VANILLIN MIT PHENOLISCHEN VERBINDUNGEN Ezfn.mpdl.mpg.de/data/Reihe_B/27/ZNB-1972-27b-0707.pdf · 8....
7
This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution 4.0 International License. Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschung in Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung der Wissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht: Creative Commons Namensnennung 4.0 Lizenz. VANILLIN MIT PHENOLISCHEN VERBINDUNGEN 707 gleichen Ladung nicht in Reaktion treten können. In beiden Fällen wird also ein Angriff am Ring ver- hindert. Mit dem HMO-Modell, das sich bei unserer Pro- blemstellung als nützlich erwiesen hat, können nun weitere Pflanzenphenole berechnet werden. Es lassen sich mit den ermittelten Ladungsdichten mögliche Reaktionszentren angeben. Eine experimentelle Überprüfung der Ergebnisse kann durchgeführt wer- den. Experimenteller Teil Eingesetzte Lösungen: 0,1 M Substanz, 0,1M Vanil- lin, 5 ml konz. H2 SO4 /l00ml Lösung; Lösungsmittel: Methanol. 5 ml Substanz-, 5 ml Vanillin- und 0,4 ml H2S04- Lösung wurden auf 20 ml aufgefüllt und mit dem Spektralphotometer SP 800 A von Unicam vermessen. Die Extinktionswerte beziehen sich auf eine Schicht- dicke von 0,5 cm. Phenol, Brenzcatechin und Hydrochinon zeigen keine Färbung. 1.3.5-Trimethoxybenzol, 1.2.4-Tri-, 1.2.4.5-Tetra- hydroxybenzol, Dinitro- und Nitrophloroglucin wurden nach Literaturangaben dargestellt. Eine vorzügliche 1 W. HEIMANN, K. HERRMANN u. G. FEUCHT, Z. Lebensmit- tel-Unters. u. -Forsch. 145, 20, 199 [1971]. 2 W. G. BERGER u. K. HERRMANN, Z. Lebensmittel-Unters, u. -Forsch. 146, 266, 275 [1971]. 3 R. RIBEREAU-GAYON, Les Composes Phenoliques Des Vegetaux, S. 49, Dunod, Paris 1968. 4 J. L. GOLDSTEIN u. T. SWAIN, Phytochem. 2, 375 [1963]. 5 T. SWAIN u. J. L. GOLDSTEIN, in: J. B. PRIDHAM, Methods in Polyphenol Chemistry, S. 137, Pergamon Press, Oxford 1963. Arnax [nm] E Phloroglucin 485 1,33 Catechin 498 0,72 Phloroglucincarbonsäure 483 0,56 Nitrophloroglucin 515 0,46 1,2,4-Trihydroxybenzol 534 0,10 Resorcin 515 0,04 2-Methylresorcin 522 0,025 1.2.4.5-Tetrahydroxybenzol 536 0,015 Pyrogallol 510 0,010 Reinigung des 1.2.4-Trihydroxybenzols gelang über Polyamid mit dem Fließmittel Benzol — Methanol —Eis- essig (10 : 2 : 1). Zu den HMO-Berechnungen Parameter nach A. STREITWIESER 12 C = 0 ho = 1,0, he =o = 1,0 C-OH ho = 2,5, hc- -o=0,8 -O- ho = 3,0 NO 2 ho = 1,0, = 2,0, &NO = 0,7 COOH ho = 2,4, he = 0,8, kc-0 = 0,8 CH 3 hcH3 = -0,1 Beim C-Atom der Carboxylgruppe ist ein indukti- ver Effekt nach D e w a r berücksichtigt worden. 0 R. GIULIANO, Ann. Chim. applic. 29, 86 [1939]. 7 J. C. PEW, J. Amer. chem. Soc. 73, 1678 [1951]. 8 J. L. GOLDSTEIN u. T. SWAIN, Phytochem. 2, 376 [1963]. 9 F. WENZEL, Mh. Chem. 34,1915 [1913]. 10 G. BRIEGLEB, Elektronen —Donator—Acceptor-Komplexe, Springer, Berlin-Göttingen-Heidelberg 1961. 11 H. HAGEMANN, Dissertationsarbeit, TU Hannover 1971. 12 A. STREITWIESER, JR., Molecular Orbital Theory for Orga- nic Chemists, John Wiley, New York-London 1961.
Transcript of VANILLIN MIT PHENOLISCHEN VERBINDUNGEN Ezfn.mpdl.mpg.de/data/Reihe_B/27/ZNB-1972-27b-0707.pdf · 8....

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
VANILLIN MIT PHENOLISCHEN VERBINDUNGEN 7 0 7
gleichen Ladung nicht in Reaktion treten können. In beiden Fällen wird also ein Angriff am Ring ver-hindert.
Mit dem HMO-Modell, das sich bei unserer Pro-blemstellung als nützlich erwiesen hat, können nun weitere Pflanzenphenole berechnet werden. Es lassen sich mit den ermittelten Ladungsdichten mögliche Reaktionszentren angeben. Eine experimentelle Überprüfung der Ergebnisse kann durchgeführt wer-den.
