
Untersuchung zur Acidierung des Harns mittels alimentärer ... · Das MMA-Syndrom tritt in der...
207
Aus dem Institut für Tierernährung der Tierärztlichen Hochschule Hannover ___________________________________________________________________________ Untersuchung zur Acidierung des Harns mittels alimentärer Calciumchlorid-Gabe bei tragenden Sauen INAUGURAL-DISSERTATION Zur Erlangung des Grades eines Doktors der Veterinärmedizin (Dr. med vet.) durch die Tierärztliche Hochschule Hannover vorgelegt von BJÖRN RÖCKER aus Garlstedt Hannover 2006
Transcript of Untersuchung zur Acidierung des Harns mittels alimentärer ... · Das MMA-Syndrom tritt in der...

Aus dem
Institut für Tierernährung der Tierärztlichen Hochschule Hannover
___________________________________________________________________________
Untersuchung zur Acidierung des Harns mittels
alimentärer Calciumchlorid-Gabe bei tragenden Sauen
INAUGURAL-DISSERTATION
Zur Erlangung des Grades eines
Doktors der Veterinärmedizin
(Dr. med vet.)
durch die Tierärztliche Hochschule Hannover
vorgelegt von
BJÖRN RÖCKER
aus Garlstedt
Hannover 2006

Wissenschaftliche Betreuung: Univ. Prof. Dr. M. Coenen 1. Gutachter: Univ. Prof. Dr. M. Coenen 2. Gutachter: Univ. Prof. Dr. M. Wendt Tag der mündlichen Prüfung: 2. Juni 2006

Meinen Eltern und
allen Menschen, die mich auf meinem Weg fördernd begleitet haben.


Inhaltsverzeichnis
Inhaltsverzeichnis
1. EINLEITUNG 9
2. LITERATURÜBERSICHT 11
2.1. DER MASTITIS- METRITIS- AGALAKTIE- KOMPLEX (MMA- KOMPLEX) 11 2.1.1 Nomenklatur 11 2.1.2. Verbreitung und Inzidenz des MMA-Komplexes 12 2.1.3. Symptomatik des MMA-Komplexes 12 2.1.4. Ätiologie und Pathogenese des MMA-Komplexes 17 2.1.4.1 Mastitis und Agalaktie 18 2.1.4.2. Metritis 20 2.1.5. Prophylaxemaßnahmen zur Verhinderung des MMA-Sydroms 23
2.2. ZUSAMMENHANG ZWISCHEN DEM PUERPERALEN KRANKHEITSKOMPLEX UND DER UNSPEZIFISCHEN FORM DER HARNWEGSINFEKTION BEI DER SAU 26 2.3. HARNWEGSINFEKTIONEN BEIM SCHWEIN 29
2.3.1. Prävalenz der unspezifischen Form der Harnwegsinfektion der Sau 30 2.3.2. Das Keimspektrum der unspezifischen Form der Harnwegsinfektion 34
2.4. DIE ALIMENTÄRE BEEINFLUSSUNG DER HARNACIDITÄT 35 2.4.1. Der Zusammenhang zwischen der Harnansäuerung und der Harnwegsgesundheit 35 2.4.2. Mechanismen der alimentären Harnansäuerung 36 2.4.3. Der Einfluss der Ernährung auf den Harn-pH-Wert 39
2.5. DIE AUSWIRKUNGEN VON VERÄNDERUNGEN IM SÄURE-BASEN-HAUSHALT AUF DEN KNOCHENSTOFFWECHSEL 42 2.6. DAS KNOCHENGEWEBE 43
2.6.1. Die Funktion des Knochengewebes 43 2.6.2. Die Struktur des Knochengewebes 43 2.6.3. Die zelluläre Grundlage des Knochengewebes 44 2.6.4. Die Knochenformation 45 2.6.5. Der Knochenabbau 47 2.7. Das Parathormon 48
2.8. BIOCHEMISCHE VERFAHREN ZUR UNTERSUCHUNG DES KNOCHENSTOFFWECHSELS 49
2.8.1. Knochenspezifische alkalische Phosphatase (bAP) 50 2.9. DAS CALCIUM 51
2.9.1. Die Verteilung und Funktion von Calcium im Körper 51 2.9.2. Die Aufnahme von Calcium 52 2.9.3. Die Ausscheidung von Calcium 54
2.10. DER PHOSPHOR 55 2.10.1. Die Funktion und Regulation des Phosphorgehaltes im Organismus 55 2.10.2. Die Absorption von Phosphor 55 2.10.3. Die renale Ausscheidung von Phosphor 56

Inhaltsverzeichnis
3. MATERIAL UND METHODEN 57
3.1. DAS ZIEL DES VERSUCHES 57 3.2. ZEITRAUM UND ORT DER VERSUCHSDURCHFÜHRUNG 58 3.3 DIE VERSUCHSTIERE 58 3.4 DIE AUFSTALLUNG DER VERSUCHSTIERE 59 3.5. DIE GRUPPENEINTEILUNG DER VERSUCHSTIERE 61 3.6. DAS FUTTER UND DIE FÜTTERUNG 62
3.6.1. Das Futter 62 3.6.2. Die Futtermittelprobengewinnung 63 3.6.3. Die Futtermittelanalysen 63 3.6.4. Die Ergebnisse der Futtermittelanalysen 67 3.5.5. Die Fütterung 68
3.6. DIE BLUTPROBEN 70 3.6.1. Die Entnahme der Blutproben 70 3.6.2. Die Aufbereitung und Aufbewahrung der Blutproben 71 3.6.3. Die Analyse der Blutproben 71 3.6.4. Analyse der Plasmaproben: 72
3.7. DIE HARNPROBEN 77 3.7.1. Die Entnahme der Harnproben 77 3.7.2. Die Aufbereitung und Aufbewahrung der Harnproben 77 3.7.3. Die Analyse der Harnproben 77
3.8. DIE STATISTISCHE AUSWERTUNGEN DER ANALYSENERGEBNISSE 79
4. ERGEBNISSE 80
4.1 DIE DATEN DER VERSUCHS- UND KONTROLLTIERE 80 4.2. DIE FUTTERAUFNAHME DER SAUEN 81 4.3 DIE DATEN ZUM ABLAUF DER GEBURTEN UND ZU DEN NEUGEBORENEN FERKELN 82 4.4. DIE ERGEBNISSE DER PLASMA- BZW. BLUTANALYSEN 84
4.4.1. Der pH-Wert im Blut der Sauen 84 4.4.2. Die Konzentration an Hydrogenkarbonat (HCO3
-) im Blut der Sauen 86 4.4.3. Der Vergleich der Konzentrationsverläufe vom Gesamtcalcium im Plasma und dem ionisierten Calcium im Blut der Sauen 88 4.4.4. Der Phosphorgehalt im Plasma der Sauen 90 4.4.5. Der Natriumgehalt im Plasma der Sauen 92 4.4.6. Der Chloridgehalt im Plasma der Sauen 94 4.4.7. Die Kaliumkonzentration im Blut der Sauen 96 4.4.8. Kreatininkonzentrationen im Plasma und im Harn 98
4.5. AUSGEWÄHLTE PARAMETER ZUR ÜBERWACHUNG DES CALCIUMHAUSHALTES 99 4.5.1. Die Parathormonkonzentration im Plasma der Sauen 99 4.5.2. Die Konzentration der knochenspezifischen alkalischen Phosphatase im Plasma der Sauen 101
4.6. DIE ERGEBNISSE DER ANALYSEN DER SPONTANHARNPROBEN 103 4.6.1. Der pH-Wert im Harn der Sauen 103 4.6.2 Die relative Mineralstoffausscheidung über den Harn 104
4.7. DIE ERGEBNISSE DER BLUTGASANALYSE DER NEUGEBORENEN FERKEL 106

Inhaltsverzeichnis
5. DISKUSSION 108
5.1. KRITIK DER METHODEN 108 5.1.1. Die Fütterung der Sauen und die Zugabe des CaCl2 zur Ration der Versuchstiere 108 5.1.2. Die Bestimmung des pH-Wertes im Harn der Sauen 109 5.1.3. Die Blutentnahme 110 5.1.4. Die Bestimmung der Anionen-Kationen-Bilanz (AKB) und der Elektrolyt-Bilanz (EB) im Futter 111 5.1.5. Der Vergleich der Blut- und Harnparameter zur Feststellung der renalen Exkretion 112
5.2. DISKUSSION DER ERGEBNISSE 113 5.2.1. Die Akzeptanz des Futters nach der Zumischung von Calciumchlorid (CaCl2) 113 5.2.2. Der pH-Wert im Harn der Sauen 114 5.2.3. Die Vorhersage des Harn-pH-Wertes im Harn anhand der Anionen- Kationen-Bilanz (AKB) im Futter 115 5.2.4. Die Vorhersage des Harn-pH-Wertes anhand der Elektrolytbilanz (EB) im Futter 118 5.2.5. Der Mechanismus der Harnansäuerung nach der Zugabe von CaCl2 120 5.2.6. Der pH-Wert und die Bikarbonatkonzentration im Blut der Sauen 122 5.2.7. Die Veränderungen der Calciumbilanz und des Knochenstoffwechsels der Sauen 124 5.2.8. Die Konzentrationen der anderen Mineralstoffe im Blut und im Harn der Sauen 133 5.2.9. Die Entwicklung der Kreatininkonzentrationen im Plasma und im Harn 135 5.2.10. Der Einfluss des CaCl2-Einsatzes auf die Ferkel und den Ablauf der Geburten 136 5.2.11. Die Inzidenz des MMA-Syndroms während des Versuches 138 5.2.12. Die praktische Anwendung von CaCl2 zur MMA-Prophylaxe 140
6. ZUSAMMENFASSUNG 143
7. SUMMARY 146
8. ABKÜRZUNGSVERZEICHNIS 149
9. LITERATURVERZEICHNIS 151
10. TABELLENANHANG 187


Einleitung
9
1. Einleitung
Das Mastitis-Metritis-Agalaktie-Syndrom (MMA-Syndrom) stellt aus tiermedizinischer Sicht
eines der Hauptprobleme im Bereich der Ferkelerzeugung dar. Seit Jahrzehnten führt die welt-
weite Verbreitung dieses Krankheitskomplexes in zahlreichen Sauenbeständen zu erheblichen
wirtschaftlichen Einbußen. Betroffene Bestände fallen dabei meist durch schlechtere Auf-
zuchtleistungen im Bereich der Ferkelproduktion und negativ beeinflusste Fruchtbarkeits-
kennzahlen der Sauenherde auf, da die Tiere in der auf den Krankheitsverlauf folgenden
Belegung oft unter Fertilitätsstörungen leiden.
Das MMA-Syndrom tritt in der Sauenpopulation mit einer Inzidenz von 14% bis zu 77% auf
(LERCH 1987) und als Faktorenkrankheit wird dessen Auftreten von zahlreichen endogenen
und exogenen Faktoren beeinflusst. Zu diesen Faktoren zählen neben den betriebs-
individuellen Haltungsbedingungen auch die alimentäre Versorgung der Tiere mit ent-
sprechendem Futter und ausreichend Wasser. Eine wichtige pathogenetische Rolle spielen in
diesem Zusammenhang einige bakterielle Erreger, die sowohl bei Tieren nachgewiesen
wurden, die an dem MMA-Komplex litten, wie auch bei Tieren, die an der unspezifischen
Form der Harnwegsinfektion des Schweines erkrankt waren. Somit besteht offensichtlich ein
Zusammenhang zwischen dem Auftreten einer Infektion der Harnwege und der Prävalenz des
MMA-Syndroms. Dieser Zusammenhang der beiden genannten Erkrankungen wird speziell
bei der Sau durch die enge anatomische Beziehung zwischen den Organen des Harntraktes
und denen des Geschlechtsapparates gefördert. Ausgehend von dieser Prämisse existieren
mehrere wirksame prophylaktische Maßnahmen, die das Ziel verfolgen durch eine Keim-
reduktion die Auswirkungen und die Prävalenz der Harnwegsinfektion möglichst zu
begrenzen, um somit sekundär auch die Prävalenz des MMA-Komplexes erheblich zu
verringern.
Eine Möglichkeit dieser Prophylaxe gegen das MMA-Syndrom zur Reduktion der ent-
sprechenden Keimflora in dem potentiellen Erregerreservoir der Harnwege besteht in der
alimentären Ansäuerung des Harnes, um in der Harnblase ein relativ keimfeindliches Milieu
zu schaffen und so die Keimbelastung des Tieres und seiner unmittelbaren Umgebung zu
reduzieren. In dieser Untersuchung wird zum Zweck der Acidierung des Harns gekapseltes
Calciumchlorid (CaCl2) eingesetzt. Aus diesem Grund ist es Gegenstand dieser Unter-
suchung die Wirkung dieses Stoffes auf den Organismus und die damit verbundenen
Regelmechanismen zu erforschen und näher zu erörtern. In erster Linie geht es hierbei darum
zu untersuchen, wie sich dieser Eingriff in den Säure-Basen-Haushalt des Körpers auf die

Einleitung
10
Mineralstoff-Homöostase und den damit in Verbindung stehenden Knochenstoffwechsel
auswirkt bzw. welche Veränderungen sich in diesen sehr fein abgestimmten Regel-
mechanismen ergeben. Dabei sind gerade Veränderungen des Parathormon-Spiegels im Blut
und dessen Auswirkungen auf die Calciumhomöostase des Organismus von Bedeutung. Ein
erhebliches Interesse gilt in dieser Untersuchung ebenfalls der Reaktion des Knochengewebes
bzw. den Veränderungen im Knochenstoffwechsel, die durch die vermehrte Calciumauf-
nahme und die dadurch erzeugte Acidierung hervorgerufen werden.
Ein weiteres Ziel dieser Arbeit ist es, aufgrund der erzielten Ergebnisse den Zeitraum für eine
effektive Prophylaxe gegen das MMA-Syndrom über die Zulage von Calciumchlorid in
gekapselter Form näher einzugrenzen.

Literaturübersicht
11
2. Literaturübersicht
2.1. Der Mastitis- Metritis- Agalaktie- Komplex (MMA- Komplex)
2.1.1 Nomenklatur
In der Literatur werden für den puerperalen Krankheitskomplex der Sau aufgrund seiner viel-
fältigen Erscheinungsformen sehr unterschiedliche Bezeichnungen gewählt. Die klassische
Erscheinungsform dieses Krankheitsbildes bestehend aus einer Mastitis, einer Metritis und
einer Agalaktie wird von mehreren Autoren (BÄCKSTRÖM et al. 1984; BERNER 1987;
SCHÖNING u. PLONAIT 1990; WAWRON 1995) als MMA-Syndrom bzw. als MMA-
Komplex bezeichnet. In vielen Fällen tritt der puerperale Krankheitskomplex der Sau jedoch
nicht in seiner klassischen Erscheinungsform auf, sondern die drei einzelnen Symptome
Mastitis, Metritis und Agalaktie zeigen sich in unterschiedlichen Ausprägungsformen oder
einzelne Anteile dieses Symptomkomplexes fehlen sogar gänzlich. Deshalb finden in der
Literatur für die Beschreibung des puerperalen Krankheitskomplexes der Sau verschiedene
Begriffe Verwendung, welche die Gewichtung der einzelnen Symptome entsprechend ihrem
klinischen Erscheinungsbild, ihrer Ätiologie und ihrer Pathogenese der jeweiligen Autoren
wiederspiegeln.
Hier werden einige Beispiele für die unterschiedlichen Bezeichnungen für den puerperalen
Krankheitskomplex der Sau von verschiedenen Autoren aufgeführt:
MMA-Syndrom bzw. MMA-Komplex (BÄCKSTRÖM et al. 1984; BERNER 1987;
SCHÖNING u. PLONAIT 1990; WAWRON 1995)
Puerperale Septikämie und Toxämie
(VANDEPLASSCHE et al. 1960; SCHULZE u. BOLLWAHN 1962; SCHULZE u. VON
MICKWITZ 1966; BOSTEDT et al. 1998)
Agalaktie
(GLAWISCHNIG 1967, 1969; NACHREINER u. GINTHER 1974; ROSS et al. 1981;
GÖRANSSON 1990)
Agalaktia toxaemica (RINGARP 1960)

Literaturübersicht
12
Hypogalaktie, Peripartales Hypogalaktiesyndrom, Periparturient Hypogalactic
Syndrome (VAN BEERS-SCHREURS 1996; IBEN 1999)
Puerperale Mastitis (BERTSCHINGER et al. 1990; AWAD-MASALMEH et al. 1990)
Coliforme Mastitis (BERTSCHINGER 1999)
Peripartales Urogenitalsyndrom (PUGS), Swine Urogenital Disease (SUGD)
(BILKEI et al. 1995b)
2.1.2. Verbreitung und Inzidenz des MMA-Komplexes
Der puerperale Krankheitskomplex der Sau stellt ein weltweites gesundheitliches Problem in
der Schweinehaltung dar, denn dieses Syndrom findet in Ferkelerzeugerbetrieben auf der
ganzen Welt Verbreitung.
Zu der Inzidenz des MMA-Komplexes werden allerdings sehr unterschiedliche Angaben
gemacht. JORSAL (1986) beschreibt eine Befallsintensität von lediglich 10% der Muttertiere,
während BOLLWAHN et al. (1989) davon berichten, dass in Problembetrieben bis zu 40%
der Sauen an dem MMA-Syndrom erkranken. Die Inzidenz des MMA-Komplexes unterliegt
in der Sauenpopulation dabei sehr großen betriebsindividuellen Schwankungen. LERCH
(1987) stellte eine Schwankungsbreite im Zusammenhang mit dem Auftreten des puerperalen
Krankheitskomplexes der Sau von 14% bis zu 77% der Muttertiere fest. Da es sich bei dem
MMA-Komplex um eine Faktorenerkrankung handelt, wird die Inzidenz der Krankheit von
zahlreichen endogenen und exogenen Faktoren beeinflusst, die im weiteren Verlauf dieses
Kapitels noch näher erörtert werden.
2.1.3. Symptomatik des MMA-Komplexes
Die Symptome des puerperalen Krankheitskomplexes im Sinne des MMA-Syndroms treten
bei einem Teil der befallenen Sauen schon während der Geburt auf (MARTIN et al. 1967),
der Grossteil der befallenen Tiere zeigt aber erst innerhalb von 48 bis 72 Stunden post partum
(GLOCK 1983; BOSTEDT et al. 1998; BERTSCHINGER 1999) die typischen Anzeichen
dieser Erkrankung.
Die betroffenen Sauen fallen in erster Linie bei der täglichen Kontrolle durch eine
eingeschränkte oder in einigen Fällen sogar sistierende Futteraufnahme (CERNE et al. 1984;
MAASS et al. 1993) auf. Bei der näheren Betrachtung zeigen diese Tiere eine allgemeine

Literaturübersicht
13
Schwäche, die sich im weiteren Verlauf bis hin zur Apathie (BERTSCHINGER 1999)
steigern kann.
Bei einer klinischen Allgemeinuntersuchung ist in den meisten Fällen eine erhöhte Herz- und
Atemfrequenz (HAGN 1995; BERTSCHINGER 1999) zu verzeichnen und oft sind die
Skleralgefäße injiziert und die Lidbindehäute gerötet (BOSTEDT et al. 1998). Teilweise
treten zyanotische Ohren und fleckenförmige, rötliche Verfärbungen und ödematöse
Veränderungen im Bereich des Unterbauches (EHRENTRAUT 1968; GLOCK 1983) auf, die
als Zeichen einer zunehmenden Kreislaufschwäche gewertet werden können.
Bei der rektalen Kontrolle der Körperinnentemperatur weisen die erkrankten Tiere meistens
eine Erhöhung dieses Wertes auf. Zum Maß dieser Erhöhung der Körpertemperatur finden
sich in der Literatur jedoch unterschiedliche Angaben. Vom puerperalen Krankheitskomplex
betroffene Sauen, bei denen die Mastitis im Vordergrund steht, weisen demnach mit + 42°C
eine höhere rektale Körpertemperatur auf, als Tiere, die hauptsächlich an einer Endometritis
leiden und bei denen meistens eine rektale Körpertemperatur von ca. + 40°C (GLOCK 1983)
gemessen wurde. Aufgrund dieser Differenzen im Bereich der rektalen Körpertemperatur gibt
es ebenfalls keine deutliche Grenze, von welcher an ein Tier als krank einzustufen ist bzw. ab
welcher Erhöhung dieser Temperatur eine medikamentöse Behandlung einzuleiten ist.
In einigen Arbeiten zu diesem Thema werden Sauen ab einer rektalen Körpertemperatur von
+ 39,5°C (MIDDLETON-WILLIAMS et al. 1977; LENSCH 1982) als erkrankt angesehen,
während andere Autoren (SCHÖNING u. PLONAIT 1990) ein erkranktes Tier ab einer
Temperatur von + 39,4°C für behandlungswürdig ansehen, und MAAS et al. (1993)
empfehlen in Problembeständen sogar schon ab einer rektalen Temperatur von + 39,3°C eine
antibiotische Therapie, ungeachtet jeglicher Krankheitssymptome.
Andererseits konnte gezeigt werden (ULMER-SHAKIBAEI 1995), dass sogar dauerhaft
erhöhte Körpertemperaturen von bis zu + 41,3°C bei völlig ungestörten laktierenden Sauen
ohne jegliche Krankheitserscheinungen auftreten können. Dieses Phänomen wird als stoff-
wechselbedingte Laktationshyperthermie bezeichnet und ist von fieberhaften Puerperal-
erkrankungen zu differenzieren, da eine Reduktion der Körpertemperatur in diesem Fall
lediglich durch eine Futterrestriktion, nicht aber durch die Anwendung von Antipyretika oder
Antibiotika zu erreichen ist.
Bei der weiteren speziellen Untersuchung des Gesäuges der erkrankten Tiere fallen ad-
spektorisch umschriebene oder diffuse Rötungen auf und palpatorisch ist oberflächlich eine
erhöhte Hauttemperatur festzustellen. Im Gewebe der Gesäugekomplexe selbst lassen sich
umschriebene oder diffuse Verhärtungen und entzündliche Ödeme palpieren, die von einer

Literaturübersicht
14
mehr oder weniger stark ausgeprägten Schmerzhaftigkeit begleitet werden (BOSTEDT et al.
1998; BERTSCHINGER 1999). Diese Symptome entwickeln sich gewöhnlich zuerst an den
kaudalen Gesäugekomplexen (EHRENTRAUT 1968; PLONAIT 1997; BOSTEDT et al.
1998) und schreiten von hier aus nach kranial fort. Bei einzelnen erkrankten Tieren erstrecken
sie sich allerdings von vornherein über die gesamte Gesäugeleiste (GLAWISCHNIG 1967).
Um dieses schmerzhaft veränderte Gesäuge zu schützen und weitere Schmerzen zu ver-
meiden, die z.B. durch saugende Ferkel verursacht werden, nehmen diese Tiere oft eine
sternoabdominale Lage ein (RINGARP 1960; HEINRITZI u. HAGN 1999) und verwehren
somit den Ferkeln den Zugang zum Gesäuge (PLONAIT 1997).
Bei der Überprüfung der Funktionsfähigkeit des Gesäuges fallen die betroffenen Gesäuge-
komplexe durch eine mangelhafte oder sogar gänzlich sistierende Milchproduktion auf
(EHRENTRAUT 1968). In den meisten Fällen ist aber nicht nur die Milchmenge verändert,
sondern auch die Beschaffenheit des Sekretes aus den betroffenen Gesäugekomplexen wird
teilweise erheblich in Mitleidenschaft gezogen. Aus den erkrankten Drüsenkomplexen lässt
sich dann rahmig bis eitrig veränderte Milch ermelken, der teilweise Fibrin- oder Blut-
bestandteile (BERTSCHINGER 1999) beigemengt sind.
Der pH-Wert dieses Sekretes verändert sich in vielen Fällen aus dem leicht sauren
physiologischen Bereich von pH 6,4 ± 0,2 (STRAUSS-ELLERMANN 1985) in den
alkalischen Bereich bis auf einen Wert von pH 8,0 (SCHÖNING 1986).
Die Ferkel der Sauen, die an dem puerperalen Krankheitskomlex leiden, zeigen oft ein sehr
unruhiges Verhalten, indem sie sich suchend durch die Abferkelbucht bewegen, um erneut zu
versuchen ihren Hunger am Gesäuge der Mutter zu stillen. Dabei bleiben diese Saugversuche
meist aufgrund der Hypo- bzw. in schwereren Fällen aufgrund der Agalaktie des Muttertieres
vergeblich (GLOCK 1983; PLONAIT 1997; BERTSCHINGER 1999). Außerdem versuchen
die erkrankten Sauen durch das Einnehmen einer sternoabdominalen Lage ihr schmerzhaft
verändertes Gesäuge vor den Ferkel zu schützen, was diesen dann zusätzlich den Zugang zu
ihrer Nahrungsquelle erschwert. Aus dieser Nahrungskarenz resultiert eine mangelhafte
Energieversorgung, die im weiteren Verlauf zu einer Hypothermie der Ferkel (IBEN 1997,
1999) führen kann. Diese geschwächten Ferkel haben oft ein gesträubtes Haarkleid und ihre
Flanken sind als Folge der Unterernährung eingefallen. Außerdem kommt es im Zuge der
fortschreitenden Exsikkose bei diesen Tieren zur Hautfaltenbildung an beiden Seiten des
Thorax. Wenn dieser Zustand länger andauert bzw. die Ferkel nicht separat mit künstlicher
Sauenmilch versorgt werden, können sie in einem hypoglykämischen Koma verenden
(PLONAIT 1997; BERTSCHINGER 1999).

Literaturübersicht
15
Die Ferkelverluste werden in dieser Periode noch zusätzlich durch vermehrte Erdrückungs-
verluste erhöht, da als Folge des puerperalen Krankheitskomplexes das Allgemeinbefinden
des Muttertieres zu diesem Zeitpunkt so stark beeinflusst sein kann, dass die Wachsamkeit
und die Beweglichkeit der Sau stark beeinträchtigt werden (MIDDLETON-WILLIAMS et al.
1977). Dies führt dazu, dass die Ferkelverluste in Form von Erdrückungen durch ein
unachtsames oder unkontrolliertes Ablegeverhalten der Sau zusätzlich gesteigert werden, da
die Ferkel die Nähe und Wärme ihrer Mutter suchen (WEARY et al. 1996), um so die durch
das Energiedefizit bedingte Hypothermie auszugleichen.
So zeigen BACKSTRÖM et al. (1984) im Rahmen einer großen Feldstudie, dass die
perinatalen Ferkelverluste in Würfen von Sauen, die am MMA-Syndrom erkrankt waren,
innerhalb der ersten Woche post partum um das Dreifache erhöht sind gegenüber den Würfen
von gesunden Sauen. In ihrer weiteren Entwicklung stehen diese Ferkel aufgrund der
verminderten oder sogar fehlenden Kolostrumaufnahme gegenüber Tieren aus Würfen von
gesunden Muttertieren ebenfalls zurück, da sie wegen der unzureichenden Versorgung mit
maternalen Antikörpern und der noch nicht ausreichend ausgebildeten Kompetenz des
eigenen Immunsystems über keinen adäquaten Immunstatus (HAMMERBERG et al. 1989;
BERTSCHINGER 1999) gegenüber Durchfallerregern und anderen in dieser Wachstums-
phase krankmachenden Faktoren (GLAWISCHNIG 1969; WAWRON 1997) verfügen.
Deshalb sind sie in dieser wichtigen Phase der Entwicklung vielen Erkrankungen schutzlos
ausgeliefert, was letztlich häufig zum Kümmern der betroffenen Tiere führt (MIDDLETON-
WILLIAMS et al. 1977; STRAUB 1990) und somit nicht nur weitere gesundheitliche
Beeinträchtigungen der Tiere selbst folgen lässt, sondern auch erhebliche wirtschaftliche
Schäden für den Ferkelerzeuger verursacht.
Die Endometritis puerperalis als weiteren Bestandteil des MMA-Syndroms zeigt sich dem
Betrachter meist durch deutlichen Vaginalausfluss, der sowohl vom Geruch als auch von
seiner Beschaffenheit erheblich verändert sein kann (GLAWISCHNIG 1969; BERNER 1984;
HEINRITZI u. HAGN 1999). Ein solches abfließendes Exsudat ist aber kein eindeutiger
Hinweis auf eine Endometritis, sondern es kann sich dabei je nach Herkunftsort ebenfalls um
eine Vestibulitis, eine Vaginitis oder aber bezogen auf die Harnblase auch um eine Cystitis
oder eine Kristallurie handeln. Des weiteren besteht auch die Möglichkeit einer Kombination
aus mehreren dieser Phänomene. Die sichere Diagnose Endometritis kann erst nach einer
umfangreichen vaginalen Untersuchung unter zur Hilfenahme eines Spekulums gestellt
werden (BERNER 1984). Im Zuge einer Endometritis verzögert sich in vielen Fällen der
Zervixschluss, was zu zervikovaginalen Exsudatsansammlungen führen kann (BOSTEDT et

Literaturübersicht
16
al. 1998). Diese Veränderungen an den Geschlechtsorganen wirken sich im weiteren Verlauf
in vielen Fällen negativ auf deren reibungslose Funktionalität aus, was sich oft erst bei der
nächsten Belegung bzw. der daraus erzielten mäßigen Trächtigkeitsrate zeigt. Somit hat der
puerperale Krankheitskomplex der Sau auch einen negativen Einfluss auf die
Fertilitätsergebnisse der betroffenen Sauen und darüber hinaus auf die Umrauscherquote des
Betriebes. Also wirkt sich diese Erkrankung auch auf diesem Wege negativ auf das
wirtschaftliche Betriebsergebnis aus.
Laut einer Untersuchung von HERRMANSSON et al. (1978) waren im Blut von Sauen, die
akut an dem MMA-Komplex erkrankten, der Hämatokrit und die Hämoglobinkonzentration
signifikant erniedrigt und die Werte für Calcium, Glukose und Magnesium lagen bei diesen
Tieren ebenfalls unterhalb der Normwerte. Die Aktivität der Aspertat-Amino-Transferase und
die Serumkortisolkonzentration waren dagegen signifikant erhöht gegenüber den Werten der
nicht erkrankten Kontrollgruppe. Im Blut der befallenen Tiere konnte außerdem eine
Leukopenie nachgewiesen werden. PLONAIT (1997) bestätigt in seiner Untersuchung ein
Absinken der Gehalte an Calcium und Glukose im Blut der an MMA erkrankten Tiere,
zusätzlich stellte er eine Erniedrigung an Prolaktin und Thyroxin fest, während er erhöhte
Konzentrationen an Harnstoff, Östradiol und Kortisol messen konnte. Außerdem wurde in
dieser Versuchsreihe eine beschleunigte Senkungsreaktion der Blutkörperchen und ein
erhöhter Gehalt an Serumgesamteiweiß beobachtet. WAWRON (1995) stellte allerdings in
ähnlichen Untersuchungen ein Absinken der Gesamteiweißkonzentration im Serum fest.
BUSCH et al. (1998) verzeichneten zusätzlich verringerte Eisengehalte im Serum erkrankter
Sauen und erhöhte Werte für PGF2α. NACHREINER et al. (1972) stellten in diesem
Zusammenhang erhöhte Phosphorwerte bei Sauen fest, denen sie intravenös und
intramammär Endotoxin von E. coli injiziert hatten.
In der neueren Literatur wird davon berichtet, dass sich die klinischen Symptome des
puerperalen Krankheitskomplexes zunehmend milder darstellen, so dass es lediglich zu
geringgradigen bis mäßigen Erhöhungen der Körpertemperatur kommt, die in Verbindung mit
einer schlechten Futteraufnahme und teilweise auch mit Exsudatabgängen aus der Scheide
(VAN BEERS-SCHREURS et al. 1996) auftreten. Über schlechte Zunahmen und erhöhte
Verluste bei den Ferkeln im Zuge einer verminderten Milchproduktion der Sauen wird ebenso
berichtet (KLOPPENSTEIN et al. 1998, 1999), obwohl die Muttertiere dabei vom
Allgemeinbefinden und von der Futteraufnahme keine weiteren Anzeichen einer Erkrankung
erkennen liessen und für eine Mastitis ebenfalls keinerlei klinische Anzeichen vorhanden
waren.

Literaturübersicht
17
Bei den bakteriologischen Untersuchungen von Milchproben und Blutuntersuchungen von
Sauen, bei denen nach der Geburt die Futteraufnahme reduziert war und deren Ferkel an
einem Energiedefizit litten, konnten im Vergleich zu gesunden Muttertieren keine
signifikanten Unterschiede bezogen auf diese Befunde festgestellt werden (VAN BEERS-
SCHREURS et al. 1996). Die Ursache für die verminderte Milchproduktion bleibt in dieser
Arbeit also ungeklärt.
2.1.4. Ätiologie und Pathogenese des MMA-Komplexes
In der Literatur stimmen zahlreiche Autoren darin überein, dass es sich bei dem puerperalen
Krankheitskomplex der Sau bzw. dem MMA-Syndrom um ein multifaktorielles Geschehen
handelt (EHRENTRAUT 1968; TRABITSCH 1988; AWAD-MASALMEH et al. 1990;
HENRITZI u. HAGN 1999; BOSTEDT 1999). Dabei wird meist eine Besiedelung des Uterus
und der Milchdrüse mit fakultativ pathogenen Erregern beobachtet, die durch eine Reihe von
Hilfsfaktoren in die Lage versetzt werden, diesen Krankheitskomplex auszulösen. Als
prädisponierende Hilfsfaktoren fungieren dabei ein mangelhaftes Stallmanagment, eine
allgemein schlechte Hygienesituation, eine zu rohfaserarme Fütterung, eine unzureichende
Wasserversorgung, ungenügende Geburtshygiene und das Auftreten von Nachgeburts-
verhaltungen oder einer fehlenden Kontraktionsbereitschaft des Uterus post partum
(MEREDITH 1991).
Eine große pathogenetische Bedeutung wird in diesem Zusammenhang bestehenden
Harnwegsinfektionen zu geschrieben (PEJSAK et al. 1982; BERNER 1984; BERNER u.
JÖCHLE 1988), da nach heutigem Wissensstand eine aus diesem Bereich aszendierende
Infektion des Uterus zu einer entsprechenden Endometritis führen kann. Bei einer sub partum
bestehenden Infektion der Harnwege ist laut WENDT (1998) fast immer mit einer
Verschleppung bzw. Einwanderung dieser Keime in den Uterus zu rechnen.
Deshalb soll im weiteren Verlauf auf die einzelnen Teilbereiche des MMA-Komplexes, die
daran beteiligten Keime, den Einfluss von Harnwegsinfektionen auf diese Erkrankung und auf
eventuelle Prophylaxemaßnahmen näher eingegangen werden.

Literaturübersicht
18
2.1.4.1 Mastitis und Agalaktie
Die Mastitis spielt als ein Teilbereich des puerperalen Krankheitskomplexes der Sau eine ent-
scheidende Rolle und hat auf dessen Verlauf nach Meinung von BERTSCHINGER
(1984;1999) einen größeren Einfluss als die Endometritis. Er berichtet in diesem Zusammen-
hang davon, dass viele Mastitiden nach adspektorischer und palpatorischer Untersuchung
häufig nicht erkannt werden, da den Milchdrüsen beim Schwein oft starke Fettgewebs-
schichten aufgelagert sind und post partum in vielen Fällen erhebliche ödematöse
Schwellungen im Bereich des Gesäuges auftreten. Auch BOLLWAHN und MEERMEIER
(1989) bestätigten diese Aussage, indem sie bei 23% von 1000 Sauen, die unter Normal-
bedingungen geschlachtet wurden, Symptome einer chronischen Mastitis feststellten. Zu
ähnlichen Ergebnissen kamen auch ROSS et al. (1981), da sie in klinisch unauffälligen
Gesäugekomplexen im Zuge einer histologischen Untersuchung Befunde für das Vorliegen
von Mastitiden fanden. Dabei handelte es sich meist um kleine, im Durchmesser oft nur 1 – 2
cm große Herde, aus denen sie E. coli als Mastitiserreger isolieren konnten. Leider erschwert
der anatomische Aufbau der Milchdrüse beim Schwein den sicheren Nachweis einer Mastitis
zusätzlich, da jeder Gesäugekomplex der Sau aus jeweils zwei (KLOPFENSTEIN et al. 1999)
bzw. in einigen Fällen sogar aus drei (HABERMEHL 1984; BERTSCHINGER 1999)
voneinander getrennten Drüsenkomplexen aufgebaut ist, die unabhängig voneinander
erkranken können und bei denen sich lediglich die Ausführungsgänge vereinigen. Im Falle
der Milchprobenentnahme führt dies dazu, dass das entnommene Sekret überwiegend oder
sogar ausschließlich aus den gesunden Subkomplexen stammt, da in dem erkrankten Gewebe
die Milchproduktion stark reduziert ist bzw. sogar völlig sistiert (BERTSCHINGER 1984,
1999). Unter diesen Vorraussetzungen ist der Nachweis einer Mastitis durch eine
bakteriologische Untersuchung des entnommenen Sekretes nicht immer zuverlässig
(WEGMANN u. BERTSCHINGER 1984; PERSSON 1997). Deshalb führt in vielen Fällen
erst eine Sektion zur gesicherten Diagnose der Mastitis beim Schwein.
Bei der mikrobiologischen Untersuchung wiesen BERTSCHINGER et al. (1977) mit
absteigender Häufigkeit E. coli, Klebsiella pneumoniae, Enterobacter cloacae, Pseudomonas
aeroginosa, Staphylococcus epidermidis und sowohl α- als auch β-hämolysierende
Streptokokken in entzündlich veränderten Gesäugekomplexes nach. Ähnliche Ergebnisse
konnten auch AWAD-MASALMEH et al. (1990) vorweisen, die neben den Milchproben
auch Zervixtupferproben von Sauen untersuchten, die an dem MMA-Syndrom litten.

Literaturübersicht
19
Sie isolierten bei 65,7% der Tiere E. coli, bei 41,8% Staphylokokken und bei 34,3 % β-
hämolysierende Streptokokken. Auch andere Autoren verweisen auf die hervorstechende
Bedeutung von E. coli als Mastitiserreger der coliformen Keimpalette (HAIMBERGER 1977;
CHAREONSIRISUTHIGUL et al. 1979; PEDERSEN et al. 1984; SCHÖNING u. PLONAIT
1990; ROSE et al. 1996; BERTSCHINGER 1999).
Bei der Entwicklung der Hypogalaktie scheinen diese coliformen Erreger ebenfalls eine
wichtige Rolle zu spielen, da bei ihrem Zerfall wie auch bei anderen Vertretern des gram-
negativen Keimspektrums Endotoxine (Lipopolysaccharide,LPS) freigesetzt werden. Diese
entfalten in der Blutbahn dann eine prolaktinantagonistische Wirkung (MARTINEAU et al.
1992), die sich negativ auf die Milchproduktion der Sauen auswirkt. Im Bereich des Gesäuges
wird durch Effekte der Endotoxine auf das endokrine System und das Immunsystem eine
Entzündungsreaktion vorangetrieben (NACHREINER et al. 1972; NACHREINER u.
GINTHER 1974). Einige Keime wurden allerdings nicht nur bei Tieren nachgewiesen, die an
dem MMA-Komplex erkrankt waren, sondern auch bei Tieren die keinerlei Symptome dieses
Krankheitsbildes zeigten. In diesem Zusammenhang gelang PERSSON (1997) die Isolation
von an- und α-hämolysierenden Streptokokken sowohl aus gesunden als auch aus erkrankten
Gesäugekomplexen, wobei β-hämolysierende Streptokokken in dieser Untersuchung lediglich
in den veränderten Gesäugekomplexen entdeckt wurden.
Auf zellulärer Ebene sprechen manche Autoren (WEGMANN u. BERTSCHINGER 1984;
WEGMANN 1985) von einem Vorliegen einer Mastitis bei frisch laktierenden Sauen, wenn
die Milch einen Zellgehalt von über 5 x 106 Zellen/ml aufweist, wobei der Anteil an
neutrophilen Granulozyten über 70% betragen sollte. Bei Drüsenkomplexen, die aufgrund
einer sehr geringen Ferkelzahl dieser Sau nicht besaugt werden, haben diese Werte allerdings
eine geringe Aussagekraft, da hier der erhöhte Zellgehalt eher als ein Zeichen des
physiologisches Versiegen des Milchflusses zu werten ist. Das Sekret aus palpatorisch auf-
fälligen Gesäugekomplexen zeigt in vielen Fällen einen erhöhten Zellgehalt, auch wenn
bakteriologische Untersuchungen dieses Sekretes einen negativen Befund erbringen
(PERSSON 1997).
Eine einheitliche Meinung der Autoren über die Entstehung einer solchen Mastitis geht aus
der Literatur leider nicht hervor. Laut BERTSCHINGER (1984) wandern die Erreger über
den Strichkanal aszendierend in das Gesäuge ein. Diese Variante wird durch Verun-
reinigungen der Liegefläche der Sau mit Erregern verursacht, die aus dem Darm oder aus dem
infizierten Urogenitaltrakt des Muttertieres stammen und so über die Außenhaut der
Zitzenkuppe in den Strichkanal eindringen können. Hautverletzungen in diesem Bereich

Literaturübersicht
20
durch Bissverletzungen der Ferkel oder ähnliche Ursachen wirken sich förderlich auf diesen
Prozess aus (DONE 1980; PERSSON 1997).
Andere Autoren halten den hämatogenen Weg der Besiedelung des Gesäuges für die wahr-
scheinlichste Ursache bezogen auf das Auftreten einer Mastitis (BERNER 1987; BOSTEDT
1999). Als Herkunftsort der krankmachenden Erreger wird aber auch von ihnen der Uro-
genitaltrakt und der Darm angesehen. Es wird vermutet, dass durch ein Herabsetzen bzw.
sogar durch ein Aufheben der Darmschranke eine Streuung von Erregern aus dem Darm in
die Milchdrüse und in andere beteiligte Organe stattfindet, obwohl diese Erreger primär kein
invasives Verhalten zeigen. AWAD- MASALMEH et al. (1990) stützen diese These, indem
sie aus Tupferproben, die aus der Milchdrüse und aus der Zervix entnommen haben, Keime
isolieren konnten, die ebenfalls im Kot dieser Tiere zu finden waren.
In der Frage nach der Ätiologie der Mastitis, die im puerperalen Krankheitskomplex der Sau
eine zentrale Position einnimmt, bleiben somit noch einige Fragen ungeklärt bzw. diese
können nicht eindeutig beantwortet werden.
2.1.4.2. Metritis
Bei dem MMA-Komplex handelt es sich, wie schon beschrieben, um eine Faktoren-
erkrankung bei der eine Reihe von Hilfsfaktoren in Verbindung mit einer bakteriellen
Infektion des Uterusinhaltes zu einer Endometritis führen (TRABITSCH 1988). Somit bildet
das Vorhandensein von Infektionserregern, die als meist fakultativ pathogene Erreger auf eine
gewisse Erregerkonzentration bzw. auf andere infektionsbegünstigende Faktoren angewiesen
sind, die Grundlage für den puerperalen Krankheitskomplex der Sau. VANDEPLASSCHE
(1981) verweist ebenfalls auf eine große Bedeutung einer Infektion des Uterus, da er bei
Sauen mit gestörtem Puerperium in allen Fällen eine abnorme bakterielle Besiedelung des
Uterus nachgewiesen hat und zusätzlich zeigen konnte, dass verschiedene begünstigende
Hilfsfaktoren ohne diese Keimbesiedelung nicht zu einer Endometritis führen. DE WINTER
et al. (1992) konnten durch die Inokulation von E.-coli-Suspensionen in den Uterus
metöstrischer Sauen typische eitrige Endometritiden erzeugen, die dem klinischen Bild der
puerperalen Endometritis sehr ähnlich waren. Damit weisen sie auf eine besondere patho-
genetische Bedeutung dieses Erregers für die Ätiologie der Endometritis hin. AMTSBERG
(1984) hat bei fast 500 Sauen, die an dem puerperalen Krankheitskomplex erkrankt waren,
Zervixtupferproben genommen. Bei der mikrobiologischen Analyse dieser Proben isolierte er
25 verschiedene Bakterienarten und konnte unter diesen mit großer Häufigkeit E. coli, an-, α-

Literaturübersicht
21
und β-hämolysierende Streptokokken, Staphylococcus epidermidis, Staphylococcus hyicus
und Proteus spp. nachweisen. Ähnliche Ergebnisse konnten auch AWAD-MASALMEH et al.
(1990) vorweisen, die neben den Zervixtupferproben allerdings auch Milchproben von an
dem MMA-Syndrom leidenden Sauen untersuchten. Sie isolierten bei 65,7% der Tiere E. coli,
bei 41,8% Staphylokokken und bei 34,3 % β-hämolysierende Streptokokken. Einige dieser
Keime wurden aber nicht nur in Proben nachgewiesen, die von Sauen stammten, welche am
puerperalen Krankheitskomplex erkrankt waren, sondern konnten in ähnlicher Häufigkeit
auch aus Proben von gesunden Tieren isoliert werden. So wiesen TYRELL (1978) und
AMTSBERG (1984) an- und α-hämolysierende Streptokokken und Staphylococcus
epidermidis in nahezu gleicher Verteilung aus Zervixtupferproben sowohl bei am MMA-
Syndrom erkrankten wie auch bei gesunden Tieren nach. Somit lässt sich festhalten, dass die
Bedeutung von Streptokokken und Staphylokokken in der Ätiologie und der Pathogenese des
MMA-Syndroms noch nicht gänzlich geklärt ist.
Ähnlich stellt sich die Situation für die Beteiligung von Mykoplasmen am MMA-Geschehen
dar. Denn MOORE et al. (1966) wiesen diese Erreger in Gewebeproben erkrankter Sauen
nach und in der neueren Literatur werden diese immer wieder als potentiell infektiöse
Faktoren aufgeführt. Andere Autoren (BERTSCHINGER 1977; ROSS et al. 1981 und
AWAD-MASALMEH et al. 1990) konnten eine Beteiligung von Mykoplasmen am
puerperalen Krankheitskomplex der Sau allerdings nicht bestätigen und für den tatsächlichen
Nachweis dieser Erreger aus Milch- oder Zervixtupferproben sind in ihren Arbeiten keine
Belege vorhanden.
Als weitere Erreger, die mit dem Auftreten vom MMA-Komplex in Verbindung gebracht
werden, betrachten WENDT et al. (1998) und EGGEMANN et al. (2000) die Chlamydien.
Diese wurden oft aus Zervixtupfern von Sauen mit Fertilisationsstörungen im Zuge einer
PCR-Analyse entdeckt und es wurde gezeigt, dass die Anzahl serologisch positiv auf
Chlamydien reagierender Sauen pro Bestand nicht nur im Zusammenhang mit Fruchtbar-
keitsstörungen sondern auch mit der Prävalenz des MMA-Syndroms steht.
Für einige Autoren besteht ein klarer Zusammenhang zwischen einer Hypo- bzw. Agalaktie,
dem Auftreten von subklinischen Mastitiden und einer vorliegenden Infektion im Uro-
genitalbereich (AKKERMANS u. POMPER 1980; PETERSEN 1982; BERNER 1984).
Im Gegensatz dazu sehen andere Autoren keinen näheren Zusammenhang zwischen einer
derartigen Infektion und der Ausprägung des puerperalen Krankheitskomplexes der Sau
(BÖNING et al. 1976; BERTSCHINGER 1977; MORCOC et al. 1983; IBEN 1999).

Literaturübersicht
22
PLONAIT (1997) konnte durch die experimentelle intrauterine Infektion keine
reproduzierbaren typischen MMA-Krankheitsbilder erzeugen. Außerdem fand er keine
zwingend auftretenden histologischen Befunde für eine Endometritis bei Tieren mit
vaginalem Ausfluss und konnte somit keine deutliche Beziehung zwischen vaginalem
Ausfluss, einer bakteriellen Infektion des Uterus und dem Auftreten einer mastitisbedingten
Hypogalaktie in Zusammenhang mit Störungen des Allgemeinbefindens darstellen.
MIDDLETON-WILLIAMS et al. (1977) fanden bei ihren pathomorphologischen Unter-
suchungen häufig eine oberflächliche Infiltration von neutrophilen Granulozyten in der
Uterusschleimhaut. Diese Befunde traten aber sowohl bei kranken wie auch bei gesunden
Tieren gleichermaßen auf. Deshalb wird dieser Zustand von ihnen als physiologische
Reaktion der Gebärmutter nach der Geburt beschrieben, woraus eine relativ geringe
Bedeutung dieser Entzündungsreaktion für das Auftreten des MMA-Syndroms resultiert.
STELZER et al. (1997) stellten im Gegensatz dazu allerdings erhöhte Erkrankungsraten und
signifikant höhere Ferkelverluste in der frühen postpartalen Phase bei Sauen fest, die schon
ante partum Vaginalausfluss zeigten. Ferner gelten Verletzungen und Exkoreationen im
Bereich des Genitaltraktes, die im Zuge obstetrischer Eingriffe (GLOCK 1983) auftreten
können, als prädisponierende Faktoren für später eintretende Infektionen (HOSPES et al.
1993) in dieser Körperregion. Außerdem wurden puerperale Endometritiden in Verbindung
mit verlangsamter Involution des Uterus und einer Retention der Lochialflüssigkeit
(EHRENTRAUT 1968; GLOCK 1983) beobachtet, die häufig nach einer verlängerten
Geburtsdauer beim Schwein auftraten (BÖNING et al. 1976; HEINRITZI u. HAGN 1999).
Diese Aussage bekräftigen ebenfalls BÄCKSTRÖM et al. (1984) und BOSTEDT et al.
(1998), die ein vermehrtes Auftreten des MMA-Syndroms bei Tieren verzeichneten, die sehr
große Würfe zur Welt brachten. Die Anzahl an totgeborenen Ferkel korreliert ihrer Ansicht
nach auch mit einem verstärkten Auftreten des puerperalen Krankheitskomplexes der Sau.
Somit kann abschließend zusammengefasst werden, dass Entzündungen im Bereich der
Gebärmutter eines der Hauptsymptome des MMA-Komplexes darstellen und deshalb in
diesem Zusammenhang von großer Bedeutung sind.

Literaturübersicht
23
2.1.5. Prophylaxemaßnahmen zur Verhinderung des MMA-Sydroms
Die Prophylaxemaßnahmen gegen das MMA-Syndrom der Sau verhindern diese Erkrankung
zwar nicht vollständig, senken aber deren Prävalenz und führen dazu, dass die Krankheit
selbst bzw. einzelne Symptome bedeutend milder verlaufen. Zur Senkung der Krankheits-
anfälligkeit für den puerperalen Krankheitskomplex der Sau wird zu hygienischen und
haltungstechnischen Maßnahmen, zur Chemoprophylaxe und zu prophylaktischen Fütterungs-
empfehlungen geraten.
Eine Vorraussetzung auch für andere prophylaktische Maßnahmen stellen die Haltungs-
bedingungen der Sauen dar. So fördert z.B. die Möglichkeit zur freien Bewegung im Verlauf
der Trächtigkeit, wie sie u.a. in der Gruppenhaltung gegeben ist, die Vitalität der Sauen und
wirkt sich somit positiv auf deren Puerperium aus (SCHADE 2000). Außerdem steigert die
Bewegung die Darmperistaltik und verhindert so Kotverhaltungen und Toxinresorptionen
(RINGRAP 1960). SANDSTEDT und SJORGREN (1982) stellten beim Vergleich
verschiedener Haltungsformen fest, das sich eine Weidehaltung über den Zeitraum der
Trächtigkeit sehr positiv auf die Inzidenz (2,6% bei Weidehaltung gegenüber 15,9% bei reiner
Stallhaltung) des MMA-Komplexes auswirkte. Es wurde schon früh erkannt, dass sich in den
letzten Tagen der Trächtigkeit Änderungen der Haltungsgewohnheiten negativ auswirken
(RINGRAP 1960; SCHULZE u. BOLLWAHN 1962; PLONAIT et al. 1986). Deshalb wird
eine gruppenweise Belegung der Abferkelabteile nach vorangegangener Reinigung und
Desinfektion im Rein-Raus-Verfahren empfohlen (MORGENTHUM u. BOLDUAN 1987).
Nur über den Zeitraum einer Gewöhnung an diese neue Umgebung machen die einzelnen
Autoren sehr unterschiedliche Angaben. LUTTER (1983) hält drei Tage zur Eingewöhnung
für ausreichend, während von LERCH (1987) zehn Tage propagiert werden und andere sogar
einen Zeitraum von drei Wochen für angemessen halten (MORGENTHUM u. BOLDUAN
1987). Gerade für Jungsauen, die neu in den Bestand eingegliedert werden, ist eine aktive
Immunisierung über den frühzeitigen Kontakt mit älteren Zuchtsauen von Bedeutung, um sich
so mit der Keimflora des Bestandes auseinander zu setzen (BOLLWAHN 1980).
Im Zuge der alimentären Prophylaxe gegen das MMA-Syndrom ist nicht nur auf ein
ausreichendes Angebot an einwandfreiem Tränkewasser während der Trächtigkeit zu achten,
sondern es muss auch dessen ausreichende Aufnahme gewährleistet sein. Dadurch kann
nämlich chronischen Harnwegsinfektionen und in diesem Zusammenhang auch sekundär
einem gehäuften Auftreten von Endometritiden entgegengewirkt werden. Im Zeitraum um die

Literaturübersicht
24
Geburt herum sollte den Tieren zusätzlich zu der Wasserversorgung über die Selbsttränke
Wasser in ausreichender Menge über den Trog verabreicht werden (FINKENSIEP 1993).
Die Prophylaxe gegen den MMA-Komplex im Bereich der Fütterung beginnt schon in der
Phase der Frühträchtigkeit, um eine optimale Körperkondition über die gesamte Trächtigkeit
zu erhalten. In der Gravidität sind daher voluminöse und Rfa-reiche Futtermittel von
Bedeutung, um die Darmperistaltik und somit auch die Chymuspassage anzuregen
(KAMPHUES u. BEENING 1998). Außerdem bereiten diese die Tiere optimal auf die
größeren Futtermengen vor, die während der Laktation aufgenommen werden müssen. In der
prophylaktischen Wirksamkeit gegen Gesundheitsstörungen und Probleme, die gehäuft im
peripartalen Zeitraum auftreten, ist allerdings die Fütterung im letzten Stadium der
Trächtigkeit entscheidend (KAMPHUES u. BEENING 1998). Deshalb wird in der Praxis
versucht durch die Kombination von mehreren fütterungstechnischen Maßnahmen eine
wirksame Prophylaxestrategie zu entwickeln. Zu diesen zählt die Restriktion der Futtermenge
kurz vor der Geburt bzw. am Tage der Geburt. Dabei konnte festgestellt werden, dass eine
restriktive Fütterung der Sauen ante partum (vom 110. Trächtigkeitstag an) mit einer
verkürzten Dauer der Geburt, einer verminderten Zahl an Totgeburten, sowie geringeren
Ferkelverlusten an den ersten Tagen post partum und einer geringeren Prävalenz an MMA-
Erkrankungen einherging (BILKEI u. BÖLCSKEI 1993). Andere Autoren, die mit einer
restriktiven Fütterung schon drei Wochen vor dem errechneten Geburtstermin beginnen,
konnten die Prävalenz der Puerperalerkrankungen sogar von 59% auf 8% senken
(SANDSTEDT u. SJORGEN 1982). Eine Futterrestriktion von diesem Ausmaß wird
allerdings in anderen Arbeiten als nicht unproblematisch eingestuft, da es einerseits zu einer
negativen Beeinflussung der Chymuspassage und in einer Phase hohen Bedarfs zu kurz-
fristigen Energie- und Nährstoffunterversorgungen kommen kann (KAMPHUES u.
BEENING 1998) und andererseits können diese Tiere dazu neigen stereotype Verhaltens-
weisen zu entwickeln (LAWRENCE u. TERLOUW 1993; WELDON et al. 1994). Eine
Überfütterung in der letzten Phase der Trächtigkeit ist aber wegen der Gefahr einer
Verminderung der Darmperistaltik zu vermeiden, da in diesem Fall eine Resorption von
Endotoxinen möglich wäre, die das MMA-Risiko erhöht (CEREZA et al. 1986;
MARTINEAU et al. 1992; KAMPHUES et al. 1998).
Als weitere fütterungstechnische Prophylaxemaßnahme zur Vermeidung von Obstipationen
und zur Verringerung des MMA-Risikos wird die Erhöhung des Rohfasergehaltes in der
Ration für die hochträchtigen und sehr frisch laktierenden Sauen empfohlen (GÖTZE 1939;
RINGRAP 1960; EHRENTRAUT 1968; LUTTER 1983; MORGENTHUM u. BOLDUAN

Literaturübersicht
25
1987; GÖRANSSON 1989). Die Rohfaserbestandteile neigen zu einer starken Aufquellung,
indem sie Wasser unter der Bildung von Hydrokolloiden binden. Durch diesen Vorgang
nimmt das Ingestavolumen zu und führt zu einem Dehnungsreiz, der die Darmperistaltik
anregt und so zu einer beschleunigten Chymuspassage führt. Außerdem entstehen beim
mikrobiellen Abbau der Rohfaser osmotisch wirksame Bestandteile, die einen laxierenden
Effekt haben (LÖSCHER 1994). Bei der Fütterung einer rohfaserreichen Ration wurden im
Vergleich zur Verfütterung einer herkömmlichen Ration geringere Zunahmen an Körper-
masse während der Gravidität und geringere Gewichtsverluste bei steigender Futteraufnahme
in der anschließenden Laktation beobachtet (LOPEZ et al. 1988). Außerdem sollen die Sauen
eine höhere Lebenserwartung haben, wenn sie derartig gefüttert werden (POLLMANN et al.
1981; POND et al. 1985; CARTER et al. 1987; NELSON et al. 1992a).
Das gleiche Ziel wie bei der Verfütterung einer rohfaserreichen Ration wird mit der Zugabe
von Natriumsulfat (Na2SO4) verfolgt. Denn das Glaubersalz gehört zu der Gruppe der schwer
resorbierbaren salinischen Laxatien (LÖSCHER 1994) und verbleibt somit nach oraler Gabe
zum größten Teil im Darmlumen. Dort ist es dann osmotisch wirksam und bindet vermehrt
Wasser, so dass ähnlich wie bei der Verfütterung von rohfaserreichen Futtermittel ein
Dehnungsreiz entsteht, über den die Darmperistaltik anregt wird. Diese Verfütterung von
Glaubersalz an hochträchtige Sauen ist in der Praxis weit verbreitet und führt nach 14-20 h
zum Abgang von relativ wässrigem Faeces.
Eine weitere Maßnahme aus dem fütterungstechnischen Bereich zur Verhinderung
puerperaler Erkrankungen besteht in der Verfütterung von probiotischen Substanzen.
Probiotika sind Kombinationspräparationen aus Bakterien und Hefen, die nach oraler
Verabreichung bioregulatorisch in die Darmbesiedelung eingreifen, indem sie die Darmflora
stabilisieren (GEDEK 1993). Dabei sollen lebend verabreichte Mikroorganismen wie z.B.
Bacillus-, Lactobacillus- und Streptococcus- Arten (Milchsäurebildner) verhindern, dass eine
schädliche Begleit- und Restflora im Darm die Übermacht gewinnt, wenn die intestinale
Hauptflora durch Stress, Magen-Darm-Infekte, den Geburtsverlauf oder eine länger
andauernde Therapie mit Antibiotika reduziert wurde (WIESSNER u. GOLBS 1991). Als
Futterzusatzstoffe finden meistens solche Mikroorganismen Verwendung, die die Innenseite
des Darmes mit einem Biofilm überziehen, indem sie sich in dem Mucus verankern, mit dem
das Darmrohr in diesem Bereich ausgekleidet ist. Durch eine direkte Konkurrenz um die
Nahrung und um die Adhäsionsmöglichkeiten an der Darmwand sollen so Infektionen
verhindert werden (SHERMAN et al. 1987). Eine weitere Möglichkeit der alimentären
Prophylaxe besteht in der Verabreichung von Substanzen, die innerhalb des Körpers zu einer

Literaturübersicht
26
Verschiebung des pH-Wertes führen, um dann sekundär auch den pH-Wert im Harn abzu-
senken. Die prophylaktischen Maßnahmen und die ihnen zugrunde liegenden Mechanismen
werden in den nachfolgenden Kapiteln näher erläutert.
Von einem prophylaktischen Einsatz von Antibiotika zur Keimreduzierung ist in diesem Fall
abzuraten, da diese Maßnahmen zwar kurzfristig zu guten Erfolgen führen, aber nicht für den
langfristigen Einsatz geeignet sind. Bei der prophylaktischen Verwendung werden aus
Kostengründen oft zu geringe Dosierungen verwendet, so dass sich auf Dauer massive
Resistenzen entwickeln können. Der vorschriftsmäßige Einsatz mit ausreichender Dosierung
der entsprechenden Medikamente ist nur im Einzelfall interessant, wenn andere pro-
phylaktische Maßnahmen versagen, da beim Masseneinsatz die finanzielle Belastung zu stark
wäre. Diese Lösung eignet sich also nur für das Einzeltier oder aber in absoluten Problem-
betrieben für eine größere Zahl von Tieren.
2.2. Zusammenhang zwischen dem puerperalen Krankheitskomplex und der unspezifischen Form der Harnwegsinfektion bei der Sau
Das Auftreten des puerperalen Krankheitskomplexes der Sau steht in einem recht engen
Zusammenhang mit dem Vorhandensein einer meist unspezifischen Form der Harnwegs-
infektion. Dieses Phänomen ist durch die anatomische Lage der Harnorgane bzw. im
Besonderen der Harnblase und den Organen des Geschlechtsapperates zu erklären. Dabei
fungiert die Harnblase als Keimreservoir, von dem aus die Erreger aszendierend den Genital-
trakt besiedeln. Im Zeitraum des Puerperiums, in dem der Verschluss des Uterus und die
Mechanismen der Keimabwehr herabgesetzt sind, funktioniert dieser Weg der Infektion sehr
gut, so dass in dieser Zeit das Krankheitsbild einer Endometritis leicht zu erzeugen ist. Ein
anderer Infektionsweg, der den Weg über das Gesäuge nimmt, kommt zustande, indem die
Tiere beim Absetzen des Harnes, der mit Erregern angereichert ist, die Liegeflächen
kontaminieren. Beim Ablegen der Sau kommt dann das Gesäuge mit dieser kontaminierten
Liegefläche in Kontakt und die Erreger können ebenfalls aszendierend über den Strichkanal
das Gesäuge bzw. einzelne Gesäugekomplexe besiedeln, um hier das Krankheitsbild einer
Mastitis zu erzeugen. Puerperale Erkrankungen, die mit stark erhöhter Körpertemperatur
einhergingen, wurden somit verstärkt bei Sauen diagnostiziert, die vorher an einer
Harnwegsinfektion litten. Außerdem konnten bei diesen Tieren Erreger, die zuvor im
entzündeten Harntrakt gefunden wurden, später im Puerperium aus Proben isoliert werden,
die aus den Genitalorganen oder sogar aus veränderten Gesäugekomplexen entnommen

Literaturübersicht
27
wurden (BERNER 1971, 1988). Chronische Zystitiden werden von WENDT (1998) ebenfalls
als wichtiger und prädisponierender Faktor für das Ausbrechen des MMA-Syndroms gesehen,
indem von diesen eine aszendierende Infektion des puerperalen Uterus und über die
bakterielle Kontamination der Liegeflächen mit Urin eine Besiedelung des Gesäuges mit
Erregern ausgehen, die eine Mastitis verursachen können. BERNER (1984) kam nach seinen
Untersuchungen zu dem Ergebnis, dass es bei Sauen, die an einer Infektion der Harnorgane
litten, in Abhängigkeit von den exogenen und endogenen Faktoren, den Arten der Erreger und
deren Infektionsdosis nach der Geburt in 13% der Fälle zu keiner Erkrankung im Sinne des
MMA-Komlexes kam, in 58% der Fälle kam es zu einer leichten bis mittelschweren Form
und bei 29% der Tiere entwickelte sich eine akute und schwerwiegende Ausprägung des
puerperalen Krankheitskomplexes. Außerdem fand BERNER (1971) heraus, dass der
Keimgehalt im Harn bei Sauen, die schon weit vor der Geburt an einer Infektion ihrer
Harnwege litten, in den letzten Wochen der Gravidität erheblich ansteigt. In einigen Fällen
wurde beobachtet, dass die Keimzahl bei solchen Tieren kurz vor, während oder aber kurz
nach der Abferkelung in die Höhe schnellte. Diese zeitliche Nähe der unspezifischen
Harnwegsinfektion zur Geburt erklärt BERNER (1988) mit einigen Faktoren, die zeitlich mit
der Geburt in Verbindung stehen und welche die Pathogenese einer Cystitis begünstigen. So
kommt es während der Spätgravidität und auch kurz nach der Geburt zur Erschlaffung der
Blasenmuskulatur, was einerseits dazu führt, dass ein erhöhtes Restharnvolumen in der
Harnblase verbleibt und andererseits die Funktion des M. sphinkter vesicae eingeschränkt ist.
Außerdem kommt es in der Spätgravidität seltener zur Blasenentleerung, woraus sich eine
geringere Keimverdünnung ergibt, und, durch das erhöhte Restharnvolumen bedingt, steht
den Keimen immer ein geeignetes Milieu zur Verfügung. Außerdem tritt eine
Resistenzminderung der Mukosa, teils bedingt durch Traumatisierungen im Urogenitalbereich
während der Geburt, teils bedingt durch die allgemein herabgesetzte Immunitätslage des
Muttertieres um den Geburtszeitraum herum ein. Zusätzlich begünstigen Nachgeburts-
verhaltungen, abgehende Lochien und ein mangelhafter Schamschluss eine aszendierende
Keimbesiedlung des Urogenitaltraktes. STIRNIMANN (1984) stellte in seinen
Untersuchungen fest, dass nur 18% der Infektionen der Harnwege während der Trächtigkeit
vorliegen, in die ersten zwei Wochen nach der Geburt fallen dagegen 40% der Erkrankungen
der Harnwege. BOLLWAHN et al. (1984) fanden vor allem bei Sauen mit einem gestörten
Puerperium ebenfalls eine postpartale Häufung an Bakteriurien und Zystitiden. Auch
subklinische Harnwegsinfektionen, bei denen keinerlei klinische Symptome vorliegen, aber
die einen positiven Befund bei der bakteriologischen Untersuchung der Harnproben

Literaturübersicht
28
erbrachten, wirken sich laut THORNTON et al. (1998) förderlich auf eine höhere
Erkrankungsrate am MMA-Syndrom und somit auf höhere Ferkelverluste bis zum Absetz-
termin aus. BOTH et al. (1980) zeigen in ihren statistischen Untersuchungen, dass ein relativ
enger Zusammenhang zwischen einer Infektion des Harnapparates und Störungen des
Puerperiums bzw. der Fruchtbarkeit in Sauenbeständen besteht. In Sauenherden mit
Puerperalstörungen lag die Prävalenz für Harnwegsinfektionen bei 44%, in Betrieben ohne
Auffälligkeiten im Puerperium lag diese Zahl dagegen deutlich niedriger bei 3%. Sogar 23%
der gesunden Tiere wiesen in den Problemherden erhöhte Keimgehalte im Harn auf, während
dieser Befund in den Kontrollbeständen lediglich bei 4% der gesunden Sauen erhoben werden
konnte. Diese Tiere stellen für BERNER (1984) so genannte „Indikatortiere“ dar, die eine
bestandsweise auftretende Kumulation bestimmter fakultativ pathogener Keime anzeigen und
somit als Vorboten für das Auftreten von Endometriden und Mastitiden fungieren.
PETERSEN (1979) bezeichnet das Vorliegen einer Harnwegsinfektion deshalb als
„Vorfeldsyndrom“ des MMA-Krankheitskomplexes und gibt die Empfehlung, vor der
Abferkelung eine Spontanharnprobe zu untersuchen, um Tiere zu identifizieren, die für das
Auftreten des MMA- Syndroms prädisponiert sind. Darüber hinaus gibt sie zu bedenken, dass
diese Sauen eine mögliche Infektionsquelle für gesunde Tiere darstellen.
Bekräftigt werden diese Aussagen auch durch die Arbeit von BERNER und JÖCHLE (1988),
die Proben von sterilen Zuchtsauen mit unspezifischen Infektionen der Geschlechtsorgane
untersuchten und dabei nahezu die gleichen Erreger isolierten wie bei einer unspezifischen
Infektion des Harnapparates. In den meisten Fällen handelte es sich bei diesen Infektionen um
so genannte Monoinfektionen. Im isolierten Keimspektrum nahm E. coli (50%) den ersten
Rang ein, gefolgt von Streptokokken (39,1%) und Staphylokokken (15,1%). Bei den Tieren,
die wegen einer unspezifischen Genitalinfektion an Fruchtbarkeitsproblemen mit Sterilitäts-
folge litten, wurden in 89% der Fälle gleichzeitig eine unspezifische Infektion der Harnwege
festgestellt. In 32% der Fälle wurden sogar die gleichen Erreger isoliert.
Somit kann abschließend festgestellt werden, dass ein Vorliegen einer unspezifischen
Infektion des Harnapparates in einem engen Zusammenhang mit dem späteren Ausbruch des
puerperalen Krankheitskomplexes der Sau steht und dass deshalb eine Verminderung dieser
Infektionen der Harnwege auch eine prophylaktische Wirkung gegen das Auftreten des
MMA-Komplexes hat. Deshalb soll im weiteren Verlauf auf die Problematik der Harnwegs-
infektionen und deren Vermeidung näher eingegangen werden.

