
Ultraviolett-Mutabilität bei Bacteriam...
7
This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution 4.0 International License. Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschung in Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht: Creative Commons Namensnennung 4.0 Lizenz. Adsorptionsmethode überhaupt eine einfache Mög- lichkeit, den Sitz und die Austrittsstellen bestimm- ter „positiver" Stoffwechselprodukte bei geeigne- ten Bakterien nachzuweisen. Durch Versuche an „gewaschenen" Bakterien im Vergleich zu „un- gewaschenen", ähnlich wie sie zum Studium der toxischen Wirksamkeit gerade von Tetanus-Bak- 7 Vgl. H. S c h m i d t , Grundlagen der spezifischen Therapie, Berlin 1940, S. 847. terien schon vielfach durchgeführt worden sind T , müßte ein solcher Nachweis möglich sein. Die Untersuchungen wurden 1944 im Virchow- K rankenhaus Berlin und im AEG-For- schungsinstitut Berlin durchgeführt. Für die Überlassung der anaeroben Bakterienstämme dan- ken wir Hrn. Prof. Z e i ß 1 e r , Bakteriol. Institut des Krankenhauses Hamburg-Altona. Die Targesin-Prä- parate wurden uns von der Firma G o e d e c k e - Ber- lin zur Verfügung gestellt. Ultraviolett-Mutabilität bei Bacteriam prodigiosum Von REINHARD WALTER KAPLAN Aus dem Kaiser-Wilhelm-Institut für Züchtungsforschung, Erwin-Baur-Institut, Voldagsen (Z. Naturforschg. 3b, 29—35 [1948J; eingegangen am 13. Oktober 1947) Die Bestrahlung von Zellensuspensionen des Bact. prodigiosum mit dem Gesamtspek- trum der Hg-Lampe ergab einen bei niedrigen Energiedosen linear mit der Dosis ab- fallenden Logarithmus der Überlebendenzahl, der bei höheren Dosen wegen der sich auswirkenden teilweisen Verklumpung der Zellen von der semilogarithmischen Lineari- tät abbiegt. Im Gebiet semilogarithmischer Tötung steigt auch die Häufigkeit abwei- chender Kolonien (m) linear mit der Dosis an, durch die Zellverklumpung fällt die Variantenrate höherer Dosen schwankend wieder ab. Die mutativen Abweichungen be- stehen in der Hauptsache in Zwergwuchs, ferner in schwächerer oder stärkerer Farbstoff- bildung sowie anomaler Farbstoffverteilung in den Kolonien. Nur die Variation Rot nach Weiß (r — w) blieb unbeeinflußt. Aus abweichenden Kolonien geimpfte Tochter- platten zeigten an, daß die eine Varianten-Kolonie bildende Stammzelle eine von den normale Kolonien bildenden Zellen abweichende erbliche Beschaffenheit besaß. Bei Abfilterung der Wellenlängen unter etwa 3000 Ä ergab sich keine Beeinflussung der Keimzahl der Suspension und ebenfalls keine Veränderung der Variantenrate. Damit waren dem früher aus der Untersuchung der spontanen Variation r — w gewonnenen Indizium zwei weitere Anzeichen dafür hinzugefügt, daß der Erbvariationsmechanismus des Bact. prodigiosum dem Genmutationsprozeß höherer Organismen vergleichbar sei. Dies wurde gestützt durch Vergleich der für die Erzeugung von 1% Mutationen not- wendigen Energiemenge beim Bacterium und Sphaerocarpus-Spermatozoid sowie den Trichophyton-Sporen und Antirrhinum-Pollen. Unter der Annahme, daß die Keimtötung den Letalmutationen entspricht, ergab sich ein Verhältnis von vitalen zu letalen Genen von 1/155 = 0,65%. Der Unterschied zwischen diesem und dem gleichen Verhältnis von 7—8% bei Drosophila kann durch die einfachere Bakterienstruktur verstanden werden. Aus der maximal möglichen Zunahme der w-Mutationen von 1% bei der Höchstdosis und der Annahme, daß an dieser Mutation nur ein bis wenige Gene beteiligt sind, ergibt sich eine untere Grenze der Genzahl für Prodigiosum von etwa 1200, falls die w-, ro- und Letal gene etwa gleiche UV-Empfindlichkeit besitzen. 1. P r o b l e m s t e l l u n g I n einer vorhergehenden Mitteilung 1 wurde über Versuche am Bact. prodigiosum berichtet, deren Ziel es war, den Mechanismus der spontanen genetischen Veränderung näher kennenzulernen, bei welcher sich Zellen, die zu roten Kolonien 1 R. W. K a p l a n , Spontane Mutabilität bei Bact. prodigiosum. Z. Naturforschg. 2b, 308 [1947]. (r) auswachsen, in solche, welche weiße Kolonien (w) bilden, und umgekehrt, verwandeln. Insbe- sondere wurde dieser Vorgang mit dem Mutations- prozeß der höheren Organismen verglichen, wozu als Methode die biophysikalische Analyse ge- wählt wurde, da uns diese die Natur des sich ändernden Erbfaktors besser verrät als die Bastardanalyse, welche wir ja bei Bakterien wegen
Transcript of Ultraviolett-Mutabilität bei Bacteriam...

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
Adsorptionsmethode überhaupt eine einfache Mög-lichkeit, den Sitz und die Austrittsstellen bestimm-ter „positiver" Stoffwechselprodukte bei geeigne-ten Bakterien nachzuweisen. Durch Versuche an „gewaschenen" Bakterien im Vergleich zu „un-gewaschenen", ähnlich wie sie zum Studium der toxischen Wirksamkeit gerade von Tetanus-Bak-
7 Vgl. H. S c h m i d t , Grundlagen der spezifischen Therapie , Berl in 1940, S. 847.
terien schon vielfach durchgeführt worden sindT, müßte ein solcher Nachweis möglich sein.
