
Duftstoffe: Die Sprache der Pflanzen. Signalrezeption, Biosynthese und Ökologie
12
366 | © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Chem. Unserer Zeit, 2006, 40, 366 – 377 Signalrezeption, Biosynthese und Ökologie Duftstoffe: Die Sprache der Pflanzen B IRGIT S CHULZE | C HRISTIAN KOST | G EN - ICHIRO A RIMURA | WILHELM B OLAND DOI: 10.1002/ciuz.200600387 Zwischen der Limabohne (Phaseolus lunatus, im Hintergrund) und unterschiedlichen Insekten, die entweder Fraßfeinde oder Verteidiger der Pflanze sind, kommt es zu komplexen Interaktionen. Die zwei typischen Herbivoren rechts unten (ein Rüsselkäfer und zwei Kurzfühlerschrecken) induzieren die Emission charakteristischer Duftstoffe, welche hier durch ein typisches GC-Spektrum des Limabohnenduftes dargestellt sind. Diese Duftmoleküle lösen die Sekretion extrafloralen Nektars an den blattständigen Nektarien aus (im Kreis rechts oben; die 4 kurzen ‚Stielchen‘). Dieser Nektar wieder- um lockt potentielle Pflanzenverteidiger wie Ameisen und Wespen an, die dann die Anzahl der angreifenden Herbivoren reduzieren.
-
Upload
birgit-schulze -
Category
Documents
-
view
214 -
download
0
Transcript of Duftstoffe: Die Sprache der Pflanzen. Signalrezeption, Biosynthese und Ökologie

366 | © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Chem. Unserer Zeit, 2006, 40, 366 – 377
Signalrezeption, Biosynthese und Ökologie
Duftstoffe: Die Sprache der PflanzenBIRGIT SCHULZE | CHRISTIAN KOST | GEN-ICHIRO ARIMURA | WILHELM BOLAND
DOI: 10.1002/ciuz.200600387
Zwischen der Limabohne (Phaseolus lunatus, im Hintergrund) und unterschiedlichen Insekten, die entweder Fraßfeinde oder Verteidiger der Pflanzesind, kommt es zu komplexen Interaktionen. Die zwei typischen Herbivoren rechts unten (ein Rüsselkäfer und zwei Kurzfühlerschrecken) induzierendie Emission charakteristischer Duftstoffe, welche hier durch ein typisches GC-Spektrum des Limabohnenduftes dargestellt sind. Diese Duftmolekülelösen die Sekretion extrafloralen Nektars an den blattständigen Nektarien aus (im Kreis rechts oben; die 4 kurzen ‚Stielchen‘). Dieser Nektar wieder-um lockt potentielle Pflanzenverteidiger wie Ameisen und Wespen an, die dann die Anzahl der angreifenden Herbivoren reduzieren.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 366

Chem. Unserer Zeit, 2006, 40, 366 – 377 www.chiuz.de © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | 367
K O M M U N I K A T I O N V O N P F L A N Z E N | B I O C H E M I E
Pflanzen können ihren Feinden nicht aus-weichen, weil sie ortsgebunden sind. Um sichdennoch gegen Fraßfeinde zu verteidigen, haben sie im Laufe der Evolution eine Fülle effektiver Schutzmechanismen entwickelt. Besonders raffiniert sind Strategien, bei denen die Pflanze durch die Produktion vonDuftstoffen Raubinsekten rekrutiert, welchedann die Pflanze vor ihren Feinden schützen.Über ihren Duft sind Pflanzen sogar dazu in der Lage, untereinander zu kommunizierenund sich so gegenseitig vor Angreifern zuwarnen.
Das Spektrum pflanzlicher Verteidigungsstrategienreicht von physikalischen Fraßbarrieren wie Dornen,
Haaren oder Blattwachsen über toxische Sekundärmeta-bolite, die angreifende Herbivore direkt töten oder ab-schrecken, bis hin zu verdauungshemmenden Proteinenoder Verbindungen, welche die Qualität des gefressenenBlattmaterials reduzieren.
Neben dieser allgemein bekannten direkten Verteidi-gung verfügen Pflanzen auch über indirekte Abwehrstrate-gien, wobei über bestimmte Signale die natürlichen Fraß-feinde der Herbivoren angelockt werden. Die „herbeigeru-fenen Beschützer“ reduzieren dann die Anzahl der Pflan-zenschädiger und üben somit einen Schutzeffekt auf diePflanze aus. Ein solches Signal kann ein charakteristischesDuftstoffbouquet sein, dessen Biosynthese durch dieFraßaktivität des Herbivoren in der Pflanze induziert wird(Abbildung 1, Pfad A). Anhand der charakteristischen che-
A B B . 1 | KO M M U N I K AT I O N I M P F L A N Z E N R E I C H
Ein Befall mit Herbivoren löst in einerPflanze A) die Emission von Duftstoffenoder B) die Sekretion extrafloralen Nek-tars aus, die dann die natürlichen Feindeihrer Angreifer anlocken und so einen in-direkten Schutzeffekt auf die Pflanze aus-üben. C) Die induzierte Duftemission ver-mittelt aber auch innerhalb derselbenPflanze, wie auch zwischen arteigenenNachbarpflanzen die Sekretion extraflo-ralen Nektars oder die Expression vertei-digungsrelevanter Gene. D) Ob auch dieDuftstoffe artfremder Pflanzen einen ähn-lichen Effekt haben, ist noch unklar.E) Auch wurzelschädigende Herbivoreinduzieren eine Emission flüchtiger Ver-bindungen, die ihre Feinde (z. B. Faden-würmer) anlocken. F) Zusätzlich könnenWurzelherbivore die Duftstoffemission inoberirdischen Pflanzenteilen auslösen undso die indirekte Verteidigung über Duftbeeinflussen. G) Inwieweit auch eineunterirdische Kommunikation zwischenPflanzen eine Rolle spielt ist noch nichtgeklärt.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 367

368 | © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.chiuz.de Chem. Unserer Zeit, 2006, 40, 366 – 377
mischen Zusammensetzung dieses Gemisches können räu-berische Insekten oder Parasitoide nicht nur die Duftstoffaussendende Pflanze, sondern sogar den Herbivoren sowiedas Ausmaß des Befalls erkennen [1].
Indirekte Verteidigungsmechanismen sind induzierbar.Sie sind also nicht permanent aktiv, sondern sie werdenerst im Bedarfsfall ausgelöst, zum Beispiel durch eine Her-bivore. Der Vorteil der Induzierbarkeit liegt im sparsamenEinsatz metabolischer Aktivitäten, die sonst für die konti-nuierliche Produktion von Verteidigungsmetaboliten auf-gewendet werden müssten. Ferner können Schadorganis-
men gegen eine nur temporär wirksame Verteidigung, diesich auch noch auf mehrere Verbindungen und Strategienstützt, kaum Resistenzmechanismen aufbauen. Für den an-zulockenden Räuber ist durch die Induzierbarkeit ein deut-licher Zusammenhang zwischen der Anwesenheit von Beu-te und dem emittierten Duftbouquet gegeben, ein perma-nent produziertes Signal wäre dagegen wegen des fehlen-den Kontrasts keine brauchbare Lokalisierungshilfe für dasRaubinsekt.
