2 Zytologische Grundlagen der FEM Modellierungtheuer/2 Zytoplasma - 2 -- 02-04-28a.pdf ·...
13
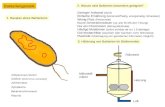
Zytologische Grundlagen der FEM-Modellierung Seite 2 - 1 2 Zytologische Grundlagen der FEM Modellierung Lebende Zellen sind hochorganisierte dynamische Einheiten, in denen wichtige zelluläre Makromoleküle spontan zu komplexen Proteinstrukturen aggregieren und wieder dissoziieren können [12]. Dabei kommt es zu häufigen lokalen Än- derungen des Aggregatzustandes. Die geschilderten Tatsachen führen zu, in den klassischen Ingenieurwissenschaften wenig bekannten, biologischen Material- eigenschaften. Abbildung 2-1 enthält die 3D-Computerrekonstruktion einer Lungenepitelzelle. Die Abbildung zeigt den Zellkern (blau), Zytoplasma (hell- blau), die Plasmamembranen (schwarz), Zellorganellen (grün) und Erythrozy- ten (rot). Im Allgemeinen haben Eukariontenzellen folgende Organellen: Endosplasmatisches Retikulum Golgi-Apparat Mitochondrien Lyosomen Peroxisomen Zellkern Die Organellen nehmen ca. 50% des Zellvolumens ein. Die restlichen 50% des Zellvolumens bilden das Zytosol. Die wichtigsten Proteinsynthese-Prozesse finden im Zytosol statt. Eukariontenzellen bestehen zu 70% aus Wasser, zu 18% aus Proteinstrukturen, zu 3% aus Pospholipiden. Die Zelltrockenmasse be- steht zu mehr als 50% aus Protein. Proteine sind aus einer Vielfalt unterschied- licher Aminosäuren aufgebaut. Der Zellkern ist das ausgeprägteste Organ der Zelle. Er enthält das von einer Lipiddoppelschicht eingeschlossene Kernplasma und einen oder mehrere Nuc- leoli. Das Kernplasma besteht aus dem Kernfasernetzwerk und der Kernflüs-
Transcript of 2 Zytologische Grundlagen der FEM Modellierungtheuer/2 Zytoplasma - 2 -- 02-04-28a.pdf ·...

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
1
2 Zytologische Grundlagen der FEM Modellierung
Lebende Zellen sind hochorganisierte dynamische Einheiten, in denen wichtige
zelluläre Makromoleküle spontan zu komplexen Proteinstrukturen aggregieren
und wieder dissoziieren können [12]. Dabei kommt es zu häufigen lokalen Än-
derungen des Aggregatzustandes. Die geschilderten Tatsachen führen zu, in den
klassischen Ingenieurwissenschaften wenig bekannten, biologischen Material-
eigenschaften. Abbildung 2-1 enthält die 3D-Computerrekonstruktion einer
Lungenepitelzelle. Die Abbildung zeigt den Zellkern (blau), Zytoplasma (hell-
blau), die Plasmamembranen (schwarz), Zellorganellen (grün) und Erythrozy-
ten (rot). Im Allgemeinen haben Eukariontenzellen folgende Organellen:
Endosplasmatisches Retikulum
Golgi-Apparat
Mitochondrien
Lyosomen
Peroxisomen
Zellkern
Die Organellen nehmen ca. 50% des Zellvolumens ein. Die restlichen 50% des
Zellvolumens bilden das Zytosol. Die wichtigsten Proteinsynthese-Prozesse
finden im Zytosol statt. Eukariontenzellen bestehen zu 70% aus Wasser, zu
18% aus Proteinstrukturen, zu 3% aus Pospholipiden. Die Zelltrockenmasse be-
steht zu mehr als 50% aus Protein. Proteine sind aus einer Vielfalt unterschied-
licher Aminosäuren aufgebaut.
Der Zellkern ist das ausgeprägteste Organ der Zelle. Er enthält das von einer
Lipiddoppelschicht eingeschlossene Kernplasma und einen oder mehrere Nuc-
leoli. Das Kernplasma besteht aus dem Kernfasernetzwerk und der Kernflüs-

