CH 13 Review
17
1 Chapter 13 Chem. 555/Biol. 545 PRINCIPLES OF BIOCHEMISTRY THE CITRIC ACID CYCLE: (Krebs/Tricarboxylic [TCA] Cycle) LECTURE OBJECTIVES 1. Understand the central position of the PDH reaction in overall metabolism. Pyruvate Dehydrogenase (PDH) is a polyfunctional enzyme complex that utilizes FIVE different cofactors to carry out oxidative decarboxylation of Pyruvate . **Commited step in Pyruvate Catabolism** 2. Know the sequence of reactions involved in the conversion of pyruvate to acetyl CoA: the structure of the pyruvate dehydrogenase (PDH) complex; the role of each enzyme and coenzyme in its action. PDH Complex: consists of 3 e nzymes and 5 cofactors • 3 enzymes – E1 = Pyruvate dehydrogenase – E2 = Dihydrolipoamide acetyltransferase – E3 = Dihydrolipoamide dehydrogenase • 5 Cofactors – Thiamine pyrophosphate – FAD – Lipoic acid – NAD + – Coenzyme A (SH-CoA) STEP ONE Enzyme: Pyruvate dehydrogenase Cofactor: Thiamine PP (TPP) - Decarboxylate s Pyruvate - The C atom flanked by the S and N acts as a carbanion which attacks the carbonyl C of Pyruvate leads to loss of CO 2 forming 1 st product in overall reaction Hydroxyethy lthiamine pyropho sphate (HETPP): First product of overal l reaction
-
Upload
nanastasia-welnick -
Category
Documents
-
view
225 -
download
0
Transcript of CH 13 Review

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 1/17
Chapter 13 Chem. 555/Biol. 545 PRINCIPLES OF BIOCHEMISTRY
THE CITRIC ACID CYCLE: (Krebs/Tricarboxylic [TCA] Cycle)
LECTURE OBJECTIVES
1. Understand the central position of the PDH reaction in overall metabolism.
Pyruvate Dehydrogenase (PDH) is a polyfunctional enzyme complex that utilizes FIVE different cofactors to car
out oxidative decarboxylation of Pyruvate.
**Commited step in Pyruvate Catabolism**
2. Know the sequence of reactions involved in the conversion of pyruvate to acetyl CoA: the structure of t
pyruvate dehydrogenase (PDH) complex; the role of each enzyme and coenzyme in its action.
PDH Complex: consists of 3 enzymes and 5 cofactors
• 3 enzymes
– E1 = Pyruvate dehydrogenase
– E2 = Dihydrolipoamide acetyltransferase
– E3 = Dihydrolipoamide dehydrogenase
• 5 Cofactors
– Thiamine pyrophosphate
– FAD
– Lipoic acid
– NAD+
– Coenzyme A (SH-CoA)
STEP ONEEnzyme: Pyruvate dehydrogenase
Cofactor: Thiamine PP (TPP)
- Decarboxylates Pyruvate
- The C atom flanked by the S and N acts as a carbanion which attacks the carbonyl C o
Pyruvate leads to loss of CO2forming 1st
product in overall reaction
Hydroxyethylthiamine pyrophosphate (HETPP): First product of overall reaction

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 2/17
- It is the acetaldehyde-adduct of the cofactor TPP
- It is decarboxylated Pyruvate
STEP 2
Enzyme: Pyruvate dehydrogenase
Cofactor: Lipoamide
- Prosthetic group- Oxidizing agent that converts HETPP (the acetaldehyde TPP adduct) Acetyl dibydrolipoamide
STEP 3Enzyme: Dihydrolipoamide acetyltransferase

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 3/17
Cofactor: Coenzyme A
- Second Substrate
The disulfhydryl (reduced) form of Lipoamide must be reoxidized back to the disulfide (oxidized) form for tenzyme complex to continue operating.
STEP 4
Enzyme: Dihydrolipoamide dehydrogenase
- The FAD group of E3 Oxidizes the reduced Lipamide of E2 forming FADH2
Cofactor: FAD
- Prosthetic group that serves as the OXIDANT to convert reduced Dihydrolipoamide back
the active oxidized disulfide form Lipoamide
STEP 5Enzyme: Dihydrolipoamide dehydrogenase
Cofactor: NAD+
- 3rd substrate
- Oxidizes the FADH2 prosthetic group of E3 back to FAD regenerating the original holoenzyme acompleting the catalytic cycle
Overview of Products and Substrates
Step 1: Step 3: Step 5:
Substrate 1: Pyruvate Substrate 2: Coenzyme A Substrate 3: NAD+
Product 1: CO2 Product 2: Acetyl CoA Product 3: NADH
(reductant) (oxidized)
(oxidant) (reduced)

