
Kognitive Neurophysiologie des Menschen - boris.unibe.ch · wissenschaftliche Artikel zu Themen der...
89
© 2012 W. Skrandies, Aulweg 129, D-35392 Giessen http://geb.uni-giessen.de/geb/volltexte/2008/6504/
Transcript of Kognitive Neurophysiologie des Menschen - boris.unibe.ch · wissenschaftliche Artikel zu Themen der...

KOGNITIVE
NEUROPHYSIOLOGIE DES
MENSCHEN
HUMAN COGNITIVE
NEUROPHYSIOLOGY
© 2012 W. Skrandies, Aulweg 129, D-35392 Giessenhttp://geb.uni-giessen.de/geb/volltexte/2008/6504/

ImpressumHerausgeber: Wolfgang Skrandies
© 2012 W. Skrandies, Aulweg 129, D-35392 [email protected]
Editorial Board:M. Doppelmayr, SalzburgA. Fallgatter, TübingenT. Koenig, BernH. Witte, Jena
ISSN 1867-576X
ii Human Cognitive Neurophysiology 2012, 5 (1)

Kognitive Neurophysiologie des Menschen wurde im Jahr 2008 gegründet. Hier sollenwissenschaftliche Artikel zu Themen der kognitiven Neurophysiologie des Menschen er-scheinen Sowohl Beiträge über Methoden als auch Ergebnisse der Grundlagen- und klinischenForschung werden akzeptiert. Jedes Manuskript wird von 3 unabhängigen Gutachtern beurteiltund so rasch wie möglich publiziert werden.Die Zeitschrift ist ein elektronisches ”Open Access”-Journal, ohne kommerzielle Interessen;http://geb.uni-giessen.de/geb/volltexte/2008/6504/.
Eine dauerhafte Präsenz der Zeitschrift im Internet wird durch die Universität Giessengewährleistet.
Human Cognitive Neurophysiology was founded in 2008. This journal will publish contribu-tions on methodological advances as well as results from basic and applied research on cogni-tive neurophysiology. Both German and English manuscripts will be accepted. Each manuscriptwill be reviewed by three independent referees.This is an electronic ”Open Access”-Journal with no commercial interest, published athttp://geb.uni-giessen.de/geb/volltexte/2008/6504/.
Online presence is guaranteed by the University of Giessen.
2012, 5 (1) Kognitive Neurophysiologie des Menschen iii

Instructions for Authors
Only original and unpublished work will be considered for publication unless it is explicitly statedthat the topic is a review. All manuscripts will be peer-reviewed. Both German and Englishversions are acceptable. After publication, the copyright will be with the editor of the journal.Usage of published material for review papers will be granted. Manuscripts (as WORD or TEXfiles ) should be sent to [email protected].
Organization of manuscripts: The title page with a concise title should give the authors’ names,address(es), and e-mail address of the corresponding author. The manuscript should includean abstract in English (maximum 300 words). Organize your work in the sections Introduction,Methods, Results, Discussion, and Literature. Please also supply a short list of keywords thatmay help to find your publication.
Illustrations: All figures should be submitted as jpeg or Coreldraw files. Please supplyfigure legends that explain the content of the figures in detail. Since this is an electronic journalcolor figures will be published free-of-charge.
The Literature should only include papers that have been published or accepted for publication.The reference list should be in alphabetical order by author. In the text, references should becited by author(s) and year (e.g. Johnson, Hsiao, & Twombly, 1995; Pascual-Marqui, Michel, &Lehmann, 1994; Zani & Proverbio, 2002).
Examples of reference formatJohnson, K., Hsiao, S., & Twombly, L. (1995). Neural mechanisms of tactile form recognition. In
M. Gazzaniga (Ed.), The Cognitive Neurosciences (p. 253-267). Cambridge, Mass.: MITPress.
Pascual-Marqui, R., Michel, C., & Lehmann, D. (1994). Low resolution electromagnetic tomog-raphy: a new method for localizing electrical activity in the brain. International Journal ofPsychophysiology , 18, 49-65.
Zani, A., & Proverbio, A. (Eds.). (2002). The Cognitive Electrophysiology of Mind and Brain.San Diego: Elsevier.
iv Human Cognitive Neurophysiology 2012, 5 (1)

Inhalt — Contents
Inhalt — Contents
D. Eckstein, T. Koenig, M. Wyss, & W. J. Perrig — Monitoring the Time Course ofPerception without Awareness . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
M. Wagner & W.-J. Kuo — Population-Adapted Averaged Head Templates . . . . . . . 22M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften und der ana-
lytischen Philosophie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting . . . . . . . 61M. Doppelmayr – Neurobiologie der Psychotherapie (Buchbesprechung) . . . . . . . . 82Announcements — Ankündigungen . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84
2012, 5 (1) Kognitive Neurophysiologie des Menschen v

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Abstract
D. Eckstein, T. Koenig, M. Wyss, & W. J. Perrig (Bern, Switzerland) — Monitoring the Time Courseof Perception without Awareness: A Comparison of Mirror Masked Words and NonwordsMirror masked words are embedded into a context that makes them appear as senseless patterns or as
strings of unfamiliar letters. Thus, mirror masked words can be shown for several hundreds of milliseconds
without being recognised as words. We sought to further investigate effects of nonsconscious reading by
monitoring event-related brain potentials (ERPs) while participants observed mirror masked letter strings.
ERPs were recorded while participants observed mirror masked words and nonwords. Data of 15 partici-
pants was segmented into periods of quasi-stable field topography (microstates). Microstates for masked
words and nonwords were compared using randomization tests, statistical parametric scalp maps and Low
Resolution Electromagnetic Tomography (LORETA).
ERPs to masked words and nonwords showed significant topographic differences between 136 and 256
ms, indicating that stimuli were nonconsciously discriminated. A LORETA model localised sources of
activation discriminating between masked words and nonwords in left operculum, the right superior pari-
etal lobe and right superior temporal gyrus indicating higher current density for nonwords than for words
in these areas.
ERPs of mirror masked stimuli can indicate unconscious discrimination even in cases where behavioural
priming is unreliable. This approach might be useful for investigating differences in early, nonconscious
stages of word perception.
Keywords: Reading; Language; Subliminal perception; ERPs; LORETA
Monitoring the Time Courseof Perception without
Awareness: A Comparison ofMirror Masked Words and
Nonwords
D. Eckstein∗1,4, T. Koenig∗2,4, M. Wyss,∗3 & W.J. Perrig∗1,4 1Department of Psychology,
University of Bern, 3000 Bern 9, 2Department ofPsychiatric Neurophysiology, University Hospital ofPsychiatry, ,3Teacher Training University of CentralSwitzerland (PHZ), Lucerne, 4Center for Cognition,
Learning, and Memory, University of Bern, Bern,Switzerland
Introduction
In the past few years, research on perceptionwithout awareness has obtained consider-able attention in cognitive science. Effectivemasking techniques have been developedwith which stimuli can be presented sublimi-nally. Priming experiments with subliminallypresented words and letter strings have beensuccessfully used to investigate processingof linguistic properties of words (cf. Frost,Forster, & Deutsch, 1997; Grainger, Colé,& Segui, 1991; Kinoshita & Lupker, 2003),
2012, 5 (1) Kognitive Neurophysiologie des Menschen 1

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
effects of attitudes and emotional content(cf. Bargh, 1992; Hassin, Uleman, & Bargh,2006; Wentura, 2002) and effects on memory(e.g., Jacoby & Whitehouse, 1989). The usualmethod of masking is to embed to-be-maskedstimuli in a rapid visual stream of patternmasks. This combination of short stimulusduration and overlapping visual percepts ofstimulus and masks leads to the visual expe-rience that the stimulus is invisible. However,this method only works within a narrow rangeof possible presentation times that excludeawareness of stimuli. It has also been notedthat the onset of a mask that follows a stimulushas the effect of interrupting any bottom-upprocessing of the stimulus up to that point(Humphreys, Besner, & Quinlan, 1988; Ko-vacs, Vogels, & Orban, 1995; Rolls & Tovee,1994). Because of its tight limits, the timingmust ideally be empirically determined on anindividual basis for every participant in a givenexperiment.
Another difficulty arises from the fact thatsubliminal perception must be indirectly in-ferred from participants’ reactions. Earlier at-tempts to use direct measures (whereby par-ticipants were asked to indicate which word ina list appeared beforehand) have suffered fromunreliability, which explains why indirect mea-surements based on priming are commonlyused. The actual paradigmatic measurementof subliminal perception consists in assessingpriming by subliminal primes on reactions tosucceeding visible probes. These priming ef-fects are not as large, and thus more difficultto replicate than priming effects obtained withvisible primes. One way to increase the ef-fect size is to familiarise participants with thetest material used in the task. Indeed, ef-
fects can be quite robust when using this strat-egy (cf. Greenwald, Draine, & Abrams, 1996;Greenwald, Klinger, & Schuh, 1995), which ex-plains why existing fMRI studies on subliminalperception have been based on this approach(e.g., Dehaene, Naccache, Cohen, & Rivière,2001; Dehaene, Naccache, Le Clec'H, & LeBihan, 1998; Naccache & Dehaene, 2001).However, it has become evident that frequentlyrepeating the same set of stimuli with fixed re-sponse mappings leads to automatisation of allprocesses involved in responding. As a conse-quence, processes of perception, memory andresponse preparation become difficult to sep-arate (Damian, 2001; Kunde, Kiesel, & Hoff-mann, 2003). This issue is further complicatedby the fact that the threshold of consciousnessdecreases with repeated presentation (Wol-ford, Marchak, & Hughes, 1988), which canlead to paradoxical priming effects, e.g. ef-fects that appear to be related to the mean-ing of stimuli but are instead driven by low-levelperceptual properties of the stimuli (Abrams &Greenwald, 2000; Greenwald, Abrams, Nac-cache, & Dehaene, 2003; Kouider & Dupoux,2004).
To overcome these methodological difficul-ties of delicate timing issues or small effectsizes, we developed a masking method thatdoes not rely on temporal masking, but in-stead uses spatial masking (Perrig & Eckstein,2005). This masking technique, which we callmirror maskingüses each letter’s mirror imageto mask the word. As can be seen in Figure 1,letters are merged with their inverted counter-part at the letters' base line. In previous exper-iments of ours, mirror masking has proven ef-fective in hiding prime words (that were shownfor 750 ms) from awareness: Across a range
2 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
of unpublished and published experiments withover 200 participants, 85% of participants re-ported not having perceived any of the primewords after the experiment (Perrig & Eckstein,2005). The masking effect is in principle dueto a visual neglect driven by attentional diver-sion, thus resembling the phenomenon of 'inat-tentional blindness' (Mack & Rock, 1998). Thefact that the mirror masked letters form newsymmetrical patterns resembling an unknownalphabet further enhances the masking effect.Indeed, when participants start seeing wordsin the patterns, they are usually unable to de-scribe the masking at first, because the wordstend to be seen as ’popping out’ of the pat-tern. Due to its different mode of functioning,mirror masking has two outstanding advan-tages compared to pattern masking: (a) Re-ports of conscious perception do not dependon response criterion levels, because wordsare seen as popping out as a whole, and (b)presentation time is in principle not critical.
We have previously observed replicablepriming of mirror masked primes with the wordstem completion task and, less reliably, withthe lexical decision task. In word stem comple-tion, where participants are asked to completea 3-letter word onset with two or three letterssuch that a valid word is formed, masked primewords that were only shown once in an exper-iment were reliably completed more often thancontrol words, and this effect remained stableover stimulus onset asynchronies (SOAs) of 1to 3 seconds (Perrig & Eckstein, 2005). Sub-sequent studies suggested that priming is notdue to single letters or word-parts, becausecompletion of words was not facilitated by sub-strings of primes (Eckstein, Sturzenegger, &Perrig, in preparation). However, it is possible
that perception of words does not reach thesemantic level, as semantic priming was foundto depend on participants’ reading ability (cf.Eckstein, Norris, Davis, & Henson, 2009).Our previous studies thus indicate that mirrormasked words are unconsciously processedat a nonsemantic lexical level. Therefore, theycan be used as a tool to investigate the mech-anisms of nonconscious perceptual and lexicalword perception that is not accompanied byawareness.
In all experiments so far, behavioural prim-ing by masked words was weak. This is ausual finding in this field, when one usesnon-repeated word presentation (cf. Damian,2001). The present study was conducted be-cause ERP measures have been found to bemore sensitive to nonconscious priming thanbehavioural priming (e.g., Holcomb, Reder,Misra, & Grainger, 2005; Kiefer, 2002; Misra &Holcomb, 2003). The main idea was to locatetemporal and topographic differences in ERPsrecorded while participants observed maskedwords and nonwords. This approach is usuallydifficult to do because ERPs of the maskedstimulus interfere with ERPs of the succeedingnonmasked stimulus in priming studies. In-stead, we were able to present mirror maskedwords for 500 ms, which allowed us for on-linemonitoring of processes occurring while partic-ipants fixed the masked letter strings. Becausewe could not assume to find the same ERPcomponents for nonconscious perception asthe ones reported for conscious word percep-tion (cf. Compton, Grossenbacher, Posner,& Tucker, 1991; Grossi & Coch, 2005; Hino-josa, Martin-Loeches, & Rubia, 2001; Nobre &McCarthy, 1995; Schendan, Ganis, & Kutas,1998; Sereno, Rayner, & Posner, 1998, for
2012, 5 (1) Kognitive Neurophysiologie des Menschen 3

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Figure 1: Example of the mirror masked term ’mirror nonwords’. Construction of mirrored letters isillustrated in the upper part of the figure. In the lower part, examples of mirrored nonwords (left, fromtop: ”skofe”, ”neibs”, ”weethce”, ”ruiohs”, ”ararlon”, ”druimed” and ”orulkce”) and words (right, fromtop: ”berth”, ”autumn”, ”liver”, ”success”, ”heaven”, ”irritation” and ”chance”) are reproduced. Note:The original language used was German.
prelexical processing; Cohen et al., 2000;Martin-Loeches, Hinojosa, Gomez-Jarabo,& Rubia, 1999; Nobre, Allison, & McCarthy,1994; Rudell & Hua, 1997; Skrandies, 1998for discrimination of word-forms or retrievalfrom word lexicon; Brown, Hagoort, & Chwilla,2000; Dietrich et al., 2001; Holcomb, et al.2005; Kutas & Hillyard, 1980; McCarthy, No-bre, Bentin, & Spencer, 1995; Wirth et al.,2006 for semantics, but see Penolazzi, Hauk,& Pulvermüller, 2007), we used the microstateand the TANOVA methods, which are bothapt for explorative determination of significantdifferences in ERP topographies between timesegments and conditions (McCarthy et al.,1995). Therefore, comparison with findingsstemming from research with visible words,according which differences at about 200 mswould be expected with lexical differences be-
tween stimuli, can only be done with extremecaution.
Analysis was based on comparisons of ERPscalp topographies across conditions in differ-ent time segments, using the microstate ap-proach. The time course of ERP topographiescontains periods of relative stability duringwhich change across time is minimal. Theseperiods of quasi stable ERP topography havebeen called ’microstates’ and are thoughtto correspond to basic steps of informationprocessing (Lehmann, 1990). Because it issensible to collapse across similar data froma statistical point of view, on- and offset timesof microstates were used to define time win-dows of analysis. Using a bootstrap method,comparison of microstate topographies acrossconditions was then used to locate differencesin active cerebral generators.
4 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
The experiment consisted in priming taskusing lexical decision, whereby each visibleprobe (word or nonword) letter string was pre-ceded by a mirror masked (word or nonword)letter string shown for 500 ms. This was doneto ensure that participants were looking at themasked stimuli when they appeared on thescreen, although their attention was directedtowards the succeeding probes. On the basisof a previous unpublished experiment, we ten-tatively expected priming effects for word-wordcompared to nonword-word trials which wouldindicate lexical processing of the primes. Inorder to rule out other reasons for such prim-ing, orthographic and semantic relatedness ofmasked and visible stimuli was minimised andevery stimulus was shown only once during theexperiment.
Materials and Methods
Subjects
Participants were 8 male and 11 female vol-unteers aged 21 to 38 years (M = 28.5, SD= 5.7). They were all native German speak-ers with normal or corrected-to-normal vision,without any neurological or neuropsychologi-cal disorder and without psychoactive medica-tion. 16 participants were self-reported right-handers and 3 were ambidextrous. All partic-ipants were naïve to the experimental hypoth-esis, had never been in contact with the mirrorword masking paradigm and were naïve withrespect to priming paradigms. The study wasapproved by the University’s ethics committeeand informed consent was obtained from allparticipants.
Stimuli
180 non-associated word pairs were selectedfrom a set of German nouns, most of whichdescribed concrete and emotionally neutralconcepts. Word length varied between 3 and8 letters and between 1 and 3 syllables. Or-thographic similarity of words in word pairswas very low as measured by an index oforthographic similarity described in Weber,1970. This set of word pairs was dividedinto four groups of 45 word pairs that werebalanced with respect to word length, numberof syllables, orthographic similarity and wordfrequency (Institut für Deutsche Sprache,1991-2007), based on occurrences in SwissNewspaper texts. Nonwords were constructedby concatenating letters that were randomlyselected from each letter position of the ex-perimental word set. 180 nonwords wereconstructed on the basis of the first word inthe word pairs, and another 180 nonwordswere constructed in the same way based onthe second word in the word pairs. Hence,letter frequencies of words and nonwordswere identical at each letter position. Pairingsof masked and nonmasked stimuli were con-trolled. The four groups of word pairs werecounterbalanced over participants and thefour possible pairing conditions (masked wordvs. nonmasked probe word, masked nonwordvs. nonmasked probe word; masked word vs.nonmasked probe nonword, masked nonwordvs. nonmasked probe nonword). Additional 15nonwords and 5 words were used for practice.
Procedure
Participants were comfortably seated ina darkened, acoustically and electrically
2012, 5 (1) Kognitive Neurophysiologie des Menschen 5

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
shielded recording chamber. A 15”’ liquiddisplay VGA screen and an IBM compatiblepersonal computer was used for stimulus dis-play. Participants were seated at a distanceof about 50 cm from the screen, their headspositioned by a head-rest mounted on a smalltable in front of them. Left and right keys ofa button box were used with the index fin-gers of the left and right hand for collectionof responses. The experimental program waswritten in MEL 2.0 (Schneider, 1988).
10 practice trials and 180 experimental trialswere shown. Each trial had a fixed sequencestarting with a 1s blank screen which was fol-lowed by a 500 ms mirror masked stimulus,then a 500 ms blank screen, and finally a 500ms nonmasked probe stimulus. The responsewindow started at probe onset and lasted forone second. Inter-trial interval duration wasone second. Participants were instructed tofixate the centre of the screen, where an ab-stract pattern (the masked letter string) wouldindicate the imminent onset of the letter stringprobe. They were asked to press as quickly aspossible the key corresponding to the type ofprobe shown. Right and left button box keyswere alternatively assigned to word and non-word probes across participants. Participantswere also instructed to keep eye movementsand eye blinks to a minimum during trials.
All stimuli were presented at the centre ofthe screen in white lettering on a black back-ground. The mirrored letter strings were pre-sented in mirror masked lower-case letterswritten in 'boxie17' font, subtending a verticalvisual angle of about 1°and a horizontal vi-sual angle of about 4°. Probe letter stringswere presented in upper-case in 'FG-16' (MELfont) which had the same height as the mir-
rored font. Sequence of the 180 trials wasrandomized. Each participant saw 45 stimu-lus pairs in each of the four conditions of word-nonword combinations, whereby word groupswere rotated between conditions and partici-pants, and nonwords were randomly chosenfrom the nonword set. Each word and non-word in the stimulus set was shown maximallyonce during the experiment. A short break wasgiven after 60 and 120 trials.
At the end of the experiment, participant’sawareness of the masked words was as-sessed with three consecutive questions: Hadthey noticed anything unusual in the patterns?Had they seen single letters in them? And, hadthey seen words in them? These questionswere used to determine which of the partici-pants were not aware of the words throughoutthe whole experiment.
Electrophysiological Recording
EEGs were recorded by 74 Ag/AgCl scalpelectrodes mounted on an elastic cap accord-ing to the international 10-10 system. Torecord eye blinks, two additional electrodeswere placed below the eyes. Cz was usedas recording reference. Electrode impedanceswere held below 10 kOhm. Signals were am-plified, bandpass filtered at 0.5-70 Hz, digi-tized at 250 Hz and continuously stored for of-fline analysis. Stimulus onsets were recordedwith a separate channel appearing in the EEGrecordings.
EEG preprocessing and averaging
From the raw EEG data, electrooculograph-ical signals (EOG) were computed as bipo-lar derivations between F9 and F10 (horizon-
6 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
tal EOG) and the channels below the eyeagainst Fp1 and Fp2 (vertical EOG). All 74scalp channels were recomputed to averagereference. After applying a 50 Hz notch filterand a 1.5 to 30 Hz bandpass filter, the analy-sis epochs were selected, starting at the onsetof each stimulus and lasting for 500 ms. (After500 ms, eye movements were more frequent).These epochs were DC corrected (baseline re-moval) and submitted to a semi-automatic ar-tifact detection that rejected epochs with chan-nels showing peak to peak amplitudes largerthan 100 uV within 100 ms and artifacts iden-tified by visual inspection. Over participants,the rate of rejection due to artifacts was about10%. For each subject, separate mean ERPswere computed for masked nonwords, maskedwords, nonmasked nonwords and nonmaskedwords. For each of these four conditions, agrand-mean across all participants was com-puted.
Identification of microstates
Identification of microstates was based onGlobal Field Power (GFP) of the grand-meanERPs for masked and nonmasked stimuli,respectively (Lehmann & Skrandies, 1980).Separate microstate analyses were donebased on the post-hoc observation that theGFP time course and the topographies ofthe grand-mean ERPs were substantially dif-ferent. GFP is a momentary, global index oftopographic strength and is defined as the spa-tial standard deviation. Periods of high GFPassumingly correspond to near-synchronousactivity of neural populations, and changesof ERP topography typically occur in localtroughs of the GFP (Lehmann, 1986). Wetherefore used the troughs of the GFP curve to
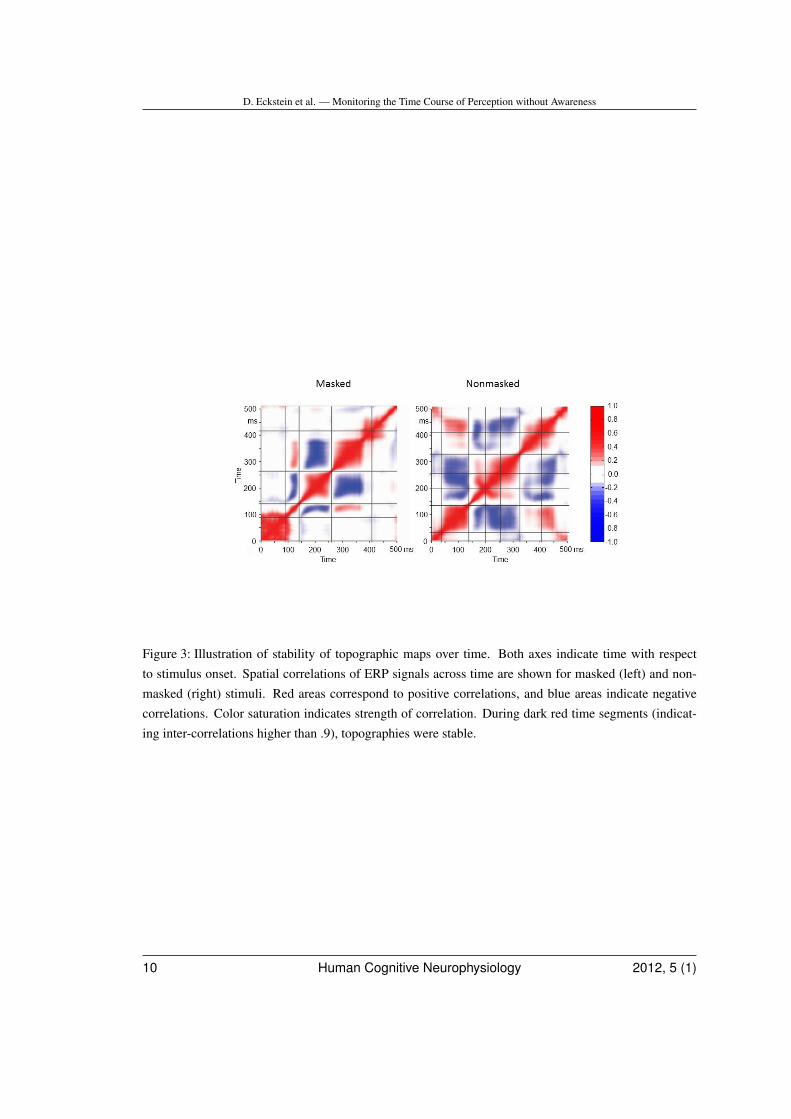
determine on- and offsets of the microstatesused for further analysis. Microstates that hadan on- or offset at the beginning or ending ofthe 0 to 500 ms post-stimulus analysis periodwere not further analysed. For illustration pur-poses, stability of microstates between GFPtroughs was further quantified by computingthe matrix of spatial correlations across time(Kochi, Koenig, Strik, & Lehmann, 1996).
Comparison of the ERPs
Statistical comparisons of word and nonwordERPs across subjects were done separatelyfor masked and nonmasked stimuli. Separateanalyses were conducted for comparison ofmicrostate amplitude and topography in eachmicrostate time window. For each subject andcondition, individual mean microstate topogra-phies were first computed. Differences be-tween microstate amplitudes associated withwords and nonwords were tested with pairedtwo-tailed t-tests over subjects on GFP (statis-tical parametric scalp maps).
For the comparison of microstate topog-raphy between word and nonword condi-tions, a randomization statistic was appliedusing the 'TANOVA' program (available on-line at www.unizh.ch/keyinst/ NewLORETA/LORETA01.htm). Using Global Map Dissimi-larity (Lehmann & Skrandies, 1980) as a GFP-independent difference measure between con-ditions, TANOVA applies a randomization test(Edgington, 1980; Manly, 1997) to establishthe exact probability of the observed differenceby assuming a null hypothesis of zero topo-graphic dissimilarity. This procedure has beenused in earlier studies (Kondakor, Pascual-Marqui, Michel, & Lehmann, 1995; Lehmannet al., 2005; Strik, Fallgatter, Brandeis, &
2012, 5 (1) Kognitive Neurophysiologie des Menschen 7