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
2
sigkeit. Die Kernfaserschicht ist wahrscheinlich mit in der inneren Kernmemb-
ran eingebetteten Verbindungsproteinen verankert und trägt entscheidend zu
der Kernform und der Kernsteifigkeit bei. Die Kernhülle ist eine ca. 40 nm di-
cke Doppelschichtmembran. Der Stoffwechsel zwischen Zytoplasma und
Kernplasma erfolgt über Poren in der Kernmembran. Der mittlere Durchmesser
der Kernmembranporen beträgt ca. 100 nm. Im Bereich der Kernmembranporen
sind die beiden Doppelschichtmembranen durch verstärkende Membranprotei-
ne miteinander verbunden. Wesentlichen Einfluß auf die Zellsteifigkeit hat die
Form und Steifigkeit des Zellkerns. Die Kernsteifigkeit wird von dem Kernge-
rüst und der doppelschichtigen Kernmembran bestimmt. Dabei spielen die vie-
len Porenverstärkungen in der Kernmembran und das Netzwerk der Lamina ei-
ne entscheidende Rolle. Bei vielen Eukariontenzellen hat die Kernfaserschicht
eine mittlere Dicke von 20 bis 40 nm. Das Chromatin säumt die Innenseite der
inneren Membrandoppelschicht. Dem Kernfasernetzwerk wird eine ordnende
Rolle bei der Positionierung und Formgebung der Chromosomen zugesprochen
[12]. Nucleoli enthalten hohe Mengen an Proteinen und RNA. Sie werden nicht
von einer Membran begrenzt. Im Nucleolus lassen sich drei Regionen definie-
ren:
Lagune
Pars granulosa 15 nm dicke ribosomale Vorläufer
Pars fibrosa 5 nm Ribonucleoproteinfaser
Die Größe des Nucleolus schwankt bei Eukariontenzellen in erheblichem Um-
fang und hat bei aktiven, proteinsynthetisierenden Zellen ein relatives, tempo-
räres Maximum.
Die meisten Tumorzellen weisen normalen Zellen gegenüber strukturelle und
funktionelle Unterschiede auf. Es sind funktionelle Unterschiede der Plasma-
membran und strukturelle Unterschiede des Zytoskelettes sowie der extrazellu-
lären Matrix. Maligne Zellen weisen in der Regel einen verstärkten Stoffwech-

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
3
seltransport durch die Plasmamembran auf. In der Lipiddoppelschicht maligner
Zellen kann oft eine verstärkte Blasenbindung konstatiert werden [29]. Die
prägnanteste strukturelle Charakteristik maligner Zellen ist eine geänderte Or-
ganisation der zellulären und extrazellulären Filament-Aggregatanordnung. Es
wird angenommen, daß durch den Verlust der mikrotubulären Kon-
trollmechanismen die Aggregation zytoplasmatischer und extrazellulärer Fila-
mentstrukturen gestört ist [12]. Es kommt zu einer diffusen, wenig gebündelten
Anordnung zellulärer Aktin-Filamente.
Aktinfaserstränge haben eine formerhaltende und formbestimmende Rolle.
Der zelluläre Kraftfluß in normalen Zellen erfolgt weitgehend über gebündelte
Aktinstränge. Eine Störung der Aktinbündelung und der makromolekularen
Verbindungsproteine führt zu einem Verlust der funktionellen Steifigkeit der
zytoplasmatischen Skelettordnung. Von entscheidender Bedeutung für die Mo-
dellierung zellulärer Strukturen ist die Steifigkeit und Viskosität der Zelle als
auch die formbestimmende Anordnung der extrazellulären Matrix.
Die Zellsteifigkeit und damit die Zellform wird hauptsächlich von der Steifig-
keit der Zytoskelettfilamente, ihrer Vernetzungsform und Bündelung sowie ih-
rer Verankerung an den Proteineinlagerungen in der Plasmamenbran bestimmt.
Das Zytoplasma ist aus dem Zytoskelett und der Zellflüssigkeit aufgebaut. Es
ist eine dynamische Anordnung hochorganisierter Proteinketten, in denen Zell-
flüssigkeit eingelagert ist. Es gibt drei wichtige zelluläre Skelettfilamente:
-Mikrofilamente ( besonders Aktinfilamente ) Durchmesser ca. 7 nm
-Mikrotubuli Durchmesser ca. 25 nm
-Intermediärfilamente Durchmesser ca. 10 nm
In Abbildung 2-2 ist die Elementdiskretisierung für das 2D-FEM Berech-
nungsmodell einer Lymphknoten-Metastasenzelle des Mammakarzionoms dar-

