
Vorlesung SoSe 2016 “Zellbiologie und Physiologie der ... · In addition, gibberellic acid (GA)...
14
1 Vorlesung SoSe 2016 “Zellbiologie und Physiologie der Pflanzen” Prof. Dr. I Finkemeier Prof. Dr. A. von Schaewen Prof. Dr. J. Kudla 02.05.2016 „Und sie bewegen sich doch…“ Pflanzliche Bewegungen Tropismen – Wachstumsbewegungen Nastien – osmotische Bewegungen Taxien – freie Ortsbewegungen (begeißelte Einzeller) Stoffgradient Alge positiv negativ Reaktion: chemotaktisch Entwicklung von frei beweglichen und festsitzenden Vielzellern…
Transcript of Vorlesung SoSe 2016 “Zellbiologie und Physiologie der ... · In addition, gibberellic acid (GA)...

1
Vorlesung SoSe 2016
“Zellbiologie und Physiologie der Pflanzen”
Prof. Dr. I Finkemeier
Prof. Dr. A. von Schaewen
Prof. Dr. J. Kudla
02.05.2016 „Und sie bewegen sich doch…“
Pflanzliche Bewegungen
Tropismen – Wachstumsbewegungen
Nastien – osmotische Bewegungen
Taxien – freie Ortsbewegungen (begeißelte Einzeller)
Stoffgradient
Alge
positiv
negativ
Reaktion: chemotaktisch
Entwicklung von frei beweglichen und festsitzenden Vielzellern…

2
Exkurs: Die Endosymbionten-Theorie
Welche Faktoren favorisieren Endosymbiosen?
The hydrogen hypothesis for the first eukaryoteMartin W, Müller M (1998) Nature, 392: 37-41.
A model for the origin of mitochondria and hydrogenosomes that directly accounts for their common ancestry. It differs from classical formulations of the endosymbiont hypothesis primarily in the premises that first, the host of mitochondrial symbiosis was not a fermenting heterotroph but rather an
autotrophic Archaebacterium that strictly depended upon molecular hydrogen (H2) for survival
(as many contemporary autotrophs do) and that, second, the symbiont was the common ancestor of mitochondria and hydrogenosomes and was a facultatively anaerobic α-proteobacterium that gratuitously supplied molecular hydrogen to the host as a waste product of its (own) metabolism.
A hypothetical symbiosis between these two cells is sketched involving gene transfer from symbiont to host that derives a single, bipartite, heterotrophic cell that possesses an archaebacterial genetic apparatus, a eubacterial glycolytic pathway, and a facultatively anaerobic, symbiotically-derived organelle in its cytosol.
Further reading (Reviews)Tracing the Thread of Plastid Diversity through the Tapestry of LifeC. F. Delwiche, The American Naturalist 1999
Mergers and acquisitions: Malaria and the great chloroplast heistG. I. McFadden, Genome Biology 2000
One, two, three: Nature´s toolbox for building plastidsB. Stoebe, U.-G. Maier, Protoplasma 2002
H2

3
Metabolische Abhängigkeit
als treibende Kraft
Maximize surface area (to improve H2 uptake)
Need to feed the partner (before complete enclosure)
Independence (of geological H2 source)
Anaerobic co-existence (as requirement)
Gene transfer
H2
H2
(Mitochondrium)
Vorteil: Kann alle heute existierenden Formen erklären (aber NICHT schlüssig die Herkunft von Peroxisomen!)
α-Proteo- withArchae-bacterium
⇒ Eukaryotic cell
TropismenPflanzliche Tropismen sind echte Wachstumsbewegungen.
Sie beruhen auf Zellstreckung (in bestimmten Zonen), die durch das Phytohormon Auxin vermittelt wird.
Dabei wird die Richtung durch den auslösenden Reiz bestimmt (⇒ gerichtete Bewegung).
