH3T3 fibroblasts (Movilla et al., 1999). In view of these results, it is evident
that the Dbl-homology domain of Vav proteins is sufficient to reorganize the actin
cytoskeleton, a process that may be required for Vav2-mediated alveolar morphogenesis.
Rho proteins constitute a subgroup of the Ras superfamily of small GTPases that are key
regulators of the actin cytoskeleton and of several cellular processes like gene transcription,
membrane trafficking, growth, movement and morphogenesis (reviewed in Van Aelst and
D’Souza-Chorey, 1997; Hall, 1998; Mackay and Hall, 1998). Rho small GTPases switch from
an inactive GDP-bound state to an active GTP-bound state; this process is accelerated by
guanine nucleotide exchange factors containing a Dbl-homology domain. Activated small
GTPases trigger actin polymerization to form stress fibers, focal adhesion complexes,
lamellipodia, membranes ruffles and filopodia. An increasing amount of genetic studies in
invertebrates provide ample evidence that GEFs and Rho proteins play a central role in several
morphogenic events during the development of a multicellular organism. It has been shown
that the putative GDP/GTP exchanger DRhoGEF2 and the small GTPase DRho1 are part of a
signaling pathway that determines cell shape changes during gastrulation of Drosophila

63
(Barrett et al., 1997; Häcker et al., 1998). The recently described Drosophila Trio GEF or its
C. elegans homolog UNC-73 have been shown to regulate axon guidance and cell migration
in the central nervous system (Steven et al., 1998; Awasaki et al., 2000; Liebl et al., 2000;
Newsome et al., 2000; Bateman et al., 2000). Still life, another Drosophila RhoGEF, has
been identified in a mutational screen for motor activity defects, and its absence determines
reduced locomotion as well as male infertility (Sone et al., 1997). Furthermore, Rho proteins
have also been shown to control dorsal closure at late stages of Drosophila embryogenesis
(Magie et al., 1999), directed cell movement in the Drosophila ovary (Murphy et al., 1999)
and planar polarity in eyes and wing hair (Eaton et al., 1995; Eaton et al., 1996; Strutt et al.,
1997). In mammals, the complexity of the small GTPase superfamily, and therefore the
potential redundancy of its members, has made the study of their role during embryogenesis
difficult. In humans, mutations of the FGD1 (faciogenital dysplasia) gene, which encodes the
Vav-related guanine nucleotide exchanger FGD1, cause an X-linked autosomal
developmental disorder (known as Aarskog-Scott syndrome or faciogenital dysplasia)
involving skeletal and genito-urinary anomalies (Pasteris et al., 1994; Olson et al., 1996;
Zheng et al., 1996). In view of the accumulating evidence of a role for GEFs and Rho proteins
in epithelial development, it is conceivable that Rho small GTPases are downstream effectors
of Vav2 in the mammary gland. Matrigel assays with EpH4 cells overexpressing
constitutively active or dominant negative forms of Rho, Rac and Cdc42 small GTPases may
elucidate their contribution to alveolar morphogenesis.
4.6 Vav2 Is a Specific Effector of Neuregulin Signals for Alveolar Morphogenesis
EpH4 mammary epithelial cells express endogenous c-Met and ErbB receptors and are
competent to elicit biological responses following stimulation by HGF/SF or neuregulin, the
ligands for such receptors (Niemann et al., 1998). EpH4 cells exhibit ductal growth when
cultured on Matrigel in the presence of HGF/SF, while they form alveoli-like structures upon
neuregulin treatment. Therefore, they provide a versatile system to study different
developmental events of the mammary gland in vitro. A Dbl-defective mutant of Vav2 did not

64
affect cell proliferation but completely blocked neuregulin-induced morphogenesis in a
dominant-negative manner (Fig. 11). In contrast, this catalytically-inactive Vav2 mutant did
not interfere with the morphogenic stimulus of HGF/SF for ductal growth (Fig. 12). These
results suggest that Vav2 is specifically involved in the morphogenic but not the mitogenic
signaling cascades of neuregulin/ErbB-2, whereas other intracellular effectors may mediate
the stimulatory effects of HGF/SF and c-Met. It is known that one such effector is the
multiadaptor protein Gab1 (Grb2-associated binder; Holgado-Madruga et al., 1996). Gab1
specifically couples to activated c-Met (Weidner et al., 96; Schaeper et al., 2000), thus
transmitting unique HGF/SF signals for essential developmental processes during embryonic
life; genetic analysis revealed that both Gab1- and c-Met-deficient mice exhibit similar
phenotypes (Bladt et al., 1996; Sachs et al., 2000). In the last years, several multiadaptor
proteins have been described as specific substrates of receptor tyrosine kinases and key
transducers of their biological function. DOS, a Gab1-related multiadaptor protein of
Drosophila, mediates retinal development downstream of the receptor tyrosine kinase
Sevenless (Herbst et al., 1996; Raabe et al., 1996). IRS-1 and IRS-2 are major substrates of
the insulin and insulin-like growth factor receptors (Sun et al., 1991; Sun et al., 1995). Mice
lacking IRS-1 are mildly insulin-resistant due to compensation by IRS-2, but show a dramatic
impairment of glucose uptake in skeletal muscle (Araki et al., 1994; Tamemoto et al., 1994;
Yamauchi et al., 1996; Bruning et al., 1997). Disruption of the IRS-2 gene results in diabetes
(Withers et al., 1998), thus clearly showing distinct and critical roles for IRS-1 and IRS-2 in
different tissues that are targeted by insulin. Recently, novel Dok proteins have been
characterized as interaction partners of c-Ret and, like the receptor, induce neurite outgrowth
in PC12 cells (Grimm et al., 2001). The present work supports a putative role of Vav2 in
mediating neuregulin signals for mammary alveolar morphogenesis. Moreover, all these
studies provide substantial evidence that specific biological responses may result from
specific signaling pathways involving particular adaptors and hence, they put forward an
alternative mechanism for the generation of the unique developmental responses of receptor
tyrosine kinases.

65
Recent studies show that activation of Vav2 and RhoA also inhibits HGF/SF-induced cell
scattering upon overexpression in Madin-Darby canine kidney cells (MDCK; Kodama et al.,
2000). This observation suggests a negative role of Vav2 in branching morphogenesis and
raises the possibility of a dual role in vivo. Vav2 may therefore mediate neuregulin-induced
alveolar morphogenesis while simultaneously repressing HGF/SF-induced ductal growth.
Other groups, in contrast, report that activation of RhoG is essential for differentiation and
neurite outgrowth in PC12 cells following NGF treatment (Katoh et al., 2000). In the present
work, catalytically active Vav2 as GDP/GTP exchanger is strictly required for neuregulin-
induced alveolar morphogenesis. Future experiments in more physiological systems are
required to clarify the regulatory role of GEF factors and their targets in different biological
responses.
4.7 Possible Molecular Mechanisms Involving Vav2 as Critical Effector of Alveolar
Morphogenesis
This work provides evidence that Vav2 and ErbB-2 may be functionally associated in vivo to
promote alveolar morphogenesis of mammary epithelium. However, it does not resolve the
issue whether Vav2 is a direct or indirect substrate of the ErbB-2 kinase. A TrkErbB2-Vav2
fusion protein becomes morphogenic upon NGF stimulation (Fig. 7); nevertheless, it is
possible that an additional kinase is recruited to this hybrid protein via Vav2. Indeed,
activation of all Vav proteins by phosphorylation through Src tyrosine kinases has been
reported (Crespo et al., 1997; Schuebel et al., 1998; Movilla et al., 1999). In addition, recent
studies show that the PDGF receptor stimulates Vav2 through tyrosine phosphorylation by Src
(Chiariello et al., 2001). Recently, in vitro kinase assays were here performed with
immunoprecipitated TrkErbB-2 receptor from lysates of stimulated cells, together with
recombinant Vav2 protein as substrate. In line with the aforementioned report, these data
suggests that Vav2 is not directly phosphorylated by ErbB-2 in vitro (data not shown). It is
thus possible that the recruitment and activation of Src by ErbB-2 represents an intermediate
step resulting in the engagement of Vav2 for morphogenesis. However, activation of Src has

66
largely been implicated in the etiology of ErbB2-overexpressing breast tumors but not in
normal mammary development (Muthuswamy and Muller, 1994; Muthuswamy and Muller,
1995a); in fact, targeted overexpression of activated c-Src in the mammary epithelium results
in epithelial hyperplasia and impaired lobulo-alveolar development followed by severe
lactational failure (Webster et al., 1995). Latest experiments show that phosphorylated Vav2
interacts with the SH2 domain of Src (data not shown). This interaction is indeed likely, since
Vav2 contains a Src-binding consensus motif involving the regulatory tyrosine Y172;
however, it requires previous phosphorylation of Vav2. It is therefore possible that ErbB-2
phosphorylates Vav2 first on some of its various, non-regulatory tyrosine residues, thus
creating docking sites for Src other than the predicted consensus motif, and then Src futher
phosphorylates and activates Vav2. In support of this hypothesis, none of the known
autophosphorylation sites of ErbB-2 corresponds to the optimal Src consensus binding motif
(Songyang et al., 1993), though direct interaction between Src and ErbB-2 was here observed
in the yeast system and has also been reported by others (Muthuswamy and Muller, 1995).
Similarly, the direct binding site for Src on the PDGFβ receptor (DGHEYpYIpYVDP; Mori
et al., 1993) does not match the predicted motif; instead, it resembles the amino acid sequence
of ErbB-2 encompassing tyrosine Y3 (pYLVP), which was mapped in the yeast system as part
of the Src binding site (Table 2). However, such sequence is not present in Vav2, thus raising
the possibility of a broader range of phosphotyrosine-containing sequences that may be
recognized by the SH2 domain of Src. Alternatively, it has been proposed that Src
phosphorylates its own binding sites on the EGF receptor (Olayioye et al., 1999); such a
model would account for simultaneous binding of Src and activation of Vav2. Taken together,
these latest experiments favor that the indirect recruitment of Vav2 to ErbB-2 is of
physiological relevance. Nevertheless, further research that addresses the role of direct and
indirect binding of Vav2 to ErbB-2 may contribute to understand the mechanisms of Vav2
recruitment and activation in neuregulin signaling.