Die Unte rsuchungen wurden 1944 im V i r c h o w -K r a n k e n h a u s B e r l i n und im A E G - F o r -s c h u n g s i n s t i t u t B e r l i n durchgeführ t . F ü r die Über lassung der anaeroben Bakter iens tämme dan-ken wir Hrn . P ro f . Z e i ß 1 e r , Bakter iol . Ins t i tu t des Krankenhauses Hamburg-Altona. Die Targes in -Prä -p a r a t e wurden uns von der F i rma G o e d e c k e - Ber-lin zur V e r f ü g u n g gestell t .
Ultraviolett-Mutabilität bei Bacteriam prodigiosum V o n REINHARD WALTER KAPLAN
Aus dem Kaiser-Wilhelm-Inst i tu t f ü r Züchtungsforschung , Erwin-Baur- Ins t i tu t , Voldagsen
(Z. N a t u r f o r s c h g . 3b, 29—35 [1948J; e ingegangen am 13. Oktober 1947)
Die Bes t rah lung von Zellensuspensionen des Bact. prodigiosum mit dem Gesamtspek-t rum der Hg-Lampe ergab einen bei n iedr igen Energiedosen l inear mit der Dosis ab-fallenden Logar i thmus der Überlebendenzahl , der bei höheren Dosen wegen der sich auswirkenden tei lweisen Verk lumpung der Zellen von der semilogari thmischen Linear i -tä t abbiegt. Im Gebiet semilogari thmischer Tö tung s te igt auch die Häuf igkei t abwei-chender Kolonien (m) l inear mit der Dosis an, durch die Ze l lverk lumpung fä l l t die Var ian tenra te höherer Dosen schwankend wieder ab. Die muta t iven Abweichungen be-stehen in der Hauptsache in Zwergwuchs, ferner in schwächerer oder stärkerer Farbstoff-bi ldung sowie anomaler Fa rbs to f fve r t e i lung in den Kolonien. Nur die Variat ion Rot nach Weiß (r — w) blieb unbeeinflußt . Aus abweichenden Kolonien geimpfte Tochter-pla t ten zeigten an, daß die eine Varianten-Kolonie bildende Stammzelle eine von den normale Kolonien bildenden Zellen abweichende erbliche Beschaffenheit besaß. Bei Abfi l terung der Wel lenlängen un te r e twa 3000 Ä ergab sich keine Beeinflussung der Keimzahl der Suspension und ebenfal ls keine Ve rände rung der Var ian tenra te . Damit waren dem f r ü h e r aus der Unte r suchung der spontanen Var ia t ion r — w gewonnenen Indizium zwei wei tere Anzeichen d a f ü r h inzugefüg t , daß der Erbvar ia t ionsmechanismus des Bact. prodigiosum dem Genmutat ionsprozeß höherer Organismen vergleichbar sei. Dies wurde gestütz t durch Vergleich der f ü r die E r z e u g u n g von 1% Mutationen not-wendigen Energ iemenge beim Bacter ium und Sphaerocarpus-Spermatozoid sowie den Trichophyton-Sporen und Ant i r rh inum-Pol len . Unter der Annahme, daß die Keimtötung den Letalmutat ionen entspricht , e rgab sich ein Verhä l tn i s von vi talen zu letalen Genen von 1/155 = 0,65%. Der Unterschied zwischen diesem und dem gleichen Verhältnis von 7—8% bei Drosophila kann durch die e infachere B a k t e r i e n s t r u k t u r vers tanden werden. Aus der maximal möglichen Zunahme der w-Mutationen von 1% bei der Höchstdosis und der Annahme, daß an dieser Mutation nur ein bis wenige Gene betei l igt sind, ergibt sich eine untere Grenze der Genzahl f ü r Prodigiosum von etwa 1200, falls die w-, ro-und Letal gene etwa gleiche UV-Empfindlichkeit besitzen.
1. P r o b l e m s t e l l u n g
In einer vorhergehenden Mitteilung1 wurde über Versuche am Bact. prodigiosum berichtet, deren
Ziel es war, den Mechanismus der spontanen genetischen Veränderung näher kennenzulernen, bei welcher sich Zellen, die zu roten Kolonien
1 R. W. K a p l a n , Spontane Mutabil i tät bei Bact. prodigiosum. Z. Naturforschg. 2b, 308 [1947].
(r) auswachsen, in solche, welche weiße Kolonien (w) bilden, und umgekehrt, verwandeln. Insbe-sondere wurde dieser Vorgang mit dem Mutations-prozeß der höheren Organismen verglichen, wozu als Methode die biophysikalische Analyse ge-wählt wurde, da uns diese die Natur des sich ändernden Erbfaktors besser verrät als die Bastardanalyse, welche wir ja bei Bakterien wegen

unserer Unkenntnis von deren Sexualvorgängen nicht durchführen können. Denn die Kreuzungs-analyse läßt nur eine Lokalisation des Erbfaktors in der Zelle bzw. den Chromosomen zu, erfaßt aber im Gegensatz zu jener Methode die innere stoffliche Struktur des Gens nicht. Die einer Ein-trefferfunktion entsprechende Abhängigkeit der Variantenrate von der Lagerzeit der Zellen sowie deren Temperaturabhängigkeit machen es wahr-scheinlich, daß die Ursache der Variation r — w und umgekehrt die Veränderung eines oder weni-ger, den mutablen Genen entsprechenden molekül-artigen Erbfaktoren darstellt. Im folgenden soll über Versuche berichtet werden, die diese Art der genetischen Analyse des Bact. prodigiosum weiterführen, und zwar mit Hilfe der ultraviolet-ten Strahlen als Mittel zur Auslösung von Erb-variationen.