Experimenteller Teil
Eingesetzte Lösungen: 0,1 M Substanz, 0,1M Vanil-lin, 5 ml konz. H2SO4/l00ml Lösung; Lösungsmittel: Methanol.
5 ml Substanz-, 5 ml Vanillin- und 0,4 ml H2S04-Lösung wurden auf 20 ml aufgefüllt und mit dem Spektralphotometer SP 800 A von Unicam vermessen. Die Extinktionswerte beziehen sich auf eine Schicht-dicke von 0,5 cm.
Phenol, Brenzcatechin und Hydrochinon zeigen keine Färbung.
1.3.5-Trimethoxybenzol, 1.2.4-Tri-, 1.2.4.5-Tetra-hydroxybenzol, Dinitro- und Nitrophloroglucin wurden nach Literaturangaben dargestellt. Eine vorzügliche
1 W . HEIMANN, K . HERRMANN u . G . FEUCHT, Z . L e b e n s m i t -tel-Unters. u. -Forsch. 145, 20, 199 [1971].
2 W . G . BERGER u . K . HERRMANN, Z . L e b e n s m i t t e l - U n t e r s , u . -Forsch. 146, 266, 275 [1971] .
3 R. RIBEREAU-GAYON, Les Composes Phenoliques Des Vegetaux, S. 49, Dunod, Paris 1968.
4 J . L . GOLDSTEIN u . T . SWAIN, P h y t o c h e m . 2 , 3 7 5 [ 1 9 6 3 ] . 5 T . SWAIN u . J . L . GOLDSTEIN, i n : J. B . PRIDHAM, M e t h o d s
in Polyphenol Chemistry, S. 137, Pergamon Press, Oxford 1963.
Arnax [nm]
E
Phloroglucin 485 1,33 Catechin 498 0,72 Phloroglucincarbonsäure 483 0,56 Nitrophloroglucin 515 0,46 1 ,2 ,4-Trihydroxybenzol 534 0,10 Resorcin 515 0,04 2-Methylresorcin 522 0,025 1.2.4.5-Tetrahydroxybenzol 536 0,015 Pyrogallol 510 0,010
Reinigung des 1.2.4-Trihydroxybenzols gelang über Polyamid mit dem Fließmittel Benzol — Methanol —Eis-essig (10 : 2 : 1).
Zu den HMO-Berechnungen Parameter nach A. S T R E I T W I E S E R 1 2
C = 0 ho = 1,0, he =o = 1,0 C - O H ho = 2,5, hc- - o = 0 , 8 - O - ho = 3,0 NO2 ho = 1,0, = 2,0, &NO = 0,7 COOH ho = 2,4, he = 0,8, kc-0 = 0,8 CH3 hcH3 = - 0 , 1
Beim C-Atom der Carboxylgruppe ist ein indukti-ver Effekt nach D e w a r berücksichtigt worden.
0 R. GIULIANO, Ann. Chim. applic. 29, 86 [1939] . 7 J. C. PEW, J. Amer. chem. Soc. 73, 1678 [1951] . 8 J . L . GOLDSTEIN u . T . SWAIN, P h y t o c h e m . 2 , 3 7 6 [ 1 9 6 3 ] . 9 F. WENZEL, Mh. Chem. 34 ,1915 [1913] .
10 G. BRIEGLEB, Elektronen —Donator—Acceptor-Komplexe, Springer, Berlin-Göttingen-Heidelberg 1961.
11 H. HAGEMANN, Dissertationsarbeit, TU Hannover 1971. 12 A. STREITWIESER, JR., Molecular Orbital Theory for Orga-
nic Chemists, John Wiley, New York-London 1961.

7 0 8 W. KÖHNLEIN UND F. MÖNKEHAUS
Experimentelle Hinweise auf intramolekulare Energieleitung in der hybriden D N A von Bacillus subtilis nach Bestrahlung
mit langwelligem UV-Licht Experimental Evidence for Intramolecular Energy Transfer in Hybrid DNA
of B. subtilis after Irradiation with Long Wavelength UV
W . KÖHNLEIN u n d F . MÖNKEHAUS
Institut für Strahlenbiologie der Universität Münster
(Z. Naturforsch. 27 b, 708—713 [1972] ; eingegangen am 20. März 1972)
Investigations on hybrid DNA of B. subtilis after irradiation with long wavelength UV (313 nm) indicate that single strand breaks are produced in the T-strand. These lesions are not observed in normal DNA. Since the action of diffusible photoproducts could be excluded the occurrence of single strand breaks in the T-strand may be due to intramolecular energy transfer. Measuring the breakage rates for the T- and B-strand by employing analytical ultracentrifugation it was found that up to 5% of the energy absorbed in the B-strand can be transferred to the T-strand and causes there single strand breaks. The breakage rates per e r g - m m - 2 for single strand breaks after long wavelength UV are determined to be AT = 0 . 5 - 1 0 - 4 and AB = 10-10~ 4 breaks/106 Dalton for the T- and B-strand of hybrid DNA respectively.
