Analyse der Struktur-Funktionsbeziehungen in OPR3 · modifizierten OPR1-Proteine im Peroxisom...
121
Universität Hohenheim Institut für Physiologie und Biotechnologie der Pflanzen Prof. Dr. A. Schaller Analyse der Struktur-Funktionsbeziehungen in OPR3 Diplomarbeit vorgelegt von Daniela Schwörer Fakultät Naturwissenschaften - Studiengang Biologie Betreuung durch Dr. Annick Stintzi Hohenheim, September 2007
Transcript of Analyse der Struktur-Funktionsbeziehungen in OPR3 · modifizierten OPR1-Proteine im Peroxisom...

Universität Hohenheim
Institut für Physiologie und Biotechnologie der Pflanzen
Prof. Dr. A. Schaller
Analyse der Struktur-Funktionsbeziehungen in OPR3
Diplomarbeit vorgelegt von Daniela Schwörer Fakultät Naturwissenschaften - Studiengang Biologie
Betreuung durch Dr. Annick Stintzi
Hohenheim, September 2007


Für meine Eltern

Zusammenfassung I
Zusammenfassung
In der vorliegenden Arbeit wurden Strukturanalysen zweier Proteine der
Oxophytodiensäure Reduktase Familie, OPR3 und OPR1, durchgeführt. Durch
zielgerichtete Mutationen sollte untersucht werden, welche Strukturen für die
Substratspezifität der Proteine verantwortlich sind. OPR3 ist in der Lage, sowohl
cis(+)OPDA als auch cis(-)OPDA umzusetzen, wohingegen OPR1 lediglich
cis(-)OPDA umsetzen kann. Es sollte ein OPR1 Protein durch zielgerichtete
Mutationen so modifizieren werden, dass es cis(+)OPDA umsetzen kann. Hierfür
wurden verschiedene Konstrukte der OPR1 cDNA hergestellt, bei welchen die
Aminosäuren Tyrosin an Position 74 in Phenylalanin getauscht und die Aminosäure
Tyrosin an Position 244 in Histidin ausgetauscht wurden. Außerdem wurde C-
Terminal ein peroxisomales SRL-Tag kloniert, welches die Lokalisation im
Peroxisomen sicherstellen sollte. Opr3-defiziente Arabidopsis thaliana wurden mit
den Konstrukten transformiert. Wenn die männliche Fertilität von opr3-defizienten
Pflanzen wiederhergestellt werden kann, so zeigt dies, dass die Aktivität von OPR3
durch die punktmutierten OPR1 Proteine komplementiert werden kann. Dies war
nicht der Fall, woraufhin weitere Untersuchungen klarstellen sollten, ob die
modifizierten OPR1-Proteine im Peroxisom vorhanden waren. Allerdings konnten
auch diese kein eindeutiges Ergebnis liefern.
Des Weiteren wurden punktmutierte OPR3-Proteine untersucht, die an Stellen die
für eine Dimerisierung entscheidend sind (Glutamat an Position 291 wurde durch
Lysin ersetzt) und eine Phosphorylierung bei Dimerisierung ausbilden (Tyrosin an
Position 364 wurde durch Phenylalanin ersetzt). Hierbei sollte untersucht werden,
ob die punktmutierten Proteine, die in vitro als Monomere kristallisieren auch in vivo
nicht dimerisieren können. Eine daraus resultierende eventuelle dauerhafte Aktivität
sollte untersucht werden. Die männliche Sterilität der opr3-defizienten Pflanzen
konnte durch die Mutierten OPR3-Proteine komplementiert werden, aber es zeigte
sich kein veränderter Phänotyp. Um eine dauerhafte Aktivität von OPR3
nachzuweisen, müssen weitere Experimente durchgeführt werden.
Außerdem wurde die Möglichkeit einer reversiblen Phosphorylierung zur Regulation
von OPR3 mit Hilfe von Western Blot Analysen untersucht. Hierbei wurden
Verwundungsexperimente an Tomaten und Arabidopsis thaliana durchgeführt. Es
stellte sich heraus, dass OPR3 nicht nur durch Verwundung induziert wird, sondern
auch, wenn eine benachbarte Pflanze verwundet wird und flüchtige Botenstoffe
(volatile organic compounds, kurz VOCs) freigesetzt werden. LeOPR3 scheint
sowohl induziert, als auch durch Phosphorylierung inaktiviert zu werden, während

Zusammenfassung II
bei Arabidopsis zwar eine Induktion von OPR3 zu erkennen ist, aber keine
eindeutige Phosphorylierung des Proteins festgestellt werden konnte.

Inhaltsverzeichnis III
INHALTSVERZEICHNIS
ZUSAMMENFASSUNG I
1. EINLEITUNG 1
1.1 Der Oktadekanoidweg 3
1.2 Das OPR3 Protein– Lokalisation und Aufgabe 5
1.3 Die opr3-Mutante 7
1.4 Strukturvergleich von OPR3 und OPR1 8
1.5 Volatiles – Kommunikation unter Pflanzen 12
1.6 Ziel dieser Arbeit 14
2. MATERIAL UND METHODEN 15
2.1 Organismen 15
2.1.1 Pflanzen 15
2.1.2 Bakterien 15
2.2 Vektoren und Plasmide 15
2.3 Oligonukleotide 17
2.4 Enzyme 18
2.5 Antikörper 19
2.5.1 Primäre Antikörper 19
2.5.2 Sekundäre Antikörper 19
2.6 Chemikalien und Verbrauchsmaterial 20
2.7 Laborgeräte 21
2.8 Antibiotika und Medien 22
2.8.1 Stammlösungen Antibiotika 22
2.8.2 Medien 22

Inhaltsverzeichnis IV
2.9 Allgemeine Methoden mit Pflanzen 23
2.9.1 Anzucht von Tomaten 23
2.9.2 Anzucht von Arabidopsis thaliana 23
2.9.3 Transformation von Arabidopsis thaliana mit der „floral dip“ Methode 23
2.9.4 Pollenkeimungstest 24
2.10 Molekularbiologische Techniken mit DNA 25
2.10.1 Polymerase-Ketten-Reaktion (PCR) 25
2.10.2 Auftrennung und Sichtbarmachen von DNA durch Gelelektrophorese 27
2.10.3 Restriktionsverdau 28
2.10.4 DNA-Gelextraktion 28
2.10.5 Klonierung 28
2.10.6 Transformation durch Elektroporation 29
2.10.7 DNA Extraktion 29
2.10.8 Zielgerichtete Mutagenese 32
2.10.9 DNA-Sequenzierung 34
2.11 Molekularbiologische Techniken mit Proteinen 35
2.11.1 Gradientenzentrifugation / Zellorganellfraktionierung 35
2.11.2 Katalaseaktivität 36
2.11.3 Proteinexpression und Aufreinigung 36
2.11.4 Gesamtproteinextraktion 39
2.11.5 Bradfordtest 39
2.11.6 Diskontinuierliche Polyacrylamidgelelektrophorese (SDS-PAGE) 40
2.11.7 Proteinfärbung 42
2.11.8 Western Blot 44
2.11.9 Antikörperaufreinigung 45
2.11.10 Immunologischer Nachweis der Proteine 47
3. ERGEBNISSE 49
3.1 Mutationen der OPR3 cDNA 49
3.2 Mutationen der OPR1 cDNA 50
3.3 Klonierung in pGreen 0229 54
3.4 Identifizierung transgener Pflanzen 56
3.5 Analyse der Mutation E291K 58
3.6 Ist die Männliche Fertilität in OPR1+SRL, Y244H und Y74F/Y244H wiederhergestellt?
60

Inhaltsverzeichnis V
3.7 Lokalisation der mutierten OPR1-Proteine 62
3.7.1 Gradientenzentrifugation 62
3.7.2 Katalaseaktivität 64
3.7.3 Western Blot auf OPR1 65
3.8 Wird OPR3 durch reversible Phosphorylierung reguliert? 67
3.8.1 OPR3 wird durch Green Leave Volatiles Induziert 71
3.8.2 LeOPR3 wird bei Induktion durch Green Leave Volatiles Phosphoryliert 73
3.8.3 Im Arabidopsis Wildtyp wird OPR1 durch die Induktion von OPR3 reprimiert 74
3.8.4 In opr1 Pflanzen ist OPR3 konstitutiv aktiv 76
4. DISKUSSION 79
4.1 OPR3 kann nicht durch punktmutierte OPR1Proteine ersetzt werden 79
4.2 Arabidopsis zeigt keinen Veränderungen durch punktmutierte OPR3 Proteine 81
4.3 OPR3 wird durch VOCs induziert 82
5. ANHANG VII
5.1 Ergebnisse der Pollenkeimungstests VII
5.1.1 Wildtyp VII
5.1.2 Arabidopsis opr3-Mutante VII
5.1.3 Arabidopsis opr3-Mutante komplementiert mit OPR3 cDNA VIII
5.1.4 Arabidopsis opr3-Mutante komplementiert mit E291K VIII
5.1.5 Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL IX
5.1.6 Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL-Y244H X
5.1.7 Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H/Y74F XII
5.2 Sequenz des AtOPR3 Promotors XIV
5.3 Aminosäure und Basensequenz von LeOPR1 XV
5.4 Aminosäure und Basensequenz von LeOPR3 XVII
5.5 Sequenzen von OPR3-Mutanten XIX
5.5.1 OPR3 E291K XIX
5.5.2 OPR3 Y364F XIX
5.6 Sequenzen von OPR1Mutanten XX
5.6.1 OPR1+SRL XX
5.6.2 OPR1 Y74F XX
5.6.3 OPR1 Y244H XXI

Inhaltsverzeichnis VI
5.6.4 OPR1 Y74F/Y244H XXI
6. VERZEICHNISSE XXII
6.1 Literatur XXII
6.2 Abkürzungen und Einheiten XXVII
6.3 Abbildungen und Tabellen XXIX
DANKE XXXIV
EHRENWÖRTLICHE ERKLÄRUNG

1. Einleitung 1
1. Einleitung
Jeder Organismus muss, wenn er überleben will, gewisse Anforderungen in seiner
Umwelt erfüllen. Sind Umgebung oder Bedingungen suboptimal, ist der
Organismus Stress ausgesetzt. Viele dieser Stresssituationen lassen sich umgehen
oder lösen, indem zum Beispiel ein Tier einen Schattenplatz aufsucht, wenn es ihm
in der Sonne zu heiß ist, oder ein Hund sich durch Hecheln Kühlung verschafft.
Pflanzen aber sind Standort und Umgebung ausgeliefert. Daher mussten Pflanzen
im Laufe der Evolution Möglichkeiten entwickeln, um mit veränderten
Umweltbedingungen fertig zu werden. Hierfür stehen der Pflanze andere
Möglichkeiten zur Verfügung als Menschen oder Tieren. Sie empfängt
Umweltsignale, die dann intra- und interzellulär weitergegeben werden um
schließlich spezifische Reaktionen hervorzurufen, die die Pflanze an die
veränderten Umweltbedingungen anpasst. Hierfür hat eine Pflanze ein großes
Spektrum an Pflanzenhormonen zur Verfügung, die eine interzelluläre
Kommunikation ermöglichen. Ein bekannter Mechanismus hierbei ist die Bildung
von Jasmonsäure bei Verwundung durch Fraßfeinde. Jasmonsäure (vgl. Abb. 1) ist
bereits in den 60er Jahren entdeckt worden, aber erst in der allerjüngsten Zeit als
Pflanzenhormon anerkannt worden.
Abb. 1: Struktur der Jasmonsäure
Jasmonsäure kommt nicht nur in Arabidopsis thaliana, der Modellpflanze, die in
dieser Arbeit zum Großteil der Experimente herangezogen wird, sondern ubiquitär
im Pflanzenreich vor (Meyer et al., 1984). Sie ist verantwortlich für die Regulation
verschiedenster Prozesse (Creelman und Mullet, 1997). Zu den Jasmonsäure-
regulierte Entwicklungsprozesse gehören die Differenzierung des männlichen
Gametophyten, die Verlängerung der Antherenfilamente, die Öffnung der Stoma
während der Anthese und den Reifungsprozess von Pollenkörnern. (McConn and
Browse, 1996;Sanders et al., 2000; Stintzi and Browse, 2000; Ishiguro et al., 2001;
Park et al., 2002). Außerdem hat Jasmonsäure eine Wirkung auf die Seneszenz,
Wurzelentwicklung und Fruchtbildung der Pflanzen (Devoto und Turner, 2003). In
hohen Dosen wirkt sie auf das Wachstum der Pflanzen hemmend (Koda, 1997).
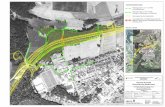
1. Einleitung 2
Weiterhin kommt ihr eine große Aufgabe als Signalmolekül in Antwort auf Stress zu
(Berger 2002). Abbildung 2 zeigt im linken Bereich die Wirkungen von Jasmonaten
auf Stressoren und im rechten Teil auf die Entwicklung der Pflanzen. Am meisten
ist aber von den Funktionen der Jasmonsäure durch Mutanten in Arabidopsis
bekannt, welche über so genannte loss-of-function Mutationen nicht mehr in der
Lage sind Jasmonsäure zu bilden oder zu erkennen.
Abb. 2: Die Wirkungen von Jasmonaten auf Stressoren und ihre Beteiligung an der
pflanzlichen Entwicklung (Wasternack, 2005)

1. Einleitung 3
1.1 Der Oktadekanoidweg
Der Oktadekanoidweg führt zur Bildung von Jasmonsäure. Sie wird ausgehend von
aus der Chloroplastenmembran freigesetzten α-Linolensäure (18:3) gebildet, die
von 13-Lipoxygenase (LOX) katalysiert in Hydroperoxylinolensäure umgewandelt
wird. Diese wird von der Allenoxid Synthetase (AOS) zu einem instabilen Epoxid,
welches sofort von der Allenoxid Cyclase (AOC) zyklisiert und somit zu
Oxophytodiensäure (OPDA) wird. Geschieht diese sofortige Umwandlung in
cis(+)OPDA nicht, entstehen racemische Formen von OPDA, die nicht als Vorläufer
der Jasmonsäure fungieren können. Cis(+)OPDA wird von einer NADPH2-
abhängigen OPDA Reduktase (OPR3) im Peroxisom an der 10,11-Doppelbindung
reduziert. Schließlich entsteht OPC 8:0, aus der nach 3 Zyklen β-Oxidation
Jasmonsäure, das Endprodukt des Biosynthesewegs entsteht (Review Schaller F.,
2005).
Abb. 3: Der Oktadekanoidweg der Jasmonysnthese (nach Vick und Zimmermann,
adaptiert nach Wasternack und Hause, 2002)

1. Einleitung 4
Es wurden fünf Isoformen der OPDA Reduktasen (OPRs) in Arabidopsis gefunden,
in Tomate drei, im Reis sogar 13 (Review: Schaller et al., 2005). Abbildung 4 zeigt
die Verwandtschaft der 3 OPR Isoformen aus Tomate und Arabidopsis.
AtOPR3
AtOPR1AtOPR2
10001000
10001000
999999
LeOPR3
LeOPR1LeOPR2
0.10.1
AtOPR3
AtOPR1AtOPR2
1000100010001000
1000100010001000
999999999999
LeOPR3
LeOPR1LeOPR2
0.10.1
Abb. 4: Verwandtschaftsvergleich der OPR Isoformen von Tomate und Arabidopsis.
Die OPRs aus Tomate und OPR1, 2, 3, 4 und 5 (Abu-Romman, Stintzi und Schaller
A., unveröffentlicht) aus Arabidopsis thaliana katalysieren die Reduktion von α,β-
ungesättigten Kohlenstoffverbindungen (Schaller F. et al., 2000; Strassner et al.,
1999). Zu dieser Gruppe gehört auch cis(+)OPDA, welche das physiologische
Substrat von OPR3 ist. Doch nur OPR3 ist in der Lage in vivo cis(+)OPDA zu OPC
8:0 umzusetzen (Stintzi und Browse, 2000). Keine der anderen Isoformen kann
diese Reaktion im Oktadekanoidweg ersetzen. OPR3 kann zwar auch cis(-)OPDA
in vitro umsetzen, allerdings bildet sich daraus nicht die aktive Form
(3R,7S)-Jasmonsäure, sondern die inaktive (3S,7R)-Jasmonsäure. Eine andere
Isoform der Oxophytodiensäure Reduktase, das OPR1, ist nur in der Lage
cis(-)OPDA umzusetzen und somit nicht für den Oktadekanoidweg geeignet
(Schaller F., 1998).
Jasmonsäure hat auf den Oktadekanoidweg eine positive Rückkopplung. Wenn
Jasmonsäure produziert wird, wird zugleich die Bildung von am Oktadekanoidweg
beteiligten Proteinen stimuliert. Ebenso ist OPDA ein Signalmolekül im
Oktadekanoidweg, das unter anderem die Bildung von im Oktadekanoidweg
vorgeschalteten Enzymen stimuliert. (Stintzi et al., 2001)

1. Einleitung 5
1.2 Das OPR3 Protein– Lokalisation und Aufgabe
Abb. 5: Vergleich der Sequenzen der 3 Isoformen von OPR in Tomaten (Le) mit
den Isoformen aus Arabidopsis thaliana (At)
AtOPR1 --MENGEAKQS--VPLLTPYKMGRFNLSHRVVLAPLTRQRSYGNVPQPHAAIYYSQRTTPGGFLITEATGVSDTAQGYQDTPGIWTKEHVEAWKPIVDAVHAKGGIFFCQ 106
AtOPR2 MEMVNAEAKQS--VPLLTPYKMGRFNLSHRVVLAPLTRQKSYGSVPQPHAILYYSQRTSPGGFLIAEATGVSDTAQGYPDTPGIWTKEHVEAWKPIVDAVHAKGGIFFCQ 108
LeOPR1 MENKVVEEKQVDKIPLMSPCKMGKFELCHRVVLAPLTRQRSYGYIPQPHAILHYSQRSTNGGLLIGEATVISETGIGYKDVPGIWTKEQVEAWKPIVDAVHAKGGIFFCQ 110
AtOPR3 ----MTAAQGNSNETLFSSYKMGRFDLSHRVVLAPMTRCRALNGVPNAALAEYYAQRTTPGGFLISEGTMVSPGSAGFPHVPGIYSDEQVEAWKQVVEAVHAKGGFIFCQ 106
LeOPR3 ----MASSAQDGNNPLFSPYKMGKFNLSHRVVLAPMTRCRALNNIPQAALGEYYEQRATAGGFLITEGTMISPTSAGFPHVPGIFTKEQVREWKKIVDVVHAKGAVIFCQ 106
1.......10........20........30........40........50........60........70........80........90.......100.......110
AtOPR1 IWHVGRVSNSGFQPNGKAPISCSDKPLMPQIRSNGI---DEALFTPPR--RLGIEEIPGIVNDFRLAARNAMEAGFDGVEIHGANG-YLIDQFMKDTVNDRTDEYGGSLQ 210
AtOPR2 IWHVGRVSNRGFQPRRQAPISCTGKPIMPQMRANGI---DEARFTPPR--RLSIEEIPGIVNDFRLAARNAMEAGFDGVEIHGAHG-YLIDQFMKDKVNDRTDEYGGSLQ 212
LeOPR1 IWHVGRVSNKDFQPNGEDPISCTDRGLTPQIMSNGI---DIAHFTRPR--RLTTDEIPQIVNEFRVAARNAIEAGFDGVEIHGAHG-YLIDQFMKDQVNDRSDKYGGSLE 214
AtOPR3 LWHVGRASHAVYQPNGGSPISSTNKPISENRWRVLLPDGSHVKYPKPR--ALEASEIPRVVEDYCLSALNAIRAGFDGIEIHGAHG-YLIDQFLKDGINDRTDQYGGSIE 213
LeOPR3 LWHVGRASHEVYQPAGAAPISSTEKPIS-NRWRILMPDGTHGIYPKPR--AIGTYEISQVVEDYRRSALNAIEAGFDGIEIHGAHG-YLIDQFLKDGINDRTDEYGGSLA 212
.......120.......130.......140.......150.......160.......170.......180.......190.......200.......210.......220
AtOPR1 NRCKFPLEIVDAVAKEIGPDRVGIRLSPFADYMESGDTNPGALGLYMAESLNKYG------ILYCHVIEARMKTMGEVHACPHT--------LMPMRKAFKGTFISAGGF 306
AtOPR2 NRCKFALEVVDAVAKEIGPDRVGIRLSPFADYMESGDTNPEALGLYMVESLNKYG------ILYCHMIEPRMKTVGEIAACSHT--------LMPMREAFKGTFISAGGF 308
LeOPR1 NRCRFALEIVEAVANEIGSDRVGIRISPFAHYNEAGDTNPTALGLYMVESLNKYD------LAYCHVVEPRMKTAWEKIECTES--------LVPMRKAYKGTFIVAGGY 310
AtOPR3 NRCRFLKQVVEGVVSAIGASKVGVRVSPAIDHLDATDSNPLSLGLAVVDMLNKLQDVNGLKLAYLHVTQPRYHAYGQTESGRQGSDEEEAKLMKSLRMAYKGTFMSSGGF 323
LeOPR3 NRCKFITQVVQAVVSAIGADRVGVRVSPAIDHLDAMDSNPLSLGLAVVERLNKIQLHSGSKLAYLHVTQPRYVAYGQTEAGRLGSEEEEARLMRTLRNAYQGTFICSGGY 322
.......230.......240.......250.......260.......270.......280.......290.......300.......310.......320.......330
AtOPR1 TREDGNEAVSKGRTDLVAYGRWFLANPDLPKRFQVDAPLNKYDRPTFYTSDPVVGYTDYPFLESTA-------- 372
AtOPR2 TREDGNEAVAKGRTDLVAYGRWFLANPDLPKRFQLDAPLNKYNRSTFYTSDPVVGYTDYPSLESTA-------- 374
LeOPR1 DREDGNRALIEDRADLVAYGRLFISNPDLPKRFELNAPLNKYNRDTFYTSDPIVGYTDYPFLETMT-------- 376
AtOPR3 NKELGMQAVQQGDADLVSYGRLFIANPDLVSRFKIDGKLNKYNRKTFYTQDPVVGYTDYPFLA------PSSRL 391
LeOPR3 TRELGIEAVAQGDADLVSYGRLFISNPDLVMRIKLNAPLNKYNRKTFYTQDPVVGYTDYPFLQGNGSNGPLSRL 396
.......340.......350.......360.......370.......380.......390.......400....

1. Einleitung 6
Während die ersten drei Enzyme des Oktadekanoidwegs im Plastiden lokalisiert
sind, ist OPR3 im Peroxisom (Siehe Abbildung 6). Ein Blick auf das C-terminale
Ende der OPR3-Sequenz zeigt, dass hier ein SRL-Tag vorhanden ist (Abbildung 5),
das für die Lokalisation im Peroxisom verantwortlich ist (Strassner et al. 2002). Hier
findet auch die β−Oxidation statt. OPDA muss also von den Plastiden ins
Peroxisom gelangen, um dort umgesetzt werden zu können (Stintzi und Browse
2000). Abbildung 6 verdeutlicht die Lokalisation der Proteine, die im
Oktadekanoidweg integriert sind und weitere Synthesewege, an denen
Jasmonsäure (JA) beteiligt ist.
Abb. 6: Lokalisation der Proteine des Oktadekanoidwegs, Jasmonsäurebiosynthese
und Metabolismus.

1. Einleitung 7
Der Umsatz von cis(+)OPDA zu cis(+)OPC-8:0 ist die Reduktion der 10,11-
Doppelbindung im Cyclopentenonring (siehe auch Abbildung 7). Hierbei wird der
FMN Cofaktor oxidiert, der von NADPH reduziert wird und somit wieder zur
Verfügung steht. Die Reduktion durch OPR3 ist also NADPH-abhängig.
OPR3O
COOH
O
COOH11
10
cis (+) OPDA OPC-8:0cis(+)cis(+)FMNH2 FMN
NADPH + H+NADP+
OPR3O
COOH
O
COOH
O
COOH11
10
O
COOH11
10
cis (+) OPDAcis (+) OPDA OPC-8:0cis(+)cis(+)OPC-8:0cis(+)cis(+)FMNH2 FMN
NADPH + H+NADP+
Abb. 7: Umsatz von cis(+)OPDA durch NADPH-abhängiges OPR3 zu cis(+)OPC-
8:0
1.3 Die opr3-Mutante
OPR3 kommt eine Aufgabe in der männlichen Fertilität zu. Für die männliche
Fertilität in Arabidopsis thaliana ist das Vorhandensein von Jasmonsäure unbedingt
nötig. In der opr3-Mutante sind zeitlichen Abläufe der Pollenfreisetzung gestört, der
Pollen wird zu spät freigesetzt. Zu diesem Zeitpunkt sind die Stigmapapillen nicht
mehr empfänglich für eine Bestäubung (Sanders et al., 1999, Sanders et al., 2000).
Allerdings kann bei opr3-Pflanzen durch Applikation von exogener Jasmonsäure
die Fertilität wiederhergestellt werden. Durch eine exogene Applikation von OPDA
kann dies nicht erreicht werden. OPDA kann Jasmonsäure als chemisches Signal
für die Antheren- und Pollenreifung nicht ersetzen (Stintzi and Browse 2000). Alle
Mutationen im Oktadekanoidweg, die die Bildung von Jasmonsäure und den darauf
folgenden Signalweg (Xie et al. 1998) unterbrechen, haben eine männliche Sterilität
zur Folge. Die in dieser Arbeit verwendeten Mutanten sind durch T-DNA Insertion
hergestellt worden.
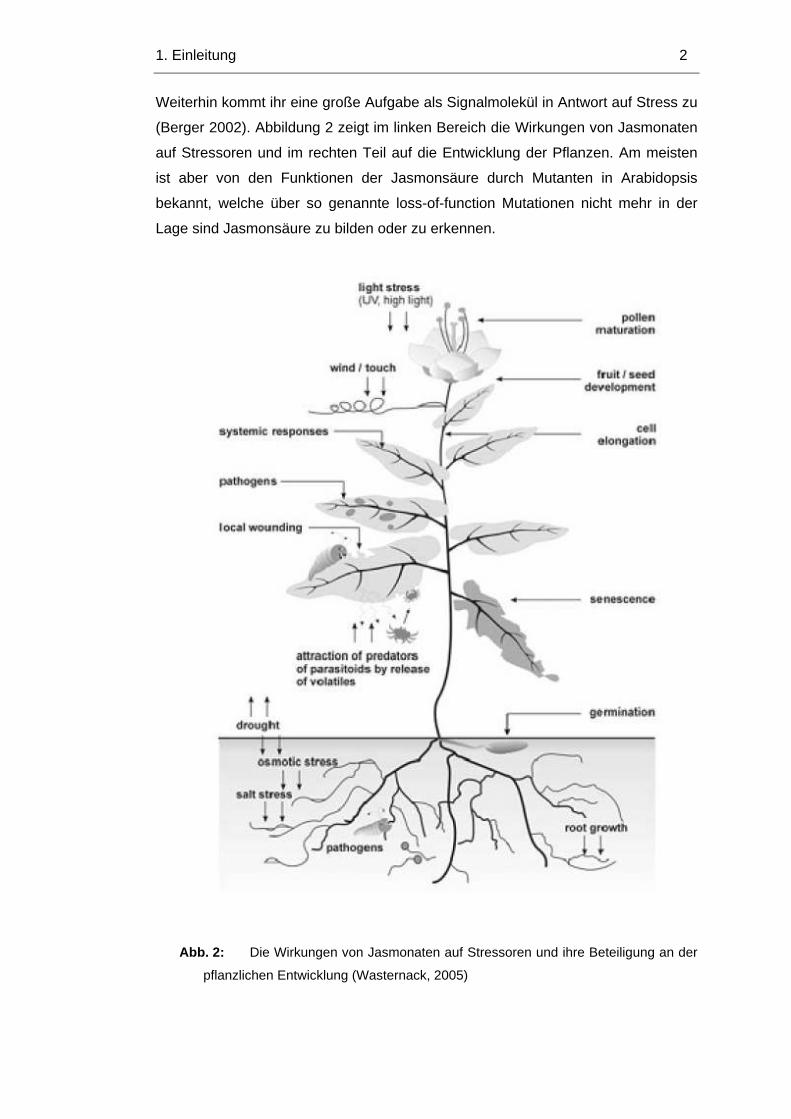
1. Einleitung 8
1.4 Strukturvergleich von OPR3 und OPR1
Um die Stereoselektivität von OPR1 besser zu verstehen, soll hier genauer auf die
Strukturen von OPR3 und OPR1 eingegangen werden. OPR3 und OPR1 haben
eine 53%ige Sequenzidentität (Abbildung 8 und Stintzi und Browse, 2000), aber
ihre Strukturen sind tatsächlich über weite Strecken identisch.
Abb. 8: 3D-Struktur von OPR1.
OPR1 (Abbildung 8) und OPR3 bilden ein 8 strängiges αβ-Fass (Breithaupt et al.,
2000 und 2006). Die Autoren konnten zeigen, dass 8 parallel verlaufende
β−Faltblätter ein Fass bilden, das von α−Helices umgeben ist. Das Aktive Zentrum
ist in allen OPR Isoformen hoch konserviert. OPR1 und OPR3 unterscheiden sich
allerdings in den β−Loops, die aus dem Protein herausragen. Exponiert hierbei ist
in OPR1 der β3-Loop (siehe Abbildung 9). Wie schon eingangs erwähnt, ist nur
OPR3 in der Lage cis(+)OPDA umzusetzen, wodurch eine aktive Form der
Jasmonsäure entsteht. Allerdings kann es in vitro auch cis(-)OPDA umsetzen. Das
nahe verwandte Protein OPR1 hingegen kann nur cis(-)OPDA umsetzen. Dies liegt
an einer strukturellen Besonderheit von OPR1. Hier ragt der β3-Loop weiter heraus
als in OPR3 und bildet somit eine Art Deckel über dem aktiven Zentrum. Dieser ist
dafür verantwortlich, dass nur cis(-)OPDA das aktive Zentrum von OPR1 erreichen
kann, nicht aber cis(+)OPDA. In OPR3 ist dieser Loop etwas kleiner, wodurch die
Höhlung um das aktive Zentrum größer wird, und sowohl cis(+) als auch cis(-
)OPDA das aktive Zentrum erreichen und umgesetzt werden können.

1. Einleitung 9
Abb. 9: Strukturvergleich von OPR3 (gelb) und OPR1(blau)
Exponierte Aminosäuren am β3-Loop sind die Tyrosine an den Positionen 246 und
78 (Breithaupt et al., 2006 und Abbildung 10). Tauscht man an OPR3 Proteinen die
korrespondierenden Aminosäuren Histidin an Position 244 und Phenylalanin an
Position 74 gegen die Tyrosine von OPR1 aus, so ist OPR3 in vitro nur noch mit
einer sehr geringen Effizient in der Lage cis(+)OPDA umzusetzen (F. Schaller und
T.Clausen, unveröffentlicht).
Abb. 10: Detaildarstellung des Aktiven Zentrums von OPR1 mit gebundenem
Substrat
Ein weiterer Unterschied zwischen den Strukturen ist der β6-Loop. In OPR1 ist
diese Struktur sehr flexibel, was dazu führt, dass sie in der Röntgenkristallstruktur
nicht visualisierbar ist (Malone et al., 2005, Breithaupt et al, 2006 und Abbildung 9,
blau). In OPR3 hingegen zeigt sie sich der β6-Loop als unflexible fingerartige
Struktur (Breithaupt et al., 2006 und Abbildung 9, gelb).

1. Einleitung 10
Röntgenkristallstrukturen haben gezeigt, dass OPR3 als Homodimer kristallisiert
(Breithaupt et al., 2006 und Abbildung 11). Hierbei lagert sich die fingerartige
Struktur des β6-Loop eines OPR3 Protomers in das aktive Zentrum eines anderen
Protomers. Genauso lagert sich der β6-Loop des zweiten Protomers spiegelbildlich
in das Aktive Zentrum des ersten. OPR3 inhibiert sich also durch Dimerisierung
selbst (Breithaupt et al., 2006).
Abb. 11: Röntgenkristallstruktur von OPR3. Der Fingerartige β6-Loop des einen
Protomers (rot) interagiert in der substratbindenden Tasche des anderen Protomers
(gelb) und umgekehrt. Nach Breithaupt et al., 2006
Die Dimerisierung wird stabilisiert durch exponierte Aminosäuren. Direkt an der
Spitze des β6-Loop befindet sich ein Glutamat (Position 291), das direkt oberhalb
des Cofaktors FMN in der Substrat bindenden Tasche des aktiven Zentrums bindet
(siehe Abbildung 12). Hierdurch wird das Aktive Zentrum blockiert. Eine
Punktmutation dieser Aminosäure hat gezeigt, dass ein Ersetzen von Glutamat
durch Lysin dazu führt, dass OPR3 als Monomer kristallisiert. Außerdem konnte
gezeigt werden, dass das mutierte OPR3 Protein E291K einen sechsfach höheren
Substratumsatz hat als das nicht-mutierte OPR3. Eine Inhibierung der Aktivität
durch Homodimerisierung konnte somit nachgewiesen werden (Breithaupt et al.,
2006).