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Pascual-Marqui, 1998). In microstates whereTANOVA indicated a topographic differenceat p <.05, further analyses were performed:Firstly, t-maps were computed, thresholdedat p < .05 (uncorrected) and displayed forcomparison with other studies (note that theset-values were not used for further statisticalinterference, since the null-hypothesis wasrejected by the TANOVA). Secondly, sinceevidence for a topographic difference impliesthat there must have been differences inthe underlying sources, a distributed sourcelocalization procedure was employed to lo-cate these putative intracerebral sources ofsignificant differences.
Source localization was based on low reso-lution electromagnetic tomography (LORETA,Pascual-Marqui et al., 1999; Pascual-Marqui,Michel, & Lehmann, 1994) applied to indi-vidual, normalized (maximum GFP = 1) mi-crostate topographies of both included stim-ulus conditions. This version computed theelectric current density in the cortical areas ofthe digitized brain atlas of the Montreal Neu-rological Institute (MNI) at ˜7 mm resolution(2394 voxels). In those microstates wherethe TANOVA indicated significant differencesof ERP topography between conditions, voxel-by-voxel t-statistics were used to identify thosevoxels that could putatively account for the dif-ferences observed on the scalp. For illustra-tion purposes, the highest threshold of the t-statistics was set at an alpha level of 5%.
Results
Behavioral Data
Four participants saw at least one word in themasked letter strings and were therefore ex-
cluded from all subsequent analyses. All otherparticipants reported that they did not noticeanything unusual in the patterns and that theyhad seen no letters or words in them. Whenasked about the mirror masked strings, someof these participants guessed that they werestrings of an unknown writing, as for instancehieroglyphics. All analyses were performedon the data of these remaining 15 participantswho were ignorant of the information hidden inthe mask. Response accuracy was reasonablyhigh, with 95% correct responses for words(SD = 3%) and 92% correct responses for non-words (SD = 5%). Average reaction times werefaster for words, M = 579 ms (SD = 55 ms),than for nonwords, M = 639 ms (SD = 75 ms),t(14) = 7.09, p < .001. No effect of priming wasfound when comparing masked-string/visiblestring pairs that where congruent vs. incon-gruent in lexicality (e.g., word-word and non-word/nonword vs. nonword-word pairs; effectof type of masked string, F(1,14) = 0.62, p >.20 and interaction of type of masked stringwith type of visible string, F(1,14) = 1.95, p =.18).
Masked Words and Nonwords
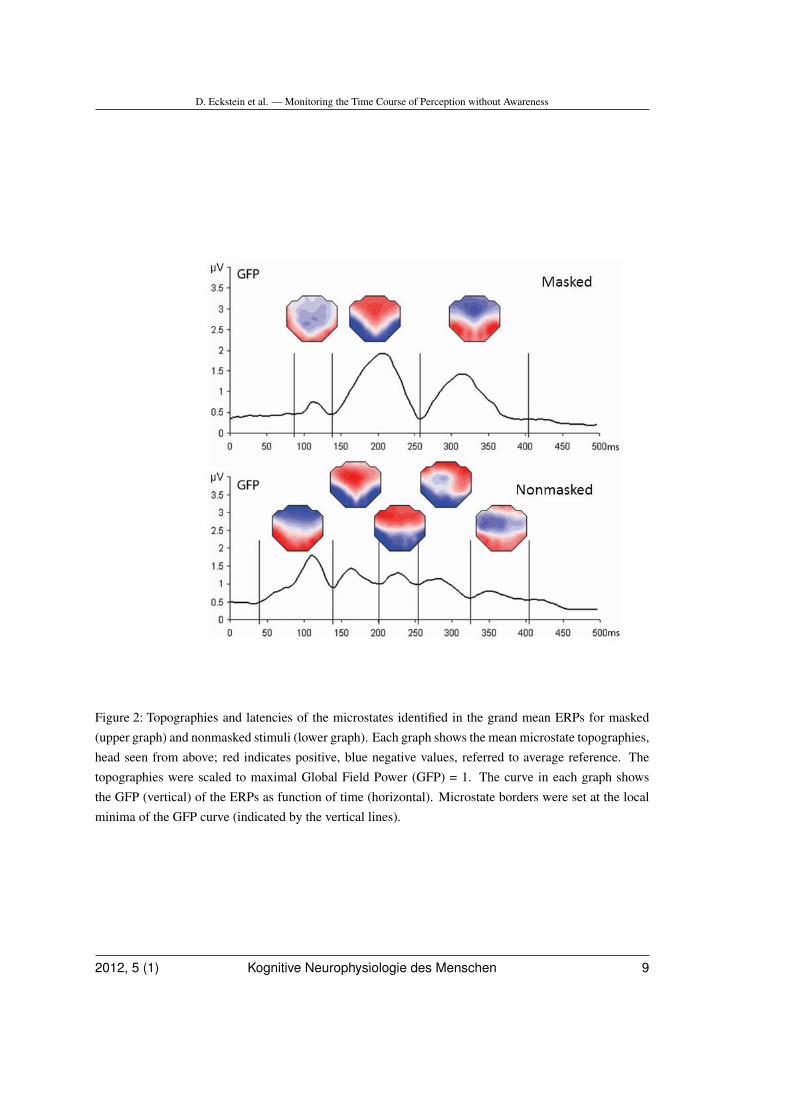
Segmentation of grand mean ERPs yielded3 microstates. Microstate topographies, mi-crostate latencies and GFP time course formasked stimuli are shown in upper part ofFig. 2. The stability of microstates was furtherquantified with spatiotemporal correlation ma-trices of ERPs averaged across subjects andconditions (Fig. 3, left).
The first microstate was a P100 topographywith a bilateral occipital positivity (88-136 ms).This was followed by a N200 that displayedan occipital bilateral negativity and a fronto-
8 Human Cognitive Neurophysiology 2012, 5 (1)
D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Figure 2: Topographies and latencies of the microstates identified in the grand mean ERPs for masked(upper graph) and nonmasked stimuli (lower graph). Each graph shows the mean microstate topographies,head seen from above; red indicates positive, blue negative values, referred to average reference. Thetopographies were scaled to maximal Global Field Power (GFP) = 1. The curve in each graph showsthe GFP (vertical) of the ERPs as function of time (horizontal). Microstate borders were set at the localminima of the GFP curve (indicated by the vertical lines).
2012, 5 (1) Kognitive Neurophysiologie des Menschen 9
D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Figure 3: Illustration of stability of topographic maps over time. Both axes indicate time with respectto stimulus onset. Spatial correlations of ERP signals across time are shown for masked (left) and non-masked (right) stimuli. Red areas correspond to positive correlations, and blue areas indicate negativecorrelations. Color saturation indicates strength of correlation. During dark red time segments (indicat-ing inter-correlations higher than .9), topographies were stable.
10 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
central positivity (136-256 ms). A third mi-crostate was characterized by a bilateral pari-etal positivity and a fronto-central negativity(256-404 ms).
Comparison of GFP differences betweenword and nonword stimulus microstates indi-cated no significant GFP differences betweenwords and nonwords. The randomization testof topographic dissimilarity however indicatedsignificant differences of microstate topogra-phies in the 136-256 ms microstate (p = .034,cf. Fig. 4).
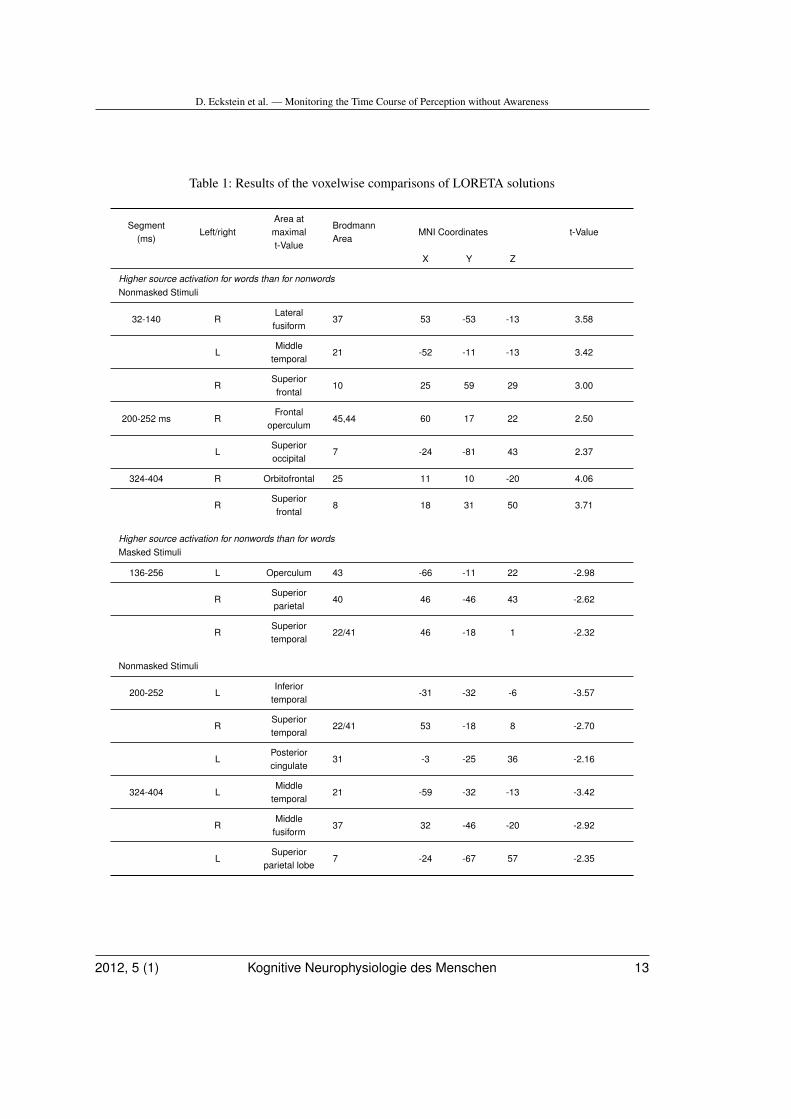
Results of the voxelwise comparisons ofLORETA solutions in microstates with word-nonword topographic differences are shown inFig. 5 and Table 1 (see also the supplementarymaterial). According to the LORETA model,differences between masked words and non-words between 136 and 256 ms were relatedto higher current density for nonwords than forwords in left operculum, right superior parietallobe and right superior temporal lobe (first rowin Figure 5).
Nonmasked words and nonwords
Grand mean ERPs and segmentation isshown in Figures 2 and 3. Five microstateswere identified: A first microstate was a P100with a bilateral occipital positivity (32-140 ms),which was followed by a N200 that displayeda strong occipital bilateral negativity and afronto-central positivity (144-200 ms). Then,two microstates followed that lasted from 200to 252 ms and from 256 to 320 ms, duringwhich the posterior negativity expanded overcentral and left parietal sites. A fifth microstatefrom 324 to 404 ms was characterized by largenegativity extending over midline and frontalelectrodes.
Comparison of GFP differences betweenword and nonword stimulus microstates indi-cated no significant GFP differences betweenwords and nonwords. The randomization testof topographic dissimilarity indicated signifi-cant differences of microstate topographies inthe 32-140 ms microstate (p = .027), the 200-252 ms microstate (p = .008) and the 324-404 ms microstate (p = .005). T-maps of themicrostates with significant topographic differ-ences are shown in Fig. 4.
According to the LORETA model, differ-ences between words and nonwords in the32-140 ms segment were localized in right lat-eral fusiform gyrus, left mediotemporal gyrusand right superior frontal gyrus. During the200-252 ms segment, differences were foundin right temporal lobe, left temporal lobe, leftposterior cingulate, right frontal operculumand left superior occipital lobe. Between 324and 404 ms, differences were found in rightorbitofrontal area, right superior frontal lobe,left middle temporal gyrus and right middlefusiform gyrus (Table I).
Discussion
This study investigated ERP time course ofmirror masked words and nonwords that werenot recognised by participants. Sequences ofstable microstates were analysed, which arethought to reflect successive stages of wordprocessing that correlate with successive acti-vation of functional brain areas (Koenig, Kochi,& Lehmann, 1998; Koenig & Lehmann, 1996;Michel et al., 2001; Pegna, Khateb, Michel, &Landis, 2004).
Based on findings of earlier studies, differ-ences between words and nonwords shouldappear in the 100-200 ms segment with vis-
2012, 5 (1) Kognitive Neurophysiologie des Menschen 11

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Figure 4: Upper row: Mean microstate maps, scaling as in Fig. 3. Lower row: Statistical parametricscalp maps for those microstates that showed significant topographic differences in the randomizationtests. Blue areas indicate larger values in the nonword condition; red areas indicate larger values in theword condition. Increasing steps of color intensity indicate p values below .10, .05 and .01, respectively.
Figure 5: Glass brain images of voxels showing differences between words and nonwords (p < .05), darkervoxels indicate higher t values.
12 Human Cognitive Neurophysiology 2012, 5 (1)
D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Table 1: Results of the voxelwise comparisons of LORETA solutions
Segment(ms)
Left/rightArea atmaximalt-Value
BrodmannArea
MNI Coordinates t-Value
X Y Z
Higher source activation for words than for nonwordsNonmasked Stimuli
32-140 RLateralfusiform
37 53 -53 -13 3.58
LMiddle
temporal21 -52 -11 -13 3.42
RSuperiorfrontal
10 25 59 29 3.00
200-252 ms RFrontal
operculum45,44 60 17 22 2.50
LSuperioroccipital
7 -24 -81 43 2.37
324-404 R Orbitofrontal 25 11 10 -20 4.06
RSuperiorfrontal
8 18 31 50 3.71
Higher source activation for nonwords than for wordsMasked Stimuli
136-256 L Operculum 43 -66 -11 22 -2.98
RSuperiorparietal
40 46 -46 43 -2.62
RSuperiortemporal
22/41 46 -18 1 -2.32
Nonmasked Stimuli
200-252 LInferior
temporal-31 -32 -6 -3.57
RSuperiortemporal
22/41 53 -18 8 -2.70
LPosteriorcingulate
31 -3 -25 36 -2.16
324-404 LMiddle
temporal21 -59 -32 -13 -3.42
RMiddle
fusiform37 32 -46 -20 -2.92
LSuperior
parietal lobe7 -24 -67 57 -2.35
2012, 5 (1) Kognitive Neurophysiologie des Menschen 13

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
ible words. Indeed, ERP differences of vis-ible words vs. nonwords were found in aP100 microstate located on the left parieto-occipital lobe which is known to reflect font,letter and n-gram differences (Compton et al.,1991; Nobre et al., 1994; Sereno et al., 1998),and is associated with foci wandering from theoccipital lobe to posterolateral fusiform gyrus(Binder & Price, 2000; Fujimaki et al., 1999;Gernsbacher & Kaschak, 2003; Petersen, Fox,Snyder, & Raichle, 1990; Pugh et al., 1996;Tarkiainen, Helenius, Hansen, Cornelissen,& Salmelin, 1999) and sometimes also com-bined with frontal areas (Hauk, Davis, Ford,Pulvermuller, & Marslen-Wilson, 2006). Justafter the 200 ms point which appears to bea marker of lexical access, significant word-nonword ERP differences were found in a seg-ment ranging from 200 to 252 ms. The differ-ence map resembled regressor maps associ-ated with word frequency reported by Hauk etal. (2006). Word frequency is closely related toword familiarity, which might have been used todiscriminate words from nonwords in our study.The late differences in the 324 to 404 ms seg-ment were reasonably in agreement with find-ings related to postlexical and semantic pro-cessing of words that are associated with pos-terior negativity for words vs. nonwords, whichis seen with frontal lobe, middle temporal gyrusand medial fusiform activation.
The topographic ERP-differences withmasked strings were different. Significantdifferences between stable ERP topographieswhen participants observed masked wordscompared to when they observed maskednonwords were found in a time segment from136 to 256 ms after stimulus onset. Assumingthe study participants validly were not aware
of the words in the masked patterns, theseERP differences suggest that some word-related properties of masked stimuli wereunconsciously processed. This segment of132 to 256 ms after stimulus onset was usedfor further analysis to identify the processesdifferentiating between masked words andnonwords. First, ERPs averaged over thewhole time segment of interest were anal-ysed. Difference maps formed a characteristicpattern of focused temporo-parietal positivityon the left hemisphere (Fig. 4). This patternbore resemblance with left-lateralised pos-itivity around the temporo-occipital junctionrelated to orthography which is discussed tobe associated with N170 (Bentin, Mouchetant-Rostaing, Giard, Echallier, & Pernier, 1999;Salmelin, Service, Kiesila, Uutela, & Salonen,1996, although this was reversed for nonwordsvs. words in our case), and parietal positivityrelated to word frequency and lexicality (P3,Proverbio, Vecchi, & Zani, 2004). Hence,there were some similarities of masked stim-ulus word-nonword differences and ERPs ofvisible stimuli as reported in the literature.
In a second step, LORETA distributedsource solution for this left lateralized positivityfor masked words vs. nonwords was com-puted (cf. Fig. 5). Results suggested that thedifference in topographies was due to strongercurrent density for nonwords than for words intwo right hemisphere areas (superior temporallobe, superior parietal lobe) and in one lefthemisphere area (posterior operculum nearthe Sylvian fissure). Activation in middle andsuperior temporal lobe has frequently beenfound with word vs. nonword comparisons,although the activity tends to be left lateralisedor bilateral (Cohen et al., 2000; Nobre et al.,
14 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
1994; Salmelin et al., 1996; Schendan et al.,1998). Why the modelled temporal sourcesare localized in the right hemisphere is difficultto explain on the basis of the present results.It is not unusual to assume that languageprocessing is bilateral but that activity is oftenstronger in the left hemisphere (cf. Damasio,Tranel, Grabowski, Adolphs, & Damasio, 2004;Scott & Wise, 2004). As one possible explana-tion, we suggest that unconscious processingof words might recruit language areas in adifferent fashion than visible words, leadingto measurable activation in the right hemi-sphere (e.g., Grossi & Coch, 2005; Holcomb &Grainger, 2006). Another possible explanationmight be that the activity results from an inhi-bition of word processing due to the masking.Such inhibition has been observed previouslyin masking studies (Eckstein, Kubat, & Perrig,2011; Wentura & Frings, 2005). The other twoareas, i.e. parietal lobe and posterior opercu-lum, have also occasionally been reported todifferentiate between words and nonwords orpseudowords (Compton et al., 1991; Hauk etal., 2006; Salmelin et al., 1996). It is howeverunusual to find higher activation for nonwordsthan words, especially in language areas inthe left hemisphere. Given that the primeswere not readable for the participants, it ispossible that this difference in activity justrepresents a difference in resource allocationor in timing. That is, higher allocation of re-sources was needed to convert substrings ofnonword primes to phonemes than for wordprimes and such a decoding probably tooklonger for nonwords than for words, whichwould explain the late timing. The sources ofactivation found with the LORETA model couldthus tentatively reflect word recognition pro-
cesses that are known to occur in the 136 to256 ms time range or earlier components thatwere delayed, but we infer this with caution, asthe temporal and parietal activation appearedto be in the right but not in the left hemisphere.
Although LORETA source models are notsuited to make direct inferences as to whichprocesses are responsible for word-nonwordERP differences, it is possible to compareLORETA solutions for different conditions inorder to identify similarities and differencesin modelled brain activations. Therefore, themasked stimuli LORETA solution for ERPs av-eraged over the 136 to 256 ms time seg-ment was compared with the nonmasked stim-uli LORETA solution for ERPs averaged overthe 200 to 252 ms time segment (cf. Fig. 4).There was a small area of overlap in right su-perior temporal lobe which discriminated be-tween words and nonwords for masked as wellas for nonmasked stimuli (maximal t valuesat MNI coordinates 46/-18/1 and 53/-18/8 formasked and nonmasked stimuli, respectively).We tentatively suggest that this overlap repre-sents a common process that was active in thistime range, whereas the non-overlap of otherareas indicate that most processes differenti-ating between words and nonwords were notshared when observing masked compared tononmasked stimuli in this specific task.
In summary, differences were found be-tween ERPs of masked words and ERPs ofmasked nonwords, and the source of thesedifferences was located in an area that wasalso found as source with visible words andnonwords. Compared to visible stimuli how-ever, differences were small. Given that mirrormasked patterns look like nonsense patternsor unknown hieroglyphs, ERPs measured
2012, 5 (1) Kognitive Neurophysiologie des Menschen 15

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
while participants see mirror masked wordsprimarily reflect processes that lead to theconscious experience of seeing nonsense pat-terns. It is therefore not surprising that most ofthe ERP differences found between words andnonwords are different for masked comparedto nonmasked stimuli. The task used did notrequire lexical processing of the mirror maskedstimuli. Nevertheless, some of the processesinvolved in perception of masked stimuli ap-peared to differentiate between words andnonwords, indicating that some aspects ofwords are automatically processed even ifthere is no direct requirement for processing.
This study has shown that it is possible tomonitor the time course of nonconscious pro-cessing of words vs. nonwords. Becausewords were presented only once during theexperiment, we can exclude stimulus learningeffects as a source of nonconscious percep-tion. Hence, our results evidence that maskedwords were spontaneously ’read’ up to a cer-tain level, although they were consciously per-ceived as meaningless patterns. Further re-search will be needed in order to identify thestages of word processing involved in noncon-scious processing of mirror masked words. Wehave done a few ERP and behavioural experi-ments that indicate that phonological process-ing is involved in the effect of mirror maskedpriming. It might be also appropriate to con-duct some comparative studies to determine ifthe same processes occur with mirror mask-ing (which is based on attentional distraction)compared to temporal pattern masking (whichis based on perceptual thresholding). At themoment however, mirror masking appears tobe a potential alternative to temporal masking,especially when there is need for longer pre-
sentation durations.
Acknowledgements
D. Eckstein and T. Koenig contributed in equalparts to this study. The authors thank AntjeHeinrich, Rik Henson and Susanne Jaeggi forvaluable comments on an earlier version of thisarticle. Part of this work was funded by SwissNational Foundation Fellowship No PA001 -113106/1.
References
Abrams, R. L., & Greenwald, A. G. (2000).Parts outweigh the whole (word) in uncon-scious analysis of meaning. PsychologicalScience, 11, 118-124.
Bargh, J. A. (1992). Does subliminality mat-ter to social psychology? Awareness of thestimulus versus awareness of its influence.In R. F. Bornstein & T. S. Pittman (Eds.), Per-ception without awareness. Cognitive, Clini-cal, and Social Perspectives (pp. 236-255):New York: Guilford.
Bentin, S., Mouchetant-Rostaing, Y., Giard,M. H., Echallier, J. F., & Pernier, J. (1999).ERP manifestations of processing printedwords at different psycholinguistic levels:time course and scalp distribution. Journalof Cognitive Neuroscience, 11, 235-260.
Binder, J., & Price, C. J. (2000). Functionalneuroimaging of language. In R. Cabeza &A. Kingstone (Eds.), Handbook of FunctionalNeuroimaging of Cognition (Vol. 1), pp. 187-251. Cambridge: MIT Press.
Brown, C. M., Hagoort, P., & Chwilla, D. J.(2000). An event-related brain potential
16 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
analysis of visual word priming effects. Brain& Language, 72, 158-190.
Cohen, L., Dehaene, S., Naccache, L.,Lehericy, S., Dehaene-Lambertz, G.,Henaff, M. A., & Michel, F. (2000). Thevisual word form area: spatial and temporalcharacterization of an initial stage of readingin normal subjects and posterior split-brainpatients. Brain, 123, 291-307.
Compton, P. E., Grossenbacher, P., Pos-ner, M. I., & Tucker, D. M. (1991). Acognitive−anatomical approach to atten-tion in lexical access. Journal of CognitiveNeuroscience, 3, 304-312.
Damasio, H., Tranel, D., Grabowski, T.,Adolphs, R., & Damasio, A. (2004). Neuralsystems behind word and concept retrieval.Cognition, 92, 179-229.
Damian, M. F. (2001). Congruity effectsevoked by subliminally presented primes:Automaticity rather than semantic process-ing. Journal of Experimental Psychology:Human Perception and Performance, 27,154-165.
Dehaene, S., Naccache, L., Cohen, L., Le Bi-han, D. , Mangin, J.-F., Poline, J. -B., &Rivière, D. (2001). Cerebral mechanismsof word masking and unconscious repetitionpriming. Nature Neuroscience, 4, 752-758.
Dehaene, S., Naccache, L., Le Clec'H,G., Koechlin, E., Mueller, M., Dehaene-Lambertz, G., van de Moortele, P.-F., &Le Bihan, D. (1998). Imaging unconscioussemantic priming. Nature, 395, 597-600.
Dietrich, D. E., Waller, C., Johannes, S.,Wieringa, B. M., Emrich, H. M., & Munte, T.
F. (2001). Differential effects of emotionalcontent on event-related potentials in wordrecognition memory. Neuropsychobiology,43, 96-101.
Eckstein, D., Kubat, M., & Perrig, W. J. (2011).Visible homonyms are ambiguous, sublim-inal homonyms are not: A close look atpriming. Consciousness and Cognition, 20,1327-1343.
Eckstein, D., Norris, D., Davis, M. H., & Hen-son, R. N. (2009). Invisible is better: de-crease of subliminal priming with increasingvisibility. Psyche, 15, 39-59.
Eckstein, D., Sturzenegger, M., & Perrig, W.J. (in preparation). The fate of orthographicinformation: Lexical decisions are sensitiveto masked words and masked nonwords.
Edgington, E. S. (1980). Validity of Random-ization Tests for One-Subject Experiments.Journal of Educational Statistics, 5, 235-251.
Frost, R., Forster, K. I., & Deutsch, A. (1997).What can we learn from the morphology ofHebrew? A masked-priming investigationof morphological representation. Journal ofExperimental Psychology: Learning, Mem-ory and Cognition, 23, 829-856.
Fujimaki, N., Miyauchi, S., Putz, B., Sasaki, Y.,Takino, R., Sakai, K., & Tamada, T. (1999).Functional magnetic resonance imagingof neural activity related to orthographic,phonological, and lexico-semantic judg-ments of visually presented characters andwords. Human Brain Mapping, 8, 44-59.
Gernsbacher, M. A., & Kaschak, M. P. (2003).Neuroimaging studies of language produc-
2012, 5 (1) Kognitive Neurophysiologie des Menschen 17

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
tion and comprehension. Annual Review ofPsychology, 54, 91-114.
Grainger, J., Colé, P., & Segui, J. (1991).Masked morphological priming in visualword recognition. Journal of Memory andLanguage, 30, 370-384.
Greenwald, A. G., Abrams, R. L., Naccache,L., & Dehaene, S. (2003). Long-term se-mantic memory versus contextual memoryin unconscious number processing. Jour-nal of Experimental Psychology: Learning,Memory, & Cognition, 29, 235-247.
Greenwald, A. G., Draine, S. C., & Abrams, R.L. (1996). Three cognitive markers of un-conscious semantic activation. Science, 273(5282), 1699-1702.
Greenwald, A. G., Klinger, M. R., & Schuh, E.S. (1995). Activation by marginally percepti-ble ("subliminal") stimuli: Dissociation of un-conscious from conscious cognition. Jour-nal of Experimental Psychology: General,124, 22-42.
Grossi, G., & Coch, D. (2005). Automatic wordform processing in masked priming: An ERPstudy. Psychophysiology, 42, 343-355.
Hassin, R. R., Uleman, J. S., & Bargh, J. A.(2006). The New Unconscious. Oxford, UK:Oxford Univ Press.
Hauk, O., Davis, M. H., Ford, M., Pulvermuller,F., & Marslen-Wilson, W. D. (2006). Thetime course of visual word recognition as re-vealed by linear regression analysis of ERPdata. NeuroImage, 30, 1383-1400.
Hinojosa, J. A., Martin-Loeches, M., & Rubia,F. J. (2001). Event-related potentials and se-
mantics: an overview and an integrative pro-posal. Brain & Language, 78, 128-139.
Holcomb, P. J., & Grainger, J. (2006). Onthe time course of visual word recognition:An event-related brain potential investigationusing masked repetition priming. Journal ofCognitive Neuroscience, 18, 1631-1643.
Holcomb, P. J., Reder, L., Misra, M., &Grainger, J. (2005). The effects of prime vis-ibility on ERP measures of masked priming.Cognitive Brain Research, 24, 155-172.
Humphreys, G. W., Besner, D., & Quinlan, P. T.(1988). Event perception and the word rep-etition effect. Journal of Experimental Psy-chology: General, 117, 51-67.
Institut für Deutsche Sprache, M. (1991-2007).COSMAS I/II (Corpus Search, Managementand Analysis System), from http://www.ids-mannheim.de/cosmas/
Jacoby, L. L., & Whitehouse, K. (1989). Anillusion of memory: False recognition influ-enced by unconscious perception. Journalof Experimental Psychology: General, 118,126-135.
Kiefer, M. (2002). The N400 is modulatedby unconsciously perceived masked words:Further evidence for an automatic spreadingactivation account of N400 priming effects.Cognitive Brain Research, 13, 27-39.
Kinoshita, S., & Lupker, S. J. E. (2003).Masked Priming: the state of the art : NewYork: Taylor & Francis.
Kochi, K., Koenig, T., Strik, W. K., & Lehmann,D. (1996). Event-related potential P300microstate topography during visual one-
18 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
and two-dimensional tasks in chronicschizophrenics. European Archives ofPsychiatry & Clinical Neuroscience, 246,288-296.
Koenig, T., Kochi, K., & Lehmann, D. (1998).Event-related electric microstates of thebrain differ between words with visual andabstract meaning. Electroencephalography& Clinical Neurophysiology, 106, 535-546.
Koenig, T., & Lehmann, D. (1996). Microstatesin language-related brain potential mapsshow noun-verb differences. Brain & Lan-guage, 53, 169-182.
Kondakor, I., Pascual-Marqui, R. D., Michel, C.M., & Lehmann, D. (1995). Event-related po-tential map differences depend on the pres-timulus microstates. Journal of Medical En-gineering & Technology, 19, 66-69.
Kouider, S., & Dupoux, E. (2004). Partialawareness creates the "illusion" of sublim-inal semantic priming. Psychological Sci-ence, 15, 75-81.
Kovacs, G., Vogels, R., & Orban, G. A. (1995).Cortical correlate of pattern backward mask-ing. Proceedings of the National Academyof Sciences of the United States of America,92, 5587-5591.
Kunde, W., Kiesel, A., & Hoffmann, J. (2003).Conscious control over the content of uncon-scious cognition. Cognition, 88, 223-242.
Kutas, M., & Hillyard, S. A. (1980). Read-ing senseless sentences: brain poten-tials reflect semantic incongruity. Science,207(4427), 203-205.
Lehmann, D. (1986). Spatial analysis of EEGand evoked potential data. In F. H. Duffy
(Ed.), Topographic Mapping of Brain Elec-trical Activity (pp. 29-61). Boston: Butter-worths.
Lehmann, D. (1990). Brain Electric Mi-crostates and Cognition: The Atoms ofThought. In E. R. John (Ed.), Machinery ofthe Mind (pp. 209-244). Boston: Birkhauser.
Lehmann, D., Faber, P. L., Galderisi, S., Her-rmann, W. M., Kinoshita, T., Koukkou, M., &Koenig, T. (2005). EEG microstate durationand syntax in acute, medication-naive, first-episode schizophrenia: a multi-center study.Psychiatry Research, 138, 141-156.
Lehmann, D., & Skrandies, W. (1980).Reference-free identification of compo-nents of checkerboard-evoked multichannelpotential fields. Electroencephalography &Clinical Neurophysiology, 48, 609-621.
Mack, A., & Rock, I. (1998). Inattentional blind-ness. Cambridge, MA: MIT Press.
Manly, B. F. J. (1997). Randomization, Boot-strap and Monte Carlo Methods in Biology.London: Chapman & Hall.
Martin-Loeches, M., Hinojosa, J. A., Gomez-Jarabo, G., & Rubia, F. J. (1999). The recog-nition potential: An ERP index of lexical ac-cess. Brain & Language, 70, 364-384.
McCarthy, G., Nobre, A. C., Bentin, S., &Spencer, D. D. (1995). Language-relatedfield potentials in the anterior-medial tem-poral lobe: 1. Intracranial distribution andneural generators. The Journal of Neuro-science, 15, 1080-1089.
McCarthy, G., Nobre, A. C., Bentin, S., &Spencer, D. D. (1995). Language-related
2012, 5 (1) Kognitive Neurophysiologie des Menschen 19

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
field potentials in the anterior-medial tempo-ral lobe: I. Intracranial distribution and neu-ral generators. Journal of Neuroscience, 15,1080-1089.
Michel, C. M., Thut, G., Morand, S., Khateb,A., Pegna, A. J., Grave de Peralta, R., &Landis, T. (2001). Electric source imagingof human brain functions. Brain Research -Brain Research Reviews, 36, 108-118.
Misra, M., & Holcomb, P. J. (2003). Event-related potential indices of masked repeti-tion priming. Psychophysiology, 40, 115-130.
Naccache, L., & Dehaene, S. (2001). Uncon-scious semantic priming extends to novelunseen stimuli. Cognition, 80, 223-237.
Nobre, A. C., Allison, T., & McCarthy, G.(1994). Word recognition in the human infe-rior temporal lobe. Nature, 372 (6503), 260-263.
Nobre, A. C., & McCarthy, G. (1995).Language-related field potentials in theanterior-medial temporal lobe: II. Effects ofword type and semantic priming. Journal ofNeuroscience, 15, 1090-1098.
Pascual-Marqui, R. D., Lehmann, D., Koenig,T., Kochi, K., Merlo, M. C., Hell, D., &Koukkou, M. (1999). Low resolution brainelectromagnetic tomography (LORETA)functional imaging in acute, neuroleptic-naive, first-episode, productive schizophre-nia. Psychiatry Research, 90, 169-179.
Pascual-Marqui, R. D., Michel, C. M., &Lehmann, D. (1994). Low resolution elec-tromagnetic tomography: a new methodfor localizing electrical activity in the brain.
International Journal of Psychophysiology,18, 49-65.
Pegna, A. J., Khateb, A., Michel, C. M., & Lan-dis, T. (2004). Visual recognition of faces,objects, and words using degraded stimuli:where and when it occurs. Human BrainMapping, 22, 300-311.
Penolazzi, B., Hauk, O., & Pulvermüller, F.(2007). Early semantic context integrationand lexical access as revealed by event-related brain potentials. Biological Psychol-ogy, 74, 374-388.
Perrig, W. J., & Eckstein, D. (2005). Uncon-scious word-stem completion priming in amirror-masking paradigm. Consciousness &Cognition, 14, 257-277.
Petersen, S. E., Fox, P. T., Snyder, A. Z., &Raichle, M. E. (1990). Activation of extrastri-ate and frontal cortical areas by visual wordsand word-like stimuli. Science, 249(4972),1041-1044.
Proverbio, A. M., Vecchi, L., & Zani, A. (2004).From orthography to phonetics: ERP mea-sures of grapheme-to-phoneme conversionmechanisms in reading. Journal of Cogni-tive Neuroscience, 16, 301-317.
Pugh, K. R., Shaywitz, B. A., Shaywitz, S. E.,Constable, R. T., Skudlarski, P., Fulbright, R.K., & Gore, J. C. (1996). Cerebral organi-zation of component processes in reading.Brain, 119, 1221-1238.
Rolls, E. T., & Tovee, M. J. (1994). Processingspeed in the cerebral cortex and the neuro-physiology of visual masking. Proceedingsof the Royal Society: Biological Sciences,257, 9-15.
20 Human Cognitive Neurophysiology 2012, 5 (1)