I. Einleitung
Wird Thymin in der DNA durch das Basen-analog Bromuracil (BU) 1 ersetzt, so beobachtet man in verschiedenen Systemen eine erhöhte Empfind-lichkeit der substituierten DNA gegenüber UV-Be-strahlung. Diese Steigerung der Empfindlichkeit ist stark von der gewählten Wellenlänge abhängig 2~4 . In einer Reihe von Arbeiten 5 - 8 wurden verschie-dene Mechanismen zur Erklärung der beobachteten Empfindlichkeitssteigerung vorgeschlagen und unter-sucht.
Während in normaler DNA nach UV-Bestrahlung Pyrimidindimere die biologisch wichtigsten Photo-produkte sind, ist in BU-substituierter DNA die Ent-stehung von Einzelstrangbrüchen von entscheidender Bedeutung 9.
Nimmt man an, daß in hybrider DNA der T-Strang wie in TT-DNA 1 und der B-Strang wie in BB-DNA inaktiviert wird, so ist eine Dosiseffekt-kurve für die TB-DNA zu erwarten, die dem arith-metischen Mittel der Dosiseffektkurven von TT- und BB-DNA entspricht. Experimentell wurde jedoch von verschiedenen Autoren gefunden 10' n , daß die biologische Aktivität stärker als nach der Hypothese der unabhängigen Inaktivierung des T- und B-Stran-ges abnimmt. Dies deutet darauf hin, daß die An-
Sonderdruckanforderungen an Dr. W. KÖHNLEIN, Insti-tut für Strahlenbiologie, D-4400 Münster, Hittorfstr. 17.
Wesenheit des BU in einem Strang zu einer zusätz-lichen Inaktivierung des thyminhaltigen Stranges während der Bestrahlung führt.
Wir bezeichnen diesen Vorgang als „Intramole-kulare Energieleitung". Es ist das Ziel der vor-liegenden Arbeit, quantitativ zu untersuchen, ob durch intramolekulare Energieleitung auch Einzel-strangbrüche im T-Strang hybrider DNA gebildet werden, wobei die Bestrahlungsdosis im biologisch interessanten Bereich liegen sollte. Dabei ist natür-lich auszuschließen, daß diese Einzelstrangbrüche durch die Wirkung reaktiver diffusibler Photopro-dukte entstanden sind. Es ist ferner zu vermeiden, daß durch die benutzte Strahlung schon primär Einzelstrangbrüche in TT-DNA und somit auch im T-Strang von TB-DNA erzeugt werden. Aus diesem Grund wird langwelliges UV (313 nm) als Strahlen-quelle benutzt, das keine Veränderungen in TT-DNA hervorruft. Schließlich ist darauf zu achten, daß das biologische System, mit dem diese Untersuchungen durchgeführt werden sollen, folgende Bedingungen erfüllt:
1. Die biologische Funktion der in vitro bestrahl-ten DNA sollte in vivo getestet werden können.
2. Die BU-Inkorporation in die DNA muß einfach und reproduzierbar sein. Die Ausbeute an hybri-der und doppelt markierter DNA soll hoch sein. Es darf kein BU im T-Strang der hybriden DNA sein.

INTRAMOLEKULARE ENERGIELEITUNG IN DNA VON BACILLUS SUBTIL1S 7 0 9
3. Die BU-DNA sollte möglichst homogen bezüglich des Mol.-Gew. sein.
4. Die BU-DNA sollte möglichst homogen bezüglich der Dichte sein. Die BU-Substitution soll mög-lichst vollständig sein.
Ein System, das alle 4 Bedingungen erfüllt, ist nur schwer realisierbar. Die transformierende DNA von Bacillus subtilis erfüllt die Bedingungen 1, 2 und 4 sehr gut. Es wurden daher zunächst Unter-suchungen über intramolekulare Energieleitung an Bacillus subtilis DNA durchgeführt.
II. Materialien und Methoden
1. Bakterien Zur Herstellung von normaler und BU-haltiger DNA
wurden ausschließlich thyminauxotrophe Mutanten von Bacillus subtilis verwendet, nämlich die Mutante B. subtilis 23 (thy-) und die Mutante B. subtilis 19-8 (thy~, try", met"). Für Transformationsexperimente wurden als Akzeptor sowohl B. subtilis 25 (try-, his-) als auch die polyauxotrophe Mutante B. subtilis M 172 (ade-, his-, try-, met-, leu-) verwendet.
2. Puffer und Medien Als Lösungsmittel für Bakterien und DNA dienten
Standardsalinecitrat (SSC, 0,15M NaCl; 0,015M Na-Zitrat) und die Verdünnung 1/10 SSC. Für CsCl-Dichtegradientenzentrifugation wurde die DNA zu-nächst in Studier-Puffer (10~4M Tris, 10~3 M NaCl, pH 7) auf die gewünschte Konzentration verdünnt.
Zum schnellen Wachstum der Bakterien wurde ein Vollmedium benutzt (3% Difco Nutrient Broth). Zum Einbau von BU in die DNA wurde ein definiertes Minimalmedium nach V O G E L und B O N N E R 12 verwen-det, das noch folgende Zusätze pro Liter enthält: 500 mg vitaminfreies Casein (Difco), 5 mg Yeast (Difco), 25mg Bromdeoxyuridin bzw. 10mg Thymin, 20 mg Desoxycytidin und je 50 mg der erforderlichen Aminosäuren.
