
Die Abgrenzung von Biospezies auf biologisch-ethologischer ... · Zool. Jb. Syst. Bd. 9G, S....
132
Zool. Jb. Syst. Bd. 9G, S. 133-264 (1969) Aus dem Institut fur Allgemeine Zoologie der Johannes Gutenberg-Universitat Maind Die Abgrenzung von Biospezies auf biologisch-ethologischer und morphologischer Grundlage am Beispiel der Gattung Ischyropsalis C. L. KOCH 1839 (Opiliones, Is~hyropsalididae)~) Von JOCHEN MARTENS Mit 68 Abbildungen Mcinem Freund Dr. VLADIM~R SILHAVY CSC gewidmet Summary Attempts are macle to reclassify the gcnus Isehyropsal~s C L KOCIIand to ascei tain the phylogenetic relationships of thc single species. This attempt is based on the modern deflnition of species, according to wliich onlysuch groups of indivldvals are considered as species which form a natural propagation commuuity ("biospecies"). ThereIore it was first endeavourcd to find the limits of several biospecies with biological-ethological criteri'is Researches regarding the mating behaviour disclosed that J secondary aeu character("brist1e") on tlic first inembcr of male chelicerae is a species marking character. 'Phis discovery permits brospecific classification of other popula- tions which possess the same character bnt which Iiave a different marlring ratio. More morpho- logical species marking characters can now bc made available and tr~nsfcrred to populations not possessing the bristle character. This type ol species is cnlled inorphospecies 'L'heir position as a biospecies can be demonstrated only when it occurs with reldted species in the same area (sym- patricor even syntopic) Tnhaltsverzeichnis .... .................. I. Einleitung und Problemstellung .> 135 11. lfaterial und Methode (Liste der validen Arten) . 1 .... : .......... 136 111. Biologisch-ökologischer Seil ......................... 138 1. Vorkommen, Biotopanspriiche ...................... 138 . 2. Höhenverbreitung ............................ 140 3. Vorkommen in Höhlen .......................... 141 4. Syntopisches Vorkommen zweier Arten .................. 143 5. ~hä&logie ............. .; ; .,, ............... 144 I 6. Nahrungjpnd Nahrungsaufnahme ... 'i" ;d,ll .,:. " , . . . . . . . . . . 147 I Q:) Kopulaii p8verhalten . . . . . V / . li ..... l ....... I. , . 149 114 ~a 1 1 : I !I 1) Teil einer Dissertation, Naturwissenschaftliche adkultät der I 1111
Transcript of Die Abgrenzung von Biospezies auf biologisch-ethologischer ... · Zool. Jb. Syst. Bd. 9G, S....

Zool. Jb . Syst. Bd. 9G, S. 133-264 (1969)
Aus dem Institut fur Allgemeine Zoologie der Johannes Gutenberg-Universitat Maind
Die Abgrenzung von Biospezies auf biologisch-ethologischer und morphologischer Grundlage am Beispiel der Gattung Ischyropsalis C. L. KOCH 1839
(Opiliones, Is~hyropsal ididae)~)
Von JOCHEN MARTENS
Mit 68 Abbildungen
Mcinem Freund Dr. VLADIM~R SILHAVY CSC gewidmet
Summary
Attempts are macle to reclassify the gcnus Isehyropsal~s C L KOCII and to ascei tain the phylogenetic relationships of thc single species. This attempt is based on the modern deflnition of species, according to wliich onlysuch groups of indivldvals are considered as species which form a natural propagation commuuity ("biospecies"). ThereIore i t was first endeavourcd to find the limits of several biospecies with biological-ethological criteri'is Researches regarding the mating behaviour disclosed tha t J secondary aeu character("brist1e") on tlic first inembcr of male chelicerae is a species marking character. 'Phis discovery permits brospecific classification of other popula- tions which possess the same character bnt which Iiave a different marlring ratio. More morpho- logical species marking characters can now bc made available and t r~nsfcr red to populations not possessing the bristle character. This type ol species is cnlled inorphospecies 'L'heir position as a biospecies can be demonstrated only when it occurs with reldted species in the same area (sym- patricor even syntopic)
T n h a l t s v e r z e i c h n i s
. . . . . . . . . . . . . . . . . . . . . . I. Einleitung und Problemstellung .> 135
11. lfaterial und Methode (Liste der validen Arten) . 1 . . . . : . . . . . . . . . . 136
111. Biologisch-ökologischer Seil . . . . . . . . . . . . . . . . . . . . . . . . . 138
1. Vorkommen, Biotopanspriiche . . . . . . . . . . . . . . . . . . . . . . 138
. 2. Höhenverbreitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . 140
3. Vorkommen in Höhlen . . . . . . . . . . . . . . . . . . . . . . . . . . 141
4. Syntopisches Vorkommen zweier Arten . . . . . . . . . . . . . . . . . . 143
5. ~hä&logie . . . . . . . . . . . . . .; ; .,, . . . . . . . . . . . . . . . 144 I
6. Nahrungjpnd Nahrungsaufnahme . . . ' i " ;d,ll . , : . " , . . . . . . . . . . 147
I Q:) Kopulaii p8verhalten . . . . . V / . li . . . . . l . . . . . . . I . , . 149 114 ~a 1 1 :
I !I
1) Teil einer Dissertation, Naturwissenschaftliche adkultät der I 1 1 1 1

IV . Morphologischer Teil . . . . . . . . . . . . . . . . . . . . . . . . . . . . 150
1 . Morphologie und systematische Wertigkcit dcs Genitalapparatcs und Organe . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . a ) Der Genitalapparat
. . . . . . . . . . . . . . . . . . . b) Das abdominale Scutuin . . . . . . . . Entwicl<lung des Scutum walirend der Ontogenese
. . . . . . . . . . . . . . . . . C) Die Cheliceren -Bewehrung
. . . . . . . . d) Die Apopliyse und das Drüscnfeld der$-Chelicere
. . . . . . . . . . . . . . . . . . . 2 Die isolierenden Afechanisn~en
peripherer . . . . . 150
. . . . . 150
. . . . . 153
. . . . . 155
. . . . . 160
. . . . . 165
. . . . . 166
. . . . . . . . . . . . . . 3 . Vergleich syrltopischer Formen nnd Estrapolatiori 169
V . Chorologie und Speziation . . . . . . . . . . . . . . . . . . . . . . . . . . 173
1 . Verbreitiing der Gattung und der Artengruppen . . . . . . . . . . . . . . . 173
R ) Das Gattungsareal . . . . . . . . . . . . . . . . . . . . . . . . . . 173
b) Die Areale der drei Artengruppen . . . . . . . . . . . . . . . . . . . . 175
2 . Verbreitungsgeschichte und Evolution der Arten . . . . . . . . . . . . . . 177
a) Die hellwig.i.Gruppe . . . . . . . . . . . . . . . . . . . . . . . . . . 178 b) Die kolluri-Gruppe . . . . . . . . . . . . . . . . . . . . . . . . . . 178
-östlich-alpines Teilareal . . . . . . . . . . . . . . . . . . . . . . . . 178 westliches Teilareal . . . . . . . . . . . . . . . . . . . . . . . . . . 181
. . . . . . . . . . . . . . . 3 . Evol~itiue Trends in der Gattung Ischyropsulis 181
:L) Dic Chcliceren-Länge . . . . . . . . . . . . . . . . . . . . . . . . . 181 . . . . . . . . . . . . . . . . . . . . . . . b) Die Cheliccren-Bewehrung 182
. . . . . . . . . . . . . . . . . . . . . . . . . c) Die Cheliceren-Bürste 182 . . . . . . . . . . . . . . . . . . . . . . . . d) Das abdominale Scutnm 183
V1 . Systeinatischer Seil . . . . . . . . . . 1 . Zur G attungsgruppierung . . . . . . 2 . Die Situation in der Spezies-Gliederung
3 . Die Taxa . . . . . . . . . . . . . . a) Die Artengruppen . . . . . . . . b) Die Arten . . . . . . . . . . . .
Die hellwigi-Gruppe . . . . . . . . I . hellwigi hellwigi (PATYZER) . . . . I . hellwigi lucantei SIMON . . . . .
. . . . . . Die dentipulpis-Gruppe . . . . . . I dentipalpis CANESTI~IXI
. . . . . . . . I adamii CANEST~XINI . . . . . . . . . . I carli LESSERT . . . . . . . . . . I luteipes SIMON
I . manicata 1, . Koca . . . . . . . . . . . . . . . . . I nodifera SJNON
J . pyrenaea SIION . . . . . . . . . . . . . . . . . . . , I robusta SIMON

Die Abgrenzung von Biospezies usw. 135
. . . . . . . . . . . . . . . . . . . . . . . . . . Die kollari-Gruppe 236 . . . . . . . . . . . . . . . . . . . . . . . . . I. kollari C. L. KOCH 236
. . . . . . . . . . . . . . . . . . . . . . . . . . . 1. hadzii ROEWER 242 . . . . . . . . . . . . . . . . . . . . . . . . . I. muellneri H A ~ ~ A N N 245
. . . . . . . . . . . . . . . . . . . . . . . . 1. slran.di KRI \TOCHV~L 249 . . . . . . . . . . . . . . . . . . . . . . . . . I. magdalenae SIMON 252
. . . . . . . . . . . . . . . . . . . . . . . . . . . . I. dispar SIMOH 255
V I I. Liste der noiiiinellen Arten (Verweis auf die validen Arten) . . . . . . . . . . . 259
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . IX. Sumniary 261
I. Einleitung und Problemstellung
Gegenwartig setzt sich immer mehr die Anschauung durch, daß Artcn (Spezies) nicht allein mittels morphologischer Merkmale definiert und gegen andcre Arteii ab- gegrenzt wcrdcn durfen. Vielmehr soll zur Kennzeichnung das entscheidende Ge- wicht auf dic biologischen Merkmale gelegt werden. Diese Forderung kommt in der heutc allgemein anerkannten Art-Definition zum Ausdruck, die nur solche Indivi- duell-Gruppen als Art bestimmt, dic eine naturliche Fortpflanzuiigsgemeinschaft bei freier Partnerwahl darstclleii (ausfuhrliche Diskussion zuletzt bei GUNTHER 1962, MAYR 196311967). Mit diesem Konzept wird zwar prinzipiell die morphologisch- typologische Arbeitsweise verlassen, auf der die allermeisten bis jetzt aufgestellten Spezies beruhen. Sie kann sich aber dennoch nur in seltenen Fallen von der bisher geubten Praxis losen, da es meistens nicht moglich ist, dic mit dieser biologisehcn Definition angcstrcbtc Arbeitswcisc konsequent durchzufuhren. Diese Schwierigkcit '
mindert anscheinend den Wcrt dcr Definition. Indes musscn die bisherigen Morpho- spezics den Biospezies nicht unbedingt widersprechen, denn „jcde Art ist auch Aus- druck formativer Krafte, sie hat eine Gestalt und nennt Lebenserscheinungen ihr eigen" (HERRE 1967). Somit läßt sich nahezu jede Art auch nach morphologischen Gcsichtspunkten umgrcnzcn, und viele nach diesen Kriterien aufgestellte Arten haben dcr kritischen Überprufuilg standgehalten. Mit dcr Definition der Biospezies wcrden also nicht dic morphologisch kcnnzeichneriden Merkmale verworfen, sondern mit ihr wird versucht, die Beziehungen zwischen biologischem „ArtseinU und den „Artkcnn- zeichen" aufzudeckcn.
Artsein und Artkennzeichcn implizieren dic Frage nach dcn Isolationsmeehanis- mcn, die sympatrisch oder gar syi~topisch lebende Individuen zwcier oder mehrerer nahverwandter Arten vor Gcnaustausch bewahren. Sic sind bei Arthropoden unter- schiedlich verwirklicht, lassen sich aber im allgemeinen auf ethologische, biochemi- sche und morphologische Mechanismen zuruckfuhren, die biswcilen kombiniert scin konnen.

Diese isolierenden Mechanismen, die als die eigentlichen artspezifischen Kenn- zeichen angesehen werden können, werden, durch vielfältige morphologische Merk- male ergänzt, die die Biospezies zusätzlich charakterisieren. Uni diese artkennzeich- nenden Merkmale in Tiergruppen erkennen zu können, in denen die herkönimlichen Kennzeichen - wie jene der Genitalmorphologie - versagen, kann die Kenntnis des Fortpflanzungsverhaltens bisweilen niitzlich sein, denn in ihm dürfen am ehesten isolierende Mechanismen vermutet werden. Lassen sich nämlich morphologische Merkmale finden, die in enger Beziehung zum Bortpflanzuiigsverhalten stehen und die bestimmte Verhaltensweisen sogar bedingen, darf uiiterschiedliche Ausprägung dieses Merkma'ls als Ausdruck von Artverschiedenheit angesehen werden. Zweifellos stellt eine'solche Extrapolation ein Wagnis dar (HERRE 1964), doch ~verdeii sich die relativ geringen Uiisicherheiten eliminieren lassen, wenn weitere morphologische, biologische und chorologische Merkmale hinzugezogeii werden.
Mit der vorstehend angedeuteten Arbeitsweise wurde Yersucht, die iii den Ge- birgen Mittel-, Süd- und Ost-Europas lebenden Populationen des Genus Ischyropsulis C. L. KOCH 1839 (Opiliones, Ischyropsalididae) auf ihre biospezifische Zusammeii- gehörigkeit zu prüfen. Das erschien um so reizvoller, als über die spezifischen Cha,rak- tere der bis jetzt aufgestellteil, zahlreichen iiominelleii Arteii keine begründeten An- schauungeii vorlagen, ferner die Biologie dieser auffälligen, aber im allgemeinen a,ls sehr selten geltenden Weberknechte bis in allerjiiiigste Zeit nahezu unbekannt war und schließlich bereits die oberflächliche Betrachtung der Cheliceren der Nanncheii einiger Arten ein sekundäres Geschlechtsmerkmal erkennen ließ, voll dem vermutet werden durfte, daß es bei bestimmten Verhaltensweiseii eine Rolle spielt.
Der zuiiächst a,iigestellte Versuch, die Gattung mit großem Material an Ha,nd der bisher ver~vendeteii Spezies-Merkmale zu revidieren, erwies sich bald als unmög- lich und blieb erfolglos. Jedoch führte die Uiitersuchuiig des Kopulation~verha~ltens zweier Arten zur Entdeckuiig eines bei Arachniden bisher uiibekarinteii Werbeverhal- tens, dessen Verlauf von dem bereits erwähiiteii sekuiidäreii Geschle~htsmerkma~l der Mäiinchen abhängt. Die mit diesen1 Kriterium era,rbeitete Neugliederung basiert überdies auch auf ökologischen und'chorologischen Merkmalen. Aus der iiun iiber- sichtlicheii Gliederung der Gattuiig (früher 56, jetzt 15 Arteii) ergeben sich Schlüsse für Speziationsvorgänge und für die Evolution der Arten innerhalb der Gattung.
11. Material und Methode
Um Einheitlichkeit in der B e n e n n u n g d e r A r t e n zu gewährleisten und um die Über- sicht zu erleichtern, wird eine Liste der validen Art-Nainen vorangestellt, wie sie sich aus den nachstehenden Untersuchungen ergeben haben. Eine Liste der Synonyine findet sich bei jedem Artkapitel (Synonyine der validen Arten) und eine weitere am Schluß dieser Arbeit (S. 250, nomi- nelle Arten mit Ver~veis auf die validen Arten).

Die Abgrenzung von Hiospezies icsw.
Ischyropsulis C. 1J. Koca 1839
1. hellwzgi-Gruppe I. hellwigz (PANZER 1794)
I. hellwzgi hellwigz (PANZER 1794) I. hellwzgz lucantez SIMON 1879
2. dentzpulpzs-Gruppe I . d e n t % p ~ l p i ~ ~ C ~ ~ E S T ~ l N 1 1872 I. udumzi CANESSRINI 1 873 I. curlz DE LESSERT 1905 I. lutezpes SIMON 1872 I . munzcuta L. KOCH 1865 I . nod~feru SII~ON 1879 I. pyrenueu SIMON 1872 I. robustu SIMON 1872
3. kollurz-Gruppe I . lcollurz C. L. KOCH 1839 . I. hudzzz ROFWER 1950 I. muellnert HAMMANN 1898 1. stranclt KRATOCHV~L 1936 I. nzagdalenue SIMON 1881 I. dzspar S~nfoni 1872
Mateiial ful Znc l i t u n d e t h o l o g i s c h e B e o b a c h t u n g e n sammelte ich selbst auf h'lhl- ie~chen Reisen in Ueutscliland (h hellwzg~, carlz), Osterieich (h hellwzgz, Icollarz), Ii'rankreich (lutez- pes, pyrenaea, h. lucantez) und Spanien (nodzfern, h lucccnte~) Lebende Tiere erhlelt ich uberdies von den Heilen J. G itiiilcix (h hellwzgz) und A. A u s o u s i , ~ (kollarz) - Eigene Nachsuche an den Loci typici von J alfkenr (.lena/ltosentc~l), I. kratochvzlz und I troglodytes (beide nilf dei Insel Kr eta) blieb eifolglos (vgl. Kap V 1 a)
Als Zu c h tgcf A R e dienten Petiisclialen nntei s~hiedlichei Große, die hirr Stabilisieiung Iioher J,uftfeucl-itigkeit init einer 1-2 cm hohen Cipssclii~bt ailsgegossen ~vilrden. Dse Gefaße u~iirrlen in einein Thermostat bei10 "C Dauer tempei~tur iinteigebracht (l'irma RUBARTIT 8.z Co ).
Cef u t t e r t \suide -von Art LU Ai t veiscliieden -mi t Cehausesclinecl<en, Asseln, Stnben-
1 fliegen und Drosophsla Besondeis Drosophzla wurde von den meisten Arten sehr gern angenommen.
Die Z e ic l in i ingen zum Systematischen Teil wurden mit einem gioßen Leitz schen Zei- chenspiegel angefeitigt (Leihgabe der DEG an Prof. Dr. R. BRAUN).
Fur die s y s t e i i i a t i s c h e B e a r b e i t u n g standen 586 adulte Individuen aller Arten zur Verfugung. Sie stammen zum Teil aus den eigenen Aufsainmlungen, zum großeren Teil aus folgen- den Museen und Piivatsanimliingen: Naturhistorisches Museum, 1,issabon; Sammlung RAMBLA, Barcelona; Museum d'IIistoire N;~tuielle, Genkve; Natuihistorisches R'luseum, Basel; Sainmlung D ~ e s c o , Paris; Sammlung THALLK, Innsbiuck; Sammlung S c i r u ~ ~ , c n / A u s o ~ s i ~ ~ , Salzburgl Bischofshofen, jetzt Natur-Miisciim und Forschungs-lnstitut Senckenberg; Naturhistorisches Museum, Wien; Sammlung C l tun~r t , Wien; Sammlung B R I G N ~ L I , Roma; Natur-Museum und Forschungs-Institut Senclrenberg, Franlifurt (mit der Sammlung ROEWER), Zoologisches Staats-

institut und hluseuin, Hamburg; Zoologisches Museuin der Universität, Berlin; Sammlung PAN, Warszawa; Sammlung SILHBVY, StafeE; Sammlung H A D ~ I , Ljubljana; Term6szettudominyi Muzeum, Budapest; Samemlung L o a s ~ , Budapest. - Leider war es trotz vieler Bemühungen nicht möglich, das Material der Sammlung SIMON, Paris, einzubeziehen.
FLir Ü b e r m i t t l u n g v o n M a t e r i a l und von Auskünften danke ich den Herren A. Ari- SOBSKY (Bischofshofen), Prof. Dr. R. B K ~ \ U K (Mainz), dott. P. BRIGNOLI (Roms), Dr. L. FORCART (Basel), Dr. H. GISIS (i, Genf), Prof. Dr. B. GAUCICLER (Xürnberg), J. GRUBER (Wien), Prof. Dr. I. H A D ~ I (Ljubljana), K.-H. HARYIS (~üb ingen) , Privatdozent Dr. 0. KRAUS (Frankfurtlhf.), S. Lö- SER (Wuppertal), 13. PIEPEN (Fulda), E. PRETNER (Postojna), Dr. V. SILHAVI CSc (S tak t ) , mag. W. STARKGA (Warszawa), Dr. K. THALER (Innsbruck), Privatdozent Dr. H. U. THIELE (Iioln) und Frau Prof. Dr. M. RAMBLA (Barcelona), ferner Doz. Dr. E. KULLYANN und Dr. V. SBORDONI.
Die arachnologische Abteilung der Senckenbergiscken Naturforschenden Gesellschaft ermöglichte durch vielseitige Hilfe, den systeinatischen Teil der Arbeit in umfassender Weise zu gestalten, wofür auch hier gedankt sei.
Besonderen Dank schulde ich meinen Freunden und Kollegen, die mich auf den zahlreichen S a m m e l e s k u r s io n e n innerhalb Europas begleitet und 'mir die Arbeit sehr erleichtert haben, vor allem Prof. Dr. R. BRAUN, Dr. V. SILHAVY CSc, Frl. D. UHRIG, 8 . AUSOBSI~Y, J. GRUBER, Ei.-H. H A R ~ , 0. V. HELT~ERSEN, H. PAULUS, H. PIEPER. In den Pyrenaen führten mich M. BER- ~ r i s und D. BOUTET zu Höhlen, die von Ischyropsalis besiedelt sind, wofür auch hier gedankt sei. Meine Tätigkeit im Vogelsberg wurde dankensn~erterweise von Herrn Prof. Dr. SCITERF unterstützt, der mir regelmäßig in der Außenstelle ,,I<ünanzhaus" des Zoologischen Institutes Gießen Auf- na,hme gewährte. FIerr Dr. U. Haaci<cx übernahin die Bestimmung kavernikoler Diplopoden aus den Pyrenäen. - Für Diskussionen über noinenklatorische und systematisch-evolutionstheo- retische Fragen danlze ich den.Herren Privatdozenten Dr. 0. KRAUS und Dr. A. W. STEFFAN und für Gespräche über okologische und glazial-historische Fragen in den Alpen den Herren 9. Ansossitu, I<.-H. H ~ ~ t a r s und Dr. K. THALER. *
Wein besonderer Dank gilt meinem Lehrer, Herrn Prof. Dr. R. BRAUN, der mich in arach- nologische Grundproblcine einführte, diese Arbeit mit ständigem Interesse verfolgte und sie durch fachliclie Diskussionen unterstützte und forderte.
. 111. Biologisch-ökologischer Teil
1. Vorkon in le i i , B i o t o p a n s p r ü c h e
Alle Arteii der Gattung Ischyropsalis sind stenök, so daß sich Gemeiilsamkeiteii in den Biotop-Ansprüchen aufzeigen lasseil. Relativ breite ökologische Valenz kenn- zeichnet jene Arten, die heute eil1 vergleichsweise ausgedehntes Area81 einnehmen (hellwigi, manicutn, luteipes); Arten mit kleinen Arealen scheinen in ihrer Valenz viel begrenzter zu sein (z. B. leben die Arten strandi, hadzii, muellneri, dispur und magda- lenue rein ka,veriiikol). Außer der historischen Entwicklung der Areale sind folgende Faktoren wichtig.
1. Als wesei~.tlicher begrenzender Faktor ist die Feuchtigkeit des Substrates und damit die relative Luftfeuchtigkeit in deii Mikrobiotopeil anzunehmen: Lebensmög- lichkeiten bestehen nur dort, wo durch hohe Niederschläge oder oberflächlich ab- fließendes Wasser ständige Durchfeuchtung des Untergrundes gewährleistet ist.


Insolation ausgesetzte Habitate werden dann besiedelt, wenn die unter 1. nnd 2. genannten Bedingungen erfullt sind; das z?igen diese beiden Beispiele:
a) I . kollari lebt in Teilen der Alpen zwischen 2000 uiid 2500 m, wo nur noch Polsterpflaiizen streckenweise gedeihen. Beschattung des Biotops fehlt.
b) I . lutezpes fand ich am Port d'Aula (Arikge, Fraiikreich) in 1690 m uber der (natürlichen?J Baumgrenze. Diese Popula-tioii lebt in stark beweideter Mattenvege- tatioi~ immer dort unter Steinen, wo sich kleine Rinnsale befinden.
Die Abhangigkcit des Siedluiigsgebietes von Feuchtigkeit und Temperatur de- nionstrieren zwei weitere Beispiele:
C) I. kollari siedelt im Stubachtal (N-Abdachung der Hohen Tauern) - soweit heute bekannt - von 1460 bis 2080 m, also in der subalpineil Stufe: In kuhlfeuchten, N-exponierten Blockhalden - von dichtem Moosfilz und Parnwuchs uberwuchert und von lockerem Zirben-Fichteil-Larchenwald beschattet - findet kollari sein Prae- ferendum und ist hier stellenweise der dominierende Weberknecht (Abb. 1). Über der Baumgrenze lebt diese Art vereinzelt vor allem dort, wo durch Humusablagerun- geil feuchte llikrobiotope entstehen. Der hochstgelegeiie Fund (2080 m) gelang in einem verarmten Vaccinio-Rhododendretum (1 subad., HARMS leg.19. VIII. 1967).
d) I. h. hellwigz lebt in der Gipfelregion des Vogelsberges (Taufstein, 770 m) rela- tiv haufig in einem kleinen Streifeil naturiiaheiz, sich selbst uberlassenen Waldes rings um die Gipfelkuppe (miozaner Basalt). Dieses Waldstuck besteht uber~viegend aus Bergahorn und Buche mit dichtem Unterwuchs ans Rubus, Sambucus, Dryopterzs. Die steilen, von Moos uiid Farn uberwucherten Blockhalden scheiiien von Ischyropsa- lzs gemieden zu werden, denn hier gelang kein einziger Fund: Das lockere Block- werk, das frei von Detritus ist, dient nicht als Feuchtigkeitsspeicher. Dagegen ist hellwzgz regelmaßig auf der ebenen bis leicht geneigten Plache zu finden, die am Fuß der Blockhalde anschließt (Abb. 2). Die Tiere sitzen hier unter morschen Stämmen und in Stubben der abgestorbenen Bäume, die von Dryopteris und Rubus uberwuchert sind. Der Boden besteht aus tiefgrundigem Lehm.
Durch die Bevorzugung von Mikrohabitaten mit niedrigen Temperaturen und hoher Luftfeuchte sind alle Arten zum Leben iii Hohlen przjadaptiert (vgl. Kap.
1
111, 3). I Z u s a m m e n f a s s u n g : Alle Arten der Gattung sind stenök; sie besiedeln nur Mikrohabitate
. . mit niedrigen Durchschnittstemperaturen, ausgeglichenem taglichen Semperatulgang und hoher
relativer Luftfeuchtigkeit. - . t z + ,
2. H ö h e n v e r b r e i t u n g Alle Arten sind als Gebirgsbewohner anzusehen. Sie sind aber nicht ausschließlich
auf Gebirgslagen beschränkt, sondern besiedeln auch das Flachlaiid, soweit die wich- tigsten ökologischen Voraussetzungen gegeben sind. - Die niedrigstgelegenen Fund- orte von I. h. hellwigi sind Merdink bei Winterswijk (Niederlande; etwa 50 m) und


wanken, Steiiler Alpen) zugebilligt werden. Die Augen sind bei dieser Art zwar noch vorhanden, aber sehr klein, fast rudimentar ausgepragt. Sie mreist (wie auch strandi) Reduktionen am abdominalend-Scutuni auf. Entsprechende Ruckbildungen zeichnet I . muellnen aus (Dinarische Gebirge), die nur ausnahmsweise im Freiland vorzu- kommen scheint. Diese Umbilduilgeii, also Augenreduktion, variable Auflosungs- zustande des abdoniiiialen Scutum, Reduktioii der Sklerotisierung des Exoskelettes, fehlen der nicht ausschließlich kaveriiikoleii I. pyrenueu.
JUBERTHIE (1964) führt sieben weitere Arten an, die vorwiegend kavernikol sein und Rück- bildungen an den Augen erkennen lassen sollen. Dabei stützt er sich vor allem auf ROEWER (1950), der diese Arten zum Teil erstbeschrieb (pente l ica , goodnighti , troylodytes, n icaea , corcyraea, navar- rens is , r a v a s i n i i H ~ D ~ I ) . Die Blitteilucgen von ROEITER sind durchweg fehlerhaft, und diese.nomi- nellen Arten sind auf längst bekannte Spezi~s zu beziehen.
Die Vertreter von Höhlen-Populatioiieii aller weiteren Arten unterscheiden sich in keinem Merkmal von Iiidividuen epigäisch lebender Populationen, so daß ange- nommen werden muß, daß sie dieses Habitat erst seit relativ kurzer Zeit besiedeln. Zu ~onderei~t~vicklungeii kam es bisher nicht.
Die Areale höhleiilebender Populationen sind mit jenen der freilebenden iden- tisch, erscheinen aber bisweilen vertikal getrennt. In tiefen Lagen, die heute aus ökologischen Gründen nicht besiedelt werden köiiiien (Trockenheit), verden Höhlen besetzt, in höheren Lageii der Gebirge aber (auch) das Freiland:
a) Die untere vertikale Arealgreiize der alpinen I. kollari liegt bei 1500 m, die obere bei nahezu 2600 m. Der einzige bekannt gewordene Fundort im Tiefland ist eine Höhle (Lurgrotte bei Graz, 350:m).
' b) I. luteipes galt in den Pyrenäen bisher als überwiegend kavernikol, wie auch ~
aus DRESCO (1932) ersichtlich ist. Ich selbst fand eiiie Freiland-Population (zusammen mit H. P a u ~ u s ) am Port d'Aula (Arikge, 1690 m ; 3. IX. 1967). Die ökologischen Vor- aussetzungen am Fuß der Pyrenäen, z. B. in der Umgebung der Höhle Touasse (bei St. Girons, Ariege), die von lute-es besiedelt ist, erwiesen sich am 2. IX. 1967 als sehr ungünstig (trockener Wald).
Deutung: Die Höhlen-Populationen Gon I . kolluri in der Lurgrotte und von I . luteipes in niederen Lagen der Pj~renäen können nur als Reliktsiedlungen einer noch iii früher postglaziader Zeit kontiiiuierlichen Verbreitung angesehen werden. In der postglazialeil Wärmezeit wurden die Areale arborealer Arten in Gebirgslagen zurück- gedrängt. Während dieser klimatisch bedingten Arealregression boten sich für die hygrophilen und kalt-stenothermen Ischyropsalis-Arten Höhlen als Ausmeichbiotope an, und allein die dort siedelnden Populationen vermochten zu persistieren. - In dem geschilderten Fall von I . kollari (Lurgrotte [350 m] - Subalpin-Stufe der Alpen [etwa ab 1500 m]) tritt das besonders kraß hervor.
Die strenge Kavernikolie von I. pyrenaea ist schwieriger zu deuten, denn diese Art lebt sicher nicht länger in Höhlen als manche Populationen der sympatrischen I.

Die Abgrenzung von Biospezies usw. 143
lutezpes. Vielleicht erlosch das epigaische Areal dieser spezialisierten Art in derselbe11 Warmezeit, imi der das von lutezpes sehr eingeengt wurde. Offensichtlich gelang es pyrenaea fast nicht, erneut epigaische Siedlungen zu bilden. Bisher wurden erst zwei Individuen in Preilandhabitaten gefunden (DRESCO 1967 c).
4. S y n t o ' p i s c h e s V o r k o m m e i l zwe ie r A r t e n
Das Gattungsareal ist heute zwar recht gut bekannt, morphologisch differente, sympatrisch lebende Formen wurden bisher aber nur ausnahmsweise festgestellt. D; alle Arten irn wesentlichen dieselben ökologischen Praeferenda haben, könnten sjTm- patrische Arteii sogar syntopisch vorkommen und dieselben Mikrobiotope einnehmen. Solche Feinheiten der'Verbreitung sind den heutigen Musealsammlungen nicht ZU
entnehmen; auch die Literatur ist hier nicht verläßlich. Auf meinen eigenen Sanlnlel- reisen widmete ich syiitopischem Vorkommen zweier Arten besondere Aufn~erksam-, keit: " .
a) I. hellwigi hellwigi und I. kollari
Fundgebiet: Stubachtal, Hohe Tauerri (Österreich, Land Salzburg).
I. kollari lebt hier in1 subalpinen Nadelwald (Rhodoreto-Vaccinietum cembre- sum) bis zum oberen Rand der Legföhren-Zone (1460-2080 m). I m lichten Wald ist diese Art stellenweise häufig. In'einem engen Bereich dieser Höhenstufe kommt nun auch I. h. hellwigi vor; von ihr wurdeii im einzelnen gefunden:
19 (1490 m, AUSOBSKY & HURDES leg. 3. IX . 1966); 1 totes Ex., ld(1475 und 1480 m, AUSOBSKY & MARTENS leg. 7. X. 1966).
I. h. hellwigi erreicht in dieser Höhenstufe die obere Grenze der Vertikalverbrei- tung. Diese umfaßt im Land Salzburg Vorkommen in den talnahen Wa'ldgesellschaf- ten ab 640 m und die untere Subalpin-Stufe (bis etwa 1500 rn) (AUSOBSKY indl.). So ist im Stubachtal eine Kontaktzone und ein kleiner uberschneiduiigsbereich niit syntopischem Vorkommeii beider Arten anzunehmen. An beiden Sammeltagen wur- den Individuen beider Arten in unmitt~elbarer Nachbarschaft gefunden, teilweise*iiur 30 m voneinander entfernt.
b) I. hellujigi lucantei und I. nodifera
Rundgebiet: Sierra de Araler, Nava,rra (Spanien).
Bereits aus Pundangaben von SIMON (1879) (sub I. superba und nodifera) ist zu entnehmen, daß beide Arten nicht nur sympatrisch, sondern sogar syntopisch vor- kommen können. Nachsuche am Locus typicus, Wälder bei St. Jean de Luz (Prank- reich, Basses-Pyrknees), blieb erfolglos (7. IX. 1967). Doch gelang es in der Sierra de Aralor, nur 50 km SW, ein syntopisches Vorkommeil beider Arten zu finden. Bereits RAMBLA (1946) meldete diese Arten (sub superba und adamii) aus mehreren Höhlen dieses waldreichen Gebirges. - Ich selbst sammelte in unterholzreichem Eicheiirvald bei Lecumberri, 650 m:

hellwzgz lucuntei: I$ (leg. 23. IX. 1967); 33, Junge und viele Gelege (leg. 25. IX. 1967) ;
nodzfera: 2 3 (leg. 23. IX. 1967). Die Fundstelleii lagen nur 10 bzmr. 40 m aus- einander. Alle Tiere und die Gelege befailden sich unter morschen Stammen und moosuberwucherten Stubben.
Im folgenden liegt keine echte Syiitopie vor:
C) I. hellwiyi lucuntei und I. pyrenueu
Pundgebiet: Grottes de Betharrai~~, Basses-Pyrt5iiees (Fraiikreich).
In diesem weitläufigen Höhlensystem lebt I. pyrenuea ( S I ~ ~ O N 1913); bei einem Besuch fand ich das bestätigt (4. IX. 1967). Aus dieser Höhle wird aber auch hellwiqi lucu?ztei gemeldet (SIMON 1879, sub lucantei). Damals wurde lediglich ein Jungtier gefunden, das D n ~ s c o (196613) abbildet; eine Bestätigung durch weitere Exemplare gelang bisher nicht. So dürfte es sich lediglich um ein Tier handeln, das zufällig in diese von pyrenaeu besetzte Höhle geraten ist. Das stimmt mit den bisherigen Er- fahrungen überein: Höhlen werden immer nur von einer Ischyropsulis-Art besiedelt. Das betonen alle Autoren, und ich fand es in Österreich und Frankreich bestä,tigt. Das gilt selbst danii, wenn die von verschiedenen Arten besetzten Höhlen nur wenige Kilometer auseinander liegen. Das erklärt sich zwanglos aus der Stenökie a'ller Arte11 und der Nischeiiarmut eines Hö1.ilensystems. In ihm kann sich immer nur eine der Arten behaupten.
Indes sollen in der Höhle La Magdalena (bei Galdames; N-Spanien) zwei Arten vorkomnien: nodiferu in den Hallen nahe dem Eingang, muqdalenue im hinteren Teil der Höhle (SIMO'I 1881). Diese Mitteilung bedarf der Bestätigung.
5. P h ä n o l o g i e
.Nur von drei Formen (Spezies bzw. Subspezies) liegen aus englokalisierten Ge- bieten so viele Daten vor, daß verläßliche Aussagen möglich sind. a) I. hellwigi hellwigi
Fuiidgebiet: Taufstein in1 Vogelsberg, 770 m ; 15 Daten aus den Jahren 1958, 1963-1967 (Abb. 3).
Der erste Adultus wurde am 19. VII. (1967), der letzte am 10. X. (1967) gefuii- den; Reifezeit erstreckt sich also über 3 Monate. Ein Gelege vom 22. IV. (1966) (noch Schneeflecken an sonneiigeschützten Stellen) weist darauf hin, daß die Gelege über- wintern. Jungtiere können somit ab Ende April erwartet werden, die letzten wurden am 20. IX. (1963) gefunden. Ob ihnen vereinzelt Überwinterung gelingt oder ob sie niit Beginn der Frostperiode absterben, läßt sich noch nicht entscheiden. - Wie stark jährliche und loka,le Unterschiede des Klimas auf den Zyklus einwirken, zeigt folgendes: Alle Jungtiere vom September stammen aus einem Jahr (1963); 1966

Die Abgrenzung von Biospezies usw. 145
Anzahl
12 /
Monate
Abb. 3. Phänologie von Ischyropsalis hellwigi hellwigi (Vogelsberg, Taufstein, '770 m).
scheint es mindestens ab 6. IX. keine Jungen mehr gegeben zu haben (6. IX.: 6 ad., 24. IX. 11 ad. gesammelt), doch fand ich am 6. IX. desselben Jahres am Geiselstein, nur 3 km nw. vom Taufstein (wie dieser eine Halde miozäner Basalttrümmer), noch ein Jungtier (3. oder 4. Stadium). Der schmale Grat der Halde erstreckt sich in NS- Richtung, so daß durch diese ungünstige Lage und die dichte Beschattung nie volle Sonneneinstrahlung möglich ist. Der Zyklus dieser Population scheint retardiert zu sein. - Alle übrigen Funddaten (dieser Subspezies) fügen sich in das Schema ein. Lediglich im nordwestlichen, bereits stark atlantisch beeinflußten Teil des Areals treten Adulti bereits im Juni auf (Sauerland), ebenso auch im Wienerwald.
b) I. hellwigi lucantei
Pundgebiet: Sierra de Aralar, Navarra (Spanien), 650 m ; 2 Daten aus 1967 (Abb. 4).
Der Zyklus dieser sehr eng lokalisierten Subspezies weicht von dem der Nominat- form ab, offenbar bedingt durch Einflüsse des atlantischen Klimas. - Zwischen dem 23. und 26. IX. (1967) fand ich 3 8 , Jungtiere des 2. und 3. Stadiums und etwa 25 verschieden weit entwickelte Gelege. Daraus läßt sich (mit Vorsicht) ableiten: Die Hauptreifezeit liegt im Juli/August; Ende September waren keine $2 mehr auffindbar. Die Jungtiere schlüpfen im September/Oktober, überwintern in verschiedenen Al- tersstadien und setzen die Entwicklung im folgenden Frühjahr fort.
C) I. lcollari
Pundgebiet: Täler der Nordabdachung der Hohen Tauern, Österreich, 1480 bis 2150 m ; 10 Daten aus 1966 und 1967 (Abb. 5).
In der schneefreien Zeit wurden Adulti Mitte Mai, dann wieder vom ersten Drit- tel August bis zum Beginn der ~ehneebedeckun~ gefunden. Der hohe Anteil subadul-
10 Zool. Jb. Syst. Bd. 96

ter Jungtiere im Sommer (16. VIII. 1967: von 18 iuv. 15 subad., uberdies 3 ad.) geht spater zugunsten der Adulti immer mehr zuruck, bis diese im ersten Drittel Oktober stark dominieren (7. X. 1966: 11 ad., 3 iuv.). In dieser Sommer/Herbst-Rcifezeit findet die Eiablage statt. Fur die ubrige Zeit des Jahres kann gefolgert werden, daß ein großer Teil der Adulti und die wenigen bis zuin Einsetzen der Sehiiccfallc ver- bliebenen Jungen uberwintern. Das belegen 2 ? 1 iuv. vom 16. V. (1966). Zu Beginn der Fruhjahrsaktivitat legen die $! Eier, und die letzten Jungtiere vom Vorjahr reifehauteil sich. Im Juni/Juli gibt es fast nur frischgeschlupfte Jungtiere uiid zur
Anrahl
1 2 1
6
4
2
I I I 111. I V V V1 V1 I V l l l IX X X I XI1
Monoie
Abb. 4. Phänologie von Ischyropsalis hellwigi lucantei (Sierra de Aralar, Navarra, Spanien,:650:m).
Anzahl
od. ,"V
Gelege
l Od
s
m I 8 ° C
L a
10 U a I -J
0 , :
E 2
6
4
2
I I I 1 1 1 I" V "I . V 1 1 V l l l IX X X I XI1
Monde
Abb. 5. Phanologie von Ischyropsalis kollart an der Nordabdachung der Hohen Sauern (Land Saliburg, ~s terre ic l i ; Kapruner Tal, Stubachtal, Ilabachtal; 1480-2150 m).

Die Abgrenzung von Biospezies usw. 147
Wende Juli/August die ersten Adulten. - Dieser Zyklus (gültig zunächst nur für die Nordabdachung der Hohen Tauern) gibt die Verhältiiisse wahrscheinlich simplifiziert wieder, denn wegen der extremen Bedingungen des Hochgebirgsklimas ist mit lokale11 Verschiebungen zu rechnen. Möglicherweise besitzen Populationen mancher Stand- orte sogar mehrjährige Zyklen, die sich bei stichprobenhafteii Aufsammlungen nicht erkennen la,sseil.
Zusa inmenf a s s u n g : Alle drei Spezies bzw. 'subspezies haben ihre Hauptreifezeit in1 Somm,er und Herbst. Die beiden Subspezies, von lzellzviyi unterscheide11 sich dadurch: daß ver- schiedene Stadien Überwintern, - bei h. hellwigi die Gelege, von h. lucantei die Jungen. - Von kol- lari überwintern auch Adulti.
6. N a h r u n g u n d N a h r u n g s a u f n a h m e
Genauere Kenntnis über die Nahrungswahl besitzen wir lediglich über I. hell- wigi und von einigen Höhlenpopulationeii der Arten luteipes, pyrenaea und kollari.
I. h. hellwigi ist als strenger Nahrungsspezialist aiizuseheii, der sich (fast) aus- schließlich von Molluskeil ernährt (VERHOEFF 1900, SCHÜLER 1950 [sub ~ ~ z c G L ~ G ~ ] , MARTENS 1965). Die massigen Cheliceren (Abb. 6 U. 7) crlauben auch die Überwältigung festschaliger Beutetiere. Xeroniorphe Cernuella neglecta w'erderi voii kräftigen Indivi- duen mühelos aufgebrochen, und von juiigeii Clausiliiden, denen die Mündungsarma- tur noch fehlt, werden die TVinduiigen abgetragen, so daß nur die Spindel zurück- bleibt. Dabei packt eine der Cheliceren-Klaueii die Schale am Mündungsrand, wobei die Mündung nach oben zeigt. Die andere Chelicere schneidet kleine Stücke aus dem Weichkörper. Zieht sich die Schnecke in hintere Umgänge der Schale zurück, greifen beide Zangen den Münduiigsrand und brechen Stücke der Schale ab, bis der Weich- körper wieder greifbar ist. - Iii Gefangenschaft wurde andere Nahrung verweigert, lediglich Mehlkäfer-Larven und -Puppen nach mehrtägigem Hungern angenommen. - Diese Ergebiiisse wurdeii a'n mitteleuropä'ischen Populationen gewonnen (Siebeii- gebirge: VERHOEFF; Salzburg: S C H ~ L L E R ; Wienerwald: GRUBER in litt. ; Vogelsberg, Böhmerwald, Villacher Alpe, Stubachtal: MARTENS). Die Befunde dürfen nicht ver- allgemeinert werden, denn die westeuropäische Rasse h. lucnntei scheint in der Nah- rungswahl vielseitiger zu sein. In der Höhle Oueil de Neez (RebAnacq, Basses-Pyr6- nees) lebt eine individuenarme Population, die sich möglicher~veise von Diplopoden ernährt. 2$ 2 (leg. ~ M A R T E ~ V S & PAULUS, 6. IX. 1967) hielten sich an kleinen Fleder- ma8usguano-Flecken auf, die von Diplopodeii besiedelt wurden (Syphloblaniulus'tro- globius und Polydesrnus g~~llzcu~; HAACKER det.). Weitere Höhlenbewohner, die als Beute in Frage kommeii, entdeckten wir nicht. In Gefangenschaft nahmen diese 4 Individuen sofort Gehäuseschiiecken an. - 3 $ einer Freiland-Population (Sierra
I de Aralar, Spanien; 23. IX. 1967) verschmähteiz Molluskeil, ließen sich aber mit Drc- sophila über Monate am Leben erhalten.
1 I. kollari (mit viel schlankeren Cheliceren a81s hellwigi) bevorzugt iii der Lur- grotte (Graz, Österreich) offenbar die Assel 1TKesoniscus alpicola, einen weitgehend
I 10:


Die Abgrenzung von Biospezies u m . 149
werden; am Eingang halten sich Trichopteren und brachycere Dipteren auf. In der Höhle von Estelas (Ariege, 1. IX. 1967) fand ich das bestätigt. Als Nahrungsgrund- lage dieser sehr großen Art spielen in dieser Höhle vor allem die Trogloxenen eine Rolle (Trichopteren, Dipteren), denn die echten Höhlen-Insekten sind viel zu selten und zu klein. Zudem wird in diese Höhle kein organisches Material in größeren Men- gen eingespült, da kein Bach hindurchfließt.
I. luteipes besitzt ein weites Nahrungsspektrum, was bereits aus der breiten öko- logischen Valenz dieser Art hervorgeht. Sie lebt in feuchtem Moos an kleinen Rinn- salen in schattigen Wäldern, auf Almwiesen und in Höhlen der Pyrenäen. Individuen aus der Höhle Touasse (Aribge) fraßen sehr gern Drosophila.
Z u s a m m e n f a s s u n g : Aus mechanischen Gründen bevorzugen lang- und schlank-cheli- cerige Arten wenig sklerotisierte Arthropoden, kurz-chelicerige fressen überdies solche mit hartem Exoskelett. Nahrungsspezialist ist (nach heutiger Kenntnis) allein I. h. hellwigi.
7. K o p u l a t i o n s v e r h a l t e n
Das Paaruiigsverhalten ist von I. strundi (JUBERTHIE 1964), 1.' hellzoigi hellwigi (MARTENS 1967, 1969), I. hellwigi lucuntei und von I . lutezpes (MBRTENS 1969) be- schrieben worden.. Eine genaue Darstellung kann hier nicht gegeben werden, fol- gende Einzelheiten sind aber an dieser Stelle wichtig:
I. hellwigi und I . luteipes leiten die Paarung mit einer typischen Balz ein, die bei diesen Arten differiert. D i e3 tragen auf dem Cheliceren-Grundglied disto-dorsal bzw. -medial ein Drüsenfeld, das äußerlich durch ein scharf umrissenes Feld kurzer Bor- sten gekennzeichnet ist (vgl. Abb. 22 und Abb. im Systematischen Teil). Während des Balzverhaltens bietet da s3 dem 9 da's auf dem Drüsenfeld austretende Sekret an. Wenn das kopulationswillig ist, hebt es das Prosoma etwas an, so da,ß die weich- häutigen Mundwerkzeuge frei liegen. D a s 3 bringt darauf das Haarfeld, die „Bürsteu, und somit das austretende Sekret mit den Mundwerkzeugen des Q in Berührung. Die Cheliceren und die vorderen Beinpaare beider Partner verschränken und verklam- mern sich, worauf das 8 die Kopulation vollzieht. - .Werbe- und Kopulationsverha,l- ten von I. hellwigi hellwigi und I . hellwigi lucantei unterscheiden sich nicht, dagegen läuft die Paarungseinleitung bei I. luteipes ,,überfallartig" ab, doch wird ebenfalls die Bürste mit den Mundwerkzeugen des $2 in Berührung gebracht.
Lage und Form der Bürste sind a.rtkonstant. Wie ihre Funktion bei der Paarung zeigt, muß ihre unterschiedliche Ausprägung reproduktive Isolation signalisieren: Hier liegt ein morphologisches Merkma,l vor, das in enger Beziehung zum Fortpflan- zungs-Verhalten steht und eine bestimmte Verha'ltensweise bedingt (vgl. Diskussion in Kap: IV 1 d).
Derartige intraspezifische Sekretdarbietungen sind innerhalb der Arthropoda bei Insekten in wenigen Fällen, bei Arachniden aber überhaupt noch nicht gefunden worden.

IV. Morphologischer Teil
1. Morphologie u n d sys t ema t i s che Wer t i gke i t des G e n i t a l a p p a r a t e s u n d pe r iphe re r Organe
a) Der Genitalapparat
Es ist bekannt, daß die Genitalorgane vieler Arthropoden-Gruppen artspezi- fisch gebaut sind und von Art zu Art viel größere Diskontinuitäten aufweisen können als die äußere Mo~phologie der entsprechenden Spezies. Der Genitalapparat von Ischyropsalis-Arten wurde erst wenige Male so abgebildet, daß art- oder gruppen- charakteristische Merkmale zu ersehen sind ( 5 1 ~ ~ ~ ~ 1 1 9 5 6 , JUBERTHIE 1963). Das liegt vor allem daran, daß die Genitalien aller Arten denkbar einfach gebaut sind und bei oberflächlicher Betrachtung keine Artunterschiede aufzuweisen scheinen.
$-Geni t a l a p p a r a t (Penis) (Abb. 8): Im einzelnen sind zu unterscheiden: Der Trui icu s p eiiis (Nr. 9 der Abb. 8): ein schlanker sklerotisierter Hohlkörper, in den Muskeln hineinziehen; er läuft distal in den hakigen S t y l u s (1) aus. Der Truncus ist in der basalen Hälfte erweitert und weist an der Basis eine Auskehlung (11) auf, sein Distalende ist zur Glans hin erweitert. Der Truncus ist dorso-distal am stärk- sten sklerotisiert; diese Sklerotisierung ist basad schwächer ausgeprägt, wodurch eine schwache F lüge lung (8) der Distalhälfte des Truncus entsteht. Die Flügelung geht ohne Absatz in die Glans über, die ventral durch einen zweilappigen oder ein- heitlichen und dann fa,st immer median gekielten Sk l e r i t (2) abgesetzt ist. Dieser trägt kräftige Beborstung: zwei p a ramedia i ie (3) Felder; je ein laterales (4), sehr kleines nahe dem Kiel der Blügeluilg ist von ventral kaum sichtbar, und auf der Dorsalseite der Glans befindet sich je ein l a t e r a l e s Feld (5). Unterhalb des Sklerits der Glaiis iniindet der T r i ch t e r (61, eine Öffnung unterschiedlicher Weite meist oberhalb oder in einer weichhäutigen Geleiikzoiie (7), an der die Glans ventrad ab- gewii~kelt werden kann. 1
Q-Geiii t a l a p p a r a t (Legeröhre, Receptacula seminis) (Abb. 9): Die Legeröhre ist bei allen Arte11 sehr kurz, uilgegliedert und ihre Öffnung voll kurzen Borsten um- stellt. Die Re cep t a cu l a s eminis sind paarig und liegen im distalen Teil der Lege- röhre lateral. Sie bestehen aus je eiiier tütenförmigen häutigen Membran (Tüte) (I), aus der eine wechselnde Anzahl von S ch läuchen tritt (2). Die Schlauche verengen sich zur Basis uiid vereinigen sich zu einem gemeinsa,inen K a n a l (3). Sie besitzen eine feste Membran, meist schwach runzelig skulpturiert, mit unterschiedlich weitem Lumen (4). Die Receptacula sind in ma,ssige Biiidegewebspolster eiilgebettet und iii der unbeschädigten Legeröhre nicht erkennbar. Über die fuiiktioilsmorphologi- schell Zusammeiihänge zwischen Sameiiübertragung, Morphologie der Legeröhre und der Receptacula und der Besamung der Eier ist nichts bekannt.