1. Einleitung 11
Abb. 12: Aktives Zentrum von dimerisiertem OPR3. In gelb und dunkelrot: jeweils
ein Protomer von OPR3. Markiert (roter Kreis) sind Aminosäuren, die für die
Dimerisierung wichtig erscheinen.
Eine weitere für die Dimerisierung wichtige Aminosäure ist Tyrosin an Position 364.
In der Röntgenkristallstruktur ist sichtbar, dass dieses Tyrosin ein Sulfat über
Wasserstoffbrücken bindet (Siehe Abbildung 12). Hierbei bindet das Sulfat auch an
2 Argininreste des einen Protomers und an einen weiteren Argininrest des anderen
Protomers, wodurch die Dimerisierung stabilisiert wird. Dieses Sulfat imitiert eine
Phosphatrestbindung eines phosphorylierten Tyrosinrestes. Wenn Tyrosin gegen
Phenylalanin ausgetauscht wird, wodurch die an Tyrosin vorhandene
Hydroxylgruppe fehlt, kristallisiert OPR3 als Monomer (Breithaupt et al., 2006). Da
OPR3 jetzt nicht mehr dimerisieren kann, scheint eine dauerhafte Aktivität gegeben
zu sein. Dies wirft die Hypothese auf, ob OPR3 in vivo möglicherweise auch durch
reversible Phosphorylierung reguliert werden kann.
Bereits vorausgegangene Experimente konnten zeigen, dass OPR3 Substrat einer
Tyrosin-spezifischen Protein-Kinase ist (Schaller A., unveröffentlicht). Western Blot
Analysen mit einem Phosphotyrosin-spezifischen monoclonalen Antikörper (Anti
PT-66) haben bewiesen, dass OPR3 effizient Phosphoryliert wird, wenn es mit
einer Tyrosin-Kinase in E.coli coexprimiert wird (Abbildung 13 und Stratagene,
TKB1). Es ist bekannt, dass bei einer Verwundung einer Pflanze der Spiegel an
Jasmonsäure ansteigt, ebenso steigt der Transkriptspiegel der Gene, die für
Proteinen kodieren, die in der Biosynthese von Jasmonsäure beteiligt sind (Stintzi
et al., 2001).
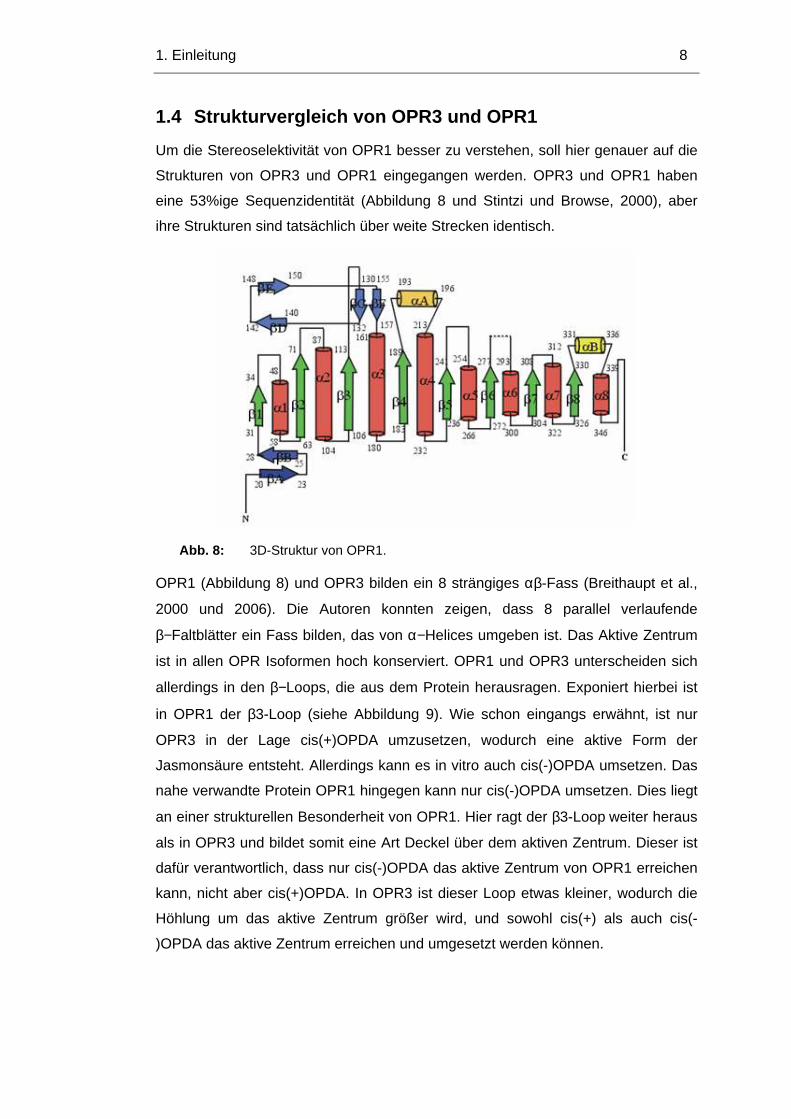
1. Einleitung 12
Dennoch ist die Kinetik der Transkriptinduktion nicht schnell genug, um für die
schnelle Akkumulation der Jasmonsäure nach Verwundung zu genügen. Dies
suggeriert, dass weitere Regulationsmechanismen für den anfänglichen Anstieg
von Jasmonsäure verantwortlich sind. Eine Möglichkeit wäre reversible
Phosphorylierung und Dephosphorylierung.
Abb. 13: In vivo Phosphorylierung eines OPR3-Proteins durch eine
Typrosinspezifische Protein Kinase im Expressionsvektor in E.coil. (Schaller,
unveröffentlicht)
1.5 Volatiles – Kommunikation unter Pflanzen
Flüchtige organische Substanzen (engl.: Volatile organic compounds, kurz VOCs)
sind Stoffe, die eine Pflanze gezielt freisetzt und die der Kommunikation dienen –
sei es nun spezifisch und dauerhaft um befruchtende Insekten anzulocken oder ad
hoc um Feinde eines fressenden Insekts herbeizurufen. Sie werden durch den
Herbivorbefall induziert und freigesetzt, aber auch eine mechanische Verwundung
ist zur Induktion ausreichend. Solche tritrophe Interaktionen sind in wenigstens
zwölf Pflanzenfamilien beschrieben worden (Frey et al., 2004).
Bei der Untersuchung der Wundreaktion in Maispflanzen auf Befall von Raupen
konnte festgestellt werden, dass durch in Raupenregurgitat enthaltene Enzyme
Linolensäure mit Glutamat zu Volicitin reagiert. Die Pflanze kann also reine
mechanische Verwundung von der Verwundung durch das Fressen einer Raupe
unterscheiden. Das entstehende Volicitin wird freigesetzt und lockt spezifisch einen
Fressfeind der Raupe an, während in der Pflanze selber der Oktadekanoidweg in
Gang gesetzt wird, um Jasmonsäure zu bilden und die systemische Wundantwort
gestartet wird. (Alborn et al., 1997). Auch Jasmonsäurederivate wie cis-Jasmon
können bei Verwundung der Pflanze freigesetzt werden und als VOCs wirken um
somit natürliche Feinde von biotischen Stressoren anzulocken (Farmer EE, 2003).
130 k
75 k
55 k
45 k
35 k
Coomassie Anti PT-66
OPR3 OPR3-(P) OPR3 OPR3-(P)

1. Einleitung 13
VOCs sind also unter anderem das Resultat einer Coevolution mit Insekten
(Dobson und Bergstrom 2002). Wie Pflanzen über VOCs mit Insekten
„kommunizieren“ legt die Vermutung nahe, dass sie ebenso untereinander
Informationen austauschen (Dicke et al., 2003). Zu den bekannten VOCs gehören:
Mono- und Sesquiterpene wie (E)-β-Ocimene und (S)-Linalool, Aromaten wie Indol
oder Methylsalicylsäure, und Oxylipine, die so genannten „green leaf volatiles“
(kurz GLVs) wie (Z)-3-Hexenal und (Z)-3-Hexenylacetat.
Green leaf volatiles (GLVs) sind C-6-Aldehyde, Alkohole und ihre Ester, die durch
den Hydroperoxid-Lyase-Weg des Oxylipin-Metabolismus hergestellt werden.
Pflanzen beginnen mit der Bildung von GLVs, nachdem sie mechanische
Verletzung an ihren Blättern erfahren haben, oder biotischem oder abiotischem
Stress ausgesetzt sind. Man vermutet, dass die Bildung von GLVs von einem
Schritt der Lipid-Hydrolyse reguliert wird, die freie Fettsäuren für den Syntheseweg
zur Verfügung stellt. Neuste Studien haben gezeigt, das GLVs eine physiologische
Bedeutung haben, außerdem konnte gezeigt werden, dass sowohl in der
Signalübertragung zwischen Pflanzen als auch in der Kommunikation mit anderen
Organismen, die die Signale aufnehmen können (Pichersky et al., 2006).
Wird eine Mais-Pflanze verletzt, so setzt diese GLVs frei. Unverwundete Mais-
Keimlinge werden daraufhin induziert, schnell Jasmonsäure zu produzieren und
Sesquiterpene auszuschütten. Außerdem produzieren Mais-Keimlinge signifikant
mehr Jasmonsäure und flüchtige Sesquiterpene nach mechanischer Verwundung,
wenn sie vorher GLVs benachbarter Pflanzen ausgesetzt waren (Engelberth et al.,
2004).
Bei Nicotiana attenuata (Baldwin et al., 2007) konnte jedoch kein Anstieg des
Jasmonsäurelevels festgestellt werden, nachdem die Pflanzen GLVs verwundeter
Pflanzen ausgesetzt waren. Transkriptom Analysen konnten zeigen, dass der
Spiegel an verschiedenen, der Jasmonsäure im Oktadekanoidweg vorgeschalteten
Proteinen einen erhöhten Transkriptionsspiegel gegenüber Kontrollpflanzen
aufwiesen, insbesondere solche, die in der Abwehrreaktion beteiligt sind wie OPR3
(Paschold et al., 2006). Obwohl also ein Transkript von OPR3 und vorgeschalteten
Proteinen im Oktadekanoidweg vorhanden ist, wird keine Jasmonsäure gebildet.
Eine Inaktivierung von OPR3 oder der vorgeschalteten Proteine, beispielsweise
durch reversible Phosphorylierung, wäre eine Möglichkeit die Bildung von
Jasmonsäure zu verhindern.

1. Einleitung 14
1.6 Ziel dieser Arbeit
Ziel der vorliegenden Arbeit ist eine strukturelle Analyse durch zielgerichtete
Mutationen von OPR3 und OPR1. Beide unterscheiden sich strukturell hauptsächlich
im β3-Loop, der bei OPR1 über das Aktive Zentrum ragt und somit den Zutritt von
cis(+)OPDA erschwert. In OPR3 ist dieser Loop kleiner, so dass cis(+)OPDA das
aktive Zentrum erreichen kann. Es wurde versucht, durch Aminosäureaustausch in
OPR1 die gleiche Substratspezifität wie in OPR3 herzustellen. Hierzu wurden
Aminosäuren des β3-Loops in OPR1 durch die korrelierenden Aminosäuren aus OPR3
ersetzt. Dadurch sollte der Zugang von cis(+)OPDA zum aktiven Zentrum ermöglicht
werden. Opr3-defiziente Arabidopsis thaliana wurden mit den punktmutierten OPR1-
Proteinen transformiert. Wenn durch die Mutationen an OPR1 die gleiche
Substratspezifität wie OPR3 hergestellt werden kann, so ist die männliche Fertilität in
opr3-defizienten Arabidopsis thaliana wieder hergestellt.
Außerdem ist bekannt, dass OPR3 Proteine als Homodimere kristallisieren und sich
dadurch in vitro selbst inhibieren. Bisher ist unklar ob sich die Proteine auch in vivo
durch Homodimerisierung inhibieren und somit eine Regulation von OPR3 stattfindet
und welche Aminosäuren an der Dimerisierung beteiligt sind. Um dies zu untersuchen
wurden opr3-defiziente Arabidopsis thaliana mit Punktmutierten OPR3 cDNAs
komplementiert. Diese enthielten Punktmutationen im Dimerisierungsloop Lβ6, und an
einem Tyrosin in der Nähe des aktiven Zentrums, dessen Hydroxylgruppe zusammen
mit 3 Argininresten eine Phosphorylierung ausbildet um die Dimerisierung zu
stabilisieren. Von beiden punktmutierten Proteinen konnte bereits gezeigt werden,
dass sie in vitro als Monomere kristallisieren. Es sollte untersucht werden, ob in vivo
eine Dimerisierung ebenfalls verhindert wird, somit eine konstitutive Aktivität besteht
und wie diese sich auf die Pflanze auswirkt.
Des Weiteren wurde untersucht ob eine Phosphorylierung von OPR3 in vivo existiert,
und falls denn so, wann sie eintritt. Zu diesem Zweck wurden sowohl Tomaten als
auch Arabidopsis-Pflanzen verwundet und der Spiegel der Phosphorylierungen durch
Western Blot Analysen untersucht.

2. Material und Methoden 15
2. Material und Methoden
2.1 Organismen
2.1.1 Pflanzen
Arabidopsis thaliana Wassilewskija
Arabidopsis thaliana Wassilewskija opr3 TDNA Insert Mutante
Arabidopsis thaliana Wassilewskija opr1 TDNA Insert Mutante
Solanum lycopersicum Castlemart
2.1.2 Bakterien
Escherichia coli OPR3-His
Escherichia coli OPR1-His
Escherichia coli OPR3-His mit TKB1 Vektor
Escherichia coli DH10B (Invitrogen)
Escherichia coli XL-1 blue (Stratagene)
Agrobacterium tumefaciens GV 3101 mit pSoup
2.2 Vektoren und Plasmide
pCR®2.1-Topo® Vektor für schnelle und effiziente Ligation von PCR-
Produkten mit T/A Überhang. (Abb. 14)
pGreen 0229 mit einem Fusionsprotein DsRED-OPR3 unter der Kontrolles des
Promotors von OPR3 (Abb. 15)
pSoup Vektor für Pflanzentransformation via Agrobakterien. Ermöglicht
pGreen-Replikation mit Hilfe des Replikationsproteins RepA

2. Material und Methoden 16
Abb. 14: Karte des pCR®2.1 Topo® Vektor
Abb. 15: Aufbau des Vektors pGreen 0229 OPR3 prom DsRED-OPR3
2130 bp
Sac
I(14
96)
Sac
II(1
487)
Not
I(14
75)
Xba
I(14
68)
Spe
I(14
62)
Bam
HI(
1456
)S
maI
(145
2)P
stI(
1448
)E
coR
I(14
38)
Eco
RV
(143
4)H
indI
II(1
426)
Cla
I(14
21)
Sal
I(14
11)
Xho
I(14
05)
Apa
I(14
00)
Kpn
I(13
94)
BglII
pSa-oriColEI ori
nptI
PvuI
BspHI
BspHI
pGreenII plasmid backbone2495 bp
nos-BarRB LBlacZ
BglII(1956)
BglII(3)
T-DNA (1959)
NotI
SacII SpeI Bam NotI1180 bp
PstI KpnI KpnIT 282
AtOPR3 prom AtOPR3 cDNARFP 7502130 bp
Sac
I(14
96)
Sac
II(1
487)
Not
I(14
75)
Xba
I(14
68)
Spe
I(14
62)
Bam
HI(
1456
)S
maI
(145
2)P
stI(
1448
)E
coR
I(14
38)
Eco
RV
(143
4)H
indI
II(1
426)
Cla
I(14
21)
Sal
I(14
11)
Xho
I(14
05)
Apa
I(14
00)
Kpn
I(13
94)
BglII
pSa-oriColEI ori
nptI
PvuI
BspHI
BspHI
pGreenII plasmid backbone2495 bp
nos-BarRB LBlacZ
Sac
I(14
96)
Sac
II(1
487)
Not
I(14
75)
Xba
I(14
68)
Spe
I(14
62)
Bam
HI(
1456
)S
maI
(145
2)P
stI(
1448
)E
coR
I(14
38)
Eco
RV
(143
4)H
indI
II(1
426)
Cla
I(14
21)
Sal
I(14
11)
Xho
I(14
05)
Apa
I(14
00)
Kpn
I(13
94)
Sac
I(14
96)
Sac
II(1
487)
Not
I(14
75)
Xba
I(14
68)
Spe
I(14
62)
Bam
HI(
1456
)S
maI
(145
2)P
stI(
1448
)E
coR
I(14
38)
Eco
RV
(143
4)H
indI
II(1
426)
Cla
I(14
21)
Sal
I(14
11)
Xho
I(14
05)
Apa
I(14
00)
Kpn
I(13
94)
BglII
pSa-oriColEI ori
nptI
PvuI
BspHI
BspHI
pGreenII plasmid backbone2495 bp
nos-BarRB LBlacZ nos-BarRB LBlacZ
BglII(1956)
BglII(3)
T-DNA (1959)
NotI
SacII SpeI Bam NotI1180 bp
PstI KpnI KpnIT 282
AtOPR3 prom AtOPR3 cDNARFP 750

2. Material und Methoden 17
2.3 Oligonukleotide
Primer wurden von Operon Biotechnologies (Köln) bezogen.
Primer Sequenz (5’ → 3’-Orientierung)
Primer M13 M13 reverse CAG GAA ACA GCT ATG AC
M13 forward GAC CGG CAG CAA AAT G
OPR3
primer
5mutLeOPR3 (34) Spe I
ACT AGT ATG GCG TCT TCA GCT CAA GAT
3mutLeOPR3 (35) Pst I
CTG CAG TCA CAG ACG CGA TAA CGG TCC A
OPR1
primer
5mutLeOPR1 (36)
Spe I
ACT AGT ATG GAA AAT AAA GTC GTT GAA GA
3mutLeOPR1 (37) Pst I
CTG CAG TCA CAG ACG CGA CGA CGG
TGTCATGGTTTCTAGAAATGGA
OPR1_876R (876) ATC GAT GGT ACC CAA CGA TTG GCT TCC
ATG CCT CCA
Mutations-
primer
LeOPR1Y74F-For TAT CTG AGA CTG GCA TAG GGT TTA AAG
ATG TAC CTG GTA TAT GG
LeOPR1Y74F-Rev
CCA TAT ACC AGG ATC TTT AAA CCC TAT
AGT CTC AGA TA
LeOPR1Y244H-For TAT CCC CAT TTG CGC ATC ATA ATG AAG
CAG GGG AC
LeOPR1Y244H-Rev
GTG CCC TGC TTC ATT ATG ATG CGC AAA
TGG GGA TA

2. Material und Methoden 18
ß-Tubulin
(TuB4)
TuB4F TGA GGA AGG AAG CTG AGA AC
TuB4R ATA CAC TCG TCA GCG TTT TC
OPR3
Promotor
1612 GCT AAT GAC CTA TCA ATG TAC
2.4 Enzyme
Calf intestine alkaline Phosphatase (CIAP) Fermentas
Ligase (T4-DNA-Ligase) Fermentas
Polymerase
(T4-DNA-Polymerase)
Fermentas
Proteinase K Fermentas
RNAse A Fermentas
Restriktionsenzyme Fermentas
Taq-DNA-Polymerase PEQLAB,
Erlangen, 5 Units/µl

2. Material und Methoden 19
2.5 Antikörper
2.5.1 Primäre Antikörper
Anti-AtOPR1 polyklonaler Antikörper aus dem Kaninchen
Anti-AtOPR3 polyklonaler Antikörper aus dem Kaninchen
beide Antiseren wurden freundlicherweise
von Dr. Florian Schaller zur Verfügung
gestellt.
Anti-AOS Serum polyklonales Antikörperserum aus dem
Kaninchen
Anti Phosphotyrosin PT-66 monoklonaler Antikörper aus der Maus
Lot:086K4751 (Sigma)
2.5.2 Sekundäre Antikörper
Anti- Rabbit IgG Peroxidasekonjugierter Antikörper aus der
Ziege (Calbiochem)
Anti-Mouse IgG Peroxidasekonjugierter Antikörper aus der
Ziege (Calbiochem)

2. Material und Methoden 20
2.6 Chemikalien und Verbrauchsmaterial
Soweit nicht anders angegeben, wurden Chemikalien von Roth (Karlsruhe) und Merck
(Darmstadt), Medien, Vitamine, Antibiotika und Agar von Duchefa (Haarlem),
Plastikverbrauchsmaterialien wie Pipettenspitzen und Reaktionsgefäße von der Firma
Sarstedt (Nümbrecht) und Petrischalen von Greiner bio-one (Kremsmünster) bezogen.
Acrylamid-Rotiphorese Gel 40 Roth
6-Aminohexansäure Merck
AmmoniumPer(oxid)Sulfat (APS) Roth
Bradford Reagenz (Protein Assay) Roth
BugBuster Novagen
Cetyltrimethylammoniumbromid (CTAB) Roth
Dimethylsulfoxid (DMSO) Roth
dNTPs MBI-Fermentas
Ethylendiamintetraacetat (EDTA) Fluka AG
Ethidiumbromid Roth
Formaldehyd Roth
GeneRuler™1kb DNA bzw100bp DNA Ladder. Fermentas
Kanamycin A Monosulfat Duchefa
LB Broth Low Salt Duchefa
ß-Mercaptoethanol Serva
Methyljasmonat Serva
Miracloth Calbiochem
Natriumdodecylsulfat (SDS) Fluka AG
Natriumthiosulfat Roth
Natiumorthovanadate Baack
PageRulerTM Prestained Protein Ladder Fermentas
Rifampicin Duchefa
Röntgenfixierer Kodak
Röntgenentwickler Kodak
Silwet L-77 Lehle Seeds, Round Rock
Tetramethylethylendiamin (TEMED) Roth
Tetracyclin Duchefa
Tris(hydroxymethyl)-aminomethan (Tris) Roth
Triton-X 100 Roth
Tween 20 Fluka AG
Tricine Roth
Xylen Cyanol Serva

2. Material und Methoden 21
2.7 Laborgeräte
Blotting-Apparatur LKB-Bromma/Pharmacia
Computerprogramme CaryWinUV (Varian Instruments, Walnut Creek)
Digitalkamera Nikon, Coolpix995
Digitalkamera für Mikroskop
und Stereomikroskop
Spot RT, Visitron Systems
Elektroporator EQUIBIO, Easyjec T
Elektrophorese-Kammer
für Agarosegele
EasyCast Electrophoresis Systems Owl Scientific, Inc.
Elektrophorese-Kammer
für Proteingele
BioRad
Fraktionatoren H.Hölzel (Hörlkofen bei Erding)
Gradientenmischer für Lineare Gradienten
H.Hölzel (Hörlkofen bei Erding)
PCR-Maschine BioRad MyCycler
Rotoren Eppendorf F 45-30-11
Eppendorf A 4-81
HB-5, Sorvall
F14, Sorvall
SW 27, Sorvall
AH 650, Sorvall
Schüttler Multitron HT Infors
Stereomikroskope Zeiss SV11
Hund SM33
UV-Vis Spektrophotometer Carry100Bio, Varian
Eppendorf bioPhotometer 6131
Zentrifugen Ultracentrifuge OTD-COMBI, Sorvall
5402, Eppendorf
Microspin 24, Sorvall
5810R, Eppendorf
RC 5B Plus, Sorvall

2. Material und Methoden 22
2.8 Antibiotika und Medien
2.8.1 Stammlösungen Antibiotika
Ampicillin 100 mg/ml in ddH2O,
Endkonzentration 100 µg/ml
Kanamycin 50 mg/ml in ddH2O,
Endkonzentration 50 µg/ml
Tetracyclin 12,5 mg/ml in 70 % EtOH
Endkonzentration 12,5 µg/ml
Alle Antikörper wurden sterilfiltriert und bei -20°C gelagert.
2.8.2 Medien
LB-Medium 2 % LB Broth (w/v)
für LB-Platten zusätzlich 1,2 % (w/v) Agar
SOC- Medium 2 % Bacto Tryptone (w/v)
0,5 % Bacto Yeast Extrakt (w/v)
10 mM NaCl
2,5 mM KCl
10 mM MgCl2
10 mM MgSO4
20 mM Glucose
pH 6.8 - 7.0 mit NaOH

2. Material und Methoden 23
2.9 Allgemeine Methoden mit Pflanzen
2.9.1 Anzucht von Tomaten
Tomatensamen wurden bei 70°C über Nacht (ÜN) inkubi ert. Anschließend wurden sie
mit 70 % EtOH gewaschen und für 3 h in 10 % Trinatriumphosphat inkubiert. Nach
mehrmaligem Waschen (5 mal 10 min) der Samen mit ddH2O wurden sie im
Gewächshaus bei einer Photoperiode von 12 h angezogen.
2.9.2 Anzucht von Arabidopsis thaliana
Trockene Samen wurden zur Behandlung gegen Thripsen für 2-3 Stunden bei -70°C
gelagert, bevor sie für 48-72 Stunden in 0,1% Agar bei 4°C stratifiziert wurden und bei
einer Photoperiode von 12 h angezogen wurden.
2.9.3 Transformation von Arabidopsis thaliana mit der „floral dip“
Methode
Saccharose-Lösung 5 % Saccharose
0,03 % Silwet L-77
Jasmonsäure zum Besprühen 0,03 % Methyljasmonsäure
0,1 % Tween 20
Basta 35 mg/l
5 l pro Pflanzkasten
4 ml LB-Flüssigkultur eines Agrobaterium tumefaciens Stammes, der das zu
transformierende Konstrukt enthielt, wurde 2 Tage bei 28 °C im Wasserbadschüttler
angezogen und anschließend für 2 weitere Tage mit einem tausendstel Volumen in
400 ml LB-Medium angeimpft.

2. Material und Methoden 24
Bei einer OD600 von 1 wurde die Kultur bei 3400 x g für 30 min abzentrifugiert, der
Überstand abgenommen und das Pellet in 300 ml Saccharoselösung aufgenommen.
Der primäre Sproß der zu transformierenden opr3-Arabidopsis-Pflanzen wurde
abgeschnitten um die Bildung von Sekundärsprossen zu fördern. Sobald diese einen
Blütenstand von 2-10 cm erreicht hatten wurde die Transformation durchgeführt.
Hierzu wurden alle Blühorgane für einige Sekunden in die Bakteriensuspension
getaucht. Um eine Transformation der noch nicht blühenden Sekundärsprosse zu
ermöglichen, wurden einige Tropfen der Bakteriensuspension auf die Rosette getropft.
Die transformierten Pflanzen wurden gut gewässert und für 2 Tage mit einer
Plastikhaube luftdicht verpackt. Die hohe Luftfeuchtigkeit erhöht die Transformations-
rate.
Im Falle einer opr3-Mutanten Pflanze, die männlich steril ist, wurden die
transformierten Pflanzen wurden alle 2 Tage mit Jasmonsäurelösung besprüht um die
Fertilität der Pollen wieder herzustellen. Nach der Fruchtreife wurden alle Samen
geerntet und auf Basta getränkter Erde ausgelegt. Samen, die das Konstrukt
enthielten besaßen eine Resistenz gegen Basta und konnten wachsen. Sobald die
Keimlinge 2 bis 4 Blätter besaßen wurden sie auf normale Erde umgesetzt.
2.9.4 Pollenkeimungstest
Keimungsmedium 17% Saccharose
2 mM CaCl2
1,65 mM H3BO4
pH 7.0 mit NaOH
0,6% Agar
In 90 mm Petrischalen gießen
und erkalten lassen
Von je 6 Blüten im Stadium 13 (Bowman et al., 1991) einer Pflanze wurden, bis auf die
Antheren alle Blütenorgane entfernt. Die Pollen wurden durch Tupfen auf dem
Pollenkeimungsmedium verteilt und für 24 Stunden im Dunkeln gelassen. Ca 300
Pollen jeder Blüte wurden unter dem Mikroskop gezählt auf Keimung untersucht.

2. Material und Methoden 25
2.10 Molekularbiologische Techniken mit DNA
2.10.1 Polymerase-Ketten-Reaktion (PCR)
5x PCR Puffer 15 mM Magnesium Chlorid
100 mM Ammoniumsulfat
0,08% (v/v)Triton-X 100
20% (v/v) Dimethylsulfoxid (DMSO)
50 mM Tris/HCl pH 8.3
0,4% (v/v) Tween® 20
dNTPs 10 mM dATP, dGTP, dCTP, dTTP
Taq 5 Units / µl
Die Standard-PCR wurde in einem Volumen von 25 µl durchgeführt. Bestandteile
waren je: 5 µl 5x PCR-Puffer, 0,5 µl 10 mM dNTP’s, 1 µl thermostabile Taq-DNA-
Polymerase und je 0,5 µl 10 µM Forward- bzw Reverse-Primer. Die Menge an
Template-DNA variierte je nach Ausgangsmaterial. Von Plasmid-DNA wurden 1 µl
einer 1:10 Verdünnung, bei genomischer DNA 1 µl unverdünnt verwendet.
Je nach Primerkombination wurden verschiedene Annealingtemperaturen gewählt. Die
Elongationszeit wurde entsprechend der Länge des zu amplifizierenden Produktes
gewählt. Die DNA-Polymerase amplifiziert etwa 1 kb/min.
Die verschiedenen Bedingungen sind in Abb. 16 und Tab. 1 dargestellt.

2. Material und Methoden 26
95°C 95°C2 min 30 sec 72°C 72°C
xx min 7 min
xx °C40 sec
10°C1x 25-35x 1x ∞
Abb. 16: Temperatur und Dauer der einzelnen Schritte einer PCR
Tab. 1: Verwendete Annealingtemperaturen und Elongationszeiten in Abhängigkeit der
Spezifischen Primerkombinationen
Primerkombination Annealingtemperatur Elongationszeit Zyklen
OPR3 komplett 34/35 58°C 75 sec 30
OPR1 komplett 36/37 58°C 75 sec 30
Tubulin
(Ladekontrolle)
TuB4F/TuB4R 50°C 40 sec 35
Nachweis der OPR1 Konstrukte
OPR1+SRL 1612/OPR1_876R 58°C 120 sec 35
OPR1Y74F 1612/Y74Frev 58°C 120 sec 35
OPR1Y244H 1612/Y244Hrev 58°C 120 sec 35
Nachweis der OPR3 Konstrukte
OPR3 1612/ 35 58°C 120 sec 35
E291K 1612/ 35 58°C 120 sec 35
Y364F 1612/ 35 58°C 120 sec 35
PCR Produkte wurden vor der weiteren Verwendung mit dem QIAquick® PCR
Purification Kit (Qiagen) aufgereinigt.

2. Material und Methoden 27
2.10.2 Auftrennung und Sichtbarmachen von DNA durch
Gelelektrophorese
TAE-Puffer 40 mM Tris,
20 mM Essigsäure,
1 mM EDTA
14 µg/l Ethidiumbromid
pH 8.0 mit HCl
DNA-Ladepuffer
Bromphenolblau (10-fach)
50 % Glycerin,
1 mM EDTA,
1 Spatelspitze Bromphenolblau
für 20 min autoklaviert, und bei RT
aufbewahrt
DNA-Ladepuffer
Xylen-Cyanol (10-fach)
50 % Glycerin
1 % Xylen Cyanol
für 20 min autoklaviert und bei RT
aufbewahrt
Größenstandard GeneRuler™ 1 kb bzw.100 bp DNA
Ladder (Fermentas)
1% TAE Agarosegel 1 g Agarose elektrophoresis grade
(Duchefa) auf 100 ml TAE Puffer
Um DNA-Fragmente ihrer Größe nach aufzutrennen wurden sie nach Zugabe von 1/10
Volumen eines 10x Ladepuffers auf 1%ige TAE-Agarosegele geladen. Für Fragmente
einer Größe von unter 500 bp wurde Xylen-Cyanol Puffer, für größere Fragmente
wurde Bromphenolblau verwendet. Die Auftrennung erfolgte bei 80-120 V für 30-60
Minuten. Das Gel wurde anschließend unter UV-Licht betrachtet und fotografisch
dokumentiert.

2. Material und Methoden 28
2.10.3 Restriktionsverdau
Restriktionsverdaue wurden meistens in einem Volumen von 20 µl durchgeführt. Der
Ansatz beinhaltete 1 µg Plasmid-DNA, 1/10 bis 1/5 des Endvolumens an
entsprechendem Restriktionsenzympuffer, und 0,5 µl Restriktionsenzym (10 U/µl).
Nach einer 2 - 2,5stündigen Inkubation bei 37°C wu rden der komplette Ansatz zur
Kontrolle des Verdaus gelelektrophoretisch aufgetrennt.
2.10.4 DNA-Gelextraktion
In TAE-Agarosegelen aufgetrennte DNA-Fragment wurden aus dem Gel
herausgeschnitten und mit dem QIAquick® Gel Extraktions Kit (Qiagen) nach den
Anweisungen des Herstellers extrahiert.
2.10.5 Klonierung
2.10.5.1 Klonierung in pCR 2.1 TOPO Vektor (Invitro gen)
Aufgereinigte DNA-Fragmente wurden zuerst in den pCR2.1-Topo®-Vektor (siehe
http://www.invitrogen.com/content/sfs/manuals/topota_man.pdf) nach dem Protokoll
des Herstellers (Invitrogen) kloniert.
2.10.5.2 Klonierung in pGreen 0229
Für eine Ligation von Vektor und Insert müssen diese mit Restriktionsenzymen so
geschnitten worden sein, dass die Überhänge sich aneinander lagern können. Eine
Dephosphorylierung des Vektors mittels Alkalischer Phosphatase verhindert eine
Selbstligation. Dafür wurden zu dem Plasmid 1 µl CIAP zugegeben und der Ansatz für
1 h bei 37°C inkubiert. Die Inaktivierung der Phosp hatase erfolgte für 10 min bei 75°C.
Für eine Ligation wurde ein molares Vektor-Insert Verhältnis von 1:5 gewählt. Die
Ligationsansätze besaßen ein Endvolumen von 15-20 µl und enthielten neben Vektor
(meinst 1 µl) und Insert, (1-6 µl) 1x Ligase-Puffer mit ATP und 1 U T4-DNA-Ligase. Die
Ligation wurde entweder in der PCR-Maschine (8 h bei 16°C, 60 min bei 30°C, 10 min
bei 65°C) oder bei Raumtemperatur (RT; Inkubation f ür 2 h) durchgeführt

2. Material und Methoden 29
2.10.6 Transformation durch Elektroporation
Zu 40 µl elektrokompetenten Bakterien E.coli DH10B (Invitrogen) oder XL-1 blue
(Stratagene), die auf Eis aufgetaut wurden, wurde 1 µl des Ligationsansatzes
pipettiert, vorsichtig gemischt und eine Minute auf Eis ruhen gelassen. Der Ansatz
wurde in vorgekühlte Elektroporationsküvetten mit 1 mm Elektrodenabstand überführt.
Die Elektroporation wurde mit den Parametern 2500 V, 15 µF, 335 Ω durchgeführt,
sofort 960 µl SOC Medium zugefügt und für 1h bei 37°C unter Schütteln inkubiert.
Anschließend wurden die Bakterien auf einem Selektionsmedium ausplattiert und bei
37°C über Nacht inkubiert. Die Inkubationstemperatu r bezieht sich auf die
Transformation von E. coli XL1-Blue bzw E. coli DH10b. Bei der Transformation von
Agrobacterium tumefaciens erfolgten alle Inkubationen bei 28°C.
2.10.7 DNA Extraktion
2.10.7.1 Plasmid DNA Extraktion aus E.coli
Lösung I 25 mM Tris pH 7.5 mit HCl
50 mM Glucose
50 mM EDTA
Lösung II 0,4 N NaOH
4 % SDS
getrennt ansetzen und kurz vor
Gebrauch 1:1 mischen
Lösung III 5 M Kaliumacetat
pH 5.5 mit Eisessig
Lösung IV 3 M Natriumacetat
pH 5.2 mit Eisessig

2. Material und Methoden 30
TE Puffer 10 mM Tris, pH 8.0 mit HCl
1mM EDTA
Pankreatische RNase 10 mg/ml in TE
10 min bei 85°C inkubieren zum
Inaktivieren von DNasen
TE Puffer gesättigtes Phenol,
Chloroform,
Phenol/Chloroform 1:1
Transgene Bakterien wurden in ca. 4 ml LB Medium mit entsprechendem
Selektionsantibiotikum unter Schütteln bei 37°C übe r Nacht angezogen. 1,5 ml der
Kultur wurden in ein Eppendorf gegeben und abzentrifugiert (20.000 x g, 2 min, RT).
Der Überstand wurde vollständig entfernt und das erhaltene Bakterienpellet in 100 µl
Lösung I resuspensiert. Anschließend wurden 150 µl Lösung II zugegeben, vorsichtig
gemischt und 5 min auf Eis stehen gelassen. Nach Zugabe von 150 µl Lösung III
wurde erneut gut gemischt und für weitere 5 min auf Eis inkubiert. Das Gemisch wurde
5 min bei 4°C und 20.000g abzentrifugiert. Der erha ltene Überstand (400 µl) wurde
vorsichtig abpipettiert und die DNA mit dem doppelten Volumen (800 µl) eiskalten
Ethanol für 15-30 min auf Eis gefällt. Nachdem für 15 min bei 4°C und 20.000 g
zentrifugiert wurde, wurde der Überstand vollständig verworfen und das Pellet in 200
µl TE Puffer gelöst. Zum Verdau der RNA wurde außerdem 1 µl pankreatische RNase
zugesetzt und bei 37°C für 30 min inkubiert. Die Re inigung der Plasmid-DNA erfolgte
durch Ausschütteln mit 200 µl Phenol/Chloroform und nochmals mit 200 µl Chloroform,
wobei nach beiden Extraktionen für jeweils 2 min zentrifugiert wurde. Hierbei wurde
nur die Oberphase in ein neues Eppendorfgefäß überführt wurde. Es folgte eine
erneute Fällung mit dem 2,5 fachen Volumen eiskalten Ethanols (550µl) und 20µl
Natriumacetat (Lösung IV) auf Eis für 15 min. Nach 15-minütiger Zentrifugation bei 4°C
und 20.000 g wurde der Überstand vollständig verworfen, das Pellet mit 70%
eiskaltem Ethanol (500 µl) gewaschen und bei 37°C g etrocknet. Abschließend wurde
das Pellet in 30 µl TE Puffer gelöst und bei -20°C aufbewahrt.