Literaturübersicht
29
2.3. Harnwegsinfektionen beim Schwein
Eine Infektion der Harnwege stellt bei Schweinen und gerade bei älteren Sauen eine häufige
Erkrankungsform dar, bei der Glomerulonephritiden, interstitielle Nephritiden, Tubulo-
nephrosen, Zystitiden und Perizystitiden beobachtet werden. Eine große Bedeutung hat in
diesem Zusammenhang die Cystitis, da das Milieu der Harnblase für viele Erreger gute
Bedingungen zum Überleben bzw. zur Vermehrung bietet. Deshalb fungiert die Harnblase oft
als Erregerreservoir, von dem aus die anderen Harnorgane besiedelt werden (SOFRENOVIC
u. BOLLWAHN 1963; WENDT 1992). Harnwegsinfektionen stellen bei notgeschlachteten
und verendeten Sauen die häufigste Abgangsursache dar und sind ein wichtiger Faktor in der
Ätiologie und der Pathogenese des puerperalen Krankheitskomplexes der Sau (BERNER
1988).
In der Literatur werden eine spezifische und eine unspezifische Form der Harnwegsinfektion
unterschieden (SMITH 1983). Die spezifische Form, die durch Eubacterium suis hervor-
gerufen wird, verursacht meist schwerwiegende aszendierende Erkrankungen im gesamten
Harntrakt (WENDT 1992). In der heutigen Literatur wird Eubacterium suis allerdings als
Actinobaculum suis bezeichnet (LAWSON et al. 1997). Der Eber fungiert bei dieser
Erkrankung meist ohne selbst an ihr zu erkranken als Keimreservoir, da er die Erreger in
seinem Präputialdivertikel beherbergt und sie beim Deckakt auf die Sau überträgt. Bei der Sau
kommt es von den Schleimhäuten des Genitaltraktes zu einer aszendierenden Infektion des
gesamten Harnapparates (WENDT et al. 1993). Die spezifische Harnwegsinfektion verläuft
subakut bis chronisch (JONES 1980; WALDMANN 1987) und die Tiere magern im Verlauf
der Erkrankung sehr stark ab. Es können allerdings auch plötzliche Todesfälle durch
intravesikales Verbluten auftreten.
Bei der unspezifischen Form der Harnwegsinfektion handelt es sich wie beim MMA-
Komplex dagegen um ein multifaktorielles Geschehen, bei dem ubiquitär vorkommende
Umweltkeime als fakultativ pathogene Erreger von anderen endogenen und exogenen
Faktoren dazu befähigt werden eine Infektion auszulösen. Die unspezifische Form der Harn-
wegsinfektion betrifft dabei häufig nur die Harnblase des Schweins (Waldmann 1987) und
verläuft oft ohne eindeutige klinische Symptome. Deshalb stellt diese Form der Harnwegs-
infektion in der Schweinepraxis ein weitaus größeres Problem dar als die spezifische Form
der Harnwegsinfektion (STIRNIMANN 1984; VERTER und UECKER 1990).

Literaturübersicht
30
2.3.1. Prävalenz der unspezifischen Form der Harnwegsinfektion der Sau
Zur Prävalenz der unspezifischen Infektion der Harnwege beim Schwein werden in der
Literatur sehr unterschiedliche Angaben gemacht. Berner (1981 a und b) stellt diese
Erkrankung in seiner Untersuchung bei 25,4% der Schlachtsauen fest, wobei sie bei 17,5%
der Tiere mit ungeklärter Herkunft und bei 42% der Sauen aus Betrieben mit Harnwegs-
problemen auftrat. In Ferkelerzeugerbetrieben stellte er bei 24,8% der Muttertiere im Bereich
des Harnapparates eine Infektion fest. Hier schwankte die Befallshäufigkeit zwischen 11,2%
und 47,8%. VESPER (1991), die 14 willkürlich ausgewählte Ferkelerzeugerbetriebe unter-
suchte, stellt eine durchschnittliche Befallsstärke von 18,7% fest, wobei die Häufigkeit des
Auftretens einer Harnwegsinfektion in einer Bandbreite zwischen 3,8% und 36,4% vertreten
war. Petersen (1979) bringt die Prävalenz von unspezifischen Harnwegsinfektionen mehr mit
der Situation des Einzelbetriebes in Verbindung. Dabei spricht er bei einer Prävalenz von
unter 10% lediglich von einem Einzeltierproblem. Eine Befallshäufigkeit von 20-30% wird
von ihm als Schwerpunktproblem bezeichnet, während seiner Meinung nach ein Bestands-
problem erst ab einer Prävalenz von 40% vorliegt. Einige Autoren zeigen unter der Prämisse,
dass es sich hier um eine Faktorenkrankheit handelt (BUSSE et al. 1982; MADEC u. DAVID
1983; VESPER 1991), dass sich exogene Faktoren wie eine unbefriedigende Hygienesituation
im Betrieb, belastende Haltungsformen oder Bewegungsstörungen der Tiere maßgeblich auf
die Häufigkeit des Auftretens der unspezifischen Form der Harnwegsinfektion auswirken.
Außerdem sind für das Krankheitsgeschehen das Alter und die Anzahl der Würfe der Sauen
von Bedeutung. STIRNIMANN (1984) stellte in diesem Zusammenhang fest, dass bei
Altsauen (≥ 6 Würfen) die Befallshäufigkeit von Harnwegsinfektionen mit 47% deutlich über
dem Wert bei Erstlingssauen mit 13% lag. BECKER et al. (1985) zeigten eine kontinuierliche
Zunahme der Prävalenz von Harnwegsinfektionen von relativ jungen Sauen (≤ 3 Würfe) mit
17,5% bis rauf auf 32% (-60%) bei Altsauen (7-10 Würfe).
Als weitere endogene Faktoren sind die ausreichende Funktion der körpereigenen Abwehr-
mechanismen zur Vermeidung von Harnwegsinfektionen und die Beschaffenheit des Harnes
als bakterielles Medium zu nennen. Der anatomische Aufbau des Harntraktes bildet dabei die
Grundlage für die Abwehr einer unerwünschten Keimbesiedelung durch eine aszendierende
Infektion aus dem äußeren Geschlechtstrakt. So wurden in einigen Untersuchungen ein
erhöhter Keimgehalt am Orificium urethrae externum festgestellt, wenn der Vulvaschluss
aufgrund von Verletzungen gestört war (MADEC u. DAVID 1984; BERNER 1988). Im
Bereich der Urethra wirkt nicht nur deren Länge (KIVISTO et al. 1977) sondern auch die so

Literaturübersicht
31
genannte „Urethral High Pressure Zone“ bestehend aus glatter und quergestreifter Muskulatur
(OSBORNE et al. 1979), die sich in der Mitte der Urethra befindet, einer aszendierenden
Keimbesiedelung entgegen. Wenn die Funktion dieser Zone oder die des Musculus sphinkter
vesicae z.B. durch Verletzungen nach der Geburt oder nach dem Deckakt gestört ist, wird
dadurch eine ascendierende bakterielle Migration gefördert (BERNER 1988).
Um eine Infektion auszulösen, müssen die Erreger sich zunächst einmal in der Schleimhaut
verankern und sich dort vermehren bzw. Kolonien bilden (REID u. SOBEL 1987). Als Teil
der körpereigenen Abwehr besiedeln apathogene Keime nach dem Prinzip der „bakteriellen
Interferenz“ den äußeren Bereich des Harnapparates und stellen somit eine Konkurrenz für
die aszendierenden Keime dar (OSBORNE et al. 1979; REID u. SOBEL 1987). Gegen eine
dauerhafte Erregeradhäsion schützt sich der Körper durch eine kontinuierliche Erneuerung
des Harntraktepithels, da im Zuge der Desquamation Keime aus den Harntrakt entfernt
werden (DAVIS et al.1977; BIBERSTEIN 1990). Außerdem werden die Epithelien von
Mukopolysacchariden (Glykosaminoglykane) überzogen, die das Anheften von Erregern
verhindern, indem sie die Epithelzellrezeptoren maskieren (SHROM et al. 1977; PARSONS
et al. 1978; REID u. SOBEL 1987; LILLY u. PARSONS 1990; WENDT 1992). Außerdem
werden in der Schleimhaut sekretorische Immunglobuline produziert (OSBORNE et al.
1979), die von PARSONS et al. (1975) sogar als wichtigste Barriere gegen das Anheften von
Erregern angesehen werden. Dabei lassen sich sowohl bei gesunden Sauen wie auch bei
Tieren, die an einer Harnwegsinfektion litten, Immunglobuline vom Typ IgG, IgA und SIgA
nachweisen (BOURNE et al. 1973; WAGNER 1990). Bei E.-coli bedingten Infektionen oder
A.-suis-Pyelonephritiden lassen sich sogar spezifische Antikörper nachweisen
(STIRNIMANN 1984; LANGFELDT et al. 1990).
Die physiologische Funktionsweise der Harnorgane hat ebenfalls den Effekt des köpereigenen
Abwehrmechanismus zur Vermeidung von Harnwegsinfektionen, da die von innen nach
außen gerichteten Muskelbewegungen einen mechanischen Spüleffekt erzeugen, dessen
Effektivität von einer ausreichenden Menge an gebildetem Harn, der Miktionsfrequenz und
der Vollständigkeit der Entleerung der Harnblase bzw. dem Restharnvolumen abhängt
(OSBORNE et al. 1979). NORDEN et al. (1968) zeigten in ihren Untersuchungen, dass durch
diesen Spüleffekt in Verbindung mit den vorher genannten Abwehrmechanismen 99,9 % der
Keime aus dem Harntrakt entfernt werden können. GREGORY et al. (1971) kontaminierten
Harnblasen von Hunden mit E. coli und fanden heraus, dass diese durch den Spüleffekt
innerhalb von 72 Stunden aus der Blase eliminiert werden.

Literaturübersicht
32
Als weiterer endogener Faktor beeinflusst der Harn selbst bzw. dessen Zusammensetzung das
Auftreten von Harnwegsinfektion. Seine physikalischen Eigenschaften wie der pH-Wert, die
Osmolarität und das spezifische Gewicht sind dabei von entscheidender Bedeutung. Für den
physiologischen Harn-pH-Wert beim Schwein wird ein relativ weites Spektrum an Werten
angegeben, wobei folgende Werte als physiologisch gelten, pH 5-8 (PETERSEN 1979), pH
6,1-7,3 (v.VOPELIUS-FELDT 1984), pH 6,5-7,5 (MADEC u. DAVID 1984) und pH 5,27-
8,11 (RUHRMANN et al. 1986). Der pH-Wert beeinflusst maßgeblich die Eignung des
Harnes als bakterielles Medium, da aus einer Verschiebung des pH-Wertes eine
Verlangsamung des Generationsintervalles bzw. ein Sistieren des Bakterienwachstums
resultieren kann, oder im Extremfall entwickelt sich sogar ein bakteriozides Medium. In
verschiedenen Untersuchungen wurde gezeigt, dass der Optimalbereich bezogen auf den pH-
Wert für das Wachstum von E. coli im Harn zwischen pH 6,0 und pH 7,0 angesiedelt ist. Bei
Abweichungen des pH-Wertes sowohl in alkalischer als auch in saurer Richtung verlangsamt
sich das Bakterienwachstum, bis es schließlich oberhalb von pH 7,5 und unterhalb von pH 5,0
zum Erliegen kommt (ASSCHER et al. 1966). Veränderungen der Wachstumsintensität sind
in diesem Bereich schon bei Verschiebungen des pH-Wertes von 0,2 Einheiten zu
registrieren. Außerhalb des Wachstumsbereiches genügen dagegen schon sehr kleine
zusätzliche Verschiebungen des pH-Wertes in den sauren pH-Bereich um 0,1 – 0,2 Einheiten
und es erfolgt ein Absterben der Bakterien (LEES 1978). Dabei sind in diesem
Zusammenhang nicht die pH-Werte im bakterioziden Bereich von Bedeutung, sondern eher
jene, die zu einer Stase des Bakterienwachstums führen. Denn diese bakteriostatischen pH-
Wert-Bereiche sind schon unter physiologischen Bedingungen zu erreichen. Diese wirken
sich so negativ auf das Bakterienwachstum aus, dass sie zusammen mit den schon
beschriebenen körpereigenen Abwehrmechanismen, vor allem mit dem Spüleffekt,
ausreichen, um eine Infektion der Harnwege zu verhindern bzw. deren Prävalenz zu
minimieren (YEAW 1944).
Der pH-Wert ist dabei allerdings in enger Verbindung mit der Osmolarität des Harns zu
betrachten. ASSCHER et al. (1966) verwiesen in ihrer Arbeit im menschlichen Urin auf die
Abhängigkeit zwischen dem pH-Wert und der Osmolarität in der Gestaltung eines
bakteriellen Mediums, indem sie zeigten, dass bei einer Osmolarität des Harns von unter 600
mOsm/kg der pH-Wert unter pH 5 gesenkt werden musste, um das Wachstum von E. coli im
Harn zu stoppen. Bei einer Osmolarität von über 600 mOsm/kg reichte dagegen ein Absenken
des pH-Wertes auf pH 5,5 aus, um den gleichen Effekt zu erzielen. Im Kaninchenharn wirkte
in den meisten Fällen allein eine Osmolarität von 800 – 1000 mOsm/kg bakteriozid für E. coli

Literaturübersicht
33
(Mulholland et al. 1969). Somit zeigt sich, dass das Keimwachstum im Medium Harn
maßgeblich von dessen pH-Wert und dessen Osmolarität beeinflusst wird. Dabei ist zu
beachten, dass je weiter sich einer dieser beiden Faktoren aus dem Optimalbereich entfernt,
sich umso weniger der jeweils andere Faktor aus diesem Bereich entfernen muss, um das
Keimwachstum zu hemmen bzw. im Extremfall sogar bakteriozid zu wirken (Lees et al.
1978).
Als weitere Eigenschaft ist das spezifische Gewicht des Harnes interessant, das von der
Konzentration der gelösten Stoffe im Harn abhängt (PLONAIT 1980) und damit von der
Diurese bzw. des Resorptions- und Konzentrationsvermögen der Nieren bestimmt wird.
Deshalb wird auch für diesen Parameter ein relativ weiter physiologischer Schwankungs-
bereich angegeben. Für die Harndichte werden somit Werte von 1005-1035 mg/dl
(BAUMGARTNER u. KRUZIG 1983) bzw. 1009-1025 mg/dl (v. VOPELIUS-FELDT 1984)
angegeben.
Weiteren Aufschluss über die Beschaffenheit des Harnes liefert die mikroskopische
Untersuchung des durch Zentrifugation gewonnenen Harnsedimentes. Dieses enthält
Epithelzellen, Bakterien, Leukozyten, Erythrozyten und Kristalle. Epithelzellen befinden sich
häufig im Harn, da es im Harntrakt zur permanenten Epithelnachbildung und somit zur
Abschilferung des Epithels kommt. Deshalb sind im Harn Plattenepithelien aus dem äußeren
Genitale und der Urethra, Übergangsepithelien aus der Blase, dem Ureter und dem
Nierenbecken und Nieren- bzw. Tubulusepithelien zu finden (HEINTZ u. ALTHOF 1984).
Das Auftreten von Leukozyten im Harn spricht für einen entzündlichen Prozess im Bereich
des Harntraktes, obwohl es seinen Ursprung auch in einer mechanischen Irritation der
Schleimhaut haben kann (SCHMIDL 1979b; BOLLWAHN u. ARNHOFER 1989).
Erythrozyten sind im Harn normalerweise nicht zu finden und stellen deshalb immer einen
pathologischen Befund dar, weil sie auf Verletzungen der harnableitenden Organe oder
schwerwiegende entzündliche Prozesse an den Nieren hinweisen.
Kristalle sind häufig und teilweise in größeren Mengen im Sauenharn zu finden (MADEC u.
DAVID 1983). Dabei kann man morphologisch amorphe Phosphate, Ammonium-
Magnesium-Phosphate (Struvit), Oxalate und Urate unterscheiden, wobei die Phosphate mit
63% am häufigsten nachgewiesen werden (MADEC u. DAVID 1983). Dieses Phänomen wird
als Kristallurie bezeichnet (WENDT et al. 1996). Das Auftreten von Kristallen unterliegt
dabei tierindividuellen Einflüssen und ist außerdem vom pH-Wert und von der Konzentration
des Harns abhängig, die wiederum vom Mineralstoffgehalt im Futter und von der Wasser-
aufnahme der Tiere bestimmt wird (LAPPE 1995). Während gesteigerte Calcium- bzw.

Literaturübersicht
34
Phosphorkonzentrationen im Harn und ein alkalischer pH-Wert dieses Mediums die
Kristallbildung fördern, wird dieser Vorgang von höheren Gehalten an Natrium und Kalium
gehemmt. Calcium und Phosphor fallen im Harn in erster Linie dann aus, wenn das
Löslichkeitsprodukt aufgrund einer erhöhten Harnkonzentration überschritten wird, die meist
als Folge reduzierten Trinkwasseraufnahme auftritt (WENDT et al. 1996).
2.3.2. Das Keimspektrum der unspezifischen Form der Harnwegsinfektion
In den meisten Fällen handelt es sich bei dem Krankheitsbild der unspezifischen Harnwegs-
infektion um so genannte Monoinfektionen, an der lediglich eine Keimart beteiligt ist. Am
häufigsten sind als Erreger E. coli bzw. coliforme Keime (58,1-75,9%) vertreten, an zweiter
Stelle kommen die Streptokokken (4,2-19,8%), gefolgt von Korynebakterien (4,2%) (heute
Actinobaculum suis), Staphylokokken (3,8- 13,4%), Proteus (1,6-4,2%) und Klebsiellen (0,3-
2,2%). Neben diesen Monoinfektionen treten in einigen Fällen auch Mischinfektionen auf, an
denen ebenso häufig E. coli, Streptokokken und Staphylokokken beteiligt sind (BERNER
1981; BUSSE et al. 1982; BECKER et al. 1985; CARR u. WALTON 1992b). Als häufigster
Erreger wurde somit Escherichia coli, ein gramnegatives Stäbchen, das durch Begeißelung
zur Bewegung befähigt ist, nachgewiesen. Die meisten Stämme von E. coli tragen außerdem
Fimbrien, die auch als Pili bezeichnet werden. Beim Schwein wurden mannosesensitive Typ-
1-Fimbrien nachgewiesen (CARR u. WALTON 1992a), die bei 41% der in der Harnblase
angesiedelten Stämme vorkommen. Diese Stämme zeigten ebenfalls eine positive
Hämagglutination mit Schweineerythrozyten (CARR u. WALTON, 1992a). Die Typ-1-
Fimbrien ermöglichen es diesen Isolaten von E. coli an die Schleimschicht aus Glyko-
proteinen, die dem Blasenepithel aufgelagert ist, anzuheften und im weiteren Verlauf das
Epithel selbst zu schädigen (CARR u. WALTON, 1992a).

Literaturübersicht
35
2.4. Die alimentäre Beeinflussung der Harnacidität
2.4.1. Der Zusammenhang zwischen der Harnansäuerung und der Harnwegsgesundheit
Auf die entscheidende Rolle des pH-Wertes im Harn für die Entstehung bzw. die Vermeidung
von Infektionen im Bereich des Harnapparates wurde schon in vorhergehenden Kapiteln hin-
gewiesen. In diesem Kapitel sollen die alimentären Möglichkeiten behandelt werden, mit
denen es gelingt den pH-Wert so zu verändern, dass die Prävalenz einer Harnwegsinfektion
minimiert wird. KASS (1957) konnte durch den Zusatz von Methionin zur Nahrung den
Harn-pH-Wert auf pH 4,5- 5 absenken und somit den Keimgehalt im Harn um mehrere
Zehnerpotenzen reduzieren. Beim Schwein erzielte JÜRGENS (1991) eine Senkung des pH-
Wertes um 2,16 pH-Einheiten mit Ammoniumchlorid bzw. um 2,45 pH-Einheiten mit DL-
Methionin, indem er diese Substanzen dem Schweinefutter untermischte. So senkte er den
Harn-pH-Wert unter pH 6,1- 6,3 und konnte die Keimausscheidung ebenfalls erheblich
verringern. Dieser Effekt blieb dabei allerdings zeitlich auf die Dauer des Einsatzes der
ansäuernden Substanzen beschränkt. Mit einer Mischung aus Ammoniumchlorid, Methionin
und Phosphorsäure erreichte KROHN (1993) eine Absenkung des Harn-pH-Wertes auf pH
5,8. Nach ihrer Aussage ist mit einem Futter-Basenexzess von -397 mmol/kg TS eine
Absenkung des Harn-pH-Wertes beim Schwein auf pH 5,4 möglich. Von KROHN (1993)
wurden in ihren Untersuchungen bei kurzzeitiger Acidierung sogar Einzelwerte von pH 4 im
Harn gemessen. Zum Zweck der Harnansäuerung wurden in einigen Versuchen auch
organische Säuren wie Ascorbinsäure, Propionsäure oder Zitronensäure eingesetzt. Diese
eigneten sich allerdings nicht besonders für diesen Verwendungszweck, da durch ihren Zusatz
lediglich eine pH-Wert-Reduktion um 0,49- 0,97 pH-Einheiten zu erreichen war (SCOTT
1971; JÜRGENS 1991; FINKENSIEP 1993). Diese Absenkungen der pH-Werte waren damit
so gering, dass keine nennenswerte Keimreduktion erreicht werden konnte, wodurch der
Einsatz dieser organischen Säuren in diesem Anwendungsgebiet zweifelhaft erscheint.

Literaturübersicht
36
2.4.2. Mechanismen der alimentären Harnansäuerung
Zur Verständlichkeit des Mechanismus der alimentären Harnansäuerung ist es von
Bedeutung, sich die Grundlagen der Regulation des Säure-Basen-Haushaltes noch einmal vor
Augen zu führen. Denn die Konstanthaltung des Blut-pH-Wertes bei ca. pH 7,4 (Referenz-
wert für das Schwein: pH 7,42, KRAFT u. DÜRR 1995) ist für den Organismus von großer
Wichtgkeit, da u.a. die Molekülformen von Proteinen und somit die physiologische Struktur
der Zellbestandteile sowie die optimale Wirksamkeit der Enzyme von diesem konstanten
Wert abhängig sind (CHAN 1974). Der Regulation zur Schaffung eines konstanten Blut-pH-
Wertes liegen drei Mechanismen zu Grunde. Diese sorgen für die Entfernung von
Kohlendioxid (CO2) und Protonen (H+), welche laufend als Produkte des Stoffwechsels
gebildet werden (MÜLLER-PLATHE 1982; FORTH 1992) und somit aus dem Körper
entfernt werden müssen.
Mechanismen zur Konstanthaltung des Blut-pH-Wertes:
1.) Bindung oder Abgabe von H+ durch Puffersysteme
2.) Regulation des Partialdruckes von CO2 durch die Atmung
3.) Regulation der renalen H+- bzw. HCO3--Ausscheidung
Veränderungen im Säure-Basen-Haushalt des Körpers werden entweder durch die Aufnahme
von Säure- bzw. Basenäquivalenten mit der Nahrung hervorgerufen oder entstehen durch
Störungen der Regulationssysteme im metabolischen oder respiratorischen Bereich, die z.B.
durch eine Beeinträchtigung der Ventilation, der Bildung abnormer Mengen endogener
Säureäquivalente oder durch renale und extrarenale Verluste von Säure- oder Basen-
äquivalenten verursacht werden können.
1.) Regulation des Säure-Basen-Haushaltes über körpereigene Puffersysteme
In der Sparte der körpereigenen Puffersysteme spielt das Bikarbonatpuffersystem eine ent-
scheidende Rolle. Denn die HCO3--Ionen nehmen im Organismus H+- Ionen auf und die
daraus entstandene Verbindung zerfällt sofort wieder in die zwei neu erzeugten Stoffe CO2
und H2O. Während das entstandene Wasser im Körper verbleibt, gelangt das Kohlendioxid
über den Blutkreislauf zur Lunge, in der es daraufhin abgeatmet wird. Bei einer vermehrten
Anflutung von H+-Ionen im Körper sorgen dessen Regelmechanismen für eine Verstärkung

Literaturübersicht
37
der Atmung und somit gleichzeitig für eine vermehrte Abatmung von CO2 (THEWS 1995)
und dadurch sekundär für eine Entsorgung der überschüssigen H+- Ionen.
Das Phosphatpuffersystem, welches aus dem primären (Säure) und dem sekundären (kor-
respondierende Base) Phosphat besteht, spielt im extrazellulären Medium aufgrund seiner
geringen Konzentration eine untergeordnete Rolle. Das anorganische Phosphat ist aber an der
H+-Ausscheidung über den Harn beteiligt (SCHEID 1994).
Bei der Regulation des intrazellulären pH-Wertes haben Proteine eine große Bedeutung, denn
sie binden H+-Ionen reversibel an die endständigen Amino- und Carboxylgruppen sowie an
die Seitengruppen. Der Imidazolring des Histidins, die Sulfhydrylgruppe des Cysteins und die
terminale NH2-Gruppe sind hier Beispiele für die physiologische Wirksamkeit (SCHEID
1994; THEWS 1995).
2.) Regulation der Säure-Basen-Haushaltes über die Atmung
An der Aufrechterhaltung eines konstanten pH-Wertes im Körper ist die Lunge maßgeblich
beteiligt, da diese den Körper durch die Abatmung von Kohlendioxid zu einem offenen
System macht und auf diese Weise das Blut von der äquivalenten Menge an H+-Ionen befreit.
Außerdem kann der Organismus durch die Regulation der Atmung Störungen im Bereich des
Säure-Basen-Haushaltes in gewissem Maße kompensieren, indem bei einer zunehmenden
Acidität des Blutes die Atmung verstärkt wird, während diese bei einer Zunahme des
Basengehaltes zum Teil eingeschränkt werden kann. Somit ist es dem Organismus möglich
über die Stärke der Ventilation regulartorisch auf den physiologischen pH-Wert einzuwirken
(THEWS 1995).
3.) Regulation des Säure-Basen-Haushaltes über die Nieren
Die Nieren sind ein sehr wichtiges Organ zur Regulation des Säure-Basen-Haushaltes des
Organismus, da sie durch ihre Filtrationsfunktion, durch Vorgänge der Reabsorption und der
Sekretion den Körper ähnlich wie die Lunge zu einem offenen System machen, das mit seiner
Umwelt in einen regen Stoffaustausch steht. Ihnen kommen somit über die tubuläre H+-
Sekretion (primär-aktiv oder sekundär-aktiv im Austausch gegen Na+) zwei bedeutende

Literaturübersicht
38
Funktionen zu, erstens die Ausscheidung nichtflüchtiger („fixer“) Säuren und zweitens die
Rückresorption des filtrierten Bikarbonats (MÜLLER-PLATHE 1982).
Im Organismus anfallende H+-Ionen werden an ihrem Entstehungsort abgepuffert und müssen
daraufhin, um die Puffersysteme zu regenerieren, aus dem Körper verbracht werden
(SILBERNAGL 1994). Im Fall einer Acidose wird deshalb in den Nieren vermehrt
sekundäres Phosphat filtriert, um im Tubulussystem sezernierte H+-Ionen aufzunehmen. Das
so entstandene primäre Phosphat wird nun als Träger einer sauren Valenz über den Harn
ausgeschieden, da das Tubulusepithel für diese Verbindung impermeabel ist (HIERHOLZER
u. FROMM 1995). Die indirekte Ausscheidung von H+-Ionen über die Niere erfolgt in Form
von Ammonium-Ionen (NH4+), deren Harnkonzentration während einer Acidose auf das 10-
fache ansteigen kann (HIERHOLZER u. FROMM 1995). Unter physiologischen
Bedingungen werden Ammonium-Ionen, die im Aminosäurestoffwechsel anfallen, in der
Leber mit äquimolaren Mengen an Bikarbonat zu Harnstoff umgewandelt, welcher dann über
die Nieren ausgeschieden wird. Im Falle einer metabolischen Acidose fallen allerdings so
viele NH4+-Ionen an, dass diese in der Leber an Glutamin gekoppelt und so zu den Nieren
transportiert werden. Hier erfolgt eine Hydrolyse, aus der NH4+ und Glutamat hervorgehen,
welches seinerseits weiter in NH4+ und α-Ketoglutarat2- gespalten wird. Das so entstandene
NH4+ dissoziiert in den Tubulusepithelzellen zu NH3 und H+, die sich nach der Diffusion in
die Tubuli in deren Lumen erneut zum NH4+ verbinden und dann in dieser Form
ausgeschieden werden. Das α-Ketoglutarat wird in Verbindung mit 2 H+-Ionen, die auf
diesem Wege zusätzlich eliminiert werden, zu Glukose verstoffwechselt (SILBERNAGL
1994). Das filtrierte Bikarbonat, das die wichtigste Puffersubstanz im Organismus darstellt,
wird schon bei physiologischen pH-Werten zu 90% im proximalen Bereich des
Tubulussystems reabsorbiert (HIERHOLZER u. FROMM 1995). Verschiebt sich aber die
Stoffwechselsituation in den Bereich einer Acidose werden hier auch noch die letzten 10%
dieses Stoffes reabsorbiert. Dieser Vorgang, der im distalen Anteil des Tubulussystems
lokalisiert ist, sorgt so dafür, dass mit dem Harn kein Bikarbonat mehr ausgeschieden wird. In
der Stoffwechselsituation einer Alkalose kommt es dagegen in diesem Bereich zu einer
nettomäßigen Sekretion von Bikarbonat (HIERHOLZER u. FROMM 1995).
Im Falle einer Acidose erfolgt die Senkung des pH-Wertes im Harn somit durch eine
gesteigerte Säureausscheidung und eine ebenfalls gesteigerte Rückresorbtion von Bikarbonat.
Dabei sind für die Säureausscheidung mengenmäßig nicht die frei ausgeschiedene H+-Ionen
sondern die „titrierbaren Säuren“ und die Ammoniumionen von Bedeutung (HIERHOLZER
u. FROMM 1995).

Literaturübersicht
39
2.4.3. Der Einfluss der Ernährung auf den Harn-pH-Wert
Mit der Nahrung werden Stoffe bzw. komplexe Verbindungen aufgenommen, deren Verstoff-
wechselungen zu Veränderungen des physiologischen Säure-Basen-Status führen. In diesem
Fall sind die dargestellten Regulationsmechanismen des Säure-Basen-Haushaltes gefordert,
um den physiologischen Zustand aufrecht zu erhalten bzw. wieder herzustellen.
Die einzelnen Bestandteile der Nahrung beeinflussen den Säure-Basen-Haushalt recht unter-
schiedlich, deshalb sollen an dieser Stelle einige Beispiele dazu gezeigt werden.
Eine Metabolisierung der neutralen Kohlenhydrate und Triglyceride führt zu keiner
Veränderung im Säure-Basen-System, da bei ihrem Abbau zu Kohlendioxid und Wasser
weder Säuren noch Basen entstehen (CHAN 1974; HALPERIN u. JUNGAS 1983).
Kohlenhydrate + O2 → CO2 + H2O
Triglyzeride + O2 → CO2 + H2O
Lediglich bei einer Abweichung von der physiologischen Stoffwechselsituation (Hypoxie,
Ketose usw.) werden diese Stoffe nur unvollständig abgebaut und können somit zur
Säurebelastung des Organismus beitragen.
Glukose + O2 → Laktat- + H+
Triglyzeride + O2 → Azetoazetat- + H+
Bei der Umsetzung der Eiweißbestandteile der Nahrung hängt die endogene Protonen-
freisetzung von der Art der Proteinquelle ab. Werden z.B. schwefelhaltige Aminosäuren
(Methionin, Cystein) oxidiert, entstehen dabei Kohlendioxid, Wasser, Harnstoff, Sulfat und
zwei Protonen. Diese 2 H+-Ionen tragen somit zur Säurebelastung des Körpers bei
(PATIENCE et al. 1987).
Met./Cys. + O2 → CO2 + H2O + Harnstoff + SO42- + 2H+

Literaturübersicht
40
Umsetzbare organische Anionen (Zitrat, Laktat usw.), die in vielen Futtermitteln enthalten
sind, gehen meist eine Verbindung mit anorganischen Kationen (Na+, K+ usw.) ein. Bei der
metabolischen Umsetzung dieser Verbindungen entsteht Bikarbonat, das als Puffersubstanz
eine alkalisierende Wirkung hat (HARRINGTON u. LEHMANN 1970).
R-COO - - K+ + O2 → CO2 + H2O + K+ + HCO3-
Die Metabolisierung von Nahrungsbestandteilen tierischer Herkunft, die aus organischen
Kationen in Verbindung mit anorganischen Anionen bestehen, führt zur Säurebelastung des
Organismus, da bei diesen Umsetzungsvorgängen Protonen freigesetzt werden
(HARRINGTON u. LEHMANN 1970).
R- NH2 + Cl- + O2 → Harnstoff + CO2 + H2O + H+ + Cl-
Die Menge an Protonen, die bei der Freisetzung von organisch gebundenem Phosphat aus
Phosphorlipiden und phosphorhaltigen Proteinen den Organismus belastet, ist abhängig vom
vorherrschenden pH-Wert (HARRINGTON u. LEHMANN 1970).
Bei pH 7,4: Phosphorproteine/lipide + H2O → ROH + 0,8 HPO42- + 0,2 H2PO4
- + 1,8 H+
Kommt es dagegen zu einer Verschiebung des pH-Wertes in den alkalischen Bereich,
verschiebt sich der prozentuale Anteil des anorganischen Phosphates in Richtung PO43-,
während sich der Gehalt an anorganischen Phosphat bei einer pH-Wert-Verschiebung in den
sauren pH-Bereich in Richtung H3PO4 verändert.
Karbonate, Hydroxide und Oxide der Alkali- und Erdalkalimetalle haben dagegen einen
direkten alkalisierenden Effekt auf den Säure-Basen-Haushalt des Körpers (HARRINGTON
u. LEHMANN 1970; BUFFINGTON 1989; KIENZLE 1991).
Die Aufnahme von anorganischem Phosphat wirkt sich ebenfalls pH-abhängig auf den Säure-
Basen-Haushalt aus. Denn je nach pH-Wert liegt es als primäres oder sekundäres Phosphat
vor, so dass die renale Ausscheidung zusammen mit einem bzw. mit zwei Alkaliionen erfolgt,
wodurch die Höhe der Säurebelastung des Körper variiert (KIENZLE 1991).

Literaturübersicht
41
Eine direkte Säurebelastung geht dagegen von den Bestandteilen der anorganischen Säuren
(HCL, H3PO4 usw.) aus, die über die Nahrung aufgenommen werden.
Erdalkalichloride (CaCl2, MgCl2) beeinflussen den Säure-Basen-Haushalt des Organismus auf
eine etwas andere Art, die sich aus dem Absorptionsverhalten der einzelnen Bestandteile
ergibt. Das Chlorid wird fast vollständig aus dem Verdauungstrakt resorbiert (ca. 90%;
MÄNNER u. BRONSCH 1987; CHING et al. 1989), während die Resorptionsrate für den
jeweils anderen Bestandteil sehr gering ist. Diese liegt nämlich für Magnesium bei 20- 40%
und für Calcium sogar nur bei 10- 20% (MÄNNER u. BRONSCH 1987; CHING et al. 1989).
Da durch diese unterschiedliche Aufnahme der einzelnen Bestandteile der Verbindungen die
Elektronenneutralität erheblich gestört ist, kommt es zur Abgabe v.a. von Bikarbonat in den
Darm, um die Neutralität wieder herzustellen. Durch diesen Stoffaustausch geht somit dem
Körper wichtige Puffersubstanz verloren, was sekundär zu einer Säurebelastung des
Organismus führt.
Alle hier aufgeführten metabolischen Abbauvorgänge, die zu einer acidotischen Belastung
des Säure-Basen-Haushaltes beitragen, führen sekundär zu einer Ansäuerung des Harns, da
der Organismus, wie schon in vorhergehenden Kapiteln beschrieben wurde, versucht über die
Niere diesen Säureüberschuss auszugleichen. Die Verschiebung des pH-Wertes im Harn ist
wiederum, wie ebenfalls schon erläutert wurde, mit negativen Folgen für die dort ansässigen
Erreger verbunden. Somit können einige dieser metabolischen Abbauvorgänge prophylaktisch
gegen das Auftreten von unspezifischen Harnwegsinfektionen oder in Zusammenhang mit
diesen zur MMA-Prophylaxe eingesetzt werden, wie es in der hier vorliegenden Unter-
suchung durch die Zumischung von Calciumchlorid zum Sauenfutter geschehen ist.
Aber Verschiebungen im Säure-Basen-Haushalt des Organismus wirken sich nicht nur an den
Organen aus, die mit der Regulation dieses Bereiches enger verbunden sind, sondern beein-
flussen den Körper gesamthaft, so dass sich auch Veränderungen in anderen Organsystemen
oder in Teilen dieser Systeme ergeben.
In diesem Zusammenhang gilt hier den Veränderungen des Knochenstoffwechsels ein
besonderes Interesse. Deshalb soll im weiteren Verlauf näher auf das Knochengewebe, dessen
Stoffwechsel und seine Verbindung zum Säure-Basen-Haushalt eingegangen werden.

Literaturübersicht
42
2.5. Die Auswirkungen von Veränderungen im Säure-Basen-Haushalt auf den Knochenstoffwechsel
In der Literatur wird beschrieben, dass eine Säurebelastung des Säure-Basen-Haushaltes eine
Nettoabgabe von Calcium aus dem Körper bewirkt (LEHMANN et al. 1966 und 1967;
LICATA et al. 1981; BRESLAU et al. 1988), indem die Exkretion von Calcium über die
Nieren ansteigt (LEHMANN et al. 1967; BUSHINSKY et al. 1982), da aufgrund einer
erhöhten extrazellulären Calciumkonzentration sekundär die Reabsorbtion von Calcium in
den Tubuli gehemmt wird (KRIEGER et al. 2000). Die Absorption von Calcium aus dem
Magendarmtrakt bleibt dagegen jedoch unverändert (GAFTER et al. 1980). Im Gegensatz
dazu konnte die renale Exkretion von Calcium durch die Zugabe einer alkalisierenden
Substanz signifikant gesenkt werden (LUTZ 1984; SEBASTIAN et al. 1994). Etliche Autoren
(ARNETT et al. 1986; GOLDHABER et al.1987; BUSHINSKY 1989, 1995; RABADJIJA et
al. 1990; CHABALA et al. 1991; BUSHINSKY et al. 1992, 1993, 1995; KRIEGER et al.
1992; SPRAGUE et al. 1994) zeigten, dass im Fall einer metabolischen Acidose vermehrt
Calcium aus dem Knochen freigesetzt wird, da das fehlende Calcium aus dem
Knochengewebe als größtem Calciumspeicher im Organismus mobilisiert werden muss
(WIDDOWSON u. DICKERSON 1964). Diese Calciummobilisation ist das Resultat
verstärkter Osteoklastenaktivität im Knochen in Verbindung mit einer verminderten
Arbeitsintensität der Osteoblasten (KRIEGER et al. 1992; SPRAGUE et al. 1994;
BUSHINSKY 1995; BUSHINSKY et al. 1995). Eine Schlüsselrolle nimmt in diesem
Zusammenhang die Konzentration an Bikarbonat ein, da im Fall einer metabolischen Acidose
mit sinkender Bikarbonatkonzentration mehr Calcium freigesetzt wird als bei einer
respiratorischen Acidose mit gleichbleibendem Bikarbonat-spiegel (BUSHINSKY 1989;
BUSHINSKY et al. 1992). Bei konstantem pH-Wert wurde bei sinkender
Bikarbonatkonzentration eine steigende Mobilisation von Calcium aus dem Knochengewebe
beobachtet (BUSHINSKY et al. 1992). Durch eine metabolischen Alkalose konnten
BUSHINSKY et al. (1996) dagegen die Kollagensynthese der Osteoblasten steigern und die
Aktivität der Osteoklasten senken.
Abschließend lässt sich also feststellen, dass sich eine Verschiebung des Säure-Basen-
Haushaltes, die einen metabolischen Ursprung hat, auf den Knochenstoffwechsel auswirkt, da
eine Abweichung in den sauren pH-Bereich zu einer Resorption und somit zu einer
Freisetzung von Calcium führt, während aus einer Verschiebung in den alkalischen pH-
Bereich ein Aufbau von Knochensubstanz resultiert und somit einen Verbrauch von
Calciumionen darstellt.

Literaturübersicht
43
2.6. Das Knochengewebe
2.6.1. Die Funktion des Knochengewebes
Seine Hauptfunktion erfüllt der Knochen als passiver Teil des Bewegungsapparates, indem er
den Muskelaktivitäten und Bewegungsabläufen als mechanisches Fundament dient, welches
den Sehnen und Bändern einen festen Ansatz bietet. Um sich den daraus resultierenden
Belastungen möglichst optimal anzupassen, finden im Knochengewebe ständig aufbauende
und abbauende Stoffwechselprozesse statt. Deshalb handelt es sich hier aus physiologischer
Sicht um ein dynamisches Gewebe, in dem Hormone und andere Faktoren für einen
geregelten Ablauf zwischen Aufbau und Resorption sorgen.
Weitere bedeutende Aufgaben erfüllt das Knochengewebe mit der Hämatopoese, der
Regulation des Calciumstoffwechsels und der Speicherung von anorganischen Ionen. Diese
Aufgaben werden dabei von den ständig ablaufenden Auf- und Abbauprozessen beeinflusst
(DUCY et al. 2000).
2.6.2. Die Struktur des Knochengewebes
Außen wird der Knochen fast vollständig von einer bindegewebige Hülle, dem Periost
(Knochenhaut), überzogen, das zur Versorgung des Knochens ein dichtes Geflecht an Blut-
und Lymphgefäßen und zahlreiche sensible Nervenfasern enthält (LIEBICH 1993).
Das Knochengewebe selbst besteht außen aus der Substantia compacta bzw. Substantia
corticalis (80% des Knochengewebes) und im Inneren aus der Substantia spongiosa (ca. 20%
des Knochengewebes). Die Substantia compacta ist zu 80- 90% mineralisiert und weist einen
typischen Aufbau im Sinne des Haverschen Systems auf, während die Substantia spongiosa
lediglich zu 5- 20% mineralisiert ist und überwiegend aus poröser Knochenmatrix besteht
(MARKS u. HARMEY 1996). Die Substantia compacta bildet die Metaphyse der Röhren-
knochen und überzieht als dünne Substantia corticalis deren Enden. Im Inneren der
Knochenenden befindet sich die Substantia spongiosa als ein Netzwerk aus kleinen Knochen-
bälkchen, deren Struktur so ausgerichtet ist, dass Druck- und Zugkräfte, die auf den Knochen
wirken, optimal abgefangen werden. Die Substantia spongiosa zeichnet sich im Gegensatz zu
der Substantia compacta in ihrer Funktion als Calciumspeicher des Organismus durch eine
große metabolische Aktivität aus (CHRISTENSON 1997).

Literaturübersicht
44
2.6.3. Die zelluläre Grundlage des Knochengewebes
Um seine differenzierten Funktionen zu erfüllen, besteht das Knochengewebe aus vier
verschiedenen Zelltypen: Den Osteoblasten, den Osteozyten, den Osteoklasten, und den
„bone lining cells“, die hier allerdings eine untergeordnete Rolle spielen und somit nicht
weiter von Interesse sind.
A) Osteoblasten
Die Osteoblasten entstehen aus pluripotenten mesenchymalen Stammzellen und wandeln sich
nach einem dreiphasigen Entwicklungsprozess zu Osteozyten. In der ersten Phase dieser
Entwicklung (Proliferationsphase) wird Prokollagen vom Typ I synthetisiert, in der zweiten
Phase (Reifungsphase) erhöht sich dann die Aktivität der alkalischen Phosphatase und in der
dritten Phase (Mineralisierungsphase) werden von ihnen schließlich calciumbindende
Proteine ausgeschieden (STEIN et al. 1990). Die Osteoblasten sind verantwortlich für die
Synthese der organischen Bestandteile der Knochengrundsubstanz (JUNQUEIRA u.
CARNEIRO 1991) und sezernieren die Vorstufen des Typ-I-Kollagens (95% der Knochen-
matrix) und andere nicht-kollagene Proteine (5% der Knochenmatrix) (CHARLES et al.
1994). Außerdem regulieren die Osteoblasten die Mineralisation der Knochenmatrix
(JUNQUEIRA u. CARNEIRO 1991), wobei hauptsächlich Calcium und Phosphat in Form
von Hydroxyapatitkristallen (Ca10(PO4)6(OH)2) in das Osteoid eingelagert werden (MARKS
u. HERMEY 1996).
B) Osteozyten
Die Osteozyten entstehen wie beschrieben aus den Osteoblasten und sind verantwortlich für
das Fortbestehen des Knochens, denn sie können Osteoid bilden und dieses im begrenzten
Maße auch resorbieren. Über den jeweiligen Funktionszustand der Osteozyten gibt dabei ihr
morphologischer Aufbau Auskunft. Wenn in erster Linie Knochenmatrix produziert wird,
besitzen sie eine für Osteoblasten typische Zellorganellausstattung. Im Falle der Resorption
von Knochengrundsubstanz sind sie jedoch mit lysosomalen Vakuolen und anderen
phagozytosetypischen Zellbestandteilen ausgerüstet. Obwohl sich jeder Osteozyt in einer
eigenen Lakune in der Knochenmatrix befindet, steht er dennoch über filopodienartige
Zellfortsätze in Kontakt zu benachbarten Osteozyten, der Knochenmatrix, den versorgenden

Literaturübersicht
45
Blutgefäßen und der internen und externen Oberfläche des Knochens (DOTY 1981; LIEBICH
1993; CHRISTENSON 1997).
D) Osteoklasten
Osteoklasten sind große, vielkernige Riesenzellen, die durch den Zusammenschluss mehrerer
Vorläuferzellen der Monozyten-Makrophagen-Linie entstehen. Die Vereinigung dieser
einzelnen Zellen ist ein Reaktionsmechanismus, der durch verschiedene resorptionsfördernde
Faktoren ausgelöst wird. Zu diesen zählen das 1,25-dihydroxy-Vitamin D3 (TAKEDA et al.
1999), Parathormon (PTH) (LIU et al. 1998), Prostaglandin E2 (PGE2) (SAKUMA et al.
2000) und Interleukin II (IL- II). Zur Differenzierung der Osteoklasten ist allerdings eine Zell-
zu-Zell-Verbindung zwischen den Osteoblasten und den Vorläuferzellen der Osteoklasten
notwendig (UDAGAWA et al. 1999), da die Osteoklasten im Gegensatz zu den Osteoblasten
selbst keine Rezeptoren für das Parathormon besitzen (LIU et al. 1998; OKADA et al. 2002).
Da die Osteoklasten zur Hauptsache die Funktion des Knochenabbaues erfüllen, besitzen sie
in ihrem Inneren zahlreiche mit Enzymen gefüllte Vakuolen. Als beteiligte Enzyme wurden
u.a. die tartratresistente, saure Phosphatase (TRAP), Matrixmetalloproteinase (MMP) und
Kathepsine (v.a. Kathepsin B, Kathepsin K und Kathepsin L) bei Ratten, Kanninchen und
Menschen nachgewiesen (GOTO et al. 1994; INAOKA et al. 1995; SATO et al. 1997;
HALLEEN et al. 1999). Da zur Lösung der Mineralstoffe aus dem Knochen ein saurer pH-
Wert notwendig ist (SUNQUIST u. MARKS 1995), befinden in der Resorptionszone der
Membran der Osteoklasten Protonenpumpen, die H+-Ionen in diesen Bereich befördern, um
so für eine ausreichende Absenkung des pH-Wertes zu sorgen (BLAIR et al. 1989; BEKKER
u. GAY 1990; VÄÄNÄNEN et al. 1990).
2.6.4. Die Knochenformation
Für die Knochenformation sind die Osteoblasten das zentrale Element, da diese nach ihrer
Entwicklung aus den mesenchymalen Stammzellen für die Synthese der Vorstufen des
Kollagens vom Typ I und für die Regulation der Mineralisierung des Knochengewebes
verantwortlich sind (JUNQUEIRA u. CARNEIRO 1991). In der folgenden Tabelle wird eine
Übersicht der Signalstoffe, die die Formation des Knochens regulieren, und ihrer
Wirkungsweisen aufgeführt.
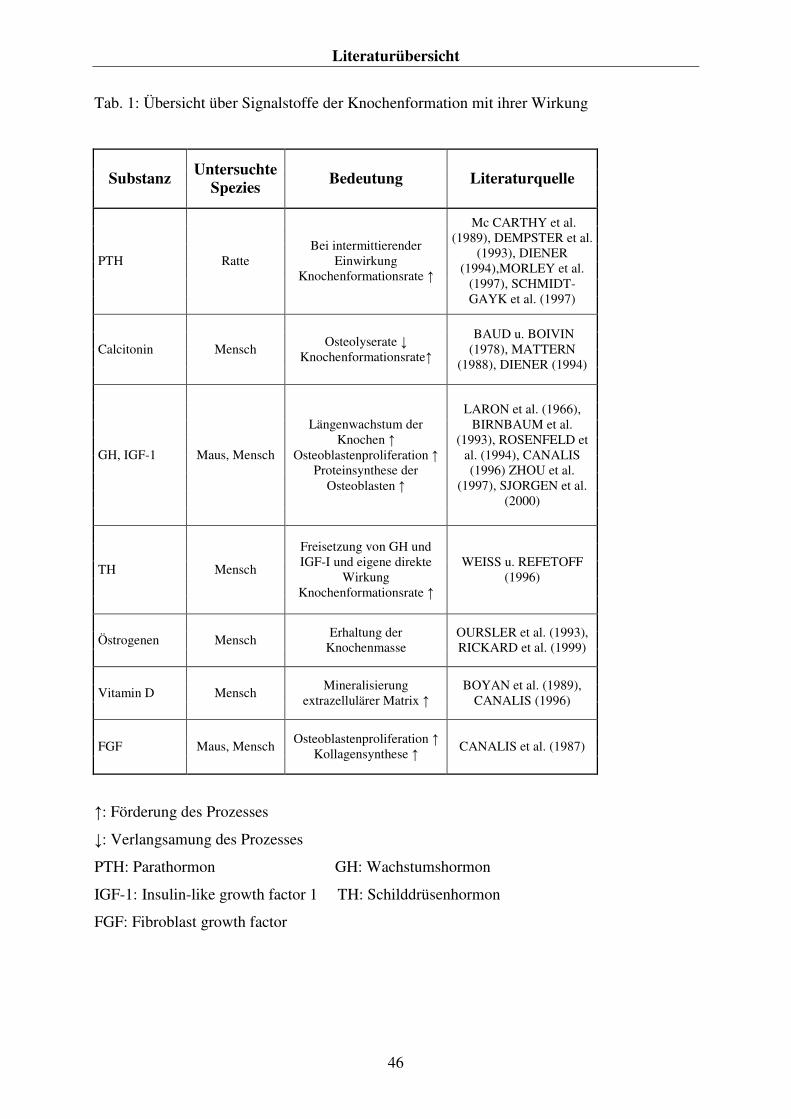
Literaturübersicht
46
Tab. 1: Übersicht über Signalstoffe der Knochenformation mit ihrer Wirkung
Substanz Untersuchte
Spezies Bedeutung Literaturquelle
PTH Ratte Bei intermittierender
Einwirkung Knochenformationsrate ↑
Mc CARTHY et al. (1989), DEMPSTER et al.
(1993), DIENER (1994),MORLEY et al.
(1997), SCHMIDT-GAYK et al. (1997)
Calcitonin Mensch Osteolyserate ↓
Knochenformationsrate↑
BAUD u. BOIVIN (1978), MATTERN
(1988), DIENER (1994)
GH, IGF-1 Maus, Mensch
Längenwachstum der Knochen ↑
Osteoblastenproliferation ↑ Proteinsynthese der
Osteoblasten ↑
LARON et al. (1966), BIRNBAUM et al.
(1993), ROSENFELD et al. (1994), CANALIS (1996) ZHOU et al.
(1997), SJORGEN et al. (2000)
TH Mensch
Freisetzung von GH und IGF-I und eigene direkte
Wirkung Knochenformationsrate ↑
WEISS u. REFETOFF (1996)
Östrogenen Mensch Erhaltung der
Knochenmasse OURSLER et al. (1993), RICKARD et al. (1999)
Vitamin D Mensch Mineralisierung
extrazellulärer Matrix ↑ BOYAN et al. (1989),
CANALIS (1996)
FGF Maus, Mensch Osteoblastenproliferation ↑
Kollagensynthese ↑ CANALIS et al. (1987)
↑: Förderung des Prozesses
↓: Verlangsamung des Prozesses
PTH: Parathormon GH: Wachstumshormon
IGF-1: Insulin-like growth factor 1 TH: Schilddrüsenhormon
FGF: Fibroblast growth factor

Literaturübersicht
47
2.6.5. Der Knochenabbau
Zur Resorption des Knochengewebes sind mehrere Teilschritte notwendig. Im ersten Schritt
erfolgt eine lokale Ansäuerung des Knochengewebes durch die Abgabe von H+- Ionen, Cl--
Ionen und lysosomalen Enzymen. Zu diesen Enzymen zählt z.B. die tartratresistente, saure
Phosphatase, außerdem werden zusätzlich einige Proteine sezerniert. Durch diese Vorgänge
wird das kristalline Hydroxyapatit gelöst und abtransportiert (FALLON et al. 1984; BARON
et al. 1985). In den weiteren Schritten der Knochenresorption wird das organische Knochen-
material abgebaut, was durch den Einsatz von Proteasen geschieht. Als wichtigste Vertreter
dieser Gruppe sind das Kathepsin K und die Matrix-Metalloproteinase 9 (MMP- 9) zu nennen
(TEZUKA et al. 1994; INAOKA et al. 1995; DRAKE et al. 1996; LITTLEWOOD- EVANS
et al. 1997).
Tab. 2: Übersicht über Signalstoffe des Knochenabbaus mit ihrer Wirkung
Substanz Untersuchte
Spezies Bedeutung Literaturquelle
PTH Ratte Bei permanenter
Einwirkung Knochenresorptionsrate ↑
Mc CARTHY et al. (1989), DEMPSTER et
al. (1993), DIENER (1994),MORLEY et al.
(1997), SCHMIDT-GAYK et al. (1997)
Glukokortikoide Maus, Mensch
Knochenformationsrate ↓ Osteoblastogenese ↓ Osteoblasten- und
Osteozytenapoptose ↑
CANALIS (1996), WEINSTEIN et al.
(1998)
TGFβ1
TGFβ2 Maus Knochenformationsrate ↓
OURSLER et al. (1993), SERRA et al. (1999)
↑: Förderung des Prozesses
↓: Verlangsamung des Prozesses
PTH: Parathormon TGFβ: Transforming growth factor beta

Literaturübersicht
48
2.7. Das Parathormon
Das Parathormon (PTH) wird als eine aus 115 Aminosäuren bestehende inaktiven Vorform in
den Hauptzellen der Nebenschilddrüse synthetisiert. Innerhalb der Hauptzellen wird daraus
dann durch eine proteolytische Spaltung die biologisch aktive Form gebildet, welche die
Aminosäuren 1-84 beinhaltet. Für die hormonelle Wirkung an den Zielzellen sind besonders
die Aminosäuren an Position 25-34 von Bedeutung, da sie an das aminoterminale Ende der
extrazellulären Rezeptorbereiche anbinden. Die Aminosäuren an Position 1- 7 sorgen darauf-
hin für die Aktivierung eines Second messengers innerhalb der Zielzelle (ERDMANN et al.
1998; LUCK et al. 1999; GRAUSCHOPF et al. 2000; GREENBERG et al. 2000).
Im Blutkreislauf hat das zirkulierende Parathormon eine Halbwertzeit von weniger als drei
Minuten, bevor es zum größten Teil (60-70%) in den Kupferschen Sternzellen und den
Hepatozyten der Leber oder in den Tubuluszellen der Niere (20-30%) oder zu einem geringen
Anteil auch in anderen Organen abgebaut wird (BRINGHURST et al. 1988).
Die Sekretion des Parathormons in der Nebenschilddrüse steuert ein negativer Feedback-
mechanismus, in dem eine hohe Konzentration an extrazellulären Calciumionen die Sekretion
hemmt, während eine niedrige Konzentration diese fördert. Der schnelle Metabolismus dieses
Hormons sorgt dafür, dass die Sekretionsrate der Nebenschilddrüse maßgeblich für die Menge
des für eine Rezeptorbindung zur Verfügung stehenden Hormons verantwortlich ist. Somit
können kurzfristige Veränderungen der Konzentration an Calciumionen durch Anpassung der
Sekretionsrate der Nebenschilddrüse wieder angeglichen werden. Gesteuert wird dies durch
Ca-sensible Rezeptoren, deren Expression in den Hauptzellen dieser Drüse sehr hoch ist und
die als Hauptvermittler bei der hemmenden Wirkung einer erhöhten extrazellulären
Konzentration an Calciumionen auf die Parathormonsekretion fungieren (DIAZ et al. 1999).
In verschiedenen Studien an Mäusen wurden unterschiedliche Parathormonrezeptoren
klassifiziert. Der PTH1- Rezeptor wird sehr stark im Bereich des Knochengewebes und der
Niere exprimiert und übermittelt somit die Wirkung des Parathormons an die wichtigsten
Zielorgane (KARAPLIS et al. 1994; LANSKE et al. 1996; VORTKAMP et al. 1996). In der
Niere von Ratten wurden Ca-sensitive Rezeptoren im basolateralen Bereich der Epithelzellen
des dicken kortikalen Mittelstückes des Nephrons gefunden (RICCARDI et al. 1998). In
diesem Bereich und in den distalen gewundenen Abschnitten der Nierenkanälchen wird die
Reabsorption von Calcium über das Parathormon gesteuert (HEBERT et al. 1997). Bei einer
niedrigen extrazellulären Konzentration an Calcium steigert das Parathormon in der Niere
nicht nur die Reabsorption von Calcium, sondern beeinflusst zusätzlich noch die glomeruläre

Literaturübersicht
49
Filtrationsrate, damit weniger Calcium mit dem Primärharn ausgeschieden wird. Beim
Menschen wird über die renale Synthese von Calcitriol auch indirekt eine Stimulation der
intestinalen Absorptionsrate von Calcium bewirkt.
Im Knochengewebe wirkt das Parathormon über Rezeptoren, die auf den Osteoblasten
vorhanden sind. Es reguliert über diese Rezeptoren die Anzahl und die Größe dieser
Knochenzellen und reduziert ihre Synthese an Osteoprotegerin, was sich wiederum indirekt
auf die Osteoklastogenese auswirkt (KANZAWA et al. 2000). Eine direkte Wirkung auf die
Osteoklasten konnte bis zum heutigen Zeitpunkt nicht festgestellt werden, so dass deren
Bildung und Funktion durch Parathormon lediglich durch die Sekretion von Osteoprotegerin
über die Osteoblasten zu beeinflussen ist (LUI et al. 1998; OKADA et al. 2002). Es wird
beschrieben, dass das Parathormon in-vitro eine hemmende Wirkung auf die Synthese und die
Abgabe von Matrix-Proteinen, wie z.B. Typ-I-Kollagen, Osteocalcin und die alkalische
Phosphatase der Osteoblasten hat (HOWARD et al. 1998; BOGDANOVIC et al. 2000). Als
Ergebnisse von in vivo durchgeführten Versuchen kam lediglich heraus, dass initial eine
erhöhte Osteoblastenapoptoserate besteht (STANISLAUS et al. 2000) und dass in den
Osteoblasten von Mäusen und Ratten vermehrt Matrixmetalloproteinasen vorhanden sind
(McCLELLAND et al. 1998; ZHAO et al. 1999). Für die physiologische Konzentration an
PTH im Plasma von Sauen, die sich im letzten Drittel der Trächtikeit befinden, sind aus der
Literatur leider keine Angaben bekannt.
2.8. Biochemische Verfahren zur Untersuchung des Knochenstoffwechsels
Da das Skelett ein dynamisches Gewebe darstellt, in dem ständig Auf- und Abbauprozesse
ablaufen, um so die Gestalt der einzelnen Knochen optimal ihren Funktionen anzupassen oder
um Defizite bzw. Überschüsse im Mineralstoffhaushalt des Körpers auszugleichen, existieren
verschiedene Methoden zur Überwachung dieser Stoffwechselprozesse. So genannte
Knochenmarker, die bei allen Umbauvorgängen am Knochen in den Blutkreislauf ausge-
schüttet werden, sind für biochemischen Nachweisverfahren zur Untersuchung des Knochen-
stoffwechsels sehr geeignet. Bei diesen Knochenmarkern handelt es sich u.a. um Enzyme der
im Knochen befindlichen Zellen und um Matrixkomponenten, die während der Umbau-
vorgänge an den Blutkreislauf abgegeben werden (PRICE 1988).
Dabei werden zwei verschiedene Arten unterschieden, zum Einen so genannte Formations-
marker, die charakteristisch für den Knochenaufbau sind. Als Beispiele werden hierfür von
SEIBEL et al. (1993) das Osteocalcin, die Propeptide des Typ-I-Kollagens und die

Literaturübersicht
50
knochenspezifische alkalische Phosphatase, auf die im weiteren Verlauf dieser Arbeit noch
näher eingegangen wird, genannt. Zum Anderen existieren so genannte Resorptionsmarker,
wie das carboxyterminale Telopeptid des Typ-I-Kollagens (ITCP) oder die tartratresistente
saure Phosphatase (TRAP), die für die Abbauprozesse innerhalb des Knochengewebes stehen
(SEIBEL et al. 1993). Die Konzentration dieser Knochenmarker im Blut wird dabei
maßgeblich von der Arbeitsweise und Arbeitsintensität der Osteoblasten, Osteozyten und
Osteoklasten bestimmmt (SEIBEL et al. 1993), so dass über die Kombination aus diesen
beiden Markertypen Rückschlüsse auf die im Knochengewebe ablaufenden Prozesse und
damit auf die momentane Stoffwechsellage des Knochens gezogen werden können
(DELMAS 1995). In dem hier vorliegenden Versuch wurde der Knochenstoffwechsel anhand
der knochenspezifischen alkalischen Phosphatase (bAP) überwacht, auf die deshalb im
nächsten Abschnitt näher eingegangen wird.
2.8.1. Knochenspezifische alkalische Phosphatase (bAP)
Die alkalischen Phosphatasen sind Enzyme, die beim Menschen von Zellen unterschiedlicher
Organe gebildet werden. Zu diesen Organen zählen die Leber, die Nieren, der Darmtrakt, die
Plazenta und das Knochengewebe, wobei die beiden Isoformen der Leber und des
Knochengewebes ihre Existenz demselben Gen verdanken. Ihre Unterscheidungsmerkmale
werden erst bei der organspezifischen posttranslationellen Glykosilierung gebildet
(SANECKI et al., 1990). Die knochenspezifische Phosphatase (bAP) kann durch chemische
Präzipitation mit Lezithin, durch Hitzeisolierung oder durch Gelelektrophorese isoliert und
anschliessend im Serum oder Plasma nachgewiesen werden (ROSALKI u. FOO, 1984). Als
Marker für die Knochenformation bzw. die Bildung von Knochengewebe eignet sich die
knochenspezifische alkalische Phosphatase sehr gut, da sie beim Knochenaufbau in großen
Mengen von den Osteoblasten gebildet wird und somit einen Indikator für deren Aktivität
darstellt (WHYTE, 1983; EPSTEIN, 1988; DELMAS 1991). Aus der Literatur gehen leider
für den Parameter „knochenspezifische alkalische Phosphatase“ keine physiologischen Werte
für die Tierart Schwein hervor.

Literaturübersicht
51
2.9. Das Calcium
2.9.1. Die Verteilung und Funktion von Calcium im Körper
Da in diesem Versuch Calciumchlorid (CaCl2) eingesetzt wird, um eine metabolische Acidose
zu erzeugen und so in den Säure-Basen-Haushalt des Körpers einzugreifen, soll im folgenden
Abschnitt näher auf den Stoffwechsel und die Funktion von Calcium im Organismus einge-
gangen werden. Calcium (Ca2+) ist im Körper sowohl intrazellulär wie auch extrazellulär
vorhanden und erfüllt dort diverse Funktionen z.B. als Co-Faktor für Gerinnungsfaktoren,
Adhäsionsmoleküle und andere Proteine sowie für die Kontrolle der neuronalen Erregbarkeit
oder als Second messenger bei der intrazellulären Signalübertragung. Die intrazelluläre
Calciumkonzentration verfügt dabei über einen relativ großen Toleranzbereich (POZZAN et
al. 1994), während das Calcium im extrazellulären Medium ständig in einer konstanten
Konzentration verfügbar sein muss. Deshalb wird der Spiegel an extrazellulären ionisiertem
Calcium in sehr engen Grenzen von homöostatischen Mechanismen, welche die
Nebenschilddrüsen, die Calcitonin-sezernierenden C-Zellen der Schilddrüse, die Nieren, den
Knochen und das Intestitium einschließen, überwacht und reguliert (BROWN 1991;
BRINGHURST et al. 1998). Konzentrationsabweichungen werden von Calciumrezeptoren
wahrgenommen, die als Schlüsselelement dieses komplexen Regelmechanismus die
Gegenregulation einleiten (BROWN 1991).
Als Reservoir für Calciumionen fungiert das dem Körper als Stützgerüst dienende Knochen-
gewebe (BRINKHURST et al. 1998). Hier bilden die Calciumsalze zusammen mit
Phosphorsalzen als mineralische Komponente das Hydroxyapatit, das der Grundsubstanz des
Knochens die nötige Stabilität verleiht.
Dabei steuert die extrazelluläre Calciumkonzentration selbst die Freisetzung von Calcium aus
dem Knochen, indem eine hohe Konzentration die Bereitstellung und die Aktivität der
Osteoblasten fördert (QUARLES 1997; YAMAGUCHI et al. 1999) und die Arbeit der
Osteoklasten hemmt (KANATANI et al. 1999; ZAIDI et al.1999). Außerdem wird in dieser
Situation die Sekretion von Calcitonin in der Nebenschilddrüse angeregt (BROWN 1991; MC
GEHEE et al. 1997; BRINGHURST et al. 1998), um die Arbeit der Osteoklasten zu stoppen
und die Reabsorption von Calcium in der Niere zu senken.
Der physiologische Calciumspiegel im Plasma beim Schwein liegt nach Angaben von KOLB
(1989) zwischen 2,3 und 2,8 mmol/l und hängt dabei seiner Meinung nach in seinem
absoluten Wert vom Alter, dem Geschlecht, der Leistung und dem Reproduktionsstadium des

Literaturübersicht
52
Tieres ab. HEINRITZI und PLONAIT (2001) geben in ihren Ausführungen zur
Calciumkonzentration im Plasma etwas höhere Werte mit einer Schwankungsbreite von 2,40
– 3,00 mmol/l beim Schwein an.
2.9.2. Die Aufnahme von Calcium
Die aktive Ca2+- Absorption aus der Nahrung erfolgt zum Hauptteil im vorderen Dünndarm
(RADDE et al. 1980; FOX et al. 1985; KAUNE et al. 1992). Möglicherweise findet auch im
Ileum eine aktive Ca2+- Absorption statt, deren quantitativer Umfang allerdings erheblich
geringer ausfällt (RADDE et al. 1980; BRONNER 1990). In anderen Untersuchungen wurde
im Gegensatz dazu im Ileum eine Nettosekretion an Calciumionen nachgewiesen (FAVUS
1985; KARBACH 1991). Im Dickdarm sind sowohl aktive wie auch passive Transport-
vorgänge zu finden, obwohl hier kaum mehr als 10% der Calciumgesamtabsorption statt-
finden (BRONNER u. PANSU 1999). Der intestinale Calciumtransport setzt sich prinzipiell
aus zwei Komponenten zusammen. Er besteht einerseits aus einer nicht-sättigbaren,
parazellulären Komponente, die als Diffusionsprozess quantitativ von einem elektrischen
bzw. chemischen Gradienten zwischen mukosalem und serosalem Kompartiment und der
Dichtigkeit der Gewebeschranke abhängt, und andererseits aus einer sättigbaren, transzel-
lulären Variante, die auf aktiven Transportmechanismen beruht (NELLANS 1990;
KARBACH 1991; BRONNER u. PANSU 1999).
A) Parazellulärer Calciumtransport
Der nicht-sättigbare, parazelluläre Transport kann bei entsprechender Konzentration an
Calciumionen im gesamten Bereich des Dünndarms in etwa dem gleichen Ausmaß stattfinden
(PANSU et al. 1983). Der Anteil, den der parazelluläre Transport bezogen auf die Gesamt-
resorption an Calcium einnimmt, hängt hauptsächlich von der Lokalisation des untersuchten
Darmabschnittes und somit von der jeweiligen Aufenthaltsdauer, der vorherrschenden trans-
muralen Potentialdifferenz, sowie dem Gehalt an Calciumionen in der aufgenommenen
Ration und deren Bioverfügbarkeit ab (BRONNER u. PANSU 1999). Bei einem hohen
Gehalt an Calciumionen in der Nahrung spielt der parazelluläre Transport dieser Ionen eine
wichtigere Rolle (BRONNER u. PANSU 1999) und kann auf etwa 50% der Gesamtresorption
an Calcium ansteigen (NELLANS 1988).

Literaturübersicht
53
B) Transzellulärer Calciumtransport
Der transzelluläre Calciumtransport lässt sich in drei Teilbereiche unterteilen, nämlich die
Aufnahme über die Bürstensaummembran, den Transport durch die Epithelzelle und die
Ausschleusung an der Basolateralmembran.
Calciumaufnahme an der Bürstensaummembran
Innerhalb der Epithelzellen, die den Darm auskleiden, liegt der Gehalt an Calciumionen um
den Faktor 1000 bis 10000 niedriger als im Darmlumen (SCHRÖDER et al. 1996), außerdem
besteht eine Potentialdifferenz von ca. 50 mV, weshalb diese Ionen anhand des elektro-
chemischen Gradienten in die Zelle eingeschleust werden können ohne Energie aufzuwenden
(FULLMER 1992). Allerdings können die Ca2+-Ionen aufgrund ihrer positiven Ladung nicht
einfach durch die ungeladene Lipiddoppelschicht der Zellmembran hindurch diffundieren,
was spezifische Transportmechanismen wie bestimmte Ionenkanäle oder Ionentransporter
erforderlich macht.
Intrazellulärer Calciumtransport
Die Calciumionen werden in den Epithelzellen an das Protein Calbindin gebunden und
passieren so in dieser komplexen Form die Zelle von der luminalen zur basolateralen Seite.
Diese Art der Durchquerung der Zelle ist schätzungsweise um das 70-fache schneller als die
freie Diffusion von Calcium (BRONNER et al. 1986; BRONNER u. PANSU 1999). Die
Bildung des Calbindins wird in den Enterozyten durch die biologische Wirkung des
Calcitriols induziert (BAR u. WASSERMANN 1974; BRONNER u. FREUND 1975;
EDELSTEIN et al. 1978). Unter physiologischen Bedingungen gemessene Konzentrationen
an Calbindin reichen aus, um große Mengen an Calciumionen durch die Zelle zu schleusen,
ohne die physiologische intrazelluläre Konzentration an freien Calciumionen zu verändern
(KAUNE 1992). Es wird allerdings angenommen, dass dieser Schritt den limitierenden Faktor
des transepithelialen Calciumtransportes darstellt (SCHRÖDER et al. 1996). Zusätzlich
spielen endozytotische Vesikel ebenfalls eine Rolle bei der Absorption von Calciumionen
(NEMERE et al. 1987).