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
4
gestellt. Die Abbildung zeigt das Kernfilamentgerüst und das zytoplasmatische
Skelett mit Mikrotubuli, Intermediärfilamenten und Aktinfilamenten.
Zusätzlich zu den drei Hauptgruppen zellulärer Proteinfilamente enthalten die
meisten Eukariontenzellen weitere makromolekulare Proteinstrukturen. Diese
können Proteinfilamente bündeln und Verbindungen zwischen Filamentsträn-
gen und Proteineinlagerungen in der Plasmamembran herstellen. Es gibt kon-
traktionsauslösende Verbindungsproteine, wie Myosin und Nicht-
Muskelmyosin, gelbildende Verbindungsproteine (Filamin und Actinin), poly-
mersisationshemmdende Proteine (Profilin), Bündelungsproteine (Fimbrin) und
Fragmentierungsproteine (Villin und Gelsolin). Aktinfilamente sind in vielen
zellulären Strukturen anzutreffen. Sie werden über unterschiedliche Vernet-
zungsproteine miteinander verbunden. Die Strukturform aktinbindender Prote-
ine beeinflußt den Aggregationszustand des Zytoplasmas. Aktin-Filamente be-
stehen aus zwei, in Helix-Anordnung aneinandergekoppelte Makromoleküle
globulärer Form. Die Ankopplungskräfte beruhen auf Wasserstoffbrücken und
bestimmen typischen Helixanordnungen oder Faltanordnungen. So entstandene
Aktinstränge können durch zusätzliche Verbindungsproteine miteinander ver-
bunden sein. Dabei wird die Steifigkeit erheblich gesteigert. Aktinmakromole-
küle haben eine durchschnittliche Länge von ca. 6 nm. Aktinstränge ordnen
den zellulären und extrazellulären Kraftfluß. Die zytoplasmatische Aktinfila-
mentstruktur bestimmt die jeweilige zytoplasmatische Viskosität.
Die dreidimensionale Anordnung zellulärer Proteinmakromoleküle werden
durch nichtkovalente Wechselwirkungen der Polipeptidketten stabilisiert. Viele
Proteine sind aus globulären Untereinheiten gebildet. Diese sind durch kurze
Polypeptidketten miteinander verbunden. Makromolekulare Bindungskräfte
halten Proteinaggregatstrukturen zusammen. Die in der Fachliteratur mehrfach
zitierte Anhäufung von Aktinfilamenten im Bereich der Lipiddoppelschicht-
Innenmembran [12 bis 20] wird durch eine entsprechende FEM-Diskretisierung
der Aktinelemente (Abbildung 2-2) berücksichtigt.

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
5
Abbildung 2-1
3D-Computerrekonstruktion einer Lungenepitelzelle. Die Abbildung zeigt den Zell-
kern (blau), Zytoplasma (hellblau), die Plasmamembranen (schwarz), Zellorganellen
(grün) und Erythrozyten (rot).

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
6
Abbildung 2-2
Elementdiskretisierung für das 2D-FEM Berechnungsmodell einer Lymphknoten-
Metastasenzelle des Mammakarzinoms. Die Abbildung zeigt das Kernfilamentgerüst
und das zytoplasmatische Skelett mit Mikrotubuli, Intermediärfilamenten und Ak-
tinfilamenten.
Zytoplasmatisches Filament Kernfilament