Eine induzierbare Verteidigung setzt eine hoch diffe-renzierte und empfindliche Wahrnehmung der Umwelt vo-raus. Da von der Rezeption des Reizes bis zur de-novo-Syn-these der Duftstoffe oder anderer Verteidigungsmetaboliteeine gewisse Zeit verstreicht, in der die Pflanze nur durchihre basalen Schutzmechanismen verteidigt ist, ist eineschnelle und verlässliche Signalweiterleitung für das Über-leben der Pflanze essentiell. Um die Effizienz einer solchenVerteidigung zu optimieren, sollte eine Pflanze schon zu Be-ginn zwischen einer einfachen mechanischen Beschädi-gung, wie sie von einen herabfallenden Ast, Wind oder ei-
S I G N A L K A S K A D E N B E I D E R P F L A N Z L I C H E N V E R T E I D I G U N G |Elizitoren aus dem Regurgitat von Insektenlarven (N-Acylaminosäu-rekonjugate oder Glucoseoxidase) und kontinuierliche mechanischeVerwundung lösen an der Zellmembran Signalkaskaden aus, die dieBiosynthese der pflanzlichen Phytohormone 12-Oxophytodiensäure(OPDA), Jasmonsäure (JA), Salicylsäure (SA) und Ethylen nach sichziehen.
N-Acylglutamine bewirken durch Vermittlung eines Rezeptors oderdurch Einlagerung in die Lipiddoppelschicht der Zellmembran eine Membrandepolarisation und den Einstrom von Ca2+-Ionen. Dieser Reiz aktiviert unter anderem eine Phospholipase A2 (PL), dieLinolensäure aus Phospholipiden freisetzt und als Substrat für eineLipoxygenase (LOX) zugänglich macht. Die so gebildete 13-Hydro-peroxyoctadecatriensäure wird über ein instabiles Allenoxid zurOPDA cyclisiert. Während OPDA in den Chloroplasten gebildet wird,finden die Reduktion der Doppelbindung am Ringsystem im Cyto-plasma und die anschließende β-Oxidation der Seitenkette in denPeroxisomen statt. Die de-novo synthetisierte JA ist cis-disubstitu-tiert, lagert aber schon in der Pflanze (vermutlich enzymkatalysiert)zur biologisch weniger aktiven trans-JA um [7].
Die Glucoseoxidase löst die Bildung reaktiver Sauerstoffspezies(H2O2) aus. Diese können mehrfach ungesättigte Fettsäuren unspe-zifisch funktionalisieren. Fettsäurehydroperoxide bilden die Aus-gangstoffe für die Biosynthese von fettsäureabgeleiteten Phytohor-monen, den Oxylipinen, deren wichtigster Vertreter JA und OPDAsind. Andere Phytohormone wie die Salicylsäure (SA) und das über1-Aminocyclopropan-1-carbonsäure (ACC) aus Methionin gebildeteEthylen modulieren die Wirkung der JA. Das Zusammenspiel derPhytohormone führt zu einer veränderten Genexpression und somitzu spezifischen Verteidigungsreaktionen.
A B B . 2 | C H E M I S C H E S T I M U L I
Drei N-Acylgluta-minderivate ausdem Regurgitatvon Insekten-larven.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 368

Chem. Unserer Zeit, 2006, 40, 366 – 377 www.chiuz.de © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | 369
K O M M U N I K A T I O N V O N P F L A N Z E N | B I O C H E M I E
nem Hagelkorn verursacht werden kann, und einem amBlatt fressenden Insekt sicher unterscheiden können.
Spezifische Verbindungen und mechanischeSchäden aktivieren die pflanzliche Verteidigung
Die einfachsten Reize sind physikalische Stimuli, die durchBerührung und Verwundung von Zellen ausgelöst werden.Viel spezifischer sind dagegen chemische Elizitoren aus demSpeichel oder dem Vorderdarminhalt (Regurgitat) eines fres-senden Herbivoren. Solche Elizitoren können Enzyme seinwie zum Beispiel β-Glucosidasen aus dem Speichel der Kohl-weißlinglarven (Pieris brassicae), aber auch niedermole-kulare Verbindungen [2]. So konnten aus dem Regurgitatvieler Raupenarten Konjugate aus ungesättigten Fettsäurenund den Aminosäuren Glutamin oder Glutamat isoliert wer-den. Als erster Vertreter dieser Verbindungsklasse wurdedas N-(17-Hydroxylinolenoyl)-L-glutamin identifiziert undwegen seiner Fähigkeit in Maispflanzen (Zea mays) eindem Raupenfraß ähnliches Duftspektrum auszulösen „Voli-citin“ genannt (von lateinisch „volatilis“ flüchtig und „eli-cere“ erregen). Bis heute konnte eine ganze Reihe von un-terschiedlich funktionalisierten N-Acylglutaminen im Re-gurgitat verschiedenster Raupenarten identifiziert werden(Abbildung 2) [1]. In Maispflanzen wurde sogar ein Volicitinbindendes Protein charakterisiert, welches an der Reizer-kennung beteiligt sein soll [3]. Die simultane Wahrnehmungeines Verwundungsreizes zusammen mit insektenspezifi-schen Verbindungen ermöglicht es den angegriffenen Pflan-zen, gezielt auf ihren jeweiligen Fraßfeind zu reagieren. DieMechanismen der Signalerkennung unterscheiden sich da-bei zwischen verschiedenen Pflanzenarten. So lösen zumBeispiel die N-Acylglutamine in der Limabohne (Phaseoluslunatus) keine Duftproduktion aus; dies gilt auch für β-Glu-cosidasen, die nur im Wechselspiel von Kohlweißlingsrau-pen und Kohlarten elizitierend wirken [2].
Elizitoraktive Verbindungen können jedoch auch überihre physikalischen Eigenschaften wirken. So besitzen dieN-Acylglutamine einen amphiphilen Charakter, der ihre Ein-lagerung in Biomembranen unterstützt und so durch Ver-änderung der Membraneigenschaften Membrandepolarisa-
tionen auslösen kann. Die Depolarisation setzt sich überdie Gesamtheit der Blattspreite fort [4] und kann nachge-schaltete Signalkaskaden auch ohne Beteiligung von Re-zeptoren aktivieren.