67
Previous studies suggest that activation of the MAPK/ERK (mitogen-activated protein
kinase/extracellular signal regulated protein kinase) pathway is a necessary step in neuregulin-
induced alveolar morphogenesis (Niemann et al., 1998). Indeed, MAP kinases are stimulated
by all ligand-activated combinations of ErbB receptors (Ben-Levy et al., 1992; Graus-Porta et
al., 1995; Karunagaran et al., 1996; Pinkas-Kramarski et al., 1996; Dankort et al., 1997;
Pinkas-Kramarski et al., 1998). The MAPK pathway is triggered via recruitment of Grb2 or
Shc/Grb2 complexes by the activated receptors (reviewed in Schlessinger and Bar-Sagi, 1994
and references therein). It has been shown that Grb2 constitutively binds Sos (Son of
Sevenless) via its SH3 domains. Association of the SH2 domain of Grb2 with activated
receptor tyrosine kinases leads to recruitment of the GDP/GTP exhanger Sos to the
membrane, where it can switch the membrane-anchored Ras GTPase to the active GTP-bound
form. Activation of Ras triggers a signaling cascade of kinases involving Raf (also termed
MAP/ERK kinase kinase MEKK, or MAPKKK), MAPK/ERK kinase (MEK, also termed
MAPKK) and finally the MAPK, which ultimately regulates transcription (reviewed in
Schaeffer and Weber, 1999). Tyrosines Y2 and Y4 of the ErbB-2 multidocking site directly
bind Grb-2 and Shc, respectively (Ricci et al., 1995; Dankort et al., 1997), and
Grb2/Sos/ErbB-2 complexes have been detected in breast cancer cells (Janes et al., 1994).
Moreover, different patterns of MAPK activation by α- and β-isoforms of NRG-1 and NRG-2
were reported (Pinkas-Kramarski et al., 1998); it has been suggested that duration of coupling
to MAPK pathway contributes to the signaling specificity by several ErbB heterodimers, and
the presence of ErbB-2 in receptor combinations contributes to prolong MAPK activation
(Karunagaran et al., 1996). Overexpression of constitutively active Vav proteins does not
activate MAPK; instead, Vav proteins strongly activate the c-Jun N-terminal kinase/stress-
activated protein kinase (JNK/SAPK; Crespo et al., 1996; Abe et al., 2000; data not shown).
Activation of JNK/SAPK by Vav proteins depends on the integrity of the Dbl-homology
domain, as it requires activation of the small GTPase Rac1. Nevertheless, a connection
between Vav2 and Ras has been suggested by synergism for cellular transformation (Schuebel
et al., 1998). It has been shown that Vav indirectly enhances Ras signaling; Vav-mediated

68
activation of small GTPases leads to subsequent activation of p21-activated protein kinases
(PAKs), which in turn activate Raf and MEK (Bustelo, 2000 and references therein). It is still
unclear whether Vav2 engages similar phosphorylation cascades. The possible requirement of
JNK/SAPK activation in neuregulin-induced alveolar morphogenesis therefore requires
attention.
It is clear that activation of the transcription factor Stat5a (signal transducer and activator of
transcription 5a) is critical in lobulo-alveolar morphogenesis. High levels of activated Stat5a
are found in the mammary gland at late pregnancy and during lactation. Inactivation of the
stat5a gene is accompanied by failure in terminal mammary differentiation but normal
production of milk proteins during pregnancy (Liu et al., 1997). Moreover, activation and
function of Stat5a during alveolar morphogenesis seems to be located downstream of ErbB-4
and prolactin signaling (Jones et al., 1999; Ihle and Kerr, 1995). Phosphorylation and
activation of Stat5 following neuregulin treatment has been observed in NIH3T3 fibroblasts
overexpressing both ErbB-2 and ErbB-4 (Olayioye et al., 1999); this observation indicates
that either heterodimers of ErbB2/erbB4, homodimers of ErbB-4 or both are responsible for
Stat5 activation. Since transgenic mice overexpressing dominant-negative ErbB-2 and ErbB-4
receptors in the mammary gland also show lactational failure, it is likely that heterodimers of
ErbB-2 and ErbB-4 control the activity of Stat5a in vivo. The activity of Stat5a in EpH4 cells
that overexpress Vav2 proteins has not been evaluated.
4.8 Conclusions
The results presented in this work suggest that Vav2 is a specific interacting partner of the
ErbB subgroup of receptor tyrosine kinases. This particular affinity of Vav2 for ErbB
receptors has a functional outcome: Vav2 can mediate specific signals from these receptors to
elicit alveolar morphogenesis of mammary epithelial cells. The enzymatic activity of Vav
proteins as GDP/GTP exchangers may provide a direct link to cytoskeletal rearrangements, an
essential step in morphogenic events. In vivo, Vav2 and ErbB-2 co-localize and are associated

69
in mammary alveolar epithelium during pregnancy, while neuregulin is simultaneously
synthesized in the mammary stroma. Therefore, this work supports a model whereby
neuregulin activates ErbB-2 in mammary epithelium, which then recruits Vav2 to trigger
unique signaling cascades that lead to lobulo-alveolar morphogenesis of the gland during
pregnancy.
As discussed above, there are some mechanistic points of ErbB-2/Vav2 signaling that still
need to be clarified. In addition, data on the function of Vav2 in mammary development in
vivo are required. Genetic ablation of the vav2 gene in mice revealed defective immune
response to thymus-independent antigens, and the additional loss of vav led to a severe defect
in B cell maturation (Doody et al., 2001; Tedford et al., 2001); however, vav2-deficient mice
apparently lack an overt epithelial phenotype, and it is not clear whether the mammary glands
from vav2 null mice undergo normal alveolar morphogenesis during pregnancy. An
explanation for this lack of an evident phenotype in epithelia is that Vav3, the other epithelial
Vav protein, may compensate for Vav2 function in tissues where both Vav2 and Vav3 are co-
expressed, as is the case of the mammary epithelium. Thus, generation of vav2/vav3 double
knockout mice may shed light on the physiological function of these guanine nucleotide
exchange factors in mammalian epithelial morphogenesis.

70
REFERENCES
Abe, K., K. L. Rossman, B. Liu, K. D. Ritola, D. Chiang, S. L. Campbell, K. Burridge, C. J. Der (2000). Vav2 is
an activator of Cdc42, Rac1, and RhoA. J. Biol. Chem. 275, 10141-10149.
Adams, J. C. and F. M. Watt (1993). Regulation of development and differentiation by the extracellular matrix.
Development 117, 1183-1198.
Adamson, E. D. (1990). Developmental activities of the epidermal growth factor receptor. Curr. Top. Dev. Biol.
24,1-29.
Aghazadeh, B., W. E. Lowry, X. Y. Huang, M. K. Rosen (2000). Structural basis for relief of autoinhibition of
the Dbl homology domain of proto-oncogene Vav by tyrosine phosphorylation. Cell 102, 625-633.
Akiyama, T., S. Matsuda, Y. Namba, T. Saito, K. Toyoshima, T. Yamamoto (1991). The transforming potential
of the c-erbB-2 protein is regulated by its autophosphorylation at the carboxyl-terminal domain. Mol. Cell
Biol. 11, 833-842.
Araki, E., M. A. Lipes, M. E. Patti, J. C. Bruning, B. Haag, R. S. Johnson, C. R. Kahn (1994). Alternative
pathway of insulin signalling in mice with targeted disruption of the IRS-1 gene. Nature 372, 186-190.
Aroian, R. V., M. Koga, J. E. Mendel, Y. Ohshima, P. W. Sternberg (1990). The let-23 gene necessary for
Caenorhabditis elegans vulval induction encodes a tyrosine kinase of the EGF receptor subfamily. Nature
348, 693-699.
Ausubel, F., R. Brent, R. Kingston, D. Moore, J. Smith, K. Struhl (1994). Current Protocols in Molecular
Biology. John Wiley & Sons, Inc.
Awasaki, T., M. Saito, M. Sone, E. Suzuki, R. Sakai, K. Ito, C. Hama (2000). The Drosophila trio plays an
essential role in patterning of axons by regulating their directional extension. Neuron 26, 119-131.
Barcellos-Hoff, M. H., J. Aggeler, T. G. Ram, M. J. Bissell (1989). Functional differentiation and alveolar
morphogenesis of primary mammary cultures on reconstituted basement membrane. Development 105,
223-235.

71
Bardelli, A., P. Longati, D. Gramaglia, M. C. Stella, P. M. Comoglio (1997). Gab1 coupling to the HGF/Met
receptor multifunctional docking site requires binding of Grb2 and correlates with the transforming
potential. Oncogene 15, 3103-3111.
Barrett, K., M. Leptin, J. Settleman (1997). The Rho GTPase and a putative RhoGEF mediate a signaling
pathway for the cell shape changes in Drosophila gastrulation. Cell 91, 905-915.
Bartel, P. L. and S. Fields (1995). Analyzing protein-protein interactions using two-hybrid system. Methods
Enzymol. 254, 241-263.
Bateman, J., H. Shu, D. Van Vactor (2000). The guanine nucleotide exchange factor trio mediates axonal
development in the Drosophila embryo. Neuron 26, 93-106.
Baulida, J., M. H. Kraus, M. Alimandi, P. P. Di Fiore, G. Carpenter (1996). All ErbB receptors other than the
epidermal growth factor receptor are endocytosis impaired. J. Biol. Chem. 271, 5251-5257.
Bchini, O., A. C. Andres, B. Schubaur, M. Mehtali, M. LeMeur, R. Lathe, P. Gerlinger (1991). Precocious
mammary gland development and milk protein synthesis in transgenic mice ubiquitously expressing
human growth hormone. Endocrinology 128, 539-546.
Behrens, J., J. P. von Kries, M. Kuhl, L. Bruhn, D. Wedlich, R. Grosschedl, W. Birchmeier (1996). Functional
interaction of beta-catenin with the transcription factor LEF-1. Nature 382, 638-642.
Ben-Levy, R., H. F. Paterson, C. J. Marshall, Y. Yarden (1994). A single autophosphorylation site confers
oncogenicity to the Neu/ErbB-2 receptor and enables coupling to the MAP kinase pathway. EMBO J. 13,
3302-3311.
Berdichevsky, F., C. Gilbert, M. Shearer, J. Taylor-Papadimitriou (1992). Collagen-induced rapid
morphogenesis of human mammary epithelial cells: the role of the alpha 2 beta 1 integrin. J. Cell Sci. 102,
437-446.
Billadeau, D. D., S. M. Mackie, R. A. Schoon, P. J. Leibson (2000). The Rho family guanine nucleotide
exchange factor Vav-2 regulates the development of cell-mediated cytotoxicity. J. Exp. Med. 192, 381-
392.