Literaturübersicht
54
Calciumextrusion an der Basolateralmembran
An der Basolateralmembran der Enterozyten müssen die Calciumionen entgegen eines
Konzentrationsgradienten in den extrazellulären Raum ausgeschleust werden, deshalb wird
bei diesem Teilabschnitt der Calciumabsorption Energie verbraucht.
Es werden verschiedene Mechanismen mit unterschiedlichen Transportkapazitäten
beschrieben, die Calcium aus den Enterozyten ausschleusen. DI POLO und BEAUGE (1983)
beschreiben einen Na+/Ca2+ Ionentauscher, der die notwendige Energie aus dem elektro-
chemischen Gradienten des Austauscherions bezieht. CARAFOLI (1987) führt einen
Uniporter an, der bei entsprechend hoher Konzentration durch eine erleichterte Diffusion dem
elektrischen Gradienten entgegenwirkt. Außerdem nennt er ATP-betriebene Ionenpumpen,
deren Antriebsenergie aus der Aufspaltung der energiereichen ATP-Verbindung stammt und
die aufgrund ihrer hohen Affinität zum Calcium zur Feinregulation der intrazellulären
Konzentration an Calciumionen eingesetzt werden (CARAFOLI 1987). Die spezifischen
Uniporter und Antiporter, die zwar eine geringere Affinität, aber dafür eine hohe
Transportkapazität aufweisen, treten bei einer hohen intrazellulären Konzentration in den
Vordergrund (DI POLO u. BEAUGE 1983; DENTON u. MC CORMACK 1985).
2.9.3. Die Ausscheidung von Calcium
Überschüssiges Calcium entsorgt der Körper beim Menschen und bei den Säugetieren über
die Niere. Im Falle einer erhöhten Calciumkonzentration im Blut wird in der Schilddrüse
Calcitonin sezerniert, welches in der Niere an den spezifischen Rezeptoren andockt. Dadurch
wird die Rückresorption der in das Tubulussystem sezernierten Calciumionen verringert.
Diese Reabsorption erstreckt sich dabei praktisch über das gesamte Nephron. Ca. 60% des
Calciums werden im proximalen Tubulus und ca. 30% im dicken, aufsteigenden Teil der
Henleschen Schleife passiv (aufgrund der transepithelialen Potentialdifferenz) parazellulär
reabsorbiert. Im distalen Konvolut wird Calcium transzellulär aktiv reabsorbiert, indem durch
luminale Kanäle passiv in die Tubulusepithelzellen einströmt und dann durch ATP-
angetriebene Carrier primär aktiv oder im Austausch gegen Natrium sekundär aktiv aus
diesen Zellen in die Blutgefäße ausgeschleust wird. Dieser aktive Ca2+-Transport wird über
das PTH gesteuert (v. ENGELHARDT u. BREVES 2000).

Literaturübersicht
55
2.10. Der Phosphor
2.10.1. Die Funktion und Regulation des Phosphorgehaltes im Organismus
Neben dem schon beschriebenen Calcium stellt gerade auch der Phosphor ein sehr wichtiges
Element im Stoffwechsel des Knochengewebes dar. Dieses wird ebenso in Verbindung mit
dem Calcium in Form von Hydroxyapatid (Ca10(PO4)6(OH)2) im Knochen gespeichert und
verleiht diesem somit die nötige Stabilität. Das Knochengewebe fungiert damit ebenfalls als
größter Speicher für dieses Element im Körper, da hier ca. 80% (v. ENGELHARDT u.
BREVES 2000) des Phosphors gespeichert werden. Die extrazelluläre Flüssigkeit enthält als
Phosphorpool, aus dem die Verbrauchsstellen mit diesem Element versorgt werden, dagegen
lediglich ca. 1% des im Körper befindlichen Phosphors. Bedeutende Funktionen erfüllt der
Phosphor z.B. als essentielles Element der Phosphorlipide zum Aufbau von Membranen oder
als Faktor für zahlreiche enzymatische Abläufe und Proteinfunktionen oder als Bestandteil
des Adenosintriphosphates, der wichtigsten Energiequelle des Körpers, und nicht zuletzt als
Teil des Hydroxyapatids zur Stabilisierung des Knochengewebes und als Teil der Nuklein-
säuren zur Speicherung und Übertragung genetischer Informationen. Aufgrund dieser
wichtigen Funktionen und des sehr geringen und damit sehr empfindlichen extrazellulären
Gehaltes wird die Phosphorkonzentration im Blut in relativ engen Grenzen reguliert. Beim
Schwein erstreckt sich die physiologische Bandbreite der Phosphorkonzentration im Plasma
laut HEINRITZI und PLONAIT (2001) von 2,10 – 3,30 mmol/l. Gesteuert werden diese
Werten über die Höhe der gastrointestinalen Absorption, der Ein- und Ausschleusung dieses
Elementes in das Knochengewebe sowie durch die Intensität der renalen Exkretion.
2.10.2. Die Absorption von Phosphor Den benötigten Phosphor nimmt der Körper dazu oral mit der Nahrung auf und dieser wird
dann als anorganische Phosphatverbindung hauptsächlich in den Darmabschnitten des
Jejunums und des Ileums absorbiert (WALLING 1977). Bei dieser Art der Resorption werden
primäre und sekundäre Phosphate (H2PO4- und HPO4
2-) mit Hilfe eines Natrium-Co-
transporters über die Bürstensaummembran in die Darmepithelzellen aufgenommen. Aus
diesen Zellen gelangt das Phosphat dann über eine carriervermittelte, erleichterte Diffusion in
das Interstitium. Durch das in der Niere gebildete Calcitriol kann bei einem Phosphormangel

Literaturübersicht
56
die Phosphorresorption gesteigert werden, indem dann vermehrt Na+-Phosphat-Cotransporter
in die Bürstensaummembran eingebaut werden (v. ENGELHARDT u. BREVES 2000).
2.10.3. Die renale Ausscheidung von Phosphor In den Nieren nimmt das Gleichgewicht zwischen glomerulärer Filtration und tubulärer
Reabsorptionsrate die Funktion der Regeleinheit für die Phosphorhomöostase wahr
(MIZGALA u. QUAMME 1985). Die Zellen, in denen diese Regulationsprozesse statt-
finden, sind die proximalen Tubuluszellen (DREZNER 2002), wo sich Carrier befinden, die
Phosphat zusammen mit Natrium im Verhältnis 1:3 aus dem Harn in die Tubuluszellen
befördern. Um bei einem Phosphorüberschuss, einer Acidose oder einer Hypocalcämie die
Phosphorretention zu senken, wirken als Regelfaktoren gegen einen Phosphorüberschuss
Glykokortikoide, TGF α, Calcitonin, PTH und das PTH related protein (PTHrP), indem sie
die Transportkapazität des Carriers verringern bzw. letztlich zum lysosomalen Abbau des
Carriers beitragen. Im Gegensatz dazu wird die Reabsorption von Phosphat bei einem
Phosphormangel u.a. von IGF-1, Thyroid Hormon, einer Alkalose und dem Phosphormangel
selbst begünstigt, indem bei dieser Konstellation die Carriertransportkapazität erhöht wird
bzw. ein erhöhter Einbau dieser Carrier in den Tubulusepithelzellen stattfindet (DREZNER
2002).

Material und Methoden
57
3. Material und Methoden
3.1. Das Ziel des Versuches
Es ist bekannt, dass durch eine forcierte Gabe von Calciumchlorid eine Beeinflussung des
Säure-Basen-Haushaltes im Organismus hervorgerufen wird und dass weiterführend über
diesen Weg eine Acidierung des Harnes, die als Folge der Regulation des Säure-Basen-
Haushaltes auftritt, erreicht werden kann. Diese Acidierung des Harnes hat bekanntlich
aufgrund des engen Zusammenhanges zwischen dem Bestehen einer Harnwegsinfektion und
dem Auftreten des MMA-Komplexes der Sau eine prophylaktische Wirkung gegen das Auf-
treten dieser Erkrankung.
Das Ziel der hier vorliegenden Studie bestand darin, näher zu erörtern, welche Auswirkungen
von den Veränderungen des Säure-Basen-Haushaltes im Organismus ausgehen, die durch eine
forcierte Gabe von Calciumchlorid (CaCl2) hervorgerufen werden. Dabei wurden verstärkt die
Interaktionen des Säure-Basen-Haushaltes mit der Mineralstoff-Homöostase und dem
Knochenstoffwechsel überprüft, da deren Regelmechanismen in sehr enger Verbindung
stehen bzw. sogar mit einander verknüpft sind.
Zu diesem Zweck sollen folgende Hypothesen geprüft werden:
a) Der Parathormon-Spiegel im Blut, der Veränderungen in der Ausschüttung dieses Hormons
anzeigt, wird als Folge der Acidierung durch die forcierte Gabe von Calciumchlorid
modifiziert und ist somit verbunden mit entsprechenden Veränderungen der Calcium-
Homöostase.
b) Die Acidierung, die über die forcierte Verabreichung von Calciumchlorid erreicht wird,
wirkt sich direkt oder indirekt auf den Knochenstoffwechsel aus und beeinflusst hier
maßgeblich die Freisetzung von Calcium. Die Reaktion der Knochenzellen kann dabei anhand
des Gehaltes der so genannten Knochenmarker im Blut bestimmt werden. Als Knochen-
marker wird in dieser Arbeit die knochenspezifische alkalische Phosphatase (bAP) analysiert.

Material und Methoden
58
c) Zur Überwachung des Mineralstoffhaushaltes und der renalen Mineralstoffausscheidung
wurden die Konzentrationen an Natrium, Chlorid, Kalium, anorganischem Phosphor und
Kreatinin im Blut und im Harn der Sauen analysiert.
Die Überprüfung dieser Hypothesen bildete die Basis für eine objektive Bewertung des
Einsatzes von Calciumchlorid zur Prophylaxe gegen das Auftreten von Harnwegsinfektionen
bei der Sau. Diese Wirkweise könnte somit sekundär auch prophylaktisch gegen eine
Erkrankung der Sauen am MMA-Komplex nützlich sein, da eine Infektion der Harnorgane als
Vorläufersyndrom dieses Krankheitskomplexes gilt. Außerdem sollte mit den hier erzielten
Ergebnissen das zeitliche Ausmaß solcher prophylaktischen Maßnahmen näher eingegrenzt
werden.
3.2. Zeitraum und Ort der Versuchsdurchführung
Die Probenentnahme des hier beschriebenen Versuches wurde vom November 2003 bis zum
Mai 2004 im Sauenbestand des Lehr- und Forschungsgutes der Stiftung der Tierärztlichen
Hochschule Hannover in Ruthe durchgeführt und die Analysen der einzelnen Parameter
erfolgte sowohl paralell wie auch im Anschluss an diesen Zeitraum im Institut für
Tierernährung der Stiftung der Tierärztlichen Hochschule Hannover.
3.3 Die Versuchstiere
Zur Durchführung dieses Versuches wurden 29 Sauen, die mindestens einmal geferkelt
hatten, aus der Sauenherde des Lehr- und Forschungsgutes der Stiftung der Tierärztlichen
Hochschule Hannover in Ruthe ausgewählt. Diese Tiere stammten ausnahmslos aus dem
Hybridzuchtprogramm der BHZP und hatten im Durchschnitt ein Alter von 2,51 ± 0,95
Jahren, wobei die älteste Sau ein Alter von 4,64 Jahren hatte, während die jüngste Sau gerade
ein Alter von 1,29 Jahren aufwies. Zum Zeitpunkt des Versuchsbeginns, ca. der 80.
Trächtigkeitstag, wurden die Tiere von den übrigen Sauen separiert, gewogen und ihre
Körperkondition wurde beurteilt. Danach erfolgte, wie im weiteren Verlauf beschrieben wird,
die Aufstallung der Tiere unter Versuchsbedingungen im Wartestallbereich. Am 108.
Trächtigkeitstag wurden die Sauen dann erneut gewogen und ihre Körperkondition wurde
wiederum beurteilt, um sie danach in den Abferkelbereich umzustallen. Dabei betrug die
mittlere Körpermasse am 80. Trächtigkeitstag 218,73 kg ± 20,19 kg, wobei die schwerste Sau

Material und Methoden
59
255,5 kg auf die Waage brachte und das leichteste Tier lediglich 192,3 kg wog. Am 108.
Trächtigkeitstag betrug die mittlere Körpermasse 238,85 kg ± 20,11 kg und das schwerste
Tier wog zu diesem Zeitpunkt 282,5 kg und die leichteste Sau hatte ein Körpergewicht von
205,8 kg.
3.4 Die Aufstallung der Versuchstiere
Die Aufstallung der Tiere vor dem 80.Trächtigkeitstag (vor Versuchsbeginn)
In dem Zeitraum zwischen der Belegung der Sauen und dem Versuchsbeginn, d.h. vor dem
80. Trächtigkeitstag, waren die Tiere in einer großen Gruppe in einem Freiraumlaufstall des
Wartestallabteils aufgestallt. Der zentrale Laufbereich dieses Stalles war mit Betonspalten-
boden ausgelegt, während die beidseitig angeordneten und durch Trennwände abgeteilten
Liegekojen über einen isolierten, planbefestigten Betonboden verfügten. Dieser Stallbereich
wurde im Flüssigmistverfahren ohne Einstreu betrieben. Die Wasserversorgung war über
Nippeltränken sichergestellt, die sich am Rand der Lauf-flächen befanden und hier an den
Trennwänden der Liegekojen befestigt waren. Die individuelle Futterzuteilung erfolgte in
diesem Aufstallungsbereich über eine Transponderfütterungsanlage mit integrierter
Selektionsmöglichkeit.
Die Aufstallung der Tiere vom 80.-108. Trächtigkeitstag
In dem Zeitraum vom 80.–108. Trächtigkeitstag waren die Tiere weiterhin im Wartestallabteil
untergebracht, allerdings wurden sie zum Beginn des Versuches (80. Trächtigkeitstag) in
einen separaten Bereich dieses Abteils umgestallt. Hier waren sie jetzt bis zum 108. Trächtig-
keitstag in Kastenständen mit Selbstfangvorrichtung aufgestallt. In der vorderen Hälfte dieser
Kastenstände bestand der Boden aus einer isolierten, planbefestigten Betonfläche, während
die hintere Hälfte und der Gang bzw. der Freilaufbereich hinter den Kastenständen mit Beton-
spaltenboden ausgelegt war. Dieses Aufstallungssystem wurde ebenfalls im Flüssig-
mistverfahren ohne Einstreu betrieben. Die Wasserversorgung war hier über Tränkeventile
sichergestellt, die die Schweine selbst bedienen konnten, so dass dann Wasser in den Trog der
Tiere gelangte. Die Futterzuteilung der einzelnen Tiere erfolgte in dieser Aufstallungsform
von Hand.

Material und Methoden
60
Die Aufstallung der Tiere vom 108. Trächtigkeitstag bis zum Versuchsende (8.Tag p.p.)
Am 108. Trächtigkeitstag, also ca. eine Woche vor dem errechneten Abferkeltermin, wurden
die Sauen vom Wartestallbereich nach dem Rein-Raus-Prinzip in ein frisch gereinigtes und
desinfiziertes Abferkelabteil umgestallt. Der Sauenstall verfügte über drei solcher Abferkel-
abteile, in denen sich jeweils acht Abferkelbuchten befanden. In der Abferkelbucht selbst
waren die Sauen in einem diagonal angeordneten Kastenstand aufgestallt. Im vorderen Drittel
der Bucht war der Boden als isolierter und planbefestigter Betonboden ausgeführt, während
die restlichen zwei Drittel teilweise aus Gussrosten oder aus Kunststoffspaltenböden
bestanden. Auch dieser Stallbereich wurde ohne Einstreu auf der Basis von Flüssigmist und
zwar abteilweise nach dem Badewannen-Prinzip bewirtschaftet.
Wasser erhielten die Sauen auch hier über Tränkenippel, die sich oberhalb der Tröge
befanden. Außerdem konnte den Tieren über eine separate Rohrleitung zusätzlich Wasser
über den Trog verabreicht werden. Für die Ferkel befand sich im hinteren Teil der
Abferkelbucht eine separate Selbsttränke, die speziell auf deren Bedürfnisse zugeschnitten
war. Die Futterzuteilung für die Sauen lief während des normalen Betriebes dieser Abteile
über Volumendosierer, an denen per Hand die individuelle Futtermenge pro Tier reguliert
werden konnte. Diese Volumendosierer wurden über einen Rohrkettenförderer aus dem
Vorratssilo mit Trockenfutter versorgt. Während der Fütterung konnten dann alle
Volumendosierer mit der individuellen Futtermenge über einen Hebelmechanismus entleert
werden. Während des Versuches wurde den Sauen allerdings die individuell abgewogene
Futtermenge von Hand vorgelegt. Den Ferkeln konnte im Bedarfsfall über Edelstahlschalen
künstliche Sauenmilch verabreicht werden. Prästarter wurde den Ferkeln rationiert über
spezielle Futterautomaten angeboten, die sich im hinteren Teil der Bucht befanden und täglich
zweimal von Hand befüllt wurden.
Die Sauen verblieben auch nach dem Ablauf des Versuches (8. Tag post partum) bis zum
Ende der 3-4 Wochen dauernden Säugezeit in diesem Abteil. Danach wurden die Sauen in das
separate Deckzentrum umgestallt, während die Ferkel im Rein-Raus-Prinzip in ein sauberes
und desinfizertes Flatdeckabteil zur weiteren Aufzucht wechselten. Das Abferkelabteil wurde
dann mit dem Hochdruckreiniger komplett gewaschen und danach desinfiziert, bevor es
erneut im Rein-Raus-Prinzip belegt wurde.

Material und Methoden
61
3.5. Die Gruppeneinteilung der Versuchstiere
Die an diesem Versuch beteiligten Tiere sind auf zwei Gruppen verteilt worden. Zum Einen
bildeten 14 Sauen die Versuchsgruppe, in der zusätzlich zu dem jeweiligen Alleinfutter
Calciumchlorid verabreicht wurde, und zum Anderen bestand die entsprechende Kontroll-
gruppe aus 15 Tieren, die zum Vergleich lediglich mit dem entsprechenden Alleinfutter
versorgt wurden. Bei der Verteilung der Tiere auf diese beiden Gruppen ist auf eine möglichst
gleichmäßige Gruppengestaltung geachtet worden, so dass die Unterschiede im Bereich des
Tiermaterials weitestgehend minimiert wurden. Als ein Kriterium dieser Einteilung wurde der
Ernährungszustand zum Zeitpunkt des Versuchsbeginns herangezogen. Deshalb wurden die
Tiere am 80. Trächtigkeitstag gewogen und einer Beurteilung ihrer Körperkondition
unterzogen.
Ein weiteres Kriterium für die Verteilung der Tiere stellte das Alter bzw. die Anzahl der
Würfe dar, die die jeweilige Sau vor dem Beginn des Versuches schon zur Welt gebracht
hatte. Des weiteren waren die Daten aus dem Sauenplaner, über den die Sauenherde verwaltet
wurde, ausschlaggebend für die möglichst gleichmäßige Verteilung der Tiere. Als Eckdaten
galten hier Berichte über Erkrankungen in den vorangegangenen Säugezeiten, speziell aus
dem Bereich des MMA-Komplexes. Aber auch andere Produktionsdaten wie die Anzahl der
lebend geborenen Ferkel und die Anzahl der abgesetzten Ferkel fanden hier Berück-
sichtigung, um eine möglichst gleichmäßige Verteilung der Sauen auf die beiden Gruppen zu
realisieren.

Material und Methoden
62
3.6. Das Futter und die Fütterung
3.6.1. Das Futter
Das Alleinfutter für tragende bzw. laktierende Sauen
Für die Fütterung der Sauen in diesem Versuch kamen zwei verschiedene Arten von
Alleinfutter zum Einsatz, bei denen es sich um Mischfutter handelte, das auf dem
Versuchsgut der Stiftung der Tierärztlichen Hochschule Hannover in Ruthe als hofeigene
Mischung selbst hergestellt wurde.
Bestandteile der verschiedenen Alleinfutter:
Alleinfutter für Alleinfutter für tragende Sauen laktierende Sauen Winterweizen - 25,1 % Wintergerste 68,9 % 46,0 % Weizenkleie 17,3 % 5,0 % Sojaschrott 10,9 % 20,0 % Mineralstoffe 2,9 % 3,9 % Sojaöl 0,7 % 2,0 %
Trockenschnitzel:
Die Trockenschnitzel, die den Sauen einmal täglich gegen ca. 11 Uhr verabreicht wurden,
waren zu Pellets von einer Größe von 1 cm Durchmesser und bis zu 3,5 cm Länge gepresst.
Diese lösten sich durch geringe Feuchtigkeit unter Quellung der Zuckerrübenschnitzel sofort
auf und sollten durch diese Volumenzunahme im Magen der Sauen ein Sättigungsgefühl
hervorrufen, um diese so zu beruhigen.
Calciumchlorid (CaCl2) in gekapselter Form:
Bei dem in der Untersuchung verwendeten Calciumchlorid handelte es sich um das Produkt
Calci-Cap 75® (Fa. Soda, Feed ingredients Division, Monaco), das laut den Angaben des
Herstellers aus 75 % Calciumchlorid (27 % Calcium und 47 % Chlorid) und aus 25 %
Ummantelung, bestehend aus pflanzlichen Fettsäuren, zusammengesetzt war.

Material und Methoden
63
3.6.2. Die Futtermittelprobengewinnung
Um Proben für die später folgenden Futtermittelanalysen zu gewinnen, wurde beim Abwiegen
der einzelnen Futterportionen jeweils ein aliquater Anteil entnommen und aus diesen Anteilen
wurde dann eine repräsentative Sammelprobe erstellt. Bis zur jeweiligen Analyse lagerten
diese Sammelproben kühl, trocken und luftdicht verschlossen in Plastikbeuteln.
Auf diese Weise fand die Entnahme der Proben für die Futtermittelanalysen von allen
verwendeten Futtermitteln statt.
3.6.3. Die Futtermittelanalysen
Die Trockensubstanz- und Rohnährstoffgehalte der einzelnen Futtersammelproben wurden
nach den Vorschriften der WEENDER-Futtermittelanalyse (nach VDLUFA Methodenbuch
III; NAUMANN und BASSLER 1997) ermittelt. Bei allen Analysen dieses Verfahrens wurde
eine Doppelbestimmung durchgeführt.
Trockensubstanz (TS):
Zur Bestimmung der Trockensubstanz wurden die Futterproben für mindestens 8 Stunden bei
einer Temperatur von 105°C bis zum Erreichen der Gewichtskonstanz in einem
Trockenschrank getrocknet.
.
Rohasche (Ra):
Die Bestimmung des Rohaschegehaltes erfolgte durch eine 6 stündige Veraschung der Proben
in einem gewichtskonstanten Tiegel in einem 600°C heißen Muffelofen.
Rohprotein (Rp):
Zur Analyse des Rohproteingehaltes in den Futterproben kam es zur Anwendung des
Kjeldahlverfahrens, bei dem durch die Verwendung konzentrierter Schwefelsäure (H2SO4)
der in der Probe enthaltene Stickstoff in die Ammoniumform oxidiert wurde. Durch die
Zugabe von 30%iger Natronlauge wurde aus dieser Verbindung Ammoniak freigesetzt, der in
eine vorgelegte 30%ige Borsäure überdestilliert wurde (Kjeldahl-Gerät, Fa. Gerhardt, Bonn),
und somit konnte anhand einer Titration mit 0,1 N HCl der Stickstoffgehalt bestimmt werden.
Anhand des Stickstoffgehaltes konnte nun mit der folgenden Formel der Gehalt an Rohprotein
in der Probe errechnet werden. Rp = N x 6,25

Material und Methoden
64
Rohfett (Rfe):
Die Bestimmung des Gehaltes an Rohfett wurde nach einem Säureaufschluss der Probe mit
30%igen Salzsäure innerhalb einer 6-stündigen Extraktion in einem Soxhlet-Apparat mit
Hilfe von Petroläther durchgeführt, worauf eine Destillation in einem Rotationsverdampfer
(Fa. Büchi, Zürich, Schweiz) folgte, bevor die Probe für 12 Stunden bei 80°C getrocknet
wurde.
Rohfaser (Rfa):
Der Rohfasergehalt der Probe wurde festgestellt, indem die Probe jeweils für 30 Minuten in
1,25 %iger Schwefelsäure (H2SO4) und in 1,25%iger Natronlauge im Rohfaseraufschlussgerät
(Fibertec System 1020, Hot Extractor, Fa. Foss, Hägganäs, Schweden) gekocht wurde. Der
Rückstand wurde mit heißem, destillierten Wasser gewaschen und für 6 Stunden in einem
Trockenschrank bei einer Temperatur von 105°C getrocknet. Danach wurde dieser im
Muffelofen für 3 Stunden bei 500°C verascht und nochmals gewogen. Über die Subtraktion
des Rohaschegewichtes vom Trockengewicht konnte dann der Rohfasergehalt errechnet
werden.
N-freie Extraktstoffe:
Der Gehalt an N-freien Extraktstoffen wurde nach der folgenden Formel errechnet.
NfE = TS – (Ra + Rp + Rfe + Rfa)
Organische Substanz:
Die Menge der organischen Substanz im Futter ergab sich aus der Differenz zwischen der
Trockensubstanz und des Gehaltes an Rohasche.
oS = TS – Ra
Stärke:
Der Stärkegehalt der Futterproben wurde per Doppelbestimmung (Blindwert und Hauptwert)
nach einer Säurehydrolyse und der Zugabe von Carrez-I- und Carrez-II-Lösung zum Zwecke
der Klärung durch eine polarimetrische Messung der optischen Drehung der Lösung
(Polartronic E, Fa. Schmidt und Haensch, Köln) durchgeführt.

Material und Methoden
65
Zucker:
Der Zuckergehalt im Futter wurde nach der Methode von LUFF-SCHOORL (1976) bestimmt.
Die dabei verbrauchte Menge an Natriumthiosulfat war äquivalent zum Zuckergehalt.
Bestimmung der Mengen- und Spurenelemente:
Nassveraschung:
Zur Bestimmung der einzelnen Spuren- und Mengenelemente wurde eine Aschelösung nach
dem Prinzip der Nassveraschung hergestellt, indem 3 g der Probe mit 15 ml eines
Veraschungsgemisches aus 65%iger Salpetersäure und 70%igen Perchlorsäure im Verhältnis
4:1 gemischt und langsam eingekocht worden sind. Nach der Abkühlung wurde nochmals
kurz mit 5 ml 7,5%iger Salzsäure aufgekocht, um die anorganischen Anteile in Salze zu
überführen. Danach wurde der Inhalt mit tridestilliertem Wasser über einen Schwarzbandfilter
(Schwarzband Rundfilter, Fa. Schleicher & Schuell, Dassel) in einen 50 ml Messkolben
überspült und dieser bis zur Eichmarkierung aufgefüllt.
Calcium und Magnesium:
Die Bestimmung des Gehaltes an Calcium und Magnesium erfolgte aus der Aschelösung, der
eine 0,5 %ige Lanthanchloridlösung zugesetzt wurde, um Störionen zu maskieren. Nachdem
die Aschelösung daraufhin nach der Methode von SLAVIN (1969) verdünnt wurde, erfolgte
die Bestimmung des Gehaltes an Calcium und Magnesium mittels Atomabsorptions-
spektroskopie (Unicam Solar 969, Fa. Unicam, Offenbach).
Natrium und Kalium:
Die Gehalte an Natrium und Kalium in der Aschelösung wurden mit Hilfe eines
Flammenphotometers (M 8D-Acetylen, Fa. Lange, Berlin) im Flammenemissionsverfahren
nach SCHUHKNECHT und SCHINKEL (1963) analysiert.
Phosphor:
Zur Bestimmung des Phosphorgehaltes wurde eine Farblösung aus 500 µl der Aschelösung
und 10 ml von einem Reagenz bestehend aus gleichen Anteilen an Salpetersäure,
Ammoniumvanadat- und Ammoniummolybdatlösung gemischt. Nach der Auffüllung auf
Volumen von 50 ml mit destilliertem Wasser wurde nach 30 Minuten bei 365 nm mit Hilfe
eines Spektralphotometers (Cads 100, Fa. Lange, Berlin) kolormetrisch nach GERICKE und

Material und Methoden
66
KURMIES (1952) die Extinktion gemessen. Mit dem Wert dieser Extinktion konnte
daraufhin der Phosphorgehalt im Futter errechnet werden.
Chlorid:
Zur Bestimmung des Chloridgehaltes wurden 5 g der zu analysierenden Probe mit 50 ml
destilliertem Wasser in einen Messzylinder eingefüllt und dann wurde mit einem Chlorid-
Analyser (Typ 925, Fa. Ciba Corning, Gießen) nach dem Prinzip einer Fällungstitration diese
Analyse durchgeführt. Dazu wird von zwei Silberelektroden des Gerätes eine konstante
Menge an Silberionen in die Probelösung abgegeben, die sich mit den Chloridionen in der
Lösung zu Silberchlorid verbinden. Wenn alle Chloridionen sich in einer solchen Verbindung
befinden, endet schließlich die Titration. Anhand der Menge der abgegebenen Silberionen
konnte dann die Chloridmenge in der Lösung bestimmt werden.
Kupfer, Mangan, Zink und Eisen:
Die Messung der Anteile dieser Elemente erfolgte mit Hilfe der Atomabsorptions-
spektroskopie (Unicam Solar 969, Fa. Unicam, Offenbach) direkt aus der Aschelösung.
Selen:
Die Bestimmung des Selengehaltes in den verwendeten Futtermitteln erfolgte nach einer
separaten Nassveraschung mit Hilfe der Atomabsorptionsspektrometrie im Hydridsystem.
Dazu wurde 1 g der Probensubstanz mit 15 ml Veraschungsgemisch aus 70 %iger Perchlor-
säure und 65 %iger Salpetersäure in einem Mischungsverhältnis 1:4 vermischt. Danach wurde
der Kolben erwärmt bis die Probe fast eingedampft war. Nach kurzer Abkühlung wurden 5 ml
verdünnter Salzsäure hinzugefügt und nochmals aufgekocht. Nachdem die Probe dann erneut
abgekühlt war, wurde sie mit 10 ml halbkonzentrierter Salzsäure versetzt und für 30 Minuten
in ein siedendes Wasserbad überführt. In diesem Reaktionsschritt wurde das bei der
Veraschung entstandene Selen V und VI vollständig zu Selen IV reduziert, da nur dieses im
weiteren Verlauf zum Selenhydrid reagieren kann. Bei der Atomabsorptionsspektrometrie
(Unicam Solar 969, Fa. Unicam, Offenbach) reagierte das Selen IV nun mit dem Wasserstoff
zum Selenhydrid. Diese Verbindung wurde in der Flamme thermisch zersetzt und die
Konzentration des Selens konnte atomabsorptionsspektrometrisch gemessen werden. Die
Messung erfolgte gegen eine wässrige Eichreihe mit den Selen-Konzentrationen 10, 20 und
40 µg/l.
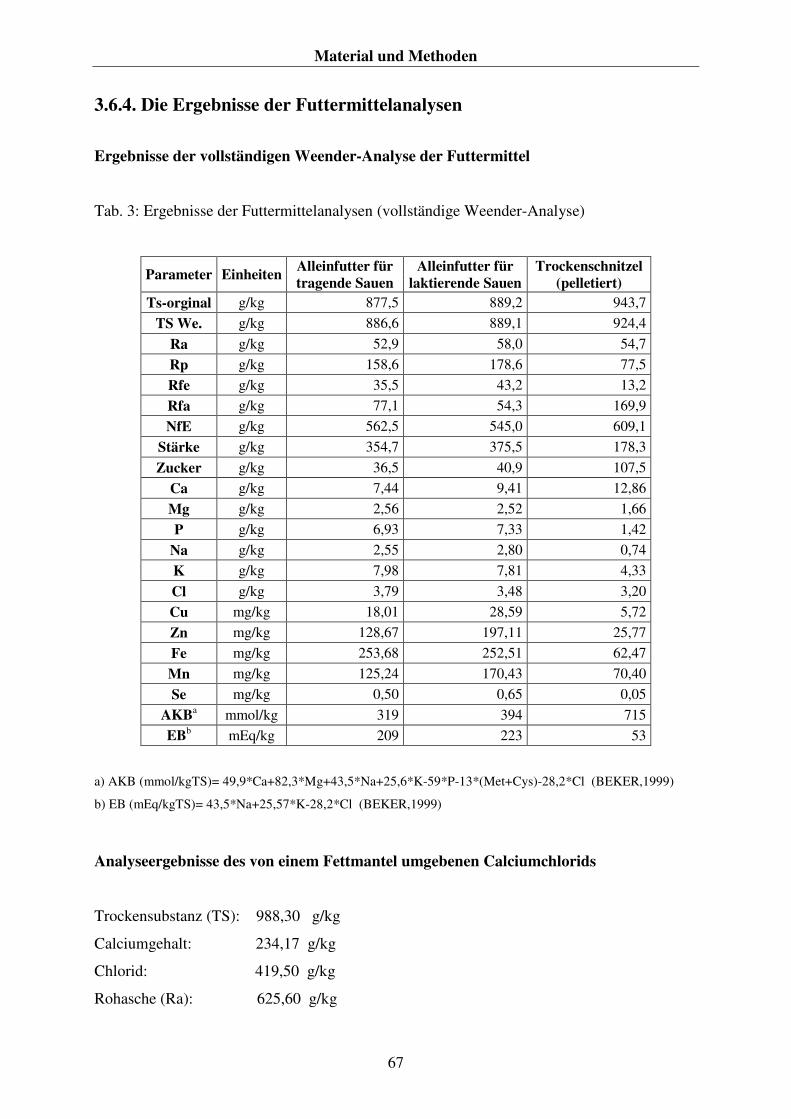
Material und Methoden
67
3.6.4. Die Ergebnisse der Futtermittelanalysen
Ergebnisse der vollständigen Weender-Analyse der Futtermittel
Tab. 3: Ergebnisse der Futtermittelanalysen (vollständige Weender-Analyse)
Parameter Einheiten Alleinfutter für tragende Sauen
Alleinfutter für laktierende Sauen
Trockenschnitzel (pelletiert)
Ts-orginal g/kg 877,5 889,2 943,7
TS We. g/kg 886,6 889,1 924,4
Ra g/kg 52,9 58,0 54,7
Rp g/kg 158,6 178,6 77,5
Rfe g/kg 35,5 43,2 13,2
Rfa g/kg 77,1 54,3 169,9
NfE g/kg 562,5 545,0 609,1
Stärke g/kg 354,7 375,5 178,3
Zucker g/kg 36,5 40,9 107,5
Ca g/kg 7,44 9,41 12,86
Mg g/kg 2,56 2,52 1,66
P g/kg 6,93 7,33 1,42
Na g/kg 2,55 2,80 0,74
K g/kg 7,98 7,81 4,33
Cl g/kg 3,79 3,48 3,20
Cu mg/kg 18,01 28,59 5,72
Zn mg/kg 128,67 197,11 25,77
Fe mg/kg 253,68 252,51 62,47
Mn mg/kg 125,24 170,43 70,40
Se mg/kg 0,50 0,65 0,05
AKBa mmol/kg 319 394 715
EBb mEq/kg 209 223 53
a) AKB (mmol/kgTS)= 49,9*Ca+82,3*Mg+43,5*Na+25,6*K-59*P-13*(Met+Cys)-28,2*Cl (BEKER,1999)
b) EB (mEq/kgTS)= 43,5*Na+25,57*K-28,2*Cl (BEKER,1999)
Analyseergebnisse des von einem Fettmantel umgebenen Calciumchlorids
Trockensubstanz (TS): 988,30 g/kg
Calciumgehalt: 234,17 g/kg
Chlorid: 419,50 g/kg
Rohasche (Ra): 625,60 g/kg

Material und Methoden
68
3.5.5. Die Fütterung
Fütterung vor Versuchsbeginn (vor dem 80. Trächtigkeitstag)
Vor dem Beginn des Versuches bzw. vor dem 80. Trächtigkeitstag wurden alle Sauen mit
dem Alleinfutter für tragende Sauen gefüttert. Dazu erhielten die Sauen eine auf ihr
Trächtigkeitsstadium und ihre Körperkondition individuell zugeschnittene Menge von diesem
Alleinfutter, welches sie beim Betreten der Futterstation über eine Transponderohrmarke
abrufen konnten. Die tägliche Menge an Alleinfutter wurde dabei auf 10 einzelne
Futterportionen verteilt. Außerdem erhielten die Tiere in diesem Zeitraum einmal täglich je
250g Trockenschnitzel, die einfach zur freien Aufnahme auf die planbefestigten Areale dieses
Stallabteils verteilt wurden.
Fütterung vom 80.–108. Trächtigkeitstag (im Wartestall)
Im Zeitraum vom 80.-108. Trächtigkeitstag waren die Sauen in Kastenständen im Warte-
stallabteil untergebracht und wurden mit Alleinfutter für tragende Sauen gefüttert. Dieses
Futter wurde vorher in der individuellen Menge, die jeder Sau pro Mahlzeit zugeteilt worden
war, abgewogen und portionsweise in Plastikbeutel abgefüllt. Die Vorlage dieses Futters
erfolgte in zwei Mahlzeiten, die erste Hälfte der Tagesration erhielten die Sauen morgens
gegen 630 Uhr und die zweite Hälfte wurde am Nachmittag gegen 1600 Uhr gefüttert. Die
Futtervorlage erfolgte immer von Hand und wurde vom Betreuungspersonal zu den
entsprechenden Zeiten durchgeführt. Die Tiere der Kontrollgruppe erhielten lediglich das für
sie bestimmte Alleinfutter für tragende Sauen in der individuellen Dosierung. Bei den Tieren
der Versuchsgruppe wurde zusätzlich zu jeder Portion des Alleinfutters für tragende Sauen
noch 14 g (US) des von einem Fettmantel umgebenen Calciumchlorids dazugegeben. Diese
Zugabe von Calciumchlorid erfolgte nach dem Abwiegen der tierspezifischen Portion an
Alleinfutter und wurde dann unter diese Portion gemischt und ebenfalls bis zur Fütterung in
dem Plastikbeutel aufbewahrt. Die Menge des individuell verabreichten Futters richtete sich
nach dem Trächtigkeitsstadium der jeweiligen Sau und dem Zustand der Körperkondition des
Tieres. Der Zeitraum vom 80. bis zum 108. Trächtigkeitstag wurde dazu in drei Teilabschnitte
aufgeteilt, in denen die individuelle Futtermenge jeweils gesteigert wurde. Der erste
Teilabschnitt reichte vom 80. Trächtigkeitstag abends bis zum 90. Trächtigkeitstag morgens,
während sich der zweite Abschnitt vom 90. Trächtigkeitstag abends bis zum 99.

Material und Methoden
69
Trächtigkeitstag abends erstreckte und der dritte Abschnitt somit am 100. Trächtigkeitstag
morgens begann und mit der Umstallung in den Abferkelbereich am 108. Trächtigkeitstag
nach der morgendlichen Fütterung endete. Die entsprechenden individuellen Futtermengen,
die pro Mahlzeit verfüttert wurden, sind den Tabellen (Tab. 26 und 27) im Kapitel
„Tabellenanhang“ zu entnehmen. Zusätzlich zu dem Alleinfutter für tragende Sauen erhielt
jede Sau täglich 250 g Trockenschnitzel, die ca. gegen 1100 Uhr von Hand in den Trog jedes
Tieres gegeben wurden. Da es sich bei dieser Fütterung um eine rationierte Form handelte,
waren nach der Futteraufnahme der Tiere keine Restmengen an Futter mehr vorhanden. Somit
konnte davon ausgegangen werden, dass die gesamte Menge des vorgelegten Futters
aufgenommen wurde.
Fütterung vom 108. Trächtigkeitstag bis zum Ende des Versuches (8. Tag p.p.)
Am 108. Trächtigkeitstag erfolgte die Umstallung der Sauen in ein Abferkelabteil und damit
auch die Umstellung der Fütterung. Ab dem Nachmittag des 108. Trächtigkeitstages erhielten
die Tiere das Alleinfutter für laktierende Sauen. Dieses Futter wurde vorher in der
individuellen Menge, die jede Sau pro Mahlzeit bekommen sollte, abgewogen und portions-
weise in Plastikbeutel abgefüllt. Die Vorlage dieses Futters erfolgte in zwei Mahlzeiten, die
erste Hälfte der Tagesration erhielten die Sauen morgens gegen 630 Uhr und die zweite Hälfte
wurde am Nachmittag gegen 1600 Uhr gefüttert. Die Futtervorlage erfolgte dabei immer von
Hand und wurde vom Betreuungspersonal zu den entsprechenden Zeiten durchgeführt. Die
Tiere der Kontrollgruppe erhielten lediglich das für sie bestimmte Alleinfutter für laktierende
Sauen in der individuellen Dosierung. Bei den Tieren der Versuchsgruppe wurde vom 108.
Trächtigkeitstag bis zur Abferkelung zusätzlich zu jeder Portion des Alleinfutters für
laktierende Sauen noch 14 g (US) des von einem Fettmantel umgebenen Calciumchlorids
dazugegeben. Diese Zugabe von Calciumchlorid erfolgte nach dem Abwiegen der
tierspezifischen Portion an Alleinfutter und wurde dann unter diese Portion gemischt und
ebenfalls bis zur Fütterung in dem Plastikbeutel aufbewahrt. Im Zeitraum vom 108. – 112.
Trächtigkeitstag erhielt jede Sau die gesamte für sie vorgesehene Menge an Alleinfutter für
laktierende Tiere. Ab dem 112. Trächtigkeitstag wurde diese Menge dann auf 1 kg
Alleinfutter pro Tier und Mahlzeit reduziert. Diese Reduktion wurde beibehalten, bis aus dem
Gesäuge der jeweiligen Sau Milch im Strahl zu ermelken war, was als sicheres Zeichen der
nahenden Geburt gilt. Ab diesem Zeitpunkt kurz vor der Geburt und jeweils eine Mahlzeit
nach der Abferkelung erhielten die Sauen lediglich Wasser und kein Futter. In der

Material und Methoden
70
darauffolgenden Zeit bis zum Versuchsende erhielten alle Sauen lediglich das Alleinfutter für
laktierende Sauen. Dabei wurde die individuelle Futtermenge pro Mahlzeit der Wurfleistung
und der Tierkondition angepasst und von Tag zu Tag langsam gesteigert.
Die individuellen Futtermengen, die jedes einzelne Tier zu den verschiedenen Fütterungszeit-
punkten erhalten hat, sind den Tabellen (Tab. 28 und 29) des Tabellenanhangs zu entnehmen.
3.6. Die Blutproben
3.6.1. Die Entnahme der Blutproben
Blutproben wurden jeder Sau und je einem Ferkel pro Wurf zu folgenden Zeiten entnommen:
Blutentnahmetag Blutentnahmezeitpunkt Tiermaterial_____
80. Trächtigkeitstag 800 – 830 Uhr Sauen
108. Trächtigkeitstag 800 – 830 Uhr Sauen
Am Tag der Geburt sobald die Sau nach dem Ende der Sauen
Geburt (Abgang der Nachgeburt) steht (+ 1 Ferkel/ Wurf)
2. Tag post partum 800 – 830 Uhr Sauen
4. Tag post partum 800 – 830 Uhr Sauen
6. Tag post partum 800 – 830 Uhr Sauen
8. Tag post partum 800 – 830 Uhr Sauen
Zur Durchführung der Blutentnahme wurden die Sauen zunächst von einer Hilfsperson mit
einer Oberkieferschlinge fixiert und dann das Blut aus der Vena jugularis externa entnommen.
Dazu wurden Heparin-Lithium-beschichtete Monovetten der Firma Sarstedt aus Nümbrecht
mit einem Volumen von 9 ml und sterile Einmalkanülen vom Fabrikat TSK-SUPRA (Fa.
Ehrhardt-Söhne GmbH, Geislingen) in der Größe 1,50 mm x 100 mm verwendet. Bei jeder
Blutentnahme wurden so jeweils 3 der beschriebenen Monovetten befüllt.
Den neugeborenen Ferkeln wurden zur Blutentnahme auf dem Rücken liegend von einer
Hilfsperson die Gliedmaßen fixiert, so dass dann eine Blutprobe aus der Vena cava cranialis
entnommen werden konnte. Dazu wurden die gleichen Monovetten wie bei der Blutentnahme
der Sauen verwendet. Allerdings kamen hier sterile Einmalkanülen mit der Bezeichnung
NEOLUS der Firma TERUMO EUROPE N.V. (Leuven, Belgien) in der Größe 0,8 mm x 40

Material und Methoden
71
mm zum Einsatz. Den Ferkeln wurde allerdings lediglich die Blutmenge einer Monovette
entnommen.
3.6.2. Die Aufbereitung und Aufbewahrung der Blutproben
Blutproben für die Blutgasanalyse:
Der Inhalt einer Monovette pro Blutentnahme bei den Sauen und die Blutprobe der Ferkel
wurden sofort nach der Blutentnahme in eine geschlossene Styroporkiste verpackt, die
ursprünglich der Anlieferung des Spermas zur künstlichen Besamung in den Sauenbestand
diente. In diesem Behältnis wurden die Proben auf gefrorenen „COOL-PACKS“ bei einer
Temperatur von + 2 bis + 4 °C gelagert, um so später für die Blutgasanalyse genutzt zu
werden.
Blutproben für die Plasmagewinnung:
Der Inhalt der anderen zwei Monovetten, die einer Sau pro Blutentnahme entnommen worden
waren, wurden zur Plasmagewinnung genutzt. Zu diesem Zweck wurden diese Proben mit
einer Zentrifuge (Typ Heraeus Christ HC 121, Fa. Heraeus Instruments GmbH, Osterode am
Harz) 20 Minuten lang bei 3000 U/Min zentrifugiert, was einer Beschleunigung von 1900 G
entsprach. Danach wurde das gewonnene Plasma in produktionssterile Reaktionsgefäße mit
Deckel und einem Volumen von 2 ml pipettiert. Diese Plasmaproben wurden dann bis zur
weiteren Analyse bei einer Temperatur von – 20 °C eingefroren.
3.6.3. Die Analyse der Blutproben Die Blutproben für die Blutgasanalyse, die nicht zentrifugiert wurden, wurden spätestens eine
Stunde nach der Blutentnahme mit Hilfe des Blutgasanalysegerätes (Typ Rapid lab 860, Fa.
Chrion Diagnostics, Fernwald) analysiert. Zu diesem Zweck wurde eine Glaskapillare mit
Probenmaterial gefüllt und in die entsprechende Öffnung des Gerätes gesteckt. Aus dieser
Probe wurden jetzt die in der folgenden Tabelle (Tab. 4) aufgeführten Parameter mit Hilfe der
entsprechenden Verfahren analysiert.
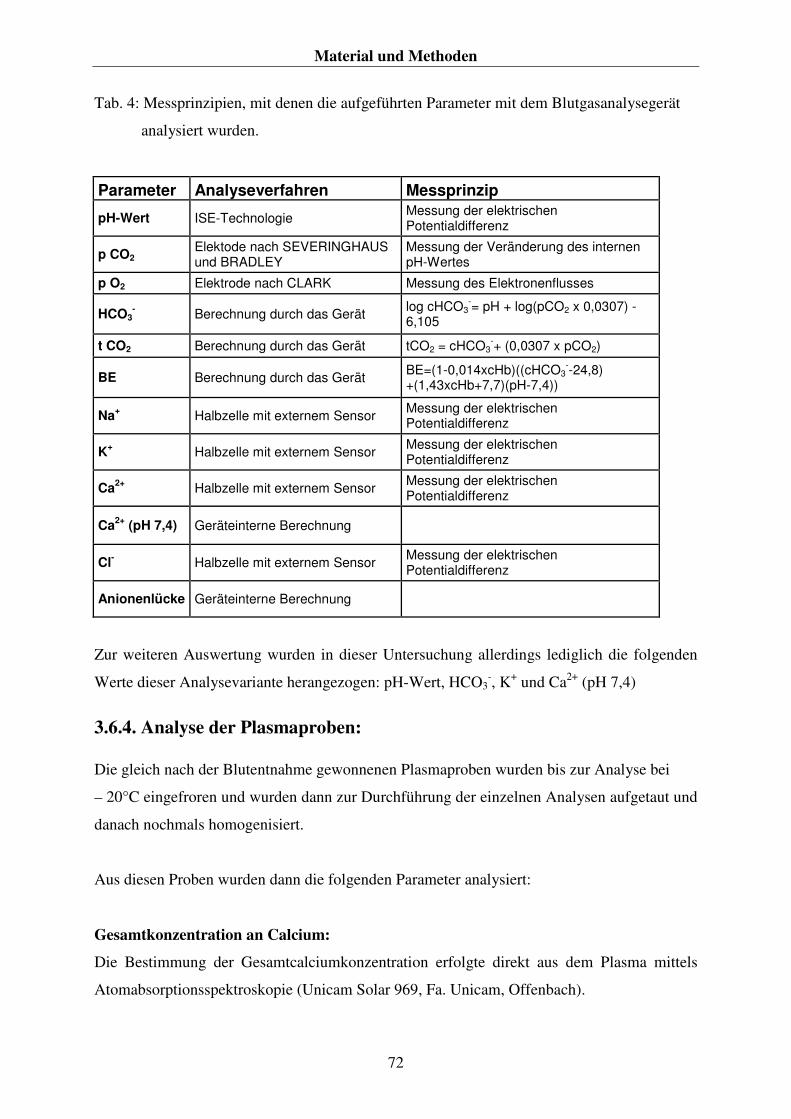
Material und Methoden
72
Tab. 4: Messprinzipien, mit denen die aufgeführten Parameter mit dem Blutgasanalysegerät
analysiert wurden.
Parameter Analyseverfahren Messprinzip
pH-Wert ISE-Technologie Messung der elektrischen Potentialdifferenz
p CO2 Elektode nach SEVERINGHAUS und BRADLEY
Messung der Veränderung des internen pH-Wertes
p O2 Elektrode nach CLARK Messung des Elektronenflusses
HCO3- Berechnung durch das Gerät
log cHCO3-= pH + log(pCO2 x 0,0307) -
6,105
t CO2 Berechnung durch das Gerät tCO2 = cHCO3-+ (0,0307 x pCO2)
BE Berechnung durch das Gerät BE=(1-0,014xcHb)((cHCO3
--24,8)
+(1,43xcHb+7,7)(pH-7,4))
Na+ Halbzelle mit externem Sensor
Messung der elektrischen Potentialdifferenz
K+ Halbzelle mit externem Sensor
Messung der elektrischen Potentialdifferenz
Ca2+
Halbzelle mit externem Sensor Messung der elektrischen Potentialdifferenz
Ca2+
(pH 7,4) Geräteinterne Berechnung
Cl- Halbzelle mit externem Sensor
Messung der elektrischen Potentialdifferenz
Anionenlücke Geräteinterne Berechnung
Zur weiteren Auswertung wurden in dieser Untersuchung allerdings lediglich die folgenden
Werte dieser Analysevariante herangezogen: pH-Wert, HCO3-, K+ und Ca2+ (pH 7,4)
3.6.4. Analyse der Plasmaproben: Die gleich nach der Blutentnahme gewonnenen Plasmaproben wurden bis zur Analyse bei
– 20°C eingefroren und wurden dann zur Durchführung der einzelnen Analysen aufgetaut und
danach nochmals homogenisiert.
Aus diesen Proben wurden dann die folgenden Parameter analysiert:
Gesamtkonzentration an Calcium:
Die Bestimmung der Gesamtcalciumkonzentration erfolgte direkt aus dem Plasma mittels
Atomabsorptionsspektroskopie (Unicam Solar 969, Fa. Unicam, Offenbach).

Material und Methoden
73
Bei diesem Analyseverfahren wird die Probe fein zerstäubt durch eine Acetylen-Luft-Flamme
gesaugt, wodurch die Elemente, die in ihr enthalten sind, in einen atomaren Zustand überführt
werden. Diese so entstandenen Atome absorbieren jeweils Strahlungsenergie einer
charakteristischen Wellenlänge, die die Elektronen auf der äußeren Schale dieser Atome
benötigen, um von ihrem Grundzustand in einen energetisch angeregten Zustand zu gelangen.
Durch diese energetische Absorption dieser Wellenlänge entsteht eine charakteristische
Resonanzlinie. Im Atomabsorptionsspektrometer sind Hohlkathodenlampen vorhanden, die
das zu analysierende Element enthalten und so das dazugehörige Linienspektrum emittieren.
Nachdem die Probe nun aus dem Strahlengang die entsprechende Strahlung absorbiert hat,
wird mit einem Empfängersystem mit Sekundärverstärker die Extinktion gemessen. Der Wert
der Extinktion ist proportional zur Menge des in der Probe enthaltenen Elementes.
Phosphor:
Die Menge an anorganischem Phosphat, welches in den Plasmaproben vorhanden war, wurde
mit Hilfe eines UV-Testes (Nobi-Flow Phosphor-UV, Fa. Nobis Labordiagnostica GmbH,
Endingen) gemessen und in mmol/l umgerechnet.
Diese Testvariante basiert auf einer Komplexbidung des Phosphates mit Ammoniummolybdat
zu einem Ammoniummolybdat-Phosphor-Komplex in schwefelsaurer Lösung. Dieser
Stoffkomplex absorbiert Strahlung im Wellenbereich des UV-Lichtes. Somit ist eine
Extinktion zu messen, die in Verbindung mit einer Standardmessung zur Berechnung des
Phosphatgehaltes in der zu analysierenden Probe dient.
Natrium:
Der Gehalt an Natrium in den Plasmaproben wurde ebenfalls wie bei der Analyse der
Futterproben mit Hilfe eines Flammenphotometers (M 8D-Acetylen, Fa. Lange, Berlin) im
Flammenemissionsverfahren nach SCHUHKNECHT und SCHINKEL (1963) analysiert.
Im Unterschied zu den Futterproben ging hier aber kein Veraschungsverfahren voraus,
sondern der Gehalt an Natrium wurde direkt aus den Plasmaproben bestimmt.
Chlorid:
Zur Bestimmung des Chloridgehaltes wurden 5 ml der zu analysierenden Plasmaprobe
abgemessen und in einen 50 ml fassenden Messzylinder eingefüllt, der dann bis zur
Eichmarke mit destilliertem Wasser aufgefüllt wurde. Im Anschluss wurde diese Lösung eine
Stunde lang geschüttelt. Es folgte eine Zentrifugation eines entsprechenden Aliquots, um

Material und Methoden
74
daran entsprechend die Messung durch zu führen. Die Bestimmung des Chloridgehaltes mit
einem Chlorid-Analyser (Typ 925, Fa. Ciba Corning, Gießen) läuft nach dem Prinzip einer
Fällungstitration ab. Dazu geben zwei Silberelektroden des Gerätes eine konstante Menge an
Silberionen in die Probelösung ab, die sich mit den Chloridionen in der Lösung zu
Silberchlorid verbinden. Wenn alle Chloridionen sich in einer solchen Verbindung befinden,
endet die Titration. Anhand der Menge der abgegebenen Silberionen konnte so die
Chloridmenge in der Lösung bestimmt werden.
Kreatiningehalt im Plasma:
Die Bestimmung des Kreatiningehaltes im Plasma wurde durch einen enzymatischen Farbtest
(Zweipunkt-Kinetik) nach der PAP-Methode durchgeführt (THOMAS 1992).
Dazu mussten zunächst zwei Reagenzlösungen hergestellt werden.
Zur Herstellung der Reagenzlösung I wurde die Enzymmischung R2, die 20 U/ml Kreatinase,
5 U/ml Sarcosinoxidase, 1 U/ml Peroxidase, 10 U/ml Ascorbatoxidase und 10 µmol/l
Kaliumhexacyanoferrat enthielt, in 50 ml einer Pufferlösung bestehend aus 150 mmol/l
Phosphatpuffer (pH 7,9) und 1 mmol/l ADPS gelöst.
Zur Herstellung der Reagenzlösung II wurde die Enzymmischung R4 , die aus 0,2 mmol/l 4-
Aminoantipyrin und 20 U/ml Kreatinase bestand, in 10 ml des Puffers, der auch zur
Herstellung der Reagenzlösung I verwendet wurde, gelöst.
Die Messung der Extinktion wurde unter der Verwendung eines 546 nm Filters bei einer
Messtemperatur von 25°C durchgeführt.
Zur Beschickung der Küvetten wurde folgendes Pipettierschema angewendet:
Leerwert Standard Analyse
Standard - 50µl -
Probe - - 50µl
Reagenzlösung I 500µl 500µl 500µl
Den Inhalt der Küvetten wurde gut gemischen und 5 Minuten bei 25°C inkubiert.
Reagenzlösung II 100µl 100µl 100µl

Material und Methoden
75
Den Inhalt der Küvetten wurde erneut gut gemischt und nach genau 2 Minuten wurde der
Wert für E1 ablesen und nach genau 2 weiteren Minuten konnte der Wert für E2 ermittelt
werden.
Die Berechnung des Kreatiningehaltes im Serum erfolgte dann nach folgender Formel:
( E2 – E1)Analyse c ( mg/ dl) = ------------------------------ x 2 mg/dl ( E2 – E1)Standard
Parathormon:
Die Parathormonkonzentration im Plasma wurde mit einem Enzyme-Linked-Immuno-
Sorbent-Assay (ELISA) (Porcine Intact PTH ELISA Kit, Immutopics, Inc., San Clemente,
Canada) ermittelt. Zur Durchführung dieser Analyse wurden jeweils 50 µl des Proben-
materials bzw. der Standardlösung oder der Kontrolllösung in die entsprechenden Kavitäten
der Microtiterplatte pipettiert und jeweils 50 µl der Arbeits-Anti-Körper-Lösung, die zu
gleichen Anteilen aus Biotin-markierten porzinen Antikörpern und aus HRP-konjugierten
porzinen Antikörpern bestand, dazugegeben. Danach wurde diese Platte mit einem
Plattenverschliesser verschlossen und mit Alufolie abgedeckt. In diesem Zustand erfolgte
dann für drei Stunden eine Inkubation bei Raumtemperatur auf einem horizontalen Rotor bei
180-220 U/Min. Nachdem nun die Alufolie und der Plattenverschliesser entfernt worden
waren, wurde aus jeder Kavität der Inhalt abgesaugt und fünfmal mit einer Waschlösung
gewaschen und erneut ausgesaugt. Dann wurden 100 µl des ELISA-HRP-Substrates dazu
pipettiert, die Mikrotiterplatte wurde erneut wie schon beschrieben verschlossen und
nochmals für 30 Minuten unter den gleichen Bedingungen inkubiert. Nach Ablauf dieser
Inkubationszeit wurde der Plattenverschluss entfernt und in einem Mikrotiterablesegerät die
Absorption bei einer Wellenlänge von 620 nm gemessen. Sofort danach wurde in jede Kavität
50µl einer ELISA-Stop-Lösung pepittiert und die Platte eine Minute bei der vorgegebenen
Drehzahl auf den Rotor verbracht. Nach diesem Schritt wurde nochmals die Absorption
allerdings bei einer Wellenlänge von 450 nm gemessen. Anhand der vorher erstellten
Eichkurven aus den mitgelieferten Standards konnte dann aus beiden Absorptionsraten mit
den dazugehörigen Eichkurven der Gehalt an PTH in der Probe bestimmt werden. Das
Ergebnis wurde in pg/ml angegeben.
Absorption der Probe Konzentration Probe = --------------------------------------- x Konzentration der Standardlsg. Absorption der Standardlösung

Material und Methoden
76
Knochenspezifische alkalische Phosphatase (bAP):
Die Bestimmung der Konzentration der knochenspezifischen Phosphatase erfolgte mit einem
Enzym-Immunassay (METRATM BAP EIA kit, Fa. QUIDEL CORPORATION, San Diego,
USA). Diese Bestimmung des bAP-Gehaltes funktioniert nach dem Prinzip eines
Immunassays, bei dem die in der zu analysierenden Probe enthaltenen bAP-Anteile an einen
mit monoklonalen Anti-bAP-Antikörper beschichteten Mikrotiterstreifen gebunden werden.
Diese gebundenen bAP-Anteile konnten dann über ihre Enzymaktivität mit Hilfe eines p-
Nitrophenylphosphat (pNPP)-Substrats bestimmt werden.
Zur Durchführung dieser Analyse wurden in die Vertiefung der mit von Mäusen stammenden
monoklonalen Anti-bAP-IgG-Antikörper beschichteten Teststreifen 125 µl einer Testpuffer-
substanz gegeben, die aus Magnesiumchlorid, Zinksulfat, Tensid und Natriumazid bestand.
Daraufhin wurden 20 µl der Standardlösung bzw. der zu untersuchenden Probensubstanz
hinzu pipettiert, um dann für 3 Stunden bei + 20 – 28°C inkubiert zu werden. Im weiteren
Verlauf wurden die Vertiefungen entleert und viermal mit einer einfach konzentrierten
Waschpufferlösung gewaschen. Im nächsten Schritt wurden in die gewaschenen Vertiefungen
150 µl einer Substratlösung gegeben, die als Substrat das p-Nitrophenylphosphat enthielt.
Nach einer weiteren Inkubationszeit von 30 Minuten bei +20-28°C wurden 100 µl einer
Stopplösung dazugegeben, die Natronlauge (NaOH) enthielt. Im Folgenden wurde nun die
Absorbanz bei 405 nm bestimmt. Die Auswertung der jeweiligen Konzentrationen an bAP in
den Plasmaproben erfolgte anhand einer Standardmesskurve, die vorher unter der
Verwendung einer Standardlösung, welche dem Testkit beigefügt war, erstellt wurde. Die
bAP-Konzentrationen im Plasma wurden in U/l angegeben.

Material und Methoden
77
3.7. Die Harnproben
3.7.1. Die Entnahme der Harnproben
Zur Gewinnung der Harnproben wurde der Mittelstrahlurin beim spontanen Absetzen des
Harnes aufgefangen (Spontanharnprobe). Diese Form der Harnentnahme wurde am 90.
Trächtigkeitstag und am 111. Trächtigkeitstag jeweils beim Auftreiben der Sauen vor der
morgendlichen Fütterung durchgeführt. Zum Auffangen des Harnes wurden durchsichtige und
weithalsige Kunststofflaschen mit einem Fassungsvermögen von 500 ml benutzt.
3.7.2. Die Aufbereitung und Aufbewahrung der Harnproben
Der Harn, der so durch das Auffangen einer Spontanharnprobe gewonnen wurde, ist
direkt nach der Messung des pH-Wertes in Milchprobenröhrchen aus Kunststoff umgefüllt
und dann in diesen Röhrchen bis zur weiteren Analyse bei – 20°C eingefroren worden.
3.7.3. Die Analyse der Harnproben
pH-Wert Bestimmung im Harn:
Die Messung des pH-Wertes im Harn erfolgte direkt nach der Gewinnung einer
Spontanharnprobe mit einem Labor-pH-Meter (Typ 23, Fa. Knick, Berlin), an das als
Messsonde eine Schott-pH-Einstabmesskette (Typ Blue Line 18 pH, Fa. Schott Glas, Mainz)
angeschlossen war.
Kreatiningehalt:
Die Bestimmung des Kreatiningehaltes im Harn wurde durch einen enzymatischen Farbtest
(Zweipunkt-Kinetik) nach der PAP-Methode durchgeführt (THOMAS 1992), die ebenfalls
zur Bestimmung des Kreatiningehaltes im Plasma verwendet worden ist. Dieses Verfahren
wurde im Kapitel 3.6.4. „Analyse der Plasmaproben“ näher beschrieben. Im Vergleich zum
Plasma musste der Harn zur Durchführung dieser Analyse allerdings vorher mit destilliertem
Wasser im Verhältnis 1:20 verdünnt werden.

Material und Methoden
78
Mineralstoffbestimmung im Harn:
Calcium:
Die Bestimmung der Calciumkonzentration erfolgte auch im Harn wie bei der Analyse der
Plasmaproben direkt aus dem Probenmaterial mittels Atomabsorptionsspektroskopie (Unicam
Solar 969, Fa. Unicam, Offenbach).
Phosphor:
Die Menge an anorganischen Phosphat, das in den Harnproben vorhanden war, wurde
ebenfalls wie bei der Analyse der Plasmaproben mit Hilfe eines UV-Testes (Nobi-Flow
Phosphor-UV, Fa. Nobis Labordiagnostica GmbH, Endingen) gemessen und in mmol/l
umgerechnet.
Natrium :
Der Gehalt an Natrium wurde wie bei der Analyse der Futterproben ebenfalls mit Hilfe des
Flammenphotometer (M 8D-Acetylen, Fa. Lange, Berlin) im Flammenemissionsverfahren
nach SCHUHKNECHT und SCHINKEL (1963) analysiert.
Chlorid:
Der Chloridgehalt in den Harnproben ist direkt aus der Probe bestimmt worden. Dazu wurden
ähnlich wie bei der Chloridanalyse im Plasma 100 µl der Harnprobe abpipettiert und mit dem
Chlorid-Analysator 925 (Fa. Ciba Corning, Gießen) wurde nach dem Prinzip der
kolorimetrischen Titration der Chloridgehalt gemessen.