Lange Zeit glaubte man, dass die offensichtlichen Un-terschiede in der Reaktion der Pflanze auf mechanische Ver-wundung und Raupenfraß allein durch die Anwesenheit be-stimmter Elizitoren des Raupenregurgitats zu erklären sei-en. So löst eine kurze Folge von Verletzungen, zum Beispieldurch Nadelstiche, in der Limabohne keine Duftprodukti-on aus. Mit Hilfe einer programmierbaren „mechanischenRaupe“ konnte den Pflanzen, ganz analog zum natürlichenFraßvorgang, eine lang anhaltende kontinuierliche Folgevon Verletzungen zugefügt werden (Abbildung 3). DieseFolge kleiner Verletzungen (alle fünf Sekunden) ist nachwenigen Stunden völlig ausreichend, um eine Duftstoffab-gabe im beschädigten, aber auch in weiter entfernten Blät-tern hervorzurufen. Unterschiede zur natürlichen, durchRaupenfraß ausgelösten Duftemission sind lediglich in derquantitativen Zusammensetzung des Bouquets sichtbar. Dabei wurde auch ersichtlich, dass die abgegebene Mengebestimmter Duftstoffe, darunter (Z)-3-Hexenylacetat und Linalool, der beschädigten Blattfläche und der Schadens-dauer proportional ist. Allerdings gehorchen nicht alleflüchtigen Verbindungen dieser Gesetzmäßigkeit. Zum Bei-spiel bleibt die Menge an emittiertem Methylsalicylat ober-halb einer gewissen Reizschwelle konstant.
Es zeigt sich also, dass Pflanzen schon an der Art der Be-schädigung ihrer Blätter ein bedrohliches Ereignis erken-nen können [5]. Diese Information wird durch die chemi-schen Signale aus dem Speichel von Raupen moduliert undführt in der Summe zu einem charakteristischen Abwehr-muster.
Nachgeschaltete Zellsignale der pflanzlichen Abwehr
Rezeptor-Ligand-Wechselwirkungen, aber auch eine Mem-brandepolarisation in den geschädigten Zellschichten führtzur Aktivierung nachgeschalteter Signalwege, die die In-formation in die Zelle weiterleiten. Dabei handelt es sich oft Abb. 3 Die „me-
chanische Raupe“:Links: Ein Metall-stift bewegt sichauf und ab undverursacht einekontinuierlichemechanische Verwundung im einem Lima-bohnenblatt. Rechts: Durch ei-nen Schrittmotorkann der Metall-stift computerge-steuert in zwei Di-mensionen bewegtund so eine defi-nierte Fläche ver-letzt werden.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 369

370 | © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.chiuz.de Chem. Unserer Zeit, 2006, 40, 366 – 377
um den Einstrom von Ca2+-Ionen durch spannungsabhän-gige Kanäle. Während bei einer einmaligen mechanischenVerwundung kein Einstrom von Ca2+-Ionen in die Zellen zubeobachten ist, wird dies durch Raupenfraß erreicht (Ab-bildung 4) [4]. Offensichtlich findet die Unterscheidungzwischen verschiedenen Reiztypen bereits auf der Ebeneder primären Signalweiterleitung statt.
Reaktive Sauerstoffspezies (ROS) wie H2O2 oder Super-oxidradikale stehen ebenfalls am Anfang der Signalkette.Sie werden nach Verletzung durch die Aktivität bestimmterEnzyme wie Superoxiddismutasen oder NADPH-Oxidasengeneriert. Bereits geringe Mengen ROS fungieren als Signalfür die Biosynthese von Abwehrmetaboliten oder lösen inmikrobiell infizierten Zellen den so genannten program-mierten Zelltod (analog zur Apoptose in tierischen Zellen)aus, der die Ausbreitung von Pathogenen verhindert [2].Darüber hinaus wirken ROS auch direkt gegen Herbivoremit stechend-saugenden Mundwerkzeugen, so etwa Blatt-läuse [2]. Durch unselektive Oxidation mehrfach ungesät-tigter Fettsäuren aus Membranlipiden begünstigen ROS dieEntstehung von Fettsäurehydroperoxiden, die in die Bio-synthese von Fettsäureoxidationsprodukten, den so ge-nannten Oxylipinen, einfließen können [6].
Reaktive Sauerstoffspezies, insbesondere H2O2, könnenin der Pflanze auch durch Enzyme im Sekret der Herbivor-en erzeugt werden. So enthält das Regurgitat des Baum-wollkapselbohrers (Helicoverpa zea) eine Glucoseoxidase,welche die Bildung von H2O2 durch Oxidation von Gluco-se katalysiert. Durch dieses Enzym kann das Insekt in dieSignalkaskade der Pflanze eingreifen. Da ROS für die Pflan-ze eher ein Zeichen für Pathogenbefall als für Herbivoriesind, täuscht die Raupe die Pflanze. Diese aktiviert durchdas Phytohormon Salicylsäure typische Abwehrstrategiengegen Pathogenbefall, die oft antagonistisch mit der Ab-wehr von Herbivoren verschaltet sind [1].
Die Rolle von Phytohormonen für die Pflanzenverteidigung
Die weitere Regulation pflanzlicher Abwehrreaktionen wirdvon Pflanzenhormonen übernommen. Der wohl wichtigsteVertreter stressinduzierter Phytohormone ist die Jasmon-
säure, die in mehreren Schritten aus 13-Hydroperoxyocta-decatriensäure entsteht [7]. Interessant ist hier die auffälli-ge Ähnlichkeit zwischen der pflanzlichen Biosynthese derJasmonsäure und der menschlichen Arachidonsäurekaska-de, bei der ebenfalls aus mehrfach ungesättigten Fettsäurenhochwirksame cyclopentanoide Gewebshormone (z. B.Prostaglandine) gebildet werden.
Die Jasmonsäurebiosynthese ist ein Schlüsselschritt beider Signaltransduktion in der Pflanze. Viele pflanzliche Ab-wehrreaktionen wie etwa die Biosynthese von Phytotoxi-nen, aber auch die Duftstoffemission können durch Jas-monsäure ausgelöst werden. Auch Metabolite und Konju-gate der Jasmonsäure (z. B. Konjugate der Jasmonsäure mitAminosäuren) sind als Signalstoffe aktiv. Vor allem das Jas-monsäure-Isoleucin-Konjugat besitzt eine von Jasmonsäureunabhängige Aktivität [7]. Analoga solcher Verbindungensind hochaktive Elizitoren der Duftstoffbiosynthese der Li-mabohne und repräsentieren als chemisch leicht modifi-zierbare Moleküle ein ideales Werkzeug zur experimentel-len Manipulation der Biosynthese von pflanzlichen Sekun-därstoffen [8].