D. Eckstein et al. — Monitoring the Time Course of Perception without Awareness
Rudell, A. P., & Hua, J. (1997). The recognitionpotential, word difficulty, and individual read-ing ability: on using event-related potentialsto study perception. Journal of Experimen-tal Psychology: Human Perception & Perfor-mance, 23, 1170-1195.
Salmelin, R., Service, E., Kiesila, P., Uutela,K., & Salonen, O. (1996). Impaired visualword processing in dyslexia revealed withmagnetoencephalography. Annals of Neu-rology, 40, 157-162.
Schendan, H. E., Ganis, G., & Kutas, M.(1998). Neurophysiological evidence for vi-sual perceptual categorization of words andfaces within 150 ms. Psychophysiology, 35,240-251.
Schneider, W. (1988). Micro ExperimentalLaboratory: An integrated system for IBMPC compatibles. Behavior Research Meth-ods, Instruments, & Computers, 20, 206-217.
Scott, S. K., & Wise, R. J. S. (2004). The func-tional neuroanatomy of prelexical process-ing in speech perception. Cognition, 92, 13-45.
Sereno, S. C., Rayner, K., & Posner, M. I.(1998). Establishing a time-line of wordrecognition: evidence from eye movementsand event-related potentials. Neuroreport,9, 2195-2200.
Skrandies, W. (1998). Evoked potential corre-lates of semantic meaning - A brain mappingstudy. Cognitive Brain Research, 6, 173-183.
Strik, W. K., Fallgatter, A. J., Brandeis, D.,& Pascual-Marqui, R. D. (1998). Three-
dimensional tomography of event-relatedpotentials during response inhibition: ev-idence for phasic frontal lobe activation.Electroencephalography & Clinical Neuro-physiology, 108, 406-413.
Tarkiainen, A., Helenius, P., Hansen, P. C.,Cornelissen, P. L., & Salmelin, R. (1999).Dynamics of letter string perception in thehuman occipitotemporal cortex. Brain, 122,2119-2132.
Weber, M. R. (1970). First-graders' use ofgrammatical context in reading. In H. Levin& J. P. Williams (Eds.), Basic Studies onReading (pp. 147-163): New York: BasicBooks.
Wentura, D. (2002). Ignoring ""brutal"" willmake ""numid"" more pleasant but ""uyuvu""more unpleasant: The role of a priori pleas-antness of unfamiliar stimuli in affectivepriming tasks. Cognition and Emotion, 16,296-298.
Wentura, D., & Frings, C. (2005). Repeatedmasked category primes interfere with re-lated exemplars: New evidence for negativesemantic priming. Journal of ExperimentalPsychology: Learning, Memory and Cogni-tion, 31, 108-120.
Wirth, M., Horn, H., Koenig, T., Federspiel, A.,Müller, T., Meier, B., & Strik, W. K. (2006).Semantic Processing Deficits in Schizophre-nia. Neuropsychobiology, 54, 10 (abstract).
Wolford, G., Marchak, F., & Hughes, H. (1988).Practice effects in backward masking. Jour-nal of Experimental Psychology: HumanPerception & Performance, 14, 101-112.
2012, 5 (1) Kognitive Neurophysiologie des Menschen 21

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Abstract
M. Wagner & W.-J. Kuo (Hamburg, Germany & Taipei, Taiwan) – Population-Adapted AveragedHead Templates for EEG and MEG Source AnalysisIn electroencephalography or magnetoencephalography source localization studies where individual sub-
ject’s magnetic resonance images (MRIs) are not available, averaged MRIs can be used instead. For pop-
ulations of subjects with head shapes that are both homogeneous and sufficiently distinct from the head
shape of an existing averaged MRI, the use of a population-adapted, averaged MRI is desirable. Examples
for such populations are non-Caucasian or pediatric subjects. A population-adapted, averaged MRI can
be created based on individual MRIs of subjects representing the population of interest. The extents of the
bounding box containing the brain, together with the locations of the anterior and posterior commissure
landmarks are measures facilitating this process. Based on their average values for the individual MRIs, it
is possible to average these individual MRIs. Alternatively and also based on these measures, an existing
averaged MRI can be adapted to better match the population’s head shape. Based on individual MRIs of
62 Chinese subjects, the procedure is demonstrated, including the creation of standard electrode locations
and realistically shaped boundary element method head models.
Keywords: Magnetic resonance imaging; Electroencephalography; Magnetoencephalography; Source
localization; Averaged MRI; Talairach Co-registration
Population-AdaptedAveraged Head Templates for
EEG and MEG SourceAnalysis
M. Wagner ∗1, W.-J. Kuo∗2
1Compumedics Germany GmbH,Heußweg 25, 20255 Hamburg, Germany,
2Institute of Neuroscience,National Yang-Ming University, Taipei, Taiwan
Introduction
In electroencephalography (EEG) andmagnetoencephalography (MEG) source anal-ysis, medical image data such as magneticresonance images (MRIs) can be utilized ina variety of ways. Localization accuracy isincreased by means of individual, realisticallyshaped head models (Fuchs, Drenckhahn,Wischmann, & Wagner, 1998; Mosher, Leahy,& Lewis, 1999). Source analysis results canbe displayed in their anatomical context byoverlaying them onto structural brain magessuch as MRI (Fuchs, Wagner, Wischmann,Ottenberg, & Dössel, 1994). Furthermore,anatomical features such as the gray matter
22 Human Cognitive Neurophysiology 2012, 5 (1)

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
may be used to constrain source analysis(Dale & Sereno, 1993; Wagner, Fuchs, Wis-chmann, Ottenberg, & Dössel, 1995).
Often, however, individual MRI data are notavailable or shall deliberately not be used. Thelatter can for example be the case in groupstudies, where performing source analyses inindividual anatomical space and later poolingthe results is more laborious than performingsource analyses in a common, representativeanatomical space. In such cases, it is goodpractice to use averaged MRI datasets instead(Fuchs, Kastner, Wagner, Hawes, & Eber-sole, 2002; Pascual-Marqui, Esslen, Kochi, &Lehmann, 2002): An averaged MRI dataset(also commonly called an MRI template)shows anatomical features that are commonto a population of subjects, without suggestinga level of detail (cortical folds) that could onlybe obtained by using the subject’s own MRI.An averaged MRI still makes it possible to de-rive and use realistically shaped head models,view source results in their anatomical context,and constrain source analysis (albeit not to thelevel of cortical folds).
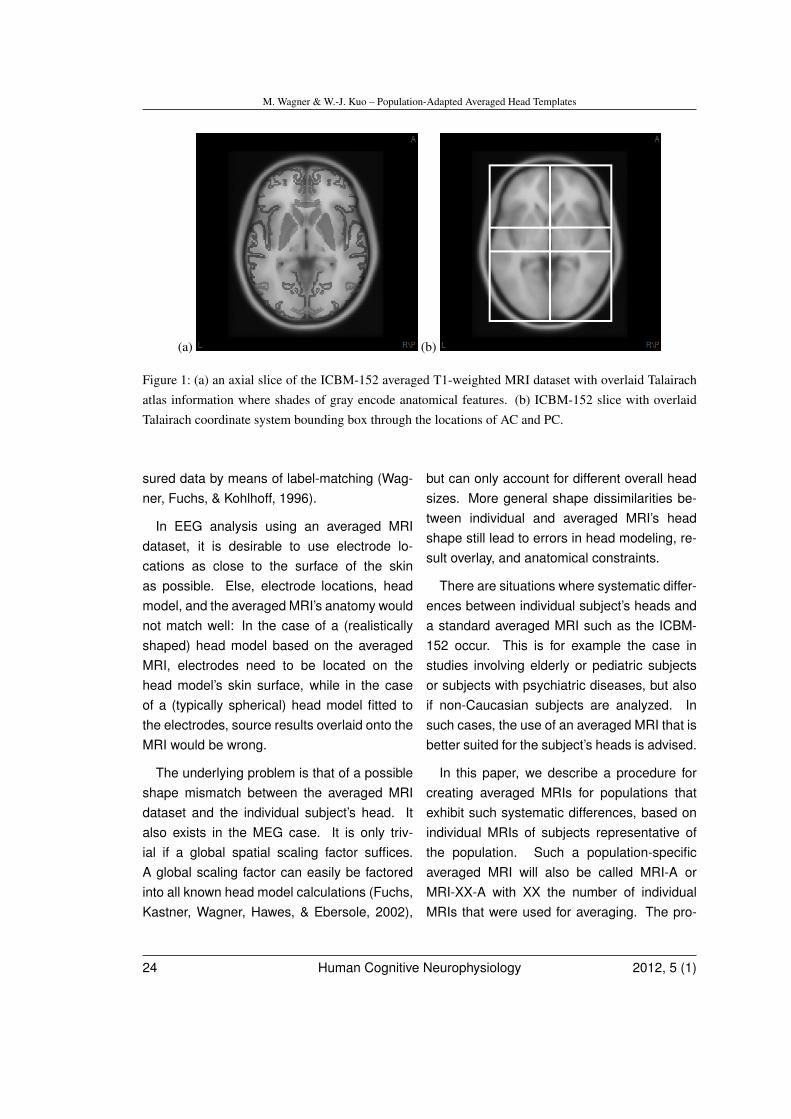
Today, a commonly used averaged MRI isthe ICBM-152 dataset (Mazziotta et al., 2001),also called the Montreal Neurological Insti-tute (MNI) brain. It represents an averageCaucasian head shape. A consequence ofthe widespread use of the ICBM-152 is theavailability of additional, co-registered three-dimensional (3-D) maps detailing e.g. tis-sue type probabilities (Mazziotta, Toga, Evans,Fox, & Lancaster, 1995) or cytoarchitectonicatlas information (Lancaster et al., 2000; Fig-ure 1a). This availability makes it straightfor-ward to guide source analysis based on tissuetypes, e.g. to constrain sources to the cortical
gray matter, or to relate source analysis resultsto atlas information.
Another important reference frame is theTalairach atlas (Talairach & Tournoux, 1988),which is based on axial cross-sections of asingle subject’s brain. It includes detailedanatomical labels. The proportional grid-based coordinate system introduced in thispublication has become the de-facto standardfor reporting locations in the brain. The Ta-lairach coordinate system is based on twolandmarks, the anterior commissure (AC),which serves as the origin, and the posteriorcommissure (PC), together with the extentsof the bounding box containing the brain.The mid-sagittal and the axial plane throughAC and PC, together with the coronal planesthrough AC and PC, respectively, subdividethis bounding box into twelve cuboid-shapedcompartments. Using a linear transformationper compartment, any brain, including theICBM-152, can be co-registered with the Ta-lairach reference frame (Desco et al., 2001,Lancaster et al., 2007). Figure 1b shows theTalairach bounding box overlaid onto an axialslice of the ICBM-152, while Figure 2 shows a3-D rendering of the bounding box.
In order to bring EEG electrode or MEGsensor locations into the anatomical referenceframe of an averaged MRI, a landmark-basedtransformation can be applied (Fuchs, M., Wis-chmann, Wagner, & Krüger, 1995). Alterna-tively, and only available in the EEG case, elec-trode locations based on the 10-20 system(Jasper, 1958) or one of its extensions (Cha-trian, 1985; Oostenveld & Praamstra, 2001)can be determined by measuring distanceson the averaged MRI’s segmented skin andrelating their locations to the actually mea-
2012, 5 (1) Kognitive Neurophysiologie des Menschen 23
M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
(a) (b)
Figure 1: (a) an axial slice of the ICBM-152 averaged T1-weighted MRI dataset with overlaid Talairachatlas information where shades of gray encode anatomical features. (b) ICBM-152 slice with overlaidTalairach coordinate system bounding box through the locations of AC and PC.
sured data by means of label-matching (Wag-ner, Fuchs, & Kohlhoff, 1996).
In EEG analysis using an averaged MRIdataset, it is desirable to use electrode lo-cations as close to the surface of the skinas possible. Else, electrode locations, headmodel, and the averaged MRI’s anatomy wouldnot match well: In the case of a (realisticallyshaped) head model based on the averagedMRI, electrodes need to be located on thehead model’s skin surface, while in the caseof a (typically spherical) head model fitted tothe electrodes, source results overlaid onto theMRI would be wrong.
The underlying problem is that of a possibleshape mismatch between the averaged MRIdataset and the individual subject’s head. Italso exists in the MEG case. It is only triv-ial if a global spatial scaling factor suffices.A global scaling factor can easily be factoredinto all known head model calculations (Fuchs,Kastner, Wagner, Hawes, & Ebersole, 2002),
but can only account for different overall headsizes. More general shape dissimilarities be-tween individual and averaged MRI’s headshape still lead to errors in head modeling, re-sult overlay, and anatomical constraints.
There are situations where systematic differ-ences between individual subject’s heads anda standard averaged MRI such as the ICBM-152 occur. This is for example the case instudies involving elderly or pediatric subjectsor subjects with psychiatric diseases, but alsoif non-Caucasian subjects are analyzed. Insuch cases, the use of an averaged MRI that isbetter suited for the subject’s heads is advised.
In this paper, we describe a procedure forcreating averaged MRIs for populations thatexhibit such systematic differences, based onindividual MRIs of subjects representative ofthe population. Such a population-specificaveraged MRI will also be called MRI-A orMRI-XX-A with XX the number of individualMRIs that were used for averaging. The pro-
24 Human Cognitive Neurophysiology 2012, 5 (1)

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Figure 2: 3-D rendering of the Talairach bounding box with the AC and PC landmarks. The mid-sagittalplane through AC and PC, the axial plane through AC and PC, and the two coronal planes through ACand PC, respectively, define twelve compartments used for the piece-wise linear transformation betweenindividual subject and Talairach coordinates.
posed procedure was designed to accomplishthe following aims:
1) The averaged MRI should clearly showprominent features such as the brain, the skull,and the skin layers, equalizing within-grouphead shape variations. Therefore, individ-ual MRIs need to be spatially normalized be-fore averaging, so that matching features co-localize.
2) Furthermore, each individual MRI shouldhave the same impact on the resulting aver-age, independent of its overall intensity. Thisimplies that MRI intensities have to be normal-ized before averaging.
3) The co-registration of averaged MRIs tothe Talairach atlas as well as to averaged MRIsof different subject populations and to indi-vidual MRIs should be straightforward. Thismakes the Talairach landmarks and bounding
boxes a natural choice as the basis of co-registration.
4) Group-intrinsic head shapes and brain di-mensions should be preserved in the resultingaverage, making it necessary to spatially nor-malize individual MRIs to the average popula-tion’s brain dimensions before averaging.
5) As an alternative to or extension of creat-ing an averaged MRI from the individual MRIsused, it should be possible to spatially nor-malize an existing averaged MRI such as theICBM-152 to match the population of inter-est. The result is a version of the ICBM-152that exhibits a head shape and size match-ing the population of interest. This population-specific ICBM-152-based dataset will also becalled an adapted ICBM-152 dataset or ICBM-152-A. For some populations (e.g. in pediatricstudies), other existing templates besides the
2012, 5 (1) Kognitive Neurophysiologie des Menschen 25

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
ICBM-152 dataset might be preferable candi-dates for adaptation.
6) In preparation of an EEG or MEG study,the resulting averaged MRI dataset should beused to create a population-specific realisti-cally shaped head model, and used as a ba-sis for calculating the 10-20 electrode systempositions on the shape of its segmented skin.
7) Finally, creating the averaged MRI shouldrequire little user interaction so that it can eas-ily be applied for a given population.
Existing approaches for creating averagedMRI datasets are summarized in (Toga &Thompson, 2001). Most approaches achievethis by means of a template and use high-parametric, non-linear transformations (Kleinet al., 2009), which allow a mapping of sub-cortical structures between individual andaveraged MRI, but introduces the problemof choosing the right template to match thepopulation-of-interest. The specific goals ofthis study in the context of EEG and MEGanalysis gives the mapping of sub-corticalstructures a lower priority, with the benefit thata template-free ansatz can be pursued, whereimage data are normalized onto the averageof their Talairach bounding boxes.
In order to demonstrate the procedure, 62individual MRIs of ethnic Chinese subjectshave been processed and an averaged Chi-nese MRI-62-A dataset as well as an adaptedICBM-152-A dataset have been created
Previous studies (Tang et al., 2010; Cheeet al., 2009; Kochunov et al., 2003) have al-ready established that significant differencesbetween Chinese and Caucasian head shapesexist. One previous study also created anaveraged Chinese MRI dataset (Tang et al.,2010) but used a different co-registration tech-
nique, which involved matching all MRIs ontoone MRI randomly selected from the popula-tion of interest as an intermediate step. Fur-thermore, this study did not demonstrate atlasco-registration nor the derivation of head mod-els and electrode layouts.
Methods
MRI Acquisition
The subject sample consisted of 62 subjects(37 females and 25 males), with an averageage of 22.4 years and a standard deviation(SD) of 2.1 years. At this age, all subjectshad already attained their adult head shapes(Friede, 1981). All subjects were right-handedethnic Chinese with no history of neurologicaldisorders. Handedness was verified using theEdinburgh Inventory (Oldfield, 1971). All sub-jects negated when asked for psychiatric dis-eases. Written consent of the participants wasobtained before MRI scanning, with the proto-col approved by the Institutional Ethics and Ra-diation Safety Committees of National Yang-Ming University, Taipei, Taiwan.
Whole-head T1-weighted MRI datasets of allsubjects were acquired at the Laboratory ofCognitive Neural Science, Yang-Ming Univer-sity, Taipei, Taiwan. A magnetization-preparedrapid gradient-echo (MP-RAGE) sequence runon a 3T TrioTim scanner (Siemens, Erlan-gen, Germany) was used for the acquisition.The pixel dimensions and slice thickness were1 mm each. The matrix size was 224x256, re-sulting in a field-of-view (FOV) of 224x256 mm.The number of slices was 192.
26 Human Cognitive Neurophysiology 2012, 5 (1)

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Landmark and Bounding BoxDetermination
MRI data were loaded into the Curry 7 soft-ware (Compumedics, Charlotte, NC, USA). InCurry, the locations of AC and PC were de-termined manually. An additional mid-sagittallandmark (MS) in a more superior slice al-lowed, together with AC and PC, to definethe mid-sagittal plane. Based on this in-formation, the 3-D bounding box containingthe brain was gauged, comprising the fol-lowing distance measures: AC-Anterior, PC-Posterior, AC-Superior, AC-Inferior, AC-Left,and AC-Right, in addition to the AC-PC dis-tance already obtained. The averages of thesemeasures were computed and compared totheir equivalents in the ICBM-152 templateand the Talairach atlas. A two-tailed z-test wasused to assess whether the observed differ-ences are significant.
Averaged MRI Dataset and AdaptedICBM-152
Each MRI was then re-sliced according to thecoordinate axes spanned by AC, PC, and themid-sagittal plane, and its twelve sub-volumesdefined by AC, PC, and the bounding box di-mensions were individually, linearly and con-tiguously transformed. For details on this pro-cess, please refer to the Appendix. The av-erage measures obtained in the previous stepwere used as the dimensions of the targetbounding box. The outcome was 62 volumet-ric images with identical locations of AC andPC landmarks, identical mid-sagittal planes,and identical bounding boxes enclosing thebrains. In these images, all brains occu-pied the same coordinate space. Then, MRI
intensities were linearly scaled so that the99th intensity percentile was the same acrossdatasets. The 99th percentile was chosen asa noise-insensitive measure of the maximum.Finally, an averaged 3-D image was computedwhich will be called the MRI-62-A in the re-mainder of the paper.
In the same way as described above forthe individual MRIs, the ICBM-152 was trans-formed such that its AC-PC distance as well asits Talairach bounding box extents matched theaverage values of the subject population, yield-ing an ICBM-152-A dataset. Figure 3 showsthis procedure in a flowchart.
Electrode Locations and Head Models
For the original and the adapted ICBM-152as well as for the MRI-62-A, the locations ofthe nasion, left and right pre-auricular point,and inion landmarks were determined. Basedon these landmarks and the skin surface seg-mented from MRI, electrode locations for theextended 10-20 system were computed by theCurry software (Wagner, Fuchs, & Kohlhoff,1996). Computations included measuring andsubdividing distances on the outer surface ofthe segmented skin, replicating the proce-dure for manual electrode placement (Jasper,1958; Chatrian, 1985). As a result, for thesethree datasets, 86 electrode locations were ob-tained, identified by their labels. These 86electrode locations included ten sub-temporallocations, which could not be reliably esti-mated for the ICBM-152 and ICBM-152-A be-cause of their limited inferior extensions. Forthe remaining 76 electrodes, the average andstandard deviation of the distances betweenelectrodes of the same label were determinedacross all three datasets.
2012, 5 (1) Kognitive Neurophysiologie des Menschen 27

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Figure 3: Flowchart for creating the MRI-A and ICBM-152-A datasets.
In a next step, realistically shaped three-compartment boundary element method(BEM) head models were created using theautomatic BEM geometry setup algorithmimplemented in the Curry software (Wagner,Fuchs, Drenckhahn, Wischmann, Köhler, &Theißen, 1997). Triangle side lengths for theinner skull, outer skull, and the skin boundarywere 5 mm, 7 mm, and 8 mm, respectively.As a byproduct of this algorithm, trianglemeshes representing the skin and the cortexare obtained.
Results
Chinese vs. Caucasian Brain Dimensions
Table 1 lists averages, standard deviations,and medians of the AC-PC distances, bound-ing box extents, and bounding box volumes ob-tained for the 62 individual MRIs and comparesthem with their counterparts for the Talairachatlas and the ICBM-152. Comparing the 62Chinese subjects with the Talairach atlas, allbounding box extents as well as the width-to-length and height-to-length ratios turn out tobe significantly different; the bounding box vol-ume and the height-to-width ratio, however, arenot. A comparison with the ICBM-152 dimen-sions shows significant differences for all butthe mediolateral dimensions and the height-to-length ratio.
For a trained user, it took about two min-utes per dataset to define the landmarks and
28 Human Cognitive Neurophysiology 2012, 5 (1)
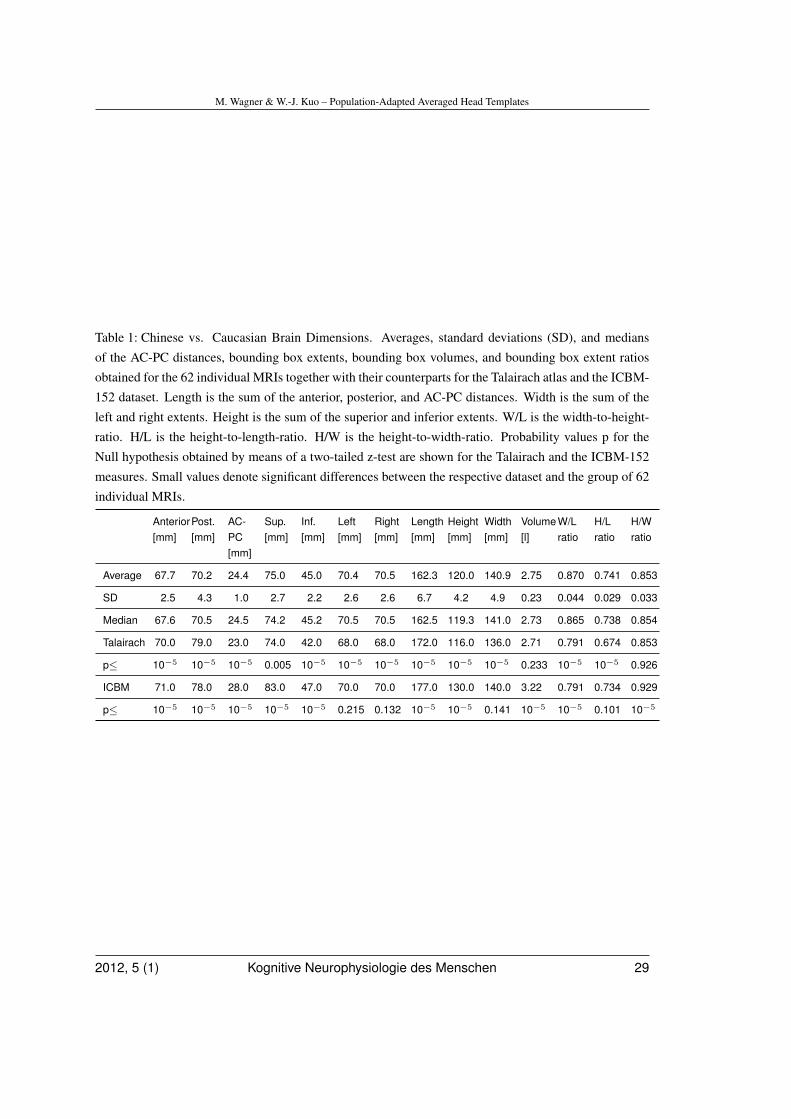
M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Table 1: Chinese vs. Caucasian Brain Dimensions. Averages, standard deviations (SD), and mediansof the AC-PC distances, bounding box extents, bounding box volumes, and bounding box extent ratiosobtained for the 62 individual MRIs together with their counterparts for the Talairach atlas and the ICBM-152 dataset. Length is the sum of the anterior, posterior, and AC-PC distances. Width is the sum of theleft and right extents. Height is the sum of the superior and inferior extents. W/L is the width-to-height-ratio. H/L is the height-to-length-ratio. H/W is the height-to-width-ratio. Probability values p for theNull hypothesis obtained by means of a two-tailed z-test are shown for the Talairach and the ICBM-152measures. Small values denote significant differences between the respective dataset and the group of 62individual MRIs.
Anterior[mm]
Post.[mm]
AC-PC[mm]
Sup.[mm]
Inf.[mm]
Left[mm]
Right[mm]
Length[mm]
Height[mm]
Width[mm]
Volume[l]
W/Lratio
H/Lratio
H/Wratio
Average 67.7 70.2 24.4 75.0 45.0 70.4 70.5 162.3 120.0 140.9 2.75 0.870 0.741 0.853
SD 2.5 4.3 1.0 2.7 2.2 2.6 2.6 6.7 4.2 4.9 0.23 0.044 0.029 0.033
Median 67.6 70.5 24.5 74.2 45.2 70.5 70.5 162.5 119.3 141.0 2.73 0.865 0.738 0.854
Talairach 70.0 79.0 23.0 74.0 42.0 68.0 68.0 172.0 116.0 136.0 2.71 0.791 0.674 0.853
p≤ 10−5 10−5 10−5 0.005 10−5 10−5 10−5 10−5 10−5 10−5 0.233 10−5 10−5 0.926
ICBM 71.0 78.0 28.0 83.0 47.0 70.0 70.0 177.0 130.0 140.0 3.22 0.791 0.734 0.929
p≤ 10−5 10−5 10−5 10−5 10−5 0.215 0.132 10−5 10−5 0.141 10−5 10−5 0.101 10−5
2012, 5 (1) Kognitive Neurophysiologie des Menschen 29

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
bounding box dimensions required.
Averaged MRI Dataset and AdaptedICBM-152 Dataset
Figure 4 shows the MRI-62-A dataset basedon 62 Chinese subjects and the ICBM-152-A. Due to the normalization of each individ-ual dataset to the Talairach bounding box, theMRI-A is least blurry at the locations of AC andPC. The overall head shape of the ICBM-152-Ais the same as for the MRI-62-A. For the MRI-62-A, the skull areas around the brain are noteverywhere as sharply delineated as for theICBM-152-A.
Electrode Locations and Head Models
A rendering of the three sets of extended10-20 system electrodes obtained for theMRI-62-A, ICBM-152, and ICBM-152-Adatasets is shown in Figure 5.
Table 2 shows the average distances andtheir standard deviations together with themedian distances between electrodes of thesame label for the three pairs of datasets.Electrode location differences are largest be-tween the MRI-62-A and the ICBM-152. Theyare smallest between the MRI-62-A and theICBM-152-A, with a mean displacement ofless than 5 mm.
The realistic BEM head model represent-ing the MRI-62-A dataset consists of 12,132triangles connecting 6,072 nodes. The ICBM-152-A BEM contains 12,264 triangles and6,138 nodes. Its inferior extension is smallerthan for the MRI-62-A model. Figure 6 showsboth BEMs together with cortical trianglemeshes. These cortical triangle meshes donot show individual gyri and sulci, due to the
non-individual MRIs they were created from.
Discussion and Conclusion
The proposed procedure meets the aims pre-sented in the introduction: As a demonstra-tion of the proposed procedure, an averagedMRI dataset representing a normal, adult Chi-nese population called the MRI-62-A datasethas been created. The MRI-62-A datasetshows prominent features such as the brain,the skull, and the skin layers. Group-intrinsichead shapes and brain dimensions have beenpreserved. Its co-registration to the Talairachatlas as well as to other MRIs can be per-formed using the same Talairach-based ap-proach as used for averaging. A population-specific ICBM-152-based dataset called theICBM-152-A has been created. It exhibits ahead shape and brain size matching the popu-lation of interest. On the basis of the MRI-62-A and the ICBM-152-A, realistic head modelsand standard electrode layout shave been cre-ated.
With about two minutes of user interac-tion per individual MRI (all other calculationswere automated), the process of creating apopulation-adapted averaged MRI is straight-forward.
The size differences between Chinese andCaucasian brains are significant, with Chinesebrains being e.g. comparatively shorter thanboth the Talairach atlas brain and the ICBM-152. These findings agree with the ones re-ported in (Tang et al., 2010), although it shouldbe mentioned, that due to the brain-strippingapproach employed in that study, measuresare not directly comparable.
30 Human Cognitive Neurophysiology 2012, 5 (1)
M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Figure 4: Upper row: MRI-62-A dataset based on 62 Chinese subjects. Lower row: ICBM-152-A datasetwith the average Chinese subject’s brain dimensions.
Table 2: Average distances and their standard deviations together with the median distances between elec-trodes of the same label for pairs of datasets. Electrode locations were obtained according to the extended10-20 electrode system procedures.
MRI-62-A vs.ICBM-152[mm]
MRI-62-A vs.ICBM-152-A[mm]
ICBM-152-A vs.ICBM-152 [mm]
Average 11.0 4.7 7.6
SD 3.7 1.8 3.0
Median 10.8 4.8 7.6
2012, 5 (1) Kognitive Neurophysiologie des Menschen 31