2. Material und Methoden 31
2.10.7.2 Plasmid DNA Extraktion aus Agrobacterium tumefaciens
Die Plasmidextraktion aus Agrobakterien zur Überprüfung des Konstruktes wurde wie
die oben beschriebene Plasmidextraktion aus E. coli durchgeführt, allerdings nur bis
zur ersten ethanolischen Fällung. Das erhaltene Pellet wurde in 30 µl TE-Puffer gelöst
und konnte mit Restiktionsenzymen verdaut werden.
2.10.7.3 Isolierung genomischer DNA aus Arabidopsis thaliana
DEX-Puffer 0,14 M Sorbitol
0,22 M Tris pH 8.0 mit HCl
0,222 M EDTA
0,8 M NaCl
0,8% CTAB
1% Sarcosine (w/v)
Autoklavieren, bei RT aufbewahren.
Vor Gebrauch 20 µl ß-Mercaptoethanol
pro ml frisch zugeben.
Bei -70°C gelagerte Blattproben wurden schnell zerr ieben, mit 100 µl DEX-Puffer
versetzt und bis zum tauen des Materiales weiter zerrieben. Es wurden 100 µl
Chloroform zugegeben und nach zehnsekündigem Mischen für 30 Minuten bei 65°C
inkubiert. Nach einer Zentrifugation bei 16 000 x g für fünf Minuten (RT) wurde der
Überstand in ein neues Eppendorf-Gefäß überführt und die DNA mit 100 µl
Isopropanol gefällt. Nach einer weiteren 5-minütigen Zentrifugation bei 16 000 x g
wurde das Pellet mit 70%igem Ethanol gewaschen, getrocknet und in 30 µl TE-Puffer
resuspendiert. Die Lagerung der DNA erfolgt bei –20°C.

2. Material und Methoden 32
2.10.8 Zielgerichtete Mutagenese
Die Ortspezifische Mutagenese dient der gezielten Mutation, Deletion oder Insertion
einzelner Basen eines Genes. Dies erfolgt während einer PCR-Reaktion mit Hilfe
spezifischer Primer, die an die Region um die zu mutierenden Basen binden, jedoch
an der Stelle der einzuführenden Mutation die geänderte Baseabfolge enthalten. Die
Reaktionen wurden mit dem Stratagen QuikChange® Site-directed Mutagenesis Kit
durchgeführt. Abbildung 17 verdeutlicht die einzelnen Schritte der Reaktion.
Als DNA Matrize dient hierbei keine genomische DNA, sondern zirkuläre Plasmid-
DNA, welches das zu mutierende Gen enthält. Laut Protokoll sollten hierfür zwischen
10 und 50 ng eingesetzt werden, die effizientere Mutagenese fand aber mit 50 ng statt,
daher wurden alle folgenden Reaktion mit 50 ng durchgeführt.. Das gesamte Plasmid
wurde amplifiziert. Hierfür wurde eine „proof-reading“ DNA Polymerase verwendet
(PfuTurbo DNA Polymerase). Um die Matrize später von den neuen, mutierten
Plasmiden zu unterscheiden, muss Plasmid-DNA eines dam+ E. coli Stammes
verwendet werden. Diese enthalten methylierte GATC-Abschnitte im Genom, die vom
Restriktionsenzym Dpn I (10 Units/µl) verdaut werden können. Nach der PCR
Reaktion fehlt den neu amplifizierten Plasmide diese Methylierung und sie werden
nicht verdaut.
Die für die Mutagenese verwendeten Primer müssen verschiedene Parameter erfüllen:
Sie sollten zwischen 25 und 45 Basen lang sein, mit einer Schmelztemperatur über
78°C.
Die gewünschte Mutation sollte sich in der Mitte der Primer befinden, so dass noch
mindestens 10-15 Basen an beiden Seiten des Primers an die Matrize binden
können.
Beide Primer müssen die gewünschte Mutation beinhalten und sich über ihre
gesamte Länge überlagern.

2. Material und Methoden 33
Die Menge der Primer liegt bei 125 ng pro Reaktion. Die Mutagenese PCR läuft über
weniger Zyklen als eine normale PCR je nachdem, ob nur Punktmutationen
vorgenommen werden (12 Zyklen), ob Aminosäuren ausgetauscht werden (16) oder
ob Deletionen oder Insertionen vorgenommen werden (18). Im vorliegenden Fall
wurden Reaktionen mit 15 und 17 Zyklen durchgeführt, wobei eine höhere Zyklenzahl
eine effizientere Mutagenese zur Folge hatte. Mehr Zyklen sind, laut Anleitung von
Stratagen, nicht effizienter, da durch die Amplifikation des gesamten Plasmides keine
geschlossenen Plasmide entstehen, die also auch nicht als Matrize dienen können.
Dementsprechend ist die Vermehrung der Plasmide nicht exponentiell wie bei einer
normalen PCR, sondern linear.
Abb. 17: Prinzip der Zielgerichteten Mutation

2. Material und Methoden 34
Die Reaktionsparameter im Einzelnen:
5 µl 10X PCR Puffer (aus Kit)
10-50 ng Zirkuläre DNA
Je 125 ng 3’ und 5’ Primer
1 µl dNTP Mix (aus Kit, Konzentration Betriebsgeheimnis)
1 µl PfuTurbo DNA Polymerase (2,5 U/µl, aus Kit)
Mit sterilfiltriertem ddH2O auf 50 µl auffüllen
Die Elongationstemperatur betrug nicht, wie bei einer normalen PCR 72°C sondern
nur 68°C, die Elongationszeit wurde entsprechend de r Größe des Plasmides gewählt.
Für ein 5 kb großes Plasmid, wie in den hier durchgeführten Experimenten, benötigt
man eine 5-minütige Elongation bei 68°C.
Direkt nach der PCR wurden die Reaktionsgefäße auf Eis gekühlt und sofort 1 µl Dpn I
Restriktionsenzym zugegeben. Der Verdau wurde für eine Stunde bei 37°C
durchgeführt. 1 µl der nach dem Verdau noch verbleibenden Plasmide wurden mittels
Elektroporation (siehe Abschnitt 2.10.6) in kompetente E.coli XL1-Blue transformiert.
Zur Überprüfung der Mutationen wurden die Plasmide sequenziert.
2.10.9 DNA-Sequenzierung
Die Sequenzierung der DNA Fragmente wurde von der Firma 4baselab in Reutlingen
übernommen. Dazu wurden 50 ng DNA eingereicht.

2. Material und Methoden 35
2.11 Molekularbiologische Techniken mit Proteinen
2.11.1 Gradientenzentrifugation / Zellorganellfrakt ionierung
Tricinepuffer 0,15 M Tricine
1 mM EDTA
10 mM Kaliumchlorid
1 mM Magnesiumchlorid
10 mM Natriumsulfat
0,1 % BSA (w/v)
12% Saccharose
pH 7.5 mit NaOH konz.
Saccharosegradient 30% Saccharose (w/v)
65% Saccharose (w/v)
Für große Mengen Pflanzenmaterial (3-10 g) wurden jeweils 12 ml der 30%igen und
65%igen Saccharoselösungen in die zwei Kammern eines Gradientenmischers
gegeben und in ein 36 ml Zentrifugenröhrchen (Beckmann) geschichtet.
Für Mengen unter 3 g Blattmaterial wurden jeweils 2 ml der Saccharoselösungen
verwendet und der Gradient in ein 5 ml Zentrifugenröhrchen (Sorvall) geschichtet. Die
so gegossenen Gradienten wurden über Nacht bei 4°C gelagert.
Arabidopsis-Pflanzen wurden geerntet, abgewogen und im Verhältnis 1:2 mit
Tricinepuffer und einer Spatelspitze Seesand homogenisiert. Diese wurde durch 4
Lagen Miracloth gefiltert und der Überstand (3-12 ml für 36 ml Gradient, 100-600 µl für
5 ml Gradient) vorsichtig auf einen Gradienten geschichtet. Die Auftrennung der
Zellorganellen erfolgte mit einem Swingout Rotor in der Ultrazentrifuge bei 61 000 x g
für 3 Stunden bei 4 °C unter Vakuum. Der Gradient w urde von unten mit einer Kanüle
angestochen und Fraktionen in Eppendorfgefäßen aufgefangen. Die Größe der
Fraktionen betrug bei den 36 ml Gradienten jeweils 2 ml, bei den 5 ml Gradienten
jeweils 250 µl. Der Saccharosegehalt der einzelnen Fraktionen wurde mit Hilfe eines
Refraktometers bestimmt.

2. Material und Methoden 36
2.11.2 Katalaseaktivität
Kaliumphosphatpuffer 100 mM Kaliumdihydrogenphosphat
100 mM Dikaliumhydrogenphosphat
werden gemischt, bis pH 7.0 erreicht ist.
10 mM H2O2
Die Messung der Katalaseaktivität dient zur Lokalisation von Peroxisomen in
Fraktionen eines Gradienten. Je 20 µl der Fraktionen wurden zu 980 µl
Kaliumphosphatpuffer mit frisch zugesetztem H2O2 gegeben. Der Abbau von H2O2
wurde über 20 min spektrophotometrisch bei 240 nm gemessen. Über die Steigung
der erhaltenen Kurve ließ sich auf die relative Aktivität der Katalase schließen.
2.11.3 Proteinexpression und Aufreinigung
Binding-Puffer 50 mM Natriumdihydrogenphosphat/
Dinatriumhydrogenphosphat
werden gemischt, bis pH 8.0 erreicht ist.
300 mM Natriumchlorid
10 mM Imidazol
4 mM Benzamidin
Lyse-Puffer Bug Buster Mix (Novagene)
mit 1 Spatelspitze DNAseI

2. Material und Methoden 37
Elutionspuffer 50 mM Natriumdihydrogenphosphat
50 mM Dinatriumhydrogenphosphat
wurden gemischt, bis pH 8.0 erreicht war.
300 mM Natriumchlorid
200 mM Imidazol
4 mM Benzamidin
Je eine Kolonie eines Bakterienstammes, der mit einem Expressionsplasmid mit
OPR1, OPR3 bzw TKB1 transformiert war, wurde in einem selektiven LB-Medium als
Vorkultur über Nacht bei 37°C angeimpft.
Die Hauptkulturen (200 ml Medium mit Antibiotika entsprechend der Vorkultur im 1l
Erlenmeyerkolben) wurden mit 2 ml der Vorkultur angeimpft und bis zu einer OD600=1
bei 37°C im Schüttler bei 200 rpm inkubiert. Die Ku ltur wurde auf 30 °C
heruntergekühlt und die Expression durch die Zugabe von 1ml 100mM IPTG induziert.
Nach 4 Stunden wurde ein 300 µl Aliquot genommen und die Zellen abzentrifugiert.
Zu den sedimentierten Bestandteilen der Expressionskulturen wurde die 5-fache
Menge (w/v) BugBuster Mix zugegeben und das Pellet durch Pipettieren
resuspendiert. Die Suspension wurde danach für 20 Minuten auf einem
Eppendorfschüttler (150 rpm) bei Raumtemperatur inkubiert, 20 Minuten bei 16000 x g
und 4 °C zentrifugiert, der Überstand in ein neues Gefäß überführt und sofort bei 4 °C
aufgereinigt. Wurde die Aufreinigung nicht sofort durchgeführt, so wurde der
Aufschluss bei -80 °C gelagert.
Die Aufreinigung der Proteine erfolgte mit Nickel-Nitrilotriessigsäure (Ni-NTA)Agarose
Säulchen. 2,3 ml Ni-NTA-Agarose wurden in eine leere Säule gegeben und der
Lagerungspuffer ablaufen gelassen. Anschließend wurde drei Mal mit je 8 ml Binding
Puffer gewaschen. Auf die Säule wurde der Protein-Extrakt aus dem BugBuster
Aufschluss gegeben und 1 Stunde bei 4 °C im Überkop fschüttler durchmischt.
Nachdem die His-Tags der Proteine an die Säulenmatrix binden konnten, wurde der
Bindungpuffer ablaufen gelassen (Probe Durchfluss). Die Säule wurde anschließend
dreimal mit je 4 ml Bindingpuffer (Wasch1; Wasch2, Wasch3) gewaschen. Das
gebundene Protein wurde in drei Elutionsschritten mit je 750 µl Elutionspuffer (Elution1,
Elution2, Elution3) von der Säule eluiert.
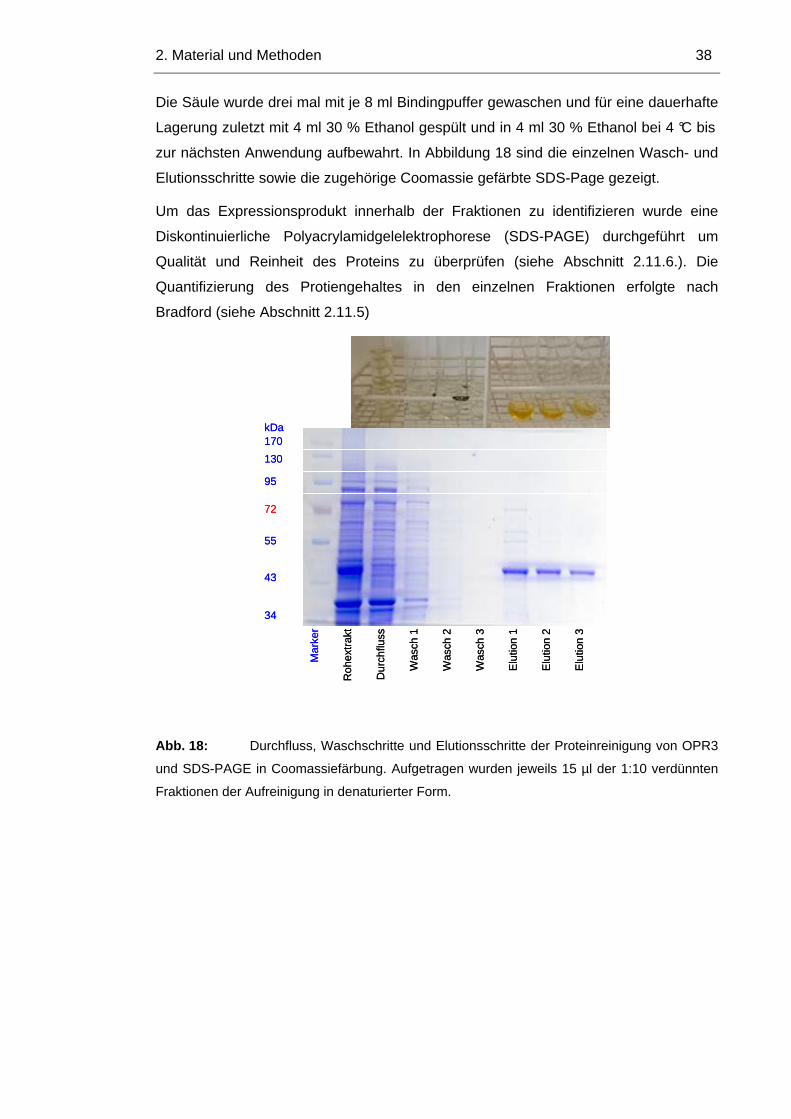
2. Material und Methoden 38
Die Säule wurde drei mal mit je 8 ml Bindingpuffer gewaschen und für eine dauerhafte
Lagerung zuletzt mit 4 ml 30 % Ethanol gespült und in 4 ml 30 % Ethanol bei 4 °C bis
zur nächsten Anwendung aufbewahrt. In Abbildung 18 sind die einzelnen Wasch- und
Elutionsschritte sowie die zugehörige Coomassie gefärbte SDS-Page gezeigt.
Um das Expressionsprodukt innerhalb der Fraktionen zu identifizieren wurde eine
Diskontinuierliche Polyacrylamidgelelektrophorese (SDS-PAGE) durchgeführt um
Qualität und Reinheit des Proteins zu überprüfen (siehe Abschnitt 2.11.6.). Die
Quantifizierung des Protiengehaltes in den einzelnen Fraktionen erfolgte nach
Bradford (siehe Abschnitt 2.11.5) M
arke
r
Roh
extr
akt
Dur
chflu
ss
Was
ch 1
Was
ch 2
Was
ch 3
Elu
tion
1
Elu
tion
2
Elu
tion
3
kDa170
130
95
72
55
43
34
Mar
ker
Roh
extr
akt
Dur
chflu
ss
Was
ch 1
Was
ch 2
Was
ch 3
Elu
tion
1
Elu
tion
2
Elu
tion
3
kDa170
130
95
72
55
43
34
Abb. 18: Durchfluss, Waschschritte und Elutionsschritte der Proteinreinigung von OPR3
und SDS-PAGE in Coomassiefärbung. Aufgetragen wurden jeweils 15 µl der 1:10 verdünnten
Fraktionen der Aufreinigung in denaturierter Form.

2. Material und Methoden 39
2.11.4 Gesamtproteinextraktion
Proteinextraktionspuffer 100mM NaCl
50 mM Tris HCl
0,5 %Triton-X 100
10 mM ß-Mercaptoethanol
10 µl/ml Protease Inhibitor Mix
10 µM Phenylarsineoxid
(in DMSO)
1 mM Natriumorthovanadate
20 mM Natriumfluorid
In flüssigem Stickstoff gefrorenes Blattmaterial wurde in einem vorgekühltem Mörser
zu einem feinen Staub gemörsert. Ca 200 mg des Materials wurden in 300 µl
Proteinextraktionspuffer aufgenommen, kurz gemischt und sofort auf Eis gestellt.
Anschließend wurden die Proben 5 min bei 4 °C und v oller Drehzahl (13.000 x g)
abzentrifugiert und der Überstand in ein neues Eppendorfgefäß überführt.
2.11.5 Bradfordtest
Der Bradfordtest erlaubt eine Bestimmung der Gesamtproteinmenge einer Probe
durch kolorimetische Messung. 800 µl einer 1:100 Verdünnung der Proteinextrakte
wurden zu 200 µl Bradfordreagenz pipettiert, für 15 min bei RT inkubiert und bei 595
nm gemessen. Für eine höhere Genauigkeit wurden jeweils 2 Proben pipettiert und
arithmetisch gemittelt. Die Konzentrationsbestimmung erfolgte anhand einer
Kalibrierungskurve, die mit Hilfe einer Verdünnungsreihe im Bereich von 0,1-20 µg/ml
BSA erstellt wurde.

2. Material und Methoden 40
2.11.6 Diskontinuierliche Polyacrylamidgelelektroph orese
(SDS-PAGE)
Sammelgelpuffer (4-fach) 0,5 M Tris-HCl (pH 6,8)
0,4 % SDS
Trenngelpuffer (4-fach) 1,5 M Tris/HCl (pH 8,8)
0,4 % SDS
APS 10 % (w/v) AmmoniumPer(oxid)Sulfat
Trenngel 10 %ig,
für 2 Gele (1, 5 mm)
4,5 ml Acrylamid-Stammlösung
(Rotiphorese®Gel 40:1 Acyrlamid :
Bisacrylamid)
4,5 ml Trenngelpuffer (4-fach)
9 ml H2O
60 µl 10 % APS
13,5 µl TEMED
Die Komponenten wurden gemischt,
ca. 2/3 hoch in Gelkassetten
gegossen, mit H2O überschichtet und
ca. 30 min polymerisieren gelassen
Sammelgel 4,5 %ig,
für 2 Gele (1,5 mm)
0,75 ml Acrylamid-Stammlösung
1,7 ml Sammelgelpuffern
(4-fach)
4,25 ml H2O
20 µl 10 % APS
12 µl TEMED

2. Material und Methoden 41
Vor dem Auftragen des Sammelgels
wurde das Wasser vollständig
entfernt. Ein Kamm mit 10 oder 15
Taschen wurde eingeschoben und
das Gel weitere 30 min polymerisieren
gelassen.
SDS-Probenpuffer (4-fach) 200 mM Tris/HCl (pH 6,8)
400 mM Dithiotreitol
8 % (w/v) SDS
0,4 % (w/v) Bromphenolblau
40 % Glycerin (v/v)
SDS -Laufpuffer (10-fach) 0,25 M Tris
1,9 M Glycin
1 % (w/v) SDS
Protein Marker PageRuler™ Prestained Protein
Ladder (Fermentas)
Die Proteinextrakte wurden mit Proteinextraktionspuffer auf die kleinste erhaltene
Proteingesamtmenge verdünnt und mit 4x SDS Probenpuffer im Verhältnis 3:1
gemischt und in einem Wasserbad bei 100°C für 5 min denaturiert. Nach der
Denaturierung wurden die Proben kurz abzentrifugiert und sofort auf Eis gestellt. Es
wurden jeweils 40 µg Gesamtprotein der Proben pro Tasche geladen.

2. Material und Methoden 42
2.11.7 Proteinfärbung
2.11.7.1 Coomassie Färbung
Coomassie-Färbelösung 2,5 g/l Coomassie Brilliant Blue R 250
450 ml Methanol
100 ml Essigsäure
mit ddH2O auf 1 l auffüllen
Coomassie-Entfärbelösung 300 ml Methanol
100 ml Essigsäure
mit ddH2O auf 1 l auffüllen
Zur Überprüfung, ob die Proteinmengen der einzelnen Proben einander entsprechen,
wurden jeweils 2 Gele aufgetragen, von denen eines in Coomassie Färbelösung für 1-
2 h unter schütteln angefärbt wurde. Mit Entfärbelösung wurde so lange entfärbt, bis
kein blauer Hintergrund mehr zu sehen war. Das Gel wurde mit ddH2O gewässert und
eingescannt. Sowohl Färben als auch Entfärben wurde durch vorsichtiges Erwärmen
auf 70 °C beschleunigt.
2.11.7.2 Silberfärbung
Sol. I 50 % Methanol
12 % Essigsäure
0,05 % Formaldehyd
Sol. II 50 % Ethanol
Sol. III 200 mg/l Na2S2O3

2. Material und Methoden 43
Sol. IV 2 g/l Silbernitrat
0,75 ml/l Formaldehyd
Sol. V 60 g/l Na2CO3
0,5 ml/l Formaldehyd
4 mg/l Na2S2O3
Sol. VI 50 % Methanol
12 % Essigsäure
Sol. VII 50 % Methanol
Das Polyacrylamid-Gel wurde 20 min mit Sol. I fixiert, 3 x 10 min in Sol. II gewaschen
und mit Sol. III für 1 min vorbehandelt. Nach dem Waschen mit Wasser (3 x 20 sec)
wurde das Gel in Silbernitratlösung (Sol. IV) für 10 min inkubiert und nochmals in
Wasser geschwenkt (2 x 20 sec). Nach Entwicklung des Gels mit Sol. V wurde
abermals mit Wasser gewaschen. Diese Entwicklungsreaktion wurde anschließend
durch eine 5-minütige Inkubation mit Sol. VI abgestoppt. Als Letztes wurde das Gel mit
Sol. VII für 20 min inkubiert und bei 4°C in Wasser aufbewahrt. Zu
Dokumentationszwecken wurde das Gel eingescannt.
2.11.7.3 Ponceau-Färbung
Ponceau-Lösung(10 x) 2 g Ponceau S,
(Sigma-Aldrich, Steinheim)
30 g Sulfosalicylsäure
30 g Trichloressigsäure
mit ddH2O auf 100 ml auffüllen

2. Material und Methoden 44
Auf Nitrocellulose Membran geblottete Proteine konnten durch Färben mit Ponceau
sichtbar gemacht werden. Hierzu wurde die Membran für 10 Minuten in Ponceau-
Lösung gelegt und anschließend ca. 5 Minuten mit Wasser entfärbt, bis die
gewünschten Banden sichtbar waren.
2.11.8 Western Blot
Kathodenlösung 40 mM 6-Aminohexansäure
20 % Methanol
Anodenlösung I 0,3 M Tris-HCl (pH 10,4)
20 % Methanol
Anodenlösung II 25 mM Tris-HCl (pH 10,4)
20 % Methanol
Um bestimmte Proteine spezifisch nachzuweisen zu machen wurden die Proteine, die
auf einer SDS-PAGE aufgetrennt wurden mittels des Semi dry Western blot
Verfahrens auf eine Nitrocellulose Membran transferiert. Hierzu wurden jeweils 6
Blätter Whatmanpapier in Anodenlösung I, je 3 Blätter in Anodenlösung II und je 6
Blätter in Kathodenlösung inkubiert. Die Nitrocellulosemembran (PROTEAN Blotting
membrane) wurde kurz in ddH2O gewässert und dann ebenfalls in Anodenlösung I
inkubiert. Die Graphitplatten der Western Blot Apparatur wurden mindestens 30 min in
ddH2O gewässert und vor dem Aufbau abgetrocknet. Der Aufbau erfolgte
entsprechend Abbildung 19. Der elektrophoretische Transfer wurde bei 100 mA pro
Blot für 90 min durchgeführt.

2. Material und Methoden 45
Abb. 19: Schematischer Aufbau eines Western Blots
2.11.9 Antikörperaufreinigung
Blockierungslösung 1 x TBS (pH 7,4)
0,1 % Tween 20
6 % Milchpulver (für AntiOPR3
und AntiOPR1)
bzw.
3% BSA (für PT-66)
Bindung 1 M Tris/HCl pH 7.5
Elution Glycin/HCl pH 2.8
Stabilisierung Tris/HCl pH 9.0

2. Material und Methoden 46
Um nur bestimmte Proteine auf einer Nitrocellulosemembran sichtbar zu machen,
benötigt man spezifische Antikörper. Diese wurden aus Serum aufgereinigt. Etwa
100 µg gereinigtes Protein wurde auf eine 10%iges Acrylamidgel geladen und bei 120
V 1,5 h aufgetrennt. Das Gel wurde dann, wie bereits im Abschnitt 2.10.8 Western Blot
erörtert auf eine Nitrocellulosemembran (WESTRAN CLEAR SIGNAL) geblottet und
mit Ponceau angefärbt. Ein ca. 0,5 x 2 cm großes Stück der Membran, das nur die
gewünschte Proteinbande enthielt, wurde ausgeschnitten, in kleine Teile zerschnitten
und in ein 2 ml Eppendorfgefäß überführt.
2 ml Blockierungslösung (6% Milch in TBS Tween bei AntiOPR3 und AntiOPR1, 3%
BSA in TBS Tween für PT-66) wurden zugegeben und für mindestens 2,5 h bei RT
oder üN bei 4°C auf einen Überkopfschüttler inkubie rt. Es folgten 3 Waschschritte mit
2 ml TBS Tween für je 3 min. 200 µl Serum und 20 µl 1 M Tris/HCl pH 7.5 wurden
zugegeben und für weitere 1,5 h bei RT unter Schütteln inkubiert. Das Serum wurde
zurückbehalten und mit 0,1% NaN3 stabilisiert. Vor der Elution wurde die
Nitrocellulosemembran erneut 3 mal 3 min mit TBS Tween gewaschen und einmal 3
min mit ddH2O.
Die Elution erfolgte mit 200 µl Glycin/HCl pH 2.8 unter schütteln für 1 min. Das Eluat
wurde sofort in ein frisches Eppendorfgefäß gegeben, in dem bereits 40 µl Tris/HCl pH
9.0 vorgelegt waren. Die Membran wurde erneut mit 200 µl Glycin/HCl eluiert (E1). Die
Elution wurde wiederholt (E2). Zur Stabilisierung der so gereinigten Antikörper wurden
jeweils 1 mg/ml BSA und 0,02 % Timerosal zugegeben.
Da OPR3 und OPR1 sehr ähnlich sind, wurde ein zusätzlicher Reinigungsschritt für
die Seren von AntiOPR3 und AntiOPR1 durchgeführt. Hierbei wurde das Serum von
OPR1 zuerst mit einer Nitrocellulose gereinigt, auf die OPR3 Protein gebunden war.
Diese wurde anschließend verworfen. Ebenso wurde mit dem Serum von AntiOPR3
eine Aufreinigung gegen OPR1 Protein durchgeführt. Nach dem Verwerfen der
Membran wurde mit dem oben beschriebenen Protokoll fort gefahren.

2. Material und Methoden 47
2.11.10 Immunologischer Nachweis der Proteine
20x TBS-Puffer (Tris buffered
Saline)
10 mM Tris-HCl pH7.4
137 mM NaCl
2,68 mM KCl
Blockierungslösung 1 x Tris buffered Saline (TBS)
pH 7.4
0,1 % Tween 20
6 % Milchpulver (für AntiOPR3
und AntiOPR1)
bzw. 3% BSA (für PT-66)
Waschlösung TBS/Tween 1 x TBS pH 7.4
0,1 % Tween 20
ECL-Lösung
(„Enhanced
Chemiluminescence“)
SuperSignal® West Dura
Extended Duration Substrate
(Pierce, Rockford)
Röntgenentwickler
Röntgenfixierer
Nach dem elektrophoretischen Transfer der Proteine auf eine Nitrocellulosemembran
wurde diese für mindestens 2 h bei RT unter Schütteln in Blockierungslösung zur
Blockierung der freien Bindungsstellen inkubiert. Die Detektion der gesuchten Proteine
erfolgte mittels der jeweiligen primären Antikörper, welche in Blockierungslösung
entsprechend verdünnt wurden (1:1000 für AntiOPR3, 1:2000 für AntiOPR1, jeweils in
6% Milchpulver in TBS/Tween und 1:4000 für Anti-Phosphotyrosine PT-66 in 3% BSA
in TBS/Tween) und bei 4°C über Nacht und unter Schü tteln inkubiert wurden.

2. Material und Methoden 48
Es folgten 3 Waschschritte mit TBS/Tween für jeweils 5 Minuten bevor die Membranen
mit dem jeweiligen sekundären Antikörper inkubiert wurden. Dieser wurde ebenfalls in
Blockierungslösung verdünnt (jeweils 1:10000, für AntiOPR3 bzw. AntiOPR1
sekundärer AntiRabbit-Antikörper, für Anti-Phosphotyrosin PT-66 sekundärer
AntiMouse-Antikörper). Zur Detektion des gebundenen zweiten Antikörpers wurde
nach erneutem dreimaligem Waschen mit TBS/Tween und einem Waschschritt mit
TBS 200 µl frisch angesetzte ECL Lösung zugegeben. Das an den sekundären
Antikörper gekoppeltes Enzym HRP (horseradish peroxidase) katalysiert die
Umsetzung von Luminol in seine oxidierte Form, bei der eine Lumineszenz entsteht.
Abbildung 20 verdeutlicht diesen Vorgang. Nachdem die Nitrocellulosemembran
luftblasenfrei in eine Folie eingepackt und in eine Röntgenkassette gelegt wurde,
konnte die Lumineszensreaktion mit Hilfe eines Röntgenfilmes sichtbar gemacht
werden kann.
Abb. 20: Schema der Antikörperdetektion

3. Ergebnisse 49
3. Ergebnisse
3.1 Mutationen der OPR3 cDNA
Es ist bekannt, das OPR3 als Homodimer kristallisiert (Breithaupt et al., 2006). Die
Kristallisierungsanalyse zeigt eine fingerartige Struktur des β6-Loops, der sich nahezu
perfekt passend in das Aktive Zentrum eines anderen Protomers lagert, während der
β6-Loop dieses Protomers wiederum das aktive Zentrum des anderen blockiert. Auf
Grund dessen ist die Interaktion des Substrates cis(+)OPDA mit dem aktiven Zentrum
nicht mehr möglich und eine Katalyse kann nicht mehr stattfinden.
Des Weiteren ist bekannt, dass die Hydroxylgruppe des Tyrosins an Stelle 364 in vitro
eine Sulfatgruppe bindet. Wird Tyrosin durch Phenylalanin ersetzt, findet die
Dimerisierung nicht mehr statt und OPR3 kristallisiert als Monomer. Tyrosinreste
können durch Tyrosinkinasen phosophoryliert werden. Die Sulfatgruppe in der OPR3
Kristallstruktur könnte ein phosohoryliertes Tyrosin imitieren. Daher wurde die
Hypothese aufgestellt, dass die Aktivität von OPR3 durch reversible Phoyphorylierung
an Position 364 reguliert werden kann.
Es wurde daher untersucht, ob eine Mutation der Dimerisierungsschleife zu einer
dauerhaften Aktivität des OPR3 Proteins führt und wie diese sich auf die Pflanze
auswirkt. Hierzu wurde im OPR3 Protein das Glutamat an Stelle 291 durch Lysin
ersetzt (E291K), sowie das Tyrosin an Stelle 364 durch Phenylalanin ersetzt (Y364F).
Kristallstruktur Analysen hatten bereits gezeigt, dass beide OPR3-Mutanten als
Monomere kristallisieren (Breithaupt et al., 2006).
Alle OPR3 cDNAs waren bereits in einem Expressionsvektor vorhanden, mussten
aber mittels PCR mit den Restriktionsenzymschnittstellen für Spe I und Pst I versehen
werden um für eine Agrobakterien-vermittelte Transformation in Arabidopsis in pGreen
kloniert werden zu können. Hierfür wurden die Primer 5mutLeOPR3 (34) und
3mutLeOPR3 (35) verwendet (Siehe Abb. 21). Die PCR Produkte dieser Reaktionen
wurden in Topo-Vektoren kloniert und die Sequenz auf die verschiedenen Mutationen
überprüft. Hierbei wurden jeweils 2 Proben eingeschickt. Die Analyse der OPR3-
Sequenzen zeigte bei beiden Proben, dass beide Schnittstellen vorhanden waren und
keine Aminosäuren ausgetauscht waren. Mit der ersten der beiden Proben wurde
weiter gearbeitet.