Die Abgrenzung von Biospezies usw.
Abb. 8. Penis, am Beispiel von Ischyropsalis muellneri. - links: von ventral; Mitte: von lateral; rechts: von dorsal. - 1 Stylus; 2 ventraler Sklerit; 3 Borstenfeld des ventralen:Sklerits; 4 late- rales Borstenfeld; 5 dorsales Borstenfeld; 6 Trichter; 7 Gelenkzone; 8 laterale Flügelung; 9 Trun- cus; 10 lateraler Kiel; I 1 basale Auskehlung.
Systematische Wertigkeit der Merkmale des Genitalapparates
Die Benutzung der $- und 9-Genitalmorphologie für systematisch-phylogeneti- sehe Uiitersuchungen an Arthropoden hat sich als gewiiiiibringend erwiesen (s. zuletzt KRAUS 1966), so daß sich hier prinzipielle Darlegungen erübrigen. Hinsichtlich der Opiliones sind aber Hinweise angebracht.
$-Genitalapparat (Penis): Offenbar wurde er erstmals von WOOD (1868)l) in einer Bearbeitung der Weberknechte der USA dargestellt und zur Artabgrenzung
1) Den Hinweis auf dieses Zitat verdanke ich Herrn Kollegen Dr. K. THALER.

152 J. MARTENS
verwendet. HANSEN & S~RENSEN (1904) untersuchten die Penes von Arten mehrerer Familien (auch Ischyropsalididae) und gaben klare Abbildungen. ROEWER verzich- tete ganz auf genitalmorphologische Kriterien und baute sein System der Opiliones allein mittels ä'ußerer Körpermerkmale auf. In neuerer Zeit gebrauchte erst SILHAVI (1938) den $-Geiiitalapparat zur Artgliederung und demonstrierte seinen hohen Trennwert an äußerlich sehr ähnlichen Arten des Genus Opilio (Phalangiidae). In- zwischen haben Merkmale des Penis entscheidende Bedeutung zur Spezies-Abgren- zung und -Gruppierung gewonnen, vor allem in Gruppen mit äußerlich sehr einheit- licher Morphologie. In manchen Verwandtschaftskreisen fehlen jedoch diese markan- ten Unterschiede von Art zu Art, so daß allein die Strukturen dieses Organs zur sicheren Abgrenzung nicht ausreichen. Betrachtet man die Areale dieser meist sehr nahe verwandten Arten, erweist sich, daß sie vikariieren. Es handelt sich offenbar um Formen, die den Prozeß der Artaufspaltung erst in erdgeschichtlich jüngster Zeit durchlaufen oder noch nicht einmal abgeschlossen haben. Aus der europäischen Fauna sind hier die Genera Astrobunus, Homalenotus (beide Sclerosomatidae), Para- nemastoma (Nemastomatidae), Qraecophalunyium, Gyas (beide Phalangiidae) und mit gewissen Einschränkungen auch Ischyropsalis zu nennen. Bei den Arten dieser Gat- tung sind die Penis-Strukturen einfach und nur in engen Grenzen variabel und somit auch bei nicht nahverwandten Arten sehr ähnlich. Da dem Schaft jegliche Armatur fehlt und lediglich die Glans geringfügig gegliedert ist, sind nur die Proportion des Schafts und der ventrale Sklerit der Glans von Art zu Art geringfügig verschieden. Auch die Beborstnng des Sklerits ist bei a,llen Arten weitgehend einheitlich. Somit erscheint der 8-Genitalapparat uniform und merkmalsarm. Das bedeutet aber nicht, daß er zur Umgrenzung von Taxa unbrauchbar wäre. Es erwies sich, da0 trotz der Einförmigkeit drei scharf trennende Alternativ-Merkmale (sensu BECK 1965) vor- handen sind. Der ventrale Sklerit der Glans ist entweder zweilappig-plan oder lanzett-
Abb. 9. Receptaculum seminis, am Beispiel von Ischyropsalis dispar (vgl. Abb. 68a). - 1. hautige Membran (Tüte); 2. Schlauche; 3. gemeinsamer Ausfuhrkanal; 4. Lumen des Schlauches.

Die Abgrenzung von Biospezies usw. 153
förmig mit medianem Kiel oder lanzettförmig-p1an:Diese Merkmale trennen drei Artengruppen (vgl. Kap. V1 3a und Abb. 32).
9-Genitalapparat (Receptacula seminis): Die Legeröhre scheint nach bish'crigen Erfahrungen zur Speziesgliederung ungeeignet. Sie ist zwar innerhalb größerer Grup- pen sehr unterschiedlich gebaut, weist aber von Art zu Art gar keine oder nur äußerst geringe Unterschiede auf. Vergleichsweise größere Bedeutung kommt den Recepta- cula seminis zu. In der Gattung Ischyropsalis ist deren interspezifische Variabilität jedoch auf die Zahl der Schläuche (Abb. 9) und auf Proportions-Unterschiede be- schränkt. Da aber die Zahl der Schläuche der paarigen Rezeptakel bereits bei e inem Individuum variieren kann, muß ihr systematisch kennzeichnender Wert gering er- scheinen. Da diese Strukturen aber bisher noch von keiner Gattung der Opilioiies vollzählig dargestellt worden sind, werden hier die Rezeptakel aller Arten abgebildet (Kap. VI).
b) Das abdominale Scutum In der Gattung Ischyropsalis sind prinzipiell zwei Formen des abdominalen '
Scutum verwirklicht. Entweder befindet sich auf den ersten funf abdominalen Seg- menten ein einheitlicher Schild (Scutum) oder die fünf einzelnen Teile dieses Schildes sind frei gegeneinander beweglich und lediglich durch Gelenk -(Intersegmental-) Häute miteinander verbunden. Bei Zwischenstadien sind lediglich zwei bis vier der Einzelsklerite miteinander verschmolzen.
Für die thorako-abdominalen Scuta-Bildungen der Weberknechte führte H A D ~ I (1942) Termini ein. Sie bezeichnen den Grad der Verschmelzung der abdominalen Sklerite untereinander und ihren0zusammentritt mit der Cephalothorax-Platte. - Pur die weiteren Ausfuhrungen sind wichtig:
1. S c u t u m magnum: Großschild aus der Cephalothorax-Platte und dem ab- dominalen Scutum der Sklerite I-V. (Bei den Ischyropsalididae ist dieser Zustand nicht verwirklicht, wohl aber bei den Nachbarfamilien Nemastomatidae und Trogu- lidae.)
2. S c u t u m pa rvum: Cephalothorax-Platte nicht mit dem Scutum der Ter- gite I-V verschmolzen, durch Gelenkhaut getrennt (nur bei den Ischyropsalididae, etwas modifiziert auch bei den Arten der Tribus Eupnoi).
3. S c u t u m l amina tum: abdominale Sklerite I-V nicht zu eiiiem Scutum verschmolzen, frei gegeneinander und gegen die thorakale Platte beweglich (nur bei den Ischyropsalididae).
Als weitere Termini führe ich ein: 4. S c u t u m in t e rmed ium: Von den abdominalen Skleriten I-V sind 2, 3
oder 4 ganz oder teilweise miteinander verschmolzen. Mit der thorakalen Platte treten sie nie zusammen. Dieses Stadium ist zwischen jenem des Scutum parvum und dem des Scutum laminatum einzustufen (nur bei den Ischyropsalididae verwirklicht).

5. S cu t um diss ec t um: Die Einzelsklerite des Scutum laminatum sind ein- fach bis mehrfach zerteilt, so daß pro Segment eine Querreihe oder eine ungeordnete Gruppe von Einzelskleriten erscheint (bei einigen Arten der Gattungen Taracus und Sabacon veiwirklicht, beide Ischyropsalididae; ferner bei Jungtieren von Ischyro- psalis).
Die Geschlechter einer Art können unterschiedliche Scutum-Bildungen zeigen; bei Ischyropsalis kommen folgende Kombinationen vor (Abb. 10):
1. $9 mit Scutum parvum; bei 8, also über der Hälfte der Arten. 2. $ mit Scutum parvum, 9 mit Scutum laminatum; bei 5 Arten, bei zwei kom-
men auch andere Scuta vor (3). 3. $ mit Scutum intermedium, mit Scutum laminatum; bei 3 Arten, bei zwei
kommen auch andere Scuta vor (8). 4. $9 mit Scutum laminatum; bei 2 Arten, bei einer kommt auch ein anderes
Scutum vor (8). Nie weist das 9-Scutum einen höheren Verschmelzungsgrad als das$-Scutum auf.
Systematische Wertigkeit der Merkmale des Scutum Die von Art zu Art und bisweilen innerhalb einer Art unterschiedliche Ausprä-
gung des Scutum erlaubt es, dieses Merkmal zur akzessorischeii Kennzeichnung der Taxa heranzuziehen. Wenn man von den wenigen Fällen absieht, in denen das $ so- wohl ein Scutum intermedium als auch ein Scutum laminatum trageii kann,'& dieses Merkmal bei beiden Geschlechterii außerordentlich konstant. Da jedoch nur vier verschiedene Kombii~ationen der drei bei Ischyropsalis vorkommenden Scuta ver- wirklicht sind (Abb. IO), ist der tatsächliche Trennwert gering. Keinesfalls kann ich
Abb. 10. Scuta-Kombinationen ($ und ?) bei Ischyropsalzs. - (links$, rechts ?) a $ und 9 Scuturn parvum: b $ Scutum parvunl, 9 Scutum laminatum; c $ Scutum intermedium, Scutum lamina- t u m ; d $ und Scutum laminatum.

Die Abgrenzung von Biospezies usw. 155
aber der Meinung von DRESCO (1967 b, C) beipflichten, der, gestützt auf die Ausfüh- rungen von H A D ~ I (1942) über variable Scuta zweier Arten, dieses Merkmal zur Spe- zies-Charakterisierung ablehnt.
~ntwicklung des Scutum wä,hrend der Ontogenese
H A D ~ I (1942) folgert allein aus der Häufigkeit des Scutum parvum bei den 6, daß dieses ein ursprüngliches Merkmal darstelle, das die $2 vor allem aus biologisch- ökologischen Gründen aufgegeben hätten: eine größere „Freiheitu des Abdomens . während der Trächtigkeit. Zur Entscheidung, ob das Scutum parvum oder dieEinzel- sklerite (Scutum laminatum) den primären Zustand darstellen, kann die ontogeneti- sche Entwicklung dieser Struktur beitragen. H A D ~ I (1942) leistete bereits Vorarbeit, ha,tte aber von keiner Art alle Entwicklungsstadien zur Verfügung, uberdies war ihm die Spezies-Zuordnung meist nicht möglich.
Hier lagen alle 6 Jugendstadien von h. hellwiyi vor, ferner 5 von luteipes und 4 von kollnri. Diese Arten durchlaufen 6 Jugendstadien, 10 % der Individuen von luteipes aber nur 5 (JUBERTHIE 1964). Aus dem Ei schlupft eine Nymphe, die menig beweglich ist und sich bereits nach 2-3 Stunden häutet.
Eidonomie der Jugendstadien
I . h. hellwigi (Abb. 11): Material aus Nachzuchten vieler Tiere vom Taufstein/ Vogelsberg (Deutschland).
1. S t a d i u m (Nymphe, 3 Ex.): Kutikula der Nymphe ohne jede Sklerotisierung und somit auch ohne Scutuin; bei den unmittelbar nach dem Schlüpfen schon häu- tuiigsbereiten Tieren ist sie mühelos abhebbar. Durch die Kutikula sind die beiden steifen Borsteii auf den1 2. Thorakal-Tergit und Behaarung der Beine des 2. Stadiums deutlich sichtbar.
2. S t a d i u m (9 Ex.): Keine Sklerite auf den abdominalen Areae, doch trägt jedes der abdominalen Segmente I-V11 ein paramedianes Paar „Spaltsiiiiiesorgaile" (sensu HAD~I) , umgeben von einem sklerotisierten Hof; Cephalothorax-Platte vor- ha,ilden. Sklerit des 1. Thora'ka'l-Segments ist mit ihr verschmolzen; 2. Thorakal- Segment durch 2 kleine Borsteii keiliitlich, an deren Basis ein kleiner sklerotisierter Hof.
3. S t a d i u m (9 Ex.): Alle abdominalen Segmente dorsal mit einer Borsteii- Q~ierreihe; Sklerotisations-Zeiltreii treten stärker hervor. Variabilität: Bei 8 Ex. ist die aus zwei Zentren bestehende Sklerotisations-Zone auf den Segmenten 1-111 zu einem einheitlichen Sklerit verschmolzen; bei 1 Ex. sind die Zentren dcr Sklerite I bis 111 und V verschmolzen, IV und die auf V folgenden zweigeteilt.
4. S t a d i u m (10 Ex.): Bei 8 Ex. besitzen die Segmente 1-111 und die auf V fol- genden eine einheitliche Skleritspange, IV ist zweiteilig; bei 2 Ex. sind alle Skleritc im progressiven einteiligen Stadium.

5. S t a d i u m (6 Ex.): Bei 3 Ex. ist Sklerit IV zwcigeteilt, bei 3 Ex. einheitlich, doch lassen von diescii noch 2 Ex. distale und caudale Einschnürung dieses Sklcrits erkenncn.
6. S t a d i u m (subadult, 4 Ex.): Bei 2 Ex. ist Sklerit IV noch zweiteilig, bei 1 Ex. hängen beide Einzelteilc zusammen, sind aber median perforiert; bei einem Ex. ist Sklerit IV, wie alle anderen, einheitlich.
7. S t a d i u m (adult): $9 regelmäßig niit Scutum parvum. Z s s a i n m e n f a s s u n g : I. h. hellwigi legt init der Häutung zum 2. Stadiuin auf den abdomi-
nalen Segmenten I-V11 je ein paariges Sklerotisations-Zentrum an. Bei den folgenden Stadien verschmelzen diese Zentren von distal nach caildal fortschreitend zu einer einheitlichen Spange. Es fallt auf, daß die Zentren des Segments 1V bis zum Subadnlt-Stadium geteilt bleiben körinen. Da,s Scutum parvum wird erst mit der Adult-Häutung angelegt. Nie verwachsen bei Jugend- stadien Skleritspangen verschiedener Seginentc z u einem Scutuin intermediuin.
I . luteipes (Abb. 1.2): Material vom Co1 de la Rkpublique, Haute-Loire, und Grottc de Touasse, Ariege (Frankreich).
1. S t a d i u m (Nymphe): Lag nicht vor; gegenüber hellwigi sind jedoch keine Ab- weichungen dieses uiispezialisierteii Stadiums zu erwarten.
2. S t a d i u m ( 3 Ex.): Auf den abdominalen Segmenten I-V11 bestehen 2 Sklero- tisations-Zentren, jedoch bei allen Ex. auf Segment I1 bereits eine mediane Spange, deren beide Einzclelemente abcr unterschiedlich stark verschmolzen sind.
Abb. 11. I. hellwzgt hellwagt. - Ontogenetische Entwicklung des Scutum (abdoniiriale Sklerite I-V) und der freien Sklerite V1 und VII . - a Stadium 11; b und e Stadium 111; d und e Stadium I V ; f Stadium V ; g Stadium V I ; h Stadiuin V11 (adult).

Die Abgrenzung von Biospezies usw. 157
3. S t a d i u m (1 Ex.): Auf den Segmenten 1-11 je eine Spange, alle ubrigen Zen- treii nocfi frei.
4. S t a d i u m (1 Ex.): Auf dcn Segcmpnten 1-111 je eine Spange, alle ubrige~i Zciitrcn noch frei.
5. S t a d i u m (2 Ex.): Auf den Segmenten I-V11 je eine Skleritspangc, gebildet aus den bciden Eiiizelzentren (= Scutum Iaminatum).
6. S t adiinm (3 Ex.): Segmente I-VII mit jc ciner Skleritspange; wie Stadiun15.
7 . S t a d ium (adult): 89 regelniaßig mit Scutum parvuiri.
Z i i s ammenf a s s u n g Das relativ geringe Mdterial (10 iuv.), d'~s mogliche Variabilitat nicht erkennen IdRt, zesgt, daß auch bei luletpcs die paiamedianen Paare der Sl\lerotisations- Zentren von distal nach c'~irda1 vornnschreitend verschniel~en Einzelne Spangen \~erwc~clisen vor der Reifehautung nicht.
I I. kollari: Material aus dem Stubachtal, Hohe Tauerii (Österreich) (Abb. 13).
1. S t a d i u m (Nymphe): Lag nicht vor. I 2. S t a d i u m (4 Ex.): Segnicnte I-V11 mit je einein paramcdianen Paar Skleroti-
satioiis-Zentren um je ein „Spaltsiiiiies-Organ".
3. S t a d i u m : Lag nicht vor. Bei einem hauturigsbereiten Tier des 2. Stadiums sind die nicderliegcnden Borsten der abdominalen Segniente zu erkennen, die dicsem Stadium noch fehlen.
4. S t a d i u m (3 Ex.): Variabilitat im Verschmelzungs-Grad der beiden Skleroti- sationszentrcn der Segmente: Bei 2 Ex. sind Skleritc 1-IV einheitlich, lcdiglich 1V distal und caudal gefurcht; bei einem Ex. hangen dic Zentren der Segmentc 11 und IV nur lose zusammen.
5. S t a d i u m (2 Ex.): Fortschreitende Tendenz, dic bciden Einzel-Zentren zu verschmelzen; bci einem Ex. Sklerite I-IV einheitlich, bci dem 2. Ex. auch Sklerit V ilahezin einheitlich.
0 0 0 . *. . 0 a b C d e
0
Abb. 12. I . lute%pes. - Ontogenetische Entwicklung des Scutum (abdominale Sklerite I-V) und der freien Sklerite V1 und VII. - a Stadium 11; b Stadiuni 111; C Stadium IV; d Stadium V und VI; e Stadium V11 (adult).

6. S t a d i u m (subadult, 11 Ex.): Diese relativ große Serie variiert sehr in der Sklerit-Auspragung; kaum ein Tier gleicht dem anderen. Jmmer eine Spange bilden die Zentren 1-111, alle folgenden sind cntwcdcr zweigeteilt oder - in ~rcchsclnder Folge - verschmolzeii. Maximal sind die Skleritc 1-V11 einhcitlich. Die Serie zeigt nicht alle moglichcn Variationcii, doch fällt auf, da6 die Sklcritc der einzcliien Seg- mente nie miteinander verschmolzcii sind, - bis auf eine Ausnal~me (Abb. 13 k).
7. S t a d i u m (adult): 8 mit Scutum parvum, $! mit Scutum lami~iatum.
Zusdiri i i icnfdssirng Es laWt sich erkennen, daH anch bei 1. kollara die distalen Zentren uiid nachfo-lgend, - von Hautung zii I-Iautung voranschreitend - die caridalen Sk le r~ t sp~~ngen verschmolzen werden. Bci altcren Jugendstadien sind die Sklcrite sehr unterschiedlich auf die einzelnen Seginentc ver teilt. Tm Siibatlult-Stadium verwachsen benachbarte Skle~rte ausnahms- weise. so daR cin Scatilm intcrn~ediirii~ entsteht.
Vergleich uiid Folgeruiigeii
Bei Arte11 der Nachbarfamilie Ncrriastomatidae vcrschmelzeii die Skleritspangcn der abdomiiialcn Scgmcnte bereits bei scibadulten Tiereii (z. B. Paranemastoma), sie tragen also ein Scutum parvum. Scuta des Cephalothorax und des Abdomens sind durch eine weiche Membran getrennt. Das Scutum magnum der adulten Neinastoma- tiden (ccphalothoraka,lc und abdon~inalc Platte verschmolzen) bildet keiiic Ischyropsa- . . - - amm a a D . 0 .' d- -L . 0 - 0 0 . - I - . . . . n . e . e . .
o - o ~ - - ~ - 0 - r - " 0 0 - a m - L - * - . m - .. a . 0 - - - . n .* * * 0
0 .* ea l m *. 0 f 9 h I k I m n
Abb. 13. I. l io l lar~. - Ontogcnet~schc Rnt\\icklung des Scirt~irn (abdominale Sltlerlte I-V) und dei freien Sklciite V1 und V11 - a Stnrl~um 11, b tind c Stadiunr J V , d und e Stadiui~i V , /, b ~ s TZ
Stadlum V (subddult).

Die Abgrenzung von Biospezies usw. 159
lis-Art mehr aus, vielmehr wird von ihnen ein typisches Jugendmerkmal der Nemasto- matidae in das Adultstadium übernommen. Die Gattung (und die ganze Familie) zeichnet sich in dieser Hinsicht durch Neotenie aus. Bei den 9 vieler Ischyro~sulis- Arten wird die Entwicklung noch mehr retardiert. Sie behalten das bereits in fruhen Jugendstadien auftretende Scutum laminatum ständig bei. Diese Tendenz, die post- embryonale Entwicklung vorzeitig zu beenden, ist nicht abgeschlossen. Das beweisen die höchst variablen Scuta intermedia von I. hudzii (Abb. 14), I. rnuellneri (Abb. 15) und I . strandi.
Im phylogenetischen Sinne darf also von einer „Auflösung" des Scutum ge- sprochen werden. Die gemeinsamen Vorfahren der Nemastomatidae und Trogulidae einerseits und der Ischyropsalididae andererseits besaßen zweifellos ein Scutum magnum. Die Ischyropsalididae bilden" es heute nicht mehr aus, denn wahrend der Ontogenese verschmelzen die als Einzelsklerite angelegten Elemente vielfach gar nicht mehr zu einem einheitlichen Schild. In dieser Hinsicht machen Scuta inter- media und laminata den Prozeß der „Ver?chmelzung" zum Scutum magnum nicht mehr durch.
Man kann einwenden, daß das Scutum laminatum nicht den abgeleiteten, son- dern den ursprunglichen Zustand darstelle, der bei altertunilichen Arachniden auch heute noch vorkomme. Das Scutum parvum ware dann als progressiver Zustand zu
Abb. 14. $-Scuta von I. hadzii. - a bis d nach Angaben von H A D ~ I (1942); e nach einem Es. vom Obir, Österreich (zwischen den Beispielen c und d einzureihen).
Abb. 15. $-Scuta von I. nzuellneru. - a Castitljiva jania; b Radmannsdorf; c Ins'el Korfu; d Jod- loch und Berg Sever/Herzegowina; e wie U ; f laina Priivec. I

werten. Dagegeu spricht, daß Scuta laminata (und als weitere Progression Scuta dis- secta) innerhalb der Opiliones nur bei den Ischyropsalididae vorkommen. Wollte man diese Bildungen als primar ansehen, wurde man unterstellen, daß Arten des Genus Ischyropsulzs in diesem Merkmal noch urspri~nglichere Verhaltliisse aufwiesen als Gruppen der Opiliones, die in vieleii Merkmalen als schr ursprunglich gelten und die das System anfuhren. Sie tragen nie Scuta laminata, sondern Scuta magna oder gar Scuta completa (Großsehild aus Ccphalothorax innd deii abdominalen Skleriten I bis
f G//ed veniro-
2 G//ed medan
haltnng). Abb. 16. Schema der Cheliceren-Bewehrung. - Querschn~tt durch das 1. und 2. Glied ( ~ n Ruhe-
VII). So kaiiii das Scutum laminatum der Ischyropsalididae allenfalls als tertiärer Zustand gewertet werden ( H A D ~ I 1942): Der primäre (Einzelsklerite) ist bei rezenten Opiliones nicht mehr anzutreffen, der sekundäre (Großsehild) ist weitverbreitet, und der tertiäre (Scutum parvum, laminaturn und dissectum) kommt nur in einer Gruppe vereinzelt vor.
C. Die Cheliceren-Bewchrung.
Alle Arten zeichnen sich durch weit uber korperlai~ge Chelieeren aus, deren erstes Glied, weniger auch das zweite, mit Dornen, Tuberkeln und Borsten besetzt sind. Alle bisherigen Autoren maßen dieser variablen Bewehrung entscheidende Be- deutung als Artkennzeichen zu. - Die bisweilen verwirrend erscheinende Bewehrung des Grundgliedes läßt sieh auf ein einfaches Schema zuruckfuhren (Abb. 16):
1. do r sa l e Re ihe ; eine meist streng ausgerichtete Reihe großer Dornen auf der Dorsalseite.
2. v e n t r o - l a t e r a l e Re ihe ; meist streng ausgerichtete Reihe großer, distad gerichteter Dorne.

Die Abgrenzung von Biospezies usw. 161
3. ven t ro -med ia l e Reihe ; wie 2., aber meist nur schwach entwickelt.
In und an diese drei meist markanten Reihen schieben sich kleine Dornen- Elemente der Lateral- und weniger auch der Medial-Seite, so daß die Anordnung meist unübersichtlich ist.
Die Ventralseite (ventro-median) dei Grundgliedes ist immer unbewehrt. In die Furche, die die beiden ventralen Dornen-Reihen bilden, wird in Ruhehaltung das 2. Glied eingeklappt (Abb. 16). Die Grundglieder einiger Arten sind unbewehrt. Le- diglich die Anlagestellen der normalerweise größten Dornen sind durch eine Borste gekennzeichnet. Es muß zunächst offenbleiben, ob diese habituell sehr ähnlichen Arten als nächstverwandt anzusehen sind oder ob Konvergenzen vorliegen.
Die Bewehrung ist bei$ und $? entweder einheitlich, so daß sich die Geschlechter sehr ähneln. In vielen Fällen ist aber die Bedornung der 9-Chelicere stärker; das fällt vor allem dann auf, wenn die $-Cheliceren glatt sind.
Eine biologische Bedeutung der Bewehrung wurde bisher nicht gefunden; nichts deutet darauf hin, daß ihr eine zukommt. Im Gegenteil weist die hohe individuelle Variabilität in Größe, Lage und Anordnung der Dornen darauf hin, daß dieser Merk- malskomplex neutralen Selektionswert besitzt.
Die Variabilität der Bedornung wird vor allem von H A D ~ I und DRESCO hervor- gehoben. Die intra- und interspezifische Variabilität dieser Polymorphie ist jedoch unbekannt und ihr Wert für die Spezies-Abgrenzung somit fraglich. Es war bis jetzt keine Aussage darüber möglich, ob Merkmale dieser Bewehrung in Form eines Klin geographisch variieren, ob sie in bestimmten Populationen Diskontinuitäten auf- weisen und somit Artverschiedenheiten andeuten, und es war deshalb auch nicht ersichtlich, ob zwei Arten sympatrisch oder gar syntopisch vorkommen können.
Um Einblick in die Variabilität der Cheliceren-Bewehrung zu erhalten, wurden untersucht:
1. Unterschiede an den Cheliceren eines Individuums;
2. Unterschiede zwischen Individuen einer streng lokalisierten Höhlen-Popu- lation ;
3. Unterschiede an den Cheliceren genitalmorphologisch sehr einheitlicher Indi- viduen ;
4. Unterschiede im $-Genital-Apparat bei Individuen mit einheitlicher Be- wehrung ;
5. Schließlich wurde nach fließenden Übergängen der Bewehrung und der Pro- portion des Grundgliedes in engumschriebenen Arealen gesucht.
ad 1 (Abb. 17): Die Grundglieder sind dorsal unterschiedlich bewehrt (Teilfigur a). Es ist nicht sicher, ob sich die großten Elemente (schwarz hervorgehoben) ent- sprechen. Sie sind zwar annahernd gleich groß, aber lagevcrschieden. Ebenso vari- ieren die interkalierten kleinen (ebenfalls schwarz markierten) Dornen; ihre gegen-
11 Zool. Jb. Syst. Bd. 96 a

Abb. 17. Variabiltiät der dorsalen Bewehrung der rechten und linken Chelicere (Grundglied) eines Individuums, von lateral. - a Monte Baldo (vgl. Abb. 63 b); b Lurgrotte (vgl. Abb. 56i).
Abb. 18. Variabilitat der dorsalen Bewehrung der rechten Chelicere (Grundglied von lateral, 8). - Individuen aus einer isolierten Hohlen-Population (Lurgrotte; vgl. Abb. 56).

Die Abgrenzung von Biospezies usw. 163
seitige Zuordnung ist nicht moglich. Entsprechendes ist Teilfigur b zu entnehmen. Die Unterschiede sind weniger ausgepragt, da die Bedornung schwacher ist.
ad 2 (Abb. 18): Folgegernaß variiert auch bei einer streng isolierten Hohleil- Population die Dorsalbewehrung dcs Grund-Gliedes erheblich. Homologisierung der Dorne bleibt immcr unsicher, da sich in die Dorsalreihe manche der lateralen Dorne einschieben.
ad 3 (Abb. 19): Die genital-morphologisch eiiihcitlichcn Individuen aus cngum- schriebcnem Areal (NO-Spaiiie~i) wcisen am Gruridglicd dorsal alle Übergknge von starker bis fchlcnder Bewehrung auf.
ad 4 (Abb. 20): Individuen ubereinstimmender Dorsalbewehrung, aber vcrschie- dener geographischer Herkunft (S-Italien, Schweiz, Frankreich, NO-Spanien, Portu- gal) diffcriercn im 8-Genitalapparat.
ad 5 (Abb. 21): Individuen aus verschiedenen engumschriebenen alpinen Popu- lationen konncn bei einheitlicher Gcnitalmorphologic nicht nur in der dorsalen Be- wehrung, sondern auch - gleitend - in der Proportion des Grundgliedes differieren.
Systematische Wertigkeit der Merkmale der Cheliceren-Bcwehrung
Merkmale der dorsalen Bewehrung dcs Grundgliedes erlauben (ohne vorherige Kentnnis von Artgrenzen) nicht cinmal, Populationen einander zuzuordnen, gc- schweige denn Arten abzugrenzcn, da 1. die Variabilität am Einzeltier bereits hoch ist, 2. die Variabilität dicses Merkmals bei englokalisierten Populationen hoch scin
Abb. 19. Variabilitat der dorsalen Bewehrung der rechtenchelicere (Grundglied von lateral,&. - Individuen aus einem engumschriebenen Areal in NO-Spanien (vgl. Abb. 49a, b, c, d, e, f).
11'

Abb. 20. Nahezu fehlende Variabilität der dorsalen Bewehrung der rechten Chelicere (Grundglied von lateral, 8). - Individuen aus disjunkten Arealen. - a S-Italien (vgl. Abb. 39a); b Österreich (vgl. Abb. 42d); c Frankreich(vg1. Abb. 44c); d Spanien (vgl. Abb. 49a); e Portugal(vg1. Abb. 54a.)
Abb. 21. Gleitende Anderung der Cheliceren-Pioportion (rechtes Giundglied Yon lateial, a-c: 3, d: $2). - Individuen aus alpinen Populationeil. - a vgl. Abb. 66b; b vgl. Abb. 6Gd; c vgl. Abb. 56e; d vgl. Abb. 63f.

Die Abgrenzung von Biospezies usw. 165
kann, 3. bei einheitlicher Genitalmorphologie Extreme in der Bewehrung auftreten, 4. bei Unterschieden in der Genitalmorphologie die Bewehrung einheitlich sein kann, 5. benachbarte Populationen fließende Übergänge in Bewehrung und Proportion des Grundgliedes aufweisen konnen.
Ein primär auf die Morphologie der Cheliceren gegründetes Spezies-Konzept (ROEWER, H A D ~ I , DRESCO) kann daher keineswegs dem der Biospezies entsprechen. Die Cheliceren-Bewehrung signalisiert keine Artgrenzen.
d) Die Apophyse und das Drüsenfeld der $-Chelicere
Apophyse : Das 1. Glied der $-Chelicere ist distal bei den meisten Arten auf der Dorsal- oder Dorsomedial-Seite angesch.cvolleii oder zu einer stumpfen Apophyse aus- gezogen (vgl. Abbildungen in Kap. V1 3 b). Sie ist bei den meisten unterschiedlich geformt, aber innerhalb einer Spezies nur wenig variabel. Die Diskontinuitäten von Art zu Art sind gering.
Ähnliche (wahrscheinlich homologe) Bildungen finden sich bei Arten der Gat- tung Sabucon und der einzigen Art der Gattung Ceratolasma (beide Ischyropsalididae) und bei nahezu allen Arten der Familie Nema'stomatidae, ferner bei den Trogulidae (Gattung Dicranolasrna). Die$ der Arten dieser Gruppen besitzen absolut uiid relativ wesentlich kleinere Cheliceren-Grundglieder; auf ihnen steht dorsal die meist la,iige und deutlich vom Grundglied abgesetzte Apophyse. Auf ihr oder aii ihrer Basis liegen Austrittsstellen für Drüsei~sekret (vgl. Kap. I11 7).
Auf oder in der Nä'he dieser Apophyse tragen die $ einiger Arten der Gattung Ischyropsalis einen dichten Besatz kurzer und feiner Härchen, die zu einem scharf begrenzten Feld zusainmentreteii (Abb. 22). HAD?I (1928) bemerkte es zuerst (bei hellwigi, sub pectinifem) und iiaiinte das Haarfeld „Bürsteu. ROEWER (1950) iiber- nahm diesen Terminus 1 er wird auch hier beibehalten. Gleichbedeutend benutze ich - hinweisend auf die anatomischeil Verhältnisse - „Drüsenfelda. Diese Bürste ist nicht
Abb. 22. ,,Bürstenu ain Distalende des 8-Cheliceren-Grundgliedes (von medial). - a hellzoigi; b lizanicata; c robustu; d car l i ; e a d a m i i ; f nod i f e ra ; g py renuea ; h lu te ipes; i dent ipalpis .

bei allen Arten vorhanden, zumindest nicht in dieser gut kenntlichen Form. Ein- zelne Haare und Borsten befinden sich am Distalende des 1. Cheliceren-Gliedes bei allen Arten. Lage und Form der Bürste erwiesen sich bei geographisch engumschrie- benen Populationen als bemerkenswert konstant, Das gilt auch dann, wenn die Cheli- Ceren bei den Individuen solcher Populationen sehr variieren (vgl. Abb. 19). Hier scheint ein sekundäres Geschlechtsmerkmal vorzuliegen, das für eine sinnvolle Spe- zies-Gliederung herangezogen werden kann. Einschränkend muß aber betont werden, daß es nur den $ und, wie gesagt, nicht allen Formen zukommt. Sein Trennwert er- scheint deshalb zunächst begrenzt. - Die Bürstenhaare frisch in Alkohol fixierter Tiere sind regelmäßig von einem weißen, koagulierten Sekret verklebt. Diese sehr auffällige Erscheinung wurde bisher übersehen, bis THALER (1966) darauf hinwies. Sie findet sich gleichermaßen an den$-Cheliceren der Arten der obengenannten Grup- pen. Zwar bemerkte bereits H A D ~ I (1928) den weitlumigen Ausführporus für das Drüsensekret (bei Paranemastoma quadrzpunctatum), deutete ihn aber als „akzessori- sches Stigma".
Systematische Wertigkeit der Merkmale an Apophyse und Drüsenfeld
Die Diskontinuität des Merkmals „Apophyseu ist bei Individuen unterschied- licher geographischer Herkunft, deren Artselbständigkeit vermutet werden darf, bis- weilen nur sehr gering. Diesem Merkmal ist zur Speziesabgrenzung nur akzessorischer Wert beizumessen.
Dem Merkmal „Drüsenfeld" kommt eine hervorragende Bedeutung in der Se- xualbiologie zu (vgl. Kap. I11 7). Es ist somit berechtigt, überwiegend dieses Merkmal für die Spezies-Gliederung heranzuziehen. Die mit ihm abgegrenzten Arten dürften der heutigen Definition der Spezies als Fortpflanzungsgemeinschaft weitgehend ent- sprechen.
Ergebnisse des Kap. I V 1: Die bisher in der Spezies-Systematik der Gattung Ischyropsalis verwendeten und einige bisher nicht beachtete Merkmale haben sich als weitgehend variabel und zur Ermittlung und Abgrenzung von Biospezies als un- brauchbar erwiesen (Genitalapparat, abdominales Scutum, Cheliceren-Bewehrung, Apophyse des Cheliceren-Grundgliedes). Reproduktive Isolation scheint lediglich die „Bürste" zu signalisieren. Die Funktion der Bürste (bzw. des Drüseiifeldes) während des Paarungsverhaltens bestätigt das (vgl. Kap. I11 7).
2. Die isoliereiideiz Mechanismen
Vorstehend wurde erläutert, daß sich das morphologisch-typologische Artkon- zept in der Gattung Ischyropsalis nicht mit dem Konzept der Biospezies decken kann. Die von Autor zu Autor schwankende Bewertung einzelner Merkmale führte zu dif-

Die Abgrenzung von Biospezies usw. 167
ferenten Gliederungen. Es gab bisher kein zuverlässiges Kriterium, nach dem eine objektive Wertung der Merkmale hätte erfolgen können.
Die Bewertung morphologischer Merkmale zum zwecke systematischer Gliede- rungen beruht vor allem auf den Erfahrungen des Kenners der Gruppe, der sie, wie HERRE (1964) ausführt, (1) von einem Schluß von bekannten Spezies-Unterschieden [hier: ~ios~ezies-dnterschieden] auf Spezies-Grenzen in neuen Fällen gewinnt, oder (2) durch eine Analyse die Formbesonderheiten und die innerartliche Ausformung der Merkmale bei verwandten Arten ermittelt und so zu einer begründeten Bewertung noch unbekannter Formeigenheiten gelangt. Beide Verfahren sind Extrapolationen, die immer ein „Wagnisu darstellen. Dies um so mehr, da in beiden Fällen zwischen ,,ArtseinH und „Artkennzeichenn, also zwischen den biologischen Eigenheiten der Art und somit ihrer Umgrenzung als Fortpflanzungsgemeinschaft einerseits und der morphologischen Kennzeichnung dieser Einheit andererseits, nicht getrennt werden kann. Das heißt, daß zunächst ungewiß bleiben muß, ob die erfaßten Kennzeichen jeweils der Ausdruck biologischer und damit spezifischer (= artlicher) Verschieden- heit sind.
Im vorliegenden Fall - Abgrenzung der Ischyropsalis-Arten - ist Verfahren (I), bei dem von bekannten Art-Unterschieden auf weitere Art-Grenzen geschlossen wird, vorläufig nicht anwendbar, da sichere Kriterien zur Abgrenzung bisher nicht bekannt sind. Anhaltspunkte lassen sich durch Verfahren (2) erschließen, also durch die Unter- suchung der Variabilität einer als Art vermuteten Gruppe von Populationen. Da aber von der Voraussetzung ausgegangen werden muß, daß in der Gattung Ischyropsalis bisher noch in keinem Falle (!) zwei oder mehr Arten ausreichend begründet gegen- einander abgegrenzt worden sind, ist auch dieses Verfahren nur bedingt verwendbar. Man könnte allenfalls aus Diskontinuitäten in der morphologischen Ausprägung sympatrisch (besser syntopisch) lebender Individuen auf Art-Verschiedenheit schlie- ßen. Sympatrisches, vor allem aber syntopisches Vorkommen mehrerer Arten gehört in dieser Gattung jedoch zu den seltenen Ausnahmen (vgl. Kap. I11 4). Ganz im Gegenteil zeichnen sich die meisten Arten durch strenge Arealvikarianz aus. So ist die Artabgrenzung auf rein morphologischer Grundlage in diesem Falle nahezu un- möglich.
Es bleibt die Möglichkeit, alle differenten Formen auf ihre reproduktive Isola- tion zu prüfen - ein Vorgehen, das sich aus technischen Gründen und der Schwierig- keit der Materialbeschaffung verbietet. Es würde jedoch genügen, das Kopulations- verhalten weniger Formen zu prüfen. Sollten sich Unterschiede im Verhalten oder in vorher nicht beachteten morphologischen Strukturen auffinden lassen, die mit dem Sexualverhalten korreliert sind und denen isolierende Punktion zugeschrieben werden muß, dürften diese Merkmale als Ausdruck von Artverschiedenheit gewertet werden. Schlüsse auf andere Formen wären dann leichter möglich.

Zunächst müssen die Mechanismeii erwähnt werden, die die freie Gendurch- mischung von Populationen verschiedener Spezies-Zugehörigkeit verhindern. Vor der Paarung greifen folgende ein:
(1) chemische, indem einPartner auf einen Sexuallockstoff des anderen anspricht;
(2) ethologische, indem bestimmte Verhaltensweisen notwen.dig sind, um Syn- chronisation der Geschlechter und damit die Kopulation zu erreichen;
(3) mechanische, indem die Genitalapparate von 8 und einen so hohen Grad von Kompliziertheit erreicht haben, daß zum 9 nur ein ganz bestimmter $-Apparat paßt („Schlüssel-Schloß-Prinzip"). Es scheint aber, daß dieser mechanischenschranke - sollte sie in manchen Gruppen tatsächlich bestehen - nur akzessorische Bedeut~ing zukommt. Tatsächlich ist den meist vorgeschalteten ethologischen Schranken größe- res Gewicht beizumessen;
(4) ökologische, indem verwandte Arten lediglich durch unterschiedliche Vor- zugsbiotope, durch Nahrungsspezialisierung oder durch verschiedene Reifezeiten vor Genaustausch bewahrt bleiben.
Nur Merkmal (3) ist morphologisch definiert und - an lebendem Material als artisolierend erkannt - auch an weiteren Populationen nachzuprüfen, ohne daß biologische Merkmale bekannt sein müßten.
Auch bei einem Schluß von einem ethologisch-biologischen Merkmal, das mit morphologischen Kriterien korreliert ist, auf Nachbarpopulationen mit unbekannten ethologisch-biologischen Merkmalen haben wir es mit einer Extrapolation zu tun, ihre Zuverlässigkeit läßt sich abgrenzen. Wenn ein Lethologisches, Genaustausch unter- bindendes Merkmal bekannt ist, das mit morphologischen Strukturen korreliert ist, scheint es berechtigt, in diesen Strukturen abweichende Individuen ebenfalls als Vertreter eigener Arten anzusehen. Uberprüfung auch des ethologischen Nerkmals der erschlossenen (mit sehr hoher Wahrscheinlichkeit) Biospezies ist wünschenswert, aber nicht unbedingt erforderlich. Zweifellos ist auch dieses Vorgehen ein „Wagnisu (HERRE 1964), doch werden sich mit dieser Methode gewonnene Ergebnisse durch morphologische, ökologische und chorologische Befunde erhärten lassen. Bei den vorliegende11 Verhältnissen scheint allein dieses Vorgehen ein Höchstmaß an Objek- tivität zu gewährleisten.
Zwar können in Einzelfällen auch die Merkmale (I), (2) und (4) für sich allein Möglichkeiten zur Abgrenzung bieten, eignen sich aber nicht zur Extrapolation an konserviertem Material und somit zur allgemeinen Verwendung. Es muß also, um eine Forderung von HERRE (1964) zu wiederholen, in der Biospezies die Morpho- Spezies gesucht werden, da wohl jede Biospezies durch jeweils eigene Besonderheiten der Gestalt oder Punktion gekennzeichnet ist. Die Biospezies läßt sich durch ihre formative Leistung erfassen.

Die Abgrenzung von Biospezies usw. 169
Das heißt im~ vorliegenden F-a11, zu prufen, ob wahrend der Paarung, in deren Verlauf am ~ehesteii isolierende Mechanismen vermutet werden durfen, Verhaltens- weisen auftreten, die von Strukturen der außereii oder inneren Morphologie (z. B. der Kopulations-Organe) abhängig sind. Es darf dann erwartet werden, daß diese Merk- male bei einer Art nicht oder hochstens geringfugig variieren. Nachbararten sollten das Merkmal diskontinuierlich, also in Form eines Merkmalsspruiiges aufweisen.
Zu diesem Ergebnis kam bereits GRASSHOFF (1968). Er fand bei bekaiiiiteii Bio- spezies der Gattung Araneus (Araneae) artkennzeichnende morphologische Merkmale am Genitalapparat und extrapolierte die Ergebnisse auf Individuen unbekannter Biospezies-Zugehorigkeit. Diese erschlossenen Morpho-Spezies konnen nahezu sicher als Biospezies gelten.
Folgender ~ r b e i t s g a n ~ bietet sich an: .
1. Das Paaruiigsverhalten einer (besser mehrerer).differenter Form(eii), deren Artselbständigkeit vermutet werden darf,'muß untersucht werden. Es läßt sich dann ,
entscheiden, ob Strukturen vorhanden sind, die als Ausdruck reproduktiver Isolation aufgefaßt werden können.
Diese Voraussetzung ist erfüllt. In Kap. I11 7 und ausführlich in ~ ! A R . T E N S (1969) ist das Kopulationsverhalten dreier differenter Formen beschrieben worden. Das Biirstenfeld der $-Chelicere wurde als trennendes Merkmal erkannt.
2. Es muß geprüft werden, ob dieses Merknial bei syiitopisch lebenden Formen unterschiedlich ausgeprägt ist. Sie haben dann als verschiedene Arte11 zu gelten.
3. Sind Unterschiede im Merkmal „Bürstea gefunden morden, können mit ihm weitere Kennzeichen erschlosseii und auf Populationen unbekakiter Artzugehörig- keit übertragen weiden. .
3. Vergleich syn top i s che r Fo rn i en u n d E x t r a p o l a t i o n
Die einzigen bisher bekannten Syntopien wurden in Ka,p. 111 4~ beschrieben. Hier werden die unterschiedlichen Merkmale dieser Formen, die als Arten gelten müssen, gegenübergestellt.
Die Tabelle zeigt, daß in der Form und der Bewehrung des Cheliceren-Grund- gliedes, in Form und Lage der Bürste und in Form des Sklerits der Glans peiiis wesent- liche Unterschiede zwischen den syntopischeii Arten bestehen. Gering sind die Unter- schiede des Scutum und des Truncus penis. Da jedoch allein die Bürste (das Drüsen- feld) als Ausdruck reproduktiver Isolation und somit von Artverschiedenheit ange- sehen werden kann (vgl. Kap. I11 7 und IV 2), muß einer weiteren Aufgliederung dieses Merkmal zugrunde gelegt werden. Für die bürstentragenden Arten ergibt sich folgende Aufteilung; außer der Bürste sind weitere kennzeichnende Merkmale be- reits berücksichtigt.