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 4/17
The Citric Acid Enzymes:-Localized in the mitochondria
Citric Acid CycleFor each acetyl group that enters the pathway:
- 2 molecules of CO2 are released - mobile coenzymes NAD+
and Q are reduced
- 1 molecule of GDP (or ADP) are phosphorylated - oxaloacetate is re-formed
Step Substrate Enzyme Reaction Type Product
1 Acetyl-CoA Citrate synthase Condensation HS-CoA, H+
2 Citrate Aconitase Dehydration
3 Isocitrate Isocitrate dehydration Oxidation
decarboxylation
NADH, CO2
4 α-Ketoglutarate α-Ketoglutarate
dehydrogenase complex
Oxidative
decarboxylation
NADH, CO2
5 Succinyl CoA Succinyl-CoA synthetase Substrate-level
phosphorylation
GTP (or ATP), HS-
CoA6 Succinate Succinate dehydrogenase
Complex
Oxidation QH2
7 Fumarate Fumarase Hydration
8 Malate Malate dehydrogenase Oxidation NADH, H+
Cycles per Glucose: 2 GTP(ATP) per cycle: 1 NADH per cycle: 3 Location: mitochondria
1. Citrate Synthase: condensation of Acetyl CoA with Oxaloacetate to form Citrate
Release of
Second CO2
Release of First CO2
Acceptor Molecule Feeder Molecule

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 5/17
Acetyl CoA + Oxaloacetate + H2O Citrate + HS-CoA + H+
First Metabolically Irreversible Reaction
- Driven by the high free energy of the thioester of Acetyl CoA
Only Reaction in TCA Cycle in which C-C bon formation occurs.
Acceptor Molecule: Oxaloacetate (4-C)Feeder Molecule: Acetyl CoA (2-C)
alcohol that cannot be oxidized under biological conditions
2. Aconitase: first causes the removal of water from Citrate to form cis-aconitate. Stereo specific r
addition of water forms Isocitrate.
Citrate↔ Isocitrate
**NOTE**
Primary alcohol: (one carbon bound)
- Oxidizes to a carboxylic acid or aldehye
Secondary alcohol: (two carbon bound)
- Oxidizes to a ketone
Tertiary alcohol: (three carbons bound)
- Not easily oxidized
- Aconitase removes H2O from Citrate to form the C=C bond of ci
Aconitate

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 6/17
- Stereospecific re-addition of H2O to cis-Aconitate forms 2 R,3S
Isocitrate.
- Aconitase is a Metabolically Reversible reaction.
3. Isocitrate Dehydrogenase: oxidizes Isocitrate to an unstable -keto form that spontaneous
decarboxylates by β-elimination, releasing CO2, forming
-Ketoglutarate.
Isocitrate + NAD+ α-Ketoglutarate + NADH + CO2
**Second METABOLICALLY IRREVERSIBLE reaction of the TCA Cycle.**
**1st
of 4 Oxidation-Reduction Reactions of the TCA Cycle**
forming an -keto group.
#2. Oxalosuccinate spontaneously converts to -KG releasing CO2 via a β-elimination reaction.- Oxalosuccinate is the intermediate in this dehydrogenation reaction and is enzyme-bound
- It is a -keto tricarboxylic acid
Citrate is a
non- oxidizable tertiary alcohol .
Isocitrate is an
oxidizable
secondary alcohol.