72
Bladt, F., D. Riethmacher, S. Isenmann, A. Aguzzi, C. Birchmeier (1995). Essential role for the c-met receptor in
the migration of myogenic precursor cells into the limb bud. Nature 376, 768-771.
Bocchinfuso, W. P. and K. S. Korach (1997). Mammary gland development and tumorigenesis in estrogen
receptor knockout mice. J. Mammary. Gland. Biol. Neoplasia. 2, 323-334.
Brisken, C., S. Park, T. Vass, J. P. Lydon, B. W. O'Malley, R. A. Weinberg (1998). A paracrine role for the
epithelial progesterone receptor in mammary gland development. Proc. Natl. Acad. Sci. U. S. A 95, 5076-
5081.
Brisken, C., S. Kaur, T. E. Chavarria, N. Binart, R. L. Sutherland, R. A. Weinberg, P. A. Kelly, C. J. Ormandy
(1999). Prolactin controls mammary gland development via direct and indirect mechanisms. Dev. Biol.
210, 96-106.
Brisken, C., A. Heineman, T. Chavarria, B. Elenbaas, J. Tan, S. K. Dey, J. A. McMahon, A. P. McMahon, R. A.
Weinberg (2000). Essential function of Wnt-4 in mammary gland development downstream of
progesterone signaling. Genes Dev. 14, 650-654.
Britsch, S., L. Li, S. Kirchhoff, F. Theuring, V. Brinkmann, C. Birchmeier, D. Riethmacher (1998). The ErbB2
and ErbB3 receptors and their ligand, neuregulin-1, are essential for development of the sympathetic
nervous system. Genes Dev. 12, 1825-1836.
Bruning, J. C., J. Winnay, B. Cheatham, C. R. Kahn (1997). Differential signaling by insulin receptor substrate 1
(IRS-1) and IRS-2 in IRS-1-deficient cells. Mol. Cell Biol. 17, 1513-1521.
Bustelo, X. R., J. A. Ledbetter, M. Barbacid (1992). Product of vav proto-oncogene defines a new class of
tyrosine protein kinase substrates. Nature 356, 68-71.
Bustelo, X. R. (2000). Regulatory and signaling properties of the Vav family. Mol. Cell Biol. 20, 1461-1477.
Carraway, K. L., J. L. Weber, M. J. Unger, J. Ledesma, N. Yu, M. Gassmann, C. Lai (1997). Neuregulin-2, a
new ligand of ErbB3/ErbB4-receptor tyrosine kinases. Nature 387, 512-516.
Chang, H., D. J. Riese, W. Gilbert, D. F. Stern, U. J. McMahan (1997). Ligands for ErbB-family receptors
encoded by a neuregulin-like gene. Nature 387, 509-512.

73
Chapman, R. S., P. C. Lourenco, E. Tonner, D. J. Flint, S. Selbert, K. Takeda, S. Akira, A. R. Clarke, C. J.
Watson (1999). Suppression of epithelial apoptosis and delayed mammary gland involution in mice with a
conditional knockout of Stat3. Genes Dev. 13, 2604-2616.
Chiariello, M., M. J. Marinissen, J. S. Gutkind (2001). Regulation of c-myc expression by PDGF through Rho
GTPases. Nat. Cell Biol. 3, 580-586.
Chodosh, L. A., H. P. Gardner, J. V. Rajan, D. B. Stairs, S. T. Marquis, P. A. Leder (2000). Protein kinase
expression during murine mammary development. Dev. Biol. 219, 259-276.
Cohen, B. D., P. A. Kiener, J. M. Green, L. Foy, H. P. Fell, K. Zhang (1996). The relationship between human
epidermal growth-like factor receptor expression and cellular transformation in NIH3T3 cells. J. Biol.
Chem. 271, 30897-30903.
Coleman-Krnacik, S. and J. M. Rosen (1994). Differential temporal and spatial gene expression of fibroblast
growth factor family members during mouse mammary gland development. Mol. Endocrinol. 8, 218-229.
Crespo, P., X. R. Bustelo, D. S. Aaronson, O. A. Coso, M. Lopez-Barahona, M. Barbacid, J. S. Gutkind (1996).
Rac-1 dependent stimulation of the JNK/SAPK signaling pathway by Vav. Oncogene 13, 455-460.
Crespo, P., K. E. Schuebel, A. A. Ostrom, J. S. Gutkind, X. R. Bustelo (1997). Phosphotyrosine-dependent
activation of Rac-1 GDP/GTP exchange by the vav proto-oncogene product. Nature 385, 169-172.
Crovello, C. S., C. Lai, L. C. Cantley, K. L. Carraway (1998). Differential signaling by the epidermal growth
factor-like growth factors neuregulin-1 and neuregulin-2. J. Biol. Chem. 273, 26954-26961.
Cunha, G. R. and Y. K. Hom (1996). Role of mesenchymal-epithelial interactions in mammary gland
development. J. Mammary. Gland. Biol. Neoplasia. 1, 21-35.
Cunha, G. R., P. Young, Y. K. Hom, P. S. Cooke, J. A. Taylor, D. B. Lubahn (1997). Elucidation of a role for
stromal steroid hormone receptors in mammary gland growth and development using tissue recombinants.
J. Mammary. Gland. Biol. Neoplasia. 2, 393-402.
Dankort, D., B. Maslikowski, N. Warner, N. Kanno, H. Kim, Z. Wang, M. F. Moran, R. G. Oshima, R. D.
Cardiff, W. J. Muller (2001). Grb2 and Shc adapter proteins play distinct roles in Neu (ErbB-2)-induced
mammary tumorigenesis: implications for human breast cancer. Mol. Cell Biol. 21, 1540-1551.

74
Dankort, D. L., Z. Wang, V. Blackmore, M. F. Moran, W. J. Muller (1997). Distinct tyrosine
autophosphorylation sites negatively and positively modulate neu-mediated transformation. Mol. Cell
Biol. 17, 5410-5425.
Doody, G. M., D. D. Billadeau, E. Clayton, A. Hutchings, R. Berland, S. McAdam, P. J. Leibson, M. Turner
(2000). Vav-2 controls NFAT-dependent transcription in B- but not T-lymphocytes. EMBO J. 19, 6173-
6184.
Doody, G. M., S. E. Bell, E. Vigorito, E. Clayton, S. McAdam, R. Tooze, C. Fernandez, I. J. Lee, M. Turner
(2001). Signal transduction through Vav-2 participates in humoral immune responses and B cell
maturation. Nature Immunol. 2, 542-547.
Dunbar, M. E. and J. J. Wysolmerski (1999). Parathyroid hormone-related protein: a developmental regulatory
molecule necessary for mammary gland development. J. Mammary. Gland. Biol. Neoplasia. 4, 21-34.
Eaton, S., P. Auvinen, L. Luo, Y. N. Jan, K. Simons (1995). CDC42 and Rac1 control different actin-dependent
processes in the Drosophila wing disc epithelium. J. Cell Biol. 131, 151-164.
Eaton, S., R. Wepf, K. Simons (1996). Roles for Rac1 and Cdc42 in planar polarization and hair outgrowth in the
wing of Drosophila. J. Cell Biol. 135, 1277-1289.
Emerman, J. T. and D. R. Pitelka (1977). Maintenance and induction of morphological differentiation in
dissociated mammary epithelium on floating collagen membranes. In Vitro 13, 316-328.
Erickson, S. L., K. S. O'Shea, N. Ghaboosi, L. Loverro, G. Frantz, M. Bauer, L. H. Lu, M. W. Moore (1997).
ErbB3 is required for normal cerebellar and cardiac development: a comparison with ErbB2-and
heregulin-deficient mice. Development 124, 4999-5011.
Falls, D. L., K. M. Rosen, G. Corfas, W. S. Lane, G. D. Fischbach (1993). ARIA, a protein that stimulates
acetylcholine receptor synthesis, is a member of the neu ligand family. Cell 72, 801-815.
Faraldo, M. M., M. A. Deugnier, M. Lukashev, J. P. Thiery, M. A. Glukhova (1998). Perturbation of beta1-
integrin function alters the development of murine mammary gland. EMBO J. 17, 2139-2147.

75
Fata, J. E., Y. Y. Kong, J. Li, T. Sasaki, J. Irie-Sasaki, R. A. Moorehead, R. Elliott, S. Scully, E. B. Voura, D. L.
Lacey, W. J. Boyle, R. Khokha, J. M. Penninger (2000). The osteoclast differentiation factor
osteoprotegerin-ligand is essential for mammary gland development. Cell 103, 41-50.
Fazioli, F., U. H. Kim, S. G. Rhee, C. J. Molloy, O. Segatto, P. P. Di Fiore (1991). The erbB-2 mitogenic
signaling pathway: tyrosine phosphorylation of phospholipase C-gamma and GTPase-activating protein
does not correlate with erbB-2 mitogenic potency. Mol. Cell Biol. 11, 2040-2048.
Fedi, P., J. H. Pierce, P. P. Di Fiore, M. H. Kraus (1994). Efficient coupling with phosphatidylinositol 3-kinase,
but not phospholipase C gamma or GTPase-activating protein, distinguishes ErbB-3 signaling from that of
other ErbB/EGFR family members. Mol. Cell Biol. 14, 492-500.
Fields, S. and O. Song (1989). A novel genetic system to detect protein-protein interactions. Nature 340, 245-
246.
Fischer, K. D., A. Zmuldzinas, S. Gardner, M. Barbacid, A. Bernstein, C. Guidos (1995). Defective T-cell
receptor signalling and positive selection of Vav-deficient CD4+ CD8+ thymocytes. Nature 374, 474-
477.
Fischer, K. D., Y. Y. Kong, H. Nishina, K. Tedford, L. E. Marengere, I. Kozieradzki, T. Sasaki, M. Starr, G.
Chan, S. Gardener, M. P. Nghiem, D. Bouchard, M. Barbacid, A. Bernstein, J. M. Penninger (1998). Vav
is a regulator of cytoskeletal reorganization mediated by the T-cell receptor. Curr. Biol. 8, 554-562.
Fixman, E. D., M. A. Naujokas, G. A. Rodrigues, M. F. Moran, M. Park (1995). Efficient cell transformation by
the Tpr-Met oncoprotein is dependent upon tyrosine 489 in the carboxy-terminus. Oncogene 10, 237-249.
Foley, J., P. Dann, J. Hong, J. Cosgrove, B. Dreyer, D. Rimm, M. Dunbar, W. Philbrick, J. Wysolmerski (2001).
Parathyroid hormone-related protein maintains mammary epithelial fate and triggers nipple skin
differentiation during embryonic breast development. Development 128, 513-525.
Fowler, K. J., F. Walker, W. Alexander, M. L. Hibbs, E. C. Nice, R. M. Bohmer, G. B. Mann, C. Thumwood, R.
Maglitto, J. A. Danks, a. et (1995). A mutation in the epidermal growth factor receptor in waved-2 mice
has a profound effect on receptor biochemistry that results in impaired lactation. Proc. Natl. Acad. Sci. U.
S. A 92, 1465-1469.