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
7
Das Ausstrecken der Mikrovilli erfolgt durch die Aggregation von Aktin-
Filamenten. Durch den Abbau von Aktin-Filamenten kommt es zu einer
Schrumpfung und Rückbildung der Mikrovilli.
Mikrotubuli sind 25 nm dicke Tubulinmakromoleküle in Hohlkörperanord-
nung. Dabei sind Protofilamente um einen zentralen Bereich zylindrisch ange-
ordnet. Mikrotubuli sind
aus 13, zentrisch angeord-
neten Tubulin-Polipeptiden
aufgebaut. Diese sind durch
zahlreiche Zusatzproteine
miteinander stabilisiert und
an angrenzende Organellen
und Membranen gebunden.
Ohne zusätzliche Vernetzungsproteine und Verankerung an angrenzende
Membranen und Organellen sind Mikrotubuli biegeweiche Hohlzylinder. In
Abbildung 2-3 wird die zytoplasmatische Organisation des Mikrotubuli-Netzes
gezeigt.
In der Interphase verlaufen Mikrotubuli zentrisch, ausgehend von dem Zellzent-
rum (Centriol) durch das Zytoplasma. Dabei kann eine Konzentration von Mi-
krotubuli um den Zellkern herum beobachtet werden. Es ist wahrscheinlich, daß
Mikrotubuli für die Organisation der gesamten intrazellulären und extrazel-
lulären Skelettfilamente verantwortlich sind [12]. Das Zytoplasma der Eukari-
ontenzellen enthält im Durchschnitt 50% nichtaggregiertes Tubulin. Durch die
Zugabe bestimmter Wirkstoffe aggregieren Tubulinmoleküle zu äußerst labilen
mikrotubulären Makrostrukturen.
Durch die Einwirkung bestimmter Wirkstoffe dissoziieren Mikotubuli-
Aggregate spontan. Andere Wirkstoffe blockieren die Aggregation mikrotubu-
Abb. 2-2.1 Zytoplasmatische Mikrotubulistruktur

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
8
lärer Strukturen oder fördern die Aggregation tubulärer Strukturen. Durch die
Zugabe von Conchilin in das Zytoplasma einer sich teilenden Zelle werden die
Mikrotubuli der mitotischen Spindel aufgelöst. Die Zellteilung wird blockiert.
Die Zelle verharrt in der Mitose. Die Entfernung des Wirkstoffes erlaubt die
Rückbildung der mitotischen Spindel und den Fortgang der Zellteilung. In der
Mitose bewirkt Vinblastin bei vielen Zellen eine Blockierung der Mikrotubuli-
Aggregation. Dadurch verharrt die Zelle in Mitose und der Teilungsprozess ist
blockiert. Ein anderer Wirkstoff Taxon bewirkt eine verstärkte Aggregation von
Mikrotubuli. Es entstehen steifere mitotische Spindeln. Dadurch wird die sich
teilende Zelle auch in der Mitose blockiert. Sie stirbt. Schweres Wasser hat die
gleiche Wirkung. Kommt es zu einer Störung der Bildung von mitotischen MT-
Spindeln werden hauptsächlich schnellteilende Zellen geschädigt.
Man konnte beobachten, daß bei einigen Eukariontenzellen die Dissoziation
mikrotubulärer Makrostrukturen bei niedrigen Temperaturen schneller verläuft
[12]. Wahrscheinlich werden Mikrotubuli durch hohe Ca2+ Konzentrationen
abgebaut und bei geringen Calciumkonzentrationen wieder aufgebaut [12]. Bei
einigen Eukariontenzellen werden Mikrotubuli bei Unterkühlung schneller ab-
gebaut und bei Erwärmung wieder aufgebaut. Die Modellierung zellulärer
Strukturen muß sowohl der äußerst komplexen Anordnung unterschiedlicher
Makromolekül-Strukturen Rechnung tragen als auch die häufigen Gel-Sol-Gel
Übergänge zytoplasmatischer Komponenten berücksichtigen.
Intermediärfilamente sind makromolekulare Proteinstrukturen mit durchschnitt-
lichen Größen von ca. 10 nm im Durchmesser. Anders als Mikrotubuli und
Mikrofilamente können Intermediärfilament nach ihrer Aggregation nur schwer
oder gar nicht dissoziieren. Es gibt Kreatinfilamente, Neurofilamente (Neurone)
und Vimentinfilamente (Fibroblasten, einige Gliazellen und Muskelzellen). Ne-
ben Mikrotubuli und Aktinfilamenten haben auch Intermediärfilamente struktu-
relle, spannungsübertragende Funktionen. Die Zellsteifigkeit und damit die
Zellform wird hauptsächlich von der Steifigkeit der Zytoskelettfilamente, ihrer
Vernetzungsform und Bündelung sowie ihrer Verankerung an den Proteineinla-