Tropismen können u.a. ausgelöst werden durch:
• Licht (Phototropismus)
• Schwerkraft (Gravitrotropismus)
• Taktile Reize (Thigmotropismus) z.B. Rankenbewegung
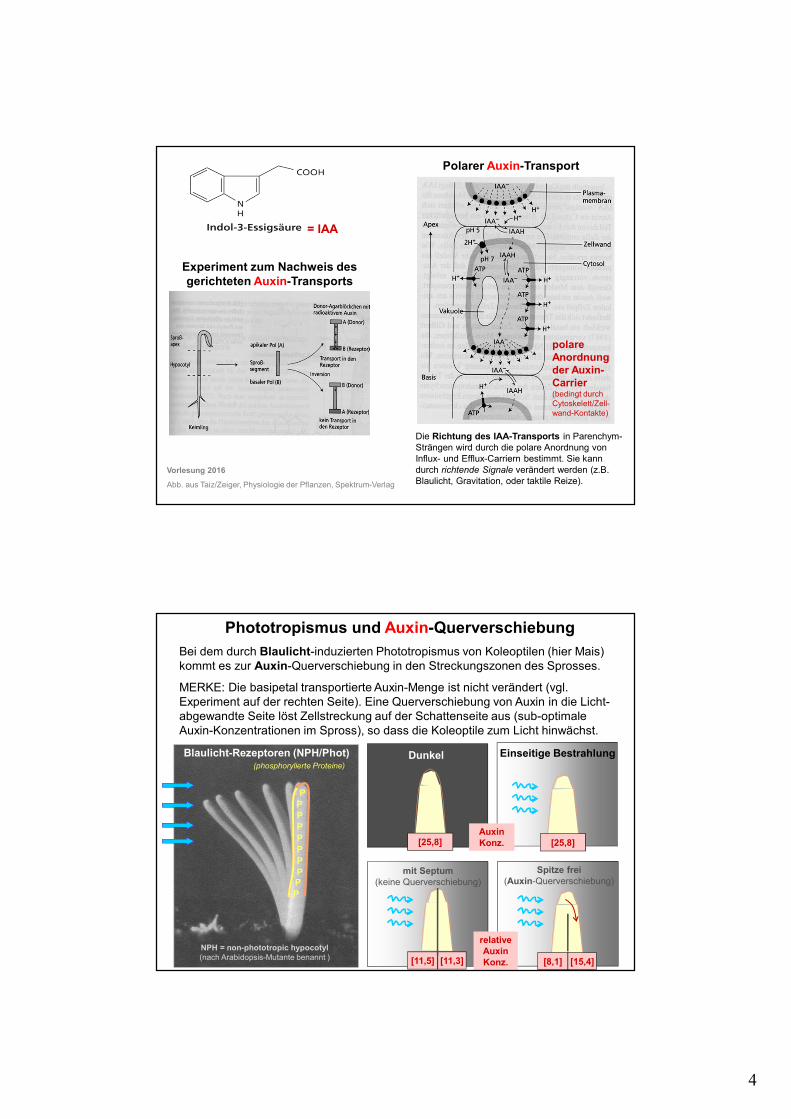
4
Basipetaler Auxin-Transport
Experiment zum Nachweis des gerichteten Auxin-Transports
polare Anordnung der Auxin-Carrier (bedingt durch Cytoskelett/Zell-wand-Kontakte)
Vorlesung 2016
Abb. aus Taiz/Zeiger, Physiologie der Pflanzen, Spektrum-Verlag
Die Richtung des IAA-Transports in Parenchym-Strängen wird durch die polare Anordnung von Influx- und Efflux-Carriern bestimmt. Sie kann durch richtende Signale verändert werden (z.B. Blaulicht, Gravitation, oder taktile Reize).
= IAA
Polarer Auxin-Transport
Phototropismus und Auxin-Querverschiebung
Bei dem durch Blaulicht-induzierten Phototropismus von Koleoptilen (hier Mais) kommt es zur Auxin-Querverschiebung in den Streckungszonen des Sprosses.