76
Freeman, M. (1998). Complexity of EGF receptor signalling revealed in Drosophila. Curr. Opin. Genet. Dev. 8,
407-411.
Freshney, N. W., S. D. Goonesekera, L. A. Feig (1997). Activation of the exchange factor Ras-GRF by calcium
requires an intact Dbl homology domain. FEBS Lett. 407, 111-115.
Gassmann, M., F. Casagranda, D. Orioli, H. Simon, C. Lai, R. Klein, G. Lemke (1995). Aberrant neural and
cardiac development in mice lacking the ErbB4 neuregulin receptor. Nature 378, 390-394.
Gorska, A. E., H. Joseph, R. Derynck, H. L. Moses, R. Serra (1998). Dominant-negative interference of the
transforming growth factor beta type II receptor in mammary gland epithelium results in alveolar
hyperplasia and differentiation in virgin mice. Cell Growth Differ. 9, 229-238.
Graus-Porta, D., R. R. Beerli, N. E. Hynes (1995). Single-chain antibody-mediated intracellular retention of
ErbB-2 impairs Neu differentiation factor and epidermal growth factor signaling. Mol. Cell Biol. 15,
1182-1191.
Graus-Porta, D., R. R. Beerli, J. M. Daly, N. E. Hynes (1997). ErbB-2, the preferred heterodimerization partner
of all ErbB receptors, is a mediator of lateral signaling. EMBO J. 16, 1647-1655.
Grimm, J., M. Sachs, S. Britsch, S. Di Cesare, T. Schwarz-Romond, K. Alitalo, W. Birchmeier (2001). Novel
p62dok family members, dok-4 and dok-5, are substrates of the c-Ret receptor tyrosine kinase and
mediate neuronal differentiation. J. Cell Biol. (in press).
Guo, L., L. Degenstein, E. Fuchs (1996). Keratinocyte growth factor is required for hair development but not for
wound healing. Genes Dev. 10, 165-175.
Guy, P. M., J. V. Platko, L. C. Cantley, R. A. Cerione, K. L. Carraway (1994). Insect cell-expressed p180erbB3
possesses an impaired tyrosine kinase activity. Proc. Natl. Acad. Sci. U. S. A 91, 8132-8136.
Hacker, U. and N. Perrimon (1998). DRhoGEF2 encodes a member of the Dbl family of oncogenes and controls
cell shape changes during gastrulation in Drosophila. Genes Dev. 12, 274-284.
Hall, A. (1998). Rho GTPases and the actin cytoskeleton. Science 279, 509-514.
Harari, D., E. Tzahar, J. Romano, M. Shelly, J. H. Pierce, G. C. Andrews, Y. Yarden (1999). Neuregulin-4: a
novel growth factor that acts through the ErbB-4 receptor tyrosine kinase. Oncogene 18, 2681-2689.

77
Hart, M. J., A. Eva, D. Zangrilli, S. A. Aaronson, T. Evans, R. A. Cerione, Y. Zheng (1994). Cellular
transformation and guanine nucleotide exchange activity are catalyzed by a common domain on the dbl
oncogene product. J. Biol. Chem. 269, 62-65.
Hazan, R., B. Margolis, M. Dombalagian, A. Ullrich, A. Zilberstein, J. Schlessinger (1990). Identification of
autophosphorylation sites of HER2/neu. Cell Growth Differ. 1, 3-7.
Herbst, R., P. M. Carroll, J. D. Allard, J. Schilling, T. Raabe, M. A. Simon (1996). Daughter of sevenless is a
substrate of the phosphotyrosine phosphatase Corkscrew and functions during sevenless signaling. Cell
85, 899-909.
Holgado-Madruga, M., D. R. Emlet, D. K. Moscatello, A. K. Godwin, A. J. Wong (1996). A Grb2-associated
docking protein in EGF- and insulin-receptor signalling. Nature 379, 560-564.
Holmes, W. E., M. X. Sliwkowski, R. W. Akita, W. J. Henzel, J. Lee, J. W. Park, D. Yansura, N. Abadi, H.
Raab, G. D. Lewis, a. et (1992). Identification of heregulin, a specific activator of p185erbB2. Science
256, 1205-1210.
Horseman, N. D., W. Zhao, E. Montecino-Rodriguez, M. Tanaka, K. Nakashima, S. J. Engle, F. Smith, E.
Markoff, K. Dorshkind (1997). Defective mammopoiesis, but normal hematopoiesis, in mice with a
targeted disruption of the prolactin gene. EMBO J. 16, 6926-6935.
Hubbard, S. R., M. Mohammadi, J. Schlessinger (1998). Autoregulatory mechanisms in protein-tyrosine kinases.
J. Biol. Chem. 273, 11987-11990.
Huelsken, J. and W. Birchmeier (2001). New aspects of Wnt signaling pathways in higher vertebrates. Curr. Op.
Genet. Dev. (in press).
Huelsken, J., R. Vogel, B. Erdmann, G. Cotsarelis, W. Birchmeier (2001). beta-Catenin controls hair follicle
morphogenesis and stem cell differentiation in the skin. Cell 105, 533-545.
Ihle, J. N. and I. M. Kerr (1995). Jaks and Stats in signaling by the cytokine receptor superfamily. Trends Genet.
11, 69-74.
Imbert, A., R. Eelkema, S. Jordan, H. Feiner, P. Cowin (2001). Delta N89 beta-catenin induces precocious
development, differentiation, and neoplasia in mammary gland. J. Cell Biol. 153, 555-568.

78
Jackson, D., J. Bresnick, C. Dickson (1997). A role for fibroblast growth factor signaling in the lobuloalveolar
development of the mammary gland. J. Mammary. Gland. Biol. Neoplasia. 2, 385-392.
Jallal, B., J. Schlessinger, A. Ullrich (1992). Tyrosine phosphatase inhibition permits analysis of signal
transduction complexes in p185HER2/neu-overexpressing human tumor cells. J. Biol. Chem. 267, 4357-
4363.
Janes, P. W., R. J. Daly, A. deFazio, R. L. Sutherland (1994). Activation of the Ras signalling pathway in human
breast cancer cells overexpressing erbB-2. Oncogene 9, 3601-3608.
Jhappan, C., A. G. Geiser, E. C. Kordon, D. Bagheri, L. Hennighausen, A. B. Roberts, G. H. Smith, G. Merlino
(1993). Targeting expression of a transforming growth factor beta 1 transgene to the pregnant mammary
gland inhibits alveolar development and lactation. EMBO J. 12, 1835-1845.
Jones, F. E., D. J. Jerry, B. C. Guarino, G. C. Andrews, D. F. Stern (1996). Heregulin induces in vivo
proliferation and differentiation of mammary epithelium into secretory lobuloalveoli. Cell Growth Differ.
7, 1031-1038.
Jones, F. E. and D. F. Stern (1999). Expression of dominant-negative ErbB2 in the mammary gland of transgenic
mice reveals a role in lobuloalveolar development and lactation. Oncogene 18, 3481-3490.
Jones, F. E., T. Welte, X. Y. Fu, D. F. Stern (1999). ErbB4 signaling in the mammary gland is required for
lobuloalveolar development and Stat5 activation during lactation. J. Cell Biol. 147, 77-88.
Joseph, H., A. E. Gorska, P. Sohn, H. L. Moses, R. Serra (1999). Overexpression of a kinase-deficient
transforming growth factor-beta type II receptor in mouse mammary stroma results in increased epithelial
branching. Mol. Biol. Cell 10, 1221-1234.
Karunagaran, D., E. Tzahar, N. Liu, D. Wen, Y. Yarden (1995). Neu differentiation factor inhibits EGF binding.
A model for trans-regulation within the ErbB family of receptor tyrosine kinases. J. Biol. Chem. 270,
9982-9990.
Karunagaran, D., E. Tzahar, R. R. Beerli, X. Chen, D. Graus-Porta, B. J. Ratzkin, R. Seger, N. E. Hynes, Y.
Yarden (1996). ErbB-2 is a common auxiliary subunit of NDF and EGF receptors: implications for breast
cancer. EMBO J. 15, 254-264.