Material und Methoden
79
3.8. Die statistische Auswertungen der Analysenergebnisse
Die statistische Auswertung der durch Analysen oder Berechnungen gewonnenen Werte
erfolgte zur Erfassung dieser Werte mit Excel® ( Microsoft Office 2000) und die eigentlichen
statistischen Berechnungen mit einem Statistikprogramm STATISTICA 97® (Version 5).
Zu diesem Zweck wurden folgende statistische Methoden angewendet:
- Mittelwertbestimmung (Mw) bei der Zusammenfassung mehrerer Einzelwerte
- Berechnung der Standardabweichung (SD) als Maß der Streuung der Einzelwerte
- Überprüfung der Werte auf die Normalverteilung mit Hilfe des Kolmogorov-Smirnov-
Testes
- Ein- bzw. zweifaktorielle Varianzanalysen mit Messwiederholung für den Vergleich
der Varianz der Mittelwerte
- Zur Überprüfung der signifikanten Effekte wurde als post-hoc-test der Least
Significant Difference-Test ( LSD-Test) durchgeführt.
Zur Einteilung der Signifikanzengrenzen galten folgende Werte:
- ein Wert von p<0,05 gilt als schwach signifikant,
- ein Wert von p<0,01 gilt als signifikant,
- ein Wert von p<0,001 gilt schließlich als hochsignifikant.
Die Darstellung von Mittelwert und Standardabweichung erfolgt im Text und in den Tabellen
wie folgt: Mw ± SD.
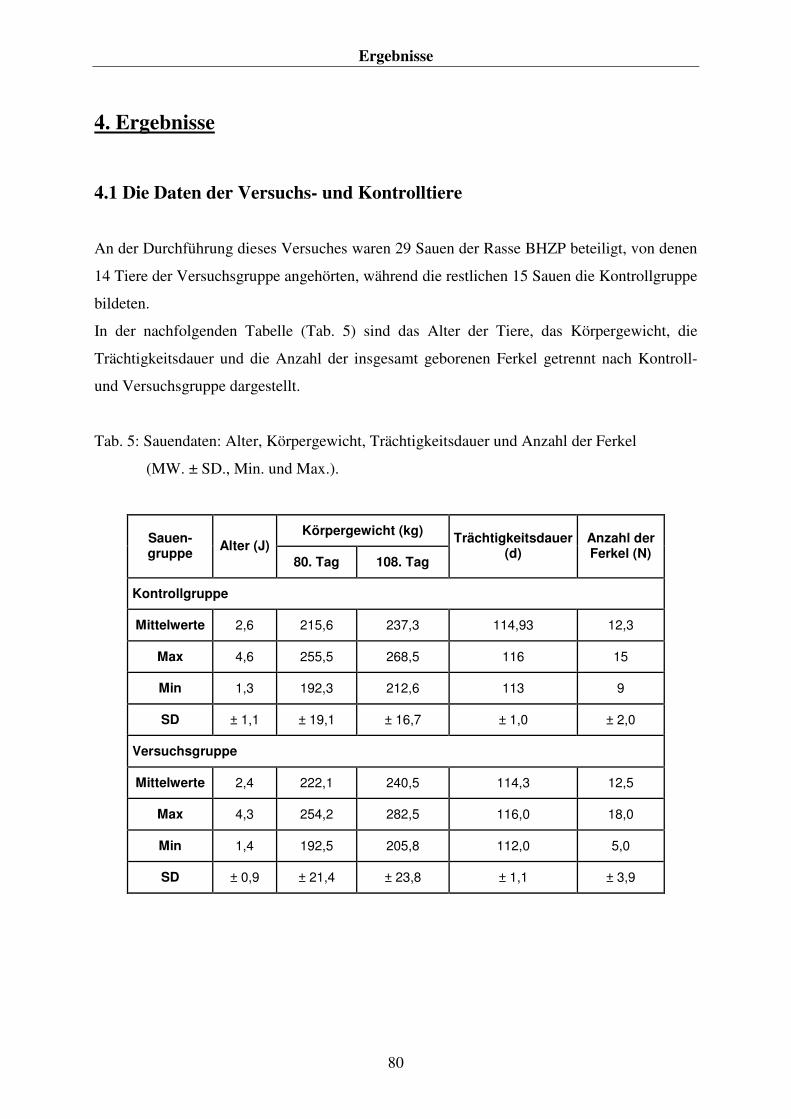
Ergebnisse
80
4. Ergebnisse
4.1 Die Daten der Versuchs- und Kontrolltiere
An der Durchführung dieses Versuches waren 29 Sauen der Rasse BHZP beteiligt, von denen
14 Tiere der Versuchsgruppe angehörten, während die restlichen 15 Sauen die Kontrollgruppe
bildeten.
In der nachfolgenden Tabelle (Tab. 5) sind das Alter der Tiere, das Körpergewicht, die
Trächtigkeitsdauer und die Anzahl der insgesamt geborenen Ferkel getrennt nach Kontroll-
und Versuchsgruppe dargestellt.
Tab. 5: Sauendaten: Alter, Körpergewicht, Trächtigkeitsdauer und Anzahl der Ferkel
(MW. ± SD., Min. und Max.).
Körpergewicht (kg) Sauen- gruppe
Alter (J)
80. Tag 108. Tag
Trächtigkeitsdauer (d)
Anzahl der Ferkel (N)
Kontrollgruppe
Mittelwerte 2,6 215,6 237,3 114,93 12,3
Max 4,6 255,5 268,5 116 15
Min 1,3 192,3 212,6 113 9
SD ± 1,1 ± 19,1 ± 16,7 ± 1,0 ± 2,0
Versuchsgruppe
Mittelwerte 2,4 222,1 240,5 114,3 12,5
Max 4,3 254,2 282,5 116,0 18,0
Min 1,4 192,5 205,8 112,0 5,0
SD ± 0,9 ± 21,4 ± 23,8 ± 1,1 ± 3,9

Ergebnisse
81
Das Alter der Tiere betrug am 80. Trächtigkeitstag zum Zeitpunkt des Versuchsbeginns im
Mittel 2,51 ± 0,95 Jahre, wobei das jüngste Tier gerade 1,3 Jahre und das Älteste 4,6 Jahre alt
war. Die Tiere der Kontrollgruppe waren im Mittel 2,6 ± 1,1 Jahre alt, während die Sauen der
Versuchsgruppe einen Mittelwert von 2,4 ± 0,9 Jahren für ihr Alter aufwiesen.
Gruppenspezifische Unterschiede, die von statistischer Relevanz wären, bestanden somit
nicht zwischen der Kontroll- und Versuchsgruppe (p>0,05).
Am 80. Trächtigkeitstag wogen die Sauen im Mittel 218,7 kg ± 20,2 kg und das Körper-
gewicht stieg bis zum 108. Trächtigkeitstag auf einen Mittelwert von 238,9 kg ± 20,1 kg an.
Der Mittelwert für die Trächtigkeitsdauer betrug 114,6 Tage und variierte zwischen minimal
112 Tagen und maximal 116 Tagen. Zwischen den gruppenspezifischen Werten von 114,9 ±
1,0 Tagen für die Versuchsgruppe und 114,9 ± 1,1 Tagen für die Tiere der Kontrollgruppe
bestehen keinerlei signifikante Unterschiede (p>0,05).
Der Mittelwert für die Anzahl der insgesamt geborenen Ferkel betrug 12,4 Ferkel pro Wurf.
Der entsprechende Wert der Kontrollgruppe lag mit 12,3 Ferkel pro Wurf leicht darunter,
während die Versuchsgruppe mit 12,5 Ferkeln pro Wurf im Mittel diesen Mittelwert leicht
überstieg. Signifikante Unterschiede zwischen den Werten dieser Gruppen bestanden in
diesem Fall aber wiederum nicht (p>0,05).
4.2. Die Futteraufnahme der Sauen
Während des Zeitraumes der Versuchsdauer vom 80. Trächtigkeitstag an bis zum 8. Tag nach
der Abferkelung wurde von allen Tieren, die in diesen Versuch involviert waren, die
angebotene Futtermenge vollständig verzehrt, so dass keinerlei Restfuttermengen aufgetreten
sind. Lediglich in der Intensität der Steigerung der Futtermenge, die den Sauen in den ersten
Tagen nach der Geburt zugeteilt wurde, gab es geringe tierindividuelle Unterschiede, da diese
Menge auf das Futteraufnahmevermögen des jeweiligen Tieres abgestimmt wurde. Somit
konnten für die weitere Auswertung der Analyseergebnisse die verabreichten Futter-
mittelinhaltsstoffe mit den verzehrten Stoffen gleichgesetzt werden.
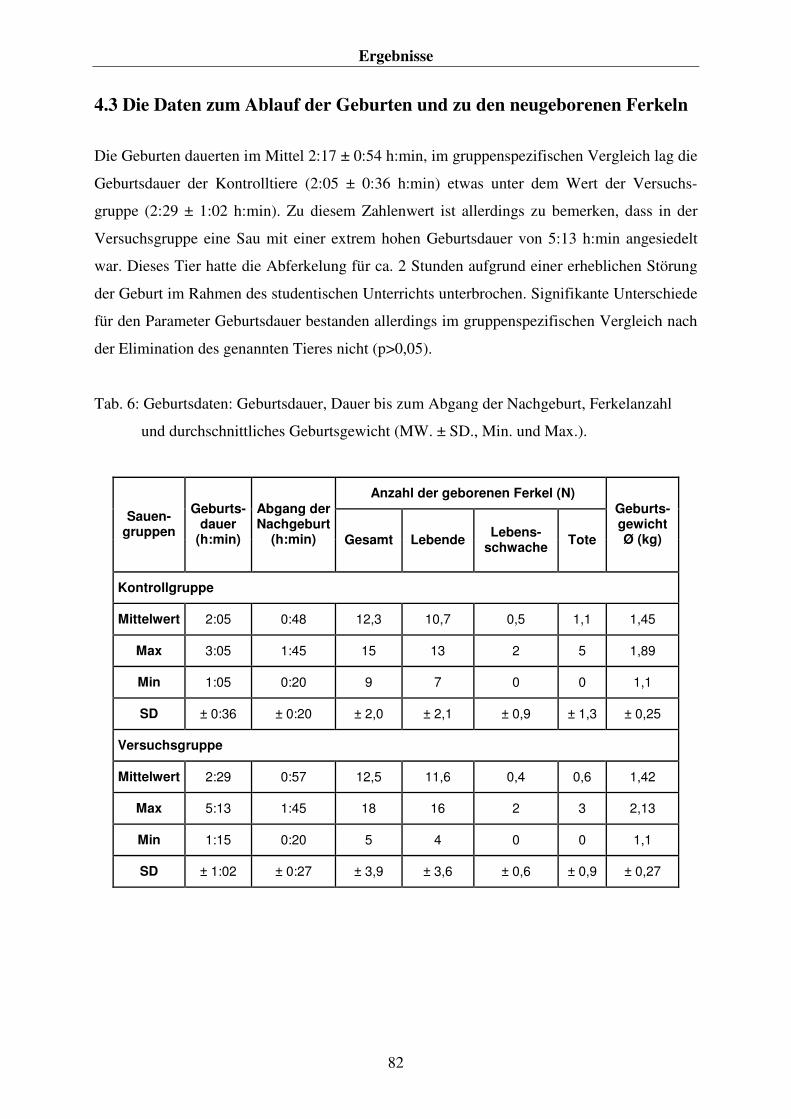
Ergebnisse
82
4.3 Die Daten zum Ablauf der Geburten und zu den neugeborenen Ferkeln
Die Geburten dauerten im Mittel 2:17 ± 0:54 h:min, im gruppenspezifischen Vergleich lag die
Geburtsdauer der Kontrolltiere (2:05 ± 0:36 h:min) etwas unter dem Wert der Versuchs-
gruppe (2:29 ± 1:02 h:min). Zu diesem Zahlenwert ist allerdings zu bemerken, dass in der
Versuchsgruppe eine Sau mit einer extrem hohen Geburtsdauer von 5:13 h:min angesiedelt
war. Dieses Tier hatte die Abferkelung für ca. 2 Stunden aufgrund einer erheblichen Störung
der Geburt im Rahmen des studentischen Unterrichts unterbrochen. Signifikante Unterschiede
für den Parameter Geburtsdauer bestanden allerdings im gruppenspezifischen Vergleich nach
der Elimination des genannten Tieres nicht (p>0,05).
Tab. 6: Geburtsdaten: Geburtsdauer, Dauer bis zum Abgang der Nachgeburt, Ferkelanzahl
und durchschnittliches Geburtsgewicht (MW. ± SD., Min. und Max.).
Anzahl der geborenen Ferkel (N)
Sauen- gruppen
Geburts- dauer
(h:min)
Abgang der Nachgeburt
(h:min) Gesamt Lebende Lebens-
schwache Tote
Geburts-gewicht Ø (kg)
Kontrollgruppe
Mittelwert 2:05 0:48 12,3 10,7 0,5 1,1 1,45
Max 3:05 1:45 15 13 2 5 1,89
Min 1:05 0:20 9 7 0 0 1,1
SD ± 0:36 ± 0:20 ± 2,0 ± 2,1 ± 0,9 ± 1,3 ± 0,25
Versuchsgruppe
Mittelwert 2:29 0:57 12,5 11,6 0,4 0,6 1,42
Max 5:13 1:45 18 16 2 3 2,13
Min 1:15 0:20 5 4 0 0 1,1
SD ± 1:02 ± 0:27 ± 3,9 ± 3,6 ± 0,6 ± 0,9 ± 0,27

Ergebnisse
83
Für die Dauer des Zeitraumes von der Geburt des letzten Ferkels bis zum Abgang der Nach-
geburt bestanden zwischen den Gruppen keine signifikanten Unterschiede (p>0,05). Bei den
Tieren der Kontrollgruppe wurde die Nachgeburt im Mittel nach 48 min ± 20 min tendenziell
etwas schneller ausgestoßen als bei den Tieren der Versuchsgruppe mit einem Mittelwert von
57 min ± 27 min. In beiden Gruppen waren die Einzelwerte jedoch in den gleichen Grenzen
von minimal 20 min bis maximal 1:45 h:min angesiedelt.
Als weitere Parameter für den Geburtsablauf werden die Gesamtzahl der Ferkel sowie die
Anzahl der lebend, lebensschwach (<800 g KGW.) und tot geborenen Ferkeln (inkl. Mumien
und missgebildete Ferkel) pro Wurf dargestellt (Tab. 6).
Im Mittel brachten die Sauen insgesamt 12,4 Ferkel pro Wurf zur Welt, von denen 11,2
Ferkel lebend und 0,8 tot geboren wurden und 0,5 Ferkel wurden aufgrund ihres Geburts-
gewichtes als lebensschwach eingestuft. Aus dem tabellarisch (Tab. 6) dargestellten Gruppen-
vergleich lassen sich keinerlei signifikante Unterschiede (p>0,05) zwischen den beiden
Tiergruppen errechnen. Der größte Wurf in diesem Versuch zählte insgesamt 18 Ferkel,
während beim kleinsten Wurf lediglich 5 Ferkel zur Welt gebracht wurden. Beide Extrema
waren bei den Tieren der Versuchsgruppe zu finden.
Die statistische Auswertung des durchschnittlichen Geburtsgewichtes von 1,44 kg pro Ferkel
zeigte im Gruppenvergleich keine signifikanten Unterschiede (p>0,05) zwischen den Würfen
der Kontrollgruppe mit 1,45 kg pro Ferkel und den Würfen der Sauen der Versuchsgruppe mit
1,42 kg pro Ferkel. Das größte durchschnittliche Wurfgewicht wurde mit 2,13 kg bei einem
Wurf aus der Versuchsgruppe ermittelt, hierbei handelte es sich allerdings auch um den
kleinsten Wurf des Versuches mit lediglich 5 Ferkeln.
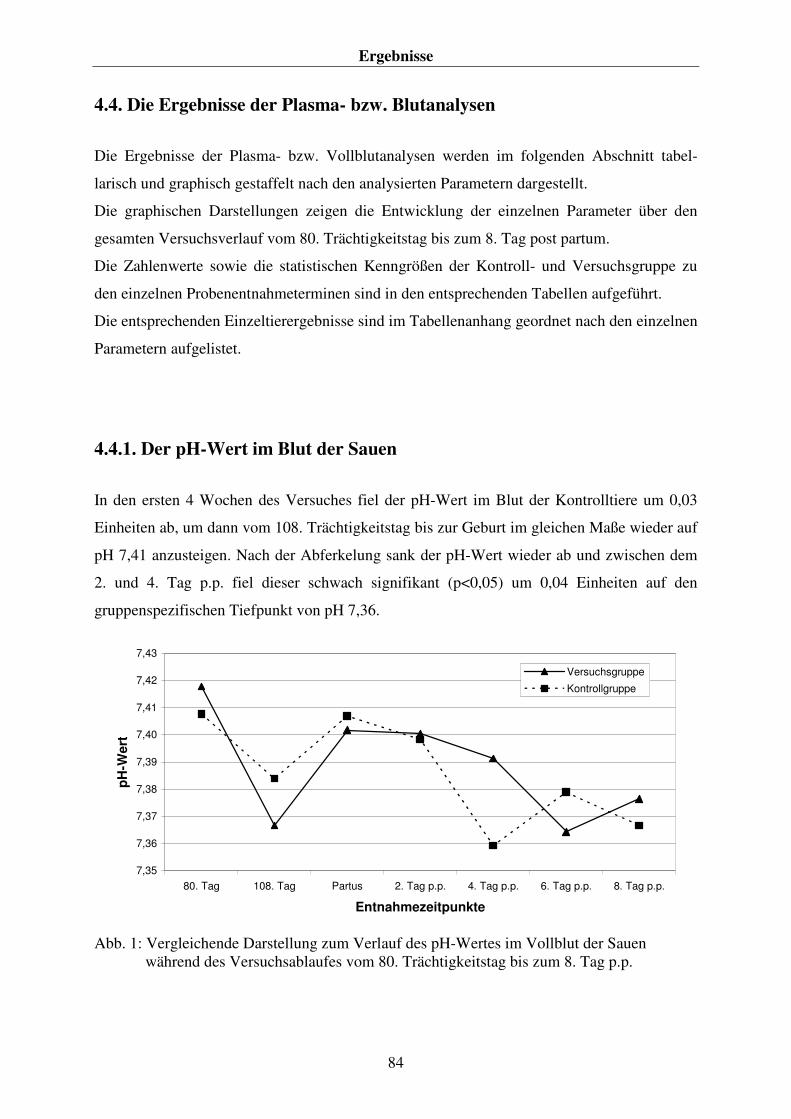
Ergebnisse
84
4.4. Die Ergebnisse der Plasma- bzw. Blutanalysen
Die Ergebnisse der Plasma- bzw. Vollblutanalysen werden im folgenden Abschnitt tabel-
larisch und graphisch gestaffelt nach den analysierten Parametern dargestellt.
Die graphischen Darstellungen zeigen die Entwicklung der einzelnen Parameter über den
gesamten Versuchsverlauf vom 80. Trächtigkeitstag bis zum 8. Tag post partum.
Die Zahlenwerte sowie die statistischen Kenngrößen der Kontroll- und Versuchsgruppe zu
den einzelnen Probenentnahmeterminen sind in den entsprechenden Tabellen aufgeführt.
Die entsprechenden Einzeltierergebnisse sind im Tabellenanhang geordnet nach den einzelnen
Parametern aufgelistet.
4.4.1. Der pH-Wert im Blut der Sauen
In den ersten 4 Wochen des Versuches fiel der pH-Wert im Blut der Kontrolltiere um 0,03
Einheiten ab, um dann vom 108. Trächtigkeitstag bis zur Geburt im gleichen Maße wieder auf
pH 7,41 anzusteigen. Nach der Abferkelung sank der pH-Wert wieder ab und zwischen dem
2. und 4. Tag p.p. fiel dieser schwach signifikant (p<0,05) um 0,04 Einheiten auf den
gruppenspezifischen Tiefpunkt von pH 7,36.
7,35
7,36
7,37
7,38
7,39
7,40
7,41
7,42
7,43
80. Tag 108. Tag Partus 2. Tag p.p. 4. Tag p.p. 6. Tag p.p. 8. Tag p.p.
Entnahmezeitpunkte
pH
-Wert
Versuchsgruppe
Kontrollgruppe
Abb. 1: Vergleichende Darstellung zum Verlauf des pH-Wertes im Vollblut der Sauen während des Versuchsablaufes vom 80. Trächtigkeitstag bis zum 8. Tag p.p.
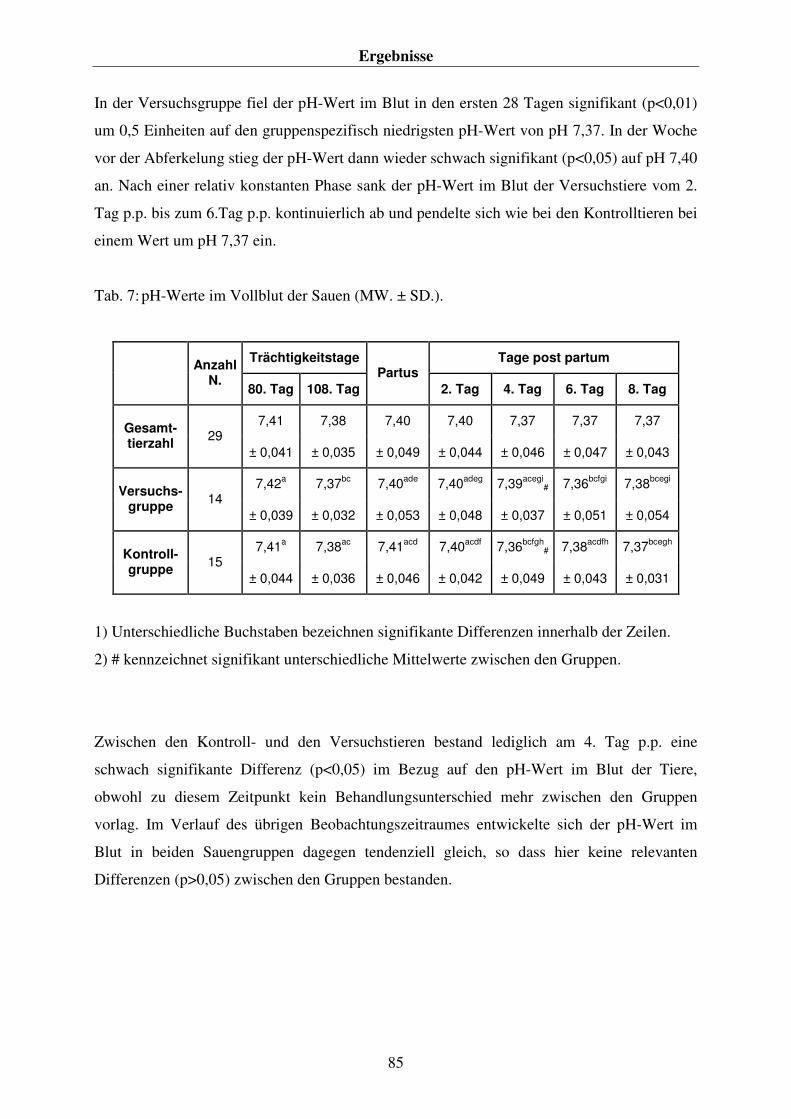
Ergebnisse
85
In der Versuchsgruppe fiel der pH-Wert im Blut in den ersten 28 Tagen signifikant (p<0,01)
um 0,5 Einheiten auf den gruppenspezifisch niedrigsten pH-Wert von pH 7,37. In der Woche
vor der Abferkelung stieg der pH-Wert dann wieder schwach signifikant (p<0,05) auf pH 7,40
an. Nach einer relativ konstanten Phase sank der pH-Wert im Blut der Versuchstiere vom 2.
Tag p.p. bis zum 6.Tag p.p. kontinuierlich ab und pendelte sich wie bei den Kontrolltieren bei
einem Wert um pH 7,37 ein.
Tab. 7: pH-Werte im Vollblut der Sauen (MW. ± SD.).
Trächtigkeitstage Tage post partum
Anzahl
N. 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
7,41 7,38 7,40 7,40 7,37 7,37 7,37 Gesamt-tierzahl
29
± 0,041 ± 0,035 ± 0,049 ± 0,044 ± 0,046 ± 0,047 ± 0,043
7,42a
7,37bc
7,40ade
7,40adeg
7,39acegi
# 7,36bcfgi
7,38bcegi
Versuchs-gruppe
14
± 0,039 ± 0,032 ± 0,053 ± 0,048 ± 0,037 ± 0,051 ± 0,054
7,41a
7,38ac
7,41acd
7,40acdf
7,36bcfgh
# 7,38acdfh
7,37bcegh
Kontroll-gruppe
15
± 0,044 ± 0,036 ± 0,046 ± 0,042 ± 0,049 ± 0,043 ± 0,031
1) Unterschiedliche Buchstaben bezeichnen signifikante Differenzen innerhalb der Zeilen.
2) # kennzeichnet signifikant unterschiedliche Mittelwerte zwischen den Gruppen.
Zwischen den Kontroll- und den Versuchstieren bestand lediglich am 4. Tag p.p. eine
schwach signifikante Differenz (p<0,05) im Bezug auf den pH-Wert im Blut der Tiere,
obwohl zu diesem Zeitpunkt kein Behandlungsunterschied mehr zwischen den Gruppen
vorlag. Im Verlauf des übrigen Beobachtungszeitraumes entwickelte sich der pH-Wert im
Blut in beiden Sauengruppen dagegen tendenziell gleich, so dass hier keine relevanten
Differenzen (p>0,05) zwischen den Gruppen bestanden.

Ergebnisse
86
4.4.2. Die Konzentration an Hydrogenkarbonat (HCO3-) im Blut der Sauen
Die Zahl der auszuwertenden Tiere war für diesen Parameter auf 25 Sauen begrenzt, von
denen 13 Tiere der Kontrollgruppe angehörten, während die übrigen 12 Sauen die
Versuchsgruppe bildeten. Für 4 Sauen (je 2 Tiere pro Gruppe) konnte aufgrund eines Fehlers
innerhalb des Blutgasanalysegerätes nicht zu jedem der Probeentnahmetermine ein Wert für
die Hydrogenkarbonatkonzentration im Blut ermittelt werden.
25,00
25,50
26,00
26,50
27,00
27,50
28,00
28,50
29,00
29,50
30,00
80. Tag 108. Tag Partus 2. Tag p.p. 4. Tag p.p. 6. Tag p.p. 8. Tag p.p.
Entnahmezeitpunkte
Hyd
rog
en
karb
on
at
(mm
ol/l)
Versuchsgruppe
Kontrollgruppe
Abb. 2: Vergleichende Darstellung des Verlaufes der Hydrogenkarbonatkonzentration
(HCO3-) im Vollblut der Sauen über den Zeitraum des Versuchsverlaufes.
Die Hydrogenkarbonatkonzentration (HCO3-) im Vollblut der Kontrolltiere stieg in den ersten
4 Wochen des Versuchs schwach signifikant (p<0,05) um 1,63 mmol/l an. Bis zum
Geburtszeitpunkt folgte dann ein leichter Konzentrationsabfall auf 27,78 mmol/l, der
allerdings über keine statistische Relevanz verfügte (p>0,05). In den ersten 8 Tagen der
Säugephase pendelten sich diese Werte zwischen 28,03 mmol/l und 28,99 mmol/l ein.
In der Versuchsgruppe erfolgte in den ersten 28 Tagen der Beobachtungszeit lediglich ein
tendenzieller Anstieg des Hydrogencarbonatgehaltes um 0,48 mmol/l, bevor die
Konzentration in der folgenden Woche bis zur Abferkelung hochsignifikant (p<0,001) von
27,9 mmol/l auf 25,62 mmol/l abfiel. In den ersten 2 Tagen nach der Abferkelung stieg dieser
Wert dann wieder hochsignifikant (p< 0,001) auf 29,28 mmol/l.

Ergebnisse
87
Am 4. Tag post partum wurde in dieser Tiergruppe schließlich die höchste mittlere
Konzentration an HCO3- im Blut von 29,85 mmol/l erreicht. Bis zum 6. Tag post partum sank
die Konzentration zunächst wieder schwach signifikant (p<0,05) um 1,62 mmol/l ab, um sich
dann am Ende der Beobachtungszeit bei 29,43 mmol/l einzupendeln.
Tab. 8: Hydrogenkarbonatkonzentration (HCO3-) im Vollblut der Sauen (mmol/l),
(MW. ± SD.).
Trächtigkeitstage Tage post partum
Anzahl
N. 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
27,02 28,10 26,74 29,06 29,40 28,13 28,93 Gesamt-tierzahl
25
± 2,46 ± 1,43 ± 1,98 ± 1,62 ± 1,79 ± 1,72 ± 1,95
27,42a
27,90ac
25,62bde
# 29,27bdfg
29,85bdfgh
28,23acfgij
29,43bdfghj
Versuchs-gruppe
12
± 1,93 ± 1,12 ± 1,75 ± 1,58 ± 1,97 ± 1,41 ± 1,39
26,65a
28,28bc
27,78ac
# 28,85bc
28,99bc
28,03bc
28,46bc
Kontroll-gruppe
13
± 2,89 ± 1,68 ± 1,61 ± 1,70 ± 1,57 ± 2,01 ± 2,30
1) Unterschiedliche Buchstaben bezeichnen signifikante Differenzen innerhalb der Zeilen.
2) # kennzeichnet signifikant unterschiedliche Mittelwerte zwischen den Gruppen.
Zum Zeitpunkt der Abferkelung war der Wert für die Hydrogenkarbonatkonzentration von
25,62 ± 1,75 mmol/l der Versuchsgruppe signifikant (p<0,01) unter dem Wert der Kontroll-
gruppe von 27,78 ± 1,61 mmol/l angesiedelt. An den anderen Analyseterminen während des
Versuches bestanden allerdings in diesem Zusammenhang keine relevanten Unterschiede
zwischen den Sauen der beiden Gruppen.
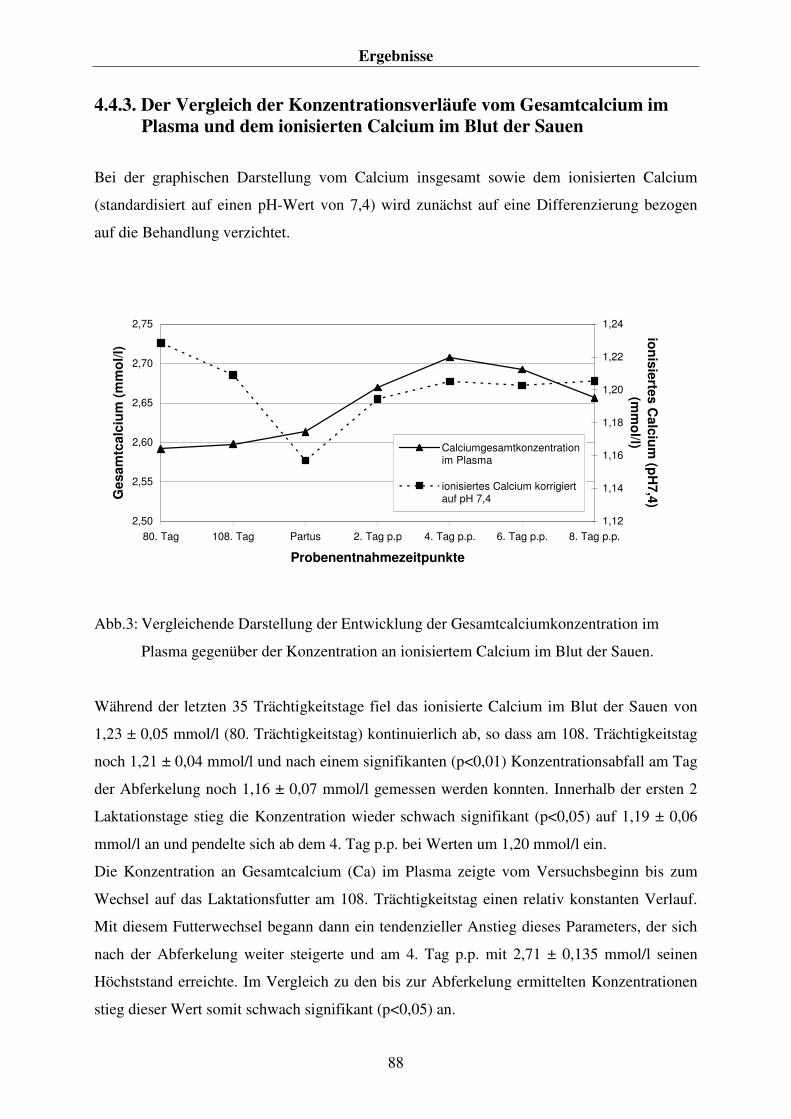
Ergebnisse
88
4.4.3. Der Vergleich der Konzentrationsverläufe vom Gesamtcalcium im Plasma und dem ionisierten Calcium im Blut der Sauen
Bei der graphischen Darstellung vom Calcium insgesamt sowie dem ionisierten Calcium
(standardisiert auf einen pH-Wert von 7,4) wird zunächst auf eine Differenzierung bezogen
auf die Behandlung verzichtet.
2,50
2,55
2,60
2,65
2,70
2,75
80. Tag 108. Tag Partus 2. Tag p.p 4. Tag p.p. 6. Tag p.p. 8. Tag p.p.
Probenentnahmezeitpunkte
Gesam
tcalc
ium
(m
mo
l/l)
1,12
1,14
1,16
1,18
1,20
1,22
1,24
ion
isie
rtes C
alc
ium
(pH
7,4
)
(mm
ol/l)
Calciumgesamtkonzentrationim Plasma
ionisiertes Calcium korrigiertauf pH 7,4
Abb.3: Vergleichende Darstellung der Entwicklung der Gesamtcalciumkonzentration im
Plasma gegenüber der Konzentration an ionisiertem Calcium im Blut der Sauen.
Während der letzten 35 Trächtigkeitstage fiel das ionisierte Calcium im Blut der Sauen von
1,23 ± 0,05 mmol/l (80. Trächtigkeitstag) kontinuierlich ab, so dass am 108. Trächtigkeitstag
noch 1,21 ± 0,04 mmol/l und nach einem signifikanten (p<0,01) Konzentrationsabfall am Tag
der Abferkelung noch 1,16 ± 0,07 mmol/l gemessen werden konnten. Innerhalb der ersten 2
Laktationstage stieg die Konzentration wieder schwach signifikant (p<0,05) auf 1,19 ± 0,06
mmol/l an und pendelte sich ab dem 4. Tag p.p. bei Werten um 1,20 mmol/l ein.
Die Konzentration an Gesamtcalcium (Ca) im Plasma zeigte vom Versuchsbeginn bis zum
Wechsel auf das Laktationsfutter am 108. Trächtigkeitstag einen relativ konstanten Verlauf.
Mit diesem Futterwechsel begann dann ein tendenzieller Anstieg dieses Parameters, der sich
nach der Abferkelung weiter steigerte und am 4. Tag p.p. mit 2,71 ± 0,135 mmol/l seinen
Höchststand erreichte. Im Vergleich zu den bis zur Abferkelung ermittelten Konzentrationen
stieg dieser Wert somit schwach signifikant (p<0,05) an.
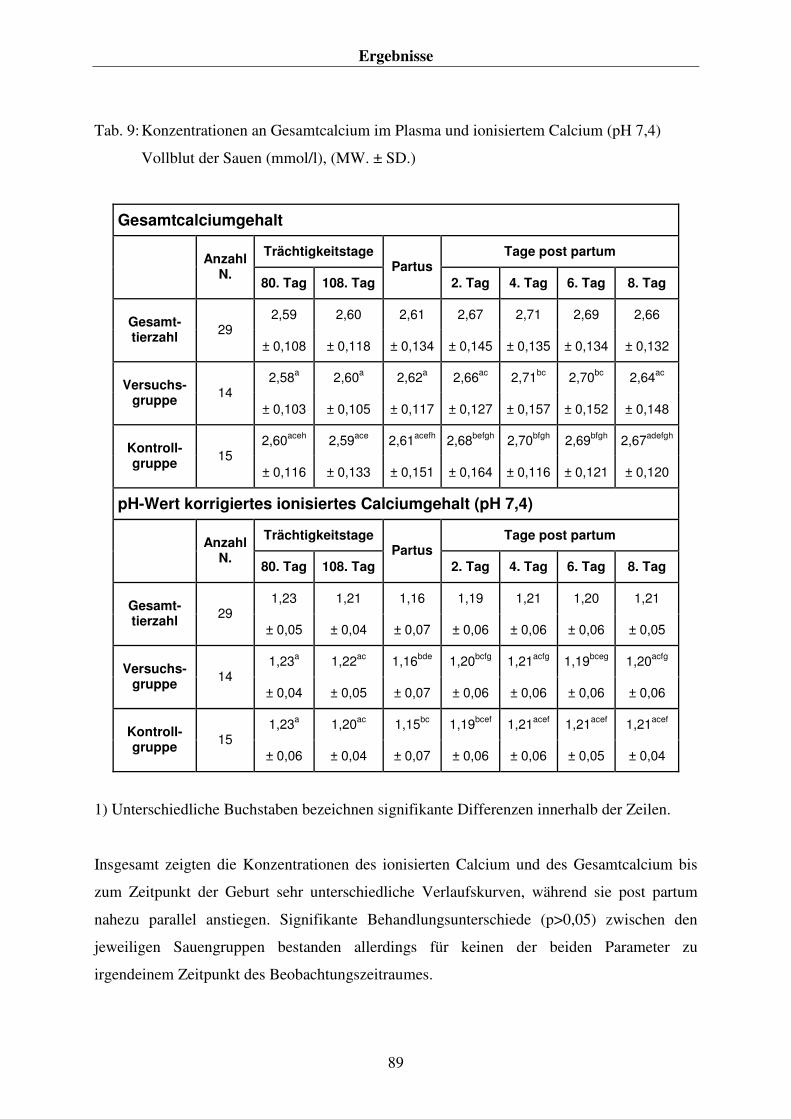
Ergebnisse
89
Tab. 9: Konzentrationen an Gesamtcalcium im Plasma und ionisiertem Calcium (pH 7,4)
Vollblut der Sauen (mmol/l), (MW. ± SD.)
Gesamtcalciumgehalt
Trächtigkeitstage Tage post partum
Anzahl
N. 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
2,59 2,60 2,61 2,67 2,71 2,69 2,66 Gesamt-tierzahl
29
± 0,108 ± 0,118 ± 0,134 ± 0,145 ± 0,135 ± 0,134 ± 0,132
2,58a
2,60a
2,62a
2,66ac
2,71bc
2,70bc
2,64ac
Versuchs-gruppe
14
± 0,103 ± 0,105 ± 0,117 ± 0,127 ± 0,157 ± 0,152 ± 0,148
2,60aceh
2,59ace
2,61acefh
2,68befgh
2,70bfgh
2,69bfgh
2,67adefgh
Kontroll-gruppe
15
± 0,116 ± 0,133 ± 0,151 ± 0,164 ± 0,116 ± 0,121 ± 0,120
pH-Wert korrigiertes ionisiertes Calciumgehalt (pH 7,4)
Trächtigkeitstage Tage post partum
Anzahl
N. 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
1,23 1,21 1,16 1,19 1,21 1,20 1,21 Gesamt-tierzahl
29
± 0,05 ± 0,04 ± 0,07 ± 0,06 ± 0,06 ± 0,06 ± 0,05
1,23a
1,22ac
1,16bde
1,20bcfg
1,21acfg
1,19bceg
1,20acfg
Versuchs-gruppe
14
± 0,04 ± 0,05 ± 0,07 ± 0,06 ± 0,06 ± 0,06 ± 0,06
1,23a
1,20ac
1,15bc
1,19bcef
1,21acef
1,21acef
1,21acef
Kontroll-gruppe
15
± 0,06 ± 0,04 ± 0,07 ± 0,06 ± 0,06 ± 0,05 ± 0,04
1) Unterschiedliche Buchstaben bezeichnen signifikante Differenzen innerhalb der Zeilen.
Insgesamt zeigten die Konzentrationen des ionisierten Calcium und des Gesamtcalcium bis
zum Zeitpunkt der Geburt sehr unterschiedliche Verlaufskurven, während sie post partum
nahezu parallel anstiegen. Signifikante Behandlungsunterschiede (p>0,05) zwischen den
jeweiligen Sauengruppen bestanden allerdings für keinen der beiden Parameter zu
irgendeinem Zeitpunkt des Beobachtungszeitraumes.

Ergebnisse
90
4.4.4. Der Phosphorgehalt im Plasma der Sauen
In den ersten 4 Wochen des Versuches erhöhte sich die Phosphorkonzentration im Plasma
aller Sauen leicht. In der Woche vor der Abferkelung stiegen diese Werte dann in beiden
Sauengruppen (Kontrollgruppe um 0,37 mmol/l und Versuchsgruppe um 0,38 mmol/l) hoch-
signifikant (p<0,001) an.
2,00
2,10
2,20
2,30
2,40
2,50
2,60
2,70
80. Tag 108. Tag Partus 2. Tag p.p. 4. Tag p.p. 6. Tag p.p. 8. Tag p.p.
Entnahmezeitpunkte
Ph
osp
ho
rko
nzen
trati
on
(m
mo
l/l)
Versuchsgruppe
Kontrollgruppe
Abb. 4: Vergleichende Darstellung über den Verlauf der Phosphorkonzentration im Plasma der Sauen für den Zeitraum des Versuches. (MW ± SD. siehe Tab. 10).
Am Tag der Abferkelung wurde sowohl bei den Kontrolltieren mit 2,61 mmol/l wie auch in
der Versuchsgruppe mit 2,54 mmol/l die höchste Phosphorkonzentration im Verlauf des
Versuches erreicht. In der postpartalen Phase sank diese Konzentration in beiden Tiergruppen
bis zum 6.Tag p.p. kontinuierlich ab. In den ersten 2 Tagen nach der Geburt fiel die
Phosphorkonzentration bei allen Sauen signifikant (p<0,01) um 0,28 mmol/l. Bei den
Kontrolltieren folgte dann in den nächsten 2 Tagen ein weiteres schwach relevantes Absinken
(p<0,05) dieser Werte, während in der Versuchsgruppe die weitere Konzentrations-
verringerung ihre Signifikanz verlor. Vom 6. Tag p.p. bis zum Ende des Versuches stieg die
Phosphorkonzentration in beiden Tiergruppen wieder schwach signifikant (p<0,05) auf im
Mittel 2,34 ± 0,28 mmol/l an.

Ergebnisse
91
Tab. 10: Phosphorgehalt im Plasma der Sauen (mmol/l), (MW. ± SD.).
Trächtigkeitstage Tage post partum
Anzahl
N. 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
2,09 2,20 2,57 2,30 2,14 2,12 2,34 Gesamt-tierzahl
29
± 0,29 ± 0,30 ± 0,35 ± 0,44 ± 0,36 ± 0,35 ± 0,28
2,03a
2,16ac
2,54bde
2,26bcfg
2,15acfgh
2,13acfgh
2,37bdegi
Versuchs-gruppe
14
± 0,28 ± 0,28 ± 0,34 ± 0,39 ± 0,39 ± 0,29 ± 0,24
2,15a
2,24ac
2,61bd
2,33ace
2,12acfg
2,10acfgh
2,30acegi
Kontroll-gruppe
15
± 0,30 ± 0,33 ± 0,36 ± 0,50 ± 0,33 ± 0,41 ± 0,32
1) Unterschiedliche Buchstaben bezeichnen signifikante Differenzen innerhalb der Zeilen.
Die Ergebnisse der Phosphoranalyse blieben von den Behandlungsunterschieden der beiden
Gruppen unbeeinflusst, so dass zu keinem Zeitpunkt des Versuches signifikante Unterschiede
zwischen den Werten der Kontrollgruppe und denen der Versuchsgruppe bestanden (p>0,05).
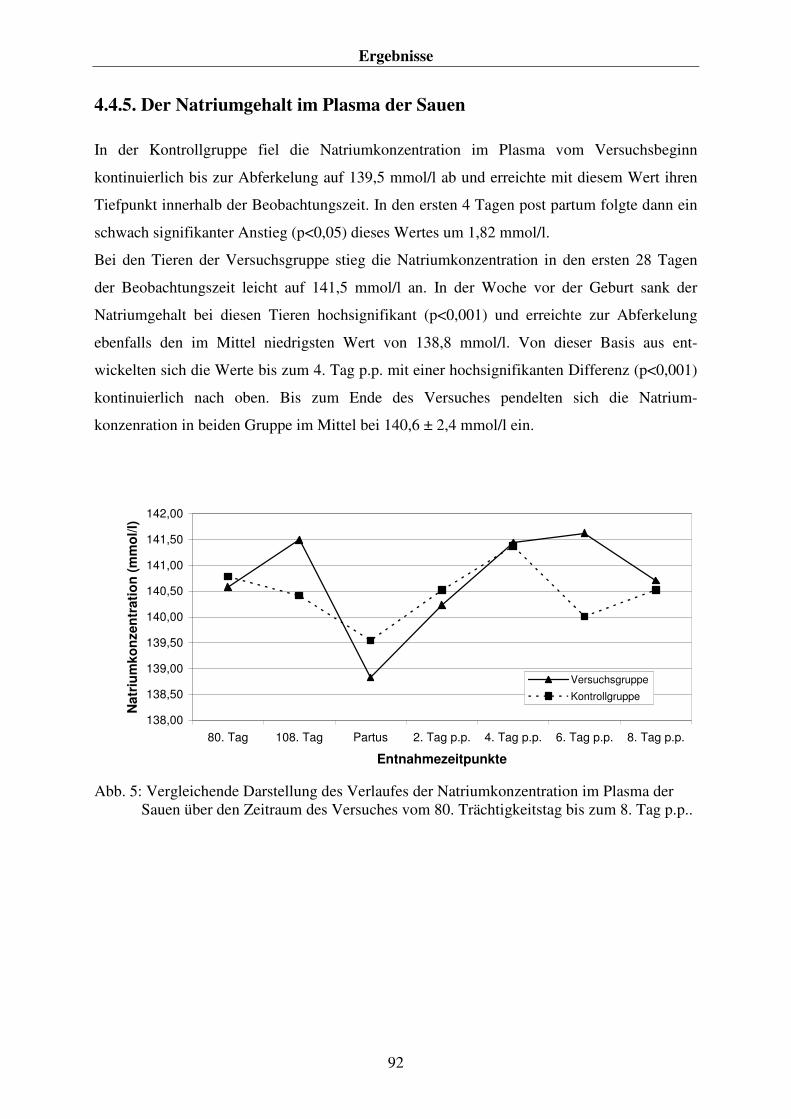
Ergebnisse
92
4.4.5. Der Natriumgehalt im Plasma der Sauen
In der Kontrollgruppe fiel die Natriumkonzentration im Plasma vom Versuchsbeginn
kontinuierlich bis zur Abferkelung auf 139,5 mmol/l ab und erreichte mit diesem Wert ihren
Tiefpunkt innerhalb der Beobachtungszeit. In den ersten 4 Tagen post partum folgte dann ein
schwach signifikanter Anstieg (p<0,05) dieses Wertes um 1,82 mmol/l.
Bei den Tieren der Versuchsgruppe stieg die Natriumkonzentration in den ersten 28 Tagen
der Beobachtungszeit leicht auf 141,5 mmol/l an. In der Woche vor der Geburt sank der
Natriumgehalt bei diesen Tieren hochsignifikant (p<0,001) und erreichte zur Abferkelung
ebenfalls den im Mittel niedrigsten Wert von 138,8 mmol/l. Von dieser Basis aus ent-
wickelten sich die Werte bis zum 4. Tag p.p. mit einer hochsignifikanten Differenz (p<0,001)
kontinuierlich nach oben. Bis zum Ende des Versuches pendelten sich die Natrium-
konzenration in beiden Gruppe im Mittel bei 140,6 ± 2,4 mmol/l ein.
138,00
138,50
139,00
139,50
140,00
140,50
141,00
141,50
142,00
80. Tag 108. Tag Partus 2. Tag p.p. 4. Tag p.p. 6. Tag p.p. 8. Tag p.p.
Entnahmezeitpunkte
Natr
ium
ko
nzen
trati
on
(m
mo
l/l)
Versuchsgruppe
Kontrollgruppe
Abb. 5: Vergleichende Darstellung des Verlaufes der Natriumkonzentration im Plasma der
Sauen über den Zeitraum des Versuches vom 80. Trächtigkeitstag bis zum 8. Tag p.p..

Ergebnisse
93
Tab. 11: Natriumgehalt im Plasma der Sauen (mmol/l), (MW. ± SD.).
Trächtigkeitstag Tage post partum
Anzahl
N 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
140,7 140,9 139,2 140,4 141,4 140,8 140,6 Gesamt-tierzahl
29
± 1,33 ± 2,25 ± 2,43 ± 2,68 ± 2,03 ± 2,38 ± 2,40
140,6a
141,5ac
138,8bde
140,2ace
141,4acf
141,6acf
# 140,7acf
Versuchs-gruppe
14
± 1,34 ± 1,34 ± 1,81 ± 2,25 ± 0,85 ± 1,06 ± 2,06
140,8a
140,4a
139,6ab
140,5ab
141,4ac
140,0ab
# 140,5ab
Kontroll-gruppe
15
± 1,37 ± 2,81 ± 2,91 ± 3,10 ± 2,75 ± 2,99 ± 2,75
1) Unterschiedliche Buchstaben bezeichnen signifikante Differenzen innerhalb der Zeilen.
2) # kennzeichnet signifikante unterschiedliche Mittelwerte zwischen den Gruppen.
Zwischen den Sauen der Kontrollgruppe und denen der Versuchsgruppe bestand am 6. Tag
post partum, also 6 Tage nach dem Ende der differenzierten Fütterung, ein schwach
signifikanter Unterschied (p<0,05) für die Natriumkonzentration im Plasma. In der übrigen
Zeit des Versuches bestanden dagegen keine statistisch relevante Unterschiede zwischen den
Gruppen und die Natriumkonzentration entwickelte sich tendenziell bei allen Sauen gleich.

Ergebnisse
94
4.4.6. Der Chloridgehalt im Plasma der Sauen
Die Chloridkonzentration war zum Beginn des Versuches in beiden Tiergruppen um 108,5
mmol/l angesiedelt und fiel bei den Tieren der Kontrollgruppe tendenziell bis zum 108.
Trächtigkeitstag auf 107,7 mmol/l. Bis zum Tag der Abferkelung fielen diese Werte in der
Kontrollgruppe dann signifikant (p<0,01) im Mittel um 2,3 mmol/l und erreichten zu diesem
Zeitpunkt ihren niedrigsten Stand. Im postpartalen Beobachtungszeitraum bis zum 8. Tag p.p.
erfolgten dann keine relevanten Konzentrationsveränderungen mehr und es wurden Werte von
106,5 mmol/l bis 106,9 mmol/l im Plasma der Kontrolltiere gemessen.
105,00
105,50
106,00
106,50
107,00
107,50
108,00
108,50
109,00
109,50
110,00
80. Tag 108. Tag Partus 2. Tag p.p. 4. Tag p.p. 6. Tag p.p. 8. Tag p.p.
Entnahmezeitpunkte
Ch
lori
dko
nzen
trati
on
(m
mo
l/l)
Versuchsgruppe
Kontrollgruppe
Abb. 6: Vergleichende Darstellung des Verlaufes der Chloridkonzentration (Cl-) im Plasma der Sauen während des Versuches vom 80. Trächtigkeitstag bis zum 8. Tag p.p..
In der Versuchsgruppe stieg die Chloridkonzentration im Plasma in den ersten 4 Wochen der
Versuches um 0,7 mmol/l an, um sich in der verbleibenden Woche bis zur Abferkelung
hochsignifikant (p<0,001) um 3,4 mmol/l zu verringern und mit 106,0 mmol/l die geringste
mittlere Konzentration im Versuchsverlauf zu erreichen. In den ersten 4 Tagen der Säuge-
phase war dann ein leichter tendenzieller Anstieg der Chloridkonzentration zu verzeichnen,
die sich im weiteren Verlauf zwischen 106,5 bis 107,0 mmol/l einpendelte.

Ergebnisse
95
Tab. 12: Chloridkonzentration im Plasma der Sauen (mmol/l), (MW. ± SD.)
Trächtigkeitstage Tage post partum
Anzahl
N. 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
108,6 108,6 105,7 106,3 106,8 107,0 106,5 Gesamt-tierzahl
29
± 2,08 ± 2,15 ± 2,58 ± 2,82 ± 2,96 ± 2,35 ± 2,47
108,7a
109,4a# 106,0
b 106,1
b 107,0
b 107,0
b 106,5
b
Versuchs-gruppe
14
± 2,23 ± 1,16 ± 2,32 ± 2,28 ± 2,35 ± 1,36 ± 2,18
108,4a
107,7ac
# 105,4bde
106,5bce
106,5bce
106,9ace
106,5bce
Kontroll-gruppe
15
± 1,99 ± 2,55 ± 2,85 ± 3,31 ± 3,50 ± 3,06 ± 2,80
1) Unterschiedliche Buchstaben bezeichnen signifikante Differenzen innerhalb der Zeilen.
2) # kennzeichnet signifikante unterschiedliche Mittelwerte zwischen den Gruppen.
Zwischen der Kontrollgruppe und den Tieren der Versuchsgruppe bestand lediglich am 108.
Trächtigkeitstag ein schwach signifikanter Unterschied (p<0,05), da die Versuchstiere zu
diesem Zeitpunkt eine um 1,7 mmol/l höhere Konzentration an Chlorid im Plasma vorwiesen.
Über den gesamten Versuchsverlauf zeigten beide Gruppen ansonsten jedoch eine tendenziell
gleiche Konzentrationsentwicklung des Chloridgehaltes im Plasma, so dass an den anderen
Analysezeitpunkten keine relevanten Differenzen (p>0,05) zwischen den Gruppen bestanden.
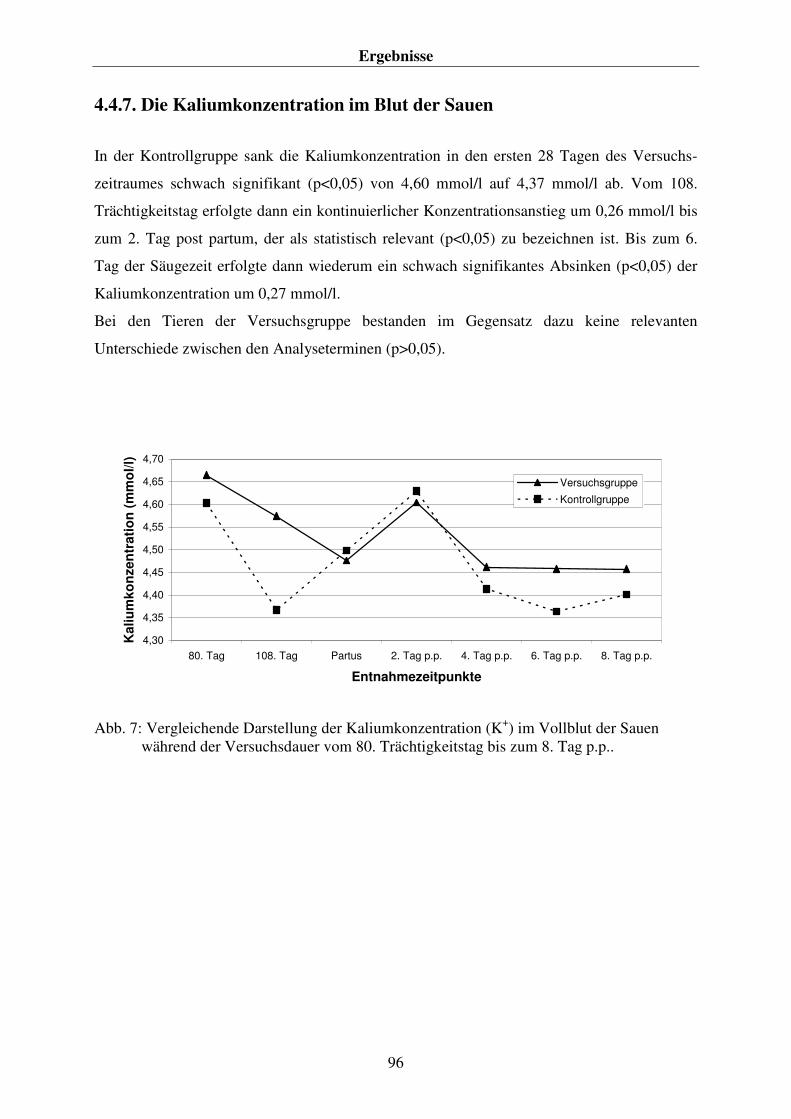
Ergebnisse
96
4.4.7. Die Kaliumkonzentration im Blut der Sauen
In der Kontrollgruppe sank die Kaliumkonzentration in den ersten 28 Tagen des Versuchs-
zeitraumes schwach signifikant (p<0,05) von 4,60 mmol/l auf 4,37 mmol/l ab. Vom 108.
Trächtigkeitstag erfolgte dann ein kontinuierlicher Konzentrationsanstieg um 0,26 mmol/l bis
zum 2. Tag post partum, der als statistisch relevant (p<0,05) zu bezeichnen ist. Bis zum 6.
Tag der Säugezeit erfolgte dann wiederum ein schwach signifikantes Absinken (p<0,05) der
Kaliumkonzentration um 0,27 mmol/l.
Bei den Tieren der Versuchsgruppe bestanden im Gegensatz dazu keine relevanten
Unterschiede zwischen den Analyseterminen (p>0,05).
4,30
4,35
4,40
4,45
4,50
4,55
4,60
4,65
4,70
80. Tag 108. Tag Partus 2. Tag p.p. 4. Tag p.p. 6. Tag p.p. 8. Tag p.p.
Entnahmezeitpunkte
Kaliu
mko
nzen
trati
on
(m
mo
l/l)
Versuchsgruppe
Kontrollgruppe
Abb. 7: Vergleichende Darstellung der Kaliumkonzentration (K+) im Vollblut der Sauen
während der Versuchsdauer vom 80. Trächtigkeitstag bis zum 8. Tag p.p..
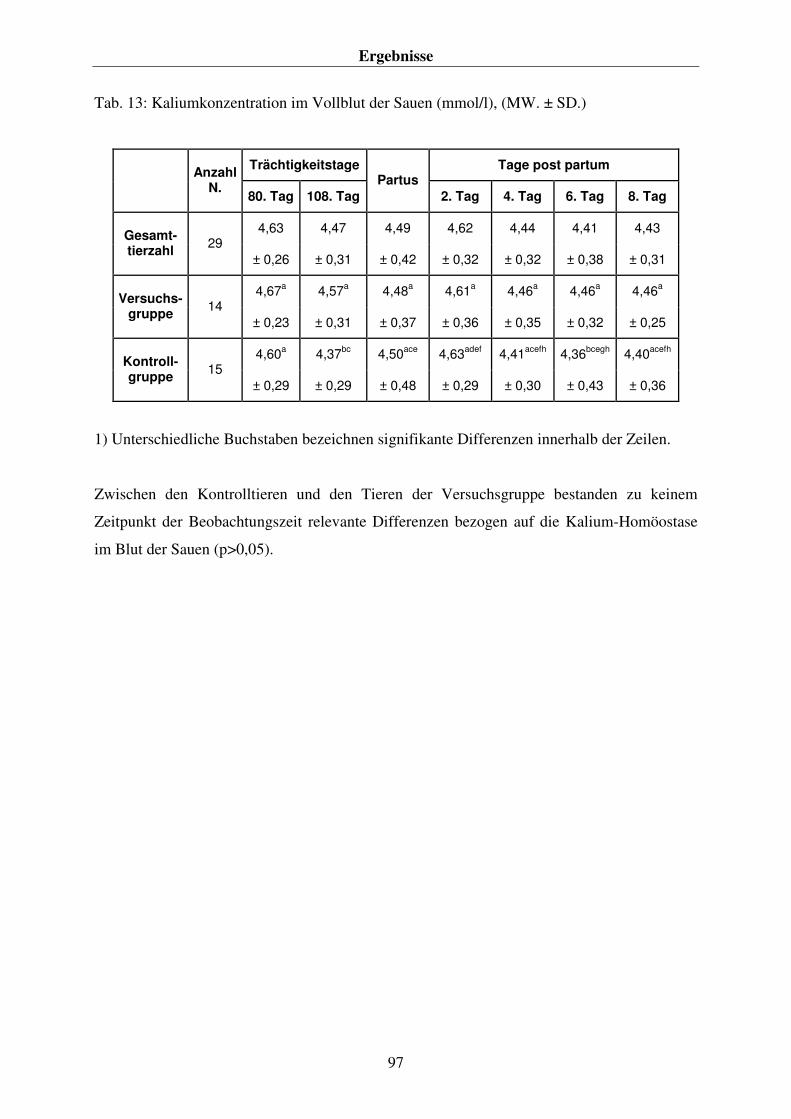
Ergebnisse
97
Tab. 13: Kaliumkonzentration im Vollblut der Sauen (mmol/l), (MW. ± SD.)
Trächtigkeitstage Tage post partum
Anzahl
N. 80. Tag 108. Tag
Partus
2. Tag 4. Tag 6. Tag 8. Tag
4,63 4,47 4,49 4,62 4,44 4,41 4,43 Gesamt-tierzahl
29
± 0,26 ± 0,31 ± 0,42 ± 0,32 ± 0,32 ± 0,38 ± 0,31
4,67a
4,57a
4,48a
4,61a
4,46a
4,46a
4,46a
Versuchs-gruppe
14
± 0,23 ± 0,31 ± 0,37 ± 0,36 ± 0,35 ± 0,32 ± 0,25
4,60a
4,37bc
4,50ace
4,63adef
4,41acefh
4,36bcegh
4,40acefh
Kontroll-gruppe
15
± 0,29 ± 0,29 ± 0,48 ± 0,29 ± 0,30 ± 0,43 ± 0,36
1) Unterschiedliche Buchstaben bezeichnen signifikante Differenzen innerhalb der Zeilen.
Zwischen den Kontrolltieren und den Tieren der Versuchsgruppe bestanden zu keinem
Zeitpunkt der Beobachtungszeit relevante Differenzen bezogen auf die Kalium-Homöostase
im Blut der Sauen (p>0,05).

Ergebnisse
98
4.4.8. Kreatininkonzentrationen im Plasma der Sauen .
Am 90. Trächtigkeitstag liegt der Mittelwert für die Kreatininkonzentration im Plasma der
Kontrollgruppe bei 213,9 ± 44,2 µmol/l, wobei die gemessenen Einzelwerte von minimal
154,7 µmol/l bis maximal 302,8 µmol/l angesiedelt sind. In der Versuchsgruppe verteilen sich
diese Werte von minimal 152,1 µmol/l bis maximal 277,6 µmol/l, so dass ein Mittelwert von
202,4 ± 38,0 µmol/l vorliegt.
Tab. 14: Kreatininkonzentrationen im Plasma der Sauen (µmol/l) (MW±SD)
Anzahl N.
90. Trächtig-keitstag
111. Trächtig-keitstag
2,36 2,34 Gesamt-tierzahl
29 ± 0,47 ± 0,47
2,29 2,31 Versuchs-gruppe
14 ± 0,43 ± 0,39
2,42 2,37 Kontroll- gruppe
15 ± 0,50 ± 0,54
Am 111. Trächtigkeitstag variieren diese Werte für die Kreatininkonzentration im Plasma bei
den Kontrolltierenen zwischen 131,7 µmol/l und 308,5 µmol/l und der Mittelwert beträgt zu
diesem Zeitpunkt 209,5 ± 47,7 µmol/l. Am 111. Trächtigkeitstag liegt in der Versuchsgruppe
ein Mittelwert von 204,2 ± 34,5 µmol/l für diesen Parameter vor, wobei sich die Einzelwerte
von minimal 145,0 µmol/l bis maximal 275,8 µmol/l erstrecken. Signifikante Differenzen
bezogen auf die Kreatininkonzentration im Plasma bestehen somit weder innerhalb der
Gruppen zwischen den beiden Analyseterminen noch zwischen den Tiergruppen zu den
entsprechenden Terminen.
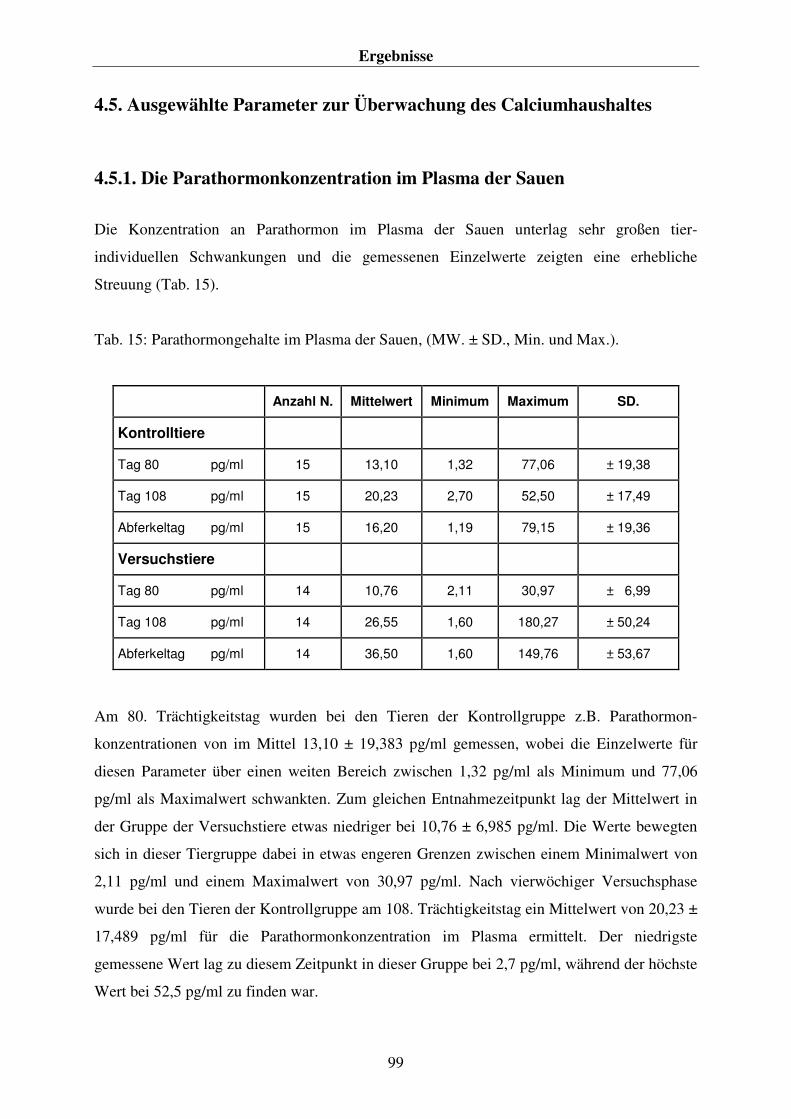
Ergebnisse
99
4.5. Ausgewählte Parameter zur Überwachung des Calciumhaushaltes
4.5.1. Die Parathormonkonzentration im Plasma der Sauen
Die Konzentration an Parathormon im Plasma der Sauen unterlag sehr großen tier-
individuellen Schwankungen und die gemessenen Einzelwerte zeigten eine erhebliche
Streuung (Tab. 15).
Tab. 15: Parathormongehalte im Plasma der Sauen, (MW. ± SD., Min. und Max.).
Anzahl N. Mittelwert Minimum Maximum SD.
Kontrolltiere
Tag 80 pg/ml 15 13,10 1,32 77,06 ± 19,38
Tag 108 pg/ml 15 20,23 2,70 52,50 ± 17,49
Abferkeltag pg/ml 15 16,20 1,19 79,15 ± 19,36
Versuchstiere
Tag 80 pg/ml 14 10,76 2,11 30,97 ± 6,99
Tag 108 pg/ml 14 26,55 1,60 180,27 ± 50,24
Abferkeltag pg/ml 14 36,50 1,60 149,76 ± 53,67
Am 80. Trächtigkeitstag wurden bei den Tieren der Kontrollgruppe z.B. Parathormon-
konzentrationen von im Mittel 13,10 ± 19,383 pg/ml gemessen, wobei die Einzelwerte für
diesen Parameter über einen weiten Bereich zwischen 1,32 pg/ml als Minimum und 77,06
pg/ml als Maximalwert schwankten. Zum gleichen Entnahmezeitpunkt lag der Mittelwert in
der Gruppe der Versuchstiere etwas niedriger bei 10,76 ± 6,985 pg/ml. Die Werte bewegten
sich in dieser Tiergruppe dabei in etwas engeren Grenzen zwischen einem Minimalwert von
2,11 pg/ml und einem Maximalwert von 30,97 pg/ml. Nach vierwöchiger Versuchsphase
wurde bei den Tieren der Kontrollgruppe am 108. Trächtigkeitstag ein Mittelwert von 20,23 ±
17,489 pg/ml für die Parathormonkonzentration im Plasma ermittelt. Der niedrigste
gemessene Wert lag zu diesem Zeitpunkt in dieser Gruppe bei 2,7 pg/ml, während der höchste
Wert bei 52,5 pg/ml zu finden war.

Ergebnisse
100
Die Versuchstiere hatten am 108. Trächtigkeitstag im Mittel einen Parathormonspiegel von
26,55 ± 50,244 pg/ml. Diese hohe Standardabweichung resultierte aus dem weiten Spektrum,
über das die Einzelwerte von minimal 1,6 pg/ml bis maximal 180,27 pg/ml verteilt waren.
Am Tag der Geburt hatten die Tiere der Kontrollgruppe einen Mittelwert von 16,2 ± 19,364
pg/ml im Plasma, dabei lag der Minimalwert bei 1,19 pg/ml und der Maximalwert bei 79,15
pg/ml. Im Mittel war die PTH-Konzentration in der Versuchsgruppe zu diesem Zeitpunkt mit
36,50 ± 53,666 pg/ml etwas höher und schwankte tierindividuell zwischen minimal 1,60
pg/ml und maximal 149,76 pg/ml. Somit war in beiden Gruppen eine sehr breite Variation
gegeben.
Da für beide Gruppen für den 80. Trächtigkeitstag eine relativ gleiche Ausgangssituation für
die Parathormonkonzentration vorlag, wird in der folgenden Abbildung (Abb. 8) die relative
Steigerung der PTH-Werte vom 80. bis zum 108. Trächtigkeitstag bzw. bis zum Tag der
Abferkelung gruppenspezifisch dargestellt.
0
2
4
6
8
10
12
14
rela
tive S
teig
eru
ng
(X
-fach
e)
Versuch 108. T.
Kontrolle 108. T.
Versuch Partus
Kontrolle Partus
Abb. 8: Relative Steigerung der Parathormon-Konzentration im Plasma der Sauen im
Vergleich zu den entsprechenden Werten vom 80. Trächtigkeitstag (X-fache).
In der Kontrollgruppe stieg die PTH-Konzentration am 108. Trächtigkeitstag im Mittel
zunächst auf das 4,93-fache des Ausgangswertes vom 80. Trächtigkeitstag an, während sie bis
zum Tag der Abferkelung wieder auf das 2,77-fache des Ausgangswertes abfiel.
In der Gruppe der Versuchstiere stieg die Konzentration an PTH dagegen vom 80. bis zum
108. Trächtigkeitstag relativ auf das 2,55-fache des Ausgangswertes an, um sich danach bis
zum Abferkeltermin weiter auf das 3,48-fache des Ausgangswertes zu erhöhen. Die relativen
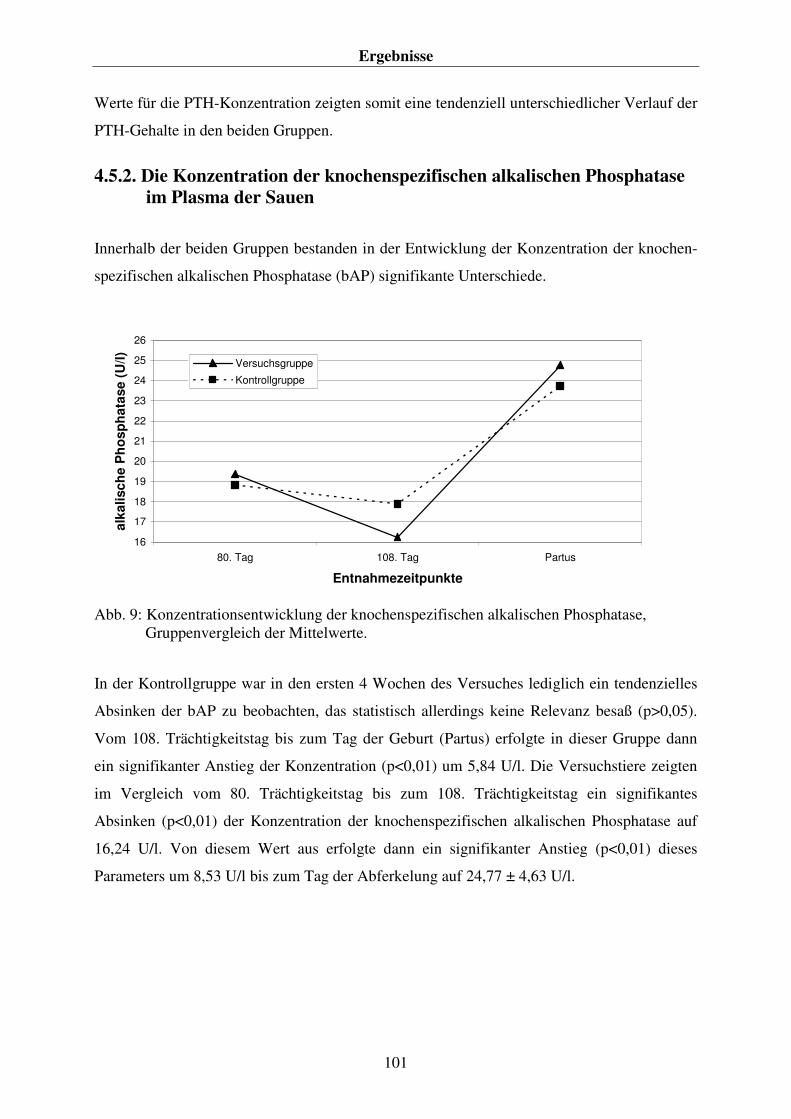
Ergebnisse
101
Werte für die PTH-Konzentration zeigten somit eine tendenziell unterschiedlicher Verlauf der
PTH-Gehalte in den beiden Gruppen.
4.5.2. Die Konzentration der knochenspezifischen alkalischen Phosphatase im Plasma der Sauen
Innerhalb der beiden Gruppen bestanden in der Entwicklung der Konzentration der knochen-
spezifischen alkalischen Phosphatase (bAP) signifikante Unterschiede.
16
17
18
19
20
21
22
23
24
25
26
80. Tag 108. Tag Partus
Entnahmezeitpunkte
alk
alisch
e P
ho
sp
hata
se (
U/l)
Versuchsgruppe
Kontrollgruppe
Abb. 9: Konzentrationsentwicklung der knochenspezifischen alkalischen Phosphatase, Gruppenvergleich der Mittelwerte.
In der Kontrollgruppe war in den ersten 4 Wochen des Versuches lediglich ein tendenzielles
Absinken der bAP zu beobachten, das statistisch allerdings keine Relevanz besaß (p>0,05).
Vom 108. Trächtigkeitstag bis zum Tag der Geburt (Partus) erfolgte in dieser Gruppe dann
ein signifikanter Anstieg der Konzentration (p<0,01) um 5,84 U/l. Die Versuchstiere zeigten
im Vergleich vom 80. Trächtigkeitstag bis zum 108. Trächtigkeitstag ein signifikantes
Absinken (p<0,01) der Konzentration der knochenspezifischen alkalischen Phosphatase auf
16,24 U/l. Von diesem Wert aus erfolgte dann ein signifikanter Anstieg (p<0,01) dieses
Parameters um 8,53 U/l bis zum Tag der Abferkelung auf 24,77 ± 4,63 U/l.