MERKE: Die basipetal transportierte Auxin-Menge ist nicht verändert (vgl. Experiment auf der rechten Seite). Eine Querverschiebung von Auxin in die Licht-abgewandte Seite löst Zellstreckung auf der Schattenseite aus (sub-optimale Auxin-Konzentrationen im Spross), so dass die Koleoptile zum Licht hinwächst.
Blaulicht-Rezeptoren (NPH/Phot)
[8,1] [15,4]
[25,8]
Einseitige BestrahlungDunkel
[25,8]Auxin Konz.
[11,5] [11,3]
relative Auxin Konz.
mit Septum(keine Querverschiebung)
Spitze frei(Auxin-Querverschiebung)
(phosphorylierte Proteine)
PPPPPPPP
PP
NPH = non-phototropic hypocotyl(nach Arabidopsis-Mutante benannt )

5
Bakterielle Photolyasen
Arabidopsis CRY
Arabidopsis Nph1/Phot2
Vorlesung 2016
Zur Erinnerung:
Blaulicht-Rezeptoren = Cryptochrome (CRY, NPH, Phot)
Struktur: Proteine mit je 2 Chromophoren
Pterin/Flavin FAD MTHFFAD
Redox-Partner?
e-ET
FMN FMN Kinase
bis zu 8x
ATP
Ser-PCys-S
Signal-Domänen
ET = Elektronentransport
C14 Fettsäure (Myristoylgruppe) C16 Fettsäure (Palmitoylgruppe)
N-Myristoylierung (an Glycin in Pos. 2, nach Prozessieren des ersten Met) vermittelt schwache, reversible Interaktionen mit Membranen (und anderen Proteinen). Über Myristoylierung lässt sich der Transport und die Lokalisation von Proteinen steuern. Bei Pflanzen spielt Myristoylierung eine wichtige Rolle bei der Signal-Transduktion (als Reaktion auf äußere Stimuli & Sress).
Phytochrom kann die phototrope Reaktionsfähigkeit erhöhen, ohne selbst phototrop wirksam zu sein!
Der wachsende Spross reagiert i. d. R. positiv, die Wurzel (wenn überhaupt) negativ phototrop.
Daneben gibt es den Dia-Phototropismus, d.h. Blätter orientieren sich quer zum Licht - bei extremer Strahlung u.U. sogar senkrecht (in Lichtachse).
Wirkungsspektrum für die phototrope Reaktion des Lucerne-Hypokotyls (UV-A und Blaulicht)
blauUV-B
Phototropismusdurch Blaulicht vermittelteWachstumsänderung von Organ-Flanken in Streckungszonen von Spross und Wurzel (positiv/negativ).
Phy
UV-A
Hypocotyl

6
1. Aktivierung von H+-ATPasen � Ansäuerung der Zellwand
2. Aktivierung von Expansin � Lockerung der Zellwand
3. Neusynthese von Zellwandmaterial
Auxin-induzierte Zellstreckung
Phototropismus
„auxin responsive
element“ (ARE)
Auxin aktiviert Signalkaskaden
„frühe“ (innerhalb Minuten) und „späte“
(im Stundenbereich) Antworten.
Die Promotorbereiche Auxin-induzierter Gene enthalten sogen. “auxin responsive elements“ (ARE) und sind reprimiert (ARF via AUX/IAA).
Extrazellulär bindet Auxin (an ABP1) wodurch nach Andocken an TMK-Rezeptoren in der PM sehr schnell messbare Reaktionen erfolgen (z.B. Aktivierung von H+-ATPasen in der PM).
Bei intrazellulärer Akkumulation bindet Auxin an TIR1, ein Auxin-Bindeprotein des cytosolischenE3/SCF (F-box)-Komplexes. Dies führt zur Ubiquitinierung von Aux/IAA-Repressoren(induzierter Abbau) und Transkription Auxin-induzierter Gene wie MHA2 (eine H+-ATPase) und ZMK1 (ein K+-Kanal an der PM).
Weitere Auxin-induzierte Gene kodieren für Proteine, die zur Manifestation der Auxin-Antwort beitragen (z.B. Enzyme, die zur Neusynthese von Zellwandmaterial benötigt werden).