Zur Transformation wurden die von B O T T und W I L S O N 13 beschriebenen Medien verwendet.
3. Präparation von normaler- und BU-DNA Normale DNA, aus im Vollmedium kultivierten Bak-
terien, wurde nach M A R M U R 14 gewonnen. Die Mar-murmethode wurde jedoch dahingehend abgeändert, daß die DNA nach Lysis der Bakterien und nach Be-handlung mit RNase und Pronase nur einmal mit Chloroform — Octanol (2/1) geschüttelt und nur ein-mal mit Äthylalkohol gefüllt wurde.
Die BU-DNA wurde im wesentlichen nach der von K Ö H N L E I N und H U T C H I N S O N 7 beschriebenen Methode hergestellt, die hier noch einmal kurz wiedergegeben sei: Bakterien werden im Vollmedium bis zur expo-nentiellen Wachstumsphase bei 37 °C kultiviert, in angereichertes thyminhaltiges Minimalmedium ver-
dünnt, nach etwa 3 Generationen zentrifugiert und in BU-haltigem Minimalmedium weiter kultiviert (37 °C). Nach etwa 21/2 Generationen im BU-Medium werden die Bakterien durch Zentrifugieren konzentriert und die DNA wie beschrieben extrahiert. Nach Auflösen der DNA in 1/10 SSC wurde ein weiterer Reinigungs-schritt durch präparative Ultrazentrifugation im CsCl-Dichtegradienten durchgeführt (Beckman-Modell L2-65B; Festwinkelrotor 65; 45 000 UpM; 20 Stdn.). Die verschiedenen DNA-Fraktionen (TT, TB, BB) wurden dann mit Hilfe einer registrierenden Zeiss-Mikroküvet-teneinrichtung15 getrennt aufgefangen und ausgiebig gegen 1/10 SSC dialysiert.
4. Transformationsexperimente Die Transformationsexperimente wurden nach der
von B O T T und W I L S O N 13 angegebenen Methode durch-geführt.
5. Bestrahlungsexperimente und Dosimetrie Aus dem Spektrum eines Quecksilberhochdruckbren-
ners (Philips HOQ 700) wurde das kurzwellige UV-Licht durch geeignete Interferenz-Reflektionsfilter (Schott & Gen. Mainz, UV-R 280) in Kombination mit UV-Kantenfiltern (Schott & Gen. Mainz, WG 5 und WG 6) ausgeblendet. Dadurch wurde erreicht, daß fast ausschließlich UV-Licht der Wellenlänge 313 nm für die Bestrahlung zur Verfügung stand.
Die Dosisleistung der Lampe wurde mit Hilfe der Kalium-Eisenoxalat-Aktinometrie bestimmt16 und be-trug am Ort der Probe 25,5 erg/mm2,sec. Die DNA-Proben wurden in einer Konzentration von 15 — 20 /ig/ ml bei einer Schichtdicke von 3 — 5 mm bestrahlt. In der gewählten Anordnung war eine homogene Be-strahlung der Proben gewährleistet. Nach bestimmten Dosen wurden jeweils 0,2 ml entnommen. Ein Teil wurde nach entsprechender Verdünnung für Transfor-mationsexperimente benutzt, der Rest für die Unter-suchungen im CsCl-Dichtegradienten verbraucht.
6. Denaturierung der DNA Zur Untersuchung der Einzelstrangbrüche wurde die
DNA in 0,1 M NaOH bei Anwesenheit von 0,5% Form-aldehyd denaturiert. Nach 5 Min. bei 50 °C wird die DNA-Lösung mit der erforderlichen Menge von 1 M KH2P04 neutralisiert. Diese Methode gestattet eine schnelle und schonende Denaturierung mit reproduzier-barer Neutralisation auf einen pH-Wert von 7,3 — 7,5.
7. Experimente an der Analytischen Ultrazentrifuge Für die analytische Ultrazentrifugation wurde eine
Anlage der Firma Heraeus-Christ (Typ AZ9100) ver-wendet, die mit UV-Optik und elektronischem Scanner ausgerüstet ist. Der Scanner schreibt wahlweise Trans-mission oder Extinktion des Zentrifugenzelleninhalts auf. Es wurden 12 mm Kel-F Mittelstücke verwendet. Sedimentationsexperimente wurden mit 15 — 20//g/ml DNA bei 20 000 UpM bzw. 30 000 UpM und 20 °C durchgeführt. Die gemessenen Sedimentationskonstan-ten wurden auf Normalbedingungen umgerechnet.

7 1 0 W. KÖHNLEIN UND F. MÖNKEHAUS
Dichtegradientenzentrifugation wurde bei 44 700 UpM und 25 °C durchgeführt. Die DNA-Konzentration be-trug im allgemeinen 1—3 //g/Band.
Nach 20 — 24 Stdn. war der Gleichgewichtszustand erreicht.