Ergebnisse
102
Tab. 16: Konzentration der knochenspezifischen Phosphatase im Plasma der Sauen (U/l), (MW ± SD)
Trächtigkeitstag Gruppe Anzahl (N)
80. Tag 108.Tag
Partus
19,19 16,82 24,41 Gesamt- tierzahl
29
± 5,11 ± 3,31 ± 4,77
19,37 16,24 24,77 Versuchs-
gruppe 14
± 4,41 ± 2,27 ± 4,63
18,84 17,89 23,73 Kontroll- gruppe
15
± 6,37 ± 4,59 ± 5,12
Zwischen den Tieren der Kontrollgruppe (15 Tiere) und den Tieren der Versuchsgruppe (14
Tiere) bestand allerdings während des Versuches zu keinem Zeitpunkt ein signifikanter
Unterschied im Gehalt der bAP (p>0,05).

Ergebnisse
103
4.6. Die Ergebnisse der Analysen der Spontanharnproben
4.6.1. Der pH-Wert im Harn der Sauen
Die Auswertung der Ergebnisse der pH-Wert-Analysen im Harn der Sauen hat folgende
signifikante Unterschiede ergeben:
1. in beiden Gruppen zwischen dem 90. und 111. Trächtigkeitstag (p<0,01)
2. am 90. Trächtigkeitstag zwischen der Kontroll- und der Versuchsgruppe (p<0,01)
3. am 111. Trächtigkeitstag zwischen der Kontroll- und der Versuchsgruppe (p<0,01)
Tab. 17: pH-Werte im Harn der Sauen, (MW ± SD, Min. und Max.)
Anzahl N. Mittelwert Minimum Maximum SD.
Kontrolltiere
pH-Wert 90. Tag 15 7,05 6,40 7,65 ± 0,343
pH-Wert 111. Tag 15 7,31 6,80 8,05 ± 0,311
Versuchstiere
pH-Wert 90. Tag 14 5,96 5,25 6,70 ± 0,446
pH-Wert 111. Tag 14 6,73 5,75 7,35 ± 0,456
Die pH-Werte im Harn der Kontrolltiere schwankten am 90. Trächtigkeitstag zwischen einem
Minimalwert von pH 6,4 und einem Maximalwert von pH 7,65, so dass der mittlere pH-Wert
bei pH 7,05 ± 0,34 angesiedelt war. Am 111. Trächtigkeitstag lag der Mittelwert in dieser
Gruppe bei pH 7,31 ± 0,311 und das an diesem Tag gemessene pH-Wert-Spektrum erstreckte
sich von minimal pH 6,8 bis maximal pH 8,05. Die Differenz zwischen diesen beiden
Zeitpunkten war somit signifikant (p<0,01).
Am 90. Trächtigkeitstag wurden im Harn der Versuchstiere pH-Werte in der Spannbreite von
pH 5,25 bis pH 6,7 gemessen. Bis zum 111. Trächtigkeitstag kam es in dieser Gruppe zu
einem signifikanten Anstieg (p<0,01) des pH-Wertes im Harn, so dass zu diesem Zeitpunkt
ein mittlerer pH-Wert von pH 6,73 ± 0,46 vorlag und die Einzelwerte zwischen minimal pH
5,75 bis maximal pH 7,35 angesiedelt waren.

Ergebnisse
104
4.6.2. Die relative Mineralstoffausscheidung über den Harn
Die Abgabe der Mineralstoffe über den Harn wurde in diesem Versuch durch den Quotienten
des entsprechenden Elementes und Kreatinin berechnet. Die Ergebnisse der statistischen
Analyse dieser Daten sind hier tabellarisch aufgeführt (Tab. 18).
Tab. 18: Konzentrationsquotienten Mineralstoffe/Kreatinin zum Vergleich der relativen
Ausscheidung der Mineralstoffe (Ca, P, Na, Cl) über den Harn.
(Mineralstoffe (mg/dl) / Kreatinin (mg/dl) im Harn (MW±SD)).
Im Harn der Sauen am 90. Trächtigkeitstag (MW±SD)
Gruppe Anzahl (N) Calcium/ Kreatinin
Phosphor/ Kreatinin
Natrium/ Kreatinin
Chlorid/ Kreatinin
0,20 0,33 0,42 1,36 Alle 29
± 0,36 ± 0,39 ± 0,33 ± 0,83
0,16 0,20#
0,34
2,02#
Versuch 14
± 0,11 ± 0,15 ± 0,24 ± 0,70
0,24 0,44#
0,49
0,74#
Kontroll 15
± 0,49 ± 0,50 ± 0,39 ± 0,28
Im Harn der Sauen am 111. Trächtigkeitstag (MW±SD)
Gruppe Anzahl (N) Calcium/ Kreatinin
Phosphor/ Kreatinin
Natrium/ Kreatinin
Chlorid/ Kreatinin
0,21 0,36 0,98 2,43 Gesamt 29
± 0,19 ± 0,30 ± 0,63 ± 1,01
0,25 0,23#
0,75
3,04#
Versuch 14
± 0,22 ± 0,16 ± 0,47 ± 0,36
0,18 0,49#
1,18
1,86#
Kontroll 15
± 0,15 ± 0,35 ± 0,70 ± 1,10
1) # kennzeichnet signifikante unterschiedliche Mittelwerte zwischen den Gruppen.

Ergebnisse
105
Zwischen den Konzentrationsquotienten von Calcium bestanden an beiden Analyseterminen
zwischen der Kontrollgruppe und der Versuchsgruppe im Mittel keine signifikanten
Differenzen (p>0,05) und auch innerhalb der Gruppen fehlte zwischen den beiden Analyse-
terminen jegliche statistisch relevante Differenz (p>0,05). Es fiel allerdings auf, dass beide
Parameter über eine sehr große relative Streuung der Einzelwerte verfügten, die eine
statistische Auswertung erheblich erschwerte.
Die relative Ausscheidung von Phosphor über den Harn wurde dagegen schwach signifikant
(p<0,05) von der zusätzlichen CaCl2-Gabe beeinflusst und führte zu einer schwach signifikant
(p<0,05) reduzierten Phosphorausscheidung der Versuchstiere von 0,203 ± 0,154 am 90.
Trächtigkeitstag und 0,233 ± 0,159 am 111. Trächtigkeitstag, während die Kontrolltiere am
90. Tag eine relative Ausscheidung von 0,438 ± 0,505 und am 111. Tag von 0,486 ± 0,349 für
Phosphor zeigten. Aber auch für diese Werten konnte von der relativ großen Standardab-
weichung eine breite Streuung der Einzelwerte abgeleitet werden. Zwischen den beiden
Analyseterminen waren weder in der Kontrollgruppe noch in der Versuchgruppe signifikante
Differenzen zu errechnen (p>0,05).
Bei der Betrachtung der relativen Ausscheidung von Natrium über den Harn zeigte sich in
beiden Gruppen ein deutlicher Unterschied zwischen dem 90.Trächtigkeitstag mit 0,42 ± 0,33
und dem 111. Trächtigkeitstag mit 0,98 ± 0,63, dieser Unterschied verfügte allerdings
aufgrund der großen relativen Streuung der Einzelwerte über keine statistische Relevanz
(p>0,05). Bei den Kontrolltieren stieg die relative Ausscheidung von Natrium zum 111.
Trächtigkeitstag auf 1,18 ± 0,70 an und lag somit zu diesem Zeitpunkt relativ weit über dem
Wert der Versuchstiere mit 0,75 ± 0,47. Eine statistisch signifikante Differenz war allerdings
auch hier nicht zu ermitteln (p>0,05). Am 90. Trächtigkeitstag bestanden ebenfalls lediglich
tendenzielle Unterschiede zwischen der Kontroll- und der Versuchsgruppe, die statistisch
widerum nicht abzusichern waren (p>0,05).
Der Chlorid-Kreatinin-Quotient war in der Gruppe der Versuchstiere etwa um den Faktor 2
deutlich erhöht gegenüber den Kontrolltieren und belegte somit eine gesteigerte renale
Chloridausscheidung der Versuchstiere. Außerdem lag in dieser Gruppe zwischen dem 90.
und dem 111. Trächtigkeitstag eine erhebliche Steigerung der Chloridsekretion von 2,02 ±
0,70 auf 3,04 ± 0,36 vor und auch in der Kontrollgruppe stieg die relative Chloridaus-
scheidung im gleichen Zeitraum von 0,74 ± 0,28 auf 1,86 ± 1,10. Bei diesen Anstiegen der
relativen Chloridausscheidung im Vergleich zur Kreatininausscheidung handelte es sich
allerdings lediglich um ein tendenzielles Geschehen, das aufgrund einer erheblichen Streuung
der Einzelwerte über keine statistische Relevanz verfügte (p>0,05). Zwischen den beiden
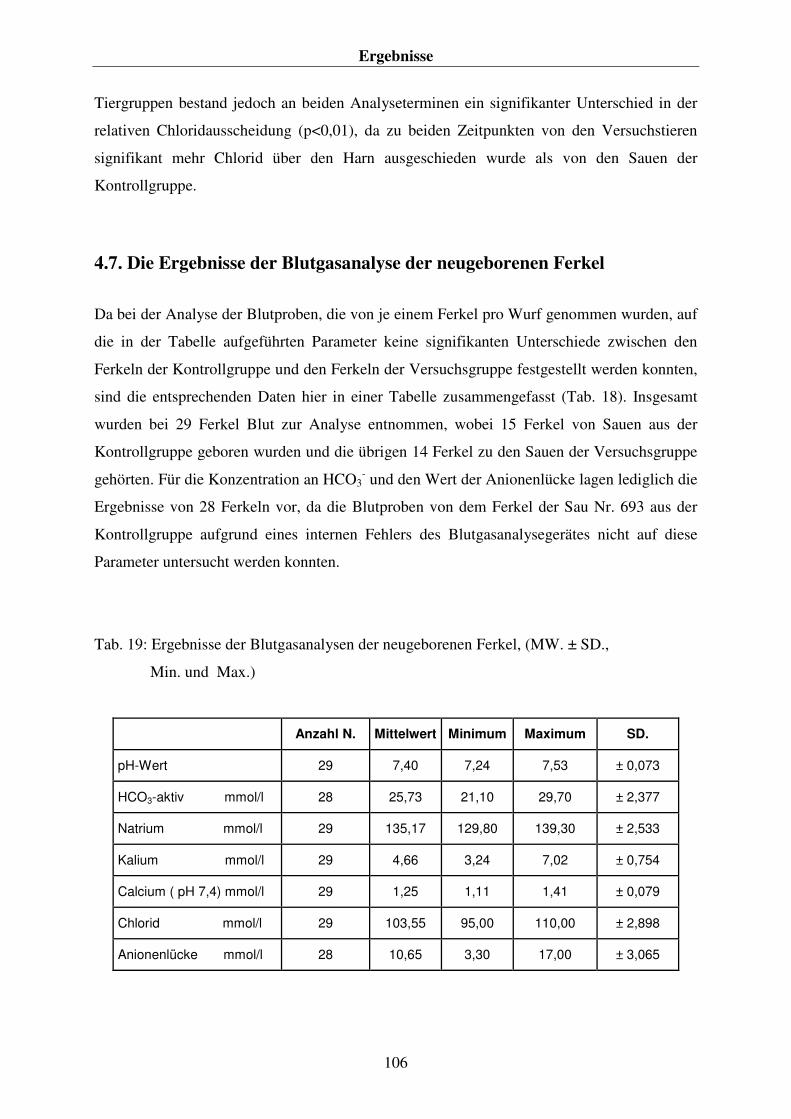
Ergebnisse
106
Tiergruppen bestand jedoch an beiden Analyseterminen ein signifikanter Unterschied in der
relativen Chloridausscheidung (p<0,01), da zu beiden Zeitpunkten von den Versuchstieren
signifikant mehr Chlorid über den Harn ausgeschieden wurde als von den Sauen der
Kontrollgruppe.
4.7. Die Ergebnisse der Blutgasanalyse der neugeborenen Ferkel
Da bei der Analyse der Blutproben, die von je einem Ferkel pro Wurf genommen wurden, auf
die in der Tabelle aufgeführten Parameter keine signifikanten Unterschiede zwischen den
Ferkeln der Kontrollgruppe und den Ferkeln der Versuchsgruppe festgestellt werden konnten,
sind die entsprechenden Daten hier in einer Tabelle zusammengefasst (Tab. 18). Insgesamt
wurden bei 29 Ferkel Blut zur Analyse entnommen, wobei 15 Ferkel von Sauen aus der
Kontrollgruppe geboren wurden und die übrigen 14 Ferkel zu den Sauen der Versuchsgruppe
gehörten. Für die Konzentration an HCO3- und den Wert der Anionenlücke lagen lediglich die
Ergebnisse von 28 Ferkeln vor, da die Blutproben von dem Ferkel der Sau Nr. 693 aus der
Kontrollgruppe aufgrund eines internen Fehlers des Blutgasanalysegerätes nicht auf diese
Parameter untersucht werden konnten.
Tab. 19: Ergebnisse der Blutgasanalysen der neugeborenen Ferkel, (MW. ± SD.,
Min. und Max.)
Anzahl N. Mittelwert Minimum Maximum SD.
pH-Wert 29 7,40 7,24 7,53 ± 0,073
HCO3-aktiv mmol/l 28 25,73 21,10 29,70 ± 2,377
Natrium mmol/l 29 135,17 129,80 139,30 ± 2,533
Kalium mmol/l 29 4,66 3,24 7,02 ± 0,754
Calcium ( pH 7,4) mmol/l 29 1,25 1,11 1,41 ± 0,079
Chlorid mmol/l 29 103,55 95,00 110,00 ± 2,898
Anionenlücke mmol/l 28 10,65 3,30 17,00 ± 3,065

Ergebnisse
107
Der pH-Wert im Blut entsprach in seinem Mittelwert für alle Ferkel zusammen mit pH 7,4 ±
0,073 dem physiologischen Wert, von dem sich die gruppenspezifischen Mittelwerte von pH
7,399 ± 0,071 für die Versuchstiere und pH 7,40 ± 0,073 für die Kontrollgruppe nur minimal
unterschieden (p>0,05).
Die gruppenspezifischen Mittelwerte der Versuchsgruppe (1,236 ± 0,0765 mmol/l) und der
Kontrollgruppe (1,254 ± 0,0856 mmol/l) des auf einen pH-Wert von pH 7,4 korrigierten
Gehalt an ionisierten Calcium waren in sehr engen Grenzen um den gemeinsamen Mittelwert
von 1,25 ± 0,079 mmol/l angesiedelt (p>0.05).
Die für den Gehalt an Kalium gemessenen Werte waren zwar mit einem Minimalwert von
3,24 mmol/l und einem Maximalwert von 7,02 mmol/l über einen weitläufigeren Bereich
verteilt, aber auch für diesem Parameter bestanden keine signifikanten Unterschiede (p>0.05)
zwischen den Tiergruppen.
Die mittleren Konzentrationen an Natrium (135,17 ± 2,533 mmol/l) und Chlorid (103,55 ±
2,898 mmol/l) zeigten ebenfalls keine relevanten Differenzen (p>0,05) zwischen den
Gruppen. Außerdem bestanden weder für die Hydrogenkarbonatkonzentration (HCO3-) mit
einem Mittelwert von 25,73 ± 2,377 mmol/l noch für die Anionenlücke mit im Mittel 10,65 ±
3,065 mmol/l signifikante Behandlungsunterschiede zwischen den Ferkeln der beiden Sauen-
gruppen (p>0,05).

Diskussion
108 108
5. Diskussion
In der hier vorliegenden Forschungsarbeit wurde den Versuchstieren Calci Cap®, ein CaCl2,
das von einem Mantel aus pflanzlichen Fettsäuren umgeben ist, zur Prophylaxe gegen das
MMA-Syndrom verabreicht. Seine prophylaktische Wirkung entfaltet diese Substanz durch
eine Ansäuerung des Harns, der sich somit zu einem bakteriostatischen oder im Extremfall
sogar zu einem bakterioziden Medium entwickelt und somit seine Funktion als Keimreservoir
verliert. Dieses Reservoir bildet in vielen Fällen den Ausgangspunkt für das Auftreten der un-
spezifischen Form der Harnwegsinfektion beim Schwein, die als so genanntes Vorläufer-
syndrom für den MMA-Komplex bezeichnet wird. Auf diesem Wege der Harnacidierung
kann somit sekundär auch die Inzidenz des MMA-Komplexes verringert werden.
Gegenstand der hier vorliegenden Forschungsarbeit war es, die von dem Calci Cap® aus-
gehenden Beeinflussungen des Organismus näher zu untersuchen.
Dabei waren folgende Aspekte von besonderem Interesse:
1. Die Anzahl der geborenen Ferkel, die Geburtsgewichte, der Ablauf der Geburten und
die Gesundheit dieser Ferkel.
2. Das Ausmaß der Acidierung über den Verlauf des Versuchszeitraumes.
3. Die Reaktion des Knochenstoffwechsels bzw. der entsprechenden Parameter wie z.B.
die Konzentrationen an PTH und bAP im Plasma.
4. Die renale Exkretion von Calcium nach der acidogenen Verschiebung der Elektrolyt-
bilanz im Alleinfutter der Sauen.
5.1. Kritik der Methoden
5.1.1. Die Fütterung der Sauen und die Zugabe des CaCl2 zur Ration der Versuchstiere
Die Zuteilung der entsprechenden Mengen der Alleinfuttermittel erfolgte in diesem Versuch
nicht pauschal, sondern wurde mit dem Ziel der Optimierung der Körperkondition zum Zeit-
punkt der Geburt durchgeführt, so dass die tierindividuellen Futtermengen in den ersten 20
Tagen des Versuches einer leichten Schwankungsbreite unterlagen, um Konditionsmängel der
einzelnen Tiere auszugleichen.

Diskussion
109 109
Die Zulage des CaCl2 erfolgte dagegen einheitlich mit zweimal täglich 14 g (US) zu den ent-
sprechenden Fütterungszeiten. Somit erfolgte diese Zuteilung weder bezogen auf die tier-
individuelle Körpermasse noch in Bezug auf die speziell pro Tier und Tag verabreichte
Menge an Alleinfutter.
Die Zuteilung von 250 g Trockenschnitzel-Pellets pro Tier und Tag vom 80. bis zum 108.
Trächtigkeitstag wurde ebenfalls einheitlich ohne Berücksichtigung der Körpermasse oder der
Körperkondition der Einzeltiere durchgeführt. Unter diesen Bedingungen bestanden dem zur
Folge tierindividuelle Schwankungen in den pro kg Körpermasse und Tag aufgenommenen
Stoffmengen, die teilweise zur Beeinflussung der Analyseparameter beitragen können und
deshalb bei der Beurteilung dieser Ergebnisse berücksichtigt werden müssen.
5.1.2. Die Bestimmung des pH-Wertes im Harn der Sauen
Die Messung des pH-Wertes im Harn der Tiere erfolgte in spontan abgesetzten Harnproben,
um so eine Katheterisierung der Sauen zu vermeiden. Zum einen wurde diese Variante der
Harnprobengewinnung aus Tierschutzgründen gewählt und zum anderen besteht bei der
Katheterisierung der Harnblase immer die Gefahr den Urogenitaltrakt mit fakultativ
pathogenen Erregern zu infizieren. Eine solche Infektion kann im weiteren Verlauf durch
Urease-bildende Bakterien zu Veränderungen des pH-Wertes im Harn führen und somit die
Messergebnisse negativ beeinflussen. Es konnte leider in diesem Versuch nicht beantwortet
werden, ob ein derartiger Effekt auf die gemessenen Harn-pH-Werte stattgefunden hat, da vor
und während des Versuches auf eine Kontrolle des bakteriologischen Harnstatus der
einzelnen Tiere verzichtet wurde. Aus diesem Grund kann ein solcher Einfluss hier leider
nicht ausgeschlossen werden. FINKENSIEP (1993) bemerkt in seinen Untersuchungen zum
Thema der Harnprobenentnahme, dass qualitativ keine Unterschiede zwischen den Harn-
proben bestanden, die durch Katheterisierung der Harnblase gewonnen wurden und
Harnproben, die durch das Auffangen von spontan abgesetztem Harn gesammelt wurden.
Eine Verfälschung der Messergebnisse nach dem Auffangen der spontan abgesetzten Proben
war in diesem Versuch nicht zu erwarten, da die Messung des pH-Wertes unmittelbar nach
dem Absatz des Harns durchgeführt wurde und die entsprechenden Proben dann relativ
schnell bzw. mit einer Zeitverzögerung von ca. 1 Stunde bei – 18°C eingefroren wurden.
In diesem Zusammenhang zeigte SCHUKNECHT (1991) in ihren Untersuchungen an un-
konserviertem Katzenharn, dass bei pH-Werten von unter pH 6,5 die Aufbewahrung der
Proben bei Zimmertemperatur zu keinerlei Veränderungen geführt hat, während Harnproben

Diskussion
110 110
mit einem pH-Wert von pH 8,0 nach 16 Stunden unter derartigen Bedingungen einen Anstieg
des pH-Wertes von maximal einer Einheit zeigten. Da diese Zeitabstände in der hier
vorliegenden Arbeit aber auf keinen Fall erreicht wurden, ist eine derartige Beeinflussung der
Ergebnisse hier nicht zu erwarten.
5.1.3. Die Blutentnahme
Bei der Entnahme der Blutproben wurde auf eine möglichst komplikationslose und stressarme
Durchführung dieses Vorganges geachtet. Da die Tiere dieses Vorhaben im Rahmen des
Versuchsplanes jedoch mehrfach über sich ergehen lassen mussten, setzte gerade bei den
ferkelführenden Sauen ein gewisser Lerneffekt zur Abwehr der zur Blutentnahme nötigen
Fixation ein. Somit waren diese Tiere einer geringfügig erhöhten Stressbelastung ausgesetzt.
In diesem Zusammenhang stellten OLDIGS und HINSCH (1987) als Folge stressbedingter
Erhöhungen der Laktatwerte Veränderungen im Säure-Basen-Haushalt und den Blutgas-
werten bei der Beprobung des Göttinger Miniaturschweins fest. Diese Form der Beein-
flussung der Ergebnisse ist daher auch in dieser Arbeit nicht auszuschließen. Da allerdings in
dem hier vorliegenden Versuch die Bedingungen in beiden Gruppen gleich waren, wird die
tendenzielle Entwicklung der Ergebnisse durch diese zusätzliche Stressbelastung nicht
beeinträchtigt.
Zwischen dem Zeitpunkt der Entnahme und dem Zeitpunkt der Analyse der Blutproben liegt
aufgrund des Transportes in das Analyselabor ein Abstand von ca. 1 Stunde, in der die Proben
in einer Styroporkiste auf Kühlakkus bei einer Temperatur von +2-4°C gelagert wurden.
ASSAL et al. (1980) untersuchten Veränderungen im Schweineblut, die sich bei 24-stündiger
Lagerung bei Zimmertemperatur (21-24°C) ergeben. Unter diesen Bedingungen wurden
Veränderungen des pH-Wertes um etwa -0,01 Einheiten, des pCO2 um + 0,55 mmHg und des
BE um -0,03 mmol/l nach einer Lagerung von 60 Minuten festgestellt. Da unter den
Aufbewahrungsbedingungen des hier vorliegenden Versuches diese Prozesse aufgrund der
niedrigeren Temperaturen noch langsamer verlaufen seien müssten, können derartige
Beeinflussungen der hier analysierten Proben vernachlässigt werden.

Diskussion
111 111
5.1.4. Die Bestimmung der Anionen-Kationen-Bilanz (AKB) und der Elektrolyt-Bilanz (EB) im Futter
Die Berechnung der AKB und der EB im Futter erfolgte nach den von BEKER (1999)
benutzten Formeln. In der nachfolgenden Tabelle (Tab. 20) sind die entsprechenden Kationen
und Anionen aufgeführt, die zur Berechnung dieser beiden Werte herangezogen wurden.
Tab. 20: Kationen und Anionen, die zur Berechnung der AKB bzw. der EB nach
BEKER (1999) benötigt werden.
Aus diesen aufgeführten Faktoren werden die AKB und EB in den entsprechenden Futterrationen berechnet (BEKER 1999)
Kationen Anionen
AKB (mmol/kg) Ca Mg Na K P S
(Met. + Cys.) Cl
EB (mEq/kg) Na K Cl
Dabei wurde deutlich, dass die einzelnen Kationen und Anionen entsprechend ihrer
Berücksichtigung unterschiedliche Einflüsse auf die Werte der AKB bzw. EB ausüben.
In die Berechnung der AKB flossen dem entsprechend als Kationen die Werte für Calcium,
Magnesium, Natrium und Kalium und als Anionen die Gehalte an Phosphat, Schwefel und
Chlorid mit ein. Die Werte für den Schwefel wurden in der hier vorliegenden Studie wie auch
in anderen Arbeiten z.B. beim Schwein (FINKENSIEP 1993; KROHN 1993) oder bei der
Katze (SCHUKNECHT 1991) durch die Summe der schwefelhaltigen Aminosäuren
Methionin und Cystein ersetzt. Da in der Futtermittelanalyse der hier vorliegenden Arbeit die
Gehalte dieser Aminosäuren nicht ermittelt wurden, mussten ihre Konzentrationen im Futter
der Sauen durch die Berechnung anhand eines Futterrationsprogramms für Schweine (Institut
für Tierernährung der TIHO Hannover) geschätzt werden. Für die Berechnung der EB wurden
dagegen ausschließlich die Gehalte an Natrium und Kalium als Kationen und als Anion
lediglich der Chloridgehalt herangezogen. Somit ist die Differenz in der Auswirkung der
zusätzlichen Gabe von zweimal täglich 14 g (US) CaCl2 auf die AKB bzw. auf die EB durch
die unterschiedlich Berücksichtigung der einzelnen Kationen und Anionen zu erklären. Die in
der nachfolgenden Tabelle (Tab. 21) aufgeführten Werte für die AKB bzw. EB der einzelnen
Fütterungsabschnitte zeigen, dass diese CaCl2-Gabe sich rechnerisch auf die EB, bei deren
Berechnung lediglich das Chlorid als Anion berücksichtigt wird, gravierender auswirkt als auf

Diskussion
112 112
die AKB, für deren Berechnung das Chlorid nur einen Teil der Ainion-Bilanz ausmacht und
außerdem für die Kationen-Bilanz noch zusätzlich das Calcium Berücksichtigung findet.
Tab. 21: Die Werte für die AKB und die EB der einzelnen Fütterungsabschnitte für die Ration
der Kontroll- und Versuchstiere.
Die nach BEKER (1999) berechneten Werte für die AKB und die EB der entsprechenden Fütterungsabschnitte
Futterration an den entsprechenden
Trächtigkeitstagen 80.-108. Tag
a 108. Tag – Partus
post partum
AKB (mmol/kg TS) 367 394 394 Kontroll-gruppe
EB (mEq/kg TS) 190 223 223
AKB (mmol/kg TS) 359 389 394 Versuchs-
gruppe EB (mEq/kg TS) 22 94 223
a) Am 108. Trächtigkeitstag erfolgt mittags die Umstallung der Sauen in den Abferkelbereich und somit die
Futterumstellung vom Alleinfutter für tragende Sauen auf das Alleinfutter für laktierende Sauen (siehe Tab.3).
Die Berechnungen der AKB und der EB in den verschiedenen Rationen zu den
entsprechenden Fütterungszeitpunkten wurde anhand der Mittelwerte der verabreichten
Futtermenge am 90. und 111. Trächtigkeitstag und für die ersten Tage der Laktationsphase
durchgeführt, da die Sauen nicht mit einer einheitlichen Menge der entsprechenden Allein-
futter versorgt wurden, sondern die Futtermengenzuteilung tierindividuell mit dem Ziel der
Optimierung der Körperkondition zum Zeitpunkt der Geburt variierte. Diese Vor-
raussetzungen müssen allerdings bei der Interpretation der Ergebnisse berücksichtigt werden.
5.1.5. Der Vergleich der Blut- und Harnparameter zur Feststellung der renalen Exkretion
Da die Sauen dieses Versuches nicht in Bilanzständen sondern in einem konventionellen
Sauenstall aufgestallt waren, konnte die pro Tier und Tag abgesetzte Harnmenge nicht
bestimmt werden. Um aber dennoch Tendenzen der renalen Ausscheidung der Mineralstoffe
erfassen zu können, wurde deren relative Ausscheidung zur ausgeschiedenen Kreatininmenge
bestimmt. Zu diesem Zweck wurden in dieser Arbeit die Kreatininkonzentrationen im Plasma
und im Harn der Sauen bestimmt. Da in beiden Gruppen die Bedingungen identisch waren, ist

Diskussion
113 113
von einer tendenziellen gruppenspezifischen Beeinflussung nicht auszugehen. Für die
Interpretation der Ergebnisse müssen diese suboptimalen Bedingungen allerdings
berücksichtigt werden.
5.2. Diskussion der Ergebnisse
5.2.1. Die Akzeptanz des Futters nach der Zumischung von Calciumchlorid (CaCl2)
Die Alleinfuttermittel für trächtige und für laktierende Sauen, die beide als hofeigene
Mischung auf dem Versuchsgut selbst hergestellt wurden, sind von allen Tieren während des
gesamten Versuches komplett verzehrt worden. Die Zumischung von je 14g (US) gecoatetem
Calciumchlorid pro Mahlzeit zu der tierindividuellen Alleinfuttermenge bei den Sauen der
Versuchsgruppe hat die Futteraufnahme in diesem Versuch nicht negativ beeinflusst, da auch
diese Tiere das angebotene Futter restlos verzehrten. In anderen Arbeiten, in denen Calcium-
chlorid an Schweine verfüttert wurde, wird dagegen teilweise von einer verminderten
Akzeptanz des Futters berichtet. YEN et al. (1981) erklärten diese reduzierte Futteraufnahme
mit der durch das CaCl2 hervorgerufenen metabolischen Acidose, da in ihrem Versuch die
Futteraufnahme durch die Zugabe von NaHCO3 wieder normalisiert werden konnte. Im
Vergleich zu dem hier vorliegenden Versuch, in dem die Sauen während der Trächtigkeit
restriktiv gefüttert wurden, fütterten YEN et al. (1981) die männlichen Mastschweine, die in
ihrem Versuch Verwendung fanden, ad libitum. In neueren Untersuchungen (DE ROUCHEY
et al. 2003) wird dagegen nicht von einer Beeinflussung der Futteraufnahme durch den Zusatz
von Calciumchlorid berichtet. Da die Akzeptanz eines Futters maßgeblich von dessen
Schmackhaftigkeit abhängig ist, wurde in dem hier durchgeführten Versuch gecoatetes CaCl2
eingesetzt, das von einem Mantel aus pflanzlichen Fettsäuren umgeben ist. Durch diese
Ummantelung werden die Geschmacksrezeptoren vor dem sehr bitteren Geschmack des
CaCl2 geschützt und somit auch dessen Akzeptanz erhöht. Gerade bei der von YEN et al.
(1981) angewandten ad-libitum-Fütterung können schon geringe geschmackliche Ver-
änderungen des verabreichten Futters zu einer erheblichen Beeinflussung der Futteraufnahme
führen. Bei dem in dieser Arbeit eingesetzten gecoateten Calciumchlorid bzw. durch die
eingesetzte Menge dieses Stoffes konnten wie beschrieben allerdings keine negativen Aus-
wirkungen auf die Futteraufnahme festgestellt werden.

Diskussion
114 114
5.2.2. Der pH-Wert im Harn der Sauen
Die statistische Auswertung der pH-Werte im Harn der Sauen hat gezeigt, dass es schon nach
einer 10-tägigen Zugabe von 14g CaCl2 (US) pro Mahlzeit möglich war, den pH-Wert im
Harn signifikant (p<0,01) auf einen mittleren pH-Wert von pH 5,96 zu reduzieren. Im
weiteren Verlauf des Versuches kommt es dann in beiden Gruppen bis zum 111. Trächtig-
keitstag zu einem signifikanten Anstieg (p<0,01) des pH-Wertes. Aber auch nach diesem
Anstieg des Harn-pH-Wertes blieb weiterhin ein signifikanter Unterschied (p<0,01) zwischen
den Tieren der Versuchsgruppe und den Tieren der Kontrollgruppe bestehen. Durch die
Zugabe von CaCl2 konnte der pH-Wert im Harn der Sauen der Versuchsgruppe also am 90.
Trächtigkeitstag signifikant um 1,19 Einheiten gesenkt werden, während am 111. Trächtig-
keitstag lediglich eine Absenkung um 0,58 Einheiten zu erreichen war. Das Ansteigen der
pH-Werte im Harn der Sauen in beiden Gruppen und die Verringerung der pH-Wert-
Differenz zum 111. Trächtigkeitstag kann durch die geänderte Fütterung der Sauen erklärt
werden, da am 108. Trächtigkeitstag die Sauen in den Abferkelbereich umgestallt wurden und
gleichzeitig ein Futterwechsel vom Alleinfutter für tragende Sauen auf das Alleinfutter für
säugende Sauen stattgefunden hat und die Zulage der Trockenschnitzel-Pellets beendet
wurde. Der allgemeine Anstieg der pH-Werte im Harn ist somit mit der auf diese Weise
entsprechend geänderten Fütterung der gesamten Tiergruppe zu erklären. Die Verringerung
der Differenz der gruppenspezifischen pH-Werte im Harn ist die Folge der prozentual
kleineren Menge an zugesetztem CaCl2 bezogen auf die entsprechend gesteigerte Menge an
Alleinfutter in der Versuchsgruppe. Diese Verringerung der Differenz wird dabei durch die
Veränderungen der acidierenden Potenz der Futterrationen im Laufe der entsprechenden
Versuchsphasen hervorgerufen, die in Tabelle 21 in Kapitel 5.1.4. aufgeführt werden.
Im Vergleich zu anderen Stoffen, die zur Ansäuerung des Harns verwendet werden, sind die
ansäuernden Effekte, die in dieser Arbeit durch den Einsatz des CaCl2 erreicht werden
konnten, im mittleren Bereich angesiedelt. Dies stellten auch BUDDE und CRENSHAW
(2003) fest, indem sie nach der Zugabe von 9,67 g CaCl2 pro kg (TS) Mischfutter zur
Verfütterung an Mastschweine eine Reduktion des pH-Wertes im Harn auf pH 5,71 messen
konnten. JÜRGENS (1991), der verschiedene Stoffe auf ihre Eignung als Harnansäuerer bei
Zuchtsauen untersuchte, erzielte durch die Verabreichung von organischen Säuren
(Propionsäure, Ascorbinsäure und Zitronensäure) negative pH-Wert-Änderungen von 0,49 bis
0,97 pH-Einheiten, während er durch die Applikation von 0,5 g DL-Methionin/kg KGW mit
0,91 pH-Einheiten bzw. 1mval Ammoniumchlorid/kg KGW mit 1,39 pH-Einheiten höhere

Diskussion
115 115
Absenkungen des pH-Wertes im Harn erreichte. Durch eine Verdoppelung der Konzentration
dieser beiden Stoffe bewirkte JÜRGENS (1991) schließlich sogar ein Absinken des pH-
Wertes um 2,16 bzw. 2,45 pH-Einheiten. FINKENSIEP (1993) erzielte durch die Zulage von
20 g DL-Methionin pro kg Mischfutter eine Reduktion des pH-Wertes im Harn von pH 6,37
auf pH 5,42 (0,95 pH-Einheiten). ARNHOFER (1987) erhöhte den Anteil an tierischem
Eiweiß in der Ration auf 25%, worauf er ein Absinken des pH-Wertes im Harn um 0,74 pH-
Einheiten feststellen konnte. FINKENSIEP (1993) berichtet außerdem von einer spontanen
Reduktion des pH-Wertes um den Zeitpunkt der Geburt, da er am Tag der Geburt eine
signifikante Säuerung des Harns auf einen pH-Wert von im Mittel pH 5,91 (im Extremfall
sogar auf pH 5,27) beobachten konnte und innerhalb von 2 Tagen post partum wieder die
Ausgangswerte dieses Parameters erreicht wurden.
Die Fragen, inwieweit eine Erhöhung in der Dosierung von CaCl2 zu einer Steigerung der
Absenkung des pH-Wertes im Harn beiträgt bzw. ob und in welcher Höhe diesem Prozess
Grenzen gesetzt sind, bleiben in der hier durchgeführten Arbeit leider ungeklärt und sollten
durch weiterführende Untersuchungen näher erörtert werden. Unbeantwortet bleibt in diesem
Zusammenhang auch die Frage, ob ab einer gewissen Dosierung die Akzeptanz der Ration
abnimmt oder ob das gecoatete Calciumchlorid auch dann keinen Einfluss auf die
Futteraufnahme hat.
5.2.3. Die Vorhersage des pH-Wertes im Harn anhand der Anionen- Kationen-Bilanz (AKB) im Futter
Bei der Zugabe von Stoffen, die zur Ansäuerung des Harns führen, ist es wichtig schon bei
der Anmischung der Futterration die Größe des acidierenden Effektes abschätzen zu können,
um diesen Effekt durch die Höhe der Zugaben zu steuern. In diesem Zusammenhang konnte
in verschiedenen Untersuchungen zum Thema Harnansäuerung durch die Verwendung von
Futterzusatzstoffen gezeigt werden, dass ein signifikanter Zusammenhang zwischen der AKB
(X) der eingesetzten Futterration und den bei diesen Versuchen ermittelten mittleren Harn-
pH-Werten (y) besteht. Dieser Zusammenhang lässt sich am besten durch folgende Gleichung
beschreiben: y = 3*10-6x2 + 0,0031x + 6,19; r = 0,95**; n = 24 (BEKER 1999).
Außerdem besteht ein linearer Zusammenhang zwischen der AKB im Futter und dem pH-
Wert im Harn, der in folgender Gleichung beschrieben wird: y = 6,39 + 0,0028x ; r = 0,92** ;
n = 24 (BEKER 1999). Dabei bleibt allerdings unberücksichtigt, dass durch die begrenzte
Säureausscheidungskapazität der Nieren ab einer bestimmten AKB keine weitere Absenkung

Diskussion
116 116
des pH-Wertes im Harn mehr zu erreichen ist (SCHEUNERT und TRAUTMANN 1987).
Beim Schwein können bei stark negativer AKB somit mittlere pH-Werte im Harn von bis zu
pH 5,4 erreicht werden (BEKER 1999). Unter Verwendung der genannten quadratischen
Gleichung errechnete BEKER (1999) eine AKB von ca. – 66 mmol/kg TS in der von ihr
verwendeten Ration, um den pH-Wert im Harn unter pH 6,0 zu senken.
Andere Autoren (JÜRGENS 1991; FINKENSIEP 1992; KROHN 1993) benutzen eine lineare
Gleichung (y = 6,53 + 0,0016 x) und errechnen damit eine AKB von mindestens – 331
mmol/kg TS in ihrer Ration, um im Harn einen pH-Wert von unter pH 6,0 zu erzeugen.
BEKER (1999) fasste diese beiden Gleichungen zusammen und errechnete daraus folgenden
Zusammenhang y = 2*10-6x2 + 0,0029x + 6,26; r = 0,93** ; n = 29 (siehe Abb. 10).
5,0
5,5
6,0
6,5
7,0
7,5
8,0
-500 -400 -300 -200 -100 0 100 200 300 400
y=6,26+0,003*x+0,000002*x2
AKB(mmolAKB(mmol//kgTSkgTS))
pHpH
a)a)
b)b)
c)c)
d)d)
a) gemessener Harn-pH-Wert der Versuchstiere am 90. Trächtigkeitstag (MW±SD)
b) gemessener Harn-pH-Wert der Versuchstiere am 111. Trächtigkeitstag (MW ± SD)
c) gemessener Harn-pH-Wert der Kontrolltiere am 90. Trächtigkeitstag (MW±SD)
d) gemessener Harn-pH-Wert der Kontrolltiere am 111. Trächtigkeitstag (MW ± SD)
Abb. 10: Abhängigkeit des pH-Wertes im Harn von der AKB im Futter nach BEKER (1999)
und die in dem hier vorliegenden Versuch berechneten Werte der AKB mit den
jeweils gemessenen pH-Werten im Harn der Sauen (a - d).
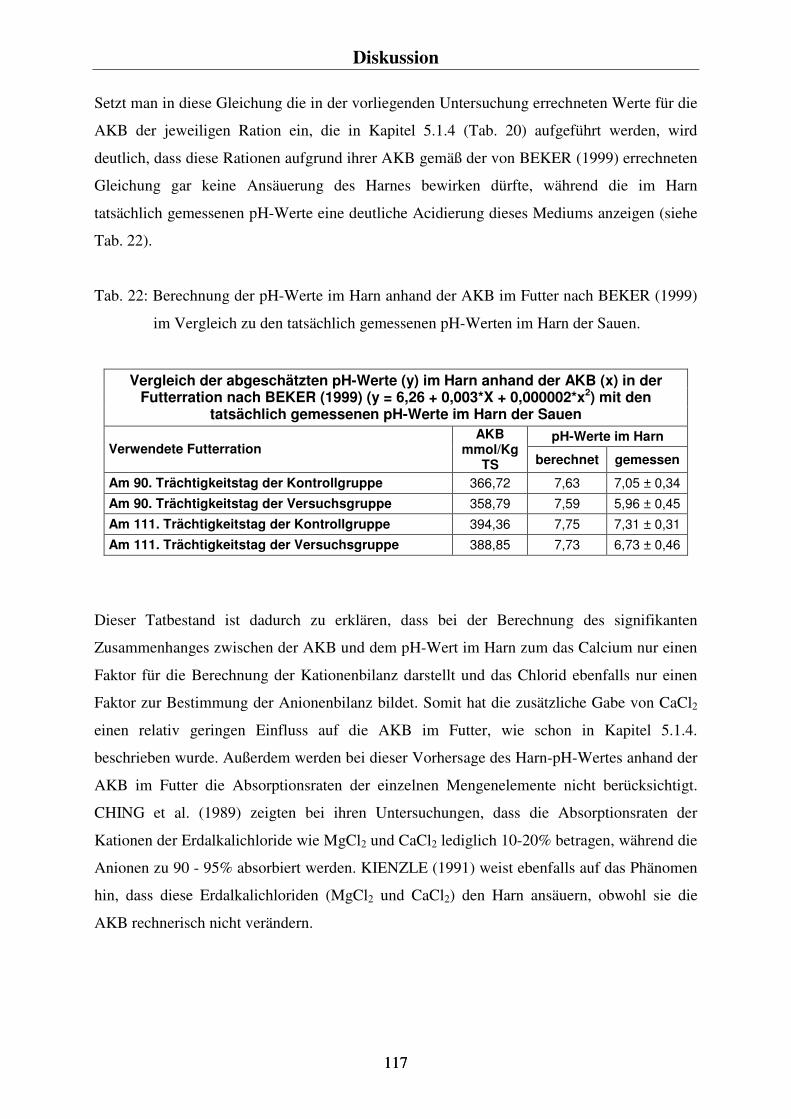
Diskussion
117 117
Setzt man in diese Gleichung die in der vorliegenden Untersuchung errechneten Werte für die
AKB der jeweiligen Ration ein, die in Kapitel 5.1.4 (Tab. 20) aufgeführt werden, wird
deutlich, dass diese Rationen aufgrund ihrer AKB gemäß der von BEKER (1999) errechneten
Gleichung gar keine Ansäuerung des Harnes bewirken dürfte, während die im Harn
tatsächlich gemessenen pH-Werte eine deutliche Acidierung dieses Mediums anzeigen (siehe
Tab. 22).
Tab. 22: Berechnung der pH-Werte im Harn anhand der AKB im Futter nach BEKER (1999)
im Vergleich zu den tatsächlich gemessenen pH-Werten im Harn der Sauen.
Vergleich der abgeschätzten pH-Werte (y) im Harn anhand der AKB (x) in der Futterration nach BEKER (1999) (y = 6,26 + 0,003*X + 0,000002*x2) mit den
tatsächlich gemessenen pH-Werte im Harn der Sauen
pH-Werte im Harn Verwendete Futterration
AKB mmol/Kg
TS berechnet gemessen
Am 90. Trächtigkeitstag der Kontrollgruppe 366,72 7,63 7,05 ± 0,34
Am 90. Trächtigkeitstag der Versuchsgruppe 358,79 7,59 5,96 ± 0,45
Am 111. Trächtigkeitstag der Kontrollgruppe 394,36 7,75 7,31 ± 0,31
Am 111. Trächtigkeitstag der Versuchsgruppe 388,85 7,73 6,73 ± 0,46
Dieser Tatbestand ist dadurch zu erklären, dass bei der Berechnung des signifikanten
Zusammenhanges zwischen der AKB und dem pH-Wert im Harn zum das Calcium nur einen
Faktor für die Berechnung der Kationenbilanz darstellt und das Chlorid ebenfalls nur einen
Faktor zur Bestimmung der Anionenbilanz bildet. Somit hat die zusätzliche Gabe von CaCl2
einen relativ geringen Einfluss auf die AKB im Futter, wie schon in Kapitel 5.1.4.
beschrieben wurde. Außerdem werden bei dieser Vorhersage des Harn-pH-Wertes anhand der
AKB im Futter die Absorptionsraten der einzelnen Mengenelemente nicht berücksichtigt.
CHING et al. (1989) zeigten bei ihren Untersuchungen, dass die Absorptionsraten der
Kationen der Erdalkalichloride wie MgCl2 und CaCl2 lediglich 10-20% betragen, während die
Anionen zu 90 - 95% absorbiert werden. KIENZLE (1991) weist ebenfalls auf das Phänomen
hin, dass diese Erdalkalichloriden (MgCl2 und CaCl2) den Harn ansäuern, obwohl sie die
AKB rechnerisch nicht verändern.

Diskussion
118 118
5.2.4. Die Vorhersage des Harn-pH-Wertes anhand der Elektrolytbilanz (EB) im Futter
Als weitere Variante zur Vorhersage des pH-Wertes, der im Harn durch die Verfütterung
bestimmter Futtermittel erzeugt wird, nennt BEKER (1999) die Bestimmung der Elektrolyt-
bilanz (mEq/kg TS) der entsprechenden Ration.
Den mittleren zu erwartenden pH-Wert (y) berechnet man, indem man die Elektrolytbilanz (x)
in die folgende lineare Gleichung y = 6,319 * 0,007*x einsetzt. In der folgenden Abbildung
(Abb. 11) werden graphisch der lineare Verlauf dieses signifikanten Zusammenhanges und
einige von BEKER (1999) untersuchte Elektrolyt-Bilanzen mit den dazugehörigen pH-
Werten im Harn und zusätzlich die in dem hier vorliegenden Versuch tatsächlich im Harn der
Sauen gemessenen pH-Werte aufgeführt.
EB(mEq/kgTS)
5,0
5,5
6,0
6,5
7,0
7,5
8,0
-100 -60 -20 20 60 100 140 180 200
y=6,319+0,007*x
pHpH
a)a)
b)b)
c)c)
d)d)
a) gemessener Harn-pH-Wert der Versuchstiere am 90. Trächtigkeitstag (MW ± SD)
b) gemessener Harn-pH-Wert der Versuchstiere am 111. Trächtigkeitstag (MW ± SD)
c) gemessener Harn-pH-Wert der Kontrolltiere am 90. Trächtigkeitstag (MW ± SD)
d) gemessener Harn-pH-Wert der Kontrolltiere am 111. Trächtigkeitstag (MW ± SD)
Abb. 11 : Abhängigkeit des pH-Wertes im Harn von der Elektrolyt-Bilanz im Futter
(BEKER 1999) und die im hier vorliegenden Versuch berechneten EB-Werte mit
den jeweils dazu gemessenen pH-Werten im Harn der Sauen.

Diskussion
119 119
Die in dem hier vorliegenden Versuch errechneten Elektrolyt-Bilanzen, die schon in Kapitel
5.1.4. (Tab. 20) aufgeführt wurden, werden in die Berechnung nach BEKER (1999) eingesetzt
und so die im Harn zu erwartenden pH-Werte berechnet. Die auf diese Weise berechneten
pH-Werte sind im Vergleich zu den in diesem Versuch tatsächlich im Harn gemessenen pH-
Werten in der nachfolgenden Tabelle (Tab. 23) aufgeführt.
Tab. 23: Berechnung der pH-Werte im Harn anhand der EB im Futter nach BEKER (1999)
im Vergleich zu den tatsächlich gemessenen pH-Werten im Harn der Sauen.
Vergleich der abgeschätzten pH-Werte (y) im Harn anhand der EB (x) in der Futterration nach Beker (1999) y = 6,319 + 0,007*x) mit den tatsächlich
gemessenen pH-Werten im Harn der Sauen
pH-Werte im Harn Verwendete Futterration
EB mEq/kg
TS berechnet gemessen
Am 90. Trächtigkeitstag der Kontrollgruppe 189,54 7,64 7,05 ± 0,34
Am 90. Trächtigkeitstag der Versuchsgruppe 21,58 6,47 5,96 ± 0,45
Am 111. Trächtigkeitstag der Kontrollgruppe 223,43 7,88 7,31 ± 0,31
Am 111. Trächtigkeitstag der Versuchsgruppe 94,05 6,97 6,73 ± 0,46
Die Ergebnisse dieser Berechnungvariante der zu erwartenden pH-Werte im Harn zeigen
wiederum keine Übereinstimmungen mit den pH-Werten, die in diesem Versuch tatsächlich
gemessen wurden, wenn die Sauen die entsprechende Futterration gefüttert bekamen.
Allerdings werden nach dieser Berechnungsmethode über die EB im Futter bessere
Ergebnisse erzielt als mit der Berechnung über die AKB in der Ration. Dies ist durch die
Berechnung der EB zu erklären. Das Chlorid bildet in dieser Berechnung das einzige Anion,
das in diese Berechnung einbezogen wird, während das Calcium als Kationen keinen Faktor
in dieser Berechnungsformel bildet, wie schon in Kapitel 5.1.4. dargestellt wurde. Da das
Chlorid, das im Darm wie geschildert zu >90% resorbiert wird, somit Berücksichtigung
findet, während das Calcium, das nur zu einem geringen Teil (ca. 10%) resorbiert wird, in die
Berechnung nicht einfliesst, lassen sich über die EB in diesem Fall relativ gute Vorhersagen
der pH-Werte treffen. Aber für eine brauchbare Vorhersage des zu erwartenden pH-Wertes im
Harn ist diese Berechnungsart unter den hier gegebenen Bedingungen ebenfalls nicht
empfehlenswert.

Diskussion
120 120
5.2.5. Der Mechanismus der Harnansäuerung nach der Zugabe von CaCl2
Die Ansäuerung des Harns durch die Zugabe von CaCl2 beruht auf einer unterschiedlichen
Absorptionsrate der beiden Bestandteile dieses Erdalkalichlorids. Während der Calciumanteil
lediglich zu 10-20% absorbiert wird (MÄNNER u. BRONSCH 1987), wird das oral aufge-
nommene Chlorid mit einer Resorptionsrate von ca. 95% fast vollständig vom Körper aufge-
nommen. Aus dieser Differenz der Resorption resultiert eine Störung der Elektronen-
neutralität im Darm, zu deren Ausgleich, angetrieben von der transmuralen Potential-
differenz, Bikarbonat (HCO3-) in das Darmlumen sezerniert wird. Somit geht dem Körper
durch diese Bikarbonatabgabe eine bedeutende Puffersubstanz verloren und es entsteht eine
metabolische Acidose. Zur Konstanterhaltung des pH-Wertes im Blut wird nun durch die
Steigerung der Reabsorption von Bikarbonat in den Nierentubuli die Acidose ausgeglichen.
Zu diesem Zweck werden in den Nierentubuli zusätzlich zu den 90% des filtrierten
Bikarbonates, die schon unter physiologischen Bedingungen reabsorbiert werden, im Fall
einer metabolischen Acidose auch noch die übrigen 10% aus dem Harn zurück in den Körper
reabsorbiert. Somit wird im Fall einer metabolischen Acidose kein bzw. kaum noch
Bikarbonat über den Harn ausgeschieden (HIERHOLZER u. FROMM 1995), so dass dessen
puffernde Wirkung im Harn wegfällt und eine entsprechende Ansäuerung des Harns erreicht
wird.
Die erhöhte orale Chloridaufnahme der Versuchstiere führt ebenfalls zur Ansäuerung des
Harns, da dieses Element unabhängig von dessen Konzentration im Futter nahezu vollständig
resorbiert wird. Um die Chloridkonzentration im Blut bzw. im Körper konstant zu halten,
wird das überschüssige Chlorid auf dem Wege der Exkretion über die Nieren mit dem Harn
wieder ausgeschieden. In dem hier vorliegenden Versuch zeigt sich dies dadurch, dass sich
die Chloridkonzentration im Plasma der Sauen der Versuchsgruppe lediglich am 108.
Trächtigkeitstag schwach signifikant (p<0,05) von dem Chloridgehalt im Plasma der
Kontrolltiere unterscheidet, in dem sie diesen Wert um 1,7 mmol/l übersteigt, während zu
allen anderen Analyseterminen kein signifikanter Unterschied zwischen den beiden
Tiergruppen besteht. In diesem Zusammenhang ist auch die signifikant erhöhte
Chloridkonzentrarion im Harn der Versuchstiere zu erklären, die auch von PATIENCE und
CHAPLIN (1997) nach dem Einsatz von CaCl2 festgestellt wurde. BUDDE und
CRENSHAW (2003) weisen in ihrer Arbeit daraufhin, dass eine vermehrte Chloridaufnahme
durch eine gesteigerte renale Exkretion von Chlorid in Verbindung mit NH4+-Ionen
kompensiert wird. Diese vermehrte Exkretion von NH4+-Ionen beruht nach der gesteigerten

Diskussion
121 121
Ausscheidung der Chloridionen (Anionen) auf einer transmuralen Potentialdifferenz, zu deren
Ausgleich nun die entsprechenden Kationen (NH4+-Ionen) ausgeschieden werden müssen.
Auf diesem Weg erfolgt letztlich wie bei der Verwendung von tierischen Eiweißen, DL-
Methionin oder Ammoniumchlorid die Ansäuerung des Harns mit dem Unterschied, dass
beim Einsatz dieser Stoffe eine Verstoffwechselung der Verbindungen erfolgt, nachdem sie
aus dem Intestinum absorbiert wurden. So entsteht z.B. aus Methionin Schwefelsäure,
Brenztraubensäure und Ammoniak, und aus Ammoniumchlorid entsteht Harnstoff und
Salzsäure, die auch bei der Metabolisierung des tierischen Eiweißes freigesetzt werden
(HARRINGTON u. LEHMANN 1970; PATIENCE et al. 1987). Das bei diesen
Umsetzungsprozessen entstandene NH4+ wird unter physiologischen Bedingungen in der
Leber mit der äquimolaren Masse an Bikarbonat zu Harnstoff umgewandelt, der dann über die
Nieren ausgeschieden wird. Im Falle einer metabolischen Acidose fallen allerdings so viele
NH4+-Ionen an, dass diese in der Leber an Glutamin gekoppelt werden. Dieses wird zu den
Nieren transportiert und dort zu NH4+ und Glutamat hydrolisiert. Das Glutamat dissoziiert
weiter in NH4+ und α-Ketoglutarat2-, welches im weiteren Verlauf unter dem Verbrauch von 2
H+-Ionen zu Glucose umgesetzt wird. Auf diesem Weg wird somit zusätzlich zur
Abpufferung der metabolischen Acidose und damit zur Konstanterhaltung des Blut-pH-
Wertes beigetragen. Die entstandenen NH4+-Ionen dissoziieren innerhalb der Epithelzellen
der Nierentubuli zu NH3 und H+, um dann in das Tubuluslumen zu diffundieren und sich hier
erneut zum NH4+ zu verbinden. Aus diesem Grund kann im Falle einer metabolischen
Acidose der Gehalt an Ammonium-Ionen im Harn um das 10fache ansteigen. In der hier
vorliegenden Untersuchung wurde die Konzentration an NH4+ im Harn allerdings nicht
bestimmt, aber im Vergleich zu anderen Arbeiten (PATIENCE u. CHAPLIN 1997; BUDDE
u. CRENSHAW 2003) müsste der Gehalt an NH4+ im Harn der Versuchstiere signifikant
angestiegen sein.

Diskussion
122 122
5.2.6. Der pH-Wert und die Bikarbonatkonzentration im Blut der Sauen
In dem hier vorliegenden Versuch war es möglich durch die Zugabe von zweimal täglich 14 g
(US) Calciumchlorid bei den Tieren der Versuchsgruppe innerhalb von 28 Tagen den pH-
Wert im Blut der Sauen signifikant (p<0,01) zu senken, so dass am 108. Trächtigkeitstag ein
Blut-pH-Wert von pH 7,37 erreicht wurde. In diesem Zeitraum des Versuches war der Harn-
pH-Wert der Versuchsgruppe (pH 5,96 ± 0,45) signifikant erniedrigt gegenüber dem Wert der
Kontrolltiere (pH 7,05 ± 0,34). Nach der Umstellung auf das Alleinfutter für laktierende
Sauen stieg dann in der letzten Woche der Trächtigkeit der Blut-pH-Wert der Versuchstiere
schwach signifikant (p<0,05) an und erreichte zum Zeitpunkt der Abferkelung wieder den
neutralen pH-Wert von pH 7,40 ± 0,05. In diesem Zeitabschnitt steigerte sich ebenfalls der
pH-Wert im Harn dieser Sauen signifikant (p<0,01), der dabei aber weiterhin im sauren pH-
Bereich von pH 6,73 ± 0,46 angesiedelt war. Zur gleichen Zeit erfolgte bei den Kontrolltieren
ein ebenso signifikanter Anstieg des Harn-pH-Wertes, während der pH-Wert im Blut lediglich
tendenziell etwas anstieg. Von verschiedenen anderen Autoren (YEN et al. 1981; PATIENCE
et al. 1987b, PATIENCE und 1997) wurde ebenfalls eine Absenkung des pH-Wertes im Blut
von Schweinen nach dem Einsatz von säuernden Substanzen beschrieben. Nach ihren
Aussagen trat diese Verschiebung des pH-Wertes im Blut in Form einer metabolischen
Acidose allerdings erst dann ein, wenn die Regelmechanismen des Säure-Basen-Haushaltes
nicht mehr ausreichten bzw. wenn die Säureausscheidungskapazität der Nieren überschritten
wurde (BEKER 1999). Dieses Vorgehen stellt eine Schutzfunktion des Körpers dar, da er für
seine optimale Funktion auf die Konstanterhaltung des pH-Wertes im Blut angewiesen ist und
schon bei einer Verschiebung dieses Parameters auf Werte von weniger als pH 6,3 bzw. mehr
als pH 7,7 der Tod eintreten kann (SELDIN u. GIEBISCH 1989). Die Umstellung auf das
Laktationsfutter führte dann in der letzten Trächtigkeitswoche aufgrund einer veränderten
Mineralstoffzusammensetzung zu einer geringeren acidotischen Belastung des Organismus,
so dass die metabolische Acidose durch die Säureausscheidungskapazität der Nieren noch
ausgeglichen werden konnte, während die Ansäuerung des Harns erhalten blieb. DE
ROUCHEY et al. (2003) beobachteten in diesem Zusammenhang bei einer kontinuierlichen
Steigerung der EB im Sauenfutter von 0 bis 500 mEq/kg einen ebenfalls kontinuierlichen
Anstieg des pH-Wertes im Blut von pH 7,33 bis pH 7,43. In der hier vorliegenden Arbeit hat
die zusätzliche Gabe von CaCl2 dazu geführt, dass die Konzentration an Bikarbonat im Blut
der Versuchstiere nach der Futterumstellung auf das Laktationsfutter hochsignifikant
(p<0,001) von 27,9 mmol/l auf 25,62 mmol/l abfiel, um nach der Geburt und somit auch nach

Diskussion
123 123
dem Ende des Calciumchlorideinsatzes wieder hochsignifikant (p<0,001) auf 29,28 mmol/l
anzusteigen. Im gleichen Versuchabschnitt ereigneten sich in der Kontrollgruppe hingegen
keine signifikanten Konzentrationsveränderungen des Bikarbonatspiegels im Blut. Zum
Zeitpunkt der Abferkelungen bestand somit zwischen den beiden Tiergruppen ein
signifikanter Unterschied (p<0,01) bezogen auf den Bikarbonatspiegel im Blut der Tiere.
BUDDE und CRENSHAW (2003) berichten ebenfalls von einem schwach signifikanten
Absinken (p<0,05) der Konzentration an Bikarbonat im Blut der Tiere in Verbindung mit
einer Absenkung des pH-Wertes im Blut, was sie damit erklären, dass die metabolische
Acidose in diesem Versuch durch die Regelmechanismen des Körpers nicht mehr
ausgeglichen werden konnte. DE ROUCHEY et al. (2003) beobachten bei der
kontinuierlichen Steigerung der EB im Sauenfutter von 0 bis 500 mEq/kg TS einen
hochsignifikanten (p<0,001) und ebenfalls kontinuierlichen Anstieg der Konzentration an
HCO3- im Blut von 19,0 bis 28,8 mmol/l. SELDIN und GIEBISCH (1989) verweisen darauf,
dass der pH-Wert und die Konzentration an HCO3- im Blut nach dem Prinzip der
HENDERSEN-HASSELBACH-GLEICHUNG zusammenhängen. Die Entwicklung des pH-
Wertes im Blut ist somit eng mit dem Verlauf der Konzentration an Bikarbonat verbunden, da
es im Körper die wichtigste Puffersubstanz darstellt (HIERHOLZER u. FROMM 1995). Im
Falle einer metabolischen Acidose steigert der Körper die Reabsorptionsrate an Bikarbonat im
proximalen Tubulusabschnitt der Nieren von ca. 90% unter physiologischen Bedingungen auf
nahezu 100%, um so diesen acidotischen Zustand auszugleichen. Im Zuge dieser
Resorptionssteigerung konnte während einer metabolischen Acidose ein signifikantes
Absinken der Konzentration an HCO3- im Harn gemessen werden (BUDDE u. CRENSHAW
2003) bzw. es wurde kein Bikarbonat mehr über den Primärharn ausgeschieden
(HIERHOLZER u. FROMM 1995).
In diesem Versuch wurde durch die Zugabe von CaCl2 in den ersten 4 Wochen zunächst eine
metabolische Acidose erzeugt, die zu einer massiven Reabsorption von Bikarbonat führte.
Durch die Umstellung auf das Laktationsfutter konnte dann keine metabolische Acidose mehr
erreicht werden, was anscheinend eine Verringerung der Reabsorption von Bikarbonat aus
dem Primärharn bewirkte. Diese Entwicklung lässt sich einerseits vom Verlauf des
Bikarbonatspiegels im Blut und andererseits anhand der gemessenen pH-Werte im Harn der
Versuchstiere ableiten.

Diskussion
124 124
5.2.7. Die Veränderungen der Calciumbilanz und des Knochenstoffwechsels der Sauen
Durch die Substitution von täglich zweimal 14 g (US) CaCl2 wird den Sauen der Versuchs-
gruppe oral eine größere Menge an Calcium verabreicht als den Tieren der Kontrollgruppe (in
den ersten 28 Tagen z.B. ca. 150% der Kontrollgruppe), somit nehmen in diesem Zeitab-
schnitt die Kontrolltiere 7,44 g und die Sauen der Versuchsgruppe 11,05 g Calcium mit jedem
Kilogramm an Trockensubstanz des Alleinfutters auf (siehe Abb. 12). Bei der Auswertung der
Calciumkonzentration im Blut der Tiere macht sich diese Differenz der oralen Zufuhr
allerdings nicht bemerkbar, da während des Versuches zwischen den beiden Tiergruppen
weder für die Gesamtkonzentration an Calcium noch für die Konzentration an ionisiertem
Calcium signifikante Unterschiede bestehen. Somit scheint die differenzierte orale Calcium-
aufnahme keinen relevanten Effekt auf die Calciumhomöostase im Blut der Sauen auszuüben.
Zu erklären ist dieser Sachverhalt mit dem Resorptionsverhalten des Darmes, da beim
Schwein lediglich 30 – 40% des mit der Nahrung aufgenommenen Calcium im Dünndarm
resorbiert werden (GIESEMANN et al. 1998), während das übrige Calcium zusammen mit
dem endogenen Calcium, das über die Galle in den Darm sezerniert wird, fäkal ausge-
schieden wird. Außerdem wird zu jeder Zeit zum Ausgleich der aus dem Darm
aufgenommenen Calciummenge überschüssiges Calcium renal über den Harn abgeführt. Die
renal ausgeschiedene Menge an Calcium ist dabei unter physiologischen Bedingungen von
der endogenen über den Darm ausgeschiedenen Menge abhängig und beträgt ca. 13% der
endogen ausgeschiedenen Calciummenge. Im Endeffekt beträgt somit die Nettoaufnahme von
Calcium nur ca. 20 - 25% der oral aufgenommenen Menge dieses Elementes (MÄNNER u.
BRONSCH 1987). Außerdem beobachteten JAMBOR u. PROCHAZKA (1977) in ihrer
Untersuchung, dass nach einer Ansäuerung der Futterration die fäkal ausgeschiedene
Calciummenge signifikant angestiegen ist, obwohl der Anteil an endogenem Calcium im Kot
auf 2- 5% abgesunken war, während im Vergleich dieser Anteil an endogenem Calcium bei
der Verfütterung einer neutralen Ration bei über 29% angesiedelt war. Sie folgerten daraus,
dass durch eine Verschiebung der AKB in der Ration zu Gunsten der Anionen die
Ausnutzung des oral aufgenommenen Calcium abnimmt. Diese Ergebnisse lassen den Schluss
zu, dass der acidierende Effekt, der bei den Versuchstieren erreicht wurde, bei diesen Sauen
zu einer erheblichen Verminderung der Calciumresorption geführt hat und dass das Calcium,
das diese Tieren zusätzlich oral verabreicht bekamen, somit schlechter ausgenutzt wurde. Mit
diesem Sachverhalt lässt sich auch der in diesem Versuch fehlende Effekt der höheren

Diskussion
125 125
Calciumaufnahme auf den Gehalt an ionisiertem Calcium im Blut und die
Gesamtkonzentration an Calcium im Plasma der Tiere erklären (siehe Abb. 12).
Abb. 12: Darstellung zum Verbleib der oral aufgenommenen Calciummenge.
Die Gesamtkonzentration an Calcium im Plasma setzt sich aus ca. 10% komplexgebundenem
Calcium, ca. 40% proteingebundenem Calcium und ca. 50- 60% aus ionisierten Calcium
zusammen (MÄNNER u. BRONSCH 1987). Das ionisierte Calcium stellt dabei die
physiologisch bedeutende Komponente des Calciumgesamtgehaltes dar (MÄNNER u.
BRONSCH 1987). In dem hier vorliegenden Versuch fiel in beiden Tiergruppen während der
ersten 28 Tage der Versuchsphase die Konzentration an ionisiertem Calcium lediglich
tendenziell ab, um dann in der letzten Woche vor der Geburt signifikant (p<0,01) abzusinken.
Somit sank die Konzentration an ionisiertem Calcium im Blut der Sauen kontinuierlich vom
Anfang des Versuches am 80. Trächtigkeitstag bis zur Abferkelung. BOSTEDT (1978)
registrierte in seinem Versuch unmittelbar vor der Geburt ebenfalls diese starke Depression
der Konzentration an ionisiertem Calcium im Blut, die teilweise bis zum 2. Tag p.p. anhielt.
HANSARD et al. (1966) berichteten zu diesem Thema von einem erhöhten Verbrauch an
Calciumionen am Ende des letzten Drittels der Trächtigkeit, da aufgrund des schnellen
Gewichtszuwachses der Feten in diesem Zeitabschnitt der Trächtigkeit ca.1,98 g Calcium pro
Tag für die Entwicklung des fetalen Knochengewebes bei einem Wurf von 8 Ferkeln
verbraucht werden. Dieser Bedarf an Calcium für die Entwicklung des fetalen
Calciumhomöostase Calciumhomöostase Konzentration im Blut Konzentration im Blut (Keine signifikanten Differenzen) (Keine signifikanten Differenzen)
Orale Orale Calciumaufnahme Calciumaufnahme
a)Kontrolltiere a)Kontrolltiere (7,44 g/ (7,44 g/ kgTS kgTS ) )
a=100% a=100% b)Versuchstiere b)Versuchstiere (11,05 g/ (11,05 g/ kgTS kgTS ) )
b=149% b=149%
Ansatz: Ansatz: a) a) Maternales Maternales Gewebe Gewebe
( ( 8g/ 8g/ kgKGW kgKGW ) ) b) Fetales Gewebe b) Fetales Gewebe
(ca. 35mg/ (ca. 35mg/ kgKM kgKM /d) /d) Keine relevanten gruppen Keine relevanten gruppen - - spezifischen Differenzen spezifischen Differenzen
Renale Renale Exkretion Exkretion (0,21 (0,21 ± ± 0,19mg/dl(Ca/Crea)) 0,19mg/dl(Ca/Crea)) Keine Keine relevanten relevanten gruppen gruppen - - spezifischen spezifischen Differenzen Differenzen
Faekale Fäkale Ausscheidung Ausscheidung Ausmaß unbekannt? Ausmaß unbekannt? Erwartet gemäß der Erwartet gemäß der oralen Calciumaufnahme: oralen Calciumaufnahme: Kontrolltiere 100% Kontrolltiere 100% Versuchstiere 149% Versuchstiere 149%

Diskussion
126 126
Knochengewebes wird aus dem Pool an ionisiertem Calcium aus dem Blut der Muttertiere
gedeckt. HANSARD et al. (1966) zeigten weiterhin, dass im Verlauf der Trächtigkeit am 70.
Trächtigkeitstag 6%, am 105. Tag der Trächtigkeit 17,8% und am 114.Tag sogar 21,3% des
von der Sau aus dem Futter resorbierten Calcium diaplazentar auf die Feten übertragen
werden. GUEGUEN und PEREZ (1981) haben in ihren ähnlich gelagerten Versuchen bei
einem Wurfgewicht von 14 kg eine Einlagerung von 3,7 g Calcium und 2,2 g Phosphor pro
Tag gemessen. Somit besteht in der Phase vom 80. Trächtigkeitstag bis zum Zeitpunkt der
Abferkelung ein erhöhter und im Verlaufe der Trächtigkeit ansteigender Bedarf an ionisierten
Calcium, der in dem hier vorliegenden Versuch mit einem durchschnittlichen Wurfgewicht
von 14,75 kg und einer durchschnittlichen Ferkelzahl von 12,42 Ferkeln pro Wurf ein
größeres Ausmaß gehabt haben dürfte als in den beiden genannten Studien zu diesem Thema.
Da zwischen den beiden Tiergruppen weder für das Wurfgewicht noch für die Ferkelzahl
nenneswerte Differenzen bestanden, kann daraus der Schluss gezogen werden, dass für den
fetalen Calciumansatz ebenfalls kein Unterschied zwischen den Tiergruppen bestand (siehe
Abb. 12). Die gleichen Verhältnisse galten übrigens auch für den Ansatz des maternalen
Gewebes während der Trächtigkeit, da die Körpermassenentwicklung ebenfalls in beiden
Gruppen nahezu gleich war. Somit sorgt der gestiegene Calciumbedarf im Laufe der
Trächtigkeit für das zunächst tendenzielle und in der letzten Woche vor der Abferkelung
sogar signifikante Absinken der Konzentration an ionisiertem Calcium im Blut der Sauen. In
den ersten 2 Tagen nach der Abferkelung stieg die Konzentration an ionisiertem Calcium
wieder signifikant an und erreichte dann im Verlauf der ersten Laktationswoche nahezu die
Ausgangswerte aus der Zeit vor dem Versuch, da zu dieser Zeit dem Körper der Muttertiere
kein ionisiertes Calcium zum Aufbau des fetalen Knochengewebes mehr entzogen wurde.
Dieser postpartale Konzentrationsanstieg an ionisiertem Calcium verlief außerdem nahezu
parallel zur Entwicklung der Gesamtkonzentration an Calcium im Plasma, die im Verlauf des
Versuches nach der Abferkelung kontinuierlich angestiegen ist und somit postpartal
signifikant über den Ausgangswerten zum Anfang des Versuches angesiedelt war. Nach den
Aussagen von BOSTEDT (1987) kann dies als Zeichen einer im Vergleich zur Trächtigkeit
besseren Calciumverdauung gewertet werden. GIESEMANN et al. (1998) beobachteten in
ihrer Untersuchung in der Säugezeit der Sauen ebenfalls einen signifikanten Anstieg der
Verwertung von Calcium und Phosphor, wobei sie davon berichteten, dass dieser Anstieg
vom Alter der Tiere beeinflusst wurde, da dieser bei älteren Sauen deutlicher ausgeprägt war
als bei den jüngeren Tieren.