Die zunächst angegangene Frage war die nach der Abhängigkeit der Variantenrate von der UV-Dosis des Hg-Gesamtspektrums. Als Varianten wurden diesmal nicht nur die Übergänge der Kolonienfärbung von Rot nach Weiß beachtet, sondern auch alle anderen erkennbaren Farb-änderungen, vor allem aber auch Wuchsabwei-chungen, besonders verzwergte Kolonien. Als zweite Frage wurde die Abgrenzung des wirk-samen Wellenlängenbereichs versucht. Die bisher von anderen Autoren gewonnenen Erfahrungen über die UV-Tötung2-3 der Bakterien und evtl. UV-Mutabilität bei höheren Organismen4 ließ vermu-ten, daß, wie dort, das aktive Gebiet unter etwa 3000 A liegt, welches sich leicht durch ein Glas-filter auslöschen läßt.
2. M e t h o d i k Als Strahlenquel le wurde eine von ihrem Glaskol-
ben bef re i t e HgQ-500-Quecksilberdampflampe von Osram mit zugehör iger Drossel verwendet . In den Versuchen I, II , I I I und IVa ist das Gesamtspek t rum e inges t rah l t worden, im letzten Versuch IVb wurden mit einer Fens te rg lasp la t t e die Strahlen un te r e twa 3000 A weitgehend abgefi l ter t . In 50 bzw. 80 cm ( I I I ) Abstand unter der Lampe stand ein Becherglas mit ebenem Boden von 5,2 cm Durchmesser , in dem sich die zu best rahlende Zellensuspension befand (10 cm3
bei I und III , 5 cm3 bei I I und 7,5 cm3 bei IVa und I V b ) . Während der Bes t rah lung wurde die Intensi-t ä t mit einem Sperrschicht-Photoelement gemessen;
2 A. B a c h e m u. M. A. D u s h k i n , A s tudy of bacter ia l sensi t ivi ty to u l t ravio le t radiat ion. BioL Bull. 69, 109 [1935].
3 P. E. L e a u. R. B. H a i n e s , The bactericidal action of u l t rav io le t l ight. J . of Hyg. 40. 162 [1940].
die W e r t e wurden nach Eichung mit einer Spezial-Vakuumthermozel le * (nach Prof . H a s e , Gehrden) un te r Berücks ich t igung der Exposit ionszeit , der be-s t rah l ten Suspensionsfläche und der gesamten Zell-zahl in der Suspension in erg/Zel le umgerechnet.
Die Suspensionsdichte wurde in einer selbsther-gestel l ten Zählkammer ausgezähl t sowie aus der Koloniezahl in den Kontrol lschalen festgestel l t . Beide Zählungen st immten ausreichend gut überein. Wegen der sehr ger ingen E indr ingungs t i e fe der Strahlen war es notwendig, mit einem aus einem Wechsel-stromsummer gefertigten Rührer fü r dauernde, schnelle und gute Durchmischung der Suspension zu sorgen, wodurch eine stat ist isch gleichmäßige Ver te i lung der eingestrahlten Energie auf alle Zellen erreicht wurde. Zur Gewinnung der einzelnen Dosen der je 5—7 Ex-posi t ionsminuten dauernden Versuche wurde nach je-weils einer gewissen Zeit (meist 1 Min.) die Be-s t r ah lung mit tels einer Blende unterbrochen und 0,5 cm3 der Suspension entnommen, diese Menge mit 5 cm3 sterilem Leitungswasser verdünnt und so weitere Verdünnungen in Zehnerpotenzen hergestel l t . Von den in Vorversuchen als güns t ig f ü r die Auszählung fes tges te l l ten Verdünnungen wurde je ein Tropfen auf eine Pet r i schale mit Nährboden (Agar 2%, Glyce-r in 2,5%, Zi t ronensäure 0.5%, NH 4H 2P0 4 0,3%, MgSO, 0,03%, F e ( N H 4 ) ( S 0 4 ) 2 0,03%, NaCl 0,03%, KOH bis pn = 7,0) gebracht und mit dem Glasspatel brei tge-str ichen. Natür l ich wurden die Ster i l i tä tsbedingungen beim Arbei ten beachtet . Die beimpften Schalen wur -den 2 Tage bis zum Abschluß des Wachstums der an-wachsenden Kolonien bei 30 ° C bebrütet .
Die Zahl der überlebenden Keime pro Tropfen, deren Logar i thmus als Maß f ü r die abtötende Wi r -k u n g der Strahlendosis in der Tabelle not ier t ist, e rgab sich aus der Anzahl Kolonien auf der dichtest bewachsenen, noch gut zählbaren Plat te , dividier t durch die Verdünnung . F e r n e r wurde die Anzahl der weißen Kolonien in allen gut zählbaren, also nicht zu dicht bewachsenen Pla t ten fes tges te l l t sowie die An-zahl al ler sonstigen, von der Norm abweichenden Kolonien (m).
3. V a r i a n t e n k o l o n i e n
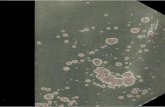
Bei den Varianten handelt es sich zum größten Teil um verzwergte Kolonien von normaler, z.Tl. aber auch von hellerer oder dunklerer Farbe. Ein kleinerer Teil der Abweichenden hatte bei nor-maler Größe eine hellere, selten eine dunklere rote Farbe oder auch abweichende Farbvertei-lung um das Koloniezentrum (helleren oder dunk-leren Nabel). Ein anderer Anteil zeichnete sich durch schlierenartig unregelmäßige Verteilung
4 W. N o e t h 1 i n g u. II. S t u b b e , Die Aus-lösungen von Genmutat ionen durch kurzwel l iges Licht., Z. ind. Abstammungs- Vererbungsl . 72, 378 [19361.
* Hrn. Prof . H a s e sei auch an dieser Stelle f ü r die Über lassung dieses Meßgerätes gedankt.