8. Bestimmung des Molekulargewichts aus der Band-breite im CsCl-Dichtegradienten
Nach MESELSON et al.17 kann aus der Standard-abweichung (o) der im Dichtegradienten erhaltenen DNA-Bänder das Mol.-Gew. berechnet werden. Zur Bestimmung von o trägt man In c/c0 = / (r —r0)2 für ein Band auf und erhält eine Gerade mit der Stei-gung — 1 : 2 g2 und daraus das Mol.-Gew. Ergibt sich bei der gewählten Auftragung eine nach oben konkave Kurve, so deutet dies auf Molekulargewichtshetero-genität hin. Man kann dann das Gewichtsmittel des Mol.-Gew. (M w) für sämtliche im beobachteten Band vorhandenen Moleküle mit Hilfe einer ebenfalls von MESELSON et al.17 angegebenen Gleichung berechnen.
M In Cj/d (r—r0)2 • Ci (r) A r IC{(r)Ar (1)
wobei
ß= , R T v (d£>/dr) r0 w2 r0
mit v = partielles spezifisches Volumen der DNA; R = universelle Gaskonstante; T = absolute Temperatur; w = Winkelgeschwindigkeit; (dg/dr) ro = Dichtegradient an der Stelle des Bandmaximums; r0 = Lage des Band-maximums.
9. Berechnung der Bruchraten aus der Änderung der Molekular gewichtsverteilung
Zur Bestimmung der Bruchraten wird auf die von CHARLESBY 18 angegebene Theorie zurückgegriffen. Da-nach gilt, daß bereits nach 5 Brüchen im Mittel pro ursprünglicher Polynukleotidkette das Verhältnis Mw / Mno (M no = Zahlenmittel der Ausgangsverteilung) un-abhängig von der Anfangsverteilung ist und gleich dem einer anfänglichen Zufallsverteilung wird. Dies ist für bakterielle DNA sicher erfüllt. Dann gilt für alle Be-strahlungsdosen die Beziehung
Mw = 2 M n . (2)
Weiter gilt nach CHARLESBC 18 ganz allgemein MM-i-MM-^w-ip, (3)
wobei p = Bruchwahrscheinlichkeit; w = Gewicht des Nukleotids. Mit
p = a-D ,
wobei a — Bruchrate; D = Dosis, erhält man schließlich für die Bruchrate pro Nukleotid
d(Mw-1) d D
III. Ergebnisse
1. Test auf Homogenität des Materials
Die Auftriebsdichten der denaturierten T-Stränge der verschiedenen DNA-Fraktionen (normale-, TT-, TB-DNA) war in allen Fällen gleich und betrug (1,722 + 0,001 )g/cm3. Damit kann ein Bromuracil-gehalt von größer als 1% im T-Strang der hybriden DNA ausgeschlossen werden. In Abb. 1 ist das Ab-
OD
a — 2-w (4)
Abb. 1. Scanneraufzeichnung eines CsCl-Dichtegradienten mit BU-DNA. m = Meniskus; TT, TB und BB repräsentieren die 3 DNA-Fraktionen (44.700 U p M ; 25 ° C ; Q = 1,753 g/
cm 3 ) .
sorptionsprofil der BU-DNA im CsCl-Dichtegradien-ten nach Erreichen des Gleichgewichtszustands dar-gestellt. Aus der Verteilung der DNA in den ver-schiedenen Bändern erkennt man folgendes:
1. Die Bänder von TT- und BB-DNA sind sym-metrisch und haben etwa gleiche Bandbreite und somit auch das gleiche Molekulargewicht.
2. Der Dichteunterschied beträgt 0,1 g/cm3. Das ent-spricht 100% Substitution durch BU.
3. Das Band der hybriden DNA ist breiter und er-scheint in vielen Aufnahmen als schlecht aufge-löstes Doppelband. Diese Dichte-Inhomogenität ist eine Folge des unterschiedlichen Thymingehal-tes in den komplementären Strängen der DNA.
4. TT- und BB-Band sind homogen bezüglich der Dichte.
Sedimentationsexperimente mit nativer TT-, TB-und BB-DNA sowie mit denaturierter TT- und BB-DNA ergeben Molekulargewichte, die mit denen aus der Standardabweichung bestimmten in etwa über-einstimmen. Eine Ausnahme macht lediglich native TB-DNA aus den in 3. erwähnten Gründen.
Da zum Nachweis der intramolekularen Energie-leitung nur denaturierte TB-DNA untersucht wird, können die T- und B-Stränge als homogen bezüglich der Dichte betrachtet werden.

I N T R A M O L E K U L A R E ENERGIELEITUNG IN DNA VON BACILLUS SUBTIL1S 7 1 1
Aus den durchgeführten Messungen folgt die er-wartete Molekulargewichtsinhomogenität. Bei der in II., 8 erwähnten Auftragung ergibt sich auch eine Abweichung von der Geraden. Man sollte daher zur Bestimmung von Mw die Gl. (1) benutzen. Ein Ver-gleich der exakten Methode mit der linearen Nähe-rung zeigt jedoch, daß sich nur geringe Unterschiede ergeben. Es ist daher zulässig, bei der Bestimmung der Mw-Werte die lineare Näherung zu benutzen.