79
Katoh, H., H. Yasui, Y. Yamaguchi, J. Aoki, H. Fujita, K. Mori, M. Negishi (2000). Small GTPase RhoG is a
key regulator for neurite outgrowth in PC12 cells. Mol. Cell Biol. 20, 7378-7387.
Katzav, S., D. Martin-Zanca, M. Barbacid (1989). vav, a novel human oncogene derived from a locus
ubiquitously expressed in hematopoietic cells. EMBO J. 8, 2283-2290.
Katzav, S., J. L. Cleveland, H. E. Heslop, D. Pulido (1991). Loss of the amino-terminal helix-loop-helix domain
of the vav proto-oncogene activates its transforming potential. Mol. Cell Biol. 11, 1912-1920.
Klapper, L. N., H. Waterman, M. Sela, Y. Yarden (2000). Tumor-inhibitory antibodies to HER-2/ErbB-2 may
act by recruiting c-Cbl and enhancing ubiquitination of HER-2. Cancer Res. 60, 3384-3388.
Kleinberg, D. L. (1997). Early mammary development: growth hormone and IGF-1. J. Mammary. Gland. Biol.
Neoplasia. 2, 49-57.
Kleinman, H. K., M. L. McGarvey, J. R. Hassell, V. L. Star, F. B. Cannon, G. W. Laurie, G. R. Martin (1986).
Basement membrane complexes with biological activity. Biochemistry 25, 312-318.
Kodama, A., T. Matozaki, A. Fukuhara, M. Kikyo, M. Ichihashi, Y. Takai (2000). Involvement of an SHP-2-Rho
small G protein pathway in hepatocyte growth factor/scatter factor-induced cell scattering. Mol. Biol. Cell
11, 2565-2575.
Kokai, Y., J. A. Cohen, J. A. Drebin, M. I. Greene (1987). Stage- and tissue-specific expression of the neu
oncogene in rat development. Proc. Natl. Acad. Sci. U. S. A 84, 8498-8501.
Korach, K. S. (1994). Insights from the study of animals lacking functional estrogen receptor. Science 266, 1524-
1527.
Kramer, R., N. Bucay, D. J. Kane, L. E. Martin, J. E. Tarpley, L. E. Theill (1996). Neuregulins with an Ig-like
domain are essential for mouse myocardial and neuronal development. Proc. Natl. Acad. Sci. U. S. A 93,
4833-4838.
Kratochwil, K. (1986). Tissue combination and organ culture studies in the development of the embryonic
mammary gland. Dev. Biol. (N. Y. 1985. ) 43,15-33.
Lacey, D. L., E. Timms, H. L. Tan, M. J. Kelley, C. R. Dunstan, T. Burgess, R. Elliott, A. Colombero, G. Elliott,
S. Scully, H. Hsu, J. Sullivan, N. Hawkins, E. Davy, C. Capparelli, A. Eli, Y. X. Qian, S. Kaufman, I.

80
Sarosi, V. Shalhoub, G. Senaldi, J. Guo, J. Delaney, W. J. Boyle (1998). Osteoprotegerin ligand is a
cytokine that regulates osteoclast differentiation and activation. Cell 93, 165-176.
Lee, K. F., H. Simon, H. Chen, B. Bates, M. C. Hung, C. Hauser (1995). Requirement for neuregulin receptor
erbB2 in neural and cardiac development. Nature 378, 394-398.
Lenferink, A. E., R. Pinkas-Kramarski, M. L. van de Poll, M. J. van Vugt, L. N. Klapper, E. Tzahar, H.
Waterman, M. Sela, E. J. van Zoelen, Y. Yarden (1998). Differential endocytic routing of homo- and
hetero-dimeric ErbB tyrosine kinases confers signaling superiority to receptor heterodimers. EMBO J. 17,
3385-3397.
Levkowitz, G., H. Waterman, E. Zamir, Z. Kam, S. Oved, W. Y. Langdon, L. Beguinot, B. Geiger, Y. Yarden
(1998). c-Cbl/Sli-1 regulates endocytic sorting and ubiquitination of the epidermal growth factor receptor.
Genes Dev. 12, 3663-3674.
Levkowitz, G., S. Oved, L. N. Klapper, D. Harari, S. Lavi, M. Sela, Y. Yarden (2000). c-Cbl is a suppressor of
the neu oncogene. J. Biol. Chem. 275, 35532-35539.
Li, M., X. Liu, G. Robinson, U. Bar-Peled, K. U. Wagner, W. S. Young, L. Hennighausen, P. A. Furth (1997).
Mammary-derived signals activate programmed cell death during the first stage of mammary gland
involution. Proc. Natl. Acad. Sci. U. S. A 94, 3425-3430.
Li, L., (2001). (submitted)
Licitra, E. J. and J. O. Liu (1996). A three-hybrid system for detecting small ligand-protein receptor interactions.
Proc. Natl. Acad. Sci. U. S. A 93, 12817-12821.
Liebl, E. C., D. J. Forsthoefel, L. S. Franco, S. H. Sample, J. E. Hess, J. A. Cowger, M. P. Chandler, A. M.
Shupert, M. A. Seeger (2000). Dosage-sensitive, reciprocal genetic interactions between the Abl tyrosine
kinase and the putative GEF trio reveal trio's role in axon pathfinding. Neuron 26, 107-118.
Lin, C. Q. and M. J. Bissell (1993). Multi-faceted regulation of cell differentiation by extracellular matrix.
FASEB J. 7, 737-743.
Liu, X., G. W. Robinson, K. U. Wagner, L. Garrett, A. Wynshaw-Boris, L. Hennighausen (1997). Stat5a is
mandatory for adult mammary gland development and lactogenesis. Genes Dev. 11, 179-186.

81
Lopez-Barahona, M., I. Fialka, J. M. Gonzalez-Sancho, M. Asuncion, M. Gonzalez, T. Iglesias, J. Bernal, H.
Beug, A. Munoz (1995). Thyroid hormone regulates stromelysin expression, protease secretion and the
morphogenetic potential of normal polarized mammary epithelial cells. EMBO J. 14, 1145-1155.
Lopez-Lago, M., H. Lee, C. Cruz, N. Movilla, X. R. Bustelo (2000). Tyrosine phosphorylation mediates both
activation and downmodulation of the biological activity of Vav. Mol. Cell Biol. 20, 1678-1691.
Luetteke, N. C., T. H. Qiu, R. L. Peiffer, P. Oliver, O. Smithies, D. C. Lee (1993). TGF alpha deficiency results
in hair follicle and eye abnormalities in targeted and waved-1 mice. Cell 73, 263-278.
Luetteke, N. C., T. H. Qiu, S. E. Fenton, K. L. Troyer, R. F. Riedel, A. Chang, D. C. Lee (1999). Targeted
inactivation of the EGF and amphiregulin genes reveals distinct roles for EGF receptor ligands in mouse
mammary gland development. Development 126, 2739-2750.
Lund, L. R., J. Romer, N. Thomasset, H. Solberg, C. Pyke, M. J. Bissell, K. Dano, Z. Werb (1996). Two distinct
phases of apoptosis in mammary gland involution: proteinase-independent and -dependent pathways.
Development 122, 181-193.
Lydon, J. P., F. J. DeMayo, C. R. Funk, S. K. Mani, A. R. Hughes, C. A. Montgomery, G. Shyamala, O. M.
Conneely, B. W. O'Malley (1995). Mice lacking progesterone receptor exhibit pleiotropic reproductive
abnormalities. Genes Dev. 9, 2266-2278.
Mackay, D. J. and A. Hall (1998). Rho GTPases. J. Biol. Chem. 273, 20685-20688.
Magie, C. R., M. R. Meyer, M. S. Gorsuch, S. M. Parkhurst (1999). Mutations in the Rho1 small GTPase disrupt
morphogenesis and segmentation during early Drosophila development. Development 126, 5353-5364.
Mann, G. B., K. J. Fowler, A. Gabriel, E. C. Nice, R. L. Williams, A. R. Dunn (1993). Mice with a null mutation
of the TGF alpha gene have abnormal skin architecture, wavy hair, and curly whiskers and often develop
corneal inflammation. Cell 73, 249-261.
Marchionni, M. A., A. D. Goodearl, M. S. Chen, O. Bermingham-McDonogh, C. Kirk, M. Hendricks, F. Danehy,
D. Misumi, J. Sudhalter, K. Kobayashi, a. et (1993). Glial growth factors are alternatively spliced erbB2
ligands expressed in the nervous system. Nature 362, 312-318.

82
Margolis, B., P. Hu, S. Katzav, W. Li, J. M. Oliver, A. Ullrich, A. Weiss, J. Schlessinger (1992). Tyrosine
phosphorylation of vav proto-oncogene product containing SH2 domain and transcription factor motifs.
Nature 356, 71-74.
Matsui, Y., S. A. Halter, J. T. Holt, B. L. Hogan, R. J. Coffey (1990). Development of mammary hyperplasia and
neoplasia in MMTV-TGF alpha transgenic mice. Cell 61, 1147-1155.
Meyer, D. and C. Birchmeier (1994). Distinct isoforms of neuregulin are expressed in mesenchymal and neuronal
cells during mouse development. Proc. Natl. Acad. Sci. U. S. A 91, 1064-1068.
Meyer, D. and C. Birchmeier (1995). Multiple essential functions of neuregulin in development. Nature 378,
386-390.
Miettinen, P. J., J. E. Berger, J. Meneses, Y. Phung, R. A. Pedersen, Z. Werb, R. Derynck (1995). Epithelial
immaturity and multiorgan failure in mice lacking epidermal growth factor receptor. Nature 376, 337-341.
Moores, S. L., L. M. Selfors, J. Fredericks, T. Breit, K. Fujikawa, F. W. Alt, J. S. Brugge, W. Swat (2000). Vav
family proteins couple to diverse cell surface receptors. Mol. Cell Biol. 20, 6364-6373.
Mori, S., L. Ronnstrand, K. Yokote, A. Engstrom, S. A. Courtneidge, L. Claesson-Welsh, C. H. Heldin (1993).
Identification of two juxtamembrane autophosphorylation sites in the PDGF beta-receptor; involvement in
the interaction with Src family tyrosine kinases. EMBO J. 12, 2257-2264.
Morini, M., S. Astigiano, M. Mora, C. Ricotta, N. Ferrari, S. Mantero, G. Levi, M. Rossini, O. Barbieri (2000).
Hyperplasia and impaired involution in the mammary gland of transgenic mice expressing human FGF4.
Oncogene 19, 6007-6014.
Morris, J. K., W. Lin, C. Hauser, Y. Marchuk, D. Getman, K. F. Lee (1999). Rescue of the cardiac defect in
ErbB2 mutant mice reveals essential roles of ErbB2 in peripheral nervous system development. Neuron
23, 273-283.
Movilla, N. and X. R. Bustelo (1999). Biological and regulatory properties of Vav-3, a new member of the Vav
family of oncoproteins. Mol. Cell Biol. 19, 7870-7885.
Murphy, A. M. and D.J.Montell (1996). Cell type specific roles for Cdc42, Rac and RhoL in drosophila
oogenesis. J. Cell Biol. 133, 617-630.