3. Ergebnisse 50
Die Analyse einer der E291K OPR3 Sequenzen wies zwei Fehler in der DNA Sequenz
auf, die zum Aminosäureaustausch führten, diese wurde verworfen, die andere
Sequenz dagegen zeigte beide Schnittstellen und die Mutation von Glutamat nach
Lysin an Stelle 291. Mit dieser Probe wurde weiter gearbeitet. Die Sequenzen der
OPR3 cDNA mit der Punktmutation Y364F waren ebenfalls beide fehlerfrei und zeigte
sowohl die Mutation von Tyrosin 364 zu Phenylalanin, als auch beide Schnittstellen.
Auch hier wurde die Erste der beiden Proben zur weiteren Arbeit verwendet.
OPR3 E291K Y364F
Primer 34/35
Abb. 21: Ausschniktt eines Ethidiumbromid gefärbten 1% Agarosegels. PCR der OPR3
Konstrukte OPR3, E291K, Y364F mit Primer 34 und 35, 1191 bp, Annealingtemperatur
58°C, Elongationszeit 75 sec.
3.2 Mutationen der OPR1 cDNA
Anhand der Sequenzanalysen von OPR3 und OPR1 ist bekannt, dass OPR3 und
OPR1 in 53 % ihrer Aminosäuresequenz identisch sind (Stintzi und Browse, 2000 und
Abbildung 21). Auch ihre Struktur ist nahezu identisch. Beide haben die Struktur eines
(αβ)8-Barells und überlagern annähernd, das aktive Zentrum beider Enzyme ist hoch
konserviert. (Breithaupt et al., 2000 und 2006 und Abbildung 9 in Einleitung)
Es wurde gezeigt, dass OPR1 stereoselektiv ist und nur cis(-)OPDA umsetzt. OPR3
hingegen reduzierten sowohl cis(+)OPDA als auch cis(-)OPDA. Allerdings kann nur
cis(+)OPDA als Vorstufe in aktive Jasmonsäure umgesetzt werden (Schaller F. et al.,
1998). Dieser Unterschied in der Substratspezifität wird in der Beschaffenheit des β6-
Loops vermutet. Exponiert sind die Aminosäuren Y246 und Y78, die im OPR1 Protein
eine Verengung der Höhlung um das Aktiven Zentrums zur Folge haben, wohingegen
ihre Korrespondierenden Aminosäuren im OPR3 Protein (H244 und F74) eine größere
Höhlung um das Aktive Zentrum lassen. Dies hat vermutlich eine höhere Selektivität
der Substrate bei OPR1 zur Folge, wohingegen OPR3 ein breites Spektrum an
Substraten zulässt (Breithaupt et al., 2006 und Abbildung 10 in Einleitung).

3. Ergebnisse 51
Aufgrund dieser Tatsache stellte sich die Frage, ob man durch den Austausch der
korrespondierenden Aminosäuren im β6-Loop die Substratspezifität von OPR1 in die
von OPR3 umwandeln könnte. Um nicht nur das selbe Substratspektrum, sondern
auch die selbe Lokalisation des mutierten Proteins zu gewährleisten musste mittels
PCR noch ein Zieltag kloniert werden, welches dafür sorgt, dass OPR1, ebenso wie
OPR3, im Peroxisom und nicht wie sonst cytosolisch lokalisiert (Strassner et al.,
2002). Das Peroxisomentag selber besteht nur aus der Aminosäurenabfolge SRL, für
die Komplementation erschien es aber sinnvoll, die an AtOPR3 noch vorhandenen
zusätzlichen Aminosäuren Prolin (P) und Serin (S) ebenfalls zu klonieren (siehe
Abbildung 22, C-terminales Ende der Sequenzen).
Abb. 22: Sequenzvergleich von OPR1, OPR2 und OPR3 aus Tomaten (Le) und
Arabidopsis thaliana (At)
Die in OPR1 zu mutierenden Aminosäuren sind hellblau markiert, die in OPR3 zu
mutierenden Aminosäuren in gelb. Das in Dunkelrot markierte SRL Zieltag für
Peroxisomen wurde ebenfalls an OPR1 kloniert.
AtAt
At
AtAt
At
α7β7 β8 α8α6
AtAt
At
β5 β6α4 α5
Dimerization
AtAt
At
β3 β4α3βC βEβD βF
Substrate specificity
αB
αB
PTS1
βA βB β1 α1 β2 α2Y74
F74
Y244
H244
E291
Y364
AtAt
At
AtAt
At
α7β7 β8 α8α6
AtAt
At
β5 β6α4 α5
Dimerization
AtAt
At
β3 β4α3βC βEβD βF
Substrate specificity
αB
αB
PTS1
βA βB β1 α1 β2 α2βA βB β1 α1 β2 α2Y74
F74
Y244
H244
E291
Y364

3. Ergebnisse 52
Somit ergeben sich die folgenden Konstrukte:
LeOPR1-PSSRL Y74F (Tyrosin 74 wurde durch Phenylalanin ersetzt)
LeOPR1-PSSRL Y244H (Tyrosin 244 wurde durch Histidin ersetzt)
LeOPR1-PSSRL Y74F/Y244H (beide Tyrosine ausgetauscht)
Im Folgenden werden die Mutanten mit Y74F, Y244H und Y74/Y244H bezeichnet.
Außerdem wurde nicht mutierte OPR1 cDNA mit dem Peroxisomentag PSSRL
versehen, um zu überprüfen ob die Anwesenheit des OPR1 Proteins im Peroxisom
bereits ausreichend ist um die männliche Sterilität zu komplementieren. Diese Mutante
wird im folgenden OPR1+SRL bezeichnet.
Die Punktmutationen, die zum spezifischen Aminosäureaustausch führten wurden mit
Zielgerichteter Mutagenese durchgeführt. Hierzu wurde zuerst durch eine PCR-
Reaktion die OPR1 cDNA mit den Restriktionsenzymschnittstellen für Spe I und Pst I
versehen, sowie das C-terminale Tag PSSRL kloniert. Dabei wurden die Primer
5mutLeOPR1 (36) und 3mutLeOPR1 (37) verwendet (Siehe Abbildung 23). Die so
gewonnenen cDNAs wurden in Topo®-Vektoren kloniert. Zum einen ist Topo ein
prädestiniertes Plasmid um PCR Produkte zu klonieren, zum anderen diente dieses
Plasmid in der Mutagenese PCR als Matrize.
Primer 36/37
OPR1+SRL
Abb. 23: Ethidiumbromid gefärbtes 1% Agarosegel. Links: PCR der OPR3 Konstrukte
OPR3, E291K, Y364F mit Primer 34 und 35, 1191 bp, Rechts: PCR der OPR1-cDNA
mit Primern 36 und 37, 1131 bp. Annealingtemperatur 58°C, Elongationszeit 75 sec.
Die zielgerichtete Mutagenese PCR wurde in jeweils 4 Ansätzen durchgeführt, wobei
je 2 mal 10 bzw. 50 ng DNA Matrize eingesetzt wurden und mit beiden Ansätzen 15
bzw. 17 Zyklen durchgeführt wurden. Nachfolgend dargestellt (Abbildung 24) sind die
Mutagenese-Primer (blau) und ihre Bindung an die OPR1-Matrize (schwarz) mit dem
jeweiligen Basenpaar. Unterstrichen ist das Basentriplett, das mutiert wurde um die
gewünschte Aminosäure auszutauschen. Die komplette Sequenz von OPR1 mit den
Mutationsprimern ist im Anhang zu finden.

3. Ergebnisse 53
Abb. 24: DNA Sequenzauschnitt von OPR1 (schwarz) mit den jeweils bindenden
Mutageneseprimern (blau). Geänderte Basentriplets, die zum Aminosäureaustausch
führen sind unterstrichen
Nach der Mutagenese PCR wurden den Ansatz 1 U Dpn I zugegeben um die noch
vorhandenen nicht mutierten Template Plasmide zu verdauen. Die noch vorhandenen
Plasmide, die eine Mutation enthalten sollten, wurden in E. coli XL-1 blue transformiert
und auf LB-Platten mit Kanamycin (50 µg/ml) ausgestrichen. Von je 3 Kolonien wurden
Flüssigkulturen angeimpft und Plasmid DNA isoliert. Um die Mutation zu überprüfen
wurden die Plasmide sequenziert. Bei der Mutation Y244H wurden 3 Sequenzen
gefunden, die die Mutation enthielten, allerdings war bei einer keine Pst I Schnittstelle
zu finden. Diese wurde verworfen, mit der ersten Sequenz wurde weiter gearbeitet. Bei
der Mutation von Y74F musste die Mutagenese PCR 2 mal wiederholt werden, da
keine positiven Plasmide gefunden wurden. Es wurden jeweils 3 Plasmid DNAs zum
Sequenzieren geschickt. Die letzte Mutagenese PCR ergab dann 3 positive
Sequenzen mit beiden Schnittstellen. Die Doppelmutation Y74F/Y244H, die auf einer
positiv getesteten Y244H-Sequenz durchgeführt wurde - hier wurde die Mutagenese
PCR nochmals mit den Primern für Y74F durchgeführt - zeigte nur eine positive
Sequenz, bei der ebenfalls beide Schnittstellen vorhanden waren, bei den anderen
hatte die Mutation keinen Erfolg. Die Sequenzanalysen der Plasmide, die zur
Klonierung verwendet wurden sind im Anhang zu finden
LeOPR1Y74F- For 5' - tatctgagactggcataggg ttt aaagatgtacctggtatatgg - 3' ||||||||||||||||||||| |||||||||| |||||||||||
OPR1-Matrize 212- tatagactctgaccgtatcccat gtttctacatggaccatatacc -256 ||||||||||||||||||||| |||||||||| ||||||||||| LeOPR1Y74F-Rev 3'-atagactctgaccgtatcccaaa tttctacatggaccatatacc-5' LeOPR1Y7244H-For 5'-tatccccatttgcgcatcat aatgaagcaggggac-3' ||||||||||||||||| ||||||||||||| |||| OPR1-Matrize 718- tatccccatttgcgcattat aatgaagcaggggac -754 ||||||||||||||||| ||||||||||||| |||| LeOPR1Y244H-Rev 3'-ataggggtaaacgcgtagta ttacttcgtcccctg-5'

3. Ergebnisse 54
3.3 Klonierung in pGreen 0229
Plasmide, die die gesuchte Mutation enthielten wurden mit Spe I und Pst I verdaut und
anschließend auf ein Agarosegel aufgetragen (Siehe Abbildung 25). Da der Topo®-
Vektor eine weitere Schnittstelle für Pst I außerhalb der Multicloning site enthält, die
ein 1,2 kb großes Fragment ergibt, welches dieselbe Größe wie das gesuchte Insert
hat, musste der Vektor zunächst mit Nae I verdaut werden. Dieses Restriktionsenzym
schneidet im 1,2 kb großen PstI/PstI Fragment. Die gesuchten Inserts mit der Größe
von 1131 bzw. 1191 bp wurden aus dem Gel herausgeschnitten und mittels QIAquick®
Gel Extraktions Kit (Qiagen) aus dem Gel extrahiert.
Abb. 25: Ausschnitt eines 1% Agarosegels, gefärbt mit Ethidiumbromid.
Restriktionsverdau der Mutation-tragenden Topo® Plasmide mit Nae I, Spe I und Pst I.
OPR3 (links, OPR3, E291K, Y364F) Konstrukte haben eine Größe von 1191 bp, OPR1
Konstrukte (rechts, OPR1+SRL, Y74F, Y244H, Y74F/Y244H) haben eine Größe von
1131 bp. Der Topo®-Vektor ohne Insert hat eine Größe von ca. 3900 bp. Der Verdau
wurde bei 37°C für 2 Stunden durchgeführt.
Wie Abbildung 15 (Siehe Abschnitt 2.2. Vektoren und Plasmide) zeigt, konnte die in
pGreen vorhandene DNA des DsRED-OPR3 mit den Restiktionsenzymen Spe I und
Pst I aus dem Vektor herausgeschnitten werden, gleichzeitig wurde so eine Ligation
mit den Inserts E291K, Y364F, OPR1+SRL, Y74F, Y244H und Y74F/Y244H
ermöglicht, die mit den selben Enzymen verdaut wurden. Im pGreen Vektor war
bereits ein 2130 bp großes Fragment des OPR3 Promotors kloniert, womit
sichergestellt war, dass die mutierten OPR3 und OPR1-cDNAs auch unter Kontrolle
eines OPR3 Promotors standen. Abbildung 26 zeigt den Aufbau des zu
transformierenden Vektors. Als Kontrolle wurde zusätzlich eine nicht mutierte OPR3
cDNA zur Komplementation vorbereitet, um zu überprüfen, ob die Transformation an
sich bereits Auswirkungen auf die Pflanzen hat.
OPR3 E291K Y364F OPR1+SRL Y74F Y244H Y74F/Y244H
Insert 1191 bp
bzw. 1131 bp
Spe I Pst I Fragment
2700 bp

3. Ergebnisse 55
Abb. 26: Schematischer Aufbau des Transformationsplasmides mit OPR3-Promotor,
Konstrukt (grün hinterlegt), nos-Terminator (T), Resistenzgenen für BASTA (nos-bar) und
Kanamycin (npt) und Origin of replication für die Replikation in Agrobakterium (pSA ori) und
E.coli (ColEI ori).
Nach erfolgreicher Klonierung der Konstrukte in pGreen wurden diese in
Agrobacterium tumefaciens GV3101 transformiert. Dieser enthält bereits das Plasmid
pSoup, das eine Replikation von pGreen ermöglicht. Die Agrobakterium-vermittelte
Transformation überträgt nicht das gesamte Plasmid ins Arabidopsis-Genom, sondern
nur eine T-DNA, die von der Linken Grenze (LB) und der Rechten Grenze (RB)
begrenzt wird (Hellens et al., 2000). Vor der Transformation mussten die
Agrobakterien noch auf das Vorhandensein des Konstruktes überprüft werden. Hierzu
wurde Plasmid DNA mittels Minipräp extrahiert und durch eine PCR das Konstrukt
nachgewiesen (Siehe Abbildung 27). Erfolgreich transformierte Agrobakterienstämme
wurden zur Transformation in Pflanzen angezogen.
Primer 34/35
OPR3 E291K Y364F OPR1+SRL Y74F Y244H Y74F/Y244H
Primer 36/37
Abb. 27: 1% Agarosegel mit Ethidiumbromid gefärbt. PCR auf Konstrukte in
Agrobakterien. OPR3 Konstrukte (links, OPR3, E291K, Y364F) haben eine Größe von
1191 bp, OPR1 Konstrukte (rechts, OPR1+SRL, Y74F, Y244H, Y74F/Y244H) haben eine
Größe von 1131 bp. Annealingtermperatur: 58°C, Elong ationszeit 120 sec.
TOPR3 Promotor Konstrukt npt ColEI ori pSA ori nos-Bar
BspH I Bgl II Not I Sac II SpeI Pst I Knp I Knp I Bgl II BspH I PvuI
LBRB
T-DNA
2130 bp 1131 - 1171 bp 282 bp

3. Ergebnisse 56
3.4 Identifizierung transgener Pflanzen
Da opr3-defiziente Arabidopsis thaliana Pflanzen männlich steril sind, müssen die
transformierten Pflanzen durch die exogene Gabe von Methyl-Jasmonsäure zur
Samenbildung gebracht werden. Pflanzen wurden hierfür alle 2 Tage mit einer
0,03%igen Methyl-Jasmonsäurelösung besprüht. Zur Identifizierung transgener
Pflanzen wurden je 15.000 der durch Jasmonsäurebesprühung gewonnenen Samen
auf BASTA-getränkter Erde ausgelegt. Samen, die das Konstrukt enthielten, waren
gegen BASTA resistent (die T-DNA enthält das nos-bar-Gen, das die BASTA-
Resistenz vermittelt) und konnten nach der Kotyledonenbildung weiter wachsen.
Samen, die das Konstrukt nicht enthielten konnten sich nicht über das
Kotyledonenstadium hinaus entwickeln. Es wurden jeweils zwischen 5 und 10
resistenten Pflanzen gefunden. Da die Transformationsrate normalerweise zwischen
0,1 und 0,7% liegt ist dies eine sehr geringe Transformationsrate. Hierfür gibt es 2
mögliche Erklärungen: Entweder konnte der weibliche Gametophyt nicht befruchtet
werden, da die Jasmonsäurebehandlung zu diesem kritischen Zeitpunkt nicht
erfolgreich genug war um männlich fertile Pollen herzustellen, oder der weibliche
Gamethophyt konnte nicht von den Agrobakterien transformiert werden.
Die Samen der Transformanten des Konstrukt Y364F allerdings zeigten eine
ungewöhnliche hohe Keimungsrate von annähernd 50% Resistenz gegen BASTA
obwohl in keinem der Keimlinge - es wurden Stichprobenartig 100 Pflanzen getestet -
das Konstrukt nachgewiesen werden konnte. Vermutlich war unter den transformierten
Pflanzen eine BASTA-Resistente, deren Samen keimen konnten, ohne transformiert
worden zu sein. Eine Identifikation von eventuell transformierten Pflanzen war daher
nicht möglich.
Aus den resistenten Pflanzen wurde aus einem Blatt DNA extrahiert und mittels PCR
auf das Vorhandensein des Konstruktes getestet. Die Primer zur Überprüfung auf
Anwesenheit des Konstruktes waren 5’ im OPR3 Promotor (1612), bei OPR3
Konstrukten 3mutLeOPR3 (35), bei OPR1 Konstrukten hingegen OPR1_876R (876).
Um die Anwesenheit und Menge der DNA zu kontrollieren wurden die Primer TuB4F
und TuB4R verwendet. Die Ergebnisse sind in Abbildung 28 dokumentiert.

3. Ergebnisse 57
OPR1+SRL Y244H Y74F/Y244H1 2 3 4 5 + – 1 2 3 4 5 6 7 + – 1 2 3 4 5 6 7 + –
1612/876
TuB4
Abb. 28: 1% Agarosegel in Ethidiumbromidfärbung. PCR Analyse der BASTA-
resistenten Pflanzen auf das jeweilige Konstrukt. Als Positive Kontrolle (+) diente das
jeweilige Transformationskonstrukt, Als Negative Kontrolle (–) diente Wildtypische
DNA. Als Ladekontrolle wurde eine PCR mit den Primern für Tubulin TuB4F und
TuB4R durchgeführt, positive Kontrolle hierfür war wildtypische DNA, negative
Kontrolle das Transformationskonstrukt. Annealingtemperatur: 50°C, Elongationszeit
40 sec. Oben: Analyse der OPR3 Konstrukte OPR3 (Komplementationskontrolle) und
E291K mit den Primern 1612 und 35 .Unten: Analyse der OPR1 Konstrukte
OPR1+SRL Y244H und die Doppelmutation Y74F/Y244H mit Primern 1612 und
OPR1_876R. Annealingtemperatur: 58°C, Elongationszeit: 120 sec.
Die Analyse der Pflanzen, die mit OPR1 Konstrukte transformiert wurden ergab für das
Konstrukt OPR1+SRL 4 positive Pflanzen, für das Konstrukt Y244H wurden 7 positive
Pflanzen und für die Doppelmutation Y74F/Y244H 5 positive Pflanzen gefunden. Die
Analyse der Pflanzen, die mit OPR3 Konstrukten transformiert wurde ergab für die
Transformationskontrolle OPR3 2 positiv getestete Pflanzen, für das Konstrukt E291K
wurden ebenfalls 2 positive Pflanzen gefunden.
Von den Konstrukten Y364F und Y74F konnten keine positiven Linien identifiziert
werden. Um Rückschlüsse über eine Transformation und ihre Auswirkungen ziehen zu
können müssen diese erneut transformiert werden.
OPR3 E291K 1 2 3 4 5 – + 1 2 3 4 5 6 + –
1612/35
TuB4
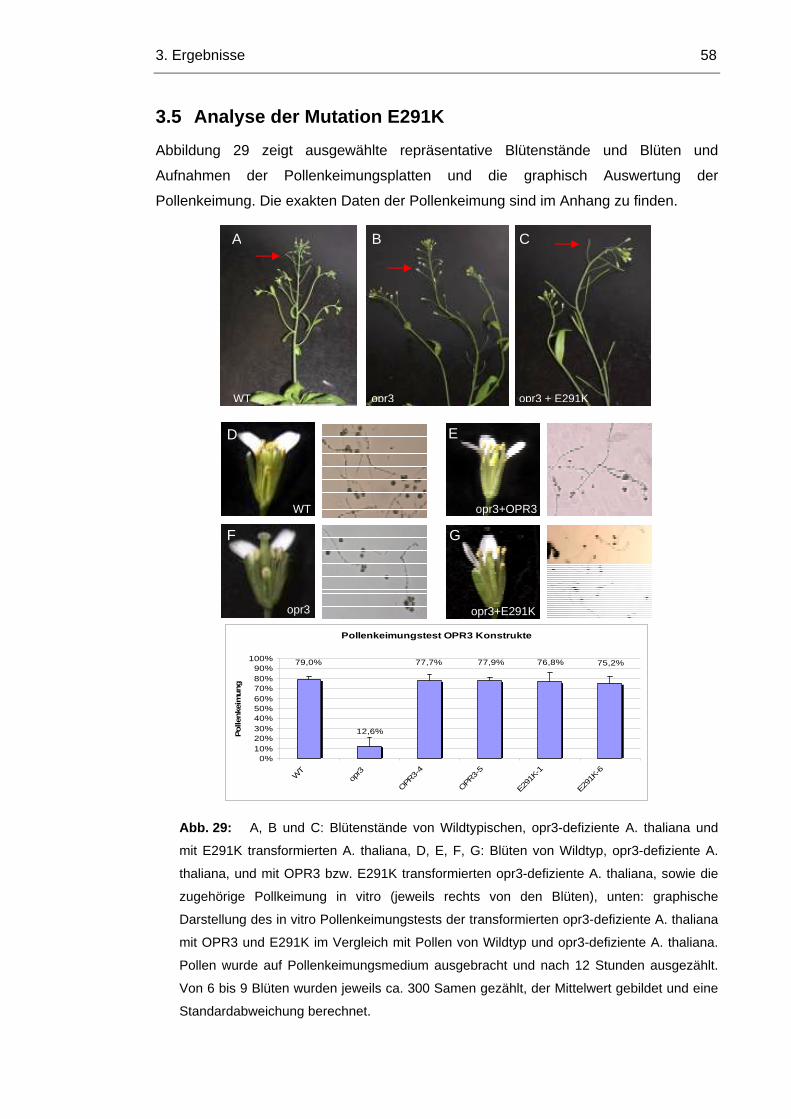
3. Ergebnisse 58
3.5 Analyse der Mutation E291K
Abbildung 29 zeigt ausgewählte repräsentative Blütenstände und Blüten und
Aufnahmen der Pollenkeimungsplatten und die graphisch Auswertung der
Pollenkeimung. Die exakten Daten der Pollenkeimung sind im Anhang zu finden.
WT
opr3 opr3+E291K
opr3+OPR3
Pollenkeimungstest OPR3 Konstrukte
77,9%77,7%79,0% 76,8% 75,2%
12,6%
0%10%20%30%40%50%60%70%80%90%
100%
WT
opr3
OPR
3-4
OPR
3-5
E291
K-1
E291
K-6
Pol
lenk
eim
ung
Abb. 29: A, B und C: Blütenstände von Wildtypischen, opr3-defiziente A. thaliana und
mit E291K transformierten A. thaliana, D, E, F, G: Blüten von Wildtyp, opr3-defiziente A.
thaliana, und mit OPR3 bzw. E291K transformierten opr3-defiziente A. thaliana, sowie die
zugehörige Pollkeimung in vitro (jeweils rechts von den Blüten), unten: graphische
Darstellung des in vitro Pollenkeimungstests der transformierten opr3-defiziente A. thaliana
mit OPR3 und E291K im Vergleich mit Pollen von Wildtyp und opr3-defiziente A. thaliana.
Pollen wurde auf Pollenkeimungsmedium ausgebracht und nach 12 Stunden ausgezählt.
Von 6 bis 9 Blüten wurden jeweils ca. 300 Samen gezählt, der Mittelwert gebildet und eine
Standardabweichung berechnet.
WT opr3 opr3 + E291K
A B C
D E
F G

3. Ergebnisse 59
Wie bereits an den Blütenständen (siehe Abbildung 29 A, B und C) zu erkennen ist
und durch Pollenkeimungstest bestätigt wird, komplementiert die Mutation E291K die
männliche Sterilität. 29 A zeigt eine Wildtypische Pflanze, die bereits einige elongierte
Schoten ausgebildet hat (Pfeil) und somit Fertilität aufweist. 29 B zeigt eine opr3-
mutante Pflanze, die keine verlängerten Schoten hat. In Abbildung 29 C ist ein
Blütenstand einer E291K-komplementierten Pflanze abgebildet, bei dem ebenfalls
elongierte Schoten zu finden sind. Die männliche Sterilität konnte also durch die
Transformation mit E291K wieder hergestellt werden.
Detailaufnahmen der Blüten zeigen in 29 D (Arabidopsis Wildtyp), E (opr3-defiziente
Arabidopsis thaliana mit OPR3 Konstrukt transformiert) und G (opr3-defiziente
Arabidopsis thaliana mit E291K Konstrukt transformiert) fertile Blüten. Die Antheren
sind verlängert und Pollen ist freigesetzt worden. Die Pollenkeimung in vitro liegt bei
zwischen 75,2% und 76,8% der Pollen bei E291K-komplementierten Pflanzen, 77,7%
bis 77,9 % bei OPR3-komplementierten Pflanzen und 79% beim Wildtyp. Abbildung 29
F hingegen zeigt eine männlich sterile opr3-defiziente Pflanze. Die Filamente sind
nicht verlängert, es wurde kein Pollen freigesetzt. Die Pollenkeimung liegt bei 12,6%.
Eine dauerhafte Aktivität von OPR3 durch die Mutation E291K, die eine Dimerisierung
und somit Inaktivität des Proteins verhindert, ist phänotypsch nicht feststellbar. Diese
könnte eine dauerhafte Ausschüttung von Jasmonsäure zu Folge haben, durch die
anderer Jasmonsäure-induzierter Gene aktiv wären, ohne dass eine Verwundung
geschehen ist. Eine Native SDS-PAGE mit anschließender Western Blot Analyse
könnte Aufschluss darüber geben, ob OPR3 weiterhin dimerisieren kann oder nicht.
Die Pflanzen befanden sich zur Fertigstellung dieser Arbeit allerdings erst in der T1
Generation, so dass Western Blot Analysen noch nicht möglich waren, da eine
größere Menge an Pflanzenmaterial benötigt wird.
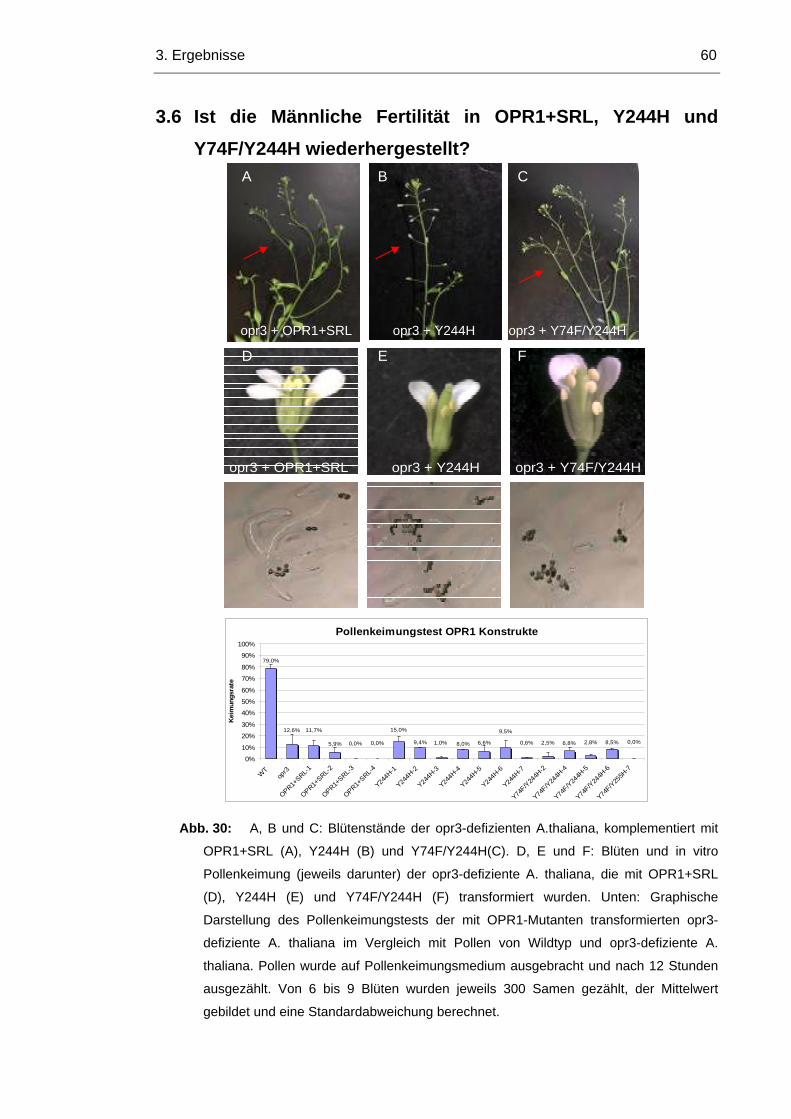
3. Ergebnisse 60
3.6 Ist die Männliche Fertilität in OPR1+SRL, Y244H un d
Y74F/Y244H wiederhergestellt?
opr3 + OPR1+SRL opr3 + Y244H opr3 + Y74F/Y244H
Pollenkeimungstest OPR1 Konstrukte
9,4% 8,0%0,0%0,0%5,9%
11,7%
79,0%
12,6%
1,0% 2,5%0,6% 0,0%8,5%2,8%6,8%
15,0%
6,6%
9,5%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
WT
opr3
OPR1+SRL-
1
OPR1+SRL-
2
OPR1+SRL-
3
OPR1+SRL-
4
Y244H
-1
Y244H
-2
Y244H
-3
Y244H
-4
Y244H
-5
Y244H
-6
Y244H
-7
Y74F/Y
244H
-2
Y74F/Y
244H
-4
Y74F/Y
244H
-5
Y74F/Y
244H
-6
Y74F/Y
255H
-7
Kei
mun
gsra
te
Abb. 30: A, B und C: Blütenstände der opr3-defizienten A.thaliana, komplementiert mit
OPR1+SRL (A), Y244H (B) und Y74F/Y244H(C). D, E und F: Blüten und in vitro
Pollenkeimung (jeweils darunter) der opr3-defiziente A. thaliana, die mit OPR1+SRL
(D), Y244H (E) und Y74F/Y244H (F) transformiert wurden. Unten: Graphische
Darstellung des Pollenkeimungstests der mit OPR1-Mutanten transformierten opr3-
defiziente A. thaliana im Vergleich mit Pollen von Wildtyp und opr3-defiziente A.
thaliana. Pollen wurde auf Pollenkeimungsmedium ausgebracht und nach 12 Stunden
ausgezählt. Von 6 bis 9 Blüten wurden jeweils 300 Samen gezählt, der Mittelwert
gebildet und eine Standardabweichung berechnet.
opr3 + OPR1+SRL opr3 + Y244H opr3 + Y74F/Y244H
D E F
A B C

3. Ergebnisse 61
Abbildung 30 zeigt ausgewählte repräsentative Blütenstände und Blüten und
Aufnahmen der Pollenkeimungsplatten, sowie die graphische Auswertung der
Pollenkeimung im Vergleich mit Wildtypischen und opr3-defizienten Arabidopsis-
Pflanzen. Die genauen Datensätze zur Pollenkeimung jeder einzelnen Mutante finden
sich im Anhang. Abbildungen 30 A, B und C zeigen jeweils Blütenstände von opr3-
mutanten Pflanzen, die mit OPR1+SRL (A), Y244H (B) und Y74F/Y244H(C)
transformiert wurden. An keiner sind elongierte Schoten zu finden, die auf eine
männliche Fertilität schließen lassen. Die Abbildungen 30 D, E und F zeigen
Detailaufnahmen der jeweiligen Blüten. Keine der mit OPR1-Konstrukten
komplementierten Pflanzen zeigt eine Verlängerung der Filamente und eine Öffnung
der Antheren und eine Freisetzung von Pollen. In vitro Pollenkeimungstest haben
gezeigt, dass sich die Pollenkeimung nicht wesentlich von der einer opr3-Mutante
unterscheidet – opr3-Mutanten haben etwa 12 % Pollenkeimung, die Transformation
mit OPR1+SRL zeigte Pollenkeimung zwischen 0 und 11,7%. Y244H zeigte zwischen
0,6 und 15% Pollenkeimung und die Doppelmutation Y74F/Y244H 0 - 8,5%.
Eine Transformation von opr3-Arabidopsis Pflanzen mit den Konstrukten OPR1+SRL,
Y244H und Y244H/Y74F ist also nicht ausreichend um die männliche Fertilität wieder
herzustellen. Dies zeigt, dass die Anwesenheit von OPR1 im Peroxisom alleine noch
nicht genügt um die Abwesenheit von OPR3 zu komplementieren, da für die
Fortführung des Oktadekanoidweges cis(+)OPDA umgesetzt werden muss. OPR1
kann nur cis(-)OPDA umsetzen. Strukturelle Daten deuten darauf hin, dass Tyrosin an
den Stellen 246 und 78 als eine Art Torhüter vor dem Aktiven Zentrum fungieren,
wodurch der Zugang von cis(+)OPDA zum Aktiven Zentrum verhindert wird. Demnach
wäre zu erwarten, das durch den Austausch der beiden Aminosäuren der Umsatz von
cis(+)OPDA möglich werden sollte. Wie die Daten in Abbildung 30 aber zeigen ist die
männliche Fertilität nicht wieder hergestellt worden. Der Zugang von cis(+)OPDA zum
Aktiven Zentrum wird vermutlich noch durch weitere Aminosäuren im β6-Loop über
dem aktiven Zentrum reguliert. Alle männlich sterilen Pflanzen wurden erneut mit
Jasmonsäure besprüht um die männliche Fertilität wieder herzustellen und Samen zu
erhalten.