Vorlesung 2016 Abb. verändert nach Taiz/Zeiger, Physiologie der Pflanzen, Spektrum-Verlag
Aux/IAA
ARF ARF
Aux/IAA
ARF ARF (auxin response factor)
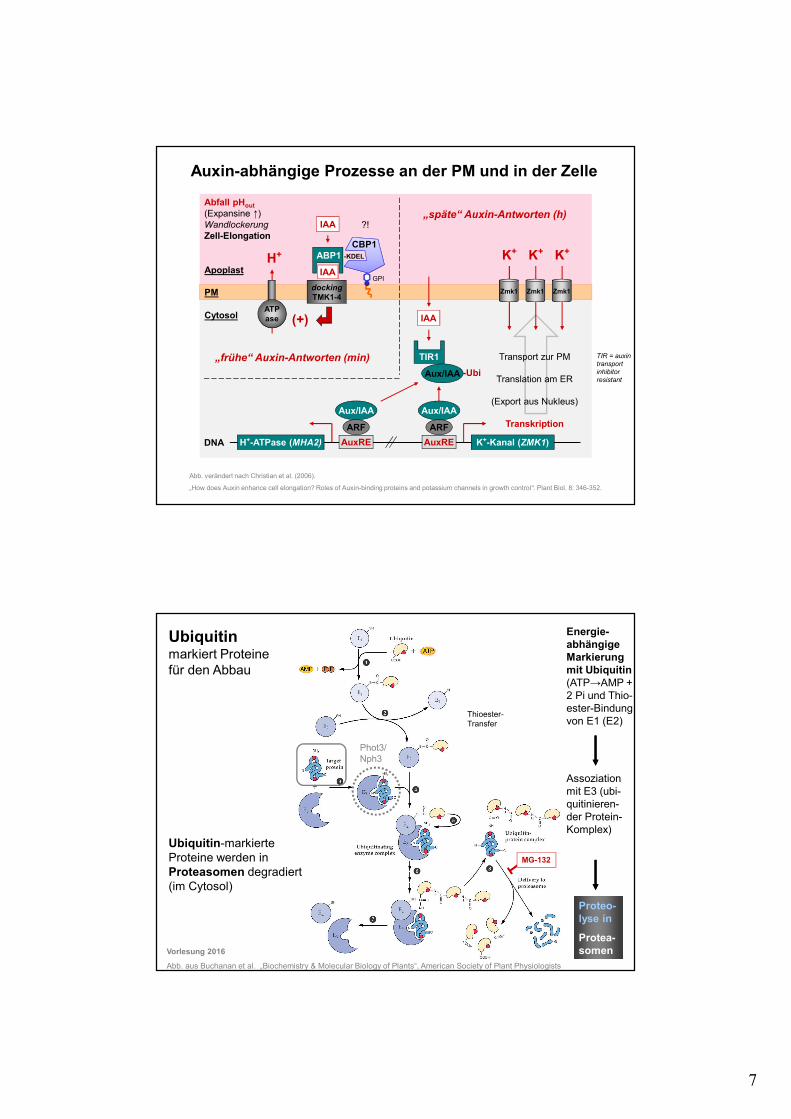
7
Auxin-abhängige Prozesse an der PM und in der Zelle
Abb. verändert nach Christian et al. (2006).
„How does Auxin enhance cell elongation? Roles of Auxin-binding proteins and potassium channels in growth control“. Plant Biol. 8: 346-352.
PMdocking
TMK1-4
ABP1 -KDEL
IAA
„frühe“ Auxin-Antworten (min)
„späte“ Auxin-Antworten (h)
IAA
K+ K+
Zmk1 Zmk1
AuxRE
ARF
TIR1
AuxRE
ARF
Aux/IAA
DNA
Aux/IAA -Ubi
K+-Kanal (ZMK1)H+-ATPase (MHA2)
Aux/IAA
Cytosol
Apoplast
(+)
Transport zur PM
Translation am ER
(Export aus Nukleus)
Transkription
Abfall pHout
(Expansine ↑) WandlockerungZell-Elongation
H+
ATPase
IAA
TIR = auxin transport inhibitor resistant
CBP1CBP1
GPI
?!