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Figure 5: Extended 10-20 system electrodes for the ICBM-152 (light gray), the ICBM-152-A (dark gray),and the MRI-62-A (black), together with the skin surface of MRI-62-A.
32 Human Cognitive Neurophysiology 2012, 5 (1)
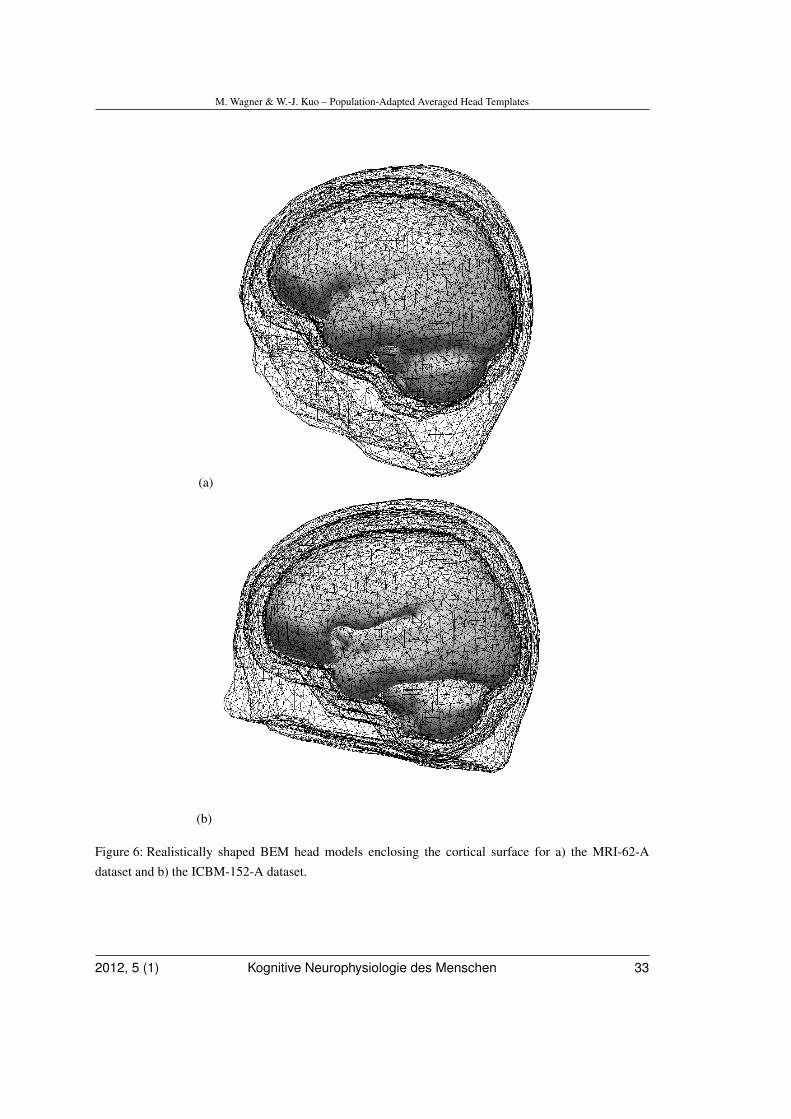
M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
(a)
(b)
Figure 6: Realistically shaped BEM head models enclosing the cortical surface for a) the MRI-62-Adataset and b) the ICBM-152-A dataset.
2012, 5 (1) Kognitive Neurophysiologie des Menschen 33

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
In a population-specific EEG or MEG study,finding significantly different brain dimensionsbetween the population-of-interest and an ex-isting, candidate MRI template should motivatethe creation and use of an MRI-A or an ICBM-152-A dataset instead of just using the tem-plate.
An important practical question is, whichof the two adapted MRIs, the MRI-A or theICBM-152-A, is better suited for being used ina given study:
Both adapted datasets can be linked to ad-ditional atlas data because their Talairach pa-rameters are known.
Due to the bounding box-based head shapenormalization before averaging, the quality ofthe MRI-A dataset is not uniform: it is highestaround the AC and PC landmarks and wherethe brain touches the planes of the boundingbox, but and lower in other areas, such as thedorsolateral prefrontal cortex (Brodmann area9). This is not the case for the ICBM-152-A,because a different, template-based approachwas used for normalizing individual MRIs in thecourse of the ICBM-152 creation.
Due to the larger field-of-view of the indi-vidual MRIs used for averaging in this study,sub-temporal electrode locations according tothe extended 10-20 system can be computedfor the MRI-62-A but not for the ICBM-152-A.For the same reason, the MRI-62-A BEM hasa wider inferior coverage, which commendsthe use of the MRI-62-A for studies involv-ing sub-temporal electrodes. For the other 76electrode locations, a mean difference of lessthan 5 mm between both adapted datasets isstill lower than the electrode location errorsbrought about by using caps with labeled elec-trodes, so that in such a setup no clear recom-
mendation can be spelled out.
Of course, the quality of individual MRIs fora given population may vary, so that using theICBM-152-A may be a welcome alternative if alow number of individual MRIs are available orif these are of low quality.
Furthermore, it should not be forgotten thatthe ICBM-152 has by now become part of thecommunity’s viewing habits, so that presentingone’s results in that context may make themeasier to perceive. This is of course not true, ifthe subject population is pediatric. In this case,either an existing pediatric template (Sanchez,Richards, & Almli 2011) may be adapted us-ing a process analogous to the one describedabove for the creation of the ICBM-152-A, orthe MRI-A should be used.
Whichever dataset is chosen for a particularstudy, the possibility to use electrode locationsand a head model that are fit for the populationof interest, to overlay source results onto ananatomical image with matching head shape,and the possibility to use adapted atlas datato constrain source analysis and report resultsrecommend using an adapted averaged MRIfor studies involving non-Caucasian or pedi-atric populations.
While it has been shown that brain exten-sions and standard electrode locations varysignificantly between the ICBM-152 and thepopulation-specific MRIs, this is only a hintthat differences in source localization are alsoto be expected. A follow-up study will exam-ine the effect of the different MRIs and theirderived head models and electrode locationsonto source localizations.
34 Human Cognitive Neurophysiology 2012, 5 (1)

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Acknowledgments
M. Wagner thanks C.W. Ponton for the initialimpetus to work on MRI averaging. This workwas supported in part by the National ScienceFoundation of Taiwan under Grant 98-2517-S-004-001-MY3 and Grant 100-2321-B-009-001.
References
Chatrian, G.E. (1985). Ten percent electrodesystem for topographic studies of sponta-neous and evoked EEG activity. Am J Elec-troencephalogr Technol, 25, 83-92.
Chee, M.W. et al. (2009). Cognitive functionand brain structure correlations in healthyelderly East Asians, NeuroImage, 46, 257-269.
Dale, A.M. & Sereno, M.I. (1993). Improvedlocalization of cortical activity by combiningEEG and MEG with MRI cortical surface re-construction: a linear approach. J. CognNeurosci, 5, 162-176.
Desco, M. et al. (2001). Multimodality im-age quantification using Talairach grid. ProcSPIE, 4322, 1385-1392.
Friede, H. (1981). Normal development andgrowth of the human neurocranium and cra-nial base. Scand J Plast Reconstr Surg; 15,163-169.
Fuchs, M., Drenckhahn, R., Wischmann,H.-A., & Wagner, M. (1998). An improvedboundary element method for realisticvolume-conductor modelling. IEEE TransBiomed Eng, 45, 980-997.
Fuchs, M., Kastner, J., Wagner, M., Hawes,S., & Ebersole, J.S. (2002). A standardizedboundary element method volume conduc-tor model. Clin Neurophysiol, 113, 702-712.
Fuchs, M., Wagner, M., Wischmann, H.-A., Ot-tenberg, K., & Dössel, O. (1994). Possi-bilities of functional brain imaging using acombination of MEG and MRT. In C. Pantev
2012, 5 (1) Kognitive Neurophysiologie des Menschen 35

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
(Ed.), Oscillatory Event-Related Brain Dy-namics (pp. 435-457). New York: PlenumPress.
Fuchs, M., Wischmann, H.-A., Wagner, M.,& Krüger, J. (1995). Coordinate systemmatching for neuromagnetic and morpho-logical reconstruction overlay. IEEE TransBiomed Eng, 42, 416-420.
Jasper, H.H. (1958). The ten twenty electrodesystem of the international federation. Elec-troencephalogr Clin Neurophysiol, 10, 371-37.
Klein, A., et al. (2009). Evaluation of 14 non-linear deformation algorithms applied to hu-man brain MRI registration. Neuroimage,46, 786-802.
Kochunov, P. et al. (2003). Localizedmorphological brain differences betweenEnglish-speaking Caucasians and Chinese-speaking Asians: new evidence of anatomi-cal plasticity. Neuroreport, 14, 961-964.
Lancaster, J.L. et al. (2007). Bias betweenMNI and Talairach coordinates analyzed us-ing the ICBM-152 brain template. Hum BrainMapp, 1028, 1194-1205.
Lancaster, J.L. et al. (2000). Automated Ta-lairach atlas labels for functional brain map-ping. Hum Brain Mapp, 10, 120-131.
Mazziotta, J.C. et al. (2001). A probabilisticatlas and reference system for the humanbrain: International Consortium for BrainMapping (ICBM). Phil Trans R Soc Lond B,356, 1293-1322.
Mazziotta, J.C., Toga, A.W., Evans, A., Fox, P.,& Lancaster, J. (1995). A probabilistic at-las of the human brain: theory and rationale
for its development. The International Con-sortium for Brain Mapping (ICBM). NeuroIm-age, 2, 89-101.
Mosher, J.C., Leahy, R.M., & Lewis, P.S.(1999). EEG and MEG: forward solutionsfor inverse methods. IEEE Trans BiomedEng, 46, 245-259.
Oldfield, R.C. (1971). The assessment andanalysis of handedness: The Edinburgh in-ventory, Neuropsychologia, 9, 97-113.
Oostenveld, R. & Praamstra, P. (2001). Thefive percent electrode system for high-resolution EEG and ERP measurements,Clin Neurophysiol, 112, 713-719.
Pascual-Marqui, R.D., Esslen, M., Kochi, K.,& Lehmann, D. (2002). Functional imag-ing with low-resolution brain electromagnetictomography (LORETA): a review. MethodsFind Exp Clin Pharmacol, 24 (supplementC), 91-95.
Sanchez, C.E., Richards, J.E., & Almli, C.R.(2011). Neurodevelopmental MRI braintemplates for children from 2 weeks to4 years of age. Dev Psychobiol, DOI:10.1002/dev.20579.
Talairach, J. & Tournoux, P. (1988). Co-planarStereotaxic Atlas of the Human Brain: 3-Dimensional Proportional System - an Ap-proach to Cerebral Imaging. New York, NY:Thieme Medical Publishers.
Tang, Y. et al. (2010). The construction ofa Chinese MRI brain atlas: A morphomet-ric comparison study between Chinese andCaucasian cohorts. NeuroImage, 51, 33-41.
36 Human Cognitive Neurophysiology 2012, 5 (1)

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Toga, A.W., & Thompson, P.M. (2001). Therole of image registration in brain mapping.Image And Vision Computing, 19, 3-24.
Wagner, M., Fuchs, M., Wischmann, H.-A.,Ottenberg, K., & Dössel, O. (1995). Cor-tex segmentation from 3D MR images forMEG reconstructions. In C. Baumgartner,L. Deecke, G. Stroink, & S. J. Williamson(Eds.), Biomagnetism: fundamental re-search and clinical applications (pp. 352-356). Amsterdam: Elsevier Science/IOSPress.
Wagner, M., Fuchs, M., & Kohlhoff, H. (1996).Automatic Generation of the 10-20 System.Brain Topogr, 8, 409.
Wagner, M., Fuchs, M., Drenckhahn, R.,Wischmann, H.-A., Köhler, T., & Theißen, A.(1997). Automatic generation of BEM andFEM meshes. NeuroImage, 5 (4 supple-ment 1), S389.
2012, 5 (1) Kognitive Neurophysiologie des Menschen 37

M. Wagner & W.-J. Kuo – Population-Adapted Averaged Head Templates
Appendix
Transformation from MRI Space to TemplateSpace
The co-registration of a specific image“dataset” (identified by the letter d) into acommon “template” space (identified by theletter t) is described below. In the scope of thispaper, the interpretation of what the “dataset”and “template” exactly are differs, dependingon which part of the process we are lookingat: When averaging individual MRIs to createan MRI-A dataset, d denotes the individualMRIs while t refers to the average spacebased on measures obtained for all individualMRIs. When creating an ICBM-152-A dataset,d refers to the ICBM-152 dataset while, asabove, t refers to the average space basedon all individual MRIs. When transforming theMRI-A or the ICBM-152-A to Talairach atlasspace (or vice versa), d refers to the former,while t represents the latter.
The locations of AC and PC in templatespace are tAC and tPC . tAC is always in theorigin, and tPC is always on the negative yaxis, so that
tAC =
0
0
0
and tPC =
0
−tAC−PC
0
where tAC−PC is the AC-PC distance. The
positive x axis points right, y points anterior,and z points up. The bounding box dimen-sions (see Figure 2) are given by the AC-Anterior distance tAC−A, the PC-Posterior dis-tance tPC−P , the already mentioned AC-PCdistance tAC−PC , the AC-Superior distance tS ,the AC-Inferior distance tI , the AC-Left dis-tance tL, and the AC-Right distance tR.
If t denotes Talairach space, the boundingbox dimensions (see Table I) are tAC−A =
70mm, tPC−P = 79mm, tAC−PC = 23mm,
tS = 74mm, tI = 42mm, tL = 68mm, andtR = 68mm.
Analogously, dAC−A, dPC−P , dAC−PC , dS ,
dI , dL, and dR describe the bounding boxmeasures in dataset space. The locations ofAC and PC in dataset space are dAC and dPC .
Additionally, the location of a superior mid-sagittal location, dMS serves to unambiguouslyidentify the orientation of the z axis.
The transformation from dataset space intotemplate space consists of three steps: trans-lation, rotation, and scaling. In a first step, atranslation maps dAC , onto tAC (the origin).Next, a rotation maps dPC onto the negativey axis and dMS onto the y-z plane. The scal-ing step ensures that the bounding box dimen-sions co-localize:
x→
x tLdL
for x ≤ 0
x tRdR
for x > 0
y →
(y + dAC−PC)×tPC−P
dPC−P− tAC−PC
for y ≤ −dAC−PC
y tAC−PC
dAC−PCfor − dAC−PC < y ≤ 0
y tAC−A
dAC−Afor y > 0
z →
z tIdI
for z ≤ 0
z tSdS
for z > 0
This scaling step ensures a contiguous map-ping without any jumps at the compartmentboundaries.
38 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Abstract
M. Ruchsow (Göppingen) — Personale Identität aus Sicht der Neurowissenschaften und der ana-lytischen PhilosophieThe problem of personal identity is discussed very controversially in analytical philosophy and contempo-
rary neuroscience. Descartes’ approach was a dualism of substances (res cogitans and res extensa). This
solution was rejected by Locke and other empiricists, because substances can never be the object of pos-
sible experience. Locke proposed (autobiographic) memory as warrant of (diachronic) personal identity.
Alternatively, the criterion of bodily continuity was suggested, with body, brain or “relevant parts” of the
brain facilitating diachronic identity. As Parfit showed, both naturalistic (neuroscientific) criteria (auto-
biographic memory and “enough of the brain”) are contradictory. Taylor’s concept of narrative identity
might be a continuative approach.
Keywords: Personal Identity, Introspection, Narrative Self
Personale Identität aus Sichtder Neurowissenschaften
und der analytischenPhilosophie
M. Ruchsow, FachkrankenhausChristophsbad, Faurndauer Str. 6-28, 73035
Einleitung
Mit Beginn der Neuzeit geht der Glaube aneinen sinnvollen, identitätsstiftenden Kos-mos zunehmend verloren. Der kanadischePhilosoph Charles Taylor sieht diese Entwick-lung durchaus ambivalent und spricht, MaxWebers Diktum von der „Entzauberung derWelt“ aufnehmend, von einem „Unbehagen ander Moderne“. Aus Taylors Sicht wurde mehr
individuelle Freiheit dadurch errungen, dasswir uns von einem älteren Moralhorizont gelösthaben. Damit einhergehend gerieten auch diealten Ordnungsgefüge in Misskredit und dieVorstellung, dass die „hierarchical order inthe universe was reflected in the hierarchiesof human society“. Gleichzeitig verliehen diealten Ordnungen aber auch „meaning to theworld and to the activities of social life“ (Taylor,1991, S. 3). Die neuzeitliche Entwicklung führtalso zu einem Zuwachs an individueller Frei-heit, andererseits aber auch zu einem Verlustan Orientierung bzw. sozialer Freiheit (d.h.Sittlichkeit im Sinne Hegels). Das Konzept dersozialen Freiheit bzw. Sittlichkeit greift denantiken Polisgedanken auf, demzufolge dieBürger „had such a deep subjective attach-ment to their polis that their social membershipcould be said to constitute a central part oftheir own identities“ (Neuhouser, 2000, S. 34).
Das Problem der personalen Identität ist
2012, 5 (1) Kognitive Neurophysiologie des Menschen 39

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
also eine neuzeitliche Fragestellung, die dengeschilderten historischen Prozess zur Vo-raussetzung hat. Typische Vertreter dieserneuzeitlichen Konstellation sind Descartesund Locke, für die Personen „desengagierte“neutrale Beobachter einer weitgehend mech-anistisch zu erklärenden äußeren Wirklichkeitsind (vgl. Taylor, 1989, S. 143 ff.).
Nach einer kurzen Präzisierung, was in dervorliegenden Arbeit unter „Identität“ zu verste-hen ist, wird zunächst Descartes’ Personbe-griff als substantielle Einheit von res cogitansund res extensa vorgestellt. Die Ablehnungvon Substanzen als möglichen Gegenständenvon Erfahrung durch den Empiristen Lockeführt u.a. zum Problem der diachronen Iden-tität, das er durch das Erinnerungskriteriumzu lösen versucht. Im weiteren Verlauf wirdsich zeigen, dass sowohl das Kriterium derpsychischen Kontinuität (Erinnerung) als auchdas der physischen Kontinuität (des Kör-pers oder des Gehirns) widersprüchlich ist.Als ein möglicherweise weiterführender Lö-sungsansatz wird dann Taylors Konzept dernarrativen Identität erläutert.
1. Identitätsbegriffe
Der Begriff der Person ist historisch wie sys-tematisch mit den beiden eng zusammenhän-genden Problemkreisen der
1. Identität von Personen und der
2. Autonomie von Personen verbunden (vgl.Krebs, Kambartel & Jantschek, 1995, S.89 ff.).
Hinsichtlich 1.) ist zu unterscheiden zwischennumerischer und qualitativer Identität.
Von numerischer oder logischer Identitätsprechen wir, wenn folgende Kriterien erfülltsind:
a.) Symmetrie (x = y →y = x),
b.) Transitivität (x = y ∧ y = z →x = z) und
c.) Reflexivität (x = x) (vgl. Gillitzer, 2001, S.24)
Zwei Dinge sind numerisch identisch (alsoder Zahl nach ein einziges Ding), wenn sie hin-sichtlich aller ihrer Eigenschaften gleich sind.
Demgegenüber bezeichnen wir zwei ver-schiedene Gegenstände als qualitativ iden-tisch, wenn sie sich lediglich ähnlich, alsonur in einer bestimmten Eigenschaft (odermehreren) gleich sind. Numerische Identitätist somit ein Grenzfall qualitativer Identität(Gleichheit hinsichtlich aller Eigenschaften vs.Gleichheit hinsichtlich einer/mehrerer Eigen-schaften; Tugendhat & Wolf, 1983, S. 168f.).
Hinsichtlich 2.) bleibt festzuhalten, dassder Begriff der qualitativen Identität auch ineiner erweiterten Bedeutung gebraucht wird.Er bezieht sich dann auf den autonomenLebensentwurf von Personen und die damiteinhergehenden existentiellen Fragen: „Werbin ich?“ und: „Wer will ich sein?“ Durchihre Antworten orientieren sich Personen gle-ichzeitig in einem „moralischen Raum“ bzw.verorten sich auf einer „moralischen Land-karte“ (Taylor, 1989, S. 25 ff.).
In der vorliegenden Arbeit wird der Termi-nus „numerische Identität“ verwendet, wenndie Zahl der zugrundeliegenden Entitäten rel-evant ist, also die Frage, ob von einer odermehreren Personen geredet wird. Der Begriffder „qualitativen Identität“ wird ausschließlichin der erweiterten Bedeutung verwendet, alsowenn die qualitativen Aspekte von Personalität
40 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
und insbesondere die moralische Dimensionvon Lebensentwürfen im Vordergrund stehen(vgl. Ruchsow & Hermle, 2007).
Es wird außerdem zwischen dem Problemder
I synchronen und dem der
II diachronen Identität unterschieden,
also der Identität einer Person zu einem bes-timmten Zeitpunkt und der Identität im Zeitver-lauf.
Im Rahmen seines Substanzendualismuswar für Descartes die Lösung von I.) beson-ders vordringlich, während II.) aufgrund derUnveränderlichkeit von Substanzen bzw. derUnsterblichkeit der Seele(nsubstanz) unprob-lematisch war. Für Locke ergab sich nachPreisgabe des Substanzbegriffs genau dieumgekehrte Problemkonstellation: II.) wardeutlich schwieriger zu lösen als I.).
2. Descartes
2.1.Descartes’ Personbegriff
Descartes gebraucht den Begriff der Per-son eher selten (Thiel, 2011, S. 36). WennDescartes von „persona“ bzw. „personne“spricht, versteht er darunter „die Vereinigungvon (Menschen-) Körper und (Menschen-)Geist“. Kemmerling unterscheidet in diesemZusammenhang drei Ichbegriffe:
1. Den Ausdruck „das Ich“, also die sub-stantivierte „ich“-Redeweise, verwendetDescartes, um sich auf die eigene Seelebzw. den eigenen Geist zu beziehen,während „ich“ sich
2. sowohl auf den eigenen Körper, als auch
3. auf die Vereinigung von eigenem Körperund Geist bezieht, d.h. die Person (Kem-merling, 2005, S. 123).
Den drei Ichbegriffen sind dementsprechenddrei Weisen der Erkenntnis zugeordnet:
1. Die (eigene) Seele („das Ich“) kann nurdurch den Intellekt erkannt werden. Derreine Intellekt wird nach Descartes durchmetaphysische Gedanken trainiert, dieuns mit dem Begriff des Geistes vertrautmachen sollen.
2. Bei der Erkenntnis des (eigenen) Kör-pers (d.h. Ausdehnung, Gestalt undBewegung) muss der Intellekt von der(bildlichen) Vorstellungskraft unterstütztwerden. Die Vorstellungskraft wird vorallem durch mathematische Studiengeschult; sie gewöhnen uns daran, deut-liche und klare Begriffe von Körpern zubilden.
3. Die Person, d.h. die Vereinigung vonKörper und Geist, wird weder durch denreinen Intellekt, noch durch den von derVorstellungskraft unterstützten Intellektadäquat erfasst. Nur „das gewöhnlicheLeben und das alltägliche Gesprächlehren uns, die Vereinigung von Körperund Geist zu begreifen, und zu diesemZwecke solle man sich tunlichst jedwedermetaphysischen und mathematischenBetätigung enthalten (Kemmerling, 2005,S. 124).“
An verschiedenen Stellen betont Descartesdiese enge Verbindung von Körper undGeist, die seinen Personbegriff ausmacht.So schreibt er in der Sechsten Meditation,dass sich seine Seele nicht einfach in seinem
2012, 5 (1) Kognitive Neurophysiologie des Menschen 41

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Körper befinde wie ein Schiffer in seinemSchiff, sondern aufs innigste mit ihm vereintsei und ihn gleichsam durchdringe (Descartes,1641/2009, S. 88). Wiederholt weist er daraufhin, dass Körper und Geist eine „substantielleEinheit“ bzw. „essentielle Einheit“ bilden (Per-ler, 2006, S. 213). Geist und Körper sind „ingewissem Sinn“ unvollständige Substanzen.Zu vollständigen (funktionsfähigen) werdensie erst in der essentiellen Einheit. Descartesunterscheidet dementsprechend zwischen„ens per se“ und „ens per accidens“. Wennzwei Substanzen zufällig miteinander verbun-den sind („ens per accidens“) „und alle ihreFunktionen auch unabhängig voneinanderausüben können, bilden sie zusammen eineakzidentelle Einheit - ein bloßes Konglom-erat, das nicht mehr ist als die Summe seinerbeiden Bestandteile. Wenn hingegen zweiSubstanzen derart miteinander verbundensind, dass sie nicht alle ihre Funktionen unab-hängig voneinander ausüben können, bildensie zusammen eine essentielle Einheit - et-was Komplexes, was mehr ist als die Summeseiner Bestandteile. Genau dies ist bei derVerbindung von Körper und Geist der Fall“(Perler, 2006, S. 213).
Die enge Verbindung von res cogitans undres extensa in der Person zeigt sich insbeson-dere im Bereich der Sinneswahrnehmung undder Schmerzempfindung, die weder der Geistnoch der Körper allein ausüben können. Beider Analyse der Sinneswahrnehmung unter-scheidet Descartes wiederum „drei Grade“oder Stufen voneinander:
1. Eine rein körperliche Stufe, die sich beiTieren und Menschen findet, und durch di-rekte Einwirkung eines Objektes auf denKörper eine Nervenreizung verursacht.
2. Eine körperlich-geistige Stufe.
3. Eine rein geistige Stufe, die genau dannvorliegt, wenn ein Urteil über das äußere,auf den Körper einwirkende Objektgebildet wird.
Im Kontext von Descartes’ Personbegriff ist diezweite Stufe von besonderer Bedeutung, „dienur aufgrund der essentiellen Einheit von Kör-per und Geist möglich ist“ (Perler, 2006, S.214).
2.2. Die aristotelisch-scholastischeTradition
Descartes’ Personbegriff muss vor dem Hin-tergrund der aristotelisch-scholastischen Tra-dition gesehen werden, der zufolge „einePerson eine natürliche Substanz ist, die ausForm und Materie besteht: Die Seele ist dieForm, der Körper die Materie“ (Perler 2006, S.210). Im Rahmen seines hylemorphistischenAnsatzes betrachtete Aristoteles die Seele alsLebensprinzip, das ein Stück Materie erst zueinem funktionierenden, lebendigen Körpermacht. Dieser Ansatz hat den Vorteil, dass diePerson von vornherein als Einheit (im Sinnesynchroner Identität) aufgefasst werden kann,jedoch den Nachteil - insbesondere für diechristlichen Kommentatoren des Mittelalters -dass die Seele nicht unsterblich ist, sondernmit dem Körper untergeht.
Die spätmittelalterlichen Aristoteliker wiez.B. Thomas von Aquin versuchten dieserKonsequenz u.a. durch eine differenzierteTheorie des Intellekts bzw. der Seele zu ent-gehen. Demnach bildeten nur die „niederen“Teile der Seele das Lebensprinzip und gehenmit dem Tod des individuellen menschlichen
42 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Körpers zugrunde. Der Intellekt als der „er-habene“ Teil der menschlichen Seele überlebtden Tod und geht nicht mit dem Körper unter.Durch eine solche Erklärung wurde versucht,die Einheit der Person und die Unsterblichkeitder Seele zusammen zu denken (vgl. Perler,2006, S. 211).
Eine besonders einflussreiche Kritik ander aristotelisch-scholastischen Lehre for-mulierte Der Renaissancephilosoph PietroPomponazzi in seiner Schrift „De Immortal-itate Animae“ („Über die Unsterblichkeit derSeele“) aus dem Jahr 1516. Er zeigt dort, dassdie Aristoteliker, insbesondere Thomas vonAquin, die Seele einerseits als sehr eng mitdem Körper verbunden dachten, andererseitsaber als eine Entität, die auch unabhängigvom Körper existieren kann. Die Seele ist alsoeinmal etwas, wodurch etwas lebendig ist (ein„quo est“) ein andermal etwas, was an sichexistiert (ein „quod est“). Nach Pomponazzilässt sich dieser Widerspruch im Rahmeneiner aristotelisch-scholastischen Theorie derPerson nicht auflösen.
Unklar ist, ob Descartes Pompanazzis Textgelesen hat, auf jeden Fall war er mit der darananschließenden Kontroverse vertraut (vgl.Perler, 2006, S. 212). Descartes versucht, dasvon Pompanazzi formulierte Problem durchAnnahme eines Substanzendualismus (vonres cogitans und res extensa) zu lösen. Aufdiese Weise versucht er die enge Einheit vonSeele und Körper mit der Unsterblichkeit derSeele zusammen zu denken. Dabei soll
1. sein Personbegriff, verstanden als sub-stantielle Einheit von Körper (res extensa)und Seele (res cogitans), den hylemor-phistischen Ansatz der aristotelisch-scholastischen Tradition ersetzen.
2. sein Substanzbegriff die Unsterblichkeitder Seele garantieren, da sich Sub-stanzen nach klassischem Verständnisnur hinsichtlich ihrer akzidentellen Eigen-schaften ändern können.
Für Descartes ist die Seele zudem - im Gegen-satz zum Körper - eine besondere („reine“)Substanz, da sie unteilbar sei (d.h. unzer-störbar = unsterblich). So betont er „dasswir einen Körper nur als teilbar, einen Geistdagegen nur als unteilbar einsehen können:Denn die Hälfte eines Geistes können wir nichtbegreifen, wie wir es doch bei jedem beliebigkleinen Körper können; so dass wir ihre Na-turen nicht nur als verschieden, sondern sogarals gewissermaßen entgegengesetzt erken-nen“ (Descartes, 1641/2009, S. 14). Aus derTeilbarkeit (= Sterblichkeit) des Körpers folgtalso keineswegs die Vernichtung des Geistes(= Sterblichkeit der Seele), wie Descarteseinige Zeilen später deutlich herausstellt. Je-doch muss er den endgültigen Beweis derUnsterblichkeit der Seele - zumindest imRahmen der „Meditationen“ - eingestanden-ermaßen schuldig bleiben. Darüber hinausist das Argument für die Unteilbarkeit (= Un-sterblichkeit) der Seele nur dann stichhaltig,wenn Descartes entweder zeigen kann, dass
1. die Zerstörung der Seele durch Teilungdie einzige Art ihres Zugrundegehens istoder
2. alle anderen Arten des Zugrundegehensdie menschliche Seele nicht betreffen(Perler, 2006, S. 179 f.).
2.3. Cartesianischer Dualismus
Descartes’ dualistische Substanzenlehre isteine Voraussetzung seines Personbegriffs.
2012, 5 (1) Kognitive Neurophysiologie des Menschen 43