Diskussion
127 127
Die Exkretion von Calcium war in dem hier vorliegenden Versuch in beiden Gruppen
ebenfalls nahezu gleichwertig und ohne signifikante Unterschiede (siehe Abb. 12). Aber unter
der Prämisse, dass infolge der Acidierung die Absorptionsrate für Calcium verringert wird, ist
in dem hier vorliegenden Versuch die Nettoabsorption von Calcium in der Versuchs-
tiergruppe nicht wesentlich gesteigert worden, obwohl diesen Tieren pro Mahlzeit zusätzlich
14 g (US) CaCl2 verabreicht wurden. GIESEMANN et al. (1998) zeigten in ihren Studien
ebenfalls, dass trotz unterschiedlicher Aufnahme von Calcium bei den Sauen die über den
Harn ausgeschiedene Menge dieses Stoffes nicht beeinflusst wurde.
BUDDE und CRENSHAW (2003) wiesen dagegen auf eine hochsignifikant (p<0,001)
gesteigerte Calciumexkretion in Verbindung mit einer acidotischen Belastung des Säure-
Basen-Haushaltes hin und wurden in dieser Aussage durch andere Autoren (LEMANN et al.
1967; NEWELL u. BEAUCHENE 1975; PATIENCE u. CHAPLIN 1997) bestätigt.
Allerdings wurde im Gegensatz zu der hier vorliegenden Arbeit in all diesen Untersuchungen
nicht mit Sauen gearbeitet, die sich im letzten Drittel der Trächtigkeit befanden und somit wie
beschrieben einen erhöhten Calciumbedarf haben, was abgesehen von der metabolischen
Acidose für sich allein schon zu einer katabolen Situation des Knochenstoffwechsels und
somit zur Freisetzung von Calciumionen führt.
Zur Regulation des Calciumstoffwechsels wird das kontinuierliche Absinken der
Konzentration an ionisiertem Calcium vom 80. Trächtigkeitstag bis zur Abferkelung im Blut
der Sauen von so genannten Calcium-sensitiven Rezeptoren wahrgenommen (DIAZ et al.
1999), die daraufhin in den Hauptzellen der Epithelkörperchen die vermehrte Produktion
einer Vorstufe des Parathormons auslösen. Diese Vorstufe wird noch innerhalb der Haupt-
zellen durch eine hydrolytische Spaltung in das aktive Parathormon umgewandelt. Diese
biologisch aktive Form des Parathormons entfaltet daraufhin seine Wirkung über spezifische
Rezeptoren im Knochengewebe und in den Nieren, um den niedrigen Calciumgehalt im Blut
wieder anzuheben. Deshalb steigert es im Tubulussystem der Nieren die Rückresorption von
Calcium und verringert zusätzlich die glomeruläre Filtrationsrate (HEBERT et al. 1997), so
dass letztendlich weniger Calcium über den Primärharn ausgeschieden wird. Außerdem
fördert das PTH in den Nieren die Bildung von 1,25-Dihydroxycholecalciferol (1,25-
(OH)2D), das im Darm die aktive Absorption von Calcium steigert, was zu der schon
beschriebenen besseren Verfügbarkeit des oral aufgenommenen Calciums während der
Laktation beiträgt. Im Knochengewebe übt das Parathormon über spezielle PTH-Rezeptoren,
die auf den Osteoblasten vorhandenen sind, eine direkte Wirkung auf den Knochen-
stoffwechsel aus. Das Parathormon reguliert die Größe und Anzahl der Osteoblasten

Diskussion
128 128
(KANZAWA et al. 2000) und verringert zusätzlich die Osteoprotegerin-Synthese in diesen
Knochenzellen, was dann sekundär den Knochenabbau über die Osteoklasten fördert (LUI et
al. 1998; OKADA et al. 2002). Somit konnte bei in-vitro-Versuchen eine hemmende Wirkung
von PTH auf die Synthese und Abgabe von Matrixproteinen wie z.B. Typ-I-Kollagen,
Osteocalcin und alkalische Phosphatase der Osteoblasten festgestellt werden
(BOGDANOVIC et al. 2000), die in Verbindung mit der gesteigerten Osteoklastentätigkeit
insgesamt zu einem Abbau von Knochengewebe führt (BUSHINSKY 1995; BUSHINSKY et
al. 1995; KRIEGER et al. 1992). Betrachtet man in diesem Zusammenhang die Entwicklung
der PTH-Konzentrationen im Plasma der Tiere, fallen zunächst die sehr großen tier-
individuellen Konzentrationsschwankungen auf, die von minimal 1,19 pg/ml bis maximal
180,27 pg/ml reichen. Diese überdimensionale Streuung der Einzelwerte machte eine
statistische Auswertung zur Herausstellung gruppenspezifischer Unterschiede nahezu un-
möglich, so dass man sich in diesem Zusammenhang mit tendenziellen Konzentrations-
entwicklungen des Parathormons zufrieden geben muss. Betrachtet man die relative
Steigerung der PTH-Konzentration im Plasma im Vergleich zu den Ausgangswerten, die am
80. Trächtigkeitstag gemessen wurden, wird das starke Ansteigen dieses Wertes in den ersten
4 Wochen bei den Kontrolltieren auf das 4,93-fache des Ausgangswertes deutlich, während in
der Versuchsgruppe lediglich eine Steigerung um das 2,55-fache erfolgte. Aufgrund dieser
Werte müsste der Abbau von Knochengewebe somit bei den Kontrolltieren tendenziell stärker
ausgeprägt gewesen sein als in der Versuchsgruppe.
Dabei bleibt allerdings zu berücksichtigen, dass der acidierende Effekt der CaCl2-Substitution
sich ebenfalls auf den Knochenstoffwechsel auswirkt und für einen Abbau von Knochen-
gewebe sorgt. Bei Lämmern und bei Ratten hatte eine längere Verfütterung von acidierenden
Rationen ebenfalls ein Ansteigen der Knochenresorption und ein Absinken des Knochen-
aufbaues zur Folge (KRAUT et al. 1986; DAMIR et al. 1991). Die Schlüsselrolle dieser
Knochenresorption infolge einer Acidose übernimmt die Konzentration an Bikarbonat
(BUSHINSKY et al. 1992), da selbst bei gleichem pH-Wert aber sinkender Bicarbonat-
konzentration die Ca-Mobilisation ansteigt (BUSHINSKY et al. 1996). In der letzten Woche
vor der Abferkelung sank die Konzentration an Bikarbonat im Blut der Versuchstiere
signifikant unter den Wert der Kontrollgruppe, so dass bei den Tieren der Versuchsgruppe
über die sinkende Bikarbonatkonzentration der Knochenabbau gesteigert wurde. SELDIN und
GIEBISCH (1989) sehen diesen Knochenabbau außerdem als Reaktion des Körpers zur
Abpufferung der extrazellulären Flüssigkeit und BROSNAN und BROSNAN (1982)
beziffern den Verbrauch von Säure auf 9,2 mol, um 10 mol Ca2+ aus dem Knochen

Diskussion
129 129
herauszulösen. Da der Verbrauch an Calcium in beiden Gruppen identisch war und das
zusätzlich oral verabreichte Calcium aufgrund der acidotischen Stoffwechselsituation der
Versuchstiere nicht genutzt werden konnte, war für beide Tiergruppen nahezu die gleiche
Menge an Calcium verfügbar. Zum relativ geringeren Parathormonanstieg im Plasma der
Versuchstiere kommt es, da in dieser Tiergruppe die katabole Ausrichtung des
Knochenstoffwechsels schon durch das signifikante Absinken des Blut-pH-Wertes in den
ersten 28 Tagen des Versuches erreicht wurde. Durch die puffernde Wirkung des
Knochenabbaues stieg der pH-Wert im Blut der Versuchstiere wieder an, da beim Abbau des
Knochens H+-Ionen verbraucht wurden. In der Woche vor der Abferkelung wurde der Abbau
von Knochensubstanz und damit auch die Freisetzung von Calcium durch den signifikanten
Konzentrationsabfall an Bikarbonat im Blut der Versuchstiere aufrechterhalten. Bei den
Tieren der Kontrollgruppe, bei denen weder eine signifikante Acidose noch ein signifikantes
Absinken des Bikarbonatspiegels beobachtet wurde, ging die Erhaltung der
Calciumhomöostase dagegen den klassischen Weg über die Ca-sensitiven Rezeptoren, die
aufgrund des abfallenen Gehaltes an ionisiertem Calcium die vermehrte Bildung von
Parathormon auslösten.
Zur Überwachung des Stoffwechsels im Knochengewebe wurde die Plasmakonzentration der
knochenspezifischen alkalischen Phosphatase analysiert, bei der es sich um einen so
genannten Knochenformationsmarker handelt. Diese Phosphatase wird von den Osteoblasten
produziert und verankert sich dann in deren Plasmamembran. Nach einiger Zeit löst sie sich
aus dieser Verbindung und ist somit als Knochenformatiosmarker im Plasma nachweisbar
(CHRISTENSON 1997), bevor sie in der Leber eliminiert wird (DELMAS 1991).
In diesem Versuch war bei den Sauen der Versuchsgruppe vom 80. bis zum 108.
Trächtigkeitstag ein signifikantes Absinken der Konzentration dieses Knochenformations-
markers zu verzeichnen, während die Kontrolltiere in diesem Zusammenhang lediglich einen
tendenziellen Konzentrationsabfall zeigten. Dieser signifikante Konzentrationsabfall der
knochenspezifischen alkalischen Phosphatase lässt auf eine Verminderung der Osteoblasten-
tätigkeit bzw. der Osteoblastenanzahl schließen und stützt somit die Aussage, dass die
metabolischen Acidose, die bei den Sauen der Versuchsgruppe mit der Substitution von CaCl2
erzeugt wurde, zu einer Resorption von Calcium aus dem Knochen geführt hat. Nachdem am
108. Trächtigkeitstag auf das Alleinfutter für laktierende Sauen umgestellt wurde, erfolgte
dann allerdings sowohl in der Kontrollgruppe als auch bei den Tieren der Versuchsgruppe ein
signifikanter Anstieg der Konzentration der knochenspezifischen alkalischen Phosphatase im
Plasma der Sauen, was als Zeichen einer gesteigerten Osteoblastentätigkeit bzw. einer

Diskussion
130 130
vermehrten Anzahl aktiver Osteoblasten gewertet werden kann. Daraus lässt sich eigentlich
schließen, dass zu diesem Zeitpunkt in beiden Gruppen anabole Stoffwechselvorgänge im
Bereich des Knochengewebes abliefen. Tatsächlich wurde den Tieren beider Gruppen nach
der Umstellung auf das Laktationsfutter auch eine größere Menge an Calcium und anderen
Mineralstoffen zugeführt, da einerseits die Mineralstoffkonzentration dieses Futters höher war
als im Alleinfutter für tragende Sauen und weil andererseits nach dieser Futterumstellung
auch größere Mengen an Alleinfutter verabreicht wurden.
Zu diesem Zeitpunkt der Trächtigkeit erscheint aufgrund eines erhöhten Calciumbedarfs bzw.
der signifikant abgefallenen Konzentration an ionisiertem Calcium im Blut jedoch eine
vermehrte Einlagerung an Calcium in das Knochengewebe zweifelhaft. Allerdings deuteten
die Entwicklungen der Parathormonkonzentration im Plasma der Sauen, die zur Geburt hin
ebenfalls etwas abfielen, auf eine bessere Calciumverfügbarkeit hin. Gruppenspezifische
Unterschiede bezogen auf den Anstieg der Konzentration der bAP im Plasma bestanden
zwischen den Tieren der Versuchsgruppe und den Kontrolltieren nicht, so dass die Zugabe
von CaCl2 hier keinen Einfluss auf die Konzentration dieses Knochenformationsmarkers im
Plasma ausübte. SCHONEWILLE et al. (1994) stellten bei einer Untersuchung an
Milchkühen ebenfalls fest, dass bei diesen Tieren die AKB einer Ration keinen Einfluss auf
die Konzentration der knochenspezifischen alkalischen Phosphatase im Serum der Tiere hatte.
LIESEGANG et al. (2005) beobachteten dagegen eine stetige und hochsignifikante
Konzentrationsverringerung (p=0,001) dieses Knochenformationsmarkers schon ab der 15.
Woche der Trächtigkeit bis zum Ende der ersten Laktationswoche und berichteten von einem
hochsignifikanten Absinken (p<0,001) der Konzentration der knochenspezifischen
alkalischen Phosphatase sofort nach der Abferkelung. SATO et al. (2002) stellten in ihrer
Untersuchung fest, dass es bei der Analyse der knochenspezifischen alkalischen Phosphatase
bei der Kuh um den Zeitpunkt der Geburt sehr schwierig war, die knochenspezifische
alkalische Phosphatase von der Form der alkalischen Phosphatase zu unterscheiden, die im
Euter zu dieser Zeit in großer Menge gebildet wird. Außerdem waren sie der Meinung, dass
die knochenspezifische alkalische Phosphatase positiv von der im Euter gebildeten
alkalischen Phosphatase beeinflusst wurde. Ob dieser Zusammenhang eventuell auch beim
Schwein besteht, bleibt unbeantwortet. Aber dieser Zusammenhang könnte eine weitere
Erklärung für den signifikanten Anstieg dieses Parameters im Plasma der Tiere aus beiden
Gruppen zum Zeitpunkt der Abferkelung sein, da auch bei der Sau die Milchproduktion des
Gesäuges kurz vor der Geburt stark ansteigt.

Diskussion
131 131
Mit dem Calciumhaushalt eng verbunden ist der Phosphorhaushalt, allerdings ist dieser
weniger strikt geregelt. Somit kann die Entwicklung der Phosphorkonzentration im Plasma
der Sauen ebenfalls als ein Produkt des katabolen Knochenstoffwechsels angesehen werden ,
der sich aufgrund der präpartalen Calciumdrainage entwickelt hat. In dem hier durchgeführten
Versuch zeigte sich dieser Zusammenhang darin, dass der signifikante Abfall der
Konzentration an ionisiertem Calcium und der hochsignifikante Anstieg der Phosphor-
konzentration praktisch zeitgleich verliefen. In der Woche vor der Abferkelung war somit bei
allen Sauen ein hochsignifikanter Anstieg der Phosphorkonzentration zu verzeichnen. In den
ersten 2 bis 4 Tagen der Laktationsphase, in der sowohl die Konzentration an ionisiertem
Calcium wie auch der Gesamtgehalt dieses Elementes anstiegen, fielen auch die
Phosphorwerte im Plasma wieder signifikant ab und erreichten in der Folgezeit wieder ihr
präpartales Ausgangsniveau. Phosphor wird in einer katabolen Stoffwechselsituation des
Knochen-gewebes freigesetzt, indem von den Osteoklasten Hydroxyapatitkristalle
(Ca10(PO4)6(OH)2) aus dem Ostoid herausgelöst werden. Ausgelöst wird dieser katabole
Knochenstoffwechsel durch den Abfall des ionisierten Calcium im Blut, da dieses zur
Bildung des fetalen Knochengewebes verbraucht wird. GUEGUEN und PEREZ (1981)
zeigten, dass zu dieser Zeit der Trächtigkeit bei einem Wurfgewicht von 14 kg 3,7 g Calcium
am Tag in das fetale Gewebe eingelagert werden, während die tägliche Einlagerungsmenge an
Phosphor lediglich 2,2 g beträgt. Somit wird von dem aus dem maternalen Knochengewebe
freigesetzten Phosphor relativ wenig auf die Feten übertragen, was den Phosphorgehalt im
Plasma demnach ansteigen lässt. BOSTEDT (1978) konnte in seinen Untersuchungen zur
ante- und postpartalen Entwicklung der Elektrolytkonzentrationen im Serum von Sauen sehr
ähnliche Beobachtungen zu den Veränderungen der Phosphorkonzentration in diesem
Zeitraum machen und berichtete von einem signifikanten Anstieg (p<0,01) dieser
Konzentration zum Zeitpunkt der Geburt und ein ebenfalls signifikantes Absinken (p<0,05 –
0,01) des Phosphorgehaltes bis zum Ende der 3. Laktationswoche. Aus diesen Ergebnissen
kann somit für den hier vorliegenden Versuch festgehalten werden, dass die
Phosphorkonzentration im Plasma nicht von der Ansäuerung durch die Zugabe von CaCl2
beeinflusst wurde, sondern dass für die in beiden Tiergruppen über den gesamten
Versuchszeitraum sehr ähnlich verlaufende Konzentrationsentwicklung maßgeblich von den
Regelmechanismen zur Aufrechterhaltung der Konzentration an ionisiertem Calcium in den
letzten Wochen der Trächtigkeit bzw. zum Zeitpunkt der Geburt abhängig war. PATIENCE
und CHAPLIN (1997) berichteten in ihrer Forschungsarbeit nach dem Einsatz von CaCl2 in

Diskussion
132 132
der Mastschweinefütterung zum Zwecke der Acidierung ebenfalls davon, dass diese
Fütterung keinen Einfluss auf den Phosphorgehalt im Blut der Tiere ausgeübt hat.
Die relative renale Exkretion von Phosphor im Vergleich zum Kreatinin wurde bei den Sauen
der Versuchgruppe durch den Einsatz von CaCl2 schwach signifikant (p<0,05) reduziert, was
durch den relativ geringeren Parathormongehalt im Plasma dieser Tiere im Vergleich zu den
Kontrolltieren bedingt war, da durch den Einfluss des Parathormons die renale
Phosphorausscheidung gesteigert wird. Dies geschieht, indem das Parathormon die Bildung
von Na+-P-Symportcarriern einschränkt bzw. deren lysosomalen Abbau in den luminalen
Membranen der Tubulusepithelzellen der proximalen Tubuli fördert und somit die tubuläre
Resorption von Phosphor in Form von Phosphat hemmt. Von anderen Autoren (NIMMO et
al. 1981; GIESEMANN et al. 1998) wurde nach dem Einsatz von CaCl2 dagegen keine
Veränderung der Phosphorausscheidung über die Nieren beobachtet, da in diesen Studien sich
die Kontrolltiere in keiner katabolen Stoffwechselsituation des Knochenstoffwechsels
befanden und somit bei diesen Tieren kein erhöhter Parathormonspiegel vorhanden war.
Ein weiterer Effekt der erhöhten Calciumaufnahme der Versuchstiere, der in der hier
vorliegenden Arbeit leider nicht untersucht wurde, ist die fördernde Wirkung dieser forcierten
Aufnahme auf die Gastrinsekretion im Magen. So berichteten LUTHMAN et al. (1977) bei
einer Untersuchung an Schafen von einem signifikant erhöhten Gastrinspiegel im Plasma der
Tiere in der letzten Phase der Trächtitigkeit infolge einer erhöhten Calciumaufnahme.
Deshalb halten sie eine Beteiligung des Gastrins an der Regulation der Calciumhomöostase
für möglich. RÜMENAPF et al. (1998) konnten zwar keine Beweise für den Zusammenhang
zwischen Gastrin, Calcitonin und PTH belegen, sie beobachteten aber ebenfalls eine
vermehrte Neigung zur Osteopenie bei der Abwesenheit von Gastrin und eine Zunahme der
Knochenmasse und des Knochenaufbaues. Das Gastrin fördert die Magensaftsekretion und
damit die Bildung von Magensäure, außerdem begünstigt es die Magenentleerung und steigert
die Darmmotilität. Da es bei Sauen, die an dem MMA-Syndrom leiden oft zu Stoff-
wechselstörungen kommt, wäre es in einer weiteren Studie deshalb sehr interessant die
Wirkung einer forcierten Calciumaufnahme auf die Gastrinsekretion und weiterführend die
Wirkung dieser Substanz auf den Knochenstoffwechsel zu prüfen.

Diskussion
133 133
5.2.8. Die Konzentrationen der anderen Mineralstoffe im Blut und im Harn der Sauen
Durch die Zugabe von CaCl2 stieg in den ersten 4 Wochen des Versuches die Chlorid-
konzentration im Plasma der Versuchstiere tendenziell kontinuierlich an, während diese in der
Kontrollgruppe kontinuierlich absank. Somit bestand am 108. Trächtigkeitstag ein schwach
signifikanter Konzentrationsunterschied (p<0,05) von 1,7 mmol/l zwischen den Versuchs-
und den Kontrolltieren. Im weiteren Verlauf des Versuches nach der Futterumstellung auf das
Laktationsfutter fiel die Chloridkonzentration im Plasma bei allen Tieren bis zur Abferkelung
deutlich ab, um sich danach in der postpartalen Phase in beiden Gruppen wieder bei einem
Chloridgehalt von 106,5 bis 107,0 mmol/l einzupendeln. Vom Beginn der CaCl2-Substitution
am 80. Trächtigkeitstag bis zu deren Ende zum Zeitpunkt der Abferkelung lag die Chlorid-
konzentration im Plasma der Versuchstiere in dieser Untersuchung somit tendenziell immer
über den Werten der Kontrolltiere. PATIENCE und CHAPLIN (1997), die ebenfalls CaCl2 an
Schweine zur Verschiebung der AKB in der Ration verfütterten, beobachteten in ihren
Untersuchungen einen signifikanten Anstieg der Chloridkonzentration im Plasma der Tiere,
wie er in der hier durchgeführten Untersuchung lediglich am 108. Trächtigkeitstag erreicht
wurde. Außerdem stellten sie fest, dass sich die Chloridresorption unter diesen Bedingungen
nicht veränderte. Im Harn stieg die Konzentration an Chlorid in der Arbeit von PATIENCE
und CHAPLIN (1997) signifikant an. Auch BUDDE und CRENSHAW (2003), die ebenfalls
CaCl2 zur Acidierung eingesetzt haben, verzeichneten einen signifikanten Anstieg der
Plasmakonzentration an Chlorid. Außerdem wiesen sie in ihrer Arbeit daraufhin, dass eine
vermehrte Chloridaufnahme durch eine gesteigerte renale Exkretion von Chlorid in
Verbindung mit NH4+-Ionen kompensiert wird. Diese vermehrte Exkretion von NH4
+-Ionen
beruht nach der gesteigerten Ausscheidung der Chloridionen (Anionen) auf einer
transmuralen Potentialdifferenz, zu deren Ausgleich nun Kationen (NH4+-Ionen)
ausgeschieden werden müssen. Auch in der hier vorliegenden Untersuchung war die relative
Ausscheidung an Chlorid im Bezug auf die Kreatininexkretion der Versuchstiere signifikant
(p<0,01) gesteigert gegenüber den Kontrolltieren. Dieses Verhältnis ist dadurch zu erklären,
dass im Gegensatz zum Calcium das Chlorid unabhängig von der Konzentration dieses
Mineralstoffes im Futter nahezu vollständig im Darm resorbiert wurde und dass die CaCl2-
Substitution auf diesem Wege zu einer gesteigerten Chloridkonzentration im Blut geführt hat.
Das überschüssige Chlorid versuchte der Körper deshalb zur Konstanthaltung der
Mineralstoffhomöostase über die renale Exkretion von Chlorid wieder aus dem Organismus

Diskussion
134 134
auszuschleusen, woraufhin die Chloridkonzentration im Harn signifikant (p<0,01)
angestiegen ist.
Die Natriumkonzentrationen im Plasma zeigte nahezu den gleichen Verlauf wie die
Konzentration an Chlorid während der CaCl2-Substitution. Während die Natrium-
konzentration der Kontrolltiere vom Anfang des Versuches bis zur Abferkelung
kontinuierlich absank, stieg dieser Parameter bei den Sauen der Versuchsgruppe in den ersten
28 Tagen des Versuches tendenziell an, um dann in der Woche vor der Geburt
hochsignifikant auf den niedrigsten Wert innerhalb des Versuches von 138,8 mmol/l abzu-
fallen. In den ersten 4 Laktationstagen stieg die Natriumkonzentration im Blut der Tiere dann
in beiden Gruppen sehr gleichförmig an. Über die Zeit der zusätzlichen Calciumchloridgabe
wurde die Natriumkonzentration allerdings nicht signifikant beeinflusst, so dass sich zu
keinem Zeitpunkt signifikante Konzentrationsdifferenzen zwischen der Kontroll- und der
Versuchsgruppe ergaben. Auffallend war jedoch der relativ parallele Konzentrationsverlauf
von Natrium und Chlorid. Dieser kann allerdings mit dem Ausgleich einer Potentialdifferenz
erklärt werden, da die negativ geladenen Chlorid-Anionen aus dem zusätzlichen CaCl2 eine
negative Potentialdifferenz aufbauen, die der Körper durch die positiv geladenen Natium-
Kationen auszugleichen versucht. PATIENCE und CHAPLIN (1997) beobachteten in ihrer
Untersuchung zur Acidierung durch die Zugabe von CaCl2 allerdings keine signifikanten
Auswirkungen auf die Natriumkonzentration im Blut der Tiere.
Die CaCl2-Substitution rief ebenfalls keine statistisch relevanten Unterschiede der Kalium-
konzentration im Blut der Sauen hervor, wovon auch PATIENCE und CHAPLIN (1997)
berichteten. Lediglich am 108. Trächtigkeitstag zeigte die Kaliumkonzentration der Versuchs-
tiere einen tendenziell höheren Wert als bei den Kontrolltieren, der sich ebenfalls wie beim
Natrium durch den Ausgleich einer Potentialdifferenz erklären lässt.
In der Exkretion von Natrium zeigten sich in diesem Versuch keine relevanten Unterschiede
zwischen den Tieren der Versuchsgruppe und den Kontrolltieren. Da allerdings in diesem
Versuch alle Tiere mit den gleichen Mengen an Natrium versorgt wurden, waren in diesem
Zusammenhang auch keine Unterschiede zu erwarten. BUDDE und CRENSHAW (2003)
beobachteten ebenfalls keine Beeinflussung der Natriumexkretion durch den Einsatz von
CaCl2.

Diskussion
135 135
5.2.9. Die Entwicklung der Kreatininkonzentrationen im Plasma und im Harn
Die Kreatininkonzentrationen im Harn und im Plasma wurden in diesem Versuch bestimmt,
um eine Bezugsgrösse für die teilweise stark schwankenden Harnkonzentrationen der
einzelnen Mineralstoffe zu bekommen. Dieser Methode bedienten sich auch RUHRMANN et
al. (1986), um mit dieser Diuresekorrektur die stark schwankenden Harnkonzentrationen der
verschiedenen Mineralstoffe vergleichen zu können. Dieses Verfahren setzt allerdings voraus,
dass ein ausreichender Zusammenhang zwischen der Kreatininkonzentration und der
Konzentration des entsprechenden Mineralstoffes besteht. Die jeweiligen Korrelations-
koeffizienten sind hierzu der Arbeit von RUHRMANN et al. (1986) zu entnehmen.
Die im Plasma der Sauen gemessenen Kreatininkonzentrationen zeigen weder zwischen den
beiden Tiergruppen noch zwischen den jeweiligen Analyseterminen signifikante
Unterschiede. Die jeweils berechneten Mittelwerte von 208,62 ± 41,55 µmol/l am 90.
Trächtigkeitstag und von 206,86 ± 41,55 µmol/l am 111. Trächtigkeitstag liegen allerdings
über dem von WENDT (1992) angegebenen Grenzwert für gesunde Sauen von 150 µmol/l.
Da in dem hier vorliegenden Versuch allerdings nahezu alle gemessenen Einzelwerte der
Kreatininkonzentration im Plasma über diesem Grenzwert liegen, ist nicht davon auszugehen,
dass bei allen Tieren eine negative Beeinflussung der Nierenfunktion vorlag, die z.B. durch
eine Infektion der Harnorgane und hier im Besonderen der Nieren ihren Ursprung haben
könnte. Ob allerdings Einzeltiere, die besonders hohe Kreatininwerte im Plasma aufwiesen,
an einer solchen Infektion litten und die aufgrund eines subklinischen Verlaufes dieser
Erkrankung weiter klinisch nicht auffällig waren, bleibt in dieser Arbeit ungeklärt, da vor der
Versuchsdurchführung auf eine bakteriologische Untersuchung der Harnproben verzichtet
wurde.

Diskussion
136 136
5.2.10. Der Einfluss des CaCl2-Einsatzes auf die Ferkel und den Ablauf der Geburten
Die Ergebnisse dieser Untersuchung zeigten, dass die zusätzliche Gabe von CaCl2 vom 80.
Trächtigkeitstag an bis zur Abferkelung weder einen nennenswerten Einfluss auf die Dauer
der Geburten noch auf die Zeitspanne von der Geburt des letzten Ferkels bis zum Abgang der
Nachgeburt hatte. Die im Mittel um 24 min verlängerte Geburtsdauer bei den Tieren der
Versuchsgruppe hatte ihren Ursprung in der Einbeziehung einer extrem verlängerten Geburts-
dauer von 5:13 h:min der Sau Nr. 667. Dieses Tier hat nach der Geburt von 3 Ferkel die
Abferkelung für ca. 2 Stunden unterbrochen, da im Rahmen des studentischen Unterrichtes
bei einem benachbarten Tier von 6 Studenten versucht wurde eine sonographische Unter-
suchung durchzuführen. Nachdem diese Störung des Geburtsablaufes vorüber war, hat die
Sau Nr. 667 die Abferkelung nach ca. 2,5 Stunden ohne Störungen fortgesetzt.
Der Zeitraum von der Geburt des letzten Ferkels bis zum Abgang der Nachgeburten zeigte in
beiden Tiergruppen eine nahezu identische Ausdehnung von minimal 20 min und maximal
1:45 h:min. Somit existierten auch in diesem Zusammenhang keine bedeutenden
Unterschiede zwischen den beiden Sauengruppen.
Der Einfluss der CaCl2-Substitution auf die Anzahl der insgesamt geborenen Ferkel, wie auch
der lebend, lebensschwach und tot geborenen Ferkel war in dieser Untersuchung ebenfalls
nicht signifikant. Obwohl die Sauen der Versuchsgruppe mit 11,57 lebend geborenen Ferkeln
pro Wurf und nur 0,36 lebensschwachen und 0,57 toten Ferkeln pro Wurf tendenziell bessere
Würfe zur Welt brachten als die Kontrolltiere mit 10,73 lebend, 0,53 lebensschwach und 1,07
tot geborenen Ferkeln. Auf diese tendenzielle Entwicklung der Anzahl an lebend geborenen
Ferkeln pro Wurf bei Sauen, denen mehr Calcium verabreicht wurde, verwiesen auch
MAHAN und FETTER (1982) in ihrer Arbeit. Die statistische Auswertung ihrer Forschungs-
ergebnisse erbrachte aber in diesem Zusammenhang wie in der hier vorliegenden Arbeit keine
signifikanten Unterschiede.
Auch die Entwicklung der Ferkel scheint von der unterschiedlichen Calciumversorgung der
Muttertiere unbeeinflusst zu bleiben. Die Auswertung der durchschnittlichen Geburtsgewichte
der Ferkel mit im Mittel 1,45 ± 0,25 kg in der Kontrollgruppe und 1,42 ± 0,27 kg in der
Versuchsgruppe ergab jedenfalls keinen signifikanten Unterschied zwischen diesen beiden
Gruppen. MAHAN und FETTER (1982) berichteten aus ihrer Arbeit dazu ebenfalls von einer
unveränderten Entwicklung des Geburtsgewichtes der Ferkel trotz vermehrter Calciumgabe.
Selbst der Mineralstoffhaushalt der Ferkel zeigte durch den Zusatz von gecoatetem Calcium-
chlorid zum Sauenfutter keine signifikante Beeinflussung in diesem Versuch. Somit konnten

Diskussion
137 137
zwischen den Ferkeln der Sauen aus der Versuchsgruppe und den Ferkel der Kontrollgruppe
bei der Blutgasanalyse weder in Bezug auf den pH-Wert oder die Konzentration an HCO3-
noch für die Konzentration an Mineralstoffen (Ca2+, Na, K und Cl) irgendwelche
signifikanten Unterschiede festgestellt werden. Diese Beobachtungen machten auch NIMMO
et al. (1981), die in ihrer Arbeit ebenfalls bei der Verfütterung von Rationen mit
verschiedenen Konzentrationen an Calcium keine Veränderungen dieser Werte bei den
Ferkeln der einzelnen Sauengruppen feststellen konnten.
Der Einsatz von gecoatetem Calciumchlorid scheint somit in der angewandten Menge
während der Trächtigkeit den Stoffaustausch zwischen dem Muttertier und den Ferkel nicht
zu beeinflussen, so dass diese Behandlung weder negative noch positive Effekte auf die
Anzahl der aufzuchtfähigen Ferkel und deren Entwicklung zum Zeitpunkt der Geburt ausübt.
Durch die prophylaktische Wirkung gegen das MMA-Syndrom, das die Aufzuchtleistungen
der befallenen Tier erheblich verringern kann, hat die CaCl2-Substitution allerdings im
Verlauf der Säugezeit einen relativ positiven Einfluss auf die Entwicklung dieser Ferkel und
somit ebenfalls auf die Leistungsfähigkeit der Sauen. Deshalb wäre in diesem Zusammenhang
in weiterführenden Untersuchungen die Entwicklung der Ferkel bis zum Absetztermin
interessant und bei den Sauen wären die Trächtigkeitsergebnisse der darauf folgenden
Belegung von Interesse, ob in diesem Zusammenhang eventuell ein positiver Einfluss auf
diese Parameter durch den CaCl2-Zusatz erreicht werden könnte.

Diskussion
138 138
5.2.11. Die Inzidenz des MMA-Syndroms während des Versuches
In dem hier durchgeführten Versuch zeigten 3 Sauen nach der Abferkelung die ersten
Anzeichen des MMA-Syndroms, dabei handelte es sich um die Sau Nr. 667 aus der Versuchs-
gruppe und die Tiere Nr. 597 und Nr. 703 aus der Kontrollgruppe. Diese Tiere fielen dem
betreuenden Personal am 1. (Nr. 667 und 597) bzw. am 2. Tag (Nr. 703) nach der
Abferkelung durch eine verzögerte Aufnahme der zugeteilten Futtermenge auf. Bei der
Kontrolle der Körperinnentemperatur zeigten alle Tiere Werte von ≥39,5°C auf und erhielten
daraufhin die in diesem Schweinebestand übliche medikamentelle Behandlung. Diese bestand
aus der Gabe von 0,4 mg Meloxicam pro kg KGW (Präparat: Metacam® Boehringer
Vetmedica GmbH, Ingelheim) 2 mal im Abstand von 24 Stunden zur Fiebersenkung und als
Antibiotikum 3 mal im Abstand von 24 Stunden Marbofloxacin (Präparat: Marboxyl® 10%,
Fa. Vetiquinol GmbH, Ravensburg) in einer Dosierung von 2 mg pro kg KGW. Zusätzlich
wurde diesen Sauen bei der Erstbehandlung ein Oxytocin-Depotpräparat mit dem Wirkstoff
Carbetocin (Präparat: Longacton®, Fa. VetCom-pharma GmbH, Tettnang) in einer Dosierung
von 0,14 mg pro Sau verabreicht. Bei allen betroffenen Tieren fiel nach dieser Behandlung
die Körpertemperatur auf <39,5°C und die Sauen zeigten wieder ein ungestörtes
Allgemeinbefinden, während die Futteraufnahme dieser Tiere allerdings nur langsam wieder
anstieg.
Das Tier mit der Nr. 667 aus der Versuchsgruppe hatte mit 05:13 h:min mit Abstand die
längste Geburtsdauer im Versuch, da dieses Tier aufgrund einer Störung während der Geburt
diese für ca. 2 Stunden unterbrochen hatte. Mit dieser stark verlängerten Geburtsdauer gehörte
dieses Tier bezogen auf das MMA-Syndrom zur Risikogruppe, da eine verlängerte Geburt das
Befallsrisiko erheblich steigern kann (BÖNING et al. 1976; HEINRITZI u. HAGN 1999).
Die Sau Nr. 597 aus der Kontrollgruppe gehörte ebenfalls zur MMA-Risikogruppe, da dieses
Tier von ihren 13 Ferkeln lediglich 8 lebend zur Welt brachte, während die übrigen 5 Ferkel
tot geboren wurden. Nach Ansicht von BÄCKSTRÖM et al. (1984) und BOSTEDT et al.
(1998) korreliert das Auftreten des MMA-Syndroms relativ eng mit der Anzahl der tot
geborenen Ferkel und mit der Größe des zur Welt gebrachten Wurfes.
Bei der Sau Nr. 703 aus der Kontrollgruppe wurde die Nachgeburt mit 01:45 h:min nach der
Geburt des letzten Ferkels im Vergleich relativ spät ausgestoßen. Dieses Tier zeigte dann
etwa 2 Tage nach der Abferkelung einen leicht eitrigen Ausfluss als Zeichen einer
puerperalen Endometritis und gehörte damit nach EHRENTRAUT (1968) und GLOCK
(1983) ebenfalls zur MMA-Risikogruppe. Die betroffenen Tiere zeigten lediglich in den

Diskussion
139 139
Tagen nach der Abferkelung eine erhöhte Körpertemperatur und eine langsamer ansteigende
Futteraufnahme. An den Gesäugen wurden dagegen keine Veränderungen festgestellt. Ihre
Ferkel wurden allerdings trotzdem zur Vorbeuge in den ersten Tagen der Laktation mit
künstlicher Sauenmilch versorgt. Bei der Betrachtung der Ergebnisse der übrigen analysierten
Parameter fielen diese Tiere dagegen weder positiv noch negativ auf.
Die Inzidenz des MMA-Syndroms lag in der Versuchsgruppe mit einer befallenen Sau von 14
Tieren bei 7,14%, während in der Kontrollgruppe mit zwei erkrankten Sauen von 15 Tieren
eine Inzidenz von 13,33% erreicht wurde. In anderen Arbeiten werden verschiedene Angaben
zu diesem Thema gemacht. So spricht JORSAL (1986) lediglich von einer Befallshäufigkeit
von 10% der Sauen, während Bollwahn et al. (1989) diese in Problembetrieben auf bis zu
40% beziffern und LERCH (1987) von einer Schwankungsbreite für diesen Wert von 14% bis
sogar auf 77% ausgeht. Die relativ geringe Inzidenz des MMA-Syndroms in dem hier
vorliegenden Versuch lässt sich durch weitgehende Optimierung der Faktoren wie z.B.
Fütterung, Wasserversorgung und Haltung erklären, die bei suboptimaler Gestaltung das
Auftreten des MMA-Komplexes erheblich begünstigen können. Ob und in wie weit die
unterschiedliche Befallshäufigkeit der Versuchtiere im Vergleich zu den Kontrolltieren auf
den Einsatz des gekapselten CaCl2 zurückzuführen war oder ob es sich dabei um einen
Zufallsbefund gehandelt hat, bedarf der weiteren Abklärung durch den Einsatz dieses Zusatz-
stoffes in einer größeren Tierpopulation. Aber die durch die Zugabe dieses Stoffes erreichte
signifikante Absenkung der Harn-pH-Werte deutete im Vergleich zu den genannten anderen
Forschungsarbeiten zu diesem Thema daraufhin, dass man durch den Einsatz von CaCl2
primär die Prävalenz von unspezifischen Harnwegsinfektionen verringern könnte, was sich
sekundär vor dem Hintergrund des Zusammenhanges der unspezifischen Form der
Harnwegsinfektion und dem späteren Auftreten des MMA-Komplexes auch auf eine
signifikante Absenkung der Prävalenz dieser Erkrankung auswirken könnte, wenn in dem
jeweiligen Bestand dieses Vorläufersyndrom des MMA-Komplexes eine Rolle spielt.

Diskussion
140 140
5.2.12. Die praktische Anwendung von CaCl2 zur MMA-Prophylaxe
Die Intention dieser Arbeit war die Überprüfung der Interaktion einer temporären Acidierung
und einer forcierten Calciumaufnahme, die von der Substitution von täglich zweimal 14 g
(US) Calciumchlorid ausgehen, das zur Prophylaxe gegen das MMA-Syndrom vom 80.
Trächtigkeitstag bis zur Abferkelung eingesetzt wurde. Die zur MMA-Prophylaxe notwendige
Absenkung des Harn-pH-Wertes wurde durch diese Substitution schon nach wenigen Tagen
erreicht, indem im Harn ein pH-Wert von im Mittel pH 5,96 ± 0,45 erzeugt werden konnte.
Die MMA-Prophylaxe nimmt hier den Weg über die Keimreduktion im Harn bzw. die
Verhinderung des Auftretens der unspezifischen Form der Harnwegsinfektion bei der Sau, die
nach heutigem Kenntnisstand als Vorläufererkrankung des MMA-Syndroms gilt.
ARNHOFER (1987) erreichte eine Verminderung der Keimzahl im Harn bei Sauen mit
signifikanter Bakteriurie, in dem er den pH-Wert im Harn unter pH 6,65 absenken konnte.
Allerdings bleibt dazu zu bemerken, dass diese Absenkung lediglich für den Zeitraum der
Ansäuerung erreicht werden konnte und nach dem Absetzen der säuernden Substanz die
Ausgangskeimzahlen wieder erreicht bzw. teilweise sogar überschritten wurden. JÜRGENS
(1991) reduzierte die Keimbelastung im Harn sogar signifikant um 1,5 Zehnerpotenzen durch
die Verabreichung von 2mval Ammoniumchlorid/kg KGW, indem er so den pH-Wert im
Harn von pH 7,29 bis auf pH 5,14 absenken konnte oder durch die Zugabe von 1g DL-
Methionin sogar eine pH-Wert-Absenkung von pH 7,24 auf pH 4,79 erreichte. JÜRGENS
(1991) kommt aufgrund seiner Untersuchungsergebnisse zu dem Schluss, dass unterhalb eines
pH-Wertes im Harn von pH 6,1 - 6,3 mit einer wirksamen Keimreduktion in diesem Medium
zu rechnen ist. In der hier vorliegenden Arbeit wurde am 90. Trächtigkeitstag bei den Tieren
der Versuchsgruppe im Mittel ein pH-Wert im Harn von pH 5,96 ± 0,45 gemessen, durch den
nach den Aussagen von JÜRGENS (1991) eine sichere Reduktion der Keimflora erreicht
werden müsste. In der hier vorliegenden Untersuchung wurde dagegen auf eine Bestimmung
der Keimzahl im Harn verzichtet. Es bleibt aber zu beachten, dass sich der Einsatz von
Substanzen, die zur Ansäuerung des Harnes eingesetzt werden, auf einen rein
phrophylaktischen Einsatz gegen die Besiedelung des Harntraktes mit einer schädigenden
Keimflora beschränkt, da durch deren Einsatz lediglich eine Verringerung des Keimgehaltes
erreicht werden kann, aber keine Elemination der Erreger stattfindet. Die Abtötung dieser
Keime bleibt somit allein dem Antibiotikaeinsatz vorbehalten.
Nach den Angaben von BERNER (1981) stellt E. coli mit 58,1-75,9% der nachgewiesenen
Keime, den hauptverantwortlichen Keim für die unspezifische Form der Harnwegsinfektion

Diskussion
141 141
beim Schwein dar. Somit müssten die erreichten Harn-pH-Werte zu einer erheblichen Keim-
reduktionen beitragen, da der Optimalbereich für das Wachstum von E. coli im pH-Bereich
zwischen pH 6 und pH 7 angesiedelt ist (ASSCHER et al. 1966). Die in dem hier
vorliegenden Versuch eingesetzte Menge an Calciumchlorid müsste deshalb ausreichen, um
pH-Werte im Harn zu erzeugen, die dazu in der Lage sind das Keimwachstum im Harntrakt
zu unterdrücken bzw. erheblich zu reduzieren. Außer diesem gewünschten Effekt, der einen
maßgeblichen Beitrag zur MMA-Prophylaxe leistet, führt die Gabe von Calciumchlorid zu
keiner negativen Beeinflussung der Sauen, da weder die Futteraufnahme noch die gemessenen
physiologischen Vorgänge im Körper der Muttertiere irgendwelche Anzeichen für eine
negative Beeinflussung gezeigt haben. Auch Auswirkungen auf die neugeborenen Ferkel oder
deren Entwicklung konnten in dieser Untersuchung nicht festgestellt werden. Aus diesem
Grund scheint die Gabe von CaCl2 eine probate Methode zur Prophylaxe gegen das Auftreten
des MMA-Komplexes bei der Sau zu sein.
Ob eine Erhöhung der verabreichten Dosierung an CaCl2 in diesem Fall zu einer Verbes-
serung der prophylaktischen Wirkung beitragen würde oder ob davon negative
Beeinflussungen der hier analysierten Parameter ausgehen würden, bleibt in dieser
Untersuchung allerdings unbeantwortet.
Über den Zeitraum des Einsatzes von CaCl2 kann man im praktischen Einsatz allerdings
verschiedene Ansichten vertreten. Aus fütterungstechnischer Sicht und aus Gründen des
Arbeitsablaufes wäre hier der Einsatz von CaCl2 ab der Umstallung der Sauen in den
Abferkelbereich zu propagieren, wenn diese Umstallung mindestens 8 – 10 Tage vor der
Abferkelung erfolgt. Da die Tiere mit der Umstallung in diesen neuen Stallbereich in der
Regel auch auf ein anderes Fütterungssystem, das speziell für diesen Bereich ausgelegt ist,
umgestellt werden, wäre mit diesem System bei entsprechender Einmischung des CaCl2 die
Futtervorlage in diesem Bereich zu automatisieren. Dabei sollte allerdings der genannte
Zeitraum der Umstallung vor der Abferkelung nicht unterschritten werden, um eine
ausreichende Vorlaufzeit des prophylaktischen Effektes zu gewährleisten. Kann diese
Zeitspanne der Umstallung vor der Geburt nicht eingehalten werden bzw. wird ein früherer
Einsatz gewünscht, wie er in diesem Versuch praktiziert wurde, wäre die separate Gabe des
CaCl2 oder die Anfertigung eines „Top-Dressings“, bei dem das CaCl2 einer kleinen Menge
an Alleinfutter oder anderen separat verabreichten Futterstoffen zu gemischt wird, das Mittel
der Wahl. Allerdings ist bei dieser Variante in den meisten Haltungssystemen für tragende
Sauen eine separate Vorlage per Hand nötig. Mit diesem zusätzlichen Arbeitsaufwand kann
der Zeitraum dieser Prophylaxemethode allerdings entsprechend ausgedehnt werden und

Diskussion
142 142
somit wie in dem hier beschriebenen Versuch auch effektiver gestaltet werden. Von einem
andauernden Einsatz von ansäuernden Substanzen während der gesamten Trächtigkeit kann
an dieser Stellen aufgrund der Auswirkungen auf das Knochengewebe und den
Calciumhaushalt nur abgeraten werden. Nach der Geburt sollte der Einsatz des CaCl2 in der
ersten Woche des Puerperiums allerdings weiter fortgesetzt werden, da in dieser Zeit eine
erhöhte Gefahr der Keimbesiedelung des Harn- und Geschlechtstraktes zu erwarten ist. Der
prophylaktische Einsatz von CaCl2 bleibt also, soweit dieses zu realisieren ist, auf das letzte
Drittel der Trächtigkeit und die erste Woche der Laktation beschränkt. Eine Erfolgskontrolle
der CaCl2-Substitution kann im praktischen Einsatz relativ einfach in Form einer Messung des
Harn-pH-Wertes durchgeführt werden. Dazu sollte zunächst der Harn-pH-Wert vor dem
Einsatz kontrolliert werden, um dann im Verlauf nach ca. 1 Woche durch erneute Messung
die entsprechende pH-Wert-Absenkung zu registrieren.

Zusammenfassung
143 143
6. Zusammenfassung
Björn Röcker
Untersuchung zur Acidierung des Harns mittels alimentärer Calciumchlorid-Gabe bei
tragenden Sauen.
In dieser Studie wurden die Auswirkungen einer Calciumchlorid-Gabe (CaCl2), die zur
Acidierung des Harns eingesetzt wurde, auf die Stoffwechselvorgänge wie den Säure-Basen-
Haushalt, den Knochenstoffwechsel und die Mineralstoffhomöostase bei tragenden Sauen
untersucht.
Der Versuchsgruppe, die aus 14 Sauen bestand, wurde dazu vom 80. Trächtigkeitstag bis zur
Abferkelung je zweimal täglich 14 g (US) Calciumchlorid (Calci Cap®) in das entsprechende
Alleinfutter gemischt. Die Kontrollgruppe, denen kein zusätzliches CaCl2 verabreicht wurde,
bestand aus 15 Tieren. Vom 80.-108. Trächtigkeitstag wurde ein Alleinfutter für tragende
Sauen (EB 208mEq/kgTS) und vom 108. Tag bis zum Versuchsende am 8. Tag p.p. ein
Alleinfutter für laktierende Sauen (EB 223mEq/kgTS) verfüttert.
Von jeder Sau wurden jeweils am 80. und 108. Trächtigkeitstag, sowie gleich nach der
Abferkelung und weiterführend jeden 2. Tag bis zum 8. Tag p.p. Blutproben gezogen, sowie
am 90. und 111. Trächtigkeitstag Spontanharnproben aufgefangen. Außerdem wurde jeweils
einem Ferkel pro Wurf gleich nach der Geburt eine Blutprobe abgenommen.
In diesen Proben wurden folgende Parameter analysiert:
- Blut: pH-Wert, Hydrogenkarbonat, Gesamtcalcium, ionisiertes Calcium, anorganisches
Phosphat, Chlorid, Natrium, Kalium, Kreatinin, Parathormon und die knochen-
spezifische alkalische Phosphatase.
- Harn: pH-Wert, Calcium, anorganisches Phosphat, Chlorid, Natrium und Kreatinin.
Um einen Einfluss der zusätzlichen CaCl2-Gabe auf den Ablauf der Geburten und die
Entwicklung der Ferkel zu prüfen, wurden außerdem die Geburtsdauer, die Zeitspanne bis
zum Abgang der Nachgeburten, die Anzahl der insgesamt geborenen Ferkel, sowie der
lebend, lebensschwach und tot geborenen Ferkel und das Gewicht dieser Ferkel bestimmt.

Zusammenfassung
144 144
Dabei zeigten sich folgende Veränderungen nach der zusätzliche CaCl2-Gabe:
1.) Der mittlere Harn-pH-Wert der Versuchstiere lag am 90. Trächtigkeitstag (pH 5,96
± 0,45) signifikant (p<0,01) unterhalb des pH-Wertes der Kontrolltiere (pH 7,05 ±
0,34). Bis zum 111. Trächtigkeitstag kam es nach der Futterumstellung in beiden
Gruppen zu einem signifikanten Anstieg der Harn-pH-Werte, so dass zu diesem
Zeitpunkt bei den Versuchstieren ein mittlerer pH-Wert von pH 6,73 ± 0,46 und in der
Kontrollgruppe von pH 7,31 ± 0,31 gemessen wurden. Der signifikante gruppen-
spezifische Unterschied (p<0,01) blieb allerdings bestehen.
2.) Auf die Entwicklung des mittleren Blut-pH-Wertes hatte die zusätzliche CaCl2-
Substitution keinen statistisch relevanten Einfluss (p>0,05).
3.) Am Tag der Abferkelung lag die Konzentration an Hydrogenkarbonat (HCO3-) im
Blut der Versuchstiere mit einer Differenz von 2,16 mmol/l schwach signifikant
(p<0,05) unterhalb Kontrollgruppe. Während der übrigen Versuchszeit zeigen sich
dagegen keine signifikanten Konzentrationsunterschiede (p>0,05) zwischen den
Tiergruppen.
4.) Die Zugabe von 2 mal täglich 14 g CaCl2 zeigte keinen signifikanten Einfluss
(p>0,05) auf die Konzentrationen von ionisiertem Calcium im Blut und Gesamt-
calciumkonzentration und anorganischem Phosphat im Plasma, sowie auf die
Plasmakonzentrationen von Natium und Chlorid und den Gehalt an Kalium im
Vollblut.
5.) Der Parathormonspiegel variierte sehr stark zwischen minimal 1,19 pg/ml bis
maximal 180,27 pg/ml. Der Vergleich der relativen Steigerung des Parathormon-
gehaltes zum gruppenspezifischen Ausgangswert zeigte, dass dieser Wert in der
Kontrollgruppe vom 80. Trächtigkeitstag bis zum 108. Tag auf das 4,93-fache an-
stieg, um dann vor der Geburt auf das 2,77-fache abzusinken. Bei den Versuchstieren
wurde dagegen am 108.Tag das 2,55-fache des Ausgangswertes erreicht und bis zur
Abferkelung erfolgte eine weitere Steigerung auf das 3,48-fache.

Zusammenfassung
145 145
6.) Bei der Auswertung der Analysedaten zur Konzentration der knochenspezifischen
alkalischen Phosphatase ergaben sich keine relevanten gruppenspezifischen
Differenzen (p>0,05).
7.) Am 90. und am 111. Trächtigkeitstag war die relative renale Ausscheidung an
Phosphor (P/Krea) der Kontrolltiere schwach signifikant (p<0,05) gegenüber der
Versuchsgruppe erhöht, während bei den Sauen der Versuchsgruppe dagegen an
beiden Terminen die relative Ausscheidung von Chlorid signifikant (p<0,01) über den
Werten der Kontrollgruppe lag. Die Ausscheidung von Calcium und Natrium über den
Harn wurde dagegen von der zusätzlichen CaCl2-Gabe nicht signifikant beeinflusst.
8.) Durch die Zugabe von CaCl2 wurden weder der Geburtsverlauf noch der Abgang
der Nachgeburten beeinflusst und auch die Anzahl der insgesamt geborenen
Ferkeln wie auch der lebend, lebensschwach und tot geborenen Ferkeln zeigte
keinen relevanten Unterschied. Die Geburtsgewichte und die Ergebnisse der
Blutgasanalyse konnten ebenfalls keine Differenzen der Ferkel aufzeigen, deren
Ursprünge in der unterschiedlichen Behandlung der Muttertiere zu suchen wären.
In diesem Versuch konnte eine Acidierung des Harns mit Harn-pH-Werten von unter pH 6,5
schon nach wenigen Tagen mit der zusätzlichen Gabe von zweimal täglich 14 g (US) erreicht
werden. Diese Acidierung bildet somit die Grundlage für eine ausreichende prophylaktische
Wirkung gegen eine Keimbesiedelung bzw. eine Infektion der Harnorgane. Da derartige
Infektionen als Vorläufersyndrom für das spätere Auftreten des MMA-Komplexes gelten,
kann somit sekundär auch eine prophylaktische Wirkung gegen diese Erkrankung, die ein
erhebliches Problem in der Sauenhaltung und Ferkelproduktion darstellt, erreicht werden.
Dieser Acidierungseffekt und die forcierte orale Calciumaufnahme haben dabei keine
negativen Auswirkungen auf die in dieser Untersuchung überprüften Stoffwechselvorgänge
im Körper der Tiere, so dass die Methode der Infektionsprophylaxe in der Praxis durchaus
ihre Berechtigung hat.

Summary
146 146
7. Summary
Björn Röcker
Investigation of acidifying the urine by alimentary calciumchloride-
administration among pregnant sows.
In this study the effects of a calcium chloride administration (CaCl2) utilized in order to
acidify the urine on the metabolic processes like the acid-base-balance, the bone-metabolism
and the mineral homeostasis of sows were investigated.
The experimental group consisted of 14 sows receiving 14 g of calcium chloride (Calci
Cap®) with their sole feed twice a day from day 80. of pregnancy until farrowing. The control
group consisted of 15 animals, which were not administrated additional CaCl2. From day 80
through 108 of pregnancy a sole feed of the pregnant sows (EB 208mEq/kgTS) was applied
and from day 108 through the end of this investigation a sole feed for lactating sows (EB.
223mEq/kgTS) was applied.
Blood samples were taken from every sow on day 80 and 108 of pregnancy, right after
farrowing as well as every second day until day 8 p.p.. Spontaneous urine samples were
collected on day 90 and 111 of pregnancy. Moreover a blood sample was taken from one
piglet of each litter right after delivery.
In these samples the following parameters were analyzed:
blood: pH-value, hydrogen carbonate, total calcium, ionized calcium, inorganic phosphate,
chloride, sodium, creatinine, pth and bone-specific alkaline phosphatases.
urine: pH-value, calcium, inorganic phosphate, chloride, sodium and creatinine.
In order to examine the impact of an additional intake of CaCl2 on the course of the delivery
and the development of the piglets the following parameters were stipulated in addition: The
duration of delivery, the period of time until the expulsion of the afterbirths, the quantity of

Summary
147 147
born piglets in total as well as the piglets classified as born alive, weak and dead plus the
weight of these piglets.
The following results were obtained analyzing these parameters:
1) The medium urine pH-value of the test animals was (pH 5.96 ± 0.45) significantly
(p<0.01) lower on day 90 of pregnancy than the pH-value of the control animals (pH
7.05 ± 0.34). Until day 111 of pregnancy after the feed change the urine pH-values
rose in both groups significantly so that at this time the medium pH-value of the test
animals was measured at pH 6.73 ± 0.46 and that of the control group at pH 7.31 ±
0.31. The significant group-specific difference (p<0,01) endured.
2) The additional substitution of CaCl2 had no statistically relevant effects (p>0,05) on
the development of the medium blood pH-value
3) On the day of farrowing the concentration of HCO3- in the blood of the experimental
animals was with a difference of 2,16 mmo/l faintly significantly (p<0.05) beneath
that of the control group. During the other time of the experiment no significant
differences of concentration (p>0,05) arose between the animal groups.
4) The administration of 14 g CaCl2 twice a day indicates no significant effects (p>0,05)
on the concentration of ionized calcium in the blood as well as on the total calcium
concentration and inorganic phosphate in the plasma. The same holds true for the
plasma concentration of sodium and chloride and the content of potassium in the
blood.
5) The value of PTH varied strongly from minimum 1.19 pg/ml to maximum 180.27
pg/ml. The comparison of the relative augmentation of the PTH level in contrast to the
group specific initial value reflects that this value multiplies by 4.93 for the control
animals from day 80 through day 108 of pregnancy. At the time of delivery this value
drops to the 2.77-fold. In contrast, the test animals attain a 2.55-fold of the initial
value on day 108 of pregnancy which rises further to a 3.48-fold until farrowing.

Summary
148 148
6) As to the analysis of the data concerning the concentration of the bone-specific
alkaline phosphatase there is no relevant group-specific disparity (p>0,05).
7) On day 90 and 111 of pregnancy relative renal excreation of phosphorus (P/Crea) of
the control animals is faintly significantly higher (p<0.05) compared to the values of
the test animals whereas the values of the relative excision of chloride of the test sows
exceeds those of the control group significantly (p<0,01) on both dates. However, the
urine excision of calcium and sodium was not significantly influenced by the
additional CaCl2 administeration.
8) By the administration of CaCl2 neither the course of the delivery or the expulsion of
the afterbirths were influenced. The same holds true for the number of piglets born
alive, weak or dead. The birth weight and the results of the analysis of blood gas of
the piglets also reveals no disparities originating in the differentiated treatment of the
females.
In this study an acidifying of urine with urine pH-values inferior to pH 6.5 already be
reached after a few days with the additional administering of 14 g (US) of CaCl2 twice a
day. A bacteriostatic effect on the germ flora of the urine organs can be assumed with a
urine pH-value of that kind. As the germ flora is considered as an origin of the later
occurrence of the MMA-complex a reduction in prevalence of this syndrome can be
obtained. The acidifying effect and the forced up oral calcium ingestion do not have a
negative impact on the metabolic processes in the animals’ body tested in this experiment,
so that this method of infection prophylaxis has in praxis absolutely legitimate claim
inpractice.