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 7/17
4. α-Ketoglutarate Dehydrogenase Complex: Oxidizes and Decarboxylates
Ketoglutarate
α-Ketoglutarate + HS-CoA + NAD+ Succinyl CoA + NADH + CO2
- Multi-enzyme complex similar to Pyruvate dehydrogenase complex
- -Ketoglutarate is an -Keto acid
- Products: CO2, NADH and Succinyl CoA.
- Succinyl CoA: high energy Thioester that participates in substrate-level phosphorylation in the next reaction
**Second Oxidative Decarboxylation**
**Third Metabolically Irreversible Reacion**
5. Succinyl- CoA Synthetase: Catalyzes the conversion of succinyl CoA to succinate
Succinyl CoA + GDP (or ADP) + Pi ↔ Succinate + GTP (or ATP) + HS-CoA
α - Ketoglutarate
dehydrogenase complex

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 8/17
- conserves energy of a
thioester to form GTP(a
nucleotide diphosphate)
- Once called
“Tkn”
- Substrate-level
phosphorylation
- Pi becomes
incorporated during the
energy transfer to
convert GDP to GTP
Proposed Mechanism of
Succinyl-CoA Synthase
Step 1 – Phosphate displaces SH-CoA
to form the mixed acid anhydride,succinyl phosphate.
Step 2 – The phosphoryl group is
transferred to a histidine residue of the
enzyme forming a relatively stable
covalent phosphoenzyme intermediate.
Step 3 – Succinate is released, and the
phosphoenzyme transfers its phosphory
group to GDP forming GTP.
8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 9/17
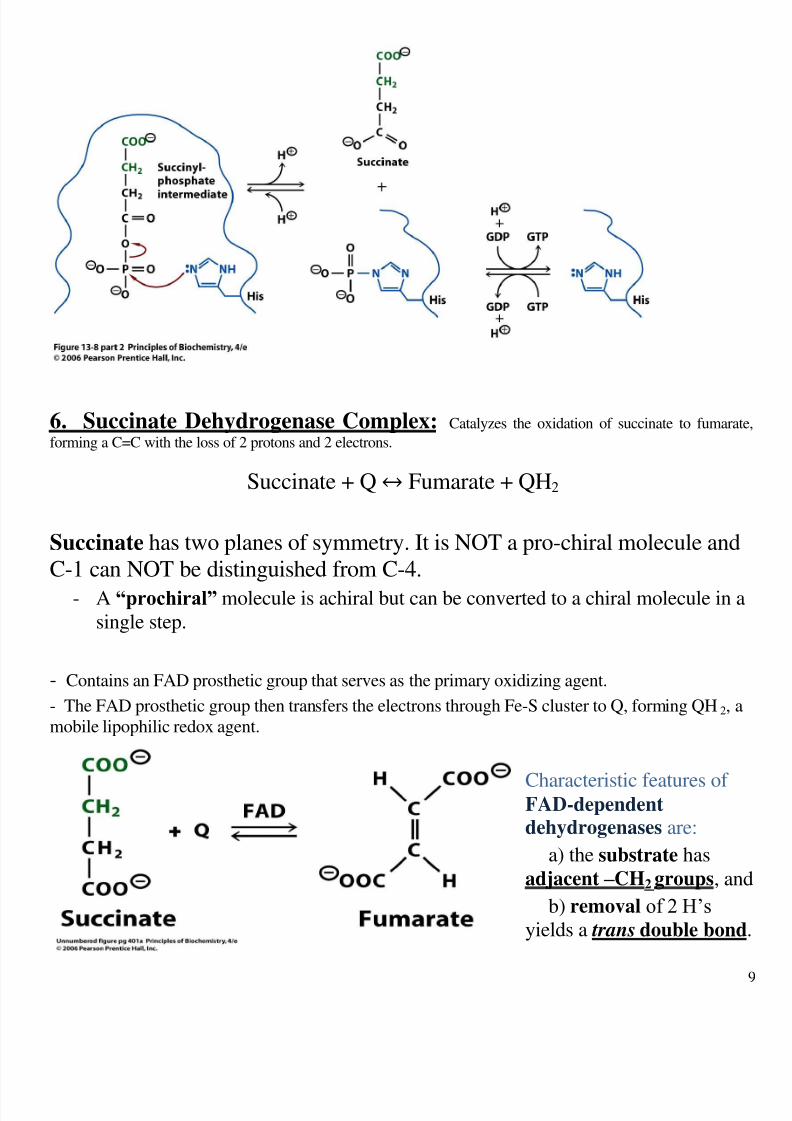
6. Succinate Dehydrogenase Complex: Catalyzes the oxidation of succinate to fumara
forming a C=C with the loss of 2 protons and 2 electrons.
Succinate + Q↔ Fumarate + QH2
Succinate has two planes of symmetry. It is NOT a pro-chiral molecule and
C-1 can NOT be distinguished from C-4.
- A “prochiral” molecule is achiral but can be converted to a chiral molecule in a
single step.
- Contains an FAD prosthetic group that serves as the primary oxidizing agent.
- The FAD prosthetic group then transfers the electrons through Fe-S cluster to Q, forming QH2, a
mobile lipophilic redox agent.
Characteristic features of FAD-dependent dehydrogenases are:
a) the substrate has
adjacent – CH2 groups, an
b) removal ’
yields a trans double bond