83
Muthuswamy, S. K. and W. J. Muller (1994). Activation of the Src family of tyrosine kinases in mammary
tumorigenesis. Adv. Cancer Res. 64, 111-123.
Muthuswamy, S. K. and W. J. Muller (1995a). Activation of Src family kinases in Neu-induced mammary tumors
correlates with their association with distinct sets of tyrosine phosphorylated proteins in vivo. Oncogene
11, 1801-1810.
Muthuswamy, S. K. and W. J. Muller (1995b). Direct and specific interaction of c-Src with Neu is involved in
signaling by the epidermal growth factor receptor. Oncogene 11, 271-279.
Nemanic, M. K. and D. R. Pitelka (1971). A scanning electron microscope study of the lactating mammary gland.
J. Cell Biol. 48, 410-415.
Newsome, T. P., S. Schmidt, G. Dietzl, K. Keleman, B. Asling, A. Debant, B. J. Dickson (2000). Trio combines
with dock to regulate Pak activity during photoreceptor axon pathfinding in Drosophila. Cell 101, 283-
294.
Nguyen, A. V. and J. W. Pollard (2000). Transforming growth factor beta3 induces cell death during the first
stage of mammary gland involution. Development 127, 3107-3118.
Nguyen, L., M. Holgado-Madruga, C. Maroun, E. D. Fixman, D. Kamikura, T. Fournier, A. Charest, M. L.
Tremblay, A. J. Wong, M. Park (1997). Association of the multisubstrate docking protein Gab1 with the
hepatocyte growth factor receptor requires a functional Grb2 binding site involving tyrosine 1356. J. Biol.
Chem. 272, 20811-20819.
Niemann, C., V. Brinkmann, E. Spitzer, G. Hartmann, M. Sachs, H. Naundorf, W. Birchmeier (1998).
Reconstitution of mammary gland development in vitro: requirement of c-met and c-erbB2 signaling for
branching and alveolar morphogenesis. J. Cell Biol. 143, 533-545.
Niranjan, B., L. Buluwela, J. Yant, N. Perusinghe, A. Atherton, D. Phippard, T. Dale, B. Gusterson, T. Kamalati
(1995). HGF/SF: a potent cytokine for mammary growth, morphogenesis and development. Development
121, 2897-2908.
O'Neill, T. J., A. Craparo, T. A. Gustafson (1994). Characterization of an interaction between insulin receptor
substrate 1 and the insulin receptor by using the two-hybrid system. Mol. Cell Biol. 14, 6433-6442.

84
Olayioye, M. A., D. Graus-Porta, R. R. Beerli, J. Rohrer, B. Gay, N. E. Hynes (1998). ErbB-1 and ErbB-2
acquire distinct signaling properties dependent upon their dimerization partner. Mol. Cell Biol. 18, 5042-
5051.
Olayioye, M. A., I. Beuvink, K. Horsch, J. M. Daly, N. E. Hynes (1999). ErbB receptor-induced activation of
stat transcription factors is mediated by Src tyrosine kinases. J. Biol. Chem. 274, 17209-17218.
Olayioye, M. A., R. M. Neve, H. A. Lane, N. E. Hynes (2000). The ErbB signaling network: receptor
heterodimerization in development and cancer. EMBO J. 19, 3159-3167.
Olson, M. F., N. G. Pasteris, J. L. Gorski, A. Hall (1996). Faciogenital dysplasia protein (FGD1) and Vav, two
related proteins required for normal embryonic development, are upstream regulators of Rho GTPases.
Curr. Biol. 6, 1628-1633.
Ormandy, C. J., A. Camus, J. Barra, D. Damotte, B. Lucas, H. Buteau, M. Edery, N. Brousse, C. Babinet, N.
Binart, P. A. Kelly (1997). Null mutation of the prolactin receptor gene produces multiple reproductive
defects in the mouse. Genes Dev. 11, 167-178.
Pandey, A., A. V. Podtelejnikov, B. Blagoev, X. R. Bustelo, M. Mann, H. F. Lodish (2000). Analysis of receptor
signaling pathways by mass spectrometry: identification of vav-2 as a substrate of the epidermal and
platelet-derived growth factor receptors. Proc. Natl. Acad. Sci. U. S. A 97, 179-184.
Park, M., M. Dean, C. S. Cooper, M. Schmidt, S. J. O'Brien, D. G. Blair, G. F. Vande-Woude (1986).
Mechanism of met oncogene activation. Cell 45, 895-904.
Partanen, A. M. (1990). Epidermal growth factor and transforming growth factor-alpha in the development of
epithelial-mesenchymal organs of the mouse. Curr. Top. Dev. Biol. 24, 31-55.
Pasteris, N. G., A. Cadle, L. J. Logie, M. E. Porteous, C. E. Schwartz, R. E. Stevenson, T. W. Glover, R. S.
Wilroy, J. L. Gorski (1994). Isolation and characterization of the faciogenital dysplasia (Aarskog-Scott
syndrome) gene: a putative Rho/Rac guanine nucleotide exchange factor. Cell 79, 669-678.
Pawson, T. (1995). Protein modules and signalling networks. Nature 373, 573-580.
Pawson, T. and J. D. Scott (1997). Signaling through scaffold, anchoring, and adaptor proteins. Science 278,
2075-2080.

85
Peles, E. and Y. Yarden (1993). Neu and its ligands: from an oncogene to neural factors. Bioessays 15, 815-824.
Peles, E., R. Ben Levy, E. Tzahar, N. Liu, D. Wen, Y. Yarden (1993). Cell-type specific interaction of Neu
differentiation factor (NDF/heregulin) with Neu/HER-2 suggests complex ligand-receptor relationships.
EMBO J. 12, 961-971.
Pepper, M. S., J. V. Soriano, P. A. Menoud, A. P. Sappino, L. Orci, R. Montesano (1995). Modulation of
hepatocyte growth factor and c-met in the rat mammary gland during pregnancy, lactation, and involution.
Exp. Cell Res. 219, 204-210.
Pierce, D. F., M. D. Johnson, Y. Matsui, S. D. Robinson, L. I. Gold, A. F. Purchio, C. W. Daniel, B. L. Hogan,
H. L. Moses (1993). Inhibition of mammary duct development but not alveolar outgrowth during
pregnancy in transgenic mice expressing active TGF-beta 1. Genes Dev. 7, 2308-2317.
Pinkas-Kramarski, R., L. Soussan, H. Waterman, G. Levkowitz, I. Alroy, L. Klapper, S. Lavi, R. Seger, B. J.
Ratzkin, M. Sela, Y. Yarden (1996). Diversification of Neu differentiation factor and epidermal growth
factor signaling by combinatorial receptor interactions. EMBO J. 15, 2452-2467.
Pinkas-Kramarski, R., R. Eilam, I. Alroy, G. Levkowitz, P. Lonai, Y. Yarden (1997). Differential expression of
NDF/neuregulin receptors ErbB-3 and ErbB-4 and involvement in inhibition of neuronal differentiation.
Oncogene 15, 2803-2815.
Pinkas-Kramarski, R., M. Shelly, B. C. Guarino, L. M. Wang, L. Lyass, I. Alroy, M. Alimandi, A. Kuo, J. D.
Moyer, S. Lavi, M. Eisenstein, B. J. Ratzkin, R. Seger, S. S. Bacus, J. H. Pierce, G. C. Andrews, Y.
Yarden, M. Alamandi (1998). ErbB tyrosine kinases and the two neuregulin families constitute a ligand-
receptor network. Mol. Cell Biol. 18, 6090-6101.
Pollard, J. W. and L. Hennighausen (1994). Colony stimulating factor 1 is required for mammary gland
development during pregnancy. Proc. Natl. Acad. Sci. U. S. A 91, 9312-9316.
Prigent, S. A. and W. J. Gullick (1994). Identification of c-erbB-3 binding sites for phosphatidylinositol 3'-kinase
and SHC using an EGF receptor/c-erbB-3 chimera. EMBO J. 13, 2831-2841.
Qian, X., W. C. Vass, A. G. Papageorge, P. H. Anborgh, D. R. Lowy (1998). N terminus of Sos1 Ras exchange
factor: critical roles for the Dbl and pleckstrin homology domains. Mol. Cell Biol. 18, 771-778.

86
Raabe, T., J. Riesgo-Escovar, X. Liu, B. S. Bausenwein, P. Deak, P. Maroy, E. Hafen (1996). DOS, a novel
pleckstrin homology domain-containing protein required for signal transduction between sevenless and
Ras1 in Drosophila. Cell 85, 911-920.
Reichmann, E., R. Ball, B. Groner, R. R. Friis (1989). New mammary epithelial and fibroblastic cell clones in
coculture form structures competent to differentiate functionally. J. Cell Biol. 108, 1127-1138.
Ricci, A., L. Lanfrancone, R. Chiari, G. Belardo, C. Pertica, P. G. Natali, P. G. Pelicci, O. Segatto (1995).
Analysis of protein-protein interactions involved in the activation of the Shc/Grb-2 pathway by the ErbB-2
kinase. Oncogene 11, 1519-1529.
Riese, D. J., T. M. van Raaij, G. D. Plowman, G. C. Andrews, D. F. Stern (1995). The cellular response to
neuregulins is governed by complex interactions of the erbB receptor family. Mol. Cell Biol. 15, 5770-
5776.
Riese, D. J. and D. F. Stern (1998). Specificity within the EGF family/ErbB receptor family signaling network.
Bioessays 20, 41-48.
Riethmacher, D., E. Sonnenberg-Riethmacher, V. Brinkmann, T. Yamaai, G. R. Lewin, C. Birchmeier (1997).
Severe neuropathies in mice with targeted mutations in the ErbB3 receptor. Nature 389, 725-730.
Robinson, G. W., A. B. Karpf, K. Kratochwil (1999). Regulation of mammary gland development by tissue
interaction. J. Mammary. Gland. Biol. Neoplasia. 4, 9-19.
Rodrigues, G. A. and M. Park (1993). Dimerization mediated through a leucine zipper activates the oncogenic
potential of the met receptor tyrosine kinase. Mol. Cell Biol. 13, 6711-6722.
Ruan, W. and D. L. Kleinberg (1999). Insulin-like growth factor I is essential for terminal end bud formation and
ductal morphogenesis during mammary development. Endocrinology 140, 5075-5081.
Sachs, M., K. M. Weidner, V. Brinkmann, I. Walther, A. Obermeier, A. Ullrich, W. Birchmeier (1996).
Motogenic and morphogenic activity of epithelial receptor tyrosine kinases. J. Cell Biol. 133, 1095-1107.
Sachs, M., H. Brohmann, D. Zechner, T. Muller, J. Hulsken, I. Walther, U. Schaeper, C. Birchmeier, W.
Birchmeier (2000). Essential role of Gab1 for signaling by the c-Met receptor in vivo. J. Cell Biol. 150,
1375-1384.