Lokalität Name Merkmale lcollari h. hellwigi h. lucantei nodifera
Lokalitat Österreich: Stubachtal Spanien: Sierra de Aralar
I. Cheliceren- Glied: Forrri B u i s t e
P e n i s l'ruiicus
Sklerit
schlank -
Haripti eihen stark vortretend, viele Einzeldorne 8 : parvum 9: Iaminatiim
schlank, basal ver- dickt plan, gelappt, stark beborstet
massig, robust sehi schlank sehr groß, dorsal, auf dem ganzen klein, mediodistal, dorsalen Diittel des Grundgliedes ellipsoid 14auptreihen wenig vortietend, fast fehlend, doisal win- viele Einzeldorne dorsal und hige Tubeikeln lateral
69: parvum 8 9 : parvunl
sehr sclilank schlank, basal verdickt
plan, ungelappt, wenig beborstet ungelappt, gekielt, stark beborstet
1.. manicata; mit großer Bürste dorso-distal, am Distalende auf die Mcdialscite übergreifend; Cheliceren-Grundglied distal apophisenartig aufgebläht (Abb. 22b) (Karpatcn-Länder).
2. adamii; kleine Bürste medio-distal; dorsal eine Längsreihe aus 3 kräftigen Dornen, winzige Apophyse disto-medial (Abb. 22 e) (Halbinsel-Italien).
3. carli; Bürste ähnlich adamii, ebenso dorsale Bewehrung; nicdrige und stumpfe Apophysc dorso-distal (Abb. 22 d) (Mittel- und West-Alpen).
4. dentipalpis; Bürste medio-distal, klein; dorsal eine Längsreihe aus 3 schlankcn Dornen; Apophysc dorsad halbkugelig vorgcwölbt (Abb. 22i) (mittlere Süd-Alpen).
5. luteipes; Bürste am Distalrand dorso-nicdial; punktförmig klein, 1)rcier-Reihe dcr dorsalen Bcwehrung nicht prägnant, markantc stumpf-kegelförmigc Apophysc dorso-distal (Abb. 22h) (franz. Zentralmassiv, Ost- u ~ i d Mittcl-Pyrenäen).
6. nodifera; große ellipsoide Bürste medio(!)-distal; dorsale Reihe der Beweh- rung stark variiercrid, wenn vorhasidcii, dann immer 3 Einzcldornc; dorsale Apophyse spitz, stark nach distal vorgezogcn (Abb. 22f) (Kantabrisehe Gebirge).
7 . pyrenaea; sehr klcine Bürste dorso-distal; dorsale ßcwehrtiilg rcduziert, fast fehlend; dorsale Apophyse stumpf (Abb. 22g) (Höhlcn dcr fraiiz. Zentral-Pyrenäen).
8. robusta; große ellipsoide Bürste medial; dorsale Bewehruilg dreizahlig; Apo- physe medio-distal vorgewölbt (Abb. 22c) (Portugal).
9. hellwigi; sehr ausgedehntc Bürste auf dem dorso-distalen Drittel; dorsale Be- wchrung aus vielen Einzeldornen, nur wenige markant hervortretend; winzige medio-

Die Abgrenzung von Biospezies usw. 171
distale Apophyse (Abb. 22 a) (2 Areale mit etwas differenten Populationen in Südost-/ ,
Mittel-Europa und in den W-PyrenäenIKantabrischen Gebirgen).
Erschließung weiterer Merkmale Für die vorstehend aufgeführten Arten ist das Merkmal „BürsteLL für die Er-
kennung und Kennzeichnung ausschlaggebend. Es besteht nun die Möglichkeit, mit seiner Hilfe weitere Merkmale so zu umgrenzen, zu „eichenu, daß sich auch jene Popu- lationen Arten zuordnen lassen, denen das Merkmal „Bürsteu fehlt. Die mit diesem Merkmal erschlossenen Kriterien werden nachfolgend in der Reihenfolge ihrer Wer- tigkeit aufgeführt.
1. Apophyse des J -Che l ice ren-Grundgl iedes . - Den lage- und größen- konstanten Bürstenfeldern ist eine Aufwölbung bzw. stumpfe Apophyse zugeord~iet, die in den meisten Fällen die Bürste trägt. Die Apophyse ist formkonstant, kann je- doch bei manchen Arten (z..B. nodifera) geringfügig variieren. Diese Struktur bietet also ein Merkmal, das sich hervorragend zur Extrapolation eignet. Sein Wert wird nur dadurch etwas gemindert, daß die interspezifischeil Unterschiede bisweilen nur gering sind (z. B. luteipeslnodifera).
Extrapolation: Der Apophyse des Grundgliedes (d) ist nach der Bürste zentrale Bedeutung zur Abgrenzung noch unbekannter Arten zuzuschreiben.
2. Bewehrung des Che l ice ren-Grundgl iedes . - Es wurde abgeleitet (vgl. Kap. IV lc ) , daß die Cheliceren-Bewehrung für sich allein, also ohne vorherige Kenntnis von Artgrenzen, für eine sinnvolle Artgliederung nicht brauchbar ist. Die Cheliceren-Bewehrung der vorstehend erschlossenen Arten, die nahezu sicher Bio- spezies darstellen, Weist indes von Spezies zu Spezies deutliche oder aber nur so geringe Unterschiede auf, daß sie nur an einer großen Serie erkannt werden können. Die große Zahl und die unterschiedliche Lage der Dornen-Elemente macht die de- skriptive Darstellung schwierig; es muß deshalb auf die Abbildungen (Kap. V1 3 b) ver- wiesen werden. Sie lassen erkennen, daß die markanten Elemente der dorsalen, ven- tro-medialen und der veiztro-lateralen Reihe entweder weitgehend konstant (z. B. hellwigi, manicuta), wenig variabel (luteiyes, carli, adamii) oder extrem va'riabel sind (nodifera).
E x t r a p o 1 a t i o n: Man kann davon a,usgeheii, daß Popula,tionen engumschrie- bener Areale, deren Individuen keine oder nur geringe Variabilität der Chelicereii- Bewehrung aufweisen, einer Spezies zugehören. Syntopische Arten weisen gerade an diesem Cheliceren-Merkmal erhebliche interspezifische Unterschiede auf. Das Merk- mal versagt bei Arten mit stark variabler Bewehrung und ist deshalb für die Er- schließung neuer Artgrenzen nur bedingt brauchbar.
3. G e n i t a l a p p a r a t des 3 (Penis). - Für die bisher behandelten Arten hat der Genitalapparat keine wesentliche Bedeutung für die Artabgrenzung. Das liegt einerseits daran, daß seine Strukturen einfach und wenig variabel sind (vgl. Kap. IV

1 a), andererseits liegt das am Alter der einzelnen Arten. Die großenteils vikariierenden Arten sind noch jung und Artunterschiede haben sich nicht oder nur sehr schwach am Genitalapparat manifestiert. Unterschiede sind am Truncus (Proportion) und an der Form des ventralen Sklerits der Glans erkennbar. Der Sklerit ist entweder schmal lanzettlich, breit auslaufend oder kurz gerundet. Dieses ~ e r k & l kann aber bereits bei einer Art erheblich variieren (z. B. carli und adamii).
E x t r a p o l a t i o n : Ohne Kenntnis von Artgrenzen ist den Merkmalen des 8- Genitalapparates für die Erschließung weiterer Arten nur geringer Wert beizumessen. - 9-Genitalapparat vgl. Kap. IV 1a .
4. abdomina l e s Scu tum. - Bei den bisher behandelten Arten hat es sich als konstant erwiesen, doch fehlen fast gänzlich Unterschiede von Art zu Art. Achtmal treten Scuta parva bei beiden Geschlechtern auf, zweimal Scutum parvum beim $ und Scutum laminatum beim 9.
E x t r a p o 1 a t i o n: Wegen zu geringer interspezifischer Unterschiede nahezu un- geeignet, um unbekannte Artgrenzen festzulegen.
5. Verbre i tung . - Neben den morphologischen Unterschieden bestehen von Art zu Art unterschiedliche chorologische Merkmale. Die vorstehend begründeten 9 Arten vikariieren weitgehend (vgl. Abb. 24, 25). Die Zuordnung zu einer bestimmten Spezies ist allein mit den chorologischen Merkmalen nahezu immer eindeutig möglich.
E x t r a p ola t i o n: Zur Erschließung von Artgrenzen sind chorologische Merk- male gut geeignet.
Z u s a m m e n f a s s u n g : Von vier morphologischen Rferkmalen eignen sich zwei (Apophyse und Bewehrung des 8-Cheliceren-Grundgliedes) gut, zwei weitere (8-Genitalapparat und abdomi- nales Scutum) nur bedingt zur Erschließung neuer Artgrenzen; gut eignen sich überdies Merkmale der geographischen Verbreitung.
Extrapolation
Die bisher nicht erfaBten Populationen (Individuen ohne Bürste) lassen sich mit den vorstehend begründeten Merkmalen zusammenfassen.
I. Winziges, reliktäres Areal in den Kantabrischen Gebirgen N-Spaniens. 10. dispar;$-Grundglied völlig glatt, höchstens dorsal drei Tuberkel angedcutet ;
dorso-distal charakteristische Apophyse, deren distale Böschung plan; Truiicus peiiis extrem lang und schlank; $9 mit Scutum parvum (Abb. 67).
11. magdalenae; $-Grundglied mit schwacher Bewehrung dorsal; dorso-distale Apophyse schwach zweizipfelig; Truncus penis untersetzt; $ Scutum parvum, $2 Scu- tum laminatum (Abb. 65).
11. Vergleichsweise viel größeres Areal in den 0-Alpen und auf der Balkan- Halbinsel.
12. kollari; $-Grundglied mit kräftiger Bewehruilg, ungeordnet, ohne markante Einzelelemente; ohne deutliche Apophyse, doch bei sehr kräftigen Individuen eine

Die Abgrenzung von Biospezies usw.
schwache dorso-mediale Aufblähung; $ Scutum parvum, 9 Scutum laminatum (Abb. 56) (mittlere und nördliche 0-Alpen). -
(Merkmale des Penis besitzen bei dieser und den folgenden Arten fast keinen Trennwert und werden somit nicht erwähnt.)
13. strandi; $- und 9-Grundglied dorsal meist exzessiv bedornt, ohne 'distale Aufwölbung oder Apophyse; $ Scutum intermedium, 9 Scutum laminatum ( ~ b b . 63) (östliche S-Alpen).
14. hadzii; Bewehrung des Grundgliedes rechtwinklig abstehend, kräftige Ein- zeldorne dorsal; disto-medial keulig verdickt, $ Scutum intermedium, ? Scutum laminatum (Abb. 59) (Karawanken, Steiner Alpen).
15. muellneri; $-Grundglied nahezu glatt, höchstens mit winzigen Tuberkeln; sehr kleine Apophyse medio-distal; $ Scutum intermedium oder laminatum, Scu- tum laminatum (Abb. 61) (Dinarische Gebirge bis N-Griechenland).
Die vorstehend charakterisierten Arten stellen Morphospezies dar. Nach den Erfahrungen, die bei der Untersuchung der bürstentragenden Arten gemacht wurden (hellwigi- und dentipalpis-Gruppe), handelt es sich mit hoher Wahrscheinlichkeit zu- gleich um Biospezies, die sich mit biologischen Merkmalen umgrenzen lassen würden.
V. Chorologie und Speziation
1. V e r b r e i t u n g der G a t t u n g u n d der A r t e n g r u p p e n a) Das Gattungsareal
Die Gattung Ischyropsalis ist auf Europa beschränkt. Ihre Arten besiedeln haupt- sächlich die Mittel- und Hochgebirge West-, Mittel-, Süd- und Teile Ost-Europas.Die drei in Japan und Korea vorkommenden Arten: abez SATO & SUZUKI 1938, yezoensis SUZUKI 1958 und coreana SUZUKI 1966 weichen von den europäischen Arten in gra- vierenden Merkmalen der Cheliceren, des $-Genitalapparates und des Exoskelettes so stark ab, daß fur sie eine eigene Gattung, Nipponopsalis MARTENS & SUZUKI 1966, errichtet werden mußte.
Die Verbreitungskarte (Abb. 23) läßt folgende Angaben des Schrifttums unbe- rücksichtigt:
1. D a n e m a r k . - HAA-SEN (1885) nennt einen einzigen Fund eines Ischyropsalis-Jungtieres in Kord-Jutland, das spater verlorenging. NEINERTZ (1964) erwahnt diese Angabe in der Bearbei- tung der danischen Opiliones nicht und halt es fur wahrscheinlich, daß dieser Fund auf Verschlep- pung ( I ) beruht (in litt.). - Ein Vorkommen von I. hellwzgz ist in Jutland moglich, bedarf jedoch der Bestatigung, denn der Fundort liegt von der Nordgrenze des hellwigz-Areals weit abgesprengt (Karte nlit diesem Fundort in MARTENS 1965).
2. B u l g a r i e n . - ROEWER (1950) beschreibt I. balcanaca aus dem Balkan-Gebirge, ohne genaue Fundorts-Angabe. Bestatigung blieb bisher aus, obwohl die Weberknecht-Fauna dieses Gebirges seitdem gut untersucht worden ist. Uberdies ist balcanica synonym manzcata, deren Areal-Sudgrenze wesentlich weiter nordlich im Karpaten-Becken verlauft.

Abb. 23. Das Areal der Gattung Ischyropsalis.
3. Griechen land . a) P e l o p o n n e s und A t t i k a . - Die Typenserie von I . moreana sammelte der Autor an-
geblich selbst bei Tripolitza (= Tripolis); sie enthält 15 pyrenaea und 3 9 ~odi fera . Herkunft dieser Tiere von der Peloponnes halte ich für ausgeschlossen. ,Sie stammen zweifellos aus den Pyrenäen bzw. den Kantabrischen Gebirgen, dem Verbreitungsgebiet beider Arten.
Aus der Umgebung von Vytina stammt angeblich I . goodnighti; die Typenserie besteht aus 1 5 9 dispar. Auch in diesem Fall halte ich Herkunft aus S-Griechenland für nahezu undenkbar.
I . pentelica kommt angeblich aus einer Höhle des Pentelikon-Gebirges bei Athen, von ROEWER selbst gesammelt. Die beiden Jungtiere der Typenserie (nicht 1 5 9 , wie irrtümlich mit- geteilt) gehören zu carli; die charakteristische Cheliceren-Apophyse des subadulten 5 zeigt das deutlich. Aus zoogeographischen Gründen ist es auszuschließen, daß das Material vom ange- gebenen Ort stammt.
. b) Ägä i sche Inse ln . - I . turki wird von Ägina gemeldet; die Typenserie enthält 1 5 9 magdalenae. Die Tiere stammen mit Sicherheit nicht von Ägina, zumal die Insel im August, dem angeblichen'Sammelmonat, keine ökologischen Bedingungen aufweist, d@ einer Ischyropsalis-Art genügen könnten. Ganz bestimmt gibt es diese nicht in einer dem vollen Sonnenlicht ausgesetzten griechischen Tempelruine, dem angeblichen Locus typicus. Davon überzeugte ich mich im Herbst 1963 selbst.
Von Kreta werden zwei Arten gemeldet: I . troglodytes aus der Topolia-Höhle in den Weißen Bergen auf W-Kreta und I : kratochvili aus dem Omalos-Katavothron, ebenfalls in den Weißen Bergen. Die Typenserie von troglodytes (5 iuv., nicht 2 5 39, wie irrtümlich mitgeteilt) ist zu kollari zu stellen, kratochvili zu adamii. Beide Fundangaben halte ich aus zoogeographischen Gründen für nicht richtig, zumal es mir im April 1965 trotz intensiver Suche nicht gelungen ist, an den Loci typici Ischyropsalis aufzufinden.

Die Abgrenzung von Biospezies usw. 175
4, S i z i l i e n , K o r s i k a . - Von diesen Inseln beschreibt ROEWER (1950) I. caporiaccoi und corsica. Tatsächlich handelt es sich um Exemplare von luteipes bzw. nodifera. Die Fundortangaben halte ich aus zoogeographischen Gründen nicht für richtig. Indes ist es durchaus möglich, daß' auf beiden Inseln Ischyropsalis vorkommt - doch dann gewiß,nicht diese Arten.
5. f r a n z ös is c h e R i v i e r a. - I . nicaea wurde nach einem magdalenae beschrieben. Das Areal dieser Art liegt in NW-Spanien. Sehr wahrscheinlich liegt auch hier eine Fundort-Verwechs-
,
lung vor.
6. D e u t s c h l a n d . - Die Arten I. alfkeni (Jena/Rosental) und kaestneri (Rauhe ~ l b / ~ c h - senhausen) wurden nach Individuen von muellneri bzw. manicata,beschrieben. Ganz bestimmt liegen hier Fundorts-Verwechslungen vor, denn die Areale beider Arten liegen in O-"bzw. SO- Europa. - Die Nachsuche am Locus typicus von alfkeni verlief negativ.
7. Ö s t e r r e i c h . - I . pestae (Leithagebirgelsonnenberg), angeblich vom Autor selbst ge- sammelt, ist ein 8 lutetpes. Die Weberknecht-Fauna des Leithagebirges wurde besonders von GRUBER (1960) intensiv untersucht, doch wurde nie Ischyropsalis gefunden. Auch eigene Nach- suche blieb erfolglos.
b) Die Areale der drei Artengruppen
Nach genital- und cheliceren-morphologischen Merkmalen sind drei Artengrup- pen zu unterscheiden, die in Kap. V1 3 näher begründet werden.
a) hellwigi-Gruppe (Abb. 24). - Ihr Areal ist identisch mit jenem der beiden Subspezies h. hellwigi (Mittel-Europa) und h. lucantei (W-Europa).
Bbb. 24. Das Areal der Aellwigi-Gruppe. - West-Areal: I . hellzoigi lucantei; Ost-Areal: I. hellwigi hellwigi.

0 250 500 I km I
Abb. 23. Das Areal der dentipalpis-Gruppe. - Arealuberschneidungen sind kreuzschraffiert. a robusta; b nodzfera; c lc pyrenaea; dld luteipes; e dentipalpis; f carli; g adamii; h mantcata.
Abb. 26. Das Areal der kollarz-Gruppe. - ala ~nagdalenae; b dispar; c strand?,; d kollarz; e hadzti; f nzuellnerz. - .

Die Abgrenzung von Biospezies usw. 177
b) dentipalpis-Gruppe (Abb. 25). - Das Areal erstreckt sich bei weitgehender Vikarianz der Teilareale vom Karpaten-Becken bis N-Portugal. Sympatrie ist ledig- lich bekannt von nodzferalrobusta (alb), luteipes/pyrenaea (c/d) und carli/dentipalpis
(elf). c) kollari-Gruppe (Abb. 26). - Es bestehen zwei wcit isolierte Tcilareale - ein
alpin/west-balkanisches und ein vergleichsweise sehr viel kleineres iberisches. Die alpinen Areale vikariieren streng; für die beiden iberischen ist das nicht gesichert, aber \vahrscheinlich.
2. Ve rb re i t ungsgesch i ch t e u n d E v o l u t i o n de r A r t e n
Der vorstehend aufgezeigte eigentümliche Verbrcitungsmodus der drei Gruppen jewcils nächstverwandter Arten, der nicht mit ökologischen Faktoren erklärt werden kann, läßt nach dem Alter der einzelnen Taxa und ihrer Geschichte in der jüngsten geologischen Vcrgangenheit fragen.
Habitucll sehr ähnlichen Arten, deren Areale vikariieren, wird kein hohes Alter zugebilligt. Bisweilen läßt sich ihr Status - „schon Spezies" oder „noch Subspezies" - gar nicht sicher feststellen; ganz abgcsehen davon, daß das moderne Artkonzept a,uf vikariierciide Formen nur bcdingt anwendbar ist.
Es ist deshalb zu fragen, ob es berechtigt ist, vikariante Arten, deren nahe Ver- wandtschaft außer Zweifel steht, überhaupt als eigene (Bio-)Spezies anzusehen. Ebenso berechtigt wäre es, von einem Artenkreis (sensu RENSCH) bzw. von einer Supcrspezies zu sprechen, die MAYR (1967: 526) definiert als „eine monophyletische Gruppe von vollständig oder im wesentlichen allopatrischen Arten, die zu distinkt sind, um als einzige Art aufgefaßt zu werden". Die artlichen Komponenten der Supcrspezies sind die „SemispeziesLC; sie haben „einige, aber noch nicht alle Merk- male von Speziesrang erworben". Die dentipalpis- und die kollari-Gruppe (mit jeweils rnehrcren Arten) genügen zumindest vom Verbreitungsbild her der Definition der Superspezies. Doch beweisen die relativ großen Zonen sympatrischen Vorkommens jcweils zweier Arten der dentipalpis-Gruppe (Abb. 25), daß bereits vollständige Spe- ziation stattgefunden hat. Für die allopatrischen Arten der kollari-Gruppe ist das prinzipiell nicht sicher zu entscheiden, doch spricht die große rnorphologischc Diver- genz der einzelnen Arten für diese Annahmc.
Es ist bekannt, daß die europäische l'acna (und ~1ora)'während des Pleistozäns tiefgreifende Wandlungen erfuhr. Als die Vergletscherung weitcr Teile Europas zu- nahm, wurden die Faunenelemente unter den Waldbcwohnern, zu denen alle Ischy- ropsalis-Arten gehören, auf die südlichcn Halbinseln abgedrängt, soweit das nordi- sche und das nach Norden vordringende alpine Eis sie nicht vernichtete. Andererseits wurdcn durch die resulticrcnden Arealzerreißungen in hohem Maße evolutive Pro- zesse ausgelöst oderlund beschleunigt, die zur Differenzierung einer Vielzahl von Rassen und Arten führten. Ließen sich also bei Ischyropsalis Speziations-Vorgänge
1 2 2001. Jb . Syst. Bd. 96

nachweisen, die auf Glazialeinflüsse zurückgehen, ließe sich das Alter zumindest einiger Arten erschließen.
Die hellwigi-Gruppe
Diese Art besitzt mit den beiden disjunkteii Teilarealen (Abb. 24) das größte Ver- breitungsgebiet aller Arten der Gattung. Die Disjunktion ist durch Glazialeinflüsse leicht deutbar. Die ursprünglich sicher über weite Teile Mittel- und W-Europas ver- breitete Art besiedelte zumindest während der letzten (Würm-)Vereisung zwei Refu- gien. Eine Population lebte in den Waldgebieten der Balkanhalbinsel, von wo sie postglazial die 0-Alpen und Teile Mittel-Europas wieder besiedelte, den Rhein aber offenbar nicht überwinden konnte. Von dieser östlichen Rasse sind keine Funde westlich des Rheins bekannt. Ein zweites Refugium befand sich in Iberien. Mit dem erneuten Vordringen des Waldes besetzte diese Populatioii Teile der Pyrenaen und der Kantabrischeii Gebirge, erreichte aber nicht das Zentralmassiv, wo heute allein luteipes siedelt. Die Isolation wirkte vielleicht nur während des letzten Glazials; sie führte zu zwei schwach differenzierten Subspezies, die reproduktiv nicht isoliert sind (MARTENS 1969).
Die kollari-Gruppe (östlich-alpines Teilareal)
Die alpinen Arten der kollari-Gruppe (kollari, hadzii, strandi) vikariieren streng (Abb. 26; vgl. auch Kap. V 2b). Sie sind aus Gründen der Genital- und Cheliceren- Morphologie als nächstverwandt anzusehen. I . kollari, mit dem größten Areal, be- siedelt große Teile der 0-Alpen, wird aber in südlichen Randmassiven durch strandi (Bergamasker Alpen, Monte Baldo, Monti Lessini, Alpi Veneti) und hadzii (Kara- wanken, Steiner Alpen) substituiert.
Es ist bekannt, vor allem durch HOLDHAUS (1954), daß viele Südalpeii-Massive während des ganzen Pleistozäns weitgehend uiivergletschert geblieben sind und für viele Elemente der Fauna und Flora zu „Massifs, de refuge" wurden (sensu CHODAT & PAMPANINI, nach HOLDHAUS), in denen die Eiszeiten überdauert werden konnten. Alle angeführten Südalpen-Züge gehören zu diesen Refugialgebieteil. Dort lebt in der oberen Waldzone und oberhalb der Waldgrenze eine große Anzahl streng lokalisierter Tierarten; allein der Anteil der terrikoleii Käfer ist bis heute genaner bekannt. Es liegt nahe, auch in deii Areale11 der Arten des kollari-Komplexes der S-Alpen eine entsprechende Genese zu seheil. Demnach wurden während der eiszeitlichen Deva- stierung der Zentral-Alpeii die dort siedelnden Ischyropsalis-Arten zur Abvanderung gezwungen bzw. vernichtet, lediglich auf den klimatisch begünstigten Raiidposten konnten sich die dort ansässigen Populationen behaupten. Aber auch dort wareii die Lebensbedingungen zu Zeiten starker Vereisung der Hauptalpen für viele Arten des Waldes sehr ungünstig, und es ist fraglich, ob den steiiöken Ischyropsalis-Arten noch Lebensmöglichkeiten gegeben wareii, HOLDHAUS (1954) führt aus, daß „die in deii

Die Abgrenzung von Biospezies iisw. 179
Gebirgswaldern des Gebictes [gemeint sind die Massifs de refuge] lcbcnde tcrrikole Coleoptercnfauna in ihrer Gesamtheit bctrachtet . . . wahrend der Eiszcit in sehr empfindlicher Weise gcstort wurde . . . Jm Vergleich mit der subalpinen Terrikol- fauna ist die Fauna dcr Grotten viel weniger verarmt, und man gewinnt den Ein- druck, daß die Blindkäfer in den Grotten bcsscre Moglichkeiten hattcn, die Eiszeit zu uberdaucrn, als im Preicn im Waldbodcn". Weriii zwischen der Ökologie terrikoler Blindkafer und jencr der Ischyropsalos-Artcn auch nur mit Vorbehalt Vcrgleiche ge- zogen werden durfcn, so ist doch auffallig, daß allc naher erlauterten Ischyropsalis- Funde der kollura-Gruppe in dcn sudalpinen Massifs de refixge nur aus Hohlen stammen- Sichere Preilandfunde (soweit sic nicht kolluri selbst bctreffcn) sind unbekannt. Selbst wenn man unterstcllt, da8 Hohleiibiotopc jcwcils schncller und grundlicher zu ex- ploriercn sind als iimliegelidc Freilaiidhabitatc, bcstatigt das doch indirekt die An- nahmt von HOJ,VHAUS uber die terrikolen Blindkafer. Die Populationcn von I . strundi des Moiitc Baldo und der ostlicheii Monti Lessini, zwei der bekaiintcstcii Massifs de refugc, leben gebietsweise noch heutc untcr ganz extremen Bediiigungcn. In der liohlc Spluga de ia Preta (1445 m), die von strandi besiedelt ist, herrschen in1 August nur 4-7 "C (JUBERTHIE 1964). Deli kleinen Populationen ist bis heute nc~~ncnswcrte Arealausdchnung nicht gelungcn; bci so cngcr Eiiii~ischung darf sie auch iiicht er- wa,rtct werden. Vur sehr lange Besiedlung der 1-lohlen sprechen auch morphologische Befundc, vor allcm bei strancli. Dic Augcn sind bei den meisten Iridividueii vollig rcduzicrt, das Exoskelett (nicht das der Chcliceren) ist nur schwach sklerotisieit uiid somit nicht tief schwarz (vgl. Kap. 111 3). JUBERTHIE (1964: 123) weist uberdies auf die von Hohlc zu Hohle stark schwankende Cheliceren-Bewchrung dcr Iiidividuen dicser Kleinst-Populationcn hin. - Die auffalligcii abgcleiteten Merkmale der bciden Arten der S-Alpcn (strando und hadzoo) wciseu auf relativ langes Hohlenlcbcii und somit auf hohes Siedlungsalter im jetzigen Areal hin.
Dagegen muß das Verbreitungsbild von lcollari so iiitcrpretiert werden, daß diese Art von einem außcralpincii Refugium (oder von deren mehreren) Teile des pra- (oder inter-)glazialen Areals wiederbesicdelte. Zwar gab es auch am Alpen-Nordrand klcincre Refugien, wo kollaro heutc vorkommt, doch war dcrcii La,ge noch ungunstiger, da ihncn Wald vollig fehlte. Pur Tierc dcs subalpinen Nadelwaldes gab es hier keine Moglichkcit dce Überdauerns, und HOLDHAUS (1. C.) zieht „aus dcm volligei~ Fehlen cndemischcr Artcn der IValdfainna in dcii iiordiichcn Kalkalpcn" dcn Schluß, daß „wahrend der Fiszcit dic Bedingungen für dic Erhaltung typisch subalpiner Ei~demi- tcii in den nordlichen Kalkalpen i~iclit vorhanden waren". Ebcnso unwahrschciillich ist cs auch, duß die Rcfugicn der recht nordlich gelegciicn Gurktalcr und Lavant- talcr Alpcn fur so großc Arthropoden-Arten ausreichende 1,ebensmoglichkciteii boten. (Es ist hier von untcrgeordnet~r Bcdcutung, daß aus diesem Gcbiet noch gar kein Fund ciiier Art der kollurt-Gruppe bekannt ist.) Bcide Gcbirgszugc weiseil iiidcs zahlrciche alpin-cndernische Carabidcii auf, und neucrdlngs gclarig der Nachweis
12

einer nahezu auf diese Alpennlassive beschränkten Subspezies der Gattung Nemastoma (N. bidentatum relictum GRUBER & MARTEKS 1968, Nemastomatidae), der ein höheres als nur Riß/Würm-Interglazialalter zugebilligt werden muß, denn die morphologi- schen und ökologischen Differenzierungen gegenüber der Nominatform sind erheblich.
Die kollari-Fundorte an der Cima Undici (Val Saguna, 2050 m, THALER leg.; nördliche Lessinische Alpen) und am Rolle-Paß (südliche Dolomiten) liegen unmittel- ba'r am Rande von Massifs de refuge, von denen eine Arealerweiterung hätte ausgehen können. Die strenge ökologische Einmischung der Glazialüberdauerer scheint diese Annahme aber zu verbieten.
So darf als nahezu sicher angesehen werden, daß kollari als „Rückwanderer auf weite Distanz" (sensu HOLDHAUS) anzusehen ist, der von einem oder mehreren außer- alpinen Refugien wieder in die Alpen einwanderte. Inzwischen sind individuenreiche Populationen in den Hohen Tauern (vor allem im Stubachtal, AUSOPSKY & HURDES leg.) gefunden worden, wo Refugialgebiete mit Sicherheit nicht bestanden und diese Annahme unterstützen.
Die Rückwanderung von kollari erfolgte auf jeden Fall in die eiszeitlich stark, devastierten Gebiete postglazial. Würm-Überwinterung in den heutigen Verbrei- tungsgebieten nehmen aber SCHMÖLZER (1962) nach Funden in den westlichen Ziller- taler Alpen (Wolfendorn) und auf der Plose (Dolomiten) (sub helvetica) und THALER (1963) nach Funden bei Luilz (OÖ) (sub danubia) an. Das erscheint zunächst für jene Fundorte zutreffend, die in der Nähe von ehemaligen Nunatakern liegen, also von Berggipfeln, die auch zu Zeiten maximaler Vereisung über die Oberfläche des Eises ragten. Es ließ sich nämlich nachweisen, daß Vertreter einiger Tiergruppen an sol- chen Extrem-Standorten teilweise alle Glazialzeiten überdauern konnten (JANET- SCHEK 1956, SCHM~LZER 1962). Das gilt für den Wolfendorn (höchste Ischyropsalis- Bunde fast 2600 m) und wahrscheinlich auch für die Plose (höchster Fundort 2500 m). Diese hochgelegenen Standorte allein rechtfertigen die Annahme'einer Würm-Über- dauerung auf Nunatakern für Ischyropspalis wohl kaum, solange nicht nachweisbar ist, daß kollari in den tiefergelegeneil subalpinen Nadelwäldern fehlt oder doch we- sentlich seltener ist als in der alpinen Stufe. Nach Erfahrungen vor allem im Stubach- tal ( ~ o h e Tauern) bilden die Nadelwälder der Subalpin-Stufe den Vorzugsbiotop von kollari, nicht die hochalpinen Grasheiden, was SCHMOLZER (1962) unterstellt. Solange die Vertikalverbreitung und die ökologischen Praeferenda von koilari in jenen Gebie- ten nicht genauer beka,nnt sind, die in Nunatak-Zonen liegen, gibt es für eine Würm- Überdauerung keinen deutlichen Hinweis.
Nach vorstehenden Erörterungen ist die Speziatioii der alpinen Arten des kollari- Komplexes während des Pleistozäns erfolgt. Manche Populatioilen überdauerten an Ort und Stelle, und durch Isolation erfolgte divergente Entwicklung selbst an be- nachbarten Standorten. Die vergleichsweise weitverbreitete Art kollari wanderte postglazial (wieder) in die Alpen ein.

Die Abgrenzung von Biospezies usw. 181
Westliches Teilareal :
Die reliktäre, vom Hauptareal der Gruppe weit abgesprengte Verbreitung der beiden Arten dispar und magdalenae in wenigen Höhlen der Kantabrischen Gebirge (Abb. 26) und die stark apomorphe Ausprägung einiger Merkmale (Cheliceren-Länge und -Bewehrung; Sklerotisierung des Exoskelettes, Penis-Bau) deuten auf hohes, wohl präglaziales Alter dieser bei'den Arten hin. Der vielfach belegte Refugial-Charak- ter Iberiens bestätigt das. - Ihr wahrscheinlich nächster Verwandter ist I. muellneri. Diese Art zeichnet sich ebenfalls durch viele Apomorphien aus und ist offensichtlich auf Höhlen der Dinarischen Alpen beschränkt. Dieses Areal entspricht dem nörd- lichen Teil des . mediterranen . Sekundär-Refugiums (sensu DE LATTIN 1967).
Zusammenfassung: In der Gattung Ischyropsalis lassen sich glazialbedingte Differen- zierungs-Prozesse aufzeigen, die auf Einengung präglazialer Artareale, inter- und postglaziale Arealerweiterungen und -Überschneidungen zurückzuführen sind. Die ältesten Arten leben in Teilen der Pyrenäe?, der Kantabrischen Gebirge, in den S-Alpen und in den Dinarischen Gebirgen, also in Gebieten, die auch alte Arten vieler anderer Tiergruppen beherbergen. Während der Isola- tion entwickelten sie abgeleitete morphologische (lange Cheliceren, reduzierte Augen und Skleroti- sation) und ökologische Merkmale (obligatorisches Höhlenleben). - Sehr wahrscheinlich würm- glazialen Ursprungs ist die Disjunktion der beiden schwach differenzierten Subspezies von I. hell- wigi. Die Arten der dentipalpis-Gruppe sind großenteils älter. Ihre Areale haben sich teilweise überlagert, und die morphologischen Unterschiede von Art zu Art sind bisweilen beträchtlich.
3. E v o l u t i v e T rends i n der G a t t u n g I schyropsa l i s Die 15 Arten lassen sich zu drei monophyletischen Gruppen zusammenfassen
(vgl. Kap. V1 3a). Einzelmerkmale undIoder Merkmalskomplexe sollten gestatten, Trends in der morphologischen Entwicklung der Arten der Gattung aufzuzeigen und die Beziehungen der Arten innerhalb der drei Gruppen zu erkennen. Bei einem sol- chen Verfahren müssen die Merkmale in ursprüngliche und abgeleitete getrennt werden. Auf diese Weise kann die ursprüngliche Gestalt eines hypothetischen Vor- fahren der ~ a t t u n i abgeleitet werden. Im folgenden wird aber lediglich versucht, die ursprünglichen Merkmale bei den Arten der dentipalpis- und der kollari-Gruppe auf- zufinden. So können die abgeleiteten Zustände erkannt und auffällige Merkmals- kongruenzen verwandtschaftlich fernstehender Arten (Homoiologien) gegenüberge- stellt werden.
a) Cheliceren-Länge Die sehr langen Cheliceren sind als Exzessiv-Bildungen anzusehen, die bei den
Opiliones in mehreren Verwandtschaftskreisen auftreten. Die betreffenden Arten be- sitzen jeweils auch nahe Verwandte, die normal proportionierte Cheliceren aufweisen. Stark verlängerte Cheliceren gelten als apomorphes (abgeleitetes) Stadium. Innerhalb der Gattung sind verschiedene Grade der Spezialisierung erkennbar. Einerseits gibt es Arten mit relativ kurzen und gedrungenen, andererseits solche mit sehr langen und

schlanken Cheliceren (bezogen auf die Länge des Cephalothorax). Schlank gilt als apomorph, gedrungen als plesiomorph.
b) Cheliceren-Bcwchrung
Mit der Änderung der Cheliceren-Proportion (kurz/plump - lang/schmal) wird auch die Bewehrung (in beiden Gruppen unabhängig) umgestaltet. Jene Arten der dentipalpis- und kollurz-Gruppe, die an den Anfang der zwei Rcihen zu stellen sind, besitzen wenig spezialisicrte Bewehrung. Die drei Hauptreihen des Grundgliedes (dor- sal, ventro-lateral, ventr.0-medial) bestehcn aus vielen kleinen Elementen, die sich nur wenig von der lateralcn Bewehrung unterscheiden. Bei progressiven Stadien sind die lateralen Dorncn reduziert, die Eiemeiite der Ilauptreihen vcrgracr t und stark hervortretend. Das Endstadium kanii iii exzessiver Bewehrung oder aber in volliger Doriienreduktiou bestehen.
dentzpalpis-Grupp (8) Am Anfang dcr Rcihc stchcii municatu (Abb. 46) und adamii (Abb. 39). Bei bei-
den Arten ist die dorsale Bemrehrung des Grniidgliedes zwar deutlich hervorgehoben, docli bestehen die beiden veiitralcn Reihen aus vielen Dorn-Elemciitcn. Kleine Tu- berkel schieben sich teilweise in die Hauptreihen ein. Bci luteipes (Abb. 44) ist die Beweliruiig der J,ateralseite fast vollig reduziert, die drei Hauptrcihen erscheinen konzciitriert und bctont. Bei curlz ist die dorsale Rcihc auf drei besonders große Doriicn beschrankt. Dicse Reduktion kennzeichnet auch nodifera (Abb. 49), bei der ubcrdics die beidcn ventralen Reihen weitgehend verschwunden sind. In manchen Populatioiien treten aber auch keine dorsalen Dorneii mehr auf, nur Borsten kcnn- zeichiien die Anlagestelle~i. Bci pyrenaecc (Abb. 51) kommt nur dieser Zustand vor.
Icollurt-Gruppe (3) Zwei Tcndeiize~i sind feststellbar. Einerseits wird die Große dcr Dor~l-Elemente
der drei Hauptreihen extrem gesteigert, aiidcrerseits vollig ruckgebildet. - Recht ursprunglichc Verhaltnissc zeigt kollari (Abb. 56) mit wenig ausgeprägtcn Haupt- reihcii; lateral stehcn Tuberkel, die iri die ventro-laterale und in dic dorsale Reihe eiiiruclreii. Bci hadztz (Abb. 59) werdci~ die drei Hauptreihen scharf akzentuiert und bei shancli (Abb. 63) erreiche11 die einzelnen Dornen cxtrcme Große. - Die zweitc Richtung fuhrt zu volliger Rediilitioii der Bcwchrung. Niedrige Hocker besitzt das Gr~iiiclglicd von rnuqclulenae (Abb. 65); ailzuschlicßcii ist dispur (Abb. 67), der die Bc~~~cliruiig fast fehlt, und schlicßlich rnuellnerz (Abb. 61), deren Grundglied (beim $) nur vcrstreute Tuberkel und Borsten aufweist.
C) Dic Cheliceren-Burste
Von den 15 Arten besitze11 9 ciiie Burste. Diese Arten weisen (außer hellwigi) alle eiiicii einlappigeil und gekielteri Sklerit der Glans peiiis auf. Die Bürste erstreckt sich

Die Abgrenzung von Biospezies usw. 183
über das dorso-distale Drittel des Grundgliedes oder ist (mit allen Übergangsstufen) auf eine winzige Region am medio-distalen Ende beschränkt. Diese reduzierte Form genügt gerade noch den funktionellen Erfordernissen während des Kopulations- Verhaltens (vgl. Kap. I11 7 U ~ ~ M A R T E K S 1969). Da diesesMerkmal sowohl sehr deut- lich ausgeprägt, offensichtlich aber auch völlig fehlen kann, ist zunächst nicht zu entscheiden, ob es sich um ein altes Merkmal handelt, das einige Arten verloren haben, oder ob eine Neuerwerbung vorliegt. Da jedoch bei zwei nahverwandten Familien (Trogulidae, Nemastomatidae) diese Stryktur ebenfalls auftritt, muß sie als relativ ursprünglich angesehen werden. Zudem ist unwahrscheinlich, daß ein so kompliziertes Merkmal iii einem engen Verwandtschaftskreis mehrfach entstanden ist. Es sollte geprüft werden, ob hier nicht eil1 altes Arachniden-Merkmal vorliegt, das innerhalb der Opiliones nur von einigen Gattungen der Dyspnoi (innerhalb der Unterordnung Palpatores) bewahrt wurde. Möglicherweise ist dieses Organ Drüsenbildungen des entsprechenden Körpersegmentes bei anderen Ordnungen der Arachnida (Araneae, Chelonethi) homolog. Andererseits besteht aber auch die Möglichkeit einer Neubil- dung innerhalb der Dyspnoi.
Ich betrachte eine große Bürste als ursprüngliches Merkmal, eine kleine und im Extremfall fehlende als abgeleiteten Zustand. Diese Annahme wird durch das „Prin- zip der wechselseitigen Erhellung" (KRAUS) gestützt, denn Arten ohne Bürste weisen auch andere abgeleitete Merkmale auf.
d) Das abdominale Scutum Es wurde gezeigt (vgl. IV l b ) , daß das Scutum magnum als plesiomorphe, Scu-
tum parvum, intermedium, laminatum und dissectum als apomorphe Ausprägungen gewertet werden müssen.
dentipalpis-Gruppe ' Die 8 tragen immer Scutum parvum, ebenfalls die 9 mit der auffälligen Aus-
nahme von manicata und dentipulpis, von denen sich marticata sollst durch viele plesiomorphe Merkmale auszeichnet (Größe der Bürste, Bewehrung der Cheliceren). Eine Reihung ist in dieser Gruppe also nicht möglich, doch erhält die Vermutung, da5 die Arten der dentipalpis-Gruppe die uysprünglichereiz Merkmale aufweisen, durch das generelle Auftreten des Scutum parvum eine weitere Stütze.
kollari-Gruppe Aufgliederung der Arten nach dem Bau des Scutum:
1. $9 Scutum parvum: dispar 2. 8 Scutum parvum 9 Scutum laminatum: rnagdalenae, kollari

3. $ Scutum intermedium: muellneri (part.), strandi, hadzii Scutum laminatum
4. 39 Scutum laminatum: muellneri (part.)
Die am stärksten abgeleitete Ausprägung (3, 4) weisen jene Arten auf, deren Chelicercn-Länge und -Bedornung ebcnfalls als apomorph charakterisiert worden sind. Dagegcn bilden dispar und magdakenae anschcincnd eine Ausnahme. Der Cheli- ceren-Bau beider Arten ist als abgeleitet zu betrachten, nicht jedoch der des Scutum. Dennoch ist es bei bciden Arten apomorph, jedoch nicht durch Auflosung in Einzel- skleritc, sondern durch Verringerung der Sklerotisierung. Es resultiert ein Zustand, der jenem der Phalangiidae nahe kommt: Ein Scutum parvum, dessen Teile (cephalo- thorakales und abdominales Schild) häutig dunn sind. HADBI (1942) nennt diesen Schild „Scutum tenue".
Diskuss ion
Inncrhalb dcr denttpalpts- und kollart-Gruppe sind Tendenzen der Merkmals- entwicklung feststcllbar, die in beiden Gruppen nahezu parallel ablaufen. Bei ver- gleichsweise sehr konstanten $-Genitalstrukturen zeigen die Arten bcider Gruppen einen Trend zur Vcrlangcrung der Cheliccren, zur Reduktion der Cheliceren-Bcweh- rung, ferncr eine Tendenz zur Verkleinerung der Cheliccren-Burste bis auf ein winzi- ges, kaum mehr erkcnnbares Feld (denttpalpts-Gruppe). Die Endglieder beider Reihen stimmen in den Merkmalen der peripheren Gestalt so uberein, daß diese Arten „auf den ersten Blick" gar nicht unterschreden werden konnen. Lediglich die Alternativ- merkmale an den $-Genitalstrukturen (Sklerit der Glans pcnis ungelappt-gekielt oder zweilappig-plan) und das Burstenrudiment der Art der denttpalpts-Gruppe weiscn bciden Arten vcrschiedene Gruppcnzugehcirigkeit und eine seit langem eigcnstandige Entwicklung zu (vgl. Abb. 51 mit Abb. 65/67). - Dic ortho-selektiven Faktoren, die diese Evolution begunstigt haben, sind vorlaufig unbekannt.
VI. Systematischer Teil
1. Zur G a t t u n g s g r u p p i e r u n g
Die drei Familien Trogulidae, Nemastomatidae und Ischyropsalididae der Sub- ordo Palpatorcs werden als Cohors Dyspnoi zusammcngefaßt und mit dem Merkmal ,,gegitterte Stigmen" charakterisiert. Viele Vertreter dieser Familien zeichiien sich überdies durch Drusenbildungen am $-Cheliceren-Grundglied aus (Ischyropsalididae: Ischyropsalis, Sabacon, Ceratolasma (s. U.); Trogulidae: Dicranolasma; Nemastoma- tidae).
Die Trogulidae und Nemastomatidae bilden wahrscheinlich Schwestergruppen. Darauf weisen die einheitliche Ausprägung des Scutum magnum und die ähnliche

Die Abgrenzung von ~ i o s ~ e z i t i s usw. 185
Gestaltung der Sternalflachen. Dagegen zeichnen sich die I~chyropsali~didae durch die Sonderbildung des Scutum parvum aus (bzw. Scutum laminatum, intermedium oder dissectum). Allein dieses Merkmal trennt die Familie von den ubrigen'Dyspnoi streng und besitzt somit familien-diagnostischen Wert. Keineswegs kennzeichnend sind die kraftigen, uber korperlangen Cheliceren, wie sie die Arten der Genera Ischy- ropsalzs, Taracus und i\rzpponopsalzs aufweisen, denn die Gattungcn Sabacon, Ruax- phzlos und Ceratolasma besitzen kleine, normal proportionierte Cheliceren. Cerato- lasma zeichnet sich uberdies, wie die meisten Troguliden und einige nordamerikani- sche Nemastoinatidae (Dendrolasma; die urspruiigliche Placierung bei dcn Troguliden ist falsch!) durch bewehrten Carapax-Brontalrand aus (Abb. 31), so daß auch dieses Merkmal (fur die Trogulidae) nicht mehr als familientrennend angesehen werden kann, was KAESTNER (1965) noch annimmt. Als cinziges, die Tschyropsalididae kenn- zeichnendes Mcrkmal bleibt der Bau des dorsalen Scutum, das die Stufe des Scutum parvum nie uberschreitet. Ob mit diesem Merkmal aber eine monophyletische Einheit umrisscn wird, muß zunachst offen bleiben und sogar fraglich erscheinen. Bei den Gattungen der Familie sind namlich die Bauelemente der Sternalflache, die bei den Nemastomatidae und Trogulidae bemerkenswert einheitlich sind, sehr unterschied- lich gebaut. Dem Bau der Sternalflache ist zur Kennzeichnung von Taxa uber dem Artniveau Bedeutung beizumessen, denn er erweist sich von Gruppe zu Gruppe als bemerkenswert konstant.
Tatsachlich liegen bei den drei lang-chelicerigen, in der Holarktis vikariierenden Genera Ischyropsalzs (Europa), Taracus (N-Amerika) und Nzpponopsalzs (Japan und Korea), deren nahe Verwandtschaft auf Grund außerer Mcrkmale zunachst vermutet werden durfte, sehr uiitcrschiedliche Verhaltiiisse vor. .
Ischyropsalzs (Abb. 27):. großes Labium, vom Sternum (unter dem Operculum . genitale) durch weichhautige Membran getrennt; Sternum groß, nicht mit den Basen der Coxen verwachsen; Lobus der Coxa I mit 2 Skleriten, Coxen 11-IV mit kleinen Loben.
Taracus (Abb. 28): kleineres Labium, vom Stcrnum durch Membran getrennt, Sternum mit den Basen der Coxae 1-111 fest verwachsen, Lobus der Coxa I nur mit einem großen Sklcrit, Coxa I1 mit kleincm Lobus.
Nzpponopsalu (Abb. 29): kleines Labium, Lobus der Coxa I mit einem Sklerit, Coxae 11-IV ohne Loben; Stcrnum mit den basalen Teilen der Coxae 1-111, teil- weise auch der Coxa IV, fest verwachsen.
Die von Gattung zu Gattung sehr unterschiedliche, bei den Arten jeder Gattung aber sehr einheitlichc Auspragung der Sternalflachen stutzt die Annahme einer nahen Verwandtschaft dieser drei Gruppen keineswegs, vielmehr muß angeiiommcn werden, daß sie sich scit geologisch relativ langer Zeit selbstandig entwickelt haben. Genital- morphologisch weisen Ischyropsalzs und Taracus zwar vergleichbare Strukturen auf, doch ist der Penis zu wenig gegliedert, um von dieser Ähnlichkeit auf enge Verwandt-

schaft sehlicßcii zu köiine~i. ~ i c iG lans penis allcr drei Nipponopsalts-Artci~ weicht hingcgen stark ab, so daß auch diese Struktur die Eigcnstandigkeit der Gruppe er- weist (Abbildungen zuletzt bei MARTENS & SUZUI~I 1966). So sind die laiigcn Cheli- ccreli der Arten der drei Genera als Homoiologieii zu werten, also als uiiabhangig voneinander ciitstandcne Strukturell iii Gruppen, die iii einem wcitlaufigen Ver- nrandtschaftsverhaltnis zueiiiandcr steheii.
Überraschenderweise gehort die einzige Art dcr Gattang Cerc~tolasma, C. tri- cantha GOODNIGJIT & GOOUNIGHT 1942 (N-Amerika, Oregon) (Abb. SI), die die Aii- toreii zu dpn Trogulidae stellten, zu den Is~hyropsalididae.~) Nach Merkmalen dcr Sternalflache ist diese Art und Gattu~ig init Ischyropsalts nachst verwaiidt.
.
Cerutolasma (Abb. 30): relativ sehr großes T,abium, vom Sterliuin durch eine Membran getrennt; Steriium relativ niedrig, nicht mit den Coxen verwachsen; Lobus der Coxa I mit cincm Sklerit, Coxae 11- IV mit je einem klcincn Lobus.
Auch dic$-Geiiitalstrukturci~ weisen auf engen Ziisammciihang init Ischyropsalts hin. Pririzipiell weicht nur dcr Stylus ab, der nicht gekrummt, soiidcrii gcstrcckt ist. Auch die Chclicereil-Druse ist bci Ceratolusma vorhanden (bei Turacus und Ntppono- psalzs fehlt sie!). Das Sekretionsfcld, das tcilweisc auf einer Apophyse liegt (Abb. 31b). ,st latcrad exponicrt.
1) Den Hinweis veidanke ich Hcirn Icollegen J. C ~ u a r n , Wien.