Neben der Jasmonsäure sind weitere Phytohormoneoder hormonähnliche Signalstoffe an der Feinregulationpflanzlicher Verteidigungsreaktionen beteiligt. Salicylsäuregilt vorwiegend als Signal für die Abwehr von Pathogenenund wird als Gegenspieler der Jasmonsäure angesehen. Dasbei Herbivorie gleichzeitig ausgesendete flüchtige HormonEthylen dagegen kann sowohl synergistisch (überwiegend),als auch antagonistisch zur Jasmonsäure wirken. Die Phyto-hormone bilden ein regulatorisches Netzwerk und beein-flussen sich wechselseitig durch Aktivierung oder Hem-mung ihrer Biosynthese oder vielmehr durch Modulation ih-rer Wirkung auf die Expression verteidigungsrelevanter Ge-ne [1]. Die Komplexität dieser Signalverschaltung wirddurch verschiedene Oxylipine weiter erhöht. Dazu zählenHydroxy- und Epoxyfettsäuren, α,β-ungesättigte Aldehydeund Ketone oder die nichtenzymatisch entstehenden cyclo-pentanoiden Phytoprostane. Diesen wird ebenfalls eineFunktion als Signalmolekül zugeschrieben, die bislang abernur unzureichend verstanden ist. Erst das Zusammenspielvon Phytohormonen, die frühen Signalen (Ca2+-Ionen, ROS)
Abb. 4 Ca2+-Ein-strom in die Pflan-zenzellen nachHerbivorie (links)und mechanischerVerletzung(rechts). Das Blattder Limabohnewurde mit einemFarbstoff (Fluo-3-AM) behandelt,der in Gegenwartvon Ca2+-Ionengrün fluoresziert.Man erkennt, dassdie Zellen amRand der Fraßzo-ne des Herbivorenerhöhte Konzen-trationen an Ca2+-Ionen aufweisen,während eine ein-fache mechani-sche Verletzungkaum zu einer Zunahme an Ca2+-Ionen führt. DieChloroplasten er-scheinen in dieserDarstellungsweiserot.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 370

Chem. Unserer Zeit, 2006, 40, 366 – 377 www.chiuz.de © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | 371
K O M M U N I K A T I O N V O N P F L A N Z E N | B I O C H E M I E
nachgeordnet sind, Transkriptionsfaktoren und noch un-bekannten Elementen ermöglicht der Pflanze die Emissioneines für den jeweiligen Herbivoren charakteristischen Duft-bouquets.
Zusammensetzung der DuftstoffbouquetsFür ihr Duftbouquet bedient sich die Pflanze eines großenRepertoires flüchtiger Verbindungen, die sich drei großen
Klassen zuordnen lassen: Terpene, Acetogenine und aro-matische Verbindungen (Abbildung 5).
Für die Biosynthese der Terpenvorstufen Isopentenyl-diphosphat (IPP) und Dimethylallyldiphosphat stehen Pflan-zen zwei verschiedene Biosynthesewege zur Verfügung [9].Der „klassische“ Weg nutzt Mevalonsäure, die aus drei Ein-heiten Acetyl-CoA gebildet wird. Der Mevalonatweg läuft imCytoplasma ab und dient hauptsächlich der Biosynthese
A B B . 5 | D I E B I OS Y N T H E S E W EG E F Ü R D I E P F L A N Z L I C H E N D U F T S TO F F K L A S S E N …
A B B . 6 | T E R PE N S Y N T H A S E N A L S M U LT I PRO D U K T E N Z Y M E
… Terpene, Aceto-genine und Aro-maten. DMAPP:Dimethylallyldi-phosphat; DXP:Desoxyxylulose-phosphat; GA3P:Glycerinaldehyd-3-phosphat; HPL:Hydroperoxidlya-se; IPP: Isopen-tenyldiphosphat;JA: Jasmonsäure;LOX: Lipoxygen-ase; MeJA: Methyl-jasmonat; TPS:Terpensynthase.
Über eine kationische Zwischenstufe sind Terpen-synthasen in der Lage, aus einer einzigen Vorstufeein komplexes Terpengemisch zu synthetisieren.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 371
372 | © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.chiuz.de Chem. Unserer Zeit, 2006, 40, 366 – 377
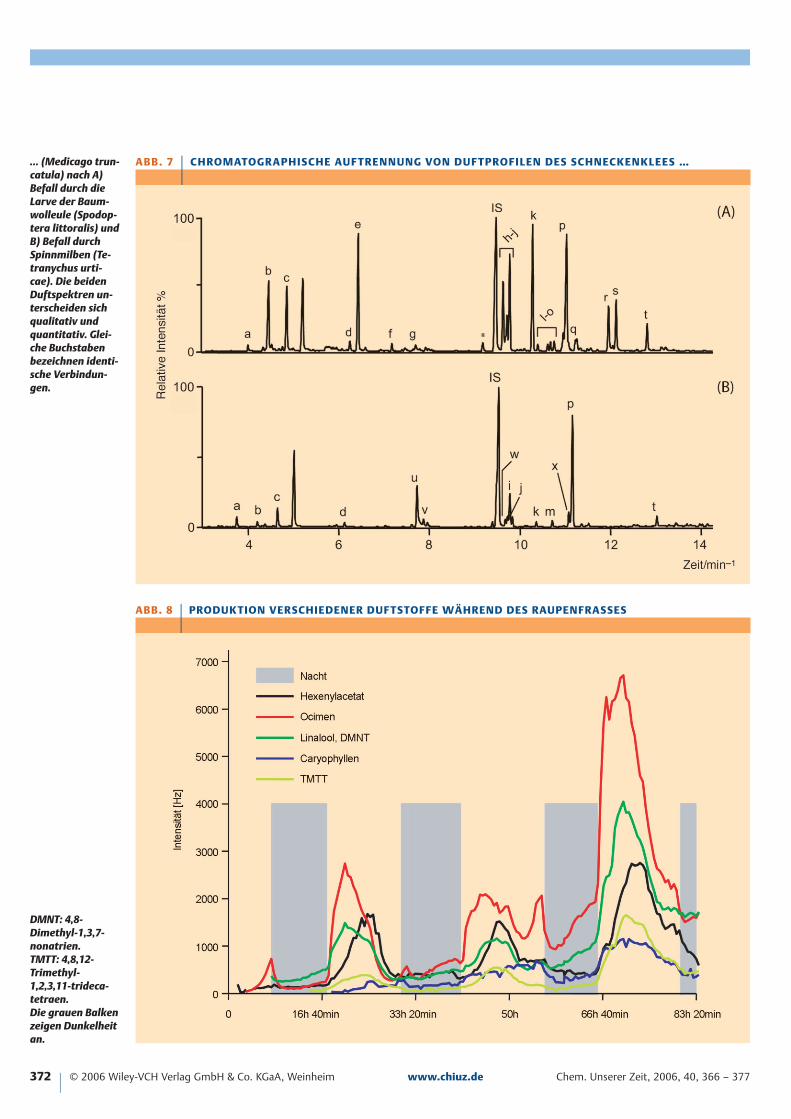
A B B . 7 | C H RO M ATO G R A PH I S C H E AU F T R E N N U N G VO N D U F T PRO F I L E N D E S S C H N EC K E N K L E E S …
ABB . 8 | PRODUK TION VER SCHIEDENER DUF TSTOFFE WÄHREND DES RAUPENFRASSES
… (Medicago trun-catula) nach A)Befall durch dieLarve der Baum-wolleule (Spodop-tera littoralis) undB) Befall durchSpinnmilben (Te-tranychus urti-cae). Die beidenDuftspektren un-terscheiden sichqualitativ undquantitativ. Glei-che Buchstabenbezeichnen identi-sche Verbindun-gen.