2. Veränderungen in der biologischen Aktivität UV-bestrahlter BU-DNA
Die verschiedenen DNA-Fraktionen (TT, TB und BB) wurden mit UV-Licht (313 nm) bestrahlt und die Transformationsaktivität mit kompetenten Bak-terien getestet. In Abb. 2 ist die Abnahme der bio-
1.0
0.5
&0.05
0.01
Abb. 2. Relative Transformationsaktivität der verschiedenen BU-DNA-Fraktionen (TT, TB und BB) in Abhängigkeit von der Bestrahlungsdosis, (— — ) hypothetische Inaktivie-
rungskurve für TB-DNA (siehe Text) .
logischen Aktivität gegen die UV-Dosis aufgetragen. Die nach der Hypothese der unabhängigen Inakti-vierung des T- und des B-Stranges zu erwartende Kurve für TB-DNA ist ebenfalls eingezeichnet. Man erkennt, daß die TT-DNA selbst nach hohen UV-Dosen noch 50% der ursprünglichen Aktivität be-sitzt. Weiter sieht man, daß die tatsächliche Trans-formationsaktivität der hybriden DNA stärker ab-nimmt, als nach der Hypothese zu erwarten ist. Man findet also auch bei der Bestrahlung mit langwelli-gem UV eine zusätzliche Schädigung des T-Stranges in hybrider DNA. Dies deutet auf intramolekulare Energieleitung hin.
3. Physikalische Veränderungen in UV-bestrahlter BU-DNA
Hybride DNA wurde nach UV-Bestrahlung (313 nm) denaturiert und im neutralen CsCl-Gradienten zentrifugiert. In Abb. 3 sind die erhaltenen Absorp-
Abb. 3. Scanneraufzeichnung eines CsCl-Dichtegradienten mit unbestrahlter und bestrahlter TB-DNA nach Denaturie-rung (0,5 • 104 erg/mm2 ) . T und B symbolisieren die entspre-chenden Einzelstränge (44.700 U p M ; 25 ° C ; Q0 = 1,775 g/
cm 3 ) .
tionsprofile bestrahlter und unbestrahlter Proben dargestellt. Man erkennt deutlich, daß in den be-strahlten Proben nicht nur das B-Band, sondern auch das T-Band breiter ist als in der Kontrolle. Abb. 4 zeigt die Auswertung einer Bestrahlungs-
1.0
0.5
C/co
0.1
0.05
0 50 100 (r-r0)2 [cm2• 10*] *-
Abb. 4. Auftragung von In c /c 0 gegen (r—r 0 ) 2 für die T-Bänder aus hybrider DNA nach Bestrahlung mit langwelli-gem UV. T — r0 = Abstand vom Bandmaximum; c /c 0 = relative Konzentration. Die Proben wurden mit folgenden Dosen be-strahlt: unbestrahlt o ; 0,3 • 104 erg/mm2 A ; 1,5-104 erg/
mm2 • ; 1,7 • 104 erg/mm2 • ; 4 • 104 erg/mm2 - f .
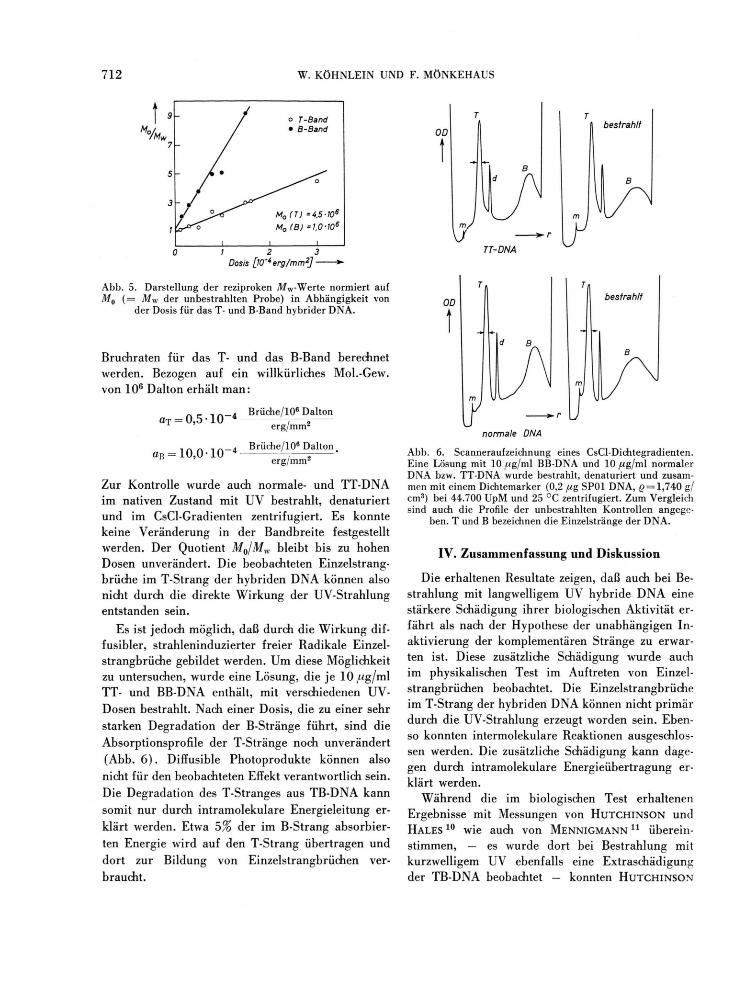
serie. Mit steigender Dosis erfolgt eine Verbreite-rung des T- wie auch des B-Bandes. In Abb. 5 sind die reziproken Mw-Werte normiert auf die M^-Werte der unbestrahlten Proben gegen die Dosis aufgetragen. Mit Hilfe der Gl. (4) können nun die
7 1 2 W. KÖHNLEIN UND F. MÖNKEHAUS
t M°/»w
7 o T-Band • B-Band
- A * -— 3
/ M0 (T) = 4,5-106
[pfCo
1 M0 (B) = 1.0-106
i i 1 2 3
Dosis [10'4 erg/mm2] —
Abb. 5. Darstellung der reziproken M w -Werte normiert auf M0 ( = Mw der unbestrahlten Probe) in Abhängigkeit von
der Dosis für das T- und B-Band hybrider DNA.