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Baker und Morris unterscheiden zwischenDescartes’ Dualismus („Descartes’ Dualism“)und dem von ihnen sogenannten Cartesian-ischen Dualismus („Cartesian Dualism“), derim wesentlichen auf einer Legendenbildung(„Cartesian Legend“) im Bereich der Analytis-chen Philosophie beruht (Baker und Morris,2002). Zentraler Bestandteil des Cartesian-ischen Dualismus ist die Fähigkeit zur Intro-spektion („Cartesian Introspection“), die Bakerund Morris folgendermaßen charakterisieren:„Physical objects are public and observable,though fallibly, via the senses; mental ob-jects are private and (quasi-) observable viathe infallible faculty of introspection“ (Bakerund Morris, 2002, S. 11). Auch die scharfeKontrastierung von „physical objects“ und„mental objects“ gehört zur CartesianischenLegendenbildung: „There are two worlds, theone populated by physical objects, the otherby mental objects.“ Physikalische Objektekönnen rein mechanistisch erklärt werden(„physical objects are essentially (bits of)clockwork“), während mentale „Gegenstände“als Bewusstseinszustände aufzufassen sind(„mental objects are essentially (states of)consciousness“; Baker und Morris, 2002, S.11). Diese „Two-Worlds View“ führt dann zumProblem der Interaktion von physikalischenGegenständen (wie z.B. dem eigenen Körper)und Mentalzuständen, bzw. zur Asymmetrievon Erster- und Dritter-Person-Perspektive.So macht es zum Beispiel Sinn zu sagen„Ich spüre meine Schmerzen“, wohingegender Satz „Ich spüre seine Schmerzen“ sinn-los ist. Meine Mentalzustände sind privat,während mir die der anderen Menschen nurindirekt zugänglich sind (z.B. über ihr Ver-halten). Im Rahmen der Cartesianischen
Legendenbildung sind somit die „doctrines of’the epistemological transparency of thought’[...] and of ’Cartesian Privacy’“ von entschei-dender Bedeutung (Baker und Morris, 2002,S. 19), wobei die Doktrin der epistemologis-chen Transparenz besagt, dass „introspectionis essentially and universally infallible“ (Ryle,1949, S. 14). „If I think, hope, remember, will,regret, hear a noise, or feel a pain, I must, ipsofacto, know that I do so“ (Ryle, 1949, S. 158).
Die Doktrin der Cartesianischen Privatheitführt demgegenüber nicht nur zur bere-its beschriebenen Asymmetrie von erster-und dritter Person, sondern ermöglicht aucheinen direkten, unmittelbaren bzw. nicht-inferentiellen Zugang zu meinen innerenZuständen („by acquaintance“; s. unten).So habe ich z.B. (aus der Ersten-Person-Perspektive) unmittelbaren Zugang zu meinenSchmerzen, ohne das aus meinem Verhaltenerst irgendwie erschließen zu müssen.
2.4. Erkenntnistheorie
Descartes’ Überlegungen zum Personbegriffführten uns zu seinem substanzdualistischenAnsatz, der wiederum aus seinen erkenntnis-theoretischen Überlegungen resultierte. In-sofern lässt es sich auch bei Abhandlungenüber Descartes’ Sicht der Person kaum ver-meiden, auf seinen epistemologischen Ansatznäher einzugehen.
Descartes’ Erkenntnistheorie ist imwesentlichen motiviert durch die Suche nacheinem sicheren und unbezweifelbaren Funda-ment des Wissens angesichts der Möglichkeiteines systematischen Skeptizismus. Er be-ginnt mit der alltäglichen Erfahrung, dass unsunsere Sinne täuschen und dementsprechenddie meisten unserer Überzeugungen falsch
44 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
sein können. Es ist gar nicht notwendig, „zuzeigen, dass meine Meinungen allesamt falschsind, [...]; sondern weil schon allein die Ver-nunft dazu rät, dass dem nicht völlig Sicherenund Unzweifelhaften die Zustimmung nichtweniger gründlich entzogen werden muss alsdem offenbar Falschen, wird es schon ausre-ichen, alles zurückzuweisen, worin ich auchnur irgendeinen Grund zum Zweifeln antreffe“(Descartes, 1641/2009, S. 20).
In einem zweiten Schritt radikalisiertDescartes die Möglichkeit des Zweifels, indemer feststellt, „dass der Wachzustand niemalsaufgrund sicherer Anzeichen vom Traumunterschieden werden kann“ (Descartes,1641/2009, S. 21).
Als dritten Zweifelsgrund nennt Descartesdie Möglichkeit, dass mich Gott bzw. einböser Dämon täuscht, so dass ich mich frage,ob „es überhaupt keine Erde, keinen Himmel,kein ausgedehntes Ding, keine Gestalt, keineGröße, keinen Ort gibt - und all dies mir trotz-dem genau so wie jetzt zu existieren scheint?“(Descartes, 1641/2009, S. 22).
Bekanntermaßen findet Descartes’ system-atischer Zweifel zu Beginn der Zweiten Med-itation im Cogito, d.h. in der Selbstgewis-sheit des Ichs sein Ende, indem „festgestelltwerden muss, dass dieser Grundsatz Ich bin,ich existiere, sooft er von mir ausgesprochenoder durch den Geist begriffen wird, notwendigwahr ist“ (Descartes, 1641/2009, S. 28). Ausdem Cogito Argument leitet Descartes eineHierarchisierung des Wissens ab. Das Wis-sen über unsere Mentalzustände (mittels Intro-spektion) ist gewisser und grundlegender alsdas Wissen über äußere Gegenstände (mit-tels Sinneswahrnehmung): „Wenn ich etwaaufgrund dessen, dass ich ihn berühre oder
sehe, urteile, dass der Erdboden existiert, soist daraus sicherlich noch viel mehr abzuleiten,dass mein Geist existiert: Es ist nämlich vielle-icht möglich, dass ich urteile, den Boden zuberühren, obwohl gar kein Erdboden existiert,nicht jedoch, dass ich dieses Urteil fälle, undmein Geist, der dieses Urteil fällt, nicht sei [...]“(Descartes, 1644/2005, S. 19 ff.).
Diese Priorisierung introspektiven Wissensführte dazu, Descartes’ erkenntnistheoretis-che Position als
• fundamentalisch und
• internalistisch zu charakterisieren.
Descartes’ Fundamentalismus („foundational-ism“) zeigt sich vor allem in seiner Auffassung,dass das mittels Introspektion gewonneneWissen unmittelbarer, sicherer und zuverläs-siger ist, als das mittels Sinneswahrnehmung,was impliziert, dass ersteres als das Funda-ment für letzteres aufzufassen ist: Descartes„sees self-knowledge as providing a founda-tion for the thinker’s general body of knowl-edge“ (Gertler, 2011, S. 33).
Der (epistemologische) Internalismus lässtsich folgendermaßen charakterisieren:
• „Epistemic internalists (generally) accept,and externalists deny, that knowledge re-quires that one has accessible reasons forone’s belief, whereas externalism rejectsthis access requirement.“
• „[...] knowledge must be supported byreasons that are within the thinker’s mind“(Gertler, 2011, S. 31 f.).
Legt man die klassische WissensdefinitionPlatons als wahre, gerechtfertigte Meinung(vgl. Platon, Theätet, 201d-206b) zugrunde,
2012, 5 (1) Kognitive Neurophysiologie des Menschen 45

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
dann unterscheiden sich (epistemologische)Internalisten und Externalisten primär hin-sichtlich des Rechtfertigungsbegriffs; derInternalist findet die Rechtfertigungsgründefür seine Wissensansprüche ausschließlich„im Kopf“, während dies für den Externalistennicht gilt. Da Descartes beide internalistischenKriterien erfüllt, wird er von vielen als „the archinternalist“ (z.B. Gertler, 2011, S. 32), aberauch als ein paradigmatischer Vertreter eines„klassischen“ Fundamentalismus gesehen.Als „klassisch“ wird ein Fundamentalismusbezeichnet, wenn er postuliert, dass
• „das Fundament aller Rechtfertigung [vonWissensansprüchen; Ergänzung: M.R]aus unfehlbaren und unanfechtbarenÜberzeugungen besteht“,
• wobei „diese basalen Überzeugungen sobeschaffen sind, dass sie selbst einenGrund für die Einsicht in ihre Wahrheitliefern“ (Grundmann, 2008, S. 284).
Internalismus und Fundamentalismus sindMöglichkeiten, einem grundsätzlichen erken-ntnistheoretischen Trilemma zu entgehen, dasin der Antike als Agrippas Trilemma, und in dergegenwärtigen Diskussion als Münchhausen-Trilemma bekannt ist (Albert, 1991, S. 15).Möchte ich z.B. Überzeugung A rechtfertigen,kann ich das mit Hilfe einer weiteren Überzeu-gung B tun, wenn sich A aus B inferenziellerschließen lässt. Wodurch wird jedoch Bgerechtfertigt? Es gibt drei Möglichkeiten, diealle inakzeptabel sind:
1. Unendlicher Regress: B wird durch C, Cdurch D gerechtfertigt, usw. ad infinitum.
2. Rechtfertigungszirkel: B, der Grund für A,
wird wiederum durch A selbst gerechtfer-tigt.
3. Abbruch der inferenziellen Begründun-gen: B, unser Grund für A, wird nichtweiter gerechtfertigt, sondern (willkürlich)als evident, intuitiv einleuchtend, basal,etc. deklariert.
Es ist intuitiv einleuchtend, dass Descartes’ in-ternalistischer Fundamentalismus Möglichkeit3 favorisiert. Descartes’ Idee, dass Perso-nen mittels Introspektion einen privilegiertenZugang zu ihren Mentalzuständen haben undmit diesen unmittelbar (d.h. nicht-inferentiell)bekannt sind (im Sinne von „acquaintance“),ist nicht nur von philosophiehistorischem Inter-esse. Auch in den gegenwärtigen Diskussio-nen ist Descartes’ Grundidee, den Personbe-griff mit der Fundierung von Erkenntnistheoriezu verbinden, von großer Relevanz und wirdkontrovers diskutiert.
2.5. Moderne Versionen desinternalistsischen Fundamentalismus
Viele Philosophen des „Wiener Kreises“ undaus der Frühphase der analytischen Philoso-phie vertraten Versionen des internalistischenFundamentalismus. So war Russell der Auf-fassung - ähnlich wie Descartes (vgl. dasZitat aus den „Prinzipien“, siehe oben, Ab-schnitt 2.4) - dass zwar an der Existenz derGegenstände der Außenwelt gezweifelt wer-den kann, nicht aber an der Realität unsererSinneswahrnehmungen: „We have seen that itis possible, without absurdity, to doubt whetherthere is a table at all, whereas it is not possibleto doubt the sense-data“ (Russell, 1912, S.74). Sinneswahrnehmungen („sense-data“)sind als Mentalzustände aufzufassen, von
46 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
denen wir unmittelbare Kenntnis („acquain-tance“) besitzen: „[...] Russell claims thatawareness of sense data is direct, whereasawareness of tables is indirect.“ Das führtdazu, dass „sense data ’stand between’ youand the table, so to speak, that your aware-ness of the table is only indirect. Sense dataplay this mediating role in every example ofperceptual awareness, [...]“ (Gertler, 2011, S.89).
Parrallel zu Russell hat auch Moore ab 1910eine Theorie der Sinnesdaten vertreten. Aucher plädiert dafür, zwischen Wahrnehmungund den von dieser unabhängig existieren-den Gegenständen zu unterscheiden: „[...] inevery sensation or idea we must distinguishtwo elements, (1) the ’object’, or that in whichone differs from another; and (2) ’conscious-ness’, or that which all have in common - thatwhich makes them sensations or mental facts“(Moore, 1903/1993, S. 37).
In analoger Weise sind auch für CarnapSätze der Realwissenschaften (Physik, Psy-chologie, etc.) rückführbar auf das wirklichGegebene. Er schreibt: „Jeder Satz der Wis-senschaft muss sich bei logischer Analyse alssinnvoll bewähren. Dabei wird entweder ge-funden, dass es sich um eine Tautologie oderum eine Kontradiktion (Negation einer Tautolo-gie) handelt; dann gehört der Satz zum Ge-biet der Logik einschließlich der Mathematik.Oder der Satz ist eine gehaltvolle Aussage,d.h. weder tautologisch noch kontradiktorisch;dann ist es ein empirischer Satz. Er ist zurück-führbar auf das Gegebene und daher grund-sätzlich als wahr oder falsch entscheidbar“(Carnap, 1931, S. 25 f.).
Bereits im „Logischen Aufbau der Welt“(1928) entwickelte Carnap ein Konstitution-
ssystem, das die Rückführung aller wis-senschaftlichen Begriffe auf „unmittelbarGegebenes“ erlauben sollte. Unmittelbargegeben sind Carnap zufolge „die Bewusst-seinsvorgänge oder Erlebnisse des Ichs“.Diese bezeichnet er als „Elementarerleb-nisse“; sie sollen „ihrem Wesen nach unz-erlegbare Einheiten“ darstellen, die „einfachso hinzunehmen“ sind, „wie sie sich geben“(Carnap, 1928/1998, S. S. 86 ff.).
Eine (zeitgenössische) Version der „Ac-quaintance Theory of Self-Knowledge“ wirdz.B. von Fumerton vertreten. Zum unmittel-baren, d.h. nicht-inferentiellen Wissen schreibter. „My suggestion is that one has a noninfer-entially justified belief that P when one has thethought that P and one is acquainted with thefact that P“ (Fumerton, 1995, S. 75).
Die „Acquaintance Theory of Self-Knowledge“ lässt sich somit folgendermaßenzusammenfassen:
• Aufgrund unmittelbarer Kenntnis („ac-quaintance“) sind wir mit unseren Sinnes-daten vertraut, z.B. dass wir Schmerzenhaben.
• Aufgrund unmittelbarer Kenntnis sind wirzu dem Urteil „Ich habe Schmerzen“, bzw.„Schmerz ist vorhanden“ gelangt.
• Aufgrund unmittelbarer Kenntnis sind wiruns der Korrespondenz zwischen un-serem Schmerzempfinden und unseremUrteil „Schmerz ist vorhanden“ bewusst(Gertler, 2011, S. 101).
Die Methode der Introspektion ist nicht nur fürdie Erkenntnistheorie und die Philosophie desGeistes von großem Interesse, sondern auchfür die modernen Neurowissenschaften. Dies
2012, 5 (1) Kognitive Neurophysiologie des Menschen 47

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
soll im nächsten Abschnitt kurz referiert wer-den.
Introspektion und NeurowissenschaftenDie Frage, ob die Introspektion eine zuver-
lässige und damit zulässige wissenschaftlicheMethode ist, wird seit Beginn des 20stenJahrhunderts sehr unterschiedlich bewertet.Der Behaviorismus und sein Nachfolger, derkognitive Funktionalismus, „declared waron introspective psychology.“ (Schwitzgebel,2010, Zugriff: 03.02.2012) Es wurden imWesentlichen vier Einwände vorgebracht:
1. „Introspective claims are unreliable be-cause they are not regularly replicated inothers.
2. Subjects confabulate (make up stories)about what is going on in themselveswhen they need to do so to make senseof behaviour.
3. Introspection has access only to a tinyfraction of what is going on in oneself cog-nitively.
4. It is impossible for introspection to accessbrain states“ (Brook & Mandik, 2004, S.390).
Die Unzuverlässigkeit introspektiver Berichte(Einwand 1.) ist Gegenstand von Wittgen-steins Privatsprachenargument, das im folgen-den Abschnitt 2.6. ausführlich diskutiert wer-den wird.
Brooks und Mandiks Einwände 2. und3. fanden ab Anfang der 1960er Jahrezunehmend empirische Bestätigung in Experi-menten, in denen die Methode der Introspek-tion selbst zum Untersuchungsgegenstandwurde. Es zeigte sich insbesondere, dass
viele höherstufige kognitive Prozesse intro-spektiv nicht zugänglich sind, wodurch sich die„anti-introspectivist view“ unter kognitiven Psy-chologen deutlich verstärkte. Mandler (1975,S. 245) fasst die Resultate dieser Experimentefolgendermaßen zusammen: „There are manysystems that cannot be brought into con-sciousness, and probably most systems thatanalyze the environment in first place havethat characteristic. In most of these cases,only the products of cognitive and mentalactivities are available to consciousness.“
Der Bostoner Philosoph Daniel Dennett(1991, S. 122) kommt zu einer ähnlichenEinschätzung. Wir können nicht direktwahrnehmen, „[...] what happens on ourretinas, in our ears, on the surface of our skin.What we actually experience is a product ofmany processes of interpretation - editorialprocesses, in effect.“ Dies hat Implikationenfür die Bedeutung der Introspektion und denCartesianischen Fundamentalismus insge-samt: „There is no single, definitive ’stream ofconsciousness’, because there is no centralHeadquarters, no Cartesian Theatre where ’itall comes together’ for the perusal of a Cen-tral Meaner. Instead of such a single stream(however wide), there are multiple channels inwhich specialist circuits try, in parallel pande-moniums, to do their various things, creatingMultiple Drafts as they go“ (Ebd., S. 253 f.).
Aus Brooks und Mandiks Einwand 4. ziehtder Philosoph und Kognitionsforscher PaulChurchland den gegenteiligen Schluss, näm-lich dass Personen mit „[...] sufficient neu-roscientific education can introspect his orher brain states as brain states“ (zit. nach:Mandik, 2006, S. 66). Churchland vertrittseine „Introspection Thesis“ im Rahmen eines
48 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
eliminativen Materialismus, der unser laien-haftes psychologisches Vokabular („folk psy-chology“) für überflüssig und irreführend hält.Eine elaborierte zukünftige Kognitions- undNeurowissenschaft muss nicht mehr auf du-biose mentale Entitäten rekurrieren, sondernspricht stattdessen von neuronalen Zustän-den. Möchten wir an uns vertrauten Begrif-fen wie z.B. „Introspektion“ festhalten, dannreferieren diese nicht mehr auf Psychischessondern notwendigerweise auf Physisches.Churchlands eliminativer Materialismus bzw.Naturalismus ist von verschiedenen Seitennachhaltig kritisiert worden (z.B: Keil, 1993),was im Rahmen dieser Arbeit jedoch nichtvertieft werden kann.
Mit dem zunehmenden Einfluss derKognitions- und Neurowissenschaften kames zu einer Kehrtwende; die Methode derIntrospektion wurde wieder „salonfähig“, ins-besondere im Bereich der Emotionsforschung,des „Mental Imagery“ und der sogenannten„Consciousness Studies“, die alle essentiellauf „subjective reports of experience“, alsoverbale „introspektive“ Berichte von Proban-den, angewiesen sind. Dies hat die paradoxeKonsequenz, „that neuroscience, the most sci-entific of approaches to human nature to date,has been forced to fall back onto a techniquerejected as unscientific over 100 years ago!“(Brook & Mandik, 2004, S. 390)
In vielen empirischen Studien gibt es aberauch die Tendenz, „subjective reports of expe-rience“ bewusst zu umgehen und durch objek-tivere Methoden zu ersetzen wie z.B. (seman-tisches) Priming oder gleich auf bildgebendeVerfahren auszuweichen. Damit wird zwar der„subjektive Faktor“ Introspektion vermieden,aber in Kauf genommen, dass neurowis-
senschaftliche Befunde nicht auf eindeutigeWeise mit entsprechenden Mentalzuständenkorreliert und insofern interpretationsbedürftigsind (vgl. Ruchsow, Hermle & Kober, 2010).Insofern scheinen neurowissenschaftlicheStudien häufig zwischen der Skylla unzuver-lässiger introspektiver Probandenberichte undder Charybdis objektiver aber uneindeutigerMessergebnisse wählen zu müssen.
Wie gezeigt, geht der internalistische Fun-damentalismus davon aus, dass unsere Men-talzustände wesentlich privat und unmittelbarzugänglich sind, zwei Annahmen, die in dennächsten beiden Abschnitten kritisch unter-sucht werden sollen.
2.6. Privatheit
Wittgensteins Überlegungen zur Privatheitbzw. zu den Möglichkeiten einer Privat-sprache in den Paragraphen 243 bis 315der „Philosophischen Untersuchungen“ (PU)lassen sich als kritische Auseinandersetzungmit dem Cartesianismus lesen. Unter einerPrivatsprache versteht Wittgenstein folgen-des: „Die Wörter dieser Sprache sollen sichauf das beziehen, wovon nur der Sprechendewissen kann; auf seine unmittelbaren, pri-vaten Empfindungen. Ein Anderer kann dieseSprache also nicht verstehen.“ (Wittgenstein,1953/1984, § 243 = PU 243)
Die Grundannahme einer derartigen pri-vaten Sprache lässt sich folgendermaßenzusammenfassen.:
P1 „Sensations are private occurences“(Schroeder, 2001, S. 196). Meine Zahn-schmerzen sind aus diesem Grundanderen Personen nicht unmittelbarzugänglich. Ebenso wenig kann ich mir
2012, 5 (1) Kognitive Neurophysiologie des Menschen 49

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
sicher sein, ob eine andere Person ähn-liche Zahnschmerzen wie ich empfindenkann.
2.6.1. „Das Rad gehört nicht zur Mas-chine“ (PU 271)
Aus (P1) ergibt sich eine skeptische Konse-quenz, die Wittgenstein in PU 272 formuliert:„Das Wesentliche am privaten Erlebnis isteigentlich nicht, dass Jeder sein eigenes Ex-emplar besitzt, sondern dass keiner weiß, obder Andere auch dies hat, oder etwas an-deres. Es wäre also die Annahme möglich- obwohl nicht verifizierbar - ein Teil derMenschheit habe eine Rotempfindung, einanderer Teil eine andere.“ Für Wittgensteinhaben private psychische Ereignisse keinerleiFunktion, vergleichbar einem überflüssigenZahnrad in einer Maschine: „Hier möchte ichsagen: das Rad gehört nicht zur Maschine,das man drehen kann, ohne dass Anderessich mitbewegt“ (PU 271).
Für Wittgenstein sind private mentaleEreignisse nicht nur überflüssig, sondernim Rahmen einer öffentlichen Sprache nichtkommunizierbar. Mentalzustände sind wieein Käfer in einer Schachtel, zu der nur derBesitzer der Schachtel Zugang hat. DerKäfer ist also wesentlich „privat“. Wittgensteinanalysiert diese Situation folgendermaßen:„Niemand kann je in die Schachtel des An-deren schaun; und jeder sagt, er wisse nurvom Anblick seines Käfers, was ein Käferist. - Da könnte es ja sein, dass Jeder einanderes Ding in seiner Schachtel hätte. Ja,man könnte sich vorstellen, dass sich einsolches Ding fortwährend veränderte. [...] dieSchachtel könnte auch leer sein“ (PU 293).
Sind Schmerzen, Käfer, etc aber wesentlichprivat, dann ergibt sich als Konsequenz:„Wenn man die Grammatik des Ausdrucks derEmpfindung nach dem Muster von ’Gegen-stand und Bezeichnung’ konstruiert, dannfällt der Gegenstand als irrelevant aus derBetrachtung heraus“ (PU 293).
2.7. Unmittelbarkeit
Eine ausführliche Kritik an der Vorstellungunmittelbaren nicht-inferentiellen Wissensfindet sich bereits im 1. Kapitel von Hegels„Philosophie des Geistes“ (Hegel, 1807/1973,S. 82 ff.). In der analytischen Philosophieverlief die Rezeptionsgeschichte Hegels sehrwechselhaft, um nicht zu sagen: dialektisch.Nach einer kurzen hegelianischen Phase der„Gründungsväter“ der analytischen Philoso-phie, Moore und Russell, verstanden sichdiese als dezidierte Anti-Hegelianer. DieserAnti-Hegelianismus im Selbstverständnis deranalytischen Philosophie ging so weit, dassdie oft gestellte Frage, warum sich die an-alytische Philosophie eigentlich „analytisch“nenne, folgendermaßen beantwortet werdensollte: „’analytisch’ bedeutet ursprünglichgenau ’anti-hegelisch’“ (Welsch, 2011, S. 71).
Seit etwa Mitte der 1950er Jahre zeichnetsich jedoch eine Wiederkehr Hegels im Bere-ich der neueren analytischen Philosophie ab.Hier ist (neben John McDowell und RobertBrandom) insbesondere der amerikanischePhilosoph Wilfrid Sellars zu nennen, derseine Kritik an der Möglichkeit unmittelbarenWissens unter dem Stichwort „Mythos desGegebenen“ („myth of the given“) auch als„Méditations Hegeliènnes“ verstanden wissenwollte (Sellars, 1956/1997, S. 45).
Nach Sellars lässt sich die Idee, dass „truth
50 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
bearers“ durch „truth makers“ verifiziert (bzw.falsifiziert) werden, nicht aufrecht erhalten:„Having a sense-impression is, by itself, an ex-ample neither of knowledge nor of consciousexperience“ (Sellars, 1956/1997, S. 4), daes sich bei Sinnesdaten nicht um (proposi-tionales) und damit wahrheitsfähiges Wissenhandelt. In anderen Worten: „[...] one mightthink one is acquainted with certain sorts ofobjects (sense data), determinate properties(this particular shade of yellow), generic uni-versals (being yellow, being colored), and,crucially, facts (my being in pain now, some-thing’s being yellow). None of the items onthis list are the kinds of things that can be trueor false [...]. The object that is yellow, the yel-lowness of the object, that fact that the objectis yellow are all neither true nor false“ (Fumer-ton, 2009, Zugriff: 22.01.2012). Der Mythosdes Gegebenen („Myth of the Given“) bestehtnach Sellars in der Idee „that observation [...]is constituted by certain self-authenticatingnonverbal episodes, the authority of which istransmitted to verbal and quasi-verbal perfor-mances [...]“ (Sellars 1956/1997, S. 77). DieVorstellung einer Fundierung unseres (propo-sitionalen) Wissens durch mentale Episodenlässt sich aber nicht halten: „For empiricalknowledge, like its sophisticated extension,science, is rational, not because it has afoundation but because it is a self-correctingenterprise which can put any claim in jeopardy,though not all at once“ (Sellars 1956/1997, S.79). Bei dem Versuch, Wahrnehmungs-berichte, also epistemisches bzw. proposi-tionales Wissen auf nicht-epistemisches bzw.nicht-propositionales Wissen zu reduzieren,handelt es sich nach Sellars vielmehr umeinen naturalistischen Fehlschluss: „Now the
idea that epistemic facts can be analyzedwithout remainder - even ’in principle’ - intonon-epistemic facts whether phenomenal orbehavioral, public or private [...] is, I believe, aradical mistake - a mistake of a piece with theso-called ’naturalistic fallacy’ in ethics“ (Sellars1956/1997, S. 19).
Aber nicht nur Descartes’ Introspektions-begriff, sondern auch der diesem zugrundeliegende Substanzbegriff war sehr umstrittenund wurde von empiristischen Philosophenwie z.B. John Locke abgelehnt.
3. Locke
3.1. Kritik am Substanzbegriff
Ähnlich wie Descartes wollte auch Lockeeinen radikalen Neuanfang in der Philosophiemachen. Er betrachtete sich als Hilfsarbeiter(„Under-Labourer“), der das Ziel hat, „[...]clearing Ground a little, and removing someof the Rubbish that lies in the way to Knowl-edge [...]“. Durch Leidenschaften, Erziehung,Gewohnheiten und Vorurteile wird unser Wis-sen systematisch verzerrt und verfälscht.Insofern ist auch eine radikale Sprachkritikerforderlich, denn „Vague and insignificantForms of Speech and Abuse of Languagehave so long passed for Mysteries of Sci-ence; And hard or misapply’d Words, withlittle or no meaning, have, by Prescriptionsuch a Right to be mistaken for deep Learningand height of Speculation, that it will not beeasy to persuade either those who speak orthose who hear that they are but the Coversof Ignorance and hindrance of true Knowl-edge“ (Locke, 1694/1979, S. 10). Das Zieldieser Demontage ist die Fundierung unseresWissens auf der Grundlage der Erfahrung,
2012, 5 (1) Kognitive Neurophysiologie des Menschen 51

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
die nach Locke die einzige Quelle sicherermenschlicher Erkenntnis ist: „Let us then sup-pose the Mind to be, as we say, white Papervoid of all Characters, without any Ideas. [...]Whence has it all the materials of Reason andKnowledge? To this I answer, in one word,From Experience [...]“ (Locke 1694/1979, S.104). Insofern ist für Locke auch die Frage, obes eine cartesianische Seelensubstanz gibtzu verneinen, da der Begriff „Substanz“ keinGegenstand möglicher Erfahrung ist.
Durch die Ablehnung des Substanzbegriffsergab sich jedoch eine neue Konstellationhinsichtlich der Identität von Personen. FürDescartes war das Problem der diachronenIdentität von eher untergeordneter Bedeutung,da sein Substanzbegriff die Unsterblichkeit derSeele (= Persistenz der Person) garantierte.Die Erklärung der synchronen Identität warjedoch weitaus problematischer, da sie sichdem Problem der Interaktion von res extensaund res cogitans stellen, bzw. den Begriff dersubstantiellen Einheit inhaltlich füllen musste.Für Locke ergab sich eine inverse Prob-lematik: die synchrone Identität ist leichter zudenken, während sich die transtemporale Ein-heit der Person (und damit die Unsterblichkeitder Seele) ohne Substanzbegriff nur schwerverstehen lässt.
3.2. Diachrone personale Identität
Locke definiert Person folgendermaßen: „[...]we must consider what Person stands for;which, I think, is a thinking intelligent Being,that has reason and reflection, and can con-sider itself as itself, the same thinking thing,in different times and places; which it doesonly by that consciousness which is insepa-rable from thinking, and, as it seems to me,
essential to it“ (Locke 1694/1979, S. 335).„Consciousness“ liefert auch das gesuchte(naturalistische bzw. neurowissenschaftliche)Kriterium diachroner Identität: „For, since con-sciousness always accompanies thinking, and’tis that which makes every one to be, whathe calls self, and thereby distinguishes himselffrom all other thinking things, in this alone con-sists personal Identity, i.e. the sameness of arational Being: And as far as this conscious-ness can be extended backwards to any pastAction or Thought, so far reaches the Identityof that Person“ (Locke 1694/1979, S. 335). Inder Nachfolge Lockes wurde „consciousness“meist mit „memory“ bzw. „Erinnerung“ über-setzt und das Kriterium diachroner Identitätdementsprechend als Erinnerungskriteriumbezeichnet. Für Locke ist wichtig, dass mansich nicht nur allgemein an eine bestimmteWahrnehmungssituation erinnert, sonderndass die damalige Wahrnehmung sozusagen„im selben Bewusstsein“ wie die jetzige erfolgt:„For as far as any intelligent Being can repeatthe Idea of any past Action with the sameconsciousness it has of any present Action;so far it is the same personal self“ (Locke1694/1979, S. 336).
Locke war der erste, der den Begriff derPerson im Gegensatz zur Scholastik und zumCartesianismus allein durch den des Bewusst-seins bestimmte (Thiel, 1997, S. 163; Gillitzer,2001, S. 111). Die diachrone Einheit derPerson konstituiert sich im Selbstbewusstseinund ist damit epistemisch ausschließlich inder Ersten-Person-Perspektive zugänglich.Quante nennt dies die „These der erstper-sönlichen Natur personaler Einheit“ (kurz:Erstpersönlichkeitsthese; Quante, 2007, S.57).
52 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Darüber hinaus vertritt Locke eine „Unab-hängigkeitsthese“, die besagt, dass person-ale Einheit „unabhängig von der Identität einerselbstbewusstseinstranszendenten Substanz“ist (Quante, 2007, S. 58 f.).. In Lockes Worten:„For it being the same consciousness thatmakes a Man be himself to himself, personalIdentity depends on that only, whether it be an-nexed only to one individual Substance, or canbe continued in a succession of several Sub-stances. [...] The same consciousness unit-ing those distant Actions into the same Person,whatever Substances contributed to their Pro-duction“(Locke 1694/1979, S. 336).
Drittens ist Locke der Meinung, dass per-sonale Einheit ein komplexes Phänomen istund sich (z.B. durch das Erinnerungskriterium)auf informative Weise weiter analysieren lässt.Quante nennt dies „die These der internenKomplexität personaler Einheit - kurz: dieKomplexitätsthese“ (Quante, 2007, S. 58).
4. Reid und Butler
Lockes Lösungsansatz wurde insbesonderevon Reid und Butler kritisiert. Reid zeigtanhand des „Brave Officer Paradox“, dassLockes Erinnerungskriterium nicht stichhaltigist: „Suppose a brave officer to have beenflogged when a boy at school for robbing anorchard, to have taken a standard from theenemy in his first campaign, and to have beenmade a general in advanced life; suppose,also, which must be admitted to be possible,that, when made a general, he was consciousof his taking a standard, but had absolutelylost the consciousness of his flogging“ (Reid,1785/1975, S. 114). Reids Kritik lässt sichpointiert folgendermaßen zusammenfassen:„Identity is transitive; memory continuity is not“
(Olson, 2010, Zugriff: 22.01.2012).
Dieser Transitivitätseinwand lässt sich je-doch dadurch entkräften, dass man LockesErinnerungsrelation im Sinne einer psychol-ogischen Verbundenheit („connectedness“)ersetzt durch die schwächere Relation derpsychologischen Kontinuität („continuity“), fürdie gilt: „Zwischen dem Bewusstseinszus-tand einer Person zum Zeitpunkt t0 und demBewusstseinszustand einer Person zum Zeit-punkt t1 besteht genau dann psychologischeKontinuität, wenn es zwischen diesen beidenBewusstseinszuständen ein stetiges Bandvon Bewusstseinszuständen gibt, die untere-inander in der Relation der psychologischenVerbundenheit stehen. Gibt es ein solchesBand, dann handelt es sich bei den beidenBewusstseinszuständen zu t0 und t1 um dieBewusstseinszustände ein und derselbenPerson“ (Quante, 2007, S. 47).
Butler wies darauf hin, dass Erinnerungenstets eine Person implizieren, die die Erin-nerungen hat. Das Erinnerungskriterium setztalso personale Identität voraus und kann dieseniemals konstituieren: „And one should re-ally think it self-evident, that consciousess ofpersonal identity presupposes, and thereforecannot constitute, personal identity, any morethan knowledge, in any other case , can con-stitute truth, which it presupposes“ (Butler,1736/1975, S. 100).
Butlers Zirkularitätseinwand lässt sich durchdie Annahme sogenannter „q-memories“entkräften, ein Vorschlag, der auf SydneyShoemaker zurückgeht. Nach Shoemakersind q-memories durch einen fehlenden im-pliziten Subjektbezug gekennzeichnet, sodass sie nicht-zirkulär personale Identität kon-stituieren können: „One way of characterizing
2012, 5 (1) Kognitive Neurophysiologie des Menschen 53