DMNT: 4,8-Dimethyl-1,3,7-nonatrien.TMTT: 4,8,12-Trimethyl-1,2,3,11-trideca-tetraen.Die grauen Balkenzeigen Dunkelheitan.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 372

Chem. Unserer Zeit, 2006, 40, 366 – 377 www.chiuz.de © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | 373
K O M M U N I K A T I O N V O N P F L A N Z E N | B I O C H E M I E
von Sesquiterpenen. Der zweite Biosyntheseweg, der Des-oxyxylulosephosphat als Zwischenstufe verwendet, ist inden Chloroplasten lokalisiert, in denen Mono- und Diter-pene synthetisiert werden. Allerdings ist ein Austausch vonIPP zwischen diesen beiden Kompartimenten möglich. Acetogenine sind fettsäureabgeleitete Verbindungen undentstehen aus Fettsäurehydroperoxiden, die enzymatischüber eine Hydroperoxidlyase in Alkohole und Aldehyde ge-spalten werden.
Auch flüchtige Metabolite der Jasmonsäure (Methyljas-monat und (Z)-Jasmon) sind im Duft vieler Pflanzen ver-treten. Aromatische Substanzen wie Methylsalicylat oder In-dol bilden die dritte Verbindungsklasse der Duftstoffe undentstammen dem Stoffwechsel aromatischer Aminosäuren,also dem Shikimatweg.
Die Terpene bilden die größte Gruppe von Naturtstof-fen und zeichnen sich durch eine besonders hohe Struk-turvielfalt aus. Um diese Komplexität mit einfachen Mittelnzu erreichen, bedienen sich die Pflanzen eines genialenTricks. Im Gegensatz zu den „klassischen“ Enzymen, die jeweils nur ein definiertes Produkt bilden, sind viele Terpensynthasen Multiproduktenzyme, das heißt sie können aus einer Vorstufe eine ganze Palette unterschied-licher Strukturen generieren. Durch Abspaltung des Di-phosphats aus dem IPP-Starter entsteht eine kationischeZwischenstufe, die sich im katalytischen Zentrum des Enzyms zu ganz unterschiedlichen Terpengrundkörpern cyclisieren und umlagern kann (Abbildung 6) [10]. Auf die-se Weise kann ein aufwändiges Duftmuster mit einem Mi-nimum an enzymatischer Ausstattung entstehen. Fernerkann das Duftprofil durch Mutation einzelner Aminosäurenim aktiven Zentrum der Terpensynthasen modifiziert wer-den, so dass hier im Zuge der Evolution eine enorme Struk-turvariabilität durch nur wenige Punkmutationen generiertwerden konnte.
Zahlreiche Faktoren beeinflussen die Duftantwort der Pflanze
Pflanzenarten unterscheiden sich zunächst durch ihren cha-rakteristischen Eigenduft (sofern vorhanden). Ferner lösenHerbivoren, die sich ja in der Art des Fraßschadens und derchemischen Zusammensetzung ihres Speichels unterschei-den, ganz unterschiedliche Duftmuster in den attackiertenPflanzen aus [11]. Während zum Beispiel Schmetterlings-larven mit ihren beißenden Mundwerkzeugen großflächigBlattmaterial entfernen, sind Milben Schädlinge, die einzel-ne Zellen anstechen und deren Inhalt aussaugen. Undtatsächlich unterscheidet sich bei der Limabohne [12] undanderen Pflanzen das emittierte Duftspektrum in einigenFällen qualitativ, stets aber quantitativ, je nachdem ob Spinn-milben (Tetranychus urticae) oder Raupen (Spodopteraspp.) den Schaden verursacht hatten. Als Beispiel dient hierdas Duftprofil des Schneckenklees (Medicago truncatula),das durch Larven der Baumwolleule (Spodoptera littoralis)oder durch Spinnmilben ausgelöst worden ist (Abbildung 7)[13].
Weitere Faktoren, die die Biosynthese von Duftstoffenbeeinflussen, sind abiotische Umweltbedingungen wie derStickstoffgehalt des Bodens oder die Wasserverfügbarkeit.Außerdem unterliegt die Duftemission einem ausgeprägtenTag-Nacht-Rhythmus. So emittiert die Limabohne nach Be-fall durch Larven von S. littoralis ein tageslichtabhängiges,rhythmisches Duftmuster, dessen Intensität mit dem Fort-gang des Fraßschadens und damit trotz verminderter Blatt-fläche zunimmt (Abbildung 8). Wird die Raupe nach eini-ger Zeit entfernt, klingt die Duftproduktion während un-
G LOSSA R |Direkte Verteidigung: Ein Verteidigungsmechanismus, der einen negativen Einfluss aufden angreifenden Fraßfeind ausübt (z.B. Dornen, Pflanzentoxine).
Elizitor: Eine (meist chemisch definierte) Verbindung, die im Zielorganismus eine Ab-wehrreaktion hervorrufen kann.
Herbivor: Pflanzenfresser.
Indirekte Verteidigung: Verteidigungsstrategie, die durch die Anlockung der natür-lichen Feinde des herbivoren Angreifers funktioniert.
Karnivor: Fleischfresser, hier Insektenfresser.
Parasitoid: Ein Parasitoid ist ein Tier, in der Regel ein Insekt, welches sich in oder an einem andern Organismus entwickelt, von diesem ernährt und den Wirt zum Abschlussder Parasitierung tötet (z. B. Schlupfwespen (Ichneumonidae)).
Trophische Ebenen: Unterschiedliche Stufen einer Nahrungskette. Ein Karnivor steht aufeiner höheren trophischen Stufe als ein Herbivor.
Regurgitat: Speichelsekret oder Vorderdarminhalt des fressenden Herbivoren, der beimFraßprozess mit dem Blatt in Kontakt kommt und von der Pflanze als Erkennungssignalgenutzt wird.
Abb. 9 In einem Zwei-Arm-Olfaktometer bewegt sich das Insekt innerhalb einerGlasröhre, wobei es an einer Y-förmigen Abzweigung die Wahl zwischen zwei Rich-tungen hat. Am Ende dieser beiden Arme befinden sich zwei verschiedene Duftquel-len, etwa eine unbehandelte und eine von Herbivoren beschädigte Pflanze. Der Duftbeider Pflanzen wird über einen Luftstrom durch die Arme der Glasröhre geleitet.An der Entscheidung des Insektes zugunsten eines bestimmten Arms des Olfakto-meters kann man erkennen, welche Düfte oder Duftkomponenten attraktiv, ab-schreckend oder neutral auf das Insekt wirken.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 373

374 | © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.chiuz.de Chem. Unserer Zeit, 2006, 40, 366 – 377
gefähr 24 Stunden aus. Tageszeitabhängige Variationen imDuftmuster sind auch von anderen Pflanzen bekannt undkönnen sowohl quantitativer als auch qualitativer Natursein. Tabakpflanzen (Nicotiana tabacum) emittierennachts ein anderes Duftbouquet als tagsüber. Der nachtsproduzierte Duft hält nachtaktive Motten von der Eiablageab, da die Tiere durch den Duft abgestoßen werden,während das tagsüber emittierte Duftspektrum die natür-lichen Feinde der Herbivoren anlockt [1].