Bruchraten für das T- und das B-Band berechnet werden. Bezogen auf ein willkürliches Mol.-Gew. von 106 Dalton erhält man:
« r- -. Brüche/106 Dalton aT = 0,5 10 4 .---•• - -erg/mm''
n n - , ^ - a Brüche/106 Dalton an = 10,0-10 4 —,—; ' erg/mm2
Zur Kontrolle wurde auch normale- und TT-DNA im nativen Zustand mit UV bestrahlt, denaturiert und im CsCl-Gradienten zentrifugiert. Es konnte keine Veränderung in der Bandbreite festgestellt werden. Der Quotient M0/Mw bleibt bis zu hohen Dosen unverändert. Die beobachteten Einzelstrang-brüche im T-Strang der hybriden DNA können also nicht durch die direkte Wirkung der UV-Strahlung entstanden sein.
Es ist jedoch möglich, daß durch die Wirkung dif-fusibler, strahleninduzierter freier Radikale Einzel-strangbrüche gebildet werden. Um diese Möglichkeit zu untersuchen, wurde eine Lösung, die je 10 / 'g/ml TT- und BB-DNA enthält, mit verschiedenen UV-Dosen bestrahlt. Nach einer Dosis, die zu einer sehr starken Degradation der B-Stränge führt, sind die Absorptionsprofile der T-Stränge noch unverändert (Abb. 6 ) . Diffusible Photoprodukte können also nicht für den beobachteten Effekt verantwortlich sein. Die Degradation des T-Stranges aus TB-DNA kann somit nur durch intramolekulare Energieleitung er-klärt werden. Etwa 5% der im B-Strang absorbier-ten Energie wird auf den T-Strang übertragen und dort zur Bildung von Einzelstrangbrüchen ver-braucht.
TT-DNA
bestrahlt
bestrahlt
normale DNA
Abb. 6. Scanneraufzeichnung eines CsCl-Dichtegradienten. Eine Lösung mit 10 /ug/ml BB-DNA und 10 /ug/ml normaler DNA bzw. TT-DNA wurde bestrahlt, denaturiert und zusam-men mit einem Dichtemarker (0,2 /ug SP01 DNA, £ = 1,740 g/ cm3) bei 44.700 UpM und 25 ° C zentrifugiert. Zum Vergleich sind auch die Profile der unbestrahlten Kontrollen angege-
ben. T und B bezeichnen die Einzelstränge der DNA.
IV. Zusammenfassung und Diskussion
Die erhaltenen Resultate zeigen, daß auch bei Be-strahlung mit langwelligem UV hybride DNA eine stärkere Schädigung ihrer biologischen Aktivität er-fährt als nach der Hypothese der unabhängigen In-aktivierung der komplementären Stränge zu erwar-ten ist. Diese zusätzliche Schädigung wurde auch im physikalischen Test im Auftreten von Einzel-strangbrüchen beobachtet. Die Einzelstrangbrüche im T-Strang der hybriden DNA können nicht primär durch die UV-Strahlung erzeugt worden sein. Eben-so konnten intermolekulare Reaktionen ausgeschlos-sen werden. Die zusätzliche Schädigung kann dage-gen durch intramolekulare Energieübertragung er-klärt werden.
Während die im biologischen Test erhaltenen Ergebnisse mit Messungen von HUTCHINSON und HALES 10 wie auch von MENNIGMANN 11 überein-stimmen, — es wurde dort bei Bestrahlung mit kurzwelligem UV ebenfalls eine Extraschädigung der TB-DNA beobachtet — konnten HUTCHINSON

INTRAMOLEKULARE ENERGIELEITUNG IN DNA VON BACILLUS SUBTIL1S 7 1 3
und HALES 10 keine Molekulargewichtsveränderun-gen im T-Strang hybrider DNA beobachten. Das ist möglicherweise so zu erklären, daß dort nur 3 DNA-Proben für solche Messungen zur Verfügung stan-den und die Bänder selbst der unbestrahlten Proben ziemlich breit und undeutlich waren. In den hier ge-schilderten Experimenten wurden wesentlich mehr DNA-Proben untersucht. Auch war die Homogenität des Materials größer. Weiter besteht die Möglich-keit, daß die photochemischen Prozesse in der BU-DNA bei langwelligem UV verschieden sind von denen bei Bestrahlung mit kurzwelligem UV. Unter-suchungen des Circular-Dichroismus der BU-DNA 19
können so interpretiert werden, daß an der lang-welligen Seite des Hauptabsorptionsbandes ein n — n*-Übergang stattfindet, der in normaler DNA nicht beobachtet wird 20. Weitere Bestrahlungsexperi-mente mit kurzwelligem UV sollten hier eine Klä-rung bringen und sind vorgesehen.