weißer zwischen roten Partien in der Kolonie aus. Zur vorläufigen Orientierung über die Erb-lichkeit dieser Differenzen gegenüber den norma-len Kolonien wurden stichprobenmäßig aus Kolo-nien der verschiedenen Typen, unter Zwischen-schaltung einer Verdünnung, Tochterplatten aus-gespatelt. Die darauf anwachsenden Kolonien waren bei den Zwergen immer von der Farbe der Elternkolonie; die Größe war nur bei dichterem Bewuchs zwergig, am Rande und an lockeren Stellen normal, was eine Hemmung des Wuchses nur bei Nährstoffmangel oder durch gewisse Stoff-wechselprodukte im Substrat anzeigt, keine Wuchs-minderung unter allen Bedingungen. Aus diesem Grunde wurden zur Feststellung der Mutanten-rate auch nur Platten verwendet, die genügend dicht bewachsen waren, und die Randpartie mit großen Kolonien ausgelassen. Als Index fü r mu-tierte Kolonien wurde nicht die absolute Größe angesehen, sondern eine Verzwergung bei nor-maler Größe der gleichdicht wachsenden Nach-barn.
Die Farbmutanten von Normalwuchs behielten ihre Abweichung in den Tochterplatten ebenfalls, selten fanden sich auch Tochterplatten, bei denen nur ein mehr oder weniger großer Teil den Ab-weichertyp bewahrte, andere Bruchteile jedoch den alten Farbtyp oder einen neuen, helleren oder dunkleren, repräsentierten. Unklar verhielten sich die Tochterplatten der Schlieren-Kolonien: sie be-standen aus einem großen Anteil normalfarbiger und einem sehr erheblichen Bruchteil (z. Tl. bis etwa zur Hälfte) weißer Kolonien, z. Tl. mit roten Sektoren; sie enthielten jedoch nur wenige Schlierenkolonien vom Elterntyp. Demnach stam-men auch die Schlierenkolonien von einer erblich abgeänderten Zelle ab. Jedoch ist wegen des wei-teren Verhaltens des abgeänderten Faktors im Verlauf des Kolonienwachstums noch nicht zu sagen, ob es sich um eine Art sehr mutables Gen
. (kurzlebiges Allel) handeln kann, oder ob viel-leicht andere, noch unbekannte Vorgänge (z. B. irgendwelche Entmischungen von Erbeinheiten oder dergl.) die Beschaffenheit der Tochterkolo-nien verursachen. Hierzu ist ein weiteres Studium nötig.
4. Q u a n t i t a t i v e E r g e b n i s s e u n d F o l -g e r u n g e n
Der Überlebenden-Logarithmus (s. Tab.) er-weist sich bei näherem Zusehen als im Anfangs-
teil etwa linear abhängig von der Strahlendosis!), was ja schon exakt von anderer Seite mehrfach nachgewiesen ist (s. z.B. Anm.3). Die Abweichun-gen von der exakten Kurve im vorliegenden Fall sind bei niederen Dosen durch die nicht sehr ge-naue Verdünnungs- und Tropfenimpfmethode hervorgerufen. Sie spielen für die Feststellung der Mutationsraten keine Rolle. Bei höheren Dosen weichen die gefundenen Kolonienzahlen ( + in der Tab.) jedoch so stark von der anfänglichen Linearität ab, daß hier ein neuer Faktor ein Aus-biegen der Kurve erzeugt haben muß. Sehr wahr-scheinlich ist diese Ursache in der auch im Mikro-skop erkennbaren, seltenen Verklumpung der Zellen zu suchen, die beim Durchlaufen der ersten 4—6 Zehnerpotenzen keine bzw. kaum merkliche Wirkung hat, die bei hohen Dosen aber zu einem Abbiegen der Kurve führ t , wie weiter unten näher ausgeführt wird. Der zufälligen Größe oder Zahl dieser Klumpen im Impftropfen entsprechen die Schwankungen der Keimzahl.
Wie die Tabelle ausweist, hat im angewandten Dosisbereich keine nachweisbare Vermehrung der Mutation r — w stattgefunden. Eine Homogenitäts-prüfung der Häufigkeiten bei den verschiedenen Dosen ergibt in Versuch I : x«= H A P = 6,5%; I I : 4 = 6,7, P = 1 6 % ; I I I : 4 = 18, P = 0 , 6 % ; IVa: 4 = 5,8, P = 12%; IVb : 4 = 1,0, P = 6 0 % * . Mit Ausnahme von I I I ist also eine Abhängigkeit der w-Mutationen von der Dosis nicht nachweisbar. Eine Betrachtung der Einzelzahlen von I I I zeigt aber, daß auch dort von einer klaren Dosisabhängig-keit keine Rede sein kann, da nur der 5. Wert mit 0,68% w herausfällt, während bei höheren Dosen wieder höhere Prozentsätze gefunden wurden. Der Homogenitätstest unter Ausschluß dieses Wertes ergibt denn auch x? = 7,7, P = 17%, also keine Dosisabhängigkeit. Die Ursache für das Herausfallen des 5. Punktes ist wohl in einem im
(ai— niPi)2 * Dabei ist y ; — 2 das übliche Maß der
Gesamtabweichung der Versuchswer te von der zu p rü fenden Hypothese, im Fal l des Homogenitäts-tes tes also von der durchschnit t l ichen Häuf igkei t p = S ß j / E n ; der Mutanten {at = Mutantenzahl, ni = Zahl ausgewer t e t e r Kolonien im Versuch i). Aus ergibt sich dann die Wahrscheinlichkeit P dafür , daß in weiteren Versuchen die Abweichung überschrit-ten wird. Tafeln h ie r fü r bei P ä t a u , Z.Vererbungs-lehre 80, 558 [1942]. Bei I u. I I wurden die Dosis-klassen mit 0 bzw. 1 wj-Kolonie mit den vorhergehen-den vere in ig t .