3. Ergebnisse 62
3.7 Lokalisation der mutierten OPR1-Proteine
3.7.1 Gradientenzentrifugation
Da eine Komplementation des Phänotyps durch die verschiedenen OPR1-Konstrukte
nicht erfolgreich war, musste zunächst noch überprüft werden, ob das mutierte OPR1-
Protein überhaupt in den Peroxisomen vorhanden war. Hierzu mussten Peroxisomen
isoliert werden. Zunächst wurden Pflanzen geerntet und mit Tricinepuffer
homogenisiert. 100µl und 600 µl des Filtrat dieses Zellaufschlusses wurde
anschließend auf einen 5 ml 65-30%igen Saccharosegradienten aufgetragen. Durch
Ultrazentrifugation werden hier die Zellorganellen entsprechend ihrer Dichte
aufgetrennt. Die Gradienten wurden in einen Fraktionator eingespannt, von unten
angebohrt und Fraktionen von je 250 µl aufgefangen. Der Zuckergehalt der einzelnen
Fraktionen wurde mit Hilfe eines Refraktometers bestimmt. In den Fraktionen
zwischen 58 % und 50 % Saccharose waren die Peroxisomen zu erwarten (Strassner
et al., 2002).
100 µl 600 µl
OPR1+SRL-1, Y244H-1, Y74F/Y244H-1 OPR1+SRL-1, Y244H-1, Y74F/Y244H-1
Abb. 31: 5 ml Gradienten der verschiedenen Linien, beladen mit 100 µl (links) und 600
µl (rechts) der Überstände von OPR1+SRL-1, Y244H-1 und Y74F/Y244H-1.

3. Ergebnisse 63
Jeweils 96 Samen einer positiven Linie von Y244H und OPR1+SRL wurden ausgesät
und nach Ausbilden von 4 Blättern mit BASTA besprüht, da nach der 2. Mendelschen
Regel (Spaltungsregel) nur 3:1 Pflanzen der Nachkommen in der T2-Generation das
Konstrukt tragen. Pflanzen, die das Konstrukt nicht trugen wurden so eliminiert. Die
Keimungsrate war sehr gering, so dass schlussendlich nur zwischen 20 und 5
Pflanzen jeder Linie überhaupt kleine Rosetten ausbildeten. Die übrigen Pflanzen
gingen entweder ein oder zeigten nur wenige kleine Rosettenblätter und bildeten
bereits Blütenstände aus, wie dies bei Hitze- und Lichtstress der Fall ist. Alle Rosetten
überlebender grünen Pflanzen wurden zur Gradientenzentrifugation geerntet
(Abbildung 31). Außerdem wurde eine positive Pflanze der Doppelmutation
Y74F/Y244H geerntet, von der noch keine Samen in der T2 Generation existierten.
.
66% 63,5% M 61% 57,5% 55% 52,5% 51% 48,5% 44% 41,5% 38,5% 35% M 30,5% 21% 18,5%
13096
72
56
43
34
26
kDa96
72
56
43
34
26
17
M 65% 63% 60% 58% 55,5% 52,5% 50% 46,5% 42% 38% M 36% 32,5% 25,5% 14%
96
72
56
43
34
26
17
kDa13096
72
56
43
34
26
Abb. 32: Coomassie- und Silber-Färbung der SDS-Gele der 2 Gradienten von Y244H.
Oben: 100µl beladener Gradient, Unten 600 µl beladener Gradient. Die Fraktion von
18,5% im 100µl Gradienten fehlt in der Silber-Färbung, da diese Fraktion nicht
vollständig gefüllt und deswegen im ersten Gel bereits aufgebraucht worden war.

3. Ergebnisse 64
Jeweils 20 µl jeder Fraktion wurden mit 5 µl SDS-Probenpuffer gemischt, denaturiert
und auf einem 10%igem SDS-Gel aufgetrennt um anschließend mit Coomassie- und
Silber-Färbung Proteine sichtbar zu machen. Hierdurch konnte gezeigt werden, dass
die Proteinmengen so gering sind, dass in den Fraktionen zwischen 58 % und 50 %
Saccharose nur geringe Mengen an Proteinen um 70 kDa sichtbar sind.
Abbildung 32 zeigt hier stellvertretend Färbungen der Gele der beiden Gradienten von
Y244H-1, die mit 100 µl (oben) und 600 µl (unten) des Filtrats des Zellaufschlusses
beladen wurden. Obwohl in den Gradienten eine deutliche grüne Bande auszumachen
ist die die Chloroplasten enthält und in den entsprechenden Fraktionen ebenfalls eine
deutliche grüne Färbung zu sehen ist, sind in den selben Taschen im SDS-Gel nur
dünne Proteinbanden auszumachen. Im dünneren Bereich des Gradienten (oberhalb
30%) sind große Mengen an Proteinen zu sehen, die vermutlich aus den
aufgeschlossenen Chloroplasten und zerstörten Zellorganellen stammen. Die
einzelnen Fraktionen wurden mittels Western Blot analysiert, außerdem wurde
versucht mit einem spektrophotometischen Aktivitätstest die Fraktionen zu
identifizieren, die Peroxisomen enthalten.
3.7.2 Katalaseaktivität
Katalase ist ein Enzym, welches in den Peroxisomen lokalisiert ist und die Umsetzung
von H2O2 zu Wasser und Sauerstoff katalysiert. Diese Umsetzung lässt sich
spektrophotometrisch bestimmen, indem man die Adsorption von Wasserstoffperoxid
bei 240 nm misst (Strassner et al., 2002). Bei Versuchen mit Pflanzenmaterial, das in
Tricinepuffer aufgenommen wurde, aber nicht auf einen Gradienten geladen worden
war, konnten eindeutige Aktivitätsspektren erzielt werden. Experimente haben gezeigt,
dass sowohl in den Fraktionen zwischen 58% und 50% Saccharose, als auch in den
obersten Fraktionen eines Gradienten eine Katalaseaktivität festzustellen ist (eigene
Daten nicht gezeigt; Strassner et al, 2002). Die Aktivität im oberen Bereich des
Gradienten rührt von beim Zellaufschluss gespaltenen Peroxisomen und somit
freigesetzter Katalase, die sich dort ansammelt. Bei den oben gezeigten Gradienten
allerdings konnte aufgrund der geringen Pflanzenmenge und der dadurch bedingten
geringen Zellorganell- und Proteinmenge in keiner Fraktion eine eindeutige Aktivität
festgestellt werden (Daten nicht gezeigt).

3. Ergebnisse 65
3.7.3 Western Blot auf OPR1
Durch die geringe Menge an Pflanzenmaterial (ca. 2 g pro Linie) von transformierten
Pflanzen waren sowohl die Detektion der Peroxisomen durch Katalaseaktivitätstest,
als auch die Protein-Detektion in der Coomassie- und Silber-Färbung äußerst
schwierig (siehe Abbildung 32). Daher wurden Western Blot Analysen mit
verschiedenen Antikörpern durchgeführt, die ein Bild davon geben sollen, welche
Fraktionen des Gradienten welche Organellen enthalten. AOS ist ein Enzym des
Oktadekanoidweges, welches in den Chloroplasten lokalisiert ist. Ein Western Blot mit
AntiAOS-Serum (Siehe Abbildung 33, oben) wurde daher als Kontrolle hinzugezogen.
OPR1 soll nach erfolgreicher Transformation, zusätzlich zur normalen Lokalisation im
Cytosol, im Peroxisom vorhanden sein.
kDa
56
43
34
65% M 63% 60% 58% 55,5% 52,5% 50% 46,5% 42% 38% 36% M 32% 25% 14% OPR1kDa
56
43
34
AntiAOS
AntiOPR1
Abb. 33: Vergleich der beiden Western Blots des Gradienten von Y244H-1(beladen mit
600µl) mit Anti-AOS (oben) und Anti-OPR1 (unten) und OPR1 Protein als Kontrolle
Western Blot Analysen zeigen, dass in den oberen Fraktionen (25-14 % Saccharose)
OPR1 Protein vorhanden ist, welches entweder aus aufgeschlossenen Peroxisomen
oder auch aus dem Cytosol stammt (siehe Abb. 33, unten). Im Normalfall ist OPR1
Cytosolisch (Strassner et al., 2002). Im Gradienten würde es sich oberhalb der
Fraktionen sammeln, in denen sich Plastiden befindet.

3. Ergebnisse 66
Der Western Blot, der mit Anti AOS detektiert wurde zeigt hingegen Banden in den
Fraktionen zwischen 55,5% und 50% Saccharose sowie zwischen 36% und 14%
Saccharose. Die Banden zwischen 55,5% und 50% Saccharose sind höchst
wahrscheinlich unspezifische Bindungen, da es sich bei Anti AOS nur um
ungereinigtes Serum handelt. Endogenes OPR1 sollte sich in diesen Fraktionen finden
lassen, ist aber vermutlich aufgrund der geringen Menge an Pflanzenmaterial nicht
detektierbar. Die AOS-Signale zwischen 36% und 25% hingegen scheinen den
Plastiden abzugrenzen. Dies sind auch die Fraktionen, die eine grüne Färbung zeigen.
Oberhalb dieser Banden sammeln sich Proteine aus aufgeschlossenen Zellorganellen,
die ebenfalls Signale zeigen. Nach wie vor ist unklar, ob das veränderte OPR1 Protein
sich nicht im Peroxisomen befindet, oder ob es nur nicht oder zu wenig exprimiert wird.

3. Ergebnisse 67
3.8 Wird OPR3 durch reversible Phosphorylierung regu liert?
Wie bereits erwähnt wurde, kristallisiert OPR3 als Homodimer bei dem das jeweilige
Aktive Zentrum eines Protomers von einer fingerartigen Struktur blockiert wird, die aus
dem anderen Protomer herausragt. In der Kristallstruktur imitiert ein Sulfat-Ion in der
Nähe des Tyrosins 364 die Phosphatgruppe eines Phosphorylierten Tryrosinrestes
und bildet Wasserstoffbrücken zwischen beiden Untereinheiten aus, die zur
Stabilisierung des Dimers beitragen. Tauscht man Tyrosin 364 durch Phenylalanin
aus, wodurch die Hydroxylgruppe verloren geht, kann keine Phosphorylierung mehr
stattfinden, und das Protein kristallisiert als Monomer. (Breithaupt et al., 2006)
Bereits vorausgegangene Experimente konnten zeigen, das OPR3 Substrat einer
Tyrosin-spezifischen Protein Kinase ist (Schaller A., unveröffentlicht). Western Blot
Analysen mit einem Phosphotyrosin-spezifischen monoclonalen Antikörper (Anti PT-
66) haben bewiesen, dass OPR3 effizient Phosphoryliert wird, wenn sie mit einer
Tyrosin-Kinase in E.coli coexprimiert werden.
All diese Vorkenntnisse weisen darauf hin, dass das Momomer-Dimer Gleichgewicht
von OPR3 in vivo durch eine reversible Phosphorylierung erfolgen könnte. Daher soll
in dieser Arbeit untersucht werden ob OPR3 in vivo phosphoryliert werden kann. Es ist
bekannt, dass bei einer Verwundung einer Pflanze der Spiegel an Jasmonsäure
ansteigt, ebenso steigt der Transkriptspiegel der Gene, die in der Biosynthese von
Jasmonsäure beteiligt sind. Dennoch ist die Kinetik der Transkriptinduktion nicht
schnell genug um für die schnelle Akkumulation der Jasmonsäure nach Verwundung
zu genügen was suggeriert, dass weitere Regulationsmechanismen für den
anfänglichen Anstieg von Jasmonsäure verantwortlich sind. Ein Beispiel wäre
reversible Phosphorylierung und Dephosphorylierung.
Es wurden daher Verwundungsexperimente durchgeführt, bei denen wildtypische
Tomaten Pflanzen zu verschiedenen Zeitpunkten nach Verwundung geerntet wurden
(meist 5, 10, 20, 30, 60 und 180 Minuten nach Verwundung) zu denselben Zeitpunkten
wurden Pflanzen geerntet, die nicht verwundet wurden, um als Kontrolle zu dienen.
Gesamtproteinextrakte der geernteten Pflanzen wurden mit SDS-Page und Westen
Blot Analysen mit Antikörpern auf OPR3 und Phosphorylierung (Anti PT-66)
untersucht.

3. Ergebnisse 68
Als Kontrollen dienten hierbei gereinigtes OPR3 Protein und gereinigtes Protein aus
einem E.coli Expressionsstamm der eine Tyrosin-spezifischen Protein Kinase enthält
und somit OPR3 phosphoryliert. In den folgenden Western Blot Analysen ist
phosphoryliertes OPR3 Protein als Kontrolle aufgetragen und als TKB1 bezeichnet.
K0’ K120’ M vw10’ vw30’ vw60’ vw180’ TKB1 OPR3kDa13072554334
Abb. 34: Western Blot des ersten Verwundungsexperiment an Tomaten mit Anti PT-66.
Aufgetragen wurde Gesamtproteinextrakt. Unverwundete Kontrollpflanzen wurden zu
Beginn des Experimentes geerntet und nach 120 Minuten (K0’ und K120’), verwundete
Pflanzen nach 10, 30, 60 und 180 Minuten. Kontrollpflanzen und verwundete Pflanzen
befanden sich im selben Raum. Als Kontrolle wurden OPR3 Protein und OPR3 Protein
aus dem Expressionsvektor TKB1 aufgetragen, dass eine Tyrosinkinase enthält und
OPR3 somit Phosphoryliert(Im Bild bezeichnet als TKB1).
Abbildung 34 zeigt den Western blot eines Verwundungsexperiments mit
Tomatenpflanzen, der mit Anti PT-66 detektiert wurde. Sowohl in den Verwundeten
(Spuren vw10’, vw30’, vw60’, vw180’), als auch in den Kontrollpflanzen (K0’ und
K120’) sind dieselben Banden an phosphorylierten Proteinen zu erkennen. Auch zu
den verschiedenen Zeitpunkten ist keine Veränderung zu erkennen. Es liegt hier also
offenbar keine induzierbare Phosphorylierung vor. Es wurden weitere Experimente mit
mehr Zeitpunkten durchgeführt um der Spiegel an OPR3 zu verschiedenen
Verwundungszeiten zu messen.
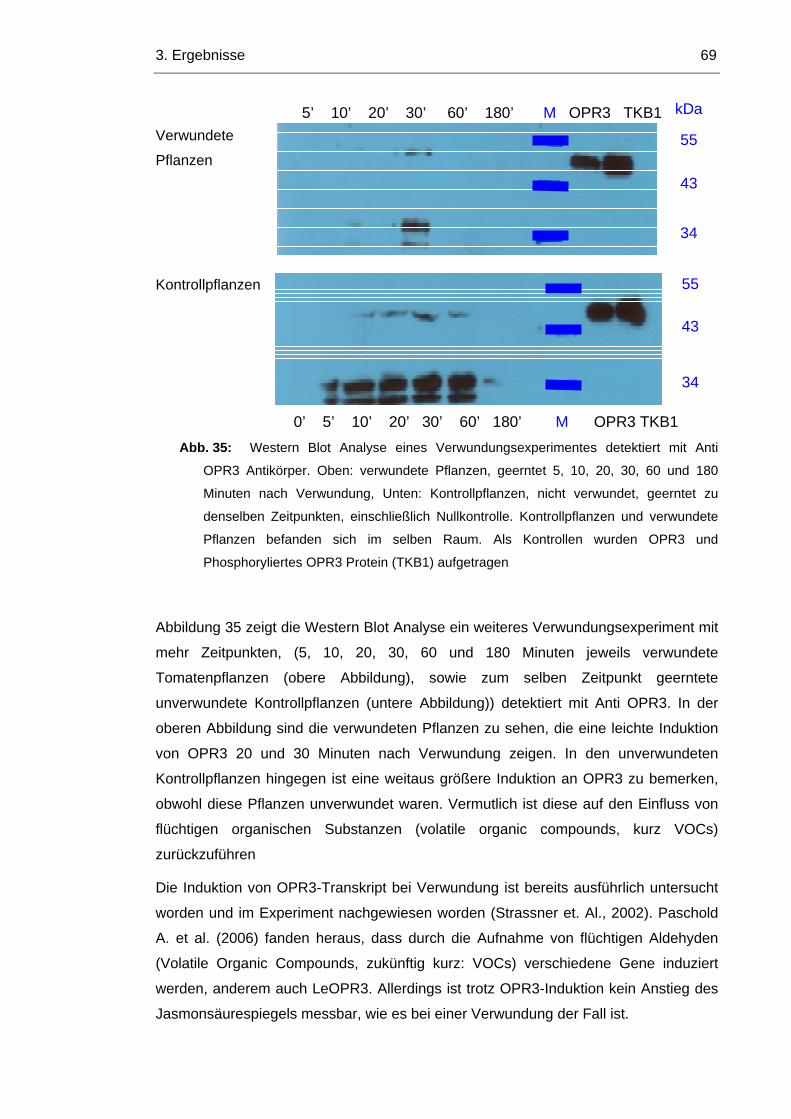
3. Ergebnisse 69
Verwundete
Pflanzen
Kontrollpflanzen
Abb. 35: Western Blot Analyse eines Verwundungsexperimentes detektiert mit Anti
OPR3 Antikörper. Oben: verwundete Pflanzen, geerntet 5, 10, 20, 30, 60 und 180
Minuten nach Verwundung, Unten: Kontrollpflanzen, nicht verwundet, geerntet zu
denselben Zeitpunkten, einschließlich Nullkontrolle. Kontrollpflanzen und verwundete
Pflanzen befanden sich im selben Raum. Als Kontrollen wurden OPR3 und
Phosphoryliertes OPR3 Protein (TKB1) aufgetragen
Abbildung 35 zeigt die Western Blot Analyse ein weiteres Verwundungsexperiment mit
mehr Zeitpunkten, (5, 10, 20, 30, 60 und 180 Minuten jeweils verwundete
Tomatenpflanzen (obere Abbildung), sowie zum selben Zeitpunkt geerntete
unverwundete Kontrollpflanzen (untere Abbildung)) detektiert mit Anti OPR3. In der
oberen Abbildung sind die verwundeten Pflanzen zu sehen, die eine leichte Induktion
von OPR3 20 und 30 Minuten nach Verwundung zeigen. In den unverwundeten
Kontrollpflanzen hingegen ist eine weitaus größere Induktion an OPR3 zu bemerken,
obwohl diese Pflanzen unverwundet waren. Vermutlich ist diese auf den Einfluss von
flüchtigen organischen Substanzen (volatile organic compounds, kurz VOCs)
zurückzuführen
Die Induktion von OPR3-Transkript bei Verwundung ist bereits ausführlich untersucht
worden und im Experiment nachgewiesen worden (Strassner et. Al., 2002). Paschold
A. et al. (2006) fanden heraus, dass durch die Aufnahme von flüchtigen Aldehyden
(Volatile Organic Compounds, zukünftig kurz: VOCs) verschiedene Gene induziert
werden, anderem auch LeOPR3. Allerdings ist trotz OPR3-Induktion kein Anstieg des
Jasmonsäurespiegels messbar, wie es bei einer Verwundung der Fall ist.
kDa
55
43
34
55
43
34
0’ 5’ 10’ 20’ 30’ 60’ 180’ M OPR3 TKB1
5’ 10’ 20’ 30’ 60’ 180’ M OPR3 TKB1

3. Ergebnisse 70
Es scheint als würde die Verwundungsreaktion nur bis zu einem bestimmten Punkt
ablaufen und dort innehalten. Grundlage der folgenden Experimente war die
Vermutung, dass OPR3 durch die Aufnahme von VOCs zwar induziert wird, aber
eventuell durch eine vorübergehende Phosphorylierung inaktiv ist, wodurch die
Pflanze bei einer wirklichen Verwundung schneller eine Abwehrreaktion starten kann.
Zwischen 50 und 100 Tomatenpflanzen wurden in regelmäßigen Kreisen aufgestellt.
Jeweils in der Mitte der Kreise und um sie herum wurde eine Tomatenpflanze gestellt,
die mit einer Gefäßklemme an mehreren Blättern verwundet wurde. Die Verwundung
wurde jeweils so gesetzt, dass sie die Blatthauptader durchquerte. Diese Pflanzen, die
VOCs absondern, sind die Emitterpflanzen, unverwundete Pflanzen, die in
unmittelbarer Nähe stehen sind in diesem Fall Empfänger-Pflanzen, anhand derer die
OPR3 Induktion durch VOCs nachgewiesen werden soll. Zusätzlich wurden Blätter
von Tomaten in kleine Stücke zerschnitten, zerdrückt und zwischen den Töpfen
verteilt, um eine optimale Verteilung der so freigesetzten VOCs zu garantieren. Des
Weiteren sind Pflanzen in abgetrennten Räumen aufgestellt, die weder verwundet
werden, noch VOCs ausgesetzt sind, um als Kontrollen zu dienen. Bei Experimenten
mit Arabidopsis Pflanzen wurden diese in regelmäßigen Rechtecken von 9 oder 16
Pflanzen zusammengestellt, wobei immer bei einer bis vier Pflanzen pro Gruppen die
Rosettenblätter mit Gefäßklemmen verwunden wurden. Auch hier wurde darauf
geachtet, dass die Verwundung die Blattader durchquert.
Eventuellem Luftzug wurde vorgebeugt indem eine Stellage aus Brettern und Kisten
um den Versuchsaufbau gestellt wurde. Ein komplettes Abschotten der Pflanzen in
beispielsweise einem Plexiglasbehälter ist nicht sinnvoll. Durch Photosynthese wird
der CO2-Gehalt im abgeschlossenen Behälter schnell reduziert. Wenn die Pflanzen
ihren CO2-Kompensationspunkt erreicht haben, erhöht sich die Zahl der geöffneten
Stomata und erhöht somit die Aufnahme von VOCs wodurch eine inadäquate
Aufnahme von VOCs gegenüber der natürlichen stattfindet und somit eine ungleich
höhere oder womöglich sogar andere Reaktion hervorruft (Paschold A. et al., 2006).
Sowohl von den Emitterpflanzen, als auch von den nicht verwundeten Receiver-
Pflanzen wurden zu bestimmten Zeitpunkten von je 6-8 Pflanzen die jüngsten Blätter
geerntet und sofort in flüssigem Stickstoff eingefroren. Außerdem wurden weitere
Pflanzen in abgetrennte Räume gestellt, und ebenfalls zu bestimmten Zeitpunkten
geerntet um als Kontrolle zu dienen. Die so gewonnenen Pflanzenproben wurden
mittels Western Blot analysiert. Alle gezeigten Ergebnisse sind mindestens dreimal
wiederholt worden, in den meisten Fällen konnte ein biologisches Replikat erstellt
werden um die Ergebnisse zu verifizieren.
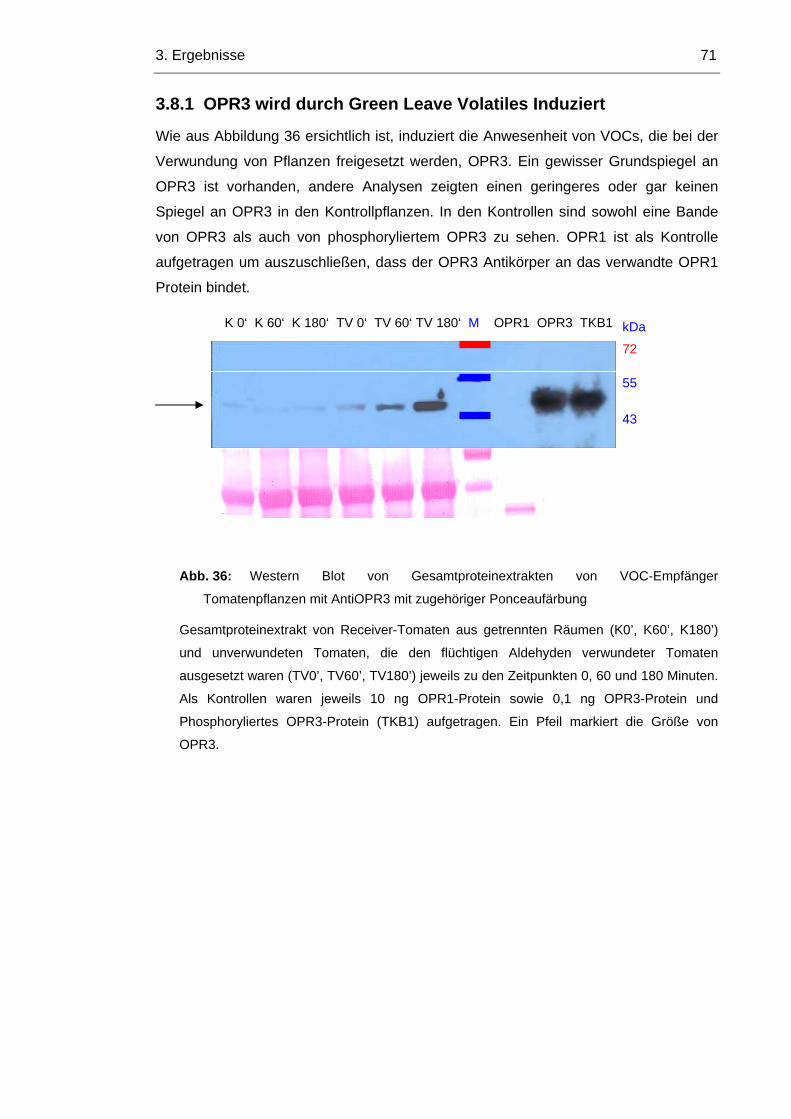
3. Ergebnisse 71
3.8.1 OPR3 wird durch Green Leave Volatiles Induzie rt
Wie aus Abbildung 36 ersichtlich ist, induziert die Anwesenheit von VOCs, die bei der
Verwundung von Pflanzen freigesetzt werden, OPR3. Ein gewisser Grundspiegel an
OPR3 ist vorhanden, andere Analysen zeigten einen geringeres oder gar keinen
Spiegel an OPR3 in den Kontrollpflanzen. In den Kontrollen sind sowohl eine Bande
von OPR3 als auch von phosphoryliertem OPR3 zu sehen. OPR1 ist als Kontrolle
aufgetragen um auszuschließen, dass der OPR3 Antikörper an das verwandte OPR1
Protein bindet.
kDa
72
55
43
K 0‘ K 60‘ K 180‘ TV 0‘ TV 60‘ TV 180‘ M OPR1 OPR3 TKB1
Abb. 36: Western Blot von Gesamtproteinextrakten von VOC-Empfänger
Tomatenpflanzen mit AntiOPR3 mit zugehöriger Ponceaufärbung
Gesamtproteinextrakt von Receiver-Tomaten aus getrennten Räumen (K0’, K60’, K180’)
und unverwundeten Tomaten, die den flüchtigen Aldehyden verwundeter Tomaten
ausgesetzt waren (TV0’, TV60’, TV180’) jeweils zu den Zeitpunkten 0, 60 und 180 Minuten.
Als Kontrollen waren jeweils 10 ng OPR1-Protein sowie 0,1 ng OPR3-Protein und
Phosphoryliertes OPR3-Protein (TKB1) aufgetragen. Ein Pfeil markiert die Größe von
OPR3.

3. Ergebnisse 72
vw5’ vw10’ vw20’ vw30’ vw45’ vw60’ vw120’ vw180’ M OPR3 TKB1kDa
72
55
43
34
Abb. 37: Western Blot von Gesamtproteinextrakten verwundeter Tomatenpflanzen mit
AntiOPR3
Gesamtproteinextrakte von verwundeten Tomatenpflanzen zu den Zeitpunkten 5, 10, 20,
30, 45, 60, 120 und 180 Minuten, im Bild bezeichnet durch vw5’, vw10’, vw20’, vw30’,
vw45’, vw60’, vw120’ und vw180’. Als Kontrollen wurden aufgetragen jeweils 0,1 ng OPR3-
Protein und phosphoryliertes OPR3-Protein. Ein Pfeil markiert die Höhe von OPR3.
Abbildung 37 zeigt, dass OPR3 durch Verwundung bereits nach 10 Minuten induziert
wird, mit einem geringen Rückgang nach 30 Minuten und einem erneuten Maximum
bei 60 Minuten. Nach 180 Minuten ist bereits kein OPR3 mehr vorhanden, während
bei der Induktion durch VOCs nach 180 Minuten das Maximum erreicht ist.
2 weitere Banden lassen sich am unteren Rand des Blots ausmachen, bei einer Höhe
zwischen 30 und 40 kDa. Diese könnten Degradationsbanden von OPR3 seien, da sie
im selben Maße zunehmen wie OPR3, allerdings um einen gewissen Zeitfaktor nach
hinten versetzt, oder Degradationsbanden anderer Proteine, die durch die
Verwundungsreaktion induziert werden und OPR3 ähnlich sind, wie z.B. andere
Proteine aus der OPR Familie, allerdings sind sie viel kleiner als OPR3.
Die Induktion bei Verwundung ist eine schnellere, aber zeitlich begrenztere Induktion
als durch VOCs. Wenn man bedenkt, dass die Induktion durch VOCs nur eine
Vorsichtsmaßnahme für eventuell folgende Verwundungen ist, macht dies durchaus
Sinn, da die Bildung von OPR3 sozusagen auf Vorrat für eine nur möglicherweise viel
später eintretende Verwundung erfolgt.
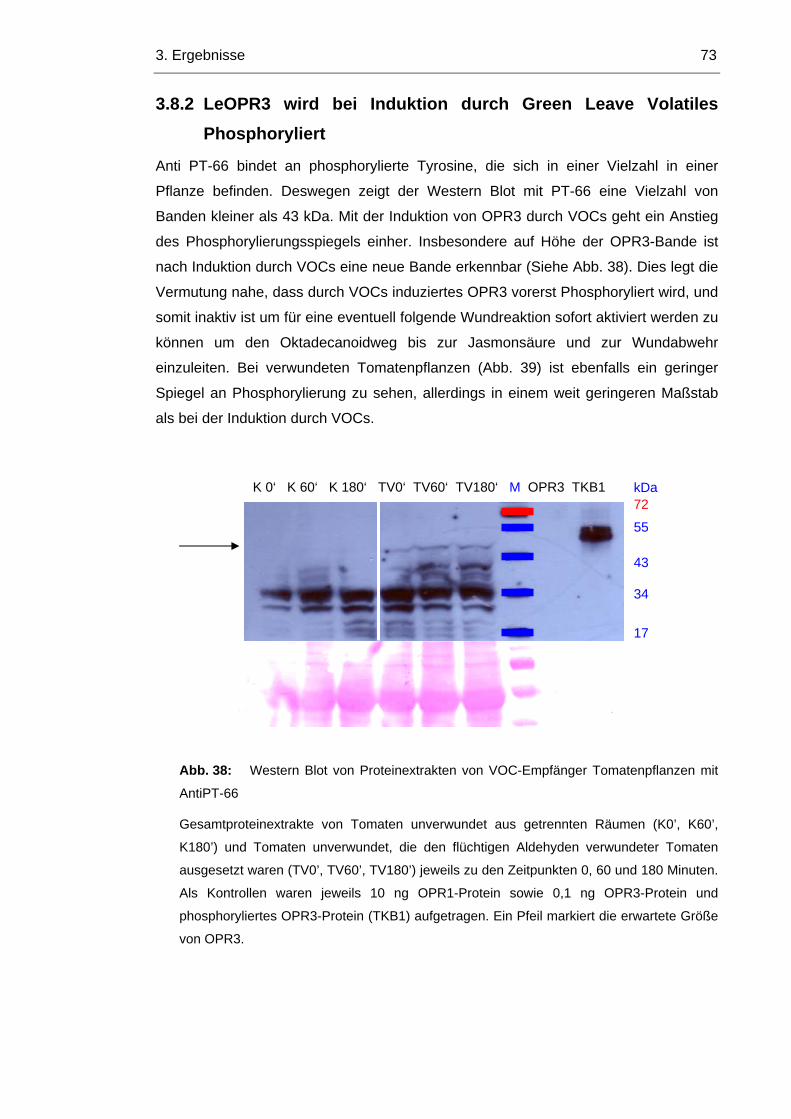
3. Ergebnisse 73
3.8.2 LeOPR3 wird bei Induktion durch Green Leave Volatiles
Phosphoryliert
Anti PT-66 bindet an phosphorylierte Tyrosine, die sich in einer Vielzahl in einer
Pflanze befinden. Deswegen zeigt der Western Blot mit PT-66 eine Vielzahl von
Banden kleiner als 43 kDa. Mit der Induktion von OPR3 durch VOCs geht ein Anstieg
des Phosphorylierungsspiegels einher. Insbesondere auf Höhe der OPR3-Bande ist
nach Induktion durch VOCs eine neue Bande erkennbar (Siehe Abb. 38). Dies legt die
Vermutung nahe, dass durch VOCs induziertes OPR3 vorerst Phosphoryliert wird, und
somit inaktiv ist um für eine eventuell folgende Wundreaktion sofort aktiviert werden zu
können um den Oktadecanoidweg bis zur Jasmonsäure und zur Wundabwehr
einzuleiten. Bei verwundeten Tomatenpflanzen (Abb. 39) ist ebenfalls ein geringer
Spiegel an Phosphorylierung zu sehen, allerdings in einem weit geringeren Maßstab
als bei der Induktion durch VOCs.
K 0‘ K 60‘ K 180‘ TV0‘ TV60‘ TV180‘ M OPR3 TKB1 kDa72
55
43
34
17
Abb. 38: Western Blot von Proteinextrakten von VOC-Empfänger Tomatenpflanzen mit
AntiPT-66
Gesamtproteinextrakte von Tomaten unverwundet aus getrennten Räumen (K0’, K60’,
K180’) und Tomaten unverwundet, die den flüchtigen Aldehyden verwundeter Tomaten
ausgesetzt waren (TV0’, TV60’, TV180’) jeweils zu den Zeitpunkten 0, 60 und 180 Minuten.
Als Kontrollen waren jeweils 10 ng OPR1-Protein sowie 0,1 ng OPR3-Protein und
phosphoryliertes OPR3-Protein (TKB1) aufgetragen. Ein Pfeil markiert die erwartete Größe
von OPR3.