Die Abgrenzung von Biospezies usw. 187
29 ,30
Abb. 27-30. Schemata der Sternalflächen von (27) Ischyropsalis ; (28) Taracus; (29) Nipponopsalis; (30) Ceratolasnza. - Feine Punktierung: Loben der Pedipalpen-Cosen; grobe Punktierung: Unter- lippe; Schraffur: häutige Teile des Lobus der Coxa I ; schwarze Flächen in der Schraffur: Sklerite des Lobus der Coxa I ; Wellenlinien: Sternum; PaCx: Coxa des Pedipalpus; Cx I: Coxa I etc.; Lb: Oberlippe; Op: Operculum genitale, das durchsichtig dargestellt i s t ; schwarze Punkte an den Basen der Cosen: Loben (Laden) der Coxen.
2. Die S i t u a t i o n i n der Spezies-Gliederung(Ischyropsa1is)
,,Schwerer wiegend scheint ein anderes, oft geäußertes Bedenken zu sein, nemlich dieses, daß in einem solchen Verzeichniss keinerlei Bürgschaft dafür geboten werde, daß die unter dein betreffenden Namen aufgeführten Objekte wirklich diesen Namen mit Recht verdienen.'' - BERTICAU 1880 (Verh. naturhist. Ver. Rheinl. Westf., 37: 215).
Die systematische Bearbeitung der Gattung besorgten vor allem SIMON (1872 bis 1913), ROEWER (1914, 1923, 1950) und HADZI (1926127, 1942,1954). - Acht der von SIMOV beschriebenen Taxa der franko-ibeiischen Fauna haben sich als valid erwiesen. Größte Bedeutuiig haben die Arbeiten von ROEWER. Seine Ge~amtbearbei tun~ der Weltfauna der Opiliones (1923) nennt 22 Arteii, bei der letzten Revision der Ischyro- psalididae (1950) bestehen 32 iiomiiielle Arteii (eine, ravusinii, wurde von ihm über- sehen) und 31 weitere werden von ihm erstbeschrieben (eine, hispunica, noch 1953). - H A D ~ I (1942, vor allem 1954) setzt sich kritisch mit den Arbeiten von ROEWER und dessen Artkonzept auseinander und liefert erstmals proportionsgerechte Zeichnungen der Cheliceren. - Neuerdings versucht DRESCO (1966, 1967) die Taxonomie der west-

europäischen Arten zu klären. Da er sich nicht vom typologischen Artkonzept seiner Vorgänger trennen kann, müssen die ~ r ~ e b n i s s e mit Zurückhaltung betrachtet wer- den. Sie stimmen nicht mit den hier vorgelegten überein. - Die in den vorstehenden Kapiteln dargelegten Ergebnisse rechtfertigen es, lediglich 15 Arten anzuerkennen.
3. Die T a x a a) Die Artengruppen
Das sekundäre 8-Geschlechtsmerkmal „Bürsteu läuft parallel mit Merkmalen 3m 8-Genitalapparat. Der ventrale Sklerit der Glans penis der bürstentragenden Arten ist immer ungelappt, entweder median gekielt oder plan (Abb. 32; vgl. auch die Ab- bildungen in den Art-Kapiteln). Bei Arten ohne deutliche Bürste ist der Sklerit zwei- lappig und ungekielt (Abb. 32b). Mit diesem Alternativ-Merkmal ergibt sich eine klare Gliederung in drei Gruppen jeweils nächstverwandter Arten. Diese Artengrup- pen haben Untergattungsrang; auf Benennung soll jedoch verzichtet werden, da die Gattung klein und dementsprechend übersichtlich ist.
a) hellwigi-Gruppe. - 1 Spezies, 2 Subspezies; h. hellwigi und h. lucantei. Mit relativ ursprünglichen Merkmalen; Sklerit der Glans penis klein, plan, mit wenigen
Abb. 31. Ceratolasma tricantha GOODNIGHT & GOODNIGHT, $. - a Habitus von lateral (Beine nicht dargestellt); b rechtes Cheliceren-Grundglied von lateral; c Glans penis von ventral; d-f Pe- nis von ventral, dorsal und lateral.
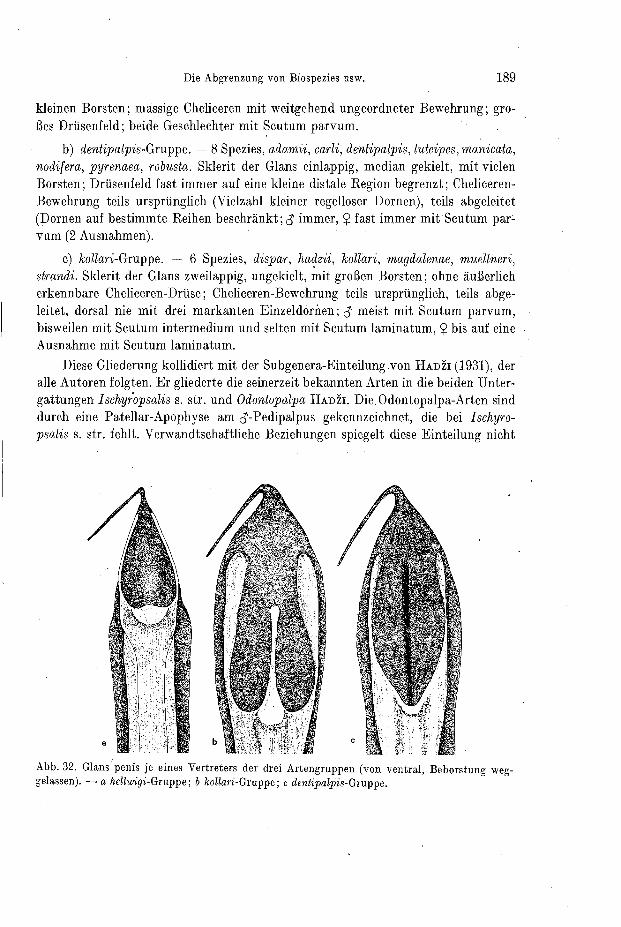
Die Abgrenzung von Biospezies usw. 189
kleinen Borsten; massige Cheliceren mit weitgehend ungeordneter Bewehrung; gro- ßes Drüsenfeld; beide Geschlechter mit Scutum parvum.
b) dentipalpis-Gruppe. - 8 Spezies, adarnii, carli, dentipalpis, luteipes, manicata, nodifera, pyrenaea, robusta. Sklerit der Glans einlappig, median gekielt, mit vielen Borsten; Drüsenfeld fast immer auf eine kleine distale Region begrenzt; Cheliceren- Bewehrung teils ursprünglich (Vielzahl kleiner regelloser Dornen), teils abgeleitet (Dornen auf bestimmte Reihen beschränkt; $ immer, $2 fast immer mit Scutum par- vum (2 Ausnahmen).
C) kollari-Gruppe. - 6 Spezies, dispar, hadzii, lcollari, rnagdalenae, muellneri, strandi. Sklerit der Glans zweilappig, ungekielt, mit großen Borsten; ohne äußerlich erkennbare Cheliceren-Drüse; Cheliceren-Bewehrung teils ursprünglich, teils abge- leitet, dorsal nie mit drei markanten Einzeldornen; $ meist mit Scutum parvum, bisweilen mit Scutum intermediuni und selten mit Scutum laminatum, Q bis auf eine Ausnahme mit Scutum laminatum.
Diese Gliederung kollidiert mit der Subgenera-Einteilung von HADHI (1931), der ~ alle Autoren folgten. Er gliederte die seinerzeit bekannten Arten in die beiden Unter- gattungen Ischykysalis s. str. und Odontopalpa HADHI. Die Odontopalpa-Arten sind durch eine Patellar-Apophyse am 8-Pedipalpus gekennzeichnet, die bei Ischyro-
I psalis s. str. fehlt. Verwandtschaftliche Beziehungen spiegelt diese Einteilung nicht
Sbb. 32. Glans penis je eines Vertreters der drei Artengruppen (von ventral, Beborstung weg- gelassen). - a hellwigi-Gruppe; b kollari-Gruppe; c dentipalpis-Gruppe.
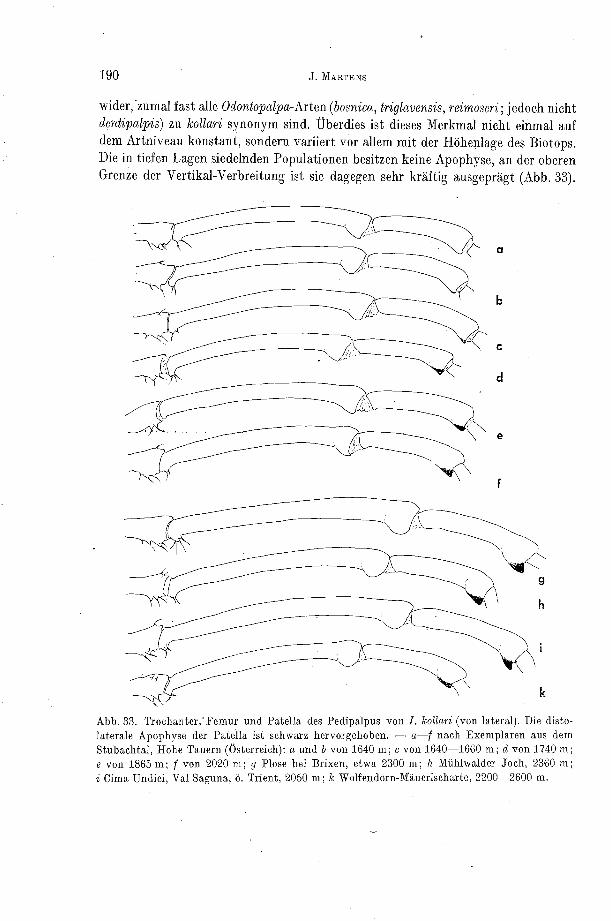
widcr,~zumal fast alle Odontopalpa-Arten (bosnica, triglavensis, reimoseri; jedoch nicht dentzpulpis) zu lcollari synonym sind. Überdies ist dieses Merkmal nicht einmal auf dem Artiiiveau konstant, sondern variiert vor allem mit der Hohenlage des Biotops. Die in tiefen Lagen siedelnden Populationen besitzen keine Apophyse, an der oberen Grenze der Vertikal-Verbreitung ist sie dagegen sehr kräftig ausgeprägt (Abb. 33).
Abb. 33. Tiochantcr,'Fernur lind Patella des Pedipalpus von I. kollart (von lateial). Die d~s to - laterale Apopliysc dei Patella ist schvara hcrvo~gchoben. - U-f nach Exempl,~ren aiis dem Stubaclital, IIohe 'I'duern (Osterrelch) a und b von 1640 m; c von 1640-1660 m ; cl von 1740 nl, e von 1865 m ; f von 2020 m; g Plose bei B~ ixen , etwa 2300 m , h Muhl~~raldei Joch, 2360 rn; z C ~ m a Undici, Val Saguna, o. Trient, 2050 rn; Ic Wolfendorn-Mauerlsck~~~~tc, 2200-2600 111.

Die Abgrenzung von Biospezies usw. 191
b) Die Arten
Vorbemerkung. - In den '~ r tka~ i t e l i 1 werden die wichtigste Literatur und alle Synonyma aufgeführt. Die Synonymie wird begründet. 'In Diagnose und Be- schreibung werden die kennzeichnenden Merkmale erörtert und ihre, Variabilität be- sprochen. Die Verbreitung jeder Art wird möglichst genau dokumentiert mittels selbst untersuchten Materials und deutbarer Literaturangaben. - Das Material, das den mit „V" bezeichneten Literaturstellen zugrunde lag, wurde vollständig oder teil- weise revidiert; „Nu bezieht sich auf Namennennungen, „D" mit nachgestellten Ge-
'
schlechtsangaben macht auf diagnostischen Text aufmerksam. -
Die Meßstrecken sind Abb. 34 zu entnehmen; mitgeteilt werden überdies: 9. Länge des Femur 11, 10. Länge des Metatarsus 11, 11. Länge des Penis, 12. Anzahl
. der Pseudogelenke des Metatarsus und 13. Anzahl der sekundären Gelenke des Tar; sus. - In Klammern stehen die arithmetischen Mittel, die Indices geben die Zahl der
I gemessenen Individuen an.
Von folgenden nominellen Arten wurde holo-, para- oder syntypisches (I), topotypisches (2), lectotypisches (3) oder klassisches Material (4) untersucht: adamii (3), alfkeni (I), amseli (I), archeri (I), asturica (I), austriaca (I), balcanica (I), bosnica (I), cancroides (4), caporiaccoi (I), carli (4), corcyraea (I), corsica (I), dacica (I), danubia (I), frankenbergeri (I), franzi (I), goodnighti (I), h. helvetica (I), h. milleri (I), hispanica (I), janetscheki (I), kaestneri (I), knirschi (I), kollari (1, 4), kratochvili (I), lusitanica (I), manicata (3), moreana (I), muellneri (I), navarrensis (I), nicaea (I), pectinifera (I), pentelica (I), pestae (I), plicata (I), pyrenaea (2), ravasinii (I), reimoseri (I), robusta (2), segregata (I), strandi (I), strasseri (I), styriaca (I), taunica (1, 3), tirolensis (I), triglavensis (I), troglodytes (1) und turki (I).
Abb. 34. Die Meßstrecken. - 1 ~ e ~ h a l o t h o r a s : ~ ä n ~ e ; 2 Cephalothorax-Breite; 3 Scutum-Länge; 9 4 Scutum-Breite; 5 Länge des 1. Cheliceren-Gliedes; 6 Länge des 2. Cheliceren-Gliedes; 7 Breite
des 2. Chelicereii-Gliedes; 8 Länge des 3. Cheliceren-Gliedes. - Die Meßstrecken 9-13 sind dem Test auf S. 191 zu entnehmen.

Tabelle zur Bestimmung der Arten
Dichotome Bestimmungsschlüssel haben den Zweck, den1 Nichtkenner einer Gruppe schiielle und sichere Determination vorliegenden Materials zu ermogliehen. Die Beiiutzuiig des Schlüssels ist um so sicherer, je mehr Alternativmerkmale er ver- wendet. Sind die einzelnen Merkmale jedoch schwer objektivierbar und zudem varia- bel, ist die Bestimmung von Stichproben schwierig, da diese nie alle Merkmale der betreffenden Spezies in sich vereiiiigeii konneii. Als Ausweg wird hier Gewicht auf die chorologischen Merkmale gelegt. Sie keiinzeichneii die Ischyropsalis-Arten ebenso sicher wie morphologische Kriterien.
y . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ventraler Sklerit der Glans penis einlappig, plan oder mit medianem IGel (Abb. 32a, c) . . . . . . . . . . . . . . . . . . . . . . . . . . . . ventraler Sklerit der Glans penis zweilappig, plan (Abb. 32b) . . . . . . . Sklerit der Glans penis plan, mit wenigen kleinen Borsten (Abb. 35); distales Drittel des Cheliceren-Grundgliedes dorsal mit Bürste (Abb. 35), nicht auf die Medialseite übergreifend . . . . . . . . . . . . . . . . . . . . . Sklerit der Glans penis gekielt, mit großen Borsten (z. B. Bbb. 39), Bürste klein, auf disto-dorsales oder disto-mediales Feld beschränkt (z. B. Abb. 22 d bis f), wenn Bürste auf ganzem disto-dorsalen Drittel des Gliedes, dann auf die Medialseite übergreifend (Abb. 22b) . . . . . . . . . . . . . . . . . Bürste des Cheliceren-Grundgliedes auf dem distalen Drittel dorsal, auf die Rledialseite übergreifend (Abb. 22b) (Karpaten-Länder) . . . . . . . . . Bürste des Cheliceren-Grundgliedes am Distalende dorsal bis medial, oft punktförmig klein, dann ganz an den Distalrand vorgeschoben (z. B. Abb.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22d-f) Cheliceren-Grundglied dorso-distal mit stumpfer, keulig aufgetriebener Apo-
. . . . . . . . . . . . . . . . . . . . . . . . physe (z. B. Abb. 42) Cheliceren-Grundglied dorso-distal ohne nennenswerte Auftreibung (z. B.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . Abb. 39) Cheliceren-Grundglied dorsal mit 3 kräftigen Dornen (Zentral-Alpen) (z. B. hbb . 37,43) . . . . . . . . . . . . . . . . . . . . . . . . . . . . Cheliceren-Grundglied dorsal mit mehr als 3 kräftigen Dornen oder nahezu
. . . . . . . . . . . . . . . . . . . . unbewehrt (z. B. Abb. 49,51) . . . . . . . . . . . . . Patella des Pedipalpus mit distaler Apophyse
Patella des Pedipalpus ohne Apophyse (vgl. Abb. 42) . . . . . . . . . . Bürste des Cheliceren-Grundgliedes medio-distal, relativ klein, nahezu rund
. . . . . . . . . . . . . . . . . . . . (Abb. 22e) (Halbinsel-Italien) Bürste des Cheliceren-Grundgliedes medio-distal, relativ groß, längsoval
. . . . . . . . . . . . . . . . . . . . . . . . (Abb. 22c) (Portugal) Cheliceren-Grundglied schlank, Bewehrung schwach, bisweilen nahezu feh- lend (z. B. Abb. 51) . . . . . . . . . . . . . . . . . . . . . . . . . Cheliceren-Grundglied untersetzt, mit kräftiger, vor allem dorsaler Beweh- rung (z. B. Abb. 44) . . . . . . . . . . . . . . . . . . . . . . . . .
9 dentipalpis carli
adamii
robusta
10
I1

Die Abgrenzung von Biospezies usw.
Augen sehr klein(Abb.53),Beine einfarbig dunke1,Bürste dorso-medialunmittel- bar amDistalrand desGrundgliedes,punktförmig(Höhlen der Zentral-Pyrenäen) Augen groß (Abb. 50), Beine kontrastreich hellgelb und dunkelbraun gerin- gelt, Bürste medial, oval, vom Distalrand des Grundgliedes meist abgesetzt (Abb. 22f) (Kantabrische Gebirge, N-Portugal) . . . . . . . . . . . . .
pyrenaea
nodifera
Dorsal-Bewehrung des Cheliceren-Grundgliedes nicht streng in einer Linie, mehr als dreizählig, Bürste punktförmig, dorso-distal, Beine nicht kontrast- reich hellgelb und dunkelbraun geringelt (franz. Zentralmassiv, 0- und Zen- tral-Pyrenaen) (vgl. Abb. 44) . . . . . . . . . . . . . . . . . . . . . Dorsal-Bewehrung des Cheliceren-Grundgliedes streng ausgerichtet, drei- zahlig, Burste langsoval, medial (Abb. 22f), Beine kontrastreich hellgelb und dunkelbraun geringelt (Kantabrische Gebirge, N-Portugal) . . . . . . . .
luteipes
nodifera
Cheliceren-Grundglied untersetzt, regellose kraftige Bedornung, Einzelreihen . . . . . . . . . . . . . . . . . wenig hervorgehoben (z. B. Abb. 56)
Cheliceren-Grundglied sehr lang und schlank, entweder nahezuaunbewehrt (Abb. 67) oder mit exzessiv hervorgehobenen Einzelreihen (Abb. 63) . Dorsale Bewehrung des Cheliceren-Grundgliedes rechtwinklig abstehend (Abb. 59), mit Scutum intermedium (Karawanken, Steiner Alpen) . . . . . Dorsale Bewehrung des Cheliceren-Grundgliedes nach vorn geneigt (Abb. 56),
hadzii
kollari . . . . . . . . . . . . . . . . . . . . mit Scutum parvum (0-Alpen)
Dorsale ~ e w e h r u n g des Cheliceren-Grundgliedes exzessiv entwickelt (Abb. 63) (S-Alpen) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Cheliceren-Grundglied nahezu unbewehrt (z. B. Abb. 61) . . . . . . . . .
strandi 15
Penis extrem lang und schmal (Abb. 67), Augen groß (Abb. 68) (Kantabri- . . . . . . . . . . . . . . . . . . . . . . . . . . . sehe Gebirge)
Penis untersetzt, nicht extrem lang und schmal (z. B. Abb. 65), Augen klein . . . . . . . . . . . . . . . . . . . . . . . . . . (Abb. 62 und 66)
dispar
Cheliceren-Grundglied ohne auffällige dorso-distale Apophyse (Abb. GI), Scutum intermedium oder laminatum (Dinarische Gebirge) . . . . . . . Cheliceren-Grundglied dorso-distal mit auffälliger Apophyse (Abb. 65), Scu-
muellneri
tum parvum (Kantabrische Gebirge) . . . . . . . . . . . . . . . . magdalenae
18 25
. . . . . . mit Scutum parvum (abdominale Sklerite I-V verschmolzen) mit Scutum laminatum (abdominale Sklerite I-V frei) . . . . . . . . . Dorsale Bewehrung des Cheliceren-Grundgliedes mit 3 (selten 4)* nahezug leich großen markanten Einzeldornen (Abb. 42) . . . . . . . . . . . . . . . Dorsale Bewehrung des Cheliceren-Grundgliedes mit mehr als 3 verschieden großen Einzeldornen (z. B. Abb. 51) : . . . . . . . . . . . . . . . . . 21
carli 20 robusta adamii 22 24
abdominales Scutum glatt, ohne Tuberkel-Querreihen . . . . . . . . . . abdominales Scutum mit Tuberkel-Querreihen . . . . . . . . . . . . . Tuber oculorum sehr groß (Abb. 55) (Portugal) . . . . . . . . . . . . . Tuber oculorum kleiner (Abb. 40) (Halbinsel-Italien) . . . . . . . . . . Cheliceren-Grundglied sehr lang und schlank (z. B. Abb. 51) . . . . . . . Cheliceren-Grundglied untersetzt klobig (z. B. Abb. 35) . . . . . . . . . Dorsale Bewehrung des Grundgliedes vor allem an der basalen Hälfte, distale Hälfte nahezu unbewehrt (Abb. 51) . . . . . . . . . . . . . . . . . . Dorsale Bewehrung des Grundgliedes gleichmäßig verteilt (Abb. 49 und 67) .
pyrenaea 23
3 Zod. Jb. Syst Bd. 96

Dorsale Bewehrung des Grundgliedes kräftig entwickelt, der distale Dorn oft am größten, aber nie kleiner als die caudad folgenden (Abb. 49), Beine kon- trastreich hell und dunkel geringelt (Kantabrische Gebirge, N-Portugal) . . Dorsale Bewehrung des Grundgliedes schwach entwickelt, distaler Dorn nie am größten, immer kleiner als die caudad folgenden (Abb. 67), Beine nicht kontrastreich geringelt (Kantabrische Gebirge) . . . . . . . . . . . . . Tuber oculorum klein (Abb. 36), Cephalothorax kräftig gewölbt (Abb. 36) . Tuber oculorunl groß (Abb. 45), Cephalothoras schwach gewölbt (Abb. 45) . Cheliceren-Grundglied sehr schlank, sehr schwach oder exzessiv bedornt (z. B.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . Abb. 61,63) Cheliceren-Grundgliedrobust, untersetzt, kräftig bedornt (2. B. Abb. 56) . . Cheliceren-Grundglied sehr schlank, exzessiv bedornt (Abb. 63) (S-Alpen) . . Cheliceren-Grundglied sehr schlank, schwach bedornt (Abb. 61 und 65) . . . Dinarische Gebirge . . . . . . . . . . . . . . . . . . . . . . . . . Kantabrische Gebirge . . . . . . . . . . . . . . . . . . . . . . . . Tuber oculorum kräftig entwickelt (Abb. 57) . . . . . . . . . . . . . . Tuber oculorum schwach entwickelt (Abb. 60) . . . . . . . . . . . . . Dorsale Bewährung des Grundgliedes distad kleiner werdend (Abb. 56), Linse der Augen klein (Abb. 5'7) (0-Alpen) . . . . . . . . . . . . . . . Dorsale Bewehrung des Grundgliedes distad mit einem kräftigen ~ o r n en-
. . . . . . . . . dend (Abb. 46), Linse groß (Abb. 47) (Karpaten-Länder) Dorsale Bewehrung des Grundgliedes rechtwinklig abstehend (Abb. 59) (Ka- rawanken, Steiner Alpen) . . . . . . . . . . . . . . . . . . . . . . Dorsale Bewehrung des Grundgliedes nach distal abgewinkelt (Abb. 37) . .
nodifera
dispar hellwigi luteipes
26 28 strandi. 2 7 nzuellneri magdalenae 29 30
kollari
nzanicata
hadzii denlipalpis
Die he l lwig i -Gruppe (Abb. 24,35-36) I. hellwigi (PANZER 1794)
1794 Phalangium Hellwigii PASZER, Fauna Germanica, 13: 8 (D??).
Die Spezies wird in zwei Subspezies gegliedert, beide zeichnen sich durch folgende Merkmale aus.
Diagnose : Einzige Art der hellwigi-Gruppe, charakterisiert durch dorsale Lage der Bürste und spärliche Beborstung der Glans peiiis.
Beschre ibung . - Cheliceren;$-Basalglied(Abb. 35a-C, k): Burste von der kleinen disto-medialen Apophyse bis zum ersten kräftigen Dorn auf der distalen Hälfte des Gliedes; caudad weitere Dornen abnehmender Länge, zwischen und neben ihnen Warzen- und tuberkel-artige Erhebungenj auch medio- und latero-basal; Ven- tralseite unbewehrt, jedoch von einer medio- und latero-basalen Reihe von je 5-7 Dornen eingefaßt. - 2. Glied (Abb. 35a, k): am „Knieu wenige mit Haaren besetzte Tuberkel, in einer medialen Reihe bis zur Basis des unbeweglichen Fingers ziehend; unbewehrte Partien (beider Glieder) netzartig gerunzelt und spärlich mit kurzen Haaren besetzt. - 9: Bewehrung wie beim $, doch Cheliceren schlanker wirkend, da ohne dorsale Bürste (Abb. 35 d-f, 1).
P e d ip a lpus ($9): alle Glieder schlank und dünn, lediglich Femur distal schwach konisch angeschwollen.

Die Abgrenzung von Biospezies usw. 195
P e de s (Laufbeine): relativ kurz und robust, ohne Bewehrung, schwarz. Cepha lo tho rax u n d Tube r ocu lorum (Abb. 36): hinter dem Tuber ocu-
lorum (an den Ansatzstellen der Chelicerenmuskulatur) starke Aufwölbung des Ce- phalothorax, dann steiler Absturz zum 2. Thorakal-Tergit; Tuber oculorum median tief gefurcht, Augen nahezu getrennt, Augenöffnung klein.
S cu t um: $9 regelmäßig mit derbem Scutum parvum ; trapezoid, längste Seite distal, ohne Bewehrung, jedoch fein bekörnelt und mit winzigen Borsten.
Abb. 35. I. hellujigi (PANZER). a-f und k-1 1. bzw. 2. rechtes Cheliceren-Glied von'lateral, dar- über jeweils das distale Ende des I. Gliedes von medial. a-i I. h. hellwigi ( P A N Z E R ) . a Burscheid (Deutschland, 8); b Rekawinkel (Österreich,$); cszakonyfalu (Ungarn,d); d und e wie c (9); f Tauf- stein (Deutschland, ?); g-h Glans penis: g Schafstein (Deutschland); lz wie C ; i Penis, Lusen (Deutschland); k-n I. h. lucantei SIMON. k Sierra Urbaso (Spanien, 8); 1 angeblicher Fund'ort ,,Ari&geU (Frankreich, 9); m Glans penis, wie k ; 9% Penis, wie k.
13'

Pen i s (Abb. 35g-i, m-n): sehr schlank und schmal, kaum bauchig erweitert; die basalen zwei Drittel des Truncus ventral mit breiter Rinne; distale Hälfte mit stark sklerotisiertem Saum. - Ventraler Skleri-t der Glans herzförmig, lateral mit wenigen kurzen Borsten besetzt, median schwach eingesenkt, jedoch ohne Rinne.
R e c e p t a c u l u m semin is (Abb. 36a-c): zehn sehr dünne Schläuche, distal2 bisweilen verästelt (6 Legeröhren präpariert).
V a r i a b i l i t ä t : das spezifische Merkmal „BürsteL' bemerkenswert konstant in Lage und Ausdehnung; Größe der Cheliceren-Bewehrung der drei Hauptreihen schwankend, Lagemuster jedoch weitgehend konstant; Stylus penis immer nach links gewendet (bis auf eine Ausnahme [Abb. 35h], dort rechts orientiert).
Ischyropsalis hellwigi hellwigi ( P A N Z E R 1794) 1794 Phalangizinz Hellwigii PANZER, Fauna Germanica, 13: 8 (D??). 1834 Phalangium Helwigii, - HAHN, Die Arachniden, 2: 5 T. 37 A-D (D$). 1834 Phalav~giurn Helwigii, - WIDER, Handzeichnungen von Arachniden. - Unveröffent-
lichtes MS, im Besitz des Si\IF. 1841 I. hellwigi, - C. L. KOCH, in HAHN & KOCH, Die Arachniden, 8: 17-18 T. 258 Abb. 603
(D). 1848 I. Herbstii C. L. KOCH, Die Arachniden, 16 (114): 68 T. 561 Abb. 1545 (D??).
mm 1 "1 1 I
I ' I 1
d a c
Abb. 36. I. hellwigi (PANZER). - Cephalothorax und Receptacula seminis. a-c Receptacula sem3nis: a Taufstein (Deutschland) ; b Szakonyfalu (Ungarn; beide I. h. hellwigi); c angeblicher Fundort ,,Ari&geU (Frankreich; I. h. lucantei); d Cephalothorax von rechts-lateral, wie a.

Die Abgrenzung von Biospezies usw.
1865 I . Helwigii, - L. KOCH, Z. Ferd. Tirol. Vorarlb., 3 (12): 164 (D). 1865 I. Herbstii, - L. KOCH, Z. Ferd. Tirol. Vorarlb., 3 (12): 164 (D). 1869 I . Helwigii, - L. KOCH, 2001. Mitt. tirol. vqrarlb. Landesmus.: 16 (N). 1885 Ischyropsalis sp., - HANSEN, 2001. Dan., 4: 99 (N). 1900 I . Helujigii, - VERHOEFF, 2001. Anz., 23 (608): 106 (N).
V 1923 I . taunica MÜLLER; in ROEWER, Weberknechte d. Erde,: 683 Abb. 850 (D$?). v 1923 I . plicata ROEWER, Weberknechte d. Erde: 692 Abb. 865 (D$?).
1923 I. herbstii, - ROEWER, Weberknechte d. Erde: 686 (part.: Laibach). V 1924 I . helwigii, - MÜLLER, 2001. Jb . Anat. 45: 414 T. 19 Abb. 5 (D$?) (part.: Altkönig). v 1924 I . taunica, - MÜLLER, 2001. Jb . Anat., 45: 418 Abb. 511 (D). V 1926127 I . pectinifera H A D ~ I , Bull. Ass. Mus. Slov., 718 B: 30 T. 7 Abb. 174-186, T. 8 Abb.
187-190 (D J). V 1928 1. pectinifera, - H A D ~ I , 2001. Anz., 77 (112): 15 (D J).
1930 I . helwigii, - B ~ T T N E R , Jb. Ver. Naturk. 192811930: 1-4 (N). 1930 I. taunica. - ZAVREL, 2001. Anz., 89: 121 Abb. 1-2 (D). 1934 I . taunica, - KRATOCHV~L, Acta SOC. scient. nat. moraviae, 9 (5): 14, T. 1 Abb. 1 8
(D$?). v 1938 I . frankenbergeri BARTOS, 2001. Anz., 123: 155 Abb. 1-7 (D?). v 1942 I . taunica, - H A D ~ I , Razprave Acad. Ljubljana, 21: 38 Abb. 13 (J). v 1960 I. taunica, - ROEWER, Senckenbergiana, 31 (112): 18 T. 2 Abb. 14 a-b, T. 3 Abb. 14c
bis d (D J $2). v 1950 I . pectinifera, - ROEWER, Senckenbergiana, 31 (112): 21 T. 2 Abb. 16 a (D J). V 1950 I . frankenbergeri, - ROEWER, Senckenbergiana, 31 (112): 21 T. 7 Abb. 42 c (D 9). V 1950 I . franzi ROEWER, Senckenbergiana, 31 (112): 22 T. 7 Abb. 44c, d (D 9). V 1950 I . plicata, - ROEWER, Senckenbergiana, 31 (112): 24 T. 3 Abb. 16 a-d (D$?). V 1950 I . plicata, - S C H ~ L L E R , Mitt. naturw. Arbeitsgem. Haus d. Natur, 1: 41 Abb. 1-3.
1950 I . crassichelis ROEWER, Senckenbergiana, 31 (112): 25 T. 10 Abb. 62 a-b (D 8). 1954 I . taunica, - H A D ~ I , Diss. Acad. scient. art. Slov., 2: 189 Abb. 9-19 (D 3). 1954 I . (hellwigi?) taunica, - RENSCH, Natur und Heimat, 14: 1 , l Abb. (N). 1956 I . taunica, - PFEIFER, Mitt. 2001. MUS. Berlin, 32: 80 (N).
V 1956 I . taunica, - SILHAVI, Fauna CSR, 7: 154, T. 24 Abb. 303-306 (D$?). v 1958 I . sp., - ~ I L H A V I , Vlast. Sbornik Vys., 2: 99 Abb. 1-5.
1961 I . taunica, - RAFALSI~I, Pozn. towar. przyj. nauk, 25: 14. .
V 1965 I . hellwigi, - MARTENS, Natur U. Museum, 95: 143 Abb. 1-4. .
196613 I . taunica, - DRESCO, Bull. Mus. nat. Hist. nat., (2) 38: 801 Abb. 6, 8 b , 11. V 1966 I . hellwigi, - THALER, Ber. ?Tat.-Med. Ver. Innsbruck, 54: 155. v 1967 I . hellwigi, - MARTENS, Naturwissenschaften, 54: 346.
Begründung der Synonymie: PANZERS ,,Phalangium Hellwigii" wurde am Nordrand der deutschen Mittelgebirge auf dem
,,Elema (heute Elm) bei Destedt nahe Braunschweig gefunden; die Abbildung läßt auf ein schließen. Im 19. Jahrhundert wurde die Art mehrfach erwähnt, aber lediglich von HAHN (1834), W 1 ~ ~ ~ ( 1 8 3 4 ) u n d C. L. KOCH (1841 ; 1848, sub herbstii) abgebildet. Offenbar ha t R o E w E R ( ~ ~ ~ ~ , 1923, 1950) diese Abbildungen falsch interpretiert, denn er versteht unter ,,hellwigiU eine lang- und schlank-chelicerige Art, deren J-Cheliceren-Grundglied nahezu unbewehrt ist. Dagegen weisen die Zeichnungen der alten Autoren eindeutig eine gestaucht-kurzchelicerige Art aus. H A D ~ I (19261 1927) schließt sich zwar der Auslegung von ROEWER an, doch bemerkt bereits er (wie ROEWER

später selbst, 1950), daß HAHN (1834, sub hellwigi) und MULLER (1923, auch 1924; sub taunica) sich auf dieselbe Art beziehen. Die Identität von hellwigi (sensn PANZER) 1gßt sich durch die chorologischen Verhältnisse eindeutig klären. Da im engeren Mittel-Europa außerhalb der Alpen (wozu der Locus typicus von hellwigi zählt), immer nur eine Art gefunden wurde, die den Be- schreibungen der alten Autoren völlig entspricht, kann sich der Name hellwigi nur auf diese Art beziehen (über hellwigi sensu ROEWER siehe unter I. muellneri). - Dasselbe gilt von I. taunica. Das Typen-Material wich von jenen Individuen, die MÜLLER unter dem Namen „hellu;igi" von ROETVER zum Vergleich erhalten hatte, erheblich ab, so daß eine Neubeschreibung gerechtfertigt schien ( M ~ L L E R , inündl. Mitteilung). - Auch I. herbsti wurde von ROEWER falsch gedeutet. E r versteht unter dieseln Namen eine Art mit markanter, dreizähliger Bewehrung des Cheliceren- Grundgliedes. Das kann aber nicht zutreffen, wie die Original-Abbildung von C. L. KOCH (1848) beweist; zudem fehlen in Krain, dein Herlrunftsland uon herbsti, Arten mit Dreier-Bewehrung des Grundgliedes völlig. - I. pectinifera, plicata und crassichelis wurden bereits von H A D ~ I (1954) zu taunica (und somit zu hellwigi) gestellt. Auch I. frankenbergeri und franzi sind auf hellwigi zu beziehen (vidi Typ.!).
DRESCO ( 1 9 6 7 ~ ) faßt hellwzgi wie ROEWER auf, obgleich die Synonymie von ~ ~ ~ R T E N S (1965) bereits geklart wurde. Das Material, das SIMON (1872) unter dem Namen „hellwigi" vorlag und das er auch abbildet (im Vergleich zu pyrenaea), gehort eindeutig zu muellneri und nicht zu hell- wzgi, n ie DRESCO unterstellt. Hier hat bereits L. KOCH geirrt, von dem SIMON dieses Exemplar erhielt.
D iagnose : Areae des Scutuni durch Furcheil deutlich gegeneinander abgesetzt, Truncus penis unterhalb der Glans nur schwach geflügelt (Abb. 35g-i).
Va r i ab i l i t ä t : an den Cheliceren gering, auch bei Individuen weit auseinander- liegeilder Fundorte (vgl. Abb. 3Sa-c).
Ve rb re i t ung u n d u n t e r s u c h t e s Ma te r i a l : vergleichsweise großes Areal in Mittel-Europa; es erstreckt sich von M-Jugoslawien (Sarajewo) über Österreich, On- garn (erst ein Fundort), Polen, die Tschechoslowakei, Deutschland und die Nieder- lande (erst ein Fundort) (vgl. Puiiktkarte Abb. 24).
Es wurden 1028, 79 9 und etwa 50 Jungtiere untersucht. J u g o s l a w i e n . - Agram ( 1 8 , SMF/RII/228); Moravika bei Rijeka ( 1 8 , Zool. Inst. War-
schau, BIELAWSL~I leg. 9. VIII. 1957); ohne genaueu Fundort, jedoch a,us Krain ( 1 8 Syntypus I. pectinifera, Zool. Inst. Ljubljaila); Friedrichsteiner Wald s. Gottschee (1 iuv. Blus. Wien, GRUBER leg. VI. 1968). - U n g a r n . - Höhle bei Szakonyfalu/W-Ungarn ( 9 8 4 9, X. 1956 - IV. 1957; 6 8 , 7 9, 1X.-X. 1958, alle LOKSA leg., coll. Lo~rs .~) . - Ö s t e r r e i c h . - Steiermark. Deutsch- landsberg, SW-Hang der Anhohe Mitteregg, 450 m ( 1 8 , 1 ?, Mus. Wien, STROUHAL leg. 24. VII. 1954); Poßruck, 3 kin s. Schloßberg, 420 111 (1 iuv., Mus. Wien. GRUBER leg. 27. X. 1967); - Kärnten. Karawanken, Obir, 1400 n1(1$, Mus. Wien, SCHEERPELTZ leg. 24.126. VI. 1950); Drau- tal oberhalb Villach ( I 8 SMF/R 1119498, ROEWER leg. VIII. 1927); Vellachtal, Seifengraben se. Oberhammer, 800 m (1 iuv. Mus. Wien, STROCHAL leg. 12. VIII . 1957); Karawanken, Lobnig- Graben, 1100 m (1 8, Mus. Wien, SCHEERPELTZ leg. 12. VIII . 1967); Koralpe, S-Hang, St. Loren- Zen, hlagdalensberg (1 iuv., Mus. Wien, FRANZ leg.); 8-Hang bei St. Vinzenz (1 iuv. Mus. Wien, FRANZ leg. 14. VIII. 1965); Villacher AlpeIDobratsch, 1900 m (1 iuv., MARTENS leg. 2. VI. 1966); Hohe Tauern, Glocknerhaus (3 0 , Typenserie I. franzi, SMF/R 1113861, ROEWER leg. VIII. 1927);

Die Abgrenzung von Biospezies usw. 199
Wörther See, Ufer ( 2 8 , 1 9 , Typenserie I. plicata, SMFIR 111227); - Nieder-Österreich. Wiener Wald, Rekawinkel ( 9 3 , 6 Q, 3 iuv., Mus. Wien, GRUBER leg. IX. 1960 - VII. 1966); mittlerer Rotgraben, W. Klosterneuenburg (1 9, Mus. Wien, GRuBERieg. 26. VIII . 1962); Reisalpe, Gscheid, W. Kalte Kuchl(1 9, Plus. Wien. GRUBER leg. 26. VIII . 1962); Reisalpe. - Ober-Österreich. Sau- wald, Engelhartszell, Kesselbachtal(1 ?, Alus. Wien, GRUBER leg. 25. IX. 1957). - Land Salzburg. Trockene Klammen bei Elsbethen (3 8 , 2 9, coll. S C H ~ L L E R , jetzt SMF, alle S C H Ü L L E R ~ ~ ~ . IX. 1934 - X. 1951); Saalfelden, Eialtenbachtal (1 9, Op. 661234 jetzt S l lF , HURDES leg. 27. VIII. 1966); Luiigau, Rotgülden, 1530 in ( 1 8 , Op. 67/181a, jetzt SMF, A U S O B ~ I ~ Y leg. 7. X. 1961); Stu- bachtal, Sprengriedl (1 9, Op. 66/26Ga, jetzt S l fF , AUSOBSI~Y & HURDES leg. 27. VIII. 1966; 2 3 , AUSOBSI~Y 81 MARTENS leg. 7. X. 1966). - Tirol. Innsbruck ( 1 8 , lfus. Wien, REIS leg.); Innsbruck, Sellraintal, Egghöfe bei St. Quirin ( I 9, coll. HAESER, GAMS leg. IX. 1961); Achen- see (1 9, Xus. Wien, GANGLBAUER leg.); Oberperfuß-Ranggerköpfl, 1400-1600 ni (9 8 , 5 0 , 2 iuv., coll. THALER, THALER leg. X. 1961-V. 1965; vgl. THALER 1965); Valsa Tal, Tscheich- Berg, etwa 1600 m (1 9, coll. KRAUS, K R - ~ U S leg. 15. VIII . 1966). - T s c h e c h o s l o w a k e i . - Rimov bei TTebiC (1 iuv., coll. SILHAVY 06234, SILH-AVY leg. VIII . 1949, vgl. SILHAVY 1958); Höhle B ic i Skala bei Brünn ( I 9, coll. SILHAVY 02038, KRATOCHV~L leg. VII. 1938); Morava-Beskydy, LysA hora (1 $, coll. SILHAYY 02087, MILLER leg. VIII . 1952); hfariansk6Laznelllarienbad (1 $2, coll. S I L I I ~ V Y 02026, SILHAVI leg. 28. VIII. 1961, vgl. SILHAVY 1966); AltvaterIPraded, Schlucht des Oppafalles ( 1 3 , coll. SILHAVY, WOLDRICH leg. VII. 1928, vgl. ZAVREL 1930). - Po len . - RiesengebirgeIIhkonoSe, Sneink J a m i (1 9, Typus I. frnnkenbergi, coll. SILHAYY 01017, FRASI<ENBERGER leg. VII. 1937, vgl. BARTOS 1938); Bystrzyca ISlodzka, lfieldtygorze ( 1 8 , coll. Zool. Inst. Warszawa, RIEDEL leg. 18./20. VII. 1962); Landeck, Grafschaft Glatz ( 1 8 , Rlus, Berlin). - D e u t s c h l a n d . - Böhmerwald, Lusen, 1000-1200 in ( 2 8 , 2 ?), Rachel, 1000 m ( I P), Arber, 950 nl ( 1 8 , I ?), alle ~ IARTENS leg. 13.-20. VIII. 1964, vgl. MARTENS 1965); Odenwald ( 1 8 , SMF 10749, HEINZ leg. 15. VIII . 1958, vgl. MARTENS 1965); Rhön, Schafstein ( 1 8 , PIEPER leg. 3. IX. 1963, vgl. B ~ A R T E N S 1965), Lettengraben, e. Wüstensachsen, 800 m (1 iuv., PIEPER leg. 4. VIII. 1966); Xilseburg (1 $, LBKGER leg. 29. VII. 1966); Stellberg ( I 9, PIEPER leg. 27. VII. 1966); Vogelsberg, Taufstein (31 ad. I1 iuv. BRAUN, iMI in~~ws , PIEPER, SILHBVY, THIELE, UHRIG leg. 1958-1967 ; Einzeldaten z. T. in M'IARTENS 1965); Taunus. - Schlangenbad, Altkönig, Hohemark, Nünster ( 3 3 , 4 9, 2 iuv., V. HEYDEN, MCLLER, ZUR STRASSEN, PERL, HESSE leg. 1820-1956; Einzeldaten bei KYRTENS 1965), zwischen Frauenstein und Georgenborn ( 1 8 , HABCI<EIZ leg. 1967); Bürge1 bei Jena (3 iuv. HOFFXAXN leg. 1952); Lauchagrund bei Jena (3 iuv. H O F F ~ ~ ~ A X N leg. 1952); Wuppertal-Beyenburg (3 8 , 4 9, LÖSER leg. XII. 1965 - VII. 1966); Paffrath, Wüstenhammer, Barmer Talsperre, Eifgenthal (108 , lO 9, LÖSER leg. VII1.-X. 1965).
Weitere Fundorte: J i i gos l amien . - TrebeviC bei Sarajevo/Bosnien ( H A D ~ I 1954); Kuma, Zlatica ( H A D ~ I 1942). - Ö s t e r r e i c h . - „Dachsteinu (DRESCO 1967a); PodcetrtekIWindisch- Laiidsberg ( H A D ~ I 1934); Lichtmeßgraben bei AdmontIEnns-Tal (Typus I. crassichelis). - T s c h e c h o s l o w a k e i . - Mittelgrnnd/Elbe (BARTOS 1938). - Po len . - G6ra Sobotka (R-AFALSKI 1961). - De U t s c h l a n d. - Unterfranken (C. IZOCH 1872); Umgebung vnn Erlangen (PFEIFER 1956); Erzgebirge, Fichtelberg-Gebiet, oberes Tal der kleinen Blittweida, Lauenhainer Mühle im Zschopautal ( B ~ T T K E R 1928130); Odenwald, Hirschborn (WIDER, MS unveröffentlicht); Bärental bei Heidelberg ( J ~ S D E R mdl.), Spessart (HAHN 1834), Siebengebirge (VERHOEFF 1900); Sauerland, Naturschutzgebiet Stimmstamm (RENSCH 1955); Böhmerwald, Dreisesselberg (GAUCKLER in litt.); Dreitannenriegel (GAUCI~LER in litt.); Münsterland, Wolbeck (FARWICK Rs RADE 1878), Herford (BARNER 1955), Elin bei Destedt (PBNZER 1794); Taunus, Feldberg (MÜLLER 1924), Naunhof, se. Leipzig(H1~sscH in l i t t . ) ; Thüringer Wald, Inselsberg (HOFFMANN in litt.). - N i e d e r l a n d e . - lfeerdink bei Winterswijk (SPOEI~ in litt.). - D ä n e m a r k . - Unsicherer Fundort, Hallund, Jütland (HANSEN 1885).

Vor kommen: I . hellwigi hellwigi ist eine relativ euryöke Form, die vom Tief- land (Niederlande, Ca. 50 m), den Mittelgebirgen und der Montan-Stufe der Alpen (gelegentlich) bis in die Subalpin-Stufe (Dobratsch, 1900 m) vorkommt. Größte Sied- lungsdichte scheint in den Kammlagen der Mittelgebirge und in der Montan-Stufe der Alpen zu bestehen. Von dieser Form sind zwei Höhlenpopulationen bekannt (Szakonyfalu/Ungarn und Bici Skala bei Brünn/Tschechoslowakei). - Vgl. Kap. I11 1 und 3.
Ischyropsalis hellwigi lucantei SIMON 1879 1879 I. lucantei SIMON, Arachnides de France, 7: 273 T. 14 Abb. 2 (D iuv.). 1881 I. superbus SIMON, Ann. Soc. espan. Hist. Nat., 10: 129 (D). 1914 I. lueantei, - ROEWER, Arch. Naturgesch., 80 A: 116 (D). 1914 I. superba, - ROEWER, Arch. Naturgesch., 80 A: 117 (D). 1923 I. superba, - ROEWER, Weberknechte d. Erde,; 689 (D 9). 1923 I. lucantei, - ROEWER, Weberknechte d. Erde,: 689 (D). 1935 I. superba, - $OEWER, Arch. Zool. GBn. ExpBr., 78: 83 (N). 1946 I. superba, - RAMBLA, Cons. sup. Invest. cient. est. pir., 58 Abb. 10 (D$). 1950 I. lucantei, - ROEWER, Senckenbergiana, 31 (112): 42 T. 6 Abb. 36c, d (D).
V 1950 I. superba, - ROEWER, Senckenbergiana, 31 (112): 42 T. 6 Abb. 38c, d (D). 1966131. superba, - DRESCO, Bull. Mus. Hist. Nat. (2) 38: 801 Abb. 1-5, 7-8a, 9a, 10,,
12-28 (D $9). B e g r ü n d u n g d e r S y n o n y m i e : DRESCO (1966b) wies nach, daß lueantei SIMON 1879 auf
einem Jungtier basiert; die zugehörigen Adulti wurden unter dem Namen I. superba SIMON 1881 bekannt. I. lucantei wurde von ROEWER (1914, 1923, 1950) zitiert, immer mit Bezug auf das eine Typus-Exemplar. Dieser Sachverhalt rechtfertigt nicht das Vorgehen von DRESCO (1966b), den Namen lueantei zum Nomen oblitum zu erklären und zu verwerfen: Gemäß den IRZN hat ein Name nur dann als Nomen oblitum zu gelten, wenn er als älteres Synonym länger als 50 Jahre unbenutzt geblieben ist. Das trifft hier nicht zu. Auch das Argument von DRESCO (1966b), man solle keine Namen aufrechterhalten, die sich auf Jungtiere stützen, ist nicht stichhaltig, da die Jungtiere in diesem Fall mit Sicherheit den Adulti zugeordnet werden können. Somit muß der älteste Name, lucantei SIMON 1879, verwendet werden.
D i ag n o s e: Areae des Scutum nahezu furchenlos ineinanderübergehend, Trun- cus penis unterhalb der Glans relativ stark geflügelt (vgl. Abb. 35).
Va r i ab i l i t ä t : Sie entspricht jener der Nominatform (vgl. Abb. 35). Ve rb re i t ung u n d u n t e r s u c h t e s Mater ia l : Diese Subspezies besiedelt
(nach heutiger Kenntnis) ein vergleichsweise sehr kleines Areal in den W-Pyrenäen und in den östlichsten Kantabrischen Gebirgen (vgl. Punktkarte Abb. 24). Das vom Siedlungsgebiet der Nominatgorm weit abgesprengte Areal erscheint reliktär (über seine Entstehung vgl. Kap. V 2).
Es wurden 6 3 , 4 9 und 10 Jungtiere untersucht. F r a n k r e i c h . - Basses-Pyrhnees. Grotte Oueil de NeezlRBbenacq (23 , 2 9, MARTENS lk
PAULUS leg. 6. IX. 1967); Fundort unsicher: „ArikgeCL (20 9, SMF/RII/5027). - S p a n i e n . - Sierra Urbaso, Cueva Cristinos (1 $); Sierra de Aralar, Lecumberri, 650 m (3 $, 10 iuv., MARTENS & PAULUS leg. 23.125. IX. 1967).

Die Abgrenzung von Biospezies usw.
Weitere Fundorte: F r a n k r e i c h . - Hautes-Pyrenees: St. Jean de Luz (SIYON 1881), St. Pb- de-Bigorre, Grotte de la Palle (DRESCO 1966b), Gedre, Peyrouse. Grotte de Betharram (SIXON 1879), - Basses-Pyrbnbes: Lescun, Ahusky, Aussurucq, Co1 dlAngacica, Alqay, Ravin d'hspeigt, '
Olhette, Urrugne (alle DRESCO 1966b). - S p a n i e n . - Sierra de Aralar, Cueva Txorrote (RAM- BLB 1946), Cueva de Legorras, Cigoitia (ROEWER 1938), Cueva de Armina, Berriatua (D iz~sco 1966 b).
Vo r k oni m en: Die ökologischen Praeferenda werden im wesentlichen jenen der Nominatform entsprechen. Diese Subspezies dürfte also hauptsächlich in schattigen und feuchten Wäldern vorkommen; das bestätigen die eigenen Funde (Sierra de Aralar: Eichenhochwald mit dichtem ~ronendach , wenig Unterhplz, jedoch dichter Bewuchs mit Gras und Stauden, Moos). Die Vertikalverbreitung erstreckt sich offen- sichtlich nahezu von Meereshöhe (St. Jean de Luz, As-in)'bis mindestens 650 m (Sierra de Aralar), wahrscheinlich werden aber noch höhere Gebirgslagen eingenom- men und auch Höhlen regelmäßig besiedelt; DRESCO (196613) nennt 9 Höhlenvor- kommen, von denen eins mii~destens 60 Jahre besteht: SIMON (1907) nennt die Grotte Oueil de Nbez bei Rebenacq, HENROT & NEGRE bestätigten das Vorkommen 1943, und ich selbst fand dort 1967 ebenfalls eine kleine ~ o ~ u l a t i o n .
A n h a n g : Meßstrecken (in mnl) (Erklärung der Rfeßstrecken vgl. Abb. ,34) von I. h. hellwigi und I. h. lucantei.
$ Cx-L ( I ) Sc-L (3) L1 (5) B2 (7) Fe I1 (9) Pen. (11) TaGe (13) Cx-L (1) Sc-L (3) L1 (5) B, ( 7 ) Fe11 (9) Mt Pg (12)
Cx-B (2) Sc-B (4) L2 (6) L, (8) Mt 11 (10) Mt-Pg (12)
CX-B (2) Sc-B (4) L2 (6) L, (8) MT11 (10) TaGe (13)
Die dentzpulpzs-Gruppe I. dentzpulpis CANESTRINI 1872 (Abb. 37-38)
1872 I. dentzpalpzs C~NESTRIKI, Ann. Mus. Civ. Stor. nat., 2 (1): 9 T. 1 Abb. 5 (D). 1879 I. dentzpalpzs, - SIMON, Arachn. de France, 7: 275 (D$). 1914 I . dentqalpss, - ROEWER, Arch. Naturgesch., 80 A: 111 (D$).
v 1916 I. helvetzca ROEWER, Arch. Naturgesch., 82 A (2): 152 Abb. 43 (D 9). 1923 I. denttpalpis, - ROEWER, Weberknechte d. Erde, : 686 (D$).
v 1923 I. helvetzca, - ROEWER, Weberknechte d. Erde, 691 Abb. 863, 864 (D$?; part.: nurdie ?-Diagnose ist auf hellietzca/denttpalpts zu beziehen).