87
Sakakura, T. (1987). Mammary embryogenesis. In The mammary gland: Development, regulation and function.
(eds. M. C. Neville and C. W. Daniel), pp. 37-66. Plenum Press, New York.
Sandgren, E. P., J. A. Schroeder, T. H. Qui, R. D. Palmiter, R. L. Brinster, D. C. Lee (1995). Inhibition of
mammary gland involution is associated with transforming growth factor alpha but not c-myc-induced
tumorigenesis in transgenic mice. Cancer Res. 55, 3915-3927.
Sawer, R. J. and J. F. Fallon (1983). Epithelial-mesenchymal interactions in development. Praeger Publishers,
New York.
Schaeffer, H. J. and M. J. Weber (1999). Mitogen-activated protein kinases: specific messages from ubiquitous
messengers. Mol. Cell Biol. 19, 2435-2444.
Schaeper, U., N. H. Gehring, K. P. Fuchs, M. Sachs, B. Kempkes, W. Birchmeier (2000). Coupling of Gab1 to c-
Met, Grb2, and Shp2 mediates biological responses. J. Cell Biol. 149, 1419-1432.
Schlessinger, J. and D. Bar-Sagi (1994). Activation of Ras and other signaling pathways by receptor tyrosine
kinases. Cold Spring Harb. Symp. Quant. Biol. 59, 173-179.
Schroeder, J. A. and D. C. Lee (1998). Dynamic expression and activation of ErbB receptors in the developing
mouse mammary gland. Cell Growth Differ. 9, 451-464.
Schuebel, K. E., X. R. Bustelo, D. A. Nielsen, B. J. Song, M. Barbacid, D. Goldman, I. J. Lee (1996). Isolation
and characterization of murine vav2, a member of the vav family of proto-oncogenes. Oncogene 13, 363-
371.
Schuebel, K. E., N. Movilla, J. L. Rosa, X. R. Bustelo (1998). Phosphorylation-dependent and constitutive
activation of Rho proteins by wild-type and oncogenic Vav-2. EMBO J. 17, 6608-6621.
Sebastian, J., R. G. Richards, M. P. Walker, J. F. Wiesen, Z. Werb, R. Derynck, Y. K. Hom, G. R. Cunha, R. P.
DiAugustine (1998). Activation and function of the epidermal growth factor receptor and erbB-2 during
mammary gland morphogenesis. Cell Growth Differ. 9, 777-785.
Segatto, O., F. Lonardo, J. H. Pierce, D. P. Bottaro, P. P. Di Fiore (1990). The role of autophosphorylation in
modulation of erbB-2 transforming function. New Biol. 2, 187-195.

88
Segatto, O., G. Pelicci, S. Giuli, G. Digiesi, P. P. Di Fiore, J. McGlade, T. Pawson, P. G. Pelicci (1993). Shc
products are substrates of erbB-2 kinase. Oncogene 8, 2105-2112.
Sibilia, M. and E. F. Wagner (1995). Strain-dependent epithelial defects in mice lacking the EGF receptor.
Science 269, 234-238.
Sibilia, M., J. P. Steinbach, L. Stingl, A. Aguzzi, E. F. Wagner (1998). A strain-independent postnatal
neurodegeneration in mice lacking the EGF receptor. EMBO J. 17, 719-731.
Silberstein, G. B. (2001). Postnatal mammary gland morphogenesis. Microsc. Res. Tech. 52, 155-162.
Sone, M., M. Hoshino, E. Suzuki, S. Kuroda, K. Kaibuchi, H. Nakagoshi, K. Saigo, Y. Nabeshima, C. Hama
(1997). Still life, a protein in synaptic terminals of Drosophila homologous to GDP-GTP exchangers.
Science 275, 543-547.
Songyang, Z., S. E. Shoelson, M. Chaudhuri, G. Gish, T. Pawson, W. G. Haser, F. King, T. Roberts, S.
Ratnofsky, R. J. Lechleider, a. et (1993). SH2 domains recognize specific phosphopeptide sequences. Cell
72, 767-778.
Songyang, Z., S. E. Shoelson, J. McGlade, P. Olivier, T. Pawson, X. R. Bustelo, M. Barbacid, H. Sabe, H.
Hanafusa, T. Yi, a. et (1994). Specific motifs recognized by the SH2 domains of Csk, 3BP2, fps/fes,
GRB-2, HCP, SHC, Syk, and Vav. Mol. Cell Biol. 14, 2777-2785.
Sonnenberg, A., C. J. Linders, P. W. Modderman, C. H. Damsky, M. Aumailley, R. Timpl (1990). Integrin
recognition of different cell-binding fragments of laminin (P1, E3, E8) and evidence that alpha 6 beta 1
but not alpha 6 beta 4 functions as a major receptor for fragment E8. J. Cell Biol. 110, 2145-2155.
Soriano, J. V., M. S. Pepper, T. Nakamura, L. Orci, R. Montesano (1995). Hepatocyte growth factor stimulates
extensive development of branching duct-like structures by cloned mammary gland epithelial cells. J. Cell
Sci. 108, 413-430.
Stein, D., J. Wu, S. A. Fuqua, C. Roonprapunt, V. Yajnik, P. D'Eustachio, J. J. Moskow, A. M. Buchberg, C. K.
Osborne, B. Margolis (1994). The SH2 domain protein GRB-7 is co-amplified, overexpressed and in a
tight complex with HER2 in breast cancer. EMBO J. 13, 1331-1340.

89
Steven, R., T. J. Kubiseski, H. Zheng, S. Kulkarni, J. Mancillas, A. Ruiz-Morales, C. W. Hogue, T. Pawson, J.
Culotti (1998). UNC-73 activates the Rac GTPase and is required for cell and growth cone migrations in
C. elegans. Cell 92, 785-795.
Streuli, C. H., N. Bailey, M. J. Bissell (1991). Control of mammary epithelial differentiation: basement
membrane induces tissue-specific gene expression in the absence of cell-cell interaction and
morphological polarity. J. Cell Biol. 115, 1383-1395.
Streuli, C. H., C. Schmidhauser, N. Bailey, P. Yurchenco, A. P. Skubitz, C. Roskelley, M. J. Bissell (1995a).
Laminin mediates tissue-specific gene expression in mammary epithelia. J. Cell Biol. 129, 591-603.
Streuli, C. H., G. M. Edwards, M. Delcommenne, C. B. Whitelaw, T. G. Burdon, C. Schindler, C. J. Watson
(1995b). Stat5 as a target for regulation by extracellular matrix. J. Biol. Chem. 270, 21639-21644.
Strutt, D. I., U. Weber, M. Mlodzik (1997). The role of RhoA in tissue polarity and Frizzled signalling. Nature
387, 292-295.
Sun, X. J., P. Rothenberg, C. R. Kahn, J. M. Backer, E. Araki, P. A. Wilden, D. A. Cahill, B. J. Goldstein, M. F.
White (1991). Structure of the insulin receptor substrate IRS-1 defines a unique signal transduction
protein. Nature 352, 73-77.
Sun, X. J., L. M. Wang, Y. Zhang, L. Yenush, M. G. Myers, E. Glasheen, W. S. Lane, J. H. Pierce, M. F. White
(1995). Role of IRS-2 in insulin and cytokine signalling. Nature 377, 173-177.
Sweeney, C. and K. L. Carraway (2000). Ligand discrimination by ErbB receptors: differential signaling through
differential phosphorylation site usage. Oncogene 19, 5568-5573.
Tamemoto, H., T. Kadowaki, K. Tobe, T. Yagi, H. Sakura, T. Hayakawa, Y. Terauchi, K. Ueki, Y. Kaburagi, S.
Satoh, a. et (1994). Insulin resistance and growth retardation in mice lacking insulin receptor substrate-1.
Nature 372, 182-186.
Tarakhovsky, A., M. Turner, S. Schaal, P. J. Mee, L. P. Duddy, K. Rajewsky, V. L. Tybulewicz (1995).
Defective antigen receptor-mediated proliferation of B and T cells in the absence of Vav. Nature 374,
467-470.

90
Tedford, K., L. Nitschke, I. Girkontaite, A. Charlesworth, G. Chan, V. Sakk, M. Barbacid, K. D. Fischer (2001).
Compensation between Vav-1 and Vav-2 in B cell development and antigen receptor signaling. Nature
Immunol. 2, 548-555.
Threadgill, D. W., A. A. Dlugosz, L. A. Hansen, T. Tennenbaum, U. Lichti, D. Yee, C. LaMantia, T. Mourton,
K. Herrup, R. C. Harris, a. et (1995). Targeted disruption of mouse EGF receptor: effect of genetic
background on mutant phenotype. Science 269, 230-234.
Timpl, R. (1996). Macromolecular organization of basement membranes. Curr. Opin. Cell Biol. 8, 618-624.
Tonner, E., M. C. Barber, M. T. Travers, A. Logan, D. J. Flint (1997). Hormonal control of insulin-like growth
factor-binding protein-5 production in the involuting mammary gland of the rat. Endocrinology 138,
5101-5107.
Tzahar, E., G. Levkowitz, D. Karunagaran, L. Yi, E. Peles, S. Lavi, D. Chang, N. Liu, A. Yayon, D. Wen, a. et
(1994). ErbB-3 and ErbB-4 function as the respective low and high affinity receptors of all Neu
differentiation factor/heregulin isoforms. J. Biol. Chem. 269, 25226-25233.
Tzahar, E., H. Waterman, X. Chen, G. Levkowitz, D. Karunagaran, S. Lavi, B. J. Ratzkin, Y. Yarden (1996). A
hierarchical network of interreceptor interactions determines signal transduction by Neu differentiation
factor/neuregulin and epidermal growth factor. Mol. Cell Biol. 16, 5276-5287.
Tzahar, E., R. Pinkas-Kramarski, J. D. Moyer, L. N. Klapper, I. Alroy, G. Levkowitz, M. Shelly, S. Henis, M.
Eisenstein, B. J. Ratzkin, M. Sela, G. C. Andrews, Y. Yarden (1997). Bivalence of EGF-like ligands
drives the ErbB signaling network. EMBO J. 16, 4938-4950.
Van Aelst, L. and C. D'Souza-Schorey (1997). Rho GTPases and signaling networks. Genes Dev. 11, 2295-2322.
van Genderen, C., R. M. Okamura, I. Farinas, R. G. Quo, T. G. Parslow, L. Bruhn, R. Grosschedl (1994).
Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in
LEF-1-deficient mice. Genes Dev. 8, 2691-2703.
Vonderhaar, B. K. (1987). Local effects of EGF, alpha-TGF, and EGF-like growth factors on lobuloalveolar
development of the mouse mammary gland in vivo. J. Cell Physiol 132, 581-584.