3. Ergebnisse 74
vw5’ vw10’ vw20’ vw30’ vw45’ vw60’ vw120’ vw180’ M OPR3 TKB1kDa55
43
34
Abb. 39: Western blot von Proteinextrakten verwundeter Tomatenpflanzen mit Anti PT-
66
Gesamtproteinextrakte von verwundeten Tomatenpflanzen zu den Zeitpunkten 5, 10, 20,
30, 45, 60, 120 und 180 Minuten, im Bild bezeichnet durch vw5’, vw10’, vw20’, vw30’,
vw45’, vw60’, vw120’ und vw180’. Als Kontrollen wurden aufgetragen jeweils 0,1 ng OPR3-
Protein und phosphoryliertes OPR3-Protein (TKB1). Ein Pfeil markiert die Höhe von OPR3.
3.8.3 Im Arabidopsis Wildtyp wird OPR1 durch die In duktion von
OPR3 reprimiert
Um sicher zu gehen, dass es sich bei den angezeigten Signalen des OPR3
Antikörpers auch wirklich um ein OPR3 Protein handelt und nicht um eine
Kreuzreaktion mit einem OPR1 Protein, wurden zusätzliche Western Blot Analysen mit
Antikörpern gegen OPR1 und OPR3 Proteine durchgeführt. Hierfür wurde ein neues
Verwundungsexperiment geplant, bei dem zusätzlich klar gestellt werden sollte, ob
VOCs spezifisch intrafamiliär wirken, oder ob auch VOCs von Artfremden Pflanzen
eine Reaktion auslösen können. Hierfür wurden Arabidopsis-Pflanzen als VOC-
Empfänger gewählt, wohingegen Tomaten und Arabidopsis-Pflanzen in 2 getrennten
Experimenten als VOC Emitter Pflanzen verwundet wurden. In beiden Versuchsreihen
wurden die VOC Empfänger Pflanzen nach 0, 30, 60 und 90 Minuten geerntet.
Verwundete Emitter Pflanzen wurden nach 60 und 90 Minuten geerntet. Des Weiteren
wurde eine Arabidopsis opr3-Mutante Pflanze ebenfalls verwundet und nach 60
Minuten geerntet.

3. Ergebnisse 75
Anti OPR1
AntiOPR3
Anti PT-66
Abb. 40: Western Blots von Arabidopsis-Pflanzen, die VOCs von Tomaten (TV) und
Arabidopsis-Pflanzen (AV) ausgesetzt waren, geerntet nach je 0, 30, 60 und 90 Minuten.
Als Kontrolle dienten verwundete opr3 Pflanzen, die nach 60 Minuten geerntet wurden,
sowie verwundete Arabidopsis-Pflanzen nach 60 und 90 Minuten Verwundung, außerdem
0,1 ng aufgereinigtes OPR3 und 10 ng OPR1 Protein bzw. 0,1 ng Phosphoryliertes OPR3
Protein (TKB1). Detektion mit AntiOPR1 (Oben), AntiOPR3 (Mitte) und Anti PT-66 (Unten)
kb
72
55
43
34
TV
0’
TV
30’
TV
60’
TV
90’
AV
0’
AV
30’
AV
60’
AV
90’
Mar
ker
opr3
vw
60’
vw60
’
vw90
’
OP
R3
OP
R1
kb
72
55
43
TV
0’
TV
30’
TV
60’
TV
90’
AV
0’
AV
30’
AV
60’
AV
90’
Mar
ker
opr3
vw
60’
vw60
’
vw90
’
OP
R3
OP
R1
kDa
72
55
43
34
TV
0’
TV
30’
TV
60’
TV
90’
AV
0’
AV
30’
AV
60’
AV
90’
Mar
ker
opr3
vw 6
0’
vw60
’
vw90
’
TK
B1
OP
R3

3. Ergebnisse 76
Abbildung 40 zeigt die Gesamtproteinextrakte beider Versuchsanordnungen. Es zeigt,
dass OPR1 60 bis 90 Minuten, nachdem in die Pflanze entweder verwundet, oder von
VOCs erreicht wurde, im Western Blot nicht mehr nachweisbar ist. Im Gegenzug steigt
der Spiegel an OPR3 bei der Induktion durch VOCs bereits nach 30 Minuten deutlich
an. Dieses Experiment lässt darauf schließen, dass sowohl OPR1 als auch OPR3
durch eine Verwundung bzw. eine Reaktion auf VOCs reguliert werden. Weitere
Experimente mit opr1- und opr3-mutanten Pflanzen sollen weiteren Aufschluss über
diesen Zusammenhang geben.
Von denselben Proteinextrakten wurden nun auch eine Western Blot Analyse mit Anti-
PT 66 durchgeführt, um Aufschluss über die Phosphorylierung von OPR3 zu erlangen.
Wie allerdings Abbildung 40 (unten) zeigt sind zwar auf Höhe des OPR3 Proteins
Signale zu sehen, die aber in der opr3-Mutante ebenfalls zu finden sind. Es kann sich
hierbei also nicht um das gesuchte OPR3 Protein handeln.
3.8.4 In opr1 Pflanzen ist OPR3 konstitutiv aktiv
Wie bereits in den vorangegangenen Experimenten erwähnt, sollte sicher gestellt
werden, dass das von Anti OPR3 detektierte Protein wirklich OPR3 ist und nicht eine
Kreuzreaktion mit OPR1 ein Signal gibt. Deswegen wurde ein weiteres VOC
Experimente mit Arabidopsis opr1-Mutanten Pflanzen durchgeführt. Hierbei stellte sich
heraus dass durch die Abwesenheit von OPR1 OPR3 konstitutiv aktiv ist (siehe Abb.
41 oben). Sowohl in den Kontrollpflanzen, als auch in allen verwundeten und durch
VOCs induzierten Pflanzen ist bereits vom Nullpunkt an ein hoher Spiegel an OPR3 zu
beobachten. Bei einer Induktion durch VOCs erhöht und verringert sich dieser Spiegel
noch im selben Maße, wie es auch bei einer normalen Verwundungsreaktion der Fall
ist. Überraschenderweise ist 120 Minuten nach der ersten VOC-Freisetzung in den
Empfänger Pflanzen OPR3 nicht mehr detektierbar, wohingegen bei Abbildung 36
noch ein Maximum an OPR3 bei 180 Minuten nach dem Empfang von VOCs zu sehen
war.
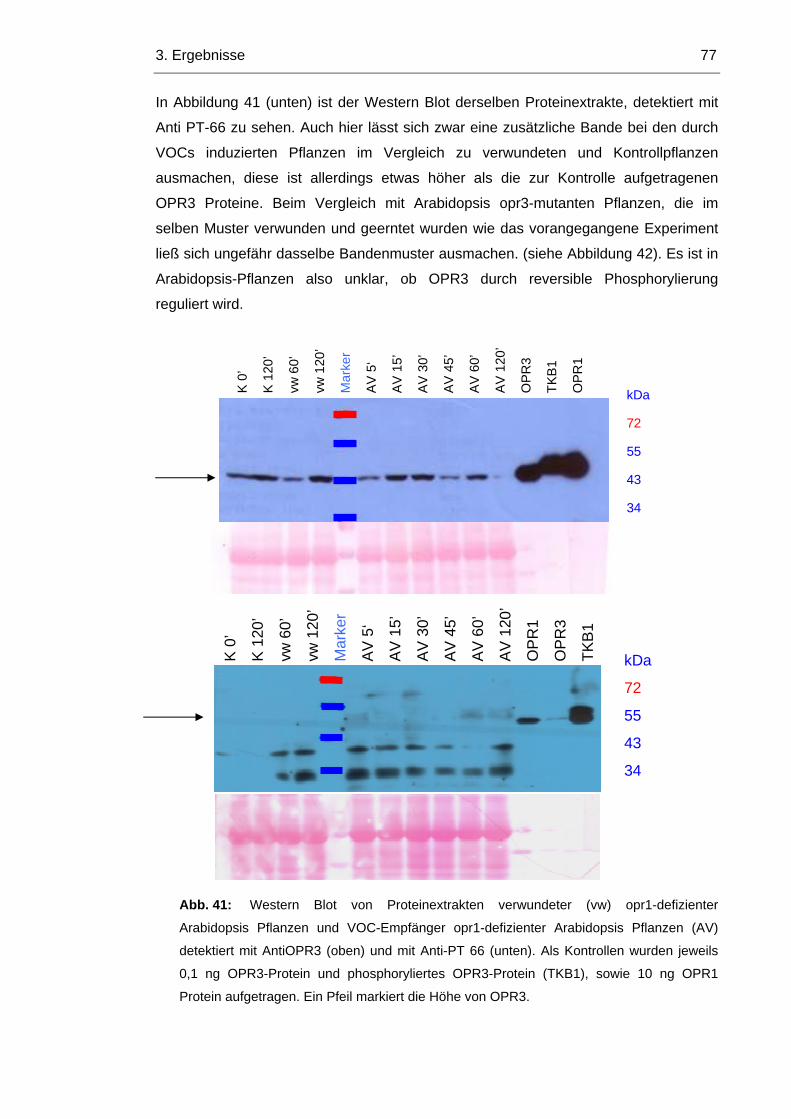
3. Ergebnisse 77
In Abbildung 41 (unten) ist der Western Blot derselben Proteinextrakte, detektiert mit
Anti PT-66 zu sehen. Auch hier lässt sich zwar eine zusätzliche Bande bei den durch
VOCs induzierten Pflanzen im Vergleich zu verwundeten und Kontrollpflanzen
ausmachen, diese ist allerdings etwas höher als die zur Kontrolle aufgetragenen
OPR3 Proteine. Beim Vergleich mit Arabidopsis opr3-mutanten Pflanzen, die im
selben Muster verwunden und geerntet wurden wie das vorangegangene Experiment
ließ sich ungefähr dasselbe Bandenmuster ausmachen. (siehe Abbildung 42). Es ist in
Arabidopsis-Pflanzen also unklar, ob OPR3 durch reversible Phosphorylierung
reguliert wird.
K 0
’
K 1
20’
vw60
’
vw12
0’
Mar
ker
AV
5‘
AV
15’
AV
30’
AV
45’
AV
60’
AV
120
’
OP
R3
TK
B1
OP
R1
kDa
72
55
43
34
K 0
’
K 1
20’
vw 6
0’
vw 1
20’
Mar
ker
AV
5‘
AV
15’
AV
30’
AV
45’
AV
60’
AV
120
’
OP
R1
OP
R3
TK
B1
kDa
72
55
43
34
Abb. 41: Western Blot von Proteinextrakten verwundeter (vw) opr1-defizienter
Arabidopsis Pflanzen und VOC-Empfänger opr1-defizienter Arabidopsis Pflanzen (AV)
detektiert mit AntiOPR3 (oben) und mit Anti-PT 66 (unten). Als Kontrollen wurden jeweils
0,1 ng OPR3-Protein und phosphoryliertes OPR3-Protein (TKB1), sowie 10 ng OPR1
Protein aufgetragen. Ein Pfeil markiert die Höhe von OPR3.

3. Ergebnisse 78
K 0
’
K 1
20’
vw 6
0’
vw 1
20’
AV
5‘
AV
15’
AV
30’
AV
45’
AV
60’
AV
120
’
Mar
ker
OP
R3
OP
R1
TK
B1
kDa
72
55
43
34
Abb. 42: Western Blot von Proteinextrakten verwundeter (vw) opr3-defizienter
Arabidopsis Pflanzen und VOC-Empfänger opr1-defizienter Arabidopsis Pflanzen (AV)
detektiert mit Anti-PT 66. Als Kontrollen wurden jeweils 0,1 ng OPR3-Protein und
phosphoryliertes OPR3-Protein (TKB1) sowie 10 ng OPR1 Protein aufgetragen. Ein Pfeil
markiert die Höhe von OPR3.

4. Diskussion 79
4. Diskussion
4.1 OPR3 kann nicht durch punktmutierte OPR1 Proteine
ersetzt werden
Die Genfamilie der Oxophytodiensäure Reduktasen ist eine weit verbreitete Familie in
Pflanzen. In Tomate wurden bisher drei, in Arabidopsis fünf Isoformen identifiziert, die
eine Reduktion von α.β−ungesättigten Kohlenstoffverbindungen katalysieren. OPR3 ist
der am weitesten entfernte Verwandte der Familie und die einzige Isoform, die in der
Lage ist, die Reduktion von cis(+)OPDA zu katalysieren. OPR1 hingegen kann in vitro
nur cis(-)OPDA reduzieren. Die hohe Übereinstimmung in der Aminosäuresequenz von
OPR3 und OPR1 und ihre ähnlichen Substrate in vitro legen nahe, dass OPR1 durch
zielgerichtete Mutationen dasselbe Substratspektrum erreichen könnte, wie OPR3.
Hierfür wurden OPR1 cDNAs an 2 Stellen punktmutiert. Diese Punktmutationen waren
im β3-Loop, der in der Struktur von OPR1 eine Art Deckel über das Aktive Zentrum
bildet und somit den Zutritt von cis(+)OPDA, des physiologischen Substrates von
OPR3, verhindert. Um die Lokalisation des mutierten Proteins im Peroxisom, dem
Wirkungsort von OPR3 sicherzustellen, wurde außerdem durch PCR ein SRL-Tag an
das C-terminale Ende von OPR1 kloniert.
Dennoch konnte eine opr3-defiziente Arabidopsis thaliana Pflanze nicht durch die
punktmutierten OPR1 Proteine komplementiert werden. Eine Komplementation durch
die Punktmutationen ist also nicht ausreichend um die Substratspezifität von OPR3
herzustellen. Um sicher zu gehen, dass die Proteine im Peroxisom lokalisiert sind und
nicht aufgrund einer Fehllokalisation keine Aktivität zeigen, wurden Zellorganellen mit
Hilfe eines Saccharosegradienten aufgetrennt und untersucht. Aufgrund der geringen
Menge an Pflanzenmaterial konnte allerdings nicht eindeutig geklärt werden, ob sich
die mutierten Proteine im Peroxisom befanden oder nicht.
Um diesen Sachverhalt eindeutig zu klären müssen erneut Samen der transformierten
Pflanzen ausgesät und die Lokalisation der Proteine durch Zellorganellfraktionierung
untersucht werden. Eine weitere Möglichkeit der Untersuchung der subzellulären
Lokalisation wäre eine Detektion durch eine in situ Protein Lokalisierung mit
fluoreszierenden Antikörpern gegen OPR1 (Sauer M. et al., 2006). Hierfür würde sich
allerdings eine opr1-defiziente Pflanze eignen, da OPR1 normalerweise im Cytoplasma
lokalisiert ist und eine Lokalisation im Peroxisom somit überstrahlt werden könnte.
Sollte wider erwarten das Protein nicht im Peroxisom vorhanden sein müsste ein neues
Konstrukt zur Transformation hergestellt werden.

4. Diskussion 80
Allerdings konnte bereits gezeigt werden, dass das SRL-Tag für eine Lokalisation
eines Proteins im Peroxisom ausreichend ist (Mullen, 1997). Ferner sollten neue
Transformationen von Arabidopsis opr3-Mutanten mit den verschiedenen Konstrukten
OPR1+SRL, Y74F, Y244H, und Y74F/Y244H erfolgen, um mehr unabhängige Linien
zu erhalten. Die Insertion der OPR1 enthaltenden T-DNA erfolgt zufällig im Genom. Es
ist möglich, dass weitere Linien einen Phänotyp zeigen.
Eventuell ist OPR1 nicht ausreichend exprimiert um eine ausreichende Menge an
Protein zu produzieren, die den Oktadekanoidweg vervollständigen kann. Es ist zwar
ein 2130 bp großes Fragment des OPR3 Promotors in der T-DNA vorhanden, das
ausreichend groß sein sollte um die Transkription von OPR1 zu gewährleisten, aber
eventuell ist durch das Vorhandensein nur einer Genkopie im diploiden Organismus die
Funktion nicht ausreichend. Die T2 Generationen der jeweiligen transformierten
Pflanzen, in denen zwei Kopien des mutierte Genes vorhanden sind, sollten hierüber
Aufschluss geben. Eventuell könnten weitere Konstrukte mit einem 35S-Promotor statt
des OPR3 Promotors Aufschluss über sowohl die Aktivität, als auch, bei einer erneuten
Zellorganellfraktionierung, über die Lokalisation der mutierten OPR1-Proteine geben.
Sollten auch diese Pflanzen männliche Sterilität zeigen, so müssen noch weitere
Aminosäuren im β3-Loop mutiert werden. Hierbei gibt es mehrere Möglichkeiten.
Entweder tauscht man die Aminosäuren aus, die in der Nähe der bereits mutierten
Tyrosine an Position 244 und 74 sind, oder man versucht den β3-Loop bereits im
Ansatz zu mutieren, um einen anderen Winkel des Loops zu erreichen, wodurch der
Zugang des Substrates zum aktiven Zentrum erleichtert wird.

4. Diskussion 81
4.2 Arabidopsis zeigt keinen Veränderungen durch
punktmutierte OPR3 Proteine
Durch Röntgenkristallanalysen konnte gezeigt werden, dass OPR3 als Homodimer
kristallisiert. Zielgerichtete Mutationen in der Dimerisierungsschleife des Glutamats an
Position 291 in Lysin (E291K) haben gezeigt, dass das so mutierte Protein in vitro nicht
mehr dimerisieren kann (Breithaupt et al., 2006). Eine Komplementation von opr3-
defizienten Arabidopsis thaliana mit dem punktmutierten Protein war erfolgreich, die
männliche Fertilität konnte wieder hergestellt werden.
Für eine weitere Mutation, Tyrosin an Position 364 wurde durch Phenylalanin ersetzt
(Y364F), konnten allerdings keine positiven transformierten Pflanzen identifiziert
werden. Eine erneute Transformation von opr3-defizienten Arabidopsis thaliana ist
notwendig um Rückschlüsse auf die Wiederherstellung der männlichen Fertilität ziehen
zu können.
Mit den T2 Generationen beider Transformanten müssen dann eine Vielzahl an
Experimenten durchgeführt werden, um Rückschlüsse ziehen zu können, ob OPR3 in
vivo durch die eingebrachten Mutationen nicht mehr dimerisieren kann. Durch native
SDS-PAGE lässt sich klären, ob das mutierte OPR3 als Monomer oder als Dimer
vorliegt.
Durch die dauerhafte Aktivität von OPR3 könnte auch konstant Jasmonsäure gebildet
werden. Ein solch erhöhter Jasmonsäurespiegel, ohne dass eine Pflanze verwundet
oder den bereits erwähnten Volatile organic compounds (VOCs) ausgesetzt wurde, ist
messbar. Wie in der Einleitung erwähnt, steuert Jasmonsäure eine Vielzahl an
Prozessen in der Pflanze. Durch eine dauerhafte Aktivität von OPR3 könnte auch
beständig Jasmonsäure gebildet werden und somit die Transkripte der durch
Jasmonsäure induzierten Gene dauerhaft vorhanden sein. Diese lassen sich durch
Microarray-Analysen im Vergleich mit wildtypischen Pflanzen und den transgenen
Pflanzen nachweisen. Auch ein erhöhter Jasmonsäurespiegel gegenüber dem Wildtyp
sollte durch die konstante Aktivität nachweisbar sein.
Sollten entgegen der Ergebnisse der Kristallisierung der beiden mutierten Proteine
OPR3 in vivo dennoch dimerisieren können, müssten Doppelmutationen der beiden
Proteine hergestellt werden um somit auszuschließen, dass nur eine der beiden
Mutationsstellen für eine erfolgreiche Dimerisierung ausreichend ist.

4. Diskussion 82
4.3 OPR3 wird durch VOCs induziert
Bei Verwundungsexperimenten und Experimenten, bei denen Tomatenpflanzen VOCs
von verwundeten Tomaten ausgesetzt waren, konnte gezeigt werden, dass OPR3
sowohl durch Verwundung, als auch durch VOCs induziert wird. Es ist also nicht nur
eine Transkript-Induktion von OPR3 wie bei Nicotiana attenuata (Paschold et al.,
2006), sondern auch ein Vorhandensein des Proteins nachweisbar.
Western Blots mit einem Phosphotyrosin spezifischem Antikörper (Anti PT-66) zeigten,
dass von der Größe des OPR3 Proteins eine zusätzliche Bande im Vergleich zu nicht
mit VOC induzierten Kontrollpflanzen sichtbar ist. Allerdings ist nicht eindeutig
nachweisbar, dass sich diese Bande auch auf OPR3 bezieht, oder ob ein anderes
Protein derselben Größe durch den Empfang von VOCs phosphoryliert wird.
Bei Arabidopsis Pflanzen hingegen ist zwar, wie in Tomatenpflanzen, dieselbe
Induktion von OPR3 durch Verwundung und VOCs sichtbar, allerdings konnte keine
eindeutige Phosphorylierung festgestellt werden. Der Vergleich von wildtypischen,
opr3-defizienten und opr1-defizienten Arabidopsis thaliana erbrachte ebenfalls keinen
Aufschluss darüber, welches Protein phosphoryliert wird. Eine zweidimensionale SDS-
Page könnte Aufschluss geben, ob OPR3 oder OPR1 durch die Induktion mit VOCs
phosphoryliert werden.
Es stellt sich natürlich die Frage, welchen Sinn ein Vorhandensein des OPR3-Proteins
hat, wenn keine Jasmonsäure gebildet wird (Paschold et al., 2006). Die Autoren
vermuten, dass durch die Transkription von Genen, die in einer Abwehrreaktion bei
Befall durch Herbivore beteiligt sind, die Fitness der Pflanze steigt, indem sie bei einem
eigenen Befall schneller Jasmonsäure ausschütten kann. Somit werden andere Gene
schneller induziert und eine Abwehrreaktion setzt schneller ein.
In Langzeitexperimenten ließe sich überprüfen, ob Pflanzen, die längere Zeit VOCs
ausgesetzt sind eine schnellere Abwehrreaktion und somit auch eine höhere Fitness
haben, als Pflanzen, die ohne vorherigen VOC-Einfluss verwundet wurden.
Es wäre interessant zu erfahren, an welchen Stellen OPR3 phosphoryliert wird und
durch welche Mechanismen diese Phosphorylierung vermittelt wird. Aktivitätstest eines
phosphorylierten OPR3 Proteins können Aufschluss darüber geben, wann eine
Phosphorylierung Einfluss auf die Aktivität hat.

4. Diskussion 83
Um einen Rückschluss auf Phosphorylierung von OPR3 im Zusammenhang mit der
Aktivität zu erhalten, empfehlen sich die mit den E291K und Y364F transformierten
Arabidopsis-Pflanzen für gleichartige Experimente. Die Mutation Y364F ist an einer
Stelle, die eine Dimerisierung durch Phosphorylierung stabilisieren soll. Wenn im
Wildtyp eine Dimerisierung durch eine Phosphorylierung auftritt und OPR3 somit
inaktiviert ist, sollte es bei dieser Mutante weder zu Phosphorylierung noch zur
Inaktivität kommen. Pflanzen die VOCs ausgesetzt sind, sollten also
unphosphoryliertes OPR3 bilden und somit auch Jasmonsäure, obwohl sie nicht
verwundet sind – gesetzt den Fall, dass Jasmonsäure nicht schon vorher durch eine
dauerhafte OPR3 Aktivität gebildet worden ist.
Des Weiteren sollten auch Experimente mit Emitter Pflanzen durchgeführt werden, die
nicht mit einer Gefäßklemme nur mechanisch verwundet wurden, sondern auf
natürliche Weise verwundet werden, beispielsweise durch Frass von Manduca sexta,
um zu überprüfen, ob sich die Reaktion der Empfänger-Pflanzen ändert. Möglich wäre,
dass sich der Phosphorylierungsspiegel ändert, wenn die Verwundung natürlicher Art
ist. Alborn et al. (1997) konnten zeigen, dass durch eine Verwundung von Mais durch
Raupenfrass Volicitin gebildet wird, was zur Freisetzung von VOCs führt. Die Pflanze
kann allerdings aufgrund der enzymatischen Aktivität des Raupenregurgitats
unterscheiden, ob sie mechanisch oder durch einen Fraßfeind verwundet wird. So
scheint die Verwundung durch Fraß einen höheren Stellenwert einzunehmen, als eine
reine mechanische Verwundung.

5. Anhang VII
5. ANHANG
5.1 Ergebnisse der Pollenkeimungstests
5.1.1 Wildtyp
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen
1 106 282 388 72,7%
2 74 327 401 81,5%
3 66 259 325 79,7%
4 66 303 369 82,1%
5 65 215 280 76,8%
6 66 233 299 77,9%
7 48 253 301 84,1%
8 68 227 295 76,9%
9 75 282 357 79,0%
Mittelwert 79,0%
Standard-abweichung 3,4%
5.1.2 Arabidopsis opr3-Mutante
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 94 8 102 7,8% 2 31 12 43 27,9% 3 112 29 141 20,6% 4 152 7 159 4,4% 5 94 2 96 2,1%
Mittelwert 12,6%
Standard-abweichung 11,2%
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 129 22 151 14,6% 2 127 6 133 4,5% 3 156 7 163 4,3% 4 223 36 259 13,9% 5 186 53 239 22,2% 6 180 34 214 15,9%
Mittelwert 12,6%
Standard-abweichung 7,0%
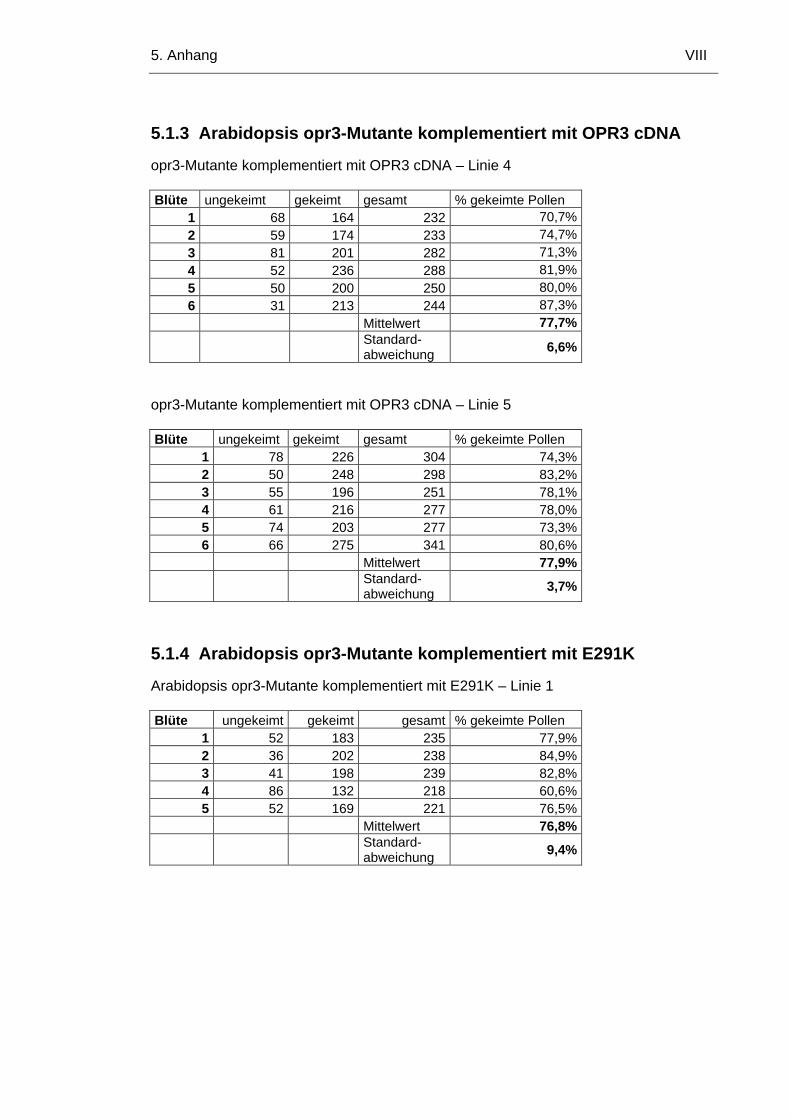
5. Anhang VIII
5.1.3 Arabidopsis opr3-Mutante komplementiert mit OPR3 cDNA
opr3-Mutante komplementiert mit OPR3 cDNA – Linie 4
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 68 164 232 70,7% 2 59 174 233 74,7% 3 81 201 282 71,3% 4 52 236 288 81,9% 5 50 200 250 80,0% 6 31 213 244 87,3%
Mittelwert 77,7%
Standard-abweichung 6,6%
opr3-Mutante komplementiert mit OPR3 cDNA – Linie 5
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 78 226 304 74,3% 2 50 248 298 83,2% 3 55 196 251 78,1% 4 61 216 277 78,0% 5 74 203 277 73,3% 6 66 275 341 80,6%
Mittelwert 77,9%
Standard-abweichung 3,7%
5.1.4 Arabidopsis opr3-Mutante komplementiert mit E291K
Arabidopsis opr3-Mutante komplementiert mit E291K – Linie 1
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 52 183 235 77,9% 2 36 202 238 84,9% 3 41 198 239 82,8% 4 86 132 218 60,6% 5 52 169 221 76,5%
Mittelwert 76,8%
Standard-abweichung 9,4%

5. Anhang IX
Arabidopsis opr3-Mutante komplementiert mit E291K – Linie 6
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 56 161 217 74,2% 2 68 200 268 74,6% 3 89 222 311 71,4% 4 56 231 287 80,5% 5 36 209 245 85,3% 6 63 134 197 68,0%
Mittelwert 75,2%
Standard-abweichung 7,2%
5.1.5 Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL – Linie 1
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 343 31 374 8,3% 2 164 23 187 12,3% 3 274 56 330 17,0% 4 156 31 187 16,6% 5 694 76 770 9,9% 6 183 15 198 7,6%
Mittelwert 11,9%
Standard-abweichung
4,1%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL – Linie 2
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 111 4 115 3,5% 2 96 8 100 8,0% 3 177 12 189 6,3%
Mittelwert 5,9%
Standard-abweichung 4,1%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL – Linie 3
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 172 0 172 0,0% 2 128 0 128 0,0% 3 217 0 217 0,0% 4 136 0 136 0,0%
Mittelwert 0,0%
Standard-abweichung
0,0%
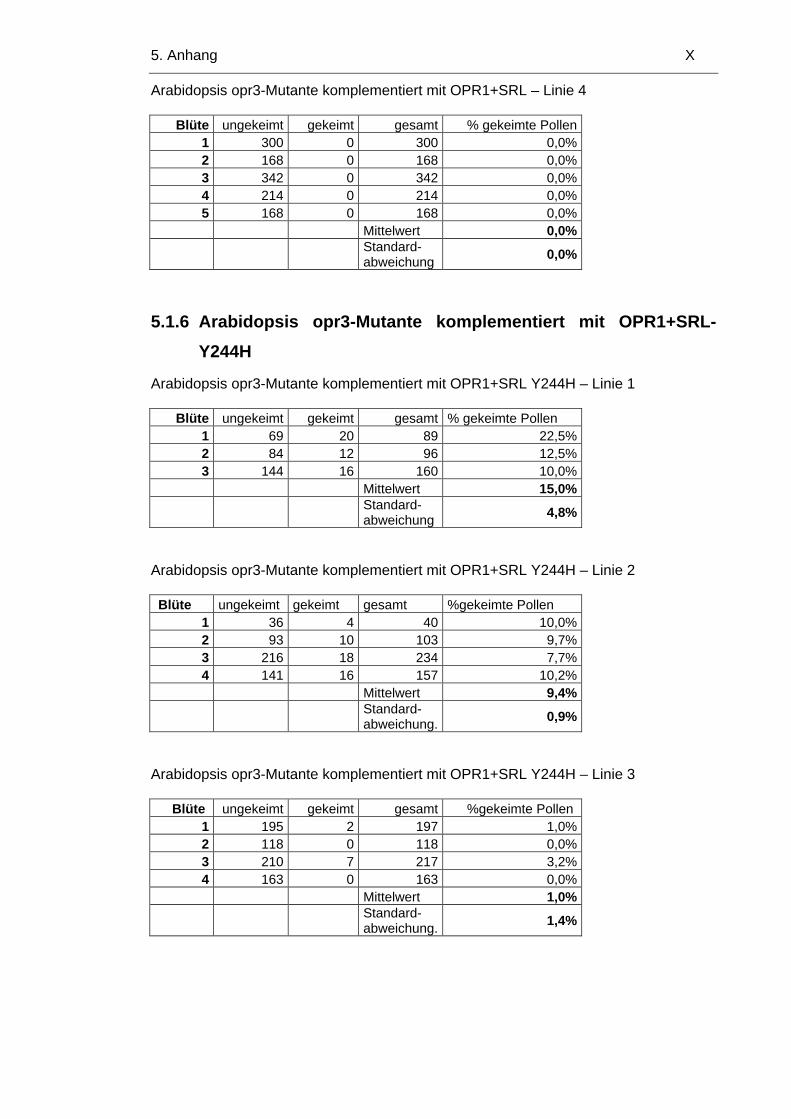
5. Anhang X
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL – Linie 4
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 300 0 300 0,0% 2 168 0 168 0,0% 3 342 0 342 0,0% 4 214 0 214 0,0% 5 168 0 168 0,0%
Mittelwert 0,0%
Standard-abweichung
0,0%
5.1.6 Arabidopsis opr3-Mutante komplementiert mit O PR1+SRL-
Y244H
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 1
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 69 20 89 22,5% 2 84 12 96 12,5% 3 144 16 160 10,0%
Mittelwert 15,0%
Standard-abweichung
4,8%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 2
Blüte ungekeimt gekeimt gesamt %gekeimte Pollen 1 36 4 40 10,0% 2 93 10 103 9,7% 3 216 18 234 7,7% 4 141 16 157 10,2%
Mittelwert 9,4%
Standard-abweichung. 0,9%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 3
Blüte ungekeimt gekeimt gesamt %gekeimte Pollen 1 195 2 197 1,0% 2 118 0 118 0,0% 3 210 7 217 3,2% 4 163 0 163 0,0%
Mittelwert 1,0%
Standard-abweichung. 1,4%