1950 T. dentipalpis, - ROEWER, Senckenbergiana, 31 (112): 36 (D). v 1950 I. helvetica helvetica, - ROEWER, Senckenbergiana, 31 (112): 20 T. 3 Abb. 19a-d(D$?;
part.: nur die ?-Diagnose ist auf helveticaldentipplpis zu beziehen). v 1959 I. dentipalpis, - DRESCO, Ann. Sp616ol., 14(3/4): 383 Abb. 18-26 (D$?).
B e g r ü n d u n g d e r S y n o n y m i e : Die Art wurde nach einem Exemplar aus den Pennini- schen Alpen beschrieben, aber erst von DRESCO (1959) wiedergefunden. Die Überprüfung zweier dieser Tiere, die mir Herr Kollege DRESCO ermöglichte, ergab, daß die beiden bisher bekannten Individuen von helvetica (P) offensichtlich zu dentipalpis gehören. Endgültige Entscheidung wird aber erst dann möglich sein, wenn von „helvetica" auch $ vorliegen.
0 D iagn o s e: Eine Art der dentipalpis-Gruppe, gekennzeichnet durch folgende Besonderheiten: latero-distale Patellarapophyse ($), Scutum magnum (3) bzw. lami- natum (?), sehr schlanke und spitz auslaufende Bewehrung des Cheliceren-Grund- gliedes ($9).
B es c h r ei b u n g. - C h eli c er en (nach Material aus dem Tessin): $-Basalglied (Abb. 37): schlank, doch dorso-distal zu einer breiten, dorsal sanft gerundeten Apo- physe auslaufeild, frontal mit rundem Borstenfeld; dorsal 6 Dornen, die drei vor- deren am größten; ventro-lateral 6 und ventro-medial 5 Dornen unterschiedlicher Größe. - 2. Glied schlank, unregelmäßige grobe Bekörnelung am Knie und an der Frontalseite. - 9: allgemeine Merkmale wie beim $, doch ohne die charakteristi- sche Apophyse und Bürste.
P e d i p a l p u s (Abb. 37g-h): Patellades $ latero-distal mit kräftiger Apophyse, die dem 9 fehlt.
P e des (Laufbeine): kräftig, mittellang.
Cepha lo tho rax u n d Tube r ocu lorum (Abb. 38): Cephalothorax auffallend flach, zerstreut fein bekörnelt; 2. Thorakaltergit mit 9-13 unterschiedlich großen Dornen, die größten (mittleren) plump kegelförmig; die Augen weit voneinander ge- trennt, Tuber oculorum nur schwach ausgebildet, Linse klein.
Scu tum: $ mit Scutum magnum, 9 mit Scutum laminatum; Areae nur fein punktiert, ohne Borsten oder Höcker-Querreihen.
Pen i s (Abb. 37e-f): Truncus schlank, auch an der Basis nicht bauchig, im distalen Drittel am schmalsten; Sklerit der Glans lang und schmal, Seitenränder nahezu parallel verlaufend; ohne deutlichen medianen Kiel (!); Beborstung kräftig, Trichter eng.
R e c e p t a c u l a semin is (Abb. 38): 4-5 relativ kurze Rezeptakel mit rauher Oberfläche (2 Legeröhren präpariert).
Va r i ab i l i t ä t : Das geringe Material läßt sichere Aussagen nicht zu. Individuen von verschiedenen Fundorten differieren in Form des ~ e ~ h a l o t h o r a x und Größe der Augen (Abb. 38) ; Form und Bewehrung der Cheliceren weitgehend übereinstimmend, vgl. Abb. 37.

a-d i------- f h
Abb. 37. I . dent ipalpis CANESTRINI. a-d 1. bzw. 2. rechtes Cheliceren-Glied von latera,l, darüber jeweils das distale Ende des 1. Gliedes von medial. - a Frasco (Schweiz, 8 ) ; b Sonogno (Schweiz, 9); c „Sessin" (Sch\reiz, Holotypus I. helzjetica, 9 ) ; d Rätikon (Österreich, P); g-h Patella des Pedipalpus von lateral: g wie a (8 ) ; 12 wie b (9) ; e Glans penis, wie a ; f Penis, wie a.

Abb. 38. I. dentipalyis CANESTRINI. - Cephalothorax und Receptaculum seminis, a-b Cephalo- thoraces von rechts-lateral: a Rätikon (Österreich); b Sonogno (Schweiz); C-d: Receptacula semi- nis: c wie a : d wie b.
Be zi e hu ii g en: Innerhalb der dentipalpis-Gruppe steht dentipalpis isoliert, enger Anschluß an Arten der Gruppe ist nicht möglich: Lage und Form von Apo- physe und Bürste der $-Chelicere, Form der Cheliceren-Bewehrung, Scutum lamina- tum des 9.
Verb re i t ung u n d u n t e r s u c h t e s Mater ia l : Bisher sind nur drei isolierte Fundorte aus den österreichischen, schweizerischen und italienischen Alpen bekannt.
Es wurden 1 $ und 3 untersucht.
Schwe iz . - Tessin, Höhle C i di Vecc bei Frasco ( 1 8 coll. DRESCO, FERRINI leg. 15. IV. 1957, vgl. DRESCO 1959), Höhle Fiadariö da1 Ragn bei Sonogno (1 9 coll. DRESCO, FERRINI leg. 3. VIII . 1957, vgl. DRESCO 1959); ,,Tessin1' ( I 9 Holotypus I. helvetica SMF/RI/1282). - Ö s t e r - re ich . - Vor,arlberg, Rätikon, Seehöhle in der Sulzfluh, 2300 m (1 9 coll. HAUSER, HAUSER leg. 24. V. 1961).
Weitere Fundorte: I t a l i e n . - Aosta-Tal, Gressoney-Saint Jean (CANESTRIXI 1872); Borna de la Glace, oberhalb von Chabavdey (Gozo 1908). -Schweiz . - Tessin, Höhle Frigee bei Frasco (DRESCO 1959).
Vor kommen: Untersuchungen über die Ökologie liegen nicht vor; dentipalpis besiedelt offensichtlich Höhlen und Freilandhabitate.
k n h a n g : Korpermaße (in mm) (Erklärung der Abkürzungen vgl. Abb. 34) (Exemplare ans dem Tessin, FERRINI leg.):
$. Cx-L ( 1 ) 1 ,5 Cx-B ( 2) 2,5; Sc-L ( 3) 2,5; SC-B ( 4) 2,G; L, ( 5 ) 3 8 ; L, ( 6 ) 5,2 ; B, ( 7 ) ' 0 . 8 ; L, ( 8 ) 1 ,9 ; Fe11 ( 9) 7,3; Mt I1 (10) 9,6 ; Pen. (11) 3,8; Mt Pg (12) 2 ; Ta Ge (13) 40.

Die Abgrenzung von Biospezies usw. 205
9 Cx-L ( 1) 1,G; CX-B ( 2) 2,4 ; Sc-L ( 3) -; Sc-B ( 4) -;
LI ( 5) 4,3; L, ( 6) 5,7; B, ( 7) 0,9; L, ( 8) 2,2; Fe11 ( 9) 6,9; Mt11 (10) 8,6; - (11) -; MtPg (12) 3 ; TaGe (13) 38.
I. adamii CANE~TRINI 1873 (Abb. 39-40) 1873 I. a d a m i i CANESTRINI, Atti SOC. venet.-trent. Sci. Nilt., (1) 2: 50 ( D P?). 1875 I. adarnii , - CANESTRTNI, Atti SOG. venet.-trent. Sci. Nat. (1) 4: 3 (N).
v 1914 1. adami i , - ROEWER, Arch. Naturgesch., 80 A (3): 109 Abb. 5.(D). v 1923 I. a d a m i i , - ROEWEK, Weberknechte d. Erde, 685 Abb. 854 (D 9). v 1923 I. carli , - ROEWER, Weberknechte d. Erde, 687 (part.: „l'irol, RosengartenLLi 2 $2, nec.
1s 39) . 1930 I. a p u a n u s C A ~ ~ O R I ~ ~ C C O , Mon. Zool. Ital. ,41 (5): 126 (D). 1943 I. prope a p u a n u s , - SROSS~\RELLI, 13011. Soc. ent. Ital., 75 (5): 50 Abb. A-B (D$?). 1948 1. earli , - LERMA, Studi spel. fauna Italia medrid., 57 (9): 163 Abb. 2 (D$).
V 1950 1. carli , - ROEWER, Senckenbergiana, 31 (112): 19 (part.: Serie R 1115025: „Tirol, Rosen- garten; 2 ?, nec 1 $ 1 ?).
v 1950 1. herbstii, - R o ~ w ~ a , Senckenbergiana, 31 (112): 25 T. 6 Abb. 34 C-d (part.; Serie SMF/R 111234; D P).
1950 I. adami i , - R o ~ w e i t , Senckenbergiana, 31 (112): 33 T. 5 Abb. 31 C-d (D $, nec Y ) . v 1950 I. amseli ROEWER, Senckenbergiana, 31 (112): 33 T. 3 Abb. 18 a-b (D$). V 1950 1. kratochvili ROEWER, Senckenbergiana, 31 (112): 32 T. 7 Abb. 40 C-d (D $2).
1950 I. a p u a n a , - ROEWER, Senclrenbergiana, 31 (112): 36 (D). v 1960 I. lute-es, - Unr~rscrrisit, Mem. Mus. Civ. Stor. Nat., 8: 102 (N).
B e g r ü n d u n g d e r S y n o n y m i e : Nach dem vorliegenden Material wird I-lalbinsel-ltalien nur von einer Art besiedelt. Die $ sind an den Cheliceren sicher zu erkennen (Apophyse, Bürste), so daß die Zeichnungen von TROSSAI~ELLI (1943, sub a p u a n u s ) und von LERMA (1948, sub carli) zugeordnet werden liönnen. - I. a p u a n a , vorn Autor nicht abgebildet, ist synonym a d a m i i , denn TROS~ARELLI bildet die Chelicercn cines $ ab, das a,us der NB,he des Locirs typicus s tammt (Alpi apuane). Sie stirninen mit anderem S- und mittelitalienischen Material völlig iiberein. - R o e w ~ n (1950) erklärt e i n d (SMF/RI/976, nec q), dessen Fundort nicht bekannt ist, zum Lectotypus. Die Angabe ,,~CULCZYNSI<I ded. ex Typus" erscheint zweifelhaft, denn nach freundlicher Auskunft von inag. W. STAREGA (Warszawa) ist in den Katalogen der Sammlung Uu~czvr j sh r keinerlei Ischyropsalis-Typenmaterial verzeichnet. - I. amsel i und I. lcratoehvili tragen alle Merkmale von a d a m i i nnd sind in die S y n o n y m i ~ zu stellen.
Diagnose: Eine Art der dentipalpis-Gruppe mit folgenden Besonderheitcn: keine deutliche Apophyse auf dem $-Cheliceren-Grundglied, Burste ciförmig, medio- distal; dorsale Bewehrung dreizählig, Areae des Scutum mit je einer Tuberkcl-Quer- reihe ($9).
Beschreibung. - Cheliceren (nach dem Lectotypus SMl?/RI/976); $-Basal- glied (Abb. 39): dorsal 3 große, in einer Linie ausgerichtete Dornen, zwischen diesen, jedoch nicht in einer Reihe, mchrere Tuberkel; ventro-lateral 2 große und unregel- mäßig gerciht kleine und ventro-medial 10 Dornen unterschiedlicher Größe; lateral und weniger medial Tuberkcl verstreut; disto-mediodorsal eine stumpf gerundcte, kurze Apophyse, medial mit einer annähernd runden Bürste besetzt. - 2. Glied

Abb. 39. I. adumit CANESTRINI. a-n 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber je- weils das distale Ende des 1. Gliedes von medial. - a Calabrien, Sta. Eufemia (Italien, wie alle folgenden Abb.; 8 ) ; b wie a ; C Aspromonte ( 3 ) ; d wie C : e genauer Fundort unbekannt (SIKFIRIII 12019,S); f Lectotypus, Fundort unbekannt (8); g Sardinien (Holotypus I. amseli, 8 ) ; h Tivoli (9); i Abbruzzen, Grotta del Azco ( P ) ; k Grotta di Cunardo (Q); 1 wie k ; m Basilicata, Lagonegro (9); n Asprornonte, Piani di Melia (9). o-s Glandes oenis: o wie C ; p wie a ; q wie f ; r wie e ; s wie g; t Penis, wie C.

Die Abgrenzung von Biospezies usw. 207
(Abb. 39a-b, e): am „Knie" einige niedrige Tuberkel. - P: wie die Bewehrung des$ doch ohne Apophyse und Burste; habituell nur geringer Unterschied zwischen den Cheliceren beider Geschlechter.
Pcd ipa lpus ($Q): ohne morphologische Besonderheiten.
P e d e s (Laufbeine): relativ kurz und robust, dunkclbraun, Fcmora basal mit hellgelbem Ringfleck.
Cepha lo tho rax u n d Tube r oculorum (Abb. 40b): Aufwolbung des Ce- phalothorax maßig stark, vgl. Abb.; 2. Thorakaltcrgit mit eincr Querreihc von 8 z. T. sehr kleinen knotigen Dornen; Augenhugel groß, median stark gefurcht, Linse der Augen deutlich vorgcwolbt.
Scu tum: $9 regclmaßig mit Scutum parvum; alle Arcae und die freien Tcrgite mit einer Qucrreihe kleincr, abcr sehr deutlicher Tuberkel, diese mit je einer kleinen Borste.
Penis (Abb. 390-t): Truncus im distalen Drittel am schmalsten, untcrhalb der Glans eine schwache Erwciterilng ; Sklerit der Glans sehr variabcl mit allen Übergangen von breit herzformig bis schmal lanzcttlich, relativ klein; Öffnung dcs Trichters immer eng.
Rccep t a c u l a scniiiiis (Abb. 40a): eine praparierte Lcgcrohrc (Calabrien, Scilla/Melia) auf der einen Seite mit 7, auf der andcren mit 6 langen und sehr schlaii- kcn Rezcptakeln.
Va r i ab i l i t a t : betrifft vor allem die Chelicercn-Bewehrung; dorsale Reihe mit 3 oder 4 markanten Dornen, bisweilen rechts und links untersehicdlich (Abb. 39i); wcsentlich variabler sind Große und Zahl der Dorn-Elemcntc der bcidcn ventralen Reihen, vgl. Abb. 39; nahezu keine Variabilitat von Lage und Form der Apophyse und der Burste.
B czichungcn: Als nachst vcrwandt ist I. carlz anzuschen; diese Art und udumzi gehen wahrscheinlich auf eine gemeinsame Stammform zuruck. Die Areale grenzen aneinander und sind nur durch die Po-Ebene getrennt.
a b Abb. 40. 1. adanzzt C A N ~ S T R I ~ V I . - n Receptaculiiin seminis (Scilla/Melis), 6 Ccphnlothora.: von rechtis-lateral (Aspiomontc).

Verb re i t ung u n d u n t e r s u c h t e s Ma te r i a l : Halbinsel-Italien und offen- sichtlich auch Sardinien (ein Fund); die nördlichsten Fundorte liegen in den Alpi Apuani (Foce di Mosceta; Grotta Carpanello bei PignoneILa Spezia), die südlichsten in Calabrien (Aspromonte) (vgl. Abb. 25 und Abb. 41).
Es wurden 7 3 , 13 und 1 Jungtier untersucht.
I t a l i e n . - Abruzzen, Grotta del Azco (I Q, Mus. Wien); Tivoli(3 9, SMFIR 111234); Cala- brien, Aspromonte (2 9, Mus. Wien, PAGANETTI leg. 1906); Asproinonte, Piani di Melia (1 Q, Alus. Wien, RUFFO leg., vgl. KRIT~CHER 1960); Scilla, 4-8 km von AIelia (2 $2, Zool. Inst. Warszawa, RSEDEL leg. 3. IV. 1964); Sta. Eufemia ( 2 3 2 9, Mus. Budapest, PAGANETTI-HUMLER leg. 1913); Basilicata, Lagonegro (1 $2, Mus. Hamburg, leg. 10. X. 1923); Lazio, Ienne, Pozzo della Creta Rossa (1 iuv. coll. BRIGNOLI, SBORDONI leg. 27. XI. 1966); Sardinien, Aft. Eccas ( 1 8 Holotypus I. anzseli, SRIF/R 111311, A m s ~ ~ l e g . ) . - Ohne Fundorts-Angabe(18 Lectotypus, SMF/R 1/976).
Weitere Fundorte: Matese, Grotta di Campo Rotondo (LERMA 1948); Golf di Squillace (CBNESTRINI 1873; Locus typicus); ,,Calabrien" (CANESTRSNI 1876); Alpi Apuane, Höhle am Foce di Mosceta (CAPORI~LCCO 1930, Locus typicus I. apuana); La Spezis, Pignoile, Grotta Car- panello (TROSSARELLI 1943).
Fundorts-Angabe sicher falsch: Italien, Trentino-Tiroler Etschland, Rosengarten (2 9 „Alle- typus" I. carli, SMFIR 1116025); Griechenland, Insel Kreta, Omolos-Katavothron (1 9 Holotypus I . kratochcili, ShlF/R 111548).
Vorkommen: Untersuchungen zu Ökologie liegen nicht vor; in S-Italien werden offensichtlich die Wälder der Gebirge bevorzugt besiedelt. Möglicherweise verträgt adamii höhere Temperaturen als die meisten anderen Ischyropsalis-Arten.
Abb. 41. Fundpunkte einiger Arten der dentipalpis-Gruppe. -. 1 robusta; 2 luteipes; 3 carli; 4 adamzi: mit Pfeil sind die Fundorte von dentipalpzs bezeichnet.

Die Abgrenzung von Biospezies usw.
A n h a n g : Meßstrecken (in mm) (Erklärung der Abkürzungen vgl. Abb. 34):
$ CX-L (1) 1,6- 2,4 (1,g4); Sc-L (3) 2,O- 2,4 (2,2,); LI (5) 3,l- 3,7 (3,341; B2 (7) 0,9- 1 9 1 (0,964); Fe I1 (9) 3,6- 3,7 (3,67,); Pen. (11) 2,9- 3,05 (2,95,); TaGe (13) 29 -37
9 Cx-L (1) 1,8 -2,3 (2,05„); SC-L (3) 2,4 -3,O (2,75„); L1 (5) 3,3 -3,7 (3,5„); B2 (7) 0,85-1,2 (1,0611); Fe I1 (9) 3,9 -5,9 (4,59„); MtPg (12) 4-5
CX-B (2) SC-B (4) L2 (6) L3 (8) Mt I1 (10) MtPg (11)
CX-B (2) SC-B (4) L2 (6) L, (8) Mt I1 (10) TaGe (13)
I . carli LESSERT 1905 (Abb. 42-43)
1875 I . manicata, - PAVESI, Atti. SOC. Ital. Sci. Nat., 18: 254 (N). 1905 Ischyropsalis Carli LESSERT, Rev. Suisse Zool., 13: 658 Abb. 15-17 (D Q). 1914 I . carli, - ROEWER, Arch. Naturgesch., 80 A (3): 111 Abb. 7 (D 9). 1914 I . herbstii, - ROEWER, Arch. Naturgesch., 80 A (3): 110 (D$??, part.: Como).
v 1917 I . carli , - LESSERT, Cat. Invert. Suisse, 9: 53 Abb. 34-37 (D $9). V 1923 I . carli , - ROEWER, Weberknechte d. Erde,: 687 Abb. 856 (DQ, necdQ (part.; Serie
„Tirol/Rosengarten" zu adarnii). 1923 I . Herbstii , - ROEWER, Weberknechte d. Frde,: 686 (D; part.: Como).
v 1923 I. herbsti, - SCHENKEL, Verh. Naturforsch. Ges. Basel, 34: 124 (N). v 1936 I. carli, - SCHENKEL, Rev. Suisse Zool., 43: 312 (N). v 1950 I . carli , - ROEWER, Senckenbergiana, 31 (112): 19 T. 7 Abb. 41 a-d (part.: nicht die
Serie SMF/RII/5025). v 1950 I . herbstii, - ROEWER, Senckenbergiana, 31 (112): 25 T. 6 Abb. 34 C-d ( D d , nec 9)
(part.: nur die Serie SMFIR 1111265 von Como). v 1950 I. pentelica ROEWER, Senckenbergiana, 31 (112): 30 T. 8 Abb. 49 a-d.
1959 I . carli, - DRESCO, Ann. SpBlkol., 14 (314): 382. 1960 I . carli, - DRESCO & DEROUET, Ann. SpBlkol., 15 (1): 112. 1966a I . carli, - DRESCO, Bull. Mus. Nat. Hist. Nat., (2) 38: 598, Abb. 22, 26. 1966 I . carli, - STRINATI, Faune cavernicole de la Suisse: 332 (N).
B e g r ü n d u n g d e r S y n o n y m i e : Diese überwiegend west- und südalpine Art interpre- tieren die meisten Autoren einheitlich. - Der Name h e ~ b s t i i wurde erstmals von ROEWER (1914) für Individuen aus den italienischen S-Alpen und Mittel-Italien gebraucht. Sie zeichnen sich durch dorsale Dreier-Bewehrung des 1. Cheliceren-Gliedes aus. Diese Zuordnung ist sicher falsch, denn Locus typicus von herbstii ist ,,KrainU, wo Arten mit solcher Cheliceren-Bewehrung nicht vorkommen; herbstii ist synonymhellwigi (vgl. diese Art). - Die Typenserie von pentelica (SMF/ R 111233) besteht aus einem subadulten 3 und einem Jungtier (also nicht 18, 1.9, wie ROEWER irrtümlich mitteilt). Bewehrung und dorso-distale Apophyse der Chelicere des subad. $ weisen eindeutig auf Zugehörigkeit zu carli. Die Fundorts-Angabe (Attika, Höhle im Pentelikon) ist sicher unrichtig - vgl. Kap. V l a . Die Nennung von manicata für das Tessin ( P A ~ E S I 1875) inuß mit der da.maligen Unkenntnis der Verbreitung der Arten erklärt werden, denn manicata ist auf die Karpaten-Länder beschränkt.
14 Zool. Jb. Syst. Bd. 96

Diagnose: Eine Art der dentipalpis-Gruppe, gekennzeichnet durch folgende Besonderheiten: Apophyse auf dem$-Cheliceren-Grundglied niedrig und stumpf, des- sen dorsale Bewehrung (fast) immer dreizählig($?), Areae des Scutum ohne Tuberkel- Querreihe ($?).
B es chr eibung. - Cheli Ceren (nach Material von Wolfenschießen, Schweiz) ; $-Basalglied (Abb. 42a): dorsal eine Reihe aus 3 sehr großen Dornen, der distale am größten; ventro-lateral 5, ventro-medial 8 Dornen unterschiedlicher Größe; lateral (basal) und medial nur wenige verstreute Tuberkel; distales Ende des Grundgliedes schwach nach ventral abgewinkelt. Dadurch entsteht der Eindruck einer sehr flachen dorso-distalen Apophyse. - 2. Glied mit minimaler Behöckerung am „Knieu. - $2: die allgemeinen Merkmale wie beim $, doch ohne Apophyse und Burste.
P e d i p a l p u s (69): normal, glatt, mit feinen Haaren besetzt. P e des (Laufbeine): relativ kurz und robust, dunkelbraun, lediglich die Tarsen
heller, Femora basal mit hellem Ringfleck. Cepha lo tho rax u n d T u b e r oculorum (Abb. 43c): Ansatzstellen der Cheli-
ceren-Muskulatur (vor allem hinter dem Tuber oculorum) wenig erhaben, 2. Thorakal-
h i k I m
Abb. 42. I. earli LESSERT. a-g 1. bzw. 2. rechtes Cheliceren-Glied von lateral, dariiber jeweils das distale Ende des 1. Gliedes von medial. - a Wolfenschießen (Schweiz, 3 ) ; b Adelboden (Schweiz, 3 ) ; c Porlezza(Italien, 3); d Lugano (Italien,$); e Como (Italien,$); f Groß-Verniunt (Österreich, 3 ) ; g SUnser See (Osterreich, 9). h Penis, wie e ; i-m Glandes penis: 2. wie b ; k wie a ; lw iee ; mwief.

Die Abgrenzung von Biospezies usw. 211
Tergit mit 4-8 kleinen Tuberkeln ; Tuber oculorum groß, median tief gefurcht; große, weit vorstehcnde Linsen.
S c u t u m : d ? regelmäßig mit Scutum parvum; die einzelnen Areae, wie auch die freien Tergitc, ohne Tuberkel-Querreihen, lediglich fein bekornelt.
Penis : Porm des Truncus im wesentlichen wie bei adamn, jedoch in der basalen Halfte bauehigcr (Abb. 42h), Sklerit der Glans lanzettformig, unterschiedlich groß, medianer Kiel deutlich; Öffnung des Trichters eng bis sehr weit (Abb. 42i-m).
R e c e p t a c u l a semin is (Abb. 43a-b): Schlauche relativ kurz und robust, nur schwach gekrummt, jedoch das distalc Ende bisweilen abgcwinkelt; Anzahl der Schlauche 5-6, bci einem Ex. rcchts und links verschieden (Damuls, Sünser See) (2 Legerohren prapariert).
Va r i ab i l i t a t : Merkmale der außeren Morphologie einheitlich, dorsale Beweh- rung des Cheliceren-Grundgliedes ($9) immer markant dreizahlig, falls weitere Dorn- Elemente in dieser Reihe, dann klcin und wenig hervortretend (z. B. Abb. 42c); dorso-distale Aufwolbung des $-Grundgliedes immer vorhanden; Lage und Porm der Burste einheitlich. Stark schwankend dagegen die beiden ventralen Dornen-Rcihen (Einzelheiten s. Abb.); Trichter der Glans penis von englumig bis breit ausladend variierend.
Bez iehungen: vgl. Ausfuhrungen bei I. carlz.
V e r b r e i t u n g u n d u n t e r s u c h t e s Ma te r i a l : Das Areal liegt in den W-Alpen (nicht westlich des Genfer Sees, soweit bis heute bekannt) und in den westlichen Zentral-Alpen dcr Schweiz, Österreichs (bis jetzt fünf Funde in Vorarlbcrg und Tirol), Deutschlands (Allgau) und Italiens. - Vgl. Punktliarte Abb. 4113.
Es wurdcn 143 ,15 9 und 10 Jungtiere untersucht. O s t e r r e i c h . - Vorarlberg; Damuls, Sunser See, 1800 ni (3 0 , 1 6 , 2 iuv. coll. THALFR,
THALER leg. 6.-8. VI. 1963); Groß-Vermunt, 1960 m (1 3 Mus. Wien, Gitushn leg. 8. VJ. 1966);
Abb. 43. I. carlz L ~ s s c n ~ a-b Receptdcula seminis: a Grisons (Schweiz); b Sunser Sce (Ostcr- reich); c Ceplialothorax von rechts-lateral (Grchß-Vermunt, Osterreich).
J
14*

W-Fuß der Mittagsfluh, zwischen Au und Schnepfau, 800 m (1 iuv. Mus. Wien, S ~ R O U H A L leg. 15. IX. 1960). - Schwe iz . - Mte. Ceneri, N-Hang, zwischen Bellinzona und Lugano (1 iuv. Mus. Wien, STROUIIAL leg. 20. IX. 1960); Tessin, Castel di Chiasso (2 9 Mus. Basel, ded. PONTANA 1902); Kanton Bern, Adelboden, rechte Seite des Engstligenbaches, 1200-1500 nl ( 2 6 , 3 Mus. Basel, SCIIENI~EL leg.; vgl. SCHENIIEL 1923); Kanton Bern, Gadmen-Tal(46, 1 9 Mus. Basel, SCHENKEL leg. 1932; vgl. SCHENKEL 1936); Kanton Bern, Beatenberg (1 8 Mus. Basel, SCIIENKEL leg. 1936); Kanton Nidwalden, Wolfenschießen, Schwarzwald bei Altzellen, 1300 in (1 8, 2 $! Mus. Basel. SCHENI~CL leg. VIII.; vgl. SCHENKEL 1923); Lugano ( 1 6 Mus. Gcnf. GHIDINI leg.; vgl. LESSERT 1917); Porlezza (1 $ Mus. Genf, GFTIDINI leg.); Grisons (1 6 Mus. Genf). - D e u t s c h l a n d . - Allgau; Umgebung von Balderschwang (2 $, 3 iuv., MARTENS leg. 21.-27. VII. 19G8), Oberstdorf, Wasserfalle des Paltenbaches (1 iuv., MARTENS leg. 26. VII. 1968). - I t a l i e n . - Como ( I $ SMF/R I112G5); Grotta di Cunardo (2 Mus. Wien, leg. 19. X. 1958).
Weitere Fundorte: Ö s t e r r e i c h . - Tirol; Ötztal, Obergurgl (SHALER in litt.); Ötztal, Um- hausen/Stuibenfall (KULLMANN leg., THALER in litt.). - Schwe iz . - Scliuls-Fettsn, Passug bei Chur, Churwalden (alle LESSER-T 1905); Tessin, Grotta del Mago (PAVESI 1875, sub manicata); Cuvel La S-chala, Grisons; St. Gallen, Wildenmannlislöchli und Hohle bei Freienbauh (alle STRI- N ~ T I 1966). - I t a l i e n . - Castel San Pietro (LESSERT 1917); Varzo (DRESCO & DRESCO-DEROUET 1960). -Fundorts-Angabe sicher falscli: Griechenland, Attika, Ilöhle irn Pentelikon (2 iuv. Sypen- serie I. pentelica, SMF/R 111239).
Vorkommen: I. carli besiedelt in mittleren Lagen der Alpen von (soweit heute bekannt) 800 m (Mittagsfluh, Voralberg) bis nahe 2000 m (Groß-Vermunt, Silvretta) bevorzugt Wald-Biotope; z. B. Rand eines lichten Bucheii-Fichten-Waldstückes an einer versumpften Wiese, Bachiiähe (1400 m) und Buchenhochwald (bei Balder- schwang im Allgäu); Waldgebüsch mit Pinus mugo, Rhododendron, Vaccinium myr- tillus (bei Groß-Vermunt; GRUBER in litt.) oder auch auf Almwiesen nahe kleinen Riniisalen (Damüls, Sünser See; THALER in litt.). Den einen der beiden Höhlenfuude meldet D ~ e s c o & DEROUET (1960): Grotte Est „sotto Tugliagga" bei Varzo (Prov. Novara; ferner existiert eine Population in der Grotta di Cunardo [wo genau?]). - Die Reifezeit liegt offensichtlich bereits im Brühsommer (Juni, Juli). Ende Juli wurden (im Allgäu) neben Adulten auch Juiigtiere verschiedener Stadien und Gelege gefunden.
A n h a n g ; Korpermaße (in min) (Erklarung der Abkurzungen vgl. Abb. 34):
C? Cx-L ( I ) Sc-L (2)
L1 (5) B2 (7) Fe I1 (9) Pen. (11)
P Cx-L (1) Sc-L (3)
LI (5) B2 (7) Fe I1 (9) MtPg (12)
2,3- 2,F (2,4,); 2,3-2,7 (2,591; I
4,l- 496 (4,391; 1,5- 1 , s (1,7,); ~ 4,9- 5,6 (5,391; 1
4- 5 ; TaGe (13) 32-36. 1 2,7- 3 2 (2,g12); 2,6- 3,6 (3,0„); 1 4,4- 5 , l (4,712); 1,7- 2,1 (1,85,2); 1 4,5- 5,3 (4,8810); 33-35. I

Die Abgrenzung von Biospezies usw. 213
I. luteipes SIMON 1872 (Abb. 44-45) 1866 Lhermia spinipes L u c ~ s , Ann. Soc. ent. France, (4) 6: 44 (Nomen nudum). 1872 Ischiropsalis luteipes SIMON, Ann. Soc. ent. France., (5) 2: 484 T. 16 Abb. 2, 5 (D 9). 1879 I . luteipes, - SIMON, Arachn. d. France, 7: 268 T. 13 Abb. 12 T. 14 Abb. 2 (D $9). 1907 I . luteipes, - SIMON, Arch. Zool. Exper. Gen., (4) 6 (1): 552 (N.) 1911 I . luteipes, - SIMON, Arch. Zool. Exper. Gen., (5) 9 (2): 203 (N). 1914 I . luteipes, - ROEWER, Arch. Naturgesch., 80 A (3): 112 Abb. 8 (D $9). 1923 I . luteipes, - ROEWER, Weberknechte d. Erde.: 687 Abb. 857 (D$?). 1926 I . luteipes, - JEANNEL, Faune cavernicole de la France: 159. 1950 I . luteipes, - ROEWER, Senckenbergiana, 31 (112): 38 T. 4 Abb. 25 a-d (DS.?).
V 1950 I . caporiaccoi ROEWER, Senckenbergiana, 31 (112): 33 T. 4 Abb. 23 a-b (DS.). V 1950 I. pestae ROEWER, Senckenbergiana, 31 (112): 28 T. 5 Abb. 26 a-b (DS).
1952 I . luteipes, - DRESCO, Not. Biospeleol., 7: 101 (N). 1961 I . luteipes, - JUBERTHIE, Ann. Speleol., 16 (4): 383 (N). 1964 I . luteipes, - JUBERTHIE, Ann. SpBl6ol., 19: 23, 54, 106a, 114, 119, 143 Abb. 13, 14a ,
34b, 37a, 42a (N). 1966 I . luteipes, - DRESCO, Bull. Mus. Nat. Hist. Nat., (2) 38 (5): 598 Abb. 23-25, 27-28
(N, D). B e g r ü n d u n g d e r S y n o n y m i e : Diese im französischen Zentralmassiv und in vielen Höh-
len der Pyrenäen häufige Art wurde bis auf zwei Ausnahmen immer einheitlich interpretiert. Die Typus-Exemplare von I . caporiaccoi (3) und I. pestae (3) weisen alle Kennzeichen von lutei- pes auf. Zudem sind die Angaben über die Fundorte sicher unrichtig, da beide Loci typici weit außerhalb des luteipes-Areals liegen.(jener von caporiaccoi in Sizilien, der von pestae in Österreich).
D i agn o s e: Eine Art der dentipalpis-Gruppe, gekennzeichnet durch folgende Besonderheiten: kräftige, mehr als dreizählige dorsale Cheliceren-Bewehrung (Grund- glied, $?) ; eine massige, stumpf-konische dorso-distale Apophyse auf dem Grundglied (3) und eine punktförmige dorso- bis medio-distale Bürste (J).
Beschreibung. - Cheliceren (nach Material vom Co1 de le Rhpublique, Haute-Loire); $-Basalglied (Abb. 44c): relativ schlank, jedoch dorso-distal zu einer stumpf-konischen, auch nach medial übergreifenden Apophyse erweitert; dorsal 3 kräftige Dornen, basad ein kleinerer anschließend; ventro-lateral 4 große, ventro- medial 5 Dornen unterschiedlicher Größe; eine punktförmig kleine Bürste am Distal- ende dorsal, etwas nach medial verschoben. - 2. Glied: am „Knieu medial und fron- tal kleine Tuberkel, jeweils mit einer Borste besetzt. - 9: die allgemeinen Merk- male wie beim $, doch ohne Apophyse und Bürste, dadurch viel schlanker wirkend.
P e d ip a lpus : robust, ohne inorphologische Besonderheiten.
Pedes (Laufbeine): relativ kurz und robust; Femora an der Basis mit hell- gelbem Ringfleck, distales Ende des Metatarsus und der Tarsus gelb, übrige Bein- glieder dunkelbraun.
Cepha lo tho rax u n d T u b e r oculorum (Abb. 45b): Cephalothorax kraftig gewölbt, fein und dicht bekörnelt; 2. Thorakaltergit mit 2-4 medianen Tuberkeln, lateral winzige Körnchen beigestellt; Tuber oculorum bemerkenswert groß, median

Abb. 44. I. luteipes SIMON. a-f 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber jeweils das distale Ende des 1. Gliedes von medial. - a Moulis (Fraiilrreich, 3); b Grotte de Enlenne, Aribge (Frankreich, 8 ) ; c Co1 dc la Republiyue, Haute-Loire (Frankreich, 8 ) ; d richtiger Fundort unbekannt (Holotypus i. caporiaccoi, 8 ) ; e richtiger Fundort unbekannt (Holotypus I. pestae, 8 ) ; f wie U, (P). g-k Glandes penis: g wie d ; h wie C; i wie b ; k wie e ; 1 Penis, wie e.
Abb. 45. I. luteipes SIMON. a Receptaculum seininis (Moulis, Frankreich); b Cephalothorax von rechts-lateral (Grotte de Touasse, Frankreich).

Die Abgrenzung von Biospezies usw. 215
bis fast auf die Hohe des Cephalothorax gefurcht. Linsen der Augen groß und weit vorgewolbt.
Scu tum: J? regelmaßig mit Scutum parvum, Areae des Scutum mit je einer lockeren Querreihe kraftiger Tuberkel, diese mit je einer winzigen Borste, die Reihe bisweilen median unterbrochen. - Ontogenetische Entwicklung des Scutum vgl. Kap. IV l b .
Pen is : Basale Halfte des Truncys(Abb. 441) breit, jedoch nicht bauchig, kurz unterhalb der Glans am schmalsten ; Sklerit der Glans schmal, bisweilen lanzenspitzen- formig ausgeweitetlAbb. 44g-k), kraftig beborstet; Trichter eng.
Re cep t a cu lum s emin is (Abb. 45a): Mit relativ dunnen, in sich geschwun- genen Schlauchen, (meist ?) 4 Stuck (1 Legerohre prapariert).
V a r i ab i l i t a t : Einzelheiten der Cheliceren-Bewehrung von einem Exemplar zum anderen sehr unterschiedlich, doch ist immer ein bestimmter Anlageplan ver- wirklicht (Abb. 44a-f): dorsal eine Reihe aus 4-6 kraftigen, unterschiedlich großen Dornen, von denen meist 3 besonders hervorragen, lateral sind kleinere Dornen- Elemente in diese Hauptreihe eingeschoben; dorso-distale Apophyse immer einheit- lich, Burste in der Große, jedoch nicht in der Lage, geringfugig variierend; Sklerit der Glans penis im wesentlichen einheitlich, wenn auch lateral unterschiedlich stark ausgeschwungen (vgl. Abb. 44g-k).
Bez iehungen: I. lutezpes gehort zu den kleinwuchsigen Arten der dentzpalpis- Gruppe, deren vikariante Areale sich vom Karpaten-Bogen bis Portugal erstrecken. Nimmt man die außere Morphologie als Maßstab, dann bestehen die engsten Be- ziehungen zu carlz. Das Areal dieser Art schließt ostlich an das von lutezpes an. Bei carlz ist die Cheliceren-Apophyse viel schwacher ausgebildet, die Burste hat dieselbe Lage, ist aber etwas großer. - Sichere Trennung beider Arten (auch 9) ist mittels der dorsalen Bewehrung des Grundgliedes moglich; sie ist bei carlz auf 3 markante Einzel- Dornen beschrankt, bei lutezpes sind es immer mehr als 3, zudem unterschiedlich groß und nicht streng in einer Linie ausgerichtet.
Ve rb re i t ung u n d u n t e r s u c h t e s Ma te r i a l : Das Areal von lutezpes ist wahrscheinlich disjunkt (vgl. Abb. 25 und die Punktkarte Abb. 41) ; die Populationen des franzosischen Zentralmassivs sind offenbar von jenen der Mittel- und W-Pyrenaen isoliert. Im Zentralmassiv sind Nachweise bekannt aus den Departements Puy-de- Dome, Cantal, Haute-Loire, Lot, Ardbche und Corrbze; in den franzosischen Pyrenaen wurde sie in den Departements Hautes-Pyrenees, Haute-Garonne, Arikge und Py- renees-Orientales gefunden. Nachweise aus den spanischen Pyrenaen stehe4 aus, doch fand ich (zusammen mit H. PAULUS) lutezpes am Port d'Aula (Aribge) unweit der franzosisch-spanischen Grenze; der einzige sichere Fund aus Spanien stammt aus dem katalonischen Bergland (Sierra de Montseny). Nennungen fur Italien (z. B. ROEWER 1923, KRITSCHER 1960) beruhen meist auf Verwechselungen mit adamzi.

Es wurden 143 ,14 und 21 Jungtiere untersucht. F r a n k r e i c h . - Haute-Loire, Co1 de la RBpublique = Co1 du Grand Bois (3 3 1 9, 7 iuv.
MARTENS & PAULUS leg. 28. VIII. 1967); Correze, Egletons (1 iuv. MARTENS & PAULUS leg. 30. IX. 1967); Arihge, Grotte de Touasse bei St. Girons (53 , 10 ? , 5 iuv. MARTENS & PAULUS leg. 2. IX. 1967); Ariege, Port #Aula, 1690 m (8 iuv., 18-Chelicere MARTENS & PAULUS leg. 3. IX. 1967); Grotte de Moulis (13 , 1 9 M A T J A ~ I ~ leg. X. 1955); Vallee de Bethmale bei St. Girons (1 9 Zool. Inst. Warszawa, RIEDEL leg. 22. 111. 1964); Grotte de Enlenne ( 3 3 Zool. Inst. Warszawa, BOUIL- LON, CHODOROWSKI & RIEDEL leg. 12. V. 1964). - S p a n i e n . - Catalonien, Santa Fe, Montseny (1 9 leg. 6. IV. 1955).
Fundorts-Angabe sicher falsch: Österreich, Leitha-Gebirge, Sonnenberg (1 3 Holotypus I. pestae SME/R 1119493, angeblich ROEWER leg. VIII. 1934). - Insel Sizilien, Umgebung von Mes- sina (1 3 Holotypus I. caporiaecoi SMF/R II/5030). -
Da I. ZzLteipes in beiden Arealteilen allgemein ver.breitet ist, erübrigt sich die Aufzählung' der weiteren Fundorte (vgl. DRESCO 1952).
Vorkommen: „Diese Art lebt in Frankreich im Zentralmassiv und auf der Nordseite der Pyrenäen zwischen 100 und 2000 m (DRESCO 1952). Sie kann sich im feuchten Moos aufhalten, im Innern alter Baumstümpfe oder unter loose liegenden Steinen, in den Stollen stillgelegter Bergwerke, in Höhlen und besonders an Sicker- stellen kleiner Wasserläufe" (JUBERTHIE 1961). Ich fand luteipes auch auf Almwiesen über der (natürlichen?) Baumgrenze in 1690 m (Port #Aula, Ariege). - Die Art ist an den meisten Standorten häufig, was bereits SIMON (1879) betont. In der nahrungs- reichen Höhle Touasse (Aribge) zählte ich am 2. IX. 1967 über 70 adulte Individuen, die gut sichtbar an den Wänden saßen; vgl. Skizze bei JUBERTHIE (1961).
A n h a n g ; Korpermaße (in mm) (Erklärung der Meßstrecken vgl. Abb. 34):
3 Cx-L (1) 1,8- 2,3 (2,06,); CX-B (2) 2,4- 2,9 (2,6,); Sc-L (3) 2,2- 2,5 (2,38,); SC-B (4) 2,5- 2,9 (2,72,); L1 (5) 33- 4,2 (3,7Ol2); L, (6) 4,75-5,7 (5,145); B2 (7) 9 - 1 , ( L3 (8) 1,9- 2,1 (2,005); Fe 11 (9) 4,3- G,8 (5,3G5); Mt I1 (10) 4,4-7,4 (5,82,); Pen. (11) 3,G (3,6,); Mt Pg (12) 3-4 TaGe (13) 31-35 .
? CX-L (I) 7 1 (l,754); CX-B (2) 2,G- 2,8 (2,68,); SC-L (3) 2,3- 2,6 (2,42,); SC-B (4) 2,5- 2,7 (2,62,);
LI (5). 3,3- 3,7 (3,554); L2 (6) 4,8- 5,O (4,974); B2 (7) 0,g- 1,05 (1,004); L3 (8) 1,9- 2,o (1,924); Fe11 (9) 5,2- 5,5 (5,32,); Mt11 (10) 5,9- 6,0 (5,82,); MtPg (12) 2-4 TaGe (13) 32-33
I. rnunicuta L. KOCH 1865 (Abb. 46-48) 1865 Isehyropsalzs man>cata L. KOCH, Z. Ferdinandeum Tirol Vorarlberg, (3) 12: 164 (Kurz
diagnose; ,,Siebenburgen"). 1904 I. manicata, - KULZYNSKI, Ann. Mus. Hungar., 2: 78 T. 9, Abb. 5-6.
V 1914 I. manzcata, - ROEWER, Arch. Naturgesch., 80 A (3): 105 Abb. 2 a-b (D 39) .

Die Abgrenzung von Riospezies usw.
V 1916 I. dacica ROEWER, Arch. Naturgesch., 82 A (2): 153 (D$$!). V 1923 I. marzicata, - ROEWER, Weberknechte d. Erde: 684 Abb. 851 a-d. (D$$!). V 1923 I. helvetica, - ROEWER, Weberknechte d. Erde: 691 Abb. 863, 864 ( ~ a r t . : ~ nur die
Abb. 864 ist auf manicata zu beziehen) (D 8). V 1934 I. manicata, - KRATOCHV~L, Acta Soc. Sc. nat. morav., 9 (5): 1 3 T. 1 Abb. 17 (D 9). V 1934 I. helvetica milleri K n ~ r o c ~ v i ~ , Acta Soc. nat. morav., 9 (5): 1 3 T. 1 Abb. 14 (D$$!).
1935 I. daciea, - ROEWER, Arch. Zool. Expkr. Gener., 78 (1): 84 (N). . . 1936 I. helvetica helvetica, - KRATOCIIV~L, Festschrift STRAND, 1: 248 (N).
V 1950 I. dacica, - SILHAVY, Polia ent., 13: 101 T. 1 Abb. 1-5 T. 2 (D). V 1950 I. kästneri ROEWER, Senckenbergiana,, 31 (112): 19 T. 9 Abb. 58 C-d (D 9). :
V 1950 I. helvetica helvetica, - ROEWER, Senckenbergiana, 31 (112): 20 T.3 Abb. 19 a-d (part.: nur die Abb. a-b sind auf flanicata zu beziehen; Serie SMFlRIIl229).
V 1950 I. helvetica milleri, - ROEWER, Senckenbergiana, 31 (112): 20 T. 3 Abb. 20c. V 1950 I. austriaea ROEWER, Senckenbergiana, 31 (112): 22 T. 8 Abb. 53 C-d (D $2). v 1950 I. manicata, - ROEWER, Senckenbergiana, 31 (112): 29 T. 4 Abb. 21 a-&(D$?). V 1950 I. dacica, - ROEWER, Senckenbergiana, 31 (112): 30 T. 4 Abb. 24 a-d (D$$!). .
V 1950 I. balcanica. R O E ~ E R , Senckenbergiana, 31 (112): 30 T. 9 Abb. 45 C-d (D P). 1954 I. milleri, - FIAD~I, Acad. Scient. Art. Sloven. ('4) 2: 193 (N). 1956 I. manicata, - SILIIAVY, Fauna CSR, 7: 156 T. 24 Abb. 299 (N). 1956 I. helvetica, - SILHAVY, Fauna CSR, 7: 157 (N).
V 1956 I. dacica - S I L H A ~ Y , Fauna CSR, 7: 157, T. 24 Abb. 298, 300-310 Farbtafel 3 Abb. 9 bis 10 (D 8 9).
1961 I. dacica - RAFALSI<I, Poz. Tow. nauk, 25: 14 (N). 1961 I. manicata, - R A F A I ~ S ~ I , Poz. Tow. nauk. 25: 1 3 (N). 1964 I. dacica, - AVRAM, Rev. roum. Biol., s6r. Zool., 9 (4): 245 Abb. 1-4 (D 8 9 ) .
v 1966 I. manicata, - STARLSGA, Fragmenta faun., 1 3 (6): 148 (N). 1966aI. manicata, -' D i t ~ s c o , Bull. Mus. Hist. Nat., (2) 38 (5): 598 Abb. 21 (N).
B e g r ü n d u n g d e r S y n o n y inie: ROEWER (1914-1950) nahm an, daß beide Geschlechter von maiiicata Scutnm parvum tragen. Das trifft aber nur für das $ zu; die 9 besitzen immer ein Scutum laminatnm. Unter dieser irrigen Voraussetzung beschrieb ROEWER (1916) I. dacica ('8 mit Scutum parvum, $! mit laminatum). Das zahlreiche hier vorliegende Material läßt keineswegs die Annahme zu, daß in den Karpaten-Ländern zwei (nahe) verwandte Arten vorkommen; alle Individuen sind in den wesentlichen Merkmalen (Scutum, Bürste, Penis) einheitlich. Bereits S T A R ~ G A (1966) hielt dacica und manicata für synonym. - Die Typenserien von I. helvetica bc- stehen aus Individuen zweier Arten. Der Rolotypus muß dentipulpis heißen (9, Tessin; SMF/RI/ 1282). Die „Allotypus"-Serie ( 1 8 1 9 , nec 18 3 9 ; Tirol, Rosengarten; SMli'/RI1/229) besteht hingegen aus nzanicata. Beide Individuen tragen alle typischen Kennzeichen dieser Art. Die Fund- orts-Angabe erscheint ebenfalls nicht richtig (vgl. „VerbreitungM). - Die unzulänglichen Abbil- dungen ROEWERS (1923) veranlaßten R s ~ r o c r r v i ~ (1934) die Subspezies h. milleri aufzustellen, die von H A D ~ I (1954) eingehend diskutiert und sogar als eigene Art angesehen wurde. Die Typen- serie ( 1 8 2 $! in der Sammlung SILHAVI) umfaßt normale I. manicata ohne besondere Eigen- schaften. - Die nominellen Arten I. kaestnera (SMF/RII/9496), balcanica (SMF/R11/312) und austriaea (SMFlR111232) sind ebenfalls zu manicata zu stellen; die Typenserien weisen gegenübcr manicata keine Abweichungen auf. Die Fundorts-Angaben müssen revidiert werden (vgl. „Ver- breitung").
Diagnose: Eine Art der dentipalpis-Gruppe, gekennzeichnet durch folgende Besonderheiten: kräftige, mehr als dreizählige dorsale Cheliceren-Bewehrung (Grund-

glied, $.Y), Grundglied dorso-distal keulig verdickt, dort dorsal und teilweise medial eine ausgedehnte Biirste ($), Scutum laminatum (9).
Beschreibung. - Cheliceren (nach dem $-Holotypus von I. dacica); $- Basalglied (Abb. 46 e): dorsal mit einer Längsreihe von 5 unterschiedlich großen, gegeneinader versetzten Dornen, ventro-lateral mit 3 großen, ventro-medial mit 5 Dornen, die basalen am kleinsten; zwischen dieser sehr auffälligen Bewehrung lateral und medial kleine Höcker und tuberkelartige Erhebungen in die 3 Hauptzüge eingeschoben. - Distales Viertel dorsal keulig aufgetrieben und kaum merklich nach ventral abgewinkelt, dorso-medial in eine flache, stumpf auslaufende Apophyse aus- gezogen, auf der sich die Bürste befindet, diese läuft dorsal bis zum dorso-distalen Dorn. - 2. Glied: lediglich am Knie einige niedrige Tuberkel. - 9: die allgemeinen Merkmale wie beim $, doch ohne Burste und die dorso-distale Auftreibung, dadurch 8-Grundglied sehr schlank erscheinend.
Abb. 46. I. mariicata L. KOCH. a-1 1. bzw. 2. rechtes Cheliceren-Glied von lateral, daruber je- weils das distale Ende des 1. Gliedes von medial. - a Batrina Bilea (Rumanien, $); b ,,TatraM ($); c Zilina/Sillein (&SR, 8); d richtiger Fundort unbekannt (,,AllotypusU I. helvetica, 8); e Kronstadt (Rumänien; Holotypus I. dacaca, 6); f richtiger Fundort unbekannt (SMF/RII/229, wie d ; 9 ) ; g richtiger Fundort unbekannt (Holotypus I. kaestnerz, 9) ; h richtiger Fundort unbe- kannt (Holotypus I. austriaca, 9); i und k „Tatraa (9) ; 1 richtiger Fundort unbekannt (Holotypus I. balcanica, 9); m-n Glandes penis: rn „Tatrau; n wie d ; o Penis, wie d.