Kommunikation zwischen Pflanze und InsektAngesichts dieser hoch variablen Duftprofile wird offen-sichtlich, welche enorme sinnesphysiologische Leistunghinter dem Erkennungsprozess der angelockten Insektensteckt. Diese müssen nicht nur dazu in der Lage sein, an-hand der wahrgenommenen Duftstoffe Rückschlüsse aufWirtspflanzen oder die Anwesenheit von Beutetieren zuziehen, sondern diese Informationen auch aus einer kom-plexen Duftlandschaft herausfiltern [14].
In einem so genannten Olfaktometer kann das Verhal-ten der Insekten gegenüber unterschiedlich behandeltenPflanzen getestet werden (Abbildung 9). Unzählige Olfak-tometerexperimente haben ergeben, dass nicht nur Raub-insekten das Duftspektrum nutzen, um die Identität desfressenden Herbivoren zu erkennen, sondern auch Herbi-voren, um ihre Nahrungspflanzen zu identifizieren. Pflanz-liche Duftstoffe können auch abschreckend, das heißt alsdirekte Verteidigung, auf Herbivore wirken. So bevorzugtenBlattkäfer unbeschädigte Blätter der Limabohne gegenüberBlättern, die mit Jasmonsäure behandelt wurden und in Fol-ge dessen eine erhöhte Duftemission aufwiesen [15]. An-hand des emittierten Dufts erkennen die Käfer den erhöh-ten Verteidigungszustand der Pflanze und wenden sichdann bevorzugt unbeschädigten Blättern zu.
Spezialisierte Insekten, die nur ein sehr begrenztesWirtsspektrum nutzen, orientieren sich eher an arttypi-schen Verbindungen. So werden auf Kohlarten spezialisierteHerbivore von den typischen schwefelhaltigen Verbindun-gen der Kohlpflanzen angelockt. Herbivore mit einem brei-ten Spektrum an Wirtspflanzen reagieren vor allem auf Sub-stanzen, die generell in Pflanzendüften enthalten sind, sozum Beispiel (Z)-3-Hexenylacetat oder Methylsalicylat. Die-se Insekten scheinen sich dabei weniger an einzelnen cha-rakteristischen Komponenten des Pflanzenduftes zu orien-
tieren, sondern eher an den Verhältnissen der unter-schiedlichen Duftkomponenten zueinander [14].
Ein ähnliches Prinzip nutzen Raubinsekten um ihre Beu-te zu lokalisieren. Angelockt von herbivorinduzierten Pflan-zendüften können diese dabei sogar zwischen Duftspek-tren, die von verschiedenen Herbivoren ausgelöst wurden,unterscheiden. Auch hier scheint diese Fähigkeit stark vomGrad der Spezialisierung des Insektes auf eine bestimmteBeute oder auch vom Ernährungsstatus des jeweiligen Raub-insektes abhängig zu sein [11]. Ein wichtiger Faktor, derdas Ansprechen der Insekten auf bestimmte Duftstoffe be-einflusst, ist aber auch ihre vorangegangene Erfahrung.Durch die Assoziation mit Erfolgserlebnissen, wie das Auf-finden von Beute oder die Duftumgebung der schlüpfendenLarven, lernen Insekten bereits in ganz frühen Stadien aufbestimmte chemische Stoffe zu reagieren.
Ökologische Bedeutung der pflanzlichen Duftsprache
Ein Großteil der bisherigen Untersuchungen zur Signalver-arbeitung und duftvermittelten Interaktion zwischen Pflan-zen und Insekten wurden unter Laborbedingungen durch-geführt. Nur an wenigen Pflanzenarten, beispielsweise anWildem Tabak (Nicotiana attenuata), wurde die Bedeu-tung dieser Interaktionen auch in Freilandversuchen be-stätigt.
Um zum Beispiel die Rolle des Oxylipinsignals für dieAktivierung der pflanzlichen Abwehr zu erforschen, wur-den gentechnisch veränderte Tabakpflanzen erzeugt, dienur noch geringe Mengen an Jasmonsäure herstellen konn-ten. In Freilandversuchen lieferten diese Pflanzen interes-sante Einblicke in das Verhalten herbivorer Insekten: Dergentechnisch veränderte Tabak mit „ausgeschalteter“ Ab-wehrreaktion, wurde nun von Zwergzikaden (Empoascasp.) als Nahrungsressource genutzt. Die Tatsache, dassPflanzen mit intakter Verteidigung nicht von diesen Zikadenbesucht werden, deutet an, dass Herbivore permanentPflanzen auf ihre Verteidigungsfähigkeit überprüfen [16].Die Beobachtung unterstreicht nachdrücklich die Notwen-digkeit und den Wert von Freilanduntersuchungen mit trans-genen Pflanzen.
Neueste Untersuchungen machen deutlich, dass auchdie Limabohne in ihrem natürlichen Umfeld vom Aussendeneiner Duftnachricht profitiert. Die Pflanze verlässt sich da-
Abb. 10 Links undMitte: Die Lima-bohne (Phaseoluslunatus) trägt anihren Blattachselnextraflorale Nek-tarien (Pfeile), dieein zuckerhaltigesSekret abscheidenund damit Amei-sen oder andereVerteidiger an-locken. Rechts: Ameisenverteidigen ihreWirtspflanze vorAngreifern. Hier haben sie eine Heuschreckeerbeutet.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 374

Chem. Unserer Zeit, 2006, 40, 366 – 377 www.chiuz.de © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | 375
K O M M U N I K A T I O N V O N P F L A N Z E N | B I O C H E M I E
bei jedoch nicht nur auf ihre Duftstoffe, sondern verfügt zu-sätzlich über eine zweite Form der indirekten Verteidigung,nämlich der Sekretion von extrafloralem Nektar. Hierbeiwird von speziellen Strukturen – den so genannten ex-trafloralen Nektarien – eine wässrige, hauptsächlich ausZuckern und Aminosäuren bestehende Lösung ausgeschie-den. Diese lockt Ameisen und andere Raubinsekten aus derUmgebung an, die die Pflanze dann vor Angreifern schüt-zen (Abbildung 10 und Abbildung 1, Pfad B). Die Aktivie-rung dieser beiden indirekten Verteidigungsarten führt zueiner reduzierten Herbivorierate sowie einer erhöhten Pro-duktion an Blüten und Früchten, was auf einen ökologi-schen Nutzen beider Verteidigungsstrategien hinweist [17].Die Sekretion des Nektar kann sogar über die Gasphasedurch den Duft verletzter Pflanzen ausgelöst werden.