K+
Zmk1
Proteo-lyse in
Protea-somen
Ubiquitin markiert Proteine für den Abbau
Ubiquitin-markierte Proteine werden in Proteasomen degradiert (im Cytosol)
Energie-abhängige Markierungmit Ubiquitin (ATP→AMP + 2 Pi und Thio-ester-Bindung von E1 (E2)
Assoziation mit E3 (ubi-quitinieren-der Protein-Komplex)
Thioester-Transfer
MG-132
Vorlesung 2016
Abb. aus Buchanan et al. „Biochemistry & Molecular Biology of Plants“, American Society of Plant Physiologists
Phot3/Nph3

8
ANALOG: PIF4-vermittelte Schattenvermeidung
Stamm & Kumar (2010)The phytohormone signal network regulating elongation growth during shade avoidance. J. Exp. Bot. 61: 2889-2903.
Fig. 2. In white light (high R:FR), PIF4-mediated transcriptional activation is inhibited. Part of the PIF4 protein pool is sequestered in inactive complexes with DELLA proteins, whereas another part is bound by the FR absorbing active PhyB (Pfr B), which leads to the degradation of PIF4 via the ubiquitin–proteasome pathway. In FR light-enriched environments, e.g. due to canopy shade, Pfr B is converted to its R-absorbing inactive form (Pr B), which abolishes its ability to bind PIF4. Thus, PIF4 can accumulate, allowing transcription of shade-induced genes. In addition, gibberellic acid (GA) levels increase in response to shade, which leads to the degradation of DELLA proteins, releasing an additional pool of PIF4 protein. In prolonged shade, HFR1 accumulates and forms heterodimers with PIF4 that do not bind to DNA any more, thus preventing an exaggerated response.
hellrotes Licht überwiegt ⇒Pfr-Form von PhyBim Zellkern
dunkelrotes Licht überwiegt⇒Pr-Form von PhyB
bleibt im Cytosol
Maiswurzeln werden auf die Seite gelegt. Nach 90 min ist eine Krümmung in Richtung des Erdmittelpunktes zu sehen. Diese Bewegung heißt aber nicht Geo-, sondern Gravitropismus.
Modifizierte Plastiden (Statolithen) in speziellen Zellen (Statocyten) der Wurzelhaube lagern sich am basalen Ende der Zellen ab. Dies führt zu einer Umleitung des Auxin-Stroms in der Wurzelspitze und damit zu ungleichem Streckungswachstum der Wurzelflanken.
Gravitropismus und Statolithen-Hypothese
Vorlesung 2016
Abb. aus Campbell, „Biologie“, Spektrum-Verlag
Im Spross wird die Schwerkraft ebenfalls perzipiert! (z.B. in spez. Stärke-Scheiden der Rinde)
g
t = 0 min
t = 90 min
Mais-Wurzel

9
Modellvorstellung zur Auxin-Umverteilung in der Wurzelspitze bei der gravitropen Reaktion
In der Wurzel wirkt eine Erhöhung der Auxin-Konzentration supra-optimal, d.h. hemmend auf die Zellstreckung, und zwar in der dem Erdmittelpunkt zugewandten Flanke (wahrscheinlich durch Ethylen-Bildung).
Statolithen-Hypothese:
Schwerkraftwahrnehmung erfolgt u.a. durch einseitige Belastung des Cytoskeletts und wird durch Stärke-haltige Amyloplasten (und andere Organellen, wie das ER) vermittelt, die am Aktin-Cytoskelett verankert sind.
Schwerkraftwahrnehmung kann daher auch in Zellen ohne spezielle Statolithen erfolgen! Wahrscheinlich öffnen Ca2+-Kanäle in der Plasmamembran über Zugkräfte am Cytoskelett (vermittelt durch Masse-reiche Organellen). Dadurch erhöht sich die intrazelluläre Ca2+-Konzentration auf der apikalen Seite (Ca2+ signaling: „PM central control“-Modell).