Versuch Dosis D t in
1 0 " 4 erg/Zelle
I I
I I I
IVa
I V b (Glas-filter)
0 (Kontr.) 4,4 8,8
13.5 18,0 22,5 26,8 81,0 0 (Kontr.) 8,55
12,4 16.4 28.5 41,2
0 (Kontr.) 2,42 4,91 7,17 9,37
13,60 15,65
0 (Kontr.) 7,1
15,0 22,8
6,8 14,0 32,0
Tötung / Keimzahl
^ \ Tropfen
8,2 6,8 5.1 3 . 3 2.4 2,6+ 2,8+ 2,0+ 8.2 5,2 4.1 4,1+ 3,6+ 2,9+
8.2 8,0 7.2 6.3 6,1 3,8 2,8 8,1 6,2 3.7 2.5 8.08 8,0 8.04
w-Rate m-Rate aJni = %
39/1493 14/76H 42/1336 13/334 : 16/528 21/485 17/8U0
0/95
40/1443 34/1697 33/1621
1/162 : 22/1041 22/1503:
44/1699 21/1129 22/1504 • 43/2665 10/1457 11/670 : 12/590 :
33/1339 : 35/1819:
7/572 : 5/395 :
38/1452: 31/1055: 43/1403 :
:2,62 : 1,83 : 3,15 : 3,90 :3,03 4,34 2,12
= 0
: 2,78 : 2,01 :2,04 :0,62 : 2,20 : 1,46
: 2,58 : 1,86 : 1,46
1,61 0,68
: 1,64 :2,03
2,46 :2,t>5 1,22
: 1,27 2,62 3,22 3,06
22/1477 1.50 34/714 : 4,75 67/1336 5,04 30/313 - 9,60 38/500 = 7,60 42/485 - - 8,67+ 73/bOO = 9,15+
7/95 = 7,37+ 26/1443 = 1,80 86/1697 = 5,07
108/1621 = 6,67 12'1K2 = 7,42+
105/1041 = 10,1 + 106/1503 = 7,05+ 21/1699 - 1,24 24/1129 - 2,12 47/1504 3,13
107/2665 4,02 78/1457 — 5.34 44/670 = 6,55 55/590 = 9,33 14/1339 = 1,05 70/1319 5,31 49/572 - 8,58 41/395 — 10,38 19/1452 = 1,31 13/1055 - 1,23 16/1403 1,14
Tab. Ergebnisse der UV-Bes t rah lungen von Bact. prodigiosum. at = Var ian tenzahl unter nt geprüf ten Kolonien in Dosis-Nummer i.
Impf tropfen befindlichen Klumpen mit (wegen ge-meinsamer Abstammung) überwiegend r-Zellen zu suchen. Die Erhöhung der Keimzahl durch diesen Klumpen ist auch deutlich im etwas zu hohen Überlebenden-Logarithmus von 6,1 gegen erwartungsgemäß etwa 5,2 zu erkennen.
Betrachten wir die gefundenen Häufigkeiten der andern registrierten Varianten (m ) , so fällt die Abhängigkeit von der Strahlendosis sofort ins Auge. Die Variantenraten steigen etwa linear mit der Dosis an und biegen ungefähr bei der Dosis, bei welcher sich an der Überlebendenkurve die Verklumpung deutlich auswirkt, zu einem un-regelmäßig abfallenden Verlauf um ( + in der Tab.). Dieses Umbiegen und schwankende Abfal-len dürfte wohl ebenfalls eine Auswirkung des Zusammenhaftens von Zellen in Klumpen sein.
Bei niederen Dosen sterben und mutieren die Einzelzellen, was zunächst die Mutantenrate an-steigen läßt, während die Überlebendenkurve fällt. Bei höheren Dosen wird die Zahl der Überleben-den-Einzelzellen allmählich geringer, so daß zu-
nehmend geringere Verdünnungen ausreichen, um eine nicht zu dicht besiedelte Agarplatte für die Auszählung der Varianten zu erhalten. Die ge-ringere Verdünnung erhöht aber die Wahrschein-lichkeit, Zellklumpen in den Impftropfen zu be-kommen. In diese Klumpen vermag der wirksame Strahlenanteil wegen seiner starken Absorption in den Zellen nur oberflächlich einzudringen. Es wird als nur eine Außenschicht von Zellen ver-ändert und abgetötet, im Inneren bleibt ein mehr oder weniger großer Teil wegen dieser Schutz-schicht unerreicht und unbeeinflußt, auch bei Dosen, die praktisch alle Zellen der Außenschicht abgetötet haben. Beim Ausspateln werden die Klumpen zerdrückt und die Zellen vereinzelt, so daß bei hohen Dosen der größte Teil der an-wachsenden Kolonien aus den unversehrten Zel-len des Klumpenzentrums hervorgeht. Damit sinkt die Variantenrate wieder ab unter Schwankungen, die von der variablen Größe und Zahl der Klum-pen im Impftropfen verursacht werden. Die Über-lebendenkurve dagegen strebt asymptotisch unter

entsprechenden Schwankungen einem konstanten Wert zu.
Für den Vergleich mit dem UV-Mutations-mechanismus der höheren Organismen kann also nur der untere Teil der aufsteigenden Varianten-kurve verwendet werden, denn nur er gibt die Dosisabhängigkeit des an Einzelzellen von Prodi-giosum ablaufenden Variationsprozesses wieder.