Abkürzungsverzeichnis
149 149
8. Abkürzungsverzeichnis
AKB Anionen-Kationen-Bilanz ap ante partum ATP Adenosin-Triphosphat bAP knochenspezifische alkalische Phosphatase BE Basenüberschuss BHZP Bundeshybridzuchtprogramm
Ca2+ Calcium-Ionen
CaCl2 Calciumchlorid
Cl- Chlorid
CO2 Kohlendioxid Cys. Cystein d Tag EB Elektrolytbilanz E. coli Escherichia coli GH Wachstumshormon
H+ Wasserstoff-Ion
H2O Wasser
H2SO4 Schwefelsäure HCL Salzsäure
HCO3- Hydrogenkarbonat
HRP Horseradish peroxidase IGF Insulin-like Growth Factor Ig Immunglobulin IL-2 Interleukin 2 J Jahre
K+ Kalium-Ion LPS Lipopolysaccharide Max Maximalwert Met. Methionin
MgCl2 Magnesiumchlorid Min Minimalwert MMA Metrits-Mastits-Agalaktie MMP Matrix-Metalloproteinase MW Mitterwert N Anzahl
Na+ Natrium-Ion
Na2SO4 Natriumsulfat
NaHCO3 Nariumhydrogenkarbonat

Abkürzungsverzeichnis
150 150
NaOH Natronlauge Nfe N-freie Extrakte
NH4+ Ammonium-Ion
O2 Sauerstoff OS Organische Substanz
pCO2 Partialdruck an Kohlendioxid PCR Polimerase-Kettenreaktion
PGE2 Prostaglandin E2
PGF2α Prostaglandin F2α
PO43- Phosphat
pp post partum PTH Parathormon PUGS Peripartales Urogenitalsyndrom Ra Rohasche Rfa Rohfaser Rfe Rohfett Rp Rohprotein SD Standardabweichung
SO42- Sulfat
SUGD Swine Urogenital Disease TH Schilddrüsenhormon TRAP Tartratresistente saure Phosphatase TS Trockensubstanz US Ursprüngliche Substanz

Literaturverzeichnis
151 151
9. Literaturverzeichnis AKKERMANS, J. P., W. POMPER (1980): The significance of bacteriuria with reference to disturbances in fertility. In: 6th Int. Pig Vet. Soc. Congr., Kopenhagen 1980, Proc., S.44 AMTSBERG, G. (1984): Ergebnisse bakteriologischer Zervixtupferuntersuchungen bei puerperal erkrankten Sauen. Tierärztl. Umsch. 39, 479-484 ARNETT, T.R., D.W. DEMPSTER (1986): Effect of pH on bone resorption by rat osteoclasts in vitro. Endocrinology 119, 119-124 ARNHOFER, G. (1987): Untersuchungen über den Einfluss von Fütterung, Haltung und Behandlung auf die Harninhaltsstoffe der Zuchtsau. München, Univ., Tierärztl. Fak., Diss. ASSAL, A.N., J. CHRISTIANSEN, J.S.D. POULSEN (1980): Acid-base status of porcine blood during storage. Nordisk Veterinaermedicin 32, 9-16 ASSCHER, A.W., M. SUSSMANN, W.E. WATERS, R.H. DAVIS, S. CHICK (1966): Urine as a medium for bacterial growth. Lancet, 1037-1041 AWAD-MASALMEH, M., W. BAUMGARTNER, A. PASSERNIG, R. SILBER, F. HINTERDORFER (1990): Bakteriologische Untersuchungen bei an puerperaler Mastitis (MMA-Syndrom) erkrankter Sauen verschiedener Tierbestände Österreichs. Tierärztl. Umsch. 45, 526-535 BÄCKSTRÖM, L., A.C. MORKOC, J. CONNOR, R. LARSON, W. PRICE (1984): Clinical study of mastitis-metritis-agalactia in sows in Illinois. J. Am. Vet. Med. Assoc. 185, 70-73

Literaturverzeichnis
152 152
BAR, A., R.H. WASSERMAN (1974): Duodenal calcium binding protein in the chick: A new bioassay for vitamin D. J. Nutr. 104, 1202-1207 BARON, R., L. NEFF, D. LOUVARD, P. J. COURTOV (1985): Cell-mediated extracellular acidification and bone resorption: Evidence for a low pH in resorbing lacunaeand localization of a 100 kD lysosomal protein at the osteoclast ruffled border. J. Cell. Biol. 101, 2210-2222 BARZEL, U.S., J. JOWSEY (1969): The effects of chronic acid and alkali administration on bone turnover in adult rats. Clin. Sci. 36, 517-524 BAUD, C.A., G. BOIVIN (1978): Effects of hormones on osteocyte function and perilacunar wall structure. Clin. Orthop. 136, 270-281 BAUMGARTNER, W., P. KRUZIK (1983): Labordiagnostik in der Klauentierpraxis. Tierärztl. Umsch. 38, 503-506 BECKER, H.A., R. KURTZ, G. VON MICKWITZ (1985): Chronische Harnwegsinfektion beim Schwein, Diagnose und Therapie. 1. Mitteilung: Diagnose, klinische Bedeutung und Verbreitung. Prakt. Tierarzt 66, 1006-1011 BEKER, S. (1999): Einstellung des Harn-pH-Wertes bei Sauen. München, Univ., Tierärztl. Fak., Diss. BEKKER, P.J., C.V. GAY (1990): Biochemical characterization of electrogenic vacuolar proton pump in purified chicken osteoblast plasma membrane vesicles. J. Bone Miner. Res. 5, 557-567 BERNER, H. (1971): Die Bedeutung chronischer Erkrankungen der Harnwege bei der Entstehung von Puerperalstörungen und Mastitiden der Muttersau. Dtsch. tierärztl. Wochenschr. 78, 241-245

Literaturverzeichnis
153 153
BERNER, H. (1981): Untersuchungen zum Vorkommen von Harnwegsinfektionen bei Muttersauen. Tierärztl. Umsch. 36, a) 162-171 und b) 250-255 BERNER, H. (1984): Die Bedeutung von Harnwegsinfektionen für die Entstehung der puerperalen Endometritis beim Schwein. Tierärztl. Umsch. 39, 450-458 BERNER, H. (1988): Cystitis in der MMA-Diagnostik. Prakt. Tierarzt 69, 124-130 BERNER, H., W. JÖCHLE (1988): The role of urogenital infections on infertility and sterility in sows. In: 10th Int. Pig Vet. Soc. Congr., Rio de Janeiro 1988, Proc., S. 306 BERTSCHINGER, H.U. (1984): Neue Aspekte der Pathogenese der puerperalen Mastitis. Tierärztl. Umsch. 39, 458-461 BERTSCHINGER, H.U. (1999): Coliforme Mastitis. In: STRAW, B.E., S. D. ALLAIRE, W.L. MENGELING, D.J. TAYLOR (Hrsg.): Diseases of swine. 8. Auflage, Iowa State University Press, Ames, Iowa, United States BERTSCHINGER, H.U., E. BÜRGI, V. ENG, P. WEGMANN (1990): Senkung der Inzidenz von puerperaler Mastitis bei der Sau durch Schutz des Gesäuges vor Verschmutzung. Schweiz. Arch. Tierheilk. 132, 557-566 BERTSCHINGER, H.U., J. POLENZ, I. HEMLEP (1977): I.Untersuchungen über das Mastitis-Metritis-Agalaktie-Syndrom (Milchfieber) der Sau. II. Bakteriologische Befunde bei Spontanfällen. Schweiz. Arch. Tierheilk. 119, 223-233 BIEBERSTEIN, E.L. (1990): Agents associated with the urinary tract. Chapt. 31: The urinary tract as a microbial habitat; urinary tract infections. Rev. Vet. Microbiol. 2, 228-242

Literaturverzeichnis
154 154
BILKEI, G., A. BÖLCSKEI (1993): Die Auswirkungen der Fütterung im letzten Trächtigkeitsmonat auf die perinatalen Parameter bei verschiedener Körperkondition und Parität der Muttersauen. Tierärztl. Umsch. 48, 629-632, 635 BILKEI, G., A. BÖLCSKEI, T. GOOS (1995b): Pathogene Befunde aus dem Urogenitaltrakt ausgemerzter Muttersauen aus einem Großbestand. Tierärztl. Prax. 23, 37-41 BIRNBAUM, R.S., K.M. WIREN, R.R. BOWSHER (1993): Changes in IGF-I expression and aecretion during osteoblast proliferation and differentiation. J. Bone Min. Res. 8 (Suppl. 1), 970a BLAIR, H.C., S.L. TEITELBAUM, R. GHISELLI, S. GLUCK (1989): Osteoclastic bone resorption by a polarized vacuolar proton pump. Science 245, 855-887 BÖNING, J., M. FRITSCH, B. BÖNING, H.J. KUBSCH (1976): Untersuchungen über das Auftreten von Puerperalerkrankungen industriemäßig gehaltener Sauen. Monatsh. Veterinärmed. 31, 124-129 BOGDANOVIC, Z., Y.F. HUANG, M. DODIG, H.S. CLARK, A.C. LICHTLER, B.E. KREAM (2000): Parathyroid hormone inhibits collagen synthesis and the activity of rat collal transgenes mainly by cAMP- mediated pathway in mouse calvariae. J. Cell Biochem. 77, 149-158 BOLLWAHN, W. (1980): Geburtsstörungen, Puerperalerkrankungen und Mastitiden. In: SCHULZE, W., K. BICKHARDT, W. BOLLWAHN, G.V. MICKWITZ, H. PLONAIT (Hrsg.): Klinik der Schweinekrankheiten. Verlag Schaper, Hannover, 273-288 BOLLWAHN, W., H. v. VOPELIUS- FELDT, A. v. VOPELIUS- FELDT, G. ARNHOFER (1984): The clinical value of bacteriuria in sows. In: 8th Int. Pig Vet. Soc. Congr., Genth 1984, Proc., S. 149 BOLLWAHN, W., G. ARNHOFER (1989): Die Bedeutung exogener Faktoren für die Zusammensetzung des Harns der Zuchtsauen. Tierärztl. Prax. 17, 43-46

Literaturverzeichnis
155 155
BOLLWAHN, W., D. MEERMEIER (1989): Häufigkeit und Diagnostik chronischer Gesäugeveränderungen bei Zuchtsauen (Schlachtbefunde). Berl. Münch. tierärztl. Wochenschr. 102, 223-227 BOSTEDT, H. (1978): Das Schwein in der ante- und postpartalen Periode. I. Mitteilung: Untersuchungen über Elektrolytkonzentrationen (Ca, anorganisches Phosphat, Mg) im Blutserum sowie über den Gehalt an Fe und Cu in Blutserum und Milch. Berl. Münch. tierärztl. Wochenschr. 91, 21-24 BOSTEDT, H. (1999): Pathogenese und klinisches Bild des MMA-Komplexes. In: bpt-Kongress, Nürnberg 1999, Proc., S. 137-139 BOSTEDT, H., G. MAIER, K. HERFEN, R. HOSPES (1998): Klinische Erhebungen bei Sauen mit puerperaler Septikämie und Toxämie. Tierärztl. Prax. 26, 332-338 BOTH, G., K. MÖLLER, F.W. BUSSE (1980): Zur Frage der Beziehungen zwischen Fruchtbarkeitsstörungen und Harninfektionen beim Schwein. 1. Mitteilung: Untersuchung an Harnproben mittels bakteriologischer Teststreifen. Tierärztl. Umsch. 35, 468-473 BOURNE, F.J., J. CURTIS, L.S. WAH (1973): Urinary immunglobulins in the adult pig. Immunol. 24, 1-12 BRESLAU, N.A., L. BRINKLEY, K. HILL, C.Y.C. PAK (1988): Relationship of animal protein-rich diet to kidney stone formation and calcium metabolism. J. Clin. Endocrinol. Metab. 66, 140-146 BRINKHURST, F.R., M.B. DEMAY, H.M. KRONENBERG (1998): Hormones and disorders of mineral metabolism. In: WILSON, J.D., D.W. FORSTER, H.M. KRONENBERG, P.D. LARSEN (Hrsg.): Williams Textbook of endocrinology, 9. Auflage, Verlag Saunders, Philadelphia, USA, S. 1155-1209 BRONNER, F., T. FREUND (1975): Intestinal CaBP: A new quantitative index of vitamin D deficiency in the rat. Am. J. Physiol. 229, 689-694

Literaturverzeichnis
156 156
BRONNER, F., D. PANSU, W.D. STEIN (1986): An analyses of intestinal calcium transport across the rat intestine. Am. J. Physiol. 250, G561-G569 BRONNER, F. (1990): Intestinal calcium transport: The cellular pathway. Miner. Electrol. Metab. 16, 94-100 BRONNER, F., D. PANSU (1999): Nutritional aspects of calcium absorption. J. Nutr. 129, 1: 9-12 BROSNAN, J.T., M.E. BROSNAN (1982): Dietary protein, metabolic acidosis, and calcium balance. In: DRAPER, H.H. (Hrsg.): Advances in nutrition research. Plenum Press, New York, S. 77-105 BROWN, E.M. (1991): Extracellular Ca2+ sensing, regulation of parathyroid cell function, and role of Ca2+ and other ions as extracellular (first) messenger. Physiol. Rev. 71, 371-411 BUDDE, R.A., T.D. CRENSHAW (2003): Chronic metabolic acid load induced by changes in dietary electrolyte balance increased chloride retention but did not compromise bone in growing swine. J. Anim. Sci. 81, 197-208 BUFFINGTON, C.A. (1989): Ernährung und Struviturolithiasis bei Katzen. Vet. Int. 1, 2-16
BUSCH, M.E., P. BAEKBO, P.H. ANDERSEN, J. DAHL (1998): Blood parameters of possible diagnostic value in the MMA complex. In: 15th Int. Pig Vet. Soc. Congr., Birmingham 1998, Proc., S. 223 BUSHINSKY, D.A. (1989): Net calcium efflux from live bone during chronic metabolic, but not respiratory, acidosis. Am. J. Physiol. 256, F 836-F842

Literaturverzeichnis
157 157
BUSHINSKY, D.A. (1995): Stimulated osteoclastic and suppressed osteoblastic activity in metabolic but not respiratory acidosis. Am. J. Physiol. 268, C80-C88 BUSHINSKY, D.A., M.J. FAVUS, A.B. SCHNEIDER, P.K. SEN, L.M. SHERWOOD, F.L. COE (1982): Effects of metabolic acidosis on PTH and 1,25(OH)2D3 response to low calcium diet. Am. J. Physiol. Renal Fluid Electrolyte Physiol. 243, F570-F575 BUSHINSKY, D.A., N.S. KRIEGER, D.I. GEISSER, E.B. GROSSMAN, F.L. COE (1983): Effects of pH on bone calcium and proton fluxes in vitro. Am. J. Physiol. 245, F204-F209 BUSHINSKY, D.A., J.M. GOLDRING, F.L.COE (1985): Cellular contribution to pH-mediated calcium flux in neonatal mouse calvariae. Am. J. Physiol. 248, F785-F789 BUSHINSKY, D.A., R. LEVI-SETTI, F.L. COE (1986): Ion microprobe determination of bone surface elements: Effects of reduced medium pH. Am. J. Physiol. 269, F1090-F1097 BUSHINSKY, D.A., N.E. SESSLER (1992): Critical role of bicarbonate in calcium release from bone. Am. J. Physiol. 263, F510-F515 BUSHINSKY, D.A., N.E. SESSLER, N.S. KRIEGER (1992): Greater unidirectional calcium efflux from bone during metabolic, compared with respiratory, acidosis. Am. J. Physiol. 262, F425-F431 BUSHINSKY, D.A., B.C. LAM, R. NESPECA, N.E. SESSLER, M.D. GRYNPAS (1993a): Decreased bone carbonate content in response to metabolic, but not respiratory, acidosis. Am. J. Physiol. 265, F530-F536 BUSHINSKY, D.A., W. WOLBACH, N.E. SESSLER, R. MOGILEVSKY, R. LEVI-SETTI (1993b): Physiochemical effects of acidosis on bone calcium flux and surface ion composition. J. Bone Miner. Res. 8, 93-102

Literaturverzeichnis
158 158
BUSHINSKY, D.A., E.L. NILSSON (1995): Additive effects of acidosis and parathyroid hormone on mouse osteoblastic and osteoclastic function. Am. J. Physiol. 269, C1364-C1370 BUSHINSKY, D.A., S.B. SMITH, K.L. GAVRILOV, L.F. GAVRILOV, J. LI, R. LEVI-SETTI (2002): Acute acidosis-induced alteration in bone bicarbonate and phosphate. Am. J. Physiol. Renal Physiol. 283, F1091-F1097 BUSSE, F.W., K. MÖLLER, G. BOTH, C. COMMICHAU (1982): Zur Frage der Beziehungen zwischen Fruchtbarkeitsstörungen und Harnwegsinfektionen beim Schwein. 3. Mitteilung: Keimspektrum und Sensibilität gegen Antibiotika. Tierärztl. Umsch. 37, 703-710 CANALIS, E. (1996): Regulation of bone remodelling. In: FAVUS, M.J. (Hrsg.): Primer of the metabolic bone diseases and disorders of mineral metabolism, 3d edition. Lippincott-Raven Publishers, Philadelphia, New York, S. 29-34 CANALIS, E., J. LORENZO, W.H. BURGESS, T. MACIAG (1987): Effects of endothelial cell growth factor on bone remodelling in vitro. J. Clin. Invest. 79, 52-58 CARAFOLI, E. (1987): Intracellular calcium homeostasis. Annu. Rev. Biochem. 56, 395-433 CARR, J., J.R. WALTON (1992a): The characterisation of E. coli isolates from porcine urogenital tract. In: 12th Int. Pig Vet. Soc. Congr., Den Haag 1992, Proc., S. 262 CARR, J., J.R. WALTON (1992b): The microflora of porcine urinary tract in cases of cystitis and pyelonephritis. In: 12th Int. Pig Vet. Soc. Congr., Den Haag 1992, Proc., S. 347 CARTER, D.I., J.D. CRENSHAW, P.M. SWANTEK, R.L. HARROLD, R.C. ZIMPRICH (1987): The effect of fiber intake by gravid swine during three consecutive parities on sow and litter performance. J. Anim. Sci. 65 (Suppl. 1), 89

Literaturverzeichnis
159 159
CEREZA, J., V. ROSELL, M. NIEVAS, et al. (1986): MMA syndrome in sows. Hormonal levels and antibiotic therapy. In: 9th Int. Pig Vet. Soc. Congr., Barcelona 1986,Proc., S. 94 CERNE, F., I. JERKOVIC, C. DEBELJAK (1984): Influence of Finadyne® on some clinical signs of MMA. In: 8th Int. Pig Vet. Soc. Congr., Genth 1984, Proc., S.290 CHABALA, J.M., R. LEVI-SETTI, D.A. BUSHINSKY (1991): Alteration in surface ion composition of cultured bone during metabolic, but not respiratory, acidosis. Am. J. Physiol. 261, F76-F84 CHAN, J.C.M. (1974): The influence of dietary intake on endogenous acid-production. Nutr. Metabol. 16, 1-9 CHAREONSIRISUTHIGUL, T., G. KIRPAL, G. AMTSBERG (1979): Untersuchungen zur Biochemie, Serologie, Pathogenität und Antibiotikaresistenz an E.coli-Stämmen von gesunden und an MMA erkrankten Sauen. Berl. Münch. tierärztl. Wochenschr. 92, 409-416 CHARLES, P., L. MOSEKILDE, L. RISTELI, J. RISTELI (1994): Assessment of bone remodeling using biochemical indicators of type I collagen synthesis and degradation: Relation to calcium kinetics. Bone and Mineral 24, 81-94 CHING, S.V., M.J. FETTMAN, D.W. HAMAR, L.A. NAGODE, K.R. SMITH (1989): The effect of chronic dietary acidification using ammonium chloride on acid-base and mineral metabolism in the adult cat. J. Nutr. 119, 902-915 CHRISTENSON, R.H. (1997): Biochemical markers of bone metabolism: An overview. Clin. Biochem. 30, 573-593 DAMIR, H.A., D. SCOTT, N. LOVERIDGE, W. BUCHAN, J. MILNE (1991): The effects of feeding diets containing either NaOH3 or NH4Cl on indices of bone formation and resorption and mineral balance in the lamb. Exp. Physiol. 76, 725-732

Literaturverzeichnis
160 160
DAVIS, C.P., E. BALISH, K. MIZUTANI, D.T. UEHLING (1977): Bladder response to Klebsiella infection. A scanning electron microscopy study. Invest. Urol. 15, 227-231 DELMAS, P.D. (1991): Clinical use of biochemical markers of bone remodelling in osteoporosis. In: CHRISTIANEN C., K. OVERGAARD (Hrsg.): Osteoporosis 1990, Osteopress ApS, Copenhagen, S. 450-458 DELMAS, P.D. (1995): Biochemical markers for bone turnover. Acta Orthop. Scand. (Suppl. 266) 66, 176-182 DENTON, R.M., J.G. MC CORMACK (1985): Ca2+ transport by mammalian mitochondria and its role in hormone action. Am. J. Physiol. 249, E543-E545 DE ROUCHEY, J.M., J.D. HANCOCK, R.H. HINES, K.R. CUMMINGS, D.J. LEE, C.A. MALONEY, D.W. DEAN, J.S. PARK, H. CAO (2003): Effects of dietary electrolyte balance on the chemistry of blood and urine in lactating sows and sow litter performance. J. Anim. Sci. 81, 3067-3074 DE WINTER, P.J.J., M. VERDONCK, A. DE KRUIF, L.A. DEVRIESE, F. HASEBROUCK (1992): Endometritis and vaginal discharge in the sow. Anim. Reprod. Sci. 28, 51-58 DIAZ, R., G. EL- HAJJ FULEIHAN, E.M. BROWN (1999): Regulation of parathyroid function. In: FRAY, J.G.S. (Hrsg.): Handbook of Physiology, Band III, Oxford Univ. Press, New York, USA, S. 607-662 DIENER, M. (1994): Allgemeine und spezielle Endokrinologie. Script für Studierende der Veterinärmedizin an der Universität in Zürich DI POLO, R., L. BEAUGE (1983): The calcium pump and sodium-calcium exchange in squid axons. Annu. Rev. Physiol. 45, 313-324

Literaturverzeichnis
161 161
DONE, J.T. (1980): Zitzenmängel beim Schwein. Dtsch. tierärztl. Wochenschr. 87, 437-439 DOTY, S.B. (1981): Morphological evidence of gap junctions between bone cells. Calcif. Tissue Int. 33, 509-512 DRAKE, F.H., A. DODDS, I.E. JAMES, J.R. CONNOR, C. DEBOUCK, S. RICHARDSON, E. LEE- RYKACZEWSKI, L. COLEMAN, D. RIEMAN, R. BARTHLOW, G. HASTINGS, M. GOWEN (1996): Cathepsin K, but not cathepsins B, L, or S, is abundantly expressed in human osteoclasts. J. Biol. Chem. 271, 12511-12516 DREZNER, M. K. (2002): Phospherus homeostasis and related disorders. In: BILEZIKIAN, J.P., L.G. RAIZ, G.A. RODAN (Hrsg.): Principles of bone biology, 2nd edition., Academic Press, San Diego, USA, S. 321-338 DUCY, P., T. SCHINKE, G. KARSENTY (2000): The osteoblast: A sophisticated fibroblast under central surveillance. Science 289, 1501-1504 EDELSTEIN, S., D. NOFF, L. SINAI, A. HARELL, J.B. PUSCHETT, E.E. GOLUB, F. BRONNER (1978): Vitamin D metabolism and expression in rats fed on low-calcium and low-phosphorus diets. Biochem. J. 170: 227-233 EGGEMANN, G., M. WENDT, L.E. HOELZLE, C. JÄGER, R. WEISS, K. FAILING (2000): Zum Vorkommen von Chlamydien-Infektionen in Zuchtsauenbeständen und deren Bedeutung für das Fruchtbarkeitsgeschehen. Dtsch. tierärztl. Wochenschr. 107, 3-10 EHRENTRAUT, W. (1968): Die puerperale Septikämie des Schweines und ihr Einfluss auf die Ferkelaufzucht. Monatsh. Veterinärmed. 23, 936-940 ENGELHARDT, v. W., G. BREVES (2000): Physiologie der Haustiere Enke Verlag, Stuttgart

Literaturverzeichnis
162 162
EPSTEIN, S. (1988): Serum and urinary markers of bone remodelling: Assessment of bone turnover. Endocr. Rev. 9, 437-449 ERDMANN, S., H. BURKHARDT, K. VON DER MARK, W. MÜLLER (1998): Mapping of a carboxyl- terminal active site of parathyroid hormone by calcium imaging. Cell Calcium 23, 413-421 FALLON, M.D. (1984): Bone resorbing fluid from osteoclasts is acidic: An in vitro micropunture study. In: CONN, C.V., J.R. FUJITA, J.R. POTTS et al. (Hrsg.): Endocrine control of bone and calcium metabolism. Elsevier, Amsterdam, Holland, S. 144-146 FAVUS, J.M. (1985): Factors that influense absorption and secretion of calcium in the small intestine and colon. Am. J. Physiol. 248, G147-G157 FINKENSIEP, A. (1993): Untersuchungen über den Einfluss des Harnstatus auf das Puerperalsyndrom der Sau unter Berücksichtigung der Fütterung und des Trinkwasserverbrauches. Hannover, Tierärztl. Hochsch., Diss. FORTH, W. (1992): Pharmakologie und Toxikologie. Wissenschaftsverlag Mannheim FOX, J., E.M.W. MAUNDER, V.A. RANDER, A.D. CARE (1985): Vitamin D-dependent rickets type I in pigs. Clin. Sci. 69, 541-548 FULLMER, C.S. (1992): Intestinal calcium absorption: Calcium entry. J. Nutr. 122, 644-650 GEDEK, B. (1993) : Probiotika zur Regulierung der Darmflora. Tierärztl. Umsch. 48, 97-104

Literaturverzeichnis
163 163
GIESEMANN, M.A., A.J. LEWIS, P.S. MILLER, M.P. AKHTER (1998): Effects of the reproductive cycle and age on calcium and phosphorus metabolism and bone integrity of sows. J. Anim. Sci. 76, 796-807 GLAWISCHNIG, E. (1967): Zum Problem der Agalaktie und der damit verbundenen schwierigen Ferkelaufzucht. Tierärztl. Umsch. 22, 174-179 GLAWISCHNIG, E. (1969): Die Agalaktie der Sau. Wien. tierärztl. Monatsschr. 56, 27-30 GLOCK, H. (1983): Zur Therapie des Mastitis-Metritis-Agalaktie-Komplexes ( MMA) des Schweines und damit zusammenhängender Fruchtbarkeitsstörungen. Prakt. Tierarzt 69, 511-516 GÖRANSSON, L. (1989): The effect of feed allowance in late pregnancy on the occurrence of agalactia post partum in the sow. J. Vet. Med. 36, 505-513 GÖRANSSON, L. (1990): The effect of protein source in late pregnancy feed on the occurrence of agalactia post partum in the sow. Acta vet. scand. 31, 117-120 GÖTZE, R. (1939): Klinische und experimentelle Prüfungen der Sulfonamidpräparate Prontosil, Prontabil und Astrosol. II. Mitteilung: Schwangerschaftsstörungen und infizierte Geburten beim Schwein. Dtsch. tierärztl. Wochenschr. 47, 697-700 GOLDHABER, P., L. RABADJIJA (1987): H+ stimulation of cell-mediated bone resorption in tissue culture. Am. J. Physiol. 253, E90-E98 GOTO, T., T. KIYOSHIMA, R. MOROI, T. TSUKUBA, Y. NISHIMURA, M. HIMENO, K. YAMAMOTO, T. TANAKA (1994): Localisation of the cathepsins B,D and L in the rat osteoclast by immuno-light and –electron microscopy. Histochemistry 101, 33-40

Literaturverzeichnis
164 164
GRAUSCHOPF, U., H. LILIE, K. HONOLD, M. WOZNY, D. REUSCH, A. ESSWEIN, W. SCHÄFER, K.P. RUCKNAGEL, R. RUDOLPH (2000): The N-terminal fragment of human parathyroid hormone receptor I constitutes a hormone binding domain and reveals a distinct disulfide pattern. Biochemistry 101, 33-40 GREENBERG, Z., A. BISELLO, D.F. MIERKE, M. ROSENBLATT, M. CHOREV (2000): Mapping the bimolecular interface of the parathyroid hormone (PTH) PTH-1 receptor complex: Spatial proximity between Lys (27) (of the hormone principal binding domain) and leu (261) (of the first extracellular loop) of the human PTH-1 receptor. Biochemistry 39, 8142-8152 GREGORY, J.G., A.J. WEIN, T.C. SANSONE (1971): Bladder resistence to infection. J. Urol. 105, 220 GUENGUEN, L., J.M. PEREZ (1981): A re-evaluation of recommended dietary allowances of calcium and phosphorus for pigs. Proc. Nutr. Soc. 40, 273-278 HABERMEHL, K.-H. (1984): Haut und Hautorgane. In: NICKEL, R., A. SCHUMMER, E. SEIFERLE (Hrsg.): Anatomie der Säugetiere, Band 3 3. Auflage. Paul Parey Verlag, Berlin, Hamburg, S. 508-515 HAGN, J. (1995): Untersuchungen zur Wirksamkeit und Verträglichkeit eines neu entwickelten Cephalosporins (Cefquinom) am Beispiel Mastitis-Metritis-Agalaktie erkrankter Sauen in Problembeständen. München, Univ., Tierärztl. Fak., Diss. HAIMBERGER, B. (1977): Klinische und bakteriologische Untersuchung von Sauen im Puerperium mit besonderer Berücksichtigung des Genitaltraktes, des Harnapparates und der Milch. Hannover, Tierärztl. Hochsch., Diss.
HALLEEN, J.M., S. RAISANEN, J.J. SALO, S.V. REDDY, G.D. ROODMAN, T.A. HENTUNEN, P.P. LEHENKARI, H. KAIJA, P. VIHKO, H.K. VÄÄNÄNEN (1999): Intracellular fragmentation of bone resorption products by reactive oxygen species generated by osteoclastic tartrate-resistant acid phosphatase. J. Biol. Chem. 274, 22907-22910

Literaturverzeichnis
165 165
HALPERIN, M.L., R.L. JUNGAS (1983): Metabolic production and renal disposal of hydrogen ions. Kidney Int. 24, 709-713 HAMMERBERG, C., G.G. SCHURIG, D.L. OCHS (1989): Immunodeficiency in young pigs. Am. J. Vet. Res. 50, 868-874 HANSARD, S.L., H. ITHO, J.C. GLENN, D.M. THRASHER (1966): Placental transfer and fetal utilization of calcium in developing swine. J. Nutr. 89, 335 HARRINGTON, J.T., J. LEHMANN (1970): The metabolic production and disposal of acid and alkali. Med. Clin. Of North Amer. 54, 1543-1553 HEBERT, S.C., E.M. BROWN, H.W. HARRIS (1997): Role of the Ca2+- sensing receptor in divalent mineral ion homeostasis. J. Expl. Biol. 200, 295-302 HEINRITZI, K., J. HAGN (1999): Untersuchungen zur Wirksamkeit und Verträglichkeit des neu entwickelten Cephalosporins Cefquinom bei an puerperaler Septikämie und Toxämie erkrankter Sauen. Tierärztl. Prax. 27, 114-121 HEINRITZI, K., H. PLONAIT (2001): Blutkrankheiten. In: WALDMANN, K.-H., M. WENDT (Hrsg.): Lehrbuch der Schweinekrankheiten. 3., durchgesehene Auflage, Blackwell Wissenschaftsverlag, Berlin, Wien, S. 169-196 HEINTZ, R., S. ALTHOF (1984): Das Harnsediment. Georg Thieme Verlag, Stuttgart, Hamburg HERMANSSON, I., S. EINARSSON, L. EKMAN, K. LARSSON (1978): On the agalactia post partum in the sow. A hematological and blood chemical study in affected and healthy sows. Nord. Vet. Med. 30, 474-481

Literaturverzeichnis
166 166
HIERHOLZER, K., M. FROMM (1995): Funktion der Nieren. In: SCHMIDT, R.F., G. THEWS (Hrsg.): Physiologie des Menschen. Springer Verlag, Berlin, Heidelberg, New York, S. 737-776 HOSPES, R., H. BOSTEDT, A. SOBIRAJ (1993): Analyse der geburtshilflichen Situation beim Schwein aus klinischer Sicht. Tierärztl. Prax. 21, 209-216 HOWARD, N.J., J. ELLIOTT, T.J. MARTIN, M.T. GILLESPIE (1998): Osteotropic agents regulate the expression of osteoclast differentiation factor and osteoprotegerin in osteoblastic stromal cells. Endocrinology 139, 4743-4746 IBEN, B. (1997): Reduction of neonatal mortility in pigs. Reprod. Domest. Anim. 32, 113 IBEN, B. (1999): Ein Beitrag zur Hypogalaktieprophylaxe der Muttersau. Prakt. Tierarzt 80, 222-228 INAOKA, T., G. BILBE, O. ISHIBASHI, K. TEZUKA, M. KUMEGAWA, T. KOKUBO (1995): Molecular cloning of human cDNA for cathepsin K: Novel cysteine proteinase predominantly expressed in bone. Biochem. Biophys. Res. Commun. 206, 89-96 JAMBOR, V., Z. PROCHAZKA (1977): Einfluß einer unterschiedlichen Anionen- und Kationenkonzentration in der Diät auf die Ausnutzung von Kalzium und Phosphor bei Schweinen. Arch. Tierernähr. 27, 701-709 JONES, J.E.T. (1980): Zystitis und Pyelonephritis bei Sauen. Dtsch. tierärztl. Wochenschr. 87, 443-445 JORSAL, S.E. (1986): Epidemiology of the MMA-syndrome. A field survey in Danish sow herds. In: 9th Int. Pig Vet. Soc. Congr., Barcelona 1986, Proc., S. 93

Literaturverzeichnis
167 167
JÜRGENS, C. (1991): Einfluss von Futterzusatzstoffen auf den pH-Wert und die Inhaltsstoffe des Harnes bei Zuchtsauen. Hannover, Tierärztl. Hochsch., Diss. JUNQUEIRA, L.C., J. CARNEIRO (1991): Knochen und Knochenverbindungen. In: JUNQUEIRA, L.C., J. CARNEIRO (Hrsg.): Histologie. Zytologie, Histologie und mikroskopische Anatomie des Menschen (Unter Berücksichtigung der Histophysiologie). Springer-Verlag, Berlin, Heidelberg, New York, S. 190-218 KAMPHUES, J., D. HEBELER, C. RADE (1998): Der Verdauungskanal als Quelle einer forcierten Endotoxin-/LPS- Belastung von Schweinen. Dtsch. tierärztl. Wochenschr. 105 (9), 359-360 KAMPHUES, J., J. BEENING (1998): Peripartale Fütterungsmaßnahmen bei Sauen. Prakt. Tierarzt 79, 76 KANATANI, M., T. SUGIMOTO, M. KANZAWA, S. YANO, K. CHIHARA (1999): High extracellular calcium inhibits osteoclast-like cell formation by directly acting on the calcium-sensing receptor existing in osteoclast precursor cells. Biochem. Biophys. Res. Commun. 261, 144-148 KANZAWA, M., T. SUGIMOTO, M. KANATANI, K. CHIHARA (2000): Involvement of osteoprotegerin/osteoclastogenesis inhibitory factor in the stimulation of osteoclast formation by parathyroid hormone in mouse bone cells. Eur. J. Endocrinol. 142, 661-664 KARAPLIS, A.C., A. LUZ, J. GLOWACKI, R.T. BRONSON, V.L.J. TYBULEWICZ, H.M. KRONENBERG, R.C. MULLIGAN (1994): Lethal skeleton dysplasia from targeted disruption of the parathyroid hormone-related peptide gene. Genes Develop. 8, 277-289 KARBACH, U. (1991): Segmental heterogeneity of cellular and paracellular calcium transport across the rat duodenum and jejunum. Gastroenterology 100, 47-58 KASS, E.H. (1957): Bacteriuria and the diagnosis of infections of the urinary tract. Arch. Intern. Med. 100, 709-714

Literaturverzeichnis
168 168
KAUNE, R. (1992): Die Regulation des intestinalen aktiven Calciumtransportes durch Calcitriol. Hannover, Tierärztl. Hochsch., Habil.-Schr. KIENZLE, E. (1991): Ernährung und Urolithiasis bei Haussäugetieren Übers. Tierernähr. 19, 157-199 KIVISTO, A.K., H. VASENIUS, M. SUNDHOLM (1977): Canine bacteriuria. J. Small Anim. Pract. 18, 707 KLOPPENSTEIN, C., M. BIGRAS-POULIN, G.P. MARTINEAU (1998): Neonatal growth, sow rectal temperature and milk composition. In: 15th Int. Pig Vet. Soc. Congr., Birmingham 1998, Proc., S. 231 KLOPPENSTEIN, C., C. FARMER, G. P. MARTINEAU (1999): Diseases of the mammary glands and lactation problems. In: STRAW, B.E., S. DALLAIRE, W.L. MENGELING, D.J. TAYLOR (Hrsg.): Diseases of swine. 8. Auflage, Iowa State University Press, Ames, Iowa, United States KOLB, E. (1989): Lehrbuch der Physiologie der Haustiere. Gustav Fischer Verlag, Stuttgart, New York KRAFT, W., U. DÜRR (1995): Klinische Labordiagnostik in der Tiermedizin. Schattauer Verlagsgesellschaft, Stuttgart, New York KRIEGER, N.S., N.E. SESSLER, D.A. BUSHINSKY (1992): Acidosis inhibits osteoblastic and stimulates osteoclastic activity in vitro. Am. J. Physiol. 262, F442-F448 KRIEGER, N.S., W.R. PARKER, K.M. ALEXANDER, D.A. BUSHINSKY (2000): Prostaglandins regulate acid-induced cell-mediated bone resorption. Am. J. Physiol. Renal Physiol. 279, F1077-F1082 LANGFELDT, N., M. WENDT, G. AMTSBERG (1990): Vergleichende Untersuchungen zum Nachweis von Corynebacterium-suis-Infektionen beim Schwein mit Hilfe der indirekten Immunofluoreszenz und der Kultur. Berl. Münch. Tierärztl. Wochenschr. 103, 273-276

Literaturverzeichnis
169 169
LANSKE, B., A.C. KARAPLIS, A. LUZ, A. VORTKAMP, A. PIRRO, M. KARPERIEN, L.H.K. DEFIZE, C. HO, R.C. MULLIGAN, A.B. ABOU- SAMRA, H. JÜPPNER, G.V. SEGRE, H.M. KRONENBERG (1996): PTH/PTHrP receptor in early development and indian hedgehog-regulated bone growth. Science 273, 663-666 LAPPE, F. (1995): Einfluss der Mineralstoffversorgung auf die Entwicklung von Kristallurie und Harnwegsinfektionen bei Sauen. Hannover, Tierärztl. Hochschule, Diss. LARON, Z., A. PERTZELAN, S. MANNHEIMER (1966): Genetic pituitary dwarfism with high serum concentration of growth hormone: A new inborn error of metabolism? Isr. J. Med. Sci. 2, 152-155 LAWRENCE, A.B., E.M.C. TERLOUW (1993): A review of behavioral factors involved in the development and continued performance of stereotypic behaviours in pigs. J. Anim. Sci. 71, 2815 LAWSON, P.A., E. FALSON, E. AKERVALL, P. VANDAMME, M.D. COLLINS (1997): Characterization of some Actinomyces-like isolates from human clinical specimens: Reclassification of Actinomyces suis (soltys and spratling) as Actinomyces suis comb. nov. and description of Actinobaculum schaali sp. nov.. Int. J. syst. Bacterial. 47, 899-903 LEES, G.E., C.A. OSBORNE, J.B. STVENS (1979): Urine: A medium for bacterial growth Vet. Clin. Of North Amer. 9, 611-616 LEHMANN, J., J.R. LITZOW, E.J. LENNON (1966): The effects of chronic acid loads in normal man: Further evidence for the participation of bone mineral in the defense against chronic metabolic acidosis. J. Clin. Invest. 45 (10), 1608-1614 LEHMANN, J., J.R. LITZOW, E.J. LENNON (1967): Studies of mechanism by which chronic metabolic acidosis augments urinary calcium excretion in man. J. Clin. Invest. 46 (8), 1318-1328

Literaturverzeichnis
170 170
LENSCH, J. (1982): Clamoxyl, ein neues Antibiotikum zur Therapie des MMA-Komplexes der Sauen aus Sicht der tierärztlichen Landpraxis. Prakt. Tierarzt 63, 723-726 LERCH, A. (1987): Ursachen und Bekämpfung des MMA-Komplexes bei Muttersauen. Wien. tierärztl. Wochenschr. 102, 73-75 LICATA, A.A., E. BOU, F.C. BARTTER, F. WEST (1981): Acute effects of dietary protein on calcium metabolism in patients with osteoporosis. J. Gerontol. 36, 14-19 LIEBICH, H.-G. (1993): Das Knochengewebe (Textus osseus). In: LIEBICH, H.-G. (Hrsg.): Funktionelle Histologie, 2. Auflage, Schattauer Verlagsgesellschaft, Stuttgart, S. 64-73 LIESEGANG, A., J. RESTELI (2005): Influence of different calcium concentrations in the diet on bone metabolism in growing dairy goats and sheep. J. Anim. Physiol. Anim. Nutr. (Berl.) 89 (3-6), 113-119 LILLY, J.D., C.L. PARSONS (1990): Bladder surface glycosaminoglycans is a human epithelial permeability barrier. Surg. Gynecol. Obstetr. 171, 493-496 LITTLEWOOD-EVANS, A., T. KOKUBO, O. ISHIBASHI, T. INAOKA, B. WLODARSKI, J.A. GALLAGHER, G. BILBE (1997): Localisation of cathepsin K in human osteoclasts by in situ hybridization and immunohistochemistry. Bone 20, 81-86 LIU, B.Y., J. GUO, B. LANSKE, P. DIVIETI, H.M. KRONENBERG, F.R. BRINGHURST (1998): Conditionally immortalized murine bone marrow stromal cells mediate parathyroid hormone dependent osteoclastogenesis in vitro. Endocrinology 139, 1952-1964

Literaturverzeichnis
171 171
LÖSCHER, W. (1994): Pharmaka mit Wirkung auf periphere Mediatoren. In: LÖSCHER, W., F.R. UNGEMACH, R. KROKER (Hrsg.): Grundlagen der Pharmakotherapie bei Haus- und Nutztieren. Paul Parey Verlag, Berlin, Hamburg, S. 60- 65 LUCK, M.D., P.H. CARTER, P.J. GARDELLA (1999): The (1-14) fragment of parathyroid hormone (PTH) activates intact and amino-terminally truncated PTH-1 receptors. Mol. Endocrinol. 13, 670-680 LUTHMANN, J., J. PERSSON, G. NILSSON, R. LINDBERG (1977): The effect of a high calcium-high energy diet on calcium metabolism and plasma gastrin levels in pregnant ewes. Zentralbl. Veterinärmed. A. 24 (6), 486-495 LUTTER, K. (1983): Aktueller Kenntnisstand zur Verhütung und Bekämpfung des MMA-Komplexes der Sau. Tierzucht 37, 463-468 LUTZ, J. (1984): Calcium balance and acid-base status of women as affected by increased protein intake and by sodium bicarbonate ingestion. Am. J. Clin. Nutr. 39, 281-288 MAASS, P., W. BUSCH, H. PHILIPP (1993): Untersuchungen zur Behandlung des Metritis-Mastitis-Agalaktie (MMA)-Syndroms bei Sauen mit Voren® in Kombination zu einer antibiotischen und symptomatischen Basistherapie. Tierärztl. Umsch. 48, 346-356 MADEC, F., F. DAVID (1984): Urinary problems in sow herds. In: 8th Int. Pig Vet. Soc. Congr., Genth 1984, Proc., S. 148 MADEC, M., F. DAVID (1983): Urinary disorders in sow herds: Diagnosis, consequences, environmental and physiological factors. Journees Rech. Porcine en France 15, 431-446

Literaturverzeichnis
172 172
MÄNNER, K., K. BRONSCH (1987): Mineralstoffe. In: SCHEUNERT, A., A. TRAUTMANN (Hrsg.): Lehrbuch der Veterinär-Physiologie. 7. Auflage, Paul Parey Verlag, Berlin, S. 93-119 MAHAN, D.C., A.W. FETTER (1982): Dietary calcium and phosphorus levels for reproducing sows. J. Anim. Sci. 54, 285 MARKS, S.C., D.C. HERMEY (1996): The structure and development of bone. In: BILZEKIAN, J.P., L.G. RAISZ, G.A. RODAN (Hrsg.): Principles of bone biology. Academic Press, San Diego, London, Boston, New York, Sidney, Tokio, Toronto, S. 3-15 MARTIN, C.E., B.E. HOOPER, C.H. ARMSTRONG, H.E. AMSTUTZ (1967): A clinical and pathologic study of the mastitis-metritis-agalactia syndrome of sows. J. Am. Vet. Med. Assoc. 151, 1629-1634 MARTINEAU, G.P., B.B. SMITH, B. DOIZE (1992): Pathogenesis, prevention and treatment of lactational insufficiency in sows. Vet. Clin. North Am. Food Anim. Pract. 8 (3), 661-684 MATTERN, H. (1988): Der Knochen. In: KRÜCH, F. (Hrsg.): Pathophysiologie. Physiologische und pathologische Grundlagen innerer Erkrankungen. Urban & Schwarzenberg, München, Wien, Baltimore, S. 692-701 MC CARTHY, T.L., M. CENTRELLA, E. CANALIS (1989): Parathyroid hormone enhances the transcript and polypeptide levels of insulin-like growth factor I in osteoblast-enriched cultures from fetal rat bone. Endocrinology 124, 1247-1253 MC CLELLAND, P., J. ONYIA, R. MILES, Y. TU, J. LIANG, A. HARVEY, S. CHANDRASEKHAR, J. HOCK, J. BIDWELL (1998): Intermittent administration of parathyroid hormone (1-34) stimulates matrixmetalloproteinase-9 (MMP-9) expression in rat long bone. J. Cell. Biol. 70, 391-401

Literaturverzeichnis
173 173
MC GEHEE, D.S., M. ALDERSBERG, K.P. LIU, S. HSUING, M.J. HEATH, H. TAMIR (1997): Mechanisms of extracellular Ca2+ receptor-stimulated hormone release from sheep thyroid parafollicular cells. J. Physiol. 502, 31-44 MERIDITH, M.J. (1991): Non specific bacterial infections of the genital tract in female pigs. Pig Vet. J. 27, 110-121
MIDDLETON-WILLIAMS, D.M., J. POHLENZ, G. LOTT-STOLZ, H.U. BERTSCHINGER (1977): Untersuchungen über das Mastitis-Metritis-Agalaktie-Syndrom (Milchfieber) der Sau. I. Pathologische Befunde bei Spontanfällen. Schweiz. Arch. Tierheilk. 119, 213-222 MIZGALA, C.L., G.A. QUAMME (1985): Renal handling of phosphate. Physiol. Rev. 65, 431-466 MOORE, R.W., H.E. REDMOND, C.E. LIVINGSTON JR. (1966): Mycoplasma as the etiology of a metritis-mastitis-syndrome in sows. Vet. Med. Small. Anim. Clin. 61, 883-887 MORGENTHUM, R., G. BOLDUAN (1987): Zur Beeinflussung der Dauer des Geburtsvorganges durch Lebendmasse und Diätfaktoren bei Sauen. Monatsh. Veterinärmed. 43, 194-196 MORKOC, A., L. BÄCKSTRÖM, L. LUND, A.R. SMITH (1983): Bacterial endotoxin in blood of dysgalactic sows in relation to microbial status of uterus, milk and intestine. J. Am. Vet. Med. Assoc. 183, 786-789 MORLEY, P., J.F. WHITFIELD, G.E. WILLICK (1997): Anabolic effects of parathyroid hormone on bone. Trends Endocrinol. Metab. 8, 225-231 MÜLLER, C., O. PLATHE (1982): Der Säure-Basen-Haushalt und die Blutgase. In: BREMER, H., H. BÜTTNER, D. STAMM (Hrsg.): Klinische Chemie in Einzeldarstellungen. 2. Aufl., Thieme Verlag, Stuttgart, S. 127-143

Literaturverzeichnis
174 174
MULHOLLAND, S.G., J.R. PEREZ, J.Y. GILLENWATER (1969): The antibacterial properties of urine. Invest. Urol. 6, 569-581 NACHREINER, R.F., M.C. GARCIA, O.J. GINTHER (1972): Clinical, hematological, and blood chemical changes in swine given endotoxin (Escherichia coli) during the immediate postpartum period. Am. J. Vet. Res. 33, 2489-2499 NACHREINER, R.F., O.J. GINTHER (1974): Induction of agalactia by administration of endotoxin ( Escherichia coli) in swine. Am. J. Vet. Res. 35, 619-622 NELLANS, H.N. (1988): Contributions of cellular and paracellular calcium transport in rat ileum: Effects of dietary calcium. Am. J. Physiol. 253, E726-E737 NELLANS, H.N. (1990): Intestinal calcium absorption: Interplay of paracellular and cellular pathways. Miner. Electrolyte Metab. 16, 101-108 NELSON, D.A., M.G. HOGBERG, E.R. MILLER, M.S. ALLEN (1992a): Wheat straw and soybean hull additions to sow gestation diets during two consecutive parities. Michigan State University Swine Rpt. 92, 10 NEMERE, I., A.W. NORMAN (1982): Vitamin D and intestinal cell membranes. Biochem. Biophys. Acta 694, 307-327 NEMERE, I., G. THEOFAN, A.W. NORMAN (1987): Rapid action of 1,25-dihydroxyvitamin D3 on calcium transport in perfused chick duodenum: effect of inhibitors. J. Bone Miner. Res. 2, 99-107 NEWELL, G.K., R.E. BEAUCHENE (1975): Effects of dietary calcium level, acid stress and age on renal, serum and bone responses of rats. J. Nutr. 105, 1039-1047

Literaturverzeichnis
175 175
NIMMO, R.D., E.R. PEO, J.D. CRENSHAW, B.D. MOSER, A.J. LEWIS (1981): Effect of level of dietary calcium-phosphorus during growth and gestation on calcium-phosphorus balance and reproductive performance of first-litter sows. J. Anim. Sci. 52, 1343-1349 NORDEN, C.W., G.M. GREEN, E.H. KASS (1968): Antibacterial mechanisms of the urinary bladder. J. Clin. Invest. 47, 2689-2700 OKADA, Y., I. MOROMOTO, K. URA, K. WATANABE, S. ETO, M. KUMEGAWA, L. RAISZ, C. PILBEAM, T. TANAKN (2002): Cell-to-cell adhesion via intercellular adhesion molecule-1 and leukocyte function-associated antigen-1 pathway is involved in 1-alpha,25(OH)2D3, PTH and IL-1-alpha-induced osteoclast differentiation and bone resorption. Endocr. J. 49, 483-495 OLDIGS, B., W. HINSCH (1987): Blut- und Serumwerte des Göttinger Miniaturschweines; 6. Mitteilung: Der Einfluß endogener und exogener Faktoren auf die Merkmale des Kohlenhydratstoffwechsels, des Säure-Basen-Haushaltes und auf die Blutkörperchensenkung. J. Vet. Med. A. 34, 627-638 OSBORNE, C.A., J.S. KLAUSNER, G.E. LEES (1979): Urinary tract infections: Normal and abnormal host defense mechanisms. Vet. Clin. Of North Amer. 9 ( 4), 587-605 OURSLER, M.J., J.P. LANDERS, B.L. RIGGS, T.C. SPELSBERG (1993): Oestrogen effects on osteoblasts and osteoclasts. Annals of Medicine 25, 361-371 PANSU, D., C. BELLATON, C. ROCHE, F. BRONNER (1983): Duodenal and ileal calcium absorption in the rat and effects of vitamin D. Am. J. Physiol. 244, G695-G700 PARSONS, C.L., C. GREENSPAN, S.G. MULHOLLAND (1975): The primary antibacterial defense mechanism of the bladder. Invest. Urol. 13, 72-76 PARSONS, C.L., S.H. SHROM, P. HANNO, S.G. MULHOLLAND (1978): Bladder surface mucin. Examination of possible mechanisms for its antibacterial effects. Invest. Urol. 16, 196-200

Literaturverzeichnis
176 176
PATIENCE, J.F., R.E. AUSTIC, R.D. BOYD (1987): Acid-base-homeostasis in swine: Nutritional perspective. Feedstuffs 6, 13-18 PATIENCE, J.F., R.K. CHAPLIN (1997): The relationship among dietary undetermined anion, acid-base-balance and nutrient in swine. J. Anim. Sci. 75, 2445-2452 PEDERSEN, A., K. KROVACEK, H. EKWALL (1984): Virulence factors in strains of Escherichia coli isolated from mastitic milk from agalactic sows. In: 8th Int. Pig Vet. Soc. Congr., Genth 1984, Proc., S. 286 PERSSON, A. (1997): Clinical assessment of udder health status of sows at time of weaning with special seference to bacteriology and cytology in milk. J. Vet. Med. A 44, 143-158 PETERSEN, B. (1979): Die Harnuntersuchung bei der Sau, eine einfache Methode zur Erkennung von Problemtieren. Prakt. Tierarzt 60, 1092-1100 PETERSEN, B. (1982): Value of methods of early recognition of puerperal and fertility disorders in the sow. In: 7th Int. Pig Vet. Soc. Congr., Mexico City 1982, Proc., S. 231 PLONAIT, H., A. WILMS-SCHULZE KUMP, G. SCHÖNING (1986): Prophylaxis of the MMA-syndrome by antibacterial medication and restricted feeding. In: 9th Int. Pig Vet. Soc. Congr., Barcelona 1986, Proc., S. 96 PLONAIT, H. (1980): Labordiagnostik für die tierärztliche Praxis Paul Parey Verlag, Berlin, Hamburg. PLONAIT, H. (1997): Geburt, Puerperium und perinatale Verluste. In: PLONAIT, H., K. BICKHARDT (Hrsg.): Lehrbuch der Schweinekrankheiten. 2. Auflage, Paul Parey Verlag, Berlin, S. 309-339 POLLMANN, D.S., D.M. DANIELSON, M.A. CRENSHAW, E.R. PEO, JR (1981): Long-term effects of dietary additions of alfalfa and tallow on sow reproductive performance. J. Anim. Sci. 51, 294

Literaturverzeichnis
177 177
POND, W.G., J.T. YEN, V.H. VAREL (1985): Effects of level and source of dietary fiber in gestation on reproductive performance and nutrient digestability in gilts. Nutr. Rep. Intern. 32 (3), 505 POZZAN, T., R. RIZZUTO, P. VOLPE, J. MELDOLESI (1994): Molecular and cellular physiology of intracellular calcium store. Physiol. Rev. 74, 595-636 PRICE, J.S. (1998): Biochemical markers of bone metabolism in horses: Potentials and limitations? Vet. J. 156, 163-165 QUARLES, L.D. (1997): Cation-sensing receptors in bone: A novel paradigm for regulating bone remodelling? J. Bone Miner. Res. 12, 1971-1974 RABADJIJA, L., E.M. BROWN, S.L. SWARTZ, C.J. CHEN, P. GOLDHABER (1990): H+-stimulated release of prostaglandin E2 and cyclic adenosine 3´,5´-monophosphoric acid and their relationship to bone resorption in neonatal mouse calvaria cultures. Bone 11, 295-304 RADDE, I.C., D. DAVIS, J. SHEEPERS, H.G. MC KERCHER (1980): Bidirectional transmucosal 45Ca and 32P fluxes across the small intestine of young piglet: Relationship to intestinal Ca2+-Mg2+-ATPase activity and postnatal age. In: DE LUCA H.F., C.S. ANAST (Hrsg.): Pediatric diseases related to calcium. Blackwell Scientific Publications, Oxford, S. 153-163 REID, G., J.D. SOBEL (1987): Bacterial adherence in the pathogenesis of urinary tract infection: A review. Rev. Inf. Dis. 9, 470-487 RICCARDI, D., A.E. HALL, N. CHATTOPADHYAY, J.Z. XU, E.M. BROWN, S.C. HEBERT (1998): Localization of the extracellular Ca2+/polyvalent cation-sensing protein in rat kidney. Am. J. Physiol. 174, F611-622 RICKARD, D.J., M. SUBRAMANIAM, T.C. SPELSBERG (1999): Molecular and cellular mechanisms of estrogen action on the skeleton. J. Cell. Biochem. 33, 123-132

Literaturverzeichnis
178 178
RINGARP, N. (1960): Clinical and experimental investigations into a postparturient syndrome with agalactia in sows. Acta. Agr. Scand. Suppl. 7, 115-118 ROSALKI S.B., A.Y. FOO (1984): Two new methods for separating and quantifying bone and liver alkaline phosphatase isoenzymes in plasma. Clin. Chem. 30, 1182-1186 ROSE, M., U. SCHNURRBUSCH, K. HEINRITZI (1996): The use of cefquinome in the treatment of pig respiratory diseases and MMA-syndrome. In: 14th Int. Pig Vet. Soc. Congr., Bologna 1996, Proc., S. 317 ROSENFELD, R.G., A.L. ROSENBLOOM, J. GUEVARA-AGUIRRE (1994): Growth hormone (GH) insensitivity due to primary GH receptor deficiency. Endocr. Rev. 15, 369-390 ROSS, R.F., A.P. ORNING, R.D. WOODS, B.J. ZIMMERMANN, D.F. COX, D.L. HARRIS (1981): Bacteriologic study of sow agalactia. Am. J. Vet. Res. 42, 949-956 RÜMENAPF, G., O.P. SCHWILLE, R.G: ERBEN, M. SCHREIBER, B. BERGE, W. FRIES, A. SCHMIEDL, S. KOROMA, W. HOHENBERGER (1998): Gastric fundectomy in the rat: Effects on mineral and bone metabolism, with emphasis on the Gastrin-Calcitonin-Parathyroid Hormone-Vitamin D Axis. Calcif. Tissue Int. (1998) 63, 433-444 RUHRMANN, A., H. HAMBITZER, E. BENT (1986): Referenzwerte verschiedener Harninhaltsstoffe bei Sauen Dtsch. tierärztl. Wochenschr. 93, 115-120 SANECKI, R.K., W.E. HOFFMANN, J.L. DORNER (1990): Purification and comparison of corticosteroid-induced and intestinal isoenzymes of alkaline phosphatase in dogs. Am. J. Vet. Res. 51, 1964-1968 SANSTEDT, H., U. SJORGEN (1982): Preventive measures against a high prevalence of mastitis-metritis-agalactia syndrome in sows. Svensk-Veterinartidning 34, 487-490

Literaturverzeichnis
179 179
SATO, T., M. DEL CARMEN OVEJERO, P. HOU, A.M. HEEGAARD, M. KUMEGAWA, N.T. FOGED, J.M. DELAISSE (1997): Identification of the membrane-type matrix metalloproteinase MTI-MMP in osteoclasts. J. Cell. Sci. 110, 589-596 SATO, J., K. OKADA, S. FUKUDA, R. SATO, J. YASUDA, Y. NAITO (2002): Serum activities of tartrate-resistant acid phosphatase and bone specific alkaline phosphatase as indices of bone metabolism in the cow. J. Vet. Med. Sci. 64(7), 653-655 SCHADE, C. (2000): Feldstudie zu Auswirkungen einer Ad-Libitum-Fütterung in der Gravidität auf die Entwicklung von Futteraufnahme, Körpermasse, Ernährungszustand und Rückenspeckdicke sowie auf die Reproduktionsleistung und Gesundheit von Sauen. Hannover, Tierärzt. Hochsch., Diss. SCHEID, P. (1994): Säure-Basen-Gleichgewicht In: KLINKE, R., S. SILBERNAGL (Hrsg.): Lehrbuch der Physiologie. Georg Thieme Verlag, Stuttgart, S. 189-216 SCHEUNERT, A., A. TRAUTMANN (1987) Lehrbuch der Veterinär-Physiologie. 7. Aufl. Paul Parey Verlag, Berlin. SCHMIDL, M. (1979b): Diagnostik von Erkrankungen der Nieren und der ableitenden Harnwege. Tierärztl. Umsch. 34, 862-866 SCHMIDT-GAYK, H., R. BOUILLON, H.J. ROTH (1997): Measurement of vitamin D metabolites (calcidiol and calcitriol) and their clinical significance. Scan. J. Clin. Lab. Invest. 57 (Suppl. 237), 35-45 SCHÖNING, G. (1986): Symptomatik und Verlauf des MMA-Syndroms der Sauen bei diätetischer und antibakterieller Metaphylaxe. Berlin, Frei Univ., Tierärztl. Fak., Diss. SCHÖNING, G., H. PLONAIT (1990): Metaphylaxe und Therapie des MMA-Syndroms der Sauen mit Baytril®. Dtsch. tierärztl. Wochenschr. 97, 5-10

Literaturverzeichnis
180 180
SCHONEWILLE, J.T., A.T. VAN KLOOSTER, A. DIRKZWAGER, A.C. BEYNEN (1994): Stimulatory effect of an anion (chloride)-rich ration on apparent calcium absorption in dairy cows. Livestock Prod. Sci. 40, 233-240 SCHRÖDER, B., C.SCHLUMBOHM, R. KAUNE, G. BREVES (1996): Role of calbindin-D9k in buffering cytosolic free Ca2+ ions in pig duodenal enterocytes. J. Physiol. 492, 3, 715-722 SCHUKNECHT, A. (1991): Untersuchungen zum Einfluß der Fütterung auf den Harn-pH-Wert und die renale Mineralstoffausscheidung bei der Katze. Hannover, Tierärztl. Hochsch., Diss. SCHULZE, W., W. BOLLWAHN (1962): Zu den Erkrankungen der Sauen vor, während und nach der Geburt. Dtsch. tierärztl. Wochenschr. 69, 641-644 SCHULZE, W., G. VON MICKWITZ (1966): Puerperalstörungen bei Schweinen und ihre therapeutischen Beeinflussungsmöglichkeiten. Dtsch. tierärztl. Wochenschr. 73, 557-560 SCOTT, D. (1971): Renal excretion of acid and base in the pig. Quart. J. exp. Physiol. 56, 169-177 SEBASTIAN, A., S.T. HARRIS, J.H. OTTAWAY, K.M. TODD, R.C. MORRIS (1994): Improved mineral balance and skeletal metabolism in postmenopausal women treated with potassium bicarbonate. N. Engl. J. Med. 330, 1776-1781 SEIBEL, M.S., H.W. WOITGE, R. ZIEGLER (1993): Biochemische Grundlagen des Knochenstoffwechsels. Klin. Lab. 39, 717-727 SELDIN, D.W., G. GIEBISCH (1989): The regulation of acid-base-balance. Raven Press, New York, NY.

Literaturverzeichnis
181 181
SERRA, R., A. KARAPLIS, P. SOHN (1999): Parathyroid hormone-related peptide (PTHrP)-dependent and independent effects of transforming growth factor beta (TGF-beta) on endochondral bone formation. J. Cell. Biol. 145, 783-794 SHERMAN, P., N. FLEMING, J. FORSTNER, N. ROOMI, G. FORSTNER (1987): Bacteria and mucus blanket in experimental small bowel bacterial over growth. Am. J. Path. 129, 527-534 SHROM, S.H., C.L. PARSONS, S.G. MULHOLLAND (1977): Role of urothelial surface mucoprotein in intrinsic bladder defense. Urology 9, 526-533 SILBERNAGL, S. (1994): Die Funktion der Nieren In: KLINKE, R., S. SILBERNAGL (Hrsg.): Lehrbuch der Physiologie. Georg Thieme Verlag, Stuttgart, New York, S. 259-283 SJǾRGEN, K., Y.M. BOHLOOLY, B. OLSSON, B. COSCHIGANO, J. TORNELL, S. MOHAN, O.G. ISAKSSON, G. BAUMANN, J. KOPCHICK, C. OHLSSON (2000): Disproportional skeletal growth and markedly decreased bone mineral content in growth hormone receptor -/- mice. Biochem. Biophys. Res. Commun. 267, 603-608 SMITH, W.J. (1983): Cystitis in sows. Pig News & Inform. 4, 279-281 SPRAGUE, S.M., N.S. KRIEGER, D.A. BUSHINSKY (1994): Greater inhibition of in vitro bone mineralization with metabolic than respiratory acidosis. Kidney Int. 46, 1199-1209 STANISLAUS, D., V. DEVANARAYAN, J. HOCK (2000): In vivo regulation of apoptosis in metaphyseal trabecular bone of young rats by synthetic human parathyroid hormone (1-34) fragment. Bone 27, 209-218 STEIN, G.S., J.B. LIAN, T:A. OWEN (1990): Relationship of cell growth to the regulation of tissue-specific gene expression during osteoblast differentiation. FASEB J. 4, 3111-3123

Literaturverzeichnis
182 182
STELZER, P., G. BILKEI, E. CLAVADETSCHER, O. BIRO, T. TAKACS, I. KOTAI, H. BILKEI (1997): Scheidenausfluß ante partum als Risikofaktor für postpartale Erkrankungen der Sau und frühpostnatale Ferkelverluste. Berl. Münch. tierärztl. Wochenschr. 110, 320-323 STIRNIMANN, J. (1984): Akute Harnwegsentzündungen bei der Muttersau. Schweiz. Arch. Tierheilk. 126, 597-605 STRAUB, O.C. (1990): Folgen der puerperalen Mastitis auf das Wachstum der Ferkel. Tierärztl. Umsch. 45, 132 STRAUSS-ELLERMANN, H. (1985): Klinische und pathomorphologische Befunde an der nicht laktierenden Milchdrüse des Schweines und deren Bedeutung für die folgende Laktation. Berlin, Freie Univ., Tierärztl. Fak., Diss. SUNDQUIST, K., S. MARKS (1995): Bafilomycin A1 inhibits bone resorption and tooth eruption in vivo. J. Bone Miner. Res. 9, 1575-1582 TAKEDA, S., T. YOSHIZAWA, Y. NAGAI, H. YAMATO, S. FUKUMOTO, K. SEKINE, S. KATO, T. MATSUMOTO, T. FUJITA (1999): Stimulation of osteoclast formation by 1,25-dihydroxyvitamin D requires its binding to vitamin D receptor (VDR) in osteoblastic cells: Studies using VDR knockout mice. General and Comparative Endocrinology 140, 1005-1008 TEZUKA, K., K. NEMOTO, Y. TEZUKA, T. SATO, Y. IKEDA, M. KOBORI, H. KAWASHIMA, H. EGUCHI, Y. HAKEDA, M. KUMEGAWA (1994): Identification of matrix metalloproteinase 9 in rabbit osteoclasts. J. Biol. Chem. 269, 15006-15009 THEWS, G. (1995): Säure-Basen-Status des Blutes In: SCHMIDT, R.F., G. THEWS (Hrsg.): Physiologie des Menschen. Springer-Verlag, Berlin, Heidelberg, New York, S. 615-623 THOMAS, L. (1992): Kreatinin. In: THOMAS, L. (Hrsg.): Labor und Diagnose. 4. Auflage, Medizinische Verlagsgesellschaft, Marburg, S. 447-455

Literaturverzeichnis
183 183
THRONTON, E.J., R.J. WILSON, I. CONNAUGHTON, K. MOORE (1998): Effect of subclinical urogenital infection on reproductive performance in the sow. In: 15th Int. Pig Vet. Soc. Congr., Birmingham 1998, Proc., S. 236 TRABITSCH, R. (1988): Versuche zur Auslösung des MMA-Syndroms bei Zuchtsauen durch Veränderung der Fütterungs- und Haltungsbedingungen. Wien. tierärztl. Monatsschr. 75, 74-79 TYRELL, D. (1978): Vergleichende klinische und bakteriologische Untersuchungen an erkrankten und gesunden Sauen im Frühpuerperium. Hannover, Tierärztl. Hochsch., Diss. ULMER-SHAKIBAEI, C. (1995): Untersuchungen zur Ätiologie und Differentialdiagnose der Laktationshyperthermie der Sau. Berlin, Freie Univ., Tierärztl. Fak., Diss. VÄÄNÄNEN, H.K., E.K. KARHUKORPI, K. SUNQUIST, B. WALLMARK, I. ROININEN, T. HENTUNEN, J. TUUKKANEN, P. LAKKAKORPI (1990): Evidence fort he presence of a proton pump on the vacuolar H+-ATPase type in the ruffled borders of osteoclasts. J. Cell. Biol. 111, 1305-1311 VAN BEERS-SCHREURS H.M.G., A. DIRKZWAGER, I.A.J.M. EIJCK (1996): Periparturient hypogalactic syndrome in sows. A field study. In: 14th Int. Pig Vet. Soc. Congr., Bologna 1996, Proc., S. 616 VANDEPLASSCHE, M., L. GEURDEN, M. VAN DEN WYNGAERT, G. SNOECK, A. DE VOS (1960): Puerperale Septikämie und Toxämie des Schweines. Dtsch. tierärztl. Wochenschr. 67, 375-377 VERTER, W., E. UECKER (1990): Zur Bedeutung und Diagnostik von Harnwegsinfektionen beim Schwein. Monatsh. Veterinärmed. 45, 234-237 VESPER, C. (1991): Untersuchungen zum Vorkommen von Harnwegsinfektionen in Zuchtsauenbeständen unter besonderer Berücksichtigung von Eubacterium suis. Hannover, Tierärztl. Hochsch., Diss.

Literaturverzeichnis
184 184
VOPELIUS-FELDT, A.v. (1984): Ergebnisse physiko-chemischer Untersuchungen von Schweineharn. Ein Beitrag zur Zystitis-Diagnostik. München, Univ., Tierärztl. Fak., Diss. VORTKAMP, A., K. LEE, B. LANSKE, G.V. SEGRE, H.M. KRONENBERG, C.J. TABIN (1996): Regulation of rate of cartilage differentiation by Indian hedgehog and PTH- related protein. Science 273, 613-622 WAGNER, S. (1990): Die Immunreaktion bei der durch Escherichia coli bedingten chronischen Harnwegsinfektion des weiblichen Schweines. München, Tierärztl. Fak., Diss. WALDMANN, K.H. (1987): Die Pyelozystitis der Zuchtsau. Tierärztl. Prax. 15, 263-267 WALDMANN, K-H., M. WENDT, K. BICKHARDT (1991): Kreatinin-Clearance als Grundlage klinischer Nierenfunktionsbestimmungen beim Schwein. Tierärztl. Prax. 19, 373-380 WALLING, M.W. (1977): Intetsinal calcium and phosphate transport: Differential responses to vitamin D3 metabolites. Am. J. Physiol. 233, E488-E494 WAWRON, W. (1995): Verhalten einiger Indikatoren der unspezifischen Immunität bei Sauen aus Zuchtbetrieben mit unterschiedlichen Raten der MMA-Syndrommorbidität. Tierärztl. Umsch. 50, 584-590 WAWRON, W. (1997): Einfluss von Immunitäts-stimulierenden Präparaten auf das Vorkommen der Puerperalerkrankungen bei Schweinen. Dtsch. Tierärztl. Wochenschr. 104, 220-223 WEARY, D.M., E.A. PAJOR, B.K. THOMPSON, D. FRASER (1996): Risky behaviour by piglets: A trade off between feeding and risk of mortality by maternal crushing? Anim. Behaviour 51, 619-624

Literaturverzeichnis
185 185
WEGMANN, P., H.U. BERTSCHINGER (1984): Sequential cytological and bacteriological examination of the secretion from sucked and unsucked mammary glands with or without mastitis. In: 8th Int. Pig Vet. Soc. Congr., Genth 1984, Proc., S. 287 WEGMANN, P. (1985): Zur Pathogenese der Colimastitis beim Mutterschwein. Zürich, Univ., Veterinärmed. Fak., Diss. WEINSTEIN, R.S., R.L. JILKA, A.M. PARFITT, S.C. MANOLAGAS (1998): Inhibition of osteoblastogenesis and promotion of apoptosis of osteoblasts and osteocytes by glucocorticoids. Potential mechanisms of their deleterious effects on bone. J. Clin. Invest. 102(2), 274-282 WEISS, R.E., S. REFETOFF (1996): Effect of thyroid hormone on growth: Lessons from the syndrome of resistance to thyroid hormone. Endocrinol. Metab. Clin. North Am. 25, 719-730 WELDON, W.C., A. LEWIS, G.F. LOUIS, J.L: KOVAR, M.A. GIESEMANN, P.S. MILLER (1994): Post partum hypophagia in primiparous sows: I. Effects of gestation feeding level on feed intake, feeding behavior and plasma metabolite concentrations during lactation. J. Anim. Sci. 72, 387 WENDT, M. (1992): Untersuchungen zur Diagnostik und zur Charaterisierung von Harnwegsinfektionen der Sau unter besonderer Berücksichtigung von Eubacterium suis. Hannover, Tierärztl. Hochsch., Habil.-Schr. WENDT, M. (1998): Urinary system dysorders of pigs. In: 15th Int. Pig Vet. Soc. Congr., Birmingham 1998, Proc., S. 195-201 WENDT, M., J. SOBESTIANSKY, W. BOLLWAHN (1993): Zur Behandlung der Eubacterium-suis-Infektion bei Ebern. Berl. Münch. tierärztl. Wochenschr. 106, 221-224 WENDT, M., F. LAPPE, K. BICKHARDT (1996): Untersuchungen zur Kristallurie bei Sauen. Dtsch. Tierärztl. Wochenschr. 103, 506-510

Literaturverzeichnis
186 186
WHYTE, M.P. (1983): Alkaline phosphatase and the measurement of bone formation in clinical disorders of bone and mineral metabolism. In: FRAME C., J.T. POTTS (Hrsg.): Clinical disorders of bone and mineral metabolism. Excerpta Medica, Amsterdam, S. 108-111 WIDDOWSON, E.M., J.W.T. DICKERSON (1964): Chemical composition of the body. In: COMAR, C.L., F. BRONNER (Hrsg.): Mineral metabolism. Acadimic, New York, S. 1-247 WIESNER, E., S. GOLBS (1991): Probiotika. In: WIESNER, E., R. RIBBECK (Hrsg.): Wörterbuch der Veterinärmedizin. 3. Auflage, Gustav Fischer Verlag, Jena, Stuttgart, S. 1187 YAMAGUCHI, T., N. CHATTOPADHYAY, E.M. BROWN (1999): G protein-coupled extracellular Ca2+ ( Ca0
2+)-sensing receptor ( CaR): Roles in cell signalling and control of diverse cellular functions. Adv. Pharmacol. 47, 209-253 YEAW, R.C. (1944): The effect of pH on the growth of bacteria. J. Urol. 44, 699-713 YEN, J.T., W.G. POND, R.L. PRIOR (1981): Calcium chloride as a regulator or feed intake and weight gain in pigs. J. Anim. Sci. 52, 778-782 ZAIDE, M., O.A. ADEBANJO, B.S. MOONGA, L. SUN, C.L. HUANG (1999): Emerging insights into the role of calcium ions in osteoclast regulation. J. Bone. Miner. Res. 14, 669-674 ZHAO, W., M. BYRNE, B. BOYCE, S. KRANE (1999): Bone resorption induced by parathyroid hormone is strikingly diminished in collagenase- resistant mutant mice. J. Clin. Invest. 103, 517-524 ZHOU, Y., B.C. XU, H.G. MAHESHWARI, L. HE, M. REED, M. LOZYKOWSKI, S. OKADA, L. CATALDO, K. COSCHIGAMO, T.E. WAGNER, G. BAUMANN (1997): A mammalian model for Laron syndrome produced by targeted disruption of the mouse growth hormone receptor/binding protein gene (the Laron mouse). Proc. Natl. Acad. Sci. USA 94, 13215-13220
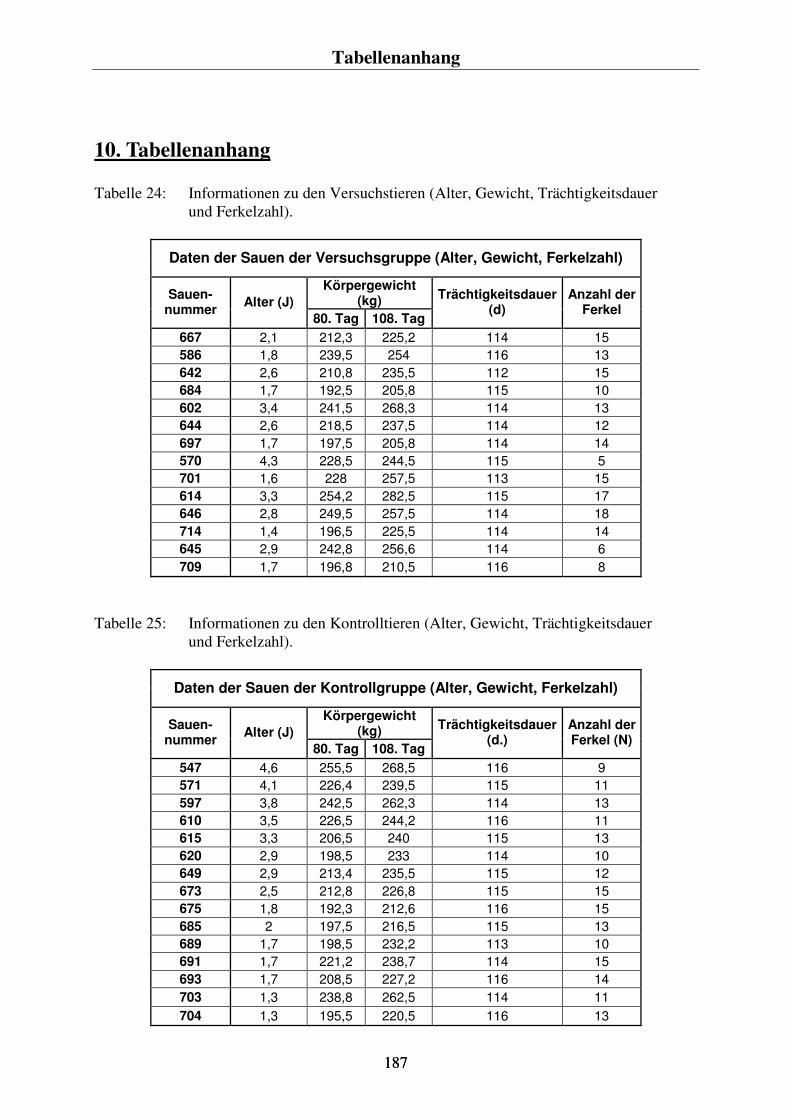
Tabellenanhang
187 187
10. Tabellenanhang Tabelle 24: Informationen zu den Versuchstieren (Alter, Gewicht, Trächtigkeitsdauer
und Ferkelzahl).
Daten der Sauen der Versuchsgruppe (Alter, Gewicht, Ferkelzahl)
Körpergewicht (kg) Sauen-
nummer Alter (J)
80. Tag 108. Tag
Trächtigkeitsdauer (d)
Anzahl der Ferkel
667 2,1 212,3 225,2 114 15
586 1,8 239,5 254 116 13
642 2,6 210,8 235,5 112 15
684 1,7 192,5 205,8 115 10
602 3,4 241,5 268,3 114 13
644 2,6 218,5 237,5 114 12
697 1,7 197,5 205,8 114 14
570 4,3 228,5 244,5 115 5
701 1,6 228 257,5 113 15
614 3,3 254,2 282,5 115 17
646 2,8 249,5 257,5 114 18
714 1,4 196,5 225,5 114 14
645 2,9 242,8 256,6 114 6
709 1,7 196,8 210,5 116 8
Tabelle 25: Informationen zu den Kontrolltieren (Alter, Gewicht, Trächtigkeitsdauer
und Ferkelzahl).
Daten der Sauen der Kontrollgruppe (Alter, Gewicht, Ferkelzahl)
Körpergewicht (kg) Sauen-
nummer Alter (J)
80. Tag 108. Tag
Trächtigkeitsdauer (d.)
Anzahl der Ferkel (N)
547 4,6 255,5 268,5 116 9
571 4,1 226,4 239,5 115 11
597 3,8 242,5 262,3 114 13
610 3,5 226,5 244,2 116 11
615 3,3 206,5 240 115 13
620 2,9 198,5 233 114 10
649 2,9 213,4 235,5 115 12
673 2,5 212,8 226,8 115 15
675 1,8 192,3 212,6 116 15
685 2 197,5 216,5 115 13
689 1,7 198,5 232,2 113 10
691 1,7 221,2 238,7 114 15
693 1,7 208,5 227,2 116 14
703 1,3 238,8 262,5 114 11
704 1,3 195,5 220,5 116 13
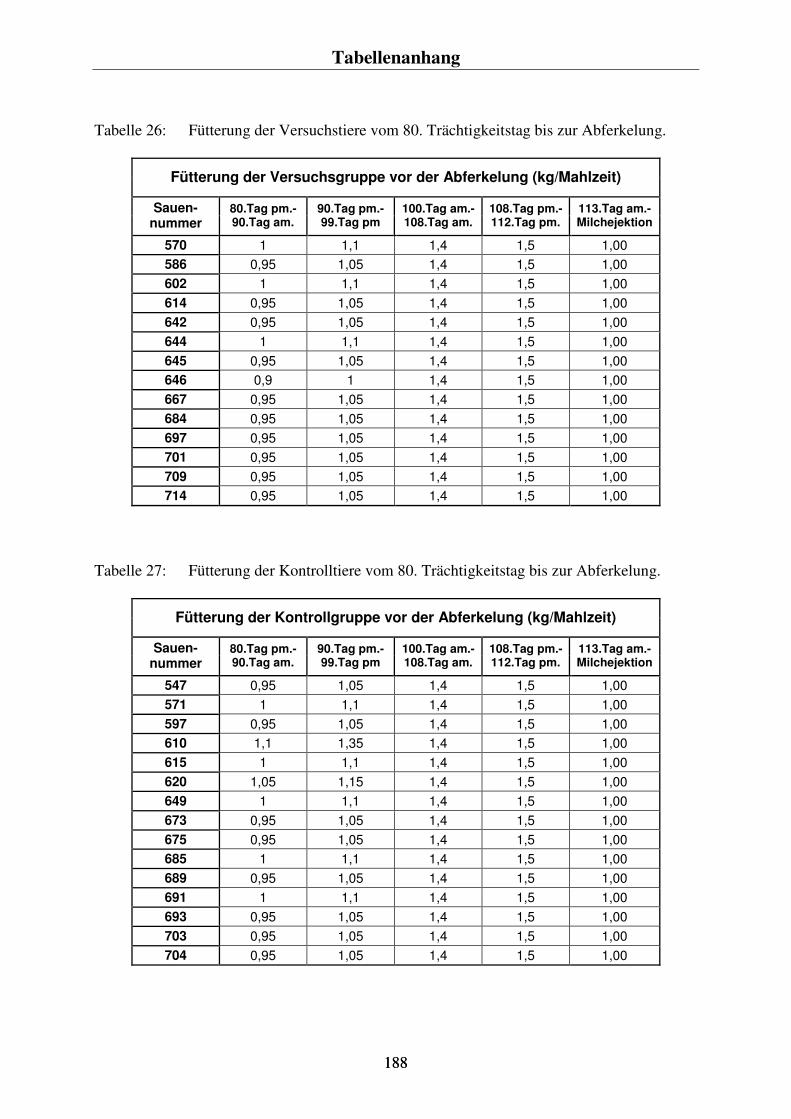
Tabellenanhang
188 188
Tabelle 26: Fütterung der Versuchstiere vom 80. Trächtigkeitstag bis zur Abferkelung.
Fütterung der Versuchsgruppe vor der Abferkelung (kg/Mahlzeit)
Sauen-nummer
80.Tag pm.-90.Tag am.
90.Tag pm.-99.Tag pm
100.Tag am.-108.Tag am.
108.Tag pm.-112.Tag pm.
113.Tag am.-Milchejektion
570 1 1,1 1,4 1,5 1,00
586 0,95 1,05 1,4 1,5 1,00
602 1 1,1 1,4 1,5 1,00
614 0,95 1,05 1,4 1,5 1,00
642 0,95 1,05 1,4 1,5 1,00
644 1 1,1 1,4 1,5 1,00
645 0,95 1,05 1,4 1,5 1,00
646 0,9 1 1,4 1,5 1,00
667 0,95 1,05 1,4 1,5 1,00
684 0,95 1,05 1,4 1,5 1,00
697 0,95 1,05 1,4 1,5 1,00
701 0,95 1,05 1,4 1,5 1,00
709 0,95 1,05 1,4 1,5 1,00
714 0,95 1,05 1,4 1,5 1,00
Tabelle 27: Fütterung der Kontrolltiere vom 80. Trächtigkeitstag bis zur Abferkelung.
Fütterung der Kontrollgruppe vor der Abferkelung (kg/Mahlzeit)
Sauen-nummer
80.Tag pm.-90.Tag am.
90.Tag pm.-99.Tag pm
100.Tag am.-108.Tag am.
108.Tag pm.-112.Tag pm.
113.Tag am.-Milchejektion
547 0,95 1,05 1,4 1,5 1,00
571 1 1,1 1,4 1,5 1,00
597 0,95 1,05 1,4 1,5 1,00
610 1,1 1,35 1,4 1,5 1,00
615 1 1,1 1,4 1,5 1,00
620 1,05 1,15 1,4 1,5 1,00
649 1 1,1 1,4 1,5 1,00
673 0,95 1,05 1,4 1,5 1,00
675 0,95 1,05 1,4 1,5 1,00
685 1 1,1 1,4 1,5 1,00
689 0,95 1,05 1,4 1,5 1,00
691 1 1,1 1,4 1,5 1,00
693 0,95 1,05 1,4 1,5 1,00
703 0,95 1,05 1,4 1,5 1,00
704 0,95 1,05 1,4 1,5 1,00