Die Abgrenzung von Biospezies usw. 219
P e d ip a lp u s ($9): Glatt und schlank, ohne Bcsonderheiten. Pedcs (Laufbeine): Relativ kurz und kräftig, Femora basal mit hellgelbem
Ringfleck. Cepha lo tho rax u n d Tube r oculorum (Abb. 47b): Ccphalothorax kräftig
gewolbt, fein granuliert und vcrcinzelt mit Borsten besetzt; 2. Thorakaltergit mit 6 kleinen Tuberkeln unterschiedlicher Große; Tuber oculorum groß, median stark gefurcht, Linse stark vorgewolbt.
Abb. 47. I. manzcata L. KOCH. a Receptaculum seminis („Ungarnc'); b Cephalotlioras von rechts-lateral (Kronstadt).
Scu t um: $ regelmaoig mit Scutum parvum, die cinzclnen Areae fein granuliert, mit ciner Querrcihc niedriger Tuberkel; Q rcgelmaßig mit Scutum laminatum (inner- halb der dentipalpzs-Gruppe nur noch bei dentipalpis selbst!); bei einem Q (als Aus- nahme) die abdominalen Sklcrits I und I1 vcrwachsen.
Pen i s (Abb. 46m-0): Truncus untersetzt, in der basalen Hälfte schwach bau- chig, unterhalb der Glans cinc schwache Erwciteruiig; Sklerit der Glans schmal lan- zettformig, mit kraftiger Beborstung; Trichter weitlumig.
Rccep t aeu lum scminis (Abb.47a):4relativ kurzc, insich gekrummte Schlauchc (1 Lcgerohrc prapariert).
Vari ab i l i t a t : AVI~AM (1964) fand bci rumanischen Tieren unterschiedliche Dornen-Zahlen der ventro-medialen ($2-9, Q 5-8) und der ventro-lateralen (2-6) Reihe des Cheliceren-Grundgliedes und des 2. Thorakaltcrgites ([2] 4-9). Uicse in- dividuellen Schwankungen lassen abcr dennoch ciiien Anlagcplan erkcnnen: Die durch wenige große Dornen betontc Dorsalreihc wird durch eiiigesehobcne Tuberkel dcr Lateralreihe aufgelockert und „unubersiehtlich". Sehr einheitlich und bczeichncnd ist die distale Erweiterung des Grundgliedes, Form und Lage der Burstc. Individuelle Krilmmungen des Truncus pcnis, die AVI~AM (1. C.) darstellt, kommen bei allen Arten vor; sie verandern die spezifischen Charaktere nicht.
Beziehungen: Inncrhalb der dentzpalpzs-Gruppe nimmt manicata einen iso- lierten Platz ein; das wird durch die eigenartige Bursten-Form, die reichliche Beweh- rung dcs Cheliceren-Grundgliedcs und das Scutum laminatum des 9 deutlich.

Verb re i t ung u n d u n t e r s u c h t e s Mater ia l : I. manicata besiedelt ein vergleichsweise großes Areal in der ~ S S R (Slowakei), in S-Polen, in der Sowjet-Union (Karpato-Ukraine) und in Rumänien; es umfaßt die Beskiden und den Karpaten- Bogen. Im heutigen Ungarn kommt manicata wahrscheinlich nicht vor, doch befindet sich im Museum Budapest ein Jungtier mit der Fundorts-Angabe „KöszegU (= Güns, W-Ungarn). Diese Angabe erscheint zweifelhaft; sie sollte bestätigt werden. Das gilt auch für die Serie mit der Angabe „bei Budapest" (SMB/RI/973, sub hellwigi). In Abb. 48 sind sie nicht berücksichtigt. - Es muß mit Nachdruck betont werden, daß manicata (nach heutiger Kenntnis) die einzige Art ist, die den Karpaten-Bogen be- siedelt. Alle anderen Angaben, die sich nachprüfen ließen, erwiesen sich als falsch (vgl. auch Abb. 25).
Es wurden 35$,50 und 20 Jungtiere untersucht. Polen. - Rogoina, Kreis Rybnik (1 9 coll. Zool. Inst. Warszawa, GUZY leg. 1884);
Bieszczady-Gebirge/= W-Teil der Waldkarpaten, Berg Tarnica, 1320-1348 m (1 9, 1 iuv. coll. Zool. Inst. Warszawa, DZIADOSZ & STAREGA leg. 9. IX. 1962); Berg Polonia Wetlihska, 1253 m

Die Abgrenzung von Biospezies usw. 221
(1 9 coll. Zool. Inst. Warszawa, RIEDEL leg. 23. V. 1962); Cisna, Kreis Lesko;Berg Rozki, 600 bis 650 m (1 iuv. coll. Zool. Inst. Warszawa, DZIADOSZ & STAREGA leg. 12. IX. 1962); ebendort (1 iuv: STAREGA leg. 1. V. 1965); Ustrzyki Gorne, Kreis U s t r z ~ k i Dolne, Wolosatka-Tal(1 9 Zool. Inst. Wazszawa, DZIADOSZ leg. 30. V. 1963); Roztoki Gorne, Kreis Lesko, Berg Jaslo, 800-1000 m (1 $2 coll. Zool. Inst. Warszawa, Exkursion PAN leg. 16. VII. 1963). - CSSR, - Slowakei, ZilinaISillein (23 , 4 9, 2 iuv. coll. SILHAVY, MILLER leg. 20. IV. 1936 - 4. XI. 1937); Mezipetoky (13,1$2 coll. SILHAVI 06227, BARTOS leg. VIII. 1938); RohaCe (3 iuv. coll. SILHAVY 06226, leg. VIII. 1951); Stub. Teplice bei %lina/Sillein (13 , 2 ? Typenserie I. helvetiea milleri, coll. S I L H A ~ Y 06222, leg. MILLER 1934); ebendort (1 iuv. coll. SILHAVI 06233, MILLER leg. 19. VI. 1935); Vrutky bei SilinalSillein (1 3 ccll. SILHAVY 06231, MILLER leg. 10. IX. 1936); Rajeckk Teplice bei Zilina/~illein (1 $2 coll. SILHAVI 06230, MILLER leg. 5. IX. 1937); Vys. Tatry (1 $2 coll. SILHAVY 06232, ~ I L H A V I leg.); Kamenica nad Cirokon/Vihorlat-Gebirge (1 $.,I 9 coll. SILHAVI 02 046, SILHA& leg. 3. VII. 1947, vgl. SILHAVY 1950); Körmöcb&nya/= Kremnitz, Skalka ( 1 3 Mus. Budapest); Lepusny völgy (3 ad. Ms. Budapest); Goldbrunni völgy (1 9, 5 3). - P o l e n 1 C S'S R. - „TatraG', ohne nähere Angaben (2 8 , 5 9 Mus. Hamburg, Berlin, Wien; SMFIR 11975, Neotypus $ SMFIR 11978). - R u m ä n i e n . - Kronstadt (63 , 4 $2, 1 iuv. Typenserie I. daciea, SMFIR I / 1283); Brasse/= Kronstadt, Ördögszikla (13 , Mus. Budapest); Umgebung von Kronstadt ( 2 6 , 2 9 SMF/R 111235); Transsylvanische Alpen (3 3 , 2 9, SMFIR II/5032), Sinaia (1 9 SMFIR 111225); Rodna-Gebirge, Gura Fintinii, Kreis Baia Mare ( 2 6 , 1 9 coll. Zool. Inst. Warszawa, DZIADOSZ leg. 30. VIII. 1965); Kreis Gheorgeni, Schlucht Cheile Bicazului bei Lacu Rosu, 900 m (1 9, coll. Zool. Inst. Warszawa, DZIADOSZ leg. 9. VII. 1965); Bezirk Maramureg, Gutiiului-Gebirge, Berg Ignig, 950-1050 m (1 9 coll. Zool. Inst. Warszawa, DZIADOSZ leg. 23. VII. 1956); Bezirk Cluj/= ,
Klausenburg, Bihar-Gebirge, Baica, am Ariegul-Fluß (1 Ex. coll. Zool. Inst. Sofia, BERON leg. 7. IX. 1963); Kelemen Lgs, Dragus ( 1 6 Mus. Budapest); Kelemen nav., Pisori Popi (1 $2 Ms. Budapest); Cojocna/= Kolosz/=Salzgrub (1 3 , 1 iuv. Mus. Budapest); Tusnadfürdö (1 $ Mus. Budapest); Torjai szanatorium (1 ? Mus. Budapest); Köresmezö (1 $2 Mus. Budapest).
Weitere Fundorte: Polen. - Cieszyn, Barania Gbra, dol. Czarnej Wiselki; Nowy Targ, Tatry, Jaskinia .Groby; Jaskinia Lodowa W Ciemniaku; Jaskinia Dziura Wyznia; Wodowiow, Babia Gbra; Ustrzyki dolne, Bieszczady, Tarnica (alle RAFALSKI 1961). - R u m ä n i e n . - Top- lita, Rastolita, Stinceni; Ciuc, Madaraz, Bäile-Tugnad (beide KOLOSVARY 1963); Bezirk Kron- stadt/=Brasov, Risnov, Moeciu; Banat, Godineeti, Regita. (beide AVRAM 1964). - So W j e t union. - Ukraine, Bogdan (KRATOCHVIL 1934, sub. I hellwigi).- Landeszugehörigkeit nicht geklärt: Batrina Bilea (1 3 , 3 Q Mus. Budapest); Madarasi Hargita (2 3 , 1 9, 4 iuv. Mus. Budapest); Ra- tosnya (1 $, 4 ? Mus. Budapest); Fongia (1 3 Mus. Budapest); Batrina Barlang (5 iuv. Mus. Buda- pest) (weitere Fundorte nennt ROEWER [1935]).
Fundorts-Angabe sicher falsch-: Ungarn, bei Budapest (1 $, SMFIR 11973); Deutschland, Raahe Alb, Waldtal bei ochsenhausen (2 Q, Typenserie I. kaestneri, SMFIR 1119496); Bulgarien, Balkan-Gebirge (1 $, Holotypus I. baleaniea, SMFIR TI/ 312); ~ u ~ h s l a w i e n , Bosnien (1 3, SMF/ R 11979); I t a l i e n , S-Tirol, Rosengarten (SMFIR 111229, Allotypus I. h. helveliea).
Vorkommen: Die spärlichen Angaben lassen darauf schließen, daß rnanicata feuchte, vor Insolation geschützte Mikrobiotope in den mittleren bis höheren Lagen der Gebirge einnimmt. Höhlen-Populationen werden vor allem aus Rumänien ge- meldet (vgl. AVRAM 1964).
A n h a n g ; Korpermaße (in mm) (Erklirung der Abkürzungen vgl. Abb. 34).
8 Cx-L (1) 1 ,s -1,9 ( 6 ) ; Cx-B (2) 2,2 - 2,6 (2,34„); Sc-L (3) 2,0 - 2,4 (2,24„); Sc-B (4) 2,2 - 2,7 (2,40„);

J„
B2 Fe 1T Pen. Ta Ce
? Cx-L L, B, Fe I1 Mt Pg
L2 L3 Mt I1 MtPg '
CX-B
JJ,
L3
Mt 11 TaGe
I. nodifera SIMON 1879 (Abb. 49-50) O
1879 Ischyropsalis nodifera SIMON, Arachnides d. Prance. 7 : 270 T. 24 Abb. 4 ( D 9, iuv. nec
$9). 1879 I . sharpi SIMON, Ann. Soc. ent. France, ( 2 ) 9 : 129 ( D iuv.). 1881 J . nodifera, - SIMON, Ann. Soc. esp. Hist. nat . 10: 128 (Synonyrnierung von sharpi
und nodifera). 1907 I . nodifera, - SIMON, Arch. xool. Expbr. G6n., ( 4 ) 6 ( 9 ) : 552 ( N ) . 1911 I..nodifera, - S r m o ~ , Arch. xool. Exper. Gen., ( 5 ) 9 ( 2 ) : 177 (N). 1913 I . petiginosa SIMON, Arch. zool. Expbr. GBn., 52: 384 (D$?) . 1914 I . nodifera, - ROEWER, Arcb. Naturgesch., 80 A ( 3 ) : 155 Abb. I 1 ( D ) . 1914 I . petiginosa, .- ROEIVER Arch. Naturgesch., 80 A (3) : 119 (U). 1923 I . nodifera, - ROEWER, Weberknechte d. Erde: G88 Abb. 860 ( D $ ? , Abb. stellt ein
Jungtier dar, nec 9). 1923 I . pecliginosa, - ROEWER, Weberknechte d. Erde: 690 (D$?) . 1936 1. nodifera, - MELLO-LEITÄO, Treb. Miis. Ciknc. Nat. Barcelona, 11: 12 (N). 193G I . pectiginosa, - MELLO-LAJTAO, Treb. Mus. Ciknc. Nat. Barcelona, 11: 12 (N). 1946 I . adanaii, - RA~CT~J,A, Aport. est. Fauna Flora vasconavar.: 56 Abb. 7 ( D 9, eindeutige
Abbildung). 1950 1. nodifera, - ROEWER, Senckenbergiana, 31 (112): 40 T. 4 Abb. 22 a-d (D$?) .
V 1950 I. naoreana R o e w m , Senckenbergiana, 3 1 (112): 3 1 T . G Abb. 33 a-d ( D $ ? , part.: nur die 3 9 sind auf nodifera zu beziehen).
v 1950 1. asturica Roswen, Senckenbergiana, 31 (112): 48 T. 8 Abb. 51 C-d ( D 9). V 1950 I. lusitanica, - ROEWER, Senckenbergiana, 31 (112): 50 T . 5 Abb. 30 a-d (part.: nur
die Serien aus Oviedo und Guipnzcoa sind auf nodifera zu beziehen). V 1950 I . janetschelci Roswisn, Senckenbergiana, 3 1 (112) 22 T . 7 Abb. 43 c-d ( D 9). v 1950 I . corsica ROEWER, Senckenbergiana, 31 (112): 34 '1'. 6 Abb. 39 C-d (D 9).
1950 I . pectiginosa, - R o ~ ~ v e i t , Senclrenbergiana, 31 (112): Abb. 48 T . 5 Abb. 29a, c-d
( D C??). V 1963 1. haspanaca ROEWER, Abh. naturwiss. Ver. Bremen, 33, ( 2 ) : 208 Abb. 4 ( D $ ) V 1953 I . pyrenaea, - R o c w ~ i t , Abh. naturwiss. Ver. Bremen, 33 ( 2 ) : 207 (N, SMF/RII /
11105). V 1963 1. lusrtnnrca, - ROEWER, Abh. naturwiss. Ver. Biemen, 33 ( 2 ) : 208 (N, SMF/RIT/
11 106). 1961 I . nodafera, - KRAUS, Senckenbergiana biol., 42 ( 4 ) . 348 (N). 1961 I . pectzgrnosa, - K ~ t ~ u s , Senckenbergiana biol., 42 (4) : 349 ( N ) . 1967 a I . nodzfera, - D ~ c s c o , Ann. Mus: Nat. u is t . Nat., ( 2 ) 39 (1): 173 Abb. 1-21 (D$?) .

Die Abgrenzung von Biospezies usw. 223
1967a I. sharpz, - DRESCO, Ann. Mus. Nat. Hist. Nat., (2) 39 (1): 174 (N). v 1967 I. 2 pyrenaea, - R ~ M B L A , Rev. Biologia, 6 (112): 12 (D 6). v 1967 I. 2 lusztanzca, - R ~ M B L L , Rev. Biologia, 6 (112). 12 (D 9).
1967 b I. petzgznosa, - DRESCO, Bull. Mus. Nat. Hist. Nat., (2) 39 (3): 506 Abb. 1-19
B e g r ü n d u n g d e r S y n o n y inie: I. nodifera und I. sharpi beziehen sich auf eine Art; das erkannte bereits SIMON (1881), und als erster revidierender Autor entschied er sich für den Xamen nodifera. - Die Beschreibung des „J" von SIMON (1879) bezieht sich auf ein 9 ; das Iäßt die Er- läuterung der Cheliceren-Bewehrung mit Sicherheit erkennen. Diese ist namlich bei den ? wesent- lich stärker als bei den 8, denen sie.bismeilen sogar völlig fehlt. Die Abbildung des ,,Q" stellt ein Jungtier dar (Scutun~ laminatum!). - Die ursprüngliche Beschreibung (ohne Abbildungen) von petiginosa reicht zur Wiedererkennung nicht aus, denn es existieren in PT-Spanien mehrere Arten, deren $-Cheliceren-Grundglied völlig oder nahezu unbewehrt ist. Erst die Zeichnungen von DRESCO (19G7b) brachten die Gewißheit, daß lediglich nodifera vorliegt und keineswegs eine valide Art neben nodiferu anzuerkennen ist. Glans penis sowie Form und Lage der Bürste von ~zodifera und petigirmsa (nach der Definition von DRESCO) stimmen völlig überein, so daß es unver- ständlich ist, daß DRESCO (19G7b) beide Arten lediglich auf Grund minimaler Unterschiede der J-Cheliceren-Bewehrung (!) aufrechterhält. Er schreibt (1967a: 175): „Dans ce cas, le $ de l'espkce [gemeint ist nodifera], dont l'article basal des chklickres ne comporte pas de grands denti- cules et en est parfois totalenlent dkpourvu, . . ." Dagegen charakterisiert DRESCO (1967 b, wie auch SIMON 1913) pefiginosa ebenfallsUdurch fehlende Bewehrung der $-Chelicere. Der Holotypus weist aber einen deutlichen dorso-distalen Dorn auf (DRESCO 1967b, Abb. 3-6; entspricht etwa der umstehenden Abb. 49e). - Die Erstbeschreibungen von ROEWER (nzoreuna, asturica, janetscheki, corsica, hispanica) müssen mit der großen Variabilität der Cheliceren-Bewehring der Spezies nodifera erklärt werden. Alle hier vorliegenden Typen-Exemplare besitzen die kennzeich- nenden Merlrmale der Art.
D i a g n o s e : Eine Art der dentzpalpzs-Gruppe, gekennzeichnet durch folgende Besonderheiten: mediale, relativ große, ellipsoide Burste ($), stumpf-kegelformige Apophyse am Grundglied dorso-distal (8) und lange, kontrastreich hell-dunkel ge- ringelte Beine ($9).
B e s c h r e i b u n g . - C h e l i c e r e n (nach dem $-Holotypus von I. hispanica); $-Basalglied (Abb. 49a): dorsal eine Reihe aus 3 kraftigen, teilweise nach vorn ge- bogenen Dornen, ventro-laterale Reihe aus 2 und die ventro-mediale aus 4 kleinen Dornen, Medialseite mit wenigen Tuberkeln, das Glied im ubrigen auffallig glatt ; das dorso-distale Ende in eine mediad geneigte, stumpfe Apophyse ausgezogen; die lang- lieh-lanzettliche Burste streng auf die Medialseite beschrankt. Sie reicht etwa von der Insertionsstelle des 2. Gliedes bis auf die Hohe des 1. dorsalen Dornes. - 2. Glied: am „Knieu mit kraftigen Hockern besetzt, medial eine Tuberkelreihe vom Knie bis zur Ansatzstelle des festen Fingers. - 9 (nach einem Ex. aus Espinaina, Picos de Europa): vom $ durch viel auffalligere Bewehrung stark abweichend; dorsal 3 kraf- tige Dornen, hinter dem 1. (von distal) ein kleiner eingeschobeii, hinter dem 3. (großen) 3 weitere sehr kleine; ventro-lateral5, die mittleren am großten, ventro-medial6 Dor- nen; Distalende des Gliedes schwach erweitert, jedoch ohne Apophyse, schwach ventrad abgebogen.. C'

Abb. 49. I. laodafera SIMOS. a-in 1. bzw. 2. rechtes Cheliceren-Glied von lateral, daruber jeweils das distale Ende des 1. Gliedes von medial. - a Sierra de Ancares (Spanien, Holotypus I. hzs- panzea, 3 ) ; b „Orense-Pontevedra" (Spanien, 3) ; c richtiger Fundort unbekannt (SNF/RII/5041, 3) ; d Serra do Geres (Portuga1,d); e Espinama, Picos de Europa (Spanien, 3); f Puente de Lozera,

Die Abgrenzung von Biospezies usw. 225
P e dip a lp u s (J?): normal, glatt, ohne morphologische Besonderheiten.
P e des (Laufbeine): relativ lang, doch sehr variierend (vgl. Maße) ; sehr auffällig und bezeichnend die Färbung: das Femur hellgelb, distal mit dunklem Ringfleck, Patella dunkelbraun, Tibia basal mit sehr schnialem, dunklem Ringfleck, anschlie- ßend eine hellgelbe und eine mittelbraune Zone, distal ein dunkelbrauner Ringfleck; Metatarsus und Tarsus gelb bis bräunlich-gelb.
Cepha lo tho rax u n d Tube r oculorum (Abb. 50e): Cephalothorax hinter dem Tuber oculorum stark gewölbt; 2. Thorakaltergit meist mit 4-6 großen Dornen; Tuber oculorum kräftig entwickelt, median tief gefurcht, Linse stark vorgewölbt.
Scutum: $9 regelmäßig mit Scutum parvum; die einzelnen Areae und die freien Tergite mit je einer Querreihe kräftiger Warzen, die mittleren am größten; bei kleinen Individuen sind nur diese vorhanden, meist als Paar.
Abb. 50. I. nodzfera SIMON. a-d Receptacula seminis: a Oviedo, Asturien; b und c Asturien (beachte unterschiedliche Zahl der Schlauche); d wie C, jedoch von lateral, um „Sekundar-Rezep- takel" zu zeigen. e Cephalothorax von rechts-lateral.
Galicien (Spanien, $) ; g wie e, 9) ; h Oviedo, Asturien (Spanien, 9) ; i wie d (9) ; k richtiger Fundort unbekannt (Holotypus I. janetscheki, 9); 1 richtiger Fundort unbekannt (Holotypus I. eorsica, 9); m richtiger Fundort unbekannt (Paratypoid I. moreana, 9); n Penis, wie U ; o-s Glandes penis: ~ w i e a ; ~ w ~ e b ; q w i e d ; r w i e e ; s w i e f .
15 Zool. Jb. Syst. Bd. 96

Pen i s (Abb. 49 11-s): Die basalen zwei Drittel des Truncus schwach erweitert, aber nicht bauchig; Sklerit der Glans breit-lanzettlich, mit großem Borstenfeld; Öff- nung des Trichters eng.
R e c e p t a c u l a m semiiiis (Abb. 50a-d): 4-5 in sich gekrümmte Schläuche; in Länge uiid Breite unterschiedlich, doch relativ kurz, distales Ende etwas aufge- bogen; bei einzelne11 Iiidividueil treten ,,Sekundär-~ezeptakel" auf (vgl. Abb. 51 d), die an der Basis der Schläuche entspringe11 uiid sich zu einem massigen, skleroti- sierten ICnoteii erweitern (4 Legeröhren präpariert).
V a r i a b i l i t ä t : Ihr unterliegen vor allem die Bewehrung und die Proportioii der Cheliceren. Beiin$ köiinen die dorsale und die beiden ventralen Reihen der Bewehrung kräftig ausgeprägt sein oder völlig fehlen; Übergänge sind vorhanden (vgl. ~ b b . 49a bis f) . Lage und Form der Bürste und der dorso-distalen Apophyse sind (bis auf ge- ringe individuelle Verschiedenheiten) einheitlich. Am stärksten bewehrt sind dieIndi- vidueii aus dem westlicheil Teil des Areals (N-Portugal, Galicieii; Abb. Q9a-b); schwach ist die Bewehrung bei den Tieren der Picos de Europa (mittleres NO-Spaiiieii; Abb. 49e; DRESCO [1967b] nennt diese Population „petiginosa"). Mittelstellung nehmen Iiidividueii aus dem östlichen Teil des Areals ein (Sierra de Aralar, W-Pore- iiäeii). - Die Bewehrung der 9-Cheliceren ist immer kräftig, in Einzelheiten jedoch individuell verschieden. Der distale dorsale Dorn ist (fast) immer am größten, nie je- doch kleiner als die caudad folgeiideii.
Be zi e h u iigen: I. nodzfera niinmt innerhalb der dentipalpis-Gruppe eine iso- lierte Stellung ein, die hauptsachlich aii der Form und an der Lage der Burste und an dem betrachtlichen Sexualdimorphismus kenntlich ist. Die Entwicklung dieser Art muß seit langem selbständig erfolgt sein.
V e r b r e i t u n g u n d u n t e r s u c h t e s Mater ia l : Das Areal erstreckt sich von den westlichsten Ausläufern der Pyrenäen (Dep. Basses-Pyreii6es) uber die Kette der Kantabrischen Gebirge bis nach N-Portugal (Serra do Geres). Zwar fehlen Funde aus Galicieii weitgehend, doch liegt das allein an mangelnder Sammeltatigkeit. - Vgl. Abb. 25 und Abb. 52.
Es wurden 158 , 12 9 und 7 Jungtiere untersucht.
Span ien . - Sierra de Anacares, Cabalia vella, Aufstieg zum Campo de Brego ( 1 8 I-lolo- typus I. hispanica SMF/R II/11108, FRANZ leg. 20. VII. 1952; vgl. R O E W E I ~ 1953); Galicien, Weg von Puente de Lozera ( 2 8 Sl'iF/R II/11107, F i t ~ ~ z leg.; vgl. ROEWER 1953); Galicien, Porto- mouro, n. Santiago, Sierra de Ancares, Pyomedos, Lozera (4 $!, 3 iuv SMF/RII/11106; vgl. ROEWER 1953); Orense-Pontevedra, Cerdedo (1 8 SB'iF/RII/11105, F I~ANZ leg. 31. VII. 1952; ~
vgl. ROEWER 1963); Asturien, Oviedo, Höhlen (1 9 Holotypus I. asturica SMF/RII/5042); Bis- caya, San Sebastian (2$ SMF/RII/5029); Bicos de Europa, Espinama und Las Ilces, 1100 bis 1350 m ( 2 8 , 19 , 1 iuv. MARTENS leg. 12.-16. IX. 1967); Siei-ra de Aralar, Lecumberri und Paß von Azpiroz (28 , 3 iuv., MARTENS leg. 23.-25. 1X. 1967). - P o r t u g a l . - Bezirk Braga, Serra do Geres, Terras de Bouro ( 1 8 , 1 9 Mus. Lissabon, leg. 10. VIII. 1945; vgl. RAMBLA 1967).

Die Abgrenzung von Biospezies usw. 227
Weitere Fundorte nennt vor allem DRESCO (1967 a, b). Da I. nodzfera in den Kantabrischen Gebirgen als allgemein verbreitet angesehen werden kann, erubrigt sich erneute Aufzahlung.
Fundorts-Angaben sicher falsch. Frankreich, Grottes de Betharram ( 4 6 SMF/RII/5041; diese Hohle ist von pyrenaea besiedelt); Griechenland, Peloponnes/= Morea, Tripolis/Tripolitza (3 9 Paratypoide I . nzoreana, SMF/RII/5026; das Ilolotypns-8 ist pyrenaea); Insel Korsika (1 P Holotypus I. corszca SMF/RI1/9495); Spanien, Prov. Huesca, Cueva de Molino (SIPION 1907).
Vorkommen: I. nodtfera lebt in Wäldern mit ausreichend feuchtem Untergrund vor allem an quelligei~ Stellen im Detritus und unter altem Holz. Ein Fundort in den Picos dc Enropa (Espinama, 1350 m ; IX. 1967) an einem Gerinne in Bucheiiwald an sudosi-exponierter Stelle war offensichtlich weithin isoliert, da der trockene Wald- boden (Kalkuntergrund) sonst keine Siedluiigsmoglichkeit bot; an dieser kleinen Plache dichte Vegetation von Rubus, Rosa, Juncz~s, Carex, Gramineen, Moos. - Im ganze11 Areal werden vielfach Hohlen besiedelt; manche Populationen durften heute vollig von der Außenwelt abgeschnitteii sein, da die unmittelbare Umgebung des Hohleneinganges kcine gunstigeii Biotope bietet, z. B. die Cueva Cullalvera bei Ra- males de la Victoria.
A n h a n g ; Korperinaße (in inm) (Erlclarung der Ablrnrzungen vgl. Abb. 34):
0' Cs-L (1) 1,65- 2,2 ( 1 8 4 ) ; Cx-B (2) 2,5 - 2,9 (2,65„); SC-L (3) 2,4 - 3,O (2,58„); Sc-B (4) 2,3 - 2,9 (2,76„); LI (5) 3,3- 4,3 (3,90„); (6) 4,55- 5,6 (5,1812); B2 (7) 0,9 - 1 ,1 (0,94„); L3 (8) L 7 - 2,5 (1,gGl2); Fe11 (9) 5,3 -11,4 (8,191,); i?ItII (10) 6,O -14,4 (9,75„); Pen. (11) 3,3 - 4,2 (3,85,); MtPg (12) 2 - 3 TaGe (13) 35 -43.
? C L (\) 1,9 - 2,4 (2,08,); Cx-ß 12) 2,8 - 3,9 (3,25,): SC-L (3) 2,7 - 3,7 (3,08,); ' Sc ß (4) 2,9 - 4 , l (3,21a); LI (5) 3,8 - 5,45 (4,4G,); L, (Gj 5,3 - 7,5 (6,02,); ß, (7) O b - 1 2 (1,128); L, (8) 2,1 - 2,6 (2,358); Fe11 (9) 5,2 -12,G (7,27,); Mt11 (10) 6,3 -15,s (8,86,); MtPg (12) 2 - 3 TaGe (13) 35 -40.
I . pyrenaea SIMON 1872 (Abb. 51-53)
1872 Ischaropsalzs Helzuzgz var. Pyrenaea SIMON, Ann. Soc. ent. Fiance (5), 2: 483 T. 16 Abb. 4
(D 4). 1875 I . pyrenaea, - BCDEL & S I ~ ~ O N , Journ. Zool., 4: 366. 1879 I. pyrenaea, - SIMON, Arachnaicles d. France, 7. 272 T. 21 Abb. 13-14 T. 24 Abb. 1 ,
1 a (D G?). 1910 I . pyrenaea, - SIMON, Arch. Zool. Exper. Gen., 45: 65 (N). 1911 I . pyrenaea, - SIMON, Arch. Zool. EspBr. Gen., 49: 203 (N). 1913 I . pyrenaea, - SIMON, Aicli. Zool. Expor. Gen., 52: 383 (N). 1914 1. pyrenaea, - R o ~ w c n , Arch. Naturgesch., 80 A (3): 114 Abb. 10 a-d (D 6 9 ) . 1923 I . pyrenaea, - ROEWER, Weberlznechte d. Erde: 688 Abb. 859 a-d (DS?) . 1926 I . pyrenaea, - JEANNEL, Faune cavernicole de la France: 159 (N). 1935 I . pyrenaea, - ROEWER, Arch. 2001. Expei. Gen., 78 (1): 83 (N) .
15*

1935 I. nodifera, - ROEWER, Arch. Zool. Expbr. Gbn., 78 (1): 82 (Nennung nach Material vom Locus typicus).
v 1950 I. pyrenaea, - ROEWER, Senckenbergiana, 31 (112): 40 T. 3 Abb. 17 a-d (Dd? ; part.: die Serien „Spanien, San Sebastian, Tolosa" sind hochstwahrscheiiilich, jene aus der Grotte von Bbtharram /SMF/RII/5041/ mit Sicherheit auf nodzfera zu beziehen; Fundort- verwechselung!).
V 1950 I. moreana ROEWER, Senckenbergiana, 31 (112): 31 T. 6 Abb. 33 a-d (D$.?, part.: nur das 8 und Abb. a-b sind auf pyrenaea zu beziehen).
1961 I. pyrenaea, - JUBERTHIE, Ann. SpBIbol., 16 (4): 387 Abb. 5, 8 (Dd?) . 1961 I. pyrenaea, - KRAUS, Senckenbergiana biol., 42 (4): 348 (N).
V 1967c I. pyrenaea, - DRESCO, Bull. Mus. Nat. Hist. Nat. (2), 39 (4): 665 Abb. 1-13,18-26a
(D 8 9 ) . 1967c I. pyrenaea venasquensis DRESCO, Bull. Mus. Nat. Hist. Nat. (2), 39(4): 670Abb. 14-17,
26 b (D 8) .
B e g r ü n d u n g d e r S y n o n y m i e : S I ~ I O N (1872) beschrieb das Material aus der Höhle von Estelas (Arikge) als Varietät von ,,hellwigi" (L. KOCH hatte irrigerweise unter diesem Namen Individuen von I. muellneri an SIMON gesandt; vgl. SIMON 1872 Abb. 4). Die 9-Cheliceren von muellneri und pyrenaea sind recht ähnlich, so daß die ~r t -Birecht igung von pyrenaea zunächst fraglich erscheinen mußte. Später wurde diese große und gut kenntliche Art immer einheitlich interpretiert, nur ROEWER stellte für 18 den Namen moreana zur Verfügung.Dieses8 besitzt alle wesentlichen pyrenaea-Merkmale. (Die Paratypoid-? gehören zu nodifera). - D R E S ~ O (1967 C) be- schreibt nach einem 8 (Freiland-Fund) die Subspezies venasquensis. Dazu ist zu bemerken: Der Fundort von „venasquensis" liegt in1 Areal der Nominatform. Da aber per definiti-onem Subspezies geographisch vikariieren, kann diesem Individuum nicht der Rang einer Subspezies zukommen. Die Trennmerkmale an der dorso-distalen Apophyse des Grundgliedes liegen im Bereich der indi- viduellen Variabilität; die Unterschiede sind geringer als jene zwischen den Abb. 51s und b (fide DRESCO 1967 C). Ich betrachte „venasquensis" als Synonynl der Nominatform.
D i ag n o s e: Eine Art der dentipalpis-Gruppe, gekennzeichnet 'durch folgende Be- sonderheiten: schlanke, nahezu glatte ($) bzw. vor allem an der basalen Hälfte schwach bewehrte (9) Cheliceren-Grundglieder, eine kräftige dorso-distale Apophyse und eine punktförmige medio-distale Bürste ($).
B es ehr eibung. - Cheliceren (nach topotypischem Material) ; 8-Basalglied (Abb. 51 a-b): schlank, kurz über der besonders dunnen Basis etwas erweitert, von dort bis zur distalen Apophyse gleichbleibend; alle 3 Hauptreihen der Bewehrung nur durch winzige Tuberkel angedeutet, lateral selbst diese nahezu fehlend; dorso- distale Apophyse stumpf kegelformig; Bürste an der distalen Basis der Apophyse; punktförmig klein. - 2. Glied: schlank, besonders frontal und medial am „Knie" mit kleinen Tuberkeln. - 9: noch schlanker als das des $, da ohne distale Apophyse, Distalende merklich nach ventral abgewinkelt und sehr schwach erweitert; spärlich, vor allem in der basalen Hälfte, mit sehr schlanken Dornen besetzt, von denen die basalen am größten sind; dorsal 5-7 (oft bei einem Individuum rechts und links verschieden); ventro-lateral, -medial je 6-8 Dornen unterschiedlicher Größe, basal konzentriert.

Die Abgrenzung von Biospezies usw.
Ped ipa lp i i s : schlank, Patella des 6 distal schwach erweitert. Pedes (Laufbeine): sehr lang und schlank, Basen der Pemora hellgelb, alle übri-
gen Glieder dunkelbraun, doch Metatarsus und Tarsus abgesetzt gelblich. Cepha lo thorax u n d T u b e r oculorum (Abb. 53b): Cephalothorax niedrig,
sehr fein granuliert; 2. Thorakaltergit mit 13-15 relativ kleinen Dornen unterschied-
Abb. 51. I. pyrenaea SIMON. - a-f 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber je- weils das distale Ende des 1. Gliedes von medial. - a Grotte de l'Estelas, Ariege (Frankreich; Locus typicus, J ) ; b richtiger Fundort unbekannt (Bolotypus I. moreana, 8); c Buco di Valenna, Piemonte (Italien, J); d wie a (P); e Grotte de Saleche, Aribge (Frankreich, 9); f wie C ( 9 ) ; g-i Glandes penis: g wie C ; h wie a ; i wie b ; k-1 Penis: k wie b; 1 wie C.

licher Großc, die mittleren an1 großten; ohne Tuber oculorum im eigentlichen Sinne: die fur eine Art der denltpulpts-Gruppe ungewohnlich kleinen Augen weit voncin- andcr (ctwa 4-5fachcr 0 der Linse) und durch eiiie breitc und tiefe Furche getrennt.
S c u t ~ ~ r n : $? regelmaßig mit Scutum parvum (DRESCO 1967c belcgt das fur 418, 409). Drc~sco (1967~) erwahnt fur ein$ „lcs tergitcs dc l'abdorneii s6parCs", also cin Scutuiri laminatum (vidi!). Auch das aus den Cottischen Alpen tragt Scutum 1;~ini- iiatum. URESCO (1. C.) bildet uberdics cin 9 ab, dessen abdominales Tergit V1 tcil- weise mit dcm Sciittrm parvum vcrschmolzeii ist.
Pen i s : Basale Halfte des Truncus schwach bauchig, Truncus kurz uilterhalb der Glans am schmalsteri, von dort zur Glaiis leicht verbrcitcrt (Abb. 51k-1); Sklerit der Glans (Abb. 51h-i) breit la11zenspitzei1-formig, lateral biswcileii' kraftig ausge- schwu~igcii; Öffiinng dcs Trichters maßig weit, Partie unterhalb der Glaiis hatitig- niciribraiios; 1 $ aus den Cottischcn Alpcil jedoch mit relativ kleinem und ciforinigem Sklcrit uiid sehr engcr Trichteroffnaiig (Abb. 51 g).
Recep t acu lum semin is (Abb. 53a): mit 4 langen und schlanken Schlauchen (1 Legerohrc prapariert).
Va r i a b i l i t a t : Bewehrung des $- und 9-Grundglicdes schwankend, beim $ bis- wcileii bis auf winzige Itndimeiite reduziert oder aber dcutlich ausgepragt (vgl. Abb.), bcim 9 oft Bewehrui~g der rcchtcii und linkeil Cheliccre verschieden, Anlageplan der Bedoriiuiig jcdoch einheitlich und charakteristisch, namlich beim $ nur wenigc Tu- berkel, beim ? schlanke Uoriicn vor allem ventral und dorso-basal; Konstaiiz dcr Form der Apophyse und der Ausdchnu~ig und Lage der Burstc. Die beidcii einzigen Individucn aus dcn Cottischcn Alpen sind weseiitlich kleiner als Tiere aus den Pyrc- naeii (vgl. Abb. 51). *
Abb. 52. Die Fundpunkte von I .pyrenaea (West-Areal; irmiaiidet, jeweils mehrere Fundorte zu- sammengefallt), I. daspar (gioßer Punkt mlt Pfeil), I. rnagdalenae (kleine L'irnkte mit Pfeil) und I. nodafera (ohne Hervorhebung).

Die Abgrenzung von Biospezies usw. 231
Bez iehungen: Im westlichen Areal-Teil von pyrenaea (bzw. anschließend in den Kantabrischen Gebirgen) siedeln 4 weitere, habituell sehr ähnliche Arten (Cheli- Ceren, Körpergröße), deren Beziehungen zu pyrenaea teilweise schwierig zu erschließen sind. Von diesen gehören dispar und magdalenae in die kollari-Gruppe (Sklerit der Glans zweilappig), also trotz äußerer Ähnlichkeit mit pyrenaea in. einen ganz anderen Verwaiidtschaftsbereich. Nicht sehr verwandt ist das mit pyrenaea vikariierende nidifera (vgl. Abb. 25 und Abb. 52), denn Lage, Formund Ausdehilung der Bürste sind sehr unterschiedlich. Das sehr viel kleinere, zunächst sehr unähiilich erscheinende, mit pyrenaea sympatrisch vorkommende luteipes weist aber auffä'llige Gemeinsam- keiten mit dieser Art auf. Apophyse und Bürste der &-Chelicere sind äußerst ähnlich, woraus auf zumindest weitläufige Beziehungen geschlossen werden darf. Die unter- schiedliche Cheliceren-Bewehrung spricht nicht dagegen. In der dentipalpis-Gruppe wird sie nämlich bei fortschreitender Verlängerung der Chelicereii regelmäßig redu- . ziert (vgl. die kurz-gedrungenen Cheliceren von luteipes, Abb. 44, mit den langen und schlanken von pyrenaea, Abb. 51; s. a. S. 215, „Beziehungenu). - Es besteht die Möglichkeit, daß der erst kürzlich entdeckten, durch geringe Größe stark abweichen- den westalpinei~ Population der Rang einer eigenen Subspezies oder gar Spezies zu- gebilligt werden muß. Die Entscheidung muß zurückgestellt werden, da bisher erst zwei adulte Individuell vorliegen.
Ve rb re i t ung u n d u n t e r s u c h t e s Mater ia l : I. pyrenaea nimmt zwei kleine, reliktär wirkende Areale ein: Teile der französischen Pyrenäen und der (italienischen) Cottischen Alpen. Fundorte in den Pyrenäen sind aus den Departements Ariege, Haute-Garoilne, Hautes-Pyr6ii6es und Basses-Pyr6n6es bekannt (fast ausschließlich in Höhlen) (Abb. 52). Von der spanischen Pyreiiäen-Seite sind (höchstwahrscheinlich nur zufällig) noch keine Nachweise bekannt. Meldungen aus anderen Gegenden Spa- niens, vor allem aus den Kantabrischen Gebirgen (vgl. ROEWER 1935), beruhen sämt- lich auf Verwechselungeii mit habituell ähnlichen Arten und haben sich in keinem Falle bestätigen lassen - vgl. auch DRESCO 1967 c und Abb. 25.
Abb. 53. I . pyrenaea SIMON. - a Receptaculum seminis; b Cephalothorax von rechts-lateral(beide vom Locus typicus).

Es wurden 5 8 , 8 $? und 7 Jungtiere untersucht. F r a n k r e i c h . - Ariege, Grotte de 1'Estelas bei Cazavet; Locus typicus ( 1 6 Zool. Inst.
Warszawa PAN, RIEDEL leg. 22. 11. 1964; 2 8 , 6 9, 2 iuv. MARTENS & PAULUS leg. I . IX. 1967); Haute-Garonne, Grotte de Saleich/Espugne (1 9 MATJASIC leg. 1955); Hautes-PyrBnBes, Grottes de BBtharram (3 ~ ~ ~ . . M A R T E N S & PAULUS leg. 4. IX. 1967). - I t a l i e n . - Cottische Alpen, Piemonte, Crissolo, Buco di Valenza ( 1 3 coll. BRIGNOLI, ACTIS leg. 7. V. 1959; 1Qcoll. BRIGNOLI, VIGNA leg. 24. VIII. 1967); Crissolo, Grotta Rio Martino (2 iuv. MORSI leg. 16. 111. 1969).
W e i t e r e F u n d o r t e : F r a n k r e i c h . - Ariege: Saint-Girons, Grotte d'Aubert (JEANNEL 1926); Cazavet, Grotte Foulquier; Salau, Aven unterhalb des Co1 du Salau; Cazavet, Gouffre de 1'Haiou; Galey, Puits de Pruhaut; Saint-Julien-du-Castillonnais, Puits de la Mate; Buzan, Grotte de la Cabane des Gardes (alle DRESCO 1 9 6 7 ~ ) ; Moulis, Grotte de LiquB (SIMON 1910); - Haute-Garonne: Milhas, Grotte de Paloumere; Milhas, Puits de Burtetch; Herran, Puits de las Templas; Portet d'Aspet, Grotte de Ruisec; Francazal, Grotte de Pas du Loup; Luchon, Port de VBnasque (DRESCO 1 9 6 7 ~ ) ; - Basses-PyrBnBes: Les Eaux-Chaudes, Grotte des Eaux-Chaudes (SIMON 1911); Arudy, Grotte de Malarode (ROEWER 1935); - Hautes-PyrBnBes: Rieulhes, Grotte de la Palle; Saint-PB-de-Bigorre, Grotte de la Bouhadere; Aragnout, Grotte des Cabanes d'Arti- gusses; Loudanvielle, Vallon dlAygues Tortes (alle DRESCO 1 9 6 7 ~ ) . - Fundorts-Angabe sicher falsch: Griechenland, Morea (= Peloponnes), Tripolis/=Tripolitza ( 1 3 Holotypus I. moreana,
SMF/RII/5026; die Paratypoid-99 sind auf nod%fera zu beziehen).
Vorkommen: I. pyrenueu scheint eine rein troglobionte Art zu sein, obwohl sie keine ausgeprägten Höhlentier-Merkmale besitzt. Viele (alle?) der Populationen sind von der Außenwelt (mindestens während des größten Teiles des Jahres) abgeschlossen, da außerhalb der Höhlen vielfach ungünstige ökologische Bedingungen herrschen. Das gilt z. B. für die Höhle von Estelas, dem Locus typicus. - Neuerdings meldet DRESCO (1967 C) zwei Preilandfunde, der eine in 2400 m. Wahrscheinlich kommen also in hohen Lagen der Pyrenäen auch (individuenarme) freilebende Populationen vor, die bisher übersehen worden sind. Es dürften ähnliche Verhältnisse wie bei luteipes herrschen (vgl. Kap. I11 3 und I11 6).
A n h a n g ; Korpermaße (in mm) (Erklarung der Meßstrecken vgl. Abb. 34); Individuen aus den Pyrenaen:
d Cx-L (1) 1,7 - 1,9 (1,854); Cx-B (2) 2,5 - 2,7 (2,65,); Sc-L (3) 2,6 - 3,2 (2,92,); SC-B (4) 2,7 - 3,2 (2,9,); LI (5) 4,7 - 5 3 (4,95,); L, (6) 6 4 - 7,1 (666,); B, (7) 037- 1 ,0 (0,92,); L, (8) 2,3 - 2,5 (2,36,); Fe11 (9) 8,2 - 9,O (8,6,); Mt11 (10) 0 4 - 1 (10,66,); Pen. (11) 4,3 - 4,75 (4,51,); MtPg (12) 4 ; TaGe (13) 36-42.
? Cx-L (1) 1,9 - 2 , l ( 9 ) Cx-B (2) 2,9 - 3,3 (3,163); SC-L (3) 2,9 - 3,3 (3,133); SC-B (4) 3,O 3,6 (3,36,); LI (5) 4,3 5,9 ( 4 1 4 ) L, (6) 6,1 - 6,7 (6,624); B, (7) I,O7-1,2 ( 1 1 , ) ; L, (8) 2,4 - 2,6 (2,5,); Fe11 (9) 8,9 - 9,8 (9,2,); Mt11 (10) 9,6 -13,3 (11,13,); MtPg (12) 3 - 5 ; TaGe (13) 41.

Die Abgrenzung von Biospezies usw. ' 233
I. robusta SIMON 1872 (Abb. 54-55) 1872 Ischiropsalis robusta SIMOK, Ann. Soc. ent. France (6), 2: 230 T. 12 Abb. 13 ( D d , nec 9). 1879 I. robusta, - SINOX, Arachnides d. France. 7 : 276 (N). 1914 I . robusta, - ROEWER, Arch. Naturgesch., 80 A (3): 118 Abb. 13 (D$., nec 9). 1923 I . robusta, - ROEWER; Weberknechte d. Erde: 690, Abb. 862 (D$.,nec 9).
,,
v 1923 I . lusitanica ROEWER, Weberknechte d. Erde: 692 Abb. 866 (D 9 , nec 3). 1936 I. robusta, - MELLO-LEITÄO, Treb. Mus. Cienc. Nat. Barcelona, 11 (9) (ser. ent.): 12
(part.: nur Gerez). 1950 I. robusta, - ROETVER, Senckenbergiana, 31 (112): 46 T. 6 Abb'. 3 c-d (D$., nec 9).
v 1960 I . lusitanica, - ROEWER, Senckenbergiana, 31 (112): 60 T. 6 Abb. 30 a-d ( D 9, nec $; D 9 & Serien aus Asturien sind auf andere Arten zu beziehen)..
v 1961 I. lusitanica, - KRAUS, Senckenbergiana biol., 42 (4): 349 Abb. 21-24 (D$.). 1966s I. robusta, - DRESCO, Bull. Mus. Nat. Hist. Nat. (2), 38 (5): 592 Abb. 3-20 (D$.?). 1967 I . robusta, - RAMBLA, Rev. Biol., 6 (112): 11 (N).
'
V 1967 I. luteipes, - RAXBLA, Rev. Biol., 6 (112): 11 (N).
B e g r ü n d u n g d e r S y n o n y m i e : Das Typus-? (nec 8, wie KRAUS [I9611 bereits verbes- serte) von lusitanica ist synonym robusta. Abweichungen gegenüber robusta-9, zu denen $. vor- liegen, bestehen nicht. I . nodifera, die ebenfalls in Portugal vorkommt, ist auch im ?-Geschlecht nicht mit robusta (und somit lusitanica) zu verwechseln. - Da KRAUS kein Material von robusta zur Verfügung hatte, mußte er das vorliegende 3 zwangsläufig für lusitanica halten. Irrig ist seine Synonymierung von adamii (sensu RANRLA 1946) mit lusitanicu (= robusta), denn RBD~BLA hielt nodifera-9 (aus der Sierra de Aralar) für adamii. Diese Art ist jedoch auf Halbinsel-Italien be- schränkt. RAMBLA (1967) ordnet portugiesische robusta-? der Art luteipes zu, die in Portugal nicht vorkommt. Dadurch wird erneut unterstrichen, daß die artspezifischen Merkmale (der iberischen Arten) bisher nicht erarbeitet worden waren.
Diagnose: Eine Art der dentipalpis-Gruppe, gekennzeichnet durch folgende Besonderheiten: eine relativ große, mediale Bürste ($), einen relativ und absolut sehr großen Tuber oculorum ($9).
Beschre ibung . - Cheliceren (nach SMP 11690); d-Basalglied (Abb. 54b): kurz und gedrungen, dorsal mit 3 mittelgroßen, nach vorn geneigten Dornen, ventro- lateral 3 und ventro-medial 5 kräftige Dornen; lateral und medial wenige kleine Tu- berkel; Apophyse nur schwach entwickelt, Distalende des Grundgliedes dorsal auf- gebläht, über der Insertionsstelle des 2. Gliedes stumpf ausgezogen. Bürste relativ groß, eiförmig, medial (!) auf der distalen Apophyse. - 2. Glied: schlank, am „Knieu nahezu unbewehrt. - $2: entspricht in den wesentlichen Merkmalen dem $, doch ohne distale Aufblähung und Burste.
P e d i p a l p u s ($9): ohne Besonderheiten. 6
Pedes (Laufbeine): kurz, Bemora basal hell geringelt, übrige Beinglieder (bis auf die Tarsen und Metatarsen) dunkelbraun bis schwarz.
Cepha lo tho rax u n d T u b e r oculorum(Allb. 55): Cephalothoraxkräftigge- wölbt und stark granuliert; 2. Thorakal-Tergit mit 2-4 Tuberkeln (DRESCO [1966a] zeichnet bis 8); Tuber oculorum sehr groß, innerhalb der ganzen Gattung bei robusta am mächstigsten entwickelt; median gefurcht, die beiden Augen jedoch nicht isoliert.

S c u t u m : 89 regelmäßig mit Scutuin parvum; die fein granulierten Areae uiid die freieii Tergite mit je einer Querreihe niedriger Tuberkel, auf den beiden erstell Areae bisweilen aber fehleiid.
P e n i s : Truiicus auch distal relativ breit, basale Hälfte erweitert, aber nicht bauchig; basaler Teil des Sklerits der Glans sehr schmal lanzettlich, distale Hälfte breit gerundet; Trichter mit weiter Öffnuiig.
R e c e p t a c u l u m semii i i s : Schläuche relativ kurz und breit, weitlumig; die eiiie präparierte Legeröhre einerseits mit 5, aiidererseits mit 6 Schläucheii.
V a r i a b i l i t ä t : Da,s geringe Material erla~ibt keine Aussageil; bezüglich der Cheli- cereii vgl. Abb. 54.
Abb. 54. I. robusta SIXOY. - a-e 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber je- weils das distale Ende des I. Gliedes von niedial'(Bürste). - U Joane (Portugal, 6); b Sierra de Estrela (Portugal,$); c Coimbra (Portugal, Holotypus I. lus i tanica , 9); d Quinta da Igreja (Portu- gal, 9); e wie a (P) ; f Penis, wie b ; g Glans penis, wie b.