Nach den bisherigen Erfahrungen an Tieren5
und Pflanzen6 folgen die durch UV-Strahlen er-zeugten Mutationen einer Eintrefferkurve. Die durch UV-Bestrahlung hervorgerufene Verände-rung der Erbvariationsrate bei Bact. prodigiosum war also daraufhin zu prüfen, ob ihre Dosisab-hängigkeit ebenfalls durch solche Eintreffervor-gänge verstanden werden kann. Hierzu wurde ein statistischer Vergleich der in den einzelnen Ver-suchen erhaltenen Variantenhäufigkeiten ht = a j t ^ mit der den unteren Teil der Eintrefferfunktion repräsentierenden Kurve p,- = c -f ,u D. durchge-führt (at- = Variantenzahl unter ni geprüften Kolonien in Dosis-Nummer i; pt — zu erwartende Variantenwahrscheinlichkeit, c = Variantenpro-zentsatz in der Kontrolle, p. = Mutationskon-stante, D i = eingestrahlte Energiedosis). Die deutlich von der Verklumpung beeinflußten Ver-suchspunkte ( + ) wurden ausgeschieden. Zu-nächst mußten die auf Grund des empirischen Zahlenmaterials wahrscheinlichsten Werte von c und n gesucht werden, wozu sich die „method of maximum likelihood"7 anbot. Unter der zu-treffenden Annahme kleiner p-Werte (Poisson-Verteilung) ergab sie die 2 Gleichungen:
(la,b) Wegen der Umständlichkeit der Errechnung der beiden Parameter c und n hieraus wurden zu-nächst die Näherungsformeln
^ (2 at D.) (2 n.) (2 tf.) (2 ni Z>.)
{2n.D:i)—{2n.Dy
U n d ~ ( 2 ^ ) — J i ( 2 n . D ) c = ^ (2b) verwendet.
Aus den für die Einzelversuche I—IVa erhalte-nen IT-Werten wurde ein gewogenes Mittel
5 I. S e l l - B e l e i t e s u. A. C a t c h , Mutations-aus l . sung durch u l t rav io le t tes Licht bei Drosophila I. Z. ind. Abstammungs-, Vererbungs l . 80, 551 [1942].
H. = 0,425-10—2 Mutationen/10-4 erg gebildet und mit diesem die Einzel-c nach (2b) errechnet. Die für die Einzelversuche erhaltenen cl = 1,65%, T n = 1,53%, 7 m = 1,19% und c 1 V a = 1,64% sowie
das gemeinsame £ = 0,425% wurden dann zur Errechnung der nach der Eintrefferhypothese zu erwartenden Mutantenwahrscheinlichkeiten pi =Tc + pl Di für alle Versuchspunkte der ganzen Versuchsreihe I—IVa verwendet und diese p. mit Hilfe der x2"Methode auf Übereinstimmung mit den entsprechenden, empirischen Mutanten-häufigkeiten h t geprüft. So wurde für Versuch I ein x® = 8,5 mit einer Überschreitungswahrschein-lichkeit P = 7,5%, fü r II ein x? = 0,70, P = 40%, für I I I ein x* = 2,5, P= 77%, für IVa ein x3= 4,7, P— 9% und für alle Versuche zusammen ein 16,6 m i t P = 28% gefunden. Die P-Werte zeigen an, daß die Eintrefferhypothese mit den beobachteten Mutantenhäufigkeiten in allen Versuchen ver-träglich ist. Da bei Verwendung der exakten For-meln ( l a , b) sich noch höhere P-Werte, also noch bessere Übereinstimmung mit der Eintreffer-funktion, ergeben hätte, konnte auf eine Prüfung mittels der aus diesen Formeln erhaltenen c und \l verzichtet werden.
Nachdem durch Untersuchung der Dosis-Effekt-Funktion ein Unterschied zwischen den durch Ultraviolettlicht ausgelösten Mutationen höherer Organismen5-6 und dem Variationspro-zeß des Bact. prodigiosum nicht aufgefunden wer-den konnte, ergab sich die weitere Frage, ob diese Bakterienvarianten durch das gleiche oder ein anderes Wellenlängengebiet als das bei höheren Organismen wirksame ausgelöst würde. Da ein UV-Monochromator ausreichender Leistung nicht zur Verfügung stand, und da andererseits das fü r den Genmutationsprozeß wirksame UV-Ge-biet der Wellenlängen unter etwa 3000 A sich durch gewöhnliches Glas leicht abfiltrieren läßt, wurde bei Versuch IVb in den Strahlengang eine Glasplatte eingeschaltet, die nur Wellenlängen oberhalb etwa 3000 A durchließ. Zum Vergleich diente der Versuch IVa mit der gleichen Zellen-suspension und der gleichen Versuchsanordnung, wobei nur das Filterglas weggelassen war. Die von der 1,4 mm dicken Glasplatte durchgelassene
0 L . I. S t a d 1 e r u . F . U. U b e r , Genetic effects of u l t r av io l e t rad ia t ion in Mays IV. J . Genetics 27, 84 [1942],
T R. A. F i s h e r , Stat is t ical methods. London 1934.

Gesamtspektralenergie wurde mit der Vakuum-thermozelle gemessen und betrug 60% ; das beson-ders für sichtbares Licht empfindliche Sperr-schichtphotoelement, welches die Intensitäts-schwankungen während des Versuchs kontrol-lierte, zeigte 90% Durchlässigkeit an. Die in der Tabelle mitgeteilten Zahlenwerte fü r I V a und IVb zeigen eindeutig, daß selbst bei so hohen Dosen wie 32-10 - 4 erg/Zelle weder eine merk-liche Abtötung noch eine erweisbare Veränderung der Variantenrate durch das X-Gebiet oberhalb 3000 A stattgefunden hat. Damit ist die Bakterien-variation durch UV auf das gleiche spektrale Ge-biet beschränkt, welches für die Bakterientötung2
und die Genmutationen4-0'8'10 schon bekannt sind. Ob für den ersten Vorgang ebenso wie für die beiden letzten2-4'6'8'10 das Wirkungsspektrum mit dem Absorptionsspektrum der Thymonucleinsäure übereinstimmt, wird zwar durch den Versuch IV wahrscheinlich gemacht, kann aber nur durch Anwendung monochromatischen Lichtes bei der Analyse sicher entschieden werden.