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 10/17
• Succinate Dehydrogenase is a complex of several polypeptides.
• It contains FAD as a prosthetic group, as well as Iron-Sulfur (Fe-S
clusters.
• The SDH Complex is located in the Inner Mitochondrial Membrane
7. Fumerase: Catalyzes the near-equilibrium conversion of fumarate to malate through the stereospecif
trans addition of water to the double bond of fumarate.
Fumarate + H2O↔ L-Malate
- Causes stereospecific addition of H2O to the trans double bond of Fumarate forming L-Malate
8. Malate Dehydrogenase: Oxidation of malate to regenerate oxaloacetate, with formation o
molecule of NADH.
L-Malate + NAD+ ↔ Oxaloacetate + NADH + H
+
- Uses NAD+ kn
- This NAD n O, nn “ ” - At equilibrium the reaction lies far to the LEFT; the level of L-Malate is much higher than the level
Oxaloacetate.
- Because of this, Oxaloacetate is the limiting substrate in the TCA Cycle.

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 11/17
IV. Energy yield of ATP per acetyl-CoA unit.
Net reaction of the Citric Acid Cycle:
Acetyl CoA + 3 NAD+
+ Q + GDP + Pi + 2 H2O HS-CoA + 3 NADH + QH2 + GTP + 2 CO2 + 2 H+
The complete oxidation of 1 molecule of acetyl CoA by the citric acid cycle and subsequent reactions is associatwith the production of about 10 molecules of ATP.
Reaction Energy-yielding product ATP equivalents
Isocitrate dehydrogenase (Step 3) NADH 2.5
α-Ketoglutarate dehydrogenase complex (Step 4) NADH 2.5
Succinyl-CoA synthetase (Step 5) GTP or ATP 1.0
Succinate dehydrogenase complex ( Step 6) QH2 1.5
Malate dehydrogenase (Step 8) NADH 2.5
Total 10.0
The TCA cycle- is a mechanism for oxidizing Acetyl CoA to CO2 using NAD
+and Q as oxidants
- Does not cause NET degradation of any cycle intermediates
--Energy is conserved in the form of Reduced Coenzymes: 3 NADH, 1 QH2 and 1 GTP (Nucleoti
Triphosphate)
--NADH and QH2 are reoxidized in the Electron Transport Scheme and produce much more ATP
Oxidative Phosphorylation.
Glycolysis converts glucose to 2 molecules of pyruvate with a net gain of 2 molecules of ATP. There are
molecules of NADH produced in the reaction catalyzed by glyceraldehyde-3-phosphate dehydrogenase. = 7 ATP
The conversion of both pyruvate molecules to acetyl CoA by the pyruvate dehydrogenase complex yields 2 NADmolecules, which correspond to about 5 additional molecules of ATP.