Da nach THORSETT 9 die Einzelstrangbrüche in weitaus stärkerem Maß als etwa Basenschäden für den Verlust der biologischen Aktivität verantwort-lich sind, ist es hier sicher erlaubt, die nach Be-strahlung mit langwelligem UV beobachtete zusätz-liche Schädigung in TB-DNA durch die, infolge
1 Abkürzungen: BU = 5 Bromuracil; BU-DNA = DNA, die an Stelle von Thymin Bromuracil enthält. BU-DNA setzt sich aus den folgenden 3 Fraktionen zu-sammen: TT = unmarkierte D N A ; TB = hybride DNA mit BU in einem Strang; BB = doppelsträngig substi-tuierte DNA. Aus thyminhaltigem Medium gewonnene DNA wird als normale DNA bezeichnet. AFW = Gewichtsmittel des Mol.-Gew. Mn — Zahlenmittel des Mol.-Gew.
2 F. HUTCHINSON, Biochim. biophysica Acta [Amsterdam] 9 1 , 5 2 7 [ 1 9 6 4 ] .
3 S . A . R A P A P O R T , V i r o l o g y 2 2 , 1 2 5 [ 1 9 6 4 ] . 4 W . GÖHDE, S t u d i a B i o p h y s i c a 1 2 , 1 5 1 [ 1 9 6 8 ] . 5 A. WACKER, Prog, in Nucleic Acid. Research 1, 369
[ 1 9 6 3 ] , 6 G . H O T Z U. H . REUSCHL, M o l e c . G e n . G e n e t i c s 9 9 , 5
[ 1 9 6 7 ] . 7 W . KÖHNLEIN U. F . HUTCHINSON, R a d i a t . R e s . 3 9 , 7 4 5
[ 1 9 6 9 ] . 8 W . D . RUPP U. W . H . PRUSOFF, B i o c h e m . b i o p h y s i c . R e s .
C o m m u n . 1 8 , 1 4 5 , 1 5 8 [ 1 9 6 5 ] .
intramolekularer Energieleitung hervorgerufenen, Einzelstrangbrüche im T-Strang zu erklären, zumal für den biologischen wie auch den physikalischen Test Bestrahlungsdosen im gleichen Dosisbereich verwendet wurden.
Durch welchen Mechanismus die beobachtete intramolekulare Energieleitung beschrieben werden kann, ist noch nicht restlos geklärt. Sicher spielen die Wasserstoffbrücken und der v a n d e r W a a l s -Kontakt der komplementären DNA-Stränge dabei eine wichtige Rolle. Untersuchungen über die UV-Degradation von BU-DNA in Abhängigkeit von der Konformation (LLON und KÖHNLEIN, unveröffent-lichte Resultate) unterstützen diese Annahme.
Um die hier gefundenen Resultate auch an einem anderen System, das die in der Einleitung erwähn-ten 4 Forderungen besser erfüllt, zu prüfen, wurden ergänzende Untersuchungen zur intramolekularen Energieleitung mit der DNA des Bacillus subtilis Phasen PBSH durchgeführt 21.
Wir danken Dr. F. H U T C H I N S O N für wertvolle An-regungen und Frau G. S T R I C K E R für ausgezeichnete technische Assistenz. Diese Arbeit wurde mit großzügi-ger Unterstützung der Deutschen Forschungsgemein-schaft durchgeführt.
9 G. THORSETT, Ph.D. Thesis, Yale 1969. 1 0 F . HUTCHINSON U. H . B . HALES, J . m o l e c u l a r B i o l . 5 0 . 5 9
[1970]. 11 H. D. MENNIGMANN, Molec. Gen. Genetics 99, 76 [1967]. 1 2 H . VOGEL U. D . BONNER, J . b i o l . C h e m i s t r y 9 7 , 2 1 8
[ 1 9 5 6 ] . 13 K. F. B o r r u. G. A. WILSON, Bacteriol. Rev. 32, 370
U968] . 14 j. MARMUR, J. molecular Biol. 3, 208 [1961] . 1 5 W . GÖHDE U. W . KÖHNLEIN, Z e i s s I n f o r m a t i o n e n , i m
Drude. 1 6 C . G . H A T C H A R D U. C . A . PARKER, P r o c . R o y . S o c . [ L o n -
don] A 235 ,518 [1956], 1 7 M . MESELSON, F . W . STAHL U. J . V I N O G R A D , P r o c . na t .
Acad. Sei. USA 43, 581 [1957] . 18 A. CHARLESBY, Proc. Roy. Soc. A 224, 120 [1954]. 19 F. HUTCHINSON, persönliche Mitteilung 1969. 2 0 D . M . G R A Y U. I . RUBENSTEIN, B i o p o l y m e r s 6 , 1 6 0 5
[1968]. 2 1 F . MÖNKEHAUS U. W . KÖHNLEIN, Z . N a t u r f o r s c h .