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
9
gerungen in der Plasmamenbran bestimmt. Wesentlichen Einfluß auf die Zell-
steifigkeit hat auch die Steifigkeit und die Form des Zellkerns. Die Kernsteifig-
keit wird von dem Kerngerüst und der doppelschichtigen Kernmembran be-
stimmt. Dabei spielen die vielen Porenverstärkungen in der Kernmembran und
das Netzwerk der Lamina eine entscheidende Rolle.
Von entscheidender Bedeutung für die Modellierung zellulärer Strukturen sind
die Steifigkeit, die Viskosität und die formbestimmende Anordnung der extra-
zellulären Matrix. Diese besteht aus Filamentnetzwerken in denen extrazellulä-
re Flüssigkeit eingelagert ist. Viele extrazelluläre Filamentstränge sind mit
Verbindungsproteinen der zellulären Lipiddoppelschicht verbunden und bilden
mit dem Zytoskelett einen geschlossenen Kraftschluß. Es wird angenommen,
daß sich die Orientierung der extrazellulären Filamentbündel an dem zytoplas-
matischen Mikrotubuli-Netzwerk ausrichtet. Desgleichen vermutet man eine
reziproke Beeinflussung der Aktinbündel im Zytoplasma durch die extrazellulä-
re Matrix.
Im Golgi-Apparat werden zytoplasmatische und extrazelluläre Proteinstrukturen
in ihre endgültige Form gebracht. Dabei erfahren die Initialproteine spezifische
kovalente Bindungsänderungen. Die Änderunsmechanismen sind noch weitge-
hend unbekannt.
Für die Modellierung der zu analysierenden Zellen müssen detaillierte Angaben
zu den formbestimmenden Parameter der entsprechenden subzellulärer Struktu-
ren (Orientierung und Stärke der Aktinbündel, Mikrotubulianordnungen, Ver-
bindungsproteine und Intermediärfilamente) vorliegen. Desgleichen sind Anga-
ben zu der Steifigkeit und Viskosität der extrazellulären Filamentanordnungen
erforderlich.
Die Mikofilamente für das Berechnungsmodell in Abb. 2-2 sind dreiecksförmig
angeordnet und haben eine durchschnittliche Länge von ca. 65 nm . Im Bereich
der Kernmembran sind den Kernfaser-Elementen (Feder-Dämpfungselemente)
steifere Materialwerte zugeordnet. Damit wird der Kernmembrandoppelschicht
mit der an ihr anhaftenden, versteifenden Lamina Rechnung getragen. Im FEM-