5. Anhang XI
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 4
Blüte ungekeimt gekeimt gesamt %gekeimte Pollen 1 262 22 284 7,7% 2 57 5 62 8,1% 3 153 11 164 6,7% 4 112 11 123 8,9% 5 46 4 50 8,0% 6 73 7 80 8,8%
Mittelwert 8,0%
Standard-abweichung 0,5%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 5
ungekeimt gekeimt gesamt %gekeimte Pollen 1 30 0 30 0,0% 2 62 6 62 9,7% 3 12 0 12 0,0% 4 27 1 28 3,6%
Mittelwert 6,6%
Standard-abweichung 4,5%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 6
ungekeimt gekeimt gesamt %gekeimte Pollen 1 36 6 42 14,3% 2 59 7 66 10,6% 3 212 32 244 13,1% 4 0 100 0,0%
Mittelwert 9,5%
Standard-abweichung 6,3%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 7
ungekeimt gekeimt gesamt %gekeimte Pollen 1 276 0 276 0,0% 2 428 2 430 0,5% 3 273 1 274 0,4% 4 421 7 428 1,6% 5 370 2 372 0,5% 6 318 2 320 0,6% Mittelwert 0,6%
Standard-abweichung 0,5%

5. Anhang XII
5.1.7 Arabidopsis opr3-Mutante komplementiert mit O PR1+SRL
Y244H/Y74F
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 2
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 66 2 68 2,9% 2 113 7 120 5,8% 3 88 7 95 7,4% 4 73 2 75 2,7% 5 17 0 17 0,0% 6 126 5 131 3,8% 7 74 0 74 0,0% 8 78 0 78 0,0% 9 15 0 15 0,0%
Mittelwert 2,5%
Standard-abweichung 2,8%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 4
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 57 6 63 9,5% 2 65 5 70 7,1% 3 124 3 127 2,4% 4 107 10 117 8,5% 5 101 9 110 8,2% 6 60 5 65 7,7% 7 73 7 80 8,8% 8 100 2 102 2,0%
Mittelwert 6,8%
Standard-abweichung
2,9%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 5
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 83 6 89 6,7% 2 268 5 273 1,8% 3 378 2 380 0,5% 4 219 5 224 2,2%
Mittelwert 2,8%
Standard-abweichung 1,4%
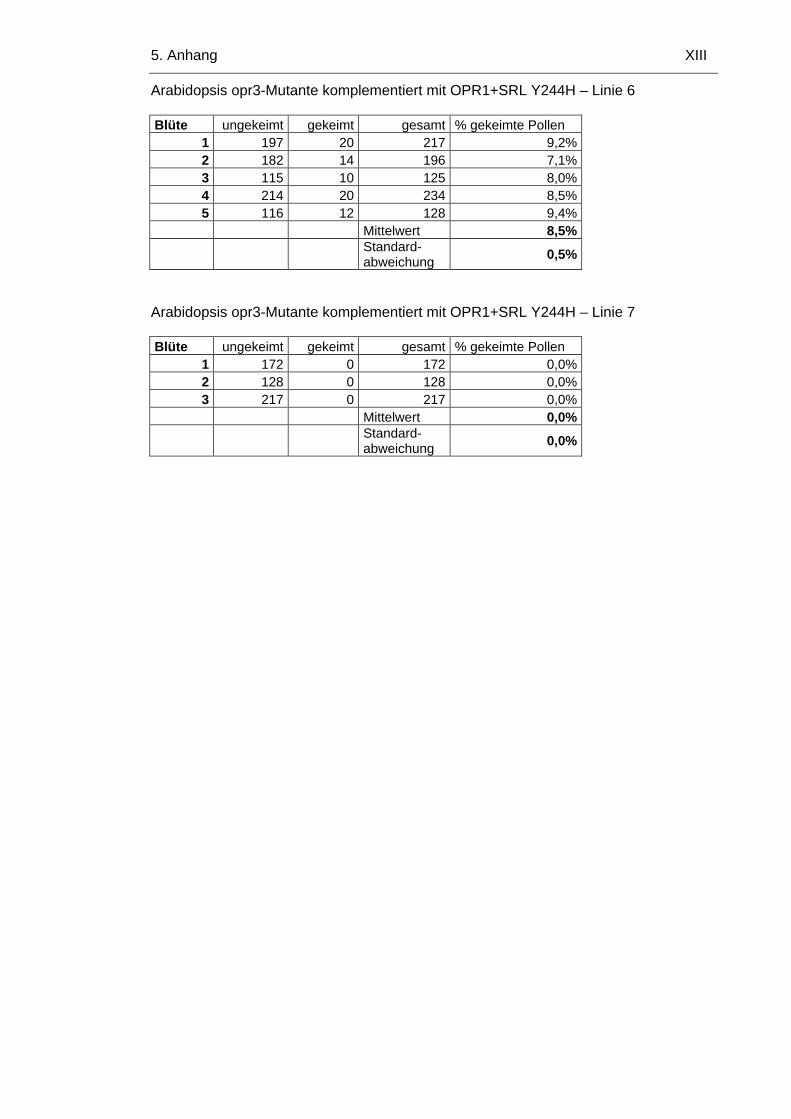
5. Anhang XIII
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 6
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 197 20 217 9,2% 2 182 14 196 7,1% 3 115 10 125 8,0% 4 214 20 234 8,5% 5 116 12 128 9,4%
Mittelwert 8,5%
Standard-abweichung
0,5%
Arabidopsis opr3-Mutante komplementiert mit OPR1+SRL Y244H – Linie 7
Blüte ungekeimt gekeimt gesamt % gekeimte Pollen 1 172 0 172 0,0% 2 128 0 128 0,0% 3 217 0 217 0,0%
Mittelwert 0,0%
Standard-abweichung
0,0%

5. Anhang XIV
5.2 Sequenz des AtOPR3 Promotors
NotI SacII Primer 19 OPR3PR-1 gGCGGCCGCGGttcgattgat tacatcctca gcgtgac
1 ttcgattgat tacatcctca gcgtgacaaa tccaaattca ca ataaatca tataagcaat 61 acagcttttc gattacatct taaaccacct tagtgaggtt ct ataactaa aatggttgaa 121 ttatattaaa ctaaacaaac aatgttttct aaaaatggta tg cccaaaaa tggtttacga 181 aaaatagact caaaaataca aatgtcattg gcgaaaaata aa tgacaatt aatgaatata 241 caaataatag ataaaaagca aaatgaaata aattttcaat gt ttcattta gttaaagata 301 gtgtatcttg aagatcataa cgtattttag attatgtacc tg atttataa gtggcttata 361 aatcacctac cattttacca gtaaccgatc taatctacac ac aaatatca tcttttactc 421 ttataagatt ttaaccatat ttttagatct cataagaatc aa tgcaattt catataaaac 481 atcaaaataa ttctataaat taaaattttc tcaagataat at ttaatttt atttataaat 541 ctcttaatta gccatctttc tacatttatc aaaagatatt ag ctatctgt tctcataggc 601 aatatgataa aaaaaaaaac tttttccaag ttaggggcac aa aaagagtt tccctgggac 661 ttgggctgag aataatgaag tggctgggaa atgattagtt aa ggatggct tgatgaattc 721 tcaatattct gttgagggaa caaaatatac taggaaggat gc aagtccag tttggtctaa 781 accattgtac gaaccaaaaa caatcaataa agtaactcag gg ctcacaaa tggtcgataa 841 tatttctaaa gtttcagttc tagaggatga gcaaaaggat tt gatgatgg gagatgtact 901 agaggtacaa gacaagagtc tcactgaagg gttgaatttc ag atatttta aatcacgttt 961 ggagtaacaa aaaacaaatg tgagaaggag ctcaattagc tc caactgaa tctagtgtta 1021 ttagggatac tttggaaact ccaattcaat ctttaagatg ta ggattgat gaattcatgg 1081 aaatcggcag atccatctat gaatttagta tggtggtgtt tc ctaggaat tgacaataca 1141 attttacttg gtgcaaagtg tcttcggagg tgtccctttc tt tttcactc ggaattggaa 1201 gttttaatat tggcaataaa aaatatcctt tcaagaagga ta aagtgtca ggcatttgaa 1261 acagataatt attcagagta agttttgatg gtacagtccc ct gaagaatg actcgctttc 1321 tctaattttt tagaagatct tgaattgcta tgtgtctcct tt tcttgctt tcttctattt 1381 catgtttatg taatactaag gcaaactatt tagcacaata tt ctaaactt ttattttctg 1441 aagttatttt ttaaaccctt tccctcctaa ttggacaacc aa tcatggaa ttagttttta 1501 cgtaatgttt taaactcact atttcacctt tattctgacc tg ttaatgga agacaactat 1561 gactttatac attgaaatgt tgtcaacctg ttatatatta at gttaacat agctaatgac 1621 ctatcaatgt acgtaaagcg aatgtcaatc atattcgtat cc gttaattt catttaggaa 1681 atcaggctca ttgccaatcc actaagaaac atctctgata ga tccaaata ataaaaataa 1741 tataatatga agcaaaatag aacgaaatat ctcacgcgga cc aaactttg aactttatta 1801 taaaacaaat cagcacgtga aagtaatggt cggttcggtc ca agatgcca agaactcctc 1861 cggtccatcc atcaacctaa ccttttttgg atataaaatt ta gcaaattc catttccacg 1921 accacacaac aacaacacat ccactcgaat tttctatttc cg aataacaa aaccggtaaa 1981 ctcatcacaa acttaagtta ctttgtagtt tgtgttttgt cc ttttatgt aatgaagaag 2041 atccaactaa ctactactca atatttttgt attgaccgtt ga tttctcag atctgacttt 2101 tacttctcct tcttccagat cggcggagac AAGAGGA AGAAGGTCTA GCCGCCTCTG TGATCAgg Primer 20 OPR3PR-R2120 SpeI

5. Anhang XV
5.3 Aminosäure und Basensequenz von LeOPR1
Farbig markiert sind Anlagerungsstellen der Mutageneseprimer, sowie die
auszutauschenden Basenpaare und daraus resultierenden ausgetauschten
Aminosäuren in der Proteinsequenz.
1: M E N K V V E E K Q V D K I P L M S P C
ATGGAAAATAAAGTCGTTGAAGAGAAACAAGTAGACAAGATCCCTCTAATGAGCCCTTGT
1 ---------!---------!---------!---------!--- ------!---------! 60
1: K M G K F E L C H R V V L A P L T R Q R
AAAATGGGAAAGTTTGAGTTATGTCATAGAGTTGTATTGGCACCATTAACAAGGCAAAGA
61 ---------!---------!---------!---------!--- ------!---------! 120
1: S Y G Y I P Q P H A I L H Y S Q R S T N
TCTTATGGTTATATTCCTCAACCACATGCTATACTTCATTACTCACAAAGAAGTACAAAT
121 ---------!---------!---------!---------!--- ------!---------! 180
LeOPR1Y74F-For tatctgagactggcatagggtttaaagat
1: G G L L I G E A T V I S E T G I G Y78 K D
GGTGGCCTTCTAATAGGAGAGGCCACAGTAATATCTGAGACTGGCATAGGGTACAAAGAT
181 ---------!---------!---------!---------!--- ------!- TTT-----! 240
LeOPR1Y74F-Rev F74
gtacctggtatatgg
1: V P G I W T K E Q V E A W K P I V D A V
GTACCTGGTATATGGACAAAAGAGCAAGTGGAGGCTTGGAAACCAATTGTAGATGCAGTT
241 ---------!---------!---------!---------!--- ------!---------! 300
1: H A K G G I F F C Q I W H V G R V S N K
CATGCTAAAGGAGGAATCTTCTTTTGCCAAATTTGGCATGTTGGTAGAGTTTCCAACAAA
301 ---------!---------!---------!---------!--- ------!---------! 360
1: D F Q P N G E D P I S C T D R G L T P Q
GATTTTCAGCCCAATGGAGAGGATCCTATCTCCTGCACAGACAGAGGACTAACACCTCAA
361 ---------!---------!---------!---------!--- ------!---------! 420
1: I R S N G I D I A H F T R P R R L T T D
ATTCGTTCCAATGGCATAGATATTGCACACTTTACACGACCTAGACGGTTGACAACAGAT
421 ---------!---------!---------!---------!--- ------!---------! 480
1: E I P Q I V N E F R V A A R N A I E A G
GAAATTCCTCAAATTGTTAACGAATTTCGAGTTGCTGCTAGAAACGCAATTGAAGCTGGA
481 ---------!---------!---------!---------!--- ------!---------! 540

5. Anhang XVI
1: F D G V E I H G A H G Y L I D Q F M K D
TTTGATGGGGTTGAGATCCACGGAGCTCATGGCTATCTAATTGATCAGTTTATGAAAGAT
541 ---------!---------!---------!---------!--- ------!---------! 600
1: Q V N D R S D K Y G G S L E N R C R F A
CAAGTTAACGATCGAAGTGATAAATATGGAGGGTCTTTAGAGAATCGTTGTAGATTTGCA
601 ---------!---------!---------!---------!--- ------!---------! 660
ta
1: L E I V E A V A N E I G S D R V G I R I
CTTGAAATAGTGGAAGCAGTTGCAAATGAGATTGGATCTGACCGAGTTGGTATAAGGATA
661 ---------!---------!---------!---------!--- ------!---------! 720
tccccatttgcgcatcataatgaagcaggggac LeOPR1Y244H-For
1: S P F A H Y246N E A G D T N P T A L G L Y
TCCCCATTTGCGCAT TATAATGAAGCAGGGGACACGAACCCGACTGCTTTGGGACTTTAC
721 ---------!----- CAT-!---------!---------!---------!---------! 780
H244 LeOPR1Y244H-Rev
1: M V E S L N K Y D L A Y C H V V E P R M
ATGGTGGAATCGTTGAACAAGTATGATCTCGCGTATTGCCATGTGGTTGAGCCTAGGATG
781 ---------!---------!---------!---------!--- ------!---------! 840
1: K T A W E K I E C T E S L V P M R K A Y
AAAACAGCTTGGGAAAAAATTGAATGTACTGAAAGCCTTGTACCGATGAGGAAGGCATAT
841 ---------!---------!---------!---------!--- ------!---------! 900
1: K G T F I V A G G Y D R E D G N R A L I
AAAGGTACTTTTATAGTAGCTGGTGGTTACGATAGAGAAGATGGAAACAGAGCTTTGATT
901 ---------!---------!---------!---------!--- ------!---------! 960
1: E D R A D L V A Y G R L F I S N P D L P
GAAGATCGAGCTGATCTTGTTGCGTATGGACGTTTATTCATATCTAATCCAGATTTACCA
961 ---------!---------!---------!---------!--- ------!---------! 1020
1: K R F E L N A P L N K Y N R D T F Y T S
AAGCGATTTGAGCTAAATGCTCCTCTTAACAAGTATAACAGAGACACATTTTATACTTCT
1021 ---------!---------!---------!---------!--- ------!---------! 1080
1: D P I V G Y T D Y P F L E T M T *
GATCCAATTGTTGGCTATACTGATTATCCATTTCTAGAAACCATGACATGA
1081 ---------!---------!---------!---------!--- ------!- 1131

5. Anhang XVII
5.4 Aminosäure und Basensequenz von LeOPR3
Farbig markiert sind die in OPR1 eingesetzten Basenpaare und daraus resultierenden
Aminosäuren in der Proteinsequenz.
1: M A S S A Q D G N N P L F S P Y K M G K
ATGGCGTCTTCAGCTCAAGATGGAAACAATCCCCTTTTCTCTCCTTACAAGATGGGCAAG
1 ---------!---------!---------!---------!--- ------!---------! 60
1: F N L S H R V V L A P M T R C R A L N N
TTCAATCTATCCCACAGGGTAGTATTGGCTCCGATGACAAGGTGCAGAGCACTGAATAAT
61 ---------!---------!---------!---------!--- ------!---------! 120
1: I P Q A A L G E Y Y E Q R A T A G G F L
ATTCCACAGGCGGCGCTAGGGGAGTATTACGAGCAGAGAGCGACGGCCGGTGGATTTCTG
121 ---------!---------!---------!---------!--- ------!---------! 180
1: I T E G T M I S P T S A G F74 P H V P G I
ATCACTGAAGGCACTATGATTTCTCCGACTTCAGCTGGGTTTCCTCATGTGCCAGGGATT
181 ---------!---------!---------!---------!--- ------!---------! 240
1: F T K E Q V R E W K K I V D V V H A K G
TTCACAAAGGAACAAGTAAGGGAATGGAAGAAAATAGTTGATGTAGTGCATGCAAAGGGT
241 ---------!---------!---------!---------!--- ------!---------! 300
1: A V I F C Q L W H V G R A S H E V Y Q P
GCTGTCATATTTTGTCAGCTGTGGCATGTTGGTCGTGCATCTCATGAAGTGTATCAACCT
301 ---------!---------!---------!---------!--- ------!---------! 360
1: A G A A P I S S T E K P I S N R W R I L
GCTGGAGCTGCACCAATATCATCCACTGAGAAGCCTATATCAAATAGGTGGAGAATTCTA
361 ---------!---------!---------!---------!--- ------!---------! 420
1: M P D G T H G I Y P K P R A I G T Y E I
ATGCCTGATGGAACTCATGGGATTTATCCAAAACCAAGAGCAATTGGAACCTATGAGATC
421 ---------!---------!---------!---------!--- ------!---------! 480
1: S Q V V E D Y R R S A L N A I E A G F D
TCACAAGTTGTTGAAGATTATCGCAGGTCGGCCTTGAATGCTATTGAAGCAGGTTTCGAT
481 ---------!---------!---------!---------!--- ------!---------! 540
1: G I E I H G A H G Y L I D Q F L K D G I
GGTATTGAAATCCATGGAGCTCACGGTTACTTGATTGATCAATTCTTGAAAGATGGGATC
541 ---------!---------!---------!---------!--- ------!---------! 600

5. Anhang XVIII
1: N D R T D E Y G G S L A N R C K F I T Q
AATGACCGGACAGATGAGTATGGTGGATCACTAGCCAACCGGTGCAAATTCATCACACAG
601 ---------!---------!---------!---------!--- ------!---------! 660
1: V V Q A V V S A I G A D R V G V R V S P
GTGGTTCAAGCAGTAGTCTCAGCAATAGGAGCTGATCGCGTAGGCGTTAGAGTTTCACCA
661 ---------!---------!---------!---------!--- ------!---------! 720
1: A I D H244L D A M D S N P L S L G L A V V
GCAATAGAT CATCTTGATGCCATGGACTCTAATCCACTCAGCCTTGGCTTAGCAGTTGTT
721 ---------!---------!---------!---------!--- ------!---------! 780
1: E R L N K I Q L H S G S K L A Y L H V T
GAAAGACTAAACAAAATCCAACTCCATTCTGGTTCCAAGCTTGCCTATCTTCATGTAACA
781 ---------!---------!---------!---------!--- ------!---------! 840
1: Q P R Y V A Y G Q T E A G R L G S E E E
CAGCCACGATACGTAGCATATGGGCAAACTGAAGCAGGCAGACTTGGCAGTGAAGAGGAA
841 ---------!---------!---------!---------!--- ------!---------! 900
1: E A R L M R T L R N A Y Q G T F I C S G
GAGGCTCGTTTAATGAGGACTTTGAGGAACGCGTATCAGGGGACATTCATTTGCAGTGGT
901 ---------!---------!---------!---------!--- ------!---------! 960
1: G Y T R E L G I E A V A Q G D A D L V S
GGATACACTAGGGAACTAGGAATTGAGGCTGTGGCACAAGGTGATGCTGATCTCGTGTCA
961 ---------!---------!---------!---------!--- ------!---------! 1020
1: Y G R L F I S N P D L V M R I K L N A P
TATGGTCGTCTTTTCATCTCTAATCCTGATTTGGTTATGAGAATCAAGCTAAATGCACCT
1021 ---------!---------!---------!---------!--- ------!---------! 1080
1: L N K Y N R K T F Y T Q D P V V G Y T D
CTAAATAAGTATAACAGGAAGACATTCTATACTCAAGATCCAGTTGTGGGATACACAGAT
1081 ---------!---------!---------!---------!--- ------!---------! 1140
1: Y P F L Q G N G S N G P L S R L *
TACCCTTTCCTTCAAGGAAATGGAAGCAATGGACCGTTATCGCGTCTGTGA
1141 ---------!---------!---------!---------!--- ------!- 1191

5. Anhang XIX
5.5 Sequenzen von OPR3-Mutanten
5.5.1 OPR3 E291K
Spe I Primer 5’mut LeOPR3 ccgccACTAG T ATGGCGTCT TCAGCTCAAG AT 1 ccgccACTAG TATGGCGTCT TCAGCTCAAG ATGGAAACAA TCCCCTTTTC TCTCCTTACA 61 AGATGGGCAA GTTCAATCTA TCCCACAGGG TAGTATTGGC TCCGATGACA AGGTGCAGAG 121 CACTGAATAA TATTCCACAG GCGGCGCTAG GGGAGTATTA CGAGCAGAGA GCGACGGCCG 181 GTGGATTTCT GATCACTGAA GGCACTATGA TTTCTCCGAC TTCAGCTGGG TTTCCTCATG 241 TGCCAGGGAT TTTCACAAAG GAACAAGTAA GGGAATGGAA GAAAATAGTT GATGTAGTGC 301 ATGCAAAGGG TGCTGTCATA TTTTGTCAGC TGTGGCATGT TGGTCGTGCA TCTCATGAAG 361 TGTATCAACC TGCTGGAGCT GCACCAATAT CATCCACTGA GAAGCCTATA TCAAATAGGT 421 GGAGAATTCT AATGCCTGAT GGAACTCATG GGATTTATCC AAAACCAAGA GCAATTGGAA 481 CCTATGAGAT CTCACAAGTT GTTGAAGATT ATCGCAGGTC GGCCTTGAAT GCTATTGAAG 541 CAGGTTTCGA TGGTATTGAA ATCCATGGAG CTCACGGTTA CTTGATTGAT CAATTCTTGA 601 AAGATGGGAT CAATGACCGG ACAGATGAGT ATGGTGGATC ACTAGCCAAC CGGTGCAAAT 661 TCATCACACA GGTGGTTCAA GCAGTAGTCT CAGCAATAGG AGCTGATCGC GTAGGCGTTA 721 GAGTTTCACC AGCAATAGAT CATCTTGATG CCATGGACTC TAATCCACTC AGCCTTGGCT 781 TAGCAGTTGT TGAAAGACTA AACAAAATCC AACTCCATTC TGGTTCCAAG CTTGCCTATC 841 TTCATGTAAC ACAGCCACGA TACGTAGCAT ATGGGCAAAC TAAAGCAGGC AGACTTGGCA E 291 in K 901 GTGAAGAGGA AGAGGCTCGT TTAATGAGGA CTTTGAGGAA CGCGTATCAG GGGACATTCA 961 TTTGCAGTGG TGGATACACT AGGGAACTAG GAATTGAGGC TGTGGCACAA GGTGATGCTG 1021 ATCTCGTGTC ATATGGTCGT CTTTTCATCT CTAATCCTGA TTTGGTTATG AGAATCAAGC 1081 TAAATGCACC TCTAAATAAG TTTAACAGGA AGACATTCTA TACTCAAGAT CCAGTTGTGG 1141 GATACACAGA TTACCCTTTC CTTCAAGGAA ATGGAAGCAA ACCTGGCAAT AGCGCAGACA 1201 CTGACGTCcc gcc CCTGGCAAT AGCGCAGACA CTGACGTC ccgcc Pst I Primer 3’mut LeOPR3
5.5.2 OPR3 Y364F
Spe I Primer 5’mut LeOPR3 ccgccACTAG T ATGGCGTCT TCAGCTCAAG AT 1 ccgccACTAG TATGGCGTCT TCAGCTCAAG ATGGAAACAA TCCCCTTTTC TCTCCTTACA 61 AGATGGGCAA GTTCAATCTA TCCCACAGGG TAGTATTGGC TCCGATGACA AGGTGCAGAG 121 CACTGAATAA TATTCCACAG GCGGCGCTAG GGGAGTATTA CGAGCAGAGA GCGACGGCCG 181 GTGGATTTCT GATCACTGAA GGCACTATGA TTTCTCCGAC TTCAGCTGGG TTTCCTCATG 241 TGCCAGGGAT TTTCACAAAG GAACAAGTAA GGGAATGGAA GAAAATAGTT GATGTAGTGC 301 ATGCAAAGGG TGCTGTCATA TTTTGTCAGC TGTGGCATGT TGGTCGTGCA TCTCATGAAG 361 TGTATCAACC TGCTGGAGCT GCACCAATAT CATCCACTGA GAAGCCTATA TCAAATAGGT 421 GGAGAATTCT AATGCCTGAT GGAACTCATG GGATTTATCC AAAACCAAGA GCAATTGGAA 481 CCTATGAGAT CTCACAAGTT GTTGAAGATT ATCGCAGGTC GGCCTTGAAT GCTATTGAAG 541 CAGGTTTCGA TGGTATTGAA ATCCATGGAG CTCACGGTTA CTTGATTGAT CAATTCTTGA 601 AAGATGGGAT CAATGACCGG ACAGATGAGT ATGGTGGATC ACTAGCCAAC CGGTGCAAAT 661 TCATCACACA GGTGGTTCAA GCAGTAGTCT CAGCAATAGG AGCTGATCGC GTAGGCGTTA 721 GAGTTTCACC AGCAATAGAT CATCTTGATG CCATGGACTC TAATCCACTC AGCCTTGGCT 781 TAGCAGTTGT TGAAAGACTA AACAAAATCC AACTCCATTC TGGTTCCAAG CTTGCCTATC 841 TTCATGTAAC ACAGCCACGA TACGTAGCAT ATGGGCAAAC TAAAGCAGGC AGACTTGGCA 901 GTGAAGAGGA AGAGGCTCGT TTAATGAGGA CTTTGAGGAA CGCGTATCAG GGGACATTCA 961 TTTGCAGTGG TGGATACACT AGGGAACTAG GAATTGAGGC TGTGGCACAA GGTGATGCTG 1021 ATCTCGTGTC ATATGGTCGT CTTTTCATCT CTAATCCTGA TTTGGTTATG AGAATCAAGC 1081 TAAATGCACC TCTAAATAAG TTTAACAGGA AGACATTCTA TACTCAAGAT CCAGTTGTGG Y 364 in F 1141 GATACACAGA TTACCCTTTC CTTCAAGGAA ATGGAAGCAA TGGACCGTTA TCGCGTCTGT 1201 GATCCATTTC TAGAAACCAT GACACCGTCG TCGCGTCTGT GACTGCAGGG CGG Primer 3’mut LeOPR3 A CTGTGGCAGC AGCGCAGACA CT GACGTCccgcc Pst I

5. Anhang XX
5.6 Sequenzen von OPR1Mutanten
5.6.1 OPR1+SRL
Spe I Primer 5’mut LeOPR1 ccgccACTAG T ATGGAAAAT AAAGTCGTTG AAGA 1 ccgccACTAG TATGGAAAAT AAAGTCGTTG AAGAGAAACA AGTAGACAAG ATCCCTCTAA 61 TGAGCCCTTG TAAAATGGGA AAGTTTGAGT TATGTCATAG AGTTGTATTG GCACCATTAA 121 CAAGGCAAAG ATCTTATGGT TATATTCCTC AACCACATGC TATACTTCAT TACTCACAAA 181 GAAGTACAAA TGGTGGCCTT CTAATAGGAG AGGCCACAGT AATATCTGAG ACTGGCATAG 241 GGTACAAAGA TGTACCTGGT ATATGGACAA AAGAGCAAGT GGAGGCTTGG AAACCAATTG 301 TAGATGCAGT TCATGCTAAA GGAGGAATCT TCTTTTGCCA AATTTGGCAT GTTGGTAGAG 361 TTTCCAACAA AGATTTTCAG CCCAATGGAG AGGATCCTAT CTCCTGCACA GACAGAGGAC 421 TAACACCTCA AATTCGTTCC AATGGCATAG ATATTGCACA CTTTACACGA CCTAGACGGT 481 TGACAACAGA TGAAATTCCT CAAATTGTTA ACGAATTTCG AGTTGCTGCT AGAAACGCAA 541 TTGAAGCTGG ATTTGATGGG GTTGAGATCC ACGGAGCTCA TGGCTATCTA ATTGATCAGT 601 TTATGAAAGA TCAAGTTAAC GATCGAAGTG ATAAATATGG AGGGTCTTTA GAGAATCGTT 661 GTAGATTTGC ACTTGAAATA GTGGAAGCAG TTGCAAATGA GATTGGATCT GACCGAGTTG 721 GTATAAGGAT ATCCCCATTT GCGCATTATA ATGAAGCAGG GGACACGAAC CCGACTGCTT 781 TGGGACTTTA CATGGTGGAA TCGTTGAACA AGTATGATCT CGCGTATTGC CATGTGGTTG 841 AGCCTAGGAT GAAAACAGCT TGGGAAAAAA TTGAATGTAC TGAAAGCCTT GTACCGATGA 901 GGAAGGCATA TAAAGGTACT TTTATAGTAG CTGGTGGTTA CGATAGAGAA GATGGAAACA 961 GAGCTTTGAT TGAAGATCGA GCTGATCTTG TTGCGTATGG ACGTTTATTC ATATCTAATC 1021 CAGATTTACC AAAGCGATTT GAGCTAAATG CTCCTCTTAA CAAGTATAAC AGAGACACAT 1081 TTTATACTTC TGATCCAATT GTTGGCTATA CTGATTATCC ATTTCTAGAA ACCATGACAC 1141 CG TCG TCG CGT CTG TGA CTGCAGggcgg AGG TAAAGATCTT TGGTACTGTG GC AGC AGC GCA GAC ACT GACGTC ccgcc P S S R L * Pst I
Primer 3’mut LeOPR3
5.6.2 OPR1 Y74F
Spe I ccgccACTAG T ATGGAAAAT AAAGTCGTTG AAGA 1 ccgccACTAG TATGGAAAAT AAAGTCGTTG AAGAGAAACA AGTAGACAAG ATCCCTCTAA 61 TGAGCCCTTG TAAAATGGGA AAGTTTGAGT TATGTCATAG AGTTGTATTG GCACCATTAA 121 CAAGGCAAAG ATCTTATGGT TATATTCCTC AACCACATGC TATACTTCAT TACTCACAAA 181 GAAGTACAAA TGGTGGCCTT CTAATAGGAG AGGCCACAGT AATATCTGAG ACTGGCATAG 241 GG TTTAAAGA TGTACCTGGT ATATGGACAA AAGAGCAAGT GGAGGCTTGG AAACCAATTG Y 74 in F 301 TAGATGCAGT TCATGCTAAA GGAGGAATCT TCTTTTGCCA AATTTGGCAT GTTGGTAGAG 361 TTTCCAACAA AGATTTTCAG CCCAATGGAG AGGATCCTAT CTCCTGCACA GACAGAGGAC 421 TAACACCTCA AATTCGTTCC AATGGCATAG ATATTGCACA CTTTACACGA CCTAGACGGT 481 TGACAACAGA TGAAATTCCT CAAATTGTTA ACGAATTTCG AGTTGCTGCT AGAAACGCAA 541 TTGAAGCTGG ATTTGATGGG GTTGAGATCC ACGGAGCTCA TGGCTATCTA ATTGATCAGT 601 TTATGAAAGA TCAAGTTAAC GATCGAAGTG ATAAATATGG AGGGTCTTTA GAGAATCGTT 661 GTAGATTTGC ACTTGAAATA GTGGAAGCAG TTGCAAATGA GATTGGATCT GACCGAGTTG 721 GTATAAGGAT ATCCCCATTT GCGCATTATA ATGAAGCAGG GGACACGAAC CCGACTGCTT 781 TGGGACTTTA CATGGTGGAA TCGTTGAACA AGTATGATCT CGCGTATTGC CATGTGGTTG 841 AGCCTAGGAT GAAAACAGCT TGGGAAAAAA TTGAATGTAC TGAAAGCCTT GTACCGATGA 901 GGAAGGCATA TAAAGGTACT TTTATAGTAG CTGGTGGTTA CGATAGAGAA GATGGAAACA 961 GAGCTTTGAT TGAAGATCGA GCTGATCTTG TTGCGTATGG ACGTTTATTC ATATCTAATC 1021 CAGATTTACC AAAGCGATTT GAGCTAAATG CTCCTCTTAA CAAGTATAAC AGAGACACAT 1081 TTTATACTTC TGATCCAATT GTTGGCTATA CTGATTATCC ATTTCTAGAA ACCATGACAC 1141 CG TCG TCG CGT CTG TGA CTGCAGggcgg AGG TAAAGATCTT TGGTACTGTG GC AGC AGC GCA GAC ACT GACGTC ccgcc P S S R L * Pst I