Die Abgrenznng w n Biospezies usw. 235
Bez iehungen: Die rneistcn Übcreinstimmungen mit robusta weist die osteuro- paische Art mantcata auf. Auch bci ihr ist die distale Apophysc dcs Grundgliedes kculig aufgeblaht, und die Burstcn beidcr Arten sind ähnlich. Bei manicala ist sie zwar ausgedehnter, da sic sich auch auC die Dorsalscite erstrcclit, doch entsprechen sich Lage und Forni dcs mcdialcii Teiles. Auch dic Forrn des Sklerits der Glans ist verglc~chbar. Offeiibar haben beide Artcn viclc Merkmale des gcmcinsamcn Vorfahrcn bewahrt. Nahc Verwandtschaft beider Arten nahm bercits SIMON (1872) aii; cr Cr-. laiiterte jedoch nicht, woraus er sie ersah.
Abb. 55. I. robustu S~n~oiv . - U Recept~~culuiii seiniiiis (Holotypus I . lusttan?ca); b Cephalothor ,~~ von rechts-lateial (Sierra clc I3str8ln)
V c r b r e i t u n g u n d u n t c r s u c h t c s Ma te r i a l : I. robustu nimmt (nach den bisherigen noch sehr IuckenhaFten Kenntnissen) ein relativ lilcincs Arcal inPortugal ein; alle lcundortc licgen in dcr nordlichen Landeshalfte, z. T. in den Greiizgcbirgen zu Galicicn, so daß Funde in Spanicn crwartct werden durfen; vgl. Puiiktkarte Abb. 41 uiid Abb. 25.
Es wurden 2 8 und 3 Q uiitcrsucht.
P o l t u g a l - Coimbra ( 1 Y Holotypus I. lusztunaca, SMF/K1/226); Bezirk Guaida, Sierra de Esttella, 1000 nl ( 1 8 SMF 11690, F I ~ A N ~ , leg. 22. Vl1. 1965, vgl. I<Ru~\s 1961); ßeziik ßraga, Joane, Vila Nova de Parn~l icdo ( 1 6 , 1 $' Mus. Lissabon, vgl. RAMULA 1967), Bezirk Vila Real. Sedielos, lt4gna (1 Mus. Lissnbon, vgl. R a w n r , ~ 1967).
Weitere Fiindoite ßeziik Glldrdrli, Sieirt~ de Estiella, Seia (RLMBLA 1967), Tr,~s-os-Montes, Serta (le Geles, Locils typicus (SIMON 1872); U e ~ i r k Beird, Brussnco, Bezirk Minho, zwischen Porto und Sa Joana de Fez (beide D i i ~ s c o 1966).
Vorkomm cn: Die Typcnserie wurde in einer I-Iohlc gcsammclt („peu pro- foiidc, mais ii6anmoiiis ohscure"); weitere Mitteilungcii zur Ökologic bestehen nicht.
A n h a n g ; Korpermaße (in min) (Erltlarung der Meßstreckcn vgl. Abb. 34):
8 Cx-1, (I) 1,G - 1,8 (1,7,); Cx-B (2) 2 , l - 2,5 (2,3,); Sc-L (3) 2,3 - 2,4 (2,35,); SC-ß (4) 2,4 - 2,7 (2,55,); LI (5) 2,9 - 3 , l (3,0,); L2 (6) 3,9 - 4 5 (4,2,); B2 (7) 0,75- 0,95 (0,86,); L, (8) L 4 - 1,5 (l,52);

' FeI I (9) " Pen. (11)
? Cx-L ( I ) Sc-L (3)
L, (5) B2 . (7) Fe I I (9) MtPg (12)
(3,921; Mt I1 (2,551) ; Mt Pg ( , 833) ; Cx-B (2L.33); Sc-B (3,383); L2
(0,9631; L3 ( G ) ; Mt I1
Ta Ge
490 - 4 2 (4,121; 3 ; TaGe (13) 26-27. 2,s - 3 , l (2,96,); 2,9 - 3,3 (3,131; 4,55-5,l (4,88,); 1,8 (1,831; 4,4 - 4,8 (433,);
26 -33.
Die ko l la r i -Gruppe
I. kollari C. L. KOCH 1839 (Abb. 56-58) 1839 I. kollari C. L. KOCH, Übersicht d. Arachnidens., 2: 24 (D). 1841 I. kollari, - C. L. KOCH, in HAIIN, Die Arachniden, 8: 19 T. 258 A6b. 604 (D 9).
,1865 I. kollari, - L.. KOCH, Z. Ferdinan. Tirol, Vorarlb., (3) 12: 164 (D). 1879 I. Kollari, - SIMON, Arachnides d. France, 7: 276 (N). 1879 I. kollari, - THORELL, Ann. Mus. civ. Stor. natur., 8: 452 (D generis; Bestimmung von
I . kollari als Generotypus). 1914 I. bosnica ROEWER, Arch. Naturgesch., 80 A: 107 Abb. 3 a-d (D$?).
v 1923 I. bosnica, - R o E w ~ n , Weberknechte d. ~ r d e , 684 Abb. 852 (D.$?). v 1931 I. (Odorztopalpa) triglavensis H A D ~ I , Prirod. Razprave, 1: 107 Abb. 4-7 (D$?). v 1933 I. helwigii, - S C H ~ L L E R , Mitt. Höhlen. Karstforsch., 1933: 44 (N).
1942 I. triglavensis, - I-IAD~I, Razprave.Akad. Ljubljana, 2: 9 Abb. 2 C-d (D). v 1950 I. danubia ROEWER, Senckenbergiana, 31 (112): 21 T. 9 Abb. 55 C-d (D 9). V 1950 I. strasseri ROEWER, Senckenbergiana, 31 (112): 22 T. 8 Abb. 52 C-d (D iuv., nec 9). V 1950 I. tirolensis ROEWER, Senckenbergiana, 31 (112): 23 T. 6 Abb. 32 a-b (D 8). V 1950 1. styriaca ROEWER, Senckenbergiana, 31 (112): 23 T. 9 Abb. 60 C-d (D 9). v 1950 I. reirnoseri ROEWER, Senckenbergiana, 31 (112): 23 T. 1 Abb. 9 a-d (D $9). v 1950 I. triglavensis, - ROEWER, Senckenbergiana, 31 (112): 24 T. 8 Abb. 2 b-d.
1950 L. spinichelis ROEWER, Senckenbergiana, 31: 25 T. 10 Abb. 65 C-d (D P). 1950 I. nivalis ROEWER, Senckenbergiana, 31: 26 T. 10 Abb. 61 C-d (D 9). 1950 I. scutata ROEWER, Senckenbergiana, 31: 26 T. 1 0 Abb. 63 C-d (D$, nec 9).
v 1950 I. segregata ROEWER, Senckenbergiana, 31: 28 T. 10 Abb. 64 C-d (D iuv., nec 9). v 1950 I. bosnica, - ROEWER, Senckenbergiana, 31: 29 T. 1 Abb. 7 a-d (D$?). V 1950 I. troglodytes ROEWER, Senckenbergiana, 31: 32 T. 8 Abb. 50 a-d (D iuv., nec $9). v 1952 I. helwigi, - VORNATSCHER, Höhle, 3: 12 (Lurgrotte; N). v 1954 I. (Odontopalpa) triglavensis, - HADEI, Razprave, 2: 143 Abb. 1-4 (D $9).
1954 I. nivalis, - H A D ~ I , Razprave, 2: 150 Abb. 5 (D V 1954 I. danubia, - H A D ~ I , Razprave, 2: 152 Abb. 6 (D 9).
1954 I. scutata, - H A D ~ I , Razprave, 2: 156 Abb. 8 (D$). 1954 I. sp. aff. janetscheki, - I ~ A D ~ I , Razprave, 2: 174 Abb. 14 (D 8). 1954 I. rnanicata, - HADEI, Razprave, 2: 176 Abb. 15 (D$). 1954 I. sp. aff. tirolensis, - H>\D?I, Razprave, 2: 179 Abb. 16 (D iuv.).
V 1963 I. cf. danubia, - THALER, Ber. naturwiss.-med. Ver. Innsbruck, 53: 278 (N).
B e g r ü n d u n g d e r S y n o n y m i e : I. kollari wurde nach Material aus der Gegend von Gastein (Land Salzburg, Österreich) beschrieben, in der Originalmitteilung jedoch nicht abge- bildet. Wenig spater bringt derselbe Autor (C. L. KOCH 1841) unter dem Namen kollari eine Farb- abbildung eines ?, das ein Scutum laminatum trägt. Nach heutiger recht genauer Kenntnis

Die Abgrenzung von Biospezies usw. 237
kommen in den Hohen Sauern (und damit in der Umgebung von Gastein) zwei Isehyropsalis- Arten vor, vo,n denen die eine, hellwigi, in beiden Geschlechtern Scutumgarvum besitzt. Diese ' bildet C. L. KOCH zusammen mit kollari ab. Die 9 der anderen Art, kolla:i, tragen immer ein Scutum laminatum. Somit ist sicher (was auch aus anderen Details der Abbildung zu ersehen ist), auf welche Art der Name kollari zu beziehen ist. Überdies befindet sich in der Sammlung ROEWER eine Serie von I. kollari (SMF/RI/974; 1 3 , 1 iuv., nec 2 iuv.), die mit ,,Typus, REIMOSER ded." bezeichnet ist. Diese völlig ausgeblichenen Tiere können zumindest als Syntypen angesehen werden (vgl. Abb. 56a). SIMON (1879) und THORELL (1879) benutzten den Namen kollari zuletzt; ROEWER hielt kollari und hellwigi für Synonyme und erwähnte die Art nur noch in den Listen. Obwohl dieser älteste verfügbare Name länger als 50 Jahre unbenutzt geblieben ist, bestehen dennoch keine Bedenken, ihn zu benutzen: Bei keiner der späteren Benennungen kann davon &us- gegangen werden (vgl. Synonymie-Liste), daß diese in „allgemeinen Gebrauch" gekommen wären. Sonlit besteht keine Veranlassung, lcollari etwa gemäß I R Z N Art. 23b als Nomen oblitum zu eerten. Durch die Wiederverwendung von kollari wird kein schutzbedürftiger späterer Name be- eiiiträchtigt.
Neben den vielen neuen nominellen Arten von ROEWER ist auch triglavensis H A D ~ I auf kollari zu beziehen. Der I-Iolotypus weist gegenüber topotypischen kollari einige Unterschiede auf, die aber die Abtrennung einer eigenen Art keineswegs rechtfertigen.
Diagnose: Eine Art der kollari-Gruppe, gekennzeichnet durch folgende Be- sonderheiten: kräftige, aus vielen Einzeldornen bestehende Bewehrung des Grund- gliedes ($?), Scutum parvum ($) 6z+. Scutum laminatum (?).
Beschreibung. - Cheliceren (nach dem Syntypus SMF/RI/974); $-Basal- glied (Abb. 56a): liräftig untersetzt, dorsal eine Längsreihe aus 9 Zähnchen, nicht in gerader Linie ausgerichtet; die proximalen am größten, lateral über das ganze Glied zum Teil winzige Tuberkel verstreut; ventro-lateral eine streng ausgerichtete Reihe von 11 Zähnchen unterschiedlicher Größe; ventro-medial 7, von diesen das 4. und 6. (von basal) am größten; Medialseite bis auf 2 kleine basale Höcker unbewehrt; dorso-distal keine deutliche Aufwölbung oder Apophyse und ohne Bürste - vgl. aber „Variabilitätu. - 2. Glied: schlank; am Knie sehr stark bewehrt, medial eine' Reihe von 5 und dorsal (frontal) ein dichtes Feld kräftiger Zähne. - $2 (Hohe Tau- ern, Stubachtal): mit den wesentlichen Kennzeichen des$, doch dorsal nur 9 Dornen, vor allem lateral von kleinen Tuberkeln begleitet; die beiden ventralen Reihen mit 7-9 Dornen unterschiedlicher Größe, medial wenige kleine Tuberkel regellos ver- streut. - 2. Glied: wie das des $.
Ped ipa lpus : Beim $ Femur und Patella im allgemeinen distal mit leichter An- schwellung, vor allem ventral betont; über die Patellarapophyse vgl. „Variabilität".
ohne diese Anschwellungen. - Alle Glieder dunkelbraun, an den Gelenkstellen aufgehellt.
P e des (Laufbeine): mittellang, robust. Cephalothoirax u n d T u b e r oculorum (Abb. 57e): Ansatzstellen der Cheli-
ceren-Muskulatur (hinter dem Tuber oculorum) nur schwach erhaben; 2. Thorakal- Tergit mit 8-12 Dornen, klein; Tuber oculorum doppelt so breit wie lang, niedrig, median stark gefurcht; Linse klein.

Abb. 66. I. lcollari C. L. I~OCI-I. - a-p 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber, jeweils das distale Ende des 1. Gliedes von riiedial. - a Gastein (Österreich; Syntypus SI\IF/RI/ 974,J) ; b Bruiieclr (Italien, J ) ; c Pustertal, Gruppechtspitze über Terenten (Italien, 8 ) ; cl Gröd- nertal (Österreich, J ) ; e Ciina Unclici, Val Saguna (Italien, 8); f Rolle-Paß (Italien; I-Iolotypus I. tirolensis, J ) ; g Plose bei Brisen (Italien, J ) ; h Brenner (Italien, J ) ; i Lnrgrotte (Österreich, 3);

Die Abgrenzung von Biospezies usw. 239
k Schladining (Österixich; Holotypus I. styriaca, 9 ) ; 1 Seeriegelhöhle (Österreich, P ) ; m ~ a i n s a u (Österreich, 9 ) ; n wie i (9) ; o Gruildlsee (Österreich, 9 ) ; p genauej Fundort unbekannt (Holotypus I. clanubia, 9); q-r Penes: p wie g ; r wie e ; s-y Glandes penis: s, t und ZL wie i ; V wie e ; ZU wie g; X wie f ; y Taubenloch bei 1Circhbei.g aiil Wechsel (Österreich).
Scu tum: $ fa'st immer mit nahezu quadratischeiii Scutuni parvum, Ecken a'b- gerundet; aii den Nahtstellen der.eiiizeliieii Are? (seichte Furchen, schwache Puiik- tieruiig) laterale hussparungeii; hreae fein beköriielt, Ait je einer Borsten-Querreihe besetzt. - Es komint vor, daß die Areae 4 und 5; bisweilen auch 3, nicht völlig mit dem übrigen Scutum verschmolzeil sind. - 9 regelmäßig mit Scutum laminatum; die abd~mina~len Skleritplättchen der Segmente 3-6 siiid gelegeiitlich iilediaii hal- biert (E~empla~re aus der Lurgrotte) - vgl. Kap. IV l b .
Pen i s (Abb. 56g-Y): Sklerit der Gla,ns rundlich bis läiiglich, ininier z~eila~ppig ; mit kräftiger Beborstuiig; ba'sale Halfte des Truncus etwas yerbreitert, schnialste Stelle im distalen Drittel; laterale slilerotisierte Randleisteii unterhalb der Gla,iis schwach erweitert; Truncus basal mit tiefer Eiiikerbuiig.
d b b . 57. I . kollari C. L. KOCH. - a-d 'Receptacula. seminis: .a Stubachtal (Gegenseite init 4 Schläuchen); h Plose bei Brisen; c Bruneck (Gegenseite mit 4 Schlauchen); d wic a ; e Cephalo- thorax ~ o n rechts-lateral (Stubachtal).

Recep tacu lum seminis (Abb. 57a-d): 4-5 relativ lange und schlanke Schläuche, sehr stark gekrümmt oder auch nahezu gerade; Distalende bisweilen zwei- zipfelig (6 Legeröhren präpariert).
Va r i ab i l i t ä t : Anlage der Cheliceren-Bewehrung immer einheitlich, im ein- zelnen aber sehr variabel, die drei Hauptreihen kräftig, aber die Dornen nie so lang
". wie das Grundglied breit und nie zu einer Linie ausgerichtet. - Das Grundglied ist . . besonders bei kräftigen Individuen dorso-distal bisweilen zu einer flachen „Apo-
physe" aufgetrieben und mit kurzen Haaren besetzt, ähnlich der Bürste der Arten der dentipalpis-'Gruppe; koaguliertes ~heliceren-sekret wurde nie bemerkt. - Der Pedi- palpus des $ kann an der Patella lateral eine mehr oder weniger deutlich ausgeprägte Apophyse aufweisen. Alle Arten mit diesem Merkmal vereinigte HADHI (1931) in der Untergattung Odontopalpa; bis auf dentipa1pis.erwiesen sich aber alle synonym kol- lari. Das Merkmal tritt vor allem iil jenen kollari-Populationen auf, die'die mittlere und obere Zone des vertikalen Arealgürtels besiedeln. Das läßt sich mit Material aus dem Stubachtal (Hohe Tauern) belegen (vgl. Abb. 33). Exemplare aus 1640 m, also nahe der unteren vertikalen Arealgrenze (tiefster Fund bisher bei 1460 m), weisen keine oder eine kaum angedeutete Apophyse auf, bei Individuen von 1740 und 1865 m ist sie bereits deutlich und bei 2020 m sehr stark ausgeprägt. Da kollari im Stubachtal bis mindestens 2080 m vorkommt (Fund eines Jungtieres), dürfen Individuen mit noch markanterer Apophyse erwartet werden. Solche sind von der Plose bei Brixen (2500 m), dem Mühlwalder Joch (2360 m), der Cima Undici (2050 m) und vom Triglav (2200 m) bekannt. Diese Korrelation „Höhenlage des Biotops - Ausprägung der Apophyse" ist nicht immer 'verwirklicht. Es gibt Populationen hoher Gebirgslagen, die nur sehr schwach ausgebildete oder gar keine Patellar-Apophysen aufweisen (z. B. Brenner, Wolfendorn-Mäuerlscharte, zwischen 2200 und 2600 m). Die Variabilität des Sklerits der Glans penis ist Abb. 56 zu entnehmen.
Beziehungen: Innerhalb des kollari-Komplexes sind die alpinen Arten als nächstverwandt anzusehen; ihre Entstehung ist in Kap. V 2 behandelt.
Ve rb re i t ung u n d u n t e r s u c h t e s K a t e r i a l : I. kollarz besiedelt fast die ganzen Ost-Alpen, erreicht jedoch offensichtlich nicht die westliche Grenze dieses Alpenteiles (die Linie Bodensee-Rheintal-Splugenpaß-Comer See) (Abb. 26 und 58). Die folgenden Fundorte konnen das bis heute bekannte Areal umreißen: im W das Brenner-Gebiet und der Schlern, im S die Cima Uiidici, der Rolle-Paß, das Triglav- Massiv und das Bachergebirge (= Pohorje), im 0 die Lurgrotte bei Graz, das Tauben- loch bei Kirchberg am Wechsel und der Hochschneeberg, im N der Ötscher und der Kasberg. Ob zwischen Drau und Mur tatsächlich eine Verbreitungslücke besteht, bleibt zu untersuchen (bisher je ein Fund vom Speikkogel/Gleinalpe und aus der Peba-Klamm bei Lienz). ,
Es wurden 91 $, 75 und 58 Jungtiere untersucht.

Die Abgrenzung von Biospezies usw. 241
Sbb. 58. Die Fundorte der alpinen Arten der kollari-Gruppe. - 1 I. strandi; 2 I. hadzii; ohne Be- zeichnung: I. kollari.
Jugoslawien. - Slowenien, Triglav, Schneefeld des Kredarico, 2200 m (1 $ Holotypus I. trsglavenszs, Zool. Inst. Ljubljana, SELISKAR leg. 31. V. 1930); Triglav, Pod Sternarjem nahe Skrlatica (1 0, Zool. Inst. Ljubljana, D. H A D ~ I leg. 1952, vgl. H A D ~ I 1954) ; - (1 s t e r r e ic h. - Steiermark, Lurgrotte bei Graz (35$,17 $2, 2 iuv. Mus. Wien, SMF; ABEL, BRAUN, KULLIANY, MARTENS, SCHMID, SCHUTZ, VORNATSCHER leg.); Taubenloch bei Kirchberg am Wechsel (1 $ Mus. Wien); Rax-Alpe (4 9 , 3 iuv. SMF 10274; 1 9 SMF 9547; 1 9, 4 iuv. SMF 11362); Veitsch (1 9, Mus. Wien); Almberger Eis- und Tropfsteinhohle/Grundlsee (1 9 Mus. ,Wien, A u ~ ~ l e g . 7. X. 1961), Se. Totes Gebirge, Sinterhaus, Brettstein (1 $ Mus. Wien, VORNATSCHER leg.); Dachstein-Massiv, GjaidsteinIGuttenberghaus ( 1 8 SMF 10271); Ramsau,soberhalb Schladming (1 9 Holotypus I. styriaca, ROEWER leg. VIII. 1927); Ramsau, Aufstieg durch die Feisterscharte, etwa 2100 m (1 3, 1 9 Typenserie I. reimoseri, SMF/RII/230, REIMOSER & ROEWER leg. VIII. 1927); Admont (SMF/ RIIl10273). Nieder-Österreich; Lunz, Herdengelhohle (1 iuv. Mus. Wien, leg. 30. V. 1963; 2 iuv. Mus. Wien, VORNATSCHER leg. 7. VII. 1963); Lunz, zwischen Obersee und Pauschenalm, Durren- stein und Ybbstaler Hutte ( 2 8 , 6 9, 6 iuv. 'THALER [& PLATTNER] leg. VIII. 1960 - 9. VI. 1962; vgl. THALER 1963); Durrenstein, 1870 m ( I$ coll. MALICLY, MALICICY leg. 2. VI1.-15. IX. 1966); Gipfel des Ötscher bei Gaming, etwa 1900 m (1 9 Mus. Wien, RESSL leg. 21. VI. 1959). - Ober- Osterreich; Gosau, am ostlichen Berghang im Wald (1 iuv. SMF/RII/9499, ROEWER leg. VIII. 1942, vgl. ROEWER 1950). -Land Salzburg; Tennengebirge, Sommereckkessel-Hohle (1 $ Schü/ Op. 599, jetzt SMF, SCHULLER leg., vgl. SCHULLER 1933); Lungau, Lignitztal, Lignitzalm, 1600 m (1 iuv. Op. 67/176b, jetzt SMF, AUSOBSKY leg. 6. X. 1967); Kleinarltal, Tappenkarsee, 1760 m (1 iuv. Op. 671105 C, jetzt SMF, AUSOBSICY leg. 5. IX. 1967); Gasteiner Tal (1 $, 1 iuv. Mus. Wien KOLLAR leg. 1855); Kaprun, s. Salzburgerhütte, 1990 m (1 $2 Op. 661236, jetzt SMF, HURDES leg. 13. VIII. 1966); Stubachtal, 1460-2080 m (16$,15 ? ,30 iuv., uberwiegend SMF; AUSOBSKY, HARMS, V. HELVERSEN, HURDES, MARTENS, PAULUS, UHRIG leg. 3. IX. 1966-19. VIII. 1967); Habachtal, sw. Fazeneck, 2150 m ( 1 3 Op. 671297 HUTZ leg. 24. X. 1967); Osterhorngruppe Kühlloch-Trattberg, etwa 1450 m (1 iiiv. LOVCIIC leg. 19. VII. 1959). - Osttirol; Lienz, Peba- Klamm (13, KOFLER leg. 20. VIII. 1965). - Tirol; Brenneralpen, Wolfendorn-Mäuerlscharte,
16 Zool. Jb. Syst. Bd. 96

2200-2600 m ( 4 8 , 4 Q, 4 iuv. coll. THALER, THALER leg. 13. VII. -11. IX. 1962); Windlöcher ( 1 8 , 1 9 ~ ~ ~ / ~ 1 1 / 1 1 2 1 1 ) ; Seeriegelhöhle(33, 4 9 SMF 11213), Gasslhöhle(2 Q SMF); - I t a l i e n . - Trentino-Tiroler Etschland, Pustertal, Gruppechtspitze über Terenten (1 Q PEEZ leg. 12. VII. 1955); Mühlwalder Joch n. Bruneck ( 5 8 , 6 9 PESKOLLER leg.); Weißenbach n. Bruneck, 2360 in
(1 9 SCHULZE leg. IX. 1962); Plose bei Brixen (1 8 , 3 Q Mus. Wien HOLDHAUS leg. ; 4 8 , 2 coll. THALER, THALER leg. 23. VI. 1963); Grödner Tal (1 $ SMF 1482); Schlern (1 Q, 6 iuv. coll. THALER, THALER leg. 27.128. VII. 1966); Dolomiten, Sella-Joch (1 iuv. SMFIRII18487, STRASSER leg.); Rolle-Paß ( 1 3 Mus. Berlin, Holotypus I. tirolensis, NÜLLER leg. 27. VI. 1934); Cima Undici, Borgo, Val Saguna e. Trient, Ca. 2050 m (1 $ coll. THALER, THALER leg. 25. IX. 1965); Dolomiten, Höhlen (1 3, 1 Q SMF 12041).
Weitere Fundorte: J u g o s l a w i e n . - Slowenien; Bachergebirge (= Pohorje), RibniSka koCa, 980 m (18 ~ ~ A T J A ~ I ~ leg., vgl. H A D ~ I 1954). - Ö s t e r r e i c h . - Steiermark; Gleinalpe, Speikkogel, 1900 m (1 Ex. leg. 5. VI. 1947, FRANZ & GUSHOLD 1954); Seckauer Zinken, Kar der Goldlacke, etwa 1900 m (FRANZ & GUNHOLD 1954); Mitteralm bei Aflenz, Schneedoline; Totes Gebirge, Umgebung Steyrer See; Hochschwab, Gipfel des Ebenstein, etwa 2200 m ; Eisenerzer Alpen, Zeyritzkampel (alle FRANZ & GUNHOLD 1934). - Nieder-Österreich; Hochschneeberg (Locus typicus von I. nivalis). - Ober-Österreich; Kasberg, Doline am Kamm gegen Schwalben- mauer; Traunstein, Gipfelregion (beide ROEWER 1950). - Fundorte sicher falsch, auf der Ver- ljreitungskarte Abb. 58 nicht berücksichtigt: Bosnien ( 2 3 SMF/RI/971, Typenserie I. bosnica); Nieder-Österreich, Wachau, Pechlarn [= Pöchlarn] (1 9 Holotypus I. danubia, SMF/RI1/2894).
V o r k o m m e n : vgl. Kap. I11 1, I11 2, I11 3, I11 4 und I11 5.
A n h a n g ; Körpermaße (in mm) (Erklärung der Meßstrecken vgl. Abb. 34):
8 Cx-L (1) 1,9 - 2,5 (2,07„); Cx-B (2) 2,2 - 3,3 (2,7lS4); SC-L (3) 2,3 - 3,2 (2,85„); SC-B (4) 2,2 - 3,3 (2,78„); L, (5) 3 8 - 5,9 (4,387,); L2 (6) 5,2 - 7,4 (6,07„); B, (7) 0385- 134 (1,0275); L, (8) 2,0 - 3,0 (2,2460); Fe11 (9) 5,O - 7,7 (5,83„); Mt11 (10) 5,7 - 9 , l (6,40„); Pen. (11) 2,6 - 3,7 (3,46„); Mt Pg (12) 3-5. TaGe (13) 32-36.
? Cx-L (1) 1 ,8 - 2,O ( 9 5 ) ; Cx-B (2) 2,O - 3,O (2,58,,); LI (5) 3,4 - 4,75 (4,24„); L, (6) 4,75- 6 2 (5,83„); B2 (7) 0 - 1 5 ( 0 ) ; L, (8) 1 ,8 - 294 (2,1859); Fe11 (9) 4,9 - 5,9 ( 5 4 ) ; Mt11 (10) 5,3 - 6,7 (6,0403); Mt Pg (12) 3 - 5 TaGe (13) 34 -38.
I. hadzii ROEWER 1950 (Abb. 59-60)
1851 Phalangium cancroides SCHRIIDT, Ber. Mitth. Freund. Naturwiss. Wien, 7: 61 (D; praeocc. durch MULLER 1776).
V 1926127 I. cancrotdes, - HADJI, Bull. Mus. Slov., 718: 26 T. 6 Abb. 146-157 (D 9). 1926127 I. manicata, - H A D ~ I , Bull. Mus. Slov., 718: 128 T. 6 Abb. 158-165 T. 7 Sbb . 166
bis 173 (D 8 ) . V 1928 I. cancroides, - H A D ~ I , Zool. Anz., 77: 14 (D 9).
1928 I. manicata, - H A D ~ I , Zool. Anz., 77: 15 (D$). 1942 I. cancroides, - H A D ~ I , Razprave Akad. Ljubljana, 2: 28 Abb. 10-12 (D$?). 1950 I. hadzii ROEWER (Nom. novum), Senckenbergiana, 31: 52 (N).
v 1964 I. hadzii, - GRUBER, Zeitschr. Arbeitsgem. Öster. Ent., 16: 2 (N).

Die Abgrenzung von Biospezies usw. 243
B e g r ü n d u n g d e r S y n o n y m i e : „Phala~zgium cancroides" wurde nach einem 9 aus der Knochenhöhle Ziavka „Gegenüber der Steineralpe, velka planina, an der Kreuzeralpe, am nörd- lichen Abhange des Mokritzberges" beschrieben. H A D ~ I ergänzte die Diagnose (1926127, 1928) nach dem Origiiialmaterial (Q), hielt aber die zugehörigen $ für manicata. - ROEWER (1950) führte das Nomen novum ein, da Phalangium cancroides bereits durch MÜLLER (1776) aufgestellt worden war.
Diagnose: Eine Art der kollari-Gruppe, gekennzeichnet durch folgende Be- sonderheiten: rechtwinklig abstehende Bewehrung des Cheliceren-Grundgliedes (d?), ein Scutum intermedium (8) bzw. laminatum 9).
Beschre ibung: - Cheliceren;$-Basalglied(Abb. 59a-b): dorsal 2-4große, rechtwinklig abstehende oder sogar caudad gekrümmte Dornen, bisweilen kleine Tuberkel in diese Reihe eingeschoben; ventro-lateral mit 6-7 sehr langen Dornen, die basalen am kleinsten; ventro-medial ausgerichtet 6-8 kleine, ebenfalls nahezu rechtwinklig abstehende Dornen; Distalende besonders nach medial zu einem flachen Höcker oder stumpfen Buckel aufgewölbt, dorso-medial mit schütterem Borsten- besatz. - 2. Glied: ,,Knieu spärlich niedrig bedornt, frontal in eine Borstenreihe übergehend. - 9: mit den allgemeinen Merkmalen des 8 , doch ohne die dorso- distale Aufwölbung.
P e d ip a lpus (J?): schlank, Patella ohne Andeutung einer Apophyse.
P e de s (Laufbeine): relativ lang, ohne strukturelle Besonderheiten.
..C 20 I w m
d.* ' W
Abb. 59. I. hadzii ROEWER. - a-C 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber je- weils das distale Ende des I. Gliedes von medial. - a Kärnten (Österreich, $); b Obir (Österreich, 8); c wie a (9); d-e Glandes penis: d wie a ; e wie b ; f Penis, wie b.
16"

Cepha lo tho rax u n d Tube r oculorum (Abb. 60b): Cephalothorax flach; 2. Thorakal-Tergit mit 8 kräftigen Dornen unterschiedlicher Größe; ohne Tuber ocu- lorum im eigentlichen Sinne, sondern beide Augen isoliert; Durchmesser der Linse sehr klein.
S c u t u m (Abb. 14, S. 159): $ offenbar nie mit vollständigem Scutum parvum, sondern von der abdominalen Platte eine oder mehrere Areae ganz oder teilweise ab- getrennt, also ein Scutum intermedium; $2 regelmäßig mit Scutum laminatum.
Penis: basale Hälfte des Truncus schwach bauchig erweitert, distales Drittel am schmalsten, zur Basis der Glans schwach erweitert; Truncus relativ lang.
Recep t acu lum seminis (Abb. 60a): Schläuche relativ kurz und dick, nur sanft geschwungen; die einzige untersuchte Legeröhre rechts und links mit je 4 Schläuchen.
Va r i ab i l i t ä t : an dem kleinen Material nicht zu erkennen; bez. des Scutum vergleiche Abb. 14, S. 159.
Beziehungen: I. hadzii gehört zu jenen Arten der kollari-Gruppe, die Teile der Glazialperioden in ihrem heutigen Siedlungsgebiet verbracht haben und sich dort divergent entwickelten. Enge verwandtschaftliche Beziehungen bestehen vor allem zu I. strandi (vgl. Kap. V 2).
Ve rb re i t ung u u d u n t e r s u c h t e s Mater ia l : I. hadzii ist ein Endemit weni- ger so-alpiner Gebirgszüge; Nachweise sind bisher nur aus den Steiner Alpen (= Sann- taler Alpen) und den Karawanken bekannt; von diesen gelang einer in Österreich (Obir) (Abb. 26 und 58).
Es wurden 2 8 , 2 und 1 Jungtier untersucht. Jugos lawien : Krain, ohne genauen Fundort (1 9 aus der Sammlung SCHMIDT, 2001. Inst.
Ljubljana); ViSevnik Jama (1 iuv. Zool. Inst. Ljubljana, PRETNER leg. 1953). - Österreich. - Kärnten, Obir ( 1 3 Mus. Wien, MANDL leg. VIII. 1953, vgl. GRUBER 1964); ,,Höhlen in Kärnten" ( 1 3 , l i;! Mus. Berlin 433). - Weitere Fundorte: Jugoslawien. - Slowenien, Höhle Erjevaci bei LuEe; Höhle TrbiSka zijalka bei LuEe, Savinjska dolina; Höhle Medvedja am Mokrica (Lotus typicus); Höhle Vjetrenica im Velika planina; Höhle Rofce im Velika planina; Höhle RjavEeva, Sannijska dolina (alle H A D ~ I 1942).
lateral (Obir).

Die Abgrenzung von Biospezies usw. 245
Vorkommen : I. hudzii wurde bisher nur in „meist weder langen noch tiefen Höhlen gefunden" (1-IAD:I); ob dennoch epigäische Populationen existieren, müssen weitere Untersuchungen zeigen.
Anhang : Körpermaße (in mm) (Erklärung der Meßstrecken vgl. Abb. 34):
6 Cx-L (I ) 1,3 - 2,0 ( 1 6 5 ) Cx-B (2) 2,4 (2,421; . L1 (5) 3,93 - 4,15 (4,05,); L, (6) 5,5 - 5,65 (5,57,);
B2 (7) 0,8 - 0,85 (0,82,); L3 (8) 2,1 - 2,15 (2,12,); Fe11 (9) 6,5 - 7,O (6,75,); Mt11 (10) 10,4 (lW1) ; Pen. (11) 3,9 (3,921 ; Mt Pg (12) 3 ; TaGe (13) 44.
? Cx-L (1) 1,51; Cx-B (2) 2,5,; LI (5) 4 2 (4,221; L2 (6) 5,9 - 6,O (5,952); B, (7) 0 3 (0,9,); L3 (8) 2,4,; Fe11 (9) 6,0,; Mt I1 (10) 8,6,; Mt Pg (12) 3 ; TaGe (13) 44.
I . muellneri HAMANN 1898 (Abb. 61-62) 1872 Isehiropsalis Helwigi, - SIMON, Ann. Soc. ent. France, (5) 2: 483 T. 16 Abb. 4 (D).
v 1898 Ischyropsalis Müllneri BAMANN, 2001. Anz., 21 (570): 535 (D$?). v 1914 I. helwigii, - ROEWER, Arch. Naturgesch., 80 A (3): 104 Abb. 1 a-d (D $9). v 1914 I . corcyraea ROEWER, Arch. Naturgesch., 80 A (3): 108 Abb. 5 a-b (D $9). v 1923 I . helwigii, - ROEWER, Weberknechte d. Erde,: 683 Abb. 849 a-d (D $9; Verbreitnngs-
angaben sind nur teilweise auf muellneri zu beziehen). v 1923 I . corcyraea, - ROEWER, Weberkncchtc d. Erde,: 685 Abb. 853 a-b (D$9). v 1924 I. helwigii, - MÜLLER, 2001. Jb. Anat., 45: 414 (part.) T. 19 Abb. 1-3, 6-7 T. 20
Abb. 12-15 (D). v 1926127 I . nziillneri, - HAD%I, Glasnik/=Bull. Assoc. Mus. Slov., 718 B: 23 T. 6 Abb. 131 bis
145 (D $9). v 1926127 I . müllneri, - I-IADZI, Glasnik/=Bull. Assoc. Mus. Slov. 718 B: 20 T. 5 Abb. 116 bis
123 (D $9). 1928 I . helwigii, - K A S T N E ~ ~ , in D A ~ L , Tierm. Deutschl. 8: 25 Abb. 42-44, 47-48. (D $9).
V 1928 1. helwigii, - H A D ~ I , Zool. Anz., 77: 12 (D $9). 11 1928 1. müllneri, - H A D ~ I , Zool. Anis., 77: 1 3 ( D $9). V 1934 I . helwigii, - K n i ~ r o c ~ v i ~ , Acta Soc. SC. nat. IKorav., 9 (5): 14 T. 1 Abb. 15-16 (D $9). V 1942 I. helwigii, - H A D ~ I , Raisprave, 21: 19 Abb. 4, 7 a-b, 8-9 (D). V 1950 I . helzvigii, - ROEII~EII, Senckcnbergiana, 31 (112): 17 T. 7 Abb. 45 a-d (D$?). V 1950 I . nzüllneri, - RQEWER, Senckenbergiana, 31 (112): 26 T. 5 Abb. 28 a-d (DS?). V 1950 I. corcyraea, - ROELVEH, Senckenbergiana, 31 (112): 28 T. 7 Abb. 46 a-b T. 8 Abb. 46c
bis d (D $9). V 1950 I. alflceni ROEWER, Senckenbergiana, 31 (112): 19 'r. 8 Abb. 47a, b (D?, ncc$). V 1954 I. helwigii müllneri, - H A D ~ I , Acad. Sc. art. slov,. 2: 191 (D). V 1956 I. helwigii, - SILIIAVY, Fauna CSR, 7: 151 '1'. 24 Abb. 293-294, 302 Farbtafel 4
Abb. 1 3 (D$?). 1964 I . helwigi müllneri, - JUBERTHIE, Ann. Spklkol., 19 (1): 27 Abb. 37 c (N).
V 1965 I. muellneri, - MARTENS, Natur U. Museum, 95 (4): 144 (N).
B e g r ü n d u n g d e r S y n o n y m i e : Bezüglich der irrtümlichen Verwendung des Namens hellwigi durch SIMON vgl. I . pyrenaea (S. 228). - ROEWER (1914, und in spätcren Bearbeitungen)

benutzte den Nanlen hellwlgz fur die sehr lang- und schlankchelicerige Ai t muellnerz, da ihm 1) die Originalbeschreibung von muellnerz bis 1950 unbekannt geblieben was und er 2) bei der Inter- pretation von hellwagn offensichtlich nicht die Original-Abbildungen von I ~ A H N (1834), Wmcn (1834) und C. L. l i o c ~ r (1841, 1848) beachtet hat, die alle eine gedrungen-chelicerige Art dar- stellen (vgl bei hellwzgz). Erst H,i»i1(1926/27 und 1928) revalidierte den vergessenen Namen und nannte, da auch er die Art-Verschiedenheit von lzellwagz und muellnerz nicht erkannte, slowenische Hohlen-Popi~l~~tionen nauellnerz; Individuen aus der Sammlung SCHMIDY, ,m denen er gering- fugwe Unterschiede zii sehen glaubte, gab ei <~ l s hellzvzg~ bekmnt . Spates degradierte ei beide zu Sabspc~ies von hellwiqr, beschrankte sich abei 1954 auf die Anerkennung von „Variantenu - Die Ubeip~utiing der Typen von corcyraea zeigte, daß muellnera vorliegt. Die cntspiechenden Ab- bildirngen sind grob scheinatisiert und treffen nicht die tdts'%~tilirhen Veilialtnisse.
Diagnose : Einc Art der kollurz-Gruppe, gckennzciehnct durch folgciide Be- sonderhciteii: nahezu glatte, sehr schlaiikc Chclieercn (J), eine winzige, medio-distale Apopliyse am Grundglied ($), ein Sentum intermedium oder laminatum (J), 9 immer mit Sciitum laminatum.
B es eh r ci burig. - Chelicer eil (~iaeh topotypisehcm Matcrial) ; J-ßasalglicd (Abb. 61): relativ lang und schr diiiiii, Distalendc schwach erweitert und leicht ven- trad abgebogen, dorso-mcdial iii winzige, spitze Apophyse auslaufciid; Bewehrung sehr schwach, die dorsale n i ~ d dic ventro-laterale Rcihe lediglich durch Kornchen a i~g~deute t . - 2. Glied: ain „KiiieU niedrige Tuberkel, medial und frontal in Reihen
Abb. 61. I. muellneri HAMANN. - a-f I . bzw. 2. rechtes Cheliceren-Glied von l a t e~a l , darunter jeweils clas distale Ende des I . Gliedes vor1 iiledial. - a Castitljiva jama (Jugoslawien; Locus typicus, 6); b Ra,dmannsdorf (Jiigoslawien, 8); c Insel ESorfir (Griechenland, 8); d wie a (Syn- typus, 9) ; e wie b (P); f wie c (Paratypoid I. corcyraea, 9 ) ; g-h Glandes penis: g wie a,; h richtiger Fundort unbekannt (SMF/R 112607); i Penis, wie h.

Die Abgrenzung von Biospezies usw. 247
kleiner Höcker auslaufend. -- 9: Proportionen wie beim $, jedoch mit stärkerer Bewehrung, die drei Hauptreihen aus vielen kleinen Dornen, kleine Tuberkel auch lateral, vor allem an der basalen Hälfte.
P e d ip a lpu s: robust, bei $9 distal schwach tropfenförmig erweitert. P e d e s (Laufbeine): phalangiidenhaft lang, Basis der Pemora hellgelb geringelt,
Coxen sehr warzig; Metatarsus und Tarsus hellgelb, a,lle übrigen Glieder mittel- bis dunkelbraun.
Cepha lo tho rax u n d T u b e r ocu lorum (Abb. 62c): Cephalothorax nur schwach aufgewölbt, fein granuliert; 2. Thorakaltergit mit 8 sehr großen distad ge- richteten Dornen; ohne Tuber oculorum im eigentlichen Sinne, sondern Augen iso- liert und weit voneinander entfernt; Linse klein.
S c u t u m (Abb. 15, S. 159): 3 mit Scutum intermedium oder laminatum; immer mit Scutum laminatum. Bei d e n 8 kommen alle Übergänge von einer abdomi- nalen Platte aus den Skleriten 1-111 (folgende Sklerite frei) bis zu völlig isolierten Eiiizelskleriten I-V111 vor. Die Ablösung der einzelnen Areae vom (rezent nicht mehr auftreteiiden) Scutum parvum unterliegt folgender~egel : Areae I wird durch laterale Einschnürung undIoder mediane Perforation von der Platte aus den Areae 1-111 gelöst (Abb. 15a-c). Nachfolgend wird die Platte aus den Areae I1 und I11 ebenso zerlegt (Abb. 15d-e), so daß ein Scutum lamiiiatum resultiert (Abb. 15f).
Pen i s (Abb. 61g-i): Truncus kräftig und gedrungen, basales Drittel erweitert, aber nicht bauchig; die distal stark sklerotisierte Ventralseite erscheint von ventral als schwache Flügelung; ventraler Sklerit sehr breit und kurz; Borsten relativ kurz; Trichter weit.
Recep t acu lum semin is (Abb. 62a-b): Schläuche kurz und dick, mäßig oder stark gekrümmt; die 2 untersuchten Legeröhren beidseitig mit je 4 Schläuchen.
Va r i ab i l i t ä t : Die Bewehrung des $-Cheliceren-Grundgliedes kann bisweilen stärker sein, als auf den Abb. 61a-c angegeben; Bewehrung des ?-Gliedes vgl. Abb. -'Variabilität des Scutum vgl. „Scutunic'.
Abb. 62. I . muellneri HAMANN. - a-b Receptacula seminis: a Insel Korfu (Syntypus I. corcyraea) b Radmannsdorf; c Cephalothorax von rechts-lateral, wie b.

Beziehungen: In den Kantabrischen Gebirgen kommen die habituell sehr ähnlichen Arten der kollari-Gruppe dispar und magdalenae vor. Bei ihnen ist aber die Apophyse des S.-Grundgliedes anders gebaut. Dennoch bestehen zu diesen beiden Arten die engsten phylogenetischen Beziehungen. Innerhalb der alpinen mittel-/ südost-europäischen Arten der kollari-Gruppe steht muellneri völlig isoliert.
Ve rb re i t ung u n d u n t e r s u c h t e s Mater ia l : Unter allen Arten der kollari- Gruppe besitzt muellneri das größte Areal. Es erstreckt sich auf der Balkanhalbinsel etwa von Ljubljana als schmaler Streifen bis N-Griechenland (nur Korfu als Fundort belegt!). Die meisten Nachweise stammen aus dem nördlichsten Teil des Areals (gute Durchforschung der Höhlen!), aus dem mittleren und südlichen Teil liegen nur ver- einzelte Funde vor; die beiden Nachweise von Korfu sollten bestätigt werden (Abb. 26). - Im Schrifttum werden viele weitere Fundorte genannt, die außerhalb des oben umrissenen Gebietes liegen (deutsche Mittelgebirge, 0-Alpen bis Monte Rosa, Tatra, Siebenbürgen [ROEWER 1923, sub hellwigi] ; Nieder-Österreich, Gaming [RoE- WER 1950, sub hellwigi]; Thüringen, Jenaer Berge [ROEWER 1950, sub alfkeni]). Die Angaben haben sich in keinem Falle bestätigen lassen; das Belegmaterial ist in den meisten Fällen falsch bestimmt worden, bisweilen liegen offensichtliche Ver- wechslungen der Fundorte vor.
Es wurden 29$, 16 9 und 1 Jungtier untersucht.
J u g o s l a w i e n . - Slowenien, Castitljiva jama (2 9 Syntypen, Mus. Berlin 8057, HAMANN & MÜLLER leg.; ferner 1 1 3 , 6 9 aus den Mus. Wien, Hamburg, Berlin, coll. SILH~IVI 25928, SMF/ RI1977); Radmannsdorf/Krain (2 $, 4 9 Mus. Wien); Jodloch/Krain (1 6 Mus:Wien); Berg Babji ZobIKrain (1 $ SMF/R 1116440); Höhle PrSivec nad Bohinjskim jezerom (2 $, 2 coll. SILHAVY 15543, ZVAN leg.); ,,KrainX, ohne genauere Angaben ( 7 8 , 3 9 Mus. Hamburg 1248, Rtus. Berlin 986, SMF/RII/2116, SMF/RI/972); - Kroatien, Berg Sever zwischen Dubrownik und Rotor (2 3 coll. SILHAVY 03113, K R A T O C H ~ ~ L leg.); - Herzegowina, Bjelalnica-Gebirge (1 $, vgl. H A D ~ I 1942). - G r i e c h e n l a n d . - Insel Korfu (2 9, 1 iuv. [nec 13, 19 , 1 iuv.] Typenserie I. corcyraea, SR!F/RI/980; ferner 13 Mus. Wien).
Weitere Fundorte: J u g o s l a ~ v i e n . - Slowenien; LuSe; Höhle Medvedja; Smarna Gora bei Ljubljana ( H A D ~ I 1942)
Fundorts-Angabe sicher falsch bzw. Landeszugehörigkeit nicht zu ermitteln: CSSR, Preßburg (1 $Mus. Wien). - Österreich, Gaming ( 1 3 SDIF/RII/2607). - Deutschland, Jena, Rosental(1 9 Holotypus I. alfkeni, SMFiRIIi9497). -Italieil/Schweiq Monte R o s a ( l 3 Mus. Berlin 4406). - Popradi t6 (1 9 Mus. Budapest, LENDL leg. 1886).
Vorkommen: Alle näher erläuterten Funde stammen aus Höhlen; ob es auch epigäische Populationen gibt, ist nicht bekannt. In der Höhle Castitljiva luknja, dem Locus typicus, lebt I. muellneri in einem Temperaturbereich von 7,8" bis 5,2 "C (Ju- BERTHIE 1964).
Anhang . Körpermaße (in mm) (Erklärung der Meßstrecken vgl. Abb. 34): 8 Cx-L (1) 1,4 - 2,O (1,671,); Cx-B (2) 2,2 - 2,5 (2,38„);
LI (5) 5,1 - 5,4 ( 5 2 ~ ) ; L2 (6) 6,4 - 7,1 (6,68191; B2 (7) 0,75- 0,85 (0,81„); L, (8) 2,2 - 2,5 (2,3515);

Die Abgrenzung von Biospezies usw. 249
Fe11 (9) 6,8 - 9,0 (8,32„); Mt11 (10) 10,6 ~ 1 2 , 5 (11,6717); Pen. (11) 3,7 - 4 , 4 ( 4 1 3 ) MtPg (12) 1 - 3 ; TaGe(13)36-48 .
9 Cx-L (1) 1,6 - 2,1 ( 1 1 ) Cs-B (2) 2,4 - 3,0 (2,6OlS); L1 (5) 4,s - 6,O ( 5 , 1 1 , ) L, (6) 6,0 - 7,7 (7,36„); B2 (7) 0,9 - 1,05 (0,98„); . L3 (8) 2,4 - 2,8 (2,60„); Fe11 (9) 6,7 - 9,2 (8,04„); I l t I I (10) 10,4 -12,2 (ll,Oll);
Pg (12) 2 - 3 ; TaGe (13) 45 -48.
I . strandi KR.~TOCHV~L 1936 (Abb. 63-64) V 1936 1. strandi I<RATOCHV~L, Festschrift STRAKD, 1 : 249 Abb. 1-2 (D 9). v 1942 I. ruvasini i H A D ~ I , Razprave, 2: 11 Abb. 5-6 (D Q). , . ? 1947 I. ruf fo i C A P O R I ~ ~ C O , Mein. Rtus. Civ. Verona, 1 : 139 (D iuv.). . "
v 1950 I. strnncli, - ROEWER, Senckenbergiana, 31 (112): 34 T. 9 'Abb. 59 C-e (D 9). 1950 I. ru f fo i , - ROEWER, Senckenbergiana, 31 (112): 35 (D iuv.).
v 1950 I. knirsclzi ROE\VER, Senckenbergiana, 31 (112): 27 T. 9 Abb. 57 a-d (D$?). V 1934 I. sp. aff. strasseri, - H A D ~ I , Razprave, P 180, 193 Abb. 17-18 (D 9).
1963 I. s trandi , - JUBERTHIE, Bnn. Spelkol., 18 (3): 326 Abb. 1-10 (D$.?). 1963 I. ru f fo i , - JUBERTHIE, Ann. Spelkol., 18 (3): 330 (iV). 1964 I. s trandi , - JUBERTHIE, Ann. Sp616ol., 19 (1): 25 4bb . 3, 12, 37 d, 38b (D).
B e g r ü n d u n g d e r S y n o n y niie: Dem E-Iolotypus-? fehlen Tuber oculoruin und Augen vollig, so daß sich s lrundi damals eindeutig gegen alle Arten abgrenzen ließ. I. raljasinii unter- scheidet sich von s trandi , wie der Autor selbst betont, lediglich durch den Besitz von Augen. Neuerdings beschrieb JUBERTHIE (1963) Individuen von s trandi , die Augenrudimente aufweisen, und ich selbst fand in falsch bestimmtem Saminlungsmaterial (SRIF, Mus. Wien) Esenlplare vom Locus typicus mit deutlich ausgeprägten Augen. Beide Arten sind also sicher synonym. - JU- BERTHIE (1963) halt den Typus von ru f fo i mit hoher Wahrscheinlichkeit für ein Jungtier von s.trandi; k+airsclzi gehört bestimmt zu strandi.
Diagnose : Eine Art der kolluri-Gruppe, gekennzeichnet durch folgende Be- sonderheiten: sehr lange, exzessiv bedornte Cheliceren-Grundglieder ($9); fehlende, höchsteiis rudiilieiitär entwickelte Augen und ein Scutum intermedium (8) bzw. laminatuni ($2).
Beschre ibung . - Chelicer en (nach topotypischem Material); $-Basalglied: schlank uiid sehr lang (struncli hat die läilgsteii Chelicereil aller Ischyropsalis-Arten!), besonders dorsa'l sehr große, meist vorgeneigte Dornen, dorsal 5-10, ventro-lateral 7-10, ventro-medial 9-11, oft a,m linken und rechten Glied unterschiedlich aus- geprägt (vgl. Abb. 17a, S. 162); lateral und sehr spärlich auch medial kleine Tuberkel eingestreut, teilweise in die Hauptreiheil eii~bezogeii; dorso-distal mit schwacher An- schwellung; doch ohne Apophyse. - 2. Glied: am „Knieu frontal niedrige Tuberku- lierung. - - 9 : keine Unterschiede zm7ischeii $- und 9-Cheliceren (!).
P e d ip a lp U s: der des $ bisweilen mit einer apophysenartigen Vorwölbung am Distalende der Patella.
P e des (Laufbeine): sehr lang, dunkelbraun, nur Metatarsus und Tarsus hell gelblich.