Tabellenanhang
189 189
Tabelle 28: Fütterung der Versuchstiere in den ersten 8 Tagen nach der Abferkelung.
Laktationsfuttermenge der Versuchstiere nach der Abferkelung (kg/Mahlzeit)
Tage nach der Abferkelung Sauen-nummer
lebende Ferkel (N) 1 2 3 4 5 6 7 8
570 5 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,25
586 11 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
602 13 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
614 14 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
642 14 1,0 1,0 1,25 1,5 1,75 2,25 2,5 2,5
644 12 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
645 4 1,0 1,0 1,25 1,25 1,5 1,75 2,0 2,25
646 16 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
667 13 1,0 1,0 1,25 1,25 1,5 1,75 2,0 2,25
684 10 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
697 13 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
701 15 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
709 8 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
714 14 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
Tabelle 29: Fütterung der Kontrolltiere in den ersten 8 Tagen nach der Abferkelung.
Laktationsfuttermenge der Kontrolltiere nach der Abferkelung (kg/Mahlzeit)
Tage nach der Abferkelung Sauen-nummer
lebende Ferkel (N) 1 2 3 4 5 6 7 8
547 7 1,0 1,0 1,25 1,5 1,5 2,0 2,25 2,5
571 9 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
597 8 1,0 1,0 1,0 1,25 1,25 1,5 1,75 2,0
610 9 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
615 13 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
620 9 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
649 11 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
673 13 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
675 13 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
685 12 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
689 9 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
691 13 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
693 12 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5
703 11 1,0 1,0 1,0 1,25 1,25 1,5 1,75 2,0
704 12 1,0 1,0 1,25 1,5 1,75 2,0 2,25 2,5

Tabellenanhang
190 190
Tabelle 30: Geburtsdaten der Versuchsgruppe (Geburtsdauer, Dauer bis zum Abgang der Nachgeburt, Ferkelzahlen und durchschnittliche Geburtsgewichte).
Geburtsdaten der Tiere der Versuchsgruppe
Anzahl der geborenen Ferkel (N) Sauen-
nummer
Geburts- dauer
(h:min)
Nachgeburt (h:min) Gesamt Lebende
Lebens-schwache
Tote
Geburts-gewicht Ø
(kg)
570 03:35 00:30 5 5 0 0 1,61
586 02:00 00:45 13 11 1 1 1,15
602 02:02 01:45 13 13 0 0 1,51
614 02:35 01:35 17 14 0 3 1,19
642 01:45 00:55 15 14 0 1 1,29
644 02:05 00:50 12 12 0 0 1,22
645 01:15 00:40 6 4 1 1 2,13
646 03:30 01:05 18 16 0 2 1,1
667 05:13 01:37 15 13 2 0 1,41
684 01:30 00:55 10 10 0 0 1,34
697 02:45 00:40 14 13 1 0 1,41
701 02:35 01:25 15 15 0 0 1,63
709 02:05 00:20 8 8 0 0 1,68
714 01:55 00:25 14 14 0 0 1,27
Tabelle 31: Geburtsdaten der Kontrollgruppegruppe (Geburtsdauer, Dauer bis zum Abgang der Nachgeburt, Ferkelzahlen und durchschnittliche Geburtsgewichte).
Geburtsdaten der Tiere der Kontrollgruppe
Anzahl der geborenen Ferkel (N) Sauen-
nummer
Geburts- dauer
(h:min)
Nachgeburt
(h:min) Gesamt Lebende Lebens-
schwache Tote
Geburts-gewicht Ø
(kg)
547 01:55 00:35 9 7 0 2 1,25
571 01:50 00:50 11 9 0 2 1,63
597 01:13 00:57 13 8 0 5 1,28
610 01:05 00:40 11 9 0 2 1,71
615 02:10 00:55 13 13 0 0 1,66
620 01:50 00:40 10 9 0 1 1,36
649 01:40 00:30 12 11 0 1 1,62
673 02:52 00:55 15 13 2 0 1,1
675 02:50 00:45 15 13 2 0 1,18
685 03:05 00:20 13 12 0 1 1,23
689 01:30 00:30 10 9 0 1 1,89
691 02:10 01:10 15 13 2 0 1,38
693 02:10 00:45 14 12 2 0 1,27
703 02:10 01:45 11 11 0 0 1,79
704 02:50 00:55 13 12 0 1 1,39

Tabellenanhang
191 191
Tabelle 32: pH-Werte im Nativblut der Sauen aus der Versuchsgruppe, entsprechend den
Blutentnahmeterminen zugeordnet.
pH-Werte im Vollblut der Sauen der Versuchsgruppe
Trächtigkeitstage Tage post partum Sauen- nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
570 7,341 7,320 7,410 7,430 7,398 7,334 7,379
586 7,439 7,342 7,333 7,346 7,373 7,437 7,333
602 7,430 7,414 7,458 7,415 7,375 7,381 7,367
614 7,400 7,385 7,477 7,350 7,366 7,304 7,331
642 7,399 7,347 7,371 7,496 7,360 7,300 7,353
644 7,433 7,367 7,417 7,436 7,387 7,444 7,417
645 7,466 7,400 7,364 7,422 7,432 7,412 7,415
646 7,394 7,350 7,444 7,365 7,373 7,357 7,276
667 7,431 7,413 7,383 7,360 7,375 7,388 7,373
684 7,411 7,343 7,330 7,322 7,473 7,266 7,510
697 7,392 7,397 7,415 7,390 7,402 7,368 7,383
701 7,398 7,357 7,484 7,394 7,366 7,369 7,377
709 7,507 7,377 7,417 7,420 7,451 7,382 7,411
714 7,409 7,321 7,322 7,459 7,347 7,358 7,346
Tabelle 33: pH-Werte im Nativblut der Sauen der Kontrollgruppe, entsprechend den
Blutentnahmeterminen zugeordnet.
pH-Werte im Vollblut der Sauen der Kontrollgruppe
Trächtigkeitstage Tage post partum Sauen- nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
547 7,379 7,378 7,406 7,349 7,319 7,328 7,350
571 7,284 7,393 7,429 7,408 7,398 7,378 7,406
597 7,415 7,405 7,416 7,393 7,367 7,340 7,359
610 7,391 7,466 7,403 7,479 7,374 7,378 7,346
615 7,393 7,349 7,348 7,456 7,340 7,323 7,349
620 7,406 7,324 7,385 7,413 7,317 7,349 7,365
649 7,433 7,409 7,369 7,338 7,354 7,397 7,424
673 7,452 7,380 7,408 7,374 7,433 7,382 7,356
675 7,459 7,387 7,402 7,393 7,341 7,350 7,336
685 7,459 7,351 7,532 7,417 7,418 7,439 7,371
689 7,451 7,332 7,382 7,383 7,258 7,416 7,310
691 7,406 7,387 7,395 7,372 7,353 7,378 7,405
693 7,389 7,382 7,349 7,465 7,441 7,479 7,411
703 7,402 7,429 7,411 7,365 7,359 7,403 7,357
704 7,397 7,386 7,469 7,371 7,317 7,343 7,355
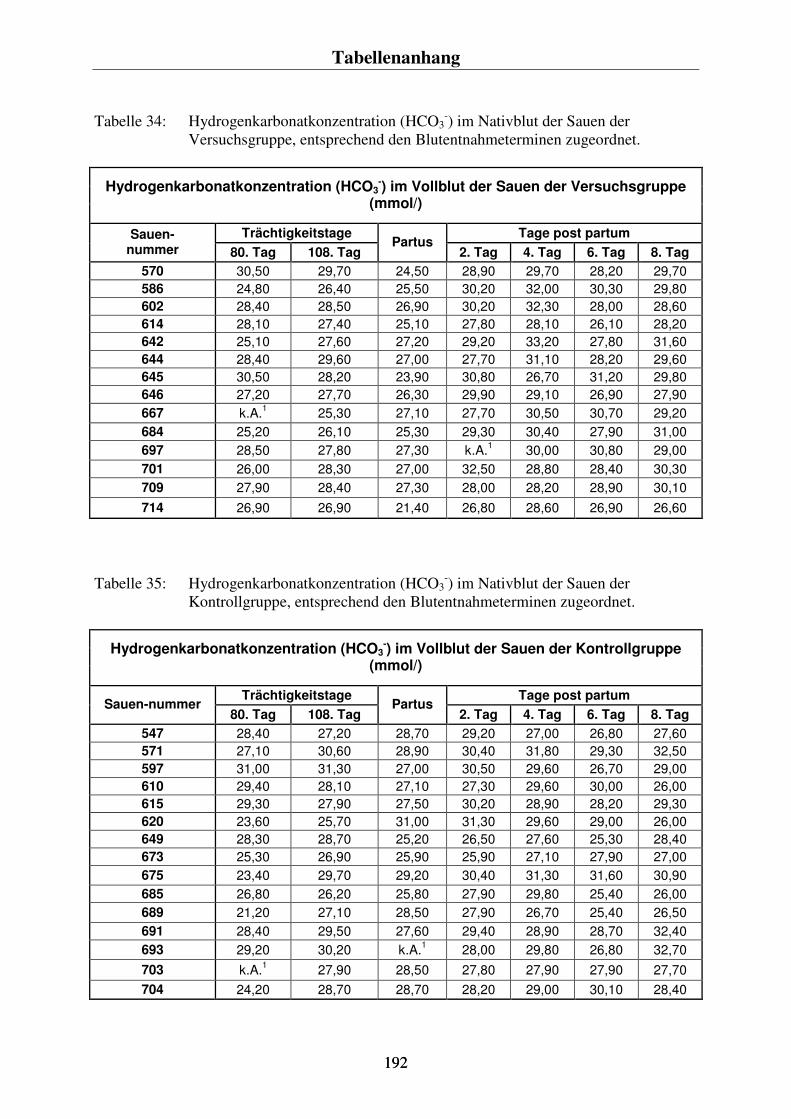
Tabellenanhang
192 192
Tabelle 34: Hydrogenkarbonatkonzentration (HCO3
-) im Nativblut der Sauen der Versuchsgruppe, entsprechend den Blutentnahmeterminen zugeordnet.
Hydrogenkarbonatkonzentration (HCO3-) im Vollblut der Sauen der Versuchsgruppe
(mmol/)
Trächtigkeitstage Tage post partum Sauen- nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
570 30,50 29,70 24,50 28,90 29,70 28,20 29,70
586 24,80 26,40 25,50 30,20 32,00 30,30 29,80
602 28,40 28,50 26,90 30,20 32,30 28,00 28,60
614 28,10 27,40 25,10 27,80 28,10 26,10 28,20
642 25,10 27,60 27,20 29,20 33,20 27,80 31,60
644 28,40 29,60 27,00 27,70 31,10 28,20 29,60
645 30,50 28,20 23,90 30,80 26,70 31,20 29,80
646 27,20 27,70 26,30 29,90 29,10 26,90 27,90
667 k.A.1 25,30 27,10 27,70 30,50 30,70 29,20
684 25,20 26,10 25,30 29,30 30,40 27,90 31,00
697 28,50 27,80 27,30 k.A.1 30,00 30,80 29,00
701 26,00 28,30 27,00 32,50 28,80 28,40 30,30
709 27,90 28,40 27,30 28,00 28,20 28,90 30,10
714 26,90 26,90 21,40 26,80 28,60 26,90 26,60
Tabelle 35: Hydrogenkarbonatkonzentration (HCO3
-) im Nativblut der Sauen der Kontrollgruppe, entsprechend den Blutentnahmeterminen zugeordnet.
Hydrogenkarbonatkonzentration (HCO3-) im Vollblut der Sauen der Kontrollgruppe
(mmol/)
Trächtigkeitstage Tage post partum Sauen-nummer
80. Tag 108. Tag Partus
2. Tag 4. Tag 6. Tag 8. Tag
547 28,40 27,20 28,70 29,20 27,00 26,80 27,60
571 27,10 30,60 28,90 30,40 31,80 29,30 32,50
597 31,00 31,30 27,00 30,50 29,60 26,70 29,00
610 29,40 28,10 27,10 27,30 29,60 30,00 26,00
615 29,30 27,90 27,50 30,20 28,90 28,20 29,30
620 23,60 25,70 31,00 31,30 29,60 29,00 26,00
649 28,30 28,70 25,20 26,50 27,60 25,30 28,40
673 25,30 26,90 25,90 25,90 27,10 27,90 27,00
675 23,40 29,70 29,20 30,40 31,30 31,60 30,90
685 26,80 26,20 25,80 27,90 29,80 25,40 26,00
689 21,20 27,10 28,50 27,90 26,70 25,40 26,50
691 28,40 29,50 27,60 29,40 28,90 28,70 32,40
693 29,20 30,20 k.A.1 28,00 29,80 26,80 32,70
703 k.A.1 27,90 28,50 27,80 27,90 27,90 27,70
704 24,20 28,70 28,70 28,20 29,00 30,10 28,40
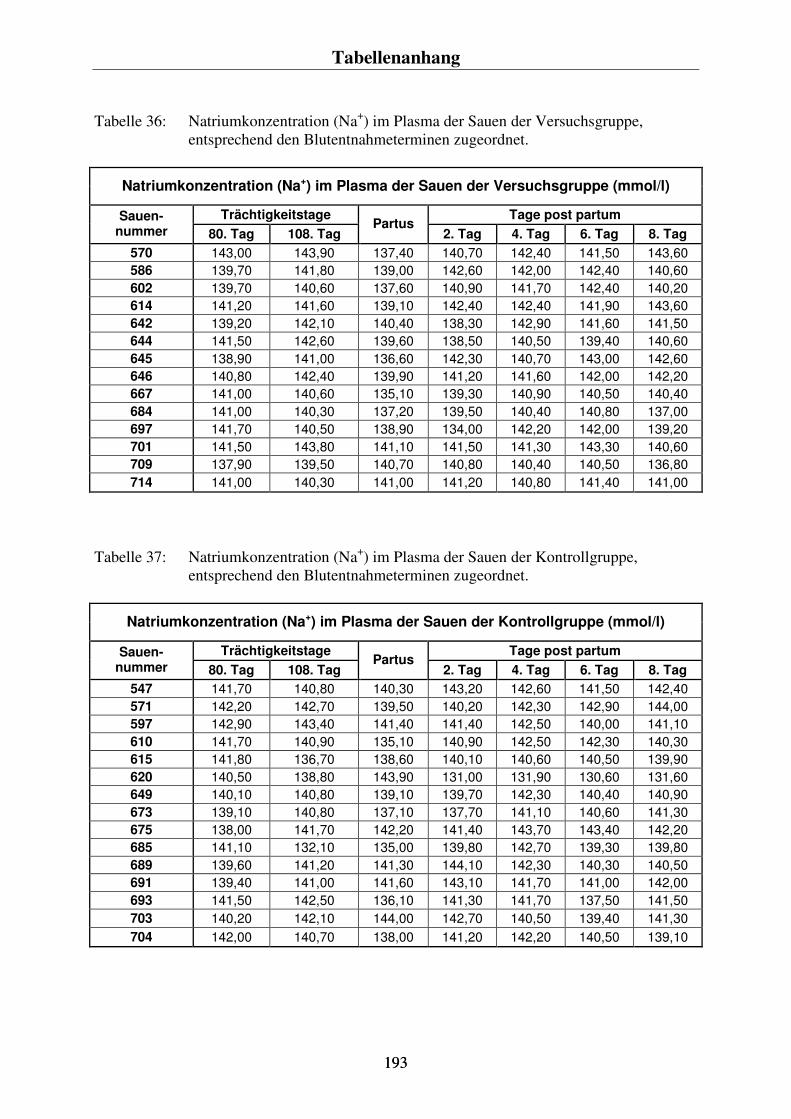
Tabellenanhang
193 193
Tabelle 36: Natriumkonzentration (Na+) im Plasma der Sauen der Versuchsgruppe,
entsprechend den Blutentnahmeterminen zugeordnet.
Natriumkonzentration (Na+) im Plasma der Sauen der Versuchsgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen-nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
570 143,00 143,90 137,40 140,70 142,40 141,50 143,60
586 139,70 141,80 139,00 142,60 142,00 142,40 140,60
602 139,70 140,60 137,60 140,90 141,70 142,40 140,20
614 141,20 141,60 139,10 142,40 142,40 141,90 143,60
642 139,20 142,10 140,40 138,30 142,90 141,60 141,50
644 141,50 142,60 139,60 138,50 140,50 139,40 140,60
645 138,90 141,00 136,60 142,30 140,70 143,00 142,60
646 140,80 142,40 139,90 141,20 141,60 142,00 142,20
667 141,00 140,60 135,10 139,30 140,90 140,50 140,40
684 141,00 140,30 137,20 139,50 140,40 140,80 137,00
697 141,70 140,50 138,90 134,00 142,20 142,00 139,20
701 141,50 143,80 141,10 141,50 141,30 143,30 140,60
709 137,90 139,50 140,70 140,80 140,40 140,50 136,80
714 141,00 140,30 141,00 141,20 140,80 141,40 141,00
Tabelle 37: Natriumkonzentration (Na+) im Plasma der Sauen der Kontrollgruppe,
entsprechend den Blutentnahmeterminen zugeordnet.
Natriumkonzentration (Na+) im Plasma der Sauen der Kontrollgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen-nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
547 141,70 140,80 140,30 143,20 142,60 141,50 142,40
571 142,20 142,70 139,50 140,20 142,30 142,90 144,00
597 142,90 143,40 141,40 141,40 142,50 140,00 141,10
610 141,70 140,90 135,10 140,90 142,50 142,30 140,30
615 141,80 136,70 138,60 140,10 140,60 140,50 139,90
620 140,50 138,80 143,90 131,00 131,90 130,60 131,60
649 140,10 140,80 139,10 139,70 142,30 140,40 140,90
673 139,10 140,80 137,10 137,70 141,10 140,60 141,30
675 138,00 141,70 142,20 141,40 143,70 143,40 142,20
685 141,10 132,10 135,00 139,80 142,70 139,30 139,80
689 139,60 141,20 141,30 144,10 142,30 140,30 140,50
691 139,40 141,00 141,60 143,10 141,70 141,00 142,00
693 141,50 142,50 136,10 141,30 141,70 137,50 141,50
703 140,20 142,10 144,00 142,70 140,50 139,40 141,30
704 142,00 140,70 138,00 141,20 142,20 140,50 139,10
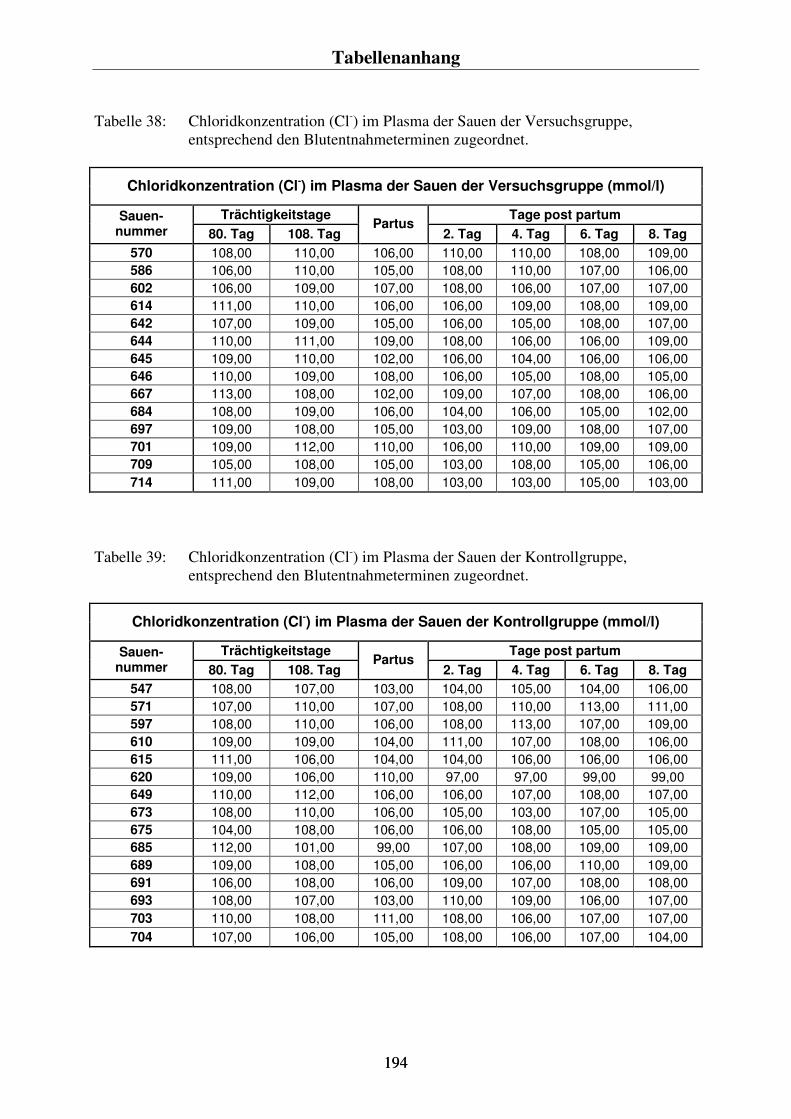
Tabellenanhang
194 194
Tabelle 38: Chloridkonzentration (Cl-) im Plasma der Sauen der Versuchsgruppe,
entsprechend den Blutentnahmeterminen zugeordnet.
Chloridkonzentration (Cl-) im Plasma der Sauen der Versuchsgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen-nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
570 108,00 110,00 106,00 110,00 110,00 108,00 109,00
586 106,00 110,00 105,00 108,00 110,00 107,00 106,00
602 106,00 109,00 107,00 108,00 106,00 107,00 107,00
614 111,00 110,00 106,00 106,00 109,00 108,00 109,00
642 107,00 109,00 105,00 106,00 105,00 108,00 107,00
644 110,00 111,00 109,00 108,00 106,00 106,00 109,00
645 109,00 110,00 102,00 106,00 104,00 106,00 106,00
646 110,00 109,00 108,00 106,00 105,00 108,00 105,00
667 113,00 108,00 102,00 109,00 107,00 108,00 106,00
684 108,00 109,00 106,00 104,00 106,00 105,00 102,00
697 109,00 108,00 105,00 103,00 109,00 108,00 107,00
701 109,00 112,00 110,00 106,00 110,00 109,00 109,00
709 105,00 108,00 105,00 103,00 108,00 105,00 106,00
714 111,00 109,00 108,00 103,00 103,00 105,00 103,00
Tabelle 39: Chloridkonzentration (Cl-) im Plasma der Sauen der Kontrollgruppe,
entsprechend den Blutentnahmeterminen zugeordnet.
Chloridkonzentration (Cl-) im Plasma der Sauen der Kontrollgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen-nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
547 108,00 107,00 103,00 104,00 105,00 104,00 106,00
571 107,00 110,00 107,00 108,00 110,00 113,00 111,00
597 108,00 110,00 106,00 108,00 113,00 107,00 109,00
610 109,00 109,00 104,00 111,00 107,00 108,00 106,00
615 111,00 106,00 104,00 104,00 106,00 106,00 106,00
620 109,00 106,00 110,00 97,00 97,00 99,00 99,00
649 110,00 112,00 106,00 106,00 107,00 108,00 107,00
673 108,00 110,00 106,00 105,00 103,00 107,00 105,00
675 104,00 108,00 106,00 106,00 108,00 105,00 105,00
685 112,00 101,00 99,00 107,00 108,00 109,00 109,00
689 109,00 108,00 105,00 106,00 106,00 110,00 109,00
691 106,00 108,00 106,00 109,00 107,00 108,00 108,00
693 108,00 107,00 103,00 110,00 109,00 106,00 107,00
703 110,00 108,00 111,00 108,00 106,00 107,00 107,00
704 107,00 106,00 105,00 108,00 106,00 107,00 104,00

Tabellenanhang
195 195
Tabelle 40: Kaliumkonzentration (K+) im Nativblut der Sauen der Versuchsgruppe,
entsprechend den Blutentnahmeterminen zugeordnet.
Kaliumkonzentration (K+) im Vollblut der Sauen der Versuchsgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen-nummer
80. Tag 108. Tag Partus
2. Tag 4. Tag 6. Tag 8. Tag
570 4,58 5,10 4,87 4,23 4,39 4,86 4,23
586 4,85 4,33 4,37 4,19 4,46 4,22 4,59
602 4,62 4,60 4,61 4,30 4,26 4,13 4,82
614 4,78 4,13 4,78 4,74 4,57 4,77 4,69
642 4,56 4,45 4,79 4,61 3,86 3,93 4,33
644 4,71 4,60 4,35 4,62 4,16 4,66 4,62
645 4,26 4,37 4,91 4,47 5,17 4,07 4,74
646 4,81 4,56 3,89 4,38 4,53 4,48 4,40
667 4,50 5,18 4,65 5,00 4,77 4,32 4,41
684 4,94 4,93 4,68 4,68 5,05 4,65 4,41
697 4,32 4,30 4,20 5,59 4,20 4,43 3,95
701 5,12 4,64 4,70 4,44 4,32 4,55 4,70
709 4,55 4,32 3,78 4,60 4,40 4,31 4,21
714 4,71 4,53 4,09 4,62 4,33 5,04 4,31
Tabelle 41: Kaliumkonzentration (K+) im Nativblut der Sauen der Kontrollgruppe,
entsprechend den Blutentnahmeterminen zugeordnet.
Kaliumkonzentration (K+) im Vollblut der Sauen der Kontrollgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen-nummer
80. Tag 108. Tag Partus
2. Tag 4. Tag 6. Tag 8. Tag
547 4,87 4,06 4,22 4,45 4,50 4,31 4,62
571 4,51 4,23 4,48 4,27 4,07 3,94 4,52
597 4,15 4,39 4,39 4,63 4,22 4,06 4,10
610 4,48 4,65 3,89 4,74 4,41 4,15 4,63
615 4,44 4,35 4,62 4,76 4,53 4,61 3,92
620 4,77 4,13 4,84 5,04 4,14 4,22 4,87
649 4,13 4,61 4,22 5,10 4,68 4,99 4,98
673 4,62 4,66 4,18 4,79 4,74 4,28 4,09
675 5,06 4,68 4,29 4,36 4,36 4,08 4,57
685 4,45 4,18 4,70 4,37 4,29 4,38 3,90
689 4,87 4,96 4,58 4,66 5,17 5,40 4,87
691 4,59 4,10 3,94 4,08 3,98 3,77 4,40
693 4,83 4,05 4,96 4,85 4,59 4,29 4,40
703 4,27 4,44 4,33 4,50 4,23 4,83 4,01
704 5,00 4,02 5,84 4,84 4,30 4,15 4,14
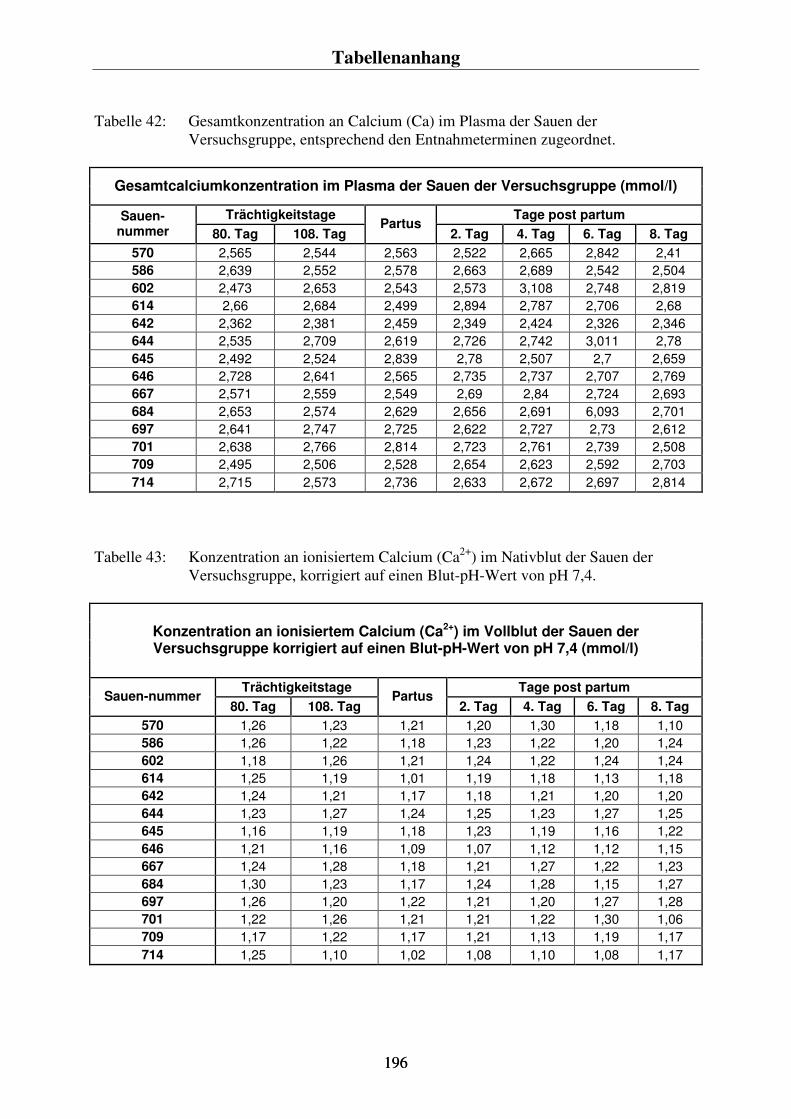
Tabellenanhang
196 196
Tabelle 42: Gesamtkonzentration an Calcium (Ca) im Plasma der Sauen der
Versuchsgruppe, entsprechend den Entnahmeterminen zugeordnet.
Gesamtcalciumkonzentration im Plasma der Sauen der Versuchsgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen- nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
570 2,565 2,544 2,563 2,522 2,665 2,842 2,41
586 2,639 2,552 2,578 2,663 2,689 2,542 2,504
602 2,473 2,653 2,543 2,573 3,108 2,748 2,819
614 2,66 2,684 2,499 2,894 2,787 2,706 2,68
642 2,362 2,381 2,459 2,349 2,424 2,326 2,346
644 2,535 2,709 2,619 2,726 2,742 3,011 2,78
645 2,492 2,524 2,839 2,78 2,507 2,7 2,659
646 2,728 2,641 2,565 2,735 2,737 2,707 2,769
667 2,571 2,559 2,549 2,69 2,84 2,724 2,693
684 2,653 2,574 2,629 2,656 2,691 6,093 2,701
697 2,641 2,747 2,725 2,622 2,727 2,73 2,612
701 2,638 2,766 2,814 2,723 2,761 2,739 2,508
709 2,495 2,506 2,528 2,654 2,623 2,592 2,703
714 2,715 2,573 2,736 2,633 2,672 2,697 2,814
Tabelle 43: Konzentration an ionisiertem Calcium (Ca2+) im Nativblut der Sauen der
Versuchsgruppe, korrigiert auf einen Blut-pH-Wert von pH 7,4.
Konzentration an ionisiertem Calcium (Ca2+) im Vollblut der Sauen der Versuchsgruppe korrigiert auf einen Blut-pH-Wert von pH 7,4 (mmol/l)
Trächtigkeitstage Tage post partum Sauen-nummer
80. Tag 108. Tag Partus
2. Tag 4. Tag 6. Tag 8. Tag
570 1,26 1,23 1,21 1,20 1,30 1,18 1,10
586 1,26 1,22 1,18 1,23 1,22 1,20 1,24
602 1,18 1,26 1,21 1,24 1,22 1,24 1,24
614 1,25 1,19 1,01 1,19 1,18 1,13 1,18
642 1,24 1,21 1,17 1,18 1,21 1,20 1,20
644 1,23 1,27 1,24 1,25 1,23 1,27 1,25
645 1,16 1,19 1,18 1,23 1,19 1,16 1,22
646 1,21 1,16 1,09 1,07 1,12 1,12 1,15
667 1,24 1,28 1,18 1,21 1,27 1,22 1,23
684 1,30 1,23 1,17 1,24 1,28 1,15 1,27
697 1,26 1,20 1,22 1,21 1,20 1,27 1,28
701 1,22 1,26 1,21 1,21 1,22 1,30 1,06
709 1,17 1,22 1,17 1,21 1,13 1,19 1,17
714 1,25 1,10 1,02 1,08 1,10 1,08 1,17
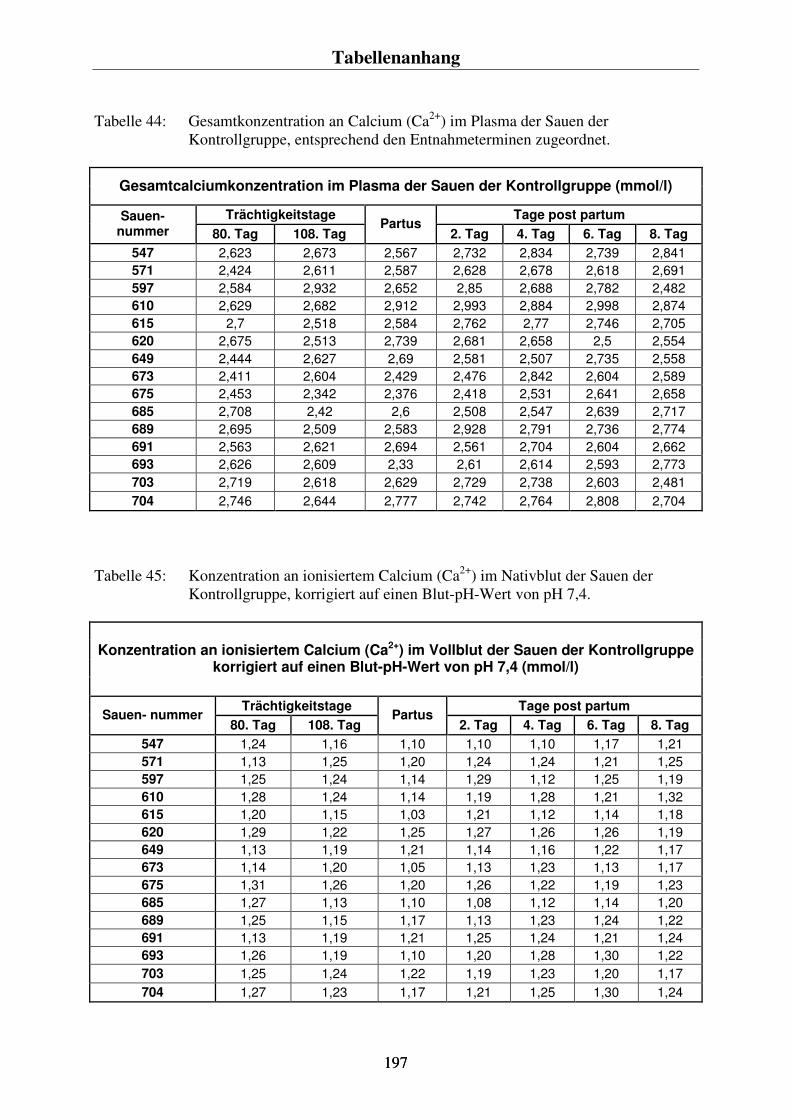
Tabellenanhang
197 197
Tabelle 44: Gesamtkonzentration an Calcium (Ca2+) im Plasma der Sauen der
Kontrollgruppe, entsprechend den Entnahmeterminen zugeordnet.
Gesamtcalciumkonzentration im Plasma der Sauen der Kontrollgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen- nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
547 2,623 2,673 2,567 2,732 2,834 2,739 2,841
571 2,424 2,611 2,587 2,628 2,678 2,618 2,691
597 2,584 2,932 2,652 2,85 2,688 2,782 2,482
610 2,629 2,682 2,912 2,993 2,884 2,998 2,874
615 2,7 2,518 2,584 2,762 2,77 2,746 2,705
620 2,675 2,513 2,739 2,681 2,658 2,5 2,554
649 2,444 2,627 2,69 2,581 2,507 2,735 2,558
673 2,411 2,604 2,429 2,476 2,842 2,604 2,589
675 2,453 2,342 2,376 2,418 2,531 2,641 2,658
685 2,708 2,42 2,6 2,508 2,547 2,639 2,717
689 2,695 2,509 2,583 2,928 2,791 2,736 2,774
691 2,563 2,621 2,694 2,561 2,704 2,604 2,662
693 2,626 2,609 2,33 2,61 2,614 2,593 2,773
703 2,719 2,618 2,629 2,729 2,738 2,603 2,481
704 2,746 2,644 2,777 2,742 2,764 2,808 2,704
Tabelle 45: Konzentration an ionisiertem Calcium (Ca2+) im Nativblut der Sauen der
Kontrollgruppe, korrigiert auf einen Blut-pH-Wert von pH 7,4.
Konzentration an ionisiertem Calcium (Ca2+) im Vollblut der Sauen der Kontrollgruppe korrigiert auf einen Blut-pH-Wert von pH 7,4 (mmol/l)
Trächtigkeitstage Tage post partum Sauen- nummer
80. Tag 108. Tag Partus
2. Tag 4. Tag 6. Tag 8. Tag
547 1,24 1,16 1,10 1,10 1,10 1,17 1,21
571 1,13 1,25 1,20 1,24 1,24 1,21 1,25
597 1,25 1,24 1,14 1,29 1,12 1,25 1,19
610 1,28 1,24 1,14 1,19 1,28 1,21 1,32
615 1,20 1,15 1,03 1,21 1,12 1,14 1,18
620 1,29 1,22 1,25 1,27 1,26 1,26 1,19
649 1,13 1,19 1,21 1,14 1,16 1,22 1,17
673 1,14 1,20 1,05 1,13 1,23 1,13 1,17
675 1,31 1,26 1,20 1,26 1,22 1,19 1,23
685 1,27 1,13 1,10 1,08 1,12 1,14 1,20
689 1,25 1,15 1,17 1,13 1,23 1,24 1,22
691 1,13 1,19 1,21 1,25 1,24 1,21 1,24
693 1,26 1,19 1,10 1,20 1,28 1,30 1,22
703 1,25 1,24 1,22 1,19 1,23 1,20 1,17
704 1,27 1,23 1,17 1,21 1,25 1,30 1,24

Tabellenanhang
198 198
Tabelle 46: Phosphorkonzentration im Plasma der Sauen der Versuchsgruppe,
entsprechend geordnet nach den Entnahmeterminen.
Phosphorkonzentration im Plasma der Sauen der Versuchsgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen- nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
570 2,035 1,829 2,668 1,718 1,800 1,917 1,839
586 2,275 2,148 2,688 2,295 1,946 2,172 2,472
602 1,669 1,938 2,104 2,005 1,972 1,792 2,194
614 2,188 2,694 2,240 1,344 2,392 2,282 2,475
642 2,478 1,985 2,564 2,494 2,181 2,115 2,351
644 1,945 2,387 2,320 2,576 2,134 2,181 2,448
645 1,823 1,872 2,907 2,486 1,924 2,482 2,185
646 1,677 2,225 2,532 2,013 1,647 1,996 2,465
667 1,800 2,229 2,149 2,528 2,372 2,402 2,033
684 2,067 2,357 2,954 2,546 3,163 2,535 2,377
697 2,156 2,344 2,177 2,031 1,966 1,790 2,637
701 1,633 1,633 2,544 2,551 1,744 1,646 2,548
709 2,388 2,254 2,434 2,464 2,438 2,538 2,721
714 2,339 2,323 3,275 2,628 2,454 2,029 2,438
Tabelle 47: Phosphorkonzentration im Plasma der Sauen der Kontrollgruppe, entsprechend
geordnet nach den Entnahmeterminen.
Phosphorkonzentration im Plasma der Sauen der Kontrollgruppe (mmol/l)
Trächtigkeitstage Tage post partum Sauen- nummer 80. Tag 108. Tag
Partus 2. Tag 4. Tag 6. Tag 8. Tag
547 2,202 2,181 2,556 2,384 2,538 1,945 2,202
571 2,079 2,177 2,829 2,146 1,437 1,345 1,973
597 1,633 1,633 1,633 1,633 1,633 1,633 1,633
610 1,604 1,862 2,061 1,973 2,038 2,077 2,221
615 1,850 2,059 2,651 2,538 2,565 2,412 2,724
620 2,325 2,262 3,001 1,946 2,272 1,896 2,671
649 2,428 2,494 2,674 2,607 2,071 2,737 2,388
673 2,348 2,587 2,665 2,685 2,578 2,751 2,621
675 2,244 2,544 2,860 2,480 2,333 2,312 2,440
685 2,186 2,050 2,842 2,497 2,217 2,088 2,085
689 2,515 2,464 2,907 3,600 1,930 2,401 2,697
691 2,080 2,761 2,721 1,770 1,881 1,494 1,942
693 2,166 2,135 2,840 2,433 2,260 2,114 2,468
703 1,863 1,800 2,402 1,720 1,881 1,937 2,113
704 2,653 2,586 2,471 2,562 2,205 2,336 2,387
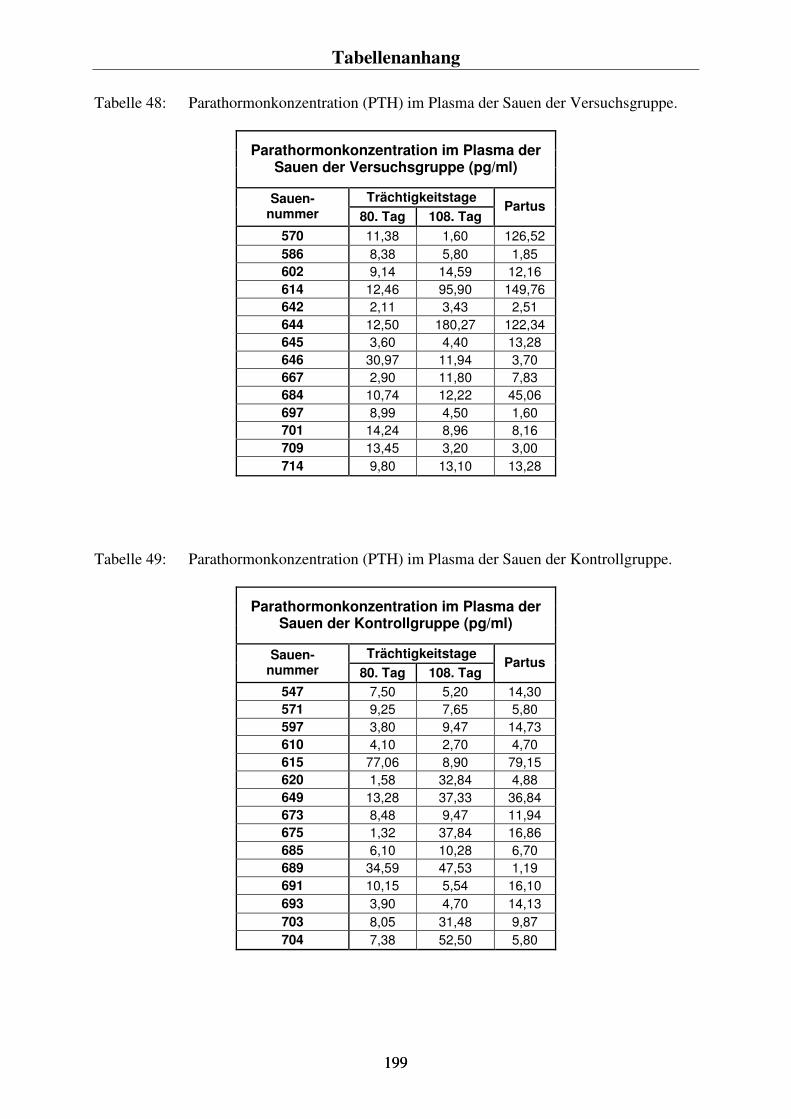
Tabellenanhang
199 199
Tabelle 48: Parathormonkonzentration (PTH) im Plasma der Sauen der Versuchsgruppe.
Tabelle 49: Parathormonkonzentration (PTH) im Plasma der Sauen der Kontrollgruppe.
Parathormonkonzentration im Plasma der Sauen der Kontrollgruppe (pg/ml)
Trächtigkeitstage Sauen- nummer 80. Tag 108. Tag
Partus
547 7,50 5,20 14,30
571 9,25 7,65 5,80
597 3,80 9,47 14,73
610 4,10 2,70 4,70
615 77,06 8,90 79,15
620 1,58 32,84 4,88
649 13,28 37,33 36,84
673 8,48 9,47 11,94
675 1,32 37,84 16,86
685 6,10 10,28 6,70
689 34,59 47,53 1,19
691 10,15 5,54 16,10
693 3,90 4,70 14,13
703 8,05 31,48 9,87
704 7,38 52,50 5,80
Parathormonkonzentration im Plasma der Sauen der Versuchsgruppe (pg/ml)
Trächtigkeitstage Sauen- nummer 80. Tag 108. Tag
Partus
570 11,38 1,60 126,52
586 8,38 5,80 1,85
602 9,14 14,59 12,16
614 12,46 95,90 149,76
642 2,11 3,43 2,51
644 12,50 180,27 122,34
645 3,60 4,40 13,28
646 30,97 11,94 3,70
667 2,90 11,80 7,83
684 10,74 12,22 45,06
697 8,99 4,50 1,60
701 14,24 8,96 8,16
709 13,45 3,20 3,00
714 9,80 13,10 13,28

Tabellenanhang
200 200
Tabelle 50: pH-Werte in den Spontanharnproben der Sauen der Versuchsgruppe.
pH-Werte im Harn der Sauen der Versuchsgruppe
Trächtigkeitstage Sauen- nummer 90. Tag 111. Tag
667 5,65 6,30
586 6,70 7,00
642 5,60 6,90
684 5,95 6,40
602 6,10 6,80
644 6,35 6,85
697 5,60 7,25
570 6,35 7,35
701 6,20 6,45
614 6,60 6,65
646 6,05 6,25
714 5,25 5,75
645 5,55 7,30
709 5,50 6,90
Tabelle 51: pH-Werte in den Spontanharnproben der Sauen der Kontrollgruppe.
pH-Werte im Harn der Sauen der Kontrollgruppe
Trächtigkeitstage Sauen- nummer 90. Tag 111. Tag
703 6,40 7,15
620 6,75 7,00
675 6,85 7,30
689 6,65 7,35
704 6,70 7,15
571 7,65 8,05
691 6,90 7,30
693 7,45 7,35
597 7,50 7,55
610 7,20 7,35
547 7,15 7,40
615 7,05 6,85
685 7,10 6,80
649 7,20 7,55
673 7,15 7,55

Tabellenanhang
201 201
Tabelle 52: Plasmakonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am
90. Trächtigkeitstag der Versuchstiere.
Plasmaparameter der Versuchstiere am 90. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
570 193,00 362,04 328,00 10,281 6,302
586 192,00 341,56 343,00 10,577 7,047
602 260,00 348,03 319,00 9,912 5,168
614 199,00 349,05 348,00 10,661 6,777
642 177,00 339,80 339,00 9,467 7,673
644 278,00 351,52 332,00 10,160 6,025
645 220,00 351,45 361,00 9,988 5,644
646 207,00 369,39 349,00 10,934 5,192
667 169,00 362,31 334,00 10,305 5,573
684 185,00 355,01 331,00 10,633 6,402
697 191,00 358,51 335,00 10,585 6,677
701 255,00 349,01 328,00 10,573 5,058
709 152,00 346,72 322,00 10,000 7,394
714 164,00 353,80 348,00 10,882 7,243
Tabelle 53: Harnkonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am
90. Trächtigkeitstag der Versuchstiere.
Harnparameter der Versuchstiere am 90. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
570 17191,0 353,83 29,01 50,96 25,98
586 3816,0 78,75 9,84 11,50 12,65
602 9409,0 170,40 12,50 14,92 10,88
614 2370,0 39,05 10,91 4,11 0,18
642 3866,0 121,87 23,34 17,10 17,54
644 3427,0 66,24 8,92 0,01 11,36
645 19986,0 286,87 9,17 0,24 8,89
646 19716,0 245,02 16,33 46,25 27,91
667 4064,0 77,58 15,42 5,43 17,71
684 3172,0 99,40 26,17 4,21 15,01
697 1452,0 56,61 9,59 3,57 6,17
701 27227,0 825,22 210,04 34,00 10,88
709 11636,0 211,57 11,42 0,01 5,12
714 2316,0 66,91 13,75 4,80 5,16

Tabellenanhang
202 202
Tabelle 54: Plasmakonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am 90. Trächtigkeitstag der Kontrolltiere.
Plasmaparameter der Kontrolltiere am 90. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
547 218,00 347,83 320,00 10,513 6,818
571 288,00 348,03 360,00 9,715 6,440
597 210,00 346,64 344,00 10,357 5,058
610 263,00 357,40 334,00 10,537 4,967
615 171,00 359,73 362,00 10,822 5,728
620 188,00 356,16 345,00 10,721 7,201
649 168,00 363,83 335,00 9,796 7,518
673 182,00 359,73 354,00 9,663 7,271
675 259,00 360,81 323,00 9,832 6,951
685 155,00 374,18 346,00 10,854 6,769
689 202,00 348,03 311,00 10,802 7,789
691 303,00 342,19 314,00 10,273 6,442
693 195,00 339,68 338,00 10,525 6,709
703 200,00 355,30 298,00 10,898 5,769
704 206,00 351,52 332,00 11,006 8,218
Tabelle 55: Harnkonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am 90. Trächtigkeitstag der Kontrolltiere.
Harnparameter der Kontrolltiere am 90. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
547 1301,0 12,14 10,92 0,17 1,60
571 2577,0 21,30 12,00 4,59 11,04
597 8317,0 35,38 19,59 6,26 7,96
610 11197,0 89,33 22,25 10,41 16,71
615 1616,0 13,02 9,92 3,74 0,71
620 595,0 9,70 9,84 12,56 12,65
649 15137,0 88,43 15,08 0,01 7,47
673 17346,0 87,24 59,00 2,27 14,19
675 1289,0 14,20 13,75 10,86 11,47
685 1347,0 11,47 13,58 3,90 1,78
689 4213,0 42,60 7,61 0,01 16,69
691 1220,0 11,36 6,93 0,01 9,58
693 2962,0 29,87 9,84 3,26 30,20
703 22847,0 70,53 14,34 7,40 48,73
704 1735,0 14,20 10,42 2,46 15,51

Tabellenanhang
203 203
Tabelle 56: Plasmakonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am
111. Trächtigkeitstag der Versuchstiere.
Plasmaparameter der Versuchstiere am 111. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
570 214,00 352,62 337,00 10,196 5,664
586 192,00 359,67 354,00 10,228 6,651
602 246,00 345,69 339,00 10,633 6,003
614 173,00 354,98 348,00 10,758 8,343
642 222,00 346,65 346,00 9,543 6,146
644 231,00 351,52 352,00 10,858 7,393
645 195,00 363,28 346,00 10,116 5,796
646 179,00 365,79 359,00 10,585 6,892
667 207,00 363,50 331,00 10,257 6,902
684 218,00 364,30 321,00 10,317 7,299
697 202,00 359,67 335,00 11,010 7,259
701 276,00 355,00 332,00 11,086 5,058
709 145,00 355,00 338,00 10,044 6,982
714 162,00 356,19 356,00 10,313 7,194
Tabelle 57: Harnkonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am
111. Trächtigkeitstag der Versuchstiere.
Harnparameter der Versuchstiere am 111. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
570 3717,0 143,87 47,01 27,82 8,12
586 3866,0 112,85 38,42 11,99 11,30
602 1568,0 52,04 7,37 9,97 5,84
614 1993,0 60,35 19,50 3,91 2,67
642 3965,0 137,24 31,17 16,40 11,13
644 2207,0 79,27 21,84 0,01 9,42
645 5549,0 200,81 16,67 0,01 4,80
646 8027,0 255,78 35,00 14,52 1,78
667 3568,0 110,49 18,67 5,00 17,20
684 2874,0 126,59 53,01 1,37 19,05
697 2788,0 106,15 50,26 5,40 8,44
701 7620,0 235,08 6,07 45,11 6,01
709 2586,0 82,47 25,67 10,22 5,76
714 5171,0 179,29 24,75 4,28 4,98
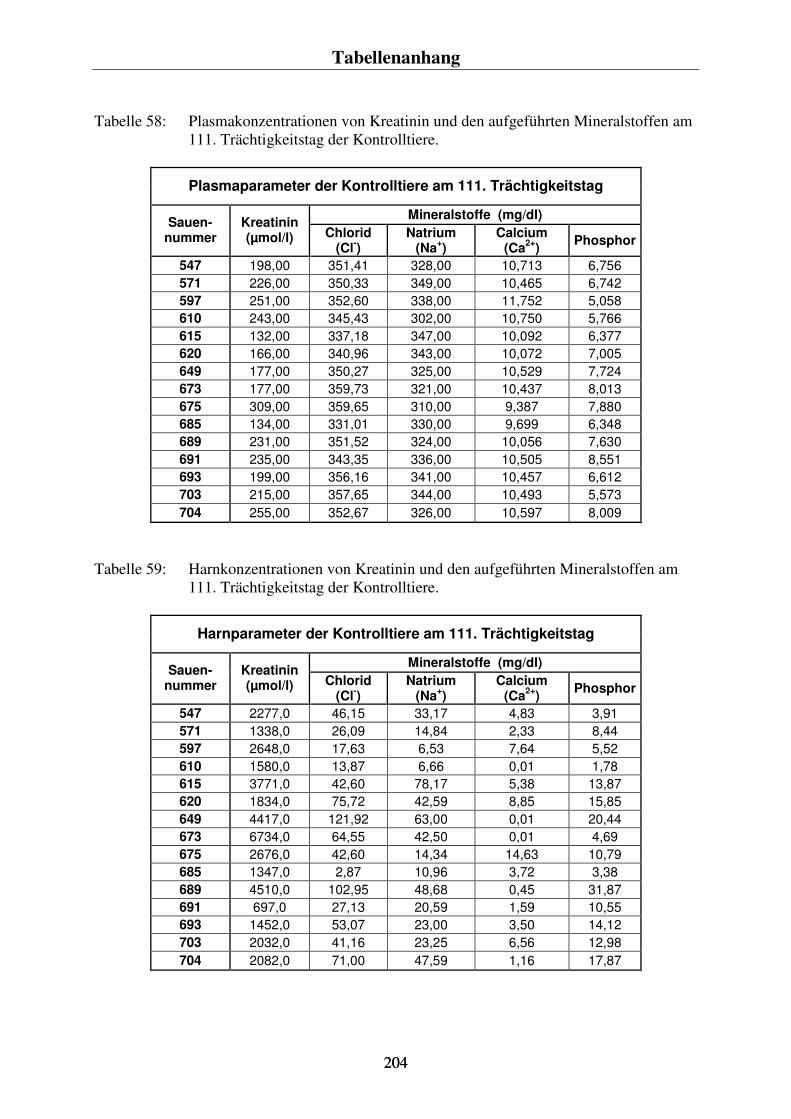
Tabellenanhang
204 204
Tabelle 58: Plasmakonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am
111. Trächtigkeitstag der Kontrolltiere.
Plasmaparameter der Kontrolltiere am 111. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
547 198,00 351,41 328,00 10,713 6,756
571 226,00 350,33 349,00 10,465 6,742
597 251,00 352,60 338,00 11,752 5,058
610 243,00 345,43 302,00 10,750 5,766
615 132,00 337,18 347,00 10,092 6,377
620 166,00 340,96 343,00 10,072 7,005
649 177,00 350,27 325,00 10,529 7,724
673 177,00 359,73 321,00 10,437 8,013
675 309,00 359,65 310,00 9,387 7,880
685 134,00 331,01 330,00 9,699 6,348
689 231,00 351,52 324,00 10,056 7,630
691 235,00 343,35 336,00 10,505 8,551
693 199,00 356,16 341,00 10,457 6,612
703 215,00 357,65 344,00 10,493 5,573
704 255,00 352,67 326,00 10,597 8,009
Tabelle 59: Harnkonzentrationen von Kreatinin und den aufgeführten Mineralstoffen am
111. Trächtigkeitstag der Kontrolltiere.
Harnparameter der Kontrolltiere am 111. Trächtigkeitstag
Mineralstoffe (mg/dl) Sauen-
nummer Kreatinin (µmol/l) Chlorid
(Cl-)
Natrium (Na
+)
Calcium (Ca
2+)
Phosphor
547 2277,0 46,15 33,17 4,83 3,91
571 1338,0 26,09 14,84 2,33 8,44
597 2648,0 17,63 6,53 7,64 5,52
610 1580,0 13,87 6,66 0,01 1,78
615 3771,0 42,60 78,17 5,38 13,87
620 1834,0 75,72 42,59 8,85 15,85
649 4417,0 121,92 63,00 0,01 20,44
673 6734,0 64,55 42,50 0,01 4,69
675 2676,0 42,60 14,34 14,63 10,79
685 1347,0 2,87 10,96 3,72 3,38
689 4510,0 102,95 48,68 0,45 31,87
691 697,0 27,13 20,59 1,59 10,55
693 1452,0 53,07 23,00 3,50 14,12
703 2032,0 41,16 23,25 6,56 12,98
704 2082,0 71,00 47,59 1,16 17,87
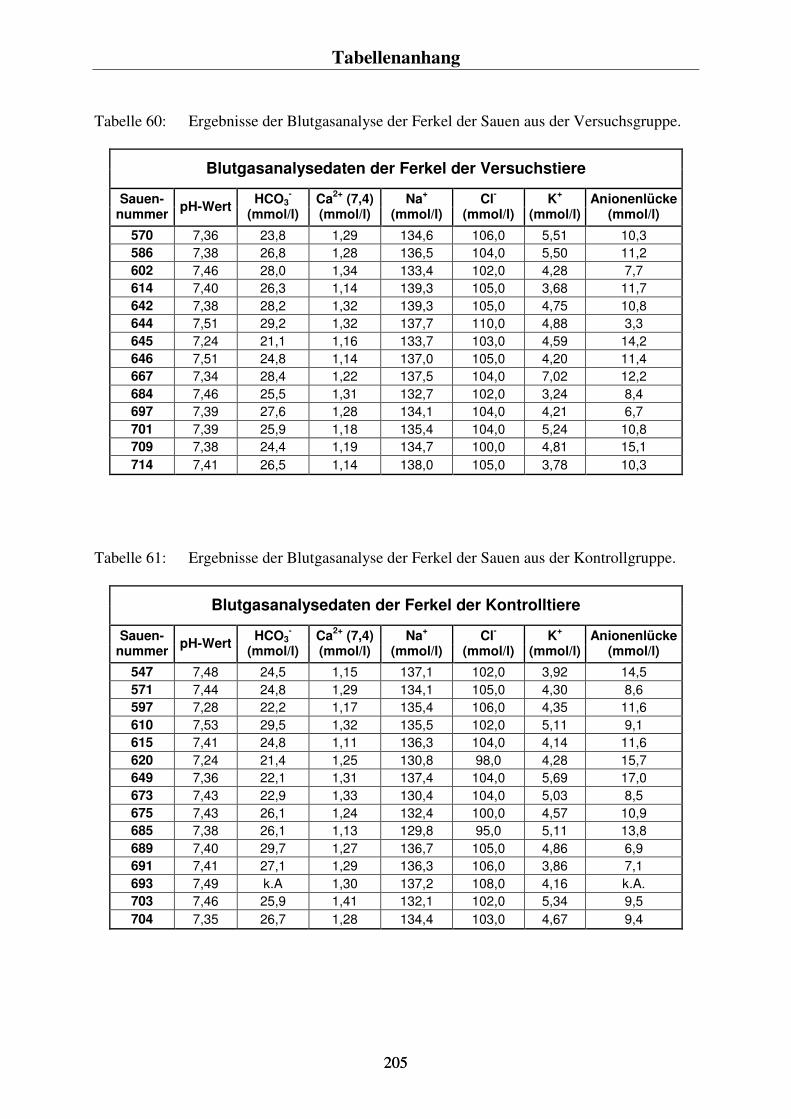
Tabellenanhang
205 205
Tabelle 60: Ergebnisse der Blutgasanalyse der Ferkel der Sauen aus der Versuchsgruppe.
Blutgasanalysedaten der Ferkel der Versuchstiere
Sauen-nummer
pH-Wert HCO3
-
(mmol/l) Ca
2+ (7,4)
(mmol/l) Na
+
(mmol/l) Cl
-
(mmol/l) K
+
(mmol/l) Anionenlücke
(mmol/l)
570 7,36 23,8 1,29 134,6 106,0 5,51 10,3
586 7,38 26,8 1,28 136,5 104,0 5,50 11,2
602 7,46 28,0 1,34 133,4 102,0 4,28 7,7
614 7,40 26,3 1,14 139,3 105,0 3,68 11,7
642 7,38 28,2 1,32 139,3 105,0 4,75 10,8
644 7,51 29,2 1,32 137,7 110,0 4,88 3,3
645 7,24 21,1 1,16 133,7 103,0 4,59 14,2
646 7,51 24,8 1,14 137,0 105,0 4,20 11,4
667 7,34 28,4 1,22 137,5 104,0 7,02 12,2
684 7,46 25,5 1,31 132,7 102,0 3,24 8,4
697 7,39 27,6 1,28 134,1 104,0 4,21 6,7
701 7,39 25,9 1,18 135,4 104,0 5,24 10,8
709 7,38 24,4 1,19 134,7 100,0 4,81 15,1
714 7,41 26,5 1,14 138,0 105,0 3,78 10,3
Tabelle 61: Ergebnisse der Blutgasanalyse der Ferkel der Sauen aus der Kontrollgruppe.
Blutgasanalysedaten der Ferkel der Kontrolltiere
Sauen-nummer
pH-Wert HCO3
-
(mmol/l) Ca
2+ (7,4)
(mmol/l) Na
+
(mmol/l) Cl
-
(mmol/l) K
+
(mmol/l) Anionenlücke
(mmol/l)
547 7,48 24,5 1,15 137,1 102,0 3,92 14,5
571 7,44 24,8 1,29 134,1 105,0 4,30 8,6
597 7,28 22,2 1,17 135,4 106,0 4,35 11,6
610 7,53 29,5 1,32 135,5 102,0 5,11 9,1
615 7,41 24,8 1,11 136,3 104,0 4,14 11,6
620 7,24 21,4 1,25 130,8 98,0 4,28 15,7
649 7,36 22,1 1,31 137,4 104,0 5,69 17,0
673 7,43 22,9 1,33 130,4 104,0 5,03 8,5
675 7,43 26,1 1,24 132,4 100,0 4,57 10,9
685 7,38 26,1 1,13 129,8 95,0 5,11 13,8
689 7,40 29,7 1,27 136,7 105,0 4,86 6,9
691 7,41 27,1 1,29 136,3 106,0 3,86 7,1
693 7,49 k.A 1,30 137,2 108,0 4,16 k.A.
703 7,46 25,9 1,41 132,1 102,0 5,34 9,5
704 7,35 26,7 1,28 134,4 103,0 4,67 9,4

Danksagung
206 206
Danksagung
Mein aufrichtiger Dank für die Überlassung dieses Themas sowie für die Unterstützung und
die gute Betreuung bei der Erstellung dieser Arbeit, die Beantwortung meiner wissenschaft-
lichen Fragen und die Hilfe bei der statistischen Auswertung der Ergebnisse gilt Herrn Prof.
Dr. Manfred Coenen.
Ein besonderer Dank gilt außerdem dem Lehr- und Forschungsgut der Stiftung der
Tierärztlichen Hochschule in Ruthe für die Möglichkeit die dort ansässige Sauenherde zur
Durchführung dieses Versuches nutzen zu dürfen.
Weiterhin möchte ich Herrn Dr. C. Sürie und Herrn H. Mohwinkel vom Lehr- und
Forschungsgut in Ruthe für die Unterstützung bei der Koordination und Umsetzung meines
Versuches danken.
Meinen besonders herzlichen Dank möchte ich Herrn A. Wischnewski als Betreuer des
Schweinestalles und Herrn J. Minks als Versuchstechniker des Lehr- und Forschungsgutes
aussprechen, die mich tatkräftig bei der praktischen Durchführung dieses Versuches
unterstützt haben und sogar vor einer Probenentnahme an Weihnachten nicht zurück
schreckten.
Für den fachlichen Rat und die praktische Unterstützung bei der Aufbereitung und den
verschiedenen Analyseverfahren der Blut- und Harnproben möchte ich Herrn Peter Rust, Frau
Katrin Meyer sowie den übrigen Mitgliedern des Laborteams des Institutes für Tierernährung
der Stiftung der Tierärztlichen Hochschule Hannover danken.
Meinen Arbeitgebern Dr. Christian Niewöhner und Ralf Bischoff bin ich dafür dankbar, dass
sie mir oft im Praxisalltag den Rücken frei gehalten haben und mir jede Möglichkeit zur
Weiterführung meiner Arbeit einräumten.
Meinen Kommilitonen Herrn Dr. Henning Schenk und Herrn Dr. Guido Martens, sowie Frau
Simone Kaiser gilt ebenfalls mein Dank, da sie immer ein offenes Ohr für meine kleinen
Probleme hatten und mir mit Rat und Tat zur Seite standen.

Danksagung
207 207
Den größten Dank möchte ich allerdings meinen Eltern und Geschwistern aussprechen, weil
sie mir die Durchführung dieser Forschungsarbeit erst ermöglicht haben und auch vorher auf
meinem bisherigen Lebensweg immer unterstützt und gefördert haben.


