So sind durch die Feststellungen der UV-Dosis-Effekt-Funktion und der Begrenzung des Wir-kungsspektrums auf das mittlere UV unterhalb 3000 A zu dem früher mitgeteilten Indizium der spontanen Variation1 zwei weitere Indizien dafür beigebracht worden, daß die bei Bact. prodigio-sum stattfindenden Erbveränderungen wenigstens z. Tl. durch Faktoren hervorgebracht werden, die mit den Genen der höheren Organismen in ihrer Abänderungs- und damit in ihrer Bauweise ver-gleichbar sind. Diese Arbeitshypothese kann noch weiter gestützt werden durch Berechnungen über die für den Mutationsprozeß notwendigen abso-luten Energiemengen pro Zelle bei verschiedenen, in der Literatur besprochenen Organismen. Wir fanden für unser Bacterium eine Mutationskon-stante n = 0,43%/10-4 erg/Zelle, also für 1% Mutationen 2,4-10-4 erg/Zelle. Die Spermatozoi-den von Sphaerocarpus8 ergeben etwa 25% Mu-tanten bei 2-105 erg/cm2, was bei einer Größen-ordnung dieser Zellen von etwa 10-7 cm2 etwa 8-10- 4 erg/Spermatozoid für 1% Mutationen er-gibt. Bedenkt man die rohe Rechnung, die wahr-scheinlich verschiedenen Absorptionsverhältnisse
8 E. K n a p p u. H. S c h r e i b e r , Quant i ta t ive Analyse der mutat ionsauslösenden W i r k u n g mono-chromatischen UV-Lichtes. Naturwiss . 27, 304 [1939].
0 F. M. U b e r , Ul t ravio le t spect rophotometry of Zea mays pollen with the quartz microscope. Amer. J . Bot. 26, 799 [1939].
im Bacterium und Spermatozoid sowie die Ver-schiedenartigkeit beider Zellen, ausgelöste Muta-tionen sichtbar zu manifestieren, so ergibt sich eine überraschende Übereinstimmung der Größen-ordnungen. Sie dürfte ihre Ursache wohl in der Ähnlichkeit des Primärvorgangs der Erbände-rung beider Zelltypen haben sowie in dem nicht sehr erheblichen Verlust an Strahlungsenergie beim Durchgang durch die den Erbfaktoren vor-gelagerten Plasmaschichten. Denn diese plasma-tische Filterschicht ist wegen der Kleinheit des Bacteriums bzw. der praktischen Nacktheit des Spermatozoidenkernes in beiden Fällen nur dünn. Wesentlich dicker ist sie in Sporen- oder Pollen-körnern, da in diesen die Strahlung erst die stark absorbierende Zellwand sowie das Zytoplasma durchsetzen muß, bis sie den Kern erreicht. Nach den Absorptionsmessungen am Mais-Pollen9 ge-nügt schon die Zellwand + 16 n Plasmaschicht, um von den wirksamen Strahlen etwa 90% zu absorbieren. Diese Absorptionsunterschiede sowie wohl auch die verschiedenen Manifestationsfähig-keiten fü r Mutationen und evtl. unterschiedliche Strahlenempfindlichkeit der Chromosomen (Nu-cleinsäuregehalt!) dürften sich in dem höheren Energieverbrauch pro bestrahlter Zelle ausdrük-ken, den wir bei den Sporen von Trichophyton10
als etwa 1,3 erg/Spore und beim Pollen von Antirrhinum4 als etwa 0,7 erg/Pollenkorn für 1% Mutationen finden.
Betrachten wir zum Schluß noch den Abtötungs-vorgang auf Grund der vorliegenden Zahlen, so können wir unter der sicher zutreffenden An-nahme der exponentiellen Tötungsfunktion N =
(lgVo-lgV.) Ar
0 e—TD die TötungskonstanteT = 2,30 — ,T , Keimzahl ^ _ »
errechnen (lgA0 = lg T r 0 f e n ' bei Dosis D = 0, lg iV. desgleichen bei Dosis D.). Wir erhalten als Durchschnitt aller Versuche T = 0,66 Zelltötun-gen/erg pro Zelle. Sehen wir die Tötung einer Bakterienzelle (d.h. Unfähigkeit zur Kolonien-bildung) durch ein UV-Quant als Letalmutation an, so würde dieser Prozeß etwa t/y. = 155-mal häufiger vorkommen als eine Mutation, die als Abänderung der Koloniebeschaffenheit sichtbar wird. Damit müssen also Gene, deren Änderung zur Zell-Letalität führt, ebenfalls in etwa 155-faeher Anzahl gegenüber den „sichtbaren" Genen
10 C. W. E m m o n s u. H o l l a e n d e r , The action of u l t rav io le t radiat ion on dermatophytes. I I . Amer. J. Bot. 26, 467 [1939].