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
the difference between quasi-rememberingand remembering is by saying that the formeris subject to a weaker previous awarenesscondition than the latter. Whereas someone’sclaim to remember a past event implies that hehimself was aware of the event at the time ofits occurrence, the claim to quasi-remember apast event implies only that someone or otherwas aware of it“ (Shoemaker, 1970/2003,S. 24). Jedoch muss sich auch ein Vertei-diger von Lockes Erinnerungskriterium, derq-memories als basal ansetzt, fragen lassen,„auf welcher Grundlage er von der Menge derQuasi-Erinnerungen zu der Menge der Erin-nerungen kommen möchte“ (Quante, 2007, S.90).
5. Das physischeKontinuitätskriterium
Quante und Olson plädieren aufgrund dergeschilderten Schwierigkeiten des Erin-nerungskriterium für ein anderes naturalis-tisches Kriterium diachroner Identität, das ph-ysisches Kontinuitätskriterium. Nach Quantelässt sich Butlers Zirkularitätseinwand effektivdurch die Annahme aus dem Weg räumen,„dass der Träger der mentalen Episoden nichtdie Person, sondern ein [...] menschlicherOrganismus ist“ (Quante, 2007, S. 90). Zudiesem Schluss kommt auch Olson, der füreinen „somatic approach“ bzw. „animalism“plädiert: „There appears to be a thinkinganimal located where you are. It also ap-pears that you are the thinking thing - theonly one - located there“. Olsons Überlegungist auch durch die absurde Konsequenz desErinnerungskriteriums motiviert, dass alleUnterbrechungen des Bewusstseinsstroms
z.B. durch Schlaf, Narkosen, Ohnmachten,etc. dazu führen „that I have never existed atany time when I was completely unconscious“(Olson, 2010, Zugriff: 22.01.2012).
Auch der New Yorker Philosoph ThomasNagel definiert personale Identität anhanddes physischen Identitätskriteriums. Die Mini-malanforderung an ein physikalisches Korrelatpersonaler Identität besteht darin, „bearer ofmental states and the cause of their continuity“zu sein (Nagel, 1986, S. 40). Hypothetischnimmt Nagel an, dass am ehesten das intakteGehirn diesen Anforderungen Genüge tut. Einalternativer Kandidat ist der menschliche Kör-per, der eng mit den Hirnfunktionen verknüpftist, jedoch „[...] the brain is the only part ofme whose destruction I could not possiblysurvive. The brain, but not the rest of theanimal, is essential to the self“. Zusammen-fassend kann (mit milder Übertreibung) gesagtwerden: „ [...] I am my brain [...]“ (Nagel, 1986,S. 40). Das Gehirn ist die notwendige undhinreichende Bedingung personaler Identität(Northoff, 2001, S. 131-132), da es die biol-ogische Grundlage aller mentalen Vorgängewie Erinnerungen aber auch des „Bewusst-seinsstroms“ ist und somit ein Fortbestehender Person nach (Teil-)verlust des Gehirnsunmöglich erscheint.
6. Parfit
Mit Hilfe eines Gedankenexperiments weistParfit nach, dass Nagels Slogan „I am mybrain“ nicht ausreicht, um numerische Identitätzu garantieren: „Suppose next that I needsurgery. All of my brain cells have a defectwhich, in time, would be fatal. But a surgeoncan replace all these cells. He can insert newcells that are exact replicas of the existing
54 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
cells except that they have no defect“ (Parfit,1984, S. 474). Parfit unterscheidet zwei Oper-ationsprozeduren: In der ersten („Case One“)werden alle Nervenzellen meines Gehirnsin einer Serie von 100 Operationen schrit-tweise entfernt und durch neue ersetzt. Beider anderen Operationsart („Case Two“) wirdin einem Schritt mein ganzes Gehirn aus-getauscht. Bei der ersten Operation ist dieKontinuität meiner Person gegeben, bei derzweiten jedoch nicht: „ There is a real dif-ference between these cases. In Case One,each of the new parts of my brain is for a timejoined to the rest of my brain. This enableseach new part to become part of my brain.When the first new part is inserted, and joinedto the rest of my brain, it wins the title to beas much part of my brain as the old parts.When the second new part is inserted, it toobecomes a part of my brain. This is true ofevery new part, because there is a time whenthis part is joined to what then counts as therest of my brain.
In Case Two, things are different. There areno times when each part is joined to the rest ofmy brain. Because of this, the new parts do notcount as parts of my brain. My brain ceases toexist“ (Parfit, 1984, S. 474 f.). Die erste Art derOperation werde ich (als Person) überleben,die zweite jedoch nicht, obwohl das Endergeb-nis in beiden Fällen dasselbe ist. Folglich istder Slogan „I am my brain“ nicht hinreichend,um numerische Identität zu gewährleisten.
Zusammenfassend bleibt festzuhalten, dassweder Descartes’ noch Lockes Ansatz einetragfähige Lösung des Problems der per-sonalen Identität darstellt. Descartes’ Sub-stanzbegriff lässt sich im Rahmen des zeit-genössischen naturalistischen Paradigmas
nicht halten. Die Aufgabe des Substanzbe-griffs führt in der Folgezeit zu einer Ver-lagerung der Diskussion von der synchronenzur diachronen Identität. Aber auch die (natu-ralistischen ) Kriterien der Erinnerung und derphysischen Kontinuität, die im Zusammenhangder diachronen Identität formuliert wurden,sind mit einer Vielzahl von Schwierigkeitenbelastet.
Im folgenden sollen Überlegungen deskanadischen Philosophen Charles Taylor zurnarrativen Identität vorgestellt werden, dersich nicht nur an der analytischen Philosophie,sondern insbesondere auch an kontinentaleu-ropäischen Traditionslinien orientiert.
7. Taylor
7.1. Selbstinterpretation
Taylors Überlegungen zur personalen Identitätknüpfen in vielen Punkten an Wittgenstein an,mit dem er sich sehr intensiv auseinanderge-setzt hat (z.B. Taylor, 1972; 1988a; 1995;2010). Taylor ist jedoch nicht nur einem an-alytischen Ansatz verpflichtet, sondern stehtu.a. in den Traditionslinien von Hermeneu-tik, Anthropologie und Deutschem Idealismus(hier insbesondere Hegel). Von Axel Honnethwurde er als ein Parteigänger der Hermeneutikim Felde der analytischen Wissenschaftstheo-rie bezeichnet (in: Taylor, 1988b, S. 296).
Für Taylor sind Personen wesentlich „self-interpreting animals“ (Taylor, 1985a, S. 45 ff.)und „dialogical selves“ (Taylor, 1995, S. 230).Sie sind also durch ihre Fähigkeit zur Selbstin-terpretation und zum sprachlich-dialogischenAustausch mit anderen Menschen gekennze-ichnet. Ähnlich wie für Wittgenstein ist für Tay-lor Sprache ein intrinsisch soziales Phänomen:
2012, 5 (1) Kognitive Neurophysiologie des Menschen 55

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
„The matter talked about is no longer justfor me or for you, but for us“ (Taylor, 1985a,S. 260). Insofern verfehlen wir völlig dasWesentliche, „if we remain with the monolog-ical model of the subject, and think of all statesof awareness, knowledge, belief, attending to,as ultimately explicable as states of individu-als“ (Taylor, 1985a, S. 265).
7.2. Sprache
Ausgehend von dieser sozialen und dialogis-chen Dimension der Sprache kritisiert Tay-lor die zeitgenössischen Sprachauffassungenals naturalistisch, worunter er Theorien ver-steht, die von einem Beobachter über einenbeobachteten Gegenstand entwickelt werden,an dem er nicht beteiligt ist.
Taylor stellt dieser naturalistischen eineromantisch-expressivistische Sprachauffas-sung gegenüber, die er auch als Herder-Humboldt-Hamann-Theorie bzw. als „triple-Htheory“ bezeichnet (Taylor, 1985a, S. 256).Die Expressivität von Sprache verdeutlichtauch deren dialogischen Charakter, indem sieeine bestimmte Art der Beziehung zu einemGegenüber ausdrückt. Sprechen heißt dann„to put it ’out there’, to have it out before us, tobe ’up front’ about it“ (Taylor, 1985a, S. 264).Sprache erzeugt etwas, „what one might call apublic space, or a common vantage point fromwhich we survey the world together“ (Taylor,1985a, S. 259). Etwas ausdrücken bedeutet„to place it in public space, and thus to bringus together qua participants in a common actof focussing“ (Taylor, 1985a, S. 260).
7.3. Hermeneutik
Wie Gadamer ist auch Taylor der Auffas-sung, dass Menschen immer schon in einebestimmte Sprachgemeinschaft, bestimmteTraditionen und ein bestimmtes Verständ-nis der Welt hineingeboren sind, das dieBasis ihres Verstehens bildet („Geworfen-heit“ im Sinne Heideggers). Intersubjektivitätgeht der Subjektivität voraus, „we are awareof the world through a ’we’ before we arethrough an ’I’“ (Taylor, 1985b, S. 40). Insofernbefindet sich jeder in einem hermeneutis-chen Zirkel, weil die Interpretationen unsererUmwelt selbst schon Resultat vorgängigerInterpretationen sind (Breuer, 2000, S. 23).Verstehen hat immer den Doppelcharakterder Teilnahme an einem Sinngeschehen,das dem Subjekt vorausgeht und sein Be-greifen ermöglicht und einem Sichverstehen(Angehrn, 2004, S. 774): Wie ich die Weltauffasse, ist davon abhängig, wie ich den Sinnmeines Daseins interpretiere, wie umgekehrtmein Selbstverständnis nicht losgelöst vonmeiner Sicht auf die Dinge zustande kommtbzw. in Honneths Worten: „Soziales Handelnist ohne Bezugnahme auf das situationsge-bundene Selbstverständnis der handelndenSubjekte gar nicht angemessen aufzufassen;der Erklärung einer Handlung muss daher einhermeneutisches Verstehen der jeder unmit-telbaren Beobachtung entzogenen Perspek-tive des Handelnden unbedingt vorhergehen“(in: Taylor, 1988b, S. 299).“
Taylor unterscheidet dementsprechend zweiInterpretationsebenen: Auf einer präreflexivenEbene ist Personen ein Selbst- und Weltver-ständnis vorgegeben, das primär und zumgrößten Teil implizit und nicht artikuliert ist (vgl.Rosa, 1998, S. 200). Dem reflexiven Bewusst-
56 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
sein wird diese gesamte erste (präreflexive)Interpretationsebene dann zum Gegenstandeiner zweiten Interpretationsebene, in der jeneauf ihre Bedeutungen und ihren Sinn befragtund sprachlich artikuliert wird.
7.4. Moralische Topographien
Ein „selbstinterpretierendes Tier“ zu seinheißt für Taylor u.a. auch, sich der Frage zustellen „Who I desire to be?“ (Rosa, 1998,S. 98) und sich durch ihre Beantwortung ineinem moralischen Raum zu verorten. UnsereIdentität als Personen wird aus seiner Sichtentscheidend durch fundamentale Wertungenmitbestimmt, so dass sich ein unauflöslicherZusammenhang von Personalität und derKonzeption des Guten ergibt.
Taylor unterscheidet (mit zunehmenderWertigkeit) zwischen „Lebensgütern“ („LifeGoods“), „Hypergütern“ („Hypergoods“) und„konstitutiven Gütern“ („Constitutive Goods“).Konstitutive Güter sind Moralquellen, d. h. „itis a something the love of which empowersus to do and be good“ (Taylor, 1989, S. 93).Dabei leistet das konstitutive Gut mehr als nurdie Bestimmung des Inhalts der Moraltheorie.„Love of it is what empowers us to be good.And hence also loving it is part of what itis to be a good human being.“. Moralthe-orie gebietet also nicht nur, in bestimmterWeise zu handeln und bestimmte moralischeEigenschaften an den Tag zu legen, sondernschreibt auch vor, zu lieben, was gut ist: „Tolove the constitutive good [...] is to be stronglymotivated in just that way which is definedas part of doing the good [...]. That is whybeing good involves loving something and notjust doing something“ (Taylor, 1989, S. 533 f.,Anm. 2).
Lebensgüter sind demgegenüber Ziele,Prinzipien und Ideale, die Menschen in ihremLeben zu verwirklichen suchen, während Hy-pergüter Leitwerte darstellen, die Kohärenzund eine einheitliche Orientierung in unserLeben zu bringen vermögen
7.5. Narrative Identität
Nach Taylor müssen wir das eigene Leben imSinne einer narrativen Darstellung begreifen.Es genügt nicht, dass ein Individuum über eineidentitätsstiftende moralische Landkarte ver-fügt bzw. sich im moralischen Raum orien-tiert. Die Ausbildung und der Erhalt person-aler Identität verlangt vielmehr, dass es in derLage ist, „sich auf dieser Landkarte zu posi-tionieren, d. h. seinen jeweiligen aktuellen Ortim Hinblick auf die verzeichneten Güter (undÜbel) zu bestimmen sowie einen Sinn für dieBewegungsrichtung des eigenen Lebens undHandelns vor dem Hintergrund dieser Güterzu entwickeln“ (Rosa, 1998, S. 166). Person-ale Identität hat eine unhintergehbare zeitlicheStruktur, der die narrative Darstellung der eige-nen Lebensgeschichte am ehesten gerechtwird. Personsein erschöpft sich nicht in derBestimmung dessen, was jemand ist, sondernumfasst auch immer, was jemand gewordenist (oder hätte werden können) bzw. was erin Zukunft werden wird. Der Mensch ist einGeschichten erzählendes Wesen: „It has oftenbeen remarked that making sense of one’s lifeas a story is also, like orientation to the good,not an optional extra; that our lives exist also inthis space of questions, which only a coherentnarrative can answer. In order to have a senseof who we are, we have to have a notion of howwe have become, and of where we are going“(Taylor, 1989, S. 47).
2012, 5 (1) Kognitive Neurophysiologie des Menschen 57

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Kritisch anzumerken bleibt, dass Narratio-nen ebenfalls ein intaktes (autobiographis-ches) Gedächtnis voraussetzen und sichdamit letztendlich als eine Variante desErinnerungskriteriums entpuppen. Sie kön-nen somit kaum die Identität der Persongarantieren, sondern haben diese vielmehrzur Voraussetzung. Taylors großes Verdi-enst besteht jedoch darin, auf die sozialeDimension personaler Identität hingewiesenzu haben. Sein Ansatz müsste durch Über-legungen des Sozialkonstruktivismus ergänztwerden (z.B. Smith, 2010; Bakhurst, 2011),was jedoch den Rahmen dieser Arbeit spren-gen würde.
Literatur
Albert, H. (1991). Traktat über kritische Ver-nunft. Tübingen: Mohr Siebeck.
Angehrn, E. (2004). Hans-Georg Gadamer- Das Projekt einer PhilosophischenHermeneutik. In: A. Beckermann & D.Perler (Hrsg.), Klassiker der PhilosophieHeute. Stuttgart: Reclam, 771-791.
Baker, G., & Morris, K. J. (2002). Descartes’Dualism. London, New York: Routledge, 2.Auflage.
Bakhurst, D. (2011). The Formation of Rea-son, Malden/MA: Wiley-Blackwell.
Breuer, I. (2000). Charles Taylor. Hamburg:Junius.
Brook, A., & Mandik, P. (2004). The Philoso-phy and Neuroscience Movement. Analyse& Kritik, 26, 382-397.
Butler, J. (1736/1975). Of Personal Identity. InJ. Perry (Hrsg.), Personal Identity. Berkeley,Los Angeles, London: University of Califor-nia Press, 99-105.
Carnap, R. (1928/1998). Der logische Aufbauder Welt. Hamburg: Meiner.
Carnap, R. (1931). Die alte und die neueLogik. Erkenntnis, 1, 12-26.
Dennett, D. C. (1991). Consciousness Ex-plained. London: The Penguin Press.
Descartes, R. (1641/2009). Meditationen. Mitsämtlichen Einwänden und Erwiderungen,Hamburg: Meiner.
Descartes, R. (1644/2005). Die Prinzipien derPhilosophie, Hamburg: Meiner.
Fumerton, R. (1995). Metaepistemology andScepticism. Lanham, MD: Rowman & Littlefield.
Fumerton, R. (2009). Knowledge by Ac-quaintance vs. Description. In E. N.Zalta (Hrsg.),The Stanford Encyclope-dia of Philosophy (Summer 2009 Edi-tion). URL = <http://plato.stanford.edu/archives/sum2009/entries/ knowledge-acquaindescrip/>.
Gertler, B. (2011). Self-knowledge. London,New York: Routledge.
Gillitzer, B. (2001). Personen, Menschenund ihre Identität. Stuttgart, Berlin, Köln:Kohlhammer.
Grundmann, T. (2008). Analytische Ein-führung in die Erkenntnistheorie. Berlin,New York: de Gruyter.
58 Human Cognitive Neurophysiology 2012, 5 (1)

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Hegel, G. W. F. (1807/1973). Phänomenologiedes Geistes. Frankfurt/M.: Suhrkamp.
Keil, G. (1993). Kritik des Naturalismus. Berlin,New York: de Gruyter.
Kemmerling, A. (2005). Ideen des Ichs:Studien zu Descartes’ Philosophie. Frank-furt/M.: Klostermann, 2. Auflage.
Krebs, A., Kambartel, F., & Jantschek, T.(1995) Person. In J. Mittelstraß (Hrsg.), En-zyklopädie Philosophie und Wissenschafts-theorie. Stuttgart, Weimar: J. B. Metzler, 89-92.
Locke, J. (1694/1979): An Essay ConcerningHuman Understanding. Oxford: ClarendonPress, 2. Auflage.
Mandik, P. (2006). The Introspectability ofBrain States as Such. In: B. L. Keeley(Hrsg.), Paul Churchland. Cambridge/UK:Cambridge University Press, 66-87.
Mandler, G. (1975). Consciousness: Re-spectable, Useful and Probably necessary.In: R. Solso (Hrsg.), Information Process-ing and Cognition: The Loyola Symposium.Hillsdale/NJ: Erlbaum, 229-254.
Moore, G. E., (1903/1993). The Refutation ofIdealism. In G. E. Moore, Selected Writings,London, New York: Routledge, 23-44.
Nagel, T. (1986). The View from Nowhere. Ox-ford: Oxford University Press.
Neuhouser, F. (2000). Foundations of Hegel’sSocial Theory. Cambridge/MA: Harvard Uni-versity Press.
Northoff, G. (2001). Personale Identität undoperative Eingriffe in das Gehirn. Pader-born: mentis.
Olson, E. T. (2010). Personal Identity.In E. N. Zalta (Hrsg.): The StanfordEncyclopedia of Philosophy. URL =<http://plato.stanford.edu/archives/win2010/entries/identity-personal/>.
Parfit, D. (1984). Reasons and Persons. Ox-ford: Clarendon Press.
Perler, D. (2006). Descartes. München: C. H.Beck, 2. Auflage.
Quante, M. (2007). Person. Berlin, New York:de Gruyter.
Reid, T. (1785/1975). Of Mr. Lockes Accountof our Personal Identity. In J. Perry (Hrsg.)Personal Identity. Berkeley, Los Angeles,London: University Press of California, 111-118.
Rosa, H. (1998). Identität und KulturellePraxis: Politische Philosophie nach CharlesTaylor. Frankfurt/M., New York: Campus.
Ruchsow, M., & Hermle, L. (2007). Zur En-twicklung des Begriffs der Person in Psychi-atrie und Philosophie. Der Nervenarzt, 78,505-510.
Ruchsow, M., Hermle, L., & Kober, M. (2010).MRT als Lügendetektor und Gedanken-leser? Kritische Bestandsaufnahme undReflexion. Der Nervenarzt, 81, 1085-1091.
Russell, B. (1912). Problems of Philosophy.New York: Henry Holt & Co.
Ryle, G. (1949). The Concept of Mind. Lon-don: Hutchinson.
Schroeder, S. (2001): Private Language andPrivate Experience. In H.-J. Glock (Hrsg.):Wittgenstein. A Critical Reader. Oxford:Blackwell, 174-198
2012, 5 (1) Kognitive Neurophysiologie des Menschen 59

M. Ruchsow — Personale Identität aus Sicht der Neurowissenschaften
Schwitzgebel, E. (2010). Introspection.In: E. N. Zalta (Hrsg.): The Stan-ford Encyclopedia of Philosophy. URL= <http://plato.stanford.edu/archives/fall2010/entries/ introspection />.
Sellars, W. (1956/1997). Empiricism and thePhilosophy of Mind. Cambridge/MA: Har-vard University Press.
Shoemaker, S. (1970/2003). Persons andTheir Pasts. In S. Shoemaker (Hrsg.), Iden-tity, Cause, and Mind. Oxford: Oxford Uni-versity Press, 19-48.
Smith, C. (2010). What is a Person? Chicago,London: University of Chicago Press.
Taylor, C. (1972). The Opening Argumentsof the Phenomenology. In A. MacIntyre(Hrsg.), Hegel. A Collection of Critical Es-says. New York: Doubleday, 151-187.
Taylor, C. (1985a). Human Agency and Lan-guage. Philosophical Papers, Band 1. Cam-bridge/UK: Cambridge University Press.
Taylor, C. (1985b). Philosophy and the Hu-man Sciences. Philosophical Papers, Band2. Cambridge/UK: Cambridge UniversityPress.
Taylor, C. (1988a). Wittgenstein, Empiricism,and the Question of the „Inner“: Com-mentary on Kenneth Gergen. In S. B.Messer, L. A. Sass, & R. L. Woolfolk (Hrsg.),Hermeneutics and Psychological Theory:Interpretive Perspectives on Personality;Psychotherapy, and Psychopathology, NewBrunswick/NJ: Rutgers University Press,52-58.
Taylor, C. (1988b). Negative Freiheit? Frank-furt/M.: Suhrkamp.
Taylor, C. (1989) Sources of the Self. The Mak-ing of Modern Identity. Cambridge/MA: Har-vard University Press.
Taylor, C. (1991) The Malaise of Modernity.Concord, Ontario: Anansi Press.
Taylor, C. (1995). Philosophical Arguments.Cambridge/MA: Harvard University Press.
Taylor, C. (2010). Language not Mysterious?In: B. Weiss & J. Wanderer (Hrsg.), ReadingBrandom. London, New York: Routledge,32-47.
Thiel, U. (1997). Individuation und Identität.In U. Thiel (Hrsg.): John Locke. Essayüber den menschlichen Verstand. Berlin:Akademie Verlag, 149-168.
Thiel, U. (2011). The Early Modern Subject.Self-Consciousness and Personal Identityfrom Descartes to Hume. Oxford, New York:Oxford University Press.
Tugendhat E., & Wolf, U. (1983). Logisch-semantische Propädeutik. Stuttgart:Reclam.
Welsch, W. (2011). Immer nur der Mensch?Berlin: Akademie Verlag.
Wittgenstein, L. (1953/1984). PhilosophischeUntersuchungen. Werkausgabe Band 1.Frankfurt/M.: Suhrkamp.
60 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
Abstracts of the 20thGermanEEG/EP Mapping Meeting,
Giessen, October 14-16, 2011
The influence of expertise on neural pro-cesses when anticipating action effectsin tennis N. Balser (1), S. Pilgramm (1,2),M. Bischoff (1,2), B. Lorey (1,2), K. Zentgraf(3), R. Stark, A (2). M. Williams (4) & J.Munzert (1) (1) Institut für Sportwissenschaft,Justus-Liebig-Universität Gießen, D-35394Gießen, Germany; (2) Bender Institute of Neu-roimaging, Justus-Liebig-Universität Gießen,D-35394 Gießen, Germany; (3) Institut fürSportwissenschaft, Universität Bern, CH-3012Bern, Switzerland; (4) Faculty of Health Sci-ences, University of Sydney, AUS-NSW 2141Sydney, AustraliaIt has recently been proposed that the actionobservation network (AON) (Zentgraf et al.,2011) plays an important role in the anticipa-tion of action effects. The cerebellum could beparticularly crucial for predicting action conse-quences because of its role in the control ofexecuted movements (Miall, 2003) and its in-clusion in the AON (Cross et al., 2009). To in-vestigate whether expertise in sport influencesthe activation of this network during an an-ticipation task, 16 expert tennis players and16 novices observed videos of tennis strokes.While lying in an fMRI scanner, participantsobserved videos showing an opponent’s fore-hand strokes that were occluded at the mo-ment of ball-racket contact. The participants’task was to anticipate the direction of the ob-served strokes without having any information
about the ball flight. This anticipation task wascontrasted with observing a player bouncing aball with no need to anticipate. Results showeda significantly better anticipation performancein expert tennis players compared to novices,t(30) = 4.19, p < .001. The functional imagingdata showed enhanced activation in the mainareas of the AON when contrasting the antic-ipation and nonanticipation tasks. More pre-cisely, the premotor cortex, the inferior frontalgyrus, and the superior parietal lobe showedgreater activation when anticipation of strokedirection was contrasted with mere observa-tion. Compared with novices, experts showedgreater activation in the cerebellum during theanticipation condition. An additional covariatetennis experiencerevealed that the cerebellaractivation in the experts was higher for playerswho had longer tennis experience. Our find-ings support the theory that the AON is usedto anticipate the effects of observed actions(Zentgraf et al., 2011). The stronger activationin the cerebellum suggests that experts relyon internal models when predicting the conse-quences of an action.
Inhibitory control assessed through simul-taneous EEG-fMRI imaging: a pilot studySarah Baumeister (1), Sarah Hohmann (1),Isabella Wolf (1), Michael Plichta (2), StefanieRechtsteiner (1), Maria Zangl (5), MatthiasRuf (3), Christine Niemeyer (4), Peter Kirsch(4), Herta Flor (5), Andreas Meyer-Lindenberg(2), Martin Holtmann (1,6), Vasil Kolev (1,7),Juliana Yordanova (1,7), Tobias Banaschewski(1), Daniel Brandeis (1,8) (1) Department ofChild and Adolescent Psychiatry and Psy-chotherapy, CIMH Mannheim; (2) Depart-ment of Psychiatry and Psychotherapy, CIMHMannheim; (3) Department Neuroimaging,
2012, 5 (1) Kognitive Neurophysiologie des Menschen 61

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
CIMH Mannheim; (4) Department of ClinicalPsychology, CIMH Mannheim; (5) Departmentof Cognitive and Clinical Neuroscience, CIMHMannheim; (6) Child and Adolescent Psychi-atry, Ruhr-University Bochum, Germany; (7)Institute of Neurobiology, Bulgarian Academyof Sciences, Sofia, Bulgaria; (8) Departmentof Child and Adolescent Psychiatry, Universityof Zürich
Inhibitory control shows individual and ge-netic variation and is impaired in AttentionDeficit Hyperactivity Disorder (ADHD). Neuro-feedback is an emerging nonpharmacologicaltreatment for ADHD (Holtmann et al., 2009),but how changes in the inhibitory pathwaycontribute to the treatment outcome is still un-clear. Advanced imaging with combined EEG(electroencephalography) and fMRI (func-tional magnetic resonance imaging) shouldoptimally characterize individual variationand treatment-induced changes of inhibitorycontrol.
In this pilot study we explored the feasibilityof simultaneous recording of 64 channel EEGand 3T (Siemens TIM Trio) fMRI to probe con-flict processing and inhibitory control using aFlanker/NoGo task (Meyer-Lindenberg et al.,2006). The task requires a left or right but-ton press, corresponding to the central arrow,when flankers are either other arrows or boxes.The button press has to be inhibited whenflankers are „XX“.
The fMRI analysis (N=23) revealed robustNoGo vs. Neutral effects, with increased acti-vation to NoGo stimuli in an inhibitory networkinvolving bilateral insula and the ACC (p<.05,FWE corrected). After gradient, ballistocardio-gram and ICA corrections, the correspondingEEG analysis (N=17) revealed a significant
(p<.01 at Cz, compared to neutral flankers)NoGo positivity lasting from about 460 to660ms. Furthermore, we found a significantly(p<.05 at Cz) enhanced N2 in the NoGocompared to the Neutral condition.Simultaneous EEG-fMRI during a Flanker/NoGotask showed landmark effects of inhibitory con-trol in both imaging modalities with a simple7 min paradigm suitable for adults as well asADHD children. In an ongoing study we nowexamine whether inhibitory control is impairedin ADHD compared to control groups, andimproves after neurofeedback training.Supported by the SFB 636 and NewMedsprojects.
Manipulierte audiovisuelle Pointlight Dis-plays beeinflussen die Nutzung internerModelle zur Prädiktion M. Bischoff (1,2), B.Lorey (1,2), S. Pilgramm (1,2), K. Zentgraf(3,2), R. Stark (2), D. Vaitl (2) & J. Munz-ert (1,2) (1) Institut für Sportwissenschaft,Justus-Liebig-Universität Giessen, D-35394,Deutschland; (2) Bender Institute of Neu-roimaging, Justus-Liebig-Universität Giessen,D-35394, Deutschland; (3) Institut fürSportwissenschaft, Universität Bern, CH-3012, Schweiz Um Handlungen und Hand-lungseffekte zu verstehen, werden im Hirn desBeobachters interne Modelle der Bewegungmit der wahrgenommenen Bewegung ver-glichen. Solche Handlungsrepräsentationenliegen in verschiedenen Sinnesmodalitätenvor, die Verarbeitung interner Modelle solltealso durch Manipulation multisensorischerWahrnehmung beeinflussbar sein. Wird beider visuellen Handlungswahrnehmung zusät-zliche auditive Information verwendet und wiewirkt sich widersprüchliche auditive Informa-tion aus?
62 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
Ein Tischtennis-Spieler, der seinen Gegen-spieler während dessen Schlagausführungbeobachtet, nutzt auch den Zeitpunkt desSchläger-Ball-Kontakts zur Einschätzung desSchlages. Wird die visuelle Bewegungsinfor-mation reduziert in Form von Pointlight Dis-plays (PLD) und werden Ball sowie Schlägerdabei ausgeblendet, dann zeigt der Ton denSchläger-Ball-Kontakt an und vervollständigtim Vergleich zu einer äquivalenten aber ton-losen Bewegung das interne Modell.
Sechsundzwanzig Beobachter wurden mitfunktioneller MRT untersucht, währendsie PLDs eines gegnerischen Tischtennis-Spielers bezüglich der Richtung des resul-tierenden Ballflugs einschätzten (die Alterna-tive war „Cross oder Longline“). In den Videoswaren nur die Marker für beide Schultern, denrechten Ellenbogen und das rechte Handge-lenk des Spielers zu sehen, sowie zwei statis-che Markierungen der gegenüberliegendenPlattenkante. Der Schläger-Ball-Kontakt kon-nte allein durch ein entsprechendes Tonsignalmarkiert werden. Dieser Ton wurde entwedergar nicht präsentiert, zum Bewegungsbe-ginn – also offensichtlich zu früh, 120ms vordem eigentlichen Zeitpunkt – unmerklich zufrüh – oder zum Zeitpunkt des Schläger-Ball-Kontakts.
Die Probanden zeigten eine erhöhte Per-formanz, wenn der Ton zeitlich kongruentgesetzt war. Region-of-Interest Analysen derfMRT-Daten ergaben als sensitiv für die Kon-gruenz multisensorische Integrationsareale imtemporo-okzipitalen Gyrus temporalis mediusund im anterioren intraparietalen Sulcus. DieRekrutierung interner Modelle im ventralenprämotorischen Kortex und Gyrus frontalisinferior (BA44) zeigte bei kongruenter audiovi-
sueller Stimulation höhere Aktivierung als beider unmerklich inkongruenten. Interne Mod-elle sind also multisensorisch repräsentiertund als funktionelle Grundlage der Prädiktionzugänglich für multisensorische Kongruenzef-fekte.
Functional and clinical abnormalities inpatientes with panic disorder and theirtreatment options S. Deppermann (1), S.Sickinger (1), N. Vennewald (2), A.J. Fallgat-ter (1), P. Zwanzger (2) & A.-C. Ehlis (1) (1)Münster, (2) TübingenRepetitive transcranial magnetic stimulation(rTMS) has already been suggested as analternative treatment method in a variety ofpsychological disorders such as major de-pression. Another frequent disabling diseaseis panic disorder (PD). PD is characterizedby the sudden onset of unexpected panicattacks often resulting in the avoidance of amultitude of situations (agoraphobia). Theneurobiological correlate of these symptomsis believed to be a dysfunction of the fearnetwork including hyperactivity of the amyg-dala and decreased activity of the prefrontalcortex. Accordingly, a model of inadequatetop-down governance in anxiety disorders hasbeen established. Whereas functional mag-netic resonance imaging (fMRI) data couldshow that cognitive behavioral therapy (CBT)is capable of normalizing both exaggeratedamygdala activity and prefrontal hypofunction,the effects of rTMS as an additional therapeu-tic tool have not been investigated much sofar. Even though CBT is an effective treatmentmethod it has the disadvantage of a delayedonset of its effect and is not always sufficientso that additional psychopharmacologicalmedication might be needed. Therefore the
2012, 5 (1) Kognitive Neurophysiologie des Menschen 63