5. Anhang XXI
5.6.3 OPR1 Y244H
Spe I ccgccACTAG T ATGGAAAAT AAAGTCGTTG AAGA 1 ccgccACTAG TATGGAAAAT AAAGTCGTTG AAGAGAAACA AGTAGACAAG ATCCCTCTAA 61 TGAGCCCTTG TAAAATGGGA AAGTTTGAGT TATGTCATAG AGTTGTATTG GCACCATTAA 121 CAAGGCAAAG ATCTTATGGT TATATTCCTC AACCACATGC TATACTTCAT TACTCACAAA 181 GAAGTACAAA TGGTGGCCTT CTAATAGGAG AGGCCACAGT AATATCTGAG ACTGGCATAG 241 GGTACAAAGA TGTACCTGGT ATATGGACAA AAGAGCAAGT GGAGGCTTGG AAACCAATTG 301 TAGATGCAGT TCATGCTAAA GGAGGAATCT TCTTTTGCCA AATTTGGCAT GTTGGTAGAG 361 TTTCCAACAA AGATTTTCAG CCCAATGGAG AGGATCCTAT CTCCTGCACA GACAGAGGAC 421 TAACACCTCA AATTCGTTCC AATGGCATAG ATATTGCACA CTTTACACGA CCTAGACGGT 481 TGACAACAGA TGAAATTCCT CAAATTGTTA ACGAATTTCG AGTTGCTGCT AGAAACGCAA 541 TTGAAGCTGG ATTTGATGGG GTTGAGATCC ACGGAGCTCA TGGCTATCTA ATTGATCAGT 601 TTATGAAAGA TCAAGTTAAC GATCGAAGTG ATAAATATGG AGGGTCTTTA GAGAATCGTT 661 GTAGATTTGC ACTTGAAATA GTGGAAGCAG TTGCAAATGA GATTGGATCT GACCGAGTTG 721 GTATAAGGAT ATCCCCATTT GCGCATCATA ATGAAGCAGG GGACACGAAC CCGACTGCTT Y 244 in H 781 TGGGACTTTA CATGGTGGAA TCGTTGAACA AGTATGATCT CGCGTATTGC CATGTGGTTG 841 AGCCTAGGAT GAAAACAGCT TGGGAAAAAA TTGAATGTAC TGAAAGCCTT GTACCGATGA 901 GGAAGGCATA TAAAGGTACT TTTATAGTAG CTGGTGGTTA CGATAGAGAA GATGGAAACA 961 GAGCTTTGAT TGAAGATCGA GCTGATCTTG TTGCGTATGG ACGTTTATTC ATATCTAATC 1021 CAGATTTACC AAAGCGATTT GAGCTAAATG CTCCTCTTAA CAAGTATAAC AGAGACACAT 1081 TTTATACTTC TGATCCAATT GTTGGCTATA CTGATTATCC ATTTCTAGAA ACCATGACAC 1141 CG TCG TCG CGT CTG TGA CTGCAGggcgg AGG TAAAGATCTT TGGTACTGTG GC AGC AGC GCA GAC ACT GACGTC ccgcc P S S R L * Pst I
5.6.4 OPR1 Y74F/Y244H
Spe I ccgccACTAG T ATGGAAAAT AAAGTCGTTG AAGA 1 ccgccACTAG TATGGAAAAT AAAGTCGTTG AAGAGAAACA AGTAGACAAG ATCCCTCTAA 61 TGAGCCCTTG TAAAATGGGA AAGTTTGAGT TATGTCATAG AGTTGTATTG GCACCATTAA 121 CAAGGCAAAG ATCTTATGGT TATATTCCTC AACCACATGC TATACTTCAT TACTCACAAA 181 GAAGTACAAA TGGTGGCCTT CTAATAGGAG AGGCCACAGT AATATCTGAG ACTGGCATAG 241 GG TTTAAAGA TGTACCTGGT ATATGGACAA AAGAGCAAGT GGAGGCTTGG AAACCAATTG Y 74 in F 301 TAGATGCAGT TCATGCTAAA GGAGGAATCT TCTTTTGCCA AATTTGGCAT GTTGGTAGAG 361 TTTCCAACAA AGATTTTCAG CCCAATGGAG AGGATCCTAT CTCCTGCACA GACAGAGGAC 421 TAACACCTCA AATTCGTTCC AATGGCATAG ATATTGCACA CTTTACACGA CCTAGACGGT 481 TGACAACAGA TGAAATTCCT CAAATTGTTA ACGAATTTCG AGTTGCTGCT AGAAACGCAA 541 TTGAAGCTGG ATTTGATGGG GTTGAGATCC ACGGAGCTCA TGGCTATCTA ATTGATCAGT 601 TTATGAAAGA TCAAGTTAAC GATCGAAGTG ATAAATATGG AGGGTCTTTA GAGAATCGTT 661 GTAGATTTGC ACTTGAAATA GTGGAAGCAG TTGCAAATGA GATTGGATCT GACCGAGTTG 721 GTATAAGGAT ATCCCCATTT GCGCATCATA ATGAAGCAGG GGACACGAAC CCGACTGCTT Y 244 in H 781 TGGGACTTTA CATGGTGGAA TCGTTGAACA AGTATGATCT CGCGTATTGC CATGTGGTTG 841 AGCCTAGGAT GAAAACAGCT TGGGAAAAAA TTGAATGTAC TGAAAGCCTT GTACCGATGA 901 GGAAGGCATA TAAAGGTACT TTTATAGTAG CTGGTGGTTA CGATAGAGAA GATGGAAACA 961 GAGCTTTGAT TGAAGATCGA GCTGATCTTG TTGCGTATGG ACGTTTATTC ATATCTAATC 1021 CAGATTTACC AAAGCGATTT GAGCTAAATG CTCCTCTTAA CAAGTATAAC AGAGACACAT 1081 TTTATACTTC TGATCCAATT GTTGGCTATA CTGATTATCC ATTTCTAGAA ACCATGACAC 1141 CG TCG TCG CGT CTG TGA CTGCAGggcgg AGG TAAAGATCTT TGGTACTGTG GC AGC AGC GCA GAC ACT GACGTC ccgcc P S S R L * Pst I

6. Verzeichnisse XXII
6. VERZEICHNISSE
6.1 Literatur
ALBORN HT, TURLINGS TCJ, JONES TH, STENHAGEN GH, LOUGHRIN JH, TUMLINSON JH. (1997) An Elicitor of Plant Volatiles from Beet Armyworm Oral Secretion
Science 276: 945-949
BALDWIN IT, HALITSCHKE R, PASCHOLD A, VON DAHL CC, PRESTON CA.(2006) Volatile signaling in plant-plant interactions: "talking trees" in the genomics era.
Science 311 812 - 815.
BERGER S, BELL E, MULLET JE. (1996) Two methyl jasmonate-insensitive mutants show altered expression of AtVsp in response to methyl jasmonate and wounding.
Plant Physiology 11: 525 - 531
BERGER S (2002) Jasmonate-related mutants of Arabidopsis as tools for studying stress signaling.
Planta 214, 497 - 504.
BREITHAUPT C, STRASSNER J, BREITINGER U, HUBER R, MACHEROUX P, SCHALLER A, CLAUSEN T. (2001) X-ray structure of 12-oxophytodienoate reductase 1 provides structural insight into substrate binding and specificity within the family of OYE.
Structure 9: 419 - 429
BREITHAUPT C, KURZBAUER R, LILIE H, SCHALLER A, STRASSNER J, HUBER R, MACHEROUX P, CLAUSEN T (2006) Crystal structure of 12-oxophytodienoate reductase 3 from tomato: Self-inhibition by dimerization
PNAS 103(39) 14337 - 14342
CREELMAN RA, MULLET JE. (1997) Biosynthesis and action of jasmonates in plants.
Plant Physiol Plant Mol Biol 48:355 – 381.
DEVOTO A AND TURNER JG. (2003) Regulation of Jasmonate-mediated Plant Responses in Arabidopsis.
Annals of Botany 92, 329 – 337.
DICKE M, AGRAWAL AA, BRUIN J (2003) Plants talk, but are they deaf?
Trends Plant Sci 8:403 - 405.

6. Verzeichnisse XXIII
DOBSON, H.E.M. AND BERGSTROM, G. (2000) The ecology and evolution of pollen odors.
Plant Syst. Evol. 222 63 – 87.
ENGELBERTH J, ALBORN HT, SCHMELZ EA, TUMLINSON JH. (2004) Airborne signals prime plants against insect herbivore attack.
Proc Natl Acad Sci USA 101(6):1781 - 1785.
FARMER, E.E., ALMERAS , E., AND KRISHNAMURTHY, V. (2003) Jasmonates and related oxylipins in plant responses to pathogenesis and herbivory.
Curr. Opin. Plant Biol. 6, 372 – 378
FARMER EE, RYAN CA. (1990) Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves.
Proc Natl Acad Sci USA 87(19) 7713 - 7716.
FOX BG, MALONE TE, JOHNSON KA, MADSON SE, ACETI D, BINGMAN CA, BLOMMEL PG, BUCHAN B, BURNS B, CAO J, CORNILESCU C, DORELEIJERS J, ELLEFSON J, FREDERICK R, GEETHA H, HRUBY D, JEON WB, KIMBALL T, KUNERT J, MARKLEY JL, NEWMAN C, OLSON A, PETERSON FC, PHILLIPS GN JR, PRIMM J, RAMIREZ B, ROSENBERG NS, RUNNELS M, SEDER K, SHAW J, SMITH DW, SREENATH H, SONG J, SUSSMAN MR, THAO S, TROESTLER D, TYLER E, TYLER R, ULRICH E, VINAROV D, VOJTIK F, VOLKMAN BF, WESENBERG G, WROBEL RL, ZHANG J, ZHAO Q, ZOLNAI Z. (2005) X-ray structure of Arabidopsis At1g77680, 12-oxophytodienoate reductase isoform 1.
Proteins 61(1) 206 - 208.
FREY M, SPITELLER D, BOLAND W, GIERL A. (2004) Transcriptional activation of Igl, the gene for indole formation in Zea mays: a structure-activity study with elicitor-active N-acyl glutamines from insects.
Phytochemistry 65(8)1047 - 1055
GRANT M, LAMB C. (2006) Systemic immunity.
Curr Opin Plant Biol. 9(4) 414 - 420.
ISHIGURO S, KAWAI-ODA A, UEDA J, NISHIDA I, OKADA K. (2001) The DEFECTIVE IN
ANTHER DEHISCENCE1 Gene Encodes a Novel Phospholipase A1 Catalyzing the Initial Step of Jasmonic Acid Biosynthesis, Which Synchronizes Pollen Maturation, Anther Dehiscence, and Flower Opening in Arabidopsis.
Plant Cell 13, 2191 – 2209.
KODA Y. (1997) Possible involvement of jasmonates in various morphogenic events.
Physiologia Plantarum 100, 639 – 646.

6. Verzeichnisse XXIV
MALONE TE, MADSON SE, WROBEL RL, JEON WB, ROSENBERG NS, JOHNSON KA, BINGMAN CA, SMITH DW, PHILLIPS GN JR, MARKLEY JL, FOX BG. (2005) X-ray structure of Arabidopsis At2g06050, 12-oxophytodienoate reductase isoform 3
Proteins 58(1) 243 - 245
MATSUI K. (2006) Green leaf volatiles: hydroperoxide lyase pathway of oxylipin metabolism.
Curr Opin Plant Biol. 9(3):274 - 280
MCCONN M AND BROWSE J (1996) The Critical Requirement for Linolenic Acid Is Pollen Development, Not Photosynthesis, in an Arabidopsis Mutant.
Plant Cell 8, 403 – 416.
PARK JH, HALITSCHKE R, KIM HB, BALDWIN IT, FELDMANN KA AND FEYEREISEN R (2002) A knock-out mutation in allene oxide synthase results in male sterility and defective wound signal transduction in Arabidopsis due to a block in jasmonic acid biosynthesis.
Plant J. 31, 1 – 12.
PASCHOLD A, HALITSCHKE R, BALDWIN IT. (2007) Co(i)-ordinating defenses: NaCOI1 mediates herbivore- induced resistance in Nicotiana attenuata and reveals the role of herbivore movement in avoiding defenses.
Plant J. 51(1):79 - 91
PASCHOLD A, HALITSCHKE R, BALDWIN IT. (2006) Using 'mute' plants to translate volatile signals.
Plant J. 45(2): 275 - 291.
PICHERSKY E, NOEL JP, DUDAREVA N. (2006) Biosynthesis of plant volatiles: nature's diversity and ingenuity.
Science. 311(5762):808 - 811.
SAMBROOK J, FRITSCH EF, MANIATIS T Molecular cloning. A Laboratory Manual.
SANDERS PM, BUI AQ, WETERINGS K, MCINTIRE KN, HSU YC, LEE PY, TRUONG MT, BEALSTP, GOLDBERG RB (1999) Anther developmental defects in Arabidopsis thaliana male-sterilemutants.
Sex Plant Reprod. 11, 297 – 322.
SANDERS PM, LEE PY, BIESGEN C, BOONE JD, BEALS TP, WEILER EW AND GOLDBERG RB (2000) The Arabidopsis DELAYED DEHISCENCE1 Gene Encodes an Enzyme in the Jasmonic Acid Synthesis Pathway.
Plant Cell 12, 1041 – 1061.

6. Verzeichnisse XXV
STINTZI A, WEBER H, REYMOND P, BROWSE J, FARMER EE (2001) Plant defense in the absence of jasmonic acid: The role of cyclopentenones
Proc Natl Acad Sci USA.98(22):12837 - 12842.
SCHALLER F, BIESGEN C, MÜSSIG C, ALTMANN T, WEILER EW (2000) 12-Oxophytodienoate reductase 3 (OPR3) is the isoenzyme involved in jasmonate biosynthesis.
Planta 210, 979 – 984.
SCHALLER F, HENNIG P, WEILER EW. (1998)12-Oxophytodienoate-10,11-reductase: occurrence of two isoenzymes of different specificity against stereoisomers of 12-oxophytodienoic acid
Plant Physiol. 118(4), 1345 - 1351.
SCHALLER F. (2001) Enzymes of the biosynthesis of octadecanoid-derived signalling molecules.
J. Exp. Botany 52, 11 – 23.
SCHALLER F, SCHALLER A, STINTZI A. (2005) Biosynthesis and Metabolism of Jasmonates. J Plant Growth Regul 23: 179 - 199.
SHIOJIRI K, KISHIMOTO K, OZAWA R, KUGIMIYA S, URASHIMO S, ARIMURA G, HORIUCHI J, NISHIOKA T, MATSUI K, TAKABAYASHI J. (2006) Changing green leaf volatile biosynthesis in plants: an approach for improving plant resistance against both herbivores and pathogens.
Proc Natl Acad Sci U S A. 103(45):16672 - 16676.
STINTZI A, BROWSE J. (2000). The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis.
PNAS 97: 10625 - 10630.
STINTZI A, WEBER. H., REYMOND P, BROWSE J UND FARMER EE. (2001) Plant defense in the absence of jasmonic acid: The role of cyclopentenones.
PNAS 98: 12837 - 12842.
STRASSNER J, FURHOLZ A, MACHEROUX P, AMRHEIN N, SCHALLER A. (1999) A homolog of old yellow enzyme in tomato. Spectral properties and substrate specificity of the recombinant protein.
J Biol Chem. 274: 35067 - 35073

6. Verzeichnisse XXVI
STRASSNER J, SCHALLER . F., FRICK UB, HOWE GA, WEILER EW, AMRHEIN N, MACHEROUX P, SCHALLER A. (2002) Characterization and c-DNA-microarray expression analysis of 12-oxodienoate reductases reveals differntial roles for octadecanoid biosynthesis in the local versus the systemic wound response.
The Plant Cell 32: 585 - 601.
TAKAHASHI F, YOSHIDA R, ICHIMURA K, MIZOGUCHI T, SEO S, YONEZAWA M, MARUYAMA K, YAMAGUCHI-SHINOZAKI K, SHINOZAKI K. (2007)The mitogen-activated protein kinase cascade MKK3-MPK6 is an important part of the jasmonate signal transduction pathway in Arabidopsis.
Plant Cell. 19(3):805 - 818.
TON J, D'ALESSANDRO M, JOURDIE V, JAKAB G, KARLEN D, HELD M, MAUCH-MANI B, TURLINGS TC. (2007)Priming by airborne signals boosts direct and indirect resistance in maize.
Plant J. 49(1):16 - 26.
TURNER JG, ELLIS C, DEVOTO A. (2002) The jasmonate signal pathway.
Plant Cell, 14: 153 - 164.
VICK BA UND ZIMMERMANN DC. (1984) Biosynthesis of Jasmonic acid by several plant species.
Plant Physiol. 75: 458 - 46
WASTERNACK C. (2005) Introductory Remarks on Biosynthesis and Diversity in Actions.
J Plant Growth Regul 23: 167 - 169.
WASTERNACK C, HAUSE B. (2002). Jasmonates and octadecanoids: Signals in plant stress responses and development.
Prog Nucl Acid Res Mol Biol 72:165 – 221.
XIE DX, FEYS BF, JAMES S, NIETO-ROSTRO M, TURNER JG (1998) COI1: An Arabidopsis
Gene Required for Jasmonate-Regulated Defense and Fertility.
Science 280, 1091 – 1094

6. Verzeichnisse XXVII
6.2 Abkürzungen und Einheiten
°C Grad Celsius
A Ampere
Abb. Abbildung
AOS Allenoxidsynthase
AOC Allenoxidcyclase
APS Ammoniumperoxodisulfat
At Arabidopsis thaliana
ATP Adenosin 5'-Triphosphat
bp Basenpaare
c centi (10P-2)
cDNA komplementäre DNA
CIP alkalische Phosphatase aus Kälberdarm
Col Columbia
CTAB Cethyltrimethyl-Ammoniumbromid
DEX DNA-Extraktionspuffer
DMSO Dimethylsulfoxid
DNA Desoxyribonukleinsäure
DNase Desoxyribonuklease
dNTPs Desoxy-Nukleotidtriphosphat
DTT Dithiothreit
ECL Enhanced Chemiluminescence
EDTA Ethylendiamintetraessigsäure
EtOH Ethanol
Forw forward
F Farad
g Erdbeschleunigung (9,81 m/s²)
g Gramm
GLVs Green Leave Volatiles – flüchtige organische Substanzen
h Stunde
Ig G Immunglobulin G
IPTG Isopropyl-ß-D-1-thiogalactopyranosid
kb Kilobasenpaar
kDa Kilo Dalton
l Liter
LB Luria Bertani

6. Verzeichnisse XXVIII
LB Left Border
Le Lycopersicum esculentum bzw. Solanum lycopersicum
LOX Lipoxygenase
µ mikro (10-6)
m milli (10-3)
m Meter
min Minute
mRNA messenger Ribonukleinsäure
M Molar
NADPH Nicotinsäureamid-Adenosin-Dinucleotid-Phosphat, reduziert
Ω Ohm
OPDA Oxophytodiensäure
PCR Polymerase-Kettenreaktion
RB Right Border
rev reverse
RNA Ribonuklein-Säure
RT Raumtemperatur
SDS Natriumdodecylsulfat
TAE Tris-Acetat-EDTA-Puffer
T-DNA Transfer-DNA
TE Tris-EDTA-Puffer
TBS Tris-Buffered Saline
TEMED Tetrametylethylendiamin
Tween20 Polyoxyethylensorbitanmonolaurat
U Units/Enzymeinheiten
ÜN über Nacht
V Volt
VOCs Volatile Organic Compounds
v/v Volumen pro Volumen
Ws Wassilewskija
WT Wildtyp
w/v Gewicht pro Volumen

6. Verzeichnisse XXIX
6.3 Abbildungen und Tabellen
Tab. 1: Verwendete Annealingtemperaturen und Elongationszeiten in
Abhängigkeit der Spezifischen Primerkombinationen 27
Abb. 1: Struktur der Jasmonsäure ............................................................................ 1
Abb. 2: Die Wirkungen von Jasmonaten auf Stressoren und ihre Beteiligung an der
pflanzlichen Entwicklung (Wasternack, 2005) ................................................................ 2
Abb. 3: Der Oktadekanoidweg der Jasmonysnthese (nach Vick und Zimmermann,
adaptiert nach Wasternack und Hause, 2002) ............................................................... 3
Abb. 4: Verwandtschaftsvergleich der OPR Isoformen von Tomate und Arabidopsis
.................................................................................................................... 4
Abb. 5: Vergleich der Sequenzen der 3 Isoformen von OPR in Tomaten (Le) mit
den Isoformen aus Arabidopsis thaliana (At) ................................................................. 5
Abb. 6: Lokalisation der Proteine des Oktadekanoidwegs, Jasmonsäurebiosynthese
und Metabolismus. ......................................................................................................... 6
Abb. 7: Umsatz von cis(+)OPDA durch NADPH-abhängiges OPR3 zu cis(+)OPC-
8:0 ............................................................................................................ 7
Abb. 8: 3D-Struktur von OPR1.................................................................................. 8
Abb. 9: Strukturvergleich von OPR3 (gelb) und OPR1(blau).................................... 9
Abb. 10: Detaildarstellung des Aktiven Zentrums von OPR1 mit gebundenem
Substrat ........................................................................................................ 9
Abb. 11: Röntgenkristallstruktur von OPR3. Der Fingerartige β6-Loop des einen
Protomers (rot) interagiert in der substratbindenden Tasche des anderen Protomers
(gelb) und umgekehrt. Nach Breithaupt et al., 2006..................................................... 10
Abb. 12: Aktives Zentrum von dimerisiertem OPR3. In gelb und dunkelrot: jeweils
ein Protomer von OPR3. Markiert (roter Kreis) sind Aminosäuren, die für die
Dimerisierung wichtig erscheinen................................................................................. 11
Abb. 13: In vivo Phosphorylierung eines OPR3-Proteins durch eine
Typrosinspezifische Protein Kinase im Expressionsvektor in E.coil. (Schaller,
unveröffentlicht) ............................................................................................................ 12
Abb. 14: Karte des pCR®2.1 Topo® Vektor ......................................................... 16

6. Verzeichnisse XXX
Abb. 15: Aufbau des Vektors pGreen 0229 OPR3 prom DsRED-OPR3............... 16
Abb. 16: Temperatur und Dauer der einzelnen Schritte einer PCR ...................... 26
Abb. 17: Prinzip der Zielgerichteten Mutation ....................................................... 33
Abb. 18: Durchfluss, Waschschritte und Elutionsschritte der Proteinreinigung von
OPR3 und SDS-PAGE in Coomassiefärbung. Aufgetragen wurden jeweils 15 µl der
1:10 verdünnten Fraktionen der Aufreinigung in denaturierter Form............................ 38
Abb. 19: Schematischer Aufbau eines Western Blots........................................... 45
Abb. 20: Schema der Antikörperdetektion............................................................. 48
Abb. 21: Ausschniktt eines Ethidiumbromid gefärbten 1% Agarosegels. PCR der
OPR3 Konstrukte OPR3, E291K, Y364F mit Primer 34 und 35, 1191 bp,
Annealingtemperatur 58°C, Elongationszeit 75 sec. .. .................................................. 50
Abb. 22: Sequenzvergleich von OPR1, OPR2 und OPR3 aus Tomaten (Le) und
Arabidopsis thaliana (At) .............................................................................................. 51
Abb. 23: Ethidiumbromid gefärbtes 1% Agarosegel. Links: PCR der OPR3
Konstrukte OPR3, E291K, Y364F mit Primer 34 und 35, 1191 bp, Rechts: PCR der
OPR1-cDNA mit Primern 36 und 37, 1131 bp. Annealingtemperatur 58°C,
Elongationszeit 75 sec. ................................................................................................ 52
Abb. 24: DNA Sequenzauschnitt von OPR1 (schwarz) mit den jeweils bindenden
Mutageneseprimern (blau) ........................................................................................... 53
Abb. 25: Ausschnitt eines 1% Agarosegels, gefärbt mit Ethidiumbromid.
Restriktionsverdau der Mutation-tragenden Topo® Plasmide mit Nae I, Spe I und Pst I.
OPR3 (links, OPR3, E291K, Y364F) Konstrukte haben eine Größe von 1191 bp, OPR1
Konstrukte (rechts, OPR1+SRL, Y74F, Y244H, Y74F/Y244H) haben eine Größe von
1131 bp. Der Topo®-Vektor ohne Insert hat eine Größe von ca. 3900 bp. Der Verdau
wurde bei 37°C für 2 Stunden durchgeführt. ......... ....................................................... 54
Abb. 26: Schematischer Aufbau des Transformationsplasmides mit OPR3-
Promotor, Konstrukt (grün hinterlegt), nos-Terminator (T), Resistenzgenen für BASTA
(nos-bar) und Kanamycin (npt) und Origin of replication für die Replikation in
Agrobakterium (pSA ori) und E.coli (ColEI ori). ............................................................ 55
Abb. 27: 1% Agarosegel mit Ethidiumbromid gefärbt. PCR auf Konstrukte in
Agrobakterien. OPR3 Konstrukte (links, OPR3, E291K, Y364F) haben eine Größe von
1191 bp, OPR1 Konstrukte (rechts, OPR1+SRL, Y74F, Y244H, Y74F/Y244H) haben
eine Größe von 1131 bp. Annealingtermperatur: 58°C, Elongationszeit 120 sec. ....... 55

6. Verzeichnisse XXXI
Abb. 28: 1% Agarosegel in Ethidiumbromidfärbung. PCR Analyse der BASTA-
resistenten Pflanzen auf das jeweilige Konstrukt. Als Positive Kontrolle (+) diente das
jeweilige Transformationskonstrukt, Als Negative Kontrolle (–) diente Wildtypische
DNA. Als Ladekontrolle wurde eine PCR mit den Primern für Tubulin TuB4F und
TuB4R durchgeführt, positive Kontrolle hierfür war wildtypische DNA, negative
Kontrolle das Transformationskonstrukt. Annealingtemperatur: 50°C, Elongationszeit
40 sec. Oben: Analyse der OPR3 Konstrukte OPR3 (Komplementationskontrolle) und
E291K mit den Primern 1612 und 35 .Unten: Analyse der OPR1 Konstrukte
OPR1+SRL Y244H und die Doppelmutation Y74F/Y244H mit Primern 1612 und
OPR1_876R. Annealingtemperatur: 58°C, Elongationsze it: 120 sec........................... 57
Abb. 29: A, B und C: Blütenstände von Wildtypischen, opr3-defiziente A. thaliana
und mit E291K transformierten A. thaliana, D, E, F, G: Blüten von Wildtyp, opr3-
defiziente A. thaliana, und mit OPR3 bzw. E291K transformierten opr3-defiziente A.
thaliana, sowie die zugehörige Pollkeimung in vitro (jeweils rechts von den Blüten),
unten: graphische Darstellung des in vitro Pollenkeimungstests der transformierten
opr3-defiziente A. thaliana mit OPR3 und E291K im Vergleich mit Pollen von Wildtyp
und opr3-defiziente A. thaliana. Pollen wurde auf Pollenkeimungsmedium ausgebracht
und nach 12 Stunden ausgezählt. Von 6 bis 9 Blüten wurden jeweils ca. 300 Samen
gezählt, der Mittelwert gebildet und eine Standardabweichung berechnet. ................. 58
Abb. 30: A, B und C: Blütenstände der opr3-defizienten A.thaliana, komplementiert
mit OPR1+SRL (A), Y244H (B) und Y74F/Y244H(C). D, E und F: Blüten und in vitro
Pollenkeimung (jeweils darunter) der opr3-defiziente A. thaliana, die mit OPR1+SRL
(D), Y244H (E) und Y74F/Y244H (F) transformiert wurden. Unten: Graphische
Darstellung des Pollenkeimungstests der mit OPR1-Mutanten transformierten opr3-
defiziente A. thaliana im Vergleich mit Pollen von Wildtyp und opr3-defiziente A.
thaliana. Pollen wurde auf Pollenkeimungsmedium ausgebracht und nach 12 Stunden
ausgezählt. Von 6 bis 9 Blüten wurden jeweils 300 Samen gezählt, der Mittelwert
gebildet und eine Standardabweichung berechnet. ..................................................... 60
Abb. 31: 5 ml Gradienten der verschiedenen Linien, beladen mit 100 µl und 600 µl
der Überstände von OPR1+SRL-1, Y244H-1 und Y74F/Y244H-1............................... 62
Abb. 32: Coomassie- und Silber-Färbung der SDS-Gele der 2 Gradienten von
Y244H. Oben: 100µl beladener Gradient, Unten 600 µl beladener Gradient. Die
Fraktion von 18,5% im 100µl Gradienten fehlt in der Silber-Färbung, da diese Fraktion
nicht vollständig gefüllt und deswegen aufgebraucht war. ........................................... 63

6. Verzeichnisse XXXII
Abb. 33: Vergleich der beiden Western Blots des Gradienten von Y244H-1
(beladen mit 600µl) mit Anti-AOS (oben) und Anti-OPR1 (unten) und OPR1 Protein als
Kontrolle 65
Abb. 34: Western Blot des ersten Verwundungsexperiment an Tomaten mit Anti
PT-66. Aufgetragen wurde Gesamtproteinextrakt. Unverwundete Kontrollpflanzen
wurden zu Beginn des Experimentes geerntet und nach 120 Minuten (K0’ und K120’),
verwundete Pflanzen nach 10, 30, 60 und 180 Minuten. Kontrollpflanzen und
verwundete Pflanzen befanden sich im selben Raum. Als Kontrolle wurden OPR3
Protein und OPR3 Protein aus dem Expressionsvektor TKB1 aufgetragen, dass eine
Tyrosinkinase enthält und OPR3 somit Phosphoryliert(Im Bild bezeichnet als TKB1). 68
Abb. 35: Western Blot Analyse eines Verwundungsexperimentes detektiert mit Anti
OPR3 Antikörper. Oben: verwundete Pflanzen, geerntet 5, 10, 20, 30, 60 und 180
Minuten nach Verwundung, Unten: Kontrollpflanzen, nicht verwundet, geerntet zu
denselben Zeitpunkten, einschließlich Nullkontrolle. Kontrollpflanzen und verwundete
Pflanzen befanden sich im selben Raum. Als Kontrollen wurden OPR3 und
Phosphoryliertes OPR3 Protein (TKB1) aufgetragen................................................... 69
Abb. 36: Western Blot von Gesamtproteinextrakten von VOC-Empfänger
Tomatenpflanzen mit AntiOPR3 mit zugehöriger Ponceaufärbung.............................. 71
Abb. 37: Western Blot von Gesamtproteinextrakten verwundeter Tomatenpflanzen
mit AntiOPR3................................................................................................................ 72
Abb. 38: Western Blot von Proteinextrakten von VOC-Empfänger
Tomatenpflanzen mit AntiPT-66................................................................................... 73
Abb. 39: Western blot von Proteinextrakten verwundeter Tomatenpflanzen mit Anti
PT-66 .................................................................................................... 74
Abb. 40: Western Blots von Arabidopsis-Pflanzen, die VOCs von Tomaten (TV)
und Arabidopsis-Pflanzen (AV) ausgesetzt waren, geerntet nach je 0, 30, 60 und 90
Minuten. Als Kontrolle dienten verwundete opr3 Pflanzen, die nach 60 Minuten
geerntet wurden, sowie verwundete Arabidopsis-Pflanzen nach 60 und 90 Minuten
Verwundung, außerdem 0,1 ng aufgereinigtes OPR3 und 10 ng OPR1 Protein bzw. 0,1
ng Phosphoryliertes OPR3 Protein (TKB1). Detektion mit AntiOPR1 (Oben), AntiOPR3
(Mitte) und Anti PT-66 (Unten) ..................................................................................... 75
Abb. 41: Western Blot von Proteinextrakten verwundeter (vw) Arabidopsis opr1-
mutanter-Pflanzen und VOC-Empfänger Arabidopsis-opr1-mutanter Pflanzen (AV)
detektiert mit AntiOPR3 (oben) und mit Anti-PT 66 (Unten) Als Kontrollen wurden

6. Verzeichnisse XXXIII
jeweils 0,1 ng OPR3-Protein und phosphoryliertes OPR3-Protein (TKB1), sowie 10 ng
OPR1 Protein aufgetragen. Ein Pfeil markiert die Höhe von OPR3. ............................ 77
Abb. 42: Western Blot von Proteinextrakten verwundeter (vw) Arabidopsis opr3-
mutanter-Pflanzen und VOC-Empfänger Arabidopsis-opr1-mutanter Pflanzen (AV)
detektiert mit Anti-PT 66. Als Kontrollen wurden jeweils 0,1 ng OPR3-Protein und
phosphoryliertes OPR3-Protein (TKB1) sowie 10 ng OPR1 Protein aufgetragen. Ein
Pfeil markiert die Höhe von OPR3................................................................................ 78

Danke
An Herrn Professor A. Schaller für die Möglichkeit meine Diplomarbeit am Institut für
Physiologie und Biotechnologie der Pflanzen durchführen zu können und für die
Vergabe dieses interessanten Themas.
An Herrn Professor A. Pfitzner der freundlicherweise die Zweitkorrektur übernommen
hat.
An Dr. Annick Stintzi für die engagierte Betreuung und Unterstützung und für die vielen
Diskussionen während meiner Diplomarbeit.
An Mama und Papa, die meinen Studienweg stets unterstützt haben, sowohl finanziell
als auch moralisch, im letzten Jahr immer an meiner Seite waren, mir Beistand
geleistet haben und mich während meiner Lern- und Schreibphasen ertragen haben.
An Sebastian für Aufmunterung, Anstachelung, Hilfe bei Word und anderen
Absurditäten, kleine Gesten und Momente, Filmabende, Kochabende, Krupuk – kurz:
Dafür, dass er immer das Richtige zum passenden Zeitpunkt getan hat.
Des Weiteren danke ich:
Frau Brehm, die mein Biologisches Interesse entdeckt und gefördert hat und
einen großen Anteil daran hat, dass ich das Studium der Biologie in Angriff
genommen habe
Jedem Einzelnen vom Institut 260 und vom Gewächshausteam für
Hilfestellung, Kaffeeklatsch und Korrekturlesen, insbesondere Jutta, Saeid,
Klaus, Benni und Kristin
Charlie und Chrissie für ihre Ablenkungsmanöver
Ekki und Martin für Diskussionen, Tipps und Tricks
Corinna, Anne, Felix, Peter, Steffen, Oli, Benni und all meinen anderen
Kommilitonen für die vielen „Kippen-und-Kaffee-Pausen“ und dafür, dass sie
mich jammern ließen
Sandra für Bombus - ohne dich hätte es kein zweites Semester gegeben
Opeth, Queen und Incubus für nächtliche Motivation
Arabidopsis, dass sie sich stressen ließ
Dem Meer, denn es trägt den Blick,
den Sternen, denn es lohnt sich nach ihnen zu greifen
und dem Wind, denn er treibt das Boot immer weiter.
„Die Wissenschaft ist etwas noch nicht ganz gefundenes und
nie ganz aufzufindendes“ Wilhelm von Humboldt

Ehrenwörtliche Erklärung
Ich versichere hiermit ehrenwörtlich, dass ich die vorliegende Arbeit selbständig
und nur unter Benutzung der angegebenen Literatur und Hilfsmittel angefertigt
habe. Wörtlich übernommene Sätze und Satzteile sind als Zitat belegt, andere
Anlehnungen hinsichtlich Aussage und Umfang unter Quellenangabe kenntlich
gemacht.
Die Arbeit hat in gleicher oder ähnlicher Form noch keiner Prüfungsbehörde
vorgelegen.
Leonberg, den 20. September 2007
_________________________________
Daniela Schwörer