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 12/17
Fate of Acetyl-CoA’s carbon Units
Carbon atoms from Acetyl CoA (red) are not lost as CO2 in the first turn of the cycle.
At Succinate, because of its 2 planes of symmetry, atoms from Acetyl CoA becom
randomized.
Only in the second cycle do labeled C-atoms from Acetyl CoA become oxidized to CO2
The 2 C atoms that enter the cycle as the acetyl group on acetyl CoA are not t
same carbon atoms that are lost as CO2.
o The carbon balance in the overall reaction pathway: fo
each 2-C group from acetyl CoA that enters the cycle,
carbon atoms are released during 1 complete turn of th
cycle.
o The 2 C atoms of acetyl CoA become half of thsymmetric 4 carbon dicarboxylic acid (succinate) in the 5
step of the cycle.
o The 2 halves of this symmetric molecule are chemicall
equivalent so carbons arising from acetyl CoA becom
evenly distributed in molecules formed from succinate.
For each 2-C acetyl-CoA unit that enters the TCA cycle:-2 CO2 molecules generated - b ’ y
-3 NADH (+ 3 H+), 1 QH2, and 1 GTP (equiv. To 1 ATP) generated.
The ’ y-CoA are not oxidized in the TCA cycle, but are transferred to redox cofactors for subseque
oxidation in the electron-transport scheme (ETS).
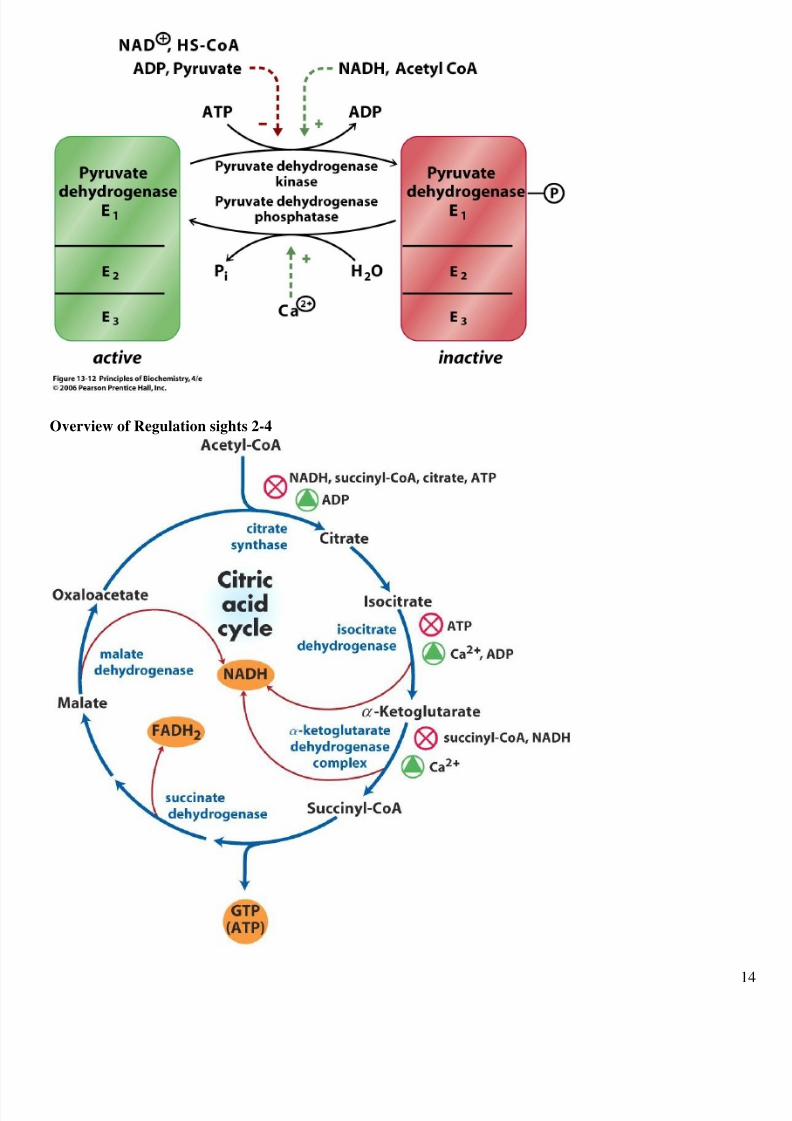
Regulation of the Citric Acid Cycle
The Rate of the Cycle is controlled at 4 sites:
1) The Pyruvate Dehydrogenase Complex, which controls the supply of Acetyl CoA.
2) Citrate Synthase
3) Isocitrate Dehydrogenase
4) The -Ketoglutarate Dehydrogenase Complex

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 13/17
The principal allosteric inhibitors are NADH and ATP, both of which signal energy excess.
1) Regulation of PDH Complex
A. By Allosteric Modulation
Increased levels of Acetyl CoA and NADH inhibit E2 & E3. Feedback Regulation
Increased levels of CoA-SH and NAD+ activate E2 & E3. Feed Forward Regulation
B. By Covalent Modification (Mammalian PDH Complex)
Phosphorylation INACTIVATES E1.Dephosphorylation ACTIVATES E1.
The Substrates make the complex ACTIVE by inhibiting PDH Kinase: NAD+,
HS-CoA, ADP and Pyruvate
The Products INACTIVATE the complex by activating PDH Kinase: NADH
and Acetyl CoA
8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 14/17
Overview of Regulation sights 2-4