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
aim of this study was to investigate the ap-plication of sham controlled prefrontal rTMSduring CBT in order to enhance the impactof the therapy. To quantify and hence beingable to evaluate the effects of rTMS in terms ofhelping to normalize the prefrontal hypoactivitythe neuroimaging method functional near in-frared spectroscopy (fNIRS) was chosen. Onemajor advantage of this method over fMRI isits compliance with especially PD patients.During the NIRS recording patients performedan emotional (emotional stroop/Westphalparadigm) as well as a cognitive task (verbalfluency). The results are not only ground-breaking regarding the future of the treatmentof PD but also demonstrate the benefits offNIRS as an imaging method to assess highercortical structures when other methods suchas fMRI are less convenient.
Frontal Midline Theta in Simple ReactionTime Tasks: Comparing Predictable andUnpredictable Stimulus Onset M. Doppel-mayr & E. Weber FB Psychologie, UniversitätSalzburg, Hellbrunnerstr 34, 5020 Salzburg,ÖsterreichIn several types of sport, fast reaction timesare of upmost importance. To gain a better un-derstanding of the cognitive processes that oc-cur in the prestimulusinterval of reactions (asfor example the start of a 100m sprint), we in-vestigated changes of the EEG frontal midlinetheta (FmT), which is known to be related toworking memory, action regulation, and sus-tained attention.EEG and simple reaction times of 18 voluntaryparticipants have been recorded during threedifferent RT- tasks. In all tasks participantswere instructed to react (push a button) as fastas possible whenever a circle was presented
on a monitor. The interstimulusinterval (ISI) ofthe presentation has been set either to 3 sec-onds, to 6 seconds or varying between 3 and6 seconds (in 125 msec steps).As expected the results indicated the fastestRT for the 3 sec ISI presentation, followed bythe 6 sec ISI. The varying ISI yielded the slow-est RT. EEG data indicated significant differ-ences in the timecourse of FmT activity at fron-tocentral and central locations. While for the6 second ISI as well as for the varying ISIFmTremained on a relatively stable level dur-ing the last second preceding the task, in the3 sec ISI condition a specific preparation pat-ternëmerged. In this task FmT increased sig-nificantly from a very low level to a level ex-ceeding the FmT activity of the other task con-ditions. Taken together these data indicate thatthe predictability of a stimulus leads to varyingpreparatory attentional processes that can bedetected by FmT.
The neuronal basis of the development ofhumans into speaking and cultural beingsDuncker, H.-R. Institut für Anatomie und Zellbi-ologie der Universität Gießen During the phy-logenetic development of humankind, both thestructures and functions of the human bodywere subjected to evolutionarily-driven biolog-ical modifications. The body has enlarged,has developed new proportions with respect toits extremities, and has dramatically increasedfunctional capabilities. Each successive Homospecies exhibits an enlarged cerebrum, whichhas developed as a consequence of new ter-tiary association areas, which make up 50By migration out of the tropical rain forest, theirancestral living area, into the enlarging EastAfrican savannahs, developing humans couldno longer make use of their inherited feed-
64 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
ing manners and behaviour. As such, theywere forced to invent all necessary activitiesfor their nourishment and defence, includingtheir social communication in the extended sa-vannahs. The inventions of single individu-als were adopted by most members of thecommunity, including the successful handingdown of these new abilities to descendants.Through these developments, social commu-nities drove the formation of cultural communi-ties, with common use of, and communicationabout, these abilities and activities, which arecollected into, and thereby enlarged, their his-torical development.
All these new capabilities and communicationabilities could not be integrated into the geneticcodes of the developing humans, but could bestored as memory in the newly-developed ter-tiary association areas of the cerebrum. Forhanding down of these capabilities to descen-dants, developing humans depended on theimitation of these activities and actions, by ob-serving the activities of their parents and othercommunity members. These imitations, whichare used occasionally by higher apes, were in-tensively used by developing humans to handdown abilities to descendants. However, theresults of the first imitations by minors hadto be intensively socially controlled by parentsand other community members, and the devel-oping minors had to repeat and rehearse theimitated actions until they mastered the neces-sary level of precision. These activities werethen neuronally fixed for automatic future ap-plication. Apart from these activities, a gesticu-latory communication developed, perhaps outof gestural pointing at or towards important ob-jects or events. The gesticulatory communica-tion incorporated primarily the language char-
acteristic of including the functional meaning ofthese objects or events for the communicatinghumans, which are represented together withthe objects or events in the cerebral memoryof the persons.This gesticulatory communication has devel-oped – very slowly and in a stepwise fashion– in combination with linguistic sounds. Onlywith the appearance of modern humans, withtheir ability to produce all vowels, has a fullcapacity for spoken communication developed.By the long phylogeny of mammals in the sub-terranean or nocturnal habitat, their auditorysystems became the most important sensorysystem for social and emotional interaction. Inthe language communication of modern hu-mans, a direct coupling of the brains of inter-locutors occurs: The speaker’s production ofcortical excitations for pronouncing words andsentences induces in the brain of the listenercomparable excitations, together with specificassociations. These capabilities of acousticcommunication depend not only on the mutualunderstanding of interlocutors, but also onethe handing down of complex thoughts to allof the members of the community, and to theirminors. All cultural developments of modernhumans depend on these abilities, in particu-lar, the development the diversity of differentoccupations, as well as thinking in highly de-veloped cultural communities.
Recording of laser evoked potentials dur-ing preferential stimulation of mechano-insensitive nociceptors unmasked by pe-ripheral nerve block M. Dusch (1*), J. vander Ham (1*), B. Weinkauf (1), J. Benrath (1),M. Schmelz (1), R. Rukwied (1), M. Ringkamp(2), R-D. Treede (3), U. Baumgärtner (3) (1)Dept. of Anaesthesiology, Medical Faculty
2012, 5 (1) Kognitive Neurophysiologie des Menschen 65

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
Mannheim, Heidelberg University, Germany;(2) Dept. of Neurosurgery, Johns Hopkins Uni-versity, Baltimore, Maryland; (3) Chair of Neu-rophysiology, Centre for Biomedicine and Med-ical Technology Mannheim, Medical FacultyMannheim, Heidelberg University, GermanyIn order to investigate preferentially mechano-insensitive (MIA) nociceptors in healthy volun-teers, laser radiant heat stimuli were applied tothe lateral proximal leg after peripheral nerveblock of the lateral femoral cutaneous nerve(LFCN). Laser-evoked potentials (LEP) wererecorded in healthy volunteers from partiallyanesthetized skin areas.Nerve block of the LFCN was performed in 10healthy male subjects with Ropivacain 1In the area of differential sensitivity LEP re-sponses were delayed compared to unaffectedskin (225 ±10.4 ms (SEM), vs. 185 ±7.8 ms)and LEP amplitudes were reduced (13.8 ±1.3µV vs. 25.6 ±1.6 µV). Reaction times of painresponses to the laser stimuli were increased(654 ms vs. 406 ms, p<0.01) and pain magni-tude reduced (1/10 vs. 5/10 NRS; p<0.01).The increase of LEP latency suggests thatmechano-insensitive heat-sensitive Aδ noci-ceptors (MIA type II) have a slower conductionvelocity or a higher response latency ascompared to mechano-sensitive Aδ nocicep-tors (type II MSA). Alternatively, recruitmentof widely branched slowly conducting andmechano-insensitive branches of Aδ nocicep-tors can explain our finding. We concludethat recording of LEP during laser stimulationof areas with differentially anesthetized skinis useful to study a mechanically-insensitive,heat-sensitive subpopulation of Aδ nocicep-tors.
Alternative approaches in estimating re-
gional individuality indices and reliabilityin complex cognition: FMRI and ERP-dataThorsten Fehr Center for Cognitive Sciences;University of BremenIndividual differences, but also the reliabilityof psychophysiological measurements are im-portant upcoming topics in the neurosciences.Usually, experimental approaches are directedto reduce information to describe neural cor-relates of complex mental processes in amore comprehensible and putatively sufficientway. However, individual aspects potentiallydue to individual learning history and mentalstrategy run the risk of being inappropriatelyneglected. Furthermore, retest-reliability inindividual physiological parameters has notbeen examined in both experimental intra-and inter-session arrangements. The presentapproach utilizes the STROOP-paradigm asone of the most consistent and reliable experi-mental procedures in psychological sciences.Based on previously published data, there wasan adapted form of the STROOP-task appliedin an EEG- and fMRI-study to estimate indi-vidual intra- and inter-session reliability of be-havioural and physiological data. Behaviouraldata showed the expected interference effectfor incongruent as compared to congruentand baseline conditions reflected in longerresponse times for the incongruent condition.Different individuality indices on the basis ofCartesian analytical geometric will be intro-duced and applied on functional neuroimagingdata. Reliability- and individuality-estimationsof ERP- and fMRI-data will be compared,illustrated, and critically discussed.
Topographic correlates of semantic prim-ing differentiate Alzheimer’s and semanticdementia M. Grieder (1), R.M. Crinelli (2), T.
66 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
Dierks (1), L.-O. Wahlund (2), T. Koenig (1), M.Wirth (3) (1) Dept. of Psychiatric Neurophys-iology, University Hospital of Psychiatry, Uni-versity of Bern, Switzerland; (2) Karolinska In-stitute, Dept. NVS, Division of Clinical Geri-atrics, Stockholm, Sweden; (3) Jagustlab, He-len Willis Neuroscience Institute, University ofCalifornia Berkeley, Berkeley, CA, USA
With the progressing course of Alzheimer’sdisease (AD), deficits in declarative memoryincreasingly restrict the patients’ daily activi-ties. Besides episodic memory impairments,semantic memory is affected by this demen-tia subtype. In contrast, patients suffering fromsemantic dementia (SD) show isolated seman-tic memory impairments. With the aim to es-tablish sensitive biological markers for the dif-ferentiation of symptom dimensions in demen-tia subtypes, the present study compared ADand SD patients with healthy controls. In par-ticular, the automatic and controlled seman-tic word processing was investigated by com-bining the recording of event related poten-tials (ERP) with the performance of a seman-tic priming (SP) task. Concretely, the task re-quired word/non-word (prime and target) lexi-cal decisions on sequentially presented wordpairs, consisting of semantically related or un-related prime-target combinations. Besidesthe analysis of the target onset reaction times(RT), early automatic and later controlled ERPcomponents were investigated. Both patientgroups showed general slowed RTs for alltask conditions (related and unrelated wordpairs), whereby the SD performed consider-able slower than the AD. However, the sizeof the SP effect was comparable in all partic-ipant groups. The ERP results indicate thatearly automatic components are only slightly
altered in AD and SD patients whereas thelate controlled components are affected mostdominantly in the SD group. Taken together,these results could reflect that the semanticdatabase itself is likely to remain preserved inAD and to a less extent also in SD patients,who suffer from deteriorated retrieval of se-mantic representations. Taken together, theautomatic spread of activation in the semanticnetwork appeared to be preserved in both pa-tient groups, while the late ERP componentsgave a clear indication of neurophysiologicalalterations in AD and SD. Together with subse-quent studies, these novel findings might im-prove a profound understanding of the demen-tia subtypes and their early detection.
EEG recorded during gross-motor behav-ior and sleep Hoedlmoser, K. (1), Birklbauer,J. (2), Rigler, S. (1), Mueller, E. (2), & Sch-abus, M. (1) (1) Laboratory for Sleep and Con-sciousness Research, Department of Psychol-ogy, University of Salzburg, Austria; (2) De-partment of Sport Science and Kinesiology,University of Salzburg, AustriaThere is now compelling evidence that sleepcontributes to the consolidation of proceduraltypes of memory and to motor learning in par-ticular. Concerning offline processing of fine-motor skills during sleep there is already pro-found knowledge. However, data on gross-motor tasks are still missing. Therefore, theaim of our study was to investigate sleep ef-fects on performance in a real-life gross-motortask: riding an inverse steering bicycle. Wetested 20 healthy male subjects (20 - 29 yrs;M=24.3, SD=2.03) by a between-subject de-sign. Each subject either participated in a 2hrmidday nap or a restful wake condition. In bothconditions participants were previously trained
2012, 5 (1) Kognitive Neurophysiologie des Menschen 67

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
on the gross-motor learning task and tested forperformance pre- and post-sleeping/resting.Gross-motor performance was decreased af-ter nap but not after resting wakefulness. Fastsleep spindle density (13-15Hz) was nega-tively related to the change of performanceover nap, whereas slow sleep spindle density(11-13Hz) was generally (baseline and exper-imental nap) negatively related to initial gross-motor performance. Together these findingssuggest that fast sleep spindles during shortmidday naps interfere with gross-motor con-solidation and individual’s general gross mo-tor learning ability is reflected in interindividualdifferences of slow sleep spindle activity. Ad-ditionally to our sleep-related hypotheses, weinvestigated EEG alpha activity during ridingthe inverse in comparison to a normal steer-ing bicycle in a subsample of our subjects(N=5). These analyses revealed a strong taskrelated alpha synchronization more prominentduring inverse compared to normal steering bi-cycling. As the inverse steering bicycle pro-vides an inversion-evoked cognitive load dueto inhibition of highly automated movementpatterns (riding a normal steering bicycle), theobserved task related power increase in the al-pha band with respect to a reference can beconsidered as a cortical correlate of the inhibi-tion of already well known, automated memorytraces.
Recording EEG and fMRI simultaneouslyduring a reward anticipation task: Pilot re-sults for a study with ADHD children SarahHohmann (1), Michael Plichta (2), IsabellaWolf (1,3), Stefanie Rechtsteiner (1), SarahBaumeister (1), Maria Zangl (5), ChristineNiemeyer (4), Matthias Ruf (3), Peter Kirsch(4), Herta Flor (5), Andreas Meyer-Lindenberg
(2), Tobias Banaschewski (1), Martin Holt-mann (1,6), Daniel Brandeis (1,7) (1) De-partment of Child and Adolescent Psychiatryand Psychotherapy, Central Institute of Men-tal Health; (2) Department of Psychiatry andPsychotherapy, CIMH; (3) Department Neu-roimaging, CIMH; (4) Department of ClinicalPsychology, CIMH; (5) Department of Cog-nitive and Clinical Neuroscience, CIMH; (6)Department of Child and Adolescent Psychia-try – University of Bochum; (7) Department ofChild and Adolescent Psychiatry - Universityof Zürich
Recording EEG during an fMRI - session isincreasingly used to study cognitive and mo-tivational states in basic and clinical neuro-science. We used this combined measure-ment technique to learn more about the hu-man reward system. Simultaneous recordingsrequire modifications of the original fMRI re-ward task to improve signal to noise ratio forEEG. One aim of the present study was thusto find out whether the fMRI core results wouldprove robust in the modified paradigm, whileresolving potential technical problems whichmay occur when combining both methods inchallenging motivational paradigms. Based onprevious work of Kirsch et al. (2003) we modi-fied a reward anticipation task for the use withEEG, which is known to reliably activate ventralstriatal brain structures. EEG data and fMRIimages were simultaneously acquired in a 3TMRI Siemens TIM Trio scanner from 20 healthyparticipants (20 to 26 years) using an event re-lated design. Money gained of the reward taskwas paid after the fMRI-Session.
We found a stronger BOLD signal in the ven-tral striatum during anticipation of reward. Therobust activation of the brain reward system in
68 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
our modified monetary incentive task confirmsthat neither simultaneous fMRI-EEG, nor thecorresponding task modifications diminishedsensitivity in detecting ventral striatal BOLD-responses. The analyses revealed meaning-ful ERPs with significant differences betweenboth conditions (win money/no money) devel-oping during early (<1s) as well as late (>1s)stages of reward anticipation.Showing that simultaneous combination ofboth methods produced good data qualityconcerning electrophysiology and hemody-namic response future work will concentrateon data fusion of both techniques to fully uti-lize the complementary information brought byEEG to fMRI and vice versa. In future studieswith ADHD-children we will apply this task tostudy effects of neurofeedback-training on thebrain reward system regarding neurobiologicaland neurophysiological alterations.
Acute and chronic response of electroen-cephalographic activity to exercise duringnormoxia and normobaric hypoxia: a pi-lot study Hülsdünker, T. (1), Gutmann, B. (1),Mierau, A. (1), Mierau, J. (1), Hollmann, W.(2), Strüder, H.K.(1) (1): Institute of Movementand Neurosciences, German Sport University(Cologne, Germany), (2): Institute for Cardiol-ogy and Sports Medicine, German Sport Uni-versity (Cologne, Germany)It has been suggested, that exercise train-ing under normobaric hypoxia may be supe-rior to normoxic training for the performanceof endurance athletes. So far, most of theprevious research has focused on vascular,metabolic and muscular changes associatedwith exercising under hypoxia without takinginto account that changes may also occur inthe central nervous system. Therefore, the
aim of the current study was to examine theeffects of acute exercise before and after train-ing under normoxic versus hypoxic conditionson Electroencephalography (EEG). Three en-durance untrained male volunteers performedone-legged cycle training for 30 minutes, threetimes per week, for four weeks. One leg wastrained under normoxic and the other leg undernormobaric hypoxic (12.5Before training, up-per alpha power increased by after normoxicbut decreased by after hypoxic exercise in theparietal region. These changes were attenu-ated after training in normoxia but even morepronounced after training in hypoxia. Our re-sults suggest that exercise effects on EEGare strongly influenced by hypoxia. Training-induced adaptations to normoxic versus hy-poxic exercise may have opposite directionshowever, with the same meaning (here: in-creased activity in the parietal cortex).
Do lateralized readiness potentials help toelucidate extraversion-related differencesin the magnitude of the psychological re-fractory period (PRP) effect? Rebekka In-dermühle, Stefan Troche, and Thomas Ramm-sayer Department of Psychology, University ofBern, Bern, SwitzerlandPsychological refractory period (PRP) refersto a major bottleneck of information process-ing that becomes evident when participantsare required to respond to two signals (S1and S2) presented in rapid succession. Typi-cally, the response to S2 becomes increasinglydelayed with decreasing stimulus onset asyn-chrony (SOA) between S1 and S2. The ma-jor goal of the present study was to answerthe question of whether extraversion-relatedindividual differences can be identified at thisstage of premotor information processing. For
2012, 5 (1) Kognitive Neurophysiologie des Menschen 69

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
this purpose, 63 introverts and 63 extravertsperformed a PRP task and stimulus-locked(S-LRP) and response-locked (LRP-R) lateral-ized readiness potential latencies were deter-mined. To quantify the PRP effect at the be-havioral level, regression analyses were per-formed separately for each participant withSOA as independent variable and responsetime to S2 as dependent variable. Thus, theresulting regression coefficient b representsan estimate of the magnitude of the individ-ual PRP effect. While at the behavioral level,a reliably more pronounced PRP effect couldbe confirmed for introverts compared to ex-traverts, extraversion-related individual differ-ences could be revealed neither for S-LRP norfor LRP-R latencies. Our findings suggest thatexperimentally induced overload of the infor-mation processing system by means of a PRPtask appears to differentially affect introvertsand extraverts. LRP analyses, however, didnot prove to be sensitive for the identificationof these behavioral differences at the electro-physiological level.This work was supported by the Swiss Na-tional Science Foundation (105314-120096).
Enhancement of delta- and theta EEGpower during perception of visual ambigu-ous stimuli. Ksenia Khalaidovski, ChristinaSchmiedt-Fehr, Canan Basar-Eroglu & BirgitMathes. Institute of Psychology and CognitionResearch, University of Bremen, Grazer Str.4,D-28359 Bremen, GermanyAmbiguous patterns make up a well-knownclass of visual phenomena in which one in-variant stimulus pattern is perceived in twodifferent, mutually exclusive ways. When sub-jects indicate a perceptual reversal by pressinga button a reversal-related slow positive wave,
which is prominent in the delta-band (0.5-3Hz), can be detected [1]. This delta compo-nent, by its functional and topographical simi-larity to the P300, was interpreted as reflectingthe conscious recognition of the perceptualchange. However, focused attention neededduring perceptual reversals should also reflectin an enhanced theta (4-7 Hz) response. ThisEEG study aimed to investigate the reversal-related delta and theta activity using the stro-boscopic alternative motion (SAM) paradigm,a multistable stimulus that during constantviewing induces internally generated changesin the perceived direction of motion of flasheddots. Reaction times from an unambiguouscontrol condition were used to estimate thestimulus onset after which the percept haspresumably changed. Reversal-related deltaas well as theta EEG power was enhancedwhen compared to a time period during whichthe percept was stable. As expected, thetheta response exhibited a frontal maximum.Further, the topography of the theta responseobserved following exogenously triggeredperceptual reversals differed from internallygenerated perceptual changes. These resultssuggest that the state of enhanced attentionmay be reflected by reversal-related activity inthe delta and theta band. These emphasizethe influence of top-down regulated processesupon multistable perception [2]. [1] C. Basar-Eroglu et al. (1993), Int J Neurosci 73, 139-51[2] B. Mathes et al. (2006), Neurosci Lett 402,145-49
EEG-Analysis with Matrix-Wavelets A. KleinPhysiologisches Institut, Aulweg 129, 35392GießenScalar wavelet analysis of EEG-recordingsposes a considerable problem due to the
70 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
amount of data produced. In particular, theresulting wavelet transforms are not obviouslyrelated to each other, and it would be verydesirable if known or suspected relationshipsbetween channels could be modelled withinthe transform itself, not necessitating any ad-ditional steps. This requirement is met by theMatrix-Wavelet-Transform which operates onvector-valued instead of scalar-valued signalsusing matrix-valued wavelet functions, andwhich guarantees that any information con-tained in the original signal is still available foranalysis in the computed transform.The possibilities of this novel method, for ex-ample the analysis of centroids of activity, di-mensionality of the signal, and time-frequencycentroids, are presented and compared withresults that were obtained with more traditionalmethods.
Neuronal correlates of focused attentionin pistol shooting sports Christoph Krein-bucher, Andrea Geipel, Denise Beckmann-Waldenmayer & Jürgen Beckmann Instituteof Sport Psychology, Technische UniversitätMünchenIn shooting sports the last few seconds of theshot preparation are crucial for a successfulshot outcome. It is not yet clear if certain fre-quencies in different regions of the brain aremandatory for achieving peak performance.Therefore, neuronal correlates of focused at-tention in sport pistol shooting ought to be re-searched further. Moreover, the investigationshould reveal whether an increase in focusedattention can be achieved through the imple-mentation of sport specific attentional trainingand if there are also changes related to thecortical activity.The participants included thirteen male and
seven female right handed youth athletes (M= 16.79, SD = 2.68) from the German shoot-ing federation (DSB). While the experimen-tal group (N = 11) received attentional train-ing based on scientifically substantiated re-search and augmented by hypnotherapeuticelements, participants in the control group (N= 9) engaged in progressive muscle relaxation.Both cohorts received six 90 minute trainingsessions. A 64-channel EEG was used be-fore and after the trainings were conducted toevaluate the efficacy of the attention trainingmeasures. In each of them a competition wasdesigned in which the pistol shooters were re-quired to shoot 60 shots on a target in 90 min-utes. The shot release was determined withthe help of an optical sensor fixed directly onthe weapon. The sensor’s analyzing softwareenabled the differentiation between good andbad shots based on an assessment of sev-eral shooting relevant criteria during the aimingprocess. In addition to also assessing the par-ticipant’s overall shot outcome, an assessmentof the athlete’s distractibility from the shoot-ing tasks was conducted by evaluating the ath-lete’s perception of subliminal stimuli. Self-report questionnaires were also used to evalu-ate the participant’s level of attention.
Results from baseline measures of the brainwaves revealed that an increase of theta ac-tivity in frontal areas is only present in badshots. Thus, there is evidently an optimal levelof the theta frequency range. In good shots,there is a higher alpha activity in parietal re-gions two seconds before shot release than inbad shots. These findings indicate an inhibi-tion of verbal and analytical processes whiletrying to become highly focused before pullingthe trigger. Further calculations are needed to
2012, 5 (1) Kognitive Neurophysiologie des Menschen 71

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
assess the degree to which a change in brainactivity occurs through the implementation ofthe attention training and how this correlateswith behavioral states.
Change detection enhances the P3 andfronto-parietal delta response during delay-dependent working memory B. Mathes (1,2),J. Schmiedt (1), C. Schmiedt-Fehr (1,2), C.Pantelis (3) & C. Basar-Eroglu (1,2) (1) Uni-versity of Bremen, Institute of Psychology andCognition Research, Bremen, Germany, (2)Centre for Cognitive Science, Germany (3)Melbourne Neuropsychiatry Centre, Universityof Melbourne, Melbourne, AustraliaBoth, recognition of previously encoded stim-uli and change detection enhance the P3. Thecurrent study contrasted these opposite influ-ences on the P3 using a delayed matching-to-sample task, which included requirements ofcommonly used working memory as well aschange detection tasks. Mean P3 amplitudeand delta activity was analysed from partic-ipants who classified probe stimuli as eitherbeing identical or modified. The P3 ampli-tudes were larger for modified than for identi-cal probes, with both eliciting a larger P3 thanduring encoding. Enhanced single-trial ampli-tude and trial-by-trial consistency of delta ac-tivity contributed to the larger P3 for the mod-ified probe. Fronto-parietal phase coherenceof delta activity was also more enhanced formodified than for identical probes. The resultsindicate that the P3 reflects an adaptable pro-cess of attentional resource allocation to opti-mize memory-guided decision-making.
Brain oscillatory changes after acute ex-ercise in pre-school children during eyes-closed versus eyes-open resting condi-
tions Mierau, A. (1), Hülsdünker, T. (1),Mierau, J. (1), Hense, A. (2), Hense, J. (2),Strüder, H.K. (1) (1) Institute of Movementand Neurosciences, German Sport Univer-sity (Cologne, Germany); (2) Children’s SportSchool NRW (Cologne, Germany)
The effects of acute exercise on brain corti-cal activity have been reported in some pre-vious studies however, only for adults. Tothe best of our knowledge, there have beenno studies which investigate such effects inpre-school children although this age-groupis characterized by a particularly high plas-ticity of the central nervous system. 8 male,pre-school children participated in the study(5.6±0.5 yrs; 122.4±4.2 cm; 22.1±2.7 kg). Ina balanced cross over design participants ei-ther completed an exercise session (EXE) of45 min or a control condition (CON) wherethey were allowed to paint and/or to talk totheir parents. The exercise session consistedin three out of five different movement games(10 min each) that were introduced in a coun-terbalanced order and a soccer match (15min) at the end of the session. Heart rate(HR) was recorded continuously before, dur-ing and after exercise as a measure of physi-cal demand. 15-20 min before (PRE) and af-ter (POST) one minute resting EEG record-ings were completed with the eyes closedand the eyes open in a counterbalanced or-der between subjects. EEG data was Fouriertransformed to provide estimates for abso-lute power in eight frequency bands (delta togamma) based on the individual peak alphafrequency and analyzed in four regions acrossthe scalp. Average HR during the exercise ses-sion was 150.2±15.8 beats/min. Lower-1 al-pha power was reduced for CON compared to
72 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
EXE (p<0.05) in the eyes open condition. Fur-thermore, beta1 and beta2 power were signifi-cantly lower in the frontal cortex for EXE com-pared to CON (p<0.001). The results of thepresent study show that acute exercise can in-duce significant changes in brain cortical ac-tivity of 5-year old children. These changesare characterized by decreased frontal activ-ity accompanied by a reduction of global corti-cal arousal however, only when visual input ispresent.
Statistical methods for the identification ofsignificant directed interactions betweenEEG signals T. Milde, K. Schwab, L. Leistritz& H. Witte Institut für Medizinische Statistik, In-formatik und DokumentationThe Granger Causality Index (GCI) and Par-tial Directed Coherence (PDC) are two of themost common methods for the identification ofdirected interactions between EEG signals. Toidentify statistically significant interactions it isnecessary to know the probability distributionsof these interaction measures under the null-hypothesis of absent interactions. As desireddistributions are mostly unknown nonparamet-ric statistical methodologies must be used toidentify significant interactions. Fundamentalsof GCI- and PDC-analysis are presented us-ing multivariate AR-models. Statistical analy-sis of resulting values via bootstrapping in themulti-trial case and via surrogate data is pre-sented. The focus is on the comparison be-tween different groups and tasks. It can beshown that topographic representations of sig-nificant directed interactions allow conclusionson interacting brain regions. Benefits of thesetechniques are demonstrated for applicationsin pain research and in clinical child psychol-ogy.
Supported by the German Federal ResearchCouncil, Wi 1166/10-2.
Bimodal processing of audio-visual stim-uli during face-to-face communicationHorst M. Müller, Verena Winter, HendrikWesselmeier SFB 673 „Alignment in Commu-nication“ and EC 277 „Cognitive InteractionTechnology“ (CITEC), University of Bielefeld,33501 Bielefeld, GermanyWhile speaking in a face-to-face dialogue, lipreading supports the language comprehensionprocess substantially. Besides this bimodalbinding process during natural language com-prehension, perception of voice and lip move-ment of the interlocutor is not synchronic. In aconversation, for instance standing two metresapart from each other, an approx. 6 ms delayof the voice exists. Due to this fact, auditorysensation after visual sensation is a commoncircumstance, but auditory sensation before vi-sual sensation should be unusual to us. How-ever, in comparison to hearing only, an audio-visual signal should have the most ecologicalvalidity. In this study, we investigated 1) howvarying the temporal synchronity of the audi-tory signal to the visual signal evokes differentERP wave forms and 2) how bimodal audio-visual processing will influence the amplitudeof the N1/P2 component. Therefore, 25 sub-jects perceived audio-visual sequences of upto four seconds in length. The auditory signallaged for 0 ms, 40 ms, 80 ms, 200 ms, 360 msand preceded 40 ms, 80 ms, 120 ms and 280ms in relation to the visual signal while record-ing the EEG. In addition, the bimodal effectswere measured against a physical devianceof the stimuli as an additional control condi-tion. In a behavioral task the naturalness ofthe stimuli was rated by a questionnaire and
2012, 5 (1) Kognitive Neurophysiologie des Menschen 73