Cepha lo tho rax u n d Tube r oculorum (Abb. 64b): Cephalothorax hinter ,
den Augen (wenn vorhanden!) deutlich erhaben, zum 1. Thorakaltergit steil abfallend; 2. Thorakaltergit mit 8-14 (JUBERTHIE 1963) kräftigen distad gerichteten Dornen, die hier vorliegenden Individuen nur mit 8-12; Cephalothorax stark granuliert und mit einzelnen größeren borstentragenden Tuberkeln besetzt; Ausprägung des Tuber oculorum sehr variabel: Der Holotypus ohne Spur eines Tuber oculorum oder von Augenrudimenten; andere Individuen mit sehr kleinen Augen (Abb. 64b), Tuber ocu- lorum jedoch nur angedeutet; e i n3 auf der rechten Seite mit deutlich ausgebildetem Auge, links jedoch völlig fehlend; wenn beide Augen vorhanden, da1111 meist unter- schiedlich stark entwickelt.
h b b . 63. I. s trandi KRATOCHV~L. - a-g I. bzm. 2 . rechtes Cheliceren-Glied von lateral. - a und b Monte Baldo (Italien, $ bzw. 9); c Val Seriana, Bergamasker Alpen (Italien, 9); d wie a (Holo- typus I. knirschi , 8); e wie a ( ~ a r a t i ~ o i d I. knirschi , 9); f Monte Baldo, Buce della Preta (Italien, 9); g Csnsiglio, Venetianer Alpen (Italien; Holotypus I. ravasini i , 9); h Penis, wie a ; i Glans penis, wie a.
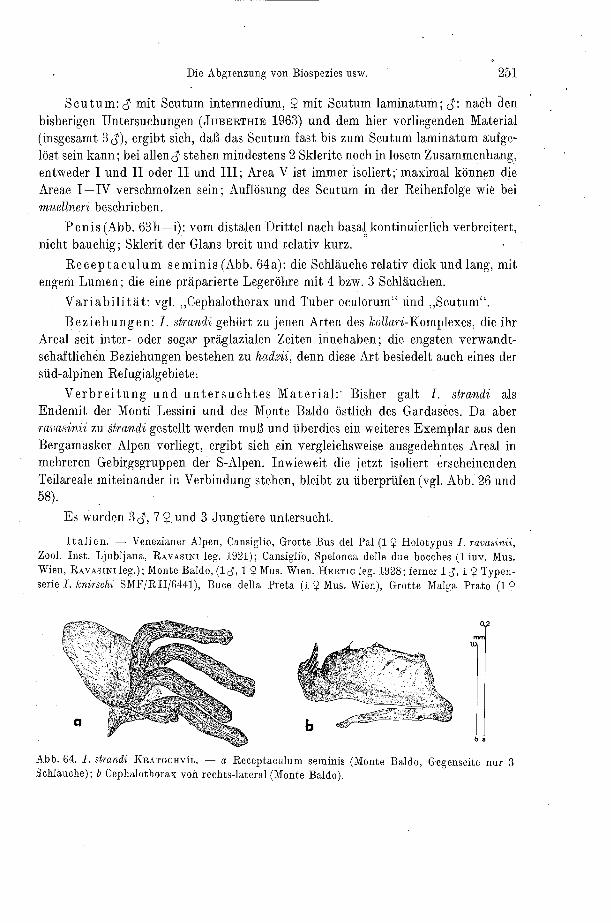
Die Abgrenzung von Biospezies usw. 251
S c u t um: $ mit Scutum intermedium, mit Scutum laminatum ; $: nach den ~
,
bisherigen Untersuchungen (JURERTHIE 1963) und dem hier vorliegenden Material (insgesamt 3 $), ergibt sich, daß das Scutum fast bis zum Scutum laminatum aufge- löst sein kann; bei allen$ stehen mindestens 2 Sklerite noch in losem Zusammenhang, entweder I und I1 oder I1 und 111; Area V ist immer 'isoliert;. maximal können die Areae I-IV verschmol'zen sein; Auflösung des Scutum iii der Reihenfolge wie bei muellneri beschrieben.
Pen i s (Abb. 63h-i): vom distalen Drittel nach basal kontinuierlich verbreitert, nicht bauchig; Sklerit de; Glans breit und relativ kurz.
R e c e p t a c u l u m semiiiis (Abb. 64a): die Schläuche relativ dick und lang, mit engem Lumen; die eine präparierte Legeröhre mit 4 bzw. 3 Schläuchen.
Va r i ab i l i t ä t : vgl. „Cephalothorax und Tuber oculorum" und „Scutum".
B eziehuiig en: I. strandi gehört zu jenen Arten des kollari-Komplexes, die ihr Areal seit inter- oder sogar präglazialen Zeiten innehaben; die engsten verwandt- schaftlichen Beziehuiigeii bestehen zu hadzii, denn diese Art besiedelt auch eines der süd-alpinen Refugialgebiete:
Ve rb re i t ung u n d u n t e r s u c h t e s Mater ia l : ' Bisher galt I. strandi als Endemit der Monti Lessini und des Monte Baldo östlich des Gardasees. Da aber ravasinii zu strandi gestellt werden muß und überdies ein weiteres Exemplar aus den Bergamaska Alpen vorliegt, ergibt sich ein vergleichsweise ausgedehntes Areal in mehreren Gebirgsgruppen der S-Alpen. Inwieweit die jetzt isoliert erscheinenden Teilareale miteinander in Verbindung stehen, bleibt zu überprüfen (vgl. Abb. 26 und 58).
Es wurden 3 6 , 7 ?,und 3 Jungtiere untersucht.
I t a l i e n . - Venezianer Alpen, Cansiglio, Grotte Bus del Pa1 (1 Holotypus I. ravasinii, Zool. Inst. Ljubljana, RAVASINI leg. 1921); Cansiglio, Spelonca delle due bocches (1 iuv. Mus. Wien, RAVASINI leg.); Monte Baldo, (1 8, 1 9 Mus. Wien. HERTIG leg. 1928; ferner 1 $, 1 9 Typen- serie I. knirsclzi SMF/RII/G441), Buce della Preta (1 ? Mus. Wien), Grotte hfalga Prato (1 ?
Abb. 64. I. strandi KRATOCHV~L. - a Receptaculum seminis (Monte Baldo, Gegenseite nur 3 Schlauche); b Cephalothorax von rechts-lateral (Monte Baldo).

Tlolotypiis coll SILHBVY 011284), i?lonti Lessini, Grotta Spluga, S Mauro di Salina, Provinz Verond (1 8, 1 9, 2 iuv coll BRIGNOLI, VIGNA leg. 24. J11 1968), Bergamasker Alpen, Val Serlana, Grotte Bus del Tacoi (1 9 coll. Zoo1 Inst L~ubI~ , lna , vgl H A D ~ I 1954, sub aff strasserz). - Wei- tere Fundor te Monte Baldo, Grotta dei Cervi, 1425 m , Monti J,esslni, Cogolo del Zavatin, 1475 m; Bus de la Fanta, 1475 nl , Buso de Ia neve dc 1d Gaibaria, 1691 rn, Buso del16Arena, 1524 m ; Spluga de In P ~ e t a , 1445 m , Covoli di Velo, 875 111, Crrottd del Monte Caobo, 925 m ; Buso de la D\'ef de la Col, 1574 nl (alle J u s ~ n r i r r ~ 1963, 19G4)
Voi l rominen: I. strand% wurde bisher n u r in Hohlen gefunden (vgl. Ilap. V 2b).
A n h a n g ; Korpermaße (in 111111) (Erkl<~rung der Meßstrecken vgl. Abb. 34):
2 CS-L (1) 1,7 - 1,9 (1,82); CX-U (2) 2,2 - 2,35 L, (5) 5,1 (5,lz); "2 (6) 6,5 B2 (7) 0 , 9 5 1,O (1,O2), 113 (8) 2,35- 2,4 F e l l (9) 6,9 (6 ,g~) ; Mt I1 (10) 8,8 - 9,0 Pen. (11) 4 , l - 4,4 (4,25,), Mt Pg (12) 2; TaGe
? Cx-1, (1) 1,s - 1,9 (1,85); Cs-B (2) 2,2 - 2,3 11, (5) 5,1 - G,O (5,341; L2 (6) (6,5) - 7,7 B2 (7) 1,O - 1,05 (1,024; L, (8) 2,5 - 2,75 ~e (9) 6,2 - 7,O (($73,); Mt11 (10) 8 , l - 9,5 R'ItPg (12) 2 - 3 TaGc (13) 38 -40.
I . mugdulenue S l n ~ o ~ 1881 (Abb. 65-66) 1881 Ischyropsalis Magdalenae SIMON, Ann. Soc. esp. Hist. Nat., 10: 130 (D 9). J914 1. madalenae (sic!), - ROEWER, Arch. Naturgescli., 80 A (3): 118 (D 9). 1923 T. madulenae, - ROEWER, Wcberlmechte d . Erde: 690 (J) 9). 194G I . helzoigzi, - Riinrijr,~, Aport. est, fauna Hora vasco-navar.,: 52 Abb. 2-4 (D P). 194G I. dispar, - R A ~ B L A , Aport. est. fauna flora vasco-rin\~ar.,: 53 Abb. 5-6 (D$). 1950 I. naudelenae (sic!), - ROEWER, Senckenbergiana, 31 (112): 46 T. 6 Abb. 35 c-d (D 9).
v 1950 1. turki R o ~ w s i z , Senckenbergiana, 31 (112): 31 T. 5 Abb. 27a-d (D$?). v 1950 1. n,uvarrensis ROETVEII, Senckenbergiana, 31 (112): 44 T. 9 Abb. 56 C-d (D 9). v 1950 I. nicaea ROEWEI~, Senckenbergiana, 31 (112): 36 T. 8 Abb. 48 a-b (D P).
B e g r ü n d u n g d e r S y n o n y m i e : Die Original-Beschreibung (ohne Abbildungen) basiert aiif einein 9, sie langt zur Wiedcrerkennung der Spezies nicht aus. ROEWER (1950) bildet zwar die Cheliceren des Typus ab., doch sind die Skizzen sehr schematisiert. Die Übercinstimrnung des Typus (einschließlich der Rez~ignahmen von ROETVER 1914, 1923 und 1950) und dem übrigen, in sich einheitlichen Matcrin,l vorstehender Liste ist nicht ganz gesichcrt, denn SIMON (1881) -
spricht von „Plaque et seginents abdoinineaus", also einein Scutum parvum.Alle vorliegenden 9 tragen aber cin Scuturii la,ininatuin (ebenso das 9 , das RAMRLA darstellt). Indes können die einzelnen SBLerite sehr hell sein, so daß bisweilen ein Scutum parvunl vorgetäuscht wird. - Sicherheit in dieser nomenlrlatorischen Frage ist nur durch die Überprüfung des Typus zu erlan- gen (Mus. Paris)l). Die vorgeschlagene Zuordnung ist aber sehr.wahrscheinlich richtig, da magda- lenae von der einzigen habituell ähnlichen Art N-Spaniens bereits nach äußeren Merkmalen (Tuber ocitlorurn) sicher getrennt werden kann (vgl. Abb. 66 und 68). - Die Typenserien von turki (d?), navarrensis (P) und nicaea (9) gehören zu einer Art; alle wesentlichen Merkmale stimmen mit den relativ guten Abbildungen von magdalenue überein, die RAMBLA (1946) bietet (sub dispar und hellwigi).
I
1) Trotz wiederholter Bemuhungen war es mir nicht moglich, diesen zu untersuchen. I
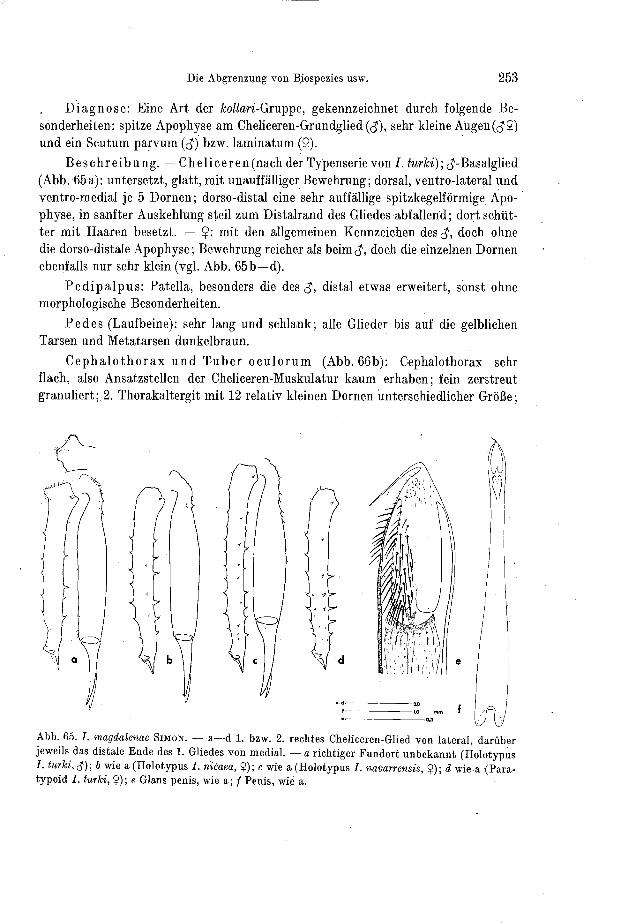
Die Abgrenzung von Bjospezies usw. 253
Diagnose: Eine Art der kollari-Gruppe, gekennzeichnet durch folgende Be- sonderheiten: spitze Apophyse am Cheliceren-Grundglied ($), sehr kleine Augen($$) und ein Scutum parvum (8) bzw. laminatum (Y) .
Beschreibung. - Cheliceren(nach der Typenserie von I. turki); $-Basalglied (Abb. 65a): untersetzt, glatt, mit unauffälliger Bewehrung; dorsal, ventro-lateral und ventro-medial je 5 Dornen; dorso-distal eine sehr auffällige spitzkegelförmige Apo- physe, in sanfter Auskehlung steil zum Distalrand des Gliedes abfallend; dort schüt- ter mit Haaren besetzt. - 9: mit den allgemeinen Kennzeichen desd, doch ohne die dorso-distale Apophyse; Bewehrung reicher als beim$, doch die einzelnen Dornen ebenfalls nur sehr klein (vgl. Abb. 65b-d).
Ped ipa lpus : Patella, besonders die des $, distal etwas erweitert, sonst ohne morphologische Besonderheiten.
Pedes (Laufbeine): sehr lang und schlank; alle Glieder bis auf die gelblicheil Tarsen und Metatarsen dunkelbraun.
Cepha lo tho rax u n d Tube r oculorum (Abb. 66b): Cephalothorax sehr flach, also Ansatzstellen der Cheliceren-Muskulatur kaum erhaben; fein zerstreut granuliert; 2. Thorakaltergit mit 1 2 relativ kleinen Dornen unterschiedlicher Größe;
Abb. 65. I. magdalenae S I M O N . - a-d 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber jeweils das distale Ende des 1. Gliedes von medial. - a richtiger Fundort unbekannt (Holotypus I. turki , 8); b wie a (Holotypus I. nicaea, 9); c wie a (Holotypus I. navarrensis, 9); d wie a (Para- typoid I. turki , ?); e Glans penis, wie a ; f Penis, wie a.

Tubcr oculorum sehr nicdrig, median bis auf die Hohe des Cephalothorax gefurcht, Augcn weit voneinander entfernt, etwa 3facher Augen-Durchmesser, Linse klein.
S cu turn: das einzige vorliegeiide $ mit ciiicm Scutum parvum, von dem Area V jedoch weitgehend gctrennt ist; allc Arcae (und die freien Tergite) mit einer Qucr- reihc langcr und steifer Borsten, jeweils auf eincni winzigen Tuberkel; 9 mit Scutum lamiiiatum, dessen Sklerite sehr klein, ebcnfalls mit einer Borstenreihe; $- und 9- Scutum fein granuliert.
Pen is : Trulicus (Abb. 65f): von der Basis bis zum distalen Drittel kontinuierlich verschmalert, von dort etwas zur Glans verbreitert; Sklerit der Glans (Abb. 65e) schmal und relativ lang, Trichtcr weit.
Recep t acu lum semin is (Abb. 66a): kurze und dicke Schlauche mit engem Lumen, stark in sich gekrummt; die einzige präparierte Legerohrc rechts und links mit je 4 Schlauchen.
Var iab i l i t a t : Schwierig zu beurteilen, da nur ein $ vorliegt; RAMBLA (1 946, sub dzspar) zeichnet ein$, dessen basales Chcliccrcn-Glicd völlig unbewehrt, lediglich mit cinigcri Borsten bcsctzt ist; kcinc Unterschiede in der dorso-distalen Apophyse; Proportion und Verteilung der Bewehrung der $2-Cheliccren vgl. Abb. 65b-d.
Bez iehung e 11: Die drei habituell sehr ähnlichen Artcn muellneri, magdalenae und dzspar der kollarz-Gruppe besiedeln zwei sehr disjunkte Areale in den Dinarischen Gebirgen dcr Balkanhalbinscl (muellneri) bzw. in den Kantabrischen Gebirgen (mag- dalenae, dzspar). Diese Arten sind zwar genitalmorphologisch recht verschieden (bc- sonders muellnerz/magdalenae gegcnuber dzspar), was außer der disjunkten Vcrbrei- tung in typischcn Rcfugialgebieten auf ihr hohes Alter hinweist, doch gehen sie (innerhalb der kollari-Gruppe) wahrscheinlich auf eineil gemeinsamen Vorfahren zu- ruck. Die synapomorphc Ausformung der 8- und auch der $2-Cheliccren deutet vor allem darauf hin.
Dic $ dieser drei Arten sind an der Form der Cheliceren-Apophyse leicht zu dif- fercnzicren, denn bei muellnerz ist sie sehr klein und liegt medial, bei den beiden anderen Arten ist sie großer und liegt dorsal, bei magdalenae sehr spitz (sie bildet einen
Abb. 66. I . rnaqdaler~ae S~mois. - a Receptaculuiri seminis (I-Iolotypus I. navarrenszs); b Crphalo- thorax von rechts-lateral (Holotypus I. turki).

Die Abgrenzung von Biospezies usw. 255
Winkel von weniger als 90°), bei dispar stumpf (Winkel über 90"). - Während die von magd~lenae und dispar leicht zu trennen sind (Augen, Scutum), unterscheiden sich magdalenae und muellneri im wesentlichen nur durch die Bewehrung des 2. Tho- rakaltergits (magdalenae kleine Dornen, muellneri sehr große).
Ve rb re i t ung u n d u n t e r s u c h t e s Ma te r i a l : I. magdalenae besiedelt ein (nach heutiger Kenntnis) sehr kleines Areal in den Kantabrischen Gebirgen N-Spa- niens. Der Holotypus stammt aus der Höhle Magdalena bei Galdames, weitere Indi- "
viduen liegen aus Höhlen der Sierra de Aralar vor, die RAXRLA (1946, sub dispar und hellwigi) gut abbildet. Aus derselben Region („Navarrau) wird der Holotypus von navarrensis gemeldet (synonym magdalenae). - Ganz offensichtlich unrichtig sind die Fundorts-Angaben „NizzaU (nicaea) und „Insel Ägina, Griechenland" (turlci); - vgl. Abb. 26 und 52.
Es wurden 1 und 3 9 untersucht.
1 S p a n i e n . - , ,Navarract (1 Holotypus 1. navarrensis. SMF/RII/5033). - Weitere Fund- orte: S p a n i e n . - Biscaya, Galdames, Höhle La Magdalena (Locus typicus, SIXON 1881); Na- varra, Sierra de Aralar, Höhlen von Aparein, Kilixketa, Putxerri und Putxerri Ko-Aizpea (vgl. RAMBLA 1946, sub hellwigi und dispar). - Fundorts-Angaben sicher falsch: Griechenland, Insel
I . Ägina ( 1 3 , 1 9 Typenserie I. turki SMF/RI1/5028, angeblich ROEWER leg. VIII. 1926); Frank-
' reich, Riviera, Nizza-Villefranche, an der Corniche-Straße (1 ? Holotypus I. nicaea, SMF/RII / 9494).
I Vorkommen: Alle Individuen, 'deren Herkunft gesichert erscheint, wurden in
I Höhlen gesammelt. Kenntnisse über die Ökologie bestehen außerdem nicht.
A n h a n g : Korpermaße (in mm) (Erklarung der Bleßstrecken vgl. Abb. 34):
$ CX-L (1) 1 9 Cx-B (2) 2,7,; SC-L (3) 2,8,; Sc-B (4) 2,4; LI (5) 5,21; L2 (6) 7,051; B2 (7) 0,851; L3 (8) 2,4, F e I I (9) 9,3,; Mt11 (10)12,8,; Pen. (11) 4,5,; MtPg (12) 3 ; TaGe (13) 49.
9 CX-L (1) 1,5 - 1,8 (1,6g3); CX-B (2) 2,7 - 2,8 (2,73,); L1 (5) 5 , l - 5,7 (5,33,); L2 (6) 6 9 - 7,5 (7,133); B, . (7) 0,87- 1,O (0,92,);
4 L3 (8) 2,2 - 2,7 (2,363); Fe11 (9) 7,9 - 8,7 (8,4,); Nt11 (10) 10,5 -11,G (11,133); MPg (12) 2 - 3 ; TaGe (13) 45 -50.
I. dispar SIMON 1872 (Abb. 67-68) 1872 Zschzropsalus dzspar S I M O ~ , Ann. Soc. ent. France, (6) 2: 277 T. 12 Abb. 11, 12 (D S, iuv.
nec $9). 1879 I. dzspar, - SIMOK, Arachnides d. France, 7: 274 (D$, iuv.). 1881 1. dispar, - SIMON, Anri. Soc. espan. Hist. Sa t . , 10: 128 (N). 1914 I . dzspar, - ROEWER, Arch. Zi'aturgesch., 80 A (3): 113 Abb. 9 a-b (D $, iuv.). 1923 1. dzspar, - ROEWER, Weberknechte d. Erde,: 687 Abb. 858 (D$, iuv., nec $9).
? 1935 1. dzspar, - ROEWER, Arch. Zool. Exper. Gen., 78 (1): 83 (N). 1936 1 dzspar, - MELLO-LEITAÖ, Treb. Mus. Cienc. Nat. Barcelona, 11 (9): 11 (N).

1950 I. dispar, - ROEWER, Senckenbergiana, 31 (112): 44 T. 2 Abb. 11 a-d (D $, iuv., nec 80).
v 1950 I. goodnighli ROEWER, Senckenbergiana, 31 (112): 31 T. 2 Abb. 13 a-d (D $9). V 1950 I. archeri ROEWER, Senckenbergiana, 31 (112): 48 T. 2 Abb. 12 a-b (Dd) .
1961 I. dispar, - I i ~ a u s , Senckenbergiana biol., 42 (4): 348 (N). 1961 I. archerz, - KRAUS, Senckenbergiana biol., 42 (4): 349 (N).
B e g r ü n d u n g d e r S y n o n y inie: Wiedererkennung nach Beschreibung und Abbildung von SIMON ist möglich, doch ist zu beachten, daß die ?-Diagnose offensichtlich auf einem Jung- tier basiert (Körper und Extremitäten „blanc testach", ,,arceaux superieurs . . . tous separbs", also ein Scutum laminatum). 1879 berichtigte SIMON den Irrtum. - Die Typenserien von good- nighti und archeri müssen zu dispar gestellt werden, denn sie weisen alle typischen Merkmale dieser leicht kenntlichen Art auf.
Diagnose : Eine Art der kollari-Gruppe, gekennzeichnet durch folgende Be- sonderheiten: ein (fast) vollig glattes, axtformiges Cheliceren-Grundglied ($), einen extrem schlanken Penis ($), einen großen Tuber oculorum ($9) und ein Scutum parvum ($9).
Beschre ibung . - Cheliceren (nach Material aus Ordulla, dem weiteren Lo- cus typicus); 3-Basalglied: glänzend glatt, bis auf einige winzige Tuberkel unbewehrt; an der Basis sehr dunn, von dort bis zur dorso-distalen Apophyse schwach, aber kontinuierlich verbreitert; Distalende etwas nach ventral abgewinkelt und dorsal zu einer stumpfen Apophyse ausgezogen, zur Insertionsstelle des 2. Gliedes plan abfal- lend, uber diese hinausragend und so eine weitere kleine medio-distale Apophyse bil- dend; ohne auffallige Dornen-Bewehrung, lediglich winzige Borsten auf Tuberkeln deuten die 3 Hauptreiheii an; die beiden Apophysen mit Besatz sehr feiner Haare. - 2. Glied: schlank, am „Knie" dorsal, medial und lateral isolierte Tuberkel. - - 9: mit den allgemeinen Kennzeichen des $, doch ohne die charakteristische dorso-distale Apophyse und mit wesentlich starkerer Bewehrung aus kräftigen, aber schlanken, dorsal nahezu rechtwinklig abstehenden Dornen, dorsal 3-7, ventro-lateral3-5 und ventro-medial 7 ; am 2. Glied kraftigere Bedornuiig als beim $; medial in einer Reihe bis zur Basis des festen Fingers reichend.
P e d ip a lp U s ($9): lang und schlank; schutter mit langen Haaren besetzt (Länge etwa wie Durchmesser des entsprechenden Gliedes).
Cepha lo tho rax u n d T u b e r ocu lorum (Abb. 68): Cephalothorax mäßig ge- wolbt, mit winzigen Borsten hinter dem Tuber oculorum; 2. Thorakaltergit mit 11 re- lativ großen, nahezu senkrecht stehenden Zahnen; Tuber oculorum und Linse der Augen groß; Tuber durch mediane Furche nahezu zweigeteilt.
P e de s (Laufbeine): lang und feingliedrig.
S C U t u m : $9 mit Scutum parvum; alle Areae (und die freieil Tergite) mit einer Querreihe dichtstehender Tuberkel, die mittleren der 1. Areae, besonders beim $, ein schlankes Dornenpaar.

Die Abgrenzung von Biospezies usw. 257
P e n i s : Truncus extrem schlank und lang; wenig unterhalb der Glans am schmal- sten, von dort bis zum basalen Viertel kontiiiuierlich verbreitert, aber nicht bauchig, dann wieder etwas schmaler; ventraler Sklerit schmal, kraftig beborstet ; Trichter eng.
R e c e p t a c u l u m s e m i n i s : 3-4 mittellange bis lange Rezeptakel (2 Legeroh- ren prapariert).
V a r i a b i l i t a t : Das vorliegende Material (nur 5 $, 4 9) weist die Art als wenig variabel aus; die Form der Apophyse des Chelicereii-Grundgliedes (die disto-frontale
Abb. 67. I . dispar SIMOX. - a-h 1. bzw. 2. rechtes Cheliceren-Glied von lateral, darüber jeweils das distale Ende des 1. Gliedes von medial. - a „San Sebastina" (Spanien; Holotypus I. archeri, 8); b Ordufia, Vizcaya (Spanien, $) ; c Ordufia, Vitoria, Prov. Vascongadas (Spanien, $) ; d richtiger Fundort unbekannt (Holotypus I. goodnighti, 8); e wie b ($2); f wie C (9); g und h wie d (Para- typoide); i-k Glandes penis: i wie d ; k wie c ; 1 Penis, wie d.
17 2001. Jb. Ssst. Bd. 96

Ebene!) ist konstant; bisweilen ist das Glied völlig dornenlos. 9-Cheliceren vgl. Abb. 67e-h.
Beziehungen: vgl. die Ausführungen bei I. magdalenae.
Ve rb re i t ung u n d u i i te rsuchtes Mater ia l : I. dispar scheint auf ein winzi- ges Areal in den Kantabrischen Gebirgen südlich Bilbao beschränkt zu sein. Sowohl die Typenserie als auch eine weitere Nennung durch SIMIN (1881) und die beiden Serien mit verläßlichen Fundorts-Angaben, die hier vorliegen, beruhen auf Material, das bei Orduiio nahe der Stadt Vitoria gesammelt wurde. Die Angabe ,,SanSebastianU von' ROEWER (sub archeri) ist ungenau und kann zur Kenntnis des Areals nicht bei- tragen. - Ganz offensichtlich unrichtig ist die Nennung für Griechenland (sub good- ~
nighti), vgl. Abb. 26 und Abb. 52.
Es wurden 5 3 , 4 9 und 1 Jungtier untersucht.
S p a n i e n . - Provinz Vizcaya; San Sebastian (2$. Typenserie I. archeri SMF/RII/5029); Orduiio (1 6, 1 9 RAMBLA ded:); Ordufio/Vitoria (1 $, 1 0 , 1 iuv. RAKBLA ded.). - Weitere Fund- orte: Provinz Vizcaya, Orduiio, Cueva de Albia (Locus typicus, SIXON 1872); Orduiio, Cueva de lia Embajada (SIMON 1881). - Fundorts-Angabe sicher falsch: Griechenland, Mores (= Pelopon- nes), Höhle am Berghang von Vityna (1 $, 2 9 Typenserie I. goodlzighti, SMF/RII/5040, angeblich ROEWER leg. 1926).
Vorkommen: Die näher erläuterten Funde stammen aus Höhlen; es ist unbe- kannt, ob auch freilebende Populationen vorkommen.
A n h a n g . Korperniaße (in mm) (Erklarung der Rleßstrecken vgl. Abb. 34):
6 CX-L (1) 1 ,5 - 1,9 ( 1 6 5 , ) Cx-B (2) 2 , l - 2,6 (2,28,); SC-L (3) 2,5 - 3 , l ( 2 0 , ) SC-B (4) 2,2 - 2,7 (2,38,); LI (5) 5,O - 5,GO (5,37,); L, (6) 6,2 - 7,O (6,53,); B, (7) 0,72- 0,85 (0,78,); L, (8) 2,05- 2,2 (2,15,); Fe11 (9) 7 , l - 8,7 (8,3,); Mt11 (10) 10,8 -13,l (11,9,); Pen. (11) 4,7 (4,7,); nrt PE (12) 2-3; TaGe (13) 44-48.
? Cx-L (1) 1,6 - 1,7 ( 1 6 ) Cx-B (2) 2 , l - 2,4 (2,27,); SC-L (3) 2,7 - 2,9 (2,77,); Sc-B (4) 2,2 - 2,s (2,33,);
Abb. 68. I. dispar SIMON. - a Receptaculum seminis (Patraypoid I. goodnightz); b-c Cephalo- thoraces von rechts-lateral: b und c Orduiio.

Die Abgrenzung von Biospezies usw.
LI (5) 5,4 - 5,s (5,57,); L, (6) 6 9 - 7,2 (7,0%); B, (7) 0,78- 0,95 (0,84,); L, (8) 2,2 - 2,4 (2,331; Fe11 (9) 7,4 - 8,3 (7,97,); J l t I1 (10) 11,7 -11,9 (11,82); Mt Pg (12) 3 ; TaGe (13) 45 -46.
VII. Liste der nominellen Arten
(Verwe i s auf d ie va l i den A r t e n )
Anmerkung: Fette Seitenzahlen beziehen sich auf die vorliegende Arbeit, Zahlen mit vor- gestelltem „R" bezeichnen die Seiten der Revision von ROEWER (1960).
abei: zur Gattung Nipponopsa l i s , R 50 adami i : 205, R 33 al fkeni : s. nauellneri, R 19 anzseli: s. adami i , R 33 apuana: s. adami i , ~ ' 3 6 archeri: s. dzspar, R 48 asturica: s. nodifera, R 48 austriaca: s. manicata , R 22 balcanica: s. manicata , R 30 bosnica: s. kollari, R 29 cancroides: s . hadzi i , R 52 caporioccoi: s. luteipes, R 33 carli: 209, R 19 corcyraea: s. mue l lner i , R 28 corsica: s. nodifera, R 34 crassichelis: s. helleaigi hellwigi, R 25 dacica: s. manicata , R 30 danub ia : s. kollari, R 21 dent ipalpis: 201, R 36 dispar: 255, R 44 frankenbergeri: s. hellwigi hellwigi, R 21 franzi: s. hellwigi helleaigi, R 22 goodnighti: s. d i spar , R 31 hadz i i : 242, R 52 hellwigi hellroigi: 196, R 18 (sub t aun ica ) hellwigi lucantei 200, R 42 (sub lucantei et
superba) helvetica helvetica: s. dent ipalpis , R 20 helvetica milleri: s. manica ta , R 20 herbsti: s. hellwigi hellwigi, R 25 hispanica: s. nodifera, R - janetscheki: s. nodifera, R 22 kaestneri: s. manicata , R 19 knirschi : s. s trandi , R 27 kollari: 236, R - kratochvili: s. a d a m i i , R 32
17*
lucantei: s. hellwigi lucante i , R 42 lus i tanica: s. robusta, R 50 luteipes: 213, R 38 naagdalenae: 252, R 46 nzanicatc~: 216, R 29 moreana: s. pyrenaea ( J ) , nod?jfera (9). R 31 nzuellneri: 245, R 26 navarrensis: s. magdalenae, R 44 nicaea: s. naagdalenae, R 36 ,
nival is : s. kollari, R 26 nodifera: 222, R 40 pectinifera: s. hellwigi hellwigi, R 21 pentelica: s. carli , R 30 pestae: s. luteipes, R 28 petiginosa: s. nodi fera , R 48 plicata: s. helleuigi hellwigi, R 24 pyrenaea: 227, R 40 pyrenaea pyrenaea: s. pyrenaea, R - pyrenaea venaspuensis: s. pyrenaea, R - racasini i : s. s trandi , R - reimoseri: s. kollari, R 23 robusta: 233, R 46 ruf fo i : s. s trandi , R 36 scutata: s. kollari, R 26 segregata: s. kollari, R 28 spinichelis: s. kollari, R 25 s trandi: 249, R 34 strasseri: s. kollari, R 22 styriaca: s. kol lar i , R 23 superba: s. hellzoigi lucantei, R 42 taunica: s. helleoigi helleoigi, R 18 t irolensis: s. kollari, R 23 triglavensis: s. kollari, R 24 troglodytes: s. kollari, R 32 t u rk i : s. magdalenae, R 31

VIIH. Zusammenfassung
1. Die Gattung Ischyropsalis C. L. KOCH wird mit bisher nicht beachteten biologisch-etho- logischen und morphologischen Merkmalen neu gegliedert, und die Verwandtschaftsbeziehungen der einzelnen Arten werden erörtert. Die 15 validen Arten sind mit sehr hoher Wahrscheinlichkeit isolierte Fortpflanzungs-Gemeinschaften (Biospezies). Für die allopatrischen Arten der Gattung läßt sich das jedoch nicht nachweisen. - Dieses Ergebnis stützt sich auf zahlreiche Einzel- untersuchungen:
2. Alle Arten der Gattung sind stenok. Sie besiedeln nur Habitate mit niedrigen Durch- schnittstemperaturen, ausgeglichenem taglichen Temperaturgang und hoher relativer Luft- feuchtigkeit.
3. Größte Siedlungsdichte besteht in Mittel- und Hochgebirgen; die höchsten Fundorte liegen zwischen 2500 und 2600 In.
4. Von allen Arten sind kavernikole Populationen bekannt. Fünf Arten (hadzii, strandi, muellneri, magdalenae, dispar) scheinen nur in Höhlen vorzukommen; diese Arten weisen typi- sche Höhlentiern~erkmale auf.
5. In zwei Fällen wurden im selben Biotop zwei Arten gefunden. Diese syntopisch lebenden Arten sind nicht eng miteinander verwandt.
6. Die Reifezeit von drei näher untersuchten Arten bzw. Rassen liegt im Sommer und Herbst. J e nach Höhenlage des besiedelten Biotops differieren die Reifezeiten der einzelnen Populationen.
7. Die Nahrung der allermeisten Arten besteht aus Arthropoden; strenger Nahrungs- spezialist scheint allein I. h. hellwigi zu sein, die sich wahrscheinlich nur von Mollusken ernährt.
8. Das Kopulationsverhalten von I. hellwigi und I. luteipes wird beschrieben. Während eines charakteristischen Werbeverhaltens bietet das 3 dem 9 ein Sekret an, das auf einem Bürsten- feld des Cheliceren-Grundgliedes austritt. Vergleichbare Verhaltensweisen sind nur von einigen Arten der Blattaria, Ensifera und von den Malachiidae (Coleoptera) bekannt.
9. Für die Strukturen des 8- und 9-Genitalapparates (Penis bzw. Receptaculum seminis) wird eine Nomenklatur eingeführt.
10. Die systematische Wertigkeit markanter Merkmale des Genitalapparates und der peri- pheren Gestalt wird untersucht (Penis, Receptaculum seminis, abdominales Scutum, Cheliceren- Bewehrung, Apophyse und Drusenfeld der 3-Chelicere). Die wichtigsten Art-Merkmale sind Form und Lage der Cheliceren-Bewehrung, der Apophyse und des Drüsenfeldes der 8-Chelicere, Pro- portion des Truncus und des ventralen Sklerits des Penis.
11. Das abdominale Scutum entwickelt sich während der Ontogenese. Bis zum Subadult- Stadium sind die Sklerite jedes Segmentes isoliert. Die abdominalen Skleritspangen I-V ver- schmelzen erst bei der Imaginalhäutung. Innerhalb der Gattung ist das Scutum parvum als phylogenetisch ältester, das Scutum laminatum als phylogenetisch jüngster Entwicklungszustand zu werten.
12. Die artisolierend wirkenden Mechanismen syntopisch vorkommender Arten werden erörtert. Es bleibt vorerst offen, ob sie in unterschiedlichem Verhalten, im Chemisnlus des Drüsen- sekretes oder in weiteren (noch unbekannten) Faktoren zu suchen sind.
13. Mit den als artkennzeichnend (biospezifisch) erkannten Merkmalen, die eine bestimmte Artengruppe auszeichnen, werden weitere Merkmale „geeicht" und auf Populationen unbekann- ter Biospezies-Zugehörigkeit übertragen. Die so „erschlossenen" Spezies stellen wahrscheinlich ebenfalls Biospezies dar.
14. Die Gattung ist auf Europa beschränkt. Die Areale der Arten, der Artengruppen und das der Gattung werden dargestellt.

Die Abgrenzung von Biospezies usw. 261
15. Die Arten der beiden polytypischen Artengruppen vikariieren in den ineisten Fallen. 16. Die Speziation läßt sich zum größten Teil auf Isolation einzelner Populationen während
der Glazia,lzeiten zurüclrführen. Postglaziale Arealüberschiebungen nächstverxvandter Arten fan- den nur ausnahinsweise statt . 3
17. In den beiden polytypischen Artengruppen lassen sich eGolutive Trends erkennen: Ver- larigerung der beiden Cheliceren-Grundgliedep, Reduktion oder exzessive Verstärkung der Chelii ceren-Bexvehrung, Auflösung des abdominalen Septum, Reduktion der Cheliciren-Bürste. .
18. Die Ischyropsalididae stehen wahrscheinlich in einem Sch~vestergruppen-Verhältnis zu den Trogulidae und Xemastomatidae.
19. Die vorliegenden Untersuchungen legen eine Einteilung der Gattung in 15 Arten nahe (früher 56). Sie werden beschrieben, die Synonymie begründet und alle vejfügbaren Daten über . .
Verbreitung, Ökologie und Biologie werden mi'tgeteilt. .,
IX. Summary
1. The genus Ischyripsalis C. L. KOCH is revised an the basis of previously unnoticed biological-ethological and morphological characteristics, and the relationships of the single species are discussed. Very likely, the 15 valid species are isolated communities (,,biospeciesU). But this will not be demonstrable for the allopatric species of the genus. - The result is based on many indi\-idual researches.
2. 911 species of the genus are stenoecious. They only settle in habitats witli lowoaverage teinperatures, balanced daily course of temperature and high humidity.
3. Highest density of settlement occurs in medium-high and high mountain chains, the highest altitudes of settlemeilt being located betmeen 2500 and '2600 m.
4. Cave-dwelling populations of all species are known. Five species (hndzii, strandi, muell- '
~zeri, magdalenae, dispnr) seem to occur only in caves. These species show typical symptoins of cave-dwelling.
5. In two cases, two species were found in the same biotop very close to each other. These syntopic living species are not closely related.
6. The maturity time of three ~vell-exainined species and/or subspecies is in summe. and autnmn. She inaturity times of the single populations differ slightly and depend an the heights of the biotops.
7. The food of most of the species consists of arthropods; the only strict food specialist seeins to be I. helltoigi helluigi, ~vhich probably feeds only on n~olluscs.
8. The copulation behaviour of I. hellzoiyi and I. luteipes is described. During a characte- ristic conrtship, the $ offers the 9 a secretion flowing out of an area of bristles (,,BürsteL') located a t the end of the basic mernber of the chelicerae. Comparable behaviour is only known in soine species of Blattaria, Ensifera and Ma,lachiidae (Coleoptera). It has so far been nnknown in Arach- nida.
9. Womenclature is introdnced for tlie $- and 9-genitalia (penis, receptaculum seiniiiis). 10. The systematic importance of prominent characteristics of the genitalia and the
esterior morphology is being examined (penis, receptaculum seminis, scutum of the abdomen, arnlament cf chelicerae, apophysis and secretion area of $-chelicerae). The inost important spe- cific characters are shape and position of the armainent of thechelicerae, of the apophysis and the secretion area of the $-chelicerae, proportion of truncus penis and the ventral sclerite of the penis.
i 11. The abdominal scutum develops during ontogenesis. The sclerites of each segment are
isolated until the subadult stage is reached. The sclerite clasps I-V of the abdomen melt together

during moult into adult stage. Within the genus, scutum parvum must be regarded as the phylo- genetic oldest, and scutum laminatum as t he phylogenetic youngest stage of development.
12. The mechanisms, isolating syntopic living species, are discussed. I t has so far not been determined.wether they consist in varying behaviour, in chemism of gland secretion or in other (still unknown) factors.
13. With the characteristics recognixed as marking species (biospecific) ~vhich make out a certain group of species, further characteristics are „gaugedU and transmitted to populations of an unkno~vn biospecific membership. Species, disclosed in such way, are probably also biospecies.
14. The genus is restricted to Europe. The distribution of the species, species groups and of the genus i s , described.
15. In most cases the species of the two polytypic species groups settle in vicarious areas. 16. To a large extent the speciation is caused by isolation of single populations during
glacial periods. Post-glacial overlap of areas of closely-related species occurred only in excep- t i o n ~ l cases.
17. In the'two polytypic species groups, trends of evolution can be observed: prolongation of the two basic members of chelicerae, reduction or escessive strengthening of chelicerae arms, dissolution of the abdominal scutum, reduction of the $-chelicerae bristle area.
18. The Ischyropsalididae probably form a sister group of the Trogulidae and Nemasto- matidae.
19. The above-mentioned researches make i t obvious tha t the genus is divided into 15 spe- cies (before 56). They are described, the Synonyms established and all available data on distribu- tion, ecology, and biology communicated.
X. Literatur
Hier werden nur die im Test zitierten Autoren aufgeführt; es erübrigt sich, das in den Synonym-Listen erwähnte Schr i f t tu~n anzuführen, da dieses bereits dort genau zitiert ist. ATTRIWI, ST. (1964), Observations sur la variabilite de l'esphce Ischyropsalis dacica ROEWER des
grottes de Roumanie (Opiliones, Palpatores, Ischyropsalis). - Rev. Roumanie Biol., sßr. Zool. 8, 245-256. Bucaresti.
BECIC, L. (1965), Über Variabilität und Wertigkeit morphologischer Merkmale bei adulten Oriba- tiden (Arachnida, Acari) am Beispiel der Gattung Rostrozetes SELLNICIC 1925. - Abh. Sen- ckenberg. naturforsch. Ges. 588, 1-64. Frankfurt am Main.
DE LATTIN, G. (1967), Grundriß der Zoogeographie. Jena. DRESCO, E. (1952), Repartition d'Ischyropsalis luteipes SIMON (Opiliones Ischyropsalidae). -
Notes biospkol. 7 , 101-104. - (1966a), Recherches sur les Opilions du genre Ischyropsalis (Fam. Ischyropsalidae). I. Les
caractkres systematiques. - 11. Ischyropsalis robusta SINON. - Bull. Mus. Nat. Hist. Nat. (2) 38, 586-602. Paris.
- (1966b), ~eche rches sur les Opilions du genre Ischyropsalis (Famille Ischyropsalidae). 111. Ischyropsalis superba SIMON et Ischyropsalis lucantei SIMON. - Bull. Mus. Nat. Hist. Nat. (2) 38, 801-816. Paris.
- (1967a), Recherches sur les Opilions du genre Ischyropsalis (Fam. Ischyropsalidae). - IV. Ischyropsalis nodifera SIXON. - Bull. Mus. Nat. Hist. Nat. (2) 39, 173-187. Paris.
- (1967 b), Recherches sur les Opilions du genre Ischyropsalis (Fam. Ischyropsalidae). V. Ischy- ropsalis petiginosa SINON. - Bull. Mus. Xat. Hist. Nat. (2) 39, 506-514. Paris.
- (1967 C), Recherches sur les Opilions du genre Ischyropsalis (Fam. Ischyropsalidae). VI. Ischy- ropsalis pyrenaea SIMON. - Bull. Mus. Kat. Hist. Nat. (2) 39, 565-679. Paris.


MARTENS, J . & S. Snzur~r (1966), Zur systematischen Stellung ostasiatischei Ischyropsalididen-Ai ten (Arachqoidea, Opiliones, Ischyropsalididae). - Annot Zool. Japon. 39, 215-221. Tokyo.
~ I ~ Y R , E. (1963/67), Animal Species and Evolution. Caiubridge Mass. - Artbegriff und Evolution. Hamburg, Berlin. Deutsche Ubertragung von G. AEBERER.
MEIIYERTL, TII. (1964), Eine zootopographische Untersuchung uber die dauischen Opilioniden. - Vidensk. Medd. fra Dknsk naturh. Foren. 126, 417-449. Kopenhagen.
MULLER, A. (1924), Zur Anatomie einiger Arten des Genus Ischyropsalts C. L. KOCII nebst vei- gleichend anatomischen Betrachtungen. - Zool. Jb . (Anatomie) 45, 405-518. Jena.
R ~ M B L A , M. (1946), Opiliones del Aralar. - Aport. est. fauna flora vasco-navarras, 45-65. Zaragoza.
- (1967), Opiliones de Portugal. - Revista Biol. 6, 1-34. Lisboa. ROEWER, C.-F. (1914), Die Familien der Ischyropsalidae und Nemastomatidae der Opiliones-
Palpatores. - Arch. Naturgesch. 80 A, 99-169. Berlin. - (1923), Die Weberknechte der Erde. G. Fischer, Jena. - (1950), Uber Ischyropsalididae und Trogulidae. Weitere Weberknechte XV. - Sencken-
bergiana 31, 11-56. Frankfurt am Main. - (1953), Mediterrane Opiliones Palpatores. - Abh. naturw. Ver. 33, 201-210. Bremen. SCIIMOLZCR, K. (1962), Die Kleintierwelt der Nunatakker als Zeugen einer Eiszeit-Uberdaueiung. - Mitt. Zool. Mus. 88, 171-400. Berlin.
SCHULLER, L. (1933) Ischyropsalts helzutgtt PANZER aus einei Salzburger Hohle. - Mitt. Hohlen. Kaistforsch. 1933, 44-45. Berlin.
-'(1950), Ein Beitrag zur Okologie ulid Biologie von Ischyropsalts pltcata ROEWER nov. spec. - Mitt. Nat. Arbeitsgem. B, 44-48. Salzburg.
SILIIAVY, V. (1938), Sur l'importnnce de la forme de l'appareil sexuel pour le systkme des Opilions et rkvision de quelques espkces europkennes du genre Optlto HERBST. - Sbornik pi'irodov. Klubu 3, 7-20. Tiebii.
- (1956), Fauna CSR, Sekaci-Opilionidea 7 , 1-272. Praha. - (1966), Fragments Opilionidologica 11. - Zpravy Cesk. spol. ent. CSAV 2,104-105. S ~ n r c ~ , E. (1872), Notice sur les Arachnides cavernicoles et hypoghs. - Ann. Soc. ent. France (5)
2, 215-244. Paris. - (1872), Notice complkmentaire sur les Arachnides cavernicoles et hypogks. - Ann. Soc. ent.
France (6) 2, 471-488. Paris. - (1879), Les Arachnides de France 7 , 116-332. Paris. - (1881), Arachnides nouveaux ou peu connus des provinces basques. - Ann. Soc. espan. Ilist.
nat . 10, 127-132. Madrid. - (1913), Biospeologica XXX. Araneae et Opiliones. Quatrieme Serie. - Arch. Zool. expkr.
gknkral 52, 359-386. Paris. THALER, K. (1963), Spinnentiere aus Lunz (Niederosterreich) nebst Bemerkungen zu einigen von . KULCZYNST,I aus Niederosterreich gemeldeten Arten. - Ber. Nat.-Med. Verein 53, 273-283.
Innsbruck. , (1966),.Fragmenta Fa~inistica Tirolensia (Diplopoda, Arachnida). - Ber. Nat.-Med. Verein
54, 151-157. Innsbruck. VERHOEFP, C. W. (1900) Zur Biologie von Ischyropsalts. - Zool. Anz. 23, 106-107. Leipzig. WOOD, H. C. (1868), On the Phalangeae of the United States of America. - Communications
Essex. Inst. 6, 10-31.
Anschrift des Verfassers: Dr. JOCHEN MSRTENS, Institut fur Allgemeine Zoologie, 65 Mainz, Saarstraße 21.


