Vorlesung 2016
Abb. aus Taiz/Zeiger, Physiologie der Pflanzen, Spektrum-Verlag
Schwerkraftwahrnehmungführt zu einer Verschiebung der Auxin-Konzentration im Gewebe ⇒ gravitropeReaktion, via Stimulation oder Hemmung der Zell-Streckung - je nach Lage des Auxin-Optimums im jeweiligen Gewebe. MERKE: In Spross und Wurzel verschieden!

10
Vorlesung 2016
Abb. aus Buchanan et al. „Biochemistry & Molecular Biology of Plants“, American Society of Plant Physiologists
Cytoskelett-gesteuerter Calzium-Kanal
„Transdukon“
oder„Signalosom“
Ein Proteinkomplex an der PM ist mit dem Cytoskelett (Aktin) verankert und dient so der Signalperzeption und -weiterleitung
Voraussetzung: Aktin-Filament
Verankerung!
Ziehen bzw. Erschlaffen
Nastien – Bewegungsrichtung ist anatomisch vorgegeben
Pflanzenbewegungen, bei denen die Richtung des auslösenden Reizes keinen Einfluss auf die Bewegungsrichtung hat: „Alles-oder-Nichts“-Reaktionen bei Überschreiten eines Schwellen-werts), sind vollständig reversibel und werden durch schnelle Turgor-Änderungen ausgelöst:
- Turgor-Erhöhung durch aktive K+-Aufnahme (K+-Pumpen!) - Erschlaffung durch Öffnen von K+-Kanälen.
Beispiele:
• Spaltöffnungsbewegung
• seismonastische Blattbewegung (von Mimosen bei Berührung, Erschütterung)
• circadiane (Tages-rhythmische) „Schlafbewegung“ von Blättern
Vorlesung 2016 Abb. aus Campbell, Biologie, Spektrum-Verlag
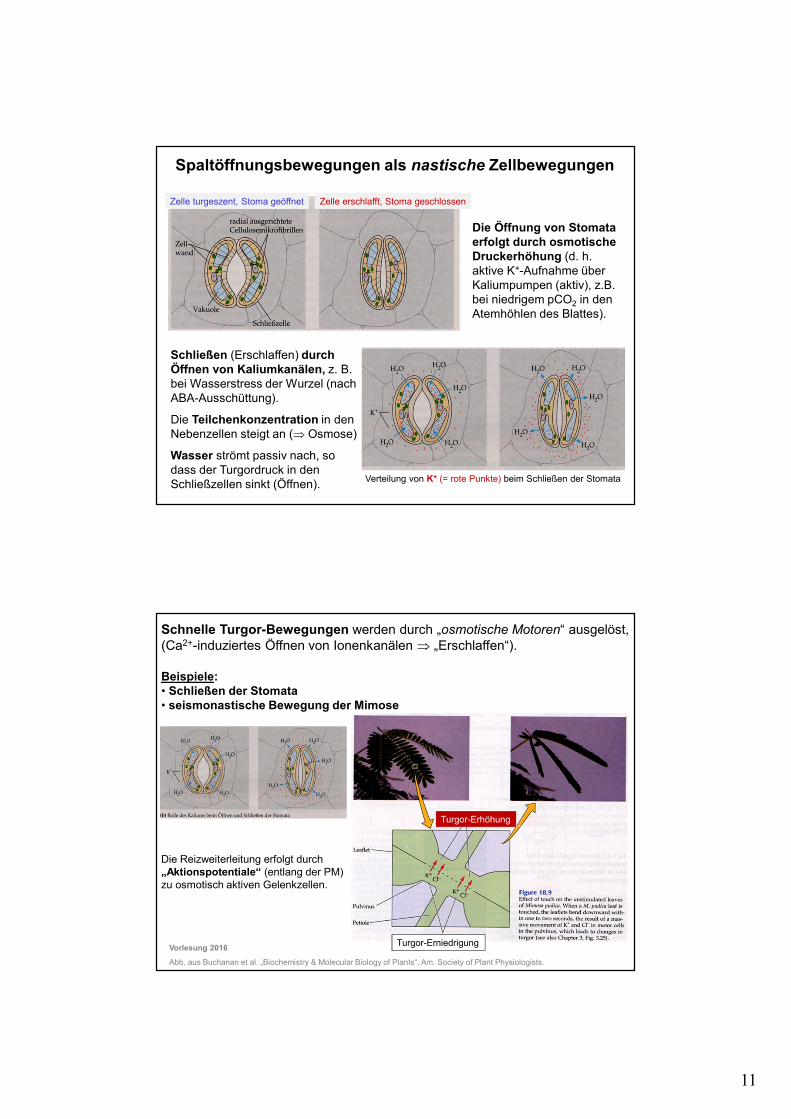
11
Spaltöffnungsbewegungen als nastische Zellbewegungen
Die Öffnung von Stomata erfolgt durch osmotische Druckerhöhung (d. h. aktive K+-Aufnahme über Kaliumpumpen (aktiv), z.B. bei niedrigem pCO2 in den Atemhöhlen des Blattes).