Kommunikation über die WurzelnDie bisherigen Untersuchungen zur duftvermittelten indi-rekten Verteidigung konzentrierten sich auf oberirdischePflanzenteile. Jüngste Untersuchungen haben jedoch ge-zeigt, dass auch Wurzeln Duftstoffe als Informationsträgernutzen, um Feinde von Wurzelfraß verursachenden Herbi-voren anzulocken (Abbildung 1, Pfad E). So produzierenMaispflanzen, deren Wurzeln zuvor von Insektenlarven be-fressen wurden, das Sesquiterpen β-Caryophyllen. Dieseslockt karnivore Fadenwürmer (Nematoden) an, welchedann die Wurzelherbivoren dezimieren [18]. Generell sinddie Interaktionen zwischen den verschiedenen trophischenEbenen im Boden von sehr komplexer Natur: Bakterien,mykorrhizierende Pilze und die Wurzeln der Nachbar-pflanzen beeinflussen den Verteidigungsstatus und damitden Metabolismus der ganzen Pflanze. Dabei können die un-terirdischen Reize die Reaktionen oberirdischer Teile so-wohl verstärken, als auch abschwächen. So bewirkt eine Be-
siedelung mit Wurzelbakterien eine Art Breitbandimmuni-sierung der Pflanze gegen viele verschiedene Pflanzen-pathogene. Wurzelfressende Insekten dagegen lösen dieDuftstoffbiosynthese in den Blättern der Pflanze aus (Ab-bildung 1, Pfad F) [19], was zu einer Störung der oberirdi-schen Pflanze-Insekt-Kommunikation führen kann. Im Ge-gensatz zu dem, was wir über die induzierbaren Verteidi-gungsreaktionen oberirdischer Pflanzenteile und ihrer Kom-munikation über Duftstoffe wissen, sind die Kenntnisse zuMechanismen und der Ökologie der viel komplexerenWechselwirkungen im Boden noch recht begrenzt.
Kommunikation zwischen PflanzenDer Informationsaustausch über flüchtige chemische Ver-bindungen ist nicht nur zwischen Pflanzen und ihren Fraß-feinden/Beschützern möglich, sondern auch zwischen denPflanzen selbst denkbar. Ob Pflanzen tatsächlich die Duft-stoffe ihrer Nachbarpflanzen wahrnehmen und sich somitauf einen eventuell bevorstehenden Befall vorbereiten kön-nen ist eine kontrovers diskutierte Frage. Begasung mit demDuftbouquet induzierter Pflanzen, aber auch mit einzelnenDuftkomponenten, führte in unbeschädigten Limabohnenzur erhöhten Expression abwehrrelevanter Gene (Abbil-dung 11). Gleichzeitig wird durch solche Duftstoffe in denEmpfängerpflanzen auch die Biosynthese von Phytohor-monen wie zum Beispiel Ethylen angeregt [1]. Bislang gibtes noch recht wenige Arbeiten, bei denen der Einfluss vonDuftstoffen auf arteigene Nachbarindividuen im Freilanduntersucht wurde [20]. Kürzlich wurde an der Limabohnebeobachtet, dass die beiden indirekten Verteidigungsstra-tegien der Pflanze, nämlich die Duftemission und Sekreti-on extrafloralen Nektars, funktionell miteinander verknüpftsind. So erhöht der Duft von angefressenen Pflanzenran-ken in unbefallenen Nachbarranken die Sekretion des ex-
A B B . 1 1 | D U F T S TO F F E I N D U Z I E R E N D I E E X PR E S S I O N V E R T E I D I G U N G S R E L E VA N T E R G E N E
Links: In einem Glasgefäß mit luftdurchlässigem Boden teilen unbeschädigte Limabohnenblätter (Empfängerblätter) ihren Gasraum mit Blättern,die entweder mechanisch beschädigt oder von Spinnmilben attackiert wurden (Emitterbätter). In beiden Blatttypen wurde anschließend das Expressionsmuster verteidigungsrelevanter Gene bestimmt. Rechts: Gelelektrophoretische Auftrennung der mit Ethidiumbromid gefärbten PCR-Produkte. SM: spinnmilbenattakierte Blätter: W: mechanisch verwundete Blätter; PR2, PR3, PR4: allgemeine Verteidigungsproteine (Chitinasen undß-1,3-Glucanasen); LOX: Lipoxygenase; TPS: Terpensynthase.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 375

376 | © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.chiuz.de Chem. Unserer Zeit, 2006, 40, 366 – 377
trafloralen Nektars (Abbildung 1, Pfad C). Die Duftstoffedienen also auch dem Signalaustausch zwischen unter-schiedlichen Pflanzenteilen, wobei die durch den erhöh-ten Nektarfluss angelockten Ameisen als eigentliche Ver-teidiger fungieren [21]. Inwieweit eine Kommunikation zwischen befressenen und unbefressenen Pflanzen der glei-chen oder einer anderen Art stattfindet (Abbildung 1, PfadD) und ob diese dann eher ober- oder unterirdisch vermit-telt wird (Abbildung 1, Pfad G), sind offene Fragen, die inzukünftigen Untersuchungen beantwortet werden müssen.
Wie gezeigt, sind Pflanzen keine passiven Lebensfor-men, sondern sie kommunizieren rege mit den sie umge-benden Organismen. Gleichzeitig können sie flexibel aufabiotische und biotische Faktoren ihrer Umwelt reagieren.Sowohl ihre Fraßfeinde als auch die Feinde ihrer Feinde ver-stehen sich darauf, die emittierten Duftmuster zu ihremVorteil zu nutzen. Im Laufe der Evolution werden beide Sei-ten immer neue Strategien entwickeln müssen, um die Ge-genseite zu „belauschen“ und ihre eigene Kommunikationzu optimieren.
Danksagung Wir danken Prof. Marcel Dicke (Wageningen Universität,Niederlande) für Abbildung 9 und Maritta Kunert und Mar-git Leitner für die Abbildungen 7 und 8.
SchlagworteDuftstoffe, pflanzliche Schutzmechanismen, Terpene, pflanz-liche Kommunikation
SummaryPlants respond with coordinated actions to threats from theirenvironment. After being attacked by a herbivorous insect,plants are able to emit a complex blend of volatiles, which at-tract carnivorous arthropods that reduce the number of her-bivores and thus benefit the plant. By modulating the emit-ted volatile spectrum, plants are able to transmit informa-tion on type and intensity of their infestation to their defen-ders. Using either various elicitors from the saliva secretion ofthe herbivore or differences in the spatio-temporal pattern ofwounding, the plant is able to discriminate between differentherbivore species. Subsequently, a complex signalling net-work ensures the specific reaction to various stresses. More-over, also plant roots are able to emit volatiles upon attack toattract the predators of root-feeding insects. New studies sug-gest that plants may also be able to communicate with eachother: By “eavesdropping” on their damaged neighbour,plants of the same or another species can use the informa-tion on a currently increased risk of being attacked by herbi-vores and, as a precaution, activate its own defences. How-ever, most of these interactions between plants and other or-ganisms have been studied mainly under laboratory condi-tions and we are only starting to understand the ecology ofthese mechanisms in nature.