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
compared to ERPs. The auditory after visualsensation caused flattened N1/P2 amplitudeswhile the odd situation with auditory before vi-sual sensation caused N1/P2 amplitudes sim-ilar to natural synchronic stimuli. The resultscorrelate with the findings from the behavioralexperiment where subjects had to judge whenan audio-visually presented stimulus was outof sync. These findings were discussed inthe framework of the lip-moving requirementsof speaking humanoid robots, which could in-crease the robustness of the machine to hu-man communication.Supported by the Deutsche Forschungsge-meinschaft (SFB 673, EC 277)
The Applicability of the Somatotopic Prin-ciple to Action Execution, Imagery, and Ob-servation T. Naumann (1), B. Lorey (1,2), S.Pilgramm (1,2), C. Petermann (4), M. Bischoff(1,2), K. Zentgraf (3,2), R. Stark (2), D. Vaitl(2) & J. Munzert (1) (1) Institute for SportsScience, Justus Liebig University Giessen, D-35394, Germany; (2) Bender Institute of Neu-roimaging, Justus Liebig University Giessen,D-35394, Germany; (3) Institute for Sports Sci-ence, University of Bern, CH-3012, Switzer-land; (4) Department of Sports Medicine, Jus-tus Liebig University Giessen, D-35394, Ger-manyJeannerod’s (2001) mental simulation theoryhypothesizes overlapping cerebral activationduring the imagery, observation, and executionof human movements. Several fMRI studieshave shown somatotopic activation patternsfor the imagery (Ehrsson et al., 2003) andobservation (Wheaton et al., 2004) of move-ments with different effectors that are roughlysimilar to the patterns found for their execu-tion. Against this background, the goal of
the first study was to compare the degree ofoverlap between both simulation states andexecution in one design. During fMRI mea-surement, participants had to imagine, ob-serve, and execute alternate, unilateral exten-sion/flexion movements with the right hand,right foot, or both effectors simultaneously.The fMRI results showed similarities betweenexecution and imagery in both the activationpattern and the dissociation of effectors inthe contralateral motor cortex (M1, PMC &SMA). Although these areas were activatedsignificantly during observation, this conditionshowed more of an overlap of both effectors. Asomatotopic dissociation was found only in theright (ipsilateral) hemisphere (BA44). To inves-tigate the relevance of somatotopy and move-ment characteristics for the degree of overlap,the second study extended the design to in-clude two additional movements. This time,participants had to imagine or observe threedifferent unilateral movements executed witheither the right hand or the right foot. The firstmovement was the rhythmic extension/flexionmovement already described, the second wasan accuracy-demanding pointing task, and inthe third task, subjects had to generate staticforce by pushing a bellows. Results showeddistinct activation sites in the contralateral mo-tor cortex for different movements during ac-tion observation. No clear somatotopic orga-nization could be found for either hand or footmovements. It is concluded that there mightbe a task-specific form of organization in ac-tion observation (Graziano, 2006), whereas aneffector-specific dissociation of different typesof movements is found for imagery.
Ehrsson, H. H. et al. (2003). Journal of Neu-rophysiology, 90, 3304-3316. Graziano, M.
74 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
(2006). Annual Review of Neuroscience, 29,105-34 Jeannerod, M. (2001). Neuroimage,14, 103-109. Wheaton, K. J., et al. (2004).Neuroimage, 22, 277-288.
Mismatch Negativity in Patients with Ma-jor Depression Christine Norra (1), Sebas-tian Waniek (2) (1) Dept. of Psychiatry, Psy-chotherapy, Preventive Medicine, Ruhr Univer-sity Bochum, (2) Dept. of Psychiatry and Psy-chotherapy, University Hospital Münster, Ger-manyMismatch negativity (MMN) is an auditoryevoked potential that has consistently detectedneural pre-attentive information processingdeficits of rare unexpected deviants mainlyin schizophrenia. Neurochemically the gluta-matergic neurotransmitter system has beenidentified as crucial for the origin of deficitsin MMN. There is also some evidence formonoaminergic modulation of MMN. Still, withregard to patients with depressive disordersthere are only a few small and rather hetero-geneous studies of MMN, so far. The aimof the study was first a comparison of MMNbetween patients with major depression and ahealthy control group, and second, a longitudi-nal examination of differential monoaminergicmedication effects on MMN.Auditory MMN was recorded using multi-channel EEG in 17 unmedicated inpatientswith an acute depressive episode and age-matched healthy controls. Patients receiveda follow-up MMN after four weeks of random-ized antidepressant treatment with selectiveserotonergic or noradrenergic re-uptake in-hibitors. In parallel, depressed symptomswere documented on a weekly basis.MMN amplitudes of scalp derived potentialswere impaired, with different alterations in the
latencies pointing to deficits of MMN gener-ation in depressed patients as opposed tohealthy controls. Brain source analysis sig-nificantly located the largest MMN impairmentin patients left frontally with increased latency.However, despite full psychopathological re-sponse patients showed no significant changeof MMN after antidepressive pharmacother-apy, irrespective of the selective nature of themonoaminergic drug.Our findings point to deficits of the auditory in-formation processing in MMN in patients withdepressive disorder, too. However, the ab-sence of any normalization of the MMN in thepost-acute phase regardless of the pharma-cological profile of monoaminergic antidepres-sants might rather refer to the trait characterof underlying pathophysiological mechanismsin depression. Further longitudinal studies arewarranted to investigate the potential seroton-ergic modulation of MMN and its clinical im-pact.
Personal Identity from the Viewpoint ofNeuroscience and (Analytical) PhilosophyM. Ruchsow Christophsbad, Faurndauer Str.6-28, 73035 GöppingenDuring the last years the controversy about di-achronic personal identity was dominated bynaturalistic approaches, proposing a (modi-fied) memory criterion (e.g. Locke), a physi-cal criterion of bodily continuity (e.g. Williams,Nagel), or a combination of both (e.g. Brand,Northoff). Contrary to this, Wittgenstein’s non-naturalistic approach shows that the meaningof a word is its use in language (Philosophi-cal Investigations [PI] § 43). With referenceto (diachronic) personal identity Wittgensteinsuccinctly remarks that „there is a great varietyof criteria for personal ’identity’“ (PI 404) which
2012, 5 (1) Kognitive Neurophysiologie des Menschen 75

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
can be characterized by family resemblance(PI 66). A detailed elaboration of Wittgen-stein’s basic idea can be found in the workof the Canadian philosopher Charles Taylorwho advocates that personal identity nowa-days is influenced by three traditions: 1.) adisengaged atomistic individualism originatingmainly from the philosophy of Enlightenment,2.) a romantic expressivism as alternative toEnlightenment philosophy, and 3.) a traditionaltheistic position. Taylor refuses disengagedatomism. In his view persons obtain identityby identification with their social institutions /social roles, which alludes to Hegel’s conceptof ethical life (Sittlichkeit) and I. Berlin’s con-cept of positive freedom. The present paperargues for a view of personality in the tradi-tion of hermeneutics and (post)analytical phi-losophy and refuses naturalistic and neurosci-entific solutions to the problem of diachronicpersonal identity.
Wavelets - what works and what doesn’t? T.Sauer Lehrstuhl für Numerische Mathematik,Heinrich-Buff-Ring 44, 35392 Gießen, Ger-manyWavelets are an extremely powerful tool to pro-cess phenomena that are local in time and fre-quency. Such effects appear frequently espe-cially in the context of EEG signals. Due tothis reason, wavelet methods, discrete ones aswell as continuous ones, can be found in moreand more toolboxes for biosignal processing.In practice, however, it happens quite oftenthat a transform is applied to a class of sig-nals for which it is neither intended nor suit-able. This can lead to very strange results andthe reasons for that are of principal nature, noteven due to incorrect or poor implementations.In this talk I will mention and explain some
of these phenomena (which, by the way, caneven appear in a similar way for the Fouriertransform); since they cannot be avoided ingeneral, it is a good idea to be at least awareof their existence.
Cortical current density oscillations in themotor cortex are correlated with muscu-lar activity during (sub-)maximal bike exer-cise Stefan Schneider (1), David Rouffet (2),Francois Billaut (2), Heiko K. Strüder (1) (1)German Sport University Cologne, Institute ofMovement and Neurosciences, Germany; (2)Victoria University Melbourne, School of Sportand Exercise Science, AustraliaDespite modern imaging techniques, assess-ing and localizing changes in brain activity dur-ing whole body moderate and intensive ex-ercise is still challenging and has not beenshown so far. Using an active EEG systemin combinations with source localization algo-rithms, in this study we aimed to asses, dis-play and localize changes in brain cortical ac-tivity at different intensities of bike exercise.Two experienced bikers (one male / one fe-male) served as subjects in this study. EEGactivity was recorded on 32 sites across themotor cortex (m1). Simultaneously muscle ac-tivity was recorded using EMG. Muscular aswell as brain activity were time locked to thepedaling cycle. Averaging 100 cycles at differ-ent Watt loads up tp 7W/kg body weight, re-vealed a high correlation between oscillationsin muscular activity and cortical current den-sity (CCD) in the lower limbs motor cortex area(MNI 0/-40/60). Results demonstrate that itis possible to assess and localize brain corti-cal activity even during maximal exercise inten-sity using highly sophisticated but also easy touse and simple techniques like EEG in com-
76 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
bination with source localization algorithms.Furthermore results confirmed the assumptionthat increasing exercise intensity is accompa-nied by an increase in cortical current densityin M1, whereas activity patterns remain com-paratively stable across different exercise in-tensities.
Multimodal imaging of complex languageprocessing - A combined NIRS-EEG studyS. Schneider (1), L. Wagels (2), A-C. Ehlis (1),L. Ernst, A.J. Fallgatter (1), A. M. Rapp (1) (1)Clinic of Psychiatry und Psychotherapy, Uni-versity of Tuebingen, 72076 Tuebingen, Ger-many; (2) Department of Clinical Psychologyand Psychotherapy, University of Tuebingen,72072 Tuebingen, GermanyModern neuroimaging methods allow for adistinct visualization of the localization, func-tion and intercommunication of the languagenetwork. The deliberate, combined applicationof different imaging methods, in particular,can benefit a holistic investigation of dynamicprocesses, on the one hand, and brain regionsand networks, on the other, that are cruciallyinvolved in language processing. The presentstudy focuses on simultaneous recordingsof electroencephalography (EEG) and near-infrared spectroscopy (NIRS) to investigateboth temporal as well as topographical as-pects of brain activation patterns associatedwith the processing of complex language, suchas metaphorical speech or coherent discourse.A group of 20 right-handed, healthy subjectsperformed two subsequent tasks while EEGand NIRS data were recorded simultaneously:First, participants had to judge the mean-ingfulness of metaphoric, literal or nonsensephrases. In the second paradigm short storieswere presented in form of sentences with ei-
ther definite articles (leading to more coherentdiscourse) or indefinite articles (leading to lesscoherent discourse) as well as a sequence ofmeaningless letter combinations as a controlcondition. Neurophysiological data revealedcondition related differences in brain activa-tion patterns for both tasks. More precisely,a combined consideration of electrophysio-logical and vascular data gave insight to thecomplex neurocognitive processes underly-ing pragmatic language comprehension. Ourresults demonstrate the utility of combinedEEG and NIRS measurements to investigateneural substrates of higher-level cognitive pro-cesses such as figural language processingand discourse comprehension.
Are effects of sleep deprivation treatmentin patients with major depression pre-dictable by event-correlated potentials?Streuer M, Son K, Juckel G, Norra C Dept.of Psychiatry, Psychotherapy and PreventiveMedicine, LWL University Hospital Bochum,Ruhr University Bochum, GermanyAs opposed to psychopharmacotherapy theantidepressive effect of sleep deprivation isimmediate and clinically striking. However,the search for neurobiological predictors likeaminergic-cholinergic dysbalance of REMsleep, alterations of circadian rhythms, effectsof vigilance or microsleep has not yet beensuccessfully terminated. The loudness depen-dence of auditory evoked potentials (LDAEP)has been found a potentially valid psychobi-ological marker for the serotonergic system.Therefore, the influence of sleep deprivationtreatment on LDAEP in depressive patientswas examined in a controlled study.In 27 inpatients with a moderate to se-vere episode of major depression taking
2012, 5 (1) Kognitive Neurophysiologie des Menschen 77

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
their individual antidepressive medication amultichannel-EEG was recorded on two con-secutive mornings. Auditory evoked potentialswith different intensities (60-100 dB) wererecorded, and LDAEP of N1 and P2 ampli-tudes was determined. 14 patients took part insupervised sleep deprivation while 13 patientsserved as controls by sleeping as usual. Inaddition, mood (BDI, HAM-D) and sleepiness(SSS) was psychometrically assessed.Self- and investigator-rated assessmentsshowed significant improvement of depres-sive mood in depressive patients after SDcompared to controls. AEP amplitudes and la-tencies were not different between the groups,and LDAEP remained intra-individually stable.Still, there was a clear tendency (p<0.069) ofa stronger LDAEP at baseline in respondersto SD compared to non-responders.Altogether this pilot study showed effectsof sleep deprivation in AEP analyses ofsubgroups with stronger LDAEP, i.e., lowerserotonergic activity, in depressive patientswho were responders to SD. The results arein line with previous studies showing that astronger LDAEP seems to be associated witha favourable treatment response to seroton-ergic antidepressants, e.g., SSRI. In orderto find the optimum indication for SD amongdepressive patients further studies of LDAEPin SD are warranted in larger patient samples.
Scalp Topography Evoked by ReadingColor Words W. Skrandies Institute of Physi-ology, Justus Liebig University, GiessenBrain imaging studies show that non-compatible Stroop stimuli activate the an-terior cingulum of the human brain, andcommonly late ERP effects are described. Inan electrophysiological study we analyzed the
processing of compatible and non-compatiblecolor words (Stroop condition) and simplecolor words (Color condition). Two groupsof 22 healthy adults each participated in twodifferent experiments. In the Stroop conditioncompatible and incompatible color words werepresented on a monitor while in the Color con-dition simple color words appeared. EEG wasrecorded from 30 channels between the inionand 5In the Stroop condition effects of stimu-lus compatibility were seen with componentsbetween 120 und 170 ms where compatiblestimuli yielded significantly larger responses.In the Color condition global field power wassignificantly smaller than in the Stroop con-dition. Components occurring at 170 and at200 ms showed significant differences in theirscalp topography. There were no topograph-ical differences at latencies smaller than 130ms. Our results illustrate that the compatibil-ity of Stroop stimuli is reflected early in fieldstrength and topography at about 120 ms la-tency. In addition, components occurring after170 ms activated different neurons in Stroopand Color condition. This suggests specifictop-down processing at the level of the visualcortex.
Topographical Correlates of Human Se-mantic Learning W. Skrandies (1) & H.Shinoda (2) (1) Institute of Physiology, JustusLiebig University, Giessen, Germany; (2) Fac-ulty of Psychology, Rissho University, Tokyo,JapanWe studied human semantic learning of Kanjisymbols in 18 healthy German adults. FortyKanji characters were presented visually inrandom order before and after a learning ses-sion of 20 minutes duration. In the learningperiod subjects acquired the meaning of 20 of
78 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
the stimuli while the other stimuli served asunlearned control condition. Learning perfor-mance was tested at the end of the experi-ment. Event-related potentials were recordedfrom 30 electrodes between the inion andFz, and were averaged offline for each con-dition separately. All subjects learned stimu-lus meaning with a recall rate of more than90Our findings show that learning of the se-mantic meaning of Kanji characters occursrapidly within 20 minutes. Learning is directlyrelated to neurophysiological changes at earlyprocessing stages. Thus learning is reflectedby systematic changes in scalp topography ofelectrical brain activity of primary visual areas.
Phase Locking Mechanisms in the AlphaRange During Retention of a Motor or Vi-suospatial Working Memory Trace E. Weber(1), B. Griesmayr (1), & P. Sauseng (2) (1) De-partment of Physiological Psychology, Univer-sity of Salzburg, A-5020, Salzburg, AustriaThe neuronal correlates of retaining a motormemory trace are examined in 30 healthy par-ticipants. Studies using transcranial magneticstimulation (Hummel et al, 2002, Sauseng etal., 2009) showed decreased cortical excitabil-ity while alpha amplitude was high. Thus, al-pha activity is assumed to be important forresponse inhibition. Concerning interregionalconnectivity, Serrien et al. (2005) found in-creased connectivity between the sensorimo-tor cortex and prefrontal cortex shortly afterpresentation of NoGo stimuli. In the presentedexperiment, a finger sequence had to be en-coded and reproduced after a retention inter-val of three seconds. In a control condition,the finger sequence only had to be recognized(but not reproduced) after the retention inter-val. In this study, power changes and phase
coupling indices have been investigated in thealpha range (8-12 Hz) for the reproduction andrecognition conditions as well as for a restingcondition. No significant differences in alphapower (8-12 Hz) between the retention inter-vals of recognition or reproduction and a rest-ing condition have been observed. However,in contrast to a resting condition, phase de-coupling of a distributed network in the Alpharange (8-10 Hz) could be found for both condi-tions. Further phase coupling analysis showedsignificant coupling mechanisms in the upperAlpha (10-12Hz) in the reproduction conditionbetween left dorsolateral premotor cortex, rightprimary motor cortex and right central andcentroparietal regions in the last second ofthe retention period. While Alpha power didnot play a major role retaining a motor mem-ory trace, phase coupling analysis uncoveredmemory-relevant network couplings. This cou-pling mechanism may be interpreted as a mo-tor memory trace which is relevant for the re-call of the respective motor response in par-ticular shortly before the execution of the ini-tially inhibited motor response. Supported bythe FWF Project P 22084
Reliability of ERP-data in the STROOP in-terference task Juliana Wiechert Departmentof Neuropsychology and Behavioral Neurobi-ology, Universität BremenThe STROOP-paradigm is one of the mostconsistent experimental approaches in psy-chological sciences. However, retest-reliabilityin individual physiological parameters has notbeen examined in both experimental intra- andinter-session arrangements. Based on previ-ously published data, an adapted form of theSTROOP-task was applied in an EEG-studyto estimate individual intra- and inter-session
2012, 5 (1) Kognitive Neurophysiologie des Menschen 79

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
reliability of behavioural and electrophysiologi-cal data. Preliminary behavioural data showedboth consistent split-half as well as re-test re-liability in 15 healthy young female study par-ticipants. There was an expected interferenceeffect in the incongruent condition reflectedin longer response times compared to bothcongruent and baseline conditions, and a fa-cilitation effect in the congruent condition re-flected in shorter response times comparedto the baseline condition. Electrophysiolog-ical data suggest larger intra- as comparedto inter-session-reliability. Topographical ERP-effects appeared to be less reliable at process-ing relevant electrode sites across sessionsas compared to electrode positions not pre-senting any condition differences. Behaviouraldata will critically be discussed in relation tothe respective electrophysiological findings inan extended sample of individuals.
Model-based strategies for time-variantanalysis of oscillatory EEG/MEG activity H.Witte, Th. Lehmann, M. Wacker, L. LeistritzInstitute of Medical Statistics, Computer Sci-ences and Documentation, Jena UniversityHospital, Friedrich Schiller University Jena,D-07740 JenaIt is shown that model-based analysis strate-gies can be beneficially used for exploringphase coupling characteristics (linear andnon-linear) and cross-frequency couplingsof and between EEG oscillations. As modelcoupled non-linear oscillators (e.g. of theDuffing type) were applied to identify thedependencies between time-variant phase-locking, n:m synchronization, and quadraticphase coupling patterns as well as their rela-tions to so-called cross-frequency couplings(amplitude-amplitude, amplitude-frequency
and frequency-frequency couplings). In a firststep the parameters of the coupled differentialequations (oscillators) are estimated by usingthe measured EEG data. In a second stepthe differential equations are solved whichresults in a corresponding modeled EEG real-ization. The comparative time-variant analysisbetween measured and simulated data re-veals the effects which can be explained bythe model. Such strategies were successfullyapplied for analysis of EEG burst patternsin sedated patients (encephalographic burstsuppression pattern) and in healthy neonates(burst-interburst patterns during quiet sleep)and can be adapted on other oscillatoryEEG/MEG activities.
Neural correlates of switching the atten-tional focus during finger movementsKristin Marie Zimmermann (1,2), MatthiasBischoff (2), Britta Lorey (2,3), Rudolf Stark(2), Joern Munzert (3) & Karen Zentgraf (1,2)(1) Institute of Sport Science, Department ofHuman Performance and Training in Sport,University of Muenster, Muenster, Germany;(2) Bender Institute of Neuroimaging, JustusLiebig University, Giessen, Germany; (3) In-stitute of Sport Science, Department of SportPsychology, Justus Liebig University, Giessen,GermanyFor more than a decade, sports-related re-search has been concerned with the effects ofvarious attentional strategies on motor perfor-mance. Thereby comparing an attentional fo-cus on moving body parts (internal focus) witha focus directed on extracorporeal movementeffects (external focus), the latter seems supe-rior in producing a benefit in performance out-come. However, the search for differences inthe underlying mechanisms of an internal and
80 Human Cognitive Neurophysiology 2012, 5 (1)

W. Skrandies — Abstracts of the 20th German EEG/EP Mapping Meeting
an external attentional focus during motor ex-ecution has just started. This study investi-gated the effects of switching from a trained toan unfamiliar attentional focus on BOLD sig-nal change by applying functional magneticresonance imaging (fMRI). While performingan overlearned finger tapping sequence (80correct sequence executions prior to scannersession), participants (M = 24.7 years; SD =2.9; gender matched) were instructed to ei-ther attend to the keys of the response box(external focus) or to their fingers (internal fo-cus). For the last half of all trials during scan-ner session, an unexpected change in atten-tional focus (from external to internal and viceversa) was verbally instructed. The instructedattentional switch led to increased responsesin the inferior frontal gyrus and the inferiorand superior parietal lobule, assumed to de-pict attentional reallocation. Moreover, chang-ing from a trained external to a novel inter-nal focus was associated with higher activationin the left primary somatosensory cortex andthe right posterior cerebellum. When switch-ing from a trained internal to an unfamiliar ex-ternal focus, increased activation was found inthe left premotor cortex, the left secondary so-matosensory cortex, bilateral anterior and pos-terior cerebellum, the vermis and the basalganglia. We suggest that the motor-related ar-eas recruited when switching to an external fo-cus of attention are reflecting a beneficial alter-ation in motor processing by enhancement ofsensorimotor integration.
2012, 5 (1) Kognitive Neurophysiologie des Menschen 81

Book Review – Buchbesprechung
M. Doppelmayr –Neurobiologie derPsychotherapie
(Buchbesprechung)
M. Doppelmayr, FB Psychologie, UniversitätSalzburg, Hellbrunnerstr. 345020 Salzburg,
In der zweiten Auflage des Buches Neurobi-ologie der Psychotherapie verfolgt Prof. Dr.G. Schiepek von der Paracelsus Medizinis-chen Universität Salzburg ein ausgesprochenschwieriges und umfangreiches Unterfan-gen. In diesem ca. 700 seitigen Sammelw-erk werden so gut wie alle neurobiologischorientierten Aspekte der Psychotherapieaufgegriffen und zumeist detailliert disku-tiert. Knapp 100 Autorinnen und Autorenhaben zu diesem Werk beigetragen, unterIhnen Persönlichkeiten wie T. Banaschewski,N. Birbaumer, H. Flor, H. Haken, U. Halsband,T. Kircher, W. Leeb, H. Markowitsch, W. Milt-ner, V. Müller, W. Singer oder P. Tass, um nureinige zu nennen.Nach einem einleitenden umfassendenÜberblick werden die unterschiedlichen Betra-chtungsweisen in acht Themenschwerpunktenaufgearbeitet. Die Themen erstrecken sich hi-erbei auf die Gebiete Messmethoden; Moleku-lare Neurobiologie von Gehirn, Immun- undEndokrinsystem; Das Gehirn als komplexesSystem; Psychische Funktionen; Neurobiolo-gie sozialer Prozesse; Psychische Störungen;Konsequenzen für Therapien; Wissenschaft-stheoretische und ethische Fragen. Diese
Übersicht zeigt zugleich den umfassendenund multidisziplinären Ansatz, der keinen derin diesem Feld notwendigen oder relevantenWissenschaftssektoren ausspart.
Für den Großteil der Leserinnen und Leserwerden vermutlich diejenigen Kapitel, dieeinen ganz klaren Bezug zu psychischenFunktionen, sozialen Prozessen oder psychis-chen Störungen haben, die Wichtigsten sein.Dieser Bereich umfasst etwas mehr als dieHälfte des Buches und schließt eine Vielzahlan Themengebieten und Störungsbilder ein.Die einzelnen Kapitel präsentieren sehrpointiert den aktuellen Stand der Forschungund sind fast ausnahmslos sehr gut und ver-ständlich geschrieben und mit entsprechen-den Tabellen oder Graphiken aufbereitet. Diehohe Relevanz der neurobiologischen Grund-lagen und Erklärungsmodelle für den Bereichder Psychotherapie wird hier klar demonstriertund abgebildet. In vielen der Kapitel wird auchganz spezifisch auf die jeweils ableitbarenpraktischen Konsequenzen der vorgelegtenBefunde hingewiesen. Abhängig vom jew-eiligen Autor werden naturgemäß die Grund-lagen und Befunde in sehr unterschiedlicherTiefe und aus unterschiedlichen Blickwinkelnpräsentiert. In seiner Gesamtheit ist dieserTeil aber ausgesprochen gut elaboriert, aktuellund zumeist hervorragend aufbereitet.
Neben dem eben beschrieben Teil gibt esnoch viele weitere Kapitel, die vornehmlichMethoden oder reine Grundlagen präsen-tieren. Wenngleich auch diese gut aufbereitetsind, aktuelle Verfahren und neue Entwicklun-gen aufzeigen, so erscheinen mir diese vomUmfang her etwas unpassend. Für Fachleutebieten diese kaum Neues und für Personendie sich nicht konkret damit beschäftigen
82 Human Cognitive Neurophysiology 2012, 5 (1)

Book Review – Buchbesprechung
erscheint es zu detailliert. Außerdem istetwas unglücklich, wenn z.B.: im Kapitelzur Elektroenzephalographie spezielle Be-funde zu Alzheimer Demenz, Depression oderSchizophrenie besprochen werden, die manin dieser Form in den spezifischen Kapiteln zuden Störungen so nicht mehr findet und somitübersieht, wenn man die Methodenkapitelnicht durcharbeitet.Zusammenfassend lässt sich sagen, dassdieses Buch eine von erstklassigen Autorenverfasste, gut gewählte, ausgewogene undinhaltlich sehr breite Palette an Wissen bi-etet. Die Kapitel sind Großteils unabhängigund bauen nicht direkt aufeinander auf, wases ermöglicht beliebige Bereiche selektivdurchzuarbeiten. Die Methoden und Grund-lagen nehmen aus meiner Sicht einen etwaszu breiten Raum ein, der besser für einevertiefte Aufarbeitung klassischer Störungs-bilder genutzt werden hätte könnte. So sindetwa dem klassischen Themenbereich ”De-pression” gerade sieben Seiten gewidmet,während Bereiche wie ”Systhemtheorie undDynamic Causal Modelling” mehr als 30Seiten umfasst. Generell aber ist diesesBuch eine hervorragende Arbeit, die sowohlTherapeutInnen, neurobiologisch orientiertenFachkräften als auch sehr interessierte Laienfaszinieren wird und mit der G. Schiepek einehilfreiche Brücke zwischen Neurobiologie undPsychotherapie schlägt.
Neurobiologie der Psychotherapie, GünterSchiepek; 2. Auflage; Schattauer Verlag, Stuttgart(2011), ISBN 978-3-7945-2674-1, 704 Seiten, 224Abbildungen, 32 Tabellen, EUR 119.-
2012, 5 (1) Kognitive Neurophysiologie des Menschen 83

Announcements — Ankündigungen
Announcements — Ankündigungen
• ISBET Meeting
The annual meeting of the International Society for Brain Electromagnetic Topography (ISBET) willtake place in Bristol, Tennessee, USA, as a Joint Meeting of ECNS - ISNIP - ISBET - EPIC fromSeptember 12-16, 2012.
Information and Registration at: http://www.ecnsweb.com/2012-conference.htm
• 21. Deutsches EEG/EP Mapping Meeting / 21th German EEG/EP Mapping Meeting
Conference language is German; English contributions will be accepted.
– 12. bis 14. Oktober 2012; Schloss Rauischholzhausen
– Schwerpunkte / Themen
* M. Falkenstein (Dortmund) ”Altern, Kognition und Elektrophysiologie”
* Dr. M. Plichta (Mannheim) Symposium über ”NIRS - Grundlagen und Anwendun-gen in der Neurophysiologie des Menschen”
– Anmeldeschluss ist der 15. Juli 2012.
– Information und Anmeldung unter: http://www.med.uni-giessen.de/physio/
84 Human Cognitive Neurophysiology 2012, 5 (1)














![Neurophysiologie Teil 2 neu 2 Power Point · 2020-03-20 · Title: Microsoft PowerPoint - Neurophysiologie Teil 2 neu 2_Power Point[Compatibility Mode] Author: sekre Created Date:](https://static.fdokument.com/doc/165x107/5ec8c83d5e733d5a8b77e0da/neurophysiologie-teil-2-neu-2-power-point-2020-03-20-title-microsoft-powerpoint.jpg)