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 15/17
Suggested Problems:
5. The disease beriberi, which results from a dietary deficiency of vitamin B1 (thiamine), is characterized by
n n y, w n y n α-ketoglutarate in the blood. How
ny n n n y n α-ketoglutarate?
Thiamine is the precursor of the coenzyme thiamine pyrophosphate (TPP), which is found in two enzyme
complexes associated with the citric acid cycle: the pyruvate dehydrogenase complex and the α-
ketoglutarate dehydrogenase complex. A deficiency of TP decreases the activities of these enzyme
complexes. Decreasing the conversion of pyruvate to acetyl CoA and of α-ketoglutarate to succinyl CoAcauses accumulation of pyruvate and α-ketoglutarate.
8. A deficiency of a citric acid cycle enzyme in both mitochondria and the cytosol of some tissues (e.g. blood
lymphocytes) results in severe neurological abnormalities in newborns. The disease is characterized by excretio
n n bny n α-ketoglutarate, succinate and fumarate. What enzyme deficiency
would lead to these symptoms?
A deficiency in the citric acid cycle enzyme fumarase would result in abnormally high concentrations of
fumarate and prior cycle intermediates including succinate and α-ketoglutarate, which could lead to
excretion of these molecules.
1. The oxidation of pyruvate to acetyl CoA and CO2 requires the cofactors coenzyme A, NAD+, lipoate, FAD
and TPP.
2. Four dicarboxylic acid intermediates of the citric acid cycle are succinate, fumarate, malate and
oxaloacetate. 3. The conversion of α-ketoglutarate also produces CO2. This is an oxidative process in which the
actual oxidant is NAD+.
4. Transformation of pyruvate to acetyl CoA is catalyzed by the pyruvate dehydrogenase complex [This
complex contains multiple copies of pyruvate dehydrogenase (E1), dihydrolipoamide acetyltransferase
(E2), and dibydrolipoamide dehydrogenase (E3).]
5. The enzyme that catalyzes the first step of the citric acid cycle is citrate synthase 6. T ny y nn α-ketoglutarate to succinyl CoA is the α-ketoglutarate
dehydrogenase complex. 7. The only step in the citric acid cycle that directly produces an energy-rich phosphoanhydride involves the
conversion of succinyl CoA to succinate
8. There are a total of five (including the conversion of pyruvate to acetyl CoA oxidation-reduction steps in
the catabolism of pyruvate via the citric acid cycle. The specific reduced coenzymes formed are NADH and
QH2 9. The competitive inhibitor malonate slows the citric acid cycle and causes succinate to accumulate.
10. Of the 8 steps in the citric acid cycle, the number of steps that produce CO2 is two.
11. Certain citric acid cycle intermediates serve as biosynthetic precursors of other materials. Two such
intermediates are α-ketoglutarate and succinyl CoA (as well as citrate and oxaloacetate) 12. A negative modulator involved in regulating each of the enzymes that catalyze the 3 metabolically irreversib
reactions of the citric acid cycle is NADH
13. The intermediate L-malate is formed by the hydration of fumarate in the cycle.14. The second CO2 producing step in the citric acid cycle involves the conversion of α-ketoglutarate to succin
CoA