91
Webster, M. A., R. D. Cardiff, W. J. Muller (1995). Induction of mammary epithelial hyperplasias and mammary
tumors in transgenic mice expressing a murine mammary tumor virus/activated c-src fusion gene. Proc.
Natl. Acad. Sci. U. S. A 92, 7849-7853.
Weidner, K. M., S. Di Cesare, M. Sachs, V. Brinkmann, J. Behrens, W. Birchmeier (1996). Interaction between
Gab1 and the c-Met receptor tyrosine kinase is responsible for epithelial morphogenesis. Nature 384, 173-
176.
Wen, D., E. Peles, R. Cupples, S. V. Suggs, S. S. Bacus, Y. Luo, G. Trail, S. Hu, S. M. Silbiger, R. Ben-Levi, R.
A. Koski, H. S. Lu, Y. Yarden (1992). Neu differentiation factor: a transmembrane protein containing an
EGF domain and an immunoglobulin homology unit. Cell 69, 559-572.
Wiesen, J. F., P. Young, Z. Werb, G. R. Cunha (1999). Signaling through the stromal epidermal growth factor
receptor is necessary for mammary ductal development. Development 126, 335-344.
Withers, D. J., J. S. Gutierrez, H. Towery, D. J. Burks, J. M. Ren, S. Previs, Y. Zhang, D. Bernal, S. Pons, G. I.
Shulman, S. Bonner-Weir, M. F. White (1998). Disruption of IRS-2 causes type 2 diabetes in mice.
Nature 391, 900-904.
Woldeyesus, M. T., S. Britsch, D. Riethmacher, L. Xu, E. Sonnenberg-Riethmacher, F. Abou-Rebyeh, R.
Harvey, P. Caroni, C. Birchmeier (1999). Peripheral nervous system defects in erbB2 mutants following
genetic rescue of heart development. Genes Dev. 13, 2538-2548.
Wysolmerski, J. J., W. M. Philbrick, M. E. Dunbar, B. Lanske, H. Kronenberg, A. E. Broadus (1998). Rescue of
the parathyroid hormone-related protein knockout mouse demonstrates that parathyroid hormone-related
protein is essential for mammary gland development. Development 125, 1285-1294.
Xie, W., A. J. Paterson, E. Chin, L. M. Nabell, J. E. Kudlow (1997). Targeted expression of a dominant negative
epidermal growth factor receptor in the mammary gland of transgenic mice inhibits pubertal mammary
duct development. Mol. Endocrinol. 11, 1766-1781.
Yamauchi, T., K. Tobe, H. Tamemoto, K. Ueki, Y. Kaburagi, R. Yamamoto-Honda, Y. Takahashi, F.
Yoshizawa, S. Aizawa, Y. Akanuma, N. Sonenberg, Y. Yazaki, T. Kadowaki (1996). Insulin signalling
and insulin actions in the muscles and livers of insulin-resistant, insulin receptor substrate 1-deficient
mice. Mol. Cell Biol. 16, 3074-3084.

92
Yang, Y., E. Spitzer, D. Meyer, M. Sachs, C. Niemann, G. Hartmann, K. M. Weidner, C. Birchmeier, W.
Birchmeier (1995). Sequential requirement of hepatocyte growth factor and neuregulin in the
morphogenesis and differentiation of the mammary gland. J. Cell Biol. 131, 215-226.
Yenush, L., R. Fernandez, M. G. Myers, T. C. Grammer, X. J. Sun, J. Blenis, J. H. Pierce, J. Schlessinger, M. F.
White (1996). The Drosophila insulin receptor activates multiple signaling pathways but requires insulin
receptor substrate proteins for DNA synthesis. Mol. Cell Biol. 16, 2509-2517.
Zhang, D., M. X. Sliwkowski, M. Mark, G. Frantz, R. Akita, Y. Sun, K. Hillan, C. Crowley, J. Brush, P. J.
Godowski (1997). Neuregulin-3 (NRG3): a novel neural tissue-enriched protein that binds and activates
ErbB4. Proc. Natl. Acad. Sci. U. S. A 94, 9562-9567.
Zhang, R., F. W. Alt, L. Davidson, S. H. Orkin, W. Swat (1995). Defective signalling through the T- and B-cell
antigen receptors in lymphoid cells lacking the vav proto-oncogene. Nature 374, 470-473.
Zheng, Y., D. J. Fischer, M. F. Santos, G. Tigyi, N. G. Pasteris, J. L. Gorski, Y. Xu (1996). The faciogenital
dysplasia gene product FGD1 functions as a Cdc42Hs-specific guanine-nucleotide exchange factor. J.
Biol. Chem. 271, 33169-33172.
Zrihan-Licht, S., B. Deng, Y. Yarden, G. McShan, I. Keydar, H. Avraham (1998). Csk homologous kinase, a
novel signaling molecule, directly associates with the activated ErbB-2 receptor in breast cancer cells and
inhibits their proliferation. J. Biol. Chem. 273, 4065-4072.

93
ABBREVIATIONS
ATP adenosine 5' triphosphate
cDNA complementary DNA
C-terminus carboxyl terminus
DMEM Dulbecco's modified Eagle medium
DMSO dimethyl sulfoxide
DNA deoxyribonucleic acid
E day of embryonic development
ECM extracellular matrix
EDTA ethylendiaminotetraacetic acid
EGF epidermal growth factor
EHS Engelbreth-Holm-Swarm murine tumor
FCS fetal calf serum
FGF fibroblast growth factor
GDP guanosine 5' diphosphate
GTP guanosine 5' triphosphate
HA hemagglutinin epitope
HEPES N-(2-hydroxyethyl)piperazine-N'-(2-ethanesulfonic acid)
HGF/SF hepatocyte growth factor/scatter factor
IgG Immunoglobulin Class G
JNK/SAPK c-Jun N-terminal kinase/stress-activated protein kinase
kDa kiloDalton
LB Luria-Bertani medium
M mol/l
MAPK mitogen-activated protein kinase
Matrigel reconstituted basement membrane from EHS
mRNA messenger RNA

94
NGF nerve growth factor
NP-40 Nonidet P-40
NRG neuregulin
OD optical density
PAGE polyacrylamide gel electrophoresis
PBS phosphate buffered saline
PCR polymerase chain reaction
PDGF platelet-derived growth factor
PEG polyethylene glycol
PLCγ1 phospholipase C gamma1
PMSF phenylmethylsulfonylfluoride
Y tyrosine residue
RNA ribonucleic acid
SDS sodium dodecylsulphate
TE tris-EDTA solution
TGF transforming growth factor
Tpr Translocated promoter region
Tris tris(hydroxymethyl)-aminomethane hydrochloride
x-Gal 5-chlor-4-brom-3-indolyl-β-galactoside
YPD yeast extract/peptone/dextrose

95
ERKLÄRUNG
Hiermit erkläre ich, dass ich die vorliegende Dissertation selbständig und nur unter
Verwendung der angegebenen Hilfsmittel erarbeitet und verfasst habe. Diese Arbeit wurde
keiner anderen Prüfungsbehörde vorgelegt.
Berlin, 5. Juli 2001.
Silvana Di cesare

96
CURRICULUM VITAE
Name Di Cesare, Silvana Patricia
Date of birth 23. September 1963
Place of birth La Plata, Argentina
Education
1968-1975 Primary school
Colegio Corazon Eucaristico de Jesus, La Plata
1976-1980 Secondary school
Colegio Corazon Eucaristico de Jesus, La Plata
1981-1989 Studies and Graduation in Biochemistry
Facultad de Ciencias Exactas, Universidad Nacional de La Plata
1992-1995 Dissertation in Human Biology: Studies of the immunoneuroendocrine
interactions in an animal model of systemic lupus erythemathosus.
Supervisor: Prof. Dr. Hugo Besedovsky
Philipps-Universität, Marburg
since 1996 Graduate student. Supervisor: Prof. Dr. Walter Birchmeier
Max-Delbrück-Centrum für Molekulare Medizin, Berlin

97
PUBLICATIONS
Weidner, K. M., S. Di Cesare, M. Sachs, V. Brinkmann, J. Behrens, W. Birchmeier (1996).
Interaction between Gab1 and the c-Met receptor tyrosine kinase is responsible for epithelial
morphogenesis. Nature 384, 173-176.
Birchmeier, W., V. Brinkmann, C. Niemann, S. Meiners, S. Di Cesare, H. Naundorf, M.
Sachs (1997). Role of HGF/SF and c-Met in morphogenesis and metastasis of epithelial cells.
In Plasminogen-related growth factors (eds. G. R. Bock and J. A. Goode). John Wiley and
Sons Ltd, West Sussex, England.
Di Cesare S., K. M. Weidner, M. Sachs, V. Brinkmann, W. Birchmeier (1996). Gab1
functionally interacts with c-Met to mediate branching morphogenesis. In Mol. Biol. Cell
(ASCB Abstracts, Supplement to Vol. 7).
Grimm, J., M. Sachs, S. Britsch, S. Di Cesare, T. Schwarz-Romond, K. Alitalo, W.
Birchmeier (2001). Novel p62dok family members, dok-4 and dok-5, are substrates of the c-
Ret receptor tyrosine kinase and mediate neuronal differentiation. J. Cell Biol. 154(2), 345-
354.
Di Cesare S., J. Huelsken, B. Erdmann, M. Sachs, J. Grimm, W. Birchmeier (2001). The
guanine nucleotide exchange factor Vav2 induces alveolar morphogenesis of mammary gland
epithelial cells. (Manuscript in preparation).