in der Prodigiosum-Zelle enthalten sein. Verglei-chen wir das so erhaltene Verhältnis „sichtba-rer" / „letaler" Gene von V155 = 0,65% bei Prodi-giosum mit dem im X-Chromosom von Droso-phila11 gefundenen von 7—8% „sichtbarer" C1B-Gene, so ergibt sich eine Differenz. Sie könnte verstanden werden auf Grund der so unterschied-lichen Organisation beider Lebewesen. Denn die Bakterienzelle hat wohl einfach nicht die „Aus-drucksfähigkeit" fü r Mutationen wegen ihres ein-facheren Baus, der auf viele Erbänderungen, welche z. B. zu Verschiebungen des Zellstoffwech-sels führen, nur mit bis zur Letalität geminder-tem Wachstum antworten kann. Bei Drosophila dagegen werden sich demgegenüber eine große Zahl Genänderungen nicht in allen den verschie-denen Zelltypen gleichartig ungünstig auswirken, wodurch eine Ausbalancierung der Vitalitätshem-mung mancher mutierter Gene durch die Arbeits-teilung der Zellen im vielzelligen Organismus er-folgen kann. Andererseits können manche Gen-änderungen wegen der Zelltypenunterschiede im Tier sich morphologisch, also sichtbar, auswir-ken, die den einzigen Zelltyp des Bacteriums nicht leicht sichtbar zu ändern vermögen und also un-erkannt bleiben. Sicherlich ist die große phylo-genetische Wandlungsfähigkeit der Vielzeller mit auf diese Fähigkeit, vitalitätshemmende Mutatio-nen durch Zellarbeitsteilung auszubalancieren, so-wie auf die größere Zahl von Möglichkeiten, Muta-tionen als sichtbare Bauunterschiede auszu-drücken, zurückzuführen.
In der ersten Mitteilung1 über die spontanen Prodigosium-VariaLtionen war die Annahme aus-gesprochen worden, daß der Übergang r — w durch Mutation nur weniger Gene, vielleicht nur eines solchen, zustande kommt. Durch die UV-Bestrahlung hatte sich eine Erhöhung der to-Häu-figkeit nicht gezeigt; falls sie überhaupt stattfin-det, ist sie von den statistischen Schwankungen überdeckt und dürfte bei der höchsten ange-wandten Dosis maximal etwa 1% betragen. Für unsere übrigen Mutationen (m-Rate) fanden wir bei der höchsten Dosis etwa 8% Erhöhung der Häufigkeit. Sie ist sicher eine „Gensummenrate", d. h. durch die Summierung der Mutationen vieler Gene zustande gekommen. Das Einzelgen hätte
11 N. W. T i m o f e e f - R e s s o w s k y u. M. D e l -b r ü c k , St rahlengenet ische Versuche über sichtbare Mutat ionen und die Mutabil i tät einzelner Gene bei D7-osophila melanogaster. Z. ind. Abstammungs-, Ver-e rbungs l . 71, 322 [1936].
bei der vorliegenden Höchstdosis demnach noch keine nachweisbare Zunahme der Mutationsrate ergeben. Dies läßt auch das gefundene Verhalten der lü-Rate verstehen, denn, wie wir sehen, wird der Übergang r — w wahrscheinlich nur durch ein spontan mutables Gen oder wenige solche hervor-gebracht. Setzen wir für das (oder die) io-Gen(e) eine in bezug auf alle anwesenden Gene durch-schnittliche Strahlenempfindlichkeit voraus, so ist die untere Grenze der an der m-Gensummenrate beteiligten Genzahl von der Größenordnung 8, in Wahrheit dürften wesentlich mehr, vielleicht 101—102 Gene an den „sichtbaren" Kolonieände-rungen beteiligt sein. Unter Berücksichtigung des oben erwähnten Verhältnisses der Letal- zu den Vitalgenen und unter der Annahme gleicher durch-schnittlicher UV-Empfindlichkeit beider Gensor-ten kommt man so zu einer unteren Grenze der in der Prodigiosum-Zelle vorhandenen Zahl von etwa 1200 durch UV beeinflußbaren Genen. Die wirkliche Zahl dürfte mehrmals größer sein, viel-leicht zwischen 103 und 104 liegen. Da für das X-Chromosom von Drosophila eine Genzahl von etwa 3300 auf Grund der Untersuchungen mit ionisierenden Strahlen geschätzt wird12, erscheint das Ergebnis der UV-Versuche bei den relativ ein-fach gebauten Zellen des Bact. prodigiosum plau-sibel. Andererseits ist die Zahl der Prodigiosum-Gene aber etwas zu hoch, wenn man die Ergeb-nisse über die Abtötung von Bacterium coli durch ionisierende Strahlen13 vergleicht, welche etwa 250 Letalgene in diesem Bacterium anzeigen. Worauf der Unterschied beruht, ist ohne weitere Versuche schwer zu entscheiden. Vielleicht liegt ein wirklicher Unterschied im Gengehalt von Bact. coli und Bact. prodigiosum vor, oder ist das Verhältnis von Letal- zu Vitalgenen in beiden wesentlich verschieden; vielleicht sind die L e a -schen 250 Letaleinheiten auch gar keine Einzel-gene, sondern (chromosomenhafte?) Gruppen sol-cher. Vielleicht (und am wahrscheinlichsten) ist aber die UV-Analyse zur genaueren Bestimmung der Genzahl schlecht geeignet wegen zu großer Unterschiede in der durchschnittlichen UV-Strahlen - Empfindlichkeit der bei Prodigiosum untersuchten Gengruppen.
12 K. G. Z i m m e r u. N. W. T i r a o f e e f - R e s -s 0 w s k y , Über einige physikal ische Vorgänge bei der Aus lösung von Genmutat ionen durch St rahlung. Z. ind. Abstammungs-, Vererbungs l . 87, 353 [1942].
13 P. E. L e a , Actions of Radiat ions on l iving cells, 1946. Nach f reundl icher Mit tei lung von Hrn. Dr . F r i e d r i c h - F r e k s a , Tübingen.













![HRN Entladungslampen 2013 - HANS RAUM · 2019. 11. 19. · HID-PV ). Nur mit "End of Life" Abschaltung (IEC61167, IEC 62035). Artikel-Nr. Lichtfarbe Watt Sockel Lichtstrom [lm] Lichtausbeute](https://static.fdokument.com/doc/165x107/60e80f5cf0249149130ab6e0/hrn-entladungslampen-2013-hans-raum-2019-11-19-hid-pv-nur-mit-end.jpg)