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 16/17
1. What is coenzyme A and what is its function in the oxidation of pyruvate via the citric acid cycle?
Coenzyme A is composed of an ADP moety, an esterified phosphate, a pantothenic acid unit, and a
mercaptoethylamine unit. Coenzyme A functions as a carrier of acyl groups such as the acetyl group
formed when pyruvate is oxidatively decarboxylated by the pyruvate dehydrogenase complex, and the
succinyl group formed when α-ketoglutarate is oxidatively decarboxylated by the α-ketoglutarate
dehydrogenase complex
3. In eukaryotes, pyruvate is produced from glucose by glycolysis in the cytosol, but the enzymes of the citric
acid cycle are in the mitochondrial matrix (except for succinate dehydrogenase, which is part of Complex II in thinner mitochondrial membrane). How are these catabolic pathways connected despite being located in differentcell compartments?
Pyruvate, produced in the cytosol by glycolysis, enters mitochondria through porins in the outer
membrane and by the aid of pyruvate translocase in the inner membrane. In the mitochondrial matrix,
pyruvate is converted to acetyl CoA, a substrate for the citric acid cycle.
True/False
1. ________ FAD is used in the citric acid cycle as an oxidizing agent2. ________ The final steps in the oxidation of glucose to CO2 occur in the citric acid cycle.
3. ________ Massive amounts of all intermediates of the citric acid cycle must be present for its efficient
operation.4. ________ In eukaryotes, the enzymes that catalyze the reactions of the citric acid cycle are found in the
cytosol.
6. ________ Pyruvate is produced in the eukaryotic cytosol but enters the mitochondrial matrix unaided.
7. ________ The eukaryotic pyruvate dehydrogenase complex is the largest multienzyme complex known8. ________ Intermediates of the citric acid cycle that are used for anabolic (synthetic) needs of the cell are
replenished by anaplerotic reactions.
9. ________ Porphyrin biosynthesis could potentially interfere with the citric acid cycle by depleting succinylCoA
10. ________ Two moles of coenzyme A are consumed per mole of pyruvate catabolized to acetyl CoA, which
subsequently consumed in the citric acid cycle.11. ________ The citric acid is controlled entirely by the availability of acetyl CoA and cycle intermediates.
12. ________ Per mole of glucose, the citric acid cycle produces as many high-energy phosphate molecules by
substrate level phosphorylation as does glycoltic catabolism of glucose to pyruvate.
13. ________ A prochiral molecule is a symmetrical molecule that can be converted to a chiral molecule bysubstitution of a single functional group.
4 ________ T α-ketoglutarate dehydrogenase complex is very much like the pyruvate dehydrogenase
complex.
15. ________ The reaction catalyzed by the succinate dehydrogenase complex is the only step in the citric acidcycle that involves substrate level phosphorylation.

8/3/2019 CH 13 Review
http://slidepdf.com/reader/full/ch-13-review 17/17
1. True. FAD is a prosthetic group of an enzyme of the succinate dehydrogenase complex that accepts hydroge
form succinate and immediately transfers them to coenzyme Q, the mobile carrier of reducing power within the
inner mitochondrial membrane.
2. True. The citric acid cycle generages 6 molecules of CO2 for each molecule of glucose catabolized.3. False. Your text points out that the cycle is a multistep catalyst. Since the process is cyclic, the intermediate
are continually recycled. However, sufficient amounts of oxaloacetate must be present to allow appropriate rate
of acetyl CoA uptake.4. False. There are 2 six-carbon intermediates, 1 five-carbon intermediate, and 5 four-carbon intermediates.
5. False. They are located in the mitochondria.6. False. Pyruvate does pas freely through porins in the outer mitochondrial membrane, but its passage, insymport with H+, through the inner mitochondrial membrane is aided by pyruvate translocase.
7. True. It is several times larger than a ribosome.
8. True.9. True. Succinyl CoA condenses with glycine to initiate porphyrin biosynthesis, but succinyl CoA can be
replenished by degradation of some amino acids.
10. False. Although conenzyme A is required in 2 different steps as pyruvate is catabolized, two moles of
coenzyme A are formed as products in the citric acid cycle, so there s no net consumption of coenzyme A.11. False. In addition to the availability of acetyl CoA and oxaloacetate to initiate citrate formation, control is
exercised through allosteric modulators and their effects on the cycle enzymes citrate synthase, isocitrate
dehydrogenase, n α-ketoglutarate dehydrogenase complex.12. True. Glycolysis produces 2 ATP per mole of glucose. From the two moles of pyruvate produced byglycolysis, the citric acid cycle produces 2 moles of GTP (or ATP)
13. True. The prochiral molecule contains a carbon of the type Caacd which can be converted to a chiral molecu
by n n α by n nn T bn wthen be Cabcd
14. True. Both are complexes of several different enzymes and utilize the same cofactors (TPP, lipoamide, FAD
CoASH, and NAD+)
15. False. It is the step catalyzed by succinyl CoA synthase that results in substrate level phosphorylation asGTP (or ATP) is formed.