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
10
Berechnungsmodell der Lymphknoten-Metastasenzelle (Abb. 2-2) werden ext-
razellulären Protein-Filamente mit stabförmigen Feder-Dämpfungselementen
simuliert. Die Elemente sind durch die Membrandoppelschicht mit dem plas-
matischen Skelett der Zelle verbunden.
Die Abbildungen 2-4 und 2-5 zeigen die FEM-Anordnung der zytoplasmati-
schen und extrazellulären Proteinfilament-Elemente bei Lymphknoten-
Metastasenzellen des Mammakarzinoms in peripherem lymphoiden Gewebe.
Im FEM-Berechnungsmodell sind den malignen Proteinfilament-Elementen
(links) reduzierte Steifigkeiten zugeordnet. Die zytologische Begründung ist der
Verlust der zytoplasmatischen und extrazellulären Skelettordnung maligner
Metastasenzellen.
In der Abbildung 2-6 ist der Kern- Deteilbereich der Elementdiskretisierung des
FEM-Berechnungsmodelles einer Lymphknoten-Metastasenzelle des Mamma-
karzinoms dargestellt.
Abbildung 2-3. Zelluläre Anordnung zytoplasmatischer Mikrotubuli. Die Ora-
nisation der Mikrotubuli verläuft radial durch das Zytoplasma, vom Zellzent-
rum (Zentriol) aus, um den Zellkern.
Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
11
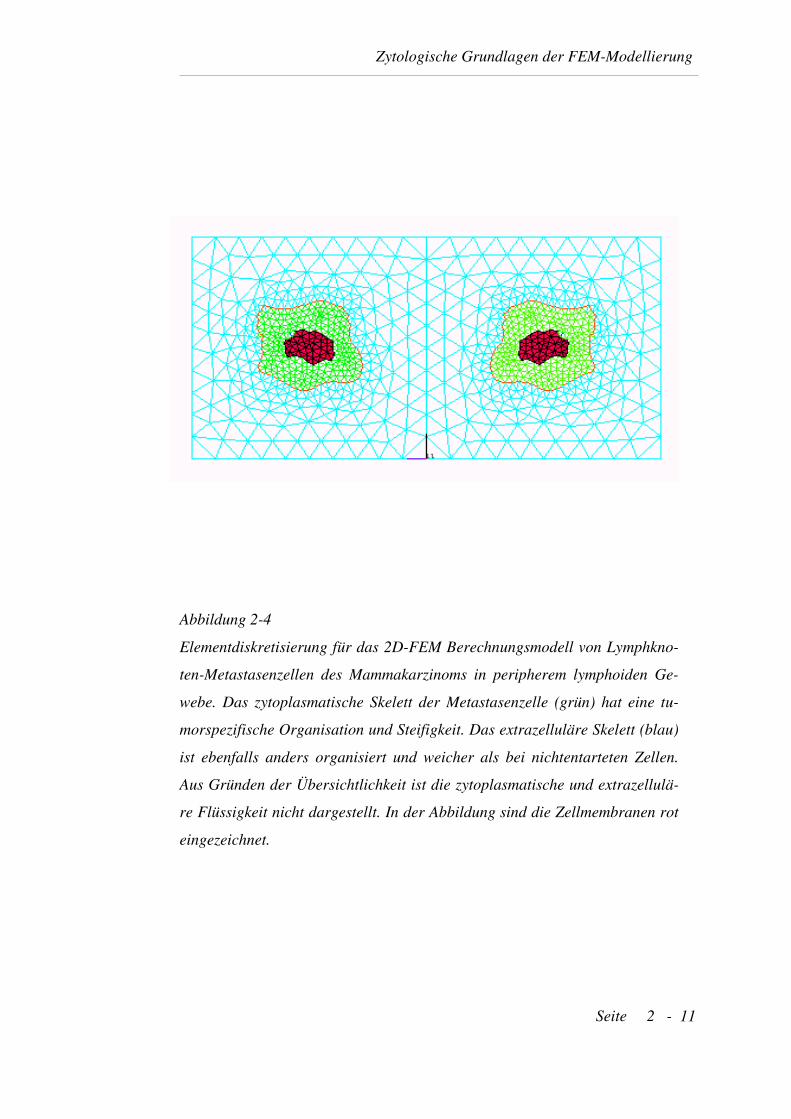
Abbildung 2-4
Elementdiskretisierung für das 2D-FEM Berechnungsmodell von Lymphkno-
ten-Metastasenzellen des Mammakarzinoms in peripherem lymphoiden Ge-
webe. Das zytoplasmatische Skelett der Metastasenzelle (grün) hat eine tu-
morspezifische Organisation und Steifigkeit. Das extrazelluläre Skelett (blau)
ist ebenfalls anders organisiert und weicher als bei nichtentarteten Zellen.
Aus Gründen der Übersichtlichkeit ist die zytoplasmatische und extrazellulä-
re Flüssigkeit nicht dargestellt. In der Abbildung sind die Zellmembranen rot
eingezeichnet.

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
12
Abbildung 2-5
Die Abbildung zeigt die Anordnung der zytoplasmatischen und extrazellulären
Proteinfilamente von Lymphknoten-Metastasenzellen des Mammakarzinoms.
Durch den Verlust der zytoplasmatischen und extrazellulären Skelettordnung
ist die funktionelle Steifigkeit der Zelle gestört. Den Zytoskelettfilamentelemen-
ten sind im Berechnungsmodell reduzierte Steifigkeiten zugeordnet. In der Ab-
bildung ist das Zytoplasma grün, das Kernplasma rot und die extrazelluläre
Flüssigkeit hellblau dargestellt.

Zytologische Grundlagen der FEM-Modellierung
Seite 2 -
13
Abbildung 2-6
Elementnetzwerk des 2D-FEM Berechnungsmodelles einer Lymphknoten-
Metastasenzelle
des Mammakarzinoms. Detailbereich des Zellkerns mit Kernlamina, Kernmemb-
ran und
zytoplasmatischem Skelett.
extrazelluläres Skelettfilament Plasmamembran Zytoskelett Kernlamina











![Soventol Fibel 082017 - apothekenkanal.de · Soventol Gel 20 mg/1 g Gel. Wirkst.: Bamipin[(RS)-lactat. Zus.-setz.: 1 g Gel enth. 20 mg Bamipin[(RS)-lactat]. Sonst. Best.-teile: Gerein.](https://static.fdokument.com/doc/165x107/5e64f7ea3480624f8923b165/soventol-fibel-082017-soventol-gel-20-mg1-g-gel-wirkst-bamipinrs-lactat.jpg)