ZusammenfassungPflanzen reagieren mit spezifischen Abwehrreaktionen auf Be-drohungen aus ihrer Umwelt. Der Angriff eines herbivoren In-sekts kann beispielsweise eine erhöhte Emission charakteris-tischer Duftstoffe auslösen. Die komplexen Mischungen sindattraktiv für die natürlichen Feinde des Herbivoren, die danndie Pflanze verteidigen. Mit dem Duftmuster übermittelt diePflanze ihren Beschützern sogar weitergehende Informationüber Art und Intensität ihres Befalls. Die Modulation des Bou-quets gründet sich auf unterschiedliche chemische Elizitorenaus dem Insektenspeichel und auf räumlich-zeitlich unter-schiedliche Muster der Verletzung. Ein komplexes Netzwerknachgeschalteter Signalkaskaden garantiert dabei eine spe-zifische Reaktion der Pflanze auf unterschiedliche Bedro-hungen. Auch die Wurzeln locken mit Duftstoffen die natür-lichen Gegenspieler von wurzelfressenden Schädlingen an und begrenzen so den verursachten Schaden. Darüber hinauskann die in den emittierten Duftstoffen kodierte Informationüber ein aktuell erhöhtes Herbivorierisiko auch von benach-barten Pflanzen der eigenen oder einen anderen Art genutztwerden. Während viele solcher pflanzlichen Reaktionen auf ihre Umwelt im Labor gut untersucht sind, beginnen wir ge-rade erst diese Zusammenhänge im natürlichen Habitat zuverstehen.
Literatur[1] G. Arimura, C. Kost und W. Boland, Biochim. Biophys. Acta Mol. Cell
Biol. Lipids. 22000055, 1734, 91.[2] L. L. Walling, J. Plant Growth Regul. 22000000, 19, 195.[3] C. L. Truitt, H. X. Wei und P. W. Paré, Plant Cell 22000044, 16, 523.[4] M. Maffei, S. Bossi, D. Spiteller, A. Mithöfer und W. Boland, Plant
Physiol. 22000044, 134, 1752.[5] A. Mithöfer, G. Wanner und W. Boland, Plant Physiol. 22000055, 137,
1160.[6] A. Mithöfer, B. Schulze und W. Boland, FEBS Lett. 22000044, 566, 1.[7] F. Schaller, A. Schaller und A. Stintzi, J. Plant Growth Regul. 22000055,
23, 179.[8] A. Mithöfer, M. Maitrejean und W. Boland, J. Plant Growth Regul.
22000044, 23, 170.[9] W. Eisenreich, F. Rohdich und A. Bacher, Trends Plant Sci. 22000011,
6, 78.[10] T. G. Köllner, C. Schnee, J. Gershenzon und J. Degenhardt, Plant Cell
22000044, 16, 1115.[11] M. Dicke, Entomol. Exp. Appl. 11999999, 91, 131.[12] R. Ozawa, G. Arimura, J. Takabayashi, T. Shimoda und T. Nishioka,
Plant Cell Physiol. 22000000, 41, 391.[13] M. Leitner, W. Boland und A. Mithöfer, New Phytol. 22000055, 167, 597.[14] T. J. A. Bruce, L. J. Wadhams und C. M. Woodcock, Trends Plant Sci.
22000055, 10, 269.[15] M. Heil, J. Chem. Ecol. 22000044, 30, 1289.[16] A. Kessler, R. Halitschke und I. T. Baldwin, Science 22000044, 305, 665.[17] M. Heil, J. Ecol. 22000044, 92, 527.[18] S. Rasmann, T. G. Köllner, J. Degenhardt, I. Hiltpold, S. Toepfer,
U. Kuhlmann, J. Gershenzon und T. C. J. Turlings, Nature 22000055, 434,732.
[19] T. M. Bezemer und N. M. van Dam, Trends Ecol. Evol. 22000055, 20, 617.[20] M. Dicke, A. A. Agrawal und J. Bruin, Trends Plant Sci. 22000033, 8, 403.[21] C. Kost und M. Heil, J. Ecol. 22000066, 94, 619-628.
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 376

Chem. Unserer Zeit, 2006, 40, 366 – 377 www.chiuz.de © 2006 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim | 377
K O M M U N I K A T I O N V O N P F L A N Z E N | B I O C H E M I E
Die AutorenBirgit Schulze wurde 1975 in Bayreuth geboren undstudierte von 1995 bis 2000 Pharmazie an derPhilipps-Universität Marburg. Nach ihrer Approba-tion arbeitete sie am Max-Planck-Institut für Bio-geochemie in Jena an der Bestimmung von Kohlen-stoffisotopen in Jahresringen von Bäumen. Im Jahr2001 wechselte sie ans Max-Planck-Institut fürchemische Ökologie zu Prof. Boland, bei dem sie imJahr 2005 ihre Dissertation über die Bedeutung vonOxylipinen als pflanzliche Signalstoffe abschloss.
Christian Kost, geboren 1975, studierte von 1995bis 2001 Biologie an der Universität Kaiserslautern.Nach einer eineinhalbjährigen Anstellung alswissenschaftlicher Mitarbeiter an der UniversitätWürzburg, begann er 2003 seine Promotion amMax-Planck-Institut für chemische Ökologie in derArbeitsgruppe von Prof. Boland. Seine Dissertationbeschäftigt sich mit freilandökologischen Aspektenzur indirekten Verteidigung der Limabohne sowiederen zwischenpflanzlicher Kommunikation.
Gen-ichiro Arimura, geboren 1972, studierte Bio-logie an der Faculty of Science, Hiroshima Univer-sity, Hiroshima, Japan. Nach dem Abschluss seinerPromotion (1998) absolvierte er Forschungsaufent-halte an der Universität Kyoto, Japan (1998-2002)sowie der University of British Columbia, Vancouver,Kanada (2002-2004). Seit 2004 ist er am Max-Planck-Institut für Chemische Ökologie und unter-sucht die Komplexität pflanzlicher Signalkaskaden.
Wilhelm Boland, Jahrgang 1950, studierte Chemiean den Universitäten Münster und Köln. DiePromotion erfolgte 1978 in Köln bei L. Jaenicke miteiner Arbeit zur Pheromonkommunikation beimarinen Braunalgen. Nach der Habilitation (1986)folgte er 1987 einem Ruf auf eine Professur fürOrganische Chemie an die Universität Karlsruhe.1994 wechselte er an die Universität Bonn auf einenLehrstuhl für Bioorganische Chemie. Seit 1997 ist erMitglied der Max-Planck-Gesellschaft und Direktorder Abteilung Bioorganische Chemie am MPI fürchemische Ökologie in Jena. Die Forschungsinteres-sen konzentrieren sich auf Interaktionen vonPflanzen und Insekten. Im Vordergrund stehen dabeidie induzierte pflanzliche Abwehr sowie diemolekulare Basis der chemischen Verteidigung vonInsekten.
KKoorrrreessppoonnddeennzzaaddrreessssee::Prof. Dr. Wilhelm BolandMax-Planck-Institut für Chemische ÖkologieHans-Knöll-Straße 8D-07745 Jena, GermanyE-Mail: [email protected]
366_CHI_387_Schulze 30.11.2006 8:38 Uhr Seite 377


