Schließen (Erschlaffen) durch Öffnen von Kaliumkanälen, z. B. bei Wasserstress der Wurzel (nach ABA-Ausschüttung).
Die Teilchenkonzentration in den Nebenzellen steigt an (⇒ Osmose)
Wasser strömt passiv nach, so dass der Turgordruck in den Schließzellen sinkt (Öffnen). Verteilung von K+ (= rote Punkte) beim Schließen der Stomata
Zelle turgeszent, Stoma geöffnet Zelle erschlafft, Stoma geschlossen
Vorlesung 2016
Abb. aus Buchanan et al. „Biochemistry & Molecular Biology of Plants“, Am. Society of Plant Physiologists.
Turgorerhöhung
Turgor-Erniedrigung
Schnelle Turgor-Bewegungen werden durch „osmotische Motoren“ ausgelöst, (Ca2+-induziertes Öffnen von Ionenkanälen ⇒ „Erschlaffen“).
Beispiele: • Schließen der Stomata• seismonastische Bewegung der Mimose
Die Reizweiterleitung erfolgt durch „Aktionspotentiale“ (entlang der PM)zu osmotisch aktiven Gelenkzellen.
Turgor-Erhöhung

12
Vorlesung 2016
Abb. aus Buchanan et al. „Biochemistry & Molecular Biology of Plants“, Am. Society of Plant Physiologists.
Aktin
Aktivierung von Spannungs-kontrollierten Ca2+-Kanälen
Blaulicht, Berührung,
etc.
Zug-regulierter Ca2+-Kanal
Spannungs-regulierter (V)
Ca2+-Kanal
Extrazelluläre Wahrnehmung führt zu
intrazellulären Verrechnungen
Mechanoperzeption
Cellulose-Mikrofibrillen haben über (Rosettenkomplexe) und transmembraneRezeptoren Kontakt zu kortikalen Mikrotubuli auf der Membran-Innenseite.
⇒ Verformung der Zellwand löst eine Reaktion des Cytoskeletts aus!
Vorlesung 2016
Abb. aus Buchanan et al. „Biochemistry & Molecular Biology of Plants“, Am. Society of Plant Physiologists.

13
Touch- or wind-induced reactions
Figure 2. Mechanical stimulation induces ion fluxes in plants.
(a) Mechanical stimulation, provided by directing a stream of air
onto the aerial parts of a plant expressing the Ca2+-dependent
luminescent protein aequorin, revealed that mechanical signals lead
to rapid and transient increases in cytosolic Ca2+. Cold shock (0°C)
was used as a control stimulus also known to elicit large scale
increases in cytosolic Ca2+ in plants.
(b) Repeated mechanical stimulation leads to attenuation of
subsequent Ca2+ responses, suggesting either downregulation of the
mechanosensor upon repeated stimulation, or exhaustion of the store
of Ca2+ being released to increase cytosolic ion levels. The recovery
after a period of no stimulation could represent refilling of the
discharged Ca2+ stores or resensitization of the mechanoreceptor
system.
(c) Model of possible plasma membrane signaling events upon
mechanical stimulation drawn from events seen during turgor-driven
root-hair and pollen-tube elongation. Membrane tension leads to
opening of a Ca2+-permeable channel. A cytosolic Ca2+ increase
then activates a proton transporter leading to cytosolic acidification
and cell-wall alkalinization. Simultaneously, at least in root hairs,
the NADPH oxidase RBOH C is activated through its Ca2+-binding
EF hand motifs leading to ROS production in the cell wall. Wall
ROS might then leak back to the cytoplasm. Cytosolic ROS, pH
decrease and Ca2+ increase are all known to elicit signaling events.
Similarly, elevated wall pH and ROS are known to rigidify the wall
matrix (pectin). ⇒ opposite of auxin, antagonist = ethylene!
Thus, this Ca2+-dependent system can directly modulate growth,
through altering wall properties, and can elicit the cytoplasmic
signaling cascades known to be linked to mechanical response.
Refraktärzeit
Review: Monshausen & Gilroy (2009) „Feeling green: Mechanosensing in Plants“, Trends in Cell Biology 19: 228-235.
Figure 4. Action of possible mechano-
responsive elements.
Plant mechanosensors probably fall into
two broad classes: those activated by
tension in the membrane, as exemplified
by the MSL family of Cl- permeable
channels, and those monitoring wall status
and/or sheer between the wall and the
plasma membrane. SAC represents the
stretch-activated Ca2+-permeable conduc-
tance identified through electrophysiology
but yet to be cloned. AtHK1 operates in an
osmosensing pathway that potentially
functions through a phospho-relay cascade
that uses the antagonistic response
regulators ARR3/4 and ARR8/9.
The Receptor-like Kinases (RLKs) such as
wall-associated kinases (WAK), Theseus
(THE1, senses inhibition of Cellulose
biosynthesis) and the lectin domain-
containing RLK provide models for how a
wall sensor might operate. They are likely
to elicit protein kinase-dependent signals
that could relay mechanical information
directly to mechano-responsive gene
expression for example, or interact with
the Ca2+-dependent signaling cascade that,
to date, remains the best characterized
mechanically induced signal transduction
event in plants.
Review: Monshausen & Gilroy (2009) „Feeling green: Mechanosensing in Plants“, Trends in Cell Biology 19: 228-235.
Mechanoperzeption (aktuelles Modell)
Wand-StatusMembran Zug

14
Berührung und andere mechanische Reize können Änderungen des Streckungswachstums auslösen
Beispiele:
• Rankenbewegung
• Kurzwüchsigkeit
Wuchshemmung durch Ethylen-
Freisetzung nach Berührung
Neben reinem Wachstum werden auch andere Entwicklungsprozesse durch mechanische Reize ausgelöst (z.B. Holz-bildung im Stängel durch Druck- oder Wind-belastung).
Auch Berührungsreize werden durch Calzium-Signale übertragen (z.B. Öffnen Spannungs-regulierter Ca2+-Kanäle nach mechanischer Verformung).
Also – Pflanzen besser nicht streicheln !
Zusammenfassung
Bewegungen bei Pflanzen
Tropismen Reaktionen zum Reiz hin (gerichtete Wachstumsbewegungen)
Nastien „Alles-oder-Nichts“-Reaktionen (ungerichtete Turgorbewegungen)
Taxien Orientierung entlang eines Stoffgradienten (bei frei schwimmenden Organismen)


















![Tricycloquinazoline (TCQ) based electron deficient ... · b]fluorene unit, and novel polymers containing the 9,9’-bisfluorenylidene unit. Our aim was to increase the chemical stability](https://static.fdokument.com/doc/165x107/5fa3dfdd979363378f3f3e32/tricycloquinazoline-tcq-based-electron-deficient-bfluorene-unit-and-novel.jpg)